Изобретение относится к биологии и медицине, а именно к генной инженерии. Предложенный способ может быть использован в фундаментальных исследованиях клеточной биологии и терапии генетических заболеваний.
Основным методом бесшовного внесения интересующей ДНК последовательности в клеточный геном является метод направленной гомологичной репарации [Capecchi М.R., Altering the genome by homologous recombination. // Science (New York, N.Y.). 1989. T. 244. №4910. C. 1288-92]. В данном методе используются донорные конструкции, содержащие выбранную для вставки ДНК последовательность между плечами гомологии - последовательностями, которые идентичны участкам, фланкирующим участок генома, в который будет проводиться вставка. Такая методика позволяет проводить точные модификации с низкой частотой: успешное встраивание последовательности в геном происходит в одной из 105-107 клеток [Capecchi М.R., Altering the genome by homologous recombination. // Science (New York, N.Y.). 1989. T. 244. №4910. C. 1288-92]. Внесение двунитевого разрыва в выбранный для редактирования участок генома увеличивает эффективность вставки конструкции с плечами гомологии на несколько порядков [Bibikova М., Carroll D., Segal D.J., Trautman J.K., Smith J., Kim Y.-G., Chandrasegaran S., Stimulation of Homologous Recombination through Targeted Cleavage by Chimeric Nucleases // Molecular and Cellular Biology. 2001. T. 21. №1. C. 289-297; Plessis A., Perrin A., Haber J.E., Dujon В., Site-specific recombination determined by I-SceI, a mitochondrial group I intron-encoded endonuclease expressed in the yeast nucleus // Genetics. 1992. T. 130. №3. C. 451-460; Rouet P., Smih F., Jasin M., Introduction of double-strand breaks into the genome of mouse cells by expression of a rare-cutting endonuclease. // Molecular and cellular biology. 1994. T. 14. №12. C. 8096-106.; Rudin N., Sugarman E., Haber J.E., Genetic and physical analysis of double-strand break repair and recombination in Saccharomyces cerevisiae. // Genetics. 1989. T. 122. №3. C. 519-34].
Наиболее популярной системой для внесения разрыва в геном является система CRISPR-Cas9, специфичность которой задается короткой последовательность гидовой РНК [Cho S.W., Kim S., Kim J.M., Kim J.-S., Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease // Nature Biotechnology. 2013. T. 31. №3. C. 230-232.; Cong L., Ran F.A., Cox D., Lin S., Barretto R., Habib N., Hsu P.D., Wu X., Jiang W., Marraffini L.A., Zhang F., Multiplex Genome Engineering Using CRISPR/Cas Systems // Science (New York, N.Y.). 2013. T. 339. №6121. C. 819.; Jinek M., East A., Cheng A., Lin S., Ma E., Doudna J., RNA-programmed genome editing in human cells // eLife. 2013. T. 2. C. e00471].
Репарация внесенного эндонуклеазой двунитевого разрыва ДНК может проходить по различным путям. Путь гомологичной репарации позволяет изменять интересующую последовательность ДНК в геноме с использованием донорной последовательности ДНК.
Эффективность редактирования по пути гомологичной репарации можно увеличить за счет привлечения к месту разрыва некоторых белков, участвующих в процессах репарации. Это было продемонстрировано для CtIP, Rad51, Mre11A, Rad52, Brex27 домена BRCA2 белка и N-концевой части ExoI белка [Sun W., Liu H., Yin W., Qiao J., Zhao X., Liu Y., Strategies for Enhancing the Homology-Directed Repair Efficiency of CRISPR-Cas Systems // The CRISPR Journal. 2022. T. 5. №1. C. 7-18].
Другой путь увеличения эффективности редактирования - привлечение к месту разрыва белков, ингибирующих конкурирующие пути репарации.
Для этого использовались FFR домен 53 ВР1 белка и RAD18 белок без SAP домена [Sun W., Liu Н., Yin W., Qiao J., Zhao X., Liu Y., Strategies for Enhancing the Homology-Directed Repair Efficiency of CRISPR-Cas Systems // The CRISPR Journal. 2022. T. 5. №1. C. 7-18.].
Еще один подход увеличения эффективности редактирования -привлечение к месту разрыва белков - участников гомологичной репарации из других организмов. Этот подход был проверен для RecT и N-концевой части UL12 белка [Sun W., Liu Н., Yin W., Qiao J., Zhao X., Liu Y., Strategies for Enhancing the Homology-Directed Repair Efficiency of CRISPR-Cas Systems // The CRISPR Journal. 2022. T. 5. №1. C. 7-18.].
Несмотря на разнообразие белков, использованных для увеличения эффективности редактирования, PALB2 белок никогда не рассматривался в качестве кандидата для увеличения эффективности редактирования генома.
Известны способы редактирования генома, в которых для увеличения эффективности редактирования используется привлечение к месту разрыва CtIP, Rad51, Mre11A белков [Tran N.-T., Bashir S., Li X., Rossius J., Chu V.Т., Rajewsky K., Kühn R., Enhancement of Precise Gene Editing by the Association of Cas9 With Homologous Recombination Factors // Frontiers in Genetics. 2019. T. 10. C. 2-5.].
Известны рибонуклеиновые комплексы для редактирования генома с увеличенной эффективностью редактирования, разработанные на основе привлечения к месту разрыва CtIP белка [RU 2749741, С1] и [RU 2750939, С1].
Эти подходы могут быть дополнены за счет использования альтернативных белков репарации.
Проблемой, на решение которой направлено данное изобретение, является расширение арсенала средств для увеличения эффективности геномного редактирования за счет гомологичной репарации в клетках млекопитающих.
Задачей настоящего изобретения является создание нового инструмента для увеличения эффективности геномного редактирования за счет гомологичной репарации в клетках млекопитающих.
Технический результат, достигаемый при осуществлении изобретения, заключается в расширении арсенала средств для увеличения эффективности геномного редактирования за счет гомологичной репарации в клетках млекопитающих.
Предлагаемый способ обеспечивает повышение эффективности геномного редактирования за счет гомологичной репарации в клетках млекопитающих.
Нами установлено, что привлечение PALB2 белка к месту разрыва ДНК позволяет увеличить эффективность системы геномного редактирования, что расширяет возможности ее применения.
Сущность изобретения заключается в следующем.
Для редактирования генома млекопитающих путем гомологичной репарации формируют разрыв генома путем использования системы CRISPR и привлекают к месту разрыва ДНК PALB2 белок или белок с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка.
Другими словами, предложено привлечение к месту разрыва ДНК PALB2 белка или белка с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка для использования с CRISPR системой для редактирования геномной ДНК.
Таким образом, нами предложено применение PALB2 белка или белка с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка для редактирования геномной ДНК путем использования CRISPR системы.
Привлечение PALB2 белка или белка с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка к месту разрыва ДНК может осуществляться с помощью измененной гидовой РНК и РНК связывающих белков.
Выбор систем для привлечения белка к месту разрыва не ограничен MS2 адаптерной системой, используемой в примерах, приведенных ниже, и может включать другие способы доставки белка к месту разрыва ДНК.
Известно, что можно проводить изменения последовательности белка без потери его функции [Ng Р.С, Henikoff S., Predicting the Effects of Amino Acid Substitutions on Protein Function // Annual Review of Genomics and Human Genetics. 2006. T. 7. №1. C. 61-80.]. Это может быть сделано за счет направленного мутагенеза аминокислотных остатков, не влияющих напрямую на его функциональность. Поэтому в данном способе возможно использование белка с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка.
Геномное редактирование за счет гомологичной репарации в клетках млекопитающих с предложенным способом происходит с использованием экзогенной последовательности ДНК, которая может быть представлена двуцепочечной или одноцепочечной последовательностью ДНК.
Внесение разрыва в геном для редактирования за счет гомологичной репарации в клетках млекопитающих происходит с использованием CRISPR белка. В примерах приведено использование spCas9 белка.
Среди белков, которые можно использовать в предлагаемом способе - spCas9, eSpCas9, SpCas9-HF1, HypaCas9, evoCas9, xCas9, HiFi Cas9, Sniper-Cas9, LZ3 Cas9 и другие варианты CRISPR белков.
PALB2 белок и адаптерная система могут быть доставлены в клетку в виде ДНК кодирующей последовательности PALB2 белка сшитого с MS2 адаптерным белком для привлечения к месту разрыва и последовательности гидовой РНК MS2 адаптерной системы; в виде мРНК для трансляции PALB2 белка сшитого с MS2 адаптерным белком и гидовой РНК MS2 адаптерной системы; в виде рибонуклеопротеидного комплекса, включающего PALB2 белок сшитый с MS2 адаптерным белком и гидовой РНК MS2 адаптерной системы.
Указанные выше способы доставки в клетку PALB2 белок аналогичны способам доставки в клетку белка с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка.
Для увеличения эффективности редактирования за счет гомологичной репарации в клетки куда доставляются PALB2 белок и адаптерная система должны быть доставлены CRISPR белок и донорная конструкция.
Доставка CRISPR белков в виде ДНК или РНК последовательностей может проходить совместно или независимо от доставки PALB2 белка и адаптерной системы.
Сборку рибонуклеопротеинового комплекса рекомендуется проводить совместно с CRISPR белком перед доставкой в клетки. Доставку донорной последовательности в виде двуцепочечной или одноцепочечной ДНК рекомендуется проводить совместно с доставкой PALB2 белка и адаптерной системы.
Специфичность CRISPR системы при использовании предложенного способа редактирования задается направляющей последовательностью РНК длиной 20 пар оснований, которая добавляется с 5' конца к последовательности гидовой РНК MS2 адаптерной системы. Рекомендуемый способ добавления направляющей РНК - это клонирование требуемой последовательности перед последовательностью ДНК, кодирующей РНК MS2 адаптерной системы по BsmBI сайтам рестрикции. Пример такой конструкции представлен на фиг. 1.
Аналогично можно проводить сшивку PALB2 белка с MS2 адаптерным белком. Для этого к кодирующей последовательности PALB2 белка методом ПЦР с перекрывающимися праймерами добавляется кодирующая последовательность MS2 адаптерного белка и сайты рестрикции. По сайтам рестрикции полученная конструкция может быть встроена в плазмиду для доставки в клетку. Пример такой конструкции представлен на фиг. 2.
Для проверки предложенного способа в эукариотических клетках была получена плазмида, экспрессирующая PALB2 белок, сшитый через короткий линкер с MS2 белком. Для считывания экспрессии конструкции к С-концу CtIP белка через разрезающуюся Т2А последовательность был добавлен красный флуоресцентный белок mRuby. В эту же плазмиду была добавлена кассета для транскрипции последовательности гидовой РНК MS2 адаптерной системы.
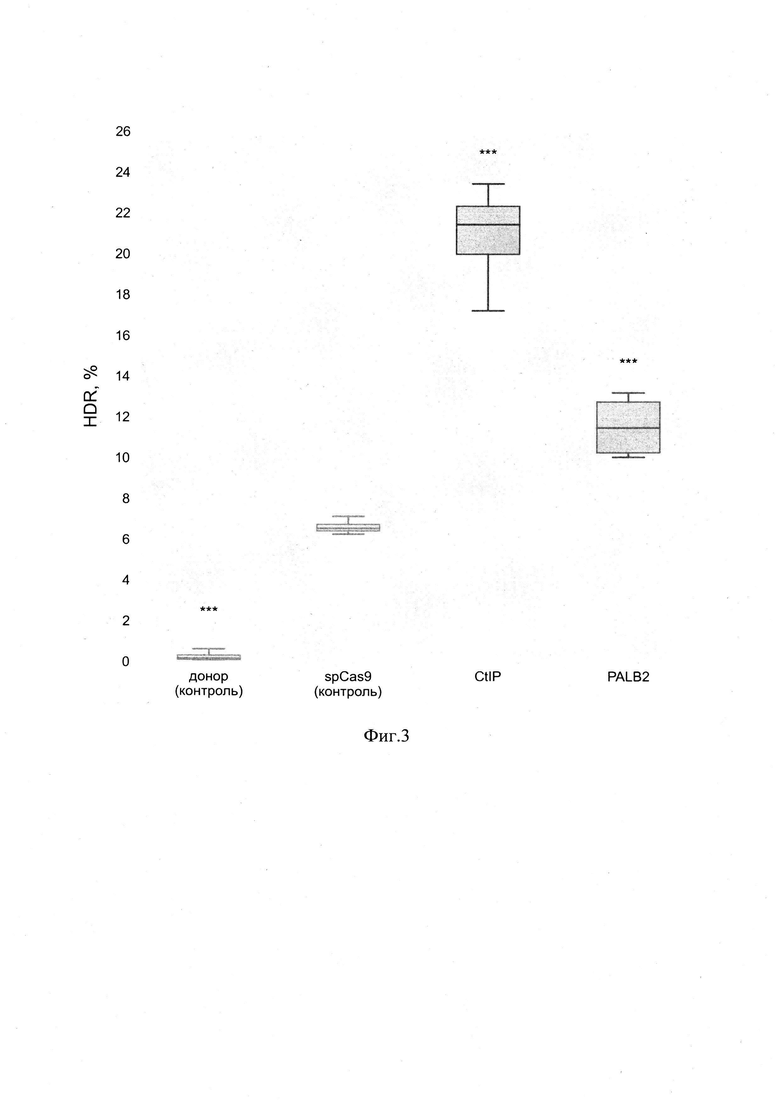
Эффективность предложенного способа была проверена на клеточной линии Hela Kyoto. В качестве места для редактирования генома был выбран TGTCCCTAGTGGCCCCACTG участок AAVS1 локуса. Редактирование генома происходило за счет вставки флуоресцентного EGFP белка. Использование флуоресцентного белка позволяет считывать эффективность редактирования за счет появления в клетках зеленого флуоресцентного сигнала, который можно детектировать методом проточной цитофлуориметрии. При проверке эффективности использовался следующий протокол. Методом лентивирусной вставки была получена моноклональная клеточная линия HelaKyoto экспрессирующая spCas9 белок. Экспрессия белка была подтверждена с помощью иммуноцитохимического окрашивания. Клетки были рассажены на планшет и трансфецированы двумя плазмидами -плазмидой, которая содержит кассету для экспрессии PALB2 белка сшитого с MS2 адаптерным белком и кассету для экспрессии последовательности гидовой РНК MS2 адаптерной системы направленной к TGTCCCTAGTGGCCCCACTG участку AAVS1 локуса, и донорной плазмидой, которая содержит EGFP последовательность для встраивания в AAVS1 локус.Через два дня после трансфекции методом цитофлуориметрического сортинга были отобраны клетки с флуоресцентным сигналом mRuby и фоновым сигналом от EGFP донора. Чтобы избавиться от фонового EGFP сигнала клетки велись на протяжении 10 дней после сортинга. После этого оставшийся процент клеток с EGFP сигналом был подсчитан методом проточной цитофлуориметрии. Эти данные позволяют делать выводы об эффективности репарации при использовании предложенного способа. В качестве контрольных точек использовались донорная EGFP плазмида без CRISPR системы, CRISPR система без PALB2 белка, CRISPR система с привлечением CtIP белка с помощью MS2 адаптерной системы. Для каждой точки эксперимент проводился в трех повторностях. Статистика рассчитывалась с использованием t-критерия Стьюдента в сравнении с контролем spCas9 (***р<0.001). Результаты эксперимента представлены на фиг. 3.
По результатам эксперимента видно, что предложенный способ значительно повышает эффективность редактирования генома за счет гомологичной репарации. Предложенный способ расширяет арсенал существующих подходов редактирования генома и облегчает их применение в фундаментальных исследованиях клеточной биологии, а также терапии генетических заболеваний.
Настоящее изобретение не предназначено для модификации генетической целостности клеток зародышевой линии человека, не предполагает использование человеческих эмбрионов.
| название | год | авторы | номер документа |
|---|---|---|---|
| Система привлечения белка к месту разрыва ДНК для увеличения эффективности редактирования генома млекопитающего | 2022 |
|
RU2808601C1 |
| Система для увеличения эффективности редактирования генома млекопитающего за счет гомологичной репарации | 2022 |
|
RU2808045C1 |
| Рибонуклеопротеиновый комплекс для редактирования генома человека путем вставки в него интересующей последовательности | 2020 |
|
RU2750939C1 |
| Рибонуклеопротеиновый комплекс для редактирования генома человека | 2020 |
|
RU2749741C1 |
| Набор для определения копийности вставки интересующей конструкции в AAVS1 локус генома человека | 2021 |
|
RU2786396C1 |
| СИСТЕМА РЕДАКТИРОВАНИЯ ГЕНОМНОЙ ДНК ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ НА ОСНОВЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩЕЙ БЕЛОК SUCAS9NLS | 2022 |
|
RU2804422C1 |
| Средство редактирования генома на основе белка LigD из бактерии Pseudomonas putida и Cas9 комплекса | 2022 |
|
RU2797049C1 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ pcDEXO-GS-SSB, СОДЕРЖАЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ, КОДИРУЮЩИЕ ХИМЕРНЫЕ БЕЛКИ SSB И ЭКЗОНУКЛЕАЗЫ АППАРАТА ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ БАКТЕРИОФАГОВ, ОБЪЕДИНЕННЫЕ ЧЕРЕЗ ГЛИЦИН-СЕРИНОВЫЙ ПОЛИЛИНКЕР | 2020 |
|
RU2763747C1 |
| Способ получения генно-модифицированных кроликов с нокаутом гена LEPR с помощью системы CRISPR/Cas9 | 2023 |
|
RU2836438C1 |
| ОРТОГОНАЛЬНЫЕ БЕЛКИ CAS9 ДЛЯ РНК-НАПРАВЛЯЕМОЙ РЕГУЛЯЦИИ И РЕДАКТИРОВАНИЯ ГЕНОВ | 2014 |
|
RU2704981C2 |


Изобретение относится к биотехнологии, в частности, к способу редактирования генома млекопитающего путем гомологичной репарации с использованием системы CRISPR, включающему формирование разрыва генома. Указанный способ предусматривает привлечение к месту разрыва белка PALB2. Изобретение позволяет расширить арсенал средств для увеличения эффективности геномного редактирования за счет гомологичной репарации в клетках млекопитающих. 3 ил.
Способ редактирования генома млекопитающего путем гомологичной репарации с использованием системы CRISPR, включающий формирование разрыва генома, отличающийся тем, что к месту разрыва привлекают PALB2 белок или белок с аминокислотной последовательностью, которая по меньшей мере на 95% идентична аминокислотной последовательности PALB2 белка.
| NGOC-TUNG TRAN et al., Enhancement of Precise Gene Editing by the Association of Cas9 With Homologous Recombination Factors, Front | |||
| Genet., Sec | |||
| Genomic Assay Technology, 2019, Volume 10, https://doi.org/10.3389/fgene.2019.00365 | |||
| Рибонуклеопротеиновый комплекс для редактирования генома человека | 2020 |
|
RU2749741C1 |
| CHARPENTIER et al., CtIP fusion to Cas9 enhances transgene integration by homology-dependent | |||

