Изобретение относится к микробиологии и может быть использовано в исследовательской работе НИИ и практической работе бактериологических лабораторий клинических учреждений, исследующих свойства микроорганизмов, взаимодействующих с эритроцитами и гемоглобином, а также как критерий отбора биотехнологически перспективных микроорганизмов, для создания лекарственных и пробиотических препаратов.
Гемоглобин эритроцитов представляет собой сложную молекулу, состоящую из небелковой части - гема, и белковой части - глобина. Основная функция гемоглобина - транспорт кислорода [Зинчук В.В. Коррекция кисло-родтранспортной функции крови при патологии сердечно-сосудистой системы / В.В. Зинчук, С.В. Гацура, К.В. Глуткина, Гродно, ГрГМУ - 2016. - 312 с.]. В процессе присоединения и отдачи кислорода железо в молекуле гемоглобина свою валентность не меняет, т.е. при присоединении кислорода или его отдаче железо не окисляется и не восстанавливается [Васильева Е.М. Биохимические особенности эритроцита. Влияние патологии // Биомедицинская химия, 2005, Т. 51, вып. 2, с. 118-126.].
При воздействии ряда факторов гемоглобин может окисляться до метгемоглобина, который не способен присоединять кислород и поэтому не может обеспечить дыхание тканей. Образование метгемоглобина происходит постоянно, ежедневно около 0,5% всего гемоглобина превращается в метгемоглобин. Метгемоглобин снова восстанавливается в гемоглобин специальным ферментом, использующим НАДН - метгемоглобинредуктазой (НАДН-цитохром Р-редуктаза). Поэтому концентрация метгемоглобина в крови в норме - меньше 2% [Фаткуллин К.В., Гильманов А.Ж., Костюков Д.В. Клиническое значении и современные методологические аспекты определения уровня карбокси- и метгемоглобина в крови // Практическая медицина, 2014, Т. 79, №3, С. 17-21.].
Однако концентрация метгемоглобина может значительно повышаться при воздействии на организм ряда факторов. Факторы, способствующие образованию метгемоглобина:
1. физические - влияние различного рода излучения [Gisele Capanema de Oliveira et al. The effect of γ-radiation on the hemoglobin of stored red blood cells: the involvement of oxidative stress in hemoglobin conformation // Ann Hematol (2013) 92:899-906];
2. химические - действие металлов переменной валентности и их оксидов [А.Н. Осипов, Г.Г. Борисенко, Ю.А. Владимиров. Биологическая роль нитрозильных комплексов гемопротеинов // Успехи биологической химии, т. 47, 2007, с. 259-292];
3. биологические - в результате нарушения ферментативной активности эритроцита и метаболизма бактерий [Глебов А.Н., Шульга Е.В., Зинчук В.В. Роль кислородсвязывающих свойств крови в развитии окислительного стресса, индуцированного липополисахаридом - Гродно, 2011, 216 с.].
Известен механизм образования метгемоглобина при взаимодействии микроорганизмов с клетками крови, на примере микроорганизмов кишечника [Трухина М. Нитраты, нитриты и пути снижения их содержания в овощах // Химия, 2001, №31, С. 35-37.].
В результате воздействия различных факторов на организм человека содержание метгемоглобина в крови может повышаться до 40% от всего гемоглобина [Фаткуллин К.В., Гильманов А.Ж., Костюков Д.В. Клиническое значении и современные методологические аспекты определения уровня карбокси- и метгемоглобина в крови // Практическая медицина, 2014, Т. 79, №3, С. 17-21.], что способствует резкому нарушению снабжения тканей кислородом и может привести к летальному исходу.
Известны способы восстановления метгемоглобина в гемоглобин в результате воздействия метгемоглобинредуктазы мембраны эритроцита, глута-тиона [Фаткуллин К.В., Гильманов А.Ж., Костюков Д.В. Клиническое значении и современные методологические аспекты определения уровня карбокси-и метгемоглобина в крови / Практическая медицина, 2014, Т. 79, №3, С. 17-21.], аскорбиновой кислоты [Казанец Е.Г. Метгемоглобинемии // Детская больница, 2009, №1, С. 38-42.], малонового альдегида [Герман С.В. Метгемоглобинемии: особенности патогенеза и клиники // Клиническая медицина, 1999, Т. 77, №4, С. 9-12.].
Задачей заявляемого технического решения является создание способа выявления у микроорганизмов протективного действия на молекулу гемоглобина.
Для решения указанной технической задачи в заявляемом способе выявления у микроорганизмов протективного действия на молекулу гемоглобина исследуемую культуру микроорганизмов выращивают в жидкой питательной среде, отделяют супернатант от микробных клеток центрифугированием, параллельно с опытной пробой готовят контрольную пробу из жидкой питательной среды, опытную и контрольную пробы пропускают через бактериальные фильтры, в каждую из проб добавляют эритроцитарную взвесь, инкубируют, центрифугируют, затем во все пробы добавляют супернатант условно-патогенного микроорганизма, способного к образованию метгемоглобина, инкубируют, лизируют эритроцитарную взвесь в опытной и контрольной пробах, замеряют оптическую плотность лизата при длине волны 630 нм, характерной для максимального пика поглощения метгемоглобина, затем рассчитывают уровень образования метгемоглобина в опытной и контрольной пробах по формуле:
Ра=[(ОДк-ОДоп)/ОДк]×100%,
где Ра - уровень протективной активности исследуемого микроорганизма (%);
ОДоп, (отн.е.) - оптическая плотность метгемоглобина в опыте,
ОДк (отн.е.) - оптическая плотность метгемоглобина в контроле.
О наличии или отсутствии протективного действия исследуемого микроорганизма на молекулу гемоглобина судят по степени образования метгемоглобина в опыте по сравнению с контролем.
Достигаемый технический результат при осуществлении изобретения состоит в создании способа выявления у микроорганизмов протективного действия на молекулу гемоглобина, который позволяет выявить новое свойство микроорганизмов - способность предотвращать образование метгемоглобина из гемоглобина, которое может быть использовано в исследовательской работе НИИ и практической работе бактериологических лабораторий клинических учреждений, исследующих свойства микроорганизмов, взаимодействующих с эритроцитами и гемоглобином, а также как критерий отбора биотехнологически перспективных микроорганизмов для создания лекарственных и пробиотических препаратов.
Аналогов изобретения в патентной и научно-технической литературе не обнаружено.
Авторами экспериментально установлено новое свойство микроорганизмов - способность предотвращать образование метгемоглобина из гемо-глобина.
Ранее был определен уровень образования метгемоглобина при воздействии условно-патогенных микроорганизмов на эритроциты [Щуплова Е.А., Фадеев С.Б. Спектральный анализ гемоглобина под действием микроорганизмов с разным уровнем антигемоглобиновой активности // Современные проблемы науки и образования. 2013. №2; URL: www.science-education.ru/108-8791]. В результате работы было обнаружено, что исследуемые условно-патогенные микроорганизмы (Staphylococcus epidermidis, Escherichia coli, S. saprophyticus и другие) способны окислять молекулу гемоглобина до метгемоглобина с разным уровнем его образования.
Для выявления протективного действия микроорганизмов на молекулу гемоглобина были проведены следующие исследования.
Культуры Corynebacterium amycolatum ICIS 53, Corynebacterium xerosis ICIS 99, Lactobacillus spp., S.aureus из коллекции Института клеточного и внутриклеточного симбиоза УрО РАН выращивали в жидкой питательной среде (для каждого вида микроорганизма использовали соответствующий тип питательной среды) в течение 24 ч при 37°C. Бульонные культуры микроорганизмов центрифугировали при 9000 g в течение 15 мин, супернатанты отделяли от микробных клеток.
Параллельно готовили контрольные пробы - жидкие питательные среды (для каждой пробы соответствующий тип питательной среды). Опытные и контрольные пробы пропускали через бактериальные фильтры (0,22 μм, Millipore). В опытные пробирки вносили 2 мл приготовленных стерильных супернатантов исследуемых микроорганизмов, в контрольные - 2 мл стерильной жидкой питательной среды и добавляли по 0,5 мл эритроцитарной взвеси в концентрации 106 эр/мл (способ приготовления взвеси: 2 мл эритроцитарной взвеси трехкратно отмывали в стерильном фосфатно-солевом буфере (ФСБ) (рН=7,4) центрифугированием в течение 5 минут при 1000 g. Далее эритроцитарную взвесь фильтровали через крупнопористые фильтры для грубых крупнодисперсных осадков и добавляли к осадку 18 мл ФСБ для получения 5% эритроцитарной взвеси (или 106 эр/мл)).
Полученные образцы инкубировали в течение 24 часов при 37°C. Далее образцы осаждали центрифугированием при 1000 g в течение 5 мин. Осадок в контрольных и опытных пробах дважды отмывали стерильным фосфатно-солевым буфером (рН=7,4) и добавляли по 2 мл супернатанта условно-патогенных микроорганизмов (S. epidermidis, Е. coli, S.saprophyticus и Candida albicans), способных к образованию метгемоглобина, предварительно пропущенных через бактериальные фильтры (0,22 μм, Millipore), инкубировали в течение 24 часов при 37°C.
Затем в опытные и контрольные образцы добавляли по 2,5 мл стерильной дистиллированной воды для получения лизата и замеряли оптическую плотность исследуемых растворов с помощью сканирующего спектрофотометра «Genesys 6» (США) в диапазоне 450-700 нм, характерного для гемовой части гемоглобина.
Для спектрального анализа молекулы гемоглобина рассчитывали показатели оптической плотности, которые строили относительно модельного спектра, содержащего оксигемоглобин. Для нивелирования разницы, обусловленной разной концентрацией гемоглобина в исследуемых пробах, спектры строили по нормированным данным, относительно поглощения оптической плотности при длине волны 576 нм [Михайлюк И.К., Разживин А.П. Метод разложения спектров на полосы по исходному спектру и набору его производных // Биофизика, - 2003 - Т. 48, вып. 3, с. 405-410.].
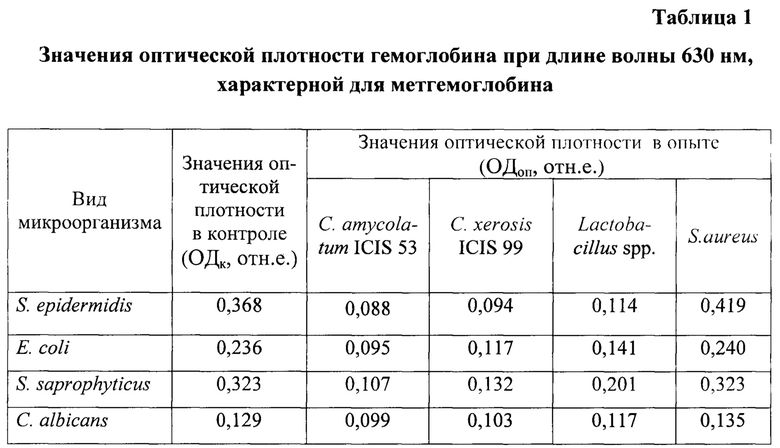
Учитывали значения оптической плотности молекулы гемоглобина в опытных и контрольных пробах при длине волны 630 нм, характерной для максимального пика поглощения метгемоглобина.
Результаты исследований приведены в таблице 1.
Как видно из таблицы 1, значения оптической плотности гемоглобина под действием условно-патогенных микроорганизмов (контроль) при длине волны 630 нм варьировали от 0,129 до 0,368 отн.е.
В опыте наблюдали существенное снижение значений оптической плотности молекулы гемоглобина после предварительной обработки эритроцитов супернатантами исследуемых микроорганизмов: С. amycolatum ICIS 53, С. xerosis ICIS 99 и Lactobacillus spp., S. aureus.
Показатели оптической плотности молекулы гемоглобина в опыте по сравнению с контролем снизились в 1,1-4,2 раза, что соответствует значениям оптической плотности в диапазоне от 0,088 до 0,323 отн.е. Необходимо отметить, что у S.aureus наблюдали повышение значений оптической плотности до 0,419 отн.е.
Далее рассчитывали уровень протективной активности (Ра) исследуемых микроорганизмов на молекулу гемоглобина по формуле:
Ра=[(ОДк-ОДоп)/ОДк]×100%,
где Ра - уровень протективной активности исследуемого микроорганизма (%);
ОДоп (отн.е.) - оптическая плотность метгемоглобина в опыте;
ОДк (отн.е.) - оптическая плотность метгемоглобина в контроле.
О наличии или отсутствии протективного действия исследуемого микроорганизма на молекулу гемоглобина судят по степени образования метгемоглобина в опыте по сравнению с контролем.
Полученные результаты представлены в таблице 2.
Как видно из таблицы 2, уровень протективной активности на молекулу гемоглобина у исследуемых микроорганизмов варьировал от -13,9 до 72,8%.
На основании полученных результатов, авторами был сделан вывод о наличии протективного действии на молекулу гемоглобина исследуемых культур микороорганизмов С. amycolatum ICIS 53, С. xerosis ICIS 99 и Lactobacillus spp. и отсутствии его у S. aureus.

С практической точки зрения, наблюдение эффекта снижения образования метгемоглобина под воздействием супернатантов условно-патогенных микроорганизмов после предварительной обработки эритроцитов суперна-тантами исследуемых микроорганизмов может быть использовано в исследовательской работе НИИ и практической работе бактериологических лабораторий клинических учреждений, исследующих свойства микроорганизмов, взаимодействующих с эритроцитами и гемоглобином, а также как критерий отбора биотехнологически перспективных микроорганизмов для создания лекарственных и пробиотических препаратов.
Способ осуществляется следующим образом:
1. Исследуемую культуру микроорганизма выращивают в 3 мл жидкой питательной среды (для каждого вида микроорганизма используется соответствующий тип питательной среды) в течение 24 часов при 37°C.
2. Бульонную культуру центрифугируют (9000 g в течение 15 минут), супернатант отделяют от микробных клеток.
3. Параллельно готовят контрольную пробу - жидкую питательную среду (для каждого вида микроорганизма соответствующий тип питательной среды).
4. Опытную и контрольную пробы пропускают через бактериальные фильтры (0,22 им, Millipore).
5. В опытную пробирку вносят 2 мл приготовленного стерильного супернатанта исследуемого микроорганизма, в контрольную - 2 мл стерильной жидкой питательной среды.
6. В опытную и контрольную пробы добавляют по 0,5 мл 5% эритроцитарной взвеси (способ приготовления взвеси: 2 мл эритроцитарной взвеси трехкратно отмывают стерильным фосфатно-солевым буфером (ФСБ) (рН=7,4) центрифугированием в течение 5 минут при 1000 g, далее эритроцитарную взвесь фильтруют через крупнопористые фильтры для грубых крупнодисперсных осадков и добавляют к осадку 18 мл ФСБ для получения 5% эритроцитарной взвеси (или 106 эр/мл)).
7. Опытную и контрольную пробы инкубируют в течение 24 ч при 37°C.
8. После инкубации опытную и контрольную пробы центрифугируют (1000g в течение 5 минут), затем осадок дважды отмывают стерильным фосфатно-солевым буфером (рН=7,4) центрифугированием (1000g в течение 5 минут).
9. В каждую из проб объемом 0,5 мл добавляют по 2 мл супернатанта условно-патогенного микроорганизма, способного образовывать метгемоглобин, предварительно пропущенного через бактериальный фильтр (0,22 μм, Millipore) и инкубируют в течение 24 ч при 37°C.
10. После инкубации в опытную и контрольную пробы добавляют по 2,5 мл стерильной дистиллированной воды для получения лизата.
11. Измеряют оптическую плотность исследуемых растворов с помощью сканирующего спектрофотометра «Genesys 6» (США) в диапазоне 450-700 нм, характерного для гемовой части гемоглобина, затем рассчитывают нормированные спектры оптической плотности молекулы гемоглобина.
12. Учитывают значения оптической плотности молекулы гемоглобина в опытной и контрольной пробах при длине волны 630 нм, характерной для максимального пика поглощения метгемоглобина.
13. Рассчитывают уровень образования метгемоглобина в опытной и контрольной пробах по формуле:
Ра=[(ОДк-ОДоп)/ОДк]×100%,
где Ра - уровень протективной активности исследуемого микроорганизма (%);
ОДоп (отн.е.) - оптическая плотность метгемоглобина в опыте;
ОДк (отн.е.) - оптическая плотность метгемоглобина в контроле.
О наличии или отсутствии протективного действия исследуемого микроорганизма на молекулу гемоглобина судят по степени образования метгемоглобина в опыте по сравнению с контролем.
Примеры конкретного выполнения способа:
Пример 1.
Из влагалища здоровой женщины выделен представитель нормофлоры Corynebacterium amycolatum ICIS 53. Для выявления у выделенной культуры микроорганизма протективного действия на молекулу гемоглобина ее выращивали в жидкой питательной среде (мясо-пептонном бульоне, Hi Media) в течение 24 часов при 37°C. Бульонную культуру С. amycolatum ICIS 53 центрифугировали при 9000 g в течение 15 мин, супернатант отделяли от микробных клеток. Параллельно готовили контроль - жидкую питательную среду (мясо-пептонный бульон, Hi Media). Опытную и контрольную пробы пропускали через бактериальные фильтры (0,22 дм, Millipore). В опытную пробирку вносили 2 мл приготовленного стерильного супернатанта С. amycolatum ICIS 53, в контрольную - 2 мл стерильной жидкой питательной среды. В опытную и контрольную пробы добавляли по 0,5 мл 5% взвеси эритроцитов (способ приготовления взвеси: 2 мл эритроцитарной взвеси трехкратно отмывали в стерильном фосфатно-солевом буфере (ФСБ) (рН=7,4) центрифугированием в течение 5 минут при 1000 g. Далее эритроцитарную взвесь фильтровали через крупнопористые фильтры для грубых крупнодисперсных осадков и добавляли к осадку 18 мл ФСБ для получения 5% взвеси эритроцитов (или 106 эр/мл)).
Опытную и контрольную пробы инкубировали в течение 24 часов при 37°C. Инкубированные смеси центрифугировали (1000 g в течение 5 минут), затем осадок дважды отмывали стерильным фосфатно-солевым буфером (рН=7,4).
К эритроцитарной взвеси добавляли по 2 мл супернатанта условно-патогенного микроорганизма - S. epidermidis, способного к образованию метгемоглобина, предварительно пропущенного через бактериальный фильтр (0,22 μм, Millipore).
Инкубировали в течение 24 часов при 37°C, периодически перемешивая. Затем в опытную и контрольную пробы добавляли по 2,5 мл стерильной дистиллированной воды для получения лизата и измеряли оптическую плотность исследуемых растворов с помощью сканирующего спектрофотометра «Genesys 6» (США) в диапазоне 450-700 нм, характерного для гемовой части гемоглобина. Для спектрального анализа молекулы гемоглобина рассчитывали показатели оптической плотности, которые строили относительно модельного спектра, содержащего оксигемоглобин. Для нивелирования разницы, обусловленной разной концентрацией гемоглобина в исследуемых пробах, спектры строили по нормированным, относительно поглощения оптической плотности при длине волны 576 нм. В опытной и контрольной пробах учитывали значения оптической плотности молекулы гемоглобина при длине волны 630 нм, характерной для максимального пика поглощения метгемоглобина. Рассчитывали уровень образования метгемоглобина в опытной и контрольной пробах по формуле:
Ра=[(ОДк-ОДоп)/ОДк]×100%,
где Ра - уровень протективной активности исследуемого микроорганизма (%);
ОДоп (отн.е.) - оптическая плотность метгемоглобина в опыте;
ОДк (отн.е.) - оптическая плотность метгемоглобина в контроле.
ОДоп=0,088
ОДк=0,323
Ра=[(0,323-0,088)/0,323]×100%=72,8%
Уровень протективной активности С. amycolatum ICIS 53 составил 72,8%. На основании полученных данных сделан вывод о наличии у исследуемой культуры С. amycolatum ICIS 53 протективного действия на молекулу гемоглобина.
Пример 2.
Со слизистой верхних дыхательных путей здорового мужчины выделен представитель нормофлоры С xerosis ICIS 99. Для выявления у выделенной культуры микроорганизма протективного действия на молекулу гемоглобина были проведены исследования согласно заявляемому способу, аналогично примеру 1.
Использовали супернатант условно-патогенного микроорганизма - Е. coli.
Результаты расчетов уровня протективной активности (Ра) исследуемого микроорганизма С. xerosis ICIS 99 следующие:
ОДоп=0,117
ОДк=0,236
Ра=[(0,236-0,117)/0,236]×100%=50,4%
Уровень протективной активности С. xerosis ICIS 99 составил 50,4%.
На основании полученных данных сделан вывод о наличии у исследуемой культуры С. xerosis ICIS 99 протективного действия на молекулу гемоглобина.
Пример 3.
Из влагалища здоровой женщины выделен представитель нормофлоры - Lactobacillus acidophilus. Для выявления у выделенной культуры микроорганизма протективного действия на молекулу гемоглобина были проведены исследования согласно заявляемому способу, аналогично примеру 1.
Использовали супернатант условно-патогенного микроорганизма - С. albicans.
Результаты расчетов уровня протективной активности (Ра) исследуемого микроорганизма L. acidophilus следующие:
ОДоп=0,117
ОДк=0,129
Ра=[(0,129-0,117)/0,129]×100%=9,3%
Уровень протективной активности L. acidophilus составил 9,3%.
На основании полученных данных сделан вывод о наличии у исследуемой культуры L. acidophilus протективного действия на молекулу гемоглобина.
Пример 4.
Из влагалища здоровой женщины выделен представитель нормофлоры - С. amycolatum. Для выявления у выделенной культуры микроорганизма протективного действия на молекулу гемоглобина были проведены исследования согласно заявляемому способу, аналогично примеру 1.
Использовали супернатант условно-патогенного микроорганизма - S. saprophytics.
Результаты расчетов уровня протективной активности (Ра) исследуемого микроорганизма С. amycolatum следующие:
ОДоп=0,102
ОДк=0,323
Ра=[(0,323-0,102)/0,323]×100%=68,4%
Уровень протективной активности С. amycolatum составил 68,4%.
На основании полученных данных сделан вывод о наличии у исследуемой культуры С. amycolatum протективного действия на молекулу гемоглобина.
Пример 5.
Со слизистой верхних дыхательных путей здорового мужчины выделен представитель микрофлоры S. aureus. Для выявления у выделенной культуры микроорганизма протективного действия на молекулу гемоглобина были проведены исследования согласно заявляемому способу, аналогично примеру 1.
Использовали супернатант условно-патогенного микроорганизма - S. epidermidis.
Результаты расчетов уровня протективной активности (Ра) исследуемого микроорганизма S. aureus следующие:
ОДоп=0,419
ОДк=0,368
Ра=[(0,368-0,419)/0,368]×100%=-13,9%
Уровень протективной активности S. aureus составил -13,9%.
На основании полученных данных сделан вывод об отсутствии у исследуемой культуры S. aureus протективного действия на молекулу гемоглобина.
Таким образом, использование предлагаемого способа позволяет выявить новое свойство микроорганизмов - способность предотвращать образование метгемоглобина из гемоглобина, которое может быть использовано в исследовательской работе НИИ и практической работе бактериологических лабораторий клинических учреждений, исследующих свойства микроорганизмов, взаимодействующих с эритроцитами и гемоглобином, а также как критерий отбора биотехнологически перспективных микроорганизмов для создания лекарственных и пробиотических препаратов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВЫЯВЛЕНИЯ У МИКРООРГАНИЗМОВ ПРОТЕКТИВНОГО ДЕЙСТВИЯ В ЭФФЕКТЕ ФЕНТОНА | 2004 |
|
RU2279079C2 |
| Штамм бактерий Bifidobacterium longum ICIS-505 - продуцент биологически активных веществ, обладающих антиперсистентной активностью в отношении условно-патогенных и патогенных бактерий и дрожжевых грибов | 2018 |
|
RU2704423C1 |
| СПОСОБ ВЫЯВЛЕНИЯ У БАКТЕРИЙ ИНГИБИТОРОВ КАТАЛАЗЫ МИКРООРГАНИЗМОВ | 2000 |
|
RU2180353C2 |
| Средство для продуцирования органических соединений, обладающих антибактериальной и антигрибковой активностью | 2021 |
|
RU2774192C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АНТИГИСТОНОВОЙ АКТИВНОСТИ МИКРООРГАНИЗМОВ | 2001 |
|
RU2203956C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АНТИГЕМОГЛОБИНОВОЙ АКТИВНОСТИ МИКРООРГАНИЗМОВ | 2004 |
|
RU2262705C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ КОЛОНИЗАЦИОННОЙ РЕЗИСТЕНТНОСТИ ЭКОЛОГИЧЕСКОЙ НИШИ ТЕЛА ЧЕЛОВЕКА | 2000 |
|
RU2175673C1 |
| СПОСОБ ВЫБОРА ШТАММОВ МИКРООРГАНИЗМОВ-ДЕСТРУКТОРОВ НЕФТИ И НЕФТЕПРОДУКТОВ | 2009 |
|
RU2426781C2 |
| СПОСОБ ВЫЯВЛЕНИЯ ПРОДУКЦИИ МИКРООРГАНИЗМАМИ ИНГИБИТОРОВ АНТИМИКРОБНОГО БЕЛКА ТРОМБОЦИТОВ (β-ЛИЗИНА) | 2007 |
|
RU2351654C2 |
| СПОСОБ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ АНТИОКСИДАНТНОЙ АКТИВНОСТИ МИКРООРГАНИЗМОВ | 2011 |
|
RU2465593C1 |
Изобретение относится к микробиологии и может быть использовано при выявлении протективного действия микроорганизмов на молекулу гемоглобина. Для этого исследуемую культуру микроорганизмов выращивают в жидкой питательной среде, отделяют супернатант от микробных клеток центрифугированием, параллельно с опытной пробой готовят контрольную пробу из жидкой питательной среды. Далее опытную и контрольную пробы пропускают через бактериальные фильтры и в каждую из проб добавляют эритроцитарную взвесь, затем инкубируют, центрифугируют и во все пробы добавляют супернатант условно-патогенного микроорганизма, способного к образованию метгемоглобина. Смеси инкубируют, лизируют эритроцитарную взвесь и измеряют оптическую плотность лизата при длине волны 630 нм и рассчитывают уровень образования метгемоглобина в опытной и контрольной пробах. О протективном действии исследуемого микроорганизма на молекулу гемоглобина судят по степени образования метгемоглобина в опыте по сравнению с контролем. Изобретение обеспечивает способ выявления протективного действия микроорганизмов на молекулу гемоглобина и может быть критерием отбора биотехнологически перспективных микроорганизмов для создания лекарственных и пробиотических препаратов. 2 табл., 3 пр.
Способ выявления у микроорганизмов протективного действия на молекулу гемоглобина, заключающийся в том, что исследуемую культуру микроорганизмов выращивают в жидкой питательной среде, отделяют супернатант от микробных клеток центрифугированием, параллельно с опытной пробой готовят контрольную пробу из жидкой питательной среды, опытную и контрольную пробы пропускают через бактериальные фильтры, в каждую из проб добавляют эритроцитарную взвесь, инкубируют, центрифугируют, затем во все пробы добавляют супернатант условно-патогенного микроорганизма, способного к образованию метгемоглобина, инкубируют, лизируют эритроцитарную взвесь в опытной и контрольной пробах, замеряют оптическую плотность лизата при длине волны 630 нм, характерной для максимального пика поглощения метгемоглобина, затем рассчитывают уровень образования метгемоглобина в опытной и контрольной пробах; о наличии или отсутствии протективного действия исследуемого микроорганизма на молекулу гемоглобина судят по степени образования метгемоглобина в опыте по сравнению с контролем.
| СПОСОБ ВЫЯВЛЕНИЯ У МИКРООРГАНИЗМОВ ПРОТЕКТИВНОГО ДЕЙСТВИЯ В ЭФФЕКТЕ ФЕНТОНА | 2004 |
|
RU2279079C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АНТИГЕМОГЛОБИНОВОЙ АКТИВНОСТИ МИКРООРГАНИЗМОВ | 2004 |
|
RU2262705C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ КОНЦЕНТРАЦИЙ ПРОИЗВОДНЫХ ГЕМОГЛОБИНА В БИОЛОГИЧЕСКИХ ТКАНЯХ | 2013 |
|
RU2517155C1 |
| ФАДЕЕВ С.Б | |||
| и др | |||
| Образование метгемоглобина при эритроцитарно-бактериальных взаимодействиях, Вестник ОГУ, 2014, 13, 174, стр | |||
| Сепаратор-центрофуга с периодическим выпуском продуктов | 1922 |
|
SU128A1 |
| ЩУПЛОВА Е.А | |||
| и др | |||
| Спектральный анализ гемоглобина под действием микроорганизмов с разным уровнем антигемоглобиновой активности, Современные проблемы науки и образования, 2013, 2., стр | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

