Область техники
Изобретение относится к области биохимии, в частности к рекомбинантному иммунотоксину, который может быть использован в медицине для терапии рассеянного склероза, способу его получения и применения. Заявляемый иммунотоксин представляет собой полипептидную цепь, которая состоит из вектора доставки (фрагмента миелин-олигодендроцитного гликопротеина) и активной части (А-цепи растительного токсина вискумина).
Уровень техники
Лечение рассеянного склероза (PC) остается одной из наиболее серьезных проблем современной медицины. Если ранее терапевтическая помощь пациентам с PC сводилась лишь к проведению симптоматического лечения, то к настоящему времени появились пути воздействия на течение болезни. К сожалению, пока не разработан препарат, который способен полностью остановить развитие заболевания. Современное лечение PC включает в себя применение препаратов, "изменяющих течение PC" (ПИТРС). Имеющиеся в распоряжении врачей препараты снижают частоту и тяжесть обострений, а также замедляют темпы накопления неврологического дефицита, что подтверждается данными магнитно-резонансной томографии головного и спинного мозга [J van der Star В, YS Vogel D, Kipp M, Puentes F, Baker D, Amor S. In vitro and in vivo models of multiple sclerosis. CNS & Neurological Disorders-Drug Targets (Formerly Current Drug Targets-CNS & Neurological Disorders). 2012 Aug 1; 11(5):570-88; Freedman, M.S., et al., Efficacy of disease-modifying therapies in relapsing remitting multiple sclerosis: a systematic comparison. Eur Neurol, 2008. 60(1): p. 1-11; Fisher E, Nakamura K, Lee JC, You X, Sperling B, Rudick RA. Effect of intramuscular interferon beta-la on gray matter atrophy in relapsing- remitting multiple sclerosis: A retrospective analysis. Multiple Sclerosis Journal. 2015 Aug 3:1352458515599072]. К ПИТРС, разрешенным к применению, относят препараты: интерферон-бета (ИНФб), глатирамера ацетат (Copaxone), митоксонтрон, финголимод, натализумаб, даклизумаб, терифлюномид, диметилфумарат, алемтузумаб [Loma, I., & Heyman, R. (2011). Multiple Sclerosis: Pathogenesis and Treatment. Current Neuropharmacology, 9(3), 409-416]. Первые два препарата наиболее активно используются при ремитирующем течении PC, однако они оказываются эффективными не во всех случаях, и тогда переходят к использованию более новых и сильных, но обладающих побочными эффектами, препаратов.
Для терапии PC на разной стадии разработки находятся как неспецифические препараты, так и препараты более направленного действия. К неспецифическим можно отнести следующие препараты, прошедшие разные стадии клинических испытаний: 1) Fingolimod (ингибитор рецепторов сфингозин-1-фосфата; останавливает выход Т- и В-клеток из вторичных лимфоидных органов, предотвращая миграцию лимфоцитов в центральную нервную систему) [Brinkmann, V., Sphingosine 1-phosphate receptors in health and disease: mechanistic insights from gene deletion studies and reverse pharmacology. Pharmacol Ther, 2007. 115(1): p. 84-105]; 2) Teriflunomide (ингибирует пролиферацию T-клеток) [Korn, Т., et al., Suppression of experimental autoimmune neuritis by leflunomide. Brain, 2001. 124(Pt 9): p. 1791-802]; 3) Tecfidera (другое название - BG-12; содержит диметилфумарат; является иммуномодулятором с неясным механизмом действия; позволяет снизить частоту рецидивов при рецидивирующе-ремиттирующей форме PC в 2 раза согласно данным III стадии клинических испытаний) [Robert J. Fox et al. Placebo-controlled Phase 3 study of oral BG-12 or glatiramer in multiple sclerosis. N Engl J Med, 2012. 367: p. 1087-1097]; 4) Roquinimex (иммуностимулятор; повышает активность натуральных киллеров, снижает ангиогенез и продукцию TNFα; препятствует формированию новых очагов воспаления в головном мозге, что было подтверждено во время II и II стадий клинических испытаний) [Wolinsky, J.S., et al., Linomide in relapsing and secondary progressive MS: part II: MRI results. MRI Analysis Center of the University of Texas-Houston, Health Science Center, and the North American Linomide Investigators. Neurology, 2000. 54(9): p. 1734-41]; 5) Laquinimod (структурный аналог Roquinimex; блокирует способность T-клеток и макрофагов перемещаться через гемато-энцефалический барьер, а также переключает фенотип с Тх-2 на Тх-3) [Yang, J.S., et al., Laquinimod (ABR-215062) suppresses the development of experimental autoimmune encephalomyelitis, modulates the Th1/Th2 balance and induces the Th3 cytokine TGF-beta in Lewis rats. J Neuroimmunol, 2004. 156(1-2): p. 3-9]; 6) Cladribine (ингибитор дезоксицитидин киназы, влияющий на ДНК репарацию и опосредованно индуцирующий гибель лимфоцитов; прошел III стадию клинических испытаний для терапии PC, после чего его внедрение было остановлено) [Thone, J. and G. Ellrichmann, Oral available agents in the treatment of relapsing remitting multiple sclerosis: an overview of merits and culprits. Drug Healthc Patient Saf, 2013. 5: p.37-47]. Из-за неспецифического действия данные препараты обладают различными серьезными побочными эффектами.
Альтернативным подходом в терапии PC является применение препаратов моноклональных антител, направленных против определенных молекул, участвующих в патогенезе заболевания [Buttmann, М. and P. Rieckmann, Treating multiple sclerosis with monoclonal antibodies. Expert Rev Neurother, 2008. 8(3): p. 433-55]. В клинической практике применяется препарат Natalizumab, представляющий собой моноклональное антитело к интегринам α4β1 [Steinman, L. (2012). The discovery of natalizumab, a potent therapeutic for multiple sclerosis. The Journal of Cell Biology, 199(3), 413-416]. Он блокирует адгезию мононуклеарных клеток на поверхности стромальных эндотелиальных клеток, что предотвращает миграцию аутореактивных лимфоцитов через гемато-энцефалический барьер и в воспаленные ткани. Эффективность этого препарата при лечении PC оказалась выше, чем эффективность интерферона бета [Ransohoff, R.M., Natalizumab for multiple sclerosis. N Engl J Med, 2007. 356(25): p. 2622-9].
Ритуксимаб (Rituximab) предназначается для терапии различных аутоиммунных заболеваний, включая PC. Rituximab - антитело класса IgG, специфичное к CD20 (мембранный белок, присутствующий на поверхности всех зрелых В-клеток). Препарат индуцирует апоптоз клеток, несущих CD20, что приводит к системному влиянию на продукцию антител, цитокиновую сеть, презентацию антигена В-клетками, активацию Т-клеток и макрофагов [Cross, А.Н., et al., Rituximab reduces В cells and T cells in cerebrospinal fluid of multiple sclerosis patients. J Neuroimmunol, 2006. 180(1-2): p. 63-70; Duddy, M. and A. Bar-Or, B-cells in multiple sclerosis. Int MS J, 2006. 13(3): p. 84-90; Hauser, S.L., et al., B-cell depletion with rituximab in relapsing-remitting multiple sclerosis. N Engl J Med, 2008. 358(7): p. 676-88; Uchida, J., et al., The innate mononuclear phagocyte network depletes В lymphocytes through Fc receptor-dependent mechanisms during anti-CD20 antibody immunotherapy. J Exp Med, 2004. 199(12): p. 1659-69; Liossis, S.N. and P.P. Sfikakis, Rituximab-induced В cell depletion in autoimmune diseases: potential effects on T cells. Clin Immunol, 2008. 127(3): p. 280-5]. Данные полученные во время второй фазы клинических испытаний показали, что введение 2000 мг препарата Rituximab значительно снижает количество очагов демиелинизации в течение 48 недель в сравнении с контрольной группой [Bar-Or, A., et al., Rituximab in relapsing-remitting multiple sclerosis: a 72-week, open-label, phase I trial. Ann Neurol, 2008. 63(3): p. 395-400]. Серьезным недостатком Rituximab является неспецифическое удаление подавляющей части популяции циркулирующих В-клеток.
Несмотря на большое количество существующих подходов к терапии PC, идея направленного подавления популяции аутореактивных лимфоцитов остается до сих пор актуальной и востребованной. Применяя подход селективной элиминации аутореактивных В-клеток, можно избежать большинства описанных ранее проблем [O'Connor PW, Oh J. Disease-modifying agents in multiple sclerosis. Handb Clin Neurol. 2014 Feb 5; 122:465-501], характерных для неспецифической терапии аутоиммунных заболеваний. Идеальными кандидатами на роль молекул адресного воздействия являются иммунотоксины, большой опыт применения которых в раковой терапии создает существенную базу для создания терапевтических препаратов аутоиммунных заболеваний [Arkfeld, D.G., The potential utility of В cell-directed biologic therapy in autoimmune diseases. Rheumatol Int, 2008. 28(3): p. 205-15; Kreitman, R.J., Immunotoxins for targeted cancer therapy. AAPS J, 2006. 8(3): p. Е532-51].
Молекулы иммунотоксинов состоят из двух ключевых фрагментов: сигнальной последовательности адресной доставки к клетке-мишени и действующей части, приводящей к гибели клетки. В качестве сигнальной последовательности могут выступать: антитела, Fab фрагменты антител, гормоны, цитокины, клеточные факторы, лиганды поверхностных Т- или В-клеточных рецепторов [Zocher, М., et al., Specific depletion of autoreactive В lymphocytes by a recombinant fusion protein in vitro and in vivo. Int Immunol, 2003. 15(7): p. 789-96; Frankel, A.E., Increased sophistication of immunotoxins. Clin Cancer Res, 2002. 8(4): p. 942-4]. Практически все иммунотоксины имеют в своем составе фрагменты антител, за исключением препарата ONTAK, который в качестве домена, отвечающего за связывание с клетками-мишенями, включает молекулу интерлейкина-2 человека. В качестве активного компонента, как правило, используют бактериальные (например, дифтерийный токсин, псевдомонадный экзотоксин А) или растительные (например, рицин) токсины.
Механизм действия иммунотоксинов заключается в следующем: вначале происходит связывание иммунотоксина со специфическим рецептором на поверхности клеток-мишеней, затем посредством рецепторно-опосредованного эндоцитоза иммунотоксин попадает внутрь клетки, после чего следует внутриклеточный транспорт и взаимодействие токсина с мишенью. В случае дифтерийного токсина и псевдомонадного экзотоксина А токсическое действие основано на модификации фактора элонгации эукариот (eEF-2), что приводит к прекращению синтеза белка. Рибосоминактивирующие белки, к которым относится рицин, также блокируют синтез белков, однако воздействуют непосредственно на рибосомы. Несмотря на различия в каталитическом механизме используемых токсинов, проникновение иммунотоксинов в клетки происходит в целом одинаково [Srivastava S., Luqman S. Immune-o-toxins as the magic bullet for therapeutic purposes // Biomed. Res. Ther. 2015. Vol.2, №1. P. 169-183;  M., Sandvig K. Ricin and Ricin-Containing Immunotoxins: Insights into Intracellular Transport and Mechanism of action in Vitro // Antibodies. 2013. Vol.2, №2. P. 236-269; Becker N., Benhar I. Antibody-Based Immunotoxins for the Treatment of Cancer // Antibodies. 2012. Vol. 1, №3. P. 39-69; Pastan I. et al. Immunotoxin treatment of cancer // Annu. Rev. Med. 2007. Vol. 58. P. 221-237]. Именно первый этап (связывание с поверхностными рецепторами) обеспечивает специфичность действия иммунотоксинов и направленную элиминацию клеток-мишеней.
M., Sandvig K. Ricin and Ricin-Containing Immunotoxins: Insights into Intracellular Transport and Mechanism of action in Vitro // Antibodies. 2013. Vol.2, №2. P. 236-269; Becker N., Benhar I. Antibody-Based Immunotoxins for the Treatment of Cancer // Antibodies. 2012. Vol. 1, №3. P. 39-69; Pastan I. et al. Immunotoxin treatment of cancer // Annu. Rev. Med. 2007. Vol. 58. P. 221-237]. Именно первый этап (связывание с поверхностными рецепторами) обеспечивает специфичность действия иммунотоксинов и направленную элиминацию клеток-мишеней.
В международной заявке WO 8906968 раскрыт способ получения и применения для лечения аутоиммунных заболеваний иммунотоксина, состоящего из полноразмерного моноклонального антитела к мембранному белку CD5, присутствующему на 85-100% зрелых Т-лимфоцитов, и А-цепи рицина. Соединение двух компонентов осуществляется за счет химической конъюгации. К недостаткам описанного метода относится получение А-цепи рицина посредством выделения рицина из растительного источника (семян клещевины), восстановления дисульфидной связи между А- и В-цепями токсина и очистки А-цепи от В-цепи и полноразмерного рицина с помощью хроматографии. Такой способ получения А-цепи рицина требует особых мер безопасности при производстве (рицин относится к биологическому оружию и является крайне токсичным для человека) и тщательного контроля полноты очистки. Кроме того, рецептор CD5 присутствует на 20-30% В-клеток у здоровых людей, что обуславливает побочное действие описанного иммунотоксина.
В решении, представленном в европейской заявке ЕР365087А1, раскрыт способ получения иммунотоксинов для терапии или профилактики аутоиммунных заболеваний (в том числе, рассеянного склероза), представляющих собой конъюгаты пептидов, распознаваемых Т-лимфоцитами, и активной части (А-цепи рицина или комплексов хелатирующих агентов с радиоактивными изотопами 90Y, 153Sm, 43Sc, 57Cu, 186Rh, 188Rh, 212Bi, 211At, 103Pd). Для терапии рассеянного склероза предлагается использовать пептиды, соответствующие фрагментам основного белка миелина 1-16, 1-37, 59-74, 68-88, 89-169, 114-122. Недостатками описанного способа являются: сложность и дороговизна химического синтеза соответствующего специфического пептида; необходимость выделения А-цепи рицина из природного источника или синтез комплекса хелатора с радиоактивным изотопом (в обоих случаях требуются особые меры безопасности при производстве); потери полупродуктов при синтезе конъюгата специфической и активной части иммунотоксина.
Благодаря научному прогрессу в области генной инженерии белков в последнее время преобладает тенденция создания рекомбинантных иммунотоксинов из-за возможности получать более гомогенные препараты высокой степени чистоты. В описанном изобретении был применен данный подход.
Раскрытие изобретения
Задачей настоящего изобретение является разработка иммунотоксина для терапии рассеянного склероза и способа его получения.
Техническим результатом заявляемого изобретения является получение иммунотоксина для терапии рассеянного склероза, обладающего высокой специфичностью при низкой токсичности. В заявляемом изобретении активный компонент для терапии рассеянного склероза характеризуется избирательным действием на аутореактивные В-лимфоциты, не затрагивая здоровые клетки. Заявляемый иммунотоксин обладает низкой токсичностью, о чем свидетельствует отсутствие токсических эффектов в проведенных токсикологических исследованиях на животных моделях. Заявляемый способ получения позволяет получать заявляемый иммунотоксин, не прибегая к дополнительным мерам безопасности. Также заявляемый способ позволяет контролировать гомогенность состава на всех стадиях синтеза.
Поставленная задача решается иммунотоксином для терапии рассеянного склероза, представляющим собой рекомбинантный белок, включающий А-субъединицу вискумина и фрагмент миелин-олигодендроцитного гликопротеина, обеспечивающий связывание с клетками-мишенями, соединенные линкером для связывания упомянутых фрагментов, при этом А-субъединица вискумина представляет собой полипептид с последовательностью аминокислот, не менее чем на 95% гомологичную SEQ ID NO: 1, а фрагмент миелин-олигодендроцитного гликопротеина представляет собой пептид с последовательностью не менее чем на 90% гомологичную SEQ ID NO: 2. Предпочтительно, когда иммунотоксин представляет собой белок с последовательностью не менее чем на 95% гомологичную SEQ ID NO: 3.
Также поставленная задача решается рекомбинантной плазмидной ДНК для экспрессии в клетках Escherichia coli гена, кодирующего заявляемый иммунотоксин, представляющая собой коммерческий вектор, модифицированный фрагментом, кодирующим заявляемый иммунотоксин, включающий следующие фрагменты:
a. фрагмент XhoI-NcoI;
b. промотор бактериофага Т7;
c. терминатор РНК-полимеразы бактериофага Т7;
d. область начала репликации ColE1/рМВ1/pBR322/pUC;
e. область, кодирующую ген фермента, обеспечивающего устойчивость к антибиотику.
При этом в качестве коммерческого вектора предпочтительно использовать плазмиды pET-22b(+), рЕТ-26b(+), рЕТ-28а-с(+).
При использовании для модификации в качестве вектора плазмиды рЕТ-28b(+), модифицированный вектор pET-28b(+)_ML1A-MOG, имеет размер 6127 п. н., в котором участки имеют следующие координаты:
a. фрагмент XhoI-NdeI вектора рЕТ-28b(+);
b. промотор бактериофага Т7;
c. терминатор РНК-полимеразы бактериофага Т7;
d. область начала репликации ColE1/pMB1/pBR322/pUC;
e. область, кодирующую ген аминогликозид-З'-фосфотрансферазы, обеспечивающую устойчивость к антибиотику канамицину;
f. уникальные сайты узнавания рестрикционными эндонуклеазами: XhoI - 158, BamHI - 233, AscI - 468, AgeI - 686, SacII - 793, NdeI - 996.
Поставленная задача также решается штаммом Escherichia coli - продуцентом заявляемого иммунотоксина, трансформированного рекомбинантной плазмидной ДНК.
Также поставленная задача решается способ получения заявляемого иммунотоксина, заключающегося в том, что культивируют штамм-продуцент с получением биомассы, содержащей тельца включения, которую затем подвергают разрушению с помощью ультразвукового дезинтегратора с последующим выделением телец включения с помощью центрифугирования. Полученные тельца включения отмывают методом последовательного растворения примесей, с получением иммунотоксина в нерастворенной форме, который затем переводят в растворенную форму с помощью раствора хаотропного агента, после чего неактивный растворенный иммунотоксин подвергают ренатурации методом разведения, диафильтрации и концентрированию с помощью тангенциальной фильтрации. При этом разрушение биомассы с помощью ультразвукового дезинтегратора производят проводят 3-4 раза по 3 минуты с 2-минутными перерывами при частоте 22 кГц и охлаждении на льду, а тельца включения отделяют от растворимых компонентов клеток с помощью центрифугирования. Очищение телец включения проводят с помощью растворов, содержащих детергенты и хаотропные агенты, обеспечивающие удаление примесей ДНК, РНК, липидов и примесных белков. Перевод иммунотоксина из нерастворимой неактивной формы в растворимую неактивную форму проводят с помощью раствора, содержащего хаотропный агент, при этом что перевод (ренатурацию) проводят с помощью разведения в растворе, содержащем аргинин и имеющем кислое значение рН, затем проводят концентрирование активного иммунотоксина после ренатурации с помощью тангенциальной фильтрации.
Предпочтительно в качестве хаотропного агента использовать растворы мочевины, тиомочевины, гуанидина гидрохлорида, а в качестве детергента использовать додецилсульфат натрия, дезоксихолат, саркосил, твин 20, твин 80, тритон Х-100.
Поставленная задача также решается фармацевтической композицией для терапии рассеянного склероза, включающая в качестве действующего вещества заявляеиый иммунотоксин в терапевтически эффективном количестве и фармацевтически приемлемую основу.
Также поставленная задача решается способом терапии рассеянного склероза, заключающийся во внутривенном введении фармацевтической композиции, содержащей заявляемый иммунотоксин в терапевтически эффективном количестве.
Краткое описание чертежей
На фиг. 1 приведена физическая карта плазмиды pET-24d(+).
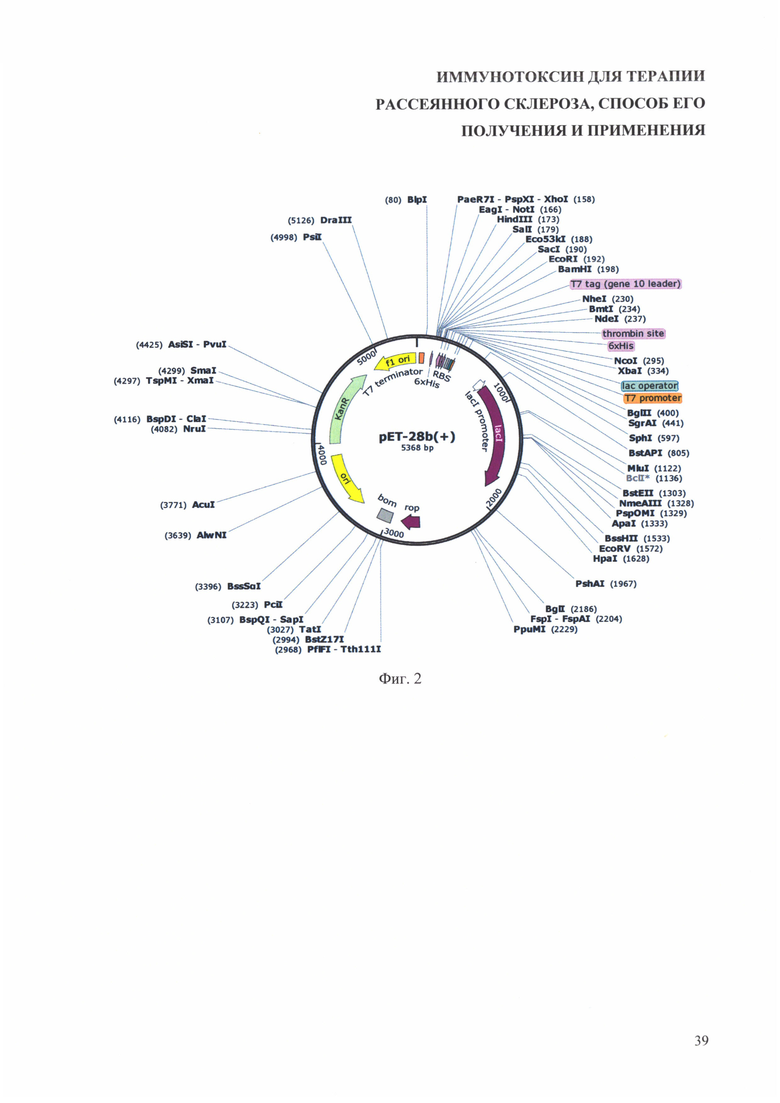
На фиг. 2 приведена физическая карта плазмиды рЕТ-28b(+).
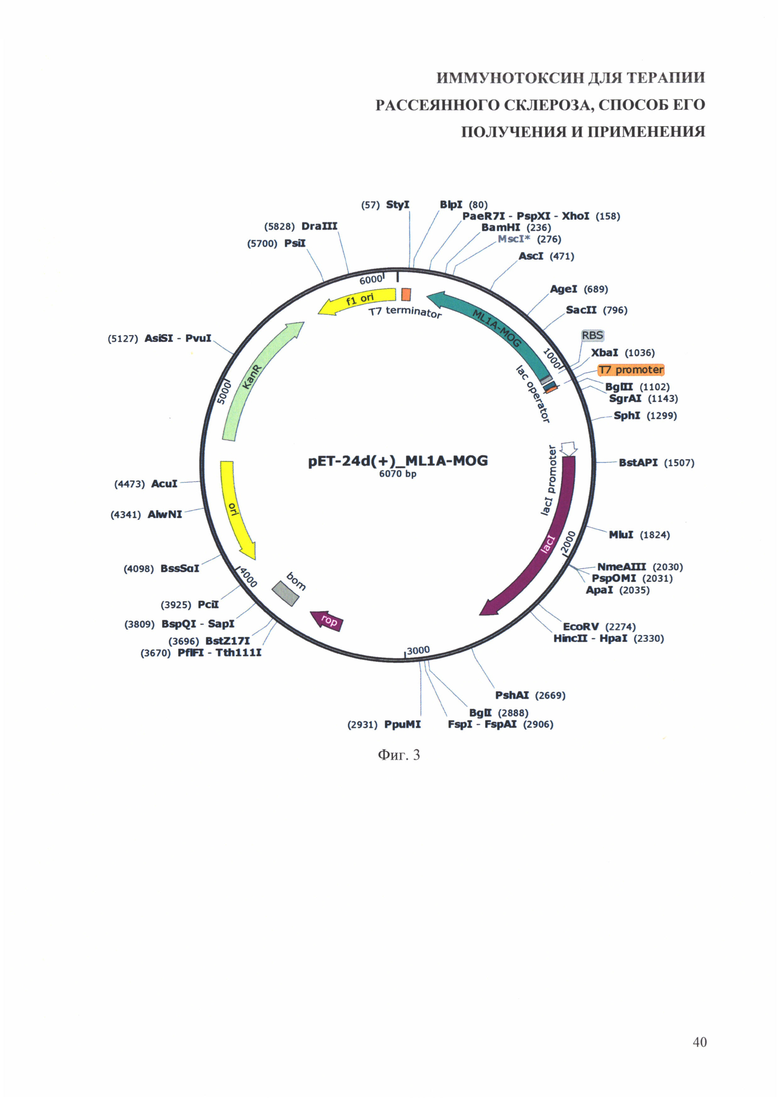
На фиг. 3 приведена физическая карта генетической конструкции рЕТ-24d(+)_ML1A-MOG.
На фиг. 4 приведена физическая карта генетической конструкции рЕТ-28b(+)_ML1A-MOG.
На фиг. 5 представлены примеры электрофореграмм субстанции иммунотоксина.
На фиг. 6 показана кривая плавления для субстанции иммунотоксина, полученной заявленным способом (зависимость производной интенсивности флуоресценции от температуры).
На фиг. 7 представлен график зависимости относительной интенсивности люминесценции (пропорциональной количеству экспрессированной люциферазы) от концентрации иммунотоксина, полученного заявленным способом.
На фиг. 8 представлен график развития признаков экспериментального аутоиммунного энцефаломиелита у экспериментальных животных после введения разных доз иммунотоксина, полученного заявленным способом.
На фиг. 9 представлена зависимость продолжительности жизни мышей с экспериментальным аутоиммунным энцефаломиелитом в экспериментальных группах после введения разных доз иммунотоксина, полученного заявленным способом.
Осуществление изобретения
Заявляемый иммунотоксин для терапии рассеянного склероза (ИТРС) в качестве вектора доставки содержит фрагмент миелин-олигодендроцитного гликопротеина (МОГ). Высокая специфичность и аффинность антител, присутствующих на мембране В-лимфоцитов, лежит в основе специфического распознавания МОГ-пептида и связывания ИТРС на поверхности аутореактивных В-клеток. Активным компонентом иммунотоксина является А-цепь растительного токсина вискумина. А-цепь вискумина обладает N-гликозидазной активностью и катализирует депуринизацию рибосом в эукариотических клетках (остатка аденина в положении 4324 в 28S рРНК большой субъединицы рибосомы), за счет чего прекращается синтез белка.
Синтез генетической конструкции (последовательность SEQ ID №3), кодирующей А-субъединицу вискумина (последовательность SEQ ID №1) и фрагмент МОГ, представляющий собой пептид с последовательностью SEQ ID №2, соединенные линкером, обеспечивающего их сшивку, которая может иметь минимальную длину (от двух аминокислот), осуществляли с использованием стандартных широко известных методов генной инженерии: клонирования, рестрикции, лигирования, трансформации [Green, M.R., Sambrook, J. (Ed.). Molecular Cloning: A Laboratory Manual (Fourth Edition). 2012. Cold Spring Harbor].
Для субклонирования полученной последовательности, кодирующей иммунотоксин используют любые коммерчески доступные плазмиды для экспрессии белков в Escherichia coli (Е. coli) под контролем промотора Т7, в которых в множественном сайте клонирования есть сайты рестрикции NcoI и XhoI, например, плазмиды pET-24d(+) (Novagen), pET-26b(+) (Novagen), pET-28a-c(+) (EMD Biosciences) (https://www.addgene.org/vector-database/query/?q_vdb=*+***%20vector_type:Bacterial). Ha фиг. 1 и фиг. 2 для примера приведены физические карты векторов pET-24d(+) и рЕТ-28b(+). На фиг. 3 и фиг. 4 приведены физические карты плазмид pET-24d(+) и рЕТ-28b(+), содержащие участок, кодирующий описанный иммунотоксин (на фигурах обозначен ML1A-MOG).
Плазмидой, несущей генетическую конструкцию, кодирующую описанный иммунотоксин, трансформируют штамм Е. coli, подходящий для экспрессии белков под контролем промотора бактериофага Т7 (экспрессирующий Т7 РНК-полимеразу, предпочтительно несущий ген lacIq или ген lysY, более предпочтительно - оба эти гена), например, Е. coli BL21(DE3) или Е. coli Lemo(DE3). После трансформации клетки Е. coli высевают на агаризованную питательную среду LB с антибиотиком, в зависимости от использованной плазмиды (например, канамицин или ампициллин) [Green, M.R., Sambrook, J. (Ed.). Molecular Cloning: A Laboratory Manual (Fourth Edition). 2012. Cold Spring Harbor].
Полученные единичные колонии трансформированных Е. coli культивируют в среде LB с антибиотиком, в зависимости от использованной плазмиды (например, канамицин или ампициллин), и полученную суспензию клеток Е. coli используют в качестве посевного материала (инокулята) для ферментера.
Для получения биомассы, содержащей ИТРС, используют автоиндуцируемую среду с добавлением антибиотика в зависимости от используемой плазмиды (например, канамицин или ампициллин). Для чего в подготовленный ферментер стерильно переносят посевной материал (среду), заполняя приблизительно 1/3 от объема ферментера (или проводят стерилизацию среды непосредственно в ферментере), асептически вносят инокулят через соответствующий порт (1/20-1/10 от объема питательной среды). Инкубируют биомассу в течение не менее чем 10 ч. После инкубации проводят проверку синтеза ИТРС иммуноферментным анализом или с помощью аналитического электрофореза. Подтверждением наличия целевого белка служит детекция сигнала (например, поглощения при определенной длине волны), определенной интенсивности при постановке иммуноферментного анализа либо обнаружение полосы, соответствующей белку определенной массы, на геле после его окрашивания. При использовании любого метода используется контрольный образец, содержащий известное количество очищенного иммунотоксина.
При проведении электрофореза в денатурирующих условиях основное пятно, соответствующее ИТРС, должно мигрировать как белок с массой 28±1 кДа. На основании предварительно построенной калибровочной кривой для площади пятна на электрофореграмме в зависимости от количества нанесенного белка ИТРС определяют его количество в анализируемых пробах.
Полученную биомассу от культуральной жидкости отделяют центрифугированием или фильтрацией. Супернатанты (культуральную жидкость) удаляют, а полученный осадок клеток до использования в следующих стадиях хранят при -20°С.
ИТРС экспрессируется в клетках Е. coli в нерастворимой форме (в виде телец включения). Для выделения телец включения биомассу клеток разрушают с помощью ультразвука, после чего тельца включения отделяют центрифугированием (остаются в осадке) и очищают от различных примесей с помощью промывки растворами определенного состава, содержащими детергенты и хаотропные агенты, обеспечивающие удаление примесей ДНК, РНК, липидов и примесных белков.
В качестве хаотропного агента могут быть использованы растворы мочевины, тиомочевины, гуанидина гидрохлорида.
В качестве детергентов могут быть использованы додецилсульфат натрия, дезоксихолат, саркосил, твин 20, твин 80, тритон Х-100.
Для выделения и отмывки телец включения используют четыре раствора (А, Б, В, Г). Все растворы готовят в деионизированной воде (I тип) и используют свежеприготовленными.
В составе раствора А содержится 1,5-2,5 М NaCl, 1-5 мМ дитиотреитол, 5-15 мМ ЭДТА и 20-100 мМ Tris-HCl или фосфатный буфер, при этом рН раствора должен составлять 8,0-9,0. Раствор Б для отмывки телец включения включает 1-3% Triton X-100, 25-150 мМ NaCl, 1-5 мМ дитиотреитол, 20-100 мМ Tris-HCl или фосфатный буфер, при этом рН раствора должна составлять рН 8,0-9,0. Раствор В сходен по составу с раствором Б, но не содержит Triton Х-100. Раствор Г для отмывки телец включения от примесных белков включает 3,5-4,5 М мочевины, 1-5 мМ дитиотреитол, 20-100 мМ Tris-HCl или фосфатный буфер, при этом рН раствора должна составлять 8,0-9,0.
Для отмывки (очистки) телец включения к полученному ранее осадку клеток приливают раствор А из расчета 1:5-1:10 (объем биомассы:объем раствора А). Осадок клеток ресуспендируют. Клетки разрушают ультразвуком при частоте 22 кГц при охлаждении на льду во избежание перегрева суспензии. Обработку ультразвуком повторяют 3-4 раза по 3 мин с 2-минутными перерывами. Затем суспензию центрифугируют при 5000-10000 g и 15±5°С для отделения телец включения от растворимых примесей разрушенных клеток. Супернатант удаляют. Обработку осадка раствором А повторяют еще 1-2 раза.
Затем к полученному осадку добавляют раствор Б из расчета 1:5-1:10 (объем осадка:объем раствора Б). Осадок ресуспендируют и гомогенизируют ультразвуком на льду 2-3 раза по 3 мин с 2-минутными перерывами. Полученную суспензию центрифугируют при 5000-10000 g и 15±5°С для осаждения телец включения. Супернатант удаляют, а осадок еще 1-2 раза отмывают раствором Б.
К полученному осадку добавляют раствор В из расчета 1:5-1:10 (объем осадка:объем раствора В). Осадок ресуспендируют и гомогенизируют ультразвуком на льду по 2-3 мин с 2-минутными перерывами. Полученную суспензию центрифугируют при 5000-10000 g и 15±5°С для осаждения телец включения. Супернатант удаляют, а осадок отмывают раствором В еще 1-2 раза (или до прекращения образования пены).
Супернатант удаляют, к осадку добавляют раствор Г из расчета 1:1-1:2 (объем осадкаюбъем раствора В). Осадок ресуспендируют и гомогенизируют ультразвуком на льду 2-3 раза по 3 мин с 2-минутными перерывами. Полученную суспензию центрифугируют при 15±5°С. Супернатант удаляют, а осадок еще 1-2 раза отмывают раствором Г.
К полученному осадку добавляют раствор для солюбилизации телец включения из расчета 1:5-1:10 (объем осадка: объем раствора).
Раствор для солюбилизации включает 5,5-6,0 М гуанидина гидрохлорид, 1-5 мМ дитиотреитол, 25-75 мМ Na-ацетатный буфер, при этом рН раствора составляет 4,5-5,5. Раствор для солюбилизации телец включения используют свежеприготовленным.
После приливания раствора для солюбилизации к отмытым тельцам включения раствор необходимо перемешать для равномерного растворения осадка (необходимо полностью отделить осадок от стенки бутыли, не вспенивая раствор). Перемешивать можно вручную или с помощью магнитной мешалки. Растворы инкубируют при +2 - +8°С в течение 1,5 ч и при периодическом перемешивании.
Из полученного опалесцирующего прозрачного вязкого раствора отделяют нерастворенные включения. Для удаления нерастворившегося белка солюбилизат фильтруют с помощью установки вакуумной фильтрации через фильтр с размерами пор 0,22 мкм в стерильные бутыли. Раствор сразу же используют для проведения рефолдинга.
Для проведения рефолдинга полученный фильтрат разводят раствором для рефолдинга. Раствор для рефолдинга включает 0,45-0,5 М аргинина, 25-75 мМ Na-ацетатный буфера, при этом рН раствора должен составлять 4,5-5,5. Полученный раствор помещают на магнитную мешалку и при комнатной температуре перемешивают в течение времени, необходимого для сворачивания иммунотоксина (20-30 ч).
Для удаления дитиотреитола и гуанидина гидрохлорида проводят диафильтрацию. Раствор для диафильтрации содержит 0,45-0,5 М аргинина, 40-60 мМ Na-ацетатный буфер, при этом рН раствора должен составлять 5,0±0,2. Перед проведением диафильтрации раствор охлаждают до +2 - +8°С. Диафильтрацию проводят с использованием установки для тангенциальной фильтрации (материал фильтра - полиэфирсульфон, отсечка по массе - 10000 Да).
Полученный раствор концентрируют в 20-25 раз и стерилизуют. Концентрирование проводят методом тангенциальной фильтрации (материал фильтра - полиэфирсульфон, отсечка по массе - 10000 Да). Стерилизацию концентрата осуществляют методом фильтрации (материал фильтра - полиэфирсульфон, размер пор - 0,22 мкм).
Заявляемый иммунотоксин предлагается в качестве активного ингредиента для приготовления фармацевтической композиции для внутривенного введения при терапии рассеянного склероза, которая может содержать эффективное количество иммунотоксина (вплоть до 50 масс%, предпочтительно вплоть до 20 масс%, более предпочтительно вплоть до 10 масс%, еще более предпочтительно до 0,1-0,5 масс%), а остальное - дополнительные фармацевтически приемлемые добавки (эксципиенты). Под фармацевтически приемлемым эксципиентами подразумеваются применяемые в сфере фармацевтики разбавители, вспомогательные агенты (целевые добавки) и/или носители, стабилизаторы, регуляторы рН, буферы, антиоксиданты, консерванты, соединения, пролонгирующие действие препарата и др., например, выбранные из группы: вода, бензалкония хлорид, натрия гидрофосфат, натрия дигидрофосфата, натрия хлорид, натрия цитрат, натрия эдетат, натрия бисульфит, натрия тиосульфат, натрия карбонат, магния стеарат, маннитол, бензалкония хлорид, хлоргексидин, фенилэтиловый спирт, борная кислота, гиэтеллоза (гидроксиэтилцеллюлоза), гидроксиметилцеллюлоза, натрия гидроксид, лимонная кислота, полиэтиленгликоль, глицерин, декстроза, декстран 40, декстран 70, поливинилпирролидон, твин-80, крахмал, лактоза и прочие фармацевтически приемлемые носители и компоненты.
Для терапии рассеянного склероза фармацевтическую композицию, включающую иммунотоксин вводят внутривенно в терапевтически эффективном количестве, при этом частоту и длительность введения выбирают в зависимости от характера и степени выраженности заболевания.
Настоящее изобретение поясняется конкретными примерами выполнения, которые не являются единственно возможным, но наглядно демонстрирует возможность достижения требуемого технического результата.
1. Получение генно-инженерной конструкции.
В качестве основы для получения плазмиды, несущей ген ML1A с фрагментом гена MOG на С-конце (pET-28b(+)_ML1A-MOG), использовали плазмиду рЕТ-22b(+) с геном ML1A. Для переклонирования в новую плазмиду с отличающейся антибиотикорезистентностью ген ML1A амплифицировали с использованием следующих праймеров (подчеркиванием выделены сайты рестрикции):
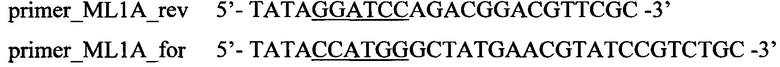
При этом на концы последовательности, кодирующей ML1A, вводили сайты рестрикции BamHI (GGATCC) и NcoI (CCATGG).
Фрагмент MOG амплифицировали с использованием праймеров:

При этом на концы последовательности, кодирующей фрагмент MOG, вводили сайты рестрикции XhoI (CTCGAG) и BamHI (GGATCC).
Полученные фрагменты длиной 778 и 92 п. н. очистили от примесей с помощью электрофореза в 2% агарозном геле с последующей экстракцией из геля с помощью набора innuPREP Gel Extraction Kit.
Для наработки достаточного количества матрицы плазмида рЕТ-28b(+) была трансформирована в штамм Е. coli DH5α. Трансформированные клетки высевали на агаризованную питательную среду LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl, 2% агар-агара) с добавлением 32 мкг/мл канамицина. Полученные колонии переносили в пробирки с 5 мл питательной среды LB с добавлением 32 мкг/мл канамицина. После инкубации в течение ночи (16 ч) при 37°С и перемешивании со скоростью 180 об/мин, клетки осадили центрифугированием в течение 5 мин при 5000 g. После этого с помощью набора irmuPREP Plasmid Mini Kit из них выделяли плазмиду рЕТ-28b(+).
Очищенные фрагменты, кодирующие ML1A и фрагмент MOG (insert_ML1A и insert_MOG), а также плазмида рЕТ-28b(+) подвергали рестрикции соответствующими ферментами: insert_ML1A - BamHI и NcoI, insert_MOG - BamHI и XhoI, pET-28b(+) -NcoI и XhoI. Затем все фрагменты лигировали с помощью ДНК-лигазы бактериофага Т4 (соотношение матрица: insert_ML1A: insert_MOG=1:2:2). Полученная лигазная смесь была трансформирована в штамм Е. coli DH5α. Трансформированные клетки высевали на агаризованную питательную среду LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl, 2% агар-агара) с добавлением 32 мкг/мл канамицина. В результате получили 23 колонии. Четыре колонии пересеяли в пробирки с 5 мл питательной среды LB с добавлением 32 мкг/мл канамицина. После инкубации в течение ночи при 37°С и перемешивании со скоростью 180 об/мин, клетки осадили центрифугированием (5 мин, 5000 g). После этого с помощью набора innuPREP Plasmid Mini Kit из них выделили плазмиды. Так как концентрация плазмид была недостаточна для проведения секвенирования, была поставлена дополнительная ПЦР с использованием универсальных праймеров T7_for (5'-TAATACGACTCACTATAGGG-3') и T7_rev (5'-GCTAGTTATTGCTCAGCGG-3'). Полученные фрагменты массой 1022 п. н. очищали в агарозном геле и передавали на секвенирование. Таким образом, в результате проведенных генно-инженерных работ получили плазмиду, несущую ген целевого белка ML1A-MOG. Полученной плазмидой трансформировали штамм Е. coli BL21 Star(DE3).
Затем с чашек Петри отбирали 8 единичных колоний Е. coli BL21 Star(DE3)_ML1A-MOG. Данные колонии в стерильных условиях переносили в пробирки с 5 мл питательной среды LB с канамицином (32 мкг/мл). Пробирки инкубировали в течение ночи (16 часов) при 34°С и 180 об/мин. Затем к полученной суспензии клеток добавляли 1,35 мл стерильного 50% раствора глицерина и перемешивали. Аликвоты по 200 мкл переносили в криопробирки и замораживали при -70°С.
Для каждого клона провели пробную экспрессию целевого белка: 25 мкл музейной культуры переносили в пробирку с 10 мл среды LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl) с канамицином (32 мкг/мл) и культивировали в течение 6 ч при 37°С и 180 об/мин, после чего добавляли ИПТГ до конечной концентрации 0,5 мМ для индукции экспрессии ML1A-MOG и инкубировали в течение 3 ч при 34°С и 130 об/мин. Аналитический электрофорез проб, отобранных до индукции экспрессии и после, показал, что ML1A-MOG продуцировался во всех клонах.
2. Выращивание посевного материала
Приготовление и стерилизация питательной среды
В стакан емкостью 500 мл вносили 210 мл воды деионизированной, добавляют навески дрожжевого экстракта (1,5 г), триптона (3,0 г), натрия хлористого (3,0 г) и перемешивают до полного растворения компонентов. Объем раствора доводили до 300 мл водой деионизированной. Приготовленный раствор переносили в шесть конических колб емкостью 250 мл (по 50 мл), закрывали ватно-марлевой пробкой и стерилизовали при 0,5-0,8 атм. в течение 60 мин в автоклаве.
В микропробирку емкостью 2 мл на весах вносили 0,75 г глицерина, 0,048 г сульфата канамицина, доводили суммарный вес водой деионизированной до 1,5 г. Полученный раствор тщательно перемешивали до полного растворения компонентов. Раствор фильтровали через фильтр с размером пор 0,22 мкм в стерильную микропробирку емкостью 2 мл. В стерильных условиях переносили по 50 мкл раствора канамицина в конические колбы с питательной средой.
Культивирование инокулята
В конические колбы с питательной средой с канамицином стерильно добавляли по 50 мкл посевного материала в глицерине, расфасованного в криопробирки (суспензия клеток штамма-продуцента Е. coli BL21 Star(DE3)_ML1A-MOG). Культивирование проводили в шейкере-инкубаторе при температуре (34±1)°С и перемешивании со скоростью 130 об/мин в течение 16 часов. Полученную суспензию клеток Е. coli использовали в качестве посевного материала (инокулята) для ферментера.
3. Получение биомассы, содержащей ИТРС
Приготовление и стерилизация неполной питательной среды
В стакан емкостью 5 л вносили 4 л воды деионизированной. Навески дрожжевого экстракта (30 г), триптона (60 г), NaCl (17,53 г), Na2HPO4*12H2O (53,72 г), KH2PO4 (20,41 г), MgSO4*7H2O (2,9578 г) и (NH4)2SO4 (3,9642 г) также переносили в стакан и растворяли на магнитной мешалке. Полученный раствор переносили в ферментер. В тот же стакан наливали еще 1,5 л воды деионизированной и также переносили в ферментер.
Все открытые выходы ферментера закрывали алюминиевой фольгой. Перекрывали все линии подачи и отбора, устанавливали заглушки на неиспользуемые порты и запускалипрограмму автоклавирования.
Приготовление раствора сахаров
Раствор готовили в стеклянном стакане емкостью 250 мл. Навеску глицерина (30 г) делали в стеклянном стакане. Вносили туда же 100 мл воды деионизированной. Раствор перемешивали с помощью магнитной мешалки. Затем переносили в полученный раствор навески глюкозы (3 г) и лактозы (12 г). После растворения Сахаров раствор стерилизовали фильтрованием через фильтр с размером пор 0,22 мкм и стерильно переносили в ферментер (сначала убеждались, что среда после автоклавирования остыла).
Приготовление раствора антибиотика
Раствор антибиотика (канамицина) готовили в полипропиленовой пробирке емкостью 15 мл. Для этого навешивали в пробирку сульфат канамицина (0,6 г) и доводили вес деионизированной водой до 12,0 г. Тщательно перемешивали раствор с помощью вортекса. После полного растворения канамицина раствор стерилизовали фильтрованием через фильтр с размером пор 0,22 мкм в стерильную пробирку емкостью 15 мл. Раствор стерильно переносили в ферментер.
Культивирование биомассы
Подсоединяли зонд для измерения температуры. Все неиспользуемые порты закрывали заглушками. Подключали воду к рубашке холодильника для возврата конденсата. Заливали глицерин в ячейку для термозонда, устанавливали термозонд. Включали прибор в сеть.
Устанавливали температуру на +37°С. Включали мотор и устанавливали перемешивание на 200 об/мин. Подавали с помощью перистальтического насоса в колбу ферментера 6 мл пеногасителя. Подсоединяли кабели зондов для определения рН и растворенного кислорода к прибору. Калибровали датчик кислорода. Устанавливали значение рН 6,9. Устанавливали аэрацию на 40%.
Асептически вносили инокулят через соответствующий порт. Инкубировали биомассу в течение 20 ч.
4. Проверка уровня экспрессии иммунотоксина (ИТРС)
Для оценки уровня экспрессии определяли содержание белка ИТРС в клетках с помощью аналитического электрофореза в денатурирующих условиях в полиакриламидном геле. Из ферментера отбирали 1-2 мл суспензии клеток в специальный коллектор. Переносили 0,1 мл суспензии в микропробирку емкостью 1,5 мл. В пробу добавляли 0,2 мл деионизированной воды и ресуспендировали пипетированием. Переносили 0,01 мл полученной суспензии в новую микропробирку и добавляли 0,01 мл буфера для нанесения образцов с β-меркаптоэтанолом. Пробу инкубировали в течение 5 мин при 95°С. Затем центрифугировали 1 мин при 5000 g. Для аналитического электрофореза в полиакриламидном геле вносили в лунки по 0,020 мл образцов и по 5 мкл смеси маркеров молекулярного веса.
После визуализации белков в геле его фотографировали в камере для документирования гелей ChemiDoc MP и проводили оценку электрофореграммы с помощью программы для обработки результатов ImageLab Software. Основное пятно, соответствующее ИТРС, должно мигрировать как белок с массой 28±1 кДа. На основании предварительно построенной калибровочной кривой для площади пятна на электрофореграмме в зависимости от количества нанесенного белка ИТРС определяли его количество в анализируемых пробах, которое должно составлять 1±0,2 мкг.
5. Отделение биомассы
Отключали аэрацию, перемешивание, контроль температуры и рН. Отключали прибор от сети. Сливали суспензию через специальный порт в пластиковую бутыль емкостью 10 л.
Биомассу от культуральной жидкости отделяли центрифугированием: переносили суспензию клеток в 6 центрифужных полипропиленовых бутылей емкостью 1000 мл и центрифугировали 20 мин при 3000 g. Супернатанты (культуральную жидкость) удаляли декантацией и отправляли на автоклавирование (после этого - в стоки). Полученный осадок клеток до использования в следующих стадиях хранили при -20°С.
После удаления суспензии клеток проводили очистку ферментера в соответствии с инструкцией производителя.
6. Выделение и очистка телец включения
Для выделения и отмывки телец включения использовали четыре раствора (А, Б, В, Г). Все растворы готовили в деионизированной воде (I тип).
Приготовление раствора А для отмывки телец включения
Раствор А (2 М NaCl, 5 мМ дитиотреитол, 5 мМ ЭДТА, 25 мМ Tris-HCl рН 8) готовили в мерном стакане емкостью 5000 мл. В стакан наливали 3000 мл деионизированной воды и растворяли навески NaCl (566,84 г), дитиотреитола (3,7418 г), гидрохлорида трис(гидроксиметил)аминометана (10,7670 г) и трис(гидроксиметил)аминометана (6,4263 г). После этого растворяли навеску ЭДТА (9,0271 г). Раствор переносили в мерный цилиндр емкостью 5000 мл, доводили объем деионизированной водой до 4850 мл и переносили обратно в стакан. Раствор А использовали свежеприготовленным.
Приготовление раствора Б для отмывки телец включения
Раствор Б (2% Triton Х-100, 50 мМ NaCl, 1 мМ дитиотреитол, 25 мМ Tris-HCl рН 8) готовили в мерном стакане емкостью 5000 мл. В стакан наливали 3000 мл холодной дистиллированной воды и растворяли навески NaCl (106,58 г), дитиотреитола (0,5630 г), гидрохлорида трис(гидроксиметил)аминометана (8,1030 г) и трис(гидроксиметил)аминометана (4,8363 г). После полного растворения солей в раствор вносили Triton Х-100 (73 мл) и тщательно перемешивали на магнитной мешалке. После растворения Triton Х-100 раствор переносили в мерный цилиндр емкостью 5000 мл, доводили объем деионизированной водой до 3650 мл и переносили обратно в стакан. Раствор Б использовали свежеприготовленным.
Приготовление раствора В для отмывки телец включения
Раствор В (1 мМ дитиотреитол, 25 мМ Tris-HCl рН 8) готовили в мерном стакане емкостью 5000 мл. В стакан наливали 3000 мл деионизированной воды и растворяли навески дитиотреитола (0,5630 г), гидрохлорида трис(гидроксиметил)аминометана (8,1030 г) и трис(гидроксиметил)аминометана (4,8363 г). После полного растворения солей раствор переносили в мерный цилиндр емкостью 5000 мл, доводили объем деионизированной водой до 3650 мл и переносили обратно в стакан. Раствор В использовали свежеприготовленным.
Приготовление раствора Г для отмывки телец включения
Раствор Г (4 М мочевина, 1 мМ дитиотреитол, 25 мМ Tris-HCl рН 8) готовили в мерном стакане емкостью 5000 мл. В стакан наливали 2500 мл деионизированной воды и растворяли навески мочевины (876,88 г), дитиотреитола (0,5630 г), гидрохлорида трис(гидроксиметил)аминометана (8,1030 г) и трис(гидроксиметил)аминометана (4,8363 г). Раствор переносили в мерный цилиндр емкостью 5000 мл, доводили объем деионизированной водой до 3650 мл и переносили обратно в стакан. Раствор Г использовали свежеприготовленным.
Очистка телец включения
В центрифужные бутыли с биомассой вносили по 400 мл раствора А. Осадок клеток ресуспендировали. Клетки разрушали ультразвуком при охлаждении на льду (3 раза по 3 мин с 2-минутными перерывами). Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин. Супернатант удаляли. К осадкам снова добавляли по 400 мл раствора А, ресуспендировали и обрабатывали ультразвуком. Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин.
Супернатант удаляли, а к осадкам добавляли по 300 мл раствора Б. Осадки ресуспендировали и гомогенизировали ультразвуком на льду (2 раза по 3 мин с 2-минутными перерывами). Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин. Супернатант удаляли, а осадок еще раз отмывали раствором Б. Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин.
Супернатант удаляли, к осадкам добавляли по 300 мл раствора В. Осадки ресуспендировали и гомогенизировали ультразвуком на льду (2 раза по 3 мин с 2-минутными перерывами). Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин.
Супернатант удаляли, к осадкам добавляли по 30 мл раствора Г. Осадки ресуспендировали и гомогенизировали ультразвуком на льду (2 раза по 3 мин с 2-минутными перерывами). Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин. Супернатант удаляли, а осадок еще раз отмывали раствором Б. Бутыли центрифугировали при 8000 g, 15°С в течение 15 мин.
Супернатант удаляли, а осадки отправляли на солюбилизацию.
Супернатанты со всех стадий объединяли для дальнейшей утилизации.
7. Солюбилизация телец включения. Приготовление раствора для солюбилизации телец включения
Приготовление раствора для солюбилизации
Раствор (6 М гуанидина гидрохлорид, 1 мМ дитиотреитол, 25 мМ Na-ацетатный буфер рН 5) готовили в пластиковой бутыли емкостью 10 л. В бутыль наливали 3380 мл деионизированной воды (I тип) (ультрачистая вода с удельным сопротивлением 18,18 МОм*см при 25°С, т.е. вода 1 типа по ASTM, NCCLS, ISO 3696, САР) и переносили навески гидрохлорида гуанидина (3467,74 г), дитиотреитола (0,9332 г) и CH3COONa⋅3H2O (20,5814 г). В бутыль опускали якорь для магнитной мешалки и переносили ее на магнитную мешалку. С помощью ледяной уксусной кислоты (около 2,9 мл) доводили значение рН раствора потенциометрически до 5,0±0,5. Раствор для солюбилизации телец включения использовали свежеприготовленным.
Солюбилизация
В центрифужные бутыли с отмытыми тельцами включения вносили по 300 мл раствора для солюбилизации. Осадок аккуратно ресуспендировали (необходимо полностью отделить осадок от стенки бутыли, не вспенивая раствор). В бутыли вносили еще по 700 мл раствора для солюбилизации. В бутыли опускали большие якори для магнитной мешалки и закрывали крышками. Растворы инкубировали при +2 - +8°С в течение 1,5 ч и при периодическом перемешивании на магнитной мешалке.
Отделение нерастворенных включений
Для удаления нерастворившегося белка солюбилизат фильтровали с помощью установки вакуумной фильтрации через фильтр с размерами пор 0,22 мкм в стерильные бутыли емкостью 1000 мл. Раствор сразу же использовали для проведения рефолдинга.
8. Получение активного ИТРС
Приготовление раствора для рефолдинга ИТРС
Раствор для рефолдинга (0,5 М аргинин, 50 мМ Na-ацетатный буфер рН 5) готовили в трех пластиковых бутылях емкостью 50 л. В бутыли переносили навески аргинина (4180,8 г) и CH3COCNa⋅3H2O (326,592 г) и заливали 43,45 л деионизированной воды (I тип). В бутыли помещали большие якори для магнитной мешалки и переносили на мешалку. После растворения солей доводили рН раствора потенциометрически до 5,0±0,2 (около 1440 мл ледяной уксусной кислоты на каждую бутыль).
Рефолдинг ИТРС
Бутыли с раствором для рефолдинга помещали на магнитную мешалку и при непрерывном перемешивании медленно вносили по 2000 мл солюбилизированных телец включения (после стадии отделения нерастворившегося белка). Процедуру повторяли для всех 3 бутылей. Растворы инкубировали в течение 20-24 ч при комнатной температуре.
9. Получение субстанции ИТРС
Приготовление раствора для диафильтрации
Раствор для диафильтрации (0,5 М аргинин, 50 мМ Na-ацетатный буфер рН 5) готовили в пластиковой апирогенной бутыли емкостью 50 л. В бутыль переносили навески аргинина (4355 г) и CH3COCNa⋅3H2O (340,2 г) и заливали 45,27 л воды для инъекций. В бутыль помещали большой якорь для магнитной мешалки и переносили на магнитную мешалку. После растворения солей доводили рН раствора потенциометрически до 5,0 (около 1500 мл ледяной уксусной кислоты). Перед проведением диафильтрации раствор охлаждали до +2 - +8°С.
Концентрирование и диафильтрация рефолдинг-среды
Раствор ИТРС после рефолдинга концентрировали на установке тангенциальной фильтрации KMPi System (мембрана - mPES, 10000 NMWL) до примерно 5 л. После этого в бутыль с концентратом добавляли 12,5 л раствора для диафильтрации. Полученный раствор снова концентрировали до примерно 5 л. Повторяли процедуру еще 3 раза. На последней стадии раствор концентрировали до 2,3±0,3 л.
Стерилизация субстанции ИТРС
Концентрат ИТРС стерилизовали в асептических условиях (локальная зона чистоты класса А) фильтрованием через полиэтиленсульфоновый фильтр с размером пор 0,22 мкм.
Подтверждение подлинности и чистоты субстанции
Подтверждение чистоты и молекулярной массы иммунотоксина проводили методом электрофоретического разделения в 13% полиакриламидном геле в невосстанавливающих условиях (без меркаптоэтанола). Визуализацию полос производили с помощью окрашивания Кумасси G-250 с последующей денсиметрией. При этом на электрофореграмме должна присутствовать основная полоса, соответствующая белку с массой 28±1 кДа, а сумма интенсивностей других полос не должна превышать 5% от суммарной интенсивности всех полос.
Для трех серий субстанции иммунотоксина, полученных заявленным способом, провели определение %-ого содержания иммунотоксина и его молекулярной массы. Примеры полученных электрофореграмм представлены на фиг. 5. Для определения молекулярной массы использовались маркеры молекулярных масс Precision Plus Protein Unstained Standards (Bio-Rad). Гели после окрашивания сканировали, определяли содержание и молекулярную массу целевого белка с помощью программы ImageLab Software (Bio-Rad). С использованием программы Microsoft Excel рассчитали стандартное отклонение и относительное стандартное отклонение. Результаты представлены в Таблице 1.

Подтверждение формирования третичной структуры проводили с помощью определения температуры плавления. Определение температуры плавления осуществляли методом Thermofluor с использованием флуоресцентного красителя SYPRO Orange. Температура плавления соответствует максимуму на кривой зависимости первой производной интенсивности флуоресценции от температуры и должна составлять 39±2°С.
На фиг. 6 приведен пример кривой плавления для субстанции ИТРС, полученной заявленным способом.
Пример, демонстрирующий возможность использования иммунотоксина в качестве активного компонента.
Из полученного указанным выше способом иммунотоксина готовили раствор препарата следующего состава:
Вспомогательные вещества:
Для этого, используя приготовленный раствор для разведения, доводили полученный раствор очищенного иммунотоксина до концентрации 0,05 мг/мл.
Раствор для разведения (рН 5,0±0,2) имеет следующий состав:
Лекарственный препарат в жидкой лекарственной форме представлял собой бесцветный прозрачный раствор без видимых частиц.
Предполагаемый способ применения в клинике: 4 однократных ежедневных внутривенных введения в дозе 2 мкг/кг. Повтор курса через месяц. Предполагаемый способ введения - внутривенная инфузия: рассчитанная разовая доза препарата отбирается из флакона с помощью шприца и растворяется в 100 мл флаконе с изотоническим раствором; препарат вводится сначала с низкой скоростью (6 мл/ч) для выявления возможной реакции гиперчувствительности, через 15 мин - 12 мл/ч, затем каждые 15 мин скорость увеличивается на 12 мл/ч до 72 мл/ч.
Исследования специфической активности
Исследование специфической активности было проведено в экспериментах in vitro и на животной модели.
Свойства иммунотоксина определяются двумя основными параметрами: проявлением каталитической активности токсина и взаимодействием с определенными рецепторами на поверхности клеток-мишеней. Каталитическая активность ИТРС обусловлена наличием в составе слитого белка А-субъединицы вискумина, которая катализирует депуринизацию остатка аденина в положении 4324 (А4324) в 28 S рРНК, что приводит к инактивации рибосом и остановке синтеза белка. В качестве модельной системы для изучения подавления трансляции была выбрана бесклеточная система экспрессии люциферазы. Люцифера - фермент, способный окислять субстрат люциферин, при этом реакция сопровождается испусканием света. Таким образом, количество модельного белка (люциферазы), синтезированного в бесклеточной системе, может быть оценено по интенсивности люминесценции. При инактивации рибосом происходит снижение детектируемого сигнала. По зависимости интенсивности люминесценции от количества ГЛФ, внесенного в реакционную смесь, можно определять ингибирующую концентрацию ИТРС (значение ИК50). ИК50 для иммунотоксина, определенное описанным методом, составляет 5±2 нМ. Пример кривой ингибирования иммунотоксином, полученным заявленным способом, трансляции белка в бесклеточной системе экспрессии приведен на фиг. 7.
Для оценки способности иммунотоксина связываться с поверхностью В-лимфоцитов была использована in vitro модель на основе гибридомы, продуцирующей специфические антитела. Для оценки связывания иммобилизованный на поверхности планшета иммунотоксин инкубировали с суспензией клеток, содержащих на своей поверхности специфические рецепторы для иммунотоксина (линия А) и не содержащих (линия В). По количеству связавшихся клеток в обоих случаях судили о специфичности иммунотоксина. В отсутствии иммунотоксина обе линии клеток практически не связывались с планшетом (в среднем 179 клеток/лунку для линии А и 209 клеток/лунку для линии В). Если поверхность была модифицирована иммунотоксином, клетки линии А связывались с планшетом (2032 клеток/лунку), а клетки линии В - нет (346 клеток/лунку).
Проверка активности препарата иммунотоксина была также проведена с использованием животной модели. Общепринятой животной моделью рассеянного склероза является экспериментальный аутоиммунный энцефаломиелит (ЭАЭ), который наиболее часто в исследованиях индуцируют у мышей или крыс. И хотя ЭАЭ не полностью отражает все аспекты развития рассеянного склероза у людей, эта модель применяется для оценки эффективности новых лекарственных средств.
Для исследования были использованы 46 мышей-самок линии C57BL/6 возраста 8 недель. Животные были разбиты на 4 группы по 10 животных и одну группу 6 животных (контрольная группа №1; мыши получали инъекции физиологичного раствора вместо индуцирующих ЭАЭ агентов и исследуемого препарата). В остальных группах ЭАЭ индуцировали путем двукратного введения (с промежутком в 1 день) коклюшного токсина РТХ (2000 нг/мышь, внутривенно в РОС) и гомогенизат спинного мозга мыши (ГСММ; 5 мг/мышь, подкожно). Одна из этих групп являлась контрольной №2 - вместо исследуемого препарата вводился носитель, а остальные группы получали разные дозы исследуемого препарата (0,05, 0,5 и 5 мкг ИТРС/мышь).
Признаки развития ЭАЭ начали появляться у животных начиная с 1 дня после начала эксперимента. Для каждой экспериментальной группы были рассчитаны средние значения оценки развития ЭАЭ (см. фиг. 8).
В группе, не получавшей исследуемый препарат ИТРС, признаки заболевания проявлялись раньше и накапливались в группе быстрее, тогда как для всех групп, получавших ИТРС можно отметить более позднее время проявления заболевания.
Также при введении животным ИТРС наблюдалось увеличение продолжительности жизни. Смерть животных из экспериментальных групп наступала из-за полного паралича и последующей дегидратации и угнетения дыхания (см. фиг. 9).
Наиболее эффективной являлась доза 0,5 мкг/мышь, так как при этой концентрации ИТРС проявление признаков ЭАЭ наступало позже, а средняя продолжительность жизни была максимальной. Полученные на животных моделях экспериментальные результаты могут быть экстраполированы на дальнейшее использование ИТРС у человека (Руководство по проведению доклинических исследований лекарственных средств. Под ред. А.Н. Миронова. Часть 1. - М., 2012).
Исследование острой токсичности
Исследование острой токсичности иммунотоксина было проведено на двух видах животных: белых нелинейных мышах (возраст 9-10 недель, масса тела 18-20 г) и белых нелинейных крысах (возраст 9-10 недель, масса тела 180-200 г). Использованные дозы иммунотоксина: 50, 200, 500, 750, 1000 мкг/кг (по 10 животных на каждую группу, 5 самцов и 5 самок), в качестве был использован физиологический раствор. Способ введения - внутривенная инъекция. Наблюдение за животными осуществлялось в течение 14 дней после введения. Оценивали общее состояние животных по состоянию шерсти и слизистых оболочек, массе тела. Через 14 дней животных подвергали эвтаназии углекислым газом, проводили паталогоанатомическое вскрытие, визуально определяли морфологическое состояние органов и слизистых покровов, определяли массовые коэффициенты органов (сердце, легкие с трахеей, тимус, печень, селезенка, почки, надпочечники, головной мозг, яичники или яички).
В ходе исследования смертность животных (как мышей, так и крыс) от препарата отсутствовала. Признаков интоксикации при осмотре не было выявлено. Сразу после введения больших доз препарата, у животных наблюдалась небольшая одышка и незначительное угнетение. Данные клинические признаки проходили в течение 1-2 часов. Масса тела и массовые коэффициенты органов животных, получавших ИТРС, не отличалась от контрольной группы. Отсутствовали видимые морфологические изменения органов и слизистых оболочек. Хроническая токсичность
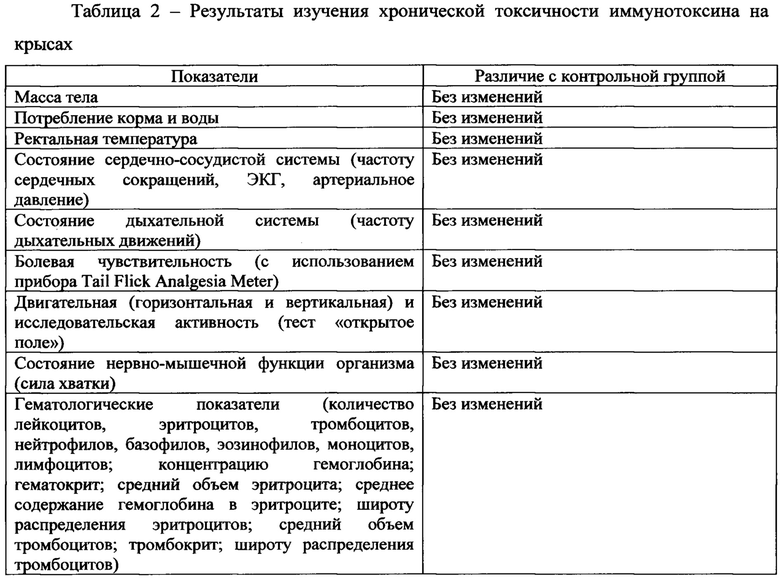
Исследование хронической токсичности иммунотоксина было проведено на двух видах животных: белых нелинейных крысах (возраст 9-10 недель, масса тела 180-200 г) и на кроликах породы «Шиншилла» (возраст 2-3 месяца, масса тела 2200-2500 г). Исследованные дозы для крыс составляли 12 и 80 мкг/кг (по 10 животных в группе, 5 самцов и 5 самок), для кроликов - 6 и 80 мкг/кг (по 6 животных в группе, 3 самца и 3 самки). Также были сформированы контрольные группы животных, получавшие физиологический раствор. Введение осуществляли внутривенно ежедневно на протяжении 30 дней.
Перечень показателей, определявшихся в ходе исследования, и наблюдавшихся изменений приведен в Таблицах 2 и 3. Смертность животных во всех экспериментальных группах отсутствовала.



Исследование иммунотоксичности
В исследовании была использована стандартная животная модель - мыши-гибриды (CBAxC57BL/6)Fl, возраст 6-8 недель, масса тела 18-20 г. Введение препарата осуществляли в режиме, приближенном к предполагающемуся в клинике (4 однократных введения, по одному каждый день); в хвостовую вену. Исследованные дозы - 200 и 400 мкг/кг (по 10 животных в группе, 5 самцов и 5 самок); контрольные группы получали физиологический раствор.
Для оценки влияния на гуморальный иммунитет определяли число антителообразующих клеток (АОК) при иммунизации Т-зависимым антигеном (эритроцитами барана). Спустя час после введения препаратов все животные были проиммунизированы внутрибрюшинной инъекцией эритроцитов барана в субоптимальной дозе 5×107 эритроцитов на животное. Подсчет числа АОК в селезенке мышей производили на 5-е сутки после иммунизации. Реакцию ставили на предметных стеклах без поддерживающей среды по Canningham. По количеству зон гемолиза эритроцитов вокруг отдельных антителообразующих клеток, определяемых под микроскопом, подсчитывали число АОК в расчете на 106 спленоцитов. Различий в количестве АОК не выявлено, что свидетельствует об отсутствии воздействия иммунотоксина на гуморальный иммунитет лабораторных животных.
Для изучения влияния на клеточный иммунитет через 1 ч после последнего введения препарата или контрольного раствора все мыши были сенсибилизированы путем подкожной инъекции эритроцитов барана в межлопаточную область. На 5 день после сенсибилизации была проведена оценка гиперчувствительности замедленного типа. Для этого всем животным в подушечку задней лапы были введены эритроциты барана (разрешающая инъекция). В другую заднюю лапу был введен физиологический раствор. Через 24 ч проводилась оценка местной воспалительной реакции по различию в массе задних лапок. Воздействия на гиперчувствительность замедленного типа к эритроцитам барана выявлено не было, что свидетельствует об отсутствии влияния иммунотоксина на клеточный иммунитет лабораторных животных.
Влияние на фагоцитарную активность перитонеальных макрофагов исследовали через 24 часа после последнего введения препарата. Для этого животных подвергали эвтаназии углекислым газом, а брюшную полость умерщвленных животных промывали культуральной средой. Концентрацию клеток перитонеального экссудата доводили до 2 млн/мл. Затем переносили по 2 мл в чашки Петри диаметром 40 мм и инкубировали в термостате в течение 1 ч при 37°С. Не прикрепившиеся клетки отмывали. В среду добавляли опсонизированные дрожжи. По завершении реакции клетки фиксировали и окрашивали по Романовскому-Гимзе. Подсчитывали под микроскопом количество фагоцитировавших и нефагоцитировавших макрофагов. Определяли процент фагоцитировавших макрофагов по отношению к общему количеству. Определяли среднее количество фагоцитированных дрожжевых клеток. У самок различий между группами выявлено не было. У самцов в ответ на введение ИТРС (200 и 400 мкг/кг) происходило увеличение фагоцитарного индекса (с 46,0±4,5 до 70,8±4,3 и 73,0±5,0, соответственно). Это может свидетельствовать об иммуностимулирующем действии препарата.
Изучение аллергизирующих свойств
Аллергизирующие свойства препарата оценивали по развитию реакции общей анафилаксии и реакции гиперчувствительности амедленного типа.
Для оценки сенсибилизирующих свойств в реакции общей анафилаксии использовали нелинейных морских свинок (возраст 4-6 недель, масса тела 240-260 г). Исследованные дозы: 10 и 100 мкг/кг (по 10 животных в группе, 5 самцов и 5 самок); животные контрольной группы получали физиологический раствор. Животным было сделано три инъекции ИТРС (первая - подкожно, две последующие - внутримышечно). Спустя 14 дней после первого введения животным была введена внутривенно разрешающая доза ИТРС. Такая же доза была введена животным контрольной группы. После этого была проведена оценка интенсивности анафилактического шока. Ни в одной из групп признаков анафилактического шока зафиксировано не было (индекс реакции 0 по шкале Weigle).
Определение гиперчувствительности замедленного типа проводили на белых нелинейных мышах (возраст 9-10 недель, масса тела 18-20 г). Исследованные дозы: 200 и 400 мкг/кг (по 10 животных в группе, 5 самцов и 5 самок); животные контрольной группы получали физиологический раствор. Мышей сенсибилизировали путем подкожного введения ИТРС (200 или 400 мкг/кг), по 1 введению в течение 5 дней. Животным контрольной группы вводили физ. раствор. Для выявления сенсибилизации через 5 дней после последней инъекции всем мышам вводили в подушечку задней лапы 50 мкл исследуемого препарата. В подушечку другой задней лапы вводили 50 мкл растворителя. Через 24 ч после этого производили эвтаназию животных и отрезали обе задние лапы. Затем производили оценку реакции ГЗТ по разнице масс контрольной и экспериментальной лапок. Достоверных различий между экспериментальными группами выявлено не было.
Таким образом, препарат в исследованиях на лабораторных животных не проявлял аллергизирующих свойств.
Исследование мутагенности
Для оценки мутагенности исследуемого препарата был проведен тест на индукцию генных мутаций. В качестве модельного объекта для мутационного теста Эймса была использована Salmonella typhimurium (штаммы ТА 97, ТА 98 и ТА 100), которая является стандартной бактериальной тест-системой. Производился учет мутаций к прототрофности по гистидину при действии соединений, индуцирующих мутации типа замены оснований или сдвига рамки считывания в геноме этого организма. В качестве негативного контроля была использована обработка растворителем. В качестве позитивного контроля использовались вещества, индуцирующие мутации у соответствующих штаммов Salmonella typhimurium. В тесте Эймса препарат не проявлял мутагенных свойств.
Для оценки мутагенности исследуемого препарата на эукариотических клетках была использована методика, рекомендованная Организацией экономического сотрудничества и развития (OECD), - «Test No. 476: In Vitro Mammalian Cell Gene Mutation Tests using the Hprt and xprt genes». Принцип метода заключается в регистрации мутаций (точечных замен, сдвига рамки считывания, делеций или вставок), приводящих к инактивации гена Hprt, кодирующему гипоксантин-гуанин-фосфорибосилтрансферазу. Активность данного фермента обуславливает цитотоксичность 6-тиогуанина, который становится токсичным только после фосфорилирования. Нормальные клетки, экспрессирующие Hprt, чувствительны к цитостатическому действию 6-тиогуанина (6-ТГ), в то время как мутантные клетки с инактивированным геном Hprt, способны пролиферировать в присутствии 6-ТГ. Тест-объектом являлись клетки СНО-К1. В данном тесте препарат также не проявлял мутагенных свойств.
Для оценки генотоксичности исследуемого препарата на эукариотических клетках была использована методика, рекомендованная Организацией экономического сотрудничества и развития (OECD) - Test No. 487: In Vitro Mammalian Cell Micronucleus Test, 2016 г. Учет микроядер в клетках представляет собой тест на генотоксичность, в ходе которого определяется доля клеток, содержащих в цитоплазме микроядра. Тест-объектом являлись клетки линий Сасо-2 и HepaRG. Различий с контролем выявлено не было.
Исследование репродуктивной токсичности
Исследование влияния на репродуктивную функцию было проведено на аутбредном стоке крыс «Вистар» (возраст 10-13 недель, масса тела 220-300 г). Исследованные дозы препарата: 8 и 40 мкг/кг; контрольные группы получали носитель. Введение: внутривенно в латеральную вену хвоста; самкам - на протяжении 15 дней до спаривания и в течение 20 дней во время беременности; самцам - на протяжении 49 дней перед спариванием. Эксперимент состоял из нескольких частей: 1) определение влияния ИТРС на генеративную функцию самок (введение препарата крысам-самкам и ссаживание с интактными самцами; регистрация беременности); 2) определение влияния ИТРС на генеративную функцию самцов (введение препарата крысам-самцам и ссаживание с интактными крысами-самками; регистрация беременности); 3) определение эмбриотоксического потенциала препарата (введение ИТРС самкам во время беременности; регистрация жизнеспособности плодов, физического развития потомства).
В ходе исследования смертности животных не было. У всех животных, получавших носитель и препарат, наблюдалась местная реакция на введение в виде цианоза хвоста разной степени выраженности. Иных проявлений интоксикации у животных не наблюдалось. В проведенном исследовании влияния препарата ИТРС на репродуктивную функцию самок, определено, что препарат, вводимый самкам внутривенно в дозе 40 мкг/кг на протяжении трех эстральных циклов перед ссадкой, оказывает негативное влияние на фертильность самок, достоверно снижая ее относительно контрольных животных (индекс фертильности снизился с 0,95 до 0,60). Показано, что препарат, вводимый самцам в период формирования гамет, не оказывал влияния на их генеративную функцию. Также показано, что потомство, рожденное от самок, получавших препарат до и во время беременности, не отличается ни по выживаемости, ни по динамике набора веса от потомства контрольных самок.
Таким образом, можно говорить о наличии репродуктивной токсичности препарата в отношении генеративной функции самок и об отсутствии репродуктивной токсичности в отношении самцов. Также можно говорить об отсутствии фетотоксического действия препарата.








Группа изобретений относится к медицине и касается иммунотоксина для терапии рассеянного склероза, представляющего собой рекомбинантный белок, включающий А-субъединицу вискумина и фрагмент миелин-олигодендроцитного гликопротеина, соединенные линкером для связывания упомянутых фрагментов. Группа изобретений также касается рекомбинантной плазмидной ДНК для экспрессии в клетках Escherichia coli гена, кодирующего указанный иммунотоксин; касается способа получения иммунотоксина; касается способа терапии рассеянного склероза, заключающегося во внутривенном введении фармацевтической композиции, содержащей указанный иммунотоксин, в терапевтически эффективном количестве. Группа изобретений обеспечивает получение иммунотоксина для терапии рассеянного склероза, обладающего высокой специфичностью при низкой токсичности. 6 н. и 13 з.п. ф-лы, 9 ил., 3 табл., 1 пр.
1. Иммунотоксин для терапии рассеянного склероза, представляющий собой рекомбинантный белок, включающий А-субъединицу вискумина и фрагмент миелин-олигодендроцитного гликопротеина, обеспечивающий связывание с клетками-мишенями, соединенные линкером для связывания упомянутых фрагментов.
2. Иммунотоксин по п. 1, характеризующийся тем, что А-субъединица вискумина представляет собой полипептид с последовательностью, не менее чем на 95% гомологичной SEQ ID NO: 1.
3. Иммунотоксин по п. 1, характеризующийся тем, что фрагмент миелин-олигодендроцитного гликопротеина представляет собой пептид с последовательностью, не менее чем на на 90% гомологичной SEQ ID NO: 2.
4. Иммунотоксин по п. 1, представляющий собой рекомбинантный белок с последовательностью, не менее чем на 95% гомологичной SEQ ID NO: 3.
5. Рекомбинантная плазмидная ДНК для экспрессии в клетках Escherichia coli гена, кодирующего иммунотоксин по п. 1, представляющая собой вектор, модифицированный фрагментом, кодирующим иммунотоксин по п. 4, включающий следующие фрагменты:
a) фрагмент XhoI-NcoI;
b) промотор бактериофага Т7;
c) терминатор РНК-полимеразы бактериофага Т7;
d) область начала репликации ColE1/pMB1/pBR322/pUC;
e) область, кодирующую ген фермента, обеспечивающего устойчивость к антибиотику.
6. Плазмидная ДНК по п. 5, характеризующаяся тем, что в качестве вектора используют pET-22b(+), рЕТ-26b(+), рЕТ-28а-с(+).
7. Штамм Escherichia coli - продуцент иммунотоксина по п. 1, трансформированный рекомбинантной плазмидной ДНК по п. 5.
8. Способ получения иммунотоксина по п. 1, характеризующийся тем, что культивируют штамм-продуцент по п. 7 с получением биомассы, содержащей тельца включения, которую подвергают разрушению с помощью ультразвукового дезинтегратора с последующим выделением телец включения с помощью центрифугирования, которые затем отмывают методом последовательного растворения примесей, с получением иммунотоксина в нерастворенной форме, который затем переводят в растворенную форму с помощью раствора хаотропного агента, после чего неактивный растворенный иммунотоксин подвергают ренатурации методом разведения, диафильтрации и концентрированию с помощью тангенциальной фильтрации.
9. Способ по п. 8, характеризующийся тем, что разрушение биомассы с помощью ультразвукового дезинтегратора производят при частоте 22 кГц и охлаждении на льду.
10. Способ по п. 9, характеризующийся тем, что ультразвуковую обработку проводят 3-4 раза по 3 минуты с 2-минутными перерывами.
11. Способ по п. 8, характеризующийся тем, что тельца включения отделяют от растворимых компонентов клеток с помощью центрифугирования.
12. Способ по п. 8, характеризующийся тем, что очищение телец включения проводят с помощью растворов, содержащих детергенты и хаотропные агенты, обеспечивающие удаление примесей ДНК, РНК, липидов и примесных белков.
13. Способ по п. 8, характеризующийся тем, что перевод иммунотоксина из нерастворимой неактивной формы в растворимую неактивную форму проводят с помощью раствора, содержащего хаотропный агент.
14. Способ по пп. 12 и 13, характеризующийся тем, что в качестве хаотропного агента используют растворы мочевины, тиомочевины, гуанидина гидрохлорида.
15. Способ по п. 12, характеризующийся тем, что в качестве детергента используют додецилсульфат натрия, дезоксихолат, саркосил, твин 20, твин 80, тритон Х-100.
16. Способ по п. 8, характеризующийся тем, что перевод иммунотоксина из растворимой неактивной формы в растворимую активную форму (ренатурацию) проводят с помощью разведения в растворе, содержащем аргинин и имеющем кислое значение рН.
17. Способ по п. 8, характеризующийся тем, что концентрирование активного иммунотоксина после ренатурации проводят с помощью тангенциальной фильтрации.
18. Фармацевтическая композиция для терапии рассеянного склероза, включающая в качестве действующего вещества иммунотоксин по п. 1 в терапевтически эффективном количестве и фармацевтически приемлемую основу.
19. Способ терапии рассеянного склероза, заключающийся во внутривенном введении фармацевтической композиции по п. 18 в терапевтически эффективном количестве.
| US 9951113 B2, 24.04.2018 | |||
| US 20160122707 A1, 05.05.2016 | |||
| НОВЫЕ IL-17-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2550272C2 |
| SHEMIAKIN IG | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| [Article in Russian], Mol Biol (Mosk) | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |

