На урожайность сельскохозяйственных культур влияют многие факторы, среди которых имеются, с одной стороны, факторы, влияющие на способность растения производить биомассу (фотосинтез, поглощение влаги и питательных веществ), а с другой стороны, факторы, влияющие на способность растения противостоять различным стрессам, наподобие биотических стрессов (насекомые, грибы, вирусы и т.п.) или абиотических стрессов (засухи, засоление почвы, нехватка питательных веществ и т.п.).
Одним значительным фактором, влияющим на создание биомассы, является фотосинтез. Фотосинтез представляет собой механизм, с помощью которого растения захватывают атмосферный углекислый газ и преобразуют его в сахар, который затем встраивается в растительные ткани, благодаря чему образуется биомасса. Фотосинтез является основным источником всей первичной продуктивности на Земле.
Большинство растений имеют фотосинтетический механизм, в котором хлоропластный фермент RuBisCo (Рибулозо-1,5-Бифосфат Карбоксилаза/Оксигеназа) является основным ферментом, захватывающим углекислый газ и преобразующим его в сахар. Эти растения, включая некоторые важные сельскохозяйственные растения, например рис, пшеницу, ячмень, картофель, рапс, относятся к так называемым С3-растениям. Одна известная проблема в фотосинтетическом механизме C3-растений состоит в том, что эффективность фиксации углерода не является оптимальной в некоторых окружающих условиях, когда часть фиксированного углерода теряется за счет альтернативной активности RuBisCo, называемой окисление.
RuBisCO способен катализировать как карбоксилирование, так и окисление рибулозо-1,5-бифосфата. Баланс между двумя данными функциями зависит главным образом от соотношения в листьях СО2/О2, которое может изменяться вслед за реакцией растений на некоторые окружающие условия. Каждая реакция карбоксилирования создает две молекулы фосфоглицерата, который входит в цикл Кальвина, с образованием в конечном итоге крахмала и сахарозы и с регенерацией рибулозо-1,5-бифосфата. Реакция окисления производит по одной молекуле фосфоглицерата и фосфогликолата. Последний рециркулирует в фосфоглицерат за счет фотодыхания (Leegood R.C. et al, 1995). На каждые две молекулы полученного фосфогликолата высвобождается одна молекула СО2, приводя к общей потере фиксированного углерода, что в конечном итоге снижает производство сахаров и биомассы. В данной реакции также теряется аммоний, и требуется его повторная фиксация через энергозатратные реакции в хлоропласте.
Сообщалось, что преодоление фотодыхания является целью для повышения максимальной эффективности фотосинтеза и повышения его производительности (Zhu et al., 2008), и было описано несколько попыток для того, чтобы снизить потерю углерода в растениях и, вследствие этого, увеличить производство сахаров и биомассы.
Kebeish et al. сообщали, что потери от фотодыхания у Arabidopsis thaliana могут быть устранены за счет введения в хлоропласты бактериального пути для катаболизма фотореспираторного промежуточного гликолата (WO 03/100066; Kebeish R. et al., 2007). Авторы впервые нацелили три субъединицы гликолатдегидрогеназы Escherichia coli в хлоропласты Arabidopsis thaliana, а затем ввели глоксилаткарболигазу Escherichia coli и редуктазу тартронового полуальдегида Escherichia coli для завершения пути, который преобразует гликолат в глицерат параллельно с эндогенным фотореспираторным путем. Данная пошаговая ядерная трансформация с пятью генами Escherichia coli приводит к растениям Arabidopsis, в которых хлоропластный гликолат преобразуется непосредственно в глицерат. Данные трансгенные растения росли быстрее, производили больше биомассы побегов и корней и содержали больше растворимых сахаров.
В PCT/EP2009/059843 раскрыт способ увеличения производства биомассы и/или производства семян и/или фиксации углерода в растениях риса, в котором растения риса трансформируют тремя субъединицами (glcD, glcE и glcF) гликолатдегидрогеназы Escherichia coli, без последующего введения глоксилаткарболигазы Escherichia coli и редуктазы тартронового полуальдегида Escherichia coli.
Целью представленного изобретения являлось использование трансляционных слияний субъединиц ферментов бактериальной гликолатдегидрогеназы (GDH) из множества субъединиц в сельскохозяйственных культурах, избегая затратного по времени и утомительного процесса многократных трансформаций или трансформации с множеством экспрессионных кассет. Бактериальные субъединицы glcD, glcE и glcF сливали с гибкими линкерами в различных расположениях и тестировали в штаммах E. Coli, испытывающих недостаток в GDH, демонстрируя, что рекомбинантные белки DEFp, EFDp и FDEp слияния из многих субъединиц GDH являются активными. Конструкции с наилучшим исполнением переносили в растения Nicotiana tabacum, риса и рапса, и трансгенные растения продемонстрировали значительно усиленный рост и улучшенную интенсивность фотосинтеза.
Представленное изобретение относится к способу увеличения производства биомассы и/или производства семян и/или фиксации углерода в растениях, включающему введение в геном клетки растения нуклеиновой кислоты, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы, при этом результатом указанного введения указанной одной нуклеиновой кислоты является вновь возникающее экспрессирование одного синтетического полипептида, имеющего ферментативную активность гликолатдегидрогеназы и при этом указанный один полипептид локализован в хлоропластах полученного растения.
В контексте изобретения, белок слияния из многих субъединиц гликолатдегидрогеназы представляет собой один полипептид, состоящий из субъединиц гликолатдегидрогеназы, которые являются существенными для активности гликолатдегидрогеназы, как правило, с пептидными линкерами между данными субъединицами.
В представленном изобретении, авторы изобретения выбрали повторяющуюся линкерную последовательность (Gly4Ser)3, подходящую для ковалентного соединения бактериальных доменов glcD, glcE и glcF в полипротеиновый формат, без ухудшения необходимых свойств, таких как правильное складывание, растворимость и активность GDH. Кроме того, линкер не должен быть подвержен протеазному расщеплению в цитозоле растения, обеспечивая возможность сверхэкспрессирования полипептидов в хлоропластах.
В контексте изобретения, биомасса представляет собой количество вещества, производимого отдельными растениями, или площадью поверхности, на которой выращивают растения. Для того, чтобы определить увеличение производства биомассы можно измерить несколько параметров. Примерами подобных параметров являются высота растения, поверхность листовой пластины, сухая масса побегов, сухая масса корней, количество семян, масса семян, размер семян и т.п. Производство семян или урожай семян можно определять на отдельное растение или на площадь поверхности, на которой выращивают растения.
Как правило, данные параметры измеряют после определенного периода выращивания в почве или на конкретной стадии выращивания, например, в конце вегетационного периода, и сравнивают между растениями, трансформированными одной или более нуклеиновыми кислотами согласно изобретению, и растениями, не трансформированными подобными одной или более нуклеиновыми кислотами.
Увеличение фиксации углерода растением можно определить посредством измерения газообмена и параметров флуоресценции хлорофилла. Подходящая методология с использованием системы LI-6400 (Li-Cor) и программного обеспечения, поставляемого производителем, описана в R. Kebeish et al., 2007 и включена в данное описание посредством ссылки.
Нуклеиновая кислота, включенная в способ изобретения, кодирует один полипептид, имеющий ферментативную активность гликолатдегидрогеназы.
Активность гликолатдегидрогеназы может быть оценена согласно Lord J.M. 1972, с использованием технологии, описанной в примере 6 представленной заявки.
В качестве альтернативы, может быть проведен комплементационный анализ с мутантами E. Coli, с дефицитом трех субъединиц, образующих активную эндогенную гликолатдегидрогеназу. Данные мутанты E. coli неспособны к росту на гликолате в качестве единственного источника углерода. Когда сверхэкспрессирование фермента у данных испытывающих недостаток мутантов восстанавливает рост бактерий на среде, содержащей гликолат в качестве единственного источника углерода, это означает, что данный фермент кодирует функциональный эквивалент гликолатдегидрогеназы E.coli. Способ и средство для комплементационного анализа описаны у Bari et al., 2004 и включены в данное описание посредством ссылки.
Молекулы нуклеиновых кислот, кодирующие один полипептид, имеющий ферментативную активность гликолатдегидрогеназы, могут быть получены посредством методик рекомбинантных ДНК (например, ПЦР) или посредством химического синтеза. Идентификация и выделение молекул подобных нуклеиновых кислот может происходить за счет использования последовательностей, или части данных последовательностей, молекул известных нуклеиновых кислот гликолатдегидрогеназы или, исходя из реальной ситуации, обратных комплементарных цепей данных молекул, например, посредством гибридизации согласно стандартным методам (см., например, Sambrook et al., 1989).
Гликолатдегидрогеназой для цели изобретения может быть любая гликолатдегидрогеназа природного происхождения, или ее любой активный фрагмент или ее любой вариант, в котором некоторые аминокислоты (предпочтительно аминокислоты 1-20, более предпочтительно 1-10, даже более предпочтительно 1-5) были замещены, добавлены или удалены таким образом, чтобы фермент сохранял свою активность гликолатдегидрогеназы.
Согласно представленному изобретению, "нуклеиновую кислоту" или "молекулу нуклеиновой кислоты" необходимо понимать, как полинуклеотидную молекулу, которая может относиться к типу ДНК или РНК, предпочтительно к типу ДНК, и в частности иметь двойную цепь. Она может иметь природное или синтетическое происхождение. Синтетические нуклеиновые кислоты генерируют in vitro. Примерами подобных синтетических нуклеиновых кислот являются кислоты, в которых кодоны, которые кодируют полипептид (полипептиды), имеющий ферментативную активность гликолатдегидрогеназы согласно изобретению, были оптимизированы в соответствии с организмом хозяина, в котором он должен экспрессироваться (например, посредством замещения кодонов такими кодонами, которые являются более предпочтительными или наиболее предпочтительными в таблицах частоты использования кодонов организма подобного хозяина или группы, к которой относится организм подобного хозяина, по сравнению с первоначальным хозяином). Способы оптимизации кодонов хорошо известны квалифицированным специалистам.
Предпочтительными белками слияния из многих субъединиц гликолатдегидрогеназы являются белки, состоящие из слияния субъединиц бактериальной гликолатдегидрогеназы, более предпочтительно белки, состоящие из слияния трех существенных субъединиц, кодируемых опероном E. coli glc (gi/1141710/gb/L43490.1/ECOGLCC). Наиболее предпочтительными являются полипептиды, которые содержат слитые аминокислотные последовательности SEQ ID №№: 2 (Glc D), 4 (Glc E) и 6 (Glc F), в которых данные аминопоследовательности могут быть связаны линкером. Соответственно, нуклеиновая кислота, включающая полинуклеотидные последовательности SEQ ID NO: 1, 3 и 5, может быть использована для осуществления представленного изобретения.
Способ изобретения охватывает введение в геном клетки растения нуклеиновой кислоты, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы, имеющий ферментативную активность гликолатдегидрогеназы, при этом указанный полипептид содержит последовательности, имеющие идентичность последовательности, составляющую по меньшей мере 60, 70, 80 или 90%, особенно по меньшей мере 95%, 97%, 98% или по меньшей мере 99% на уровне аминокислотной последовательности с SEQ ID NO: 2, 4 и 6, соответственно, причем результатом введения нуклеиновой кислоты (кислот) является вновь возникающее экспрессирование одного полипептида, имеющего ферментативную активность гликолатдегидрогеназы, и при этом указанная активность локализована внутри хлоропластов.
Способ изобретения также охватывает введение в геном клетки растения нуклеиновой кислоты, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы, имеющий ферментативную активность гликолатдегидрогеназы, при этом указанная нуклеиновая кислота содержит последовательности нуклеиновых кислот с по меньшей мере 60, 70, 80 или 90%, особенно по меньшей мере 95%, 97%, 98% или по меньшей мере 99% идентичности последовательности к нуклеотидным последовательностям SEQ ID NO: 1, 3 и 5 соответственно, причем результатом введения нуклеиновой кислоты является вновь возникающее экспрессирование по меньшей мере одного полипептида, имеющего ферментативную активность гликолатдегидрогеназы, и при этом указанная активность локализована внутри хлоропластов.
Для цели данного изобретения, "идентичность последовательности" двух родственных нуклеотидных или аминокислотных последовательностей, выраженная в виде процентного соотношения, относится к числу позиций в двух оптимально выровненных последовательностях, которые имеют идентичные остатки (x100), деленные на число сравниваемых позиций. Делеция, т.е. позиция в выравнивании, где остаток присутствует в одной последовательности, но отсутствует в другой, рассматривается как позиция с неидентичными остатками. Выравнивание двух последовательностей может быть выполнено с помощью алгоритма Needleman и Wunsch (Needleman and Wunsch 1970) в EMBOSS (Rice et al., 2000) для поиска оптимального выравнивания по всей длине последовательностей, используя настройки по умолчанию (штраф на внесение делеции 10, штраф на продолжение делеции 0,5).
После того как последовательность чужеродной ДНК становится известной, можно разрабатывать праймеры и зонды, которые специфично узнают данные последовательности в нуклеиновой кислоте (ДНК или РНК) образца с помощью методики молекулярной биологии. Например, может быть разработан метод ПЦР для идентификации генов, используемых в способе изобретения (генов gdh) в биологических образцах (таких как образцы растений, растительный материал или продукты, содержащие растительный материал). Подобная ПЦР основана по меньшей мере на двух специфичных "праймерах", при этом, например, оба узнают последовательность внутри кодирующей области gdh, используемой в изобретении (например, кодирующей области SEQ ID № 1, 3, 5), или один узнает последовательность внутри кодирующей области gdh, а другой узнает последовательность внутри соответствующей последовательности транзитного пептида или внутри регуляторных областей, таких как промотор или 3’-конец химерного гена, содержащего ДНК gdh, используемого в изобретении. Праймеры предпочтительно имеют последовательность между 15 и 35 нуклеотидами, которые в условиях оптимизированной ПЦР специфически распознают последовательность внутри химерного гена gdh, используемого в изобретении, так что специфичный фрагмент ("фрагмент интеграции" или дискриминирующий ампликон) амплифицируется из образца нуклеиновой кислоты, содержащего ген gdh, используемый в изобретении. Это означает, что в условиях оптимизировнной ПЦР амплифицируется только намеченный фрагмент интеграции, и никакая другая последовательность в геноме растения или чужеродной ДНК.
Способ изобретения охватывает также введение в геном клетки растения нуклеиновой кислоты, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы, имеющий ферментативную активность гликолатдегидрогеназы, при этом указанная одна нуклеиновая кислота гибридизирует в жестких условиях в нуклеотидную последовательность, выбранную из группы SEQ ID NO: 1, 3 и 5, при этом результатом введения нуклеиновой кислоты (кислот) является вновь возникающее экспрессирование одного полипептида, имеющего ферментативную активность гликолатдегидрогеназы, и при этом указанная активность локализована внутри хлоропластов. Жесткие условия гибридизации, как используется в данном описании, относится в частности к следующим условиям: иммобилизация релевантных последовательностей ДНК на фильтре, и предварительная гибридизация фильтров на протяжении каждых 1-2 часов в 50% формамиде, 5% SSPE, 2x растворе Денхардта и 0,1% SDS при 42°C, или 1-2 часов в 6x SSC, 2x растворе Денхардта и 0,1% SDS при 68°C. Денатурированный зонд, меченый dig или радиоактивной меткой, затем добавляли непосредственно в предгибридизационную текучую среду и проводили инкубацию на протяжении от 16 до 24 часов при соответствующей температуре, указанной выше. После инкубации фильтры затем промывали в течение 30 минут при комнатной температуре в 2x SSC, 0,1% SDS, с последующими 2 промываниями по 30 минут каждое при 68°C в 0,5 × SSC и 0,1% SDS. Авторадиограф установили посредством экспозиции фильтров в течение 24-48 часов на рентгеновской пленке (Kodak XAR-2 или аналогичной) при -70°C с усиливающим экраном. Конечно, в данном процессе могут быть использованы аналогичные условия и параметры, при этом все еще сохраняются необходимые жесткие условия гибридизации.
Терминология ДНК или белок, "включающий в себя" определенную последовательность X, как используется на протяжении всего текста, относится к ДНК или белку, включающему или содержащему по меньшей мере последовательность X, так что другие последовательности нуклеотидов или аминокислот могут быть включены в 5’-(или N-терминальный) и/или 3’-(или C-терминальный) конец, например (нуклеотидная последовательность кодирования) селектируемый маркер белок, (нуклеотидная последовательность кодирования) транзитный пептид, и/или 5’-лидерную последовательность или 3’-трейлерную последовательность. Аналогичным образом, должно быть понятно, что применение термина "содержат", "содержащие" или "содержит" на протяжении всего текста и формулы изобретения данной заявки подразумевает включение приведенного числа или стадии или группы чисел или стадий, но не исключение какого-либо другого числа или стадии или стадии или группы чисел или стадий.
Способ представленного изобретения состоит в инсталляции активности гликолатдегидрогеназы внутрь хлоропласта. Это может быть произведено либо посредством введения нуклеиновой кислоты, кодирующей активность гликолатдегидрогеназы, в ядерный геном клеток растения, при этом кодирующую последовательность белка затем сливают с нуклеиновой кислотой, кодирующей хлоропластный транзитный пептид. В качестве альтернативы, активность гликолатдегидрогеназы может быть добавлена хлоропласту с помощью непосредственной трансформации генома хлоропласта нуклеиновой кислотой (кислотами), кодирующей соответствующий фермент.
Могут использоваться общие методики для трансформирования клеток растений или тканей растений. Одна группа способов содержит бомбардирование клеток, протопластов или тканей частицами, с которыми соединены последовательности ДНК. Еще одна группа способов содержит применение, в качестве средства для переноса в растение, химерного гена, который вставляют в плазмиду Ti Agrobacterium tumefaciens или плазмиду Ri Agrobacterium rhizogenes. Могут быть использованы другие способы, такие как микроинъекции или электропорация, или прямая преципитация иным образом с использованием PEG. Квалифицированные специалисты могут выбрать любой подходящий способ и средство для трансформации клетки растения или растения.
С целью экспрессирования нуклеиновой кислоты, которая кодирует полипептид, обладающий ферментативной активностью, которая требуется для представленного изобретения в клетках растений, могут использоваться любые подходящие регуляторные последовательности. Регуляторные последовательности будут предоставлять области инициации транскрипции и трансляции, а также терминальные области, в которых инициация транскрипции может быть конститутивной или индуцируемой. Кодирующая область является функционально связанной с такими регуляторными последовательностями. Подходящие регуляторные последовательности представлены конститутивным промотором 35S. В качестве альтернативы, может быть использован конститутивный промотор убиквитина, в частности промотор убиквитина кукурузы (GenBank: gi19700915). Примеры для индуцируемых промоторов представлены светоиндуцируемыми промоторами малой субъединицы RUBISCO и промоторами "белков, связывающих светоулавливающий комплекс (lhcb)". Преимущественно, может быть использована промоторная область gos2 гена Oryza sativa, включающая 5’-нетранслируемую область гена GOS2 с интроном (de Pater et al., 1992), промоторная область рибулозо-1,5-бифосфонаткарбоксилазы малой субъединицы гена Oryza sativa (Kyozuka J. et al., 1993), или промоторная область гена актина 1 Oryza sativa (McElroy D. et al., 1990).
Согласно изобретению, можно осуществить применение, в сочетании с промотором, других регуляторных последовательностей, которые распложены между промотором и кодирующей последовательностью, такой как активаторы транскрипции ("энхансеры"), например активатор трансляции вируса табачной мозаики (TMV), описанный в заявке WO 87/07644, или вируса гравировки табака (TEV), описанный Carrington & Freed 1990, например, или интроны, такие как интрон adhl кукурузы или интрон актина 1 риса.
В качестве регуляторного терминатора или последовательности полиаденилирования, можно осуществить применение любой соответствующей последовательности бактериального происхождения, такой как, например, терминатор nos Agrobacterium tumefaciens, вирусного происхождения, такой как, например, терминатор CaMV 35S, или растительного происхождения, такой как, например, терминатор гистон, как описано в заявке EP 0633317.
В одном отдельном варианте осуществления изобретения, в результате того, что предпочтительной является трансформация ядерного генома, нуклеиновая кислота, которая кодирует транзитный пептид хлоропласта, задействует 5’ последовательности нуклеиновой кислоты, кодирующей гликолатдегидрогеназу, при этом данная последовательность транзитного пептида расположена между областью промотора и нуклеиновой кислотой, кодирующей гликолатдегидрогеназу, таким образом, чтобы позволить экспрессирование транзитного пептида/белка слияния гликолатдегидрогеназы. Транзитный пептид делает возможным направить гликолатдегидрогеназу в пластиды, более конкретно в хлоропласты, при этом белок слияния расщепляется между транзитным пептидом и гликолатдегидрогеназой, когда последний попадает в пластиду. Транзитный пептид может быть единственным пептидом, таким как транзитный пептид EPSPS (описанный в патенте США 5188642), или транзитный пептид малой субъединицы рибулозобискарбоксилазы/оксигеназы (RuBisCO ssu) растения, например, транзитный пептид хлоропласта, полученный из гена рибулозо-1,5-бифосфаткарбоксилазы из Solanum tuberosum (GenBank: ген gi21562, кодирующий белок G68077, аминокислоты 1-58), при включении соответствующим образом нескольких аминокислот N-терминальной части зрелой RuBisCO ssu (EP 189707), или направляющий пептид хлоропласта гена rbcS1 (gi21562) картофеля. Транзитный пептид может быть целым транзитным пептидом природного происхождения (дикий тип), его функциональным фрагментом, его функционально активным мутантом. Он также может представлять собой химерный транзитный пептид, в котором по меньшей мере два транзитных пептида связаны друг с другом, или в котором части различных транзитных пептидов связаны друг с другом функциональным образом. Один пример такого химерного транзитного пептида включает транзитный пептид RuBisCO ssu подсолнечника, слитый с N-терминальной частью RuBisCO ssu кукурузы, слитой с транзитным пептидом кукурузы RuBisCO ssu, как описано в патенте EP 508909.
В качестве альтернативы, полипептиды могут быть непосредственно экспрессированы в хлоропласт с использованием трансформации генома хлоропласта. Способы для интегрирования интересующих нуклеиновых кислот в геном хлоропласта являются известными в данной области, в частности способы, основанные на механизме гомологичной рекомбинации. Подходящие векторы и системы селекции являются известными квалифицированным специалистам в данной области. Кодирующие последовательности для полипептидов могут либо переноситься в отдельные векторы, либо в одну конструкцию, в которой отдельные открытые рамки считывания могут быть слиты с одной или несколькими полицистронными РНК с сайтами, связанными рибосомами, добавленными перед каждой отдельной открытой рамкой считывания для того, чтобы допустить независимую трансляцию. Пример средств и способов, которые могут использоваться для данной интеграции в геном хлоропласта, приведен, например, в WO 06/108830, содержание который включено в данное описание посредством ссылки.
Когда нуклеиновые кислоты непосредственно интегрируют в геном хлоропласта, последовательность транзитного пептида не требуется. В таком случае, инициирующий трансляцию кодон (Met) может быть добавлен к последовательности, кодирующей зрелый белок, чтобы обеспечить инициирование трансляции.
Объектом представленного изобретения также являются нуклеиновые кислоты, кодирующие белок слияния из многих субъединиц гликолатдегидрогеназы.
В отдельном варианте осуществления, нуклеиновая кислота изобретения кодирует белок слияния из многих субъединиц гликолатдегидрогеназы, который содержит аминокислотную последовательность, которая нацеливает указанный белок в хлоропласт.
В еще одном отдельном варианте осуществления, нуклеиновая кислота изобретения кодирует белок слияния из многих субъединиц гликолатдегидрогеназы, который представляет собой результат слияния субъединиц бактериальной гликолатдегидрогеназы.
В еще одном отдельном варианте осуществления, нуклеиновая кислота изобретения кодирует белок слияния из многих субъединиц гликолатдегидрогеназы, который представляет собой результат слияния трех субъединиц, кодируемых опероном glc E. coli.
В еще одном отдельном варианте осуществления, нуклеиновая кислота изобретения кодирует белок слияния из многих субъединиц гликолатдегидрогеназы, который содержит аминокислотные последовательности, имеющие по меньшей мере 60% идентичности последовательности к последовательностям SEQ ID NO: 2, 4 и 6 соответственно.
В еще одном отдельном варианте осуществления, нуклеиновая кислота изобретения кодирует белок слияния из многих субъединиц гликолатдегидрогеназы, который содержит полинуклеотидные последовательности, имеющие по меньшей мере 60% идентичности последовательности к полинуклеотидным последовательностям SEQ ID NO: 1, 3 и 5, соответственно.
Объектом представленного изобретения также являются клетки растений, ткани растений, растения и их часть или семена, включающие одну нуклеиновую кислоту, кодирующую белок слияния из многих субъединиц гликолатдегидрогеназы, и экспрессирующие внутри хлоропласта один полипептид, обладающий ферментативной активностью гликолатдегидрогеназы.
Отдельные варианты осуществления нуклеиновых кислот, введенных в клетки растений, ткани растений, растения и часть или их семена, упомянуты выше.
Представленное изобретение также относится к растениям, которые содержат трансформированные клетки, в частности растения, которые являются регенерированными из трансформированных клеток. Регенерация может быть получена посредством любого подходящего способа. Могут быть использованы следующие патенты и патентные заявки, в частности, относящиеся к способам транформирования клеток растений и регенерации растений: US 4459355, US 4536475, US 5464763, US 5177010, US 5187073, EP 267159, EP 604662, EP 672752, US 4945050, US 5036006, US 5100792, US 5371014, US 5478744, US 5179022, US 5565346, US 5484956, US 5508468, US 5538877, US 5554798, US 5489520, US 5510318, US 5204253, US 5405765, EP 442174, EP 486233, EP 486234, EP 539563, EP 674725, WO 91/02071 и WO 95/06128.
Представленное изобретение также относится к трансформированным растениям или их частям, которые получены посредством культивирования и/или скрещиванием с регенерированными растениями, и к семенам трансформированных растений, характеризующимся тем, что они содержат трансформированную клетку растения согласно изобретению.
В отдельном варианте осуществления изобретения, трансформированные растения или их части выбраны из риса, пшеницы, ячменя, картофеля, рапса, табака.
В отдельном варианте осуществления изобретения, трансгенные семена и мука, масло или пища, полученная из них, происходят из растений риса, пшеницы, ячменя, рапса или табака.
Представленное изобретение также относится к любым продуктам, таким как мука, которые получены посредством переработки растений, их частей или семян изобретения. Например, изобретение охватывает зерна, полученные при переработке семян согласно изобретению, но также муку, полученную при дальнейшей переработке семян или зерен, а также любой пищевой продукт, полученный из указанной муки.
Список последовательностей:
SEQ ID № 1: ДНК последовательность glc D Escherichia coli
SEQ ID № 2: аминокислотная последовательность, кодируемая SEQ ID № 1
SEQ ID № 3: ДНК последовательность glc E Escherichia coli
SEQ ID № 4: аминокислотная последовательность, кодируемая SEQ ID № 3
SEQ ID № 5: ДНК последовательность glc F Escherichia coli
SEQ ID № 6: аминокислотная последовательность, кодируемая SEQ ID № 5
ФИГУРЫ
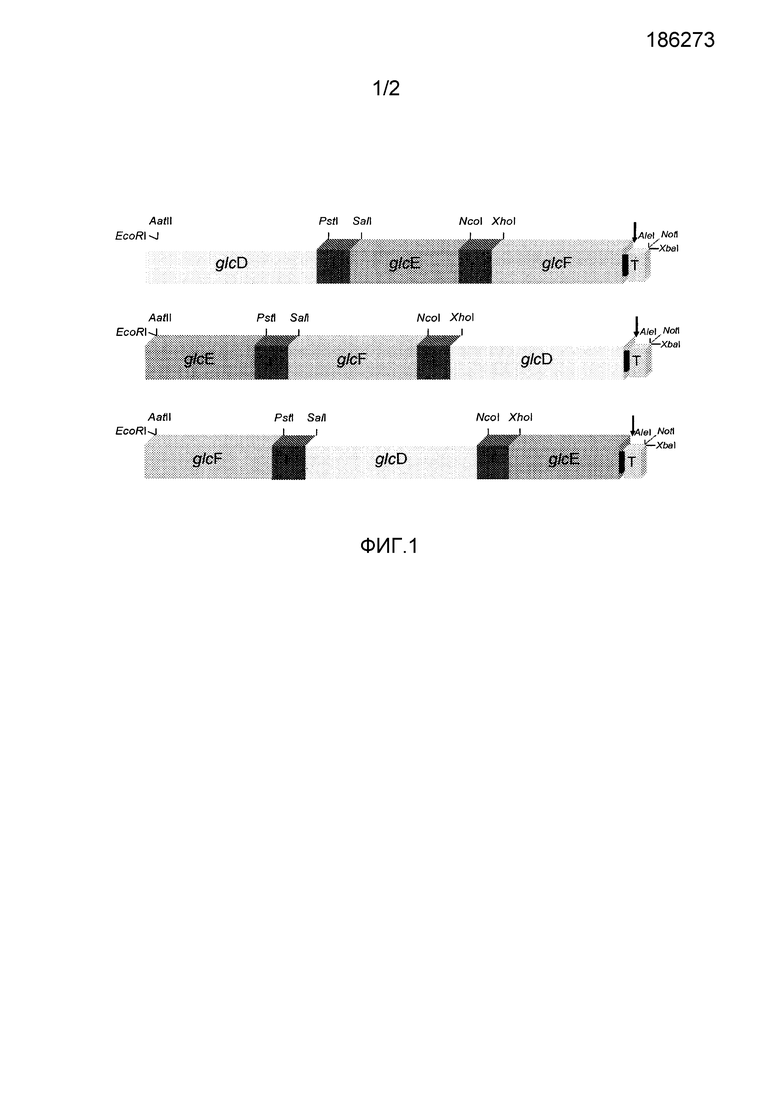
Фиг.1: Конструирование синтетических, состоящих из множества субъединиц кассет слияния. glcD, glcE и glcF: бактериальные гены, кодирующие субъединицы D, E и F GDH. I: линкер (Gly4Ser)3. T: метка His6. Стрелка: сайт расщепления энтерокиназы. Указаны введенные сайты рестрикции.
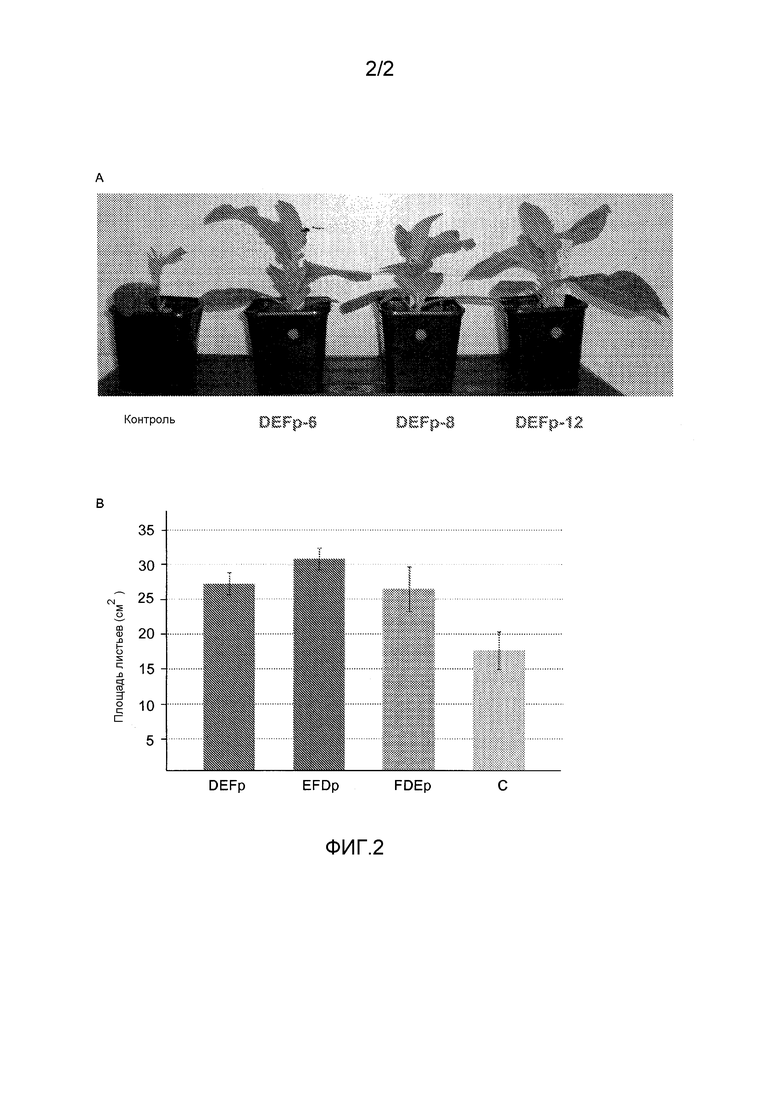
Фиг.2: Параметры роста трансгенных и нетрансгенных линий. A: Фенотип 4-недельных трансгенных растений T0, продуцирующих DEFp в хлоропласте. B: Площадь листьев 7-недельных растений табака. DEFp (n=6), EFDp (n=5), FDEp (n=4), C: растения нетрансгенного N. tabacum cv. Petit Havana SR1 (n=6).
ПРИМЕР 1: Создание синтетических, состоящих из множества субъединиц генных конструкций
Бактериальные кДНК glcD, glcE и glcF, кодирующие субъединицы D, E и F GDH, были слиты с гибким линкером, кодирующим мотив (Gly4Ser)3, в генной конструкции из множества субъединиц (Фиг.1).
Сайты рестрикции, показанные в Фиг.1, ввели, чтобы обеспечить реаранжировку субъединиц glcD, glcE и glcF и субклонирование кассет, состоящих из множества субъединиц, в бактериальные и растительные векторы экспрессии. Кроме того, внутренние сайты рестрикции (PstI/SalI и NcoI/XhoI) ввели, чтобы облегчить замену линкера (Gly4Ser)3 другими гибкими линкерами. В дополнение, генерирование кассет слияния двух генов будет возможным посредством делеции кДНК в срединном положении через сайты рестрикции SalI/XhoI.
C-терминальную метку His6 ввели, чтобы обеспечить возможность обнаружения и очистки рекомбинантных белков. Чтобы избежать возможного влияния метки His6 на ферментативную активность фермента энтерокиназы, состоящего из множества субъединиц GDH, добавили сайт расщепления перед меткой His6, обеспечивая возможность удаления C-терминальной метки.
ПРИМЕР 2: Синтез кассет слияния множества субъединиц
Разработали три кассеты слияния множества субъединиц, содержащие кДНК трех бактериальных субъединиц в трех различных компоновках glcD-glcE-glcF, glcE-glcF-glcD и glcF-glcD-glcE, и синтезировали синтетические гены, кодирующие соответствующие полипептиды DEFp, EFDp и FDEp, соответственно. Перед синтезом, кодоны синтетических генов оптимизировали для максимального результата экспрессирования в соответствии с частотой использования кодона Brassica napus. Кроме того, на основании генетического алгоритма, синтетические гены одновременно оптимизировали по большому набору конкурирующих параметров, таких как вторичная структура мРНК, скрытые участки сплайсинга, повторения кодонов и мотивов, и гомогенного содержания GC.
ПРИМЕР 3: Комплементационный анализ мутантов E. coli испытывающих недостаток в трех субъединицах, образующих активную эндогенную гликолатдегидрогеназу
Чтобы определить, способны ли DEFp, EFDp и FDEp дополнять гликолатоксидазные мутанты E. coli, провели комплементационный анализ с мутантом E. coli JA155, который несет транспозонную вставку в субъединице glcD оперона glc и неспособен к росту на гликолате в качестве единственного источника углерода. Сверхэкспрессия DEFp, EFDp и FDEp в данном мутанте сохранила рост бактерий в среде, содержащей гликолат в качестве единственного источника углерода, показывая, что все три полипротеина являются функциональными in vivo и могут дополнять субъединицу glcD активного фермента EcGO.
ПРИМЕР 4: Субклонирование кДНК DEFp, EFDp и FDEp в растительные векторы экспрессии
Чтобы in vivo оценить действие бактериального полипротеина из множества субъединиц DEFp, EFDp и FDEp на активность GDH и производство биомассы в растениях N. tabacum cv. Petit Havanna SR1, кДНК, кодирующую DEFp, EFDp и FDEp, вставляли в растительный вектор экспрессии, обеспечивая нацеливание рекомбинантного белка на хлоропласты клеток растения. Трансгенной экспрессией управляли с помощью промотора CaMV 35S с дуплицированной областью энхансера.
Синтезированную кДНК DEFp первоначально вставляли в челночный вектор pTRAkc, используя сайты рестрикции EcoRI и XbaI перед терминатором CaMV 35S, генерируя плазмиду pTRA-nptll-DEFp. Плазмида pTRA содержит область крепления каркаса гена RB7 табака (gi3522871) и кассету nptll pPCV002 (Konz и Schell, 1986) для отбора трансгенных растений на канамицине (Фиг.2). Вслед за этим, конститутивный дважды усиленный промотор CaMV 35S, нетранслируемую 5’-область гена хальконсинтазы и последовательность направляющего пептида хлоропласта гена rbcS1 картофеля амплицифировали посредством ПЦР, используя в качестве шаблона плазмиду pTRAkc-rbcs1-cTP. Амплифицированный с помощью ПЦР фрагмент затем субклонировали в pTRA-nptll-DEF, используя сайты рестрикции AscI и AatII.
Клонирование кДНК EFDp и FDEp в растительный вектор экспрессии проводили аналогичным образом. Три итоговые конструкции обозначили: pTRA-35S-rbcs-cTP:DEFp, pTRA-35S-rbcs-cTP:EFDp и pTRA-35S-rbcs-cTP:FDEp, соответственно.
ПРИМЕР 5: Трансформация и регенерация растения табака
Растительные векторы экспрессии вводили в клетки GV3101 Agrobacterium tumefaciens, используя систему электропорации Gene Pulser II (BioRad, Hercules, CA, USA) согласно инструкциям производителя. Для исследования эффекта накопления DEFp, EFDp и FDEp в устойчиво трансформированных растениях табака (N. tabacum cv. Petit Havana SR1), трансгенные растения T0 генерировали посредством трансформации листового диска рекомбинантной A. tumefaciens (Horsch et al., 1985), используя в качестве маркера селекции канамицин. Полученные растения культивировали в парнике в стандартной почве DE73 с фотопериодом естественного дневного освещения 16 часов и температурой дневного времени 22°C /ночного времени 20°C.
До 33 трансгенных растений T0 скринировали на наличие трансгена и рекомбинантного белка с помощью множественной ПЦР и анализа методом иммуноблоттинга, соответственно. 33-48% исследованных линий продуцировали DEFp, EFDp или FDEp, соответственно, с ожидаемым размером молекул, составляющим 142 кДа. Семь линий T0, показывающих наивысший уровень рекомбинантных белков (от 0,03 до 0,09% общего растворимого белка), использовали для создания поколения T1.
ПРИМЕР 6: Выделение хлоропласта и ферментные анализы
Интактные хлоропласты выделяли, используя методику, описанную Kleffmann et al., 2007. Данные препараты свободны от действия загрязняющей каталазы ифумаразы (>95% чистоты).
Функцию гликолатдегидрогеназы измеряли, как описано у Lord J.M., 1972. 100 мкг экстракта белка хлоропласта добавляли к 100 мкмоль фосфата калия (pH 8,0), 0,2 мкмоль DCIP, 0,1 мл 1% (м/о) PMS, и 10 мкмоль гликолата калия в итоговом объеме, составляющем 2,4 мл. В фиксированные временные интервалы отдельные исследования завершали, добавляя 0,1 мл 12 M HCI. После выдерживания в течение 10 мин, добавляли 0,5 мл 0,1 M фенилгидразин-HCI. Обеспечивали возможность выдерживания смеси на протяжении дальнейших 10 мин, а затем торможение вследствие образования глиоксилатфенилгидразона измеряли при 324 нм.
ПРИМЕР 7: Высвобождение СО2 из меченого гликолата в экстрактах хлоропластов
1 мкCi [1,2-14C]-гликолата (Hartmann Analytics) добавляли в 50 мкг экстракта белка хлоропласта в плотно закупоренной 15-мл пробирке. Высвобожденный СО 2 абсорбировали в 500-мкл пробирке, содержащей 0,5 M NaOH, присоединенный к внутренней стенке 15-мл пробирки. Образцы инкубировали в течение 5 часов, при этом газовую фазу в пробирке часто перемешивали с помощью шприца.
ПРИМЕР 8: Оценка фенотипа растений, экспрессирующих GDH E.coli
Рост трансгенных растений, продуцирующих рекомбинантный белок DEFp, EFDp или FDEp в хлоропласте, отслеживали посредством измерения площади листьев согласно формуле:
Площадь листьев (см2) = 3,73 × (длина × ширина/100) + 0,011 × (длина × ширина/100)2.
Трансгенные линии T0 табака, конститутивно продуцирующие DEFp, EFDp или FDEp, соответственно, показали значительное увеличение площади листьев (1,54-, 1,75- и 1,5-кратно, соответственно p<0,05), по сравнению с нетрансгенными контрольными растениями (фиг.4). В дополнение, трансгенные растения имели больше листьев, чем дикий тип SR1, и дополнительные маленькие листья, помимо огромных.
Производительность фотосинтеза трансгенных растений отслеживали посредством Licor LI-6400 с помощью измерений наблюдаемой ассимиляции CO2 и компенсационной точки. Наблюдаемая норма ассимиляции CO2 в условиях окружающей среды была значительно сильнее у трансгенных растений DEFp и EFDp по сравнению с растениями дикого типа. Кроме того, трансгенные линии DEFp обладают значительным снижением (P<0,05) компенсационных точек CO2 (54 м.д. CO2) по сравнению с контрольным (64 м.д. CO2), указывая на более высокие скорости фотосинтеза для линий T0 DEFp.
Увеличенная биомасса и пониженное фотодыхание дополнительно подтверждались в поколении T1 (Таблица 1) и T2. Кроме того, производительность дикого типа и трансгенных линий, выращенных без добавления удобрений, анализировали в поколении Т1. В данных условиях, растения табака, сверхэкспрессирующие полипротеины DEFp, EFDp и FDEp, продемонстрировали уменьшенный хлороз и производство более высокой биомассы, чем контрольный дикий тип.
удобрения в воде
Взятые вместе, эти данные указывают на то, что растения, продуцирующие бактериальный полипротеин гликолатдегидрогеназу в своих хлоропластах, обладают значительно повышенной биомассой и улучшенной скоростью фотосинтеза.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<120> Способ увеличения фотосинтетической фиксации углерода,
с использованием белка слияния из многих субъединиц гликолатдегидрогеназы
<130> BCS 10-4005
<160> 6
<170> PatentIn version 3.4
<210> 1
<211> 1500
<212> ДНК
<213> Escherichia coli
<220>
<221> Кодирующая последовательность
<222> (1)..(1497)
<223> gcl D
<400> 1
atg agc atc ttg tac gaa gag cgt ctt gat ggc gct tta ccc gat gtc 48
Met Ser Ile Leu Tyr Glu Glu Arg Leu Asp Gly Ala Leu Pro Asp Val
1 5 10 15
gac cgc aca tcg gta ctg atg gca ctg cgt gag cat gtc cct gga ctt 96
Asp Arg Thr Ser Val Leu Met Ala Leu Arg Glu His Val Pro Gly Leu
20 25 30
gag atc ctg cat acc gat gag gag atc att cct tac gag tgt gac ggg 144
Glu Ile Leu His Thr Asp Glu Glu Ile Ile Pro Tyr Glu Cys Asp Gly
35 40 45
ttg agc gcg tat cgc acg cgt cca tta ctg gtt gtt ctg cct aag caa 192
Leu Ser Ala Tyr Arg Thr Arg Pro Leu Leu Val Val Leu Pro Lys Gln
50 55 60
atg gaa cag gtg aca gcg att ctg gct gtc tgc cat cgc ctg cgt gta 240
Met Glu Gln Val Thr Ala Ile Leu Ala Val Cys His Arg Leu Arg Val
65 70 75 80
ccg gtg gtg acc cgt ggt gca ggc acc ggg ctt tct ggt ggc gcg ctg 288
Pro Val Val Thr Arg Gly Ala Gly Thr Gly Leu Ser Gly Gly Ala Leu
85 90 95
ccg ctg gaa aaa ggt gtg ttg ttg gtg atg gcg cgc ttt aaa gag atc 336
Pro Leu Glu Lys Gly Val Leu Leu Val Met Ala Arg Phe Lys Glu Ile
100 105 110
ctc gac att aac ccc gtt ggt cgc cgc gcg cgc gtg cag cca ggc gtg 384
Leu Asp Ile Asn Pro Val Gly Arg Arg Ala Arg Val Gln Pro Gly Val
115 120 125
cgt aac ctg gcg atc tcc cag gcc gtt gca ccg cat aat ctc tac tac 432
Arg Asn Leu Ala Ile Ser Gln Ala Val Ala Pro His Asn Leu Tyr Tyr
130 135 140
gca ccg gac cct tcc tca caa atc gcc tgt tcc att ggc ggc aat gtg 480
Ala Pro Asp Pro Ser Ser Gln Ile Ala Cys Ser Ile Gly Gly Asn Val
145 150 155 160
gct gaa aat gcc ggc ggc gtc cac tgc ctg aaa tat ggt ctg acc gta 528
Ala Glu Asn Ala Gly Gly Val His Cys Leu Lys Tyr Gly Leu Thr Val
165 170 175
cat aac ctg ctg aaa att gaa gtg caa acg ctg gac ggc gag gca ctg 576
His Asn Leu Leu Lys Ile Glu Val Gln Thr Leu Asp Gly Glu Ala Leu
180 185 190
aca ctt gga tcg gac gcg ctg gat tca cct ggt ttt gac ctg ctg gcg 624
Thr Leu Gly Ser Asp Ala Leu Asp Ser Pro Gly Phe Asp Leu Leu Ala
195 200 205
ctg ttc acc gga tcg gaa ggt atg ctc ggc gtg acc acc gaa gtg acg 672
Leu Phe Thr Gly Ser Glu Gly Met Leu Gly Val Thr Thr Glu Val Thr
210 215 220
gta aaa ctg ctg ccg aag ccg ccc gtg gcg cgg gtt ctg tta gcc agc 720
Val Lys Leu Leu Pro Lys Pro Pro Val Ala Arg Val Leu Leu Ala Ser
225 230 235 240
ttt gac tcg gta gaa aaa gcc gga ctt gcg gtt ggt gac atc atc gcc 768
Phe Asp Ser Val Glu Lys Ala Gly Leu Ala Val Gly Asp Ile Ile Ala
245 250 255
aat ggc att atc ccc ggc ggg ctg gag atg atg gat aac ctg tcg atc 816
Asn Gly Ile Ile Pro Gly Gly Leu Glu Met Met Asp Asn Leu Ser Ile
260 265 270
cgc gcg gcg gaa gat ttt att cat gcc ggt tat ccc gtc gac gcc gaa 864
Arg Ala Ala Glu Asp Phe Ile His Ala Gly Tyr Pro Val Asp Ala Glu
275 280 285
gcg att ttg tta tgc gag ctg gac ggc gtg gag tct gac gta cag gaa 912
Ala Ile Leu Leu Cys Glu Leu Asp Gly Val Glu Ser Asp Val Gln Glu
290 295 300
gac tgc gag cgg gtt aac gac atc ttg ttg aaa gcg ggc gcg act gac 960
Asp Cys Glu Arg Val Asn Asp Ile Leu Leu Lys Ala Gly Ala Thr Asp
305 310 315 320
gtc cgt ctg gca cag gac gaa gca gag cgc gta cgt ttc tgg gcc ggt 1008
Val Arg Leu Ala Gln Asp Glu Ala Glu Arg Val Arg Phe Trp Ala Gly
325 330 335
cgc aaa aat gcg ttc ccg gcg gta gga cgt atc tcc ccg gat tac tac 1056
Arg Lys Asn Ala Phe Pro Ala Val Gly Arg Ile Ser Pro Asp Tyr Tyr
340 345 350
tgc atg gat ggc acc atc ccg cgt cgc gcc ctg cct ggc gta ctg gaa 1104
Cys Met Asp Gly Thr Ile Pro Arg Arg Ala Leu Pro Gly Val Leu Glu
355 360 365
ggc att gcc cgt tta tcg cag caa tat gat tta cgt gtt gcc aac gtc 1152
Gly Ile Ala Arg Leu Ser Gln Gln Tyr Asp Leu Arg Val Ala Asn Val
370 375 380
ttt cat gcc gga gat ggc aac atg cac ccg tta atc ctt ttc gat gcc 1200
Phe His Ala Gly Asp Gly Asn Met His Pro Leu Ile Leu Phe Asp Ala
385 390 395 400
aac gaa ccc ggt gaa ttt gcc cgc gcg gaa gag ctg ggc ggg aag atc 1248
Asn Glu Pro Gly Glu Phe Ala Arg Ala Glu Glu Leu Gly Gly Lys Ile
405 410 415
ctc gaa ctc tgc gtt gaa gtt ggc ggc agc atc agt ggc gaa cat ggc 1296
Leu Glu Leu Cys Val Glu Val Gly Gly Ser Ile Ser Gly Glu His Gly
420 425 430
atc ggg cga gaa aaa atc aat caa atg tgc gcc cag ttc aac agc gat 1344
Ile Gly Arg Glu Lys Ile Asn Gln Met Cys Ala Gln Phe Asn Ser Asp
435 440 445
gaa atc acg acc ttc cat gcg gtc aag gcg gcg ttt gac ccc gat ggt 1392
Glu Ile Thr Thr Phe His Ala Val Lys Ala Ala Phe Asp Pro Asp Gly
450 455 460
ttg ctg aac cct ggg aaa aac att ccc acg cta cac cgc tgt gct gaa 1440
Leu Leu Asn Pro Gly Lys Asn Ile Pro Thr Leu His Arg Cys Ala Glu
465 470 475 480
ttt ggt gcc atg cat gtg cat cac ggt cat tta cct ttc cct gaa ctg 1488
Phe Gly Ala Met His Val His His Gly His Leu Pro Phe Pro Glu Leu
485 490 495
gag cgt ttc tga 1500
Glu Arg Phe
<210> 2
<211> 499
<212> Белок
<213> Escherichia coli
<400> 2
Met Ser Ile Leu Tyr Glu Glu Arg Leu Asp Gly Ala Leu Pro Asp Val
1 5 10 15
Asp Arg Thr Ser Val Leu Met Ala Leu Arg Glu His Val Pro Gly Leu
20 25 30
Glu Ile Leu His Thr Asp Glu Glu Ile Ile Pro Tyr Glu Cys Asp Gly
35 40 45
Leu Ser Ala Tyr Arg Thr Arg Pro Leu Leu Val Val Leu Pro Lys Gln
50 55 60
Met Glu Gln Val Thr Ala Ile Leu Ala Val Cys His Arg Leu Arg Val
65 70 75 80
Pro Val Val Thr Arg Gly Ala Gly Thr Gly Leu Ser Gly Gly Ala Leu
85 90 95
Pro Leu Glu Lys Gly Val Leu Leu Val Met Ala Arg Phe Lys Glu Ile
100 105 110
Leu Asp Ile Asn Pro Val Gly Arg Arg Ala Arg Val Gln Pro Gly Val
115 120 125
Arg Asn Leu Ala Ile Ser Gln Ala Val Ala Pro His Asn Leu Tyr Tyr
130 135 140
Ala Pro Asp Pro Ser Ser Gln Ile Ala Cys Ser Ile Gly Gly Asn Val
145 150 155 160
Ala Glu Asn Ala Gly Gly Val His Cys Leu Lys Tyr Gly Leu Thr Val
165 170 175
His Asn Leu Leu Lys Ile Glu Val Gln Thr Leu Asp Gly Glu Ala Leu
180 185 190
Thr Leu Gly Ser Asp Ala Leu Asp Ser Pro Gly Phe Asp Leu Leu Ala
195 200 205
Leu Phe Thr Gly Ser Glu Gly Met Leu Gly Val Thr Thr Glu Val Thr
210 215 220
Val Lys Leu Leu Pro Lys Pro Pro Val Ala Arg Val Leu Leu Ala Ser
225 230 235 240
Phe Asp Ser Val Glu Lys Ala Gly Leu Ala Val Gly Asp Ile Ile Ala
245 250 255
Asn Gly Ile Ile Pro Gly Gly Leu Glu Met Met Asp Asn Leu Ser Ile
260 265 270
Arg Ala Ala Glu Asp Phe Ile His Ala Gly Tyr Pro Val Asp Ala Glu
275 280 285
Ala Ile Leu Leu Cys Glu Leu Asp Gly Val Glu Ser Asp Val Gln Glu
290 295 300
Asp Cys Glu Arg Val Asn Asp Ile Leu Leu Lys Ala Gly Ala Thr Asp
305 310 315 320
Val Arg Leu Ala Gln Asp Glu Ala Glu Arg Val Arg Phe Trp Ala Gly
325 330 335
Arg Lys Asn Ala Phe Pro Ala Val Gly Arg Ile Ser Pro Asp Tyr Tyr
340 345 350
Cys Met Asp Gly Thr Ile Pro Arg Arg Ala Leu Pro Gly Val Leu Glu
355 360 365
Gly Ile Ala Arg Leu Ser Gln Gln Tyr Asp Leu Arg Val Ala Asn Val
370 375 380
Phe His Ala Gly Asp Gly Asn Met His Pro Leu Ile Leu Phe Asp Ala
385 390 395 400
Asn Glu Pro Gly Glu Phe Ala Arg Ala Glu Glu Leu Gly Gly Lys Ile
405 410 415
Leu Glu Leu Cys Val Glu Val Gly Gly Ser Ile Ser Gly Glu His Gly
420 425 430
Ile Gly Arg Glu Lys Ile Asn Gln Met Cys Ala Gln Phe Asn Ser Asp
435 440 445
Glu Ile Thr Thr Phe His Ala Val Lys Ala Ala Phe Asp Pro Asp Gly
450 455 460
Leu Leu Asn Pro Gly Lys Asn Ile Pro Thr Leu His Arg Cys Ala Glu
465 470 475 480
Phe Gly Ala Met His Val His His Gly His Leu Pro Phe Pro Glu Leu
485 490 495
Glu Arg Phe
<210> 3
<211> 1053
<212> ДНК
<213> Escherichia coli
<220>
<221> Кодирующая последовательность
<222> (1)..(1050)
<223> gcl E
<400> 3
atg cta cgc gag tgt gat tac agc cag gcg ctg ctg gag cag gtg aat 48
Met Leu Arg Glu Cys Asp Tyr Ser Gln Ala Leu Leu Glu Gln Val Asn
1 5 10 15
cag gcg att agc gat aaa acg ccg ctg gtg att cag ggc agc aat agc 96
Gln Ala Ile Ser Asp Lys Thr Pro Leu Val Ile Gln Gly Ser Asn Ser
20 25 30
aaa gcc ttt tta ggt cgc cct gtc acc ggg caa acg ctg gat gtt cgt 144
Lys Ala Phe Leu Gly Arg Pro Val Thr Gly Gln Thr Leu Asp Val Arg
35 40 45
tgt cat cgc ggc att gtt aat tac gac ccg acc gag ctg gtg ata acc 192
Cys His Arg Gly Ile Val Asn Tyr Asp Pro Thr Glu Leu Val Ile Thr
50 55 60
gcg cgt gtc gga acg ccg ctg gtg aca att gaa gcg gcg ctg gaa agc 240
Ala Arg Val Gly Thr Pro Leu Val Thr Ile Glu Ala Ala Leu Glu Ser
65 70 75 80
gcg ggg caa atg ctc ccc tgt gag ccg ccg cat tat ggt gaa gaa gcc 288
Ala Gly Gln Met Leu Pro Cys Glu Pro Pro His Tyr Gly Glu Glu Ala
85 90 95
acc tgg ggc ggg atg gtc gcc tgc ggg ctg gcg ggg ccg cgt cgc ccg 336
Thr Trp Gly Gly Met Val Ala Cys Gly Leu Ala Gly Pro Arg Arg Pro
100 105 110
tgg agc ggt tcg gtc cgc gat ttt gtc ctc ggc acg cgc atc att acc 384
Trp Ser Gly Ser Val Arg Asp Phe Val Leu Gly Thr Arg Ile Ile Thr
115 120 125
ggc gct gga aaa cat ctg cgt ttt ggt ggc gaa gtg atg aaa aac gtt 432
Gly Ala Gly Lys His Leu Arg Phe Gly Gly Glu Val Met Lys Asn Val
130 135 140
gcc gga tac gat ctc tca cgg tta atg gtc gga agc tac ggt tgt ctt 480
Ala Gly Tyr Asp Leu Ser Arg Leu Met Val Gly Ser Tyr Gly Cys Leu
145 150 155 160
ggc gtg ctc act gaa atc tca atg aaa gtg tta ccg cga ccg cgc gcc 528
Gly Val Leu Thr Glu Ile Ser Met Lys Val Leu Pro Arg Pro Arg Ala
165 170 175
tcc ctg agc ctg cgt cgg gaa atc agc ctg caa gaa gcc atg agt gaa 576
Ser Leu Ser Leu Arg Arg Glu Ile Ser Leu Gln Glu Ala Met Ser Glu
180 185 190
atc gcc gag tgg caa ctc cag cca tta ccc att agt ggc tta tgt tac 624
Ile Ala Glu Trp Gln Leu Gln Pro Leu Pro Ile Ser Gly Leu Cys Tyr
195 200 205
ttc gac aat gcg ttg tgg atc cgc ctt gag ggc ggc gaa gga tcg gta 672
Phe Asp Asn Ala Leu Trp Ile Arg Leu Glu Gly Gly Glu Gly Ser Val
210 215 220
aaa gca gcg cgt gaa ctg ctg ggt ggc gaa gag gtt gcc ggt cag ttc 720
Lys Ala Ala Arg Glu Leu Leu Gly Gly Glu Glu Val Ala Gly Gln Phe
225 230 235 240
tgg cag caa ttg cgt gaa caa caa ctg ccg ttc ttc tcg tta cca ggt 768
Trp Gln Gln Leu Arg Glu Gln Gln Leu Pro Phe Phe Ser Leu Pro Gly
245 250 255
acc tta tgg cgc att tca tta ccc agt gat gcg ccg atg atg gat tta 816
Thr Leu Trp Arg Ile Ser Leu Pro Ser Asp Ala Pro Met Met Asp Leu
260 265 270
ccc ggc gag caa ctg atc gac tgg ggc ggg gcg tta cgc tgg ctg aaa 864
Pro Gly Glu Gln Leu Ile Asp Trp Gly Gly Ala Leu Arg Trp Leu Lys
275 280 285
tcg aca gcc gag gac aat caa atc cat cgc atc gcc cgc aac gct ggc 912
Ser Thr Ala Glu Asp Asn Gln Ile His Arg Ile Ala Arg Asn Ala Gly
290 295 300
ggt cat gcg acc cgc ttt agt gcc gga gat ggt ggc ttt gcc ccg cta 960
Gly His Ala Thr Arg Phe Ser Ala Gly Asp Gly Gly Phe Ala Pro Leu
305 310 315 320
tcg gct cct tta ttc cgc tat cac cag cag ctt aaa cag cag ctc gac 1008
Ser Ala Pro Leu Phe Arg Tyr His Gln Gln Leu Lys Gln Gln Leu Asp
325 330 335
cct tgc ggc gtg ttt aac ccc ggt cgc atg tac gcg gaa ctt tga 1053
Pro Cys Gly Val Phe Asn Pro Gly Arg Met Tyr Ala Glu Leu
340 345 350
<210> 4
<211> 350
<212> Белок
<213> Escherichia coli
<400> 4
Met Leu Arg Glu Cys Asp Tyr Ser Gln Ala Leu Leu Glu Gln Val Asn
1 5 10 15
Gln Ala Ile Ser Asp Lys Thr Pro Leu Val Ile Gln Gly Ser Asn Ser
20 25 30
Lys Ala Phe Leu Gly Arg Pro Val Thr Gly Gln Thr Leu Asp Val Arg
35 40 45
Cys His Arg Gly Ile Val Asn Tyr Asp Pro Thr Glu Leu Val Ile Thr
50 55 60
Ala Arg Val Gly Thr Pro Leu Val Thr Ile Glu Ala Ala Leu Glu Ser
65 70 75 80
Ala Gly Gln Met Leu Pro Cys Glu Pro Pro His Tyr Gly Glu Glu Ala
85 90 95
Thr Trp Gly Gly Met Val Ala Cys Gly Leu Ala Gly Pro Arg Arg Pro
100 105 110
Trp Ser Gly Ser Val Arg Asp Phe Val Leu Gly Thr Arg Ile Ile Thr
115 120 125
Gly Ala Gly Lys His Leu Arg Phe Gly Gly Glu Val Met Lys Asn Val
130 135 140
Ala Gly Tyr Asp Leu Ser Arg Leu Met Val Gly Ser Tyr Gly Cys Leu
145 150 155 160
Gly Val Leu Thr Glu Ile Ser Met Lys Val Leu Pro Arg Pro Arg Ala
165 170 175
Ser Leu Ser Leu Arg Arg Glu Ile Ser Leu Gln Glu Ala Met Ser Glu
180 185 190
Ile Ala Glu Trp Gln Leu Gln Pro Leu Pro Ile Ser Gly Leu Cys Tyr
195 200 205
Phe Asp Asn Ala Leu Trp Ile Arg Leu Glu Gly Gly Glu Gly Ser Val
210 215 220
Lys Ala Ala Arg Glu Leu Leu Gly Gly Glu Glu Val Ala Gly Gln Phe
225 230 235 240
Trp Gln Gln Leu Arg Glu Gln Gln Leu Pro Phe Phe Ser Leu Pro Gly
245 250 255
Thr Leu Trp Arg Ile Ser Leu Pro Ser Asp Ala Pro Met Met Asp Leu
260 265 270
Pro Gly Glu Gln Leu Ile Asp Trp Gly Gly Ala Leu Arg Trp Leu Lys
275 280 285
Ser Thr Ala Glu Asp Asn Gln Ile His Arg Ile Ala Arg Asn Ala Gly
290 295 300
Gly His Ala Thr Arg Phe Ser Ala Gly Asp Gly Gly Phe Ala Pro Leu
305 310 315 320
Ser Ala Pro Leu Phe Arg Tyr His Gln Gln Leu Lys Gln Gln Leu Asp
325 330 335
Pro Cys Gly Val Phe Asn Pro Gly Arg Met Tyr Ala Glu Leu
340 345 350
<210> 5
<211> 1224
<212> ДНК
<213> Escherichia coli
<220>
<221> Кодирующая последовательность
<222> (1)..(1221)
<223> gcl F
<400> 5
atg caa acc caa tta act gaa gag atg cgg cag aac gcg cgc gcg ctg 48
Met Gln Thr Gln Leu Thr Glu Glu Met Arg Gln Asn Ala Arg Ala Leu
1 5 10 15
gaa gcc gac agc atc ctg cgc gcc tgt gtt cac tgc gga ttt tgt acc 96
Glu Ala Asp Ser Ile Leu Arg Ala Cys Val His Cys Gly Phe Cys Thr
20 25 30
gca acc tgc cca acc tat cag ctt ctg ggc gat gaa ctg gac ggg ccg 144
Ala Thr Cys Pro Thr Tyr Gln Leu Leu Gly Asp Glu Leu Asp Gly Pro
35 40 45
cgc ggg cgc atc tat ctg att aaa cag gtg ctg gaa ggc aac gaa gtc 192
Arg Gly Arg Ile Tyr Leu Ile Lys Gln Val Leu Glu Gly Asn Glu Val
50 55 60
acg ctt aaa aca cag gag cat ctc gat cgc tgc ctc act tgc cgt aat 240
Thr Leu Lys Thr Gln Glu His Leu Asp Arg Cys Leu Thr Cys Arg Asn
65 70 75 80
tgt gaa acc acc tgt cct tct ggt gtg cgc tat cac aat ttg ctg gat 288
Cys Glu Thr Thr Cys Pro Ser Gly Val Arg Tyr His Asn Leu Leu Asp
85 90 95
atc ggg cgt gat att gtc gag cag aaa gtg aaa cgc cca ctg ccg gag 336
Ile Gly Arg Asp Ile Val Glu Gln Lys Val Lys Arg Pro Leu Pro Glu
100 105 110
cga ata ctg cgc gaa gga ttg cgc cag gta gtg ccg cgt ccg gcg gtc 384
Arg Ile Leu Arg Glu Gly Leu Arg Gln Val Val Pro Arg Pro Ala Val
115 120 125
ttc cgt gcg ctg acg cag gta ggg ctg gtg ctg cga ccg ttt tta ccg 432
Phe Arg Ala Leu Thr Gln Val Gly Leu Val Leu Arg Pro Phe Leu Pro
130 135 140
gaa cag gtc aga gca aaa ctg cct gct gaa acg gtg aaa gct aaa ccg 480
Glu Gln Val Arg Ala Lys Leu Pro Ala Glu Thr Val Lys Ala Lys Pro
145 150 155 160
cgt ccg ccg ctg cgc cat aag cgt cgg gtt tta atg ttg gaa ggc tgc 528
Arg Pro Pro Leu Arg His Lys Arg Arg Val Leu Met Leu Glu Gly Cys
165 170 175
gcc cag cct acg ctt tcg ccc aac acc aac gcg gca act gcg cga gtg 576
Ala Gln Pro Thr Leu Ser Pro Asn Thr Asn Ala Ala Thr Ala Arg Val
180 185 190
ctg gat cgt ctg ggg atc agc gtc atg cca gct aac gaa gca ggc tgt 624
Leu Asp Arg Leu Gly Ile Ser Val Met Pro Ala Asn Glu Ala Gly Cys
195 200 205
tgt ggc gcg gtg gac tat cat ctt aat gcg cag gag aaa ggg ctg gca 672
Cys Gly Ala Val Asp Tyr His Leu Asn Ala Gln Glu Lys Gly Leu Ala
210 215 220
cgg gcg cgc aat aat att gat gcc tgg tgg ccc gcg att gaa gca ggt 720
Arg Ala Arg Asn Asn Ile Asp Ala Trp Trp Pro Ala Ile Glu Ala Gly
225 230 235 240
gcc gag gca att ttg caa acc gcc agc ggc tgc ggc gcg ttt gtc aaa 768
Ala Glu Ala Ile Leu Gln Thr Ala Ser Gly Cys Gly Ala Phe Val Lys
245 250 255
gag tat ggg cag atg ctg aaa aac gat gcg tta tat gcc gat aaa gca 816
Glu Tyr Gly Gln Met Leu Lys Asn Asp Ala Leu Tyr Ala Asp Lys Ala
260 265 270
cgt cag gtc agt gaa ctg gcg gtc gat tta gtc gaa ctt ctg cgc gag 864
Arg Gln Val Ser Glu Leu Ala Val Asp Leu Val Glu Leu Leu Arg Glu
275 280 285
gaa ccg ctg gaa aaa ctg gca att cgc ggc gat aaa aag ctg gcc ttc 912
Glu Pro Leu Glu Lys Leu Ala Ile Arg Gly Asp Lys Lys Leu Ala Phe
290 295 300
cac tgt ccg tgt acc cta caa cat gcg caa aag ctg aac ggc gaa gtg 960
His Cys Pro Cys Thr Leu Gln His Ala Gln Lys Leu Asn Gly Glu Val
305 310 315 320
gaa aaa gtg ttg ctt cgt ctt gga ttt acc tta acg gac gtt ccc gac 1008
Glu Lys Val Leu Leu Arg Leu Gly Phe Thr Leu Thr Asp Val Pro Asp
325 330 335
agc cat ctg tgc tgc ggt tca gcg gga aca tat gcg tta acg cat ccc 1056
Ser His Leu Cys Cys Gly Ser Ala Gly Thr Tyr Ala Leu Thr His Pro
340 345 350
gat ctg gca cgc cag ctg cgg gat aac aaa atg aat gcg ctg gaa agc 1104
Asp Leu Ala Arg Gln Leu Arg Asp Asn Lys Met Asn Ala Leu Glu Ser
355 360 365
ggc aaa ccg gaa atg atc gtc acc gcc aac att ggt tgc cag acg cat 1152
Gly Lys Pro Glu Met Ile Val Thr Ala Asn Ile Gly Cys Gln Thr His
370 375 380
ctg gcg agc gcc ggt cgt acc tct gtg cgt cac tgg att gaa att gta 1200
Leu Ala Ser Ala Gly Arg Thr Ser Val Arg His Trp Ile Glu Ile Val
385 390 395 400
gaa caa gcc ctt gaa aag gaa taa 1224
Glu Gln Ala Leu Glu Lys Glu
405
<210> 6
<211> 407
<212> Белок
<213> Escherichia coli
<400> 6
Met Gln Thr Gln Leu Thr Glu Glu Met Arg Gln Asn Ala Arg Ala Leu
1 5 10 15
Glu Ala Asp Ser Ile Leu Arg Ala Cys Val His Cys Gly Phe Cys Thr
20 25 30
Ala Thr Cys Pro Thr Tyr Gln Leu Leu Gly Asp Glu Leu Asp Gly Pro
35 40 45
Arg Gly Arg Ile Tyr Leu Ile Lys Gln Val Leu Glu Gly Asn Glu Val
50 55 60
Thr Leu Lys Thr Gln Glu His Leu Asp Arg Cys Leu Thr Cys Arg Asn
65 70 75 80
Cys Glu Thr Thr Cys Pro Ser Gly Val Arg Tyr His Asn Leu Leu Asp
85 90 95
Ile Gly Arg Asp Ile Val Glu Gln Lys Val Lys Arg Pro Leu Pro Glu
100 105 110
Arg Ile Leu Arg Glu Gly Leu Arg Gln Val Val Pro Arg Pro Ala Val
115 120 125
Phe Arg Ala Leu Thr Gln Val Gly Leu Val Leu Arg Pro Phe Leu Pro
130 135 140
Glu Gln Val Arg Ala Lys Leu Pro Ala Glu Thr Val Lys Ala Lys Pro
145 150 155 160
Arg Pro Pro Leu Arg His Lys Arg Arg Val Leu Met Leu Glu Gly Cys
165 170 175
Ala Gln Pro Thr Leu Ser Pro Asn Thr Asn Ala Ala Thr Ala Arg Val
180 185 190
Leu Asp Arg Leu Gly Ile Ser Val Met Pro Ala Asn Glu Ala Gly Cys
195 200 205
Cys Gly Ala Val Asp Tyr His Leu Asn Ala Gln Glu Lys Gly Leu Ala
210 215 220
Arg Ala Arg Asn Asn Ile Asp Ala Trp Trp Pro Ala Ile Glu Ala Gly
225 230 235 240
Ala Glu Ala Ile Leu Gln Thr Ala Ser Gly Cys Gly Ala Phe Val Lys
245 250 255
Glu Tyr Gly Gln Met Leu Lys Asn Asp Ala Leu Tyr Ala Asp Lys Ala
260 265 270
Arg Gln Val Ser Glu Leu Ala Val Asp Leu Val Glu Leu Leu Arg Glu
275 280 285
Glu Pro Leu Glu Lys Leu Ala Ile Arg Gly Asp Lys Lys Leu Ala Phe
290 295 300
His Cys Pro Cys Thr Leu Gln His Ala Gln Lys Leu Asn Gly Glu Val
305 310 315 320
Glu Lys Val Leu Leu Arg Leu Gly Phe Thr Leu Thr Asp Val Pro Asp
325 330 335
Ser His Leu Cys Cys Gly Ser Ala Gly Thr Tyr Ala Leu Thr His Pro
340 345 350
Asp Leu Ala Arg Gln Leu Arg Asp Asn Lys Met Asn Ala Leu Glu Ser
355 360 365
Gly Lys Pro Glu Met Ile Val Thr Ala Asn Ile Gly Cys Gln Thr His
370 375 380
Leu Ala Ser Ala Gly Arg Thr Ser Val Arg His Trp Ile Glu Ile Val
385 390 395 400
Glu Gln Ala Leu Glu Lys Glu
405










Изобретение относится к области биохимии, в частности к нуклеиновой кислоте, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы, а также трансгенной клетке растения, трансгенному растению и трансгенному семени, содержащим вышеуказанную нуклеиновую кислоту. Также раскрыт способ увеличения производства биомассы и фиксации углерода в растениях по сравнению с диким видом растения, включающий введение в геном клетки растения нуклеиновой кислоты, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы. Изобретение позволяет эффективно получать растение с увеличенным производством биомассы и фиксации углерода по сравнению с диким видом растения. 5 н. и 14 з.п. ф-лы, 2 ил., 1 табл., 8 пр.
1. Способ увеличения производства биомассы и фиксации углерода в растениях по сравнению с диким видом растения, включающий введение в геном клетки растения нуклеиновой кислоты, кодирующей белок слияния из многих субъединиц гликолатдегидрогеназы, в котором результатом указанного введения указанной одной нуклеиновой кислоты является экспрессирование de novo одного полипептида, имеющего ферментативную активность гликолатдегидрогеназы, где указанный один полипептид локализован в хлоропластах полученного растения и где белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотные последовательности, имеющие по меньшей мере 95% идентичность последовательностям SEQ ID NO: 2, 4 и 6 соответственно.
2. Способ по п. 1, в котором осуществляют указанное введение указанной одной нуклеиновой кислоты в ядерный геном клеток растения и в котором указанная одна нуклеиновая кислота кодирует один полипептид, содержащий фрагмент аминокислоты, который нацеливает полипептид в хлоропласт.
3. Способ по п. 1 или 2, в котором белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотные последовательности SEQ ID NO: 2, 4 и 6 соответственно.
4. Способ по любому из пп. 1-2, где нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, содержит полинуклеотидные последовательности, имеющие по меньшей мере 95% идентичность полинуклеотидным последовательностям SEQ ID NO: 1, 3 и 5 соответственно.
5. Способ по п. 4, в котором нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, содержит полинуклеотидные последовательности SEQ ID NO: 1, 3 и 5 соответственно.
6. Нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, где указанный белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотные последовательности, имеющие по меньшей мере 95% идентичность последовательностям SEQ ID NO: 2, 4 и 6 соответственно.
7. Нуклеиновая кислота по п. 6, в которой белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотную последовательность, которая нацеливает указанный белок в хлоропласт.
8. Нуклеиновая кислота по п. 6 или 7, в которой указанный белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотные последовательности SEQ ID NO: 2, 4 и 6 соответственно.
9. Нуклеиновая кислота по п. 6 или 7, где нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, содержит полинуклеотидные последовательности, имеющие по меньшей мере 95% идентичность полинуклеотидным последовательностям SEQ ID NO: 1, 3 и 5 соответственно.
10. Нуклеиновая кислота по п. 9, в которой нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, содержит полинуклеотидные последовательности SEQ ID NO: 1, 3 и 5 соответственно.
11. Трансгенная клетка растения для получения трансгенного растения с увеличенным производством биомассы и фиксацией углерода по сравнению с клеткой дикого вида растения, содержащая нуклеиновую кислоту, кодирующую белок слияния из многих субъединиц гликолатдегидрогеназы, где указанный белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотные последовательности, имеющие по меньшей мере 95% идентичность последовательностям SEQ ID NO: 2, 4 и 6 соответственно.
12. Трансгенная клетка растения по п. 11, в которой указанный белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотные последовательности SEQ ID NO: 2, 4 и 6 соответственно.
13. Трансгенная клетка растения по п. 11 или 12, где нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, содержит полинуклеотидные последовательности, имеющие по меньшей мере 95% идентичность полинуклеотидным последовательностям SEQ ID NO: 1, 3 и 5 соответственно.
14. Трансгенная клетка растения по п. 13, в которой нуклеиновая кислота, кодирующая белок слияния из многих субъединиц гликолатдегидрогеназы, содержит полинуклеотидные последовательности SEQ ID NO: 1, 3 и 5 соответственно.
15. Трансгенная клетка растения по п. 11 или 12, в которой белок слияния из многих субъединиц гликолатдегидрогеназы содержит аминокислотную последовательность, которая нацеливает указанный белок в хлоропласт.
16. Трансгенное растение с увеличенным производством биомассы и фиксацией углерода по сравнению с диким видом растения, содержащее трансгенную клетку растения по любому одному из пп. 11-15.
17. Трансгенное растение по п. 16, выбранное из риса, пшеницы, ячменя, картофеля, рапса, табака.
18. Трансгенное семя для получения растения с увеличенным производством биомассы и фиксацией углерода по сравнению с диким видом растения, содержащие трансгенную клетку растения по любому из пп. 11-15.
19. Трансгенное семя по п. 18, выбранное из риса, пшеницы, ячменя, рапса, табака.
| Четырехтактный реверсивный распределитель импульсов для управления шаговым двигателем | 1986 |
|
SU1367127A1 |
| RASHAD KEBEISH et al., Chloroplastic photorespiratory bypass increases photosynthesis and biomass production in Arabidopsis thaliana, NATURE BIOTECHNOLOGY, 2007, Vol | |||
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| AJAY K | |||
| GARG et al., Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses, Current Issue, 2002, vol | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Прибор для нанесения точек геодолитных ходов по дирекционным углам и расстояниям | 1929 |
|
SU15898A1 |
| МОЛЕКУЛЫ ДНК, КОДИРУЮЩИЕ ПЛАСТИДНЫЙ ТРАНСЛОКАТОР 2-ОКСОГЛУТАРАТ/МАЛАТ, РЕКОМБИНАНТНАЯ МОЛЕКУЛА ДНК, ТРАНСФОРМИРОВАННЫЕ БАКТЕРИЯ И ДРОЖЖИ И ПРОТЕИН, ОБЛАДАЮЩИЙ АКТИВНОСТЬЮ ПЛАСТИДНОГО ТРАНСЛОКАТОРА 2-ОКСОГЛУТАРАТ/МАЛАТ | 1995 |
|
RU2188866C2 |

