ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка заявляет приоритет предварительной заявки на патент США № 62/926313, поданной 25 октября 2019 г., и заявки на европейский патент № EP20315271.5, поданной 28 мая 2020 г., раскрытия которых включены в данный документ посредством ссылки в их полном объеме.
ПРЕДСТАВЛЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ТЕКСТОВОМ ФАЙЛЕ В ФОРМАТЕ ASCII
[0002] Содержание нижеследующего представленного текстового файла в формате ASCII включено в данный документ посредством ссылки в его полном объеме: машиночитаемая форма (CRF) перечня последовательностей (название файла: 183952032840SEQLIST.TXT, дата составления: 29 мая 2020 г., размер: 2 KБ).
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Настоящее изобретение относится к способам мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием клеткой-хозяином, а также к связанным с ними способам продуцирования и скрининга. Настоящее изобретение также относится к способам мониторинга продуцирования антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, клеткой-хозяином, а также к связанным с ними способам продуцирования и скрининга.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0004] Полиспецифические антитела, которые связывают два или более разных эпитопа на одном или разных антигенах, в последние годы стали перспективными терапевтическими вариантами в иммуноонкологии (Baeuerle, P. A. & Reinhardt, C., Cancer Res 2009, 69 (12), 4941-4). Нацеливание на несколько мишеней использовали для различных целей, таких как достижение повышенной специфичности лекарственных средств или имитация природных лигандов в путях передачи сигнала, например, в средствах для лечения гемофилии на основе одновременного связывания с парами рецепторов на поверхности одной и той же клетки (Kitazawa, T. et al. Nat Med 2012, 18 (10), 1570-4). Важным вариантом применения является концепция активатора Т-клеток (TCE), в котором одно плечо молекулы активирует Т-клетки посредством связывания с рецептором CD3/CD28, а другое плечо нацелено на опухолевый антиген для уничтожения опухоли (Krah, S. et al. N Biotechnol 2017, 39 (Pt B), 167-173; Correnti, C. E. et al. Leukemia 2018, 32 (5), 1239-1243).
[0005] IgG-подобные триспецифические антитела (tsAb) состоят из двух разных тяжелых цепей и двух разных легких цепей, обычно экспрессируемых в одной клетке-хозяине с последующей внутриклеточной сборкой цепей. Хотя в этом способе продуцирования устраняется потребность в наличии двух отдельных линий клеток и процессов очистки, в нем могут образовываться нежелательные молекулы с ошибочным спариванием в дополнение к необходимому tsAb. В отсутствие рационального дизайна и при случайной ассоциации субъединиц теоретический выход молекул с правильным спариванием составляет всего 12,5% (ФИГ. 1B). Принудительная гетеродимеризация тяжелых цепей была успешно выполнена с применением подходов на основе белковой инженерии, таких как дизайн "выступы-во-впадины" (Ridgway, J. B. et al. Protein Eng 1996, 9 (7), 617-21). Однако, когнатное спаривание легких цепей с правильными тяжелыми цепями остается сложной задачей при продуцировании tsAb.
[0006] Таким образом, сохраняется потребность в способах мониторинга или анализа продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием. Кроме того, сохраняется необходимость в способах мониторинга или анализа продуцирования антитела или производных антител, которые предусматривают несколько молекул, отличающихся по молекулярной массе (например, с различными химическими модификациями, такими как химически модифицированный цистеин или другие остатки, или с разными гликоформами).
[0007] Все приведенные в данном документе ссылочные материалы, в том числе заявки на патенты, патентные публикации и номера доступа в UniProtKB/Swiss-Prot, включены в данный документ посредством ссылки в их полном объеме, как если бы каждый отдельный ссылочный материал был специально и отдельно указан для включения в качестве ссылки.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0008] Для удовлетворения этих и других потребностей в данном документе предусмотрены способы мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием (например, линией клеток). В этих способах предусмотрена, среди прочего, высокопроизводительная аналитическая платформа, основанная на SEC-LC и MS интактных молекул в денатурирующих условиях, для идентификации и относительной количественной оценки ошибочного спаривания цепей и других молекул, связанных с IgG. Предпочтительно, эти способы позволяют проводить MS-анализ интактных связанных с mAb молекул (например, полиспецифических связывающих белков, антител, белков, являющихся продуктом слияния с Fc, фрагментов антител и т. д.), непосредственно из осветленного собранного материала клеток, за счет чего избегают трудоемкой и дорогостоящей очистки (например, аффинной хроматографии с использованием белка А) и этапов замены буфера. Таким образом, эти способы можно применять для быстрого скрининга большого количества клонов в отношении потенциальных линий клеток-продуцентов.
[0009] В некоторых вариантах осуществления в данном документе предусмотрены способы мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием, где способы включают определение посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS) количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием в среде для культивирования клеток, содержащей полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием. В некоторых вариантах осуществления полиспецифический связывающий белок предусматривает ассоциацию двух или более полипептидных цепей, содержащих по меньшей мере первую полипептидную цепь и вторую полипептидную цепь, отличную от первой полипептидной цепи, и одна или несколько молекул с ошибочным спариванием содержат две или более полипептидных цепей, содержащих по меньшей мере одну из первой и второй полипептидных цепей в ассоциации, отличной от ассоциации полиспецифического связывающего белка.
[0010] В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой полиспецифическое антитело, содержащее тяжелую цепь первого антитела, легкую цепь первого антитела, тяжелую цепь второго антитела, отличную от тяжелой цепи первого антитела, и легкую цепь второго антитела, отличную от легкой цепи первого антитела. В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой биспецифическое антитело, фрагмент антитела или белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой триспецифическое антитело, фрагмент антитела или белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего: ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две тяжелые цепи первого антитела; ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две тяжелые цепи второго антитела; ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две легкие цепи первого антитела; и ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две легкие цепи второго антитела. В некоторых вариантах осуществления полиспецифический связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих сайта,
при этом первая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
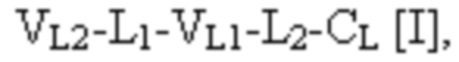
и вторая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
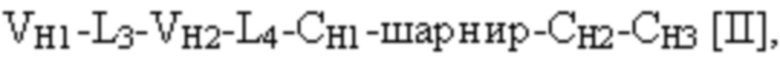
и третья полипептидная цепь связывающего белка содержит структуру, представленную формулой:
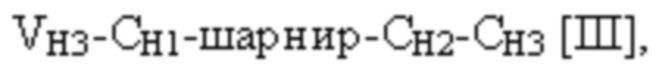
и четвертая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
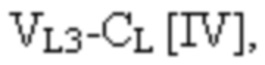
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
при этом полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи, где VH1 и VL1 образуют первый антигенсвязывающий сайт, где VH2 и VL2 образуют второй антигенсвязывающий сайт, и где VH3 и VL3 образуют третий антигенсвязывающий сайт. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего: ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой I; ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой II; ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой III; и ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой IV.
[0011] В некоторых вариантах осуществления определение количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием предусматривает деконволюцию одного или нескольких MS-спектров, полученных посредством MS. В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка по сравнению с одной или несколькими молекулами с ошибочным спариванием. В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка по сравнению с количеством одной или нескольких отдельных молекул с ошибочным спариванием. В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка по сравнению с общим количеством молекул с ошибочным спариванием. В некоторых вариантах осуществления перед определением способы дополнительно включают предоставление или получение (например, из линии клеток, которая продуцирует полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием) среды для культивирования клеток, содержащей полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием. В некоторых вариантах осуществления перед определением способы дополнительно включают отделение среды для культивирования клеток от линии клеток, которая продуцирует полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием. В некоторых вариантах осуществления перед определением среду для культивирования клеток отделяют от линии клеток посредством центрифугирования. В некоторых вариантах осуществления среду для культивирования клеток подвергают SE-UPLC без предварительного хроматографического разделения. В некоторых вариантах осуществления среду для культивирования клеток подвергают SE-UPLC без предварительной аффинной хроматографии с использованием белка А. В некоторых вариантах осуществления MS представляет собой MS интактных молекул. В некоторых вариантах осуществления MS представляет собой квадрупольную времяпролетную (QToF) MS. В некоторых вариантах осуществления SE-UPLC представляет собой SE-UPLC в денатурирующих условиях. В некоторых вариантах осуществления SE-UPLC объединена с MS непосредственно. В некоторых вариантах осуществления перед SE-UPLC-MS способы дополнительно включают приведение полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием в контакт с протеазой. В некоторых вариантах осуществления протеаза представляет собой IdeS или IdeZ. В некоторых вариантах осуществления SE-UPLC проводят с начальной скоростью потока, составляющей менее приблизительно 0,4 мл/мин. В некоторых вариантах осуществления SE-UPLC проводят с начальной скоростью потока, составляющей приблизительно 0,1 мл/мин. В некоторых вариантах осуществления SE-UPLC проводят со скоростью потока, составляющей приблизительно 0,1 мл/мин. в течение первых 25 минут, а затем со скоростью потока, составляющей приблизительно 0,4 мл/мин. (например, в течение минут 25-33). В некоторых вариантах осуществления SE-UPLC проводят с использованием изократического элюирования подвижной фазой. В некоторых вариантах осуществления подвижная фаза содержит раствор, содержащий смесь ацетонитрил:вода в соотношении 30:70. В некоторых вариантах осуществления подвижная фаза содержит муравьиную кислоту и трифторуксусную кислоту (TFA). В некоторых вариантах осуществления подвижная фаза содержит приблизительно 0,05% муравьиной кислоты и приблизительно 0,05% трифторуксусной кислоты (TFA). В некоторых вариантах осуществления определение количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием выполняют за приблизительно 33 минуты или меньше. В некоторых вариантах осуществления, В некоторых вариантах осуществления выполнение способа занимает приблизительно 33 минуты или меньше. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 300 Да, между полиспецифическим связывающим белком и одной или несколькими молекулами с ошибочным спариванием или между двумя молекулами с ошибочным спариванием. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 162 Да, между полиспецифическим связывающим белком или молекулами с ошибочным спариванием и одной или несколькими гликоформами.
[0012] В некоторых вариантах осуществления линия клеток представляет собой линию клеток млекопитающего. В некоторых вариантах осуществления линия клеток представляет собой линию клеток яичника китайского хомячка (СНО). В некоторых вариантах осуществления перед определением линию клеток культивируют с использованием среды для культивирования клеток в непрерывной культуре клеток, например, в биореакторе с перемешиванием. В некоторых вариантах осуществления перед определением линию клеток культивируют с использованием среды для культивирования клеток в периодической культуре клеток.
[0013] В некоторых вариантах осуществления в данном документе предусмотрены способы получения полиспецифического связывающего белка, включающие: (a) культивирование линии клеток, содержащей один или несколько полинуклеотидов, кодирующих полиспецифический связывающий белок, в среде для культивирования клеток в условиях, подходящих для продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием линией клеток; (b) отделение от линии клеток среды для культивирования клеток, содержащей полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием; (c) определение количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием в среде для культивирования клеток посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS); и (d) удаление по меньшей мере некоторой доли одной или нескольких молекул с ошибочным спариванием из полиспецифического связывающего белка, продуцируемого линией клеток, или определение одного или обоих из показателей качества и чистоты полиспецифического связывающего белка, продуцируемого линией клеток. В некоторых вариантах осуществления полиспецифический связывающий белок предусматривает ассоциацию двух или более полипептидных цепей, содержащую по меньшей мере первую полипептидную цепь и вторую полипептидную цепь, отличную от первой полипептидной цепи. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием содержат две или более полипептидных цепей, содержащих по меньшей мере одну из первой и второй полипептидных цепей в ассоциации, отличной от ассоциации полиспецифического связывающего белка.
[0014] В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой полиспецифическое антитело, содержащее тяжелую цепь первого антитела, легкую цепь первого антитела, тяжелую цепь второго антитела, отличную от тяжелой цепи первого антитела, и легкую цепь второго антитела, отличную от легкой цепи первого антитела. В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой биспецифическое антитело или белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой триспецифическое антитело или белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего: ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две тяжелые цепи первого антитела; ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две тяжелые цепи второго антитела; ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две легкие цепи первого антитела; и ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две легкие цепи второго антитела. В некоторых вариантах осуществления полиспецифический связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих сайта,
при этом первая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
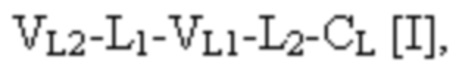
и вторая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
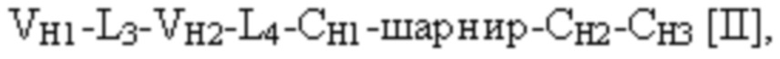
и третья полипептидная цепь связывающего белка содержит структуру, представленную формулой:
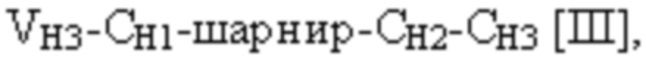
и четвертая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
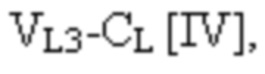
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи, где VH1 и VL1 образуют первый антигенсвязывающий сайт, где VH2 и VL2 образуют второй антигенсвязывающий сайт, и где VH3 и VL3 образуют третий антигенсвязывающий сайт. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего: ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой I; ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой II; ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой III; и ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой IV.
[0015] В некоторых вариантах осуществления определение количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием предусматривает деконволюцию одного или нескольких MS-спектров, полученных посредством MS. В некоторых вариантах осуществления определение полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием предусматривает оценку общего титра полиспецифического связывающего белка, продуцируемого линией клеток. В некоторых вариантах осуществления определение полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием предусматривает оценку общего титра молекул с ошибочным спариванием, продуцируемых линией клеток. В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка по сравнению с одной или несколькими молекулами с ошибочным спариванием. В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка по сравнению с количеством одной или нескольких отдельных молекул с ошибочным спариванием. В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка по сравнению с общим количеством молекул с ошибочным спариванием. В некоторых вариантах осуществления среду для культивирования клеток отделяют от линии клеток посредством центрифугирования. В некоторых вариантах осуществления в рамках (с) среду для культивирования клеток подвергают SE-UPLC без предварительного хроматографического разделения. В некоторых вариантах осуществления в рамках (с) среду для культивирования клеток подвергают SE-UPLC без предварительной аффинной хроматографии с использованием белка А. В некоторых вариантах осуществления MS представляет собой MS интактных молекул. В некоторых вариантах осуществления MS представляет собой квадрупольную времяпролетную (QToF) MS. В некоторых вариантах осуществления SE-UPLC представляет собой SE-UPLC в денатурирующих условиях. В некоторых вариантах осуществления SE-UPLC объединена с MS непосредственно. В некоторых вариантах осуществления после культивирования линии клеток и перед определением способы дополнительно включают приведение полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием в контакт с протеазой. В некоторых вариантах осуществления протеаза представляет собой IdeS или IdeZ. В некоторых вариантах осуществления SE-UPLC проводят с начальной скоростью потока, составляющей менее приблизительно 0,4 мл/мин. В некоторых вариантах осуществления SE-UPLC проводят с начальной скоростью потока, составляющей приблизительно 0,1 мл/мин. В некоторых вариантах осуществления SE-UPLC проводят со скоростью потока, составляющей приблизительно 0,1 мл/мин. в течение первых 25 минут, а затем со скоростью потока, составляющей приблизительно 0,4 мл/мин. В некоторых вариантах осуществления SE-UPLC проводят с использованием изократического элюирования подвижной фазой. В некоторых вариантах осуществления подвижная фаза содержит раствор, содержащий смесь ацетонитрил:вода в соотношении 30:70. В некоторых вариантах осуществления подвижная фаза содержит муравьиную кислоту и трифторуксусную кислоту (TFA). В некоторых вариантах осуществления подвижная фаза содержит приблизительно 0,05% муравьиной кислоты и приблизительно 0,05% трифторуксусной кислоты (TFA). В некоторых вариантах осуществления определение выполняют за приблизительно 33 минуты или меньше. В некоторых вариантах осуществления выполнение способа занимает приблизительно 33 минуты или меньше. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 300 Да, между полиспецифическим связывающим белком и одной или несколькими молекулами с ошибочным спариванием или между двумя молекулами с ошибочным спариванием. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 162 Да, между полиспецифическим связывающим белком или молекулами с ошибочным спариванием и одной или несколькими гликоформами.
[0016] В некоторых вариантах осуществления линия клеток представляет собой линию клеток млекопитающего. В некоторых вариантах осуществления линия клеток представляет собой линию клеток яичника китайского хомячка (СНО). В некоторых вариантах осуществления линию клеток культивируют в непрерывной культуре клеток (например, в биореакторе с перемешиванием). В некоторых вариантах осуществления линию клеток культивируют в периодической культуре клеток.
[0017] В некоторых вариантах осуществления в данном документе предусмотрены способы скрининга совокупности линий клеток в отношении продуцирования полиспецифического связывающего белка, где способы включают: определение количества полиспецифического связывающего белка, продуцируемого первой линией клеток из совокупности, в соответствии со способом по любому из приведенных выше вариантов осуществления; и определение количества полиспецифического связывающего белка, продуцируемого второй линией клеток из совокупности, в соответствии со способом по любому из приведенных выше вариантов осуществления.
[0018] В некоторых вариантах осуществления способы дополнительно включают: сравнение количества полиспецифического связывающего белка, продуцируемого первой линией клеток, с количеством полиспецифического связывающего белка, продуцируемым второй линией клеток; и основывающийся на сравнении выбор линии клеток, которая продуцировала более высокое количество полиспецифического связывающего белка. В некоторых вариантах осуществления способы дополнительно включают: определение количества одной или нескольких молекул с ошибочным спариванием, продуцируемого первой линией клеток, в соответствии со способом по любому из приведенных выше вариантов осуществления; и определение количества одной или нескольких молекул с ошибочным спариванием, продуцируемого второй линией клеток, в соответствии со способом по любому из приведенных выше вариантов осуществления. В некоторых вариантах осуществления после определения количества одной или нескольких молекул с ошибочным спариванием, продуцируемого первой и второй линиями клеток, способы дополнительно включают: сравнение количества одной или нескольких молекул с ошибочным спариванием, продуцируемого первой линией клеток, с количеством одной или нескольких молекул с ошибочным спариванием, продуцируемым второй линией клеток; и основывающийся на сравнении выбор линии клеток, при продуцировании в которой значение соотношения полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием является более высоким. В некоторых вариантах осуществления линию клеток выбирают на основе более высокого значения соотношения количества полиспецифического связывающего белка и количества одной или нескольких отдельных молекул с ошибочным спариванием. В некоторых вариантах осуществления линию клеток выбирают на основе более высокого значения соотношения количества полиспецифического связывающего белка и общего количества молекул с ошибочным спариванием.
[0019] В некоторых вариантах осуществления в данном документе предусмотрены способы мониторинга продуцирования антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, где способы включают: определение посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS) количества антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, в среде для культивирования клеток, содержащей антитело или производное антитела и одну или несколько молекул, являющихся весовыми вариантами, где антитело или производное антитела и одна или несколько молекул, являющихся весовыми вариантами, различаются по молекулярной массе. В некоторых вариантах осуществления в данном документе предусмотрены способы получения антитела или производного антитела, включающие: (a) культивирование линии клеток, содержащей один или несколько полинуклеотидов, кодирующих антитело или производное антитела, в среде для культивирования клеток в условиях, подходящих для продуцирования антитела, или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, линией клеток; (b) отделение от линии клеток среды для культивирования клеток, содержащей антитело или производное антитела и одну или несколько молекул, являющихся весовыми вариантами; (c) определение количества антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, в среде для культивирования клеток посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS); и (d) удаление по меньшей мере некоторой доли одной или нескольких молекул, являющихся весовыми вариантами из антитела или производного антитела, продуцируемых линией клеток, или определение одного или обоих из показателей качества и чистоты антитела или производного антитела, продуцируемых линией клеток.
[0020] В некоторых вариантах осуществления в данном документе предусмотрены способы скрининга совокупности линий клеток в отношении продуцирования антитела или производного антитела, где способы включают: определение количества антитела или производного антитела, продуцируемого первой линией клеток из совокупности в соответствии со способом по любому из приведенных выше вариантов осуществления; и определение количества антитела или производного антитела, продуцируемого второй линией клеток из совокупности, в соответствии со способом по любому из приведенных выше вариантов осуществления. В некоторых вариантах осуществления способы дополнительно включают: сравнение количества антитела или производного антитела, продуцируемого первой линией клеток, с количеством антитела или производного антитела, продуцируемым второй линией клеток; и основывающийся на сравнении выбор линии клеток, которая продуцировала большее количество антитела или производного антитела. В некоторых вариантах осуществления способы дополнительно включают: определение количества одной или нескольких молекул, являющихся весовыми вариантами, продуцируемого первой линией клеток, в соответствии со способом по любому из приведенных выше вариантов осуществления; и определение количества одной или нескольких молекул, являющихся весовыми вариантами, продуцируемого второй линией клеток, в соответствии со способом по любому из приведенных выше вариантов осуществления. В некоторых вариантах осуществления после определения количества одной или нескольких молекул, являющихся весовыми вариантами, продуцируемого первой и второй линиями клеток, способы дополнительно включают: сравнение количества одной или нескольких молекул, являющихся весовыми вариантами, продуцируемого первой линией клеток, с количеством одной или нескольких молекул, являющихся весовыми вариантами, продуцируемым второй линией клеток; и основывающийся на сравнении выбор линии клеток, при продуцировании в которой значение соотношения антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, являлось более высоким, или выбор линии клеток, при продуцировании в которой относительная доля одной молекулы, являющейся весовым вариантом, по отношению к общему количеству антитела или производного антитела и одной или нескольких продуцируемых молекул, являющихся весовыми вариантами, являлась более высокой. В некоторых вариантах осуществления количество антитела или производного антитела и/или одной или нескольких молекул, являющихся весовыми вариантами, в соответствии с любым из вариантов осуществления в данном документе относится к относительному количеству (например, по сравнению с одной или несколькими другими молекулами или по сравнению с общим количеством продуцируемых антител/молекул).
[0021] В некоторых вариантах осуществления антитело или производное антитела содержат остаток цистеина в положении 293 (Cys293). В некоторых вариантах осуществления антитело или производное антитела и одна или несколько молекул, являющихся весовыми вариантами, предусматривают молекулы со свободным цистеином (например, не связанным дисульфидной связью с другим цистеином антитела или производного антитела), которые были подвержены цистеинилированию, N-ацетилцистеинилированию или глутатионилированию. В некоторых вариантах осуществления антитело или производное антитела не являются N-гликозилированными (например, в Fc-области антитела). В некоторых вариантах осуществления антитело или производное антитела содержат мутацию в Fc-области, которая приводит к снижению или устранению N-гликозилирования. В некоторых вариантах осуществления антитело или производное антитела содержат мутацию N300A (согласно EU-индексу). В некоторых вариантах осуществления антитело или производное антитела являются N-гликозилированными (например, в Fc-области антитела), и способ дополнительно включает (например, перед SE-UPLC-MS) удаление N-гликозилирования антитела. В некоторых вариантах осуществления удаление N-гликозилирования антитела предусматривает обработку антитела ферментом, представляющим собой пептид:N-гликозидазу (например, PNGазу F). В некоторых вариантах осуществления определение количества антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами, предусматривает деконволюцию одного или нескольких MS-спектров, полученных посредством MS. В некоторых вариантах осуществления перед определением способы дополнительно включают предоставление или получение (например, из линии клеток, которая продуцирует антитело или производное антитела и одну или несколько молекул, являющихся весовыми вариантами) среды для культивирования клеток, содержащей антитело или производное антитела и одну или несколько молекул, являющихся весовыми вариантами. В некоторых вариантах осуществления перед определением способы дополнительно включают отделение среды для культивирования клеток от линии клеток, которая продуцирует антитело или производное антитела и одну или несколько молекул, являющихся весовыми вариантами. В некоторых вариантах осуществления перед определением среду для культивирования клеток отделяют от линии клеток посредством центрифугирования. В некоторых вариантах осуществления среду для культивирования клеток подвергают SE-UPLC без предварительного хроматографического разделения. В некоторых вариантах осуществления среду для культивирования клеток подвергают SE-UPLC без предварительной аффинной хроматографии с использованием белка А. В некоторых вариантах осуществления MS представляет собой MS интактных молекул. В некоторых вариантах осуществления MS представляет собой квадрупольную времяпролетную (QToF) MS. В некоторых вариантах осуществления SE-UPLC представляет собой SE-UPLC в денатурирующих условиях. В некоторых вариантах осуществления SE-UPLC объединена с MS непосредственно. В некоторых вариантах осуществления одна или несколько молекул, являющихся весовыми вариантами, представляют собой молекулы антитела или производного антитела, содержащие химически модифицированный остаток цистеина. В некоторых вариантах осуществления антитело или производное антитела и одна или несколько молекул, являющихся весовыми вариантами, отличаются по молекулярной массе на по меньшей мере 119 Да. В некоторых вариантах осуществления MS позволяет определять разницу в массе между цистеинилированными (сдвиг по массе 119 Да), N-ацетилцистеинилированными (сдвиг по массе 161 Да) и глутатионилированными (сдвиг по массе 305 Да) молекулами. Антитело или производное антитела и одна или несколько молекул, являющихся весовыми вариантами, отличаются по молекулярной массе на по меньшей мере 162 Да. В некоторых вариантах осуществления антитело или производное антитела являются N-гликозилированными (например, в Fc-области антитела), и одна или несколько молекул, являющихся весовыми вариантами, представляют собой гликоформы антитела или производного антитела. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 162 Да, между антителом или производным антитела и одной или несколькими молекулами, являющимися весовыми вариантами, которые представляют собой гликоформы антитела или производного антитела. В некоторых вариантах осуществления антитело или производное антитела представляют собой полиспецифическое антитело.
[0022] Следует понимать, что один, несколько или все свойства из различных вариантов осуществления, описанных в данном документе, можно комбинировать с получением других вариантов осуществления настоящего изобретения. Эти и другие аспекты настоящего изобретения будут очевидны специалисту в данной области. Эти и другие варианты осуществления настоящего изобретения дополнительно описаны с помощью нижеследующего подробного описания.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0023] На ФИГ. 1A изображено схематическое представление триспецифического связывающего белка, содержащего четыре полипептидные цепи, которые образуют три антигенсвязывающих сайта, которые связывают три белка-мишени: CD28, CD3 и HER2. Первая пара полипептидов содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией (VH1-VH2 и VL2-VL1), образующие два антигенсвязывающих сайта, которые распознают CD3 и CD28 (CODV), и вторая пара полипептидов содержит один вариабельный домен (VH3 и VL3), образующий один антигенсвязывающий сайт, который распознает HER2 (Fab). В триспецифическом связывающем белке, показанном на ФИГ. 1A, используется константная область с мутацией по типу "выступы-во-впадины", где выступ находится на второй паре полипептидов с одним вариабельным доменом.
[0024] На ФИГ. 1B изображены молекулы, полученные в результате ошибочного спаривания тяжелой цепи (слева) или легкой цепи (справа). Слева показаны молекулы, которые получаются в результате ошибочного спаривания двух тяжелых цепей Fab (вверху) или двух тяжелых цепей CODV (внизу). Справа показаны молекулы, которые получаются в результате ошибочного спаривания двух легких цепей Fab (внизу слева), двух легких цепей CODV (внизу справа) или ошибочного спаривания легких цепей Fab и CODV не с теми тяжелыми цепями (вверху справа), а также триспецифический связывающий белок с правильным спариванием (вверху слева).
[0025] На ФИГ. 2A и 2B показаны спектры масс-спектрометрии (MS) после деконволюции для интактных (ФИГ. 2A) триспецифических связывающих белков, специфичных к CD38, и их F(ab')2 фрагментов, полученных при расщеплении с помощью IdeS (ФИГ. 2B), при этом разные молекулы обозначены. Изображенные молекулы включают триспецифический связывающий белок с правильным спариванием (H1L1/H2L2), молекулы с ошибочным спариванием с двумя легкими цепями Fab (H1L1/H2L1) и молекулы с ошибочным спариванием с двумя легкими цепями CODV (H1L2/H2L2). В аннотациях показаны значения массы, полученные экспериментальным путем, по сравнению с расчетными значениями массы (в скобках).
[0026] На ФИГ. 3A и 3B показано продуцирование триспецифического связывающего белка из серии клонов. На ФИГ. 3A показано ранжирование клонов, продуцирующих триспецифический связывающий белок, специфический к CD38, по проценту tsAb с правильной массой (чистоте). На ФИГ. 3B показана продуктивность каждого клона (нормализованный титр) в том же порядке, как изображено на ФИГ. 3A (т. е. ранжированные по чистоте).
[0027] На ФИГ. 3C показаны секторные диаграммы, изображающие доли примесей, представляющих собой молекулы с ошибочным спариванием и полуантитела, а также молекул с правильным спариванием в случайно выбранной подгруппе клонов, продуцирующих триспецифический связывающий белок, специфический к CD38, полученных с использованием периодической культуры.
[0028] На ФИГ. 4A-4C показаны результаты анализа триспецифических связывающих белков посредством SEC c MS интактных молекул. На ФИГ. 4A показана иллюстративная хроматограмма основных пиков для анализа на основе SEC в денатурирующих условиях c MS интактных молекул в осветленной собранной жидкости с tsAb. На ФИГ. 4B показаны комбинированные спектры под каждым хроматографическим пиком. На ФИГ. 4C показан увеличенный масс-спектр после деконволюции для смеси триспецифического связывающего белка, специфического к CD38, и триспецифического связывающего белка, специфического к HER2, обеспечивающий разрешение молекул с ошибочным спариванием легких цепей из двух конструкций с Δмассы=302 Да.
[0029] На ФИГ. 5A показана относительная количественная оценка молекул с правильным спариванием и ошибочным спариванием, полученная для разных значений загрузки колонки (мкг белка), от 4,4 до 21,8 мкг для триспецифического связывающего белка, специфического к HER2, и от 11,0 до 55,0 мкг для триспецифического связывающего белка, специфического к CD38, демонстрирующая устойчивость и надежность этого способа для анализа образцов собранного материала с низким и высоким титром. Для триспецифического связывающего белка, специфического к HER2, значения загрузки колонки показаны как 4,4 мкг, 8,7 мкг, 13,1 мкг, 17,4 мкг и 21,8 мкг (слева направо). Для триспецифического связывающего белка, специфического к CD38, значения загрузки колонки показаны как 11,0 мкг, 22,0 мкг, 33,0 мкг, 44,0 мкг и 55,0 мкг (слева направо).
[0030] На ФИГ. 5B и 5C показаны необработанные масс-спектры и масс-спектры после деконволюции, полученные посредством SEC-LC-MS-анализа осветленного собранного материала (ФИГ. 5B) и очищенного с использованием ProA (ФИГ. 5C) триспецифического связывающего белка, специфического к CD38, демонстрирующие сопоставимость этих двух способов. В аннотациях к спектрам после деконволюции показаны значения массы, полученные экспериментальным путем, и значения % интенсивности для каждой молекулы.
[0031] На ФИГ. 5D и 5E показано ранжирование клонов, продуцирующих триспецифический связывающий белок, специфический к HER2, на основе процентного содержания tsAb с правильной массой (чистоты) (ФИГ. 5D) и продуктивности, измеренной по титру, для одних и тех же клонов (ФИГ. 5E).
[0032] На ФИГ. 6A и 6B показан выход tsAb с правильной массой в различных клонах триспецифического связывающего белка, специфического к CD38 (ФИГ. 6A), и триспецифического связывающего белка, специфического к HER2 (ФИГ. 6B), выращенных в различных условиях культивирования клеток (ambr или периодическая культура, как указано).
[0033] На ФИГ. 6C-6E показано влияние различных условий выращивания на уровни ошибочного спаривания цепей и уровни полуантител в клонах CD38 TCE, выращенных при двух различных условиях в ambr (ФИГ. 6C и 6D) и в условиях периодической культуры (ФИГ. 6E).
[0034] На ФИГ. 6F и 6G показано влияние различных условий выращивания на уровни ошибочного спаривания цепей в клонах TCE, специфических к HER2, выращенных в условиях культивирования в ambr (ФИГ. 6F) или в условиях периодической культуры (ФИГ. 6G).
[0035] На ФИГ. 7 изображен иллюстративный рабочий процесс для высокопроизводительной MS интактных молекул в соответствии с некоторыми вариантами осуществления. Помимо высокопроизводительных функциональных возможностей, дополнительные преимущества включают: (1) отсутствие необходимости в очистке белка А, что обеспечивает экономию времени и расходов; (2) обеспечение более непосредственной оценки производительности линий клеток за счет предоставления информации о потенциально потерянной биомассе в виде свободных цепей, полуантител и других субмолекул, которые могут не пройти очистку с использованием белка А; и (3) применимость для таких лишенных Fc форм, как Fab, scFv и другие фрагменты антител.
[0036] На ФИГ. 8A и 8B показаны масс-спектры интактных молекул после деконволюции, полученные для клонов клеток, продуцирующих триспецифические антитела, специфические к HIV\CD28\CD3, с наиболее высокими и наиболее низкими уровнями ошибочного спаривания цепей соответственно.
[0037] На ФИГ. 9A и 9B показана продуктивность (оцененная по титру, мкг/мл; ФИГ. 9A) и продуцирование продукта с правильным спариванием (% молекул с правильной массой; ФИГ. 9B) для 50 клонов клеток, продуцирующих триспецифические антитела, специфические к HIV\CD28\CD3, выращенных в условиях периодической культуры.
[0038] На ФИГ. 10A и 10B показаны подробные профили гликанов, полученные для молекул с эталонной массой (H1L1/H2L2) в выбранном клоне, продуцирующем триспецифическое антитело, специфическое к HIV\CD28\CD3. Клон культивировали в двух различных условиях: в центрифужной пробирке (условия периодической культуры; ФИГ. 10A) и в биореакторе ambr15 (условия периодической культуры с подпиткой; ФИГ. 10B).
[0039] На ФИГ. 11A и 11B показан результат MS-анализа собранного материала антител со сконструированным цистеином (Cys293) в отношении статуса блокировки цистеина. На ФИГ. 11A показаны результаты идентификации статуса блокировки Cys293 молекул, полученные с применением прямого MS-анализа интактных молекул. Это антитело содержало мутацию N300A, которая устраняла N-гликозилирование Fc-области. На ФИГ. 11B показаны результаты идентификации статуса блокировки Cys293, полученные с применением прямого MS-анализа интактных молекул для антител, обработанных с помощью PNGазы F для удаления N-гликанов перед анализом.
ПОДРОБНОЕ ОПИСАНИЕ
[0040] В настоящем изобретении, среди прочего, предусмотрены способы мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием, например полиспецифического антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или другого полиспецифического связывающего белка, который предусматривает ассоциацию двух или более полипептидных цепей, содержащих по меньшей мере первую полипептидную цепь и вторую полипептидную цепь, отличную от первой полипептидной цепи, и одной или нескольких молекул с ошибочным спариванием, которые содержат две или более полипептидных цепей, содержащих по меньшей мере одну из первой и второй полипептидных цепей в ассоциации, отличной от ассоциации полиспецифического связывающего белка. Аналогичные способы можно применять для продуцирования полиспецифического связывающего белка, скрининга линий клеток в отношении продуцирования полиспецифического связывающего белка или при продуцировании антител или белков, являющихся продуктом слияния с Fc, а также для продуцирования антитела или производного антитела (например, продуцируемых с одной или несколькими молекулами, являющихся весовыми вариантами) или скрининга линий клеток в отношении продуцирования антитела или производного антитела (например, продуцируемых с одной или нескольких молекул, являющихся весовыми вариантами). Предпочтительно эти способы позволяют проводить MS-анализ интактных связанных с mAb молекул непосредственно в осветленном собранном материале, за счет чего избегают трудоемкой и дорогостоящей очистки (например, аффинной хроматографии с использованием белка А) и этапов замены буфера. Таким образом, эти способы можно применять для быстрого скрининга большого количества клонов в отношении потенциальных линий клеток-продуцентов.
[0041] В нижеследующем описании изложены иллюстративные способы, параметры и т. п. Однако следует понимать, что такое описание не предусмотрено в качестве ограничения объема настоящего изобретения, а наоборот - предусмотрено в качестве описания иллюстративных вариантов осуществления.
Определения
[0042] Используемые в соответствии с настоящим изобретением следующие термины, если не указано иное, следует понимать как имеющие следующие значения. Если контекстом не требуется иное, то термины в единственном числе будут включать форму множественного числа, и термины во множественном числе будут включать форму единственного числа.
[0043] Понятно, что аспекты и варианты осуществления настоящего изобретения, описанные в данном документе, включают аспекты и варианты осуществления "содержащие", "состоящие из" и/или "фактически состоящие из".
[0044] Используемый в данном документе термин "полинуклеотид" относится к полимерам в виде однонитевой или двунитевой нуклеиновой кислоты, имеющих длину по меньшей мере 10 нуклеотидов. В определенных вариантах осуществления нуклеотиды, составляющие полинуклеотид, могут представлять собой рибонуклеотиды или дезоксирибонуклеотиды, или модифицированную форму любого типа нуклеотида. Такие модификации включают модификации оснований, как, например, бромуридин, модификации рибозы, как, например, арабинозид и 2',3'-дидезоксирибоза, и модификации межнуклеотидных связей, как, например, фосфотиоат, фосфодитиоат, фосфоселеноат, фосфодиселеноат, фосфоанилотиоат, фосфоаниладат и фосфоамидат. Термин "полинуклеотид" конкретно подразумевает однонитевые и двунитевые формы ДНК.
[0045] "Выделенный полинуклеотид" представляет собой полинуклеотид, имеющий геномное, cDNA или синтетическое происхождение или характеризующийся некоторой их комбинацией, который: (1) не ассоциирован со всем полинуклеотидом, в котором выделенный полинуклеотид встречается в природе, или его частью, (2) связан с полинуклеотидом, с которым он не связан в природе, или (3) не встречается в природе в виде части более крупной последовательности.
[0046] "Выделенный полипептид" представляет собой такой полипептид, который: (1) не содержит по меньшей мере некоторых других полипептидов, с которыми он обычно встречается, (2) фактически не содержит других полипептидов из одного и того же источника, например, из одного и того же вида, (3) экспрессируется клеткой из другого вида, (4) был отделен по меньшей мере от приблизительно 50 процентов полинуклеотидов, липидов, углеводов или других веществ, с которыми он связан в природе, (5) не связан (посредством ковалентного или нековалентного взаимодействия) с частями полипептида, с которыми "выделенный полипептид" связан в природе, (6) функционально связан (посредством ковалентного или нековалентного взаимодействия) с полипептидом, с которым он не связан в природе, или (7) не встречается в природе. Такой выделенный полипептид может кодироваться геномной ДНК, cDNA, mRNA или другой РНК синтетического происхождения или любой их комбинацией. Предпочтительно выделенный полипептид по сути не содержит полипептидов или других контаминантов, которые встречаются в его природном окружении, которые бы вызывали затруднения при его применении (терапевтическом, диагностическом, профилактическом, исследовательском или ином).
[0047] Встречающиеся в природе антитела обычно представляют собой тетрамер. Каждый такой тетрамер обычно состоит из двух идентичных пар полипептидных цепей, при этом каждая пара имеет одну полноразмерную "легкую" цепь (обычно имеющую молекулярную массу приблизительно 25 кДа) и одну полноразмерную "тяжелую" цепь (обычно имеющую молекулярную массу приблизительно 50-70 кДа). Используемые в данном документе термины "тяжелая цепь" и "легкая цепь" обозначают любой полипептид иммуноглобулина, содержащий последовательность вариабельного домена, достаточную для придания специфичности в отношении антигена-мишени. Аминоконцевая часть каждой легкой и тяжелой цепей обычно содержит вариабельный домен из приблизительно 100-110 аминокислот или больше, который обычно отвечает за распознавание антигена. Карбоксиконцевая часть каждой цепи обычно определяет константный домен, ответственный за эффекторную функцию. Таким образом, во встречающемся в природе антителе полноразмерный полипептид, представляющий собой тяжелую цепь иммуноглобулина, содержит вариабельный домен (VH) и три константных домена (CH1, CH2 и CH3), где домен VH находится на аминоконце полипептида, а домен CH3 находится на карбоксильном конце, и полноразмерный полипептид, представляющий собой легкую цепь иммуноглобулина, содержит вариабельный домен (VL) и константный домен (CL), где домен VL находится на аминоконце полипептида, а домен CL находится на карбоксильном конце.
[0048] Легкие цепи человека обычно классифицируют как легкие каппа- и лямбда-цепи, а тяжелые цепи человека обычно классифицируют как мю, дельта, гамма, альфа или эпсилон, и они определяют изотип антитела IgM, IgD, IgG, IgA и IgE соответственно. IgG имеет несколько подклассов, в том числе без ограничения IgG1, IgG2, IgG3 и IgG4. IgM имеет подклассы, в том числе без ограничения IgM1 и IgM2. IgA аналогичным образом подразделяют на подклассы, в том числе без ограничения IgA1 и IgA2. В полноразмерных легких и тяжелых цепях вариабельные и константные домены обычно соединены с помощью "J"-области из приблизительно 12 или больше аминокислот, при этом тяжелая цепь также содержит "D"-область из приблизительно 10 или больше аминокислот. См., например, Fundamental Immunology (под ред. Paul, W., Raven Press, 2-е изд., 1989), которая включена посредством ссылки во всей своей полноте для всех целей. Вариабельные области каждой пары легкая/тяжелая цепь обычно образуют антигенсвязывающий сайт. Вариабельные домены встречающихся в природе антител обычно характеризуются одинаковой общей структурой относительно консервативных каркасных областей (FR), соединенных с помощью трех гипервариабельных областей, также называемых определяющими комплементарность областями или CDR. CDR из двух цепей каждой пары обычно выровнены с помощью каркасных областей, что может обеспечивать возможность связывания со специфическим эпитопом. От аминоконца к карбоксильному концу вариабельные домены как легкой, так и тяжелой цепей обычно содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
[0049] Термин "совокупность CDR" обозначает группу из трех CDR, которые содержатся в одной вариабельной области, способной связывать антиген. Точные границы этих CDR определяли по-разному в соответствии с различными системами. Система, описанная Kabat (Kabat et al., Sequences of Proteins of Immunological Interest (Национальные институты здравоохранения, Бетесда, Мэриленд. (1987) и (1991)), не только предусматривает однозначную систему нумерации остатков, применимую к любой вариабельной области антитела, но также предусматривает точные границы остатков, определяющие три CDR. Эти CDR могут называться CDR в соответствии с Kabat. Chothia и коллеги (Chothia and Lesk, 1987, J. Mol. Biol. 196: 901-17; Chothia et al., 1989, Nature 342: 877-83) обнаружили, что определенные субфрагменты в пределах CDR в соответствии с Kabat принимают почти идентичные конформации пептидного каркаса, несмотря на наличие большого различия на уровне аминокислотной последовательности. Эти субфрагменты были обозначены как L1, L2 и L3 или H1, H2 и H3, где "L" и "H" обозначают области легкой цепи и тяжелой цепи соответственно. Эти области могут называться CDR в соответствии с Chothia, границы которых совпадают с CDR в соответствии с Kabat. Другие границы, определяющие CDR, которые совпадают с CDR в соответствии с Kabat, были описаны Padlan, 1995, FASEB J. 9: 133-39; MacCallum, 1996, J. Mol. Biol. 262(5): 732-45; и Lefranc, 2003, Dev. Comp. Immunol. 27: 55-77. Еще одни определения границ CDR могут не строго соответствовать одной из систем, приведенных в данном документе, но тем не менее будут совпадать с CDR в соответствии с Kabat, несмотря на то, что они могут быть укорочены или удлинены с учетом прогностических или экспериментальных выводов о том, что конкретные остатки или группы остатков или даже все CDR не влияют в значительной степени на связывание антигена. В способах, используемых в данном документе, можно использовать CDR, определенные в соответствии с любой из этих систем, несмотря на то, что в определенных вариантах осуществления применяют CDR, определенные в соответствии с Kabat или Chothia. Идентификация прогнозируемых CDR с помощью аминокислотной последовательности хорошо известна в данной области техники, например, в Martin, A.C. "Protein sequence and structure analysis of antibody variable domains" в Antibody Engineering, Vol. 2. под ред. Kontermann R., Dübel S., Springer-Verlag, Берлин, стр. 33-51 (2010). Аминокислотную последовательность вариабельного домена тяжелой и/или легкой цепи можно также исследовать с целью идентификации последовательностей CDR с помощью других традиционных способов, например, путем сравнения с известными аминокислотными последовательностями других вариабельных областей тяжелой и легкой цепей с определением областей гипервариабельности в последовательности. Пронумерованные последовательности можно выравнивать вручную или путем использования программы выравнивания, такой как одна из пакета программ CLUSTAL, как описано в Thompson, 1994, Nucleic Acids Res. 22: 4673-80. Молекулярные модели традиционно используют для того, чтобы правильно определить каркасные и CDR-области и таким образом скорректировать основанные на последовательности интерпретации.
[0050] Используемый в данном документе термин "Fc" обозначает молекулу, будь то в мономерной или мультимерной форме, содержащую последовательность, не являющуюся частью антигенсвязывающего фрагмента, которая получена в результате расщепления антитела или получена другими способами, и при этом она может содержать шарнирную область. Исходный иммуноглобулиновый источник нативного Fc предпочтительно происходит от человека и может представлять собой любой из иммуноглобулинов, тем не менее предпочтительными являются IgG1 и IgG2. Молекулы Fc составлены из мономерных полипептидов, которые могут быть связаны в димерные или мультимерные формы посредством ковалентной (т. е. дисульфидных связей) и нековалентной связи. Число межмолекулярных дисульфидных связей между мономерными субъединицами нативных молекул Fc варьирует от 1 до 4, в зависимости от класса (например, IgG, IgA и IgE) или подкласса (например, IgG1, IgG2, IgG3, IgA1 и IgGA2). Одним примером Fc является димер с дисульфидной связью, полученный в результате расщепления IgG папаином. Термин "нативный Fc", используемый в данном документе, является общим для мономерных, димерных и мультимерных форм.
[0051] F(ab)-фрагмент обычно содержит одну легкую цепь и домены VH и CH1 одной тяжелой цепи, где часть VH-CH1 тяжелой цепи F(ab)-фрагмента не может образовывать дисульфидную связь с другим полипептидом тяжелой цепи. Как используется в данном документе, F(ab)-фрагмент также может содержать одну легкую цепь, содержащую два вариабельных домена, разделенных аминокислотным линкером, и одну тяжелую цепь, содержащую два вариабельных домена, разделенных аминокислотным линкером, и домен CH1.
[0052] F(ab')-фрагмент обычно содержит одну легкую цепь и часть одной тяжелой цепи, которая содержит большую часть константной области (между доменами CH1 и CH2), вследствие чего межцепочечная дисульфидная связь может быть образована между двумя тяжелыми цепями с образованием молекулы F(ab')2.
[0053] Используемый в данном документе термин "связывающий белок" относится к не встречающейся в природе (или рекомбинантной, или сконструированной) молекуле, которая специфически связывается с по меньшей мере одним антигеном-мишенью. В некоторых вариантах осуществления связывающий белок содержит два или более антигенсвязывающих домена. В некоторых вариантах осуществления связывающий белок представляет собой полиспецифическое антитело, фрагмент антитела или белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления связывающий белок представляет собой биспецифическое антитело или фрагмент антитела. В некоторых вариантах осуществления связывающий белок представляет собой триспецифическое антитело или фрагмент антитела. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический связывающий белок, например, как описано ниже. В некоторых вариантах осуществления связывающий белок содержит одну или две Fc-области, слитые с одним, двумя, тремя или более антигенсвязывающими доменами или другими полипептидами (например, белком, являющимся продуктом слияния с Fc). В некоторых вариантах осуществления связывающий белок представляет собой иммуноглобулин с двойным вариабельным доменом (DVD), например, как описано в WO 2012061558. В некоторых вариантах осуществления связывающий белок содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией, например, как описано в WO 2012135345. В некоторых вариантах осуществления связывающий белок содержит четыре полипептидные цепи, которые образуют четыре антигенсвязывающих сайта, при этом две полипептидные цепи содержат структуру, представленную формулой:
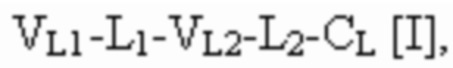
и две полипептидные цепи содержат структуру, представленную формулой:
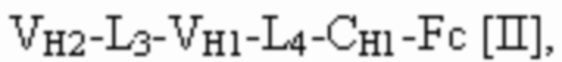
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VR2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
Fc представляет собой шарнирную область иммуноглобулина и константные домены CH2, CH3 тяжелой цепи иммуноглобулина; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры; и
где полипептиды формулы I и полипептиды формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи.
[0054] Триспецифический связывающий белок по настоящему изобретению, если не указано иное, содержит четыре полипептидные цепи, которые образуют по меньшей мере три антигенсвязывающих сайта, где первая полипептидная цепь имеет структуру, представленную формулой:
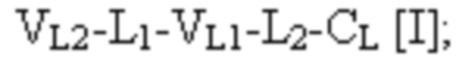
и вторая полипептидная цепь имеет структуру, представленную формулой:
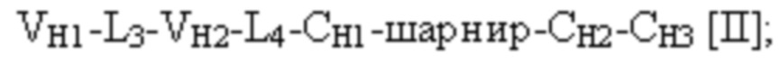
и третья полипептидная цепь имеет структуру, представленную формулой:
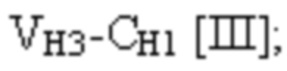
и четвертая полипептидная цепь имеет структуру, представленную формулой:
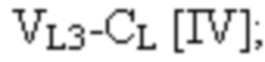
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина; и
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2;
L1, L2, L3 и L4 представляют собой аминокислотные линкеры;
и где полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи.
[0055] "Рекомбинантная" молекула представляет собой молекулу, которая была получена, экспрессирована, создана или выделена рекомбинантными способами.
[0056] В одном варианте осуществления настоящего изобретения представлены связывающие белки, характеризующиеся биологической и иммунологической специфичностью в отношении одного-трех антигенов-мишеней. В другом варианте осуществления настоящего изобретения представлены молекулы нуклеиновой кислоты, содержащие нуклеотидные последовательности, кодирующие полипептидные цепи, которые образуют такие связывающие белки. В другом варианте осуществления настоящего изобретения представлены векторы экспрессии, содержащие молекулы нуклеиновой кислоты, содержащие нуклеотидные последовательности, кодирующие полипептидные цепи, которые образуют такие связывающие белки. В еще одном варианте осуществления настоящего изобретения представлены клетки-хозяева, которые экспрессируют такие связывающие белки (т. е. содержащие молекулы нуклеиновой кислоты или векторы, кодирующие полипептидные цепи, которые образуют такие связывающие белки).
[0057] Используемый в данном документе термин "способность к обмену" обозначает способность к взаимозамене вариабельных доменов в формате связывающего белка, при этом с сохранением укладки и наивысшей аффинности связывания. "Способность к полному обмену" относится к способности менять порядок доменов как VH1, так и VH2 и, следовательно, порядок доменов VL1 и VL2 в полипептидной цепи формулы I или полипептидной цепи формулы II (т. е. обращать порядок), при этом поддерживая полную функциональность связывающего белка, о чем свидетельствует сохранение аффинности связывания. Кроме того, следует отметить, что обозначения VH и VL относятся лишь к положению домена в конкретной белковой цепи в конечном формате. Например, VH1 и VH2 могут быть получены из доменов VL1 и VL2 исходных антител и введены в положения, соответствующие VH1 и VH2 связывающего белка. Аналогично VL1 и VL2 могут быть получены из доменов VH1 и VH2 исходных антител и помещены в положения, соответствующие VH1 и VH2 связывающего белка. Таким образом, обозначения VH и VL относятся к существующему в данный момент положению, а не к первоначальному положению в исходном антителе. Следовательно, домены VH и VL являются "способными к обмену".
[0058] Используемый в данном документе термин "антиген", или "антиген-мишень", или "целевой антиген" обозначает молекулу или часть молекулы, которая может связываться связывающим белком и дополнительно может использоваться у животного для получения антител, способных связываться с эпитопом этого антигена. Антиген-мишень может иметь один или несколько эпитопов. С учетом того, что каждый антиген-мишень распознается связывающим белком, связывающий белок способен конкурировать с интактным антителом, которое распознает антиген-мишень.
[0059] Термин "моноспецифический связывающий белок" обозначает связывающий белок, который специфически связывается с одним антигеном-мишенью.
[0060] Термин "моновалентный связывающий белок" обозначает связывающий белок, который имеет один антигенсвязывающий сайт.
[0061] Термин "биспецифический связывающий белок" обозначает связывающий белок, который специфически связывается с двумя различными антигенами-мишенями.
[0062] Термин "бивалентный связывающий белок" обозначает связывающий белок, который имеет два связывающих сайта.
[0063] Термин "триспецифический связывающий белок" обозначает связывающий белок, который специфически связывается с тремя различными антигенами-мишенями.
[0064] Термин "тривалентный связывающий белок" обозначает связывающий белок, который имеет три связывающих сайта. В конкретных вариантах осуществления тривалентный связывающий белок может связываться с одним антигеном-мишенью. В других вариантах осуществления тривалентный связывающий белок может связываться с двумя антигенами-мишенями. В других вариантах осуществления тривалентный связывающий белок может связываться с тремя антигенами-мишенями.
[0065] "Выделенный" связывающий белок представляет собой связывающий белок, который был идентифицирован и отделен и/или извлечен из компонента своего природного окружения. Компоненты-контаминанты из его природного окружения представляют собой вещества, которые бы вызывали затруднения при диагностических или терапевтических применениях связывающего белка, и могут включать ферменты, гормоны и другие растворенные вещества белковой или небелковой природы. В некоторых вариантах осуществления связывающий белок будет очищенным: (1) до более чем 95% по весу антитела, как определяется способом Лоури, и наиболее предпочтительно до более чем 99% по весу, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с применением секвенатора с вращающимся стаканом, или (3) до гомогенности с помощью SDS-PAGE в восстанавливающих или невосстанавливающих условиях с применением кумасси голубого или предпочтительно серебряного красителя. Выделенные связывающие белки включают связывающий белок in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент природного окружения связывающего белка не будет присутствовать.
[0066] Используемые в данном документе термины "в значительной степени чистый" или "в значительной степени очищенный" обозначают соединение или структуру, которая является преобладающей присутствующей структурой (т. е. в расчете на моль она более многочисленна, чем любая другая отдельная структура в композиции). В некоторых вариантах осуществления в значительной степени очищенная фракция представляет собой композицию, где структура составляет по меньшей мере приблизительно 50% (в расчете на моль) от всех присутствующих макромолекулярных структур. В других вариантах осуществления в значительной степени чистая композиция будет содержать более чем приблизительно 80%, 85%, 90%, 95% или 99% от всех макромолекулярных структур, присутствующих в композиции. В еще одних вариантах осуществления структура является очищенной до необходимой гомогенности (загрязняющие структуры не могут быть выявлены в композиции с помощью традиционных способов выявления), где композиция состоит фактически из одной макромолекулярной структуры.
[0067] Термин "эпитоп" подразумевает любую детерминанту, предпочтительно полипептидную детерминанту, способную специфически связываться с иммуноглобулином или T-клеточным рецептором. В определенных вариантах осуществления эпитопные детерминанты включают химически активные поверхностные группы молекул, такие как аминокислоты, боковые цепи сахаров, фосфорильные группы или сульфонильные группы, и в определенных вариантах осуществления могут иметь специфические характеристики трехмерной структуры и/или специфические характеристики заряда. Эпитоп представляет собой область антигена, которая связывается антителом или связывающим белком. В определенных вариантах осуществления считается, что связывающий белок специфически связывает антиген, если он предпочтительно распознает свой антиген-мишень в сложной смеси белков и/или макромолекул. В некоторых вариантах осуществления считается, что связывающий белок специфически связывает антиген, если равновесная константа диссоциации составляет ≤ 10-8 M, более предпочтительно если равновесная константа диссоциации составляет ≤ 10-9 M и наиболее предпочтительно если константа диссоциации составляет ≤ 10-10 M.
[0068] Константу диссоциации (KD) связывающего белка можно определить, например, с помощью поверхностного плазмонного резонанса. В целом в анализе поверхностного плазмонного резонанса измеряют связывающие взаимодействия между лигандом (антигеном-мишенью на биосенсорной матрице) и аналитом (связывающим белком в растворе) в реальном времени с помощью поверхностного плазмонного резонанса (SPR) с применением системы BIAcore (Pharmacia Biosensor; Пискатауэй, Нью-Джерси). Анализ поверхностного плазмонного резонанса можно также выполнять посредством иммобилизации аналита (связывающего белка на биосенсорной матрице) и представления лиганда (антигена-мишени). Используемый в данном документе термин "KD" обозначает константу диссоциации взаимодействия между конкретным связывающим белком и антигеном-мишенью.
[0069] Используемый в данном документе термин "специфически связывается" обозначает способность связывающего белка или его антигенсвязывающего фрагмента связываться с антигеном, содержащим эпитоп, с Kd, составляющей по меньшей мере приблизительно 1×10-6 M, 1×10-7 M, 1×10-8 M, 1×10-9 M, 1×10-10 M, 1×10-11 M, 1×10-12 M или больше, и/или связываться с эпитопом с аффинностью, которая в по меньшей мере два раза превышает его аффинность в отношении неспецифического антигена.
[0070] Используемый в данном документе термин "линкер" обозначает один или несколько аминокислотных остатков, вставленных между доменами иммуноглобулина для обеспечения подвижности, достаточной для того, чтобы домены легкой и тяжелой цепей сворачивались в иммуноглобулины с двумя вариабельными областями в перекрестной ориентации. Линкер вставляют в переходный участок между вариабельными доменами или между вариабельным и константным доменами, соответственно, на уровне последовательности. Переходный участок между доменами можно идентифицировать, поскольку приблизительный размер доменов иммуноглобулина хорошо известен. Точное местоположение переходного участка между доменами можно определить путем определения положения пептидных отрезков, которые не образуют элементов со вторичной структурой, такой как бета-слои или альфа-спирали, что показано с помощью экспериментальных данных или что можно прогнозировать с помощью методик моделирования или прогнозирования вторичной структуры. Линкеры, описанные в данном документе, обозначены как L1, который расположен на легкой цепи между C-концом домена VL2 и N-концом домена VL1; и L2, который расположен на легкой цепи между C-концом домена VL1 и N-концом домена CL. Линкеры тяжелой цепи называют как L3, который расположен между C-концом домена VH1 и N-концом домена VH2; и L4, который расположен между C-концом домена VH2 и N-концом домена CH1.
[0071] Используемый в данном документе термин "вектор" относится к любой молекуле (например, нуклеиновой кислоте, плазмиде или вирусу), используемой для переноса закодированной информации в клетку-хозяина. Термин "вектор" подразумевает молекулу нуклеиновой кислоты, которая способна транспортировать другую нуклеиновую кислоту, с которой ее связали. Одним типом вектора является "плазмида", которая обозначает кольцевую двунитевую молекулу ДНК, в которую могут быть вставлены дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, при этом в геном вируса могут быть вставлены дополнительные сегменты ДНК. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы для млекопитающих). Другие векторы (например, неэписомные векторы для млекопитающих) могут быть интегрированы в геном клетки-хозяина после введения в клетку-хозяина, и за счет этого они реплицируются вместе с геномом хозяина. Кроме того, определенные векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называются в данном документе "рекомбинантными векторами экспрессии" (или просто "векторами экспрессии"). Как правило, векторы экспрессии, полезные в технологиях рекомбинантных ДНК, часто находятся в форме плазмид. Термины "плазмида" и "вектор" могут использоваться взаимозаменяемо в данном документе, поскольку плазмида является наиболее широко используемой формой вектора. Однако подразумевается, что в настоящее изобретение включены и другие формы векторов экспрессии, такие как вирусные векторы (например, ретровирусы, аденовирусы и аденоассоциированные вирусы с дефектной репликацией), которые выполняют эквивалентные функции.
[0072] Используемая в данном документе фраза "рекомбинантная клетка-хозяин" (или "клетка-хозяин") обозначает клетку, в которую был введен рекомбинантный вектор экспрессии. Подразумевается, что рекомбинантная клетка-хозяин или клетка-хозяин обозначают не только конкретную рассматриваемую клетку, но также и потомство такой клетки. Поскольку в последующих поколениях могут возникать определенные модификации вследствие мутации либо влияний окружающей среды, такое потомство в действительности может не быть идентичным родительской клетке, однако такие клетки по-прежнему включены в объем используемого в данном документе термина "клетка-хозяин". Широкое разнообразие систем экспрессии в клетке-хозяине можно применять для экспрессии связывающих белков, в том числе системы экспрессии на основе бактерий, дрожжей, бакуловирусов и клеток млекопитающих (а также системы экспрессии на основе фагового дисплея). Примером подходящего бактериального вектора экспрессии является pUC19. Для рекомбинантной экспрессии связывающего белка клетку-хозяина трансформируют или трансфицируют одним или несколькими рекомбинантными векторами экспрессии, несущими фрагменты ДНК, кодирующие полипептидные цепи связывающего белка, вследствие чего полипептидные цепи экспрессируются в клетке-хозяине и предпочтительно секретируются в среду, в которой клетки-хозяева культивируются, при этом из данной среды можно извлечь связывающий белок.
[0073] Используемый в данном документе термин "трансформация" обозначает изменение генетических характеристик клетки, и при этом клетка была трансформирована, если ее модифицировали с тем, чтобы она содержала новую ДНК. Например, клетка является трансформированной, если она генетически модифицирована по сравнению с ее нативным состоянием. После трансформации трансформирующая ДНК может подвергаться рекомбинации с ДНК клетки путем физической интеграции в хромосому клетки, или может временно сохраняться в виде эписомального элемента без репликации, или может реплицироваться независимо как плазмида. Считается, что клетка является стабильно трансформированной, если ДНК реплицируется при делении клетки. Используемый в данном документе термин "трансфекция" обозначает захват чужеродной или экзогенной ДНК клеткой, и при этом клетка была "трансфицирована", если экзогенную ДНК ввели внутрь от клеточной мембраны. Ряд методик трансфекции хорошо известен из уровня техники. Такие методики можно применять для введения одной или нескольких экзогенных молекул ДНК в подходящие клетки-хозяева.
[0074] Используемый в данном документе термин "встречающийся в природе" и применяемый в отношении объекта обозначает тот факт, что объект может обнаруживаться в природе и не был подвергнут манипуляциям со стороны человека. Например, полинуклеотид или полипептид, присутствующий в организме (включая вирусы), который может быть выделен из природного источника и который не был преднамеренно модифицирован человеком, является встречающимся в природе. Аналогично используемый в данном документе термин "не встречающийся в природе" обозначает объект, который не обнаруживается в природе или который был структурно модифицирован или синтезирован человеком.
[0075] Как используется в данном документе, двадцать стандартных аминокислот и их аббревиатуры соответствуют общепринятой практике. Стереоизомеры (например, d-аминокислоты) двадцати стандартных аминокислот; не встречающиеся в природе аминокислоты и аналоги, такие как α-,α-двузамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие нестандартные аминокислоты также могут быть подходящими компонентами полипептидных цепей связывающих белков. Примеры нестандартных аминокислот включают 4-гидроксипролин, γ-карбоксиглутамат, ε-N,N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В системе обозначений полипептидов, используемой в данном документе, левое направление представляет собой направление в сторону аминоконца, а правое направление представляет собой направление в сторону карбоксильного конца в соответствии со стандартной практикой и правилами.
[0076] Встречающиеся в природе остатки можно разделить на классы на основании общих свойств боковой цепи:
(1) гидрофобные: Met, Ala, Val, Leu, Ile, Phe, Trp, Tyr, Pro;
(2) полярные гидрофильные: Arg, Asn, Asp, Gln, Glu, His, Lys, Ser, Thr;
(3) алифатические: Ala, Gly, Ile, Leu, Val, Pro;
(4) алифатические гидрофобные: Ala, Ile, Leu, Val, Pro;
(5) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(6) кислые: Asp, Glu;
(7) основные: His, Lys, Arg;
(8) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(9) ароматические: His, Trp, Tyr, Phe; и
(10) ароматические гидрофобные: Phe, Trp, Tyr.
[0077] Консервативные аминокислотные замены могут включать обмен представителя одного из этих классов на другого представителя того же класса. Неконсервативные замены могут включать обмен представителя одного из этих классов на представителя другого класса.
[0078] Специалист в данной области техники сможет определить подходящие варианты полипептидных цепей связывающих белков с помощью общеизвестных методик. Например, специалист в данной области может идентифицировать подходящие фрагменты в полипептидной цепи, которые можно изменить без нарушения активности, путем нацеливания на области, которые, как предполагается, не являются важными с точки зрения активности. Альтернативно специалист в данной области техники может идентифицировать остатки и части молекул, которые являются консервативными среди схожих полипептидов. Кроме того, даже участки, которые могут быть важными с точки зрения биологической активности или с точки зрения структуры, могут подвергаться консервативным аминокислотным заменам без нарушения биологической активности или без неблагоприятного воздействия на структуру полипептида.
Способы
[0079] Определенные аспекты настоящего изобретения относятся к способам мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием. В некоторых вариантах осуществления способы включают определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием) посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS). В некоторых вариантах осуществления продукт и одну или несколько молекул с ошибочным спариванием определяют в среде для культивирования клеток. В данном документе предусмотрены иллюстративные и неограничивающие описания полиспецифических связывающих белков.
[0080] Некоторые аспекты настоящего изобретения относятся к способам мониторинга продуцирования антитела или производного антитела и одной или нескольких молекул, являющихся весовыми вариантами. В некоторых вариантах осуществления способы включают определение количества продукта (например, одной или нескольких молекул, являющихся весовыми вариантами антитела или производного антитела) посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS). В некоторых вариантах осуществления антитело или производное антитела и одну или несколько молекул, являющихся весовыми вариантами, определяют в среде для культивирования клеток. Антитело или производное антитела и одна или несколько молекул, являющихся весовыми вариантами, могут представлять собой, например, несколько молекул с различной молекулярной массой (например, с различными химическими модификациями, такими как химически модифицированный цистеин или другие остатки, или с разными гликоформами). Как показано в приведенных ниже примерах, способы, описанные в данном документе в отношении анализа антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием, можно применять для анализа антитела или производного антитела и одной или более молекул, являющихся весовыми вариантами.
[0081] В некоторых вариантах осуществления среда для культивирования клеток представляет собой осветленный собранный материал культуры клеток. Например, среда для культивирования клеток может представлять собой среду, полученную из культуры клеток и осветленную посредством тангенциальной поточной фильтрации (TFF), глубинной фильтрации и/или центрифугирования. В некоторых вариантах осуществления способы дополнительно включают отделение среды для культивирования клеток или собранного материала от линии клеток, продуцирующей продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием). В некоторых вариантах осуществления среду для культивирования клеток осветляют посредством центрифугирования. В некоторых вариантах осуществления среду для культивирования клеток подвергают SE-UPLC без предварительного хроматографического разделения. Другими словами, в некоторых вариантах осуществления среда для культивирования клеток представляет собой среду, которая не подвергалась предварительному хроматографическому разделению. В некоторых вариантах осуществления среду для культивирования клеток подвергают SE-UPLC без предварительной аффинной хроматографии с использованием белка А. Другими словами, в некоторых вариантах осуществления среда для культивирования клеток представляет собой среду, которую не приводили в контакт с белком А.
[0082] В некоторых вариантах осуществления определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием) предусматривает деконволюцию одного или нескольких MS-спектров, полученных посредством MS. Способы деконволюции MS-спектров известны из уровня техники. Неограничивающий пример программного обеспечения, применимого для деконволюции, приведен ниже.
[0083] В некоторых вариантах осуществления определяют относительное количество полиспецифического связывающего белка, например, по сравнению с количеством одной или нескольких молекул с ошибочным спариванием. Например, количество продуцируемого полиспецифического связывающего белка можно сравнивать с количеством одной или нескольких отдельных молекул с ошибочным спариванием и/или с общим количеством молекул с ошибочным спариванием (например, общим количеством всех молекул с ошибочным спариванием или их определенной подгруппой).
[0084] В некоторых вариантах осуществления MS представляет собой MS интактных молекул. В некоторых вариантах осуществления MS представляет собой квадрупольную времяпролетную (QToF) MS. Из уровня техники известно множество способов QToF MS. Иллюстративные и неограничивающие параметры для QToF MS описаны в приведенных ниже примерах.
[0085] В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 1000 Да или меньше, приблизительно 900 Да или меньше, приблизительно 800 Да или меньше, приблизительно 700 Да или меньше, приблизительно 600 Да или меньше, приблизительно 500 Да или меньше, приблизительно 400 Да или меньше, приблизительно 300 Да или меньше или приблизительно 200 Да или меньше. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 300 Да. Например, в некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 300 Да, между полиспецифическим связывающим белком и одной или несколькими молекулами с ошибочным спариванием или между двумя молекулами с ошибочным спариванием. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 162 Да. Например, в некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 162 Да, между гликоформами, например между полиспецифическим связывающим белком или молекулами с ошибочным спариванием и одной или несколькими гликоформами.
[0086] В некоторых вариантах осуществления SE-UPLC представляет собой SE-UPLC в денатурирующих условиях. В некоторых вариантах осуществления SE-UPLC объединена с MS непосредственно. Из уровня техники известны различные способы SE-UPLC. Иллюстративные и неограничивающие условия для SE-UPLC описаны в приведенных ниже примерах.
[0087] В некоторых вариантах осуществления SE-UPLC выполняют с более медленной начальной скоростью потока, за которой следует более высокая скорость потока. Не ограничиваясь теорией, считается, что более медленная начальная скорость потока позволяет более эффективно разделять компоненты собранного материала культуры клеток на основе размера, а повышение скорости потока после этого начального периода приводит к повышению общей скорости способа. В некоторых вариантах осуществления SE-UPLC выполняют с начальной скоростью потока, составляющей менее приблизительно 0,4 мл/мин., за которой следует скорость потока, составляющая приблизительно 0,4 мл/мин. или больше. В некоторых вариантах осуществления начальная скорость потока составляет менее приблизительно 0,4 мл/мин., менее приблизительно 0,3 мл/мин., менее приблизительно 0,2 мл/мин. или приблизительно 0,1 мл/мин. В некоторых вариантах осуществления начальная скорость потока относится к первым 15, 20 или 25 минутам SE-UPLC. В некоторых вариантах осуществления SE-UPLC выполняют со скоростью потока, составляющей менее приблизительно 0,4 мл/мин. в течение первых 25 минут, за которой следует скорость потока, составляющая более приблизительно 0,4 мл/мин. или больше (например, 25-33 минуты). В некоторых вариантах осуществления SE-UPLC проводят со скоростью потока, составляющей приблизительно 0,1 мл/мин. в течение первых 25 минут, а затем со скоростью потока, составляющей приблизительно 0,4 мл/мин. (например, от 25 до 33 минут).
[0088] В некоторых вариантах осуществления SE-UPLC проводят с направлением части(частей) потока, образуемого при жидкостной хроматографии (LC), в котором не происходит элюирования белка (например, 0-10 минут и/или 24-33 минуты), в отходы, за счет чего обеспечивают более быстрый анализ для скрининга и избегают контаминации источника MS.
[0089] В некоторых вариантах осуществления SE-UPLC проводят с использованием изократического элюирования подвижной фазой. Например, в некоторых вариантах осуществления подвижная фаза содержит раствор, содержащий смесь ацетонитрил:вода в соотношении 30:70. В некоторых вариантах осуществления подвижная фаза содержит муравьиную кислоту (FA) и трифторуксусную кислоту (TFA). Не ограничиваясь теорией, считается, что, хотя TFA применима для разделения посредством жидкостной хроматографии (LC), она может подавлять MS-сигнал, и, следовательно, подвижная фаза, содержащая смесь FA и TFA, может обеспечивать эффективное разделение посредством жидкостной хроматографии (LC) при сохранении адекватного соотношения сигнал-шум для MS-анализа. В некоторых вариантах осуществления подвижная фаза содержит приблизительно 0,05% муравьиной кислоты и приблизительно 0,05% трифторуксусной кислоты (TFA).
[0090] Предпочтительно способы, предусмотренные в данном документе, позволяют осуществлять быстрый анализ продуктов, что позволяет проводить высокопроизводительный скрининг (например, множества потенциальных линий клеток-продуцентов). Например, в некоторых вариантах осуществления определение выполняют за приблизительно 30 минут или меньше, за приблизительно 33 минуты или меньше, за приблизительно 35 минут или меньше, за приблизительно 40 минут или меньше, за приблизительно 45 минут или меньше или за приблизительно 60 минут или меньше.
[0091] В некоторых вариантах осуществления способы дополнительно включают перед SE-UPLC-MS приведение продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием) в контакт с протеазой. В некоторых вариантах осуществления протеаза представляет собой IdeS или IdeZ, например, для анализа фрагментов антител.
[0092] В некоторых вариантах осуществления антитело или производное антитела содержат остаток цистеина в положении 293 (Cys293). В некоторых вариантах осуществления антитело или производное антитела и одна или несколько молекул, являющихся весовыми вариантами, предусматривают молекулы со свободным цистеином (например, не связанным дисульфидной связью с другим цистеином антитела или производного антитела), которые были подвержены цистеинилированию, N-ацетилцистеинилированию или глутатионилированию. В некоторых вариантах осуществления антитело или производное антитела не являются N-гликозилированными (например, в Fc-области антитела). В некоторых вариантах осуществления антитело или производное антитела содержат мутацию в Fc-области, которая приводит к снижению или устранению N-гликозилирования. В некоторых вариантах осуществления антитело или производное антитела содержат мутацию N300A (согласно EU-индексу). В некоторых вариантах осуществления антитело или производное антитела являются N-гликозилированными (например, в Fc-области антитела), и способ дополнительно включает (например, перед SE-UPLC-MS) удаление N-гликозилирования антитела. В некоторых вариантах осуществления удаление N-гликозилирования антитела предусматривает обработку антитела ферментом, представляющим собой пептид:N-гликозидазу (например, PNGазу F).
[0093] В некоторых вариантах осуществления одна или несколько молекул, являющихся весовыми вариантами, представляют собой молекулы антитела или производного антитела, содержащие химически модифицированный остаток цистеина. В некоторых вариантах осуществления MS позволяет определять разницу в массе между цистеинилированными (сдвиг по массе 119 Да), N-ацетилцистеинилированными (сдвиг по массе 161 Да) и глутатионилированными (сдвиг по массе 305 Да) молекулами. Например, антитела и производные антител, содержащие свободный остаток цистеина (например, цистеин, не связанный дисульфидной связью с другим остатком цистеина антитела/производного антитела), и/или антитела и производные антител, содержащие дополнительный остаток цистеина, встроенный в их Fc-область, могут быть применимыми, например, для конъюгации с соединением (например, цитотоксическим лекарственным средством с образованием конъюгата антитело-лекарственное средство). Этот дополнительный цистеин является неспаренным (свободным), т. е. не задействован в дисульфидных связях с другими остатками цистеина в молекуле mAb. Предыдущая работа показала, что этот дополнительный цистеин (Cys293) может образовывать нежелательные дисульфидные связи с соседними остатками цистеина, что также известно как "перестановка дисульфидных связей", которая потенциально приводит к структурной нестабильности антитела. Неконтролируемые химические реакции сульфгидрильных групп также могут обуславливать значительный риск в отношении выполнимости производственного процесса в ходе обработки для образования ADC, что приводит к нежелательному профилю конъюгации лекарственного средства. Для избежания этих рисков необходимо обеспечить блокирование свободного цистеина с помощью дисульфидной модификации, такой как цистеинилирование и глутатионилирование, путем регулирования условий культивирования клеток. Эти модификации можно избирательно устранять с образованием свободного цистеина перед этапом конъюгации с лекарственным средством. Способы, описанные в данном документе, можно применять, например, для анализа продуцирования антитела или производного антитела, в том числе различных молекул, являющихся весовыми вариантами, таких как молекулы, содержащие химически модифицированный цистеин, такой как цистеин, блокированный дисульфидной модификацией, такой как цистеинилирование и глутатионилирование.
[0094] В некоторых вариантах осуществления антитело или производное антитела являются N-гликозилированными (например, в Fc-области антитела), и одна или несколько молекул, являющихся весовыми вариантами, представляют собой гликоформы антитела или производного антитела. В некоторых вариантах осуществления MS позволяет определять разницу в массе, составляющую приблизительно 162 Да, между антителом или производным антитела и одной или несколькими молекулами, являющимися весовыми вариантами, которые представляют собой гликоформы антитела или производного антитела. В некоторых вариантах осуществления антитело или производное антитела представляют собой полиспецифическое антитело. В некоторых вариантах осуществления антитело или производное антитела представляют собой моноклональное антитело.
[0095] Другие аспекты настоящего изобретения относятся к способам получения антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка. В некоторых вариантах осуществления способы включают культивирование линии клеток, содержащей один или несколько полинуклеотидов, кодирующих продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием), в условиях, подходящих для продуцирования продукта линией клеток; отделение от линии клеток среды для культивирования клеток, содержащей продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием); определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием) в среде для культивирования клеток посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS); и (d) удаление по меньшей мере некоторой доли одной или нескольких молекул с ошибочным спариванием из полиспецифического связывающего белка, продуцируемого линией клеток. В некоторых вариантах осуществления способы включают культивирование линии клеток, содержащей один или несколько полинуклеотидов, кодирующих продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием), в условиях, подходящих для продуцирования продукта линией клеток; отделение от линии клеток среды для культивирования клеток, содержащей продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием); определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием) в среде для культивирования клеток посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS); и (d) определение одного или обоих из показателей качества и чистоты полиспецифического связывающего белка, продуцируемого линией клеток. В некоторых вариантах осуществления способы включают культивирование линии клеток, содержащей один или несколько полинуклеотидов, кодирующих продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием), в условиях, подходящих для продуцирования продукта линией клеток; отделение от линии клеток среды для культивирования клеток, содержащей продукт (например, антитело, фрагмент антитела, белок, являющийся продуктом слияния с Fc, или полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием); определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием) в среде для культивирования клеток посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS); и (d) определение одного или обоих из показателей качества и чистоты полиспецифического связывающего белка, продуцируемого линией клеток, и удаление по меньшей мере некоторой доли одной или нескольких молекул с ошибочным спариванием из полиспецифического связывающего белка, продуцируемого линией клеток. Любой из иллюстративных продуктов, описанных в данном документе, может найти применение в этих и других способах.
[0096] Другие аспекты настоящего изобретения относятся к способам скрининга совокупности линий клеток в отношении продуцирования продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием). В некоторых вариантах осуществления способы включают определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием), продуцируемого первой линией клеток (например, с применением любого из способов, описанных в данном документе), и определение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием), продуцируемого второй линией клеток (например, с применением любого из способов, описанных в данном документе), например, отличной от первой линии клеток. Любой из иллюстративных продуктов, описанных в данном документе, может найти применение в этих и других способах.
[0097] В некоторых вариантах осуществления способы дополнительно включают сравнение количества продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием), продуцируемого первой линией клеток, с количеством продукта (например, антитела, фрагмента антитела, белка, являющегося продуктом слияния с Fc, или полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием), продуцируемого второй линией клеток; и основывающийся на сравнении выбор линии клеток, которая продуцирует большее количество продукта. В некоторых вариантах осуществления проводят скрининг 2 или более, 3 или более, 4 или более, 5 или более, 10 или более, 20 или более, 30 или более, 40 или более или 50 или более различных линий клеток. Предпочтительно способы, описанные в данном документе, являются быстрыми и применимы для высокопроизводительного скрининга.
[0098] В некоторых вариантах осуществления способы дополнительно включают определение количества одной или нескольких молекул с ошибочным спариванием, продуцируемого первой линией клеток; и определение количества одной или нескольких молекул с ошибочным спариванием, продуцируемого второй линией клеток. В некоторых вариантах осуществления, например, после определения количества одной или нескольких молекул с ошибочным спариванием, продуцируемого первой и второй линиями клеток, способы дополнительно включают сравнение количества одной или нескольких молекул с ошибочным спариванием, продуцируемого первой линией клеток, с количеством одной или нескольких молекул с ошибочным спариванием, продуцируемым второй линией клеток. В некоторых вариантах осуществления на основе сравнения выбирают линию клеток, при продуцировании в которой значение соотношения полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием является более высоким. В некоторых вариантах осуществления линию клеток выбирают на основе более высокого значения соотношения полиспецифического связывающего белка и количества одной или нескольких отдельных молекул с ошибочным спариванием и/или на основе более высокого значения соотношения количества полиспецифического связывающего белка и общего количества молекул с ошибочным спариванием.
[0099] В некоторых вариантах осуществления выбираемую линию клеток выбирают для применения в качестве линии клеток-продуцентов.
Полиспецифические связывающие белки
[0100] Определенные аспекты настоящего изобретения относятся к полиспецифическим связывающим белкам.
[0101] В некоторых вариантах осуществления полиспецифический связывающий белок предусматривает ассоциацию двух или более полипептидных цепей, содержащую по меньшей мере первую полипептидную цепь и вторую полипептидную цепь, отличную от первой полипептидной цепи. Например, в некоторых вариантах осуществления полиспецифический связывающий белок представляет собой биспецифическое антитело или биспецифический белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой триспецифическое антитело или триспецифический белок, являющийся продуктом слияния с Fc. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием содержат две или более полипептидных цепей, содержащих по меньшей мере одну из первой и второй полипептидных цепей в ассоциации, отличной от ассоциации полиспецифического связывающего белка.
[0102] В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой полиспецифическое антитело, содержащее тяжелую цепь первого антитела, легкую цепь первого антитела, тяжелую цепь второго антитела, отличную от тяжелой цепи первого антитела, и легкую цепь второго антитела, отличную от легкой цепи первого антитела. В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего: ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две тяжелые цепи первого антитела; ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две тяжелые цепи второго антитела; ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две легкие цепи первого антитела; и ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две легкие цепи второго антитела.
[0103] В некоторых вариантах осуществления полиспецифические связывающие белки содержат четыре полипептидные цепи, которые образуют три антигенсвязывающих сайта, которые специфически связываются с одним или несколькими белками-мишенями, где первая пара полипептидов, образующих связывающий белок, содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией, и где вторая пара полипептидов, образующих связывающий белок, содержит один вариабельный домен.
[0104] В некоторых вариантах осуществления полиспецифические связывающие белки содержат одну или две Fc-области, слитые с одним, двумя, тремя или более антигенсвязывающими доменами или другими полипептидами (например, белок, являющийся продуктом слияния с Fc). В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой иммуноглобулин с двойным вариабельным доменом (DVD), например, как описано в WO 2012061558. В некоторых вариантах осуществления полиспецифический связывающий белок содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией, например, как описано в WO 2012135345. В некоторых вариантах осуществления связывающий белок содержит четыре полипептидные цепи, которые образуют четыре антигенсвязывающих сайта, при этом две полипептидные цепи содержат структуру, представленную формулой:
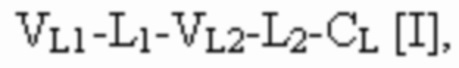
и две полипептидные цепи содержат структуру, представленную формулой:

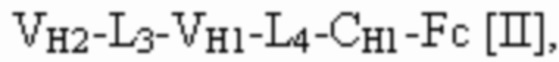
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VR2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
Fc представляет собой шарнирную область иммуноглобулина и константные домены CH2, CH3 тяжелой цепи иммуноглобулина; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры;
где полипептиды формулы I и полипептиды формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи.
[0105] В некоторых вариантах осуществления каждый из трех антигенсвязывающих сайтов связывает другую мишень (например, полипептидный антиген). В некоторых вариантах осуществления триспецифический связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих сайта, где первая полипептидная цепь содержит структуру, представленную формулой:
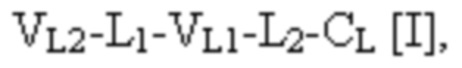
и вторая полипептидная цепь содержит структуру, представленную формулой:
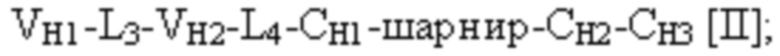
и третья полипептидная цепь содержит структуру, представленную формулой:
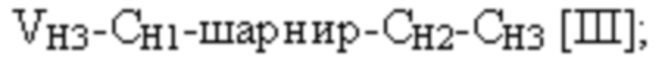
и четвертая полипептидная цепь содержит структуру, представленную формулой:
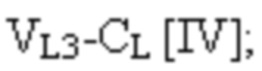
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи.
[0106] В некоторых вариантах осуществления полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи. В некоторых вариантах осуществления VH1 и VL1 образуют первый антигенсвязывающий сайт, при этом VH2 и VL2 образуют второй антигенсвязывающий сайт, и при этом VH3 и VL3 образуют третий антигенсвязывающий сайт.
[0107] В некоторых вариантах осуществления одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего: ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой I; ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой II; ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой III; и ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой IV.
[0108] В некоторых вариантах осуществления полиспецифические связывающие белки являются триспецифическими и/или тривалентными. В некоторых вариантах осуществления полиспецифические связывающие белки являются биспецифическими и/или бивалентными.
[0109] Предполагается, что любому из антигенсвязывающих сайтов, описанных в данном документе, можно найти применение в триспецифическом связывающем белке по настоящему изобретению, например, содержащем четыре полипептидные цепи, характеризующиеся структурами, описанными выше. Например, в некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт, который связывает опухолевый белок-мишень. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт, который связывает белок-мишень HIV. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, который связывает белок-мишень HIV, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает белок-мишень HIV, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт, который связывает белок-мишень HIV. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт, который связывает опухолевый белок-мишень. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт, который связывает полипептид CD38. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий сайт, который связывает полипептид HER2.
[0110] В некоторых вариантах осуществления связывающий белок по настоящему изобретению связывает один или несколько опухолевых белков-мишеней и один или несколько Т-клеточных белков-мишеней. В некоторых вариантах осуществления связывающий белок способен специфически связывать один опухолевый белок-мишень и два различных эпитопа на одном Т-клеточном белке-мишени. В некоторых вариантах осуществления связывающий белок способен специфически связывать один опухолевый белок-мишень и два различных Т-клеточных белка-мишени (например, CD28 и CD3). В некоторых вариантах осуществления первая и вторая полипептидные цепи связывающего белка образуют два антигенсвязывающих сайта, которые специфически нацелены на два Т-клеточных белка-мишени, а третья и четвертая полипептидные цепи связывающего белка образуют антигенсвязывающий сайт, который специфически связывает опухолевый белок-мишень. В некоторых вариантах осуществления белок-мишень представляет собой CD38 или HER2. Дополнительные опухолевые белки-мишени представлены ниже. В некоторых вариантах осуществления один или несколько T-клеточных белков-мишеней представляют собой один или несколько из CD3 и CD28.
[0111] Иллюстративные полиспецифические связывающие белки, которые можно применять в способах по настоящему изобретению, также включают без ограничения белки, описанные в международных публикациях №№ WO 2017074878, WO 2017180913, WO 2018151841 и WO 2019074973.
[0112] В некоторых вариантах осуществления связывающий белок представляет собой триспецифический связывающий белок. В некоторых вариантах осуществления триспецифический связывающий белок содержит антигенсвязывающий сайт, который связывает полипептид CD38, антигенсвязывающий сайт, который связывает полипептид CD28, и антигенсвязывающий сайт, который связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые содержат три антигенсвязывающих сайта, где полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи (например, как описано в данном документе). В некоторых вариантах осуществления домены VH и VL любого из антигенсвязывающих сайтов, специфических к CD38, описанных выше, представляют собой VH3 и VL3 и образуют третий антигенсвязывающий сайт, который связывает полипептид CD38. В некоторых вариантах осуществления VH1 и VL1 образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, VH2 и VL2 образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и VH3 и VL3 образуют третий антигенсвязывающий сайт, который связывает полипептид CD38.
[0113] В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий сайт, который связывает полипептид HER2, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид HER2, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, описанных выше, где домены VH3 и VL3 образуют пару, которая образует третий антигенсвязывающий сайт, который связывает полипептид HER2.
[0114] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий сайт, который связывает опухолевый белок-мишень. В некоторых вариантах осуществления опухолевый белок-мишень представляет собой полипептид CD38 (например, полипептид CD38 человека). В некоторых вариантах осуществления опухолевый белок-мишень представляет собой полипептид HER2 (например, полипептид HER2 человека). В некоторых вариантах осуществления опухолевый белок-мишень по настоящему изобретению включает без ограничения A2AR, APRIL, ATPDазу, BAFF, BAFFR, BCMA, BlyS, BTK, BTLA, B7DC, B7H1, B7H4 (также известный как VTCN1), B7H5, B7H6, B7H7, B7RP1, B7-4, C3, C5, CCL2 (также известный как MCP-1), CCL3 (также известный как MIP-1a), CCL4 (также известный как MIP-1b), CCL5 (также известный как RANTES), CCL7 (также известный как MCP-3), CCL8 (также известный как mcp-2), CCL11 (также известный как эотаксин), CCL15 (также известный как MIP-1d), CCL17 (также известный как TARC), CCL19 (также известный как MIP-3b), CCL20 (также известный как MIP-3a), CCL21 (также известный как MIP-2), CCL24 (также известный как MPIF-2/эотаксин-2), CCL25 (также известный как TECK), CCL26 (также известный как эотаксин-3), CCR3, CCR4, CD3, CD19, CD20, CD23 (также известный как FCER2, рецептор IgE), CD24, CD27, CD28, CD38, CD39, CD40, CD70, CD80 (также известный как B7-1), CD86 (также известный как B7-2), CD122, CD137 (также известный как 41BB), CD137L, CD152 (также известный как CTLA4), CD154 (также известный как CD40L), CD160, CD272, CD273 (также известный как PDL2), CD274 (также известный как PDL1), CD275 (также известный как B7H2), CD276 (также известный как B7H3), CD278 (также известный как ICOS), CD279 (также известный как PD-1), CDH1 (также известный как E-кадгерин), хитиназу, CLEC9, CLEC91, CRTH2, CSF-1 (также известный как M-CSF), CSF-2 (также известный как GM-CSF), CSF-3 (также известный как GCSF), CX3CL1 (также известный как SCYD1), CXCL12 (также известный как SDF1), CXCL13, CXCR3, DNGR-1, эктонуклеозидтрифосфатдифосфогидролазу 1, EGFR, ENTPD1, FCER1A, FCER1, FLAP, FOLH1, Gi24, GITR, GITRL, GM-CSF, Her2, HHLA2, HMGB1, HVEM, ICOSLG, IDO, IFNα, IgE, IGF1R, IL2R бета, IL1, IL1A, IL1B, IL1F10, IL2, IL4, IL4Ra, IL5, IL5R, IL6, IL7, IL7Ra, IL8, IL9, IL9R, IL10, rhIL10, IL12, IL13, IL13Ra1, IL13Ra2, IL15, IL17, IL17Rb (также известный как рецептор IL25), IL18, IL22, IL23, IL25, IL27, IL33, IL35, ITGB4 (также известный как интегрин b4), ITK, KIR, LAG3, LAMP1, лептин, LPFS2, молекулы MHC класса II, MUC-1, NCR3LG1, NKG2D, NTPDазу-1, OX40, OX40L, PD-1H, тромбоцитарный рецептор, PROM1, S152, SISP1, SLC, SPG64, ST2 (также известный как рецептор IL33), STEAP2, Syk-киназу, TACI, TDO, T14, TIGIT, TIM3, TLR, TLR2, TLR4, TLR5, TLR9, TMEF1, TNFa, TNFRSF7, Tp55, TREM1, TSLP (также известный как корецептор IL7Ra), TSLPR, TWEAK, VEGF, VISTA, Vstm3, WUCAM и XCR1 (также известный как GPR5/CCXCR1). В некоторых вариантах осуществления одна или несколько из указанных выше антигенных мишеней представляют собой антигенные мишени человека.
[0115] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий сайт, который связывает белок-мишень HIV. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит три антигенсвязывающих сайта, каждый из которых связывается с белком-мишенью HIV. В некоторых вариантах осуществления связывающие белки специфически связываются с одним или несколькими белками-мишенями HIV (например, как описано выше) и одним или несколькими белками-мишенями на T-клетке, в том числе Т-клеточным рецепторным комплексом. Примеры белков-мишеней на T-клетках включают без ограничения, среди прочего, CD3 и CD28. В некоторых вариантах осуществления белок-мишень HIV представляет собой гликопротеин 120, гликопротеин 41 и гликопротеин 160. В некоторых вариантах осуществления связывающий белок связывает один или несколько из гликопротеина 120, гликопротеина 41 и гликопротеина 160. К иллюстративным белкам HIV относятся без ограничения MPER белка gp41 HIV-1, CD4-связывающий сайт белка gp120 HIV-1, гликан в петле V3 белка gp120 HIV-1 или верхняя часть тримера, представляющего собой белок gp120 или gp160 HIV-1. Например, в некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий сайт, который связывает CD4-связывающий сайт белка gp120 HIV-1. К иллюстративным антигенсвязывающим сайтам, которые связывают белки-мишени HIV, предусмотренные для применения в данном документе, относятся без ограничения сайты, описанные в международной публикации № WO 2017/074878, такие как сайты из антител CD4BS "a", CD4BS "b", MPER, MPER_100W, V1/V2 "a", V1/V2 "b" или V3.
[0116] В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий сайт, который связывает полипептид CD28, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD28, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD28, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, и один из которых связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид CD38. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид HER2. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает опухолевый белок-мишень.
[0117] В некоторых вариантах осуществления любого из приведенных выше вариантов осуществления связывающий белок представляет собой триспецифический связывающий белок. В некоторых вариантах осуществления триспецифический связывающий белок содержит антигенсвязывающий сайт, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2), антигенсвязывающий сайт, который связывает полипептид CD28, и антигенсвязывающий сайт, который связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые содержат три антигенсвязывающих сайта, где полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи (например, как описано в данном документе). В некоторых вариантах осуществления домены VH и VL любого из антигенсвязывающих сайтов, специфических к CD28, описанных выше, представляют собой VH1 и VL1 и образуют первый антигенсвязывающий сайт, который связывает полипептид CD28. В некоторых вариантах осуществления VH1 и VL1 образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, VH2 и VL2 образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и VH3 и VL3 образуют третий антигенсвязывающий сайт, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2).
[0118] В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий сайт, который связывает полипептид CD3, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, и один из которых связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид CD38. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид HER2. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий сайт, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих сайта, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает опухолевый белок-мишень.
[0119] В некоторых вариантах осуществления связывающий белок представляет собой триспецифический связывающий белок. В некоторых вариантах осуществления триспецифический связывающий белок содержит антигенсвязывающий сайт, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2), антигенсвязывающий сайт, который связывает полипептид CD28, и антигенсвязывающий сайт, который связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые содержат три антигенсвязывающих сайта, где полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи (например, как описано в данном документе). В некоторых вариантах осуществления VH2 и VL2 образуют первый антигенсвязывающий сайт, который связывает полипептид CD3. В некоторых вариантах осуществления VH1 и VL1 образуют первый антигенсвязывающий сайт, который связывает полипептид CD28, VH2 и VL2 образуют второй антигенсвязывающий сайт, который связывает полипептид CD3, и VH3 и VL3 образуют третий антигенсвязывающий сайт, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2).
Линкеры
[0120] В некоторых вариантах осуществления длина линкеров L1, L2, L3 и L4 варьирует от нуля аминокислот (длина=0) до приблизительно 100 аминокислот или менее чем 100, 50, 40, 30, 20 или 15 аминокислот или меньше. Длина линкеров также может составлять 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислоту. Все из L1, L2, L3 и L4 в одном связывающем белке могут иметь одинаковую аминокислотную последовательность или все могут иметь различные аминокислотные последовательности.
[0121] Примеры подходящих линкеров включают, например, GGGGSGGGGS (SEQ ID NO:1), GGGGSGGGGSGGGGS (SEQ ID NO: 2), S, RT, TKGPS (SEQ ID NO:3), GQPKAAP (SEQ ID NO: 4), GGSGSSGSGG (SEQ ID NO: 5) и DKTHT (SEQ ID NO:6), а также раскрытые в международных публикациях №№ WO 2017/074878 и WO 2017/180913. Вышеперечисленные примеры не предназначены для какого-либо ограничения объема настоящего изобретения, и при этом было показано, что линкеры, содержащие выбранные случайным образом аминокислоты, выбранные из группы, состоящей из валина, лейцина, изолейцина, серина, треонина, лизина, аргинина, гистидина, аспартата, глутамата, аспарагина, глутамина, глицина и пролина, подходят для связывающих белков.
[0122] Состав и последовательность аминокислотных остатков в линкере может меняться в зависимости от типа элемента вторичной структуры, который необходимо достигнуть при помощи линкера. Например, глицин, серин и аланин являются наиболее подходящими для линкеров, характеризующихся максимальной гибкостью. Некоторые комбинации глицина, пролина, треонина и серина применимы, если необходим более жесткий и удлиненный линкер. При необходимости любой аминокислотный остаток может рассматриваться как линкер в комбинации с другими аминокислотными остатками для конструирования более длинных пептидных линкеров в зависимости от необходимых свойств.
[0123] В некоторых вариантах осуществления длина L1 в по меньшей два раза превышает длину L3. В некоторых вариантах осуществления длина L2 в по меньшей два раза превышает длину L4. В некоторых вариантах осуществления длина L1 в по меньшей мере два раза превышает длину L3, а длина L2 в по меньшей мере два раза превышает длину L4. В некоторых вариантах осуществления длина L1 составляет 3-12 аминокислотных остатков, длина L2 составляет 3-14 аминокислотных остатков, длина L3 составляет 1-8 аминокислотных остатков, и длина L4 составляет 1-3 аминокислотных остатка. В некоторых вариантах осуществления длина L1 составляет 5-10 аминокислотных остатков, длина L2 составляет 5-8 аминокислотных остатков, длина L3 составляет 1-5 аминокислотных остатков, и длина L4 составляет 1-2 аминокислотных остатка. В некоторых вариантах осуществления длина L1 составляет 7 аминокислотных остатков, длина L2 составляет 5 аминокислотных остатков, длина L3 составляет 1 аминокислотный остаток, и длина L4 составляет 2 аминокислотных остатка.
[0124] В некоторых вариантах осуществления каждый из L1, L2, L3 и L4 независимо имеет длину, составляющую ноль аминокислот, или содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO:1), GGGGSGGGGSGGGGS (SEQ ID NO: 2), S, RT, TKGPS (SEQ ID NO: 3), GQPKAAP (SEQ ID NO: 4) и GGSGSSGSGG (SEQ ID NO: 5). В некоторых вариантах осуществления каждый из L1, L2, L3 и L4 независимо содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO:1) GGGGSGGGGSGGGGS (SEQ ID NO: 2), S, RT, TKGPS (SEQ ID NO: 3), GQPKAAP (SEQ ID NO: 4) и GGSGSSGSGG (SEQ ID NO: 5). В некоторых вариантах осуществления L1 содержит последовательность GQPKAAP (SEQ ID NO: 4), L2 содержит последовательность TKGPS (SEQ ID NO:3), L3 содержит последовательность S, и L4 содержит последовательность RT.
[0125] В некоторых вариантах осуществления по меньшей мере один из L1, L2, L3 или L4 содержит последовательность DKTHT (SEQ ID NO:6). В некоторых вариантах осуществления L1, L2, L3 и L4 содержат последовательность DKTHT (SEQ ID NO:6).
Fc-области и константные домены
[0126] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит вторую полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит третью полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит вторую полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, и третью полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина.
[0127] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит полноразмерную тяжелую цепь антитела или полипептидную цепь, содержащую Fc-область. В некоторых вариантах осуществления Fc-область представляет собой Fc-область человека, например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека. В некоторых вариантах осуществления Fc-область включает шарнир, CH1, CH2, CH3 и необязательно домены CH4. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека. В некоторых вариантах осуществления Fc-область содержит одну или несколько мутаций, описанных ниже. В некоторых вариантах осуществления Fc-область представляет собой Fc-область одного из полипептидов тяжелой цепи (например, полипептида 2 или 3) связывающего белка, представленного в таблице 4. В некоторых вариантах осуществления константная область тяжелой цепи представляет собой константную область одного из полипептидов тяжелой цепи (например, полипептида 2 или 3) связывающего белка, представленного в таблице 4. В некоторых вариантах осуществления константная область легкой цепи представляет собой константную область одного из полипептидов легкой цепи (например, полипептида 1 или 4) связывающего белка, представленного в таблице 4.
[0128] В некоторых вариантах осуществления связывающий белок согласно настоящему изобретению содержит один или два варианта Fc. Термин "вариант Fc", используемый в данном документе, обозначает молекулу или последовательность, которая модифицирована по сравнению с нативным Fc, но все еще содержит сайт связывания для рецептора реутилизации FcRn (неонатального Fc-рецептора). Приводимые в качестве примера варианты Fc и их взаимодействие с рецептором реутилизации известны из уровня техники. Таким образом, термин "вариант Fc" может предусматривать молекулу или последовательность, полученную в результате гуманизации нативного Fc отличного от человеческого происхождения. Кроме того, нативный Fc содержит области, которые можно удалить, поскольку они обеспечивают структурные признаки или биологическую активность, которые не требуются для антителоподобных связывающих белков по настоящему изобретению. Таким образом, термин "вариант Fc" предусматривает молекулу или последовательность, в которой отсутствует один или несколько сайтов или остатков нативного Fc или в которой были модифицированы один или несколько сайтов или остатков Fc, воздействующих на или вовлеченных в: (1) образование дисульфидной связи, (2) несовместимость с выбранной клеткой-хозяином, (3) гетерогенность N-концов при экспрессии в выбранной клетке-хозяине, (4) гликозилирование, (5) взаимодействие с системой комплемента, (6) связывание с Fc-рецептором, отличным от рецептора реутилизации, или (7) антителозависимую клеточную цитотоксичность (ADCC).
[0129] В некоторых вариантах осуществления связывающий белок по настоящему изобретению (например, триспецифический связывающий белок) содержит мутацию по типу "выступа" на второй полипептидной цепи и мутацию по типу "впадины" на третьей полипептидной цепи. В некоторых вариантах осуществления связывающий белок согласно настоящему изобретению содержит мутацию по типу "выступа" на третьей полипептидной цепи и мутацию по типу "впадины" на второй полипептидной цепи. В некоторых вариантах осуществления мутация по типу "выступа" предусматривает замену (замены) в положениях, соответствующих положениям 354 и/или 366 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S354C, T366W, T366Y, S354C и T366W или S354C и T366Y. В некоторых вариантах осуществления мутация по типу "выступа" предусматривает замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S354C и T366W. В некоторых вариантах осуществления мутация по типу "впадины" предусматривает замену(замены) в положениях, соответствующих положениям 407 и необязательно 349, 366 и/или 368 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой Y407V или Y407T и необязательно Y349C, T366S и/или L368A. В некоторых вариантах осуществления мутация по типу "впадины" предусматривает замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V.
[0130] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную замену(замены) в положениях, соответствующих положениям 366 и необязательно 354 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366W или T366Y и необязательно S354C; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную замену(замены) в положениях, соответствующих положениям 407 и необязательно 349, 366 и/или 368 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y407V или Y407T и необязательно Y349C, T366S и/или L368A.
[0131] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную замену(замены) в положениях, соответствующих положениям 407 и необязательно 349, 366 и/или 368 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y407V или Y407T и необязательно Y349C, T366S и/или L368A; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную замену(замены) в положениях, соответствующих положениям 366 и необязательно 354 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366W или T366Y и необязательно S354C.
[0132] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную замену в положении, соответствующем положению 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотная замена представляет собой T366W; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную замену(замены) в положениях, соответствующих положениям 366, 368 и/или 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366S, L368A и/или Y407V.
[0133] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную замену(замены) в положениях, соответствующих положениям 366, 368 и/или 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366S, L368A и/или Y407V; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную замену в положении, соответствующем положению 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотная замена представляет собой T366W.
[0134] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W. В некоторых вариантах осуществления первый и/или второй Fc-области представляют собой Fc-области IgG1 человека. В некоторых вариантах осуществления первый и/или второй Fc-области представляют собой Fc-области IgG4 человека.
[0135] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, S354C, T366W и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, Y349C, T366S, L368A, Y407V и R409K. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, Y349C, T366S, L368A, Y407V и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, S354C, T366W и R409K.
[0136] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 354 и 366 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, S354C и T366W; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 349, 366, 368 и 407 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, Y349C, T366S, L368A и Y407V. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 349, 366, 368 и 407 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, Y349C, T366S, L368A и Y407V; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 354 и 366 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, S354C и T366W.
[0137] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит одну или несколько мутаций для снижения эффекторной функции, например, антителозависимого клеточного фагоцитоза, опосредованного Fc-рецепторами (ADCP), комплементзависимой цитотоксичности (CDC) и/или антителозависимой клеточной цитотоксичности (ADCC). В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где первая и вторая Fc-области представляют собой Fc-области IgG1 человека; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A и L235A. В некоторых вариантах осуществления Fc-области второй и третьей полипептидных цепей представляют собой Fc-области IgG1 человека, и при этом каждая из Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A и L235A. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где первая и вторая Fc-области представляют собой Fc-области IgG1 человека; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234, 235 и 329 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A, L235A и P329A. В некоторых вариантах осуществления Fc-области второй и третьей полипептидных цепей представляют собой Fc-области IgG1 человека, и где каждая из Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234, 235 и 329 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A, L235A и P329A. В некоторых вариантах осуществления Fc-области второй и третьей полипептидных цепей представляют собой Fc-области IgG4 человека, и каждая из Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A и L235A. В некоторых вариантах осуществления связывающий белок содержит вторую полипептидную цепь, дополнительно содержащую первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, и при этом третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A и L235A.
[0138] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, S354C, T366W и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, Y349C, T366S, L368A, Y407V и R409K. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, Y349C, T366S, L368A, Y407V и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, S354C, T366W и R409K.
[0139] В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую одну или несколько мутаций, которые ослабляют или устраняют связывание FcγI и/или FcγII. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую одну или несколько мутаций, которые ослабляют или устраняют связывание FcγI и/или FcγII, но не влияют на связывание FcRn. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 228 и/или 409 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S228P и/или R409K. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 234 и/или 235 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой F234A и/или L235A. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 228, 234, 235 и/или 409 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S228P, F234A, L235A и/или R409K. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 233-236 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой E233P, F234V, L235A, и делецию в положении 236. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные мутации в положениях, соответствующих положениям 228, 233-236 и/или 409 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные мутации представляют собой S228P; E233P, F234V, L235A, и делецию в положении 236; и/или R409K.
[0140] В некоторых вариантах осуществления Fc-область содержит одну или несколько мутаций, которые уменьшают или устраняют связывание Fc-рецептора и/или эффекторную функцию Fc-области (например, антителозависимый клеточный фагоцитоз, опосредованный Fc-рецепторами (ADCP), комплементзависимую цитотоксичность (CDC) и/или антителозависимую клеточную цитотоксичность (ADCC)).
[0141] В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую одну или несколько аминокислотных замен в положениях, соответствующих положениям 234, 235 и/или 329 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой L234A, L235A и/или P329A. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 298, 299 и/или 300 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S298N, T299A и/или Y300S.
[0142] В некоторых вариантах осуществления связывающий белок согласно настоящему изобретению содержит одну или несколько мутаций для улучшения стабильности, например, шарнирной области и/или поверхности контакта димера IgG4 (см., например, Spiess, C. et al. (2013) J. Biol. Chem. 288:26583-26593). В некоторых вариантах осуществления мутация предусматривает замены в положениях, соответствующих положениям 228 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P и R409K. В некоторых вариантах осуществления связывающий белок содержит вторую полипептидную цепь, дополнительно содержащую первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, и при этом третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где первая и вторая Fc-области представляют собой Fc-области IgG4 человека; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 228 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P и R409K. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит мутации по типу выступа и впадины и одну или несколько мутаций для улучшения стабильности. В некоторых вариантах осуществления первый и/или второй Fc-области представляют собой Fc-области IgG4 человека.
[0143] В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую одну или несколько аминокислотных замен в положениях, соответствующих положениям 234, 235 и/или 329 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой L234A, L235A и/или P329A. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 298, 299 и/или 300 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S298N, T299A и/или Y300S.
Нуклеиновые кислоты и векторы
[0144] Стандартные технологии рекомбинантной ДНК применяют для конструирования полинуклеотидов, которые кодируют полипептиды, образующие связывающие белки, встраивания этих полинуклеотидов в рекомбинантные векторы экспрессии и введения таких векторов в клетки-хозяева. См., например, Sambrook et al., 2001, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 3-е изд.). Ферментативные реакции и методики очистки можно проводить в соответствии со спецификациями производителей, которые обычно выполняют в данной области техники или как описано в данном документе. Если не предусмотрены конкретные определения, то терминология, используемая в контексте аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, а также лабораторные процедуры и их методики, описанные в данном документе, являются общеизвестными и широко используемыми в данной области техники. Аналогично традиционные методики можно использовать для вариантов химического синтеза, вариантов химического анализа, фармацевтического получения, составления, доставки и лечения пациентов.
[0145] В некоторых вариантах осуществления выделенная нуклеиновая кислота функционально связана с гетерологичным промотором для управления транскрипцией последовательности нуклеиновой кислоты, кодирующей связывающий белок. Промотор может относиться к контролирующим последовательностям нуклеиновой кислоты, которые управляют транскрипцией нуклеиновой кислоты. Первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, где первая последовательность нуклеиновой кислоты находится в функциональной связи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с последовательностью, кодирующей связывающий белок, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности. Примеры промоторов могут включать без ограничения промоторы, полученные из геномов вирусов (таких как вирус полиомы, вирус оспы кур, аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита B, вирус обезьян 40 (SV40) и т. п.), гетерологичные промоторы эукариот (такие как промотор гена актина, промотор гена иммуноглобулина, промоторы генов белков теплового шока и т. п.), CAG-промотор (Niwa et al., Gene 108(2):193-9, 1991), промотор гена фосфоглицераткиназы (PGK), индуцируемый тетрациклином промотор (Masui et al., Nucleic Acids Res. 33:e43, 2005), lac-систему, trp-систему, tac-систему, trc-систему, основные операторные и промоторные области лямбда-фага, промотор гена 3-фосфоглицераткиназы, промоторы гена кислой фосфатазы дрожжей и промотор гена альфа-фактора спаривания дрожжей. Полинуклеотиды, кодирующие по белки согласно настоящему изобретению, могут находиться под контролем конститутивного промотора, индуцируемого промотора или любого другого подходящего промотора, описанного в данном документе, или другого подходящего промотора, который легко распознает специалист в данной области техники.
[0146] В некоторых вариантах осуществления выделенную нуклеиновую кислоту встраивают в вектор. В некоторых вариантах осуществления вектор представляет собой вектор экспрессии. Векторы экспрессии могут содержать одну или несколько регуляторных последовательностей, функционально связанных с полинуклеотидом, подлежащим экспрессии. Термин "регуляторная последовательность" включает промоторы, энхансеры и другие элементы, контролирующие экспрессию (например, сигналы полиаденилирования). Примеры подходящих энхансеров могут включать без ограничения энхансерные последовательности из генов млекопитающих (таких как гены глобина, эластазы, альбумина, α-фетопротеина, инсулина и т. п.) и энхансерные последовательности из вируса эукариотических клеток (такие как энхансер SV40 на участке позднего начала репликации (100-270 п. о.), ранний промотор/энхансер цитомегаловируса, энхансер полиомы на участке позднего начала репликации, энхансеры аденовирусов и т. п.). Примеры подходящих векторов могут включать, например, плазмиды, космиды, эписомы, транспозоны и вирусные векторы (например, векторы на основе аденовируса, вируса осповакцины, вируса Синдбис, вируса кори, вируса герпеса, лентивируса, ретровируса, аденоассоциированного вируса и т. д.). Векторы экспрессии можно применять для трансфекции клеток-хозяев, таких как, например, клетки бактерий, клетки дрожжей, клетки насекомых и клетки млекопитающих. Биологически функциональные векторы на основе ДНК вируса и плазмидной ДНК, способные экспрессироваться и реплицироваться в хозяине, известны из уровня техники и могут применяться для трансфекции любой клетки, представляющей интерес.
Клетки-хозяева
[0147] Другие аспекты настоящего изобретения связаны с клеткой-хозяином (например, выделенной клеткой-хозяином), содержащей один или несколько выделенных полинуклеотидов, векторов и/или векторных систем, описанных в данном документе. Например, клетка-хозяин или линия клеток могут быть применены для продуцирования продукта по настоящему изобретению.
[0148] В некоторых вариантах осуществления выделенную клетку-хозяина по настоящему изобретению культивируют in vitro. В некоторых вариантах осуществления клетка-хозяин представляет собой клетку бактерии (например, клетку E. coli). В некоторых вариантах осуществления клетка-хозяин представляет собой клетку дрожжей (например, клетку S. cerevisiae). В некоторых вариантах осуществления клетка-хозяин представляет собой клетку насекомого. Примеры клеток-хозяев насекомого могут включать, например, клетки Drosophila (например, клетки S2), клетки Trichoplusia ni (например, клетки High Five™) и клетки Spodoptera frugiperda (например, клетки Sf21 или Sf9). В некоторых вариантах осуществления клетка-хозяин представляет собой клетку млекопитающего. Примеры клеток-хозяев, представляющих собой клетки млекопитающих, могут включать, например, клетки почки эмбриона человека (например, клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре), клетки Expi293™, клетки яичника китайского хомячка (CHO), клетки почки новорожденного хомячка (например, BHK, ATCC CCL 10), клетки Сертоли мыши (например, клетки TM4), клетки почки обезьяны (например, CV1 ATCC CCL 70), клетки почки африканской зеленой мартышки (например, VERO-76, ATCC CRL-1587), клетки карциномы шейки матки человека (например, HELA, ATCC CCL 2), клетки почки собаки (например, MDCK, ATCC CCL 34), клетки печени крысы линии Buffalo (например, BRL 3A, ATCC CRL 1442), клетки легкого человека (например, W138, ATCC CCL 75), клетки печени человека (например, Hep G2, HB 8065), клетки опухоли молочной железы мыши (например, MMT 060562, ATCC CCL51), клетки TRI, клетки MRC 5, клетки FS4, линию клеток гепатомы человека (например, Hep G2) и клетки миеломы (например, клетки NS0 и Sp2/0).
[0149] В некоторых вариантах осуществления клетку-хозяина или линию клеток по настоящему изобретению культивируют в непрерывной культуре клеток, например, в биореакторе с перемешиванием. В некоторых вариантах осуществления клетку-хозяина или линию клеток по настоящему изобретению культивируют в периодической культуре клеток. Подходящий реактор с перемешиванием и оборудование и методики для периодической культуры клеток известны из уровня техники; иллюстративные и неограничивающие описания приведены в примерах ниже. Иллюстративные условия и методики, подходящие для продуцирования любого из продуктов, описанных в данном документе, с использованием клетки-хозяина или линии клеток, описанных в данном документе (например, выше), известны из уровня техники.
[0150] Другие аспекты настоящего изобретения связаны со способом получения любого из связывающих белков, описанных в данном документе. В некоторых вариантах осуществления способ предусматривает: a) культивирование клетки-хозяина (например, любой из клеток-хозяев, описанных в данном документе), содержащей выделенную нуклеиновую кислоту, вектор и/или векторную систему (например, любое из выделенных нуклеиновых кислот, векторов и/или векторных систем, описанных в данном документе), в условиях, при которых клетка-хозяин экспрессирует связывающий белок; и b) выделение связывающего белка из клетки-хозяина. Способы культивирования клеток-хозяев в условиях, обеспечивающих экспрессию белка, хорошо известны специалисту средней квалификации в данной области. Способы выделения белков из культивируемых клеток-хозяев хорошо известны специалисту средней квалификации в данной области, в том числе, например, с помощью аффинной хроматографии (например, двустадийной аффинной хроматографии, включающей аффинную хроматографию с использованием белка A с последующей эксклюзионной хроматографией).
ПРИМЕРЫ
[0151] Приведенные ниже примеры иллюстрируют конкретные варианты осуществления настоящего изобретения и различные варианты их применения. Они приведены только для пояснительных целей, и их не следует рассматривать как каким-либо образом ограничивающие объем настоящего изобретения.
Пример 1. Идентификация и клональная вариация ошибочного спаривания цепей для асимметричных триспецифических связывающих белков
[0152] IgG-подобные полиспецифические антитела с асимметричными конструкциями в последние годы стали широко использоваться в терапевтических целях. Правильная сборка субъединиц в этом классе терапевтических средств является критической характеристикой качества (CQA), которая оказывает непосредственное влияние на биологическую активность. Таким образом, при работе по разработке на ранней стадии (например, отбор клонов) необходимо руководствоваться информацией о потенциальном ошибочном спаривании цепей для обеспечения своевременного принятия решений и снижения риска.
[0153] Антитела с ошибочным спариванием могут характеризоваться неоптимальными уровнями ожидаемой биологической активности и считаются родственными примесями. Удаление таких родственных примесей mAb требует дополнительных этапов очистки после колонки с использованием белка A (ProA), что приводит к повышению стоимости и сроков разработки способа. Следовательно, любое потенциальное ошибочное спаривание цепей должно быть идентифицировано и его мониторинг должен быть осуществлен в ходе разработки на ранней стадии, начиная с разработки линии клеток. Для этого требуется высокопроизводительная аналитическая платформа, обеспечивающая возможность скрининга большого количества образцов в отношении уровней ошибочного спаривания, с малым общим временем производственного цикла.
[0154] Масс-спектрометрию интактного белка использовали для идентификации и относительной количественной оценки молекул с ошибочным спариванием в биспецифических IgG. Сообщалось о нескольких различных подходах к MS интактных белков, основанных на разделении с использованием обращенной фазы или эксклюзионной жидкостной хроматографии для MS-анализа с денатурацией и MS-анализа нативных молекул (Wang, C. et al. MAbs 2018, 10 (8), 1226-1235; Schaefer, W. et al. MAbs 2016, 8 (1), 49-55; Schachner, L. et al. Anal Chem 2016, 88 (24), 12122-12127; Yin, Y. et al. MAbs 2016, 8 (8), 1467-1476). Эти подходы требуют очистки антитела, как правило, с использованием аффинности к белку А перед LC-MS-анализом.
[0155] В следующих примерах описывается высокопроизводительная аналитическая платформа, состоящая из эксклюзионной жидкостной хроматографии в денатурирующих условиях (SEC) в сочетании с QToF MS для MS-анализа интактных молекул mAb, таких как триспецифический связывающий белок, показанный на ФИГ. 1A. Этот способ оценки массы интактных молекул можно осуществлять непосредственно в осветленной собранной жидкости без предварительной очистки или пробоподготовки. Этот способ можно применять непосредственно в отношении собранной жидкости культуры клеток перед очисткой с использованием белка А для получения объективной информации из первоисточника о типах и распределении молекул, родственных mAb, образуемых клетками, что дает представление о производительности и характеристиках клонов и условиях выращивания. Эта аналитическая платформа позволила проводить скрининг большого количества клонов клеток CHO (яичника китайского хомячка), экспрессирующих различные конструкции tsAb в отношении уровней ошибочного спаривания и полуантител. Результаты этого способа оценки массы интактных молекул, особенно на ранних этапах разработки, облегчают отбор клонов СНО, которые продуцируют tsAb с подходящим качеством продукта.
Материалы и методы
Обеспечение экспрессии и очистки антител
[0156] Триспецифические антитела (tsAb) получали путем обеспечения совместной экспрессии всех четырех субъединиц в одной системе клеток-хозяев CHO с использованием периодической культуры в центрифужных пробирках или периодической культуры с подпиткой в биореакторной системе ambr®. Титр определяли в собранной жидкости с применением Octet с биосенсорами ProA. Образцы осветленного собранного материала культур клеток хранили в замороженном виде при температуре -80°C до проведения масс-спектрометрического анализа. Очистку tsAb от осветленных жидкостей культуры клеток проводили с применением центрифужных колонок с использованием белка А в формате 96-луночных планшетов с использованием системы обработки жидкостей Tecan с последующим измерением концентрации с помощью Octet.
Расщепление антител посредством IdeS
[0157] К 10 мкл раствора tsAb в концентрации ~2 мг/мл добавляли 20 мкл 25 мМ буфера Tris, рН 7,2. Образец смешивали с 1,5 мкл раствора фермента IdeS (66,6 единиц/мкл) от Genovis Inc. (Кембридж, Массачусетс, США) с последующей инкубацией при 37°C в течение двух часов.
Эксклюзионная хроматография (SEC) для масс-спектрометрии (MS)
[0158] Разделение посредством эксклюзионной жидкостной хроматографии (LC) проводили с использованием колонки UPLC BEH SEC, 1,7 мкм, 200  , 4,6×300 мм, контролируемой Acquity UPLC H-класса (Waters Corp., Милфорд, Массачусетс, США). Для образцов антител, очищенных с использованием ProA, в изократическом режиме в качестве подвижной фазы использовали 0,1% трифторуксусную кислоту (TFA) в смеси ацетонитрил:вода в соотношении 30:70 при скорости потока 0,2 мл/мин. в течение 15 минут, а для осветленных образцов собранного материала - 0,05% муравьиную кислоту (FA) и 0,05% TFA в смеси ацетонитрил:вода в соотношении 30:70 при скорости потока 0,1 мл/мин. (от 0 до 25 минут) и 0,4 мл/мин. (от 25 до 33 минут). Как FA, так и TFA со степенью чистоты для LC-MS были приобретены в Sigma (Сент-Луис, Миссури, США). Воду и ацетонитрил для LC-MS получали от Thermo Fisher Scientific (Уолтем, Массачусетс, США).
, 4,6×300 мм, контролируемой Acquity UPLC H-класса (Waters Corp., Милфорд, Массачусетс, США). Для образцов антител, очищенных с использованием ProA, в изократическом режиме в качестве подвижной фазы использовали 0,1% трифторуксусную кислоту (TFA) в смеси ацетонитрил:вода в соотношении 30:70 при скорости потока 0,2 мл/мин. в течение 15 минут, а для осветленных образцов собранного материала - 0,05% муравьиную кислоту (FA) и 0,05% TFA в смеси ацетонитрил:вода в соотношении 30:70 при скорости потока 0,1 мл/мин. (от 0 до 25 минут) и 0,4 мл/мин. (от 25 до 33 минут). Как FA, так и TFA со степенью чистоты для LC-MS были приобретены в Sigma (Сент-Луис, Миссури, США). Воду и ацетонитрил для LC-MS получали от Thermo Fisher Scientific (Уолтем, Массачусетс, США).
Масс-спектрометрия
[0159] Жидкостную хроматографию (LC) сочетали с масс-спектрометром Xevo G2 QToF (Waters Corp., Милфорд, Массачусетс, США) для осуществляемого в режиме онлайн сбора данных по MS интактных молекул в режиме чувствительности и в диапазоне m/z 500-4000 с с использованием следующих исходных параметров: напряжение на капилляре - 3 кВ, напряжение на конусе для отбора проб - 40 В, напряжение на конусе для извлечения - 2,5 В, температура источника - 150°C, температура десольватации - 500°C, поток газа конуса - 50 л/ч/, поток газа десольватации - 800 л/ч/, энергия столкновения - 6 В.
Анализ данных
[0160] Полученные файлы первичных данных LC-MS обрабатывали в пакетном режиме с использованием Byos® (Protein Metrics Inc.) для осуществления деконволюции и относительной количественной оценки.
Результаты
[0161] Ошибочное спаривание тяжелых и легких цепей в триспецифических антителах приводит к отклонениям массы от ожидаемой массы молекул с правильным спариванием, что легко определяется посредством масс-спектрометрии интактных белков. Единственным исключением являются молекулы с потенциальной перестановкой легких цепей, которые характеризуются той же массой, что и структура с правильным спариванием, и, таким образом, не различимы посредством масс-спектрометрии (ФИГ. 1B).
[0162] MS-анализ интактных молекул в собранной жидкости культуры клеток затруднен в связи со сложной матрицей и требует эффективного хроматографического разделения в режиме онлайн с MS-определением. В частности, молекулы tsAb подвергаются негативному влиянию подавления ионов вследствие значительного количества свободных легких цепей CODV (~ 36 кДа) и Fab (~ 23 кДа) в собранном материале и их изначально низкой эффективности ионизации в связи с их большим размером (до 210 кДа для некоторых молекул с ошибочным спариванием). Следовательно, при разработке способа на основе SE-UPLC-MS интактных молекул проводили оптимизацию параметров хроматографии, в том числе скорости потока, модификаторов подвижной фазы и содержания в ней органических соединений, размера пор колонки для SEC и уровня загрузки белка на колонку, а также параметров MS источника, для получения высококачественных данных по MS при прямом введении собранной жидкости без пробоподготовки.
[0163] Первоначальный рабочий процесс включал автоматическую очистку tsAb с использованием ProA из осветленного собранного материала с последующей SEC-LC в денатурирующих условиях в сочетании с QToF MS для получения данных по MS интактных молекул. SEC в денатурирующих условиях позволяет быстро (15 минут) и эффективно обессоливать, а также отделять tsAb от сверхэкспрессированных легких цепей, которые часто очищаются совместно с mAb.
[0164] На ФИГ. 2A показан масс-спектр после деконволюции для TCE с tsAb к CD38 с массой 174188 Да, соответствующей требуемой сборке, H1L1/H2L2. Два другие значения массы, наблюдаемые при 160683 Да и 187692 Да, соответствуют расчетным значениям массы молекул H1L1/H2L1 и H1L2/H2L2 с ошибочным спариванием легких цепей соответственно. Для дополнительного подтверждения структурной интерпретации молекул образец tsAb расщепляли посредством IdeS, цистеиновой протеазы, которая расщепляет антитела в определенном месте ниже шарнира, создавая пул фрагментов F(ab')2 и Fc/2. Как видно на ФИГ. 2B, после расщепления посредством IdeS образуются три разных Fab-фрагмента с таким же паттерном сдвига массы, что и наблюдаемый в интактных молекулах, что еще раз подтверждает интерпретацию легких цепей с ошибочным спариванием.
[0165] Разработка линии клеток для получения средств терапии на основе белков, в том числе антител, является основой развития на ранней стадии. Основная цель состоит в том, чтобы создать стабильную моноклональную линию клеток (обычно клетки СНО), которая может стабильно экспрессировать рекомбинантный белок-мишень в большом количестве и с необходимыми характеристиками качества продукта, такими как гликозилирование, гетерогенность заряда и т. д. (Chusainow, J. et al. Biotechnol Bioeng 2009, 102 (4), 1182-96). В контексте полиспецифических антител потенциальное ошибочное спаривание субъединиц является критической характеристикой качества (CQA), мониторинг которой необходимо осуществлять во время выбора пулов и конечных продуцирующих клонов.
[0166] Для определения типов и относительных уровней родственных примесей, в том числе цепей с ошибочным спариванием и полуантител, которые получают в результате очистки вместе с необходимой формой tsAb, проводили MS-анализ очищенных с использованием ProA материалов из 38 клонов клеток СНО, экспрессирующих TCE, специфические к CD38, с применением способа на основе SEC в денатурирующих условиях c MS интактных молекул. Относительные уровни различных молекул рассчитывали на основе значений интенсивности массы после деконволюции объединенных MS-спектров, полученных по хроматографическому пику tsAb. Поскольку разные родственные IgG молекулы могут значительно варьировать по массе, особенно полуантитела по сравнению со структурами полных tsAb, и, следовательно, по их эффективности ионизации, относительная количественная оценка молекул на основе MS является эффективным инструментом для сравнения и ранжирования клонов на основе их ошибочного спаривания и уровней  антител.
антител.
[0167] На ФИГ. 3A показан % чистоты (т. е. процент молекул с правильной массой) в различных клонах конструкции TCE, специфической к CD38, выращенных с применением периодической культуры клеток CHO. Между клонами наблюдали высокую степень вариабельности по чистоте, от 90% до 10%. Кроме того, относительные уровни молекул с ошибочным спариванием и полуантител значительно варьировали от клона к клону, как показано на ФИГ. 3C. Кроме того, сопоставление результатов определения чистоты со значениями продуктивности клонов (ФИГ. 3B), измеренными по титру, демонстрирует отсутствие корреляции между ними, что означает, что высокопродуктивные клоны потенциально могут обеспечивать на выходе высокий процент молекул mAb с ошибочным спариванием и неполной сборкой.
[0168] Эти результаты указывают на важность скрининга клонов tsAb в отношении уровней ошибочного спаривания и использования этой информации в сочетании с продуктивностью и другими параметрами культуры клеток для разработки линий клеток и отбора лучших клонов-продуцентов.
Пример 2. Прямой MS-анализ интактных молекул в осветленном собранном материале
[0169] Для снижения стоимости и времени выполнения скрининга клонов и оптимизации рабочего процесса параметры способа на основе SEC-MS в денатурирующих условиях оптимизировали для обеспечения возможности измерения посредством MS интактных молекул для молекул mAb непосредственно в осветленной жидкости для культивирования клеток, полученной во время разработки линии клеток, что позволяет избежать предварительной очистки или манипуляции с образцами.
[0170] MS интактных молекул в жидкости для культивирования клеток затруднен в связи со сложной матрицей, особенно в связи с присутствием неионогенных поверхностно-активных веществ, таких как Pluronic F-68, обычно добавляемых в среду для защиты клеток от гидродинамического повреждения, которое может приводить к совместному элюированию и подавлению ионов во время MS-анализа. Кроме того, молекулы tsAb подвергаются негативному влиянию подавления ионов вследствие значительного количества свободных легких цепей CODV (~ 36 кДа) и Fab (~ 23 кДа), сверхэкспрессированных в собранном материале, а также изначально низкой эффективности ионизации tsAb в связи с их большим размером (до 210 кДа для некоторых молекул с ошибочным спариванием). Хроматографическое отделение белков от поверхностно-активных веществ на стационарных фазах с обращенной фазой (RP), обычно используемых для MS интактных белков, может быть довольно сложным, если не невозможным, в связи с гидрофобностью обоих типов молекул. Кроме того, данный способ на основе RP-LC демонстрирует различное поведение в отношении избирательности для разных tsAb в зависимости от аминокислотной последовательности и не может использоваться в качестве способа платформы для нескольких конструкций триспецифических антител.
[0171] В этом примере SEC-LC в денатурирующих условиях успешно применяли для отделения на основе размера молекул антител от малых молекул и поверхностно-активных веществ в осветленных жидкостях для культивирования клеток. Параметры жидкостной хроматографии, в том числе скорость потока, модификаторы подвижной фазы и содержание в ней органических соединений, а также параметры источника MS, оптимизировали для получения высококачественных данных по MS для молекул tsAb в собранном материале, сопоставимых с данными для материала, очищенного с использованием ProA. Оптимизированный способ на основе SEC в денатурирующих условиях позволял осуществлять эффективное отделение родственные mAb молекулы, в том числе tsAb и сверхэкспрессированные легкую цепь CODV и легкую цепь Fab, друг от друга и от молекул поверхностно-активных веществ, как показано на хроматограмме основного пика на ФИГ. 4A и 4B.
[0172] Поверхностно-активные вещества элюируются последними на хроматограмме и поток направляется в отходы для избежания контаминации источника MS (ФИГ. 4A). Этот способ на основе SEC-MS является платформой, не зависящей от конкретного продукта, и позволяет проводить быстрый и высокопроизводительный скрининг большого количества клонов в отношении различных конструкций tsAb. Кроме того, способ, который разработали, можно применять в отношении других родственных mAb форм, в том числе форматов антител, лишенных Fc, таких как слитые белки на основе scFv2 и Fab (Brinkmann, U. and Kontermann, R. E. MAbs 2017, 9 (2), 182-212), которые не могут быть очищены с использованием автоматизированных систем очистки с использованием ProA.
[0173] Наименьшая разница в массе между двумя молекулами с ошибочным спариванием в конструкции TCE, специфической к CD38, составляет 810 Да, а в конструкции TCE, специфической к HER2, составляет 1095 Да, что значительно ниже разрешающей способности способа по массе. Для демонстрации способности способа обеспечивать различение молекул с ошибочным спариванием, которые являются более близкими по массе, осветленные образцы собранного материала из обеих конструкций смешивали в соотношении 1:1 для получения более сложной смеси молекул с ошибочным спариванием. Оба образца содержали молекулы H1L1/H2L1 с разницей в расчетных значениях массы, составляющей ≈302 Да, для CD38 TCE, и TCE, специфического к HER2. Как видно на ФИГ. 4C, осуществляемый в режиме онлайн SEC-MS-анализ собранного материала обеспечивал полное разрешение этих двух молекул и их гликоформ, отмеченных сдвигами по массе +162 Да.
[0174] Затем оценивали повторяемость, надежность и устойчивость MS-сигнала с использованием осветленных собранных жидкостей двух различных конструкций tsAb: TCE, специфической к HER2, и TCE, специфической к CD38, со значениями титра 435 и 1100 мкг/мл соответственно. На ФИГ. 5A показано, что относительные уровни трех различных сборок tsAb являются сопоставимыми при различных значениях загрузки колонки, достигаемых за счет повышения объема инъекции от 10 до 50 мкл. Эти результаты демонстрируют устойчивость и надежность количественной оценки на основе MS в диапазоне значений загрузки колонки 4-55 мкг. Таким образом, осуществляемый онлайн SEC-MS с фиксированным объемом инъекции осветленного собранного материала можно использовать для клонов с низкими и высокими титрами, обычно в диапазоне 100-1100 мкг/мл. Повторяемость способа, оцененная посредством LC-MS-анализа в трех повторах, при каждом уровне загрузки белка для двух конструкций tsAb на ФИГ. 4A-4C характеризовались значениями %RSD ниже 10% (таблица A).
Таблица A. Относительные уровни молекул с ошибочным спариванием и молекул с эталонной массой и результаты определения прецизионности для анализа образцов собранного материала TCE, специфического к CD38, и TCE, специфического к HER2, в трех повторностях при различных значениях загрузки колонки.
[0175] Кроме того, сравнение результатов количественной оценки ошибочного спаривания, полученных посредством SEC-LC-MS интактных молекул в осветленном собранном материале и в очищенных с использованием ProA материалах, показало близкое соответствие результатов измерения между двумя способами, что еще раз подтвердило применимость прямого MS-анализа интактных молекул в собранном материале для определения ошибочного спаривания (ФИГ. 5B и 5C).
[0176] Способ на основе SEC-MS для собранного материала применяли для высокопроизводительного скрининга 50 клонов TCE, специфического к HER2. В этом исследовании было определено, что правильно собранная молекула (175576 Да) является основной формой, в то время как молекулы с ошибочным спариванием легких цепей H1L1/H2L1 (162187 Да) и H1L2/H2L2 (188965 Да) на различных уровнях были определены во всех протестированных образцах. В тестируемых клонах полуантитела не определяли или определяли на очень низких уровнях. Аналогично TCE, специфическому к CD38, корреляции между уровнями ошибочного спаривания и продуктивностью клона не наблюдали (ФИГ. 5D и 5E).
Пример 3. Влияние условий выращивания культуры клеток на ошибочное спаривание цепей
[0177] Выполняли две различные группы исследований для изучения влияния условий выращивания клеток на ошибочное спаривание цепей и образование полуантител в tsAb.
[0178] В первом исследовании клоны TCE, специфического к CD38, выращивали в а) периодической культуре и собирали в день 10, и в b) и c) в двух разных условиях выращивания в микробиореакторе ambr®. Анализ на основе SEC c MS интактных молекул (материалов, очищенных с использованием ProA) показал вариации в распределении антител с ошибочным спариванием и полуантител в каждом клоне, выращенном в трех различных условиях (ФИГ. 6C-6E). Однако ранжирование клонов на основе общего уровня примесей в целом оставалось неизменным независимо от условий выращивания, как показано на ФИГ. 6C.
[0179] Аналогичные результаты были получены в исследовании TCE, специфического к HER2, где осветленную собранная жидкость из выбранных клонов, выращенных в условиях периодической культуры (собранный материал дня 10) и в микробиореакторе ambr®, анализировали посредством SEC-LC-MS интактных молекул в отношении их относительных уровней ошибочного спаривания. В то время как распределение молекул с ошибочным спариванием в каждом конкретном клоне отличается для условий периодической культуры и условий ambr (ФИГ. 6F и 6G), ранжирование клонов, основанное на общем количестве ошибочных спариваний, в целом было одинаковым для двух условий выращивания (ФИГ. 6B). Эти результаты подчеркивают важность и обоснованность первоначального отбора клонов на основе данных об ошибочном спаривании, несмотря на неизбежные изменения условий способа на более поздних стадиях разработки лекарственных средств.
[0180] Следует отметить, что в обоих исследованиях наблюдали более высокую вариацию выхода tsAb с правильным спариванием между клонами, выращенными в ambr®, по сравнению с периодической культурой; что характеризуется % чистоты в диапазоне 10-90% по сравнению с 47-80% для CD38 TCE, и 46-70% по сравнению с 45-58% для TCE, специфического к HER2 (ФИГ. 6A и 6B). Интересно, что в обоих случаях верхние клоны, по-видимому, функционировали более эффективно, о чем свидетельствовало повышение чистоты на 10-20%, в биореакторе ambr, чем в периодической культуре.
Выводы
[0181] Неправильно собранные молекулы представляют собой часто встречающиеся родственные примеси, при производстве полиспецифических антител, которые приводят к снижению выхода продукции и требуют дорогостоящих этапов очистки, а также надежных способов анализа для мониторинга на ранней стадии. В приведенных выше примерах была описана высокопроизводительная аналитическая платформа, основанная на SEC-LC-MS интактных молекул в денатурирующих условиях для идентификации и относительной количественной оценки ошибочного спаривания цепей и других родственных IgG молекул. Разработанный способ позволяет проводить MS-анализ интактных молекул для родственных mAb молекул непосредственно в очищенном собранном материале, за счет чего избегают трудоемкой и дорогостоящей очистки с использованием ProA и этапов замены буфера (ФИГ. 7). MS-анализ интактных молекул в виде двух различных конструкций tsAb; TCE, специфический к CD38, и TCE, специфический к HER2, из разных клонов клеток CHO показали вариации в уровне выхода tsAb с правильным спариванием от клона к клону без очевидной корреляции между продуктивностью клона (титром) и ошибочным спариванием. Кроме того, различные условия выращивания могут влиять на типы и распределение примесей, но не на ранжирование качества клонов, что подтверждает валидность исходного отбора клонов на основе данных об ошибочном спаривании, несмотря на неизбежные изменения условий способа на более поздних стадиях разработки лекарственных средств.
Пример 4. Анализ массы интактных молекул в собранном материале для триспецифических связывающих белков
[0182] В прошлом масс-спектрометрию интактных белков использовали для идентификации и относительной количественной оценки биотерапевтических средств, их вариантов и родственных примесей. Сообщалось о нескольких различных подходах к MS интактных белков, основанных на разделении фаз с использованием обращенной фазы или эксклюзионной LC, для MS-анализа в денатурирующих условиях и MS-анализа нативных молекул (Xu, L. et al. (2017) Science 358:85-90; Ridgway, J.B. et al. (1996) Protein Eng. 9:617-621; Wang, C. et al. (2018) MAbs 10:1226-1235; Schaefer, W. et al. (2016) MAbs 8:49-55). Эти подходы требуют очистки антитела, как правило, с использованием аффинности к белку А, перед LC-MS-анализом.
[0183] В приведенных выше примерах описана высокопроизводительная аналитическая платформа, состоящая из эксклюзионной жидкостной хроматографии (SEC) в денатурирующих условиях в сочетании с QToF масс-спектрометрией, для MS-анализа интактных молекул для определения ошибочного спаривания цепей в триспецифических антителах (tsAb). См. также Tousi, F. et al. (2020) Anal. Chem. 92:2764-2769. Этот способ оценки массы интактных молекул можно осуществлять непосредственно в осветленной собранной жидкости без предварительной очистки или пробоподготовки. Эта аналитическая платформа сделала возможным скрининг большого количества клонов клеток CHO (яичника китайского хомячка), экспрессирующих различные конструкции tsAb, в отношении их уровней молекул с ошибочным спариванием и полуантител. Результаты этого способа оценки массы интактных молекул, особенно на ранних этапах разработки, облегчают отбор клонов СНО, которые продуцируют tsAb приемлемого качества продукта.
[0184] В примерах 4 и 5 описаны варианты применения одного и того же аналитического способа отбора клонов для различных родственных mAb конструкций: триспецифического антитела, специфического к HIV\CD28\CD3 (пример 4) и моноклонального антитела (mAb) с конструированием, осуществленным в отношении цистеина (пример 5).
Материалы и методы
Эксклюзионная жидкостная хроматография (SEC)
[0185] Разделение посредством LC проводили с использованием колонки UPLC BEH SEC, 1,7 мкм, 200  , 4,6×300 мм, контролируемой Acquity UPLC H-класса (Waters Corp., Милфорд, Массачусетс, США). В качестве подвижной фазы в изократическом режиме использовали раствор 0,05% муравьиной кислоты (FA) и 0,05% TFA в смеси ацетонитрил:вода в соотношении 30:70 при скорости потока 0,1 мл/мин. (от 0 до 25 минут) и 0,4 мл/мин. (от 25 до 33 минут). Объем 50 мкл осветленного собранного материала вводили непосредственно в систему LC-MS с использованием автоматического пробоотборника Acquity, установленного на 4°C. Как FA, так и TFA характеризовались степенью чистоты для LC-MS от Sigma (Сент-Луис, Миссури, США). Воду и ацетонитрил для LC-MS получали от Thermo Fisher Scientific (Уолтем, Массачусетс, США).
, 4,6×300 мм, контролируемой Acquity UPLC H-класса (Waters Corp., Милфорд, Массачусетс, США). В качестве подвижной фазы в изократическом режиме использовали раствор 0,05% муравьиной кислоты (FA) и 0,05% TFA в смеси ацетонитрил:вода в соотношении 30:70 при скорости потока 0,1 мл/мин. (от 0 до 25 минут) и 0,4 мл/мин. (от 25 до 33 минут). Объем 50 мкл осветленного собранного материала вводили непосредственно в систему LC-MS с использованием автоматического пробоотборника Acquity, установленного на 4°C. Как FA, так и TFA характеризовались степенью чистоты для LC-MS от Sigma (Сент-Луис, Миссури, США). Воду и ацетонитрил для LC-MS получали от Thermo Fisher Scientific (Уолтем, Массачусетс, США).
Масс-спектрометрия (MS)
[0186] LC сочетали с QToF масс-спектрометром Xevo G2 (или G2XS) (Waters Corp., Милфорд, Массачусетс, США) для осуществляемого в режиме онлайн сбора данных по MS интактных молекул в режиме чувствительности и в диапазоне m/z, составляющем 500-4000. Параметры ионизации электрораспылением (ESI) для прибора Xevo G2 включали: напряжение на капилляре - 3 кВ, напряжение на конусе для отбора проб - 40 В, напряжение на конусе для извлечения - 2,5 В, температура источника - 150°C, температура десольватации - 500°C, поток газа конуса - 50 л/ч., поток газа десольватации - 800 л/ч. и энергия столкновения - 6 В. Параметры ESI для Xevo G2XS были следующими: напряжение на капилляре - 3 кВ, напряжение на конусе для отбора проб - 200 В, температура источника - 150°C, отклонение от источника - 150°C, температура десольватации - 250°C, поток газа конуса - 50 л/ч., поток газа десольватации - 800 л/ч. и энергия столкновения - 6 В.
Анализ данных
[0187] Полученные файлы первичных данных LC-MS обрабатывали в пакетном режиме с использованием Byos® (Protein Metrics Inc.) для деконволюции, аннотации и относительной количественной оценки.
Результаты
[0188] Очищенные образцы собранного материала культуры клеток из 50 клонов клеток СНО, экспрессирующих tsAb к HIV\CD28\CD3, анализировали с использованием описанного выше способа на основе SEC в денатурирующих условиях с MS интактных молекул. При анализе полученных файлов необработанных данных MS между протестированными клонами наблюдали высокую степень вариабельности уровней ошибочного спаривания (%), находящуюся в диапазоне от 1% до 100% ошибочного спаривания. H1L1/H2L1 (165-167 кДа) представляла собой доминирующую молекулу с ошибочным спариванием. Небольшое количество клонов содержало форму с ошибочным спариванием H1L2/H2L2. Гомодимер H2L2/H2L2 определили в нескольких клонах. Полуантитела (H1L1 и H2L2) присутствовали в некоторых клонах, но при низких уровнях. В отличие от протестированных ранее триспецифических антител, в клонах TCE, специфических к HIV, не было выявлено значительного количества ошибочного спаривания типа H1L2/H2L2. На ФИГ. 8A и 8B показаны масс-спектры интактных молекул после деконволюции для клонов, специфических к HIV\CD28\CD3, с наиболее высокими и наиболее низкими уровнями ошибочного спаривания цепей соответственно.
[0189] На ФИГ. 9A и 9B показана продуктивность клонов, измеренная по титру белка А, и относительные уровни молекул с правильной массой для 50 клонов клеток, продуцирующих триспецифические антитела к HIV\CD28\CD3, выращенных в условиях периодической культуры, во время стадии отбора клонов в качестве этапа разработки линии клеток. Результаты анализа ошибочного спаривания в сочетании с данными о продуктивности позволили отобрать клоны с наиболее высокими титрами, а также с наиболее низкими уровнями ошибочного спаривания цепей. Интересно, что несколько клонов с относительно высокой продуктивностью показали 0% tsAb с правильным спариванием, как показано в правом конце колонок на ФИГ. 9B.
[0190] Превосходное хроматографическое разделение в сочетании с использованием высокочувствительного масс-спектрометра позволило выявить крайне гетерогенный профиль гликанов на уровне интактных молекул в связи с присутствием сиалилированных N-гликанов на LC Fab в дополнение к обычным N-гликанам тяжелой цепи Fc. Это обеспечивает быстрый мониторинг профилей N-гликозилирования клонов без необходимости выполнять более сложный анализ гликанов с использованием обычно трудоемких и времязатратных способов. В качестве примера на ФИГ. 10A и 10B показан подробный профиль гликанов для молекулы с эталонной массой (H1L1/H2L2) в выбранном клоне, продуцирующем антитело к HIV\CD28\CD3, культивированном при двух различных условиях: 1 - в центрифужной пробирке (условия периодической культуры) и 2 - в биореакторе ambr15 (условия периодической культуры с подпиткой) соответственно, при этом показан сдвиг в сторону более галактозилированных и сиалилированных N-гликанов, очевидный из сдвига в сторону более высоких значений массы. В связи с крайней гетерогенностью гликанов, усиленной гликозилированием легкой цепи, молекулярная масса эталонной формы (H1L1/H2L2) и формы H1L1/H2L1 с ошибочным спариванием находится в диапазоне от 1000 Да до 2000 Да соответственно. Интерпретация гликоформ для эталонной формы интактного антитела к HIV\CD28\CD3 подробно описана в таблице 1.
Пример 5. Анализ массы интактных молекул в собранном материале для сконструированного антитела с цистеином
[0191] Этот продукт представляет собой thiomAb, т. е. антитело, содержащее дополнительный цистеиновый остаток, встроенный в его Fc-область, применимое, например, для конъюгации с соединением (например, цитотоксическим лекарственным средством с образованием конъюгата антитело-лекарственное средство). Этот дополнительный цистеин является неспаренным (свободным), т. е. не задействован в дисульфидных связях с другими остатками цистеина в молекуле mAb. Предыдущая работа показала, что этот дополнительный цистеин (Cys293) может образовывать нежелательные дисульфидные связи с соседними остатками цистеина, что также известно как "перестановка дисульфидных связей", которая потенциально приводит к структурной нестабильности антитела. Неконтролируемые химические реакции сульфгидрильных групп также могут обуславливать значительный риск в отношении выполнимости производственного процесса в ходе обработки для образования ADC, что приводит к нежелательному профилю конъюгации лекарственного средства. Для избежания этих рисков необходимо обеспечить блокирование свободного цистеина с помощью дисульфидной модификации, такой как цистеинилирование и глутатионилирование, путем регулирования условий культивирования клеток. Эти модификации можно избирательно устранять с образованием свободного цистеина перед этапом конъюгации с лекарственным средством.
[0192] В этом примере разработанный способ на основе SEC в денатурирующих условиях с MS интактных молекул для собранного материала культуры клеток использовали для быстрого скрининга клонов в отношении их статуса блокировки Cys293 во время разработки линии клеток. Первый изученный кандидат для thiomAb содержал мутацию N300A в Fc-области, которая обеспечивала устранение N-гликозилирования Fc. Отсутствие гликанов позволило идентифицировать блокировку Cys293 с помощью прямого MS-анализа интактных молекул в собранном материале без необходимости каких-либо манипуляций с образцами (ФИГ. 11A).
[0193] Однако второй кандидат для thiomAb с типичным профилем N-гликозилирования Fc характеризовался очень сложным паттерном в данных MS интактных молекул в связи с присутствием нескольких молекул с блокировкой, в том числе N-ацетилцистеинилирования (+161 Да), которое мешало сдвигу по массе в связи с присоединением остатков моносахаридов гексозы (+162 Да) к N-гликанам. Для устранения этой проблемы и облегчения интерпретации данных, осветленный собранный материал второго кандидата обрабатывали с помощью PNGазы F для удаления N-гликанов в нативных условиях. Затем дегликозилированный собранный материал подвергали SEC с MS интактных молекул, как и ранее. На ФИГ. 11B показан масс-спектр интактных молекул после деконволюции, полученный для второго кандидата после дегликозилирования, с идентифицированными тремя типами модификаций блокировки: цистеинилирование (Cys) со сдвигом массы +119 Да, N-ацетилцистеинилирование (N-acetyl Cys) со сдвигом массы +161 Да и глутатионилирование (Glu) со сдвигом массы +305 Да. Скрининг 22 клонов клеток СНО, экспрессирующих thiomAb, посредством этого способа показал 100% блокировку сконструированного цистеина во всех тестируемых клонах.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANOFI
<120> СПОСОБЫ АНАЛИЗА ОШИБОЧНОГО СПАРИВАНИЯ ЦЕПЕЙ
В ПОЛИСПЕЦИФИЧЕСКИХ СВЯЗЫВАЮЩИХ БЕЛКАХ
<130> 18395-20328.40
<140> Пока еще не назначен
<141> Одновременно с этим
<150> EP 20315271.5
<151> 2020-05-28
<150> US 62/926,313
<151> 2019-10-25
<160> 6
<170> FastSEQ для Windows версии 4.0
<210> 1
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 1
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 2
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 2
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 3
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 3
Thr Lys Gly Pro Ser
1 5
<210> 4
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 4
Gly Gln Pro Lys Ala Ala Pro
1 5
<210> 5
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 5
Gly Gly Ser Gly Ser Ser Gly Ser Gly Gly
1 5 10
<210> 6
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 6
Asp Lys Thr His Thr
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К CD38 И КОМБИНАЦИИ С АНТИТЕЛАМИ К CD3 И CD28 | 2018 |
|
RU2812910C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ, ОБЛАДАЮЩИЕ ОГРАНИЧЕННЫМ СВЯЗЫВАНИЕМ С СD3, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2813816C2 |
| ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, СВЯЗЫВАЮЩИЕ CD38, CD28 И CD3, А ТАКЖЕ СПОСОБЫ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ВИРУСНОЙ ИНФЕКЦИИ | 2019 |
|
RU2820351C2 |
| ПРЕДПОЧТИТЕЛЬНОЕ СПАРИВАНИЕ ДОМЕНОВ АНТИТЕЛ | 2018 |
|
RU2792440C2 |
| ТРИ- ИЛИ ТЕТРАСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2010 |
|
RU2570633C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| БИСПЕЦИФИЧЕСКИЕ ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2604189C2 |
| МОДИФИЦИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2832921C1 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ ДЛЯ АКТИВАЦИИ КЛЕТОК-НАТУРАЛЬНЫХ КИЛЛЕРОВ И ИХ ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2018 |
|
RU2809125C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2826991C2 |
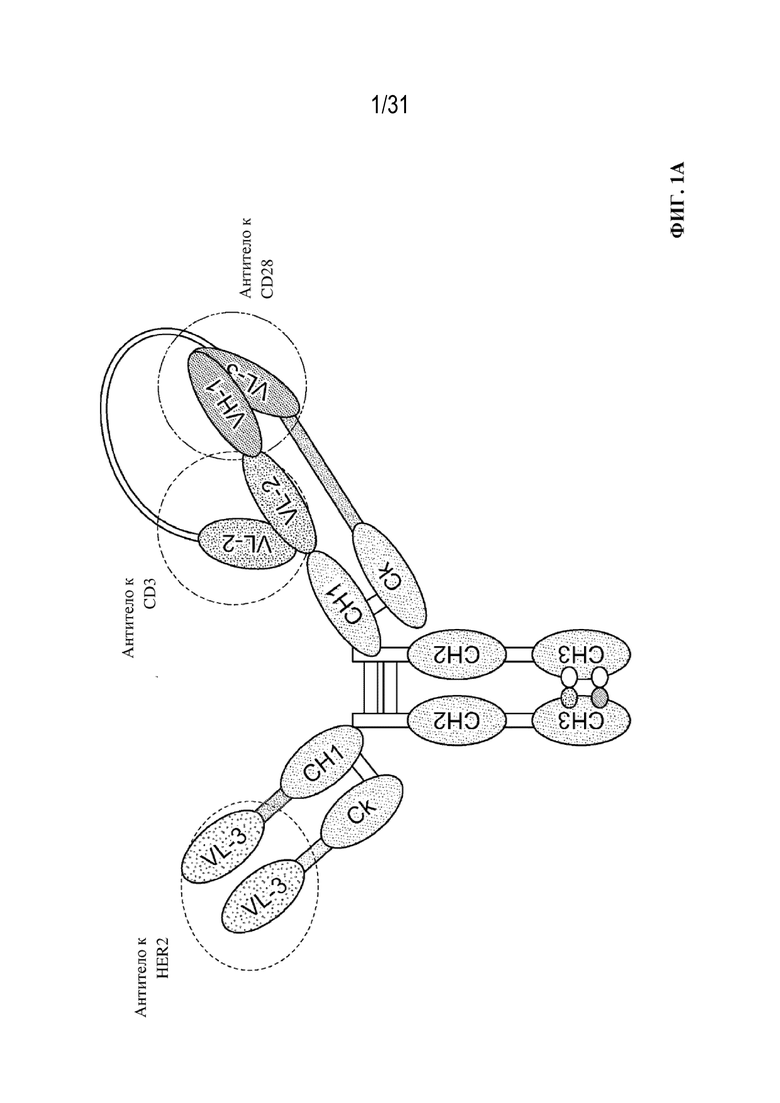
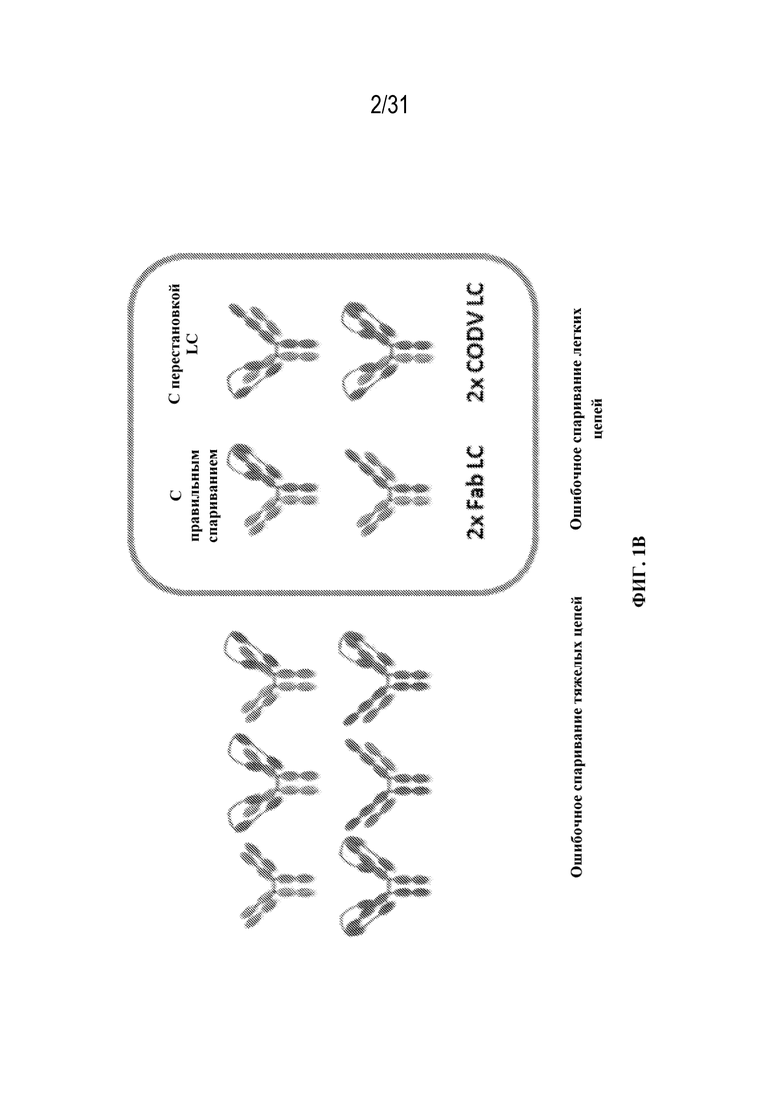
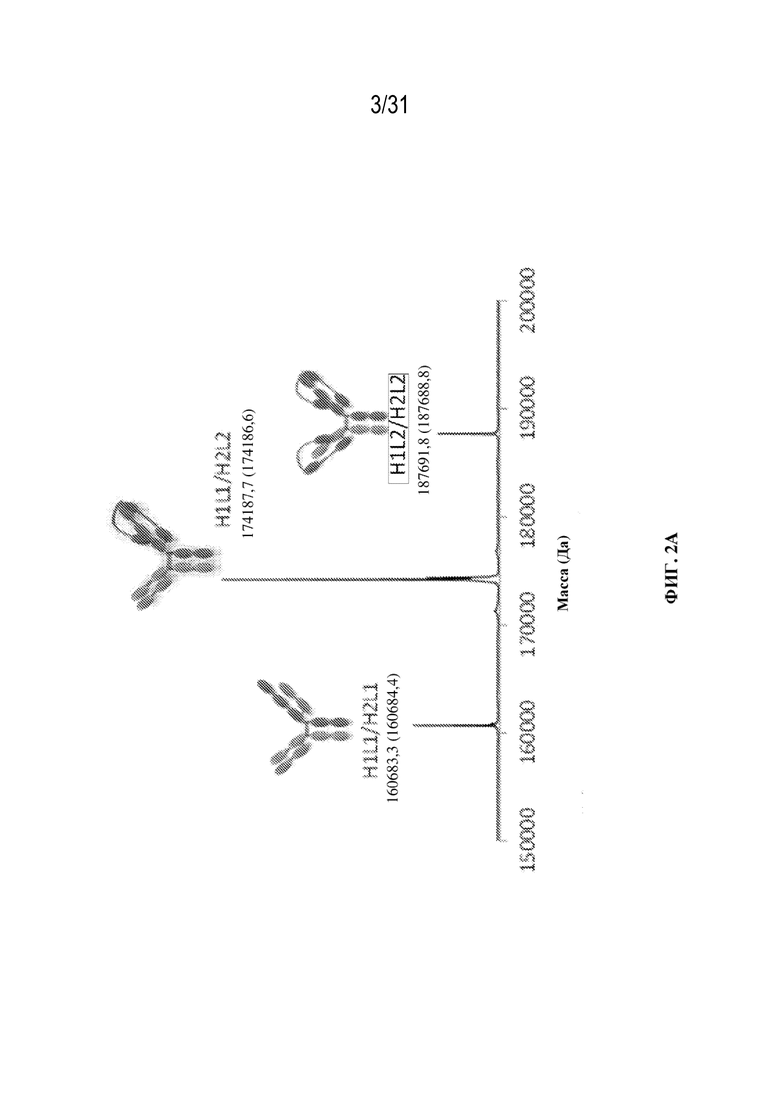
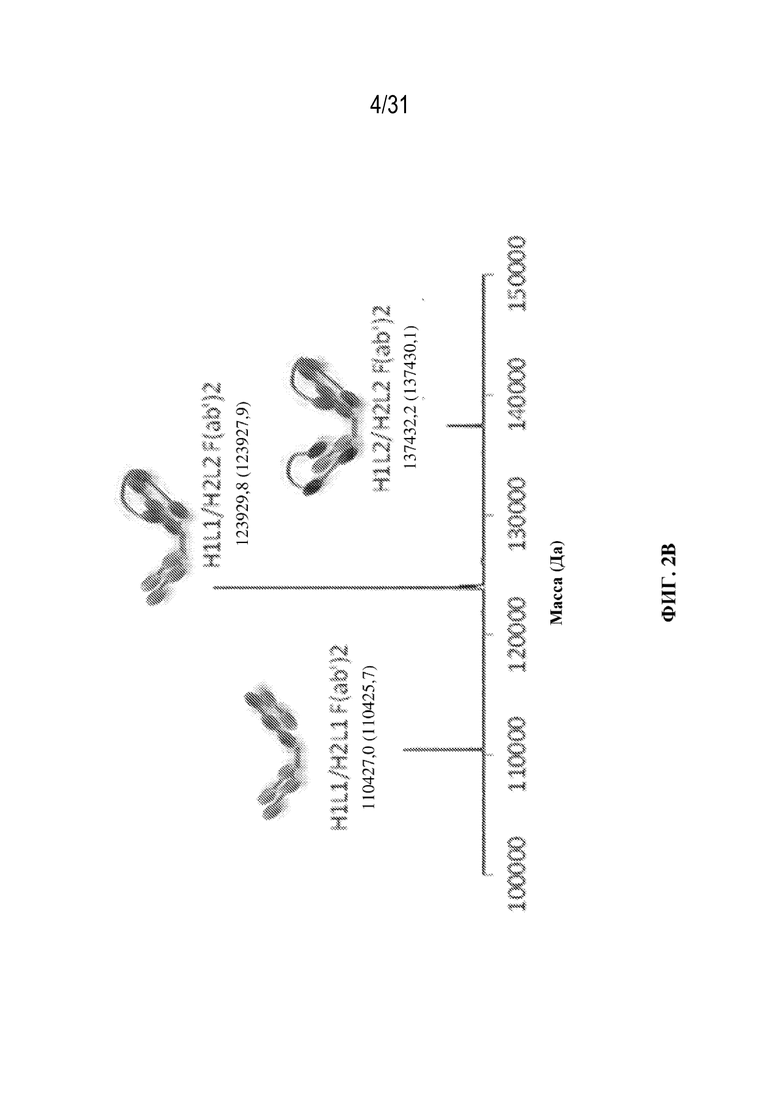
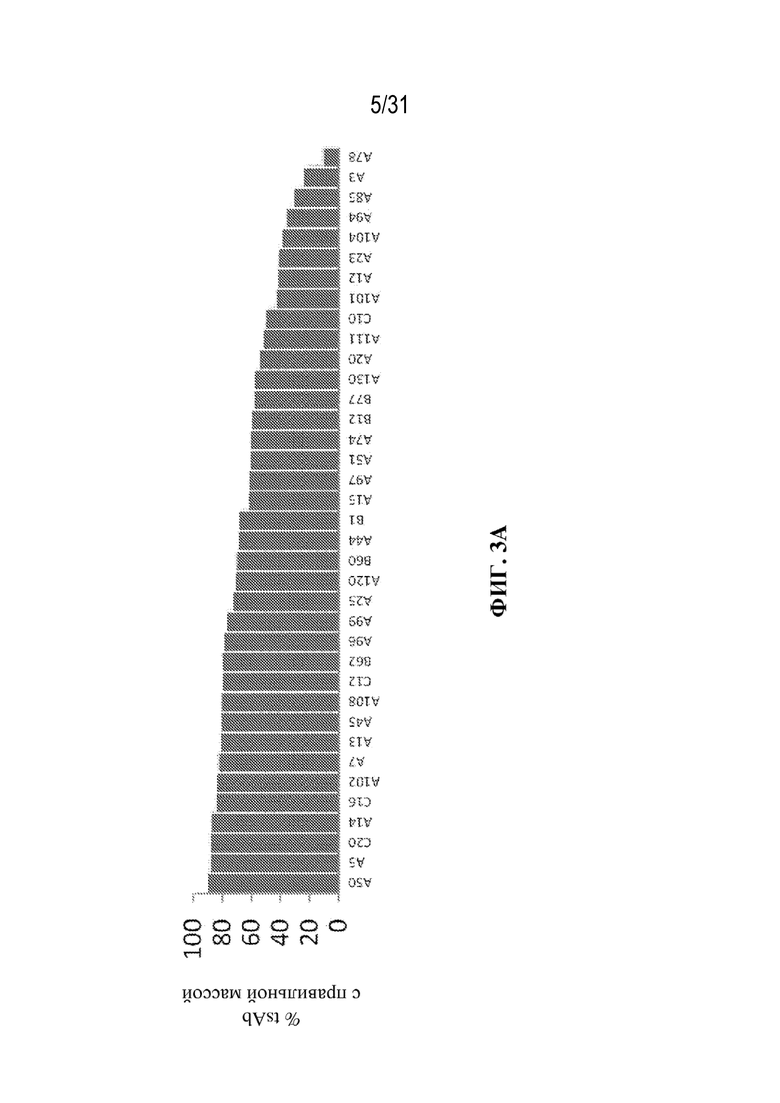
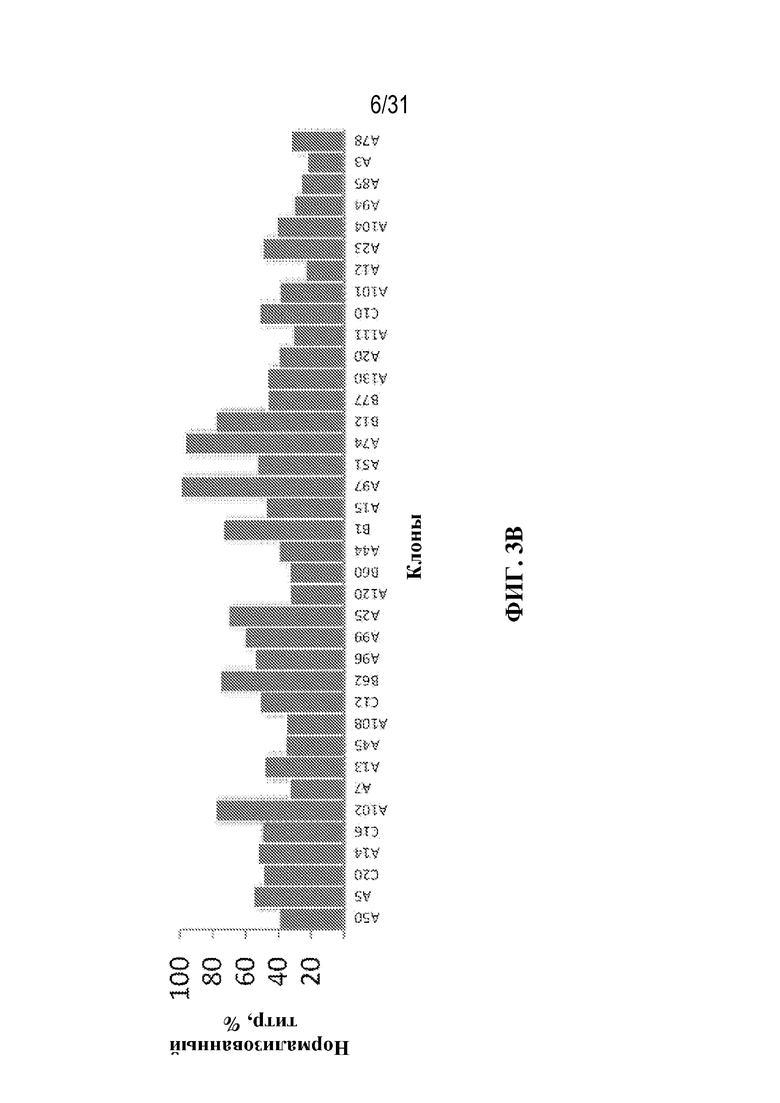
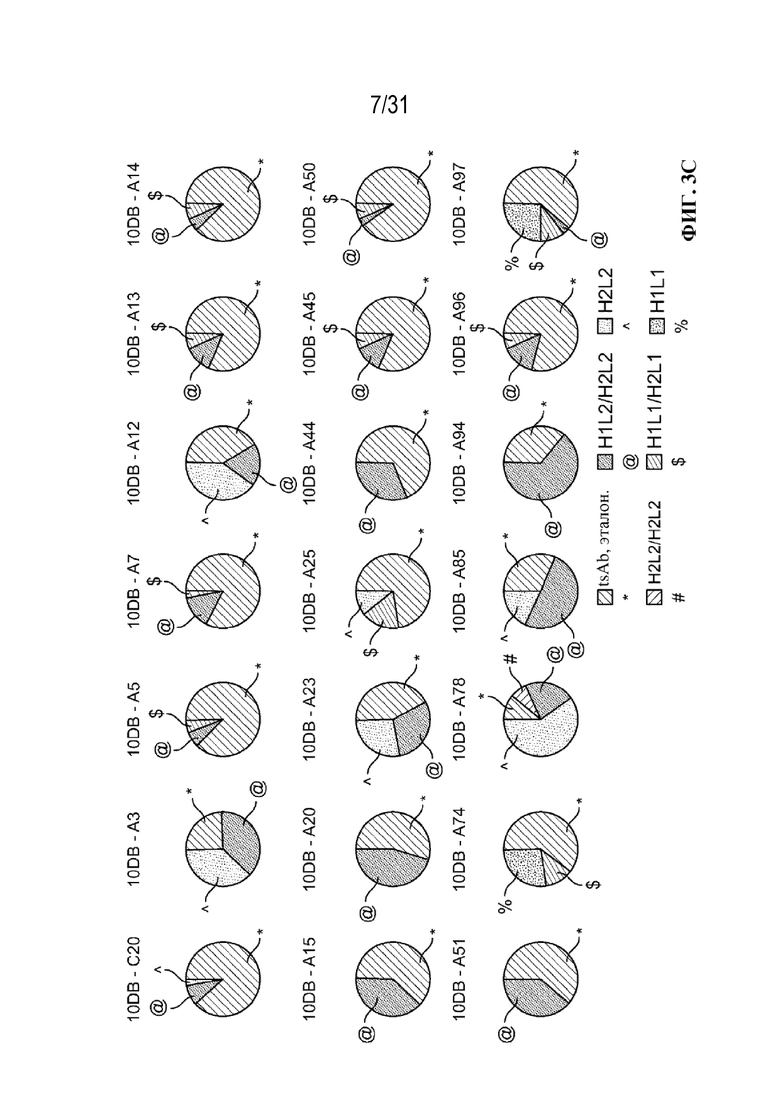
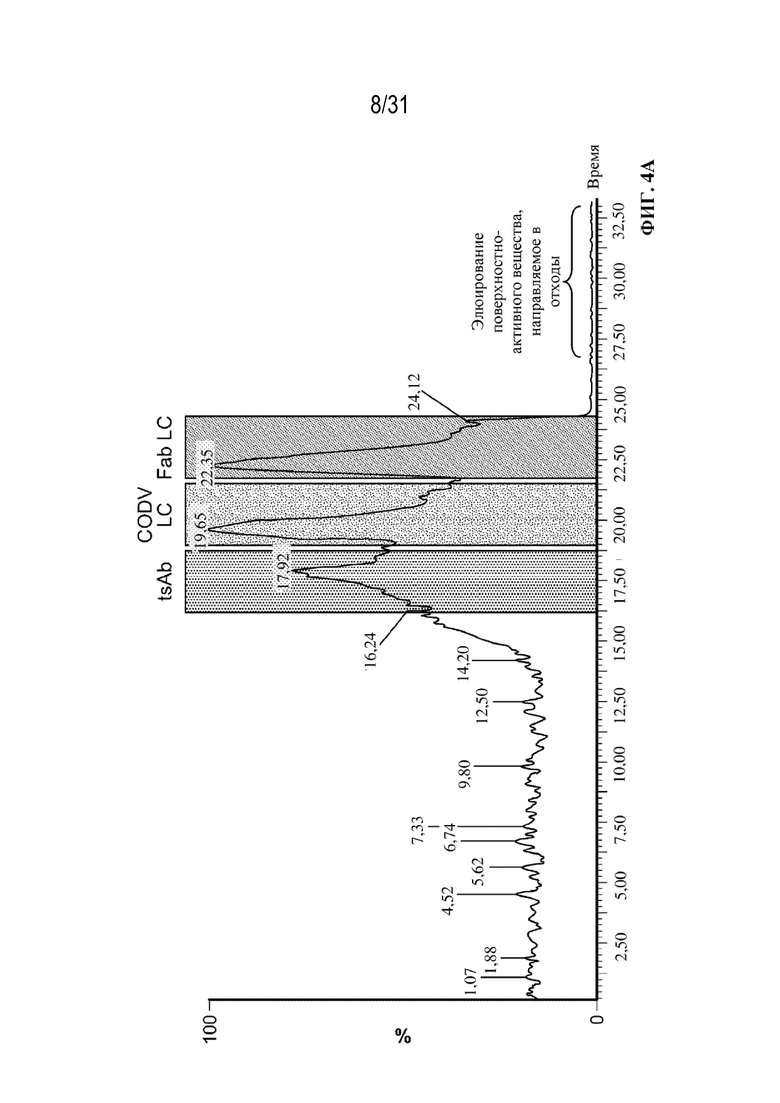
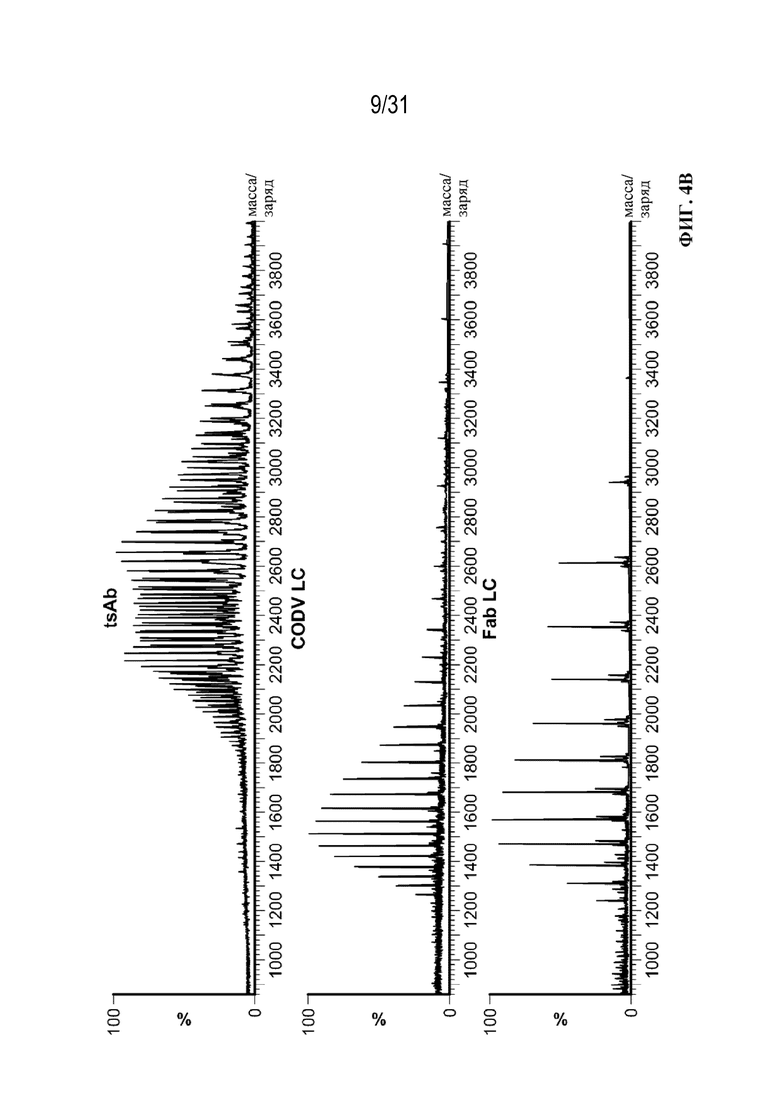
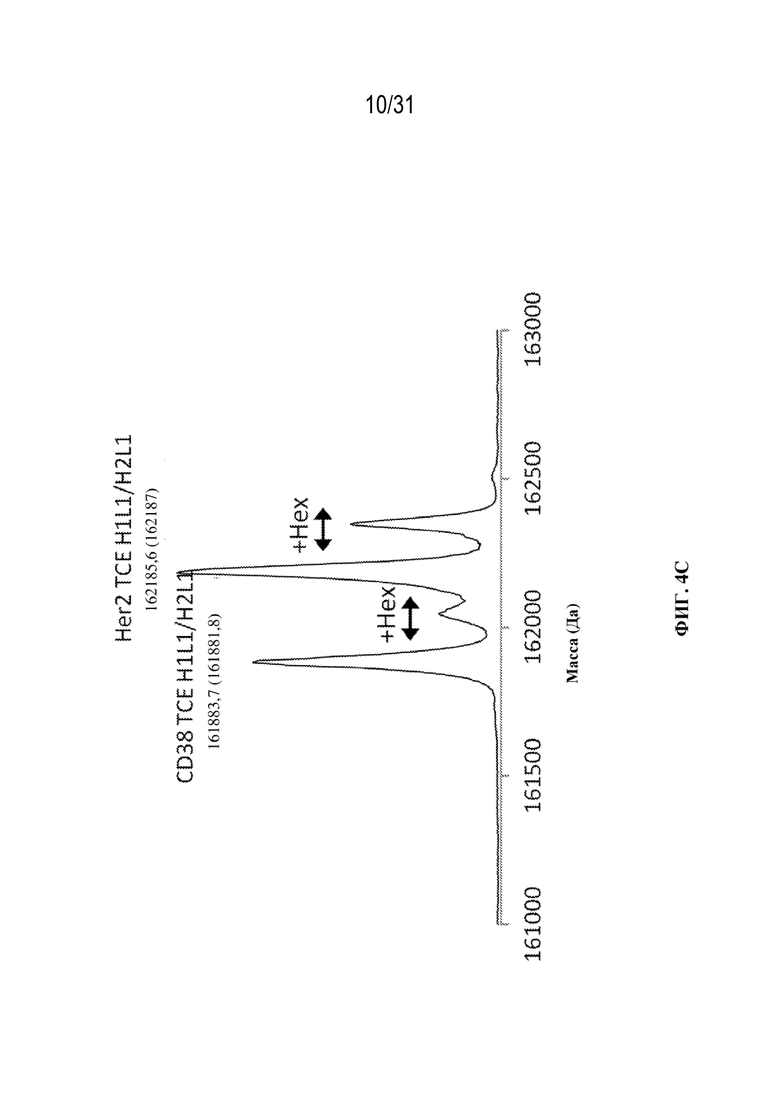
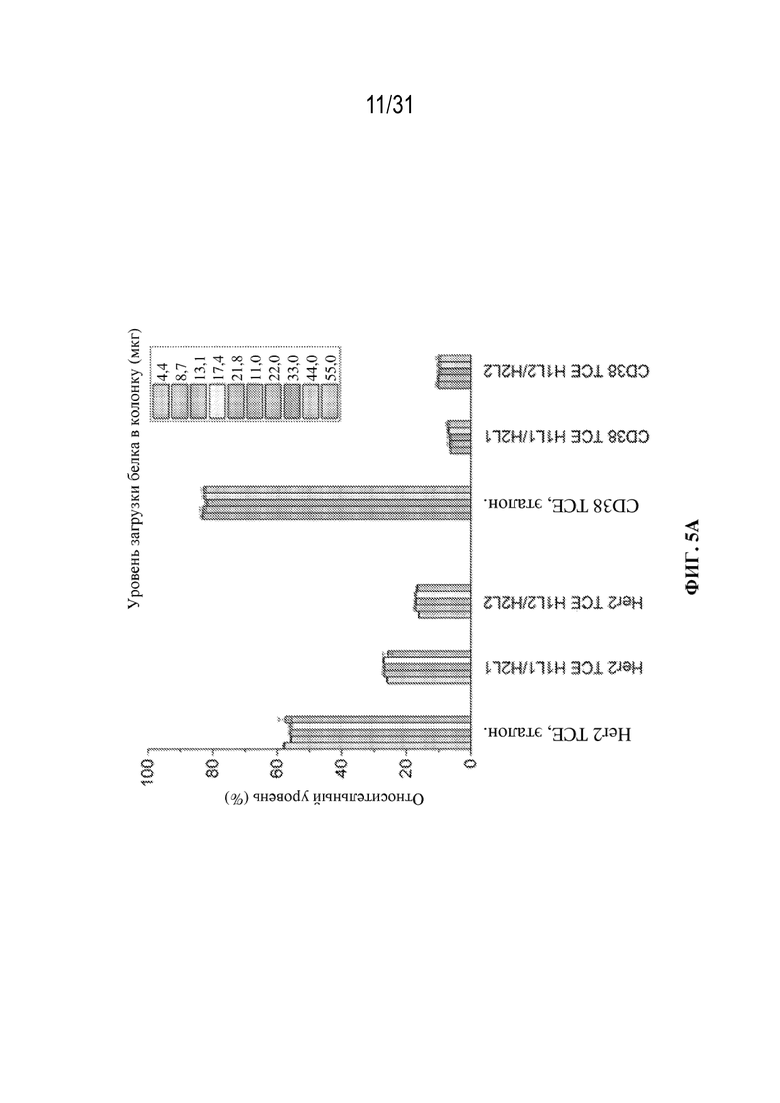
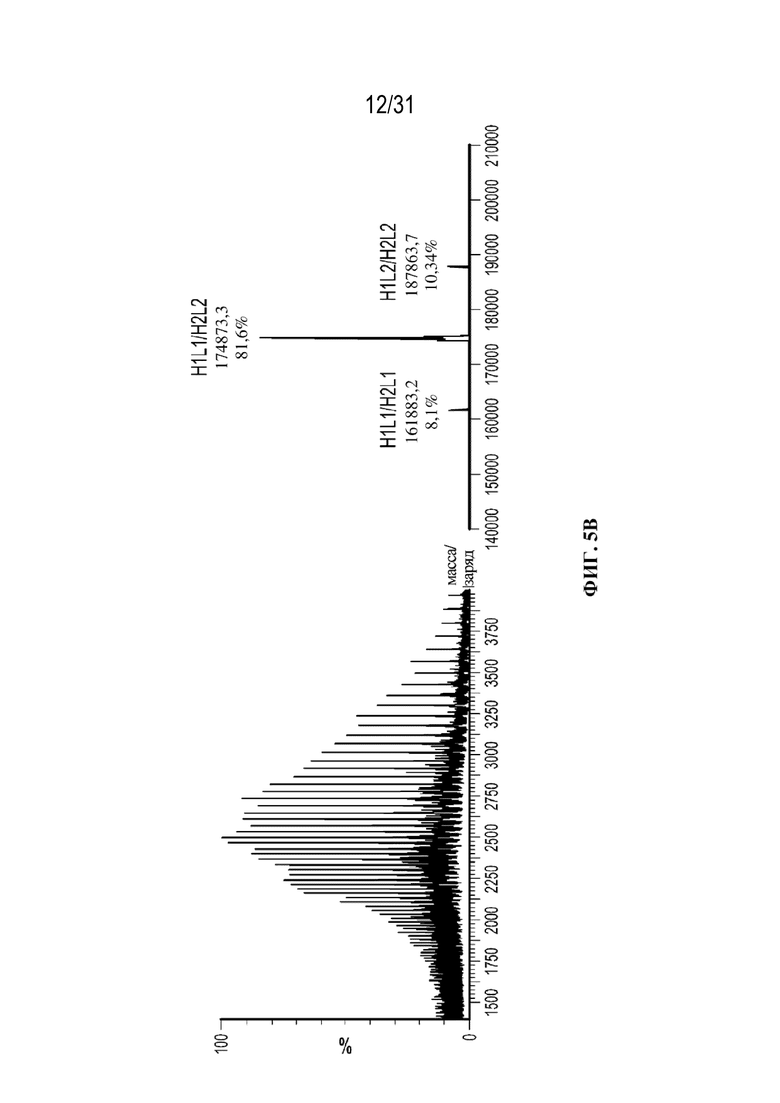
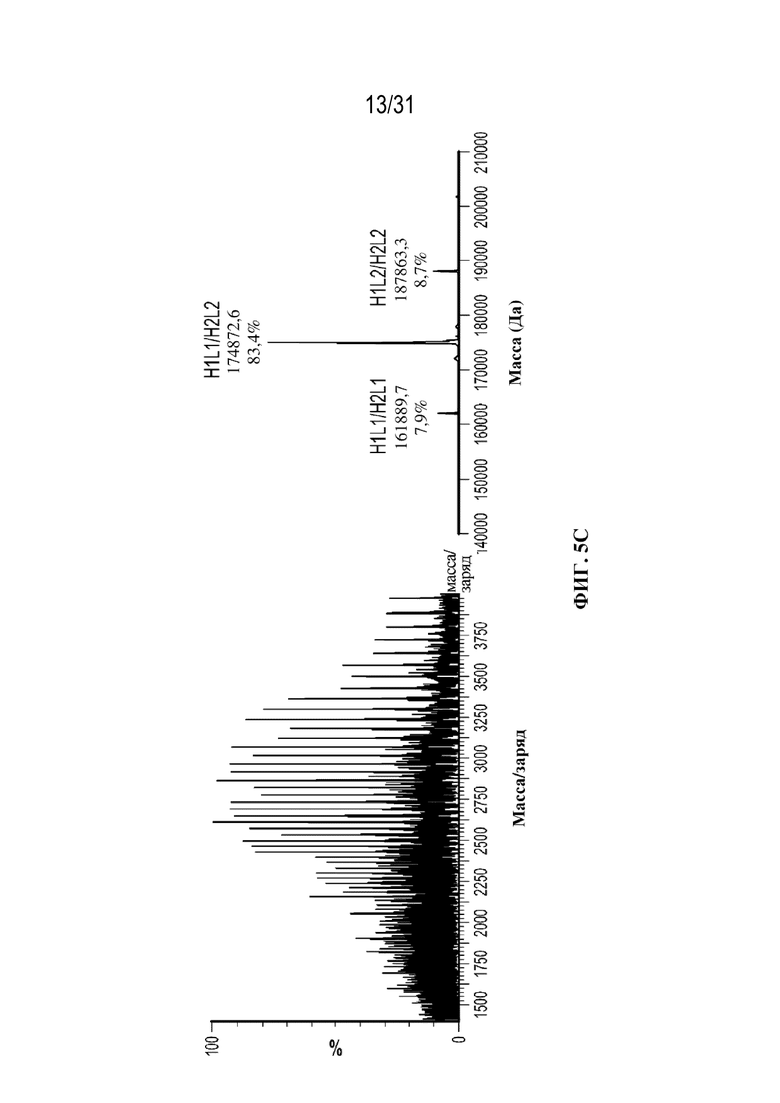
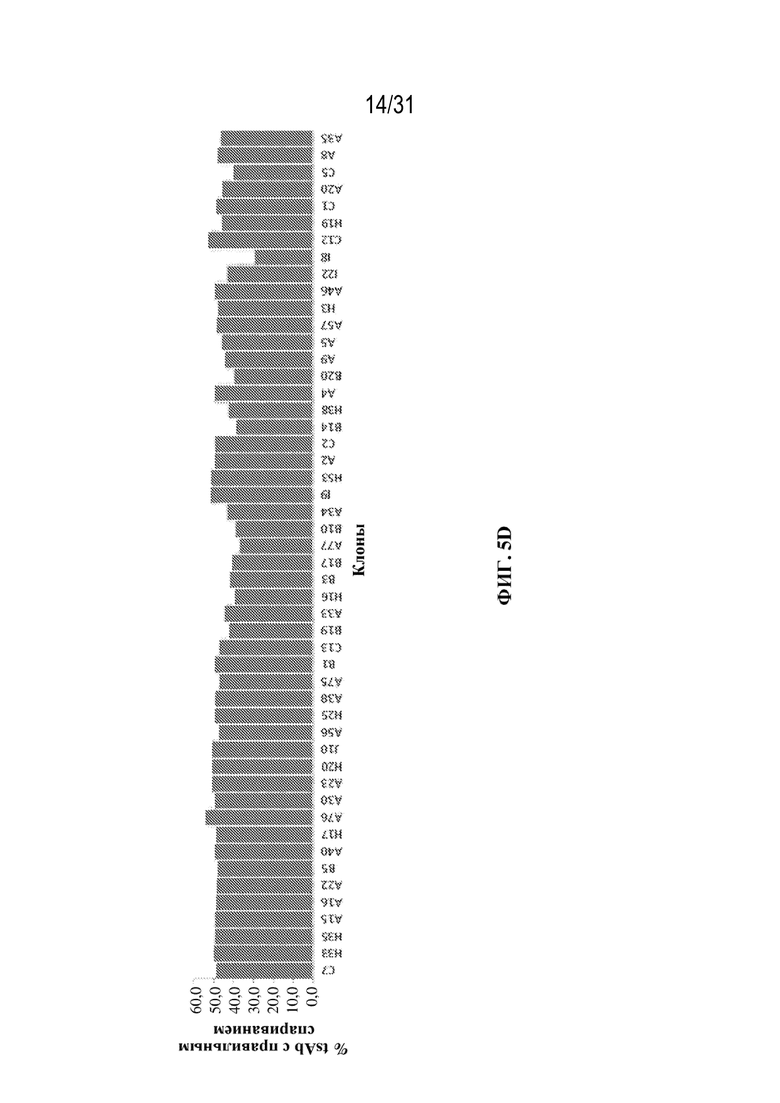

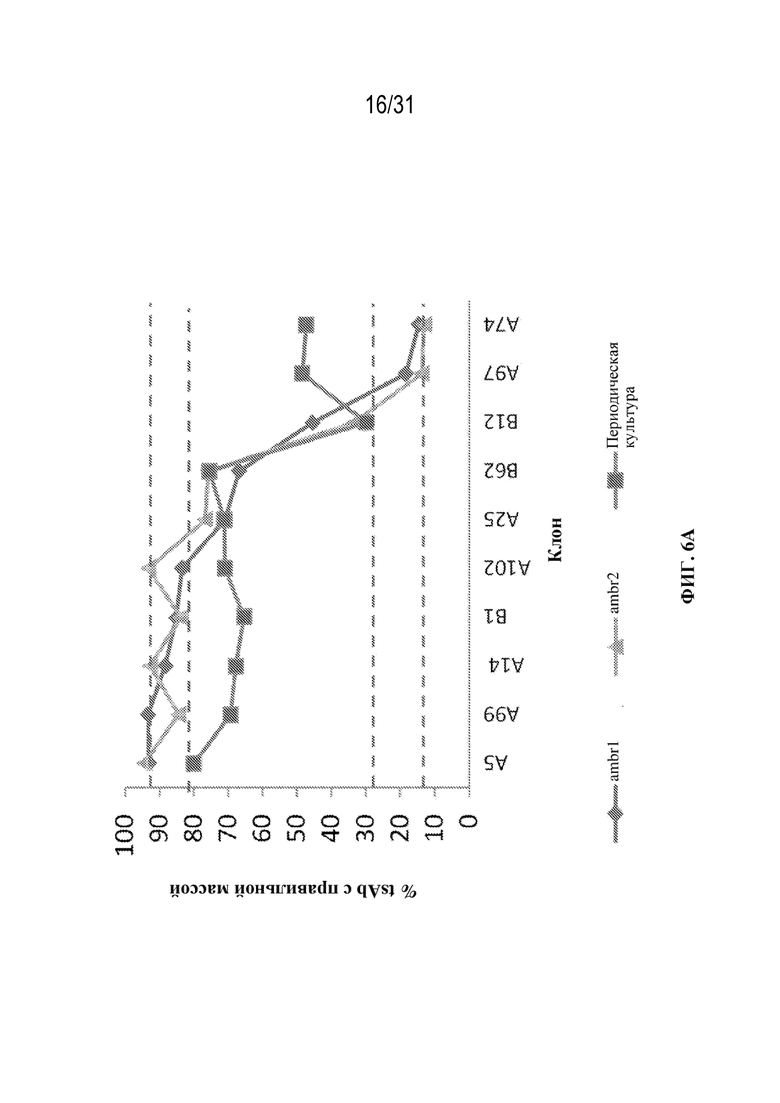
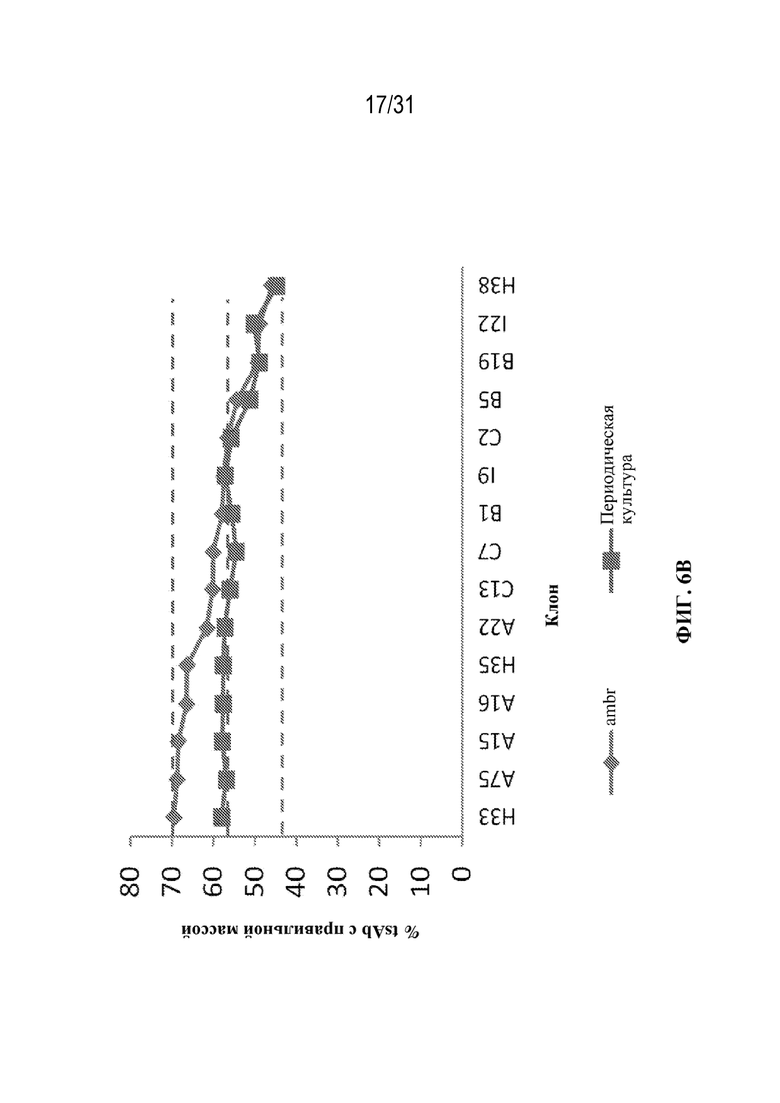
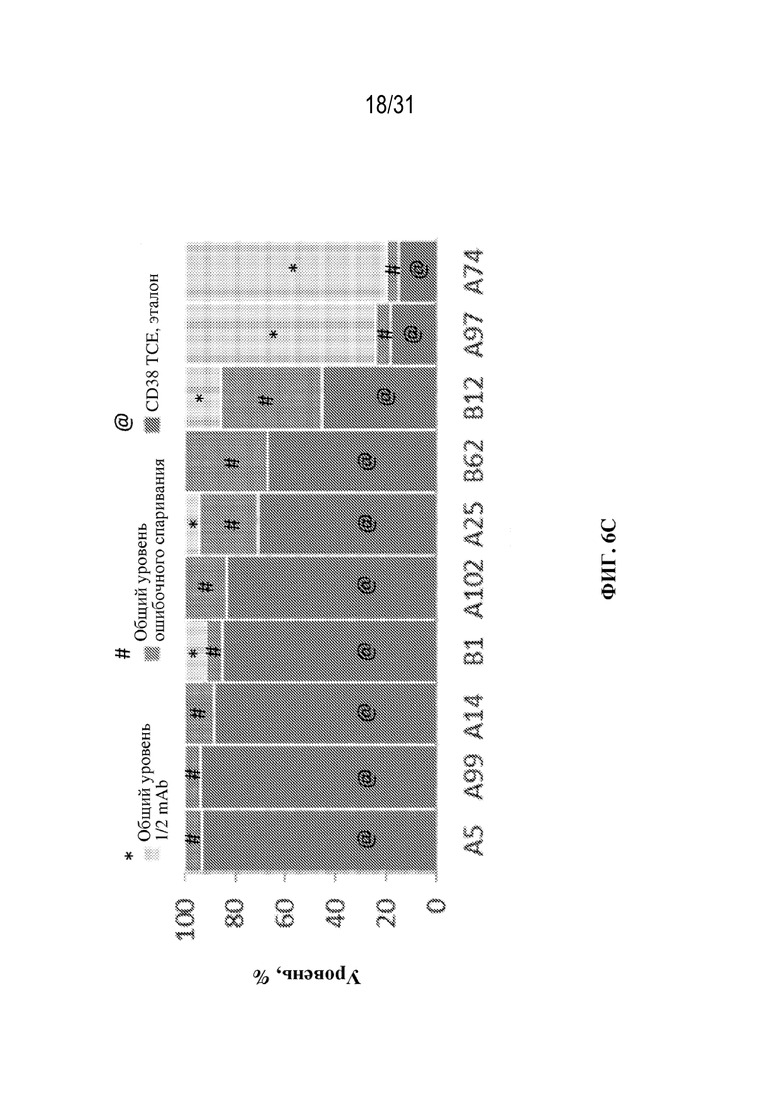
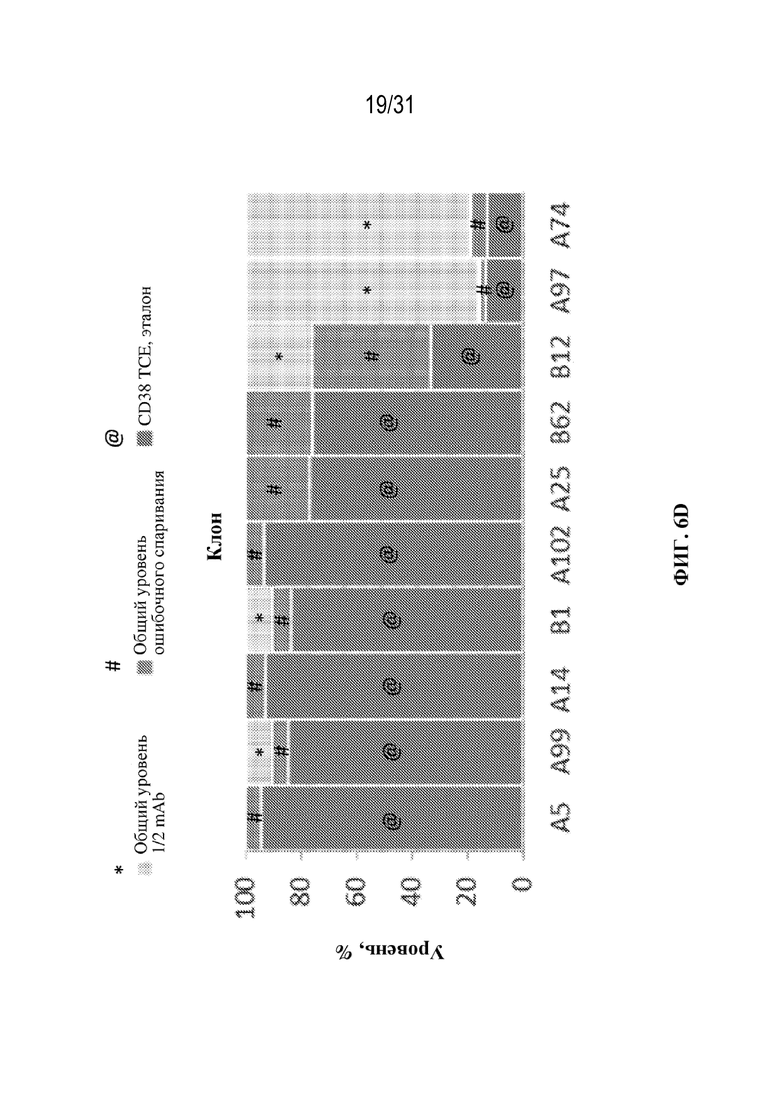
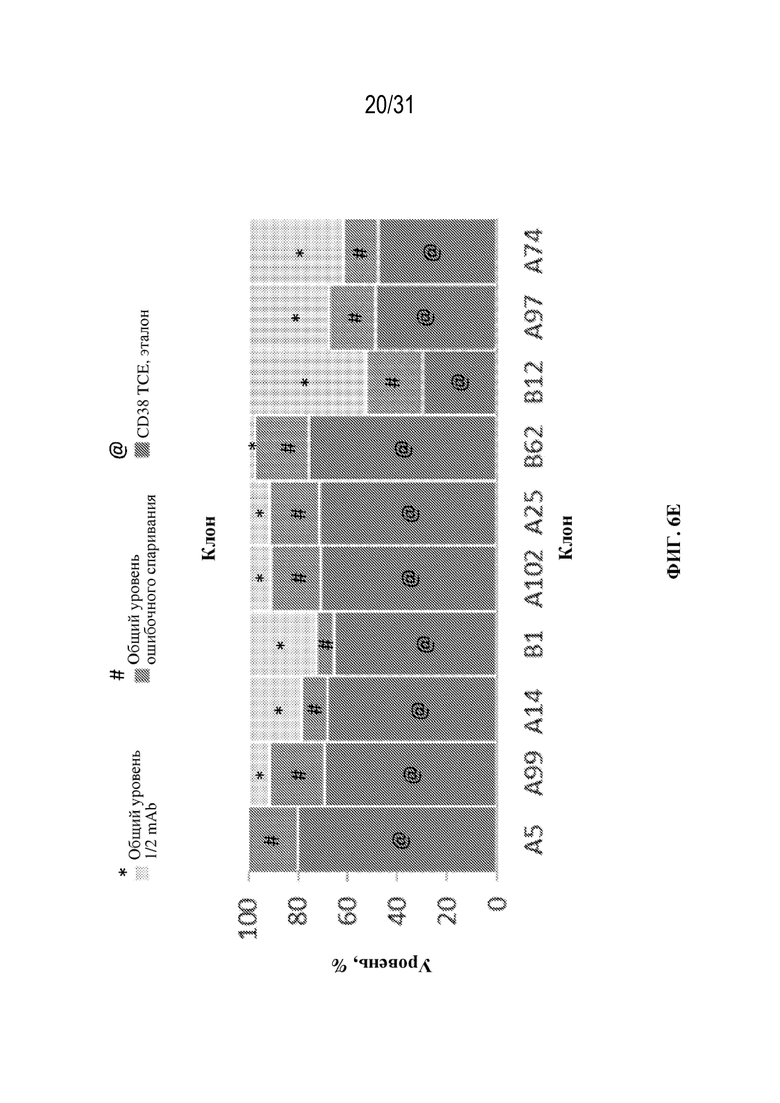

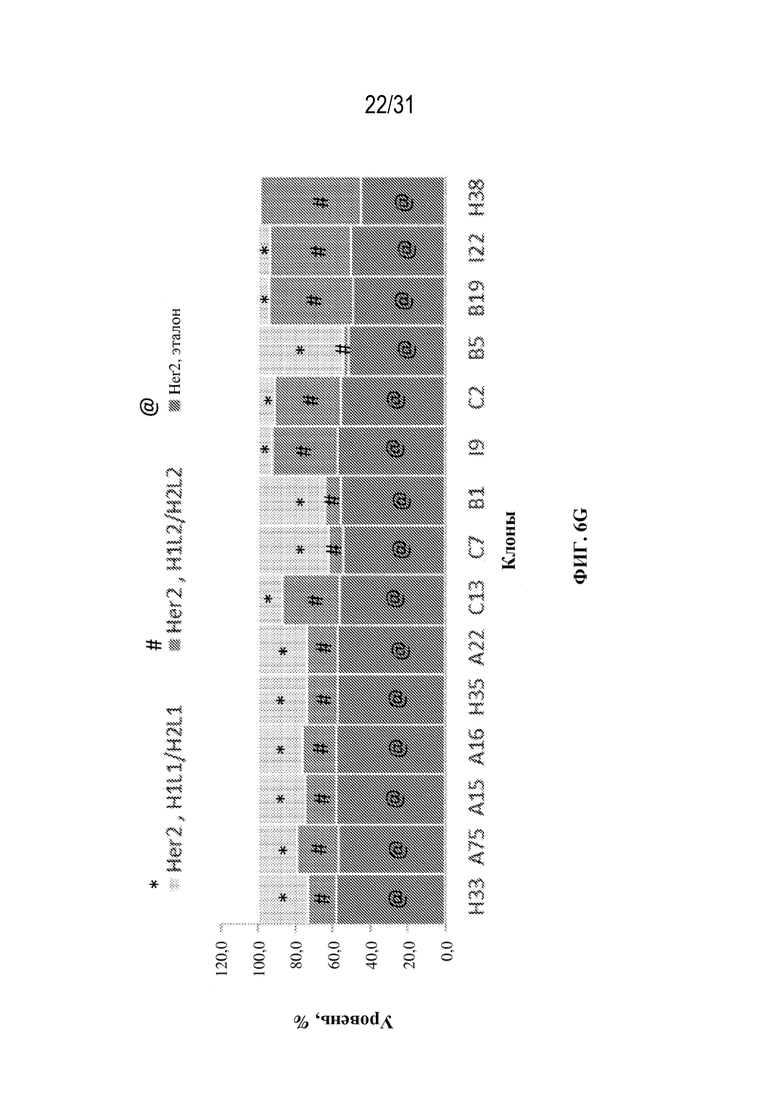
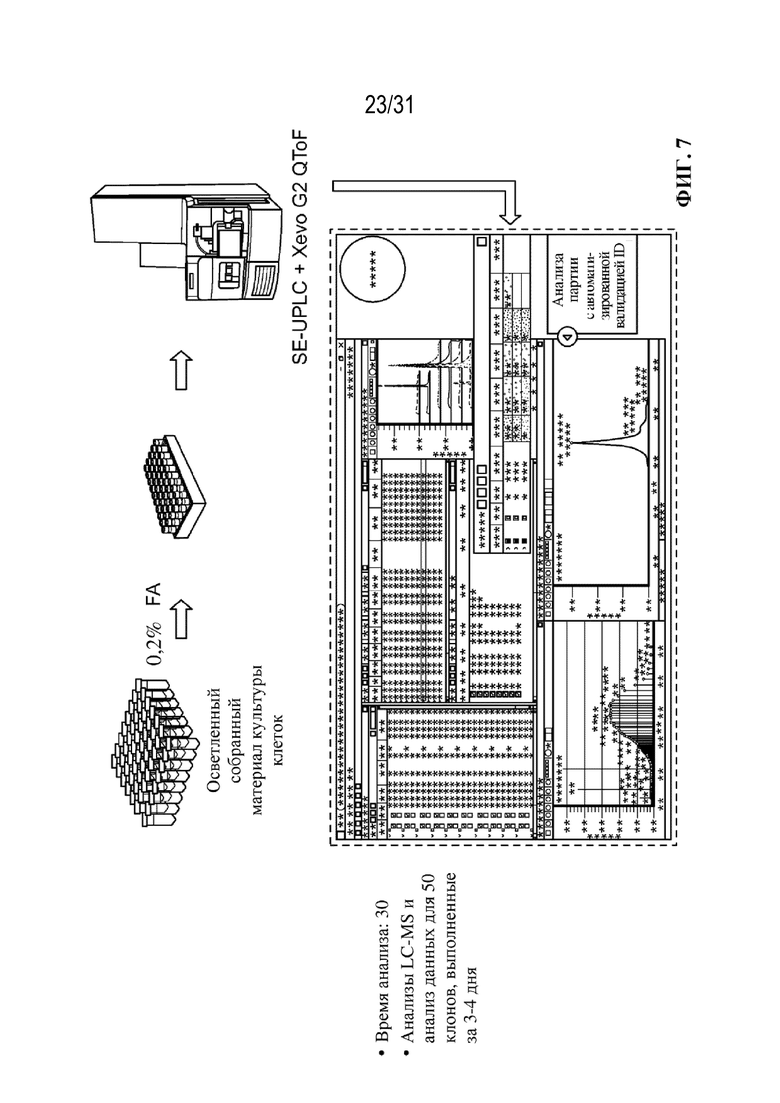

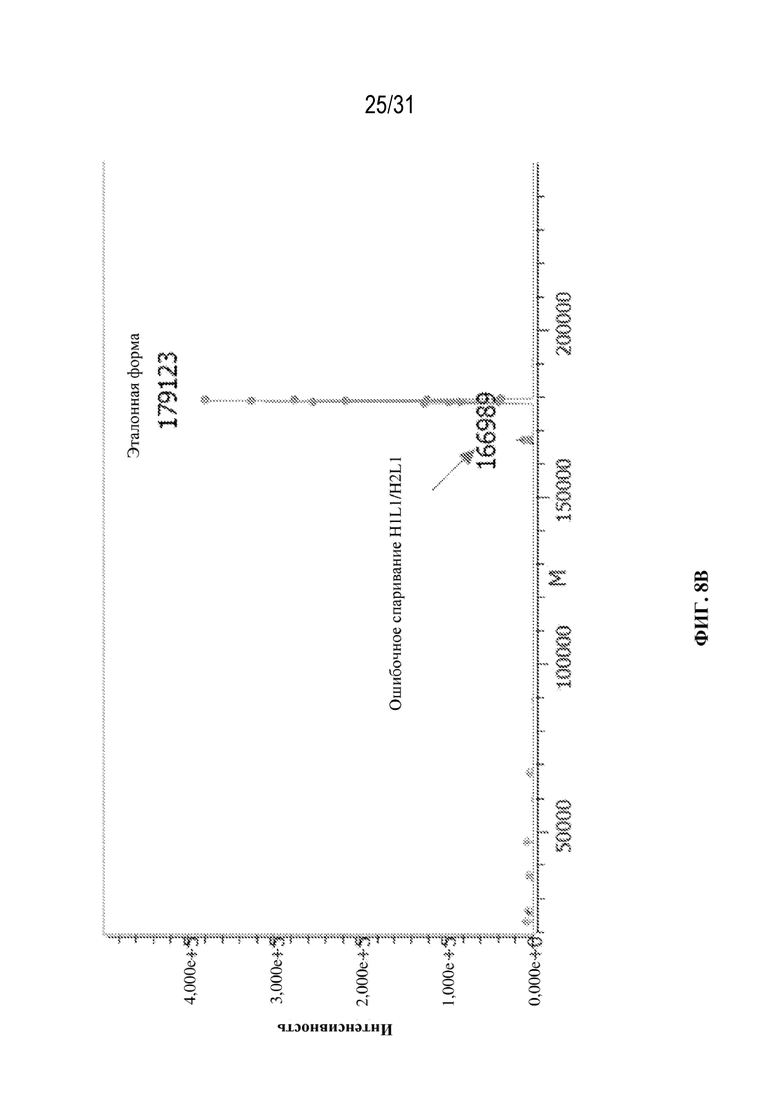

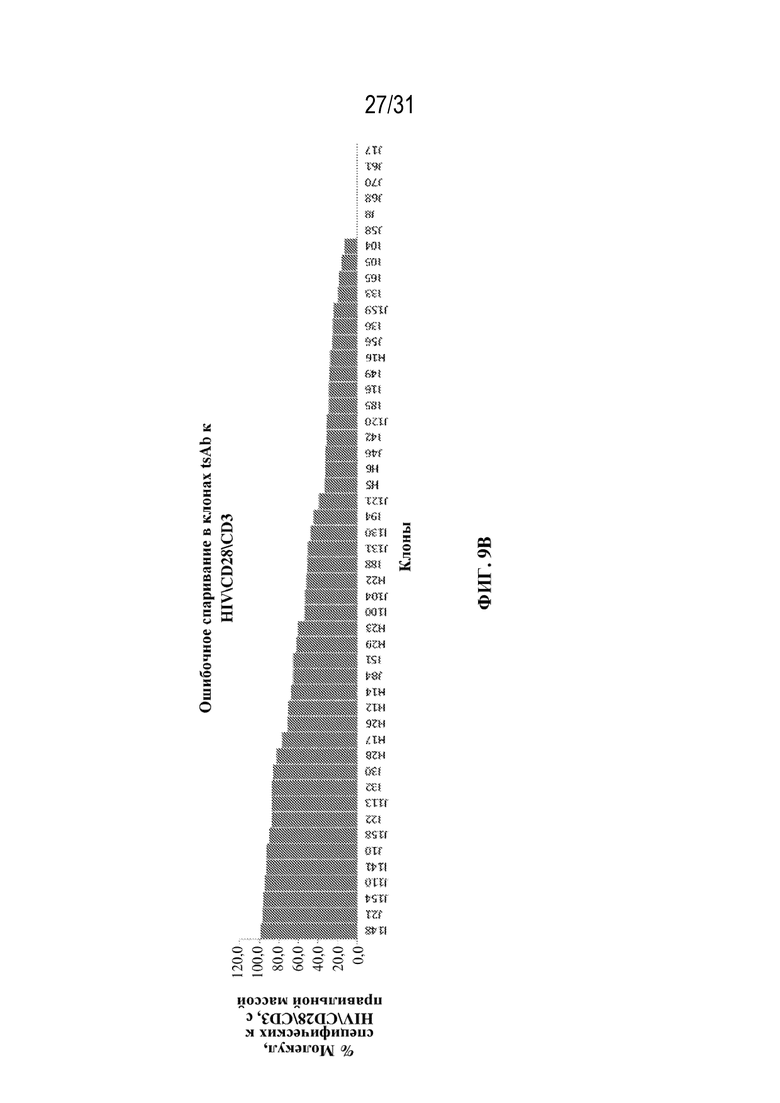
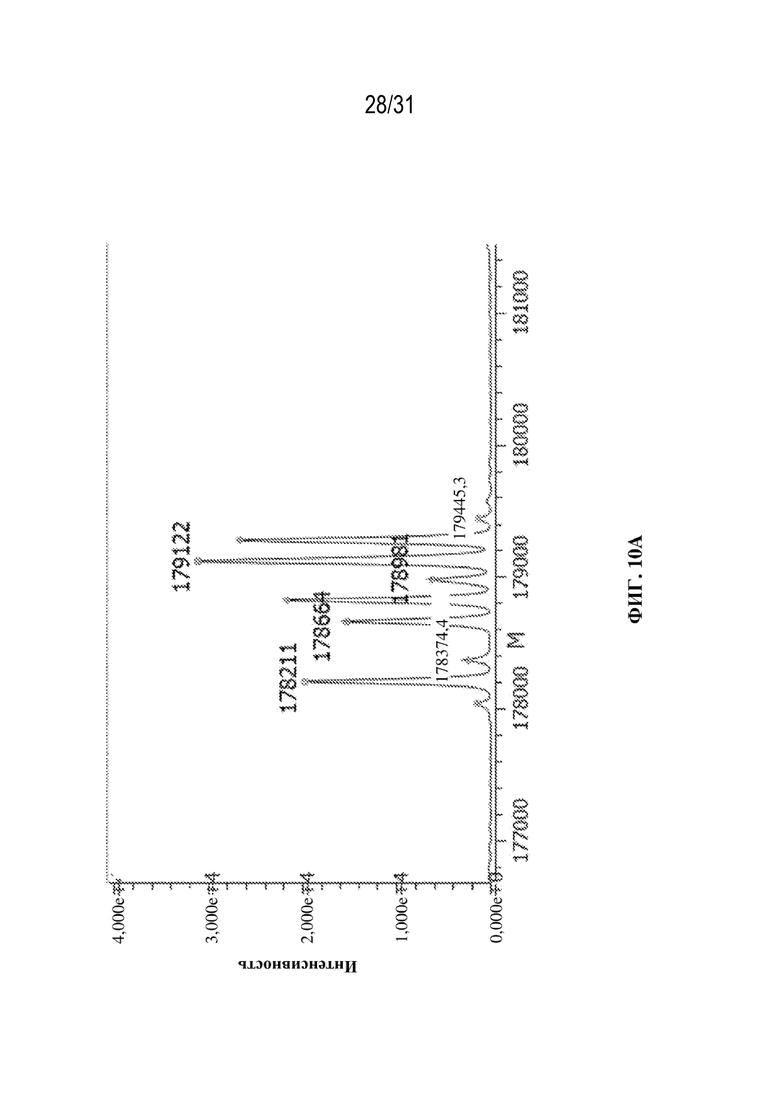
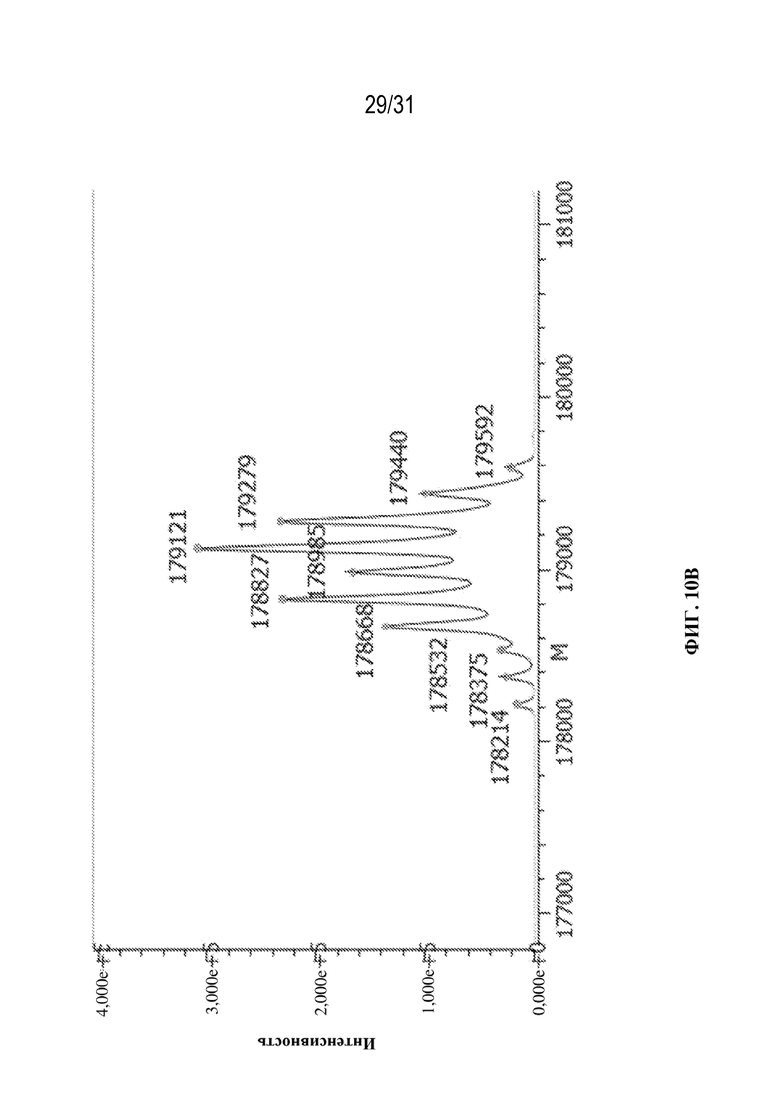
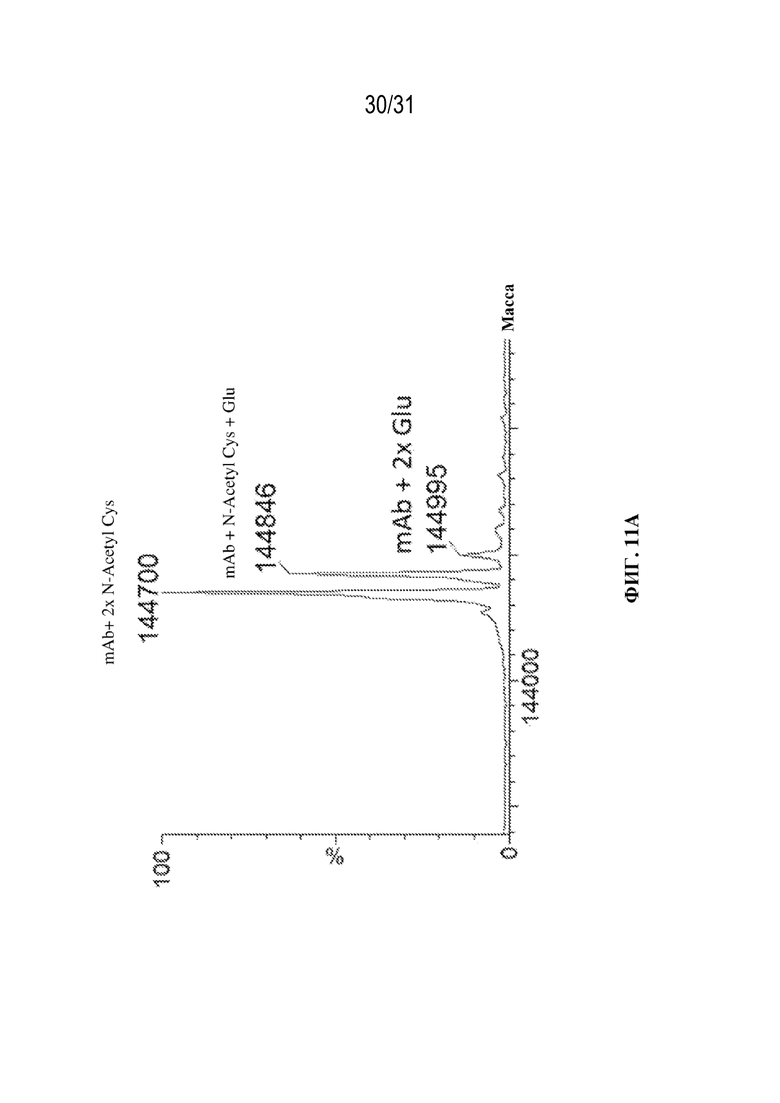
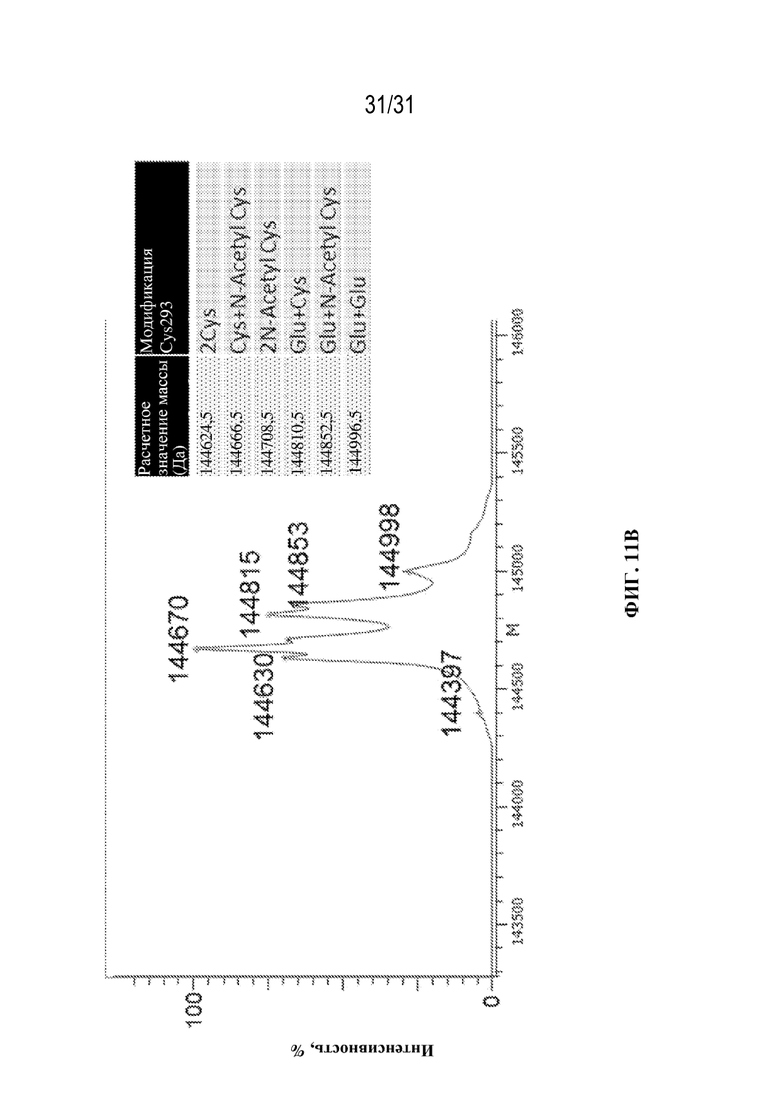
Изобретение относится к биотехнологии, в частности к способу мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием линией клеток. В некоторых вариантах осуществления способы включают определение количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием в среде для культивирования клеток посредством эксклюзионной сверхэффективной жидкостной хроматографии в денатурирующих условиях и масс-спектрометрии (SE-UPLC-MS). В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой полиспецифическое антитело, фрагмент антитела или белок, являющийся продуктом слияния с Fc. Изобретение предоставляет способ мониторинга или анализа продуцирования антитела или производных антител, которые предусматривают несколько молекул, отличающихся по молекулярной массе. 14 з.п. ф-лы, 11 ил., 2 табл., 5 пр.
1. Способ мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием, где способ включает:
определение посредством эксклюзионной сверхэффективной жидкостной хроматографии и масс-спектрометрии (SE-UPLC-MS) количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием в среде для культивирования клеток, содержащей полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием, с обеспечением таким образом мониторинга продуцирования полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием;
при этом полиспецифический связывающий белок предусматривает ассоциацию двух или более полипептидных цепей, содержащую по меньшей мере первую полипептидную цепь и вторую полипептидную цепь, отличную от первой полипептидной цепи;
при этом одна или несколько молекул с ошибочным спариванием содержат две или более полипептидных цепей, содержащих по меньшей мере одну из первой и второй полипептидных цепей в ассоциации, отличной от ассоциации полиспецифического связывающего белка;
где SE-UPLC представляет собой SE-UPLC в денатурирующих условиях.
2. Способ по п. 1, где полиспецифический связывающий белок представляет собой полиспецифическое антитело, содержащее тяжелую цепь первого антитела, легкую цепь первого антитела, тяжелую цепь второго антитела, отличную от тяжелой цепи первого антитела, и легкую цепь второго антитела, отличную от легкой цепи первого антитела, и где необязательно одна или несколько молекул с ошибочным спариванием содержат одно или несколько из следующего:
(i) ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две из тяжелых цепей первого антитела;
(ii) ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две из тяжелых цепей второго антитела;
(iii) ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две из легких цепей первого антитела; и
(iv) ассоциацию четырех полипептидных цепей полиспецифического антитела, которая содержит две из легких цепей второго антитела.
3. Способ по п. 1, где полиспецифический связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих сайта,
где первая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II],
и третья полипептидная цепь связывающего белка содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III],
и четвертая полипептидная цепь связывающего белка содержит структуру, представленную формулой:
VL3-CL [IV],
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
при этом полипептид формулы I и полипептид формулы II образуют пару перекрестно расположенных легкой цепи и тяжелой цепи, при этом VH1 и VL1 образуют первый антигенсвязывающий сайт, при этом VH2 и VL2 образуют второй антигенсвязывающий сайт, и при этом VH3 и VL3 образуют третий антигенсвязывающий сайт.
4. Способ по п. 3, где одна или несколько молекул с ошибочным спариванием предусматривают одно или несколько из следующего:
(i) ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой I;
(ii) ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой II;
(iii) ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой III; и
(iv) ассоциацию четырех полипептидных цепей связывающего белка, которая содержит две полипептидные цепи в соответствии с формулой IV.
5. Способ по любому из пп. 1-4, где определение количества полиспецифического связывающего белка и одной или нескольких молекул с ошибочным спариванием предусматривает деконволюцию одного или нескольких MS-спектров, полученных посредством MS.
6. Способ по любому из пп. 1-5, где определяют относительное количество полиспецифического связывающего белка по сравнению с одной или несколькими молекулами с ошибочным спариванием, где необязательно
(а) определяют относительное количество полиспецифического связывающего белка по сравнению с количеством одной или нескольких отдельных молекул с ошибочным спариванием; или
(b) определяют относительное количество полиспецифического связывающего белка по сравнению с общим количеством молекул с ошибочным спариванием.
7. Способ по любому из пп. 1-6, где среда для культивирования клеток представляет собой собранный материал культуры клеток, где способ необязательно дополнительно включает перед определением отделение среды для культивирования клеток от линии клеток, которая продуцирует полиспецифический связывающий белок и одну или несколько молекул с ошибочным спариванием, где необязательно перед (а) среду для культивирования клеток отделяют от линии клеток посредством центрифугирования.
8. Способ по любому из пп. 1-7, где среду для культивирования клеток подвергают SE-UPLC без
(а) предварительного хроматографического разделения, или
(b) предварительной аффинной хроматографии с использованием белка А.
9. Способ по любому из пп. 1-8, где MS представляет собой MS интактных молекул и/или квадрупольную времяпролетную (QToF) MS.
10. Способ по любому из пп. 1-9, где
(a) SE-UPLC объединена с MS непосредственно; и/или
(b) SE-UPLC проводят с начальной скоростью потока, составляющей менее приблизительно 0,4 мл/мин, где необязательно SE-UPLC проводят с начальной скоростью потока, составляющей приблизительно 0,1 мл/мин, и/или SE-UPLC проводят со скоростью потока, составляющей приблизительно 0,1 мл/мин, в течение первых 25 минут, а затем со скоростью потока, составляющей приблизительно 0,4 мл/мин.
11. Способ по любому из пп. 1-10, где SE-UPLC проводят с применением изократического элюирования подвижной фазой; где необязательно подвижная фаза содержит раствор, содержащий смесь ацетонитрил:вода в соотношении 30:70, и/или подвижная фаза содержит муравьиную кислоту и трифторуксусную кислоту (TFA), необязательно содержащая приблизительно 0,05% муравьиной кислоты и приблизительно 0,05% трифторуксусной кислоты (TFA).
12. Способ по любому из пп. 1-11, где определение выполняют за приблизительно 33 минуты или меньше.
13. Способ по любому из пп. 1-12, где MS позволяет определять разницу в массе, составляющую приблизительно 300 Да, между полиспецифическим связывающим белком и одной или несколькими молекулами с ошибочным спариванием или между двумя молекулами с ошибочным спариванием, и/или позволяет определять разницу в массе, составляющую приблизительно 162 Да, между полиспецифическим связывающим белком или молекулами с ошибочным спариванием и одной или несколькими гликоформами.
14. Способ по любому из пп. 7-13, где линия клеток представляет собой линию клеток млекопитающего и необязательно линию клеток яичника китайского хомячка (СНО).
15. Способ по любому из пп. 1-14, где перед определением линию клеток культивируют с использованием среды для культивирования клеток в непрерывной культуре в биореакторе с перемешиванием или в периодической культуре клеток.
| HABERGER M | |||
| et al., Rapid characterization of biotherapeutic proteins by size-exclusion chromatography coupled to native mass spectrometry, MAbs, 2016, vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Накладной висячий замок | 1922 |
|
SU331A1 |
| WOODS R.J | |||
| et al., LC-MS characterization and purity assessment of a prototype bispecific antibody, MAbs, 2013, vol | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ШТАНГЕН-ЦИРКУЛЬ С ВЫДВИЖНОЮ НОЖКОЮ | 1922 |
|
SU711A1 |
| YAN Y | |||
| et al., Coupling | |||

