Настоящее изобретение относится к анализу и к соответствующему набору для оценки эффективности доставки молекулы в эукариотическую клетку.
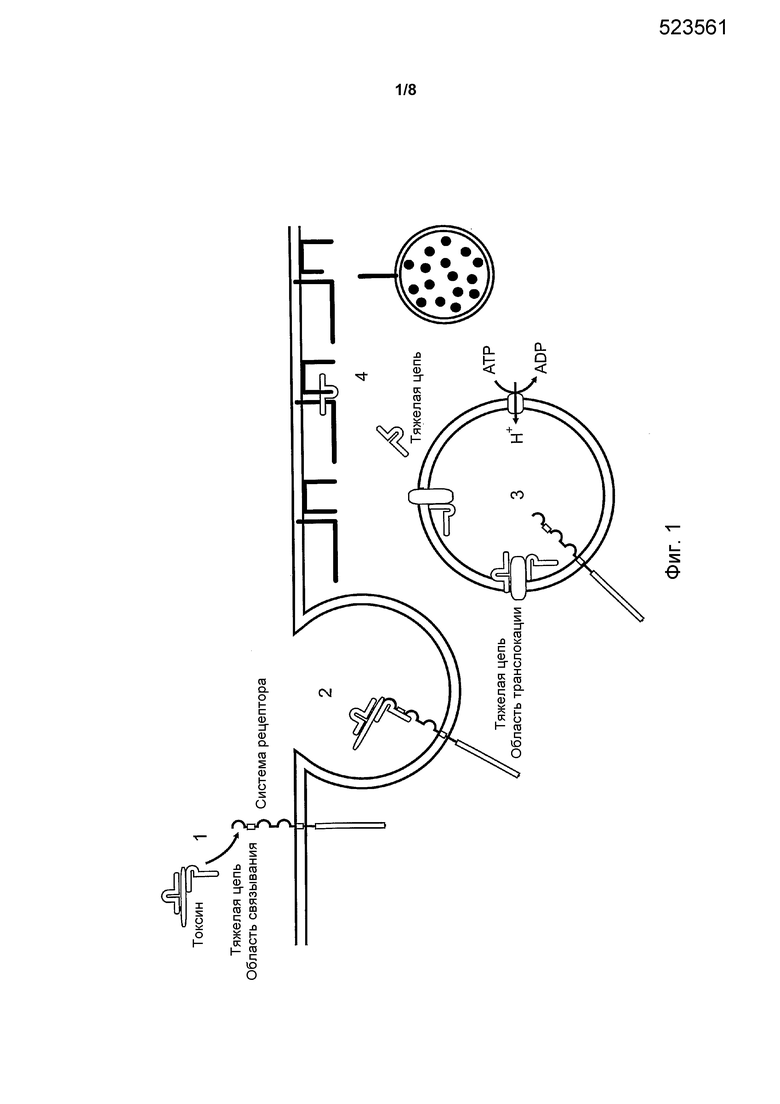
Ключевым требованием для активности любой потенциальной терапевтической молекулы является то, что она должна быть способна демонстрировать хорошую активность. Настоящее изобретение относится к молекулам, которые доходят до попадания в цитозоль эукариотических клеток через хорошо известный процесс эндоцитоза. Имея это в виду, становится важным понимать этапы, затрагивающие режим введения в клетку. Таким образом, для содействия иллюстрации ключевых этапов, связанных с этим режимом введения в клетку, следует обратиться к Фиг. 1. На этапе 1 молекула связывается с центром связывания (например, с рецептором или акцептором), присутствующим на поверхности клетки. На этапе 2 рецептор (плюс связанная молекула) становится интернализованным в клетку, этот этап обычно называется «эндоцитозом» или «формированием эндосомы». На этапе 3, вслед за интернализацией, молекула попадает в эндосомальную мембрану, и происходит высвобождение молекулы (или ее части) из внутри эндосомы, через эндосомальную мембрану и с попаданием в цитозоль эурокариотической клетки. Сразу после попадания в цитозоли (этап 4), молекула становится способной воздействовать на ее внутриклеточкую мишень (например, ингибирование молекулы-мишени, такое как протолитическое расщепление клеточного белка-мишени).
Поэтому, хорошее тестирование активности основано на точной оценке одного или более из вышеуказанных этапов.
До настоящего времени, возможное тестирование молекул, способных попадать в эукариотическую клетку, через «эндоцитоз, опосредуемый рецептором», был сфокусирован на молекулах токсина, таких как клостридиальные нейротоксины. В качестве примера, для серийно выпускаемых клостридиальных нейротоксинов (например, нейротоксина ботулина, который выпускается на рынок под такими наименованиями, как Dysport™, Neurobloc™ и Botox™), были использованы следующие анализы.
Анализ LD50 на мыши в настоящее время является единственным анализом, апробированным FDA (Food and Drug Administration, Управление по контролю за продуктами и лекарствами), для исследования высвобождения ботулинических токсинов. Анализ одновременно позволяет исследовать действие всех трех доменов ботулинического нейротоксина (т.е. связывания, транслокации и протеазы). Более подробно, анализ определяет среднюю смертельную внутрибрюшинную дозу токсина в заданный момент времени, обычно 2-4 дня спустя, после введения дозы (активность выражена в единицах LD50 для мыши). Однако, к сожалению, в анализах LD50 приходится использовать большие количества животных. Более того, единицы LD50 не являются абсолютными измерениями, поскольку они не являются биологическими константами, как таковые, они сильно зависят от условия анализа. В частности, ошибки, связанные с этим анализом, могут составлять вплоть до 60% среди различного испытательного оборудования (Sesardic et al. 2003; Biologicals 31 (4):265-276).
Анализ на периферический паралич, проводимое на мышах, которое также известен как «анализ на абдоминальный птоз, проводимый на мышах», связывает активность ботулинического токсина с уровнем вспучивания живота, наблюдаемого после того, как токсин был подкожно введен в левую бедренно-паховую область у мыши, степень паралича зависит от дозировки. Этот подход был предложен в качестве усовершенствования анализа LD50, проводимого на мышах, поскольку он основан на гуманных соображениях. Этот анализ является приблизительно в 10 раз более чувствительным, чем анализ LD50, в нем используется сублетальная доза токсина, и он является более быстрым, чем анализ LD50, поскольку он обеспечивает результат в 24-48 часов, по сравнению с 72-96 часами для типичного анализа LD50. Результаты, полученные из этого анализа, показывают исключительное совпадение со значениями LD50 (Sesardic и др., 1996). Хотя в этом анализе использовано 20% животных, используемых в анализе LD50, оно еще делает необходимым использование животных.
Такие анализы, как анализ диафрагмального нерва купола диафрагмы у мышей/крыс (которые основаны на использовании нервно-мышечных препаратах вне организма) связывают активность ботулинического нейротоксина с понижением амплитуды судорог, в ответ на действие препарата после его введения в стабилизирующую среду. Обычной конечной точкой анализа является достижение времени, требуемого для того, чтобы можно было наблюдать 50%-ное снижение амплитуды. Однако, к сожалению, анализ купола диафрагмы (например, анализ LD50) приводит к использованию больших количеств животных. В дополнение, для анализа требуется высококвалифицированный персонал, натренированный в использовании сложного и дорогого оборудования.
Анализы по расщеплению субстрата, с использованием культивированных нейронов спинного мозга, связывают активность ботулинического нейротоксина с расщеплением специфического белка, присутствующего в упомянутых нейронах. Притом, что в анализе используется меньшее количество животных, чем в анализах в живом организме (LD50, анализ на периферический паралич, проводимый на мышах) и вне живого организма (анализы на куполе диафрагмы), для анализа требуется высококвалифицированный персонал, для выполнения рассечения и выращивания культур, так как технологии рассечения и выращивания культур являются продолжительными, и они должны быть запланированы примерно за 3 недели заранее, тогда, когда они требуются. Другим недостатком является то, что измерения по расщеплению субстрата могут быть сильно изменчивыми.
Все вышеприведенные анализы имеют конкретные недостатки, в частности, связанные с вопросами жизнеобеспечения животных и/или с ограничениями для тестируемых молекул, которые связаны с нейромышечным соединением (neuromuscular junction, NMJ), последнее представляет собой клетки-мишени, с которыми связывается природный клостридиальный нейротоксин.
В WO 95/33850 описан анализ по расщеплению бесклеточного субстрата, в котором расщепленный продукт выявляют с помощью антитела, специфичного для эпитопа. Поскольку антитело является пригодным для различения между собой расщепленного субстрата и нерасщепленного белка субстрата, можно количественно определить активность клостридиального нейротоксина. Притом, что этот анализ не страдает идентифицированным выше ущербом для жизнеобеспечения животного, или от специфических недостатков для NMJ, это, несомненно, «бесклеточный» анализ и, как таковой, он пригоден лишь для оценки активности в единственном контексте расщепления белка. Следовательно, исходя из WO 95/33850, нельзя оценить активность, применительно к любому одному или более в равной степени важных этапов, а именно: связывание клетки; формирование эндосомы; или транслокация через эндосомальную мембрану. Если пойти другим путем, идентификация молекулы (согласно WO 95/33850) в качестве эффективной молекулы, расщепляющей субстрат, может обладать небольшой или вообще никакой полезной терапевтической активностью, например, из-за ее недостатков, связанных с любым одним или более свойствами: оптимальным связыванием клетки; оптимальным формированием эндосомы; и/или оптимальной функцией транслокации.
Поэтому, в данной области техники возникает необходимость в альтернативном и/или усовершенствованном анализе активности, которое обращается к одной или более идентифицированным выше проблемам. Например, возникает необходимость в гуманном анализе, что решает существующие проблемы, связанные с благополучием животных. Аналогично, существует необходимость в анализе, который не ограничен тестированием молекул, которые специфическим образом связываются с NMJ и/или с нейронными клетками. Аналогично, существует необходимость в анализе, который обеспечивает надежные результаты по активности для молекул, в частности, для анализа, который обеспечивает результаты по активности, отражающие активность препарата в живом организме.
Настоящее изобретение обращается к идентифицированным выше проблемам путем обеспечения анализа, который включает:
i) приведение эукариотной клетки в контакт с тестовой молекулой, которая подлежит оценке на способность высвобождения эндосомой, причем упомянутая эукариотная клетка содержит клеточную мембрану, включающую в себя центр связывания, присутствующий на внешней поверхности клеточной мембраны упомянутой клетки;
ii) инкубирование тестовой молекулы с упомянутой эукариотной клеткой, что позволяет, таким образом,
a) связывать тестовую молекулу и образовывать связанный комплекс с центром связывания, присутствующим на эукариотной клетке, что позволяет, таким образом, упомянутому связанному комплексу попадать в эукариотную клетку, за счет эндоцитоза;
b) формировать одну или более эндосом в упомянутой клетке, причем одна или более эндосом содержит тестовую молекулу; и
c) упомянутой тестовой молекуле попадать в цитозоль эукариотной клетки сквозь эндосомальную мембрану одной или более эндосом;
iii) удаление избыточной тестовой молекуле, которая не связана с центрами связывания, присутствующими на эукариотных клетках;
iv) после истечения заданного периода времени, детектирование количества тестовых молекул, присутствующих в одной или более эндосомах, или детектирование количества тестовых молекул, присутствующих в цитозоли упомянутой эукариотной клетки;
v) сопоставление количества тестовых молекул, выявленных на этапе iv), с контрольным значением, причем упомянутое контрольное значение отображает количество тестовых молекул, присутствующих в одной или более эндосомах, или количество тестовых молекул, присутствующих в цитозоли перед выполнением этапа iv);
vi) расчет параметра высвобождения эндосомы для тестовой молекулы, путем определения относительного изменения количества тестовых молекул, которые присутствуют в одной или более эндосомах, или путем определения относительного изменения количества тестовых молекул, присутствующих в цитозоли упомянутой эукариотной клетки.
Эукариотная клетка может быть выбрана из клетки дрожжей, клетки насекомого, клетки позвоночного животного, клетки млекопитающего, растительной клетки и грибной клетки. Примеры таких животных клеток включают в себя клетки человека, грызуна, мыши и хомяка.
В одном варианте воплощения центр связывания эукариотной клетки является пригодным либо для эндоцитоза, опосредуемого рецептором, либо для эндоцитоза, не опосредуемого рецептором. Центр связывания может представлять собой рецептор или акцептор. Примеры пептидных последовательностей, подвергнутых эндоцитозу через нерецепторные центры связывания, включают в себя пептиды, имеющие последовательности, обогащенные аргинином (например, RRRRRRRR, RRRRRRRW), и такие пептиды, как PHLIP, Pep-1, SAPE, PFVYLI и AAVALIPAVILALLAP, производные от Kaposi FGF. Считается, что эти пептиды должны подвергаться эндоцитозу в клетках через неспецифические ионные взаимодействия. Центр связывания может представлять собой центр связывания, который возникает естественным путем на поверхности эукариотной клетки. В качестве альтернативы, эукариотная клетка может представлять собой рекомбинантную эукариотную клетку, которая была модифицирована для экспрессии центра связывания, который мог не возникнуть естественным путем на поверхности упомянутой эукариотной клетки.
Этап инкубирования ii) может длиться в течение любого заданного периода времени, например, для периода времени от 5 минут до 5 дней. Типичный период времени составляет 1-12 часов, например 2-10 часов, 4-8 часов, или 6-8 часов. В течение этого периода, эукариотная клетка (т.е. внешняя поверхность клеточной мембраны) обнажена для тестовой молекулы (как правило, избыточной тестовой молекулы) с результатом, который достигается в «стабильном состоянии», причем тестовая молекула попадает во внутриклеточные эндосомы и выходит из них приблизительно при той же скорости. Этот момент времени отображает оптимальный момент времени, в который выполняются этапы (iii) и/или (iv).
Этап (iii) влечет за собой уменьшение или удаление источника тестовой молекулы, внешнего по отношению к эукариотной клетке, со снижением, таким образом, количества (или по существу, с предотвращением) тестовых молекул, попадающих в клетку. Упомянутое снижения количества тестовых молекул, попадающих в эукариотную клетку, в свою очередь, обеспечивает изменение количества тестовых молекул, попадающих в эндосомы, что, в свою очередь, приводит к изменению в количестве (или скорости) тестовой молекулы, покидающие эндосомы, и/или попадающей в цитозоль эукариотной клетки. Это является количеством (или скоростью) тестовой молекулы, покидающей структуры эндосом, которые обеспечивают основные анализы согласно настоящему изобретению, упомянутое количество (или скорость) тестовой молекулы, покидающей структуры эндосом, могут быть измерены путем изменения количества тестовых молекул, присутствующих в эндосомех, и/или путем изменения в количестве тестовых молекул, присутствующих в цитозоли. При измерении количества тестовых молекул, присутствующих в эндосомех, как правило, наблюдается снижение количества имеющихся тестовых молекул. При измерении количества тестовых молекул, присутствующих в цитозоли, можно наблюдать повышение или понижение количества тестовых молекул, присутствующих в цитозоли. В качестве примера, повышение количества тестовых молекул в цитозоли можно наблюдать, когда этап (iii) начинается перед установлением стабильного состояния эндосомального переноса тестовой молекулы. В качестве альтернативы, понижение количества тестовых молекул в цитозоли можно наблюдать, когда скорость выхода тестовой молекулы из эукариотной клетки при клеточной секреции превышает скорость эндосомального переноса тестовой молекулы из эндосом в цитозоль.
Эукариотные клетки, используемые в анализе, могут быть иммобилизованы на поверхности. Иммобилизация клеток может быть выполнена как этап предварительного анализа (т.е. предварительной иммобилизации), или может быть выполнена как часть протокола анализа. Таким образом, в одном варианте воплощения клетки, подлежащие анализу, являются иммобилизованными. Иммобилизация эукариотных клеток может быть выполнена любым стандартным средством. В качестве примера, клетки вводят в качестве затравок в аналитические планшеты при высокой плотности и позволяют им прилипнуть перед проведением анализа. В качестве альтернативы, клетки вводят в качестве затравок в аналитические планшеты и культивируются в течение нескольких дней перед использованием для обеспечения слившегося монослоя. Приклеивание клетки может быть усилено за счет использования стандартных покрытий, таких как пластины, покрытые поли-D-лизином.
В одном варианте воплощения иммобилизация эукариотных клеток может быть выполнена перед или в ходе этапа (iii), с обеспечением, таким образом, простого средства для отделения упомянутых клеток от свободной (например, несвязанной или экзогенной) тестовой молекулы. В качестве альтернативы, иммобилизация может быть выполнена после этапа (iii), например, для облегчения выполнения этапа детектирования (iv).
Этап (iii) может включать в себя этап фильтрования или этап осуществления сродства к лиганду, в ходе которого эукариотные клетки отделяют от избыточной (например, несвязанной или экзогенной) тестовой молекулы. Этап (iii) может включать в себя этап промывания, при котором избыточную (например, несвязанную или экзогенную) тестовую молекулу отмывают от эукариотной клетки, например, с использованием стандартного буферного раствора. Избыточная тестовая молекула предназначена играть роль средней тестовой молекулы, которая присутствуют в среде для количественного определения, внешней по отношению к эукариотным клеткам, и которая еще не была связана с центром связывания, присутствующим на поверхности эукариотных клеток.
Детектирование тестовой молекулы на этапе (iv), как правило, выполняют вскоре после этапа (iii). В качестве примера, типичные временные рамки для этапа (iv) составляют от 5 минут до 5 часов, вслед за этапом (iii). В одном варианте воплощения, этап (iv) выполняют в течение 15-240 минут или 30-180 минут, или 45-150, минут вслед за этапом (iii). Этап детектирования (iv) может быть повторен в течение нескольких периодов времени, например, в течение интервалов 10 минут или 15 минут или 30 минут, это позволит рассчитать скорость эндосомального высвобождения.
Этап детектирования (iv) может быть выполнен с помощью любого стандартного средства. В качестве примера, детектирование, независимое от пермеабилизации, может быть достигнуто путем мониторинга самой тестовой молекулы. Это может быть достигнуто путем маркировки тестовой молекулы флуофором. Подходящие флуофоры, которые могут быть прикреплены к тестовой молекуле, включают в себя флуоресцин, родамин, зеленый флуоресцентный белок, одностенные углеродные нанотрубки и alexa-fluor®488.
Детектирование тестовой молекулы может происходить на основе внутриклеточной локализации упомянутой тестовой молекулы, примером такой локализации в цитозоли является локализация у ядра клетки. В этом варианте воплощения тестовую молекулу помечают (например, флуофором), а также она обладает сигналом ядерной локализации. Таким образом, метка может быть выявлена в эндосоме и при выходе из эндосомы, определяемом путем мониторинга накопления метки (например, за счет флуоресценции) в ядре клетки. С этой точки зрения, само по себе ядро может быть помечено красителем, стандартные ядерные красители хорошо известны из уровня техники (например, пятно Hoescht).
Для мониторинга тестовой молекулы может быть использовано любое стандартное средство детектирования, такое как метод резонансного переноса энергии флуоресценции (Fluorescence Resonance Energy Transfer, FRET). В качестве примера, пара донор/акцептор может представлять собой голубой флуоресцентный белок (ГФБ) и желтый флуоресцентный белок (ЖФБ). Из области техники хорошо известны и другие пары донор/акцептор, пригодные для использования в FRET. В одном варианте воплощения может быть выявлена тестовая молекула, помеченная ГФБ и ЖФБ, и может быть определено количество тестовых молекул, присутствующих в эндосоме и/или в цитозоли. В одном варианте воплощения могут быть проконтролированы потери сигнала FRET. Например, из эндосомы в цитоплазму должна быть перемещена только часть тестовых молекул, и это может привести к потерям сигнала FRET, если пара донор/акцептор (например, ГФБ и ЖФБ) оказываются разделенными. Таким образом, например, тестовая молекула может содержать (или состоять из) L-цепи и области HN клостридиального нейротоксина (или его производных, или функциональных эквивалентов, например, TSI). Донорные/акцепторные метки (например, ГФБ и ЖФБ) могут быть помещены на упомянутые два компонента, например, L-цепь может быть помечена ГФБ, а компонент HN может быть помечен ЖФБ. В одном варианте воплощения сигнал FRET является детектируемым, притом что тестовая молекула, содержащая LHN, присутствует в эндосоме клетки. Функция транслокации компонента HN затем способствует высвобождению терапевтической частицы (т.е. L-цепи) в цитозоль, а сигнал FRET исчезает, поскольку метки ГФБ и ЖФБ становятся отделенными друг от друга.
Для мониторинга/детектирования тестовой молекулы также может быть использован метод резонансного переноса энергии биолюминесценции (Bioluminescence Resonance Energy Transfer, BRET). В BRET донорный флуофор пары FRET заменен люциферазой. Подходящие методологии BRET хорошо известны специалистам в данной области техники.
В одном варианте воплощения детектирование тестовой молекулы может быть основано на отличии природного pH, который существует в эндосоме, относительно pH цитозоли. В этом варианте воплощения тестовая молекула может быть помечена pH-чувствительной меткой, такой как pH-чувствительный краситель. Метку можно выбрать таким образом, чтобы она была обнаруживаемой/видимой (например, с повышенной интенсивностью) в определенном диапазоне pH. Таким образом, метка может быть выбрана таким образом, чтобы она была обнаруживаемой/видимой (например, с повышенной интенсивностью), только когда она присутствует в эндосоме, и невидимой/не обнаруживаемой (например, с низкой интенсивностью), когда она присутствует в цитозоли. В качестве альтернативы, метку можно выбрать таким образом, чтобы она становилась обнаруживаемой/видимой, только при транслокации тестовой молекулы в цитозоль. Специалистам в данной области техники известны подходящие метки (например, красители), такие как флуоресциновые pH-индикаторы (например, BCECF, BCPCF) и их производные, бензоксантеновые красители, цианиновые красители и другие мелкомолекулярные pH-индикаторы (например, комплекс европия, производные флуорена, 1,4-дигидроксифиалонитрил (1,4-DHPN), 8-гидроксипирен-1,3,6-трисульфоновая кислота (HPTS)), кислотные pH-индикаторы, такие как пиридилоксазоловый зонд желто-синий DND-160 PDMPO 59, антрафеновый сенсор DND-167 60, DND-189 61, DND-153 62, и DND 192 63, красители на основе BODIPY (например, BODIPY 68, NH2BDP 68a, DiMeNBDP 68b, EtMeNBDP 68c и DiEtNBDP 68d) и индикаторы pHrodo.
В зависимости от типа используемого средства детектирования, может быть желательным осуществить пермеабилизацию эукариотных клеток-мишеней. В качестве примера, белковая или небелковая метка (такая как микологическая метка или биотин) может быть прикреплена к тестовой молекуле, или сама тестовая молекула может по своей природе включать в себя обнаруживаемый компонент (такой как эпитоп), и последующее детектирование может быть достигнуто с использованием помеченного первичного антитела или непомеченного первичного антитела, наряду с помеченным вторичным антителом. Реагенты и методики выявления антител хорошо известны и привычны для специалистов в данной области техники. Аналогично, связывающиеся парные вещества хорошо известны из уровня техники, и, например, помеченный стрептавидин может быть использован для выявления биотинилированной контрольной молекулы. Пермеабилизация обычно включает в себя исходный этап фиксации (например, с формальдегидом, параформальдегидом, этанолом или метанолом), с последующей пермеабилизациуй с помощью подходящего агента, стандартные пермеабилизационные агенты хорошо известны из уровня техники (например, тритон X-100, дигитонин, твин 20 и/или сапонин).
В сравнительном этапе (v) применяют использование контрольного значения, которое отображает количество тестовых молекул, присутствующих в эндосомах и/или в цитозоли перед этапом детектирования (iv). Контрольное значение, как правило, определяется тем же средством/способом, посредством которого определяют количество тестовых молекул на этапе детектирования (iv). Контрольное значение, как правило, отображает количество тестовых молекул, присутствующих в эндосомах и/или в цитозоли в ходе или до этапа (iii). В качестве примера, контрольное значение может отображать количество тестовых молекул, присутствующих в эндосомах и/или в цитозоли в ходе или в конце этапа (ii), в одном варианте воплощения контрольное значение отображает количество тестовых молекул, которые присутствуют в эндосомах и/или в цитозоли, когда было установлено «стабильное состояние» скорости транслокации, а именно, когда тестовая молекула попадает во внутриклеточные эндосомы и покидает их приблизительно при одинаковой скорости.
Тестовая молекула может представлять собой «малую молекулу», терапевтическую, такую как финголимод, монастрол, памидронат, метатрексат, буспирон, немонапард, апипипипразол, бифепрунокс, SKF82958, октреотид, MK-5046, FO38-WE-05, рилменидин, SCH655842, сальвинорин, CP55940 или наночастица. Наночастицы были успешно нацелены на клетки и оказались интернализированными в них за счет конъюгирования через антитело (см., например, работу Wartlick et al., J. Drug Target. 12: 461-471). Таким образом, в одном варианте воплощения тестовая молекула может представлять собой наночастицу, связанную, среди прочего, с компонентом мишень-направляющей частицы, таким как антитело или лиганд для рецептора, присутствующего на испытуемой клетке.
В качестве альтернативы, тестовая молекула может представлять собой более крупную молекулу, такую как полипептид или белок. Конкретные примеры включают в себя токсины, такие как цитотоксические белки и не цитотоксические белки. С этой точки зрения, ссылка на полипептиды и белки включает в себя полипептиды белки, как возникшие естественным путем, так и приготовленные путем рекомбинации.
Цитотоксические белки действуют, убивая свои природные клетки-мишени. Примерами этой группы токсинов являются, среди прочего, растительные токсины, такие как рицин и абрин, и бактериальные токсины, такие как токсин дифтерии и псевдомонасовый экзотоксин A. Цитотоксические токсины, как правило, убивают свои клетки-мишени за счет торможения внутриклеточного процесса синтеза белка. Этот класс белка включает в себя перенаправленные цитотоксические белки, в которых природная способность к связыванию белка была модифицирована путем введения связующего лиганда (также известного как мишень-направляющая частица), что придает, таким образом, модифицированному белку новые свойства связывания клетки-мишени.
Напротив, не цитотоксические белки действуют на клетки-мишени, выводя из строя внутриклеточную функцию. Важно, что не цитотоксические токсины не убивают клетки-мишени, на которые они действуют. Некоторые из лучше всего известных примеров не цитотоксической протеазы включают в себя клостридиальные нейротоксины (например, ботулинический нейротоксин, который продается под такими наименованиями, как Dysport™, Neurobloc™ и Botox™), протеазы иммуноглобулина A (см., например, WO 99/032272), и протеазы антареазы (см., например, WO 2011/022357). Не цитотоксические протеазы действуют путем протеолитического расщепления и, таким образом, выводят из строя белки внутриклеточного переноса, известные как белки SNARE (например, SNAP-25, VAMP или Syntaxin), см. работу Геральд К. (2002) «Клетка и молекулярная биология» (4е издание), John Wiley & Sons, Inc. Аббревиатура SNARE происходит от термина растворимый связывающий рецептор NSF (Soluble NSF Attachment Receptor), где NSF означает N-этилмалеимид-сенситивный фактор. Белки SNARE являются основными компонентами процесса везикулярной секреции в эукариотных клетках. Таким образом, не цитотоксические протеазы действуют за счет подавления внутриклеточной секреции. Этот класс белка включает в себя перенаправленные не цитотоксические белки, в которых природная способность к связыванию белка была модифицирована путем введения связующего лиганда (также известного как мишень-направляющая частица), придавая, таким образом, модифицированному белку свойства связывания новых клеток-мишеней. Заявитель впервые разработал технологию, относящуюся к перенаправлению не цитотоксической протеазы, которая восходит к 1990-м годам (см., например, WO 94/21300, WO 96/33273 и WO 98/07864). Упомянутые перенаправленные белки называются (среди всей литературе и всего научного сообщества) мишень-направленными ингибиторами секреции (Targeted Secretion Inhibitors, TSI), ссылка на TSI включает в себя структурные эквиваленты, такие как описанные в WO 2011/018665.
Аспект настоящего изобретения, связанный с анализом «тестовой молекулы», может дополнительно включать в себя этап детектирования, непосредственно на месте, активности тестовой молекулы. Например, применительно к клостридиальному нейротоксину или TSI, анализ может дополнительно содержать этап детектирования расщепления белка SNARE. Любое такое средство детектирования может быть применено указанным образом, например, способ, описанный в WO 95/33850, или путем FRET-анализа.
Анализ согласно настоящему изобретению может быть применен отдельно, для оценки воздействия тормозящих эффектов блокирующей молекулы на систему эндосомальный перенос эукариотной клетки. Таким образом, в соответствующем аспекте настоящее изобретение обеспечивает анализ, который включает:
i) приведение эукариотной клетки в контакт с контрольной молекулой, которая связывается с центром связывания, присутствующим на поверхности упомянутой эукариотной клетки, причем упомянутая контрольная молекула образует связанный комплекс с центром связывания, попадает в эукариотную клетку за счет эндоцитоза, в ходе которого образуется эндосома, которая содержит контрольную молекулу, и при этом упомянутая контрольная молекула попадает в цитозоль эукариотной клетки сквозь эндосомальную мембрану эндосомы;
ii) инкубирование контрольной молекулы с упомянутой эукариотной клеткой, что позволяет, таким образом,
a) связывать контрольную молекулу и образовывать связанный комплекс с центром связывания, присутствующим на эукариотной клетке, что позволяет, таким образом, упомянутому связанному комплексу попадать в эукариотную клетку за счет эндоцитоза;
b) одной или более эндосом формироваться в упомянутой клетке, причем одна или более эндосом содержит контрольную молекулу; и
c) упомянутой контрольной молекуле попадать в цитозоль эукариотной клетки сквозь эндосомальную мембрану одной или более эндосом;
iii) приведение эукариотной клетки в контакт с тестовой молекулой ингибитора, которая подлежит оценке на ее способность к подавлению эндосомального высвобождения контрольной молекулы из одной или более эндосом. Упомянутое приведение в контакт эукариотной клетки с тестовой молекулой ингибитора может быть выполнено до, в ходе или после этапа (i) и/или в ходе или после этапа ii);
iv) после истечения заданного периода времени, детектирование количества контрольных молекул, присутствующих в одной или более эндосомах, или детектирование количества контрольных молекул, присутствующих в цитозоли упомянутой эукариотной клетки;
v) сопоставление количества контрольных молекул, выявленных на этапе (iv), с контрольным значением, причем упомянутое контрольное значение отображает количество контрольных молекул, присутствующих в одной или более эндосомах, или количество контрольных молекул, присутствующих в цитозоли перед выполнением этапа (iii);
vi) присвоение параметра ингибирования тестовой молекуле ингибитора путем определения относительного изменения количества контрольных молекул, которые присутствуют в одной или более эндосомах, или путем определения относительного изменения количества контрольных молекул, присутствующих в цитозоли упомянутой эукариотной клетки.
Контрольная молекула может представлять собой «малую молекулу», являющуюся терапевтической, как было упомянуто выше, такую как ингибитор PMSA (см. Liu и др., 2008) или финголимод, монастрол, памидронат, метатрексат, буспирон, немонапард, апипипипразол, бифепрунокс, SKF82958, октреотид, MK-5046, FO38-WE-05, рилменидин, SCH655842, сальвинорин, CP55940 или наночастицу.
В качестве альтернативы, контрольная молекула может представлять собой более крупную молекулу, такую как полипептид или белок. Конкретные примеры включают в себя токсины, такие как цитотоксические белки и не цитотоксические белки. С этой точки зрения, ссылка на полипептиды и белки включает в себя полипептиды и белки, как возникшие естественным путем, так и приготовленные рекомбинантным образом. Примеры подходящих цитотоксических и не цитотоксических белков были описаны здесь ранее. Что касается не цитотоксических белков, предпочтительные примеры включают в себя клостридиальные нейротоксины (такие как токсин столбняка, BoNT/A, BoNT/B, BoNT/C1, BoNT/D, BoNT/E, BoNT/F, BoNT/G или C, бутиркум, включающий в себя его природную и модифицированные версии, а также их сочетания), TSI (такие как TSI, описанные ранее), протеазы антареазы и протеазы иммуноглобулина A.
Примеры ингибиторов тестовой молекулы включают в себя блокаторы пор, такие как тусенданин, см. работу Fischer, A. Et al. (2009) PNAS, Vol. 106, No. 5, pp. 1330-1335; блокаторы протон-АТФазы, такие как бафиломицин, см. работу Bartz et al. (2011), Biochem J 435 pp. 475-487, конканамицин (Tscherne et al. (2006) J Virol, Vol. 80, No. 4, pp. 1234-1741) и продигиозины, см. работу Ohkuma et al. (1998) J Biochem Vol. 334, pp. 731-741.
Все этапы, какие были описаны ранее, со ссылкой на базовый анализ согласно настоящему изобретению, применимы в равной мере и к вышеописанному анализу тестовой молекулы ингибитора.
Этап приведения в контакт (iii) в вышеописанном анализе тестовой молекулы ингибитора может протекать в течение любого заданного периода времени, например, в течение периода времени от 5 минут до 5 дней, перед началом этапа инкубирования (ii), например, в течение периода времени от 30 минут до 12 часов, или от 30 минут до 10 часов, или от 30 минут до 8 часов, или в течение 1-8 часов, вслед за началом этапа инкубирования (ii). В качестве альтернативы, этап (iii) может быть выполнен в течение периода времени такой же длительности, что и этап (ii), или его начало может быть отложено на период времени от 5 минут до 5 дней, например, на 1-12 часов или на 2-10 часов, или на 4-8 часов, или на 6-8 часов. Как правило, этап приведения в контакт (iii) выполняют сразу после достижения «стабильного состояния» (через этап (ii)), когда контрольная молекула попадает во внутриклеточные эндосомы и покидает их приблизительно при одинаковой скорости.
Аспект настоящего изобретения, связанный с ингибитором, также может включать в себя этап удаления избыточной контрольной молекулы и/или тестовой молекулы ингибитора. Упомянутый «этап удаления» означает «этап (iiia)» применительно к аспекту анализа ингибитора. Все описанные ранее варианты воплощения этапа (iii) «удаления» применительно к аспекту анализа «тестовой молекулы» согласно настоящему изобретению применимы в равной мере к этапу (iiia) «удаления» аспекта анализа «молекулы ингибитора» согласно настоящему изобретению.
ПЕРЕЧЕНЬ ФИГУР
Фиг. 1. Ключевые этапы связаны с эндоцитозом. Четыре ключевых этапа обозначены как 1-4.
Фиг. 2. Интернализация TSI была направлена на клетки GH3, что было выражено как GHRHR. Фиг. 2A: клетки были инкубированы в отсутствии TSI. Фиг. 2B: клетки были инкубированы в присутствии 3 мкМ TSI, с мишень-направляющей частицей GHRH, в течение 60 минут.
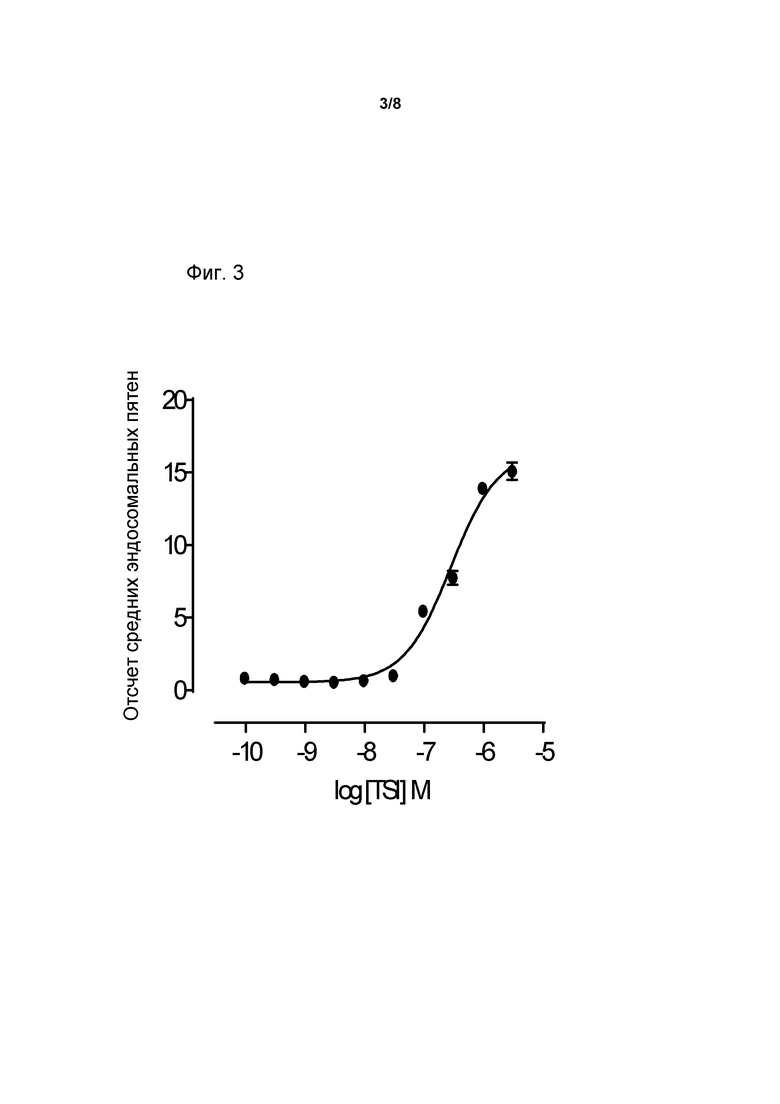
Фиг. 3. Зависимая от концентрации интернализация TSI была направлена на клетки GH3, что было выражено как GHRHR.
Фиг. 4. Интернализация TSI была направлена на крысиные питуициты. Фиг. 4A: клетки были инкубированы, в отсутствие TSI. Фиг. 4B: клетки были инкубированы, в присутствии 1 мкМ TSI, с мишень-направляющей частицей GHRH в течение 60 минут.
Фиг. 5. Зависимая от концентрации интернализация TSI с мишень-направляющей частицей GHRH в крысиные питуициты. Клетки были инкубированы с: (•) TSI, имеющий мишень-направляющую частицу GHRH; (o) соответствующий нелигандированного TSI в течение 60 минут.
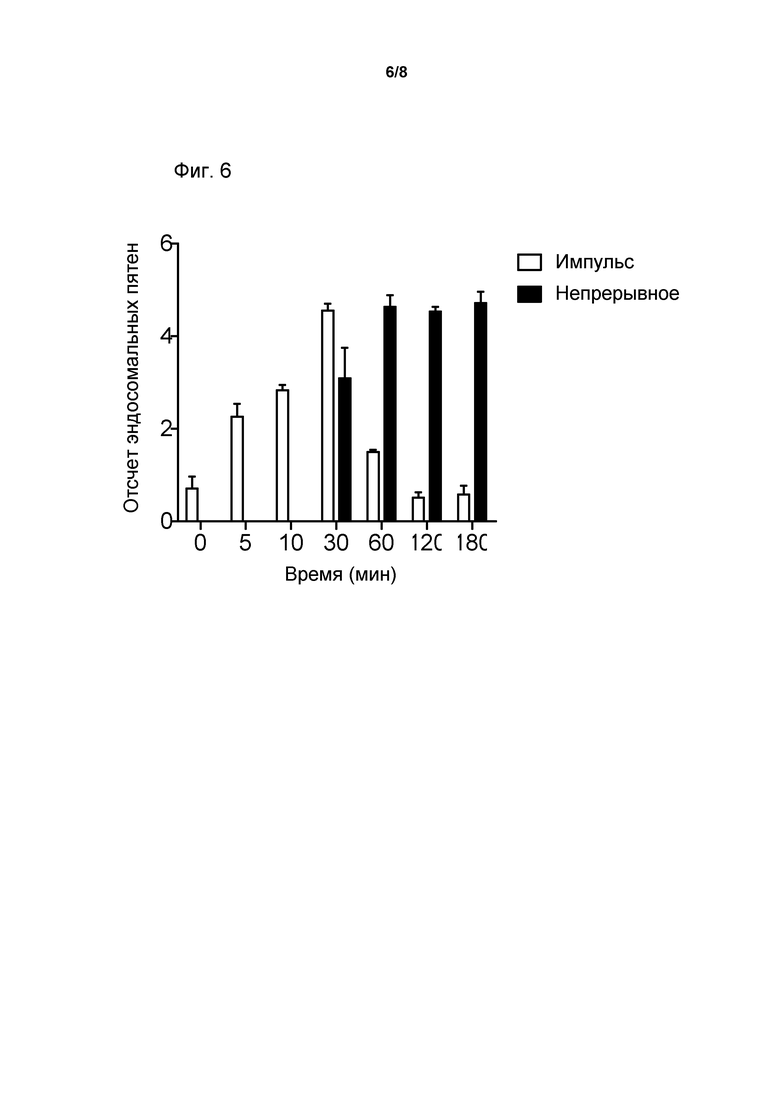
Фиг. 6. Интернализация и эндосомальный выход TSI были направлены на клетки, что было выражено как GHRHR. Режим непрерывной обработки сопоставлен с форматом обработки типа вытеснения метки.
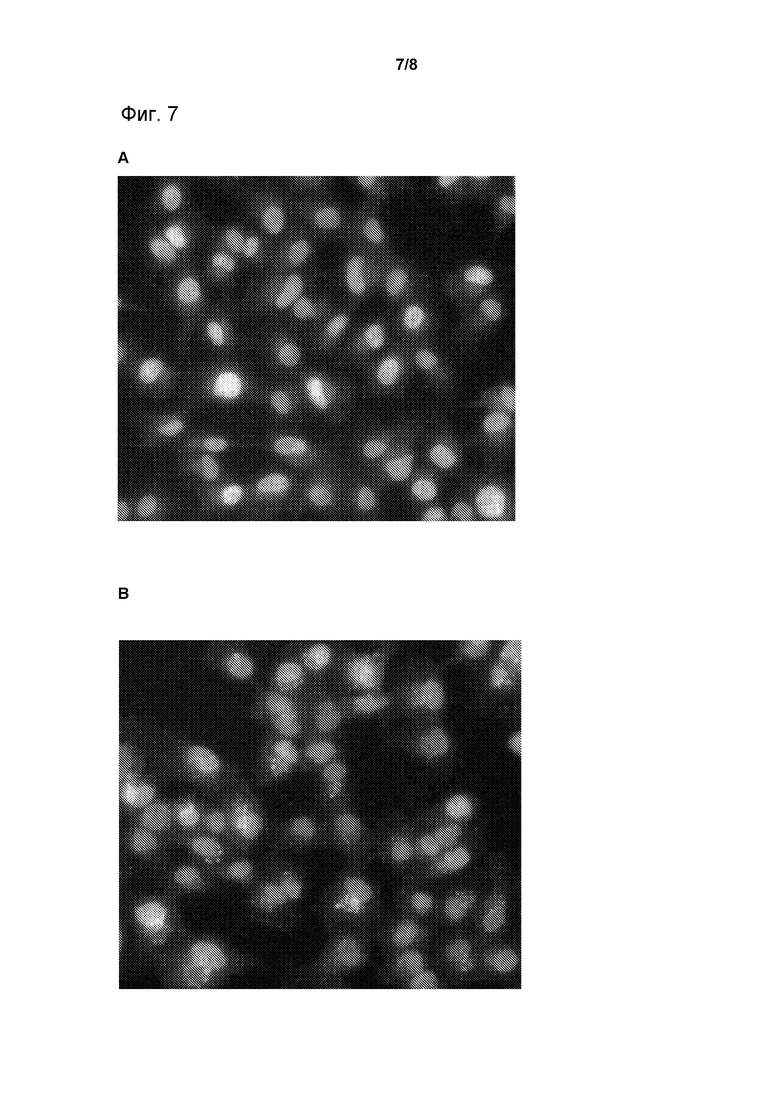
Фиг. 7. Интернализация TSI была направлена на клетки HT-1080. Фиг. 7A: клетки были инкубированы в отсутствии TSI. Фиг. 7B: клетки были инкубированы в присутствии 2 мкМ TSI с мишень-направляющей частицей EGFR в течение 60 минут.
Фиг. 8. Зависимая от концентрации интернализация TSI была направлена на клетки CHO-K1, что было выражено как GHRHR.
ПРИМЕРЫ
Пример 1
Клетки A549 вводят в качестве затравок в планшеты на 96 скважин и выращивают в течение 2 дней, для достижения ~90%-ной степени смыкания монослоя. Эпидермальный фактор роста (epidermal growth factor, EGF), помеченный как alexa-fluor®488, инкубируют с клетками в течение 60 минут. Клетки промывают и после повышения времени инкубирования, общую флуоресценцию клетки, отсчет пятен эндосом и эндосомальную флуоресценцию определяют, с использованием скрининга высокого содержания.
Пример 2
Клетки GH3, экспрессирующие рецептор высвобождения гормона роста (growth hormone-releasing hormone, GHRH) у крыс, вводят в качестве затравок в 96-скважинные планшеты, покрытые поли-D-лизин. По истечении 24 часов, TSI, содержащий область-мишень GHRH (1-44), и области транслокации и тяжелой цепи ботулинического нейротоксина серотипа C инкубируют с клетками в течение 30 минут. Клетки промывают, а TSI, связанный с поверхностью клетки, удаляют путем кислотной промывки, при повышении времени инкубирования. Клетки скрепляют параформальдегидом и подвергают пермеабилизации с использованием дигитонином. Тяжелую цепь TSI ботулинического нейротоксина C выявляют с использованием анти-LC ботулинического нейротоксина C кролика, первичного антитела и вторичного антитела, помеченного козлиным антикроликовым антителом alexa-fluor®488. Накопление в эндосомах и эндосомальный выход контролируют с использованием скрининга высокого содержания.
Пример 3
Нейроны спинного мозга эмбриона подготавливают и культивируют в 96-скважинных планшетах, покрытых матригелем. Ботулинический нейротоксин инкубируют с нейронами в течение 10 минут, после чего клетки промывают. Клетки подвергают пермеабилизации, с использованием сапонина, после повышения времени инкубирования, и LC ботулинического нейротоксина, выявленный с использованием антиботулинического нейротоксина LC антитела. Накопление в эндосомах и эндосомальный выход регулируют, с использованием скрининга высокого содержания.
Пример 4
Клетки HEK293, экспрессирующие человеческий рецептор паращитовидного гормона (parathyroid hormone, PTH1) вводят в качестве затравок в 96-скважинные планшеты, покрытые поли-D-лизином. После инкубирования, проведенного за ночь, клетки инкубируются с PTH, помеченным грибком (1-34) в течение 45 минут. Клетки промывают, а после повышения времени инкубирования, клетки фиксируют и подвергают пермеалибизации с помощью твина 20. PTH, помеченный грибком (1-34), выявляют, с использованием антигрибкового антитела, помеченного родамином. Накопление в эндосомах и эндосомальный выход контролируют, с использованием скрининга высокого содержания.
Пример 5
Клетки CHO-K1, экспрессирующие рецептор вазоактивного кишечного пептида 1, вводят в качестве затравок в 96-скважинные планшеты и оставляют инкубироваться на ночь. Клетки инкубируются по одиночке, или с повышением концентраций ингибитора эндосомального выхода в течение 60 минут, после чего биотинилированный TSI совместно инкубируют с клетками в течение 20 минут. TSI удаляют путем промывания, а закрепленные клетки, после повышения времени инкубирования, удаляют с использованием параформальдегида. Клетки подвергают пермеабилизации, с использованием дигитонина, и подвергают биотинилированию. TSI выявляют, с использованием стрептавидина и антистрептавидинового антитела, помеченного alexa-fluor® 488. Накопление TSI в эндосомах и эндосомальный выход контролируют с использованием скрининга высокого содержания. Эффект от ингибитора оценивают по сниженной скорости эндосомального выхода, по сравнению с контрольными клетками, не инкубированными с ингибитором.
Пример 6
Клетки GH3, экспрессирующие крысиный GHRH-R, были засеяны в 96-скважинные планшеты и инкубированы в течение ночи. Клетки были инкубированы в присутствии (B) и в отсутствии (A) 3 мкМ TSI с мишень-направляющей частицей GHRH в течение 60 минут. Клетки были закреплены и прозондированы с помощью кроликового антитела для тяжелой цепи TSI и козлиного «анти-кроликового» антитела Alexa Fluor® 488. Ядра были окрашены, с использованием пятна Hoescht. Ядро (A) показано бледно-серым, без выявленных пунктирных белых пятен. Ядро (B) показано окрашенным в бледно-серый цвет (Фиг. 2).
Пример 7
Клетки GH3, экспрессирующие крысиный GHRH-R, были засеяны в 96-скважинные планшеты и инкубированы в течение ночи. Клетки были инкубированы, с повышением концентрации TSI, с мишень-направляющей частицей GHRH в течение 60 минут. Клетки были закреплены и прозондированы с помощью кроликового антитела для тяжелой цепи TSI и козлиного «анти-кроликового» антитела Alexa Fluor® 488. Подсчет эндосомных пятен был проведен с использованием скрининга высокого содержания. Данные означают среднее значение ± s.e. (standard error, стандартная ошибка) от данных, полученных в трех экспериментах, выполненных троекратно (Фиг. 3).
Пример 8
Диспергированный клетки гипофиза крыс были выращены в течение 5 дней и инкубированы в присутствии (B) и в отсутствии (A) 1 мкМ TSI с мишень-направляющей частицей GHRH в течение 60 минут. Клетки были закреплены и прозондированы с помощью кроликового антитела для тяжелой цепи TSI и козлиного «анти-кроликового» антитела Alexa Fluor® 488. Ядра были окрашены, с использованием красителя Hoescht. Ядро (A) показано бледно-серым, без выявленных пунктирных белых пятен. Ядро (B) показано окрашенным в бледно-серый цвет. Белые пунктирные пятна указывают на наличие тяжелой цепи TSI в эндосомах (Фиг. 4).
Пример 9
Диспергированные клетки гипофиза крыс были выращены в течение 5 дней и троекратно инкубированы, с повышением концентрации TSI, имеющего мишень-направляющую частицу GHRH или соответствующий нелигандированный TSI, в течение 60 минут. Клетки были закреплены и прозондированы с помощью кроликового антитела для тяжелой цепи TSI и козлиного «анти-кроликового» антитела Alexa Fluor® 488. Подсчет эндосомных пятен был проведен с использованием скрининга высокого содержания. Данные означают среднее значение ± s.e. (standard error, стандартная ошибка) от данных, полученных в одном эксперименте, выполненном троекратно (Фиг. 5).
Пример 10
Клетки GH3, экспрессирующие крысиный GHRH-R, были засеяны в 96-скважинные планшеты и инкубированы в течение ночи. Клетки были троекратно инкубированы с 1 мкМ TSI, имеющим мишень-направляющую частицу GHRH, для повышения периодов времени, после которых клетки были закреплены и прозондированы с помощью кроликового антитела, для тяжелой цепи of TSI и козлиного «антикроликового» антитела Alexa Fluor® 488. Клетки были подвергнуты непрерывному инкубированию с TSI или были возбуждены импульсами с TSI в течение 30 минут, а затем промыты, до того как они были инкубированы для повышения периодов времени (30, 90, 150 мин). При этих периодах времени клетки были закреплены и впоследствии прозондированы с антителом, для тяжелой цепи D. Подсчет эндосомных пятен был проведен, с использованием скрининга высокого содержания. Данные означают среднее значение ± s.e. (standard error, стандартная ошибка) от данных, полученных в трех экспериментах, выполненных троекратно (Фиг. 6).
Пример 11
Клетки HT-1080, экспрессирующие человеческий EGFR, были просеяны в 96-скважинные планшеты и инкубированы в присутствии (B) и в отсутствии (A) 2 мкМ TSI с мишень-направляющей частицей EGFR в течение 60 минут. Клетки были закреплены и прозондированы с помощью кроликового антитела для тяжелой цепи TSI и козлиного «анти-кроликового» антитела Alexa Fluor® 488. Ядра были окрашены, с использованием красителя Hoescht. Ядро (A) показано бледно-серым, без выявленных пунктирных белых пятен. Ядро (B) показано окрашенным в бледно-серый цвет. Белые пунктирные пятна указывают на наличие тяжелой цепи TSI в эндосомах (Фиг. 7).
Пример 12
Клетки CHO-K1, экспрессирующие GHRHR, вводят в качестве затравок в 96-скважинные планшеты и инкубируют в течение ночи. Клетки были инкубированы троекратно, с повышением концентрации TSI, имеющего мишень-направляющую частицу GHRH, и с транслокацией, и области тяжелых цепей ботулинического нейротоксина серотипа A в течение 30 мин. Клетки были зафиксированы, подвергнуты пермеабилизации с дигитонином и прозондированы с помощью антитела кролика для цепи TSI и козлиного антикроликового антитела Alexa Fluor® 488. Подсчет везикул на клетку был выполнен с использованием скрининга высокого содержания. Показанные данные означают среднее значение ± s.e. (standard error, стандартная ошибка) от данных, полученных в трех экспериментах, выполненных троекратно (Фиг. 8).

Изобретение касается способа оценки способности высвобождения эндосомой тестовой молекулы, причем упомянутая тестовая молекула представляет собой клостридиальный нейротоксин или перенаправленную не цитотоксическую протеазу и обладает способностью к протеолитическому расщеплению и инактивированию белка SNARE, где способ включает: i) приведение эукариотной клетки в контакт с тестовой молекулой, которая должна быть оценена на способность высвобождения эндосомой, причем упомянутая эукариотная клетка содержит клеточную мембрану, включающую в себя центр связывания, присутствующий на внешней поверхности клеточной мембраны упомянутой клетки; ii) инкубирование тестовой молекулы с упомянутой эукариотной клеткой, что, таким образом, допускает: a) связывание тестовой молекулы и образование связанного комплекса с центром связывания, присутствующим на эукариотной клетке, что позволяет, таким образом, упомянутому связанному комплексу попадать в эукариотную клетку за счет эндоцитоза; b) формирование одной или более эндосом в упомянутой клетке, причем одна или более эндосом содержит тестовую молекулу; и c) введение упомянутой тестовой молекулы в цитозоли эукариотной клетки сквозь эндосомальную мембрану одной или более эндосом; iii) удаление избыточной тестовой молекулы, которая не связана с центрами связывания, присутствующими на эукариотных клетках; iv) детектирование количества тестовых молекул, присутствующих в одной или более эндосом, или выявление количества тестовых молекул, присутствующих в цитозоли упомянутой эукариотной клетки; v) сопоставление количества тестовых молекул, выявленных на этапе iv), с контрольным значением, причем упомянутое контрольное значение отображает количество тестовых молекул, присутствующих в одной или более эндосомах, или количество тестовых молекул, присутствующих в цитозоли, перед выполнением этапа iv); vi) расчет параметра высвобождения эндосомы для тестовой молекулы путем определения относительного изменения количества тестовых молекул, которые присутствуют в одной или более эндосом, или путем определения относительного изменения количества тестовых молекул, присутствующих в цитозоли упомянутой эукариотной клетки; причем этап (iv) содержит детектирование тестовой молекулы с использованием флуоресцентной метки. Изобретение также касается способа оценки способности тестовой молекулы ингибитора подавлять эндосомальное высвобождение контрольной молекулы с одной или более эндосом эукариотной клетки. 2 н. и 7 з.п. ф-лы, 12 пр., 8 ил.
1. Способ оценки способности высвобождения эндосомой тестовой молекулы,
причем упомянутая тестовая молекула представляет собой клостридиальный нейротоксин или перенаправленную не цитотоксическую протеазу, в которой природная способность к связыванию не цитотоксической протеазы была модифицирована путем введения связующего лиганда или мишень-направляющей частицы, и
причем упомянутая тестовая молекула обладает способностью к протеолитическому расщеплению и инактивированию белка SNARE;
где упомянутый способ включает:
i) приведение эукариотной клетки в контакт с тестовой молекулой, которая должна быть оценена на способность высвобождения эндосомой, причем упомянутая эукариотная клетка содержит клеточную мембрану, включающую в себя центр связывания, присутствующий на внешней поверхности клеточной мембраны упомянутой клетки;
ii) инкубирование тестовой молекулы с упомянутой эукариотной клеткой, что, таким образом, допускает:
a) связывание тестовой молекулы и образование связанного комплекса с центром связывания, присутствующим на эукариотной клетке, что позволяет, таким образом, упомянутому связанному комплексу попадать в эукариотную клетку за счет эндоцитоза;
b) формирование одной или более эндосом в упомянутой клетке, причем одна или более эндосом содержит тестовую молекулу; и
c) введение упомянутой тестовой молекулы в цитозоли эукариотной клетки сквозь эндосомальную мембрану одной или более эндосом;
iii) удаление избыточной тестовой молекулы, которая не связана с центрами связывания, присутствующими на эукариотных клетках;
iv) детектирование количества тестовых молекул, присутствующих в одной или более эндосом, или выявление количества тестовых молекул, присутствующих в цитозоли упомянутой эукариотной клетки;
v) сопоставление количества тестовых молекул, выявленных на этапе iv), с контрольным значением, причем упомянутое контрольное значение отображает количество тестовых молекул, присутствующих в одной или более эндосомах, или количество тестовых молекул, присутствующих в цитозоли, перед выполнением этапа iv);
vi) расчет параметра высвобождения эндосомы для тестовой молекулы путем определения относительного изменения количества тестовых молекул, которые присутствуют в одной или более эндосом, или путем определения относительного изменения количества тестовых молекул, присутствующих в цитозоли упомянутой эукариотной клетки;
причем этап (iv) содержит детектирование тестовой молекулы с использованием флуоресцентной метки.
2. Способ по п.1, в котором эукариотную клетку выбирают из клетки дрожжей, клетки насекомого, клетки позвоночного животного, клетки млекопитающего, растительной клетки и грибной клетки.
3. Способ по любому из предыдущих пунктов, в котором этап инкубирования ii) длится в течение периода времени от 5 минут до 5 дней, например 1-12 часов, или 2-10 часов, или 4-8 часов, или 6-8 часов.
4. Способ по п.1 или 2, в котором этап детектирования iv) выполняют в течение времени от 5 минут до 5 часов вслед за этапом iii), например 15-240 минут, или 30-180 минут, или 45-150 минут вслед за этапом iii).
5. Способ оценки способности тестовой молекулы ингибитора подавлять эндосомальное высвобождение контрольной молекулы с одной или более эндосом эукариотной клетки, где упомянутый способ включает:
i) приведение эукариотной клетки в контакт с контрольной молекулой, которая связывается с центром связывания, присутствующим на поверхности упомянутой эукариотной клетки, причем упомянутая контрольная молекула представляет собой клостридиальный нейротоксин или перенаправленную не цитотоксическую протеазу, в которой природная способность к связыванию не цитотоксической протеазы была модифицирована путем введения связующего лиганда, или мишень-направляющей частицы, и в которой упомянутая контрольная молекула обладает способностью к протеолитическому расщеплению и инактивированию белка SNARE,
причем упомянутая контрольная молекула образует связанный комплекс с центром связывания, попадает в эукариотную клетку за счет эндоцитоза, в ходе которого образуется эндосома, которая содержит контрольную молекулу, и при этом упомянутая контрольная молекула попадает в цитозоль эукариотной клетки сквозь эндосомальную мембрану эндосомы;
ii) инкубирование контрольной молекулы с упомянутой эукариотной клеткой, что позволяет, таким образом:
a) связывать контрольную молекулу и формировать связанный комплекс с центром связывания, присутствующим на эукариотной клетке, что позволяет, таким образом, упомянутому связанному комплексу попадать в эукариотную клетку за счет эндоцитоза;
b) формировать одну или более эндосом в упомянутой клетке, причем одна или более эндосом содержит контрольную молекулу; и
c) упомянутой контрольной молекуле попадать в цитозоль эукариотной клетки сквозь эндосомальную мембрану одной или более эндосом;
iii) приведение эукариотной клетки в контакт с тестовой молекулой ингибитора, которая подлежит оценке на ее способность к подавлению эндосомального высвобождения контрольной молекулы из эндосом;
iv) детектирование количества контрольных молекул, присутствующих в одной или более эндосомах, или детектирование количества контрольных молекул, присутствующих в цитозоли упомянутой эукариотной клетки;
v) сопоставление количества контрольных молекул, выявленных на этапе iv), с контрольным значением, причем упомянутое контрольное значение отображает количество контрольных молекул, присутствующих в одной или более эндосомах, или количество контрольных молекул, присутствующих в цитозоли, перед выполнением этапа iii);
vi) присвоение параметра ингибирования тестовой молекуле ингибитора, путем определения относительного изменения количества контрольных молекул, которые присутствуют в одной или более эндосомах, или путем определения относительного изменения количества контрольных молекул, присутствующих в цитозоли упомянутой эукариотной клетки;
причем этап (iv) содержит детектирование контрольной молекулы с использованием флуоресцентной метки.
6. Способ по п.5, причем эукариотную клетку выбирают из клетки дрожжей, клетки насекомого, клетки позвоночного животного, клетки млекопитающего, растительной клетки и грибной клетки.
7. Способ по любому из пп.5, 6, причем этап инкубирования ii) длится в течение периода времени от 5 минут до 5 дней, например в течение 1-12 часов, или 2-10 часов, или 4-8 часов, или 6-8 часов.
8. Способ по любому из пп.5, 6, причем этап детектирования iv) выполняют в течение времени от 5 минут до 5 часов вслед за этапом iii), например в течение 15-240 минут, или 30-180 минут, или 45-150 минут вслед за этапом iii).
9. Способ по любому из пп.5, 6, причем этап приведения в контакт iii) выполняют в течение времени от 5 минут до 5 дней вслед за началом этапа инкубирования ii), например в течение времени от 30 минут до 12 часов, или в течение времени от 30 минут до 10 часов, или в течение времени от 30 минут до 8 часов, или в течение времени 1-8 часов вслед за началом этапа инкубирования ii).
| KIRCHMEIER MARC J ET AL.: "Correlations between the rate of intracelluar release of endocytosed liposomal doxorubicin and cytotoxicity as determined by a new assay", JOURNAL OF LIPOSOME RESEARCH, vol | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| US 2008038296 A1, 14.02.2008 | |||
| СПОСОБ ПРОИЗВОДСТВА КОНСЕРВОВ "ТЮЛЬКА ОБЖАРЕННАЯ В ТОМАТНОМ СОУСЕ" | 2011 |
|
RU2462938C1 |

