ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
Настоящая заявка утверждает приоритет заявки на патент Китая №201810901518.0, поданной 9 августа 2018 г., которая включена в данное описание посредством ссылки во всей своей полноте.
ОБЛАСТЬ ТЕХНИКИ
Настоящая заявка в целом относится к области лекарственных средств, полученных методами генетической инженерии и на основе антител. В частности, настоящая заявка относится к биспецифическим антителам против вируса бешенства и их применению.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Бешенство представляет собой острое инфекционное заболевание, вызываемое вирусом бешенства, и поражает как людей, так и животных. Вирус бешенства передается главным образом среди собак, волков и кошек, но и другие млекопитающие, такие как еноты, скунсы, летучие мыши и лисы, также являются общеизвестными хозяевами. Животные передают данный вирус, кусая друг друга, и часто люди заражаются от укусов больных животных. На сегодняшний день не существует никакого эффективного лечения бешенства. Смертность пациентов от бешенства составляет почти 100%. Обычно пациенты умирают от дыхательной недостаточности или недостаточности кровообращения в пределах 3-6 дней. По оценкам от этого заболевания во всем мире каждый год умирают более 70000 человек и миллионы нуждаются в постконтактном лечении.
Вирус бешенства представляет собой имеющий форму пули оболочечный вирус, содержащий одноцепочечную РНК, и принадлежит семейству Rhabdoviridae и роду лиссавирусов. В геноме вируса бешенства закодированы пять вирусных белков, т.е. РНК-зависимая РНК-полимераза (L), нуклеопротеин (N), фосфорилированный белок (Р), матриксный белок (М), расположенный с внутренней стороны оболочки вирусного белка, и гликопротеин наружной поверхности (G). Гликопротеин (G-белок) вируса бешенства связывается с ацетилхолином, что определяет нейрофагоцитоз вируса бешенства. Белок G (62-67 кДа) представляет собой гликопротеин I типа, состоящий из 505 аминокислот, образует выпуклость, покрывающую наружную поверхность оболочки вирусной частицы, и показано, что он индуцирует нейтрализующие вирус антитела. Белок G имеет по меньшей мере пять нейтрализующих эпитопов. Эпитоп II представляет собой прерывистый пространственный эпитоп, включающий в себя аминокислотные остатки 34-42 и аминокислотные остатки 198-200. Эпитоп III локализован в положениях 330-338 и представляет собой линейный эпитоп. Примерно 97% обнаруженных антител распознают эпитоп II и эпитоп III. Нейтрализующее вирус бешенства антитело CR4098 связывается с эпитопом III. Некоторые из антител распознают эпитоп I и эпитоп IV. Нейтрализующее вирус бешенства антитело CR57 распознает линейный эпитоп I, т.е. положения 218-240, в котором основным связывающим доменом является последовательность KLCGVL в положении 226-231. Эпитоп IV содержит остатки 251 и 264. Еще один эпитоп представляет собой микроэпитоп, который не перекрывается с эпитопом III, отделяется от эпитопа III тремя аминокислотными остатками и имеет только два аминокислотных остатка 342-343.
Для предупреждения и лечения инфекции вирусом бешенства Всемирная организация здравоохранения (ВОЗ) рекомендует, чтобы, в случае укусов диких животных, классифицирующихся как воздействие III класса и воздействие выше II класса, субъекты получали как активную, так и пассивную иммунотерапию для получения быстрой защиты. В настоящее время разработка вакцин против вируса бешенства для активного лечения является относительно отработанной, и многочисленные вакцины против вируса бешенства поставляются на рынок внутри страны и за рубеж для активного предупреждения заражения вирусом бешенства. При этом лекарственными средствами, используемыми для пассивного лечения после контакта с вирусом бешенства, являются главным образом человеческий антирабический иммуноглобулин (HRIG) и лошадиный антирабический иммуноглобулин (ERIG). Поскольку для людей ERIG является гетерологичным белком, он иногда вызывает серьезные побочные эффекты. HRIG характеризуется высокой стоимостью, поставляется в ограниченном количестве, и его применение связано с риском возможного заражения патогенами, передающимися вместе с кровью. Существует клиническая потребность в разработке новых лекарственных средств для пассивной терапии инфекции вирусом бешенства.
В 1989 г. Schumacherl и др. получили ряд мышиных моноклональных антител к гликопротеину и нуклеопротеину вируса бешенства. Эксперименты по определению защитного действия с использованием такой терапии смесью моноклональных антител на мышах и хомяках показали, что этот способ, после пассивной иммунизации, не только демонстрировал способность полностью противостоять атаке вируса бешенства в летальной дозе, но также оказывал постконтактное защитное действие. В 1990 г. Bernhard Dietzschold и др. получили ряд человеческих моноклональных антител против вируса бешенства, используя методы слияния клеток, в которых моноклональное антитело 57 (МаЬ57) показало высокую аффинность к гликопротеину вируса бешенства, значительно нейтрализовало вирус бешенства и обеспечивало защиту мышей от атаки вируса бешенства. После получения гена антител против вируса бешенства появилась возможность приготовления человеческих моноклональных антител против вируса бешенства с использованием биореактора. В 2005 г. Goudsmit и др. и исследовательская группа под руководством Bakker опубликовали сообщение о создании двух человеческих моноклональных антител CR57 и CR4098 к G-белку вируса бешенства. Эти два моноклональных антитела смешивали непосредственно при использовании и сравнивали с HRIG. Результат показал, что действие смеси этих моноклональных антител было сопоставимо с действием HRIG при постконтактной профилактике и обеспечивало хорошую перекрестную реактивность со многими штаммами вируса бешенства, демонстрируя практическую доступность рекомбинантных человеческих моноклональных антител против вируса бешенства или даже способность заменять RIG. В настоящее время лекарственное средство CL184 на основе этих двух моноклональных антител проходит клиническое исследование, и во время клинических испытаний не обнаружено каких-либо побочных эффектов. Данное лекарственное средство является безопасным и эффективным и может заменить в клинике RIG в случае постконтактной профилактики. Однако, CL184, проходящее сейчас клинические исследования, является препаратом на основе смеси двух моноклональных антител, и его получение является относительно высокозатратным.
В данной области техники желательна разработка и применение новых биспецифических антител к G-белку вируса бешенства.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно первому аспекту настоящей заявки предложено биспецифическое антитело, содержащее два антиген-связывающих фрагмента, которые связываются с разными эпитопами G-белка вируса бешенства, и данное биспецифическое антитело обладает активностью нейтрализации вируса бешенства.
В некоторых воплощениях один антиген-связывающий фрагмент в биспецифическом антителе связывается с эпитопом I G-белка вируса бешенства, а другой антиген-связывающий фрагмент связывается с эпитопом III G-белка вируса бешенства.
В некоторых воплощениях антиген-связывающий фрагмент, который связывается с эпитопом I G-белка вируса бешенства, содержит:
HCDR1 (определяющий комплементарность участок 1 тяжелой цепи), имеющий аминокислотную последовательность RYTIN, HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG, HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP, LCDR1 (определяющий комплементарность участок 1 легкой цепи), имеющий аминокислотную последовательность TGTSSDIGAYDYVS, LCDR2, имеющий аминокислотную последовательность DATKRPS, и LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV; или
HCDR1, имеющий аминокислотную последовательность RYSIN, HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG, HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP, LCDR1, имеющий аминокислотную последовательность TGTSSDIDGYDFVS, LCDR2, имеющий аминокислотную последовательность DATKRPS, и LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV; или
HCDR1, имеющий аминокислотную последовательность GYTIN, HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG, HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP, LCDR1, имеющий аминокислотную последовательность TGTSSDLGGYDFVS, LCDR2, имеющий аминокислотную последовательность DATKRPS, и LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV;
при этом аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях антиген-связывающий фрагмент, который связывается с эпитопом III G-белка вируса бешенства, содержит:
HCDR1, имеющий аминокислотную последовательность SYGMH, HCDR2, имеющий аминокислотную последовательность TISYDGSIKDYADSVKG, HCDR3, имеющий аминокислотную последовательность GDRTGNLDY, LCDR1, имеющий аминокислотную последовательность RASQNIRNALN, LCDR2, имеющий аминокислотную последовательность DASTRQS, и LCDR3, имеющий аминокислотную последовательность QQNSEFPPT;
при этом аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 24, а аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 25; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 26, а аминокислотная последовательность вариабельной области легкой цепи является такой, как показано в SEQ ID NO: 27; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 28, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 29.
В некоторых воплощениях аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 3.
В некоторых воплощениях аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 24, а аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 25; и аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи является такой, как показано в SEQ ID NO: 3; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 26, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 27; и аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, показана в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи является такой, как показано в SEQ ID NO: 3; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 28, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 29; и аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, показана в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 3.
В некоторых воплощениях формы двух антиген-связывающих фрагментов независимо выбраны из одноцепочечного антитела (scFv) или Fab-фрагмента.
В некоторых воплощениях антиген-связывающий фрагмент, который связывается с эпитопом I G-белка вируса бешенства, представляет собой одноцепочечное антитело (scFv), а антиген-связывающий фрагмент, который связывается с эпитопом III G-белка вируса бешенства, представляет собой Fab-фрагмент. В некоторых воплощениях биспецифическое антитело содержит аминокислотную последовательность, приведенную в одной из SEQ ID NO: 32, 33 и 34. В некоторых воплощениях биспецифическое антитело содержит аминокислотные последовательности, приведенные в SEQ ID NO: 30 и SEQ ID NO: 31.
Согласно второму аспекту в настоящей заявке предложена фармацевтическая композиция, содержащая биспецифическое антитело по первому аспекту и фармацевтически приемлемый эксципиент, разбавитель или носитель.
В некоторых воплощениях фармацевтическую композицию используют для предупреждения или лечения бешенства.
Согласно третьему аспекту в настоящей заявке предложено применение биспецифического антитела по первому аспекту в изготовлении лекарственного средства для предупреждения или лечения бешенства.
Согласно четвертому аспекту в настоящей заявке предложен способ предупреждения или лечения бешенства, включающий введение субъекту, нуждающемуся в этом, биспецифического антитела по первому аспекту или фармацевтической композиции по второму аспекту.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 показан результат иммуноферментного твердофазного анализа (ELISA) способности полностью человеческих моноклональных антител C34m и C11m связываться с инактивированным вирусом бешенства.
На Фиг. 2 показан результат ELISA-анализа ингибирования полностью человеческими моноклональными антителами C34m и C11m связывания конструкции фаг-C11m с инактивированным вирусом бешенства.
На Фиг. 3 показан результат ELISA-анализа ингибирования полностью человеческими моноклональными антителами C34m и C11m связывания конструкции фаг-C34m с инактивированным вирусом бешенства.
На Фиг. 4 показан результат ELISA-анализа ингибирования полностью человеческими моноклональными антителами C11m и C34m связывания химерного антитела CR57-mIgG2a (мышиный иммуноглобулин 2а) с инактивированным вирусом бешенства.
На Фиг. 5 показан результат ELISA-анализа ингибирования полностью человеческими моноклональными антителами C11m и C34m связывания химерного антитела CR4098-mIgG2a с инактивированным вирусом бешенства.
На Фиг. 6 показан результат ELISA-анализа способности слитого белка C11m-scFv-Fc на основе одноцепочечного антитела и Fc, трех слитых на основе одноцепочечного антитела и Fc, содержащих мутантные формы C11m-scFv, и полностью человеческого моноклонального антитела C34m связываться с инактивированным вирусом бешенства.
На Фиг. 7 показан результат ELISA-анализа ингибирования связывания химерного антитела C34m-mIgG2a с инактивированным вирусом бешенства под действием слитого белка C11m-scFv-Fc на основе одноцепочечного антитела и Fc и трех слитых белков на основе одноцепочечного антитела и Fc, содержащих мутантные формы C11m-scFv.
На Фиг. 8 показан результат ELISA-анализа ингибирования связывания химерного антитела C11m-mIgG2a с инактивированным вирусом бешенства под действием слитого белка C11m-scFv-Fc на основе одноцепочечного антитела и Fc и трех слитых белков на основе одноцепочечного антитела и Fc, содержащих мутантные формы C11m-scFv.
На Фиг. 9 показан результат ELISA-анализа способности отдельных антител к G-белку вируса бешенства связываться с инактивированным вирусом бешенства.
На Фиг. 10 показан результат ELISA-анализа ингибирования связывания химерного антитела C34m-mIgG2a с инактивированным вирусом бешенства под действием биспецифического антитела S2E3-scFv-FcH+C34m-IgG1K, слитого белка S2E3-scFv-Fc на основе одноцепочечного антитела и Fc и полностью человеческого моноклонального антитела C34m.
На Фиг. 11 показан результат ELISA-анализа ингибирования связывания химерного белка S2E3-scFv-mFc с инактивированным вирусом бешенства под действием биспецифического антитела S2E3-scFv-FcH+C34m-IgG1K, слитого белка S2E3-scFv-Fc на основе одноцепочечного антитела и Fc и полностью человеческого моноклонального антитела C34m.
На ФИГ. 12 показан результат ELISA-анализа способности антител к G-белку вируса бешенства C34m и S2E3-scFv-Fc связываться с мутантными формами G-белка вируса бешенства штамма CVS-11. На ФИГ. 12А показан результат ELISA-анализа способности C34m и S2E3-scFv-Fc связываться со слитым белком GCVS11-CCD-His. На ФИГ. 12В показан результат ELISA-анализа способности C34m и S2E3-scFv-Fc связываться со слитым белком GCVS11-G229E-CCD-His. На ФИГ. 12С показан результат ELISA-анализа способности C34m и S2E3-scFv-Fc связываться со слитым белком GCVS11-I338T-CCD-His.
ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 показывает аминокислотную последовательность вариабельной области тяжелой цепи C34mVH антитела C34m.
SEQ ID NO: 2 показывает аминокислотную последовательность вариабельной области тяжелой цепи C11mVH антитела C11m.
SEQ ID NO: 3 показывает аминокислотную последовательность вариабельной области легкой цепи C34mVK антитела C34m.
SEQ ID NO: 4 показывает аминокислотную последовательность вариабельной области легкой цепи C11mVL антитела C11m.
SEQ ID NO: 5 показывает аминокислотную последовательность константной области тяжелой цепи иммуноглобулина человека (homo sapiens) CH-IgG1.
SEQ ID NO: 6 показывает аминокислотную последовательность константной области легкой цепи иммуноглобулина человека (homo sapiens) CK.
SEQ ID NO: 7 показывает аминокислотную последовательность константной области легкой цепи иммуноглобулина человека (homo sapiens) CL.
SEQ ID NO: 8 показывает аминокислотную последовательность одноцепочечного антитела C34m-scFv.
SEQ ID NO: 9 показывает аминокислотную последовательность одноцепочечного антитела C11m-scFv.
SEQ ID NO: 10 показывает аминокислотную последовательность Fc-сегмента человеческого (homo sapiens) IgG1.
SEQ ID NO: 11 показывает аминокислотную последовательность Fc-сегмента мышиного (mus musculus) IgG2a.
SEQ ID NO: 12 и SEQ ID NO: 15 показывают аминокислотные последовательности вариабельной области тяжелой цепи CR57VH и аминокислотные последовательности вариабельной области легкой цепи CR57VL моноклонального антитела CR57 к эпитопу I G-белка вируса бешенства, соответственно.
SEQ ID NO: 13 и SEQ ID NO: 16 показывают аминокислотные последовательности вариабельной области тяжелой цепи CR4098VH и вариабельной области легкой цепи CR4098VK моноклонального антитела CR4098 к эпитопу III G-белка вируса бешенства, соответственно.
SEQ ID NO: 14 показывает аминокислотную последовательность константной области тяжелой цепи мышиного (mus musculus) IgG2a.
SEQ ID NO: 17 показывает аминокислотную последовательность константной области легкой цепи мыши (mus musculus) mCL.
SEQ ID NO: 18 показывает аминокислотную последовательность константной области легкой цепи мыши (mus musculus) mCK.
SEQ ID NO: 19 показывает аминокислотную последовательность Fc-сегмента, содержащего мутацию, приводящую к образованию выступа (knob) (FcK).
SEQ ID NO: 20 показывает аминокислотную последовательность Fc-сегмента, содержащего мутацию, приводящую к образованию впадины (hole) (FcH).
SEQ ID NO: 21 показывает аминокислотную последовательность мутантной формы S1C10-scFv одноцепочечного антитела C11m-scFv.
SEQ ID NO: 22 показывает аминокислотную последовательность мутантной формы S2Al-scFv одноцепочечного антитела C11m-scFv.
SEQ ID NO: 23 показывает аминокислотную последовательность мутантной формы S2E3-scFv одноцепочечного антитела C11m-scFv.
SEQ ID NO: 24 и 25 показывают аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи мутантной формы S1C10-scFv одноцепочечного антитела C11m-scFv, соответственно.
SEQ ID NO: 26 и 27 показывают аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи мутантной формы S2Al-scFv одноцепочечного антитела C11m-scFv, соответственно.
SEQ ID NO: 28 и 29 показывают аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи мутантной формы S2E3-scFv одноцепочечного антитела C11m-scFv, соответственно.
SEQ ID NO: 30 показывает аминокислотную последовательность C34mVK-CK.
SEQ ID NO: 31 показывает аминокислотную последовательность C34mVH-IgG1K.
SEQ ID NO: 32 показывает аминокислотную последовательность слитого белка S2E3-scFv-FcH на основе одноцепочечного антитела и Fc.
SEQ ID NO: 33 показывает аминокислотную последовательность слитого белка S1C10-scFv-FcH на основе одноцепочечного антитела и Fc.
SEQ ID NO: 34 показывает аминокислотную последовательность слитого белка S2Al-scFv-FcH на основе одноцепочечного антительного и Fc.
SEQ ID NO: 35 показывает аминокислотную последовательность G-белка штамма вируса бешенства дикого типа CVS-11 (GCVS11).
SEQ ID NO: 36 показывает аминокислотную последовательность эпитопа I мутантной формы G-белка (GCVS11-G229E) штамма вируса бешенства CVS-11 (GCVS11).
SEQ ID NO: 37 показывает аминокислотную последовательность эпитопа III мутантной формы G-белка (GCVS11-I338T) штамма вируса бешенства CVS-11 (GCVS11).
SEQ ID NO: 38 показывает аминокислотную последовательность тримерного домена CCD белка 1А коронавируса человека (homo sapiens).
SEQ ID NO: 39 показывает аминокислотную последовательность слитого белка GCVS11-CCD-His.
SEQ ID NO: 40 показывает аминокислотную последовательность слитого белка GCVS11 -G229E-CCD-His.
SEQ ID NO: 41 показывает аминокислотную последовательность слитого белка GCVS11 -I338T-CCD-His.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящей заявки разработали новые биспецифические антитела против вируса бешенства с использованием методов конструирования антител. Согласно различным аспектам настоящей заявки предложены новые биспецифические антитела против вируса бешенства, полинуклеотиды, кодирующие эти биспецифические антитела, векторы, содержащие данные полинуклеотиды, клетки хозяина, содержащие эти полинуклеотиды или векторы, способы получения и очистки биспецифических антител и медицинское и биологическое применение биспецифических антител. На основании последовательностей вариабельных областей биспецифических антител, приведенных в данном описании, могут быть сконструированы полноразмерные молекулы биспецифических антител для клинического применения в качестве лекарственного средства для предупреждения или лечения бешенства.
Если не указано иное, данное изобретение может быть реализовано на практике с использованием традиционных для данной области техники методов молекулярной биологии, микробиологии, клеточной биологии, биохимии и иммунологии.
Если не указано иное, термины, использованные в настоящей заявке, имеют значения, обычно понимаемые специалистами в данной области техники.
ОПРЕДЕЛЕНИЯ
Использованный в данном описании термин «антитело» относится к молекуле иммуноглобулина, которая способна специфически связываться с мишенью посредством по меньшей мере одного антиген-распознающего сайта, расположенного в вариабельной области молекулы иммуноглобулина. Мишени включают, но не ограничиваются этим, углеводы, полинуклеотиды, липиды и полипептиды. Использованный в данном описании термин «антитело» включает в себя не только интактное (т.е. полноразмерное) антитело, но также и его антиген-связывающий фрагмент (например, Fab, Fab', F(ab')2, Fv), его вариант, слитый белок, содержащий части антитела, гуманизированное антитело, химерное антитело, диатело, линейное антитело, одноцепочечное антитело, мультиспецифическое антитело (например, биспецифическое антитело) и любые другие модифицированные форматы молекулы иммуноглобулина, содержащие желаемый специфический антиген-распознающий сайт, включая гликозилированный вариант антитела, вариант аминокислотной последовательности антитела и ковалентно модифицированное антитело.
Обычно интактное или полноразмерное антитело содержит две тяжелых цепи и две легких цепи. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (VH) и первую, вторую и третью константные области (CH1, СН2 и СН3). Каждая легкая цепь содержит вариабельную область легкой цепи (VL) и константную область (CL). Полноразмерное антитело может быть любого типа, как например, антитело IgD, IgE, IgG, IgA или IgM типа (или их подтипов), но не обязательно принадлежит какому-либо конкретному типу. Иммуноглобулины могут быть отнесены к разным типам в зависимости от аминокислотных последовательностей константных доменов своих тяжелых цепей. Как правило, иммуноглобулины принадлежат к пяти основным типам, т.е. IgA, IgD, IgE, IgG и IgM, и некоторые из этих типов могут быть дополнительно классифицированы на подтипы (изотипы), такие как IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие отдельным типам иммуноглобулинов, обозначаются как α, δ, ε, γ и μ, соответственно. Субъединичные структуры и пространственные структуры иммуноглобулинов разных типов хорошо известны.
Использованный в данном описании термин «биспецифическое антитело» означает антитело, обладающее способностью связываться с двумя разными антигенами. Например, биспецифическое антитело может состоять из двух Fc фрагментов и двух слитых с ними антиген-связывающих частей, соответственно.
Использованный в данном описании термин «антиген-связывающий фрагмент» или «антиген-связывающая часть» можно использовать взаимозаменяемо, и они относятся к части или участку молекулы интактного антитела, отвечающей(ему) за связывание с антигеном. Антиген-связывающий домен может содержать вариабельную область тяжелой цепи (VH), вариабельную область легкой цепи (VL) или обе области. Каждая из VH и VL обычно содержит три определяющих комплементарность участка, т.е. CDR1, CDR2 и CDR3.
Специалистам в данной области техники хорошо известно, что определяющие комплементарность участки (CDR, обычно включающие CDR1, CDR2 и CDR3) представляют собой участки вариабельной области, которые наиболее сильно влияют на аффинность и специфичность антитела. Последовательности CDR в VH или VL имеют два общепризнанных определения, т.е. определение согласно Kabat и определение согласно Chothia (см. например, Kabat, "Sequences of Proteins of Immunological Interest", National Institutes of Health, Bethesda, Md.(1991); Al-Lazikani et at, J. Mol. Biol., 273: 927-948 (1997); и Martin et al, Proc. Natl. Acad. Sci. USA, 86: 9268-9272 (1989)). Что касается последовательностей вариабельных областей заданного антитела, то последовательности CDR участков в VH и VL могут быть определены согласно определению по Kabat или определению по Chothia. В некоторых воплощениях настоящей заявки последовательности CDR определены согласно Kabat.
Что касается последовательностей вариабельных областей заданного антитела, то последовательности CDR участков в последовательностях вариабельных областей могут быть проанализированы рядом путей, например, с использованием он-лайн программного обеспечения Abysis (http://www.abysis.org/).http://www.abysis.org/
Что касается традиционных антител, то примеры антиген-связывающего фрагмента включают, но не ограничиваются этим, (1) Fab-фрагмент, который может представлять собой моновалентный фрагмент, имеющий цепь VL-CL и цепь VH-CH1; (2) F(ab')2 фрагмент, который может представлять собой двухвалентный фрагмент, имеющий два Fab'-фрагмента, соединенных дисульфидным мостиком в шарнирной области (т.е. димер Fab'); (3) Fv фрагмент, имеющий VL- и VH-домены в одном плече антитела; (4) одноцепочечный Fv (scFv), который может представлять собой одну полипептидную цепь, состоящую из VH-домена и VL-домена, соединенных посредством полипептидного линкера; и (5) (scFv)2, который может содержать два VH-домена, соединенных посредством пептидного линкера, и два VL-домена, которые объединены с двумя VH-доменами через дисульфидный мостик.
В конструкции биспецифического антитела «антиген-связывающая часть» включает, но не ограничивается этим, Fab-фрагмент или одноцепочечное антитело (scFv).
Использованный в данном описании термин «одноцепочечный вариабельный фрагмент (scFv)» относится к антителу одноцепочечной структуры, содержащей полипептидную цепь, состоящую из вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), которую как правило конструируют с использованием генно-инженерных методов. В этой конструкции гибкий линкер обычно располагается между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, в результате чего вариабельная область тяжелой цепи и вариабельная область легкой цепи могут сворачиваться в надлежащую конформацию, обладающую способностью связываться с антигеном.
Использованный в данном описании термин «Fab-фрагмент (антиген-связывающий фрагмент)», «Fab-часть» или тому подобное относится к фрагменту антитела, способному связываться с антигеном, образующемуся после обработки интактного антитела папаином, содержащему интактную легкую цепь (VL-CL), вариабельную область тяжелой цепи и фрагмент CH1 (VH-CH1).
Использованный в данном описании термин «моноклональное антитело» относится к антителу из по существу гомогенной популяции антител, т.е. антитела, составляющие эту популяцию, являются абсолютно идентичными за исключением наличия природных мутаций, которые могут присутствовать в небольшом количестве в отдельных антителах. Моноклональные антитела, описанные в данной заявке, в частности, включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующей последовательности в антителе, происходящем из конкретного вида или принадлежащем конкретному типу или подтипу антител, в то время как остальная часть тяжелой и/или легкой цепи идентична или гомологична соответствующей последовательности в антителе, происходящем из другого вида или принадлежащем другому типу или подтипу антител, и также включают фрагменты таких антител до тех пор, пока они проявляют желаемую биологическую активность (см. патент США №4816567; и Morrison et al, Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984)).
Использованный в данном описании термин «эпитоп», также называемый как антигенная детерминанта (AD), относится к конкретной химической группе в молекуле антигена, которая определяет специфичность этого антигена. Антиген связывается со своим антигенным рецептором на поверхности соответствующего лимфоцита через антигенный эпитоп, активируя тем самым лимфоцит и вызывая иммунный ответ. Антиген также проявляет свои иммунные эффекты посредством специфического связывания эпитопа с соответствующим антителом или сенсибилизированным лимфоцитом. Природа, количество и пространственная конфигурация антигенного эпитопа определяет специфичность антигена.
Использованный в данном описании термин «специфическое связывание» относится к реакции неслучайного связывания между двумя молекулами, например, к связыванию антитела с антигенным эпитопом.
В нуклеиновокислотных последовательностях, описанных в данной заявке, используют обозначения оснований в вырожденных положениях (помимо традиционных оснований А, Т, С и G), и они имеют те же значения, которые обычно понимаются специалистами в данной области техники. Например, R означает А или G; Y означает С или Т, М означает А или С; K означает G или Т; S означает С или G; W означает А или Т; Н означает А, или С, или Т; В означает С, или G, или Т; V означает А, или С, или G; D означает А, или G, или Т; N означает А, или С, или G, или Т.
Согласно первому аспекту настоящей заявки предложено биспецифическое антитело, содержащее два антиген-связывающих фрагмента, которые связываются с разными эпитопами G-белка вируса бешенства, при этом данное биспецифическое антитело обладает активностью нейтрализации вируса бешенства.
В некоторых воплощениях один антиген-связывающий фрагмент в биспецифическом антителе связывается с эпитопом I G-белка вируса бешенства, а другой антиген-связывающий фрагмент связывается с эпитопом III G-белка вируса бешенства.
В некоторых воплощениях антиген-связывающий фрагмент, который связывается с эпитопом I G-белка вируса бешенства, содержит:
HCDR1, имеющий аминокислотную последовательность RYTIN, HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG, HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP, LCDR1, имеющий аминокислотную последовательность TGTSSDIGAYDYVS, LCDR2, имеющий аминокислотную последовательность DATKRPS, LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV; или
HCDR1, имеющий аминокислотную последовательность RYSIN, HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG, HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP, LCDR1, имеющий аминокислотную последовательность TGTSSDIDGYDFVS, LCDR2, имеющий аминокислотную последовательность DATKRPS, LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV; или
HCDR1, имеющий аминокислотную последовательность GYTIN, HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG, HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP, LCDR1, имеющий аминокислотную последовательность TGTSSDLGGYDFVS, LCDR2, имеющий аминокислотную последовательность DATKRPS, LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV;
при этом аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях антиген-связывающий фрагмент, который связывается с эпитопом III G-белка вируса бешенства содержит:
HCDR1, имеющий аминокислотную последовательность SYGMH, HCDR2, имеющий аминокислотную последовательность TISYDGSIKDYADSVKG, HCDR3, имеющий аминокислотную последовательность GDRTGNLDY, LCDR1, имеющий аминокислотную последовательность RASQNIRNALN, LCDR2, имеющий аминокислотную последовательность DASTRQS, LCDR3, имеющий аминокислотную последовательность QQNSEFPPT;
при этом аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
В некоторых воплощениях аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 24, а аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 25; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 26, а аминокислотная последовательность вариабельной области легкой цепи является такой, как показано в SEQ ID NO: 27; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 28, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 29.
В некоторых воплощениях аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства является такой, как представлено в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 3.
В некоторых воплощениях аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 24, а аминокислотная последовательность вариабельной области легкой цепи является такой, как представлено в SEQ ID NO: 25; и аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, является такой, как представлено в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи является такой, как показано в SEQ ID NO: 3; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 26, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 27; и аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, показана в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи является такой, как показано в SEQ ID NO: 3; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 28, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 29; и аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, показана в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи показана в SEQ ID NO: 3.
В некоторых воплощениях формы двух антиген-связывающих фрагментов независимо выбраны из одноцепочечного антитела (scFv) или Fab-фрагмента.
В некоторых воплощениях антиген-связывающий фрагмент, который связывается с эпитопом I G-белка вируса бешенства, представляет собой одноцепочечное антитело (scFv), а антиген-связывающий фрагмент, который связывается с эпитопом III G-белка вируса бешенства, представляет собой Fab-фрагмент. В некоторых воплощениях биспецифическое антитело содержит аминокислотную последовательность, приведенную в одной из SEQ ID NO: 32, 33 и 34. В некоторых воплощениях биспецифическое антитело содержит аминокислотные последовательности, приведенные в SEQ ID NO: 30 и SEQ ID NO: 31.
Согласно второму аспекту настоящей заявки предложена фармацевтическая композиция, содержащая биспецифическое антитело по первому аспекту и фармацевтически приемлемый эксципиент, разбавитель или носитель.
В некоторых воплощениях фармацевтическая композиция предназначена для применения в предупреждении или лечении бешенства.
В некоторых воплощениях фармацевтическая композиция может дополнительно содержать одно или более чем одно из следующего: смазывающее вещество, такое как тальк, стеарат магния и минеральное масло; увлажняющий агент; эмульгатор; суспендирующий агент; консервант, такой как бензойная кислота, сорбиновая кислота и пропионат кальция; подсластитель и/или корригент.
В некоторых воплощениях упомянутая в данном описании фармацевтическая композиция может быть приготовлена в виде таблетки, пилюли, порошка, пастилки, эликсира, суспензии, эмульсии, раствора, сиропа, суппозитория или капсулы.
В некоторых воплощениях для доставки фармацевтической композиции по настоящей заявке может быть использован любой физиологически приемлемый путь введения, включая, но не ограничиваясь этим, пероральное введение, парентеральное введение, назальное введение, ректальное введение, внутрибрюшинное введение, внутрисосудистую инъекцию, подкожное введение, трансдермальное введение или ингаляционное введение.
В некоторых воплощениях фармацевтическая композиция для терапевтического применения может быть приготовлена для хранения в форме лиофилизированной композиции или в форме водного раствора путем смешивания агента желаемой степени чистоты с фармацевтически приемлемым носителем или эксципиентом там, где это целесообразно.
Согласно третьему аспекту настоящей заявки предложено применение биспецифического антитела по первому аспекту в изготовлении лекарственного средства для предупреждения или лечения бешенства.
Согласно четвертому аспекту настоящей заявки предложен способ предупреждения или лечения бешенства, включающий введение субъекту, нуждающемуся в этом, биспецифического антитела по первому аспекту или фармацевтической композиции по второму аспекту.
Согласно другим аспектам настоящей заявки предложена молекула нуклеиновой кислоты, кодирующая биспецифическое антитело по первому аспекту. В некоторых воплощениях молекула нуклеиновой кислоты функционально связана с регуляторной последовательностью, которая может распознаваться клеткой хозяина, трансформированной вектором.
В настоящей заявке также предложен вектор, содержащий молекулы выделенной нуклеиновой кислоты, кодирующей биспецифическое антитело по настоящей заявке, и клетка-хозяин, содержащая такую молекулу нуклеиновой кислоты или такой вектор.
Согласно другим аспектам настоящей заявки предложен способ получения биспецифического антитела по настоящей заявке. В некоторых воплощениях способ получения биспецифического антитела включает культивирование клетки хозяина для обеспечения экспрессии нуклеиновой кислоты. В некоторых воплощениях способ получения биспецифического антитела дополнительно включает извлечение биспецифического антитела из культуральной среды, содержащей клетку хозяина.
Следует понимать, что приведенное выше подробное описание предназначено только для того, чтобы предоставить специалистам в данной области техники возможность лучшего понимания настоящей заявки, и не предназначено для того, чтобы каким-либо образом являться причиной ограничений. Специалистами в данной области техники могут быть выполнены различные модификации и вариации к описанным воплощениям.
Следующие далее примеры приведены только в иллюстративных целях и не предназначены для ограничения объема настоящей заявки.
Примеры
Пример 1. Получение и проверка моноклональных антител к G-белку вируса бешенства
Авторы настоящей заявки использовали инактивированный вирус бешенства в качестве антигена для скрининга моноклональных антител и идентифицировали два человеческих моноклональных антитела с активностью нейтрализации, которым были присвоены названия C11m и C34m, соответственно. Затем авторы изобретения выполнили анализ последовательностей вариабельных областей тяжелой и легкой цепей двух моноклональных антител C11m и C34m.
Сначала получали полностью человеческие моноклональные антитела C34m и C11m к G-белку вируса бешенства, соответствующие последовательностям вариабельных областей для C11m и C34m, соответственно. Конкретно, гены, кодирующие вариабельную область тяжелой цепи антитела C34mVH (SEQ ID NO: 1) и вариабельную область легкой цепи C34mVK (SEQ ID NO: 3), клонировали в эукариотический экспрессирующий вектор (такой как pcDNA3.1, Invitrogen, Inc.), несущий гены, кодирующие константную область тяжелой цепи иммуноглобулина человека CH-IgG1 (SEQ ID NO: 5) и константную область легкой цепи CK (SEQ ID NO: 6), соответственно, получая тем самым экспрессирующий вектор для рекомбинантного антитела C34m. Помимо этого, гены, кодирующие вариабельную область тяжелой цепи антитела C11mVH (SEQ ID NO: 2) и вариабельную область легкой цепи C11mVL (SEQ ID NO: 4), клонировали в эукариотический экспрессирующий вектор (такой как pcDNA3.1, Invitrogen, Inc.), несущий гены, кодирующие константную область тяжелой цепи иммуноглобулина человека CH-IgG1 (SEQ ID NO: 5) и константную область легкой цепи CL (SEQ ID NO: 7), соответственно, получая тем самым экспрессирующий вектор для рекомбинантного антитела C11m. Полученные экспрессирующий вектор для рекомбинантного антитела C34m и экспрессирующий вектор для рекомбинантного антитела C11m использовали для трансфекции клеток почки эмбриона человека (HEK)293 (например, клеток HEK293F, Invitrogen, Inc.) с применением липосом (например, 293 fectin, Invitrogen, Inc.) или других катионных реагентов для трансфекции (например, полиэтиленимин (PEI)), соответственно. Клетки культивировали в суспензии в бессывороточных средах в течение 3-5 суток. Затем культуральные супернатанты собирали центрифугированием.
Кроме того, получали слитый белок на основе одноцепочечного антитела и Fc (scFv-Fc) к G-белку вируса бешенства. Конкретно, добавляли гибкий пептидный линкер GGGGGSGGGGSGGGGS (SEQ ID NO: 58) между вариабельными областями тяжелой и легкой цепей моноклональных антител C34m и C11m, соответственно, конструируя тем самым одноцепочечные антитела C34m-scFv (SEQ ID NO: 8) и C11m-scFv (SEQ ID NO: 9) в форме VH-линкер-VK. Гены, кодирующие одноцепочечные антитела C34m-scFv и C11m-scFv, затем клонируют в эукариотические экспрессирующие векторы (такие как pcDNA3.1, Invitrogen, Inc.), несущие гены, кодирующие Fc-сегмент человеческого IgG1 (Fc, SEQ ID NO: 10) или Fc-сегмент мышиного IgG2a (mFc, SEQ ID NO: 11), соответственно, получая тем самым экспрессирующий вектор для рекомбинантного антитела на основе одноцепочечного антитела C34m-scFv-Fc/mFc и экспрессирующий вектор для рекомбинантного антитела на основе одноцепочечного антитела C11m-scFv-Fc/mFc. Полученные экспрессирующий вектор для рекомбинантного антитела на основе одноцепочечного антитела C34m-scFv-Fc/mFc и экспрессирующий вектор для рекомбинантного антитела на основе одноцепочечного антитела C11m-scFv-Fc/mFc использовали для трансфекции клеток HEK293 (таких как HEK293F, Invitrogen, Inc.) с применением липосом (как например, 293 fectin, Invitrogen, Inc.) или других катионных реагентов для трансфекции (таких как PEI), соответственно. Клетки культивировали в суспензии в бессывороточных средах в течение 3-5 суток. Затем культуральные супернатанты собирали центрифугированием.
Кулыуральный супернатант собранных полностью человеческих моноклональных антител IgG1 или слитых белков на основе одноцепочечного антитела и Fc к G-белку вируса бешенства подвергали одностадийной очистке с использованием колонок для аффинной хроматографии с иммобилизованными белками A/G (например, Mabselect SURE, GE, Inc.). Затем консервирующие буферные растворы для рекомбинантных антител заменяли на буферы на основе PBS (забуференный фосфатом физиологический раствор) (рН 7,0), используя колонку для обессоливания (такую как Hitrap desaulting, GE, Inc.), или на другие подходящие буферы. При необходимости образцы антител можно подвергнуть стерилизации фильтрованием и затем хранить в виде аликвот при -20°С.
Пример 2. Подтверждение связывания моноклональных антител с неконкурирующими эпитопами G-белка вируса бешенства
Лунки 96-луночных планшетов для ELISA покрывали полученным инактивированным вирусом бешенства (полученным с использованием MRC-5 клеток) (1 ME (международная единица)/мл, 100 мкл/лунка) в течение ночи при 4°С. После блокирования при 37°С в течение 1 часа блокирующим раствором (буфером на основе PBS с твином (PBST), содержащим 2% молока) добавляли полностью человеческие моноклональные антитела C34m и C11m, полученные в примере 1, в эквимолярной начальной концентрации (200 нМ) и в серийных 3-кратных последовательных разведениях (всего 8 концентраций), соответственно, и инкубировали при 37°С в течение 1 часа. Планшеты для ELISA промывали PBST-буфером, добавляли конъюгированное с пероксидазой хрена (HRP) антитело к IgG человека (вторичное антитело) и инкубировали при 37°С в течение 1 часа. Затем планшеты для ELISA промывали PBST-буфером и добавляли раствор субстрата opmo-фенилендиамина (OPD) для развития окраски. После инкубирования в течение 5-10 минут развитие окраски останавливали, добавляя 1 М раствор H2SO4, и значения оптической плотности определяли, используя микропланшетный ридер на двух длинах волны 490 нм/630 нм. Результаты ELISA-анализа (ФИГ. 1) показывают, что оба полностью человеческих моноклональных антитела C34m и C11m способны связываться с иммобилизованным инактивированным вирусом бешенства с сопоставимой связывающей способностью.
Гены, кодирующие вариабельные области тяжелой цепи и вариабельные области легкой цепи полностью человеческих моноклональных антител C34m и C11m, клонировали в двухвекторные дисплейные системы pADK-S и pAG-S, соответственно (для подробного рассмотрения экспериментальных протоколов см. пример 4.1 в заявке на патент Китая №201510097117.0). Получали и очищали конструкции фаг-C34m и фаг-C11m, экспонирующие отдельные варианты Fab, и замораживали при -20°С для применения после определения титра. Лунки 96-луночных планшетов для ELISA покрывали полученным инактивированным вирусом бешенства (полученным с использованием MRC-5 клеток) (1 МЕ/мл, 100 мкл/лунка) в течение ночи при 4°С. Полностью человеческие моноклональные антитела C34m и C11m, полученные в примере 1 в эквимолярной начальной концентрации (200 нМ), разбавляли конструкциями фаг-C34m и фаг-C11m в фиксированной концентрации (1×1012 колониеобразующих единиц (КОЕ)/мл), используя 3-кратные последовательные разведения (всего 8 концентраций), соответственно, затем добавляли в 96-луночные планшеты из расчета 100 мкл/лунка и инкубировали при 37°С в течение 1 часа. Ингибирование связывания конструкций фаг-C34m и фаг-C11mc инактивированным вирусом бешенства под действием полностью человеческих моноклональных антител C34m и C11m детектировали, используя конъюгированное с HRP анти-M13-IgG (вторичное антитело). Результаты ELISA-анализа (ФИГ. 2 и 3) показывают, что полностью человеческое моноклональное антитело C11m полностью блокирует связывание конструкции фаг-C11m c инактивированным вирусом бешенства, а полностью человеческое моноклональное антитело C34m не полностью блокирует связывание конструкции фаг-C11m c инактивированным вирусом бешенства. Полностью человеческое моноклональное антитело C34m полностью блокирует связывание конструкции фаг-C34m с инактивированным вирусом бешенства, а полностью человеческое моноклональное антитело C11m не может блокировать связывание конструкции фаг-C34m с инактивированным вирусом бешенства. Эти результаты указывают на то, что полностью человеческие моноклональные антитела C34m и C11m имеют неконкурирующие связывающие эпитопы на G-белке вируса бешенства.
Пример 3. Проверка эпитопов моноклональных антител к G-белку вируса бешенства
Гены, кодирующие вариабельную область тяжелой цепи CR57VH (SEQ ID NO: 12) моноклонального антитела CR57 к эпитопу I G-белка вируса бешенства и вариабельную область тяжелой цепи CR4098VH (SEQ ID NO: 13) моноклонального антитела CR4098 к эпитопу III G-белка вируса бешенства (Bakker, А. В. et al. Novel human monoclonal antibody combination effectively neutralizing natural rabies virus variants and individual in vitro escape mutants. J. Virol., 79, 9062-9068; и патент США №9005624 B2), соответственно, клонировали в эукариотические экспрессирующие векторы (такие как pcDNA3.1, Invitrogen, Inc.), несущие ген, кодирующий константную область тяжелой цепи мышиного IgG2a (CH-mIgG2a, SEQ ID NO: 14). Ген, кодирующий вариабельную область легкой цепи CR57VL (SEQ ID NO: 15) антитела CR57, клонировали в эукариотический экспрессирующий вектор (такой как pcDNA3.1, Invitrogen, Inc.), несущий ген, кодирующий константную область легкой цепи мыши mCL (SEQ ID NO: 17), а ген, кодирующий вариабельную область легкой цепи CR4098VK (SEQ ID NO: 16) антитела CR4098 клонировали в эукариотический экспрессирующий вектор (такой как pcDNA3.1, Invitrogen, Inc.), несущий константную область легкой цепи мыши mCK (SEQ ID NO: 18). Химерные антитела, CR57-mIgG2a на основе CR57 и CR4098-mIgG2a на основе CR4098, конструировали согласно способу из примера 1, соответственно.
Лунки 96-луночных планшетов для ELISA (1 МЕ/мл, 100 мкл/лунка) покрывали полученным инактивированным вирусом бешенства (полученным с использованием MRC-5 клеток) в течение ночи при 4°С. Полностью человеческие моноклональные антитела C11m и C34m в эквимолярной начальной концентрации (200 нМ) разбавляли химерными антителами CR57-mIgG2a и CR4098-mIgG2 в фиксированной концентрации (2,5 мкг/мл), используя 3-кратные последовательные разведения (всего 12 концентраций), соответственно, добавляли в 96-луночные планшеты из расчета 100 мкл/лунка и инкубировали при 37°С в течение 1 часа. Для детекции ингибирования способности химерных антител CR57-mIgG2a и CR4098-mIgG2a связываться с инактивированным вирусом бешенства под действием полностью человеческих моноклональных антител C11m и C34m, соответственно, использовали конъюгированное с HRP антитело к IgG мыши (вторичное антитело). Результаты ELISA-анализа (ФИГ. 4, ФИГ. 5) показывают, что полностью человеческое моноклональное антитело C11m конкурирует с химерным антителом CR57-mIgG2a за связывание с G-белком вируса бешенства, но не с химерным антителом CR4098-mIgG2a, и это указывает на то, что полностью человеческое моноклональное антитело C11m специфично связывается с эпитопом I G-белка вируса бешенства. Полностью человеческое моноклональное антитело C34m конкурирует с химерным антителом CR4098-mIgG2a за связывание с G-белком вируса бешенства, но не с химерным антителом CR57-mIgG2a, и это указывает на то, что полностью человеческое моноклональное антитело C34m специфично связывается с эпитопом III G-белка вируса бешенства.
Пример 4. Скрининг мутантных форм C11m-scFv с более подходящими значениями pI
Биспецифические антитела, основанные на применении стратегии KIH («выступы-во-впадины»), лучше образуют гетеродимеры, однако предотвратить образование гомодимеров при крупномасштабных способах получения все еще трудно. Для удаления примесей после одностадийной очистки с использованием белка А в способе очистки биспецифических антител обычно используют ионообменную хроматографию (IEC). Время удерживания гомодимера на ионообменной смоле зависит от изоэлектрической точки (pI) молекулы антитела, поэтому эффективность процессов последующей обработки при получении антител можно улучшить путем изменения изоэлектрической точки.
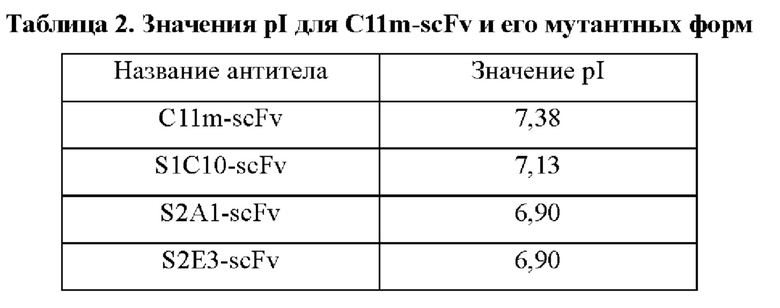
Ген, кодирующий одноцепочечное антитело C11m-scFv, полученное в примере 1, клонировали в вектор pADscFv-s (для подробного рассмотрения экспериментальных протоколов см. пример 1.3 в заявке на патент Китая №201510097117.0), получая рекомбинантный экспрессирующий вектор pADscFv-C11m-scFv. В участки CDR рекомбинантного экспрессирующего вектора pADscFv-C11m-scFv вносили мутации с использованием методов полимеразной цепной реакции (ПЦР) с перекрывающимися праймерами, конструируя тем самым библиотеку мутантных форм C11m-scFv с емкостью библиотеки более 4,6 × 106. Основные праймеры, необходимые при амплификации, показаны в Таблице 1. Скрининг библиотеки мутантных форм C11m-scFv проводили за три раунда, используя инактивированный вирус бешенства (полученный с использованием MRC-5 клеток) в качестве антигена. Окончательно идентифицировали три мутантные формы S1C10-scFv (SEQ ID NO: 21), S2Al-scFv (SEQ ID NO: 22) и S2E3-scFv (SEQ ID NO: 23) с улучшенными связывающими свойствами и ожидаемо более низкими значениями pI (http://www.bioinformatics.org/sms2/protein_iep.html). Значения pI для отдельных мутантных форм показаны в Таблице 2.
Согласно способу из примера 1 получали слитые белки на основе одноцепочечного антитела и Fc (scFv-Fc) S1C10-scFv-Fc, S2Al-scFv-Fc и S2E3-scFv-Fc, содержащие три указанные выше мутантные формы, слитые с Fc-сегментом IgG1 человека. Кроме того, для проведения функционального анализа получали химерное антитело C34m-mIgG2a на основе C34m, слитого с константной областью тяжелой цепи (CH-mIgG2a) и константной областью легкой цепи (mCK) мышиного IgG2a, и химерное антитело C11m-mIgG2a на основе C11m, слитого с константной областью тяжелой цепи (CH-mIgG2a) и константной областью легкой цепи (mCL) мышиного IgG2a. Согласно способам из примера 2 и примера 3 проводили определение способностей связываться с G-белком вируса бешенства и неконкурирующих эпитопов слитых белков на основе одноцепочечного антитела и Fc для трех мутантных форм C11m-scFv, соответственно. Результаты ELISA-анализа, показанные на ФИГ. 6, ФИГ. 7 и ФИГ. 8, демонстрируют, что способности связываться с G-белком вируса бешенства у трех мутантных форм слитых белков на основе одноцепочечного антитела и Fc значительно улучшены и сопоставимы с таковыми для полностью человеческого моноклонального антитела C34m. Слитый белок C11m-scFv-Fc на основе одноцепочечного антитела и Fc и слитые белки на основе одноцепочечного антитела и Fc для этих трех мутантных форм C11m-scFv связываются с неконкурирующим в отношении C34m-mIgG2a эпитопом на G-белке вируса бешенства. Слитый белок C11m-scFv-Fc на основе одноцепочечного антитела и Fc и слитые белки на основе одноцепочечного антитела и Fc для этих трех мутантных форм C11m-scFv конкурируют с C11m-mIgG2a за связывание с эпитопом на G-белке вируса бешенства.

Пример 5. Получение биспецифических антител
С целью конструирования гетеродимеров человеческого IgG1 на основе технологии KIH («выступы-во-впадины») создавали антиген-связывающие фрагменты для эпитопов I и III G-белка вируса бешенства в форме scFv и форме Fab, соответственно. То есть, проводили слияние антиген-связывающего фрагмента антитела C34m в форме Fab с N-концом Fc-сегмента, содержащего мутацию, приводящую к образованию выступа (FcK, SEQ ID NO: 19), и проводили слияние происходящих из мутантных форм антитела C11m антиген-связывающих фрагментов в форме scFv с N-концом Fc-сегмента, содержащего мутацию, приводящую к образованию впадины (FcH, SEQ ID NO: 20), конструируя тем самым биспецифическое антитело к G-белку вируса бешенства.
Тремя сконструированными для экспрессии эукариотическими векторами, экспрессирующими S1C10-scFv-FcH, C34mVH-IgG1K и C34mVK-CK, соответственно, с использованием липосом совместно трансфицировали клетки HEK293F, и клетки культивировали в суспензии в бессывороточной среде в течение 3-5 суток. Культуральный супернатант собирали центрифугированием. Биспецифические антитела из культурального супернатанта очищали, используя колонку для аффинной хроматографии с иммобилизованным белком A/G (например, Mabselect SURE, GE Inc.). Затем консервирующий буферный раствор для рекомбинантных антител заменяли на буфер на основе PBS (рН 7,0), используя колонку для обессоливания (например, Hitrap desaulting, GE Inc.), или на другие подходящие буферы. Обессоленный раствор белка очищали гель-проникающей хроматографией (SEC) с использованием Superdex 200 (GE), получая тем самым биспецифическое антитело S1C10-scFv-FcH+C34m-IgG1K. При необходимости образцы антител можно подвергнуть стерилизации фильтрованием и затем хранить в виде аликвот при -20°С.
Аналогичным образом получали два биспецифических антитела S2Al-scFv-FcH+C34m-IgG1K и S2E3-scFv-FcH+C34m-IgG1K.
Пример 6. Функциональная проверка биспецифических антител
Лунки 96-луночных планшетов для ELISA покрывали полученным инактивированным вирусом бешенства (полученным с использованием MRC-5 клеток) (1 МЕ/мл, 100 мкл/лунка) в течение ночи при 4°С. После блокирования в течение 1 часа при 37°С блокирующим раствором (буфером на основе PBST, содержащим 2% молока) добавляли биспецифические антитела к G-белку вируса бешенства (S1C10-scFv-FcH+C34m-IgG1K, S2Al-scFv-FcH+C34m-IgG1K, S2E3-scFv-FcH+C34m-IgG1K, S1C10-scFv-Fc, S2A1-scFv-Fc, S2E3-scFv-Fc, C11m-scFv-Fc и C34m) в эквимолярной начальной концентрации (200 нМ) и в серийных 3-кратных последовательных разведениях (всего 12 концентраций), соответственно, и инкубировали при 37°С в течение 1 часа. Планшеты для ELISA промывали PBST-буфером, добавляли конъюгированное с HRP антитело к IgG человека (вторичное антитело) и инкубировали при 37°С в течение 1 часа. Затем планшеты для ELISA промывали PBST-буфером и добавляли раствор субстрата OPD для развития окраски. После инкубирования в течение 5-10 минут развитие окраски останавливали, добавляя 1 М раствор H2SO4, и определяли значения оптической плотности, используя микропланшетный ридер на двух длинах волны 490 нм/630 нм. Результаты ELISA-анализа (ФИГ. 9) показывают, что способность биспецифических антител связываться с инактивированным вирусом бешенства оказывалась лучше таковой для слитых белков на основе одноцепочечного антитела и Fc и для полностью человеческого моноклонального антитела C34m.
Отбирали одноцепочечное антитело S2E3-scFv, имеющее низкое значение pi, и получали химерный белок S2E3-scFv-mFc, следуя методикам, приведенным в примере 1. Биспецифическое антитело S2E3-scFv-FcH+C34m-IgG1K, слитый белок S2E3-scFv-Fc на основе одноцепочечного антитела и Fc и полностью человеческое моноклональное антитело C34m в эквимолярной начальной концентрации (200 нМ) разбавляли C34m-mIgG2a и S2E3-scFv-mFc в фиксированной концентрации (2,5 мкг/мл), используя 3-кратные последовательные разведения (всего 12 концентраций), соответственно, затем добавляли в 96-луночные планшеты из расчета 100 мкл/лунка и инкубировали при 37°С в течение 1 часа. Для детекции сигналов связывания использовали конъюгированное с HRP антитело к IgG мыши (вторичное антитело). Результаты ELISA-анализа (ФИГ. 10 и ФИГ. 11) показывают, что биспецифическое антитело S2E3-scFv-FcH+C34m-IgG1K способно ингибировать связывание химерного антитела S2E3-scFv-mFc и химерного антитела C34m-mIgG2a с вакциной от вируса бешенства. То есть, биспецифическое антитело S2E3-scFv-FcH+C34m-IgG1K к G-белку вируса бешенства «работает» по двум неконкурирующим связывающим эпитопам и способно связываться как с эпитопом I, так и с эпитопом III G-белка вируса бешенства.
Пример 7. Анализ титра антител на основе теста быстрого ингибирования фокусов флуоресценции (RFFIT) на нейтрализующих антителах против вируса бешенства
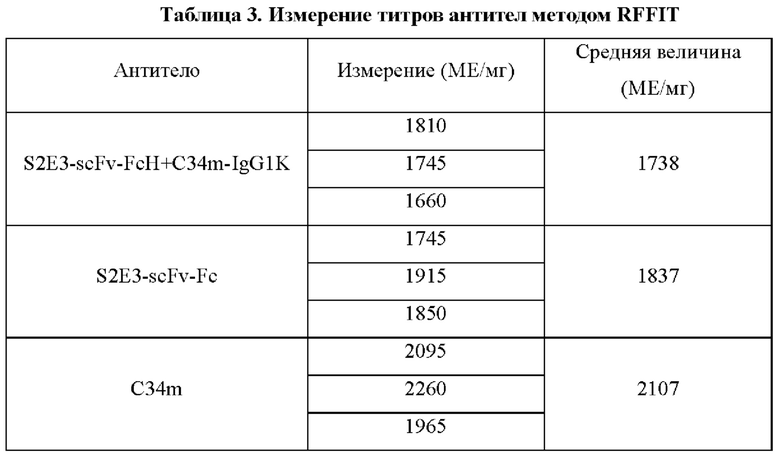
В соответствии с тестом быстрого ингибирования фокусов флуоресценции (RFFIT) на нейтрализующих антителах против вируса бешенства готовили серийные 3-кратные последовательные разведения стандартной сыворотки (0,5 МЕ/мл) и предварительно разведенных тестируемых антител (биспецифического антитела S2E3-scFv-FcH+C34m-IgG1K, слитой конструкции S2E3-scFv-Fc на основе одноцепочечного антитела и Fc, и полностью человеческого моноклонального антитела C34m, 10 мкг/мл). Стандартную сыворотку или тестируемые антитела добавляли в 96-луночные планшеты (50 мкл/лунка) в трех повторах для каждого образца. В каждую лунку добавляли подлежащий нейтрализации вирус бешенства, разведенный в соответствующем отношении. Лунки без стандартной сыворотки или тестируемого антитела служили в качестве контрольных лунок для разведений вируса, а лунки без стандартной сыворотки, тестируемого антитела или вируса бешенства служили в качестве контрольных лунок для клеток. Планшеты инкубировали при 37°С в течение 1 часа, в каждую лунку добавляли по 50 мкл клеток BSR (1 × 106 клеток/мл) и планшеты инкубировали при 37°С в инкубаторе с 5% СО2. Через 24 часа добавляли 50 мкл холодного ацетона для фиксации в течение 30 мин и затем промывали PBST-буфером. В каждую лунку добавляли по 50 мкл рабочего раствора флуоресцентного антитела против вируса бешенства и планшеты инкубировали при 37°С в течение 30 мин. После промывки PBS-буфером в каждую лунку добавляли глицерин (80%), флуоресцентное окрашивание проводили под инвертированным флуоресцентным микроскопом и титры тестируемых образцов рассчитывали согласно методу Рида-Менча.
Анализировали три антитела, используя стандартный штамм атакующего вируса бешенства CVS-11, и результаты показаны в Таблице 3. Результаты указывают на то, что биспецифическое антитело S2E3-scFv-FcH+C34m-IgG1K, слитый белок S2E3-scFv-Fc на основе одноцепочечного антитела и Fc и полностью человеческое моноклональное антитело C34m все обладают хорошей активностью нейтрализации.
Пример 8. Защита золотистых хомяков антителами против вируса бешенства от атаки вируса бешенства в летальной дозе
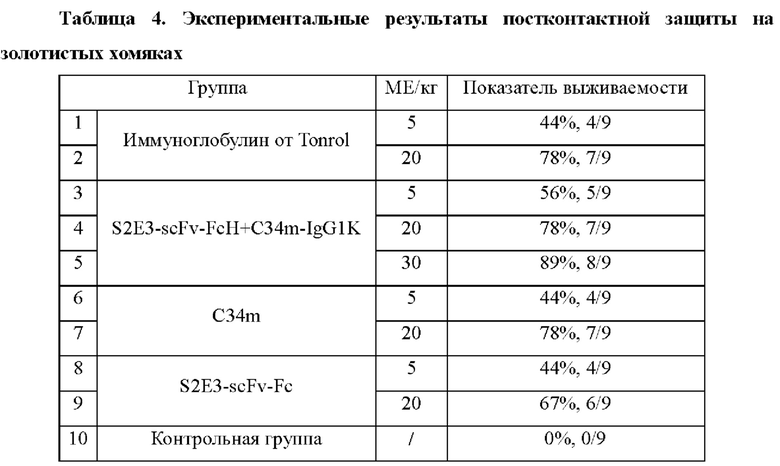
Защитное действие каждой группы антител определяли по результатам постконтактного введения иммуноглобулинов золотистым хомякам, которым путем инъекции вводили летальную дозу вируса бешенства, и сравнивали с действием имеющихся в продаже продуктов.
Золотистых хомяков подвергали атаке стандартного штамма атакующего вируса бешенства CVS-11 (6,67 lgLD50 (доза, вызывающая гибель 50% популяции)/мл; 0,2 мл/животное) с введением в их правые задние лапы. Антитела против вируса бешенства вводили в эти же места спустя 24 часа. Разделение на группы выполняли в соответствии с вводимыми дозами (МЕ/кг). При тестировании биспецифического антитела S2E3-scFv-FcH+C34m-IgG1K проводили разделение на три группы в соответствии с вводимыми дозами, составляющими 5 МЕ/кг, 20 МЕ/кг и 30 МЕ/кг. При тестированиях с использованием слитого белка S2E3-scFv-Fc на основе одноцепочечного антитела и Fc, полностью человеческого моноклонального антитела C34m и человеческого иммуноглобулина против вируса бешенства коммерческого происхождения (Tonrol Biopharmaceutical Co., Ltd.) (называемого далее как иммуноглобулин от Tonrol) проводили разделение, соответственно, на две группы в соответствии с вводимыми дозами, составляющими 5 МЕ/кг и 20 МЕ/кг. Золотистые хомяки, которым не вводили антитело против вируса бешенства, служили в качестве контрольной группы. Всего использовали 10 групп по 9 животных в группе. Каждый образец разбавляли PBS-буфером согласно результатам взвешивания золотистых хомяков, и для удобства введения окончательный объем инъекции на одно животное составлял 100 мкл.
Экспериментальные результаты показаны в Таблице 4. Все золотистые хомяки в контрольной группе умирали от бешенства в течение 2 недель после атаки, в то время как золотистые хомяки в получающей лечение группе имели более высокие значения показателей выживаемости. Защитные действия антител по настоящей заявке против вируса бешенства были сопоставимы с таковыми для инъекции иммуноглобулина от Tonrol.
Пример 9. Идентификация связывающего эпитопа для антител к G-белку вируса бешенства
Для идентификации эпитопов, распознаваемых полностью человеческим моноклональным антителом C34m, обладающим активностью нейтрализации вируса бешенства, и слитым белом S2E3-scFv-Fc на основе одноцепочечного антитела и Fc по сравнению с мутантной формой, происходящей из C11m-scFv, исследовали влияние аминокислотных замен в нейтрализующих эпитопах I-III гликопротеина вируса бешенства на связывание антител против вируса бешенства с G-белком. В сообщениях показано, что мутация в любом из эпитопов I-III G-белка вируса бешенства позволяет избежать распознавания содержащего мутацию эпитопа G-белка вируса бешенства антителом, которое связывается с данным эпитопом. В настоящем примере выбрана одноточечная мутация в аминокислотах нейтрализующих эпитопов I и III G-белка штамма вируса бешенства дикого типа CVS-11 для получения некоторой аминокислотной последовательности (Новый нейтрализующий эпитоп вируса бешенства, распознаваемый человеческим моноклональным антителом: точное картирование и анализ «ускользнувших» мутантов; Вирус бешенства: влияние на патогенность и характеристика последовательности мутаций вируса бешенства, затрагивающих антигенный сайт III гликопротеина).
Согласно традиционным методам молекулярной биологии гены внеклеточного домена, кодирующие G-белок штамма CVS-11 вируса бешенства дикого типа GCVS11 (SEQ ID NO: 35) и его мутантную по эпитопу I форму GCVS11-G229E (SEQ ID NO: 36) и мутантную по эпитопу III форму GCVS11-I338T (SEQ ID NO: 37), получают, используя ПЦР-амплификацию, и осуществляют слияние их С-концов с геном, кодирующим тримерный домен CCD белка 1А коронавируса человека (SEQ ID NO: 38), соответственно, с обеспечением хорошего образования тримеров секретируемых гликопротеинов и сохранения у них конформации и иммуногенности своего нативного белка. Полученные гены, кодирующие слитые белки, содержащие внеклеточные области G-белка вируса бешенства или двух мутантных форм (G229E и I338T) и CCD, соответственно, клонировали в эукариотические экспрессирующие векторы, несущие His-метки на С-конце (такие как pcDNA3.1, Invitrogen, Inc.), и полученные рекомбинантные экспрессирующие антиген векторы применяли соответственно для трансфекции клеток HEK293, используя липосомы (как например, 293 fectin, Invitrogen, Inc.). Клетки культивировали в суспензии в бессывороточной среде в течение 3-5 суток и культуральные супернатанты собирали посредством центробежной фильтрации. Полученный культуральный супернатант концентрировали примерно в 10 раз, используя пробирку для ультрафильтрации на центрифуге, и хранили при -80°С для последующего применения.
Лунки 96-луночных планшетов для ELISA покрывали полученным полностью человеческим моноклональным антителом C34m и слитым белком S2E3-scFv-Fc на основе одноцепочечного антитела и Fc (5 мкг/мл, 100 мкг/лунка), соответственно, при 4°С в течение ночи. После блокирования в течение 1 часа при 37°С блокирующим раствором (буфером на основе PBST, содержащим 2% молока) выполняли 3-кратные последовательные разведения (всего 11 концентраций) концентратов полученных слитых белков GCVS11-CCD-His (SEQ ID NO: 39), GCVS11-G229E-CCD-His (SEQ ID NO: 40), GCVS11-I338T-CCD-His (SEQ ID NO: 41) и добавляли в лунки 96-луночных планшетов, покрытые C34m или S2E3-scFv-Fc (100 мкл/лунка). Планшеты инкубировали при 37°С в течение 1 часа. Детектирование сигналов связывания осуществляли, используя HRP-меченные антитела к His-метке (вторичное антитело).
Результаты ELISA-анализа (ФИГ. 12А, 12В, 12С) показывают, что C34m и S2E3-scFv-Fc обладают сопоставимыми способностями связываться со слитым белком GCVS11-CCD-His (ФИГ. 12А). Способность S2E3-scFv-Fc связываться со слитым белком GCVS11-G229E-CCD-His оказывается значительно ниже, чем у C34m (ФИГ. 12В), и это указывает на то, что связывающий эпитоп для S2E3-scFv-Fc представляет собой эпитоп I G-белка вируса бешенства. Аналогично, способность C34m связываться со слитым белком GCVS11-I338T-CCD-His оказывается значительно ниже, чем у S2E3-scFv-Fc (ФИГ. 12С), и это указывает на то, что связывающий эпитоп для C34m представляет собой эпитоп III G-белка вируса бешенства.
--->
ИЗМЕНЕННЫЙ ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
для рассмотрения на национальной стадии
<110> Beijing Wisdomab Biotechnology Co., Ltd
GENRIX (Shanghai) Biopharmaceutical Co.Ltd.
hongqing GENRIX Biopharmaceutical Co., Ltd.
<120> БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ
<150> 201810901518.0
<151> 2018-08-09
<160> 64
<170> SIPOSequenceListing 1.0
<210> 1
<211> 118
<212> ПРТ
<213> Искусственная последовательность
<400> 1
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Tyr Asp Gly Ser Ile Lys Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Asp Arg Thr Gly Asn Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 2
<211> 127
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 2
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Thr Leu Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 3
<211> 107
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Ile Arg Asn Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Thr Arg Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asn Ser Glu Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 4
<211> 112
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 4
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp
85 90 95
Tyr Thr Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 5
<211> 330
<212>ПРТ
<213> Homo sapiens
<400> 5
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 6
<211> 107
<212>ПРТ
<213> Homo sapiens
<400> 6
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 7
<211> 106
<212>ПРТ
<213> Homo sapiens
<400> 7
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 8
<211> 240
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 8
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Tyr Asp Gly Ser Ile Lys Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Asp Arg Thr Gly Asn Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
130 135 140
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln
145 150 155 160
Asn Ile Arg Asn Ala Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
165 170 175
Pro Lys Leu Leu Ile Tyr Asp Ala Ser Thr Arg Gln Ser Gly Val Pro
180 185 190
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asn
210 215 220
Ser Glu Phe Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
225 230 235 240
<210> 9
<211> 254
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 9
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Thr Leu Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Gly Tyr Asn Tyr
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
245 250
<210> 10
<211> 232
<212>ПРТ
<213> Homo sapiens
<400> 10
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 11
<211> 232
<212>ПРТ
<213> Mus musculus
<400> 11
Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala
1 5 10 15
Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile
20 25 30
Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val
35 40 45
Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val
50 55 60
Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp
65 70 75 80
Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln
85 90 95
Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Asp
100 105 110
Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val
115 120 125
Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr
130 135 140
Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu
145 150 155 160
Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr
165 170 175
Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr
180 185 190
Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr
195 200 205
Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr Thr Lys
210 215 220
Ser Phe Ser Arg Thr Pro Gly Lys
225 230
<210> 12
<211> 127
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Asn Arg Tyr
20 25 30
Thr Val Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 13
<211> 119
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 13
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Ala Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Leu Tyr Asp Gly Ser Asp Lys Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Ala Val Ala Gly Thr His Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 14
<211> 330
<212>ПРТ
<213> Mus musculus
<400> 14
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 15
<211> 112
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 15
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Gly Tyr
20 25 30
Asn Phe Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp
85 90 95
Tyr Thr Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 16
<211> 107
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 16
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser Tyr Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 106
<212>ПРТ
<213> Mus musculus
<400> 17
Gly Gln Pro Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp
20 25 30
Phe Tyr Pro Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro
35 40 45
Val Thr Gln Gly Met Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Met Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu
65 70 75 80
Arg His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val
85 90 95
Glu Lys Ser Leu Ser Arg Ala Asp Cys Ser
100 105
<210> 18
<211> 107
<212>ПРТ
<213> Mus musculus
<400> 18
Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu
1 5 10 15
Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg
35 40 45
Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu
65 70 75 80
Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser
85 90 95
Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 19
<211> 232
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 19
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 20
<211> 232
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 20
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 21
<211> 254
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Ala Tyr Asp Tyr
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
245 250
<210> 22
<211> 254
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Asp Gly Tyr Asp Phe
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
245 250
<210> 23
<211> 254
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Gly Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Leu Gly Gly Tyr Asp Phe
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
245 250
<210> 24
<211> 127
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 24
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 25
<211> 112
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 25
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Ala Tyr
20 25 30
Asp Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp
85 90 95
Tyr Thr Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 26
<211> 127
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 26
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 27
<211> 112
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 27
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Asp Gly Tyr
20 25 30
Asp Phe Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp
85 90 95
Tyr Thr Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 28
<211> 127
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 28
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Gly Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 29
<211> 112
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 29
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Leu Gly Gly Tyr
20 25 30
Asp Phe Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp
85 90 95
Tyr Thr Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 30
<211> 214
<212>ПРТ
<213> Homo sapiens
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Ile Arg Asn Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Thr Arg Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Asn Ser Glu Phe Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 31
<211> 448
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 31
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Tyr Asp Gly Ser Ile Lys Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Asp Arg Thr Gly Asn Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 32
<211> 488
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Gly Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Leu Gly Gly Tyr Asp Phe
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ser
245 250 255
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
260 265 270
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
275 280 285
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
290 295 300
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
305 310 315 320
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
325 330 335
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
340 345 350
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
355 360 365
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
370 375 380
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
385 390 395 400
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
405 410 415
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
420 425 430
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
435 440 445
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
450 455 460
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
465 470 475 480
Ser Leu Ser Leu Ser Pro Gly Lys
485
<210> 33
<211> 488
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 33
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Thr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Ala Tyr Asp Tyr
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ser
245 250 255
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
260 265 270
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
275 280 285
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
290 295 300
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
305 310 315 320
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
325 330 335
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
340 345 350
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
355 360 365
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
370 375 380
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
385 390 395 400
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
405 410 415
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
420 425 430
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
435 440 445
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
450 455 460
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
465 470 475 480
Ser Leu Ser Leu Ser Pro Gly Lys
485
<210> 34
<211> 488
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 34
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Tyr Ser Arg Tyr
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Gln Gly Arg Leu Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Phe Ser Gly
100 105 110
Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser
130 135 140
Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln Ser Val
145 150 155 160
Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Asp Gly Tyr Asp Phe
165 170 175
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
180 185 190
Tyr Asp Ala Thr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
195 200 205
Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala
210 215 220
Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Asp Tyr Thr
225 230 235 240
Pro Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ser
245 250 255
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
260 265 270
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
275 280 285
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
290 295 300
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
305 310 315 320
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
325 330 335
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
340 345 350
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
355 360 365
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
370 375 380
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
385 390 395 400
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
405 410 415
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
420 425 430
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
435 440 445
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
450 455 460
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
465 470 475 480
Ser Leu Ser Leu Ser Pro Gly Lys
485
<210> 35
<211> 440
<212>ПРТ
<213>Вирусбешенства
<400> 35
Lys Phe Pro Ile Tyr Thr Ile Pro Asp Lys Leu Gly Pro Trp Ser Pro
1 5 10 15
Ile Asp Ile His His Leu Ser Cys Pro Asn Asn Leu Val Val Glu Asp
20 25 30
Glu Gly Cys Thr Asn Leu Ser Glu Phe Ser Tyr Met Glu Leu Lys Val
35 40 45
Gly Tyr Ile Ser Ala Ile Lys Val Asn Gly Phe Thr Cys Thr Gly Val
50 55 60
Val Thr Glu Ala Glu Thr Tyr Thr Asn Phe Val Gly Tyr Val Thr Thr
65 70 75 80
Thr Phe Lys Arg Lys His Phe Arg Pro Thr Pro Asp Ala Cys Arg Ala
85 90 95
Ala Tyr Asn Trp Lys Met Ala Gly Asp Pro Arg Tyr Glu Glu Ser Leu
100 105 110
His Asn Pro Tyr Pro Asp Tyr His Trp Leu Arg Thr Val Arg Thr Thr
115 120 125
Lys Glu Ser Leu Ile Ile Ile Ser Pro Ser Val Thr Asp Leu Asp Pro
130 135 140
Tyr Asp Lys Ser Leu His Ser Arg Val Phe Pro Gly Gly Lys Cys Ser
145 150 155 160
Gly Ile Thr Val Ser Ser Thr Tyr Cys Ser Thr Asn His Asp Tyr Thr
165 170 175
Ile Trp Met Pro Glu Asn Pro Arg Pro Arg Thr Pro Cys Asp Ile Phe
180 185 190
Thr Asn Ser Arg Gly Lys Arg Ala Ser Lys Gly Asn Lys Thr Cys Gly
195 200 205
Phe Val Asp Glu Arg Gly Leu Tyr Lys Ser Leu Lys Gly Ala Cys Arg
210 215 220
Leu Lys Leu Cys Gly Val Leu Gly Leu Arg Leu Met Asp Gly Thr Trp
225 230 235 240
Val Ala Met Gln Thr Ser Asp Glu Thr Lys Trp Cys Pro Pro Asp Gln
245 250 255
Leu Val Asn Leu His Asp Phe Arg Ser Asp Glu Ile Glu His Leu Val
260 265 270
Val Glu Glu Leu Val Lys Lys Arg Glu Glu Cys Leu Asp Ala Leu Glu
275 280 285
Ser Ile Met Thr Thr Lys Ser Val Ser Phe Arg Arg Leu Ser His Leu
290 295 300
Arg Lys Leu Val Pro Gly Phe Gly Lys Ala Tyr Thr Ile Phe Asn Lys
305 310 315 320
Thr Leu Met Glu Ala Asp Ala His Tyr Lys Ser Val Arg Thr Trp Asn
325 330 335
Glu Ile Ile Pro Ser Lys Gly Cys Leu Lys Val Gly Gly Arg Cys His
340 345 350
Pro His Val Asn Gly Val Phe Phe Asn Gly Ile Ile Leu Gly Pro Asp
355 360 365
Gly His Val Leu Ile Pro Glu Met Gln Ser Ser Leu Leu Gln Gln His
370 375 380
Met Glu Leu Leu Lys Ser Ser Val Ile Pro Leu Met His Pro Leu Ala
385 390 395 400
Asp Pro Ser Thr Val Phe Lys Glu Gly Asp Glu Ala Glu Asp Phe Val
405 410 415
Glu Val His Leu Pro Asp Val Tyr Lys Gln Ile Ser Gly Val Asp Leu
420 425 430
Gly Leu Pro Asn Trp Gly Lys Tyr
435 440
<210> 36
<211> 440
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 36
Lys Phe Pro Ile Tyr Thr Ile Pro Asp Lys Leu Gly Pro Trp Ser Pro
1 5 10 15
Ile Asp Ile His His Leu Ser Cys Pro Asn Asn Leu Val Val Glu Asp
20 25 30
Glu Gly Cys Thr Asn Leu Ser Glu Phe Ser Tyr Met Glu Leu Lys Val
35 40 45
Gly Tyr Ile Ser Ala Ile Lys Val Asn Gly Phe Thr Cys Thr Gly Val
50 55 60
Val Thr Glu Ala Glu Thr Tyr Thr Asn Phe Val Gly Tyr Val Thr Thr
65 70 75 80
Thr Phe Lys Arg Lys His Phe Arg Pro Thr Pro Asp Ala Cys Arg Ala
85 90 95
Ala Tyr Asn Trp Lys Met Ala Gly Asp Pro Arg Tyr Glu Glu Ser Leu
100 105 110
His Asn Pro Tyr Pro Asp Tyr His Trp Leu Arg Thr Val Arg Thr Thr
115 120 125
Lys Glu Ser Leu Ile Ile Ile Ser Pro Ser Val Thr Asp Leu Asp Pro
130 135 140
Tyr Asp Lys Ser Leu His Ser Arg Val Phe Pro Gly Gly Lys Cys Ser
145 150 155 160
Gly Ile Thr Val Ser Ser Thr Tyr Cys Ser Thr Asn His Asp Tyr Thr
165 170 175
Ile Trp Met Pro Glu Asn Pro Arg Pro Arg Thr Pro Cys Asp Ile Phe
180 185 190
Thr Asn Ser Arg Gly Lys Arg Ala Ser Lys Gly Asn Lys Thr Cys Gly
195 200 205
Phe Val Asp Glu Arg Gly Leu Tyr Lys Ser Leu Lys Gly Ala Cys Arg
210 215 220
Leu Lys Leu Cys Glu Val Leu Gly Leu Arg Leu Met Asp Gly Thr Trp
225 230 235 240
Val Ala Met Gln Thr Ser Asp Glu Thr Lys Trp Cys Pro Pro Asp Gln
245 250 255
Leu Val Asn Leu His Asp Phe Arg Ser Asp Glu Ile Glu His Leu Val
260 265 270
Val Glu Glu Leu Val Lys Lys Arg Glu Glu Cys Leu Asp Ala Leu Glu
275 280 285
Ser Ile Met Thr Thr Lys Ser Val Ser Phe Arg Arg Leu Ser His Leu
290 295 300
Arg Lys Leu Val Pro Gly Phe Gly Lys Ala Tyr Thr Ile Phe Asn Lys
305 310 315 320
Thr Leu Met Glu Ala Asp Ala His Tyr Lys Ser Val Arg Thr Trp Asn
325 330 335
Glu Ile Ile Pro Ser Lys Gly Cys Leu Lys Val Gly Gly Arg Cys His
340 345 350
Pro His Val Asn Gly Val Phe Phe Asn Gly Ile Ile Leu Gly Pro Asp
355 360 365
Gly His Val Leu Ile Pro Glu Met Gln Ser Ser Leu Leu Gln Gln His
370 375 380
Met Glu Leu Leu Lys Ser Ser Val Ile Pro Leu Met His Pro Leu Ala
385 390 395 400
Asp Pro Ser Thr Val Phe Lys Glu Gly Asp Glu Ala Glu Asp Phe Val
405 410 415
Glu Val His Leu Pro Asp Val Tyr Lys Gln Ile Ser Gly Val Asp Leu
420 425 430
Gly Leu Pro Asn Trp Gly Lys Tyr
435 440
<210> 37
<211> 440
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 37
Lys Phe Pro Ile Tyr Thr Ile Pro Asp Lys Leu Gly Pro Trp Ser Pro
1 5 10 15
Ile Asp Ile His His Leu Ser Cys Pro Asn Asn Leu Val Val Glu Asp
20 25 30
Glu Gly Cys Thr Asn Leu Ser Glu Phe Ser Tyr Met Glu Leu Lys Val
35 40 45
Gly Tyr Ile Ser Ala Ile Lys Val Asn Gly Phe Thr Cys Thr Gly Val
50 55 60
Val Thr Glu Ala Glu Thr Tyr Thr Asn Phe Val Gly Tyr Val Thr Thr
65 70 75 80
Thr Phe Lys Arg Lys His Phe Arg Pro Thr Pro Asp Ala Cys Arg Ala
85 90 95
Ala Tyr Asn Trp Lys Met Ala Gly Asp Pro Arg Tyr Glu Glu Ser Leu
100 105 110
His Asn Pro Tyr Pro Asp Tyr His Trp Leu Arg Thr Val Arg Thr Thr
115 120 125
Lys Glu Ser Leu Ile Ile Ile Ser Pro Ser Val Thr Asp Leu Asp Pro
130 135 140
Tyr Asp Lys Ser Leu His Ser Arg Val Phe Pro Gly Gly Lys Cys Ser
145 150 155 160
Gly Ile Thr Val Ser Ser Thr Tyr Cys Ser Thr Asn His Asp Tyr Thr
165 170 175
Ile Trp Met Pro Glu Asn Pro Arg Pro Arg Thr Pro Cys Asp Ile Phe
180 185 190
Thr Asn Ser Arg Gly Lys Arg Ala Ser Lys Gly Asn Lys Thr Cys Gly
195 200 205
Phe Val Asp Glu Arg Gly Leu Tyr Lys Ser Leu Lys Gly Ala Cys Arg
210 215 220
Leu Lys Leu Cys Gly Val Leu Gly Leu Arg Leu Met Asp Gly Thr Trp
225 230 235 240
Val Ala Met Gln Thr Ser Asp Glu Thr Lys Trp Cys Pro Pro Asp Gln
245 250 255
Leu Val Asn Leu His Asp Phe Arg Ser Asp Glu Ile Glu His Leu Val
260 265 270
Val Glu Glu Leu Val Lys Lys Arg Glu Glu Cys Leu Asp Ala Leu Glu
275 280 285
Ser Ile Met Thr Thr Lys Ser Val Ser Phe Arg Arg Leu Ser His Leu
290 295 300
Arg Lys Leu Val Pro Gly Phe Gly Lys Ala Tyr Thr Ile Phe Asn Lys
305 310 315 320
Thr Leu Met Glu Ala Asp Ala His Tyr Lys Ser Val Arg Thr Trp Asn
325 330 335
Glu Thr Ile Pro Ser Lys Gly Cys Leu Lys Val Gly Gly Arg Cys His
340 345 350
Pro His Val Asn Gly Val Phe Phe Asn Gly Ile Ile Leu Gly Pro Asp
355 360 365
Gly His Val Leu Ile Pro Glu Met Gln Ser Ser Leu Leu Gln Gln His
370 375 380
Met Glu Leu Leu Lys Ser Ser Val Ile Pro Leu Met His Pro Leu Ala
385 390 395 400
Asp Pro Ser Thr Val Phe Lys Glu Gly Asp Glu Ala Glu Asp Phe Val
405 410 415
Glu Val His Leu Pro Asp Val Tyr Lys Gln Ile Ser Gly Val Asp Leu
420 425 430
Gly Leu Pro Asn Trp Gly Lys Tyr
435 440
<210> 38
<211> 32
<212>ПРТ
<213> Homo sapiens
<400> 38
Val Ser Arg Leu Glu Glu Glu Met Arg Lys Leu Gln Ala Thr Val Gln
1 5 10 15
Glu Leu Gln Lys Arg Leu Asp Arg Leu Glu Glu Thr Val Gln Ala Lys
20 25 30
<210> 39
<211> 488
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 39
Lys Phe Pro Ile Tyr Thr Ile Pro Asp Lys Leu Gly Pro Trp Ser Pro
1 5 10 15
Ile Asp Ile His His Leu Ser Cys Pro Asn Asn Leu Val Val Glu Asp
20 25 30
Glu Gly Cys Thr Asn Leu Ser Glu Phe Ser Tyr Met Glu Leu Lys Val
35 40 45
Gly Tyr Ile Ser Ala Ile Lys Val Asn Gly Phe Thr Cys Thr Gly Val
50 55 60
Val Thr Glu Ala Glu Thr Tyr Thr Asn Phe Val Gly Tyr Val Thr Thr
65 70 75 80
Thr Phe Lys Arg Lys His Phe Arg Pro Thr Pro Asp Ala Cys Arg Ala
85 90 95
Ala Tyr Asn Trp Lys Met Ala Gly Asp Pro Arg Tyr Glu Glu Ser Leu
100 105 110
His Asn Pro Tyr Pro Asp Tyr His Trp Leu Arg Thr Val Arg Thr Thr
115 120 125
Lys Glu Ser Leu Ile Ile Ile Ser Pro Ser Val Thr Asp Leu Asp Pro
130 135 140
Tyr Asp Lys Ser Leu His Ser Arg Val Phe Pro Gly Gly Lys Cys Ser
145 150 155 160
Gly Ile Thr Val Ser Ser Thr Tyr Cys Ser Thr Asn His Asp Tyr Thr
165 170 175
Ile Trp Met Pro Glu Asn Pro Arg Pro Arg Thr Pro Cys Asp Ile Phe
180 185 190
Thr Asn Ser Arg Gly Lys Arg Ala Ser Lys Gly Asn Lys Thr Cys Gly
195 200 205
Phe Val Asp Glu Arg Gly Leu Tyr Lys Ser Leu Lys Gly Ala Cys Arg
210 215 220
Leu Lys Leu Cys Gly Val Leu Gly Leu Arg Leu Met Asp Gly Thr Trp
225 230 235 240
Val Ala Met Gln Thr Ser Asp Glu Thr Lys Trp Cys Pro Pro Asp Gln
245 250 255
Leu Val Asn Leu His Asp Phe Arg Ser Asp Glu Ile Glu His Leu Val
260 265 270
Val Glu Glu Leu Val Lys Lys Arg Glu Glu Cys Leu Asp Ala Leu Glu
275 280 285
Ser Ile Met Thr Thr Lys Ser Val Ser Phe Arg Arg Leu Ser His Leu
290 295 300
Arg Lys Leu Val Pro Gly Phe Gly Lys Ala Tyr Thr Ile Phe Asn Lys
305 310 315 320
Thr Leu Met Glu Ala Asp Ala His Tyr Lys Ser Val Arg Thr Trp Asn
325 330 335
Glu Ile Ile Pro Ser Lys Gly Cys Leu Lys Val Gly Gly Arg Cys His
340 345 350
Pro His Val Asn Gly Val Phe Phe Asn Gly Ile Ile Leu Gly Pro Asp
355 360 365
Gly His Val Leu Ile Pro Glu Met Gln Ser Ser Leu Leu Gln Gln His
370 375 380
Met Glu Leu Leu Lys Ser Ser Val Ile Pro Leu Met His Pro Leu Ala
385 390 395 400
Asp Pro Ser Thr Val Phe Lys Glu Gly Asp Glu Ala Glu Asp Phe Val
405 410 415
Glu Val His Leu Pro Asp Val Tyr Lys Gln Ile Ser Gly Val Asp Leu
420 425 430
Gly Leu Pro Asn Trp Gly Lys Tyr Gly Gly Gly Gly Ser Val Ser Arg
435 440 445
Leu Glu Glu Glu Met Arg Lys Leu Gln Ala Thr Val Gln Glu Leu Gln
450 455 460
Lys Arg Leu Asp Arg Leu Glu Glu Thr Val Gln Ala Lys Ala Ser Gly
465 470 475 480
Ala Ala His His His His His His
485
<210> 40
<211> 488
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 40
Lys Phe Pro Ile Tyr Thr Ile Pro Asp Lys Leu Gly Pro Trp Ser Pro
1 5 10 15
Ile Asp Ile His His Leu Ser Cys Pro Asn Asn Leu Val Val Glu Asp
20 25 30
Glu Gly Cys Thr Asn Leu Ser Glu Phe Ser Tyr Met Glu Leu Lys Val
35 40 45
Gly Tyr Ile Ser Ala Ile Lys Val Asn Gly Phe Thr Cys Thr Gly Val
50 55 60
Val Thr Glu Ala Glu Thr Tyr Thr Asn Phe Val Gly Tyr Val Thr Thr
65 70 75 80
Thr Phe Lys Arg Lys His Phe Arg Pro Thr Pro Asp Ala Cys Arg Ala
85 90 95
Ala Tyr Asn Trp Lys Met Ala Gly Asp Pro Arg Tyr Glu Glu Ser Leu
100 105 110
His Asn Pro Tyr Pro Asp Tyr His Trp Leu Arg Thr Val Arg Thr Thr
115 120 125
Lys Glu Ser Leu Ile Ile Ile Ser Pro Ser Val Thr Asp Leu Asp Pro
130 135 140
Tyr Asp Lys Ser Leu His Ser Arg Val Phe Pro Gly Gly Lys Cys Ser
145 150 155 160
Gly Ile Thr Val Ser Ser Thr Tyr Cys Ser Thr Asn His Asp Tyr Thr
165 170 175
Ile Trp Met Pro Glu Asn Pro Arg Pro Arg Thr Pro Cys Asp Ile Phe
180 185 190
Thr Asn Ser Arg Gly Lys Arg Ala Ser Lys Gly Asn Lys Thr Cys Gly
195 200 205
Phe Val Asp Glu Arg Gly Leu Tyr Lys Ser Leu Lys Gly Ala Cys Arg
210 215 220
Leu Lys Leu Cys Glu Val Leu Gly Leu Arg Leu Met Asp Gly Thr Trp
225 230 235 240
Val Ala Met Gln Thr Ser Asp Glu Thr Lys Trp Cys Pro Pro Asp Gln
245 250 255
Leu Val Asn Leu His Asp Phe Arg Ser Asp Glu Ile Glu His Leu Val
260 265 270
Val Glu Glu Leu Val Lys Lys Arg Glu Glu Cys Leu Asp Ala Leu Glu
275 280 285
Ser Ile Met Thr Thr Lys Ser Val Ser Phe Arg Arg Leu Ser His Leu
290 295 300
Arg Lys Leu Val Pro Gly Phe Gly Lys Ala Tyr Thr Ile Phe Asn Lys
305 310 315 320
Thr Leu Met Glu Ala Asp Ala His Tyr Lys Ser Val Arg Thr Trp Asn
325 330 335
Glu Ile Ile Pro Ser Lys Gly Cys Leu Lys Val Gly Gly Arg Cys His
340 345 350
Pro His Val Asn Gly Val Phe Phe Asn Gly Ile Ile Leu Gly Pro Asp
355 360 365
Gly His Val Leu Ile Pro Glu Met Gln Ser Ser Leu Leu Gln Gln His
370 375 380
Met Glu Leu Leu Lys Ser Ser Val Ile Pro Leu Met His Pro Leu Ala
385 390 395 400
Asp Pro Ser Thr Val Phe Lys Glu Gly Asp Glu Ala Glu Asp Phe Val
405 410 415
Glu Val His Leu Pro Asp Val Tyr Lys Gln Ile Ser Gly Val Asp Leu
420 425 430
Gly Leu Pro Asn Trp Gly Lys Tyr Gly Gly Gly Gly Ser Val Ser Arg
435 440 445
Leu Glu Glu Glu Met Arg Lys Leu Gln Ala Thr Val Gln Glu Leu Gln
450 455 460
Lys Arg Leu Asp Arg Leu Glu Glu Thr Val Gln Ala Lys Ala Ser Gly
465 470 475 480
Ala Ala His His His His His His
485
<210> 41
<211> 488
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 41
Lys Phe Pro Ile Tyr Thr Ile Pro Asp Lys Leu Gly Pro Trp Ser Pro
1 5 10 15
Ile Asp Ile His His Leu Ser Cys Pro Asn Asn Leu Val Val Glu Asp
20 25 30
Glu Gly Cys Thr Asn Leu Ser Glu Phe Ser Tyr Met Glu Leu Lys Val
35 40 45
Gly Tyr Ile Ser Ala Ile Lys Val Asn Gly Phe Thr Cys Thr Gly Val
50 55 60
Val Thr Glu Ala Glu Thr Tyr Thr Asn Phe Val Gly Tyr Val Thr Thr
65 70 75 80
Thr Phe Lys Arg Lys His Phe Arg Pro Thr Pro Asp Ala Cys Arg Ala
85 90 95
Ala Tyr Asn Trp Lys Met Ala Gly Asp Pro Arg Tyr Glu Glu Ser Leu
100 105 110
His Asn Pro Tyr Pro Asp Tyr His Trp Leu Arg Thr Val Arg Thr Thr
115 120 125
Lys Glu Ser Leu Ile Ile Ile Ser Pro Ser Val Thr Asp Leu Asp Pro
130 135 140
Tyr Asp Lys Ser Leu His Ser Arg Val Phe Pro Gly Gly Lys Cys Ser
145 150 155 160
Gly Ile Thr Val Ser Ser Thr Tyr Cys Ser Thr Asn His Asp Tyr Thr
165 170 175
Ile Trp Met Pro Glu Asn Pro Arg Pro Arg Thr Pro Cys Asp Ile Phe
180 185 190
Thr Asn Ser Arg Gly Lys Arg Ala Ser Lys Gly Asn Lys Thr Cys Gly
195 200 205
Phe Val Asp Glu Arg Gly Leu Tyr Lys Ser Leu Lys Gly Ala Cys Arg
210 215 220
Leu Lys Leu Cys Gly Val Leu Gly Leu Arg Leu Met Asp Gly Thr Trp
225 230 235 240
Val Ala Met Gln Thr Ser Asp Glu Thr Lys Trp Cys Pro Pro Asp Gln
245 250 255
Leu Val Asn Leu His Asp Phe Arg Ser Asp Glu Ile Glu His Leu Val
260 265 270
Val Glu Glu Leu Val Lys Lys Arg Glu Glu Cys Leu Asp Ala Leu Glu
275 280 285
Ser Ile Met Thr Thr Lys Ser Val Ser Phe Arg Arg Leu Ser His Leu
290 295 300
Arg Lys Leu Val Pro Gly Phe Gly Lys Ala Tyr Thr Ile Phe Asn Lys
305 310 315 320
Thr Leu Met Glu Ala Asp Ala His Tyr Lys Ser Val Arg Thr Trp Asn
325 330 335
Glu Thr Ile Pro Ser Lys Gly Cys Leu Lys Val Gly Gly Arg Cys His
340 345 350
Pro His Val Asn Gly Val Phe Phe Asn Gly Ile Ile Leu Gly Pro Asp
355 360 365
Gly His Val Leu Ile Pro Glu Met Gln Ser Ser Leu Leu Gln Gln His
370 375 380
Met Glu Leu Leu Lys Ser Ser Val Ile Pro Leu Met His Pro Leu Ala
385 390 395 400
Asp Pro Ser Thr Val Phe Lys Glu Gly Asp Glu Ala Glu Asp Phe Val
405 410 415
Glu Val His Leu Pro Asp Val Tyr Lys Gln Ile Ser Gly Val Asp Leu
420 425 430
Gly Leu Pro Asn Trp Gly Lys Tyr Gly Gly Gly Gly Ser Val Ser Arg
435 440 445
Leu Glu Glu Glu Met Arg Lys Leu Gln Ala Thr Val Gln Glu Leu Gln
450 455 460
Lys Arg Leu Asp Arg Leu Glu Glu Thr Val Gln Ala Lys Ala Ser Gly
465 470 475 480
Ala Ala His His His His His His
485
<210> 42
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<400> 42
Arg Tyr Thr Ile Asn
1 5
<210> 43
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<400> 43
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Arg Phe Gln
1 5 10 15
Gly
<210> 44
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<400> 44
Glu Asn Leu Asp Asn Ser Gly Thr Tyr Tyr Tyr Tyr Phe Ser Gly Trp
1 5 10 15
Phe Asp Pro
<210> 45
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<400> 45
Thr Gly Thr Ser Ser Asp Ile Gly Ala Tyr Asp Tyr Val Ser
1 5 10
<210> 46
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<400> 46
Asp Ala Thr Lys Arg Pro Ser
1 5
<210> 47
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<400> 47
Cys Ser Tyr Ala Gly Asp Tyr Thr Pro Gly Val Val
1 5 10
<210> 48
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<400> 48
Arg Tyr Ser Ile Asn
1 5
<210> 49
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<400> 49
Thr Gly Thr Ser Ser Asp Ile Asp Gly Tyr Asp Phe Val Ser
1 5 10
<210> 50
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<400> 50
Gly Tyr Thr Ile Asn
1 5
<210> 51
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<400> 51
Thr Gly Thr Ser Ser Asp Leu Gly Gly Tyr Asp Phe Val Ser
1 5 10
<210> 52
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<400> 52
Ser Tyr Gly Met His
1 5
<210> 53
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<400> 53
Thr Ile Ser Tyr Asp Gly Ser Ile Lys Asp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 54
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<400> 54
Gly Asp Arg Thr Gly Asn Leu Asp Tyr
1 5
<210> 55
<211> 11
<212> ПРТ
<213> Искусственная последовательность
<400> 55
Arg Ala Ser Gln Asn Ile Arg Asn Ala Leu Asn
1 5 10
<210> 56
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<400> 56
Asp Ala Ser Thr Arg Gln Ser
1 5
<210> 57
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<400> 57
Gln Gln Asn Ser Glu Phe Pro Pro Thr
1 5
<210> 58
<211> 16
<212>ПРТ
<213>Искусственнаяпоследовательность
<400> 58
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 59
<211> 28
<212> ДНК
<213> Искусственная последовательность
<400> 59
ccagccatgg cgcaggtgca gctggtgc 28
<210> 60
<211> 60
<212> ДНК
<213> Искусственная последовательность
<400> 60
ctggggcctg ccgcacccag yyganasyaw atbygyyawa ggtgccgccg ctggccttgc 60
<210> 61
<211> 19
<212> ДНК
<213> Искусственная последовательность
<400> 61
tgggtgcggc aggccccag 19
<210> 62
<211> 54
<212> ДНК
<213> Искусственная последовательность
<400> 62
tgctgctgat accagctcac awagyyawaa bcabcgangt cgctgctggt gccg 54
<210> 63
<211> 20
<212> ДНК
<213> Искусственная последовательность
<400> 63
gtgagctggt atcagcagca 20
<210> 64
<211> 32
<212> ДНК
<213> Искусственная последовательность
<400> 64
gatgtgcggc cgccaggacg gtaagcttgg tg 32
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| АНТИ-TSLP АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2834227C1 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛА К PCSK-9, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2782792C2 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО К SOST, И ЕЕ ПРИМЕНЕНИЯ | 2018 |
|
RU2779430C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ АНТИТЕЛА К PD-L1 И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2766590C2 |
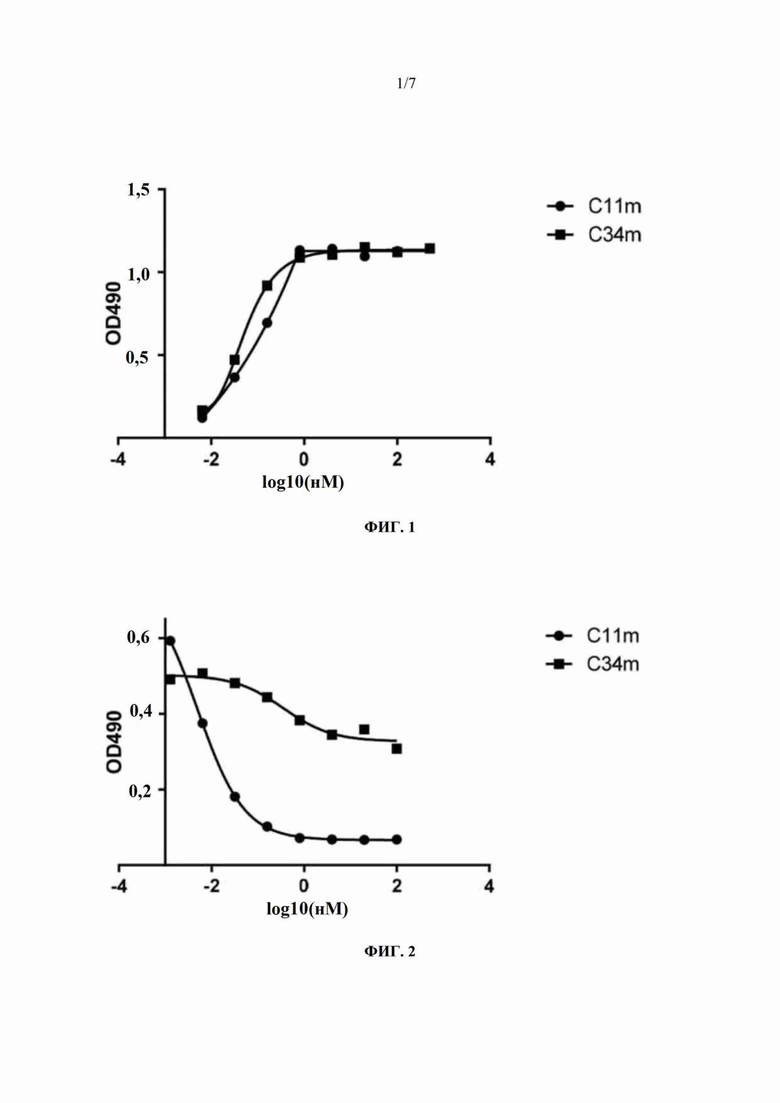
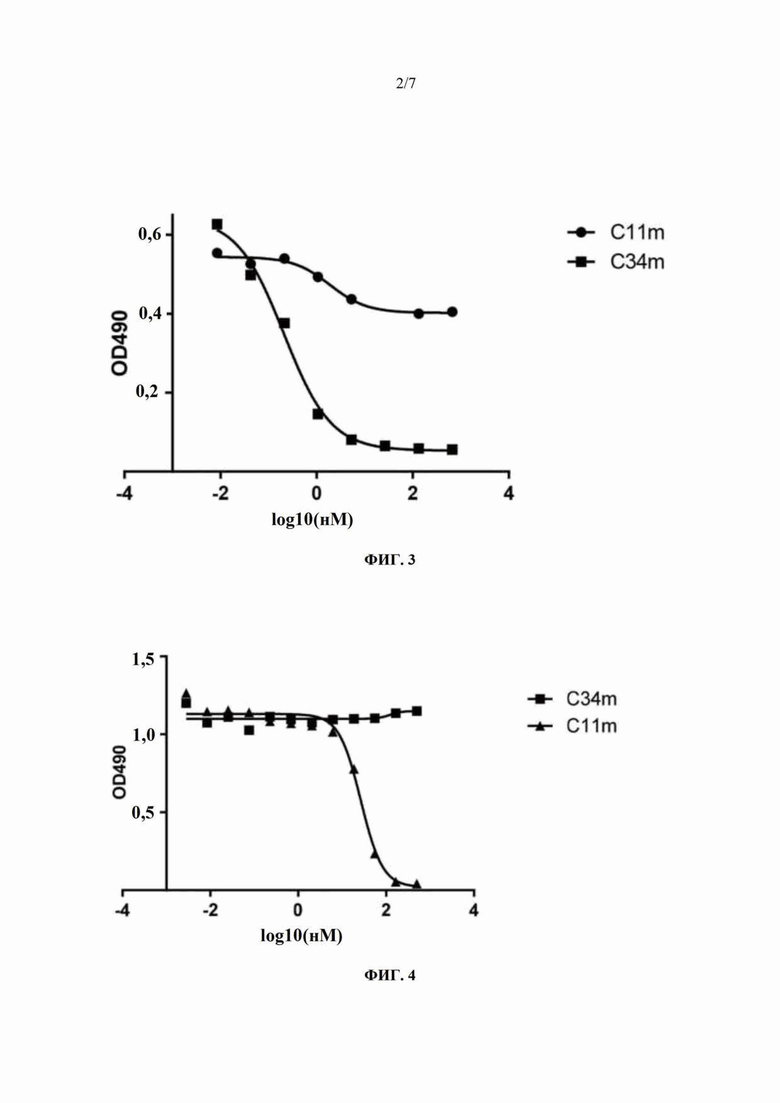
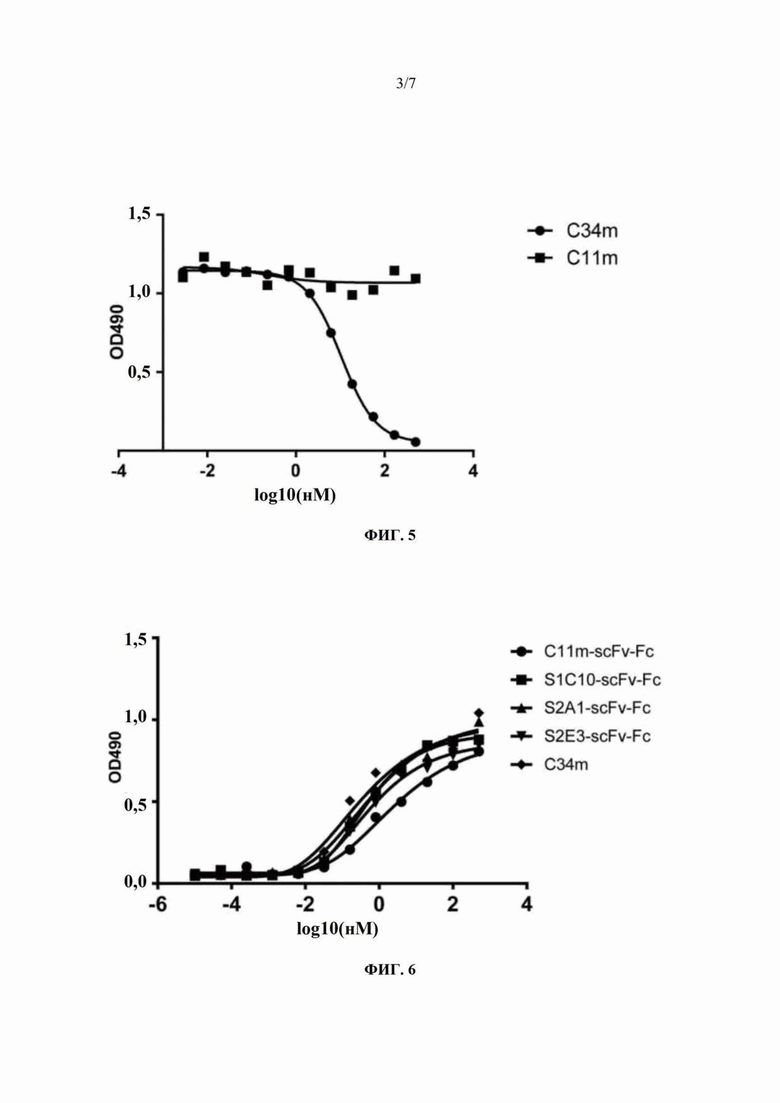
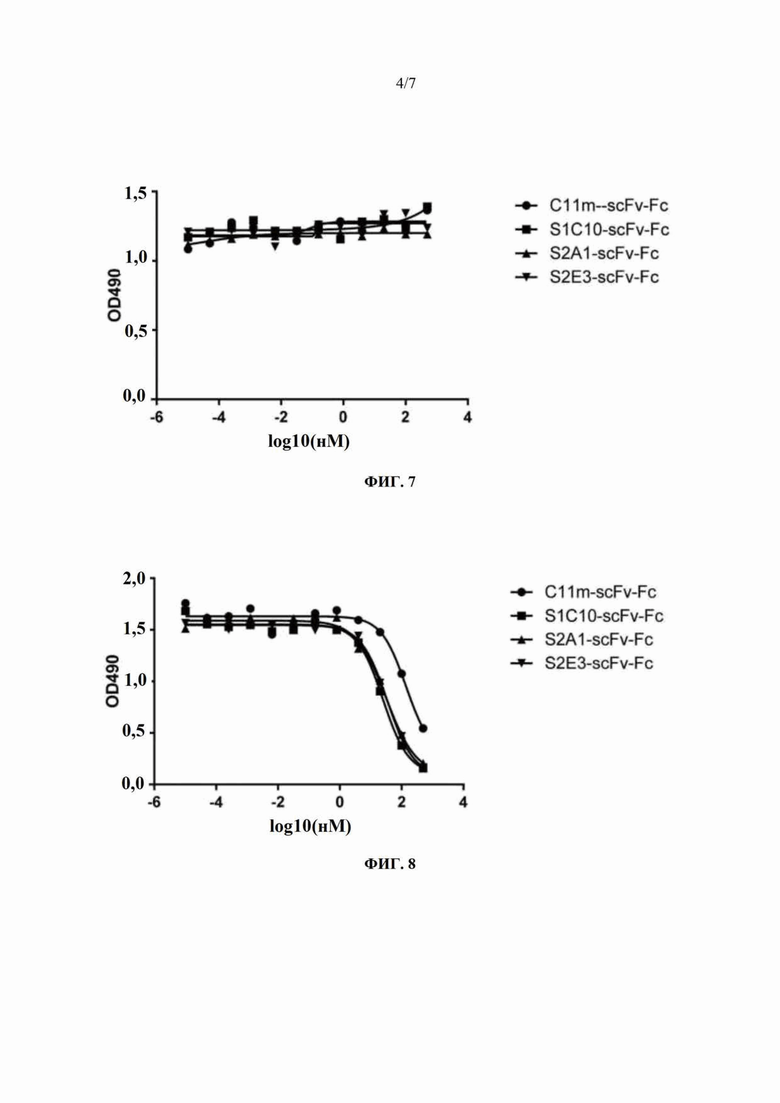

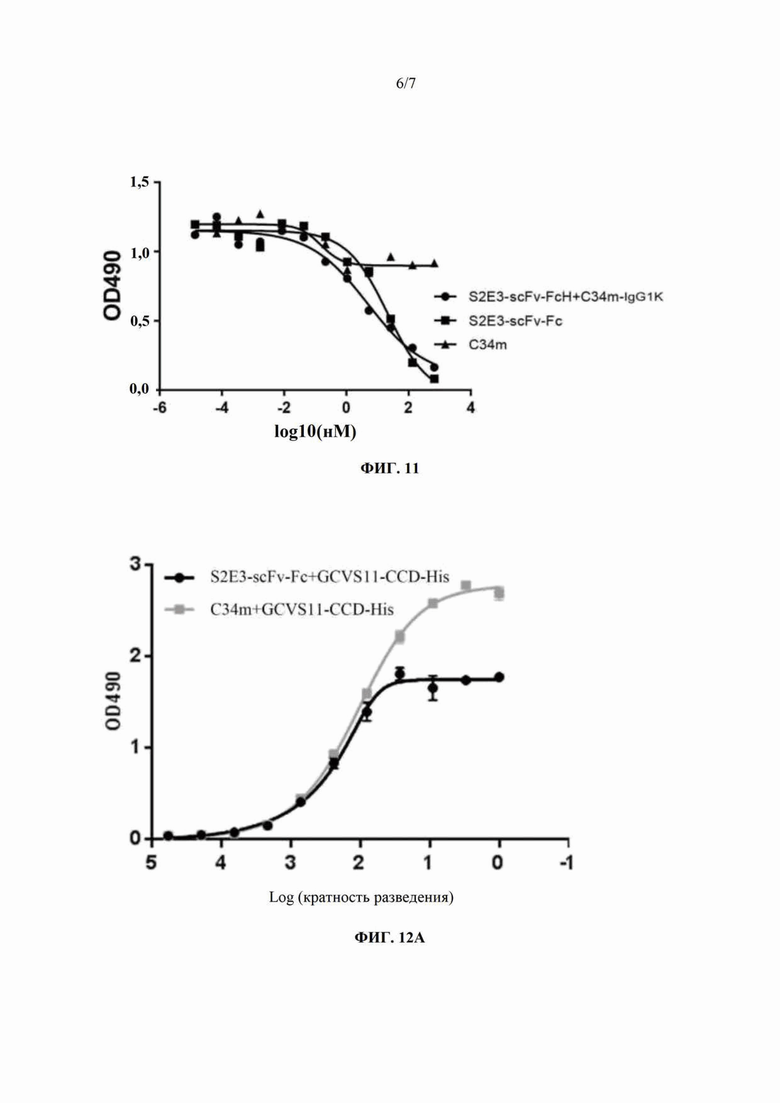
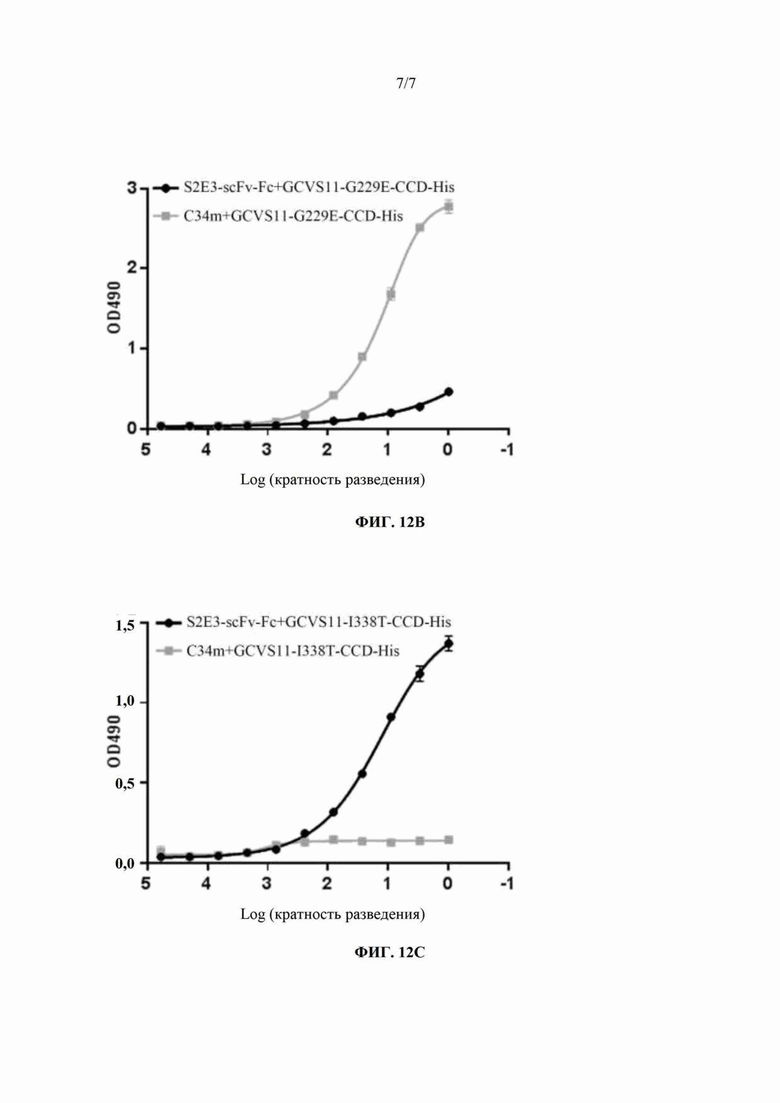
Изобретение относится к области биотехнологии. Предложено биспецифическое антитело против вируса бешенства. Биспецифическое антитело содержит два антиген-связывающих фрагмента, которые связываются с эпитопом I и эпитопом III G-белка вируса бешенства, и обладает нейтрализующей вирус бешенства активностью. Также изобретение относится к фармацевтической композиции, включающей указанное антитело, и способу предупреждения или лечения бешенства. 3 н. и 5 з.п. ф-лы, 14 ил., 4 табл., 9 пр.
1. Биспецифическое антитело, содержащее два антиген-связывающих фрагмента, которые связываются с эпитопом I и эпитопом III G-белка вируса бешенства, соответственно, где биспецифическое антитело обладает активностью нейтрализации вируса бешенства,
где антиген-связывающий фрагмент, который связывается с эпитопом I G-белка вируса бешенства, содержит:
HCDR1 (определяющий комплементарность участок 1 тяжелой цепи), имеющий аминокислотную последовательность RYTIN (SEQ ID NO: 42), HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG (SEQ ID NO: 43), HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP (SEQ ID NO: 44), LCDR1 (определяющий комплементарность участок1 легкой цепи), имеющий аминокислотную последовательность TGTSSDIGAYDYVS (SEQ ID NO: 45), LCDR2, имеющий аминокислотную последовательность DATKRPS (SEQ ID NO: 46), LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV (SEQ ID NO: 47); или
HCDR1, имеющий аминокислотную последовательность RYSIN (SEQ ID NO: 48), HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG (SEQ ID NO: 43), HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP (SEQ ID NO: 44), LCDR1, имеющий аминокислотную последовательность TGTSSDIDGYDFVS (SEQ ID NO: 49), LCDR2, имеющий аминокислотную последовательность DATKRPS (SEQ ID NO: 46), LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV (SEQ ID NO: 47); или
HCDR1, имеющий аминокислотную последовательность GYTIN (SEQ ID NO: 50), HCDR2, имеющий аминокислотную последовательность GIIPIFGTANYAQRFQG (SEQ ID NO: 43), HCDR3, имеющий аминокислотную последовательность ENLDNSGTYYYYFSGWFDP (SEQ ID NO: 44), LCDR1, имеющий аминокислотную последовательность TGTSSDLGGYDFVS (SEQ ID NO: 51), LCDR2, имеющий аминокислотную последовательность DATKRPS (SEQ ID NO: 46), LCDR3, имеющий аминокислотную последовательность CSYAGDYTPGVV (SEQ ID NO: 47); и
где антиген-связывающий фрагмент, который связывается с эпитопом III G-белка вируса бешенства, содержит:
HCDR1, имеющий аминокислотную последовательность SYGMH (SEQ ID NO: 52), HCDR2, имеющий аминокислотную последовательность TISYDGSIKDYADSVKG (SEQ ID NO: 53), HCDR3, имеющий аминокислотную последовательность GDRTGNLDY (SEQ ID NO: 54), LCDR1, имеющий аминокислотную последовательность RASQNIRNALN (SEQ ID NO: 55), LCDR2, имеющий аминокислотную последовательность DASTRQS (SEQ ID NO: 56), LCDR3, имеющий аминокислотную последовательность QQNSEFPPT (SEQ ID NO: 57);
где аминокислотные последовательности HCDR и LCDR определены согласно Kabat.
2. Биспецифическое антитело по п. 1, где
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 24, а аминокислотная последовательность вариабельной области легкой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 25; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 26, а аминокислотная последовательность вариабельной области легкой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 27; или
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 28, а аминокислотная последовательность вариабельной области легкой цепи антиген-связывающего фрагмента, который связывается с эпитопом I G-белка вируса бешенства, показана в SEQ ID NO: 29.
3. Биспецифическое антитело по п. 1, где
аминокислотная последовательность вариабельной области тяжелой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, показана в SEQ ID NO: 1, а аминокислотная последовательность вариабельной области легкой цепи антиген-связывающего фрагмента, который связывается с эпитопом III G-белка вируса бешенства, показана в SEQ ID NO: 3.
4. Биспецифическое антитело по п. 1, где формы двух антиген-связывающих фрагментов независимо выбраны из одноцепочечного антитела (scFv) или Fab-фрагмента.
5. Биспецифическое антитело по п. 4, где антиген-связывающий фрагмент, который связывается с эпитопом I G-белка вируса бешенства, представляет собой одноцепочечное антитело (scFv), а антиген-связывающий фрагмент, который связывается с эпитопом III G-белка вируса бешенства, представляет собой Fab-фрагмент.
6. Биспецифическое антитело по п. 5, которое содержит аминокислотную последовательность, приведенную в одной из SEQ ID NO: 32, 33 и 34, и/или содержит аминокислотную последовательность, приведенную в SEQ ID NO: 30 и SEQ ID NO: 31.
7. Фармацевтическая композиция для предупреждения или лечения бешенства, содержащая биспецифическое антитело по п. 1 и фармацевтически приемлемый эксципиент, разбавитель или носитель.
8. Способ предупреждения или лечения бешенства, включающий введение субъекту, нуждающемуся в этом, биспецифического антитела по п. 1.
| WO 2009147248 A2, 10.12.2009 | |||
| WO 2009147248 A2, 10.12.2009 | |||
| US 7740852 B2, 22.06.2010 | |||
| МОЛОДКИН А.В | |||
| и др., Моноклональные антитела к вирусу бешенства, Ветеринарная Патология, 2006, Номер 4 (19), с.152-160. |

