Уровень техники
Настоящее изобретение касается антител с регулируемой ассоциацией тяжелой цепи и легкой цепи, способов получения антитела с регулируемой ассоциацией тяжелой цепи и легкой цепи, способов регулирования ассоциации тяжелой цепи и легкой цепи антитела, фармацевтических композиций, включающих это антитело в качестве активного ингредиента, и т.п.
Область изобретения
Ранее сообщалось о нескольких способах получения биспецифических антител IgG-типа, имеющих константные области человека (антител IgG-типа, имеющих константную область человека, обладающую специфичностью связывания с антигеном A на одном плече и специфичностью связывания с антигеном B на другом плече). В целом биспецифические антитела IgG-типа состоят из двух типов H-цепей (то есть, H-цепи для антигена A и H-цепи для антигена В) и двух типов L-цепей (то есть, L-цепи для антигена A и L-цепи для антигена B). Когда экспрессируются такие биспецифические антитела IgG-типа, возможны 10 типов комбинаций в качестве комбинаций H2L2, поскольку экспрессируются два типа H-цепей и два типа L-цепей. Среди них существует один тип комбинации, имеющий необходимую специфичность связывания (IgG, имеющий специфичность связывания с антигеном A на одном плече и специфичность связывания с антигеном В на другом плече). Таким образом, для получения необходимого биспецифического антитела необходимо выделить один тип нужного антитела из десяти типов антител, что является очень малоэффективным и затруднительным.
Сообщалось о способах решения этой проблемы, включающих преимущественно секретирование IgG, имеющего гетерологичную комбинацию H-цепи для антигена A и H-цепи для антигена В, путем замещения аминокислот в CH3-области IgG H-цепи (патентные документы 1, 2, 3 и 4 и непатентные документы 1 и 2). Среди них сообщалось о способах, в которых используются физические препятствия в форме "выступов" и "впадин", и тех, в которых применяется отталкивание электрических зарядов.
Также сообщалось о способе эффективного получения необходимой молекулы, согласно которому используется общая L-цепь, в которой L-цепь для антигена A и L-цепь для антигена В присутствуют на одной аминокислотной последовательности (патентные документы 5 и 6). Однако, поскольку применение общей L-цепи может значительно снижать аффинность антигена, этот способ не всегда является оптимальным. Таким образом, для того, чтобы биспецифическое антитело связывалось с двумя антигенами с высокой аффинностью, ассоциируют только L-цепь и H-цепь для антигена A и только L-цепь и H-цепь для антигена В. Кроме того, сообщается о способе, позволяющем ассоциировать H-цепи и L-цепи для каждого антигена независимо от вариабельных областей, который включает замещение аминокислот в доменах CH1 и CL, которые являются константными областями, вместо аминокислот в вариабельных областях (патентные документы 2 и 7). Однако этот способ все же недостаточен для эффективного получения необходимого биспецифического антитела.
[Документы существующего уровня техники]
[Патентные документы]
[Патентный документ 1] WO 96/27011
[Патентный документ 2] WO 2006/106905
[Патентный документ 3] WO 2009/089004
[Патентный документ 4] WO 2010/129304
[Патентный документ 5] WO 98/050431
[Патентный документ 6] WO 2006/109592
[Патентный документ 7] WO 2007/147901
[Непатентные документы]
[Непатентный документ 1] Ridgway JB et al., Protein Engineering, 1996, Vol. 9, p. 617-621
[Непатентный документ 2] Merchant AM et al., Nature Biotechnology, 1998, Vol. 16, p. 677-681
Краткое описание изобретения
[Проблемы, решаемые изобретением]
Настоящее изобретение достигалось при таких условиях. Цель настоящего изобретения состоит в обеспечении антител, в которых ассоциация тяжелых цепей и легких цепей является регулируемой, способа получения антител, в которых ассоциация тяжелых цепей и легких цепей является регулируемой, и способа регулирования ассоциации тяжелых цепей и легких цепей антитела. Кроме того, в одном варианте осуществления настоящего изобретения цель настоящего изобретения состоит в обеспечении биспецифических антител, в которых ассоциация на границе CH1 и CL является регулируемой, и способа эффективного получения биспецифического антитела путем регулирования ассоциации на границе CH1 и CL.
[Средства решения проблем]
Авторами настоящего изобретения были выбраны константная область тяжелой цепи, CH1, и константная область легкой цепи (CL) в качестве областей тяжелой цепи и легкой цепи для применения в регулировании ассоциации и проводились специальные исследования регулирования ассоциации CH1 и CL. В результате авторами настоящего изобретения было обнаружено, что ассоциация CH1 и CL может подавляться замещением аминокислотных остатков, присутствующих на границе CH1 и CL, аминокислотными остатками, которые электрически взаимно отталкиваются, или аминокислотными остатками, которые не отталкиваются, и гетерологичные молекулы образуются более эффективно, чем при применении модификаций, которые только включают выступ и впадину в CH3, как было описано выше.
Таким образом, согласно открытиям, сделанным авторами настоящего изобретения, существует возможность регулирования ассоциации CH1 и CL. Кроме того, настоящее изобретение может применяться не только к регулированию ассоциации между CH1 и CL, а также к регулированию ассоциации между произвольными полипептидами.
Кроме того, авторы настоящего изобретения также подтвердили, что биспецифическое антитело согласно настоящему изобретению с регулируемой ассоциацией тяжелой цепи и легкой цепи фактически сохраняет функцию.
Как описывалось выше, авторами настоящего изобретения были успешно разработаны антигенсвязывающие молекулы, в которых ассоциация тяжелой цепи и легкой цепи является регулируемой и, таким образом, выполнено настоящее изобретение.
Настоящее изобретение касается антигенсвязывающих молекул, в которых ассоциация тяжелых цепей и легких цепей является регулируемой, способов получения антигенсвязывающей молекулы, в которой ассоциация тяжелых цепей и легких цепей является регулируемой, и способов регулирования ассоциации тяжелых цепей и легких цепей антигенсвязывающей молекулы. В частности, настоящее изобретение касается следующего:
[1] антигенсвязывающей молекулы, в которой ассоциация тяжелой цепи и легкой цепи является регулируемой, причем
один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), в тяжелой цепи и легкой цепи в антигенсвязывающей молекуле, являются аминокислотными остатками, которые электрически взаимно отталкиваются:
(a) аминокислотный остаток, включенный в константную область тяжелой цепи (CH1) в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в константную область легкой цепи (CL) в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 160, как указывается согласно нумерации EC;
[2] антигенсвязывающей молекулы [1], также характеризующейся тем, что в ней аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в п. (d), являются аминокислотными остатками, которые электрически взаимно отталкиваются:
(d) аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC;
[3] антигенсвязывающей молекулы [1] или [2], в которой аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) и (Y), как указано ниже:
(X) глутаминовая кислота (E) или аспарагиновая кислота (D); и
(Y) лизин (K), аргинин (R) или гистидин (H);
[4] любой антигенсвязывающей молекулы с [1] по [3], также характеризующейся тем, что в ней два или более аминокислотных остатков, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, являются аминокислотными остатками, которые электрически взаимно отталкиваются;
[5] антигенсвязывающей молекулы [4], в которой аминокислотные остатки, которые электрически взаимно отталкиваются, представляют собой один набор или два набора аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных в пп. (a) или (b):
(a) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 44 согласно нумерации Kabat;
[6] антигенсвязывающей молекулы [4] или [5], в которой аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в любой из наборов (X) и (Y), как указано ниже:
(X) глутаминовая кислота (E) или аспарагиновая кислота (D); и
(Y) лизин (K), аргинин (R), или гистидин (Н);
[7] антигенсвязывающей молекулы, в которой ассоциация тяжелой цепи и легкой цепи является регулируемой, причем
один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), в ассоциации тяжелой цепи и легкой цепи в антигенсвязывающей молекуле, являются аминокислотными остатками, которые электрически взаимно не отталкиваются:
(a) аминокислотный остаток, включенный в константную область тяжелой цепи (CH1) в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в константную область легкой цепи (CL) в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 160, как указывается согласно нумерации EC;
[8] антигенсвязывающей молекулы [7], также характеризующейся тем, что в ней аминокислотные остатки из набора аминокислотных остатков, показанных ниже в п. (d), представляют собой аминокислотные остатки, которые электрически взаимно не отталкиваются:
(d) аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC;
[9] антигенсвязывающей молекулы [7] или [8], в которой аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся указанные ниже пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z) и (Z) и (Z):
(X) глутаминовая кислота (E) или аспарагиновая кислота (D);
(Y) лизин (K), аргинин (R) или гистидин (H); и
(Z) аланин (A), аспарагин (N), цистеин (C), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (М), фенилаланин (F), пролин (P), серии (S), треонин (T), триптофан (W), тирозин (Y) или валин (V);
[10] любой антигенсвязывающей молекулы с [7] по [9], в которой аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, который является лизином (K); и аминокислотные остатки, включенные в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, которые все являются глутаминовой кислотой (Е);
[11] любой антигенсвязывающей молекулы с [7] по [9], в которой аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, включенные в CH1 в позиции 147 и позиции 175, как указывается согласно нумерации EC, которые являются глутаминовой кислотой (E); и аминокислотные остатки, включенные в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, которые все являются лизином (K);
[12] антигенсвязывающей молекулы [11], также характеризующейся тем, что в ней аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, является глутаминовой кислотой (E), и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC, является лизином (K);
[13] любой антигенсвязывающей молекулы с [7] по [12], в которой дополнительно два или более аминокислотных остатков, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, представляют собой аминокислотные остатки, которые электрически взаимно не отталкиваются;
[14] антигенсвязывающей молекулы [13], в которой аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой один набор или два набора аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. (а) или (b):
(a) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 44 согласно нумерации Kabat;
[15] антигенсвязывающей молекулы [13] или [14], в которой аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся указанные ниже пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z) и (Z) и (Z):
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D);
(Y) лизин (K), аргинин (R) или гистидин (Н); и
(Z) аланин (А), аспарагин (N), цистеин (С), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (М), фенилаланин (F), пролин (Р), серии (S), треонин (Т), триптофан (W), тирозин (Y) или валин (V);
[16] антигенсвязывающая молекулы по любому из пп. с [1] по [15], причем антигенсвязывающая молекула является биспецифическим антителом;
[17] способа получения антигенсвязывающей молекулы, согласно которому ассоциация тяжелой цепи и легкой цепи является регулируемой, который включает этапы с (1) по (3):
(1) модификации нуклеиновых кислот, кодирующих константную область тяжелой цепи (CH1) и константную область легкой цепи (CL), таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), электрически взаимно отталкивались:
(a) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 160, как указывается согласно нумерации EC,
(2) включения модифицированных нуклеиновых кислот в клетку-хозяин и культивирование клетки-хозяина таким образом, чтобы экспрессировались нуклеиновые кислоты, и
(3) сбора антигенсвязывающих молекул из культуры клетки-хозяина;
[18] способа получения антигенсвязывающей молекулы [17], который также включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в п. (d), электрически взаимно отталкивались:
(d) аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC;
[19] способа получения антигенсвязывающей молекулы [17] или [18], который включает на этапе (1) модификацию нуклеиновых кислот таким образом, что аминокислотные остатки, которые электрически взаимно отталкиваются, выбираются из аминокислотных остатков, включенных в группу (X) и (Y), как указано ниже:
(X) глутаминовая кислота (E) или аспарагиновая кислота (D); и
(Y) лизин (K), аргинин (R), или гистидин (H);
[20] способа получения антигенсвязывающей молекулы по любому из пп. с [17] по [19], который также включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы два или более аминокислотных остатков, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи являются аминокислотными остатками, которые электрически взаимно отталкиваются;
[21] способа получения антигенсвязывающей молекулы [20], согласно которому аминокислотные остатки, которые электрически взаимно отталкиваются, являются аминокислотными остатками любого набора, выбранного из группы, к которой относятся наборы аминокислотных остатков, показанных в пп. (a) или (b):
(a) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 44 согласно нумерации Kabat;
[22] способа получения антигенсвязывающей молекулы [20] или [21], согласно которому аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) и (Y), как указано ниже:
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D); и
(Y) лизин (K), аргинин (R), или гистидин (H);
[23] способа получения антигенсвязывающей молекулы, согласно которому ассоциация тяжелой цепи и легкой цепи является регулируемой, включая следующие этапы с (1) по (3):
(1) модификации нуклеиновых кислот, кодирующих константную область тяжелой цепи (CH1) и константную область легкой цепи (CL), таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), электрически взаимно не отталкивались:
(a) аминокислотный остаток, включенный в константную область тяжелой цепи (CH1) в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в константную область легкой цепи (CL) в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 160, как указывается согласно нумерации EC,
(2) включения модифицированных нуклеиновых кислот в клетку-хозяин и культивирование клетки-хозяина таким образом, чтобы экспрессировались нуклеиновые кислоты, и
(3) сбора антигенсвязывающих молекул из культуры клетки-хозяина;
[24] способа получения антигенсвязывающей молекулы [23], который также включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в п. (d), электрически взаимно не отталкивались:
(d) аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC;
[25] способ получения антигенсвязывающей молекулы [23] или [24], который включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки, которые электрически взаимно не отталкиваются, представляли собой аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся указанные ниже пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z) и (Z) и (Z):
(X) глутаминовая кислота (E) или аспарагиновая кислота (D);
(Y) лизин (K), аргинин (R), или гистидин (H); и
(Z) аланин (A), аспарагин (N), цистеин (C), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), пролин (P), серии (S), треонин (T), триптофан (W), тирозин (Y) или валин (V);
[26] способа получения антигенсвязывающей молекулы по любому из пп. с [23] по [25], который включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, который является лизином (К), и аминокислотные остатки, включенные в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, которые все являются глутаминовой кислотой (Е);
[27] способа получения антигенсвязывающей молекулы по любому из пп. с [23] по [25], который включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, включенные в CH1 в позиции 147 и позиции 175, как указывается согласно нумерации EC, которые являются глутаминовой кислотой (E), и аминокислотные остатки, включенные в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, которые все являются лизином (K);
[28] способа получения антигенсвязывающей молекулы по п. [27], который также включает модификацию нуклеиновых кислот таким образом, чтобы аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, являлся глутаминовой кислотой (Е), и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC, являлся лизином (K);
[29] способа получения антигенсвязывающей молекулы по любому из пп. с [23] по [28], который также включает на этапе (1) модификацию нуклеиновых кислот таким образом, чтобы два или более аминокислотных остатков, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, представляли собой аминокислотные остатки, которые электрически взаимно не отталкиваются;
[30] способа получения антигенсвязывающей молекулы по п. [29], согласно которому аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки любого набора, выбранного из группы, к которой относятся наборы аминокислотных остатков, показанных в пп. (a) или (b):
(a) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 44 согласно нумерации Kabat;
[31] способа получения антигенсвязывающей молекулы по пп. [29] или [30], согласно которому аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся указанные ниже пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z) и (Z) и (Z):
(X) глутаминовая кислота (E) или аспарагиновая кислота (D);
(Y) лизин (K), аргинин (R) или гистидин (H); и
(Z) аланин (A), аспарагин (N), цистеин (C), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), пролин (P), серии (S), треонин (T), триптофан (W), тирозин (Y) или валин (V);
[32] антигенсвязывающей молекулы, полученной согласно способу получения антигенсвязывающей молекулы по любому из пп. с [17] по [31];
[33] антигенсвязывающей молекулы [32], причем антигенсвязывающая молекула является биспецифическим антителом;
[34] способ регулирования ассоциации тяжелой цепи и легкой цепи антигенсвязывающей молекулы, который включает:
модификацию нуклеиновых кислот таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), являлись аминокислотными остатками, которые электрически взаимно отталкиваются:
(a) аминокислотный остаток, включенный в CHl в позиции 147. как указывается согласно нумерации ЕС, и аминокислотный остаток, включенный в CL в позиции 180, как указывается согласно нумерации ЕС;
(b) аминокислотный остаток, включенный в CHl в позиции 147, как указывается согласно нумерации ЕС, и аминокислотный остаток, включенный в CL в позиции 131, как, указывается согласно нумерации ЕС; и
(c) аминокислотный остаток, включенный в CHl в позиции 175, как указывается согласно нумерации ЕС, и аминокислотный остаток, включенный в CL в позиции 160, как указывается согласно нумерации ЕС;
[35] способа по п. [34], который также включает модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в п. (d), являлись аминокислотными остатками, которые электрически взаимно отталкиваются:
(d) аминокислотный остаток, включенный в CHl в позиции 213, как указывается согласно нумерации ЕС, и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации ЕС;
[36] способа по пп. [34] или [35], согласно которому аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) и (Y), как указано ниже:
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D); и
(Y) лизин (K), аргинин (R), или гистидин (Н);
[37] способа по любому из пп. с [34] по [36], согласно которому дополнительно два или более аминокислотных остатков, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, являются аминокислотными остатками, которые электрически взаимно отталкиваются;
[38] способа по п. [37], согласно которому аминокислотные остатки, которые электрически взаимно отталкиваются, являются аминокислотными остатками любого набора, выбранного из группы, к которой относятся наборы аминокислотных остатков, показанных в пп. (а) или (b):
(а) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 38, указанный согласно нумерации Kabat; или
(b) аминокислотный остаток, включенный в вариабельную область тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, включенный в вариабельную область легкой цепи в позиции 44 согласно нумерации Kabat;
[39] способа по пп. [37] или [38], согласно которому аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) и (Y), как указано ниже:
(X) глутаминовая кислота (E) или аспарагиновая кислота (D); и
(Y) лизин (K), аргинин (R), или гистидин (H);
[40] способа регулирования ассоциации тяжелой цепи и легкой цепи антигенсвязывающей молекулы, включающего:
модификацию нуклеиновых кислот таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), представляли собой аминокислотные остатки, которые электрически взаимно не отталкиваются:
(a) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, включенный в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, включенный в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 160, как указывается согласно нумерации EC;
[41] способа по п. [40], который также включает модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в п. (d), представляли собой аминокислотные остатки, которые электрически взаимно не отталкиваются:
(d) аминокислотный остаток, включенный в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, включенный в CL в позиции 123, как указывается согласно нумерации EC;
[42] способа по пп. [40] или [41], согласно которому аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся указанные ниже пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z) и (Z) и (Z):
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D);
(Y) лизин (K), аргинин (R), или гистидин (Н); и
(Z) аланин (А), аспарагин (N), цистеин (С), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (М), фенилаланин (F), пролин (Р), серии (S), треонин (Т), триптофан (W), тирозин (Y) или валин (V);
[43] способа по любому из пп. с [40] по [42], согласно которому аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотный остаток, включенный в CHl в позиции 175, как указывается согласно нумерации ЕС, который является лизином (K), и аминокислотные остатки, включенные в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации ЕС, которые все являются глутаминовой кислотой (Е);
[44] способа по любому из пп. с [40] по [42], согласно которому аминокислотные остатки, которые электрически взаимно не отталкиваются, представляют собой аминокислотные остатки, включенные в CHl в позиции 147 и позиции 175, как указывается согласно нумерации ЕС, которые являются глутаминовой кислотой (Е), и аминокислотные остатки, включенные в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации ЕС, которые все являются лизином (К);
[45] способа по п. [44], согласно которому дополнительно аминокислотный остаток, содержащийся в CHl в позиции 213, как указывается согласно нумерации ЕС, является глутаминовой кислотой (Е), и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации ЕС, является лизином (K);
[46] способа по любому из пп. с [34] по [45], согласно которому антигенсвязывающая молекула является биспецифическим антителом;
[47] композиции, содержащей антигенсвязывающую молекулу по любому из пп. с [1] по [16], [32] и [33] и фармацевтически приемлемый носитель;
[48] нуклеиновой кислоты, кодирующей антигенсвязывающую молекулу по любому из пп. с [1] по [16], [32] и [33]; и
[49] клетки-хозяина, включающей нуклеиновую кислоту [48].
Краткое описание фигур
Фиг. 1 представляет диаграмму модели границы CH1/CL.
Фиг. 2 концептуально представляет антитело с показом возможных комбинаций H-цепи и L-цепи, когда антитело получают путем смешивания одного типа H-цепи и двух типов L-цепей. Считается, что мутированные участки, имеющие большую пропорцию антитела с комбинацией E и K, как показано в квадрате, взаимодействуют электрически.
Фиг. 3 представляет графики, показывающие результаты анализа AIEX каждого из антител.
Фиг. 4 представляет графики, показывающие результаты анализа AIEX каждого из антител.
Фиг. 5 представляет графики, показывающие результаты анализа AIEX каждого из антител.
Фиг. 6 представляет график, показывающий результат анализа AIEX каждого из антител.
Фиг. 7 представляет графики, показывающие результаты анализа CIEX каждого из антител.
Фиг. 8 представляет графики, показывающие результаты анализа CIEX каждого из антител.
Фиг. 9-1 представляет графики, показывающие результаты анализа CIEX каждого из антител.
Фиг. 9-2 является продолжением Фиг. 9-1.
Фиг. 10 представляет графики, показывающие результаты анализа CIEX каждого из антител.
Фиг. 11 представляет графики, показывающие результаты анализа CIEX каждого из антител.
Фиг. 12 представляет диаграмму, которая сравнивает CH1 H-цепи путем выравнивания аминокислотных последовательностей человеческого IgA1 (SEQ ID NO: 63), IgA2 (SEQ ID NO: 64), IgD (SEQ ID NO: 65), IgE (SEQ ID NO: 66), IgG1 (SEQ ID NO: 67), IgG2 (SEQ ID NO: 68), IgG3 (SEQ ID NO: 69), IgG4 (SEQ ID NO: 70) и IgM (SEQ ID NO: 71); и CL L-цепи путем выравнивания аминокислотных последовательностей человеческого IgK (Каппа) (SEQ ID NO: 72), IgL1 (SEQ ID NO: 73), IgL2 (SEQ ID NO: 74), IgL3 (SEQ ID NO: 75), IgL6 (SEQ ID NO: 76), IgL7 (SEQ ID NO: 77) (Лямбда).
[Режим осуществления изобретения]
Настоящее изобретение касается антител, в которых ассоциация тяжелых цепей и легких цепей является регулируемой, способов получения антитела, в котором ассоциация тяжелых цепей и легких цепей является регулируемой, и способов регулирования ассоциации тяжелых цепей и легких цепей антитела.
Согласно настоящему изобретению, термин "антитело" применяется синонимично с термином "антигенсвязывающая молекула". То есть, согласно настоящему изобретению, термины "антитело" и "антигенсвязывающая молекула" применяются в самом широком смысле и включают моноклональные антитела, поликлональные антитела и варианты антител (такие, как химерные антитела, гуманизированные антитела, низкомолекулярные антитела (включая фрагменты антител, к которым могут произвольно добавляться другие молекулы) и мультиспецифические антитела), при условии, что они демонстрируют необходимую антигенсвязывающую активность или биологическую активность. Примером "антитела" или "антигенсвязывающей молекулы" согласно настоящему изобретению может быть молекула, в которой к Fab (антителу, в котором нормальной является только часть Fab) добавлен HAS-связывающий скелет. Кроме того, согласно настоящему изобретению, "антитело" также может быть полипептидом или гетеромерным мультимером. Предпочтительными антителами являются моноклональные антитела, химерные антитела, гуманизированные антитела, человеческие антитела, Fc-слитые антитела и низкомолекулярные антитела, такие, как фрагменты антител.
Антитело согласно настоящему изобретению представляет собой антитело, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой, в котором тяжелая цепь и легкая цепь, составляющие антитело, являются нужной комбинацией тяжелой цепи и легкой цепи, и в котором аминокислотные остатки в данных местах в константной области тяжелой цепи (CH1) и константной области легкой цепи являются электрически взаимно отталкивающимися аминокислотными остатками (обладающими одноименными зарядами).
Согласно настоящему изобретению, преобразование аминокислотных остатков в данных местах в константной области тяжелой цепи (CH1) и константной области легкой цепи нежелательной комбинации тяжелой цепи и легкой цепи в аминокислотные остатки, которые электрически взаимно отталкиваются (то есть, обладающие одноименным зарядом), позволяет избегать образования нежелательных комбинаций тяжелой цепи и легкой цепи путем применения этого отталкивания зарядов, и в результате может быть образована необходимая комбинация тяжелой цепи и легкой цепи.
В другом варианте осуществления антитело согласно настоящему изобретению является антителом, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой, в котором тяжелая цепь и легкая цепь, составляющие антитело, ассоциируются как нужная комбинация тяжелой цепи и легкой цепи, и в котором аминокислотные остатки в данных местах в константной области тяжелой цепи (CH1) и в константной области легкой цепи электрически взаимно не отталкиваются. Преобразование аминокислотных остатков в данных местах в константной области тяжелой цепи (CH1) и константной области легкой цепи нужной комбинации тяжелой цепи и легкой цепи в аминокислотные остатки, которые электрически взаимно не отталкиваются, позволяет образовывать нужную комбинацию тяжелой цепи и легкой цепи, например, путем применения силы притяжения электрических зарядов.
В контексте настоящего изобретения термин "полипептид" в целом относится к пептидам и белкам, длина которых составляет приблизительно десять аминокислот или более. Полипептиды изначально берут из организмов, но не обязательно ограничиваются организмами и, например, могут состоять из искусственно построенной последовательности. Они также могут быть любыми природными полипептидами, синтетическими полипептидами, рекомбинантными полипептидами и т.п. Кроме того, фрагменты вышеупомянутых полипептидов также включаются в полипептиды согласно настоящему изобретению.
В контексте настоящего изобретения фразы "регулировать ассоциацию" и "ассоциация является регулируемой" касаются регулирования для достижения нужного состояния ассоциации, более конкретно - регулирования таким образом, чтобы не образовывались нежелательные ассоциации между тяжелой цепью и легкой цепью.
В контексте настоящего изобретения термин "граница" в целом касается поверхности ассоциации, образуемой в результате ассоциации (взаимодействия), и аминокислотные остатки, образующие границу, изначально представляют собой один или несколько аминокислотных остатков, включенных в полипептидные участки, участвующие в ассоциации, и более предпочтительно являются аминокислотными остатками, приближающимися друг к другу во время ассоциации, и принимают участие во взаимодействии. Более конкретно это взаимодействие включает, например, случаи, в которых аминокислотные остатки приближаются вплотную во время ассоциации для образования водородных связей, электростатического взаимодействия или солевых мостиков друг с другом.
В контексте настоящего изобретения фраза "аминокислотные остатки, образующие границу" более конкретно касается аминокислотных остатков, включенных в полипептидный участок, составляющий границу. Например, полипептидные участки, составляющие границу, означают полипептидные участки, отвечающие за выборочное связывание между молекулами, такими, как антитела, лиганды, рецепторы или субстраты. Более конкретно в антителах такие примеры включают константные области тяжелой цепи, вариабельные области тяжелой цепи, константные области легкой цепи и вариабельные области легкой цепи.
"Модификация" аминокислотных остатков в контексте настоящего изобретения конкретно касается замещения первоначального(ых) аминокислотного(ых) остатка(ов) другим(и) аминокислотным(и) остатком(ами), удаление первоначального(ых) аминокислотного(ых) остатка(ов), добавление нового(ых) аминокислотного(ых) остатка(ов) и т.п., однако предпочтительно касается замещения одного или нескольких первоначальных аминокислотных остатков другими аминокислотными остатками.
В предпочтительном варианте осуществления антитела, согласно настоящему изобретению, антитело имеет аминокислотные остатки в данных местах в константной области тяжелой цепи (CH1) и константной области легкой цепи нежелательной комбинации тяжелой цепи и легкой цепи перед регулированием ассоциации, которые электрически отталкиваются (которые обладают одноименным зарядом).
Считается, что путем модификации аминокислотных остатков в вышеупомянутом антителе до аминокислотных остатков, которые электрически взаимно отталкиваются (обладают одноименным зарядом) ассоциация этих аминокислотных остатков ингибируется силой отталкивания электрических зарядов.
В другом предпочтительном варианте осуществления антитела согласно настоящему изобретению антитело имеет аминокислотные остатки, участвующие в ассоциации на границе полипептидов, которые электрически взаимно не отталкиваются.
Считается, что в вышеупомянутом антителе путем модификации аминокислотных остатков, участвующих в ассоциации на границе полипептидов, до аминокислотных остатков, которые электрически взаимно не отталкиваются, обеспечивается содействие ассоциации этих аминокислотных остатков, например, силой притяжения их электрических зарядов.
Таким образом, в вышеупомянутом антителе модифицированными аминокислотными остатками предпочтительно являются аминокислотные остатки, приближающиеся друг к другу при ассоциации, в полипептидных участках, образующих границу.
Аминокислотные остатки, приближающиеся во время ассоциации, могут определяться, например, путем анализа трехмерной структуры полипептида и исследования аминокислотных последовательностей полипептидных участков, образующих границу во время ассоциации полипептидов. Аминокислотные остатки на границе, которые взаимно приближаются друг к другу, предпочтительно являются мишенями "модификации" в антителе согласно настоящему изобретению.
Известно, что некоторые аминокислоты являются электрически заряженными. Как правило, лизин (K), аргинин (R) и гистидин (H) являются аминокислотами, обладающими положительным зарядом (положительно заряженными аминокислотами). Аспарагиновая кислота (D), глутаминовая кислота (E) и т.п. известны как аминокислоты, обладающие отрицательным зарядом (отрицательно заряженными аминокислотами). Кроме того, аланин (A), аспарагин (N), цистеин (C), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), пролин (P), серии (S), треонин (Т), триптофан (W), тирозин (Y), валин (V) и т.п. известны как аминокислоты, не обладающие зарядом, или неполярные аминокислоты.
Таким образом, аминокислотами, которые электрически взаимно отталкиваются (обладают одноименным зарядом), согласно настоящему изобретению, являются:
(1) аминокислоты, в которых одна из аминокислот является положительно заряженной аминокислотой, и другая аминокислота также является положительно заряженной аминокислотой, и
(2) аминокислоты, в которых одна из аминокислот является отрицательно заряженной аминокислотой, и другая аминокислота также является отрицательно заряженной аминокислотой.
Кроме того, аминокислотами, которые электрически взаимно не отталкиваются, согласно настоящему изобретению, являются:
(1) аминокислоты, в которых одна из аминокислот является положительно заряженной аминокислотой, а другая аминокислота является отрицательно заряженной аминокислотой,
(2) аминокислоты, в которых одна из аминокислот является положительно заряженной аминокислотой, а другая аминокислота является незаряженной аминокислотой или неполярной аминокислотой,
(3) аминокислоты, в которых одна из аминокислот является отрицательно заряженной аминокислотой, и другая аминокислота является незаряженной аминокислотой или неполярной аминокислотой, и
(4) аминокислоты, в которых обе аминокислоты являются незаряженными аминокислотами или неполярными аминокислотами.
Аминокислоты могут быть модифицированы в соответствии с различными способами, известными специалистам в данной области. Примерами таких способов являются, помимо прочих, сайт-специфический мутагенез (Hashimoto-Gotoh, T., Mizuno, T., Ogasahara, Y. and Nakagawa, M. (1995) An oligodeoxyribonucleotide-directed dual amber method for site-directed mutagenesis, Gene 152, 271 - 275; Zoller, M.J. and Smith, M. (1983) Oligonucleotide-directed mutagenesis of DNA fragments cloned into M13 vectors, Methods Enzymol. 100, 468-500; Kramer, W., Drutsa, V., Jansen, H.W., Kramer, B., Pflugfelder, M. and Fritz, H.J. (1984) The gapped duplex DNA approach to oligonucleotide-directed mutation construction, Nucleic Acids Res. 12, 9441 - 9456; Kramer, W. and Fritz, H.J. (1987) Oligonucleotide-directed construction of mutations via gapped duplex DNA, Methods Enzymol. 154, 350-367; Kunkel, T.A. (1985) Rapid and efficient site-specific mutagenesis without phenotypic selection, Proc. Natl. Acad. Sci. USA 82, 488-492), ПЦР-мутагенез, кассетный мутагенез и т.п.
Примерами модификации аминокислот могут быть модификации незаряженной аминокислоты или неполярной аминокислоты до положительно заряженной аминокислоты, модификация незаряженной аминокислоты или неполярной аминокислоты до отрицательно заряженной аминокислоты, модификация положительно заряженной аминокислоты до отрицательно заряженной аминокислоты и модификация отрицательно заряженной аминокислоты до положительно заряженной аминокислоты. Кроме того, модификация незаряженной аминокислоты или неполярной аминокислоты до другой незаряженной или неполярной аминокислоты, модификация положительно заряженной аминокислоты до другой положительно незаряженной аминокислоты и модификация отрицательно заряженной аминокислоты до другой отрицательно заряженной аминокислоты также включаются в модификации аминокислот согласно настоящему изобретению.
Модификация аминокислот согласно настоящему изобретению включает создание одной модификации в каждой из тяжелой и легкой цепей или создание множественных модификаций для каждой из тяжелой и легкой цепей. Кроме того, количество модификаций, добавленных к тяжелой цепи и легкой цепи, может быть одинаковым или разным.
Модификация аминокислот согласно настоящему изобретению включает создание множественных модификаций в положительно заряженных аминокислотах в тяжелой цепи или легкой цепи и создание множественных модификаций в отрицательно заряженных аминокислотах в другой цепи. Кроме того, множественные модификации в положительно заряженных аминокислотах, а также множественные модификации в отрицательно заряженных аминокислотах возможны в одной и той же тяжелой цепи или легкой цепи. В этих модификациях также могут соответствующим образом комбинироваться модификации до незаряженных аминокислот или неполярных аминокислот, а также модификации незаряженных аминокислот или неполярных аминокислот.
При модификациях согласно настоящему изобретению, например, аминокислоты в одной из цепей могут использоваться в немодифицированном виде, и в таких случаях тяжелая цепь и легкая цепь не обязательно обе должны быть модифицированы, а может быть модифицирована лишь одна из цепей
Хотя не предусмотрено особых ограничений в отношении количества аминокислотных остатков, подлежащих модификации в антителе согласно настоящему изобретению, например, при модификации константной области антитела, для того, чтобы не снижать активность связывания с антигеном и не повышать иммуногенность, предпочтение отдается модификации как можно меньшего количества аминокислотных остатков. Вышеупомянутое слово "меньшее" означает, например, количество приблизительно от 1 до 30, предпочтительно приблизительно от 1 до 20, более предпочтительно - количество приблизительно от 1 до 15, и наиболее предпочтительно - количество от 1 до 5.
В контексте настоящего изобретения термин "антитело" применяется в самом широком смысле и включает моноклональные антитела, поликлональные антитела, варианты антител (такие, как химерные антитела, гуманизированные антитела, низкомолекулярные антитела (включая фрагменты антител) и мультиспецифические антитела), при условии, что они демонстрируют необходимую биологическую активность. Кроме того, "антитело" согласно настоящему изобретению может быть полипептидом или гетеромерным мультимером. Предпочтительными антителами являются моноклональные антитела, химерные антитела, гуманизированные антитела, человеческие антитела, Fc-слитые антитела и низкомолекулярные антитела, такие, как фрагменты антител.
В контексте настоящего изобретения термин "мультиспецифическое антитело" (применяемый в представленном описании в том же значении, что и "полиспецифическое антитело") относится к антителу, которое может специфически связываться с разными типами эпитопов. Более конкретно, мультиспецифическими антителами являются антитела, обладающие специфичностью по меньшей мере к двум разным типам эпитопов и, помимо антител, распознающих разные антигены, также включаются антитела, распознающие разные эпитопы на одном антигене. (Например, если антигены являются гетерологичными рецепторами, мультиспецифические антитела могут распознавать различные домены, составляющие гетерологичные рецепторы; в альтернативном варианте, если антигены являются мономерами, мультиспецифические антитела распознают множественные сайты на мономерных антигенах). Первоначально такие молекулы связываются с двумя антигенами (биспецифические антитела; применяются согласно изобретению в том же значении, что и "антитела двойной специфичности"), однако они даже могут обладать специфичностью в отношении большего количества антигенов (например, трех типов).
Помимо описанных выше антител, к антителам согласно настоящему изобретению относятся антитела, аминокислотные последовательности которых были модифицированы аминокислотными замещениями, делециями, добавлениями и/или инсерциями или химеризацией, гуманизацией и т.п.Такие модификации аминокислотных последовательностей, такие, как аминокислотные замещения, делеции, добавления и/или инсерции и гуманизация и химеризация, могут достигаться способами, известными специалистам в данной области. Если антитела согласно настоящему изобретению получают как рекомбинантные антитела, подобным образом аминокислотные последовательности вариабельных и константных областей антитела также могут быть модифицированы аминокислотными замещениями, делециями, добавлениями и/или инсерциями или путем химеризации, гуманизации и т.п.
Антитела согласно настоящему изобретению могут быть взяты из организма любого животного, такого, как мышь, человек, крыса, кролик, коза или верблюд. Кроме того, антитела могут быть модифицированы, например, химерные антитела, в особенности модифицированные антитела, включающие аминокислотные замещения в их последовательностях, такие, как гуманизированные антитела. Антитела могут относиться к любому типу антител, такому, как продукты модификации антител, связанные с различными молекулами, фрагменты антител и низкомолекулярные антитела.
"Химерные антитела" являются антителами, полученными путем сочетания последовательностей, взятых из организмов различных животных. Примером может быть антитело, имеющее вариабельные (V) области тяжелой и легкой цепи антитела мыши и константные (C) области тяжелой и легкой цепи человеческого антитела. Химерные антитела могут быть получены известными способами. Для получения таких химерных антител, например, ДНК, кодирующая V-область антитела, может быть сшита с ДНК, кодирующей константную область человеческого антитела; полученный в результате продукт сшивания может быть вставлен в вектор экспрессии; и последовательность может быть включена в организм-хозяин для образования химерного антитела.
"Гуманизированные антитела" также называются реконструированными человеческими антителами и могут быть получены путем замены гипервариабельного участка (CDR) человеческого антитела на CDR антитела, взятого из организма отличного от человека млекопитающего, например, мыши. Способы распознавания CDR известны специалистам в данной области (Kabat et al., Sequence of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al, Nature (1989) 342: 877). Общие технологии генетической рекомбинации, подходящие для этой цели, также известны (см. публикацию европейской патентной заявки №ЕР 125023; и документ WO 96/02576). Например, CDR антитела мыши может быть определен известными способами, и ДНК может быть подготовлена таким образом, чтобы она кодировала антитело, в котором CDR сшит с каркасным участком (FR) человеческого антитела. Гуманизированное антитело затем может быть получено с применением системы, в которой используются традиционные векторы экспрессии. Такие ДНК могут быть синтезированы путем ПЦР, с применением в качестве праймеров нескольких олигонуклеотидов, построенных таким образом, чтобы они включали части, которые частично накладываются на концы участков CDR и FR (см. способ, описанный в документе WO 98/13388). FR антитела человека, соединенные через CDR, выбирают таким образом, чтобы CDR образовывали подходящий антигенсвязывающий центр. В случае необходимости аминокислоты в FR вариабельной области антитела могут быть модифицированы таким образом, чтобы CDR реконструированного человеческого антитела могли образовывать подходящий антигенсвязывающий центр (Sato, К. et al., Cancer Res. (1993) 53: 851-856). Модифицируемые аминокислотные остатки в FR включают части, которые непосредственно связываются с антигеном через нековалентные связи (Amit et al., Science (1986) 233: 747-53), части, определенным образом влияющие или воздействующие на структуру CDR (Chothia et al, J. Mol. Biol. (1987) 196: 901-17), и части, участвующие во взаимодействии между VH и VL (документ EP 239400).
Константная область тяжелой цепи антитела согласно настоящему изобретению предпочтительно является человеческой константной областью тяжелой цепи. Кроме того, примерами константных областей тяжелой цепи антитела могут быть константные области типов IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 и IgM. Константная область тяжелой цепи антитела согласно настоящему изобретению предпочтительно является константной областью типа IgG1, особенно предпочтительно - константной областью человеческого IgG1, но не ограничивается ею. Несколько аллотипических последовательностей, полученных при помощи генетического полиморфизма, описываются в публикации Sequences of Proteins, имеющих иммунологическое значение, Публикация NIH №91-3242 как константная область человеческого IgG1, и любая из них может применяться согласно настоящему изобретению.
Кроме того, константная область легкой цепи антитела согласно настоящему изобретению предпочтительно является человеческой константной областью легкой цепи. Примерами константной области легкой цепи антитела являются константные области типов IgK (Каппа), IgL1, IgL2, IgL3, IgL6 и IgL7 (Лямбда). Константная область легкой цепи антитела согласно настоящему изобретению предпочтительно является константной областью человеческого IgK (Каппа), но не ограничивается ею. Аминокислотная последовательность константной области человеческого IgK (Каппа) известна (SEQ ID NO: 72). Несколько аллотипических последовательностей, полученных при помощи генетического полиморфизма описываются в публикации Sequences of Proteins, имеющих иммунологическое значение, Публикация NIH №91-3242 как константная область человеческого IgK (Каппа) и константная область человеческого IgL7 (Лямбда), и любая из них может использоваться согласно настоящему изобретению.
Константные области антитела, в частности, константные области тяжелой цепи, могут быть модифицированы в случае необходимости с целью улучшения функции антитела или устойчивости антитела. Примерами модификаций для улучшения функции антитела являются модификации, усиливающие или ослабляющие связывание между антителом и Feγ рецептором (FcγR), модификации, усиливающие или ослабляющие связывание между антителом и FcRn, модификации, усиливающие или ослабляющие цитотоксическая активность антитела (такую, как антителозависимая клеточно-обусловленная цитотоксичность и комплементзависимая цитотоксичность) и т.п. Кроме того, также могут включаться модификации, улучшающие гетерогенность антитела и модификации, улучшающие иммуногенность и/или фармакокинетику.
Кроме того, в качестве гетерогенности C-концевой последовательности тяжелой цепи антитела IgG сообщалось об амидировании C-концевой карбоксильной группы путем делеции C-концевой аминокислоты, лизинового остатка или путем делеции двух C-концевых аминокислот, глицина и лизина (Anal. Biochem. 2007 Jan 1: 360 (1): 75-83). Таким образом, согласно настоящему изобретению, для уменьшения гетерогенности С-конца тяжелой цепи, предпочтение отдается использованию IgG, в котором C-концевой лизин или C-концевой лизин и глицин были делегированы.
Поскольку их антигенность в организме человека была ослаблена, ожидается, что химерные или гуманизированные антитела с использованием взятых из организма человека последовательностей могут использоваться при введении в организм человека з терапевтическими или другими подобными целями.
Кроме того, низкомолекулярные антитела могут использоваться в качестве антител благодаря их in vivo кинетическим характеристикам и дешевому производству с применением E. coli, растительных клеток и т.п.
Фрагменты антител представляют один тип низкомолекулярного антитела. Термин "низкомолекулярное антитело" включает антитела, включающие фрагмент антитела в качестве частичного структурного элемента. Низкомолекулярные антитела согласно настоящему изобретению не имеют особых ограничений ни по структуре, ни по способу получения, при условии, что они сохраняют антигенсвязывающую активность. Некоторые низкомолекулярные антитела обладают активностью, превышающей активность целого антитела (Orita et al, Blood (2005) 105: 562-566). В контексте данного описания "фрагменты антител" не имеют особых ограничений, при условии, что они являются частью целого антитела (например, целого IgG). Однако фрагменты антител предпочтительно включают вариабельную область тяжелой цепи (VH) или вариабельную область легкой цепи (VL), а также включают CH1 или CL. Примерами предпочтительных фрагментов антител являются: Fab, F(ab')2 и Fab'. Аминокислотные последовательности VH, VL, CH1 и CL в фрагменте антитела могут быть модифицированы путем замещения, делеции, добавления и/или инсерции. Кроме того, некоторые части CHI1 CL, VH и VL могут быть делетированы, при условии, что образованные в результате фрагменты сохраняют свою антигенсвязывающую способность, и фрагменты антител, такие, как scFv, Fab, доменное антитело (dAb) и VHH, HAS-связывающий скелет, PEG, альбумин, цитокины, токсины и т.п. (молекулы, описанные в публикации Biodrugs, 2009, 23 (2): 93-109; Methods Mol. Med., 2005, 109: 347-74; AAPS J., 2006 Aug 18, 8 (3): E532 - 51; и т.д.) также могут добавляться для повышения фармакокинетики (РК) или эффективности медикамента.
Фрагмент антитела может быть получен путем обработки антитела ферментом, например, протеазой, такой, как папаин или пепсин (см. Morimoto et al., J. Biochem. Biophys. Methods (1992) 24: 107-17; Brennan et al., Science (1985) 229: 81). В альтернативном варианте фрагменты антител также могут быть получены путем генетической рекомбинации на основе их аминокислотной последовательности.
Низкомолекулярное антитело, имеющее структуру, которая является результатом модификации фрагмента антитела, может быть получено с применением фрагментов антител, полученных путем ферментной обработки или генетической рекомбинации. В альтернативном варианте после построения гена, кодирующего целое низкомолекулярное антитело, и включения последовательности в вектор экспрессии низкомолекулярное антитело может быть экспрессировано в соответствующих клетках-хозяевах (см., например, Со et al., J. Immunol. (1994) 152: 2968-76; Better and Horwitz, Methods Enzymol. (1989) 178: 476-96; Pluckthun and Skerra, Methods Enzymol. (1989) 178: 497-515; Lamoyi, Methods Enzymol. (1986) 121: 652-63; Rousseaux et al., Methods Enzymol. (1986) 121: 663-9; Bird and Walker, Trends Biotechnol. (1991) 9: 132-7).
Предпочтительным примером антитела согласно настоящему изобретению является гетеромерный мультимер, имеющий два или более типов CH1 и два или более типов CL. Этот гетеромерный мультимер предпочтительно распознает два или более типов эпитопов, и его примером может быть полиспецифическое антитело.
Предпочтительным примером полиспецифического антитела согласно настоящему изобретению является биспецифическое антитело. Таким образом, примером предпочтительного варианта осуществления антитела согласно настоящему изобретению является биспецифическое антитело, состоящее из двух типов тяжелых цепей (первой тяжелой цепи и второй тяжелой цепи) и двух типов легких цепей (первой легкой цепи и второй легкой цепи).
В более подробном описании "биспецифических антител" согласно предпочтительным вариантам осуществления антител в соответствии с настоящим изобретением вышеупомянутая "первая тяжелая цепь" означает одну из двух тяжелых цепей (H-цепей), образующих антитело, в "вторая H-цепь" означает другую H-цепь, отличную от первой H-цепи. То есть, из двух H-цепей одна может произвольно определяться как первая H-цепь, а другая может определяться как вторая H-цепь. Подобным образом "первая легкая цепь" означает одну из двух легких цепей (L-цепей), образующих биспецифическое антитело, а "вторая L-цепь" означает другую L-цепь, отличную от первой L-цепи. Из двух L-цепей одна может произвольно определяться как первая L-цепь, а другая может определяться как вторая L-цепь. Как правило, первая L-цепь и первая H-цепь происходят из одного антитела, которое распознает определенный антиген (или эпитоп), а вторая L-цепь и вторая H-цепь также происходят из одного антитела, которое распознает определенный антиген (или эпитоп). В контексте данного описания пара L-цепь-H-цепь, образуемая первыми H-цепью и L-цепью, называется первой парой, а пара L-цепь-H-цепь, образуемая вторыми H-цепью и L-цепью, называется второй парой. Антиген (или эпитоп), используемый для получения антитела, из которого происходит вторая пара, предпочтительно отличается от антигена, используемого для получения антитела, из которого происходит первая пара. Более конкретно, антигены, распознаваемые первой парой и второй парой, могут быть одинаковыми, однако в предпочтительном варианте пары распознают различные антигены (или эпитопы). В этом случае H-цепи и L-цепи первой пары и второй пары предпочтительно имеют аминокислотные последовательности, отличающиеся одна от другой. Когда первая пара и вторая пара распознают разные эпитопы, первая пара и вторая пара могут распознавать полностью отличный антиген, или же они могут распознавать разные центры (разные эпитопы) на одном антигене. Кроме того, одна из них может распознавать антиген, такой, как белок, пептид, ген или сахар, а другая может распознавать цитотоксичные вещества, такие, как радиоактивные вещества, химиотерапевтические вещества или взятые из клеток токсины. Однако в случае потребности в создании антитела, имеющего пары, образованные специфическими комбинациями H-цепей и L-цепей, эти специфические H-цепи и L-цепи могут произвольно определяться как первая пара и вторая пара.
Что касается генов, кодирующих H-цепь или L-цепь антител перед включением мутаций в настоящее изобретение (в данном описании оно может просто указываться как "антитело согласно настоящему изобретению"), могут использоваться известные последовательности, или же они могут быть получены способами, известными специалистам в данной области. Например, они могут быть получены из библиотеки антител, или же они могут быть получены путем клонирования генов, кодирующих антитело из гибридом, вырабатывающих моноклональные антитела.
Что касается библиотек антител, многие библиотеки антител уже хорошо известны, и поскольку способы получения библиотек антител известны, специалисты в данной области смогут соответствующим образом получить библиотеки антител. Например, в отношении фаговых библиотек антител можно обратиться к литературным источникам, таким, как публикации Clackson et al, Nature 1991, 352: 624-8; Marks et al, J. Mol. Biol. 1991, 222: 581-97; Waterhouses et al, Nucleic Acids Res. 1993, 21: 2265-6; Griffiths et al, EMBO J. 1994, 13: 3245-60; Vaughan et al, Nature Biotechnology 1996, 14: 309-14; и публикация японского патента Kohyo (JP-A) H10-504970 (нерассмотренная японская публикация национальной фазы, соответствующая неяпонской международной публикации). Кроме того, могут применяться известные способы, такие, как способы, согласно которым используются эукариотные клетки, в качестве библиотек (WO 95/15393) и способы рибосомных дисплеев. Кроме того, также известны технологии получения человеческих антител путем пэннинга с применением библиотек антител человека. Например, вариабельные области человеческих антител могут быть экспрессированы на поверхности фагов как одноцепочечные антитела (scFvs) с применением способов фаговых дисплеев, и могут быть выбраны фаги, связывающиеся с антигенами. Генетический анализ выбранных фагов может определять последовательности ДНК, кодирующие вариабельные области человеческих антител, которые связываются с антигенами. Сразу после обнаружения последовательностей ДНК scFvs, которые связываются с антигенами, могут быть получены соответствующие векторы экспрессии на основе этих последовательностей для получения человеческих антител. Эти способы уже хорошо известны, и можно обратиться к источникам WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438 и WO 95/15388.
Что касается способов получения генов, кодирующих антитела из гибридом, в основном могут применяться известные технологии, которые включают использование необходимых антигенов или клеток, экспрессирующих необходимые антигены в качестве сенсибилизирующих антигенов, их использование для иммунизации в соответствии с традиционными способами иммунизации, слияние полученных таким образом иммунных клеток с известными родительскими клетками обычными способами слияния клеток, отбор вырабатывающих моноклональное антитело клеток (гибридом) обычными способами отбора, синтез кДНК вариабельных областей антитела (V-областей) из мРНК полученных гибридом с применением обратной транскриптазы и их соединение с ДНК, кодирующими необходимые константные области антитела.
Сенсибилизирующие антигены для получения вышеупомянутых генов антител, кодирующих H-цепь и L-цепь, не ограничиваются конкретными описанными ниже примерами, а включают как полные антигены, обладающие иммуногенностью, так и неполные антигены, включая гаптены и т.п., которые не демонстрируют иммуногенности. Особых ограничений относительно антигена для антител согласно настоящему изобретению не существует, и могут использоваться, например, белок полной длины или частичный пептид белка-мишени, а также вещества, состоящие из полисахаридов, нуклеиновых кислот, липидов и т.п., которые наверняка могут служить в качестве антигена. Антигены могут быть получены в соответствии со способами, известными специалистам в данной области, такими, как способы с использованием бакуловирусов (таких, как описанные в документе WO 98/46777). Гибридомы могут быть получены, например, согласно способам, описанным в публикации Milstein et al. (G. Kohler and C. Milstein, Methods Enzymol. 1981, 73: 3-46) и т.п.При низкой иммуногенности антигена он может связываться с макромолекулой, обладающей иммуногенностью, такой, как альбумин, а затем применяться для иммунизации. Кроме того, благодаря связыванию антигенов с другими молекулами в случае необходимости, они могут быть преобразованы в растворимые антигены. При применении трансмембранных молекул, таких, как рецепторы, в качестве антигенов части внеклеточных участков рецепторов могут использоваться как фрагмент, или клетки, экспрессирующие трансмембранные молекулы на их поверхности, могут использоваться как иммуногены.
Вырабатывающие антитело клетки могут быть получены путем иммунизации животных с применением соответствующих сенсибилизирующих антигенов, описанных выше. В альтернативном варианте вырабатывающие антитело клетки могут быть получены путем in vitro иммунизации лимфоцитов, которые могут вырабатывать антитела. Различные животные могут использоваться в качестве животных для иммунизации, причем обычно используют грызунов, зайцеобразных и приматов. Конкретными примерами таких животных являются мыши, крысы и хомяки как грызуны, кролики как зайцеобразные и обезьяны, включая яванского макака, макака-резуса, гамадрила и шимпанзе, как приматы.
Также известны трансгенные животные, имеющие наборы генов человеческого, и человеческие антитела могут быть получены путем использования этих животных (см. документ WO 96/34096; Mendez et al, Nat. Genet. 1997, 15: 146-56). Вместо использования таких трансгенных животных, например, могут быть получены необходимые человеческие антитела, обладающие активностью связывания против антигенов путем in vitro сенсибилизации человеческих лимфоцитов с нужными антигенами или клеток, экспрессирующих нужные антигены, с последующим слиянием сенсибилизированных лимфоцитов с клетками миеломы человека, например, U266 (см. публикацию японской патентной заявки Kokoku (JP-B) H1-59878 (рассмотренная, утвержденная японская патентная заявка, опубликованная для противопоставления)). Кроме того, нужные человеческие антитела могут быть получены путем иммунизации трансгенных животных, имеющих полный набор генов человеческого антитела с нужными антигенами (см. документы WO 93/12227, WO 92/03918, WO 94/02602, WO 96/34096 и WO 96/33735).
Иммунизацию животных осуществляют путем соответствующего разведения и суспендирования сенсибилизирующего антигена в фосфатно-буферном растворе (PBS), физиологическом растворе и т.п., с образованием эмульсии путем смешивания адъюванта в случае необходимости и ее внутрибрюшинной или подкожной инъекции животным. После этого сенсибилизирующий антиген, смешанный с неполным адъювантом Фрейнда, в предпочтительном варианте вводят несколько раз каждые 4-21 день. Выработка антитела может подтверждаться путем измерения титра антитела-мишени в сыворотке животных с применением традиционных способов.
Вырабатывающие антитело клетки, полученные из лимфоцитов или животных, иммунизированных необходимым антигеном, могут сливаться с клетками миеломы для образования гибридом с применением традиционных факторов, вызывающих слияние (например, полиэтиленгликоля) (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, 1986, 59-103). Затем в случае необходимости клетки гибридомы культивируют и выращивают и измеряют специфичность связывания, аффинность или активность антитела, выработанного из этих гибридом, применяя известные способы анализа, такие, как иммунопреципитация, радиоиммуноанализ (RIA) и твердофазный иммуносорбентный анализ (ELISA). Затем гибридомы, вырабатывающие нужные антитела, специфичность связывания, аффинность или активность которых была определена, могут быть субклонированы такими способами, как предельное разведение.
Затем гены, кодирующие нужные антитела, могут быть клонированы из гибридом или вырабатывающих антитело клеток (сенсибилизированных лимфоцитов и т.п) с применением зондов, которые могут специфично связываться с антителами (например, олигонуклеотидов, комплементарными последовательностям, кодирующим константные области антитела). Также возможно клонирование из мРНК с применением RT-PCR. Иммуноглобулины разделяют на пять разных классов, IgA, IgD, IgE, IgG и IgM. Эти классы далее разделяются на несколько подклассов (изотипов) (например, IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2; и т.п.). H-цепи и L-цепи, используемые согласно настоящему изобретению для выработки антител, не имеют особых ограничений и могут быть взяты из антител, относящихся к этим классам или подклассам; однако особенное предпочтение отдается IgG.
Также существует возможность модификации кодирующих H-цепь генов и кодирующих L-цепь генов с применением технологий генной инженерии. Генетически модифицированные антитела, такие, как химерные антитела и гуманизированные антитела, которые были искусственно модифицированы с целью снижения гетерологичной антигенности, и антитела против человека, могут быть соответствующим образом получены для таких антител, как антитела мыши, антитела крысы, антитела кролика, антитела хомяка, антитела овцы и антитела верблюда.
Химерными антителами являются антитела, состоящие из вариабельных областей H-цепи и L-цепи антитела отличного от человека млекопитающего, например, принадлежащих антителу мыши, и константных областей H-цепи и L-цепи человеческого антитела. Они могут быть получены путем сшивания ДНК, кодирующей вариабельную область антитела мыши, с ДНК, кодирующей константную область человеческого антитела, включения их в вектор экспрессии и включения вектора в клетку-хозяин для выработки антитела. Гуманизированное антитело также называется реконструированным человеческим антителом. Это гуманизированное антитело может быть синтезировано путем PCR из многих олигонуклеотидов, имеющих перекрывающиеся части на концах последовательностей ДНК, построенных для соединения гипервариабельных участков (CDR) антитела отличного от человека животного (такого, как мышь). Полученная ДНК может быть сшита с ДНК, кодирующей константную область человеческого антитела. Сшитая ДНК может быть включена в вектор экспрессии, и вектор может быть включен в клетку-хозяин для выработки антитела (см. EP 239400 и WO 96/02576). FR антител человека, которые сшиваются через CDR, выбирают в случаях, когда CDR образуют подходящий антигенсвязывающий центр. В случае необходимости аминокислоты в каркасном участке вариабельной области антитела могут быть замещены таким образом, чтобы CDR реконструированного человеческого антитела образовывали соответствующий антигенсвязывающий центр (K. Sato et al., Cancer Res. 1993, 53: 851-856).
Помимо описанных выше способов гуманизации, антитела могут быть модифицированы для улучшения их биологических свойств, например, антигенной аффинности. Такие модификации могут осуществляться с применением таких способов, как сайт-специфический мутагенез (см., например, Kunkel (1985) Proc. Natl. Acad. Sci. USA 82: 488), ПЦР-мутагенез и кассетный мутагенез. В целом мутантные антитела, биологические свойства которых были улучшены, демонстрируют гомологию и/или подобие аминокислотной последовательности 70% или выше, более предпочтительно 80% или выше, еще более предпочтительно - 90% или выше (например, 95% или выше, 97%, 98%, 99% и т.д.) по сравнению с аминокислотной последовательностью вариабельной области первоначального антитела. В контексте данного описания гомология и/или подобие последовательности определяется как соотношение аминокислотных остатков, которые являются гомологичными (одинаковые остатки) или подобными (аминокислотные остатки, относящиеся к одной группе на основе общих свойств аминокислотных боковых цепей) остаткам первоначального антитела, после максимального увеличения значения гомологии последовательности путем выравнивания последовательности и включения гэпа в случае необходимости. Как правило, природные аминокислотные остатки разделяются на группы на основе характеристик их боковых цепей:
(1) гидрофобные: аланин, изолейцин, норлейцин, валин, метионин и лейцин;
(2) нейтральные гидрофильные: аспарагин, глутамин, цистеин, треонин и серии;
(3) кислотные: аспарагиновая кислота и глутаминовая кислота;
(4) основные: аргинин, гистидин и лизин;
(5) остатки, влияющие на ориентацию цепи: глицин и пролин; и
(6) ароматические: тирозин, триптофан и фенилаланин.
Как правило, антигенсвязывающий центр антитела образуется путем взаимодействия всех шести гипервариабельных участков (участков определения комплементарности; CDR), присутствующих в вариабельных областях H-цепи и L-цепи. Известно, что даже одна из этих вариабельных областей способна распознавать и связывать антиген, хотя аффинность ниже, чем показатель вариабельных областей, содержащих все места связывания. Таким образом, в отношении генов антител согласно настоящему изобретению, кодирующих H-цепи и L-цепи, полипептиды, кодируемые этими генами, требуются только для поддержания способности к связыванию с нужным антигеном и для кодирования части фрагмента, содержащей соответствующие антигенсвязывающие центры H-цепи и L-цепи.
Более подробное объяснение представлено ниже на примере биспецифического антитела IgG-типа, имеющего два типа константных областей тяжелой цепи CHI (CH1-А и CH1-В) и два типа константных областей легкой цепи (CL-A и CL-B); однако настоящее изобретение подобным образом также может применяться к другим антителам.
В случае необходимости в получении биспецифического антитела, которое должно распознавать один эпитоп первой CH1-A и первой CL-A, а другой эпитоп - второй CH1-В и второй CL-B, теоретически существует возможность выработки 10 типов молекул антитела, когда каждый из четырех типов цепей экспрессируется для образования этого антитела.
В этом случае молекулы необходимых антител в предпочтительном варианте могут быть получены, если, например, ассоциация регулируется таким образом, чтобы подавлялась ассоциация CH1-A и CL-B и/или между CH1-B и CL-A.
Примером может быть модификация аминокислотных остатков, образующих границу между CH1-A и CL-B, до положительно заряженных аминокислотных остатков и модификация аминокислотных остатков, образующих границу между CH1-B и CL-A до отрицательно заряженных аминокислотных остатков. В результате этих модификаций непреднамеренная ассоциация между CH1-A и CL-B подавляется, поскольку аминокислотные остатки, образующие границу, оба являются положительно заряженными, и ассоциация между CH1-B и CL-A также подавляется, поскольку аминокислотные остатки, образующие границу оба являются отрицательно заряженными. Таким образом, непреднамеренная ассоциация между CH1-A и CL-B и ассоциация между CH1-B и CL-A подавляются, поскольку аминокислотные остатки, образующие границы, обладают одноименными зарядами. В результате могут быть эффективно получены антитела, имеющие преднамеренную ассоциацию между CH1-A и CL-A и преднамеренную ассоциацию между CH1-В и CL-B. Кроме того, преднамеренная ассоциация между CH1-A и CL-A активизируется, поскольку аминокислотные остатки, образующие границу, обладают разными зарядами; и преднамеренная ассоциация между CH1-B и CL-B также активизируется, поскольку аминокислотные остатки, образующие границу, обладают разными зарядами. Таким образом, могут быть эффективно получены антитела с преднамеренной ассоциацией.
Другим примером может быть модификация аминокислотных остатков, образующих границу между CH1-A и CL-B, до положительно заряженных аминокислотных остатков, при которой аминокислотные остатки, образующие границу между CL-A и CH1-B, являются взаимно незаряженными или неполярными аминокислотами. В результате этой модификации непреднамеренная ассоциация между CH1-A и CL-B подавляется, поскольку аминокислотные остатки, образующие границу оба являются положительно заряженными. С другой стороны, поскольку аминокислотные остатки, образующие границу, являются аминокислотами, которые электрически взаимно не отталкиваются, преднамеренная ассоциация между CH1-A и CL-A и преднамеренная ассоциация между CH1-B и CL-B происходят легче, чем в случае, когда аминокислоты электрически отталкиваются. Таким образом, могут быть эффективно получены антитела, имеющие преднамеренную ассоциацию между CH1-A и CL-A и преднамеренную ассоциацию между CH1-B и CL-B. В то же время, в этом примере в случае, когда аминокислотные остатки, образующие границу между CL-A и CH1-B, не являются взаимно незаряженными или неполярными аминокислотами, они могут быть модифицированы таким образом, чтобы стать взаимно незаряженными или неполярными аминокислотами.
В другом примере, когда аминокислотные остатки, образующие границу между CL-B и CH1-B, являются взаимно незаряженными или неполярными аминокислотами, один из аминокислотных остатков, образующих границу между CH1-A и CL-A, модифицируется до положительно заряженного аминокислотного остатка, а другой модифицируется до отрицательно заряженного аминокислотного остатка. В результате этой модификации, хотя преднамеренная ассоциация между CH1-A и CL-A активизируется, поскольку аминокислотные остатки, образующие границу, представляют собой комбинацию положительного заряда и отрицательного заряда, преднамеренная ассоциация между CH1-B и CL-B не подавляется, поскольку аминокислотные остатки, образующие границу, являются аминокислотами, которые электрически взаимно не отталкиваются. В результате можно эффективно получить антитело, имеющее преднамеренную ассоциацию между CH1-A и CL-A и преднамеренную ассоциацию между CH1-B и CL-B. В то же время, в этом примере в случае, когда аминокислотные остатки, образующие границу между CL-B и CH1-B, не являются взаимно незаряженными или неполярными аминокислотами, они могут быть модифицированы таким образом, чтобы стать взаимно незаряженными или неполярными аминокислотами.
Кроме того, в другом примере, когда аминокислотные остатки, образующие границу между CL-B и CH1-B, являются незаряженными или неполярными аминокислотами в CH1-B, один из аминокислотных остатков, образующих границу между CH1-A и CL-A, модифицируется до положительно заряженного аминокислотного остатка, а другой модифицируется до отрицательно заряженного аминокислотного остатка; и аминокислотные остатки, образующие границу между CL-B и CH1-B в CL-B модифицируются таким образом, чтобы обладать одноименным зарядом с модификацией, осуществленной в CH1-A. В результате этой модификации, хотя преднамеренная ассоциация между CH1-A и CL-A активизируется, поскольку аминокислотные остатки, образующие границу, представляют собой комбинацию положительного заряда и отрицательного заряда, преднамеренная ассоциация между CH1-B и CL-B не подавляется, поскольку аминокислотные остатки, образующие границу, являются аминокислотами, которые электрически взаимно не отталкиваются. В результате можно эффективно получить антитело, имеющее преднамеренную ассоциацию между CH1-А и CL-A и преднамеренную ассоциацию между CH1-В и CL-B. В то же время, в этом примере в случае, когда аминокислотные остатки, образующие границу между CL-B и CH1-B, не являются незаряженными или неполярными аминокислотами в CH1-B, они могут быть модифицированы таким образом, чтобы стать незаряженными или неполярными аминокислотами.
Кроме того, применение регулирования ассоциации согласно настоящему изобретению позволяет подавлять ассоциацию между CH1s (CHI-A и CH1-B) или ассоциацию между CL (CL-A и CL-B).
Специалисты в данной области смогут соответствующим образом определить типы аминокислотных остатков, которые приближаются вплотную во время ассоциации на границе CH1 и CL в нужном полипептиде, для которого требуется регулирование ассоциации согласно настоящему изобретению.
Кроме того, специалисты в данной области также могут соответствующим образом получить последовательности, которые могут использоваться как CH1 или CL антитела в организме, например, человека, обезьяны, мыши, кролика и т.п.путем использования открытых баз данных и т.п.Более конкретно, информация об аминокислотной последовательности CH1 или CL может быть получена способами, описанными ниже в Примерах.
Например, в отношении биспецифических антител, описанных ниже в Примерах, конкретные примеры аминокислотных остатков, приближающихся вплотную (находящихся напротив друг друга или контактирующих) на границе CH1 и CL после ассоциации, включают показанные ниже комбинации:
- лизин (K) в позиции 147, как указывается согласно нумерации EC в CH1 (например, позиция 147 в аминокислотной последовательности SEQ ID NO: 1) и находящийся напротив (контактирующий) треонин (T) в позиции 180, как указывается согласно нумерации EC в CL;
- лизин (K) в позиции 147, как указывается согласно нумерации EC в CH1 и находящийся напротив (контактирующий) серии (S) в позиции 131, как указывается согласно нумерации EC в CL;
- глутамин (Q) в позиции 175, как указывается согласно нумерации EC в CH1 и находящийся напротив (контактирующий) глутамин (Q) в позиции 160, как указывается согласно нумерации EC в CL; и
- лизин (K) в позиции 213, как указывается согласно нумерации EC в CH1 и находящийся напротив (контактирующий) глутаминовая кислота (E) в позиции 123, как указывается согласно нумерации EC в CL.
Номера, указываемые в нумерации EC согласно настоящему изобретению указываются в соответствии с нумерацией EC (последовательности белков, имеющих иммунологическое значение, Публикация NIH №91-3242). В контексте настоящего изобретения фразы "аминокислотный остаток в позиции X, как указывается согласно нумерации EC" и "аминокислота в позиции X, как указывается согласно нумерации EC" (где X означает произвольное число) также могут рассматриваться как "аминокислотный остаток, соответствующий позиции X, как указывается согласно нумерации EC" и "аминокислота, соответствующая позиции X, как указывается согласно нумерации EC".
Как указывается в описываемых ниже Примерах, необходимые антитела могут быть в предпочтительном варианте получены путем модификации этих аминокислотных остатков и осуществления способов согласно настоящему изобретению.
Соответственно, настоящее изобретение обеспечивает антитело, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой, причем один, два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), в тяжелой цепи и легкой цепи антитела, являются аминокислотными остатками, которые электрически взаимно отталкиваются:
(a) аминокислотный остаток, содержащийся в константной области тяжелой цепи (CH1) в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в константной области легкой цепи (CL) в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 160, как указывается согласно нумерации EC.
В другом варианте осуществления настоящее изобретение также обеспечивает антитело, в котором аминокислотные остатки в указанном ниже наборе аминокислотных остатков (d) являются аминокислотными остатками, которые электрически взаимно отталкиваются:
(d) аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC.
В вышеупомянутом антителе "аминокислотные остатки, которые электрически взаимно отталкиваются" или "аминокислотные остатки обладающими одноименными зарядами" предпочтительно являются выбранными из аминокислотных остатков, содержащихся, например, в любом из указанных ниже наборов (X) или (Y):
(X) глутаминовая кислота (E) или аспарагиновая кислота (D); или
(Y) лизин (K), аргинин (R), или гистидин (Н).
В вышеупомянутом антителе конкретные примеры аминокислотных остатков, которые электрически взаимно отталкиваются, включают следующие аминокислотные остатки:
- аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, является лизином (K), и аминокислотные остатки, содержащиеся в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, все являются глутаминовой кислотой (E); и,
- аминокислотные остатки, содержащиеся в CH1 в позиции 147 и позиции 175, как указывается согласно нумерации EC, являются глутаминовой кислотой (E), и аминокислотные остатки, содержащиеся в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, все являются лизином (K).
В вышеупомянутом антителе примеры аминокислотных остатков, которые электрически не отталкиваются, также включают пример, в котором аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, является глутаминовой кислотой (E), и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC, является лизином (K).
Кроме того, способы получения вышеупомянутого антитела и соответствующие настоящему изобретению способы регулирования ассоциации путем модификации аминокислотных остатков в наборах вышеупомянутых аминокислотных остатков с (a) по (d) до аминокислотных остатков, которые электрически взаимно отталкиваются, также являются предпочтительными вариантами осуществления настоящего изобретения.
Кроме того, настоящее изобретение обеспечивает антитело, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой, причем один, два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), в ассоциации тяжелой цепи и легкой цепи антитела представляют собой аминокислотные остатки, которые электрически взаимно не отталкиваются:
(a) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 160, как указывается согласно нумерации EC.
В другом варианте осуществления настоящее изобретение также обеспечивает антитело, в котором аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в п. (d), представляют собой аминокислотные остатки, которые электрически взаимно не отталкиваются:
(d) аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC.
Как указывается в Примерах, описанных ниже на Фиг. 1, каждый из аминокислотных остатков вышеупомянутых комбинаций приближается вплотную при ассоциации. Специалисты в данной области смогут определить места, соответствующие аминокислотным остаткам, описанным в вышеупомянутых пп. с (a) по (d), для необходимых CH1 или CL путем гомологичного моделирования и т.п.с применением доступного на рынке программного обеспечения и для соответствующей модификации аминокислотных остатков в этих местах.
В вышеупомянутом антителе "аминокислотные остатки, которые электрически взаимно не отталкиваются" в предпочтительном варианте являются выбранными, например, из каждого из двух наборов, выбранных из группы, к которой относятся пп. с (X) по (Z), указанные ниже, и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z) и (Z) и (Z):
(X) глутаминовая кислота (E) или аспарагиновая кислота (D);
(Y) лизин (K), аргинин (R) или гистидин (H);
(Z) аланин (A), аспарагин (N), цистеин (C), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), пролин (P), серии (S), треонин (T), триптофан (W), тирозин (Y) или валин (V).
Пример включает выбор (X) и (Y) из группы, включающей пп. с (X) по (Z), выбор глутаминовой кислоты из (X) и выбор лизина (K) из (Y); и модификацию аминокислотного остатка, содержащегося в CH1 в позиции 147, как указывается согласно нумерации EC, до глутаминовой кислоты (E) и модификацию аминокислотного остатка, содержащегося в CL в позиции 180, как указывается согласно нумерации EC, до лизина (K). В этом случае существует потребность в модификации аминокислотного остатка, содержащегося в CH1 в позиции 147, как указывается согласно нумерации EC, если аминокислотным остатком является глутаминовая кислота (E) до модификации.
В вышеупомянутом антителе конкретные примеры аминокислотных остатков, которые электрически взаимно не отталкиваются, включают аминокислотные остатки, указанные ниже:
- аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, является лизином (K), и аминокислотные остатки, содержащиеся в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, все являются глутаминовая кислота (E); и
- аминокислотные остатки, содержащиеся в CH1 в позиции 147 и позиции 175, как указывается согласно нумерации EC, являются глутаминовой кислотой (E), и аминокислотные остатки, содержащиеся в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, все являются лизином (K).
В вышеупомянутом антителе, примеры аминокислотных остатков, которые электрически не отталкиваются, также включают пример, в котором аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, является глутаминовой кислотой (E), и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC, является лизином (K).
Кроме того, способы получения вышеупомянутого антитела и соответствующие настоящему изобретению способы регулирования ассоциации путем модификации аминокислотных остатков в вышеупомянутых наборах аминокислотных остатков в пп. с (a) по (d) до аминокислотных остатков, которые электрически взаимно не отталкиваются, также являются предпочтительными вариантами осуществления настоящего изобретения.
Технология включения электрического отталкивания в границу второй константной области тяжелой цепи (CH2) или третьей константной области тяжелой цепи (CH3) для подавления нежелательной ассоциации между тяжелыми цепями, технология включения электрического отталкивания в границу вариабельной области тяжелой цепи и вариабельной области легкой цепи для подавления непреднамеренной ассоциации между тяжелой цепью и легкой цепью или технология модификации аминокислотных остатков, образующих гидрофобное ядро, присутствующее на границе вариабельной области тяжелой цепи и вариабельной области легкой цепи до полярных аминокислот, обладающих электрическим зарядом, для подавления непреднамеренной ассоциации между тяжелой цепью и легкой цепью также могут применяться к антителу согласно настоящему изобретению (см. документ WO 2006/106905).
Согласно технологии подавления непреднамеренной ассоциации между тяжелыми цепями путем включения электрического отталкивания на границе CH2 или CH3, примерами аминокислотных остатков, контактирующих на границе других константных областей тяжелой цепи, могут быть участки, соответствующие позиции 377 (позиция 356) и позиции 470 (позиция 439), позиции 378 (позиция 357) и позиции 393 (позиция 370), а также позиции 427 (позиция 399) и позиции 440 (позиция 409) в CH3-области. Нумерацию константных областей антитела можно найти в публикации Kabat et al. (Kabat, E.A., et al., 1991, Sequences of Proteins of Immunological Interest, NIH); а для нумерации константных областей тяжелой цепи номера согласно EC представлены в круглых скобках.
Технология модификации аминокислотного остатка в позиции 435, как указывается согласно нумерации EC, который является центром, касающимся связывания между IgG и Протеином A, до аминокислоты, обладающей другой силой связывания в отношении Протеина A, такой, как Arg, также может применяться к антителу согласно настоящему изобретению в комбинации с вышеупомянутыми технологиями. Путем применения этой технологии взаимодействие между H-цепью и Протеином A может быть изменено, и только гетеродимерные антитела могут быть эффективно очищены с использованием колонки с Протеином A. Эта технология также может применяться независимо без сочетания с вышеупомянутыми технологиями.
Более конкретно, например, в антителе, содержащем два типа CH3 областей тяжелой цепи, от одного до трех наборов аминокислотных остатков в CH3 области первой тяжелой цепи, которые выбирают из наборов аминокислотных остатков по указанным ниже пп. с (1) по (3), могут быть преобразованы таким образом, чтобы они электрически взаимно отталкивались:
(1) аминокислотные остатки, содержащиеся в CH3 области тяжелой цепи в позиции 356 и позиции 439, как указывается согласно нумерации EC;
(2) аминокислотные остатки, содержащиеся в CH3 области тяжелой цепи в позиции 357 и позиции 370, как указывается согласно нумерации EC; и
(3) аминокислотные остатки, содержащиеся в CH3 области тяжелой цепи в позиции 399 и позиции 409, как указывается согласно нумерации EC.
Кроме того, антитело может быть антителом, имеющим набор аминокислотных остатков во второй CH3 области тяжелой цепи, отличной от вышеупомянутой первой CH3 области тяжелой цепи, причем набор аминокислотных остатков выбран из наборов аминокислотных остатков, показанных выше в пп. с (1) по (3), и от одного до трех наборов аминокислотных остатков, соответствующих наборам аминокислотных остатков, показанным выше в пп. с (1) по (3), которые электрически взаимно отталкиваются в первой CH3 области тяжелой цепи, электрически не отталкиваются от соответствующих аминокислотных остатков в первой CH3 области тяжелой цепи.
Аминокислотные остатки, описанные выше в пп. с (1) по (3) приближаются друг к другу после ассоциации. Специалисты в данной области смогут определить места, соответствующие аминокислотным остаткам, описанных выше в пп. с (1) по (3) для нужной CH3 области тяжелой цепи или константной области тяжелой цепи, путем гомологичного моделирования и т.п. с применением доступного на рынке программного обеспечения и для соответствующей модификации аминокислотных остатков в этих местах.
В вышеупомянутом антителе "электрически отталкивающиеся" или "обладающие одноименным зарядом" означает, что, например, любые два или более аминокислотных остатков имеют аминокислотные остатки, содержащиеся в любой одной из вышеупомянутых групп (X) и (Y). С другой стороны, "электрически не отталкивающиеся" означает, что, например, антитело имеет аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся (X) и (Y), упомянутые выше и (Z) ниже, причем два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z), а также (Z) и (Z):
(Z) аланин (A), аспарагин (N), цистеин (C), глутамин (Q), глицин (G), изолейцин (I), лейцин (L), метионин (M), фенилаланин (F), пролин (P), серии (S), треонин (T), триптофан (W), тирозин (Y) или валин (V).
В предпочтительном варианте осуществления вышеупомянутого антитела первая CH3 область тяжелой цепи и вторая CH3 область тяжелой цепи могут быть сшиты дисульфидными связями.
Согласно настоящему изобретению, аминокислотный остаток, подлежащий "модификации" не ограничивается аминокислотным остатком вариабельной области антитела или константной области антитела, упомянутыми выше. Специалисты в данной области смогут определить аминокислотные остатки, образующие границу в варианте полипептида или гетеромерном мультимере путем гомологичного моделирования и т.п. с применением доступного на рынке программного обеспечения, и модифицировать аминокислотные остатки в этих местах с целью регулирования ассоциации. Гомологичное моделирование представляет собой технологию прогнозирования трехмерной структуры белка с применением доступного на рынке программного обеспечения. При построении структуры белка с неизвестной трехмерной структурой, в первую очередь ищут белок, определяемый как имеющий высокогомологичную трехмерную структуру с белком. Затем, используя эту трехмерную структуру в качестве шаблона строят структуру белка с неизвестной структурой, и структуру также оптимизируют с применением способов молекулярной динамики и т.п. для прогнозирования трехмерной структуры неизвестного белка.
Согласно технологии включения электрического отталкивания в границу вариабельной области тяжелой цепи и вариабельной области легкой цепи для подавления нежелательной ассоциации тяжелой цепи и легкой цепи, примеры аминокислотных остатков, которые контактируют на границе вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), включают глутамин (Q) в позиции 39 согласно нумерации Kabat в вариабельной области тяжелой цепи (FR2 область) и находящийся напротив (контактирующий) глутамин (Q) в позиции 38 согласно нумерации Kabat в вариабельной области легкой цепи (FR2 область). Кроме того, предпочтительным примером является лейцин (L) в позиции 45 согласно нумерации Kabat в вариабельной области тяжелой цепи (FR2) и находящийся напротив пролин (Р) в позиции 44 согласно нумерации Kabat в вариабельной области легкой цепи (FR2). Для нумерации этих центров обращались к публикации Kabat, et al. (Kabat, E.A., et al., 1991, Sequence of Proteins of Immunological Interest, NIH).
Поскольку известно, что эти аминокислотные остатки являются высококонсервативными в организмах человека и мыши (J. Mol. Recognit. 2003; 16: 113-120), ассоциация вариабельных областей антитела может регулироваться для VH-VL ассоциации антител, отличных от указываемых в Примерах, путем модификации аминокислотных остатков, соответствующих вышеупомянутым аминокислотным остаткам.
Конкретным примером является антитело, в котором два или более аминокислотных остатков, образующих границу вариабельной области тяжелой цепи и вариабельной области легкой цепи, являются аминокислотными остатками, которые электрически взаимно отталкиваются.
Более конкретно, примеры включают антитело с одним набором или двумя наборами аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. (a) или (b):
(a) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи (1) в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, содержащийся в легкой цепи (2) в позиции 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи (3) в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, содержащийся в вариабельной области легкой цепи (4) в позиции 44 согласно нумерации Kabat.
Аминокислотные остатки, описанные в вышеупомянутых пп. (a) или (b), приближаются друг к другу при ассоциации. Специалисты в данной области смогут определить места, соответствующие аминокислотным остаткам, описанным в вышеупомянутых пп. (a) или (b) в нужной вариабельной области тяжелой цепи или вариабельной области легкой цепи, путем гомологичного моделирования и т.п. с применением доступного на рынке программного обеспечения, и для соответствующей модификации аминокислотных остатков в этих местах.
В вышеупомянутом антителе "аминокислотные остатки, которые электрически взаимно отталкиваются" предпочтительно являются выбранными из аминокислотных остатков, содержащихся, например, в любом из указанных ниже наборов (X) и (Y):
(X) глутаминовая кислота (E) или аспарагиновая кислота (D); или
(Y) лизин (K), аргинин (R), или гистидин (H).
Кроме того, другой вариант осуществления антитела согласно настоящему изобретению представляет собой, например, антитело, в котором два или более аминокислотных остатков, образующие границу вариабельной области тяжелой цепи и вариабельной области легкой цепи, являются аминокислотными остатками, которые электрически не отталкиваются друг от друга. В частности, примером такого антитела может быть антитело, имеющее один набор или два набора аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных в вышеупомянутых пп. (a) и (b).
Соответствующие аминокислотные остатки, описанные в вышеупомянутых пп. (a) или (b), приближаются друг к другу при ассоциации. Специалисты в данной области смогут определить места, соответствующие аминокислотным остаткам, описанным в вышеупомянутых пп. (a) или (b), для нужной вариабельной области тяжелой цепи или вариабельной области легкой цепи путем гомологичного моделирования и т.п. с применением доступного на рынке программного обеспечения, и для соответствующего подвергания аминокислотных остатков в этих местах модификации.
В вышеупомянутом антителе "аминокислотные остатки, которые электрически взаимно не отталкиваются" означают, например, аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся вышеупомянутые пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z), а также (Z) и (Z).
Как правило, аминокислотными остатками, описанными в вышеупомянутых пп. (a) или (b), в организмах человека и мыши являются:
(1) глутамин (Q),
(2) глутамин (Q),
(3) лейцин (L) и
(4) пролин (P).
Таким образом, в предпочтительном варианте осуществления настоящего изобретения эти аминокислотные остатки подвергаются модификации (например, замещению аминокислотами, обладающими электрическим зарядом). Кроме того, типы аминокислотных остатков вышеупомянутых пп. (a) или (b) не обязательно ограничиваются вышеупомянутыми аминокислотными остатками, а могут быть также другими аминокислотами, равноценными этим аминокислотам. Например, аминокислота в позиции 38 согласно нумерации Kabat в вариабельной области легкой цепи может быть, например, гистидином (H), если речь идет об организме человека.
Согласно технологии модификации аминокислотных остатков, образующих гидрофобное ядро, присутствующее на границе вариабельной области тяжелой цепи, и вариабельной области легкой цепи до полярных аминокислот, обладающих электрическим зарядом, для подавления непреднамеренной ассоциации тяжелой цепи и легкой цепи предпочтительные примеры аминокислотных остатков, способных образовывать гидрофобное ядро на границе вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), включают лейцин (L) в позиции 45 согласно нумерации Kabat в вариабельной области тяжелой цепи (FR2) и находящийся напротив пролин (P) в позиции 44 согласно нумерации Kabat в вариабельной области легкой цепи (FR2). Для нумерации этих центров обращались к публикации Kabat, et al. (Kabat, E.A., et al., 1991, Sequence of Proteins of Immunological Interest, NIH).
Как правило, термин "гидрофобное ядро" относится к части, образуемой путем сборки боковых цепей гидрофобной аминокислоты внутри ассоциированных полипептидов. Примеры гидрофобных аминокислот включают аланин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан и валин. Кроме того, аминокислотные остатки, отличные от гидрофобных аминокислот (например, тирозина) могут принимать участие в образовании гидрофобного ядра. Это гидрофобное ядро вместе с гидрофильной поверхностью, на которой боковые цепи гидрофильной аминокислоты являются открытыми наружу, становится движущей силой, содействующей ассоциации водорастворимых полипептидов. Если гидрофобные аминокислоты двух разных доменов присутствуют на молекулярной поверхности и открыты для воздействия молекул воды, энтропия возрастает, и возрастает свободная энергия. Соответственно, два домена ассоциируются друг с другом для снижения свободной энергии и приобретения устойчивости, и гидрофобные аминокислоты на границе маскируются внутри молекулы для образования гидрофобного ядра.
Считается, что при ассоциации полипептидов образование гидрофобного ядра подавляется модификацией гидрофобных аминокислот, образующих гидрофобное ядро, до полярных аминокислот, обладающих электрическим зарядом; следовательно, считается, что подавляется ассоциация полипептидов.
Специалисты в данной области смогут определить присутствие или отсутствие гидрофобного ядра, центр (участок) образования и т.п. путем анализа аминокислотных последовательностей для необходимого полипептида. То есть, антитело согласно настоящему изобретению представляет собой антитело, характеризующееся тем, что аминокислотные остатки, способные образовывать гидрофобное ядро на границе, модифицируются до аминокислотных остатков, обладающих электрическим зарядом. Более конкретно, примеры включают антитело, в котором аминокислотные остатки, показанные ниже в пп. (1) или (2), являются аминокислотными остатками, обладающими электрическим зарядом. Боковых цепей аминокислотных остатков, показанных ниже в пп. (1) и (2), являются смежными и могут образовывать гидрофобное ядро:
(1) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи в позиции 45 согласно нумерации Kabat; и
(2) аминокислотный остаток, содержащийся в вариабельной области легкой цепи в позиции 44 согласно нумерации Kabat.
Предпочтительные примеры аминокислотных остатков, обладающих электрическим зарядом в вышеупомянутом антителе, включают глутаминовую кислоту (E), аспарагиновую кислоту (D), лизин (K), аргинин (R) и гистидин (H). Более предпочтительные примеры включают глутаминовую кислоту (E) и лизин (K).
Как правило, аминокислотными остатками, описываемыми в вышеупомянутых пп. (1) и (2), в организмах человека и мыши являются следующие:
(1) лейцин (L) и
(2) пролин (P).
Таким образом, в предпочтительном варианте осуществления настоящего изобретения эти аминокислотные остатки подвергаются модификации (например, замещению аминокислотами, обладающими электрическим зарядом). Кроме того, типы вышеупомянутых аминокислотных остатков (1) и (2) не обязательно ограничиваются вышеупомянутыми аминокислотными остатками, а также могут быть другими аминокислотами, равноценными этим аминокислотным остаткам.
К антителам согласно настоящему изобретению могут применяться и другие известные технологии. Например, для содействия ассоциации первой VH (VH1) и первой VL (VL1) и/или второй VH (VH2) и второй VL (VL2) боковая цепь аминокислоты, присутствующая в вариабельной области одной из H-цепей, может быть замещена большей боковой цепью (выступом), а боковая цепь аминокислоты, присутствующая в противоположной вариабельной области другой H-цепи, может быть замещена меньшей боковой цепью (впадиной), таким образом, чтобы выступ мог находиться во впадине, и активизировалась ассоциация VH1 и VL1 и/или VH2 и VL2; следовательно, ассоциация VH1 и VL2 и/или VH2 и VL1 также может подавляться (документ WO 1996/027011; Ridgway, J.B., et al., Protein Engineering (1996) 9, 617-621; Merchant, A.M., et al., Nature Biotechnology (1998) 16, 677-681).
Например, в случае человеческого IgG1 для того, чтобы сделать боковую цепь аминокислоты в CH3-области одной H-цепи большей боковой цепью (выступом), осуществляют модификации Y349C и T366W, а для того, чтобы сделать боковую цепь аминокислоты в CH3-области другой H-цепи меньшей боковой цепью, осуществляют модификации D356C, T336S, L368A и Y407V.
К антителам согласно настоящему изобретению могут применяться и другие известные технологии. Антитело-мишень может быть эффективно получено путем комплементарной ассоциации CH3 с применением подвергнутого инженерии путем замены цепи домена CH3, в котором часть CH3 одной H-цепи антитела заменена на последовательность, взятую из IgA, соответствующего этой части, а комплементарная часть CH3 другой H-цепи включена с последовательностью, взятой из IgA, соответствующей этой части (Protein Engineering Design & Selection, 23: 195-202, 2010).
К антителам согласно настоящему изобретению могут применяться и другие известные технологии. При получении биспецифических антител биспецифическое антитело-мишень может быть получено, например, путем обеспечения разницы в изоэлектрической точке путем различной модификации аминокислот для каждой из вариабельных областей двух типов H-цепей и использования разницы в изоэлектрической точке для очищения путем ионообменной хроматографии (документ WO 2007/114325).
Модификации согласно настоящему изобретению могут применяться для таких антител, как указанное ниже, например, антитела, имеющего структуру, в которой для содействия ассоциации первой VH (VH1) и первой VL (VL1) и/или второй VH (VH2) и второй VL (VL2) VH1 соединяют с Fc областью через первую CH1 и VL1 соединяют с первой CL, и VH2 соединяют с другой Fc областью через вторую CL и VL2 соединяют со второй CH1 (документ WO 09/80254).
Могут применяться несколько, например, две или более из вышеупомянутых известных технологий в комбинации с антителом согласно настоящему изобретению. Кроме того, антитело согласно настоящему изобретению может быть получено на основе антитела, в отношении которого применялись модификации вышеупомянутых известных технологий.
Указанные ниже соответствующие настоящему изобретению способы регулирования ассоциации обеспечивают, например, возможность эффективной выработки активных антител или полипептидов. Примерами такой активности может быть активность связывания, нейтрализующая активность, цитотоксичная активность, агонистическая активность, антагонистическая активность, ферментная активность и т.п. Агонистическая активность означает активность, вызывающую определенный вид изменений в физиологической активности через связывание антитела с антигеном, таким, как рецептор, вызывающий трансдукцию сигнала и т.п. в клетках. Примерами физиологической активности, помимо прочих, могут быть рост, выживание, дифференциация, транскрипция, мембранный транспорт, активность связывания, протеолитическая активность, фосфорилирование/дефосфорилирование, окислительно-восстановительная активность, перенос, нуклеолитическая активность, дегидратация, вызывание гибели клеток и вызывание апоптоза и т.п.
Антитела или полипептиды, распознающие необходимые антигены или связывающиеся с нужными рецепторами, могут быть эффективно получены с применением способов согласно настоящему изобретению.
Антигены согласно настоящему изобретению не имеют особых ограничений, и может применяться любой тип антигена. Примерами антигенов, помимо прочих, могут быть рецепторы или их фрагменты, антигены рака, антигены МНС и антигены дифференциации и т.п.
Примерами рецепторов согласно настоящему изобретению могут быть рецепторы, относящиеся к семейству рецепторов гемопоэтического фактора, семейству рецепторов цитокина, семейству рецепторов тирозинкиназного типа, семейству рецепторов серии/треонинкиназного типа, семейству рецепторов TNF, G-белок-связанному семейству рецепторов, GPI-заякоренному семейству рецепторов, семейству рецепторов тирозинфосфатазного типа, семейству факторов адгезии, гормональному семейству рецепторов и т.п. Информацию о рецепторах, относящихся к этим семействам рецепторов и их характеристиках можно найти в различных задокументированных источниках, например, в публикациях Cooke BA., King RJB., van der Molen HJ. ed. New Comprehensive Biochemistry Vol. 18B "Hormones and their Actions Part II" pp. 1-46 (1988) Elsevier Science Publishers BV., New York, USA; Patthy L. (1990) Cell, 61: 13-14; Ullrich A., et al. (1990) Cell, 61: 203-212; Massagul J. (1992) Cell, 69: 1067-1070; Miyajima A., et al. (1992) Annu. Rev. Immunol., 10: 295-331; Taga T. and Kishimoto T. (1992) FASEB J., 7: 3387-3396; Fantl WI., et al. (1993) Annu. Rev. Biochem., 62: 453-481; Smith CA., et al. (1994) Cell, 76: 959-962; Flower DR. (1999) Biochim. Biophys. Acta, 1422: 207-234; Miyasaka M. ed. Cell Technology, Handbook Series "Handbook for adhesion factors" (1994) Shujunsha, Токио, Япония; и др. Примерами специфических рецепторов, относящихся к вышеупомянутым семействам рецепторов, могут быть рецептор эритропоэтин (EPO) человека или мыши, рецептор гранулоцитарного колониестимулирующего фактора (G-CSF) человека или мыши, рецептор тромбопоэтина (TPO) человека или мыши, рецептор инсулина человека или мыши, рецептор Flt-3-лиганда человека или мыши, рецептор тромбоцитарного фактора роста (PDGF) человека или мыши, рецептор интерферона (IFN)-α или -β человека или мыши, рецептор лептина человека или мыши, рецептор гормона роста (GH) человека или мыши, рецептор интерлейкина (IL)-10 человека или мыши, рецептор инсулиноподобного фактора роста (IGF)-I человека или мыши, рецептор ингибирующего лейкемию фактора (LIF) человека или мыши и рецептор цилиарного нейротрофического фактора (CNTF) человека или мыши (hEPOR: Simon, S. et al. (1990) Blood 76, 31-35; mEPOR: D’Andrea, AD. et al. (1989) Cell 57, 277-285; hG-CSFR: Fukunaga, R. et al. (1990) Proc. Natl. Acad. Sci. USA. 87, 8702-8706; mG-CSFR: Fukunaga, R. et al. (1990) Cell 61, 341-350; hTPOR: Vigon, I. et al. (1992) 89, 5640-5644.; mTPOR: Skoda, RC. et al. (1993) 12, 2645-2653; hInsR: Ullrich, A. et al. (1985) Nature 313, 756-761; hFlt-3: Small, D. et al. (1994) Proc. Natl. Acad. Sci. USA. 91, 459-463; hPDGFR: Gronwald, RGK. et al. (1988) Proc. Natl. Acad. Sci. USA. 85, 3435-3439; hIFN α/β R: Uze, G. et al. (1990) Cell 60, 225-234; и Novick, D. et al. (1994) Cell 77, 391-400).
Антигены рака представляют собой антигены, экспрессируемые вследствие злокачественной трансформации клеток, и также называются опухолеспецифическими антигенами. Кроме того, нарушенные сахарные цепи, появляющиеся на поверхности клеток или на молекулах белков при озлокачествлении клеток, также являются антигенами рака, и они также называются антигенами раковых сахарных цепей. Примерами антигенов рака являются ЕрСАМ, которая экспрессируется в разных раковых клетках, включая рак легких (Proc. Natl. Acad. Sci. USA (1989) 86 (1), 27-31) (ее полинуклеотидная последовательность указывается под инвентарным номером RefSeq NM_002354.2 (SEQ ID NO: 78), а ее полипептидная последовательность указывается под инвентарным номером RefSeq NP_002345.2 (SEQ ID NO: 79)), CA19-9, CA15-3, сиалил SSEA-1 (SLX) и т.п.
Антигены МНС могут быть разделены на два широких класса: антигены МНС I класса и антигены МНС II класса: антигены МНС I класса включают HLA-A, -B, -C, -E, -F, -G, и -Н; а антигены МНС II класса включают HLA-DR, -DQ и -DP.
К антигенам дифференциации относятся CD1, CD2, CD3, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15s, CD16, CD18, CD19, CD20, CD21, CD23, CD25, CD28, CD29, CD30, CD32, CD33, CD34, CD35, CD38, CD40, CD41a, CD41b, CD42a, CD42b, CD43, CD44, CD45, CD45RO, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD51, CD54, CD55, CD56, CD57, CD58, CD61, CD62E, CD62L, CD62P, CD64, CD69, CD71, CD73, CD95, CD102, CD106, CD122, CD126 и CDwl30.
Антитела согласно настоящему изобретению могут быть биспецифическими антителами; в этом случае два антигена (или эпитопа), распознаваемые этим антителом, могут быть произвольно выбраны из вышеупомянутых рецепторов или их фрагментов, антигенов рака, антигенов МНС, антигенов дифференциации и т.п. Например, два антигена могут быть выбраны из рецепторов или их фрагментов, два могут быть выбраны из антигенов рака, два могут быть выбраны из антигены МНС, или два могут быть выбраны из антигенов дифференциации. Кроме того, каждый антиген может быть выбран из двух антигенов, произвольно выбранных, например, из рецепторов или их фрагментов, антигенов рака, антигенов МНС и антигенов дифференциации.
Кроме того, настоящее изобретение обеспечивает способ получения антитела, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой.
Предпочтительный вариант осуществления способа согласно настоящему изобретению представляет собой способ получения антитела, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой, который включает:
(1) модификацию нуклеиновых кислот, кодирующих CH1 и CL, таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), становились аминокислотными остатками, которые электрически отталкиваются друг от друга:
(a) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 160, как указывается согласно нумерации EC,
(2) включение модифицированных нуклеиновых кислот в клетки-хозяева и культивирование таким образом, чтобы клетки-хозяева экспрессировали нуклеиновые кислоты, и
(3) сбор антитела из культуры клеток-хозяев.
Другой вариант осуществления способа получения согласно настоящему изобретению включает способ получения антитела, согласно которому этап (1) вышеупомянутого способа получения также включает модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки из набора аминокислотных остатков, показанных ниже в п. (d), становились аминокислотными остатками, которые электрически отталкиваются друг от друга:
(d) аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC.
Кроме того, настоящее изобретение касается способа получения, включающего на вышеупомянутом этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки, которые электрически отталкиваются друг от друга, были выбраны из аминокислотных остатков, содержащихся в любой из групп вышеупомянутых пп. (X) и (Y).
Кроме того, настоящее изобретение касается способа получения, включающего на вышеупомянутом этапе (1) модификацию нуклеиновых кислот таким образом, чтобы два или более аминокислотных остатков, образующих границу вариабельной области тяжелой цепи и вариабельной области легкой цепи, являлись аминокислотными остатками, которые электрически отталкиваются друг от друга. Предпочтительно аминокислотные остатки, которые электрически отталкиваются друг от друга, представляют собой любой набор аминокислотных остатков, выбранных из группы, состоящей из, например, наборов аминокислотных остатков, указанных ниже в пп. (a) и (b):
(a) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, содержащийся в вариабельной области легкой цепи в позиции 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, содержащийся в вариабельной области легкой цепи в позиции 44 согласно нумерации Kabat.
Вышеупомянутые аминокислотные остатки, которые электрически отталкиваются друг от друга, предпочтительно являются выбранными из аминокислотных остатков, содержащихся в любом наборе вышеупомянутых (X) и (Y).
Еще один предпочтительный вариант осуществления способа получения согласно настоящему изобретению включает а способ получения антитела, в котором ассоциация тяжелой цепи и легкой цепи является регулируемой, который включает:
(1) модификацию нуклеиновых кислот, кодирующих CH1 и CL, таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), становились аминокислотными остатками, которые электрически не отталкиваются друг от друга:
(a) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 160, как указывается согласно нумерации EC,
(2) включения модифицированных нуклеиновых кислот в клетки-хозяева и культивирование таким образом, чтобы клетки-хозяева экспрессировали нуклеиновые кислоты, и
(3) сбор антитела из культуры клеток-хозяев.
Другой вариант осуществления способа получения согласно настоящему изобретению включает способ получения, который кроме того включает на этапе (1) вышеупомянутого способа получения, модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки из набора аминокислотных остатков, показанных ниже в п. (d), становились аминокислотными остатками, которые электрически не отталкиваются друг от друга:
(d) аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC.
Кроме того, настоящее изобретение касается способа получения, включающего, на вышеупомянутом этапе (1) модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки, которые электрически не отталкиваются друг от друга, являлись аминокислотными остатками, выбранными из каждого из двух наборов, выбранных из группы, к которой относятся вышеупомянутые пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z), а также (Z) и (Z).
Кроме того, конкретные примеры аминокислотных остатков, которые электрически не отталкиваются друг от друга на вышеупомянутом этапе (1) настоящего изобретения, включают следующие аминокислотные остатки:
- аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, который является лизином (K); и аминокислотные остатки, содержащиеся в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, которые все являются глутаминовой кислотой (E); и
- аминокислотные остатки, содержащиеся в CH1 в позиции 147 и позиции 175, как указывается согласно нумерации EC, которые являются глутаминовой кислотой (E), и аминокислотные остатки, содержащиеся в CL в позиции 180, позиции 131 и позиции 160, как указывается согласно нумерации EC, которые все являются лизином (K).
Кроме того, в другом примере, аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, является глутаминовой кислотой (E), и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC, является лизином (K).
Кроме того, настоящее изобретение касается способа получения, включающего на вышеупомянутом этапе (1) модификацию нуклеиновых кислот таким образом, чтобы два или более аминокислотных остатков, образующих границу вариабельной области тяжелой цепи и вариабельной области легкой цепи, были аминокислотными остатками, которые электрически не отталкиваются друг от друга. Предпочтительно аминокислотными остатками, которые электрически не отталкиваются друг от друга, являются, например, аминокислотные остатки любого набора, выбранного из группы, к которой относятся наборы аминокислотных остатков, указанных ниже в пп. (a) и (b):
(a) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи в позиции 39 согласно нумерации Kabat, и аминокислотный остаток, содержащийся в вариабельной области легкой цепи в позиции 38 согласно нумерации Kabat; и
(b) аминокислотный остаток, содержащийся в вариабельной области тяжелой цепи в позиции 45 согласно нумерации Kabat, и аминокислотный остаток, содержащийся в вариабельной области легкой цепи в позиции 44 согласно нумерации Kabat.
Вышеупомянутые аминокислотные остатки, которые электрически не отталкиваются друг от друга, предпочтительно представляют собой аминокислотные остатки, выбранные из каждого из двух наборов, выбранных из группы, к которой относятся вышеупомянутые пп. с (X) по (Z), и два набора выбраны из комбинаций (X) и (Y), (X) и (Z), (Y) и (Z), а также (Z) и (Z).
Кроме того, настоящее изобретение обеспечивает способ регулирования ассоциации тяжелых цепей и легких цепей антитела.
Предпочтительный вариант осуществления способа регулирования ассоциации настоящего изобретения представляет собой способ регулирования ассоциации тяжелых цепей и легких цепей антитела, включающий модификацию нуклеиновых кислот таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), становились аминокислотными остатками, которые электрически отталкиваются друг от друга:
(a) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 160, как указывается согласно нумерации EC.
Другой вариант осуществления настоящего изобретения обеспечивает способ регулирования ассоциации в антителе, который также включает модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки из набора аминокислотных остатков, показанных ниже в п. (d), были аминокислотными остатками, которые электрически отталкиваются друг от друга:
(d) аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC.
Еще один предпочтительный вариант осуществления способа регулирования ассоциации настоящего изобретения, представляет собой способ регулирования ассоциации тяжелых цепей и легких цепей антитела, включающий модификацию нуклеиновых кислот таким образом, чтобы один набор или два или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (a) по (c), становились аминокислотными остатками, которые электрически не отталкиваются друг от друга:
(a) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 180, как указывается согласно нумерации EC;
(b) аминокислотный остаток, содержащийся в CH1 в позиции 147, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 131, как указывается согласно нумерации EC; и
(c) аминокислотный остаток, содержащийся в CH1 в позиции 175, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 160, как указывается согласно нумерации EC.
Другой вариант осуществления настоящего изобретения обеспечивает способ регулирования ассоциации в антителе, который также включает модификацию нуклеиновых кислот таким образом, чтобы аминокислотные остатки из набора аминокислотных остатков, указанных ниже в п. (d), были аминокислотными остатками, которые электрически не отталкиваются друг от друга:
(d) аминокислотный остаток, содержащийся в CH1 в позиции 213, как указывается согласно нумерации EC, и аминокислотный остаток, содержащийся в CL в позиции 123, как указывается согласно нумерации EC.
В соответствии со способом регулирования ассоциации согласно настоящему изобретению, необходимое биспецифическое антитело может быть получено оптимальным и эффективным способом, как было описано выше. То есть, необходимый гетеромерный мультимер в форме биспецифического антитела может быть эффективно образован из смеси мономеров.
Фраза "модифицировать нуклеиновые кислоты" согласно вышеупомянутым способам настоящего изобретения относится к модификации нуклеиновых кислот таким образом, чтобы они соответствовали аминокислотным остаткам, включенным путем "модификаций" согласно настоящему изобретению. Более конкретно она касается модификации нуклеиновых кислот, кодирующих первоначальные (предварительно модифицированные) аминокислотные остатки, до нуклеиновых кислот, кодирующих аминокислотные остатки, подлежащие включению путем модификации. Как правило, она означает выполнение манипуляций с генами или мутационной обработки, которая в результате приводит как минимум к одной нуклеотидной инсерции, делеции или замещению в первоначальной нуклеиновой кислоте, таким образом, чтобы образовывались кодоны, кодирующие нужные аминокислотные остатки. Более конкретно, кодоны, кодирующие первоначальные аминокислотные остатки, замещаются кодонами, кодирующими аминокислотные остатки, которые подлежат включению путем модификации. Такие модификации нуклеиновых кислот могут выполняться специалистами в данной области с применением известных технологий, таких, как сайт-специфичный мутагенез и ПЦР-мутагенез.
Кроме того, настоящее изобретение обеспечивает нуклеиновые кислоты, которые кодируют антитело согласно настоящему изобретению. Кроме того, векторы, несущие нуклеиновые кислоты, также включаются в настоящее изобретение.
Нуклеиновые кислоты согласно настоящему изобретению, как правило, вставляют в соответствующие векторы, а затем включают в клетки-хозяева. Эти векторы не имеют особых ограничений, при условии, что вставленная нуклеиновая кислота устойчиво сохраняется. Например, при использовании E. coli в качестве хозяина клонирующим вектором предпочтительно является вектор pBluescript (Stratagene) и т.п., однако могут использоваться и различные векторы промышленного производства. Векторы экспрессии особенно приемлемы в качестве векторов для получения полипептидов согласно настоящему изобретению. Векторы экспрессии не имеют особых ограничений при условии, что они могут экспрессировать полипептиды в пробирках, E. coli, культивируемых клетках или отдельных организмах. Например, к предпочтительным векторам относятся вектор pBEST (Promega) для экспрессии в пробирках, pET вектор (Invitrogen) для E. coli, вектор pME18S-FL3 (инвентарный номер в GenBank AB009864) для культивируемых клеток и вектор pME18S (Mol. Cell Biol. 8: 466-472 (1998)) для отдельных организмов. Инсерция ДНК согласно настоящему изобретению в векторы может выполняться, например, с применением комплекта In-Fusion Advantage PCR Cloning Kit (Clontech).
Кроме того, настоящее изобретение обеспечивает клетки-хозяева, несущие вышеописанные нуклеиновые кислоты. Клетки-хозяева не имеют особых ограничений, и в соответствии с этой целью, могут использоваться различные клетки-хозяева, такие, как E. coli и различные клетки животных. Клетки-хозяева могут использоваться, например, в качестве вырабатывающей системы для выработки и экспрессии антитела или полипептидов согласно настоящему изобретению. In vitro и in vivo вырабатывающие системы имеются в наличии для систем выработки полипептидов. Вырабатывающие системы, в которых используются эукариотные клетки или прокариотные клетки, являются примерами in vitro вырабатывающих систем.
Эукариотные клетки, которые могут использоваться в качестве клеток-хозяев, включают, например, клетки животных, растительные клетки и грибковые клетки. К клеткам животных относятся: клетки млекопитающих, например, CHO (J. Exp. Med. (1995) 108: 945), COS, 3T3, миелома, ВНК (клетки почки новорожденного хомяка), HeLa, C127, HEK293, клетки меланомы Боуэса и Vero; клетки земноводных, такие, как ооциты Xenopus laevis (Valle, et al. (1981) Nature 291: 338-340); и клетки насекомых (например, Drosophila S2, Sf9, Sf21 и Tn5). Для экспрессии антител согласно настоящему изобретению могут соответственно использоваться клетки CHO-DG44, CHO-DX11B, COS7 и клетки ВНК. Среди клеток животных клетки CHO являются особенно предпочтительными для крупномасштабной экспрессии.
Векторы могут включаться в клетку-хозяин известными способами, например, способами с применением фосфата кальция, DEAE-декстрана, способами с использованием катионных липосом DOTAP (Boehringer-Mannheim), способами электропорации (Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons, Section 9.1-9.9), липофекции, способами с использованием липофектамина (GIBCO-BRL) или способами микроинъекции. Кроме того, введение генов для экспрессии полипептидов также может осуществляться с применением системы экспрессии Free Style 293 (Invitrogen).
К растительным клеткам относятся, например, клетки, взятые из Nicotiana tabacum, известные как вырабатывающая белок система. Культивируют каллюсы из этих клеток для выработки антител согласно настоящему изобретению.
Известными системами экспрессии белка являются системы, в которых используются грибковые клетки, включая дрожжевые клетки, например, клетки рода Saccharomyces, такие, как Saccharomyces cerevisiae и Saccharomyces pombe; и клетки нитчатых грибов, например, клетки рода Aspergillus, такие, как Aspergillus niger. Эти клетки могут использоваться в качестве хозяев для выработки антител согласно настоящему изобретению.
Бактериальные клетки могут использоваться в вырабатывающих системах, использующих прокариотные клетки. Примерами бактериальных клеток могут быть Streptococcus, Staphylococcus, E. coli, Steptomyces, Bacillus subtilis, а также описанная выше E. coli. Такие клетки могут использоваться для выработки антител согласно настоящему изобретению.
При выработке антитела с использованием клетки-хозяина согласно настоящему изобретению полинуклеотид, кодирующий антитело согласно настоящему изобретению, может быть экспрессирован путем культивирования клеток-хозяев, преобразованных вектором экспрессии, содержащим полинуклеотид. Культивирование выполняют с применением известных способов. Например, при использовании клеток животных в качестве хозяев DMEM, MEM, RPMI 1640 или IMDM могут использоваться в качестве культуральной среды, которая может дополняться сывороткой, такой, как FBS или сыворотка эмбрионов крупного рогатого скота (FCS). Также приемлемы бессывороточные культуры. Предпочтительный уровень pH в процессе культивирования составляет приблизительно от 6 до 8. Культивирование обычно осуществляют при температуре приблизительно от 30°C до 40°C в течение приблизительно 15-200 часов. В случае необходимости среду меняют, аэрируют или перемешивают.
С другой стороны, вырабатывающие системы с использованием животных или растений могут применяться в качестве систем для выработки полипептидов in vivo. Необходимый полинуклеотид вводят в организм животного или растение, полипептид вырабатывается в организме животного или растении, а затем его собирают. К "хозяевам" согласно настоящему изобретению относятся такие животные и растения.
Для секреции экспрессируемых клетками-хозяевами полипептидов в просвет эндоплазматического ретикулюма, периплазматическое пространство или внеклеточную среду в нужные полипептиды могут включаться соответствующие сигналы секреции. Эти сигналы могут быть внутренними или инородными для нужных полипептидов.
При секреции полипептидов согласно настоящему изобретению в культуральную среду полипептиды, вырабатываемые вышеупомянутым способом, могут быть собраны путем сбора среды. Если полипептиды согласно настоящему изобретению вырабатываются внутри клеток, сначала клетки подвергают лизису и эти полипептиды собирают.
Животными, используемыми для вырабатывающей системы, могут быть млекопитающие или насекомые. Могут использоваться такие млекопитающие, как козы, свиньи, овцы, мыши и крупный рогатый скот (Vicki Glaser, SPECTRUM Biotechnology Applications (1993)). В альтернативном варианте млекопитающие могут быть трансгенными животными.
Например, полинуклеотид, кодирующий антитело согласно настоящему изобретению, может быть получен как составной ген с геном, кодирующим полипептид, специфично вырабатываемый в молоке, такой, как β-казеин козы. Фрагменты полинуклеотидов, содержащие составной ген, вводят путем инъекции в эмбрионы козы, которые затем вводят обратно в организм коз. Нужное антитело может быть получено из молока, вырабатываемого трансгенными козами, рожденными от коз, которые получали эмбрионы, или от их потомства. Соответствующие гормоны могут вводиться трансгенным козам для увеличения объема молока, содержащего антитело, выработанное трансгенными козами (Ebert et al, Bio/Technology 12: 699-702 (1994)).
Насекомые такие, как шелкопряд, также могут использоваться для выработки антител согласно настоящему изобретению. Бакуловирусы, несущие полинуклеотид, кодирующий нужное антитело, могут использоваться для инфицирования шелкопряда, и нужное антитело может быть получено из жидкостей их организмов (Susumu et al, Nature 315: 592-594 (1985)).
К растениям, используемым для выработки антител согласно настоящему изобретению, относится, например, табак. В случае использования табака полинуклеотид, кодирующий нужное антитело, вставляют в растительный вектор экспрессии, например, pMON 530, а затем вектор вводят в бактерию, такую, как Agrobacterium tumefaciens. Затем бактерии используют для инфицирования табака, такого, как Nicotiana tabacum, и нужные антитела могут быть добыты из листьев табака (Ma et al, Eur. J. Immunol. 24: 131-138 (1994)).
Полученное в результате антитело может быть выделено изнутри или извне (например, из среды и молока) клетки-хозяина и очищено как по сути чистое и гомогенное антитело. Способы не ограничиваются каким-либо конкретным способом, и может применяться любой стандартный способ выделения и очистки антител. Антитела могут быть выделены и очищены путем соответствующего отбора и комбинирования, например, фракционирования сульфатом аммония или этанолом, экстрагирования кислотой, при помощи хроматографических колонок, фильтрации, ультрафильтрации, высаливания, осаждения из растворителя, экстракции растворителем, дистилляции, иммунопреципитации, электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия, изоэлектрического фокусирования, диализа, рекристаллизации и т.п.
Хроматография включает, например, аффинную хроматографию, ионообменную хроматографию, такую, как анионообменная хроматография и катионообменная хроматография, хроматографию на фосфоцеллюлозе, хроматографию с гидрофобным взаимодействием, гель-фильтрацию, хроматографию с обращением фаз, адсорбционную хроматографию, хроматографию с гидроксиапатитом и лектиновую хроматографию (Strategies for Protein Purification and Characterization: A Laboratory Course Manual. Ed Daniel R. Marshak et al, Cold Spring Harbor Laboratory Press, 1996). Эти виды хроматографии могут осуществляться с применением жидкофазной хроматографии, такой, как HPLC и FPLC. Примерами колонок для аффинной хроматографии могут быть колонки с протеином A и колонки с протеином G. Примерами колонок с протеином А могут быть Hyper D, POROS и Sepharose F.F. (Pharmacia).
Антитело может быть свободно модифицировано и пептидные компоненты могут быть удалены из него путем обработки антитела соответствующим белком, модифицирующим фермент до или после очистки антитела в случае необходимости. К таким модифицирующим белок ферментам относятся, например, трипсины, химотрипсины, лизил-эндопептидазы, протеинкиназы и глюкозидазы.
В другом предпочтительном варианте осуществления настоящее изобретение также включает способы получения антител согласно настоящему изобретению, включающие этапы культивирования клеток-хозяев согласно настоящему изобретению, как описывалось выше, и сбор антител из такой клеточной культуры.
Кроме того, настоящее изобретение касается фармацевтических композиций (лекарственных средств), включающих антитело согласно настоящему изобретению и фармацевтически приемлемый носитель. Согласно настоящему изобретению, фармацевтические композиции обычно означают лекарственные средства для лечения или профилактики или исследования и диагностики болезней.
Фармацевтические композиции согласно настоящему изобретению рецептируют способами, известными специалистам в данной области. Кроме того, антитела согласно настоящему изобретению в случае необходимости могут быть рецептированы в комбинации с другими фармацевтическими веществами. Например, они могут применяться парентерально в форме инъекции стерильного раствора или суспензии с водой или другой фармацевтически приемлемой жидкостью. Например, они могут быть рецептированы как стандартные дозы, соответствующие требованиям к приготовлению фармацевтических препаратов, путем надлежащего сочетания с фармацевтически приемлемыми носителями или средами, в частности, со стерильной водой, физиологическим раствором, растительным маслом, эмульгатором, суспензией, детергентом, стабилизатором, ароматизатором, формообразующим, носителем, консервантом, связующим веществом и т.п. В таких композициях количество активного ингредиента регулируют такими образом, чтобы доза пребывала в соответствующем заданном диапазоне.
Стерильные композиции для инъекций могут быть рецептированы с применением носителей, таких, как дистиллированная вода для инъекций, в соответствии со стандартными протоколами рецептирования.
Водные растворы для инъекций включают, например, физиологический раствор и изотонические растворы, содержащие декстрозу или другие адъюванты (например, D-сорбит, D-маннозу, D-маннит и хлорид натрия). Соответствующие солюбилизаторы, например, спирты (этанол и т.п.), полиспирты (пропиленгликоль, полиэтиленгликоль и т.п.), неионные детергенты (полисорбат 80™, НСО-50 и т.п.) могут применяться в комбинации.
К маслам относятся кунжутное и соевое масла. Бензилбензоат и/или бензиловый спирт могут применяться в комбинации в качестве солюбилизаторов. Также могут сочетаться буферы (например, фосфатный буфер и ацетат натрия), успокоительные вещества (например, новокаин), стабилизаторы (например, бензиловый спирт и фенол) и/или антиоксиданты. Приготовленные препараты для инъекций, как правило, помещают в соответствующие ампулы.
Фармацевтические композиции согласно настоящему изобретению предпочтительно вводят парентерально. Например, композиции могут быть инъекционными, трансназальными композициями, транспульмональными композициями или трансдермальными композициями. Например, такие композиции могут вводиться системно или по месту путем внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции и т.п.
Способы введения могут быть соответствующим образом выбраны с учетом возраста и симптомов пациента. Доза фармацевтической композиции, содержащей антитело или полинуклеотид, кодирующий антитело, может составлять, например, от 0,0001 до 1000 мг/кг при каждом введении. В альтернативном варианте доза может составлять, например, от 0,001 до 100,000 мг на пациента. Однако дозы не обязательно ограничиваются описанными выше диапазонами. Дозы и способы введения могут отличаться в зависимости от массы тела, возраста, симптомов пациента и т.п. Специалисты в данной области смогут выбрать соответствующие дозы и способы введения с учетом описанных выше факторов.
Аминокислоты, содержащиеся в аминокислотных последовательностях согласно настоящему изобретению, могут быть модифицированы посттрансляционно (например, модификация N-концевого глутамина до пироглутаминовой кислоты путем пироглутамилирования хорошо известна специалистам в данной области). Обычно такие посттрансляционно модифицированные аминокислоты включаются в аминокислотные последовательности согласно настоящему изобретению.
Все документы существующего уровня техники, приведенные в данном описании, включены путем ссылки.
Примеры
Далее настоящее изобретение описывается со ссылкой на конкретные Примеры, однако они не ограничивают объема настоящего изобретения. [Пример 1] Поиск центров, регулирующих границу CH1/CL Считалось, что при включении мутаций в каждый из доменов CH1 и CL биспецифического антитела и использовании электрического заряда на границе CH1/CL для регулирования границы CH1/CL специфично ассоциируются только H-цепь и L-цепь для антигена А, и только H-цепь и L-цепь для антигена В. Далее этот процесс указывается как регулирование границы CH1/CL. Проводилось исследование для поиска позиций, в которых граница CH1/CL может контролироваться, с применением модели кристаллической структуры. Аминокислоты поддерживают взаимодействие между боковыми цепями путем гидрофобного взаимодействия, электрического взаимодействия, водородного связывания и т.п.Известно, что такое взаимодействие происходит между боковыми цепями, присутствующими в диапазоне приблизительно 4 Å. Таким образом, аминокислоты были обнаружены в PDB модели 1HZH, в которой расстояние между аминокислотами, присутствующими в CH1, и аминокислотами, присутствующими в CL на границе между CH1 и CL, составляет приблизительно 4 Å. Центры, в которых находятся аминокислоты, показаны на Фиг. 1 и в Таблице 1 (перечень модифицированных центров). Номера аминокислот, показанных в Таблице 1, указаны в соответствии с нумерацией EC (последовательности белков, имеющих иммунологическое значение, Публикация NIH №91-3242). Кроме того, последующие номера аминокислот также указываются в соответствии с нумерацией EC. В представленном примере IgG1 использовали для H-цепи, a IgK (Каппа) использовали для L-цепи.
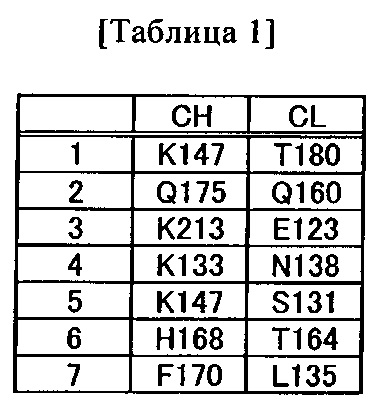
Для регулирования границы CH1/CL с использованием электрического заряда аминокислот аминокислоты, находящиеся в CH1 H-цепи или CL L-цепи (Таблица 1) замещали положительно заряженными Lys или His и отрицательно заряженными Glu или Asp.Более конкретно, подготавливали константные области: TH2 и TH11, в которых аминокислоты CH1 человеческого G1d (SEQ ID NO: 1) замещали положительно заряженным Lys; и TH1, TH3, TH4, TH9, TH10 и TH12, в которых аминокислоты CH1 человеческого G1d (SEQ ID NO: 1) замещали отрицательно заряженным Glu или Asp.Подобным образом подготавливали константные области: TL2, TL4, TL5, TL6, TL8 и TL12, в которых аминокислоты человеческого CL (SEQ ID NO: 13) замещали положительно заряженным Lys; TL11, в которых аминокислоты человеческого CL (SEQ ID NO: 13) замещали His; и TL1, TL3, TL7, TL9, TL10 и TL13, в которых аминокислоты человеческого CL (SEQ ID NO: 13) замещали отрицательно заряженным Glu или Asp.Названия, центры мутации и номера последовательностей предпочтительных константных областей сведены в Таблице 2 (перечень модифицированных центров).
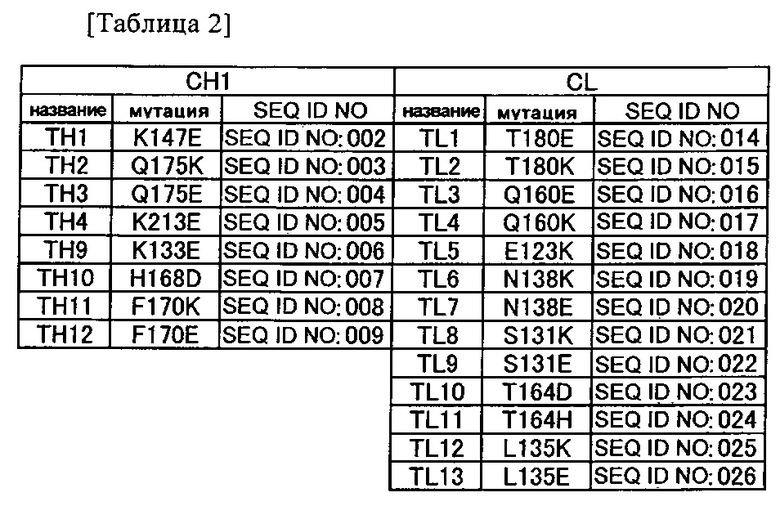
[Пример 2] Способ отбора центров, регулирующих границу CH1/CL и приготовления и анализа каждого антитела
Влияние на регулирование границы CH1/CL обнаруженных аминокислот подтверждали с применением описанных ниже способов. Отбор проводили с использованием антитела против GPC3. Сначала строили векторы экспрессии H-цепи и L-цепи. Вектор экспрессии H-цепи, имеющий вариабельную область GpH7 H-цепи (SEQ ID NO: 34) и константную область H-цепи, полученные согласно Примеру 1, и вектор экспрессии L-цепи, имеющий вариабельную область GpL16 L-цепи (SEQ ID NO: 35) и константную область L-цепи, полученные согласно Примеру 1, строили в соответствии со Справочным примером 1. Затем комбинации полученных векторов экспрессии H-цепи и L-цепи отбирали описанным ниже способом. Единичную H-цепь, в которой аминокислота в обнаруженном центре обладает положительным зарядом или отрицательным зарядом, выбирали из константных областей, полученных согласно Примеру 1. В таких случаях не обязательно вводили мутацию. Например, хотя позиция 147 ТН1 замещена Glu, мутация не вводится, поскольку аминокислота в позиции 147 G1d представляет собой Lys и изначально обладает положительным зарядом. Затем L-цеп, имеющие мутации в позициях, соответствующих мутированным позициям в CH1 выбранных H-цепей согласно Таблице 1, были выбраны из Таблицы 2. Например, при выборе TH1 для H-цепи TL1 и TL2 выбирали для L-цепи, поскольку ожидалось, что аминокислота в позиции 147 CH1 и аминокислота в позиции 180 CL должны взаимодействовать в качестве границы CH1/CL. Затем выбранные две L-цепи смешивали с выбранной одной H-цепью и антитела экспрессировали в соответствии со Справочным примером 1. И наконец, экспрессированные антитела анализировали в соответствии со Справочным примером 3 или Справочным примером 4, и модификации, эффективные для регулирования границы CH1/CL, отбирали в соответствии с соотношением экспрессии каждого антитела. Поскольку IgG состоит из комплекса из двух H-цепей и двух L-цепей, если один тип H-цепи и два типа L-цепей смешиваются и экспрессируются, должны экспрессироваться три комбинации. Например, если экспрессируются комбинации TH1 в качестве H-цепи и TL1 и TL2 в качестве L-цепей, экспрессируются три указанные ниже комбинации (Фиг. 2):
H-цепь_1:H-цепь_2:L-цепь_1:L-цепь_2=TH1:TH1:TL1:TL1 (указывается как TH1/TL1),
H-цепь_1:H-цепь_2:L-цепь_1:L-цепь_2=TH1:TH1:TL1:TL2 (указывается как TH1/TL1_TL2)
H-цепь_1:H-цепь_2:L-цепь_1:L-цепь_2=TH1:TH1:TL2:TL2 (указывается как TH1/TL2).
В случае, если ассоциация H-цепи и L-цепи не является выборочной, H-цепь и L-цепи должны экспрессироваться в соотношении TH1/TL1:TH1/TL1_TL2:TH1/TL2=1:2:1, поскольку две L-цепи присутствуют в равных количествах. Однако в случае, если H-цепь избирательно связывается лишь с одной из L-цепей на границе CH1/CL, считается, что только эта комбинация избирательно экспрессируется. Например, в случае, если аминокислота в позиции 147 CH1 и аминокислота в позиции 180 CL задействованы на границе CH1/CL, TH1 экспрессируется в качестве H-цепи, а TL1 и TL2 экспрессируются в качестве L-цепей, комбинация ТН1, в которой аминокислота в позиции 147 CH1 является Glu (отрицательно заряженным), и TL2, в которой аминокислота в позиции 180 CL L-цепи является Lys (положительно заряженным), должна избирательно экспрессироваться. Однако в случае, если аминокислота в позиции 147 CH1 и аминокислота в позиции 180 CL не взаимодействуют на границе CH1/CL, поскольку ассоциация H-цепи и L-цепи не является выборочной, они должны экспрессироваться в соотношении TH1/TL1:TH1/TL1_TL2:TH1/TL2=1:2:1. Таким образом, модификации, эффективные для регулирования границы CH1/CL (модификации, задействованные на границе CH1/CL), отбирали путем смешивания и экспрессии одного типа H-цепи и двух типов L-цепей и использования их баланса экспрессии в качестве индикатора.
Комбинации H-цепи и L-цепей сведены в Таблице 3 (также показаны комбинации H-цепи и L-цепей, используемые при экспрессии; центры мутации в H-цепи и L-цепи).
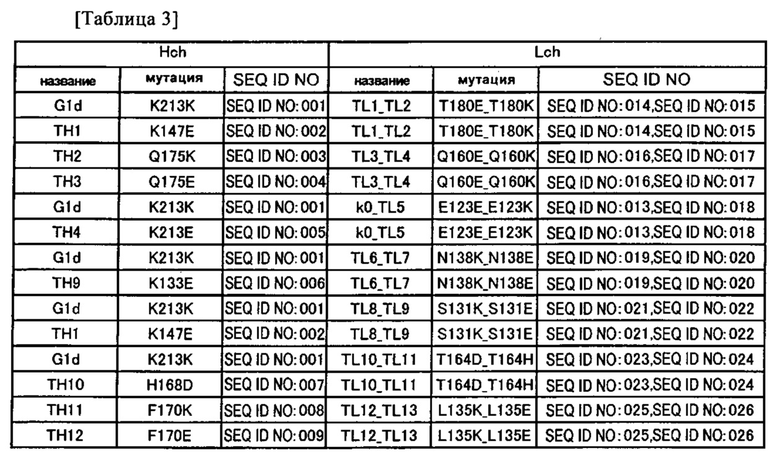
Антитела экспрессировались в соответствии с комбинациями, показанными в Таблице 3, и подтверждалось влияние на регулирование выбранной границы CH1/CL. В это время антитела одного типа H-цепи и одного типа L-цепи одновременно экспрессировали и использовали в анализах в качестве контроля. Антитела экспрессировали в соответствии со способом из Справочного примера 1.
Полученные антитела анализировали при помощи AIEX в соответствии со способом из Справочного примера 3. Данные анализа AIEX сведены на Фиг. 3. Поскольку для анализа AIEX используют анионообменную колонку, отрицательно заряженные антитела элюируются быстрее. Анализируемые данные сведены и показаны в Таблице 4. Пики указываются как пики 1, 2 и 3 в порядке уменьшения времени элюирования. Соотношение каждого пика рассчитывали при общей площади под пиком 100%.
Как показано на Фиг. 2, в случае, если позиции, введенные при мутациях, создают электрическое взаимодействие, соотношение антитела в позиции серого пика возрастает. То есть, места мутации, в которых соотношение окрашенного серым антитела превышает 25%, считаются электрически взаимодействующими.
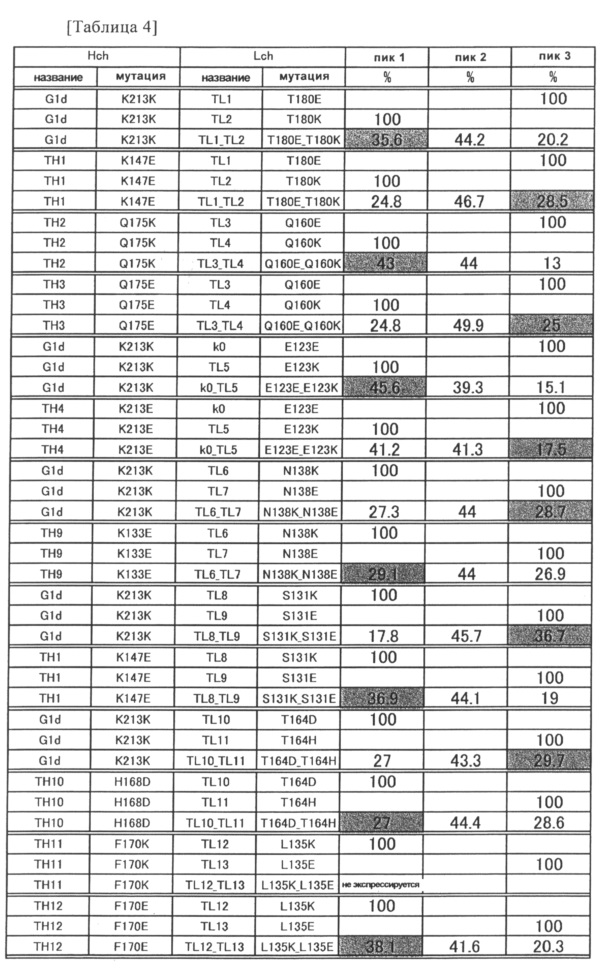
В результате такого исследования разных центров модификации позиции 147, 175 и 213 H-цепи и позиции 123, 131, 160 и 180 L-цепи были признаны эффективными для регулирования границы CH1/CL. Кроме того, было обнаружено, что модификации только позиции 147 H-цепи и позиции 123 L-цепи, указанные в документах WO 2006/106905 и WO 2007/147901, являются недостаточными для обеспечения специфической ассоциации H-цепи и L-цепи, и специфическая ассоциация возможна только при сочетании модификаций, присутствующих в представленном примере.
[Пример 3] Получение и анализ антител с комбинированными центрами модификации
Считалось, что граница CH1/CL более эффективно регулируется путем комбинирования центров К147, Q175 и К213 в CH1 и центров Е123, S131, Q160 и Т180 в CL, показанных в Примере 2, которые считались оказывающими заметное влияние на регулирование границы CH1/CL. Комбинации модификаций в полученных антителах и экспрессированные антитела сведены в Таблице 5 (Сводка центров модификации).
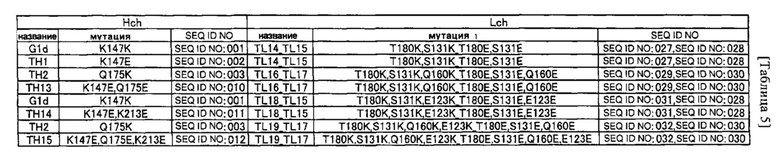
Получение векторов экспрессии H-цепи или L-цепи, введенных путем мутаций, и экспрессию антитела осуществляли в соответствии со Справочным примером 1, и анализы полученных антител осуществляли в соответствии со Справочным примером 3 или Справочным примером4. Результаты сведены в Таблице 6.
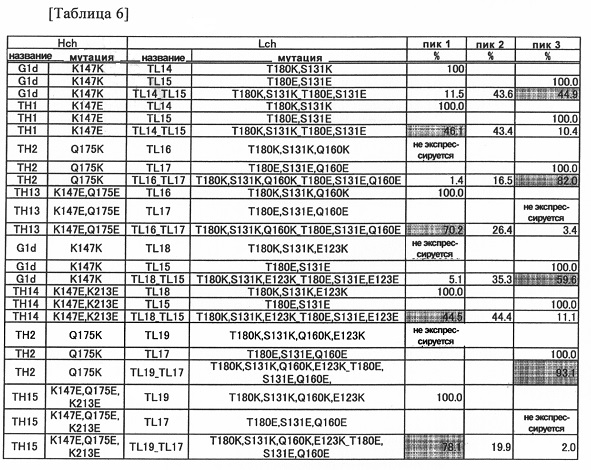
Если сравнить Таблицу 4 и Таблицу 6, станет понятно, что соотношение направленных комбинаций H-цепи и L-цепи повышается благодаря комбинированию модификаций, в отличие от введения одной модификаций. Таким образом, считается, что антитело, в котором ассоциированы только необходимые H-цепь и L-цепь, может быть эффективно получено путем комбинирования модификаций.
[Пример 4] Экспрессия и анализ биспецифических антител В Примере 3 рассматривалось получение биспецифических антител (биспецифических Ab) для ТН2, ТН13 и ТН15 H-цепей и TL16, TL17, TL19 и TL20 L-цепей, которые продемонстрировали значительное влияние на регулирование границы CH1/CL. В этом примере биспецифические антитела получали с применением антитела против IL6R и антитела против GPC3.
Константные области H-цепи (SEQ ID NO: 59) и L-цепи (SEQ ID NO: 60), распознающие антитело против IL6R, и константные области H-цепи (SEQ ID NO: 61) и L-цепь (SEQ ID NO: 62), распознающие антитело против GPC3, замещали ТН2, ТН13 и ТН15 для константных областей H-цепи и TL16, TL17, TL19 и TL20 для CL L-цепи. Кроме того, H-цепь, введенную при модификации "выступ-во-впадину" (KiH) (документ WO 96/27011) получали во избежание ассоциации между гомогенными Н-цепями. Места мутации в этих полученных антителах и экспрессированные антитела сведены в Таблице 7 (Комбинации H-цепи и L-цепи каждого биспецифического антитела).
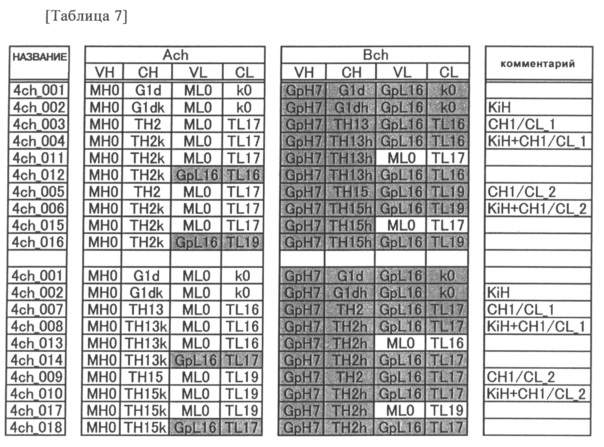
Выше в Таблице 7 "k" прибавляется после названия варианта для константных областей, в которых была введена модификация "выступ" в H-цепь, и "h" прибавляется после названия варианта, для которого константные области, в которых была введена модификация "впадина". Например, "TH1k" означает, что модификация "выступ" была введена дополнительно к мутации ТН1, а "TH1h" означает, что модификация "впадина" была введена дополнительно к мутации ТН1. Получение векторов экспрессии H-цепи или L-цепи, в которые была введена мутация, а также экспрессию антител осуществляли в соответствии со Справочным примером 1, и анализы полученных антител осуществляли в соответствии со способом анализа CIEX, показанным в Справочном примере 4.
Комбинации H-цепей и L-цепей антитела против IL6R и антитела против GPC3, которые используются в биспецифических антителах, сведены в Таблице 8.
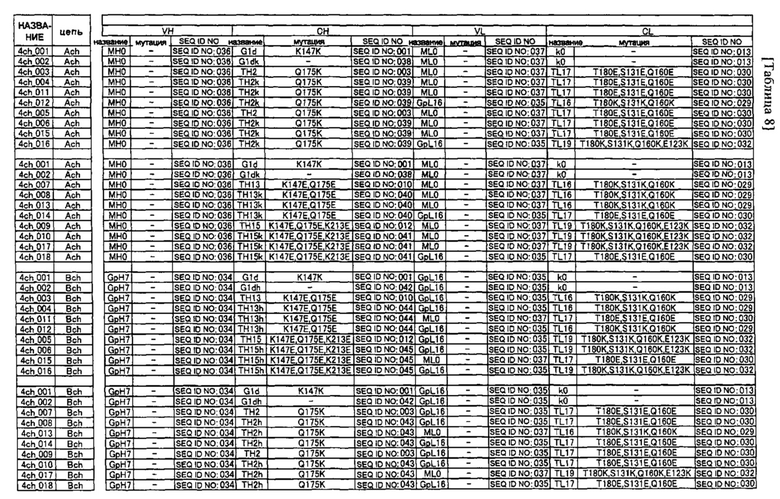
Каждая комбинация объясняется в данном описании с использованием 4ch_001, 4ch_002, 4ch_003, 4ch_004, 4ch_011 и 4ch_012 в качестве примеров. Модификации, регулирующие границу CH1/CL, сведены в Таблице 8.
4ch_001 экспрессировали с использованием H-цепей и L-цепей, в которые не вводились модификации для регулирования границы CH1/CL и модификация KiH. 4ch_002 экспрессировали с использованием H-цепей и L-цепи, в которые вводилась модификация KiH. 4ch_003 экспрессировали с использованием H-цепей и L-цепей, в которые вводились модификации для регулирования границы CH1/CL. 4ch_004 экспрессировали с использованием H-цепей и L-цепей, в которые вводились модификация KiH и модификации для регулирования границы CH1/CL. Кроме того, 4ch_011 экспрессировали с использованием H-цепей антитела против IL6R и H-цепей антитела против GPC3, в которые вводились модификация KiH и модификации для регулирования границы CH1/CL, и L-цепей антитела против IL6R, в которые вводились модификации для регулирования границы CH1/CL. 4ch_012 экспрессировали с использованием H-цепей антитела против IL6R и H-цепей антитела против GPC3, в которые вводились модификация KiH и модификации для регулирования границы CH1/CL, и L-цепей антитела против GPC3, в которые вводились модификации для регулирования границы CH1/CL. Каждое антитело экспрессировали в соответствии со Справочным примером 1 и анализировали при помощи CIEX в соответствии со Справочным примером 4; результаты сведены на Фигурах 9-1 и 9-2. Случай использования вариабельной области H-цепи антитела против IL6R обозначается MHO, а случай использования вариабельной области H-цепи антитела против GPC3 обозначается GpH7. Случай использования вариабельной области L-цепи антитела против IL6R обозначается ML0, а случай использования вариабельной области L-цепи антитела против GPC3 обозначается GpL16. Множество гетерогенных компонентов, которые считаются различными комбинациями H-цепи и L-цепи, обнаруживали путем хроматографии для 4ch_001, которую не вводят при мутациях для контроля границы CH1/CL и KiH. Для сравнения, поскольку ассоциация гомогенных H-цепей подавлялась в 4ch_002 путем мутации KiH, уменьшается количество хроматографических пиков, которые считаются примесями. Кроме того, поскольку подавлялась ассоциация H-цепей и L-цепей в 4ch_003 с использованием ТН2 и ТН13 в Н-цепях и TL16 и TL17 в L-цепях, в которые вводились модификации для регулирования границы CH1/CL, уменьшалось количество хроматографических пиков, которые считаются представляющими примеси. Кроме того, было обнаружено, что 4ch_004 с комбинацией мутаций KiH и модификаций для регулирования границы CH1/CL обычно является главным пиком. Причиной того, что хроматографические пики 4ch_004 почти совпадают с хроматографическими пиками 4ch_011, считается то, что их пики не могут быть отделены путем хроматографии из-за их подобных изоэлектрических точек (pI). Также проводились исследования на 4ch_005, 4ch_006, 4ch_015 и 4ch_016, к которым применялось другое регулирование границы CH1/CL по сравнению с 4ch_004; 4ch_007, 4ch_008, 4ch_013 и 4ch_014, в которых модификации для регулирования границы CH1/CL, введенные в 4ch_004, чередовались между антителом против GPC3 и антителом против IL6R; и 4ch_009, 4ch_010, 4ch_017 и 4ch_018, в которых модификации для регулирования границы CH1/CL, введенные в 4ch_006, чередовались между антителом против GPC3 и антителом против IL6R, с применением тех же способов, что и для 4ch_003, 4ch_004, 4ch_011 и 4ch_012. В результате гетерогенные компоненты, присутствующие на хроматограмме, были значительно уменьшены по сравнению с 4ch_001.
Из вышеуказанного становится очевидно, что биспецифические антитела могут быть эффективно получены путем комбинирования модификаций для регулирования границы CH1/CL и модификации KiH.
[Пример 5] Эффект модифицирующего регулирования границы CH1/CL с использованием различных антител
Из Примера 4 становится очевидным, что регулирование границы с использованием CH1/CL может применяться для получения биспецифических антител. Таким образом, эффект регулирования границы CH1/CL подтверждался с использованием антитела против CD3, М12 (H-цепь: SEQ ID NO: 54, L-цепь: SEQ ID NO: 57) и антитела против GPC3, GC33(2) (H-цепь: SEQ ID NO: 55, L-цепь: SEQ ID NO: 58). Как и в Примере 4, биспецифические антитела получали с использованием ТН2, ТН13 и ТН15 в Н-цепях и TL16, TL17 и TL19 в L-цепях, что продемонстрировало значительный эффект в регулировании границы CH1/CL.
Константные области H-цепи (SEQ ID NO: 54) и L-цепи (SEQ ID NO: 57) CD3, распознающие антитело Ml2, и константные области H-цепи (SEQ ID NO: 55) и L-цепи (SEQ ID NO: 58) GPC3, распознающие антитело GC33(2), замещали ТН2, ТН13 и ТН15 для CHI H-цепи и TL16, TL17 и TL19 для CL L-цепи. Кроме того, H-цепь с модификациями "выступ-во-впадину" (KiH) (Патентный документ 1) получали во избежание ассоциации между гомогенными Н-цепями. Места мутации этих полученных антител и экспрессированные антитела сведены в Таблице 9 (Сводка центров модификации).
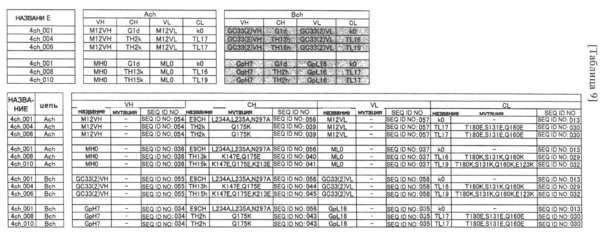
Получение векторов экспрессии H-цепей и L-цепей с мутациями и экспрессию антител осуществляли в соответствии со Справочным примером 1 и анализы полученных антител осуществляли в соответствии со способом анализа CIEX, показанным в Справочном примере 4.
Очевидно, что регулирование границы CH1/CL также применимо для получения биспецифических антител с антителом против CD3 и антителом против GPC3.
[Пример 6] Комбинация регулирования границы CH1/CL и регулирования границы вариабельной области
При получении биспецифических антител включение электрического отталкивания в вариабельные области VH и VL является известной технологией, обеспечивающей возможность специфической ассоциации заданных H-цепей м L-цепей (Патентный документ WO 2006/106905). Таким образом, для эффективной экспрессии только заданных компонентов следует вызвать отталкивание между вариабельными областями H-цепи и L-цепи, помимо регулирования границы CH1/CL. Этот процесс называется регулированием границы VH/VL. Получали МН01 (SEQ ID NO: 46), где Gln в позиции 39 согласно нумерации Kabat H-цепи антитела против IL6R был замещен Lys; МН02 (SEQ ID NO: 47), где Gln был замещен Glu; ML01 (SEQ ID NO: 50), где Gln в позиции 38 согласно нумерации Kabat L-цепи был замещен Glu; и ML02 (SEQ ID NO: 51), где Glu был замещен Lys. Кроме того, соответственно получали GpH71 (SEQ ID NO: 48), где Gln в позиции 39 согласно нумерации Kabat H-цепи антитела против GPC3 был замещен Lys; GpH72 (SEQ ID NO: 49), где Gln был замещен Glu; GpL161 (SEQ ID NO: 52), где Gln в позиции 38 согласно нумерации Kabat L-цепи был замещен Glu; и GpL162 (SEQ ID NO: 53), где Gln был замещен Lys. Получение векторов экспрессии антитела осуществляли в соответствии со способом из Справочного примера 1. Биспецифические антитела экспрессировали, используя полученные антитела. Комбинации модификаций в полученных антителах и экспрессированные антитела сведены в Таблице 10.
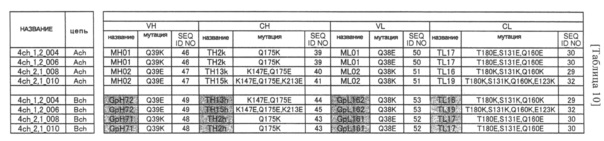
Экспрессию антитела осуществляли в соответствии со Справочным примером 1 и анализы полученных антител осуществляли в соответствии со Справочным примером 4.
Поскольку пики, наблюдаемые на хроматограммах 4ch_006 и 4ch_008, в которые были введены мутации для регулирования границы CH1/CL, которые считаются представляющими гетерогенные компоненты, исчезали в 4ch_1,2 006 и 4ch_2,1 008, в которые были вставлены мутации для регулирования границы VH/VL, стало очевидным, что только заданные компоненты могут быть эффективно получены путем применения регулирования границы VH/VL помимо регулирования границы CH1/CL (Фигуры 10 и 11). Кроме того, пики, которые считаются представляющими новые гетерогенные компоненты, не обнаруживались даже при осуществлении дополнительной мутации для регулирования границы VH/VL в 4ch_004 и 4ch_010, в которых мутации были введены только в CH1/CL, и в которых только компоненты, которые рассматривались как заданные компоненты, считались очищенными (Фигуры 10 и 11).
Из вышеуказанного становится очевидно, что добавление регулирования границы VH/VL к регулированию границы CH1/CL дополнительно способствует очистке заданных компонентов и при этом не имеет неблагоприятного воздействия на очистку, когда только заданные компоненты считаются уже очищенными.
[Пример 7] Измерение Tm антител с комбинированными центрами модификации
Модификации для регулирования границы CH1/CL могут влиять на устойчивость Fab. Таким образом, устойчивость Fab или Tm измеряли в соответствии со способом из Справочного примера 2 для комбинаций TH2/TL17, TH13/TL16 и TH15/TL19. Антитела, в которых H-цепь/L-цепь состояли из TH2/TL17, TH13/TL16 и TH15/TL19, получали с применением антитела против IL6R. Комбинации модификаций антител и экспрессированные антитела сведены в Таблице 11.
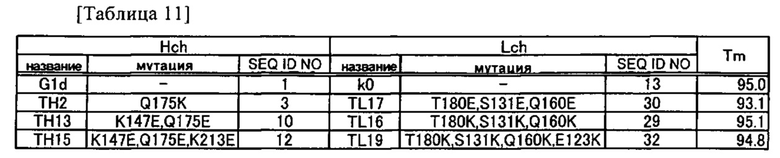
Экспрессию антител измеряли в соответствии со Справочным примером 1, и Tm (°C) каждого из полученных антител измеряли в соответствии со Справочным примером 2. Результат показывает, что значения Tm для Fab G1d/k0 без введения мутаций и для Fab TH2/TL17, TH13/TL16 и TH15/TL19 с введенными мутациями в CH1/CL, составляли 95,0°C, 93,1°C, 95,1°C и 94,8°C, соответственно. Было обнаружено, что мутации для регулирования границы CH1/CL не имеют влияния на устойчивость Fab.
[Пример 8] Влияние введения мутаций для регулирования границы CH1/CL на активность связывания
Возможность модификаций для регулирования границы CH1/CL, влияющих на связывание антигена, не может быть полностью исключена. Таким образом, для измерения аффинности к IL-6R и GPC3 измеряли активность связывания TH2/TL17, TH13/TL16 и TH15/TL19 в соответствии со способом из Справочного примера 5 (Таблица 12).
Поскольку активность связывания с IL-6R и активность связывания с GPC3 TH2/TL17, TH13/TL16 и TH15/TL19, в которые были введены мутации для регулирования границы CH1/CL, не отличалась от активности связывания природного G1d/k0 с IL-6R и GPC3, стало очевидным, что модификации для регулирования границы CH1/CL не влияют на аффинность. Кроме того, при измерении аффинности к двум антигенам, IL-6R и GPC3, в соответствии со Справочным примером 5 с применением 4ch_004, 4ch_006, 4ch_008 и 4ch_010, полученных согласно Примеру 4, было обнаружено, что их аффинность равняется показателю природного G1d/k0, показанного в Таблице 12 (Таблицы 13 и 14).
В соответствии с исследованиями, проведенными согласно Примерам с 1 по 8, стало очевидным, что только заданные компоненты могут быть эффективно очищены путем введения мутации в CH1/CL без снижения устойчивости Fab и без снижения активности связывания.
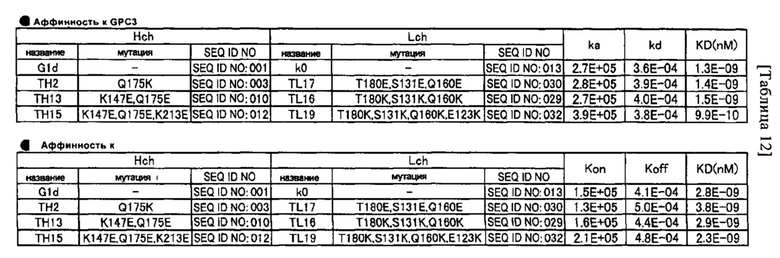
[Пример 9]
Аминокислотные последовательности человеческого IgA1 (SEQ ID NO: 63), IgA2 (SEQ ID NO: 64), IgD (SEQ ID NO: 65), IgE (SEQ ID NO: 66), IgG1 (SEQ ID NO: 67), IgG2 (SEQ ID NO: 68), IgG3 (SEQ ID NO: 69), IgG4 (SEQ ID NO: 70) и IgM (SEQ ID NO: 71) выравнивали относительно CH1 H-цепи; и аминокислотные последовательности человеческого IgK (Каппа) (SEQ ID NO: 72), IgL1 (SEQ ID NO: 73), IgL2 (SEQ ID NO: 74), IgL3 (SEQ ID NO: 75), IgL6 (SEQ ID NO: 76) и IgL7 (SEQ ID NO: 77) (Лямбда) выравнивали относительно CL L-цепи с их последующим сравнением. Результаты показаны на Фиг. 12. Модификации, обнаруженные в представленном примере, показаны стрелками. В результате введения аминокислот, обладающих разными зарядами, в H-цепь и L-цепь, таким образом, чтобы аминокислоты, показанные стрелками, отталкивались между CH1 H-цепи и CL L-цепи, как указано в представленном примере, считается, что заданные H-цепь и L-цепь могут быть специфично ассоциированы.
[Справочный пример 1] Получение векторов экспрессии антитела и экспрессия и очистка антител
Аминокислотные замещения вводили согласно способу, известному среди специалистов в данной области, с применением комплекта QuikChange для сайт-специфического мутагенеза (Stratagene), ПЦР или комплекта для ПЦР-клонирования In-fusion Advantage (Takara) и т.п., с последующим построением векторов экспрессии. Основные последовательности полученных векторов экспрессии определяли в соответствии со способом, известным среди специалистов в данной области. Антитела экспрессировали путем временной трансфекции полученных плазмид в производные от эмбриональных раковых клеток почек человека линии клеток НЕК293Н (Invitrogen) или клетки FreeStyle 293 (Invitrogen). Антитела очищали от полученного супернатанта культуры в соответствии со способом, известным среди специалистов в данной области, с применением rProtein A Sepharose™ Fast Flow (GE Healthcare). Концентрацию очищенных антител определяли путем измерения оптической плотности при 280 нм при помощи спектрофотометра и концентрацию антитела рассчитывали по полученному показателю с использованием коэффициента поглощения, рассчитанного в соответствии со способом PACE (Protein Science 1995; 4: 2411-2423).
[Справочный пример 2] Определение температуры плавления (Tm) модифицированных антител при помощи дифференциальной сканирующей калориметрии
В этом исследовании определяли температурную устойчивость путем измерения температуры плавления (Tm) антител при помощи дифференциального сканирующего калориметра; MicroCal Capillary DSC (DKSH).
Аликвоты по 500 мкл раствора каждого антитела помещали в измерительную чашку и температуру повышали с 20°C до 115°C. Наблюдали скорость повышения температуры 120°C/ч и изменения в теплоемкости.
Данные анализировали при помощи Origin7 (Light Stone) и температуру, при которой наблюдались изменения в теплоемкости, рассчитывали и определяли как значение Tm.
[Справочный пример 3] Анализ при помощи анионообменной хроматографии (AIEX)
Полученные антитела анализировали с применением способа AIEX при помощи системы Alliance (Waters). Анализы осуществляли согласно способу с применением двухжидкостного градиента с использованием TSK-геля DEAE-NPR (Tosoh) для аналитической колонки, 10 ммоль/л Tris-HCl (pH 7,5) для мобильной фазы A и 10 ммоль/л Tris-HCl и 500 ммоль/л NaCl (рН 7,5) для мобильной фазы B. Измерения осуществляли при длине волны 280 нм.
Данные анализировали при помощи Empower2 (Waters) с последующим вычислением соотношения каждого обнаруженного пика.
[Справочный пример 4] Анализ при помощи катионообменной хроматографии (CIEX)
Полученные антитела анализировали с применением способа CIEX при помощи системы Alliance (Waters). Анализы осуществляли согласно способу с применением двухжидкостного градиента с использованием WCX-10 (Dionex) для аналитической колонки, 25 ммоль/л MES (pH 6,1) для мобильной фазы A и 25 ммоль/л MES и 500 ммоль/л NaCl (pH 6,1) для мобильной фазы B. Измерения осуществляли при длине волны 280 нм.
Данные анализировали при помощи Empower2 (Waters) с последующим вычислением соотношения каждого обнаруженного пика.
[Справочный пример 5] Измерение аффинности к IL6R и GPC3
Взаимодействие между антителом-мишенью и hIL6R или GPC3 анализировали при помощи Biacore Т100 (GE Healthcare). HBS-EP+(GE Healthcare) использовали для подвижного буфера, и температура измерения составляла 25°C. Протеин A/G (Thermo Scientific) иммобилизировали на сенсорном чипе серии S СМ5 (GE Healthcare) путем аминного связывания и использовали в качестве чипа. Антитело-мишень захватывали на чип, и оно взаимодействовало с каждым антигеном, разведенным подвижным буфером. Антитела, захваченные на чипе, смывали путем реакции с 10 мМ глицина-HCl (pH 1,5) для восстановления чипа для повторного использования.
Константу диссоциации (KD) каждого антитела для антигена рассчитывали путем проведения кинетического анализа результатов измерения Biacore. Более конкретно, константу скорости ассоциации ka (л/моль/с) и константу скорости диссоциации kd (л/с) рассчитывали при помощи сенсограмм глобальной подгонки, полученных путем измерения с применением программы Biacore Evaluation в 1:1 модели связывания Ленгмюра с последующим вычислением константы диссоциации KD (моль/л) на основе этих значений. Промышленная применимость
Способ, обеспечиваемый настоящим изобретением, позволяет регулировать ассоциацию без изменения структуры, функции, активности и т.п. первоначального полипептида (антитела) и является чрезвычайно полезным, поскольку требует лишь малого количества аминокислотных замещений. Кроме того, способ также мало воздействует на антигенность.
Применение настоящего изобретения обеспечивает возможность эффективного получения биспецифических антител, которые фактически сохраняют активность.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2774414C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| ПРЕДПОЧТИТЕЛЬНОЕ СПАРИВАНИЕ ДОМЕНОВ АНТИТЕЛ | 2018 |
|
RU2792440C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2663123C2 |
| ТРИ- ИЛИ ТЕТРАСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2010 |
|
RU2570633C2 |
| ГИПЕРГЛИКОЗИЛИРОВАННЫЕ СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2014 |
|
RU2708314C2 |
































































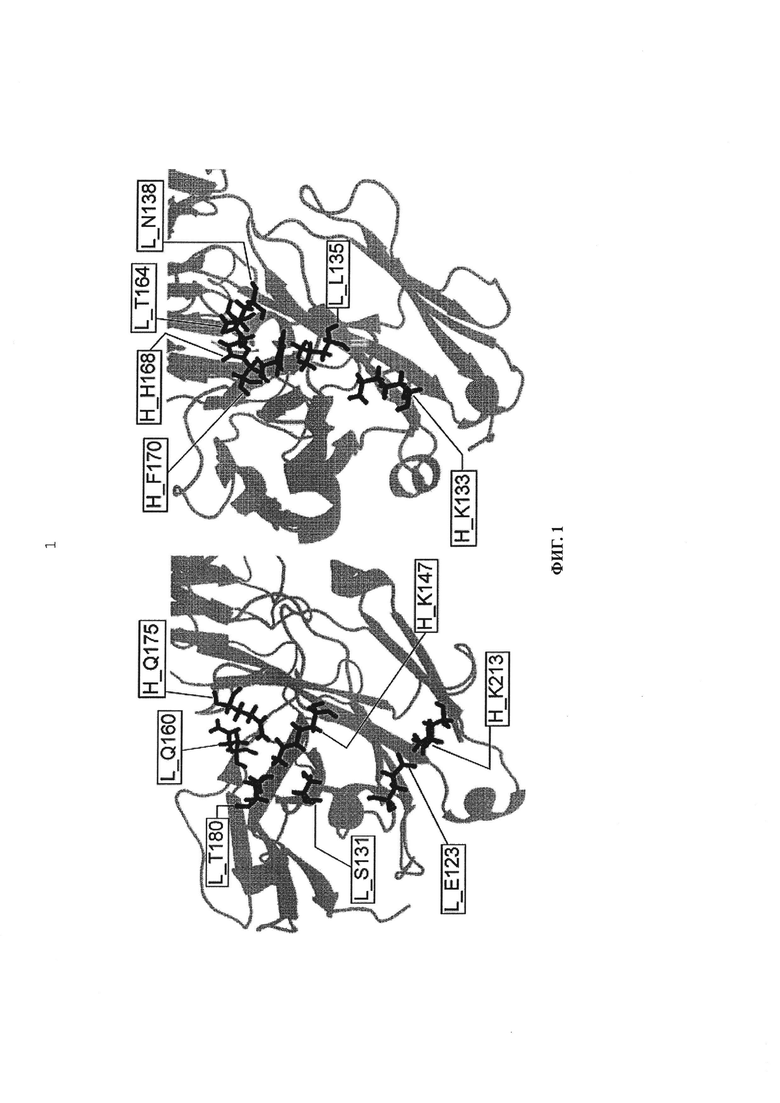
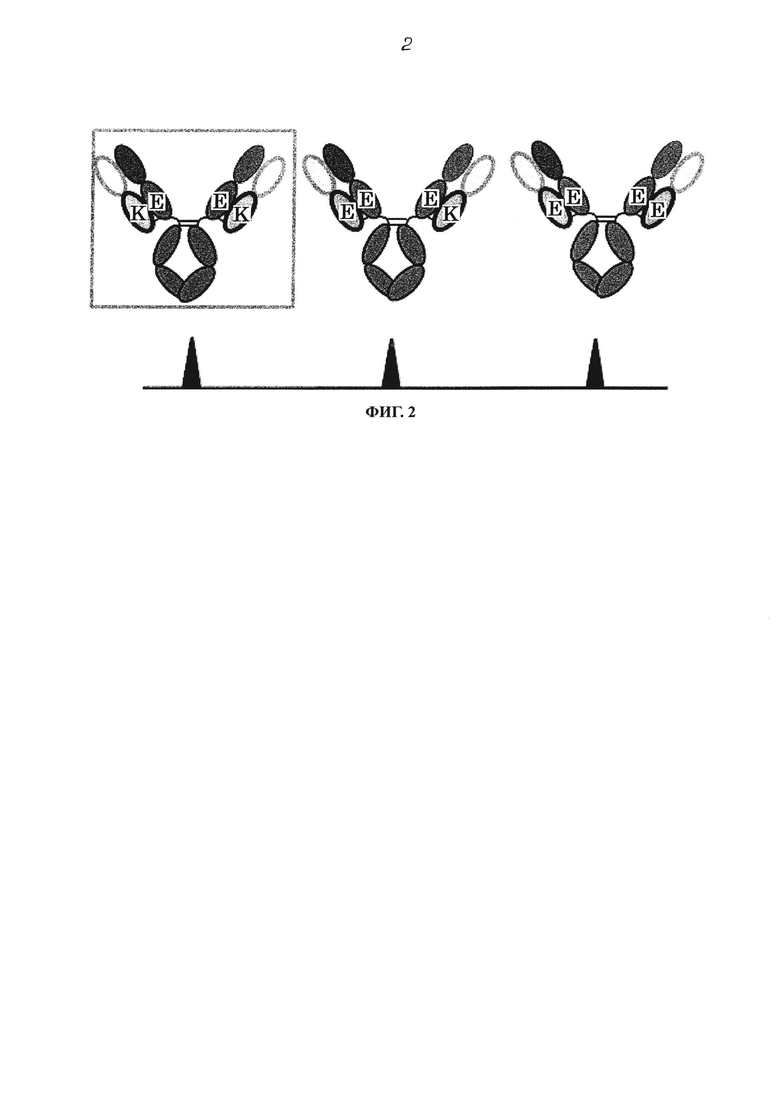
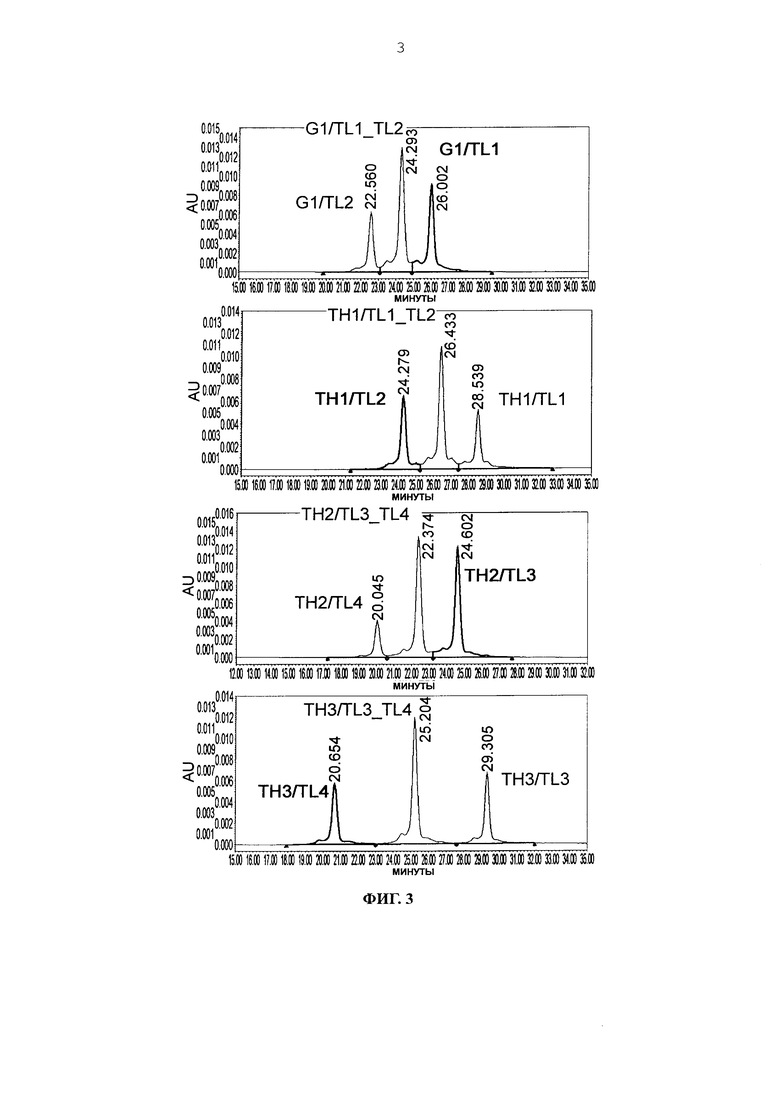
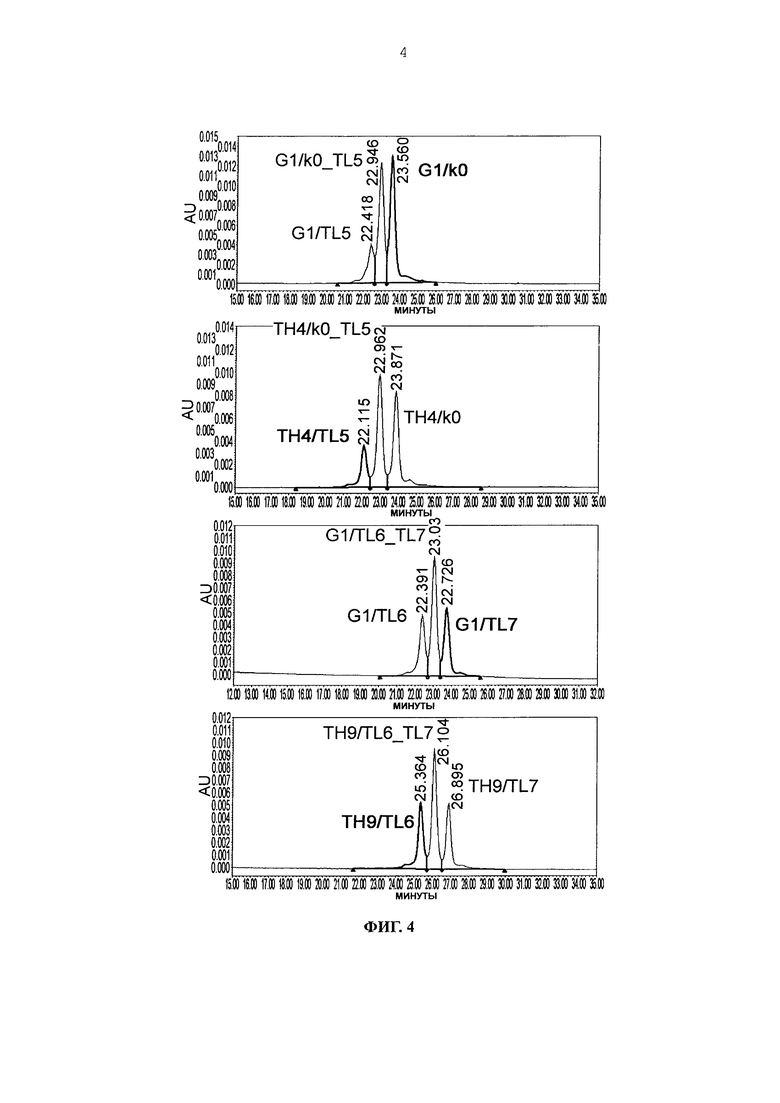
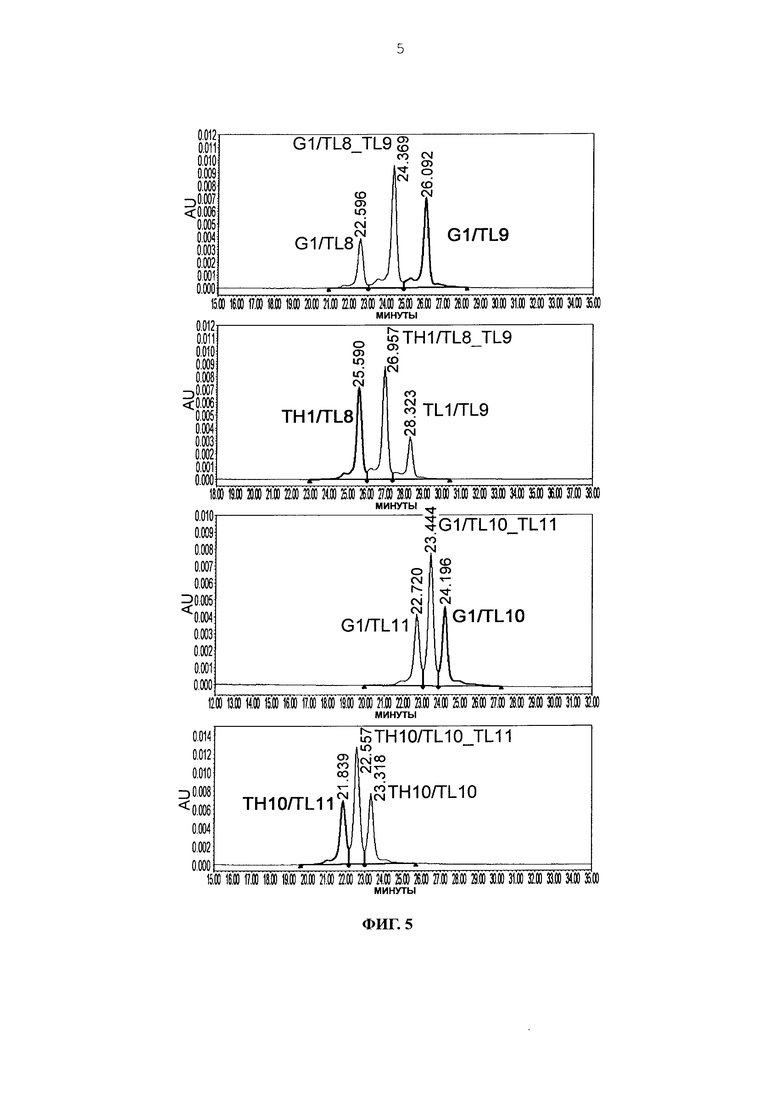
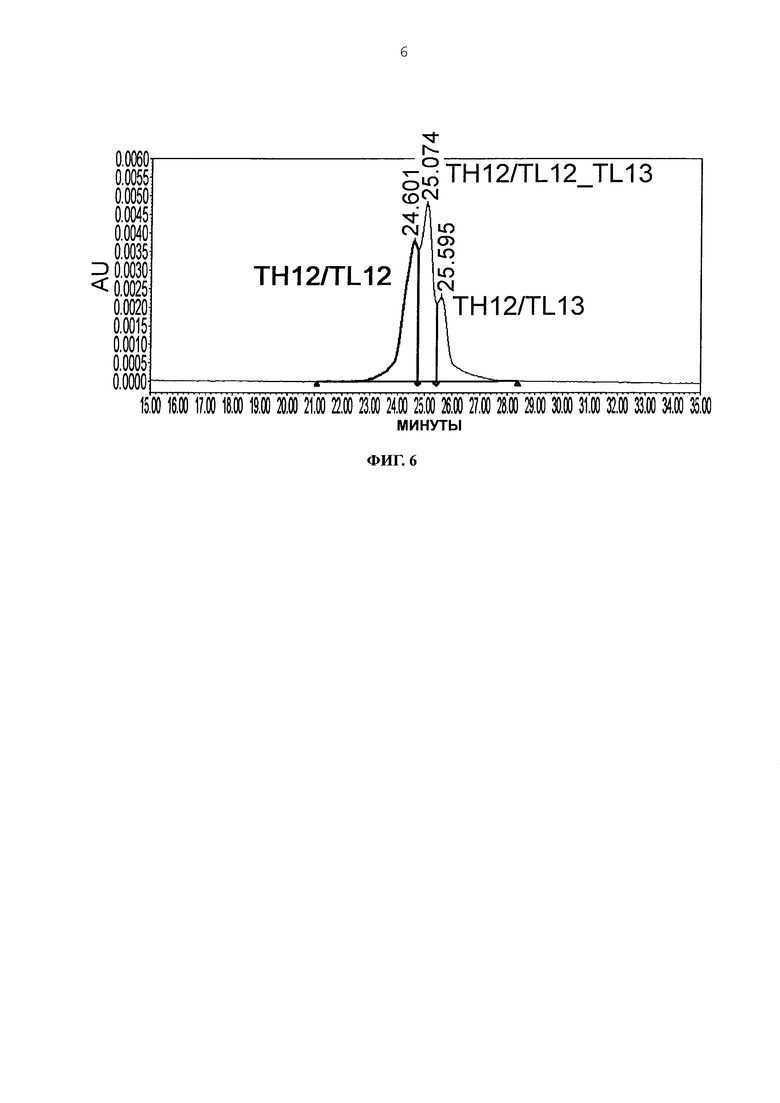
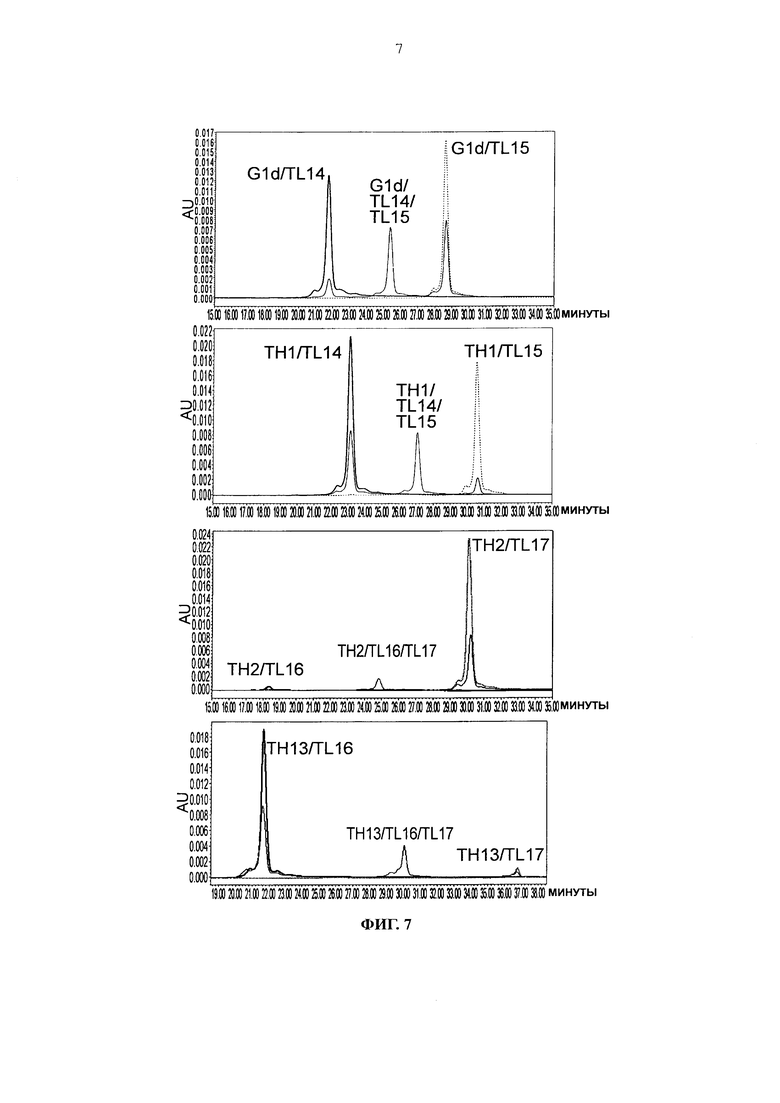
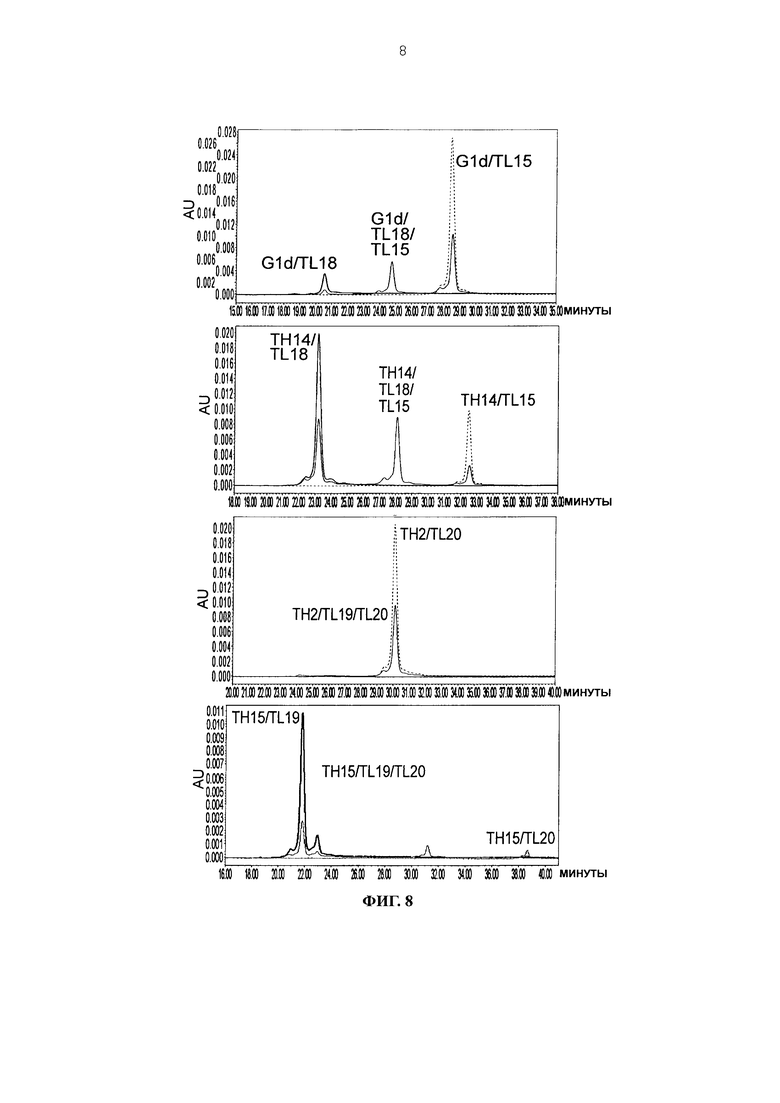


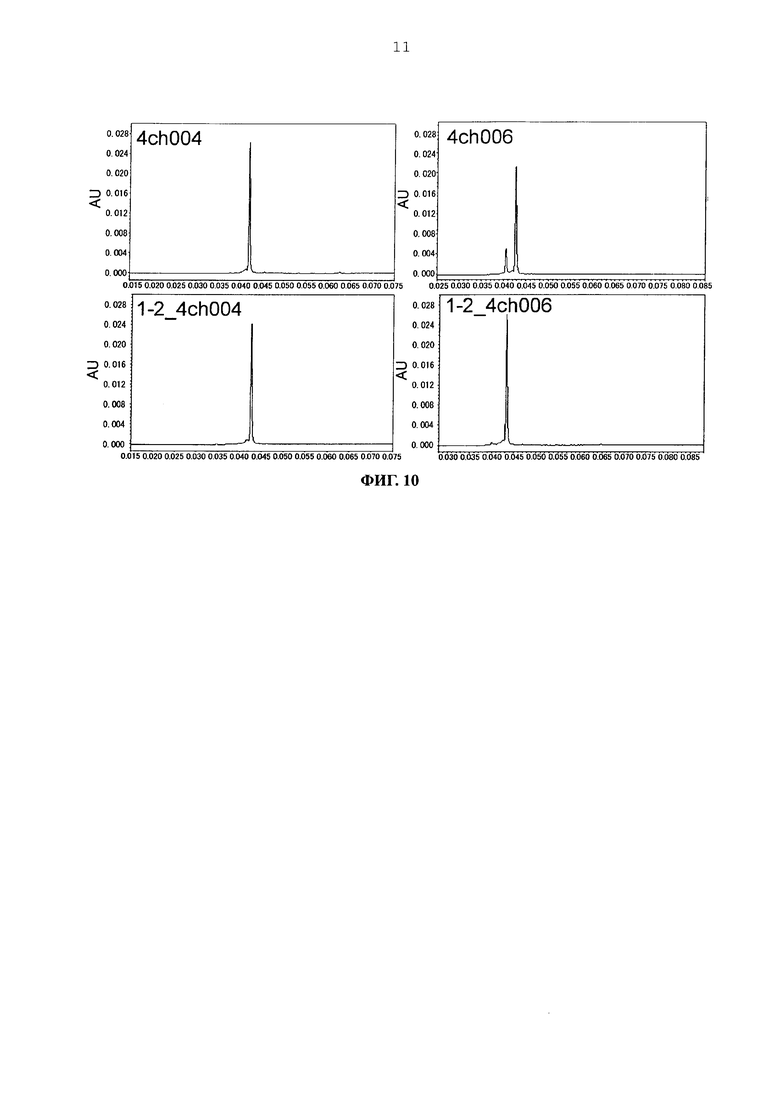
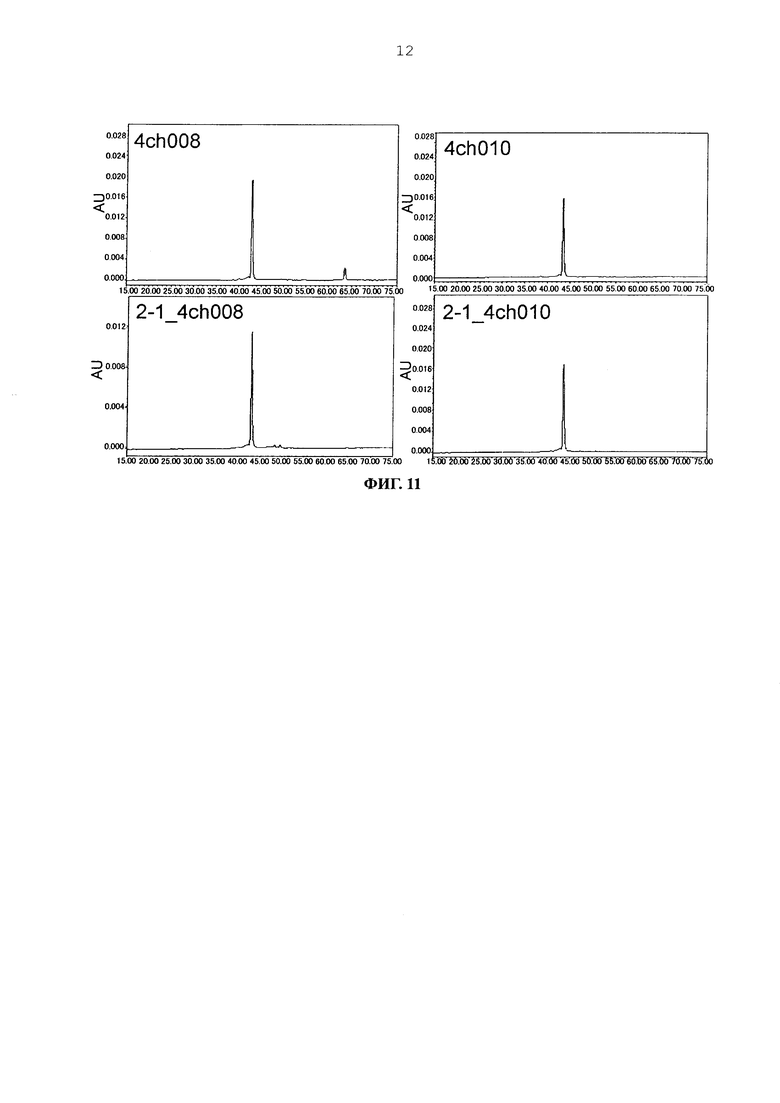

Настоящее изобретение относится к области иммунологии. Предложен способ получения биспецифического антитела, предусматривающий введение электрически взаимно отталкивающихся аминокислотных остатков в положения 147 тяжелой цепи и 180 легкой цепи, 147 тяжелой цепи и 131 легкой цепи или 175 тяжелой цепи и 160 легкой цепи (по нумерации ЕС), где тяжелая цепь представляет собой IgG1, а легкая цепь представляет собой IgK (Каппа). Данное изобретение обеспечивает подавление нежелательной ассоциации между константной области 1 тяжелой цепи (CH1) и константной области легкой цепи (CL) путем замещения аминокислот, присутствующих на границе между CH1 и CL, электрически заряженными аминокислотами. Также предложен способ регулирования ассоциации тяжелой цепи и легкой цепи биспецифического антитела такой, что нежелательные ассоциации не образуются между тяжелой и легкой цепями. 2 н. и 10 з.п. ф-лы, 12 ил., 14 табл., 9 пр.
1. Способ получения биспецифического антитела, согласно которому нежелательные ассоциации не образуются между тяжелой и легкой цепями, где тяжелая цепь представляет собой IgG1, а легкая цепь представляет собой IgK (Каппа), который включает этапы с (1) по (3):
(1) введение замен в нуклеиновые кислоты, кодирующие тяжелую и легкую цепи биспецифического антитела таким образом, что тяжелая и легкая цепи содержат один или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в пп. с (а) по (с), которые электрически взаимно отталкиваются:
(a) аминокислотный остаток, присутствующий в константной области 1 тяжелой цепи (СН1) в положении 147, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в константной области легкой цепи (CL) в положении 180, как указывается согласно нумерации ЕС;
(b) аминокислотный остаток, присутствующий в СН1 в положении 147, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в CL в положении 131, как указывается согласно нумерации ЕС; и
(c) аминокислотный остаток, присутствующий в СН1 в положении 175, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в CL в положении 160, как указывается согласно нумерации ЕС,
(2) введение нуклеиновых кислот с указанными заменами в клетку-хозяина и культивирование клетки-хозяина таким образом, чтобы экспрессировались нуклеиновые кислоты, и
(3) сбор биспецифического антитела из культуры клетки-хозяина.
2. Способ получения биспецифического антитела по п. 1, дополнительно включающий на этапе (1) введение замен в нуклеиновые кислоты таким образом, что аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в (d), электрически взаимно отталкиваются:
(d) аминокислотный остаток, присутствующий в СН1 в положении 213, как указывается согласно нумерации EU, и аминокислотный остаток, присутствующий в CL в положении 123, как указывается согласно нумерации EU.
3. Способ получения биспецифического антитела по п. 1 или 2, включающий на этапе (1) введение замен в нуклеиновые кислоты таким образом, что аминокислотные остатки, которые электрически взаимно отталкиваются, выбираются из аминокислотных остатков, включенных в группу (X) или (Y), как указано ниже:
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D); и
(Y) лизин (К), аргинин (R) или гистидин (Н).
4. Способ получения биспецифического антитела по п. 1 или 2, дополнительно включающий на этапе (1) введение замен в нуклеиновые кислоты таким образом, что два или более аминокислотных остатка, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, являются аминокислотными остатками, которые электрически взаимно отталкиваются.
5. Способ получения биспецифического антитела по п. 4, отличающийся тем, что аминокислотные остатки, которые электрически взаимно отталкиваются, являются аминокислотными остатками любого набора, выбранного из группы, к которой относятся наборы указанных ниже аминокислотных остатков, показанных ниже в (а) или (b):
(a) аминокислотный остаток, присутствующий в вариабельной области тяжелой цепи в положении 39 согласно нумерации Kabat, и аминокислотный остаток, присутствующий в вариабельной области легкой цепи в положении 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, присутствующий в вариабельной области тяжелой цепи в положении 45 согласно нумерации Kabat, и аминокислотный остаток, присутствующий в вариабельной области легкой цепи в положении 44 согласно нумерации Kabat.
6. Способ получения биспецифического антитела по п. 5, отличающийся тем, что аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) или (Y), как указано ниже:
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D); и
(Y) лизин (К), аргинин (R) или гистидин (Н).
7. Способ регулирования ассоциации тяжелой цепи и легкой цепи биспецифического антитела такой, что нежелательные ассоциации не образуются между тяжелой и легкой цепями, где тяжелая цепь представляет собой IgG1, а легкая цепь представляет собой IgK (Каппа), включающий:
1) введение замен в нуклеиновые кислоты, кодирующие тяжелую и легкую цепи биспецифического антитела таким образом, что тяжелая и легкая цепи содержат один или более наборов аминокислотных остатков, выбранных из группы, состоящей из наборов аминокислотных остатков, показанных ниже в (а)-(с), являются аминокислотными остатками, которые электрически взаимно отталкиваются:
(a) аминокислотный остаток, присутствующий в СН1 в положении 147, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в CL в положении 180, как указывается согласно нумерации ЕС;
(b) аминокислотный остаток, присутствующий в СН1 в положении 147, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в CL в положении 131, как указывается согласно нумерации ЕС; и
(c) аминокислотный остаток, присутствующий в СН1 в положении 175, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в CL в положении 160, как указывается согласно нумерации ЕС,
(2) введение нуклеиновых кислот с указанными заменами в клетку-хозяина и культивирование клетки-хозяина таким образом, чтобы экспрессировались нуклеиновые кислоты, и
(3) сбор биспецифического антитела из культуры клетки-хозяина.
8. Способ по п. 7, дополнительно включающий введение замен в нуклеиновые кислоты таким образом, что аминокислотные остатки в наборе аминокислотных остатков, показанных ниже в (d), являются аминокислотными остатками, которые электрически взаимно отталкиваются:
(d) аминокислотный остаток, присутствующий в СН1 в положении 213, как указывается согласно нумерации ЕС, и аминокислотный остаток, присутствующий в CL в положении 123, как указывается согласно нумерации ЕС.
9. Способ по п. 7 или 8, отличающийся тем, что аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) или (Y), как указано ниже:
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D); и
(Y) лизин (К), аргинин (R) или гистидин (Н).
10. Способ по п. 7 или 8, отличающийся тем, что дополнительно два или более аминокислотных остатка, образующих границу между вариабельной областью тяжелой цепи и вариабельной областью легкой цепи, являются аминокислотными остатками, которые электрически взаимно отталкиваются.
11. Способ по п. 10, отличающийся тем, что аминокислотные остатки, которые электрически взаимно отталкиваются, являются аминокислотными остатками любого набора, выбранного из группы, к которой относятся наборы аминокислотных остатков, показанных ниже в (а) или (b):
(a) аминокислотный остаток, присутствующий в вариабельной области тяжелой цепи в положении 39 согласно нумерации Kabat, и аминокислотный остаток, присутствующий в вариабельной области легкой цепи в положении 38 согласно нумерации Kabat; или
(b) аминокислотный остаток, присутствующий в вариабельной области тяжелой цепи в положении 45 согласно нумерации Kabat, и аминокислотный остаток, присутствующий в вариабельной области легкой цепи в положении 44 согласно нумерации Kabat.
12. Способ по п. 11, отличающийся тем, что аминокислотные остатки, которые электрически взаимно отталкиваются, выбраны из аминокислотных остатков, включенных в набор (X) или (Y), как указано ниже:
(X) глутаминовая кислота (Е) или аспарагиновая кислота (D); и
(Y) лизин (К), аргинин (R) или гистидин (Н).
| PADLAN E.A | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| EP 1870459 A1, 26.12.2007 | |||
| СПОСОБ ТРАНСПОРТИРОВКИ ГРУЗА ВДОЛЬ ЛЕСТНИЧНЫХ МАРШЕЙ ЗДАНИЯ | 1990 |
|
RU2009101C1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| MULLER K.M | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| HORNE C | |||
| et al | |||
| "Noncovalent association of heavy and light chains of human immunoglobulins | |||
| III | |||
| Specific interactions between VH and VL." The Journal of Immunology, 1982, 129(2): 660-664 | |||
| FEIGE M.J | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| KABAT E.A | |||
| et al., "Sequences of proteins of immunological interest." DIANE publishing, 1991, pp.647-652, 661-669 | |||
| ZHU ZHENPING et al | |||
| "Remodeling domain interfaces to enhance heterodimer formation." Protein science, 1997, 6(4): 781-788 | |||
| МОИСЕЕНКО В.М | |||
| "Моноклональные антитела в лечении злокачественных опухолей", Практическая онкология, 2003, Т | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

