РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет временной заявки США с серийным номером 61/650883, поданной 23 мая 2012 года, временной заявки США с серийным номером 61/720102, поданной 30 октября 2012 года, и PCT/IB2012/056424, поданной 14 ноября 2012 года, все из которых включены в настоящее описание в качестве ссылок в полном объеме.
УРОВЕНЬ ТЕХНИКИ
Интерлейкин-6 (IL-6) является главным провоспалительным цитокином. Он отвечает за пролиферацию и дифференцировку иммунокомпетентных и кроветворных клеток. IL-6 человека представляет собой единичный гликопротеин, состоящий из 212 аминокислот, с двумя участками N-связанного гликозилирования, и он имеет молекулярную массу приблизительно 26 кДа. Структура IL-6 содержит четыре α-спиральных домена с мотивом из четырех остатков цистеина, которые необходимы для его третичной структуры. Передача сигнала IL-6 опосредуется связыванием IL-6 либо с растворимой, либо со связанной с поверхностью альфа-цепью рецептора IL-6 (IL-6Rα), обеспечивая взаимодействие комплекса с трансмембранной субъединицей gp130 клеточной поверхности, которая опосредует внутриклеточную передачу сигнала.
IL-6 вовлечен в патогенез воспалительных заболеваний, включая воспалительные аутоиммунные заболевания, такие как ревматоидный артрит (RA), артропатия со спондилезом, системная красная волчанка (SLE), воспалительное заболевание кишечника (IBD) и болезнь Кастлмана. IL-6 также вовлечен в патогенез злокачественных опухолей, включая рак предстательной железы, диффузную крупноклеточную лимфому, множественную миелому и почечно-клеточный рак. Также описана роль IL-6 в стимуляции обусловленной опухолью анорексии, мукозита полости рта и кахексии.
Хотя связывающие IL-6 молекулы, полученные путем иммунизации не являющихся человеком животных, известны в данной области, эти молекулы, как правило, требуют обширной модификации антител способами инженерии (например, пересадка CDR и гуманизация) для снижения их иммуногенности. Более того, полученные гуманизированные варианты, как правило, имеют недостаток в виде субоптимальной аффинности связывания с IL-6-мишенью и требуют обширной модификации антител способами инженерии и созревания аффинности для восстановления аффинности связывания IL-6. Конечным результатом является то, что большинство антител против IL-6 проявляют субоптимальную аффинность связывания с мишенью IL-6.
Таким образом, учитывая важность IL-6 в патогенезе заболевания и недостатки известных антител против IL-6, в данной области существует очевидная потребность в усовершенствованных (например, минимально модифицированных способами инженерии) средствах против IL-6, которые могут ингибировать биологическую активность IL-6, и, таким образом, осуществлять лечение заболеваний, связанных с активностью IL-6.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение усовершенствует уровень техники путем предоставления связывающих молекул (например, антитела или их антигенсвязывающие фрагменты) с улучшенными профилями связывания, которые специфически связываются с IL-6 (например, IL-6 человека и не являющегося человеком примата) с высокой аффинностью связывания (например, пикомолярная аффинность связывания) и эффективно ингибируют его биологическую активность (например, связывание с рецептором IL-6). В определенных иллюстративных вариантах осуществления связывающие IL-6 молекулы по изобретению происходят из общепринятого репертуара антител вида животных семейства верблюжьих (например, лама), которых подвергали активной иммунизации антигеном IL-6. Например, происходящие из животных семейства верблюжьих связывающие IL-6 молекулы по изобретению могут содержать парные домены VH/VL или другие альтернативные каркасы, где одна или несколько гипервариабельных петель (например, H1, H2, H3, L1, L2 и/или L3) VH- или VL-доменов происходят из вида животных семейства верблюжьих. Более того, в определенных вариантах осуществления по меньшей мере одна из гипервариабельных петель имеет каноническую укладку (или комбинацию канонических укладок), которая идентична или по существу идентична укладке антитела человека. Такие связывающие молекулы проявляют высокую гомологию с человеческими (последовательность и структура) и, таким образом, они являются особенно подходящими для лечения связанных с IL-6 заболеваний или нарушений (например, воспалительное заболевание и злокачественная опухоль) вследствие их низкой иммуногенности. Неожиданно, антитела против IL-6 по изобретению проявляют высокую аффинность связывания, возможность производства и термическую стабильность без необходимости в обширной и длительной модификации антител способами инженерии и созревания аффинности, которые, как правило, требуются для известных антител против IL-6.
Таким образом, в одном из аспектов изобретение относится к связывающей молекуле, которая специфически связывается с IL-6, причем связывающая молекула содержит по меньшей мере одну CDR антитела, где CDR содержит по меньшей мере один аминокислотный остаток, который погружен в полость F229 или в полость F279 на IL-6, когда связывающая молекула связана с IL-6. В определенных вариантах осуществления связывающая молекула содержит VH-домен, причем VH-домен имеет аминокислоту в положении 98 в соответствии с Kabat, которая погружена в полость F229 на IL-6, когда антитело или фрагмент связаны с IL-6. В одном конкретном варианте осуществления аминокислота в положении 98 представляет собой триптофан. В определенных вариантах осуществления связывающая молекула содержит VL-домен, причем VL-домен имеет аминокислоту в положении 30 в соответствии с Kabat, которая погружена в полость F229 на IL-6, когда антитело или фрагмент связаны с IL-6. В одном конкретном варианте осуществления аминокислота в положении 30 представляет собой тирозин. В определенных вариантах осуществления связывающая молекула содержит VH-домен, причем VH-домен имеет аминокислоту в положении 99, в соответствии с Kabat, которая погружена в полость F279 на IL-6, когда антитело или фрагмент связаны с IL-6. В одном конкретном варианте осуществления аминокислота в положении 99 представляет собой валин.
В определенных вариантах осуществления связывающая молекула содержит VH-домен и VL-домен, причем указанный VH-домен содержит гипервариабельные петли H1, H2 и H3, где указанный полипептид VH-домена образует пару с VL-доменом, содержащим гипервариабельные петли L1, L2 и L3, где по меньшей мере одна из гипервариабельных петель H1-H3 и L1-L3 получена из общепринятого антитела вида Lama посредством активной иммунизации животного вида Lama антигеном IL-6. В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель H1, H2, L1, L2 и L3 проявляет спрогнозированную или истинную структуру канонической укладки, которая идентична или по существу идентична соответствующей структуре канонической укладки гипервариабельной петли H1, H2, L1, L2 или L3, которая встречается в антителе человека.
В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель H1 и H2 обладает спрогнозированной или истинной структурой канонической укладки, которая идентична или по существу идентична соответствующей структуре канонической укладки человека. В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель L1, L2 и L3 обладает спрогнозированной или истинной структурой канонической укладки, которая идентична или по существу идентична соответствующей структуре канонической укладки человека. В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель H1 и H2 образует комбинацию спрогнозированных или истинных структур канонической укладки, которая идентична или по существу идентична соответствующей комбинации структур канонической укладки, о которых известно, что они встречаются в VH-домене эмбрионального типа человека. В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель H1 и H2 образует комбинацию структур канонической укладки, соответствующую комбинации структур канонической укладки человека, выбранных из группы, состоящей из 1-1, 1-2, 1-3, 1-4, 1-6, 2-1, 3-1 и 3-5.
В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель L1 и L2 образует комбинацию спрогнозированных или истинных структур канонической укладки, которая идентична или по существу идентична соответствующей комбинации структур канонической укладки, о которой известно, что она встречается в доменах VL эмбрионального типа человека. В одном конкретном варианте осуществления по меньшей мере одна из гипервариабельных петель L1 и L2 образует комбинацию структур канонической укладки, соответствующую комбинации структур канонической укладки человека, выбранных из группы, состоящей из 11-7, 13-7(A,B,C), 14-7 (A,B), 12-11, 14-11, 12-12, 2-1, 3-1, 4-1 и 6-1.
В определенных вариантах осуществления связывающая молекула содержит VH-домен и VL-домен, где VH-домен и/или VL-домен связывающей молекулы обладает идентичностью последовательности 90% или более с одним или несколькими соответствующими VH- или VL-доменами человека в каркасных областях FR1, FR2, FR3 и FR4. В определенных вариантах осуществления связывающая молекула содержит VH-домен и VL-домен, и она представляет собой преобразованный в эмбриональный тип вариант исходной связывающей молекулы, где один или оба из VH-домена и VL-домена связывающей молекулы содержат всего от 1 до 10 аминокислотных замен в каркасных областях по сравнению с соответствующим VH-доменом и VL-доменом родительского не являющегося человеческим антитела. В одном конкретном варианте осуществления родительская связывающая молекула представляет собой общепринятое антитело животного семейства верблюжьих. В определенных вариантах осуществления связывающая молекула представляет собой антитело или его антигенсвязывающий фрагмент.
В определенных вариантах осуществления связывающая молекула содержит VH-домен, причем VH-домен содержит аминокислотную последовательность HCDR3, указанную в SEQ ID NO:500 [X1PDVVTGFHYDX2], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, D или Y;
X2 представляет собой любую аминокислоту, предпочтительно, Y или N; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность HCDR3 выбрана из группы, состоящей из SEQ ID NO:497-499.
В определенных вариантах осуществления VH-домен дополнительно содержит аминокислотную последовательность HCDR2, указанную в SEQ ID NO:507 [VIX1YX2X3DTYYSPSLX4S], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, D, Y или N;
X2 представляет собой любую аминокислоту, предпочтительно, D или E;
X3 представляет собой любую аминокислоту, предпочтительно, A или G;
X4 представляет собой любую аминокислоту, предпочтительно, E или K; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность HCDR2 выбрана из группы, состоящей из SEQ ID NO:501-506.
В определенных вариантах осуществления VH-домен дополнительно содержит аминокислотную последовательность HCDR1, указанную в SEQ ID NO:512 [X1X2YYX3WX4], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, T, S или P;
X2 представляет собой любую аминокислоту, предпочтительно, R или S;
X3 представляет собой любую аминокислоту, предпочтительно, A или V;
X4 представляет собой любую аминокислоту, предпочтительно, S или T; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность HCDR1 выбрана из группы, состоящей из SEQ ID NO:508-511.
В определенных вариантах осуществления связывающая молекула содержит VH-домен, содержащий аминокислотные последовательности HCDR3, HCDR2 и HCDR1, указанные в SEQ ID NO:497, 501 и 508, соответственно.
В определенных вариантах осуществления связывающая молекула дополнительно содержит VL-домен, где VL-домен содержит аминокислотную последовательность LCDR3, указанную в SEQ ID NO:524 [ASYX1X2X3X4X5X6X7], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, R или K;
X2 представляет собой любую аминокислоту, предпочтительно, N, H, R, S, D, T или Y;
X3 представляет собой любую аминокислоту, предпочтительно, F, Y, T, S или R;
X4 представляет собой любую аминокислоту, предпочтительно, N или I;
X5 представляет собой любую аминокислоту, предпочтительно, N или D;
X6 представляет собой любую аминокислоту, предпочтительно, V, N, G или A;
X7 представляет собой любую аминокислоту, предпочтительно, V или I; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность LCDR3 выбрана из группы, состоящей из SEQ ID NO:513-523.
В определенных вариантах осуществления VL-домен дополнительно содержит аминокислотную последовательность LCDR2, указанную в SEQ ID NO:535 [X1VX2X3RX4S], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, R, K, D, A или E;
X2 представляет собой любую аминокислоту, предпочтительно, S, N или T;
X3 представляет собой любую аминокислоту, предпочтительно, T, K или Y;
X4 представляет собой любую аминокислоту, предпочтительно, A, T или V; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность LCDR2 выбрана из группы, состоящей из SEQ ID NO:525-534.
В определенных вариантах осуществления VL-домен дополнительно содержит аминокислотную последовательность LCDR1, указанную в SEQ ID NO:542 [AGX1X2X3DX4GX5X6X7YVS], или вариант этой последовательности, где
X1 представляет собой любую аминокислоту, предпочтительно, A или T;
X2 представляет собой любую аминокислоту, предпочтительно, S или N;
X3 представляет собой любую аминокислоту, предпочтительно, S, E или N;
X4 представляет собой любую аминокислоту, предпочтительно, V или I;
X5 представляет собой любую аминокислоту, предпочтительно, G, Y, T или F;
X6 представляет собой любую аминокислоту, предпочтительно, G или Y;
X7 представляет собой любую аминокислоту, предпочтительно, N, D или A; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности.
В одном конкретном варианте осуществления аминокислотная последовательность LCDR1 выбрана из группы, состоящей из SEQ ID NO:538-541. В определенных вариантах осуществления связывающая молекула содержит VL-домен, содержащий аминокислотные последовательности LCDR3, LCDR2 и LCDR1, указанные в SEQ ID NO:513, 525 и 536, соответственно. В определенных вариантах осуществления связывающая молекула содержит: VH-домен, имеющий аминокислотные последовательности HCDR3, HCDR2 и HCDR1, указанные в SEQ ID NO:497, 501 и 508, соответственно; и VL-домен, имеющий аминокислотные последовательности LCDR3, LCDR2 и LCDR1, указанные в SEQ ID NO:513, 525 и 536, соответственно.
В определенных вариантах осуществления связывающая молекула содержит VH-домен, обладающий по меньшей мере 85% идентичностью последовательности с аминокислотной последовательностью, указанной в SEQ ID NO:152. В определенных вариантах осуществления связывающая молекула содержит аминокислотную последовательность VH-домена, выбранную из группы, состоящей из SEQ ID NO:127-232 и 569-571. В определенных вариантах осуществления связывающая молекула содержит аминокислотную последовательность VH-домена, которая представляет собой SEQ ID NO:152. В определенных вариантах осуществления связывающая молекула содержит VL-домен, обладающий по меньшей мере 85% идентичностью последовательности с аминокислотной последовательностью, указанной в SEQ ID NO:416. В определенных вариантах осуществления связывающая молекула содержит аминокислотную последовательность VL-домена, выбранную из группы, состоящей из SEQ ID NO:391-496. В определенных вариантах осуществления связывающая молекула содержит аминокислотную последовательность VL-домена, которая представляет собой SEQ ID NO:416. В определенных вариантах осуществления связывающая молекула содержит: VH-домен, имеющий аминокислотную последовательность, указанную в SEQ ID NO:152; и VL-домен имеет аминокислотную последовательность, указанную в SEQ ID NO:416.
В определенных вариантах осуществления связывающая молекула содержит петли H1 и H2, которые образуют комбинацию структур канонической укладки, соответствующую комбинации 3-1 структур канонической укладки человека, как в структуре антитела человека 1ACY.
В определенных вариантах осуществления связывающая молекула содержит петли L1 и L2, которые образуют комбинацию структур канонической укладки, соответствующую комбинации 6λ-1 структур канонической укладки человека, встречающейся в структуре антитела человека 3MUG. В определенных вариантах осуществления связывающая молекула содержит петли L1, L2 и L3, которые образуют комбинацию структур канонической укладки, соответствующую комбинации 6λ-1-5 структур канонической укладки человека, встречающейся в структуре антитела человека 3MUG.
В определенных вариантах осуществления связывающая молекула содержит VH-домен, причем VH-домен содержит аминокислотную последовательность HCDR3, указанную в SEQ ID NO:544 [RAGX1GX2G], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, W;
X2 представляет собой любую аминокислоту, предпочтительно, M, A, L, S или N; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность HCDR3 выбрана из группы, состоящей из SEQ ID NO:543, SEQ ID NO:566, SEQ ID NO:567 и SEQ ID NO:568.
В определенных вариантах осуществления VH-домен дополнительно содержит аминокислотную последовательность HCDR2, указанную в SEQ ID NO:554 [X1ISX2X3GX4SX5X6YX7DSVKG], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, A, P или R;
X2 представляет собой любую аминокислоту, предпочтительно, A или S;
X3 представляет собой любую аминокислоту, предпочтительно, S или G;
X4 представляет собой любую аминокислоту, предпочтительно, G или V;
X5 представляет собой любую аминокислоту, предпочтительно, A или T;
X6 представляет собой любую аминокислоту, предпочтительно, Y, N или S;
X7 представляет собой любую аминокислоту, предпочтительно, G, A или T; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность HCDR2 выбрана из группы, состоящей из SEQ ID NO:545-553.
В определенных вариантах осуществления VH-домен дополнительно содержит аминокислотную последовательность HCDR1, указанную в SEQ ID NO:562 [X1X2X3X4 X5], или вариант этой последовательности, где:
X1 представляет собой любую аминокислоту, предпочтительно, S или T;
X2 представляет собой любую аминокислоту, предпочтительно, H или Y;
X3 представляет собой любую аминокислоту, предпочтительно, A или R;
X4 представляет собой любую аминокислоту, предпочтительно, M или L;
X5 представляет собой любую аминокислоту, предпочтительно, S или Y; и
где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В одном конкретном варианте осуществления аминокислотная последовательность HCDR1 выбрана из группы, состоящей из SEQ ID NO:555-561.
В определенных вариантах осуществления VH-домен содержит HCDR3, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:543, 566, 567 и 568, и аминокислотные последовательности HCDR2 и HCDR1, указанные в SEQ ID NO:545 и 555, соответственно. В определенных вариантах осуществления связывающая молекула дополнительно содержит VL-домен, где VL-домен содержит аминокислотную последовательность LCDR3, указанную в SEQ ID NO:563, или вариант этой последовательности, где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В определенных вариантах осуществления VL-домен дополнительно содержит аминокислотную последовательность LCDR2, указанную в SEQ ID NO:564, или вариант этой последовательности, где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В определенных вариантах осуществления VL-домен дополнительно содержит аминокислотную последовательность LCDR1, указанную в SEQ ID NO:565, или вариант этой последовательности, где вариант последовательности содержит одну, две или три аминокислотных замены в указанной последовательности. В определенных вариантах осуществления VL-домен содержит аминокислотные последовательности LCDR3, LCDR2 и LCDR1, указанные в SEQ ID NO:563, 564 и 565, соответственно. В определенных вариантах осуществления связывающая молекула содержит VH-домен, имеющий аминокислотные последовательности HCDR3, HCDR2 и HCDR1, указанные в SEQ ID NO:544, 545 и 555, соответственно; и VL-домен, имеющий аминокислотные последовательности LCDR3, LCDR2 и LCDR1, указанные в SEQ ID NO:563, 564 и 565, соответственно. В определенных вариантах осуществления связывающая молекула содержит VH-домен по меньшей мере с 85% идентичностью последовательности с аминокислотной последовательностью, указанной в SEQ ID NO:86. В определенных вариантах осуществления связывающая молекула содержит VH-домен, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:39-126 и 569-571. В определенных вариантах осуществления связывающая молекула содержит VH-домен, имеющий аминокислотную последовательность, выбранную из SEQ ID NO:86, SEQ ID NO:569, SEQ ID NO:570 и SEQ ID NO:571. В определенных вариантах осуществления связывающая молекула содержит VL-домен, обладающий по меньшей мере 85% идентичностью последовательности с аминокислотной последовательностью, указанной в SEQ ID NO:350. В определенных вариантах осуществления связывающая молекула содержит VL-домен, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:303-390. В определенных вариантах осуществления связывающая молекула содержит VL-домен, имеющий аминокислотную последовательность, которая представляет собой SEQ ID NO:350. В определенных вариантах осуществления связывающая молекула содержит: VH-домен, имеющий аминокислотную последовательность, указанную в SEQ ID NO:86, SEQ ID NO:569, SEQ ID NO:570 или SEQ ID NO:571; и VL-домен, имеющий аминокислотную последовательность, указанную в SEQ ID NO:350.
В определенных вариантах осуществления связывающая молекула содержит петли H1 и H2, которые образуют комбинацию структур канонической укладки, соответствующую комбинации 1-3 структур канонической укладки человека, встречающейся в структуре антитела человека 1DFB. В определенных вариантах осуществления связывающая молекула содержит петли L1 и L2, которые образуют структуры канонической укладки, соответствующие комбинации 7λ-1 структур канонической укладки человека, встречающейся в структуре антитела человека 1MFA. В определенных вариантах осуществления связывающая молекула содержит петли L1, L2 и L3 из комбинации структур канонической укладки, соответствующей комбинации 7λ-1-4 структур канонической укладки человека, встречающейся в структуре антитела человека 3MUG.
В определенных вариантах осуществления связывающая молекула представляет собой Fab-фрагмент, который связывается с IL-6 человека с константой диссоциации (koff, измеряемая с помощью поверхностного плазмонного резонанса) менее 2×10-5 с-1. В определенных вариантах осуществления связывающая молекула связывается с антигеном IL-6 человека с субпикомолярной аффинностью связывания. В определенных вариантах осуществления связывающая молекула связывается с антигеном IL-6 человека с одноразрядной фемтомолярной аффинностью связывания. В определенных вариантах осуществления связывающая молекула содержит гипервариабельные петли, которые получены из общепринятого антитела ламы без последующего созревания аффинности. В определенных вариантах осуществления связывающая молекула ингибирует индуцируемую IL-6 пролиферацию клеток гибридомы B9 с IC50 менее 0,1 пМ.
В определенных вариантах осуществления связывающая молекула обладает температурой плавления (Tm) более 65°C. В определенных вариантах осуществления связывающая молекула представляет собой преобразованный в эмбриональный тип вариант родительского антитела животного семейства верблюжьих, причем указанный преобразованный в эмбриональный тип вариант имеет более высокую температуру плавления, чем родительское антитело животного семейства верблюжьих. В определенных вариантах осуществления связывающая молекула экспрессируется на уровне по меньшей мере 20 мг/мл после временной экспрессии в клетке HEK293. В определенных вариантах осуществления связывающая молекула характеризуется показателем EpiBase®, составляющим менее чем приблизительно 10,0, например, менее чем приблизительно 6,0. В определенных вариантах осуществления связывающая молекула ингибирует связывание IL-6 с рецептором IL-6. В определенных вариантах осуществления связывающая молекула ингибирует связывание gp130 с рецептором IL-6. В определенных вариантах осуществления связывающая молекула специфически связывается с IL-6 человека и яванского макака. В определенных вариантах осуществления связывающая молекула содержит по меньшей мере одну CDR из антитела животного семейства верблюжьих, которое специфически связывается с IL-6.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей связывающую молекулу согласно любому из предшествующих пунктов и один или несколько фармацевтически приемлемых носителей.
В другом аспекте изобретение относится к способу лечения обусловленного IL-6 заболевания или нарушения, включающему введение индивидууму, нуждающемуся в лечении, эффективного количества фармацевтической композиции по изобретению.
В другом аспекте изобретение относится к выделенной нуклеиновой кислоте, кодирующей связывающую молекулу, описанную в настоящем описании.
В другом аспекте изобретение относится к рекомбинантному вектору экспрессии, содержащему молекулу нуклеиновой кислоты по изобретению.
В другом аспекте изобретение относится к клетке-хозяину, содержащей рекомбинантный вектор экспрессии по изобретению.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фиг. 1 представлены результаты анализов пролиферации клеток, измеряющих активность антител по изобретению в отношении нейтрализации IL-6 in vitro.
На фиг. 2 представлены результаты экспериментов на мышах с ксенотрансплантатами опухоли эпителиального рака яичника, в которых определяли эффективность антител по изобретению in vivo.
На фиг. 3A-B показано, что происходящие из животного семейства верблюжьих гипервариабельные петли (L1-L3, H1 и H2) антитела 61H7 по изобретению имеют спрогнозированную каноническую укладку и комбинации канонических укладок антител человека.
На фиг. 4A-B показано, что происходящие из животного семейства верблюжьих гипервариабельные петли (L1-L3, H1 и H2) антитела 68F2 по изобретению и его варианта зародышевой линии (129D3) имеют спрогнозированные канонические укладки и комбинации канонических укладок антител человека.
На фиг. 5 представлена модель с заполнением пространства для IL-6 с наложением: (A) F229 рецептора IL-6; (B) F229 рецептора IL-6 и W98 VH 61H7, (C) F229 рецептора IL-6 и Y30 VL 68F2; и (D) F229 рецептора IL-6, W98 VH 61H7 и V99 VH 68F2, в соответствии с нумерацией Kabat.
На фиг. 6 представлена модель с заполнением пространства для двух поверхностных связывающих полостей на IL-6, важных для связывания с рецептором IL-6, с наложением остатков F229 и F279 рецептора IL-6, и остатков Y30 VL 68F2 и V99 VH 68F2, в соответствии с нумерацией Kabat (Y32 и V104 в структуре).
На фиг. 7A-B представлена термическая стабильность 68F2 и его варианта эмбрионального типа 129D3 при измерении в Biacore с иммобилизованным гликозилированным IL-6 человека в отношении (A) других вариантов антител против IL-6 эмбрионального типа по изобретению и (B) других эталонных антител. На верхней части каждой фигуры представлены кривые плавления, в то время как на нижней части приведена величина Tm для каждого антитела.
На фиг. 8 представлена стабильность в сыворотке клонов антител 68F2, 129D3 (вариант эмбрионального типа 68F2) и 103A1 (вариант 61H7). Также включено эталонное антитело GL 18.
На фиг. 9 представлены показатели низкой иммуногенности (Epibase) для антител против IL-6 по изобретению по сравнению с эталонными антителами (показанными полужирным шрифтом), включая полностью человеческое антитело адалимумаб (Humira).
На фиг. 10A-B представлено выравнивание VH и VL (A) 68F2 и (B) 61H7, где показан высокий уровень гомологии последовательности с каркасными областями их соответствующих вариантов эмбрионального типа 129D3 и 111A7. Также показано минимальное количество изменений каркасной области, внесенных в каждую молекулу (всего 13).
На фиг. 11A-B представлено выравнивание VH и VK (A) CNTO328 и (B) VH_rabbit (ALD518), где изображен высокий уровень гомологии последовательностей с каркасными областями их соответствующих вариантов эмбрионального типа CNTO136 и VH_human(ALD518). Также представлено минимальное количество изменений каркасной области, внесенных в каждую молекулу (всего 36 и 46).
На фиг. 12 представлены профили фармакокинетики для IgG1-антител 129D3 и их вариантов у яванских макаков.
На фиг. 13 представлены результаты экспериментов в модели на мышах с сывороточным амилоидом A (SAA), в которых проводили измерение эффективности антител по изобретению in vivo.
На фиг. 14 представлены результаты экспериментов на мышах с псориатическими ксенотрансплантатами, в которых проводили определение эффективности антител по изобретению in vivo.
На фиг. 15 представлены данные о росте опухоли, выявленные в экспериментах, в которых определяли эффективность in vivo антител по изобретению в модели на мышах с ксенотрансплантатом почечно-клеточного рака.
На фиг. 16 представлен график Каплана-Мейера для данных выживаемости, выявленных в экспериментах, в которых определяли эффективность антител по изобретению in vivo в модели на мышах с ксенотрансплантатом почечно-клеточного рака.
На фиг. 17 представлены данные роста опухоли, выявленные в экспериментах, в которых проводили определение эффективности антител по изобретению in vivo в модели на мышах с ксенотрансплантатом почечно-клеточного рака, в которой все средства вводили в дозе 3 мг/кг.
На фиг. 18 представлен график Каплана-Мейера для данных выживаемости, выявленных в экспериментах, в которых проводили измерение эффективности антител по изобретению in vivo в модели на мышах с ксенотрансплантатом почечно-клеточного рака, в которой все средства вводили в дозе 3 мг/кг.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Определения
Чтобы настоящее изобретение было более понятным, сначала определены определенные термины.
Как используют в рамках изобретения термин "IL-6" относится к интерлейкину-6. Нуклеотидные и полипептидные последовательности IL-6 хорошо известны в данной области. Иллюстративная аминокислотная последовательность IL-6 человека представлена в депозите GenBank GI: 10834984, и иллюстративная аминокислотная последовательность IL-6 мыши представлена в депозите GenBank GI: 13624311.
Как используют в рамках изобретения термин "антитело" относится к молекулам иммуноглобулинов, содержащим четыре полипептидных цепи: две тяжелых (H) цепи и две легких (L) цепи, связанных между собой дисульфидными связями, а также к их мультимерам (например, IgM). Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно обозначаемую как VH) и константную область тяжелой цепи. Константная область тяжелой цепи содержит три домена: CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область легкой цепи (сокращенно обозначаемую как VL) и константную область легкой цепи. Константная область легкой цепи содержит один домен (CL1). Области VH и VL могут быть далее подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), между которыми находятся области, которые являются более консервативными, называемые каркасными областями (FR).
Как используют в рамках изобретения, термин "антигенсвязывающий фрагмент" антитела включает любой природный, получаемый ферментативно, синтетический или модифицированный способами генной инженерии полипептид или гликопротеин, которые специфически связываются с антигеном с образованием комплекса. Антигенсвязывающие фрагменты антитела могут происходить, например, из полноразмерных молекул антител с использованием любых пригодных стандартных способов, таких как протеолитическое расщепление или рекомбинантные способы генной инженерии, вовлекающие манипулирование и экспрессию ДНК, кодирующей вариабельный и необязательно константный домены антитела. Неограничивающие примеры антигенсвязывающих частей включают: (i) Fab-фрагменты; (ii) F(ab')2-фрагменты; (iii) Fd-фрагменты; (iv) Fv-фрагменты; (v) одноцепочечные молекулы Fv (scFv); (vi) dAb-фрагменты; и (vii) минимальные распознающие элементы, состоящие из аминокислотных остатков, которые имитируют гипервариабельную область антитела (например, выделенная определяющая комплементарность область (CDR)). Другие модифицированные способами инженерии молекулы, такие как диантитела, триантитела, тетраантитела и миниантитела, также охватываются выражением "антигенсвязывающая часть".
Как используют в рамках изобретения, термины "вариабельная область" или "вариабельный домен" относятся к тому факту, что определенные части вариабельных доменов VH и VL значительно отличаются по последовательности среди антител, и они используются в связывании и специфичности каждого конкретного антитела к его антигену-мишени. Однако вариабельность не равномерно распределена в вариабельных доменах антител. Она сосредоточена в трех сегментах, называемых "гипервариабельными петлями" в каждом из VL-домена и VH-домена, которые образуют часть антигенсвязывающего центра. Первую, вторую и третью гипервариабельные петли домена легкой цепи V-лямбда обозначают в настоящем описании как L1(λ), L2(λ) и L3(λ), и они могут быть определены как области, содержащие остатки 24-33 (L1(λ), состоящая из 9, 10 или 11 аминокислотных остатков), 49-53 (L2(λ), состоящая из 3 остатков) и 90-96 (L3(λ), состоящая из 5 остатков) в VL-домене (Morea et al., Methods 20:267-279 (2000)). Первую, вторую и третью гипервариабельные петли домена легкой цепи V-каппа обозначают в настоящем описании как L1(κ), L2(κ) и L3(κ), и они могут быть определены как области, содержащие остатки 25-33 (L1(κ), состоящая из 6, 7, 8, 11, 12 или 13 остатков), 49-53 (L2(κ), состоящая из 3 остатков) и 90-97 (L3(κ), состоящая из 6 остатков) в VL-домене (Morea et al., Methods 20:267-279 (2000)). Первую, вторую и третью гипервариабельные петли VH-домена обозначают в настоящем описании как H1, H2 и H3, и они могут быть определены как области, содержащие остатки 25-33 (H1, состоящая из 7, 8 или 9 остатков), 52-56 (H2, состоящая из 3 или 4 остатков) и 91-105 (H3, высоковариабельная по длине) в VH-домене (Morea et al., Methods 20:267-279 (2000)).
Если нет иных указаний, термины L1, L2 и L3, соответственно, относятся к первой, второй и третьей гипервариабельным петлям VL-домена, и они охватывают гипервариабельные петли, полученные как из изотипа V-каппа, так и из изотипа V-лямбда. Термины H1, H2 и H3, соответственно, относятся к первой, второй и третей гипервариабельным петлям VH-домена, и они охватывают гипервариабельные петли, полученные из любого из известных изотипов тяжелых цепей, включая γ, ε, δ, α или μ.
Каждая из гипервариабельных петель L1, L2, L3, H1, H2 и H3 может содержать часть "определяющей комплементарность области" или "CDR", как определено ниже. Термины "гипервариабельная петля" и "определяющая комплементарность область" не являются строго синонимичными, поскольку гипервариабельные петли (HV) определяют, исходя из структуры, в то время как определяющие комплементарность области (CDR) определяют, исходя из вариабельности последовательности (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD., 1983) и границы HV и CDR могут различаться в некоторых доменах VH и VL.
CDR доменов VL и VH, как правило, могут быть определены как области, содержащие следующие аминокислоты: остатки 24-34 (CDRL1), 50-56 (CDRL2) и 89-97 (CDRL3) в вариабельном домене легкой цепи, и остатки 31-35 или 31-35b (CDRH1), 50-65 (CDRH2) и 95-102 (CDRH3) в вариабельном домене тяжелой цепи; (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Таким образом, HV может содержаться в соответствующих CDR и указание в настоящем описании на "гипервариабельные петли" доменов VH и VL следует интерпретировать как также охватывающее соответствующие CDR и наоборот, если нет иных указаний.
Более высококонсервативные вариабельные домены называют каркасной областью (FR), как определено ниже. Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR (FR1, FR2, FR3 и FR4, соответственно), по большей части принимающих конфигурацию β-слоя, соединенных тремя гипервариабельными петлями. Гипервариабельные петли в каждой цепи удерживаются близко друг к другу посредством FR и, вместе с гипервариабельными петлями из другой цепи, участвуют в образовании антигенсвязывающего центра антител. Структурный анализ антител показал взаимосвязь между последовательностью и формой участка связывания, образованного определяющими комплементарность областями (Chothia et al., J. Mol. Biol. 227: 799-817 (1992)); Tramontano et al., J. Mol. Biol, 215:175-182 (1990)). Несмотря на высокую вариабельность последовательности, пять из шести петель имеют небольшой набор конформаций главной цепи, называемых "каноническими структурами". Эти конформации, во-первых, определяются длиной петель, а, во-вторых, присутствием ключевых остатков в определенных положениях петель и каркасных областей, которые определяют конформацию посредством их укладки, образования водородных связей или способности принимать необычные конформации основной цепи.
Как используют в рамках изобретения, термины "определяющая комплементарность область" или "CDR" относятся к несоседним антигенсвязывающим центрам в вариабельной области полипептидов как тяжелой, так и легкой цепей. Эти конкретные области описаны Kabat et al., J. Biol. Chem. 252, 6609-6616 (1977) и Kabat et al., Seqences of protein of immunological interest. (1991), и Chothia et al., J. Mol. Biol. 196:901-917 (1987), и MacCallum et al., J. Mol. Biol. 262:732-745 (1996), где определения включают перекрывание подгрупп аминокислотных остатков при сравнении друг с другом. Аминокислотные остатки, которые охватывают CDR, определяемые каждой из указанных выше ссылок, указаны для сравнения. Предпочтительно, термин "CDR" представляет собой CDR, как определено Kabat на основе сравнений последовательностей.
Определения CDR
2Нумерация остатков согласно номенклатуре Chothia et al., выше
3Нумерация остатков согласно номенклатуре MacCallum et al., выше.
Как используются в рамках изобретения термины "каркасная область" или "FR-область" включают аминокислотные остатки, которые являются частью вариабельной области, но которые не являются частью CDR (например, при использовании определения CDR согласно Kabat). Таким образом, каркасная область вариабельной области имеет длину приблизительно 100-120 аминокислот, но включает только аминокислоты вне CDR. В качестве конкретного примера вариабельной области тяжелой цепи и CDR, определяемых в соответствии с Kabat et al., каркасная область 1 соответствует домену вариабельной области, охватывающему аминокислоты 1-30; каркасная область 2 соответствует домену вариабельной области, охватывающему аминокислоты 36-49; каркасная область 3 соответствует домену вариабельной области, охватывающему аминокислоты 66-94, и каркасная область 4 соответствует домену вариабельной области от аминокислоты 103 до конца вариабельной области. Каркасные области для легкой цепи сходным образом разделены каждой CDR вариабельной области легкой цепи. Аналогично, с использованием определения CDR согласно Chothia et al. или McCallum et al. границы каркасной области определяются соответствующими концами CDR, как описано выше. В предпочтительных вариантах осуществления CDR являются такими, как определено Kabat.
В природных антителах шесть CDR, присутствующих на каждом мономерном антителе, представляют собой короткие несоседние последовательности аминокислот, которые расположены определенным образом так, чтобы образовывать антигенсвязывающий центр, когда антитело принимает его трехмерную конфигурацию в водной среде. Остальные части вариабельных доменов тяжелой и легкой цепей демонстрируют меньшую межмолекулярную вариабельность в аминокислотной последовательности, и их называют каркасными областями. Каркасные области по большей части принимают конформацию β-слоя, и CDR формируют петли, которые связывают структуры β-слоя и в некоторых случаях формируют их часть. Таким образом, эти каркасные области действуют, формируя каркас, который обеспечивает размещение шести CDR в правильной ориентации посредством межцепочечных нековалентных взаимодействий. Антигенсвязывающий центр, образованный размещенными CDR, определяет поверхность, комплементарную эпитопу, на иммунореактивном антигене. Эта комплементарная поверхность обеспечивает нековалентное связывание антитела с иммунореактивным эпитопом антигена. Положение CDR может быть без труда идентифицировано специалистом в данной области.
Как используют в рамках изобретения термин "полость F229" относится к полости на поверхности IL-6 человека, которую занимает остаток фенилаланина 229 рецептора IL-6 человека в комплексе IL-6/рецептор IL-6, как указано в Boulanger et al., 2003, Science 27, 2101-2104, которая включена в настоящее описание в качестве ссылки в полном объеме.
Как используют в рамках изобретения термин "полость F279" относится к полости на поверхности IL-6 человека, которую занимает остаток фенилаланина 279 рецептора IL-6 человека в комплексе IL-6/рецептор IL-6, как указано в Boulanger et al., 2003, Science 27, 2101-2104, которая включена в настоящее описание в качестве ссылки в полном объеме.
Как используют в рамках изобретения термин "происходящий из животного семейства верблюжьих" относится к аминокислотным последовательностям вариабельной области антитела (например, последовательности каркасной области или CDR), естественным образом присутствующим в молекулах антител животных семейства верблюжьих (например, лама). Происходящие из животного семейства верблюжьих антитела могут быть получены из любого вида животных семейства верблюжьих, включая, но не ограничиваясь ими, ламу, дромедар, альпаку, викунью, гуанако или верблюда. В определенных вариантах осуществления животное семейства верблюжьих (например, лама) активно иммунизировано IL-6 (например, IL-6 человека). В определенных вариантах осуществления термин "происходящий из животного семейства верблюжьих" ограничен последовательностями антител, которые происходят из общепринятого набора антител животного семейства верблюжьих и, в частности, исключает последовательности антител, происходящие из набора антител только с тяжелой цепью (VHH) животного семейства верблюжьих.
Как используют в рамках изобретения термин "общепринятое антитело" относится к антителам любого изотипа, включая IgA, IgG, IgD, IgE или IgM. Нативные или природные "общепринятые" антитела животных семейства верблюжьих обычно представляют собой гетеротетрамерные гликопротеины, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых цепей (H). Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как количество дисульфидных связей варьирует среди тяжелых цепей различных изотипов иммуноглобулинов. Каждая тяжелая и легкая цепь также имеет расположенные с равными интервалами внутрицепочечные дисульфидные мостики. Каждая тяжелая цепь имеет не одном конце (N-конец) вариабельный домен (VH), за которым следует ряд константных доменов. Каждая легкая цепь имеет вариабельный домен (VL) на одном конце (N-конец) и константный домен (CL) на ее другом конце; константный домен легкой цепи расположен параллельно первому константному домену тяжелой цепи, и вариабельный домен легкой цепи расположен параллельно вариабельному домену тяжелой цепи. Полагают, что конкретные аминокислотные остатки образуют поверхность контакта между вариабельными доменами легкой и тяжелой цепей.
Как используют в рамках изобретения термин "специфически связывается с" относится к способности антитела или его антигенсвязывающего фрагмента связываться с IL-6 с KD по меньшей мере приблизительно 1×10-6 (например, 1×10-6 M, 1×10-7 M, 1×10-8 M, 1×10-9 M, 1×10-10 M, 1×10-11 M, 1×10-12 M, 1×10-13 M, 1×10-14 M, 1×10-15 M или более), предпочтительно, от 1×10-12 M до 1×10-15 M или более, и/или связываться с IL-6 с аффинностью, которая по меньшей мере в два раза превышает аффинность в отношении неспецифического антигена. Однако должно быть понятно, что антитело или его антигенсвязывающий фрагмент способны специфически связываться с двумя или более антигенами, которые имеют сходную последовательность. Например, антитела или их антигенсвязывающие фрагменты, описанные в настоящем описании, могут специфически связываться как с IL-6 человека, так и с IL-6 не человека (например, мыши или не являющегося человеком примата).
Как используют в рамках изобретения термин "антиген" относится к участку связывания или эпитопу, распознаваемому вариабельной областью антитела.
Как используют в рамках изобретения термин "лечить", "осуществление лечения" и "лечение" относятся к терапевтическим или профилактическим мерам, описанным в настоящем описании. В способах "лечения" используется введение индивидууму антитела или его антигенсвязывающего фрагмента по настоящему изобретению, например, индивидууму, имеющему обусловленное IL-6 заболевание или нарушение (например, воспаление или злокачественная опухоль) или предрасположенному к наличию такого заболевания или нарушения, чтобы предотвратить, излечить, замедлить, снизить тяжесть или смягчить один или несколько симптомов заболевания или нарушения или рецидив заболевания или нарушение, или чтобы продлить выживание индивидуума по сравнению с выживанием, ожидаемым в отсутствие такого лечения.
Как используют в рамках изобретения, термин "обусловленное IL-6 заболевание или нарушение" включает болезненное состояние и/или симптомы, обусловленные активностью IL-6. Иллюстративные обусловленные IL-6 заболевания или нарушения включают, но не ограничиваются ими, воспалительные заболевания (например, воспалительные аутоиммунные заболевания, такие как ревматоидный артрит и системная красная волчанка), злокачественную опухоль (например, рак предстательной железы, диффузная крупноклеточная лимфома, множественная миелома и почечно-клеточный рак) и обусловленные злокачественной опухолью нарушения (например, анорексия и кахексия).
Как используют в рамках изобретения, термин "эффективное количество" относится к количеству антитела или его антигенсвязывающего фрагмента, которое является достаточным для обеспечения лечения, прогнозирования или диагностики обусловленного IL-6 заболевания или нарушения, как описано в настоящем описании, при введении индивидууму. Терапевтически эффективное количество варьирует в зависимости от индивидуума и болезненного состояния, подвергаемого лечению, массы тела и возраста индивидуума, тяжести болезненного состояния, способа введения и т.п., которые могут быть без труда определены специалистом в данной области. Дозировки для введения могут находиться в диапазоне, например, от приблизительно 1 нг до приблизительно 10000 мг, от приблизительно 1 мкг до приблизительно 5000 мг, от приблизительно 1 мг до приблизительно 1000 мг, от приблизительно 10 мг до приблизительно 100 мг антитела или его антигенсвязывающего фрагмента по изобретению. Режимы дозирования можно корректировать соответствующим образом, чтобы обеспечить оптимальный терапевтический ответ. Эффективное количество также представляет собой количество, при котором какие-либо токсические или вредоносные эффекты (т.е. побочные эффекты) связывающего полипептида минимизированы и/или перевешиваются полезными эффектами.
Как используют в рамках изобретения термин "индивидуум" включает любого человека или не являющегося человеком животного.
Как используют в рамках изобретения, термин "поверхностный плазмонный резонанс" относится к оптическому явлению, которое позволяет анализ взаимодействий в реальном времени посредством обнаружения изменений концентрации белка на матрице биосенсора, например, с использованием системы BIAcore™ (Biacore Life Sciences division of GE Healthcare, Piscataway, NJ).
Как используют в рамках изобретения термин "KD" относится к равновесной константе диссоциации конкретного взаимодействия связывающий полипептид/антиген.
Как используют в рамках изобретения термин "константа диссоциации" относится к константе диссоциации (Koff) для конкретного связывающего взаимодействия.
II. Связывающие IL-6 молекулы
В одном из аспектов изобретение относится к связывающим молекулам (антитела или их антигенсвязывающие фрагменты), которые специфически связываются с IL-6 и ингибируют его активность. Такие связывающие молекулы, как правило, содержат по меньшей мере одну аминокислотную последовательность области CDR, указанную в таблицах 13-18 настоящего описания.
Анализ кристаллической структуры IL-6 человека в комплексе с рецептором IL-6 человека показал, что 2 остатка рецептора IL-6, F229 и F279, являются важными для взаимодействия IL-6/рецептор IL-6 (см. например, Boulanger et al., 2003, Science 27, 2101-2104, которая включена в настоящее описание в качестве ссылки в полном объеме). В комплексе IL-6/рецептор IL-6, F229 и F279 погружены в отдельные полости на поверхности IL-6. В определенных вариантах осуществления связывающие молекулы по изобретению используют эти полости на IL-6, достигая высокой аффинности связывания. В одном из конкретных вариантов осуществления связывающие молекулы по изобретению содержат область CDR антитела, где область CDR содержит аминокислотный остаток, погруженный в полость F229 или в полость F279 на IL-6, когда связывающая молекула связана с IL-6.
Как правило, связывающие молекулы по изобретению ингибируют активность IL-6 (например, путем антагонизма связыванию IL-6 с рецептором IL-6). В определенных вариантах осуществления связывающие молекулы также ингибируют связывание gp130 с рецептором IL-6. Однако в других вариантах осуществления связывающие молекулы могут связываться с IL-6 без ингибирования связывания gp130 с рецептором IL-6.
Связывающие молекулы по изобретению, как правило, обладают высокой аффинностью к IL-6 и, как правило, являются высокоэффективными в отношении ингибирования активности IL-6 in vivo и in vitro. В определенных вариантах осуществления связывающие молекулы по изобретению связываются с IL-6 человека с константой диссоциации (koff, измеряемая с помощью поверхностного плазмонного резонанса) менее чем приблизительно 1×10-4 с-1 (например, приблизительно 9×10-5, 8×10-5, 7×10-5, 6×10-5, 5×10-5, 4×10-5, 3×10-5, 2×10-5 и 1×10-5). В других вариантах осуществления связывающие молекулы по изобретению ингибируют индуцированную IL-6 пролиферацию клеток гибридомы B9 с IC50 менее 0,1 пМ. В некоторых других вариантах осуществления связывающие молекулы по изобретению конкурируют с определенным антителом, связывающимся с IL-6, где такое определенное антитело содержит последовательность VH и последовательность VL, выбранные из аминокислотных последовательностей VH и VL, указанных в таблице 13-18. В некоторых других вариантах осуществления связывающие молекулы по изобретению конкурентно связываются по меньшей мере с 30%, 40%, 50%, 60%, 70%, 80% или 90% IL-6, связываемого определенным антителом, связывающимся с IL-6. В некоторых других вариантах осуществления связывающие молекулы по изобретению конкурируют за связывание IL-6 с 20A4, 24D10, 68F2, 61H7, 129D3 или 111A7, например, конкурентно связываются по меньшей мере с 50%, 60%, 70%, 80% или 90% IL-6, связываемого одним из этих антител против IL-6. В некоторых других вариантах осуществления связывающие молекулы по изобретению конкурируют за связывание IL-6 с 17F10, 24C9, 18C11, 29B11, 28A6 или 126A3, например, конкурентно связываются по меньшей мере с 50%, 60%, 70%, 80% или 90% IL-6, связываемого одним из этих антител против IL-6.
Как правило, связывающие молекулы по изобретению также обладают высокой термической стабильностью. В определенных вариантах осуществления связывающие молекулы обладают температурой плавления (Tm), превышающей 55°C (например, по меньшей мере 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75°C или выше). В определенных иллюстративных вариантах осуществления связывающие IL-6 молекулы по изобретению представляют собой варианты эмбрионального типа, которые обладают термической стабильностью, сравнимой или превышающей стабильность их родительских происходящих из животных семейства верблюжьих аналогов. В определенных иллюстративных вариантах осуществления термическую стабильность измеряют после инкубации в подходящем буфере (например, PBS) при концентрации 100 мкг/мл в течение 1 часа. В других иллюстративных вариантах осуществления термическая стабильность связывающей IL-6 молекула представляет собой стабильность, которую проявляет полноразмерный формат IgG (например, содержащий Fc-область IgG1 или IgG4).
Связывающие молекулы по изобретению также характеризуются высокими уровнями экспрессии функционального антитела с низкими уровнями нефункциональных примесей, таких как высокомолекулярные и низкомолекулярные агрегаты. Например, связывающие IL-6 молекулы по изобретению могут характеризоваться уровнями продукции по меньшей мере 20 мг/л (например, по меньшей мере 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 мг/л или выше). В определенных иллюстративных вариантах осуществления связывающие IL-6 молекулы представляют собой варианты эмбрионального типа, которые проявляют уровень экспрессии, сравнимый или превышающий уровень экспрессии их родительских аналогов животных семейства верблюжьих. В других иллюстративных вариантах осуществления уровень экспрессии определяют с использованием формата полноразмерного IgG связывающей IL-6 молекулы по изобретению, например, посредством временной экспрессии в клетке HEK293.
Связывающие молекулы по изобретению также, как правило, характеризуются низкой спрогнозированной иммуногенностью. Например, связывающие IL-6 молекулы по изобретению обладают показателями EpiBase® (например, общие показатели DRB1) менее 15,0, менее 12,0 или менее 10,0. В определенных иллюстративных вариантах осуществления связывающие молекулы имеют показатели иммуногенности приблизительно 9,0, приблизительно 8,0, приблизительно 7,0 или приблизительно 6,0. В других вариантах осуществления показатель иммуногенности является меньшим, чем показатель иммуногенности Humira®, например, приблизительно 6,0, приблизительно 5,0 или приблизительно 4,0.
Связывающие молекулы по изобретению могут связываться с любым IL-6, включая, но не ограничиваясь ими, IL-6 человека и яванского макака. Предпочтительно, связывающие молекулы могут связываться IL-6 как человека, так и яванского макака.
i) Антитела против IL-6 или их антигенсвязывающие фрагменты
В определенных вариантах осуществления изобретение относится к антителам или их антигенсвязывающим фрагментам, которые специфически связываются с IL-6 (например, IL-6 человека) и осуществляют антагонизм связывания IL-6 с рецептором IL-6. Последовательности VH, VL и CDR иллюстративных клонов Fab по изобретению указаны в таблицах 13-18. Антитела по изобретению могут содержать любую из аминокислотных последовательностей каркасной области и/или CDR этих клонов Fab.
Антитела по изобретению могут содержать последовательность области CDR с аминокислотным остатком (например, ароматическая аминокислота, такая как триптофан или тирозин), который погружен в полость F229 на IL-6, когда антитело или фрагмент связаны с IL-6. Иллюстративные антитела содержат VH-домен с триптофаном в положении 98 и/или VL-домен с тирозином в положении 30, в соответствии с Kabat. Такие антитела обладают особенно высокой аффинностью к IL-6.
Дополнительно или альтернативно, антитела по изобретению могут содержать последовательность области CDR с аминокислотным остатком, который погружен в полость F279 на IL-6, когда антитело или фрагмент связаны с IL-6. Иллюстративные антитела включают VH-домен с валином в положении 99 в соответствии с Kabat.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат VH, содержащую 1, 2 или 3 аминокислотных последовательности CDR из VH, указанных в таблицах 13-16.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат VL, содержащую 1, 2, или 3 аминокислотных последовательности CDR из VL, указанных в таблицах 13-16.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат: VH, содержащую 1, 2 или 3 аминокислотных последовательности CDR из VH, указанных в таблицах 13-18; и VL, содержащую 1, 2 или 3 аминокислотных последовательности CDR из VL, указанных в таблицах 13-18. В предпочтительном варианте осуществления все шесть CDR происходят из одного и того же клона Fab.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат VH, указанную с таблицах 13-16.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат VL, указанную в таблицах 13-16.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат VH и VL, указанные в таблицах 13-16.
В определенных вариантах осуществления антитела против IL-6 или фрагменты по изобретению содержат VH и VL из одного клона Fab, указанные в таблицах 13-16.
В определенных вариантах осуществления изобретение относится к антителам или их антигенсвязывающим фрагментам, которые специфически связываются с IL-6, причем антитела или фрагменты содержат вариант аминокислотных последовательностей CDR, VH, и VL, указанных в таблицах 13-18.
В определенных вариантах осуществления вариант последовательности содержит аминокислотную последовательность VH и/или VL приблизительно с 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью с аминокислотными последовательностями области VH или VL, указанными в таблицах 13-16.
В других вариантах осуществления вариант последовательности содержит аминокислотную последовательность VH, VL или CDR, выбранную из таблиц 13-18, которая изменена путем внесения одной или нескольких консервативных аминокислотных замен. Консервативные аминокислотные замены включают замену аминокислоты одного класса аминокислотой того же класса, где класс определяется общими физико-химическими свойствами боковой цепи аминокислот и высокими частотами замещения в гомологичных белках, встречающихся в природе, при определении, например, с помощью стандартной матрицы обмена частотами Dayhoff или матрицы BLOSUM. Было выделено шесть общих классов боковых цепей аминокислот, и они включают: класс I (Cys); класс II (Ser, Thr, Pro, Ala, Gly); класс III (Asn, Asp, Gln, Glu); класс IV (His, Arg, Lys); класс V (Ile, Leu, Val, Met) и класс VI (Phe, Tyr, Trp). Например, замена Asp на другой остаток класса III, такой как Asn, Gln или Glu, представляет собой консервативную замену. Таким образом, спрогнозированный несущественный аминокислотный остаток в антителе против IL-6 или его антигенсвязывающем фрагменте, предпочтительно, заменен другим аминокислотным остатком того же класса. Способы идентификации аминокислотных консервативных замен, которые не устраняют связывание антигена, хорошо известны в данной области (см., например, Brummell et al., Biochem. 32:1180-1187 (1993); Kobayashi et al. Protein Eng. 12(10):879-884 (1999); и Burks et al. Proc. Natl. Acad. Sci. USA 94:412-417 (1997)).
В других вариантах осуществления вариант последовательности содержит аминокислотную последовательность VH, VL или CDR, выбранную из таблиц 13-18, которая изменена для повышения продукции и/или производства антитела, например, путем замены метионина на аланин, серин или лейцин. В некоторых других вариантах осуществления вариант последовательности содержит аминокислотную последовательность VH, VL или CDR, выбранную из таблиц 13-18, которая изменена для повышения продукции антитела, например, путем замены глутамина на глутаминовую кислоту или аспарагина на аланин или родственные аминокислоты. В иллюстративных вариантах осуществления один или несколько остатков глутамина вне областей CDR аминокислотных последовательностей VH, представленных в таблице 16, заменены остатком(ами) глутаминовой кислоты, например, один или несколько остатков глутамина в положении 1, 3, 5 или 16 или любой комбинации этих положений в SEQ ID NO:152 заменены остатком(ами) глутаминовой кислоты для повышения продукции или стабильности антитела. В одном конкретном варианте осуществления глутамин в положении 1 SEQ ID NO:152 заменен глутаминовой кислотой.
ii) Связывающие IL-6 молекулы с высокой гомологией с человеческими
В некоторых аспектах связывающие IL-6 молекулы по изобретению представляют собой антитела (или антигенсвязывающие фрагменты) с высокой гомологией с человеческими. Антитело считают имеющим "высокую гомологию с человеческим", если VH-домены и VL-домены, взятые вместе, проявляют по меньшей мере 90% идентичность аминокислотной последовательности с наиболее сходными последовательностями VH и VL человека эмбрионального типа. Антитела, обладающие высокой гомологией с человеческими, могут включать антитела, содержащие домены VH и VL нативных не являющихся человеческими антител, которые проявляют достаточно высокий % идентичности последовательности с последовательностями человека эмбрионального типа, включая, например, антитела, содержащие VH- и VL-домены общепринятых антител животных семейства верблюжьих, а также модифицированные способами инженерии, особенно гуманизированные, варианты таких антител и также "полностью человеческие" антитела.
В одном из вариантов осуществления VH-домен антитела с высокой гомологией с человеческим может проявлять идентичность аминокислотной последовательности или гомологию последовательности, составляющие 80% или более, с одним или несколькими VH-доменами человека в каркасных областях FR1, FR2, FR3 и FR4. В других вариантах осуществления идентичность аминокислотных последовательностей или гомология последовательностей между VH-доменом полипептида по изобретению и наиболее сходной последовательностью VH-домена человека эмбрионального типа могут составлять 85% или более, 90% или более, 95% или более, 97% или более, или вплоть до 99% или даже 100%.
В одном из вариантов осуществления VH-домен антитела с высокой гомологией с человеческим может содержать менее 10 (например, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1) замен аминокислотной последовательности в каркасных областях FR1, FR2, FR3 и FR4, по сравнению с наиболее сходной последовательностью VH человека.
В другом варианте осуществления VL-домен антитела с высокой гомологией с человеческим может обладать идентичностью последовательности или гомологией последовательности, составляющей 80% или более с одним или несколькими VL-доменами человека в каркасных областях FR1, FR2, FR3 и FR4. В других вариантах осуществления идентичность аминокислотной последовательности или гомология последовательности между VL-доменом полипептида по изобретению и наиболее сходной последовательностью VL-домена человека эмбрионального типа может составлять 85% или более, 90% или более, 95% или более, 97% или более или вплоть до 99% или даже 100%.
В одном из вариантов осуществления VL-домен антитела с высокой гомологией может содержать менее 10 (например 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1) замен аминокислотной последовательности в каркасных областях FR1, FR2, FR3 и FR4, по сравнению с наиболее сходной последовательностью VL человека.
Антитела с высокой гомологией с человеческими также могут содержать гипервариабельные петли или CDR, имеющие каноническую укладку человека или подобную человеческой каноническую укладку, как подробно рассмотрено ниже. В одном из вариантов осуществления по меньшей мере одну гипервариабельную петлю или CDR либо в VH-домене, либо в VL-домене, антитела с высокой гомологией с человеческим можно получать, или она может происходить, из VH- или VL-домена не являющегося человеческим антитела, например, общепринятого антитела из вида животных семейства верблюжьих, но, тем не менее, она проявляет спрогнозированную или истинную структуру канонической укладки, по существу идентичную структуре канонической укладки, которая встречается в антителах человека.
Следует отметить, что антитела с высокой гомологией с человеческими не обязательно обладают структурами канонической укладки человека или подобными человеческим структурами канонической укладки. Например, антитела приматов обладают высокой гомологией последовательности с антителами человека, но, тем не менее, часто не обладают структурами канонической укладки человека или подобными человеческим структурами канонической укладки.
В данной области является общепризнанным, что, хотя первичные аминокислотные последовательности гипервариабельных петель, присутствующих как в VH-доменах, так и в VL-доменах, кодируемых последовательностью человека эмбрионального типа, по определению, являются высоковариабельными, все гипервариабельные петли, за исключением CDR H3 домена VH, принимают только несколько отчетливых структурных конформаций, называемых канонической укладкой (Chothia et al., J. Mol. Biol. 196:901-917 (1987); Tramontano et al. Proteins 6:382-94 (1989)), которая зависит как от длины гипервариабельной петли, так и от присутствия так называемых канонических аминокислотных остатков (Chothia et al., J. Mol. Biol. 196:901-917 (1987)). Истинные канонические структуры гипервариабельных петель в интактных доменах VH или VL можно определять с помощью структурного анализа (например, рентгеновская кристаллография), но также каноническую структуру возможно спрогнозировать на основе ключевых аминокислотных остатков, которые характерны для конкретной структуры (дополнительно рассмотрено ниже). По существу, конкретный характер остатков, которые определяют каждую каноническую структуру, формирует "признак", который позволяет канонической структуре распознаваться в гипервариабельных петлях домена VH или VL неизвестной структуры; таким образом, канонические структуры могут быть спрогнозированы только на основе первичной аминокислотной последовательности.
Спрогнозированные структуры канонической укладки для гипервариабельных петель любой данной последовательности VH или VL в антителе с высокой гомологией можно анализировать с использованием алгоритмов, которые являются общедоступными от www.bioinf.org.uk/abs/chothia.html, www.biochem.ucl.ac.uk/~martin/antibodies.html и www.bioc.unizh.ch/antibody/Sequences/Germlines/Vbase_hVk.html. Эти инструменты позволяют выравнивать запрашиваемые последовательности VH или VL с последовательностями домена VH или VL человека с известной канонической структурой и осуществлять прогнозирование канонической структуры для гипервариабельных петель последовательности запроса.
В случае VH-домена, петли H1 и H2 можно оценивать как имеющие структуру канонической укладки, "по существу идентичную" структуре канонической укладки, о которой известно, что она встречается в антителах человека, если удовлетворяется по меньшей мере первое, и предпочтительно, оба, из следующих критериев:
1. Идентичная длина, определяемая количеством остатков, с наиболее сходным классом канонических структур человека.
2. По меньшей мере 33% идентичность, предпочтительно, по меньшей мере 50% идентичность, с ключевыми аминокислотными остатками, описанными для соответствующих классов канонических структур H1 и H2 человека,
(следует отметить, что для целей вышеуказанного анализа петли H1 и H2 исследуют по отдельности и каждую из них сравнивают с наиболее сходным классом канонических структур человека).
Представленный выше анализ основан на прогнозировании канонической структуры петель H1 и H2 представляющего интерес антитела. Если истинные структуры петель H1 и H2 в представляющем интерес антителе известны, например, исходя из рентгеновской кристаллографии, тогда петли H1 и H2 в представляющем интерес антителе также могут быть оценены как имеющие структуру канонической укладки, "по существу идентичную" структуре канонической укладки, о которой известно, что она встречается в антителах человека, если длина петли отличается от длины петли наиболее сходного класса канонических структур человека (как правило, на ±1 или ±2 аминокислоты), однако истинная структура петель H1 и H2 в представляющем интерес антителе соответствует структуре канонической укладки человека.
Ключевые аминокислотные остатки, встречающиеся в классах канонических структур человека для первой и второй гипервариабельных петель VH-доменов человека (H1 и H2) описаны Chothia et al., J. Mol. Biol. 227:799-817 (1992), содержание которой включено в настоящее описание в качестве ссылки в полном объеме. В частности, в таблице 3 на стр. 802 Chothia et al., которая прямо включена в настоящее описание в качестве ссылки, приведены предпочтительные аминокислотные остатки в ключевых участках для канонических структур H1, встречающихся в последовательности человека эмбрионального типа, в то время как в таблице 4 на стр. 803, также прямо включенной в настоящее описание в качестве ссылки, приведены предпочтительные аминокислотные остатки в ключевых участках для канонических структур CDR H2, встречающихся в последовательности человека эмбрионального типа.
В одном из вариантов осуществления как H1, так и H2, в VH-домене антитела с высокой гомологией с человеческим, обладают спрогнозированной или истинной структурой укладки, которая по существу идентична структуре канонической укладки, которая встречается в антителах человека.
Антитела с высокой гомологией с человеческими могут содержать VH-домен, в котором гипервариабельные петли H1 и H2 образуют комбинацию структур канонической укладки, которые идентичны комбинации канонических структур, о которых известно, что они встречаются по меньшей мере в одном домене VH человека эмбрионального типа. Было выявлено, что только определенные комбинации структур канонической укладки в H1 и H2 в действительности встречаются в VH-доменах, кодируемых последовательностью человека эмбрионального типа. В варианте осуществления H1 и H2 в VH-домене антитела с высокой гомологией с человеческим могут быть получены из VH-домена не являющегося человеком вида, например, вида животного семейства верблюжьих, но тем не менее они образуют комбинацию со спрогнозированными или истинными структурами канонической укладки, которая идентична комбинации структур канонической укладки, о которых известно, что они встречаются в последовательности человека эмбрионального типа или подвергнутого соматической мутации VH-домена. В неограничивающих вариантах осуществления H1 и H2 в VH-домене антитела с высокой гомологией с человеческим могут быть получены из VH-домена не являющегося человеком вида, например, вида животных семейства верблюжьих, и они образуют одну из следующих комбинаций канонических укладок: 1-1, 1-2, 1-3, 1-6, 1-4, 2-1, 3-1 и 3-5.
Антитело с высокой гомологией с человеческим может содержать VH-домен, который обладает высокой идентичностью последовательности/гомологией последовательности с VH человека, и который содержит гипервариабельные петли, проявляющие структурную гомологию с VH человека.
Может быть преимущественным для канонических укладок, присутствующих в H1 и H2 в VH-домене антитела с высокой гомологией с человеческим, и их комбинации, чтобы они были "исправлены" на последовательность VH человека эмбрионального типа, которая наиболее сходна с VH-доменом антитела с высокой гомологией с человеческим с точки зрения общей идентичности первичной аминокислотной последовательности. В качестве примера, если наибольшим является сходство последовательности с VH3-доменом человека эмбрионального типа, тогда может быть преимущественным для H1 и H2 образовывать комбинацию канонических укладок, которые также встречаются в природе в VH3-домене человека. Это может быть особенно важным в случае антител с высокой гомологией, которые происходят из не являющегося человеком вида, например, антител, содержащих VH- и VL-домены, которые происходят из общеизвестных антител животных семейства верблюжьих, особенно антител, содержащих гуманизированные домены VH и VL животного семейства верблюжьих.
Таким образом, в одном из вариантов осуществления VH-домен антитела против IL-6 с высокой гомологией с человеческим может обладать идентичностью последовательности или гомологией последовательности 80% или более, 85% или более, 90% или более, 95% или более, 97% или более, или вплоть до 99% или даже 100% с доменом VH человека в каркасных областях FR1, FR2, FR3 и FR4, и, кроме того, H1 и H2, в том же антителе получен из не являющегося человеческим VH-домена (например, происходящего из вида животных семейства верблюжьих), однако они образуют комбинацию со спрогнозированными или истинными структурами канонической укладки, которая является такой же, как комбинация канонических укладок, о которой известно, что она встречается в природе в том же VH-домене человека.
Например, в одном иллюстративном варианте осуществления петли H1 и H2 антитела против IL-6 по изобретению (например, 61H7) могут содержать комбинацию 1-2 структур канонической укладки человека, встречающуюся, например, в структуре антитела человека 1DFB. В другом иллюстративном варианте осуществления петли H1 и H2 антитела против IL-6 по изобретению (например, 68F2 или его вариант эмбрионального типа 129D3) могут содержать комбинацию 3-1 структур канонической укладки человека, встречающуюся, например, в структуре антитела человека 1ACY.
В других вариантах осуществления каждую из L1 и L2 в VL-домене антитела с высокой гомологией с человеческим получают из VL-домена не являющегося человеком вида (например, VL-домен, происходящий из животного семейства верблюжьих), и каждая из них обладает спрогнозированной или истинной структурой укладки, которая по существу идентична структуре канонической укладки, которая встречается в антителах человека.
Как и в случае VH-доменов, гипервариабельные петли VL-доменов как типа V-лямбда, так и типа V-каппа, могут принимать ограниченное количество конформаций или канонических структур, определяемых, частично, длиной, а также присутствием ключевых аминокислотных остатков в определенных канонических положениях.
В представляющем интерес антителе, обладающем высокой гомологией с человеческим, петли L1, L2 и L3, полученные из VL-домена не являющегося человеком вида, например, вида Camelidae, можно считать имеющими структуру канонической укладки, "по существу идентичную" структуре канонической укладки, о которой известно, что она встречается в антителах человека, если удовлетворяется по меньшей мере первое, и предпочтительно, оба, из следующих критериев:
1. Идентичная длина, определяемая количеством аминокислотных остатков, с наибольшим сходством с классом структур человека.
2. По меньшей мере 33% идентичность, предпочтительно, по меньшей мере 50% идентичность, с ключевыми аминокислотными остатками, описанными для соответствующих классов канонических структур L1 или L2 человека из репертуара либо V-лямбда, либо V-каппа,
(следует отметить, что для целей указанного выше анализа петли L1 и L2 исследуют по отдельности и каждую из них сравнивают с наиболее сходным классом канонических структур человека).
Указанный выше анализ основан на прогнозировании канонической структуры петель L1, L2 и L3 в VL-домене представляющего интерес антитела. Если истинная структура петель L1, L2 и L3 известна, например, исходя из рентгеновской кристаллографии, тогда петли L1, L2 или L3, происходящие из представляющего интерес антитела, также можно считать имеющими структуру канонической укладки, "по существу идентичную" структуре канонической укладки, о которой известно, что она встречается в антителах человека, если длина петли отличается от наиболее сходного класса канонических структур человека (как правило, на ±1 или ±2 аминокислоты), но истинная структура петель животных семейства верблюжьих соответствует канонической укладке человека.
Ключевые аминокислотные остатки, встречающиеся в классах канонических структур человека для CDR доменов V-лямбда и V-каппа человека, описаны Morea et al. Methods, 20: 267-279 (2000) и Martin et al., J. Mol. Biol., 263:800-815 (1996). Структурный набор доменов V-каппа человека также описан Tomlinson et al. EMBO J. 14:4628-4638 (1995), и структурный набор доменов V-лямбда описан Williams et al. J. Mol. Biol., 264:220-232 (1996). Содержание всех из этих документов включено в настоящее описание в качестве ссылки.
L1 и L2 в VL-домене антитела с высокой гомологией с человеческим могут образовывать комбинацию спрогнозированных или истинных структур канонической укладки, которая идентична комбинации структур канонической укладки, о которой известно, что она встречается в VL-домене человека эмбрионального типа. В неограничивающих вариантах осуществления L1 и L2 в домене V-лямбда антитела с высокой гомологией с человеческим (например, антитело, содержащее VL-домен, происходящий из животного семейства верблюжьих, или его гуманизированный вариант) могут образовывать одну из следующих комбинаций канонических укладок: 11-7, 13-7(A,B,C), 14-7(A,B), 12-11, 14-11 и 12-12 (как определено в Williams et al. J. Mol. Biol. 264:220-32 (1996) и как показано на http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVL.html). В неограничивающих вариантах осуществления L1 и L2 в домене V-каппа могут образовывать одну из следующих комбинаций канонических укладок: 2-1, 3-1, 4-1 и 6-1 (как определено в Tomlinson et al. EMBO J. 14:4628-38 (1995) и как показано на http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVK.html). Например, в одном иллюстративном варианте осуществления петли L1 и L2 антитела против IL-6 по изобретению (например, 61H7) могут содержать комбинацию 7λ-1 структур канонической укладки человека, встречающуюся, например, в структуре антитела человека 1MFA. В другом иллюстративном варианте осуществления петли L1 и L2 антитела против IL-6 по изобретению (например, 68F2 или его вариант эмбрионального типа 129D3) могут содержать комбинацию 6λ-1 структур канонической укладки человека, встречающуюся, например, в структуре антитела человека 3MUG.
В следующем варианте осуществления все три из L1, L2 и L3 в VL-домене антитела с высокой гомологией с человеческим могут обладать по существу человеческой структурой. Предпочтительно, чтобы VL-домен антитела с высокой гомологией с человеческим обладал высокой идентичностью последовательности/гомологией последовательности с VL человека, а также, чтобы гипервариабельные петли в VL-домене обладали структурной гомологией с VL человека. Например, в одном иллюстративном варианте осуществления петли L1-L3 антитела против IL-6 по изобретению (например, 61H7) могут содержать комбинацию структур канонической укладки человека 7λ-1-4, встречающуюся, например, в структуре антитела человека 1MFA. В другом иллюстративном варианте осуществления петли L1-L3 антитела против IL-6 по изобретению (например, 68F2 или его вариант эмбрионального типа 129D3) могут содержать комбинацию структур канонической укладки человека 6λ-1-5, встречающуюся, например, в структуре антитела человека 3MUG.
В одном из вариантов осуществления VL-домен антитела против IL-6 с высокой гомологией с человеческим может обладать идентичностью последовательности 80% или более, 85% или более, 90% или более, 95% или более, 97% или более или вплоть до 99% или даже 100% с VL-доменом человека в каркасных областях FR1, FR2, FR3 и FR4, и, кроме того, гипервариабельная петля L1 и гипервариабельная петля L2 могут образовывать комбинацию спрогнозированных или истинных структур канонической укладки, которая является такой же, как и комбинация канонических структур, о которой известно, что она встречается в природе в том же VL-домене человека.
Безусловно, предусматривается, что VH-домены, обладающие высокой идентичностью последовательности/гомологией последовательности с VH человека, а также структурной гомологией с гипервариабельными петлями VH человека, могут находиться в комбинации с VL-доменами, обладающими высокой идентичностью последовательностей/гомологией последовательностей с VL человека, а также структурной гомологией с гипервариабельными петлями VL человека, с образованием антител с высокой гомологией с человеческими, содержащих пары VH/VL (например, происходящие из животных семейства верблюжьих пары VH/VL) с максимальной гомологией последовательности и структуры с кодируемыми у человека парами VH/VL.
iii). Не являющиеся иммуноглобулинами связывающие молекулы
В следующем аспекте изобретение относится к не являющимся иммуноглобулинами связывающим молекулам, которые специфически связываются с IL-6. Как используют в рамках изобретения, термин "не являющиеся иммуноглобулинами связывающие молекулы" представляют собой связывающие молекулы, участки связывания которых содержат участок (например, остов или каркас), происходящий из полипептида, отличного от иммуноглобулина, но которые могут быть сконструированы (например, путем добавления последовательностей CDR-областей) так, чтобы сообщать желаемую специфичность связывания связывающей молекуле. Не являющиеся иммуноглобулинами связывающие молекулы по изобретению, как правило, содержат одну или несколько из областей CDR, указанных в таблицах 13-18, пересаженных в не являющийся иммуноглобулином полипептид.
В определенных вариантах осуществления не являющиеся иммуноглобулинами связывающие молекулы содержат участки связывания, которые происходят из представителя суперсемейства иммуноглобулинов, который не является иммуноглобулином (например, T-клеточный рецептор или белок клеточной адгезии (например, CTLA-4, N-CAM, телокин)). Такие связывающие молекулы содержат участок связывания, который сохраняет конформацию укладки иммуноглобулина и способен специфически связываться с IL-6, когда он модифицирован включением одной или нескольких областей CDR, указанных в таблицах 13-18. В других вариантах осуществления не являющиеся иммуноглобулинами связывающие молекулы по изобретению содержат участок связывания с топологией в белке, которая не основана на укладке иммуноглобулина (например, белки с анкириновыми повторами, тетранектины и фибронектины), но они тем не менее способны специфически связываться с мишенью (например, IL-6), когда они модифицирован включением одной или нескольких областей CDR, указанных в таблицах 13-18.
В одном из вариантов осуществления связывающая молекула по изобретению содержит молекулу тетранектина. Тетранектины представляют собой белки плазмы с трехвалентной структурой. Каждый мономер тримера тетранектина содержит пять различных аминокислотных петель, которые могут быть заменены или модифицированы способами инженерии так, чтобы они содержали последовательности CDR антител (например, области CDR, указанные в таблицах 13-18). Способы получения связывающих полипептидов на основе тетранектинов описаны, например, в US 20110086770, которая включена в настоящее описание в качестве ссылки в полном объеме.
В одном из вариантов осуществления связывающая молекула по изобретению содержит молекулу фибронектина. Связывающие молекулы на основе фибронектина (например, молекулы, содержащие домены фибронектина типа I, II или III), обладают CDR-подобными петлями, которые могут быть заменены или модифицированы способами инженерии так, чтобы они содержали последовательности CDR антител (например, области CDR, указанные в таблицах 13-18). Способы получения связывающих полипептидов на основе фибронектина описаны, например, в WO 01/64942 и в патентах США 6673901, 6703199, 7078490 и 7119171, все из которых включены в настоящее описание в качестве ссылок в полном объеме.
В другом варианте осуществления связывающая молекула по изобретению содержит участок связывания из аффиантитела. Аффиантитела происходят из связывающих иммуноглобулин доменов белка A стафилококков (SPA) (см., например, Nord et al., Nat. Biotechnol., 15: 772-777 (1997)). Участки связывания аффиантител, используемых в настоящем описании, можно синтезировать путем мутагенеза родственного SPA белка (например, белок Z), происходящего из домена SPA (например, домен B) и выбора мутантных родственных SPA полипептидов, обладающих аффинностью связывания с IL-6. Другие способы получения участков связывания аффиантител описаны в патентах США 6740734 и 6602977 и в WO 00/63243, каждый из которых включен в настоящее описание в качестве ссылки.
В другом варианте осуществления связывающая молекула по изобретению содержит участок связывания из антикалина. Антикалины (также известные как липокалины) являются представителями разнообразного семейства белков с бета-бочонками, функцией которых является связывание молекул-мишеней в их области бочонка/петли. Участки связывания липокалинов можно конструировать так, чтобы они связывали IL-6, путем рандомизации последовательностей петель, соединяющих цепи бочонка (см. например, Schlehuber et al., Drug Discov. Today, 10: 23-33 (2005); Beste et al., PNAS, 96: 1898-1903 (1999). Участки связывания антикалина, используемые в связывающих молекулах по изобретению, можно получать, начиная с полипептидов семейства липокалинов, которые являются мутантными в четырех сегментах, которые соответствуют положениям последовательности в линейной полипептидной последовательности, содержащим положения аминокислот с 28 по 45, с 58 по 69, с 86 по 99 и с 114 по 129 билин-связывающего белка (BBP) Pieris brassica. Другие способы получения участков связывания антикалина описаны в WO 99/16873 и WO 05/019254, каждая из которых включена в настоящее описание в качестве ссылки.
В другом варианте осуществления связывающая молекула по изобретению содержит участок связывания из богатого цистеином полипептида. Богатые цистеином домены, используемые при применении на практике настоящего изобретения, как правило, не образуют структуры альфа-спирали, бета-слоя или бета-бочонка. Как правило, дисульфидные связи обеспечивают укладку домена в трехмерную структуру. Обычно богатые цистеином домены имеют по меньшей мере две дисульфидных связи, более конкретно по меньшей мере три дисульфидных связи. Иллюстративным цистеин-богатым полипептидом является белок с A-доменом. A-домены (иногда называемые "повторами комплементарного типа") содержат приблизительно 30-50 или 30-65 аминокислот. В некоторых вариантах осуществления эти домены содержат приблизительно 35-45 аминокислот и в некоторых случаях приблизительно 40 аминокислот. В пределах 30-50 аминокислот имеется приблизительно остатков 6 цистеина. Среди шести остатков цистеина дисульфидные связи, как правило, расположены между следующими остатками цистеина: C1 и C3, C2 и C5, C4 и C6. A-домен представляет собой лиганд-связывающую часть. Остатки цистеина в домене связаны дисульфидной связью с образованием компактной, стабильной, функционально независимой части. Кластеры этих повторов составляют лигандсвязывающий домен, и различная кластеризация может обеспечить специфичность в отношении связывания лиганда. Иллюстративные белки, содержащие A-домены, включают, например, компоненты комплемента (например, C6, C7, C8, C9 и фактор I), сериновые протеазы (например, энтеропептидаза, матриптаза и корин), трансмембранные белки (например, ST7, LRP3, LRP5 и LRP6) и эндоцитарные рецепторы (например, сортилин-родственный рецептор, LDL-рецептор, VLDLR, LRP1, LRP2 и ApoER2). Способы получения белков с A-доменом с желаемой специфичностью связывания описаны, например, в WO 02/088171 и WO 04/044011, каждая из которых включена в настоящее описание в качестве ссылки.
В других вариантах осуществления связывающая молекула по изобретению содержит участок связывания из белка с повтором. Белки с повторами представляют собой белки, которые содержат последовательно расположенные копии небольших (например, от приблизительно 20 до приблизительно 40 аминокислотных остатков) структурных элементов или повторов, которые располагаются друг за другом с образованием непрерывных доменов. Белки с повторами можно модифицировать, чтобы они подходили к конкретному участку связывания на мишени, путем модификации количества повторов в белке. Иллюстративные белки с повторами включают сконструированные белки с анкириновыми повторами (т.е. DARPin) (см. например, Binz et al., Nat. Biotechnol., 22: 575-582 (2004)) или белки с богатыми лейцином повторами (т.е. LRRP) (см. например, Pancer et al., Nature, 430: 174-180 (2004)). Также определенные до настоящего времени третичные структуры элементов анкириновых повторов имеют общий характерный элемент, состоящий из бета-шпильки, за которой следует две антипараллельных альфа-спирали, и оканчивающийся петлей, соединяющей элемент повтора со следующим элементом. Домены, сконструированные из элементов анкиринового повтора, образованы посредством расположения друг за другом элементов повтора в виде вытянутой и искривленной структуры. Участки связывания LRRP из части адаптивной иммунной системы морских миног и других бесчелюстных рыб напоминают антитела, поскольку они образованы посредством рекомбинации набора генов с богатыми лейцином повторами во время созревания лимфоцитов. Способы получения участков связывания DARpin или LRRP описаны в WO 02/20565 и WO 06/083275, каждая из которых включена в настоящее описание в качестве ссылки.
Другие не являющиеся иммуноглобулинами участки связывания, которые можно использовать в связывающих молекулах по изобретению, включают участки связывания, происходящие из доменов гомологии Src (например, SH2- или SH3-домены), PDZ-доменов, бета-лактамазы, высокоаффинных ингибиторов протеаз и или небольших связанных дисульфидными связями белковых каркасов, таких как токсины скорпиона. Способы получения участков связывания, происходящих из этих молекул, описаны в данной области, см., например, Panni et al., J. Biol. Chem., 277: 21666-21674 (2002), Schneider et al., Nat. Biotechnol., 17: 170-175 (1999); Legendre et al., Protein Sci., 11:1506-1518 (2002); Stoop et al., Nat. Biotechnol., 21: 1063-1068 (2003); и Vita et al., PNAS, 92: 6404-6408 (1995). Другие участки связывания могут происходить из связывающего домена, выбранного из группы, состоящей из EGF-подобного домена, Kringle-домена, PAN-домена, Gla-домена, SRCR-домена, ингибиторного домена трипсина поджелудочной железы Кунитца/ингибиторного домена трипсина поджелудочной железы крупного рогатого скота, ингибиторного домена сериновой протеазы типа Kazal, домена трилистника (P-типа), C-домена фактора Виллебранда, анафилатоксин-подобного домена, CUB-домена, тиреоглобулин-подобного повтора типа I, A-домена класса рецепторов LDL, домена Sushi, домена Link, домена тромбоспондина типа I, иммуноглобулин-подобного домена, домена лектина C-типа, домена MAM, домена фактора Виллебранда типа A, домена соматомедина B, корового домена с четырьмя дисульфидными связями WAP-типа, C-домена типа F5/8, домена гемопексина, EGF-подобного домена ламининового типа, C2-домена и других таких доменов, известных средним специалистам в данной области, а также их производных и/или вариантов.
Неиммуноглобулиновые связывающие молекулы можно идентифицировать путем выбора или выделения связывающегося с мишенью варианта из библиотеки связывающих молекул, имеющих искусственно диверсифицированные участки связывания. Диверсифицированные библиотеки можно получать путем включения библиотеки последовательностей CDR (например, выбранных из последовательностей CDR, указанных в таблицах 13-18) и/или с помощью полностью случайных подходов (например, ПЦР с пониженной точностью, шаффлинг экзонов или направленные изменения) и/или их можно осуществлять с помощью признанных в данной области стратегий моделирования. Например, положения аминокислот, которые обычно вовлечены, когда участок связывания взаимодействует с его молекулой-мишенью, можно рандомизировать путем встраивания вырожденных кодонов, тринуклеотидов, случайных пептидов или целых петель в соответствующих положениях в нуклеиновой кислоте, которая кодирует участок связывания (см., например, публикацию США № 20040132028). Положение аминокислотной замены можно идентифицировать путем исследования кристаллической структуры участка связывания в комплексе с молекулой-мишенью. Положения-кандидаты для включения последовательностей CDR (например, выбранных из последовательностей CDR, указанных в таблицах 13-18) и/или рандомизации включают петли, плоские поверхности, спирали и связывающие полости участка связывания. В определенных вариантах осуществления аминокислоты в участке связывания, которые, вероятно, являются кандидатами для диверсификации, можно идентифицировать по их гомологии с укладкой иммуноглобулина. Например, остатки в CDR-подобных петлях фибронектина можно рандомизировать для получения библиотеки связывающих фибронектин молекул (см., например, Koide et al., J. Mol. Biol., 284: 1141-1151 (1998)). После включения последовательностей CDR (например, выбранных из последовательностей CDR, указанных в таблице 2-6) и/или рандомизации, диверсифицированную библиотеку можно подвергать процедуре селекции для получения связывающих молекул с желаемыми характеристиками связывания, например, специфическим связыванием с IL-6. Выбор можно осуществлять известными в данной области способами, такими как фаговый дисплей, дрожжевой дисплей или дисплей нуклеиновых кислот.
iv. Преобразование происходящих из животных семейства верблюжьих VH- и VL-доменов в эмбриональный тип
Общепринятые антитела животных семейства верблюжьих обеспечивают преимущественную исходную точку для получения антител, применимых в качестве лекарственных средств у человека вследствие следующих факторов (рассмотрены в US 12/497239, который включен в настоящее описание в качестве ссылки в полном объеме):
1) Высокая % гомология последовательности между VH- и VL-доменами животных семейства верблюжьих и их аналогами из человека;
2) Высокая степень структурной гомологии между CDR VH- и VL-доменов животных семейства верблюжьих и их аналогами из человека (т.е. подобные человеческим структуры канонической укладки и подобные человеческим комбинации канонических укладок).
Платформа животных семейства верблюжьих (например, лама) также обеспечивает значительное преимущество с точки зрения функционального разнообразия антител против IL-6, которые могут быть получены.
Применимость антител против IL-6, содержащих домены VH и/или VL животного семейства верблюжьих для применения у человека могут быть еще более улучшены путем "преобразования в эмбриональный тип" природных VH- и VL-доменов животных семейства верблюжьих, например, чтобы сделать их менее иммуногенными у хозяина-человека. Основной целью преобразования в эмбриональный тип является получение молекулы, в которой VH- и VL-домены проявляют минимальную иммуногенность, когда их вводят человеку, при сохранении специфичности и аффинности антигенсвязывающего участка, образованного родительскими VH- и VL-доменами.
Определение гомологии между доменом VH (или VL) животного семейства верблюжьих и доменами VH (или VL) человека является важной стадией в процессе преобразования в эмбриональный тип, как для выбора аминокислотных остатков животного семейства верблюжьих, подлежащих замене (в данном домене VH или VL), так и для выбора подходящего заменяющего аминокислотного остатка(ов).
Подход для преобразования в эмбриональный тип общепринятых антител животных семейства верблюжьих был разработан на основе выравнивания большого количества последовательностей доменов VH (и VL), как правило, подвергнутых соматической мутации доменов VH (или VL), о которых известно, что они связывают антиген-мишень, с последовательностями VH (или VL) эмбрионального типа человека, консенсусными последовательностями VH (и VL) человека, а также информации о последовательности эмбрионального типа, доступной для llama pacos.
Ниже описаны принципы, которые можно использовать (i) для выбора аминокислотных остатков "животного семейства верблюжьих" для замены в происходящем из животного семейства верблюжьих домене VH или VL или их CDR, и (ii) для выбора заменяющих аминокислотных остатков "человека" для замещения при преобразовании в эмбриональный тип любого данного домена VH (или VL) животного семейства верблюжьих. Этот подход можно использовать для получения преобразованных в эмбриональный тип вариантов последовательностей VH и VL, указанных в таблицах 13-16 настоящего описания.
Стадия 1. Выбрать семейство последовательностей человека (эмбрионального типа) и представителя этого семейства, который демонстрирует наиболее высокую гомологию/идентичность со зрелой последовательностью животного семейства верблюжьих, подлежащей преобразованию в эмбриональный тип. Общая методика для идентификации наиболее сходной последовательности человека эмбрионального типа для любого данного домена VH (или VL) животного семейства верблюжьих приведена ниже.
Стадия 2. Выбрать конкретного представителя семейства эмбрионального типа человека, используемого в качестве эталона для преобразования в эмбриональный тип. Предпочтительно, он представляет собой последовательность эмбрионального типа с наиболее высокой гомологией или другого представителя семейства последовательностей эмбрионального типа из того же семейства.
Стадия 3. Идентифицировать предпочтительные положения для преобразования в эмбриональный тип, исходя из таблицы использования аминокислот для последовательности эмбрионального типа животного семейства верблюжьих, которая является наиболее сходной с выбранной последовательностью человека эмбрионального типа.
Стадия 4. Попробовать заменить аминокислоты в эмбриональной последовательности эмбрионального типа животного семейства верблюжьих, которая отличается от наиболее сходной последовательности человека эмбрионального типа; преобразование в эмбриональный тип остатков FR является предпочтительным относительно остатков CDR.
a. Предпочтительными являются положения, которые отличаются от выбранной последовательности человека эмбрионального типа, использованной в качестве эталонной последовательности для преобразования в эмбриональный тип, где аминокислота, встречающаяся в последовательности животного семейства верблюжьих, не соответствует выбранной последовательности эмбрионального типа и не встречается в других последовательностях эмбрионального типа того же подкласса (для аминокислот FR, кодируемых как V, так и J).
b. Положения, которые отличаются от выбранного представителя семейства эмбрионального типа человека, но которые используются в других последовательностях эмбрионального типа того же семейства, также могут быть рассмотрены в процессе преобразования в эмбриональный тип.
c. Также можно рассматривать дополнительные несоответствия (например, вследствие дополнительных соматических мутаций) с выбранной последовательностью человека эмбрионального типа.
Следующий подход можно использовать для определения наибольшего сходства с последовательностью человека эмбрионального типа для данного домена VH (или VL) животного семейства верблюжьих:
До анализа процентной идентичности последовательностей между VH и VL эмбрионального типа животных семейства верблюжьих и человека сначала можно определять каноническую укладку, которая позволяет идентификацию семейства сегментов человека эмбрионального типа с идентичной комбинацией канонических укладок для H1 и H2 или L1 и L2 (и L3). Затем представитель семейства эмбрионального типа человека, который имеет наиболее высокую степень гомологии последовательности с представляющей интерес вариабельной областью животного семейства верблюжьих, может быть выбран для оценки гомологии последовательностей. Определение канонических классов гипервариабельных петель Chothia L1, L2, L3, H1 и H2 можно проводить с помощью биоинформатических инструментов, общедоступных на web-странице www.bioinf.org.uk/abs/chothia.html.page. Выходные данные программы демонстрируют ключевые требования к остаткам в файле данных. В этих файлах данных ключевые положения остатков представлены с предусмотренными аминокислотами в каждом положении. Последовательность вариабельной области антитела приведена в качестве входных данных и ее сначала выравнивают с консенсусной последовательностью антитела для приведения в соответствие со схемой нумерации Kabat. В анализе канонической укладки используется набор шаблонов ключевых остатков, установленных с помощью автоматизированного способа, разработанного Martin and Thornton (Martin et al., J. Mol. Biol. 263:800-815 (1996)). Границы индивидуальных каркасных областей могут быть определены с использованием схемы нумерации IMGT, которая является модификацией схемы нумерации Chothia (Lefranc et al., NAR 27: 209-212 (1999); imgt.cines.fr).
Когда известен конкретный сегмент V человека эмбрионального типа, в котором используется одна и та же комбинация канонических укладок для H1 и H2 или L1 и L2 (и L3), можно определять наиболее сходного представителя семейства с точки зрения гомологии последовательности. Процентную идентичность последовательностей между аминокислотными последовательностями каркасной области доменов VH и VL животных семейства верблюжьих и соответствующими последовательностями, кодируемыми последовательностью эмбрионального типа человека, можно определять с использованием биоинформатических инструментов, однако также можно использовать выравнивание последовательностей вручную. Последовательности иммуноглобулинов человека можно идентифицировать из нескольких баз данных белков, таких как VBase (vbase.mrc-cpe.cam.ac.uk/) или база данных Pluckthun/Honegger (http://www.bioc.unizh.ch/antibody/Sequences/Germlines). Для сравнения последовательностей человека с V-областями доменов VH или VL животных семейства верблюжьих можно использовать алгоритм выравнивания, такой как алгоритм, доступный через web-сайты, такие как www.expasy.ch/tools/#align, но также можно проводить выравнивание вручную для ограниченного набора последовательностей. Последовательности легкой и тяжелой цепей человека эмбрионального типа из семейств с одинаковыми комбинациями канонических укладок и с наиболее высокой степенью гомологии с каркасными областями 1, 2 и 3 каждой цепи, можно выбирать и сравнивать с представляющей интерес вариабельной областью животного семейства верблюжьих; также FR4 проверяют против областей JH и JK или JL человека эмбрионального типа.
Следует отметить, что в кровотоке общую процентную гомологию последовательностей для остатков FR1, FR2 и FR3 оценивают с использованием наиболее сходной последовательности из семейства последовательностей человека эмбрионального типа с идентичной комбинацией канонических укладок. Оценивают только остатки, отличающиеся от наиболее сходной последовательности или других представителей того же семейства с той же комбинацией канонических укладок (примечание - исключая любые кодируемые праймерами различия). Однако для целей преобразования в эмбриональный тип остатки в каркасных областях, идентичные представителям других семейств последовательностей человека эмбрионального типа, которые не имеют той же комбинации канонических укладок, можно рассматривать для преобразования в эмбриональный тип, несмотря на то, что их оценивают как "отрицательные" в соответствии с условиями, описанными выше. Это предположение основано на подходе "смешения и приведения в соответствие" для преобразования в эмбриональный тип, в котором каждую из FR1, FR2, FR3 и FR4 по отдельности сравнивают с ее наиболее сходной последовательностью человека эмбрионального типа, и, таким образом, преобразованная в эмбриональный тип молекула содержит комбинацию различных FR, как было осуществлено Qu и коллегами (Qu et la., Clin. Cancer Res. 5:3095-3100 (1999)) и Ono и коллегами (Ono et al., Mol. Immunol. 36:387-395 (1999)).
IV. Модифицированные связывающие молекулы
В определенных вариантах осуществления связывающие полипептиды по изобретению могут содержать одну или несколько модификаций. Модифицированные формы связывающих полипептидов по изобретению можно получать с использованием любых способов, известных в данной области.
i) Уменьшение риска иммуногенности
В определенных вариантах осуществления связывающие молекулы (например, антитела или их антигенсвязывающие фрагменты) по изобретению модифицированы, чтобы далее снизить их риск иммуногенности с использованием известных в данной области способов. Например, антитела, или их фрагменты можно преобразовывать в эмбриональный тип в соответствии со способами, описанными выше. Альтернативно, связывающие молекулы по изобретению могут быть химеризованными, гуманизированными и/или деиммунизированными.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент могут быть химерными. Химерное антитело представляет собой антитело, в котором различные части антитела происходят из различных видов животных, такое как антитела, имеющие вариабельную область, происходящую из моноклонального антитела животного семейства верблюжьих (например, лама), и константную область иммуноглобулина человека. Способы получения химерных антител или их фрагментов известны в данной области. См., например, Morrison, Science 229:1202 (1985); Oi et al., BioTechniques 4:214 (1986); Gillies et al., J. Immunol. Methods 125:191-202 (1989); патенты США № 5807715; 4816567 и 4816397, которые включены в настоящее описание в качестве ссылок в полном объеме. Для синтеза указанных молекул можно использовать способы, разработанные для получения "химерных антител" (Morrison et al., Proc. Natl. Acad. Sci. 81:851-855 (1984); Neuberger et al., Nature 312:604-608 (1984); Takeda et al., Nature 314:452-454 (1985)). Например, последовательность гена, кодирующую специфичность связывания молекулы антитела животного семейства верблюжьих против IL-6, можно подвергать слиянию с последовательностью из молекулы антитела человека с соответствующей биологической активностью. Как используют в рамках изобретения, химерное антитело представляет собой молекулу, в которой различные части происходят из различных видов животных, таких как животные, имеющие вариабельную область, происходящую из моноклонального антитела животного семейства верблюжьих (например, лама), и константную область иммуноглобулина человека, например, преобразованные в эмбриональный тип или гуманизированные антитела.
В другом варианте осуществления антитело или его антигенсвязывающая часть по изобретению являются гуманизированными. Гуманизированные антитела обладают специфичностью связывания, включающей одну или несколько определяющих комплементарность областей (CDR) из не являющегося человеческим антитела и каркасные области из молекулы антитела человека. Часто, каркасные остатки в каркасных областях человека заменяют соответствующим остатком из CDR донорного антитела для изменения, предпочтительно, повышения, связывания антигена. Эти замены каркасной области идентифицируют способами, хорошо известными в данной области, например, путем моделирования взаимодействий CDR и каркасных остатков для идентификации каркасных остатков, важных для связывания антигена и сравнения последовательностей, чтобы идентифицировать необычные каркасные остатки в конкретных положениях. (См., например, Queen et al., патент США № 5585089; Riechmann et al., Nature 332:323 (1988), которые включены в настоящее описание в качестве ссылок в полном объеме.) Антитела можно гуманизировать с использованием различных способов, известных в данной области, включая, например, пересадку CDR (EP 239400; публикация PCT WO 91/09967; патенты США № 5225539; 5530101 и 5585089), гиперхимеризацию или преобразование поверхности (EP 592106; EP 519596; Padlan, Molecular Immunology 28(4/5):489-498 (1991); Studnicka et al., Protein Engineering 7(6):805-814 (1994); Roguska. et al., PNAS 91:969-973 (1994)), и шаффлинг цепей (патент США 5565332).
В некоторых вариантах осуществления можно использовать деиммунизацию для дальнейшего снижения риска иммуногенности связывающих IL-6 молекул (например, антитело или его антигенсвязывающая часть). Как используют в рамках изобретения, термин "деиммунизация" включает изменение полипептида (например, антитела или его антигенсвязывающей части) для модификации T-клеточных эпитопов (см., например, WO9852976A1, WO0034317A2). Например, последовательности VH и VL из исходного специфичного к IL-6 антитела или его антигенсвязывающей части по изобретению можно анализировать и можно получать "карту" T-клеточных эпитопов человека из каждой V-области, демонстрирующую положение эпитопов относительно определяющих комплементарность областей (CDR) и других ключевых остатков в последовательности. Индивидуальные T-клеточные эпитопы из карты T-клеточных эпитопов анализируют для идентификации альтернативных аминокислотных замен с низким риском изменения активности конечного антитела. Конструируют ряд альтернативных последовательностей VH и VL, содержащих комбинации аминокислотных замен, а затем эти последовательности включают в ряд специфичных к IL-6 антител или их фрагментов для применения в способах диагностики и лечения, описанных в настоящем описании, которые затем исследуют в отношении функции. Как правило, получают и исследуют от 12 до 24 вариантов антител. Затем полные гены тяжелых и легких цепей, содержащих модифицированные V-области и содержащих C-области человека, клонируют в экспрессирующие векторы, а затем плазмиды вводят в клеточные линии для продукции целого антитела. Затем антитела сравнивают в соответствующих биохимических и биологических анализах и идентифицируют оптимальный вариант.
ii) Эффекторные функции и модификации Fc
В определенных вариантах осуществления связывающие молекулы по изобретению могут содержать константную область антитела (например, константную область IgG, например, константную область IgG человека, например, константную область IgG1 или IgG4 человека), которая опосредует одну или несколько эффекторных функций. Например, связывание компонента C1 комплемента с константной областью антитела может активировать систему комплемента. Активация комплемента является важной для опсонизации и лизиса клеточных патогенов. Активация комплемента также стимулирует воспалительный ответ и также может быть вовлечена в аутоиммунную гиперчувствительность. Кроме того, антитела связываются с рецепторами на различных клетках через Fc-область, причем участок связывания Fc-рецептора на Fc-области антитела связывается с Fc-рецептором (FcR) на клетке. Существует ряд Fc-рецепторов, которые являются специфичными к различным классам антител, включая IgG (гамма-рецепторы), IgE (эпсилон-рецепторы), IgA (альфа-рецепторы) и IgM (мю-рецепторы). Связывание антитела с Fc-рецепторами на клеточных поверхностях запускает ряд важных и разнообразных биологических ответов, включая поглощение и разрушение покрытых антителом частиц, выведение иммунных комплексов, лизис покрытых антителом клеток-мишеней клетками-киллерами (называемый антителозависимой клеточно-опосредуемой цитотоксичностью или ADCC), высвобождение медиаторов воспаления, плацентарный перенос и контроль продукции иммуноглобулинов. В предпочтительных вариантах осуществления связывающие молекулы (например, антитела или их антигенсвязывающие фрагменты) по изобретению связываются с рецептором Fc-гамма. В альтернативных вариантах осуществления связывающие молекулы по изобретению могут содержать константную область, которая лишена одной или нескольких эффекторных функций (например, активность ADCC) и/или неспособна связывать Fcγ-рецептор.
Определенные варианты осуществления изобретения включают антитела против IL-6, в которых по меньшей мере одна аминокислота в одном или нескольких доменах константной области удалена или иным образом изменена, чтобы обеспечить желаемые биохимические характеристики, такие как сниженные или усиленные эффекторные функции, способность к нековалентной димеризации, увеличенная способность локализоваться в области опухоли, уменьшенное время полужизни в сыворотке или увеличенное время полужизни в сыворотке по сравнению с целым неизмененным антителом приблизительно с той же иммуногенностью. Например, определенные антитела или их фрагменты для применения в способах диагностики и лечения, описанных в настоящем описании, представляют собой антитела, которые содержат полипептидную цепь, сходную с тяжелой цепью иммуноглобулина, но которые лишены по меньшей мере части одного или нескольких доменов тяжелой цепи. Например, в определенных антителах удаляют один целый домен константной области модифицированного антитела, например, удаляют весь CH2-домен или его часть.
В некоторых других вариантах осуществления связывающие молекулы содержат константные области, происходящие из различных изотипов антител (например, константные области из двух или более из IgG1, IgG2, IgG3 или IgG4 человека). В других вариантах осуществления связывающие молекулы содержат химерную шарнирную область (т.е. шарнирная область, содержащая участки шарнирной области, происходящие из шарнирных доменов отличающихся изотипов антител, например, верхнего шарнирного домена из молекулы IgG4 и среднего шарнирного домена IgG1). В одном из вариантов осуществления связывающие молекулы содержат Fc-область или ее часть из молекулы IgG4 человека и мутацию Ser228Pro (нумерация EU) в центральной шарнирной области молекулы.
В определенных вариантах осуществления в Fc-часть можно вносить мутацию для увеличения или снижения эффекторной функции с использованием способов, известных в данной области. Например, делеция или инактивация (посредством точковых мутаций или других способов) домена константной области может снизить связывание с Fc-рецептором циркулирующего модифицированного антитела, тем самым, повышая локализацию в опухоли. В других случаях возможно, что модификации константной области в соответствии с настоящим изобретением снижают связывание комплемента и, таким образом, уменьшают время полужизни в сыворотке и неспецифическое связывание конъюгированного цитотоксина. Другие модификации константной области можно использовать для модификации дисульфидных связей или олигосахаридных частей, которые обеспечивают увеличение локализации вследствие увеличенной антигенной специфичности или универсальности. Полученный в результате физиологический профиль, биодоступность и другие биохимические эффекты модификаций, такие как локализация в опухоли, биораспределение и время полужизни в сыворотке, можно без труда измерять и количественно определять с использованием хорошо известных иммунологических способов без излишнего экспериментирования.
В определенных вариантах осуществления Fc-домен, используемый в антителе по изобретению, представляет собой вариант Fc. Как используют в рамках изобретения, термин "вариант Fc" относится к Fc-домену, имеющему по меньшей мере одну аминокислотную замену относительно Fc-домена дикого типа, из которого происходит указанный Fc-домен. Например, когда Fc-домен происходит из IgG1-антитела человека, вариант Fc указанного Fc-домена IgG1 человека содержит по меньшей мере одну аминокислотную замену относительно указанного Fc-домена.
Аминокислотная замена(ы) в варианте Fc может быть расположена в любом положении (т.е. любое аминокислотное положение согласно EU) в Fc-домене. В одном из вариантов осуществления вариант Fc содержит замену в положении аминокислоты, расположенном в шарнирном домене или его части. В другом варианте осуществления вариант Fc содержит замену в положении аминокислоты, расположенном в CH2-домене или его части. В другом варианте осуществления вариант Fc содержит замену в положении аминокислоты, расположенном в CH3-домене или его части. В другом варианте осуществления вариант Fc содержит замену в положении аминокислоты, расположенном в домене CH4 или его части.
В связывающих молекулах по изобретению можно использовать любой известный в данной области вариант Fc, о котором известно, что он обеспечивает улучшение (например, снижение или усиление) эффекторной функции и/или связывания FcR. Указанные варианты Fc могут включать, например, любую из аминокислотных замен, описанных в международных публикациях PCT WO 88/07089 A1, WO 96/14339 A1, WO 98/05787 A1, WO 98/23289 A1, WO 99/51642 A1, WO 99/58572 A1, WO 00/09560 A2, WO 00/32767 A1, WO 00/42072 A2, WO 02/44215 A2, WO 02/060919 A2, WO 03/074569 A2, WO 04/016750 A2, WO 04/029207 A2, WO 04/035752 A2, WO 04/063351 A2, WO 04/074455 A2, WO 04/099249 A2, WO 05/040217 A2, WO 05/070963 A1, WO 05/077981 A2, WO 05/092925 A2, WO 05/123780 A2, WO 06/019447 A1, WO 06/047350 A2 и WO 06/085967 A2 или в патентах США 5648260; 5739277; 5834250; 5869046; 6096871; 6121022; 6194551; 6242195; 6277375; 6528624; 6538124; 6737056; 6821505; 6998253 и 7083784, все из которых включены в настоящее описание в качестве ссылки. В одном иллюстративном варианте осуществления связывающий полипептид по изобретению может содержать вариант Fc, содержащий аминокислотную замену в положении EU 268 (например, H268D или H268E). В другом иллюстративном варианте осуществления связывающий полипептид по изобретению может содержать аминокислотную замену в положении EU 239 (например, S239D или S239E) и/или в положении EU 332 (например, I332D или I332Q).
В определенных вариантах осуществления связывающий полипептид по изобретению может содержать вариант Fc, содержащий аминокислотную замену, которая изменяет независимые от антигена эффекторные функции антитела, в частности, время полужизни связывающего полипептида в кровотоке. Такие связывающие молекулы обладают либо увеличенным, либо сниженным связыванием с FcRn по сравнению со связывающими молекулами без этих замен, таким образом, они обладают увеличенным или сниженным временем полужизни в сыворотке, соответственно. Ожидается, что варианты Fc с увеличенной аффинностью будут иметь увеличенное время полужизни в сыворотке, и такие молекулы имеют полезные применения в способах лечения млекопитающих, где является желательным длительное время полужизни введенного антитела, например, для лечения хронического заболевания или нарушения. Напротив, ожидается, что варианты Fc со сниженной аффинностью связывания с FcRn будут иметь более короткое время полужизни, и такие молекулы также подходят, например, для введения млекопитающему, где укороченное время в кровотоке может быть преимущественным, например, для диагностической визуализации in vivo или в ситуациях, где исходное антитело имеет токсичные побочные эффекты, когда оно присутствует в кровотоке в течение длительных периодов времени. Варианты Fc со сниженной аффинностью связывания с FcRn также с меньшей вероятностью проходят через плаценту и, таким образом, также пригодны для лечения заболеваний или нарушений у беременных женщин.
Кроме того, другие применения, в которых может быть желательной сниженная аффинность связывания FcRn, включают применения, в которых является желательной локализация в головном мозге, почке и/или печени. В одном иллюстративном варианте осуществления измененные связывающие молекулы (например, антитела или их антигенсвязывающие фрагменты) по изобретению обладают сниженным транспортом через эпителий гломерул почки из сосудов. В другом варианте осуществления измененные связывающие молекулы (например, антитела или их антигенсвязывающие фрагменты) по изобретению обладают сниженным транспортом через гематоэнцефалический барьер (BBB) из головного мозга в сосудистое пространство. В одном из вариантов осуществления антитело с измененным связыванием FcRn содержит Fc-домен, имеющий одну или несколько аминокислотных замен в "FcRn-связывающей петле" Fc-домена. FcRn-связывающая петля содержит аминокислотные остатки 280-299 (в соответствии с нумерацией EU). Иллюстративные аминокислотные замены, которые изменяют активность связывания FcRn, описаны в международной публикации PCT № WO 05/047327, которая включена в настоящем описании в качестве ссылки.
В определенных иллюстративных вариантах осуществления связывающие молекулы (например, антитела или их антигенсвязывающие фрагменты) по изобретению содержат Fc-домен, имеющий одну или несколько из следующих замен: V284E, H285E, N286D, K290E и S304D (нумерация EU). В других иллюстративных вариантах осуществления связывающие молекулы по изобретению содержат Fc-домен человека с двойной мутацией H433K/N434F (см., например, патент США 8163881). В конкретном варианте осуществления связывающие молекулы по изобретению содержат одну или несколько вариабельных областей, выбранных из таблицы 16, и Fc-домен человека с двойной мутацией H433K/N434F. В другом конкретном варианте осуществления связывающая молекула по изобретению содержит одну или несколько последовательностей CDR из таблицы 17 и Fc-домен человека с двойной мутацией H433K/N434F. В другом конкретном варианте осуществления связывающая молекула по изобретению содержит VH-домен с SEQ ID NO:152 и Fc-домен человека с двойной мутацией H433K/N434F. В другом конкретном варианте осуществления связывающая молекула по изобретению содержит VH-домен SEQ ID NO:152, где глутамин в одном или нескольких положениях, например, в положении 1, 3, 5 или 16, или любой их комбинации заменен на глутаминовую кислоту(ы), и Fc-домен человека с двойной мутацией H433K/N434F. В другом конкретном варианте осуществления связывающая молекула по изобретению содержит VH-домен SEQ ID NO:152, где глутамин в положении 1 заменен на глутаминовую кислоту, и Fc-домен человека с двойной мутацией H433K/N434F.
В других вариантах осуществления связывающие молекулы для применения в способах диагностики и лечения, описанных в настоящем описании, имеют константную область, например, константную область тяжелой цепи IgG1 или IgG4, которая изменена для снижения или устранения гликозилирования. Например, связывающие молекулы (например, антитела или их антигенсвязывающие фрагменты) по изобретению также могут содержать вариант Fc, содержащий аминокислотную замену, которая изменяет гликозилирование Fc антитела. Например, указанный вариант Fc может иметь сниженное гликозилирование (например, N- или O-связанное гликозилирование). В иллюстративных вариантах осуществления вариант Fc обладает сниженным гликозилированием N-связанным гликаном, обычно находящимся в положении аминокислоты 297 (нумерация EU). В другом варианте осуществления антитело имеет аминокислотную замену вблизи мотива гликозилирования или в мотиве гликозилирования, например, в мотиве N-связанного гликозилирования, который содержит аминокислотную последовательность NXT или NXS. В конкретном варианте осуществления антитело содержит вариант Fc с аминокислотной заменой в положении аминокислоты 228 или 299 (нумерация EU). В более конкретных вариантах осуществления антитело содержит константную область IgG1 или IgG4, содержащую мутацию S228P и T299A (нумерацию EU).
Иллюстративные аминокислотные замены, которые сообщают сниженное или измененное гликозилирование, описаны в международной публикации PCT WO 05/018572, которая включена в настоящее описание в качестве ссылки. В предпочтительных вариантах осуществления антитела или их фрагменты по изобретению являются модифицированными для устранения гликозилирования. Такие антитела или их фрагменты могут быть обозначены как антитела "agly" или их фрагменты (например, антитела "agly"). Без связи с какой-либо конкретной теорией, полагают, что антитела "agly" или их фрагменты могут иметь улучшенный профиль безопасности и стабильности in vivo. Антитела Agly могут быть любого изотипа или подкласса, например, IgG1, IgG2, IgG3 или IgG4. В определенных вариантах осуществления антитела agly или их фрагменты содержат агликозилированную Fc-область IgG4-антитела, которая лишена Fc-эффекторной функции, тем самым устраняя потенциал к опосредуемой Fc токсичности для нормальных жизненно-важных органов, которые экспрессируют IL-6. В других вариантах осуществления антитела или их фрагменты по изобретению содержат измененный гликан. Например, антитело может иметь сниженное количество остатков фукозы на N-гликане в положении Asn297 Fc-области, т.е. оно является афукозилированным. Афукозилирование увеличивает связывание FcRgII на NK-клетках и потенциально увеличивает ADCC. Было показано, что диантитело, содержащее scFv против IL-6 и scFv против CD3, индуцирует уничтожение экспрессирующих IL-6 клеток посредством ADCC. Таким образом, в одном из вариантов осуществления афукозилированное антитело против IL-6 используют для нацеливания на экспрессирующие IL-6 клетки и их уничтожения. В другом варианте осуществления антитело может иметь измененное количество остатков сиаловой кислоты на N-гликане в положении Asn297 Fc-области. В данной области доступны многочисленные известные в данной области способы для получения антител "agly" или антител с измененными гликанами. Например, для получения таких антител можно использовать модифицированные способами генной инженерии клетки-хозяева (например, модифицированные дрожжи, например, клетки Picchia или CHO) с модифицированными каскадами гликозилирования (например, делеции гликозилтрансферазы).
iii) Ковалентное присоединение
Связывающие молекулы по изобретению могут быть модифицированы, например, посредством ковалентного связывания молекулы со связывающим полипептидом, так что ковалентное связывание не препятствует специфическому связыванию связывающего полипептида с его эпитопом. Например, но не ограничиваясь этим, антитела или их фрагменты по изобретению можно модифицировать путем гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидации, преобразования в производное с помощью известных защитных/блокирующих групп, протеолитического расщепления, связывания с клеточным лигандом или другим белком, и т.д. Любые из многочисленных химических модификаций можно осуществлять известными способами, включая, но не ограничиваясь ими, специфическое химическое расщепление, ацетилирование, формилирование и т.д. Кроме того, производное может содержать одну или несколько неклассических аминокислот.
Связывающий полипептид (например, антитела или их фрагменты), кроме того, может быть рекомбинантным образом слитым с гетерологичным полипептидом на N- или C-конце или химически конъюгированным (включая ковалентную и нековалентную конъюгацию) с полипептидами или другими композициями. Например, антитела против IL-6 могут быть рекомбинантным образом слитыми или конъюгированными с молекулами, пригодными в качестве меток в анализах для обнаружения, и эффекторными молекулами, такими как гетерологичные полипептиды, лекарственные средства, радионуклиды или токсины. См., например, публикации PCT WO 92/08495; WO 91/14438; WO 89/12624; патент США 5314995 и EP 396387.
Связывающие молекулы могут быть слиты с гетерологичными полипептидами для повышения времени полужизни in vivo или для применения в иммуноанализах с использованием способов, известных в данной области. Например, в одном из вариантов осуществления со связывающими молекулами по изобретению можно конъюгировать PEG для увеличения их времени полужизни in vivo. Leong, S. R., et al., Cytokine 16:106 (2001); Adv. in Drug Deliv. Rev. 54:531 (2002); или Weir et al., Biochem. Soc. Transactions 30:512 (2002).
Более того, связывающие молекулы по изобретению могут быть слиты с маркерными последовательностями, такими как пептид, способствующий их очистке или обнаружению. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид, такой как метка, предоставленная в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, Calif., 91311), среди прочих, многие из которых являются коммерчески доступными. Как описано в Gentz et al., Proc. Natl. Acad. Sci. USA 86:821-824 (1989), например, гексагистидин обеспечивает удобную очистку слитого белка. Другие пептидные метки, пригодные для очистки, включают, но не ограничиваются ими, "HA"-метку, которая соответствует эпитопу, происходящему из белка гемагглютинина вируса гриппа (Wilson et al., Cell 37:767 (1984)) и "flag"-метку.
Связывающие молекулы по изобретению можно использовать в неконъюгированной форме или их можно конъюгировать по меньшей мере с одной из множества молекул, например, для улучшения терапевтических свойств молекулы, для облегчения обнаружения мишени или для визуализации или терапии у пациента. Связывающие молекулы по изобретению могут быть меченными или конъюгированными непосредственно до или после очистки, когда проводят очистку. В частности, связывающие молекулы по изобретению можно конъюгировать с лекарственными средствами, пролекарствами, пептидами, белками, ферментами, вирусами, липидами, модификаторами биологического ответа, фармацевтическими средствами или PEG.
Кроме того, настоящее изобретение охватывает связывающие молекулы по изобретению, конъюгированные с диагностическим или лекарственным средством. Связывающие молекулы можно использовать в диагностических способах, например, для мониторинга развития или прогрессирования нарушения иммунных клеток (например, CLL) в качестве части методики клинического испытания, например, для определения эффективности данного режима лечения и/или профилактики. Обнаружение можно облегчать путем связывания связывающих молекул с поддающимся обнаружению веществом. Примеры поддающихся обнаружению веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминсцентные материалы, радиоактивные материалы, позитронно-активные металлы, используемые в различных способах позитронно-эмиссионной томографии, и нерадиоактивные парамагнитные ионы металлов. См., например, патент США 4741900 для ионов металлов, которые могут быть конъюгированы с антителами для применения в качестве диагностических средств в соответствии с настоящим изобретением. Примеры пригодных ферментов включают пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; примеры пригодных комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных материалов включают умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; пример люминесцентного материала включает люминол; примеры биолюминесцентных материалов включают люциферазу, люциферин и экворин; и примеры подходящего радиоактивного материала включают 125I, 131I, 111In или 99Tc.
Связывающие молекулы для применения в способах диагностики и лечения, описанных в настоящем описании, можно конъюгировать с цитотоксинами (такими как радиоактивные изотопы, цитотоксические лекарственные средства или токсины), терапевтическими средствами, цитостатическими средствами, биологическими токсинами, пролекарствами, пептидами, белками, ферментами, вирусами, липидами, модификаторами биологического ответа, фармацевтическими средствами, иммунологически активными лигандами (например, лимфокины или другие антитела, где полученная молекула связывается как с неопластической клеткой, так и с эффекторной клеткой, такой как T-клетка) или PEG.
В другом варианте осуществления антитело против IL-6 для применения в способах диагностики и лечения, описанных в настоящем описании, может быть конъюгировано с молекулой, которая снижает рост опухолевых клеток. В других вариантах осуществления описанные композиции могут содержать антитела или их фрагменты, связанные с лекарственными средствами или пролекарствами. Другие варианты осуществления настоящего изобретения включают применение антител или их фрагментов, конъюгированных с конкретными биотоксинами или их цитотоксическими фрагментами, такими как рицин, гелонин, экзотоксин Pseudomonas или дифтерийный токсин. Выбор того, какое конъюгированное или неконъюгированное антитело использовать, зависит от типа и стадии злокачественной опухоли, использования дополнительного лечения (например, химиотерапия или внешнее облучение) и состояния пациента. Специалисту в данной области будет понятно, что такой выбор можно без труда сделать ввиду указаний в настоящем описании.
Будет понятно, что в предшествующих исследованиях противоопухолевые антитела, меченные изотопами, успешно использовали для разрушения опухолевых клеток в моделях на животных и в некоторых случаях у человека. Иллюстративные радиоизотопы включают: 90Y, 125I, 131I, 123I, 111In, 105Rh, 153Sm, 67Cu, 67Ga, 166Ho, 177Lu, 186Re и 188Re. Радионуклиды действуют, генерируя ионизирующее излучение, которое вызывает множество разрывов в цепи ядерной ДНК, что приводит к гибели клетки. Изотопы, используемые для получения терапевтических конъюгатов, как правило, генерируют высокоэнергетические альфа- или бета-частицы, которые имеют короткую длину пути. Такие радионуклиды уничтожают клетки, рядом с которыми они находятся, например, неопластические клетки, с которыми конъюгат связался или в которые он проник. Они имеют небольшой эффект или не имеют эффекта на нелокализованные клетки. Радионуклиды являются по существу неиммуногенными.
V. Экспрессия связывающих молекул
После манипуляции с выделенным генетическим материалом для обеспечения связывающих молекул по изобретению, как указано выше, гены, как правило, встраивают в вектор экспрессии для внесения в клетки-хозяева, которые можно использовать для получения желаемого количества заявленных антител или их фрагментов.
Термин "вектор" или "вектор экспрессии" используют в настоящем описании и формуле изобретения для обозначения векторов, используемых в соответствии с настоящим изобретением в качестве носителя для введения в клетку и экспрессии желаемого гена в клетке. Как известно специалистам в данной области, такие векторы можно без труда выбирать из группы, состоящей из плазмид, фагов, вирусов и ретровирусов. Как правило, векторы, совместимые с настоящим изобретением, содержат селективный маркер, соответствующие участки рестрикции для облегчения клонирования желаемого гена, и обладают способностью проникать и/или реплицироваться в эукариотических или прокариотических клетках.
Для настоящего изобретения можно использовать многочисленные экспрессирующие векторные системы. Например, в одном классе векторов используются элементы ДНК, которые происходят из вирусов животных, таких как вирус папилломы крупного рогатого скота, вирус полиомы, аденовирус, вирус коровьей оспы, бакуловирус, ретровирусы (RSV, MMTV или MOMLV) или вирус SV40. В другом классе вовлечено применение полицистронных систем с внутренними участками связывания рибосом. Кроме того, клетки, в хромосомы которых встроилась ДНК, можно отбирать путем введения одного или нескольких маркеров, которые обеспечивают селекцию трансфицированных клеток-хозяев. Маркер может обеспечить прототрофность ауксотрофному хозяину, устойчивость к биоциду (например, антибиотики) или устойчивость к тяжелым металлам, таким как медь. Ген селективного маркера может быть либо прямо связан с последовательностями ДНК, подлежащими экспрессии, либо он может быть введен в ту же клетку путем котрансформации. Также могут потребоваться дополнительные элементы для оптимального синтеза мРНК. Эти элементы могут включать сигнальные последовательности, сигналы сплайсинга, а также промоторы транскрипции, энхансеры и сигналы терминации. В особенно предпочтительных вариантах осуществления клонированные гены вариабельной области встраивают в экспрессирующий вектор вместе с синтетическими генами константной области тяжелой и легкой цепей (предпочтительно, человека), как описано выше.
В других предпочтительных вариантах осуществления связывающие молекулы или их фрагменты по изобретению можно экспрессировать с использованием полицистронных конструкций. В таких экспрессирующих системах множество представляющих интерес продуктов генов, таких как тяжелые и легкие цепи антител, можно продуцировать с одной полицистронной конструкции. В этих системах преимущественно используется участок внутренней посадки рибосомы (IRES) для обеспечения относительно высоких уровней полипептидов по изобретению в эукариотических клетках-хозяевах. Совместимые последовательности IRES описаны в патенте США № 6193980, который включен в настоящем описании в качестве ссылки. Специалистам в данной области будет понятно, что такие экспрессирующие системы можно использовать для эффективного получения полного диапазона полипептидов, описанных в настоящей заявке.
В общем, после того, как вектор или последовательность ДНК, кодирующие антитело или его фрагмент, получены, вектор экспрессии можно вводить в подходящую клетку-хозяина. Иными словами, клетки-хозяева можно трансформировать. Введение плазмиды в клетку-хозяина можно проводить с помощью различных способов, хорошо известных специалистам в данной области. Они включают, но не ограничиваются ими, трансфекцию (включая электрофорез и электропорацию), слияние протопластов, осаждение с фосфатом кальция, слияние клеток с ДНК в оболочке, микроинъекцию и инфицирование интактным вирусом. См., Ridgway, A. A. G. "Mammalian Expression Vectors" Chapter 24,2, pp. 470-472 Vectors, Rodriguez and Denhardt, Eds. (Butterworths, Boston, Mass. 1988). Наиболее предпочтительно, введение плазмиды в хозяина проводят посредством электропорации. Трансформированные клетки выращивают в условиях, пригодных для продукции легких цепей и тяжелых цепей, и анализируют в отношении синтеза белка тяжелой и/или легкой цепи. Иллюстративные способы анализа включают твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) или анализ с помощью сортера для активированной флуоресценцией сортировки клеток (FACS), иммуногистохимию и т.п.
Как используют в рамках изобретения, термин "трансформация" используют в широком значении для обозначения введения ДНК в реципиентную клетку-хозяина, которая изменяет генотип, а затем приводит к изменению в реципиентной клетке.
Наравне с этими линиями, "клетки-хозяева" относятся к клеткам, трансформированным векторами, сконструированными с использованием способов рекомбинантных ДНК и кодирующими по меньшей мере один гетерологичный ген. В описании процессов выделения полипептидов из рекомбинантных хозяев термины "клетка" и "клеточная культура" используют взаимозаменяемо для обозначения источника антитела, если явно не указано иное. Иными словами, выделение полипептида из "клеток" может означать выделение либо посредством осаждения центрифугированием цельных клеток, либо из клеточной культуры, содержащей как среду, так и суспендированные клетки.
В одном из вариантов осуществления линия клеток-хозяев, используемая для экспрессии антител, имеет происхождение из млекопитающих; специалисты в данной области могут определить конкретные линии клеток-хозяев, которые наилучшим образом пригодны для экспрессии в них желаемого продукта гена. Иллюстративные линии клеток-хозяев включают, но не ограничиваются ими, DG44 и DUXB11 (линии клеток яичника китайского хомячка, DHFR-минус), HELA (карцинома шейки матки человека), CVI (линия клеток почки обезьяны), COS (производное CVI с T-антигеном SV40), R1610 (фибробласт китайского хомячка) BALBC/3T3 (фибробласт мыши), HAK (линия почки хомячка), SP2/O (миелома мыши), BFA-1c1BPT (эндотелиальные клетки крупного рогатого скота), RAJI (лимфоциты человека), 293 (почка человека). В одном из вариантов осуществления клеточная линия обеспечивает измененное гликозилирование, например, афукозилирование, антитела, экспрессированного из нее (например, PER.C6.RTM. (Crucell) или линии клеток CHO с нокаутом FUT8 (Potelligent.RTM. Cells) (Biowa, Princeton, N.J.)). В одном из вариантов осуществления можно использовать клетки NS0. Клетки CHO являются особенно предпочтительными. Линии клеток-хозяев, как правило, доступны от коммерчески поставщиков, American Tissue Culture Collection или из опубликованной литературы.
Продукция in vitro позволяет увеличение масштаба для получения больших количеств желаемых полипептидов. Способы культивирования клеток млекопитающих в этих условиях культивирования тканей известны в данной области и включают гомогенную суспензионную культуру, например, в аэрлифтном ректоре и в реакторе с постоянным перемешиванием или иммобилизованную или заключенную клеточную культуру, например, в полых волокнах, микрокапсулах, на агарозных микрогранулах или керамических кассетах. Если необходимо и/или желательно, растворы полипептидов можно очищать обычными способами хроматографии, например, гель-фильтрацией, ионообменной хроматографией, хроматографией над DEAE-целлюлозой и/или (иммуно)аффинной хроматографией.
Гены, кодирующие связывающие молекулы или их фрагменты по изобретению, также могут экспрессироваться в клетках, не являющихся клетками млекопитающих, таких как бактерии, или дрожжи, или клетки растений. В этом отношении будет понятно, что также можно трансформировать различные одноклеточные не являющиеся млекопитающими микроорганизмы, такие как бактерии, т.е. микроорганизмы, способные к росту в культурах или ферментации. Бактерии, подверженные трансформации, включают представителей энтеробактерий, таких как штаммы Escherichia coli или Salmonella; Bacillaceae, такие как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Кроме того, будет понятно, что при экспрессии в бактериях полипептиды могут стать частью телец включения. Полипептиды необходимо выделять, очищать, а затем собирать в функциональные молекулы.
В дополнение к прокариотам, также можно использовать эукариотические микроорганизмым. Saccharomyces cerevisiae или обычные пекарские дрожжи наиболее широко используются среди эукариотических микроорганизмов, хотя ряд других штаммов является общедоступным. Для экспрессии в Saccharomyces, обычно используют плазмиду YRp7, например, (Stinchcomb et al., Nature, 282:39 (1979); Kingsman et al., Gene, 7:141 (1979); Tschemper et al., Gene, 10:157 (1980)). Эта плазмида уже содержит ген TRP1, который обеспечивает маркер для селекции мутантного штамма дрожжей, лишенного способности расти в триптофане, например ATCC № 44076 или PEP4-1 (Jones, Genetics, 85:12 (1977)). Затем присутствие повреждения trpl в качестве характеристики генома дрожжевых клеток-хозяев обеспечивает эффективные условия для обнаружения трансформации по росту в отсутствие триптофана.
VI. Фармацевтические составы и способы введения связывающих молекул
В другом аспекте изобретение относится к фармацевтическим композициям, содержащим связывающую молекулу (например, антитело или его антигенсвязывающий фрагмент).
Способы получения и введения молекул по изобретению индивидууму хорошо известны и их без труда определят специалисты в данной области. Путь введения антител или их фрагментов по изобретению может быть пероральным, парентеральным, ингаляционным или местным. Термин "парентеральный", как используют в рамках изобретения, включает внутривенное, внутриартериальное, внутрибрюшинное, внутримышечное, подкожное, ректальное или вагинальное введение. Внутривенные, внутриартериальные, подкожные и внутримышечные формы парентерального введения, как правило, являются предпочтительными. Хотя явно предусматривается, что все эти формы введения входят в объем изобретения, иллюстративной формой для введения является раствор для инъекции, в частности, для внутривенной или внутриартериальной инъекции или капельного вливания. Обычно подходящая фармацевтическая композиция для инъекции может содержать буфер (например, ацетатный, фосфатный или цитратный буфер), поверхностно-активное вещество (например, полисорбат), необязательно стабилизатор (например, альбумин человека) и т.д. В других способах, совместимых с идеями, описанными в настоящем описании, полипептиды можно доставлять прямо в область, в которой находится нежелательная клеточная популяция, тем самым повышая экспозицию пораженной заболеванием ткани лекарственному средству. Например, высокая термическая стабильность и свойства растворимости связывающих молекул по изобретению делают их идеальными средствами для местного введения, например, посредством подкожных (Sub-Q) инъекций. Кроме того, чрезвычайно высокая аффинность и эффективность антител по изобретению позволяют использовать более низкую эффективную дозу, тем самым, упрощая подкожную инъекцию. Таким образом, связывающие молекулы особенно хорошо подходят для лечения или профилактики связанных с воспалением нарушений (например, ревматоидный артрит), злокачественных опухолей или обусловленных ими симптомов (например, кахексия или анорексия), для которых может быть желательной локализованная доставка связывающей молекулы.
Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии, и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло и инъецируемые органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая солевые и буферные среды. В настоящем изобретении фармацевтически приемлемые носители включают, но ими не ограничиваются, 0,01-0,1 M и предпочтительно, 0,05 M фосфатный буфер или 0,8% солевой раствор. Другие распространенные парентеральные носители включают растворы фосфата натрия, раствор декстрозы Рингера, раствор декстрозы и хлорида натрия, лактатный раствор Рингера или жирные масла. Внутривенные носители включают средства для восполнения жидкости и питательных веществ, средства для восполнения электролитов, такие как средства на основе раствора декстрозы Рингера и т.п. Также могут присутствовать консерванты и другие добавки, например, такие как противомикробные средства, антиоксиданты, хелатирующие средства и инертные газы и т.п. Более конкретно, фармацевтические композиции, пригодные для применения путем инъекции, включают стерильные водные растворы (в случае, когда они растворимы в воде) или дисперсии и стерильные порошки для приготовления непосредственно перед применением стерильных инъецируемых растворов или дисперсий. В таких случаях композиция должна быть стерильной и должна быть текучей до той степени, чтобы легко заполнять шприц. Она должна быть стабильной в условиях изготовления и хранения и предпочтительно, защищена от заражающего действия микроорганизмов, таких как бактерии и грибы. Носитель может представлять собой растворитель или дисперсную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, и жидкий полиэтиленгликоль и т.п.), и их подходящие смеси. Надлежащую текучесть можно поддерживать, например, с использованием покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Предупреждение действия микроорганизмов можно обеспечивать с помощью различных антибактериальных и противогрибковых средств, например, парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимеросала и т.п. Во многих случаях, было бы предпочтительно, включать в композицию обеспечивающие изотоничность средства, например, сахара, полиспирты, такие как маннит, сорбит или хлорид натрия. Пролонгированного действия инъецируемых композиций можно достигать путем включения в композицию средства, которое замедляет всасывание, например, моностеарата алюминия и желатина.
В любом случае, стерильные инъецируемые растворы можно получать путем включения активного соединения (например, само антитело или в комбинации с другими активными веществами) в требуемом количестве в надлежащем растворителе с одним или с комбинацией ингредиентов, перечисленных в настоящем документе, при необходимости, с последующей стерилизацией фильтрацией. Как правило, дисперсии получают путем включения активного соединения в стерильный носитель, который содержит основную дисперсионную среду и другие требуемые ингредиенты из тех, которые перечислены выше. В случае стерильных порошков для получения стерильных инъецируемых растворов, предпочтительными способами получения являются вакуумная сушка и сублимационная сушка, которая приводит к порошку активного ингредиента вместе с любым дополнительным желаемым ингредиентом из их ранее стерилизованного фильтрацией раствора. Препараты для инъекций перерабатывают, заполняют в контейнеры, такие как ампулы, мешки, бутылки, шприцы и флаконы, и герметизируют в асептических условиях способами, известными в данной области. Кроме того, препараты можно упаковывать и продавать в форме набора, такого как наборы, описанные в совместно рассматриваемых патентной заявке США с серийным номером 09/259337 и в патентной заявке США с серийным номером 09/259338, каждая из которых включена в настоящее описание в качестве ссылки. Такие изделия предпочтительно, имеют ярлыки или вкладыши в упаковку, указывающие на то, что соответствующие композиции пригодны для лечения индивидуума, страдающего аутоиммунными или неопластическими нарушениями или предрасположенного к ним.
Связывающие молекулы по изобретению можно составлять в широком диапазоне концентраций для фармацевтического применения. Например, связывающую молекулу можно составлять в концентрации от 5 мг/мл до 50 мг/мл (например, 5, 10, 20, 50 мг/мл). Альтернативно связывающие молекулы по изобретению могут быть адаптированы к более высоким концентрациям для местного (например, подкожного) введения. Например, связывающая молекула может быть составлена в концентрации от 50 мг/мл до 200 мг/мл, например, приблизительно 50, приблизительно 75, приблизительно 100, приблизительно 150, приблизительно 175 или приблизительно 200 мг/мл).
Эффективные дозы связывающих молекул по настоящему изобретению для лечения описанных выше состояний варьируют в зависимости от множества различных факторов, включающих средства для введения, область-мишень, физиологическое состояние пациента, то, является ли пациент человеком или животным, другие введенные лекарственные средства и то, является ли лечение профилактическим или терапевтическим. Обычно пациентом является человек, однако также можно лечить не являющихся человеком млекопитающих, включая трансгенных млекопитающих. Дозировки для введения можно титровать с использованием общепринятых способов, известных специалистам в данной области, для оптимизации безопасности и эффективности.
Для пассивной иммунизации антителом по изобретению дозировка может находиться в диапазоне, например, приблизительно от 0,0001 до 100 мг/кг, и чаще от 0,01 до 5 мг/кг (например, 0,02 мг/кг, 0,25 мг/кг, 0,5 мг/кг, 0,75 мг/кг, 1 мг/кг, 2 мг/кг и т.д.) массы тела хозяина. Например, дозировки могут составлять 1 мг/кг массы тела или 10 мг/кг массы тела, или могут находиться в диапазоне 1-10 мг/кг, предпочтительно, по меньшей мере 1 мг/кг. Подразумевается, что в объем изобретения входят дозировки, промежуточные для указанных выше диапазонов.
Индивидуумам можно вводить такие дозировки каждые сутки, раз в двое суток, раз неделю или по любой другой схеме, определенной с помощью эмпирического анализа. Иллюстративное лечение охватывает введение в виде множества дозировок в течение длительного времени, например, по меньшей мере шести месяцев. Дополнительные иллюстративные режимы лечения охватывают введение один раз в две недели, или один раз в месяц, или один раз в 3-6 месяцев. Иллюстративные схемы дозирования включают 1-10 мг/кг или 15 мг/кг каждые сутки, 30 мг/кг раз в двое суток или 60 мг/кг раз в неделю. В некоторых способах два или более моноклональных антител с отличающейся специфичностью связывания вводят одновременно, и в этом случае дозировка каждого вводимого антитела может находиться в пределах указанных диапазонов.
Связывающие молекулы по изобретению можно вводить неоднократно. Интервалы между единичными дозировками могут составлять, например, раз в сутки, раз в неделю, раз в месяц или раз в год. Интервалы также могут быть нерегулярными, определяемыми при измерении уровней в крови полипептида или молекулы-мишени у пациента. В некоторых способах дозировку корректируют для достижения определенной концентрации антитела или токсина в плазме, например, 1-1000 мкг/мл или 25-300 мкг/мл. Альтернативно антитела или их фрагменты можно вводить в составе с замедленным высвобождением, в случае которого требуется менее частое введение. Дозировка и частота варьируют в зависимости от времени полужизни антитела у пациента. Как правило, преобразованные в эмбриональный тип или гуманизированные антитела демонстрируют наиболее длительное время полужизни, за которым следуют химерные антитела и не являющиеся человеческими антитела. В одном из вариантов осуществления антитела или их фрагменты по изобретению можно вводить в неконъюгированной форме. В другом варианте осуществления антитела по изобретению можно вводить многократно в конъюгированной форме. В другом варианте осуществления антитела или их фрагменты по изобретению можно вводить в неконъюгированной форме, затем в конъюгированной форме или наоборот.
Дозировка и частота введения могут варьировать в зависимости от того, является ли лечение профилактическим или терапевтическим. В профилактических применениях композиции, содержащие антитела по настоящему изобретению или их смесь, вводят пациенту, еще не имеющему болезненного состояния, для повышения устойчивости пациента. Такое количество определяют как "профилактически эффективная доза". В этом применении точные количества вновь зависят от состояния здоровья пациента и общего иммунитета, однако, как правило, они находятся в диапазоне от 0,1 до 25 мг на дозу, особенно от 0,5 до 2,5 мг на дозу. Относительно низкую дозировку вводят с относительно нечастыми интервалами в течение длительного периода времени. У некоторых пациентов лечение продолжается до конца их жизни.
В терапевтических применениях иногда требуется относительно высокая дозировка (например, приблизительно от 1 до 400 мг/кг антитела на дозу, причем дозировки от 5 до 25 мг являются более часто используемыми для радиоиммуноконъюгатов и более высокие дозы используют для конъюгированных молекул цитотоксин-лекарственное средство) с относительно короткими интервалами до тех пор, пока прогрессирование заболевания не снизится или не завершится, и, предпочтительно, до тех пор пока у пациента не произойдет частичного или полного смягчения симптомов заболевания. После этого у пациента можно проводить профилактический режим.
В одном из вариантов осуществления индивидууму можно вводить молекулу нуклеиновой кислоты, кодирующую полипептид по изобретению (например, в векторе). Дозы для нуклеиновых кислот, кодирующих полипептиды, находятся в диапазоне приблизительно от 10 нг до 1 г, от 100 нг до 100 мг, от 1 мкг до 10 мг или 30-300 мкг ДНК на пациента. Дозы для инфекционных вирусных векторов варьируют в пределах 10-100 или более, вирионов на дозу.
Лекарственные средства можно вводить парентеральным, местным, внутривенным, пероральным, подкожным, внутриартериальным, внутричерепным, внутрибрюшинным, интраназальным или внутримышечным путем для профилактического и/или терапевтического лечения. Внутримышечная инъекция или внутривенная инфузия являются предпочтительными для введения антитела по изобретению. В некоторых способах терапевтические антитела или их фрагменты инъецируют прямо в череп. В некоторых способах антитела или их фрагменты вводят в композиции с замедленным высвобождением или с помощью устройства для замедленного высвобождения, такого как устройство Medipad™.
Средства по изобретению необязательно можно вводить в комбинации с другими средствами, которые являются эффективными для лечения нарушения или состояния, нуждающихся в лечении (например, профилактическом или терапевтическом). Предпочтительные дополнительные средства представляют собой средства, которые известны в данной области и которые стандартным образом вводят при конкретном нарушении.
Эффективные единичные дозировки для введения (т.е. терапевтически эффективные количества) меченных 90Y антител по изобретению находятся в диапазоне от приблизительно 5 до приблизительно 75 мКи, более предпочтительно, от приблизительно 10 до приблизительно 40 мКи. Эффективные единичные дозировки меченных 131I антител для однократного применения, которые разрушают костный мозг, находятся в диапазоне от приблизительно 5 до приблизительно 70 мКи, более предпочтительно, от приблизительно 5 до приблизительно 40 мКи. Эффективные единичные дозировки, которые разрушают костный мозг (т.е. могут требовать аутологичной трансплантации костного мозга) для меченных 131I антител находятся в диапазоне от приблизительно 30 до приблизительно 600 мКи, более предпочтительно, от приблизительно 50 до менее чем приблизительно 500 мКи.
В то время как был наработан существенный клинический опыт с 131I и 90Y, в данной области известны другие радиоактивные метки, и их можно использовать для сходных целей. Для визуализации используют другие радиоизотопы. Например, дополнительные радиоизотопы, которые совместимы с настоящим изобретением, включают, но ими не ограничиваются, 123I, 125I, 32P, 57Co, 64Cu, 67Cu, 77Br, 81Rb, 81Kr, 87Sr, 113In, 127Cs, 129Cs, 132I, 197Hg, 203Pb, 206Bi, 177Lu, 186Re, 212Pb, 212Bi, 47Sc, 105Rh, 109Pd, 153Sm, 188Re, 199Au, 225Ac, 211At, 213Bi. В этом отношении все из альфа-, гамма- и бета-эммитеров совместимы с настоящим изобретением. Кроме того, ввиду настоящего изобретения предполагается, что специалист в данной области может без труда определить то, какие радионуклиды совместимы с выбранным курсом лечения без излишнего экспериментирования. В связи с этим, дополнительные радионуклиды, которые уже используют в клинической диагностике, включают 125I, 123I, 99Tc, 43K, 52Fe, 67Ga, 68Ga, а также 111In. Антитела также метят различными радионуклидами для потенциального применения в направленной иммунотерапии (Peirersz et al. Immunol. Cell Biol. 65: 111-125 (1987)). Эти радионуклиды включают 188Re и 186Re, а также 199Au и 67Cu в меньшей степени. В патенте США 5460785 предоставлены дополнительные данные, касающиеся таких радиоизотопов, и он включен в настоящее описание в качестве ссылки.
Как рассмотрено выше, связывающие молекулы по изобретению можно вводить в фармацевтически эффективном количестве для лечения in vivo нарушений у млекопитающих. В связи с этим понятно, что описанные антитела или их фрагменты составляют так, чтобы облегчать введение и повышать стабильность активного вещества. Предпочтительно, фармацевтические композиции в соответствии с настоящим изобретением содержат фармацевтически приемлемый нетоксичный стерильный носитель, такой как физиологический раствор, нетоксичные буферы, консерванты и т.п. Для целей настоящей заявки, фармацевтически эффективное количество антитела по изобретению, конъюгированного или неконъюгированного с лекарственным средством, должно означать количество, достаточное для достижения эффективного связывания с мишенью и для достижения пользы, например, для смягчения симптомов заболевания или нарушения или для обнаружения клетки. В случае опухолевых клеток, полипептид, предпочтительно, способен взаимодействовать с выбранными иммунореактивными антигенами на неопластических или иммунореактивных клетках и обеспечивать увеличение гибели этих клеток. Безусловно, фармацевтические композиции по настоящему изобретению можно вводить в однократной или многократных дозах для обеспечения фармацевтически эффективного количества полипептида.
В соответствии с объемом настоящего изобретения, связывающие молекулы по изобретению можно вводить человеку или другому животному в соответствии с упомянутыми выше способами лечения в количестве, достаточном для обеспечения терапевтического или профилактического эффекта. Полипептиды по изобретению можно вводить такому человеку или другому животному в общепринятой дозированной форме, полученной путем комбинирования антитела по изобретению с общепринятым фармацевтически приемлемым носителем или разбавителем в соответствии с известными способами. Специалисту в данной области будет понятно, что форма и характер фармацевтически приемлемого носителя или разбавителя определяется количеством активного ингредиента, который подлежит комбинированию, пути введения и других хорошо известных переменных. Специалистам в данной области далее будет понятно, что особенно эффективной может оказаться смесь, содержащая один или несколько типов полипептидов в соответствии с настоящим изобретением.
VII. Способы лечения обусловленных IL-6 заболеваний или нарушений
Связывающие молекулы по изобретению пригодны для антагонизма активности IL-6. Таким образом, в другом аспекте изобретение относится к способам лечения обусловленных IL-6 заболеваний или нарушений путем введения индивидууму, нуждающемуся в этом, фармацевтической композиции, содержащей одну или несколько связывающих молекул по изобретению.
Обусловленные IL-6 заболевания или нарушения, подлежащие лечению, включают, но не ограничиваются ими, воспалительные заболевания и злокачественную опухоль.
Специалист в данной области способен с помощью стандартного экспериментирования определить, каким является эффективное нетоксичное количество антитела (или дополнительного лекарственного средства) для цели лечения обусловленного IL-6 заболевания или нарушения. Например, терапевтически активное количество полипептида может варьировать в зависимости от таких факторов, как стадия заболевания (например, стадия I относительно стадии IV), возраст, пол, медицинские осложнения (например, иммунодепрессивные состояния или заболевания) и масса тела индивидуума, и способность антитела индуцировать желаемый ответ у индивидуума. Режим дозирования можно корректировать для обеспечения оптимального терапевтического ответа. Например, несколько разделенных доз можно вводить каждые сутки, или дозу можно пропорционально снижать, как указано в зависимости от потребностей терапевтической ситуации. Однако, как правило, ожидается, что эффективная дозировка будет находиться в диапазоне приблизительно от 0,05 до 100 миллиграмм на килограмм массы тела в сутки и, более предпочтительно, приблизительно от 0,5 до 10, миллиграмм на килограмм массы тела в сутки.
VIII. Примеры
Пример 1. Получение и селекция специфичных к IL-6 антагонистических Fab
Лам иммунизировали IL-6 человека (либо продуцированным в E.coli, приобретенным в MACS Miltenyi Biotec (каталожный номер № 130-093-934), либо продуцированным в клетках почки эмбриона человека, приобретенным от Humanzyme (каталожный номер № HZ-1044)). Иммунизацию лам и сбор лимфоцитов периферической крови (PBL), а также последующую экстракцию РНК и амплификацию фрагментов антител проводили, как описано De Haard и коллегами (De Haard H, et al., JBC. 274:18218-30, 1999). После последней иммунизации проводили взятие крови и тотальную РНК экстрагировали из PBL, полученных с использованием градиента Ficoll-Paque и способа, описанного Chomczynski P et al. (Anal. Biochem. 162: 156-159, 1987). Затем экстрагированную РНК использовали для случайного синтеза кДНК и амплификации способом ПЦР V-областей тяжелой и легкой цепей (Vλ и Vκ) для конструирования содержащих Fab фагмидных библиотек, как описано De Haard H, et al. (Biol. Chem. 274, 1999).
Фаг, экспрессирующий Fab, получали в соответствии со стандартными протоколами и далее подвергали селекции на иммобилизованном IL-6 человека, либо биотинилированном и уловленном с помощью нейтравидина, либо прямо нанесенном на планшеты maxisorp. Тотальное или конкурентное элюирование связывающего IL-6 фага с трипсином проводили в соответствии со стандартными протоколами фагового дисплея.
Специфичные к IL-6 Fab далее подвергали скринингу в отношении перекрестной конкуренции с нейтрализующим IL-6 антителом, B-E8 и рецептором IL-6 с использованием конкурентного анализа на основе ELISA. Аминокислотные последовательности VH и VL иллюстративных антагонистических специфичных к IL-6 Fab, идентифицированных с использованием этого анализа, указаны в таблице 13 ниже.
Кинетику связывания специфичных к IL-6 Fab, которые были способны перекрестно конкурировать с антителом B-E8, оценивали с использованием поверхностного плазмонного резонанса (Biacore). В частности, биотинилированный (прокариотический) IL-6 улавливали на сенсорном чипе со стрептавидином biacore (CM5-SA) и различные концентрации очищенных Fab инжектировали в течение 3 минут, а затем промывали в течение 5 минут буфером. В фазе, полученной путем промывания, определяли константу диссоциации (kd), в то время как в инжектированной фазе вычисляли константу ассоциации (ka) с использованием концентрации и константы диссоциации в качестве параметров. Измеренные константы диссоциации и константы ассоциации и вычисленные аффинности Fab-антагонистов представлены в таблице 2. Fab 24C9 и 24D10 имеют константы диссоциации в диапазоне 10-5 с-1, и они обладают аффинностью 660 и 270 пМ, соответственно. Кинетику связывания антагонистических очищенных Fab (таблица 3) и периплазматических фракций экспрессирующих Fab бактерий (таблица 4) также оценивали с помощью поверхностного плазмонного резонанса (Biacore) с использованием продуцированного как бактериальными клетками, так и эукариотическими клетками, IL-6 человека, прямо нанесенного на чип CM5. Исследованные Fab имели константы диссоциации от 6×10-4 до 2×10-5 с-1.
Кинетика связывания выбранных очищенных антагонистических клонов Fab
(1/с)
(M)
Кинетика связывания выбранных очищенных антагонистических клонов Fab
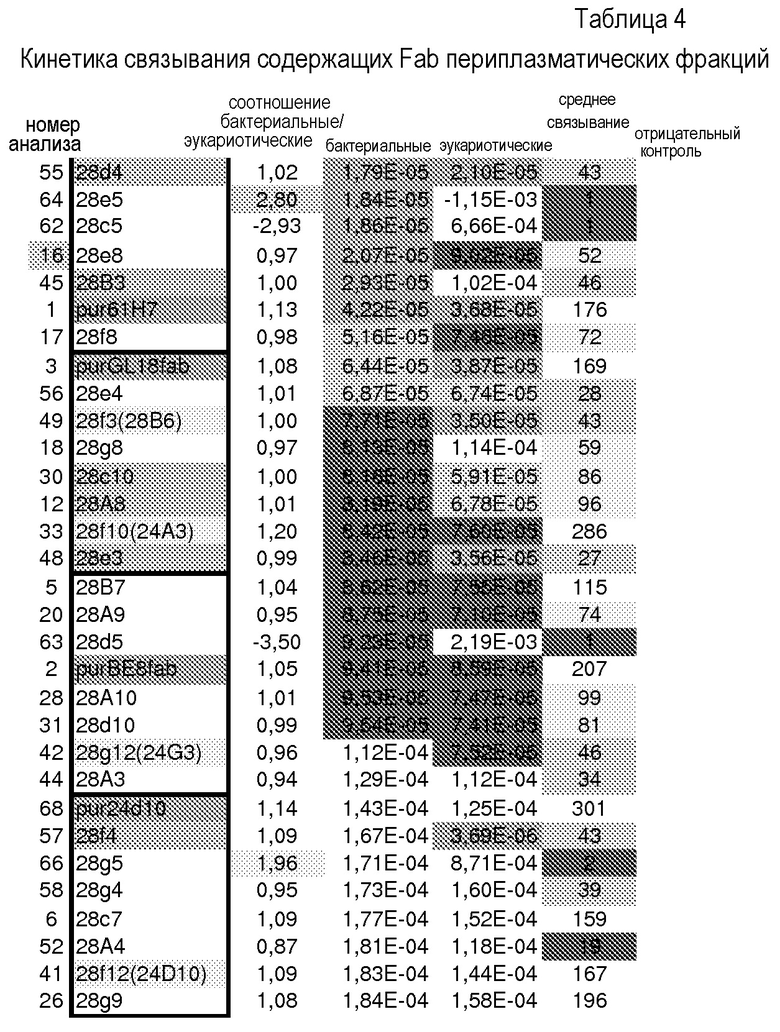
Пример 2. Шаффлинг VH /VL для повышения аффинности
Шаффлинг цепи VL использовали для повышения аффинности Fab 17F10, 18C7, 18C9, 18C11, 20A4, 29B11, 16D2 и 28A6. В этом способе тяжелую цепь этих клонов (в качестве фрагмента VHCH1) повторно вводили в первичную фагмидную библиотеку легких цепей (см. пример 1). Проводили селекцию по аффинности для отбора Fab с подвергнутыми шаффлингу цепями с увеличенной аффинностью к IL-6. Кинетику связывания Fab с подвергнутыми шаффлингу цепями оценивали с помощью поверхностного плазмонного резонанса (Biacore) с использованием продуцированных как бактериальными клетками, так и эукариотическими клетками, IL-6 человека и IL-6 яванского макака (таблицы 5-7). Аминокислотные последовательности VH и VL иллюстративных специфичных к IL-6 Fab, отобранных с помощью способа шаффлинга цепей VL, представлены в таблице 14 ниже.
Шаффлинг VH-цепей также использовали для повышения аффинности Fab 24D10. В этом способе легкую цепь 24D10 повторно включали в первичную фагмидную библиотеку тяжелых цепей (см. пример 1) и подвергали селекции с использованием анализа скорости диссоциации. При этом типе селекции фагу позволяли связываться с антигеном на субстрате в течение от 1,5 до 2 часов. На 2 раунде после 15 промывания посредством PBS-Tween проводили дополнительное промывание с присутствием избытка растворимого IL-6. Принцип состоит в том, что фаговые антитела с более низкими константами диссоциации и, таким образом, более быстро диссоциирующие, улавливаются избытом растворимой мишени и удаляются в ходе промывания. Эта методика предотвращает повторное связывание такого фага с покрытой мишенью. Время дополнительной стадии промывания увеличивали с увеличением количества проведенных раундов, и также увеличивали температуру до 37°C, чтобы отобрать более стабильные варианты Fab. Кинетику связывания Fab с шаффлингом антител и эталонных антител BE8 и GL18 оценивали с помощью поверхностного плазмонного резонанса (Biacore) с использованием продуцированного как бактериальными, так и эукариотическими клетками IL-6 человека (таблицы 8 и 9). Аминокислотные последовательности VH и VL иллюстративных специфичных к IL-6 Fab, отобранных способом шаффлинга VH-цепи, указаны в таблице 14, ниже.
Кинетика связывания очищенных Fab 17F10, 18C11, 18C7 и 20A4, подвергнутых шаффлингу цепей
68нМ
3,3E-05
8,0E-05
Кинетика связывания периплазматических фракций, содержащих Fab 29B11, подвергнутый шаффлингу легкой цепи
Кинетика связывания периплазматических фракций, содержащих Fab 28A6, подвергнутые шаффлингу цепей
Кинетика связывания периплазматических фракций, содержащих Fab 24D10, подвергнутые шаффлингу цепей
Кинетика связывания очищенных Fab 24D10, полученных шаффлингом цепей
Пример 3 Кристаллическая структура Fab 61H7 в комплексе с IL-6
i) Охарактеризация комплекса IL-6/Fab 61H7
Эксклюзионную хроматографию проводили на системе ВЭЖХ Alliance 2695 (Waters) с использованием колонки с силикагелем KW803 (Shodex), которую элюировали с помощью 50 мМ Tris-HCl pH 7,5, 150 мМ NaCl при скорости потока 0,5 мл/мин. Обнаружение проводили с использованием детектора с рассеянием света под тремя углами (Mini-DAWN™ TREOS, Wyatt technology, Sanata Barbara, США). Определение молекулярной массы проводили с помощью программного обеспечения ASTRA V (технология Wyatt).
ii) Кристаллизация
Первоначальный скрининг кристаллизации комплекса IL-6/Fab 61H7 проводили с помощью коммерческих наборов Structure screen 1 и 2, Proplex screen и Stura Footprint screen (Molecular Dimensions Ltd). Капли формировали при соотношении 1:1 (об.:об.) белка (8,45 мг/мл) и маточной жидкости в общем объеме 200 нл на 96-луночных планшетах Greiner с использованием робота Cartesian MicroSys SQ. Пригодные для дифракции кристаллы комплекса получали посредством диффузии пара с сидячей каплей при 277 K после оптимизации в 27,14% PEG MME 2K, 0,1M Na Hepes pH 7,14. Кристаллы принадлежат к пространственной группе C2 с размерами единичной ячейки: a=108,2 Å, b= 47,5 Å, и c=148,3 Å и β=97°. Они содержат один комплекс ILF/Fab на асимметричный элемент с величиной Vm 2,36 Å3/Да, которая соответствует содержанию растворителя 48%.
iii) Анализ структуры комплекса IL-6:61H7
Каноническими структурами, спрогнозированными для петель CDR mAb 61H7, являются 1 и 3 для H1 и H2, соответственно, и 7, 1 и 4 для L1, L2 и L3, соответственно, (www.bioinf.org.uk/abs/chothia.html). Наложение 61H7 VH на последовательность записи Protein Data Bank (PDB) 1dfb (происходящей из полученного от пациента антитела против белка gp41 ВИЧ-1) демонстрирует, что H1 и H2 имеют спрогнозированную каноническую укладку (см. фиг. 3A). Наложение 61H7 VL на последовательность записи Protein Data Bank (PDB) 1mfa демонстрирует, что все три CDR легкой цепи имеют спрогнозированные конформации (см. фиг. 3B). Таким образом, структурный анализ подтверждает, что VH (представитель семейства VH3) 61H7 принадлежит к комбинации 1-3 человека канонических структур H1-H2, в то время как VL (представитель семейства VL8) 61H7 принадлежит к комбинации 7λ1-4 человека канонических структур человека.
IL-6 ранее был кристаллизован и классифицирован как пучок из четырех спиралей, связанных петлями, и дополнительной мини-спирали (Somers et al., 1997, EMBO Journal 16, 989-997, которая включена в настоящее описание в качестве ссылки в полном объеме). Наложение apo IL-6 (pdb 1ALU) на IL-6 из комплекса IL-6:61H7 демонстрирует высокое соответствие между обеими моделями (rms 0,54 Å). Это подтверждает, что Fab 61H7 распознает и связывает нативную конформацию IL-6.
Кристаллическая структура IL-6 в комплексе с его рецептором и передающим сигнал рецептором gp130 продемонстрировала гексамерный комплекс (Boulanger et al., 2003, Science 27, 2101-2104, который включен в настоящее описание в качестве ссылки в полном объеме). IL-6 образует не передающий сигнал комплекс с IL-6R через участок I. Участок II представляет собой составной эпитоп, образованный бинарным комплексом IL-6 и IL-6R. Взаимодействие участка III с gp130 образует передающий сигнал комплекс.
Наложение структуры IL-6:IL-6R (pdb 1P9M) на структуру IL-6:61H7 демонстрирует высокое соответствие (rms 1,2 Å) между обеими структурами IL-6. Конформация двух петель различается. Первая петля, охватывающая остатки Asn48-Asn61, представляет собой длинную петлю, которая является неструктурированной в комплексе apo IL-6 и IL-6:61H7. Эта петля стабилизирована в структуре IL-6:IL-6R посредством связывания IL-6R. Вторая петля, конформация которой отличается, представляет собой так называемую петлю BC.
Кристаллографический асимметричный элемент содержит единичный комплекс 1:1. Общая структура комплекса IL-6:61H7 демонстрирует, что как VH (60%), так и VL (40%) обеспечивают большую поверхность взаимодействия (940 Å2). Из кристаллической структуры были определены остатки, важные для взаимодействия с IL-6. Водородные связи и солевые мостики, образованные между 61H7 Fab и цитокином, приведены в таблице 10. Взаимодействия ограничиваются CDR1 и CDR3 легкой цепи и CDR1, CDR2 и CDR3 тяжелой цепи.
Водородные связи и солевые мостики в комплексе 61H7:IL-6
SER 32[OG]
ASN 33[ND2]
TYR 34[OH]
TYR 34[OH]
SER 30
ASN 31
TYR 32
TYR 32
2,66
2,99
2,62
2,55
ASP 26[OD1]
GLU 23[OE2]
ARG 30[NH1]
ASP 26[OD2]
ASP 93[OD2]
GLY 95[O]
GLY 95[N]
GLY 95[O]
ASP 96[OD1]
ASP 96[OD1]
ASP 91
GLY 93
GLY 93
GLY 93
ASP 94
ASP 94
2,99
3,45
2,91
3,06
3,04
2,94
ARG 182[NH2]
SER 22[N]
GLU 23[OE1]
GLU 23[]
SER 22[OG]
SER 22[N]
SER 31[OG]
ARG 33[N]
SER 31
ARG 33
2,96
2,73
GLN 75[O]
GLN 183[OE1]
ALA 53[N]
GLY 54[N]
GLY 56[N]
TYR 59[ OH ]
ALA 52a
GLY 53
GLY 55
TYR 58
3,66
3,23
2,78
3,54
GLU 80[OE1]
GLU 80[OE1]
GLU 80[OE2]
MET 184[O]
ALA 100[O]
ALA 96
2,94
GLN 183[ NE2]
ASP 93[OD2]
ASP 91
2,99
ARG 182[NH2]
Наложение комплекса IL-6:61H7 и комплекса IL-6:IL-6R демонстрирует, что существует пространственное препятствие между Fab 61H7 и IL-6R. В основном пространственное столкновение с IL-6R происходит в VL. Эпитопы IL-6R и 61H7 находятся очень близко друг к другу. Для подтверждения того, существует ли перекрывание между обоими эпитопами, остатки в пределах 4,0 Å от IL-6R и остатки в пределах 4,0 Å от Fab 61H7 картировали и подвергали поиску в отношении перекрывания между эпитопами. Перекрывание между обоими эпитопами является довольно небольшим и в основном образовано паратопом VH. Перекрывание сосредоточено вокруг полости, которую занимают как петля HCDR3 61H7, так и молекула IL-6R. Этот участок связывания IL-6R на IL-6 был обозначен как участок I (Boulanger et al., 2003, Science 27, 2101-2104, который включен в настоящее описание в качестве ссылки в полном объеме). Образующий полость участок I занят гидрофобной боковой цепью Phe 229 IL-6R. Эту аминокислоту Boulanger et al. называют остатком горячей точки, поскольку исследования с мутагенезом показали ее критическую роль во взаимодействии между рецептором и цитокином. Мутация этого остатка на валин или серин полностью устраняла связывание IL-6R с IL-6 (Kalai et al., 1997, Blood, 1319-1333, обе из которых включены в настоящее описание в качестве ссылок в полном объеме). Trp98 в центре петли CDR3 тяжелой цепи Fab 61H7 занимает ту же полость, что указывает на то, что он, таким образом, взаимодействует с критическим эпитопом в IL-6, блокируя его взаимодействие с IL-6R (показано на фиг. 5). Вероятно, Trp98 является ключевым остатком для сверхвысокой аффинности Fab61H7 в отношении IL-6.
Пример 4. Кристаллические структуры Fab 68F2 и 129D3 в комплексе с IL-6
i) Получение, сбор данных и определение структуры кристалла IL-6:68F2
8 мг mAb 68F2 (4 мг/мл) в фосфатно-солевом буфере Дульбекко (d-PBS), pH 7,2, подвергали замене буфера на буфер для расщепления, содержавший 20 мМ цистеин-HCl на центрифужной колонке для обессоливания Zeba TM (Pierce Fab Preparation Kit Thermo Scientific). Образец инкубировали с иммобилизованным папаином (Pierce Thermo Scientific) и расщепляли в течение 6 часов при 37°C. Фрагменты Fc отделяли от Fab-фрагментов с использованием аффинной матрицы с Fc человека CaptureSelect (BAC BV Unilever), уравновешенной в d-PBS. Fab-фрагменты выделяли из проточной жидкости и фрагменты Fc элюировали с использованием 0,1 M глицина, pH 2,0. Концентрацию белка определяли с помощью УФ-спектрометрии, исходя из поглощения при 280 нм. 4,6 мг (>50%) очищенного Fab 68F2 выделяли и концентрировали до 1,53 мг/мл на Amicon-Ultra (предел 10 кДа).
2,5 мг rh IL-6 (Immunotools) инкубировали с 2,6 мг Fab 68F2 в фосфатно-солевом буфере Дульбекко (d-PBS), pH 7,2, в течение 1 часа при 4°C, а затем концентрировали до 1 мл на Amicon-Ultra (пороговое значение 10 кДа). Затем комплекс IL-6:68F2 отделяли от избытка свободного IL-6 с помощью хроматографии с гель-фильтрацией на колонке Superdex75 в d-PBS и, наконец, концентрировали до 8,1 мг/мл на концентраторе Amicon-Ultra (пороговое значение 10 кДа). Очистку комплекса оценивали с помощью SDS-PAGE.
Эксклюзионную хроматографию проводили на системе ВЭЖХ Alliance 2695 (Waters) с использованием колонки с силикагелем KW803 (Shodex), которую элюировали с помощью 50 мМ Tris-HCl pH 7,5, 150 мМ NaCl при скорости потока 0,5 мл/мин. Обнаружение проводили с использованием детектора с рассеянием света под тремя углами (Mini-DAWN™ TREOS, Wyatt technology, Santa Barbara, США). Определение молекулярной массы проводили с помощью программного обеспечения ASTRA V (технология Wyatt). Первоначальный скрининг кристаллизации комплекса IL-6:68F2 проводили с помощью коммерческих наборов Structure screen 1 и 2, Proplex screen и Stura Footprint screen (Molecular Dimensions Ltd). Капли формировали при соотношении 1:1 (об.:об.) белка (8,1 мг/мл) и маточной жидкости в общем объеме 200 нл на 96-луночных планшетах Greiner с использованием робота Cartesian MicroSys SQ. Пригодные для дифракции кристаллы комплекса получали посредством диффузии пара с сидячей каплей при 277 K через 9 месяцев в 25% PEG 4K, 0,15M (NH4)2SO4, 0,1M MES pH 5,5.
Кристаллы для сбора данных переносили в маточную жидкость с 7,5% этиленгликолем и быстро замораживали в жидком азоте. Данные дифракции собирали в стандартных криогенных условиях на линии пучка ID14-4 с использованием детектора ADSC Quantum 4 на синхротроне ESRF (Grenoble, Франция), обрабатывали с использованием XDS и масштабировали с помощью XSCALE. Кристаллическую структуру IL-6 в комплексе с Fab 68F2 определяли из экспериментов по нативной дифракции при одной длине волны путем молекулярного вытеснения с помощью 129D3 и структуры IL-6 с использованием MOLREP (таблица 4). Уточнение проводили с помощью BUSTER. Кристалл IL-6:129D3 получали сходным образом.
iii) Анализ структуры комплекса IL-6:68F2 и IL-6:129D3
Кристаллическая структура комплекса IL-6:68F2 имеет разрешение 2,9 Å. Детализацию модели проводили до R-фактора 26,7% и Rfree-фактора 29,6% с приемлемой стереохимией (что означает, что более 95% остатков принимают допустимые конформации). Кристаллическая структура комплекса IL-6:129D3 имеет разрешение 2,8 Å. Детализацию модели проводили до R-фактора 28,5% и Rfree-фактора 31,3% с приемлемой стереохимией.
Проводили наложение кристаллизованных Fab 68F2 и 129D3 (r.m.s.d. 1,4 Å), VH-домена (r.m.s.d. 0,5 Å) и VL-домена (r.m.s.d. 0,4). Это наложение демонстрирует, что не существует значимых отличий между родительским 68F2 и преобразованными в эмбриональный тип Fab-структурами 129D3.
Канонические структуры, спрогнозированные для петель CDR mAb 68F2 и его варианта 129D3 эмбрионального типа, представляют собой 3 и 1 для H1 и H2, соответственно, и 6λ, 1 и 5 для L1, L2 и L3, соответственно. Канонические укладки тяжелой цепи были спрогнозированы с помощью сервера на www.bioinf.org.uk/abs/chothia.html, канонические укладки легкой цепи определяли вручную. Эталонный Fab (PG16) для сравнения канонических укладок легкой цепи, выявляли вручную путем поиска на Antibody Structure Summary Page (www.bioinf.org.uk/abs/sacs/antibody_structure_summary_page.html). Наложение VL 68F2 и 129D3 на VL PG16 (запись Protein Data Bank (PDB) 3MUG) демонстрирует, что все три CDR принимают спрогнозированные конформации (см. фиг. 4A). Наложение VH 68F2/129D3 на эталонную последовательность записи PDB 1ACY демонстрирует, что H1 и H2 принимают спрогнозированные канонические укладки (см. фиг. 4B). Таким образом, структурный анализ подтверждает, что VL (представитель семейства VL2) 68F2/129D3 принадлежит к комбинации 6λ-1-5 человека канонических структур L1-L3, в то время как VH (представитель семейства VH4) 68F2/129D3 принадлежит комбинации 3-1 канонических структур H1-H2 человека.
Кристаллографический асимметричный элемент содержит единый комплекс 1:1. Общая структура комплекса IL-6:68F2 демонстрирует, что VH (50%) и VL (50%) в равной степени обеспечивают крупную поверхность взаимодействия (1156 Å2). Поверхность взаимодействия немного превышает поверхность взаимодействия 61H7 с IL-6 (940 Å2). Аналогично комплексу 61H7:IL-6, только петля L2 не прямо вовлечена во взаимодействие с IL-6.
Поверхность контакта между тяжелой и легкой цепью соответствует 1764,7 Å2 для структуры 68F2 и 1768,5 Å2 для структуры 129D3. Эта площадь поверхности контакта была сравнима с площадями, измеренными для антител 61H7 (1580,6 Å2) и 27B3 (1700,3 Å2). Все площади поверхности вычисляли с помощью программы PISA web-службы EMBL.
Из кристаллической структуры определяли остатки, важные для взаимодействия с IL-6. Водородные связи и солевые мостики, образованные между Fab 68F2 и цитокином, приведены в таблице 11. Взаимодействия ограничены CDR1 и CDR3 легкой цепи и CDR1, CDR2 и CDR3 тяжелой цепи.
Шаффлинг VL клона 20A4 привел к клону 68F2, который имеет в 10× лучшую аффинность, чем родительский 20A4. Оба антитела отличаются в основном их CDR L1 и L2. Петля CDR2 легкой цепи не вносит вклад в связывание IL-6, таким образом, увеличение аффинности должно быть объяснено мутациями в петле L1. Два из трех остатков, которые участвуют в важных взаимодействиях с IL-6, не являются консервативными в клоне 20A4: Asn26, который образует водородную связь с Ser76 IL-6, представляет собой Ser в 20A4; и Thr31, который образует две водородных связи с IL-6, представляет собой Gly в родительской L1 20A4. Добавление трех новых водородных связей при изменении этих двух остатков, вероятно, объясняет некоторое увеличение аффинности, наблюдаемое после шаффлинга VL. Наиболее важно, изменения в L1, возможно, стабилизируют и/или размещают Y30 (нумерация Kabat) надлежащим образом в полости F229, обеспечивая дополнительную высокую эффективность.
Водородные связи и солевые мостики в комплексе 68F2:IL-6
TYR 32[OH]
THR 31[N]
THR 31[O]
TYR 30
THR 29
THR 29
3,43
3,55
3,42
SER 176[O]
SER 76[OG]
GLN 75[N]
ASN 97[OD1]
ASN 95
3,73
ARG 179[NH2]
TYR 33[OH]
ARG 32[NE]
ARG 32[NH2]
TYR 33
ARG 32
ARG 32
2,16
3,20
3,28
ASP 34[OD1]
ASP 34[OD2]
ASP 34[OD2]
ASP 54[OD2]
ASP 56[OD1]
ASP 58[OD1]
ASP 58[OD2]
THR 59[O]
TYR 60[OH]
TYR 60[OH]
ASP 52
ASP 54
ASP 56
ASP 56
THR 57
TYR 58
TYR 58
3,37
2,90
3,86
3,42
2,52
3,69
3,15
LYS 171[NZ]
SER 37[OG]
HIS 164[NE2]
LYS 171[NZ]
ARG 168[NH2]
ARG 168[NE]
LYS 171[NZ]
VAL 104[O]
VAL 99
3,69
ARG 179[NH1]
ARG 32[NH2]
ARG 30
3,28
ASP 34[OD2]
ASP 58[OD1]
ASP 58[OD2]
ASP 56
ASP 56
3,86
3,42
HIS 164[NE2]
LYS 171[NZ]
ASP 102[OD2]
ASP 97
2,77
ARG 30[NH2]
Наложение комплекса IL-6:68F2 и комплекса IL-6:IL-6R демонстрирует, что существует пространственное препятствование между Fab 68F2 и IL-6R, как наблюдали для комплекса 61H7:IL-6. Однако, в противоположность комплексу 61H7:IL-6, именно VH 68F2 в основном обеспечивает пространственное столкновение с IL-6R в комплексе 68F2:IL-6. Fab 68F2 взаимодействует с IL-6 точно в том же участке, что и IL-6R. Более того, 68F2 не перекрывается с участками связывания gp130 и, таким образом, конкурирует специфически и только с IL-6R.
Наложение комплексов IL-6:IL-6R и IL-6:68F2 указывает на то, что эпитопы IL-6R и 68F2 расположены очень близко друг к другу. Остатки, принадлежащие обоим эпитопам, были картированы на IL-6 и перекрывание было определено. Перекрывание эпитопа 68F2 с IL-6R является практически полным. Участок связывания IL-6R на IL-6 назван участком I (Boulanger et al., 2003, Science 27, 2101-2104). Образующий полость участок I занят гидрофобной боковой цепью Phe 229 IL-6R. Эту аминокислоту Boulanger et al называют остатком горячей точки, поскольку исследования с мутагенезом показали ее критическую роль во взаимодействии между рецептором и цитокином. Мутация этого остатка на валин или серин полностью устраняла связывание IL-6R с IL-6 (Kalai et al., 1997, Blood, 1319-1333). Исследование полости, описанной в качестве участка I в структуре IL-6:68F2, показало, что ее занимает петля CDR1 легкой цепи Fab 68F2. В частности, Tyr32 (положение 30 согласно нумерации Kabat) в CDR1 легкой цепи играет ключевую роль в связывании этого участка (фиг. 5C).
Другим ключевым остатком во взаимодействии между IL-6 и IL-6R является Phe279 IL-6R. Этот остаток соответствует 20% (129 Å2) от общей поверхности контакта при связывании (по сравнению с 28% (174 Å2) для Phe 229), что делает его вторым из наиболее важных взаимодействий. Подобно Phe229, Phe279 также связывается в полости, образованной на поверхности IL-6. Эту полость также занимает остаток 68F2, более конкретно Val104 (положение 99 согласно нумерации Kabat) петли CDR3 тяжелой цепи (фиг. 6).
IL-6 ранее был кристаллизован и классифицирован как пучок из четырех спиралей, связанных петлями, и дополнительной мини-спирали (Somers et al., 1997, EMBO Journal 16, 989-997). Кристаллическая структура IL-6 в комплексе с его рецептором и передающим сигнал рецептором gp130 также была решена (Boulanger et al., 2003, Science 27, 2101-2104, который включен в настоящее описание в качестве ссылки в полном объеме). IL-6 образует не передающий сигнал комплекс с IL-6R через участок I. Участок II представляет собой составной эпитоп, образованный бинарным комплексом IL-6 и IL-6R. Взаимодействие участка III с gp130 образует передающий сигнал комплекс.
Сравнение структуры комплекса IL-6:61H7 со структурой IL-6:68F2 демонстрирует, что, хотя оба mAb конкурируют с IL-6R за связывание с участком I на IL-6, два антитела связывают IL-6 в двух различных эпитопах. 68F2 связывается на стороне IL-6 в форме бочонка и конкурирует исключительно за связывание IL-6R, в то время как 61H7 взаимодействует в большей степени в основании IL-6 в форме бочонка, конкурируя как с IL-6R, так и с gp130. Интересно, что уникальный перекрывающийся эпитоп на IL-6 представляет собой полость, которая заполнена остатком горячей точки Phe229 IL-6R (фиг. 5). Это указывает на то, что связывание с этой полостью является ключевым для высокой эффективности, наблюдаемой для 68F2 и 61H7.
Пример 5. Структурный анализ функций W98 в HCDR3 61H7
Поразительной особенностью чрезвычайно высокомощных антител, описанных в настоящем описании, является их способность занимать полость на IL-6, с которой связывается F229 IL-6R (в настоящем обозначаемую как полость F229). Для 61H7 (и ее преобразованных в эмбриональный тип вариантов, например, 111A7), полость F229 занимает остаток триптофана 98 (W98) HCDR3. Для оценки функциональной важности W98 для связывания 61H7 с IL-6 получали мутантов 111A7, в которых в положение 98 VH была внесена мутация на все возможные аминокислоты с фоновыми M100A или M100L. Кинетику связывания периплазматических фракций бактерий, содержащих мутантные Fab, исследовали с использованием поверхностного плазмонного резонанса (Biacore) и определяли константу диссоциации для каждого мутанта. Результаты анализа мутаций представлены в таблице 12. Эти данные отчетливо демонстрируют, что триптофан (W) в положении 98 является наилучшей возможной аминокислотой для достижения наилучшей константы диссоциации. Мутация W98 всегда была вредоносной для связывания 111A7 с IL-6.
Константа диссоциации для Fab 61H7 с мутациями VH в положениях 98 и 100
Пример 6. Преобразование в эмбриональный тип VH и VL клонов Fab 61H7 и 68F2
Последовательности VH и VL клонов 68F2 и 61H7 выравнивали против последовательностей VH и VL человека эмбрионального типа для идентификации наиболее сходных последовательностей эмбрионального типа. Процесс преобразования в эмбриональный тип проводили, как описано в WO 2010/001251, и Baca et al. (J. Biol. Chem. (1997) 272: 10678-10684) и Tsurushita et al. (J. Immunol. Methods (2004) 295: 9-19). Использовали подход библиотеки/фагового дисплея, в котором включали отличающиеся остатки FR как для остатков человека, так и для остатков ламы.
Происходящие из животного семейства верблюжьих антитела против IL-6 были примечательно сходными с человеческими по последовательности и структуре. В результате, в конечные преобразованные в эмбриональный тип варианты включали только минимальное количество изменений последовательности (путем преобразования в эмбриональный тип). Например, из 87 (VH) и 79 (VL) аминокислот в каркасные области VH и VL родительского антитела 68F2 вносили только 6 (VH) и 7 (VL) аминокислотных изменений (всего 13 аминокислотных замен), что приводит к конечной преобразованной в эмбриональный тип лидирующей последовательности (129D3) с идентичностью 93,1% и 91,1% в их соответствующих каркасах VH и VL (см. выравнивание на фиг. 10A). Аналогично, из 87 (VH) и 79 (VL) аминокислот в каркасные области VH и VL родительского антитела 61H7 вносили только 8 (VH) и 5 (VL) аминокислотных замен (всего 13 изменений аминокислот) с получением конечной преобразованной в эмбриональный тип лидирующей последовательности (111A7) с 90,8 и 93,7% идентичностью в их соответствующих каркасных областях VH и VL (см. выравнивание на фиг. 10B).
Напротив, для известных в данной области антител против IL-6 требуется обширная модификация способами инженерии (пересадка CDR) и изменения последовательности (обратные мутации) для получения варианта, пригодного для терапевтического применения. Например, для эталонного антитела против IL-6 мыши CNTO-328 требовалось 14 (VH) и 22 (VL) изменений аминокислот (всего 36) для получения гуманизированного варианта CNTO136 только с 84% и 72,5% гомологией в его каркасных областях VH и VL (для выравнивания см. фиг. 11A). Для другого эталонного антитела кролика против IL-6 ALD518 требовалось всего 46 изменений каркасной области (26 в VH и 20 в VL) для получения конечных гуманизированных вариантов только с 70,5% и 74% гомологией последовательности с родительским антителом. Таким образом, для антител против IL-6, очевидно, требуется только минимальная модификация способами инженерии, и она приводит к молекулам, последовательность которых является более сходной с человеческой.
Более того, небольшое количество остатков FR, подлежащих изменению, дает возможность включать изменения в остатках CDR в процесс преобразования в эмбриональный тип. Такие мутации CDR можно использовать для удаления аминокислоты, обеспечивающей вариабельность при продукции (участок гликозилирования, окисление, изомеризация и т.д.) или для изменения остатков CDR на аминокислоты, встречающиеся в отличающемся варианте антитела, на эмбриональный тип.
Фаговый дисплей с использованием жестких условий селекции использовали для селекции дополнительных функциональных Fab. Индивидуальные клоны подвергали скринингу в отношении константы диссоциации и наилучшие последовательности секвенировали для определения идентичности с последовательностью человека. Аминокислотные последовательности VH и VL иллюстративных преобразованных в эмбриональный тип специфичных к IL-6 Fab указаны в таблицах 15 и 16 ниже.
Последовательности CDR-области из всех идентифицированных доменов VH и VL, которые являются вариантами Fab 129D3 и 111A7, сравнивали и определяли аминокислотные консенсусные последовательности CDR. Варианты CDR 129D3 и 111A7 и полученные консенсусные последовательности CDR указаны в таблицах 17 и 18 ниже.
Пример 7. Анализ эффективности in vitro
Эффективность in vitro клонов 129D3, 68F2, 61H7, 133A9, 133H2, 133E5 и 132E7 определяли с использованием клеточного биоанализа нейтрализации с использованием клеточной линии B9, по существу как описано в Helle et al. 1988, Eur. J. Immunol 18; 1535-1540. Клетки B9 происходят из линии B-клеточной гибридомы B13.29, которой для выживания и пролиферации необходим IL-6, и которая отвечает на очень низкие концентрации IL-6 человека. Анализ проводили с использованием 10 пг/мл (или 0,5 пМ) IL-6 человека и серию концентраций очищенных клеток 129D3, B-E8, CAT6001, CNTO136, 61H7, UCB124.g1. B9 высевали в не содержащую IL-6 среду в количестве 5000 клеток/200 мкл в лунки с плоским дном в присутствии IL-6, с антителами и без них. Пролиферацию измеряли путем добавления [3H]тимидина на 64-72 ч. Результаты (представленные на фиг. 1 и в таблице 23) демонстрируют, что все клоны обладают высокой эффективностью, причем 129D3 имеет IC50 в этом анализе менее 0,1 пМ. Интересно, что, как показано на фиг. 1, IC50 клона 129D3 была лучшей относительно эталонных антител CNTO136, CAT6001, UCB124 и B-E8.
Эффективность in vitro клонов 133E5, 133A9, 133H2, 111B1, 104C1, 129D3, 68F2, 61H7 также определяли с использованием клеточного биоанализа нейтрализации с использованием клеточной линии 7TD1, по существу как описано в Van Snick et al. PNAS; 83, :9679 (1986), которая включена в настоящее описание в качестве ссылки в полном объеме. Клетки 7TD1 представляют собой клеточную линию гибридомы мыши, образованную путем слияния клеточной линии миеломы мыши Sp2/0-Ag14 с клетками селезенки из мыши C57BL/6, иммунизированной липополисахаридами Escherichia coli за трое суток до слияния. Рост клеточной линии 7TD1 зависит от IL-6 и устранение IL-6 приводит к гибели клеток посредством апоптоза. Анализ проводили с использованием 75 пг/мл IL-6 человека и серии концентраций очищенных 133E5, 133A9, 133H2, 111B1, 104C1, 129D3, 68F2, 61H7, B-E8, GL18LB, CNTO136 и hu1U.
В кратком изложении, клетки 7TD1 (7,10E3) инкубировали в течение 2-4 ч в среде RPMI1640 +10%FCS, а затем добавляли IL-6 (конечная концентрация 75 пг/мл) в микропланшеты для титрования (конечный объем 200 мкл). Клетки инкубировали в течение 3 суток при 37°C, а затем проводили промывание PBS и добавляли 60 мкл раствора субстрата (п-нитрофенил-N-ацетил-β-D-глюкозаминид (Sigma N-9376); 7,5 мМ субстрата в 0,05 M цитрате Na pH 5; 0,25 об.% triton X-100) на 4 ч. Ферментативную реакцию останавливали с помощью 90 мкл стоп-раствора (100 мМ глицин + 10 мМ EDTA pH 10,4) и измеряли OD при 405 нм. Результаты (представлены в таблице 24) демонстрируют, что все клоны обладают высокой эффективностью в этом анализе с IC50 менее 1 пМ.
Пример 8. Анализ на мышах с ксенотрансплантатом эпителиального рака яичника
Эффективность in vivo клона 129D3 определяли с использованием модели на мышах с ксенотрансплантатом. Эпителиальные клетки IGROV-1 (5×106) инъецировали подкожно мышам “nude”. Через трое суток мышам начинали вводить 129D3 или CNT0328 раз в две недели в дозировке 4 или 20 мг/кг. В каждой группе было 5 мышей и в контрольной группе было 10 мышей. Процентную выживаемость мышей в каждой исследуемой группе определяли каждую неделю, и результаты наносили на график в качестве кривой выживаемости. Результаты (представленные на фиг. 2) демонстрируют, что 129D3 проявляет эффективность in vivo, которая является лучшей, чем у эталонного антитела против IL-6 CNT0328.
Пример 9. Анализ иммуногенности
Области VH и VL антител против IL-6 по изобретению оценивали в отношении присутствия потенциальных иммуногенных последовательностей (например, предполагаемые ограниченные по HLA класса II эпитопы, также известные как эпитопы TH) и сравнивали с показателями иммуногенности для различных коммерчески доступных эталонных антител с использованием способа определения профиля Epibase®.
Определение профиля проводили на уровне аллотипа для 18 DRB1, 6 DRB3/4/5, 13 DQ и 5 DP, т.е. всего для 42 рецепторов HLA класса II. Идентифицировали антитела с прочным и умеренным связыванием DRB1, DRB3/4/5, а также антитела с прочным связыванием эпитопов DQ и DP. Подсчет эпитопов проводили по отдельности для антител, связывающихся с DRB1 с высокой и умеренной аффинностью. Пептиды, связывающиеся с несколькими аллотипами одной и той же группы, подсчитывали как один. Приблизительный показатель, выражающий иммуногенный риск в наихудшем случае, вычисляли следующим образом: показатель=Σ (подсчитанный эпитоп×частота аллотипа). Иными словами, количество эпитопов, присутствующих в случае конкретного аллотипа HLA, умножают на частоту аллеля данного аллотипа. Для данной последовательности продукты суммировали для всех аллотипов DRB1, использованных в исследовании, которые присутствуют в 2% или более популяции европиоидов.
Показатели в отношении DRB1 для антител против IL-6 по изобретению и репрезентативных эталонных антител представлены на фиг. 9. Общие показатели в отношении DRB1 представляли собой составные показатели для VH и VL каждого антитела и низкие показатели указывают на низкую иммуногенность антитела. Таким образом, на фиг. 9 продемонстрировано, что антитела против IL-6 по изобретению являются в равной или меньшей степени иммуногенными, чем эталонные антитела против IL-6, а также другие коммерчески доступные антитела (например, Humira и Remicade).
Пример 10. Возможность производства
VH и VL преобразованных в эмбриональный тип версий 68F2 повторно клонировали в векторы экспрессии pUPE для тяжелой цепи и легкой цепи, соответственно, для временной экспрессии полноразмерных IgG1-антител. После временной экспрессии в клетках HEK293E, IgG1-антитела очищали с помощью белка A и количественно определяли путем измерения OD280. В таблице 19 ниже обобщенно представлены уровни продукции преобразованных в эмбриональный тип производных вместе с уровнями родительского антитела 68F2. Также измеряли эффективность (в пМ) в анализе пролиферации на основе 7TD1 для каждого антитела.
Было выявлено, что варианты 126A3, 127F1, 129D3 и 129F1, все из которых были отобраны из преобразованных в эмбриональный тип библиотек в жестких условиях, имеют сходную эффективность с 68F2 дикого типа (т.е. от 0,5 до 0,7 пМ). Более того, все преобразованные в эмбриональный тип варианты очень хорошо экспрессировались, т.е. на уровне от 24 до 28 мкг/мл. Исключением был преобразованный в эмбриональный вариант 129F2, который обеспечил выход продукции 9 мкг/мл.
Пример 11. Анализ стабильности
Для исследования термической стабильности преобразованных в эмбриональный тип и родительских версий 68F2 в формате полноразмерного IgG1 человека образцы антител инкубировали в концентрации 100 мкг/мл (в PBS) при 4, 50, 55, 60, 65, 70 и 75°C в течение 1 часа. После этого образцы медленно охлаждали в течение 15 минут до 25°C и держали при этой температуре в течение 2 часов, после чего их хранили в течение ночи при 4°C. После центрифугирования для удаления преципитатов (денатурированного антитела), концентрацию функционального антитела, оставшегося в растворе, измеряли с использованием Biacore (разведение 1/10 в PBS и 1/10 в HBSEP). Наклон кривой ассоциации, полученной после инжектирования на чип с иммобилизованным IL-6, является показателем концентрации функционального антитела.
Как показано на фиг. 7A, преобразование в производное очевидно не влияло на кривые плавления и температуры плавления для 68F2 дикого типа и его производных дикого типа. Совершенно неожиданно, температуры плавления даже оказались увеличенными. Например, 129D3 имеет Tm приблизительно 70°C, что на 3°C выше, чем у родительского антитела 68F2. Кривые плавления для варианта эмбрионального типа 129D3 и родительского антитела 68F2 также сравнивали с эталонными антителами GL18 и CNTO136 (см. фиг. 7B). Благоприятная термическая стабильность антитела SIMPLE 68F2 и особенно его преобразованного в производное варианта 129D3 (Tm 70°C) была поразительной по сравнению с Tm 65°C для CNTO136 и Tm 61°C для антитела GL18. Неожиданно, обширная модификация способами инженерии антител (например, гуманизация) и созревание аффинности in vitro, использованные для обоих эталонных антител, значительно влияли на их стабильность, в то время как полученное in vivo антитело SIMPLE и минимальная модификация способами инженерии путем преобразования в эмбриональный тип приводила к чрезвычайно высокой термической стабильности.
Стабильность в сыворотке полноразмерных версий IgG1 человека 68F2 и его преобразованного в эмбриональный тип варианта (129D3) (и преобразованного в эмбриональный тип варианта 103A1, происходящего из лидирующего антитела 61H7 SIMPLE™) сравнивали со стабильностью эталонных антител. После инкубации при 37°C в сыворотке человека функциональную концентрацию антитела измеряли на 1, 2, 4, 8, 12, 16, 24, 32 и 56 неделю и сравнивали с предварительно распределенным на аликвоты стандартом. Как представлено на фиг. 8, стабильность в сыворотке антител по изобретению была лучшей по сравнению с эталонными антителами.
Пример 12. Оптимизация CMC
Несколько остатков или мотивов не рекомендованы для производства антител качества CMC. Среди них находится присутствие метионина в петлях CDR антитела. Метионин может окисляться, что приводит к химически измененному варианту антитела с измененными свойствами, такими как аффинность, эффективность и стабильность. Таким образом, метионин, присутствующий в CDR3 111A7 (и его преобразованном в эмбриональный тип варианте 61H7) подвергали мутации на аланин (111A7MA), лейцин (111A7ML) или серин (111A7MS). Полученные оптимизированные для CMC последовательности представлены в таблице 20 ниже. Как показано в таблице 21, мутация остатка метионина имеет незначительный эффект на связывание с IL-6.
Пример 13. Исследование фармакокинетики (PK) клона 129D3 и его Fc-мутантов у яванского макака
Проводили фармакокинетический анализ клона антитела 129D3 в формате различных молекул IgG1. Анализировали следующие антитела: IgG1 дикого типа 129D3 (129D3-WT), IgG1 129D3 с мутациями M252Y/S254T/T256E в Fc-области (129D3-YTE) и IgG1 129D3 с мутациями H433K и N434F в Fc-области (129D3-HN).
Яванским макакам (3 на исследованное антитело) внутривенно инъецировали однократную дозу 5 мг/кг 129D3-WT, 129D3-YTE или 129D3-HN. Взятие образцов проводили в различные моменты времени и их исследовали в отношении концентрации mAb в плазме с помощью ELISA. В частности, микропланшет для титрования (Maxisorb Nunc) покрывали 1 мкг/мл IL-6 (Immunotools) в PBS в течение ночи при 4°C. Планшет промывали 2 раза PBS-Tween и блокировали в течение 2 часов 300 мкл PBS-1% казеин. После 2 промываний PBS-Tween вносили образцы. Все разведения проводили в 1% объединенной простой плазме (которая представляет собой объединенную плазму от 3 наивных яванских макак, см. главу 4.2). Образцам позволяли связываться в течение 2 часов при к.т. Затем планшеты промывали 5 раз посредством PBS-Tween и добавляли биотинилированные адсорбированные поликлональные антитела обезьяны против тяжелой и легкой цепи IgG при разведении в 1000 раз (Bethyl, каталожный номер no: A80-319B) и им позволяли связываться в течение 1 часа при к.т. После промывания планшетов 5 раз посредством PBS-Tween, добавляли стрептавидин, конъюгированный с HRP (Jackson Immunoresearch 016-030-084) в разведении в 300000 раз и ему позволяли связываться в течение 1 часа при к.т. Затем планшеты промывали 5 раз посредством PBS-Tween и добавляли смесь 1:1 TMB (calbiochem CL07)-s(HS)TMB weakener (SDT, #sTMB-W). Окрашиванию позволяли протекать в течение 10 минут, а затем его останавливали с помощью 0,5 M H2SO4, после чего измеряли оптическую плотность при 450 нм. Образцы анализировали четыре раза и 129D3-WT (из той же партии, которую инъецировали животным) использовали для стандартной кривой.
Соответствующие параметры PK для некомпартментного анализа представлены в таблице 22 ниже. Фармакокинетические профили для различных IgG1-антител 129D3 представлены графически на фиг. 12 (результаты представлены в качестве среднего результата для группы обезьян). Эти данные явно демонстрируют, что 129D3-YTE и 129D3-HN имеют более длительное среднее время нахождения (MRT), чем родительское антитело 129D3-WT. Более того, 129D3-YTE и 129D3-HN имеют более медленную скорость элиминации и, таким образом, по существу пролонгированное время полужизни по сравнению с 129D3-WT.
Интересно, что, хотя оба антитела содержат Fc-область IgG1 дикого типа, время полужизни 129D3-WT является значительно более длительным, чем время полужизни IgG1-антитела против IL-6 MedImmune (GL18), описанного в US201200344212. В частности, 129D3-WT имеет время полужизни приблизительно 15,6 суток по сравнению с приблизительно 8,5 суток для антитела GL18. Таким образом, увеличенное время полужизни антител по изобретению, по-видимому, является следствием свойств их соответствующих Fab-областей.
Пример 14. Модель на мышах с сывороточным амилоидом A (SAA)
Эффективность in vivo клонов 68F2 и 61H7 далее исследовали путем измерения способности этих антител блокировать индукцию сывороточного амилоида A (SAA) в ответ на инъецированный IL-6. Общие способы для проведения этого анализа указаны в WO2006/119115A2, которая включена в настоящее описание в качестве ссылки в полном объеме. В частности, мышам Balb/c внутривенно инъецировали 68F2, 61H7, эталонные антитела GL18 или CNTO136, или солевой раствор (контроль). Через четыре часа после введения антитела мышам инъецировали 0,1 мкг IL-6. Через дополнительные 16 часов проводили взятие крови от мышей и концентрацию сывороточного амилоида A определяли с помощью ELISA. Экспериментальные группы, дозировки и результаты представлены в таблице 26 настоящего описания. Эффекты дозы также графически представлены на фиг. 13. Результаты демонстрируют, что клоны антител 68F2 и 61H7 обладают эффективностью in vivo, по меньшей мере равной высокоэффективным эталонным контролям GL18 и CNTO136.
Пример 15. Гуманизированная модель псориаза у мышей с ксенотрансплантатом
Модель с ксенотрансплантатом на мышах использовали для оценки профилактической эффективности клона 68F2 в отношении развития индуцированных псориатических очагов повреждения. В частности, мышам BNX трансплантировали биоптаты кожи диаметром 5 мм полной толщины из непораженной кожи от пациентов с псориазом (1 на мышь). Через 3 недели в трансплантаты инъецировали 0,5×106 активированных PBMC. Введение мышам клона 68F2, антитела против TNF (Remicade) или бетаметазона дипропионата (положительный контроль) начинали за 1 сутки до того, как активированные клетки инъецировали в трансплантаты. Детали групп введения и режимов представлены в таблице 25.
Эффективность лечения определяли по толщине эпидермального рубца, измеряемой с помощью световой микроскопии. Значимость отличий между группами анализировали статистически с использованием дисперсионного анализа (ANOVA) с последующими апостериорными тестами среднеквадратичных отличий (LSD) для установления статистически значимых отличий между группами введения. Величина p<0,05 означала значимое различие между группами.
Результаты этих экспериментов представлены на фиг. 14 настоящего описания. Толщина эпидермального рубца в контрольной группе (группа 2) составляла 156 мкм±4 (среднее значение±s.e.m.). Введение бетаметазона (группа 1, n=3) значимо снижало толщину эпидермального рубца до 83 мкм±13 (p< 0,05) (фиг. 12 и 13). Средняя толщина эпидермального рубца в группе введения Remicade (группа 3) составила 125 мкм±12 и в группе введения 68F2 (группа 4) она составила 125 мкм±12. Эти данные демонстрируют, что клон 68F2 является настолько же эффективным, как и антитело против TNF Remicade в гуманизированной модели псориаза на мышах.
Пример 16. Модель на мышах с ксенотрансплантатом почечно-клеточного рака
Эффективность in vivo клонов 68F2 исследовали в модели на мышах с ксенотрансплантатом почечно-клеточного рака. Основные способы для проведения этого анализа представлены в WO 2008/144763, которая включена в настоящее описание в качестве ссылки в полном объеме. В кратком изложении, клетки RXF393 (2×106) инжектировали подкожно в оба бока мыши “nude”. Опухолям позволяли расти до объема от 50 до 300 мм3, до введения антитела. У 90% мышей после инъекции развивалась опухоль. 40 мышей разделяли на 5 групп по 8 мышей. В каждой группе проводили внутрибрюшную инъекцию либо PBS (контроль), либо конкретной дозы клона 68F2 (1, 3, 10 или 30 мг/кг). Мониторинг размера опухоли и выживаемости проводили два раза в неделю.
Данные о выживаемости, представленные на фиг. 15 настоящего описания, демонстрируют, что 68F2 является эффективным в отношении замедления гибели мышей относительно контроля. В частности, наблюдаемое среднее время выживания составляло 15,5 суток для группы PBS, 21 сутки для группы 1 мг/кг, 3 мг/кг и 30 мг/кг 68F2, и 27,5 суток для группы 10 мг/кг 68F2. Данные о выживаемости, представленные на фиг. 18 настоящего описания, демонстрируют, что 129D3 и 111A7, введенные в дозе 3 мг/кг, являются эффективными в отношении замедления гибели мышей относительно контроля. Данные о скорости роста опухоли, представленные на фиг. 16 настоящего описания, демонстрируют, что клон 68F2 является эффективным в отношении ингибирования роста опухоли дозозависимым образом с насыщающими эффектами в дозе 10 мг/кг. Данные о скорости роста опухоли, представленные на фиг. 17 настоящего описания, демонстрируют, что клоны 129D3 и 111A7SDMA>A являются эффективными в отношении ингибирования роста опухоли в дозе 3 мг/кг.
Дополнительные таблицы.
Аминокислотные последовательности VH и VL иллюстративных нейтрализующих Fab против IL-6
Аминокислотные последовательности VH и VL иллюстративных нейтрализующих Fab против IL-6, полученных посредством шаффлинга VH или VL
(вместо последовательности приведено название клона, если последовательность идентична VH в другом клоне)
(вместо последовательности приведено название клона, если последовательность идентична VL в другом клоне)
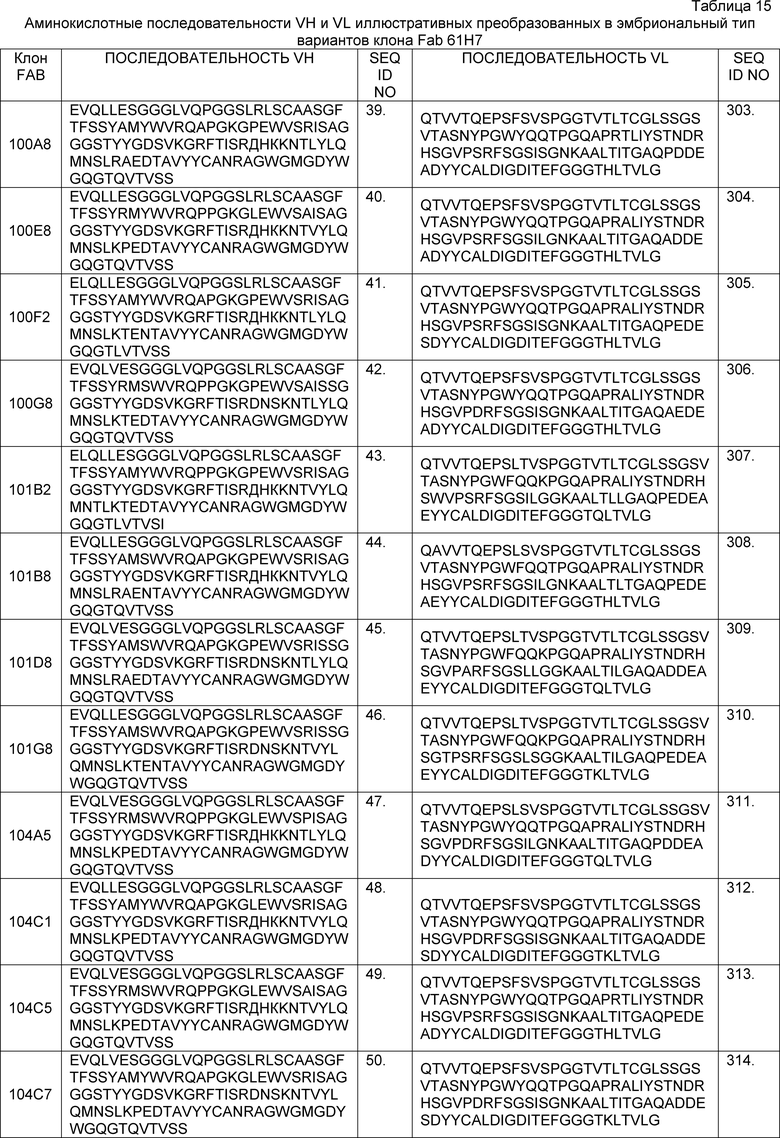
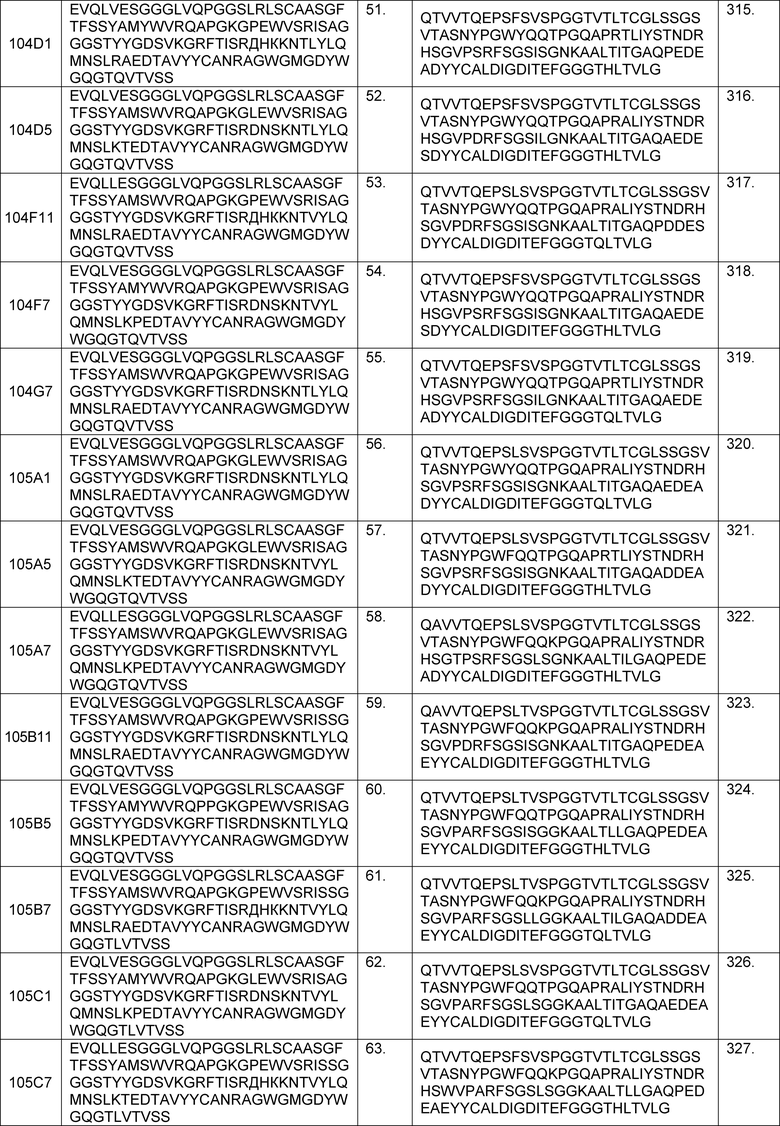
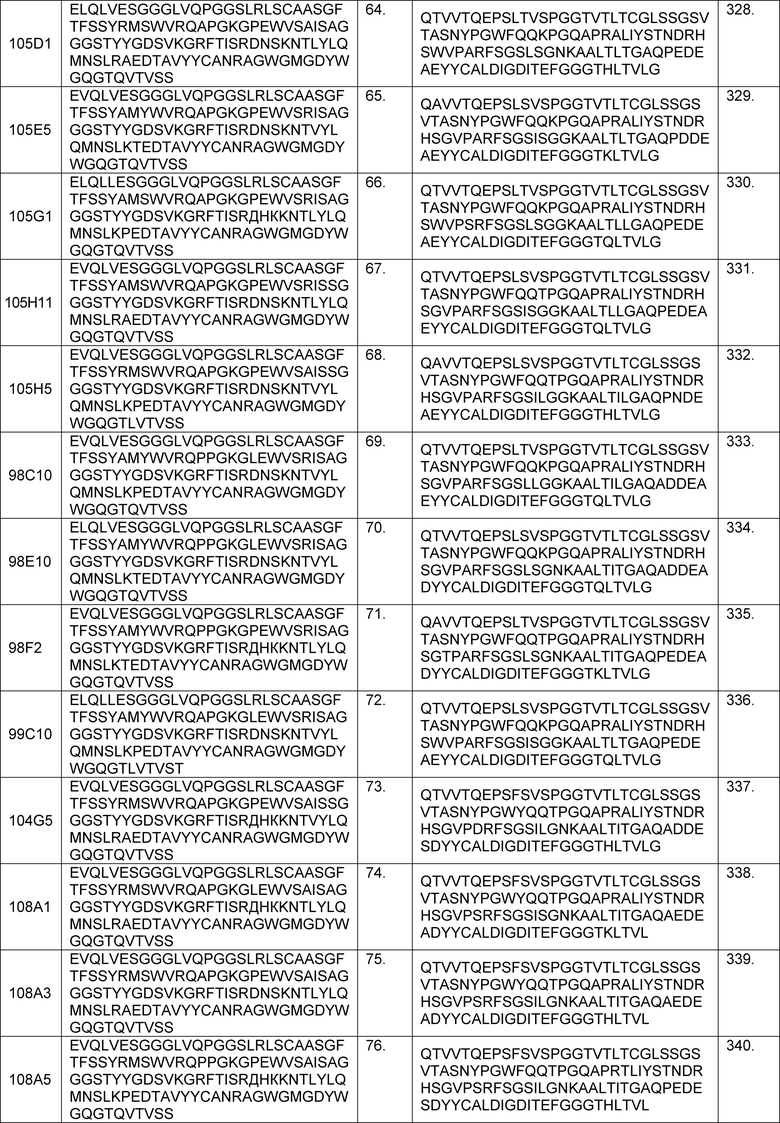
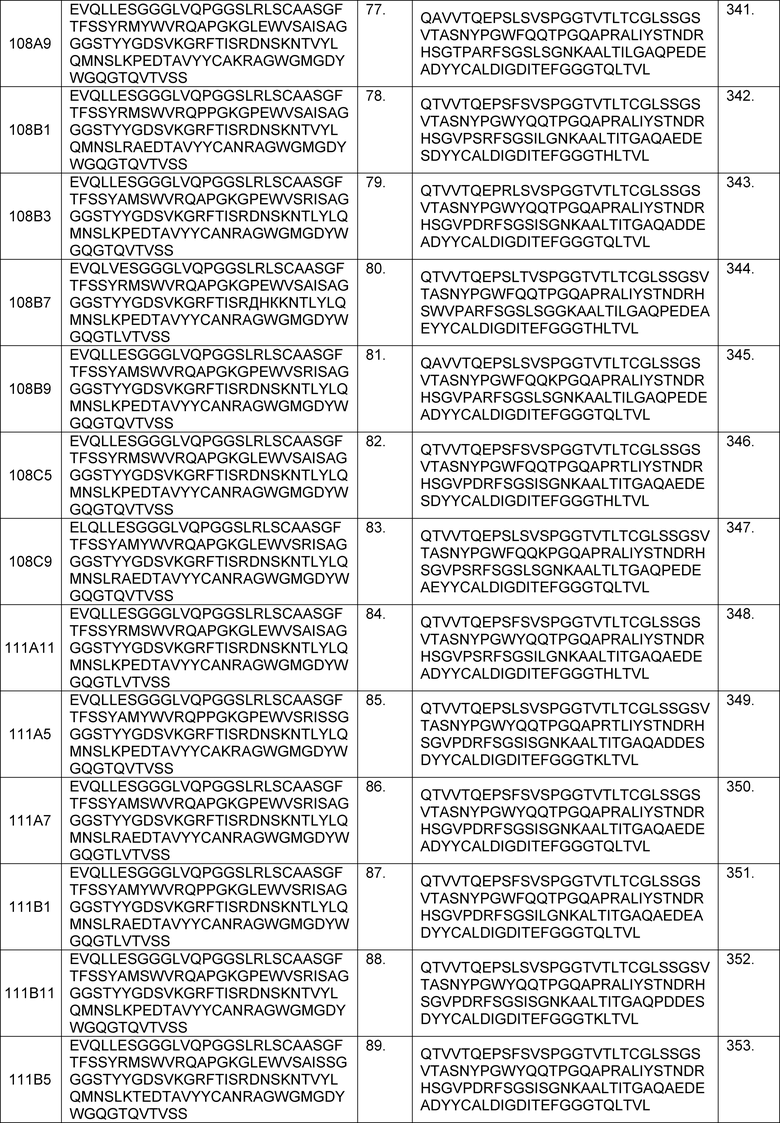
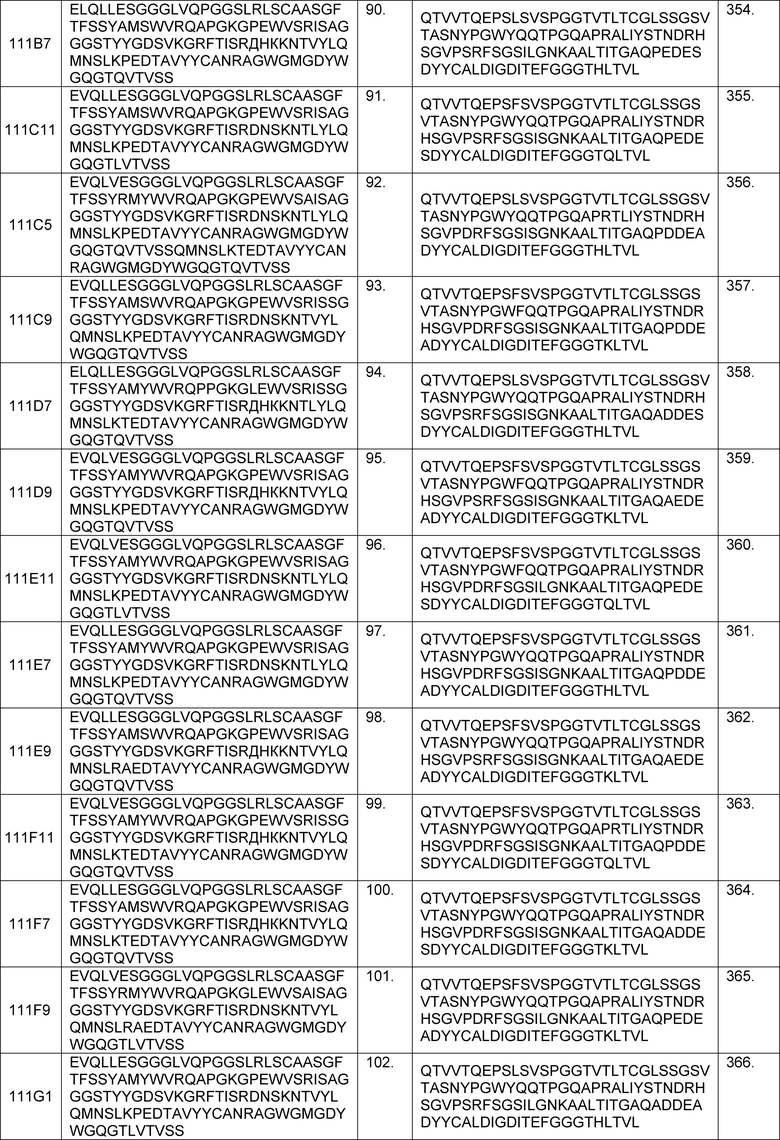
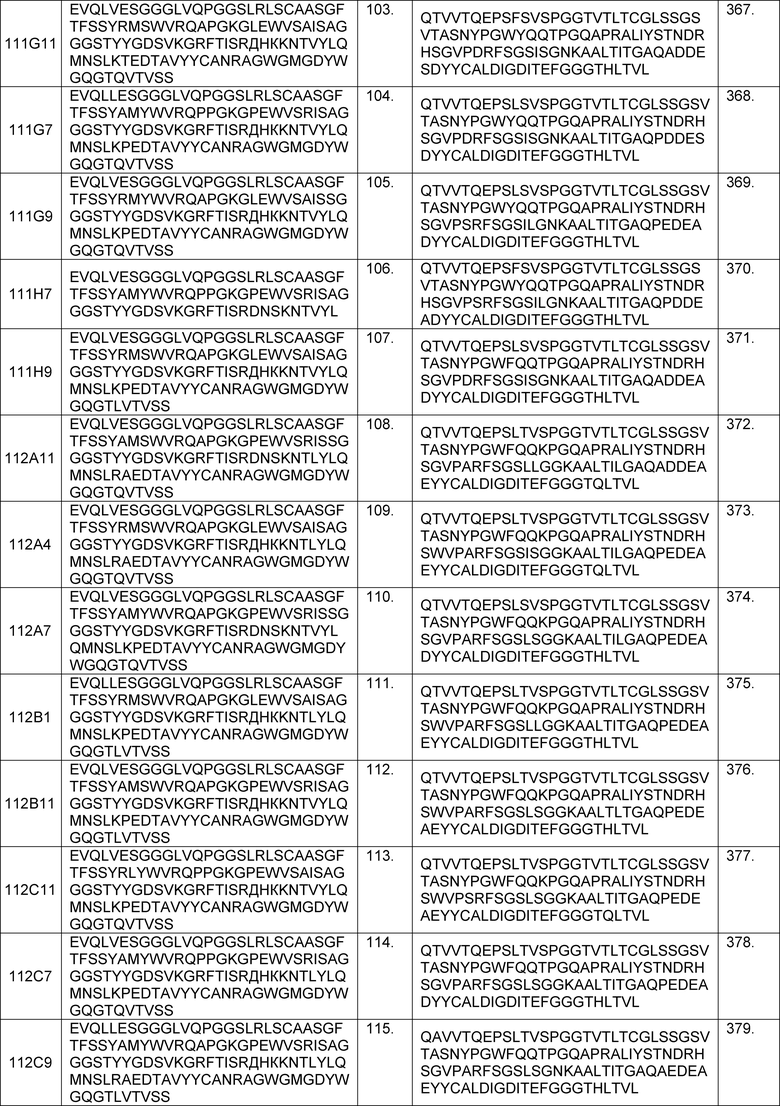
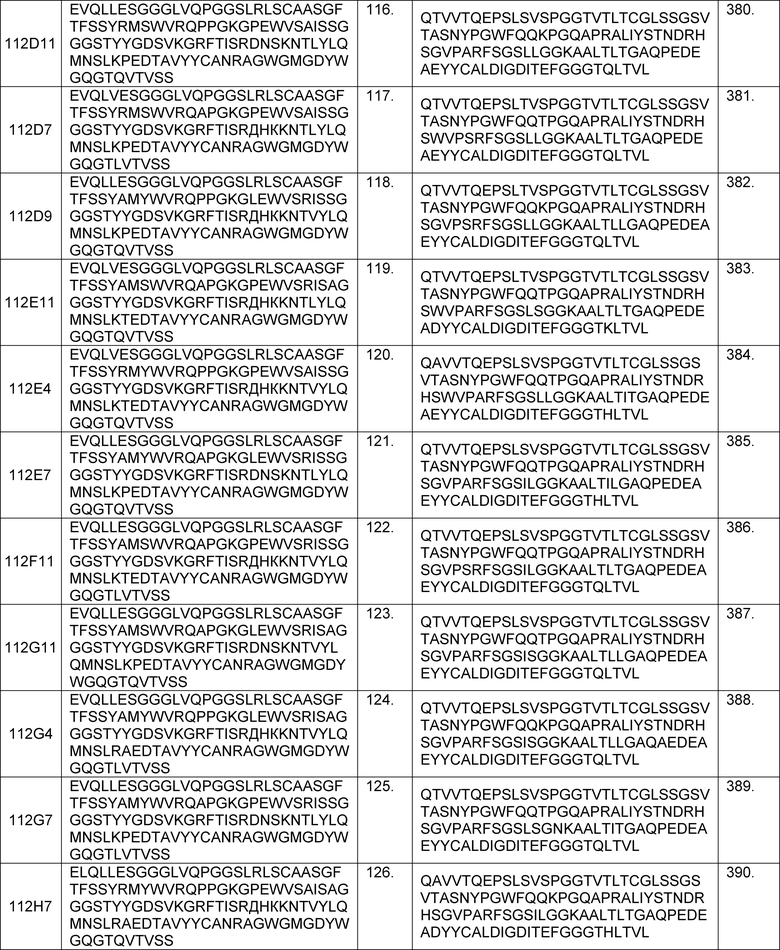
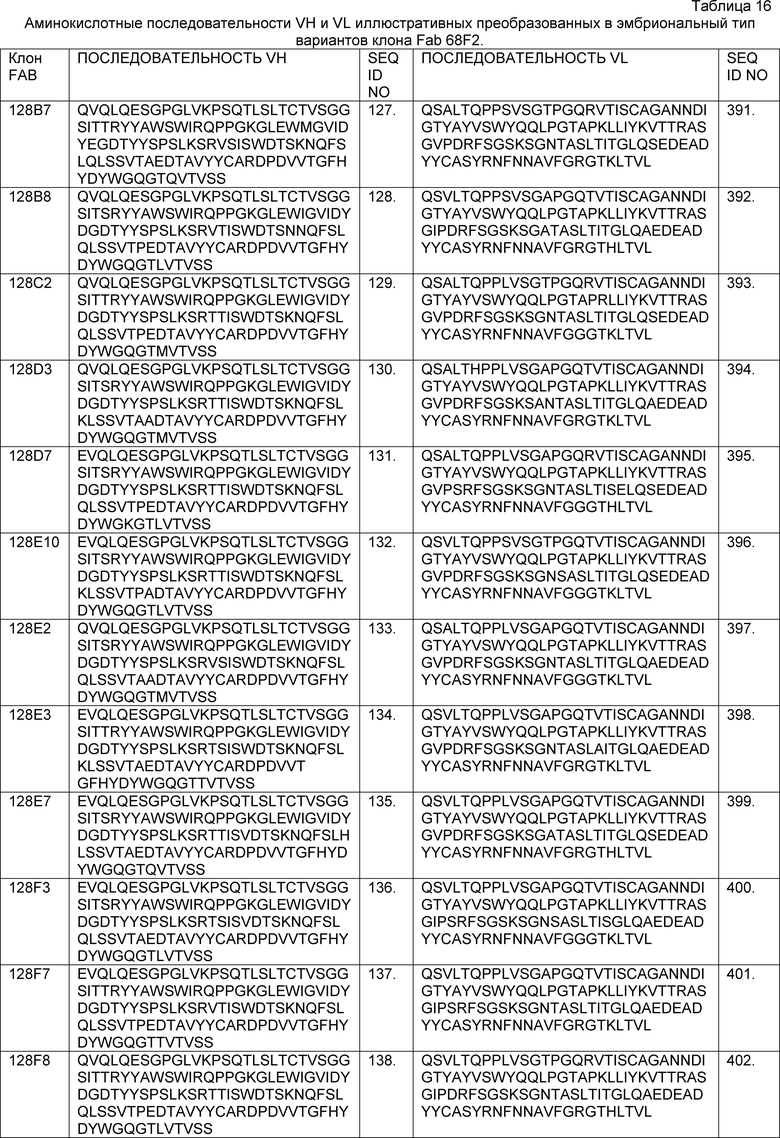
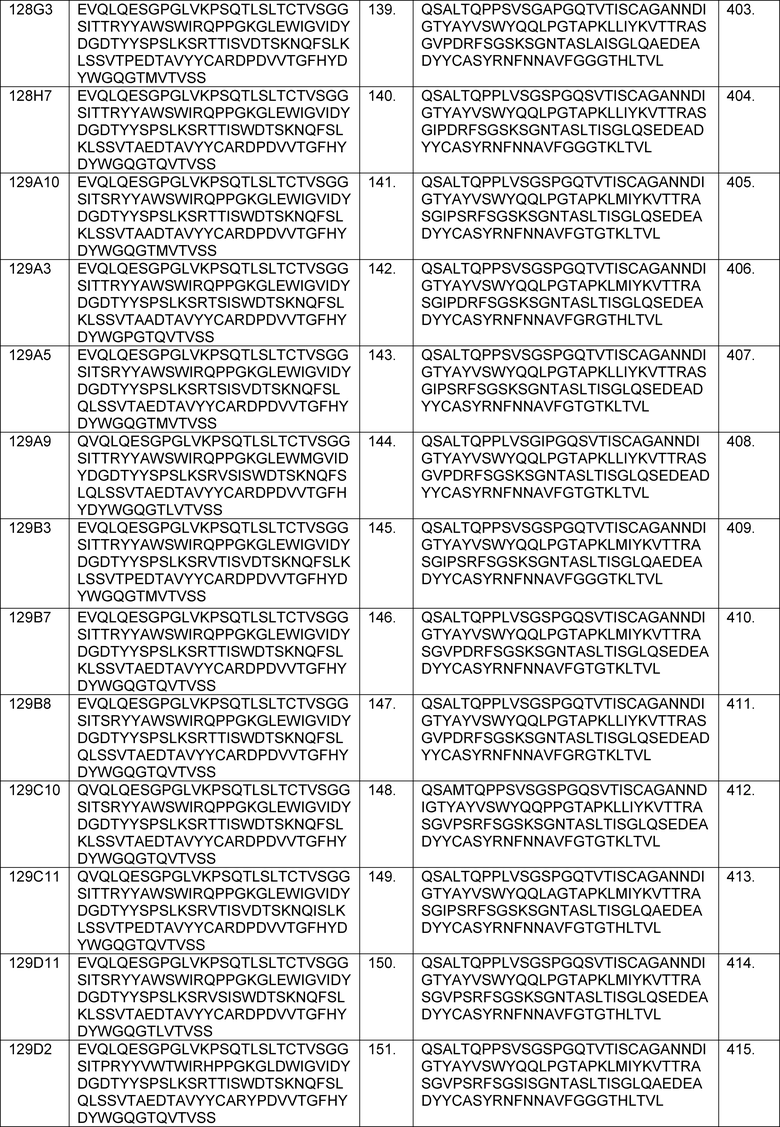
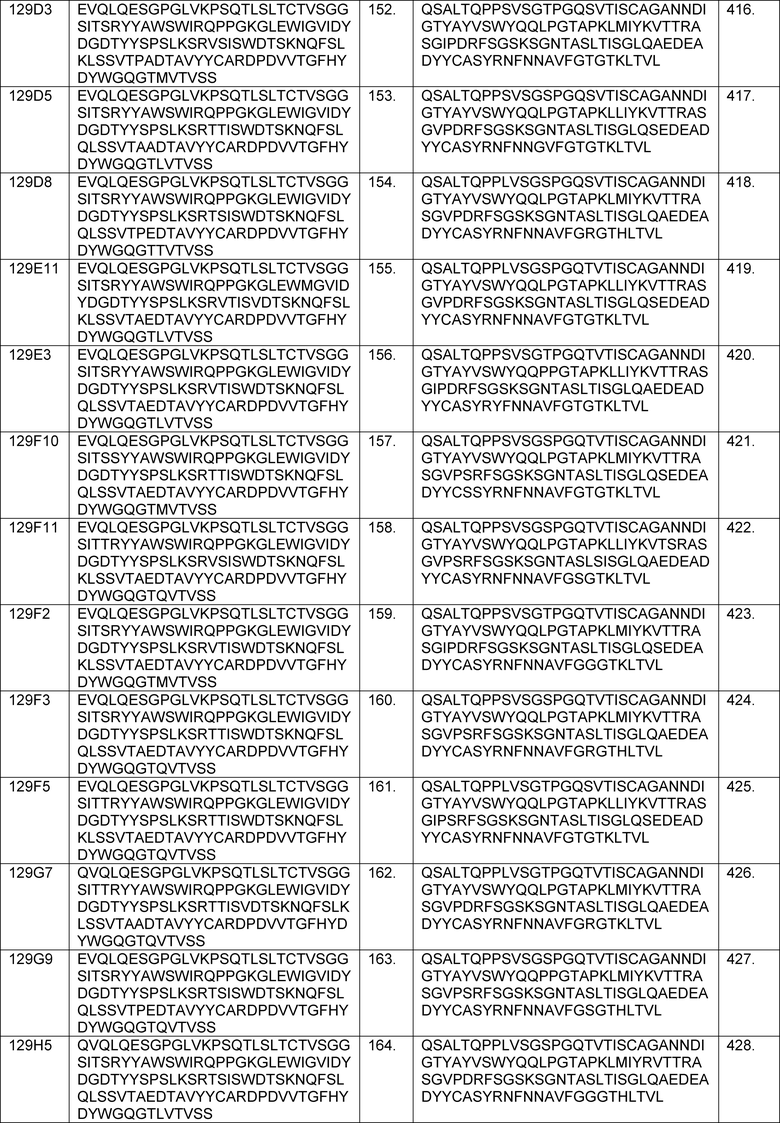
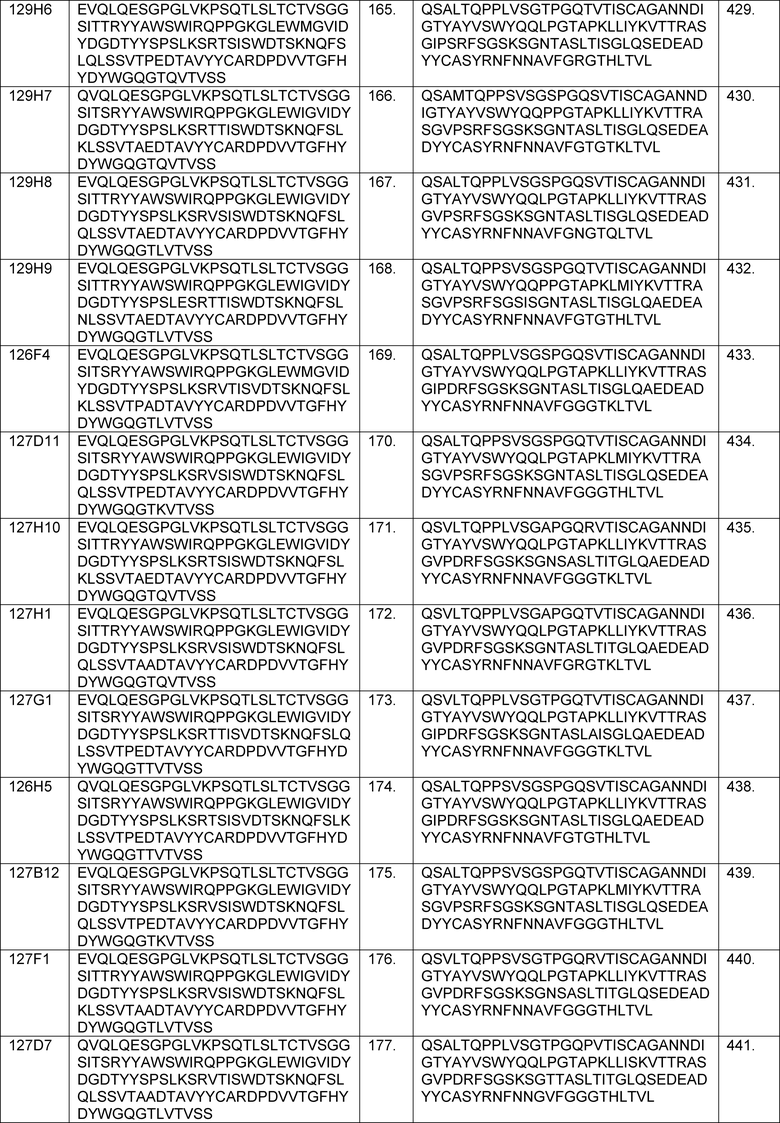
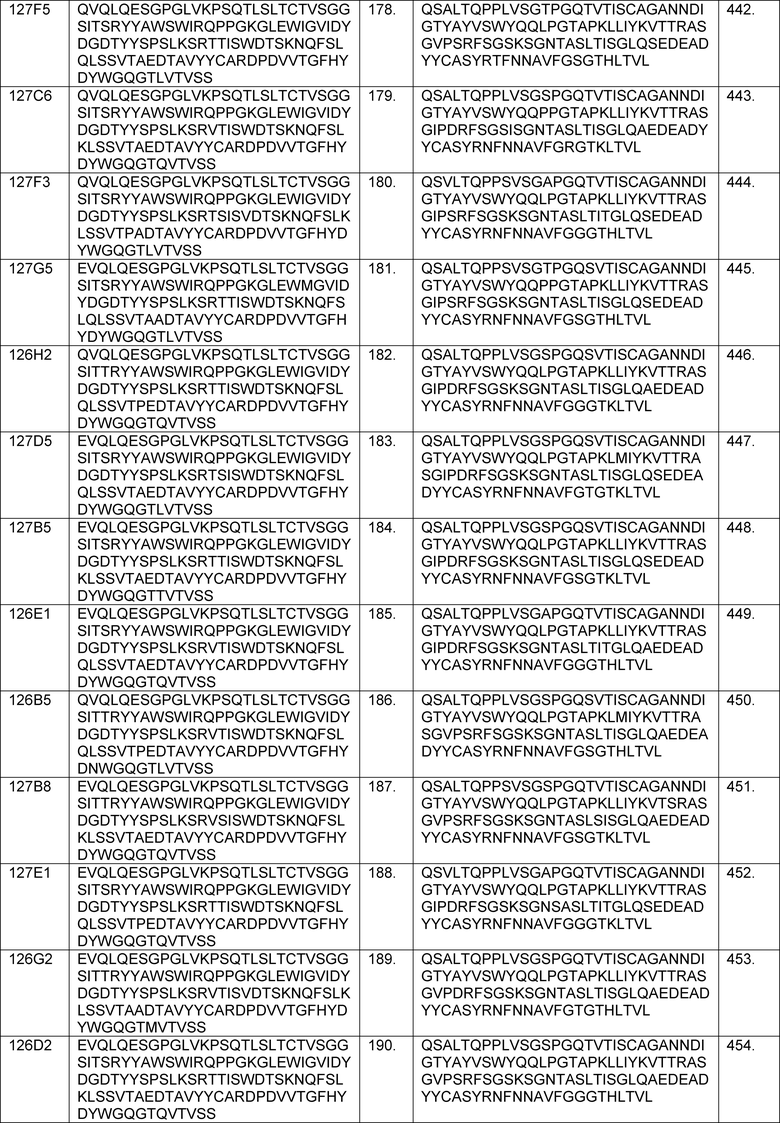
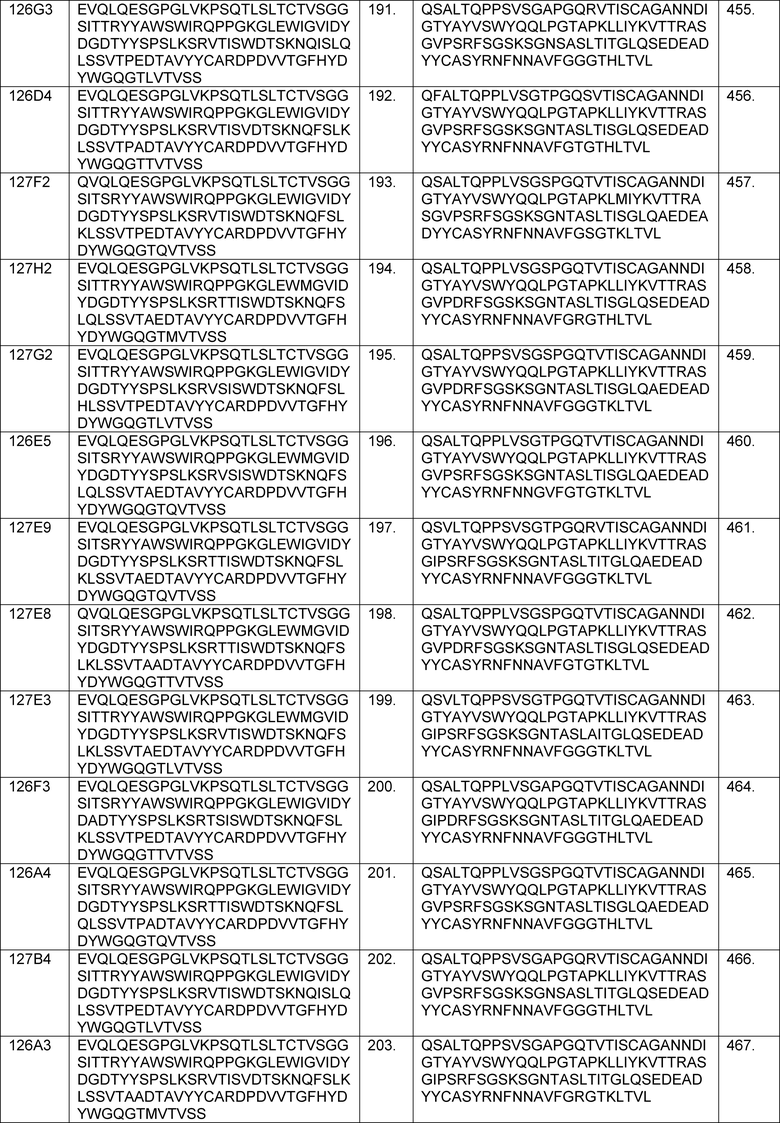
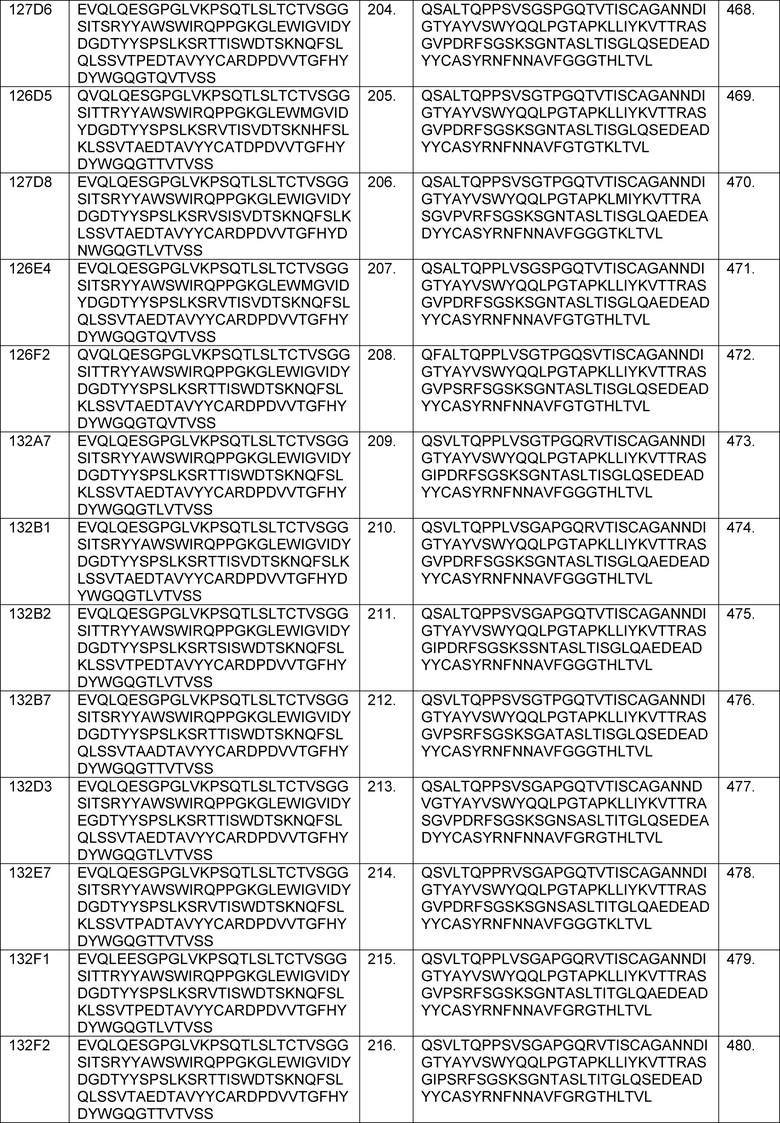
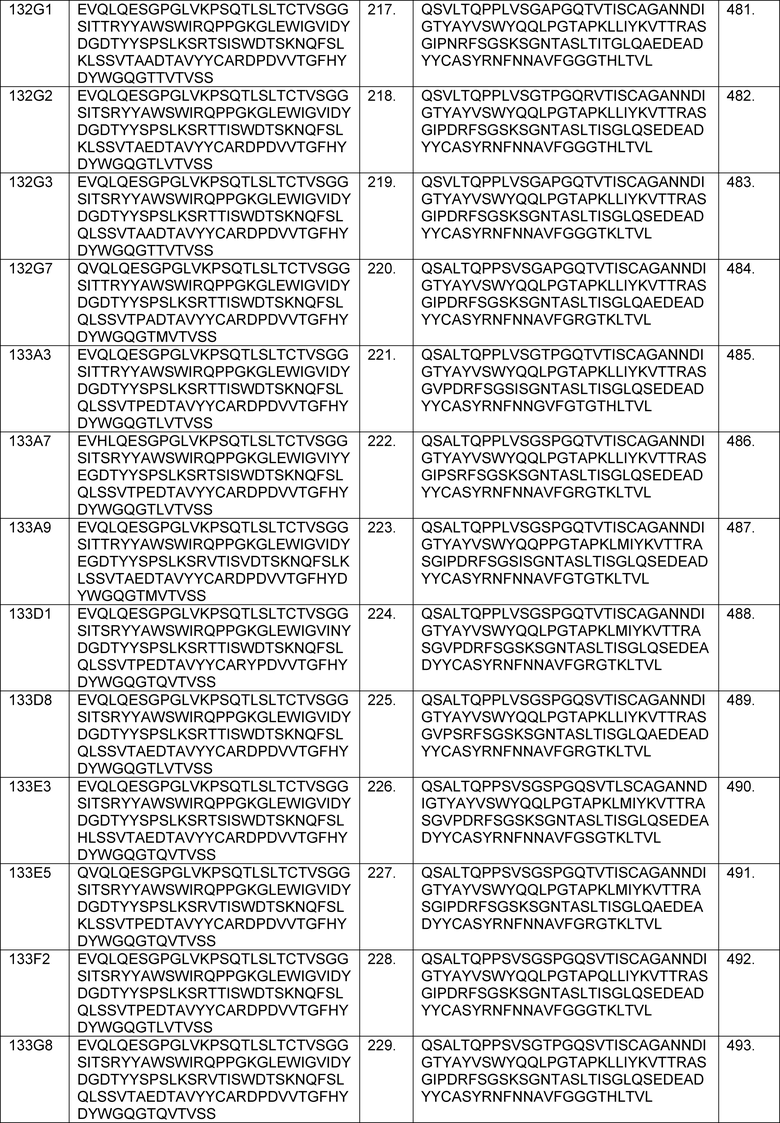
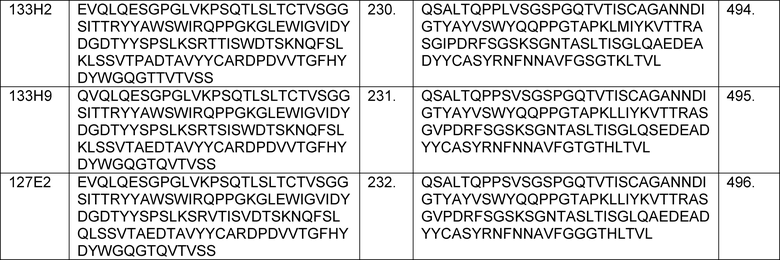
Варианты последовательностей аминокислотных последовательностей CDR Fab 129D3 и их консенсусные последовательности CDR
где:
X1= D или Y
X2= Y или N
где:
X1= D, Y или N
X2= D или E
X3= A или G
X4= E или K
где:
X1= T, S или P
X2= R или S
X3= A или V
X4= S или T
где:
X1= R или K
X2= N, H, R, S, D, T или Y
X3= F, Y, T, S или R
X4= N или I
X5= N или D
X6= V, N, G или A
X7= V или I
где:
X1= R, K, D, A ИЛИ E
X2= S, N или T
X3= T, K или Y
где:
X1= A или T
X2= S или N
X3= S, E или N
X4= V или I
X5= G, Y, T или F
X6= G или Y
X7= N, D или A
Варианты последовательности аминокислотных последовательностей CDR Fab 111A7 и их консенсусные последовательности CDR
где:
X1 = любая аминокислота или нет аминокислоты
X2 = любая аминокислота
где:
X1= A, P или R
X2= A или S
X3= S или G
X4= G или V
X5= A или T
X6= Y, N или S
X7= G, A или T
где:
X1= S или T
X2= H или Y
X3= A или R
X4= M или L
X5= S или Y
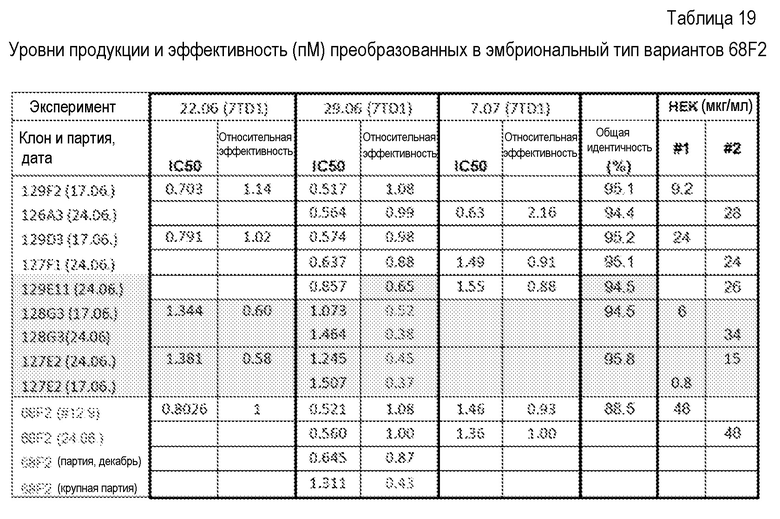
Оптимизированные CMC варианты последовательностей Fab 111A7
Кинетика связывания IL-6 для оптимизированных CMC вариантов последовательности Fab 111A7
Некомпартментный анализ PK mAb против IL-6 после однократного внутривенного введения яванскому макаку
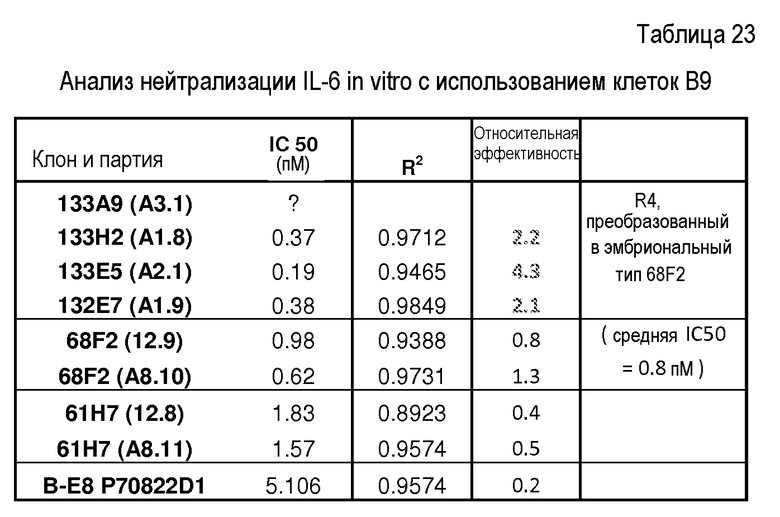
Анализ нейтрализации IL-6 in vitro с использованием клеток 7TD1
Группы и режимы введения, использованные в модели псориаза с ксенотрансплантатом
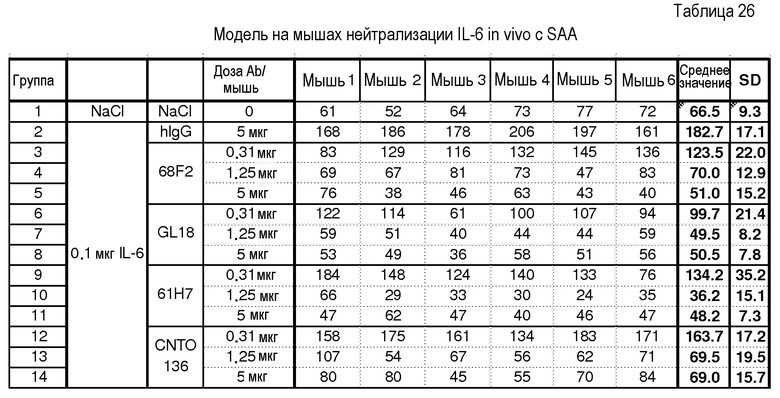
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА РЕЦЕПТОРА С-МЕТ | 2011 |
|
RU2608644C2 |
| АНТИТЕЛА ПРОТИВ CD70 | 2012 |
|
RU2604196C2 |
| Белок, связанный с гликопротеином человека, композиция, фармацевтическая композиция, вектор экспрессии | 2016 |
|
RU2773819C2 |
| Выделенный гуманизированный белок, связывающийся с гликопротеином VI человека, выделенное антитело, связывающееся с гликопротеином VI человека | 2018 |
|
RU2778884C2 |
| ГУМАНИЗАЦИЯ АНТИТЕЛ КРОЛИКА С ИСПОЛЬЗОВАНИЕМ УНИВЕРСАЛЬНОГО КАРКАСА АНТИТЕЛА | 2014 |
|
RU2652907C2 |
| ГУМАНИЗАЦИЯ АНТИТЕЛ КРОЛИКА С ИСПОЛЬЗОВАНИЕМ УНИВЕРСАЛЬНОГО КАРКАСА АНТИТЕЛА | 2018 |
|
RU2696202C1 |
| ГУМАНИЗАЦИЯ АНТИТЕЛ КРОЛИКА С ИСПОЛЬЗОВАНИЕМ УНИВЕРСАЛЬНОГО КАРКАСА АНТИТЕЛА | 2009 |
|
RU2567006C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ РЕЦЕПТОР ПРОЛАКТИНА, И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2664463C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2605327C2 |

































































































































































































































































































































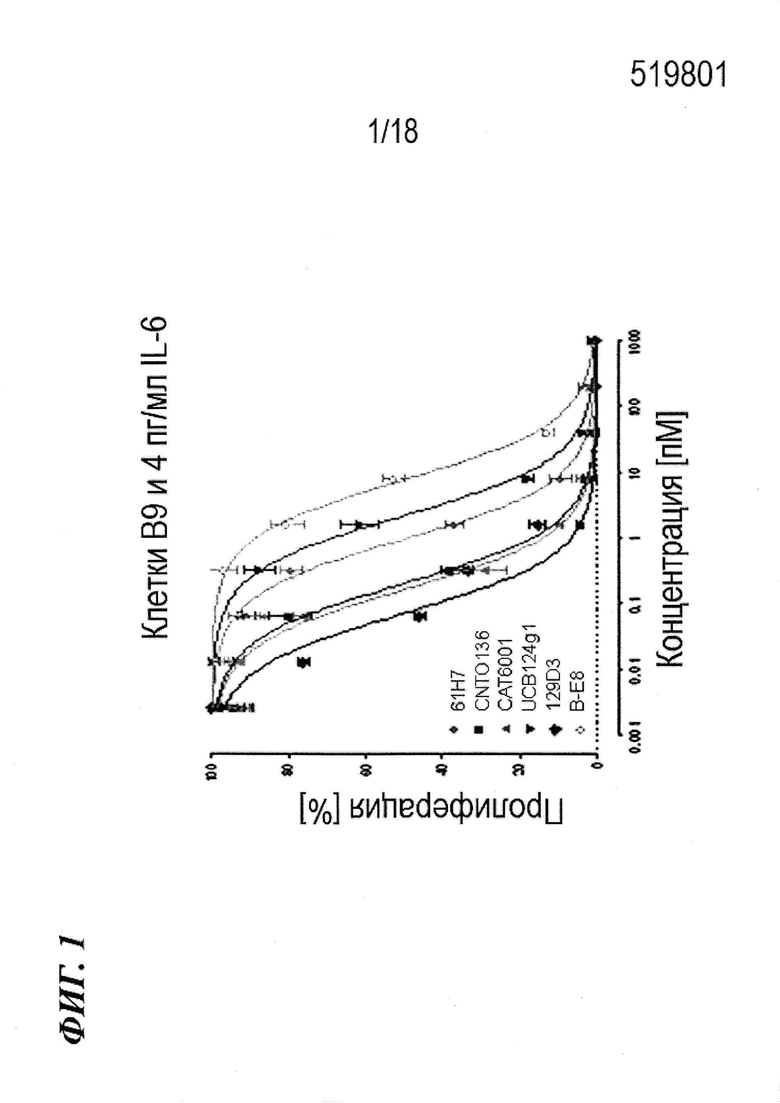
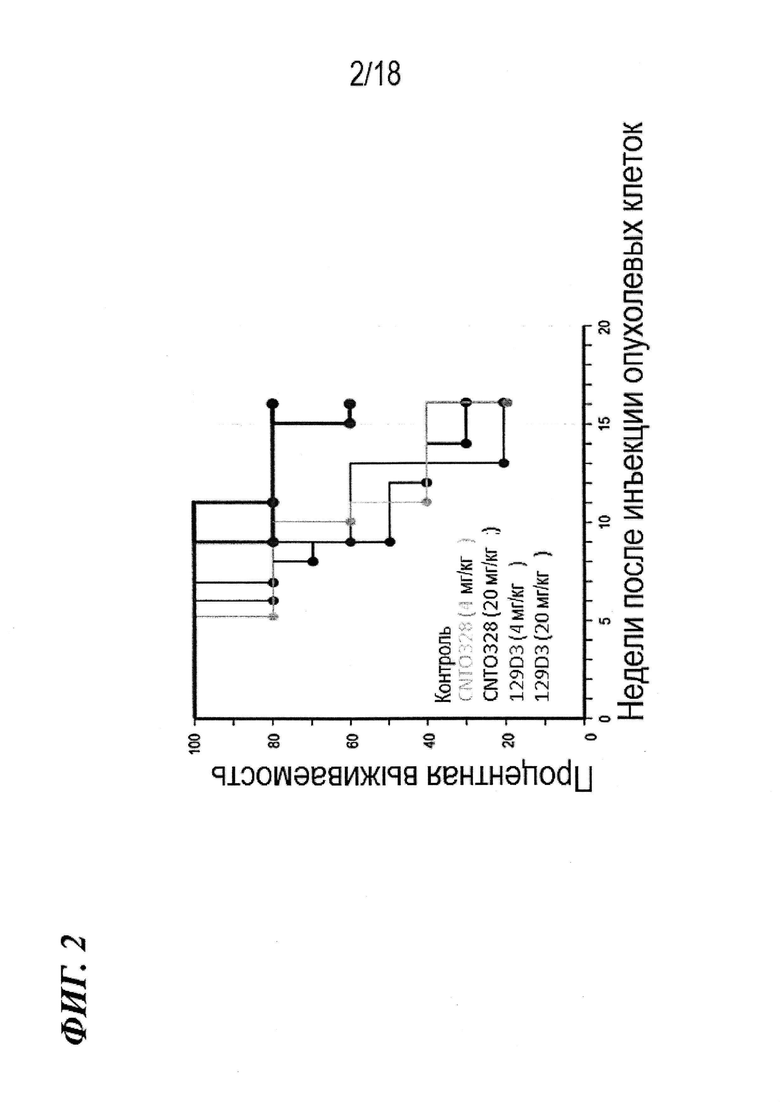
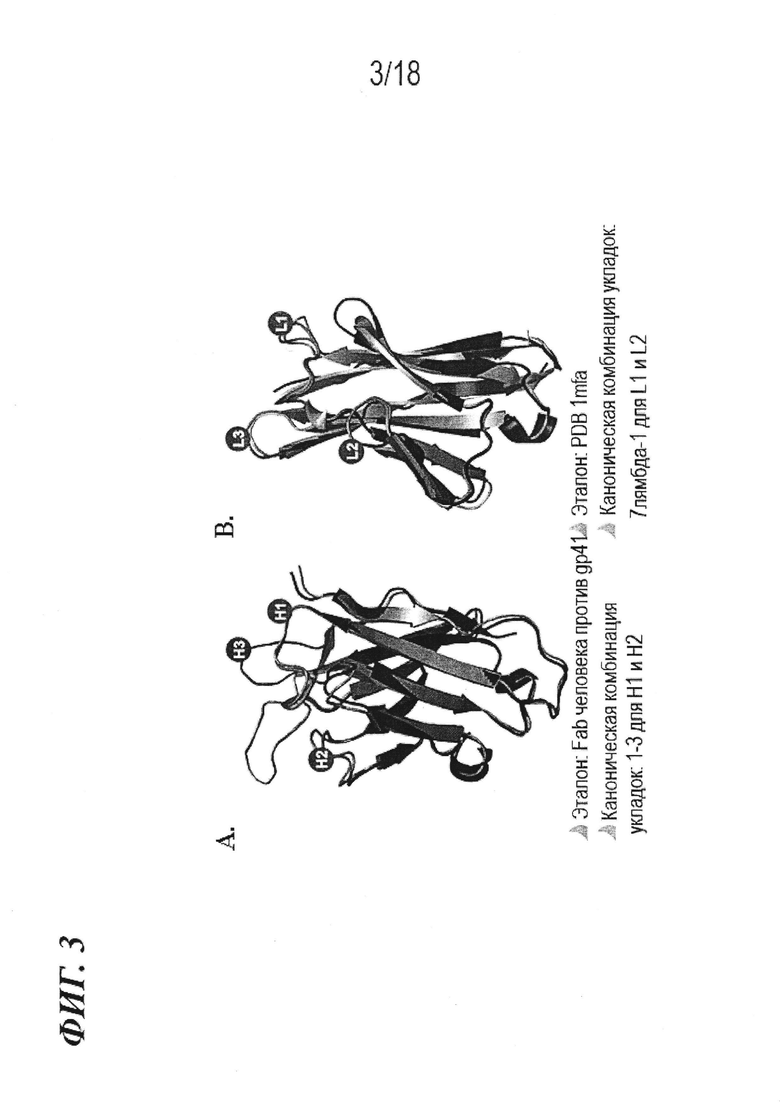
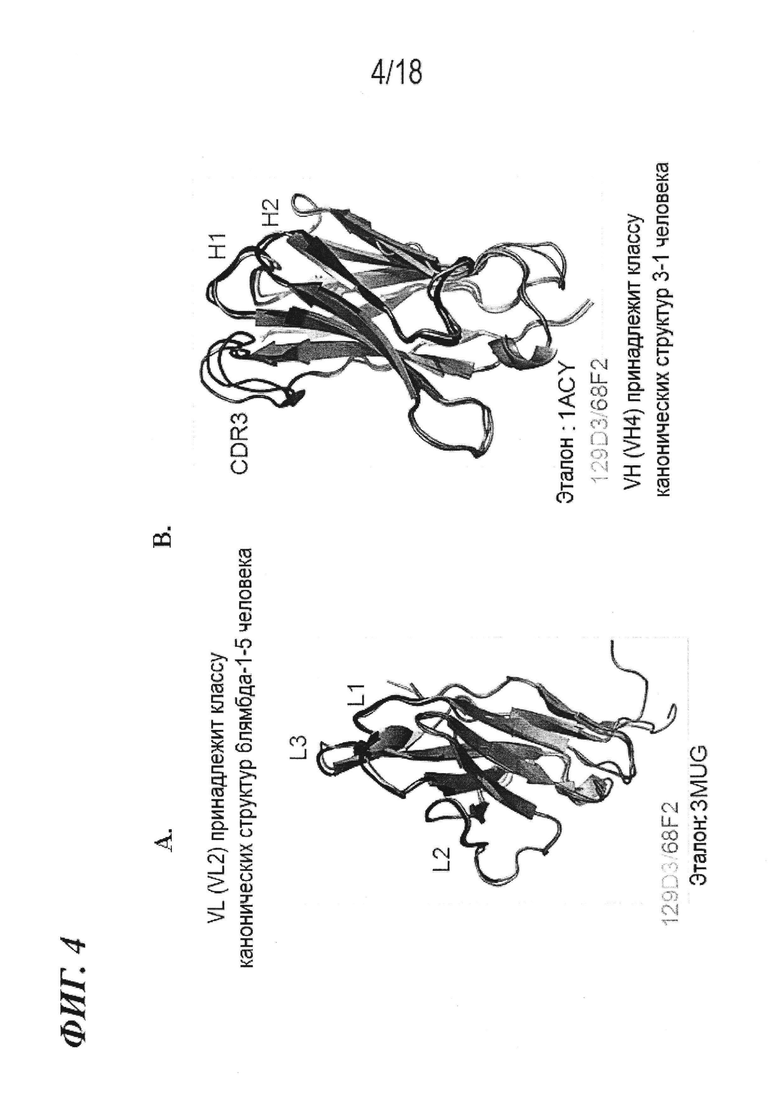
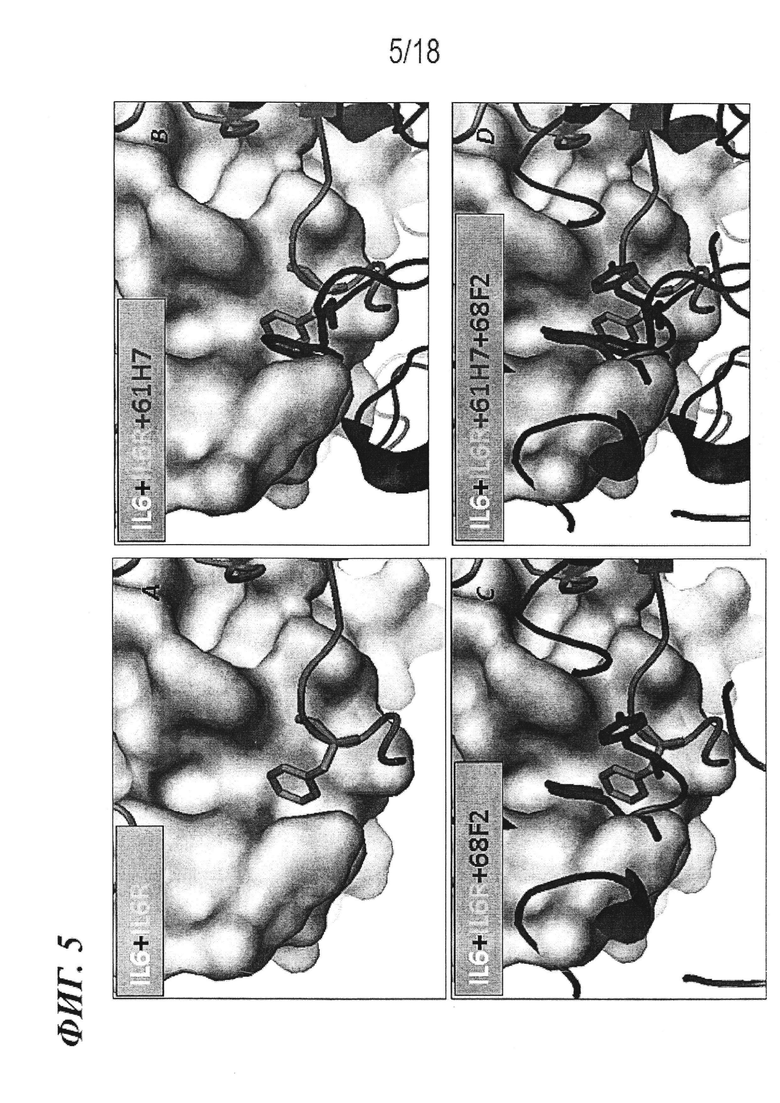
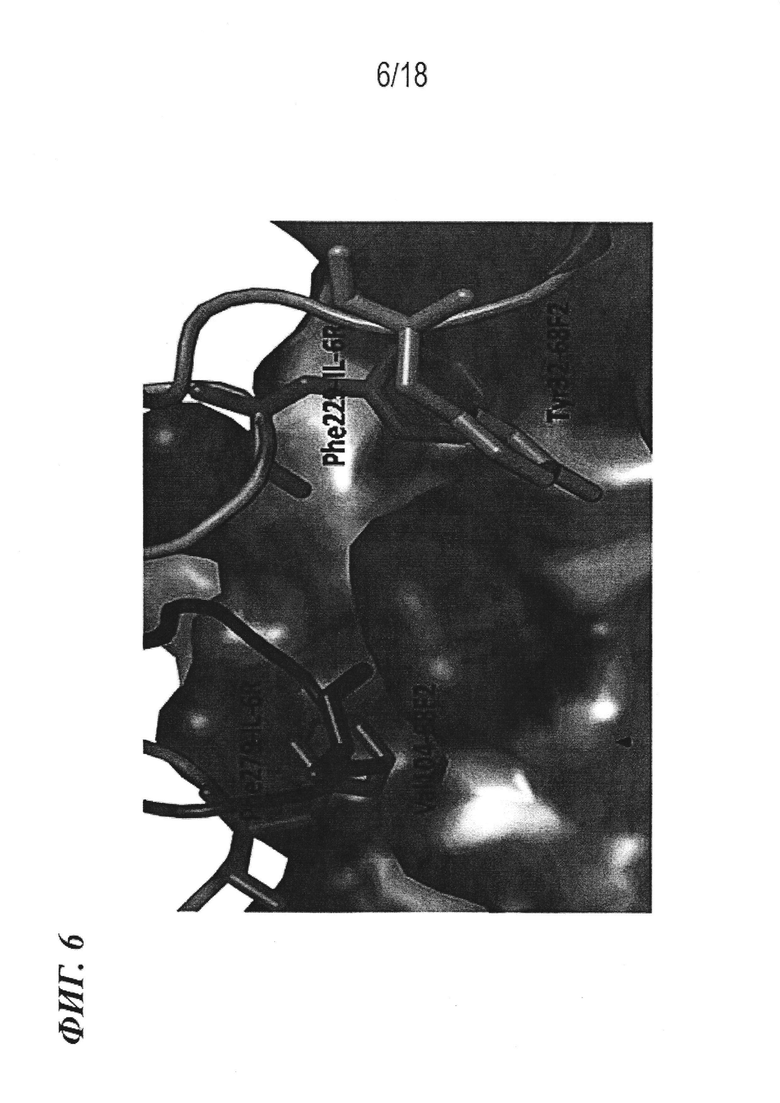
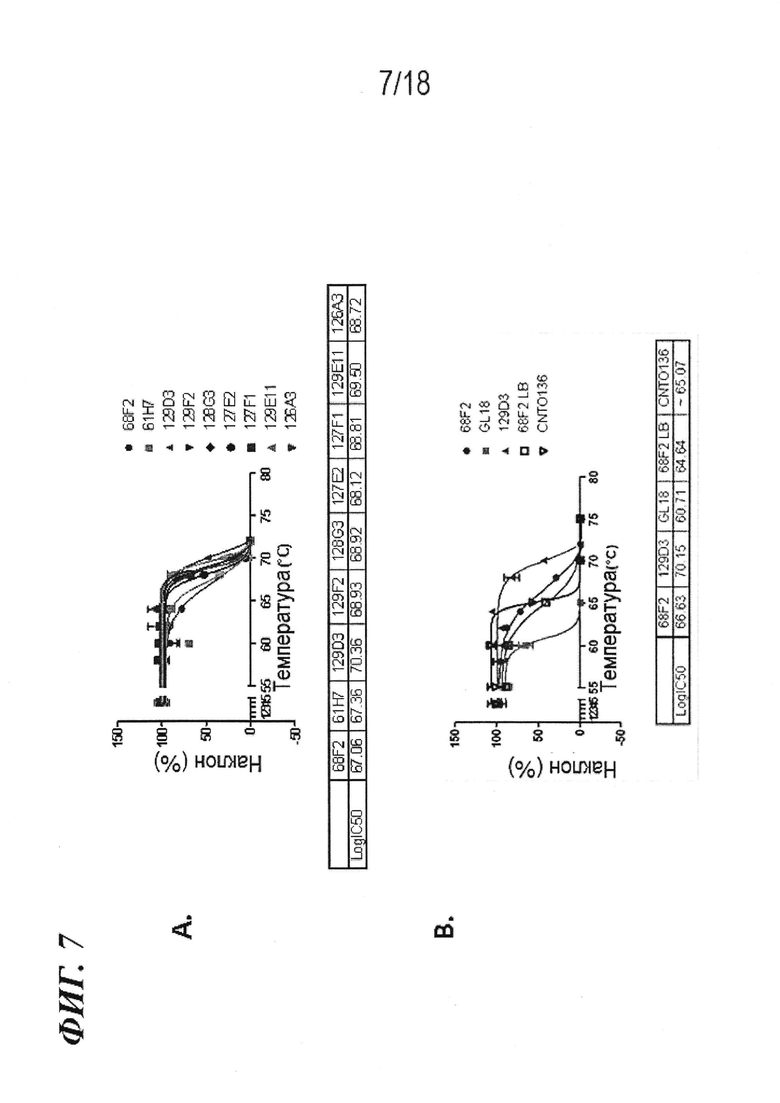
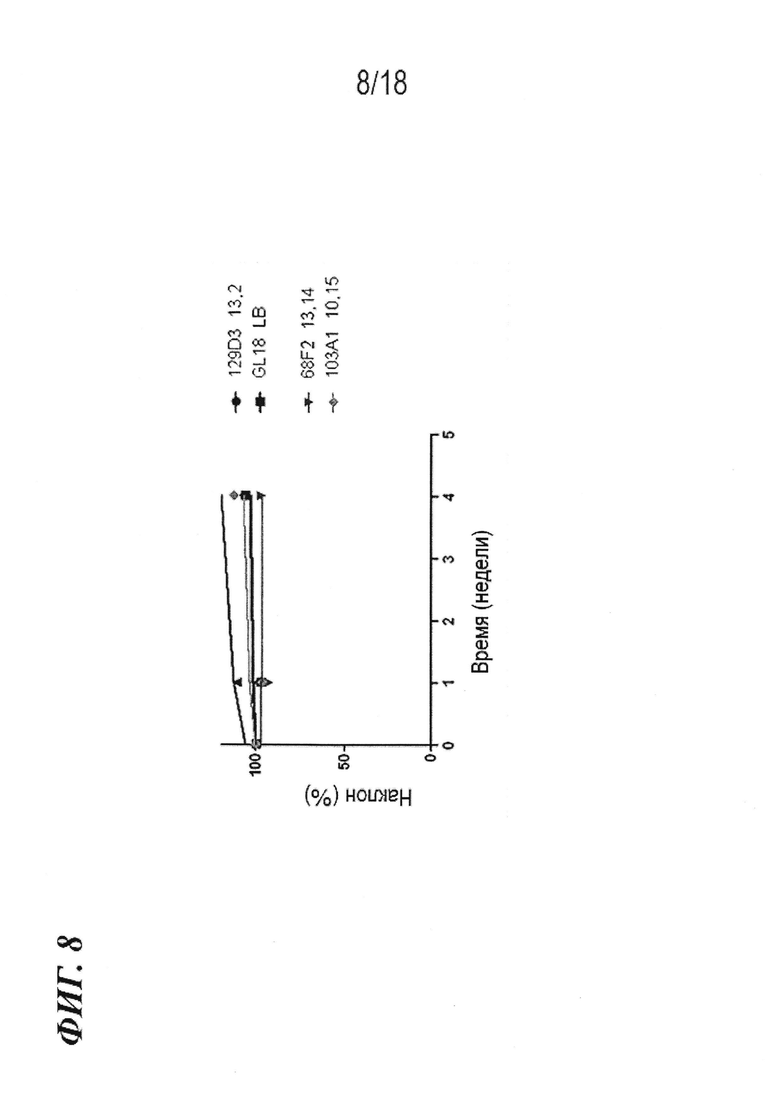

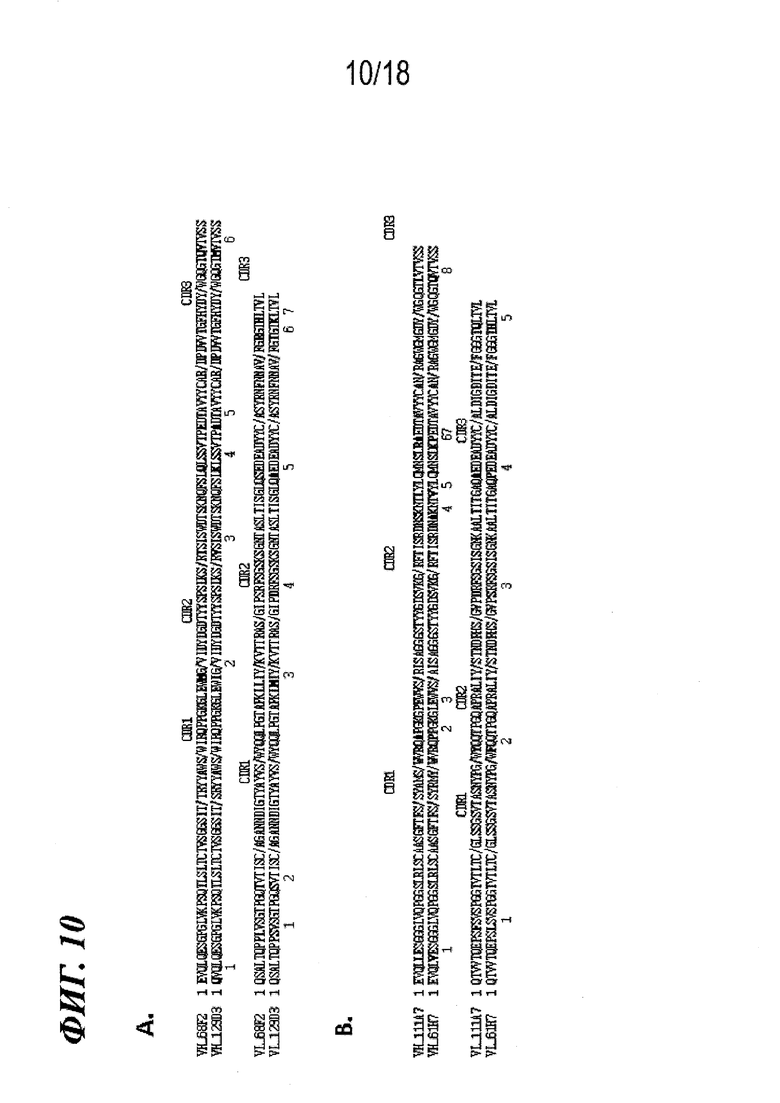
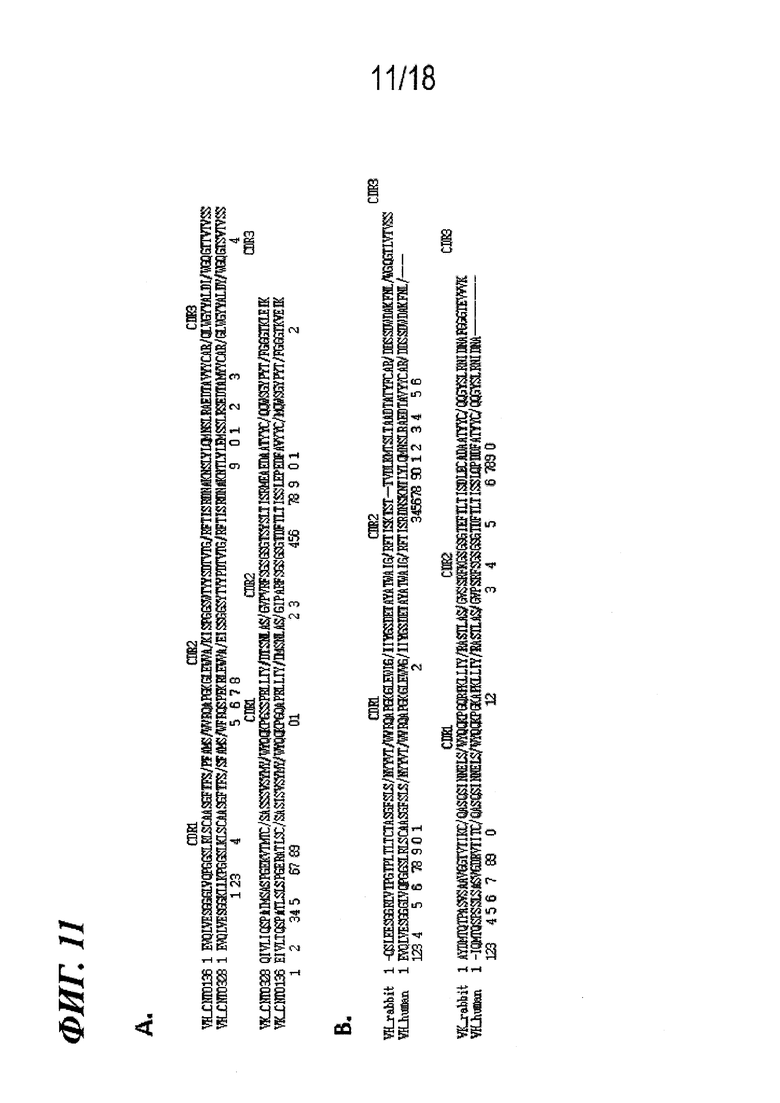
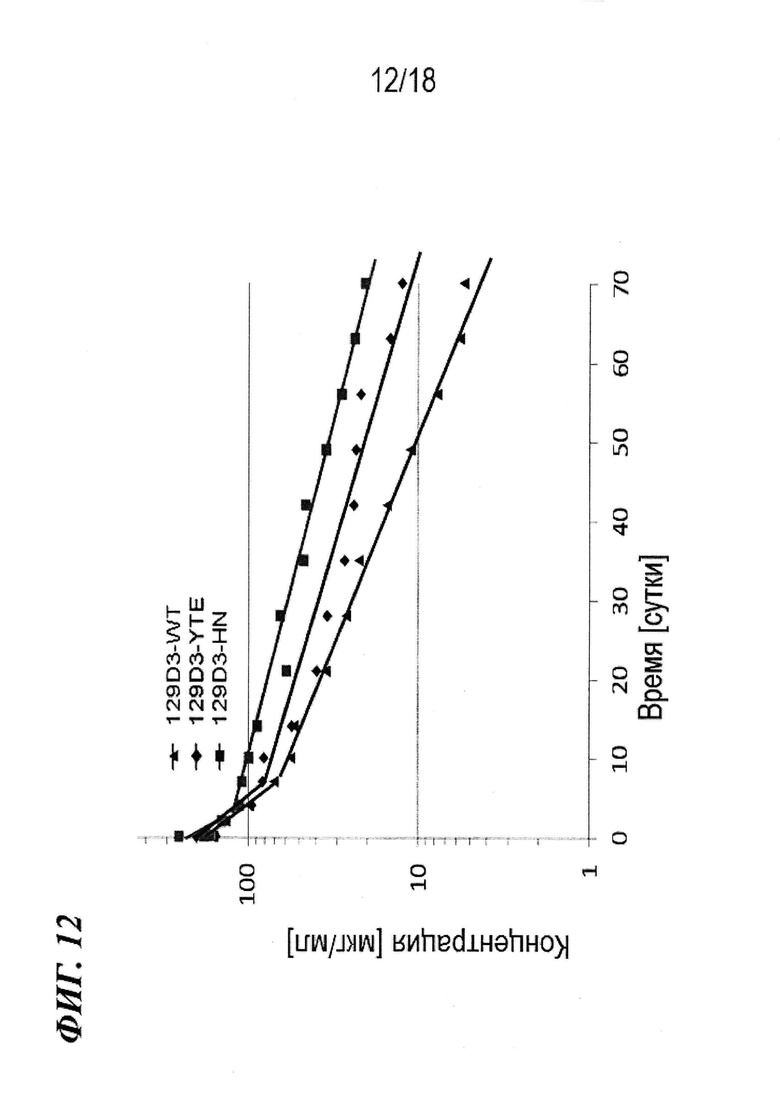
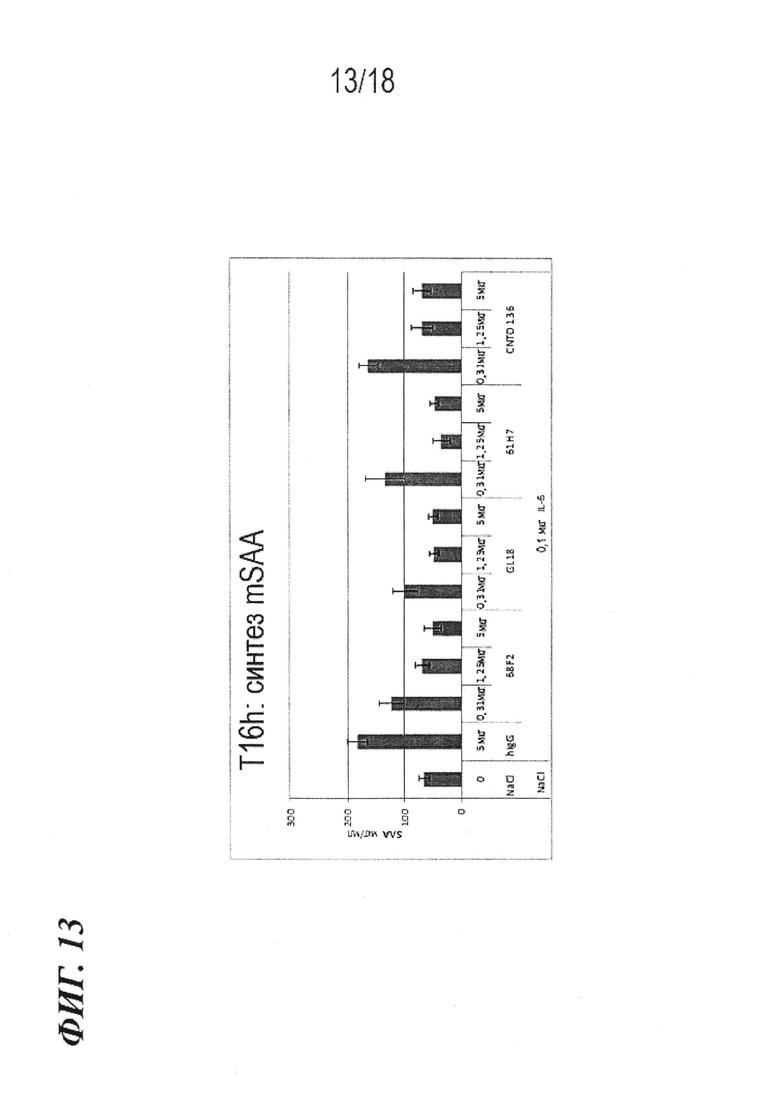

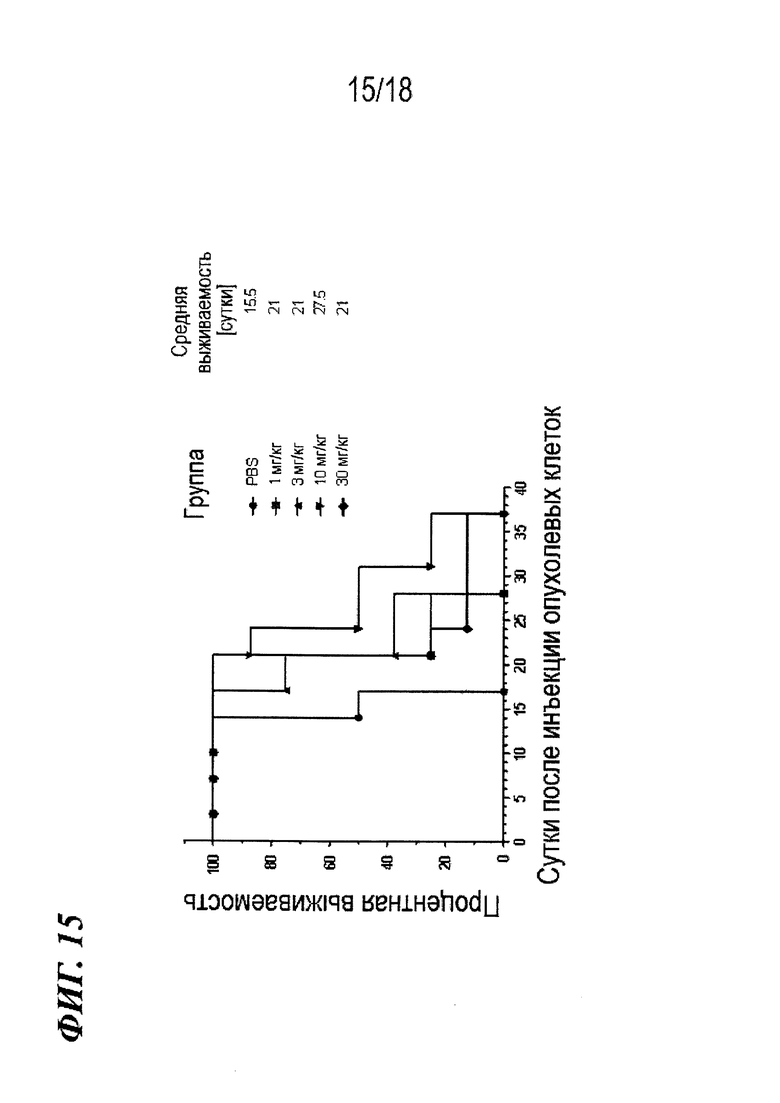
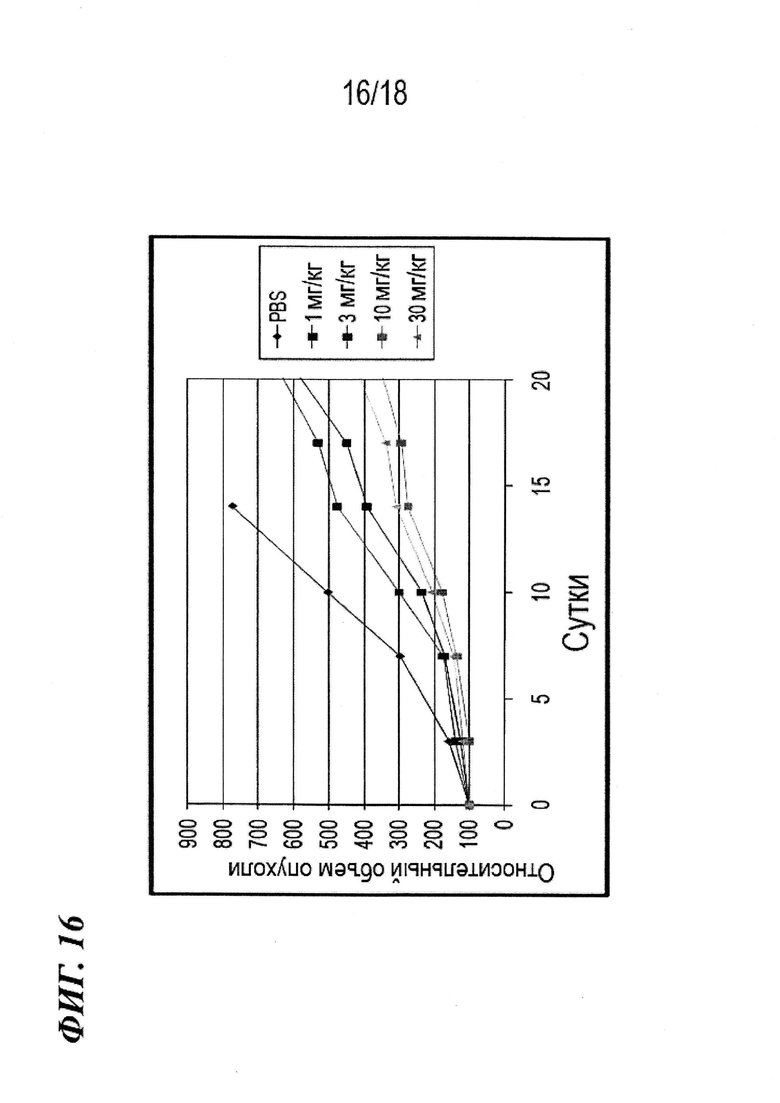
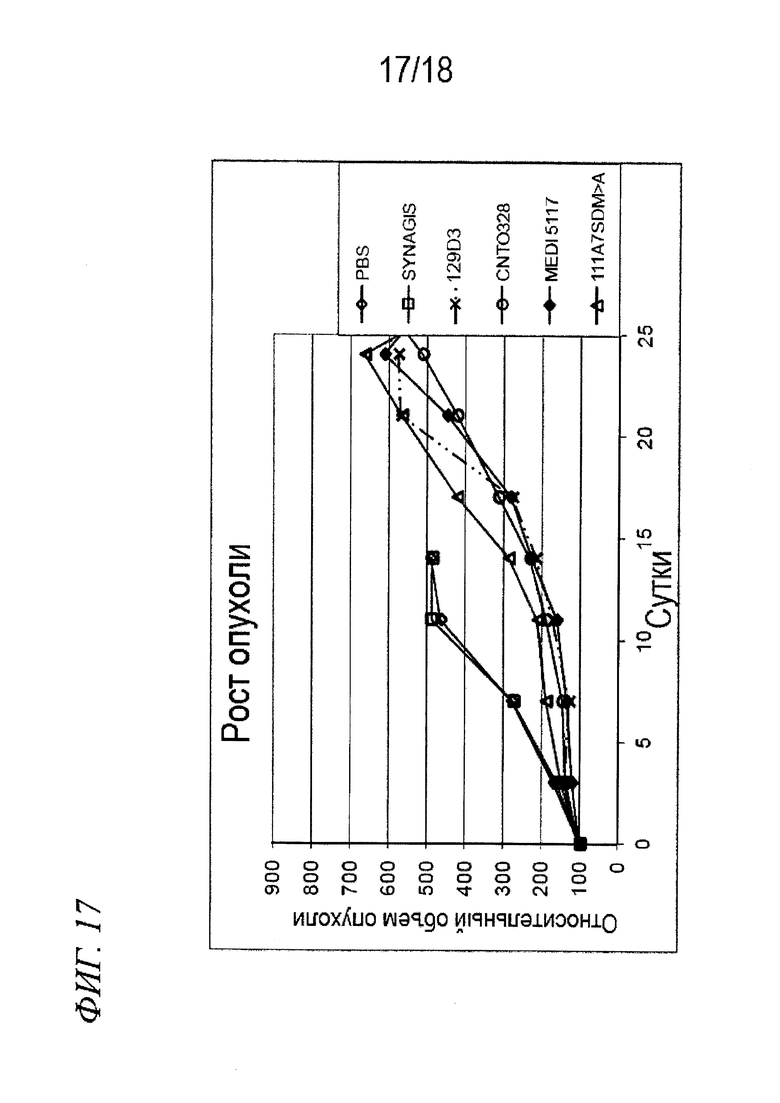
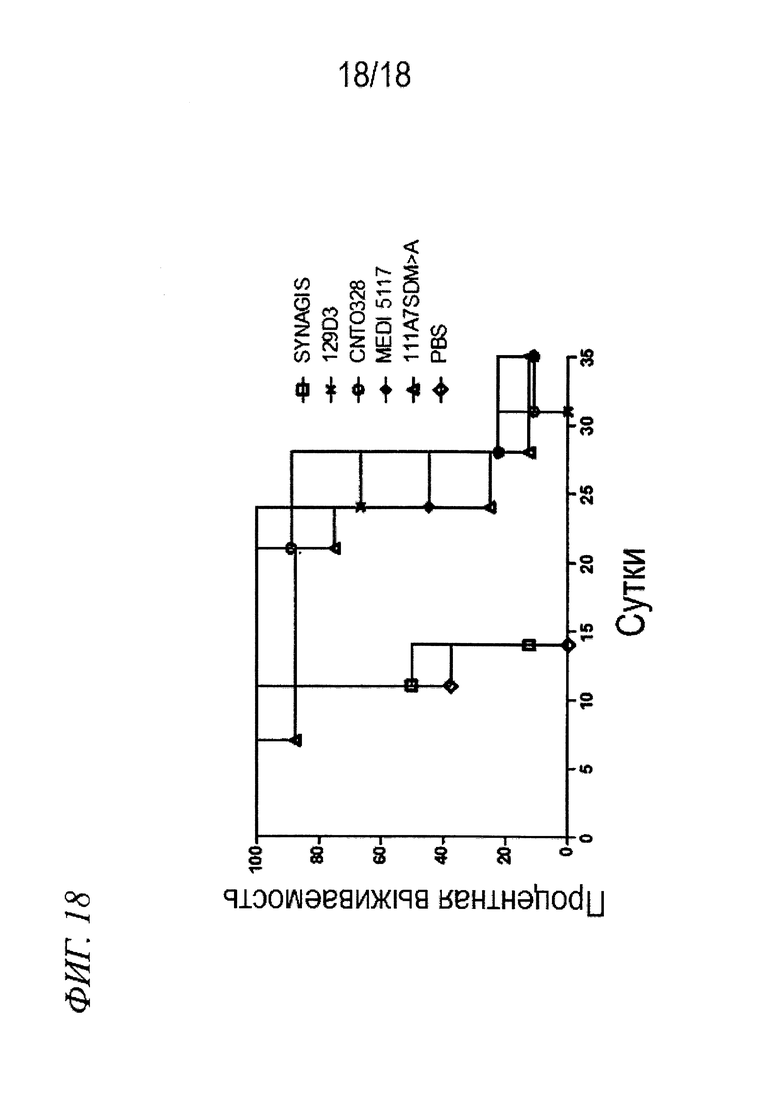
Изобретение относится к области биохимии, в частности к антителу или его антигенсвязывающему фрагменту, специфически связывающемуся с IL-6, а также к фармацевтической композиции, его содержащей. Также раскрыт способ лечения связанного с IL-6 заболевания или нарушения с использованием вышеуказанной композиции. Изобретение также относится к выделенной нуклеиновой кислоте, кодирующей вышеуказанное антитело или его антигенсвязывающий фрагмент, а также к вектору и клетке-хозяину, содержащим вышеуказанную нуклеиновую кислоту. Изобретение позволяет эффективно осуществлять лечение связанного с IL-6 заболевания или нарушения. 8 н. и 4 з.п. ф-лы, 18 ил., 26 табл., 16 пр.
1. Антитело или его антигенсвязывающий фрагмент, которое специфически связывается с IL-6, содержащее
VH-домен, содержащий
аминокислотную последовательность HCDR3 в соответствии с SEQ ID NO:497;
аминокислотную последовательность HCDR2 в соответствии с SEQ ID NO:501; и
аминокислотную последовательность HCDR1 в соответствии с SEQ ID NO:508;
и
VL-домен, содержащий
аминокислотную последовательность LCDR3 в соответствии с SEQ ID NO:513;
аминокислотную последовательность LCDR2 в соответствии с SEQ ID NO:525; и
аминокислотную последовательность LCDR1 в соответствии с SEQ ID NO:536.
2. Антитело или его антигенсвязывающий фрагмент, которое специфически связывается с IL-6, содержащее
VH-домен, содержащий
аминокислотную последовательность HCDR3 в соответствии с SEQ ID NO:543;
аминокислотную последовательность HCDR2 в соответствии с SEQ ID NO:545; и
аминокислотную последовательность HCDR1 в соответствии с SEQ ID NO:555;
и
VL-домен, содержащий
аминокислотную последовательность LCDR3 в соответствии с SEQ ID NO:563;
аминокислотную последовательность LCDR2 в соответствии с SEQ ID NO:564; и
аминокислотную последовательность LCDR1 в соответствии с SEQ ID NO:565.
3. Антитело или его антигенсвязывающий фрагмент по п.1, в котором:
VH-домен содержит аминокислотную последовательность в соответствии с SEQ ID NO:152, и
VL-домен содержит аминокислотную последовательность в соответствии с SEQ ID NO:416.
4. Антитело или его антигенсвязывающий фрагмент по п.2, в котором:
VH-домен содержит аминокислотную последовательность в соответствии с SEQ ID NO:86, и
VL-домен содержит аминокислотную последовательность в соответствии с SEQ ID NO:350.
5. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-4, которое:
a) ингибирует связывание IL-6 с рецептором IL-6;
b) ингибирует связывание gp130 с рецептором IL-6;
c) специфично связывается с IL-6 человека и яванского макака;
d) содержит по меньшей мере одну CDR из антитела животного семейства верблюжьих, которое специфично связывается с IL-6;
e) характеризуется показателем EpiBase® менее чем приблизительно 10,0;
f) экспрессируется на уровне по меньшей мере 20 мг/мл при временной экспрессии в клетке HEK293;
g) обладает температурой плавления (Tm) более 65°C;
h) ингибирует индуцируемую IL-6 пролиферацию клеток гибридомы B9 с IC50 менее 0,1 пМ;
i) связывается с IL-6 человека с константой диссоциации (koff, измеренная с помощью поверхностного плазмонного резонанса) менее 2×10-5 с-1;
j) представляет собой преобразованный в эмбриональный тип вариант родительского антитела животного семейства верблюжьих, причем указанный преобразованный в эмбриональный тип вариант имеет более высокую температуру плавления, чем родительское антитело животного семейства верблюжьих;
k) содержит по меньшей мере одну CDR из общепринятого антитела ламы без последующего созревания аффинности;
l) имеет время полужизни в сыворотке по меньшей мере 9 суток, предпочтительно, по меньшей мере 15 суток, при введении внутривенно яванскому макаку в формате нативного Fc IgG1; или
m) представляет собой преобразованный в эмбриональный тип вариант родительского нечеловеческого антитела, где антитело или антигенсвязывающий фрагмент (i) содержит VH- и VL-домен, и где один или оба из VH- и VL-доменов связывающей молекулы содержат всего от 1 до 10 аминокислотных замен в каркасных областях по сравнению с соответствующими VH- и VL-доменами родительского не являющегося человеческим антитела; и/или (ii) содержит VH- и VL-домен, где один или оба из VH-домена или VL-домена антитела или антигенсвязывающего фрагмента идентичны по последовательности на 90% или более одному или нескольким соответствующим доменам VH или VL человека в каркасных областях FR1, FR2, FR3 и FR4.
6. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-4, содержащее Fc-домен человека с двойной мутацией H433K/N434F.
7. Фармацевтическая композиция для лечения заболевания или нарушения, связанного с IL-6, содержащая антитело или антигенсвязывающий фрагмент по любому из предшествующих пунктов в эффективном количестве и один или несколько фармацевтически приемлемых носителей.
8. Способ лечения связанного с IL-6 заболевания или нарушения, включающий введение индивидууму, нуждающемуся в его лечении, эффективного количества фармацевтической композиции по п.7.
9. Применение фармацевтической композиции по п.7 в способе лечения заболевания или нарушения, связанного с IL-6, причем способ включает введение субъекту, нуждающемуся в лечении, эффективного количества фармацевтической композиции; где заболевание или расстройство, связанное с IL-6, включает: воспалительные аутоиммунные заболевания, такие как ревматоидный артрит (RA), артропатия со спондилезом, системная красная волчанка (SLE), воспалительное заболевание кишечника (IBD) и болезнь Кастлмана, рак предстательной железы, диффузная крупноклеточная лимфома, множественная миелома, почечно-клеточный рак, обусловленная опухолью анорексия, мукозит полости рта и кахексия.
10. Выделенная нуклеиновая кислота, кодирующая антитело или антигенсвязывающий фрагмент по любому из пп.1-4.
11. Рекомбинантный вектор экспрессии, содержащий нуклеиновую кислоту по п.10.
12. Клетка-хозяин для продукции антитела или антиген-связывающего фрагмента по любому из пп.1-4, содержащая рекомбинантный вектор экспрессии по п.11.
| WO 2004039826 A1, 13.05.2004 | |||
| WO 2006119115 A2, 09.11.2006 | |||
| WO 2007104529 A2, 20.09.2007 | |||
| WO 2009155180 A1, 23.12.2009 | |||
| Устройство для перемещения инструмента по поверхности сложной конфигурации | 1980 |
|
SU870029A1 |

