Область техники, к которой относится изобретение:
Изобретение относится к области биоинженерии, в частности к усовершенствованию метода тестирования активности промоторов, предназначенных для создания векторов для экспрессии генов в экстремофильных дрожжах Yarrowia lipolytica, методом транскрипционных репортеров.
Предметом изобретения является последовательность гена LacZ из Escherichia coli в составе генетической конструкции pUVLT2 (находится под контролем промотора гена митохондриального потенциал-зависимого порина VDAC). Особенностью гена LacZ в составе конструкции pUVLT2 по сравнению с аналогичной конструкцией pUVLT1, содержащей ген LacZ дикого типа, является наличие искусственно введенного дополнительного колона GGA (Gly), совмещенного с рестриктным сайтом, непосредственно после инициаторного кодона ATG (Met) в начале открытой рамки трансляции. В соответствии с «N-концевым правилом Варшавского», наличие этого дополнительного кодона придает продукту гена LacZ повышенную протеолитическую стабильность в цитоплазме Y. lipolytica. В свою очередь, улучшенная стабильность репортерного белка существенно облегчает проведение экспериментов по определению активности промоторов на основании измерения ферментативной активности β-галактозидазы in vivo.
Уровень техники:
Использование транскрипционных репортеров в настоящее время является стандартным методом молекулярного исследования, позволяющим количественно измерять активность промоторов in vivo как при проведении фундаментальных исследований, так и при создании экспрессионных систем коммерческого назначения. Ген β-галактозидазы из E. coli, предложенный в работах основоположников генной инженерии Жакоба и Моно, является наиболее традиционным и популярным среди всех известных репортеров. Он неоднократно применялся и при исследовании активности промоторов экстремофильных дрожжей Yarrowia lipolytica [1, 2]. Авторы опубликованных статей использовали спонтанно возникший мутантный вариант гена LacZ, имеющий делецию 8 кодонов на 5'-конце открытой рамки считывания. Однако ни один из авторов не обсуждал значимость этого факта и, приводя методики определения ферментативной активности репортерного белка LacZ, не рассматривал вопрос о его устойчивости к протеолизу in vivo. В работе [2] эта проблема сведена к указанию на необходимость использовать коктейль ингибиторов протеаз при гомогенизации биомассы рекомбинантного штамма.
В статье [3] описан механизм, влияющий на скорость протеосомной убиквитин-зависимой деградации в дрожжах. На основе анализа статистики установлено, что наличие на N-конце первичного продукта трансляции (вслед за инициаторным Met) остатка Gly или Pro резко повышает стабильность белка в цитоплазме S. saccharomyces. Анализ природной последовательности N-конца продукта трансляции гена lacZ из E. coli показал его неоптимальность с точки зрения этого правила: она состоит из остатков Met-Thr-Met-Ile: для инициации трансляции с образованием активного продукта могут использоваться оба указанные здесь остатка Met.
В ходе реализации изобретения было установлено, что использование гена lacZ из E. coli с нативной структурой N-конца первичного продукта трансляции приводит к образованию продукта с низкой протеолитической стабильностью, что делает невозможным достоверное определение его ферментативной активности. Создание модифицированного гена, содержащего дополнительный кодон GGA (Gly), совмещенного с сайтом рестриктазы BamHI (GGATCC) непосредственно вслед за инициаторным кодоном Met позволяет резко повысить стабильность продукта и добиться воспроизводимых результатов измерения репортерного сигнала в лизатах, несущих его в геноме рекомбинантных штаммов Y. lipolytica при использовании в качестве субстрата β-галактозидазы 3-бром, 4-хлор, 3-индолил-βD-галактопиранозида (X-gal). При реализации изобретения в качестве модельного промотора использовался промотор гена митохондриального потенциал-зависимого порина VDAC Y. lipolytica.
Характеризуя наиболее близкие аналоги заявляемого изобретения, касающиеся разработки систем экспрессии генов в Y. lipolytica, целесообразно указать на действующие в РФ патенты Юзбашевой и соавт. № 2451749 [4] и Выборной и соавт. № 2451075 [5]. В этих патентах описано получение на основе Y. lipolytica продуцентов липазы и генетической конструкции для экспозиции белка на поверхности клеточной стенки дрожжей. В этих заявках не упоминаются ни ген LacZ, ни способы измерения активности β-галактозидазы в Y. lipolytica. Таким образом, можно утверждать, что заявляемое изобретение не затрагивает каких-либо принципов, описываемых в цитируемых патентах, ни вопросов протеолитической стабилизации белков, накапливающихся в цитоплазме Y. lipolytica (предметом этих патентов являются белки, подвергающиеся секреции).
Раскрытие изобретения
1) Фрагмент, кодирующий ген LacZ из E. coli амплифицируют с праймерами Lac-for1 и Lac-rev1: размер продукта 3000 п.н. Продукт очищают и подвергают рестрикции по сайтам SalI и HindIII. Проводят клонирование в вектор pET23b, линеаризованный по сайтам SalI и HindIII. Проводят трансформацию E. coli TG1, отбирают колонии синего цвета.
2) Терминатор гена щелочной протеазы XPR клонируют путем ПЦР из генома штамма Y. lipolytica CLIB 122 с использованием двух праймеров: xprT-R и Txpr-F. Продукт размером 140 пн, рестрикцированный по сайтам XmaJI /XbaI, клонируют в вектор pUK21. Отбирают канамицин-резистентные колонии белого цвета, содержащие вставку в прямой ориентации относительно хода транскрипции α-пептида LacZ.
3) Из конструкции pET23b-LacZ выделяют фрагмент с геном LacZ путем двойной рестрикции SalI/NotI и клонируют в конструкцию, pUT, на базе pUK21, несущую терминатор XPR, линеаризованную по сайтам SalI/NotI.
4) Маркерный ген URA3 экстрагируют из конструкции pUC18-URA3 по сайтам NcoI и SalI и переносят в конструкцию на базе pUK21, несущую терминатор XPR и ген LacZ, линеаризованную по сайтам NcoI и XhoI.
5) Промотор гена митохондриального потенциал-зависимого порина из генома штамма Y. lipolytica CLIB 122 клонируют с помощью ПЦР с использованием праймеров VDAC-F и VDAC-R2. Продукт размером 1050 пн обрабатывают рестриктазами Bsp1407/ XhoI и клонируют в конструкцию на базе pUK21, несущую терминатор XPR, ген URA3 и ген LacZ, линеаризованную по сайтам KpnI и XhoI.
6) При проведении генно-инженерного конструирования используют олигонуклеотидные ДНК-праймеры следующей структуры:
Lac-for1 (SalI) GGGTCGACACCATGATTACGGATTCACTG
Lac-rev1 (HindIII) GGAAGCTTATTTTTGACACCAGACCAACTG
xprT-R (XbaI) ATATCTAGAGCCACCTACAAGCCAG
Txpr-F (Eco31I, XmaJI) CATAAAATGCGAGACCTAGGTAGGCAATTAACAG
VDAC-F-Bsp1407 GGTGTACACACATGCATATAGGATACATTT
VDAC-R-Sal GGGTCGACTGGGTTAGTACGTGTTGCGTGT
VDAC-R2-XhoI GGCTCGAGGGATCCCATGTCGACTGGGTTAGTACGTGT
7) Полученная конструкция pUVLT2 при введении в штамм Y. lipolytica PO1f обеспечивает получение трансформантов, обладающих воспроизводимо определяемой активностью β-галактозидазы в клеточных лизатах, зависимой от условий культивирования рекомбинантного штамма. Аналогичный результат не может быть получен с применением конструкции pUVLT1, аналогичной pUVLT2, но содержащей ген LacZ нативной структуры (без внесения искусственного кодона GGA (Gly) dj втором положении открытой рамки считывания вслед за иницаторным кодоном ATG (Met)).
Краткое описание графических изображений:
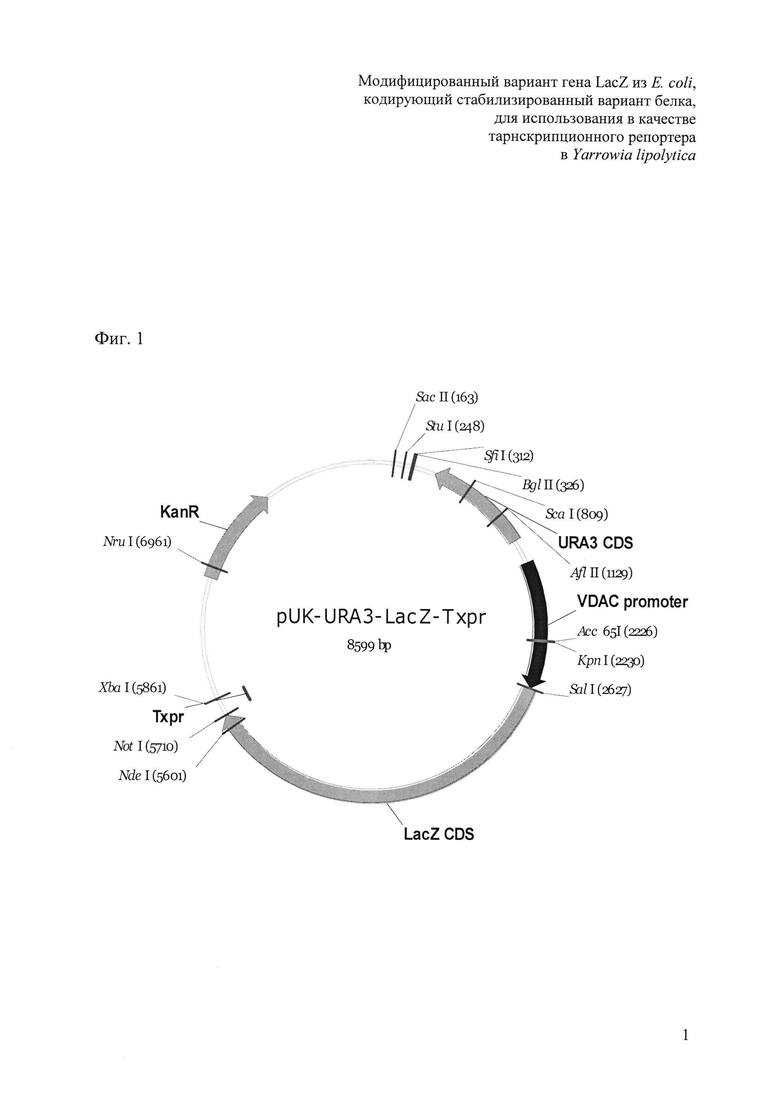
Фиг. 1. Схема функциональных элементов конструкций pUVLT1 (SEQ ID NO 1) и pUVLT2 (SEQ ID NO 2), предназначенной для введения в клетки Y. lipolytica гена lacZ из E. coli. Конструкции содержат промотор VDAC, терминатор XPR, ген lacZ из E. coli C600, маркер компенсации ауксотрофности по урацилу URA3. Поддержание конструкции в E. coli осуществляется за счет элементов из состава стандартного вектора pUK21: репликона плазмиды pMC16 и гена канамицинфосфотрансферазы (детерминанта устойчивости к канамицину).
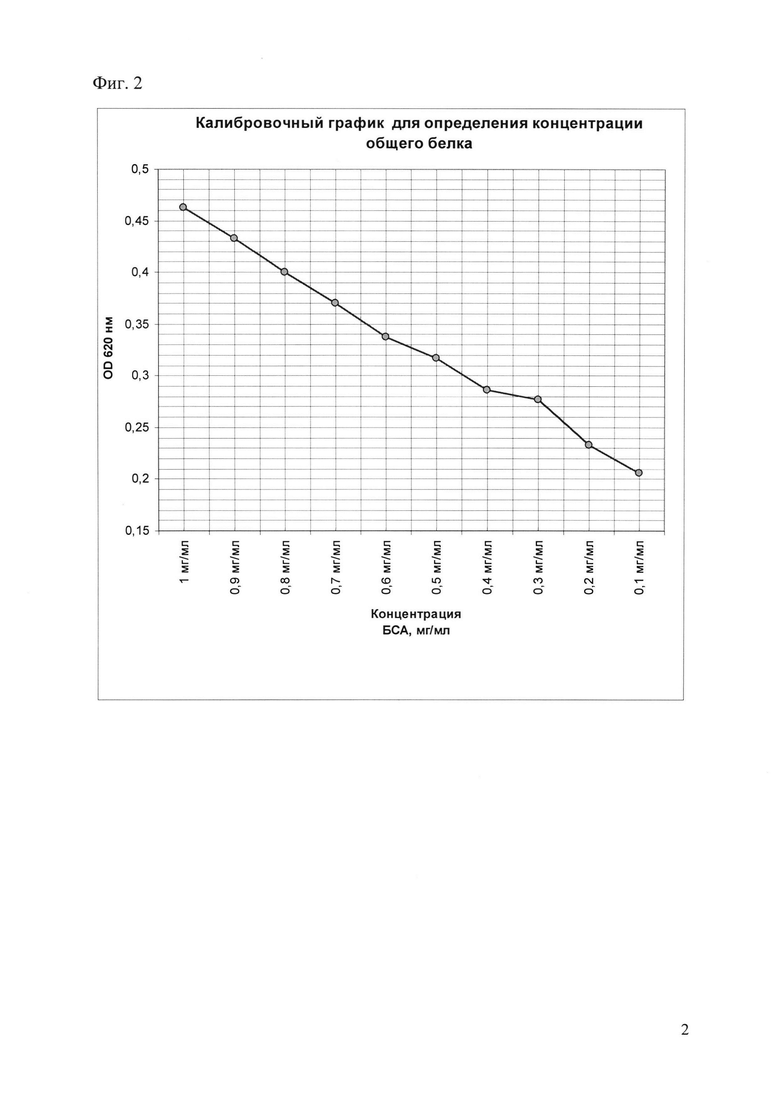
Фиг. 2. Калибровочный график для определения концентрации общего белка.
Осуществление изобретения:
1. Генно-инженерное конструирование
ДНК из биомассы штамма E. coli C600 (F2, thi-1, thr-1, leu136, lacYI, tonA21, supE44) [6], полученной культивированием на бульоне LB, выделяют с помощью набора Проба-ГС (ДНК-технология, Москва), растворяя полученный препарат в 50 мкл деионизованной воды. В качестве матрицы для проведения ПЦР используют 1 мкл препарата геномной ДНК. Реакцию проводят в 30 мкл, используя по 10 пмоль каждого из двух праймеров: Lac-for1 (SalI) GGGTCGACACCATGATTACGGATTCACTG и Lac-rev1 (HindIII) GGAAGCTTATTTTTGACACCAGACCAACTG [7]. Продукт размером 3100 пн очищают путем нанесения-десорбции на стеклянное молоко с применением набора GenJet (MBI Fermentas, Литва) и растворяют в 30 мкл деионизованной воды. 10 мкл препарата отбирают для двойной рестрикции SalI/HindIII и лигируют с ДНК вектора pET23b (Novagen)в стандартных условиях. Лигазной смесью проводят трансформацию E. coli TG1, трансформанты высевают на среду с добавлением 100 мкг/мл ампициллина и 0,01% Х-gal. Отбирают ампициллин-резистентные колонии синего цвета. Полученная конструкция получает обозначение pET23b-LacZ.
Для клонирования терминатора гена щелочной протеазы XPR используют штамм Y. lipolytica CLIB 122. ДНК из биомассы, полученной культивированием на бульоне YPD выделяют с помощью набора Проба-ГС (ДНК-технология, Москва), растворяя полученный препарат в 50 мкл деионизованной воды. В качестве матрицы для проведения ПЦР используют 1 мкл препарата геномной ДНК Y. lipolytica CLIB 122. Реакцию проводят в 30 мкл, используя по 10 пмоль каждого из двух праймеров: xprT-R (XbaI) ATATCTAGAGCCACCTACAAGCCAG и Txpr-F (Eco31I, XmaJI) CATAAAATGCGAGACCTAGGTAGGCAATTAACAG. Продукт размером 140 пн очищают путем нанесения-десорбции на стеклянное молоко с применением набора GenJet (MBI Fermentas, Литва) и растворяют в 30 мкл деионизованной воды. 10 мкл препарата отбирают для двойной рестрикции XmaJI/XbaI и лигируют с ДНК вектора pUK21 в стандартных условиях. Лигазной смесью проводят трансформацию E. coli TG1, трансформанты высевают на среду с добавлением 30 мкг/мл канамицина и 0,01% Х-gal. Отбирают канамицин-резистентные колонии белого цвета. Полученная конструкция, содержащая вставку в прямой ориентации относительно хода транскрипции α-пептида LacZ (проверяется путем контрольной рестрикции по сайтам EcoRI/XbaI - от вектора должен отделяться фрагмент длиной 140 пн), получает обозначение pUK21-T.
Маркерный ген URA3 извлекают из конструкции pUC18-Ura3 [4] в виде фрагмента длиной 860 пн путем двойной рестрикции SalI/NcoI. ДНК вектора pUK21-tXPR подвергается двойной рестрикции XhoI/NcoI. ДНК вставки и вектора объединяют и лигируют в стандартных условиях. Лигазной смесью проводят трансформацию E. coli TG1, трансформанты высевают на среду с добавлением 30 мкг/мл канамицина. Полученная конструкция получает обозначение pUT.
Из конструкции pET23b-LacZ выделяют фрагмент с геном LacZ путем двойной рестрикции SalI/NotI и лигируют с ДНК конструкции pUT, рестрицированной по тем же сайтам. Лигазной смесью проводят трансформацию E. coli TG1, трансформанты высевают на среду с добавлением 30 мкг/мл канамицина. Полученная конструкция получает обозначение pULT.
Клонирование промотора гена VDAC проводят двумя способами.
Вариант 1
В качестве матрицы для проведения ПЦР используют 1 мкл препарата геномной ДНК Y. lipolytica CLIB 122. Реакцию проводят в 30 мкл, используя по 10 пмоль каждого из двух праймеров: VDAC-F-Bsp1407 GGTGTACACACATGCATATAGGATACATTT и VDAC-R-Sal GGGTCGACTGGGTTAGTACGTGTTGCGTGT. Продукт размером 1050 пн очищают путем нанесения-десорбции на стеклянное молоко с применением набора GenJet (MBI Fermentas, Литва) и растворяют в 30 мкл деионизованной воды. 10 мкл препарата отбирают для двойной рестрикции Bsp1407/SalI и лигируют с ДНК вектора pULT, расщепленным по сайтам KpnI/SalI в стандартных условиях. Лигазной смесью проводят трансформацию E. coli TG1, трансформанты высевают на среду с добавлением 30 мкг/мл канамицина. Отбирают канамицин-резистентные колонии белого цвета. Полученная конструкция получает обозначение pUVLT1.
Вариант 2
В качестве матрицы для проведения ПЦР используют 1 мкл препарата геномной ДНК Y. lipolytica CLIB 122. Реакцию проводят в 30 мкл, используя по 10 пмоль каждого из двух праймеров: VDAC-F-Bsp1407 GGTGTACACACATGCATATAGGATACATTT и VDAC-R2-XhoI GGCTCGAGGGATCCCATGTCGACTGGGTTAGTACGTGT. Продукт размером 1050 пн очищают путем нанесения-десорбции на стеклянное молоко с применением набора GenJet (MBI Fermentas, Литва) и растворяют в 30 мкл деионизованной воды. 10 мкл препарата отбирают для двойной рестрикции Bsp1407/XhoI и лигируют с ДНК вектора pULT, расщепленным по сайтам KpnI/SalI в стандартных условиях. Лигазной смесью проводят трансформацию E. coli TG1, трансформанты высевают на среду с добавлением 30 мкг/мл канамицина. Отбирают канамицин-резистентные колонии белого цвета. Полученная конструкция получает обозначение pUVLT2.
2. Получение рекомбинантных штаммов Y. lipolytica
Введение ДНК конструкций pUVLT1 и pUVLT2 в клетки Y. lipolytica PO1f проводится методом трансформации с использованием солей Li+ и отбором трансформантов на минимальной синтетической среде, свободной от урацила. Для проведения трансформации ДНК конструкций pUVLT1 и pUVLT2 препаративно выделяют из клеток E. coli TG1 и линеаризуют с помощью рестриктазы BglI. Полученные штаммы-трансформанты троекратно пассируют на синтетической среде, свободной от урацила, для удаления клеток штамма дикого типа, не несущего конструкции. После этого по 10 клонов, несущих каждую конструкцию, тестируют на наличие активности β-галактозидазы путем высева на среду YPD, содержащую 80 мкг/мл X-gal. Учет результатов проводят после культивирования штамма при +28°С в течение 72 ч. Для дальнейшего тестирования используют клоны, обладающие выраженным синим цветом.
3. Исследование стабильности определения активности β-галактозидазы в рекомбинантных штаммах Y. lipolytica
3.1 Получение посевной культуры.
В микробиологическую пробирку объемом 20 мл, содержащую 3 мл жидкой среды YPD (1% дрожжевого экстракта, 1% пептона, 2% глюкоза), микробиологической петлей засевают материал отдельной колонии Y. lipolytica PO1f, полученной на плотной среде YPD, содержащей 80 мкг/мл X-gal. Культивирование ведут при +28°С в течение 24 ч.
3.2 Получение рабочей культуры.
3 мкл посевной культуры вносят в 3 мл среды YPD с необходимыми добавками. Культивирование ведут при +28°С в течение 18 ч. В качестве добавок вносят следующие растворы:
- 150 мкл 1 мМ K,Na-фосфатного буфера pH= 5,55 (рН среды после внесения добавки 6,1);
- 60 мкл 2,5 М K,Na-фосфатного буфера pH= 9,0; (рН среды после внесения добавки 8,1);
- 60 мкл 2,5 М Трис-сукцинатного буфера pH= 9,7 (рН среды после внесения добавки 9,0);
- 180 мкл 1 М буфера Трис-HCl pH= 8,0 (рН среды после внесения добавки 6,9).
3.3 Сбор биомассы.
Биомассу рабочих культур собирают центрифугированием в пробирках для гомогенизации Qiagen (кат. № 990381) при 9 тыс. G в течение 30 с. Промывают, суспендируя в 400 мкл 0,9% стерильного раствора NaCl, осаждают при 9 тыс. G в течение 30 с.
К осадку биомассы добавляют 400 мкл буфера Z (на 500 мл воды 4,27 г Na2HPO4, 2,75 г NaH2PO4×H2O, 0,375 г KCl, 0,125 г MgSO4×7H2O, pH=7,0) и 1 стальной шарик 7 мм (Qiagen). Гомогенизацию проводят на приборе TissueLyser II (QIAGEN) при 30 оборотах/сек в течение 1,5 мин. Гомогенаты переносят в чистые пробирки без шариков.
3.4 Определение β-галактозидазной активности с использованием в качестве субстрата X-gal.
Готовят рабочий раствор, смешивая 880 мкл буфера (1,5 М глицин, 0,33 М Трис-основание, рН 8,6) и 3520 мкл деионизованной воды. Вносят 44 мкл 1% раствора Х-gal в диметилформамиде. В лунки 96-луночного иммунологического планшета Costar вносят по 100 мкл рабочего раствора, после чего добавляют по 5 мкл гомогенатов рабочих культур. Каждый образец измеряют в трех повторностях. В качестве отрицательного контроля используют деионизованную воду объемом 5 мкл. Реакцию ведут при +37°С в течение 21 ч. Определение оптической плотности проводят на планшетном иммуноферментном фотометре (ОАО «М3» Сапфир, модель Эфос 9305) при длине волны 620 нм. Измерения проводят через 2, 4, 5. и 21 ч.
3.5 Определение содержания белка с помощью бицинхониновой кислоты.
Рабочий раствор получают, смешивая 13922 мкл реагента А и 278 мкл реагента В. В лунки 96-луночного иммунологического планшета Costar вносят по 20 мкл гомогенатов. Разведение образцов для определения белка проводят следующим образом: в лунки столбцов 2 - 5 и 7 - 10 разносят по 10 мкл буфера Z. Переносят по 10 мкл образцов из 1 в 2, перемешивают, отбирают 10 мкл образца из 2 в 3 и т.д. Из последней лунки забирают 10 мкл. Таким образом, получают разведения образцов 1, ½, ¼, 1/8, 1/16.
В лунки 96-луночного иммунологического планшета Costar разносят по 200 мкл рабочего раствора, после чего добавляют в лунки по 10 мкл разведений гомогенатов и стандарта (бычий сывороточный альбумин). Планшет инкубируют 1 ч при 37°С и измеряют на планшетном иммуноферментном фотометре (ОАО «М3» Сапфир, модель Эфос 9305) при длине волны 620 нм.
3.6 Расчет удельной ферментативной активности β-галактозидазы на мг общего белка клеточного лизата.
Каждое измерение активности β-галактозидазы и общего белка в лизате биомассы исследуемого штамма проводят в трех повторностях для каждого типа условий внешней среды. Лизаты готовят и измеряют независимо с момента засева рабочей культуры.
Концентрацию белка определяют по калибровочному графику (табл. 1, фиг. 2), построенному при использовании следующих разведений бычьего сывороточного глобулина (БСА) мг/мл: 1, 0.9, 0.8, 0.7, 0.6, 0.5, 0.4, 0.3, 0.2, 0.1.
Коэффициент молярной экстинкции суспензии индиго в воде считают равным 5,4 ммоль/л×см.
Единица активности (Ед.) - количество фермента, которое катализирует превращение одного микромоля субстрата в мин.
Среднее значение определяют путем усреднения соответствующих значений по следующей формуле:
Значение =(СРЗНАЧ(A620 B-Gal, 21 час;1:3)-СРЗНАЧ(A620 B-Gal, 4,5 часа;1:3))/(5,4 ОЕ/ммоль×990 мин×СРЗНАЧ (конц. белка мг/мл; 1:3))×1000;
Результаты определений представлены в таблице 2.
Таблица 2.
Список использованных источников
1. Gaillardin C, Ribet AM. LEU2 directed expression of beta-galactosidase activity and phleomycin resistance in Yarrowia lipolytica. Curr Genet. 1987. V. 11. № 5. P. 369-75.
2. Juretzek T, Le Dall M, Mauersberger S, Gaillardin C, Barth G, Nicaud J. Vectors for gene expression and amplification in the yeast Yarrowia lipolytica. Yeast. 2001. V. 18. № 2. P. 97-113.
3. Varshavsky A. Recent studies of the ubiquitin system and the N-end rule pathway. Harvey Lect. 2001. V. 96. P. 93-116.
4. Юзбашева Е. Ю., Юзбашев Т. В., Лаптев И. А., Ларина А. С., Синеокий С. П.. Генетическая конструкция для экспозиции белка на поверхности клеточной стенки дрожжей Yarrowia lipolytica. Патент РФ на изобретение № 2451749.
5. Выборная Т. В., Юзбашев Т. В., Соболевская Т. И., Лаптев И. А., Юзбашева Е. Ю., Синеокий С. П.. Рекомбинантный штамм дрожжей Yarrowia lipolytica - продуцент липазы. Патент РФ на изобретение № 2451075.
6. Bachmann BJ. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol Rev. 1972. V. 36. № 4. P. 525-57.
7. Смирнова М.С., Кузнецова Т.В., Леонович О.А., Гордейчук И.В., Шевелев А.Б. Метод получения библиотек серийных двухсторонних делеций с помощью ПЦР с вырожденным праймером. Патент РФ на изобретение № 2511424.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ YARROWIA LIPOLYTICA - ПРОДУЦЕНТ ИНКАПСУЛИРОВАННОЙ ФИТАЗЫ OBESUMBACTERIUM PROTEUS | 2017 |
|
RU2664476C1 |
| Рекомбинантный штамм Yarrowia lipolytica - продуцент инкапсулированной фитазы Paenibacillus sp. | 2023 |
|
RU2814490C1 |
| ИНТЕГРАТИВНАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pQ-SRUS ДЛЯ ПОЛУЧЕНИЯ ШТАММОВ ДРОЖЖЕЙ Yarrowia lipolytica, ОБЛАДАЮЩИХ СПОСОБНОСТЬЮ К ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ ГЕНОМА МИТОХОНДРИЙ ЗА СЧЁТ ЭКСПРЕССИИ ГЕНА БЕЛКА RecA БАКТЕРИАЛЬНОГО ПРОИСХОЖДЕНИЯ | 2014 |
|
RU2562869C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ YARROWIA-LIPOLYTICA - ПРОДУЦЕНТ ЛИПАЗЫ | 2011 |
|
RU2451075C1 |
| Способ репликации человеческого митохондриального генома в клетках дрожжей Yarrowia lipolytica | 2016 |
|
RU2660715C2 |
| Модифицированная нуклеотидная последовательность, кодирующая полипептид, обладающий активностью фитоен синтазы и геранилгеранилпирофосфат синтазы (варианты), ее использование при конструировании трансформантов дрожжей Yarrowia lipolytica, продуцирующих каротиноиды | 2021 |
|
RU2794980C1 |
| ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS PS107(pPIC9HAbIL-2), ЯВЛЯЮЩИЙСЯ ПРОДУЦЕНТОМ ГИБРИДНОГО БЕЛКА, СОСТОЯЩЕГО ИЗ АЛЬБУМИНА ПЛАЗМЫ КРОВИ ЧЕЛОВЕКА И ИНТЕРЛЕЙКИНА-2 ЧЕЛОВЕКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pPIC9HAbIL-2 И СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ | 2006 |
|
RU2315105C1 |
| Трансформант дрожжей Yarrowia lipolytica, продуцирующий линалоол | 2022 |
|
RU2809554C1 |
| ШТАММ ДРОЖЖЕЙ Yarrowia lipolytica - ПРОДУЦЕНТ ЛИПАЗЫ | 2007 |
|
RU2355754C1 |
| Трансформант дрожжей Yarrowia lipolytica, продуцирующий кантаксантин | 2023 |
|
RU2827562C1 |







Настоящее изобретение относится к биохимии, в частности к способу модификации гена LacZ из Escherichia coli. Для осуществления способа фрагмент ДНК, содержащий кодирующую область гена LacZ, нарабатывают с помощью полимеразной цепной реакции с использованием олигонуклеотидного праймера, позволяющего ввести дополнительный кодон GGA, кодирующий глицин, непосредственно после инициаторного кодона ATG. Настоящий способ позволяет адаптировать указанный ген для его экспрессии в клетках Yarrowia lipolytica. 2 ил., 2 табл.
Способ модификации гена LacZ из Escherichia coli, позволяющий адаптировать его для экспрессии в клетках дрожжей Yarrowia lipolytica, заключающийся в том, что фрагмент ДНК, содержащий кодирующую область гена LacZ, нарабатывают с помощью полимеразной цепной реакции с использованием олигонуклеотидного праймера, позволяющего ввести дополнительный кодон GGA, кодирующий глицин, непосредственно после инициаторного кодона ATG.
| GAILLARDIN C et.al., LEU2 directed expression of betagalactosidase activity and phleomycin resistance in Yarrowia lipolytica, Curr Genet 1987, 11(5), pp.369-75 | |||
| A THOMAS JURETZEK et.al., Vectors for gene expression and amplification in the yeast Yarrowia lipolytica, Yeast 2001, 18, pp.97-113 | |||
| РЕКОМБИНАНТНЫЙ БЕЛОК LACspCBD, ОБЛАДАЮЩИЙ БЕТА-ГАЛАКТОЗИДАЗНОЙ АКТИВНОСТЬЮ И СПОСОБНОСТЬЮ САМОПРОИЗВОЛЬНО СВЯЗЫВАТЬСЯ С ЦЕЛЛЮЛОЗОСОДЕРЖАЩИМИ СОРБЕНТАМИ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА LACspCBD, ШТАММ Escherichia coli M15 [pREP4, pLACspCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА LACspCBD, СПОСОБ ПОЛУЧЕНИЯ ИММОБИЛИЗОВАННОГО РЕКОМБИНАНТНОГО БЕЛКА LACspCBD НА ЦЕЛЛЮЛОЗЕ И СПОСОБ ФЕРМЕНТАТИВНОГО РАСЩЕПЛЕНИЯ ЛАКТОЗЫ | 2004 |
|
RU2278160C2 |
| БЕТА-ГАЛАКТОЗИДАЗА С ТРАНСГАЛАКТОЗИЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2007 |
|
RU2435856C2 |

