Настоящее изобретение относится к области технологии рекомбинантных антител, особенно к области антител с заданными свойствами. В данном документе описан способ прогнозирования времени полужизни антитела in vivo на основе времени удерживания, определенного на колонке для аффинной хроматографии на основе FcRn.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Человеческие иммуноглобулины класса G (IgG) содержат две антигенсвязывающие области (Fab), которые придают специфичность в отношении антигена-мишени, и константную область (область Fc), которая ответственна за взаимодействия с рецепторами Fc ([1, 2]). Человеческие IgG подклассов 1, 2 и 4 имеют среднее время полужизни в сыворотке 21 сутки, которое является более продолжительным, чем время полужизни любого другого известного сывороточного белка ([3]). Это продолжительное время полужизни преимущественно опосредуется взаимодействием между областью Fc и неонатальным рецептором Fc (FcRn) ([4, 5]). Это является одной из причин того, почему IgG или слитые белки, содержащие Fc, используются в качестве широко распространенного класса терапевтических средств.
Неонатальный рецептор Fc - FcRn представляет собой ассоциированный с мембраной рецептор, участвующий в гомеостазе как IgG, так и альбумина, в транспорте у матери IgG через плаценту и в фагоцитозе иммунного комплекса антиген-IgG ([6, 9]). Человеческий FcRn представляет собой гетеродимер, состоящий из гликозилированного белка, подобного главному комплексу гистосовместимости класса I (α-FcRn), и субъединицы β2 микроглобулина (β2m) ([10]). FcRn связывается с сайтом в области CH2-CH3 области Fc ([11-14]), и две молекулы FcRn могут одновременно связываться с областью Fc ([15, 16]). Аффинность между FcRn и областью Fc является pH-зависимой, демонстрируя наномолярную аффинность при эндосомальном pH 5-6 и пренебрежимо малое связывание при физиологическом pH 7,4 ([13, 17, 18]). Основополагающий механизм, придающий IgG продолжительное время полужизни, может быть объяснен тремя фундаментальными этапами. Во-первых, IgG подвергаются неспецифичному пиноцитозу разными типами клеток ([19, 20]). Во-вторых, IgG сталкиваются и связываются с FcRn в кислотной эндосоме при pH 5-6, защищая, посредством этого, IgG от лизосомальной деградации ([11, 21]). Наконец, IgG высвобождаются во внеклеточное пространство при физиологическом pH 7,4 [4]. Этот сильно зависящий от pH механизм связывания и высвобождения является критически важным для рециклирования IgG, и любое отклонение характеристик связывания при разных значениях pH может сильно влиять на время полужизни IgG в системе кровообращения ([22]).
Помимо специфичного взаимодействия области Fc с FcRn предполагали, что области Fab также способствуют связыванию с FcRn ([23-25]). Например, опосредованное Fab остаточное связывание при нейтральном pH коррелировало с фармакокинетическими свойствами набора терапевтических антител, указывая на то, что IgG с избыточным связыванием с FcFn при pH 7,3 страдают от уменьшенного времени полужизни в конечной фазе ([24]). Недавно Schlothauer et al. ([25]) описали новый способ аффинной хроматографии на основе FcRn с градиентом pH, который близко имитирует физиологические условия для диссоциации между FcRn и IgG. Кроме того, они продемонстрировали то, что IgG с идентичными областями Fc отличаются по их диссоциации от FcRn, указывая, посредством этого, на влияние области Fab на связывание с FcRn.
Однако лежащий в основе механизм того, как область Fab влияет на связывание с FcRn, все еще не выяснен.
Аналитическая аффинная хроматография на основе FcRn для функциональной характеризации моноклональных антител описана Schlothauer, Т et al. ([25]). Wang, W., et al. ([24]) описывают то, что моноклональные антитела с идентичными последовательностями Fc могут дифференциально связываться с FcRn с фармакокинетическими последствиями. Важность неонатального FcR в регуляции сывороточного времени полужизни терапевтических белков, содержащих домен Fc человеческого IgG1, описывается Suzuki, Т., et al. ([23]). Igawa, Т., et al. ([37]) описывают ослабленное устранение антител IgG путем конструирования вариабельной области. Конструирование области Fc иммуноглобулинов G для модуляции уровней антител in vivo описыается Vaccaro, C., et al. ([22]). Prabhat, P., et al. ([40]) описывают выяснение внутриклеточных путей рециклирования, приводящих к экзоцитозу рецептора Fc - FcRn, используя многофокусную микроскопию. Стратегии оценки сопоставимости фармакокинетики, фармакодинамики и иммуногенности для моноклональных антител описываются Putnam, W.S., et al. ([36]). Boswell, C.A., et al. ([38]) описывают эффекты заряда на распределение и фармакокинетику антител в тканях. Фармакокинетические характеристики и биораспределение меченных радиоактивным йодом химерных моноклональных антител TNT-1, -2 и -3 после химической модификации биотином описываются Khawli, L.A., et al. ([35]).
В WO 2013/120929 описывается аффинная хроматография на основе рецептора Fc. В US 2011/0111406 описывается способ многократного связывания антигенсвязывающих молекул с антигенами. В US 2014/0013456 описываются антитела с легкой цепью, модифицированной гистидином, и генетически модифицированные животные, не являющиеся человеком, для получения данных антител.
Недавно обсуждалось влияние области Fab на взаимодействия с FcRn [(23, 24, 25]).
Однако антитела, имеющие одинаковые области Fc, не должны просто иметь аналогичный PK (фармакокинетический) профиль. Был описан дополнительный вклад области Fab в связывание с FcRn, но лежащий в его основе механизм оставался неизвестным ([47], [24], [25]).
Помимо специфичного взаимодействия области Fc с FcRn, также предполагали, что области Fab способствуют взаимодействию FcRn-IgG ([37, 24, 25]).
В сообщении, опубликованном Li, В., et al. ([48]), описывается то, что отбор каркасной области может влиять на фармакокинетику гуманизированного терапевтического антитела через различия в заряде молекулы.
Sampei, Z., et al. ([49]) описывают идентификацию и многомерную оптимизацию асимметричного биспецифичного антитела IgG, имитирующего функцию активности кофактора фактора VIII.
Wang et al. ([24]) описывают то, что IgG с разными специфичностями в отношении мишеней и областями Fab, но идентичными последовательностями Fc могут иметь разные аффинности в отношении FcRn. Опосредованное Fab остаточное связывание при pH, близком к физиологическому, коррелировало с фармакокинетическими свойствами набора терапевтических антител, указывая на то, что IgG с избыточным связыванием с FcRn при pH 7,3 страдают от уменьшенного времени полужизни в конечной фазе.
Недавно Schlothauer et al. ([25]) описали новый способ аффинной хроматографии на основе FcRn с градиентом pH, который близко имитирует физиологические условия для диссоциации между FcRn и IgG. Кроме того, они показали то, что IgG с идентичными областями Fc отличаются по их диссоциации от FcRn in vitro, указывая, посредством этого, на влияние области Fab на взаимодействие FcRn-IgG.
Benson, J.M., et al. ([50]) описывают открытие и механизм действия устекинумаба - человеческого моноклонального антитела, нацеленного на интерлейкин-12 и интерлейкин-23, для лечения иммуноопосредованных расстройств.
Аминокислотные последовательности антитела бриакинумаб описаны в WO 2013/087911 (SEQ ID NO: 39 и SEQ ID NO: 40), антитела устекинумаб - в WO 2013/087911 (SEQ ID NO: 37 и SEQ ID NO: 38) и антитела бевацизумаб - в записи DB00112 банка лекарственных средств.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Обнаружили то, что распределение зарядов в домене Fv влияет на связывание антитело-FcRn и приводит к дополнительным взаимодействиям между антителом и FcRn. Это изменяет характеристики связывания с FcRn, особенно в отношении диссоциации комплекса антитело-FcRn при pH 7,4, уменьшая, посредством этого, зависимое от FcRn время полужизни антитела в конечной фазе.
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, включающий следующие стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом присутствие взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, определяется, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
Взаимодействие Fab антитела-FcRn представляет собой взаимодействие между областью Fab антитела с FcRn. Данное взаимодействие происходит, если оно вообще присутствует, после связывания антитела посредством FcRn. Таким образом, установление данного взаимодействия представляет собой двухэтапный процесс. На первом этапе образуется комплекс антитело-FcRn, более точно - комплекс Fc антитела-FcRn. Вторым этапом после образования комплекса Fc антитела-FcRn является установление взаимодействия Fab антитела-FcRn. Как можно видеть из этого, данные два взаимодействия, т.е. взаимодействие Fc антитела-FcRn и взаимодействие Fab антитела-FcRn, могут устанавливаться только с полноразмерным антителом.
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab-FcRn в комплексе антитело-FcRn, влияющего на время полужизни in vivo, включающий следующие стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом присутствие присутствия взаимодействия Fab-FcRn в комплексе антитело-FcRn, влияющего на время полужизни in vivo, определяется, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
Другим аспектом, как описано в данном документе, является способ определения относительного времени полужизни антитела in vivo, включающий следующие стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом антитело имеет относительное время полужизни in vivo, которое уменьшается по сравнению со стандартным/природным антителом класса IgG, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
В одном воплощении антитело класса IgG представляет собой антитело подкласса IgG1, IgG2, IgG3 или IgG4. В одном воплощении антитело класса IgG представляет собой антитело подкласса IgG1, IgG3 или IgG4. В одном воплощении антитело класса IgG представляет собой антитело подкласса IgG1 или IgG4. В одном воплощении антитело класса IgG представляет собой антитело подкласса IgG1. В одном воплощении антитело класса IgG представляет собой антитело подкласса IgG4.
Другим аспектом, как описано в данном документе, является способ определения увеличения или уменьшения времени полужизни варианта антитела in vivo относительно его родительского антитела, включающий следующие стадии:
а) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом время полужизни варианта антитела in vivo относительно его родительского антитела возрастает, если i) время удерживания варианта антитела, определенное на стадии а), дольше, чем время удерживания его родительского антитела, определенное на стадии а), и ii) время удерживания варианта антитела, определенное на стадии а), и время удерживания варианта антитела, определенное на стадии б), являются по существу одинаковыми, при этом время полужизни варианта антитела in vivo относительно его родительского антитела уменьшается, если i) время удерживания варианта антитела, определенное на стадии а), короче, чем время удерживания его родительского антитела, определенное на стадии а), и ii) время удерживания варианта антитела, определенное на стадии а), и время удерживания варианта антитела, определенное на стадии б), являются по существу одинаковыми.
Другим аспектом, как описано в данном документе, является способ отбора антитела с повышенным или пониженным временем полужизни in vivo относительно контрольного антитела, включающий следующие стадии:
а) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом, в случае отбора антитела с увеличенным временем полужизни in vivo относительно контрольного антитела, отбирается антитело, которое имеет i) время удерживания, определенное на стадии а), которое дольше, чем время удерживания контрольного антитела, определенное на стадии а), и ii) время удерживания, определенное на стадии а), которое является по существу таким же, как и время удерживания, определенное на стадии б),
при этом, в случае отбора антитела с уменьшенным временем полужизни in vivo относительно контрольного антитела, отбирается антитело, которое имеет i) время удерживания, определенное на стадии а), которое короче, чем время удерживания контрольного антитела, определенное на стадии а), и ii) время удерживания, определенное на стадии а), которое является по существу таким же, как и время удерживания, определенное на стадии б).
Другим аспектом, как описано в данном документе, является способ отбора антитела без взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом отбирается антитело, которое имеет время удерживания, определенное на стадии а), которое по существу не отличается от времени удерживания, определенного на стадии б), и отбирая, посредством этого, антитело без взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo.
Одним аспектом, как описано в данном документе, является способ получения антитела, включающий следующие стадии:
а) предоставление клетки, содержащей одну или более чем одну нуклеиновую кислоту, кодирующую антитело с увеличенным или уменьшенным временем полужизни in vivo относительно контрольного антитела, отобранное способом, описанным в данном документе, и
б) культивирование данной клетки в культивационной среде и выделение антитела из клетки или культивационной среды и получение, посредством этого, антитела.
Одним аспектом, как описано в данном документе, является способ увеличения времени полужизни антитела in vivo, включающий стадию:
- замены заряженного аминокислотного остатка в положениях 27, 55 и 94 в легкой цепи антитела на гидрофобный или нейтральный гидрофильный аминокислотный остаток (нумерация согласно Kabat) и увеличение, посредством этого, времени полужизни антитела in vivo.
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, включающий следующие стадии:
а) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn при первом значении pH с элюцией солевым градиентом,
б) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn при втором значении pH с элюцией солевым градиентом,
при этом присутствие взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, определяется, если отношение времен удерживания антитела и контрольного антитела, определенных на стадии а), существенно отличается от отношения времен удерживания антитела и контрольного антитела, определенных на стадии б).
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, включающий следующие стадии:
а) определение для варианта антитела и его родительского антитела значений KD при pH 6 с использованием поверхностного плазмонного резонанса,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли,
при этом присутствие взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, определяется, если значения KD отличаются самое большее в 10 раз, времена удерживания, определенные на стадии б), являются существенно отличными между вариантом антитела и его родительским антителом.
Одним аспектом, как описано в данном документе, является способ определения относительного времени полужизни антитела in vivo, включающий следующие стадии:
а) определение для варианта антитела и его родительского антитела значений KD при pH 6 с использованием поверхностного плазмонного резонанса,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли,
при этом антитело имеет относительное время полужизни in vivo, которое уменьшено по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания, определенное на стадии б), варианта антитела является более коротким/меньшим, чем время удерживания его родительского антитела, и
при этом антитело имеет относительное время полужизни in vivo, которое увеличено по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания, определенное на стадии б), варианта антитела является более длительным/большим, чем время удерживания его родительского антитела.
Одним аспектом, как описано в данном документе, является способ определения увеличения или уменьшения времени полужизни антитела in vivo, включающий следующие стадии:
а) определение для варианта антитела и его родительского антитела значений KD при pH 6 с использованием поверхностного плазмонного резонанса,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли,
при этом антитело имеет уменьшение времени полужизни in vivo по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания, определенное на стадии б), варианта антитела является более коротким/меньшим, чем время удерживания его родительского антитела, и
при этом антитело имеет увеличение времени полужизни in vivo по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания, определенное на стадии б), варианта антитела является более длительным/большим, чем время удерживания его родительского антитела.
В одном воплощении антитело представляет собой полноразмерное антитело.
В одном воплощении всех аспектов положительный линейный градиент pH составляет от примерно pH 5,5 до примерно pH 8,8.
В одном воплощении всех аспектов соль выбрана из хлорида натрия, сульфата натрия, хлорида калия, сульфата калия, цитрата натрия или цитрата калия.
В одном воплощении всех аспектов соль представляет собой хлорид натрия.
В одном воплощении всех аспектов первая концентрация соли составляет от 50 мМ до 200 мМ.
В одном воплощении всех аспектов первая концентрация соли составляет примерно 140 мМ.
В одном воплощении всех аспектов вторая концентрация соли составляет от 300 мМ до 600 мМ.
В одном воплощении всех аспектов вторая концентрация соли составляет примерно 400 мМ.
В одном воплощении всех аспектов времена удерживания, которые существенно отличаются на стадии а) и стадии б), отличаются по меньшей мере на 5%.
В одном воплощении всех аспектов времена удерживания, которые существенно отличаются на стадии а) и стадии б), отличаются по меньшей мере на 10%.
В одном воплощении всех аспектов времена удерживания, которые существенно отличаются на стадии а) и стадии б), отличаются по меньшей мере на 15%.
В одном воплощении всех аспектов, если времена удерживания существенно отличаются на стадии а) и стадии б), время удерживания на стадии а) больше/продолжительнее, чем на стадии б).
В одном воплощении всех аспектов, если времена удерживания существенно отличаются на стадии а) и стадии б), время удерживания на стадии б) меньше/короче, чем на стадии а).
В одном воплощении всех аспектов, если времена удерживания существенно отличаются на стадии а) и стадии б), данные времена удерживания являются пропорциональными единице, поделенной на квадратный корень концентрации соли (~1/√(с(соли))).
В одном воплощении всех аспектов родительское или контрольное антитело представляет собой антитело против IL-1R (рецептор интерлейкина-1) с SEQ ID NO: 01 (тяжелая цепь) и SEQ ID NO: 02 (легкая цепь) для подкласса IgG1 и антитело против IL-1R с SEQ ID NO: 03 (тяжелая цепь) и SEQ ID NO: 04 (легкая цепь) для подкласса IgG4.
В одном воплощении всех аспектов родительское или контрольное антитело представляет собой антитело против HER2 (рецептор 2 эпидермального фактора роста человека) с SEQ ID NO: 36 (тяжелая цепь) и SEQ ID NO: 37 (легкая цепь) для подкласса IgG1 и антитело против HER2 с SEQ ID NO: 38 (тяжелая цепь) и SEQ ID NO: 39 (легкая цепь) для подкласса IgG4.
В одном воплощении всех аспектов родительское или контрольное антитело представляет собой устекинумаб с аминокислотными последовательностями легкой и тяжелой цепи, описанными на Фиг. 5.
В одном воплощении всех аспектов колонка для аффинной хроматографии на основе FcRn содержит нековалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m).
В одном воплощении всех аспектов колонка для аффинной хроматографии на основе FcRn содержит ковалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m).
В одном воплощении всех аспектов комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m) связывается с твердой фазой.
В одном воплощении всех аспектов твердая фаза представляет собой хроматографическое вещество.
В одном воплощении всех аспектов комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m) является биотинилированным, и твердая фаза дериватизируется стрептавидином.
В одном воплощении всех аспектов бета-2-микроглобулин происходит из того же самого вида, что и неонатальный рецептор Fc (FcRn).
В одном воплощении всех аспектов бета-2-микроглобулин происходит из другого вида, чем FcRn.
В одном воплощении всех аспектов FcRn выбран из FcRn человека, FcRn яванского макака, FcRn мыши, FcRn крысы, FcRn овцы, FcRn собаки, FcRn свиньи, FcRn карликовой свиньи и FcRn кролика.
В одном воплощении всех аспектов антитело представляет собой моноспецифичные антитело или фрагмент слитого полипептида в виде антитела, или биспецифичные антитело или фрагмент слитого полипептида в виде антитела, или трехспецифичные антитело или фрагмент слитого полипептида в виде антитела, или четырехспецифичные антитело или фрагмент слитого полипептида в виде антитела.
В одном воплощении антитело представляет собой антитело класса IgG. В одном воплощении антитело представляет собой антитело подкласса IgG1, IgG2, IgG3 или IgG4. В одном воплощении антитело представляет собой антитело подкласса IgG1 или IgG4.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Объединение результатов структурного анализа взаимодействия FcRn-mAb (mAb - моноклональное антитело) приводит к заключению о том, что домен Fv и особенно вариабельный домен легкой цепи (VL) обеспечивает главное влияние на диссоциацию FcRn-mAb. Это открытие было неожиданным, так как домен Fv является удаленным от когнатного сайта связывания FcRn.
Антитела не демонстрировали различий в аффинности при pH 6,0, следовательно, область Fab, по-видимому, не имеет влияния на связывание при pH 6,0. В отличие от этого, диссоциация между FcRn и антителами подвергалась влиянию области Fab.
pH диссоциации FcRn-IgG in vitro линейно коррелировали с временем полужизни в конечной фазе in vivo. В заключение эти данные поддерживают предположение о том, что антитела, демонстрирующие более медленную диссоциацию при более высоких значениях pH, транспортируются обратно в клетку и затем деградируют вместо высвобождения обратно в кровоток.
Обнаружили то, что распределение зярядов в домене Fv влияет на связывание антитело-FcRn и приводит к дополнительным взаимодействиям между антителом и FcRn. Это изменяет характеристики связывания с FcRn, особенно в отношении диссоциации комплекса антитело-FcRn при pH 7,4, уменьшая, посредством этого, зависимое от FcRn время полужизни антитела в конечной фазе.
I. Определения
Термины в единственном числе обозначают один или два, или три, или четыре, или пять, или шесть и вплоть до 109.
Термин «примерно» обозначает интервал плюс/минус 20% от следующего далее числового значения. В одном воплощении термин «примерно» обозначает интервал плюс/минус 10% от следующего далее числового значения. В одном воплощении термин «примерно» обозначает интервал плюс/минус 5% от следующего далее числового значения.
Термин «содержащий» также включает термин «состоящий из».
Термин «изменение» обозначает мутацию (замену), вставку (присоединение), модификацию (дериватизацию) или делецию одного или более чем одного аминокислотного остатка в родительском антителе или слитом полипептиде, например, слитом полипептиде, содержащем по меньшей мере часть области Fc, связывающейся с FcRn, для получения модифицированного антитела или слитого полипептида. Термин «мутация» обозначает то, что определенный аминокислотный остаток заменяется другим аминокислотным остатком. Например, мутация L234A обозначает то, что аминокислотный остаток лизин в положении 234 в области Fc антитела (полипептиде) заменен аминокислотным остатком аланином (замена лизина на аланин) (нумерация согласно индексу EU (Европейский Союз)).
Термин «мутация аминокислоты» обозначает замену по меньшей мере одного существующего аминокислотного остатка другим отличным аминокислотным остатком (заменяющим аминокислотным остатком). Заменяющий аминокислотный остаток может представлять собой «встречающийся в природе аминокислотный остаток» и может быть выбран из группы, состоящей из аланина (трехбуквенный код: ala, однобуквенный код: А), аргинина (arg, R), аспарагина (asn, N), аспарагиновой кислоты (asp, D), цистеина (cys, С), глутамина (gln, Q), глутаминовой кислоты (glu, Е), глицина (gly, G), гистидина (his, Н), изолейцина (ile, I), лейцина (leu, L), лизина (lys, K), метионина (met, М), фенилаланина (phe, F), пролина (pro, Р), серина (ser, S), треонина (thr, Т), триптофана (trp, W), тирозина (tyr, Y) и валина (val, V). Заменяющий аминокислотный остаток может представлять собой «не встречающийся в природе аминокислотный остаток». См., например, US 6586207, WO 98/48032, WO 03/073238, US 2004/0214988, WO 2005/35727, WO 2005/74524, Chin, J.W., et al., J. Am. Chem. Soc. 124 (2002) 9026-9027; Chin, J.W. and Schultz, P.G., ChemBioChem 11 (2002) 1135-1137; Chin, J.W., et al., PICAS United States of America 99 (2002) 11020-11024 и Wang, L. and Schultz, P.G., Chem. (2002) 1-10 (все полностью включенные в данный документ посредством ссылки).
Термин «аминокислотная вставка» обозначает (дополнительное) включение по меньшей мере одного аминокислотного остатка в заданном положении в аминокислотной последовательности. В одном воплощении вставка будет представлять собой вставку одного или двух аминокислотных остатков. Вставленный(ные) аминокислотный(ные) остаток(тки) может(гут) представлять собой любой встречающийся в природе или не встречающийся в природе аминокислотный остаток.
Термин «делеция аминокислоты» обозначает удаление по меньшей мере одного аминокислотного остатка в заданном положении в аминокислотной последовательности.
Термин «антитело» в данном документе используется в широком смысле и охватывает разные структуры антитела, включающие моноклональные антитела и мультиспецифичные антитела (например, биспецифичные антитела, трехспецифичные антитела), при условии, что они представляют собой полноразмерные антитела и демонстрируют желательную активность связывания в отношении антигена и/или FcRn, но не ограничивающиеся ими.
Термин «связывание (с антигеном)» обозначает связывание антитела в анализе in vitro. В одном воплощении связывание определяется в анализе связывания, в котором антитело связывается с поверхностью, и связывание антигена с антителом измеряется поверхностным плазмонным резонансом (SPR). Связывание означает, например, аффинность связывания (KD) 10-8 М или менее, в некоторых воплощениях - от 10-13 до 10-8 М, в некоторых воплощениях - от 10-13 до 10-9 М.
Связывание может исследоваться посредством анализа BIAcore (GE Healthcare Biosensor АВ, Упсала, Швеция). Аффинность связывания определяется показателями ka (константа скорости для ассоциации антитела для комплекса антитело/антиген), kd (константа диссоциации) и KD (kd/ka).
Термин «буферное вещество» обозначает вещество, которое, при нахождении в растворе, может нивелировать измененя значения pH раствора, например, из-за добавления или высвобождения кислотных или основных веществ.
Термин «домен СН2» обозначает часть полипептида тяжелой цепи антитела, которая простирается приблизительно от положения 231 по EU до положения 340 по EU (система нумерации EU по Kabat). В одном воплощении домен СН2 имеет аминокислотную последовательность SEQ ID NO: 05: 

Термин «домен СН3» обозначает часть полипептида тяжелой цепи антитела, которая простирается приблизительно от положения 341 по EU до положения 446 по EU. В одном воплощении домен СН3 имеет аминокислотную последовательность SEQ ID NO: 06: 
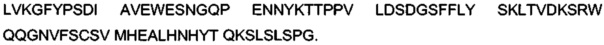
«Класс» антитела относится к типу константного домена или константной области, которыми обладает его тяжелая цепь. Существуют пять главных классов антител: IgA, IgD, IgE, IgG и IgM, и несколько из них могут дополнительно подразделяться на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называются α, δ, ε, γ и μ соответственно.
Термин «эффективное количество» агента, например, фармацевтической композиции, относится к эффективному количеству в дозировках и в течение перодов времени, необходимых для достижения желательного терапевтического или профилактического результата.
Термин «слитый с Fc полипептид» обозначает слияние связывающего домена (например, антигенсвязывающего домена, такого как одноцепочечное антитело, или полипептида, такого как лиганд рецептора) с областью Fc антитела, которое демонстрирует желательную активность связывания мишени и/или белка А, и/или FcRn.
Термин «область Fc человеческого происхождения» обозначает С-конецевую область тяжелой цепи иммуноглобулина человеческого происхождения, которая содержит по меньшей мере часть шарнирной области, домен СН2 и домен СН3. В одном воплощении область Fc тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. В одном воплощении область Fc имеет аминокислотную последовательность SEQ ID NO: 07. Однако С-концевой лизин (Lys447) области Fc может присутствовать или может не присутствовать. Если в данном документе не определено иначе, нумерация аминокислотных остатков в области Fc или в константной области осуществляется согласно системе нумерации EU, также известной как индекс EU, как описано в Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91 3242. Область Fc состоит из двух полипептидов области Fc тяжелой цепи, которые могут быть ковалентно связаны друг с другом через остатки цистеина шарнирной области, образующие межполипептидные дисульфидные связи.
Термин «FcRn» обозначает человеческий неонатальный рецептор Fc. FcRn функционирует для спасения IgG от пути лизосомальной деградации, приводя к пониженному клиренсу и увеличенному времени полужизни. FcRn представляет собой гетеродимерный белок состоящий из двух полипептидов: 50 кДа белка, подобного главному комплексу гистосовместимости класса I (α-FcRn) и 15 кДа β2-микроглобулина (β2m). FcRn с высокой аффинностью связывается с частью СН2-СН3 области Fc IgG. Взаимодействие между IgG и FcRn является строго pH-зависимым и происходит в стехиометрии 1:2 - с одним IgG, связывающимся с двумя молекулами FcRn через его две тяжелые цепи (Huber, А.Н., et al., J. Mol. Biol. 230 (1993) 1077-1083). Связывание с FcRn происходит в эндосоме при кислотном pH (pH меньше 6,5), и IgG высвобождается на нейтральной поверхности клетки (pH примерно 7,4). pH-чувствительная природа взаимодействия облегчает опосредованную FcRn защиту IgG, пиноцитированных в клетки, от внутриклеточной деградации посредством связывания с рецептором в кислотной среде эндосом. FcRn затем облегчает рециклирование IgG к поверхности клетки и последующее высвобождение в кровоток при воздействии на комплекс FcRn-IgG среды с нейтральным pH вне клетки.
Термин «связывающаяся с FcRn часть области Fc» обозначает часть полипептида тяжелой цепи антитела, которая простирается приблизительно от положения 243 по EU до положения 261 по EU и приблизительно от положения 275 по EU до положения 293 по EU, и приблизительно от положения 302 по EU до положения 319 по EU, и приблизительно от положения 336 по EU до положения 348 по EU, и приблизительно от положения 367 по EU до положения 393 по EU, и положения 408 по EU, и приблизительно от положения 424 по EU до положения 440 по EU. В одном воплощении изменен один или более чем один из следующих аминокислотных остатков согласно нумерации EU по Kabat: F243, Р244, Р245 Р, K246, Р247, K248, D249, Т250, L251, М252, I253, S254, R255, Т256, Р257, Е258, V259, Т260, С261, F275, N276, W277, Y278, V279, D280, V282, Е283, V284, Н285, N286, А287, K288, Т289, K290, Р291, R292, Е293, V302, V303, S304, V305, L306, Т307, V308, L309, Н310, Q311, D312, W313, L314, N315, G316, K317, Е318, Y319, I336, S337, K338, А339, K340, G341, Q342, Р343, R344, Е345, Р346, Q347, V348, С367, V369, F372, Y373, Р374, S375, D376, I377, А378, V379, Е380, W381, Е382, S383, N384, G385, Q386, Р387, Е388, N389, Y391, Т393, S408, S424, С425, S426, V427, М428, Н429, Е430, А431, L432, Н433, N434, Н435, Y436, Т437, Q438, K439 и S440 (нумерация по EU).
Термин «полноразмерное антитело» обозначает антитело, имеющее структуру, по существу аналогичную структуре нативного антитела. Полноразмерное антитело содержит две полноразмерные легкие цепи антитела, содержащие вариабельный домен легкой цепи и константный домен легкой цепи, и две полноразмерные тяжелые цепи антитела, содержащие вариабельный домен тяжелой цепи, первый константный домен, шарнирную область, второй константный домен и третий константный домен. Полноразмерное антитело может содержать дополнительные домены, такие как, например, дополнительный scFv или scFab, конъюгированные с одной или более чем одной из цепей полноразмерного антитела. Данные конъюгаты также охватываются термином полноразмерное антитело.
Термин «шарнирная область» обозначает часть полипептида тяжелой цепи антитела, которая соединяет домен СН1 и домен СН2, например, от примерно положения 216 до примерно положения 230 согласно системе нумерации EU по Kabat. В одном воплощении шарнирная область представляет собой укороченную шарнирную область, содержащую остатки 221-230 согласно системе нумерации EU по Kabat. Шарнирная область обычно представляет собой димерную молекулу, состоящую из двух полипептидов с идентичной аминокислотной последовательностью. Шарнирная область обычно содержит примерно 25 аминокислотных остатков и является гибкой, обеспечивая независимое перемещение антигенсвязывающих областей. Шарнирная область может быть подразделена на три домена: верхний, средний и нижний шарнирный домен (Roux, et al., J. Immunol. 161 (1998) 4083).
Термины «клетка-хозяин», «линия клетки-хозяина» и «культура клетки-хозяина» используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформантов» и «трансформированные клетки», которые включают первично трансформированные клетки и происходящее от них потомство, не придавая значения числу пассажей. Потомство может не быть полностью идентичным родительской клетке по составу нуклеиновых кислот, но может содержать мутации. В данный документ включается мутантное потомство, которое имеет такую же функцию или биологическую активность, что и исходно трансформированная клетка, подвергнутая скринингу или отобранная на них.
Термин «происходящая от» обозначает то, что аминокислотная последовательность происходит от родительской аминокислотной последовательности путем введения изменений по меньшей мере в одном положении. Таким образом, происходящая аминокислотная последовательность отличается от соответствующей родительской аминокислотной последовательности в по меньшей мере одном соответствующем положении (нумерация согласно индексу EU по Kabat для областей Fc антитела). В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, отличается по одному-пятнадцати аминокислотным остаткам в соответствующих положениях. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, отличается по одному-десяти аминокислотным остаткам в соответствующих положениях. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, отличается по одному-шести аминокислотным остаткам в соответствующих положениях. Подобным образом, происходящая аминокислотная последовательность имеет высокую идентичность аминокислотной последовательности с ее родительской аминокислотной последовательностью. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, имеет 80%-ную или большую идентичность аминокислотной последовательности. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, имеет 90%-ную или большую идентичность аминокислотной последовательности. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, имеет 95%-ную или большую идентичность аминокислотной последовательности.
Термин «полипептид человеческой области Fc» обозначает аминокислотную последовательность, которая является идентичной «нативному» полипептиду человеческой области Fc или полипептиду человеческой области Fc «дикого типа». Термин «полипептид варианта (человеческой) области Fc» обозначает аминокислотную последовательность, которая происходит от «нативного» полипептида человеческой области Fc или полипептида человеческой области Fc «дикого типа» посредством по меньшей мере одного «изменения аминокислоты». «Человеческая область Fc» состоит из двух полипептидов человеческой области Fc. «Вариант (человеческой) области Fc» состоит из двух полипептидов области Fc, при этом оба могут представлять собой полипептиды варианта (человеческой) области Fc, или один представляет собой полипептид человеческой области Fc, а другой представляет собой полипептид варианта (человеческой) области Fc.
В одном воплощении полипептид человеческой области Fc имеет аминокислотную последовательность полипептида области Fc человеческого IgG1 SEQ ID NO: 07 или полипептида области Fc человеческого IgG2 SEQ ID NO: 08, или полипептида области Fc человеческого IgG3 SEQ ID NO: 09, или полипептида области Fc человеческого IgG4 SEQ ID NO: 10. В одном воплощении полипептид области Fc происходит от полипептида области Fc SEQ ID NO: 07 или 08, или 09, или 10 и имеет по меньшей мере одну мутацию аминокислоты по сравнению с полипептидом области Fc SEQ ID NO: 07 или 08, или 09, или 10. В одном воплощении полипептид области Fc содержит/имеет от примерно одной до примерно десяти аминокислотных мутаций, и в одном воплощении - от примерно одной до примерно пяти аминокислотных мутаций. В одном воплощении полипептид области Fc имеет по меньшей мере примерно 80%-ную гомологию с полипептидом человеческой области Fc SEQ ID NO: 07 или 08, или 09, или 10. В одном воплощении полипептид области Fc имеет по меньшей мере примерно 90%-ную гомологию с полипептидом человеческой области Fc SEQ ID NO: 07 или 08, или 09, или 10. В одном воплощении полипептид области Fc имеет по меньшей мере примерно 95%-ную гомологию с полипептидом человеческой области Fc SEQ ID NO: 07 или 08, или 09, или 10.
Полипептид области Fc, происходящий от полипептида человеческой области Fc SEQ ID NO: 07 или 08, или 09, или 10, определяется изменениями аминокислот, которые в нем содержатся. Таким образом, например, термин P329G обозначает полипептид области Fc, происходящий от полипептида человеческой области Fc с мутацией пролина до глицина в положении аминокислоты 329 относительно полипептида человеческой области Fc SEQ ID NO: 07 или 08, или 09, или 10.
Для всех положений тяжелой цепи, обсуждаемых в настоящем изобретении, нумерация осуществляется согласно индексу EU. Индекс EU или индекс EU по Kabat, или индекс EU Кэбота, или схема нумерации EU относится к нумерации антитела EU (Edelman, et al., Proc. Natl. Acad. Sci. USA 63 (1969) 78-85, тем самым целиком включенная посредством ссылки). Нумерация остатков легкой цепи осуществляется согласно номенклатуре Kabat (Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91 3242).
Полипептид области Fc человеческого IgG1 имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 07).
(SEQ ID NO: 07).
Полипептид области Fc с мутациями L234A, L235A, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

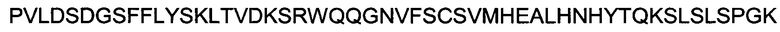 (SEQ ID NO: 11).
(SEQ ID NO: 11).
Полипептид области Fc с мутациями Y349C, T366S, L368A и Y407V, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 12).
(SEQ ID NO: 12).
Полипептид области Fc с мутациями S354C, T366W, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 13).
(SEQ ID NO: 13).
Полипептид области Fc с мутациями L234A, L235A и с мутациями Y349C, T366S, L368A и Y407V, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

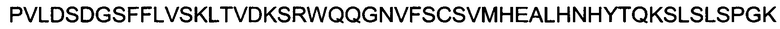 (SEQ ID NO: 14).
(SEQ ID NO: 14).
Полипептид области Fc с мутациями L234A, L235A и S354C, T366W, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 15).
(SEQ ID NO: 15).
Полипептид области Fc с мутацией P329G, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

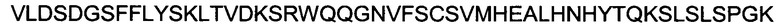 (SEQ ID NO: 16).
(SEQ ID NO: 16).
Полипептид области Fc с мутациями L234A, L235A и мутацией P329G, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 17).
(SEQ ID NO: 17).
Полипептид области Fc с мутацией P329G и мутациями Y349C, T366S, L368A, Y407V, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 18).
(SEQ ID NO: 18).
Полипептид области Fc с мутацией P329G и мутациями S354C, T366W, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:


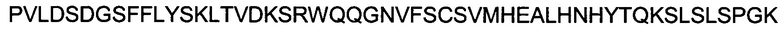 (SEQ ID NO: 19).
(SEQ ID NO: 19).
Полипептид области Fc с мутациями L234A, L235A, P329G и Y349C, T366S, L368A, Y407V, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

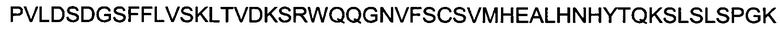 (SEQ ID NO: 20).
(SEQ ID NO: 20).
Полипептид области Fc с мутациями L234A, L235A, P329G и мутациями S354C, T366W, происходящий от области Fc человеческого IgG1, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 21).
(SEQ ID NO: 21).
Полипептид области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 10).
(SEQ ID NO: 10).
Полипептид области Fc с мутациями S228P и L235E, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 22).
(SEQ ID NO: 22).
Полипептид области Fc с мутациями S228P, L235E и мутацией P329G, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 23).
(SEQ ID NO: 23).
Полипептид области Fc с мутациями S354C, T366W, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:

 (SEQ ID NO: 24).
(SEQ ID NO: 24).
Полипептид области Fc с мутациями Y349C, T366S, L368A, Y407V, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 25).
(SEQ ID NO: 25).
Полипептид области Fc с мутациями S228P, L235E и S354C, T366W, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 26).
(SEQ ID NO: 26).
Полипептид области Fc с мутациями S228P, L235E и Y349C, T366S, L368A, Y407V, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 27).
(SEQ ID NO: 27).
Полипептид области Fc с мутацией P329G, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 28).
(SEQ ID NO: 28).
Полипептид области Fc с мутациями P329G и Y349C, T366S, L368A, Y407V, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 29).
(SEQ ID NO: 29).
Полипептид области Fc с мутациями P329G и S354C, T366W, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 30).
(SEQ ID NO: 30).
Полипептид области Fc с мутациями S228P, L235E, P329G и Y349C, T366S, L368A, Y407V, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 31).
(SEQ ID NO: 31).
Полипептид области Fc с мутациями S228P, L235E, P329G и S354C, T366W, происходящий от области Fc человеческого IgG4, имеет следующую аминокислотную последовательность:
 (SEQ ID NO: 32).
(SEQ ID NO: 32).
Выравнивание разных человеческих областей Fc показано ниже (нумерация по EU):

Термин «гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из HVR (гипервариабельная область), не являющихся человеческими, и аминокислотные остатки из человеческих FR (каркасная область). В некоторых воплощениях гуманизированное антитело будет содержать по существу все из по меньшей мере одного, и типично два вариабельных домена, в которых все или по существу все из HVR (например, CDR (область, определяющая комплементарность)) соответствуют HVR антитела, не являющегося человеческим, и все или по существу все из FR соответствуют FR человеческого антитела. Гуманизированное антитело возможно может содержать по меньшей мере часть константной области антитела, происходящую от человеческого антитела. Термин «гуманизированная форма» антитела, например, антитела, не являющегося человеческим, относится к антителу, которое подверглось гуманизации.
«Индивид» или «субъект» представляет собой млекопитающее. Млекопитающие включают одомашненных животных (например, коровы, овцы, кошки, собаки и лошади), приматов (например, люди и приматы, не являющиеся человеком, такие как обезьяны), кроликов и грызунов (например, мыши и крысы), но не ограничиваются ими. В некоторых воплощениях индивид или субъект представляет собой человека.
«Выделенное» антитело представляет собой антитело, которое было отделено от компонента его природного окружения. В некоторых воплощениях антитело очищают до более чем 95%-ной или 99%-ной чистоты, например, при определении электрофоретическими (например, SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), изоэлектрофокусировка (IEF), капиллярный электрофорез) или хроматографическими (например, гель-фильтрация или ионообменная ВЭЖХ (высокоэффективная жидкостная хроматография), или ВЭЖХ с обращенной фазой) способами. Относительно обзора способов оценки чистоты антител, см., например, Flatman, S. et al., J. Chrom. В 848 (2007) 79-87.
Термин «выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее природного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат данную молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты присутствует внехромосомно или в участке хромосомы, который отличается от его природного хромосомного участка.
Термин «моноклональное антитело» в том виде, как он используется в данном документе, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, составляющие популяцию, являются идентичными и/или связываются с тем же самым эпитопом, за исключением возможных вариантов антител, например, содержащих встречающиеся в природе мутации или возникающие во время получения препарата моноклонального антитела, причем такие варианты обычно присутствуют в минорных количествах. В отличие от препаратов поликлональных антител, которые типично включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклонального антитела направлено против одной детерминанты на антигене. Таким образом, модификатор «моноклональное» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует истолковывать как требующий получения антитела каким-либо конкретным способом. Например, моноклональные антитела, подлежащие применению согласно настоящему изобретению, можно получать целым рядом методик, включающих способ гибридомы, способы генной инженерии, способы фагового дисплея и способы с использованием трансгенных животных, содержащих все локусы человеческого иммуноглобулина или их часть, но не ограничивающихся ими, причем такие способы и другие типичные способы получения моноклональных антител описываются в данном документе.
Термин «нативные антитела» относится к встречающимся в природе молекулам иммуноглобулинов с варьирующими структурами. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины с массой примерно 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидными связями. От N- до С-конца каждая тяжелая цепь имеет вариабельную область (VH), также именуемую вариабельный тяжелый домен или вариабельный домен тяжелой цепи, с последующими тремя константными доменами (СН1, СН2 и СН3). Аналогичным образом, от N- до С-конца каждая легкая цепь имеет вариабельную область (VL), также именуемую вариабельный легкий домен или вариабельный домен легкой цепи, с последующим константным легким (CL) доменом. Легкая цепь антитела может быть приписана к одному из двух типов, именуемых каппа (κ) и лямбда (λ), на основе аминокислотной последовательности ее константного домена.
Термин «отрицательный линейный градиент pH» обозначает градиент pH, начинающийся при высоком (например, нейтральном или щелочном) значении pH и заканчивающийся при меньшем (т.е. нейтральном или кислотном) значении pH. В одном воплощении отрицательный линейный градиент pH начинается при значении pH примерно 8,8 и заканчивается при значении pH примерно 5,5.
Термин «аминокислотный остаток, не встречающийся в природе» обозначает аминокислотный остаток, отличный от аминокислотных остатков, встречающихся в природе, как перечислено выше, который может быть ковалентно связан со смежными аминокислотными остатками в полипептидной цепи. Примерами аминокислотных остатков, не встречающихся в природе, являются норлейцин, орнитин, норвалин, гомосерин. Другие примеры перечислены в Ellman, et al., Meth. Enzym. 202 (1991) 301-336. Типичный способ синтеза аминокислотных остатков, не встречающихся в природе, описан, например, в Noren, et al., Science 244 (1989) 182 и Ellman et al., выше.
Термин «фармацевтическая композиция» относится к препарату, который находится в такой форме, чтобы обеспечивать эффективную биологическую активность активного ингредиента, содержащегося в нем, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому вводилась бы композиция.
Термин «фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтической композиции, отличному от активного ингредиента, который является нетоксичным для субъекта. Фармацевтически приемлемый носитель включает буфер, эксципиент, стабилизатор или консервант, но не ограничивается ими.
Термин «плазмида» в том виде, в котором он используется в данном документе, относится к молекуле нуклеиновой кислоты, способной размножать другую нуклеиновую кислоту, с которой она связана. Данный термин включает плазмиду в виде самореплицирующейся структуры нуклеиновой кислоты, а также плазмиду, включенную в геном клетки-хозяина, в которую она была введена. Некоторые плазмиды способны управлять экспрессией нуклеиновых кислот, с которыми они связаны функциональным образом. Такие плазмиды именуются здесь «экспрессионные плазмиды».
Термин «положительный линейный градиент pH» обозначает градиент pH, начинающийся при низком (т.е. более кислотном) значении pH и заканчивающийся при большем (т.е. менее кислотном, нейтральном или щелочном) значении pH. В одном воплощении положительный линейный градиент pH начинается при значении pH примерно 5,5 и заканчивается при значении pH примерно 8,8.
Термин «рекомбинантное антитело» в том виде, как он используется в данном документе, обозначает все антитела (химерные, гуманизированные и человеческие), которые получают, экспрессируют, создают или выделяют способами генной инженерии. Это включает антитела, выделенные из клетки-хозяина, такой как клетка NS0 или СНО (яичники китайского хомяка), или из животного (например, мыши), которое является трансгенным в отношении человеческих генов иммуноглобулинов или антител, экспресируемых с использованием рекомбинантной экспрессионной плазмиды, трансфицированной в клетку-хозяина. Такие рекомбинантные антитела имеют вариабельные и константные области в реаранжированной форме. Рекомбинантные антитела, как описано в данном документе, могут подвергаться соматической гипермутации in vivo. Таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые, происходя от и будучи родственными последовательностям VH и VL человеческой зародышевой линии, могут не существовать в природе в репертуаре человеческих антител зародышевой линии in vivo.
Термин «твердая фаза» обозначает нежидкое вещество, и оно включает частицы (включая микрочастицы и шарики), сделанные из таких материалов, как полимер, металл (парамагнитные, ферромагнитные частицы), стекло и керамика; гелевые вещества, такие как диоксид кремния, оксид алюминия и полимерные гели; капилляры, которые могут быть сделаны из полимера, металла, стекла и/или керамики; цеолиты и другие пористые вещества; электроды; планшеты для микротитрования; твердые полоски и кюветы, пробирки или другие контейнеры для спектрометрических образцов. Компонент анализа в виде твердой фазы отличается от инертных твердых поверхностей тем, что «твердая подложка» содержит на ее поверхности по меньшей мере одну группировку, которая предназначена для химического взаимодействия с молекулой. Твердая фаза может представлять собой стационарный компонент, такой как чип, пробирка, полоска, кювета или планшет для микротитрования, или может представлять собой нестационарные компоненты, такие как шарики и микрочастицы. Микрочастицы также можно использовать в качестве твердой подложки для форматов гомогенного анализа. Можно использовать целый ряд микрочастиц, которые обеспечивают как нековалентное, так и ковалентное присоединение белков и других веществ. Такие частицы включают полимерные частицы, такие как полистирольные и поли(метилметакрилатные); частицы золота, такие как наночастицы золота и коллоиды золота; и керамические частицы, такие как частицы из диоксида кремния, стекла и оксидов металлов. См., например, Martin, C.R., et al., Analytical Chemistry-News & Features, May 1 (1998) 322A-327A, которая включена в данный документ посредством ссылки. В одном воплощении твердая подложка представляет собой сефарозу.
Термин «по существу одинаковые» означает то, что два значения, например, времена удерживания на колонке для аффинной хроматографии на основе FcRn двух разных антител, находятся в пределах 5% друг от друга, т.е. они отличаются меньше, чем на 5%. Например, первое время удерживания, составляющее 80 минут, и второе время удерживания, составляющее 84 минуты, являются по существу одинаковыми, тогда как время удерживания, составляющее 80 минут, и время удерживания, составляющее 85 минут, не являются по существу одинаковыми - данные времена удерживания являются отличными. В одном воплощении термин «по существу одинаковый» означает то, что два значения находятся в пределах 3,5% друг от друга, т.е. то, что они отличаются на 3,5% или менее. В одном воплощении термин «по существу одинаковый» означает то, что два значения находятся в пределах 2,5% друг от друга, т.е. то, что они отличаются на 2,5% или менее. Меньшее из двух значений берется как основа для данного расчета.
Термин «лечение» (и его грамматические вариации, такие как «лечить» или «осуществление лечения») в том виде, как он используется в данном документе, относится к клиническому вмешательству в попытке изменить природный ход заболевания индивида, которого лечат, и оно может осуществляться либо для профилактики, либо по ходу клинической патологии. Желательные эффекты лечения включают предупреждение появления или рецидива заболевания, ослабление симптомов, уменьшение любых прямых или непрямых патологических последствий заболевания, предупреждение метастазов, уменьшение скорости прогрессирования заболевания, ослабление или временное облегчение болезненного состояния и ремиссию или улучшенный прогноз, но не ограничиваются ими. В некоторых воплощениях антитела или слитые полипептиды с областью Fc, как описано в данном документе, используются для задержки развития заболевания или для замедления прогрессирования заболевания.
Термин «валентный» в том виде, как он используется в настоящей заявке, обозначает присутствие определенного числа сайтов связывания в молекуле (антитела). Термины «двухвалентный», «четырехвалентный» и «шестивалентый», как таковые, обозначают присутствие в молекуле (антитела) двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания соответственно. Биспецифичные антитела, как описано в данном документе, в одном предпочтительном воплощении являются «двухвалентными».
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с его антигеном: вариабельные домены тяжелой цепи и легкой цепи (VH и VL соответственно) антитела обычно имеют аналогичные структуры, причем каждый домен содержит четыре каркасные области (FR) и три гипервариабельные области (HVR) (см., например, Kindt, T.J. et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., N.Y. (2007), стр. 91). Один домен VH или VL может быть достаточным для придания специфичности связывания антигена. Кроме того, антитела, которые связываются с конкретным антигеном, могут быть выделены с использованием домена VH или VL из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных доменов VL или VH соответственно. См., например, Portolano, S. et al., J. Immunol. 150 (1993) 880-887; Clackson, T. et al., Nature 352 (1991) 624-628).
Термины «вариант», «модифицированное антитело» и «модифицированный слитый полипептид» обозначают молекулы, которые имеют аминокислотную последовательность, которая отличается от аминокислотной последовательности родительской молекулы. Типично такие молекулы имеют одно или более чем одно изменение, вставку или делецию. В одном воплощении модифицированное антитело или модифицированный слитый полипептид содержит аминокислотную последовательность, содержащую по меньшей мере часть области Fc, которая не встречается в природе. Такие молекулы имеют меньше, чем 100%-ную идентичность последовательности с родительским антителом или родительским слитым полипептидом. В одном воплощении вариант антитела или вариант слитого полипептида имеет аминокислотную последовательность, которая имеет от примерно 75%-ной до менее чем 100%-ной идентичности аминокислотной последовательности с аминокислотной последовательностью родительского антитела или родительского слитого полипептида, особенно от примерно 80%-ной до менее чем 100%-ной, особенно от примерно 85%-ной до менее чем 100%-ной, особенно от примерно 90%-ной до менее чем 100%-ной и особенно от примерно 95%-ной до менее чем 100%-ной. В одном воплощении родительское антитело или родительский слитый полипептид и вариант антитела или вариант слитого полипептида отличаются одним (одиночным), двумя или тремя аминокислотными остатками.
II. Способы, описанные в данном документе
Данное изобретение основано, по меньшей мере частично, на открытии того, что распределение заряда в домене Fv влияет на связывание антитело-FcRn и приводит к дополнительным взаимодействиям между антителом и FcRn. Это изменяет характеристики связывания с FcRn, особенно в отношении диссоциации комплекса антитело-FcRn при pH 7,4, уменьшая, посредством этого, FcRn-зависимое время полужизни антитела в конечной фазе.
а) Неонатальный рецептор Fc (FcRn)
Неонатальный рецептор Fc (FcRn) является важным для метаболической судьбы антител класса IgG in vivo. FcRn функционирует для спасения IgG дикого типа от пути лизосомальной деградации, приводя к пониженному клиренсу и увеличенному времени полужизни. Он представляет собой гетеродимерный белок, состоящий из двух полипептидов: 50 кДа белка, подобного главному комплексу гистосовместимости класса I (α-FcRn) и 15 кДа β2-микроглобулина (β2m). FcRn связывается с высокой аффинностью с частью СН2-СНЗ области Fc антитела класса IgG. Взаимодействие между антителом класса IgG и FcRn является pH-зависимым и происходит в стехиометрии 1:2, т.е. одна молекула антитела IgG может взаимодействовать с двумя молекулами FcRn через ее два полипептида области Fc тяжелой цепи (см., например, [16]).
Таким образом, свойства/характеристики связывания IgG с FcRn in vitro указывают на его фармакокинетические свойства in vivo в системе кровообращения.
Во взаимодействии между FcRn и областью Fc антитела класса IgG участвуют разные аминокислотные остатки домена СН2 и СН3 тяжелой цепи. Аминокислотные остатки, взаимодействующие с FcRn, расположены приблизительно между положением 243 по EU и положением 261 по EU, приблизительно между положением 275 по EU и положением 293 по EU, приблизительно между положением 302 по EU и положением 319 по EU, приблизительно между положением 336 по EU и положением 348 по EU, приблизительно между положением 367 по EU и положением 393 по EU, в положении 408 по EU и приблизительно между положением 424 по EU и положением 440 по EU. Более конкретно, во взаимодействии между областью Fc и FcRn участвуют следующие аминокислотные остатки согласно нумерации EU по Kabat: F243, P244, P245 P, K246, P247, K248, D249, T250, L251, M252, I253, S254, R255, T256, P257, E258, V259, T260, C261, F275, N276, W277, Y278, V279, D280, V282, E283, V284, H285, N286, A287, K288, T289, K290, P291, R292, E293, V302, V303, S304, V305, L306, T307, V308, L309, H310, Q311, D312, W313, L314, N315, G316, K317, E318, Y319, I336, S337, K338, A339, K340, G341, Q342, P343, R344, E345, P346, Q347, V348, C367, V369, F372, Y373, P374, S375, D376, I377, A378, V379, E380, W381, E382, S383, N384, G385, Q386, P387, E388, N389, Y391, T393, S408, S424, C425, S426, V427, M428, H429, E430, A431, L432, H433, N434, H435, Y436, T437, Q438, K439 и S440.
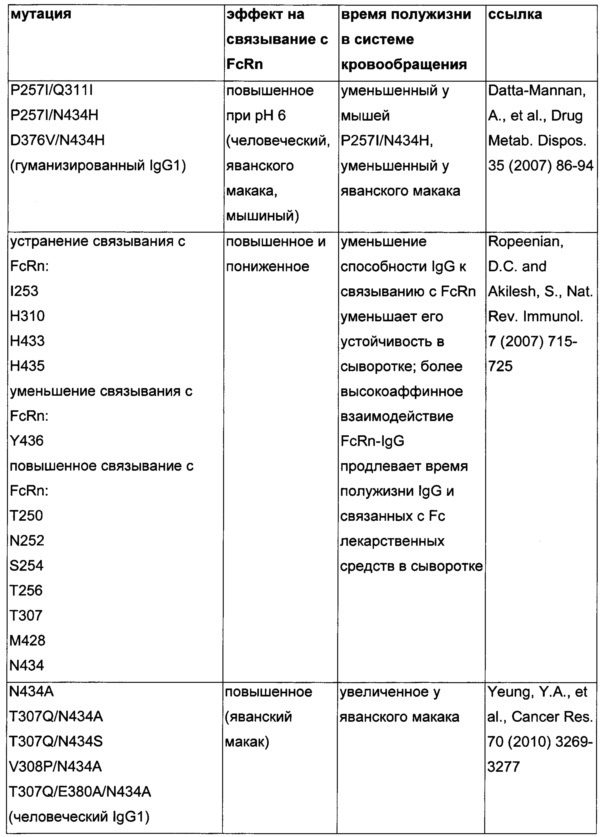
Исследования посредством сайт-направленного мутагенеза подтвердили то, что критически важными сайтами связывания в области Fc IgG для FcRn являются гистидин 310, гистидин 435 и изолейцин 253, и в меньшей степени - гистидин 433 и тирозин 436 (см., например, Kim, J.K., et al., Eur. J. Immunol. 29 (1999) 2819-2825; Raghavan, M., et al., Biochem. 34 (1995) 14649-146579; Medesan, C., et al., J Immunol. 158 (1997) 2211-2217).
Способы для увеличения связывания IgG с FcRn осуществляли посредством мутирования IgG по разным аминокислотным остаткам: треонину 250, метионину 252, серину 254, треонину 256, треонину 307, глутаминовой кислоте 380, метионину 428, гистидину 433 и аспарагину 434 (см. Kuo, Т.Т., et al., J. Clin. Immunol. 30 (2010) 777-789).
В некоторых случаях желательными являются антитела с уменьшенным временем полужизни в системе кровообращения. Например, лекарственные средства для интравитреального применения должны иметь продолжительное время полужизни в глазу и короткое время полужизни в системе кровообращения пациента. Такие антитела также имеют преимущество усиленного воздействия на сайт заболевания, например, в глазу.
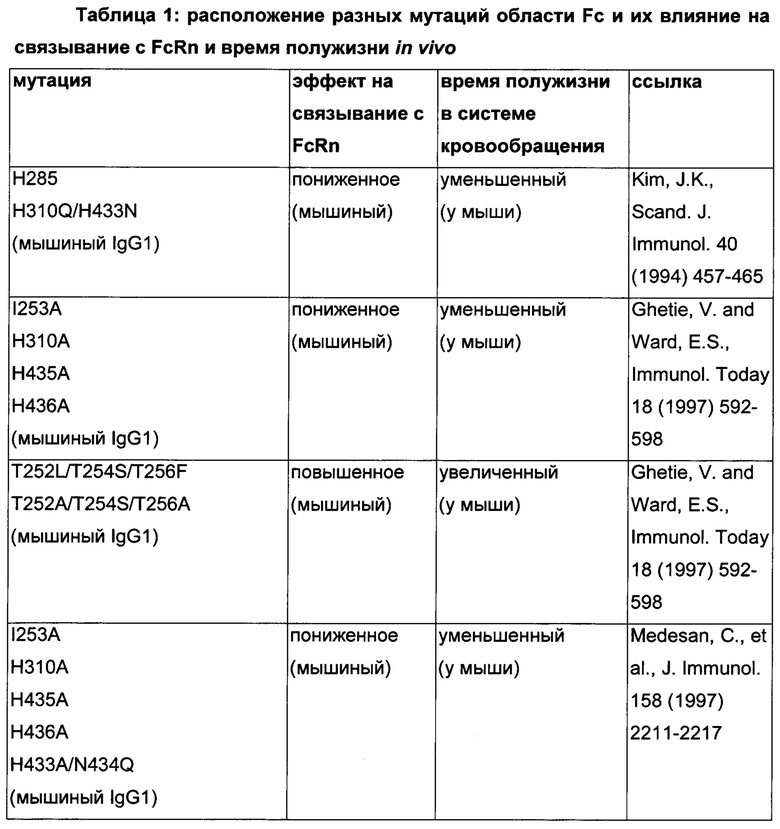
Известны разные мутации, которые влияют на связывание с FcRn и, вместе с этим, на время полужизни в системе кровообращения. Посредством сайт-направленного мутагенеза были идентифицированы критически важные остатки области Fc для взаимодействия Fc мыши с FcRn (см., например, Dall'Acqua, W.F., et al. J. Immunol 169 (2002) 5171-5180). Остатки I253, H310, H433, N434 и Н435 (нумерация EU согласно Kabat) участвуют в данном взаимодействии (Medesan, С., et al., Eur. J. Immunol. 26 (1996) 2533-2536; Firan, M., et al., Int. Immunol. 13 (2001) 993-1002; Kim, J.K., et al., Eur. J. Immunol. 24 (1994) 542-548). Обнаружили, что остатки I253, H310 и Н435 являются критически важными для взаимодействия человеческого Fc с мышиным FcRn (Kim, J.K., et al., Eur. J. Immunol. 29 (1999) 2819-2825). Dall'Acqua et al. посредством исследований взаимодействия белок-белок описали, что остатки M252Y, S254T, Т256Е улучшают связывание с FcRn (Dall'Acqua, W.F., et al. J. Biol. Chem. 281 (2006) 23514-23524). Исследования комплекса человеческий Fc-человеческий FcRn показали, что остатки I253, S254, Н435 и Y436 являются решающими для взаимодействия (Firan, М., et al., Int. Immunol. 13 (2001) 993-1002; Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604). У Yeung, Y.A., et al. (J. Immunol. 182 (2009) 7667-7671) были описаны и проверены разные мутанты по остаткам 248-259 и 301-317, и 376-382, и 424-437. Типичные мутации и их эффект на связывание с FcRn перечислены в следующей Таблице 1.
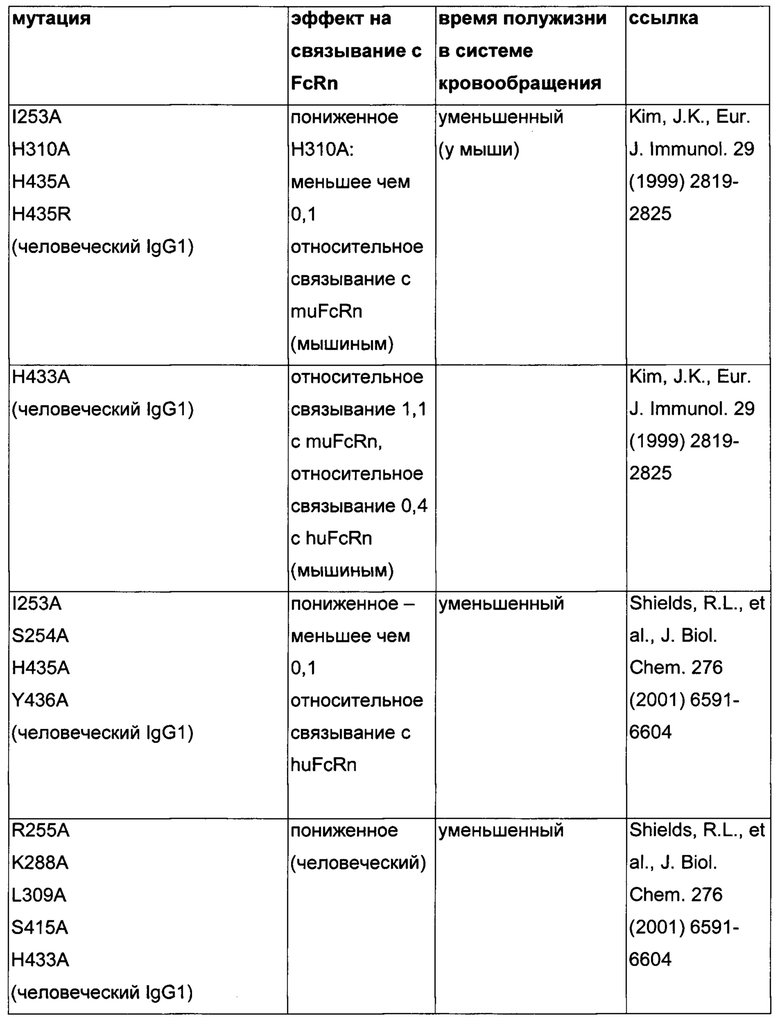
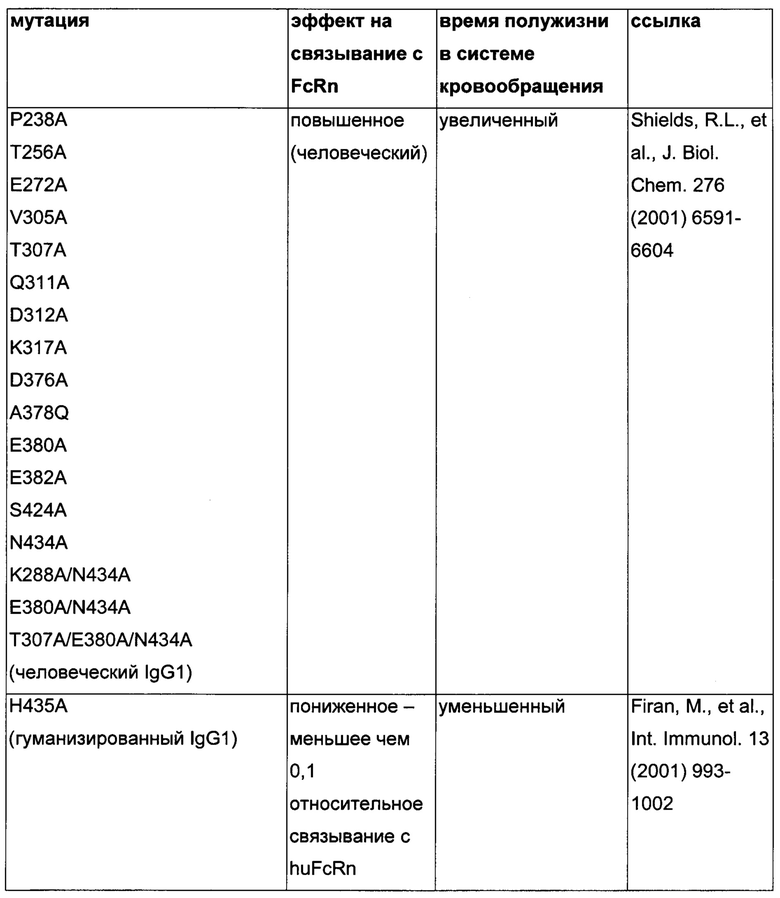
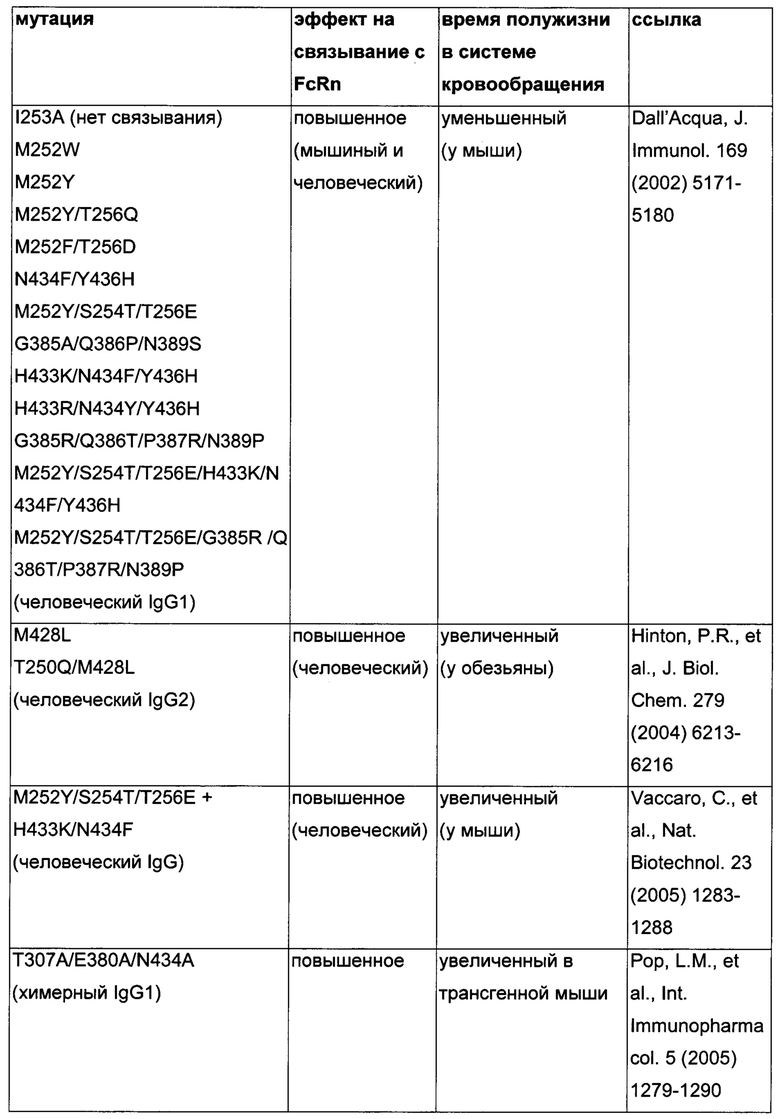
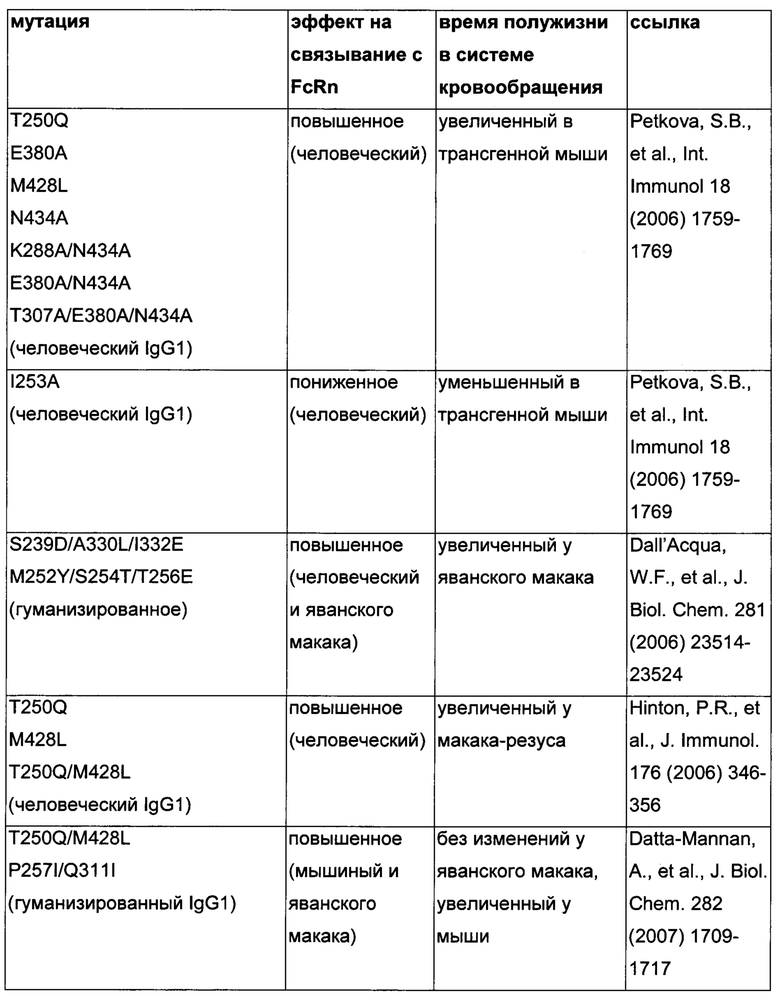

Обнаружили, что распределение заряда в домене Fv влияет на связывание антитело-FcRn и может приводить к дополнительным взаимодействиям между антителом и FcRn. Это изменяет характеристики связывания с FcRn, особенно в отношении диссоциации комплекса антитело-FcRn при pH 7,4, посредством этого влияя (уменьшая) зависимое от FcRn время полужизни антитела в конечной фазе.
Человеческий неонатальный рецептор Fc (FcRn) играет важную роль в катаболизме IgG. Свойства/характеристики связывания IgG с FcRn in vitro указывают на его фармакокинетические свойства in vivo. Такие способы in vitro имели бы большое значение во время разработки антитела, поскольку можно избежать многочисленных исследований in vivo (уменьшенное число экспериментов на животных, меньшее время и затраты).
Взаимодействия IgG-FcRn можно анализировать с использованием анализов поверхностного плазмонного резонанса (SPR) (Wang, W., et al., Drug Metab. Disp.39 (2011) 1469-1477; Datta-Mannan, A., et al., Drug Metab. Disp.40 (2012) 1545-1555; Vaughn, D.E. and Bjorkman, P.J., Biochemistry 36 (1997) 9374-9380; Raghavan, M., et al., Proc. Natl. Acad. Sci. USA 92 (1995) 11200-11204; Martin, W.L and Bjorkman, P.J., Biochemistry 38 (1999) 12639-12647).
Для оценки аффинности связывания IgG с FcRn также были описаны калориметрические способы и способы фракционирования в поле ассиметричного потока (Huber, А.Н., et al., J. Mol. Biol. 230 (1993) 1077-1083; Pollastrini, J., et al., Anal. Biochem. 414 (2011) 88-98).
Помимо наличия комплексных анализов, несколько исследований, в которых изучается корреляция между параметрами связывания с FcRn in vitro, определенными SPR, и временем полужизни антител в сыворотке in vivo, до настоящего времени не смогли продемонстрировать такую корреляцию, несмотря на улучшенные условия реакции связывания и подходящее моделирование (Gurbaxani, В., et al., Mol. Immunol. 43 (2006) 1462-1473; Gurbaxani, B.M. and Morrison, S.L., Mol. Immunol. 43 (2006) 1379-1389; Gurbaxani, В., Clin. Immunol. 122 (2007) 121-124).
Инженерия области Fc IgG1 для улучшения аффинности IgG1 к FcRn при pH 6 и при нейтральном pH при измерении технологией SPR не привела к улучшенной фармакокинетике у яванских макаков (Yeung, Y.A., et al., J. Immunol. 182 (2009) 7663-7671). Однако только умеренные увеличения в аффинности к FcRn при pH 6 у варианта IgG1 N434A без сопутствующего значимого связывания с FcRn при pH 7,4 приводили к улучшенной фармакокинетике у приматов, демонстрируя важность высвобождения от FcRn при pH 7,4 (см. Yeung, Y.A. выше).
Например, анализ SPR взаимодействия IgG-FcRn дает качественный результат, указывающий на ожидаемые или нарушенные характеристики связывания образца, но не дает ни намек на причину нарушенного связывания, ни количественную оценку количества антитела с нарушенным связыванием.
Способ аффинной хроматографии на основе FcRn с использованием элюции положительным линейным градиентом был описан в WO 2013/120929.
б) Взаимодействия FcRn-Fab, опосредованные зарядом
Известно, что конкретная манипуляция в области Fc влияет на PK параметры посредством изменения взаимодействия между областью Fc и FcRn, и ее использовали для конструирования терапевтических антител со специфическими PK свойствами [33, 34].
Несмотря на то, что влияние области Fab на взаимодействия с FcRn недавно обсуждались, когда антитела с одинаковыми последовательностями человеческой области Fc дикого типа, но с разными областями Fab демонстрировали различия в аффинности в отношении FcRn и измененную PK, механизм данного взаимодействия оставался неясным [23, 24].
Для демонстрации влияющих факторов области Fab на гомеостаз IgG, опосредованный FcRn, в качестве модельной системы использовали пару антител бриакинумаб (Ozespa™) и устекинумаб (Stelara™). И бриакинумаб, и устекинумаб представляют собой полностью человеческие моноклональные антитела IgG1. Они связываются с той же самой субъединицей р40 интерлейкина 12 (IL-12) и интерлейкина 23 (IL-23) [26], и они не являются перекрестно реагирующими с соответствующим мышиным IL-12 и IL-23 [27, 28]. Бриакинумаб и устекинумаб представляют собой антитело IgG1κ с вариабельными доменами тяжелой и легкой цепи семейств VH5 и Vκ1D зародышевой линии и антитело IgG1λ с вариабельными доменами тяжелой и легкой цепи семейств VH3 и Vλ1 зародышевой линии соответственно. Помимо разных вариабельных доменов бриакинумаб и устекинумаб демонстрируют различия в нескольких специфичных для аллотипа аминокислотах в константных доменах (см. Фиг. 5). Однако данные аминокислотные остатки находятся вне областей (когнатного) связывания с FcRn и, следовательно, могут рассматриваться как не играющие роли в PK, зависимой от FcRn [11]. Интересно то, что устекинумаб имеет (приведенное) медианное время полужизни в конечной фазе, составляющий 22 сутки [29], тогда как бриакинумаб имеет время полужизни в конечной фазе, составляющий только 8-9 суток [26, 30, 31].
в) Распределение заряда и pH-зависимый нетто заряд
Бриакинумаб демонстрирует неоднородное распределение заряда при физиологическом pH 7,4 (см., например, опубликованную кристаллическую структуру устекинумаба [27] и модель гомологии бриакинумаба). Бриакинумаб демонстрирует большую положительно заряженную область на домене Fv (см. Фиг. 1а), которая отсутствует у устекинумаба (см. Фиг. 1б). Кроме того, FcRn обладает протяженной областью с сильным отрицательным зарядом (см. Фиг. 1в), которая, однако, не участвует в когнатном связывании с областью Fc. Бриакинумаб и устекинумаб имеют расчетные изоэлектрические точки 9,7 и 9,4 соответственно. Кроме того, нетто-заряд бриакинумаба является немного более положительным во всем диапазоне pH (см. Фиг. 1г).
Аффинность связывания бриакинумаба и устекинумаба с FcRn при pH 6,0 является сравнимой, т.е. оба значения отличаются самое большее на один порядок, в одном воплощении - самое большее в 5 раз, тогда как диссоциация от FcRn является очень разной. С использованием вариантов бриакинумаба и устекинумаба можно было бы показать, что взаимодействие преимущественно является электростатическим и коррелирует с размером положительно заряженной области (см. ниже).
г) pH-зависимое взаимодействие FcRn-IgG
Посредством аффинной хроматографии на основе FcRn (см. Таблицу 2) были синтезированы и охарактеризованы десять вариантов бриакинумаба и устекинумаба в отношении их свойств связывания с FcRn (см. Таблицу 2). В данных вариантах вариабельные области были модифицированы и протестированы в отношении аффинности связывания с FcRn при pH 6 и диссоциации от FcRn с использованием поверхностного плазмонного резонанса (SPR) и аффинной хроматографии на основе FcRn (см. Таблицу 3) соответственно.
Таблица 2: систематически сконструированные варианты бриакинумаба и устекинумаба. Структурные части, подобные Fv, LC и CDR обменивали между бриакинумабом (светлый) и устекинумабом (темный): mAb 1-6. Три и пять основных аминокислот в НС (тяжелая цепь) бриакинумаба заменяли на остатки аланина (посредством сайт-направленного мутагенеза) для mAb 7 (моноклональное антитело 7) и mAb 8 соответственно. MAb 9 представляет собой бриакинумаб с тремя основными аминокислотами в CDR легкой цепи, замененными на остатки аланина, и mAb 10 представляет собой mAb 9 с дополнительной заменой пяти основных аминокислот в НС. Замена одиночных основных аминокислот показана кружками, три замененные аминокислоты изображены в виде 1 кружка, а 5 замененных аминокислот - в виде 2 кружков.
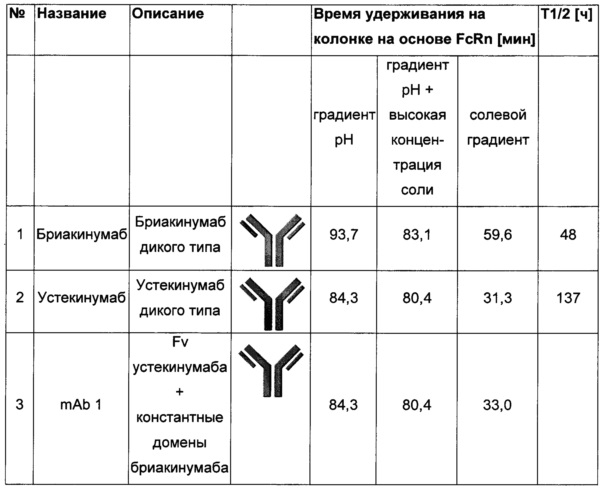
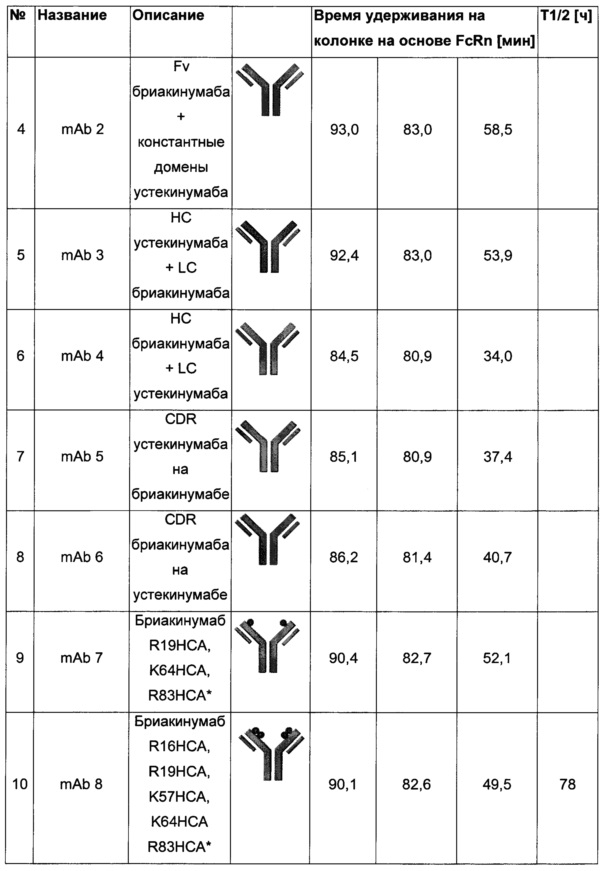
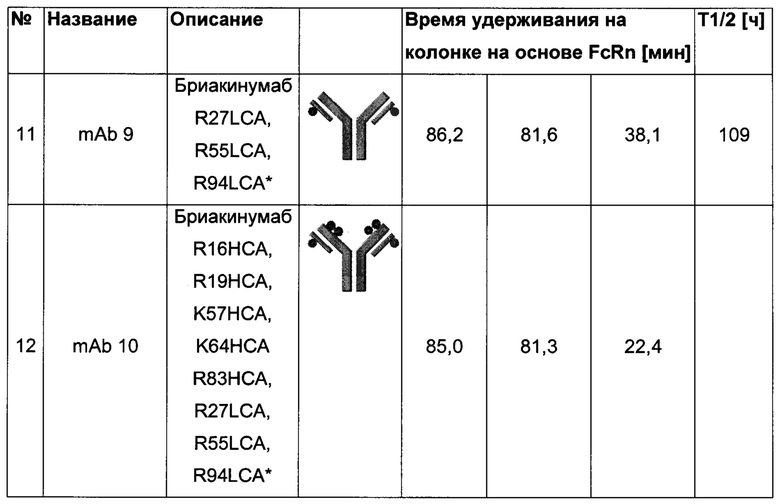
Таблица 3: аффинности связывания с FcRn и распределения заряда всех протестированных антител. Антитела сортировали согласно временам удерживания на колонке на основе FcRn. Равновесную константу диссоциации KD рассчитывали в виде стационарной аффинности и нормировали к KD устекинумаба. Сравнение относительных значений KD (устекинумаб - 1) представлено в виде среднего (n равно 3) плюс/минус стандартное отклонение (SD). Были рассчитаны изоэлектрические точки и нетто-заряды доменов Fv при pH 6,0 и pH 7,4 (SaWi-Tools). Времена удерживания на колонке на основе FcRn не коррелируют с изоэлектрической точкой или нетто-зарядом домена Fv при pH 6,0 или pH 7,4.
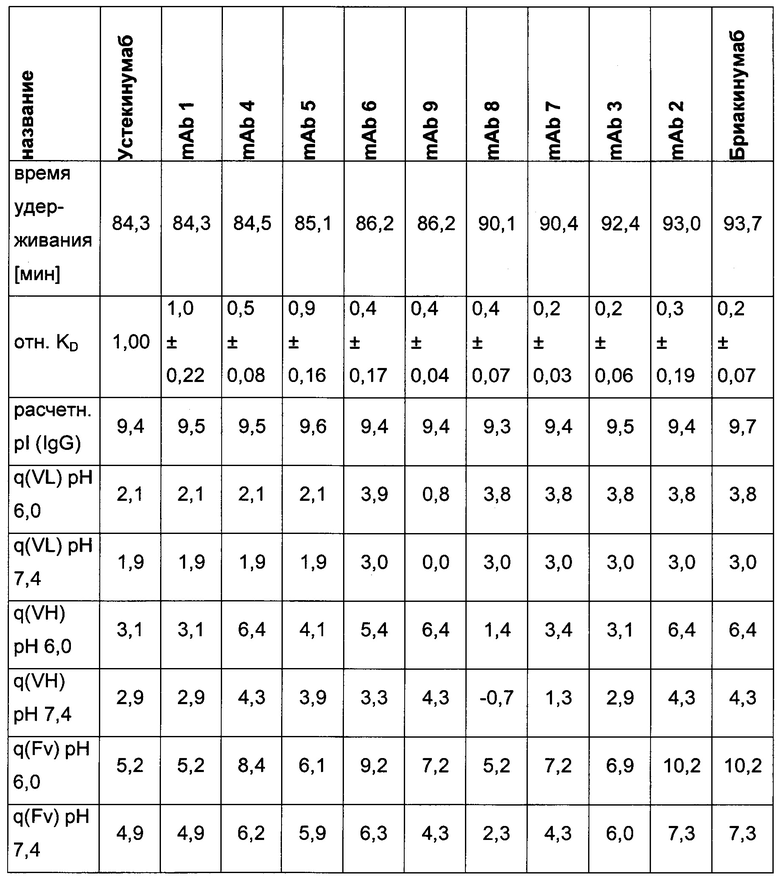
Аффинности связывания в отношении FcRn при pH 6 для одиннадцати антител попадают в узкий интервал (см. Таблицу 3). Равновесную константу диссоциации (KD) рассчитывали по отношению к устекинумабу (устекинумаб - 1,00). Бриакинумаб имел относительную KD 0,2, и девять вариантов имели варьирующие значения между бриакинумабом и устекинумабом. Таким образом, можно заключить то, что разное время полужизни в конечной фазе in vivo не вызывается разным связыванием с FcRn при pH 6,0.
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, влияющего на время полужизни антитела in vivo, включающий следующие стадии:
а) определение для варианта антитела и его родительского антитела значений KD при pH 6 с использованием поверхностного плазмонного резонанса,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли,
при этом присутствие взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, влияющего на время полужизни in vivo, определяется, если значения KD отличаются самое большее в 10 раз, и время удерживания, определенное на стадии б), между вариантом антитела и его родительским антителом является по существу отличным.
Одним аспектом, как описано в данном документе, является способ определения относительного времени полужизни антитела in vivo, включающий следующие стадии:
а) определение для варианта антитела и его родительского антитела значений KD при pH 6 с использованием поверхностного плазмонного резонанса,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли,
при этом антитело имеет относительное время полужизни in vivo, которое уменьшено по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания варианта антитела, определенное на стадии б), короче/меньше, чем время удерживания его родительского антитела, и
при этом антитело имеет относительное время полужизни in vivo, которое увеличивается по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания варианта антитела, определенное на стадии б), длиннее/больше, чем время удерживания его родительского антитела.
Одним аспектом, как описано в данном документе, является способ определения увеличения или уменьшения времени полужизни антитела in vivo, включающий следующие стадии:
а) определение для варианта антитела и его родительского антитела значений KD при pH 6 с использованием поверхностного плазмонного резонанса,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли,
при этом антитело имеет уменьшение времени полужизни in vivo по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания варианта антитела, определенное на стадии б), короче/меньше, чем время удерживания его родительского антитела, и
при этом антитело имеет увеличение времени полужизни in vivo по сравнению с его родительским антителом, если значения KD отличаются самое большее в 10 раз, и время удерживания варианта антитела, определенное на стадии б), длиннее/больше, чем время удерживания его родительского антитела.
Профили элюции двенадцати антител анализировали с использованием аффинной колонки на основе FcRn с элюцией положительным линейным градиентом pH (см. Фиг. 2). Устекинумаб и mAb1, которое несет домен Fv устекинумаба на константных частях бриакинумаба, демонстрировали неотличимые времена удерживания около 84 минут, демонстрируя то, что домен Fv влияет на взаимодействие с FcRn. Бриакинумаб, с другой стороны, элюировался при времени удерживания 94 минуты и, следовательно, имел явно отличное время удерживания по сравнению с устекинумабом. Неразличимые времена удерживания отщепленных IdeS областей Fc бриакинумаба (85,7 мин) и устекинумаба (85,2 мин) указывали на пренебрежимо малую роль области Fc. MAb 4, содержащее LC устекинумаба (LC - легкая цепь, НС - тяжелая цепь) и НС бриакинумаба, имеет время удерживания, близкое к устекинумабу, демонстрируя влияние LC на связывание с FcRn.
Варианты антител mAb 5 и mAb 6 несут CDR (части тяжелой и легкой цепи) устекинумаба на каркасе бриакинумаба и наоборот.Пересадка CDR устекинумаба на бриакинумаб (mAb 5) сдвигала время удерживания mAb 5 близко ко времени удерживания устекинумаба. Пересадка CDR бриакинумаба на устекинумаб (mAb 6) описывала/представляла профиль элюции, который был все еще близок к устекинумабу.
Сильный сдвиг времени удерживания от бриакинумаба в направлении устекинумаба наблюдали для mAb 9, которое представляет собой вариант бриакинумаба, в котором три положительно заряженных остатка в CDR легкой цепи мутировали до остатков аланина.
В mAb 7 и mAb 8 были мутированы, соответственно, три и пять положительно заряженных остатка в тяжелой цепи бриакинумаба. У данных вариантов время удерживания сдвигалось относительно бриакинумаба.
MAb 3, содержащее НС устекинумаба и LC бриакинумаба, а также mAb 2, содержащее домен Fv бриакинумаба на константных доменах устекинумаба, оба элюировались близко к бриакинумабу.
В целом, данные показывают то, что домен Fv влияет на диссоциацию от FcRn, а не на связывание с FcRn (при pH 6,0).
Времена удерживания на колонке на основе FcRn сопоставляли с изоэлектрическими точками и нетто зарядами антител. Не наблюдали корреляции между временами удерживания на колонке на основе FcRn и изоэлектрическими точками или нетто зарядами доменов Fv при лизосомальном pH 6,0 или физиологическом pH 7,4 (см. Таблицу 3). Однако измеренные времена удерживания на колонке на основе FcRn возрастали с выраженностью положительно заряженных областей, особенно вокруг вариабельных доменов легкой цепи (см. Фиг. 2).
Одним аспектом, как описано в данном документе, является способ увеличения времени полужизни антитела in vivo, включающий стадию:
- замены заряженного аминокислотного остатка в положениях 27, 55 и 94 в легкой цепи антитела на гидрофобный или нейтральный гидрофильный аминокислотный остаток (нумерация согласно Kabat) и, посредством этого, увеличения времени полужизни антитела in vivo.
Аминокислоты могут быть сгруппированы согласно общим свойствам боковой цепи:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Кислотные и основные аминокислотные остатки совместно образуют группу заряженных аминокислотных остатков.
Времена удерживания на колонке на основе FcRn также были определены в другой постановке с увеличенной ионной силой в подвижной фазе, т.е. в присутствии повышенных концентраций соли. Известно, что опосредованные зарядом взаимодействия ослабевают при условиях высокой ионной силы, тогла как гидрофобные взаимодействия типично усиливаются солью. Обнаружили, что время удерживания бриакинумаба на колонке на основе FcRn было укорочено в присутствии соли и было обратнопропорциональным квадратному корню ионной силы, как подсказывает закон экранирования заряда Дебая-Хюкеля [32]. Время удерживания устекинумаба оставалось по существу неизменным (см. Фиг. 6). Таким образом, значительная часть избыточного взаимодействия FcRn-бриакинумаб опосредована зарядом.
Обобщая приведенное выше, аффинная хроматография на основе FcRn генетически модифицированных вариантов показала то, что антитела с тем же самым доменом Fv (mAb 1 и mAb 2) и той же самой LC (mAb 3 и mAb 4) элюируются при почти идентичных временах удерживания на колонке на основе FcRn. Кроме того, пересадка CDR устекинумаба на бриакинумаб (mAb 5) сдвигает рН элюции близко к рН элюции устекинумаба. Таким образом, CDR легкой цепи обеспечивают главное влияние на связывание бриакинумаба с FcRn.
Пересадка CDR бриакинумаба на устекинумаб (mAb 6) давала профиль элюции, который был все еще близким к профилю элюции устекинумаба. Таким образом, не будучи связанными данной теорией, могло бы быть так, что взаимодействие антитело-FcRn в большей степени подвергается влиянию разрушения большой положительно заряженной области бриакинумаба, чем созданию меньшей положительно заряженной области. Как предлагается посредством моделирования MD (молекулярная динамика), прямое стабилизирующее взаимодейтвие между положительно заряженной областью Fv и отрицательно заряженной областью FcRn является легко возможным. Таким образом, взаимодействие FcRn-Fv может приводить к более медленной, чем нормальная диссоциации комплекса FcRn-IgG при физиологических условиях.
д) Корреляция pH элюции с FcRn с фармакокинетикой у трансгенных мышей с человеческим FcRn
В предыдущих исследованиях обсуждалось то, что нетто заряд является движущей силой для измененных фармакокинетических свойств посредством воздействия на электростатическое взаимодействие между антителом и отрицательно заряженными группами на поверхности эндотелиальных клеток [35, 36]. Например, Igawa et al. [37] наблюдали то, что антитела IgG4 с меньшими изоэлектрическими точками (pl) из-за генетической модификации в вариабельной области имели меньшую скорость пиноцитоза в жидкой фазе и, в свою очередь, пониженную скорость элиминации. Кроме того, Boswell et al. [38] предположили то, что различия pl для влияния на PK должны составлять по меньшей мере одну единицу.
pl бриакинумаба, устекинумаба, mAb 8 и mAb 9 варьируют от 9,3 до 9,7. Следовательно, можно предположить, что влияние на пиноцитоз в жидкой фазе является минимальным из-за различий pl. Однако более короткие времена удерживания бриакинумаба на колонке на основе FcRn при условиях высокой ионной силы (см. выше), а также больший электростатический вклад во взаимодействие FcRn-Fv по сравнению с устекинумабом при моделировании MD (см. ниже) демонстрируют то, что специфически локализованный заряд может представлять собой главный влияющий фактор на взаимодействие FcRn-IgG. Влияние заряда в домене Fv анализировали с использованием мутантов, которые имеют три (mAb 7) и пять (mAb 8) мутированных положительно заряженных остатков в НС бриакинумаба, а также бриакинумаба с тремя положительно заряженными остатками, мутированными в CDR легкой цепи (mAb 9). MAb 7 и mAb 8 демонстрируют маленькие сдвиги времени удерживания в направлении устекинумаба, подтверждающие опосредованное зарядом взаимодействие, тогда как mAb 9 демонстрирует большой сдвиг времени удерживания в направлении устекинумаба. Таким образом, специфически локализованный(ные) заряд(ды) в CDR легкой цепи сильно влияет(ют) на диссоциацию от FcRn.
Для оценки того, переводится ли эффект мутированных заряженных остатков в вариабельных доменах бриакинумаба на связывание с FcRn в модулированные PK свойства in vivo, провели исследования PK на трансгенных мышах с человеческим FcRn. Тестировали бриакинумаб и устекинумаб, наряду с двумя вариантами бриакинумаба (mAb 8 и mAb 9), которые имели времена удерживания на колонке на основе FcRn, находящиеся между бриакинумабом и устекинумабом. Процессы распределения и элиминации четырех антител согласуются с исследованиями PK других IgG (см. Фиг. 3а). Бриакинумаб демонстрировал более быстрое снижение в α-фазе, чем другие антитела. Интересно то, что бриакинумаб и устекинумаб демонстрировали время полужизни в конечной фазе 48 часов и 137 часов соответственно (см. Фиг. 3б). Варианты mAb 8 и mAb 9, которые имеют меньшие положительно заряженные области в домене Fv, имели время полужизни в конечной фазе 78 часов и 109 часов соответственно. Статистическая значимость могла быть выявлена между временем полужизни в конечной фазе бриакинумаба и устекинумаба, бриакинумаба и mAb 9, а также устекинумаба и mAb 8. Устекинумаб, mAb 9, mAb 8 и бриакинумаб элюировались в 84,3; 86,2; 90,1 и 93,7 минут, соответствуя pH элюции 7,4; 7,5; 7,7 и 7,9 соответственно. Таким образом, обнаружили, что время полужизни в конечной фазе четырех IgG линейно коррелирует со значениями pH элюции на колонке на основе FcRn in vitro (см. Фиг. 3б).
В PK экспериментах проверяли время полужизни в конечной фазе, которое рассчитывается исключительно в фазе выведения, где доминирует рециклирование FcRn [39]. Время полужизни четырех антител в конечной фазе линейно коррелирует с pH элюции на колонке на основе FcRn in vitro: чем выше pH элюции на колонке на основе FcRn, тем короче время полужизни в конечной фазе, демонстрируя, посредством этого, то, что колонка на основе FcRn представляет собой прогностический/чувствительный инструмент для определения диссоциации от FcRn in vitro. Корреляция между временем полужизни в конечной фазе и pH элюции на колонке на основе FcRn подтверждает важность быстрой диссоциации FcRn-IgG при физиологическом pH.
Не будучи связанными данной теорией, комплекс FcRn-IgG строится в эндосомах при pH 6,0, следовательно, меньшее связывание приводит к меньшему рециклированию IgG и более быстрому клиренсу. Посредством экзоцитоза комплекс FcRn-IgG высвобождается к плазматической мембране, где при физиологическом pH 7,4 в пределах короткого периода времени [40] должна происходить диссоциация IgG и FcRn. Следовательно, диссоциация при физиологическом pH также является важной для продолжительного времени полужизни [22, 40].
Таким образом, обнаружили, что диссоциация при более высоких значениях рН указывает на более медленную диссоциацию от FcRn. Не будучи связанными данной теорией, это могло бы приводить к деградации антител в лизосоме, вместо высвобождения антител обратно в кровоток.
Таким образом, обнаружили то, что заряд в домене Fv IgG влияет на время полужизни в конечной фазе путем изменения взаимодействия между IgG и FcRn. Были локализованы структурные части Fab, которые взаимодействуют с FcRn, и было продемонстрировано то, что взаимодействие опосредовано зарядом. Исследование PK выявило линейную корреляцию между диссоциацией FcRn-IgG in vitro и временем полужизни в конечной фазе in vivo.
е) Моделирование молекулярной динамики (MD) моделей FcRn-IgG
Получали модель гомологии комплекса человеческий FcRn-Fc с использованием опубликованной структуры крысиного FcRn в качестве матрицы. Положение доменов Fv бриакинумаба и устекинумаба моделировали на основе кристаллической структуры полного IgG1 (код PDB (база данных белковых структур) - 1HZH). Данные модели гомологии содержат две копии FcRn (α-FcRn с β2m) на одной полной молекуле IgG (см. Фиг. 4а). Расстояние между FcRn и доменами Fv составляет более 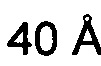 в исходной структуре и превышает дебаевский радиус приблизительно
в исходной структуре и превышает дебаевский радиус приблизительно 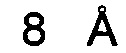 при физиологических условиях [32]. Динамику комплексов FcRn-IgG моделировали посредством моделирования молекулярной динамики на протяжении периода 100 нс в формальной воде и при физиологической ионной силе. По ходу моделирования одна из двух областей Fab достигала кончика FcRn и сохранялась в данной конформации в течение оставшегося времени моделирования (см. Фиг. 4б, в, г). Область на FcRn, для которой было обнаружено взаимодействие с доменом Fv, до настоящего времени не была описана как участвующая в связывании с IgG. Неожиданно то, что в моделированиях MD не только бриакинумаб, но также и устекинумаб принимал конформацию с Fv и FcRn, взаимодействующими друг с другом (см. Фиг. 4б, в). Обнаружили, что в обоих комплексах две разные пары доменов Fv и FcRn в асимметричной исходной структуре достигали друг друга. Обнаружили, что электростатический вклад во взаимодействие FcRn-Fv в бриакинумабе примерно в два раза больше, чем в устекинумабе (см. Фиг. 4д).
при физиологических условиях [32]. Динамику комплексов FcRn-IgG моделировали посредством моделирования молекулярной динамики на протяжении периода 100 нс в формальной воде и при физиологической ионной силе. По ходу моделирования одна из двух областей Fab достигала кончика FcRn и сохранялась в данной конформации в течение оставшегося времени моделирования (см. Фиг. 4б, в, г). Область на FcRn, для которой было обнаружено взаимодействие с доменом Fv, до настоящего времени не была описана как участвующая в связывании с IgG. Неожиданно то, что в моделированиях MD не только бриакинумаб, но также и устекинумаб принимал конформацию с Fv и FcRn, взаимодействующими друг с другом (см. Фиг. 4б, в). Обнаружили, что в обоих комплексах две разные пары доменов Fv и FcRn в асимметричной исходной структуре достигали друг друга. Обнаружили, что электростатический вклад во взаимодействие FcRn-Fv в бриакинумабе примерно в два раза больше, чем в устекинумабе (см. Фиг. 4д).
В заключение, обнаружили, что собственная гибкость плеч Fab комплексов FcRn-IgG структурно обеспечивает непосредственное стабилизирующее взаимодействие домена Fv с кончиком FcRn.
ж) Способы согласно настоящему изобретению
ж.i) Элюция линейным положительным градиентом pH при разных концентрациях соли
В данном документе описан способ, включающий следующие две стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли.
Вторая концентрация соли обычно выше/больше, чем первая концентрация соли, так что данные концентрации не являются примерно идентичными, т.е. они отличаются по меньшей мере на 10%, в одном воплощении - по меньшей мере на 20%.
При использовании данного способа можно определить присутствие взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn в простом хроматографическом способе посредством сравнения времен удерживания, полученных в присутствии разных концентраций соли (см. Фиг. 13), или посредством сравнения времени удерживания полного антитела и его области Fc. Это является важным, так как взаимодействия Fab антитела-FcRn влияют на время полужизни антитела in vivo.
Для пар антитело/контрольное антитело существуют разные связи в отношении их времен удерживания на колонке для аффинной хроматографии на основе FcRn и, к тому же, в отношении их взаимодействия с FcRn:
1) антитело и контрольное антитело имеют по существу одинаковое время удерживания на стадии а) и стадии б): в данном случае время полужизни обоих антител in vivo должен быть по существу одинаковым, т.е. на время полужизни in vivo не влияют взаимодействия Fab антитела-FcRn, или
2) антитело и контрольное антитело имеют по существу одинаковое время удерживания на стадии а), но разное время удерживания на стадии б): в данном случае время полужизни антитела in vivo короче, чем время полужизни контрольного антитела in vivo, т.е. на время полужизни in vivo влияют взаимодействия Fab антитела-FcRn.
Антитело может представлять собой вариант родительского антитела, причем в данном случае контрольное антитело представляет собой родительское антитело.
В одном случае контрольное антитело представляет собой антитело, которое имеет по существу такое же время удерживания, что и его область Fc, после отщепления IdeS или отщепления папаином.
Для предложения терапевтических схем для лечения множества заболеваний, известных в настоящее время, и также заболеваний, которые будут выявлены в будущем, существует потребность в антителах, а также в полипептидах, содержащих область Fc, с заданными свойствами.
Для задания характеристик связывания антитела с FcRn остатки, участвущие во взаимодействии с FcRn, модифицируются, и образующиеся модифицированные антитела должны быть протестированы. Если требующиеся характеристики не удовлетворяются, вновь проводится тот же самый способ.
Таким образом, было бы полезным предложить способ, который прогнозирует изменения характерных свойств модифицированного антитела на основе простого хроматографического способа, и который не требует исследований in vivo для анализа изменений характеристик у модифицированного антитела.
В некоторых случаях желательными являются антитела с увеличенным временем полужизни. Например, лекарственные средства с увеличенным временем полужизни в системе кровообращения пациента, нуждающегося в лечении, требуют уменьшенного дозирования или увеличенных интервалов дозирования. Такие антитела также имеют преимущество увеличенного воздействия на сайт заболевания, например, на опухоль.
Время полужизни in vivo коррелирует со временем удерживания на колонке для аффинной хроматографии на основе FcRn. Это особенно верно, если взаимодействие между антителом и FcRn почти целиком опосредовано остатками в области Fc антитела. Но, если остатки вне области Fc антитела, например, в Fab антитела, также взаимодействуют с FcRn, данная корреляция должна быть дополнительно подтверждена. Это может осуществляться способом, описанным в данном документе, использующим изменение времени удерживания в способе аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом рН в присутствии низкой и высокой концентраций соли, или интактного антитела и антитела с отщепленной областью Fv (равно области Fc). Если время удерживания по существу не подвергается влиянию изменения от низкой до высокой концентрации соли или отщепления области Fc, тогда отсутствует взаимодействие Fab антитела-FcRn, и большее время удерживания на колонке для аффинной хроматографии на основе FcRn коррелирует с увеличенным временем полужизни in vivo. Однако, если время удерживания подвергается влиянию, особенно если оно уменьшается посредством изменения от низких до высоких концентраций соли или посредством отщепления области Fc, тогда время полужизни in vivo коррелирует иным образом с временем удерживания на колонке для аффинной хроматографии на основе FcRn, т.е. более длительное время удерживания на колонке для аффинной хроматографии на основе FcRn коррелирует с более коротким временем полужизни in vivo из-за ослабленной диссоциации антитело-FcRn при физиологическом pH и, не будучи связанными данной теорией, повышенной лизосомальной деградации антитела.
Используемая в данном изобретении колонка для аффинной хроматографии на основе FcRn содержит матрицу и связанные с матрицей хроматографические функциональные группы, где связанная с матрицей хроматографическая функциональная группа содержит нековалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина.
Обычно исходным пунктом для способа, опсанного в данном документе, является родительское или контрольное антитело, которое характеризуется его связыванием с FcRn.
Одним аспектом, как описано в данном документе, является применение способа, как описано в данном документе, для определения присутствия взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, влияющего на время полужизни in vivo, включающего следующие стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом присутствие взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, влияющего на время полужизни in vivo, определяется, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, влияющего на время полужизни in vivo, включающий следующие стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом присутствие взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, влияющего на время полужизни in vivo, определяется, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
Варианты антител демонстрируют либо увеличенное, либо уменьшенное связывание с FcRn по сравнению с полипептидом родительского антитела или по сравнению с контрольным антителом и, таким образом, имеют модифицированное время полужизни по сравнению с родительским/контрольным антителом в сыворотке.
Обычно сначала прогнозируется, что варианты области Fc с повышенной аффинностью в отношении FcRn (т.е. с увеличенным временем удерживания на колонке на основе FcRn по сравнению с родительским антителом или контрольным антителом) имеют более продолжительное время полужизи в сыворотке по сравнению с вариантами с пониженной аффинностью в отношении FcRn (т.е. с уменьшенным временем удерживания на колонке на основе FcRn по сравнению с родительским антителом или контрольным антителом).
Данное прогнозируемое время полужизни in vivo должно быть затем подтверждено. Для этого подтверждения может быть использован способ, описанный в данном документе.
Варианты антител с увеличенным временем полужизни in vivo имеют применения в способах лечения млекопитающих, особенно людей, где желательным является продолжительное время полужизни введенного антитела, таких как лечение хронического заболевания или расстройства.
Варианты антител с пониженной аффинностью в отношении FcRn имеют применения в способах лечения млекопитающих, особенно людей, где желательным является короткое время полужизни введенного антитела или слитого полипептида, как, например, в диагностической визуализации in vivo.
Весьма вероятно, что варианты антител с пониженной аффинностью связывания с FcRn будут способны пересекать плаценту и, таким образом, могут использоваться в лечении заболеваний или расстройств у беременных женщин, особенно заболеваний или расстройств неродившихся детей. Кроме того, пониженная аффинность связывания с FcRn может быть желательной для тех лекарственных средств, которые предназначены для введения/транспорта в мозг, почку и/или печень.
Одним аспектом, как описано в данном документе, является применение способа, как описано в данном документе, для идентификации антител, которые демонстрируют ослабленный транспорт через эпителий клубочков почки из сосудистой системы.
Одним аспектом, как описано в данном документе, является применение способа, как описано в данном документе, для идентификации антител, которые демонстрируют ослабленный транспорт через гематоэнцефалический барьер из мозга в пространство сосудов.
В одном воплощении всех аспектов, как описано в данном документе, FcRn выбран из человеческого FcRn, FcRn яванского макака, мышиного FcRn, крысиного FcRn, овечьего FcRn, собачьего FcRn, свиного FcRn, FcRn карликовой свиньи и кроличьего FcRn.
В одном воплощении всех аспектов, как описано в данном документе, бета-2-микроглобулин происходит из того же самого вида, что и FcRn.
В одном воплощении всех аспектов, как описано в данном документе, бета-2-микроглобулин происходит из другого вида, чем FcRn.
В одном воплощении родительское антитело содержит по меньшей мере один связывающий домен и по меньшей мере одну область Fc. В одном воплощении родительское антитело содержит два связывающих домена и две области Fc.
В одном воплощении родительское антитело содержит по меньшей мере один связывающий домен, который специфично связывается с мишенью, которая опосредует биологический эффект (в одном воплощении - лиганд, способный к связыванию с рецептором поверхности клетки, или рецептор поверхности клетки, способный к связыванию с лигандом) и опосредует передачу негативного или позитивного сигнала в клетку. В одном воплощении родительское антитело содержит по меньшей мере один связывающий домен, специфичный в отношении антигена, предназначенного для уменьшения уровня или устранения (в одном воплощении - антиген поверхности клетки или растворимый антиген), и по меньшей мере одну область Fc.
Антитела, специфично связывающиеся с мишенью, могут быть индуцированы у млекопитающих посредством многократных подкожных или внутрибрюшинных инъекций релеватного антигена (например, очищенный антиген, клетки или клеточные экстракты, содержащие такие антигены, или ДНК, кодирующая такой антиген) и, возможно, адъюванта.
В одном воплощении антитело представляет собой полноразмерное антитело.
В одном воплощении антитело представляет собой моноклональное антитело.
В одном воплощении родительское антитело представляет собой биспецифичное антитело.
В одном воплощении родительское антитело представляет собой химерное антитело.
В одном воплощении всех предыдущих аспектов pH представляет собой градиент от примерно pH 5,5 до примерно pH 8,8.
В общем, растворимый внеклеточный домен FcRn (SEQ ID NO: 33 для человеческого FcRn) с С-концевой меткой His-Avi (SEQ ID NO: 34) соэкспрессировали с β2-микроглобулином (SEQ ID NO: 35 для человеческого бета-2-микроглобулина) в клетках млекопитающих. Нековалентный комплекс FcRn-микроглобулин биотинилировали и загружали на сефарозу, дериватизированную стрептавидином.
В одном воплощении всех аспектов, как описано в данном документе, нековалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина связывается с твердой фазой.
В одном воплощении конъюгирование нековалентного комплекса с твердой фазой осуществляется посредством химического связывания через N-концевые и/или ε-аминогруппы (лизин), ε-аминогруппы разных лизинов, карбоксильные, сульфгидрильные, гидроксильные и/или фенольные функциональные группы аминокислотного остова антитела, и/или гидроксильные группы сахаров углеводной структуры антитела.
В одном воплощении нековалентный комплекс конъюгирован с твердой фазой посредством пары специфичного связывания. В одном воплощении нековалентный комплекс конъюгирован с биотином, и иммобилизация на твердую подложку осуществляется посредством иммобилизованного на твердой подложке авидина или стрептавидина.
Пара специфичного связывания (первый компонент/второй компонент) в одном воплощении выбрана из стрептавидина или авидина/биотина, антитела/антигена (см., например, Hermanson, G.T., et al., Bioconjugate Techniques, Academic Press (1996)), лектина/полисахарида, стероида/белка, связывающего стероид, гормона/рецептора гормона, фермента/субстрата, IgG/белка А и/или G, и т.д.
В принципе, в способах, описанных в данном документе, можно использовать любое буферное вещество.
Остатки Fc, критичные для взаимодействия мышиный Fc-мышиный FcRn, были идентифицированы посредством сайт-направленного мутагенеза (см, например, Dall'Acqua, W.F., et al. J. Immunol 169 (2002) 5171-5180). Остатки I253, H310, H433, N434 и H435 (нумерация EU согласно Kabat) участвуют в данном взаимодействии (Medesan, C., et al., Eur. J. Immunol. 26 (1996) 2533; Firan, M., et al., Int. Immunol. 13 (2001) 993; Kim, J.K., et al., Eur. J. Immunol. 24 (1994) 542). Обнаружили, что остатки I253, H310 и Н435 являются критичными для взаимодействия человеческого Fc с мышиным FcRn (Kim, J.K., et al., Eur. J. Immunol. 29 (1999) 2819). Остатки M252Y, S254T, T256E были описаны Dall'Acqua et al. как улучшающие связывание с FcRn посредством исследований взаимодействия белок-белок (Dall'Acqua, W.F., et al. J. Biol. Chem. 281 (2006) 23514-23524). Исследования комплекса человеческий Fc-человеческий FcRn показали то, что остатки I253, S254, Н435 и Y436 являются решающими для взаимодействия (Firan, М., et al., Int. Immunol. 13 (2001) 993; Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604). У Yeung, Y.A., et al. (J. Immunol. 182 (2009) 7667-7671) были описаны и проверены разные мутанты по остаткам 248-259, 301-317, 376-382 и 424-437.

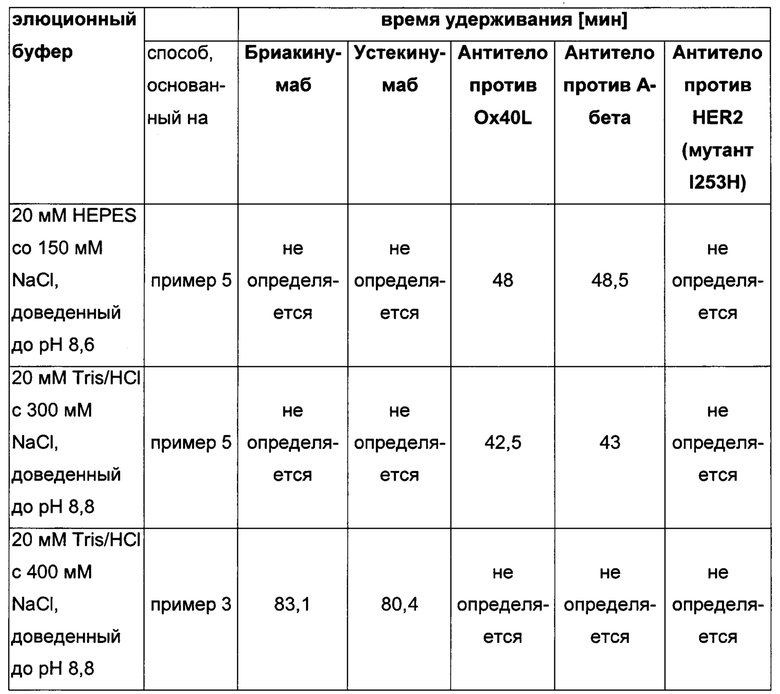
Термин мутант YTE обозначает тройной мутант M252Y/S254T/T256E.
В одном воплощении используется фармацевтически приемлемое буферное вещество, такое как, например, фосфорная кислота или ее соли, уксусная кислота или ее соли, лимонная кислота или ее соли, морфолин или его соли, 2-(N-морфолино)этансульфоновая кислота (MES) или ее соли, гистидин или его соли, глицин или его соли, трис(гидроксиметил)аминометан (TRIS) или его соли, (4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота (HEPES) или ее соли.
В одном воплощении буферное вещество выбрано из фосфорной кислоты или ее солей, или уксусной кислоты или ее солей, или лимонной кислоты или ее солей, или гистидина или его солей.
В одном воплощении буферное вещество имеет концентрацию от 10 мМ до 500 мМ. В одном воплощении буферное вещество имеет концентрацию от 10 мМ до 300 мМ. В одном воплощении буферное вещество имеет концентрацию от 10 мМ до 250 мМ. В одном воплощении буферное вещество имеет концентрацию от 10 мМ до 100 мМ. В одном воплощении буферное вещество имеет концентрацию от 15 мМ до 50 мМ. В одном воплощении буферное вещество имеет концентрацию примерно 20 мМ.
Типичный исходный раствор для положительного линейного градиента pH содержит в одном воплощении 20 мМ MES и 140 мМ NaCl, доведенные до pH 5,5.
Типичный конечный раствор для положительного линейного градиента pH содержит в одном воплощении 20 мМ TRIS и 140 мМ NaCl, доведенные до pH 8,8.
Во время применения градиента на колонку для аффинной хроматографии на основе FcRn наносится смесь исходного раствора и конечного раствора, при этом положительный линейный градиент начинается со 100% исходного раствора (т.е. чистого исходного раствора), затем доля исходного раствора уменьшается от 100% до 0%, и доля конечного раствора увеличивается от 0% до 100%, таким образом, что после применения положительного линейного градиента pH на колнку наносится 100% конечного раствора.
В одном воплощении исходный и конечный раствор содержит дополнительную соль. В одном воплощении дополнительная соль выбрана из хлорида натрия, сульфата натрия, хлорида калия, сульфата калия, цитрата натрия или цитрата калия. В одном воплощении растворы содержат от 50 мМ до 1000 мМ дополнительной соли. В одном воплощении растворы содержат от 50 мМ до 750 мМ дополнительной соли. В одном воплощении растворы содержат от 50 мМ до 500 мМ дополнительной соли. В одном воплощении раствор содержит от 50 мМ до 750 мМ дополнительной соли. В одном воплощении раствор содержит от примерно 140 мМ до примерно 400 мМ дополнительной соли.
В одном воплощении исходный и конечный раствор содержит хлорид натрия. В одном воплощении исходный и конечный раствор содержит от примерно 140 мМ до примерно 400 мМ хлорида натрия.
Обнаружили, что вид соли и буферного вещества влияет на время удерживания и разрешение. Может быть определена оптимальная концентрация соли для связывания антител с FcRn (140 мМ NaCl). Если концентрация соли выше (400 мМ), связывание с FcRn уменьшается из-за мешающего влияния взаимодействий зарядов посредством увеличения ионной силы раствора, и получается более короткое время удерживания.
Таким образом, в способе, описанном в данном документе, растворы, используемые на стадии а) и стадии б), а также градиент, нанесенный на стадии а) и стадии б), а также загрузка колонки на стадии а) и стадии б), а также размер колонки и количество аффинного хроматографического вещества на основе FcRn на стадии а) и стадии б), а также плотность аффинного лиганда на основе FcRn в аффинном хроматографическом веществе на основе FcRn на стадии а) и стадии б), а также конъюгирование FcRn с твердой фазой на стадии а) и стадии б), а также природа b2m и FcRn на стадии а) и стадии б) являются аналогичными или даже идентичными. Таким образом, в способе, как описано в данном документе, аффинная хроматография на основе FcRn на стадии а) и стадии б) проводится при идентичных условиях, за исключением концентрации соли, которая отличается между стадией а) и стадией б). В одном воплощении вторая концентрация соли больше, чем первая концентрация соли. В одном воплощении вторая концентрация соли по меньшей мере в два раза больше первой концентрации соли.
Как можно видеть из Фиг. 7, количество нанесенного антитела демонстрирует линейную корреляцию с площадью под кривой элюированного пика.
Восемь антител анализировали в виде полного антитела и после расщепления ферментом IdeS. Расщепление контролировали посредством SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и аналитической SEC (гель-фильтрация). Область Fc и Fab антитела разделяли препаративной SEC.
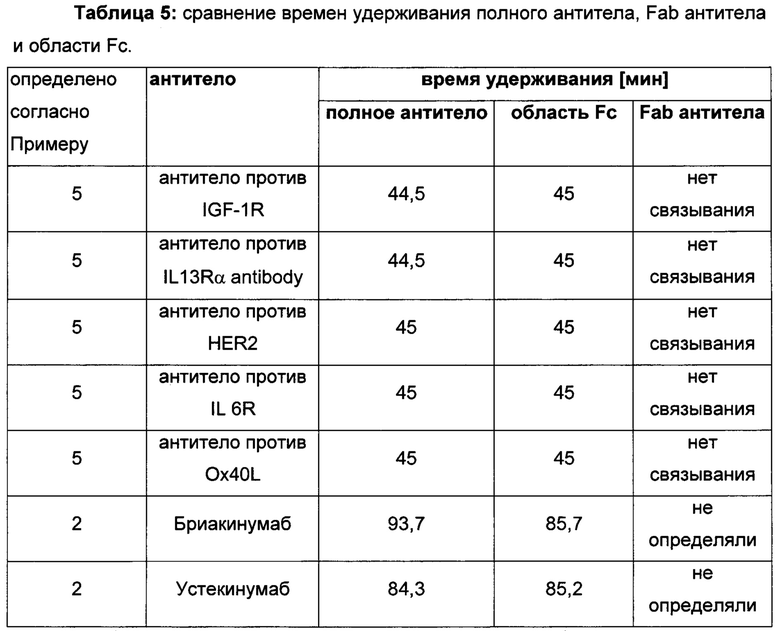
В общем, время удерживания антител, имеющих область Fc дикого типа (IgG1 или IgG2, или IgG4), варьирует от 45 до 49 мин (тестировали с 35 терапевтическими антителами против 36 антигенов, данные не показаны) при условиях Примера 5. При использовании условий Примера 2 время удерживания увеличивается до примерно 85 мин, так как градиент длиннее.

В общем, время удерживания в способах и применениях, как описано в данном документе, зависит от крутизны градиента pH и используемой концентрации соли. При использовании в качестве контроля антитела дикого типа, на более слабое связывание указывает более короткое время удерживания (более ранняя элюция), тогда как на более сильное связывание указывает более длительное время удерживания (более поздняя элюция).
Разные мутанты в области Fc IgG ведут себя по-разному на колонке FcRn, демонстрируя модифицированные времена удерживания.
Например, мутант YTE антитела против IGF-1R демонстрирует повышенное время удерживания (см. Фиг. 8).
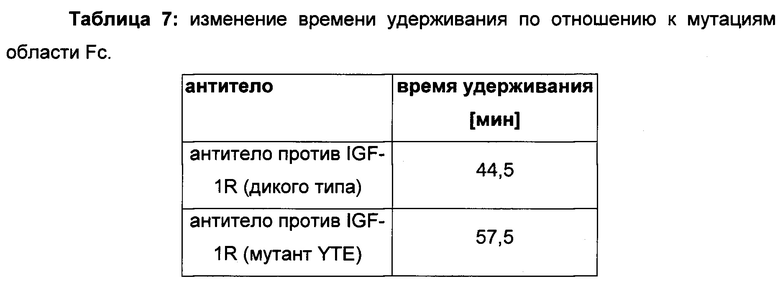
Термин «мутант YTE» обозначает тройного мутанта M252Y/S254T/T256E.
Одним аспектом, как описано в данном документе, является способ определения относительного времени полужизни антитела in vivo, включающий следующие стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом антитело имеет относительное время полужизни in vivo, которое уменьшено по сравнению со стандартным/природным антителом подкласса IgG1, IgG3 или IgG4, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
Одним аспектом, как описано в данном документе, является способ, как описано в данном документе, определения относительного времени полужизни антитела in vivo, где данный способ включает следующие этапы:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом антитело имеет относительное время полужизни in vivo, которое уменьшено по сравнению со стандартным/природным антителом подкласса IgG1, IgG3 или IgG4, если время удерживания, определенное на стадии а), и время удерживания, определенное на стадии б), являются существенно отличными.
Одним аспектом, как описано в данном документе, является способ определения увеличения или уменьшения времени полужизни варианта антитела in vivo относительно его родительского антитела, включающий следующие этапы:
а) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом время полужизни in vivo варианта антитела относительно его родительского антитела увеличивается, если i) время удерживания варианта антитела, определенное на стадии а), больше/продолжительнее, чем время удерживания его родительского антитела, определенное на стадии а), и ii) время удерживания варианта антитела, определенное на стадии а), и время удерживания варианта антитела, определенное на стадии б), являются по существу одинаковыми,
при этом время полужизни варианта антитела in vivo относительно его родительского антитела уменьшается, если i) время удерживания варианта антитела, определенное на стадии а), меньше/короче, чем время удерживания его родительского антитела, определенное на стадии а), и ii) время удерживания варианта антитела, определенное на стадии а), и время удерживания варианта антитела, определенное на стадии б), являются по существу одинаковыми.
Одним аспектом, как описано в данном документе, является применение способа, как описано в данном документе, для определения увеличения или уменьшения времени полужизни варианта антитела in vivo относительно его родительского антитела, где данный способ включает следующие этапы:
а) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания варианта антитела и его родительского антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли,
при этом время полужизни in vivo варианта антитела относительно его родительского антитела увеличивается, если i) время удерживания варианта антитела, определенное на стадии а), больше/продолжительнее, чем время удерживания его родительского антитела, определенное на стадии а), и ii) время удерживания варианта антитела, определенное на стадии а), и время удерживания варианта антитела, определенное на стадии б), являются по существу одинаковыми,
при этом время полужизни in vivo варианта антитела относительно его родительского антитела уменьшается, если i) время удерживания варианта антитела, определенное на стадии а), меньше/короче, чем время удерживания его родительского антитела, определенное на стадии а), и ii) время удерживания варианта антитела, определенное на стадии а), и время удерживания варианта антитела, определенное на стадии б), являются по существу одинаковыми.
Обнаружили, что антитела, которые демонстрировали позднюю элюцию из колонки на основе FcRn, т.е. которые имели более длительное время удерживания на колонке на основе FcRn, и которые не демонстрировали взаимодействия Fab антитела-FcRn, имели более продолжительное время полужизни in vivo (см. Пример 6).
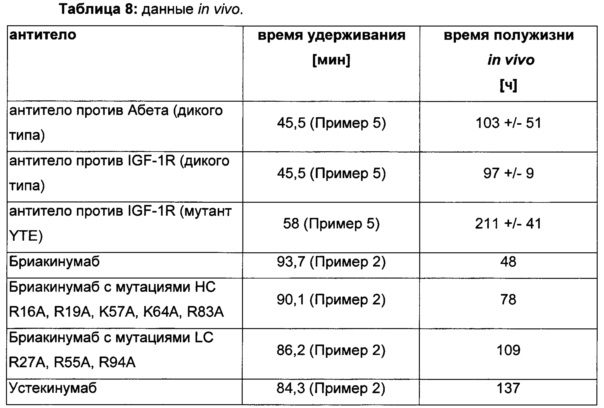
Термин «мутант YTE» обозначает тройного мутанта M252Y/S254T/T256E.
Одним аспектом, как описано в данном документе, является применение способа, как описано в данном документе, для определения времени полужизни антитела in vivo.
Набор экспериментов in vitro и in vivo, проведенных с IgG дикого типа и вариантами IgG с мутациями YTE в области Fc, позволил продемонстрировать полуколичественную корреляцию данных аффинной хроматографии на основе FcRn с данными в фармакокинетических исследованиях in vivo с использованием трансгенных мышей в отношении человеческого FcRn (Spiekerman, G.M., et al. J. Exp. Med. 196 (2002) 303-310; Dall'Acqua, W.F., et al., J. Biol. Chem. 281 (2006) 23514-23524). Мутация YTE приводит к значительно увеличенному времени полужизни и более медленному клиренсу в плазме. Более продолжительное время полужизни in vivo соответствовало более длительному времени удерживания в хроматографии на основе FcRn. Недавно показали, что вариант трастузумаба с генетически модифицированным Fc, характеризующийся увеличенным временем полужизни, имеет увеличенное связывание in vitro с FcRn, измеренное проточной цитометрией (Petkova, S.B., et al., Int. Immunol. 18 (2006) 1759-1769). Было показано, что вариант антитела IgG1 против VEGF (фактор роста эндотелия сосудов) бевацизумаба с 11-кратно улучшенной аффинностью в отношении FcRn имеет пятикратно увеличенное время полужизни у трансгенных мышах с человеческим FcRn и в три раза более продолжительное время полужизни у яванских макаков (Zalevsky, J., et al., Nat. Biotechnol. 28 (2010) 157-159).
Показали, что формат антитела не имел влияния на связывание с колонкой на основе FcRn. Это было показано для формата «выступ во впадину» и для нескольких форматов биспецифичных антител. Таким образом, описанный здесь способ может использоваться для оценки новых форматов антител.
В одном воплощении комплекс является монобиотинилированным.
В одном воплощении хроматографическое вещество, содержащее нековалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина в качестве лиганда, имеет в описанных в данном документе способах и применениях стабильность по меньшей мере 100 циклов. Циклом является градиент pH от первого значения pH до второго значения pH соответствующего способа или применения, при этом для регенерации вещества не требуется дополнительного изменения условий, чем конечные условия способа или применения. Таким образом, в одном воплощении циклом является градиент pH от примерно значения pH 5,5 до примерно значения pH 8,8.
ж.ii) Элюция антитела и его области Fc линейным положительным градинтом pH при той же самой концентрации соли
В данном документе описан способ, включающий следующие две стадии:
а) определение времени удерживания антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли,
б) определение времени удерживания области Fc антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли.
С использованием данного способа возможно определение присутствия взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn в простом хроматографическом способе посредством сравнения времен удерживания антитела и его области Fc. Область Fc, например, может быть получена ферментативным отщеплением ферментом IdeS или папаином, или может быть получена рекомбинантно. Это является важным, так как взаимодействия Fab антитела-FcRn влияют на время полужизни антитела in vivo.
Для пар антитело/область Fc антитела существуют разные связи в отношении их времен удерживания на колонке для аффинной хроматографии на основе FcRn и, подобным образом, в отношении их взаимодействия с FcRn:
1) антитело и его область Fc имеют по существу одинаковое время удерживания: в данном случае на время полужизни антитела in vivo не влияет взаимодействие Fab антитела-FcRn,
2) антитело и его область Fc имеют разные времена удерживания, и время удерживания области Fc короче, чем время удерживания антитела: в данном случае на время полужизни антитела in vivo влияет взаимодействие Fab антитела-FcRn.
В случае если рассматриваемое антитело представляет собой вариант антитела, следует рассматривать дополнительные аспекты: в варианте антитела взаимодействие Fc антитела-FcRn, а также взаимодействие Fab антитела-FcRn может быть изменено из-за введенных модификаций по отношению к родительскому антителу.
Таким образом, существуют следующие возможные связи между родительским антителом, вариантом антитела и их соответствующими областями Fc (см. Фиг. 10):
1) родительское антитело (1), вариант антитела (3) и соответствущие области Fc (2,4) имеют по существу одинаковое время удерживания: в данном случае время полужизни варианта антитела in vivo i) не подвергается влиянию взаимодействия Fab антитела-FcRn и ii) соответствует времени полужизни родительского антитела in vivo (см. Фиг. 10А),
2) вариант антитела (3) и его область Fc (4) имеют разные времена удерживания, время удерживания области Fc варианта антитела короче, чем время удерживания варианта антитела, и родительское антитело (1), область Fc родительского антитела (2) и область Fc варианта антитела (4) имеют по существу одинаковое время удерживания: в данном случае время полужизни варианта антитела in vivo i) подвергается влиянию взаимодействия Fab антитела-FcRn, и ii) короче, чем время полужизни родительского антитела in vivo (см. Фиг. 10Б),
3) родительское антитело (1) и вариант антитела (3) имеют разные времена удерживания, время удерживания области Fc варианта антитела (4) по существу такое же, как и время удерживания варианта антитела (3), и время удерживания варианта антитела дольше, чем время удерживания родительского антитела (1): в данном случае время полужизни варианта антитела in vivo i) не подвергается влиянию взаимодействия Fab антитела-FcRn и ii) дольше, чем время полужизни родительского антитела in vivo (см. Фиг. 10В),
4) родительское антитело (1) и вариант антитела (3) имеют разные времена удерживания, время удерживания области Fc варианта антитела (4) является по существу таким же, как и время удерживания варианта антитела (3), и время удерживания варианта антитела (3) короче, чем время удерживания родительского антитела (1): в данном случае время полужизни варианта антитела in vivo i) не подвергается влиянию взаимодействия Fab антитела-FcRn и ii) короче, чем время полужизни родительского антитела in vivo,
5) родительское антитело (1) и вариант антитела (3) имеют разные времена удерживания, время удерживания области Fc варианта антитела (4) отличается от времени удерживания варианта антитела (3) и также отличается от времени удерживания родительского антитела (1) и его области Fc (2), и время удерживания области Fc варианта антитела (4) находится между временем удерживания варианта антитела (3) и родительского антитела (4): в данном случае время полужизни варианта антитела in vivo i) подвергается влиянию взаимодействия Fab антитела-FcRn и ii) отличается от времени полужизни родительского антитела in vivo (см. Фиг. 10Г).
В одном случае контрольное антитело представляет собой антитело, которое имеет по существу такое же время удерживания, что и его область Fc после отщепления IdeS или отщепления папаином.
Как описано выше, взаимодействие Fab антитела-FcRn может иметь влияние на время полужизни антитела in vivo. Также, как описано выше, взаимодействие Fc антитела-FcRn может иметь влияние на время полужизни антитела in vivo. Таким образом, должны учитываться оба взаимодействия.
Например, Ropeenian и Akilesh (Nat. Rev. Immunol. 7 (2007) 715-725) описывают то, что, например, вариант Asn434Ala (N434A) гуманизированного антитела IgG1 hu4D5 (герцептин; Genentech; моноклональное антитело, специфичное в отношении ERBB2) и вариант с тройной заменой Thr307Ala/Asn434Ala/Glu380Ala (T307A/N434A/Q380A) связываются с человеческим FcRn при pH 6,0 с 3-кратно и 12-кратно большей аффинностью соответственно, чем антитело hu4D5 дикого типа. Неожиданно то, что у трансгенных по FcRn гуманизированных мышей время полужизни данных двух вариантов антител были по существу эквивалентными. Данное расхождение согласно Ropeenian и Akilesh может быть объяснено повышенной аффинностью варианта с тремя заменами в отношении FcRn при pH 7,4. Мутации области Fc, которые улучшают аффинность связывания при pH 7,4, а также при pH 6,0, действительно могут ускорять клиенс антитела in vivo, скорее чем продлевать его время полужизни.
Таким образом, следует дополнительно рассматривать, если время удерживания области Fc варианта антитела дольше, чем критическое время удерживания. Не будучи связанными данной теорией, это приводит к тому эффекту, что взаимодействие при pH 7,4 усиливается так значительно, что диссоциация комплекса антитело-FcRn при pH 7,4 уменьшается, приводя к усиленной деградации антитела (см. Фиг. 11).
Одним аспектом настоящего изобретения является способ отбора антитела, включающий следующие стадии (см. Фиг. 12):
а) определение времени удерживания i) антитела (3), ii) области Fc антитела (4), iii) контрольного антитела (1), iv) области Fc контрольного антитела (2) и v) контрольного антитела с мутацией N434A в области Fc (5) на колонке для аффинной хроматографии на основе FcRn с использованием одинаковых условий элюции,
б) отбор антитела, для которого
б-i) контрольное антитело (1), вариант антитела (3) и соответствующие области Fc (2, 4) имеют по существу одинаковое время удерживания, и время удерживания антитела короче, чем время удерживания контрольного антитела с мутацией N434A в области Fc (5), и, посредством этого, отбирая антитело, время полужизни которого in vivo i) не подвергается влиянию взаимодействия Fab антитела-FcRn и ii) соответствует времени полужизни in vivo родительского антитела (см. Фиг. 12А),
б-ii) антитело (3) и его область Fc (4) имеют разные времена удерживания, время удерживания области Fc антитела (4) короче, чем время удерживания антитела (3), и является таким же или более продолжительным, чем время удерживания контрольного антитела (1) или его области Fc (2), время удерживания контрольного антитела (1), области Fc контрольного антитела (2) и антитела (3) короче, чем время удерживания контрольного антитела с мутацией N434A в области Fc (5), и, посредством этого, отбор антитела, время полужизни in vivo которого i) подвергается влиянию взаимодействия Fab антитела-FcRn и ii) короче, чем время полужизни in vivo контрольного антитела (см. Фиг. 12Б),
б-iii) контрольное антитело (1) и антитело (3) имеют разные времена удерживания, время удерживания области Fc антитела (4) является по существу таким же, как и время удерживания антитела (3), время удерживания антитела (3) дольше, чем время удерживания контрольного антитела (1), и время удерживания антитела (3) короче, чем время удерживания контрольного антитела с мутацией N434A в области Fc (5), и, посредством этого, отбор антитела, время полужизни in vivo которого i) не подвергается влиянию взаимодействия Fab антитела-FcRn и ii) дольше, чем время полужизни in vivo контрольного антитела (см. Фиг. 12В),
б-iv) контрольное антитело (1) и антитело (3) имеют разные времена удерживания, время удерживания области Fc антитела (4) является по существу таким же, как и время удерживания антитела (3), время удерживания антитела (3) короче, чем время удерживания контрольного антитела (1), и время удерживания антитела (3) короче, чем время удерживания контрольного антитела с мутацией N434A в области Fc (5) и, посредством этого, отбор антитела, время полужизни in vivo которого i) не подвергается влиянию взаимодействия Fab антитела-FcRn и ii) короче, чем время полужизни in vivo контрольного антитела (см. Фиг. 12Г),
б-v) контрольное антитело (1) и антитело (3) имеют разные времена удерживания, время удерживания области Fc антитела (4) отличается от времени удерживания антитела (3) и также отличается от времени удерживания контрольного антитела (1) и его области Fc (2), время удерживания области Fc антитела (3) находится между временем удерживания антитела (3) и контрольного антитела (1), и время удерживания антитела (3) короче, чем время удерживания контрольного антитела с мутацией N434A в области Fc (5), и, посредством этого, отбор антитела, время полужизни in vivo которого i) подвергается влиянию взаимодействия Fab антитела-FcRn и ii) отличается от времени полужизни in vivo контрольного антитела (см. Фиг. 12Д).
В одном воплощении элюция осуществляется положительным линейным градиентом pH при постоянной концентрации соли или посредством применения линейного солевого градиента при постоянном значении pH.
В одном воплощении антитело представляет собой вариант родительского антитела, и контрольное антитело представляет собой родительское антитело. В одном воплощении вариант антитела имеет замены аминокислот в области Fab антитела и/или области Fc антитела.
ж.iii) Элюция солевым градиентом
В данном документе описан способ, включающий следующие стадии:
а) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn при первом значении pH с элюцией солевым градиентом,
б) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn при втором значении pH с элюцией солевым градиентом.
Обнаружили, что для определения того, присутствуют ли или нет взаимодействия Fab антитела-FcRn в комплексе Fc антитела-FcRn, помимо элюции градиентом pH при постоянной концентрации соли, также может быть использована элюция солевым градиентом при постоянном значении pH.
Как уже описывалось выше, взаимодействие Fab антитела-FcRn представляет собой вторичное взаимодействие, которое устанавливается, если оно вообще присутствует, после образования комплекса Fc антитела-FcRn.
Оба взаимодействия, т.е. взаимодействие Fc антитела-FcRn и Fab антитела-FcRn, представляют собой опосредованные зарядом нековалентные взаимодействия.
Одним аспектом, как описано в данном документе, является способ определения присутствия взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, включающий следующие стадии:
а) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn при первом значении pH с элюцией солевым градиентом,
б) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn при втором значении pH с элюцией солевым градиентом,
при этом присутствие взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo, определяется, если отношение времен удерживания антитела и контрольного антитела, определенных на стадии а), существенно отличается от отношения времен удерживания антитела и контрольного антитела, определенных на стадии б).
В одном воплощении первое значение pH составляет 5,5. В одном воплощении второе значение pH составляет 8,8.
В одном воплощении солевой градиент на стадии а) и стадии б) являются идентичными.
В одном воплощении солевой градиент представляет собой градиент хлорида натрия.
В одном воплощении солевой градиент составляет от 0 мМ до 250 мМ соли.
з) Бевацизумаб и мутант бевацизумаба
Другая молекула без заряженного участка в CDR выбрана для создания положительно заряженного участка в LC-CDR для подтверждения описанных выше данных о том, что положительный заряд в данном положении, в общем, влияет на аффинность связывания антител с FcRn.
Бевацизумаб был выбран, так как он имеет лишь маленький заряд в LC-CDR. Три основных аминокислотных остатка, которые были идентифицированы с использованием бриакинумаба, представляют собой остатки аргинина R27, R55 и R94. В аминокислотной последовательности бевацизумаба аспарагиновая кислота D27, лейцин L54 и треонин Т93 заменены на остатки лизина с созданием положительно заряженного участка (см. Фиг. 14).
Хроматограмма бевацизумаба дикого типа и мутанта бевацизумаба аффинной хроматографии на основе FcRn показана на Фиг. 9, и соответствующие времена удерживания перечислены в следующей Таблице 9.
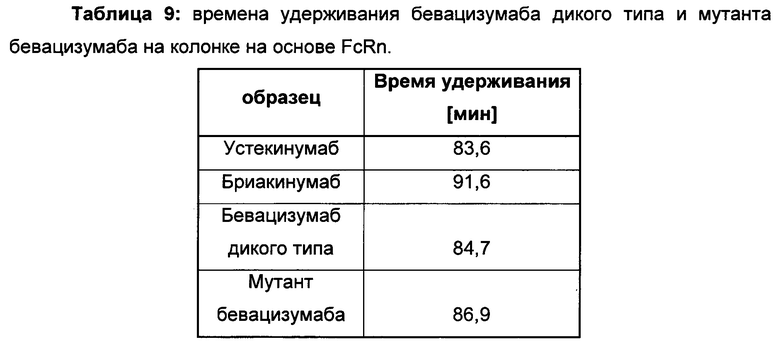
Бевацизумаб дикого типа имеет время удерживания 84,7 минут, тогда как мутант бевацизумаба элюируется через 86,9 минут. Таким образом, положительно заряженный участок в Fv бевацизумаба вызывает сдвиг времени удерживания на 2,2 минуты. Данные результаты указывают на то, что заряд в Fv IgG1, в общем, влияет на аффинность связывания с FcRn, особенно подвергается влиянию диссоциация от FcRn.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Распределение заряда и pH-зависимый нетто-заряд. Изопотенциальные поверхности белков, протонированных при pH 7,4, и с нанесением контура при 2 kBT/е; черный: положительный/отрицательный. (а) Бриакинумаб. Легкая цепь показана светло-серым, тяжелая цепь показана более темным серым. Ракурсы среднего и правого изображений связаны с ракурсом на левой панели посредством вращения по вертикальной и горизонтальной оси соответственно. (б) Устекинумаб. Легкие и тяжелые цепи закрашены светло- и темно-серым соответственно. Ракурсы являются идентичными ракурсам на (а). (в) Изопотенциальная поверхность, нанесенная в виде контура при 2 kBT/е, модели гомологии человеческого FcRn в комплексе с человеческим β2 микроглобулином (β2m). Домен Fc показан для ясности. (г) Рассчитанный на основе последовательности нетто-заряд относительно pH бриакинумаба и устекинумаба. Структуры белка получали с использованием DiscoveryStudio Pro.
Фиг. 2. рН-зависимое взаимодействие FcRn-IgG. Хроматограммы одиннадцати вариантов IgG аффинной хроматографии на основе FcRn нормировали по интенсивности для ясности. Представление молекулярной поверхности структурных моделей, протонированных при pH 7,4, накладывали на изопотенциальные поверхности с нанесением контура при 2 kBT/е. Ракурс идентичен правой панели на Фиг. 1а и сосредоточен на областях CDR. Вторая горизонтальная ось указывает рН элюции, интерполированный из измерений рН вне системы.
Фиг. 3. Эффект pH элюции на колонке на основе FcRn на фармакокинетику в трансгенных мышах с человеческим FcRn. Антитела вводили в виде одиночной в.в. (внутривенной) болюсной инъекции 10 мг/кг 6 животным на группу. Точки данных представляют собой среднее значение плюс/минус стандартное отклонение. (а) Кривые уровня в крови бриакинумаба (оранжевый), устекинумаба (зеленый), mAb8 (пурпурный) и mAb9 (голубой). (б) Корреляция между временем полужизни в конечной фазе и pH элюции на колонке на основе FcRn.
Фиг. 4. Моделирование молекулярной динамики моделей FcRn-IgG.
(а) Конформация в начале моделирования. Пунктирная линия показывает расстояние между двумя типичными аминокислотами в области Fv и в FcRn, которые достигают друг друга во время моделирования MD (молекулярная динамика), как показано на панели (в). Цвета являются идентичными Фиг. 1. (б) Конформация в конце моделирования (t равно 100 нс). Прямоугольник показывает часть молекулы, показанную на (в). (в) Подробный вид взаимодействия между доменами FcRn и Fv. Заметьте, что взаимодействующие остатки каркаса, CDR и FcRn в бриакинумабе и устекинумабе являются отличными. (г) Расстояние между остатками 245 (FcRn) и 100 (LC устекинумаба) и 29 (LC бриакинумаба) соответственно по ходу моделирования. (д) Энергии взаимодействия в конце моделирования (средние и стандартные отклонения конформаций в 96, 97, 98, 99 и 100 нс). «VDW» и «Электростатические» обозначают вандерваальсовы и электростатические вклады во взаимодействие FcRn-Fab соответственно. Структуры белков получали с использованием PyMol™ (Schrodinger LLC).
Фиг. 5. Выравнивание последовательности легкой и тяжелой цепей бриакинумаба и устекинумаба. Области VH и VL показаны курсивом; CDR отмечены звездочкой (*); решетка (#) обозначает аминокислоты в непосредственной близости (меньше 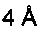 ) к FcRn в исходной структуре. Символ «
) к FcRn в исходной структуре. Символ « » отмечает остаток, мутированный до Cys, для установления дисульфидного мостика с FcRn для целей MD.
» отмечает остаток, мутированный до Cys, для установления дисульфидного мостика с FcRn для целей MD.
Фиг. 6. Зависимость от соли времен удерживания бриакинумаба и устекинумаба на аффинной колонке на основе FcRn. Бриакинумаб и устекинумаб подвергали колоночной хроматографии на основе FcRn с элюцией градиентом pH в присутствии возрастающих количеств NaCl. Данные аппроксимируются к функции обратного квадратного корня для объяснения эффекта экранирования заряда посредством растворенной соли. Времена удерживания бриакинумаба уменьшаются с 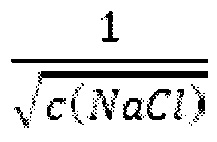 (r2 равно 0,898), тогда как времена удерживания устекинумаба по существу не подвергаются влиянию.
(r2 равно 0,898), тогда как времена удерживания устекинумаба по существу не подвергаются влиянию.
Фиг. 7. Линейность между количеством введенного антитела и площадью под кривой для хроматографии с использованием колонки на основе FcRn, как описано в данном документе.
Фиг. 8. Хроматограмма антитела дикого типа против IGF-1R и мутанта YTE на колонке на основе FcRn, как описано в данном документе.
Фиг. 9. Хроматограмма авастина дикого типа и мутанта авастина аффинной хроматографии на основе FcRn.
Фиг. 10. Схема изменения времени удерживания на колонке для аффинной хроматографии на основе FcRn, в зависимости от взаимодействий антитело-FcRn области Fc и Fab антитела; 1: родительское антитело, 2: область Fc родительского антитела, 3: вариант антитела, 4: область Fc варианта антитела; сплошная линия: полное антитело (Fab антитела плюс область Fc), пунктирная линия: только область Fc;
А: область Fc, подобная области Fc дикого типа, нет взаимодействия Fab антитела-FcRn;
Б: область Fc, подобная области Fc дикого типа, взаимодействие Fab антитела-FcRn;
В: генетически модифицированная область Fc с улучшенным связыванием с FcRn, нет взаимодействия Fab антитела-FcRn;
Г: генетически модифицированная область Fc с улучшенным связыванием с FcRn, взаимодействие Fab антитела-FcRn.
Фиг. 11. Схема, показывающая генетически модифицированное антитело с улучшенным связыванием с FcRn, взаимодействие Fab антитела-FcRn, но уменьшенное время полужизни in vivo, так как взаимодействие антитело-FcRn приводит к улучшенному клиренсу (время удерживания, превышающее критическое время удерживания).
Фиг. 12. Схема изменения времени удерживания на колонке для аффинной хроматографии на основе FcRn, в зависимости от взаимодействий антитело-FcRn области Fc и Fab антитела; 1: контрольное антитело, 2: область Fc контрольного антитела, 3: антитело, 4: область Fc антитела; сплошная линия: полное антитело (Fab антитела плюс область Fc), пунктирная линия: только область Fc.
Фиг. 13. Зависимость времени удерживания аффинной хроматографии на основе FcRn от концентрации соли и взаимодействия Fab антитела-FcRn.
Фиг. 14. Выравнивание последовательности вариабельных доменов легкой цепи бевацизумаба и варианта бевацизумаба. Идентичные и аналогичные аминокислоты показаны серым; CDR отмечены звездочкой (*).
Фиг. 15. Взаимодействие с IL-12 бриакинумаба, устекинумаба и mAb 1-6; 1: бриакинумаб, 2: устекинумаб, 3: mAb 1, 4: mAb 2, 5: mAb 3, 6: mAb 4, 7: mAb 5, 8: mAb 6.
Фиг. 16. Взаимодействие с IL-12 бриакинумаба, устекинумаба и mAb 7-10; 1: бриакинумаб, 2: устекинумаб, 3: mAb 7, 4: mAb 8, 5: mAb 9, 6: mAb 10.
Конкретные воплощения
1. Способ отбора антитела, включающий следующие стадии:
i) определение первого времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом рН в присутствии первой концентрации соли, и определение второго времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли или
ii) определение первого времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при первом значении pH, и определение второго времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при втором значении pH или
iii) определение для антитела и контрольного антитела значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли, или
iv) определение для антитела и контрольного антитела значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом, или
v) определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH, или
vi) определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при высоком значении рН, или
vii) определение для антитела и его области Fc значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли, или
viii) определение для антитела и его области Fc значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при высоком значении pH,
и, посредством этого, отбор
а) антитела, которое имеет первое время удерживания, которое по существу является таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое по существу является таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет по существу такое же время удерживания, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD области Fc самое большее в 10 раз, и которое имеет время удерживания, которое по существу является таким же, как и время удерживания его области Fc.
2. Способ отбора антитела, включающий следующие стадии:
- определение первого времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии первой концентрации соли, и определение второго времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии второй концентрации соли, и
- отбор
а) антитела, которое имеет первое время удерживания, которое по существу является таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое по существу является таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое по существу является таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
3. Способ отбора антитела, включающий следующие стадии:
- определение первого времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при первом значении pH, и определение второго времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при втором значении pH, и
- отбор
а) антитела, которое имеет первое время удерживания, которое по существу является таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое по существу является таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
4. Способ отбора антитела, включающий следующие стадии:
- определение для антитела и контрольного антитела значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли, и
- отбор
а) антитела, которое имеет первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения КD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое по существу является таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
5. Способ отбора антитела, включающий следующие стадии:
- определение для антитела и контрольного антитела значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом, и
- отбор
а) антитела, которое имеет первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
6. Способ отбора антитела, включающий следующие стадии:
- определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH и
- отбор
а) антитела, которое имеет первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
7. Способ отбора антитела, включающий следующие стадии:
- определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при высоком значении pH и
- отбор
а) антитела, которое имеет первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
8. Способ отбора антитела, включающий следующие стадии:
- определение для антитела и его области Fc значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH в присутствии высокой концентрации соли, и
- отбор
а) антитела, которое имеет первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
9. Способ отбора антитела, включающий следующие стадии:
- определение для антитела и его области Fc значения KD при pH 6 с использованием поверхностного плазмонного резонанса, и определение времени удерживания антитела и его области Fc на колонке для аффинной хроматографии на основе FcRn с элюцией линейным солевым градиентом при высоком значении pH и
- отбор
а) антитела, которое имеет первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc.
10. Способ по любому из воплощений 1-10, где данный способ предназначен для отбора антитела, которое не имеет взаимодействия Fab антитела-FcRn, влияющего на время полужизни антитела in vivo.
11. Способ по любому из воплощений 1, 6, 7, 8 и 9, где
- данный способ предназначен для отбора антитела, которое имеет относительное время полужизни in vivo, которое увеличивается по сравнению с антителом подкласса IgG1, IgG3 или IgG4, и
- далее определяется время удерживания контрольного антитела или контрольной области Fc и
- посредством этого, отбор
а) антитела, которое имеет первое время удерживания, которое дольше, чем первое время удерживания контрольного антитела, и первое время удерживания, которое является по существу таким же, как и второе время удерживания, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое дольше, чем время удерживания контрольного антитела, или
в) антитела, которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, или которое дольше, чем время удерживания контрольного антитела, или
г) антитела, которое имеет значение KD, которое отличается от значения KD его области Fc самое большее в 10 раз, и которое имеет время удерживания, которое является по существу таким же, как и время удерживания его области Fc, и которое дольше, чем время удерживания контрольного антитела.
12. Способ по любому из воплощений 1, 6, 7, 8 и 9, где
- данный способ предназначен для определения относительного увеличения или уменьшения времени полужизни антитела in vivo по сравнению с контрольным антителом, и
- дополнительно определяется время удерживания контрольного антитела или контрольной области Fc и,
- дополнительно определяется время удерживания области Fc IgG с мутацией N434A и
- посредством этого, отбор
а) антитела, которое имеет первое время удерживания, которое дольше, чем первое время удерживания контроля, который имеет первое время удерживания, и второе время удерживания, которые являются по существу одинаковыми, и которое имеет первое время удерживания, которое короче, чем время удерживания области Fc с мутацией N434A и, посредством этого, отбор антитела с относительно увеличенным временем полужизни in vivo, или
б) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое дольше, чем время удерживания контрольного антитела, и которое имеет первое время удерживания, которое короче, чем время удерживания области Fc с мутацией N434A и, посредством этого, отбор антитела с относительно увеличенным временем полужизни in vivo, или
в) антитела, которое имеет первое время удерживания, которое короче, чем первое время удерживания контрольного антитела, и которое имеет первое время удерживания и второе время удерживания, которые являются по существу одинаковыми, и, посредством этого, отбор антитела с относительно уменьшенным временем полужизни in vivo, или
г) антитела, которое имеет значение KD, которое отличается от значения KD контрольного антитела самое большее в 10 раз, и которое имеет время удерживания, которое короче, чем время удерживания контрольного антитела, и, посредством этого, отбор антитела с относительно увеличенным временем полужизни in vivo.
13. Способ по любому из воплощений 1, 2, 4, 6, 8 и 10-12, где (положительный) линейный градиент pH идет от примерно pH 5,5 до примерно pH 8,8.
14. Способ по любому из воплощений 1-13, где соль выбрана из хлорида натрия, сульфата натрия, хлорида калия, сульфата калия, цитрата натрия или цитрата калия.
15. Способ по любому из воплощений 1-14, где соль представляет собой хлорид натрия.
16. Способ по любому из воплощений 1, 2 и 10-15, где первая концентрация соли составляет от 50 мМ до 200 мМ.
17. Способ по любому из воплощений 1, 2 и 10-16, где первая концентрация соли составляет примерно 140 мМ.
18. Способ по любому из воплощений 1, 2 и 10-17, где вторая концентрация соли составляет от 300 мМ до 600 мМ.
19. Способ по любому из воплощений 1, 2 и 10-18, где вторая концентрация соли составляет примерно 400 мМ.
20. Способ по любому из воплощений 1, 3, 5, 7 и 9-19, где линейный солевой градиент идет от 0 мМ соли до 500 мМ соли.
21. Способ по любому из воплощений 1, 3, 5, 7 и 9-20, где линейный солевой градиент идет от 0 мМ соли до 250 мМ соли.
22. Способ по любому из воплощений 1, 3 и 10-21, где первое значение pH составляет примерно 5,5.
23. Способ по любому из воплощений 1, 3 и 10-22, где второе значение рН составляет примерно 7,4.
24. Способ по любому из воплощений 1, 4, 8 и 10-23, где высокая концентрация соли составляет от 250 мМ до 600 мМ.
25. Способ по любому из воплощений 1, 4, 8 и 10-24, где высокая концентрация соли составляет примерно 400 мМ.
26. Способ по любому из воплощений 1, 7 и 9-25, где высокое значение pH составляет от pH 6,5 до pH 8,8.
27. Способ по любому из воплощений 1, 7 и 9-26, где высокое значение pH составляет примерно pH 7,4.
28. Способ по любому из воплощений 1-27, где по существу разные времена удерживания отличаются по меньшей мере на 5%.
29. Способ по любому из воплощений 1-28, где по существу разные времена удерживания отличаются по меньшей мере на 10%.
30. Способ по любому из воплощений 1-29, где по существу разные времена удерживания отличаются по меньшей мере на 15%.
31. Способ по любому из воплощений 1-30, где по существу одинаковые времена удерживания отличаются меньше, чем на 5%.
32. Способ по любому из воплощений 1-31, где по существу одинаковые времена удерживания отличаются на 3,5% или менее.
33. Способ по любому из воплощений 1-32, где по существу одинаковые времена удерживания отличаются на 2,5% или менее.
34. Способ по любому из воплощений 1-33, где, если времена удерживания являются по существу отличными, данные времена удерживания являются пропорциональными единице, поделенной на квадратный корень концентрации соли (~1/√(концентрация соли)).
35. Способ по любому из воплощений 1-34, где контрольное антитело представляет собой либо антитело против IL-1R с SEQ ID NO: 01 (тяжелая цепь) и SEQ ID NO: 02 (легкая цепь) для подкласса IgG1 и антитело против IL-1R с SEQ ID NO: 03 (тяжелая цепь) и SEQ ID NO: 04 (легкая цепь) для подкласса IgG4, или антитело против HER2 с SEQ ID NO: 36 (тяжелая цепь) и SEQ ID NO: 37 (легкая цепь) для подкласса IgG1, и антитело против HER2 с SEQ ID NO: 38 (тяжелая цепь) и SEQ ID NO: 39 (легкая цепь) для подкласса IgG4.
36. Способ по любому из воплощений 1-35, где колонка для аффинной хроматографии на основе FcRn содержит нековалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m).
37. Способ по любому из воплощений 1-36, где колонка для аффинной хроматографии на основе FcRn содержит ковалентный комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m).
38. Способ по любому из воплощений 36-37, где комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m) связывается с твердой фазой.
39. Способ по воплощению 38, где твердая фаза представляет собой хроматографическое вещество.
40. Способ по любому из воплощений 36-39, где комплекс неонатального рецептора Fc (FcRn) и бета-2-микроглобулина (b2m) биотинилируется, и твердая фаза дериватизируется стрептавидином.
41. Способ по любому из воплощений 36-40, где бета-2-микроглобулин происходит из того же самого вида, что и неонатальный рецептор Fc (FcRn).
42. Способ по любому из воплощений 36-40, где бета-2-микроглобулин происходит из другого вида, чем FcRn.
43. Способ по любому из воплощений 1-42, где FcRn выбран из FcRn человека, FcRn яванского макака, FcRn мыши, FcRn крысы, FcRn овцы, FcRn собаки, FcRn свиньи, FcRn карликовой свиньи и FcRn кролика.
44. Способ по любому из воплощений 1-43, где антитело представляет собой моноспецифичное антитело или фрагмент слитого полипептида в виде антитела, или биспецифичное антитело или фрагмент слитого полипептида в виде антитела, или триспецифичное антитело или фрагмент слитого полипептида в виде антитела, или тетраспецифичное антитело или фрагмент слитого полипептида в виде антитела.
45. Способ по любому из воплощений 1-44, где антитело представляет собой полноразмерное антитело.
46. Антитело по любому из воплощений 1-45, где антитело представляет собой моноклональное антитело.
47. Способ получения антитела, включающий следующие стадии:
а) предоставление клетки, содержащей одну или более чем одну нуклеиновую кислоту, кодирующую антитело, отобранное с использованием способа по любому из воплощений 1-46,
б) культивирование клетки в культивационной среде и
в) выделение антитела из клетки или культивационной среды и, посредством этого, получение антитела.
Список ссылок
1. Edelman, G.M., Scand. J. Immunol. 34 (1991) 1-22.
2. Reff, M.E. and Heard, C., Crit. Rev. Oncol. Hematol. 40 (2001) 25-35.
3. Waldmann, T.A. and Strober, W., Prog. Allergy 13 (1969) 1-110.
4. Ghetie, V. and Ward, E.S., Annu. Rev. Immunol. 18 (2000) 739-766.
5. Chaudhury, C., et al., J. Exp. Med. 197 (2003) 315-322.
6. Brambell, F.W., et al., Nature 203 (1964) 1352-1354.
7. Brambell, F.W., Proc. Nutr. Soc. 28 (1969) 35-41.
8. Simister, N.E. and Mostov, K.E., Nature 337 (1989) 184-187.
9. Ropeenian, D.C., et al., J. Immunol. 170 (2003) 3528-3533.
10. Kuo, T.T., et al., J. Clin. Immunol. 30 (2010) 777-789.
11. Ropeenian, D.C. and Akilesh, S., Nat. Rev. Immunol. 7 (2007) 715-725.
12. Martin, W.L., et al., Mol. Cell 7 (2001) 867-877.
13. Goebl, N.A., et al., Mol. Biol. Cell 19 (2008) 5490-5505.
14. Kim, J.K., et al., Eur. J. Immunol. 24 (1994) 542-548.
15. Sanchez, L.M., et al., Biochemistry 38 (1999) 9471-9476.
16. Huber, A.H., et al., J. Mol. Biol. 230 (1993) 1077-1083.
17. Ober, R.J., et al., Proc. Natl. Acad. Sci. USA 101 (2004) 11076-11081.
18. Ober, R.J., et al., J. Immunol. 172 (2004) 2021-2029.
19. Akilesh, S., et al., J. Immunol. 179 (2007) 4580-4588.
20. Montoyo, H.P., et al., Proc. Natl. Acad. Sci. USA 106 (2009) 2788-2793.
21. Rodewald, R., J. Cell Biol. 71 (1976) 666-669.
22. Vaccaro, C., et al., Nat. Biotechnol. 23 (2005) 1283-1288.
23. Suzuki, Т., et al., J. Immunol. 184 (2010) 1968-1976.
24. Wang, W., et al., Drug Metab Dispos. 39 (2011) 1469-1477.
25. Schlothauer, Т., et al., MAbs. 5 (2013) 576-586.
26. Gandhi, M., et al., Semin. Cutan. Med. Surg. 29 (2010) 48-52.
27. Luo, J., et al., J. Mol. Biol. 402 (2010) 797-812.
28. Traczewski, P. and Rudnicka, L., BioDrugs. 26 (2012) 9-20.
29. Zhu, Y., et al., J. Clin. Pharmacol. 49 (2009) 162-175.
30. Lima, X.T., et al. Expert. Opin. Biol. Ther. 9 (2009) 1107-1113.
31. Weger, W., Br. J. Pharmacol. 160 (2010) 810-820.
32. Israelachvili, J.N., Intermolecular and Surface Forces, Academic Press Inc. (1985).
33. Petkova, S.B., et al., Int. Immunol. 18 (2006) 1759-1769.
34. Dall'Acqua, W.F., et al., J. Biol. Chem. 281 (2006) 23514-23524.
35. Khawli, L.A., et al., Cancer Biother. Radiopharm. 17 (2002) 359-370.
36. Putnam, W.S., et al., Trends Biotechnol. 28 (2010) 509-516.
37. Igawa, Т., et al., Protein Eng. Des. Sel. 23 (2010) 385-392.
38. Boswell, C.A., et al., Bioconjug. Chem. 21 (2010) 2153-2163.
39. Ropeenian, D.C., et al., Methods Mol. Biol. 602 (2010) 93-104.
40. Prabhat, P., et al., Proc. Natl. Acad. Sci. USA 104 (2007) 5889-5894.
41. Faber, C., et al., Immunotechnology. 3 (1998) 253-270.
42. Li, L., et al., BMC. Biophys. 5 (2012) 9.
43. Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604.
44. Hess, В., et al., J. Chem. Theory Comput. 4 (2008) 435-447.
45. Kortkhonjia, E., et al., MAbs. 5 (2013) 306-322.
46. Jorgensen, W.L., et al., J. Am. Chem. Soc. 118 (1996) 11225-11236.
47. Zalevsky, J., et al., Nat. Biotechnol. 28 (2010) 157-159.
48. Li, В., et al. (mAbs 6 (2014) 1255-1264).
49. Sampei, Z., et al. (PLoS One 8 (2013) e57479.
50. Benson, J.M., et al., mAbs 3 (2011) 535-545.
51. Ding, C., et al., Curr. Opin. Investig. Drugs 9 (2008) 515-522.
52. Weber, J., and Keam, S. J., BioDrugs. 23 (2009) 53-61.
Следующие примеры, графические материалы и последовательности приведены для того, чтобы помочь пониманию настоящего изобретения, истинный объем которого изложен в приложенной формуле изобретения. Понятно, что в изложенных процедурах могут быть сделаны модификации без отступления от сущности изобретения.
Материалы и методы
Антитела
Антитела, используемые в экспериментах, представляли собой устекинумаб (CNTO 1275, Stelara™), регистрационный номер CAS 815610-63-0, вариабельные домены в SEQ ID NO: 42 и 43), бриакинумаб (АВТ 874, J 695, Ozespa™, вариабельные домены в SEQ ID NO: 40 и 41), а также десять вариантов и мутантов устекинумаба и бриакинумаба, именуемые далее mAb 1-mAb 10 соответственно. Всего исследовали 12 IgG (см. Талицу 2).
Синтетические гены для устекинумаба, бриакинумаба, mAb5 и mAb6 были получены в Geneart (Life technologies GmbH, Carlsbad, CA, США). Для замены специфических аминокислот с получением mAb 1, mAb 2, mAb 7, mAb 8 и mAb 9 использовали сайт-направленный мутагенез. MAb 3 трансфицировали с использованием плазмид, кодирующих тяжелые цепи устекинумаба и легкие цепи бриакинумаба, a mAb 4 - наоборот.
Моноклональные антитела, используемые в данном документе, временно экспрессировали в клетках HEK293 (клетки эмбриональной почки человека 293) (см. ниже), и очистку проводили посредством хроматографии на белке А с использованием стандартных методик (см. ниже).
Биохимическая характеризация включала гель-фильтрацию (Waters BioSuite™ 250 7,8×300 мм, элюент: 200 мМ KH2PO4, 250 мМ KCl, pH 7,0) и анализ распределения молекулярной массы с использованием BioAnalyzer 2100 (Agilent technologies, Santa Clara, CA, США).
Фрагменты Fc получали посредством расщепления антител IdeS в пределах 30 минут при 37°С с использованием набора FabRICATOR (GENOVIS, Лунд, Швеция).
Экспрессионные плазмиды
Для экспрессии вышеописанных антител применяли варианты экспрессионных плазмид для временной экспрессии (например, в клетках HEK293-F) на основе либо организации кДНК с промотором CMV-интроном А (цитомегаловирус) или без них, либо геномной организации с промотором CMV.
Помимо экспрессионной кассеты антитела плазмиды содержали:
- репликатор, который обеспечивает репликацию данной плазмиды в Е. coli,
- ген β-лактамазы, который придает устойчивость к ампициллину в Е. coli, и
- ген дигидрофолатредуктазы из Mus musculus в качестве селектируемого маркера в эукариотических клетках.
Транскрипционная единица гена антитела состояла из следующих элементов:
- уникальный(ные) сайт(ты) рестрикции на 5'-конце
- немедленный ранний энхансер и промотор из человеческого цитомегаловируса,
- с последующей последовательностью интрона А в случае организации кДНК,
- 5' нетранслируемая область гена человеческого антитела,
- сигнальная последовательность тяжелой цепи иммуноглобулина,
- цепь человеческого антитела либо в виде кДНК, либо в виде геномной организации с организацией экзон-интрон иммуноглобулина,
- 3' нетранслируемая область с сигнальной последовательностью полиаденилирования и
- уникальный(ные) сайт(ты) рестрикции на 3'-конце.
Слитые гены, содержащие цепи антитела, получали посредством ПЦР и/или синтеза гена и собирали известными способами и методиками генной инженерии соединением соответствующих отрезков нуклеиновой кислоты, например, с использованием уникальных рестрикционных сайтов в соответствующих плазмидах. Субклонированные последовательности нуклеиновой кислоты подтверждали посредством секвенирования ДНК. Для временных трансфекций получали большие количества плазмид посредством получения плазмид из культур трансформированной Е. coli (Nucleobond АХ, Macherey-Nagel).
Методики культуры клеток
Использовали стандартные методики культуры клеток, как описано в Current Protocols in Cell Biology (2000), Bonifacino, J.S., Dasso, M., Harford, J.В., Lippincott-Schwartz, J. and Yamada, K.M. (eds.), John Wiley & Sons, Inc.
Временные трансфекции в системе HEK293-F
Антитела получали временной трансфекцией соответствующими плазмидами (например, кодирующими тяжелую цепь, а также соответствующую легкую цепь) с использованием системы HEK293-F (Invitrogen) согласно инструкции изготовителя. Вкратце, клетки HEK293-F (Invitrogen), растущие в суспензии либо во встряхиваемой колбе, либо в ферментере с перемешиванием в бессывороточной среде для экспрессии FreeStyle™ 293 (Invitrogen), трансфицировали смесью соответствующих экспрессионных плазмид и 293fectin™ или фектина (Invitrogen). Для 2 л встряхиваемой колбы (Corning) клетки HEK293-F высевали в плотности 1*106 клеток/мл в 600 мл и инкубировали при 120 об./мин, 8% CO2. Через одни сутки клетки трансфицировали при плотности клеток примерно 1,5*106 клеток/мл с примерно 42 мл смеси А) 20 мл Opti-MEM (Invitrogen) с 600 мкг общей плазмидной ДНК (1 мкг/мл), кодирующей тяжелую цепь соответственно и соответствующую легкую цепь в эквимолярном соотношении, и Б) 20 мл Opti-MEM плюс 1,2 мл 293 фектина или фектина (2 мкл/мл). Согласно потреблению глюкозы по ходу ферментации добавляли раствор глюкозы. Супернатант, содержащий секретируемое антитело, отбирали через 5-10 суток, и антитела либо непосредственно очищали из супернатанта, либо супернатант замораживали и хранили.
Очистка
Антитела очищали из супернатантов культуры клеток посредством аффинной хроматографии с использованием MabSelectSure-Sepharose™ (GE Healthcare, Швеция), хроматографии на основе гидрофобных взаимодействий с использованием бутил-сефарозы (GE Healthcare, Швеция) и гель-фильтрации на Superdex 200 (GE Healthcare, Швеция).
Вкратце, супернатанты культуры клеток после стерилизующей фильтрации захватывали на смоле MabSelectSuRe, уравновешенной буфером PBS (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, pH 7,4), промытой уравновешивающим буфером, и элюировали 25 мМ цитратом натрия при pH 3,0. Элюированные фракции антител объединяли и нейтрализовали 2 М Tris, pH 9,0. Для хроматографии на основе гидрофобных взаимодействий получали пулы антител посредством добавления 1,6 М раствора сульфата аммония до конечной концентрации 0,8 М сульфата аммония, и pH доводили до pH 5,0 с использованием уксусной кислоты. После уравновешивания бутил-сефарозной смолы 35 мМ ацетатом натрия, 0,8 М сульфатом аммония, pH 5,0, наносили на смолу антитела, промывали уравновешивающим буфером и элюировали линейным градиентом до 35 мМ ацетата натрия, pH 5,0. Фракции, содержащие антитело, объединяли и дополнительно очищали гель-фильтрацией с использованием колонки Superdex 200 26/60 GL (GE Healthcare, Швеция), уравновешенной 20 мМ гистидином, 140 мМ NaCl, pH 6,0. Фракции, содержащие антитело, объединяли, концентрировали до требующейся концентрации с использованием устройств для ультрафильтрации Vivaspin (Sartorius Stedim Biotech S.A., Франция) и хранили при -80°С.
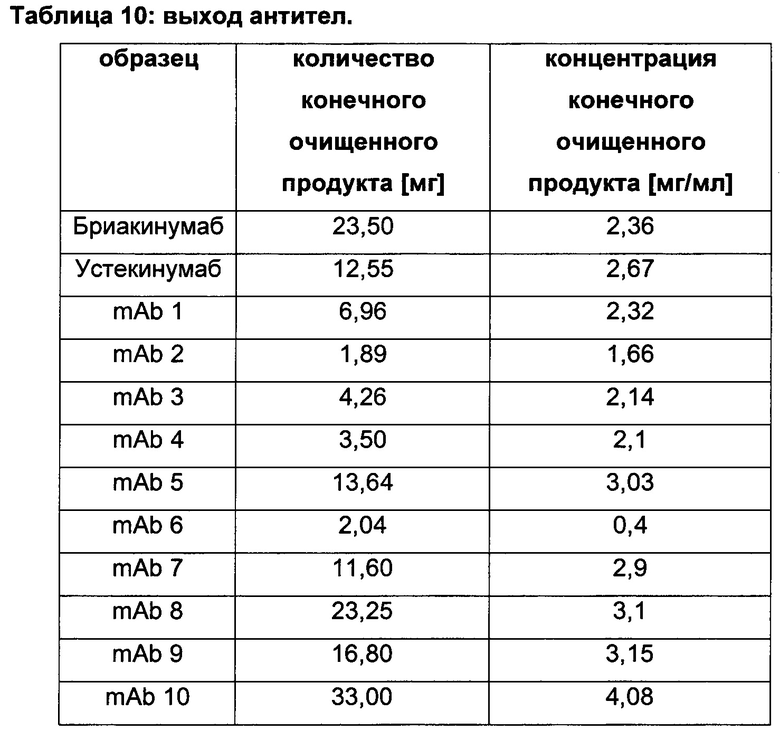
Чистоту и целостность антител анализировали после каждого этапа очистки посредством CE-SDS (капиллярный электрофорез с додецилсульфатом натрия) с использованием микрожидкостной технологии Labchip (Caliper Life Science, США). Для анализа CE-SDS готовили пять мкл раствора белка с использованием набора реактивов НТ Protein Express согласно инструкциям изготовителя и анализировали на системе LabChip GXII с использованием чипа НТ Protein Express. Данные анализировали с использованием программы LabChip GX.
Таблица 11: обзорная биохимическая характеризация всех mAb. Концентрация приведена в виде среднего значения 3 измерений. Содержание мономеров определяется интегрированием хроматограммы SEC (гель-фильтрация). Чистота видов антитела определяется посредством CE-SDS интактных mAb и после восстановления DTT (дитиотрейтол). Гидрофобность представляется относительно низко- и высокогидрофобных RS (остатки).
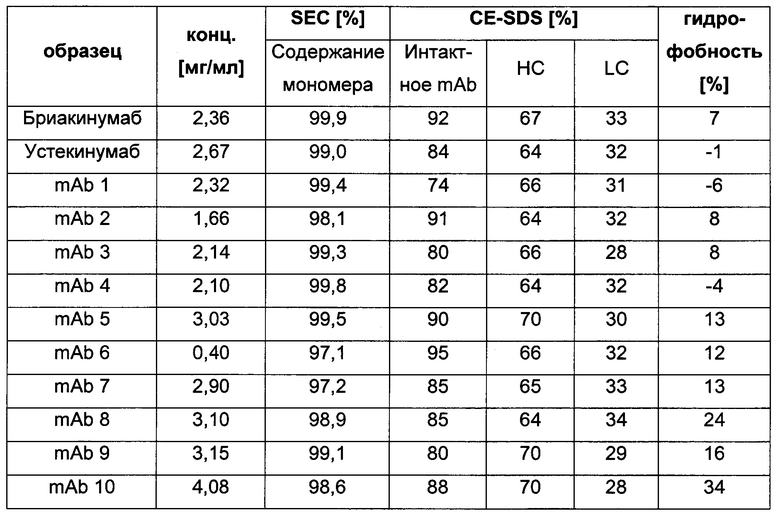
Функциональная характеризация
Функциональная характеризация включает анализ взаимодействия с мишенью (человеческий IL-12) для тестирования того, продуцировались ли бриакинумаб и устекинумаб правильно, и является ли связывание с мишенью все еще функциональным. Варианты mAb модифицировали в области Fab, и тестируется, изменяют ли данные модификации связывание с мишенью. Кроме того, анализируется взаимодействие антител, используемых в PK исследовании на мышах, с мышиным IL-12/-23 для исключения эффектов клиренса, опосредованного мишенью, в исследовании на мышах. Кроме того, измеряются уровни связывания с мышиным рецептором Fcγ I (muFcγRI), так как более сильное связывание с мышиным FcγRI могло бы приводить к более быстрому снижению в PK исследовании из-за более быстрого поглощения в антигенпрезентирующие клетки.
- Взаимодействие с человеческим IL-12
Бриакинумаб, устекинумаб и варианты имеют структурные различия в области Fab, которые могут влиять на связывание с IL-12, следовательно, результаты всех mAb представлены подробно.
- ELISA (твердофазный иммуноферментный анализ)
Кривые поглощение-концентрация вариантов с перекрестными заменами (mAb 1-6) и с модифицированным распределением заряда (mAb 7-10) показаны на Фиг. 15 и 16 соответственно. Концентрацию каждого mAb рассчитывали с использованием аппроксимации калибровочной кривой бриакинумаба с получением связывания с IL-12 относительно бриакинумаба (бриакинумаб - 100%). Оценивали, что различия связывания, меньшие или равные 30%, демонструют аналогичное связывания с IL-12 - такое же, как у бриакинумаба, различия, большие или равые 30% указывают на пониженное связывание с IL-12. Бриакинумаб, устекинумаб и mAb с обменом доменами Fv (mAb 1 и mAb 2) демонстрируют аналогичные профили связывания с IL-12. Связывание бриакинумаба, устекинумаба и mAb 2 варьирует в 20%-ном интервале, mAb 3 - в 30%-ном интервале. MAb с обменом LC (mAb 3 и mAb 4) и mAb с обменом CDR (mAb 5 и mAb 6) не связываются с IL-12.
Варианты бриакинумаба с модифицированным распределением заряда (mAb 7-9) связываются с IL-12 в интервале 30% относительно бриакинумаба, указывая на аналогичное связывание с IL-12. Только mAb 10 демонстрирует пониженное связывание с IL-12 с 63% связывания по сравнению с бриакинумабом.
- поверхностный плазмонный резонанс
SPR использовали для подтверждения результатов ELISA, специфичного в отношении мишени. Устекинумаб и бриакинумаб имеют почти идентичную константу скорости ассоциации (ka) ka (Бриакинумаб) 8*105 1/Мс относительно ka (Устекинумаб) 9*105 1/Мс). Диссоциация IL-12 и mAb является очень медленной, следовательно, расчет константы скорости диссоциации (kd) и затем равновесной константы диссоциации (KD) может отличаться от реальных значений. Несмотря на ограничение способа в данной постановке, расчетные значения могут давать общую оценку и могут использоваться для подтверждения результатов ELISA. Бриакинумаб и устекинумаб связываются с IL-12 с высокой аффинностью, и KD находится в низком нМ интервале (KD (Бриакинумаб) равно 0,2 нМ относительно KD (Устекинумаб), равного 0,07 нМ). Измеренная аффинность бриакинумаба со значениями ka, kd и KD, составляющими 8*105 1/Мс, 6*10-5 1/с и 70 пМ соответственно согласуется с литературными данными (ka - 5*105 1/Мс, kd - 5,1*10-5 1/с, KD - 100 пМ) ([51]). Высокая аффинность устекинумаба к IL-12 также описана в литературе ([52]).
В Таблице 12 обобщены расчетные кинетические параметры взаимодействия с мишенью. Моноклональные антитела с обменом доменами Fv (mAb 1 и mAb 2) и mAb с модифицированными распределениями заряда (mAb 7-10) имеют аффинности к IL-12, аналогичные бриакинумабу и устекинумабу. MAb 3 и mAb 5 не связываются с IL-12, и mAb 4 и mAb 6 демонстрируют очень слабое связывание с IL-12. Эти данные согласуются с результатами ELISA.

Аффинность FcRn-mAb при pH 6,0
KD рассчитывали относительно устекинумаба (устекинумаб - 1,0). Для оценки значений KD, аффинность mAb и FcRn оценивали как аналогичную аффинности устекинумаб-FcRn, если различия были меньшими, чем десятикратные относительно KD устекинумаб-FcRn. KD оценивали как различные, если различия KD были большими, чем десятикратные относительно KD устекинумаб-FcRn. Аффинности в отношении FcRn при pH 6,0 попадают в узкий интервал для всех mAb. Бриакинумаб имел относительный KD 0,2, а варианты имели варьирующие значения между бриакинумабом и устекинумабом, за исключением mAb 10, которое имело относительную KD 1,1.
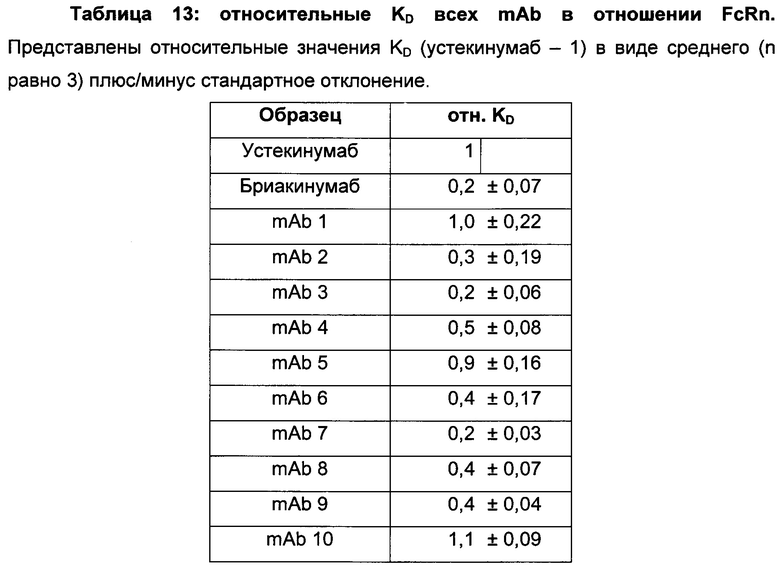
Диссоциация FcRn-mAb
Диссоциацию FcRn и mAb анализировали посредством SPR и аффинной хроматографии на основе FcRn.
- диссоциация FcRn-mAb с использованием SPR
Для оценки значений KD оценивали значения KD меньше 1 мкМ для демонстрации умеренной аффинности, от 1 до 5 мкМ - для демонстрации слабой аффинности и более 5 мкМ - для демонстрации отсутствия связывания с FcRn. Бриакинумаб и устекинумаб демонстрировали аналогичные аффинности при pH 6,0. Устекинумаб демонстрировал очень слабую аффинность при pH 6,6 и отсутствие аффинности при pH 6,8. В отличие от этого, бриакинумаб демонстрировал умеренную аффинность вплоть до pH 6,8, слабую аффинность при pH 7,0 и отсутствие связывания при pH 7,2.
Таблица 14: KD бриакинумаба и устекинумаба в отношении FcRn. KD рассчитывали с использованием буферов с возрастающими значениями pH. Значения KD, большие чем 5 мкМ, классифицировали как отсутствие связывания.
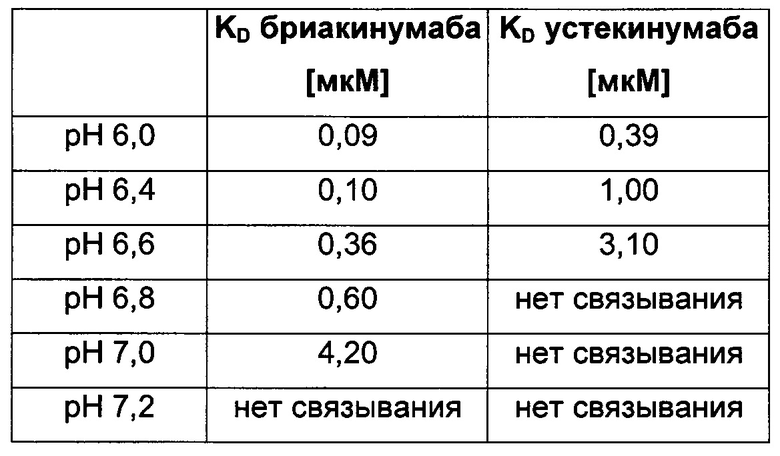
Биохимическая характеризация всех антител не продемонстрировала выраженных различий между бриакинумабом, устекинумабом и вариантами.
Получение фрагментов антител
Фрагмент F(ab')2 и фрагмент области Fc получали инкубированием в течение 30 мин при 37°С с использованием набора FabRICATOR (GENOVIS, Лунд, Швеция). Образующиеся продукты расщепления - F(ab')2 и область Fc разделяли на гель-фильтрационной колонке (SEC) (Superdex 200, GE Healthcare, Цюрих, Швейцария) с использованием хроматографической системы  Explorer (GE Healthcare, Упсала, Швеция), и фракции пиков объединяли. Стандарты молекулярной массы на той же самой колонке служили для идентификации двух продуктов расщепления на основе их времен удерживания.
Explorer (GE Healthcare, Упсала, Швеция), и фракции пиков объединяли. Стандарты молекулярной массы на той же самой колонке служили для идентификации двух продуктов расщепления на основе их времен удерживания.
Анализ поверхностным плазмонным резонансом (SPR) на основе FcRn
Характеристики связывания антител с FcRn анализировали технологией поверхностного плазмонного резонанса (SPR) с использованием установки BIAcore Т100 (BIAcore АВ, Упсала, Швеция). Данная система является надежно отработанной для исследования молекулярных взаимодействий. Она обеспечивает непрерывный мониторинг в реальном времени связываний лиганд/аналит и, таким образом, определение кинетических параметров в разных постановках анализа. Технология SPR основана на измерении показателя преломления близко к поверхности биосенсорного чипа, покрытого золотом. Изменения показателя пеломления указывают на изменения массы на поверхности, вызванные взаимодействием иммобилизованного лиганда с аналитом, инъецированным в растворе. При связывании молекул с иммобилизованным лигандом на поверхности масса возрастает, в случае диссоциации масса уменьшается. В настоящем анализе рецептор FcRn был иммобилизирован на биосенсорном чипе СМ5 для BIAcore (GE Healthcare Bioscience, Упсала, Швеция) посредством аминного связывания до уровня 400 единиц ответа (RU). Анализ проводили при комнатной темпертуре с PBS (фосфатно-солевой буферный раствор), 0,05% Tween 20, pH 6,0 (GE Healthcare Bioscience) в качестве подвижного буфера и буфера для разведения. Образцы 200 нМ нативных или окисленных антител инъецировали при скорости тока 50 мкл/мин при комнатной температуре. Время ассоциации составляло 180 с, фаза диссоциации занимала 360 с. Регенерация поверхности чипа достигалась короткой инъекцией HBS-P, pH 8,0. Оценку данных SPR проводили сравнением высоты сигнала биологического ответа в 180 с после инъекции и в 300 с после инъекции. Соответствующие параметры представляют собой максимальный уровень RU (180 с после инъекции) и позднюю стабильность (300 с после окончания инъекции).
Стационарные уровни связывания и равновесные константы диссоциации (KD) для huFcRn и IgG определяли при pH 6,0 с использованием прибора BIAcore Т100 SPR (GE Healthcare, Little Chalfont, Великобритания). Человеческий FcRn иммобилизировали на биосенсорном чипе СМ5 для BIAcore (GE Healthcare Bioscience) посредством аминного связывания до уровня 50 единиц ответа (RU). Для mAb 5 и mAb 6 использовали биосенсорный чип СМ4. Анализ проводили с использованием PBS с 0,05%-ным Tween 20 (оба от Roche Diagnostics, Мангейм, Германия), доведенного до pH 6,0, в качестве подвижного буфера и буфера для разведения при комнатной температуре. Ряд концентраций образцов готовили в интервале от 1500 нМ до 23 нМ, и каждый образец инъецировали при скорости тока 5 мкл/мин. Использовали время ассоциации и диссоциации 600 и 360 секунд соответственно. Чип регенерировали инъекцией PBS, содержащего 0,05% Tween 20 при pH 7,5. Равновесную константу диссоциации, KD, рассчитывали в виде стационарной аффинности и нормировали к KD устекинумаба.
Мыши
Для фармакокинетических исследований использовали мышей B6.Cg-Fcgrttm1Dcr Tg(FCGRT)276Dcr, дефицитных по гену α-цепи мышиного FcRn, но гемизиготных трансгенных в отношении гена α-цепи человеческого FcRn (muFcRn-/- huFcRn tg+/-, линия 276) [39]. Содержание мышей осуществляли при специфических беспатогенных условиях. Мышей приобретали в Jackson Laboratory (Bar Harbor, ME, США) (самки в возрасте 4-10 недель, масса 17-22 г во время дозирования). Все эксперименты на животных были одобрены правительством Верхней Баварии, Германия (номер разрешения 55.2-1-54-2532.2-28-10) и проведены в аккредитованном AAALAC (Ассоциация по аттестации и аккредитации содержания лабораторных животных) виварии согласно Нормативу Европейского Союза по уходу и использованию экспериментальных животных. Животных содержали в стандартных клетках, и они имели свободный доступ к пище и воде на протяжении всего периода исследования.
Фармакокинетические исследования
Одну дозу антитела в.в. (внутривенно) инъецировали через боковую хвостовую вену при уровне дозы 10 мг/кг. Мышей делили на 3 группы по 6 мышей в каждой для того, чтобы охватить в целом 9 моментов времени отбора сыворотки (в 0,08, 2, 8, 24, 48, 168, 336, 504 и 672 часа после дозы). Каждую мышь дважды подвергали ретроорбитальному кровоизвлечению, проведенному при легкой анестезии изофлураном™ (CP-Pharma GmbH, Бургдорф, Германия); третий образец крови отбирали во время умерщвления. Кровь отбирали в пробирки для сыворотки (Microvette 500Z-Gel, Sarstedt, Нюмбрехт, Германия). После 2 ч инкубации образцы центрифугировали в течение 3 мин при 9300 g для получения сыворотки. После центрифугирования образцы сыворотки хранили в замороженном состоянии при -20° до анализа.
Определение сывороточных концентраций человеческих антител
Концентрации устекинумаба, бриакинумаба, mAb 8 и mAb 9 в мышиной сыворотке определяли специфичными твердофазными иммуноферментыми анализами. Для захвата и выявления использовали биотинилированный интерлейкин 12, специфичный в отношении антител, и меченное дигоксигенином мышиное моноклональное антитело против человеческого Fc (Roche Diagnostics, Пенцберг, Германия) соответственно. Покрытые стрептавидином планшеты для микротитрования (Roche Diagnostics, Пенцберг, Германия) покрывали биотинилированным захватывающим антителом, разведенным в буфере для анализа (Roche Diagnostics, Пенцберг, Германия), в течение 1 ч. После промывки добавляли образцы сыворотки в разных разведениях, с последующей еще одной стадией инкубации в течение 1 ч. После многократных промывок связавшиеся человеческие антитела выявляли посредством последующей инкубации с выявляющим антителом, с последующим антителом против дигоксигенина, конъюгированным с пероксидазой хрена (HRP; Roche Diagnostics, Пенцберг, Германия). В качестве субстрата HRP для образования окрашенного продукта реакции использовали ABTS (2,2'-азино-ди[3-этилбензтиазолинсульфонат]; Roche Diagnostics, Германия). Поглощение образующегося реакционного продукта считывали при 405 нм с использованием контрольной длины волны 490 нм с использованием планшет-ридера Tecan sunrise (Mannedorf, Швейцария).
Все образцы сыворотки, образцы позитивного и негативного контроля анализировали в двойных повторностях и калибровали относительно эталонного стандарта.
PK анализ
Фармакокинетические параметры рассчитывали посредством некомпартментного анализа с использованием WinNonlin™ 1.1.1 (Pharsight, CA, США).
Вкратце, значения площади под кривой (AUC0-inf) рассчитывали логарифмическим трапециедальным способом из-за нелинейного уменьшения уровня антител и экстраполировали к бесконечности с использованием кажущейся терминальной константы скорости λz, с экстраполяцией от наблюдаемой концентрации в последний момент времени.
Клиренс в плазме рассчитывали как мощность дозы (D), разделенную на AUC0-inf. Кажущееся время полужизни в конечной фазе (Т1/2) выводили из уравнения Т1/2=ln2/λz.
Статистический анализ
Выпадающие сывороточные концентрации выявляли с использованием критерия выпадающих значений Налимова и исключали из дальнейшего анализа.
В качестве статистического критерия для анализа статистически значимых различий времени полужизни в конечной фазе использовали критерий подлинной значимости Тьюки (критерий Тьюки HSD).
Расчет pH-зависимого нетто заряда
pH-зависимый нетто заряд («кривые титрования») рассчитывали с использованием программы EMBOSS iep, находящейся в открытом доступе, предполагая, что все цистеины участвуют в дисульфидных мостиках.
Создание модели гомологии бриакинумаба и расчет изопотенциальных поверхностей
Модель гомологии для фрагмента Fab бриакинумаба получали с использованием программы для моделирования 9v7, используя структуру PDB (база данных белковых структур) 1AQK [41] в качестве матрицы. Изопотенциальные поверхности для Fab бриакинумаба и устекинумаба рассчитывали из данной модели (бриакинумаб) или кристаллической структуры устекинумаба (ID PDB 3НМХ) соответственно. Структуры протонировали с использованием протокола «получить белок» с использованием поля силы CHARMm в DiscoveryStudio Pro, Версия 3.5 (Accelrys Inc., Сан-Диего, США) при pH 7,4 и ионной силе 0,145 М. Электростатический потенциал рассчитывали с использованием протокола «электростатический потенциал» в DiscoveryStudio Pro, который задействует программу DelPhi [42].
Моделирование молекулярной динамики комплексов бриакинумаб- и устекинумаб-FcRn
Строили модели гомологии бриакинумаба и устекинумаба в виде полных IgG, используя DiscoveryStudio Pro, Версия 3.5, с использованием кристаллической структуры полного IgG1 (ID PDB 1HZH) без гликанов в качестве матрицы. Данное упрощение считалось подходящим, так как in vitro гликозилирование не имеет значимого эффекта на связывание с FcRn [43]. Домены Fab в данной матрице заменяли структурами Fab, описанными выше, после выравнивания их доменов CH1 и CL. Модель гомологии человеческого FcRn строили с использованием DiscoveryStudio Pro, используя комплекс крысиный FcRn-Fc (ID PDB 1I1A) в качестве матрицы. Отсутствующие остатки строили с использованием скрипта «получить белок» DiscoveryStudio Pro. Модель гомологии человеческого FcRn моделировали для обеих тяжелых цепей моделей IgG бриакинумаба и устекинумаба посредством наложения С-альфа атомов крысиных доменов Fc в пределах  около FcRn с их гомологичными партнерами на человеческую область Fc. Для моделирования MD вводили дисульфидную связь на поверхность контакта FcRn:Fc (между остатком 108 в FcRn и остатком 255 в Fc, Фиг. S1) для предотвращения диссоциации комплекса на протяжении времени моделирования. Образующиеся структуры представляют собой полный IgG, несущий две копии гетеродимера FcRn/β2mg.
около FcRn с их гомологичными партнерами на человеческую область Fc. Для моделирования MD вводили дисульфидную связь на поверхность контакта FcRn:Fc (между остатком 108 в FcRn и остатком 255 в Fc, Фиг. S1) для предотвращения диссоциации комплекса на протяжении времени моделирования. Образующиеся структуры представляют собой полный IgG, несущий две копии гетеродимера FcRn/β2mg.
Моделирования молекулярной динамики (MD) комплексов IgG-FcRn проводили с использованием программного пакета для моделирования GROMACS 4.6.2 (доступен на www.gromacs.org) [44], по существу как описано Kortkhonjia et al. [45]. Моделирования проводили параллельно на 160 процессорах компьютерного кластера, работающих в операционной системе Linux. Использовали поле силы OPLSAA [46], и структуры были полностью сольватированными с приблизительно 128000 TIP3 молекулами воды. Добавляли атомы хлора или натрия для нейтрализации общего заряда системы. Использовали усеченный октаэдр с периодическими граничными условиями с границей 7,5 ангстрем вокруг белка. Электростатические взаимодействия рассчитывали с использованием суммирования РМЕ с электростатическим порогом отсечения в реальном пространстве 1,0 нм. Потенциал Леннарда-Джонса был отсечен на уровне 1,0 нм. Для ограничения всех длин связей белка использовали LINCS, обеспечивающую шаг во времени 2 фс. Поддерживали постоянную температуру 300 К с использованием алгоритма V-rescale. После минимизации энергии (цель: максимальная сила меньше 1000 кДж/моль/нм) осуществляли 30 пс уравновешивание перед моделированием траектории на протяжении 100 нс.
Расчет энергии взаимодействия IgG-FcRn
Электростатический вклад в не приводящие к образованию связей взаимодействия между FcRn и доменом Fab, который достигает FcRn в траектории MD, рассчитывали с использованием DiscoveryStudio Pro. Для расчета энергии белок протонировали при pH 7,4, ионной силе 145 мМ и температуре 37°С с использованием тех же самых установок, как описано выше. Структуры минимизировали с использованием максимум 1000 шагов протокола «умный минимизатор» перед расчетом энергий взаимодействия с использованием протокола «рассчитать энергию взаимодействия» с полем силы CHARMm в DiscoveryStudio Pro. Использовали подразумеваемую воду и модель электростатики GBMV. Данный расчет проводили в начале траектории (0 нс) и в 96-100 нс с 1 нс интервалами.
Пример 1
Получение аффинной колонки на основе FcRn
Экспрессия FcRn в клетках HEK293
FcRn временно экспрессровали посредством трансфекции клеток HEK293 двумя плазмидами, содержащими кодирующую последовательность FcRn и бета-2-микроглобулина. Трансфицированные клетки культивировали во встряхиваемых колбах при 36,5°С, 120 об./мин (амплитуда шейкера - 5 см), 80%-ной влажности и 7% CO2. Клетки разводили каждые 2-3 суток до плотности 3-4*105 клеток/мл.
Для временной экспрессии запускали 14 л биореактор из нержавеющей стали с объемом культуры 8 л при 36,5°С, pH 7,0 плюс/минус 0,2, pO2 35% (продуванием N2 и воздухом, общий ток газа 200 мл мин-1) и скоростью мешалки 100-400 об./мин. При достижении плотности клеток 20*105 клеток/мл 10 мг пазмидной ДНК (эквимолярные количества обеих плазмид) разводили в 400 мл Opti-MEM (Invitrogen). В данную смесь добавляли 20 мл 293фектина (Invitrogen), которую затем инкубировали в течение 15 минут при комнатной температуре и затем переносили в ферментер. Начиная со следующих суток клеткам поставляли питательные вещества в непрерывном режиме: подпиточный раствор добавляли со скоростью 500 мл в сутки и глюкозу, по мере необходимости, для поддержания уровня более 2 г/л. Супернатант отбирали через 7 суток после трансфекции с использованием центрифуги с поворотными головками с 1 л бакетами: 4000 об./мин в течение 90 минут. Супернатант (13 л) осветляли посредством фильтра Sartobran Р (0,45 мкм плюс 0,2 мкм, Sartorius), и очищали из него комплекс FcRn-бета-2-микроглобулин.
Биотинилирование неонатального рецептора Fc
3 мг FcRn растворяли/разводили в 5,3 мл 20 мМ буфера на основе дигидрофосфата натрия, содержащего 150 мМ хлорид натрия, и добавляли к 250 мкл PBS и 1 таблетке полного протеазного ингибитора (целые таблетки ULTRA, Roche Diagnostics GmbH). FcRn биотинилировали с использованием набора для биотинилирования от Avidity согласно инструкциям изготовителя (Bulk BIRA, Avidity LLC). Реакцию биотинилирования осуществляли при комнатной температуре в течение ночи.
Биотинилированный FcRn диализировали против 20 мМ буфера на основе дигидрофосфата натрия, содержащего 150 мМ NaCl, pH 7,5 при 4°С в течение ночи для удаления избытка биотина.
Связывание со стрептавидин-сефарозой
Для связывания со стрептавидин-сефарозой один грамм стрептавидин-сефарозы (GE Healthcare, Великобритания) добавляли к биотинилированному и диализированному FcRn и инкубировали при 4°С в течение ночи. Сефарозой, дериватизированной FcRn, заполняли 1 мл колонку ХK (GE Healthcare, Великобритания), и колонку на основе FcRn затем уравновешивали 20 мМ буфером на основе натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), содержащим 140 мМ хлорид натрия, pH 5,5.
Пример 2
Хроматография с использованием аффинной колонки на основе FcRn и градиента pH
Сефарозой, дериватизированной рецептором, заполняли 1 мл колонку ХK (GE Healthcare), и колонку на основе FcRn затем уравновешивали 20 мМ буфером на основе 2-(N-морфолин)-этансульфоновой кислоты (MES), содержащим 140 мМ NaCl, pH 5,5.
Условия:
Образцы готовили в 20 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), 140 мМ хлориде натрия, pH 5,5. Каждый образец содержал 30 мкг mAb на инъекцию. Антитела элюировали линейным градиентом pH от pH 5,5 до 8,8 в пределах 120 минут с использованием 20 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), 140 мМ хлорида натрия, pH 5,5, и 20 мМ трис(гидроксиметил)аминометана (TRIS), 140 мМ хлорида натрия, pH 8,8, в качестве элюентов и скорости тока 0,5 мл/мин. Колоночная хроматография на основе FcRn демонстрирует связывание при кислотном pH (pH 5,5-6,0) и высвобождение при более высоких значениях pH. Для полной элюции антител pH возрастает в градиенте вплоть до pH 8,8. Хроматограммы интегрировали вручную с использованием программы Chromeleon (Dionex, Германия). Эксперименты проводили при комнатной температуре. Профиль элюции получали непрерывным измерением поглощения при 280 нм. Для определения pH элюции при определенных временах удерживания образцы отбирали каждые 5 минут, и pH измеряли вне системы.
Пример 3
Хроматография с использованием аффинной колонки на основе FcRn, условий градиента pH и высокой концентрации соли
Сефарозой, дериватизированной рецептором, заполняли 1 мл колонку ХK (GE Healthcare), и колонку на основе FcRn затем уравновешивали 20 мМ буфером на основе 2-(N-морфолин)-этансульфоновой кислоты (MES), содержащим 400 мМ NaCl, pH 5,5.
Условия:
Образцы готовили в 20 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), 400 мМ хлориде натрия, pH 5,5. Каждый образец содержал 30 мкг mAb на инъекцию. Антитела элюировали линейным градиентом pH от pH 5,5 до 8,8 в пределах 120 минут с использованием 20 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), 400 мМ хлорида натрия, pH 5,5, и 20 мМ трис(гидроксиметил)аминометана (TRIS), 400 мМ хлорида натрия, pH 8,8, в качестве элюентов и скорости тока 0,5 мл/мин. Колоночная хроматография на основе FcRn демонстрирует связывание при кислотном pH (pH 5,5-6,0) и высвобождение при более высоких значениях pH. Для полной элюции антител рН возрастает в градиенте вплоть до pH 8,8. Хроматограммы интегрировали вручную с использованием программы Chromeleon (Dionex, Германия). Эксперименты проводили при комнатной температуре. Профиль элюции получали непрерывным измерением поглощения при 280 нм. Для определения pH элюции при определенных временах удерживания образцы отбирали каждые 5 минут, и pH измеряли вне системы.
Пример 4
Хроматография с использованием аффинной колонки на основе FcRn и солевого градиента
Сефарозой, дериватизированной рецептором, заполняли 1 мл колонку XK (GE Healthcare), и колонку на основе FcRn затем уравновешивали 10 мМ буфером на основе 2-(N-морфолин)-этансульфоновой кислоты (MES), pH 7,8.
Условия:
Образцы готовили в 10 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), pH 7,8. Каждый образец содержал 30 мкг mAb на инъекцию. Антитела элюировали линейным солевым градиентом от 0 нМ до 250 нМ хлорида натрия в пределах 60 минут с использованием 10 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), pH 7,8, и 10 мМ натриевой соли 2-(N-морфолин)-этансульфоновой кислоты (MES), 250 мМ хлорида натрия, pH 7,8, в качестве элюентов и скорости тока 0,5 мл/мин. Эксперименты проводили при комнатной температуре. Профиль элюции получали непрерывным измерением поглощения при 280 нм. Хроматограммы интегрировали вручную с использованием программы Chromeleon (Dionex, Германия).
Пример 5
Хроматография с использованием аффинной колонки на основе FcRn
Сефарозой, дериватизированной рецептором, заполняли 1 мл колонку XK (GE Healthcare), и колонку на основе FcRn затем уравновешивали 20 мМ буфером на основе 2-(N-морфолин)-этансульфоновой кислоты (MES), содержащим 150 мМ NaCl, pH 5,5.
Условия:
Образцы антитела или слитого белка, содержащие 50-100 мкг белка, доводили до pH 5,5 и наносили на колонку на основе FcRn с использованием  explorer 10 XT или Dionex Summit (Dionex, Idstein, Германия). Колонку с высотой слоя 5 см затем промывали 5-10 объемами колонки уравновешивающего буфера - 20 мМ MES, 150 мМ NaCl, pH 5,5. Аффинно связанные белки, содержащие Fc, элюировали градиентом pH до 20 мМ Tris/HCl, 150 мМ NaCl, pH 8,8, за 30 объемов колонки. Для полной элюции модифицированных антител pH увеличивали в градиенте вплоть до pH 8,8. Эксперименты проводили при комнатной температуре. Профиль элюции получали непрерывным измерением поглощения при 280 нм. Время, затраченное на достижение пиком аналита, X, детектора после инъекции образца, называли временем удерживания.
explorer 10 XT или Dionex Summit (Dionex, Idstein, Германия). Колонку с высотой слоя 5 см затем промывали 5-10 объемами колонки уравновешивающего буфера - 20 мМ MES, 150 мМ NaCl, pH 5,5. Аффинно связанные белки, содержащие Fc, элюировали градиентом pH до 20 мМ Tris/HCl, 150 мМ NaCl, pH 8,8, за 30 объемов колонки. Для полной элюции модифицированных антител pH увеличивали в градиенте вплоть до pH 8,8. Эксперименты проводили при комнатной температуре. Профиль элюции получали непрерывным измерением поглощения при 280 нм. Время, затраченное на достижение пиком аналита, X, детектора после инъекции образца, называли временем удерживания.
Пример 6
Корреляция времени удерживания на колонке на основе FcRn с временем полужизни in vivo
Время полужизни in vivo измеряли в трансгенных мышах C57BL/6J с человеческим FcRn после одинократного в.в. введения 10 мг/кг (n равно 8) и сравнивали с временем удерживания на колонке на основе FcRn (см. Таблицу 15). Обнаружили, что антитела, которые демонстрировали позднюю элюцию из колонки на основе FcRn, имели более длительное время полужизни у трансгенных по FcRn мышей.
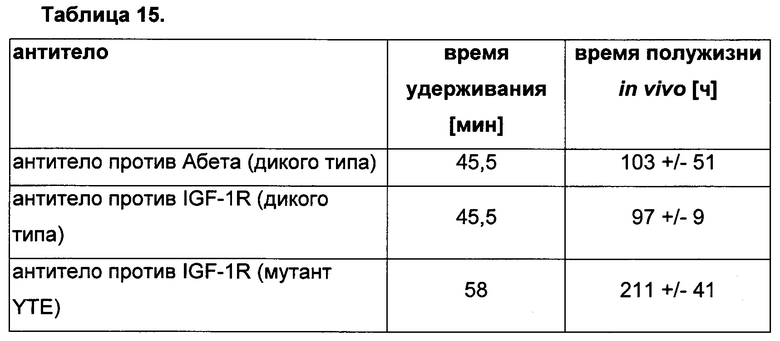
Пример 7
Очистка человеческого FcRn, мышиного FcRn и FcRn яванского макака
Осветленные супернатанты, содержащие белки, меченные гекса-His, загружали на аффинную хроматографическую смолу на основе Ni-NTA (Qiagen, Hanbrechtikon, Швейцария) при 4°С. После стадий промывки 20 мМ натрий-фосфатным буфером, содержащим 500 мМ NaCl, pH 7,4, и содержащим 20 мМ и 100 мМ имидазол соответственно, белки элюировали при скорости тока 2 мл/мин с использованием периодической элюции тем же самым буфером, содержащим 300 мМ имидазол, на хроматографической системе Prime (Amersham Pharmacia Biotech, Упсала, Швеция). Фракции объединяли и дополнительно очищали в натрий-фосфатном буфере, содержащем 500 мМ NaCl, на гель-фильтрационной хроматографии (Superdex™ 200, GE Healthcare, Цюрих, Швейцария). Очищенные белки количественно измеряли с использованием спектрофотометра Nanodrop (Nanodrop Technologies, Wilmington, DE) и анализировали посредством SDS-PAGE на 4-12% Bis-Tris гелях NuPAGE в MES буфере при денатурирующих и восстанавливающих условиях.
Пример 8
Аффинные колоночные хроматографии на основе мышиного FcRn и FcRn яванского макака
В следующей Таблице 16 приведены времена удерживания типичных человеческих антител на аффинных колонках, содержащих FcRn от яванского макака. Данные были получены с использованием следующих условий: элюционный буфер - 20 мМ TRIS/HCl, 150 мМ NaCl, pH 8,5. Дополнительное описание: см. Пример 2. Термин мутант YTE обозначает тройного мутанта M252Y/S254T/T256E.

В следующей Таблице 17 приведены времена удерживания типичных человеческих антител на мышином FcRn. Данные были получены с использованием следующих условий: 1,2 мг рецептора, связанного на 1 мл сефарозы. Элюционный буфер - 20 мМ TRIS/HCl, 150 мМ NaCl, pH 8,5. Дополнительное описание: см. Пример 2. Мутанты YTE не включены в данную таблицу, так как они не могли быть элюированы, пока pH элюционного буфера не был доведен до 9,5.

Аффинная колонка на основе FcRn яванского макака ведет себя аналогично аффинной колонке на основе человеческого FcRn в том, что касается связывания гуманизированных антител. С другой стороны, связывание гуманизированных антител с колонкой на основе мышиного FcRn сильнее, чем с аффинной колонкой на основе человеческого FcRn, что можно видеть по более долгому удерживанию.
Пример 9
Получение фрагментов антител
Фрагмент F(ab')2 и фрагмент в виде области Fc получали путем расщепления полноразмерного антитела, разведенного 1:1 100 мМ Tris, pH 8,0, посредством добавления 1 мкг цистеиновой протеазы IdeS на 50 мкг антитела и инкубации в течение 2 ч при 37°С. Образующиеся в результате продукты расщепления - F(ab')2 и Fc - разделяли на гель-фильтрационной колонке (SEC) (Superdex 200, GE Healthcare, Цюрих, Швейцария) с использованием хроматографической системы Explorer (GE Healthcare, Упсала, Швеция), и фракции пиков объединяли. Стандарты молекулярной массы на той же самой колонке служили для идентификации двух продуктов расщепления на основе их времен удерживания.
Времена удерживания полноразмерных антител заметно варьировали. В отличие от этого, времена удерживания соответствующих Fc частей всех протестированных антител фактически не отличались друг от друга (меньше 1%).
При использовании плазмина для расщепления полноразмерных антител получали такие же данные (данные не показаны).
Пример 10
Фармакокинетическое исследование у мышей с человеческим FcRn
Все процедуры проводили согласно руководствам Ассоциации по аттестации и аккредитации содержания лабораторных животных (www.aaalac.org). Исследование было разрешено Региональным советом Верхней Баварии, Германия.
Во всем данном фармакокинетическом исследовании использовали самцов и самок мышей C57BL/6J (фон); мышей, дефицитных по FcRn, но гемизиготных трансгенных в отношении человеческого FcRn (huFcRn (276) -/tg (30, 31).
Во время введения животные весили от 17 до 25 г. Соответствующее антитело давали в виде однократной внутривенной болюсной инъекции через хвостовую вену. Из-за ограниченного объема крови мышей для охвата девяти моментов времени отбора образцов требовались три группы из четырех самцов и четырех самок животных каждая, т.е. три момента времени отбора образцов на животное. Образцы крови отбирали в группе 1 в 5 мин, 24 часа и 336 часов, в группе 2 - в 2 часа, 168 часов и 504 часа, и в группе 3 - в 8 часов, 48 часов и 672 часа после введения. Образцы крови - примерно 100 мкл - получали ретробульбарной пункцией и хранили при комнатной температуре в течение 60 мин для обеспечения свертывания. Образцы сыворотки - по меньшей мере 40 мкл - получали посредством центрифугирования при 9300 g при 4°С в течение 3 мин и немедленно замораживали и хранили при -20°С до анализа.
Сывороточные концентрации человеческих терапевтических антител в мышиной сыворотке определяли твердофазным иммуноферментным анализом (ELISA) с захватом антигена, специфичным в отношении антигенсвязывающей области (Fab) введенного антитела и его вариантов. Все реактивы или образцы инкубировали при комнатной температуре на шейкере при 400 об./мин. Каждый этап промывки включал три цикла. Вкратце, покрытые стрептавидином планшеты для микротитрования покрывали биотинилированным антителом, разведенным в буфере для анализа. После промывки фосфатно-солевым буферным раствором-полисорбатом 20 (Tween 20), добавляли образцы сыворотки в разных разведениях и инкубировали в течение 1 ч. После промывки связавшиеся человеческие терапевтические антитела выявляли посредством последующей инкубации с фрагментами Fab моноклонального антитела, специфичными в отношении человеческого Fcγ, конъюгированными с дигоксигенином, которые не реагируют перекрестно с мышиным IgG. После промывки добавляли антитело против дигоксигенина, конъюгированное с пероксидазой хрена (HRP), и инкубировали в течение 1 ч. После промывки добавляли ABTS (2,2'-азино-ди[3-этилбензтиазолинсульфонат; Roche Diagnostics, Германия) в качестве субстрата HRP с образованием окрашенного продукта реакции. Поглощение образующегося продукта реакции считывали при 405 нм с контрольной длиной волны 490 нм. Все образцы сыворотки и образцы позитивного или негативного контроля анализировали в повторностях и калибровали относительно эталонного стандарта.
Фармакокинетические параметры рассчитывали посредством некомпартментного анализа с использованием программы фармакокинетической оценки WinNonlin™ (Pharsight, St. Louis, МО, США), версия 5.2.1. Вкратце, площадь под кривой концентрация/время AUC(0-672) рассчитывали по линейному трапециедальному правилу (с линейной интерполяцией) от времени 0 до бесконечности. Кажущееся время полужизни в конечной фазе (Т1/2) выводили из уравнения: T1/2=ln2/λz. Общий клиренс в организме (CL) рассчитывали как доза/AUC. Статистически значимые различия в фармакокинетических параметрах между антителом дикого типа и его вариантами определяли посредством анализа ANOVA.
Фармакокинетическое исследование у мышей C57BL/6J, дефицитных в отношении мышиного FcRn, но гемизиготных трансгенных в отношении человеческого FcRn (huFcRn (276) -/tg), продемонстрировало то, что мутация YTE усиливала фармакокинетику антитела. На уровне статистической значимости мутант YTE имел в 1,74 раза большую AUC(0-672), в 1,95 раза более медленный клиренс и в 2,2 раза более продолжительное время полужизни в конечной фазе по сравнению с антителом дикого типа (Таблица 14).
Таблица 18: фармакокинетические параметры для антитела дикого типа и его тройного мутанта YTE, полученные некомпартментным анализом сывороточных концентраций, измеренных ELISA, после однократной в.в. болюсной инъекции 10 мг/кг трансгенным мышам с человеческим FcRn. Среднее плюс/минус SD, n равно 8 на группу, анализ ANOVA значимости по сравнению с антителом дикого типа (+++, p меньше 0,001). AUC(0-672) - площадь под кривой зависимости сывороточной концентрации от времени от времени 0 до 672 ч.

Пример 11
Фармакокинетическое исследование у мышей с человеческим FcRn
Все процедуры проводили согласно руководствам Ассоциации по аттестации и аккредитации содержания лабораторных животных (www.aaalac.org). Исследование было разрешено Региональным советом Верхней Баварии, Германия.
Во всем данном фармакокинетическом исследовании использовали самцов и самок мышей C57BL/6J (фон); мышей, дефицитных по FcRn, но гемизиготных трансгенных в отношении человеческого FcRn (huFcRn (276) -/tg (30, 31).
В исследовании in vivo использовали четыре антитела: бриакинумаб, устекинумаб, mAb 8 и mAb 9.
Соответствующее антитело давали в виде однократной внутривенной болюсной инъекции (10 мг/кг). Из-за ограниченного объема крови мышей для охвата девяти моментов времени отбора образцов требовались три группы из шести животных каждая. Последним моментом отбора образцов были четыре недели после введения.
Результаты показаны на Фиг. 3.
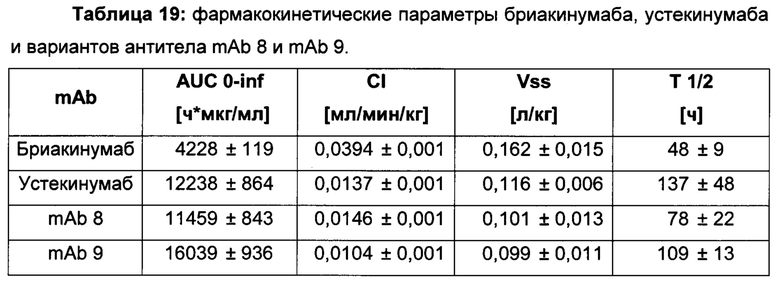
Для подтверждения того, что различия времени полужизни в конечной фазе у трансгенных мышей с человеческим FcRn были вызваны разными взаимодействиями FcRn-mAb, провели второе исследвание in vivo у мышей, нокаутных по FcRn. Для уменьшения числа мышей, используемых в данном исследовании, использовали только три антитела: бриакинумаб, устекинумаб и mAb 9.
После в.в. введения 10 мг/кг антитела клиренс всех антител значительно быстрее у мышей, нокаутных по FcRn, чем у трансгенных мышей с человеческим FcRn из-за отсутствующего рециклирования IgG, опосредованного FcRn. Разделение на альфа и бета фазу не является явно поддающимся определению из-за очень быстрого устранения антител. Может быть продемонстрировано то, что бриакинумаб имеет отличное фармакокинетическое поведение с более быстрым распределением в первые часы после введения по сравнению с устекинумабом и mAb 9. Данные открытия также наблюдали у трансгенных мышей с человеческим FcRn, указывая на то, что процесс распределения в первые часы после введения не является опосредованным FcRn.
Рассчитывали и обобщали следующие PK параметры: AUC0-inf, CI, VSS и Т1/2.
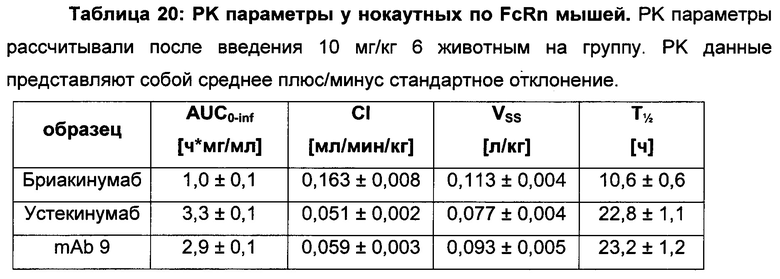
Устекинумаб и mAb 9 являются сопоставимыми в отношении AUC0-inf, CI, VSS и Т1/2. Бриакинумаб имеет меньшую AUC0-inf, более быстрый клиренс и меньшее Т1/2, чем устекинумаб и mAb 9. Расчет Т1/2 может отличаться от реального значения, так как для более точного расчета времени полужизни в конечной фазе потребовались бы моменты времени после 3 и 4 суток.
Статистический анализ времени полужизни в конечной фазе осуществляли с использованием критерия Тьюки HSD. Статистическая значимость могла быть выявлена между временем полужизни в конечной фазе бриакинумаба и устекинумаба, и устекинумаба и бриакинумаба, и mAb 9.
Образование ADA тестировали посредством выявления иммунных комплексов лекарственное средство/ADA. У нокаутных по FcRn мышей введение 10 мг/кг бриакинумаба приводило к образованию иммунных комплексов бриакинумаб/ADA примерно через 168-192 часа (7-8 суток).
Таблица 21: ADA-позитивные образцы после введения бриакинумаба у нокаутных по FcRn мышей. Сывороточные концентрации в каждый момент времени отбора образцов после в.в. введения 10 мг/кг бриакинумаба у нокаутных по FcRn мышей. ADA-позитивные образцы отмечены как * и **, описывая умеренное и сильное образование иммунных комплексов лекарственное средство/ADA соответственно.
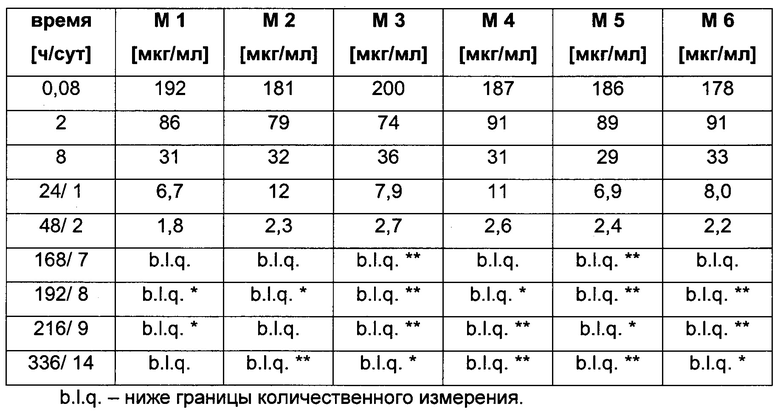
После введения mAb 9 иммунные комплексы лекарственное средство/ADA также впервые выявляли примерно через 168 часов (7 суток, Таблица 28). После введения устекинумаба у нокаутных по FcRn мышей не выявляли комплексов устекинумаб/ADA (Таблица 27). Кривые концентрация-время бриакинумаба и mAb 9 не покаывают быстрого снижения из-за образования ADA. Устекинумаб и mAb 9 имеют очень похожие кривые концентрация-время, указывающие на то, что образование ADA после введения mAb 9 не влияет на PK.
Таблица 22: сывороточные концентрации бриакинумаба у трансгеных мышей с человеческим FcRn. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу. ADA-позитивные образцы отмечены как * и ** для умеренного и сильного образования иммунных комплексов лекарственное средство/ADA соответственно.
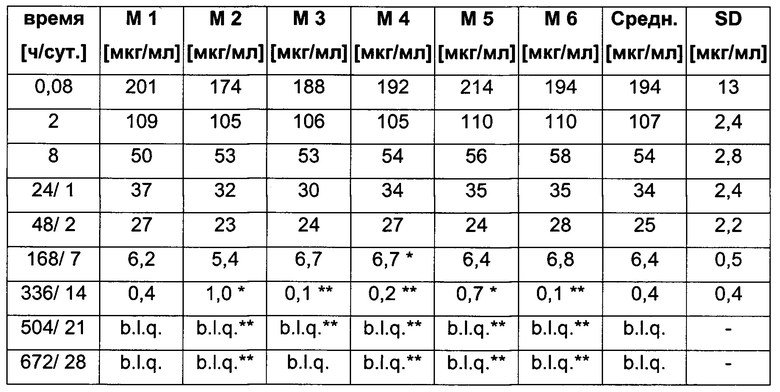
Таблица 23: сывороточные концентрации устекинумаба у трансгеных мышей с человеческим FcRn. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу. ADA-позитивные образцы отмечены как * и ** для умеренного и сильного образования иммунных комплексов лекарственное средство/ADA соответственно.
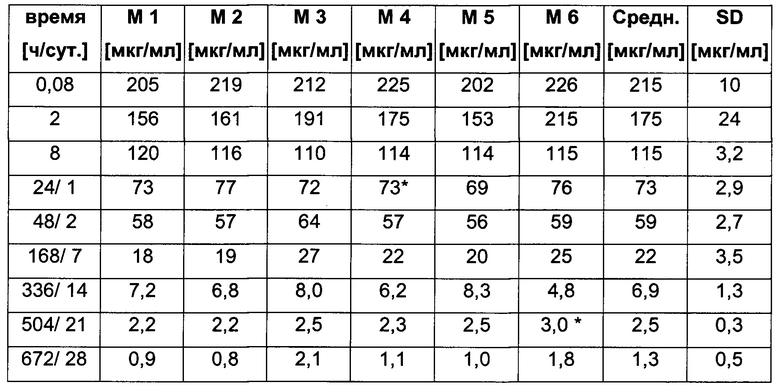
Таблица 24: сывороточные концентрации mAb 8 у трансгеных мышей с человеческим FcRn. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу. ADA-позитивные образцы отмечены как * и ** для умеренного и сильного образования иммунных комплексов лекарственное средство/ADA соответственно.
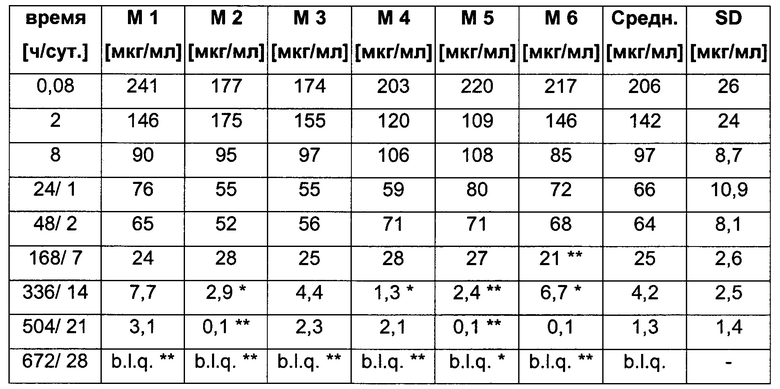
Таблица 25: сывороточные концентрации mAb 9 у трансгеных мышей с человеческим FcRn. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу. ADA-позитивные образцы отмечены как * и ** для умеренного и сильного образования иммунных комплексов лекарственное средство/ADA соответственно.
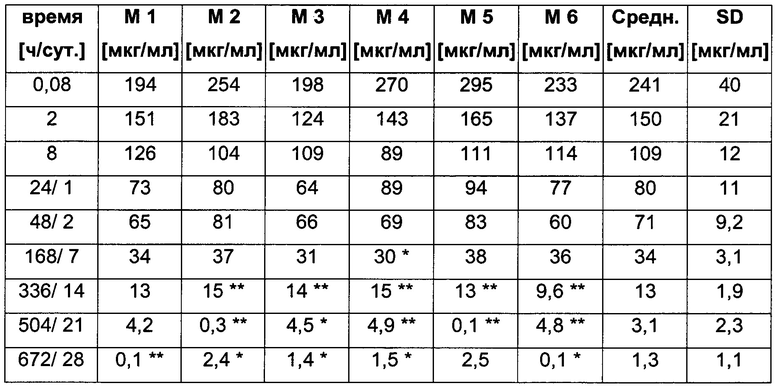
Таблица 26: сывороточные концентрации бриакинумаба у нокаутных по FcRn мышей. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу. ADA-позитивные образцы отмечены как * и ** для умеренного и сильного образования иммунных комплексов лекарственное средство/ADA соответственно.
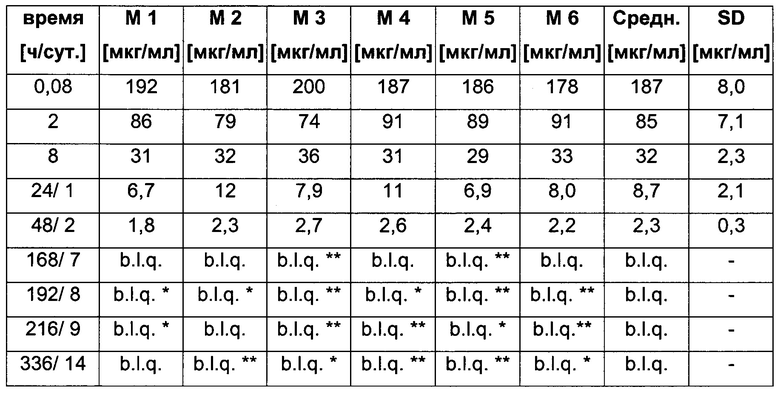
Таблица 27: сывороточные концентрации устекинумаба у нокаутных по FcRn мышей. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу.
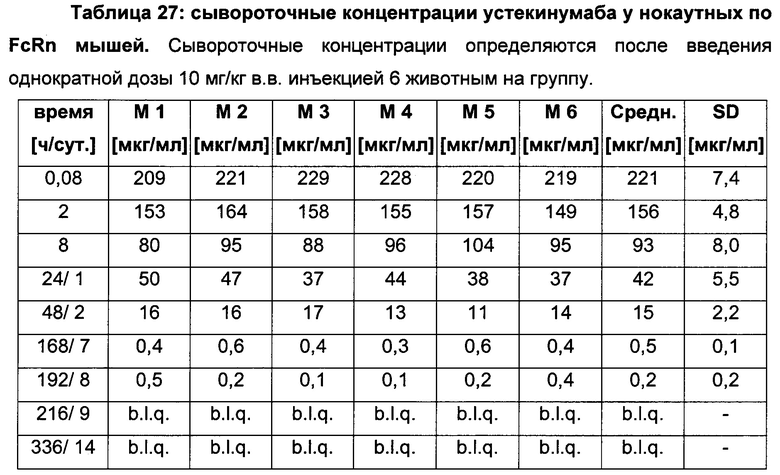
Таблица 28: сывороточные концентрации mAb 9 у нокаутных по FcRn мышей. Сывороточные концентрации определяются после введения однократной дозы 10 мг/кг в.в. инъекцией 6 животным на группу. ADA-позитивные образцы отмечены как * и ** для умеренного и сильного образования иммунных комплексов лекарственное средство/ADA соответственно.

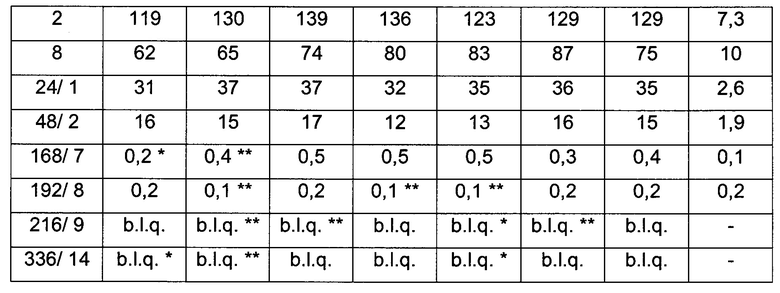
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОТБОРА АНТИТЕЛ С МОДИФИЦИРОВАННЫМ ВЗАИМОДЕЙСТВИЕМ С FcRn | 2015 |
|
RU2714153C2 |
| АФФИННАЯ ХРОМАТОГРАФИЯ С ПРИМЕНЕНИЕМ FC-РЕЦЕПТОРОВ | 2013 |
|
RU2624128C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |
| АНТИТЕЛА С РН-ЗАВИСИМЫМ СВЯЗЫВАНИЕМ АНТИГЕНА | 2011 |
|
RU2570729C2 |
| IL-8-СВЯЗЫВАЮЩИЕ АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2728430C2 |
| FC ВАРИАНТЫ АНТИТЕЛА | 2012 |
|
RU2607014C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С УЛУЧШЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С БЕЛКОМ А | 2015 |
|
RU2698969C2 |
| ГЕННО-ИНЖЕНЕРНЫЕ ПОЛИПЕПТИДЫ И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2822498C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| СПОСОБ ОТБОРА И ПОЛУЧЕНИЯ ВЫСОКОСЕЛЕКТИВНЫХ И МУЛЬТИСПЕЦИФИЧНЫХ НАЦЕЛИВАЮЩИХ ГРУПП С ЗАДАННЫМИ СВОЙСТВАМИ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ СВЯЗЫВАЮЩИЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2639287C2 |




















































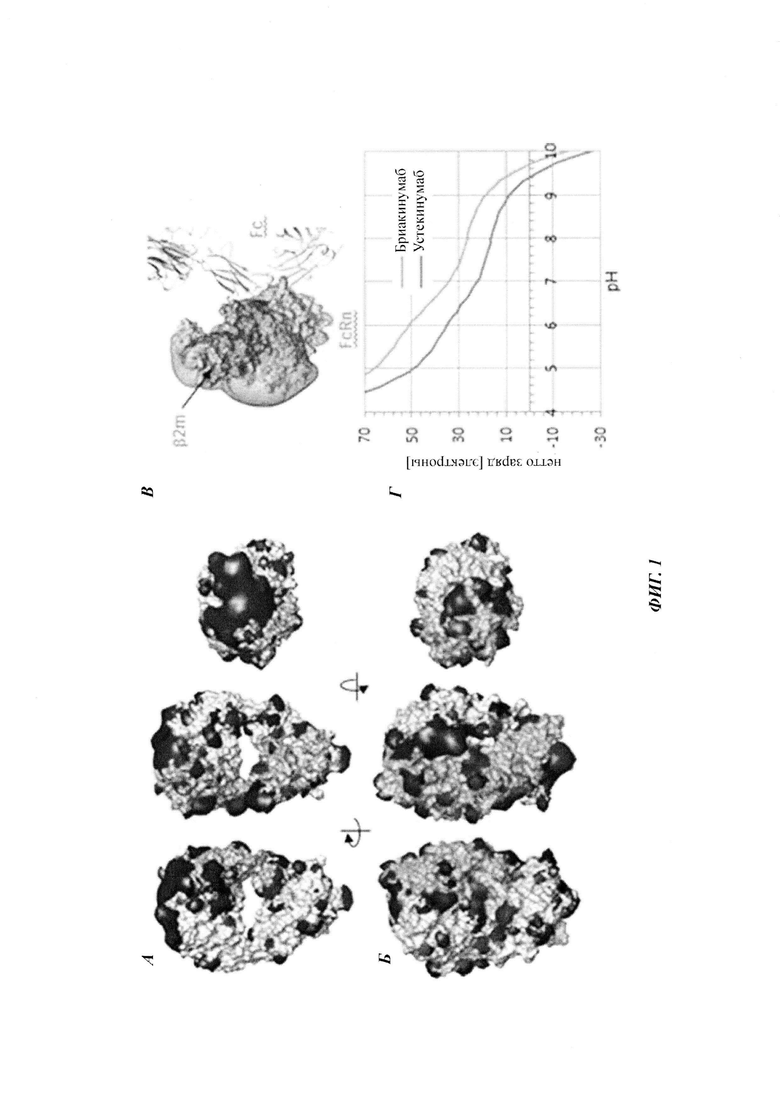

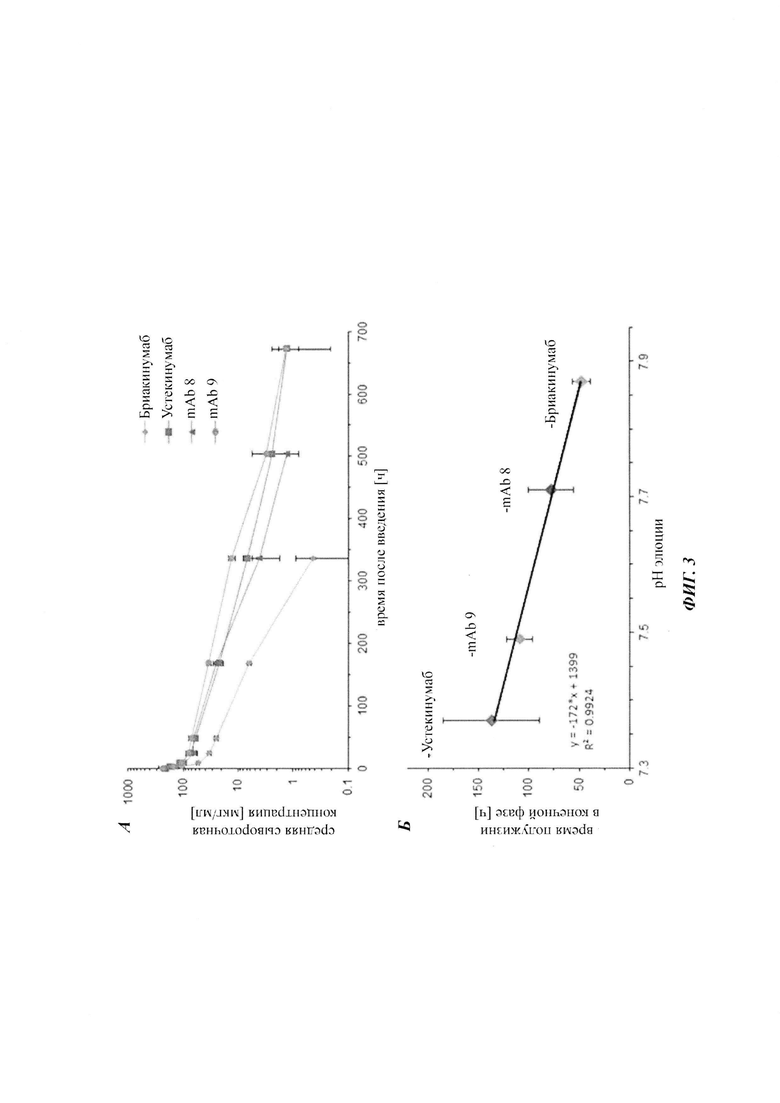
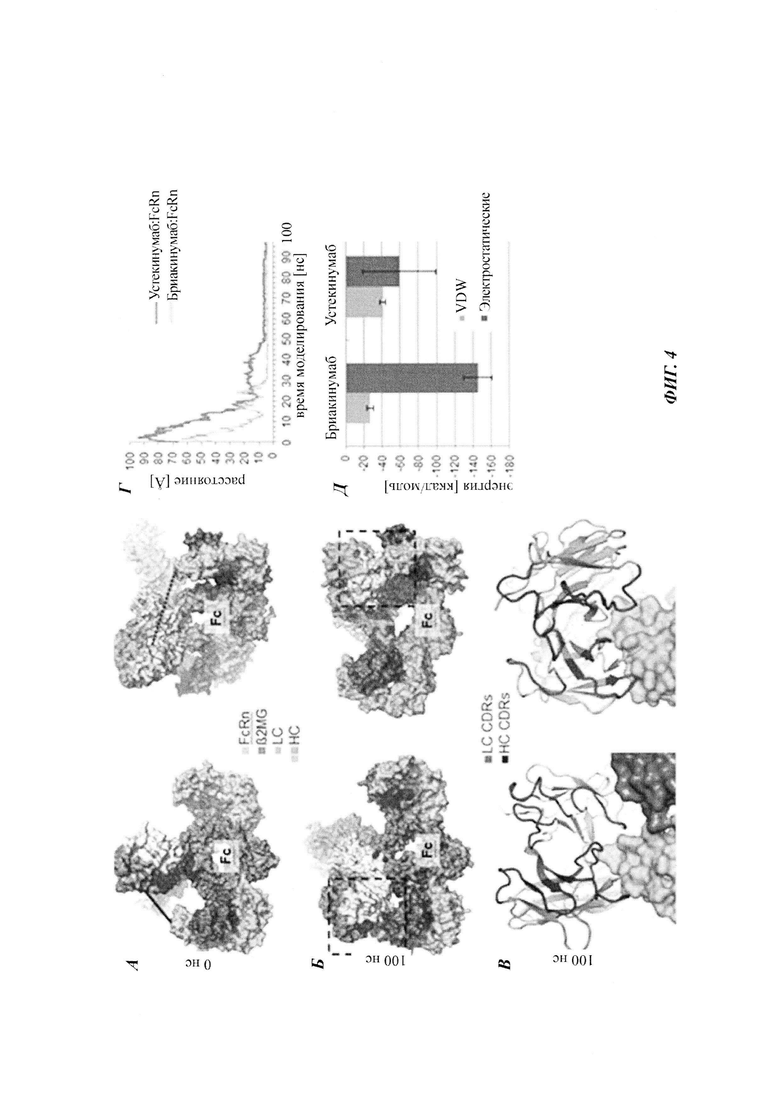


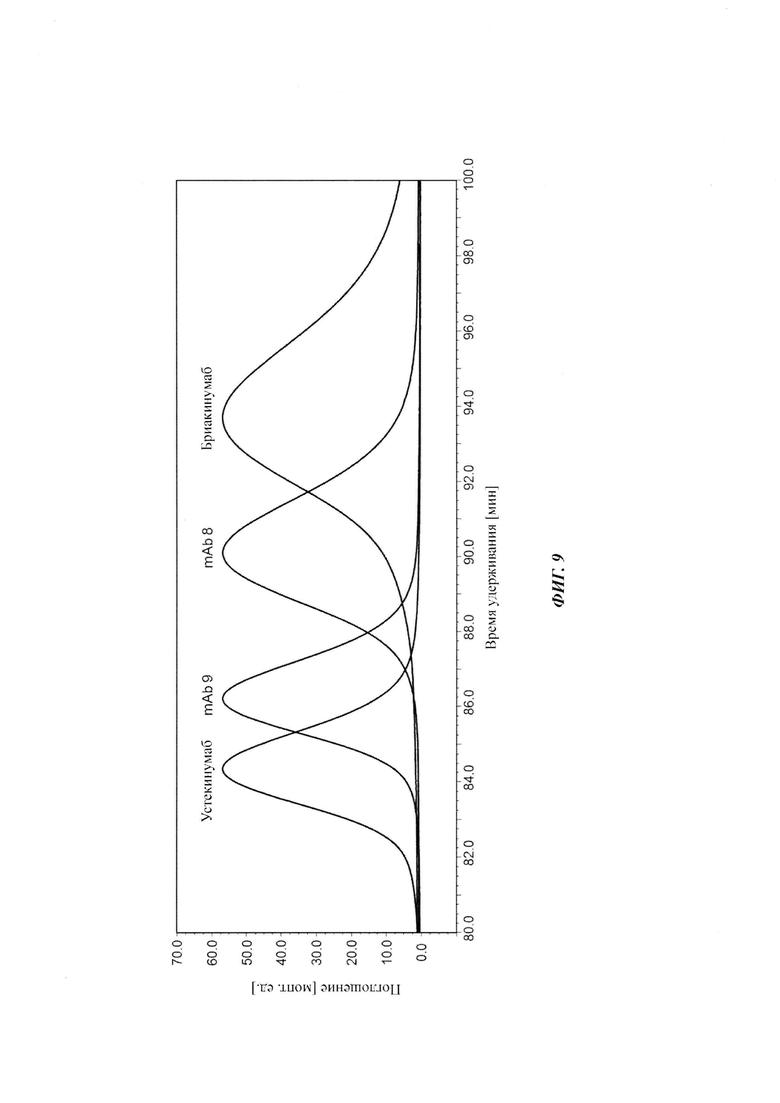
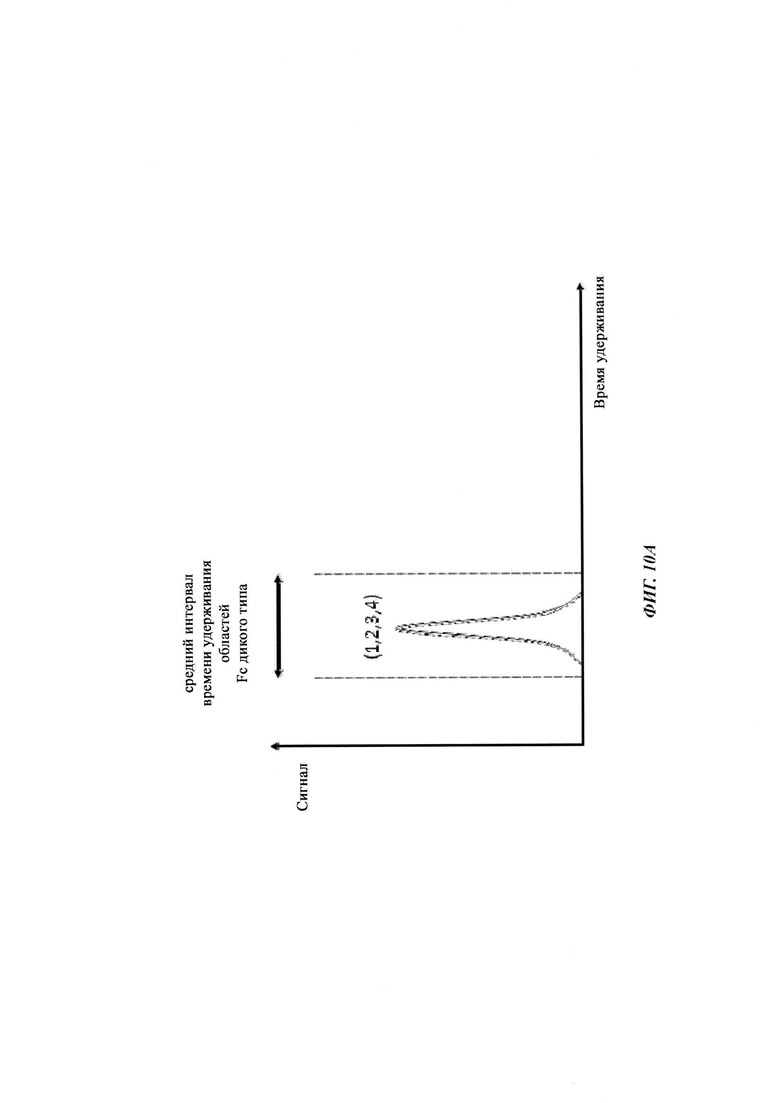
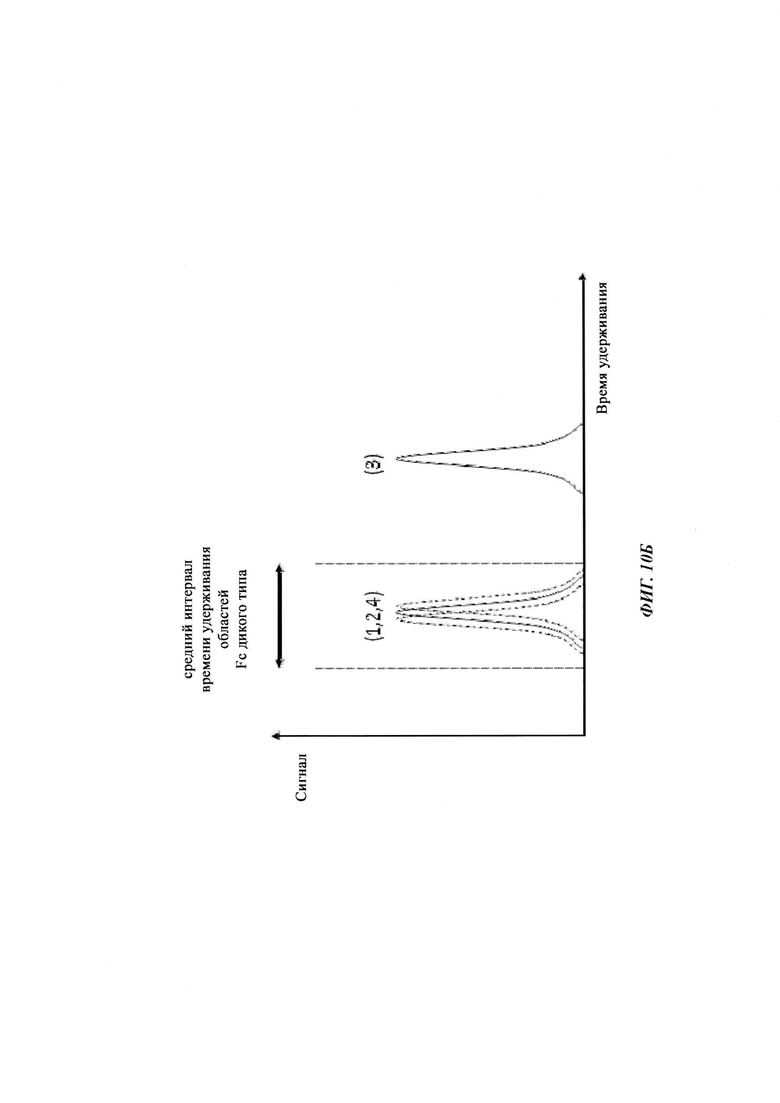
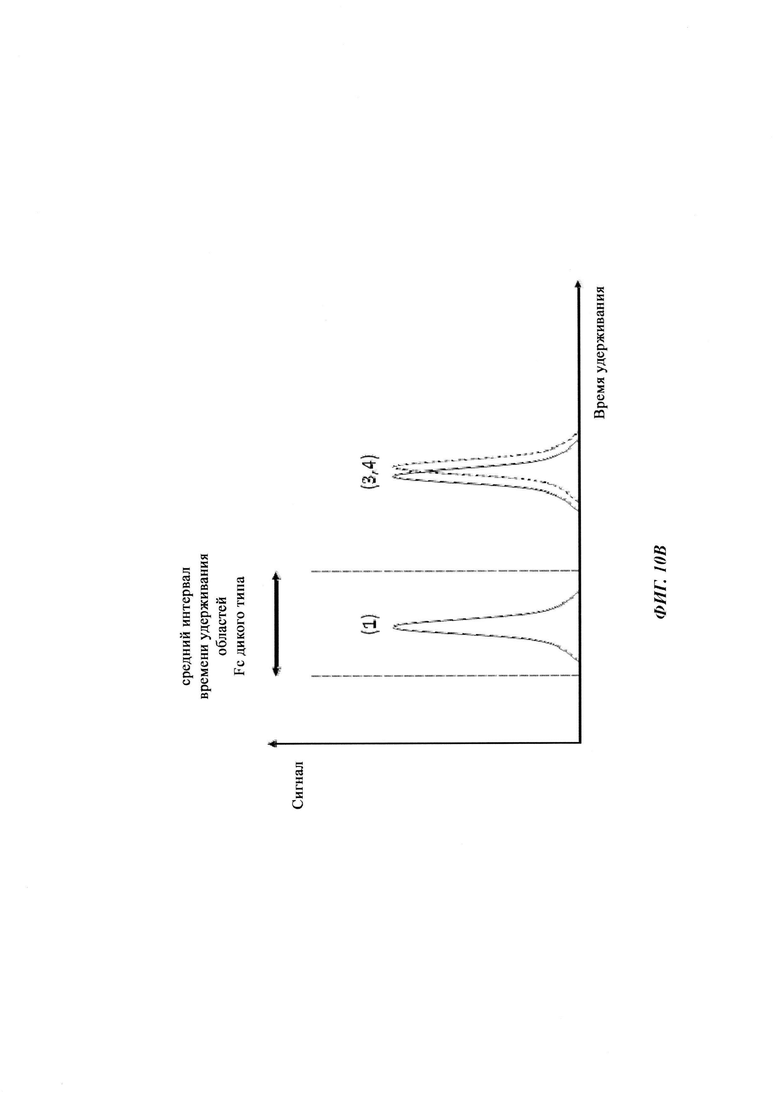
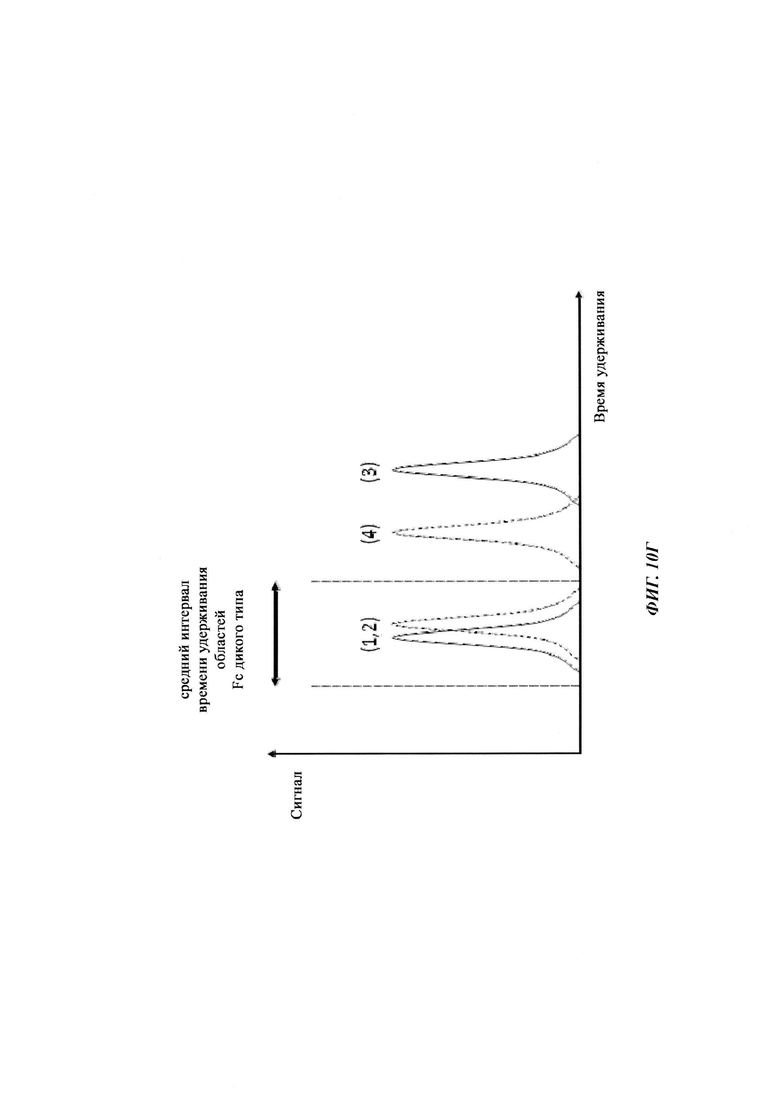
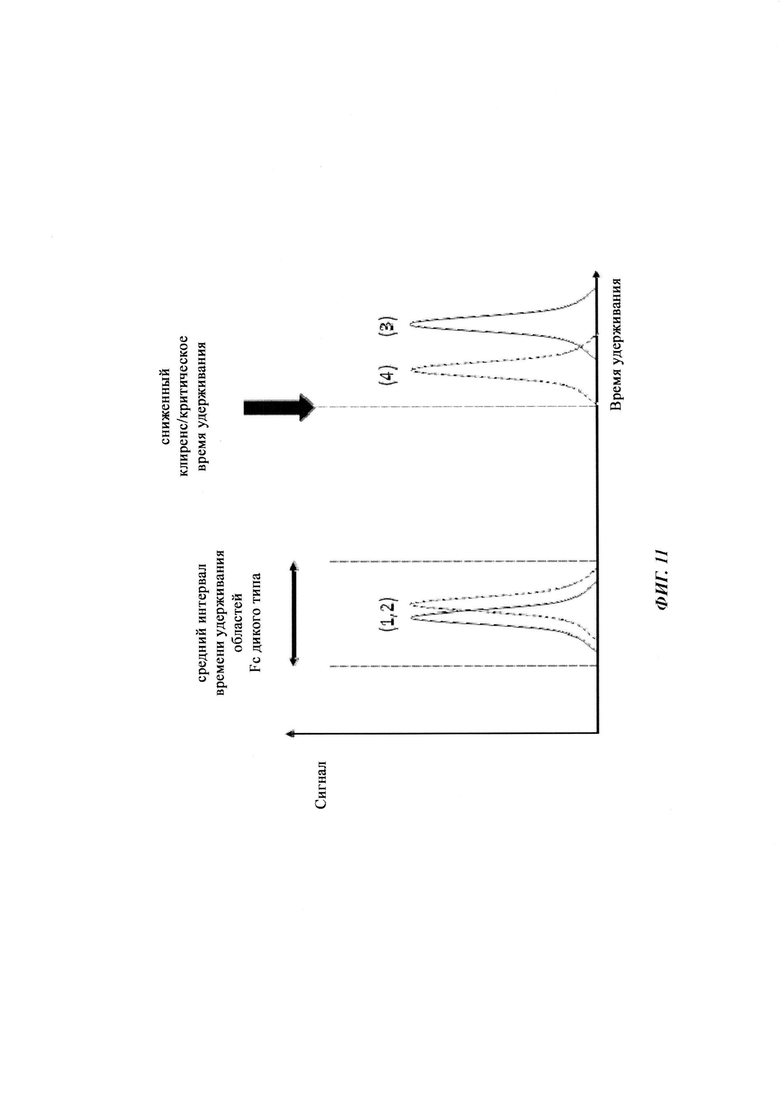
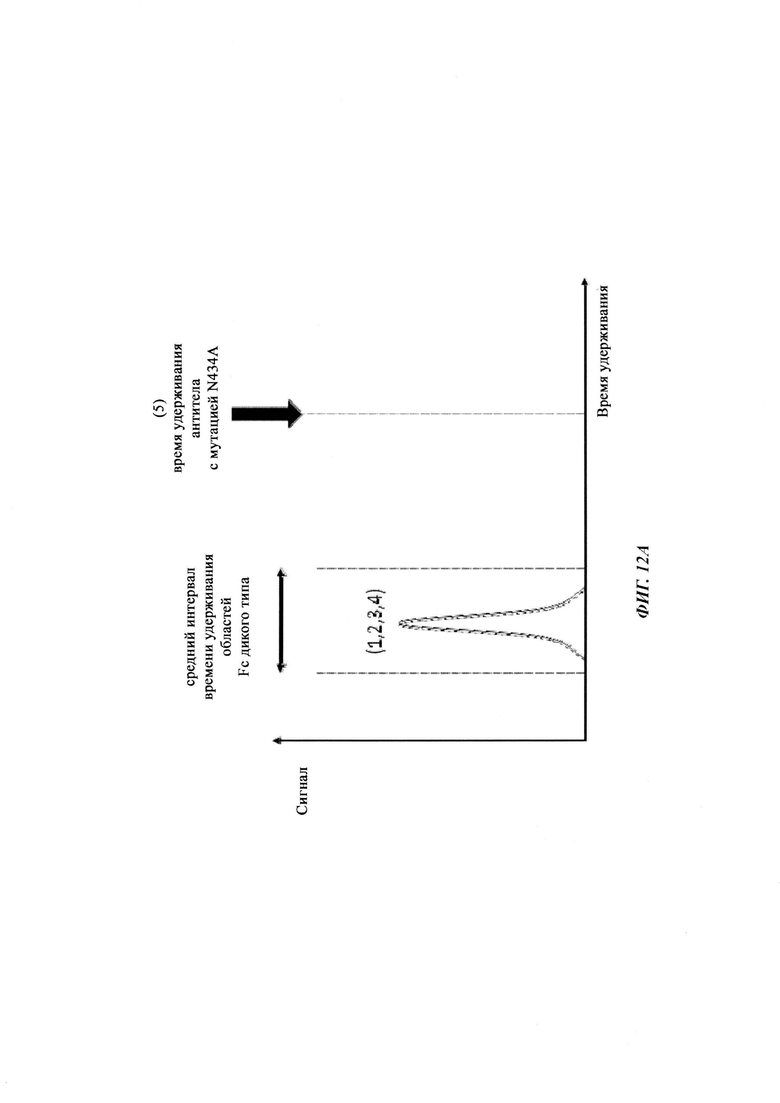
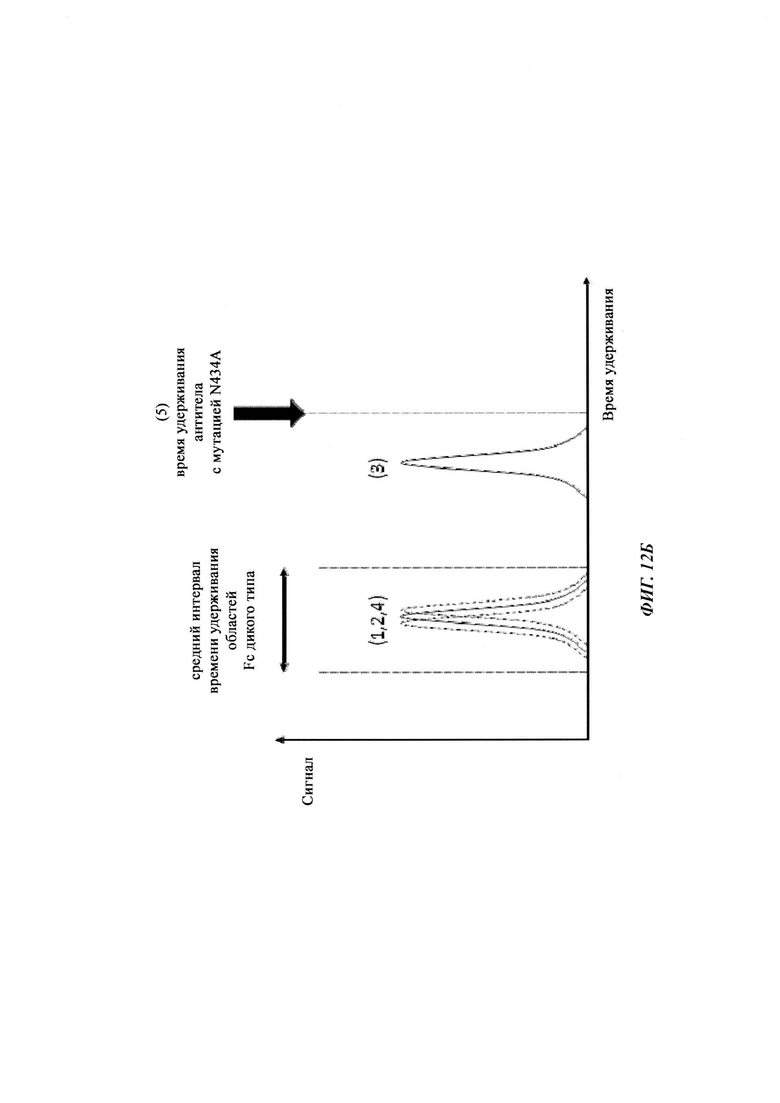
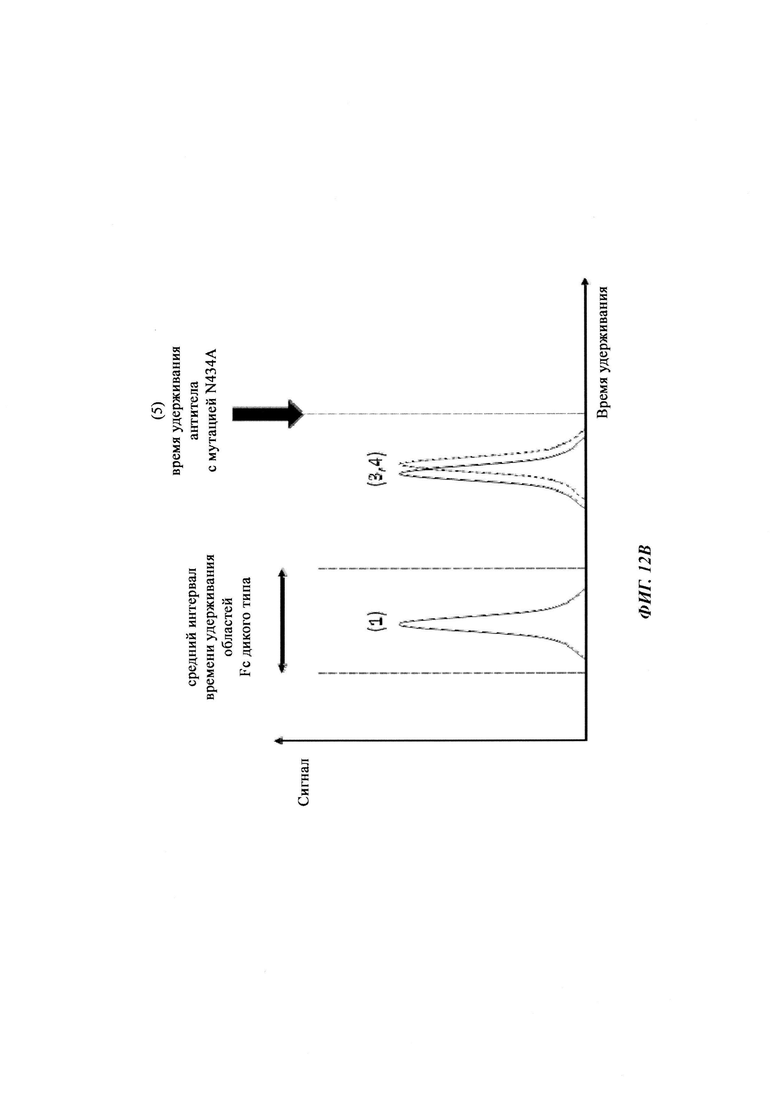
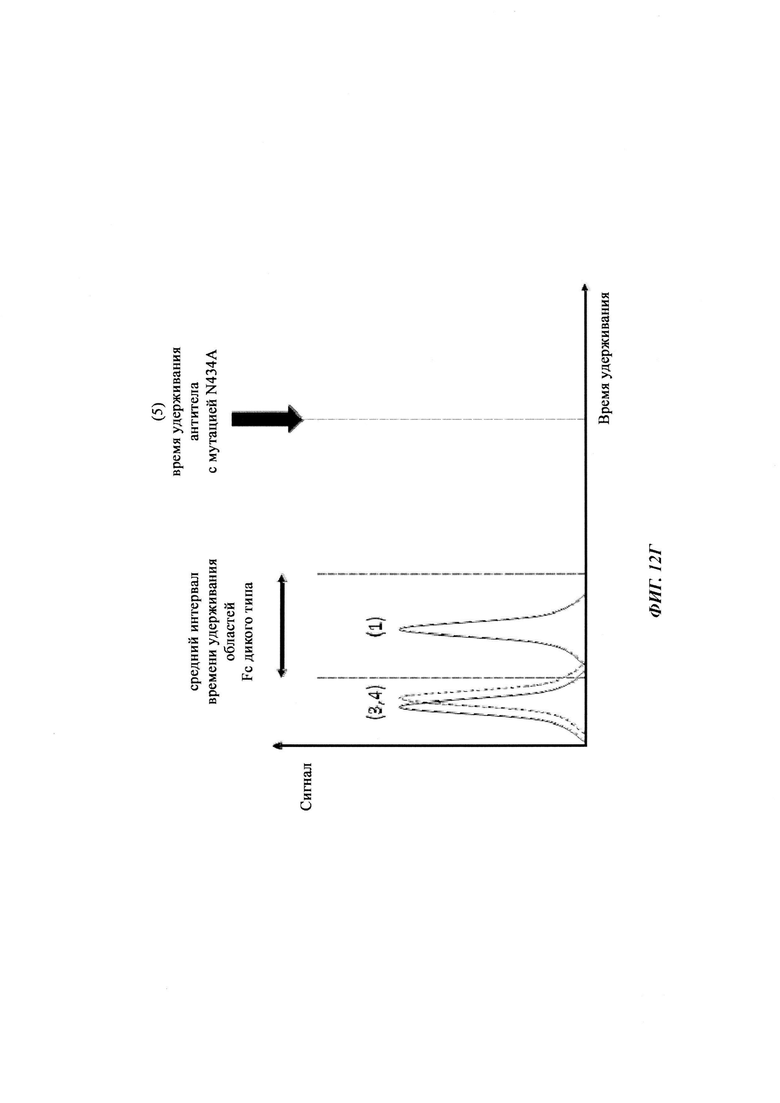
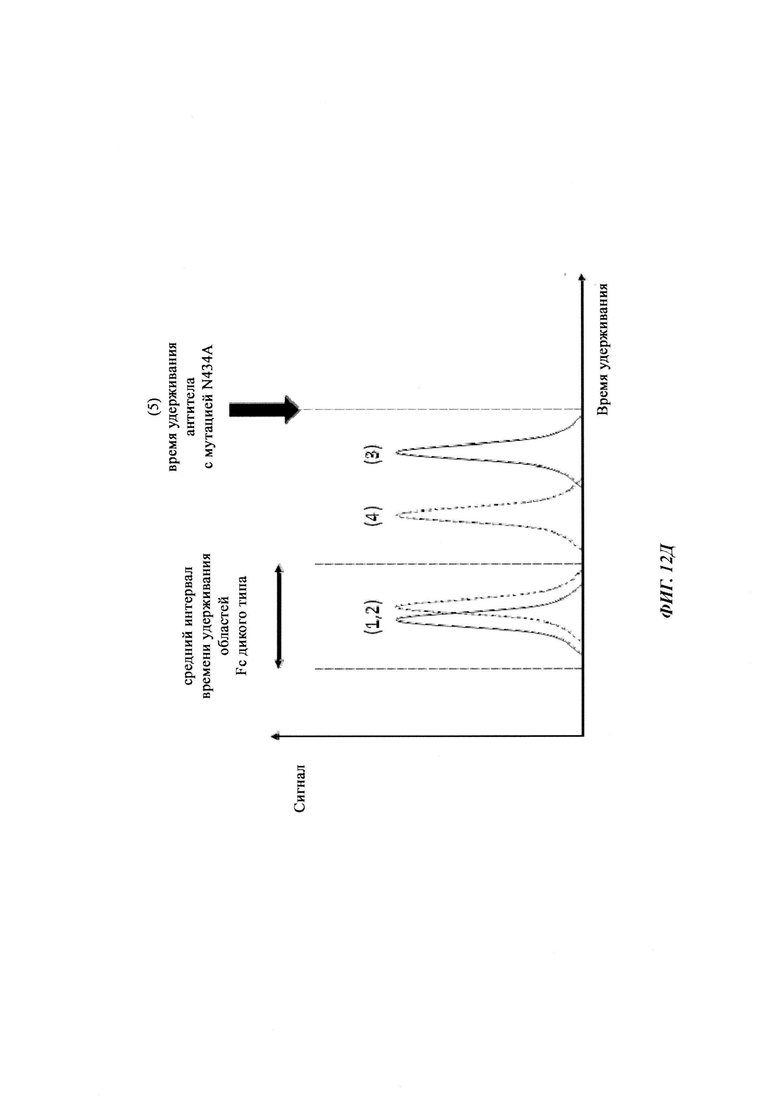
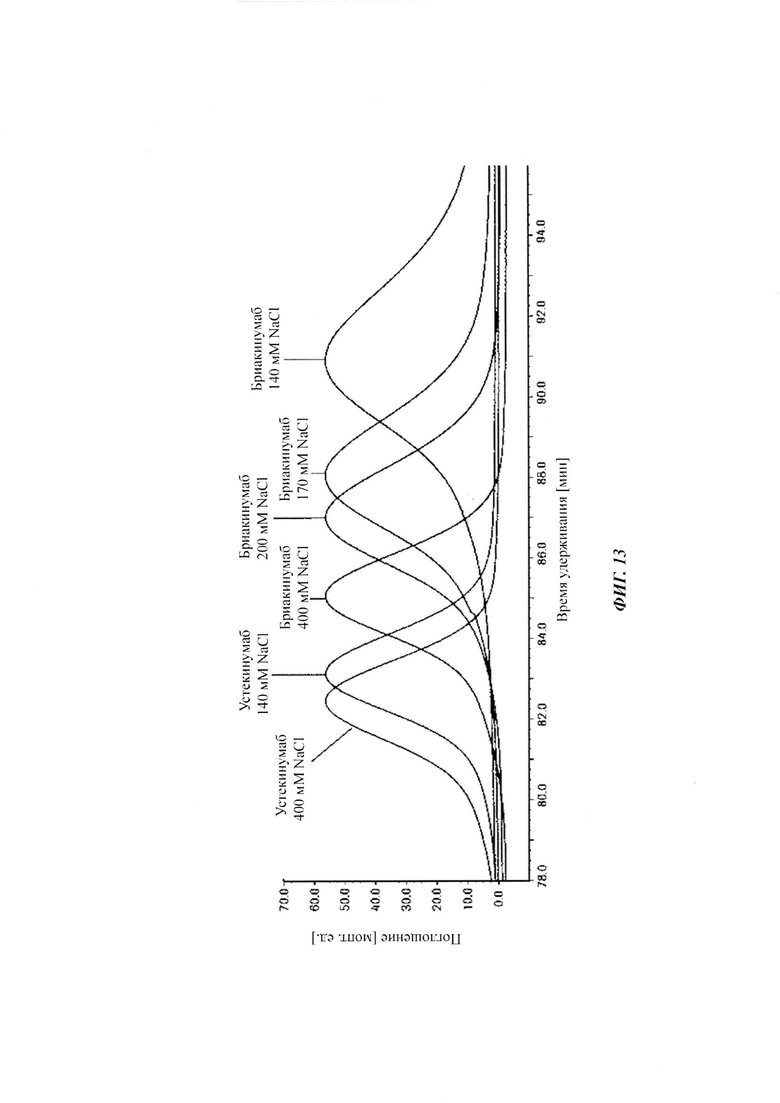

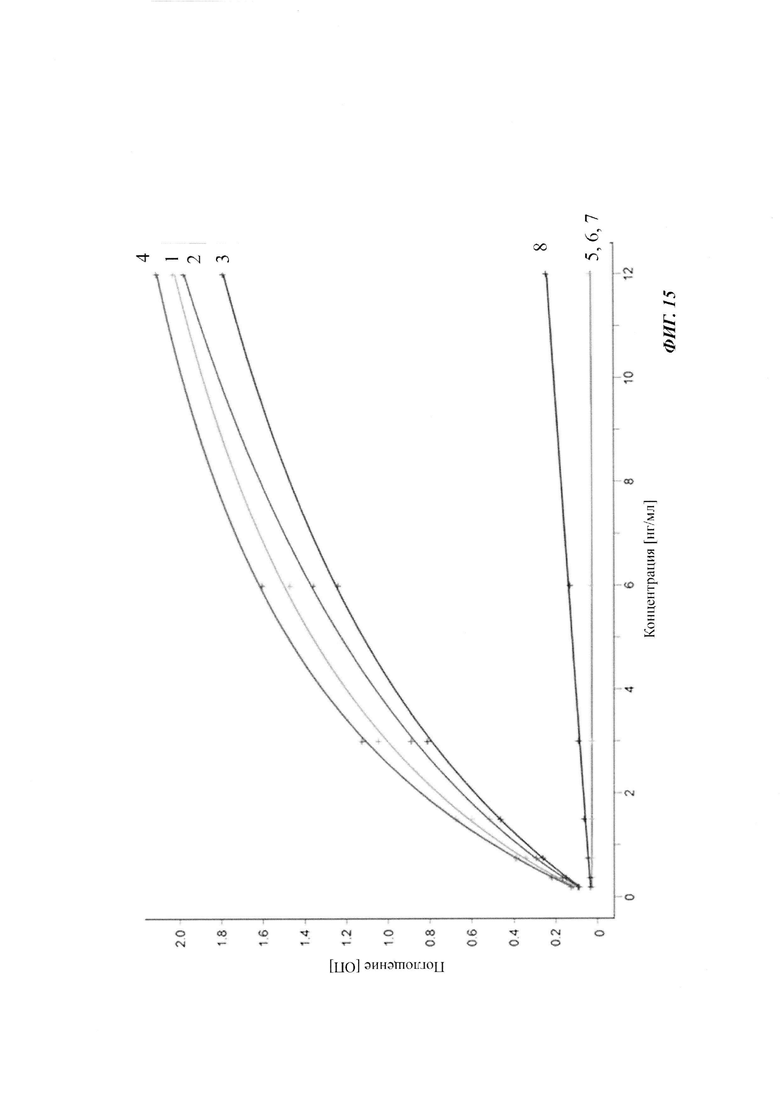
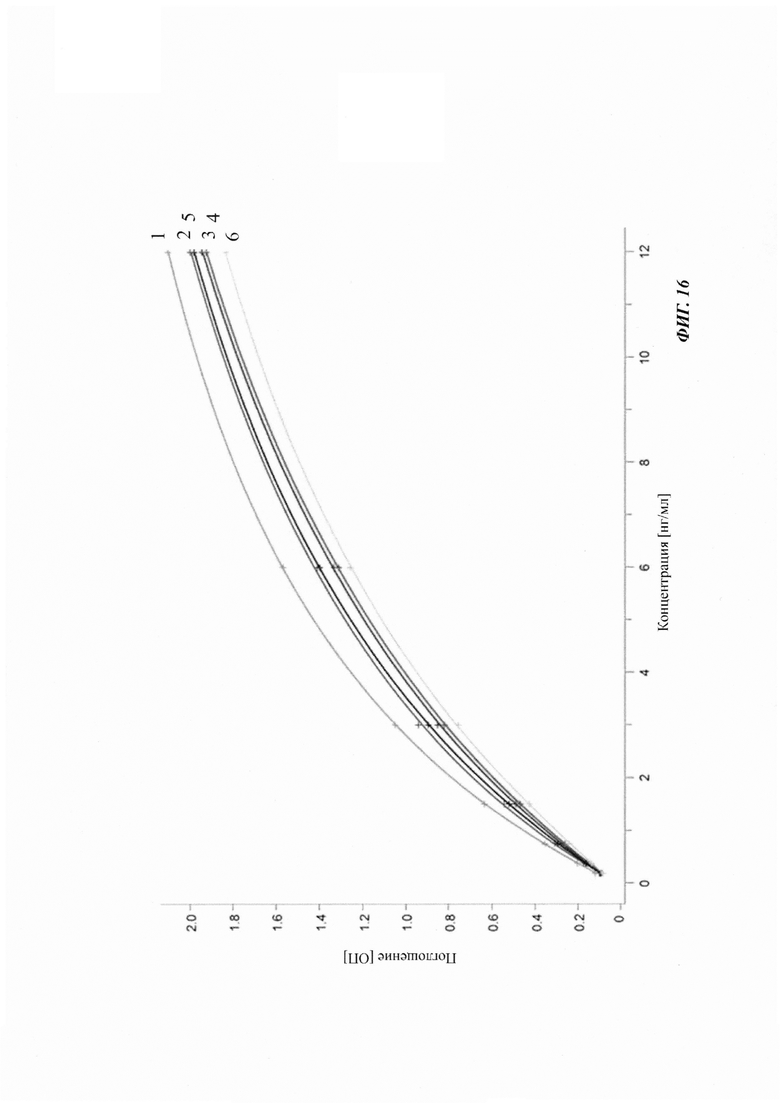
Изобретение относится к биотехнологии. Предложен способ отбора антитела с увеличенным или уменьшенным временем полужизни in vivo относительно полноразмерного контрольного антитела. Способ включает следующие стадии: а) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом рН в присутствии от 50 мМ до 200 мМ хлорида натрия; б) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом рН в присутствии от 300 мМ до 600 мМ хлорида натрия. В качестве антитела с увеличенным временем полужизни in vivo относительно контрольного антитела отбирают антитело, которое имеет время удерживания, определенное на стадии а), которое по меньшей мере на 5% больше, чем время удерживания контрольного антитела, определенное на стадии а), и время удерживания, определенное на стадии а), которое находится в пределах 5% от времени удерживания, определенного на стадии б). В качестве антитела с уменьшенным временем полужизни in vivo относительно контрольного антитела отбирают антитело, которое имеет время удерживания, определенное на стадии а), которое по меньшей мере на 5% меньше, чем время удерживания контрольного антитела, определенное на стадии а), и время удерживания, определенное на стадии а), которое находится в пределах 5% от времени удерживания, определенного на стадии б). 5 з.п. ф-лы, 16 ил., 28 табл., 11 пр.
1. Способ отбора антитела с увеличенным или уменьшенным временем полужизни in vivo относительно полноразмерого контрольного антитела, включающий следующие стадии:
а) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом рН в присутствии первой концентрации соли,
б) определение времени удерживания антитела и контрольного антитела на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом рН в присутствии второй концентрации соли,
где в случае отбора антитела с увеличенным временем полужизни in vivo относительно контрольного антитела отбирают антитело, которое имеет i) время удерживания, определенное на стадии а), которое по меньшей мере на 5% больше, чем время удерживания контрольного антитела, определенное на стадии а), и ii) время удерживания, определенное на стадии а), которое находится в пределах 5% от времени удерживания, определенного на стадии б),
где в случае отбора антитела с уменьшенным временем полужизни in vivo относительно контрольного антитела отбирают антитело, которое имеет i) время удерживания, определенное на стадии а), которое по меньшей мере на 5% меньше, чем время удерживания контрольного антитела, определенное на стадии а), и ii) время удерживания, определенное на стадии а), которое находится в пределах 5% от времени удерживания, определенного на стадии б),
где положительный линейный градиент рН составляет от рН 5,5 до рН 8,8,
где соль представляет собой хлорид натрия,
где первая концентрация соли составляет от 50 мМ до 200 мМ, и
где вторая концентрация соли составляет от 300 мМ до 600 мМ.
2. Способ по п. 1, где
- данный способ предназначен для отбора антитела, которое имеет относительное время полужизни in vivo, которое увеличивается по сравнению с антителом подкласса IgG1, IgG3 или IgG4,
- посредством этого, отбор антитела, которое имеет первое время удерживания, которое по меньшей мере на 5% больше, чем первое время удерживания контрольного антитела, и первое время удерживания, которое находится в пределах 5% от второго времени удерживания.
3. Способ по п. 1, где
дополнительно определяется время удерживания области Fc IgG с мутацией N434A и
- посредством этого, отбор
а) антитела, которое имеет первое время удерживания, которое по меньшей мере на 5% больше, чем первое время удерживания контрольного антитела, которое имеет первое время удерживания и второе время удерживания, которые находятся в пределах 5% друг от друга, и которое имеет первое время удерживания, которое по меньшей мере на 5% меньше, чем время удерживания области Fc с мутацией N434A, и, посредством этого, отбор антитела с относительно увеличенным временем полужизни in vivo, или
б) антитела, которое имеет первое время удерживания, которое по меньшей мере на 5% меньше, чем первое время удерживания контрольного антитела, и которое имеет первое время удерживания и второе время удерживания, которые находятся в пределах 5% друг от друга, осуществляют отбор антитела с относительно уменьшенным временем полужизни in vivo.
4. Способ по любому из пп. 1-3, где первая концентрация соли составляет 140 мМ.
5. Способ по любому из пп. 1-4, где вторая концентрация соли составляет 400 мМ.
6. Способ по любому из пп. 1-5, где, если времена удерживания отличаются по меньшей мере на 5%, данные времена удерживания пропорциональны единице, поделенной на квадратный корень концентрации соли (~1/√(концентрация соли)).
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| HOFFMANN-LA ROCHE AG) | |||
| Козлов И | |||
| Г | |||
| Моноклональные антитела новая эра в фармакологии и терапии //Лечебное дело | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| - n | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

