Область применения изобретения
Изобретение относится к области биотехнологии, физико-химической биологии, молекулярной биологии и направлено на разработку эффективного способа доставки биологически активных макромолекул (белков, нуклеиновых кислот различного размера и нуклеопротеидных комплексов белков с нуклеиновыми кислотами) в клетки растений. В соответствии с настоящим изобретением получены инфильтрируемые в клетки растений наноплатформы на основе частиц хитозана, функционализированные биологически активными макромолекулами, которые могут быть использованы в различных био- нанотехнологиях, например с целью их последующего применения для редактирования генома растений методом, исключающим неконтролируемое внесение чужеродного генетического материала, защиты сельскохозяйственных растений от различных патогенов, включая бактерии, оомицеты, грибы и вирусы, а также для генетической трансформации растений.
Описание изобретения
Терминология: Все термины и определения, приведенные в тексте, следует понимать в том смысле, в котором они обычно применяются в соответствующих областях науки и техники.
Некоторые определения, оговоренные специально, не являются исключениями и приводятся только в целях облегчения понимания сущности изобретения.
Эффективность доставки биомолекул - процент попадания внутрь растительной клетки целевых биомолекул, определяемый с помощью введения флуоресцирующих/флуоресцентно меченных молекул.
Эффективность геномного редактирования - процент локусов в геноме растительных клеток, в которые были внесены направленные мутации.
Биологически активные макромолекулы, биомолекулы или биополимеры - полимеры, встречающихся в природе в естественном виде, входящие в состав живых организмов.
Постоянно растущее население мира и сокращающийся фонд сельскохозяйственных пахотных земель стимулируют исследования, направленные на повышение эффективности сельского хозяйства. Стандартные средства повышения урожайности сельскохозяйственных и садовых культур включают в себя селекционные методики, основанные на выявлении растений, характеризующихся желаемыми признаками. Технологии молекулярной биологии, основанные на манипуляции генетическим материалом (нуклеиновых кислот в виде ДНК или РНК) и последующем введении этого генетического материала в растение, позволяют создавать культуры, характеризующиеся различными улучшенными экономическими, агротехническими и садово-техническими признаками.
Задача настоящего изобретения состояла в разработке технически простого универсального метода доставки биологически активных макромолекул в клетки растений, который применим для доставки разнообразных макромолекул, включая короткие РНК, ДНК, белки и комплексы белков с нуклеиновыми кислотами, с эффективностью, позволяющей регулировать различные клеточные процессы, в том числе активность клеточных генов и защитный ответ растений на инфекцию различными патогенами.
В качестве перспективного инструмента для доставки нуклеиновых кислот, белков и нуклеопротеинов в клетку в настоящее время рассматриваются полимерные системы, в том числе наночастицы и нанокапсулы [Марквичева Е.А. Хитозан и его производные в биоинкапсулировании. Хитин и хитозан: получение, свойства и применение. М.: Изд-во Наука, 2002. с. 315-326.]. Документы WO/2010/118077, WO/2009/046384, WO/2012/006439, WO/2012/006443, WO 2008/083233, WO 2011US57813A раскрывают применение наночастиц для доставки ДНК и трансформации растений. Известен также опосредованный золотыми наночастицами метод доставки сиквенс-специфических нуклеаз в растительные клетки, раскрытый в документах RU 2612156 C2 и RU 2556376 C2 (ДАУ АГРОСАЙЕНСИЗ, (US)). В отличие от известного метода, в настоящем изобретении в качестве материала для создания наночастиц для применения в доставке макромолекул в растительные клетки предложен хитозан. Хитозан широко применяют в сельском хозяйстве при разработке средств защиты растений, однако к настоящему времени не разработано способов его применения для эффективной доставки макромолекул в растительные клетки. Хитозан, в составе которого преобладают остатки глюкозамина, - природный полимер, обладающий широким спектром биологических свойств, в том числе биосовместимостью и биодеградируемостью. В последнее время хитозан рассматривается как один из перспективных материалов при создании полимерных матриц для доставки в лекарственных препаратов и ДНК в клетки животных.
Созданные на его основе системы доставки в клетки животных с внедренными в них ДНК-векторами имеют ряд преимуществ: на их поверхности можно ковалентно присоединить лиганды, которые обеспечивают высокоспецифическое взаимодействие с клеточными рецепторами; можно ввести в клетку не одну, а несколько ДНК-плазмид в одной наночастице или нанокапсуле; можно защитить нуклеиновую кислоту от расщепления ферментами; ДНК-плазмиды приобретают компактный вид, что предотвращает их механический разрыв и облегчает проникновение через мембраны клеток; такие наночастицы можно лиофильно высушить и длительно хранить в такой форме без потери активности [Gene Ther. 2004; v. 11:19:1441-1452.]. Методы введения генетического материала, ДНК, РНК, протеинов и нуклеопротеидных комплексов в растительные клетки значительно отличаются от тех, что используются для клеток животных. Основным препятствием на пути макромолекул является клеточная стенка. Ее химический состав и особенности расположения составляющих ее компонентов способствуют связыванию нуклеиновых кислот и белков различной структуры, ограничивая тем самым доступ их к мембране клетки. При использовании не вирусных векторов, например, несущей целевой ген бактериальной плазмидной ДНК, эффективность прямой трансформации оказывается очень низкой. Проводимые в 1970-1980-е годы эксперименты по введению суспензий молекул ДНК в завязи цветков или молодые цветочные побеги с помощью шприца (макроинъекции), замачиванию сухих семян растений или пыльцы в подобных растворах, применению электрофореза для усиления проникновения ДНК в ткани проростков показали, что частота образования трансформантов крайне низка (менее 2%). Альтернативными являются методы введения ДНК в протопластированные (лишенные клеточной стенки) растительные клетки, где для удаления клеточной стенки растительных клеток применяют комплекс ферментов грибного или бактериального происхождения, состоящий из пектинлиаз и целлюлаз.
В настоящее время часто используют физический способ доставки ДНК в клетку - бомбардировку ее микрочастицами (биобаллистика или сокращенно биолистика). ДНК на частицах диаметром от 0,4 до 1,2 мкм закрепляют путем обработки растворами нуклеиновой кислоты и СаСl2, или спермидина, или полиэтиленгликоля. Покрытые ДНК частицы с помощью специальных приборов разгоняют до скорости 300-600 м/с и направляют поток на изолированные клетки, фрагменты или целое растение. Для создания потока летящих частиц используют газы, образующиеся при сгорании пороха, сжатый воздух или гелий. Частицы пробивают клеточную стенку и мембраны растительной клетки. Доставленная в клетки ДНК частично переходит в свободное состояние и взаимодействует с молекулами ДНК в ядре, митохондриях или пластидах.
Метод бомбардировки микрочастицами позволяет трансформировать растения практически любых видов. Но при этом известно, что происходит она с невысокой частотой, а интеграция и экспрессия чужеродных генов может зависеть от вектора, используемого для их введения. Например, частота трансформации повышается, если используется линейная, а не кольцевая ДНК. Кроме того, при бомбардировке микрочастицами высокомолекулярные плазмиды (более 10 т.п.н.) могут фрагментироваться, поэтому уровень экспрессии чужеродных генов окажется ниже, чем в случае плазмид меньшего размера.
В качестве микрочастиц для бомбардировки растительных тканей используют сферические частицы благородных металлов - золота или вольфрама, поскольку они обладают необходимой массой и твердостью и при этом не являются токсичными.
В настоящем изобретении в качестве материала для создания частиц-носителей макромолекул используется хитозан, который, благодаря своей поликатионной природе, способен образовывать полиэлектролитные комплексы с отрицательно заряженными полимерами, такими как нуклеиновые кислоты, белки, нуклеопротеиды (Прикладная Биохимия и Микробиология 2005; т. 41:1:9-16. Biomacromolecules 2004; v. 5:928-936.). Авторы изобретения разработали оригинальную методику, позволяющую использовать хитозан для конструирования наночастиц-носителей макромолекул (ДНК, нуклеопротеидных комплексов и белков) и доставки их в растительную клетку.
В частности, с помощью раскрытого в настоящем изобретении способа доставки макромолекул авторы изобретения предлагают редактировать геном высших растений, для чего могут быть использованы элементы системы CRISPR-Cas9. Способность частиц хитозана с высокой эффективностью связываться с макромолекулами различного состава и размера крайне важна в данном случае: для работы системы CRISPR необходима особым образом процессированная CRISPR-PHК (crPHК). CRISPR - методы редактирования генома основаны на использовании ДНК-интерференции, направляющей (гидовой) РНК и нацеленности против специфических последовательностей ДНК. Ряд важных открытий, касающихся устройства систем CRISPR типа II (в частности, выяснение необходимости для ее работы белка Cas9 и дополнительной - помимо crРНК - малой РНК, названной tracrPHК), позволил в 2012 году экспериментально опробовать первую искусственно разработанную систему CRISPR типа II. Искусственные системы CRISPR-Cas могут работать не только в клетках бактерий и in vitro, но и в клетках эукариот.
Ожидаемым техническим результатом настоящего изобретения являлась разработка унифицированной процедуры доставки макромолекул в растительные клетки, расширение методического арсенала способов и средств доставки в растительную клетку макромолекул, в частности нуклеиновых кислот, белков и нуклеопротеидных комплексов, с возможностью последующего перепрограммирования растительных клеток и/или редактирования их генома.
Описание чертежей
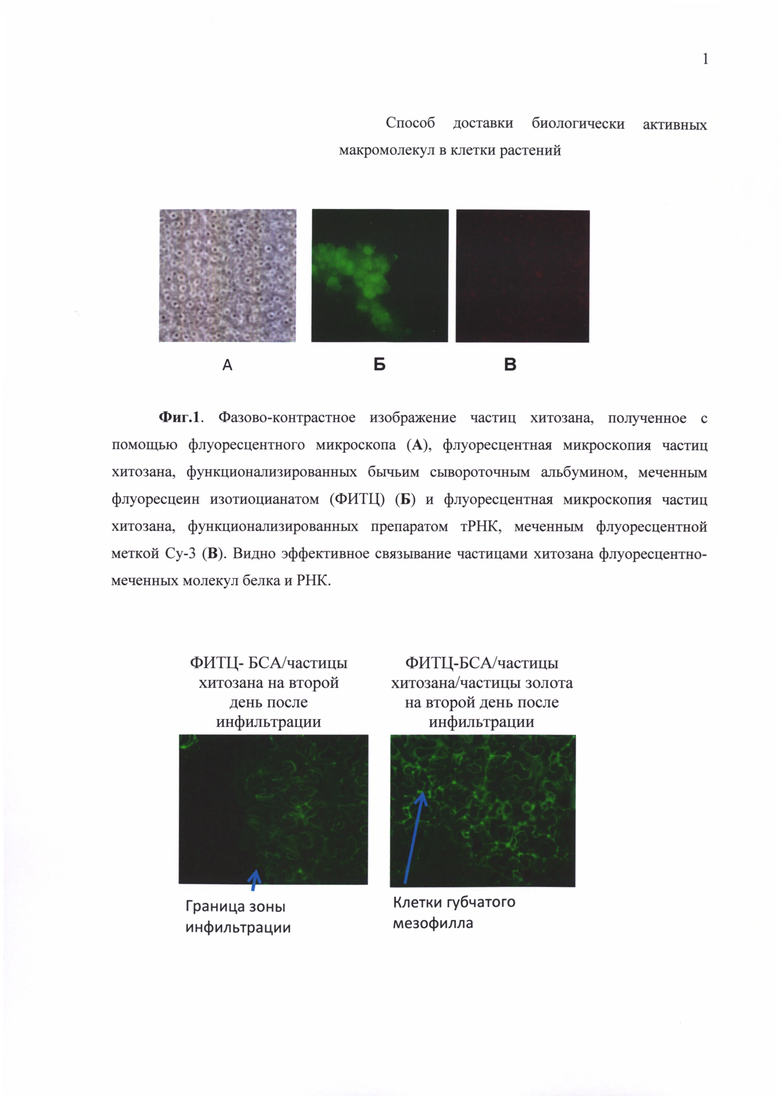
Фиг. 1. Фазово-контрастное изображение частиц хитозана, полученное с помощью флуоресцентного микроскопа (А), флуоресцентная микроскопия частиц хитозана, функционализированных бычьим сывороточным альбумином, меченным флуоресцеин изотиоцианатом (ФИТЦ) (Б) и флуоресцентная микроскопия частиц хитозана, функционализированных препаратом тРНК, меченным флуоресцентной меткой Су-3 (В). Видно эффективное связывание частицами хитозана флуоресцентно-меченных молекул белка и РНК.
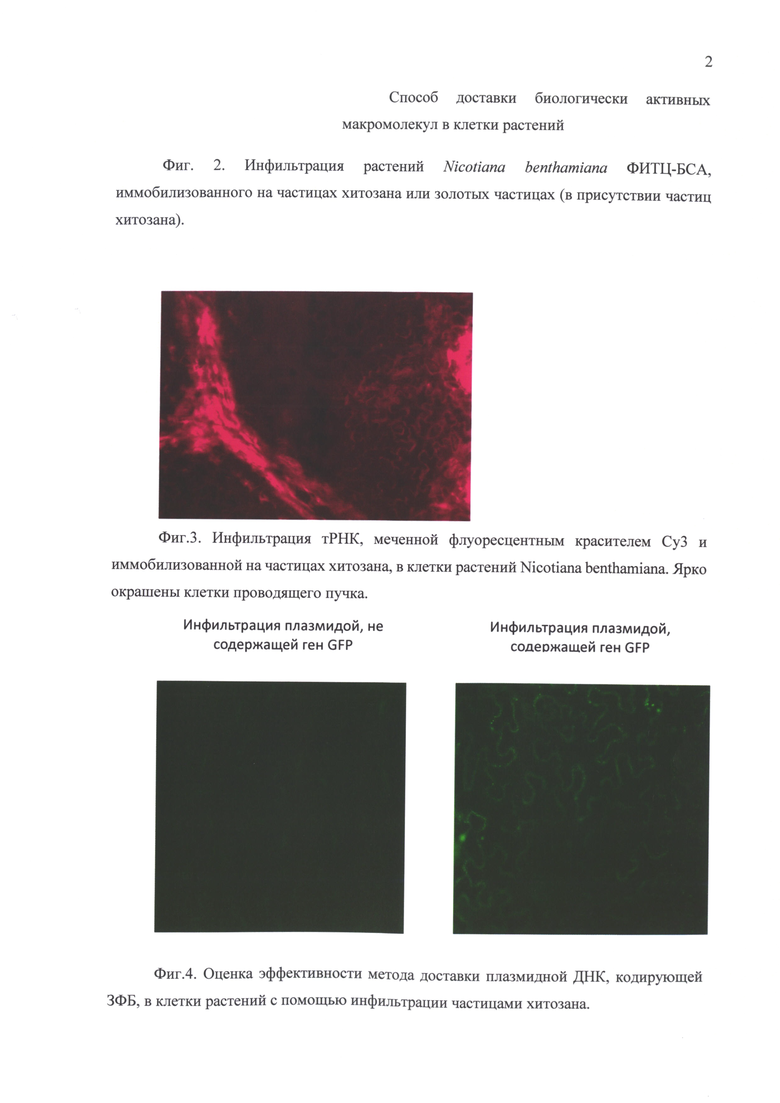
Фиг. 2. Инфильтрация растений Nicotiana benthamiana ФИТЦ-БСА, иммобилизованного на частицах хитозана или золотых частицах (в присутствии частиц хитозана).
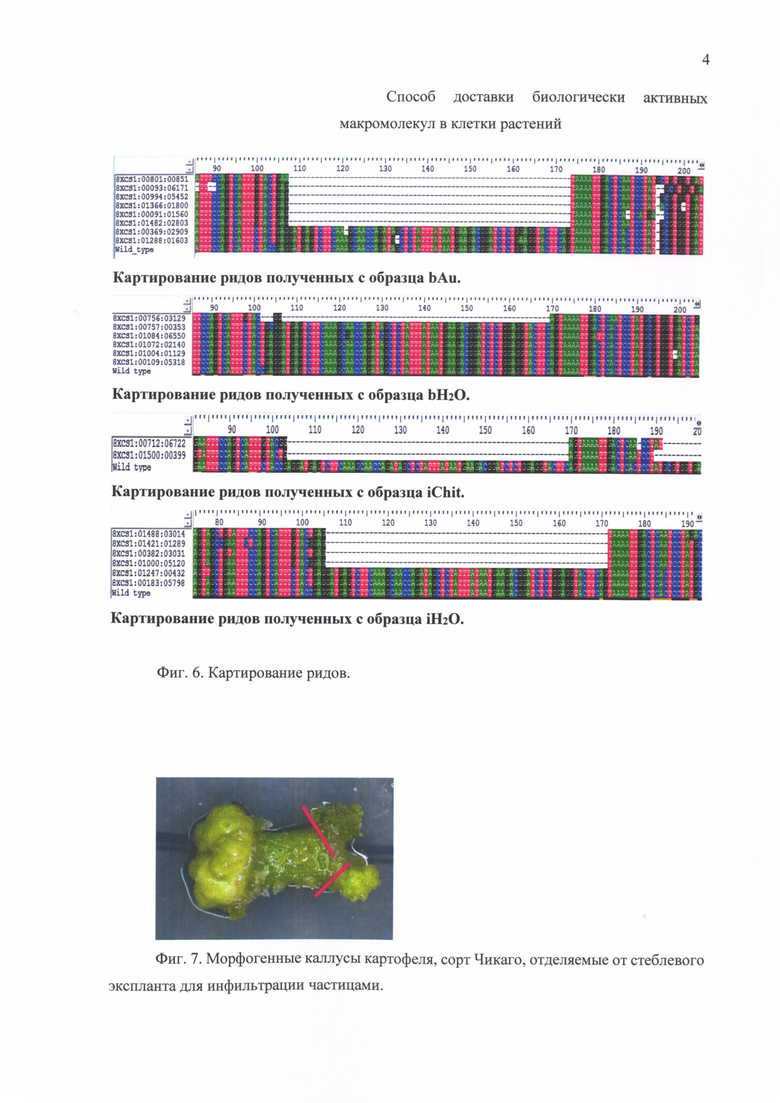
Фиг. 3. Инфильтрация тРНК, меченной флуоресцентным красителем Су3 и иммобилизованной на частицах хитозана, в клетки растений Nicotiana benthamiana. Ярко окрашены клетки проводящего пучка.
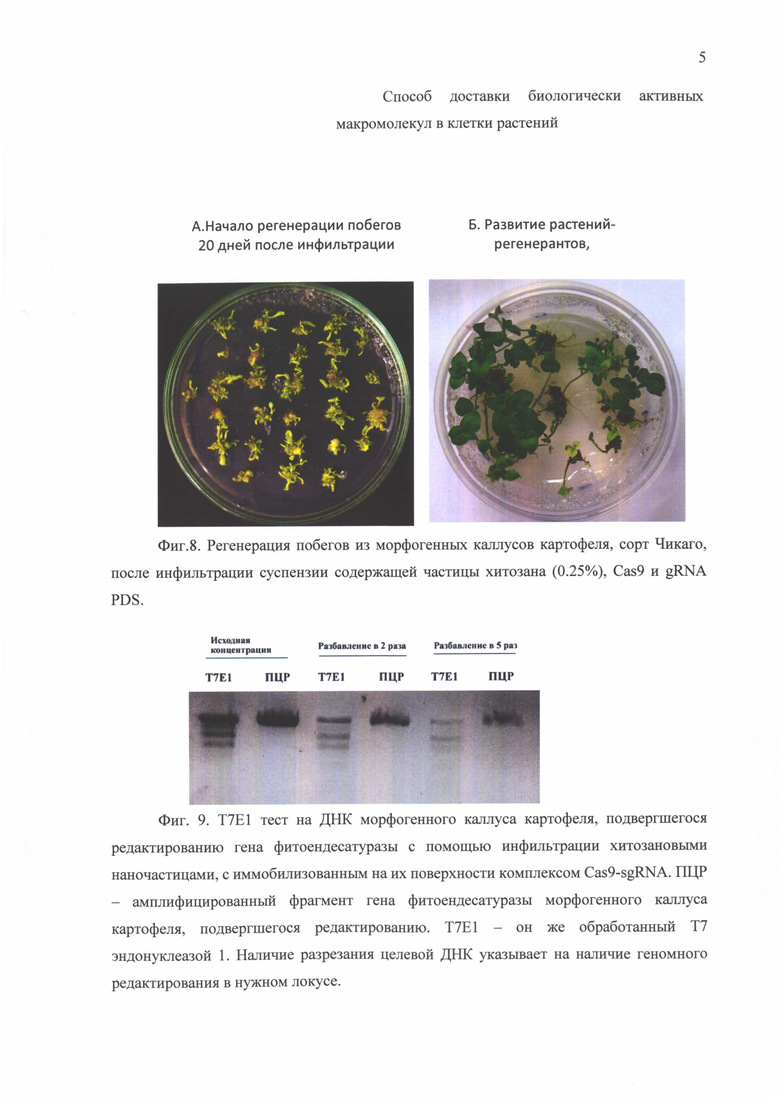
Фиг. 4. Оценка эффективности метода доставки плазмидной ДНК, кодирующей ЗФБ, в клетки растений с помощью инфильтрации частицами хитозана.
Фиг. 5. Электрофореграмма библиотеки до и после очистки.
М - маркер молекулярного веса ДНК.
1 bАu - биолистический бомбардмент золотыми наночастицами, покрытыми комплексами Cas9-sgRNA
2 bН2O - биолистический бомбардмент комплексами Cas9-sgRNA без наночастиц (- контроль)
3 iChit - вакуумная инфильтрация комплексами Cas9-sgRNA на хитозановых наночастицах
4 iH2O - вакуумная инфильтрация комплексами Cas9-sgRNA без наночастиц (-контроль)
Фиг. 6. Картирование ридов для выявления редактирования гена коилина картофеля.
Фиг. 7. Морфогенные каллусы картофеля, сорт Чикаго, отделяемые от стеблевого экспланта для инфильтрации частицами.
Фиг. 8. Регенерация побегов из морфогенных каллусов картофеля, сорт Чикаго, после инфильтрации суспензии содержащей частицы хитозана (0.25%), Cas9 и gRNA PDS.
Фиг. 9. Т7Е1 тест на ДНК морфогенного каллуса картофеля, подвергшегося редактированию гена фитоендесатуразы с помощью инфильтрации хитозановыми наночастицами, с иммобилизованным на их поверхности комплексом Cas9-sgRNA. ПЦР - амплифицированный фрагмент гена фитоендесатуразы морфогенного каллуса картофеля, подвергшегося редактированию. Т7Е1 - он же обработанный Т7 эндонуклеазой 1. Наличие разрезания целевой ДНК указывает на наличие геномного редактирования в нужном локусе.
Раскрытие изобретения
Технический результат достигается за счет оптимальной комбинации приемов и средств, применяемых при осуществлении процедуры доставки: в частности, получения частиц хитозана оптимальной величины, их связывания с макромолекулами различной природы (нуклеиновыми кислотами, нуклеопротеидными комплексами и белками в оптимальных условиях, определяемых структурой и свойствами доставляемой макромолекулы, и введения нагруженных таким образом наночастиц в растительные клетки.
Получение частиц хитозана и их функционализация.
Хитозан (Sigma, CAS Number 9012-76-4) в концентрации 1,75 мг/мл растворяли в 30 мМ Na-ацетатном буфере рН 4,5 (буфер готовили, подводя рН раствора 30 мМ СН3СОONa до 4,5 10% раствором СН3СООН) при постоянном встряхивании до полного растворения хитозана. Затем к раствору добавляли водный раствор триполифосфата натрия (ТПФ, Галреахим) в концентрации 1 мг/мл в соотношении хитозан : ТПФ, равном 5:2. Данные условия приводили к формированию гомогенного препарата частиц хитозана с диаметром 600-800 нм (фазово-контрастное изображение, полученное с помощью моторизованного инвертированного флуоресцентного микроскопа Axiovert 200М (Carl Zeiss, Германия), рис. 1a. Для некоторых методик (указаны ниже) исходный раствор частиц хитозана с рН 4,5 нейтрализовали добавлением 10% раствора NaOH до рН 6,5. К полученному раствору частиц хитозана добавляли растворы белка, РНК или ДНК (концентрации указаны в соответствующих примерах), перемешивали и озвучивали (обрабатывали ультразвуком с частотой 50/60 Hz) при температуре 25°С в течение 10 минут с использованием ультразвуковой мойки Elmasonic s300h (Германия).
Эффективность загрузки наноплатформ оценивалась методом флуоресцентной микроскопии, основываясь на количестве флуоресцирующих частиц по отношению к общему количеству частиц, видимых в световом поле (Фиг. 1). Сигналы флуоресцентных красителей (ФИТЦ и Су-3), которыми с использованием стандартных методик были мечены соответственно белки и РНК, регистрировали с помощью моторизованного инвертированного флуоресцентного микроскопа Axiovert 200М (Carl Zeiss, Германия), управляемого программным комплексом Axiovision 4.5 (Carl Zeiss Microimaging, Германия) и оборудованного объективами Plan-Neofluar с увеличениями 10х и 20х. Сигналы детектировали в зеленом и красном каналах. Параллельно регистрировали фазово-контрастное изображение каждого поля зрения. Для регистрации изображения использовали цифровую камеру ORCA II ERG-2 (Hamamatsu Photonics, Япония). Дополнительно флуоресцентно-меченные белки и РНК в клетках растений детектировались при помощи конфокального лазерного сканирующего микроскопа, модель Leica TCS SP5 (Leica Mycrosystems, Heidelberg, Germany). В клетках проводилась детекция зеленого флуоресцирующего белка (GFP). Для возбуждения свечения использовался аргоновый лазер с длиной волн 488 нм.
Оценка показала, что частицы хитозана в условиях эксперимента эффективно связывают флуоресцентно меченные белок/РНК. Меченные флуоресцентными красителями белки и РНК, связанные с частицами хитозана, показаны на фиг. 1б, в. Полученные наноплатформы сохраняли стабильность и функциональность в растворе в течение 24-48 час.
Для осуществления способа не является принципиальным, каким именно способом осуществлена функционализация частиц хитозана - например, она может быть обеспечена электростатической адсорбцией, конъюгацией с лигандом на поверхности наночастицы, покрытием наночастицы малым кофактором, который доставляемая макромолекула РНК, ДНК, белка или нуклеопротеина может распознать и связать, и непосредственной конъюгацией макромолекулы с наночастицей.
Получение экспериментальных растений. В экспериментах использовали растения Nicotiana benthamiana и картофеля Solanum tuberosum. Растения выращивали из семян/миниклубней и содержали в условиях теплицы при 16 часовом дне, с минимальной дневной температурой 28°С и минимальной температурой ночью 22°С.
Инфильтрация функционализированных частиц хитозана в клетки растений.
Инфильтрацию проводили частицами хитозана, функционализированными белком/РНК/ДНК/РНП или частицами хитозана (0,5% раствор) с добавлением раствора золотых частиц ((BioRad, Microcarrier Gold размером 0,6 мкм), функционализированными белком/РНК/ДНК/РНП. Полученные суспензии/растворы частиц предварительно озвучивали при температуре 25°С в течение 10 мин. с частотой 50/60 Hz с использованием ультразвуковой мойки Elmasonic s300h (Германия). Для инфильтрации тканей растений использовали методы механической и вакуумной инфильтрации. Механическую инфильтрацию проводили макроинъекцией с использованием инъекционного стерильного шприца. Инфильтрация вакуумированием может проводиться путем помещения ткани в эксикатор и создания в нем вакуума, после чего, при разгерметизации и резком повышении давления, происходит забрасывание нанесенной суспензии внутрь ткани. Препарат частиц хитозана, функционализированных белком/РНК/ДНК или комплексом белок-РНК, белок-ДНК, инфильтрировали в нижнюю поверхность листа через небольшое повреждение наружного эпидермиса. Площадь инфильтрации составляла не менее 1 см в диаметре. Вакуумную инфильтрацию проводили с использованием PDS-1000/Не пушки (BioRad). Для растительных образцов использовано давление 28-30 дюймов рт.ст., в соответствии с инструкцией производителя. Образцы (высечки листовой пластинки размером 5×5 мм, апикальные меристемы (около 10 меристем) или пазушные почки) помещали в камеру PDS-1000 /Не пушки, на образец наносили до 100 мкл приготовленного препарата, поднимали давление в камере до рекомендованного уровня, выдерживали в течение 60 сек, а затем снижали давление до нормального уровня. Инфильтрированные функционализированными частицами хитозана клетки/ткани растения выдерживали в климатической камере или при комнатной температуре. Оценку результатов проводили на 2-3 день после инфильтрации методами флуоресцентной микроскопии и лазерной конфокальной микроскопии.
В качестве материала для доставки наночастиц, дополнительно к вышеназванным, могут быть использованы и другие растительные ткани, например, меристемы, морфогенный каллус, пазушные почки, экспланты, зародыш, каллус, пыльца, листья, пыльники, корни, корневые кончики, цветки, семена, стручки и стебли. Для улучшения эффективности доставки дополнительно можно применять предварительную обработку растительного материала (например, частичную мацерацию клеточных стенок, инкубацию растительного материала с растворами повышенными осмотическими свойствами, к примеру раствором маннита).
Для покрытия частиц хитозана могут быть использованы макромолекулы, обеспечивающие направленную доставку в определенный субклеточный компартмент: цитозоль, ядро, тонопласты, пластиды, этиопласты, хромопласты, лейкопласты, элайопласты, протеинопласты, амилопласты, хлоропласты и полости двухслойной мембраны.
Разработанный в данной заявке способ может быть использован для доставки в клетки и ткани растений белков (бычий сывороточный альбумин, БСА; зеленый флуоресцентный белок, ЗФБ), малых РНК (тРНК, направляющей (гидовой) РНК; sgRNA и любых других), ДНК (плазмида, кодирующая ЗФБ) и рибонуклеиновых комплексов системы CRISPR-Cas (нуклеаза Cas9 совместно с гидовой РНК); для создания растений с повышенной урожайностью путем доставки в растение нуклеиновых кислот, кодирующих специфические полипептиды, для переноса в растение гена интерферона-альфа или -бета человека для получения растений, устойчивых к вирусной инфекции. Полученные результаты показывают, что разработанный способ доставки биологических макромолекул в клетки растений, основанный на инфильтрации (механической или вакуумной) препарата частиц хитозана или совместно частиц хитозана и золота с иммобилизованными на их поверхности макромолекулами, обеспечивает эффективную доставку полученных наноплатформ, что подтверждается по выявлению меченных флуоресцентными метками белков и нуклеиновых кислот в клетках растений (фиг. 1), а также результатами прямого секвенирования следующего поколения в случае доставки специфичных CRISPR/Cas9 рибонуклеопротеидных комплексов (см. примеры).
Оценка числа отредактированных клеток следующего поколения в экспериментах по редактированию генома после доставки в ткани растений Nicotiana benthamiana или Solanum tuberosum рибонуклеопротеидного комплекса системы CRISPR/Cas показала, что эффективность редактирования при обработке образцов комплексами Cas9-sgRNA на хитозановых наночастицах неожиданно высока, не менее чем 3-5 раз, а в некоторых случаях в 10 раз, превышая таковую при аналогичной обработке тканей комплексами Cas9-sgRNA в отсутствие наночастиц, что является важным новым техническим результатом.
Эксперименты по оценке количества доставляемого наночастицами хитозана материала показали, что при использовании макроинъекций или вакуумной инфильтрации для различных видов белков оно составляет от 1 до 5 мг/мл, для РНК около 15мМ и для ДНК около 1 мг/мл, до 5 мг\мл для нуклеопротеидных комплексов. При осуществлении функционализации молекулы хитозана не существует каких-либо ограничений для структуры доставляемой макромолекулы, т.е. нагрузка наночастицы может быть осуществлена любым известным в настоящее время белком животного, растительного или искусственного происхождения, любым комплексом белок\ДНК или белок\РНК, ДНК или РНК любой длины и любой степени очистки.
Предложенный в настоящем изобретении способ доставки макромолекул в ткани растений является универсальным и может быть применен для любых известных сельскохозяйственных культур (в этом списке можно назвать, например, зерновые, зернобобовые, кормовые, масличные, эфиромасличные, технические, овощные, лекарственные, цветочные, плодовые, ягодные растения, картофель, томаты, сахарная свекла, виноград, табака, морковь, кукуруза, канола, рапс, хлопчатник, пальма, арахис, соя, Oryza sp., Arabidopsis sp., Ricinus sp. сахарный тростник) и декоративных растений.
Дальнейшее повышение эффективности доставки макромолекул, ДНК, РНК и белков достигается при инкубации растительного материала в осмотических растворах. Для этого спустя 4-20 часов после проведения инфильтрации обработанный растительный материал инкубируется на высокоосмотической среде, содержащей до 500 mOsm/кг сахарозы. Подобная обработка способна еще в 4-5 раз увеличить эффективность доставки макромолекул.
Примеры
Пример 1. Доставка зеленого флуоресцентного белка (ЗФБ) в клетки растений Nicotiana benthamiana.
Пробу для инфильтрации получали, смешивая 1,75 мг/мл раствора хитозана в 30 мМ Na-ацетатном буфере (рН 4,5), водный раствор ТПФ (1 мг/мл) в объеме 50 и 20 мкл, соответственно для формирования частиц хитозана. затем к пробе добавляли 15 мкл раствора ЗФБ (TurboGFP, Евроген) с концентрацией 1 мг/мл и инкубировали в течение 15 мин. при температуре 25°С при постоянном перемешивании для иммобилизации белка на частицах хитозана. Доставку наноплатформ в клетки растений осуществляли путем механической или вакуумной инфильтрации.
В ряде опытов в пробу для инфильтрации дополнительно вносили частицы золота (BioRad, Microcarrier Gold размером 0,6 мкм). Использовали раствор частиц золота с концентрацией 30 мкг/мл. Предварительно проводили озвучивание золотых частиц при температуре 25°С в течение 10 минут ультразвуком с частотой 50/60 Hz с использованием ультразвуковой мойки Elmasonic s300h (Германия). Пробу для инфильтрации получали, смешивая раствор золотых частиц с концентрацией 30 мкг/мл, 0,5% раствор частиц хитозана, приготовленных по вышеописанной методике, и раствор ЗФБ (TurboGFP, Евроген с концентрацией 1 мкг/мл) в соотношении 40, 140 и 30 мкл, соответственно. Пробу инкубировали в течение 15 мин. при температуре 25°С при постоянном перемешивании. Доставку наноплатформ в клетки растений осуществляли путем механической или вакуумной инфильтрации. Видимой разницы в эффективности доставки, осуществляемой при помощи частиц хитозана, при добавлении золотых наночастиц и без них не обнаружено, что свидетельствует о примерно одинаковой эффективности способов. Фазово-контрастное изображение частиц хитозана при различной обработке, полученное с помощью флуоресцентного микроскопа, показано на Фиг. 1.
Пример 2. Доставка бычьего сывороточного альбумина, меченного ФИТЦ, в клетки растений Nicotiana benthamiana и Solanum tuberosum.
Предварительно проводили мечение бычьего сывороточного альбумина (БСА) ФИТЦ. Для этого непосредственно перед мечением готовили раствор БСА в концентрации 5 мг/мл в 0,1 М карбонатном буфере с рН 9,0 и на каждый 1 мл белка вносили 50 мкл исходного раствора ФИТЦ (Sigma), затем к раствору добавляли хлорид аммония (до конечной концентрации 50 мМ) и оставляли на ночь при 4°С в темноте. Меченный ФИТЦ БСА отделяли от свободного ФИТЦ с помощью гельфильтрации на Сефадексе G-50 (Pharmacea). Эффективность мечения БСА ФИТЦ была рассчитана по соответствующей формуле, определенное молярное соотношение ФИТЦ/БСА составило 1,5. Пробу для инфильтрации получали, смешивая раствор частиц хитозана, полученный согласно вышеприведенной методике (1,75 мг/мл раствора хитозана в 30 мМ ацетатном буфере (рН 4,5), раствор ТПФ (1 мг/мл) в соотношении 100 к 40 мкл) и 100 мкл раствора БСА/ФИТЦ (концентрация 3 мг/мл). Пробу инкубировали в течение 15 мин. при температуре 25°С при постоянном перемешивании. Доставку наноплатформ в клетки растений осуществляли путем механической (макроинъекция шприцом) или вакуумной инфильтрации.
В ряде опытов в пробу для инфильтрации дополнительно вносили частицы золота (BioRad, Microcarrier Gold размером 0,6 мкм). Использовали раствор частиц золота с концентрацией 30 мкг/мл. Предварительно проводили озвучивание золотых частиц при температуре 25°С в течение 10 минут ультразвуком с частотой 50/60 Hz с использованием ультразвуковой мойки Elmasonic s300h (Германия). Пробу для инфильтрации получали, смешивая 40 мкл раствора золотых частиц с концентрацией 30 мкг/мл, 140 мкл 0,5% раствор частиц хитозана, приготовленных по вышеописанной методике, и 100 мкл раствора ФИТЦ-БСА с концентрацией 3 мг/мл. Пробу инкубировали в течение 15 мин. при температуре 25°С при постоянном перемешивании. Доставку наноплатформ в клетки растений осуществляли путем механической или вакуумной инфильтрации. Видимой разницы в эффективности доставки, осуществляемой при помощи частиц хитозана, при добавлении золотых наночастиц и без них не обнаружено, что свидетельствует о примерно одинаковой эффективности способов (Фиг. 2).
Пример 3. Доставка малых РНК, меченных флуоресцентной меткой Су-3, в клетки растений Nicotiana benthamiana.
В экспериментах использовали препараты тРНК или синтезированной in vitro (как описано ниже, гидовой РНК) Предварительно проводили мечение малых РНК флуоресцентным красителем Су3 (Jena Bioscience) с использованием стандартной методики. Пробу для инфильтрации получали, смешивая раствор частиц хитозана, полученный согласно вышеприведенной методике (1,75 мг/мл раствора хитозана в 30 мМ ацетатном буфере (рН 4,5), раствор ТПФ (1 мг/мл) в соотношении 100 к 40 мкл) и 30 мкл раствора гидовой РНК (концентрация 15 мМ) или препарата тРНК (концентрация 1 мг/мл). Пробу инкубировали в течение 5 мин. при температуре 4°С при постоянном перемешивании. Доставку наноплатформ в клетки растений осуществляли путем механической или вакуумной инфильтрации. Проникновение тРНК в растительную клетку показано на Фиг. 3.
Пример 4. Доставка плазмиды (ДНК), кодирующей зеленый флуоресцентный белок, в клетки растений Nicotiana benthamiana.
Плазмидную ДНК из клеток E.coli штамма XL-blue выделяли стандартным методом с помощью набора Plasmid Miniprep (Евроген) в концентрации не менее 1мкг/мкл. Пробу для инфильтрации получали, смешивая раствор частиц хитозана, полученный согласно вышеприведенной методике (1,75 мг/мл раствора хитозана в 30 мМ ацетатном буфере (рН 4,5), раствор ТПФ (1 мг/мл) в соотношении 100 к 40 мкл) и 20 мкл раствора ДНК. Пробу инкубировали в течение 15 мин. при температуре 25°С при постоянном перемешивании. Доставку наноплатформ в клетки растений осуществляли путем механической или вакуумной инфильтрации. На Фиг. 4 наблюдается эффективная флуоресценция (экспрессия гена ЗФБ) в клетках растения после инфильтрации плазмидой, содержащей ген ЗФБ, по сравнению с контрольной плазмидой (без гена ЗФБ).
Пример 5. Редактирование гена коилина для получения растений с повышенной устойчивостью к Y вирусу картофеля.
Редактирование осуществлялось путем доставки рибонуклеопротеидного комплекса системы CRISPR/Cas, включающего направляющую (гидовую) РНК и нуклеазу Cas9 S. pyogenes в клетки растений Nicotiana benthamiana или Solanum tuberosum.
Для получения рибонуклеопротеидного комплекса использовали препарат EnGen® Cas9 NLS, S. pyogenes производства England Biolabs и гидовые РНК, синтезированные при помощи MEGAscript® Т7 Transcription Kit производства Invitrogen™. РНП получали, смешивая 1 мкл нуклеазы Cas9 с исходной концентрацией 20мкМ, 2 мкл гидовой РНК (концентрация 15 мкМ), 3 мкл 10-кратного буфера для Cas9 (10Х Cas9 Nuclease Reaction Buffer) и 14 мкл стерильной, свободной от нуклеаз Н2О. Пробу инкубировали 10 мин. при комнатной температуре и смешивали с 80 мкл предварительно озвученного раствора частиц золота или с раствором частиц хитозана, полученным как описано выше. В качестве контроля в пробу вносили 80 мкл Н2О. Доставку наноплатформ в клетки растений осуществляли путем механической или вакуумной инфильтрации.
Редактирование гена коилина оценивали методом высокопроизводительного секвенирования следующего поколения: в составе гена коилина Solanum tuberosum выявлена делеция размером около 70 нуклеотидов в районе предполагаемого сайта разрезания.
Пример 6: Секвенирование отредактированного гена коилина картофеля
Таргетная амплификация:
Амплификацию области геномного редактирования апикальных меристем, подвергшихся редактированию гена коилина, проводили с использованием праймеров F 5'-TGATGTTCCCTTCTGCTGTAGGTTGGGAA-3' и R 5'-GCGTGGTTTTGTATTGAGAAGAGTCАТА-3'. Данные праймеры были разработаны с возможностью идентификации не только точечных мутаций, но и крупных делеций. Амплификацию проводили в 50 мкл смеси используя DreamTaq PCR Master Mix 2х (Thermo Scientific, США) со следующими параметрами: первичная денатурация при 94°С в течение 2 минут; 40 циклов: 94°С, 65°С и 72°С каждый цикл по 40 секунд. Продукты амплификации были очищены с помощью Agencourt AMPure beads (Beckman Coulter, США)
Приготовление библиотеки и секвенирование:
Подготовка библиотеки для секвенирования и секвенирование были произведены в соответствие с инструкциями от производителя оборудования (Thermo Scientific, США). Определение концентрации библиотеки для секвенирования было выполнено с помощью набора Ion Library TaqMan™ Quantitation Kit (Thermo Scientific, США). Дополнительным контролем качества служило постановка электрофореза. В результате были выявлены 2 фрагмента длиной ≈360 и 300 пар нуклеотидов (Фиг. 5).
Подготовка матрицы и загрузка на чип было осуществлено с Ion Chef (Thermo Scientific, США) используя реактивы Ion 520™ & Ion 530™ Kit-Chef (Thermo Scientific, США) и Ion 520™ Chip Kit (Thermo Scientific, США). Секвенирование было произведено на приборе Ion S5 (Thermo Scientific, США).
Обработка данных последовательностей:
Полученные риды были подвержены вторичной обработке, данные секвенирования были отфильтрованы по качеству QV≥20 и были удалены риды длиной менее 200 нуклеотидов. Картирование было произведено на последовательность дикого типа (WT) используя стандартный протокол. Выравнивания представлены на Фиг. 6.
Пример 7: Количественная оценка эффективности редактирования генома растений.
Эффективность редактирования оценивали в эксперименте по редактированию гена коилина после доставки в ткани растений рибонуклеопротеидного комплекса системы CRISPR/Cas, включающего гидовую РНК и нуклеазу Cas9 S. pyogenes в клетки растений Nicotiana benthamiana или Solanum tuberosum. Эффективность оценивалась в процентах как отношение числа отредактированных последовательностей (локусов) (с отредактированным геном коилина) к неотредактированным (имеющим исходную последовательность гена). Для этого из тканей обработанных наночастицами клеток/апикальных меристем/пазушных почек/целых растений выделяется ДНК, после чего проводится, как описано в примере 6, таргетное секвенирование целевого участка ДНК, подвергнутого редактированию. После чего сравнивается число прочтений (нуклеотидных последовательностей), содержащих направленные мутации, с прочтениями, нуклеотидные последовательности которых не содержат подобных мутаций. Отношение числа прочтений, содержащих акты редактирования к общему числу прочтений и дает эффективность геномного редактирования.
При обработке золотыми наночастицами, покрытыми комплексами Cas9-sgRNA, и при вакуумной либо механической инфильтрации комплексами Cas9-sgRNA на хитозановых наночастицах эффективность трансформации находилась в пределах 10-20%. Обработка вакуумной или механической инфильтрацией комплексами Cas9-sgRNA без наночастиц (- контроль) показал эффективность доставки в пределах 0-5%.
Пример 8. Геномное редактирование морфогенных каллусов картофеля с помощью инфильтрации хитозановыми наночастицами.
Для получения морфогенных каллусов и растений-регенерантов стеблевые сегменты (0.5-1.0 см) срезали с растений in vitro картофеля и культивировали в чашках петри 21 день на среде Мурасиге и Скуга, содержащую набор витаминов по Мурасиге и Скуга (2 мг/л glycine, 0.5 мг/л nicotinic acid, 0.5 мг/л pyridoxine НСl, 0.4 мг/л thiamine НСl), 100 мг/л инозитола, 30 г/л сахарозы, 7 г/л агара, рН 5.8, а также фитогормоны Zeatin 3 мг/л, NAA 2 мг/л, GA3 1 мг/л. Экспланты культивировали в условиях освещения (16 часов 40 μmol m-2s1) при 22-25°С. Образовавшиеся морфогенные каллусы отделяли от стеблевых эксплантов (показано на Фиг. 7) и погружали в суспензию частиц. Проводили инфильтрацию хитозановыми наночастицами, с иммобилизованным на их поверхности комплексом Cas9-sgRNA к гену фитоендесатуразы, путем вакуумирования при ~0.9 bar, после чего каллусы переносили на среду для регенерации побегов указанную выше. Через 21 день после инфильтрации каллусы, формирующие побеги (Фиг. 8, А), переносили на свежую среду Мурасиге и Скуга не содержащую фитогормонов (Фиг. 8, Б) для дальнейшего развития и роста побегов и проведения их анализа. Геномное редактирование подтверждалось с помощью Т7Е1-эндонуклеазного теста, который проводился по стандартной методике, описанной на сайте производителя эндонуклеазы (https://www.neb.com/protocols/2014/08/11/determining-genome-targeting-efficiency-using-t7-endonuclease-i). Результаты теста детектировались с помощью электрофореза в агарозном геле. В среднем, один из 10 морфогенных каллусов демонстрировал наличие геномного редактирования в нужном локусе (Фиг. 8).

Изобретение относится к области биохимии, в частности к способу доставки макромолекул, выбранных из нуклеиновых кислот, белков или нуклеопротеиновых комплексов, в растительную клетку. При этом способ включает обеспечение наличия растительной клетки, имеющей клеточную стенку, покрытие поверхности наночастицы хитозана размером 600-800 нм нуклеиновой кислотой, белком или нуклеопротеиновым комплексом, приведение растительной клетки, имеющей клеточную стенку, и указанной покрытой наночастицы в контакт друг с другом и обеспечение поглощения указанной наночастицы в растительную клетку, содержащую клеточную стенку. Изобретение позволяет эффективно осуществлять доставку макромолекул, выбранных из нуклеиновых кислот, белков или нуклеопротеиновых комплексов, в растительную клетку, имеющую клеточную стенку, с возможностью последующего перепрограммирования растительных клеток и/или редактирования их генома. 16 з.п. ф-лы, 9 ил., 8 пр.
1. Способ доставки макромолекул, выбранных из нуклеиновых кислот, белков или нуклеопротеиновых комплексов, в растительную клетку, где способ включает:
обеспечение наличия растительной клетки, имеющей клеточную стенку;
покрытие поверхности наночастицы хитозана размером 600-800 нм нуклеиновой кислотой, белком или нуклеопротеиновым комплексом;
приведение растительной клетки, имеющей клеточную стенку, и указанной покрытой наночастицы в контакт друг с другом; и
обеспечение поглощения указанной наночастицы в растительную клетку, содержащую клеточную стенку.
2. Способ по п. 1, в котором покрытие наночастицы включает иммобилизацию макромолекулы за счет нековалентной адсорбции на поверхности наночастицы.
3. Способ по п. 1, в котором наночастица является мультифункционализированной наночастицей.
4. Способ по п. 1, который дополнительно включает поглощение макромолекулы внутрь наночастицы.
5. Способ по любому из пп. 1-4, который дополнительно включает обеспечение поглощения наночастицы компартментом растительной клетки.
6. Способ по любому из пп. 1-4, который дополнительно включает покрытие наночастицы пептидом, проникающим в клетку, и/или белком, обеспечивающим направленную доставку в субклеточный компартмент.
7. Способ по любому из пп. 1-6, где наночастицы вводят в растительную клетку с помощью макроинъекции, биолистики или вакуумирования.
8. Способ по любому из пп. 1-7, в котором наночастицы используют для доставки в клетку РНК, ДНК и рибонуклеиновых комплексов.
9. Способ по п. 8, где РНК выбрана из тРНК, гидовой РНК, субгеномной РНК.
10. Способ по п. 8, где рибонуклеиновые комплексы относятся к системе CRISPR-Cas.
11. Способ по п. 8, где рибонуклеиновый комплекс содержит нуклеазу Cas9 совместно с гидовой РНК.
12. Способ по любому из пп. 1-11, который дополнительно включает отбор редактированных клеток.
13. Способ по п. 12, в котором отобранные клетки являются регенерируемыми клетками.
14. Способ по п. 12, где отбор проводят с использованием методов генетического анализа.
15. Способ по п. 12, где отбор проводят с помощью методов ПЦР-РВ и/или секвенирования.
16. Способ по любому из пп. 1-15, где доставка осуществляется в растительную ткань, выбранную из меристем, морфогенного каллуса, пазушных почек, эксплантов, зародышей, каллуса, пыльцы, листьев, пыльников, корней, корневых кончиков, цветков, семян, стручков и стеблей.
17. Способ по любому из пп. 1-16, где дополнительно применяют предварительную обработку растительного материала инкубацией с растворами с повышенными осмотическими свойствами.
| ABDEL-RAZIK A.B | |||
| et al., Transformation of Thionin Genes Using Chitosan Nanoparticle into Potato Plant to Be Resistant to Fungal Infection, IOSR Journal of Biotechnology and Biochemistry (IOSR-JBB), May | |||
| - June | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| QIONG WANG et al., Establishment of a suspension cell system for transformation of Jatropha curcas using nanoparticles, Advanced Materials Research, 2012, Vol | |||
| АППАРАТ ДЛЯ РАСПРЕДЕЛЕНИЯ ТОРФА НА СУШИЛЬНОМ ПОЛЕ | 1922 |
|
SU608A1 |
| Мяльно-трепальный станок | 1921 |
|
SU314A1 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2010 |
|
RU2612156C2 |
| WO 2016094159 A1, 16.06.2016. | |||

