Область техники, к которой относится изобретение
Изобретение относится к области биоинженерии, в частности к способу направленной коррекции мутаций митохондриального генома человека с применением генетически модифицированных штаммов Yarrowia lipolytica с искусственно приданной способностью к гомологичной рекомбинации митоходриальной ДНК и введенными маркерами для фенотипической селекции наличия/отсутствия дефектов в генах тРНК-Lys и тРНК-Leu митохондриального генома. Изобретение используется для коррекции вредных мутаций митохондриального генома человека и препаративной наработки скорректированной по последовательности интактной митохондриальной ДНК в клетках дрожжей с целью ее последующего введения в мезенхимальные стромальные клетки пациента, что позволяет обеспечить лечение наиболее тяжелых распространенных митохондриальных болезней человека (например, MELAS и MERFF), нарушающих функциональность генов митохондриальных Lys- Leu-специфичных рРНК.
Уровень техники
Активное изучение митохондриального генома позволило выделить целый класс болезней, в основе которых лежат мутации генов митохондриального генома. Эти мутации способны вовлекать тРНК, рРНК или структурные гены и могут выражаться биохимически как дефекты всей электронно-транспортной цепи или как дефекты отдельных ферментов. Мутационные изменения в мтДНК могут приводить к возникновению митохондриальных наследственных болезней человека, связанных с нарушением процессов окислительного фосфорилирования и энергетического обмена в клетках. Примерно две трети (166 мутаций) патогенных точечных мутаций мтДНК локализованы в генах тРНК, они проявляются в виде разнообразных клинических синдромов. Наиболее распространены мутации в генах тРНКLeu и тРНКLys. Так, в основе синдрома MELAS (митохондриальная энцефалопатия с инсультоподобными эпизодами и лактатацидозом) лежат точечные мутации мтДНК. Примерно 80% из них - в гене мтДНК, кодирующем лейциновую тРНК митохондрий - в гене тРНК лейцина (UUR), (tRNA-Leu), где происходит однонуклеотидная замена А ~> G в 3243-м положении (А3243G tRNA leu). Эта мутация приводит к инактивации транскрипционного терминатора, заключенного внутри гена тРНК Leu. Последствием этой мутации является изменение транскрипционного соотношения рРНК и мРНК и снижение эффективности трансляции. Мутации могут возникать впервые у конкретного пациента либо наследоваться по материнской линии. Всего обнаружено 23 точечных мутаций и 4 делеции мтДНК, приводящих к MELAS [Рассел Д. Синдром MELAS. Изд.: "VSD". 2013. ISBN: 978-5-5097-2091-8]. Другая наиболее распространенная мутация – транзиция A8344G в гене тРНКLys, в 80% случаев ассоциирована с MERRF (миоклональная эпилепсия с “разорванными красными мышечными волокнами) [Мазунин И.О. и др. Мол. биол. 2010. Т. 44. № 5. C. 755–772.3]. В результате этой мутации изменяется высококонсервативный нуклеотид в псевдоуридиновой петле тРНК, что приводит к блокированию митохондриального синтеза белка.
В настоящее время ни в одной из опубликованных печатных работ, включая патентные заявки, не предложено способов направленного воздействия на митохондриальный геном человека [Di Donato, S. et al . J. Neurol. 2009. V. 256. P. 693–710; Harbauer, A.B. et al. Cell Metab. 2014. V. 4. P. 357–372; Мазунин И.О. и др. Мол. биол. 2010. Т. 44. № 5. C. 755–772; Spelbrink, J.N. et al. IUBMB Life. 2010. V. 62. P. 19–32; Тодоров, И.Н. и др. Биохимия. 2009. Т. 74. C. 1184–1194].
Этиотропные средства лечения митохондриальных болезней человека отсутствуют [Harbauer, A.B. et al. Cell Metab. 2014. V. 4. P. 357–372; Мазунин И.О. и др. Мол. биол. 2010. Т. 44. № 5. C. 755–772; Патрушев М.В. и др. Биохимия. 2014, Т. 79 (11). C. 1417 – 1428.; de Laat, P. et al. JIMD Rep. 2013. V. 8. P. 47–50.; Swalwell, H. et al. Eur J Hum Genet. 2011. V. 19(7). P. 769-75.].
Все существующие и перспективные средства лечения этих болезней основаны на купировании последствий митохондриальной недостаточности [Мазунин И.О. и др. Мол. биол. 2010. Т. 44. № 5. C. 755–772.; Carelli, V. et al. Biochim. Biophys. Acta. 2009. V. 1787. P. 518–528.; Yu-Wai-Man, P. et al. J. Med. Genet. 2009. V. 46. P. 145–158.; Rotig, A. Diabetes Metab. 2010. V. 36. P. 97–107.; A Human Mitochondrial Genome Database. 2009. www. mitomap.org.].
При этом разработка средств направленной коррекции генома митохондрий создает принципиально новые возможности для практической медицины [Никитина Л.П. и др. Забайкальский медицинский вестник. 2011. № 2. C. 184-190.; Егорова Л.А. и др. Клиницист. 2013. № 2. C. 6-13.; Рассел Д Изд.: "VSD". 2013. ISBN: 978-5-5097-2091-8.; Прохорова Н.С. и др. Таврический медико-биологический вестник 2010. Т. 13. № 4 (52). C. 146-149.].
В основном, в имеющихся патентах освещены способы медикаментозной коррекции митохондриальных дисфункций, в частности синдрома MELAS и болезни Паркинсона. В этих заявках не упоминаются ни способы воздействия на геном митохондрии, ни методы репликации генома митохондрий человека в гетерологичных системах (в том числе дрожжах). Данные изобретения направлены на медикаментозное лечение заболеваний, связанных с дисфункцией митохондрий [WO/2015/144964; US2015196500; IN515DEN2015], а также на разработку методов тестирования и поиск маркеров митохондриальных нейродегенеративных заболеваний [WO/2015/144964; CN101818193], с возможным прогнозированием заболеваний у детей [RU 2366959]. В этой части выявлена активно развивающаяся новая тенденция к использованию в качестве маркера развития митохондриальных заболеваний эпигенетических маркеров: сайтов метилирования митохондриального генома.
Известны патенты, описывающие продуценты белков и низкомолекулярных метаболитов на основе дрожжей Yarrowia lipolytica [RU 2422526; RU 2451075; RU 2451749], в том числе сверхпродуценты янтарной и лимонной кислот, обладающие измененными в функциональном отношении митохондриями. Однако, в этих патентах не описаны методы непосредственного воздействия на геном митохондрий, в частности, с использованием систем гомологичной рекомбинации. Имеются патенты [RU 2376376], в которых показаны генетические конструкции, которые способны влиять на работу митохондрий, но в этих патентах не упоминается о белке RecA или иных системах гомологичной или иной рекомбинации ДНК.
В публикации [Yoon, Y.G. и Koob, M.D. «Efficient cloning and engineering of entire mitochondrial genomes in Escherichia coli and transfer into transcriptionally active mitochondria» Nucleic Acids Res. 2003; 31(5): 1407–1415] описан эксперимент по репликации полноразмерного генома митохондрий мыши и дрожжей в E. coli. Однако по соображениям биоэтики и биобезопасности этот подход не пригоден для накопления митохондрий человека в интересах генотерапии, так как при его использовании в состав митохондриального генома пациента были бы введены дополнительные физиологически активные элементы: бактериальный репликон и маркер лекарственной устойчивости.
В патенте [FR2857977, опубл. 23.07.2003] описан способ препаративной наработки РНК для генотерапии болезней человека, основанный на использовании векторов, поддерживающихся в митохондриях дрожжей, лишенных собственного митохондриального генома.
Известен патент КНР [CN 104450768 A «Shuttle vector of targeted yeast mitochondria and application of shuttle vector» (опубл. 25.03.2015)], описывающий структуру репликативного вектора, размножающегося в митохондриях пекарских дрожжей S. cerevisiae.
Известен патент авторов И. Тарасова и Н. Энтелис из университета Старсбурга (Франция) «Vector for therapy of mitochondrial disease» (US2015361450, C12N15/85, опубл. 17.12.2015), в котором описана молекула РНК, позволяющая частично компенсировать митохондриальные болезни, снижая уровень гетероплазмии митохондриального генома в организме. Операции по введению в митохондрии ДНК в патенте не затрагивались.
Известен патент «Mitochondrial expression vector and process for the transformation of mitochondria» US2015315608, C12N15/85, опубл. 05.11.2015, в котором описывается независимый от эндогенного митохондриального генома плазмидный вектор, обеспечивающий репликацию и транскрипцию трансгенов в гигантских митохондриях клеток млекопитающих.
Известен патент I. Malcuit и A. Sorokin «Plant mitochondrial transformation method» (PCT/GB2009/002755, опубл. 28.09.2011)], описывающий способ введения РНК в митохондрии растений в сочетании с обратной транскриптазой, что позволяет нарабатывать трансгены in vivo и интегрировать их в митохондриальный геном и более ранний патент «Plant Mitochondria Transformation Method» WO_2010_061187_A2, опубл. 3.06.2010, описывающий структуру векторов, поддерживающихся в митохондриях растений, включая репликоны и эффективные промоторы. В этом же ряду находится более ранний патент CA2272788 патентообладателя Garching Innovation Gmbh (Германия) (опубл. 11.12.2000).
Таким образом, можно резюмировать, что работы по направленной коррекции дефектов генома митохондрий человека и животных нигде в мире не проводились или не привели к патентоспособным результатам. Единственным примером изобретения, направленного на коррекцию истинных причин митохондриальных заболеваний MELAS и MERFF, является патент [US 2015361450], описывающий метод доставки недостающих тРНК из цитоплазмы в митохондрии, в частности с целью подавления фенотипических проявлений мутаций в митохондриальных ДНК, воздействуя на уровень гетероплазии. Однако этот метод даже теоретически не позволяет восстановить генетическую неполноценность митохондриального генома таких пациентов и на практике не используется.
Анализируя известные в патентной и иной литературе сведения о методах терапии митохондриальных болезней человека, в том числе, прямо или косвенно использующих методы генотерапии митохондриального генома, приходится констатировать, что предметы изобретения известных патентов ограничиваются способами конструирования векторных молекул ДНК для репликации в митохондриях различных организмов (млекопитающих, растений и дрожжей), содержащих неприродные элементы. Ни один из описанных методов не позволяет реплицировать полностью нативную молекулу митохондриальной ДНК человека и вносить в неё направленные изменения типа точечных замен.
Известны также способы диагностики митохондриальных болезней человека, не касаясь возможностей их лечения. Так в патенте [CN101177702] представлен генный чип, способный распознавать участки с мутациями, связанными с синдромами MELAS и MERRF митохондриальных заболеваний. Действующие патенты [WO/2015/144964, WO/2015/108077] направлены на диагностику митохондриальных болезней человека, в том числе на разработку метода диагностики или оценки риска нейродегенератиных заболеваний [WO/2015/144964] и определение содержания белка GDF15 в качестве биомаркера для оценки тяжести митохондриальных заболеваний и эффективности их лечения [WO/2015/108077]. Все эти изобретения не затрагивают тему таких синдромов, как MELAS и MERFF, а также не используют Y. lipolytica либо иные организмы для лечения таких болезней.
Таким образом, можно утверждать, что заявляемое изобретение не затрагивает каких-либо принципов, описываемых в цитируемых патентах.
Раскрытие изобретения
Разработка методов генной инженерии для работы с геномом митохондрий является полностью новым направлением биотехнологии, не имеющим аналогов. Задачей изобретения является создание метода направленной коррекции митохондриального генома человека (в частности, устранения вредных мутаций, ассоциированных с синдромами MELAS и MERFF) в гетерологичной дрожжевой системе с целью препаративной наработки скорректированного генома и введения его в мезенхимальные стромальные клетки (МСК) человека, культивируемые in vitro. МСК с восстановленной таким образом функцией митохондрий могут быть использованы для введения пациентам, что может обеспечить частичное или полное купирование симптомов MELAS и MERFF. К преимуществам описываемого способа генотерапии человека относится возможность направленной (точечной) корректировки единственного нуклеотида из состава генома, ответственного за появление митохондриальной болезни, тогда как весь остальной геном человека остается неизменным. Существует возможность предварительного полного секвенирования митохондриального генома человека, реплицирующегося в Y. lipolytica перед введением в митохондрии МСК человека. Это позволяет добиться существенного снижения риска возникновения случайных мутаций по сравнению с альтернативным подходом: ферментативным восстановлением всей последовательности митохондриального генома человека из химически синтезированных олигонуклеотидов.
Анализируя причины практически полного отсутствия описанных в патентной и иной литературе методов направленного воздействия на геном митохондрий (включая задачу генотерапии митохондриальных болезней человека), необходимо отметить, что ни один из исследованных видов эукариот не обладает собственной системой гомологичной рекомбинации митохондриального генома. Известный из уровня техники штамм дрожжей Y. lipolytica W29, несущий конструкцию pQ-SRUS, предназначенную для введения в клетки дрожжей гена RecA из Bacillus subtilis, обладает уникальной для живых объектов способностью к интеграции ДНК-конструкций в митохондриальный геном [RU 2562869]. В рамках настоящего изобретения на базе Y. lipolytica W29 разработана генетическая система, позволяющая с помощью химической трансформации вводить в митохондрии дрожжей геном митохондрий, используя фенотипическую селекцию трансформантов по восстановлению искусственно нарушенной способности штамма расти на минимальных средах с сукцинатом в качестве единственного источника энергии. В момент трансформации митохнодриальный геном человека проникает в митохондрии дрожжей и начинает реплицироваться в них наряду с собственным митохондриальным геномом Y. lipolytica, используя имеющийся там собственный ферментативный аппарат репликации. При этом ядерный геном человека, попадая в клетки дрожжей, не реплицируется и не оказывает влияния на генотип отбираемых штаммов-трансформантов. Поэтому для получения трансформантов Y. lipolytica, поддерживающих репликацию митохондриального генома человека, не требуется предварительное физическое разделение митохондриального и ядерного генома человека.
Для создания системы фенотипической селекции трансформантов Y. lipolytica, поддерживающих репликацию митохондриального генома человека, в рамках изобретения создаются штаммы с дефектами тРНК митохондриального кодирования. Удаление гена Yalif-MT39, кодирующего тРНК-Lys, или гена Yalif-MT36, кодирующего тРНК-Leu, из митохондриального генома Y. lipolytica достигается путем замещения на ген устойчивости к гигромицину с применением искусственно введенной в митохондрии системы гомологичной рекомбинации бактериального происхождения приводит к возникновению условно летального фенотипа: утрате способности использовать метаболиты цикла Кребса (в частности, сукцинат) в качестве единственного источника энергии. Для создания таких конструкций необходимо использование специальным образом модифицированного гена гигромициновой устойчивости (аминогликозидфосфотрансферазы), по кодоновому составу элемент и типу элемента инициации трансляции в адаптированному для распознавания митохондриальной системой трансляции Y. lipolytica. В виду своеобразия системы митохондриальной системы трансляции использование для этой цели генов устойчивости, адаптированных для работы в бактериях или в ядерном геноме эукариот, невозможно.
С целью адаптации 5’-некодирующей области искусственного гена устойчивости к гигромицину для экспрессии в митохондриях Y. lipolytica была использована последовательность гена ND1 из митохондриального генома этого организма, кодирующая главную субъединицу протонной NADH-оксидазы внутренней мембраны митохондрий – один из наиболее активно экспрессирующихся компонентов митохондриального генома [Edmondson et al., Mol Genet Genomics. 2005. V. 273 P. 115-22]. Она содержит рибосом-связывающую последовательность – аналог последовательности Шайна-Дельгарно у бактерий [Paul et al., Mutat. Res. 2001. V. 5. P. 486]. При конструировании кодирующей области гена особое внимание было уделено кодонам, соответствующим Leu: во всех случаях использовался кодон TTG. В качестве Arg-специфичного кодона использовался AGG. 50% кодонов, соответствующих Trp, были заменены с TGG на TGA (терминаторный кодон стандартного генетического кода). Вследствие этого искусственный ген полностью лишен способности экспрессироваться в ядерном геноме Y. lipolytica или в бактериях: возникновение устойчивости к гигромицину достигается только при встраивании гена в геном митохондрий Y. lipolytica. Для кодирования Met в двух из четырёх позиций был использован кодон ATA (соответствует Ile в стандартном коде). В качестве терминаторного был использован кодон TAA, наиболее часто использующийся в генах митохондриального кодирования Y. lipolytica [Kolesnikova et al., RNA. 2010. V. 16. P. 926–941]. Для кодирования Thr в двух позициях использован кодон CTC, соответствующий Leu в стандартном генетическом коде.
Штаммы Y. lipolytica с искусственно введенными дефектами генов Yalif-MT39 и Yalif-MT36 (замена гена тРНК на ген гигромициновой устойчивости по принципу двуплечевой гомологичной рекомбинации с использованием белка RecA – продукта конструкции pQ-SRUS) сохраняют способность эффективно расти на полноценной питательной среде, содержащей глюкозу, используя при этом аэробный метаболизм.
Введение митохондриального генома человека в клетки Y. lipolytica, несущие ноль-мутации в генах Yalif-MT39 и Yalif-MT36, приводит к комплементации утраченной функции митохондрий – способности использовать метаболиты цикла Кребса в качестве единственного источника энергии. При этом в присутствии белка RecA одновременное введение в штаммы Y. lipolytica мутантного варианта митохондриального генома человека, несущего вредную мутацию в гене тРНК-Lys, и ПЦР-копии фрагмента этой ДНК с искусственно скорректированной мутацией, приводит к образованию in vivo (в митохондриях клеток дрожжей) варианта митохондриального генома человека со скорректированной мутацией. Такая ДНК может быть выделена из клеток Y. lipolytica и использована для введения в митохондрии МСК пациента, несущего вредную мутацию.
Осуществление изобретения:
1. Конструирование синтетического гена устойчивости к гигромицину (аминогликозидфосфотрансфераза из Tn5370, размер 332 а.о.) (Фиг. 1), по кодоновому составу и организации старта трансляции адаптированного для экспрессии в митохондриях Y. lypolytica (Фиг. 2, SEQ ID NO 3).
2. На основе искусственного гена устойчивости к гигромицину создание генетических конструкций в форме линейной двунитевой ДНК, пригодных для введения в состав митохондриального генома Y. lipolytica путем гомологичной рекомбинации с 3’-UTR оперона 1 (SEQ ID NO 1 и SEQ ID NO 2). Одновременно с приданием устойчивости к гигромицину конструкции удаление из митохондриального генома дрожжей гена тРНК-Lys или тРНК-Leu.
3. Получение трансформантов Y. lipolytica, несущих генетические конструкции на основе гена устойчивости к гигромицину (SEQ ID NO 3), в составе митохондриального генома.
4. Введение митохондриального генома человека в клетки трансформантов дрожжей с фенотипическим отбором трансформантов по комплементации дефекта в генах тРНК- Leu и тРНК-Lys. Оценка эффективности введения митохондриального генома человека в штамм дрожжей с применением метода ПЦР в реальном времени (по одной или нескольким маркерным последовательностям).
1. Разработка и получение синтетического гена устойчивости к гигромицину
Последовательность гигромицинфосфотрансферазы В - hygromycine B phosphotransferase из Tn5370 Mycobacterium smegmatis, штамм MC2 155 – детерминаниты устойчивости к гигромицину, доступна из базы данных NCBI GenBank Proteins под номером AII01819 (Фиг. 1). Соответствующая ей нуклеотидная последовательность природного гена доступна из базы данных NCBI GenBank Nucleotides под номером KM232615. При конструировании искусственного гена введение в его состав промотора не предусматривается, поскольку интегрированный в геном митохондрий трангсен эффективно транскрибируется с эффективно действующего эндогенного промотора оперона 1, находящегося выше сайта интеграции по ходу транскрипции. С целью адаптации 5’-некодирующей области искусственного гена устойчивости к гигромицину для экспрессии в митохондриях Y. lipolytica используется последовательность гена ND1 из митохондриального генома этого организма, кодирующая главную субъединицу протонной NADH-оксидазы внутренней мембраны митохондрий. Далее следует кодирующая последовательность гена, соответствующая нуклеотидной последовательности SEQ ID NO 3 (Фиг. 2). Для обеспечения высокой эффективности трансляции мРНК HygR в митохондриях и исключения экспрессии соответствующего гена вне митохондриального генома (например, при интеграции его в ядерный геном) особое внимание должно быть уделено кодонам, значение которых отличается в митохондриальном и ядерном геноме Y lipolytica. Таким кодоном, в первую очередь, является Leu: при разработке искусственного гена во всех случаях целесообразно использовать кодон TTG. В качестве Arg-специфичного кодона должен использоваться AGG. Около половины всех имеющихся кодонов, соответствующих Trp, должны быть заменены с TGG на TGA (терминаторный кодон стандартного генетического кода). Вследствие этого искусственный ген оказывается полностью лишен способности экспрессироваться в ядерном геноме Y. lipolytica или в бактериях. Для кодирования Met в двух из четырёх позиций используют кодон ATA (соответствует Ile в стандартном коде). В качестве терминаторного выбирают кодон TAA, наиболее часто использующийся в генах митохондриального кодирования Y. lipolytica. Для кодирования Thr в двух позициях используют кодон CTC, соответствующий Leu в стандартном генетическом коде. С 5’-конца искусственный ген ограничивают сайтом рестрикции BamHI, с 3’-конца – сайтом HindIII. Последовательность искусственного гена устойчивости к гигромицину представлена на Фиг. 2 (SEQ ID NO 3).
Синтез искусственного гена осуществляют полуферментативным методом. Во избежание неточностей химического синтеза длина олигонуклеотидов, введенных в реакцию лигирования, не должна превышать 50 нт. Температура гибридизации смеси олигонуклеотидов перед началом реакции лигирования – 60°С, время гибридизации – 20 мин.
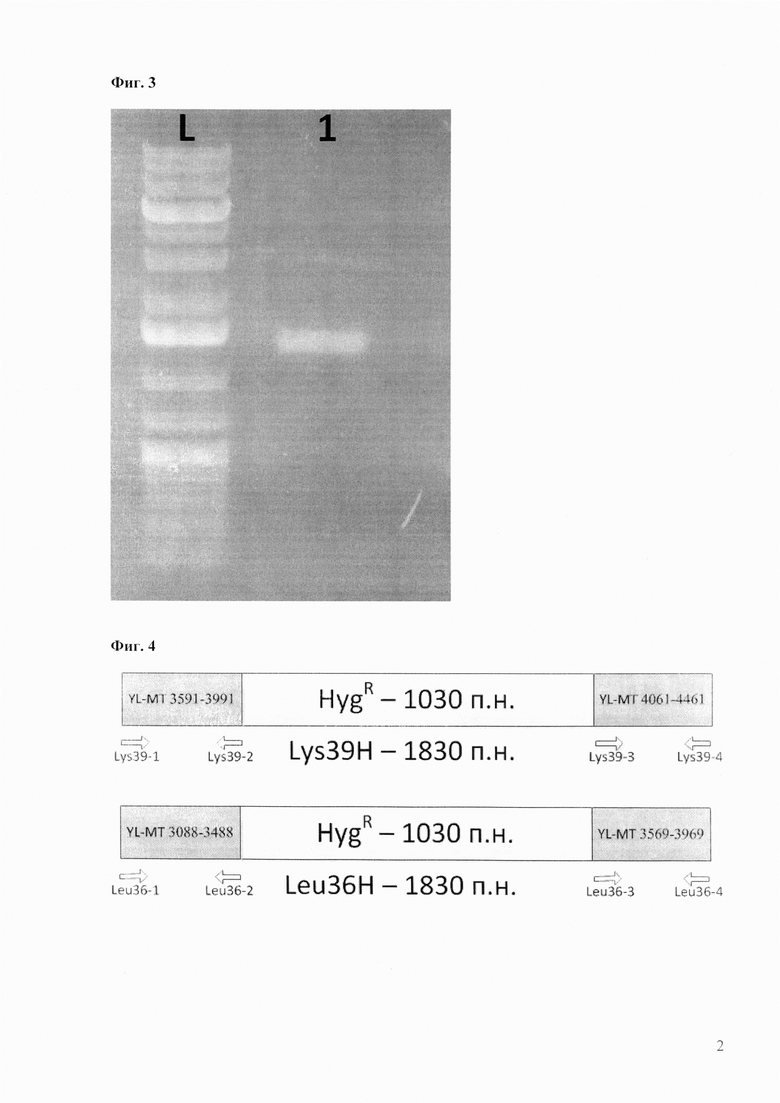
Каждый олигонуклеотид предварительно подвергают фосфорилированию 5’-конца с помощью полинуклеотидкиназы и вводят в реакцию в количестве 200 пмоль. В результате лигирования был получают продукт длиной 1030 п.н. После очистки с помощью набора GeneJet (MBI Fermentas) без использования дополнительной ПЦР-амплификации получают ~56 пмоль двуцепочечной ДНК ожидаемого размера 1030 п.н. (Фиг. 3). Её непосредственно используют для создания конструкций для трансформации Y. lipolytica в линейной форме.
2. Разработка генетических конструкций для замещения генов тРНК- Leu и тРНК-Lys на ген гигромициновой устойчивости в составе митохондриального генома Y. lipolytica
Для проведения трансформации используют ранее описанный рекомбинантный штамм Y. lipolytica W29 (pQ-SRUS), обладающий активностью рекомбиназы RecA бактериального происхождения в митохондриях [Патент RU 2562869].
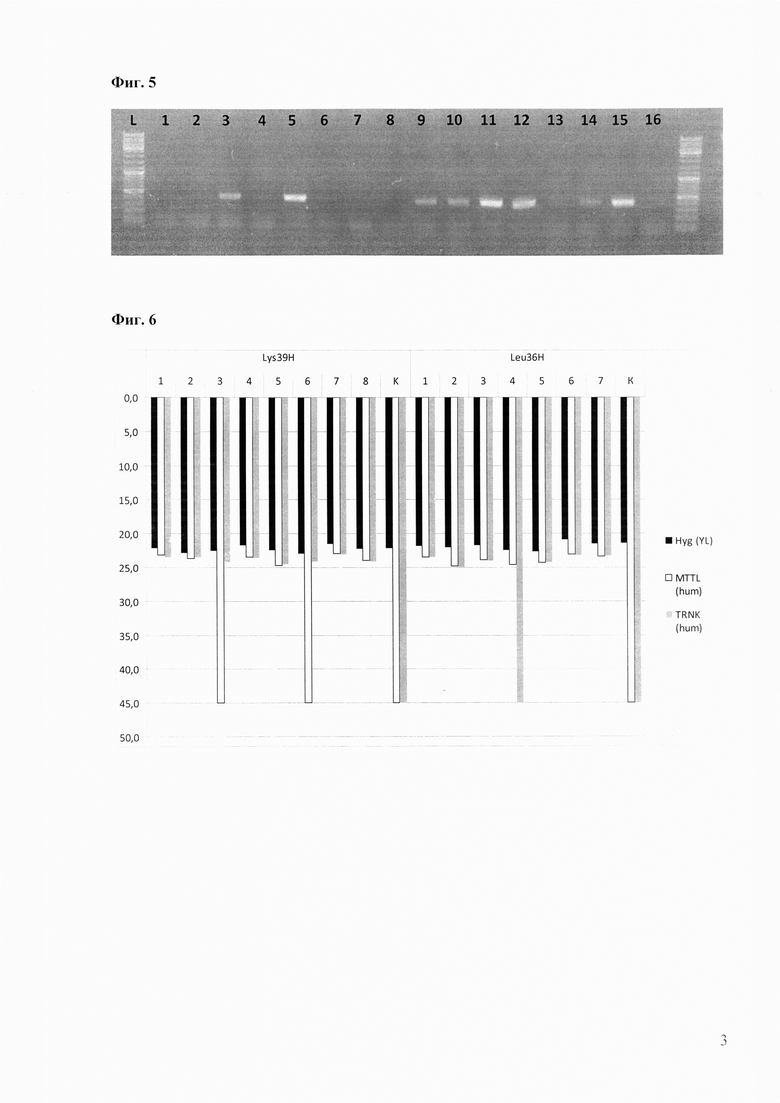
Создают две конструкции, где адресные последовательности длиной 400 п.н. для гомологичной рекомбинации с флангами генов Yalif-Mt36 и Yalif-Mt39 присоединяют к искусственному гену устойчивости к гигромицину (Фиг. 4). Конструкции Lys39H и Leu36H создают в линейной форме и препаративно нарабатывают с помощью препаративной ПЦР.
Для создания конструкции Lys39H синтезируют праймеры:
необходимые для ПЦР-клонирования области генома митохондрий 3591-3991 длиной 400 п.н. со следующей последовательностью:
1 cctttaaaag tagaatatat atataacata ataacaaaaa gtagactctg aaccatctct
61 ttgagacggt tcgagtccta tttttgttat agaagggaat aaatatatac acataaaaga
121 ataatatata agaaaaaaaa ataacgaatg tccctaactt catccttttg gattgtaagt
181 tccgataaat atatatatta tatgttcata aattaagata tatatatatc tattatcggt
241 ccttctcttt gataatttat taataatgtg ttaggtagcg ggcagggtat ttaatacttt
301 tagctctact gagttgtaga ttaattggca aatcacttaa atttgagtta agactatgaa
361 agttcgaatc tttccaattc atattaaata tattaattaa g
При этом в состав праймера Lys39-2 вводят сайт BamHI, необходимый для стыковки с последовательностью искусственного гена гигромицина HygR.
Для ПЦР-клонирования области генома митохондрий 4061-4461 длиной 400 п.н. используют праймеры:
с помощью которых прводят ПЦР-клонирование фрагмента ДНК со следующей последовательностью:
1 ctttctatta tttatattaa ataatataat tattatataa aggaccaaat agaatcgaac
61 tatcggcttg ccgaaggttc gattcctata ggaccccttt ctattattta tattaaataa
121 tataattatt tatgaaatat tcaatagata attataaaaa tttaaggtac tttttaacta
181 attgaatcga aacatcttag tttgtaaagc tatatataat tatatatagg taaaagttgt
241 atatgtaaaa aatttaagtt aaaaatagta aaaaccttta atttaacggt aaaatacata
301 attgataatt atgatacgtc aggttcgact cctacaaggt ttaataaagg atatttggct
361 gagcggttga aagcatgata tttgagctat catatataat t
Пары праймеров Lys39-1/Lys39-2 и Lys39-3/Lys39-4 вводят в реакцию ПЦР с использованием в качестве матрицы геномной ДНК типового коллекционного штамма Y. lipolytica CLIB-122.
Для создания конструкции Leu36H используют праймеры:
необходимые для ПЦР-клонирования области генома митохондрий 3088-3488 длиной 400 п.н. со следующей последовательностью:
1 attgttttgt aatcagtagc ttttggttca aatccaaaat ttaacattaa aatttataat
61 attatcctcc cataattaaa agagttgatt tttaagacct tatcgtcaaa cgtaagacgt
121 tagaacttca ctctaaaaat acgggttcaa gtcccgttaa ggttaaaaaa tataatatta
181 ttaaagtaac atagtgtaat ggtatcacat caggctcata atctgataat atttggttcg
241 aatccatttg ttgctatata atttataata ggttaataga aagtttggaa ctatagttta
301 aaggttaaaa ctaaccgctc ataacggttc aatgattgtt cgagtcaatc tggttctata
361 aaaatatata aatatatttt ttatatttat taaataataa g
При этом в состав праймера Leu36-2 вводят сайт BamHI, необходимый для стыковки с последовательностью искусственного гена HygR.
Для ПЦР-клонирования области генома митохондрий 3569-3969 длиной 400 п.н. используют праймеры:
с помощью которых проводят ПЦР-клонирование фрагмента ДНК со следующей последовательностью:
1 atagaaggga gtaataatac ttcctttaaa agtagaatat atatataaca taataacaaa
61 aagtagactc tgaaccatct ctttgagacg gttcgagtcc tatttttgtt atagaaggga
121 ataaatatat acacataaaa gaataatata taagaaaaaa aaataacgaa tgtccctaac
181 ttcatccttt tggattgtaa gttccgataa atatatatat tatatgttca taaattaaga
241 tatatatata tctattatcg gtccttctct ttgataattt attaataatg tgttaggtag
301 cgggcagggt atttaatact tttagctcta ctgagttgta gattaattgg caaatcactt
361 aaatttgagt taagactatg aaagttcgaa tctttccaat t
После очистки с применением набора GeneJet (MBI Fermentas) «плечи для рекомбинации» длиной 400 п.н. обрабатывают рестриктазой BamHI (продукты ПЦР, полученные с помощью праймеров Lys39-1/Lys39-2 и Leu39-1/Lys39-2) или HindIII (продукты ПЦР, полученные с праймерами Lys39-3/Lys39-4 и Leu39-3/Leu39-4). Фрагмент, соответствующий искусственному гену HygR, обрабатывают одновременно рестриктазами BamHI и HindIII.
Далее с целью получения конструкции Lys39H смешивают по 3 пмоль фрагментов, полученных с праймерами Lys39-1/Lys39-2, Lys39-3/Lys39-4 и фрагмента гена HygR, проводят обработку Т4 ДНК-лигазой в течение 12 часов. Лигазную смесь используют в качестве матрицы для ПЦР с праймерами Lys39-1 и Lys39-4.
1. Получение трансформантов дрожжей, несущих генетические конструкции для замещения генов тРНК- Leu и тРНК-Lys на ген гигромициновой устойчивости в составе митохондриального генома Y. lipolytica
Для проведения трансформации используют штамм Y. lipolyica W29, несущий ген рекомбиназы RecA под контролем промотора и терминатора транскрипции гена SOD2 в составе интегративной конструкции pQ-SRUS [RU 2562869].
Для получения препарата компетентных клеток штамм культивируют на полноценной жидкой среде YP состава (г/л): протеозный пептон (Difco) – 20, дрожжевой экстракт (Difco) – 10, при интенсивной аэрации при 30°С в течение 16 часов.
ДНК конструкции Lys39H нарабатывают с помощью препаративной ПЦР при помощи праймеров Lys39-1 и Lys39-4, а ДНК конструкции Leu36H - при помощи праймеров Leu36-1 и Leu36-4. Праймеры в количестве 10 пмоль каждого вносят в объем реакционной смеси 30 мкл. При проведении ПЦР используют температуру отжига 58°С. По окончании реакции продукты очищают с помощью набора GeneJet (MBI Fermentas) и определяют концентрацию с помощью спектрфотометра NanoDrop (Eppendorf). Для эксперимента используют препараты, имеющие концентрацию ДНК не менее 0,1 мкг/мкл.
Процедуру трансформации с использованием ацетата Li и полиэтиленгликоля 6000 проводят по методу, описанному ранее в работе [Davidow L.S. et al. Curr. Genet. 1985. V. 10. P. 39-48]. При этом к аликвоте клеток, содержащей 108 к.о.е., добавляют 1,2 мкг очищенной ДНК конструкции Lys39H или Leu36H. По окончании трансформации клетки, тщательно промытые деионизованной водой для удаления остатков полэтиленгликоля, высевают на полноценную агаризованную среду YPD (2% бактопептон, 2% дрожжевой экстракт Difco, 2% глюкоза, 2% бактоагар HyMedia) с добавлением гигромицина (G-418, Promega) в концентарции 23 мкг/мл.
Чашки инкубируют в термостате в течение 48 часов при 30°С. Выросшие колонии (15 колоний, полученных при трансформации ДНК конструкции Lys39H, и 13 колоний, полученных при трансформации ДНК конструкции Leu36H) пересевают на селективную среду того же состава, которая использовалась для трансформации. Полученную чашку используют в качестве исходного материала для проверки фенотипа трансформантов. Для этого каждую колонию пересевают на синтетическую агаризованную среду YNB (с добавлением витаминов и микроэлементов), содержащую в качестве единственного источника углерода и энергии 100 мМ Трис-сукцинат, рН 5,5. Для поддержания рН в среду вносят также 2 M KPi буфер; pH 5,5, приготовленный по прописи: 272 г/л безводного KH2PO4 (Amresco кат. № 0781) растворяют в 1 л воды и доводят рН с помощью рН-метрического титрования с помощью 2 M K2HPO4 (получают путем растворения 342 г/л безводного K2HPO4 (Amresco кат № 0705). Фосфатный буфер вносят в среду в объёмном соотношении 1:40 перед засевом. Из 28 проверенных колоний 12 (в том числе 7 колоний, полученных при трансформации ДНК конструкции Lys39H, и 5 колоний, полученных при трансформации ДНК конструкции Leu36H, сохраняют способность расти на минимальной среде, что свидетельствует об отсутствии перестройки митохондриального генома в результате встройки конструкций. Эти клоны в дальнейшей работе не используют. Оставшиеся 16 колоний, не обладающих способностью к росту на минимальной среде с содержанием сукцината в качестве единственного источника энергии, подвергают генотипированию.
Клетки, не обладающие способностью к росту на минимальной среде с сукцинатом в качестве единственного источника энергии, выращивают в неселективных условиях на жидкой полноценной среде YPD при интенсивной аэрации в объеме 3 мл и выделяют суммарную геномную ДНК с помощью набора Проба-ГС (ДНК-Технология, Москва). Полученные препараты геномной ДНК исследуют с помощью аналитической ПЦР с праймерами Lys39-1/Lys39-2 (8 клонов, полученных при трансформации конструкцией Lys39H), или с Leu36-1/Leu36-2 (8 клонов, полученных при трансформации конструкцией Leu36H). Результаты проверки представлены на Фиг. 5.
Результат генотипирования показывает, что два клона W29 (pQ-SRUS, Lys39H)#3 и W29 (pQ-SRUS, Leu36H)#15, сохраняют интактную структуру 5’-концевого фланга митохондриального генома, примыкающего к конструкции. Эти клоны используют для дальнейших экспериментов по введению митохондриального генома человека и восстановлению структуры и активности генов TRNL1 и TRNK/MTTK в ней.
4. Введение митохондриального генома человека в клетки трансформантов дрожжей с фенотипическим отбором трансформантов по комплементации дефекта в генах тРНК- Leu и тРНК-Lys
Для получения препарата компетентных клеток штаммы W29 (pQ-SRUS, Lys39H)#3 и W29 (pQ-SRUS, Leu36H)#15 культивируют на полноценной жидкой среде YP состава (г/л): протеозный пептон (Difco) – 20, дрожжевой экстракт (Difco) – 10, при интенсивной аэрации при 30°С в течение 16 часов и далее обрабатывают, как описано [Davidow L.S. et al. Curr. Genet. 1985. V. 10. P. 39-48].
Суммарную геномную ДНК человека для использования при трансформации дрожжей выделяют с помощью набора DNA IQ System (Qiagen) на основе магнитных микрочастиц. Используют первичную культуру фибробластов человека в количестве 1010 клеток с диплоидным набором хромосом. Для единичного эксперимента используют 10 мкг препарата геномной ДНК первичных фибробластов человека. При этом к аликвоте клеток, содержащей 108 к.о.е., добавляют ДНК в объеме 10 мкл. Трансформанты высевают на синтетическую агаризованную среду YNB (с добавлением витаминов и микроэлементов), содержащую в качестве единственного источника углерода и энергии 100 мМ Трис-сукцинат, рН 5,5 с добавлением гигромицина (G-418, Promega) в концентрации 23 мкг/мл. В качестве отрицательного контроля используют аликвоты компетентных клеток W29 (pQ-SRUS, Lys39H)#3 и W29 (pQ-SRUS, Leu36H)#15 без добавления ДНК человека. При трансформации штамма, несущего конструкцию W29 (pQ-SRUS, Lys39H)#3, получают 8 колоний, при трансформации штамма W29 (pQ-SRUS, Leu36H)#15 – 7 колоний. В отрицательных контролях колонии появляться не должны. Выросшие колонии двукратно пассируют на агаризованной синтетической среде с сукцинатом в качестве единственного источника энергии, после чего их выращивают в неселективных условиях на жидкой полноценной среде YPD при интенсивной аэрации в объеме 3 мл и выделяют суммарную геномную ДНК с помощью набора Проба-ГС (ДНК-Технология, Москва). Полученные препараты геномной ДНК исследуют с помощью ПЦР в реальном времени, используя набор производства Евроген (Москва), каталожный номер PK145S, содержащий хот-стартовый комплекс ДНК-полимеразы Taq и интеркалирующий краситель Sybr Green и три пары праймеров: HygU1 (GGGGATCCacaaaccataaaaaaaaatgCTTcaagaatccTTGttaTTGT)/HygL1 (GGAAGCTTAggcgccgggggcggtgtccggcggcccccaCAAgaactgcg) (для оценки содержания собственного митохондриального генома Y. lipolytica, маркированного геном HygR), MTTL1 (gggtttgttAagatggcagagcccggt)/MTTL2 (ggttggccatgggtatgttgttaagaa) (для выявления последовательности гена тРНК – Leu генома человека) и TRNK1 (agaaccaacacctctttacagtgaaat)/ TRNK2 (ttagttgggtgatgaggaatagtgtaa) (для выявления последовательности гена тРНК – Lys генома человека). Реакцию проводят помощью термоциклера с оптической системой детекции «ДТ-322» (ООО «НПО ДНК-Технология», Россия). Учёт результатов реакции выполняют с помощью программного обеспечения детектирующего термоциклера «ДТ-322» (Фиг. 6). В качестве отрицательных контролей используют препараты геномной ДНК родительских штаммов Y. lipolytica W29 (pQ-SRUS, Lys39H)#3 и W29 (pQ-SRUS, Leu36H)#15.
Измерения параметра Ct показывает наличие следующих средних величин для каждой группы клонов (Таблица 1).
Таблица 1. Средние значения и среднеквадратичные отклонения параметра Ct в среднем для групп трансформантов Y. lipolytica W29 (pQ-SRUS)
При этом величина Ct во всех контрольных точках составляет 45 циклов (максимально возможное значение с учетом числа выполненных циклов). Таким образом, использованный метод ПЦР в реальном времени обеспечивает возможность точно идентифицировать образцы, в которых отсутствует исследуемая мишень.
Клоны №3 и №6, полученные при трансформации геномной ДНК человека штамма Y. lipolytica W29 (pQ-SRUS, Lys39H)#3 показали отсутствие маркера MTTL человеческого митохондриального генома при наличии маркера TRNK.
Симметричная ситуация наблюдалась при трансформации штамма W29 (pQ-SRUS, Leu36H)#15. В части клонов трансформантов (например, клон №4, Фиг. 6) при трансформации происходит утрата части митохондриального генома человека, приводящая к отсутствию в генотипе маркера TRNK. Утрата селектируемого в процессе трансформации маркеров (например, MTTL в случае штамма W29 (pQ-SRUS, Leu36H)#15) не ожидается. Вероятность утраты случайно взятого локуса митохондриального генома в процессе трансформации составляет от 25% (в случае штамма W29 (pQ-SRUS, Lys39H)#3) до 14% (в случае штамма W29 (pQ-SRUS, Leu36H)#15).
Краткое описание графических изображений:
Фиг. 1. Аминокислотная последовательность маркера устойчивости к гигромицину (гигромицинфосфотрансфераза В - hygromycine B phosphotransferase) из Tn5370 Mycobacterium smegmatis, штамм MC2 155, депонированная в базе данных NCBI GenBank Proteins под номером AII01819.
Фиг. 2. Нуклеотидная последовательность искусственного гена устойчивости к гигромицину (гигромицинфосфотрансфераза В - hygromycine B phosphotransferase) из Tn5370 Mycobacterium smegmatis, адаптированная для экспрессии в митохондриальном геноме Y. lipolytica. Рамкой выделена 5’-нетранслируемая область, взятая из гена ND1 Y. lipolytica, инициаторный и терминаторный кодоны выделены жирным шрифтом и подчёркнуты. Кодоны, используемые в соответствии с уникальными особенностями генетического кода митохондрий дрожжей, показаны прописными буквами. Последовательности фланкирующих рестриктных сайтов BamHI и HindIII выделены серой заливкой.
Фиг. 3. Электрофоретический анализ фрагмента ДНК, полученного в результате лигирования фосфорилированных олигонуклеотидов. Ожидаемый размер искусственного гена устойчивости к гигромицину (гигромицинфосфотрансфераза В - hygromycine B phosphotransferase) из Tn5370 Mycobacterium smegmatis, адаптированного для экспрессии в митохондриальном геноме Y. lipolytica – 1030 п.н. (дорожка 1).
Фиг. 4. Схема интегративных конструкций Lys39H и Leu36H.
Фиг. 5. Результаты генотипирования клонов, полученных в результате трансформации Y. lipolytica W29 (pQ-SRUS) линейными конструкциями Lys39H и Leu36H, предназначенными для интеграции в геном митохондрий.
Дорожки 1-8: геномная ДНК клонов 1-15, полученных при трансформации конструкцией Lys39H и утративших способность к росту на среде с сукцинатом в качестве единственного источника энергии - ПЦР праймерами Lys39-1/Lys39-2.
Дорожки 9-16: геномная ДНК клонов 1-13, полученных при трансформации конструкцией Leu36H и утративших способность к росту на среде с сукцинатом в качестве единственного источника энергии - ПЦР с праймерами Leu36-1/Leu36-2.
Фиг. 6. Результаты определения содержания митохондриального генома человека в биомассе клонов, полученных при трансформации штаммов W29 (pQ-SRUS, Lys39H)#3) и W29 (pQ-SRUS, Leu36H)#15 суммарной ДНК человека с последующим отбором на минимальной среде с сукцинатом в качестве единственного источника углерода.
По оси абсцисс указаны номера клонов, полученных при трансформации родительских штаммов W29 (pQ-SRUS, Lys39H)#3) и W29 (pQ-SRUS, Leu36H)#15 соответственно. Литерой «К» отмечен контроль – ДНК соответствующего исходного родительского штамма. По оси ординат приведены значения порогового цикла (Ct), автоматически определенного встроенным программным обеспечением детектирующего термоциклера «ДТ-322».
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНТЕГРАТИВНАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pQ-SRUS ДЛЯ ПОЛУЧЕНИЯ ШТАММОВ ДРОЖЖЕЙ Yarrowia lipolytica, ОБЛАДАЮЩИХ СПОСОБНОСТЬЮ К ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ ГЕНОМА МИТОХОНДРИЙ ЗА СЧЁТ ЭКСПРЕССИИ ГЕНА БЕЛКА RecA БАКТЕРИАЛЬНОГО ПРОИСХОЖДЕНИЯ | 2014 |
|
RU2562869C1 |
| Трансформант дрожжей Yarrowia lipolytica, продуцирующий линалоол | 2022 |
|
RU2809554C1 |
| Модифицированная нуклеотидная последовательность, кодирующая полипептид, обладающий активностью фитоен синтазы и геранилгеранилпирофосфат синтазы (варианты), ее использование при конструировании трансформантов дрожжей Yarrowia lipolytica, продуцирующих каротиноиды | 2021 |
|
RU2794980C1 |
| Модифицированный вариант гена LacZ из E. coli, кодирующий стабилизированный вариант белка, для использования в качестве транскрипционного репортера в Yarrowia lipolytica | 2015 |
|
RU2609646C2 |
| Способ повышения продукции изолимонной кислоты у дрожжей Yarrowia lipolytica, дрожжи вида Yarrowia lipolytica, обладающие способностью к продукции изолимонной кислоты | 2018 |
|
RU2713124C2 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ ЛИПАЗЫ | 2007 |
|
RU2355754C1 |
| Штамм дрожжей Yarrowia lipolytica, продуцирующий изолимонную кислоту | 2021 |
|
RU2757603C1 |
| ДРОЖЖИ РОДА YARROWIA, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ ВНУТРИКЛЕТОЧНО НАКАПЛИВАТЬ СЛОЖНЫЕ ЭФИРЫ ЖИРНЫХ КИСЛОТ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ТАКИХ ЭФИРОВ | 2013 |
|
RU2539744C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ YARROWIA-LIPOLYTICA - ПРОДУЦЕНТ ЛИПАЗЫ | 2011 |
|
RU2451075C1 |
| ИНТЕГРАТИВНЫЙ ВЕКТОР Random-URA3-RPT ДЛЯ ПОСЛЕДОВАТЕЛЬНОГО ВВЕДЕНИЯ МНОЖЕСТВЕННЫХ КОПИЙ ГЕНЕТИЧЕСКИХ ЭЛЕМЕНТОВ В ДРОЖЖИ Yarrowia lipolytica | 2006 |
|
RU2376376C2 |




Изобретение относится к биотехнологии. Предложен способ репликации человеческого митохондриального генома в клетках дрожжей Yarrowia lipolytica. Используя метод химической трансформации, осуществляют трансформацию препарата суммарной ДНК человека в штамм Yarrowia lipolytica, содержащий ранее интегрированные конструкции pQ-SRUS и Lys39H (SEQ ID NO 1) на основе маркера устойчивости к гигромицину HygR (SEQ ID NO 3), либо штамм Yarrowia lipolytica, содержащий ранее интегрированные конструкции pQ-SRUS и Leu36H (SEQ ID NO 2). Проводят отбор трансформантов на минимальной синтетической среде, содержащей сукцинат в качестве единственного источника энергии. Определяют наличие реплицирующейся ДНК митохондриального генома человека с помощью ПЦР в реальном времени с парами праймеров, специфичных к ДНК митохондриального генома человека: MTTL1 (gggtttgttAagatggcagagcccggt)/MTTL2 (ggttggccatgggtatgttgttaagaa) для выявления последовательности гена тРНК - Leu генома человека и TRNK1 (agaaccaacacctctttacagtgaaat)/ TRNK2 (ttagttgggtgatgaggaatagtgtaa) для выявления последовательности гена тPHK-Lys генома человека. Изобретение может быть использовано для коррекции вредных мутаций митохондриального генома человека и препаративной наработки, скорректированной по последовательности интактной митохондриальной ДНК в клетках дрожжей с целью ее последующего введения в мезенхимальные стромальные клетки пациента в интересах лечения распространенных митохондриальных болезней человека, нарушающих функциональность генов митохондриальных Lys-Leu-специфичных рРНК. 6 ил., 1 табл.
Способ репликации человеческого митохондриального генома в клетках дрожжей Yarrowia lipolytica, отличающийся тем, что штамм Yarrowia lipolytica, содержащий ранее интегрированные конструкции pQ-SRUS и Lys39H (SEQ ID NO 1) на основе маркера устойчивости к гигромицину HygR (SEQ ID NO 3), либо штамм Yarrowia lipolytica, содержащий ранее интегрированные конструкции pQ-SRUS и Leu36H (SEQ ID NO 2), трансформируют препаратом суммарной ДНК человека, используя метод химической трансформации, после чего проводят отбор трансформантов на минимальной синтетической среде, содержащей сукцинат в качестве единственного источника энергии, и определяют наличие реплицирующейся ДНК митохондриального генома человека с помощью ПЦР в реальном времени с парами праймеров, специфичных к ДНК митохондриального генома человека: MTTL1 (gggtttgttAagatggcagagcccggt)/MTTL2 (ggttggccatgggtatgttgttaagaa) для выявления последовательности гена тРНК - Leu генома человека и TRNK1 (agaaccaacacctctttacagtgaaat)/ TRNK2 (ttagttgggtgatgaggaatagtgtaa) для выявления последовательности гена тPHK - Lys генома человека.
| US 20150315608 A1, 05.11.2015 | |||
| ИНТЕГРАТИВНАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pQ-SRUS ДЛЯ ПОЛУЧЕНИЯ ШТАММОВ ДРОЖЖЕЙ Yarrowia lipolytica, ОБЛАДАЮЩИХ СПОСОБНОСТЬЮ К ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ ГЕНОМА МИТОХОНДРИЙ ЗА СЧЁТ ЭКСПРЕССИИ ГЕНА БЕЛКА RecA БАКТЕРИАЛЬНОГО ПРОИСХОЖДЕНИЯ | 2014 |
|
RU2562869C1 |
| WO2009150441 A1, 17.12.2009 | |||
| СПОСОБ ОЦЕНКИ ПРОЦЕССОВ РАЗРУШЕНИЯ КОНСТРУКЦИЙ ПРИ АКУСТИКО-ЭМИССИОННОМ КОНТРОЛЕ | 2007 |
|
RU2367941C1 |
| US 20150361450 A1, 17.12.2015 | |||
| SHADEL G | |||
| S | |||
| INSIGHTS FROM MODEL SYSTEMS Yeast as a Model for Human mtDNA Replication // Am | |||
| J | |||
| Hum | |||
| Genet | |||
| Разборное приспособление для накатки на рельсы сошедших с них колес подвижного состава | 1920 |
|
SU65A1 |

