Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится, в целом, к области фармацевтических препаратов на основе антител, созданных методами генетической инженерии, и, в частности, к моноклональному антителу к человеческому IL-17 и его применению. Настоящее изобретение относится к новому моноклональному антителу к человеческому IL-17 и к применению предлагаемого антитела для лечения заболеваний, опосредованных IL-17.
Уровень техники
В настоящее время в состав семейства интерлейкина 17 (IL-17) входят шесть цитокинов, а именно IL-17A, IL-17B, IL-17C, IL-17D, IL-17E и IL-17F. Члены семейства IL-17 отличаются от других известных цитокинов последовательностью, а также характеризуются относительно низким сходством последовательности среди членов семейства (Gaffen SL, Nat. Rev. Immunol. 2009 Aug; 9(8):556-67). Функция членов семейства IL-17 преимущественно связана с модуляцией иммунного ответа.
IL-17a, гомодимерный гликопротеин с молекулярной массой 20-30 кДа, главным образом вырабатывается активированными CD4+ Т-клетками и действует как провоспалительный цитокин. Секреция IL-17 активированными Т-клетками происходит в очаге воспаления, но не во время их нахождения в системном кровотоке. IL-17 обладает множеством биологических свойств, включая усиление синтеза молекул адгезии и индукцию продукции многих воспалительных цитокинов и хемокинов в различных типах клеток, включая синовиальные клетки, хрящевые клетки, фибробласты, эндотелиальные клетки, клетки эпителия, кератиноциты и макрофаги. Наряду с этим, IL-17 индуцирует агрегацию нейтрофильных гранулоцитов в очаге воспаления, индуцируя секрецию хемокинов, стимулирует продукцию простагландинов и металлопротеаз, а также ингибирует синтез протеогликанов. Кроме того, IL-17 играет решающую роль в созревании гемопоэтических клеток-предшественников. IL-17 участвует в передаче сигнала в различных органах и тканях, включая легкое, суставной хрящ, кость, головной мозг, кроветворные клетки, почки, кожные покровы и кишечник. Таким образом, иммунная реакция, опосредованная IL-17A/Th17, имеет системный характер и приводит к воспалительной реакции, проявляющейся преимущественно в виде инфильтрации нейтрофильными гранулоцитами.
Как видно из многочисленных исследований, повышение уровня IL-17 наблюдается при различных заболеваниях, включая воспаление дыхательных путей, ревматоидный артрит (RA), остеоартрит, эрозию костей, воспалительное заболевание кишечника (IBD), отторжение аллотрансплантата, псориаз, некоторые виды злокачественной опухоли, ангиогенез, атеросклероз и рассеянный склероз (MS) (Witkowski et al, Cell. Mol. Life Sci.61:567-579, 2004). Кроме того, IL-17 играет роль в деградации коллагенового матрикса, а также в регуляции воспаления и постравматических процессов в суставах, независимо от IL-1B. Также, IL-17 и TNF-α действуют синергично, усиливая воспаление. Как было подтверждено в дальнейших исследованиях, блокирование in vivo биологической активности IL-17 с помощью антитела, специфически связывающегося с IL-17 или с растворимым рецептором IL-17, эффективно уменьшает воспаление и костную эрозию в различных животных моделях артрита. Таким образом, IL-17 представляет собой новую терапевтическую мишень в лечении RA и других аутоиммунных заболеваний. Более того, поскольку большая часть IL-17 локализуется в очаге воспаления, лекарственный препарат, нацеленный на IL-17, потенциально может оказаться безопаснее лекарственных препаратов, нацеленных на другие провоспалительные цитокины в системном кровотоке (например, на TNF).
В настоящее время Европейский союз и Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) в США одобрили препарат косентикс фармацевтической компании Novartis Pharma AG для лечения взрослых пациентов, страдающих от умеренной - тяжелой формы бляшечного псориаза. Активный ингредиент косентикса - секукинумаб, который представляет собой моноклональное антитело к IL-17. Тем не менее, в настоящей области техники все еще сохраняется потребность в создании более совершенных антител к IL-17, пригодных для лечения пациента.
Сущность изобретения
Согласно одному аспекту настоящей заявки предлагается моноклональное антитело, которое специфически связывается с человеческим IL-17A и содержит в своем составе вариабельный участок тяжелой цепи, включающий участки HCDR1, HCDR2 и HCDR3, причем HCDR1 имеет последовательность GX1X2X3X4X5Y, HCDR2 имеет последовательность NQDGX6E и HCDR3 имеет последовательность DYYDX7ISDYYIHYWYFDL; где последовательность Х1Х2Х3Х4Х5 представляет собой FTIDN, MSMSD или ITMDD, Х6 представляет собой N или D, а Х7 представляет собой V или L; и где участки HCDR определены согласно Chothia.
Согласно некоторым вариантам осуществления вариабельный участок тяжелой цепи антитела имеет аминокислотную последовательность, указанную под SEQ ID NO: 24, 25 или 26.
Согласно одному аспекту настоящей заявки предлагается моноклональное антитело, которое специфически связывается с человеческим IL-17A и содержит в своем составе вариабельный участок легкой цепи, включающий участки LCDR1, LCDR2 и LCDR3, причем LCDR1 имеет последовательность RASQNVHNRLT, LCDR2 имеет последовательность GASNLES и LCDR3 имеет последовательность QQYNGSPTT; где участки LCDR определены согласно Chothia.
Согласно некоторым вариантам осуществления вариабельный участок легкой цепи антитела имеет аминокислотную последовательность, указанную под SEQ ID NO: 21.
Согласно одному аспекту настоящей заявки предлагается моноклональное антитело, которое специфически связывается с человеческим IL-17A и содержит в своем составе вариабельный участок тяжелой цепи, включающий участки HCDR1, HCDR2 и HCDR3, и вариабельный участок легкой цепи, включающий участки LCDR1, LCDR2 и LCDR3, причем HCDR1 имеет последовательность GX1X2X3X4X5Y, HCDR2 имеет последовательность NQDGX6E, HCDR3 имеет последовательность DYYDX7ISDYYIHYWYFDL, LCDR1 имеет последовательность RASQNVHNRLT, LCDR2 имеет последовательность GASNLES и LCDR3 имеет последовательность QQYNGSPTT; где последовательность X1X2X3X4X5 представляет собой FTIDN, MSMSD или ITMDD, Х6 представляет собой N или D, а Х7 представляет собой V или L; и где участки HCDR и LCDR определены согласно Chothia.
Согласно некоторым вариантам осуществления вариабельный участок тяжелой цепи антитела имеет последовательность, указанную под SEQ ID NO: 24, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21; или вариабельный участок тяжелой цепи имеет последовательность, указанную под SEQ ID NO: 25, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21; или вариабельный участок тяжелой цепи имеет последовательность, указанную под SEQ ID NO: 26, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело представляет собой интактное антитело, по существу, интактное антитело, Fab фрагмент, F(ab')2 фрагмент или одноцепочечный Fv фрагмент.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело представляют собой полностью человеческое антитело.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело дополнительно включает константный участок тяжелой цепи, выбранный из группы, состоящей из подтипов IgG1 и IgG4, и/или константный участок легкой цепи, выбранный из группы, состоящей из подтипов каппа и лямбда.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов предлагаемое моноклональное антитело в концентрации менее 1 нМ способно ингибировать на 50% активность человеческого IL-17a в концентрации 2 нМ, где ингибирование активности измерено путем определения индуцированной человеческим IL-17А продукции IL-6 в дермальных фибробластах человека (HDFa).
Согласно одному аспекту настоящей заявки раскрыта фармацевтическая композиция, содержащая в своем составе моноклональное антитело, описанное в любом из вышеупомянутых аспектов.
Согласно одному аспекту настоящей заявки раскрыто применение моноклонального антитела, описанного в любом из вышеупомянутых аспектов, для лечения заболевания, опосредованного человеческим IL-17A.
Согласно некоторым вариантам осуществления заболевание представляет собой аутоиммунное заболевание.
Согласно некоторым вариантам осуществления заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
Согласно одному аспекту настоящей заявки раскрыто применение моноклонального антитела, описанного в любом из вышеупомянутых аспектов, для производства лекарственного препарата, предназначенного для лечения заболевания, опосредованного человеческим IL-17A.
Согласно некоторым вариантам осуществления заболевание представляет собой аутоиммунное заболевание.
Согласно некоторым вариантам осуществления заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
Согласно одному аспекту настоящей заявки раскрыт способ лечения заболевания, опосредованного человеческим IL-17A, предусматривающий введение моноклонального антитела, описанного в любом из вышеупомянутых аспектов, нуждающемуся в том субъекту.
Согласно некоторым вариантам осуществления заболевание представляет собой аутоиммунное заболевание.
Согласно некоторым вариантам осуществления заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
Краткое описание чертежей
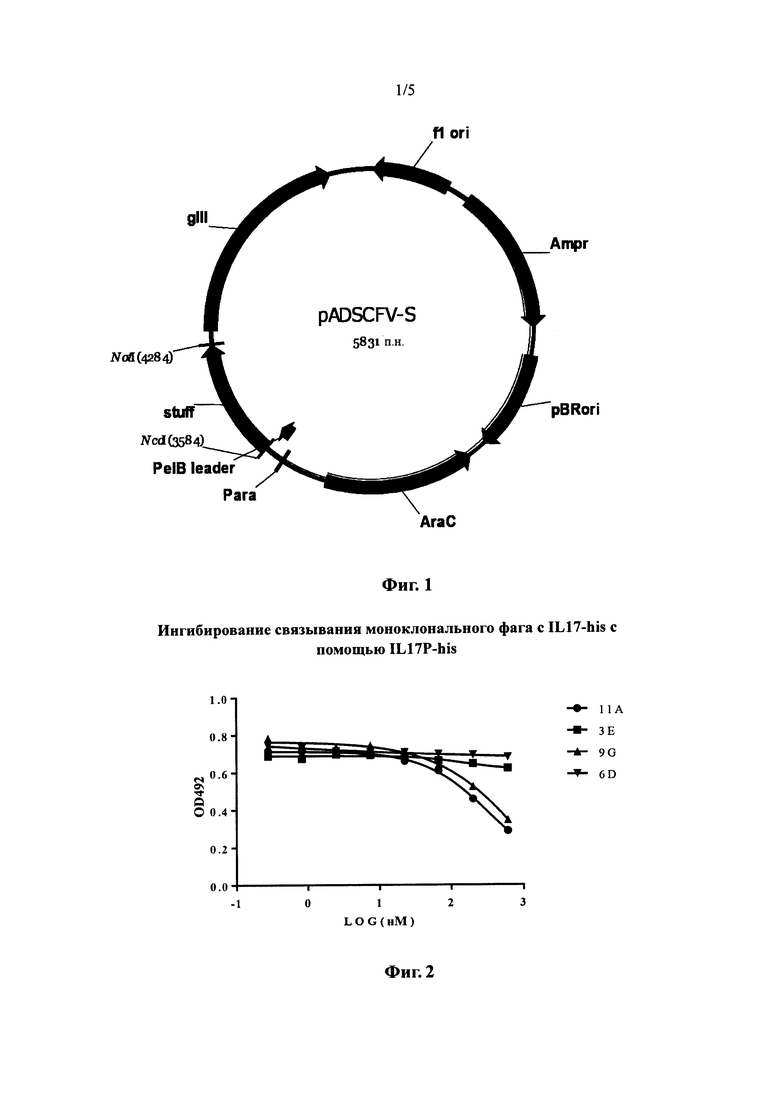
На фиг. 1 показана схема строения нового вектора для фагового дисплея pADSCFV-S (Para: арабинозный промотор, PelB leader: ген сигнального пептида PelB, NcoI: сайт рестрикции NcoI, Stuff: спейсерный фрагмент длиной 750 п. н., NotI: сайт рестрикции NotI, gIII: ген gIII фага М13, flori: сайт инициации репликации фага М13, Ampr: ген устойчивости к ампициллину, pBRori: сайт инициации репликации плазмиды pBR322, AraC: ген AraC).
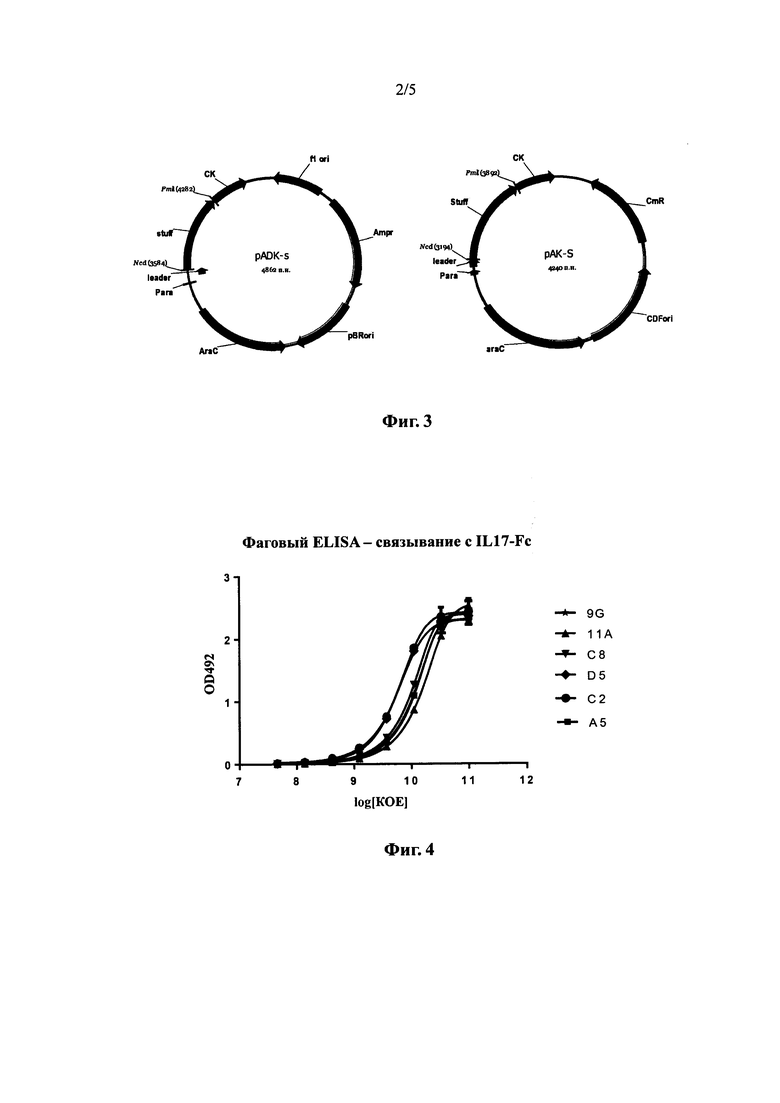
На фиг. 2 показаны результаты конкурентного связывания с IL-17 моноклонального фага и IL17R.
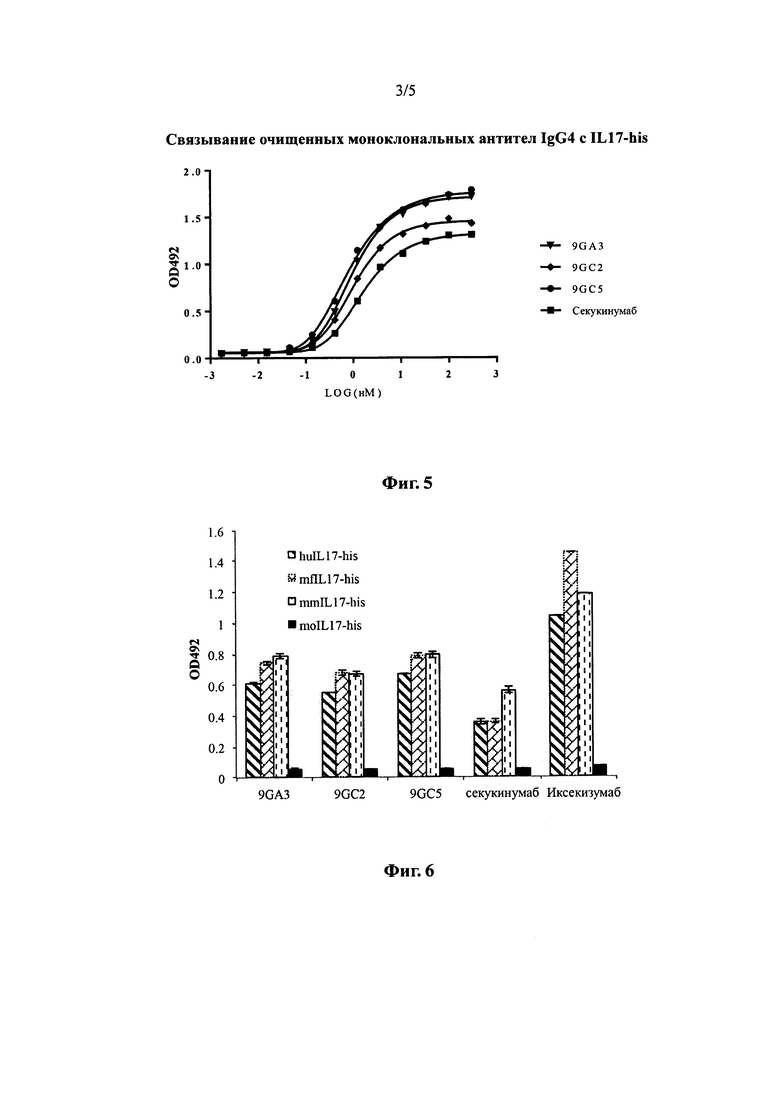
На фиг. 3 представлены схемы строения плазмид pADK-S и pAK-S (Para: арабинозный промотор, leader: ген сигнального пептида PelB, NcoI: сайт рестрикции NcoI, Stuff: спейсерный фрагмент длиной 750 п. н., PmlI: сайт рестрикции PmlI, CK: константный участок легкой каппа-цепи человеческого антитела, f1ori: сайт инициации репликации фага М13, Ampr: ген устойчивости к ампициллину, pBRori: сайт инициации репликации плазмиды pBR322, AraC: ген AraC, CmR: ген устойчивости к хлорамфениколу, CDFori: сайт инициации репликации плазмиды CDF).
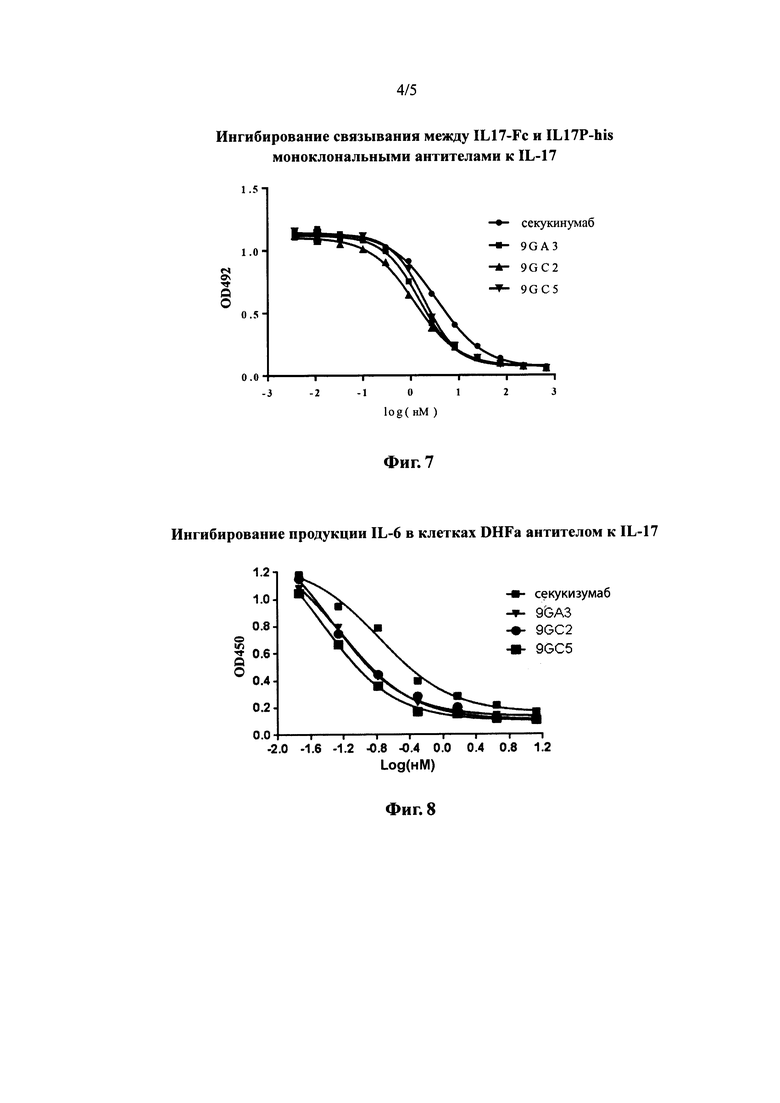
На фиг. 4 показаны результаты измерения относительной аффинности фаговых антител с помощью анализов фагового ELISA.
На фиг. 5 показаны результаты оценки с помощью анализов ELISA способности антител к IL-17 связываться с человеческим IL-17.
На фиг. 6 показаны результаты оценки с помощью анализов ELISA способности моноклональных антител к IL-17 связываться с IL-17 из разных видов животных.
На фиг. 7 показаны результаты конкурентного связывания с IL-17 моноклональных антител к человеческому IL-17 и IL17R.
На фиг. 8 показаны результаты сравнительного анализа ингибирования моноклональными антителами к IL-17 индуцированной IL-17 продукции IL-6 в клетках HDFa.
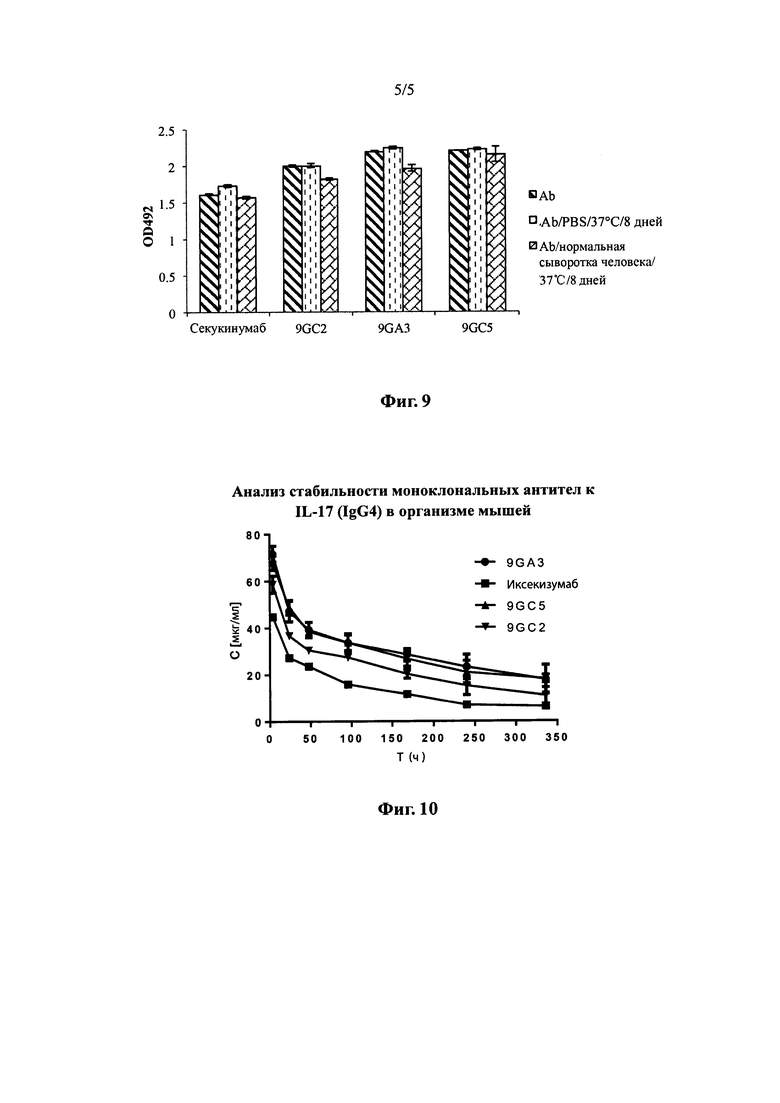
На фиг. 9 показаны результаты анализа стабильности различных моноклональных антител к IL-17 в сыворотке крови человека.
На фиг. 10 показана тенденция изменения концентрации четырех моноклональных антител к IL-17 в организме мышей.
Подробное описание изобретения
Различные изобретения, упоминаемые в настоящей заявке, частично основаны на создании фаговой библиотеки человеческих антител. Авторами настоящего изобретения был проведен скрининг моноклональных антител к человеческому IL-17 с необходимыми свойствами в созданной библиотеке антител.
Согласно различным аспектам настоящей заявки предлагается новое моноклональное антитело к человеческому IL-17 или его антиген-связывающий фрагмент; полинуклеотид, кодирующий указанное моноклональное антитело или антиген-связывающий фрагмент; вектор, содержащий в своем составе указанный полинуклеотид; клетка-хозяин, содержащая указанный полинуклеотид или вектор; способ получения и очистки указанного антитела; и медицинское применение указанного антитела или антиген-связывающего фрагмента. На основании последовательностей перспективных участков антитела, предлагаемого в настоящей заявке, молекулу интактного антитела можно составить в форме лекарственного препарата для клинического лечения аутоиммунного заболевания, опосредованного IL-17, в том числе без ограничения псориаза, ревматоидного артрита и анкилозирующего спондилита.
Если не оговорено иначе, все технические и научные термины, использованные в настоящем документе, имеют то же значение, что и в общепринятом их понимании специалистом в настоящей области техники.
В контексте настоящего документа термин «полипептид» относится к полимеру, состоящему из аминокислотных остатков, связанных посредством пептидной связи. Термин «белок» обычно относится к полипептиду относительно большого размера. Термин «пептид» обычно относится к полипептиду относительно небольшого размера (например, содержащему не более 100, 80, 60, 50, 30 или 20 аминокислотных остатков).
В контексте настоящего документа термин «антитело» относится к молекуле иммуноглобулина, которая специфически связывается с целевой молекулой посредством по меньшей мере одного антиген-распознающего паратопа в вариабельном участке молекулы иммуноглобулина. Примеры целевой молекулы включают без ограничения углевод, полинуклеотид, липид, полипептид. В контексте настоящего документа термин «антитело» включает в себя не только интактное (полноразмерное) антитело, но также и антиген-связывающий фрагмент (например, Fab, Fab', F(ab')2, Fv); вариантный, слитый белок, содержащий в своем составе часть антитела; гуманизированное антитело; химерное антитело; диатело; антитело, распознающее линейный эпитоп; одноцепочечное антитело; мультиспецифическое антитело (например, биспецифическое антитело) и любые другие модифицированные версии молекулы иммуноглобулина, содержащие антиген-распознающий участок, обладающий необходимой специфичностью, включая гликозилированный вариант антитела, вариант антитела с модифицированной аминокислотной последовательностью и ковалентно модифицированное антитело.
Интактное или полноразмерное антитело обычно содержит две тяжелые цепи и две легкие цепи. Каждая тяжелая цепь содержит вариабельный участок тяжелой цепи (VH), а также первый, второй и третий константный участок (CH1, СН2 и СН3). Каждая легкая цепь содержит вариабельный участок легкой цепи (VL) и константный участок (CL). Полноразмерное антитело может принадлежать к любому из классов, например, IgD, IgE, IgG, IgA или IgM (или подклассу любого из вышеупомянутых). Вместе с тем, антитела, предлагаемые в настоящем изобретении, не обязательно принадлежат к какому-либо определенному классу. Иммуноглобулин может быть отнесен к разным классам на основании аминокислотной последовательности константного домена тяжелой цепи антитела. В целом, существует пять классов иммуноглобулинов, а именно IgA, IgD, IgE, IgG и IgM. Некоторые из этих классов могут быть дополнительно разделены на несколько подклассов (изоформ), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей различных классов иммуноглобулинов называются α, δ, ε, γ и μ. Структуры субъединиц и трехмерные структуры различных классов иммуноглобулинов хорошо известны из уровня техники.
В контексте настоящего документа термин «антиген-связывающий фрагмент» относится к части или участку молекулы интактного антитела, отвечающему за связывание антигена. Антиген-связывающий домен может содержать вариабельный участок тяжелой цепи (VH), вариабельный участок легкой цепи (VL), или оба эти участка. Каждый из участков VH и VL обычно содержит в своем составе три определяющих комплементарность участка, а именно CDR1, CDR2 и CDR3.
Примеры антиген-связывающего фрагмента включают без ограничения (1) Fab фрагмент, который может быть моновалентным фрагментом, содержащим VL-CL цепь и VH-CH1 цепь; (2) F(ab')2 фрагмент, который может быть двухвалентным фрагментом, содержащим два Fab фрагмента, связанные дисульфидным мостиком в шарнирной области (например, димер Fab); (3) Fv фрагмент, содержащий в своем составе VL домен и VH домен одного плеча антитела; (4) одноцепочечный Fv (scFv), который может представлять собой одну полипептидную цепь, содержащую в своем составе VH домен и VL домен, соединенные пептидным линкером; и 5) (scFv)2, который может содержать в своем составе два VH домена, связанные пептидным линкером, и два VL домена, соединенные с двумя VH доменами посредством дисульфидных мостиков.
В контексте настоящего документа термин «специфически связывается с» относится к реакции неслучайного связывания между двумя молекулами, например, к связыванию антитела с эпитопом антигена.
При записи последовательностей нуклеиновых кислот, приведенных в настоящей заявке, наряду с обычными символами А, Т, С и G использованы символы вырожденных оснований. Символы, обозначающие вырожденные основания, использованы в том значении, как их обычно понимает специалист в настоящей области техники. Например, R обозначает А или G, Y обозначает С или Т, М обозначает А или С, K обозначает G или Т, S обозначает С или G, W обозначает А или Т, Н обозначает А или С или Т, В обозначает С или G или Т, V обозначает А или С или G, D обозначает А или G или Т, а N обозначает А или С или G или Т.
Согласно одному аспекту настоящей заявки предлагается моноклональное антитело, которое специфически связывается с человеческим IL-17A и содержит в своем составе вариабельный участок тяжелой цепи, включающий участки HCDR1, HCDR2 и HCDR3, причем HCDR1 имеет последовательность GX1X2X3X4X5Y, HCDR2 имеет последовательность NQDGX6E и HCDR3 имеет последовательность DYYDX7ISDYYIHYWYFDL; где последовательность Х1Х2Х3Х4Х5 представляет собой FTIDN, MSMSD или ITMDD, Х6 представляет собой N или D, а Х7 представляет собой V или L; и где участки HCDR определены согласно Chothia.
Согласно некоторым вариантам осуществления вариабельный участок тяжелой цепи антитела имеет аминокислотную последовательность, указанную под SEQ ID NO: 24, 25 или 26.
Согласно одному аспекту настоящей заявки предлагается моноклональное антитело, которое специфически связывается с человеческим IL-17A и содержит в своем составе вариабельный участок легкой цепи, включающий участки LCDR1, LCDR2 и LCDR3, причем LCDR1 имеет последовательность RASQNVHNRLT, LCDR2 имеет последовательность GASNLES и LCDR3 имеет последовательность QQYNGSPTT; где участки LCDR определены согласно Chothia.
Согласно некоторым вариантам осуществления вариабельный участок легкой цепи имеет аминокислотную последовательность, указанную под SEQ ID NO: 21.
Согласно одному аспекту настоящей заявки предлагается моноклональное антитело, которое специфически связывается с человеческим IL-17A и содержит в своем составе вариабельный участок тяжелой цепи, включающий участки HCDR1, HCDR2 и HCDR3, и вариабельный участок легкой цепи, включающий участки LCDR1, LCDR2 и LCDR3, причем HCDR1 имеет последовательность GX1X2X3X4X5Y, HCDR2 имеет последовательность NQDGX6E, HCDR3 имеет последовательность DYYDX7ISDYYIHYWYFDL, LCDR1 имеет последовательность RASQNVHNRLT, LCDR2 имеет последовательность GASNLES и LCDR3 имеет последовательность QQYNGSPTT; где последовательность X1X2X3X4X5 представляет собой FTIDN, MSMSD или ITMDD, Х6 представляет собой N или D, а Х7 представляет собой V или L; и где участки HCDR и LCDR определены согласно Chothia.
Согласно некоторым вариантам осуществления вариабельный участок тяжелой цепи антитела имеет последовательность, указанную под SEQ ID NO: 24, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21; или вариабельный участок тяжелой цепи имеет последовательность, указанную под SEQ ID NO: 25, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21; или вариабельный участок тяжелой цепи имеет последовательность, указанную под SEQ ID NO: 26, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело представляет собой интактное антитело, по существу интактное антитело, Fab фрагмент, F(ab')2 фрагмент или одноцепочечный Fv фрагмент.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело представляют собой полностью человеческое антитело.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело дополнительно включает константный участок тяжелой цепи, выбранный из группы, состоящей из подтипов IgG1 и IgG4, и/или константный участок легкой цепи, выбранный из группы, состоящей из подтипов каппа и лямбда.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов предлагаемое моноклональное антитело в концентрации менее 1 нМ способно ингибировать на 50% активность человеческого IL-17a в концентрации 2 нМ, где ингибирование активности измерено путем определения индуцированной человеческим IL-17A продукции IL-6 в дермальных фибробластах человека (HDFa).
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов предлагаемое антитело представляет собой полностью человеческое моноклональное антитело и ослабляет или устраняет по меньшей мере одну биологическую активность in vitro или in vivo, в которой участвует IL-17 или его часть, или противодействует ей.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов предлагаемое антитело характеризуется высокой аффинностью связывания с человеческим IL-17.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов предлагаемое антитело характеризуется тем, что оно способно специфически связываться с человеческим IL-17, IL-17 Масаса mulatta и IL-17 Масаса fascicularis на уровне, превышающем фоновый, но не связывается с IL-17 Mus musculus.
Согласно некоторым вариантам осуществления вышеупомянутых трех аспектов антитело содержит в своем составе вариабельный участок и константный участок, причем константный участок тяжелой цепи может принадлежать к подтипу IgG1 (SEQ ID NO: 29) или подтипу IgG4 (SEQ ID NO: 30), а константный участок легкой цепи может принадлежать к подтипу каппа (SEQ ID NO: 31) или подтипу лямбда (SEQ ID NO: 32).
Согласно еще одному аспекту настоящей заявки предлагается полинуклеотид, кодирующий моноклональное антитело или его антиген-связывающий фрагмент, описанные в вышеупомянутых аспектах, вектор, содержащий в своем составе указанный полинуклеотид, и клетка-хозяин, трансфицированная указанным вектором. Согласно некоторым вариантам осуществления клетка-хозяин представляет собой клетку линии СНО или HEK293.
Согласно одному аспекту настоящей заявки раскрыта фармацевтическая композиция, содержащая в своем составе моноклональное антитело, описанное в любом из вышеупомянутых аспектов.
Согласно некоторым вариантам осуществления фармацевтическая композиция дополнительно содержит в своем составе фармацевтически приемлемый носитель, вспомогательное вещество и/или разбавитель.
Согласно некоторым вариантам осуществления фармацевтическая композиция содержит в своем составе моноклональное антитело в терапевтически эффективном количестве.
Согласно одному аспекту настоящей заявки раскрыто применение моноклонального антитела, описанного в любом из вышеупомянутых аспектов, для лечения заболевания, опосредованного человеческим IL-17A.
Согласно некоторым вариантам осуществления заболевание представляет собой аутоиммунное заболевание.
Согласно некоторым вариантам осуществления заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
Согласно одному аспекту настоящей заявки раскрыто применение моноклонального антитела, описанного в любом из вышеупомянутых аспектов, для производства лекарственного препарата, предназначенного для лечения заболевания, опосредованного человеческим IL-17A.
Согласно некоторым вариантам осуществления заболевание представляет собой аутоиммунное заболевание.
Согласно некоторым вариантам осуществления заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
Согласно одному аспекту настоящей заявки раскрыт способ лечения заболевания, опосредованного человеческим IL-17A, предусматривающий введение моноклонального антитела, описанного в любом из вышеупомянутых аспектов, нуждающемуся в том субъекту.
Согласно некоторым вариантам осуществления заболевание представляет собой аутоиммунное заболевание.
Согласно некоторым вариантам осуществления заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
Согласно некоторым вариантам осуществления фармацевтическая активность предлагаемого в настоящей заявке антитела может быть проверена с помощью стандартного для настоящей области метода анализа, см., например, Hwang SY, Kim JY, Kim KW, Park MK, Moon Y, Kim WU, Kim HY: IL-17 induces production of IL-6 and IL-8 in rheumatoid arthritis synovial fibroblasts via NF-kappaB- and PI3-kinase/Akt-dependent pathways. Arthritis research & therapy 2004, 6(2): R120-128. Вкратце, рекомбинантный IL-17 применяют для стимуляции дермальных фибробластов человека (клеток HDFa) в присутствии разных концентраций антитела, предлагаемого в настоящей заявке. После стимуляции в течение 24 ч, отбирают культуральную жидкость и определяют содержание в ней IL-6 методом ELISA. При проведении описанного выше анализа предлагаемое в настоящей заявке антитело в отношении ингибирования продукции IL-6 предпочтительно характеризуется значением IC50 около 0,22 нМ или менее, более предпочтительно, около 0,12 нМ или менее и, еще более предпочтительно, 0,07 нМ или менее.
Согласно некоторым вариантам осуществления стабильность предлагаемого в настоящей заявке антитела в сыворотке человека может быть проверена с помощью стандартного для настоящей области метода анализа. Вкратце, образец моноклональных антител, простерилизованный методом фильтрации, разбавляют в объеме 200 мкл соответственно стерильной смеси на основе нормальной человеческой сыворотки или буфера PBS до конечной концентрации 30 мкг/мл, перемешивают и помещают в водяную баню с температурой 37°С на 8 дней (192 ч). После этого с помощью анализа ELISA оценивают связывание с человеческим IL-17 у моноклональных антител в образце с сывороткой, моноклональных антител в образце с буфером PBS и моноклональных антител из образца, хранившегося в замороженном состоянии. Сравнивают изменение значений способности к связыванию с IL-17 у моноклональных антител в различных образцах. При проведении описанной выше проверки предлагаемое в настоящей заявке антитело предпочтительно сохраняет около 87%, более предпочтительно, около 91% и, еще более предпочтительно, около 97% способности к связыванию с IL-17.
Согласно некоторым вариантам осуществления фармакокинетика предлагаемого в настоящей заявке антитела в организме мышей может быть оценена с помощью стандартного в настоящей области метода анализа. Вкратце, готовят растворы образцов моноклональных антител в буфере PBS (рН 7,0) в концентрации 0,5 мг/мл и вводят его мышам в дозе 5 мг/кг в виде однократной инъекции в хвостовую вену. В каждую группу включают восемь мышей BALB/c, в числе которых четыре самца и четыре самки. После этого осуществляют забор образцов крови из ретроорбитального синуса через 4, 24, 48, 96, 168, 240 и 336 часов после инъекции, собранные образцы хранят при -70°С. После завершения сбора образцов проводят количественное определение концентраций моноклонального антитела в образцах. Определение концентрации лекарственного средства в плазме проводят методом количественного анализа ELISA с иммобилизованным IL-17. В соответствии с моделью реакции первого порядка строят график зависимости натурального логарифма концентрации антител (lnC) от времени (t), таким образом получая угловой коэффициент k. После этого рассчитывают период полужизни (t1/2) моноклонального антитела к IL-17 у мышей по формуле t1/2=0,693/k. При проведении описанной выше проверки предлагаемое в настоящей заявке антитело имеет период полужизни (t1/2) у мышей предпочтительно около 6,7 ч, более предпочтительно, около 7,4 ч и, еще более предпочтительно, около 8,5 ч.
Согласно одному из аспектов настоящей заявки предлагается выделенная молекула нуклеиновой кислоты, кодирующая предлагаемое в настоящей заявке антитело или его легкую цепь или тяжелую цепь, вектор, содержащий в своем составе указанную молекулу нуклеиновой кислоты, клетка-хозяин, содержащая указанный вектор, и способ получения указанного антитела. Согласно некоторым вариантам осуществления молекула нуклеиновой кислоты функционально связана с регуляторной последовательностью, которую способна распознавать клетка-хозяин, трансформированная указанным вектором. Согласно некоторым вариантам осуществления способ получения антитела предусматривает культивирование указанной клетки-хозяина, обеспечивающее экспрессию указанной нуклеиновой кислоты. Согласно некоторым вариантам осуществления способ получения антитела дополнительно предусматривает выделение антитела из кулыуральной среды, в которой культивируют клетку-хозяина.
Согласно одному из аспектов настоящей заявки предлагаются IL-17 Mus musculus (SEQ ID NO: 9), IL-17 Масаса mulatta (SEQ ID NO: 10) или IL-17 Масаса fascicularis (SEQ ID NO: 11), выделенная молекула нуклеиновой кислоты, кодирующая один из них, вектор, содержащий в своем составе указанную молекулу нуклеиновой кислоты, клетка-хозяин, содержащая указанный вектор, и способ получения IL-17 Mus musculus, IL-17 Масаса mulatta или IL-17 Масаса fascicularis.
Примеры
Следующие примеры приведены здесь лишь с целью дополнительных иллюстраций настоящего изобретения, при этом не следует рассматривать их как какое-либо ограничение для настоящего изобретения.
Пример 1
Создание высококачественных фаговых библиотек антител
Метод создания библиотек антител является одним из важных способов получения человеческих моноклональных антител. В настоящее время для получения человеческих моноклональных антител разработан и успешно применяется метод создания библиотек антител, основанный на применении фагового дисплея. В этом примере наглядно показана стратегия и методология создания фаговой библиотеки антител с применением методов генетической инженерии.
1.1 Получение генов вариабельного участка тяжелой цепи и легкой цепи антитела (VH и VL)
Для создания библиотеки человеческих антител прежде всего необходимо получить гены вариабельных участков тяжелых цепей (VH) и вариабельных участков легких цепей (VL) человеческих антител. Гены вариабельных участков антител могут быть получены из лимфоцитов периферической крови здоровых субъектов или могут быть полностью синтетическими.
1.1.1 Получение генов вариабельных участков естественных человеческих антител
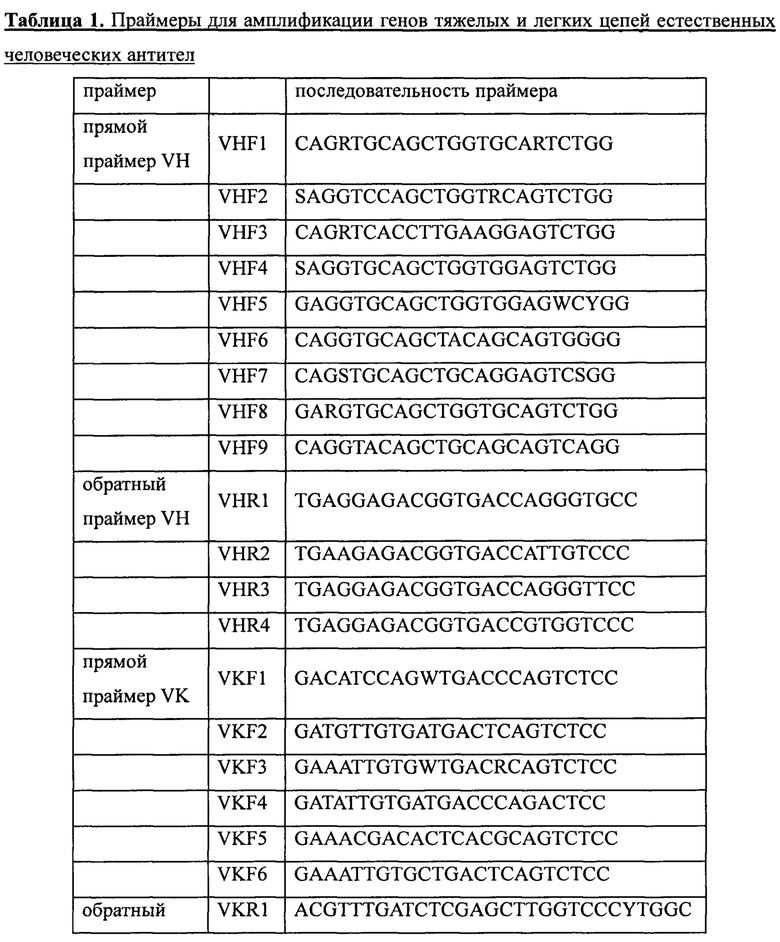
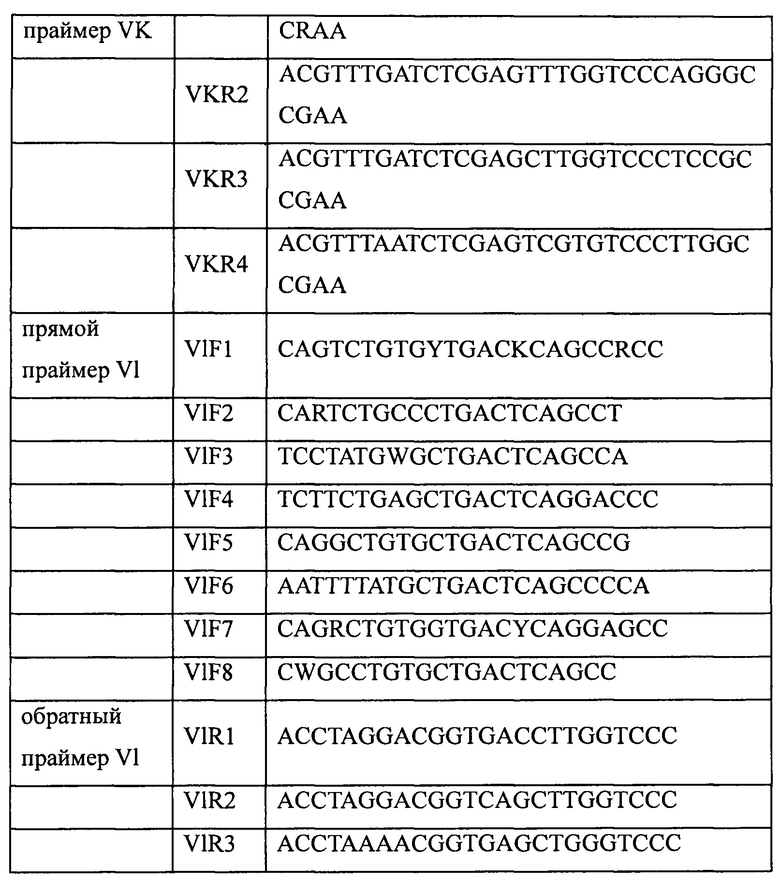
Забор крови проводили у 19 здоровых добровольцев (50 мл от каждого). После этого выделяли лимфоциты с помощью раствора для выделения лимфоцитов (MP Biomedicals Inc., кат. №0850494). РНК получали с использованием набора для выделения суммарной РНК производства Omega Inc., (кат. № R6834-01). кДНК получали с использованием набора для обратной транскрипции производства TransGen Biotech Inc. (кат. № АТ301-03). Наконец, в ПЦР-реакции использовали наборы праймеров, приведенные ниже в таблице 1, для амплификации соответственно генов вариабельных участков тяжелых цепей (VH) и генов вариабельных участков легких цепей (VL, включая Vk и Vl) антител. Выделение и очистку амплифицированных продуктов ПЦР (VH, VK или Vl) проводили с помощью стандартного электрофореза в агарозном геле, после чего хранили их для дальнейшего использования при -20°С.
1.1.2 Получение генов вариабельных участков полностью синтетических человеческих антител
Основная стратегия получения генов полностью синтетических антител предусматривает использование вырожденных праймеров для введения необходимых мутаций в участки CDR выбранного в качестве матрицы гена антитела. Для создания библиотеки полностью синтетических человеческих антител в этом примере были выбраны три матрицы вариабельных участков тяжелой цепи человеческого антитела (VH1; VH3 и VH5) и две матрицы вариабельных участков легкой цепи человеческого антитела (VK1 и Vl3).
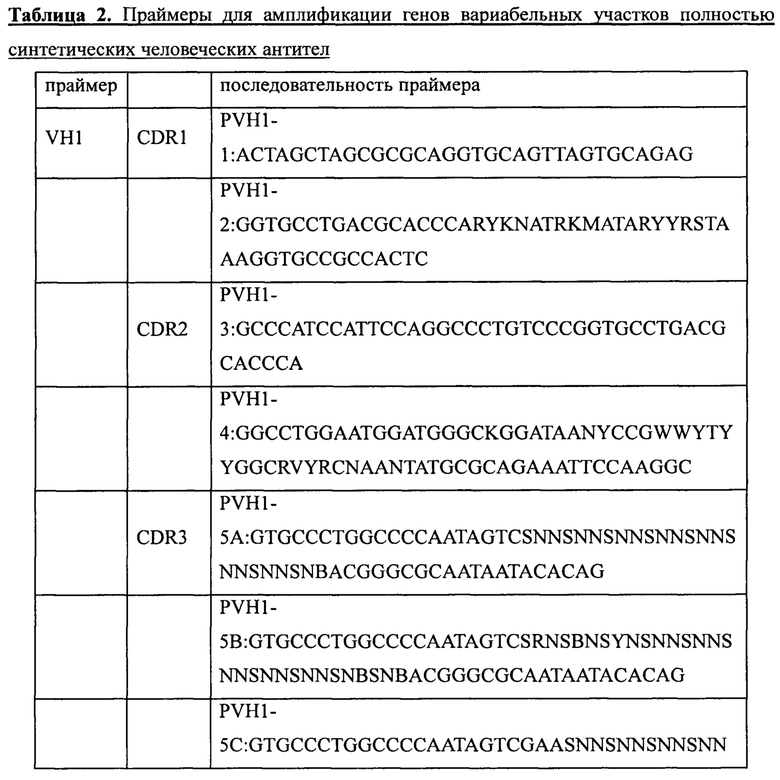
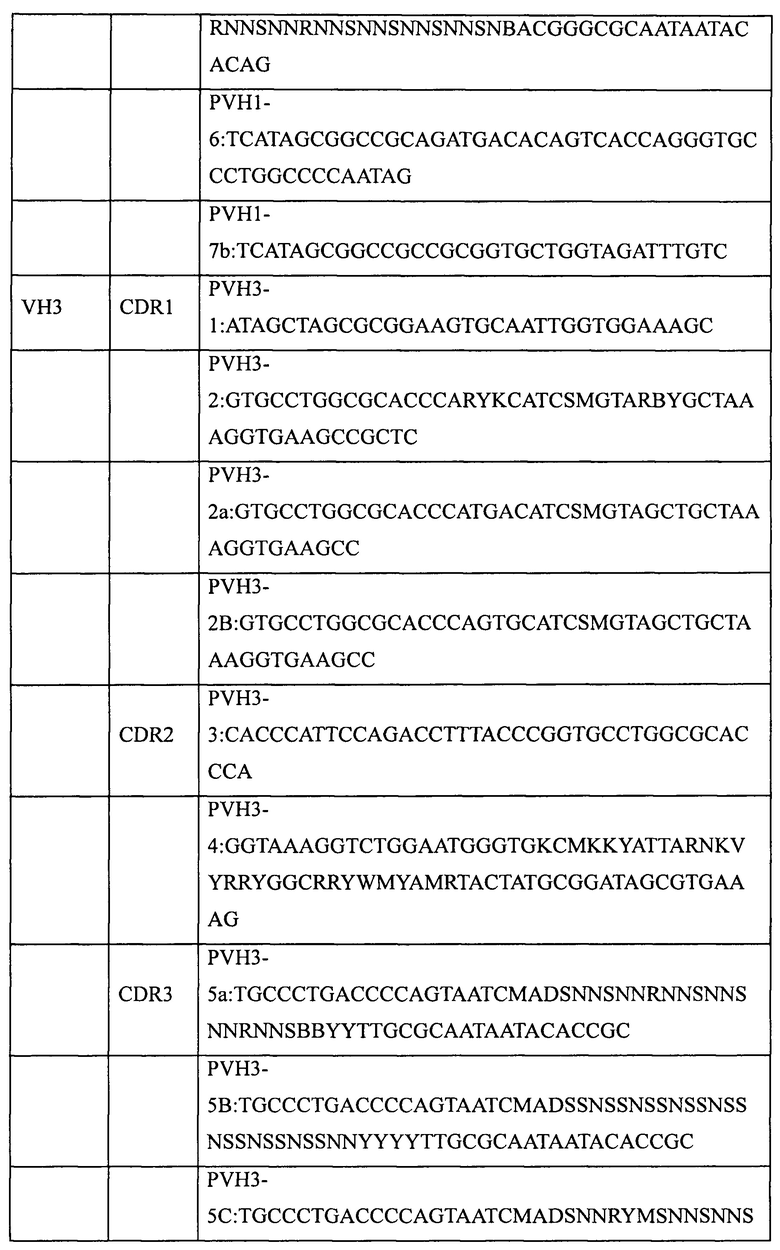
Пять генов вариабельных участков антител, а именно VH1 (SEQ ID NO: 1), VH3 (SEQ ID NO: 2), VH5 (SEQ ID NO: 3), VK1 (SEQ ID NO: 4) и Vl3 (SEQ ID NO: 5), были сконструированы и синтезированы в Ming Chen Zhi Yuan Inc. Для введения необходимых мутаций в участки CDR1, CDR2 и CDR3 пяти генов вариабельных участков были соответственно разработаны и синтезированы праймеры, приведенные в таблице 2. Необходимые мутации в соответствующие участки CDR вводили с использованием стандартного метода ПЦР и соответствующих наборов вырожденных праймеров, содержащих необходимые мутации. Затем с помощью 2-3 раундов ПЦР с перекрывающимися праймерами создавали полные гены вариабельных участков тяжелой цепи (VH1, VH3, VH5) или вариабельных участков легких цепей (VK1, VL3). Наконец, выделение амплифицированных продуктов ПЦР (VH, VK или Vl) генов вариабельных участков проводили с помощью электрофореза в агарозном геле, после чего сохраняли их для дальнейшего использования при -20°С.
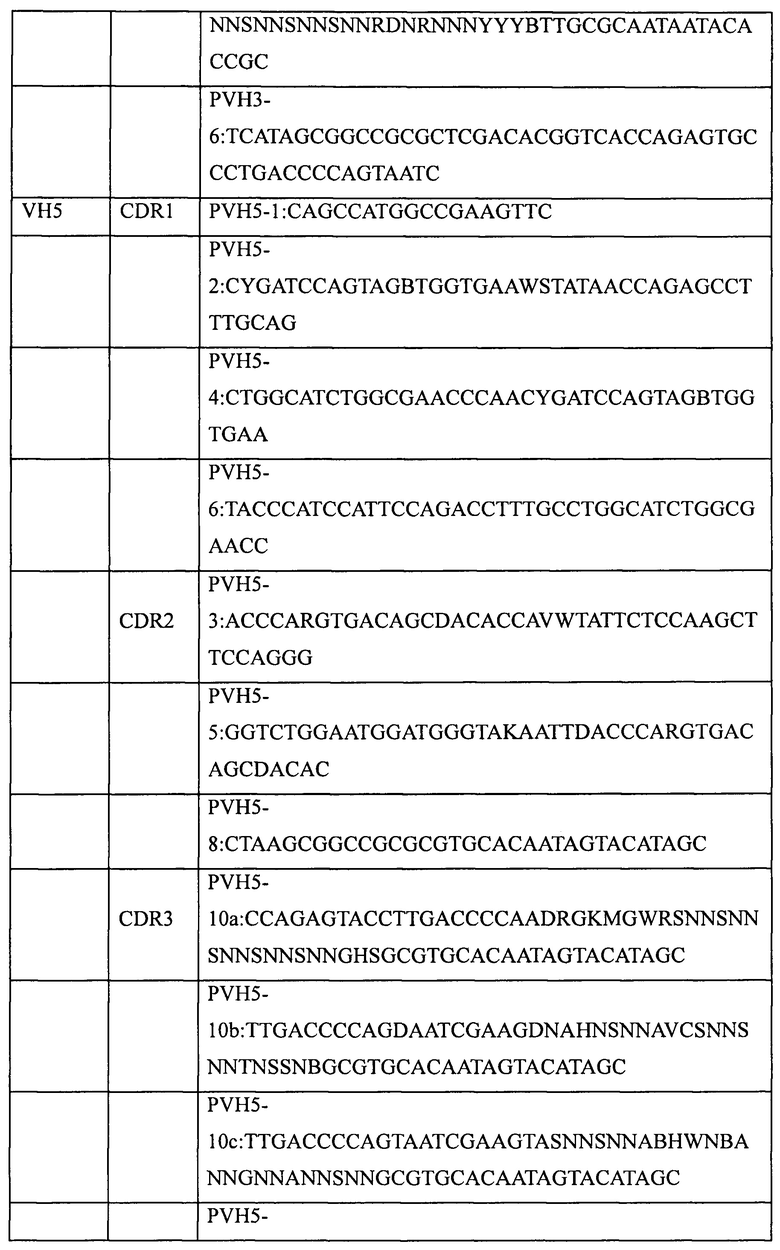
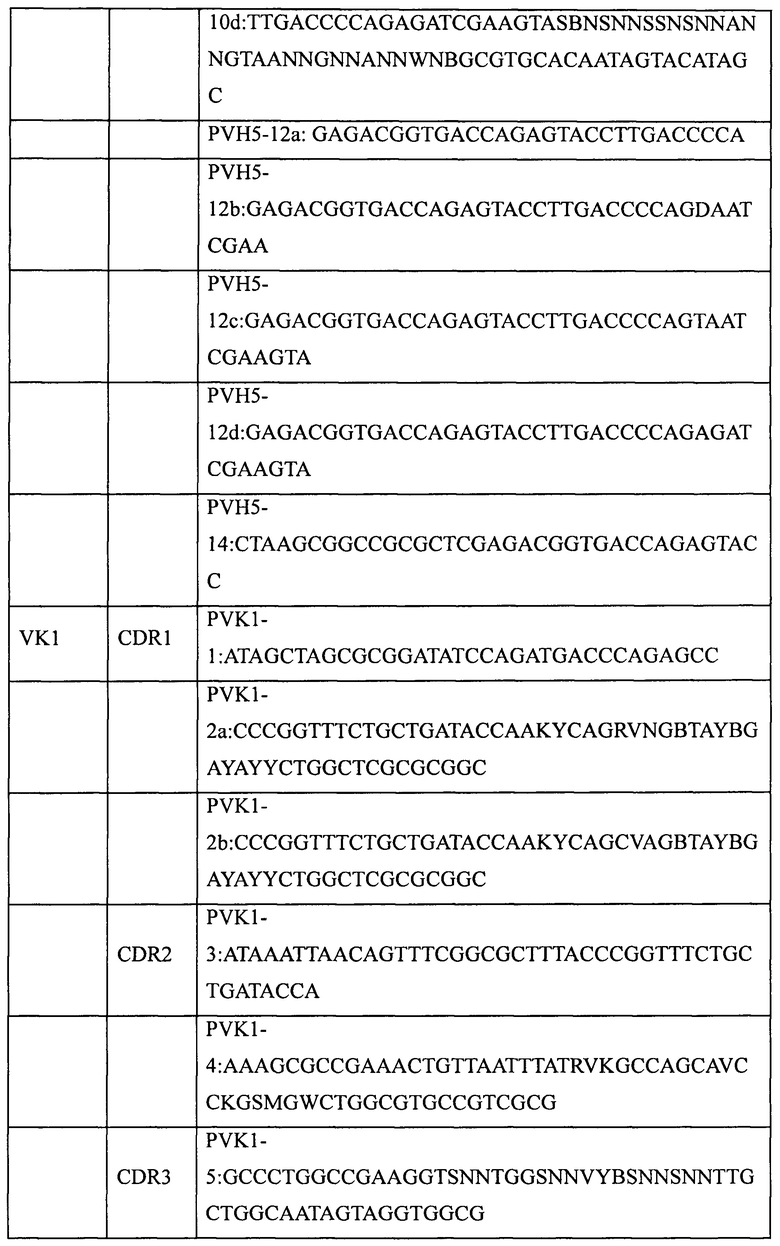
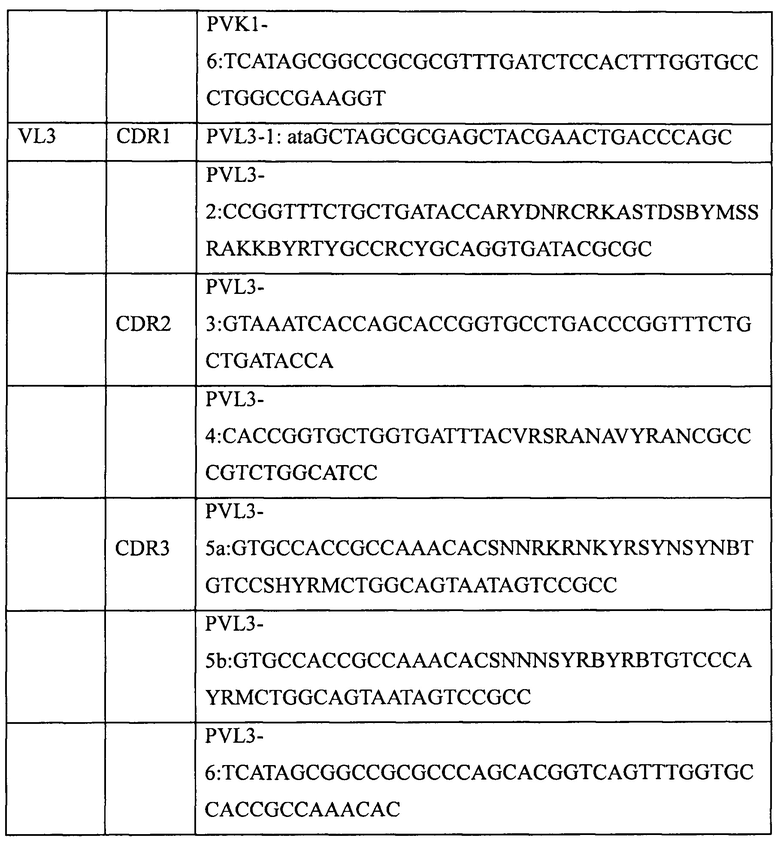
1.2 Создание генов одноцепочечных антител (одноцепочечных Fv, ScFv)
Для создания генов одноцепочечных антител (ScFv) вариабельный участок тяжелой цепи (VH) и вариабельный участок легкой цепи (VL) соединяли с помощью обычно используемого для этой цели гибкого линкера, состоящего из 15 аминокислот. Линкер имел аминокислотную последовательность GGGGSGGGGSGGGGS, которую кодирует последовательность ggtggaggcggttctggcggaggtgggagcggaggcggaggttca. Сконструированное одноцепочечное антитело имело структуру VН-линкер-VL.
С помощью способа, описанного в первом подразделе этого примера, было получено множество генов вариабельных участков тяжелой и легкой цепей, представленных в таблице 3 ниже, в том числе четыре различных гена вариабельных участков тяжелой цепи и три гена вариабельных участков легкой цепи.
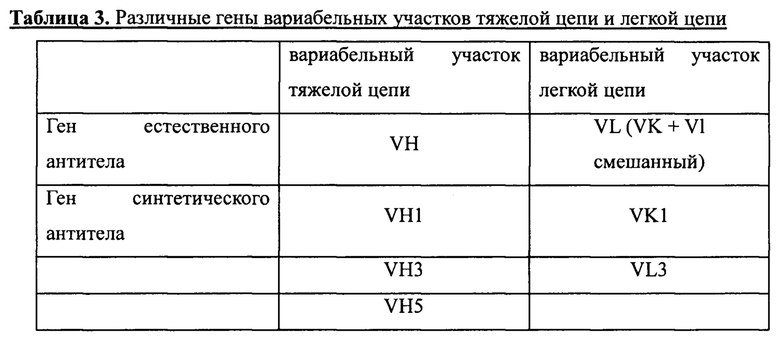
Как видно из этой таблицы, использование описанной выше стратегии конструирования одноцепочечных антител и отработанного метода ПЦР с перекрывающимися праймерами позволяет успешно объединять различные тяжелые цепи и легкие цепи. В общей сложности было создано 12 различных генов одноцепочечных антител. Эти 12 генов одноцепочечных антител, амплифицированных с помощью ПЦР, очищали и выделяли с помощью электрофореза в агарозном геле, после чего сохраняли их для дальнейшего использования при -20°С.
1.3 Создание вектора для фагового дисплея на основе арабинозного промотора
В основе традиционных векторов для фагового дисплея лежат lac промоторы (Plac). Однако использование lac промоторов из-за их особенностей, в частности фоновой экспрессии в отсутствие индуктора, отрицательно влияет на размер и разнообразие библиотеки антител. Вектор pCANTAB5E (Amersham Biosciences/GE, Inc.), который обычно используют для фагового дисплея, был изменен нами с использованием стандартных методов молекулярной биологии как описано далее.
С помощью двойного расщепления вектора pCANTAB5E ферментами AflIII и NotI была осуществлена замена Plac промотора и участка сигнального пептида g3 на фрагмент, содержащий ген AraC, арабинозный промотор (Para) и лидерную последовательность PelB, где ген AraC и Para промотор представляли собой часть вектора pBADhis (Invitrogen Inc.), а лидерная последовательность PelB (SEQ ID NO: 6) представляла собой искусственную последовательность. Затем с помощью двойного расщепления ферментами NcoI и NotI между сайтами NcoI и NotI была клонирована спейсерная последовательность длиной около 750 п.н. (SEQ ID NO: 7), в результате чего был создан конечный новый вектор для фагового дисплея pADSCFV-S (фиг. 1). Сайты NcoI и NotI в этом векторе могут облегчать клонирование гена одноцепочечного антитела (ScFv).
1.4 Получение библиотеки одноцепочечных человеческих антител и фаговой библиотеки антител
С использованием стандартных молекулярно-биологических методов и двойного расщепления ферментами NcoI и NotI было проведено соответствующее клонирование в вектор pADSCFV-S 12 генов ScFvs, получение которых описано в разделе 1.2. Продукты лигирования были использованы для трансформации методом электропорации компетентных клеток TG1, при этом для получения каждой суб-библиотеки было проведено около 20 раундов трансформации, а в общей сложности было проведено около 240 раундов трансформации. Размер каждой суб-библиотеки определяли с использованием способов разведений. Для оценки точности каждой суб-библиотеки проводили секвенирование 30-40 колоний, отобранных случайным образом из каждой суб-библиотеки. Данные о размере и точности для 12 суб-библиотек приведены в таблице 4. Суммарный размер 12 суб-библиотек достигал 1,0* 10Е9 при средней величине точности более 75%.
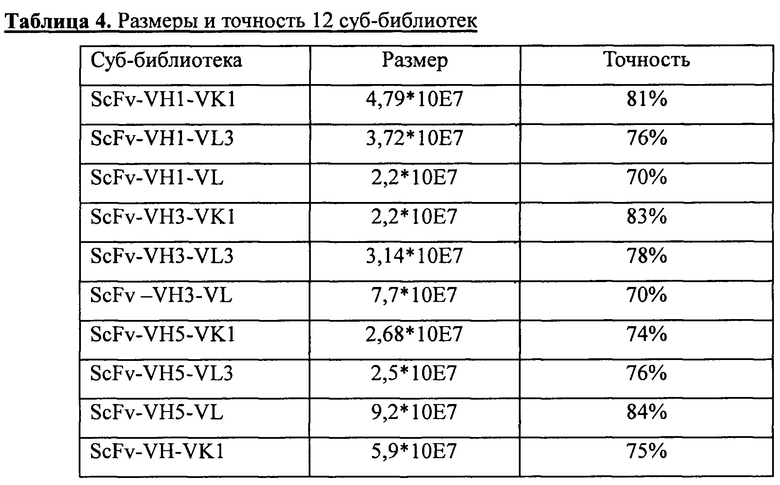
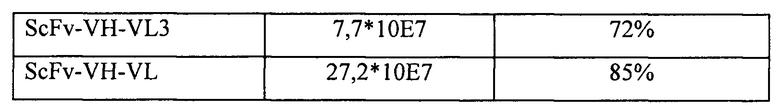
Полученные 12 суб-библиотек соответственно высевали в жидкую среду 2YTAG (А: ампициллин, 100 мкг/мл; G: глюкоза, 2%) и инкубировали при 37°С на качалке при частоте вращения 220 об./мин. до достижения клетками логарифмической фазы роста (OD600=0,8). После этого клетки инфицировали фагами-помощниками М13 (M13KO7, NEB Inc.). После инфицирования среду заменяли на жидкую среду 2YTAKA (А: ампициллин, 100 мкг/мл; K: канамицин, 70 мкг/мл; А: арабиноза, 0,001%) и для размножения фага инкубировали клетки в течение ночи при 28°С на качалке при частоте вращения 220 об./мин. Затем, для получения препаратов очищенного фага (фаг-ScFv), использовали способ осаждения с PEG/NaCl, после чего проводили их титрование. После этого препараты фаг-ScFv, полученные из 12 суб-библиотек, смешивали пропорционально с учетом размера библиотек и, таким образом, получали фаговую библиотеку человеческих антител. Конечный титр фаговой библиотеки составлял 6*10Е12 КОЕ/мл. Полученную библиотеку хранили при -70°С. Данную фаговую библиотеку антител можно было использовать для скрининга специфических человеческих антител к различным представляющим интерес антигенам.
Пример 2
Получение рекомбинантных белковых антигенов
Получение моноклональных антител к IL-17 связано с необходимостью применения нескольких различных рекомбинантных белков, включая человеческий (Homo sapiens) IL-17 (huIL17, SEQ ID NO: 8), мышиный (Mus musculus) IL-17 (moIL17, SEQ ID NO: 9), IL-17 Масаса mulatta (mmIL17, SEQ ID NO: 10), IL-17 Масаса fascicularis (mfIL17, SEQ ID NO: 11) и внеклеточный участок рецептора IL-17 человека (IL17R, SEQ ID NO: 12). Поскольку эти белки содержат модификации, связанные с гликозилированием, предпочтительной для сохранения структур и функций рекомбинантных белков является система экспрессии на основе клеток млекопитающих. Кроме того, для осуществления очистки в структуру этих рекомбинантных белков со стороны С-конца вводили гистидиновую (His) метку (SEQ ID NO: 13) или Fc фрагмент человеческого антитела (SEQ ID NO: 14).
На основе аминокислотных последовательностей различных рекомбинантных белков, представленных в базе данных Uniprot, осуществляли конструирование и последующий синтез генов (содержащих последовательность, кодирующую гистидиновую метку или Fc фрагмент) вышеупомянутых рекомбинантных белков. Синтетические гены рекомбинантных белков с использованием стандартных молекулярно-биологических методов клонировали в векторы, пригодные для экспрессии в эукариотических клетках (например, pcDNA3.1, производства Invitrogen Inc.). Затем полученные плазмиды экспрессии рекомбинантных белков вводили в клетки линии HEK293 (например, клетки HEK293F, Invitrogen Inc.) путем трансфекции с использованием липосом (например, реагента 293fectin, Invitrogen Inc.) или других средств для трансфекции (например, PEI (полиэтиленимина)). Клетки инкубировали в суспензии в бессывороточных условиях в течение 3-4 дней. После этого собирали культуральную жидкость, осаждая клетки центрифугированием.
Для получения рекомбинантных белков, слитых с гистидиновой меткой, рекомбинантные белки в культуральной жидкости подвергали дополнительной очистке на колонках для металл-хелатной аффинной хроматографии (например, HisTrap FF, производства GE Inc.). Для получения рекомбинантных белков, слитых с фрагментом Fc, для дополнительной очистки использовали колонки для аффинной хроматографии с иммобилизованным белком А или G (например, Mabselect SURE, производства GE Inc.). Затем с использованием колонок для обессоливания (например, Hitrap desaulting, производства GE Inc.) проводили замену буферного раствора для хранения рекомбинантных белков на фосфатно-солевой буфер (PBS) (рН 7,0) или другой подходящий буфер. При необходимости, образцы антител могли быть подвергнуты стерилизации фильтрованием, после чего их делили на аликвоты и хранили при -20°С.
Пример 3
Получение моноклональных антител к человеческому IL-17 методом фаговой библиотеки антител
3.1 Обогащение фаговой библиотеки антител моноклональными антителами к человеческому IL-17
Белок huIL17-his (далее именуемый «IL17-His») (5 мкг/мл) использовали как антиген для иммобилизации на планшете для ELISA (по 150 мкл в лунку, всего четыре лунки). Планшет инкубировали в течение ночи при 4°С. После этого планшет для ELISA блокировали фосфатно-солевым буфером с 0,1% Твин 20 (PBST), с добавлением 1% BSA при 37°C в течение 1 ч. Вместе с тем, фаговую библиотеку антител (фаг-scFv), получение которой описано в примере 1, блокировали путем инкубации с PBST с 1% BSA при комнатной температуре в течение 1 ч, при этом количество вносимых фаговых частиц составляло приблизительно 1012. После блокирования фаговую библиотеку антител вносили в планшет для ELISA для проведения связывания антител с антигеном и оставляли на 1 ч при 37°С. Для удаления несвязанных фаговых частиц планшет промывали PBST/PBS. Наконец, связанных фагов элюировали 0,1 М раствором глицин-HCl (рН 2,2), элюат нейтрализовали 1,5 М раствором Трис-HCl (рН 8,8).
Нейтрализованный препарат фагов использовали для инфицирования бактериальных клеток TG1, которые выращивали до логарифмической фазы. Бактерии инкубировали 30 мин. при 37°С и еще 30 мин. при 37°С на качалке при частоте вращения 150 об./мин. Для подсчета количества клеток 1% суспензии бактерий высевали, а оставшуюся часть центрифугировали при 4000 об./мин. 5 мин. После удаления супернатанта бактерии высевали на твердую среду 2YTAG (А: ампициллин, 100 мкг/мл; G: глюкоза, 2%) в чашку и инкубировали в течение ночи при 33°С для проведения последующей процедуры скрининга.
Бактерии собирали с чашек. Необходимое количество собранных бактерий высевали в жидкую среду 2YTAG (А: ампициллин, 100 мкг/мл; G: глюкоза, 2%) и инкубировали на качалке до достижения логарифмической фазы роста. Бактерии инфицировали фагом M13KO7. После инфицирования среду заменяли на жидкую среду 2YTAKA (А: ампициллин, 100 мкг/мл; K: канамицин, 70 мкг/мл; А: арабиноза, 0,001%) и инкубировали бактерии для размножения фага в течение ночи при 28°С на качалке при частоте вращения 220 об./мин. Очистку фагов для проведения последующей процедуры скрининга проводили осаждением с PEG/NaCl. В общей сложности для обогащения фаговой библиотеки было проведено 3 раунда скрининга.
3.2 Выявление отдельных колоний с последовательностями антител к IL-17
После трех раундов скрининга отбирали хорошо различимые одиночные колонии и высевали их в лунки 96-луночного планшета, содержащие среду 2YTAG, и инкубировали при 37°С на качалке при частоте вращения 220 об./мин. до достижения логарифмической фазы роста. В каждую лунку добавляли около 1010 частиц фага-помощника M13KO7. Планшет с клетками инкубировали 30 мин. при 37°С и еще 30 мин. при 37°С на качалке при частоте вращения 150 об./мин., после чего центрифугировали 10 мин. при 2000 об./мин. при комнатной температуре. После удаления супернатанта бактериальные клетки повторно суспендировали в среде 2YTAKA (А: ампициллин, 100 мкг/мл; K: канамицин, 70 мкг/мл; А: арабиноза, 0,001%) и инкубировали в течение ночи при 28°С на качалке при частоте вращения 220 об./мин.
Для иммобилизации вносили IL17-Fc в карбонатном буфере (рН 9,6) и инкубировали в течение ночи при 4°С. После трехкратной промывки PBST, для блокировки при 37°С в течение 1 ч использовали 4% раствор сухого молока в PBST. Культуральную жидкость, собранную после инкубирования в течение ночи моноклональной фаговой культуры, пропорционально разводили в PBST с добавлением 4% сухого молока, который вносили в количестве 100 мкл на лунку блокированного планшета для ELISA для связывания при 37°С в течение 1 ч. Планшет для ELISA отмывали PBST. Конъюгат пероксидазы хрена (HRP) с антителом к М13 разводили в соотношении 1:5000 и вносили в количестве 100 мкл на лунку планшета для ELISA. После инкубации в течение 1 ч при 37°С добавляли проявляющий раствор OPD и проявляли 5-20 мин. Затем реакцию проявления останавливали добавлением 1 М H2SO4 по 50 мкл в лунку. Значения оптической плотности определяли с помощью спектрофотометра для микропланшетов на двух длинах волн 492 нм/630 нм.
После проведения описанных выше процедур были получены приблизительно 300 отдельных колоний, из числа которых были выявлены четыре линии с одноцепочечными антителами (scFv), имеющими различную последовательность и относительно высокую аффинность к IL-17, получившие название 11А, 9G, 3Е и 6D.
3.3 Предварительный функциональный анализ моноклональных антител к IL-17
Четыре моноклональных линии (11А, 9G, 3Е, 6D) высевали в жидкую среду 2YTAG и инкубировали до достижения логарифмической фазы роста. Для инфицировали добавляли фаг-помощник M13KO7 в количестве, превышавшем количество бактерий примерно в 20 раз. После инфицирования среду заменяли на среду 2YTAKA и инкубировали клетки для размножения фага в течение ночи при 28°С на качалке при частоте вращения 220 об./мин. Очистку фагов проводили способом осаждения с PEG/NaCl.
Для иммобилизации вносили IL17-his в карбонатном растворе (рН 9,6) и инкубировали в течение ночи при 4°С. После трехкратной промывки PBST, для блокировки при 37°С в течение 1 ч использовали 4% раствор сухого молока в PBST. Готовили разведение препаратов четырех фагов с титром 5*10Е11 КОЕ/мл. Разведения четырех фагов использовали для приготовления разведений IL17R-his. Начальная концентрация IL17R-his составляла 20 мкг/мл. Для каждого образца готовили по семь трехкратных серийных разведений. Подготовленные разведения вносили в лунки планшета для ELISA в объеме 100 мкл на лунку и инкубировали при 37°С 1 ч. После этого планшет для ELISA промывали раствором PBST. Конъюгат HRP с вторичным антителом к М13 разводили в соотношении 1:5000 и вносили в количестве 100 мкл на лунку планшета для ELISA. После инкубации в течение 1 ч при 37°С добавляли проявляющий раствор OPD и проявляли 5-20 мин. Затем реакцию проявления останавливали добавлением 1 М H2SO4 по 50 мкл в лунку. Значения оптической плотности определяли с помощью спектрофотометра для микропланшетов на двух длинах волн 492 нм/630 нм.
Из полученных результатов (см. фиг. 2) видно, что одноцепочечные антитела 11А и 9G были способны конкурировать с IL17R-his за связывание с IL17-his, что свидетельствует о том, что 11А (SEQ ID NO: 15) и 9G (SEQ ID NO: 16) имеют такой же сайт связывания с IL-17, что и IL17R. В то же время, одноцепочечные антитела 3Е и 6D не конкурировали с IL17R-his и представляют собой нефункциональные антитела.
Пример 4
Повышение аффинности моноклональных антител к IL-17 на основе стратегии замещения легких цепей
4.1 Создание системы фагового дисплея с двумя векторами, необходимой для повышения аффинности антител
Для облегчения введения мутаций в легкие и тяжелые цепи антител были созданы три набора (всего шесть) векторов прокариотической экспрессии, которые могут сосуществовать в одной клетке Е. coli. Этот процесс описан подробнее ниже.
С использованием плазмиды pADSCFV-S, описанной в примере 1, в качестве исходной плазмиды и с помощью двойного расщепления ферментами PmlI и XbaI, вместо гена gIII в изначальном векторе в вектор pADSCFV-S был клонирован ген константного участка легкой каппа-цепи синтетического человеческого антитела (SEQ ID NO: 17), в результате чего был создан вектор pADK-S (см. фиг. 3). Этот вектор можно было использовать для клонирования и экспрессии гена легкой каппа-цепи человеческого антитела.
Подобным образом, с использованием плазмиды pADSCFV-S, описанной в примере 1, в качестве исходной плазмиды и с помощью помощью двойного расщепления ферментами PmlI и XbaI, вместо гена gIII в изначальном векторе в вектор pADSCFV-S был клонирован слитый ген, кодирующий константный участок СН1 тяжелой цепи синтетического человеческого антитела и С-концевой домен белка gIII, (SEQ ID NO: 18) или константный участок легкой лямбда-цепи человеческого антитела (SEQ ID NO: 19), в результате чего были созданы векторы pADG-S и pADL-S.
Ген устойчивости к хлорамфениколу (CmR) был получен из плазмиды pACYC184 (NEB Inc.) путем амплификации с помощью ПЦР. Сайт инициации репликации плазмиды CDF (CDFori, SEQ ID NO: 20) был синтезирован искусственным путем. Затем с помощью стандартной методики ПЦР с перекрывающимися праймерами был создан слитый ген CmR-CDoriF (по разным концам которого соответственно были добавлены сайты XbaI и AflIII). Затем, после двойного расщепления ферментами XbaI и AflIII, слитый ген CmR-CDFori клонировали соответственно в каждый из векторов pADK-S, pADG-S и pADL-S вместо участка f1ori-Ampr-pBRori, в результате чего были созданы три новые плазмиды pAK-S (см. фиг. 3), pAG-S и pAL-S.
4.2 Создание библиотеки легких цепей человеческих антител для процедуры замены легких цепей
Для облегчения проведения процедуры замены легких цепей антител, в качестве исходных плазмид были использованы плазмиды pADK-S и pADL-S для создания высококачественной библиотеки легких цепей человеческих антител. Этот процесс описан подробнее ниже.
Гены VK1 получали путем амплификации по стандартному методу ПЦР с использованием в качестве матриц ДНК из четырех суб-библиотек, содержащих гены вариабельного участка VK1, получение которых описано в примере 1, и праймеров, приведенных в таблице 5. Затем, с помощью двойного расщепления ферментами NcoI и PmlI, амплифицированные последовательности генов VK1 клонировали в вектор pADK-S на место спейсерной последовательности. Продукты лигирования вводили методом электропорации в компетентные клетки TG1, в результате чего была создана библиотека легких цепей VK1 человека. Размер библиотеки рассчитывали с использованием способов разведений. Для оценки точности каждой библиотеки легких цепей VK1 случайным образом отбирали 30 колоний для проведения секвенирования. Результаты представлены в таблице 6.
Аналогичным образом, гены VL3 получали путем амплификации с использованием в качестве матриц ДНК из четырех суб-библиотек, содержащих гены вариабельного участка VL3, получение которых описано в примере 1, и праймеров, приведенных в таблице 5. Затем с помощью двойного расщепления ферментами NcoI и PmlI амплифицированные последовательности генов VL3 клонировали в вектор pADL-S на место спейсерной последовательности. Продукты лигирования вводили методом электропорации в компетентные клетки TG1, в результате чего была создана библиотека легких цепей VL3 человека. Размер библиотеки рассчитывали с использованием способов разведений. Для оценки точности каждой библиотеки легких цепей VL3 случайным образом отбирали 30 колоний для проведения секвенирования. Результаты представлены в таблице 6. Суммарный размер двух библиотек легких цепей составлял 1,0*10Е8 при точности более 90%.
Затем две полученные суб-библиотеки легких цепей соответственно высевали в жидкую среду 2YTAG (А: ампициллин, 100 мкг/мл; G: глюкоза, 2%) и инкубировали до достижения логарифмической фазы роста. После этого клетки инфицировали фагами-помощниками М13 (M13KO7). После инфицирования среду заменяли на жидкую среду 2YTAKA (А: ампициллин, 100 мкг/мл; K: канамицин, 70 мкг/мл; G: глюкоза, 2%) и инкубировали клетки для размножения фага в течение ночи при 28°С на качалке при частоте вращения 220 об./мин. Затем для получения препаратов очищенного фага использовали способ осаждения с PEG/NaCl, после чего проводили их титрование. После этого препараты фагов из двух полученных суб-библиотек смешивали пропорционально с учетом размера библиотек и таким образом получали фаговую библиотеку (фаг-VK1+VL3) для сборки легких цепей человеческих антител. Конечный титр фаговой библиотеки составлял 5,4*10Е11 КОЕ/мл. Полученную библиотеку хранили при -70°С. Эту фаговую библиотеку можно было использовать в процедурах замены легких цепей в различных антителах.
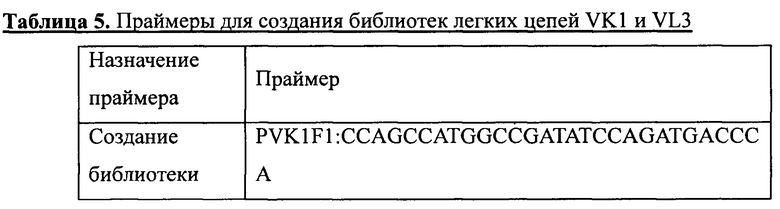
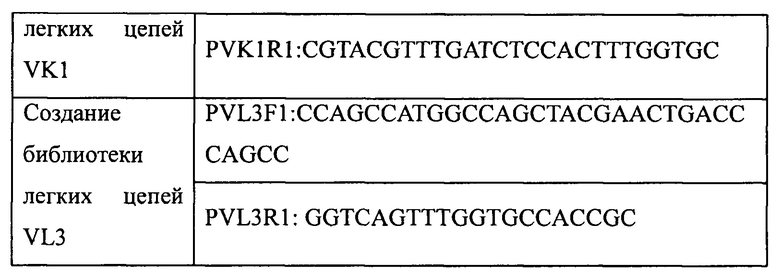

4.3 Процедуры замены легких цепей в антителах к IL-17
Легкие цепи (11AVK, 9GVK) и тяжелые цепи (11AVH, 9GVH) двух одноцепочечных антител 11А и 9G, полученных в результате скрининга из библиотеки человеческих антител, клонировали соответственно в векторы прокариотической экспрессии pADK-s и pAG-s, благодаря чему были получены плазмиды pADK-11AVK, pADK-9GVK, pAG-11AVH и pAG-9GVH, позволяющие экспрессировать соответствующие легкие и тяжелые цепи. Плазмиды с клонированными генами тяжелой цепи (pAG-HAVH, pAG-9GVH) использовали для трансформации клеток TG1. Затем клетки инфицировали фагом, обеспечивающим упаковку библиотеки легких цепей (фаг-VK1+VL3), тем самым получая библиотеки мутированных легких цепей двух антител (11AVH-VK1+VL3 и 9GVH-VK1+VL3).
После этого две библиотеки мутированных легких цепей (11AVH-VK1+VL3 и 9GVH-VK1+VL3) соответственно высевали в жидкую среду 2YTACG (А: ампициллин, 100 мкг/мл; С: хлорамфеникол, 34 мкг/мл; G: глюкоза, 2%) и инкубировали до достижения клетками логарифмической фазы роста. После этого клетки инфицировали фагами-помощниками М13 (M13KO7). После инфицирования среду заменяли на жидкую среду 2YTACKA (A: ампициллин, 100 мкг/мл; С: хлорамфеникол, 34 мкг/мл; K: канамицин, 70 мкг/мл; А: арабиноза, 0,001%) и инкубировали клетки для размножения фага в течение ночи при 28°С на качалке при частоте вращения 220 об./мин. Затем, для получения препаратов очищенной фаговой библиотеки (фаг-11AVH-VK1+VL3 и фаг-9GVH-VK1+VL3), использовали способ осаждения с PEG/NaCl, после чего проводили их титрование. Две фаговые библиотеки смешивали в равных пропорциях. Конечный титр фаговой библиотеки составлял 1,9*10Е13 КОЕ/мл. Полученную библиотеку хранили при -70°С.
Согласно стандартной методологии и метода фагового дисплея использовали IL17-His в двух раундах обогащения смешанной библиотеки фаг-11AVH-VK1+VL3 и фаг-9GVH-VK1+VL3. Затем использовали способ фагового ELISA для выявления обогащенных отдельных колоний (приблизительно 400 колоний). Колонии с высокими значениями сигнала в ELISA отбирали для секвенирования. Колонии, для которых секвенирование подтвердило правильность клонированной последовательности, использовали для получения препаратов очищенного фага. Тем временем плазмиды, с которых экспрессировались легкие цепи и тяжелые цепи двух антител (11А и 9G), были использованы для совместной трансформации клеток TG1. В качестве положительного контроля были использованы препараты очищенного фага.
4.4 Сравнение относительной аффинности моноклональных мутантных антител к IL-17 методом ELISA очищенного фага
Титры множества очищенных моноклональных фагов доводили до 1*10Е11 КОЕ/мл. Трехкратные серийные разведения готовили с использованием 4% раствора сухого молока в буфере PBST. Способность фаговых антител связывать IL-17 анализировали для отдельных разведений стандартным способом фагового ELISA с иммобилизованным IL17-Fc (2 мкг/мл). Из результатов фагового ELISA видно, что из нескольких фаговых антител, отобранных с помощью скрининга после проведения процедуры замены легких цепей на антителах 11А и 9G, все могли связываться с IL-17. Кроме того, С2 (SEQ ID NO: 21) и D5 (SEQ ID NO: 22) обладали значительно более высокой относительно аффинностью по сравнению с антителами от других колоний (см. фиг. 4). Было отмечено, что антитела С2 и D5 содержали тяжелые цепи 9GVH.
Пример 5
Создание и скрининг библиотеки мутантных последовательностей, кодирующих вариабельный участок тяжелой цепи 9G (9GVH)
В целях дополнительного повышения аффинности моноклональных антител к IL-17 была создана библиотека мутантных последовательностей, кодирующих вариабельный участок тяжелой цепи 9G. Для скрининга библиотеки мутантных тяжелых цепей была использована система фагового дисплея с двумя векторами.
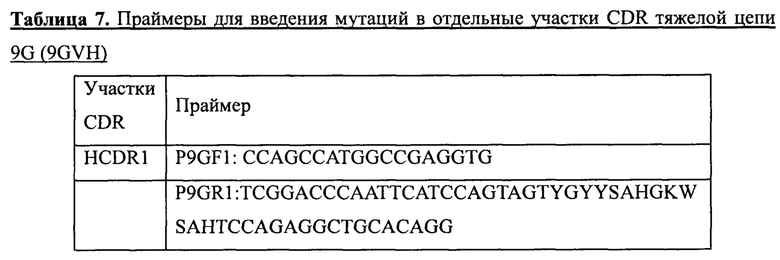
В качестве матрицы использовали ген, кодирующий вариабельный участок тяжелой цепи 9G (9GVH, SEQ ID NO: 23). Различные запланированные мутации вводили в участки CDR 9GVH по стандартному методу ПЦР с использованием вырожденных праймеров, показанных в таблице 7. После этого в процедуре сборки с целью получения мутированных генов интактных вариабельных участков тяжелой цепи 9G использовали способ ПЦР с перекрывающимися праймерами. Мутированные гены 9GVH клонировали в вектор pADG-S с использованием двойного расщепления ферментами NcoI и PmlI. Продукты лигирования были использованы для трансформации методом электропорации компетентных клеток TG1, в результате чего была создана библиотека мутантных генов 9GVH с размером приблизительно 4*10Е6. Для секвенирования было случайным образом отобрано 20 отдельных колоний. По результатам секвенирования точность библиотеки мутантных генов составляла 70% (14/20).
Затем созданную библиотеку мутантных генов 9GVH (в бактериальных клетках TG1) высевали в жидкую среду 2YTAG (А: ампициллин, 100 мкг/мл; G: глюкоза, 2%) и инкубировали до достижения клетками логарифмической фазы роста. После этого клетки инфицировали фагами-помощниками М13 (M13KO7). После инфицирования среду заменяли на жидкую среду 2YTAKA (А: ампициллин, 100 мкг/мл; K: канамицин, 70 мкг/мл; G: глюкоза, 2%) и инкубировали клетки для размножения фага в течение ночи при 28°С на качалке при частоте вращения 220 об./мин. Затем, для получения препаратов очищенного фага, упаковывающего библиотеку мутантных генов 9GVH (фаг-9GVH), использовали способ осаждения с PEG/NaCl, после чего проводили их титрование. Конечный титр фаговой библиотеки составлял 1,4*10Е13 КОЕ/мл. Полученную библиотеку хранили при -70°С.
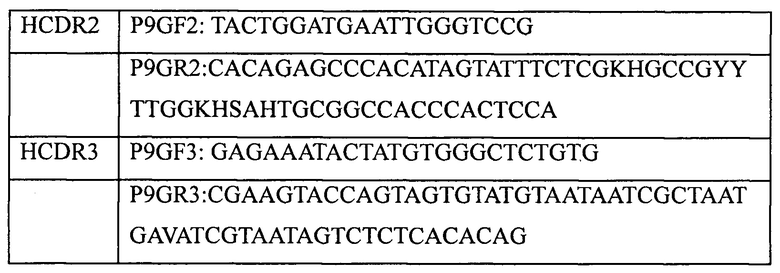
С использованием стандартного метода ПЦР два гена легкой цепи С2 и D5, скрининг которых описан в примере 4, клонировали в вектор pAK-S, который затем использовали для трансформации бактерий TG1, в результате чего были получены две линии pAK-C2/TG1 и pAK-D5/TG1. Затем клетки pAK-C2/TG1 и pAK-D5/TG1 в логарифмической фазе роста инфицировали фаговой библиотекой (фаг-9GVH), в результате чего были получены две библиотеки мутантных генов 9GVH (легкими цепями соответственно были С2 и D5).
Затем, согласно стандартным протоколам фагового дисплея, для инфицирования использовали M13KO7. Фаги экспонировали на своей поверхности две библиотеки Fab-фрагментов, содержащие мутированные цепи 9GVH, таким образом были созданы две библиотеки фагового дисплея Fab-фрагментов (фаг-С2-9GVH и фаг-D5-9GVH). Две библиотеки фагового дисплея смешивали в равных пропорциях. Было проведено два раунда скрининга смешанной библиотеки фаг-С2-9GVH и фаг-D5-9GVH с использованием IL-17. Для выявления с помощью моноклонального фагового ELISA было отобрано приблизительно 300 колоний. В итоге, для дальнейшего исследования были отобраны три мутантных линии 9GVH, которые были названы 9GA3 (SEQ ID NO: 24), 9GC2 (SEQ ID NO: 25) и 9GC5 (SEQ ID NO: 26). Кроме того, все три мутанта 9GVH содержали обладавшую наибольшей специфичностью легкую цепь С2 (SEQ ID NO: 21).
Пример 6
Экспрессия и очистка моноклональных антител
Поскольку антитела представляют собой большие белковые молекулы со сложными пост-трансляционными модификациями, для экспрессии рекомбинантных интактных антител обычно используют системы экспрессии на основе клеток млекопитающих. Кроме того, с помощью способа аффинной хроматографии с иммобилизованным белком А или G можно было легко очистить антитела до степени чистоты более 95%. В этом примере кратко описаны способы и методы получения обычных рекомбинантных антител.
Гены тяжелых и легких цепей представляющего интерес антитела с использованием стандартного молекулярно-биологического метода клонировали в векторы, пригодные для экспрессии в эукариотических клетках (например, pcDNA3.1, производства Invitrogen Inc.). Константный участок тяжелой цепи антитела мог принадлежать к подтипу IgG1 (SEQ ID NO: 29) или подтипу IgG4 (SEQ ID NO: 30), a константный участок легкой цепи мог принадлежать к подтипу каппа (SEQ ID NO: 31) или подтипу лямбда (SEQ ID NO: 32).
Затем полученные плазмиды, с которых экспрессировалась тяжелая цепь и легкая цепь, вводили совместно в клетки HEK293 (например, HEK293F от Invitrogen Inc.) или клетки СНО (например, CHO-S от Invitrogen Inc.) путем трансфекции с использованием липосом (например, реагента 293fectin от Invitrogen Inc.) или других средств для трансфекции (например, PEI). Клетки инкубировали в суспензии в бессывороточных условиях в течение 3-4 дней. После этого собирали культуральную жидкость, осаждая клетки центрифугированием. Для дополнительной очистки рекомбинантного антитела из культуральной жидкости использовали колонку для аффинной хроматографии с иммобилизованным белком А или G (например, Mabselect SURE, производства GE Inc.). Затем с использованием колонок для обессоливания (например, Hitrap desaulting, производства GE Inc.) проводили замену буферного раствора для хранения антитела на фосфатно-солевой буфер (PBS) (рН 7,0) или другой подходящий буфер (например, 0,1 М NaCl, 0,01 М цитрат натрия, рН 6,0). При необходимости, образцы антитела могли быть подвергнуты стерилизации фильтрованием, после чего их делили на аликвоты и хранили при -20°С.
Пример 7
Анализ связывания моноклональных антител к IL-17 с человеческим IL-17
Способность связываться с IL17-His четырех моноклональных антител к IL-17, 9GA3, 9GC2, 9GC5 и контрольного антитела секукинумаб (VH: SEQ ID NO: 27, VK: SEQ ID NO: 28), оценивали в ходе анализов ELISA.
IL17-His использовали для иммобилизации на планшетах в течение ночи при 4°С (0,5 мкг/мл, 100 мкл на лунку). Готовили серийные трехкратные разведения каждого моноклонального антитела с начальной концентрацией 300 нМ. Для каждого образца моноклонального антитела делали одиннадцать серийных разведений. Способность моноклональных антител к связыванию IL-17 в каждом разведении определяли с помощью конъюгата HRP с козьим антителом к человеческому IgG (см. фиг. 5).
Пример 8
Связывание моноклональных антител к IL-17 с IL-17 других видов животных
Четыре типа приготовленных IL-17 (huIL17, mIL17, mmIL17 и mfIL17) использовали для иммобилизации на 96-луночных планшетах для ELISA в течение ночи при 4°С (1 мкг/мл, 100 мкл на лунку). В планшеты добавляли блокирующий раствор (2% сухое молоко в PBST) и инкубировали планшеты при 37°С 1 ч. После этого на планшеты наносили различные моноклональные антитела к IL-17, в том числе 9GA3, 9GC2, 9GC5, секукинумаб и иксекизумаб (VH: SEQ ID NO:33, VK: SEQ ID NO: 34), и оставляли для прохождения связывания при 37°С 1 ч. Затем планшеты промывали четыре раза посредством PBST. После этого добавляли конъюгат HRP с антителом к человеческому IgG (вторичное антитело) и оставляли инкубироваться при 37°С на 1 ч. Затем планшеты промывали четыре раза посредством PBST. В лунки планшета вносили проявляющий раствор OPD и проявляли 5-10 мин. Реакцию проявления останавливали добавлением 1 М H2SO4. Значения оптической плотности при длине волны 492 нм определяли с помощью спектрофотометра для планшетов (Spectra Мах190, Molecular Device). Как видно из результатов на фиг. 6, моноклональные антитела к IL-17, предлагаемые в настоящем изобретении, и контрольные антитела были способны специфически распознавать человеческий IL-17, IL-17 Масаса mulatta и Масаса fascicularis, но не связывались с мышиным IL-17.
Пример 9
Конкурентное связывание IL-17 моноклональными антителами к человеческому IL-17 и рецептором IL-17 (IL17R)
Функциональное моноклональное антитело к IL-17 должно было быть способным блокировать связывание IL17R с IL-17 на уровне белка. В этом примере оценивали способность четырех моноклональных антител к IL-17 (9GA3, 9GC2, 9GC5 и секукинумаб) ингибировать связывание IL17R с IL-17.
IL17-Fc использовали для иммобилизации на планшетах в течение ночи при 4°С (0,5 мкг/мл, 100 мкл на лунку). Готовили серийные трехкратные разведения каждого моноклонального антитела (с начальной концентрацией 200 мкг/мл) с добавлением IL17R-his (2 мкг/мл). Для каждого образца моноклонального антитела делали десять серийных разведений. Для определения сигнала связывания IL17R-his с IL17-Fc использовали конъюгат HRP с мышиным моноклональным антителом к гистидиновой метке. Затем для анализа данных и построения графиков использовали ПО GraphPad Prism 6 (см. фиг. 7).
Клетки HDFa (взрослые дермальные фибробласты) были приобретены у Sciencell Inc. (кат. №2320). Клетки инкубировали и пересевали согласно инструкциям, предоставленным Sciencell Inc.
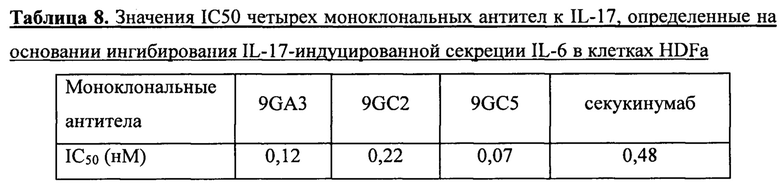
В случае оценки значений активности моноклональных антител к IL-17 с использованием клеток HDFa, клетки HDFa высевали в 96-луночные планшеты с плотностью клеток 1*10Е4 клеток в лунку. В среду FM добавляли 1% FBS, по содержанию остальных компонентов питательная среда не отличалась от полной среды FM. Клетки культивировали в течение ночи при 37°С. На следующий день среду заменяли на среду (FM+1% FBS) с добавлением 2 нМ IL17-His и моноклональных антител к IL-17 (9GA3, 9GC2, 9GC5 и секукинумаба) в разных концентрациях (0,01-10 нМ) и культивировали клетки в течение 24 ч. После этого собирали культуральную жидкость. Содержание IL-6 в культуральной жидкости каждой культуры определяли с использованием набора реагентов для количественного определения IL-6 (кат. №ЕНоо4-96), производства Excell Biology Inc. Затем для анализа данных и построения графиков использовали ПО GraphPad Prism 6 (см. фиг. 8). Значения IC50 для каждого моноклонального антитела к IL-17, определенные на основании ингибрования секреции IL-6, приведены в таблице 8.
Пример 11
Оценка стабильности моноклональных антител к IL-17 в сыворотке крови человека
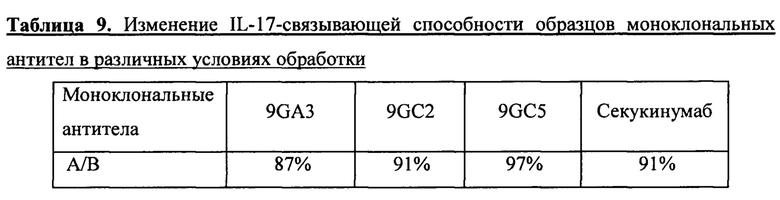
С целью предварительной оценки специфичности различных моноклональных антител к IL-17 и их стабильности в сыворотке крови была проведена оценка стабильности моноклональных антител к IL-17 в сыворотке крови человека. Для этого исследования были использованы четыре моноклональных антитела к IL-17, а именно 9GA3, 9GC2, 9GC5 и секукинумаб. Готовили растворы образцов очищенных моноклональных антител в 0,01 М цитрате натрия, 0,1 М NaCl, рН 6,0. Образцы моноклональных антител, подвергнутые стерилизации фильтрованием, разводили в 200 мкл смеси на основе стерильной сыворотки, полученной от здоровых субъектов, или в PBS до конечной концентрации 30 мкг/мл. После тщательного перемешивания образцы помещали на водяную баню с температурой 37°С на восемь дней (192 ч). Спустя восемь дней с помощью анализов ELISA проводили оценку способности связываться с человеческим IL-17 у образцов в сыворотке (А), образцов в PBS (В) и образцов моноклональных антител, хранившихся в замороженном виде (С) (см. фиг. 9). Для каждого образца моноклонального антитела проводили сравнение (А/С) изменения IL-17-связывающей способности. Как видно из данных в приведенной ниже таблице 9, четыре моноклональных антитела к IL-17 обладали хорошей стабильностью в сыворотке крови.
Пример 12
Оценка фармакокинетики (pK) in vivo в организме мышей
Для определения метаболических свойств моноклональных антител к IL-17 in vivo в организме мышей различные моноклональные антитела к IL-17 вводили в виде однократной дозы в хвостовую вену мышей с целью оценки pK. Для этого исследования были использованы четыре моноклональных антитела к IL-17 (подтип IgG4), а именно 9GA3, 9GC2, 9GC5 и иксекизумаб. Готовили растворы образцов моноклональных антител в буфере PBS (рН 7,0) с концентрацией 0,5 мг/мл. Доза введения составляла 5 мг/кг. Однократную дозу вводили в хвостовую вену. В каждую группу вошло восемь мышей BALB/c, половина из которых были самцы, а половина - самки. Забор образцов крови осуществляли через 4, 24, 48, 96, 168, 240 и 336 часов после введения (из ретроорбитального синуса), собранные образцы хранили при -70°С. После завершения сбора образцов определяли концентрации моноклональных антител.
Определение значений концентрации в крови проводили с помощью анализа количественной ELISA на иммобилизованном IL-17. В качестве стандартных образцов в количественной оценке использовали образцы четырех очищенных моноклональных антител к IL-17 (9GA3, 9GC2, 9GC5 и иксекизумаба), на основании полученных данных соответственно строили калибровочные кривые для использования в ходе количественной оценки значений концентрации моноклональных антител в крови. Тенденции изменения концентраций четырех моноклональных антител in vivo в организме мышей показаны на фиг. 10.
В соответствии с моделью реакции первого порядка строили график зависимости натуральных логарифмов концентрации антител (lnC) от времени (t), таким образом рассчитывая угловой коэффициент k. После этого по формуле t1/2=0,693/k рассчитывали значения периода полужизни (t1/2) четырех моноклональных антител к IL-17 у мышей. Результаты представлены в таблице 10.

| название | год | авторы | номер документа |
|---|---|---|---|
| Антитело против IL-4R и его применение | 2018 |
|
RU2758091C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| PD-1 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ. | 2016 |
|
RU2722451C1 |
| ГЕТЕРОДИМЕРНОЕ БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ TNFα/ПРОТИВ IL-17A, ПО СТРУКТУРЕ НАПОМИНАЮЩЕЕ ПРИРОДНОЕ АНТИТЕЛО, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2019 |
|
RU2781816C1 |
| СООТВЕТСТВУЮЩАЯ МИШЕНЬ ДЛЯ ЛЕЧЕНИЯ ФИБРОЗИРУЮЩИХ ЗАБОЛЕВАНИЙ И ЕЕ ПРИМЕНЕНИЕ | 2021 |
|
RU2830698C1 |
| АНТИТЕЛО ПРОТИВ ЛИГАНДА 1 ЗАПРОГРАММИРОВАННОЙ ГИБЕЛИ КЛЕТОК (PD-L1), ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2727914C2 |
| Мутантное полностью гуманизированное антитело против HER2 и кодирующий ген и их применение | 2015 |
|
RU2639531C1 |















Настоящее изобретение относится к области иммунологии. Предложены антитело и фрагмент антитела, способные к связыванию с человеческим IL-17A. Также рассмотрены фармацевтическая композиция и применение антитела или фрагмента антитела. Данное изобретение может найти дальнейшее применение в терапии различных заболеваний, опосредованных человеческим IL-17A. 3 н. и 7 з.п. ф-лы, 10 ил., 10 табл., 12 пр.
1. Моноклональное антитело или его антигенсвязывающий фрагмент, которое специфически связывается с человеческим IL-17A и содержит вариабельный участок тяжелой цепи, включающий участки HCDR1, HCDR2 и HCDR3, и вариабельный участок легкой цепи, включающий участки LCDR1, LCDR2 и LCDR3, причем
HCDR1 имеет последовательность GFTIDNY, HCDR2 имеет последовательность NQDGNE, HCDR3 имеет последовательность DYYDVISDYYIHYWYFDL или
HCDR1 имеет последовательность GMSMSDY, HCDR2 имеет последовательность NQDGDE, HCDR3 имеет последовательность DYYDLISDYYIHYWYFDL или
HCDR1 имеет последовательность GITMDDY, HCDR2 имеет последовательность NQDGNE, HCDR3 имеет последовательность DYYDLISDYYIHYWYFDL; и
LCDR1 имеет последовательность RASQNVHNRLT, LCDR2 имеет последовательность GASNLES и LCDR3 имеет последовательность QQYNGSPTT; и
где участки HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 определены согласно Chothia.
2. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 1, причем
вариабельный участок тяжелой цепи антитела имеет последовательность, указанную под SEQ ID NO: 24, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21; или
вариабельный участок тяжелой цепи имеет последовательность, указанную под SEQ ID NO: 25, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21; или
вариабельный участок тяжелой цепи имеет последовательность, указанную под SEQ ID NO: 26, а вариабельный участок легкой цепи имеет последовательность, указанную под SEQ ID NO: 21.
3. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1,2, причем антитело представляет собой интактное антитело, по существу интактное антитело, Fab фрагмент, F(ab')2 фрагмент или одноцепочечный Fv фрагмент.
4. Моноклональное антитело или его антигенсвязывающий фрагмент по п. 3, причем антитело представляет собой полностью человеческое антитело.
5. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-4, причем антитело дополнительно включает константный участок тяжелой цепи, выбранный из группы, состоящей из подтипов IgG1 и IgG4, и/или константный участок легкой цепи, выбранный из группы, состоящей из подтипов каппа и лямбда.
6. Моноклональное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-5, причем моноклональное антитело в концентрации менее 1 нМ способно ингибировать на 50% активность человеческого IL-17А в концентрации 2 нМ, где ингибирование активности измерено путем определения индуцированной человеческим IL-17A продукции IL-6 в дермальных фибробластах человека (HDFa).
7. Фармацевтическая композиция для лечения заболевания, опосредованного человеческим IL-17A, содержащая моноклональное антитело или его антигенсвязывающий фрагмент в эффективном количестве по любому из пп. 1-6.
8. Применение моноклонального антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6 в производстве лекарственного препарата для лечения заболевания, опосредованного человеческим IL-17A.
9. Применение по п. 8, причем заболевание представляет собой аутоиммунное заболевание.
10. Применение по п. 8, причем заболевание представляет собой псориаз, ревматоидный артрит или анкилозирующий спондилит.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К РЕЦЕПТОРУ ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2005 |
|
RU2402569C2 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |

