Настоящее изобретение относится к использованию клеток, имеющих мутантный, но функциональный белок поверхности клеток в медицинских применениях, когда избирательное истощение или обогащение популяций клеток является желательным. Мутантные, но функциональные белки поверхности клеток можно вводить в клетки способами редактирования генов, включая гомологичную репарацию двухцепочечных разрывов ДНК, в частности, в ходе редактирования генов посредством CRISPR/Cas, или с использованием редакторов оснований. Изобретение, кроме того, относится к средству и способу для избирательного истощения редактированных клеток in vivo.
Клеточная терапия является очень мощным вариантом терапии, но часто ассоциирована с тяжелыми нежелательными побочными эффектами. Трансгены и/или генетическая инженерия перенесенных клеток могут вызывать злокачественную трансформацию. Перенос CAR-T-клеток может приводить к тяжелым специфическим и неспецифическим эффектам (синдрому высвобождения цитокинов), и перенос аллогенных T-клеток может вызывать заболевание «трансплантат против хозяина» (GvHD). Успех клеточной терапии в онкологии может, вероятно, сильно стимулировать клеточную терапию по другим показаниям, включая незлокачественные заболевания. Для увеличения безопасности видов клеточной терапии, в частности, если их используют для лечения нелетальных заболеваний, является важным убедиться в том, что перенесенные клетки остаются безопасными в течение многих лет после переноса. В случаях, когда развиваются тяжелые нежелательные побочные эффекты, возможность избирательного истощения перенесенных клеток посредством «безопасного или аварийного выключателя» может значительно увеличивать безопасность клеточной терапии.
Проблемой, лежащей в основе настоящего изобретения, является предоставление системы, которая служит для постоянной маркировки и отслеживания клеток и позволяет избирательное истощение маркированных или немаркированных клеток in vitro или in vivo. Эти проблемы разрешены посредством объекта независимых пунктов формулы изобретения.
Описание
Первый аспект изобретения относится к клетке млекопитающего, экспрессирующей первую изоформу поверхностного белка, где указанная первая изоформа указанного поверхностного белка является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного поверхностного белка, для использования в медицинском лечении пациента, имеющего клетки, экспрессирующие указанную вторую форму указанного поверхностного белка.
Выражение «иммунологически отличимые» относится к первой и второй изоформе поверхностного белка, которые можно различать посредством лиганда, специфически связывающегося либо с первой, либо с второй изоформой.
В контексте настоящего описания, выражение «специфическое связывание» относится к связыванию с константой диссоциации KD ≤ 10 E -7.
В контексте настоящего описания, выражение «лиганд» относится к антителу или подобной антителу молекуле. Антитело или подобная антителу молекула может являться связанной с другой молекулой (например, иммунотоксином), или может присутствовать на поверхности клетки, в частности, иммуноцита.
В контексте настоящего описания, термин «антитело» используют в его значении, известном в данной области клеточной биологии и иммунологии; он относится к полноразмерным антителам, включая, но без ограничения, иммуноглобулин типа G (IgG), типа A (IgA), типа D (IgD), типа E (IgE) или типа M (IgM), любой антигенсвязывающий фрагмент или их одноцепочечные формы и родственные или производные конструкции. Полноразмерное антитело представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, связанные между собой посредством дисульфидных связей. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи (CH). Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначенной в настоящем описании как VL) и константной области легкой цепи (CL). Константная область легкой цепи состоит из одного домена, CL. Вариабельные области тяжелых и легких цепей содержат связывающий домен, взаимодействующий с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент классической системы комплемента.
Термин «подобная антителу молекула», в контексте настоящего описания относится к молекуле, способной к специфическому связыванию с другой молекулой или мишенью с высокой аффинностью/KD ≤ 10E-7 моль/л, в частности, KD ≤ 10E-8 моль/л. Подобная антителу молекула связывается со своей мишенью подобно специфическому связыванию антитела. Термин подобная антителу молекула включает белок с повторами, такой как сконструированный белок с анкириновыми повторами (Molecular Partners, Zürich), полипептид, происходящий из белков с повторами армадилло, полипептид, происходящий из белков с богатыми лейцином повторами, аффимер, происходящую из антитела молекулу, такую как химерный рецептор антигена (CAR) и полипептид, происходящий из белков с тетратрикопептидным повтором.
Термин подобная антителу молекула, кроме того, включает полипептид, происходящий из доменов белка A, полипептид, происходящий из домена FN3 фибронектина, полипептид, происходящий из консенсусных доменов фибронектина, полипептид, происходящий из липокалинов, полипептид, происходящий из цинковых пальцев, полипептид, происходящий из гомологичного Src домена 2 (SH2), полипептид, происходящий из гомологичного Src домена 3 (SH3), полипептид, происходящий из доменов PDZ, полипептид, происходящий из гамма-кристаллина, полипептид, происходящий из убиквитина, полипептид, происходящий из полипептида с цистеиновым узлом, и полипептид, происходящий из ноттина.
В идеале, первую и вторую изоформы поверхностного белка можно различать посредством двух лигандов, где один лиганд является способным специфически узнавать первую изоформу, и другой лиганд является способным специфически узнавать вторую изоформу. Иными словами, каждый лиганд является способным специфически связываться с одной изоформой, но не с другой изоформой. Иными словами, лиганды являются способными к установлению отличий между двумя изоформами посредством специфического связывания только одной изоформы, но не другой изоформы.
Выражение «функционально неотличимые» относится к первой и второй изоформам, которые равным образом способны осуществлять одну и ту же функцию в клетке без значительного нарушения. Иными словами, первая и вторая изоформы являются функционально в большой степени неотличимыми. В конкретных вариантах осуществления, небольшое функциональное нарушение может являться приемлемым.
В контексте настоящего описания, выражение «первая и/или вторая изоформы белка поверхности клеток» относятся к первому и второму аллелю белка поверхности клеток.
В конкретных вариантах осуществления, клетка млекопитающего представляет собой клетку человека.
В конкретных вариантах осуществления, вторая изоформа поверхностного белка относится к форме белка дикого типа (иными словами: к форме, которая обычно встречается в природе), и первая изоформа относится к изоформе, полученной посредством введения мутации в последовательность нуклеиновой кислоты, кодирующую вторую изоформу.
В конкретных вариантах осуществления, вторая изоформа поверхностного белка представляет собой нативную изоформу, и первая изоформа представляет собой генетически модифицированную изоформу, происходящую из нативной изоформы.
В контексте настоящего описания, выражение «нативный белок» относится к белку, кодируемому последовательностью нуклеиновой кислоты в геноме клетки, где эта последовательность нуклеиновой кислоты не была вставлена или подвергнута мутагенезу посредством генетической манипуляции. Иными словами, нативный белок представляет собой белок, который не является трансгенным белком или генетически модифицированным белком.
В конкретных вариантах осуществления, поверхностный белок содержит внеклеточную полипептидную последовательность, и указанная первая изоформа содержит вставку, делецию и/или замену 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 15 или 20 аминокислот по сравнению с указанной второй изоформой.
В конкретных вариантах осуществления, поверхностный белок содержит внеклеточную полипептидную последовательность и указанная первая изоформа содержит вставку, делецию и/или замену 1-20, в частности 1-5, более конкретно, 1-3 аминокислот по сравнению с указанной второй изоформой.
В конкретных вариантах осуществления, поверхностный белок содержит внеклеточную полипептидную последовательность, и указанная первая изоформа содержит вставку, делецию и/или замену 1 аминокислоты по сравнению с указанной второй изоформой.
В конкретных вариантах осуществления, вставка, делеция и/или замена расположена в участке, который не является консервативным среди различных видов млекопитающих.
В конкретных вариантах осуществления, вставка, делеция и/или замена не приводит к изменению вторичной структуры поверхностного белка.
В конкретных вариантах осуществления, вставка, делеция и/или замена локализована в участке, который является доступным для связывания лиганда, в соответствии с анализом кристаллической структуры или компьютерным прогнозированием структуры.
В конкретных вариантах осуществления, вставка, делеция и/или замена локализована в участке, который имеет уникальную топологию по сравнению с другими белками млекопитающих в соответствии с анализом кристаллической структуры или компьютерным прогнозированием структуры.
В контексте настоящего описания, выражение «уникальная топология» относится к топологии, которая присутствует только в поверхностном белке, подлежащем модификации посредством вставки, делеции и/или замены 1-20 аминокислот, а не в других белках млекопитающих, в частности, не в других поверхностных белках человека. Присутствие такой же или очень сходной топологии в других белках может затруднять получение специфического антитела, узнающего эпитоп, локализованный в указанном участке.
В конкретных вариантах осуществления, вставка, делеция и/или замена не является локализованной в участке, вовлеченном в прогнозируемое, или экспериментально установленное или подтвержденное белок-белковое взаимодействие поверхностного белка.
В конкретных вариантах осуществления, вставка, делеция и/или замена не приводит к делеции или введению дисульфидной связи для между- или внутримолекулярного взаимодействия, или гидрофобного стэкинга. В случаях, когда вставка, делеция и/или замена приводит к удалению между- или внутримолекулярного взаимодействия посредством солевого мостика, необходимо подтвердить, что это удаление взаимодействия посредством солевого мостика компенсировано новыми взаимодействиями внутри белка. В ином случае, удаления взаимодействия посредством солевого мостика следует избегать.
В конкретных вариантах осуществления, вставка, делеция и/или замена не приводит к делеции или введению участка посттрансляционной модификации белка, в частности, участка гликозилирования, который является важным для сворачивания белка.
В конкретных вариантах осуществления, вставка, делеция и/или замена приводит к делеции или введению участка посттрансляционной модификации белка, в частности, участка гликозилирования, который не является важным для сворачивания белка, таким образом, создавая новый эпитоп.
В конкретных вариантах осуществления, первую изоформу можно отличать от второй изоформы посредством связывания подобной антителу молекулы или связывания антитела.
В конкретных вариантах осуществления, первую изоформу можно отличать от второй изоформы посредством реакции иммуноэффекторной клетки, несущей антитело. В конкретных вариантах осуществления, первую изоформу можно отличать от второй изоформы посредством реакции иммуноэффекторной клетки, несущей подобную антителу молекулу. В конкретных вариантах осуществления, первую изоформу можно отличать от второй изоформы посредством реакции иммуноэффекторной клетки, несущей антитело. В конкретных вариантах осуществления, первую изоформу можно отличать от второй изоформы посредством реакции иммуноэффекторной клетки, несущей подобную антителу молекулу. В конкретных вариантах осуществления, первую изоформу можно отличать от второй изоформы посредством реакции T-клетки, в частности, активированной T-клетки, несущей химерный рецептор антигена (CAR).
В контексте настоящего описания, химерный рецептор антигена (CAR) относится к сконструированному рецептору, который прививает специфичность связывания, в частности, специфичность моноклонального антитела, T-клетке. Общепринятая форма CAR содержит внеклеточный домен, происходящий из моноклонального антитела, имеющего желательную специфичность связывания, трансмембранный домен и внутриклеточный домен (Gill and June, Imm Rev, 2014). Внеклеточный домен содержит одноцепочечный вариабельный фрагмент, содержащий вариабельные области тяжелой и легкой цепи иммуноглобулина. Нуклеотидная последовательность, кодирующая внеклеточный домен такого CAR, может происходить из клеток гибридомы, продуцирующих антитело с желательной специфичностью связывания (Gill and June, Imm Rev, 2014; Fields, Nat Prot, 2013).
В конкретных вариантах осуществления, поверхностный белок выбран из CD1a, CD1b, CD1c, CD1d, CD1e, CD2, CD3, CD3d, CD3e, CD3g, CD4, CD5, CD6, CD7, CD8a, CD8b, CD9, CD10, CD11a, CD11b, CD11c, CD11d, CDwl2, CD13, CD14, CD15, CD15u, CD15s, CD15su, CD16, CD16b, CD17, CD18, CD19, CD20, CD21, CD22, CD23, CD24, CD25, CD26, CD27, CD28, CD29, CD30, CD31, CD32, CD33, CD34, CD35, CD36, CD37, CD38, CD39, CD40, CD41, CD42a, CD42b, CD42c, CD42d, CD43, CD44, CD45, CD45RA, CD45RB, CD45RC, CD45RO, CD46, CD47, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD50, CD51, CD52, CD53, CD54, CD55, CD56, CD57, CD58, CD59, CD60a, CD60b, CD60c, CD61, CD62E, CD62L, CD62P, CD63, CD64, CD65, CD65s, CD66a, CD66b, CD66c, CD66d, CD66e, CD66f, CD68, CD69, CD70, CD71, CD72, CD73, CD74, CD75, CD75s, CD77, CD79a, CD79b, CD80, CD81, CD82, CD83, CD84, CD85a, CD85d, CD85j, CD85k, CD86, CD87, CD88, CD89, CD90, CD91, CD92, CD93, CD94, CD95, CD96, CD97, CD98, CD99, CD99R, CD100, CD101, CD102, CD103, CD104, CD105, CD106, CD107a, CD107b, CD108, CD109, CD110, CD111, CD112, CD113, CD114, CD115, CD116, CD117, CD118, CD119, CD120a, CD120b, CD121a, CD121b, CD122, CD123, CD124, CD125, CD126, CD127, CD129, CD130, CD131, CD132, CD133, CD134, CD135, CD136, CD137, CD138, CD139, CD140a, CD140b, CD141, CD142, CD143, CD144, CDw145, CD146, CD147, CD148, CDw149, CD150, CD151, CD152, CD153, CD154, CD155, CD156a, CD156b, CD156c, CD157, CD158e, CD158i, CD158k, CD159a, CD159c, CD160, CD161, CD162, CD163, CD164, CD165, CD166, CD167a, CD167b, CD168, CD169, CD170, CD171, CD172a, CD172b, CD172g, CD173, CD174, CD175, CD175s, CD176, CD177, CD178, CD179a, CD179b, CD180, CD181, CD182, CD183, CD184, CD185, CD186, CD191, CD192, CD193, CD194, CD195, CD196, CD197, CDw198, CD199, CD200, CD201, CD202b, CD203c, CD204, CD205, CD206, CD207, CD208, CD209, CD210, CDw210b, CD212, CD213a1, CD213a2, CD215, CD217a, CD218a, CD218b, CD220, CD221, CD222, CD223, CD224, CD225, CD226, CD227, CD228, CD229, CD230, CD231, CD232, CD233, CD234, CD235a, CD235b, CD236, CD236R, CD238, CD239, CD240CE, CD240DCE, CD240D, CD241, CD242, CD243, CD244, CD245, CD246, CD247, CD248, CD249, CD252, CD253, CD254, CD256, CD266, CD267, CD268, CD269 (BCMA), CD270, CD271, CD272, CD273, CD274, CD275, CD276, CD277, CD278, CD279, CD280, CD281, CD282, CD283, CD284, CD286, CD289, CD290, CD292, CDw293, CD294, CD295, CD296, CD297, CD298, CD299, CD300a, CD300c, CD300e, CD301, CD302, CD303, CD304, CD305, CD306, CD307a, CD307b, CD307c, CD307d, CD307e, CD308, CD309, CD312, CD314, CD315, CD316, CD317, CD318, CD319, CD320, CD321, CD322, CD324, CD325, CD326, CD327, CD328, CD329, CD331, CD332, CD333, CD334, CD335, CD336, CD337, CD338, CD339, CD340, CD344, CD349, CD350, CD351, CD352, CD353, CD354, CD355, CD357, CD358, CD360, CD361, CD362, CD363, CD364, CD365, CD366, CD367, CD368, CD369, CD370, CD371, легкой цепи иммуноглобулина (лямбда или каппа), белка HLA и β2-микроглобулина.
В контексте настоящего описания, HLA обозначает «человеческий лейкоцитарный антиген» и включает HLA-A, HLA.B, HLA-C, HLA-E, HLA-F, HLA-G, HLA-DM, HLA-DO, HLA-DP, HLA-DQ и HLA-DR.
В конкретных вариантах осуществления, поверхностный белок выбран из CD2, CD3, CD4, CD5, CD8, CD19, CD20, CD22, CD23, CD33, CD34, CD90, CD45, CD123, CD269 (BCMA), легкой цепи иммуноглобулина (лямбда или каппа), белка HLA и β2-микроглобулина.
В конкретных вариантах осуществления, поверхностный белок выбран из CD45, CD3, CD4, CD8a, CD8b и CD279.
В конкретных вариантах осуществления, поверхностный белок выбран из CD45, CD45RA и CD45RO.
В конкретных вариантах осуществления, поверхностный белок выбран из CD45, CD34, CD38, CD59, CD90 и CD117.
В конкретных вариантах осуществления, поверхностный белок выбран из CD45, CD19, CD20, CD22, CD22, CD23, CD38, CD138, CD268, CD269 (BCMA) и CD319.
В конкретных вариантах осуществления, поверхностный белок выбран из CD5, CD19, CD20, CD33, CD123, CD38 и CD269.
В конкретных вариантах осуществления, поверхностный белок выбран из CD45, CD19, CD4 и CD8.
В конкретных вариантах осуществления, поверхностный белок представляет собой CD45.
В конкретных вариантах осуществления, поверхностный белок представляет собой Thy1 (CD90).
В конкретных вариантах осуществления, поверхностный белок представляет собой CD19.
В контексте настоящего описания, «Thy1» обозначает «антиген 1 клетки тимуса», тета; альтернативное наименование: CD90; UniProt ID P04216 (человека).
В контексте настоящего описания, «CD45» обозначает «протеинтирозинфосфатаза, рецептор типа C (Ptprc)»; UniProt ID P08575 (человека).
В экспериментах на животных из раздела примеры, CD45 и CD90 относятся к мышиным гомологам человеческих генов и белков, соответственно. Пункты формулы изобретения 30-31 и 34-36 конкретно относятся к мышиным гомологам: «мышиному CD45» и «мышиному CD90/Thy1».
В конкретных вариантах осуществления, поверхностный белок представляет собой CD4. В конкретных вариантах осуществления, поверхностный белок представляет собой CD2. В конкретных вариантах осуществления, поверхностный белок представляет собой CD8. В конкретных вариантах осуществления, поверхностный белок представляет собой белок HLA.
В конкретных вариантах осуществления, первая изоформа поверхностного белка не кодирована в нативной геномной ДНК пациента.
В конкретных вариантах осуществления, клетка представляет собой аллогенную клетку. Аллогенная клетка относится к клетке, полученной от донора, генетически сходного с индивидуумом, которому вводят клетку. Донор может являться родственным или не родственным индивидуумом.
В конкретных вариантах осуществления, клетка представляет собой аутологичную клетку. Аутологичная клетка относится к клетке, полученной от того же самого индивидуума, которому вводят клетку.
В конкретных вариантах осуществления, первая изоформа получена посредством изменения последовательности, кодирующей указанный поверхностный белок, в нативной геномной ДНК пациента, приводящего к индукции указанной вставки, делеции и/или замены аминокислоты.
В конкретных вариантах осуществления, первая изоформа получена посредством изменения мРНК, кодирующей указанный поверхностный белок, способами редактирования РНК (Zhang, 2017). Этот способ оставляет геномную ДНК неизменной, но приводит к вставке, делеции и/или замене аминокислоты в аминокислотной последовательности поверхностного белка.
В конкретных вариантах осуществления, первая изоформа получена посредством индукции вставки, делеции и/или замены 1, 2, 3, 4 или 5 (или даже 6, 7, 8, 9, 10, 11, 12, 15 или 20) аминокислот в аминокислотной последовательности указанной второй изоформы указанного поверхностного белка.
В конкретных вариантах осуществления, первая изоформа получена посредством индукции вставки, делеции и/или замены 1-20, в частности, 1-5, более конкретно, 1-3 аминокислот в аминокислотной последовательности указанной второй изоформы указанного поверхностного белка.
В конкретных вариантах осуществления, первая изоформа получена посредством индукции вставки, делеции и/или замены 1 аминокислоты в аминокислотной последовательности указанной второй изоформы указанного поверхностного белка.
Эффект вставки, делеции и/или замены аминокислоты можно получать посредством изменения последовательности, кодирующей указанный поверхностный белок, в нативной геномной ДНК пациента (редактирование генов), или посредством изменения мРНК, кодирующей указанный поверхностный белок (редактирование РНК). Обе формы редактирования приводят к изменению аминокислотной последовательности белка.
В конкретных вариантах осуществления, эффект вставки, делеции и/или замены получают посредством
a. сравнения (выравнивания) одной или более гомологичных последовательностей различных видов млекопитающих, в частности, гомологов мыши, крысы, примата и человека, и расположения вставки, делеции и/или замены в неконсервативном участке;
b. выбора вставки, делеции и/или замены таким образом, что анализ кристаллической структуры или компьютерное прогнозирование структуры не прогнозирует изменения вторичной структуры отрезка, подвергнутого вставке, делеции и/или замене;
c. выбора вставки, делеции и/или замены таким образом, что она локализована в участке, который является доступным для связывания лиганда в соответствии с анализом кристаллической структуры или компьютерным прогнозированием структуры;
d. выбора вставки, делеции и/или замены в последовательности, не вовлеченной в прогнозируемое или экспериментально установленное белок-белковое взаимодействие поверхностного белка;
e. выбора вставки, делеции и/или замены без делеции или введения дисульфидной связи для между- или внутримолекулярного взаимодействия, или гидрофобного стэкинга; или
f. выбора вставки, делеции и/или замены без делеции или введения участка посттрансляционной модификации белка, в частности, участка гликозилирования, важного для сворачивания белка.
В конкретных вариантах осуществления, эффект вставки, делеции и/или замены получают посредством
a. сравнения (выравнивания) одной или более гомологичных последовательностей различных видов млекопитающих, в частности, гомологов мыши, крысы, примата и человека, и расположения вставки, делеции и/или замены в неконсервативном участке;
b. выбора вставки, делеции и/или замены таким образом, что анализ кристаллической структуры или компьютерное прогнозирование структуры не прогнозирует изменения вторичной структуры отрезка, подвергнутого вставке, делеции и/или замене;
c. выбора вставки, делеции и/или замены таким образом, что она локализована в участке, который является доступным для связывания лиганда в соответствии с анализом кристаллической структуры или компьютерным прогнозированием структуры;
d. выбора вставки, делеции и/или замены в последовательности, не вовлеченной в прогнозируемое или экспериментально установленное белок-белковое взаимодействие поверхностного белка;
e. выбора вставки, делеции и/или замены без делеции или введения дисульфидной связи для между- или внутримолекулярного взаимодействия, или гидрофобного стэкинга; и
f. выбора вставки, делеции и/или замены без делеции или введения участка посттрансляционной модификации белка, в частности, участка гликозилирования, важного для сворачивания белка.
В конкретных вариантах осуществления, вставку, делецию и/или замену выбирают таким образом, что она локализована в участке, который имеет уникальную топологию по сравнению с другими белками млекопитающих в соответствии с анализом кристаллической структуры или компьютерным прогнозированием структуры.
Неконсервативный участок, в контексте настоящего описания, относится к участку, который часто подвергается мутагенезу в ходе периода эволюции, как определено по множественному выравниванию последовательности (MSA) больших количеств гомологичных последовательностей. Сайт-специфическая консервативность является показателем присутствия функциональных или структурных ограничений, действующих в специфических участках, позволяющих оценку их важности в сохранении структуры или функции белка. Состояние доступности для растворителя в каждом участке, прогнозируемое или наблюдаемое для доступных экспериментально определенных структур, можно использовать для установления различий между структурно важными участками, которые часто являются высоко консервативными и углубленными, и функционально важными участками, вовлеченными в связывание лиганда, связывание субстрата, или белок-белковые взаимодействия, которые являются высоко консервативными и экспонированными. Таким образом, неконсервативный участок, в контексте настоящего описания, относится к участку, для которого показана низкая эволюционная консервативность и высокая доступность для растворителя.
Способы поиска сходных последовательностей хорошо разработаны и являются общепринятыми для выведения заключений о гомологии, и они включают широко используемый инструмент BLAST, так же как более чувствительные способы на основе профилей или скрытых Марковских моделей, такие как PSI-BLAST, HMMER, HHblits.
Множественные выравнивания последовательностей для гомологичных последовательностей можно получать посредством эффективной комбинации информации от локального и глобального выравнивания, как реализовано в T-COFFEE или, когда возможно, посредством включения структурной информации для управления конструированием MSA (MAFFT, PROMALS3D, 3D Coffee).
Способы эффективной оценки сайт-специфической консервативности по MSA основаны на измерениях энтропии по Шеннону, матрицах на основе сходства (т.е. BLOSUM) или вероятностных эволюционных моделях, подобных принципу максимального правдоподобия и эмпирическим байесовским парадигмам (Rate4Site).
Эффективные способы прогнозирования трехмерной структуры белка включают, но без ограничения, способы на основе сравнения, такие как SWISS-MODEL, MODELLER, RaptorX и IntFOLD.
В способах прогнозирования B-клеточных эпитопов используют физико-химические свойства аминокислот, т.е. гидрофобность, гибкость, полярность и экспонированная поверхность для получения вероятности для остатка являться частью либо прерывистого, либо линейного эпитопа. Эти инструменты включают, но без ограничения: Ellipro, SEPPA, BepiPred, ABCpred, DiscoTope, EpiSearch.
В конкретных вариантах осуществления, вставка, делеция и/или замена локализована в внеклеточной части указанного первого поверхностного белка, в частности, в внеклеточной петле. Мутации в внеклеточных петлях с меньшей вероятностью влияют на функцию белка.
В случаях, когда антитело или подобная антителу молекула, реакционноспособные по отношению к представляющему интерес поверхностному белку, уже существуют, информацию о том, какой эпитоп представляющего интерес поверхностного белка узнает это антитело, можно использовать для выбора участка вставки, делеции и/или замены. Тогда изоформа представляющего интерес белка, содержащая эпитоп, с которым связывается существующее антитело, может соответствовать второй изоформе, и новая сконструированная изоформа представляющего интерес белка, содержащая сконструированный эпитоп, может соответствовать первой изоформе.
Моделирование CDR описано в Messih, Bioinformatics, 2014.
В конкретных вариантах осуществления, клетку вводят до, во время или после специфической абляции клеток, экспрессирующих указанную вторую изоформу указанного поверхностного белка. В конкретных вариантах осуществления, абляцию клеток, экспрессирующих указанную вторую изоформу указанного поверхностного белка осуществляют посредством введения указанному пациенту средства, выбранного из подобной антителу молекулы, антитела, иммуноэффекторной клетки, несущей антитело или подобную антителу молекулу, и иммуноэффекторной клетки, в частности, T-клетки, несущей химерный рецептор антигена, где указанное средство является специфически реакционноспособным по отношению к указанной второй изоформе (но не к указанной первой изоформе) указанного белка поверхности клетки.
В конкретных вариантах осуществления, клетка экспрессирует антитело или подобную антителу молекулу, реакционноспособную по отношению к второй изоформе указанного поверхностного белка.
В конкретных вариантах осуществления, клетка экспрессирует химерный рецептор антигена, реакционноспособный по отношению к второй изоформе указанного поверхностного белка.
В конкретных вариантах осуществления, клетка экспрессирует антитело или подобную антителу молекулу, в частности, химерный рецептор антигена, реакционноспособный по отношению к второй изоформе поверхностного белка, и поверхностный белок выбран из CD45, CD19, CD8 и CD4, в частности, поверхностный белок представляет собой CD45.
В конкретных вариантах осуществления, клетка экспрессирует
a. первую изоформу первого поверхностного белка, где указанная первая изоформа указанного первого поверхностного белка является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного первого поверхностного белка, и
b. первую изоформу второго поверхностного белка, где указанная первая изоформа указанного второго поверхностного белка является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного второго поверхностного белка.
В конкретных вариантах осуществления, первый поверхностный белок представляет собой CD19, и второй поверхностный белок представляет собой CD45.
В конкретных вариантах осуществления, клетка млекопитающего выбрана из группы, включающей гематопоэтическую стволовую клетку (гемоцитобласт), CD4+ T-клетку, CD8+ T-клетку, T-клетку памяти, регуляторную T-клетку (Tрег), клетку естественного киллера (NK), врожденную лимфоидную клетку (ILC), дендритную клетку (DC), B-лимфоцит, ассоциированную со слизистой оболочкой инвариантную T-клетку (MAIT) и гамма-дельта-T-клетку (γδ T).
В конкретных вариантах осуществления, клетка млекопитающего представляет собой гематопоэтическую клетку. В конкретных вариантах осуществления, клетка млекопитающего представляет собой гематопоэтическую стволовую клетку. В конкретных вариантах осуществления, клетка млекопитающего представляет собой иммуноцит. В конкретных вариантах осуществления, клетка млекопитающего представляет собой T-клетку. В конкретных вариантах осуществления, клетка млекопитающего представляет собой B-клетку.
В конкретных вариантах осуществления, клетка содержит генетическое изменение, корректирующее дефект или противодействующее дефекту связанного с заболеванием гена, присутствующему у указанного пациента. Выражение «генетическое изменение, корректирующее дефект связанного с заболеванием гена» предназначено для включения вставки трансгена, генетической коррекции связанной с заболеванием мутации, делеции гена, в частности, гена, несущего связанную с заболеванием мутацию, изменение эпигенетической модификации, важной для экспрессии гена, или их комбинации. Выражение «делеция гена» предназначено для включения функциональной делеции гена, иными словами, предотвращения экспрессии гена, например, посредством вставки преждевременного стоп-кодона или посредством вставки регуляторной репрессорной последовательности.
В конкретных вариантах осуществления, клетка содержит трансген. В конкретных вариантах осуществления, трансген представляет собой последовательность нуклеиновой кислоты, кодирующую функциональную изоформу белка, затронутого связанным с заболеванием гена дефектом. Трансген не несет дефект связанного с заболеванием гена.
В конкретных вариантах осуществления, клетка имеет генетическую коррекцию связанной с заболеванием мутации. В случаях, когда клетка имеет генетическую коррекцию дефекта связанного с заболеванием гена, вызывающая заболевание мутация в нативном гене скорректирована с использованием способов редактирования генов.
В контексте настоящего описания, редактирование генов или генетическая инженерия относится к способам, осуществляющим вставку, делецию или замену последовательности нуклеиновой кислоты в геноме живого организма. Редактирование генов включает сайт-специфические двухцепочечные разрывы или сайт-специфические одноцепочечные разрывы (ники) в геномной ДНК. В качестве неограничивающего примера, редактирование генов можно осуществлять посредством HDR после опосредованных CRISPR/Cas сайт-специфических двухцепочечных разрывов или с использованием сайт-специфических редакторов оснований.
В конкретных вариантах осуществления, связанный с заболеванием гена дефект представляет собой мутацию в гене, выбранном из гена Foxp3, гена CD25, гена Stat5b, гена Stat1 и гена Itch.
В конкретных вариантах осуществления, связанный с заболеванием гена дефект представляет собой мутацию в гене Foxp3.
Конкретные варианты осуществления относятся к случаям, когда заболевание «трансплантат против хозяина» (GvHD) развивается после введения клетки млекопитающего пациенту. В этих случаях, клетку подвергают специфической абляции посредством введения пациенту антитела или подобной антителу молекулы, специфически связывающихся с указанной первой изоформой (но не с указанной второй изоформой) указанного белка поверхности клетки, или иммуноэффекторной клетки, в частности, CAR-T-клетки, специфически реакционноспособной по отношению к указанной первой изоформе (но не к указанной второй изоформе) указанного белка поверхности клетки.
В конкретных вариантах осуществления, лечение включает трансплантацию гематопоэтической стволовой клетки.
В конкретных вариантах осуществления, лечение включает трансплантацию T-клетки (иными словами: T-клеточную терапию).
В конкретных вариантах осуществления, лечение включает трансплантацию органа.
В конкретных вариантах осуществления, лечение относится к лечению генетического гематопоэтического заболевания. Неограничивающим примером генетического гематопоэтического заболевания являются талассемии.
В конкретных вариантах осуществления, лечение относится к лечению опосредованного T-клетками заболевания, в частности, генетического опосредованного T-клетками заболевания, более конкретно, подобного IPEX синдрома, ассоциированной с CTLA-4 иммунной дисрегуляции, гемофагоцитарного синдрома, синдрома ALPS или синдрома, вызванного гетерозиготными зародышевыми мутациями PTEN.
В конкретных вариантах осуществления, опосредованнон T-клетками заболевание представляет собой Х-сцепленный синдром иммунной дисрегуляции, полиэндокринопатии и энтеропатии (IPEX; OMIM http://www.omim.org/entry/304790) или подобный IPEX синдром, и указанная геномная локализация представляет собой мутацию, содержащуюся в гене, выбранном из гена Foxp3, гена CD25, гена Stat5b, гена Stat1 и гена Itch (Verbsky and Chatila, Curr Opin Pediatr. 2013 Dec;25(6):708-14). Мутации в указанных генах предотвращают экспрессию или нормальное функционирование продукта гена. Редактирование этих геномных локализаций для уничтожения мутации восстанавливает экспрессию гена и белка.
В конкретных вариантах осуществления, лечение относится к лечению иммунодефицита, в частности, синдром тяжелого комбинированного иммунодефицита (SCID).
В конкретных вариантах осуществления, дефект связанного с заболеванием гена представляет собой мутацию в гене Foxp3, и указанное заболевание представляет собой Х-сцепленный синдром иммунной дисрегуляции, полиэндокринопатии и энтеропатии (IPEX). В конкретных вариантах осуществления, дефект связанного с заболеванием гена представляет собой мутацию в Foxp3, в частности, мутацию Foxp3K276X. Эта мутация предотвращает нормальное функционирование продукта гена. Редактирование этой геномной локализации обращает мутацию к последовательности ДНК дикого типа и таким образом, восстанавливает экспрессию гена Foxp3 и белка Foxp3 (фиг. 20).
IPEX можно рассматривать как злокачественное заболевание, сравнимое с опухолю/злокачественной опухолью, поскольку оно является очень агрессивным, хотя имеет другой механизм. Абляция патогенных гематопоэтическик клеток/иммуноцитов, несущих дефект связанного с заболеванием гена, оправдана, по меньшей мере, временно.
В контексте настоящего описания, термин «ген Foxp3» относится к гену белка P3 человека с боксом головки вил, NCBI GENE ID: 50943. В экспериментах на животных, описанных в разделе примеры, осуществляли репарацию мышиной мутации Foxp3K276X посредством редактирования генов. Эта мутация воспроизводит клинически значимую мутацию Foxp3 человека (Ramsdell et al., Nature reviews. Immunology 14, 343-349 (2014); Lin et al., The Journal of allergy and clinical immunology 116, 1106-1115 (2005)).
Авторы настоящего изобретения показали, что является возможной коррекция мутации Foxp3K276X в мышиных T-клетках с использованием редактирования генов (фиг. 20, 21, 22, 26). Авторы настоящего изобретения, кроме того, показали, что подвергнутые репарации T-клетки являются функциональными и могут супрессировать другие иммуноциты (фиг. 24, 25). Эти T-клетки могут либо являться непосредственно подвергнутыми репарации, либо могут происходить из репарированных HSC или происходящих из iPS T-клеток. Авторы настоящего изобретения, кроме того, показали, что адоптивно перенесенные T-клетки размножаются в хозяевах с недостаточностью Foxp3. Вплоть до половины T-клеток становятся Tрег и супрессируют заболевания (фиг. 29). Это показывает осуществимость T-клеточной терапии для синдрома scurfy/IPEX. Авторы настоящего изобретения показали, что истощение CD4 может предотвращать заболевание (фиг. 30-32). Таким образом, клетки CD4+ представляют собой патогенные клетки. Таким образом, в принципе, терапевтическим способом может являться истощение CD4+ T-клеток и затем перенос CD4+ T-клеток с репарированным Foxp3 для восстановления иммунологического равновесия. Однако, поскольку как подвергнутые генетической репарации T-клетки, так и патогенные T-клетки с недостаточностью Foxp3, экспрессируют CD4, непрерывное истощение CD4+ T-клеток может уничтожать не только патогенные клетки, но также адоптивно перенесенные, подвергнутые генетической репарации клетки. Решением может являться одновременная коррекция гена Foxp3 и параллельное осуществление аллельной инженерии. Авторы настоящего изобретения показали осуществимость коррекции гена Foxp3 и перевода CD45.2 в CD45.1 (фиг. 20). Таким образом, патогенные клетки с недостаточностью Foxp3 можно истощать с использованием антитела, нацеленного против природного эпитопа CD45 (например, CD45.2). Если подвергнутые репарации Foxp3 клетки переводят к сконструированному аллелю CD45 (например, CD45.1), тогда перенесенные, подвергнутые генетической репарации клетки больше не подвергаются истощению. Это позволяет одновременное истощение патогенных клеток и пополнение функциональных, подвергнутых репарации клеток, пока иммунологическое равновесие не восстановится. В этой точке избирательную абляцию патогенных клеток-хозяев можно останавливать. Авторы настоящего изобретения показали, что истощение CD45 действительно существенно расширяет выживаемость мышей с недостаточностью Foxp3.
Кроме того, клетки, подвергнутые генетической коррекции Foxp3, имеют не только повторную экспрессию белка Foxp3, но также повышающую регуляцию CD25, высокоаффинного рецептора интерлейкина-2 (IL-2) in vitro и in vivo (фиг. 20, 21, 22, 26). Физиологически, Foxp3 осуществляет положительную регуляцию экспрессии CD25 в T-клетках дикого типа, и ось IL-2/CD25/Stat5 является необходимой для выживаемости регуляторных T-клеток. Таким образом, авторы настоящего изобретения постулировали, что T-клетки, повторно экспрессирующие подвергнутый генетической коррекции Foxp3, можно размножать in vivo посредством терапевтического введения IL-2 (фиг. 23). Хорошо установлено, что T-клетки можно размножать in vivo с использованием комплексов IL-2/mAb против IL-2, терапии низкими дозами IL-2, (сконструированных) вариантов IL-2, других способов размножения Tрег. На Фиг. 27 показано, что подвергнутые репарации T-клетки можно размножать in vivo с использованием комплексов IL-2/mAb против IL-2.
В конкретных вариантах осуществления, дефект связанного с заболеванием гена представляет собой мутацию, содержащуюся в гене CTLA-4, и заболевание представляет собой синдром иммунной дисрегуляции человека, ассоциированный с мутациями CTLA-4 (Schubert et al., Science Translational Medicine 5, 215ra174-215ra174 (2013); Kuehn et al., Science (New York, N.Y.) 345, 1623-1627 (2014)).
Второй аспект изобретения относится к средству, выбранному из
a. соединения, содержащего или состоящего из антитела или подобной антителу молекулы, и
b. иммуноэффекторной клетки, несущей подобную антителу молекулу, или иммуноэффекторной клетки, несущей химерный рецептор антигена,
для использования в лечении медицинского состояния. Средство является специфически реакционноспособным по отношению либо к первой, либо к второй изоформе поверхностного белка, где первая изоформа поверхностного белка является функционально неотличимой, но иммунологически отличимой от второй изоформы поверхностного белка. Средство вводят для абляции клетки, несущей изоформу, по отношению к которой средство является реакционноспособным.
В конкретных вариантах осуществления, медицинское состояние представляет собой гематопоэтическое нарушение.
В конкретных вариантах осуществления, медицинское состояние представляет собой злокачественное гематопоэтическое заболевание.
В конкретных вариантах осуществления, медицинское состояние представляет собой злокачественное гематопоэтическое заболевание, устойчивое к лечению с использованием CAR-T-клеток против CD19 (= CAR19 клеток).
В конкретных вариантах осуществления, медицинское состояние представляет собой незлокачественное гематопоэтическое заболевание.
В конкретных вариантах осуществления, медицинское состояние представляет собой аутоиммунное заболевание.
В конкретных вариантах осуществления, медицинское состояние представляет собой заболевание «трансплантат против хозяина» (GvHD).
В контексте настоящего описания, заболевание «трансплантат против хозяина» относится к медицинскому осложнению после получения в качестве реципиента трансплантированной ткани от генетически отличного индивидуума. Иммуноциты в служащей донацией ткани (трансплантате) узнают реципиента (хозяина) как чужеродного.
В конкретных вариантах осуществления, медицинское состояние представляет собой заболевание «трансплантат против хозяина», вызванное трансплантацией гематопоэтической стволовой клетки.
В конкретных вариантах осуществления, медицинское состояние представляет собой заболевание «трансплантат против хозяина», вызванное адоптивным переносом. В контексте настоящего описания, адоптивный перенос или адоптивная клеточная терапия относится к переносу клеток человека, обычно, иммуноцитов, пациенту. Клетки могут являться аутологичными или аллогенными. Способы направленной модификации, осуществляемой посредством редактирования генов, позволяют индивидуализацию перенесенного продукта клеток для репарации генетических дефектов, увеличение эффективности перенесенных клеток или снабжение клеток дополнительными желательными признаками, такими как направляющие молекулы или аварийные выключатели.
В конкретных вариантах осуществления, медицинское состояние представляет собой заболевание «трансплантат против хозяина», вызванное трансплантацией органа.
В конкретных вариантах осуществления, антитело или подобную антителу молекулу связывают с токсином, таким образом, получая иммунотоксин. В конкретных вариантах осуществления, антитело или подобную антителу молекулу связывают с сапорином.
В конкретных вариантах осуществления, средство представляет собой биспецифическое антитело или биспецифическую подобную антителу молекулу. Иными словами, средство представляет собой антитело или подобную антителу молекулу, которые могут одновременно связываться с двумя различными типами антигена.
В конкретных вариантах осуществления, средство представляет собой иммуноэффекторную клетку, несущую биспецифическое антитело или биспецифическую подобную антителу молекулу.
В конкретных вариантах осуществления, средство представляет собой иммуноэффекторную клетку, несущую
a. первое антитело или подобную антителу молекулу, специфически реакционноспособные по отношению либо к первой, либо к второй изоформе первого поверхностного белка, где указанная первая и указанная вторая изоформы указанного первого поверхностного белка являются функционально неотличимыми, но иммунологически отличимыми, и
b. второе антитело или подобную антителу молекулу, специфически реакционноспособные по отношению либо к первой, либо к второй изоформе второго поверхностного белка, где указанная первая и указанная вторая изоформы указанного второго поверхностного белка являются функционально неотличимыми, но иммунологически отличимыми.
В конкретных вариантах осуществления, первый поверхностный белок представляет собой CD19.
В конкретных вариантах осуществления, второй поверхностный белок представляет собой CD45 или CD34.
В конкретных вариантах осуществления, первый поверхностный белок представляет собой CD19, и второй поверхностный белок представляет собой CD45.
В конкретных вариантах осуществления, средство представляет собой иммуноэффекторную клетку, несущую антитело или подобную антителу молекулу, или иммуноэффекторную клетку, в частности, T-клетку, несущую химерный рецептор антигена, для использования в способе лечения злокачественного гематопоэтического заболевания.
В конкретных вариантах осуществления, средство представляет собой T-клетку, несущую химерный рецептор антигена, нацеленный против CD45, для использования в способе лечения гематопоэтического заболевания, в частности, злокачественного гематопоэтического заболевания.
CD45 является особенно хорошей мишенью для лечения гематопоэтических заболеваний, поскольку он экспрессируется на всех гематопоэтических клетках, включая злокачественные клетки. CD45 является критическим для выживаемости клеток, таким образом, для клеток будет более сложно развить устойчивость.
Другой аспект изобретения относится к комбинированному лекарственному средству, содержащему
a. первое средство в соответствии с вторым аспектом изобретения, где средство является реакционноспособным по отношению к первому поверхностному белку, и
b. второе средство в соответствии с вторым аспектом изобретения, где средство является реакционноспособным по отношению к второму поверхностному белку,
Конкретные варианты осуществления относятся к комбинированному лекарственному средству для лечения гематопоэтического заболевания.
Конкретные варианты осуществления относятся к комбинированному лекарственному средству для лечения злокачественного гематопоэтического заболевания.
Конкретные варианты осуществления относятся к комбинированному лекарственному средству для лечения незлокачественного гематопоэтического заболевания.
В конкретных вариантах осуществления, первое и второе средство представляют собой T-клетки, несущие химерный рецептор антигена.
В конкретных вариантах осуществления, первый поверхностный белок представляет собой CD19, и второй поверхностный белок представляет собой CD45.
В конкретных вариантах осуществления, первое средство представляет собой T-клетку с CAR против CD19, и второе средство представляет собой T-клетку с CAR против CD45.
В конкретных вариантах осуществления, злокачественное гематопоэтическое заболевание является устойчивым к лечению с использованием только T-клеток с CAR против CD19 (= CAR19 клеток).
Другой аспект изобретения относится к способу отслеживания in vivo клетки, экспрессирующей первую изоформу поверхностного белка, где указанная первая изоформа указанного поверхностного белка является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного поверхностного белка, включающему введение указанному пациенту лиганда, специфически реакционноспособного по отношению к указанной первой изоформе.
Альтернативный этому аспект изобретения относится к способу отслеживания клетки, экспрессирующей первую изоформу поверхностного белка, в ткани, полученной от пациента, где указанная первая изоформа указанного поверхностного белка является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного поверхностного белка, включающему введение в указанную ткань, полученную от указанного пациента, лиганда, специфически реакционноспособного по отношению к указанной первой изоформе.
В конкретных вариантах осуществления этого аспекта изобретения, ткань, полученная от пациента, относится к образцу крови. Такой образец крови можно анализировать с использованием FACS. В конкретных вариантах осуществления этого аспекта изобретения, ткань, полученная от пациента, относится к образцу органа, например, биоптату, например, образцу печени. Такой образец ткани можно анализировать с использованием гистологических способов.
Другой аспект изобретения относится к способу избирательного истощения или обогащения клетки in vivo, включающему стадии
a. получения клетки, где указанная клетка экспрессирует первую изоформу поверхностного белка, которая отличается от второй изоформы указанного поверхностного белка в отношении аминокислотного маркера, где указанная первая изоформа содержит аминокислотный маркер A, кодируемый последовательностью нуклеиновой кислоты A, и указанная вторая изоформа содержит аминокислотный маркер B, кодируемый последовательностью нуклеиновой кислоты B;
b. индукции мутагенеза от указанной последовательности нуклеиновой кислоты A до указанной последовательности нуклеиновой кислоты B в геномной ДНК указанной клетки;
c. избирательного обогащения/истощения указанной клетки на основании экспрессии указанной первой или указанной второй изоформы указанного поверхностного белка.
В контексте настоящего описания, термин «избирательное истощение клеток» относится к избирательному уменьшению общего количества или концентрации клеток, экспрессирующих конкретные маркер/аллель/изоформу.
Специалисту в данной области понятно, что в случаях, когда определенный объем содержит клетки, экспрессирующие первую изоформу, и клетки, экспрессирующие вторую изоформу, избирательное истощение клеток, экспрессирующих первую изоформу, соответствует обогащению клеток, экспрессирующих вторую изоформу.
В качестве неограничивающего примера, избирательного истощения можно достигать посредством комплементзависимой цитотоксичности (CDC), антителозависимой клеточной цитотоксичности (ADCC), конъюгата антитело-лекарственное средство (ADC) или посредством реакции клеток, в частности, иммунорецепторных клеток, несущих природный рецептор антигена или химерный рецептор антигена (CAR). Избирательное истощение можно также осуществлять посредством введения антитела или подобной антителу молекулы, которые не являются связанными с эффекторным соединением, таким как лекарственное средство или токсин.
Авторы настоящего изобретения показали, что избирательное истощение in vivo является возможным с использованием антител против CD45.2 или CD45.1, соответственно (фиг. 12). Использование антитела, нацеленного против эпитопа, разделяемого двумя популяциями клеток (например, CD45.2+ и CD45.1+ T-клетками), приводит к неизбирательному истощению обеих популяций клеток (например, истощению против CD4). В отличие от этого, истощение с использованием антитела, которое избирательно связывается с одним (например, CD45.2), но не с другим (например, CD45.1) аллелем повсеместно экспрессированного поверхностного белка (например, CD45), истощает только клетки, экспрессирующие аллель, связываемый специфическим mAb. В примере показано, что истощение CD45.2+ клеток сохраняет CD45.1+ клетки и таким образом, приводит к относительному обогащению CD45.1+ клеток. Истощение может быть более эффективным, если токсин связывают с mAb.
Преимуществом использования антитела или подобной антителу молекулы, которые не являются связанными с токсином, может являться то, что лечение с использованием иммунотоксина приводит к истощению HSC, в то время как лечение с использованием антитела или подобной антителу молекулы, не связанных с токсином, сохраняет HSC. Этот может являться желательным, поскольку гематопоэтическая система частично сохраняется.
Как показано в EP16196860.7, EP16196858.1, PCT/EP2017/059799, авторы настоящего изобретения показали, что отличие одиночной аминокислоты можно конструировать в клетке и можно отличать посредством различных лигандов, которые специфически связываются с двумя изоформами/аллелями (нативной по сравнению со сконструированной). Специфически сконструированную искусственную мутацию или редкую, но встречающуюся в природе мутацию, такую как однонуклеотидный полиморфизм (SNP), конструируют в эндогенном экспрессированном на поверхности гене для изменения его антигенности. Специалисту в данной области известно, что эту мутацию можно вводить любым способов, известным в данной области, включая HDR и редакторы оснований. Этот измененный эпитоп затем используют для избирательного истощения успешно редактированных клеток с использованием лиганда, который специфически и избирательно узнает этот искусственный эпитоп. Альтернативно, редактированным клеткам придают устойчивость к истощению посредством лиганда, который узнает природный эпитоп (и таким образом, может истощать клетки-хозяева), но не узнает измененный эпитоп, и таким образом, сохраняет перенесенные клетки.
В случаях, когда «редактированные/сконструированные клетки» (клетки, в которых первая изоформа белка поверхности клеток изменена до второй изоформы) впоследствии используют для трансплантации, в частности, адоптивного переноса, две различные изоформы можно использовать для установления отличий между перенесенными клетками и клетками-хозяевами. Это позволяет отслеживание перенесенных клеток, поскольку они являются постоянно маркированными. Отслеживание можно осуществлять с использованием меченных лигандов либо in vivo, либо ex vivo например, посредством проточной цитометрии или гистохимии, на клетках или тканях. Использование in vivo лигандов, специфических либо для перенесенных клеток, либо для клеток-хозяев, позволяет избирательное истощение либо перенесенных клеток, либо клеток-хозяев, с использованием антитела, которое связывается только с перенесенными, сконструированными клетками или с клетками-хозяевами, соответственно. Избирательного истощения можно также достигать посредством клеток, несущих природный или химерный рецептор антигена (CAR), узнающий либо перенесенные клетки, либо клетки-хозяева.
Избирательное истощение сконструированных клеток составляет важный признак безопасности посредством предоставления «безопасного или аварийного выключателя». Основная концепция аварийных выключателей и генов самоубийства описана в Jones et al., Front Pharmacol.; 5:254. doi: 10.3389. Способ авторов настоящего изобретения проще, безопаснее и более разносторонний. В принципе, любую клетку, которую адоптивно переносят, можно модифицировать, чтобы она несла измененный аллель/эпитоп в качестве комбинированного выключателя in vitro или in vivo отбора, отслеживания, безопасного выключателя и/или выключателя избирательной абляции. Неисключительные примеры включают клетки, несущие только сконструированный аллель, но не являющиеся иным образом генетически модифицированными, или клетки, несущие, кроме того, сконструированные признаки, такие как CAR-клетки. Например, перенесенные аллогенные клетки, которые используют для их эффекта трансплантат против лейкоза, могут вызывать заболевание «трансплантат против хозяина» (GvHD). Если сконструированный аллель вводят до переноса, их можно уничтожать посредством сконструированного аллеля для уменьшения/лечения GvHD (фиг. 2). Подобным образом, перенесенные аутологичные инфильтрующие опухоль лимфоциты (TIL) или специфические для патогена лимфоциты можно конструировать, чтобы они несли измененный аллель, для их уничтожения, если нежелательные побочные эффекты возникнут из-за неспецифических эффектов или слишком интенсивных специфических эффектов (фиг. 2). CAR можно получать из mAb для комбинации специфичности известного mAb с признаками клетки, например, T-клетки-киллера. Принцип перенацеливания уничтожающей (или супрессорной) активности данной (T)-клетки на специфический антиген-мишень посредством введения CAR является, в принципе, применимым к широкому ряду заболеваний, включая злокачественные новообразования, но также аутоиммунные заболевания, трансплантацию или другие гематопоэтические заболевания. Успех использования CAR19-T-клеток показывает потенциал CAR-клеток в качестве лекарственных средств. Важно, однако, что в то время как CAR19-T-клетки являются высоко эффективными для лечения некоторых CD19+ опухолей, некоторые CAR-T-клетки, реакционноспособные против множества различных молекул-мишеней, могут оказывать тяжелые, временами летальные, побочные эффекты, такие как синдром высвобождения цитокинов и/или нейротоксичность (показанная для нескольких конструкций CAR, нацеленных на CD19, но также на другие мишени, включая, но без ограничения, CD123). Таким образом, возможность контролировать/уничтожать CAR-T-клетки после переноса является важной (фиг. 2, 10, 19). В случае CAR-клеток, измененный аллель может служить аварийным выключателем. Более того, перенесенные, сконструированные клетки можно также уничтожать в случае, когда они становятся злокачественными или вызывают любой тип нежелательного специфического или неспецифического повреждения (возможно, через несколько лет). Альтернативно, вызывающие заболевание клетки-хозяева можно подвергать избирательной абляции, в то же время сохраняя аутологичные, но модифицированные клетки. В отличие от способа авторов настоящего изобретения, существующая технология ограничена абляцией перенесенных клеток, но не позволяет простой абляции клеток-хозяев. Измененная изоформа позволяет переносить, например, подвергнутые репарации генов или иным образом модифицированные аутологичные клетки во время абляции клеток-хозяев. Без переключения изоформ, введенного способом по изобретению, абляцию клеток-хозяев необходимо остановить после переноса здоровых клеток. В этом случае, в то время как заново перенесенные, подвергнутые репарации клетки размножаются, клетки-хозяева также могут размножаться и больше не могут поддаваться абляции, приводя к риску того, что вызывающие заболевание клетки-хозяева могут победить при конкуренции с подвергнутыми репарации клетками. Таким образом, придание сконструированным клеткам устойчивости к истощению способом по изобретению является высоко значимым в качестве терапевтического способа. В качестве примера, эпитоп CD19, узнаваемый клетками с CAR против CD19, можно подвергать мутагенезу в аутологичных гематопоэтических клетках таким образом, что истощающие mAb против CD19 или клетки с CAR против CD19 больше не могут связывать и разрушать сконструированные клетки, но CD19 может оставаться функциональным. Это может исключать основное затруднение с современными эффективными клетками с CAR против CD19. В то время как для CAR против CD19 получили очень высокий процент успеха уничтожения экспрессирующих CD19 гематопоэтических малигном, они согласованно приводят к уничтожению экспрессирующих CD19 здоровых клеток-хозяев. Это приводит к гипогаммаглобулинемии и таким образом, увеличенному риску инфекций. Мутантный CD19 может позволять реконструкцию иммунной системы хозяина с использованием здоровых аутологичных гематопоэтических стволовых клеток (HSC), которые могут давать начало B-клеткам, которые являются устойчивыми к клеткам с CAR против CD19. CAR 19-T-клетки могут, таким образом, постоянно предотвращать рецидив, в то время как подвергнутые редактированию устойчивые клетки могут обеспечивать естественную защиту против инфекций. Пациенты, таким образом, могут больше не зависеть от инфузий IVIG. Трансплантацию HSC можно потенциально осуществлять как частичный химеризм посредством негенотоксической предварительной подготовки, например, посредством антител (Nat Biotech, 2016). Альтернативно, клетки с CAR против CD45, узнающие эпитоп, обнаруженный в основной человеческой популяции (например, аналогичный CD45.2), можно использовать для уничтожения всех гематопоэтических клеток-хозяев, включая злокачественные или иным образом вызывающие заболевание гематопоэтические клетки. CD45 является хорошей мишенью, поскольку он экспрессируется на всех гематопоэтических клетках, включая большинство злокачественных клеток. Более того, CD45 является критическим для выживаемости лимфоцитов, таким образом, если на него нацелены CAR-T-клетки, является менее вероятным, что клетки могут осуществлять понижающую регуляцию CD45 или мутагенез CD45, чтобы избегать нацеливания CAR-T-клеток. Это может уменьшать риск рецидива. Трансплантация здоровых аутологичных гематопоэтических стволовых клеток (HSC) или других гематопоэтических клеток, несущих сконструированный эпитоп CD45 (например, CD45.1), как проиллюстрировано посредством экспериментов с переключением от CD45.2 до CD45.1, может позволить реконструировать у хозяина здоровую гематопоэтическую систему, которая больше не истощается посредством клеток с CAR против CD45. Главным преимуществом может являться то, что на все экспрессирующие CD45 злокачественные новообразования (включая, но без ограничения, T-клеточные и миелоидные злокачественные новообразования), можно нацеливаться без необходимости специфических для опухоли или типа клеток антигенов, т.е. изобретение может относиться к универсально применимой системе для лечения гематопоэтических злокачественных новообразований и других незлокачественных гематопоэтических заболеваний. Это может преодолевать основное затруднение CAR-T-клеточной терапии: идентификацию подходящих антигенов-мишеней (Klebanoff, Nat Med 2016). Это может, таким образом, существенно расширять показатели заболевания, поддающиеся CAR-T-клеточной терапии. Несмотря на то, что CAR-T-клеточная терапия является очень успешной для лечения CD19+ опухолей, другие гематопоэтические злокачественные новообразования представляют большие проблемы. В качестве примера, лечение множественной миеломы остается большой проблемой, частично обусловленной отсутствием подходящего антигена-мишени (Mikkilineni, Blood, 2017; Sadelain, Nature 2017). Кроме того, опухоли гематопоэтической системы можно лечить без необходимости аллогенных клеток, таким образом, исключая GvHD в качестве главного осложнения. Кроме того, реконструкцию можно начинать во время фазы истощения, что может сокращать время для восстановления. Важно, что мутацию, использованную для придания перенесенным клеткам устойчивости к истощению, можно позже также опять использовать для истощения этих клеток, если это станет необходимым. Зависимое от CAR-клеток истощение HSC можно потенциально использовать в качестве альтернативного способа осуществления мягкой, т.е. негенотоксической предварительной подготовки. CAR45 клетки могут также уничтожать HSC. Используемый в настоящее время способ «уничтожения и замены», применяемый для трансплантации «костного мозга» или HSC, можно осуществлять с использованием CAR45. Вместо использования токсичной химиотерапии и радиоактивного облучения, предварительную подготовку пациента можно осуществлять с использованием CAR45. Преимуществом удаления HSC и гематопоэтической системы с использованием CAR45 клеток и их замены устойчивыми HSC со сконструированным аллелем, может являться то, что опасный период времени после трансплантации можно исключать, поскольку гематопоэтическую реконструкцию можно начинать во время абляции нежелательных клеток. Таким образом, пациенты могут всегда иметь функциональную иммунную систему вместо того, чтобы проходить через длительную фазу истощения костного мозга и иммуносупрессии. Таким образом, способ авторов настоящего изобретения может исключать инфекции в качестве главных осложнений современной трансплантации HSC. Клетки с CAR, нацеленным против антигена или комбинации антигенов, для специфического ограничения клеток-мишеней HSC, можно использовать для истощения эндогенных HSC. Это может представлять собой, например, CAR против CD45 или против CD34 плюс второй антиген в способе синтетической биологии (например, с отбором И) для специфического и исключительного нацеливания CAR-клеток против HSC.
Лечение с использованием CAR45 в комбинации с замещением гематопоэтической системы HSC со сконструированным аллелем можно использовать в качестве альтернативы терапии с использованием CAR19 (поскольку все CD19+ клетки также экспрессируют CD45), или его можно использовать в качестве комбинированной терапии с использованием CAR19 плюс CAR45. Терапию с использованием CAR45 можно также использовать для рецидивирующих отрицательных по CD19 или подвергшихся мутагенезу CD19 малигном после терапии с использованием CAR19.
Этот аспект изобретения относится к универсальному способу замещения клеток. Клетки могут представлять собой гематопоэтические клетки, аутологичные или аллогенные. Если замещающие клетки представляют собой HSC, описанный способ можно использовать для лечения любого гематопоэтического злокачественного новообразования или других гематопоэтических нарушений.
Другие преимущества способа авторов настоящего изобретения по сравнению с существующими способами «аварийного выключателя» включают следующие. В способе авторов настоящего изобретения используют эндогенный белок. Нет необходимости вводить трансген или метку в клетку. Два эпитопа являются функционально идентичными, но их можно различать посредством специфически связывающихся лигандов. Способ позволяет истощение перенесенных клеток или клеток-хозяев, в зависимости от того, какой лиганд используют. Поскольку сконструированную мутацию вводят в геном, признак безопасности постоянно остается в клетках и не подвергается сайленсингу, который может произойти с введенными посредством вирусов трансгенными аварийными выключателями. Кроме того, сконструированный эпитоп может являться менее антигенным, чем искусственные большие конструкции аварийного выключателя/гена самоубийства, и будет, таким образом, с меньшей вероятностью отторгаться клетками-хозяевами. Кроме того, использование сконструированных изоформ основано на направленных мутациях и является, таким образом, вероятно, более безопасным, чем другие аварийные выключатели/гены самоубийства, которые случайным образом интегрированы в геном, обычно посредством вирусной доставки, и могут, таким образом, приводить к инсерционному мутагенезу (Cornu, Nat Med, 2017).
Специалисту в данной области известно, что для изменения белка поверхности клеток от первой изоформы до второй изоформы, можно использовать альтернативные способы вместо HDR. В качестве неограничивающего примера, переключение изоформ можно осуществлять с использованием редакторов оснований, как описано в следующей публикации: Komor et al., Nature 533, 420-424, doi:10.1038/nature17946. Этот способ может увеличивать безопасность даже более, посредством обеспечения редактирования желательной аминокислоты без необходимости разрыва дцДНК. Редакторы оснований или родственные инструменты можно доставлять в форме плазмид или миниколец (дцДНК), мРНК или RNP.
В случаях, когда переключение первой изоформы белка поверхности клеток до второй изоформы комбинируют с репарацией вызывающего заболевание гена (например, гена Foxp3), можно истощать не подвергнутые репарации клетки in vivo (т.е., после переноса хозяину) посредством истощения клеток, экспрессирующих первую изоформу. Авторы настоящего изобретения показали, что в случаях, когда как переключение изоформы, так и репарацию дефекта гена осуществляют посредством HDR, вероятность успешной репарации гена увеличена в клетках, в которых произошло переключение изоформы (фиг. 20). Комбинация переключения изоформы первого гена с генетической модификацией второго гена позволяет включать признак безопасности в генетически модифицированные клетки.
Переключение изоформы можно использовать также в качестве маркера для отслеживания редактированных, перенесенных клеток у хозяина.
В соответствии с альтернативным аспектом, настоящее изобретение относится к способу избирательного истощения или обогащения клетки в композиции нередактированных и редактированных клеток, включающему стадии
a. получения клетки, экспрессирующей первую изоформу поверхностного белка, которая отличается от второй изоформы поверхностного белка, применительно к аминокислотному маркеру, где первая изоформа содержит аминокислотный маркер A, кодируемый последовательностью нуклеиновой кислоты A, и вторая изоформа содержит аминокислотный маркер B, кодируемый последовательностью нуклеиновой кислоты B;
b. индукции в указанной клетке посредством сайт-специфической генетической манипуляции замены последовательности нуклеиновой кислоты A на последовательность нуклеиновой кислоты B, таким образом, изменения в указанной клетке экспрессии первой изоформы на экспрессию второй изоформы;
c. избирательного обогащения/истощения клетки на основании экспрессии первой или второй изоформы поверхностного белка.
В конкретных вариантах осуществления, генетическую манипуляцию, приводящую к указанной вставке, делеции и/или замене аминокислоты, осуществляют посредством гомологичной репарации после двухцепочечного разрыва, индуцированного ассоциированной с CRISPR эндонуклеазой (Cas9) и направляющей РНК, где указанная направляющая РНК является способной к гибридизации в указанной первой геномной локализации.
В контексте настоящего описания, «ассоциированная с CRISPR эндонуклеаза» относится к эндонуклеазе Cas9, известной в данной области как способствующая подобному CRISPR направляемому последовательностью разрезанию цепей ДНК. Неограничивающими примерами ассоциированной с CRISPR эндонуклеазы являются эндонуклеазы Cas9 Streptococcus pyogenes (SpyCas9), эндонуклеаза Cpf1 Francisella (FnCpf1), Acidaminococcus (AsCpf1) и бактерии Lachnospiraceae (LbCpf1), любые ортологи SpyCas9, FnCpf1, AsCpf1 или LbCpf1, или любые сконструированные варианты белков SpyCas9, FnCpf1, AsCpf1 или LbCpf1, или их ортологов. Специалисту в данной области понятно, что изобретение относится также к новым открытым или сконструированным вариантам CRISPR/Cas.
В контексте настоящего описания, термин «ортолог» относится к гену и соотвествующему ему полипептиду, входящим в вертикальное происхождение от одного предкового гена. Иными словами, ортологи генов/полипептидов имеют общего предка и разделяются при дивергенции вида на два отдельных вида. Копии одного гена в двух полученных видах затем обозначают как ортологи. Чтобы убедиться, что два гена являются ортологами, специалист в данной области может провести филогенетический анализ линии гена посредством сравнения выровненных нуклеотидных или аминокислотных последовательностей генов или полипептидов.
В контексте настоящего описания, термин «направляющая РНК» относится к синтетической РНК, способной направлять ассоциированную с CRISPR эндонуклеазу к представляющей интерес геномной локализации (где эндонуклеаза может расщеплять фосфодиэфирную связь в геномной ДНК). Специалисту в данной области понятно, что если используют эндонуклеазу Cas9, выражение «направляющая РНК» может относиться к одиночной направляющей РНК (онРНК), содержащей как последовательность, необходимую для связывания Cas9, так и определяемую пользователем «нацеливающую последовательность», или к комбинации двух молекул РНК, где одна содержит последовательность, необходимую для связывания Cas9 (tracrРНК), а другая содержит определяемую пользователем «нацеливающую последовательность» (crРНК). Если используют эндонуклеазу Cpf1, выражение «направляющая РНК» относится к одиночной молекуле РНК, содержащей как последовательность, необходимую для связывания Cpf1, так и определяемую пользователем «нацеливающую последовательность», или к нескольким направляющим РНК, транскрибируемым в форме одиночного массива crРНК (Zetsche, Nat Biotech, 2016). «Нацеливающая последовательность» является способной к гибридизации в представляющей интерес геномной локализации и таким образом, определяет геномную мишень, подлежащую модификации, и обычно содержит приблизительно 20 нуклеотидов.
Расщепление ДНК посредством Cas9 зависит от присутствия короткого смежного с протоспейсером мотива (PAM) в ДНК-мишени, ограничивающего выбор поддающихся нацеливанию последовательностей. CAS9 из Streptococcus pyogenes (SpyCas9), например, соответствует последовательности PAM 5'-NGG-3'. В конкретных вариантах осуществления, конструкция для репарации ДНК содержит мутантную последовательность PAM. Мутация делает последовательность PAM нефункциональной, но не влияет на экспрессию, стабильность или функцию белка. Использование конструкции для репарации ДНК, содержащей мутантную последовательность PAM, увеличивает эффективность HDR.
Специалисту в данной области известно, что помимо системы CRISPR, существуют альтернативные средства для сайт-специфического редактирования ДНК, а именно, использование эндонуклеаз с цинковыми пальцами, подобных активаторам транскрипции эффекторных нуклеаз (TALEN), мегануклеаз или систем на основе аргонавтов (Nat Biotechnol. 2016 Jul;34(7):768-73) или редакторов оснований (Komor et al., Nature 533, 420-424, doi:10.1038/nature17946). Изобретение относится также к использованию этих альтернативных средств для сайт-специфического редактирования ДНК.
В конкретных вариантах осуществления, первая конструкция для репарации ДНК не является субстратом для системы CRISPR, используемой на первой стадии способа (введении разрыва цепи в геномную ДНК), поскольку она не содержит последовательности PAM. Таким образом, вставленную последовательность больше нельзя разрезать после вставки посредством второго события с использованием эндонуклеазы.
В конкретных вариантах осуществления, генетическую манипуляцию, приводящую к указанной вставке, делеции и/или замене аминокислоты, осуществляют посредством обеспечения, в частности, трансфекции/электропорации, указанной клетки, редактором оснований (как описано в Komor et al., Nature 533, 420-424, doi:10.1038/nature17946 и/или Gaudelli, Nature 2017), способным менять последовательность нуклеиновой кислоты A, кодирующую аминокислотный маркер A, на последовательность нуклеиновой кислоты B, кодирующую аминокислотный маркер B, и направляющей РНК, способной направлять указанный редактор оснований на последовательность нуклеиновой кислоты A, кодирующую аминокислотный маркер A. Дизайн редактора оснований, способного менять мышиный CD45.1 на CD45.2, проиллюстрирован в разделе примеры. (фиг. 9). Это иллюстрирует осуществимость использования редакторов оснований для аллельной инженерии в качестве альтернативы доказанному способу HDR. Однако, из-за ограничений, таких как то, какие нуклеотиды в геноме поддаются редактированию оснований, авторы настоящего изобретения были неспособны сконструировать редакторы оснований для перевода CD90.2 в CD90.1 или CD90.1 в CD90.2. Кроме того, на основании дизайна исходного редактора оснований, основанного на PAM NGG (как описано в Komor et al., Nature 2016), было невозможно сконструировать редактор оснований для перевода либо CD45.1 в CD45.2, либо CD45.2 в CD45.1. В отличие от этого, получение сконструированных вариантов Cas9, узнающих измененный участок PAM, расширило специфичность на ряд нуклеотидов, на которые можно нацеливаться посредством редакторов оснований (Kim, Nat Biotech 2017). Авторы настоящего изобретения идентифицировали участок PAM для Staphylococcus aureus (SaKKH-BE3) поблизости от G, присутствующего в последовательности CD45.1, который, при мутагенезе до A, приводит к получению в результате аллеля CD45.2. Они сконструировали онРНК, которая помещает G, подлежащий переводу в A, в положение 5 в окне редактирования, поскольку SaKKH-BE3 наиболее эффективно редактирует G в положении 5 по сравнению с другими положениями в окне редактирования. Затем, авторы настоящего изобретения разработали пары направляющая РНК/редактор оснований для перевода CD45.2 в CD45.1 (Фигура 9b). Вновь сконструированные редакторы оснований аденина A/T (ABE) могут осуществлять перевод G/C. С использованием этого нового ABE является, в принципе, возможным переводить необходимый A в аллеле CD45.2 в G в аллеле CD45.1. Однако доступные в настоящее время ABE требуют PAM NGG поблизости от желательной мутации, но не присутствует такого PAM для корректного расположения ABE. Таким образом, авторы настоящего изобретения разработали гипотетический ABE на основании правил, применимых к цитидин-дезаминазным редакторам оснований. Поскольку известные цитидин-дезаминазные редакторы оснований и новые ABE осуществляют редактирование в сходном окне редактирования, разумно заключить, что этот способ может работать. Авторы настоящего изобретения идентифицировали 2 варианта, один на основе гипотетического слитого белка Cas9 SaKKH-ABE, и один на основе гипотетического слитого белка Cas9 VQR-ABE (фиг. 9b). Нельзя исключить, что фланкирующие A также будут подвергаться переводу, что может изменять эпитоп. Таким образом, перевод CD45.2 в CD45.1 с использованием редакторов оснований требует высоко специфических редакторов оснований с узким окном редактирования. Недавно опубликованные редакторы оснований имеют окна редактирования из 1-2 нуклеотидов, и таким образом, являются подходящими (Kim, Nat Biotech, 2017). Совместно, эти примеры иллюстрируют, что принцип перевода основания для аллельного редактирования является независимым от платформы и может быть осуществлен посредством опосредованных HDR способов или альтернативных способов, таких как редактирование оснований. В зависимости от типа клеток и геномного контекста желательной мутации, можно выбирать один или другой.
В контексте настоящего описания, «конструкция для репарации ДНК» относится к конструкции ДНК, которую используют в качестве матрицы для репарации повреждения цепи ДНК, в частности, двухцепочечного разрыва (DSB) в геномной ДНК, посредством HDR. Конструкция для репарации ДНК содержит гомологичные плечи и представляющую интерес трансгенную последовательность. Гомологичные плечи являются гомологичными последовательностям геномной ДНК 5' и 3' от DSB. Представляющая интерес трансгенная последовательность локализована между гомологичными плечами. В ходе репарации геномной ДНК посредством HDR, представляющую интерес трансгенную последовательность вставляют в геномную ДНК. Специалисту в данной области понятно, что конструкция для репарации ДНК может являться линейной (одноцепочечной или двухцепочечной) или кольцевой (например, плазмида, миникольцевая плазмида).
Специалисту в данной области понятно, что выражение «направляющая РНК является способной гибридизоваться с представляющей интерес геномной локализацией» относится к тому факту, что часть направляющей РНК (определенная пользователем «нацеливающая последовательность») является способной гибридизоваться с представляющей интерес геномной локализацией в условиях высокой строгости. Направляющая РНК содержит другие части, которые не являются способными гибридизоваться с представляющей интерес геномной локализацией. Посредством (частичной) гибридизации с представляющей интерес геномной локализацией, направляющая РНК направляет ассоциированную с CRISPR эндонуклеазу к представляющей интерес геномной локализации, таким образом, осуществляя DSB в представляющей интерес геномной локализации.
В конкретных вариантах осуществления, улучшающие HDR реагенты, в частности, ванилин или рукапариб, используют в способе редактирования генов. В контексте настоящего описания, ванилин обозначает 4-гидрокси-3-метоксибензальдегид, CAS No. 121-33-5. В контексте настоящего описания, рукапариб обозначает 8-фтор-2-{4-[(метиламино)метил]фенил}-1,3,4,5-тетрагидро-6H-азепино[5,4,3-cd]индол-6-он, CAS No. 283173-50-2.
Везде, где альтернативы одиночным разделяемым признакам выделяют в настоящем описании как «варианты осуществления», следует понимать, что такие альтернативы можно свободно комбинировать для формирования отдельных вариантов осуществления изобретения, описанных в настоящем документе.
Содержание заявок EP16196860.7, EP16196858.1, PCT/EP2017/059799 и EP17197820,8 приведено в настоящем документе в качестве ссылки.
Изобретение далее проиллюстрировано посредством следующих примеров и фигур, из которых можно вывести дополнительные варианты осуществления и преимущества. Эти примеры предназначены для иллюстрации изобретения, а не для ограничения его объема.
Краткое описание чертежей
На фиг. 1 проиллюстрирован принцип конструирования иммунологически отличимых вариантов белков поверхности клеток. Изоформы A и B являются способными осуществлять одинаковую функцию (являются функционально сравнимыми). В этом примере изоформа A представляет собой природный поверхностный белок, который модифицирован в вариант изоформы B.
На фиг. 2 проиллюстрировано, как иммунологически отличимые варианты клеточной поверхности увеличивают безопасность клеточной терапии у человека.
На фиг. 3 показано, что две изоформы поверхностного белка можно различать посредством моноклональных антител (mAb). Последовательности: верхний ряд: SEQ ID NO 020, нижний ряд: SEQ ID NO 021.
На фиг. 4 показан эксперимент для доказательства концепции: Перевод CD90.2 (аллель A) в CD90.1 (аллель B) в первичных клетках. Левая панель проточной цитометрии показывает чистую исходную популяцию CD90.2+ первичных T-клеток. Затем клетки модифицируют in vitro для перевода CD90.2 в CD90.1. После редактирования можно отличать 4 различных популяции: CD90.2+ гомозиготные клетки (нередактированная исходная популяция, вверху слева), CD90.1+/CD90.2+ гетерозиготные клетки (вверху справа), CD90.1+ гомозиготные клетки (внизу справа) и клетки, утратившие CD90 (внизу слева). Клетки с модифицированным аллелем в нижнем правом квадрате больше не экспрессируют исходный/оригинальный эндогенный аллель (CD90.2). Чистую популяцию CD90.1+CD90.2- клеток можно использовать для выделения корректно отредактированных клеток in vitro перед переносом пациенту.
На фиг. 5 показан второй эксперимент для доказательства концепции: Перевод CD45.2 (аллель A) в CD45.1 (аллель B) в первичных клетках. Последовательности: верхний ряд: SEQ ID NO 022, нижний ряд: SEQ ID NO 023. Как показано для перевода аллеля CD90.2 в CD90.1 (фиг. 4), клетки с модифицированным аллелем в нижнем правом квадрате больше не экспрессируют исходный/оригинальный эндогенный аллель (CD45.2). Это можно использовать для выделения чистой популяции корректно отредактированных клеток перед переносом пациенту.
На фиг. 6 показано редактирование генов в клетках EL4 с использованием рибонуклеопротеиновых частиц (RNP) Cas9. Клетки EL4 трансфицировали с использованием комплекса crРНК:tracrРНК/Cas9 и +/- матрицы для HDR 2 т.п.о. таким же образом, как для способа на основе плазмид, за исключением условий электропорации (описанных в разделе способы). Это показывает, что редактирование аллелей можно осуществлять различными способами, например, на основе плазмид или RNP, т.е. оно является независимым от платформы.
На фиг. 7 показано редактирование генов в первичных T-клетках мыши с использованием рибонуклеопротеиновых частиц (RNP) Cas9. Первичные T-клетки мыши трансфицировали с использованием комплекса crРНК:tracrРНК/Cas9 и +/- матрицы для HDR 2 т.п.о. таким же образом, как для способа на основе плазмид. Подобно фиг. 6, это показало, что редактирование аллелей для перевода аллеля A в аллель B является независимым от платформы.
На фиг. 8 показан отбор с высокой чистотой ex vivo сконструированных клеток с модифицированным аллелем. Левая панель: После модификации CD45.2 клеток в CD45.1 клетки возникают 4 популяции (сравнимо с фиг. 4). Затем проточную цитометрию использовали для очистки всех 4 популяций до очень высокой чистоты. На правой панели показана чистота после сортировки для каждой из 4 популяций. Заключение: Все 4 возможные популяции клеток можно очищать ex vivo до высокой чистоты на основании модифицированного аллеля. Корректное редактирование генов подтверждали посредством секвенирования ДНК по Сэнгеру (не показано).
На фиг. 9 показан дизайн редактора оснований для перевода CD45.1 в CD45.2 (A) и CD45.2 в CD45.1 (B). Последовательности (A): верхний ряд: SEQ ID NO 024, нижний ряд: SEQ ID NO 025 (B): верхний ряд (вариант 1 и 2): SEQ ID NO 026, нижний ряд (вариант 1 и 2): SEQ ID NO 027. A) направляющую РНК конструировали для использования с рестрицированным по PAM NNNRRT редактором оснований SaKKH-BE3. Выбранная последовательность PAM выделена черной чертой из точек (ATTTGT). Направляющая РНК выделена серой чертой, окно редактирования оснований направляющей РНК, нуклеотиды 4-7, подчеркнуто. G в положении 5 можно переводить в A с использованием этого редактора оснований. Таким образом, аллель CD45.1 можно переводить в аллель CD45.2. B) Сконструированные редакторы оснований аденина могут переводить A в G. Для перевода CD45.2 в CD45.1 идентифицировано два варианта: Вариант 1: онРНК (серая черта) сконструирована для использования с Cas9 SaKKH (PAM выделен черной чертой из точек). Это может переводить A в положении 5 окна редактирования оснований в G. Вариант 2: онРНК (серая черта) для использования с Cas9 VQR. PAM выделен пунктирной черной чертой. Это может переводить A в положении 6 окна редактирования оснований в G. Варианты 1 и 2 могут переводить аллель CD45.2 в аллель CD45.1.
На фиг. 10 показана концепция помимо необязательной абляции перенесенных клеток. Клетки отбирают у пациента, затем редактируют (ex vivo) для перевода одного аллеля в другой аллель. Успешно отредактированные клетки выбирают для инфузии чистой популяции клеток, несущих эпитоп, необходимый для избирательного истощения. Аллельная инженерия, таким образом, добавляет вариант для абляции перенесенных клеток, при необходимости.
На фиг. 11 показаны применения избирательного истощения перенесенных клеток или клеток-хозяев in vivo.
На фиг. 12 показан эксперимент для доказательства концепции для избирательной абляции in vivo. Модель для абляции in vivo: 1) Реконструкция у мышей с иммунодефицитом, лишенных всех T-клеток (Rag KO) с использованием смеси 1:1 очищенных CD45.2+ и CD45.1+ T-клеток (конгенных клеток, нередактированных клеток). 2) Опосредованное антителом истощение (неизбирательное (против CD4) по сравнению с избирательным (против CD45.2)). Антитело против CD45.2 без токсина или связанное с токсином (сапорином). 3) Анализ через 1 неделю.
На фиг. 13 показано избирательное истощение CD45.2+ клеток in vivo: Количественная оценка T-клеток в крови.
На фиг. 14 показано избирательное истощение CD45.2+ клеток in vivo: Количественная оценка относительных количеств T-клеток в лимфоидных органах. Такая же постановка эксперимента, как на фиг. 12, но с анализом лимфатических узлов (LN) и мезентериальных лимфатических узлов (мезLN).
На фиг. 15 показано избирательное истощение CD45.2+ клеток in vivo: Количественная оценка относительных количеств T-клеток в лимфоидных органах. Такая же постановка эксперимента, как на фиг. 12, но с анализом селезенки (SP).
На фиг. 16 показано избирательное истощение CD45.2+ клеток in vivo: Количественная оценка абсолютных количеств T-клеток в лимфоидных органах. Такая же постановка эксперимента, как на фиг. 12, но с анализом лимфатических узлов (LN) и мезентериальных лимфатических узлов (мезLN).
На фиг. 17 показано избирательное истощение CD45.2+ клеток in vivo: Количественная оценка абсолютных количеств T-клеток в лимфоидных органах. Такая же постановка эксперимента, как на фиг. 12, но с анализом селезенки (SP).
На фиг. 18 показан второй эксперимент для доказательства концепции для избирательной абляции in vivo клеток с редактированным аллелем. Постановка эксперимента: 1) Электропорация CD45.2 T-клеток ex vivo для модификации до CD45.1 2) Адоптивный перенос мышам с недостаточностью T-клеток (Rag KO); (без отбора по чистоте до переноса) 3) Через несколько недель контроль качества, чтобы показать редактирование аллеля 4) Опосредованное антителом истощение CD45.2 клеток, оставляющее только отредактированные клетки. Заключения: доказательство концепции, показывающее истощение in vivo CD45.2+ клеток-хозяев и нередактированных клеток, с сохранением в то же время CD45.1+ клеток с модифицированным аллелем.
На фиг. 19 показано применение изобретения: Аллельная инженерия увеличивает безопасность, например, для CAR-T-клеточной терапии. Сконструированные клетки экспрессируют первую изоформу белка поверхности клеток (например, CD45) и химерный рецептор антигена, реакционноспособный по отношению к представляющей интерес мишени, например, CD19 (CAR-19). В то время как CAR19 конструируют в T-клетках, второй аллель CD45 переводят в первый аллель CD45, чтобы позволить установление отличий перенесенных CAR-T-клеток от немодифицированных клеток-хозяев. В случае связанной с лечением токсичности, патогенные CAR-T-клетки можно подвергать избирательной абляции для остановки связанной с CAR-T токсичности.
На фиг. 20 показана генетическая коррекции клеток с scurfy и клеток, несущих мутацию Foxp3K276X человека, так же как обогащение относительной частоты подвергнутых генетической репарации клеток при отборе по суррогатному поверхностному маркеру с переключенной изоформой.
A) Выравнивание геномных последовательностей ДНК foxp3 (C57BL/6) дикого типа (SEQ ID NO 028), локуса Foxp3 с направленной мутацией Foxp3K276X (SEQ ID NO 029), которая вводит преждевременный стоп-кодон, и локуса Foxp3 мышей с scurfy (B6.Cg-Foxp3sf/J), несущих спонтанную вставку 2 п.о., приводящую к сдвигу рамки (SEQ ID NO 030). Участки связывания онРНК (зеленая линия) и последовательности PAM (черная линия). B) Способ редактирования генов тотальных CD4+ T-клеток из мышей Foxp3K276X C57BL/6. Активация и электропорация in vitro (стадия 1) с использованием плазмид, кодирующих онРНК, нацеленную на мутацию Foxp3K276X, и кольцевой плазмиды, содержащей матрицу для репарации 1 т.п.о. дикого типа (wt) Foxp3. Успешно трансфицированные клетки выделяют на основании экспрессии GFP (стадия 2). Размножение клеток in vitro для редактирования генов в присутствии rhIL-2, TGF-β, отдельно или в комбинации с ретиноевой кислотой (RA) и нейтрализующими цитокины антителами (против IL-4 и против IFNγ в течение 7 суток (стадия 3). C) Постановка эксперимента, как в B, с тотальными CD4+ T-клетками от контрольных мышей (WT) или мышей Foxp3K276X. Проточная цитометрия экспрессии CD25 и Foxp3 (с отбором живых CD4+ T-клеток). Клетки дикого типа после электропорации пустой плазмидой px458 дифференцируют в CD4+Foxp3+CD25+ T-клетки (левая панель), отсутствие дифференцировки Foxp3 в Foxp3K276X клетках после электропорации с использованием только онРНК Foxp3K276X (средняя панель) и восстановление экспрессии белка Foxp3 в Foxp3K276X клетках после электропорации с использованием онРНК Foxp3K276X и 1 т.п.о. дцДНК-матрицы для репарации Foxp3 (правая панель). Верхний ряд: индукция Foxp3 с использованием только TGF-β, нижний ряд: индукция Foxp3 с использованием TGF-β в комбинации с RA. По сравнению только с TGF-β, комбинация TGF-β и RA приводит к более высокой частоте экспрессирующих Foxp3 клеток среди клеток, имеющих интактный локус Foxp3 (т.е. клеток дикого типа и подвергнутых репарации клеток). Репрезентативные данные для 2 экспериментов с Foxp3K276X клетками и одного эксперимента с Foxp3sf/J клетками. D) Обогащение подвергнутых репарации гена, экспрессирующих Foxp3 клеток с использованием мультиплексного переключения изоформ CD45 в качестве суррогатного маркера. Постановка эксперимента, как в b, но с одновременной электропорацией плазмид, кодирующих 2 онРНК (онРНК Foxp3K276X и онРНКCD45.2_R1) и две 1 т.п.о. дцДНК-матрицы (Foxp3 дикого типа и CD45.1). Через семь суток проточная цитометрия CD45.2, CD45.1, CD25 и Foxp3 (с отбором по живым CD4+ клеткам). Верхняя панель: Предварительный отбор CD45.1- клеток (зеленая линия) и CD45.1+ клеток (красная линия). Нижняя панель: Обогащение CD25+Foxp3+ клеток в подвергнутых переключению изоформы CD45.1+ клетках. Репрезентативные данные для 2 экспериментов с Foxp3K276X клетками и одного эксперимента с Foxp3sf/J клетками.
На фиг. 21 показана репарация гена Foxp3 с использованием способа на основе плазмид и способа на основе RNP. A: CD4 T-клетки от мышей Foxp3 KO подвергали трансфекции с использованием плазмиды с онРНК, отдельно или вместе с матрицей для HDR Foxp3 дикого типа. GFP+ и GFP- клетки подвергали сортировке через 24 час после трансфекции (трансфекции плазмидой) и немедленно после сортировки клеток размножали до окончания эксперимента в присутствии коктейля для дифференцировки Foxp3. B: CD4 T-клетки от мышей Foxp3 KO подвергали трансфекции с использованием комплекса crРНК:tracrРНК/Cas9 RNP, отдельно или +/- матрицы для HDR (180 п.о. оцДНК или 2 т.п.о. плазмида). Общий пул подвергнутых трансфекции RNP клеток размножали до окончания эксперимента в присутствии коктейля для дифференцировки Foxp3.
На фиг. 22 показана успешная репарация гена Foxp3 в T-клетках in vitro. Верхний ряд: Выравнивание последовательностей геномной ДНК wt Foxp3 (C57BL/6) SEQ ID NO 031 и локуса Foxp3 с направленной мутацией Foxp3K276X SEQ ID NO 032, которая вводит преждевременный стоп-кодон. Участки связывания онРНК (зеленая линия) и последовательности PAM (черная линия). Тотальные CD4+ T-клетки из контроля wt или мышей Foxp3K276X подвергали трансфекции. Проточная цитометрия экспрессии CD25 и Foxp3 (с отбором по живым CD4+ T-клеткам). Дифференцировка клеток wt после электропорации пустой плазмидой px458 в CD4+Foxp3+CD25+ T-клетки (верхняя панель), отсутствие дифференцировки Foxp3 в Foxp3K276X клетках после электропорации с использованием только онРНКFoxp3K276X (средняя панель) и восстановление экспрессии белка Foxp3 в Foxp3K276X клетках после электропорации с использованием онРНКFoxp3K276X и 1 т.п.о. дцДНК-матрицы для репарации Foxp3 (нижняя панель). Индукция Foxp3 с использованием TGF-бета в комбинации с RA.
На фиг. 23 показан способ редактирования генов тотальных CD4+ T-клеток от мышей Foxp3K276X C57BL/6. Активация и электропорация in vitro с использованием плазмид, кодирующих онРНК, нацеленную на мутацию Foxp3K276X, и кольцевой плазмиды, содержащей матрицу для репарации 1 т.п.о. wt Foxp3. Успешно трансфицированные клетки выделяют на основании экспрессии GFP. Клетки переносят in vivo для дифференцировки Foxp3 с последующим режимом обработки IL-2.
На фиг. 24 показан клинический фенотип мышей через 14 недель после AT клеток. Мыши WT и мыши после репарации являются свободными от заболевания, перенос клеток KO приводит к воспалению кожи и алопеции. Постановка эксперимента, как на фиг. 23.
На фиг. 25 показано, что клинический фенотип (инфильтрация T-клеток) у мышей, которым вводили клетки KO, коррелирует с инфильтрацией CD4/CD3+ T-клеток в коже уха и хвоста. Отсутствие или очень небольшие количества CD4/CD3+ T-клеток обнаружены в коже уха и хвоста мышей, которым вводили клетки WT или подвергнутые репарации T-клетки. Данные изображены как % и абсолютные количества.
На фиг. 26 показано присутствие CD25/Foxp3+ Tрег клеток у мышей, которым вводили клетки WT и подвергнутые репарации клетки. Отсутствие CD25/Foxp3+ T-клеток обнаружено у мышей, которым вводили клетки KO. Данные показаны как % и MFI Foxp3. Постановка эксперимента, как на фиг. 23.
На фиг. 27 показано, что подвергнутые репарации Tрег отвечают на IL-2. Дополнительная обработка IL-2 в ходе последней недели эксперимента приводит к увеличенному присутствию CD25/Foxp3+ Tрег клеток у мышей, которым вводили клетки WT и подвергнутые репарации клетки. Отсутствие CD25/Foxp3+ T-клеток обнаружено у мышей, которым вводили клетки KO. Эти данные показывают, что клетки WT и подвергнутые репарации Tрег клетки отвечают на IL-2 в сходной степени.
На фиг. 28 показано, что клетки WT и подвергнутые репарации CD25/Foxp3 Tрег клетки экспрессируют сходные уровни различных маркеров поверхности клеток (GITR, ICOS, TIGIT, PD1 и KLRG1). Комбинацию CD25 и дополнительных маркеров (например, GITR) можно использовать для обогащения этих популяций. T-клетки с недостаточностью Foxp3 являются фенотипически отличными, например, они лишены экспрессии GITR и KLRG1.
На фиг. 29 показано, что AT CD45.1+ WT-клеток спасает мышей с недостаточностью Foxp3 и продлевает продолжительность их жизни вплоть до 8 недель (наиболее поздняя проверенная временная точка, без признаков заболевания). Высокая доля WT Tрег клеток (50%), обнаруженная у мышей Foxp3 KO, супрессирует активацию T-клеток хозяина (меньший % клеток с CD44 высоким). Это показывает, что WT T-клетки имеют способность полностью восстанавливать иммунную регуляцию и контролировать заболевание scurfy. Кроме того, микроокружение с недостаточностью Foxp3 резко стимулирует усиленное размножение T-клеток без недостаточности Foxp3.
На фиг. 30 показан эксперимент по истощению CD4 у мышей Foxp3 KO (лимфатический узел, LN). Комбинация репарации гена в T-клетках с избирательным истощением антителом: Истощают патогенные клетки, затем переносят T-клетки после генетической коррекции. Истощение CD4 в большой степени предотвращает заболевание scurfy и значительно продлевает ожидаемую продолжительность жизни. Это наблюдение поддержано данными FACS, показывающими T-клетки с CD4 низким или с отсутствием CD4 (истощение KO по сравнению с мышами KO или мышами WT без лечения).
На фиг. 31 показан эксперимент по истощению CD4 у мышей Foxp3 KO (селезенка, SP). Комбинация репарации гена в T-клетках с избирательным истощением антителом: Истощают патогенные клетки, затем переносят T-клетки после генетической коррекции. Истощение CD4 в большой степени предотвращает заболевание scurfy и значительно продлевает ожидаемую продолжительность жизни. Это наблюдение поддержано данными FACS, показывающими T-клетки с CD4 низким или с отсутствием CD4 (истощение KO по сравнению с мышами KO или мышами WT без лечения).
На фиг. 32A показано, что истощение CD4 предотвращает развитие заболевания scurfy у мышей с недостаточностью Foxp3-.
На фиг. 32B показано, что истощение CD4 в большой степени предотвращает развитие дерматита в коже хвоста мышей с недостаточностью Foxp3.
Примеры
Описание эффективной абляции генов на основе плазмид в первичных T-клетках, направленного введения точечных мутаций в первичных T-клетках, обогащение редактированных посредством HDR клеток посредством мониторирования переключения изоформ суррогатного поверхностного маркера клетки и описание генетической коррекции мышиных клеток scurfy можно обнаружить в EP16196860.7, EP16196858.1 и PCT/EP2017/059799.
Общие соображения применительно к дизайну сконструированных мутаций (конструирование аллелей):
Конструирование аллеля гена посредством введения направленной небольшой мутации, возможно, мутации одиночного нуклеотида или одиночной аминокислоты, с целью изменения специфического связывания лиганда и обеспечения связывания второго, специфического лиганда, можно использовать в качестве общего принципа для терапевтического применения. Мутацию разрабатывают таким образом, чтобы сохранить функцию сконструированного белка максимально, насколько возможно. Иммуногенность необходимо изменять, чтобы иметь возможность стимуляции образования специфических связывающих лигандов, таких как моноклональные антитела (mAb) или подобные антителам молекулы, такие как аффимеры, белки DARPIN, нанотела и т.д. Необходимо проводить скрининг таких лигандов по специфическому связыванию, и характер иммуногена требует тщательной разработки. В то же самое время, мутация составляет незначительное антигенное изменение, и таким образом, маловероятно вызывает сильную иммунологическую реакцию in vivo. Конгенные маркеры у мышей точно удовлетворяют этим критериям. Отличие между CD90.1 и CD90.2, так же как между CD45.1 и CD45.2, составляет одиночный нуклеотид, приводящий к отличию одиночной аминокислоты. В обоих случаях, это отличие можно детектировать посредством специфических mAb, с получением в результате пары mAb для каждого гена. Несмотря на то, что может происходить образование mAb против этих различий, когда конгенные клетки переносят совпадающим конгенным мышам-хозяевам, т.е. CD90.1+ клетки - CD90.2+ мышам-хозяевам или наоборот, и CD45.1+ клетки - CD45.2+ хозяевам и наоборот, не происходит иммунологического отторжения клеток. Это означает, что к этому незначительному антигенному различию иммунная система in vivo является толерантной. Совместно, эти свойства сделали клетки с конгенными маркерами очень полезным инструментом для иммунологических исследований, используемым десятилетиями. Иммунологически, эти клетки используют в качестве суррогатных клеток для аутологичных переносов (поскольку эти клетки являются генетически идентичными, за исключением одной этой мутации мутация), однако, это конгенное незначительное отличие позволяет дискриминацию подвергнутых переносу клеток от клеток-хозяев.
Выбор белка, подлежащего конструированию
Для разработки сходной системы для терапевтического применения для человека, несколько соображений необходимо принимать во внимание. Мутацию, в принципе, можно вводить в любой ген, кодирующий белок. Паттерн экспрессии представляющего интерес белка может являться соответствующим, т.е. представлять собой повсеместно распространенную или специфическую для клеток или тканей экспрессию, если желательно. На белки, экспрессированные на поверхности, можно нацеливаться наиболее непосредственно, и они могут, таким образом, в большинстве случаев, являться выбранными белками. Примеры включают, но без ограничения, белки, охарактеризованные как система кластера дифференцировки CD1-CD371) (Engel, J Immunol November 15, 2015, 195 (10) 4555-4563 и http://www.hcdm.org/). Другие представляющие интерес белки могут представлять собой повсеместно экспрессированные белки, такие как бета-2-микроглобулин или константные части HLA класса I, которые экспрессируются на всех клетках, включая неиммуногенные клетки. Альтернативно, могут представлять интерес белки, экспрессированные в специфических типах клеток, например, CD45 человека для гематопоэтических клеток, CD3 человека для T-клеток, константные области компонентов T-клеточного рецептора человека, константные области иммуноглобулинов B-клеток, таких как константные области легких цепей каппа и лямбда, или тяжелых цепей, корецепторы CD4 или CD8 человека, или маркеры B-клеток, такие как CD19, CD20, CD21, CD22, CD23, костимулирующие молекулы, такие как CD28 или CD40, CD34, на гематопоэтических стволовых клетках, или даже специфические изоформы, экспрессированные на подгруппах клеток, таких как CD45RA или CD45RO. В этом последнем случае, можно разрабатывать мутацию в вариабельной области (альтернативно, в подвергаемых сплайсингу экзонах), отличающуюся между CD45RA и CD45RO. Конструирование мутации в повсеместно экспрессированной молекуле, такой как бета-2-микроглобулин или HLA-I, может служить уникальной системой, которую можно использовать фактически в любой клетке млекопитающего, более конкретно, в любой клетке человека. Это можно использовать для отслеживания и абляции сконструированных клеток при сохранении немодифицированных клеток-хозяев. Такой признак, например, можно использовать в качестве аварийного выключателя для клеточной терапии, включающей различные типы стволовых клеток (например, стволовых клеток мышц), гепатоцитов или клеток, происходящих из индуцированных плюрипотентных стволовых клеток (iPS). Конструирование мутации в специфическом для типа клеток белке, таком как CD45, можно использовать для нацеливания на все клетки из этой специфической ткани. В случае гематопоэтической системы, модифицированный CD45 человека может служить для маркировки гематопоэтических стволовых клеток (HSC), которые используют для замены эндогенной гематопоэтической системы. Все потомство этих HSC будет нести одну и ту же мутацию, сконструированную в геноме исходных HSC. Если предшествующую гематопоэтическую систему хозяина удаляют любыми способами (например, посредством облучения, химиотерапии, истощения лигандов, таких как mAb или подобные антителам молекулы, mAb, связанных с токсином, опосредованной клетками абляции, например, опосредованной CAR-T-клетками абляции и т.д.), тогда замещающую гематопоэтическую систему, даже если она является аутологичной, можно дискриминировать по сконструированной мутации. Альтернативно, это может позволять абляцию клеток-хозяев при сохранении сконструированных клеток.
Соображения дизайна:
- Паттерн экспрессии белка (повсеместно распространенный в отличие от специфического для клеток или тканей)
- Внеклеточные белки: В большинстве случаев мутации можно конструировать во внеклеточных частях представляющих интерес белков, которые являются доступными для специфических лигандов. Мутации можно конструировать в постоянно экспрессируемых экзонах, или альтернативно, в подвергаемых сплайсингу экзонах, если специфическая для подгруппы экспрессия является желательной.
- Сохранение консервативности среди видов: консервативные аминокислоты (ак) или структуры, наиболее вероятно, являются функционально значимыми, и скорее всего, должны, таким образом оставаться незатронутыми. Мутации следует вместо этого включать в менее консервативные области.
- Химические свойства аминокислот, например, полярность, электрический заряд, гидрофобность
- Сходство аминокислот: Для сохранения функции, мутация должна переводить данную ак в родственную ак, но изменения все еще должны поддаваться детекции посредством специфических лигандов.
- Встречающиеся в природе варианты: мутации, соответствующие вариантам аминокислот, наблюдаемым в данном положении при выравнивании множества последовательностей членов семейства белков (гомологичных последовательностей из различных организмов), вероятно, не влияют на функцию и структуру белка, и могут быть выбраны для конструирования мутаций.
- Структурные соображения: Структурные данные могут помогать в рациональном дизайне потенциальных мутаций. Аминокислота, предназначенная для модификации, должна являться доступной для связывания лиганда. Мутации в петлях являются функционально относительно хорошо переносимыми и таким образом, представляют собой интерес.
- Избегание известных вызывающих заболевания мутаций.
- Принятие во внимание известных для человека генетических вариантов: функционально переносимые однонуклеотидные полиморфизмы (SNP) являются хорошими кандидатами для мутаций.
- В участки, важные для вторичных модификаций, таких как гликозилирование, не следует вводить мутации,
- В участки, важные для дисульфидных связей, не следует вводить мутации.
- В участки, важные для опубликованных взаимодействий (например, взаимодействий водородных связей, солевых мостиков, гидрофобного стэкинга), не следует вводить мутации, как внутри белка, также и для белок-белковых взаимодействий для мультибелковых комплексов или взаимодействий рецептор-лиганд.
- Принятие во внимание предшествующих успешных систем, таких как мышиные CD90.1/CD90.2 (Q-R)
- Принятие во внимание участков связывания известных связывающих лигандов, таких как известные mAb, и компьютерного моделирования определяющих комплементарность областей (CDR).
- Исключение известных «активных участков», например, каталитических участков ферментов
- Исключение структурно очень сходных доменов, присутствующих в других белках человека, поскольку это, вероятно, увеличивает риск наличия перекрестно-специфических лигандов
- Принятие во внимание иммуногенности: выбор участка, который, вероятно, приведет в результате к получению подходящего пептидного иммуногена.
Аллельная инженерия в качестве общеприменимого, независимого от платформы принципа
В EP16196860.7, EP16196858.1 и PCT/EP2017/059799 авторы настоящего изобретения показали и охарактеризовали, как платформу CRISPR/Cas можно использовать для конструирования направленной точечной мутации для аллельной инженерии с последующей избирательной абляцией. Авторы настоящего изобретения сконструировали две отдельные точечные мутации в двух отдельных генах и показали, что сконструированную мутацию можно использовать для отслеживания и/или избирательной абляции сконструированных клеток in vivo. Авторы настоящего изобретения получали гомологичную репарацию (HDR) на основе системы CRISPR/Cas9 и дцДНК-матрицы. Однако использованный пример является только одним способом введения мутации. Альтернативы включают различные типы нуклеаз, таких как белки с цинковыми пальцами, TALEN или другие природные или сконструированные системы CRISPR/Cas, такие как Cas9, Cpf-1, высокоточные нуклеазы или нуклеазы с модифицированной зависимостью от PAM (Komor, Cell, 2017). Кроме того, описанный принцип является независимым от формы доставки нуклеаз. Их можно доставлять в форме плазмиды, мРНК, рекомбинантного белка, рекомбинантного белка в комплексе с направляющей РНК (т.е. рибонуклеопротеинового комплекса, RNP), разделенной рекомбиназы или в форме интегрирующего или неинтегрирующего вируса, например, ретровируса, лентивируса, бакуловируса или другой платформы вирусной доставки. Способ доставки может включать электропорацию или другие формы, такие как липофекция, наночастица доставка, сжатие клеток или физический прокол. Кроме того, матрица для HDR может представлять собой оцДНК или дцДНК в форме матрицы - короткой оцДНК, длинной оцДНК или кольцевой или линейной, миникольцевой ДНК, плазмидной ДНК или вирусной ДНК, например, аденоассоциированного вируса (AAV). В зависимости от клетки-мишени, специфические серотипы AAV используют для доставки «груза» в клетки. Для T-клеток человека и гематопоэтических стволовых клеток человека, матрицы для HDR часто предоставляют в форме AAV6, однако, эндонуклеазу RNP вместе с короткими оцДНК-матрицами для HDR также можно успешно использовать. Таким образом, способ, которым вводят мутацию, может являться гибким.
Аллельная инженерия с использованием альтернативных способов, например, конвертеров оснований или редакторов оснований
Авторы настоящего изобретения показали, как направленный разрыв дцДНК с последующей HDR можно использовать для введения небольших, направленных, т.е. точных мутаций для аллельной инженерии. Кроме того, существуют альтернативные способы того, как разработанные точечные мутации можно конструировать в геноме живой клетки. В качестве примера, новые сконструированные химерные слитые белки позволяют прямой перевод основания-мишени ДНК в другое основание (Komor, Nature 2016; Nishida, Science 2016; Yang, Nat Comm, 2016; Ma, Nat Meth 2016). Ферменты, такие как дезаминазы, можно сливать с ДНК-связывающими модулями, такими как (но без ограничения) белки с цинковым пальцами, системы TALEN или CRISPR/Cas. Природные цитидин-дезаминазы (APOBEC1, APOBEC3F, APOBEC3G) и индуцируемая активацией дезаминаза (AID) или ортолог AID PmCDA1 (из морской миногы) могут переводить основания цитидина (C) в основания урацила (U) в ДНК. Аппарат репликации ДНК может обрабатывать U как T, если репликация ДНК произойдет до репарации U. Это приводит к переводу пары оснований C:G в T:A (Yang, Nat Comm, 2016; Komor, Nature 2016). Таким образом, несколько групп разработали сконструированные химерные белки с дезаминазами, слитыми с ДНК-связывающими модулями, которые используют для доставки дезаминазы к специфическому геномному локусу. Например, систему CRISPR/Cas можно использовать в качестве системы для доставки, при использовании сконструированного, каталитически неактивного (dCas9) варианта нуклеазы Cas9. Этот способ успешно использовали для нацеливания слитых эффекторных молекул на специфические геномные локусы. Применения включают нацеливание флуоресцентных белков на специфические локусы или доставку трансактиваторов или репрессоров транскрипции к специфическим геномным локусам для контроля специфической экспрессии гена (Wang, Ann Rev Biochem, 2016). Слияние цитидин-дезаминазы или AID с никазой Cas9 и дополнительная модификация для улучшения эффективности редактирования оснований позволяет прямой направленный перевод оснований (Komor, Nature 2016; Nishida, Science 2016). В конкретных условиях, они могут иметь преимущество над способом HDR, поскольку редактирование оснований ни индуцирует разрыв дцДНК, ни требует доставки ДНК-матрицы для HDR. Таким образом, этот альтернативный способ может приводить к меньшему количеству инделов и таким образом, может представлять собой безопасную или даже более безопасную альтернативу основанной на HDR геномной инженерии. Кроме того, доставка редактора оснований в форме мРНК или RNP может являться достаточной и может являться менее токсичной для конкретных типов клеток. Для T-клеток человека, RNP с Cas9 предоставляют успешный способ редактирования генома (Schumann, PNAS, 2015), таким образом, можно предполагать, что конвертеры оснований в форме RNP могут являться особенно хорошо подходящими для гематопоэтических клеток, включая гематопоэтические стволовые клетки (HSC) и T-клетки, однако, в принципе, перевод оснований может являться подходящим способом аллельной инженерии, применимым для любой клетки, включая клетки млекопитающих. Введенную сконструированную точечную мутацию можно затем использовать для нижестоящих применений, таких как маркировка клеток, отслеживание клеток и избирательная абляция. Редакторы оснований, доставленные в форме RNP, успешно редактировали нуклеотиды-мишени в клетках млекопитающих, эмбрионах мыши и данио рерио, и внутреннем ухе живых мышей (Rees, Nat Comm, 2017; Kim, Nat Biotech, 2017 doi:10.1038/nbt.3816). Таким образом, эти исследования показывают осуществимость специфического, не требующего дополнительной ДНК редактирования оснований. Однако, количество нуклеотидов в геноме (человека), поддающихся переводу оснований, более ограничено, чем для способа HDR, поскольку оно не только зависит от последовательности PAM (для конвертеров оснований на основе CRISPR/Cas), но подлежит дополнительным ограничениям. Несмотря на то, что цитидин-дезаминазные редакторы оснований могут переводить только C в T (или G в A), новые сконструированные редакторы оснований аденина могут переводить A в G (или T в C (Gaudelli, Nature 2017)). Однако, в дополнение к требованиям PAM, перевод оснований происходит только внутри определенного окна, определяемого специфическим дизайном и/или конструированием слитого белка. Таким образом, по сравнению с опосредованным HDR введением точечной мутации, количество поддающихся редактированию нуклеотидов является намного более ограниченным при использовании редакторов оснований (Komor, Nature 2016). Кроме того, окно перевода оснований включает несколько нуклеотидов. Например, для так называемого редактора оснований 3 (BE3), в котором используют Cas9 S. pyogenes (SpBE3, см. Komor et al., Nature 2016), это окно включает приблизительно 5 нуклеотидов. Таким образом, редактирование одиночного C до T внутри отрезка из множества смежных нуклеотидов C невозможно и может приводить к дополнительным нежелательным мутациям, которые могут изменять аминокислоту в полученном белке. Сходные окна редактирования применимы к редакторам оснований аденина. Кроме того, различные BE3, основанные на Cas9 S. aureus, SaBE3, могут приводить к поддающемуся детекции редактированию оснований в C-мишенях вне канонического окна редактирования BE3. Для преодоления этих ограничений, сконструированы более новые варианты конвертера оснований для получения более узких окон редактирования и специфичностей PAM отличных от NGG (например, NGA, NGAG, NGCG, NNGRRT и NNNRRT), однако, дизайн специфического редактора оснований для перевода одного данного нуклеотида остается проблематичным (Kim, Nat Biotech 2017, doi:10.1038/nbt.3803). Несмотря на то, что новые конвертеры оснований расширили диапазон поддающихся нацеливанию нуклеотидов в геноме млекопитающих, не каждый нуклеотид можно редактировать по желанию. Тем не менее, чтобы получить преимущество от некоторых благоприятных свойств конвертеров оснований, авторы настоящего изобретения предприняли попытку разработки редактора оснований для аллельной инженерии CD90.1 до CD90.2, CD90.2 до CD90.1, CD45.1 до CD45.2 или CD45.2 до CD45.1. Однако, с использованием исходных рестрицированных по NGG редакторов оснований, ни один из этих переводов невозможно осуществить с использованием доступных редакторов оснований. И это несмотря на тот факт, что для обоих генов, CD90 и CD45, аллельное отличие кодировано заменой G на A, которая, в принципе, должна поддаваться переводу посредством дезаминазы. Единственным решением было использование варианта BE3 на основе SaCas9, содержащего 3 мутации, ослабляющие требования этого варианта Cas9 к PAM до NNNRRT (Kleinstiver, Nature 2015; Kim, Nat Biotech, 2016 (doi:10.1038/nbt.3803). Выбор этого редактора оснований обеспечил возможность разработки редактора оснований с использованием совпадающей онРНК, который может переводить аллель CD45.1 в аллель CD45.2 (фиг. 9A). Однако этот редактор оснований может действовать только для перевода CD45.1 в CD45.2, но не перевода CD45.2 в CD45.1, авторы настоящего изобретения разработали гипотетические редакторы оснований аденина (слитые белки редакторов оснований аденина с Cas9 SaKKH или Cas9 VQR (фиг. 9B). В отличие от этого, не существовало возможных переводов аллеля CD90, даже с использованием существующего варианта редактора оснований.
Способы
Подробные способы, описывающие редактирование генов в первичных мышиных CD4+ T-клетках, редактирование генов в клетках EL-4, и способ репарации для Foxp3, можно обнаружить в EP16196860.7, EP16196858.1 и PCT/EP2017/059799.
Выделение T-клеток человека и антитела
Первичные T-клетки человека выделяли из лейкоцитарных пленок (Blutspendezentrum, Basel) от здоровых доноров с использованием градиента плотности Lymphoprep™ (Stemcell Technologies). Наивные CD4+ T-клетки предварительно обогащали с использованием набора для обогащения наивных CD4+ T-клеток человека Easysep Human naïve CD4+ T-cell enrichment kit (Stemcell Technologies) в соответствии с протоколом производителя. Альтернативно, пуповинную кровь использовали в качестве источника PBMC, без использования стадии выделения наивных T-клеток, при условии высокой частоты наивных T-клеток. Образцы до и после обогащения наивных CD4+ T-клеток окрашивали с использованием следующих антител для оценки чистоты: αCD4-FITC (OKT-4), αCD25-APC (BC96), αCD45RA-BV711 (HI100), αCD45RO-BV450 (UCHL1), αCD62L-BV605 (DREG-56), αCD3-PerCP (HIT3a) и красителя для оценки жизнеспособности Zombie-UV, все закуплены из Biolegend.
Кратко, для 1 лейкоцитарной пленки 50 мл: подготавливают 2×50 мл пробирки Falcon с фильтром и добавляют 16 мл Lymphoprep в каждую пробирку, центрифугируют при 300 g в течение 1 мин. Распределяют кровь поровну в обе 50 мл пробирки с фильтром дополняют PBS до 50 мл. Центрифугируют при 2000 об/мин (ускорение 4, замедление 1) в течение 15 мин. Удаляют некоторое количество сыворотки и отбрасывают. Осторожно объединяют белые лейкоцитарные пленки в чистой 50 мл пробирке Falcon. Добавляют стерильный PBS к обогащенной фракции PBMC до приблизительно 50 мл и центрифугируют при 300 g в течение 5 мин. Отбрасывают супернатант и ресуспендируют осадок с использованием 10 мл PBS, и дополняют до 50 мл, и центрифугируют при 300 g в течение 5 мин. Лизируют эритроциты, при необходимости, с использованием буфера для лизиса эритроцитов, перед стадией очистки.
Способ трансфекции T-клеток человека
Наивные CD4+ T-клетки или тотальные PBMC из крови или пуповинной крови использовали для трансфекции. Для активации T-клеток, 2×106 клеток рассевали в 24-луночный планшет (Corning), покрытый моноклональными антителами (mAb) против CD3 (клон гибридомы OKT3, 5 (высокая концентрация), 2,5 (средняя концентрация), 1 (низкая концентрация) мкг/мл) и против CD28 (клон гибридомы CD28. 2,5 (высокая концентрация), 1 (средняя концентрация), 0,5 (низкая концентрация) мкг/мл, оба из Biolegend), в течение 24 час при 37°C с 5% CO2 в присутствии 50 МЕ/мл рекомбинантного человеческого интерлейкина-2 (rhIL-2) (RD systems). Через 24 час T-клетки собирали и промывали с использованием PBS. 2×106 активированных T-клеток подвергали электропорации с использованием системы для трансфекции Amaxa, программы T-020 (для плазмид), или с использованием системы для трансфекции Neon® (ThermoFisher) в следующих условиях: напряжение (1600 В), ширина (10 мс), импульсы (3), наконечник 100 мкл, буфер R (для RNP). Клетки трансфицировали с использованием 6,5 мкг пустой плазмиды px458 (Addgene, плазмида номер: 48138) или комплекса crРНК:tracerRNA-Atto 550 (IDT) и Cas9 (Berkeley). После электропорации клетки рассевали в 24-луночный планшет в 650 мкл полной среды с 50 МЕ rhIL-2/мл в присутствии связанных с планшетом mAb в половине концентраций, использованных для исходной активации, т.е. против CD3 (2,5, 1,25, 0,5 мкг/мл) и против CD28 (1,25, 0,5, 0,25 мкг/мл). Экспрессию в GFP+ или Atto550+ клетках оценивали через 24 час с использованием анализатора Fortessa (BD Biosciences).
Эксперимент истощения CD45.2
CD4+ T-клетки выделяли от мышей C57BL6 (CD45.2) и конгенных C57BL6 (CD45.1) мышей с использованием набора для выделения CD4+ T-клеток мыши EasySep Mouse CD4+ T Cell Isolation Kit (Stem cell Technologies). Мышей RAG KO подвергали восстановлению с использованием соотношения 1:1 10×106 CD45.2 и CD45.1 CD4+ T-клеток донора. На те же сутки, что и переносу T-клеток, мышей также подвергали внутрибрюшинным инъекциям PBS (группа без обработки) или истощающего Ab против CD4 (клон GK1.5, 250 мкг) в течение 3 последовательных суток. Иммунотоксины CD45.2-ZAP получали посредством комбинации биотинилированного антитела CD45.2 (MW 160 кДа, Biolegend) с конъюгатом стрептавидин-SAP (2,8 молекул сапорина на стрептавидин, MW 135 кДа, Advanced Targeting Systems) в молярном соотношении 1:1, и затем разводили в PBS непосредственно перед использованием, таким же образом, как описано в исходной публикации: (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5179034/). Введение in vivo иммунотоксина или контроля с неконъюгированным антителом CD45.2 проводили посредством внутривенных инъекций. Через одну неделю, кровь, периферические лимфатические узлы (LN), мезентериальные LN (мезLN) и селезенки (SP) собирали, и клетки окрашивали с использованием следующих конъюгированных с флуорохромом mAb: против CD45.2 (104), против CD45.1 (A20), против CD4 (RM4-5), против CD3 (145-2C11), все из Biolegend. Образцы получали в BD Fortessa (BD Biosciences) и анализировали с использованием программного обеспечения FlowJo (Tree Star).
Пункты
1. Способ определения первого события гомологичной репарации (HDR) в первой геномной локализации в эукариотической клетке, где
- указанная клетка экспрессирует первую изоформу первого поверхностного белка, которая отличается от второй изоформы указанного первого поверхностного белка в отношении аминокислотного маркера, где указанная первая изоформа содержит аминокислотный маркер A, кодируемый последовательностью нуклеиновой кислоты A, и указанная вторая изоформа содержит аминокислотный маркер B, кодируемый последовательностью нуклеиновой кислоты B;
- указанная первая геномная локализация содержит указанную последовательность нуклеиновой кислоты A;
и указанный способ включает стадии
a. индукции первого двухцепочечного разрыва ДНК в указанной первой геномной локализации;
b. получения конструкции для репарации ДНК, содержащей указанную последовательность нуклеиновой кислоты B и первую пару гомологичных плеч, которые являются гомологичными последовательностям ДНК на 5' и 3' от указанной первой геномной локализации;
c. определения экспрессии указанной первой и/или указанной второй изоформы указанного первого поверхностного белка на указанной клетке и необязательно, очистки указанной клетки на основании экспрессии указанной первой и/или указанной второй изоформы указанного поверхностного белка; и
d. определения возникновения указанного первого события HDR, где экспрессия указанной второй изоформы указанного первого поверхностного белка на указанной клетке является эквивалентной возникновению указанного первого события HDR.
2. Способ по пункту 1, где возникновение указанного первого события HDR определяют по меньшей мере в двух различных экспериментальных условиях, и увеличенное соотношение экспрессии указанной второй изоформы к указанной первой изоформе в первых экспериментальных условиях по сравнению с вторыми экспериментальными условиями, показывает увеличенную эффективность HDR в указанных первых экспериментальных условиях.
3. Способ по любому из вышеуказанных пунктов, где стадии a и b проводят в культуральной среде, содержащей ванилин и/или рукапариб, в частности, в концентрации 50 мкМ - 500 мкМ ванилина и/или 0,5 мкМ - 2,5 мкМ рукапариба, более конкретно, приблизительно 300 мкМ ванилина и/или приблизительно 1 мкМ рукапариба.
4. Способ по любому из вышеуказанных пунктов, где указанная первая и указанная вторая изоформы указанного первого поверхностного белка можно отличать друг от друга посредством лиганда, в частности, антитела, где указанный лиганд является способным к дискриминирующему связыванию с указанным аминокислотным маркером A (и не с B) или с указанным аминокислотным маркером B (и не с A), соответственно.
5. Способ по любому из вышеуказанных пунктов, где указанный первый поверхностный белок представляет собой нативный белок.
6. Способ по любому из вышеуказанных пунктов, где указанный первый поверхностный белок представляет собой трансгенный белок.
7. Способ по любому из вышеуказанных пунктов, где указанную очистку осуществляют посредством активируемой флуоресценцией сортировки клеток (FACS).
8. Способ по любому из пунктов 1-6, где указанная очистка включает обогащение на основе магнитных бусин клетки, экспрессирующей указанную первую или указанную вторую изоформу указанного первого поверхностного белка.
9. Способ по любому из вышеуказанных пунктов, где указанный первый поверхностный белок представляет собой Thy1 или CD45.
10. Способ по любому из вышеуказанных пунктов, где указанный первый двухцепочечный разрыв индуцируют в указанной первой геномной локализации посредством трансфекции указанной клетки с использованием экспрессирующей конструкции ДНК, кодирующей ассоциированную с CRISPR эндонуклеазу (Cas9), и направляющей РНК, где указанная направляющая РНК является способной к гибридизации в указанной первой геномной локализации.
11. Способ по любому из вышеуказанных пунктов, где указанные гомологичные плечи содержат приблизительно 2000 пар оснований (п.о.) каждое.
12. Способ избирательного истощения или обогащения отредактированной клетки в композиции из нередактированных и редактированных клеток, где
a. указанные нередактированные клетки экспрессируют первую изоформу поверхностного белка, и указанные отредактированные клетки отредактированы способом по любому из пунктов 1-11 для экспрессии второй изоформы указанного поверхностного белка, которая отличается от указанной первой изоформы в отношении аминокислотного маркера, где указанная первая изоформа содержит аминокислотный маркер A, кодируемый последовательностью нуклеиновой кислоты A, и указанная вторая изоформа содержит аминокислотный маркер B, кодируемый последовательностью нуклеиновой кислоты B; и
b. указанная отредактированная клетка является избирательно обогащенной или истощенной на основании экспрессии указанной первой или указанной второй изоформы указанного поверхностного белка.
13. Способ избирательного истощения или обогащения клетки в композиции клеток,
включающий стадии
c. получения клетки, где указанная клетка экспрессирует первую изоформу поверхностного белка, которая отличается от второй изоформы указанного поверхностного белка в отношении аминокислотного маркера, где указанная первая изоформа содержит аминокислотный маркер A, кодируемый последовательностью нуклеиновой кислоты A, и указанная вторая изоформа содержит аминокислотный маркер B, кодируемый последовательностью нуклеиновой кислоты B;
d. индукции двухцепочечного разрыва ДНК в геномной локализации, содержащей указанную последовательность нуклеиновой кислоты A;
e. получения конструкции для репарации ДНК, содержащей указанную последовательность нуклеиновой кислоты B и пару гомологичных плеч, которые являются гомологичными последовательностям ДНК на 5' и 3' от указанной геномной локализации;
f. избирательного обогащения/истощения указанной клетки на основании экспрессии указанной первой или указанной второй изоформы указанного поверхностного белка.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Univфersitat Basel
<120> Иммунологически отличимые варианты поверхности клеток для применения
в клеточной терапии
<130> uz331
<160> 32
<170> PatentIn версии 3.5
<210> 1
<211> 20
<212> ДНК
<213> Mus musculus
<400> 1
gttttgtgag cttcaagtct 20
<210> 2
<211> 20
<212> ДНК
<213> Mus musculus
<400> 2
gtttgtgagc ttcgagtctc 20
<210> 3
<211> 20
<212> ДНК
<213> Mus musculus
<400> 3
ggctaatact tcaatttgtt 20
<210> 4
<211> 20
<212> ДНК
<213> Mus musculus
<400> 4
ttggagtgga aaacagaaaa 20
<210> 5
<211> 20
<212> ДНК
<213> Mus musculus
<400> 5
aacaaaaaac cttgatttca 20
<210> 6
<211> 20
<212> ДНК
<213> Mus musculus
<400> 6
aaacaaaaaa ccttgatttc 20
<210> 7
<211> 180
<212> ДНК
<213> Mus musculus
<400> 7
cgtcaccctc tccaaccagc cctatatcaa ggtccttacc ctagccaact tcaccaccaa 60
ggatgagggc gactactttt gtgagcttcg agtctcgggc gcgaatccca tgagctccaa 120
taaaagtatc agtgtgtata gaggtgagac tggttcccag aaagataaaa tgtccaggtt 180
<210> 8
<211> 180
<212> ДНК
<213> Mus musculus
<400> 8
cgtcaccctc tccaaccagc cctatatcaa ggtccttacc ctagccaact tcaccaccaa 60
ggatgagggc gactactttt gtgagcttcg agtctcaggc gcgaatccca tgagctccaa 120
taaaagtatc agtgtgtata gaggtgagac tggttcccag aaagataaaa tgtccaggtt 180
<210> 9
<211> 179
<212> ДНК
<213> Mus musculus
<400> 9
cgtcaccctc tccaaccagc cctatatcaa ggtccttacc ctagccaact tcaccaccaa 60
ggatgagggc gactactttt gtgagcttcg agtaagcgga gcgaatccca tgagctccaa 120
taaaagtatc agtgtgtata gaggtgagac tggttcccag aaagataaaa tgtccaggt 179
<210> 10
<211> 2000
<212> ДНК
<213> Mus musculus
<400> 10
aaatggggac ccacctgtcc taccagctgg ctgacctgta gctttcccca ccacagaatc 60
caagtcggaa ctcttggcac catgaaccca gccatcagcg tcgctctcct gctctcaggt 120
actgggcaag ggtcagggct ggcattctaa ggaatctggc ttcctcccat cccgggaagt 180
agcctctttg ccatagtctc aggggcacag gtggttggga ggtgcggggg tggggagtgg 240
ggaggagcct caacctcacc agtggtggtc tttgacatat tagaaactcc ataatggatc 300
taggaactcc tcttgctggg tggtggtggt tgtggtacac acctttaatc tcagcactca 360
ggaggcagag tcaggtggat ctgttagtct gaagccagcc tggtctacag agcaaattcc 420
aggacagcca gagctattct caagatagag aatccctttc ttgaaaaaac catttaaaaa 480
caaaaacaaa agcaacacac tcctcttgat ctcctgttct tgaaacacat tgttgggacc 540
cagaacttca gtagattgat ggaagttgga gtctgcaagt ggtggaacat cccaccaata 600
cctcaagggc gagtgcaaac cccacatccc cccagctcaa gtctactttt cctgcaggtg 660
ggaggccccg ggtctgtgtc tccccaaatt cagagaaggc actgctgtgc cagtcttgca 720
ggtgtcccga gggcagaagg tgaccagcct gacagcctgc ctggtgaacc aaaaccttcg 780
cctggactgc cgccatgaga ataacaccaa ggataactcc atccagcatg agttcagcct 840
gacccgagag aagaggaagc acgtgctctc aggcaccctt gggatacccg agcacacgta 900
ccgctcccgc gtcaccctct ccaaccagcc ctatatcaag gtccttaccc tagccaactt 960
caccaccaag gatgagggcg actacttttg tgagcttcga gtctcaggcg cgaatcccat 1020
gagctccaat aaaagtatca gtgtgtatag aggtgagact ggttcccaga aagataaaat 1080
gtccaggtta gccaggctgg ggtagccaat aaaaaaaaaa aaaaaaaaaa aaaaaaaaac 1140
aggcacctcc attacccttc ccctaactgc tggtctcctg ggaaactgct gctgtctatg 1200
tgagtggggc aagattaggg gccagaaagg gggagcttgt agtaaaagca cagttgagga 1260
aactaaatgg gaaaggcagt acagtggtga ttcttgtggt gtggaggttc tgttacagca 1320
tccggtggag ccgctaagat gagaaagcgc cagctagctg ccttgaacag ctgacacctg 1380
tctttgcccg cctgagtcct gatctcccct cctcccggca ccccttctct atccacagac 1440
aagctggtca agtgtggcgg cataagcctg ctggttcaga acacatcctg gatgctgctg 1500
ctgctgcttt ccctctccct cctccaagcc ctggacttca tttctctgtg actggttggg 1560
cccaaggaga aacaggagcc ctcgaggtcc ttcctctgca gaggtcttgc ttctcccggt 1620
cagctgactc cctccccaag tccttcaaat atctcagaac atggggagaa acggggacct 1680
tgtccctcct aaggaacccc agtgctgcat gccatcatcc cccccaccct cgcccccacc 1740
cccgccactt ctccctccat gcataccact agctgtcatt ttgtactctg tatttattcc 1800
agggctgctt ctgattattt agtttgttct ttccctggag acctgttaga acataagggc 1860
gtatggtggg taggggaggc aggatatcag tccctggggc gagttcctcc ctgccaacca 1920
agccagatgc ctgaaagaga tatggatgag ggaagttgga ctgtgcctgt acctggtaca 1980
gtcatactct gttgaaagaa 2000
<210> 11
<211> 3679
<212> ДНК
<213> Mus musculus
<400> 11
accccatcaa aatcctattt gagtgtacgg ttcggagaac ctcatttatc cggtaaatgt 60
cttttactct gctctcaggg agctgaggca ggacatcctg agatacattg ggagaggaga 120
tacagtttca ataaaataat aggttgggtg gaggtacatg cctataatgc caccactcag 180
gaaatggtgg cagcttcgtg agtttgaggc caacccaaga aacatagtga aaccctgtca 240
gtaaataagt aagcaagtat ttgagtatct actatatgct agggctgacc tggacattag 300
gggtcatctt ctgaacaaac tagtgcttga gggaggtatt tggggttttt gtttgtttaa 360
tggatctgaa tgagttccag agactggcta cacagcgata tgactgagct taacacccct 420
aaagcataca gtcagaccaa ttagacaata aaaggtatgt atagcttacc aaataaaaaa 480
attgtatttt caagagagtg tctgtctgtg tagccctggc tgttcttgaa ctcactctgt 540
agaccaggct ggcctggaaa tccatctgcc tgcctctgcc tctctgcctc tctgcctctc 600
tgcctctctc tctgcctctc tctgcctctc tctgcccctc tctgcccctc tctgcccctc 660
tctgcccctc tctgccgccc tctgccttct gccctctgcc ctctggcctc tggcctctgc 720
cctctgccct ctggcctctg gcctctgcct ctgcctcttg agtgctggaa tcaaaggtgt 780
gagctctgta ggtcttaagt tccagaagaa agtaatgaag tcacccagca gggaggtgct 840
cagggacagc acagacacac acccaggaca taggctccca cttccttggc tttctctgag 900
tggcaaagga ccttaggcag tgtcactccc taagagaagg ggataaagag aggggctgag 960
gtattcatca tgtgctccgt ggatctcaag ccctcaaggt aaatggggac ccacctgtcc 1020
taccagctgg ctgacctgta gctttcccca ccacagaatc caagtcggaa ctcttggcac 1080
catgaaccca gccatcagcg tcgctctcct gctctcaggt actgggcaag ggtcagggct 1140
ggcattctaa ggaatctggc ttcctcccat cccgggaagt agcctctttg ccatagtctc 1200
aggggcacag gtggttggga ggtgcggggg tggggagtgg ggaggagcct caacctcacc 1260
agtggtggtc tttgacatat tagaaactcc ataatggatc taggaactcc tcttgctggg 1320
tggtggtggt tgtggtacac acctttaatc tcagcactca ggaggcagag tcaggtggat 1380
ctgttagtct gaagccagcc tggtctacag agcaaattcc aggacagcca gagctattct 1440
caagatagag aatccctttc ttgaaaaaac catttaaaaa caaaaacaaa agcaacacac 1500
tcctcttgat ctcctgttct tgaaacacat tgttgggacc cagaacttca gtagattgat 1560
ggaagttgga gtctgcaagt ggtggaacat cccaccaata cctcaagggc gagtgcaaac 1620
cccacatccc cccagctcaa gtctactttt cctgcaggtg ggaggccccg ggtctgtgtc 1680
tccccaaatt cagagaaggc actgctgtgc cagtcttgca ggtgtcccga gggcagaagg 1740
tgaccagcct gacagcctgc ctggtgaacc aaaaccttcg cctggactgc cgccatgaga 1800
ataacaccaa ggataactcc atccagcatg agttcagcct gacccgagag aagaggaagc 1860
acgtgctctc aggcaccctt gggatacccg agcacacgta ccgctcccgc gtcaccctct 1920
ccaaccagcc ctatatcaag gtccttaccc tagccaactt caccaccaag gatgagggcg 1980
actacttttg tgagcttcga gtctcaggcg cgaatcccat gagctccaat aaaagtatca 2040
gtgtgtatag aggtgagact ggttcccaga aagataaaat gtccaggtta gccaggctgg 2100
ggtagccaat aaaaaaaaaa aaaaaaaaaa aaaaaaaaac aggcacctcc attacccttc 2160
ccctaactgc tggtctcctg ggaaactgct gctgtctatg tgagtggggc aagattaggg 2220
gccagaaagg gggagcttgt agtaaaagca cagttgagga aactaaatgg gaaaggcagt 2280
acagtggtga ttcttgtggt gtggaggttc tgttacagca tccggtggag ccgctaagat 2340
gagaaagcgc cagctagctg ccttgaacag ctgacacctg tctttgcccg cctgagtcct 2400
gatctcccct cctcccggca ccccttctct atccacagac aagctggtca agtgtggcgg 2460
cataagcctg ctggttcaga acacatcctg gatgctgctg ctgctgcttt ccctctccct 2520
cctccaagcc ctggacttca tttctctgtg actggttggg cccaaggaga aacaggagcc 2580
ctcgaggtcc ttcctctgca gaggtcttgc ttctcccggt cagctgactc cctccccaag 2640
tccttcaaat atctcagaac atggggagaa acggggacct tgtccctcct aaggaacccc 2700
agtgctgcat gccatcatcc cccccaccct cgcccccacc cccgccactt ctccctccat 2760
gcataccact agctgtcatt ttgtactctg tatttattcc agggctgctt ctgattattt 2820
agtttgttct ttccctggag acctgttaga acataagggc gtatggtggg taggggaggc 2880
aggatatcag tccctggggc gagttcctcc ctgccaacca agccagatgc ctgaaagaga 2940
tatggatgag ggaagttgga ctgtgcctgt acctggtaca gtcatactct gttgaaagaa 3000
tcatcgggga gggggggggg ggctcaagat gggagagctc tgctgagcct ttgtggacca 3060
tccaatgagg atgagggctt agattctacc aggtcattct cagccaccac acacaagcgc 3120
tctgccatca ctgaagaagc cccctagggc tcttgggcca gggcacactc agtaaagatg 3180
caggttcagt cagggaatga tggggaaagg ggtaggaggt gggggaggga tcaccccctc 3240
ctctaaaaca cgagcctgct gtctccaaag gcctctgcct gtagtgaggg tggcagaaga 3300
agacaaggag ccagaactct gactccagga tctaagtccg tgcaggaagg ggatcctaga 3360
accatccggt tggacccagc ttaccaaggg agagccttta ttcttctttc ccttgcccct 3420
ctgtgccagc ccctcttgct gtccctgatc ccccagacag cgagagtctt gcaacctgcc 3480
tcttccaaga cctcctaatc tcaggggcag gcggtggagt gagatccggc gtgcacactt 3540
tttggaagat agctttccca aggatcctct cccccactgg cagctctgcc tgtcccatca 3600
ccatgtataa taccaccact gctacagcat ctcaccgagg aaagaaaact gcacaataaa 3660
accaagcctc tggagtgtg 3679
<210> 12
<211> 960
<212> ДНК
<213> Mus musculus
<400> 12
cacattgttg ggacccagaa cttcagtaga ttgatggaag ttggagtctg caagtggtgg 60
aacatcccac caatacctca agggcgagtg caaaccccac atccccccag ctcaagtcta 120
cttttcctgc aggtgggagg ccccgggtct gtgtctcccc aaattcagag aaggcactgc 180
tgtgccagtc ttgcaggtgt cccgagggca gaaggtgacc agcctgacag cctgcctggt 240
gaaccaaaac cttcgcctga ctgccgccat gagaataaca ccaaggataa ctccatccag 300
catgagttca gcctgacccg agagaagagg aagcacgtgc tctcaggcac ccttgggata 360
cccgagcaca cgtaccgctc ccgcgtcacc ctctccaacc agccctatat caaggtcctt 420
accctagcca acttcaccac caaggatgag ggcgactact tttgtgagct tcgagtctca 480
ggcgcgaatc ccatgagctc caataaaagt atcagtgtgt atagaggtga gactggttcc 540
cagaaagata aaatgtccag gttagccagg ctggggtagc caataaaaaa aaaaaaaaaa 600
aaaaaaaaaa aaacaggcac ctccattacc cttcccctaa ctgctggtct cctgggaaac 660
tgctgctgtc tatgtgagtg gggcaagatt aggggccaga aagggggagc ttgtagtaaa 720
agcacagttg aggaaactaa atgggaaagg cagtacagtg gtgattcttg tggtgtggag 780
gttctgttac agcatccggt ggagccgcta agatgagaaa gcgccagcta gctgccttga 840
acagctgaca cctgtctttg cccgcctgag tcctgatctc ccctcctccc ggcacccctt 900
ctctatccac agacaagctg gtcaagtgtg gcggcataag cctgctggtt cagaacacat 960
<210> 13
<211> 160
<212> ДНК
<213> Mus musculus
<400> 13
cgtcaccctc tccaaccagc cctatatcaa ggtccttacc ctagccaact tcaccaccaa 60
ggatgagggc gactactttt gtgagcttcg agtctcaggc gcgaatccca tgagctccaa 120
taaaagtatc agtgtgtata gaggtgagac tggttcccag 160
<210> 14
<211> 120
<212> ДНК
<213> Mus musculus
<400> 14
agtttgtctc tatacacact gatactttta ttggagctca tgggattcgc gcccgagact 60
tgaagctcac aaaagtagtc gccctcatcc ttggtggtga agttggctag ggtaaggacc 120
<210> 15
<211> 180
<212> ДНК
<213> Mus musculus
<400> 15
accagcaggc ttatgccgcc acacttgacc agtttgtctc tatacacact gatactttta 60
ttggagctca tgggattcgc gcccgagact tgaagctcac aaaagtagtc gccctcatcc 120
ttggtggtga agttggctag ggtaaggacc ttgatatagg gctggttgga gagggtgacg 180
<210> 16
<211> 180
<212> ДНК
<213> Mus musculus
<400> 16
gtttcctcca cagggactga caagttttcg ctacatgact gcacaccaaa agaaaaggct 60
aatacttcaa tttgtttaga gtggaaaaca gaaaaccttg atttcagaaa atgcaacagt 120
gacaatattt catatgtact ccactgtgag ccaggtacga tgctgggcag agaagttcta 180
<210> 17
<211> 1000
<212> ДНК
<213> Mus musculus
<400> 17
caaatgggaa aaatcagccg tttatcttga tgtttccttg agttgcattt ataaaagcag 60
cacacatttc tactgaaatc tgtggtacga agaaaatatg ttatcttccc taatgaatgg 120
caaatatctg agttgataaa caaactgcca gcaagtggtg tctacacata tagatatatc 180
aaaacactaa taatgtgcac atatttgtat aattgtgcat cagctgaaat ggagataaat 240
taaaaattag gaatgtgact agatcatgaa gaagcatcag accttgggag agcttagatg 300
tcccctagcg aaatctcctg cttgcaagac tgtactccat gagtcttatt tcagagttga 360
gagggttcac atctccacac caggaaccat cacctaagag cacctctccg tttcctccac 420
agggactgac aagttttcgc tacatgactg cacaccaaaa gaaaaggcta atacttcaat 480
ttgtttagag tggaaaacag aaaaccttga tttcagaaaa tgcaacagtg acaatatttc 540
atatgtactc cactgtgagc caggtacgat gctgggcaga gaagttctat tatcagaaat 600
tattccagac gtggcttaaa tgttctttct gtagccttgt cctcctcacc caccctcagt 660
gatccgccat aaattagaat aaaataaccc tagtatctct ggcactgaaa caaattcaca 720
acgtagataa atgaaaagag caacccatgg atgataaata ttatgaaaaa ttaattaaaa 780
gaattttctt gagtactgtt atataccaga cattgttgaa agggtgatga gatgcaggca 840
ttataaaaga aagatggtat ccatagtctg tagaaaggaa gcatagtatt atccacaccg 900
gaggttgctt tctcagactt tcatacatga aaatcattgg aaagttcaag aatctgcagg 960
tagcaaggaa aacaagtgct tctaggaggg aacatgggtt 1000
210> 18
<211> 1995
<212> ДНК
<213> Mus musculus
<400> 18
atggtttaca gagcaagttt caagacagcc caggatacac agagaaaccc tgtctcaaaa 60
caaacaaaac aaaacaaaaa gcaaaaaaca caaaactaaa acaacataac aacaaaccat 120
caaaaagcag ttcctatgga ttagaaacag cctattttgt attttgtgaa ataattatac 180
ataagattgc tgttttttca tttttttctt tcttcttttt ggtatttctt tttaaattca 240
aaatgttctc attagagaaa agaatgtagt taaacatcta atgttttctg gaaaaaaaaa 300
tcaaaggggg aattctagtt atgttaatac caaagttgtg atcacagcac gtggaaggca 360
ggagtacctt atgtgttcaa agccagtcta tgctgtatct tgaattctgt ggcagccttc 420
acagtgtgag attacacctt gtttccaaaa ccccaaacag atcaacaaca agcgtgaaaa 480
cagcaaagaa attgtttttc aaatgggaaa aatcagccgt ttatcttgat gtttccttga 540
gttgcattta taaaagcagc acacatttct actgaaatct gtggtacgaa gaaaatatgt 600
tatcttccct aatgaatggc aaatatctga gttgataaac aaactgccag caagtggtgt 660
ctacacatat agatatatca aaacactaat aatgtgcaca tatttgtata attgtgcatc 720
agctgaaatg gagataaatt aaaaattagg aatgtgacta gatcatgaag aagcatcaga 780
ccttgggaga gcttagatgt cccctagcga aatctcctgc ttgcaagact gtactccatg 840
agtcttattt cagagttgag agggttcaca tctccacacc aggaaccatc acctaagagc 900
acctctccgt ttcctccaca gggactgaca agttttcgct acatgactgc acaccaaaag 960
aaaaggctaa tacttcaatt tgtttagagt ggaaaacaga aaaccttgat ttcagaaaat 1020
gcaacagtga caatatttca tatgtactcc actgtgagcc aggtacgatg ctgggcagag 1080
aagttctatt atcagaaatt attccagacg tggcttaaat gttctttctg tagccttgtc 1140
ctcctcaccc accctcagtg atccgccata aattagaata aaataaccct agtatctctg 1200
gcactgaaac aaattcacaa cgtagataaa tgaaaagagc aacccatgga tgataaatat 1260
tatgaaaaat taattaaaag aattttcttg agtactgtta tataccagac attgttgaaa 1320
gggtgatgag atgcaggcat tataaaagaa agatggtatc catagtctgt agaaaggaag 1380
catagtatta tccacaccgg aggttgcttt ctcagacttt catacatgaa aatcattgga 1440
aagttcaaga atctgcaggt agcaaggaaa acaagtgctt ctaggaggga acatgggttt 1500
ctctagaggt tggatctttt aaaacatata tttgaattca gtgaggtgga ctctttctcc 1560
tctaggcata cagctatgtg ccaggacaca cttgacaaga cagagggagg tacatacacc 1620
cagaggcgga agaggaaatt tcactctgat tgtaaacatt gtgctaacag gaaattgctg 1680
ggctaagata caaataggat gttcacagct ggaacaaata tatgttgagt taacattttt 1740
ttaaaaaatt ttaaataagt cacaatactg atttctcaac aaaattgagt atagcaatgt 1800
tatctgcagg atcttggttt attatttcac cattttcaga gacttgacct taacatgaag 1860
tcttaaacac atggtgccat aaaagaagcc catttactat gtggcatttc atatactcat 1920
gtgcaatact tacttctttc agaaaataat acaaaatgca ttagaagaaa tacattcata 1980
cctgaaagat gtcag 1995
<210> 19
<211> 4000
<212> ДНК
<213> Mus musculus
<400> 19
ttaattcacc aagtattatt gaattgttga taacagacag atatatcata aacccgagtt 60
aatgataatg aacaaaatag actagttatg gttctcatgg acttttcaat ttagttggaa 120
agatagatac caaaccaata attatcatag tataaactgg gatcttttat aggagaaagg 180
atttcaaaat aatgatgtag gggtacatga tacagttctt ctatttttca tattcctgga 240
gatactgtgc tacaacctac aatctgcttt ggtgatattt tggtatatta cctctgctat 300
atgctaatta ttcttttact tctgcttgct gagtactgaa tttgaaaccc aaaatacact 360
tatttcatca gatcattctc tgtacaaaag aatatgttaa gacatttgta tccaatgtag 420
tatcattcat tctcgatgat gattgtacta atttcctttt tttttttttc cagctgccat 480
gtttgggaac attactgtga attacaccta tgaatctagt aatcagactt ttaaggcaga 540
cctcaaagat gtccaaaatg ctaagtgtgg aaatgaggat tgtgaaaacg tgttaaataa 600
tctagaagaa tgctcacaga taaaaaacat cagtgtgtct aatgactcat gtgctccagc 660
tacaactata gatttatatg taccaccagg tgaatgtcaa tttctctcta tctgggtatt 720
tagagcaaca gaaggatttc atgtatgtgt tcctgtgctt cttcaagcta atgtggtcag 780
aaaaaagaac cgagggaaag caagtgagct ttgataagag tgtgtttcct gctgagaacc 840
ccggagagca gagagaaata cacacttatt ccttaggacc accacctggc tctcttattt 900
cttttataaa gagattccca gctgggaagt ggtgtccctc accttcaatc tcagcacttc 960
agaggcagag gcaggcagat ctcagggttc caaggcagcc atggtttaca gagcaagttt 1020
caagacagcc caggatacac agagaaaccc tgtctcaaaa caaacaaaac aaaacaaaaa 1080
gcaaaaaaca caaaactaaa acaacataac aacaaaccat caaaaagcag ttcctatgga 1140
ttagaaacag cctattttgt attttgtgaa ataattatac ataagattgc tgttttttca 1200
tttttttctt tcttcttttt ggtatttctt tttaaattca aaatgttctc attagagaaa 1260
agaatgtagt taaacatcta atgttttctg gaaaaaaaaa tcaaaggggg aattctagtt 1320
atgttaatac caaagttgtg atcacagcac gtggaaggca ggagtacctt atgtgttcaa 1380
agccagtcta tgctgtatct tgaattctgt ggcagccttc acagtgtgag attacacctt 1440
gtttccaaaa ccccaaacag atcaacaaca agcgtgaaaa cagcaaagaa attgtttttc 1500
aaatgggaaa aatcagccgt ttatcttgat gtttccttga gttgcattta taaaagcagc 1560
acacatttct actgaaatct gtggtacgaa gaaaatatgt tatcttccct aatgaatggc 1620
aaatatctga gttgataaac aaactgccag caagtggtgt ctacacatat agatatatca 1680
aaacactaat aatgtgcaca tatttgtata attgtgcatc agctgaaatg gagataaatt 1740
aaaaattagg aatgtgacta gatcatgaag aagcatcaga ccttgggaga gcttagatgt 1800
cccctagcga aatctcctgc ttgcaagact gtactccatg agtcttattt cagagttgag 1860
agggttcaca tctccacacc aggaaccatc acctaagagc acctctccgt ttcctccaca 1920
gggactgaca agttttcgct acatgactgc acaccaaaag aaaaggctaa tacttcaatt 1980
tgtttagagt ggaaaacaga aaaccttgat ttcagaaaat gcaacagtga caatatttca 2040
tatgtactcc actgtgagcc aggtacgatg ctgggcagag aagttctatt atcagaaatt 2100
attccagacg tggcttaaat gttctttctg tagccttgtc ctcctcaccc accctcagtg 2160
atccgccata aattagaata aaataaccct agtatctctg gcactgaaac aaattcacaa 2220
cgtagataaa tgaaaagagc aacccatgga tgataaatat tatgaaaaat taattaaaag 2280
aattttcttg agtactgtta tataccagac attgttgaaa gggtgatgag atgcaggcat 2340
tataaaagaa agatggtatc catagtctgt agaaaggaag catagtatta tccacaccgg 2400
aggttgcttt ctcagacttt catacatgaa aatcattgga aagttcaaga atctgcaggt 2460
agcaaggaaa acaagtgctt ctaggaggga acatgggttt ctctagaggt tggatctttt 2520
aaaacatata tttgaattca gtgaggtgga ctctttctcc tctaggcata cagctatgtg 2580
ccaggacaca cttgacaaga cagagggagg tacatacacc cagaggcgga agaggaaatt 2640
tcactctgat tgtaaacatt gtgctaacag gaaattgctg ggctaagata caaataggat 2700
gttcacagct ggaacaaata tatgttgagt taacattttt ttaaaaaatt ttaaataagt 2760
cacaatactg atttctcaac aaaattgagt atagcaatgt tatctgcagg atcttggttt 2820
attatttcac cattttcaga gacttgacct taacatgaag tcttaaacac atggtgccat 2880
aaaagaagcc catttactat gtggcatttc atatactcat gtgcaatact tacttctttc 2940
agaaaataat acaaaatgca ttagaagaaa tacattcata cctgaaagat gtcagttgga 3000
caaccttcgt gcccaaacaa attacacatg tgtagcagaa atcttatatc gcggtgtaaa 3060
actcgtcaaa aatgttataa atgtgcagac agatttgggg agtaagtata tcgtttatgt 3120
ttataaaata aataaaattt ctttttcttt tatgggttta cttttatttt tggagagtca 3180
tagaaaataa tggtatagaa ttacagaaag catatcagtg aagaaaaaag tttaaatgca 3240
ctttaaggtc aattttaaaa actaggttat acaaacctaa ctaaagttct ttttctgttg 3300
ttcaaattgt gtctgccgta agtctggtag aaacacttga aatggtttac atagattatt 3360
cccattttat tcattttctc ctcccttctt tttataacac catttgttct gtattctaac 3420
acttctttca caattatgct ctgcttctaa tagaaataaa ataagcaaag aaactaataa 3480
aactcaagca aatctacatg tttaaacata aaatacggag tatagttatc aaattttggc 3540
aatagaatta ataaccggat aaaccttatt aaatagaaag gggttctact tgggaagtca 3600
gatgtgaaat tttcaaatta aatgtaacat ttgatggtaa atgctaattt tttaaaatgg 3660
tgattaatga ataatcaacc ataaagaaga caaaagatgt cattgctcac aagcagtaaa 3720
gacattttta gaaggaagag aagagaagca gtaggaaagg aggaagggag agatagatga 3780
tagatagata atagatagat agatagatag atagatagat agatagagac agacagacag 3840
acagaaagaa agagagagac agaaacagag agacacaaaa aaagaggaaa gagaagactg 3900
ggagaaaggg aaagtaaggg atggagaaaa aagggagaga gataaaaaga agacaaaaac 3960
ataaatgaaa gcaaggaatg atgatatata tctgttatgt 4000
<210> 20
<211> 57
<212> ДНК
<213> Mus musculus
<400> 20
ttttgtgagc ttcgagtctc ggtcgcgaat cccatgagct ccaataaaag tatcagt 57
<210> 21
<211> 57
<212> ДНК
<213> Mus musculus
<400> 21
ttttgtgagc ttcaagtctc ggtcgcgaat cccatgagct ccaataaaag tatcagt 57
<210> 22
<211> 63
<212> ДНК
<213> Mus musculus
<400> 22
aaaaggctaa tacttcaatt tgtttggagt ggaaaacaaa aaaccttgat ttcagaaaat 60
gca 63
<210> 23
<211> 63
<212> ДНК
<213> Mus musculus
<400> 23
aaaaggctaa tacttcaatt tgtttagagt ggaaaacaga aaaccttgat ttcagaaaat 60
gca 63
<210> 24
<211> 48
<212> ДНК
<213> Mus musculus
<400> 24
ggctaatact tcaatttgtt tggagtggaa aacagaaaac cttgattt 48
<210> 25
<211> 48
<212> ДНК
<213> Mus musculus
<400> 25
ggctaatact tcaatttgtt tggagtggaa aacaaaaaac cttgattt 48
<210> 26
<211> 64
<212> ДНК
<213> Mus musculus
<400> 26
ggctaatact tcaatttgtt tggagtggaa aacaaaaaac cttgatttca gaaaatgcaa 60
cagt 64
<210> 27
<211> 64
<212> ДНК
<213> Mus musculus
<400> 27
ggctaatact tcaatttgtt tagagtggaa aacagaaaac cttgatttca gaaaatgcaa 60
cagt 64
<210> 28
<211> 63
<212> ДНК
<213> Mus musculus
<400> 28
gcctcaatgg acaagagctc ttgctgcatc gtagccacca gtactcaggg cagtgtgctc 60
ccg 63
<210> 29
<211> 63
<212> ДНК
<213> Mus musculus
<400> 29
gcctcaatgg actagatatc ttgctgcatc gtagccacca gtactcaggg cagtgtgctc 60
ccg 63
<210> 30
<211> 63
<212> ДНК
<213> Mus musculus
<400> 30
gcctcaatgg acaaaagagc tcttgctgca tcgtagccac cagtactcag ggcagtgtgc 60
tcc 63
<210> 31
<211> 34
<212> ДНК
<213> Mus musculus
<400> 31
gcctcaatgg acaagagctc ttgctgcatc gtag 34
<210> 32
<211> 34
<212> ДНК
<213> Mus musculus
<400> 32
gcctcaatgg actagatatc ttgctgcatc gtag 34
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУННЫЕ КЛЕТКИ, ДЕФЕКТНЫЕ ПО Suv39h1 | 2018 |
|
RU2784531C2 |
| ИНСЕРЦИЯ ТАРГЕТНОГО ГЕНА ДЛЯ УЛУЧШЕННОЙ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2017 |
|
RU2824204C2 |
| СПОСОБЫ, КОМПОЗИЦИИ И КОМПОНЕНТЫ ДЛЯ РЕДАКТИРОВАНИЯ TGFBR2 ПОСРЕДСТВОМ CRISPR-CAS9 В T-КЛЕТКАХ ДЛЯ ИММУНОТЕРАПИИ | 2018 |
|
RU2798380C2 |
| НАЦЕЛИВАНИЕ НА SRC-3 В ИММУННЫХ КЛЕТКАХ В КАЧЕСТВЕ ИММУНОМОДУЛИРУЮЩЕГО ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ РАКА | 2021 |
|
RU2828447C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ИНГИБИРОВАНИЯ ИСТОЩЕНИЯ Т-КЛЕТОК | 2018 |
|
RU2804282C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ | 2019 |
|
RU2835579C2 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| УСЛОВНО АКТИВНЫЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ МОДИФИЦИРОВАННЫХ Т-КЛЕТОК | 2015 |
|
RU2764074C2 |
| УСЛОВНО АКТИВНЫЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ МОДИФИЦИРОВАННЫХ Т-КЛЕТОК | 2016 |
|
RU2759957C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
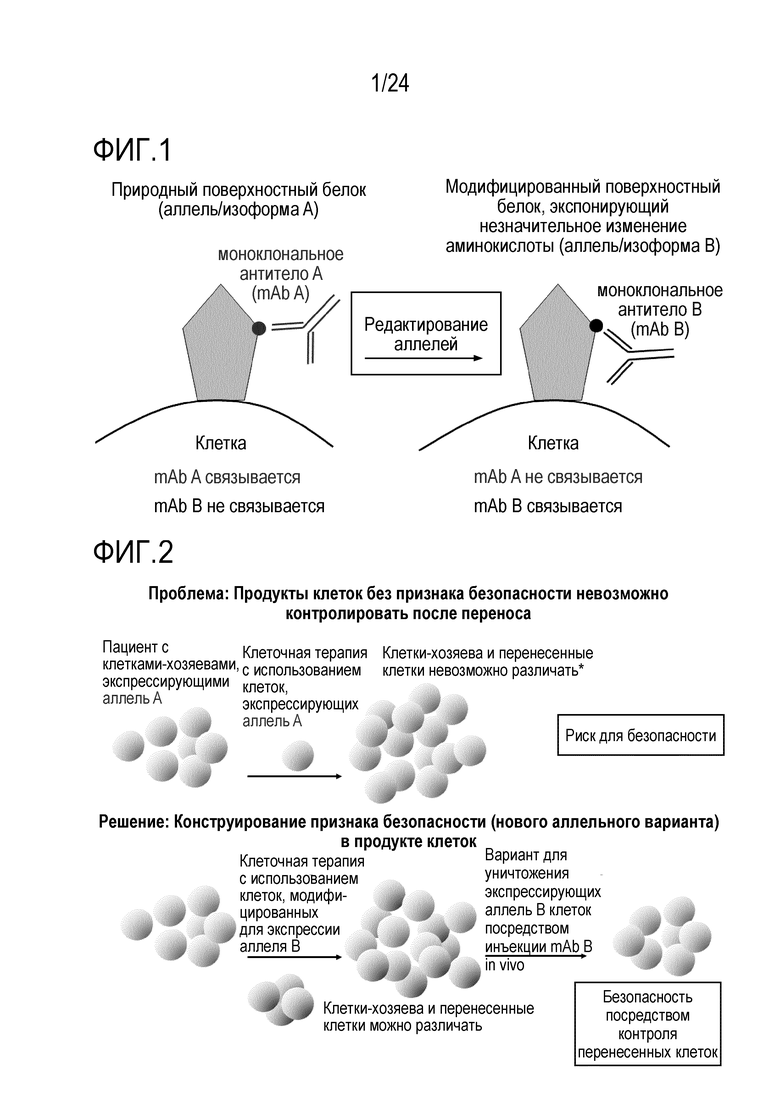
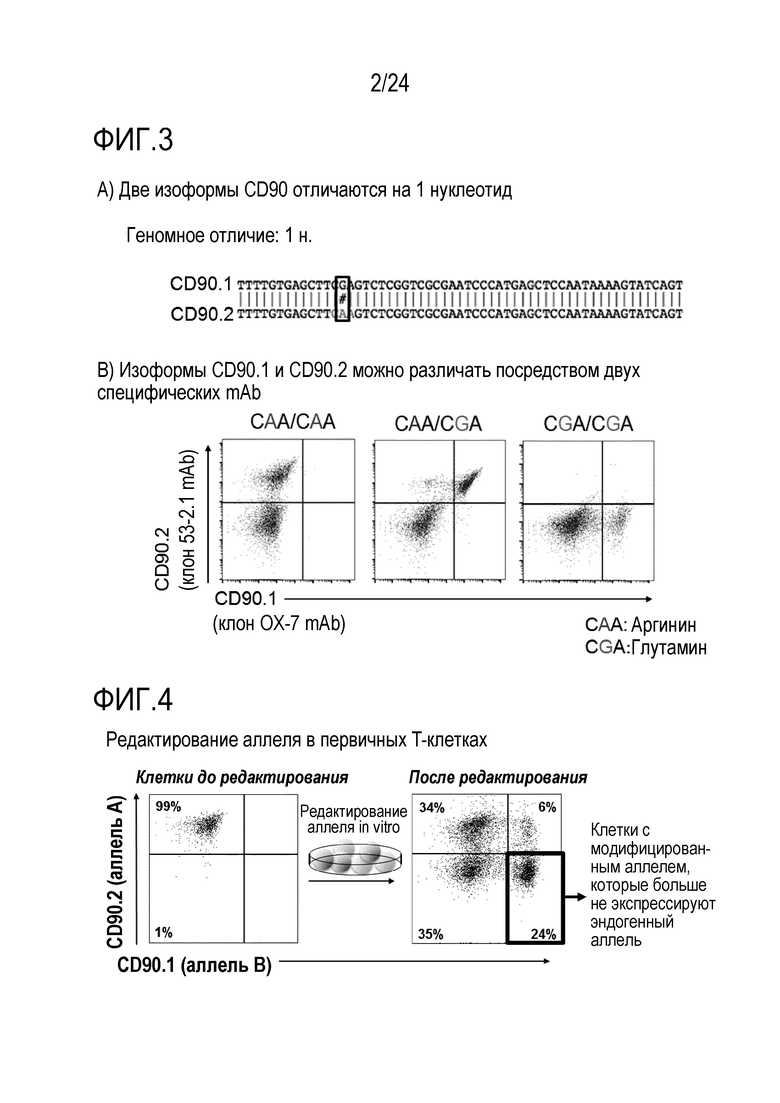

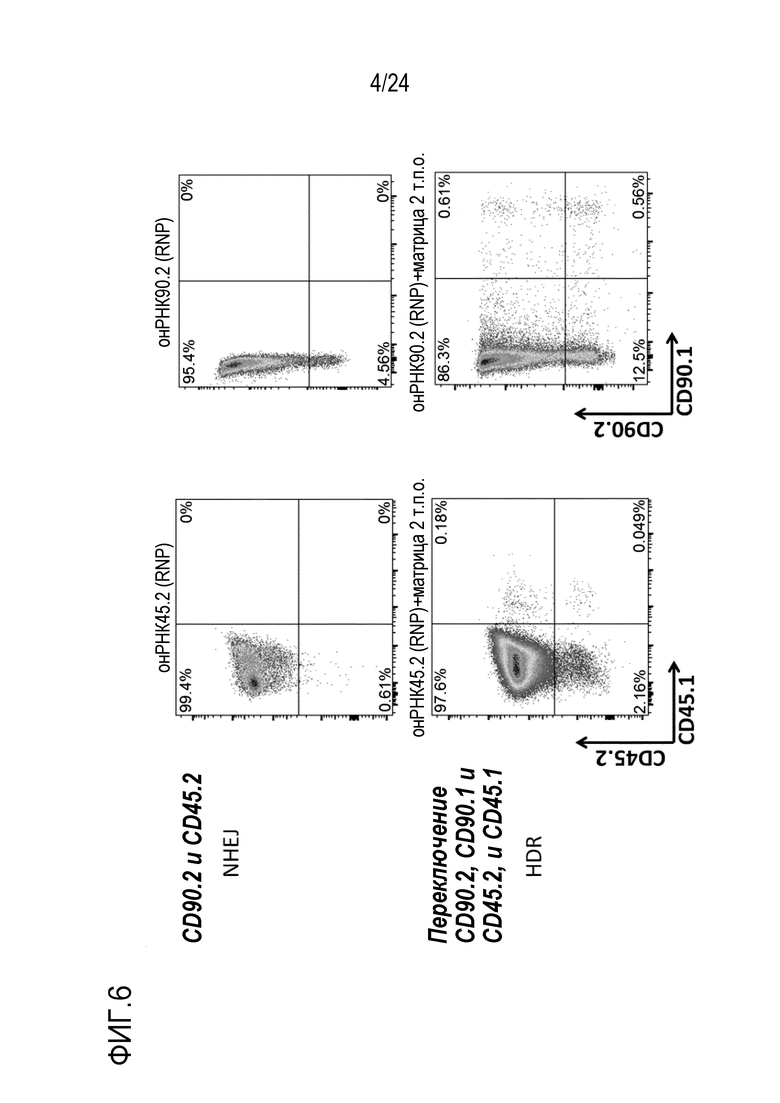
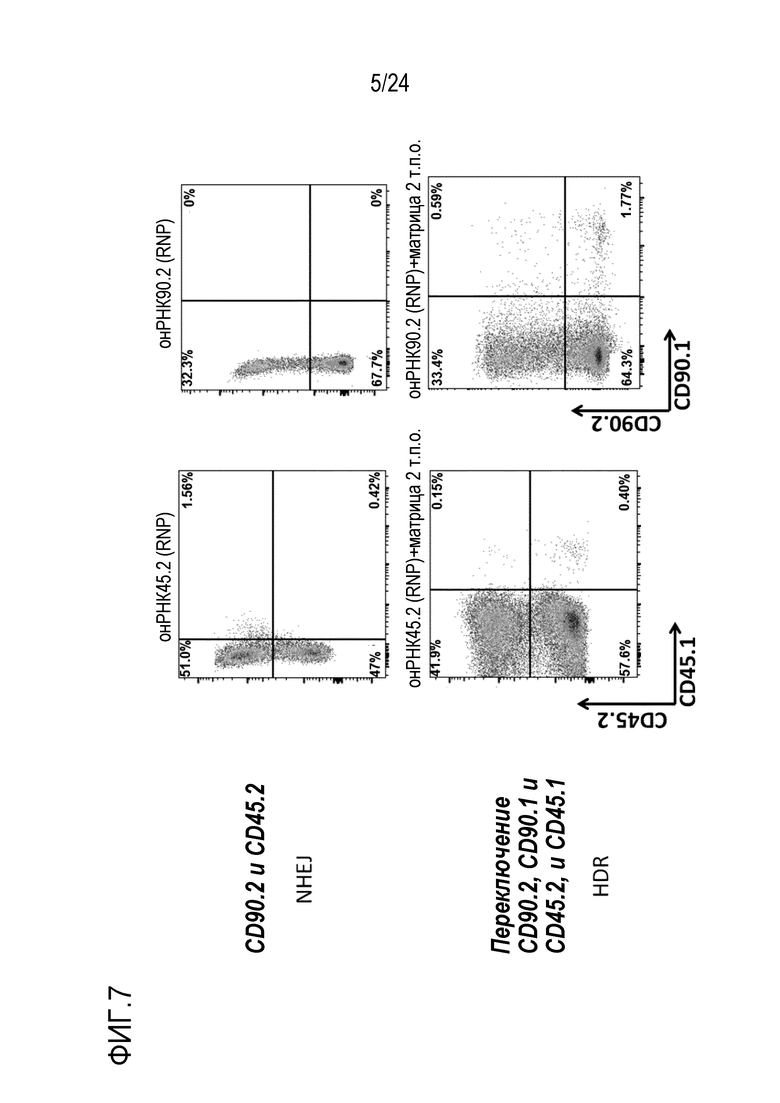

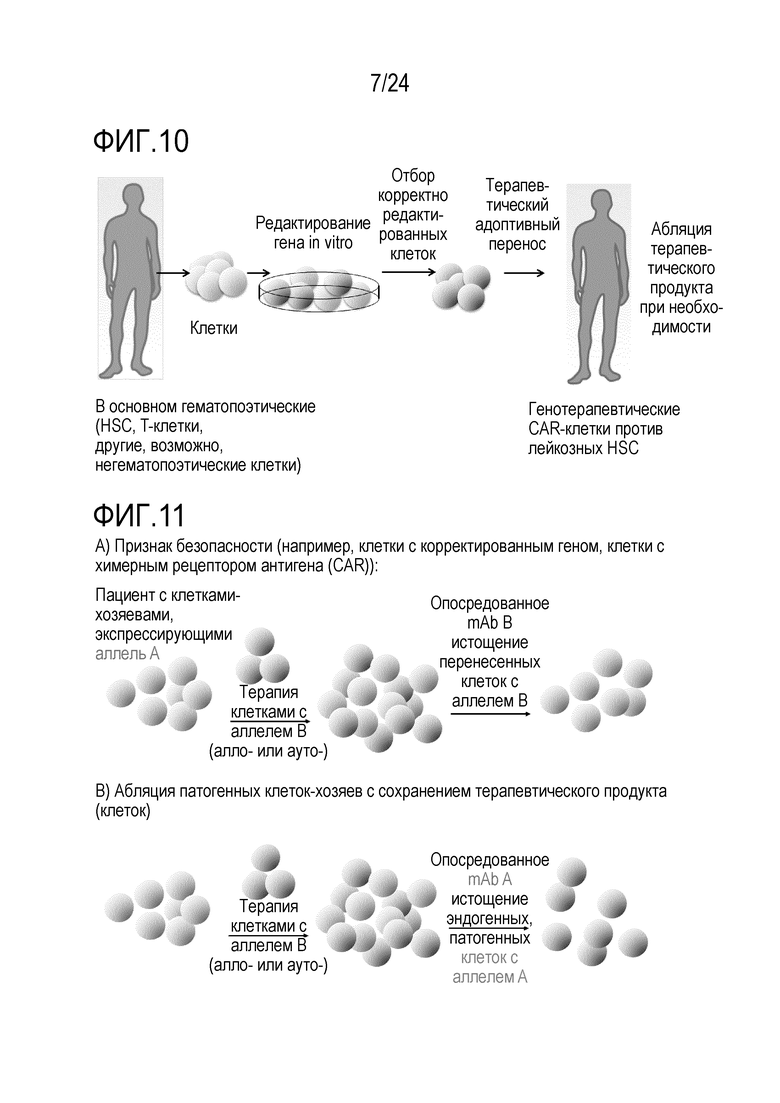

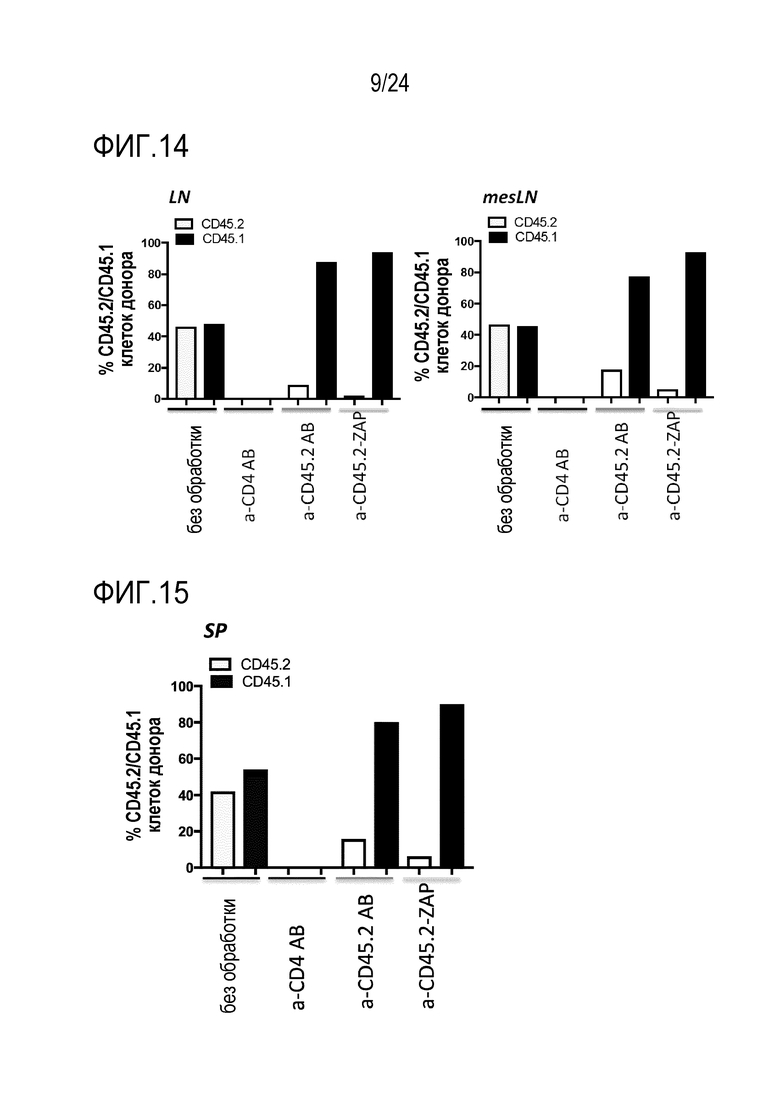
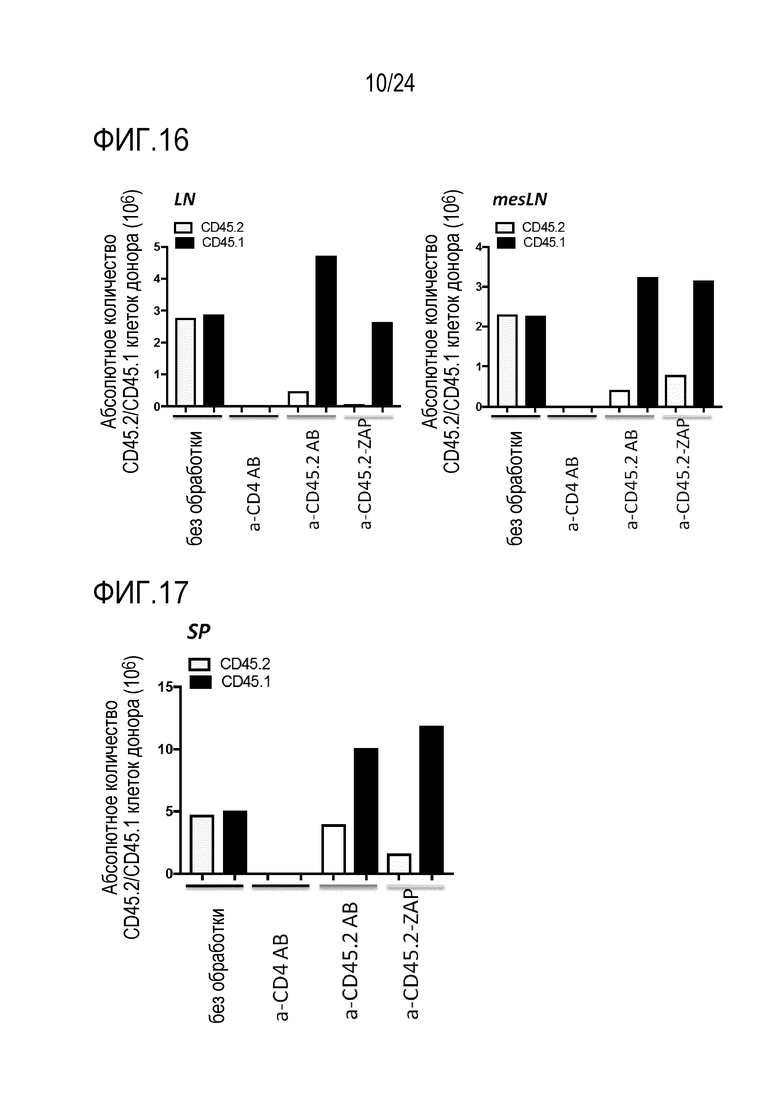
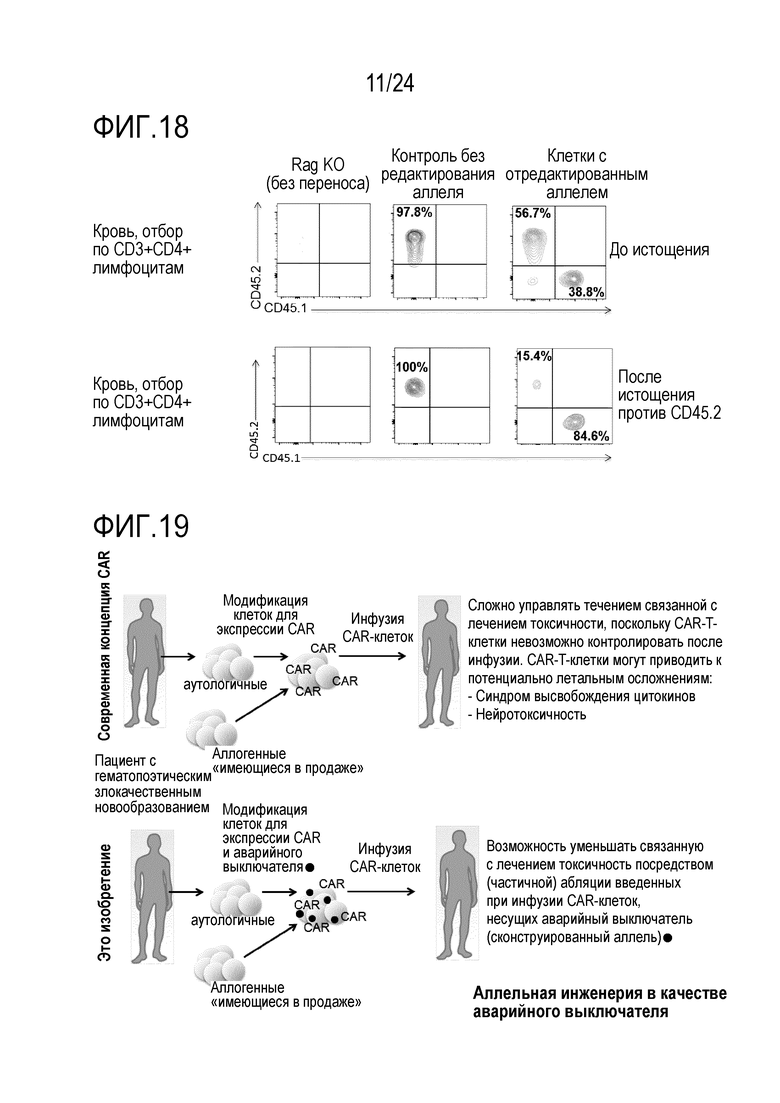

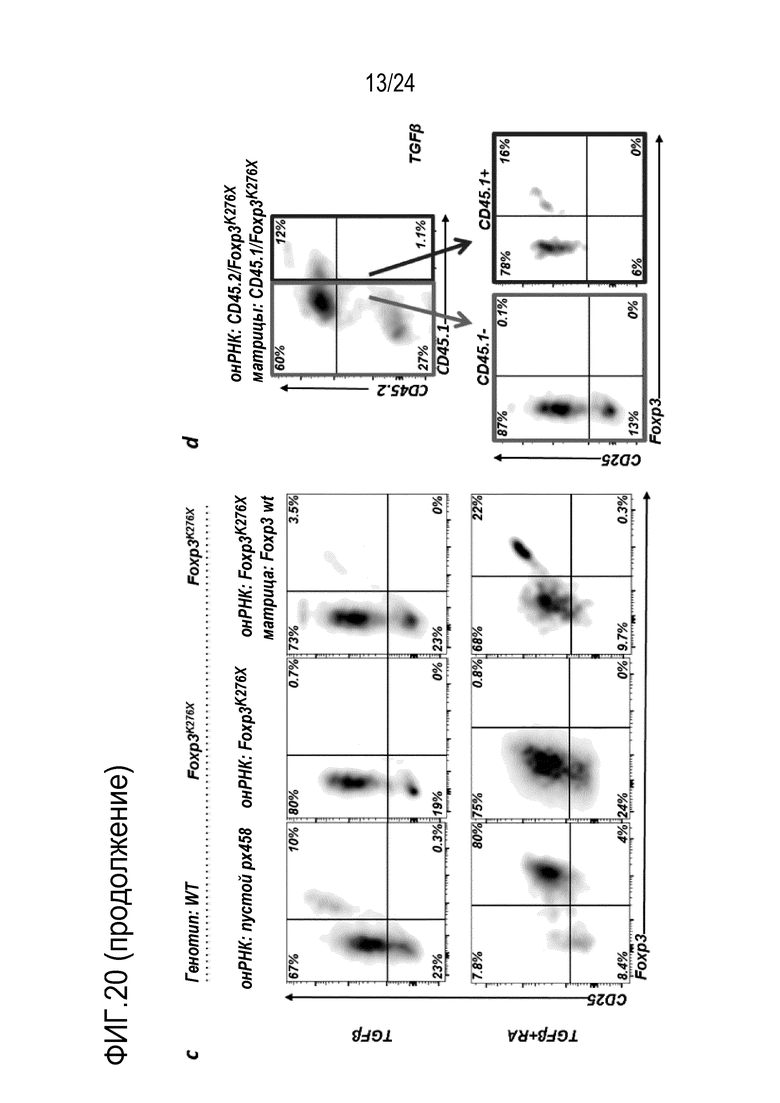
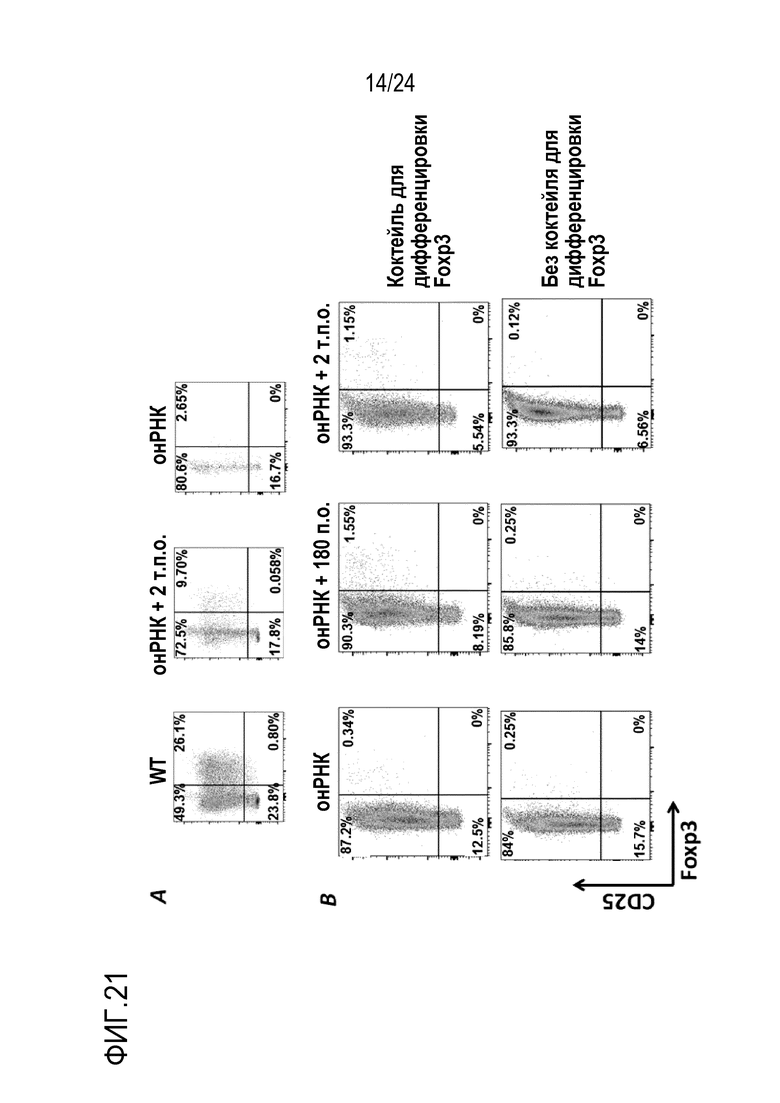




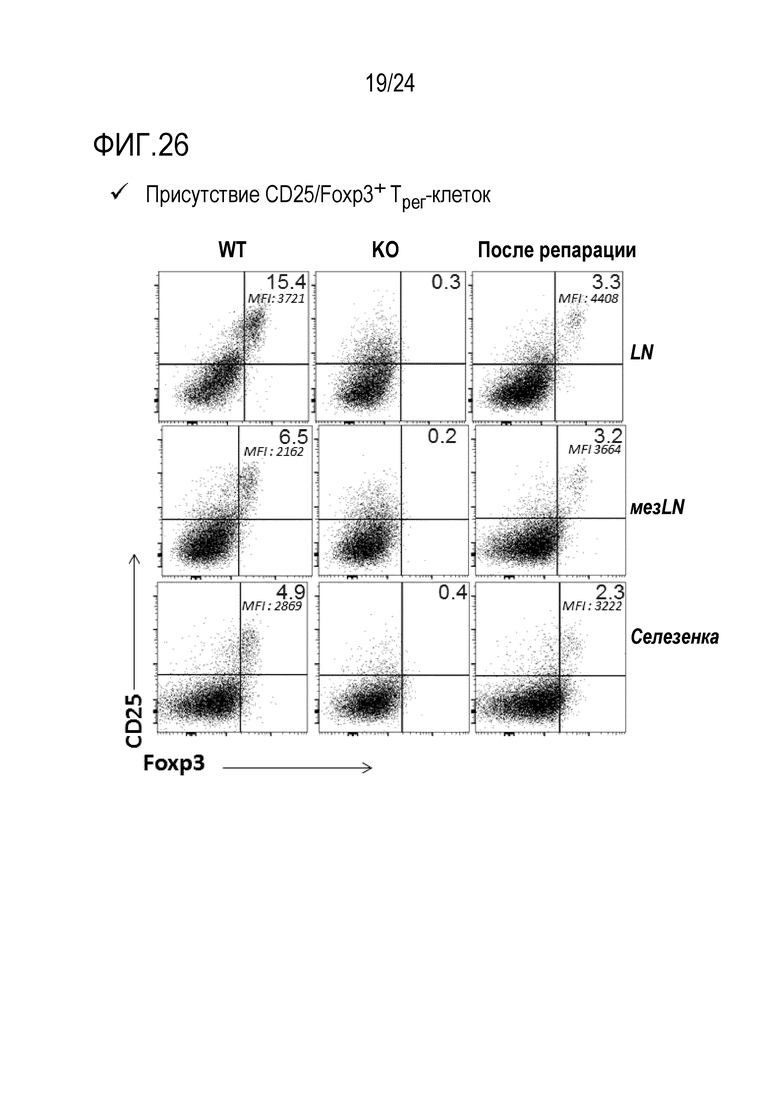


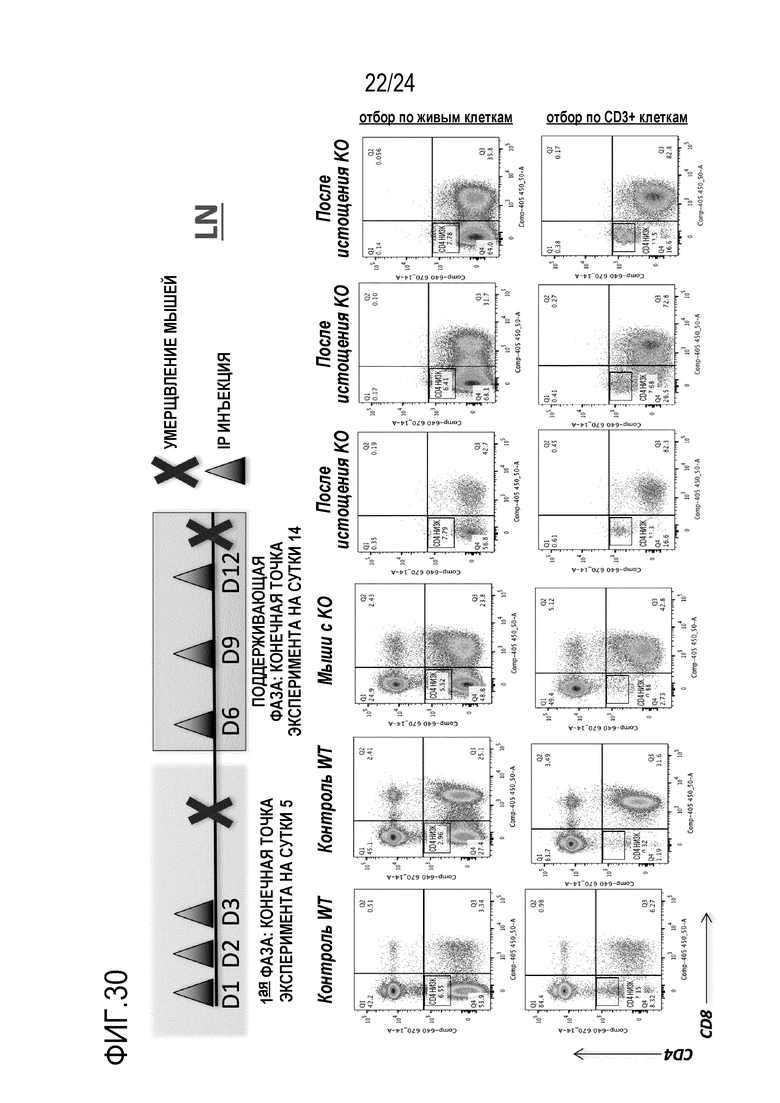


Изобретение относится к биотехнологии и генетической инженерии, в частности к способу избирательного истощения клеток, экспрессирующих вторую изоформу поверхностного белка in vivo. Для осуществления способа сначала пациенту вводят гематопоэтическую клетку человека, которая экспрессирует первую изоформу указанного поверхностного белка, которая является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного поверхностного белка. Затем пациенту вводят антитело или подобную антителу молекулу, которые являются специфически реакционноспособными по отношению ко второй изоформе указанного поверхностного белка, для абляции клетки, экспрессирующей указанную вторую изоформу. Далее происходит избирательное истощение клеток, экспрессирующих вторую изоформу указанного поверхностного белка, на основании экспрессии второй изоформы указанного поверхностного белка. Указанную клетку вводят до, во время или после специфической абляции клеток, экспрессирующих указанную вторую изоформу указанного поверхностного белка. А указанный поверхностный белок содержит внеклеточную полипептидную последовательность, и указанная первая изоформа содержит вставку, делецию и/или замену 1, 2, 3, 4 или 5 аминокислот по сравнению с указанной второй изоформой. Настоящее изобретение позволяет получить систему, которая служит для постоянной маркировки и отслеживания клеток и позволяет избирательное истощение маркированных или немаркированных клеток in vitro или in vivo. 7 з.п. ф-лы, 32 ил., 1 пр.
1. Способ избирательного истощения клеток, экспрессирующих вторую изоформу поверхностного белка in vivo, включающий
a) введение пациенту гематопоэтической клетки человека, где указанная клетка экспрессирует первую изоформу указанного поверхностного белка, которая является функционально неотличимой, но иммунологически отличимой от второй изоформы указанного поверхностного белка,
b) введение пациенту антитела или подобной антителу молекулы, которые являются специфически реакционноспособными по отношению ко второй изоформе указанного поверхностного белка, для абляции клетки, экспрессирующей указанную вторую изоформу,
c) избирательное истощение клеток, экспрессирующих вторую изоформу указанного поверхностного белка, на основании экспрессии второй изоформы указанного поверхностного белка,
где указанная клетка не является эмбриональной клеткой человека,
где указанную клетку вводят до, во время или после специфической абляции клеток, экспрессирующих указанную вторую изоформу указанного поверхностного белка,
где указанные антитело или подобная антителу молекула являются специфически реакционноспособными по отношению к указанной второй изоформе, но не к указанной первой изоформе, указанного белка поверхности клетки, и
где указанный поверхностный белок содержит внеклеточную полипептидную последовательность, и указанная первая изоформа содержит вставку, делецию и/или замену 1, 2, 3, 4 или 5 аминокислот по сравнению с указанной второй изоформой.
2. Способ по п.1, где указанную первую изоформу можно отличать от указанной второй изоформы посредством связывания подобной антителу молекулы или связывания антитела.
3. Способ по любому из предшествующих пунктов, где указанный поверхностный белок выбран из CD1a, CD1b, CD1c, CD1d, CD1e, CD2, CD3, CD3d, CD3e, CD3g, CD4, CD5, CD6, CD7, CD8a, CD8b, CD9, CD10, CD11a, CD11b, CD11c, CD11d, CDwl2, CD13, CD14, CD15, CD15u, CD15s, CD15su, CD16, CD16b, CD17, CD18, CD19, CD20, CD21, CD22, CD23, CD24, CD25, CD26, CD27, CD28, CD29, CD30, CD31, CD32, CD33, CD34, CD35, CD36, CD37, CD38, CD39, CD40, CD41, CD42a, CD42b, CD42c, CD42d, CD43, CD44, CD45, CD45RA, CD45RB, CD45RC, CD45RO, CD46, CD47, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD50, CD51, CD52, CD53, CD54, CD55, CD56, CD57, CD58, CD59, CD60a, CD60b, CD60c, CD61, CD62E, CD62L, CD62P, CD63, CD64, CD65, CD65s, CD66a, CD66b, CD66c, CD66d, CD66e, CD66f, CD68, CD69, CD70, CD71, CD72, CD73, CD74, CD75, CD75s, CD77, CD79a, CD79b, CD80, CD81, CD82, CD83, CD84, CD85a, CD85d, CD85j, CD85k, CD86, CD87, CD88, CD89, CD90, CD91, CD92, CD93, CD94, CD95, CD96, CD97, CD98, CD99, CD99R, CD100, CD101, CD102, CD103, CD104, CD105, CD106, CD107a, CD107b, CD108, CD109, CD110, CD111, CD112, CD113, CD114, CD115, CD116, CD117, CD118, CD119, CD120a, CD120b, CD121a, CD121b, CD122, CD123, CD124, CD125, CD126, CD127, CD129, CD130, CD131, CD132, CD133, CD134, CD135, CD136, CD137, CD138, CD139, CD140a, CD140b, CD141, CD142, CD143, CD144, CDw145, CD146, CD147, CD148, CDw149, CD150, CD151, CD152, CD153, CD154, CD155, CD156a, CD156b, CD156c, CD157, CD158e, CD158i, CD158k, CD159a, CD159c, CD160, CD161, CD162, CD163, CD164, CD165, CD166, CD167a, CD167b, CD168, CD169, CD170, CD171, CD172a, CD172b, CD172g, CD173, CD174, CD175, CD175s, CD176, CD177, CD178, CD179a, CD179b, CD180, CD181, CD182, CD183, CD184, CD185, CD186, CD191, CD192, CD193, CD194, CD195, CD196, CD197, CDw198, CD199, CD200, CD201, CD202b, CD203c, CD204, CD205, CD206, CD207, CD208, CD209, CD210, CDw210b, CD212, CD213a1, CD213a2, CD215, CD217a, CD218a, CD218b, CD220, CD221, CD222, CD223, CD224, CD225, CD226, CD227, CD228, CD229, CD230, CD231, CD232, CD233, CD234, CD235a, CD235b, CD236, CD236R, CD238, CD239, CD240CE, CD240DCE, CD240D, CD241, CD242, CD243, CD244, CD245, CD246, CD247, CD248, CD249, CD252, CD253, CD254, CD256, CD266, CD267, CD268, CD269, CD270, CD271, CD272, CD273, CD274, CD275, CD276, CD277, CD278, CD279, CD280, CD281, CD282, CD283, CD284, CD286, CD289, CD290, CD292, CDw293, CD294, CD295, CD296, CD297, CD298, CD299, CD300a, CD300c, CD300e, CD301, CD302, CD303, CD304, CD305, CD306, CD307a, CD307b, CD307c, CD307d, CD307e, CD308, CD309, CD312, CD314, CD315, CD316, CD317, CD318, CD319, CD320, CD321, CD322, CD324, CD325, CD326, CD327, CD328, CD329, CD331, CD332, CD333, CD334, CD335, CD336, CD337, CD338, CD339, CD340, CD344, CD349, CD350, CD351, CD352, CD353, CD354, CD355, CD357, CD358, CD360, CD361, CD362, CD363, CD364, CD365, CD366, CD367, CD368, CD369, CD370, CD371, BCMA, легкой цепи иммуноглобулина лямбда или каппа, белка HLA и β2-микроглобулина,
в частности, где указанный поверхностный белок выбран из CD2, CD3, CD4, CD5, CD8, CD19, CD20, CD22, CD23, CD33, CD34, CD90, CD45, CD123, BCMA, легкой цепи иммуноглобулина лямбда или каппа, белка HLA и β2-микроглобулина.
4. Способ по любому из предшествующих пунктов, где указанная первая изоформа не кодирована в нативной геномной ДНК пациента.
5. Способ по любому из предшествующих пунктов, где указанная первая изоформа получена путем изменения последовательности, кодирующей указанный поверхностный белок в нативной геномной ДНК пациента, посредством редактирования генов или с помощью изменения мРНК, кодирующей указанный поверхностный белок, посредством редактирования РНК.
6. Способ по любому из предшествующих пунктов, где указанная вставка, делеция и/или замена локализована во внеклеточной части указанного первого поверхностного белка, в частности во внеклеточной петле.
7. Способ по любому из предшествующих пунктов, где указанная гематопоэтическая клетка человека выбрана из группы, состоящей из гематопоэтической стволовой клетки, CD4+ T-клетки, CD8+ T-клетки, T-клетки памяти, регуляторной T-клетки (Tрег), клетки естественного киллера (NK), врожденной лимфоидной клетки (ILC), дендритной клетки (DC), B-лимфоцита, ассоциированной со слизистой оболочкой инвариантной T-клетки (MAIT) и гамма-дельта-T-клетки (γδ T).
8. Способ по любому из предшествующих пунктов, где указанная гематопоэтическая клетка человека представляет собой гематопоэтическую стволовую клетку или T-клетку.
| KAWAKAMI Y et al., Somatic gene therapy for diabetes with an immunological safety system for complete removal of transplanted cells, Diabetes, 1992, 41(8), pp.956-961 | |||
| SUSHMITA ROY et al., Donor hematopoietic stem cells confer long-term marrow reconstitution by self-renewal divisions exceeding to that of host cells, PLoS One, 2012, 7(12), e50693 |

