ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет по предварительной заявке на патент США № 62/831572, поданной 9 апреля 2019 г., и заявке на европейский патент № EP 19306261.9, поданной 2 октября 2019 г., описания каждой из которых полностью включены в настоящий документ посредством ссылки.
ПРЕДСТАВЛЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ТЕКСТОВОМ ФАЙЛЕ В ФОРМАТЕ ASCII
[0002] Содержание нижеследующего представленного текстового файла в формате ASCII включено в данный документ посредством ссылки в его полном объеме: машиночитаемая форма (CRF) перечня последовательностей (название файла: 183952032040SEQLIST.TXT, дата составления: 6 апреля, 2020 г., размер: 526 КБ).
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Настоящее изобретение относится к триспецифическим и/или тривалентным связывающим белкам, содержащим четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, которые специфически связывают один или несколько белков-мишеней, где первая пара полипептидов, образующих связывающий белок, содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией. Настоящее изобретение также относится к способам получения триспецифических и/или тривалентных связывающих белков и вариантам применения таких связывающих белков.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0004] Биотерапевтические средства на основе моноклональных антител стали важным направлением в разработке новых лекарственных средств. Технология моноклональных антител предлагает специфическое нацеливание на конкретную популяцию клеток, точную доставку в них сигналов и/или полезной нагрузки и обеспечивает длительный биологический эффект посредством функций, опосредованных Fc-фрагментом. Усилия, предпринятые в области конструирования антител, позволили разработать биспецифические антитела, в которых объединены специфичности двух моноклональных антител, для различных вариантов применения в биологических целях, расширив сферу разработки лекарственных средств на основе антител. Недавно открытые нейтрализующие антитела с улучшенной широтой спектра действия и активностью могут обеспечить больше возможных вариантов разработки биотерапевтических средств для лечения комплексных заболеваний, таких как рак, артрит и/или воспалительные заболевания.
[0005] Иммуноонкология представляет собой многообещающий развивающийся терапевтический подход для управления ходом заболевания при раке. Иммунная система представляет собой первую линию защиты от развития и прогрессирования рака. В настоящее время существует множество доказательств того, что Т-клетки способны контролировать рост опухоли и продлевать время выживания пациентов, страдающих раком, как на ранних, так и на поздних стадиях заболевания. Однако Т-клетки, специфичные в отношении опухолей, могут быть ограничены во многих отношениях, что не позволяет им контролировать заболевание.
[0006] Как часть адаптивного иммунитета человека, Т-клеточный иммунитет играет решающую роль в контроле вирусной инфекции и рака, возможно, устраняя инфицированные клетки и злокачественные клетки, что приводит к освобождению от вирусной инфекции или излечению от рака. При хронических инфекционных заболеваниях, таких как инфекция, вызванная вирусом герпеса (HSV, CMV, EBV и т. д.), HIV и HBV, вирусы сохраняют свою устойчивость у людей с помощью различных механизмов, включая подавление иммунитета, истощение Т-клеток и установление латентного периода. Тем не менее, вирусная инфекция обычно индуцирует вирусный антигенспецифический иммунитет, включающий антигенспецифические CD8 Т-клетки, которые могут легко распознавать инфицированные клетки для контроля или уничтожения посредством высвобождения цитокинов или процессов уничтожения, опосредованных цитотоксическими Т-клетками (CTL).
[0007] Таким образом, активация и/или амплификация вирусных антигенспецифических Т-клеток in vivo и/или ex vivo может обеспечить терапевтические стратегии против хронических вирусных инфекций.
[0008] Все приведенные в данном документе ссылочные материалы, в том числе заявки на патенты, патентные публикации и номера доступа в UniProtKB/Swiss-Prot, включены в данный документ посредством ссылки в их полном объеме, как если бы каждый отдельный ссылочный материал был специально и отдельно указан для включения в качестве ссылки.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0009] Для удовлетворения этих и других потребностей в данном документе предусмотрены триспецифические связывающие белки (например, антитела), которые образуют три антигенсвязывающих участка. Данные связывающие белки могут специфически связывать одну, две или три антигенные мишени или белка-мишени, таких как CD28, CD3 и опухолевый белок-мишень. Некоторые опухоли экспрессируют специфические антигены. Например, амплификацию и сверхэкспрессию HER2 можно обнаружить в молекулярных подтипах рака молочной железы, а также при карциномах желудка, яичника, легкого и предстательной железы. Для оптимальной активации Т-клеток необходимы два фактора: 1. распознавание антигена и 2. костимуляция. При использовании триспецифических связывающих HER2/CD28xCD3 белков, описанных в данном документе, сигнал 1 обеспечивается CD3-связывающим участком-агонистом, а сигнал 2 обеспечивается CD28-связывающим участком-агонистом. Триспецифический связывающий белок осуществляет рекрутинг Т-клеток к опухоли посредством связывающего участка для HER2, CD38 или связывающего участка, распознающего другой опухолевый белок-мишень, и активирует привлеченные Т-клетки посредством CD3- и CD28-связывающих участков. Происходящая в результате активация индуцирует потенциал иммунных клеток в отношении уничтожения близлежащих опухолевых клеток. Кроме того, описаны CD3-связывающие участки, характеризующиеся высокой аффинностью связывания с полипептидами CD3 человека и устраненными потенциальными недостатками при их получении (например, участками дезамидирования).
[0010] В данном документе дополнительно предусмотрены триспецифические антитела к CD38/CD28xCD3, которые разрабатывали и оценивали в отношении их способности активировать Т-клетки и последующую пролиферацию и/или амплификацию антигенспецифических Т-клеток. Эти триспецифические Ab могут эффективно расширять популяции CD4 и CD8 эффекторных клеток и клеток памяти, в том числе антигенспецифических CD8 центральных T-клеток памяти и эффекторных клеток памяти in vitro. В частности, продемонстрировали in vitro размножение специфических в отношении CMV, EBV, HIV-1, вируса гриппа CD8 центральных клеток памяти и эффекторных клеток памяти. Триспецифические антитела к CD38/CD28xCD3, описываемые в данном документе, демонстрировали новые свойства путем вовлечения CD3/CD28/CD38 с обеспечением пути передачи сигнала для стимуляции и размножения Т-клеток, что может обеспечить эффективную стратегию лечения хронических инфекционных заболеваний, таких как инфекции, вызванные HSV, CMV, EBV, HIV-1 и HBV.
[0011] Для удовлетворения этих и других потребностей в данном документе предусмотрены связывающие белки, которые связывают полипептиды CD38 (например, полипептиды CD38 человека и макака-крабоеда), включая моноспецифические, биспецифические или триспецифические связывающие белки с по меньшей мере одним антигенсвязывающим участком, который связывает полипептид CD38. Преимущественно такие связывающие белки характеризуются способностью к рекрутингу Т-клеток в непосредственной близости от раковых клеток с последующей активацией Т-клеток и стимуляцией уничтожения посредством активированных Т-клеток соседних раковых клеток с помощью механизма гранзим/перфорин, обеспечивая таким образом другой механизм действия для противоопухолевой активности антител к CD38, таких как DARZALEX® (даратумумаб). Более того, способность связывать полипептиды CD38 как человека, так и макака-крабоеда обеспечивает возможность простого тестирования связывающих белков в доклинических токсикологических исследованиях, например для оценки их профилей безопасности с целью последующего клинического применения.
[0012] В некоторых вариантах осуществления в данном документе предусмотрены связывающие белки, содержащие четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I];
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок;
где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65); и
где VH3 и VL3 образуют третий антигенсвязывающий участок.
[0013] В некоторых вариантах осуществления первый связывающий участок связывает полипептид CD28. В некоторых вариантах осуществления домен VH1 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYY (SEQ ID NO:49), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGNVNT (SEQ ID NO:50), и последовательность CDR-H3, содержащую аминокислотную последовательность TRSHYGLDWNFDV (SEQ ID NO:51), и домен VL1 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QNIYVW (SEQ ID NO:52), последовательность CDR-L2, содержащую аминокислотную последовательность KAS (SEQ ID NO:53), и последовательность CDR-L3, содержащую аминокислотную последовательность QQGQTYPY (SEQ ID NO:54). В некоторых вариантах осуществления домен VH1 содержит аминокислотную последовательность qvqlvqsgaevvkpgasvkvsckasgytftsyyihwvrqapgqglewigsiypgnvntnyaqkfqgratltvdtsistaymelsrlrsddtavyyctrshygldwnfdvwgkgttvtvss (SEQ ID NO:91), и/или домен VL1 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcqasqniyvwlnwyqqkpgkapklliykasnlhtgvpsrfsgsgsgtdftltisslqpediatyycqqgqtypytfgqgtkleik (SEQ ID NO:92).
[0014] В некоторых вариантах осуществления последовательность CDR-L1 домена VL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из QSLVHQNAQTY (SEQ ID NO:59), QSLVHENLQTY (SEQ ID NO:60), QSLVHENLFTY (SEQ ID NO:61) и QSLVHENLRTY (SEQ ID NO:62). В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHQNAQTY (SEQ ID NO:59), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLQTY (SEQ ID NO:60), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLFTY (SEQ ID NO:61), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLRTY (SEQ ID NO:62), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления домен VH2 содержит аминокислотную последовательность qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93), и/или домен VL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из divmtqtplslsvtpgqpasisckssqslvhqnaqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:95), divmtqtplslsvtpgqpasisckssqslvhenlqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:96), divmtqtplslsvtpgqpasisckssqslvhenlftylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:97), и divmtqtplslsvtpgqpasisckssqslvhenlrtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:98). В некоторых вариантах осуществления домен VH2 содержит аминокислотную последовательность qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93) или qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyassvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:302),, и/или домен VL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из divmtqtplslsvtpgqpasisckssqslvhqnaqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:95), divmtqtplslsvtpgqpasisckssqslvhenlqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:96), divmtqtplslsvtpgqpasisckssqslvhenlftylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:97), и divmtqtplslsvtpgqpasisckssqslvhenlrtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:98). В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:95. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:302, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:95. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:96. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:97. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, содержащий вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:98.
[0015] В некоторых вариантах осуществления третий антигенсвязывающий участок связывает опухолевый белок-мишень. В некоторых вариантах осуществления опухолевый белок-мишень представляет собой полипептид CD38 (например, полипептид CD38 человека). В некоторых вариантах осуществления опухолевый белок-мишень представляет собой полипептид HER2 (например, полипептид HER2 человека). В некоторых вариантах осуществления опухолевый белок-мишень по настоящему изобретению включает без ограничения A2AR, APRIL, ATPDазу, BAFF, BAFFR, BCMA, BlyS, BTK, BTLA, B7DC, B7H1, B7H4 (также известный как VTCN1), B7H5, B7H6, B7H7, B7RP1, B7-4, C3, C5, CCL2 (также известный как MCP-1), CCL3 (также известный как MIP-1a), CCL4 (также известный как MIP-1b), CCL5 (также известный как RANTES), CCL7 (также известный как MCP-3), CCL8 (также известный как mcp-2), CCL11 (также известный как эотаксин), CCL15 (также известный как MIP-1d), CCL17 (также известный как TARC), CCL19 (также известный как MIP-3b), CCL20 (также известный как MIP-3a), CCL21 (также известный как MIP-2), CCL24 (также известный как MPIF-2/эотаксин-2), CCL25 (также известный как TECK), CCL26 (также известный как эотаксин-3), CCR3, CCR4, CD3, CD19, CD20, CD23 (также известный как FCER2, рецептор IgE), CD24, CD27, CD28, CD38, CD39, CD40, CD70, CD80 (также известный как B7-1), CD86 (также известный как B7-2), CD122, CD137 (также известный как 41BB), CD137L, CD152 (также известный как CTLA4), CD154 (также известный как CD40L), CD160, CD272, CD273 (также известный как PDL2), CD274 (также известный как PDL1), CD275 (также известный как B7H2), CD276 (также известный как B7H3), CD278 (также известный как ICOS), CD279 (также известный как PD-1), CDH1 (также известный как E-кадгерин), хитиназу, CLEC9, CLEC91, CRTH2, CSF-1 (также известный как M-CSF), CSF-2 (также известный как GM-CSF), CSF-3 (также известный как GCSF), CX3CL1 (также известный как SCYD1), CXCL12 (также известный как SDF1), CXCL13, CXCR3, DNGR-1, эктонуклеозидтрифосфатдифосфогидролазу 1, EGFR, ENTPD1, FCER1A, FCER1, FLAP, FOLH1, Gi24, GITR, GITRL, GM-CSF, Her2, HHLA2, HMGB1, HVEM, ICOSLG, IDO, IFNα, IgE, IGF1R, IL2R бета, IL1, IL1A, IL1B, IL1F10, IL2, IL4, IL4Ra, IL5, IL5R, IL6, IL7, IL7Ra, IL8, IL9, IL9R, IL10, rhIL10, IL12, IL13, IL13Ra1, IL13Ra2, IL15, IL17, IL17Rb (также известный как рецептор IL25), IL18, IL22, IL23, IL25, IL27, IL33, IL35, ITGB4 (также известный как интегрин b4), ITK, KIR, LAG3, LAMP1, лептин, LPFS2, молекулы MHC класса II, MUC-1, NCR3LG1, NKG2D, NTPDазу-1, OX40, OX40L, PD-1H, тромбоцитарный рецептор, PROM1, S152, SISP1, SLC, SPG64, ST2 (также известный как рецептор IL33), STEAP2, Syk-киназу, TACI, TDO, T14, TIGIT, TIM3, TLR, TLR2, TLR4, TLR5, TLR9, TMEF1, TNFa, TNFRSF7, Tp55, TREM1, TSLP (также известный как корецептор IL7Ra), TSLPR, TWEAK, VEGF, VISTA, Vstm3, WUCAM и XCR1 (также известный как GPR5/CCXCR1). В некоторых вариантах осуществления одна или несколько из указанных выше антигенных мишеней представляют собой антигенные мишени человека.
[0016] В некоторых вариантах осуществления третий антигенсвязывающий участок связывает полипептид CD38 человека. В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYA (SEQ ID NO:13), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGQGGT (SEQ ID NO:14), и последовательность CDR-H3, содержащую аминокислотную последовательность ARTGGLRRAYFTY (SEQ ID NO:15), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSVSSYGQGF (SEQ ID NO:16), последовательность CDR-L2, содержащую аминокислотную последовательность GAS (SEQ ID NO:17), и последовательность CDR-L3, содержащую аминокислотную последовательность QQNKEDPWT (SEQ ID NO:18). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYTLTEFS (SEQ ID NO:19), последовательность CDR-H2, содержащую аминокислотную последовательность FDPEDGET (SEQ ID NO:20), и последовательность CDR-H3, содержащую аминокислотную последовательность TTGRFFDWF (SEQ ID NO:21), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSVISRF (SEQ ID NO:22), последовательность CDR-L2, содержащую аминокислотную последовательность GAS (SEQ ID NO:23), и последовательность CDR-L3, содержащую аминокислотную последовательность QQDSNLPIT (SEQ ID NO:24). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYAFTTYL (SEQ ID NO:25), последовательность CDR-H2, содержащую аминокислотную последовательность INPGSGST (SEQ ID NO:26), и последовательность CDR-H3, содержащую аминокислотную последовательность ARYAYGY (SEQ ID NO:27), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QNVGTA (SEQ ID NO:28), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:29), и последовательность CDR-L3, содержащую аминокислотную последовательность QQYSTYPFT (SEQ ID NO:30). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYSFTNYA (SEQ ID NO:31), последовательность CDR-H2, содержащую аминокислотную последовательность ISPYYGDT (SEQ ID NO:32), и последовательность CDR-H3, содержащую аминокислотную последовательность ARRFEGFYYSMDY (SEQ ID NO:33), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHSNGNTY (SEQ ID NO:34), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:35), и последовательность CDR-L3, содержащую аминокислотную последовательность SQSTHVPLT (SEQ ID NO:36).
[0017] В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFSSYG (SEQ ID NO:37), последовательность CDR-H2, содержащую аминокислотную последовательность IWYDGSNK (SEQ ID NO:38), и последовательность CDR-H3, содержащую аминокислотную последовательность ARDPGLRYFDGGMDV (SEQ ID NO:39), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QGISSY (SEQ ID NO:40), последовательность CDR-L2, содержащую аминокислотную последовательность AAS (SEQ ID NO:41), и последовательность CDR-L3, содержащую аминокислотную последовательность QQLNSFPYT (SEQ ID NO:42). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFSSYG (SEQ ID NO:43), последовательность CDR-H2, содержащую аминокислотную последовательность IWYDGSNK (SEQ ID NO:44), и последовательность CDR-H3, содержащую аминокислотную последовательность ARMFRGAFDY (SEQ ID NO:45), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QGIRND (SEQ ID NO:46), последовательность CDR-L2, содержащую аминокислотную последовательность AAS (SEQ ID NO:47), и последовательность CDR-L3, содержащую аминокислотную последовательность LQDYIYYPT (SEQ ID NO:48). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность Qvqlvqsgaevvkpgasvkvsckasgytftsyamhwvkeapgqrlewigyiypgqggtnynqkfqgratltadtsastaymelsslrsedtavyfcartgglrrayftywgqgtlvtvss (SEQ ID NO:79), и/или домен VL3 содержит аминокислотную последовательность Divltqspatlslspgeratiscrasqsvssygqgfmhwyqqkpgqpprlliygassratgiparfsgsgsgtdftltisplepedfavyycqqnkedpwtfgggtkleik (SEQ ID NO:80). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность qvqlvqsgaevkkpgasvkvsckvsgytltefsihwvrqapgqglewmggfdpedgetiyaqkfqgrvimtedtstdtaymemnslrsedtaiyycttgrffdwfwgqgtlvtvss (SEQ ID NO:81), и/или домен VL3 содержит аминокислотную последовательность eiiltqspailslspgeratlscrasqsvisrflswyqvkpglaprlliygastratgipvrfsgsgsgtdfsltisslqpedcavyycqqdsnlpitfgqgtrleik (SEQ ID NO:82). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность qvqlvqsgaevkkpgasvkvsckasgyafttylvewirqrpgqglewmgvinpgsgstnyaqkfqgrvtmtvdrssttaymelsrlrsddtavyycaryaygywgqgtlvtvss (SEQ ID NO:83), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqnvgtavawyqqkpgkspkqliysasnrytgvpsrfsgsgsgtdftltisslqpedlatyycqqystypftfgqgtkleik (SEQ ID NO:84). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность qvqlvesgggvvqpgrslrlscaasgftfssygmywvrqapgkglewvaviwydgsnkyyadsvkgrftisrdnskntlylqmnslraedtavyhcardpglryfdggmdvwgqgttvtvss (SEQ ID NO:87), и/или домен VL3 содержит аминокислотную последовательность diqltqspsflsasvgdrvtitcrasqgissylawyqqkpgkapkllifaastlhsgvpsrfsgsgsgteftltisslqpedfatyycqqlnsfpytfgqgtkleik (SEQ ID NO:88). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность qvqlvesgggvvqpgrslrlscaasgftfssygmhwvrqapgkglewvaviwydgsnkyyadsvkgrftisgdnskntlylqmnslraedtavyycarmfrgafdywgqgtlvtvss (SEQ ID NO:89), и/или домен VL3 содержит аминокислотную последовательность aiqmtqspsslsasvgdrvtitcrasqgirndlgwyqqkpgkapklliyaasslqsgvpsrfsgsgsgtdftltisglqpedsatyyclqdyiyyptfgqgtkveik (SEQ ID NO:90). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность qvqlvqsgaevkkpgasvkvsckasgysftnyavhwvrqapgqglewmgvispyygdttyaqkfqgrvtmtvdkssstaymelsrlrsddtavyycarrfegfyysmdywgqgtlvtvss (SEQ ID NO:85), и/или домен VL3 содержит аминокислотную последовательность dvvmtqsplslpvtlgqpasiscrpsqslvhsngntylnwyqqrpgqspklliykvskrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycsqsthvpltfgggtkveik (SEQ ID NO:86).
[0018] В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:156 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:156; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:157 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:157; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:158 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:158; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:159 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:159. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:160 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:160; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:161 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:161; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:162 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:162; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:163 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:163. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:164 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:164; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:165 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:165; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:166 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:166; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:167 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:167. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:168 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:168; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:169 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:169; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:170 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:170; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:171 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:171. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:172 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:172; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:173 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:173; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:174 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:174; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:175 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:175. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:176 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:176; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:177 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:177; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:178 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:178; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:179 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:179. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:181 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:181; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:182 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:182; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:183 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:183; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:184 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:184. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:185 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:185; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:186 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:186; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:187 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:187; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:188 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:188.
[0019] В некоторых вариантах осуществления третий антигенсвязывающий участок связывает полипептид HER2 человека. В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1) или GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), IYPTQGYT (SEQ ID NO:4) или IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6), SRWGGEGFYAMDY (SEQ ID NO:7) или SRWGGSGFYAMDY (SEQ ID NO:8), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9) или QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность evqlvesggglvqpggslrlscaasgfnikdtyihwvrqapgkglewvariyptngytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggdgfyamdywgqgtlvtvss (SEQ ID NO:72), Evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:73), evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:74), evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:75), или evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:76), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77) или diqmtqspsslsasvgdrvtitcrasqdvqtavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:78). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность evqlvesggglvqpggslrlscaasgfnikdtyihwvrqapgkglewvariyptngytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggdgfyamdywgqgtlvtvss (SEQ ID NO:72), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность Evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:73), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:75), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:74), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:76), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления домен VH3 содержит аминокислотную последовательность evqlvesggglvqpggslrlscaasgfnikdtyihwvrqapgkglewvariyptngytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggdgfyamdywgqgtlvtvss (SEQ ID NO:72), и/или домен VL3 содержит аминокислотную последовательность diqmtqspsslsasvgdrvtitcrasqdvqtavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:78).
[0020] В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:100 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:100; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:101 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:101; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:102 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:102; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:103 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:103. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:104 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:104; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:105 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:105; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:106 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:106; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:107 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:107. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:112 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:112; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:113 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:113; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:114 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:114; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:115 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:115. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:116 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:116; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:117 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:117; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:118 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:118; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:119 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:119. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:120 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:120; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:121 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:121; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:122 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:122; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:123 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:123. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:124 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:124; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:125 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:125; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:126 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:126; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:127 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:127. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:128 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:128; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:129 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:129; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:130 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:130; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:131 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:131. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:132 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:132; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:133 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:133; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:134 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:134; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:135 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:135. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:136 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:136; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:137 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:137; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:138 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:138; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:139 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:139. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:140 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:140; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:141 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:141; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:142 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:142; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:143 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:143. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:144 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:144; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:145 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:145; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:146 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:146; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:147 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:147. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:148 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:148; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:149 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:149; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:150 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:150; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:151 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:151. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:152 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:152; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:153 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:153; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:154 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:154; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:155 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:155. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:286 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:286; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:287 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:287; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:288 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:288; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:289 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:289. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:290 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:290; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:291 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:291; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:292 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:292; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:293 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:293. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:294 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:294; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:295 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:295; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:296 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:296; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:297 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:297. В некоторых вариантах осуществления первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:298 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:298; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:299 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:299; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:300 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:300; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:301 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:301.
[0021] В некоторых вариантах осуществления, которые могут представлять собой объединение любых других вариантов осуществления, описанных в данном документе, по меньшей мере один из L1, L2, L3 или L4 независимо имеет длину, составляющую 0 аминокислот. В некоторых вариантах осуществления каждый из L1, L2, L3 и L4 независимо имеет длину, составляющую ноль аминокислот, или содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO:69), GGGGSGGGGSGGGGS (SEQ ID NO: 70), S, RT, TKGPS (SEQ ID NO: 68), GQPKAAP (SEQ ID NO: 67) и GGSGSSGSGG (SEQ ID NO: 71). В некоторых вариантах осуществления каждый из L1, L2, L3 и L4 независимо содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO:69), GGGGSGGGGSGGGGS (SEQ ID NO:70), S, RT, TKGPS (SEQ ID NO:68), GQPKAAP (SEQ ID NO: 67) и GGSGSSGSGG (SEQ ID NO:71). В некоторых вариантах осуществления L1 содержит последовательность GQPKAAP (SEQ ID NO: 67), L2 содержит последовательность TKGPS (SEQ ID NO:68), L3 содержит последовательность S, и L4 содержит последовательность RT. В некоторых вариантах осуществления по меньшей мере один из L1, L2, L3 или L4 содержит последовательность DKTHT (SEQ ID NO:66). В некоторых вариантах осуществления L1, L2, L3 и L4 содержат последовательность DKTHT (SEQ ID NO:66).
[0022] В некоторых вариантах осуществления, которые могут представлять собой объединение любых других вариантов осуществления, описанных в данном документе, домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-IgG4-CH2-CH3 человека, и при этом каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A и L235A. В некоторых вариантах осуществления домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-IgG4-CH2-CH3 человека, и при этом каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 233-236 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой E233P, F234V, L235A и делецию в положении 236 В некоторых вариантах осуществления домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-IgG4-CH2-CH3 человека, и при этом каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 228 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P и R409K. В некоторых вариантах осуществления домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-IgG1-CH2-CH3 человека, и при этом каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 234, 235 и 329 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A, L235A и P329A. В некоторых вариантах осуществления домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-IgG1-CH2-CH3 человека, и при этом каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 298, 299 и 300 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S298N, T299A и Y300S. В некоторых вариантах осуществления домен шарнир-CH2-CH3 второй полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V; и где домен шарнир-CH2-CH3 третьей полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W. В некоторых вариантах осуществления домен шарнир-CH2-CH3 второй полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W; и где домен шарнир-CH2-CH3 третьей полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V.
[0023] В некоторых вариантах осуществления в данном документе предусмотрены выделенные молекулы нуклеиновой кислоты, содержащие нуклеотидную последовательность, кодирующую связывающий белок по любому из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления в данном документе предусмотрены векторы экспрессии, содержащие молекулу нуклеиновой кислоты по любому из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления в данном документе предусмотрены выделенные клетки-хозяева, содержащие молекулу нуклеиновой кислоты по любому из вышеуказанных вариантов осуществления или вектор экспрессии по любому из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления клетка-хозяин представляет собой клетку млекопитающего или насекомого.
[0024] В некоторых вариантах осуществления в данном документе предусмотрены фармацевтические композиции, содержащие связывающий белок по любому из вышеуказанных вариантов осуществления и фармацевтически приемлемый носитель.
[0025] В некоторых вариантах осуществления в данном документе предусмотрены способы предупреждения и/или лечения рака у пациента, включающие введение пациенту терапевтически эффективного количества по меньшей мере одного связывающего белка или фармацевтической композиции по любому из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления в данном документе предусмотрены связывающий белок или фармацевтическая композиция в соответствии с любым из указанных выше вариантов осуществления для применения в способе предупреждения и/или лечения рака у пациента, где способ включает введение пациенту терапевтически эффективного количества связывающего белка или фармацевтической композиции. В некоторых вариантах осуществления в данном документе предусмотрены связывающий белок или фармацевтическая композиция в соответствии с любым из указанных выше вариантов осуществления для применения в производстве лекарственного препарата для предупреждения и/или лечения рака у пациента.
[0026] В некоторых вариантах осуществления по меньшей мере один связывающий белок вводят совместно с химиотерапевтическим средством. В некоторых вариантах осуществления пациент представляет собой человека.
[0027] В некоторых вариантах осуществления третий антигенсвязывающий участок связывает полипептид CD38 человека, и при этом раковые клетки индивидуума или пациента экспрессируют CD38. В некоторых вариантах осуществления рак представляет собой множественную миелому. В некоторых вариантах осуществления рак представляет собой острый миелоидный лейкоз (AML), острый лимфобластный лейкоз (ALL), хронический лимфоцитарный лейкоз (CLL) или B-клеточную лимфому. В некоторых вариантах осуществления перед введением связывающего белка пациента подвергали лечению даратумумабом без периода вымывания.
[0028] В некоторых вариантах осуществления третий антигенсвязывающий участок связывает полипептид HER2 человека, и при этом раковые клетки индивидуума или пациента экспрессируют HER2. В некоторых вариантах осуществления рак представляет собой рак молочной железы, рак толстой кишки, рак желудка или немелкоклеточный рак легкого (NSCLC).
[0029] В некоторых вариантах осуществления в данном документе представлен способ размножения специфических в отношении вируса Т-клеток памяти, предусматривающий приведение специфической в отношении вируса Т-клетки памяти в контакт со связывающим белком, где связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65), и где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид CD38.
[0030] В некоторых вариантах осуществления в данном документе предусмотрен связывающий белок, который содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65), и где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид CD38, для применения при размножении специфических в отношении вируса Т-клеток памяти.
[0031] В некоторых вариантах осуществления специфическую в отношении вируса Т-клетку памяти приводят в контакт со связывающим белком in vitro или ex vivo. В некоторых вариантах осуществления приведение специфической в отношении вируса Т-клетки памяти в контакт со связывающим белком вызывает активацию и/или пролиферацию специфических в отношении вируса Т-клеток памяти.
[0032] В некоторых вариантах осуществления в данном документе представлен способ размножения Т-клеток, предусматривающий приведение Т-клетки в контакт со связывающим белком in vitro или ex vivo, где связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65), и где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид CD38.
[0033] В некоторых вариантах осуществления в данном документе предусмотрен связывающий белок, который содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65), и где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид CD38, для применения в способе размножения T-клеток.
[0034] В некоторых вариантах осуществления Т-клетка представляет собой Т-клетку памяти или эффекторную Т-клетку. В некоторых вариантах осуществления Т-клетка экспрессирует химерный антигенный рецептор (CAR) на своей клеточной поверхности или содержит полинуклеотид, кодирующий CAR.
[0035] В некоторых вариантах осуществления в данном документе предусмотрен способ лечения хронической вирусной инфекции, включающий введение индивиду или пациенту, нуждающемуся в этом, эффективного количества связывающего белка, где связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I];
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65), и где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид CD38.
[0036] В некоторых вариантах осуществления в данном документе предусмотрен связывающий белок, который содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь, и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65), и где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид CD38, для применения в способе лечения хронической вирусной инфекции, где указанный способ включает введение индивиду или пациенту, нуждающемуся в этом, эффективного количества связывающего белка.
[0037] В некоторых вариантах осуществления индивидуум или пациент представляет собой человека. В некоторых вариантах осуществления связывающий белок вводят индивидууму или пациенту в виде фармацевтического состава, содержащего связывающий белок и фармацевтически приемлемый носитель. В некоторых вариантах осуществления введение связывающего белка приводит к активации и/или пролиферации специфических в отношении вируса Т-клеток памяти у индивидуума или пациента.
[0038] В некоторых вариантах осуществления, которые могут представлять собой объединение любых других вариантов осуществления, описываемых в данном документе, Т-клетки памяти представляют собой CD8+ или CD4+ Т-клетки памяти. В некоторых вариантах осуществления Т-клетки памяти представляют собой центральные Т-клетки памяти (TCM) или эффекторные Т-клетки памяти (TEM).
[0039] В некоторых вариантах осуществления, которые могут представлять собой объединение любых других вариантов осуществления, описываемых в данном документе, вирус представляет собой вирус иммунодефицита человека (HIV), вирус гриппа, цитомегаловирус (CMV), вирус гепатита В (HBV), папилломавирус человека (HPV), вирус Эпштейна-Барр (EBV), спумавирус человека (HFV), вирус простого герпеса 1 (HSV-1) или вирус простого герпеса 1 (HSV-2).
[0040] В некоторых вариантах осуществления, которые могут представлять собой объединение любых других вариантов осуществления, описываемых в данном документе, полипептид CD28 представляет собой полипептид CD28 человека, где полипептид CD3 представляет собой полипептид CD3 человека, и где полипептид CD38 представляет собой полипептид CD38 человека.
[0041] В некоторых вариантах осуществления в данном документе предусмотрена векторная система, содержащая один или несколько векторов, кодирующих первую, вторую, третью и четвертую полипептидные цепи связывающего белка по любому из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления векторная система содержит первый вектор, кодирующий первую полипептидную цепь связывающего белка, второй вектор, кодирующий вторую полипептидную цепь связывающего белка, третий вектор, кодирующий третью полипептидную цепь связывающего белка, и четвертый вектор, кодирующий четвертую полипептидную цепь связывающего белка.
[0042] В некоторых вариантах осуществления в данном документе предусмотрены наборы, содержащие одну, две, три или четыре полипептидные цепи связывающего белка в соответствии с любым из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления наборы дополнительно содержат инструкции по применению полипептидной цепи или связывающего белка в соответствии с любым из способов или вариантов применения, описанных в данном документе, например выше.
[0043] В некоторых вариантах осуществления в данном документе предусмотрены наборы, содержащие один, два, три или четыре полинуклеотида в соответствии с любым из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления в данном документе предусмотрены наборы полинуклеотидов, содержащие один, два, три или четыре полинуклеотида из набора полинуклеотидов, содержащего (a) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:189, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:190, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:191, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:192; (b) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:193, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:194, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:195, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:196; (c) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:197, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:198, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:199, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:200; (d) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:201, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:202, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:203, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:204; (e) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:205, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:206, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:207, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:208; (f) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:209, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:210, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:211, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:212; (g) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:213, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:214, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:215, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:216; (h) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:217, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:218, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:219, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:220; (i) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:221, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:222, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:223, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:224; (j) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:225, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:226, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:227, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:228; (k) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:229, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:230, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:231, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:232; (l) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:233, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:234, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:235, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:236; (m) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:237, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:238, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:239, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:240; (n) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:241, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:242, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:243, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:244; (o) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:245, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:246, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:247, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:248; (p) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:249, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:250, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:251, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:252; (q) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:253, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:254, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:255, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:256; (r) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:257, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:258, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:259, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:260; (s) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:261, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:262, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:263, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:264; (t) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:265, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:266, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:267, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:268; (u) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:269, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:270, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:271, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:272; или (v) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:273, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:274, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:275, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:276.
[0044] Следует понимать, что один, несколько или все свойства из различных вариантов осуществления, описанных в данном документе, можно комбинировать с получением других вариантов осуществления настоящего изобретения. Эти и другие аспекты настоящего изобретения будут очевидны специалисту в данной области. Эти и другие варианты осуществления настоящего изобретения дополнительно описаны с помощью нижеследующего подробного описания.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0045] На фиг. 1A представлено схематическое изображение триспецифического связывающего белка, содержащего четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, которые связывают три белка-мишени: CD28, CD3 и HER2. Первая пара полипептидов содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией (VH1-VH2 и VL2-VL1), образующих два антигенсвязывающих участка, которые распознают CD3 и CD28, а вторая пара полипептидов содержит один вариабельный домен (VH3 и VL3), образующий один антигенсвязывающий участок, который распознает HER2. В триспецифическом связывающем белке, показанном на фиг. 1A, используют константную область с мутацией по типу "выступы-во-впадины", где выступ находится на второй паре полипептидов с одним вариабельным доменом.
[0046] На фиг. 1B представлена кратность изменения (по сравнению с исходными) значений аффинности связывания вариантов триспецифических антител к CD28/CD3/HER2, в которых используют указанные HER2-, CD3- и CD28-связывающие домены. Мутации 3233QQ - QEQ (сверху вниз) относятся к мутациям, внесенным в остатки 32-35 домена VL CD3-связывающего участка (обозначены *); остальные мутации внесли в домен VH или VL HER2-связывающего участка трастузумаба (обозначены #; нумерация согласно Кабат). В случае мутаций в HER2-связывающем участке мутацию 30Q внесли в домен VL, а остальные мутации внесли в домен VH. Значения аффинности связывания измеряли с помощью ELISA, и величины представлены относительно исходного триспецифического антитела.
[0047] На фиг. 1C представлены кривые связывания указанных триспецифических антител, связывающихся с HER2 человека, CD28 человека и CD3, определенные с помощью ELISA.
[0048] На фиг. 1D представлен предполагаемый механизм действия при активации Т-клеток, опосредованной триспецифическим антителом к HER2/CD28xCD3, и уничтожении раковых клеток HER2+.
[0049] На фиг. 2A представлено схематическое изображение триспецифического связывающего белка, содержащего четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, которые связывают три белка-мишени: CD28, CD3 и CD38. Первая пара полипептидов содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией (VH1-VH2 и VL2-VL1), образующих два антигенсвязывающих участка, которые распознают CD3 и CD28, а вторая пара полипептидов содержит один вариабельный домен (VH3 и VL3), образующий один антигенсвязывающий участок, который распознает CD38. В триспецифическом связывающем белке, показанном на фиг. 2A, используют константную область IgG4 с мутацией по типу "выступы-во-впадины", где выступ находится на второй паре полипептидов с одним вариабельным доменом.
[0050] На фиг. 2B-2E продемонстрированы измеренные с помощью ELISA значения аффинности связывания триспецифических антител IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными CD38-связывающими доменами для антигенов-мишеней CD38 человека (фиг. 2B), CD38 макака-крабоеда (фиг. 2C), CD3 человека (фиг. 2D) и CD28 человека (фиг. 2E).
[0051] На фиг. 3 продемонстрированы конкурентные SPR-анализы связывания даратумумаба и моноспецифических антител к CD38, содержащих указанные CD38-связывающие домены, с CD38. Если антитело распознавало эпитоп на CD38, который отличался от эпитопа для даратумумаба, то введение антитела приводило к усилению сигнала SPR. Если антитело распознавало эпитоп, перекрывающийся с распознаваемым даратумумабом эпитопом, введение антитела не усиливало сигнал SPR.
[0052] На фиг. 4A-4B продемонстрирована in vitro активность триспецифических антител IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными CD38-связывающими доменами в отношении уничтожения клеток множественной миеломы человека NCI-H929 (CD38+/CD28+). Анализы проводили в присутствии 5 нМ антитела изотипического контроля (фиг. 4A) или даратумумаба (фиг. 4B). В присутствии даратумумаба триспецифические антитела продолжали проявлять активность в отношении уничтожения клеток.
[0053] На фиг. 5A-5B продемонстрирована in vitro активность триспецифических антител IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными CD38-связывающими доменами в отношении уничтожения клеток лимфомы человека OCI-Ly19 (CD38+/CD28-). Анализы проводили в присутствии 5 нМ антитела изотипического контроля (фиг. 5A) или даратумумаба (фиг. 5B). Даратумумаб вызывал снижение активности триспецифических антител к CD38/CD28xCD3 в отношении уничтожения клеток.
[0054] На фиг. 6A-6J продемонстрирована характеристика размножения in vitro субпопуляции Т-клеток в PBMC, собранных от инфицированного CMV донора D в ответ на триспецифические антитела IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными альтернативными CD38-связывающими доменами. Триспецифическое антитело, лишенное CD38-связывающего домена CD38VH1, использовали в качестве отрицательного контроля (IgG4 FALA ΔCD38VH1/ΔCD28sup x ΔCD3mid). Популяции Т-клеток измеряли в указанные моменты времени (D3 относится к дню 3; D7 относится к дню 7). Указанные триспецифические антитела тестировали в указанных концентрациях, составляющих 0,2 нМ и 1 нМ. Проточную цитометрию использовали для количественного определения специфических в отношении CMV Т-клеток CD8+ (фиг. 6A-6B), специфических в отношении CMV клеток Tcm CD8+ (фиг. 6C-6D) и специфических в отношении CMV клеток Tem CD8+ (фиг. 6E-6F). Кроме того, процентное содержание специфических в отношении CMV клеток Tcm CD8+ (фиг. 6G-6H) и клеток Tem CD8+ (фиг. 6I-6J) количественно определяли в указанные моменты времени. Все протестированные триспецифические антитела способствовали пролиферации специфических в отношении CMV Т-клеток памяти CD8+ с различной активностью и кинетикой в зависимости от дозы.
[0055] На фиг. 7A-7J продемонстрирована характеристика размножения in vitro субпопуляции Т-клеток в PBMC, собранных от инфицированного CMV донора E в ответ на триспецифические антитела IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными CD38-связывающими доменами. Триспецифическое антитело, лишенное CD38-связывающего домена CD38VH1, использовали в качестве отрицательного контроля (IgG4 FALA ΔCD38VH1/ΔCD28sup x ΔCD3mid). Антитела, представленные в пояснении условных обозначений сверху вниз, представлены на графиках слева направо. Популяции Т-клеток измеряли в указанные моменты времени (D3 относится к дню 3; D7 относится к дню 7). Указанные триспецифические антитела тестировали в указанных концентрациях, составляющих 0,2 нМ, 1 нМ и 2 нМ. Проточную цитометрию использовали для количественного определения специфических в отношении CMV Т-клеток CD8+ (фиг. 7A-7B), специфических в отношении CMV клеток Tcm CD8+ (фиг. 7C-7D) и специфических в отношении CMV клеток Tem CD8+ (фиг. 7E-7F). Кроме того, процентное содержание специфических в отношении CMV клеток Tcm CD8+ (фиг. 7G-7H) и клеток Tem CD8+ (фиг. 7I-7J) количественно определяли в указанные моменты времени. Все протестированные триспецифические антитела способствовали пролиферации специфических в отношении CMV Т-клеток памяти CD8+ с различной активностью и кинетикой в зависимости от дозы.
[0056] На фиг. 8A-8J продемонстрирована характеристика размножения in vitro субпопуляции Т-клеток в PBMC, собранных от инфицированного EBV донора C в ответ на триспецифические антитела IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными альтернативными CD38-связывающими доменами. Триспецифическое антитело, лишенное CD38-связывающего домена CD38VH1, использовали в качестве отрицательного контроля (IgG4 FALA ΔCD38VH1/ΔCD28sup x ΔCD3mid). Популяции Т-клеток измеряли в указанные моменты времени (D3 относится к дню 3; D7 относится к дню 7). Указанные триспецифические антитела тестировали в указанных концентрациях, составляющих 0,2 нМ и 1 нМ. Проточную цитометрию использовали для количественного определения специфических в отношении EBV Т-клеток CD8+ (фиг. 8A-8B), специфических в отношении CMV клеток Tcm CD8+ (фиг. 8C-8D) и специфических в отношении CMV клеток Tem CD8+ (фиг. 8E-8F). Кроме того, процентное содержание специфических в отношении EBV клеток Tcm CD8+ (фиг. 8G-8H) и клеток Tem CD8+ (фиг. 8I-8J) количественно определяли в указанные моменты времени. Все протестированные триспецифические антитела способствовали пролиферации специфических в отношении CMV Т-клеток памяти CD8+ с различной активностью и кинетикой в зависимости от дозы.
[0057] На фиг. 9A-12 продемонстрирована характеристика размножения in vitro субпопуляции Т-клеток в PBMC, собранных от инфицированного EBV донора D в ответ на триспецифические антитела IgG4 FALA CD38/CD28sup x CD3mid_ENLQ DKTHT с указанными альтернативными CD38-связывающими доменами. Триспецифическое антитело, лишенное CD38-связывающего домена CD38VH1, использовали в качестве отрицательного контроля (IgG4 FALA ΔCD38VH1/ΔCD28sup x ΔCD3mid). Популяции Т-клеток измеряли в указанные моменты времени (D3 относится к дню 3; D7 относится к дню 7). Указанные триспецифические антитела тестировали в указанных концентрациях, составляющих 0,2 нМ и 1 нМ. Проточную цитометрию использовали для количественного определения специфических в отношении EBV Т-клеток CD8+ (фиг. 9A-9B), специфических в отношении EBV клеток Tcm CD8+ (фиг. 9C-9D) и специфических в отношении EBV клеток Tem CD8+ (фиг. 9E-9F). Кроме того, процентное содержание специфических в отношении EBV клеток Tcm CD8+ (фиг. 9G-10) и клеток Tem CD8+ (фиг. 11-12) количественно определяли в указанные моменты времени. Все протестированные триспецифические антитела способствовали пролиферации специфических в отношении EBV Т-клеток памяти CD8+ с различной активностью и кинетикой в зависимости от дозы.
[0058] На фиг. 13A-13D продемонстрировано изменение с течением времени (дни) объема опухоли (фиг. 13A) и массы тела (фиг. 13B) у мышей NSG, несущих опухоль ZR-75-1, которым вводили размноженные in vitro T-клетки CD3+ человека. Группы из 10 мышей обрабатывали с помощью либо несущей среды, либо триспецифического антитела Her2/CD28 x CD3 в указанных дозах. Стрелки указывают дни введения. Объем опухоли представлен как среднее значение ± SEM, мм3. Изменение массы тела представлено как изменение в %, среднее значение ± SEM. По оси X показаны дни после имплантации клеток ZR-75-1. Объем опухоли (мм3) с течением времени у отдельных мышей в каждой группе обработки представлен на фиг. 13C. Масса опухоли (мг) в каждой группы обработки представлена на фиг. 13D. **=p<0,001; ***=p<0,0003 (двухфакторный дисперсионный анализ, контроль в сравнении с 100 и 10 мкг/кг).
[0059] На фиг. 14A-14C продемонстрировано влияние обработки с использованием триспецифических антител к Her2/CD28 x CD3 на Т-клетки цельной крови. На фиг. 14A продемонстрирован анализ клеток hCD45+, CD8+, CD4+ и mCD45+ с помощью проточной цитометрии. На фиг. 14B продемонстрировано влияние обработки с использованием контрольных или триспецифических антител к Her2/CD28 x CD3 (в указанных дозах) на количество клеток hCD45+, CD8+, CD4+ и mCD45+. На фиг. 14C продемонстрировано влияние обработки с использованием контрольных или триспецифических антител к Her2/CD28 x CD3 (в указанных дозах) на соотношения клеток человека (CD4+/CD45+ и CD8+/CD45+). Для каждого параметра оси x, представленного на фиг. 14B и 14C, условия являются следующими (слева направо): контроль, триспецифическое антитело в дозе 100 мкг/кг, триспецифическое антитело в дозе 10 мкг/кг, триспецифическое антитело в дозе 1 мкг/кг и триспецифическое антитело в дозе 0,1 мкг/кг. Процентные показатели, представленные на фиг. 14B и 14C, основаны на сравнении контрольного образца с образцом, содержащим 100 мкг/кг.
[0060] На фиг. 15A-15C продемонстрировано влияние обработки с использованием триспецифических антител к Her2/CD28 x CD3 на инфильтрирующие опухоль лимфоциты (TIL), которое исследовании с помощью иммуногистохимии (IHC). Стрелки указывают на инфильтрирующие опухоль T-клетки, идентифицированные в опухолях молочной железы ZR-75-1. Верхние изображения представлены в 1-кратном увеличении; нижние изображения представлены в 20-кратном увеличении. На обоих наборах изображений окрашивание CD45 человека, CD4 человека и CD8 человека показано слева направо. Показаны опухоли мышей, обработанных с использованием контрольной несущей среды (фиг. 15A), триспецифического антитела в дозе 100 мкг/кг (фиг. 15B) или триспецифического антитела в дозе 0,1 мкг/кг (фиг. 15C).
[0061] На фиг. 16A-16C продемонстрирована количественная оценка влияния обработки с использованием триспецифических антител к Her2/CD28 x CD3 на TIL, произведенная с помощью IHC. Каждая точка представляет одну опухоль от отдельной мыши; прямоугольники представляют средние групповые значения; и планки погрешностей указывают стандартное отклонение. *=p<0,05 по сравнению с контрольной группой, получавшей несущую среду (ANOVA). Продемонстрированы количества клеток CD45+ (фиг. 16A), CD4+ (фиг. 16B) или CD8+ (фиг. 16C). На фиг. 16C показано применение способа количественной оценки площади $ для клеток CD8+ вместо алгоритма подсчета клеток в связи с чрезмерным неспецифическим сигналом в IHC микропрепарате с CD8.
[0062] На фиг. 17A-17F продемонстрирован in vitro клеточный лизис клеток-мишеней рака молочной железы HER2+ в присутствии Т-клеток CD8+ человека с помощью триспецифического антитела к Her2/CD28 x CD3, содержащего антигенсвязывающий домен дикого типа трастузумаба и антигенсвязывающий домен для CD3 без мутаций 32/35 QQ в домене VL ("ctl"), в сравнении с триспецифическими антителами к Her2/CD28 x CD3, содержащими мутации в HER2-связывающем плече и домене VL CD3-связывающего плеча (нумерация, показанная в таблице 1). Показаны значения активности касательно уничтожения клеток в отношении линий клеток с различной экспрессией HER2: HCC1954 с высокой экспрессией HER2 (фиг. 17A), BT20 со средней экспрессией HER2 (фиг. 17C) и MDA-MD-231 с низкой экспрессией HER2 (фиг. 17E). Представлены графики, на которых показано уничтожение клеток в зависимости от концентрации антител против клеток-мишеней HCC1954 (фиг. 17B), BT20 (фиг. 17D) и MDA-MD-231 (фиг. 17F), с сопоставлением связывающего белка № 2 с ctl или связывающего белка № 1 и № 5 с ctl.
[0063] На фиг. 18A и 18B обобщенно приведены средние значения EC50 (пМ) по уничтожению клеток in vitro с помощью исследуемых или контрольных триспецифических антител к Her2/CD28 x CD3 в отношении указанных линий раковых клеток молочной железы (фиг. 18A) или рака желудка (фиг. 18B). Аминокислотные последовательности указанных триспецифических антител представлены в таблице 1.
ПОДРОБНОЕ ОПИСАНИЕ
[0064] В настоящем изобретении предусмотрены триспецифические и/или тривалентные связывающие белки, содержащие четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, которые специфически связываются с одним или несколькими белками-мишенями, где первая пара полипептидов, образующих связывающий белок, содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией.
Общие определения
[0065] Используемые в соответствии с настоящим изобретением следующие термины, если не указано иное, следует понимать как имеющие следующие значения. Если контекстом не требуется иное, то термины в единственном числе будут включать форму множественного числа, и термины во множественном числе будут включать форму единственного числа.
[0066] Понятно, что аспекты и варианты осуществления настоящего изобретения, описанные в данном документе, включают аспекты и варианты осуществления "содержащие", "состоящие из" и/или "фактически состоящие из".
[0067] Используемый в данном документе термин "полинуклеотид" относится к полимерам в виде однонитевой или двунитевой нуклеиновой кислоты, имеющих длину по меньшей мере 10 нуклеотидов. В определенных вариантах осуществления нуклеотиды, составляющие полинуклеотид, могут представлять собой рибонуклеотиды или дезоксирибонуклеотиды, или модифицированную форму любого типа нуклеотида. Такие модификации включают модификации оснований, как, например, бромуридин, модификации рибозы, как, например, арабинозид и 2',3'-дидезоксирибоза, и модификации межнуклеотидных связей, как, например, фосфотиоат, фосфодитиоат, фосфоселеноат, фосфодиселеноат, фосфоанилотиоат, фосфоаниладат и фосфоамидат. Термин "полинуклеотид" конкретно подразумевает однонитевые и двунитевые формы ДНК.
[0068] "Выделенный полинуклеотид" представляет собой полинуклеотид, имеющий геномное, cDNA или синтетическое происхождение или характеризующийся некоторой их комбинацией, который: (1) не ассоциирован со всем полинуклеотидом, в котором выделенный полинуклеотид встречается в природе, или его частью, (2) связан с полинуклеотидом, с которым он не связан в природе, или (3) не встречается в природе в виде части более крупной последовательности.
[0069] "Выделенный полипептид" представляет собой такой полипептид, который: (1) не содержит по меньшей мере некоторых других полипептидов, с которыми он обычно встречается, (2) фактически не содержит других полипептидов из одного и того же источника, например, из одного и того же вида, (3) экспрессируется клеткой из другого вида, (4) был отделен по меньшей мере от приблизительно 50 процентов полинуклеотидов, липидов, углеводов или других веществ, с которыми он связан в природе, (5) не связан (посредством ковалентного или нековалентного взаимодействия) с частями полипептида, с которыми "выделенный полипептид" связан в природе, (6) функционально связан (посредством ковалентного или нековалентного взаимодействия) с полипептидом, с которым он не связан в природе, или (7) не встречается в природе. Такой выделенный полипептид может кодироваться геномной ДНК, cDNA, mRNA или другой РНК синтетического происхождения или любой их комбинацией. Предпочтительно выделенный полипептид по сути не содержит полипептидов или других контаминантов, которые встречаются в его природном окружении, которые бы вызывали затруднения при его применении (терапевтическом, диагностическом, профилактическом, исследовательском или ином).
[0070] Встречающиеся в природе антитела обычно представляют собой тетрамер. Каждый такой тетрамер обычно состоит из двух идентичных пар полипептидных цепей, при этом каждая пара имеет одну полноразмерную "легкую" цепь (обычно имеющую молекулярную массу приблизительно 25 кДа) и одну полноразмерную "тяжелую" цепь (обычно имеющую молекулярную массу приблизительно 50-70 кДа). Используемые в данном документе термины "тяжелая цепь" и "легкая цепь" обозначают любой полипептид иммуноглобулина, содержащий последовательность вариабельного домена, достаточную для придания специфичности в отношении антигена-мишени. Аминоконцевая часть каждой легкой и тяжелой цепей обычно содержит вариабельный домен из приблизительно 100-110 аминокислот или больше, который обычно отвечает за распознавание антигена. Карбоксиконцевая часть каждой цепи обычно определяет константный домен, ответственный за эффекторную функцию. Таким образом, во встречающемся в природе антителе полноразмерный полипептид, представляющий собой тяжелую цепь иммуноглобулина, содержит вариабельный домен (VH) и три константных домена (CH1, CH2 и CH3), где домен VH находится на аминоконце полипептида, а домен CH3 находится на карбоксильном конце, и полноразмерный полипептид, представляющий собой легкую цепь иммуноглобулина, содержит вариабельный домен (VL) и константный домен (CL), где домен VL находится на аминоконце полипептида, а домен CL находится на карбоксильном конце.
[0071] Легкие цепи человека обычно классифицируют как легкие каппа- и лямбда-цепи, а тяжелые цепи человека обычно классифицируют как мю, дельта, гамма, альфа или эпсилон, и они определяют изотип антитела IgM, IgD, IgG, IgA и IgE соответственно. IgG имеет несколько подклассов, в том числе без ограничения IgG1, IgG2, IgG3 и IgG4. IgM имеет подклассы, в том числе без ограничения IgM1 и IgM2. IgA аналогичным образом подразделяют на подклассы, в том числе без ограничения IgA1 и IgA2. В полноразмерных легких и тяжелых цепях вариабельные и константные домены обычно соединены с помощью "J"-области из приблизительно 12 или больше аминокислот, при этом тяжелая цепь также содержит "D"-область из приблизительно 10 или больше аминокислот. См., например, Fundamental Immunology (Paul, W., ed., Raven Press, 2nd ed., 1989), которая включена посредством ссылки во всей своей полноте для всех целей. Вариабельные области каждой пары легкая/тяжелая цепь обычно образуют антигенсвязывающий участок. Вариабельные домены встречающихся в природе антител обычно характеризуются одинаковой общей структурой относительно консервативных каркасных областей (FR), соединенных с помощью трех гипервариабельных областей, также называемых определяющими комплементарность областями или CDR. CDR из двух цепей каждой пары обычно выровнены с помощью каркасных областей, что может обеспечивать возможность связывания со специфическим эпитопом. От аминоконца к карбоксильному концу вариабельные домены как легкой, так и тяжелой цепей обычно содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
[0072] Термин "совокупность CDR" обозначает группу из трех CDR, которые содержатся в одной вариабельной области, способной связывать антиген. Точные границы этих CDR определяли по-разному в соответствии с различными системами. Система, описанная Kabat (Kabat et al., Sequences of Proteins of Immunological Interest (Национальные институты здравоохранения, Бетесда, Мэриленд. (1987) и (1991)), не только предусматривает однозначную систему нумерации остатков, применимую к любой вариабельной области антитела, но также предусматривает точные границы остатков, определяющие три CDR. Эти CDR могут называться CDR в соответствии с Kabat. Chothia и соавторы (Chothia and Lesk, 1987, J. Mol. Biol. 196: 901-17; Chothia et al., 1989, Nature 342: 877-83) обнаружили, что определенные субфрагменты в пределах CDR в соответствии с Kabat принимают почти идентичные конформации пептидного каркаса, несмотря на наличие большого различия на уровне аминокислотной последовательности. Эти субфрагменты были обозначены как L1, L2 и L3 или H1, H2 и H3, где "L" и "H" обозначают области легкой цепи и тяжелой цепи соответственно. Эти области могут называться CDR в соответствии с Chothia, границы которых совпадают с CDR в соответствии с Kabat. Другие границы, определяющие CDR, которые совпадают с CDR в соответствии с Kabat, были описаны Padlan, 1995, FASEB J. 9: 133-39; MacCallum, 1996, J. Mol. Biol. 262(5): 732-45; и Lefranc, 2003, Dev. Comp. Immunol. 27: 55-77. Еще одни определения границ CDR могут не строго соответствовать одной из систем, приведенных в данном документе, но тем не менее будут совпадать с CDR в соответствии с Kabat, несмотря на то, что они могут быть укорочены или удлинены с учетом прогностических или экспериментальных выводов о том, что конкретные остатки или группы остатков или даже все CDR не влияют в значительной степени на связывание антигена. В способах, используемых в данном документе, можно использовать CDR, определенные в соответствии с любой из этих систем, несмотря на то, что в определенных вариантах осуществления применяют CDR, определенные в соответствии с Kabat или Chothia. Идентификация прогнозируемых CDR с помощью аминокислотной последовательности хорошо известна в данной области техники, например, в Martin, A.C. "Protein sequence and structure analysis of antibody variable domains" в Antibody Engineering, Vol. 2. Kontermann R., Dübel S., eds. Springer-Verlag, Berlin, p. 33-51 (2010). Аминокислотную последовательность вариабельного домена тяжелой и/или легкой цепи можно также исследовать с целью идентификации последовательностей CDR с помощью других традиционных способов, например, путем сравнения с известными аминокислотными последовательностями других вариабельных областей тяжелой и легкой цепей с определением областей гипервариабельности в последовательности. Пронумерованные последовательности можно выравнивать вручную или путем использования программы выравнивания, такой как одна из пакета программ CLUSTAL, как описано в Thompson, 1994, Nucleic Acids Res. 22: 4673-80. Молекулярные модели традиционно используют для того, чтобы правильно определить каркасные и CDR-области и таким образом скорректировать основанные на последовательности выравнивания.
[0073] Используемый в данном документе термин "Fc" обозначает молекулу, будь то в мономерной или мультимерной форме, содержащую последовательность, не являющуюся частью антигенсвязывающего фрагмента, которая получена в результате расщепления антитела или получена другими способами, и при этом она может содержать шарнирную область. Исходный иммуноглобулиновый источник нативного Fc предпочтительно происходит от человека и может представлять собой любой из иммуноглобулинов, тем не менее предпочтительными являются IgG1 и IgG2. Молекулы Fc составлены из мономерных полипептидов, которые могут быть связаны в димерные или мультимерные формы посредством ковалентной (т. е. дисульфидных связей) и нековалентной связи. Число межмолекулярных дисульфидных связей между мономерными субъединицами нативных молекул Fc варьирует от 1 до 4, в зависимости от класса (например, IgG, IgA и IgE) или подкласса (например, IgG1, IgG2, IgG3, IgA1 и IgGA2). Одним примером Fc является димер с дисульфидной связью, полученный в результате расщепления IgG папаином. Термин "нативный Fc", используемый в данном документе, является общим для мономерных, димерных и мультимерных форм.
[0074] F(ab)-фрагмент обычно содержит одну легкую цепь и домены VH и CH1 одной тяжелой цепи, где часть VH-CH1 тяжелой цепи F(ab)-фрагмента не может образовывать дисульфидную связь с другим полипептидом тяжелой цепи. Как используется в данном документе, F(ab)-фрагмент также может содержать одну легкую цепь, содержащую два вариабельных домена, разделенных аминокислотным линкером, и одну тяжелую цепь, содержащую два вариабельных домена, разделенных аминокислотным линкером, и домен CH1.
[0075] F(ab')-фрагмент обычно содержит одну легкую цепь и часть одной тяжелой цепи, которая содержит большую часть константной области (между доменами CH1 и CH2), вследствие чего межцепочечная дисульфидная связь может быть образована между двумя тяжелыми цепями с образованием молекулы F(ab')2.
[0076] Используемый в данном документе термин "связывающий белок" относится к не встречающейся в природе (или рекомбинантной, или сконструированной) молекуле, которая специфически связывается с по меньшей мере одним антигеном-мишенью. Триспецифический связывающий белок по настоящему изобретению, если не указано иное, содержит четыре полипептидные цепи, которые образуют по меньшей мере три антигенсвязывающих участка, где первая полипептидная цепь имеет структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I];
и вторая полипептидная цепь имеет структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь имеет структуру, представленную формулой:
VH3-CH1 [III];
и четвертая полипептидная цепь имеет структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина; и
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2;
L1, L2, L3 и L4 представляют собой аминокислотные линкеры;
и где полипептид формулы I и полипептид формулы II образуют перекрестную пару легкая цепь-тяжелая цепь.
[0077] "Рекомбинантная" молекула представляет собой молекулу, которая была получена, экспрессирована, создана или выделена рекомбинантными способами.
[0078] В одном варианте осуществления настоящего изобретения представлены связывающие белки, характеризующиеся биологической и иммунологической специфичностью в отношении одного-трех антигенов-мишеней. В другом варианте осуществления настоящего изобретения представлены молекулы нуклеиновой кислоты, содержащие нуклеотидные последовательности, кодирующие полипептидные цепи, которые образуют такие связывающие белки. В другом варианте осуществления настоящего изобретения представлены векторы экспрессии, содержащие молекулы нуклеиновой кислоты, содержащие нуклеотидные последовательности, кодирующие полипептидные цепи, которые образуют такие связывающие белки. В еще одном варианте осуществления настоящего изобретения представлены клетки-хозяева, которые экспрессируют такие связывающие белки (т. е. содержащие молекулы нуклеиновой кислоты или векторы, кодирующие полипептидные цепи, которые образуют такие связывающие белки).
[0079] Используемый в данном документе термин "способность к обмену" обозначает способность к взаимозамене вариабельных доменов в формате связывающего белка, при этом с сохранением укладки и наивысшей аффинности связывания. "Способность к полному обмену" относится к способности менять порядок доменов как VH1, так и VH2 и, следовательно, порядок доменов VL1 и VL2 в полипептидной цепи формулы I или полипептидной цепи формулы II (т. е. обращать порядок), при этом поддерживая полную функциональность связывающего белка, о чем свидетельствует сохранение аффинности связывания. Кроме того, следует отметить, что обозначения VH и VL относятся лишь к положению домена в конкретной белковой цепи в конечном формате. Например, VH1 и VH2 могут быть получены из доменов VL1 и VL2 исходных антител и введены в положения, соответствующие VH1 и VH2 связывающего белка. Аналогично VL1 и VL2 могут быть получены из доменов VH1 и VH2 исходных антител и помещены в положения, соответствующие VH1 и VH2 связывающего белка. Таким образом, обозначения VH и VL относятся к существующему в данный момент положению, а не к первоначальному положению в исходном антителе. Следовательно, домены VH и VL являются "способными к обмену".
[0080] Используемый в данном документе термин "антиген", или "антиген-мишень", или "целевой антиген" обозначает молекулу или часть молекулы, которая может связываться связывающим белком и дополнительно может использоваться у животного для получения антител, способных связываться с эпитопом этого антигена. Антиген-мишень может иметь один или несколько эпитопов. С учетом того, что каждый антиген-мишень распознается связывающим белком, связывающий белок способен конкурировать с интактным антителом, которое распознает антиген-мишень.
[0081] Термин "Her2" обозначает рецептор эпидермального фактора роста человека 2 типа, который является представителем семейства рецепторов эпидермального фактора роста.
[0082] Термин "CD3" означает полипептид фактора 3 кластера дифференцировки и представляет собой поверхностный белок T-клеток, который обычно является частью Т-клеточного рецепторного (TCR) комплекса.
[0083] Термин "CD28" означает полипептид 28 кластера дифференцировки и представляет собой поверхностный белок T-клеток, передающий костимулирующие сигналы для активации и выживания Т-клеток.
[0084] "CD38" означает полипептид 38 кластера дифференцировки и представляет собой гликопротеин, обнаруживаемый на поверхности многих иммунных клеток.
[0085] Термин "рекрутер T-клеток" обозначает связывающие белки, направленные на иммунную систему хозяина, более конкретно на цитотоксическую активность T-клеток, а также направленные на опухолевый белок-мишень.
[0086] Термин "моноспецифический связывающий белок" обозначает связывающий белок, который специфически связывается с одним антигеном-мишенью.
[0087] Термин "моновалентный связывающий белок" обозначает связывающий белок, который имеет один антигенсвязывающий участок.
[0088] Термин "биспецифический связывающий белок" обозначает связывающий белок, который специфически связывается с двумя различными антигенами-мишенями.
[0089] Термин "бивалентный связывающий белок" обозначает связывающий белок, который имеет два связывающих участка.
[0090] Термин "триспецифический связывающий белок" обозначает связывающий белок, который специфически связывается с тремя различными антигенами-мишенями.
[0091] Термин "тривалентный связывающий белок" обозначает связывающий белок, который имеет три связывающих участка. В конкретных вариантах осуществления тривалентный связывающий белок может связываться с одним антигеном-мишенью. В других вариантах осуществления тривалентный связывающий белок может связываться с двумя антигенами-мишенями. В других вариантах осуществления тривалентный связывающий белок может связываться с тремя антигенами-мишенями.
[0092] "Выделенный" связывающий белок представляет собой связывающий белок, который был идентифицирован и отделен и/или извлечен из компонента своего природного окружения. Компоненты-контаминанты из его природного окружения представляют собой вещества, которые бы вызывали затруднения при диагностических или терапевтических применениях связывающего белка, и могут включать ферменты, гормоны и другие растворенные вещества белковой или небелковой природы. В некоторых вариантах осуществления связывающий белок будет очищенным: (1) до более чем 95% по весу антитела, как определяется способом Лоури, и наиболее предпочтительно до более чем 99% по весу, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с применением секвенатора с вращающимся стаканом, или (3) до гомогенности с помощью SDS-PAGE в восстанавливающих или невосстанавливающих условиях с применением кумасси голубого или предпочтительно серебряного красителя. Выделенные связывающие белки включают связывающий белок in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент природного окружения связывающего белка не будет присутствовать.
[0093] Используемые в данном документе термины "в значительной степени чистый" или "в значительной степени очищенный" обозначают соединение или структуру, которая является преобладающей присутствующей структурой (т. е. в расчете на моль она более многочисленна, чем любая другая отдельная структура в композиции). В некоторых вариантах осуществления в значительной степени очищенная фракция представляет собой композицию, где структура составляет по меньшей мере приблизительно 50% (в расчете на моль) от всех присутствующих макромолекулярных структур. В других вариантах осуществления в значительной степени чистая композиция будет содержать более чем приблизительно 80%, 85%, 90%, 95% или 99% от всех макромолекулярных структур, присутствующих в композиции. В еще одних вариантах осуществления структура является очищенной до необходимой гомогенности (загрязняющие структуры не могут быть выявлены в композиции с помощью традиционных способов выявления), где композиция состоит фактически из одной макромолекулярной структуры.
[0094] Термин "эпитоп" подразумевает любую детерминанту, предпочтительно полипептидную детерминанту, способную специфически связываться с иммуноглобулином или T-клеточным рецептором. В определенных вариантах осуществления эпитопные детерминанты включают химически активные поверхностные группы молекул, такие как аминокислоты, боковые цепи сахаров, фосфорильные группы или сульфонильные группы, и в определенных вариантах осуществления могут иметь специфические характеристики трехмерной структуры и/или специфические характеристики заряда. Эпитоп представляет собой область антигена, которая связывается антителом или связывающим белком. В определенных вариантах осуществления считается, что связывающий белок специфически связывает антиген, если он предпочтительно распознает свой антиген-мишень в сложной смеси белков и/или макромолекул. В некоторых вариантах осуществления считается, что связывающий белок специфически связывает антиген, если равновесная константа диссоциации составляет ≤ 10-8 M, более предпочтительно если равновесная константа диссоциации составляет ≤ 10-9 M и наиболее предпочтительно если константа диссоциации составляет ≤ 10-10 M.
[0095] Константу диссоциации (KD) связывающего белка можно определить, например, с помощью поверхностного плазмонного резонанса. В целом в анализе поверхностного плазмонного резонанса измеряют связывающие взаимодействия между лигандом (антигеном-мишенью на биосенсорной матрице) и аналитом (связывающим белком в растворе) в реальном времени с помощью поверхностного плазмонного резонанса (SPR) с применением системы BIAcore (Pharmacia Biosensor; Пискатауэй, Нью-Джерси). Анализ поверхностного плазмонного резонанса можно также выполнять посредством иммобилизации аналита (связывающего белка на биосенсорной матрице) и представления лиганда (антигена-мишени). Используемый в данном документе термин "KD" обозначает константу диссоциации взаимодействия между конкретным связывающим белком и антигеном-мишенью.
[0096] Используемый в данном документе термин "специфически связывается" обозначает способность связывающего белка или его антигенсвязывающего фрагмента связываться с антигеном, содержащим эпитоп, с Kd, составляющей по меньшей мере приблизительно 1×10-6 M, 1×10-7 M, 1×10-8 M, 1×10-9 M, 1×10-10 M, 1×10-11 M, 1×10-12 M или больше, и/или связываться с эпитопом с аффинностью, которая в по меньшей мере два раза превышает его аффинность в отношении неспецифического антигена.
[0097] В некоторых вариантах осуществления антигенсвязывающий домен и/или связывающий белок по настоящему изобретению "перекрестно реагирует" с полипептидами CD38 человека и макака-крабоеда, например внеклеточными доменами CD38, изоформой A CD38 человека, изоформой E CD38 человека и CD38 макака-крабоеда. Связывание связывающего белка с антигеном 1(Ag1) является "перекрестно реактивным" в отношении антигена 2 (Ag2), если значения EC50 находятся в аналогичном диапазоне для обоих антигенов. В настоящей заявке связывание связывающего белка с Ag1 является перекрестно-реактивным в отношении Ag2, если соотношение аффинности к Ag2 и аффинности к Ag1 равняется 20 или менее, при этом значения аффинности измеряют с помощью одного и того же способа в случае обоих антигенов.
[0098] Используемый в данном документе термин "линкер" обозначает один или несколько аминокислотных остатков, вставленных между доменами иммуноглобулина для обеспечения подвижности, достаточной для того, чтобы домены легкой и тяжелой цепей сворачивались в иммуноглобулины с кроссоверными двумя вариабельными областями. Линкер вставляют в переходный участок между вариабельными доменами или между вариабельным и константным доменами, соответственно, на уровне последовательности. Переходный участок между доменами можно идентифицировать, поскольку приблизительный размер доменов иммуноглобулина хорошо известен. Точное местоположение переходного участка между доменами можно определить путем определения положения пептидных отрезков, которые не образуют элементов со вторичной структурой, такой как бета-слои или альфа-спирали, что показано с помощью экспериментальных данных или что можно прогнозировать с помощью методик моделирования или прогнозирования вторичной структуры. Линкеры, описанные в данном документе, обозначены как L1, который расположен на легкой цепи между C-концом домена VL2 и N-концом домена VL1; и L2, который расположен на легкой цепи между C-концом домена VL1 и N-концом домена CL. Линкеры тяжелой цепи называют как L3, который расположен между C-концом домена VH1 и N-концом домена VH2; и L4, который расположен между C-концом домена VH2 и N-концом домена CH1.
[0099] Используемый в данном документе термин "вектор" относится к любой молекуле (например, нуклеиновой кислоте, плазмиде или вирусу), используемой для переноса закодированной информации в клетку-хозяина. Термин "вектор" подразумевает молекулу нуклеиновой кислоты, которая способна транспортировать другую нуклеиновую кислоту, с которой ее связали. Одним типом вектора является "плазмида", которая обозначает кольцевую двунитевую молекулу ДНК, в которую могут быть вставлены дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, при этом в геном вируса могут быть вставлены дополнительные сегменты ДНК. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы для млекопитающих). Другие векторы (например, неэписомные векторы для млекопитающих) могут быть интегрированы в геном клетки-хозяина после введения в клетку-хозяина, и за счет этого они реплицируются вместе с геномом хозяина. Кроме того, определенные векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называются в данном документе "рекомбинантными векторами экспрессии" (или просто "векторами экспрессии"). Как правило, векторы экспрессии, полезные в технологиях рекомбинантных ДНК, часто находятся в форме плазмид. Термины "плазмида" и "вектор" могут использоваться взаимозаменяемо в данном документе, поскольку плазмида является наиболее широко используемой формой вектора. Однако подразумевается, что в настоящее изобретение включены и другие формы векторов экспрессии, такие как вирусные векторы (например, ретровирусы, аденовирусы и аденоассоциированные вирусы с дефектной репликацией), которые выполняют эквивалентные функции.
[0100] Используемая в данном документе фраза "рекомбинантная клетка-хозяин" (или "клетка-хозяин") обозначает клетку, в которую был введен рекомбинантный вектор экспрессии. Подразумевается, что рекомбинантная клетка-хозяин или клетка-хозяин обозначают не только конкретную рассматриваемую клетку, но также и потомство такой клетки. Поскольку в последующих поколениях могут возникать определенные модификации вследствие мутации либо влияний окружающей среды, такое потомство в действительности может не быть идентичным родительской клетке, однако такие клетки по-прежнему включены в объем используемого в данном документе термина "клетка-хозяин". Широкое разнообразие систем экспрессии в клетке-хозяине можно применять для экспрессии связывающих белков, в том числе системы экспрессии на основе бактерий, дрожжей, бакуловирусов и клеток млекопитающих (а также системы экспрессии на основе фагового дисплея). Примером подходящего бактериального вектора экспрессии является pUC19. Для рекомбинантной экспрессии связывающего белка клетку-хозяина трансформируют или трансфицируют одним или несколькими рекомбинантными векторами экспрессии, несущими фрагменты ДНК, кодирующие полипептидные цепи связывающего белка, вследствие чего полипептидные цепи экспрессируются в клетке-хозяине и предпочтительно секретируются в среду, в которой клетки-хозяева культивируются, при этом из данной среды можно извлечь связывающий белок.
[0101] Используемый в данном документе термин "трансформация" обозначает изменение генетических характеристик клетки, и при этом клетка была трансформирована, если ее модифицировали с тем, чтобы она содержала новую ДНК. Например, клетка является трансформированной, если она генетически модифицирована по сравнению с ее нативным состоянием. После трансформации трансформирующая ДНК может подвергаться рекомбинации с ДНК клетки путем физической интеграции в хромосому клетки, или может временно сохраняться в виде эписомального элемента без репликации, или может реплицироваться независимо как плазмида. Считается, что клетка является стабильно трансформированной, если ДНК реплицируется при делении клетки. Используемый в данном документе термин "трансфекция" обозначает захват чужеродной или экзогенной ДНК клеткой, и при этом клетка была "трансфицирована", если экзогенную ДНК ввели внутрь от клеточной мембраны. Ряд методик трансфекции хорошо известен из уровня техники. Такие методики можно применять для введения одной или нескольких экзогенных молекул ДНК в подходящие клетки-хозяева.
[0102] Используемый в данном документе термин "встречающийся в природе" и применяемый в отношении объекта обозначает тот факт, что объект может обнаруживаться в природе и не был подвергнут манипуляциям со стороны человека. Например, полинуклеотид или полипептид, присутствующий в организме (включая вирусы), который может быть выделен из природного источника и который не был преднамеренно модифицирован человеком, является встречающимся в природе. Аналогично используемый в данном документе термин "не встречающийся в природе" обозначает объект, который не обнаруживается в природе или который был структурно модифицирован или синтезирован человеком.
[0103] Как используется в данном документе, двадцать стандартных аминокислот и их аббревиатуры соответствуют общепринятой практике. Стереоизомеры (например, d-аминокислоты) двадцати стандартных аминокислот; не встречающиеся в природе аминокислоты и аналоги, такие как α-,α-двузамещенные аминокислоты, N-алкиламинокислоты, молочная кислота и другие нестандартные аминокислоты также могут быть подходящими компонентами полипептидных цепей связывающих белков. Примеры нестандартных аминокислот включают 4-гидроксипролин, γ-карбоксиглутамат, ε-N, N,N-триметиллизин, ε-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, σ-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В системе обозначений полипептидов, используемой в данном документе, левое направление представляет собой направление в сторону аминоконца, а правое направление представляет собой направление в сторону карбоксильного конца в соответствии со стандартной практикой и правилами.
[0104] Встречающиеся в природе остатки можно разделить на классы на основании общих свойств боковой цепи:
(1) гидрофобные: Met, Ala, Val, Leu, Ile, Phe, Trp, Tyr, Pro;
(2) полярные гидрофильные: Arg, Asn, Asp, Gln, Glu, His, Lys, Ser, Thr;
(3) алифатические: Ala, Gly, Ile, Leu, Val, Pro;
(4) алифатические гидрофобные: Ala, Ile, Leu, Val, Pro;
(5) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(6) кислые: Asp, Glu;
(7) основные: His, Lys, Arg;
(8) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(9) ароматические: His, Trp, Tyr, Phe; и
(10) ароматические гидрофобные: Phe, Trp, Tyr.
[0105] Консервативные аминокислотные замены могут включать обмен представителя одного из этих классов на другого представителя того же класса. Неконсервативные замены могут включать обмен представителя одного из этих классов на представителя другого класса.
[0106] Специалист в данной области техники сможет определить подходящие варианты полипептидных цепей связывающих белков с помощью общеизвестных методик. Например, специалист в данной области может идентифицировать подходящие фрагменты в полипептидной цепи, которые можно изменить без нарушения активности, путем нацеливания на области, которые, как предполагается, не являются важными с точки зрения активности. Альтернативно специалист в данной области техники может идентифицировать остатки и части молекул, которые являются консервативными среди схожих полипептидов. Кроме того, даже участки, которые могут быть важными с точки зрения биологической активности или с точки зрения структуры, могут подвергаться консервативным аминокислотным заменам без нарушения биологической активности или без неблагоприятного воздействия на структуру полипептида.
[0107] Используемый в данном документе термин "пациент" подразумевает субъектов-людей и субъектов-животных.
[0108] Используемые в данном документе термины "лечение" или "лечить" обозначают как терапевтическое лечение, так и профилактические или превентивные меры. К нуждающимся в лечении относят тех, у которых имеется нарушение, а также тех, которые предрасположены к нарушению, или тех, у которых нужно предотвратить нарушение. В конкретных вариантах осуществления связывающие белки можно применять для лечения людей с раком или людей, подверженных раку, или для уменьшения тяжести рака у субъекта-человека. Связывающие белки также можно применять для предупреждения рака у пациента-человека. В конкретных вариантах осуществления рак представляет собой множественную миелому, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, острый миелоидный лейкоз, лимфому, рак молочной железы, такой как Her2+ рак молочной железы, лимфому из B-клеток зародышевого центра или B-клеточный острый лимфобластный лейкоз. В других вариантах осуществления связывающие белки можно применять для лечения людей с воспалительными нарушениями или людей, подверженных воспалительным нарушениям, или для уменьшения тяжести воспалительных нарушений у субъекта-человека.
[0109] Используемые в данном документе термины "фармацевтическая композиция" или "терапевтическая композиция" обозначают соединение или композицию, способные индуцировать необходимый терапевтический эффект при введении пациенту должным образом.
[0110] Используемый в данном документе термин "фармацевтически приемлемый носитель" или "физиологически приемлемый носитель" обозначает одно или несколько веществ состава, подходящие для осуществления или улучшения доставки связывающего белка.
[0111] Термины "эффективное количество" и "терапевтически эффективное количество" при использовании в отношении фармацевтической композиции, содержащей один или несколько связывающих белков, обозначают количество или дозу, достаточные для получения необходимого терапевтического результата. Более конкретно, терапевтически эффективное количество представляет собой количество связывающего белка, достаточное для ингибирования, на некоторый период времени, одного или нескольких клинически определенных патологических процессов, ассоциированных с состоянием, подлежащим лечению. Эффективное количество может меняться в зависимости от конкретного связывающего белка, подлежащего применению, а также зависит от ряда факторов и состояний, связанных с пациентом, подлежащим лечению, и тяжестью нарушения. Например, если связывающий белок подлежит введению in vivo, то факторы, такие как возраст, масса тела и состояние здоровья пациента, а также кривые зависимости "доза-эффект" и данные о токсичности, полученные в доклиническом исследовании на животных, будут входить в число таковых учитываемых факторов. Определение эффективного количества или терапевтически эффективного количества указанной фармацевтической композиции находится в пределах компетенции специалистов в данной области техники.
[0112] В одном варианте осуществления настоящего изобретения представлена фармацевтическая композиция, содержащая фармацевтически приемлемый носитель и терапевтически эффективное количество связывающего белка.
Триспецифические и/или тривалентные связывающие белки
[0113] Определенные аспекты настоящего изобретения относятся к триспецифическим и/или тривалентным связывающим белкам, содержащим четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, которые специфически связываются с одним или несколькими белками-мишенями, где первая пара полипептидов, образующих связывающий белок, содержит двойные вариабельные домены, характеризующиеся перекрестной ориентацией, и где вторая пара полипептидов, образующих связывающий белок, содержит один вариабельный домен. Любые из CDR или вариабельных доменов любого из антигенсвязывающих белков, описываемых в данном документе, могут найти применение в получении триспецифического связывающего белка по настоящему изобретению.
[0114] В некоторых вариантах осуществления каждый из трех антигенсвязывающих участков связывает другую мишень (например, полипептидный антиген). В некоторых вариантах осуществления триспецифический связывающий белок содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой:
VL2-L1-VL1-L2-CL [I],
и вторая полипептидная цепь содержит структуру, представленную формулой:
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой:
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой:
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь.
[0115] Предполагается, что любому из антигенсвязывающих участков, описанных в данном документе, можно найти применение в триспецифическом связывающем белке по настоящему изобретению, например содержащем четыре полипептидные цепи, характеризующиеся структурами, описанными выше. Например, в некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий участок, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий участок. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий участок, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий участок. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий участок, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий участок, который связывает опухолевый белок-мишень. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий участок, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий участок, который связывает опухолевый белок-мишень. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий участок, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий участок, который связывает полипептид CD38. В некоторых вариантах осуществления триспецифический связывающий белок по настоящему изобретению содержит пару доменов VH1 и VL1, которые образуют первый антигенсвязывающий участок, который связывает полипептид CD28, пару доменов VH2 и VL2, которые образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и пару доменов VH3 и VL3, которые образуют третий антигенсвязывающий участок, который связывает полипептид HER2.
[0116] В некоторых вариантах осуществления связывающий белок по настоящему изобретению связывает один или несколько опухолевых белков-мишеней и один или несколько Т-клеточных белков-мишеней. В некоторых вариантах осуществления связывающий белок способен специфически связывать один опухолевый белок-мишень и два различных эпитопа на одном Т-клеточном белке-мишени. В некоторых вариантах осуществления связывающий белок способен специфически связывать один опухолевый белок-мишень и два различных Т-клеточных белка-мишени (например, CD28 и CD3). В некоторых вариантах осуществления первая и вторая полипептидные цепи связывающего белка образуют два антигенсвязывающих участка, которые специфически целенаправленно воздействуют на два Т-клеточных белка-мишени, а третья и четвертая полипептидные цепи связывающего белка образуют антигенсвязывающий участок, который специфически связывает опухолевый белок-мишень. В некоторых вариантах осуществления белок-мишень представляет собой CD38 или HER2. Дополнительные опухолевые белки-мишени представлены ниже. В некоторых вариантах осуществления один или несколько T-клеточных белков-мишеней представляют собой один или несколько из CD3 и CD28. Иллюстративные и неограничивающие примеры полипептидов, которые можно применить в любом из триспецифических связывающих белков, описанных в данном документе, представлены в таблице 1.
[0117] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:156 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:156; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:157 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:157; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:158 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:158; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:159 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:159.
[0118] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:160 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:160; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:161 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:161; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:162 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:162; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:163 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:163.
[0119] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:164 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:164; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:165 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:165; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:166 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:166; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:167 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:167.
[0120] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:168 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:168; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:169 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:169; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:170 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:170; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:171 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:171.
[0121] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:172 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:172; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:173 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:173; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:174 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:174; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:175 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:175.
[0122] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:176 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:176; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:177 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:177; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:178 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:178; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:179 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:179.
[0123] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:181 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:181; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:182 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:182; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:183 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:183; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:184 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:184.
[0124] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:185 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:185; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:186 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:186; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:187 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:187; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:188 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:188.
[0125] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:100 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:100; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:101 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:101; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:102 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:102; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:103 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:103.
[0126] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:104 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:104; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:105 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:105; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:106 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:106; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:107 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:107.
[0127] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:112 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:112; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:113 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:113; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:114 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:114; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:115 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:115.
[0128] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:116 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:116; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:117 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:117; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:118 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:118; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:119 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:119.
[0129] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:120 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:120; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:121 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:121; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:122 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:122; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:123 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:123.
[0130] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:124 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:124; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:125 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:125; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:126 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:126; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:127 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:127.
[0131] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:128 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:128; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:129 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:129; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:130 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:130; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:131 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:131.
[0132] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:132 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:132; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:133 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:133; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:134 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:134; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:135 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:135.
[0133] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:136 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:136; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:137 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:137; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:138 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:138; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:139 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:139.
[0134] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:140 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:140; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:141 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:141; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:142 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:142; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:143 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:143.
[0135] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:144 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:144; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:145 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:145; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:146 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:146; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:147 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:147.
[0136] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:148 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:148; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:149 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:149; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:150 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:150; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:151 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:151.
[0137] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:152 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:152; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:153 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:153; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:154 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:154; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:155 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:155.
[0138] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:286 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:286; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:287 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:287; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:288 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:288; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:289 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:289.
[0139] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:290 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:290; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:291 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:291; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:292 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:292; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:293 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:293.
[0140] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:294 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:294; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:295 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:295; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:296 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:296; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:297 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:297.
[0141] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:298 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:298; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:299 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:299; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:300 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:300; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO:301 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO:301.
Связывающие участки антитела к CD38
[0142] Определенные аспекты настоящего изобретения относятся к связывающим белкам, которые содержат антигенсвязывающий участок, который связывает полипептид CD38. В некоторых вариантах осуществления полипептид CD38 представляет собой полипептид CD38 человека, также известный как ADPRC1. Полипептиды CD38 человека известны в уровне техники и включают без ограничения полипептид, представленный под номером доступа в NCBI NP_001766.2, или полипептид, полученный с помощью гена с идентификационным номером в NCBI 952. В некоторых вариантах осуществления антигенсвязывающий участок связывает полипептид CD38 человека, полипептид CD38 примата, не относящегося к человеку (например, макака-крабоеда), или полипептид CD38 человека и полипептид CD38 примата, не относящегося к человеку (например, макака-крабоеда). В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий участок, который связывает полипептид CD38, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным.
[0143] В некоторых вариантах осуществления любой из CDR и/или вариабельных доменов CD38-связывающих участков, описанных ниже, можно использовать в моноспецифическом антителе.
[0144] В других вариантах осуществления любой из CDR и/или вариабельных доменов CD38-связывающих участков, описанных ниже, можно использовать в любом связывающем участке триспецифического связывающего белка, содержащего четыре полипептида, которые образуют три антигенсвязывающих участка, например описанных выше. В определенных вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD38, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, описанных выше, где домены VH3 и VL3 образуют пару и образуют третий антигенсвязывающий участок, который связывает полипептид CD38.
[0145] В данном документе описаны различные особенности иллюстративных связывающих участков и связывающих белков. Например, в некоторых вариантах осуществления CD38-связывающий участок перекрестно реагирует с CD38 человека (например, полипептидом изоформы A и/или изоформы E CD38 человека) и CD38 макака-крабоеда. В некоторых вариантах осуществления связывающий белок, содержащий CD38-связывающий участок, индуцирует апоптоз клетки CD38+. В некоторых вариантах осуществления связывающий белок, содержащий CD38-связывающий участок, осуществляет рекрутинг Т-клетки к клетке CD38+ и необязательно активирует Т-клетку (например, посредством стимуляции и/или костимуляции TCR).
[0146] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYA (SEQ ID NO:13), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGQGGT (SEQ ID NO:14), и последовательность CDR-H3, содержащую аминокислотную последовательность ARTGGLRRAYFTY (SEQ ID NO:15); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSVSSYGQGF (SEQ ID NO:16), последовательность CDR-L2, содержащую аминокислотную последовательность GAS (SEQ ID NO:17), и последовательность CDR-L3, содержащую аминокислотную последовательность QQNKEDPWT (SEQ ID NO:18). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYA (SEQ ID NO:13), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGQGGT (SEQ ID NO:14), и последовательность CDR-H3, содержащую аминокислотную последовательность ARTGGLRRAYFTY (SEQ ID NO:15); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSVSSYGQGF (SEQ ID NO:16), последовательность CDR-L2, содержащую аминокислотную последовательность GAS (SEQ ID NO:17), и последовательность CDR-L3, содержащую аминокислотную последовательность QQNKEDPWT (SEQ ID NO:18).
[0147] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYTLTEFS (SEQ ID NO:19), последовательность CDR-H2, содержащую аминокислотную последовательность FDPEDGET (SEQ ID NO:20), и последовательность CDR-H3, содержащую аминокислотную последовательность TTGRFFDWF (SEQ ID NO:21); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSVISRF (SEQ ID NO:22), последовательность CDR-L2, содержащую аминокислотную последовательность GAS (SEQ ID NO:23), и последовательность CDR-L3, содержащую аминокислотную последовательность QQDSNLPIT (SEQ ID NO:24). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYTLTEFS (SEQ ID NO:19), последовательность CDR-H2, содержащую аминокислотную последовательность FDPEDGET (SEQ ID NO:20), и последовательность CDR-H3, содержащую аминокислотную последовательность TTGRFFDWF (SEQ ID NO:21); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSVISRF (SEQ ID NO:22), последовательность CDR-L2, содержащую аминокислотную последовательность GAS (SEQ ID NO:23), и последовательность CDR-L3, содержащую аминокислотную последовательность QQDSNLPIT (SEQ ID NO:24).
[0148] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYAFTTYL (SEQ ID NO:25), последовательность CDR-H2, содержащую аминокислотную последовательность INPGSGST (SEQ ID NO:26), и последовательность CDR-H3, содержащую аминокислотную последовательность ARYAYGY (SEQ ID NO:27); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QNVGTA (SEQ ID NO:28), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:29), и последовательность CDR-L3, содержащую аминокислотную последовательность QQYSTYPFT (SEQ ID NO:30). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYAFTTYL (SEQ ID NO:25), последовательность CDR-H2, содержащую аминокислотную последовательность INPGSGST (SEQ ID NO:26), и последовательность CDR-H3, содержащую аминокислотную последовательность ARYAYGY (SEQ ID NO:27); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QNVGTA (SEQ ID NO:28), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:29), и последовательность CDR-L3, содержащую аминокислотную последовательность QQYSTYPFT (SEQ ID NO:30).
[0149] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYSFTNYA (SEQ ID NO:31), последовательность CDR-H2, содержащую аминокислотную последовательность ISPYYGDT (SEQ ID NO:32), и последовательность CDR-H3, содержащую аминокислотную последовательность ARRFEGFYYSMDY (SEQ ID NO:33); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHSNGNTY (SEQ ID NO:34), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:35), и последовательность CDR-L3, содержащую аминокислотную последовательность SQSTHVPLT (SEQ ID NO:36). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYSFTNYA (SEQ ID NO:31), последовательность CDR-H2, содержащую аминокислотную последовательность ISPYYGDT (SEQ ID NO:32), и последовательность CDR-H3, содержащую аминокислотную последовательность ARRFEGFYYSMDY (SEQ ID NO:33); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHSNGNTY (SEQ ID NO:34), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:35), и последовательность CDR-L3, содержащую аминокислотную последовательность SQSTHVPLT (SEQ ID NO:36).
[0150] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFSSYG (SEQ ID NO:37), последовательность CDR-H2, содержащую аминокислотную последовательность IWYDGSNK (SEQ ID NO:38), и последовательность CDR-H3, содержащую аминокислотную последовательность ARDPGLRYFDGGMDV (SEQ ID NO:39); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QGISSY (SEQ ID NO:40), последовательность CDR-L2, содержащую аминокислотную последовательность AAS (SEQ ID NO:41), и последовательность CDR-L3, содержащую аминокислотную последовательность QQLNSFPYT (SEQ ID NO:42). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFSSYG (SEQ ID NO:37), последовательность CDR-H2, содержащую аминокислотную последовательность IWYDGSNK (SEQ ID NO:38), и последовательность CDR-H3, содержащую аминокислотную последовательность ARDPGLRYFDGGMDV (SEQ ID NO:39); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QGISSY (SEQ ID NO:40), последовательность CDR-L2, содержащую аминокислотную последовательность AAS (SEQ ID NO:41), и последовательность CDR-L3, содержащую аминокислотную последовательность QQLNSFPYT (SEQ ID NO:42).
[0151] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFSSYG (SEQ ID NO:43), последовательность CDR-H2, содержащую аминокислотную последовательность IWYDGSNK (SEQ ID NO:44), и последовательность CDR-H3, содержащую аминокислотную последовательность ARMFRGAFDY (SEQ ID NO:45); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QGIRND (SEQ ID NO:46), последовательность CDR-L2, содержащую аминокислотную последовательность AAS (SEQ ID NO:47), и последовательность CDR-L3, содержащую аминокислотную последовательность LQDYIYYPT (SEQ ID NO:48). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFSSYG (SEQ ID NO:43), последовательность CDR-H2, содержащую аминокислотную последовательность IWYDGSNK (SEQ ID NO:44), и последовательность CDR-H3, содержащую аминокислотную последовательность ARMFRGAFDY (SEQ ID NO:45); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QGIRND (SEQ ID NO:46), последовательность CDR-L2, содержащую аминокислотную последовательность AAS (SEQ ID NO:47), и последовательность CDR-L3, содержащую аминокислотную последовательность LQDYIYYPT (SEQ ID NO:48).
[0152] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности Qvqlvqsgaevvkpgasvkvsckasgytftsyamhwvkeapgqrlewigyiypgqggtnynqkfqgratltadtsastaymelsslrsedtavyfcartgglrrayftywgqgtlvtvss (SEQ ID NO:79), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности Divltqspatlslspgeratiscrasqsvssygqgfmhwyqqkpgqpprlliygassratgiparfsgsgsgtdftltisplepedfavyycqqnkedpwtfgggtkleik (SEQ ID NO:80). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:79, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:80. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:79, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:80.
[0153] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqlvqsgaevkkpgasvkvsckvsgytltefsihwvrqapgqglewmggfdpedgetiyaqkfqgrvimtedtstdtaymemnslrsedtaiyycttgrffdwfwgqgtlvtvss (SEQ ID NO:81), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности eiiltqspailslspgeratlscrasqsvisrflswyqvkpglaprlliygastratgipvrfsgsgsgtdfsltisslqpedcavyycqqdsnlpitfgqgtrleik (SEQ ID NO:82). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:81, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:82. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:81, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:82.
[0154] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqlvqsgaevkkpgasvkvsckasgyafttylvewirqrpgqglewmgvinpgsgstnyaqkfqgrvtmtvdrssttaymelsrlrsddtavyycaryaygywgqgtlvtvss (SEQ ID NO:83), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqnvgtavawyqqkpgkspkqliysasnrytgvpsrfsgsgsgtdftltisslqpedlatyycqqystypftfgqgtkleik (SEQ ID NO:84). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит: вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:83, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:84. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:83, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:84.
[0155] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftfssygmywvrqapgkglewvaviwydgsnkyyadsvkgrftisrdnskntlylqmnslraedtavyhcardpglryfdggmdvwgqgttvtvss (SEQ ID NO:87), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqltqspsflsasvgdrvtitcrasqgissylawyqqkpgkapkllifaastlhsgvpsrfsgsgsgteftltisslqpedfatyycqqlnsfpytfgqgtkleik (SEQ ID NO:88). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:87, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:88. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:87, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:88.
[0156] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftfssygmhwvrqapgkglewvaviwydgsnkyyadsvkgrftisgdnskntlylqmnslraedtavyycarmfrgafdywgqgtlvtvss (SEQ ID NO:89), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности aiqmtqspsslsasvgdrvtitcrasqgirndlgwyqqkpgkapklliyaasslqsgvpsrfsgsgsgtdftltisglqpedsatyyclqdyiyyptfgqgtkveik (SEQ ID NO:90). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:89, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:90. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:89, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:90.
[0157] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqlvqsgaevkkpgasvkvsckasgysftnyavhwvrqapgqglewmgvispyygdttyaqkfqgrvtmtvdkssstaymelsrlrsddtavyycarrfegfyysmdywgqgtlvtvss (SEQ ID NO:85), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности dvvmtqsplslpvtlgqpasiscrpsqslvhsngntylnwyqqrpgqspklliykvskrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycsqsthvpltfgggtkveik (SEQ ID NO:86). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:85, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:86. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:85, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:86.
[0158] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqlqqsgpelvrpgtsvkvsckasgyafttylvewikqrpgqglewigvinpgsgstnynekfkgkatltvdrssttaymhlsgltsddsavyfcaryaygywgqgttltvss (SEQ ID NO:277), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности divmtqsqkfmsasvgdrvsitckasqnvgtavawyqqqpghspkqliysasnrytgvpdrftgsgagtdftltisniqsedladyfcqqystypftfgsgtkleik (SEQ ID NO:278). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:277, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:278. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:277, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:278. В некоторых вариантах осуществления домены VH и/или VL являются гуманизированными.
[0159] В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности qvqllqsgaelvrpgvsvkisctgsgysftnyavhwvkqshvkslewigvispyygdttynqkftgkatmtvdkssstaymelarltsedsaiyfcarrfegfyysmdywgqgtsvtvss (SEQ ID NO:279), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности dvvmiqtplslpvslgdqasiscrpsqslvhsngntylnwylqrpgqspklliykvskrfsgvpdrfsgsgsgtdftlkisrveaedlgvylcsqsthvpltfgsgtqleik (SEQ ID NO:280). В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:279, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:280. В некоторых вариантах осуществления связывающий участок, который связывает CD38, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:279, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:280. В некоторых вариантах осуществления домены VH и/или VL являются гуманизированными.
[0160] В некоторых вариантах осуществления любого из приведенных выше вариантов осуществления связывающий белок представляет собой триспецифический связывающий белок. В некоторых вариантах осуществления триспецифический связывающий белок содержит антигенсвязывающий участок, который связывает полипептид CD38, антигенсвязывающий участок, который связывает полипептид CD28, и антигенсвязывающий участок, который связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический белок, содержащий четыре полипептида, которые содержат три антигенсвязывающих участка, где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь (например, как описано в данном документе). В некоторых вариантах осуществления домены VH и VL любого из антигенсвязывающих участков, связывающих CD38, описанных выше, представляют собой VH3 и VL3 и образуют третий антигенсвязывающий участок, который связывает полипептид CD38. В некоторых вариантах осуществления VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и домены VH и VL любого из антигенсвязывающих участков, связывающих CD38, описанных выше и/или в таблице 2, представляют собой VH3 и VL3 и образуют третий антигенсвязывающий участок, который связывает полипептид CD38.
[0161] Последовательности иллюстративных антигенсвязывающих участков для CD38, представлены в таблице 2. В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий участок для CD38 по настоящему изобретению, содержит 1, 2, 3, 4, 5 или все 6 последовательностей CDR антитела к CD38, которые описаны в таблице 2. В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий участок для CD38 по настоящему изобретению, содержит последовательность домена VH и/или последовательность домена VL антитела к CD38, которые описаны в таблице 2.
Таблица 2. Последовательности CD38-связывающих белков.
VH1
[0162] Дополнительно в данном документе предусмотрены антитела (например, моноспецифические антитела), содержащие любой из CD38-связывающих CDR и/или вариабельных доменов, описанных выше.
[0163] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, который связывает внеклеточный домен полипептида CD38 человека и внеклеточный домен полипептида CD38 макака-крабоеда. Иллюстративные анализы для определения того, связывает ли антигенсвязывающий участок антиген, описаны в данном документе и известны в уровне техники, включая (без ограничения) анализы ELISA, SPR и проточную цитометрию.
HER2-связывающие участки
[0164] Определенные аспекты настоящего изобретения относятся к связывающим белкам, которые содержат антигенсвязывающий участок, который связывает полипептид HER2. В некоторых вариантах осуществления полипептид HER2 представляет собой полипептид HER2 человека, также известный как NEU, NGL, ERBB2, TKR1, CD340, HER-2, MLN19 и HER-2/neu. Полипептиды HER2 человека известны в уровне техники и включают без ограничения полипептиды, представленные под номерами доступа в NCBI XP_024306411.1, XP_024306410.1, XP_024306409.1, NP_001276867.1, NP_001276866.1, NP_001276865.1, NP_001005862.1 или NP_004439.2, или полипептид, полученный с помощью гена с идентификационным номером в NCBI 2064. В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий участок, который связывает полипептид HER2, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид HER2, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, описанных выше, где домены VH3 и VL3 образуют пару, которая образует третий антигенсвязывающий участок, который связывает полипептид HER2.
[0165] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1) или GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), IYPTQGYT (SEQ ID NO:4) или IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6), SRWGGEGFYAMDY (SEQ ID NO:7) или SRWGGSGFYAMDY (SEQ ID NO:8); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9) или QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1) или GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), IYPTQGYT (SEQ ID NO:4) или IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6), SRWGGEGFYAMDY (SEQ ID NO:7) или SRWGGSGFYAMDY (SEQ ID NO:8); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9) или QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0166] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0167] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0168] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0169] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0170] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO:9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0171] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит: вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO:1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO:3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO:6); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0172] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0173] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0174] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO:4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO:8); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0175] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO:2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO:5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO:7); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO:10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO:11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO:12).
[0176] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности evqlvesggglvqpggslrlscaasgfnikdtyihwvrqapgkglewvariyptngytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggdgfyamdywgqgtlvtvss (SEQ ID NO:72), Evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:73), evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:74), evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:75), или evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:76); и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77) или diqmtqspsslsasvgdrvtitcrasqdvqtavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:78). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:72, SEQ ID NO:73, SEQ ID NO:74, SEQ ID NO:75 или SEQ ID NO:76; и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77 или SEQ ID NO:78. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:72, SEQ ID NO:73, SEQ ID NO:74, SEQ ID NO:75 или SEQ ID NO:76; и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77 или SEQ ID NO:78.
[0177] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности evqlvesggglvqpggslrlscaasgfnikdtyihwvrqapgkglewvariyptngytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggdgfyamdywgqgtlvtvss (SEQ ID NO:72), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:72, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:72, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77.
[0178] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности Evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:73), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:73, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:73, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77.
[0179] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:75), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:75, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:75, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77.
[0180] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptqgytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggsgfyamdywgqgtlvtvss (SEQ ID NO:74), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:74, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:74, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77.
[0181] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности evqlvesggglvqpggslrlscaasgfnirdtyihwvrqapgkglewvariyptnaytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggegfyamdywgqgtlvtvss (SEQ ID NO:76), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvntavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:77). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:76, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:76, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:77.
[0182] В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности evqlvesggglvqpggslrlscaasgfnikdtyihwvrqapgkglewvariyptngytryadsvkgrftisadtskntaylqmnslraedtavyycsrwggdgfyamdywgqgtlvtvss (SEQ ID NO:72), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcrasqdvqtavawyqqkpgkapklliysasflysgvpsrfsgsrsgtdftltisslqpedfatyycqqhyttpptfgqgtkveik (SEQ ID NO:78). В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:72, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:78. В некоторых вариантах осуществления связывающий участок, который связывает HER2, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:72, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:78.
[0183] В некоторых вариантах осуществления антигенсвязывающий участок для HER2 по настоящему изобретению содержит 1, 2, 3, 4, 5 или все 6 последовательностей CDR антитела к HER2 трастузумаба, 30R/55Q/102E, 30R/56A/102S, 30R/55Q/102S, 30R/56A/102E или 30Q. В некоторых вариантах осуществления антигенсвязывающий участок для HER2 по настоящему изобретению содержит последовательность домена VH и/или последовательность домена VL антитела к HER2 трастузумаба, 30R/55Q/102E, 30R/56A/102S, 30R/55Q/102S, 30R/56A/102E или 30Q.
[0184] Последовательности иллюстративных антигенсвязывающих участков для HER2, представлены в таблице 3. В некоторых вариантах осуществления антигенсвязывающий участок для HER2 по настоящему изобретению содержит 1, 2, 3, 4, 5 или все 6 последовательностей CDR антитела к HER2, которые описаны в таблице 3. В некоторых вариантах осуществления антигенсвязывающий участок для HER2 по настоящему изобретению содержит последовательность домена VH и/или последовательность домена VL антитела к HER2, которые описаны в таблице 3.
Таблица 3. Последовательности HER2-связывающих белков.
(трастузумаб)
CDR тяжелой цепи
30R
55Q
56A
(исходная)
102E
102S
(трастузумаб)
CDR легкой цепи
(исходная)
30Q
(исходная)
(исходная)
Другие связывающие опухолевые мишени участки
[0185] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит антигенсвязывающий участок, который связывает опухолевый белок-мишень. В некоторых вариантах осуществления опухолевый белок-мишень представляет собой полипептид CD38 (например, полипептид CD38 человека). В некоторых вариантах осуществления опухолевый белок-мишень представляет собой полипептид HER2 (например, полипептид HER2 человека). В некоторых вариантах осуществления опухолевый белок-мишень по настоящему изобретению включает без ограничения A2AR, APRIL, ATPDазу, BAFF, BAFFR, BCMA, BlyS, BTK, BTLA, B7DC, B7H1, B7H4 (также известный как VTCN1), B7H5, B7H6, B7H7, B7RP1, B7-4, C3, C5, CCL2 (также известный как MCP-1), CCL3 (также известный как MIP-1a), CCL4 (также известный как MIP-1b), CCL5 (также известный как RANTES), CCL7 (также известный как MCP-3), CCL8 (также известный как mcp-2), CCL11 (также известный как эотаксин), CCL15 (также известный как MIP-1d), CCL17 (также известный как TARC), CCL19 (также известный как MIP-3b), CCL20 (также известный как MIP-3a), CCL21 (также известный как MIP-2), CCL24 (также известный как MPIF-2/эотаксин-2), CCL25 (также известный как TECK), CCL26 (также известный как эотаксин-3), CCR3, CCR4, CD3, CD19, CD20, CD23 (также известный как FCER2, рецептор IgE), CD24, CD27, CD28, CD38, CD39, CD40, CD70, CD80 (также известный как B7-1), CD86 (также известный как B7-2), CD122, CD137 (также известный как 41BB), CD137L, CD152 (также известный как CTLA4), CD154 (также известный как CD40L), CD160, CD272, CD273 (также известный как PDL2), CD274 (также известный как PDL1), CD275 (также известный как B7H2), CD276 (также известный как B7H3), CD278 (также известный как ICOS), CD279 (также известный как PD-1), CDH1 (также известный как E-кадгерин), хитиназу, CLEC9, CLEC91, CRTH2, CSF-1 (также известный как M-CSF), CSF-2 (также известный как GM-CSF), CSF-3 (также известный как GCSF), CX3CL1 (также известный как SCYD1), CXCL12 (также известный как SDF1), CXCL13, CXCR3, DNGR-1, эктонуклеозидтрифосфатдифосфогидролазу 1, EGFR, ENTPD1, FCER1A, FCER1, FLAP, FOLH1, Gi24, GITR, GITRL, GM-CSF, Her2, HHLA2, HMGB1, HVEM, ICOSLG, IDO, IFNα, IgE, IGF1R, IL2R бета, IL1, IL1A, IL1B, IL1F10, IL2, IL4, IL4Ra, IL5, IL5R, IL6, IL7, IL7Ra, IL8, IL9, IL9R, IL10, rhIL10, IL12, IL13, IL13Ra1, IL13Ra2, IL15, IL17, IL17Rb (также известный как рецептор IL25), IL18, IL22, IL23, IL25, IL27, IL33, IL35, ITGB4 (также известный как интегрин b4), ITK, KIR, LAG3, LAMP1, лептин, LPFS2, молекулы MHC класса II, MUC-1, NCR3LG1, NKG2D, NTPDазу-1, OX40, OX40L, PD-1H, тромбоцитарный рецептор, PROM1, S152, SISP1, SLC, SPG64, ST2 (также известный как рецептор IL33), STEAP2, Syk-киназу, TACI, TDO, T14, TIGIT, TIM3, TLR, TLR2, TLR4, TLR5, TLR9, TMEF1, TNFa, TNFRSF7, Tp55, TREM1, TSLP (также известный как корецептор IL7Ra), TSLPR, TWEAK, VEGF, VISTA, Vstm3, WUCAM и XCR1 (также известный как GPR5/CCXCR1). В некоторых вариантах осуществления одна или несколько из указанных выше антигенных мишеней представляют собой антигенные мишени человека.
Связывающие участки антитела к CD28
[0186] Определенные аспекты настоящего изобретения относятся к связывающим белкам, которые содержат антигенсвязывающий участок, который связывает полипептид CD28. В некоторых вариантах осуществления полипептид CD28 представляет собой полипептид CD28 человека, также известный как Tp44. Полипептиды CD28 человека известны из уровня техники, и к ним относятся без ограничения полипептиды, представленные под номерами доступа NCBI XP_011510499.1, XP_011510497.1, XP_011510496.1, NP_001230007.1, NP_001230006.1 или NP_006130.1, или полипептид, полученный из гена под номером ID NCBI 940. В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий участок, который связывает полипептид CD28, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD28, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD28, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, и один из которых связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид CD38. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид HER2. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает опухолевый белок-мишень.
[0187] В некоторых вариантах осуществления связывающий участок, который связывает CD28, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYY (SEQ ID NO:49), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGNVNT (SEQ ID NO:50), и последовательность CDR-H3, содержащую аминокислотную последовательность TRSHYGLDWNFDV (SEQ ID NO:51), и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QNIYVW (SEQ ID NO:52), последовательность CDR-L2, содержащую аминокислотную последовательность KAS (SEQ ID NO:53), и последовательность CDR-L3, содержащую аминокислотную последовательность QQGQTYPY (SEQ ID NO:54). В некоторых вариантах осуществления связывающий участок, который связывает CD28, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYY (SEQ ID NO:49), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGNVNT (SEQ ID NO:50), и последовательность CDR-H3, содержащую аминокислотную последовательность TRSHYGLDWNFDV (SEQ ID NO:51); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QNIYVW (SEQ ID NO:52), последовательность CDR-L2, содержащую аминокислотную последовательность KAS (SEQ ID NO:53), и последовательность CDR-L3, содержащую аминокислотную последовательность QQGQTYPY (SEQ ID NO:54).
[0188] В некоторых вариантах осуществления связывающий участок, который связывает CD28, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности QVQLVQSGAEVVKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGSIYPGNVNTNYAQKFQGRATLTVDTSISTAYMELSRLRSDDTAVYYCTRSHYGLDWNFDVWGKGTTVTVSS (SEQ ID NO:91), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или на 100% идентична аминокислотной последовательности diqmtqspsslsasvgdrvtitcqasqniyvwlnwyqqkpgkapklliykasnlhtgvpsrfsgsgsgtdftltisslqpediatyycqqgqtypytfgqgtkleik (SEQ ID NO:92). В некоторых вариантах осуществления связывающий участок, который связывает CD28, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:91, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:92. В некоторых вариантах осуществления связывающий участок, который связывает CD28, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:91, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:92.
[0189] В некоторых вариантах осуществления любого из приведенных выше вариантов осуществления связывающий белок представляет собой триспецифический связывающий белок. В некоторых вариантах осуществления триспецифический связывающий белок содержит антигенсвязывающий участок, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2), антигенсвязывающий участок, который связывает полипептид CD28, и антигенсвязывающий участок, который связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический белок, содержащий четыре полипептида, которые содержат три антигенсвязывающих участка, где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь (например, как описано в данном документе). В некоторых вариантах осуществления домены VH и VL любого из антигенсвязывающих участков для CD28, описанных выше, представляют собой VH1 и VL1 и образуют первый антигенсвязывающий участок, который связывает полипептид CD28. В некоторых вариантах осуществления домены VH и VL любого из антигенсвязывающих участков для CD28, описанных выше и/или в таблице 4, представляют собой VH1 и VL1 и образуют первый антигенсвязывающий участок, который связывает полипептид CD28, VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и VH3 и VL3 и образуют третий антигенсвязывающий участок, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2).
[0190] Последовательности иллюстративных антигенсвязывающих участков для CD28, представлены в таблице 4. В некоторых вариантах осуществления антигенсвязывающий участок для CD28 по настоящему изобретению содержит 1, 2, 3, 4, 5 или все 6 последовательностей CDR антитела к CD28, которые описаны в таблице 4. В некоторых вариантах осуществления антигенсвязывающий участок для CD28 по настоящему изобретению содержит последовательность домена VH и/или последовательность домена VL антитела к CD28, которые описаны в таблице 4.
Таблица 4. Последовательности CD28-связывающих белков.
(sup)
Связывающие участки антитела к CD3
[0191] Определенные аспекты настоящего изобретения относятся к связывающим белкам, которые содержат антигенсвязывающий участок, который связывает полипептид CD3. В некоторых вариантах осуществления полипептид CD3 представляет собой полипептид CD3 человека, в том числе CD3-дельта (также известный как T3D, IMD19 и CD3-DELTA), CD3-эпсилон (также известный как T3E, IMD18 и TCRE) и CD3-гамма (также известный как T3G, IMD17 и CD3-GAMMA). Полипептиды CD3 человека известны из уровня техники, и к ним относятся без ограничения полипептиды, представленные под номерами доступа NCBI XP_006510029.1 или NP_031674.1, или полипептид, полученный из гена под номерами ID NCBI 915, 916 или 917. В некоторых вариантах осуществления связывающий белок, содержащий антигенсвязывающий участок, который связывает полипептид CD3, является моноспецифическим и/или моновалентным, биспецифическим и/или бивалентным, триспецифическим и/или тривалентным или полиспецифическим и/или поливалентным. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, и один из которых связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид CD38. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает полипептид HER2. В некоторых вариантах осуществления связывающий белок, который содержит антигенсвязывающий участок, который связывает полипептид CD3, представляет собой триспецифический связывающий белок, содержащий четыре полипептида, которые образуют три антигенсвязывающих участка, один из которых связывает полипептид CD28, один из которых связывает полипептид CD3, и один из которых связывает опухолевый белок-мишень.
[0192] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F (SEQ ID NO:180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления последовательность CDR-L1 домена VL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из QSLVHQNAQTY (SEQ ID NO:59), QSLVHENLQTY (SEQ ID NO:60), QSLVHENLFTY (SEQ ID NO:61) и QSLVHENLRTY (SEQ ID NO:62).
[0193] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит: вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHQNAQTY (SEQ ID NO:59), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHQNAQTY (SEQ ID NO:59), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65).
[0194] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLQTY (SEQ ID NO:60), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLQTY (SEQ ID NO:60), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65).
[0195] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLFTY (SEQ ID NO:61), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLFTY (SEQ ID NO:61), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65).
[0196] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и/или вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLRTY (SEQ ID NO:62), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO:55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO:56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO:57); и вариабельный домен легкой цепи (VL) антитела, содержащий последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHENLRTY (SEQ ID NO:62), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO:64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO:65).
[0197] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности, выбираемой из группы, состоящей из divmtqtplslsvtpgqpasisckssqslvhqnaqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:95), divmtqtplslsvtpgqpasisckssqslvhenlqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:96), divmtqtplslsvtpgqpasisckssqslvhenlftylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:97), и divmtqtplslsvtpgqpasisckssqslvhenlrtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:98). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97 и SEQ ID NO:98. В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:95, SEQ ID NO:96, SEQ ID NO:97 и SEQ ID NO:98.
[0198] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности divmtqtplslsvtpgqpasisckssqslvhqnaqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:95). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:95. В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:95.
[0199] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности divmtqtplslsvtpgqpasisckssqslvhenlqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:96). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:96. В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:96.
[0200] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности divmtqtplslsvtpgqpasisckssqslvhenlftylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:97). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:97. В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:97.
[0201] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyadsvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:93), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности divmtqtplslsvtpgqpasisckssqslvhenlrtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:98). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:98. В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:93, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:98.
[0202] В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности qvqlvesgggvvqpgrslrlscaasgftftkawmhwvrqapgkqlewvaqikdksnsyatyyassvkgrftisrddskntlylqmnslraedtavyycrgvyyalspfdywgqgtlvtvss (SEQ ID NO:302), и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность, которая на по меньшей мере 85%, на по меньшей мере 86%, на по меньшей мере 87%, на по меньшей мере 88%, на по меньшей мере 89%, на по меньшей мере 90%, на по меньшей мере 91%, на по меньшей мере 92%, на по меньшей мере 93%, на по меньшей мере 94%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98%, на по меньшей мере 99% или 100% идентична аминокислотной последовательности divmtqtplslsvtpgqpasisckssqslvhqnaqtylswylqkpgqspqsliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedvgvyycgqgtqypftfgsgtkveik (SEQ ID NO:95). В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:302, и/или вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:95. В некоторых вариантах осуществления связывающий участок, который связывает CD3, содержит вариабельный домен тяжелой цепи (VH) антитела, содержащий аминокислотную последовательность под SEQ ID NO:302, и вариабельный домен легкой цепи (VL) антитела, содержащий аминокислотную последовательность под SEQ ID NO:95.
[0203] В некоторых вариантах осуществления любого из приведенных выше вариантов осуществления связывающий белок представляет собой триспецифический связывающий белок. В некоторых вариантах осуществления триспецифический связывающий белок содержит антигенсвязывающий участок, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2), антигенсвязывающий участок, который связывает полипептид CD28, и антигенсвязывающий участок, который связывает полипептид CD3. В некоторых вариантах осуществления связывающий белок представляет собой триспецифический белок, содержащий четыре полипептида, которые содержат три антигенсвязывающих участка, где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь (например, как описано в данном документе). В некоторых вариантах осуществления домены VH и VL любого из антигенсвязывающих участков для CD3, описанных выше, представляют собой VH2 и VL2 и образуют второй антигенсвязывающий участок, который связывает полипептид CD3. В некоторых вариантах осуществления VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, домены VH и VL любого из антигенсвязывающих участков, связывающих CD3, описанных выше и/или в таблице 5, представляют собой VH2 и VL2 и образуют второй антигенсвязывающий участок, который связывает полипептид CD3, и VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает опухолевый белок-мишень (в том числе без ограничения CD38 или HER2).
[0204] Последовательности иллюстративных антигенсвязывающих участков для CD3, представлены в таблице 5. В некоторых вариантах осуществления антигенсвязывающий участок для CD3 по настоящему изобретению содержит 1, 2, 3, 4, 5 или все 6 последовательностей CDR антитела к CD3, которые описаны в таблице 5. В некоторых вариантах осуществления антигенсвязывающий участок для CD3 по настоящему изобретению содержит последовательность домена VH и/или последовательность домена VL антитела к CD3, которые описаны в таблице 5.
Таблица 5. Последовательности CD3-связывающих белков.
исходная
исходная
исходная
исходная
исходная
Исходная
где X1 представляет собой E или Q, X2 представляет собой A или L, и X3 представляет собой Q, R или F
Исходная
32/35 QQ
ENLQ
ENLF
ENLR
DNAQ
Линкеры
[0205] В некоторых вариантах осуществления длина линкеров L1, L2, L3 и L4 варьирует от нуля аминокислот (длина=0) до приблизительно 100 аминокислот или менее чем 100, 50, 40, 30, 20 или 15 аминокислот или меньше. Длина линкеров также может составлять 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислоту. Все из L1, L2, L3 и L4 в одном связывающем белке могут иметь одинаковую аминокислотную последовательность или все могут иметь различные аминокислотные последовательности.
[0206] Примеры подходящих линкеров включают, например, GGGGSGGGGS (SEQ ID NO:69), GGGGSGGGGSGGGGS (SEQ ID NO: 70), S, RT, TKGPS (SEQ ID NO: 68), GQPKAAP (SEQ ID NO: 67), GGSGSSGSGG (SEQ ID NO: 71) и DKTHT (SEQ ID NO:66), а также раскрытые в международных публикациях №№ WO2017/074878 и WO2017/180913. Вышеперечисленные примеры не предназначены для какого-либо ограничения объема настоящего изобретения, и при этом было показано, что линкеры, содержащие выбранные случайным образом аминокислоты, выбранные из группы, состоящей из валина, лейцина, изолейцина, серина, треонина, лизина, аргинина, гистидина, аспартата, глутамата, аспарагина, глутамина, глицина и пролина, подходят для связывающих белков.
[0207] Состав и последовательность аминокислотных остатков в линкере может меняться в зависимости от типа элемента вторичной структуры, который необходимо достигнуть при помощи линкера. Например, глицин, серин и аланин являются наиболее подходящими для линкеров, характеризующихся максимальной гибкостью. Некоторые комбинации глицина, пролина, треонина и серина применимы, если необходим более жесткий и удлиненный линкер. При необходимости любой аминокислотный остаток может рассматриваться как линкер в комбинации с другими аминокислотными остатками для конструирования более длинных пептидных линкеров в зависимости от требуемых свойств.
[0208] В некоторых вариантах осуществления длина L1 в по меньшей два раза превышает длину L3. В некоторых вариантах осуществления длина L2 в по меньшей два раза превышает длину L4. В некоторых вариантах осуществления длина L1 в по меньшей мере два раза превышает длину L3, а длина L2 в по меньшей мере два раза превышает длину L4. В некоторых вариантах осуществления длина L1 составляет 3-12 аминокислотных остатков, длина L2 составляет 3-14 аминокислотных остатков, длина L3 составляет 1-8 аминокислотных остатков, и длина L4 составляет 1-3 аминокислотных остатка. В некоторых вариантах осуществления длина L1 составляет 5-10 аминокислотных остатков, длина L2 составляет 5-8 аминокислотных остатков, длина L3 составляет 1-5 аминокислотных остатков, и длина L4 составляет 1-2 аминокислотных остатка. В некоторых вариантах осуществления длина L1 составляет 7 аминокислотных остатков, длина L2 составляет 5 аминокислотных остатков, длина L3 составляет 1 аминокислотный остаток, и длина L4 составляет 2 аминокислотных остатка.
[0209] В некоторых вариантах осуществления каждый из L1, L2, L3 и L4 независимо имеет длину, составляющую ноль аминокислот, или содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO:69), GGGGSGGGGSGGGGS (SEQ ID NO: 70), S, RT, TKGPS (SEQ ID NO: 68), GQPKAAP (SEQ ID NO: 67) и GGSGSSGSGG (SEQ ID NO: 71). В некоторых вариантах осуществления каждый из L1, L2, L3 и L4 независимо содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO:69), GGGGSGGGGSGGGGS (SEQ ID NO: 70), S, RT, TKGPS (SEQ ID NO: 68), GQPKAAP (SEQ ID NO: 67) и GGSGSSGSGG (SEQ ID NO: 71). В некоторых вариантах осуществления L1 содержит последовательность GQPKAAP (SEQ ID NO: 67), L2 содержит последовательность TKGPS (SEQ ID NO:68), L3 содержит последовательность S, и L4 содержит последовательность RT.
[0210] В некоторых вариантах осуществления по меньшей мере один из L1, L2, L3 или L4 содержит последовательность DKTHT (SEQ ID NO:66). В некоторых вариантах осуществления L1, L2, L3 и L4 содержат последовательность DKTHT (SEQ ID NO:66).
Fc-области и константные домены
[0211] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит вторую полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит третью полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит вторую полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, и третью полипептидную цепь, дополнительно содержащую Fc-область, связанную с CH1, при этом Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина.
[0212] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит полноразмерную тяжелую цепь антитела или полипептидную цепь, содержащую Fc-область. В некоторых вариантах осуществления Fc-область представляет собой Fc-область человека, например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека. В некоторых вариантах осуществления Fc-область включает шарнир, CH1, CH2, CH3 и необязательно домены CH4. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека. В некоторых вариантах осуществления Fc-область содержит одну или несколько мутаций, описанных ниже. В некоторых вариантах осуществления Fc-область представляет собой Fc-область одного из полипептидов тяжелой цепи (например, полипептида 2 или 3) связывающего белка, представленного в таблице 4. В некоторых вариантах осуществления константная область тяжелой цепи представляет собой константную область одного из полипептидов тяжелой цепи (например, полипептида 2 или 3) связывающего белка, представленного в таблице 4. В некоторых вариантах осуществления константная область легкой цепи представляет собой константную область одного из полипептидов легкой цепи (например, полипептида 1 или 4) связывающего белка, представленного в таблице 4.
[0213] В некоторых вариантах осуществления связывающий белок согласно настоящему изобретению содержит один или два варианта Fc. Термин "вариант Fc", используемый в данном документе, обозначает молекулу или последовательность, которая модифицирована по сравнению с нативным Fc, но все еще содержит участок связывания для рецептора реутилизации FcRn (неонатального Fc-рецептора). Приводимые в качестве примера варианты Fc и их взаимодействие с рецептором реутилизации известны в данной области техники. Таким образом, термин "вариант Fc" может предусматривать молекулу или последовательность, полученную в результате гуманизации нативного Fc отличного от человеческого происхождения. Кроме того, нативный Fc содержит области, которые можно удалить, поскольку они обеспечивают структурные признаки или биологическую активность, которые не требуются для антителоподобных связывающих белков по настоящему изобретению. Таким образом, термин "вариант Fc" предусматривает молекулу или последовательность, в которой отсутствует один или несколько участков или остатков нативного Fc или в которой были модифицированы один или несколько участков или остатков Fc, воздействующих на или вовлеченных в: (1) образование дисульфидной связи, (2) несовместимость с выбранной клеткой-хозяином, (3) гетерогенность N-концов при экспрессии в выбранной клетке-хозяине, (4) гликозилирование, (5) взаимодействие с системой комплемента, (6) связывание с Fc-рецептором, отличным от рецептора реутилизации, или (7) антителозависимую клеточную цитотоксичность (ADCC).
[0214] В некоторых вариантах осуществления связывающий белок по настоящему изобретению (например, триспецифический связывающий белок) содержит мутацию по типу "выступа" на второй полипептидной цепи и мутацию по типу "впадины" на третьей полипептидной цепи. В некоторых вариантах осуществления связывающий белок согласно настоящему изобретению содержит мутацию по типу "выступа" на третьей полипептидной цепи и мутацию по типу "впадины" на второй полипептидной цепи. В некоторых вариантах осуществления мутация по типу "выступа" предусматривает замену (замены) в положениях, соответствующих положениям 354 и/или 366 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S354C, T366W, T366Y, S354C и T366W или S354C и T366Y. В некоторых вариантах осуществления мутация по типу "выступа" предусматривает замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S354C и T366W. В некоторых вариантах осуществления мутация по типу "впадины" предусматривает замену(замены) в положениях, соответствующих положениям 407 и необязательно 349, 366 и/или 368 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой Y407V или Y407T и необязательно Y349C, T366S и/или L368A. В некоторых вариантах осуществления мутация по типу "впадины" предусматривает замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V.
[0215] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную(-ые) замену(-ы) в положениях, соответствующих положениям 366 и необязательно 354 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366W или T366Y и необязательно S354C; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную(-ые) замену(-ы) в положениях, соответствующих положениям 407 и необязательно 349, 366 и/или 368 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y407V или Y407T и необязательно Y349C, T366S и/или L368A.
[0216] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную(-ые) замену(-ы) в положениях, соответствующих положениям 407 и необязательно 349, 366 и/или 368 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y407V или Y407T и необязательно Y349C, T366S и/или L368A; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную(-ые) замену(-ы) в положениях, соответствующих положениям 366 и необязательно 354 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366W или T366Y и необязательно S354C.
[0217] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную замену в положении, соответствующем положению 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотная замена представляет собой T366W; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную(-ые) замену(-ы) в положениях, соответствующих положениям 366, 368 и/или 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366S, L368A и/или Y407V.
[0218] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотную(-ые) замену(-ы) в положениях, соответствующих положениям 366, 368 и/или 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой T366S, L368A и/или Y407V; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотную замену в положении, соответствующем положению 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотная замена представляет собой T366W.
[0219] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W. В некоторых вариантах осуществления первый и/или второй Fc-области представляют собой Fc-области IgG1 человека. В некоторых вариантах осуществления первый и/или второй Fc-области представляют собой Fc-области IgG4 человека.
[0220] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, S354C, T366W и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, Y349C, T366S, L368A, Y407V и R409K. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, Y349C, T366S, L368A, Y407V и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, S354C, T366W и R409K.
[0221] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 354 и 366 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, S354C и T366W; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 349, 366, 368 и 407 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, Y349C, T366S, L368A и Y407V. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 349, 366, 368 и 407 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, Y349C, T366S, L368A и Y407V; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 234, 235, 354 и 366 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A, L235A, S354C и T366W.
[0222] В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит одну или несколько мутаций для снижения эффекторной функции, например, антителозависимого клеточного фагоцитоза, опосредованного Fc-рецепторами (ADCP), комплементзависимой цитотоксичности (CDC) и/или антителозависимой клеточной цитотоксичности (ADCC). В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где первая и вторая Fc-области представляют собой Fc-области IgG1 человека; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A и L235A. В некоторых вариантах осуществления Fc-области второй и третьей полипептидных цепей представляют собой Fc-области IgG1 человека, и при этом каждая из Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A и L235A. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где первая и вторая Fc-области представляют собой Fc-области IgG1 человека; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234, 235 и 329 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A, L235A и P329A. В некоторых вариантах осуществления Fc-области второй и третьей полипептидных цепей представляют собой Fc-области IgG1 человека, и где каждая из Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234, 235 и 329 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A, L235A и P329A. В некоторых вариантах осуществления Fc-области второй и третьей полипептидных цепей представляют собой Fc-области IgG4 человека, и каждая из Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A и L235A. В некоторых вариантах осуществления связывающий белок содержит вторую полипептидную цепь, дополнительно содержащую первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, и при этом третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A и L235A.
[0223] В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, S354C, T366W и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, Y349C, T366S, L368A, Y407V и R409K. В некоторых вариантах осуществления вторая полипептидная цепь дополнительно содержит первую Fc-область, связанную с CH1, где первая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где первая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 349, 366, 368, 407 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, Y349C, T366S, L368A, Y407V и R409K; и где третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, где вторая Fc-область представляет собой Fc-область IgG4 человека, содержащую шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, где вторая Fc-область содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235, 354, 366 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P, F234A, L235A, S354C, T366W и R409K.
[0224] В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую одну или несколько мутаций, которые ослабляют или устраняют связывание FcγI и/или FcγII. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую одну или несколько мутаций, которые ослабляют или устраняют связывание FcγI и/или FcγII, но не влияют на связывание FcRn. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 228 и/или 409 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S228P и/или R409K. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 234 и/или 235 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой F234A и/или L235A. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 228, 234, 235 и/или 409 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S228P, F234A, L235A и/или R409K. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 233-236 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой E233P, F234V, L235A, и делецию в положении 236. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG4 человека, содержащую аминокислотные мутации в положениях, соответствующих положениям 228, 233-236 и/или 409 IgG4 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные мутации представляют собой S228P; E233P, F234V, L235A, и делецию в положении 236; и/или R409K.
[0225] В некоторых вариантах осуществления Fc-область содержит одну или несколько мутаций, которые уменьшают или устраняют связывание Fc-рецептора и/или эффекторную функцию Fc-области (например, антителозависимый клеточный фагоцитоз, опосредованный Fc-рецепторами (ADCP), комплементзависимую цитотоксичность (CDC) и/или антителозависимую клеточную цитотоксичность (ADCC)).
[0226] В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую одну или несколько аминокислотных замен в положениях, соответствующих положениям 234, 235 и/или 329 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой L234A, L235A и/или P329A. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 298, 299 и/или 300 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S298N, T299A и/или Y300S.
[0227] В некоторых вариантах осуществления связывающий белок согласно настоящему изобретению содержит одну или несколько мутаций для улучшения стабильности, например, шарнирной области и/или поверхности контакта димера IgG4 (см., например, Spiess, C. et al. (2013) J. Biol. Chem. 288:26583-26593). В некоторых вариантах осуществления мутация предусматривает замены в положениях, соответствующих положениям 228 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P и R409K. В некоторых вариантах осуществления связывающий белок содержит вторую полипептидную цепь, дополнительно содержащую первую Fc-область, связанную с CH1, при этом первая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина, и при этом третья полипептидная цепь дополнительно содержит вторую Fc-область, связанную с CH1, при этом вторая Fc-область содержит шарнирную область иммуноглобулина и константные домены CH2 и CH3 тяжелой цепи иммуноглобулина; где первая и вторая Fc-области представляют собой Fc-области IgG4 человека; и где каждая из первой и второй Fc-областей содержит аминокислотные замены в положениях, соответствующих положениям 228 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P и R409K. В некоторых вариантах осуществления связывающий белок по настоящему изобретению содержит мутации по типу выступа и впадины и одну или несколько мутаций для улучшения стабильности. В некоторых вариантах осуществления первый и/или второй Fc-области представляют собой Fc-области IgG4 человека.
[0228] В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую одну или несколько аминокислотных замен в положениях, соответствующих положениям 234, 235 и/или 329 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой L234A, L235A и/или P329A. В некоторых вариантах осуществления Fc-область представляет собой Fc-область IgG1 человека, содержащую аминокислотные замены в положениях, соответствующих положениям 298, 299 и/или 300 IgG1 человека в соответствии с EU-индексом. В некоторых вариантах осуществления аминокислотные замены представляют собой S298N, T299A и/или Y300S.
Нуклеиновые кислоты
[0229] Другие аспекты настоящего изобретения относятся к выделенным молекулам нуклеиновой кислоты, содержащим нуклеотидную последовательность, кодирующую любой из связывающих белков, описанных в данном документе. Иллюстративные и неограничивающие последовательности нуклеиновых кислот представлены в таблице 5.
[0230] Другие аспекты настоящего изобретения связаны с наборами полинуклеотидов, например, которые кодируют один или несколько полипептидов связывающего белка, описанного в данном документе. В некоторых вариантах осуществления набор полинуклеотидов по настоящему изобретению содержит один, два, три или четыре полинуклеотида из набора полинуклеотидов, который содержит: (a) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:189, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:190, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:191, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:192; (b) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:193, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:194, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:195, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:196; (c) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:197, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:198, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:199, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:200; (d) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:201, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:202, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:203, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:204; (e) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:205, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:206, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:207, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:208; (f) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:209, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:210, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:211, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:212; (g) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:213, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:214, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:215, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:216; (h) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:217, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:218, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:219, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:220; (i) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:221, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:222, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:223, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:224; (j) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:225, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:226, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:227, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:228; (k) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:229, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:230, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:231, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:232; (l) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:233, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:234, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:235, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:236; (m) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:237, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:238, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:239, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:240; (n) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:241, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:242, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:243, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:244; (o) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:245, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:246, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:247, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:248; (p) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:249, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:250, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:251, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:252; (q) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:253, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:254, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:255, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:256; (r) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:257, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:258, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:259, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:260; (s) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:261, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:262, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:263, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:264; (t) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:265, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:266, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:267, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:268; (u) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:269, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:270, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:271, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:272; или (v) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:273, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:274, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:275, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO:276.
[0231] Другие аспекты настоящего изобретения относятся к векторной системе, содержащей один или несколько векторов, кодирующих первую, вторую, третью и четвертую полипептидные цепи любого из связывающих белков, описанных в данном документе. В некоторых вариантах осуществления векторная система содержит первый вектор, кодирующий первую полипептидную цепь связывающего белка, второй вектор, кодирующий вторую полипептидную цепь связывающего белка, третий вектор, кодирующий третью полипептидную цепь связывающего белка, и четвертый вектор, кодирующий четвертую полипептидную цепь связывающего белка, например, которые показаны в полинуклеотидах таблицы 6. В некоторых вариантах осуществления векторная система содержит первый вектор, кодирующий первую и вторую полипептидные цепи связывающего белка, и второй вектор, кодирующий третью и четвертую полипептидные цепи связывающего белка. В некоторых вариантах осуществления векторная система содержит первый вектор, кодирующий первую и третью полипептидные цепи связывающего белка, и второй вектор, кодирующий вторую и четвертую полипептидные цепи связывающего белка. В некоторых вариантах осуществления векторная система содержит первый вектор, кодирующий первую и четвертую полипептидные цепи связывающего белка, и второй вектор, кодирующий вторую и третью полипептидные цепи связывающего белка. В некоторых вариантах осуществления векторная система содержит первый вектор, кодирующий первую, вторую, третью и четвертую полипептидные цепи связывающего белка. Один или несколько векторов векторной системы могут представлять собой любые из векторов, описанных в данном документе. В некоторых вариантах осуществления один или несколько векторов представляют собой векторы экспрессии. В некоторых вариантах осуществления первый, второй, третий и четвертый полинуклеотиды присутствуют в одном или нескольких векторах экспрессии, например, одном, двух, трех или четырех векторах экспрессии.
[0232] Стандартные технологии рекомбинантной ДНК применяют для конструирования полинуклеотидов, которые кодируют полипептиды, образующие связывающие белки, встраивания этих полинуклеотидов в рекомбинантные векторы экспрессии и введения таких векторов в клетки-хозяева. См., например, Sambrook et al., 2001, Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 3rd ed.). Ферментативные реакции и методики очистки можно проводить в соответствии со спецификациями производителей, которые обычно выполняют в данной области техники или как описано в данном документе. Если не предусмотрены конкретные определения, то терминология, используемая в контексте аналитической химии, синтетической органической химии и медицинской и фармацевтической химии, а также лабораторные процедуры и их методики, описанные в данном документе, являются общеизвестными и широко используемыми в данной области техники. Аналогично традиционные методики можно использовать для вариантов химического синтеза, вариантов химического анализа, фармацевтического получения, составления, доставки и лечения пациентов.
[0233] В некоторых вариантах осуществления выделенная нуклеиновая кислота функционально связана с гетерологичным промотором для управления транскрипцией последовательности нуклеиновой кислоты, кодирующей связывающий белок. Промотор может относиться к контролирующим последовательностям нуклеиновой кислоты, которые управляют транскрипцией нуклеиновой кислоты. Первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, где первая последовательность нуклеиновой кислоты находится в функциональной связи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с последовательностью, кодирующей связывающий белок, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности. Примеры промоторов могут включать без ограничения промоторы, полученные из геномов вирусов (таких как вирус полиомы, вирус оспы кур, аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита B, вирус обезьян 40 (SV40) и т. п.), гетерологичные промоторы эукариот (такие как промотор гена актина, промотор гена иммуноглобулина, промоторы генов белков теплового шока и т. п.), CAG-промотор (Niwa et al., Gene 108(2):193-9, 1991), промотор гена фосфоглицераткиназы (PGK), индуцируемый тетрациклином промотор (Masui et al., Nucleic Acids Res. 33:e43, 2005), lac-систему, trp-систему, tac-систему, trc-систему, основные операторные и промоторные области лямбда-фага, промотор гена 3-фосфоглицераткиназы, промоторы гена кислой фосфатазы дрожжей и промотор гена альфа-фактора спаривания дрожжей. Полинуклеотиды, кодирующие по белки согласно настоящему изобретению, могут находиться под контролем конститутивного промотора, индуцируемого промотора или любого другого подходящего промотора, описанного в данном документе, или другого подходящего промотора, который легко распознает специалист в данной области техники.
[0234] В некоторых вариантах осуществления выделенную нуклеиновую кислоту встраивают в вектор. В некоторых вариантах осуществления вектор представляет собой вектор экспрессии. Векторы экспрессии могут содержать одну или несколько регуляторных последовательностей, функционально связанных с полинуклеотидом, подлежащим экспрессии. Термин "регуляторная последовательность" включает промоторы, энхансеры и другие элементы, контролирующие экспрессию (например, сигналы полиаденилирования). Примеры подходящих энхансеров могут включать без ограничения энхансерные последовательности из генов млекопитающих (таких как гены глобина, эластазы, альбумина, α-фетопротеина, инсулина и т. п.) и энхансерные последовательности из вируса эукариотических клеток (такие как энхансер SV40 на участке позднего начала репликации (100-270 п. о.), ранний промотор/энхансер цитомегаловируса, энхансер полиомы на участке позднего начала репликации, энхансеры аденовирусов и т. п.). Примеры подходящих векторов могут включать, например, плазмиды, космиды, эписомы, транспозоны и вирусные векторы (например, векторы на основе аденовируса, вируса осповакцины, вируса Синдбис, вируса кори, вируса герпеса, лентивируса, ретровируса, аденоассоциированного вируса и т. д.). Векторы экспрессии можно применять для трансфекции клеток-хозяев, таких как, например, клетки бактерий, клетки дрожжей, клетки насекомых и клетки млекопитающих. Биологически функциональные векторы на основе ДНК вируса и плазмидной ДНК, способные экспрессироваться и реплицироваться в хозяине, известны в данной области техники и могут применяться для трансфекции любой клетки, представляющей интерес.
Клетки-хозяева
[0235] Другие аспекты настоящего изобретения связаны с клеткой-хозяином (например, выделенной клеткой-хозяином), содержащей один или несколько выделенных полинуклеотидов, векторов и/или векторных систем, описанных в данном документе. В некоторых вариантах осуществления выделенную клетку-хозяина по настоящему изобретению культивируют in vitro. В некоторых вариантах осуществления клетка-хозяин представляет собой клетку бактерии (например, клетку E. coli). В некоторых вариантах осуществления клетка-хозяин представляет собой клетку дрожжей (например, клетку S. cerevisiae). В некоторых вариантах осуществления клетка-хозяин представляет собой клетку насекомого. Примеры клеток-хозяев насекомого могут включать, например, клетки Drosophila (например, клетки S2), клетки Trichoplusia ni (например, клетки High Five™) и клетки Spodoptera frugiperda (например, клетки Sf21 или Sf9). В некоторых вариантах осуществления клетка-хозяин представляет собой клетку млекопитающего. Примеры клеток-хозяев млекопитающего могут включать, например, клетки почки эмбриона человека (например, клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре), клетки Expi293TM, клетки CHO, клетки почки новорожденного хомяка (например, BHK, ATCC CCL 10), клетки Сертоли мыши (например, клетки TM4), клетки почки обезьяны (например, CV1 ATCC CCL 70), клетки почки африканской зеленой мартышки (например, VERO-76, ATCC CRL-1587), клетки карциномы шейки матки человека (например, HELA, ATCC CCL 2), клетки почки собаки (например, MDCK, ATCC CCL 34), клетки печени крысы линии Buffalo (например, BRL 3A, ATCC CRL 1442), клетки легкого человека (например, W138, ATCC CCL 75), клетки печени человека (например, Hep G2, HB 8065), клетки опухоли молочной железы мыши (например, MMT 060562, ATCC CCL51), клетки TRI, клетки MRC 5, клетки FS4, линию клеток гепатомы человека (например, Hep G2) и клетки миеломы (например, клетки NS0 и Sp2/0).
[0236] Другие аспекты настоящего изобретения связаны со способом получения любого из связывающих белков, описанных в данном документе. В некоторых вариантах осуществления способ предусматривает: a) культивирование клетки-хозяина (например, любой из клеток-хозяев, описанных в данном документе), содержащей выделенную нуклеиновую кислоту, вектор и/или векторную систему (например, любое из выделенных нуклеиновых кислот, векторов и/или векторных систем, описанных в данном документе), в условиях, при которых клетка-хозяин экспрессирует связывающий белок; и b) выделение связывающего белка из клетки-хозяина. Способы культивирования клеток-хозяев в условиях, обеспечивающих экспрессию белка, хорошо известны специалисту средней квалификации в данной области. Способы выделения белков из культивируемых клеток-хозяев хорошо известны специалисту средней квалификации в данной области, в том числе, например, с помощью аффинной хроматографии (например, двухстадийной аффинной хроматографии, включающей аффинную хроматографию с использованием белка A с последующей эксклюзионной хроматографией).
Фармацевтические композиции
[0237] Терапевтические или фармацевтические композиции, содержащие связывающие белки, находятся в пределах объема настоящего изобретения. Такие терапевтические или фармацевтические композиции могут содержать терапевтически эффективное количество связывающего белка или конъюгата связывающий белок-лекарственное средство в смеси с фармацевтически или физиологически приемлемым средством для составления, выбранным в соответствии со способом введения.
[0238] Приемлемые вещества для составления являются нетоксичными для потребителя при используемых дозах и концентрациях.
[0239] Фармацевтическая композиция может содержать вещества для составления, предназначенные для модификации, поддержания или сохранения, например pH, осмолярности, вязкости, прозрачности, цвета, изотоничности, запаха, стерильности, стабильности, скорости растворения или высвобождения, всасывания или проникновения композиции. Подходящие вещества для составления включают без ограничения аминокислоты (такие как глицин, глутамин, аспарагин, аргинин или лизин), противомикробные средства, антиоксиданты (такие как аскорбиновая кислота, сульфит натрия или гидросульфит натрия), буферы (такие как борат, бикарбонат, Tris-HCl, цитраты, фосфаты или другие органические кислоты), объемообразующие средства (такие как маннит или глицин), хелатирующие средства (такие как этилендиаминтетрауксусная кислота (EDTA)), комплексообразующие средства (такие как кофеин, поливинилпирролидон, бета-циклодестрин или гидроксипропил-бета-циклодекстрин), наполнители, моносахариды, дисахариды и другие углеводы (такие как глюкоза, манноза или декстрины), белки (такие как сывороточный альбумин, желатин или иммуноглобулины), красители, ароматизаторы и разбавители, эмульгаторы, гидрофильные полимеры (такие как поливинилпирролидон), низкомолекулярные полипептиды, солеобразующие противоионы (такие как натрий), консерванты (такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота или пероксид водорода), растворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль), сахароспирты (такие как маннит или сорбит), суспендирующие средства, поверхностно-активные вещества или смачивающие средства (такие как разновидности плюроника; PEG; сложные эфиры сорбитана; полисорбаты, такие как полисорбат 20 или полисорбат 80; тритон; трометамин; лецитин; холестерин или тилоксапал), средства, повышающие стабильность (такие как сахароза и сорбит), средства, повышающие тоничность (такие как галогениды щелочных металлов, например, хлорид натрия или калия или маннит, сорбит), среды для доставки, разбавители, вспомогательные вещества и/или фармацевтические адъюванты (см., например, Remington's Pharmaceutical Sciences (18th Ed., A.R. Gennaro, ed., Mack Publishing Company 1990) и его последующие издания, которые включены в данный документ посредством ссылки для любой цели).
[0240] Оптимальную фармацевтическую композицию будет определять специалист в данной области техники в зависимости, например, от предполагаемого пути введения, формата доставки и требуемой дозы. На такие композиции могут оказывать влияние физическое состояние, стабильность, скорость высвобождения in vivo и скорость выведения in vivo связывающего белка.
[0241] Основная среда или носитель в фармацевтической композиции могут быть водными либо неводными по своей природе. Например, подходящей средой или носителем для инъекции может быть вода, физиологический солевой раствор или искусственная спинномозговая жидкость, возможно, с добавкой других веществ, общепринятых для композиций для парентерального введения. Нейтральный забуференный солевой раствор или солевой раствор, смешанный с сывороточным альбумином, представляют собой дополнительные приводимые в качестве примера среды. Другие приводимые в качестве примера фармацевтические композиции содержат буфер Tris со значением pH, составляющим приблизительно 7,0-8,5, или ацетатный буфер со значением pH, составляющим приблизительно 4,0-5,5, который может дополнительно включать сорбит или подходящий заменитель. В одном варианте осуществления настоящего изобретения композиции на основе связывающего белка могут быть получены для хранения в форме лиофилизированной массы или водного раствора путем смешивания выбранной композиции с требуемой степенью чистоты с необязательными средствами для составления. Кроме того, связывающий белок может быть составлен в форме лиофилизата с применением подходящих вспомогательных веществ, таких как сахароза.
[0242] Фармацевтические композиции согласно настоящему изобретению могут предназначаться для парентеральной или подкожной доставки. Альтернативно композиции могут предназначаться для ингаляции или для доставки через пищеварительный тракт, например, перорально. Получение таких фармацевтически приемлемых композиций находится в пределах компетенции специалиста в данной области техники.
[0243] Компоненты состава присутствуют в концентрациях, которые являются приемлемыми для места введения. Например, буферы используют для поддержания значения pH композиции на физиологическом уровне или при немного меньшем значении pH, обычно в диапазоне значений pH от приблизительно 5 до приблизительно 8.
[0244] Если предусмотрено парентеральное введение, то терапевтические композиции, предназначенные для применения, могут быть в форме апирогенного приемлемого для парентерального введения водного раствора, содержащего требуемый связывающий белок в фармацевтически приемлемой среде. Особенно подходящей средой для парентеральной инъекции является стерильная дистиллированная вода, с помощью которой связывающий белок составляют в виде стерильного изотонического раствора с подходящими консервантами. Еще один препарат может предусматривать состав на основе требуемой молекулы со средством, таким как инъецируемые микросферы, биоразлагаемые частицы, полимерные соединения (такие как полимолочная кислота или полигликолевая кислота), гранулы или липосомы, которое обеспечивает контролируемое или замедленное высвобождение продукта, который затем может быть доставлен посредством депо-инъекции. Можно также использовать гиалуроновую кислоту, и она может оказывать эффект обеспечения длительного пребывания в кровотоке. Другие подходящие средства для введения требуемой молекулы включают имплантируемые устройства для доставки лекарственных средств.
[0245] В одном варианте осуществления фармацевтическая композиция может быть составлена для ингаляции. Например, связывающий белок может быть составлен в виде сухого порошка для ингаляции. Растворы для ингаляций на основе связывающего белка можно также составлять с пропеллентом для аэрозольной доставки. В еще одном варианте осуществления растворы можно распылять.
[0246] Также предусматривается, что определенные составы можно вводить перорально. В одном варианте осуществления настоящего изобретения связывающие белки, которые вводят таким способом, можно составлять с такими носителями, которые обычно используют при составлении твердых лекарственных форм, таких как таблетки и капсулы, или без них. Например, капсулу можно разрабатывать с возможностью высвобождения активной части состава в момент попадания в пищеварительный тракт, где биодоступность повышается до максимума, а распад до попадания в системный кровоток сводится к минимуму. Можно включать дополнительные средства для облегчения всасывания связывающего белка. Также можно использовать разбавители, ароматизаторы, легкоплавкие воски, растительные масла, смазывающие вещества, суспендирующие средства, средства для улучшения распадаемости таблеток и связующие средства.
[0247] Другая фармацевтическая композиция может включать эффективное количество связывающих белков в смеси с нетоксичными вспомогательными веществами, которые являются подходящими для изготовления таблеток. Путем растворения таблеток в стерильной воде или другой подходящей среде можно получить растворы в форме со стандартной дозой. Подходящие вспомогательные вещества включают без ограничения инертные разбавители, такие как карбонат кальция, карбонат или бикарбонат натрия, лактоза или фосфат кальция; или связующие вещества, такие как крахмал, желатин или аравийская камедь; или смазывающие вещества, такие как стеарат магния, стеариновая кислота или тальк.
[0248] Дополнительные фармацевтические композиции согласно настоящему изобретению будут очевидны специалистам в данной области техники, в том числе составы, включающие связывающие белки, в форме с замедленной или контролируемой доставкой. Методики составления ряда других средств с замедленной или контролируемой доставкой, таких как липосомные носители, биоразлагаемые микрочастицы или пористые гранулы и формы для депо-инъекций, также известны специалистам в данной области техники. Дополнительные примеры препаратов с замедленным высвобождением включают полупроницаемые полимерные матрицы в форме изделий определенной формы, например, пленок или микрокапсул. Матрицы с замедленным высвобождением могут включать сложные полиэфиры, гидрогели, полилактиды, сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, поли(2-гидроксиэтилметакрилат), этиленвинилацетат или поли-D(-)-3-гидроксимасляную кислоту. Композиции с замедленным высвобождением также могут включать липосомы, которые можно получить любым из нескольких известных из уровня техники способов.
[0249] Фармацевтические композиции, подлежащие применению для введения in vivo, обычно должны быть стерильными. Этого можно достичь путем фильтрации через стерильные фильтрационные мембраны. Если композиция является лиофилизированной, то стерилизация с помощью этого способа может быть проведена либо до, либо после лиофилизации и восстановления. Композиция для парентерального введения может храниться в лиофилизированной форме или в форме раствора. Кроме того, композиции для парентерального введения, как правило, помещают в контейнер, имеющий стерильное входное отверстие, например, мешок с раствором для внутривенной инъекции или флакон с пробкой, которую можно проколоть гиподермической иглой для инъекций.
[0250] После того как фармацевтическая композиция составлена, ее можно хранить в стерильных флаконах в виде раствора, суспензии, геля, эмульсии, твердого вещества или в виде обезвоженного или лиофилизированного порошка. Такие составы можно хранить либо в готовой к применению форме, либо в форме (например, лиофилизированной), требующей восстановления перед введением.
[0251] Настоящее изобретение также охватывает наборы для получения формы для введения однократной дозы. Каждый из наборов может содержать как первый контейнер с высушенным белком, так и второй контейнер с водным составом. В объем настоящего изобретения также включены наборы, содержащие одно- и многокамерные предварительно заполненные шприцы (например, шприцы с жидкостью и шприцы с лиофилизатом).
[0252] Эффективное количество фармацевтической композиции на основе связывающего белка, подлежащей терапевтическому применению, будет зависеть, например, от терапевтического случая и целей. Специалисту в данной области техники будет понятно, что подходящие уровни доз для лечения будут, таким образом, меняться отчасти в зависимости от доставляемой молекулы, показания, в соответствии с которым будут применять связывающий белок, пути введения и метрических характеристик (массы тела, поверхности тела или размера органа) и состояния (возраста и общего состояния здоровья) пациента. Соответственно врач может подобрать дозу и видоизменить путь введения для получения оптимального терапевтического эффекта.
[0253] Частота дозирования будет зависеть от фармакокинетических параметров связывающего белка в составе, подлежащем применению. Обычно врач будет вводить композицию до тех пор, пока доза не достигнет такого уровня, при котором достигается необходимый эффект. Следовательно, композицию можно вводить в виде однократной дозы, в виде двух или более доз (которые могут содержать одинаковое количество требуемой молекулы или могут не содержать его) в течение определенного периода времени или в виде непрерывной инфузии посредством имплантируемого устройства или катетера. Дополнительное уточнение подходящей дозировки выполняется стандартным способом рядовыми специалистами в данной области техники и находится в диапазоне задач, которые они обычно выполняют. Подходящие дозировки можно определять с помощью соответствующих данных о дозозависимом ответе.
[0254] Путь введения фармацевтической композиции соответствует известным способам, например, перорально; посредством инъекции с помощью внутривенного, интраперитонеального, интрацеребрального (интрапаренхиматозного), интрацеребровентрикулярного, внутримышечного, внутриглазного, внутриартериального, интрапортального или внутриочагового путей; с помощью систем с замедленным высвобождением или с помощью имплантируемых устройств. При необходимости композиции можно вводить в виде болюсной инъекции или непрерывно путем инфузии или с помощью имплантируемого устройства.
[0255] Композицию также можно вводить местно посредством имплантации мембраны, губки или другого подходящего материала, в который необходимая молекула была поглощена или инкапсулирована. Если применяют имплантируемое устройство, то устройство можно имплантировать в любую подходящую ткань или орган, и доставка требуемой молекулы может осуществляться посредством диффузии, болюсного введения с регулируемым во времени высвобождением или непрерывного введения.
[0256] Фармацевтические композиции можно применять для предупреждения и/или лечения инфекции, вызванной HIV. Фармацевтические композиции можно применять в виде отдельного средства для терапии или в комбинации со стандартным средством для антиретровирусной терапии.
Настоящее изобретение также относится к набору, содержащему связывающий белок и другие реагенты, применимые для выявления уровней целевых антигенов в биологических образцах. Такие реагенты могут включать выявляемую метку, блокирующую сыворотку крови, образцы положительного и отрицательного контроля и реагенты для выявления. В некоторых вариантах осуществления набор содержит композицию, содержащую любой связывающий белок, полинуклеотид, вектор, векторную систему и/или клетку-хозяина, описанные в данном документе. В некоторых вариантах осуществления набор содержит контейнер и этикетку или листовку-вкладыш на контейнере или вместе с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, мешки с раствором для в/в инъекции и т. д. Контейнеры могут быть изготовлены из ряда материалов, таких как стекло или пластик. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией является эффективной для лечения, предупреждения и/или диагностирования состояния (например, инфекции, вызванной HIV), и он может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет с раствором для внутривенной инъекции или флакон с пробкой, которую можно проколоть иглой для подкожных инъекций). В некоторых вариантах осуществления на этикетке или листке-вкладыше указано, что композиция применяется для предупреждения, диагностирования и/или лечения выбранного состояния. Альтернативно или дополнительно изделие или набор может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-буферный раствор солевой раствор Рингера и раствор декстрозы. Он может дополнительно включать другие вещества, необходимые с коммерческой точки зрения и с точки зрения потребителя, в том числе другие буферы, разбавители, наполнители, иглы и шприцы.
Способы и варианты применения связывающих белков
Вирус
[0257] Некоторые аспекты настоящего изобретения относятся к способам размножения специфических в отношении вируса Т-клеток памяти. В некоторых вариантах осуществления способы предусматривают приведение в контакт специфической в отношении вируса Т-клетки памяти со связывающим белком по настоящему изобретению, например, триспецифическим связывающим белком, который содержит первый антигенсвязывающий участок, который связывает полипептид CD28, второй антигенсвязывающий участок, который связывает полипептид CD3, и третий антигенсвязывающий участок, который связывает полипептид CD38.
[0258] В некоторых вариантах осуществления специфическую в отношении вируса Т-клетку памяти приводят в контакт со связывающим белком in vitro или ex vivo.
[0259] В некоторых вариантах осуществления приведение специфической в отношении вируса Т-клетки памяти в контакт со связывающим белком вызывает активацию и/или пролиферацию специфических в отношении вируса Т-клеток памяти.
[0260] Другие аспекты настоящего изобретения относятся к способам размножения Т-клеток. В некоторых вариантах осуществления способы предусматривают приведение в контакт Т-клетки со связывающим белком по настоящему изобретению, например, триспецифическим связывающим белком, который содержит первый антигенсвязывающий участок, который связывает полипептид CD28, второй антигенсвязывающий участок, который связывает полипептид CD3, и третий антигенсвязывающий участок, который связывает полипептид CD38.
[0261] В некоторых вариантах осуществления Т-клетка представляет собой Т-клетку памяти или эффекторную Т-клетку.
[0262] В некоторых вариантах осуществления Т-клетка экспрессирует химерный антигенный рецептор (CAR) на своей клеточной поверхности или содержит полинуклеотид, кодирующий CAR.
[0263] Другие аспекты настоящего изобретения относятся к способам лечения хронической вирусной инфекции, например, у индивидуума, нуждающегося в этом. В некоторых вариантах осуществления способы предусматривают введение индивидууму, нуждающемуся в этом, эффективного количества связывающего белка по настоящему изобретению, например, триспецифического связывающего белка, который содержит первый антигенсвязывающий участок, который связывает полипептид CD28, второй антигенсвязывающий участок, который связывает полипептид CD3, и третий антигенсвязывающий участок, который связывает полипептид CD38.
[0264] В некоторых вариантах осуществления индивидуум является человеком.
[0265] В некоторых вариантах осуществления связывающий белок вводят индивидууму в фармацевтическом составе, содержащем связывающий белок и фармацевтически приемлемый носитель.
[0266] В некоторых вариантах осуществления введение связывающего белка приводит к активации и/или пролиферации специфических в отношении вируса Т-клеток памяти у индивидуума.
[0267] В любом из представленных выше способов Т-клетки памяти могут представлять собой CD8+ или CD4+ Т-клетки памяти. В любом из представленных выше способов Т-клетки памяти могут представлять собой центральные Т-клетки памяти (TCM) или эффекторные Т-клетки памяти (TEM).
Рак
[0268] Некоторые аспекты настоящего изобретения относятся к способам предупреждения и/или лечения рака у пациента. В некоторых вариантах осуществления способы включают введение пациенту терапевтически эффективного количества связывающего белка или фармацевтической композиции по настоящему изобретению.
[0269] В некоторых вариантах осуществления связывающий белок по настоящему изобретению вводят пациенту, нуждающемуся в этом, для лечения или предупреждения рака. В некоторых вариантах осуществления настоящее изобретение относится к способу предупреждения и/или лечения пролиферативного заболевания или нарушения (например, рака). В некоторых вариантах осуществления способ включает введение пациенту терапевтически эффективного количества по меньшей мере одного из связывающих белков или фармацевтических композиций на его основе, описанных в данном документе. В некоторых вариантах осуществления настоящее изобретение относится к вариантам применения по меньшей мере одного из связывающих белков или фармацевтических композиций на его основе, описанных в данном документе, для предупреждения и/или лечения пролиферативного заболевания или нарушения (например, рака) у пациента, нуждающегося в этом. В некоторых вариантах осуществления настоящее изобретение относится по меньшей мере к одному из связывающих белков или фармацевтических композиций на его основе, описанных в данном документе, для применения в получении лекарственного препарата для предупреждения и/или лечения пролиферативного заболевания или нарушения (например, рака) у пациента, нуждающегося в этом. В некоторых вариантах осуществления пациент представляет собой человека.
[0270] В некоторых вариантах осуществления по меньшей мере один связывающий белок вводят (или необходимо вводить) в комбинации с одним или несколькими средствами противоопухолевой терапии (например, любым средством противоопухолевой терапии, известным в уровне техники, таким как химиотерапевтическое средство или терапия). В некоторых вариантах осуществления по меньшей мере один связывающий белок вводят (или необходимо вводить) перед введением одного или нескольких средств противоопухолевой терапии. В некоторых вариантах осуществления по меньшей мере один связывающий белок вводят (или необходимо вводить) одновременно с одним или несколькими средствами противоопухолевой терапии. В некоторых вариантах осуществления по меньшей мере один связывающий белок вводят (или необходимо вводить) после введения одного или нескольких средств противоопухолевой терапии.
[0271] В некоторых вариантах осуществления связывающий белок содержит один или два антигенсвязывающих участка, которые связывают поверхностный белок Т-клетки, и другой антигенсвязывающий участок, который связывает внеклеточный домен полипептида HER2 человека. В некоторых вариантах осуществления связывающий белок содержит антигенсвязывающий участок, который связывает внеклеточный домен полипептида HER2 человека, антигенсвязывающий участок, который связывает полипептид CD28 человека, и антигенсвязывающий участок, который связывает полипептид CD3 человека.
[0272] В некоторых вариантах осуществления клетки рака индивидуума экспрессируют HER2. В некоторых вариантах осуществления пациента выбирают для лечения на основании того, что клетки рака экспрессируют полипептид HER2 человека. Анализы, известные в уровне техники, применимые для обнаружения экспрессии HER2 клетками рака, включают без ограничения иммуногистохимические анализы (IHC) и флуоресцентную гибридизацию in situ (FISH) .
[0273] В некоторых вариантах осуществления рак (например, HER2-положительный рак) представляет собой рак молочной железы, рак толстой кишки, рак желудка или немелкоклеточный рак легкого (NSCLC).
[0274] В некоторых вариантах осуществления связывающий белок содержит один или два антигенсвязывающих участка, которые связывают поверхностный белок Т-клетки, и другой антигенсвязывающий участок, который связывает внеклеточный домен полипептида CD38 человека. В некоторых вариантах осуществления связывающий белок содержит антигенсвязывающий участок, который связывает внеклеточный домен полипептида CD38 человека, антигенсвязывающий участок, который связывает полипептид CD28 человека, и антигенсвязывающий участок, который связывает полипептид CD3 человека.
[0275] В некоторых вариантах осуществления клетки рака индивидуума экспрессируют CD38. В некоторых вариантах осуществления клетки рака экспрессируют на своей клеточной поверхности полипептид изоформы A CD38 человека. В некоторых вариантах осуществления клетки рака экспрессируют на своей клеточной поверхности полипептид изоформы Е CD38 человека. В некоторых вариантах осуществления пациента выбирают для лечения на основании того, что клетки рака экспрессируют на своей клеточной поверхности полипептид изоформы Е CD38 человека. В некоторых вариантах осуществления клетки рака экспрессируют CD38 и CD28. В некоторых вариантах осуществления клетки рака экспрессируют CD38 и не экспрессируют CD28.
[0276] В некоторых вариантах осуществления рак (например, CD38-положительный рак) представляет собой множественную миелому, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, острый миелоидный лейкоз, лимфому, рак молочной железы, такой как рак молочной железы Her2+, рак предстательной железы, лимфому из В-клеток зародышевого центра или В-клеточный острый лимфобластный лейкоз. В определенных вариантах осуществления рак представляет собой множественную миелому. В определенных вариантах осуществления рак представляет собой острый миелоидный лейкоз (AML), острый лимфобластный лейкоз (ALL), хронический лимфоцитарный лейкоз (CLL) или B-клеточную лимфому.
[0277] В определенных вариантах осуществления рак представляет собой множественную миелому. Антитела к CD38, такие как даратумумаб, тестировали в отношении применения для лечения множественной миеломы. Однако несмотря на то, что множественная миелома считается излечимой, рецидив неизбежен практически у всех пациентов, что приводит к развитию резистентного к лечению заболевания. В некоторых вариантах осуществления рак представляет собой рецидивирующую или рефрактерную множественную миелому. В некоторых вариантах осуществления пациент был подвергнут лечению с использованием предшествующего курса лечения множественной миеломы. В некоторых вариантах осуществления связывающий белок по настоящему изобретению вводят пациенту как лечение множественной миеломы 1й, 2й или 3й линии. Не вдаваясь в теорию, полагают, что связывающий белок-антитело к CD38x-CD28x-CD3 по настоящему изобретению может быть применим в лечении множественной миеломы, например, посредством рекрутинга Т-клеток к опухолевым клеткам при участии антитела к CD38 (или антитела к CD38/CD28), активации вовлеченных Т-клеток при участии антител к CD3/CD28 и/или уничтожения опухолевых клеток с помощью механизмов на основе перфорина/гранзима. Сообщалось, что CD28 является новым маркером рака при множественной миеломе. См. Nair, J.R. et al. (2011) J. Immunol. 187:1243-1253.
[0278] Любые из связывающих белков, описываемых в данном документе, могут найти применение в способах по настоящему изобретению.
[0279] В некоторых вариантах осуществления любого из способов по настоящему изобретению перед введением связывающего белка пациента подвергали лечению даратумумабом. Как описано в данном документе, в настоящем изобретении предусмотрены CD38-связывающие белки и участки, которые не конкурируют за связывание CD38 с даратумумабом. Не ограничиваясь какой-либо теорией, полагают, что это является преимуществом, поскольку пациента, ранее подвергавшегося лечению даратумумабом, можно лечить связывающим белком по настоящему изобретению, например, без периода вымывания перед лечением.
[0280] Связывающие белки можно использовать в любом известном способе анализа, таком как анализы конкурентного связывания, прямые и непрямые сэндвич-анализы и анализы иммунопреципитации для выявления и количественного определения одного или нескольких антигенов-мишеней. Связывающие белки будут связывать один или несколько антигенов-мишеней с аффинностью, которая соответствует используемому способу анализа.
[0281] Для диагностических применений в определенных вариантах осуществления связывающие белки можно метить выявляемым фрагментом. Выявляемый фрагмент может представлять собой любой фрагмент, который способен непосредственно либо опосредованно генерировать выявляемый сигнал. Например, выявляемый фрагмент может представлять собой радиоизотоп, такой как 3H, 14C, 32P, 35S, 125I, 99Tc, 111In или 67Ga; флуоресцентное или хемилюминесцентное соединение, такое как изотиоцианат флуоресцеина, родамин или люциферин; или фермент, такой как щелочная фосфатаза, β-галактозидаза или пероксидаза хрена.
[0282] Связывающие белки также применимы для визуализации in vivo. Связывающий белок, меченный выявляемым фрагментом, можно вводить животному, предпочтительно в кровоток, и анализировать присутствие и положение меченного антитела в организме хозяина. Связывающий белок можно метить любым фрагментом, который можно выявить в организме животного с помощью ядерного магнитного резонанса, рентгенологического исследования либо с помощью других способов выявления, известных в данной области техники.
[0283] Для клинических или исследовательских применений в некоторых вариантах осуществления связывающие белки могут быть конъюгированы с цитотоксическим средством. Для нацеливания цитотоксических полезных нагрузок на конкретные опухолевые клетки использовали разнообразные антитела, связанные с цитотоксическими средствами (то есть конъюгаты антитело-лекарственное средство). Цитотоксические средства и линкеры, которые обеспечивают конъюгацию средств с антителом, известны в уровне техники; см., например, Parslow, A.C. et al. (2016) Biomedicines 4:14 и Kalim, M. et al. (2017) Drug Des. Devel. Ther. 11:2265-2276.
Терапевтические композиции на основе связывающих белков и их введение
[0284] Терапевтические или фармацевтические композиции, содержащие связывающие белки, находятся в пределах объема настоящего изобретения. Такие терапевтические или фармацевтические композиции могут содержать терапевтически эффективное количество связывающего белка или конъюгата связывающий белок-лекарственное средство в смеси с фармацевтически или физиологически приемлемым средством для составления, выбранным в соответствии со способом введения. Эти фармацевтические композиции могут найти применение в любом из способов и вариантов применения, описываемых в данном документе (например, ex vivo, in vitro и/или in vivo).
[0285] Приемлемые вещества для составления предпочтительно являются нетоксичными для получающих их пациентов при используемых дозах и концентрациях.
[0286] Фармацевтическая композиция может содержать вещества для составления, предназначенные для модификации, поддержания или сохранения, например pH, осмолярности, вязкости, прозрачности, цвета, изотоничности, запаха, стерильности, стабильности, скорости растворения или высвобождения, всасывания или проникновения композиции. Подходящие вещества для составления включают без ограничения аминокислоты (такие как глицин, глутамин, аспарагин, аргинин или лизин), противомикробные средства, антиоксиданты (такие как аскорбиновая кислота, сульфит натрия или гидросульфит натрия), буферы (такие как борат, бикарбонат, Tris-HCl, цитраты, фосфаты или другие органические кислоты), объемообразующие вещества (такие как маннит или глицин), хелатирующие вещества (такие как этилендиаминтетрауксусная кислота (EDTA)), комплексообразующие вещества (такие как кофеин, поливинилпирролидон, бета-циклодекстрин или гидроксипропил-бета-циклодекстрин), наполнители, моносахариды, дисахариды и другие углеводы (такие как глюкоза, манноза или декстрины), белки (такие как сывороточный альбумин, желатин или иммуноглобулины), красители, ароматизаторы и разбавители, эмульгаторы, гидрофильные полимеры (такие как поливинилпирролидон), низкомолекулярные полипептиды, солеобразующие противоионы (такие как ион натрия), консерванты (такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота или пероксид водорода), растворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль), сахароспирты (такие как маннит или сорбит), суспендирующие вещества, поверхностно-активные вещества или смачивающие вещества (такие как плюроники; PEG; сложные эфиры сорбита; полисорбаты, такие как полисорбат 20 или полисорбат 80; тритон; трометамин; лецитин; холестерин или тилоксапол), вещества, повышающие стабильность (такие как сахароза и сорбит), вещества, повышающие тоничность (такие как галогениды щелочных металлов - предпочтительно хлорид натрия или калия - или маннит, сорбит), среды для доставки, разбавители, вспомогательные вещества и/или фармацевтические адъюванты (см., например, Remington's Pharmaceutical Sciences (18th Ed., A.R. Gennaro, ed., Mack Publishing Company 1990) и его последующие издания, которые включены в данный документ посредством ссылки для любой цели).
[0287] Оптимальную фармацевтическую композицию будет определять специалист в данной области техники в зависимости, например, от предполагаемого пути введения, формата доставки и требуемой дозы. На такие композиции могут оказывать влияние физическое состояние, стабильность, скорость высвобождения in vivo и скорость выведения in vivo связывающего белка.
[0288] Основная среда или носитель в фармацевтической композиции могут быть водными либо неводными по своей природе. Например, подходящей средой или носителем для инъекции может быть вода, физиологический солевой раствор или искусственная спинномозговая жидкость, возможно, с добавкой других веществ, общепринятых для композиций для парентерального введения. Нейтральный забуференный солевой раствор или солевой раствор, смешанный с сывороточным альбумином, представляют собой дополнительные приводимые в качестве примера среды. Другие приводимые в качестве примера фармацевтические композиции содержат буфер Tris со значением pH, составляющим приблизительно 7,0-8,5, или ацетатный буфер со значением pH, составляющим приблизительно 4,0-5,5, который может дополнительно включать сорбит или подходящий заменитель. В одном варианте осуществления настоящего изобретения композиции на основе связывающего белка могут быть получены для хранения в форме лиофилизированной массы или водного раствора путем смешивания выбранной композиции с требуемой степенью чистоты с необязательными средствами для составления. Кроме того, связывающий белок может быть составлен в форме лиофилизата с применением подходящих вспомогательных веществ, таких как сахароза.
[0289] Фармацевтические композиции согласно настоящему изобретению могут предназначаться для парентеральной или подкожной доставки. Альтернативно композиции могут предназначаться для ингаляции или для доставки через пищеварительный тракт, например, перорально. Получение таких фармацевтически приемлемых композиций находится в пределах компетенции специалиста в данной области техники.
[0290] Компоненты состава присутствуют в концентрациях, которые являются приемлемыми для места введения. Например, буферы используют для поддержания значения pH композиции на физиологическом уровне или при немного меньшем значении pH, обычно в диапазоне значений pH от приблизительно 5 до приблизительно 8.
[0291] Если предусмотрено парентеральное введение, то терапевтические композиции, предназначенные для применения, могут быть в форме апирогенного приемлемого для парентерального введения водного раствора, содержащего требуемый связывающий белок в фармацевтически приемлемой среде. Особенно подходящей средой для парентеральной инъекции является стерильная дистиллированная вода, с помощью которой связывающий белок составляют в виде стерильного изотонического раствора с подходящими консервантами. Еще один препарат может предусматривать состав на основе требуемой молекулы со средством, таким как инъецируемые микросферы, биоразлагаемые частицы, полимерные соединения (такие как полимолочная кислота или полигликолевая кислота), гранулы или липосомы, которое обеспечивает контролируемое или замедленное высвобождение продукта, который затем может быть доставлен посредством депо-инъекции. Можно также использовать гиалуроновую кислоту, и она может оказывать эффект обеспечения длительного пребывания в кровотоке. Другие подходящие средства для введения требуемой молекулы включают имплантируемые устройства для доставки лекарственных средств.
[0292] В одном варианте осуществления фармацевтическая композиция может быть составлена для ингаляции. Например, связывающий белок может быть составлен в виде сухого порошка для ингаляции. Растворы для ингаляций на основе связывающего белка можно также составлять с пропеллентом для аэрозольной доставки. В еще одном варианте осуществления растворы можно распылять.
[0293] Также предусматривается, что определенные составы можно вводить перорально. В одном варианте осуществления настоящего изобретения связывающие белки, которые вводят таким способом, можно составлять с такими носителями, которые обычно используют при составлении твердых лекарственных форм, таких как таблетки и капсулы, или без них. Например, капсула может быть предназначена для высвобождения активной части состава в момент попадания в пищеварительный тракт, где биодоступность повышается до максимума, а распад до попадания в системный кровоток сводится к минимуму. Можно включать дополнительные средства для облегчения всасывания связывающего белка. Также можно использовать разбавители, ароматизаторы, легкоплавкие воски, растительные масла, смазывающие вещества, суспендирующие средства, средства для улучшения распадаемости таблеток и связующие средства.
[0294] Другая фармацевтическая композиция может включать эффективное количество связывающих белков в смеси с нетоксичными вспомогательными веществами, которые являются подходящими для изготовления таблеток. Путем растворения таблеток в стерильной воде или другой подходящей среде можно получить растворы в форме со стандартной дозой. Подходящие вспомогательные вещества включают без ограничения инертные разбавители, такие как карбонат кальция, карбонат или бикарбонат натрия, лактоза или фосфат кальция; или связующие вещества, такие как крахмал, желатин или аравийская камедь; или смазывающие вещества, такие как стеарат магния, стеариновая кислота или тальк.
[0295] Дополнительные фармацевтические композиции согласно настоящему изобретению будут очевидны специалистам в данной области техники, в том числе составы, включающие связывающие белки, в форме с замедленной или контролируемой доставкой. Методики составления ряда других средств с замедленной или контролируемой доставкой, таких как липосомные носители, биоразлагаемые микрочастицы или пористые гранулы и формы для депо-инъекций, также известны специалистам в данной области техники. Дополнительные примеры препаратов с замедленным высвобождением включают полупроницаемые полимерные матрицы в форме изделий определенной формы, например, пленок или микрокапсул. Матрицы с замедленным высвобождением могут включать сложные полиэфиры, гидрогели, полилактиды, сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата, поли(2-гидроксиэтилметакрилат), этиленвинилацетат или поли-D(-)-3-гидроксимасляную кислоту. Композиции с замедленным высвобождением также могут включать липосомы, которые можно получить любым из нескольких известных из уровня техники способов.
[0296] Фармацевтические композиции, подлежащие применению для введения in vivo, обычно должны быть стерильными. Этого можно достичь путем фильтрации через стерильные фильтрационные мембраны. Если композиция является лиофилизированной, то стерилизация с помощью этого способа может быть проведена либо до, либо после лиофилизации и восстановления. Композиция для парентерального введения может храниться в лиофилизированной форме или в форме раствора. Кроме того, композиции для парентерального введения, как правило, помещают в контейнер, имеющий стерильное входное отверстие, например, мешок с раствором для внутривенной инъекции или флакон с пробкой, которую можно проколоть гиподермической иглой для инъекций.
[0297] После того как фармацевтическая композиция составлена, ее можно хранить в стерильных флаконах в виде раствора, суспензии, геля, эмульсии, твердого вещества или в виде обезвоженного или лиофилизированного порошка. Такие составы можно хранить либо в готовой к применению форме, либо в форме (например, лиофилизированной), требующей восстановления перед введением.
[0298] Настоящее изобретение также относится к набору, содержащему связывающий белок и другие реагенты, применимые для выявления уровней целевых антигенов в биологических образцах. Такие реагенты могут включать выявляемую метку, блокирующую сыворотку крови, образцы положительного и отрицательного контроля и реагенты для выявления. В некоторых вариантах осуществления набор содержит композицию, содержащую любой связывающий белок, полинуклеотид, вектор, векторную систему и/или клетку-хозяина, описанные в данном документе. В некоторых вариантах осуществления набор содержит контейнер и этикетку или листовку-вкладыш на контейнере или вместе с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, мешки с раствором для в/в инъекции и т. д. Контейнеры могут быть изготовлены из ряда материалов, таких как стекло или пластик. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией является эффективной для лечения, предупреждения и/или диагностирования состояния и может иметь стерильное входное отверстие (например, контейнер может представлять собой мешок с раствором для внутривенной инъекции или флакон с пробкой, которую можно проколоть иглой для подкожных инъекций). В некоторых вариантах осуществления на этикетке или листке-вкладыше указано, что композиция применяется для предупреждения, диагностирования и/или лечения выбранного состояния. Альтернативно или дополнительно изделие или набор может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-буферный раствор солевой раствор Рингера и раствор декстрозы. Он может дополнительно включать другие вещества, необходимые с коммерческой точки зрения и с точки зрения потребителя, в том числе другие буферы, разбавители, наполнители, иглы и шприцы.
[0299] Настоящее изобретение также охватывает наборы для получения формы для введения однократной дозы. Каждый из наборов может содержать как первый контейнер с высушенным белком, так и второй контейнер с водным составом. В объем настоящего изобретения также включены наборы, содержащие одно- и многокамерные предварительно заполненные шприцы (например, шприцы с жидкостью и шприцы с лиофилизатом).
[0300] Эффективное количество фармацевтической композиции на основе связывающего белка, подлежащей терапевтическому применению, будет зависеть, например, от терапевтического случая и целей. Специалисту в данной области техники будет понятно, что подходящие уровни доз для лечения будут, таким образом, меняться отчасти в зависимости от доставляемой молекулы, показания, в соответствии с которым будут применять связывающий белок, пути введения и метрических характеристик (массы тела, поверхности тела или размера органа) и состояния (возраста и общего состояния здоровья) пациента. Соответственно врач может подобрать дозу и видоизменить путь введения для получения оптимального терапевтического эффекта.
[0301] Частота дозирования будет зависеть от фармакокинетических параметров связывающего белка в составе, подлежащем применению. Обычно врач будет вводить композицию до тех пор, пока доза не достигнет такого уровня, при котором достигается необходимый эффект. Следовательно, композицию можно вводить в виде однократной дозы, в виде двух или более доз (которые могут содержать одинаковое количество требуемой молекулы или могут не содержать его) в течение определенного периода времени или в виде непрерывной инфузии посредством имплантируемого устройства или катетера. Дополнительное уточнение подходящей дозировки выполняется стандартным способом рядовыми специалистами в данной области техники и находится в диапазоне задач, которые они обычно выполняют. Подходящие дозировки можно определять с помощью соответствующих данных о дозозависимом ответе.
[0302] Путь введения фармацевтической композиции соответствует известным способам, например, перорально; посредством инъекции с помощью внутривенного, интраперитонеального, интрацеребрального (интрапаренхиматозного), интрацеребровентрикулярного, внутримышечного, внутриглазного, внутриартериального, интрапортального или внутриочагового путей; с помощью систем с замедленным высвобождением или с помощью имплантируемых устройств. При необходимости композиции можно вводить в виде болюсной инъекции или непрерывно путем инфузии или с помощью имплантируемого устройства.
[0303] Композицию также можно вводить местно посредством имплантации мембраны, губки или другого подходящего материала, в который необходимая молекула была поглощена или инкапсулирована. Если применяют имплантируемое устройство, то устройство можно имплантировать в любую подходящую ткань или орган, и доставка требуемой молекулы может осуществляться посредством диффузии, болюсного введения с регулируемым во времени высвобождением или непрерывного введения.
Таблица 1. Полипептидные последовательности триспецифического связывающего белка
BP № 1
BP № 2
BP № 8
BP № 3
BP № 4
BP № 5
BP № 6
BP № 9
BP № 10
BP № 11
BP № 12
BP № 15
BP № 25
BP № 26
BP № 27
BP № 28
BP № 29
BP № 31
BP № 1
BP № 5'
BP № 6'
BP № 7
BP № 8
BP № 9
Таблица 6. Полинуклеотидные последовательности триспецифического связывающего белка
BP № 1
BP № 2
BP № 8
BP № 9
BP № 10
BP № 11
BP № 12
BP № 15
BP № 25
BP № 26
BP № 27
BP № 28
BP № 29
BP № 31
BP № 1
BP № 5
BP № 6
BP № 7
BP № 8
BP № 9
ПРИМЕРЫ
[0304] Приведенные ниже примеры иллюстрируют конкретные варианты осуществления настоящего изобретения и различные варианты их применения. Они приведены только для пояснительных целей, и их не следует рассматривать как каким-либо образом ограничивающие объем настоящего изобретения.
Пример 1. Разработка триспецифических антител к HER2/CD28xCD3 и вариантов CD3-связывающих участков
[0305] Иммуноонкология представляет собой многообещающий развивающийся терапевтический подход для управления ходом заболевания при раке. Иммунная система представляет собой первую линию защиты от развития и прогрессирования рака. В настоящее время существует множество доказательств того, что Т-клетки способны контролировать рост опухоли и продлевать время выживания пациентов, страдающих раком, как на ранних, так и на поздних стадиях заболевания. Однако Т-клетки, специфичные в отношении опухолей, могут быть ограничены во многих отношениях, что не позволяет им контролировать заболевание.
[0306] Для устранения ограничений T-клеток под воздействием неконтролируемых опухолей разработали новые антитела в формате триспецифических антител, изображенном на фиг. 1A, чтобы специфически активировать Т-клетки для взаимодействия с экспрессирующими HER2 раковыми клетками. Данные новые триспецифические антитела способны связываться с тремя мишенями: HER2, CD3 и CD28. HER2- и CD3-связывающие участки дополнительно оптимизировали для обеспечения связывания с высокой аффинностью и уменьшения потенциальных недостатков при их получении.
[0307] Амплификацию и сверхэкспрессию HER2 можно обнаружить в молекулярных подтипах рака молочной железы, а также при карциномах желудка, яичника, легкого и предстательной железы. Для оптимальной активации Т-клеток необходимы два фактора: (1) распознавание антигена и (2) костимуляция. При использовании триспецифических связывающих HER2/CD28xCD3 белков, описанных в данном документе, сигнал 1 обеспечивается CD3-связывающим участком-агонистом, а сигнал 2 обеспечивается CD28-связывающим участком-агонистом (см., например, фиг. 1D). Считается, что триспецифические антитела, описанные в последующих примерах, обеспечивают рекрутинг Т-клеток к опухоли посредством HER2 и активируют привлеченные Т-клетки путем связывания с CD3 и CD28. Происходящая в результате активация индуцирует потенциал иммунных клеток в отношении уничтожения близлежащих опухолевых клеток.
Материалы и методы
Получение и определение характеристик антител
[0308] Варианты триспецифических антител получали путем транзиентной трансфекции клеток Expi293 экспрессионными плазмидами. Через 5 дней после трансфекции надосадочную жидкость культуры трансфицированных клеток собирали, подвергали количественному определению и нормализовали по поглощению при 280 нм на спектрофотометре Nano Drop. Связывание надосадочной жидкости с соответствующими антигенами определяли с помощью ELISA, и показатель поглощения исходного триспецифического Ab WT к HER2 принимали равным 1,0. Кратность изменений связывания у других вариантов рассчитывали путем деления соответствующего показателя поглощения на показатель поглощения исходного Ab.
[0309] Варианты триспецифических антител очищали с использованием аффинной очистки на основе белка A с последующей очисткой посредством SEC. Связывание очищенных антител с соответствующими антигенами определяли с помощью ELISA. EC50 определяли на основании кривой связывания, построенной с помощью Graphpad Prism7.
Результаты
[0310] Получали варианты триспецифических Ab с несколькими мутациями в связывающих плечах с целью частичного устранения потенциальных недостатков при их получении, например, участков дезамидирования. Анализ связывания ELISA проводили для оценки связывания указанных триспецифических антител с каждой из трех мишеней: HER2, CD3 и CD28. На фиг. 1B триспецифические антитела к HER2/CD3xCD28 с указанными HER2- или CD3-связывающими вариантами сравнивали с исходным триспецифическим антителом. Введение некоторых наборов мутаций (например, 32/33 QQ и 33/35QQ) в домен VL CD3-связывающего участка привело к значительному снижению связывания с CD3, тогда как мутации 32/35 QQ сохраняли связывание почти на уровне дикого типа. MS-анализ пептидов показал, что связывающие участки с мутациями DNAQ в CDR-L1 (SEQ ID NO:63) все еще подвергались дезамидированию, составлявшему более 15%, тогда как мутации ENLQ (SEQ ID NO:281), ENLF (SEQ ID NO:282) и ENLR (SEQ ID NO:283) приводили к дезамидированию, составлявшему менее 5%. Важно отметить, что данные варианты также сохраняли связывание с CD3.
[0311] В дополнение к этому, кривые связывания указанных антител, связывающихся с HER2 человека, CD28 человека и CD3, представлены на фиг. 1C. Значения EC50 выбранных вариантов триспецифических антител представлены в таблице E.
Таблица E. Анализ связывания ELISA выполняли с очищенными триспецифическими антителами для определения их аффинности связывания с HER2, CD3 человека и CD28 человека.
EC50
[0312] Не ограничиваясь какой-либо теорией, как показано на фиг. 1D, полагают, что триспецифические антитела к HER2/CD3/CD28 обеспечивают рекрутинг T-клеток к раковым клеткам за счет HER2- и CD3/CD28-связывающих плечей. Дополнительно полагают, что привлеченные Т-клетки активируются с помощью CD28/CD3-связывающих плечей. Не ограничиваясь какой-либо теорией, полагают, что уничтожение раковых клеток происходит за счет механизмов, опосредованных Т-клетками (например, с участием перфорина, гранзима). Не ограничиваясь какой-либо теорией, предполагается, что аналогичные механизмы могут обеспечить возможность уничтожения других типов опухолей за счет замещения на антигенсвязывающие участки, которые распознают другие опухолевые белки-мишени.
Пример 2. Разработка триспецифических антител к CD38/CD3xCD28
[0313] Разрабатывали триспецифические антитела к CD38/CD3xCD28 и определяли их свойства связывания с полипептидами CD38, CD3 и CD28.
Материалы и методы
Получение триспецифических антител к CD38/CD28xCD3
[0314] Панель антител к CD38, к CD3 и к CD28, а также Fc-домены IgG4 человека использовали для получения триспецифических антител к CD38/CD28xCD3 в формате триспецифического антитела, изображенном на фиг. 2A.
[0315] Триспецифические связывающие белки получали путем транзиентной трансфекции клеток Expi293 4 экспрессионными плазмидами с помощью набора для трансфекции ExpiFectamine™ 293 (Thermo Fisher Scientific) в соответствии с протоколом производителя. Вкратце 25% (вес/вес) каждой плазмиды разбавляли в Opti-MEM, смешивали с предварительно разбавленным реагентом ExpiFectamine в течение 20-30 минут при комнатной температуре (к. т.) и добавляли к клеткам Expi293 (2,5×106 клеток/мл). Оптимизацию трансфекции для определения наилучшего соотношения плазмид часто использовали с целью получения триспецифического связывающего белка с хорошим выходом и чистотой.
[0316] Через 4-5 дней после трансфекции надосадочную жидкость культуры трансфицированных клеток собирали и фильтровали через фильтрующий блок с размером пор 0,45 мкм (Nalgene). Триспецифический связывающий белок в надосадочной жидкости очищали с помощью 3-стадийной процедуры. На первой стадии применяли аффинную очистку с помощью белка A, и связанное Ab элюировали с помощью "буфера для элюирования IgG" (Thermo Fisher Scientific). На второй стадии продукт подвергали диализу против PBS (pH 7,4) в течение ночи с 2 заменами буфера PBS. Перед проведением следующей стадии весь образовавшийся осадок очищали путем фильтрации через фильтрующий блок с размером пор 0,45 мкм (Nalgene). На третьей стадии применяли очистку с помощью эксклюзионной хроматографии (SEC) (Hiload 16/600 Superdex 200pg или Hiload 26/600 Superdex 200pg, GE Healthcare) для удаления агрегатов и других структур в образце. Перед объединением фракции анализировали с помощью SDS-PAGE в восстанавливающих и невосстанавливающих условиях, чтобы идентифицировать фракции, содержащие мономерный триспецифический связывающий белок. Очищенное антитело можно разделить на аликвоты и хранить при -80°C в течение длительного срока.
Анализ связывания ELISA
[0317] Аффинности связывания каждого антигена-мишени у связывающих CD38/CD28xCD3 активаторов Т-клеток измеряли с помощью ELISA. Вкратце, каждый антиген использовали для покрытия 96-луночного планшета Immuno Plate (Thermo Fisher Scientific) на протяжении ночи при 4°C с использованием каждого антигена из расчета 200 нг/лунка в PBS (pH 7,4). Покрытый планшет блокировали с использованием 5% обезжиренного молока+2% BSA в PBS в течение одного часа при к. т. с последующим трехкратным промыванием с помощью PBS+0,25% Tween 20 (Aqua Max 400, Molecular Devices). Получали серийные разведения антител (триспецифических и контрольных Ab) и добавляли в планшеты для ELISA (100 мкл/лунка в двух повторах), инкубировали при комнатной температуре (к. т.) в течение одного часа с последующим 5-кратным промыванием с помощью PBS+0,25% Tween 20. После промывания в каждую лунку добавляли вторичное антитело к Fab человека, конъюгированное с HRP (1:5000, № по кат. 109-035-097, Jackson ImmunoResearch Inc) и инкубировали при к. т. в течение 30 минут. После 5-кратного промывания с помощью PBS+0,25% Tween 20 в каждую лунку добавляли по 100 мкл субстрата для пероксидазы TMB Microwell (KPL, Гейтерсберг, Мэриленд, США). Реакцию останавливали путем добавления 50 мкл 1 M H2SO4, а OD450 измеряли с помощью SpectraMax M5 (Molecular Devices) и анализировали с помощью программного обеспечения SoftMax Pro6.3 (Molecular Devices). Окончательные данные переносили в программное обеспечение GraphPad Prism (GraphPad Software, Калифорния, США) и откладывали на графике. EC50 рассчитывали с помощью того же программного обеспечения.
Измерение связывания триспецифических антител с использованием SPR
[0318] Для полного кинетического анализа использовали антигены CD38-His человека (Cambridge Biologics, Кембридж, Массачусетс). Получение кинетических характеристик очищенных антител выполняли с помощью технологии SPR на BIACORE 3000 (GE Healthcare). Применяли анализ захвата с использованием захвата антителами, специфическими к IgG1 человека, и ориентации исследуемых антител. Для захвата белковых конструкций, содержащих Fc, использовали набор для захвата антител человека (GE Healthcare). Для захвата антигена, меченного His, использовали набор для захвата на основе антител к His (GE Healthcare). Антитело захвата иммобилизовали за счет первичных аминогрупп (11000 RU) на чипе CM5 высокой чистоты (GE Life Sciences) с помощью стандартных процедур. Анализируемое антитело захватывали при скорости потока 10 мкл/мин. со скорректированным значением RU, что должно было давать в результате максимальный сигнал связывания с аналитом, обычно составляющий 30 RU. Характеристики кинетики связывания измеряли относительно триспецифических антител. В качестве аналитического буфера использовали HBS-EP (10 мМ HEPES, pH 7,4, 150 мМ NaCl, 3 мМ EDTA и 0,005% поверхностно-активного вещества P20) при скорости потока 30 мкл/мин. Поверхности чипов подвергали регенерации с помощью регенерирующего раствора из соответствующего набора для захвата. Кинетические параметры анализировали и рассчитывали с помощью пакета программ BIA evaluation v4.1 с использованием проточной кюветы без захваченного антитела в качестве эталона и модели Ленгмюра для связывания 1:1 с массобменом.
Анализ конкурентного связывания с участием даратумумаба
[0319] Даратумумаб иммобилизовали по амино-группе на активной поверхности чипа CM5 для анализа конкурентного связывания с участием даратумумаба. Эталонную поверхность оставляли пустой и использовали для вычитания сигнала, обусловленного любым неспецифическим связыванием введенных молекул. Рекомбинантный CD38-His (Sino Biological, номер партии 10818-H08H) вводили над поверхностью с даратумумабом с последующим введением тестируемых антител. Если моноспецифическое антитело к CD38 распознавало эпитоп на CD38, отличный от распознаваемого даратумумабом эпитопа, введение антитела приводило к усилению сигнала SPR. Если антитело распознавало эпитоп, перекрывающийся с распознаваемым даратумумабом эпитопом, введение антитела не усиливало сигнал SPR.
Результаты
[0320] Аффинность связывания выбранных триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами в отношении CD38 человека определяли с помощью SPR. Константа скорости ассоциации (KOn), константа скорости диссоциации (KOff) и KD выбранных триспецифических антител представлены в таблице A. Выбранные триспецифические антитела продемонстрировали различную степень аффинности с антигеном CD38 человека.
Таблица A. Характеристики связывания выбранных триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами в отношении CD38 человека, определенные с помощью SPR.
[0321] Затем аффинность связывания выбранных триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами в отношении CD3 человека, CD28 человека, CD38 человека и CD38 макака-крабоеда определяли с помощью ELISA, как описано выше. Как показано на фиг. 2B-2E, выбранные триспецифические антитела CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами продемонстрировали различную аффинность в отношении CD38 человека (фиг. 2B) и CD38 макака-крабоеда (фиг. 2C), но аналогичную аффинность в отношении CD3 человека (фиг. 2D) и CD28 человека (фиг. 2E). Затем рассчитывали значения EC50 с помощью GraphPad Prism 7.02 с использованием модели переменного углового коэффициента с четырехпараметрической логистической кривой. Значения EC50 у выбранных триспецифических антител в отношении CD3 человека, CD28 человека, CD38 человека и CD38 макака-крабоеда представлены в таблице B. Контрольным антителом был человеческий IgG4 изотипического контроля.
Таблица B. Значения EC50 у выбранных триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами в отношении CD3 человека, CD28 человека, CD38 человека и CD38 макака-крабоеда.
[0322] Проводили конкурентный SPR-анализ для определения того, конкурируют ли антитела к CD38, hhy6284, hhy992, hhy5379 или hhy1195 (тестируемые в формате моноспецифических антител), с даратумумабом за связывание с CD38. После введения CD38 над даратумумабом (иммобилизованном на сенсорном чипе для SPR) тестируемые антитела (или даратумумаб) вводили над комплексом даратумумаб/CD38. Как показано на фиг. 3, введение Hyb6264, hhy992, Hyb5379 и Hhy1195 увеличивало сигнал SPR, что указывает на то, что данные антитела распознавали эпитопы на CD38, которые отличаются от эпитопа, который распознает даратумумаб. Как и ожидалось, введение свободного даратумумаба (контроль конкурентного связывания) не увеличивало сигнал SPR.
[0323] Связывание антител к CD38 с полипептидами CD38 человека или макака-крабоеда обобщенно представлено в таблице B2.
Таблица B2. Краткое изложение характеристик связывания антител к CD38 c CD38 человека или макака-крабоеда.
[0324] Конкурентное связывание антител к CD38 и даратумумаба также тестировали с помощью SPR-анализа. Даратумумаб иммобилизовали по амино-группе на активной поверхности чипа CM5 для анализа конкурентного связывания с участием даратумумаба. Эталонную поверхность оставляли пустой и использовали для вычитания сигнала, обусловленного любым неспецифическим связыванием введенных молекул. Рекомбинантный CD38-His (Sino Biological, номер партии 10818-H08H) вводили над поверхностью с даратумумабом с последующим введением тестируемых антител. Если антитело распознает эпитоп на CD38, отличный от распознаваемого даратумумабом эпитопа, введение антитела приведет к усилению сигнала SPR. Если антитело распознает эпитоп, перекрывающийся с распознаваемым даратумумабом эпитопом, введение антитела не будет усиливать сигнал SPR. Согласно результатам данных анализов протестированные антитела hhy992, hyb6284, hhy1195 и hhy1370 не конкурировали с даратумумабом.
Пример 3. Триспецифические антитела к CD38/CD3xCD28 содействуют лизису опухолевых клеток множественной миеломы и лимфомы человека.
[0325] Для определения того, обладают ли триспецифические антитела к CD38/CD3xCD28 активностью в отношении опухолевых клеток, использовали анализ клеточного лизиса in vitro с использованием клеток множественной миеломы и лимфомы человека.
Материалы и методы
Анализ уничтожения опухолевых клеток in vitro с использованием Т-клеток человека
[0326] Опухолевые клетки-мишени метили с помощью мембранного красителя PKH-26 (Sigma) и в течение 24 часов культивировали совместно с PBMC человека или обогащенными CD8 T-клетками в качестве эффекторных клеток при соотношении E: T, составляющем 10: 1 (E: T=3: 1 при использовании обогащенных CD8 Т-клеток), в присутствии триспецифических или соответствующих контрольных антител в указанных концентрациях. Мононуклеарные клетки периферической крови выделяли из крови нормальных людей-доноров путем разделения с помощью фиколла, а аутологичные CD8+ или все T-клетки обогащали с использованием наборов от Miltenyi Biotech (Сан-Диего, Калифорния, США). Степень клеточного лизиса у клеток-мишеней определяли за счет окрашивания с помощью набора для окрашивания погибших клеток LIVE/DEAD™ Fixable Violet Stain (Life Technologies) и измеряли по числу погибших клеток в популяции меченых клеток-мишеней путем прогона образцов на приборе LSRFortessa (BD Biosciences) с последующим анализом с применением программного обеспечения Flowjo (Treestar).
Анализ уничтожения опухолевых клеток in vitro с использованием Т-клеток человека в присутствии даратумумаба
[0327] Даратумумаб или антитела изотипического контроля в количестве 5 нМ предварительно инкубировали с опухолевыми клетками-мишенями, меченными PKH-26 (105 клеток/лунку), в течение 30 минут с последующим добавлением триспецифических TCE в указанных концентрациях и PBMC человека (E:T=10:1). Через 24 часа степень клеточного лизиса у клеток-мишеней определяли путем окрашивания с помощью набора для окрашивания погибших клеток LIVE/DEAD™ Fixable Violet Stain (Life Technologies) и измеряли по числу погибших клеток в популяции меченых клеток-мишеней путем прогона образцов на приборе LSRFortessa (BD Biosciences) с последующим анализом с применением программного обеспечения Flowjo (Treestar).
Результаты
[0328] Активность уничтожения клеток in vitro у триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами определяли с использованием линии клеток множественной миеломы человека NCI-H929, которые экспрессируют как CD38, так и CD28. Анализ проводили в присутствии даратумумаба или антител изотипического контроля в количестве 5 нМ (присутствуют в течение периода анализа). Как показано на фиг. 4A-4B, все протестированные триспецифические антитела приводили к лизису клеток зависимым от концентрации образом в присутствии и в отсутствие даратумумаба. Затем рассчитывали значения EC50 в присутствии и в отсутствие даратумумаба (таблица C). Даратумумаб снижал активность уничтожения клеток у триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с CD38-связывающими доменами CD38VH1 или CD38hhy1370, в то же время триспецифические антитела с CD38-связывающими доменами CD38hyb5739, CD38hyb6284 или CD38hhy1195 демонстрировали 3-8-кратное снижение активности уничтожения клеток в присутствии даратумумаба (таблица C).
Таблица C. Активность уничтожения in vitro у триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами в отношении линии клеток множественной миеломы человека NCI-H929 (CD38+/CD28+) в присутствии даратумумаба.
[0329] Кроме того, анализ клеточного лизиса in vitro использовали для измерения активности уничтожения клеток у выбранных триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами с использованием линии клеток лимфомы человека OCI-LY19, которые экспрессируют CD38, но не экспрессируют CD28. Анализ проводили в присутствии даратумумаба или антител изотипического контроля в количестве 5 нМ, которые присутствовали на протяжении периода анализа. Как показано на фиг. 5A-5B, все протестированные триспецифические антитела приводили к лизису клеток зависимым от концентрации образом в присутствии и в отсутствие даратумумаба. Затем рассчитывали значения ЕС50 в присутствии и в отсутствие даратумумаба (таблица D). Даратумумаб снижал активность уничтожения клеток у триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с CD38-связывающим доменом CD38VH1 в приблизительно 24 раза, в то же время триспецифические антитела с CD38-связывающими доменами CD38hhy992, CD38hyb5739, CD38hyb6284, CD38hhy1195 или CD38hhy1370 также демонстрировали снижение активности уничтожения клеток в присутствии даратумумаба (таблица D).
Таблица D. Активность уничтожения in vitro у выбранных триспецифических антител CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами в отношении линии клеток лимфомы человека OCI-LY19 (CD38 +/CD28-) в присутствии даратумумаба.
Пример 4. Триспецифические антитела к CD38/CD28xCD3 содействуют иммунному ответу, специфическому в отношении CMV
[0330] Как часть адаптивного иммунитета, Т-клеточный иммунитет играет решающую роль в контроле вирусной инфекции и рака, возможно, устраняя инфицированные клетки и злокачественные клетки, что приводит к освобождению от вирусной инфекции или излечению от рака. При хронических инфекционных заболеваниях, таких как инфекция, вызванная вирусом герпеса (HSV, CMV, EBV и т. д.), HIV и HBV, вирусы сохраняют свою устойчивость у людей с помощью различных механизмов, включая подавление иммунитета, истощение Т-клеток и установление латентного периода. Тем не менее, вирусная инфекция обычно индуцирует вирусный антигенспецифический иммунитет, включающий антигенспецифические CD8 Т-клетки, которые могут легко распознавать инфицированные клетки для контроля или уничтожения посредством высвобождения цитокинов или процессов уничтожения, опосредованных цитотоксическими Т-клетками (CTL). Таким образом, активация и/или амплификация in vivo и/или ex vivo специфических в отношении вирусного антигена Т-клеток обеспечивает терапевтические стратегии против хронических вирусных инфекций.
[0331] Разрабатывали триспецифические антитела к CD38/CD28xCD3 и оценивали в отношении их способности активировать Т-клетки и содействовать пролиферации и/или амплификации антигенспецифических Т-клеток. Эти триспецифические Ab могут эффективно расширять популяции CD4 и CD8 эффекторных клеток и клеток памяти, в том числе антигенспецифических центральных CD8 T-клеток памяти и эффекторных клеток памяти in vitro. В частности, продемонстрировали in vitro размножение специфических в отношении CMV и EBV CD8 центральных клеток памяти и эффекторных клеток памяти. Триспецифические антитела к CD38/CD28xCD3, описываемые в данном документе, демонстрировали новые свойства путем вовлечения CD3/CD28/CD38 с обеспечением пути передачи сигнала для стимуляции и размножения Т-клеток, что может обеспечить эффективную стратегию лечения хронических инфекционных заболеваний, таких как инфекции, вызванные HSV, CMV, EBV, HIV-1 и HBV.
[0332] В этом примере определяли способность триспецифических антител к CD38/CD28xCD3 содействовать активации и размножению специфических в отношении CMV Т-клеток.
Материалы и методы
Измерение пролиферации Т-клеток in vitro
[0333] Т-клетки выделяли у доноров PBMC человека путем отрицательной селекции с использованием набора для выделения всех T-клеток в магнитном поле (Miltenyi Biotec GmbH, Германия). Антитела наносили на 96-луночные планшеты для культивирования клеток при приготовлении антител в стерильном PBS и распределении по 50 мкл в каждую лунку (350 нг/лунка). Затем планшеты инкубировали при 37°C в течение по меньшей мере 2 часов, а потом промывали стерильным PBS. В покрытые антителами планшеты добавляли интактные Т-клетки (5×105 клеток/мл) и инкубировали при 37°C в течение нескольких дней. В день 4 клетки пересевали с новой средой для культивирования клеток на планшеты, только что покрытые антителами. В определенных экспериментах с 7-дневной инкубацией добавляли только свежую среду без замены на планшет, только что покрытый антителами. Клетки собирали в определенные моменты времени, и число клеток рассчитывали с использованием микроносителей для подсчета CountBright™.
Анализ пролиферации Т-клеток и определение субпопуляции Т-клеток in vitro
[0334] Мононуклеарные клетки периферической крови выделяли из крови здоровых доноров-людей, собранной компанией Research Blood Components, LLC (Бостон, Массачусетс, США). РВМС добавляли в покрытые антителами планшеты (350 нг/лунка) (5×105 клеток/мл), как описано выше, и инкубировали при 37°C в течение 3 и 7 дней. Клетки собирали в определенные моменты времени и анализировали с помощью проточной цитометрии с определением субпопуляций T-клеток: не подвергавшихся воздействию (CCR7+ CD45RO-), Tcm (CCR7+ CD45RO+), Tem (CCR7- CD45RO+), Tregs (CD4+ Foxp3+ CD25hi). CD8+ Т-клетки, специфические в отношении pp65 CMV и специфические в отношении BMLF EBV, выявляли с использованием конъюгированного с флуоресцентной меткой пентамера, ограниченного по HLA/вирусному пептиду доноров PBMC (A*02:01/NLVPMVATV, SEQ ID NO:284), (A*02:01/GLCTLVAML, SEQ ID NO:285) соответственно (ProImmune, Оксфорд, Великобритания). PBMC получали из HemaCare (Ван-Найс, Калифорния, США) от доноров с подтвержденной инфекцией CMV или EBV. PMBC от доноров, отрицательных по ограничивающему типа HLA, использовали в качестве отрицательного контроля. Окрашивание проводили в соответствии с протоколом производителя.
Количественное определение специфических в отношении CMV T-клеток
[0335] Как указано выше, мононуклеарные клетки периферической крови (PBMC) выделяли из крови доноров-людей с подтвержденным инфицированием CMV и добавляли в планшеты, содержащие триспецифическое антитело или контрольное антитело. Планшеты инкубировали при 37°C. Клетки собирали в определенные моменты времени и анализировали с помощью проточной цитометрии.
Результаты
[0336] Триспецифические антитела CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами, ΔVH1CD38 (контроль), CD38VH1, CD38hhy992, CD38hyb5739, CD38hyb6284, CD38hhy1195 и CD38hhy1370, тестировали, как описано выше, с использованием PBMC, выделенных от донора человека D, инфицированного CMV (фиг. 6A-6J) и донора-человека Е, инфицированного CMV (фиг. 7A-7J). Все протестированные триспецифические Ab к CD38 активировали специфические в отношении CMV Т-клетки и содействовали их пролиферации, что приводило к увеличению числа специфических в отношении CMV CD8+ T-клеток (клеток/лунка) с различной эффективностью и кинетикой дозозависимым образом в течение 7-дневного эксперимента (CMV донор D, фиг. 6A-6B; CMV донор E, фиг. 7A-7B). Кроме того, все протестированные триспецифические Ab к CD38 содействовали амплификации (измеряется как число клеток/лунка) специфических в отношении CMV центральных CD8+ T-клеток памяти (Tcm) (CMV донор D, фиг. 6C-6D; CMV донор E, фиг. 7C-7D) и эффекторных CD8+ Т-клеток памяти (Tem) (CMV донор D, фиг. 6E-6F; CMV донор E, фиг. 7E-7F), при этом обе субпопуляции подверглись значительной амплификации в течение 7 дней. На фиг. 6G-6J (CMV донор D) и фиг. 7G-7J (CMV донор E) представлены графики временной динамики, демонстрирующие процентное содержание специфических в отношении CMV клеток Tcm и Tem в дни 0, 3 и 7 в ходе 7-дневных экспериментов, описанных выше.
[0337] В совокупности эти данные указывают на то, что триспецифические антитела к CD38/CD28xCD3 содействуют активации и размножению специфических в отношении CMV Т-клеток, таких как специфические в отношении CMV CD8+ Т-клетки, специфических в отношении CMV эффекторные CD8+ Т-клетки памяти (Tem) и специфические в отношении CMV центральных CD8+ Т-клетки памяти (Tcm).
Пример 5. Триспецифические антитела к CD38/CD28xCD3 содействуют иммунному ответу, специфическому в отношении EBV
[0338] Далее определяли способность триспецифических антител к CD38/CD28xCD3 содействовать активации и размножению специфических в отношении вируса Эпштейна-Барр (EBV) Т-клеток.
Материалы и методы
Количественное определение специфических в отношении EBV T-клеток
[0339] Как указано выше, мононуклеарные клетки периферической крови (PBMC) выделяли из крови доноров-людей с подтвержденным инфицированием EBV и добавляли в планшеты, содержащие триспецифическое антитело или контрольное антитело. Планшеты инкубировали при 37°C в течение максимум 11 дней. Клетки собирали в определенные моменты времени и анализировали с помощью проточной цитометрии.
Результаты
[0340] Триспецифические антитела CD38/CD28sup x CD3mid_ENLQ DKTHT IgG4 FALA с альтернативными CD38-связывающими доменами, ΔVH1CD38 (контроль), CD38VH1, CD38hhy992, CD38hyb5739, CD38hyb6284, CD38hhy1195 и CD38hhy1370, также тестировали, как описано выше, с использованием PBMC, выделенных от донора С, инфицированного EBV (фиг. 8A-8J) и от донора D, инфицированного EBV (фиг. 9A-12). Все протестированные триспецифические Ab к CD38 активировали специфические в отношении EBV T-клетки и способствовали их пролиферации, что приводило к увеличению числа специфических в отношении EBV CD8+ T-клеток (клеток/лунка) с различной эффективностью и кинетикой дозозависимым образом в течение 7-дневного эксперимента (EBV донор C, фиг. 8A-8B; EBV донор D, фиг. 9A-9B). Кроме того, все протестированные триспецифические антитела к CD38 содействовали амплификации (измеряется как число клеток/лунка) специфических в отношении EBV центральных CD8+ T-клеток памяти (Tcm) (EBV донор C, фиг. 8C-8D; EBV донор D, фиг. 9C-9D) и эффекторных CD8+ Т-клеток памяти (Tem) (EBV донор C, фиг. 8E-8F; EBV донор D, фиг. 9E-9F), при этом обе субпопуляции подверглись значительной амплификации в течение 7 дней. На фиг. 8G-8J (EBV донор C) и фиг. 9G-12 (EBV донор D) представлены графики временной динамики, демонстрирующие процентное содержание специфических в отношении EBV клеток Tcm и Tem в дни 0, 3 и 7 в ходе 7-дневных экспериментов, описанных выше.
[0341] В совокупности эти данные указывают на то, что триспецифические антитела к CD38/CD28xCD3 содействуют активации и размножение специфических в отношении EBV Т-клеток, таких как специфические в отношении EBV CD8+ Т-клетки, специфические в отношении EBV эффекторные CD8+ Т-клетки памяти (Tem) и специфические в отношении EBV центральные CD8+ Т-клетки памяти (Tcm).
Пример 6. Противоопухолевые эффекты триспецифических антител к Her2/CD28 x CD3 у мышей, несущих опухоль
[0342] В данном примере триспецифическое антитело к Her2/CD28 x CD3 тестировали в отношении противоопухолевых эффектов на модели с мышами Nod scid gamma (NSG), несущими опухоль ZR-75-1, которым вводили Т-клетки, размноженные in vitro.
Материалы и методы
[0343] Мышей NSG разделили на 5 групп по 10 мышей в каждой. В день 0 клетки рака молочной железы человека ZR-75-1 с 50% матригеля имплантировали в жировую подушку молочной железы каждой мыши из расчета 5 миллионов клеток/мышь. В дни 17/18 начали размножение CD3+ Т-клеток человека. Рандомизацию мышей осуществляли в день 24, когда размер опухолей составлял приблизительно 150 мм3. В день 25 всем мышам вводили размноженные in vitro CD3+ T-клетки человека из расчета 10 миллионов клеток в 300 мкл/мышь (1QW, 1 внутрибрюшинная инъекция).
[0344] Начиная с дня 25, одна группа мышей получала дозы только среды-носителя (8% вес/объем сахарозы, 0,05% вес/объем полисорбата 80, 10 мМ гистидина, pH 5,5), в то время как другие 4 группы получали триспецифическое антитело к Her2/CD28 x CD3, и то, и другое из расчета 10 мл/кг. Группам, получавшим триспецифические антитела, вводили дозы 100, 10, 1 или 0,1 мкг/кг. Антитело или среду-носитель вводили 1QW внутривенно 2 дозами (например, в дни 25 и 32). Кровь и ткань опухоли собирали в день 38 или 39.
Результаты
[0345] Триспецифическое антитело к Her2/CD28 x CD3 (связывающий белок № 2 из таблицы 1, соответствующий SEQ ID No:104-107) сравнивали с контролем со средой-носителем в отношении их эффектов на рост опухоли молочной железы человека в модели с мышами NSG, которым вводили размноженные in vitro Т-клетки человека, описанные выше. Обработка триспецифическим антителом к Her2/CD28 x CD3 при самой высокой дозе (100 мкг/кг) приводила к наиболее значительному ингибированию роста и регрессии опухоли, хотя доза 10 мкг/кг также продемонстрировала противоопухолевые эффекты (фиг. 13A и 13D). Значительной потери массы тела не наблюдали (фиг. 13B). Объемы отдельных опухолей с течением времени для каждой группы обработки триспецифическим антителом представлены на фиг. 13C.
[0346] Затем исследовали влияние обработки триспецифическим антителом на отдельные субпопуляции иммунных клеток. Популяции клеток с CD45+ человека, CD8+ человека и CD4+ человека измеряли с помощью проточной цитометрии, так же как и клетки с CD45+ мыши (фиг. 14A-14C). Самая высокая доза (100 мкг/кг) триспецифического антитела приводила к истощению клеток с CD4+ человека, и данный эффект был дозозависимым (фиг. 14B и 14C). Введение триспецифического антитела преимущественно не влияло на число клеток с CD8+ человека.
[0347] Влияние обработки триспецифическим антителом на инфильтрирующие опухоль лимфоциты (TIL) также оценивали с помощью иммуногистохимического (IHC) окрашивания для выявления CD45, CD4 и CD8 человека. При окрашивании гематоксилином-эозином опухоли из групп с низкими дозами (триспецифическое антитело в дозе 1 мкг/кг или 0,1 мкг/кг) в целом были сопоставимы по размеру с опухолями из группы контроля со средой-носителем. Как показано на фиг. 15A-15C, число TIL человека увеличилось в группе, получавшей низкую дозу триспецифического антитела к Her2/CD28 x CD3, при этом TIL человека были редкими в группе, получавшей высокую дозу. Фотографии с результатами IHC также исследовали количественно (фиг. 16A-16C). Данные результаты указывают на значительное снижение числа CD45+ и CD8+ клеток в группах, получавших более высокие дозы триспецифических антител (100 мкг/кг и 10 мкг/кг).
[0348] По сравнению с контролем со средой-носителем опухоли из групп обработки высокими дозами триспецифических антител (100 мкг/кг или 10 мкг/кг) характеризовались редкими TIL. В группах обработки триспецифическими антителами в дозах 1 мкг/кг и 0,1 мкг/кг наблюдали от умеренного до большого количества CD45+, CD4+ или CD8+ TIL человека. Данные TIL в основном присутствовали на краях опухоли, но иногда распространялись глубже в центральную часть опухоли.
[0349] В заключение, данные результаты демонстрируют, что обработка мышей NSG, несущих опухоль молочной железы ZR-75-1, которым вводили активированные in vitro Т-клетки с применением нацеливающегося на HER2 триспецифического антитела, обеспечивающего рекрутинг Т-клеток, вводимого в 2 внутривенных дозах, составляющих 100 мкг/кг или 10 мкг/кг, приводила к значительному уменьшению объема опухоли и, одновременно, к значительному снижению TIL. При дозе триспецифического антитела 1 мкг/кг наблюдали несущественную и непоследовательную тенденцию к увеличению TIL по сравнению с контролем со средой-носителем.
Пример 7. Влияние последовательностей связывающих антиген HER2 и CD3 доменов в триспецифических антителах к Her2/CD28 x CD3 на уничтожение раковых клеток
[0350] В данном примере описано влияние последовательностей связывающих Her2 и CD3 вариабельных доменов на уничтожение клеток-мишеней. В данном примере триспецифическое антитело к Her2/CD28 x CD3 ("контроль") с антигенсвязывающим доменом из трастузумаба дикого типа и связывающим антиген CD3 доменом без мутаций 32/35 QQ в домене VL (см. пример 1) сравнивали с триспецифическими антителами к Her2/CD28 x CD3 № 1-6 из таблицы 1, соответствующими SEQ ID No:100-103, 104-107, 286-289, 290-293, 294-297 и 298-301 соответственно.
Материалы и методы
[0351] CD8+ Т-клетки выделяли из PBMC человека от здорового донора с использованием набора для выделения на основе микроносителей в магнитном поле (Miltenyi Biotec). Т-клетки использовали в качестве эффекторных клеток против линий клеток рака молочной железы, экспрессирующих различные уровни HER2, при соотношении 3:1 (эффектор : мишень). Клетки инкубировали с исследуемым или контрольным триспецифическим антителом в течение 2 дней перед проведением проточной цитометрии с использованием красителя для оценки жизнеспособности клеток (Invitrogen) и окрашивания клеток-мишеней с помощью PKH26 (Sigma). Среднее значение ЕС50 для лизиса клеток-мишеней рассчитывали у 2-3 доноров РВМС для каждого триспецифического антитела.
Результаты
[0352] Все триспецифические антитела оценивали в отношении клеточного лизиса in vitro на трех линиях клеток-мишеней HER2+ рака молочной железы: HCC1954, BT20 и MDA-MB-231. Обнаружили, что клетки рака молочной железы HCC1954 экспрессируют высокие уровни HER2, по оценке с помощью проточной цитометрии (до ~150000 рецепторов на клетку), IHC (3+) или анализа экспрессии HER2 с помощью набора HercepTest (3+) (фиг. 17A). Обнаружили, что клетки рака молочной железы BT20 экспрессируют промежуточные уровни HER2, по оценке с помощью проточной цитометрии (~ 60000 рецепторов на клетку), IHC (1+) или анализа экспрессии HER2 с помощью набора HercepTest (1+) (фиг. 17C). Обнаружили, что клетки рака молочной железы MDA-MD-231 экспрессируют низкие уровни HER2, по оценке с помощью проточной цитометрии (~ 9000 рецепторов на клетку), IHC (0+) или анализа экспрессии HER2 с помощью набора HercepTest (0) (фиг. 17E). Результаты анализов уничтожения клеток, нацеленного на клетки HCC1954, BT20 или MDA-MB-231, представлены на фиг. 17B, 17D и 17F соответственно, при сопоставлении связывающего белка № 2 с контролем или связывающих белков № 1 и № 5 с контролем. Результаты показали, что триспецифические антитела к Her2/CD28 x CD3, имеющие мутации 30R/55Q/102E в HER2-связывающем плече и мутациями 32/35 QQ в домене VL CD3-связывающего плеча, демонстрировали улучшенное уничтожение клеток-мишеней у всех трех клеточных линий, в частности при более низких концентрациях антитела.
[0353] Среднее значение EC50 (пМ) уничтожения клеток in vitro определяли для всех триспецифических антител, нацеливающихся на три указанные выше линии клеток рака молочной железы (HCC1954, BT20 и MDA-MB-231), а также на линии клеток рака желудка OE19 (высокий уровень экспрессии HER2) и GSU (промежуточный уровень экспрессии HER2). В целом, триспецифические антитела к Her2/CD28 x CD3, имеющие мутации в HER2-связывающем плече и в домене VL CD3-связывающего плеча, демонстрировали более низкую EC50 (и, таким образом, лучшее уничтожение клеток) в отношении всех трех линий клеток рака молочной железы (фиг. 18A) и обеих линий клеток рака желудка (фиг. 18B). Данные результаты показывают, что, хотя все триспецифические антитела способны индуцировать уничтожение HER2+ клеток, подвергнутые мутации триспецифические антитела стабильно демонстрировали улучшенную эффективность уничтожения клеток в отношении нескольких типов клеток-мишеней.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Sanofi
<120> ТРИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, ОТНОСЯЩИЕСЯ К НИМ
СПОСОБЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ
<130> 18395-20320.40
<140> Пока еще не назначен
<141> Одновременно с этим
<150> EP 19306261.9
<151> 2019-10-02
<150> US 62/831,572
<151> 2019-04-09
<160> 302
<170> FastSEQ для Windows версии 4.0
<210> 1
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 1
Gly Phe Asn Ile Lys Asp Thr Tyr
1 5
<210> 2
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 2
Gly Phe Asn Ile Arg Asp Thr Tyr
1 5
<210> 3
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 3
Ile Tyr Pro Thr Asn Gly Tyr Thr
1 5
<210> 4
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 4
Ile Tyr Pro Thr Gln Gly Tyr Thr
1 5
<210> 5
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 5
Ile Tyr Pro Thr Asn Ala Tyr Thr
1 5
<210> 6
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 6
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 7
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 7
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 8
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 8
Ser Arg Trp Gly Gly Ser Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 9
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 9
Gln Asp Val Asn Thr Ala
1 5
<210> 10
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 10
Gln Asp Val Gln Thr Ala
1 5
<210> 11
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 11
Ser Ala Ser
1
<210> 12
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 12
Gln Gln His Tyr Thr Thr Pro
1 5
<210> 13
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 13
Gly Tyr Thr Phe Thr Ser Tyr Ala
1 5
<210> 14
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 14
Ile Tyr Pro Gly Gln Gly Gly Thr
1 5
<210> 15
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 15
Ala Arg Thr Gly Gly Leu Arg Arg Ala Tyr Phe Thr Tyr
1 5 10
<210> 16
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 16
Gln Ser Val Ser Ser Tyr Gly Gln Gly Phe
1 5 10
<210> 17
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 17
Gly Ala Ser
1
<210> 18
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 18
Gln Gln Asn Lys Glu Asp Pro Trp Thr
1 5
<210> 19
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 19
Gly Tyr Thr Leu Thr Glu Phe Ser
1 5
<210> 20
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 20
Phe Asp Pro Glu Asp Gly Glu Thr
1 5
<210> 21
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 21
Thr Thr Gly Arg Phe Phe Asp Trp Phe
1 5
<210> 22
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 22
Gln Ser Val Ile Ser Arg Phe
1 5
<210> 23
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 23
Gly Ala Ser
1
<210> 24
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 24
Gln Gln Asp Ser Asn Leu Pro Ile Thr
1 5
<210> 25
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 25
Gly Tyr Ala Phe Thr Thr Tyr Leu
1 5
<210> 26
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 26
Ile Asn Pro Gly Ser Gly Ser Thr
1 5
<210> 27
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 27
Ala Arg Tyr Ala Tyr Gly Tyr
1 5
<210> 28
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 28
Gln Asn Val Gly Thr Ala
1 5
<210> 29
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 29
Ser Ala Ser
1
<210> 30
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 30
Gln Gln Tyr Ser Thr Tyr Pro Phe Thr
1 5
<210> 31
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 31
Gly Tyr Ser Phe Thr Asn Tyr Ala
1 5
<210> 32
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 32
Ile Ser Pro Tyr Tyr Gly Asp Thr
1 5
<210> 33
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 33
Ala Arg Arg Phe Glu Gly Phe Tyr Tyr Ser Met Asp Tyr
1 5 10
<210> 34
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 34
Gln Ser Leu Val His Ser Asn Gly Asn Thr Tyr
1 5 10
<210> 35
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 35
Lys Val Ser
1
<210> 36
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 36
Ser Gln Ser Thr His Val Pro Leu Thr
1 5
<210> 37
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 37
Gly Phe Thr Phe Ser Ser Tyr Gly
1 5
<210> 38
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 38
Ile Trp Tyr Asp Gly Ser Asn Lys
1 5
<210> 39
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 39
Ala Arg Asp Pro Gly Leu Arg Tyr Phe Asp Gly Gly Met Asp Val
1 5 10 15
<210> 40
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 40
Gln Gly Ile Ser Ser Tyr
1 5
<210> 41
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 41
Ala Ala Ser
1
<210> 42
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 42
Gln Gln Leu Asn Ser Phe Pro Tyr Thr
1 5
<210> 43
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 43
Gly Phe Thr Phe Ser Ser Tyr Gly
1 5
<210> 44
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 44
Ile Trp Tyr Asp Gly Ser Asn Lys
1 5
<210> 45
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 45
Ala Arg Met Phe Arg Gly Ala Phe Asp Tyr
1 5 10
<210> 46
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 46
Gln Gly Ile Arg Asn Asp
1 5
<210> 47
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 47
Ala Ala Ser
1
<210> 48
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 48
Leu Gln Asp Tyr Ile Tyr Tyr Pro Thr
1 5
<210> 49
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 49
Gly Tyr Thr Phe Thr Ser Tyr Tyr
1 5
<210> 50
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 50
Ile Tyr Pro Gly Asn Val Asn Thr
1 5
<210> 51
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 51
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val
1 5 10
<210> 52
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 52
Gln Asn Ile Tyr Val Trp
1 5
<210> 53
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 53
Lys Ala Ser
1
<210> 54
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 54
Gln Gln Gly Gln Thr Tyr Pro Tyr
1 5
<210> 55
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 55
Gly Phe Thr Phe Thr Lys Ala Trp
1 5
<210> 56
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 56
Ile Lys Asp Lys Ser Asn Ser Tyr Ala Thr
1 5 10
<210> 57
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 57
Arg Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr
1 5 10
<210> 58
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 58
Gln Ser Leu Val His Asn Asn Ala Asn Thr Tyr
1 5 10
<210> 59
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 59
Gln Ser Leu Val His Gln Asn Ala Gln Thr Tyr
1 5 10
<210> 60
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 60
Gln Ser Leu Val His Glu Asn Leu Gln Thr Tyr
1 5 10
<210> 61
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 61
Gln Ser Leu Val His Glu Asn Leu Phe Thr Tyr
1 5 10
<210> 62
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 62
Gln Ser Leu Val His Glu Asn Leu Arg Thr Tyr
1 5 10
<210> 63
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 63
Gln Ser Leu Val His Asp Asn Ala Gln Thr Tyr
1 5 10
<210> 64
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 64
Lys Val Ser
1
<210> 65
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 65
Gly Gln Gly Thr Gln Tyr Pro Phe Thr
1 5
<210> 66
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 66
Asp Lys Thr His Thr
1 5
<210> 67
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 67
Gly Gln Pro Lys Ala Ala Pro
1 5
<210> 68
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 68
Thr Lys Gly Pro Ser
1 5
<210> 69
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 69
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 70
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 70
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 71
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 71
Gly Gly Ser Gly Ser Ser Gly Ser Gly Gly
1 5 10
<210> 72
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 72
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 73
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 73
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 74
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 74
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Ser Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 75
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Ala Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Ser Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 76
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 76
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Ala Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 77
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 77
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 78
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 78
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Gln Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 79
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 79
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Ala Met His Trp Val Lys Glu Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Gly Gln Gly Gly Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Thr Gly Gly Leu Arg Arg Ala Tyr Phe Thr Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 80
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 80
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Gly Gln Gly Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Pro Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asn Lys
85 90 95
Glu Asp Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 81
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 81
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Val Ser Gly Tyr Thr Leu Thr Glu Phe
20 25 30
Ser Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Phe Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Ile Met Thr Glu Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Thr Thr Gly Arg Phe Phe Asp Trp Phe Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 82
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 82
Glu Ile Ile Leu Thr Gln Ser Pro Ala Ile Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ile Ser Arg
20 25 30
Phe Leu Ser Trp Tyr Gln Val Lys Pro Gly Leu Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Val Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Cys Ala Val Tyr Tyr Cys Gln Gln Asp Ser Asn Leu Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 83
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 83
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Thr Tyr
20 25 30
Leu Val Glu Trp Ile Arg Gln Arg Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asn Pro Gly Ser Gly Ser Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Val Asp Arg Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Ala Tyr Gly Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 84
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 84
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Val Gly Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Gln Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 85
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 85
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asn Tyr
20 25 30
Ala Val His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Ser Pro Tyr Tyr Gly Asp Thr Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Phe Glu Gly Phe Tyr Tyr Ser Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 86
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 86
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Pro Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 87
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 87
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr His Cys
85 90 95
Ala Arg Asp Pro Gly Leu Arg Tyr Phe Asp Gly Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 88
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 88
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Ala Ser Thr Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 89
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 89
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gly Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Phe Arg Gly Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 90
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 90
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Gly Leu Gln Pro
65 70 75 80
Glu Asp Ser Ala Thr Tyr Tyr Cys Leu Gln Asp Tyr Ile Tyr Tyr Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 91
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 91
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 92
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 92
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asn Ile Tyr Val Trp
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Gln Thr Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 93
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 93
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val
35 40 45
Ala Gln Ile Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Arg Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 94
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 94
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asn
20 25 30
Asn Ala Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 95
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 95
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 96
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 96
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 97
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 97
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Phe Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 98
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 98
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Arg Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 99
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 99
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asp
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 100
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 100
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 101
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 101
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 102
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 102
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 103
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 103
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 104
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 104
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 105
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 105
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 106
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 107
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 107
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 108
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 108
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asp
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 109
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 109
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 110
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 110
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 111
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 111
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 112
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 112
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 113
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 113
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 114
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 114
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 115
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 115
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 116
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 116
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asn
20 25 30
Asn Ala Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 117
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 117
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 118
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 118
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Ser Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 119
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 119
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 120
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 120
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asn
20 25 30
Asn Ala Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 121
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 121
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 122
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 122
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Ala Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Ser Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 123
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 123
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 124
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 124
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asn
20 25 30
Asn Ala Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 125
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 125
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 126
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 126
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Ala Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 127
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 127
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 128
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 128
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 129
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 129
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 130
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 130
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 131
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 131
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 132
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 132
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asn
20 25 30
Asn Ala Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 133
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 133
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 134
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 134
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 135
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 135
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 136
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 136
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Arg Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 137
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 137
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 138
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 138
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 139
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 139
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 140
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 140
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 141
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 141
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 142
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 142
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 143
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 143
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 144
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 144
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Phe Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 145
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 145
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 146
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 146
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 147
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 147
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 148
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 148
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Asn
20 25 30
Asn Ala Asn Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 149
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 149
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 150
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 150
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 151
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 151
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 152
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 152
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Arg Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 153
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 153
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 154
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 154
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 155
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 155
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 156
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 156
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 157
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 157
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 158
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 158
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Ala Met His Trp Val Lys Glu Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Pro Gly Gln Gly Gly Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Ala Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Thr Gly Gly Leu Arg Arg Ala Tyr Phe Thr Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 159
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 159
Asp Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Gly Gln Gly Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Pro Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asn Lys
85 90 95
Glu Asp Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 160
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 160
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 161
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 161
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 162
<211> 442
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 162
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Val Ser Gly Tyr Thr Leu Thr Glu Phe
20 25 30
Ser Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Phe Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Ile Met Thr Glu Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Thr Thr Gly Arg Phe Phe Asp Trp Phe Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu
340 345 350
Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 163
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 163
Glu Ile Ile Leu Thr Gln Ser Pro Ala Ile Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ile Ser Arg
20 25 30
Phe Leu Ser Trp Tyr Gln Val Lys Pro Gly Leu Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Val Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Cys Ala Val Tyr Tyr Cys Gln Gln Asp Ser Asn Leu Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 164
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 164
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 165
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 165
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 166
<211> 440
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 166
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Thr Tyr
20 25 30
Leu Val Glu Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Asn Pro Gly Ser Gly Ser Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Arg Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met His Leu Ser Gly Leu Thr Ser Asp Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Tyr Ala Tyr Gly Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
115 120 125
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
210 215 220
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
225 230 235 240
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
245 250 255
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
260 265 270
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
275 280 285
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
290 295 300
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
305 310 315 320
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
325 330 335
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met
340 345 350
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
355 360 365
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
370 375 380
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
385 390 395 400
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
405 410 415
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
420 425 430
Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 167
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 167
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Gln Pro Gly His Ser Pro Lys Gln Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ala Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Ile Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Ser Thr Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 168
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 168
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 169
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 169
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 170
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 170
Gln Val Gln Leu Leu Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Val
1 5 10 15
Ser Val Lys Ile Ser Cys Thr Gly Ser Gly Tyr Ser Phe Thr Asn Tyr
20 25 30
Ala Val His Trp Val Lys Gln Ser His Val Lys Ser Leu Glu Trp Ile
35 40 45
Gly Val Ile Ser Pro Tyr Tyr Gly Asp Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Thr Gly Lys Ala Thr Met Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ala Arg Leu Thr Ser Glu Asp Ser Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Arg Phe Glu Gly Phe Tyr Tyr Ser Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 171
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 171
Asp Val Val Met Ile Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Pro Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asn Trp Tyr Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Leu Cys Ser Gln Ser
85 90 95
Thr His Val Pro Leu Thr Phe Gly Ser Gly Thr Gln Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 172
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 172
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 173
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 173
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 174
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 174
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr His Cys
85 90 95
Ala Arg Asp Pro Gly Leu Arg Tyr Phe Asp Gly Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 175
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 175
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Ala Ser Thr Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 176
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 176
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 177
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 177
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 178
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 178
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gly Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Phe Arg Gly Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Gln
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 179
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 179
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Gly Leu Gln Pro
65 70 75 80
Glu Asp Ser Ala Thr Tyr Tyr Cys Leu Gln Asp Tyr Ile Tyr Tyr Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 180
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> ВАРИАНТ
<222> 6
<223> Xaa = Glu или Gln
<220>
<221> ВАРИАНТ
<222> 8
<223> Xaa = Ala или Leu
<220>
<221> ВАРИАНТ
<222> 9
<223> Xaa = Gln, Arg или Phe
<400> 180
Gln Ser Leu Val His Xaa Asn Xaa Xaa Thr Tyr
1 5 10
<210> 181
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 181
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 182
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 182
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 183
<211> 440
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 183
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Thr Tyr
20 25 30
Leu Val Glu Trp Ile Arg Gln Arg Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asn Pro Gly Ser Gly Ser Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Val Asp Arg Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Ala Tyr Gly Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
115 120 125
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
210 215 220
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
225 230 235 240
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
245 250 255
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
260 265 270
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
275 280 285
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
290 295 300
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
305 310 315 320
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
325 330 335
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met
340 345 350
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
355 360 365
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
370 375 380
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
385 390 395 400
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
405 410 415
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
420 425 430
Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 184
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 184
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Val Gly Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Gln Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 185
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 185
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Glu
20 25 30
Asn Leu Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 186
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 186
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 187
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 187
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Asn Tyr
20 25 30
Ala Val His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Ser Pro Tyr Tyr Gly Asp Thr Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Phe Glu Gly Phe Tyr Tyr Ser Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 188
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 188
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Pro Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 189
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 189
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg caccagaacg cccagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 190
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 190
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 191
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 191
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 192
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 192
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 193
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 193
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg caccagaacg cccagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 194
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 194
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 195
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 195
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca cccagggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgaaggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 196
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 196
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 197
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 197
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgacaacg cccagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 198
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 198
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 199
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 199
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca cccagggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgaaggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 200
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 200
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 201
<211> 1014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 201
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg caccagaacg cccagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaagggcc agcccaaggc cgcccccgac 360
atccagatga cccagagccc cagcagcctg tctgccagcg tgggcgacag agtgaccatc 420
acctgtcagg ccagccagaa catctacgtg tggctgaact ggtatcagca gaagcccggc 480
aaggccccca agctgctgat ctacaaggcc agcaacctgc acaccggcgt gcccagcaga 540
ttttctggca gcggctccgg caccgacttc accctgacaa tcagctccct gcagcccgag 600
gacattgcca cctactactg ccagcagggc cagacctacc cctacacctt tggccagggc 660
accaagctgg aaatcaagac caagggcccc agccgtacgg tggccgctcc cagcgtgttc 720
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 780
aacaacttct acccccgcga ggccaaagtg cagtggaagg tggacaacgc cctgcagagc 840
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 900
agcaccctga cactgagcaa ggccgactac gagaagcaca aggtgtacgc ctgcgaagtg 960
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgt 1014
<210> 202
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 202
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
agccaggtgc agctggtgga atctggcggc ggagtggtgc agcctggcag aagcctgaga 420
ctgagctgtg ccgccagcgg cttcaccttc accaaggcct ggatgcactg ggtgcgccag 480
gcccctggaa agcagctgga atgggtggcc cagatcaagg acaagagcaa cagctacgcc 540
acctactacg ccgacagcgt gaagggccgg ttcaccatca gccgggacga cagcaagaac 600
accctgtacc tgcagatgaa cagcctgcgg gccgaggaca ccgccgtgta ctactgtcgg 660
ggcgtgtact atgccctgag ccccttcgat tactggggcc agggaaccct cgtgaccgtg 720
tctagtcgga ccgccagcac aaagggccca tcggtgttcc ctctggcccc ttgcagcaga 780
agcaccagcg aatctacagc cgccctgggc tgcctcgtga aggactactt tcccgagccc 840
gtgaccgtgt cctggaactc tggcgctctg acaagcggcg tgcacacctt tccagccgtg 900
ctccagagca gcggcctgta ctctctgagc agcgtcgtga cagtgcccag cagcagcctg 960
ggcaccaaga cctacacctg taacgtggac cacaagccca gcaacaccaa ggtggacaag 1020
cgggtggaat ctaagtacgg ccctccctgc cctccttgcc cagcccctga agctgccggc 1080
ggaccctccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat cagccggacc 1140
cccgaagtga cctgcgtggt ggtggatgtg tcccaggaag atcccgaggt gcagttcaat 1200
tggtacgtgg acggcgtgga agtgcacaac gccaagacca agcccagaga ggaacagttc 1260
aacagcacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 1320
aaagagtaca agtgcaaggt gtccaacaag ggcctgccca gctccatcga gaaaaccatc 1380
agcaaggcca agggccagcc ccgcgagcct caagtgtgta ccctgccccc tagccaggaa 1440
gagatgacca agaaccaggt gtccctgagc tgtgccgtga aaggcttcta ccccagcgac 1500
attgccgtgg aatgggagag caacggccag cccgagaaca actacaagac caccccccct 1560
gtgctggaca gcgacggctc attcttcctg gtgtccaagc tgaccgtgga caagagccgg 1620
tggcaggaag gcaacgtgtt cagctgctcc gtgatgcacg aggccctgca caaccactac 1680
acccagaagt ccctgtctct gtccctgggc 1710
<210> 203
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 203
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca cccagggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgaaggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 204
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 204
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 205
<211> 1014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 205
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacaacaacg ccaacaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaagggcc agcccaaggc cgcccccgac 360
atccagatga cccagagccc cagcagcctg tctgccagcg tgggcgacag agtgaccatc 420
acctgtcagg ccagccagaa catctacgtg tggctgaact ggtatcagca gaagcccggc 480
aaggccccca agctgctgat ctacaaggcc agcaacctgc acaccggcgt gcccagcaga 540
ttttctggca gcggctccgg caccgacttc accctgacaa tcagctccct gcagcccgag 600
gacattgcca cctactactg ccagcagggc cagacctacc cctacacctt tggccagggc 660
accaagctgg aaatcaagac caagggcccc agccgtacgg tggccgctcc cagcgtgttc 720
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 780
aacaacttct acccccgcga ggccaaagtg cagtggaagg tggacaacgc cctgcagagc 840
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 900
agcaccctga cactgagcaa ggccgactac gagaagcaca aggtgtacgc ctgcgaagtg 960
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgt 1014
<210> 206
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 206
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
agccaggtgc agctggtgga atctggcggc ggagtggtgc agcctggcag aagcctgaga 420
ctgagctgtg ccgccagcgg cttcaccttc accaaggcct ggatgcactg ggtgcgccag 480
gcccctggaa agcagctgga atgggtggcc cagatcaagg acaagagcaa cagctacgcc 540
acctactacg ccgacagcgt gaagggccgg ttcaccatca gccgggacga cagcaagaac 600
accctgtacc tgcagatgaa cagcctgcgg gccgaggaca ccgccgtgta ctactgtcgg 660
ggcgtgtact atgccctgag ccccttcgat tactggggcc agggaaccct cgtgaccgtg 720
tctagtcgga ccgccagcac aaagggccca tcggtgttcc ctctggcccc ttgcagcaga 780
agcaccagcg aatctacagc cgccctgggc tgcctcgtga aggactactt tcccgagccc 840
gtgaccgtgt cctggaactc tggcgctctg acaagcggcg tgcacacctt tccagccgtg 900
ctccagagca gcggcctgta ctctctgagc agcgtcgtga cagtgcccag cagcagcctg 960
ggcaccaaga cctacacctg taacgtggac cacaagccca gcaacaccaa ggtggacaag 1020
cgggtggaat ctaagtacgg ccctccctgc cctccttgcc cagcccctga agctgccggc 1080
ggaccctccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat cagccggacc 1140
cccgaagtga cctgcgtggt ggtggatgtg tcccaggaag atcccgaggt gcagttcaat 1200
tggtacgtgg acggcgtgga agtgcacaac gccaagacca agcccagaga ggaacagttc 1260
aacagcacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 1320
aaagagtaca agtgcaaggt gtccaacaag ggcctgccca gctccatcga gaaaaccatc 1380
agcaaggcca agggccagcc ccgcgagcct caagtgtgta ccctgccccc tagccaggaa 1440
gagatgacca agaaccaggt gtccctgagc tgtgccgtga aaggcttcta ccccagcgac 1500
attgccgtgg aatgggagag caacggccag cccgagaaca actacaagac caccccccct 1560
gtgctggaca gcgacggctc attcttcctg gtgtccaagc tgaccgtgga caagagccgg 1620
tggcaggaag gcaacgtgtt cagctgctcc gtgatgcacg aggccctgca caaccactac 1680
acccagaagt ccctgtctct gtccctgggc 1710
<210> 207
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 207
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca cccagggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggctccggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaat ttctgggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 208
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 208
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 209
<211> 1014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 209
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacaacaacg ccaacaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaagggcc agcccaaggc cgcccccgac 360
atccagatga cccagagccc cagcagcctg tctgccagcg tgggcgacag agtgaccatc 420
acctgtcagg ccagccagaa catctacgtg tggctgaact ggtatcagca gaagcccggc 480
aaggccccca agctgctgat ctacaaggcc agcaacctgc acaccggcgt gcccagcaga 540
ttttctggca gcggctccgg caccgacttc accctgacaa tcagctccct gcagcccgag 600
gacattgcca cctactactg ccagcagggc cagacctacc cctacacctt tggccagggc 660
accaagctgg aaatcaagac caagggcccc agccgtacgg tggccgctcc cagcgtgttc 720
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 780
aacaacttct acccccgcga ggccaaagtg cagtggaagg tggacaacgc cctgcagagc 840
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 900
agcaccctga cactgagcaa ggccgactac gagaagcaca aggtgtacgc ctgcgaagtg 960
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgt 1014
<210> 210
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 210
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
agccaggtgc agctggtgga atctggcggc ggagtggtgc agcctggcag aagcctgaga 420
ctgagctgtg ccgccagcgg cttcaccttc accaaggcct ggatgcactg ggtgcgccag 480
gcccctggaa agcagctgga atgggtggcc cagatcaagg acaagagcaa cagctacgcc 540
acctactacg ccgacagcgt gaagggccgg ttcaccatca gccgggacga cagcaagaac 600
accctgtacc tgcagatgaa cagcctgcgg gccgaggaca ccgccgtgta ctactgtcgg 660
ggcgtgtact atgccctgag ccccttcgat tactggggcc agggaaccct cgtgaccgtg 720
tctagtcgga ccgccagcac aaagggccca tcggtgttcc ctctggcccc ttgcagcaga 780
agcaccagcg aatctacagc cgccctgggc tgcctcgtga aggactactt tcccgagccc 840
gtgaccgtgt cctggaactc tggcgctctg acaagcggcg tgcacacctt tccagccgtg 900
ctccagagca gcggcctgta ctctctgagc agcgtcgtga cagtgcccag cagcagcctg 960
ggcaccaaga cctacacctg taacgtggac cacaagccca gcaacaccaa ggtggacaag 1020
cgggtggaat ctaagtacgg ccctccctgc cctccttgcc cagcccctga agctgccggc 1080
ggaccctccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat cagccggacc 1140
cccgaagtga cctgcgtggt ggtggatgtg tcccaggaag atcccgaggt gcagttcaat 1200
tggtacgtgg acggcgtgga agtgcacaac gccaagacca agcccagaga ggaacagttc 1260
aacagcacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 1320
aaagagtaca agtgcaaggt gtccaacaag ggcctgccca gctccatcga gaaaaccatc 1380
agcaaggcca agggccagcc ccgcgagcct caagtgtgta ccctgccccc tagccaggaa 1440
gagatgacca agaaccaggt gtccctgagc tgtgccgtga aaggcttcta ccccagcgac 1500
attgccgtgg aatgggagag caacggccag cccgagaaca actacaagac caccccccct 1560
gtgctggaca gcgacggctc attcttcctg gtgtccaagc tgaccgtgga caagagccgg 1620
tggcaggaag gcaacgtgtt cagctgctcc gtgatgcacg aggccctgca caaccactac 1680
acccagaagt ccctgtctct gtccctgggc 1710
<210> 211
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 211
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacgccta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggctccggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaat ttctgggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 212
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 212
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 213
<211> 1014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 213
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacaacaacg ccaacaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaagggcc agcccaaggc cgcccccgac 360
atccagatga cccagagccc cagcagcctg tctgccagcg tgggcgacag agtgaccatc 420
acctgtcagg ccagccagaa catctacgtg tggctgaact ggtatcagca gaagcccggc 480
aaggccccca agctgctgat ctacaaggcc agcaacctgc acaccggcgt gcccagcaga 540
ttttctggca gcggctccgg caccgacttc accctgacaa tcagctccct gcagcccgag 600
gacattgcca cctactactg ccagcagggc cagacctacc cctacacctt tggccagggc 660
accaagctgg aaatcaagac caagggcccc agccgtacgg tggccgctcc cagcgtgttc 720
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 780
aacaacttct acccccgcga ggccaaagtg cagtggaagg tggacaacgc cctgcagagc 840
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 900
agcaccctga cactgagcaa ggccgactac gagaagcaca aggtgtacgc ctgcgaagtg 960
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgt 1014
<210> 214
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 214
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
agccaggtgc agctggtgga atctggcggc ggagtggtgc agcctggcag aagcctgaga 420
ctgagctgtg ccgccagcgg cttcaccttc accaaggcct ggatgcactg ggtgcgccag 480
gcccctggaa agcagctgga atgggtggcc cagatcaagg acaagagcaa cagctacgcc 540
acctactacg ccgacagcgt gaagggccgg ttcaccatca gccgggacga cagcaagaac 600
accctgtacc tgcagatgaa cagcctgcgg gccgaggaca ccgccgtgta ctactgtcgg 660
ggcgtgtact atgccctgag ccccttcgat tactggggcc agggaaccct cgtgaccgtg 720
tctagtcgga ccgccagcac aaagggccca tcggtgttcc ctctggcccc ttgcagcaga 780
agcaccagcg aatctacagc cgccctgggc tgcctcgtga aggactactt tcccgagccc 840
gtgaccgtgt cctggaactc tggcgctctg acaagcggcg tgcacacctt tccagccgtg 900
ctccagagca gcggcctgta ctctctgagc agcgtcgtga cagtgcccag cagcagcctg 960
ggcaccaaga cctacacctg taacgtggac cacaagccca gcaacaccaa ggtggacaag 1020
cgggtggaat ctaagtacgg ccctccctgc cctccttgcc cagcccctga agctgccggc 1080
ggaccctccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat cagccggacc 1140
cccgaagtga cctgcgtggt ggtggatgtg tcccaggaag atcccgaggt gcagttcaat 1200
tggtacgtgg acggcgtgga agtgcacaac gccaagacca agcccagaga ggaacagttc 1260
aacagcacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 1320
aaagagtaca agtgcaaggt gtccaacaag ggcctgccca gctccatcga gaaaaccatc 1380
agcaaggcca agggccagcc ccgcgagcct caagtgtgta ccctgccccc tagccaggaa 1440
gagatgacca agaaccaggt gtccctgagc tgtgccgtga aaggcttcta ccccagcgac 1500
attgccgtgg aatgggagag caacggccag cccgagaaca actacaagac caccccccct 1560
gtgctggaca gcgacggctc attcttcctg gtgtccaagc tgaccgtgga caagagccgg 1620
tggcaggaag gcaacgtgtt cagctgctcc gtgatgcacg aggccctgca caaccactac 1680
acccagaagt ccctgtctct gtccctgggc 1710
<210> 215
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 215
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacgccta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgaaggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaat ttctgggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 216
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 216
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 217
<211> 1014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 217
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg caccagaacg cccagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaagggcc agcccaaggc cgcccccgac 360
atccagatga cccagagccc cagcagcctg tctgccagcg tgggcgacag agtgaccatc 420
acctgtcagg ccagccagaa catctacgtg tggctgaact ggtatcagca gaagcccggc 480
aaggccccca agctgctgat ctacaaggcc agcaacctgc acaccggcgt gcccagcaga 540
ttttctggca gcggctccgg caccgacttc accctgacaa tcagctccct gcagcccgag 600
gacattgcca cctactactg ccagcagggc cagacctacc cctacacctt tggccagggc 660
accaagctgg aaatcaagac caagggcccc agccgtacgg tggccgctcc cagcgtgttc 720
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 780
aacaacttct acccccgcga ggccaaagtg cagtggaagg tggacaacgc cctgcagagc 840
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 900
agcaccctga cactgagcaa ggccgactac gagaagcaca aggtgtacgc ctgcgaagtg 960
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgt 1014
<210> 218
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 218
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
agccaggtgc agctggtgga atctggcggc ggagtggtgc agcctggcag aagcctgaga 420
ctgagctgtg ccgccagcgg cttcaccttc accaaggcct ggatgcactg ggtgcgccag 480
gcccctggaa agcagctgga atgggtggcc cagatcaagg acaagagcaa cagctacgcc 540
acctactacg ccgacagcgt gaagggccgg ttcaccatca gccgggacga cagcaagaac 600
accctgtacc tgcagatgaa cagcctgcgg gccgaggaca ccgccgtgta ctactgtcgg 660
ggcgtgtact atgccctgag ccccttcgat tactggggcc agggaaccct cgtgaccgtg 720
tctagtcgga ccgccagcac aaagggccca tcggtgttcc ctctggcccc ttgcagcaga 780
agcaccagcg aatctacagc cgccctgggc tgcctcgtga aggactactt tcccgagccc 840
gtgaccgtgt cctggaactc tggcgctctg acaagcggcg tgcacacctt tccagccgtg 900
ctccagagca gcggcctgta ctctctgagc agcgtcgtga cagtgcccag cagcagcctg 960
ggcaccaaga cctacacctg taacgtggac cacaagccca gcaacaccaa ggtggacaag 1020
cgggtggaat ctaagtacgg ccctccctgc cctccttgcc cagcccctga agctgccggc 1080
ggaccctccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat cagccggacc 1140
cccgaagtga cctgcgtggt ggtggatgtg tcccaggaag atcccgaggt gcagttcaat 1200
tggtacgtgg acggcgtgga agtgcacaac gccaagacca agcccagaga ggaacagttc 1260
aacagcacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 1320
aaagagtaca agtgcaaggt gtccaacaag ggcctgccca gctccatcga gaaaaccatc 1380
agcaaggcca agggccagcc ccgcgagcct caagtgtgta ccctgccccc tagccaggaa 1440
gagatgacca agaaccaggt gtccctgagc tgtgccgtga aaggcttcta ccccagcgac 1500
attgccgtgg aatgggagag caacggccag cccgagaaca actacaagac caccccccct 1560
gtgctggaca gcgacggctc attcttcctg gtgtccaagc tgaccgtgga caagagccgg 1620
tggcaggaag gcaacgtgtt cagctgctcc gtgatgcacg aggccctgca caaccactac 1680
acccagaagt ccctgtctct gtccctgggc 1710
<210> 219
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 219
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 220
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 220
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 221
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 221
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacaacaacg ccaacaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 222
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 222
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 223
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 223
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 224
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 224
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 225
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 225
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcgtaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 226
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 226
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 227
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 227
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 228
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 228
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 229
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 229
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 230
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 230
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 231
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 231
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 232
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 232
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 233
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 233
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgttcaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 234
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 234
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 235
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 235
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 236
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 236
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 237
<211> 1014
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 237
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacaacaacg ccaacaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaagggcc agcccaaggc cgcccccgac 360
atccagatga cccagagccc cagcagcctg tctgccagcg tgggcgacag agtgaccatc 420
acctgtcagg ccagccagaa catctacgtg tggctgaact ggtatcagca gaagcccggc 480
aaggccccca agctgctgat ctacaaggcc agcaacctgc acaccggcgt gcccagcaga 540
ttttctggca gcggctccgg caccgacttc accctgacaa tcagctccct gcagcccgag 600
gacattgcca cctactactg ccagcagggc cagacctacc cctacacctt tggccagggc 660
accaagctgg aaatcaagac caagggcccc agccgtacgg tggccgctcc cagcgtgttc 720
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 780
aacaacttct acccccgcga ggccaaagtg cagtggaagg tggacaacgc cctgcagagc 840
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 900
agcaccctga cactgagcaa ggccgactac gagaagcaca aggtgtacgc ctgcgaagtg 960
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgt 1014
<210> 238
<211> 1710
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 238
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
agccaggtgc agctggtgga atctggcggc ggagtggtgc agcctggcag aagcctgaga 420
ctgagctgtg ccgccagcgg cttcaccttc accaaggcct ggatgcactg ggtgcgccag 480
gcccctggaa agcagctgga atgggtggcc cagatcaagg acaagagcaa cagctacgcc 540
acctactacg ccgacagcgt gaagggccgg ttcaccatca gccgggacga cagcaagaac 600
accctgtacc tgcagatgaa cagcctgcgg gccgaggaca ccgccgtgta ctactgtcgg 660
ggcgtgtact atgccctgag ccccttcgat tactggggcc agggaaccct cgtgaccgtg 720
tctagtcgga ccgccagcac aaagggccca tcggtgttcc ctctggcccc ttgcagcaga 780
agcaccagcg aatctacagc cgccctgggc tgcctcgtga aggactactt tcccgagccc 840
gtgaccgtgt cctggaactc tggcgctctg acaagcggcg tgcacacctt tccagccgtg 900
ctccagagca gcggcctgta ctctctgagc agcgtcgtga cagtgcccag cagcagcctg 960
ggcaccaaga cctacacctg taacgtggac cacaagccca gcaacaccaa ggtggacaag 1020
cgggtggaat ctaagtacgg ccctccctgc cctccttgcc cagcccctga agctgccggc 1080
ggaccctccg tgttcctgtt ccccccaaag cccaaggaca ccctgatgat cagccggacc 1140
cccgaagtga cctgcgtggt ggtggatgtg tcccaggaag atcccgaggt gcagttcaat 1200
tggtacgtgg acggcgtgga agtgcacaac gccaagacca agcccagaga ggaacagttc 1260
aacagcacct accgggtggt gtccgtgctg accgtgctgc accaggactg gctgaacggc 1320
aaagagtaca agtgcaaggt gtccaacaag ggcctgccca gctccatcga gaaaaccatc 1380
agcaaggcca agggccagcc ccgcgagcct caagtgtgta ccctgccccc tagccaggaa 1440
gagatgacca agaaccaggt gtccctgagc tgtgccgtga aaggcttcta ccccagcgac 1500
attgccgtgg aatgggagag caacggccag cccgagaaca actacaagac caccccccct 1560
gtgctggaca gcgacggctc attcttcctg gtgtccaagc tgaccgtgga caagagccgg 1620
tggcaggaag gcaacgtgtt cagctgctcc gtgatgcacg aggccctgca caaccactac 1680
acccagaagt ccctgtctct gtccctgggc 1710
<210> 239
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 239
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatcaag gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca ccaacggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgacggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 240
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 240
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 241
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 241
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcgtaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 242
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 242
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 243
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 243
gaagtgcagc tggtggaatc tggcggcgga ctggtgcagc ctggcggatc tctgagactg 60
agctgtgccg ccagcggctt caacatccgg gacacctaca tccactgggt gcgccaggcc 120
cctggcaagg gactggaatg ggtggccaga atctacccca cccagggcta caccagatac 180
gccgacagcg tgaagggccg gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgtag tagatgggga 300
ggcgaaggct tctacgccat ggactattgg ggccagggca ccctcgtgac cgtgtctagt 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 244
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 244
gacatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca ggacgtgaac accgccgtgg cctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcccagc 180
agattcagcg gaagcagaag cggcaccgac ttcaccctga ccatcagctc cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag cactacacca ccccccccac atttggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 245
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 245
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgttcaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 246
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 246
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 247
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 247
caggtgcagc tggtgcagtc tggcgccgaa gtcgtgaaac ctggcgcctc cgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctacgcca tgcactgggt caaagaggcc 120
cctggccaga gactggaatg gatcggctac atctaccccg gccagggcgg caccaactac 180
aaccagaagt tccagggcag agccaccctg accgccgata caagcgccag caccgcctac 240
atggaactga gcagcctgcg gagcgaggat accgccgtgt acttctgtgc cagaacaggc 300
ggcctgaggc gggcctactt tacctattgg ggccagggca ccctcgtgac cgtgtctagc 360
gctagcacaa agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 248
<211> 654
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 248
gacatcgtgc tgacacagag ccctgccacc ctgtctctga gccctggcga gagagccacc 60
atcagctgta gagccagcca gagcgtgtcc agctacggcc agggcttcat gcactggtat 120
cagcagaagc ccggccagcc ccccagactg ctgatctatg gcgccagcag cagagccaca 180
ggcatccccg ccagattttc tggctctggc agcggcaccg acttcaccct gacaatcagc 240
cccctggaac ccgaggactt cgccgtgtac tactgccagc agaacaaaga ggacccctgg 300
accttcggcg gaggcaccaa gctggaaatc aagcgtacgg tggccgctcc cagcgtgttc 360
atcttcccac ctagcgacga gcagctgaag tccggcacag cctctgtcgt gtgcctgctg 420
aacaacttct acccccgcga ggccaaggtg cagtggaagg tggacaatgc cctgcagagc 480
ggcaacagcc aggaaagcgt gaccgagcag gacagcaagg actccaccta cagcctgagc 540
agcaccctga ccctgtccaa ggccgattac gagaagcaca aggtgtacgc ctgcgaagtg 600
acccaccagg gcctgtctag ccccgtgacc aagagcttca accggggcga gtgc 654
<210> 249
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 249
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 250
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 250
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 251
<211> 1326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 251
caggttcagc tggttcagtc tggcgccgaa gtgaagaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg tgtccggcta caccctgacc gagttcagca tccactgggt ccgacaggct 120
ccaggacaag gcttggaatg gatgggcggc ttcgatcccg aggacggcga aacaatctac 180
gcccagaaat tccagggccg cgtgatcatg accgaggaca cctctaccga caccgcctac 240
atggaaatga acagcctgcg gagcgaggat accgccatct actactgtac caccggcaga 300
ttcttcgact ggttctgggg ccagggcacc ctggttacag tctcttctgc gtcgaccaag 360
ggcccatcgg tgttccctct ggccccttgc agcagaagca ccagcgaatc tacagccgcc 420
ctgggctgcc tcgtgaagga ctactttccc gagcccgtga ccgtgtcctg gaactctggc 480
gctctgacaa gcggcgtgca cacctttcca gccgtgctcc agagcagcgg cctgtactct 540
ctgagcagcg tcgtgacagt gcccagcagc agcctgggca ccaagaccta cacctgtaac 600
gtggaccaca agcccagcaa caccaaggtg gacaagcggg tggaatctaa gtacggccct 660
ccctgccctc cttgcccagc ccctgaagct gccggcggac cctccgtgtt cctgttcccc 720
ccaaagccca aggacaccct gatgatcagc cggacccccg aagtgacctg cgtggtggtg 780
gatgtgtccc aggaagatcc cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg 840
cacaacgcca agaccaagcc cagagaggaa cagttcaaca gcacctaccg ggtggtgtcc 900
gtgctgaccg tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 960
aacaagggcc tgcccagctc catcgagaaa accatcagca aggccaaggg ccagccccgc 1020
gagcctcaag tgtataccct gcccccttgc caggaagaga tgaccaagaa ccaggtgtcc 1080
ctgtggtgtc tcgtgaaagg cttctacccc agcgacattg ccgtggaatg ggagagcaac 1140
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggacagcga cggctcattc 1200
ttcctgtact ccaagctgac cgtggacaag agccggtggc aggaaggcaa cgtgttcagc 1260
tgctccgtga tgcacgaggc cctgcacaac cactacaccc agaagtccct gtctctgtcc 1320
ctgggc 1326
<210> 252
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 252
gagatcatcc tgacacagag ccccgccatc ctgtcactgt ctccaggcga aagagccaca 60
ctgagctgta gagccagcca gagcgtgatc agcagattcc tgagctggta tcaagtgaag 120
cccggactgg cccctcggct gttgatatat ggcgcctcta cacgcgccac aggcatccct 180
gttagatttt ctggcagcgg ctccggcacc gacttcagcc tgacaattag cagcctgcag 240
cctgaggact gcgccgtgta ctactgtcag caggacagca acctgcctat caccttcggc 300
cagggcacca gactggaaat caagcgtacg gtggccgctc ccagcgtgtt catcttccca 360
cctagcgacg agcagctgaa gtccggcaca gcctctgtcg tgtgcctgct gaacaacttc 420
tacccccgcg aggccaaagt gcagtggaag gtggacaacg ccctgcagag cggcaacagc 480
caggaaagcg tgaccgagca ggacagcaag gactccacct acagcctgag cagcaccctg 540
acactgagca aggccgacta cgagaagcac aaggtgtacg cctgcgaagt gacccaccag 600
ggcctgtcta gccccgtgac caagagcttc aaccggggcg agtgt 645
<210> 253
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 253
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 254
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 254
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 255
<211> 1320
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 255
caggttcagc tgcagcagtc tggccccgaa ctcgttagac ctggcacctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cgcctttacc acctacctgg tggaatggat caagcagagg 120
cctggacagg gcctcgagtg gatcggagtg atcaatcctg gcagcggcag caccaactac 180
aacgagaagt tcaagggcaa agccacactg accgtggaca gaagcagcac cacagcctac 240
atgcacctga gcggcctgac ctctgatgac agcgccgtgt acttctgcgc cagatacgcc 300
tatggctatt ggggccaggg cacaaccctg accgttagct ctgcgtcgac caagggccca 360
tcggtgttcc ctctggcccc ttgcagcaga agcaccagcg aatctacagc cgccctgggc 420
tgcctcgtga aggactactt tcccgagccc gtgaccgtgt cctggaactc tggcgctctg 480
acaagcggcg tgcacacctt tccagccgtg ctccagagca gcggcctgta ctctctgagc 540
agcgtcgtga cagtgcccag cagcagcctg ggcaccaaga cctacacctg taacgtggac 600
cacaagccca gcaacaccaa ggtggacaag cgggtggaat ctaagtacgg ccctccctgc 660
cctccttgcc cagcccctga agctgccggc ggaccctccg tgttcctgtt ccccccaaag 720
cccaaggaca ccctgatgat cagccggacc cccgaagtga cctgcgtggt ggtggatgtg 780
tcccaggaag atcccgaggt gcagttcaat tggtacgtgg acggcgtgga agtgcacaac 840
gccaagacca agcccagaga ggaacagttc aacagcacct accgggtggt gtccgtgctg 900
accgtgctgc accaggactg gctgaacggc aaagagtaca agtgcaaggt gtccaacaag 960
ggcctgccca gctccatcga gaaaaccatc agcaaggcca agggccagcc ccgcgagcct 1020
caagtgtata ccctgccccc ttgccaggaa gagatgacca agaaccaggt gtccctgtgg 1080
tgtctcgtga aaggcttcta ccccagcgac attgccgtgg aatgggagag caacggccag 1140
cccgagaaca actacaagac caccccccct gtgctggaca gcgacggctc attcttcctg 1200
tactccaagc tgaccgtgga caagagccgg tggcaggaag gcaacgtgtt cagctgctcc 1260
gtgatgcacg aggccctgca caaccactac acccagaagt ccctgtctct gtccctgggc 1320
<210> 256
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 256
gacatcgtga tgacccagag ccagaaattc atgagcgcca gcgtgggcga cagagtgtcc 60
atcacatgta aagccagcca gaacgtgggc acagccgtgg cttggtatca gcagcagcct 120
ggccactctc ctaagcagct gatctacagc gccagcaaca gatacaccgg cgtgcccgat 180
agattcacag gatctggcgc cggaaccgac ttcaccctga ccatcagcaa catccagagc 240
gaggacctgg ccgactactt ctgccagcag tacagcacat accccttcac ctttggcagc 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 257
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 257
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 258
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 258
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 259
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 259
caggttcagc tgctgcagtc tggcgccgaa cttgtcagac ctggcgtgtc cgtgaagatc 60
agctgtacag gcagcggcta cagcttcacc aactacgccg tgcactgggt caagcagagc 120
cacgtgaagt ccctggaatg gatcggcgtg atcagcccct actacggcga caccacctac 180
aaccagaagt tcaccggcaa ggccaccatg accgtggaca agtctagcag caccgcctac 240
atggaactgg ccagactgac cagcgaggac agcgccatct acttttgcgc cagaagattc 300
gagggcttct actacagcat ggactactgg ggccagggca ccagcgtgac agtttcttct 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 260
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 260
gacgtggtca tgatccagac acctctgagc ctgcctgtgt ctctgggaga tcaggccagc 60
atcagctgca gacctagcca gtctctggtg cacagcaacg gcaacaccta cctgaactgg 120
tatctgcaga ggcccggaca gagccccaag ctgctgatct acaaggtgtc caagcggttc 180
agcggcgtgc ccgatagatt ttctggcagc ggctctggca ccgacttcac cctgaagatt 240
agcagagtgg aagccgagga cctgggcgtg tacctgtgtt ctcagagcac acacgtgccc 300
ctgacctttg gcagcggaac ccagctggaa atcaagcgta cggtggccgc tcccagcgtg 360
ttcatcttcc cacctagcga cgagcagctg aagtccggca cagcctctgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaaa gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggaaag cgtgaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgacactgag caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc tagccccgtg accaagagct tcaaccgggg cgagtgt 657
<210> 261
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 261
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 262
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 262
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 263
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 263
caggtgcagc tggtggaatc tggcggcgga gtggtgcagc ctggcagaag cctgagactg 60
agctgtgccg ccagcggctt caccttcagc agctacggca tgtactgggt gcgccaggcc 120
cctggcaaag gcctggaatg ggtggccgtg atttggtacg acggcagcaa caagtactac 180
gccgacagcg tgaagggccg gttcaccatc tcccgggaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgcg ggccgaggac acagccgtgt atcactgcgc cagagatccc 300
ggcctgcggt actttgacgg cggcatggat gtgtggggcc agggcacaac cgtgaccgtg 360
tcatctgcgt cgaccaaggg cccatcggtg ttccctctgg ccccttgcag cagaagcacc 420
agcgaatcta cagccgccct gggctgcctc gtgaaggact actttcccga gcccgtgacc 480
gtgtcctgga actctggcgc tctgacaagc ggcgtgcaca cctttccagc cgtgctccag 540
agcagcggcc tgtactctct gagcagcgtc gtgacagtgc ccagcagcag cctgggcacc 600
aagacctaca cctgtaacgt ggaccacaag cccagcaaca ccaaggtgga caagcgggtg 660
gaatctaagt acggccctcc ctgccctcct tgcccagccc ctgaagctgc cggcggaccc 720
tccgtgttcc tgttcccccc aaagcccaag gacaccctga tgatcagccg gacccccgaa 780
gtgacctgcg tggtggtgga tgtgtcccag gaagatcccg aggtgcagtt caattggtac 840
gtggacggcg tggaagtgca caacgccaag accaagccca gagaggaaca gttcaacagc 900
acctaccggg tggtgtccgt gctgaccgtg ctgcaccagg actggctgaa cggcaaagag 960
tacaagtgca aggtgtccaa caagggcctg cccagctcca tcgagaaaac catcagcaag 1020
gccaagggcc agccccgcga gcctcaagtg tataccctgc ccccttgcca ggaagagatg 1080
accaagaacc aggtgtccct gtggtgtctc gtgaaaggct tctaccccag cgacattgcc 1140
gtggaatggg agagcaacgg ccagcccgag aacaactaca agaccacccc ccctgtgctg 1200
gacagcgacg gctcattctt cctgtactcc aagctgaccg tggacaagag ccggtggcag 1260
gaaggcaacg tgttcagctg ctccgtgatg cacgaggccc tgcacaacca ctacacccag 1320
aagtccctgt ctctgtccct gggc 1344
<210> 264
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 264
gacatccagc tgacccagag ccccagcttt ctgagcgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca gggcatcagc agctacctgg cctggtatca gcagaagccc 120
ggcaaggccc ccaaactgct gatctttgcc gccagcacac tgcacagcgg cgtgcccagc 180
agattttctg gcagcggctc cggcaccgag ttcaccctga caatcagcag cctgcagccc 240
gaggacttcg ccacctacta ctgccagcag ctgaacagct tcccctacac cttcggccag 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 265
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 265
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 266
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 266
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 267
<211> 1329
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 267
caggtgcagc tggtggaaag cggcggaggc gtggtgcagc ctggcaggtc tctgagactg 60
agctgtgccg ccagcggctt caccttcagc agctacggaa tgcactgggt gcgccaggcc 120
cctggcaaag gactggaatg ggtggccgtg atttggtacg acggcagcaa caagtactac 180
gccgacagcg tgaagggccg gttcaccatc agcggcgaca acagcaagaa caccctgtac 240
ctgcagatga acagcctgcg ggccgaggac accgccgtgt actactgcgc cagaatgttc 300
agaggcgcct tcgactactg gggccagggc acactcgtga ccgtgtctag tgcgtcgacc 360
aagggcccat cggtgttccc tctggcccct tgcagcagaa gcaccagcga atctacagcc 420
gccctgggct gcctcgtgaa ggactacttt cccgagcccg tgaccgtgtc ctggaactct 480
ggcgctctga caagcggcgt gcacaccttt ccagccgtgc tccagagcag cggcctgtac 540
tctctgagca gcgtcgtgac agtgcccagc agcagcctgg gcaccaagac ctacacctgt 600
aacgtggacc acaagcccag caacaccaag gtggacaagc gggtggaatc taagtacggc 660
cctccctgcc ctccttgccc agcccctgaa gctgccggcg gaccctccgt gttcctgttc 720
cccccaaagc ccaaggacac cctgatgatc agccggaccc ccgaagtgac ctgcgtggtg 780
gtggatgtgt cccaggaaga tcccgaggtg cagttcaatt ggtacgtgga cggcgtggaa 840
gtgcacaacg ccaagaccaa gcccagagag gaacagttca acagcaccta ccgggtggtg 900
tccgtgctga ccgtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 960
tccaacaagg gcctgcccag ctccatcgag aaaaccatca gcaaggccaa gggccagccc 1020
cgcgagcctc aagtgtatac cctgccccct tgccaggaag agatgaccaa gaaccaggtg 1080
tccctgtggt gtctcgtgaa aggcttctac cccagcgaca ttgccgtgga atgggagagc 1140
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggacag cgacggctca 1200
ttcttcctgt actccaagct gaccgtggac aagagccggt ggcaggaagg caacgtgttc 1260
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtctctg 1320
tccctgggc 1329
<210> 268
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 268
gccatccaga tgacccagag ccccagcagc ctgtctgcca gcgtgggcga cagagtgacc 60
atcacctgta gagccagcca gggcatccgg aacgacctgg gctggtatca gcagaagcct 120
ggcaaggccc ccaagctgct gatctacgcc gctagctctc tgcagtccgg cgtgcccagc 180
agattttctg gcagcggctc cggcaccgac ttcaccctga caatctctgg cctgcagccc 240
gaggacagcg ccacctacta ctgtctgcaa gactacatct actaccccac cttcggccag 300
ggcaccaagg tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 269
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 269
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 270
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 270
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 271
<211> 1320
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 271
caggttcagc tggttcagtc tggcgccgaa gtgaagaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg cctctggcta cgccttcacc acctacctgg tggaatggat cagacagagg 120
cctggacagg gcctcgaatg gatgggcgtg atcaatcctg gcagcggcag caccaattac 180
gcccagaaat tccagggcag agtgaccatg accgtggaca gaagcagcac caccgcctac 240
atggaactga gcagactgag aagcgacgac accgccgtgt actactgtgc cagatacgcc 300
tacggctatt ggggccaggg aaccctggtt accgttagct ctgcgtcgac caagggccca 360
tcggtgttcc ctctggcccc ttgcagcaga agcaccagcg aatctacagc cgccctgggc 420
tgcctcgtga aggactactt tcccgagccc gtgaccgtgt cctggaactc tggcgctctg 480
acaagcggcg tgcacacctt tccagccgtg ctccagagca gcggcctgta ctctctgagc 540
agcgtcgtga cagtgcccag cagcagcctg ggcaccaaga cctacacctg taacgtggac 600
cacaagccca gcaacaccaa ggtggacaag cgggtggaat ctaagtacgg ccctccctgc 660
cctccttgcc cagcccctga agctgccggc ggaccctccg tgttcctgtt ccccccaaag 720
cccaaggaca ccctgatgat cagccggacc cccgaagtga cctgcgtggt ggtggatgtg 780
tcccaggaag atcccgaggt gcagttcaat tggtacgtgg acggcgtgga agtgcacaac 840
gccaagacca agcccagaga ggaacagttc aacagcacct accgggtggt gtccgtgctg 900
accgtgctgc accaggactg gctgaacggc aaagagtaca agtgcaaggt gtccaacaag 960
ggcctgccca gctccatcga gaaaaccatc agcaaggcca agggccagcc ccgcgagcct 1020
caagtgtata ccctgccccc ttgccaggaa gagatgacca agaaccaggt gtccctgtgg 1080
tgtctcgtga aaggcttcta ccccagcgac attgccgtgg aatgggagag caacggccag 1140
cccgagaaca actacaagac caccccccct gtgctggaca gcgacggctc attcttcctg 1200
tactccaagc tgaccgtgga caagagccgg tggcaggaag gcaacgtgtt cagctgctcc 1260
gtgatgcacg aggccctgca caaccactac acccagaagt ccctgtctct gtccctgggc 1320
<210> 272
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 272
gacatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 60
atcacctgta gagccagcca gaatgtggga acagccgtgg cctggtatca gcagaagcct 120
ggcaagagcc ccaagcagct gatctacagc gccagcaaca gatacaccgg cgtgcccagc 180
agattttctg gcagcggctc tggcaccgac ttcaccctga ccatatctag cctgcagcct 240
gaggacctgg ccacctacta ctgtcagcag tacagcacat accccttcac cttcggccag 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccacct 360
agcgacgagc agctgaagtc cggcacagcc tctgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaagtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gaaagcgtga ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgaca 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtctagcc ccgtgaccaa gagcttcaac cggggcgagt gt 642
<210> 273
<211> 1008
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 273
gacatcgtga tgacccagac ccccctgagc ctgagcgtga cacctggaca gcctgccagc 60
atcagctgca agagcagcca gagcctggtg cacgagaacc tgcagaccta cctgagctgg 120
tatctgcaga agcccggcca gagcccccag tccctgatct acaaggtgtc caacagattc 180
agcggcgtgc ccgacagatt ctccggcagc ggctctggca ccgacttcac cctgaagatc 240
agccgggtgg aagccgagga cgtgggcgtg tactattgtg gccagggcac ccagtacccc 300
ttcacctttg gcagcggcac caaggtggaa atcaaggaca aaacccatac cgacatccag 360
atgacccaga gccccagcag cctgtctgcc agcgtgggcg acagagtgac catcacctgt 420
caggccagcc agaacatcta cgtgtggctg aactggtatc agcagaagcc cggcaaggcc 480
cccaagctgc tgatctacaa ggccagcaac ctgcacaccg gcgtgcccag cagattttct 540
ggcagcggct ccggcaccga cttcaccctg acaatcagct ccctgcagcc cgaggacatt 600
gccacctact actgccagca gggccagacc tacccctaca cctttggcca gggcaccaag 660
ctggaaatca aggataagac ccacacccgt acggtggccg ctcccagcgt gttcatcttc 720
ccacctagcg acgagcagct gaagtccggc acagcctctg tcgtgtgcct gctgaacaac 780
ttctaccccc gcgaggccaa agtgcagtgg aaggtggaca acgccctgca gagcggcaac 840
agccaggaaa gcgtgaccga gcaggacagc aaggactcca cctacagcct gagcagcacc 900
ctgacactga gcaaggccga ctacgagaag cacaaggtgt acgcctgcga agtgacccac 960
cagggcctgt ctagccccgt gaccaagagc ttcaaccggg gcgagtgt 1008
<210> 274
<211> 1731
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 274
caggtgcagc tggtgcagtc tggcgccgag gtcgtgaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cacctttacc agctactaca tccactgggt gcgccaggcc 120
cctggacagg gactggaatg gatcggcagc atctaccccg gcaacgtgaa caccaactac 180
gcccagaagt tccagggcag agccaccctg accgtggaca ccagcatcag caccgcctac 240
atggaactga gccggctgag aagcgacgac accgccgtgt actactgcac ccggtcccac 300
tacggcctgg attggaactt cgacgtgtgg ggcaagggca ccaccgtgac agtgtctagc 360
gacaaaaccc atacccaggt gcagctggtg gaatctggcg gcggagtggt gcagcctggc 420
agaagcctga gactgagctg tgccgccagc ggcttcacct tcaccaaggc ctggatgcac 480
tgggtgcgcc aggcccctgg aaagcagctg gaatgggtgg cccagatcaa ggacaagagc 540
aacagctacg ccacctacta cgccgacagc gtgaagggcc ggttcaccat cagccgggac 600
gacagcaaga acaccctgta cctgcagatg aacagcctgc gggccgagga caccgccgtg 660
tactactgtc ggggcgtgta ctatgccctg agccccttcg attactgggg ccagggaacc 720
ctcgtgaccg tgtctagtga taagacccac accgccagca caaagggccc atcggtgttc 780
cctctggccc cttgcagcag aagcaccagc gaatctacag ccgccctggg ctgcctcgtg 840
aaggactact ttcccgagcc cgtgaccgtg tcctggaact ctggcgctct gacaagcggc 900
gtgcacacct ttccagccgt gctccagagc agcggcctgt actctctgag cagcgtcgtg 960
acagtgccca gcagcagcct gggcaccaag acctacacct gtaacgtgga ccacaagccc 1020
agcaacacca aggtggacaa gcgggtggaa tctaagtacg gccctccctg ccctccttgc 1080
ccagcccctg aagctgccgg cggaccctcc gtgttcctgt tccccccaaa gcccaaggac 1140
accctgatga tcagccggac ccccgaagtg acctgcgtgg tggtggatgt gtcccaggaa 1200
gatcccgagg tgcagttcaa ttggtacgtg gacggcgtgg aagtgcacaa cgccaagacc 1260
aagcccagag aggaacagtt caacagcacc taccgggtgg tgtccgtgct gaccgtgctg 1320
caccaggact ggctgaacgg caaagagtac aagtgcaagg tgtccaacaa gggcctgccc 1380
agctccatcg agaaaaccat cagcaaggcc aagggccagc cccgcgagcc tcaagtgtgt 1440
accctgcccc ctagccagga agagatgacc aagaaccagg tgtccctgag ctgtgccgtg 1500
aaaggcttct accccagcga cattgccgtg gaatgggaga gcaacggcca gcccgagaac 1560
aactacaaga ccaccccccc tgtgctggac agcgacggct cattcttcct ggtgtccaag 1620
ctgaccgtgg acaagagccg gtggcaggaa ggcaacgtgt tcagctgctc cgtgatgcac 1680
gaggccctgc acaaccacta cacccagaag tccctgtctc tgtccctggg c 1731
<210> 275
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 275
caggttcagc tggttcagtc tggcgccgaa gtgaagaaac ctggcgcctc tgtgaaggtg 60
tcctgcaagg ccagcggcta cagcttcacc aattacgccg tgcactgggt ccgacaggct 120
ccaggacaag gactggaatg gatgggcgtg atcagcccct actacggcga taccacatac 180
gcccagaaat tccagggcag agtgaccatg accgtggaca agagcagcag caccgcctac 240
atggaactga gcagactgag aagcgacgac accgccgtgt actactgcgc cagaagattc 300
gagggcttct actacagcat ggactactgg ggccagggca ccctggttac agtctcttct 360
gcgtcgacca agggcccatc ggtgttccct ctggcccctt gcagcagaag caccagcgaa 420
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 480
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct ccagagcagc 540
ggcctgtact ctctgagcag cgtcgtgaca gtgcccagca gcagcctggg caccaagacc 600
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 660
aagtacggcc ctccctgccc tccttgccca gcccctgaag ctgccggcgg accctccgtg 720
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 780
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 840
ggcgtggaag tgcacaacgc caagaccaag cccagagagg aacagttcaa cagcacctac 900
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agagtacaag 960
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1020
ggccagcccc gcgagcctca agtgtatacc ctgccccctt gccaggaaga gatgaccaag 1080
aaccaggtgt ccctgtggtg tctcgtgaaa ggcttctacc ccagcgacat tgccgtggaa 1140
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggacagc 1200
gacggctcat tcttcctgta ctccaagctg accgtggaca agagccggtg gcaggaaggc 1260
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1320
ctgtctctgt ccctgggc 1338
<210> 276
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 276
gacgtggtca tgacacagag ccctctgagc ctgcctgtga cactgggaca gcctgccagc 60
atcagctgta gacctagcca gagcctggtg cacagcaacg gcaacaccta cctgaactgg 120
tatcagcaga ggcccggaca gagccccaag ctgctgatct acaaggtgtc caagcggttc 180
agcggcgtgc ccgatagatt ttctggcagc ggctctggca ccgacttcac cctgaagatt 240
agcagagtgg aagccgagga cgtgggcgtg tactactgta gccagtctac ccacgtgcca 300
ctgacctttg gcggcggaac aaaggtggaa atcaagcgta cggtggccgc tcccagcgtg 360
ttcatcttcc cacctagcga cgagcagctg aagtccggca cagcctctgt cgtgtgcctg 420
ctgaacaact tctacccccg cgaggccaaa gtgcagtgga aggtggacaa cgccctgcag 480
agcggcaaca gccaggaaag cgtgaccgag caggacagca aggactccac ctacagcctg 540
agcagcaccc tgacactgag caaggccgac tacgagaagc acaaggtgta cgcctgcgaa 600
gtgacccacc agggcctgtc tagccccgtg accaagagct tcaaccgggg cgagtgt 657
<210> 277
<211> 114
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 277
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Thr Thr Tyr
20 25 30
Leu Val Glu Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Asn Pro Gly Ser Gly Ser Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Arg Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met His Leu Ser Gly Leu Thr Ser Asp Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Tyr Ala Tyr Gly Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 278
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 278
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Gln Pro Gly His Ser Pro Lys Gln Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ala Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Ile Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Ser Thr Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 279
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 279
Gln Val Gln Leu Leu Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Val
1 5 10 15
Ser Val Lys Ile Ser Cys Thr Gly Ser Gly Tyr Ser Phe Thr Asn Tyr
20 25 30
Ala Val His Trp Val Lys Gln Ser His Val Lys Ser Leu Glu Trp Ile
35 40 45
Gly Val Ile Ser Pro Tyr Tyr Gly Asp Thr Thr Tyr Asn Gln Lys Phe
50 55 60
Thr Gly Lys Ala Thr Met Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ala Arg Leu Thr Ser Glu Asp Ser Ala Ile Tyr Phe Cys
85 90 95
Ala Arg Arg Phe Glu Gly Phe Tyr Tyr Ser Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 280
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 280
Asp Val Val Met Ile Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Pro Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asn Trp Tyr Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Lys Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Leu Cys Ser Gln Ser
85 90 95
Thr His Val Pro Leu Thr Phe Gly Ser Gly Thr Gln Leu Glu Ile Lys
100 105 110
<210> 281
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 281
Glu Asn Leu Gln
1
<210> 282
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 282
Glu Asn Leu Phe
1
<210> 283
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 283
Glu Asn Leu Arg
1
<210> 284
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 284
Asn Leu Val Pro Met Val Ala Thr Val
1 5
<210> 285
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 285
Gly Leu Cys Thr Leu Val Ala Met Leu
1 5
<210> 286
<211> 336
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 286
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Asp Lys Thr His Thr Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
115 120 125
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln
130 135 140
Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala
145 150 155 160
Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly Val Pro
165 170 175
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly
195 200 205
Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
210 215 220
Asp Lys Thr His Thr Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys
245 250 255
Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val
260 265 270
Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln
275 280 285
Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser
290 295 300
Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His
305 310 315 320
Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
325 330 335
<210> 287
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 287
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 288
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 288
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Ala Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Ser Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 289
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 289
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 290
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 290
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 291
<211> 577
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 291
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Asp Lys Thr His Thr Gln Val Gln
115 120 125
Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile
165 170 175
Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Glu Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg
210 215 220
Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser Asp Lys Thr His Thr Ala Ser Thr Lys Gly
245 250 255
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
260 265 270
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
275 280 285
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
290 295 300
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
305 310 315 320
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
325 330 335
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
340 345 350
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
355 360 365
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
370 375 380
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
385 390 395 400
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
405 410 415
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
420 425 430
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
435 440 445
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
450 455 460
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
465 470 475 480
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
485 490 495
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
500 505 510
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
515 520 525
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
530 535 540
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
545 550 555 560
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
565 570 575
Gly
<210> 292
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 292
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 293
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 293
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 294
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 294
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Asn Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 295
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 295
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Ser Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 296
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 296
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 297
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 297
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 298
<211> 338
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 298
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Gln
20 25 30
Ser Ala Gln Thr Tyr Leu Ser Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Ser Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Gly Gln Gly
85 90 95
Thr Gln Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Val Glu Ile Lys
100 105 110
Gly Gln Pro Lys Ala Ala Pro Asp Ile Gln Met Thr Gln Ser Pro Ser
115 120 125
Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala
130 135 140
Ser Gln Asn Ile Tyr Val Trp Leu Asn Trp Tyr Gln Gln Lys Pro Gly
145 150 155 160
Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Asn Leu His Thr Gly
165 170 175
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
180 185 190
Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
195 200 205
Gln Gly Gln Thr Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
210 215 220
Ile Lys Thr Lys Gly Pro Ser Arg Thr Val Ala Ala Pro Ser Val Phe
225 230 235 240
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
245 250 255
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
260 265 270
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
275 280 285
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
290 295 300
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
305 310 315 320
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
325 330 335
Glu Cys
<210> 299
<211> 570
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 299
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Val Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Ala Thr Leu Thr Val Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser His Tyr Gly Leu Asp Trp Asn Phe Asp Val Trp Gly Lys
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ser Gln Val Gln Leu Val Glu Ser
115 120 125
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
130 135 140
Ala Ser Gly Phe Thr Phe Thr Lys Ala Trp Met His Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gln Leu Glu Trp Val Ala Gln Ile Lys Asp Lys Ser
165 170 175
Asn Ser Tyr Ala Thr Tyr Tyr Ala Ser Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asp Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Arg Gly Val Tyr Tyr
210 215 220
Ala Leu Ser Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
225 230 235 240
Ser Ser Arg Thr Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
245 250 255
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
260 265 270
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
275 280 285
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
290 295 300
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
305 310 315 320
Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
325 330 335
Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro
340 345 350
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
355 360 365
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
370 375 380
Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn
385 390 395 400
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
405 410 415
Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
420 425 430
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
435 440 445
Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
450 455 460
Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu
465 470 475 480
Glu Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
485 490 495
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
500 505 510
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
515 520 525
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
530 535 540
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
545 550 555 560
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
565 570
<210> 300
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 300
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Arg Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Gln Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Glu Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Cys Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 301
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 301
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 302
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 302
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Lys Ala
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gln Leu Glu Trp Val
35 40 45
Ala Gln Ile Lys Asp Lys Ser Asn Ser Tyr Ala Thr Tyr Tyr Ala Ser
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Arg Gly Val Tyr Tyr Ala Leu Ser Pro Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРИСПЕЦИФИЧЕСКИЕ И/ИЛИ ТРИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ПРИМЕНЕНИЕМ КРОССОВЕРНОГО ФОРМАТА С ДВОЙНЫМ ВАРИАБЕЛЬНЫМ ДОМЕНОМ (CODV) ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ, ВЫЗВАННОЙ HIV | 2020 |
|
RU2820164C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РАКОВЫМИ КЛЕТКАМИ И ОБЕСПЕЧИВАЮТ НАПРАВЛЕННЫЙ ПЕРЕНОС РАДИОНУКЛИДОВ К УКАЗАННЫМ КЛЕТКАМ | 2020 |
|
RU2833191C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К CCL2 | 2020 |
|
RU2819613C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| АНТИ-LILRB1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2801535C1 |
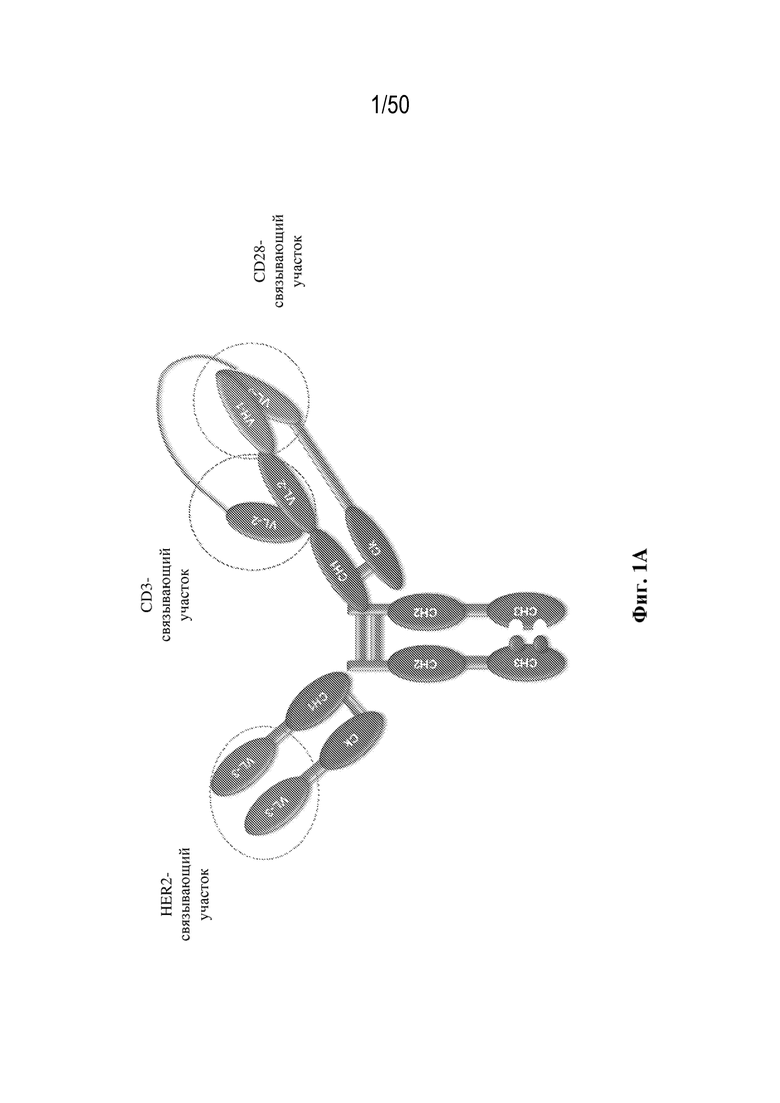
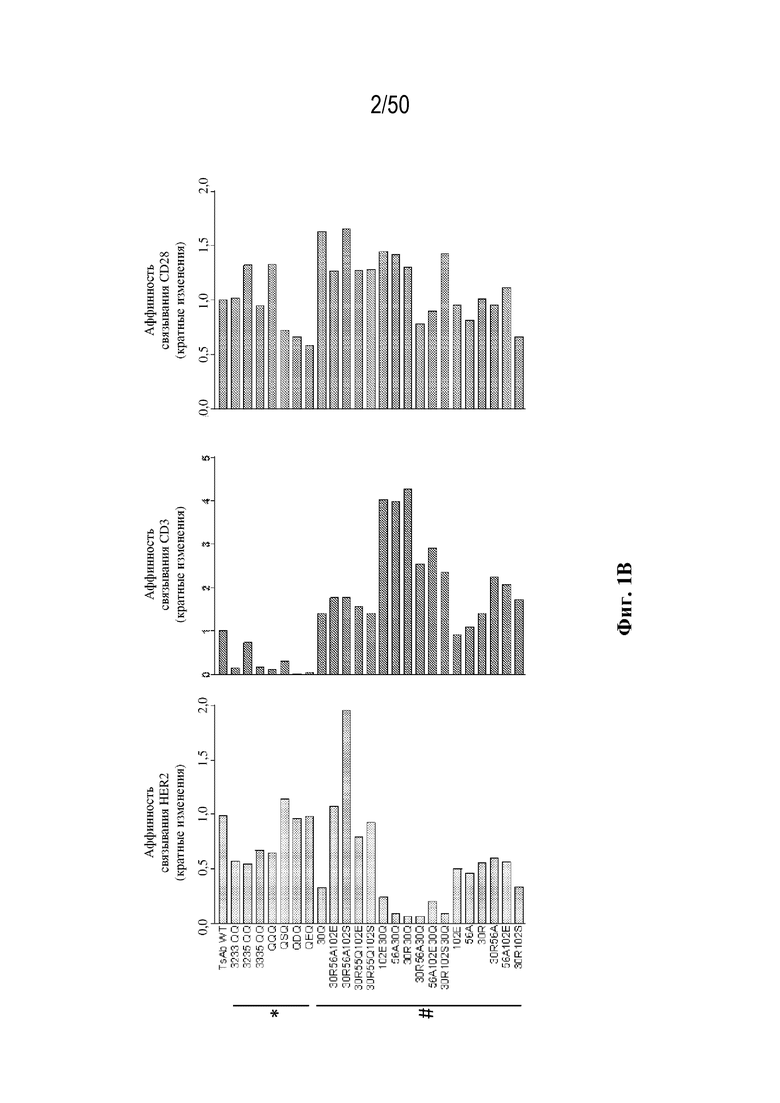
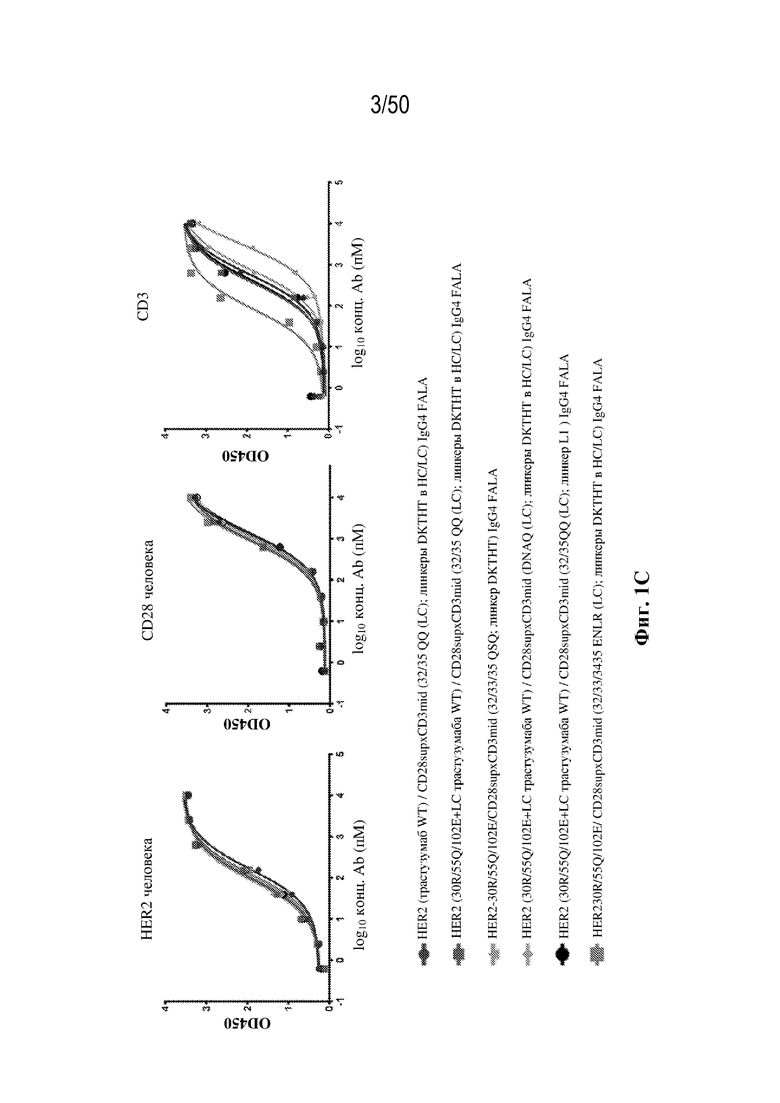

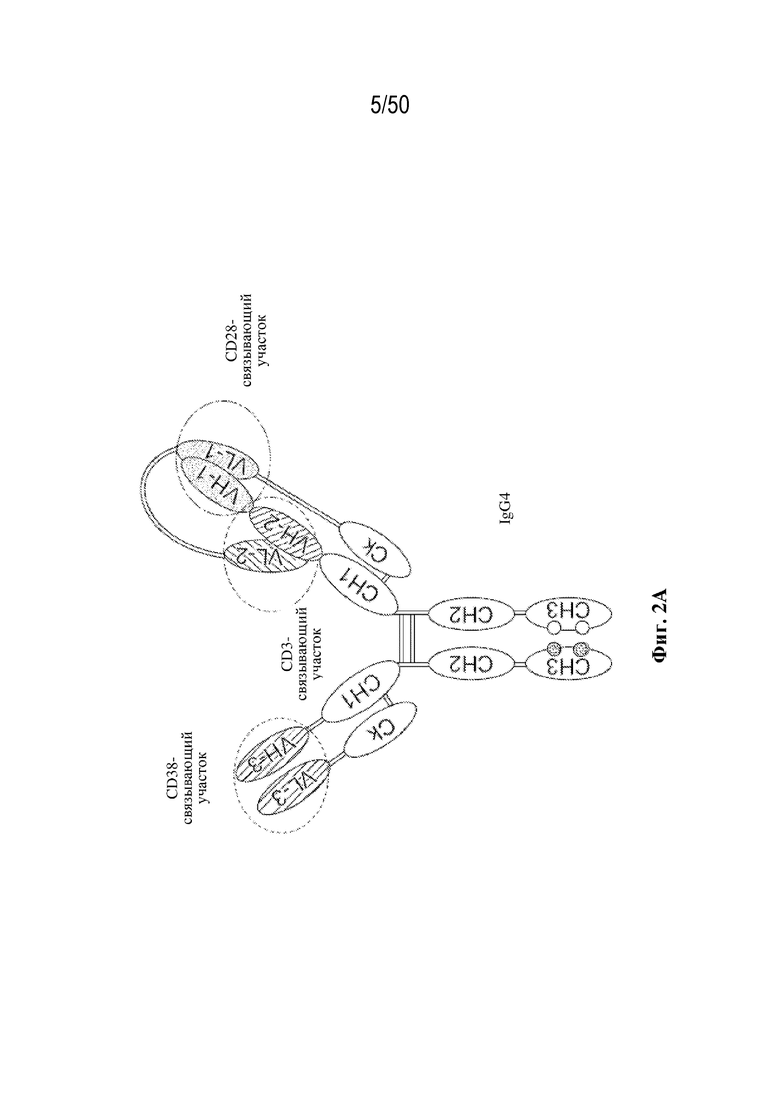
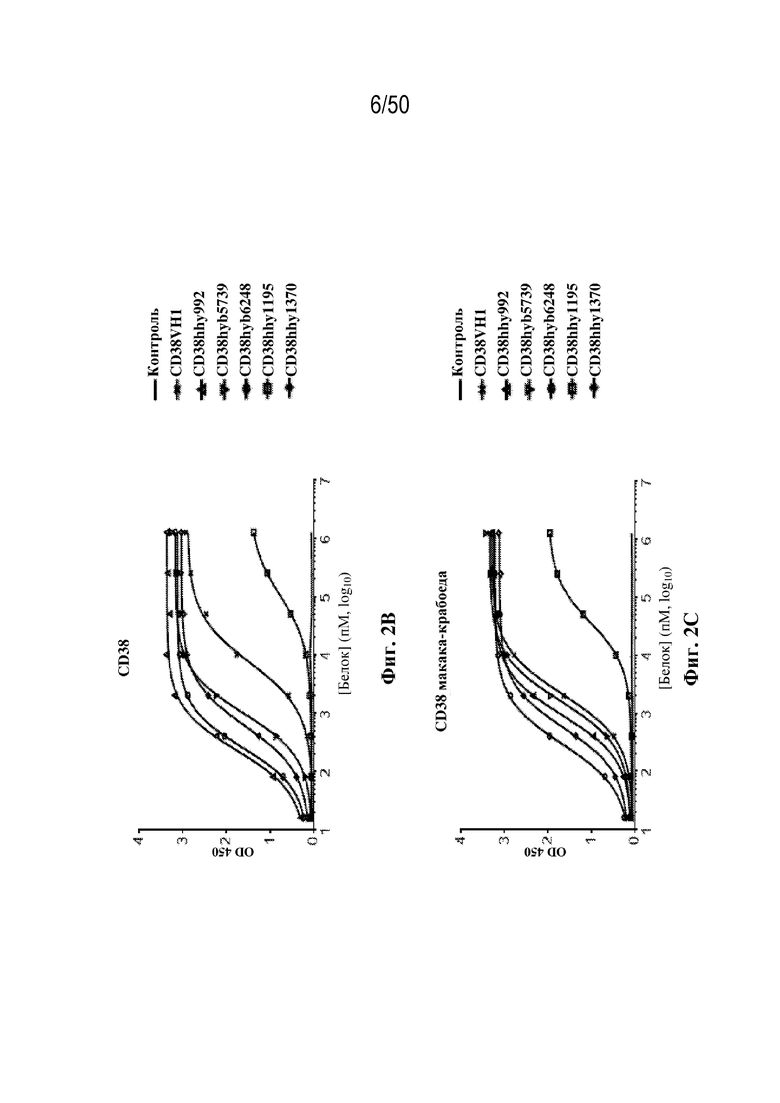
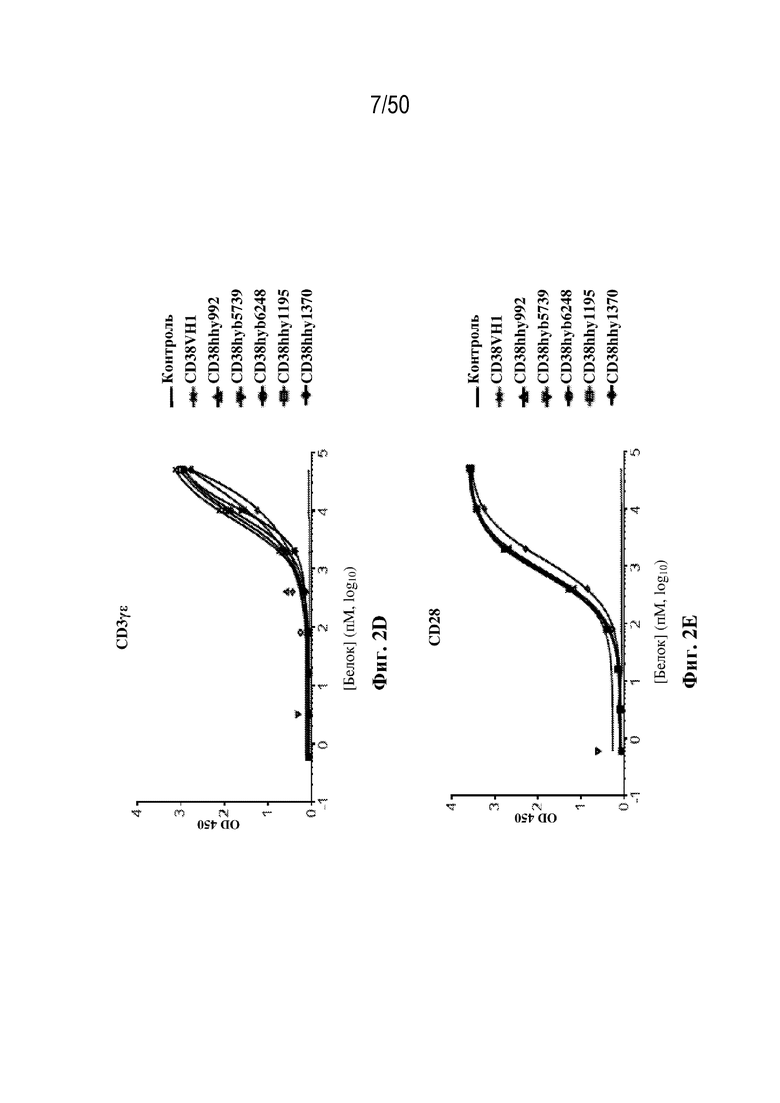
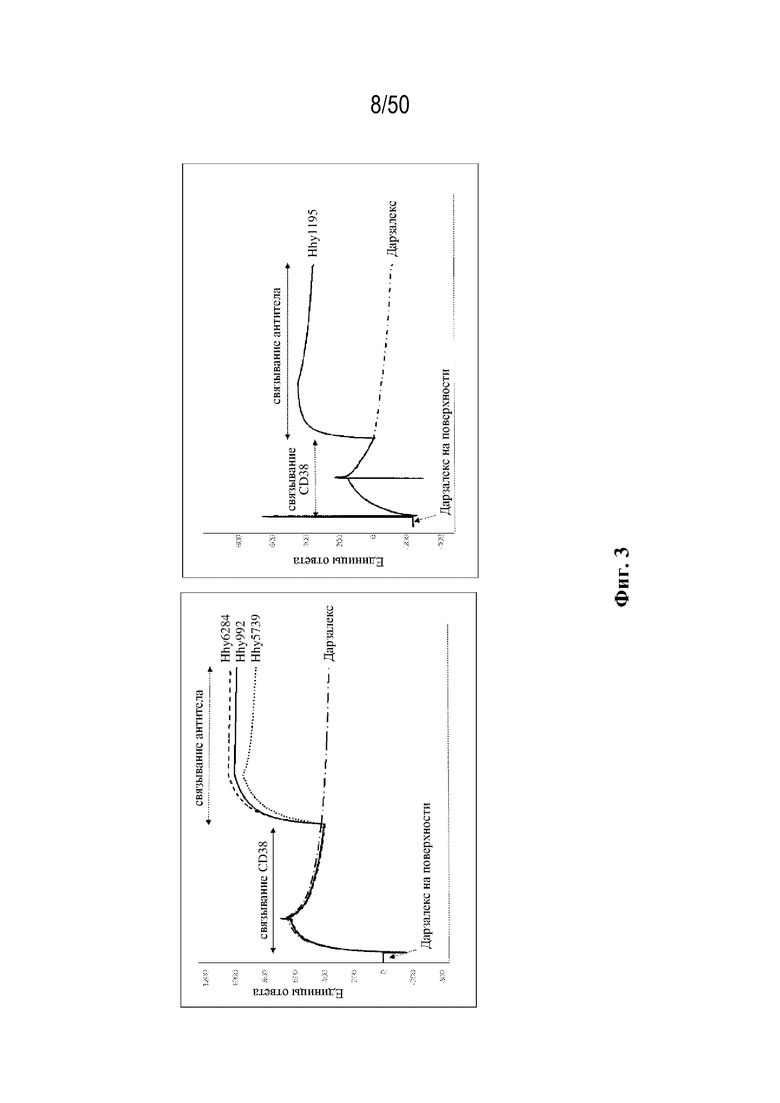
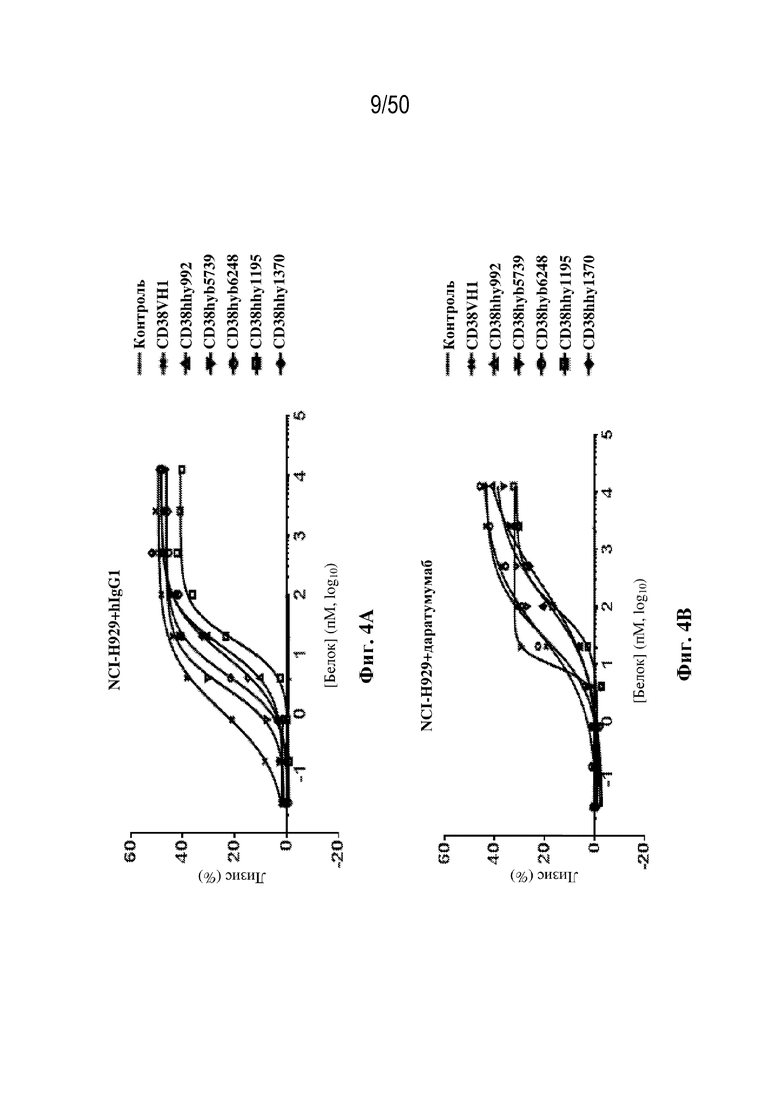
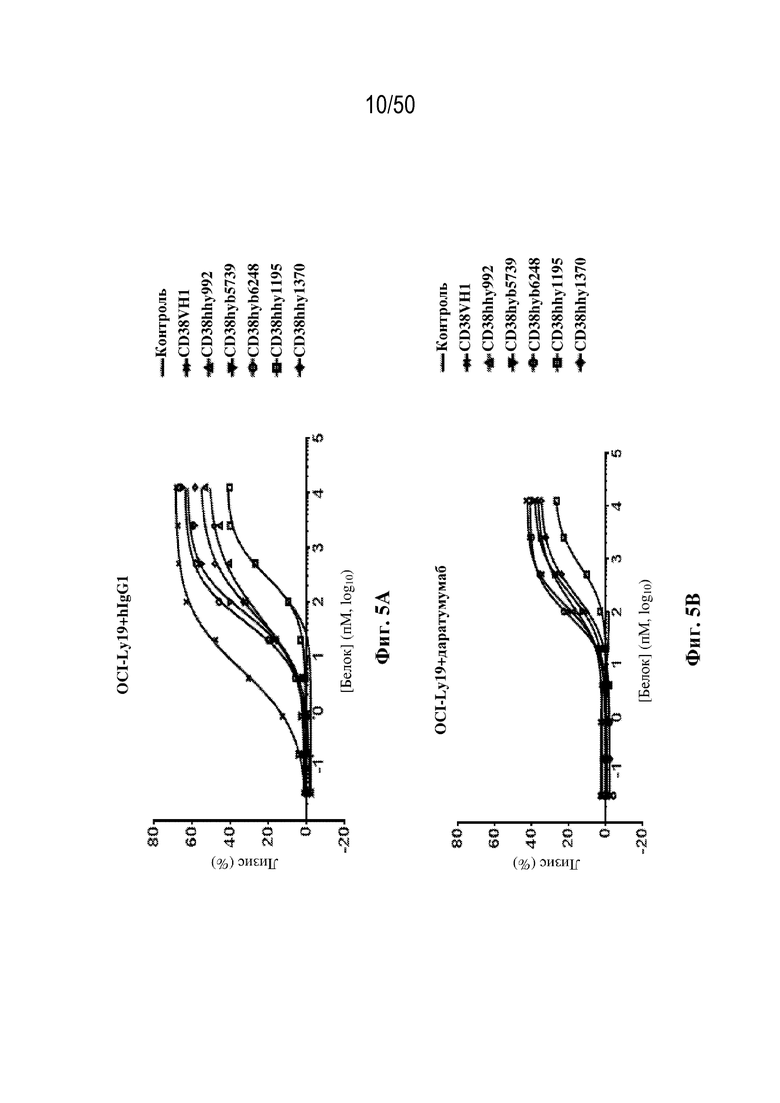
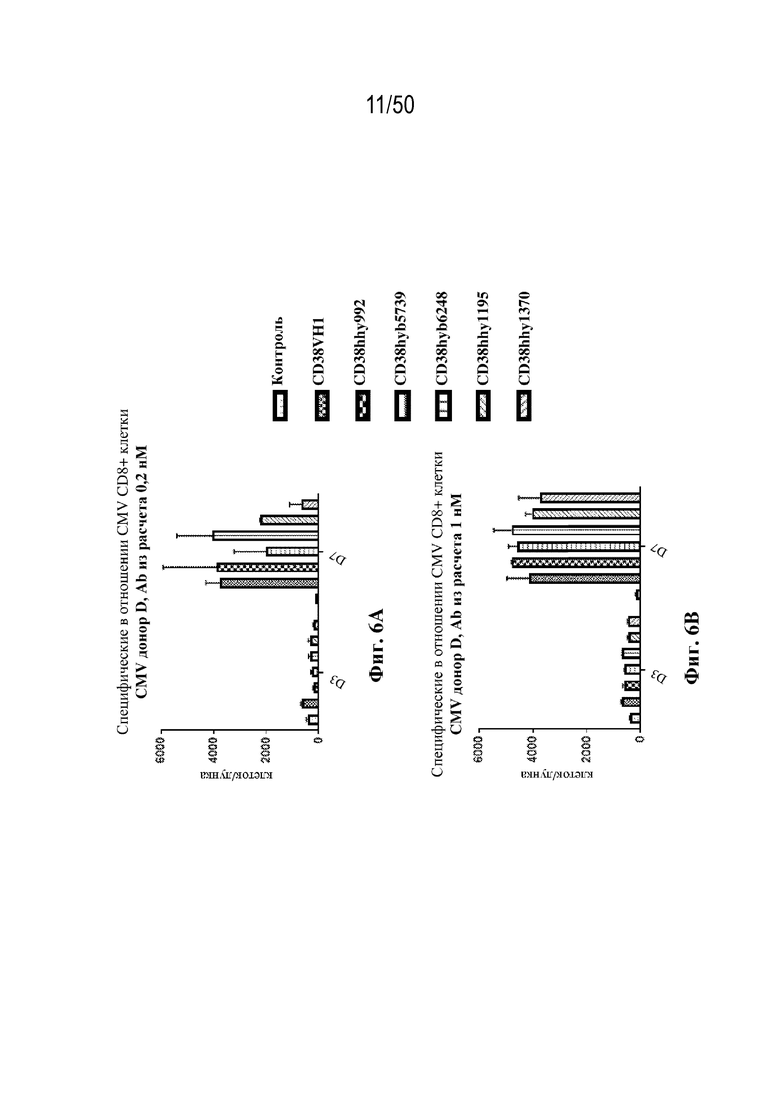
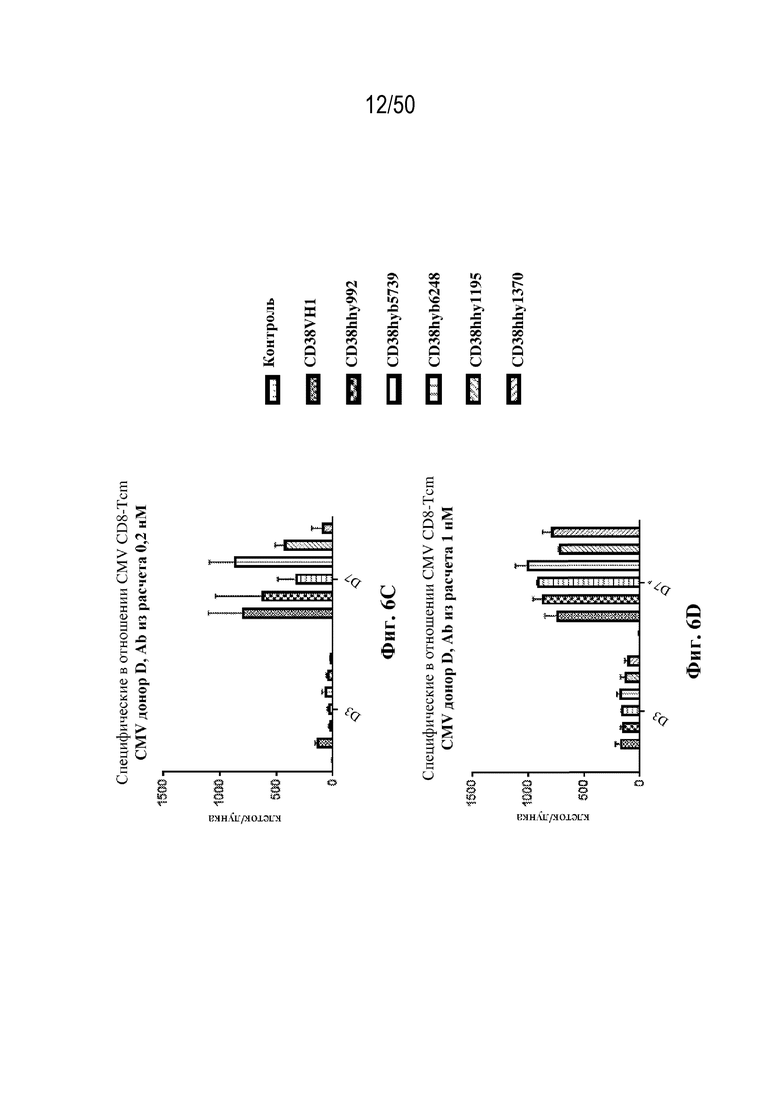
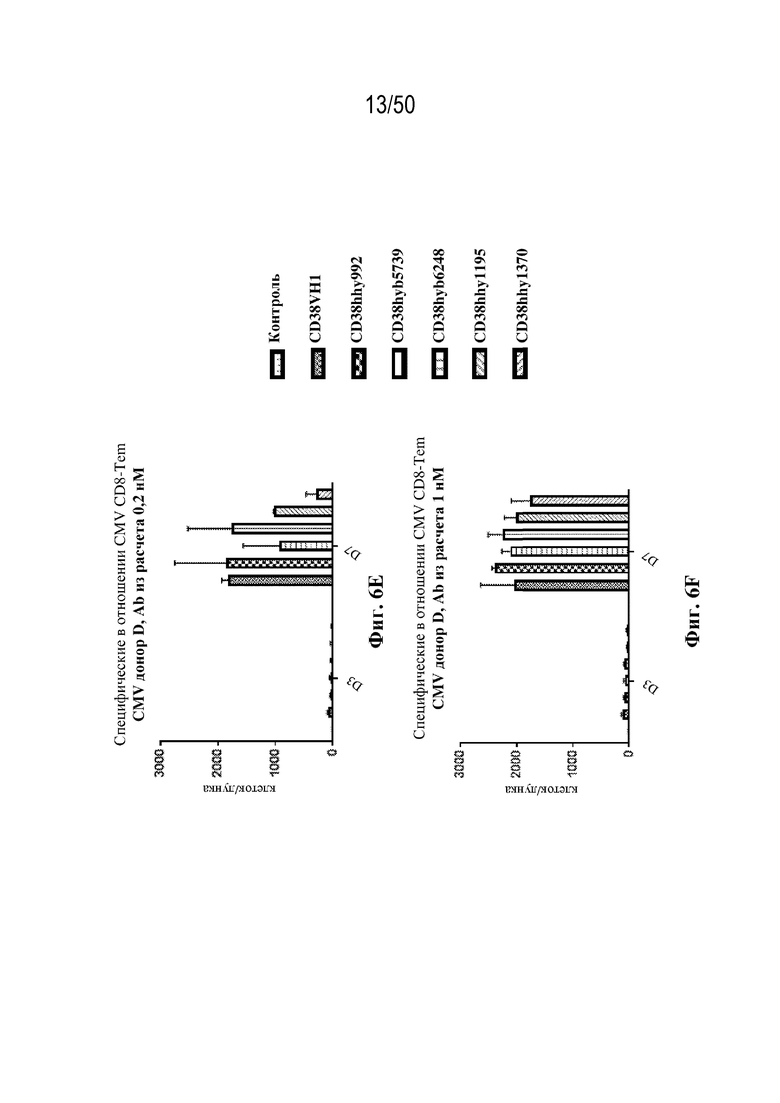
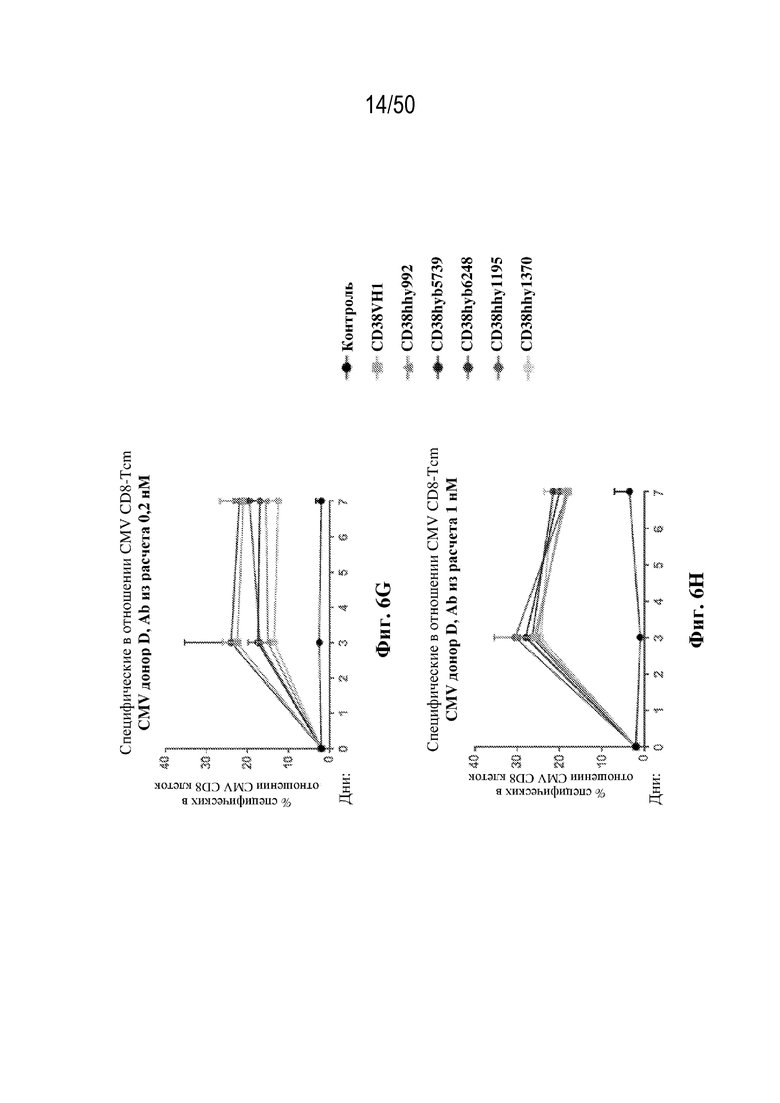
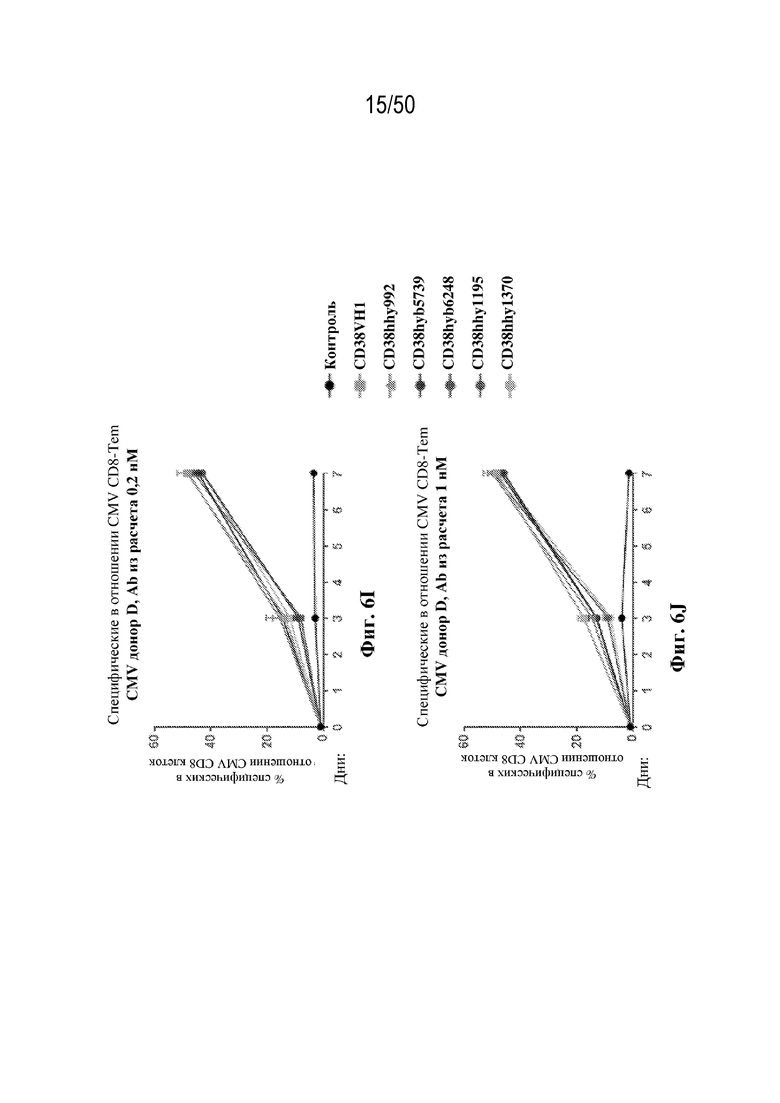
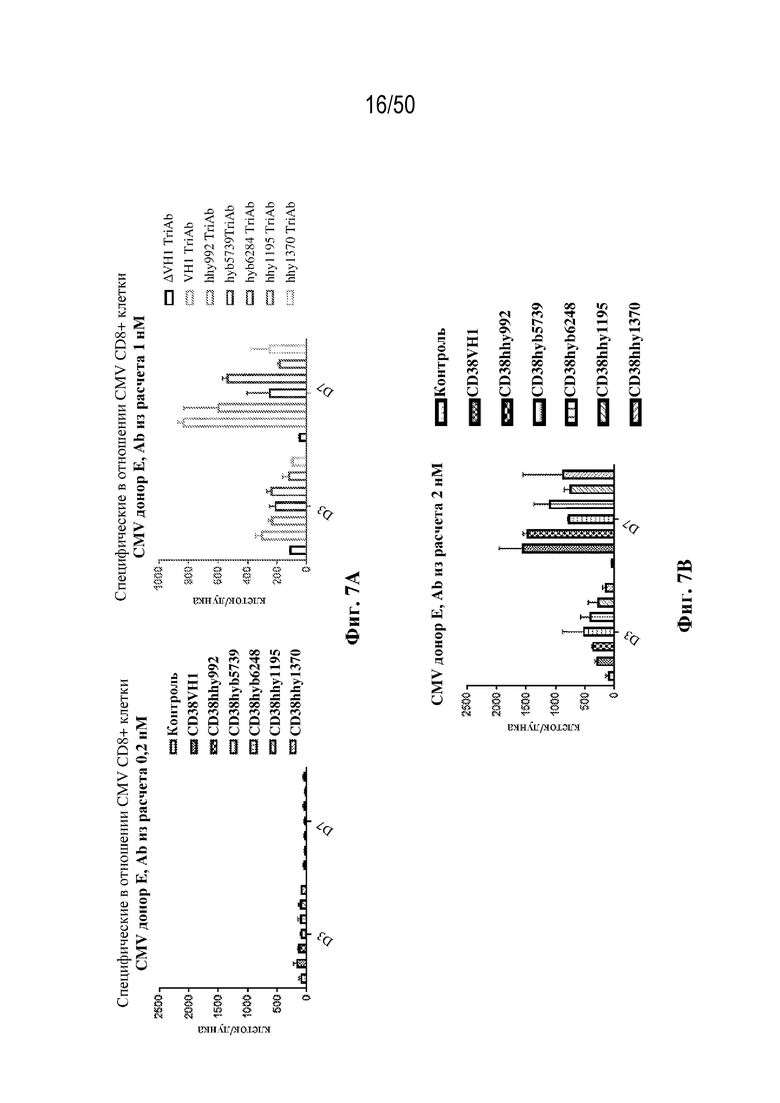
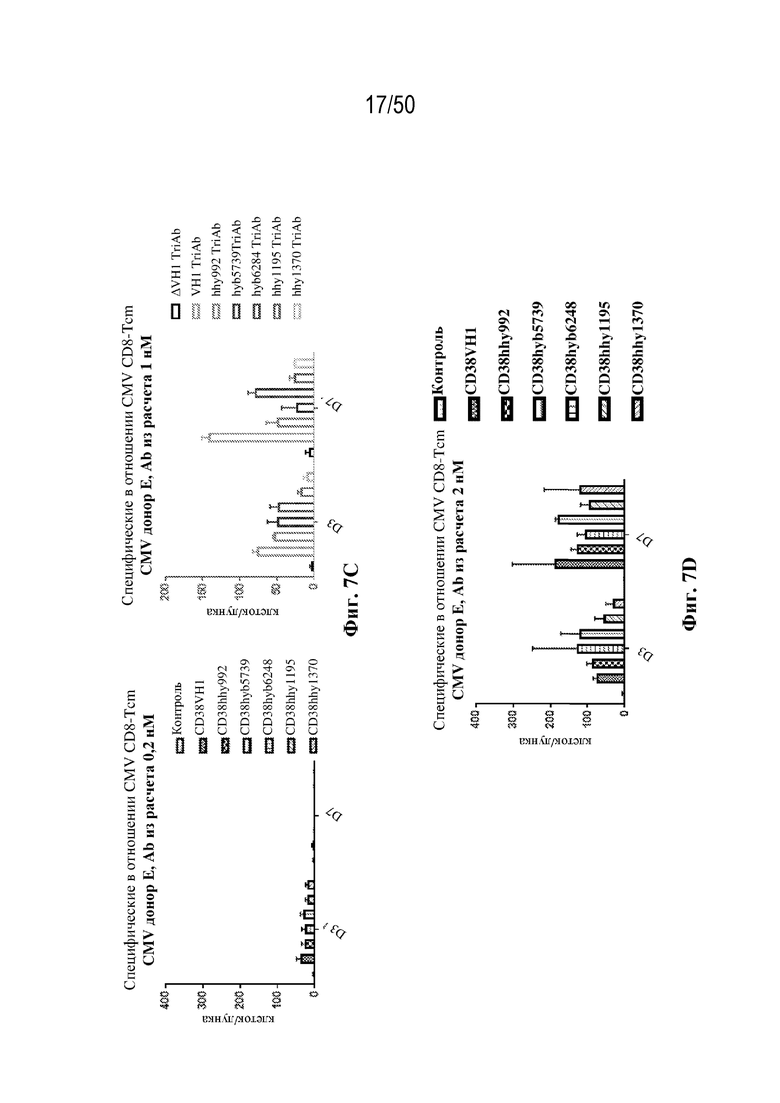
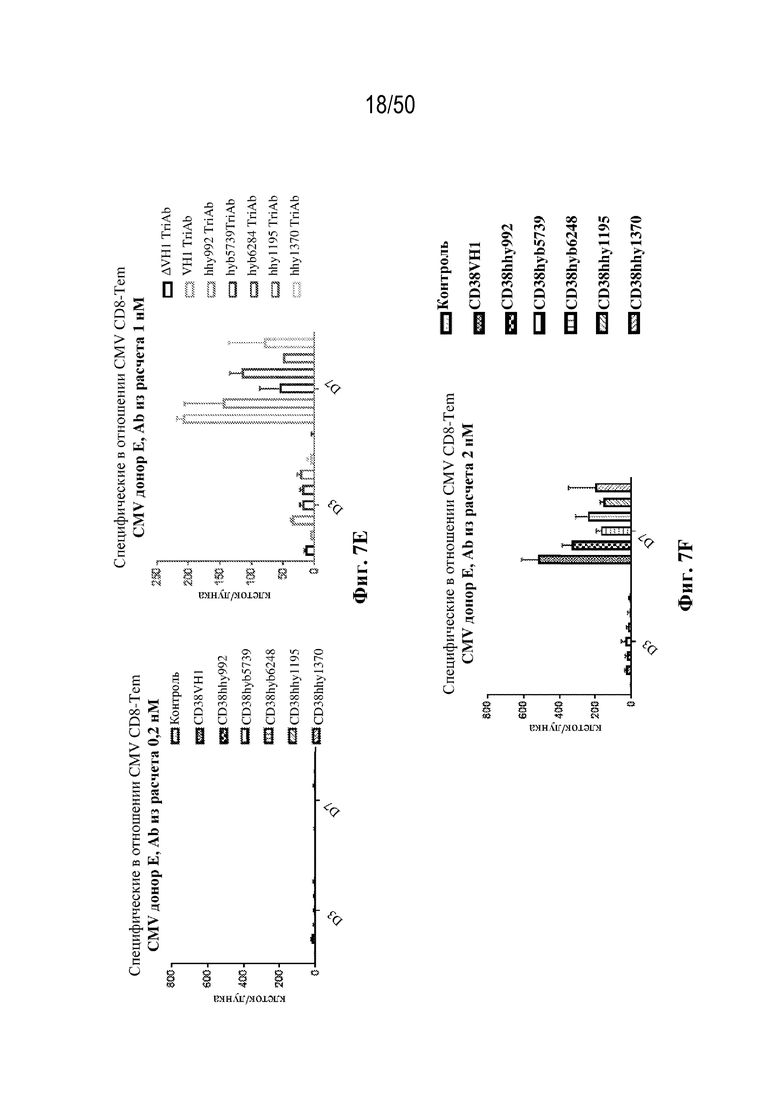
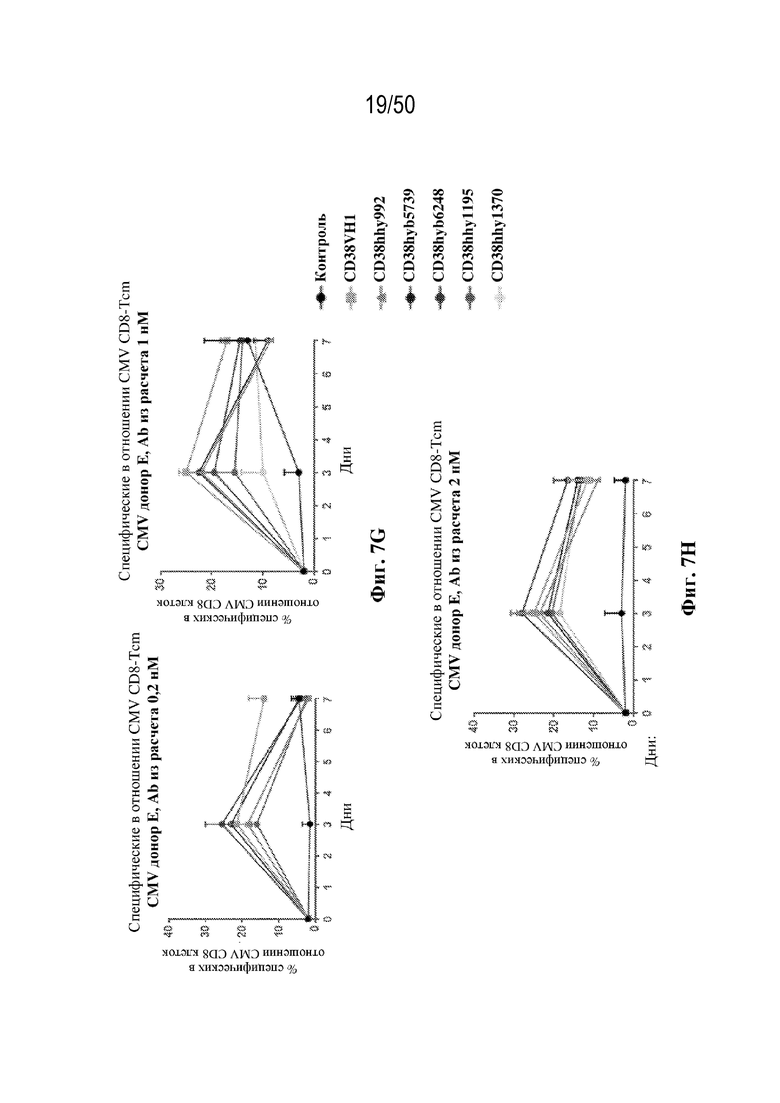
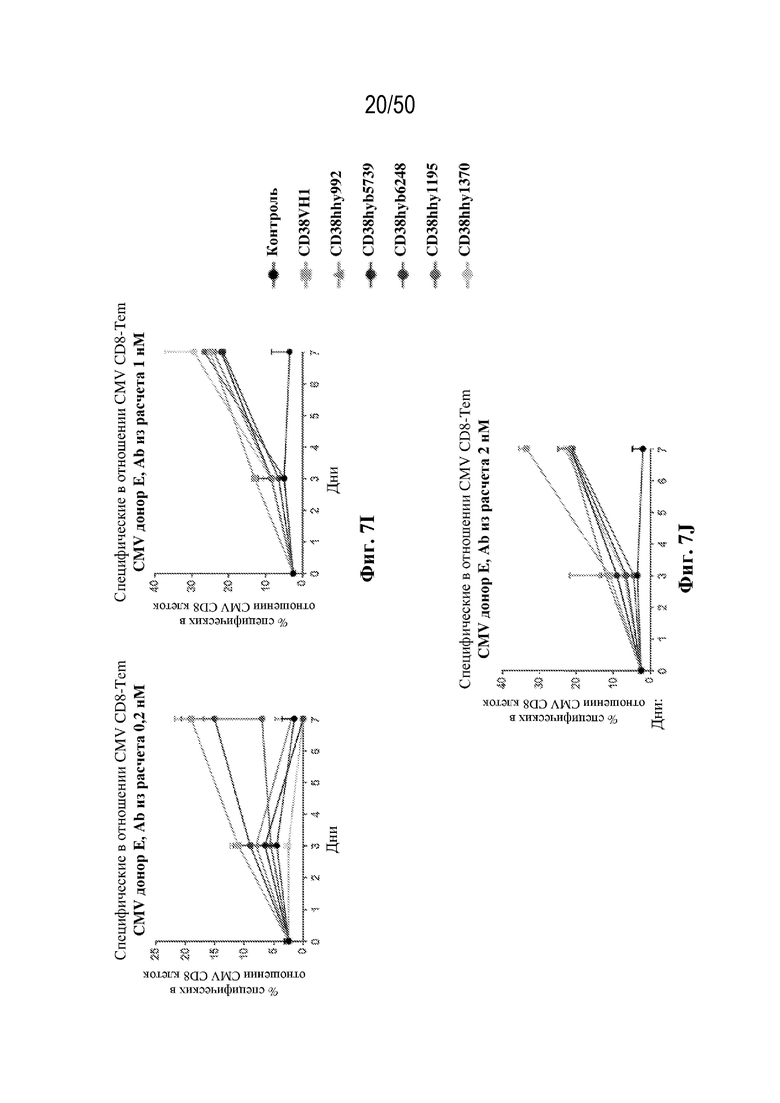
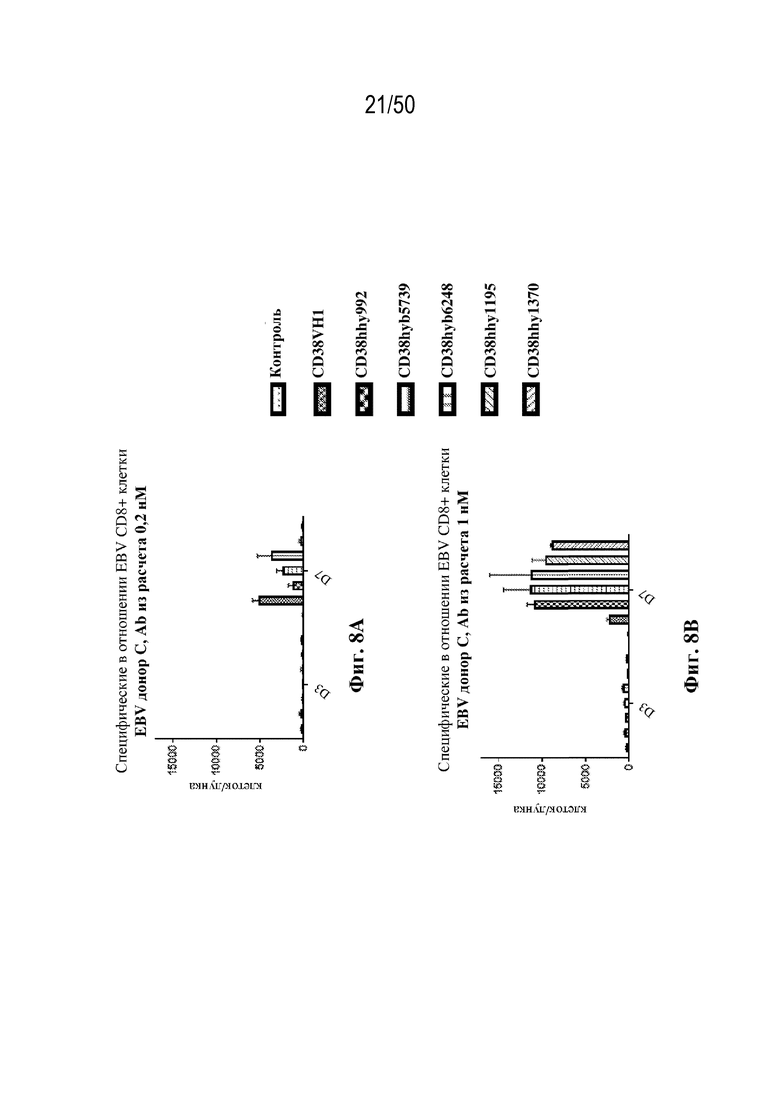
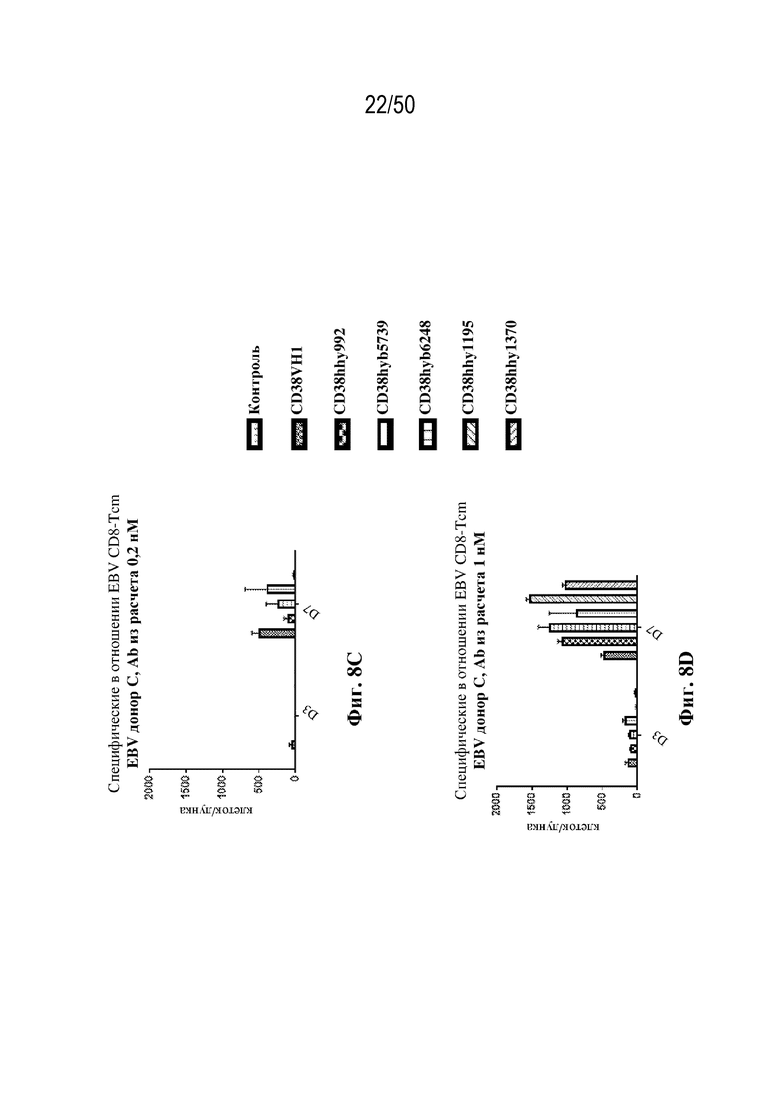
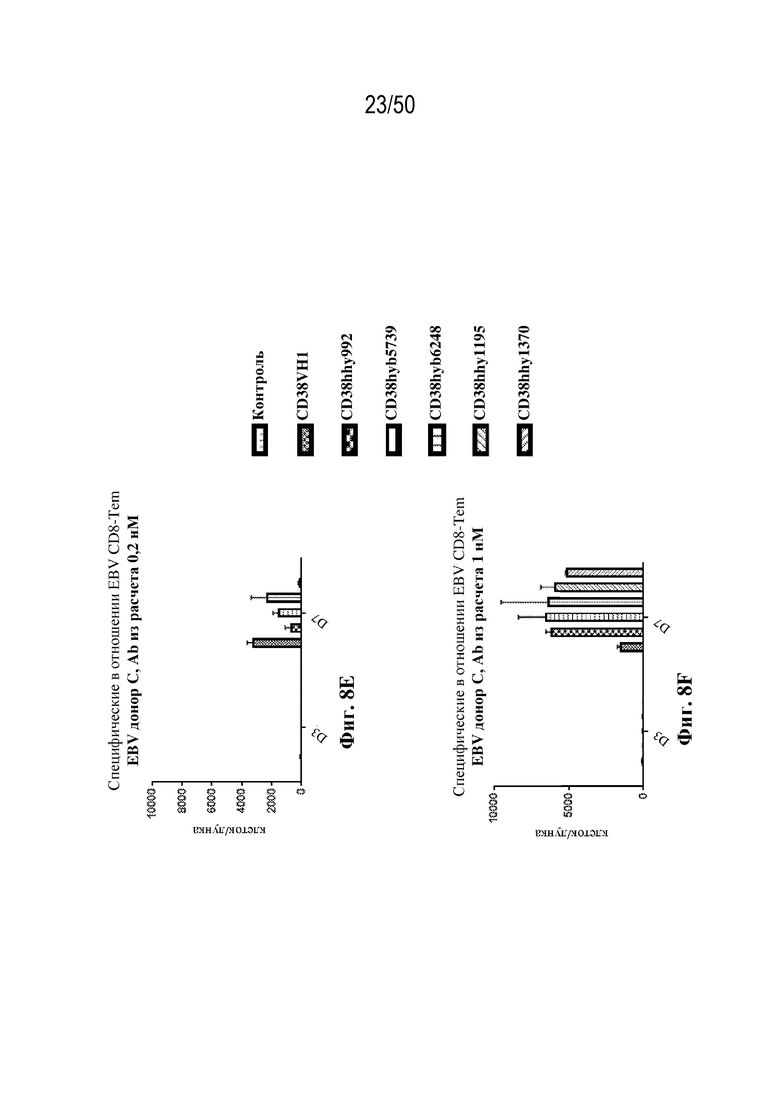
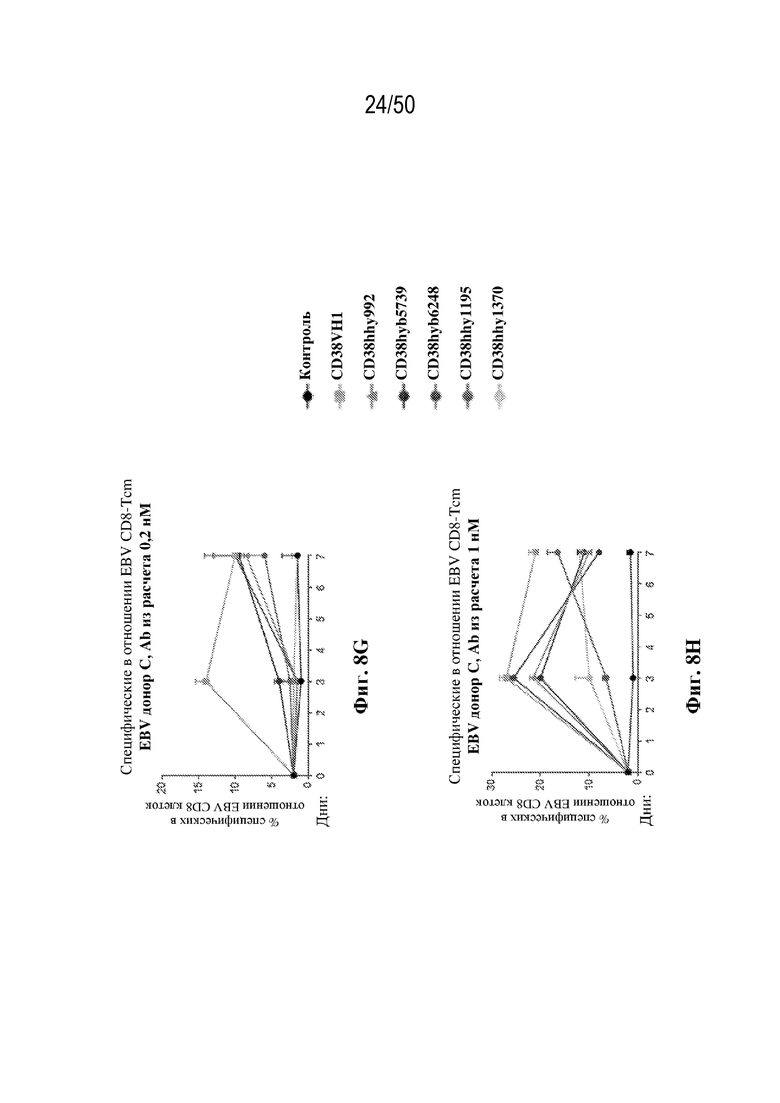
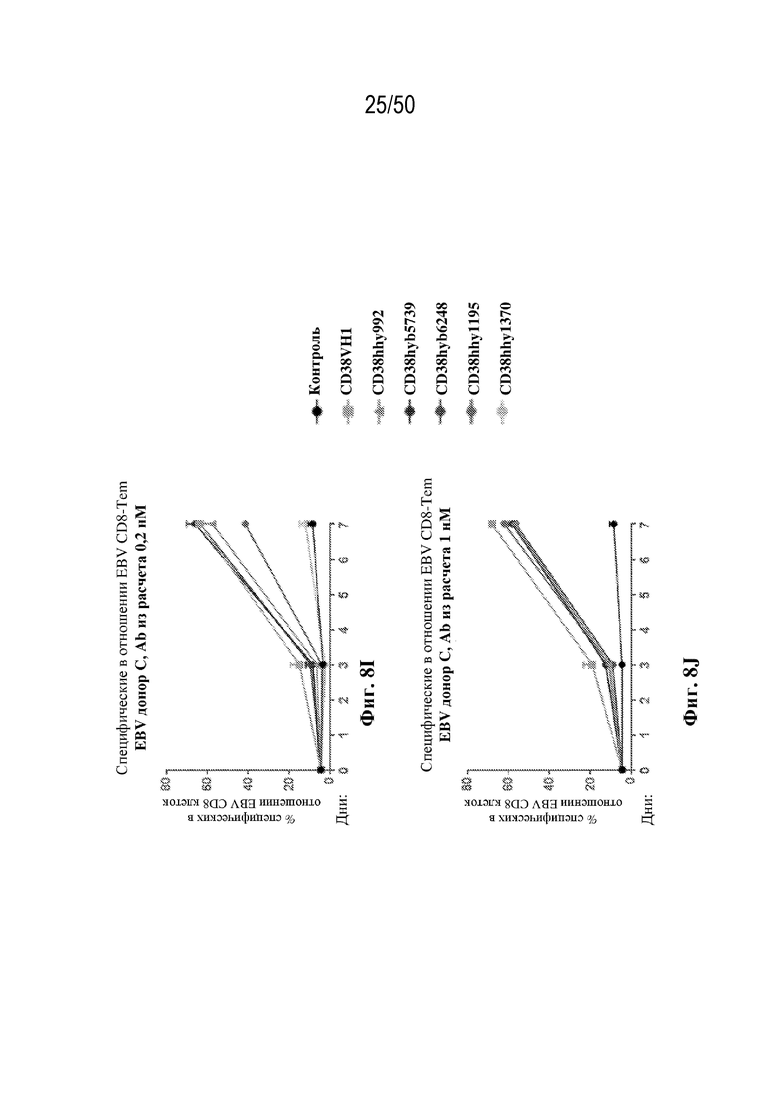
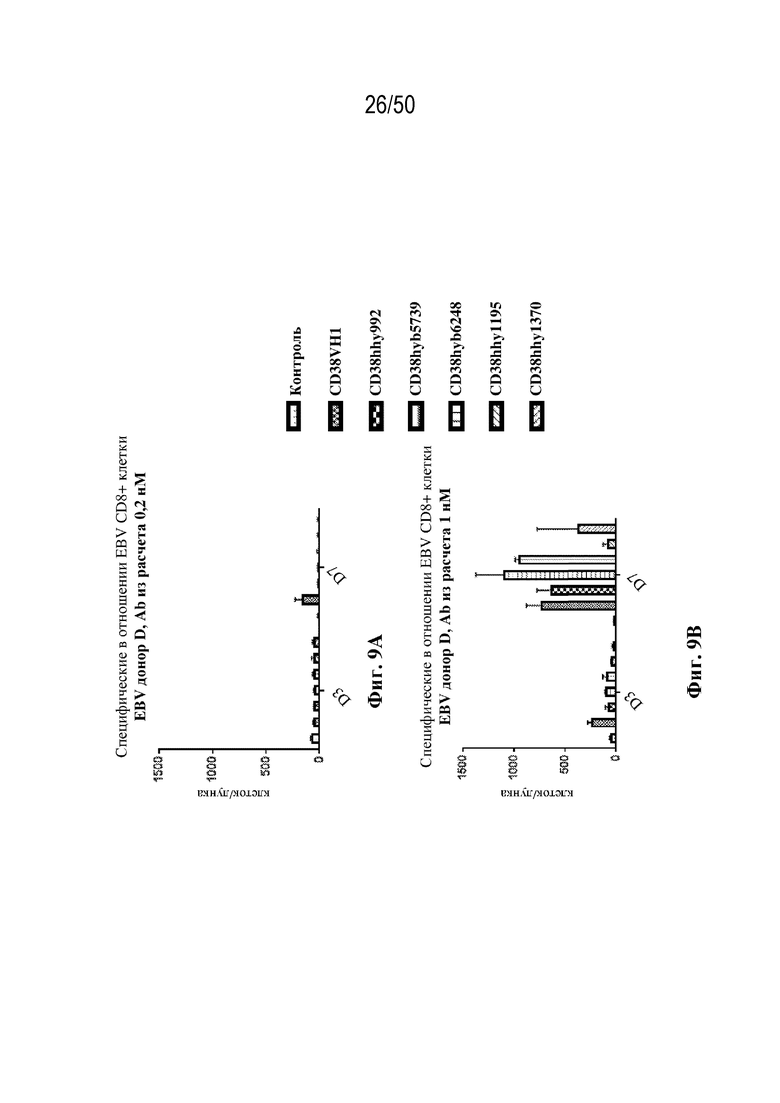
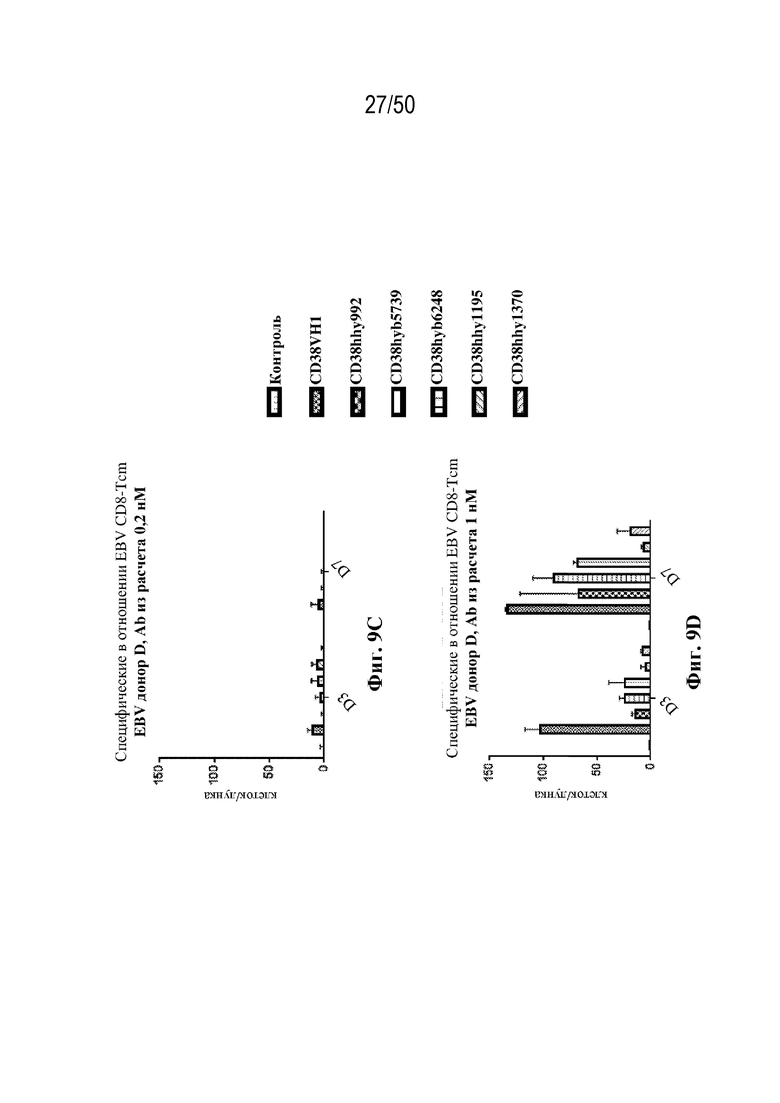
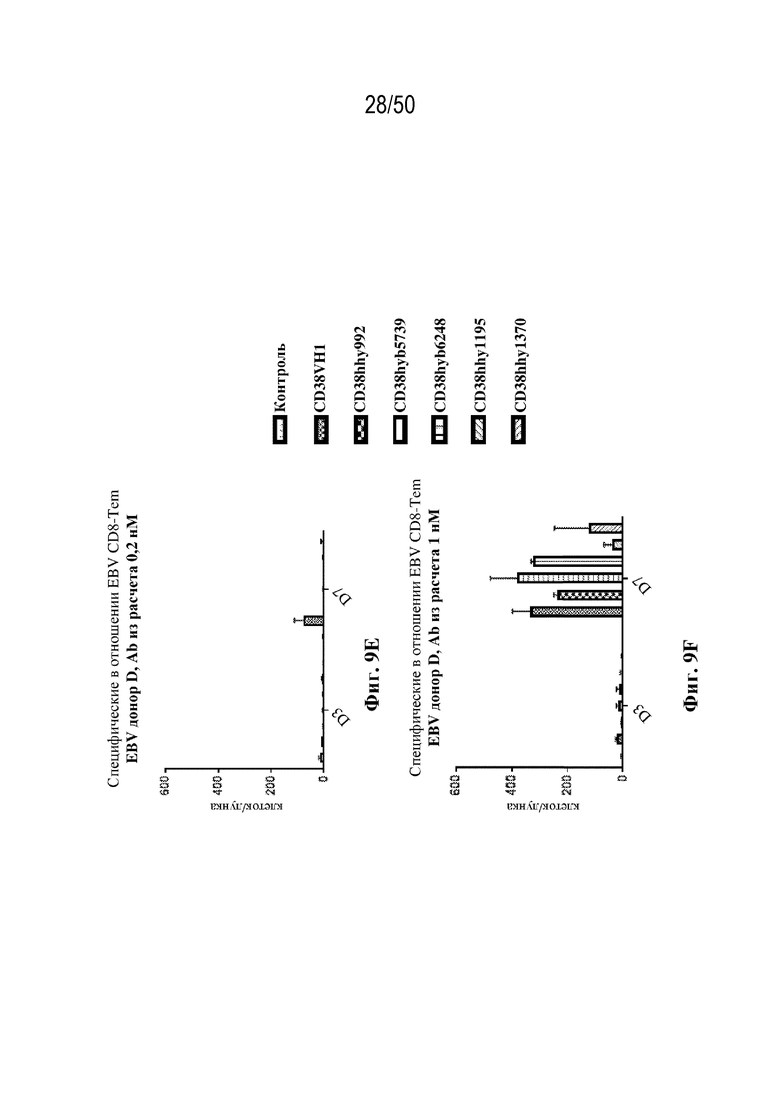
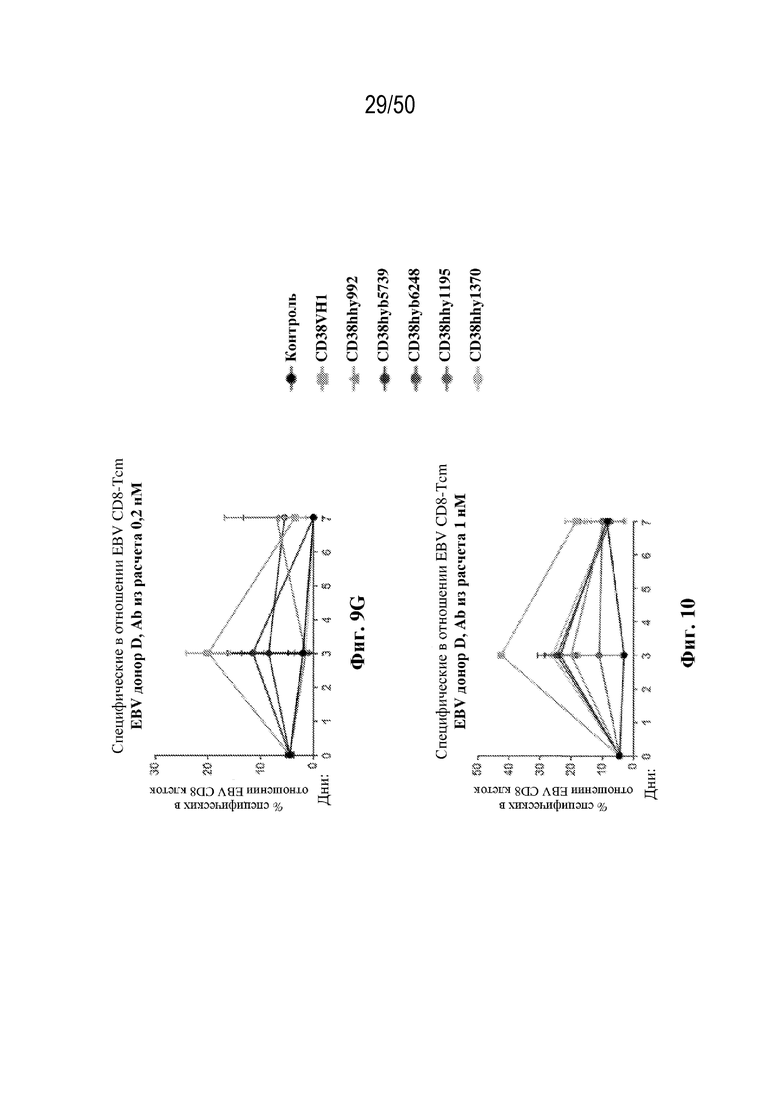
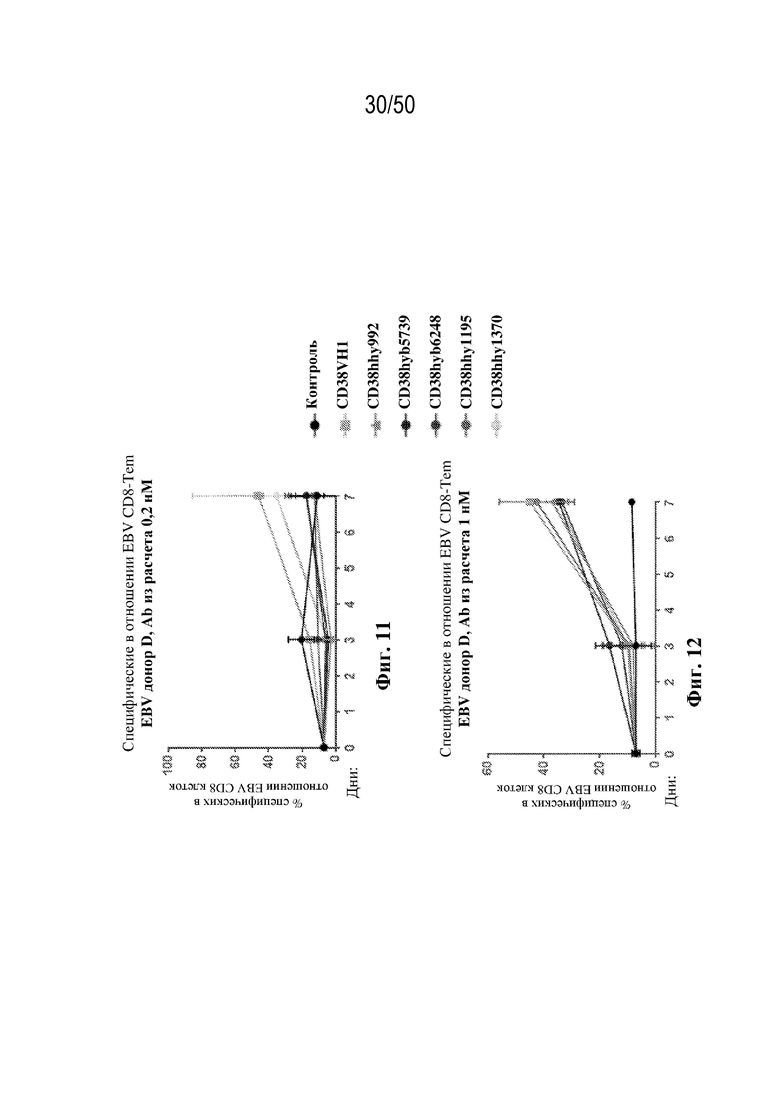
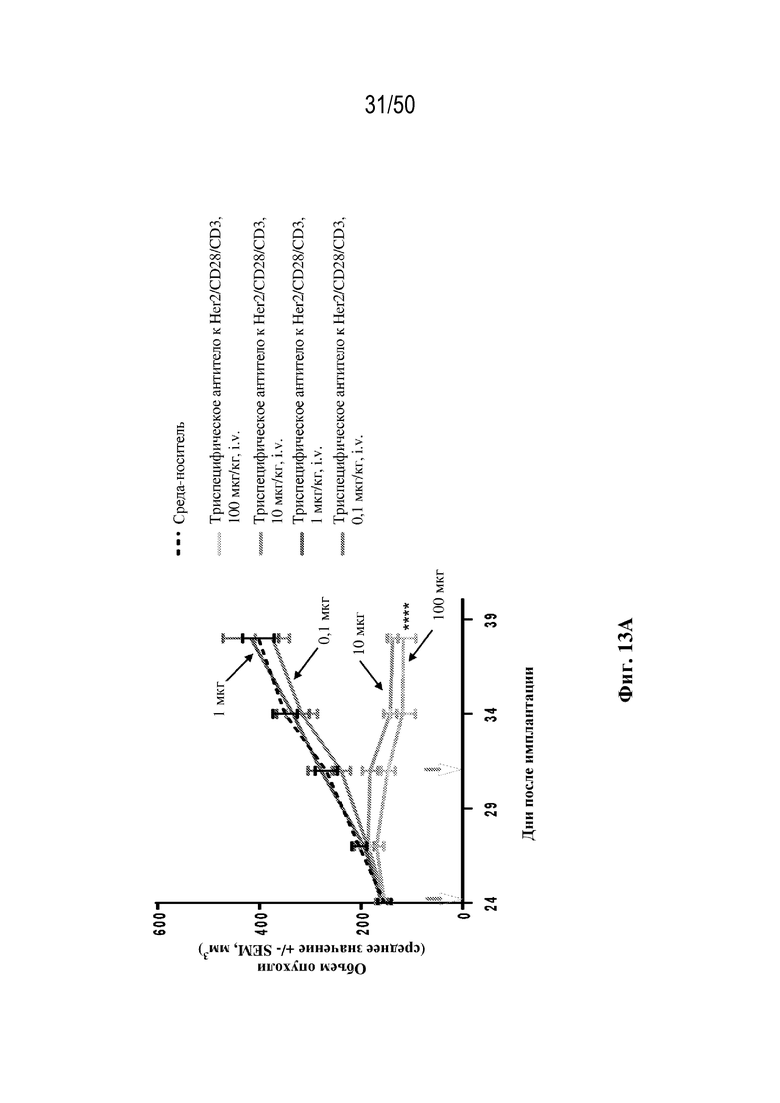
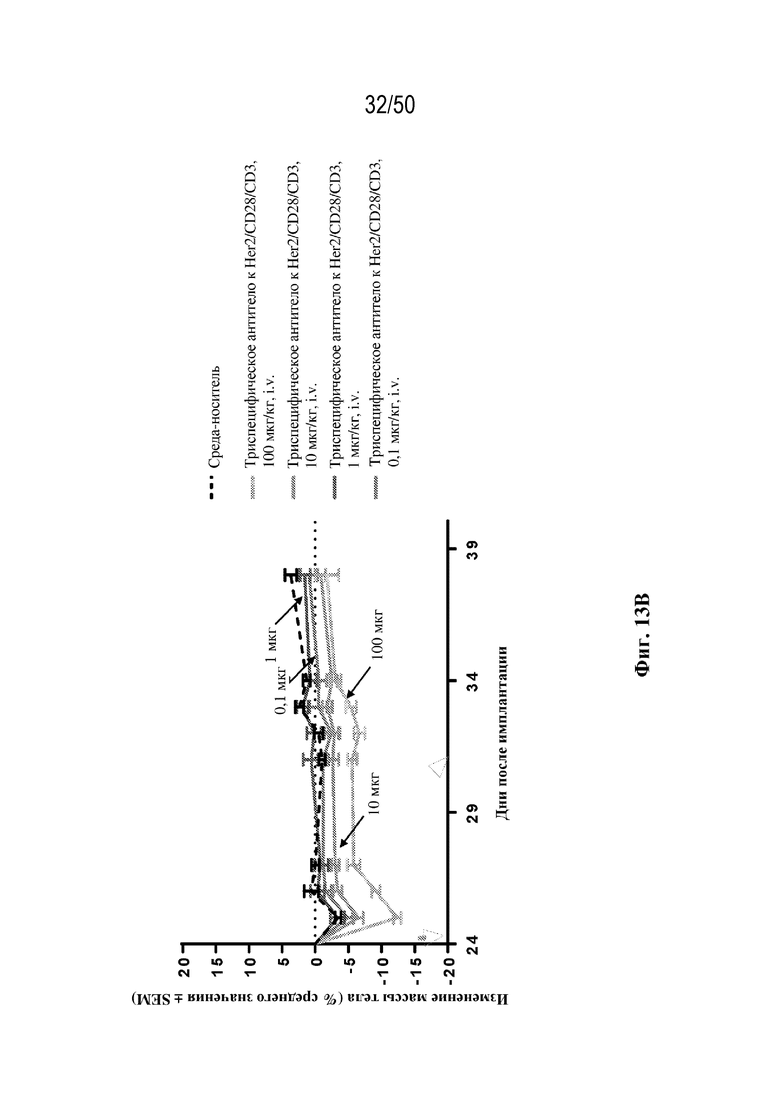
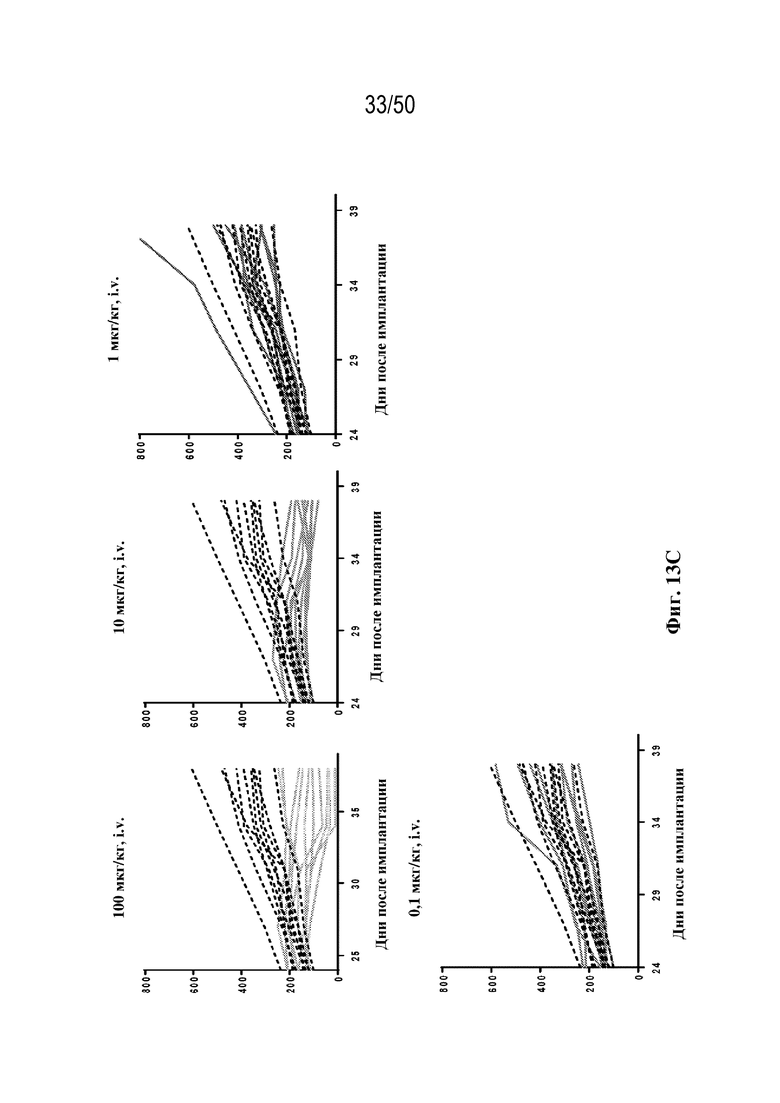
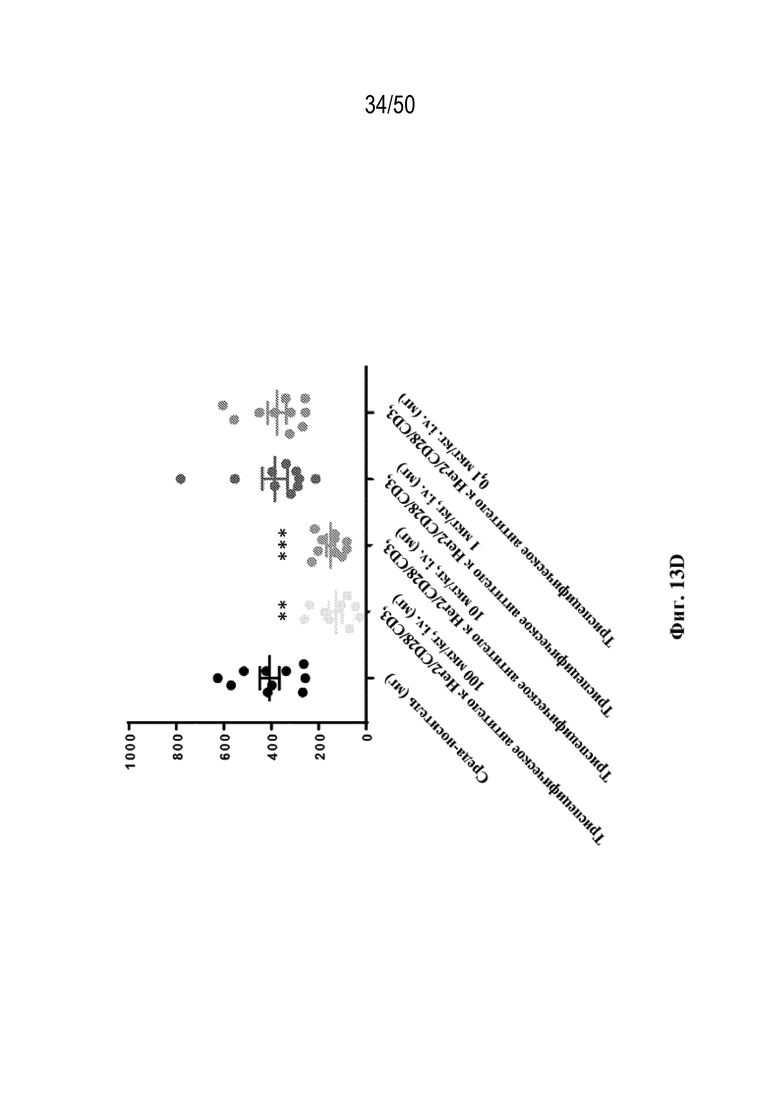
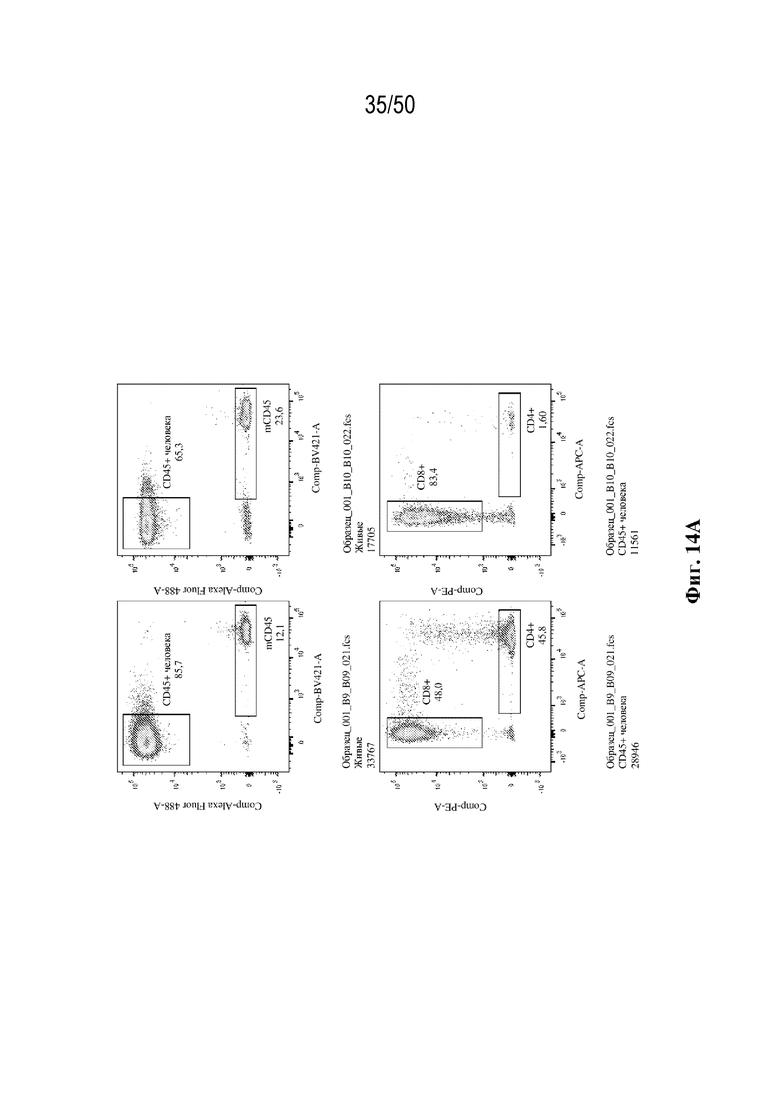



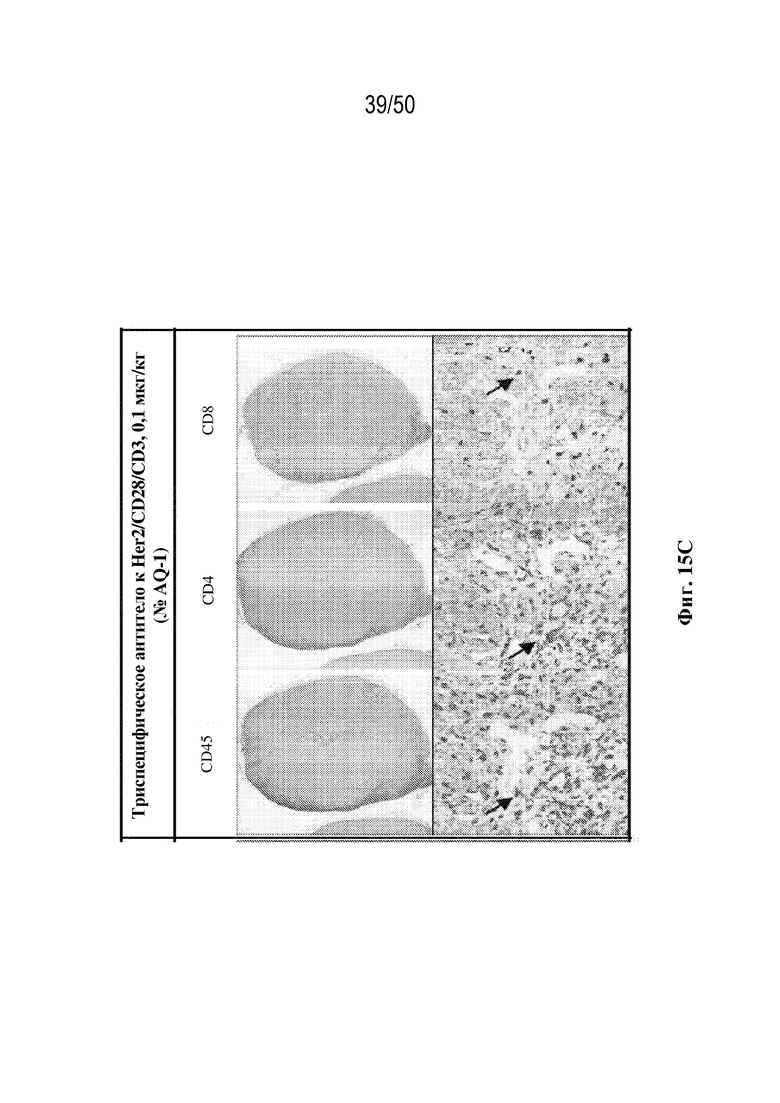
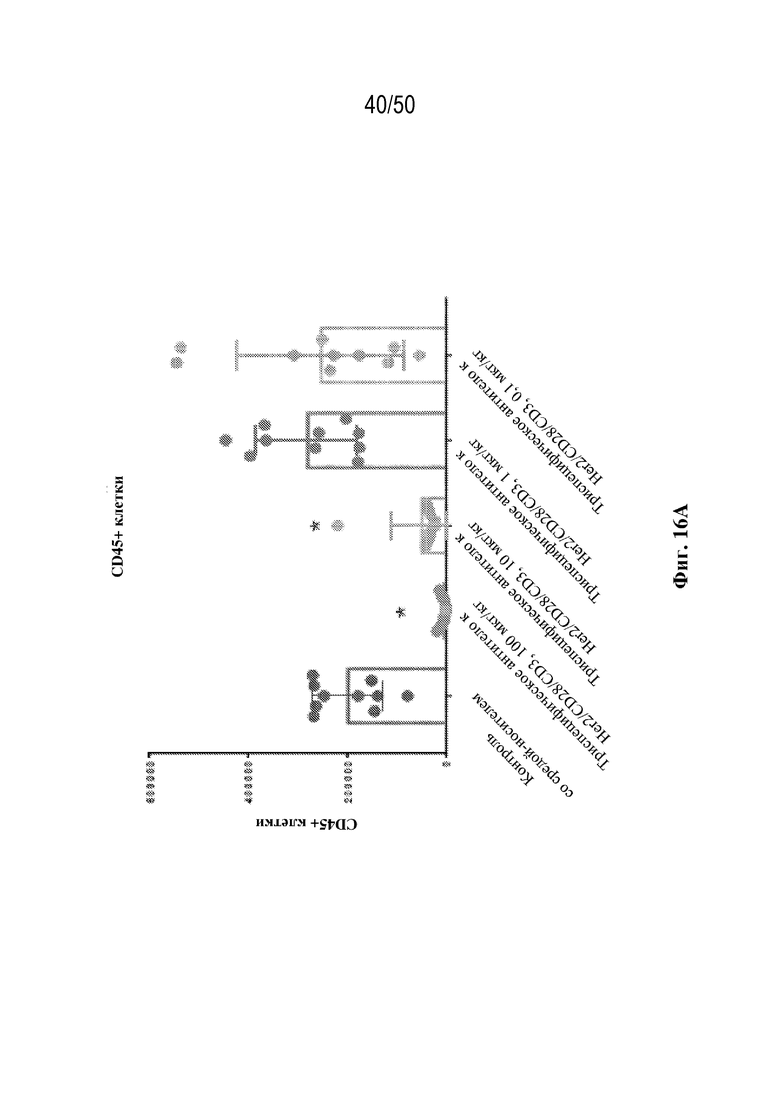
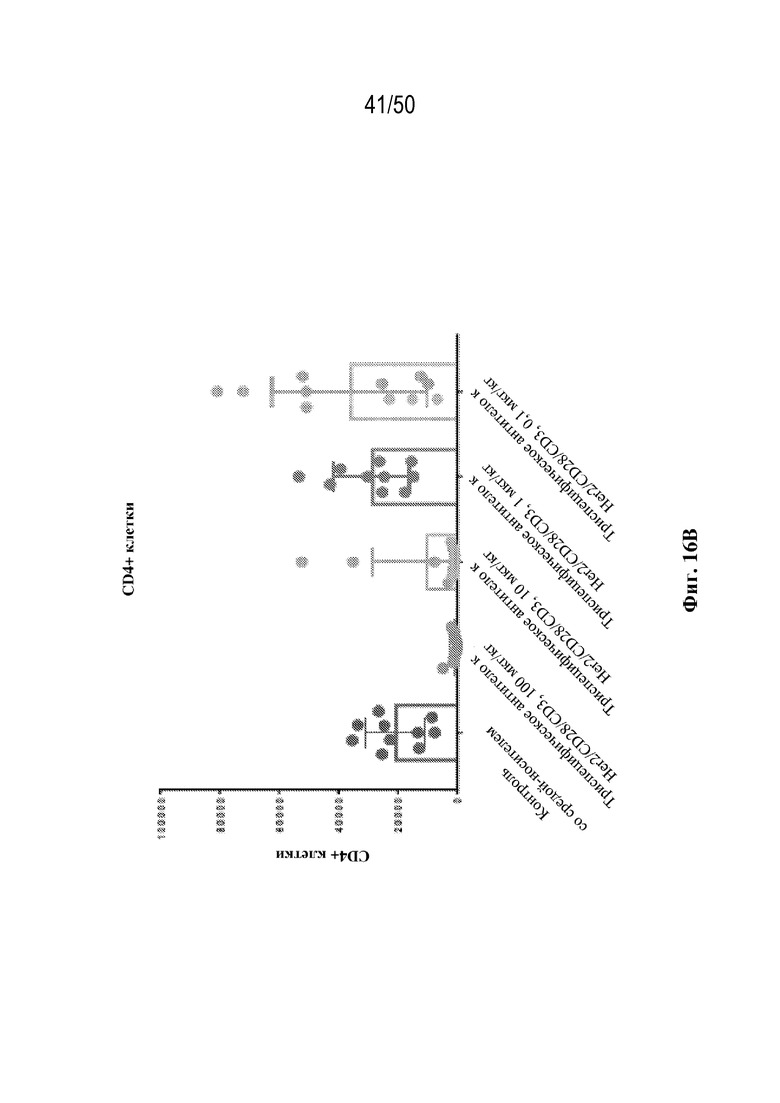
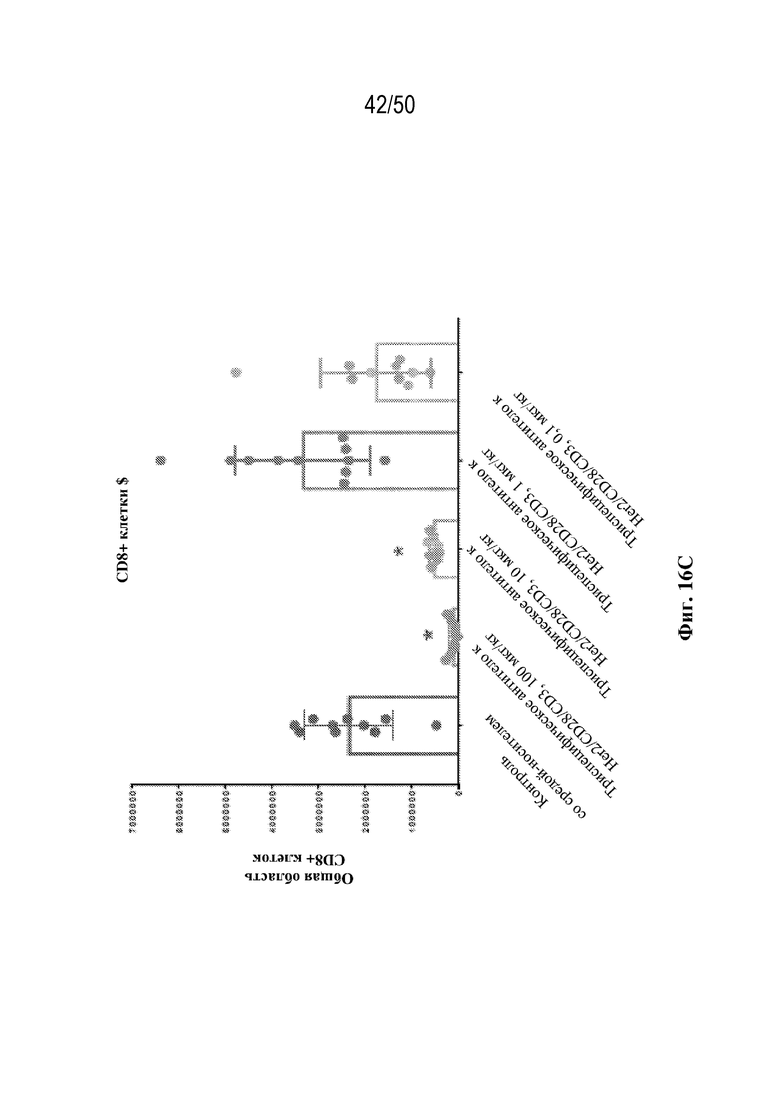

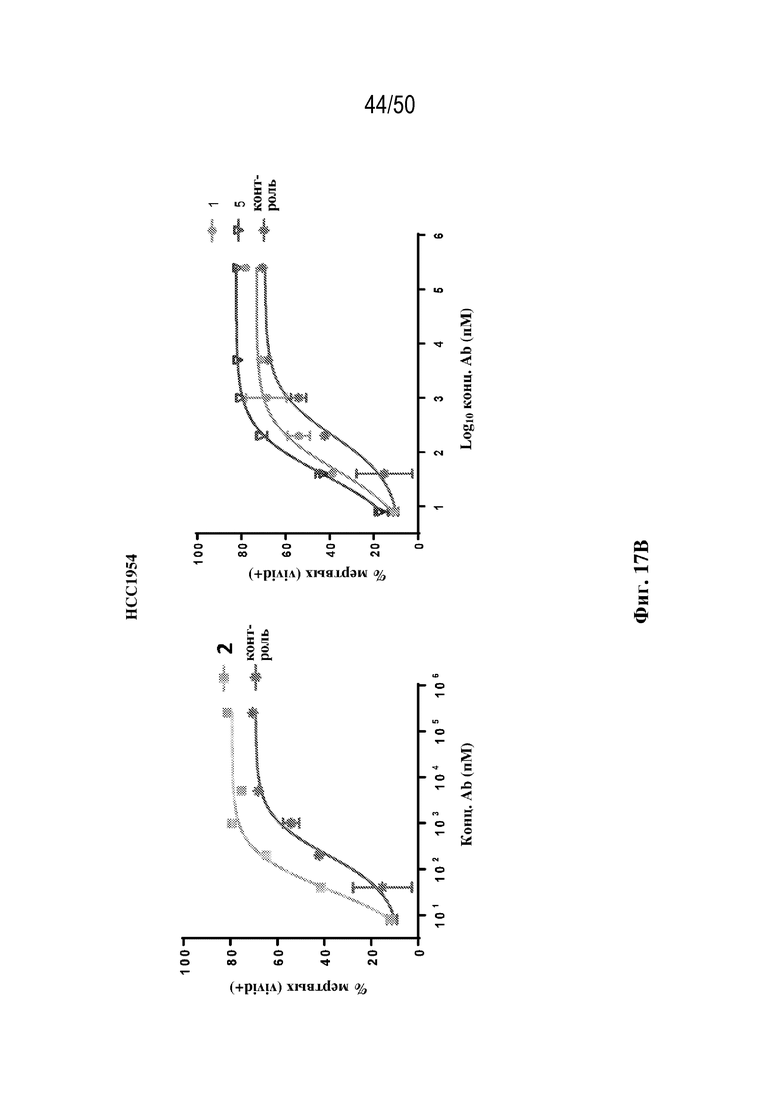
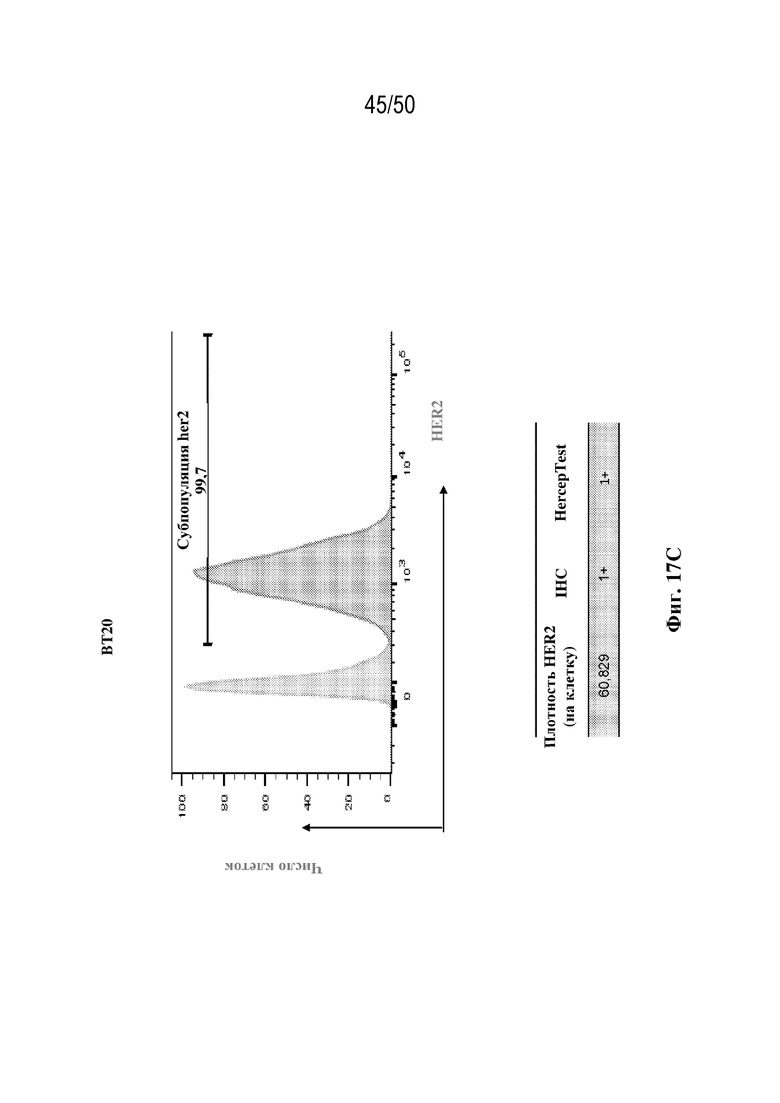
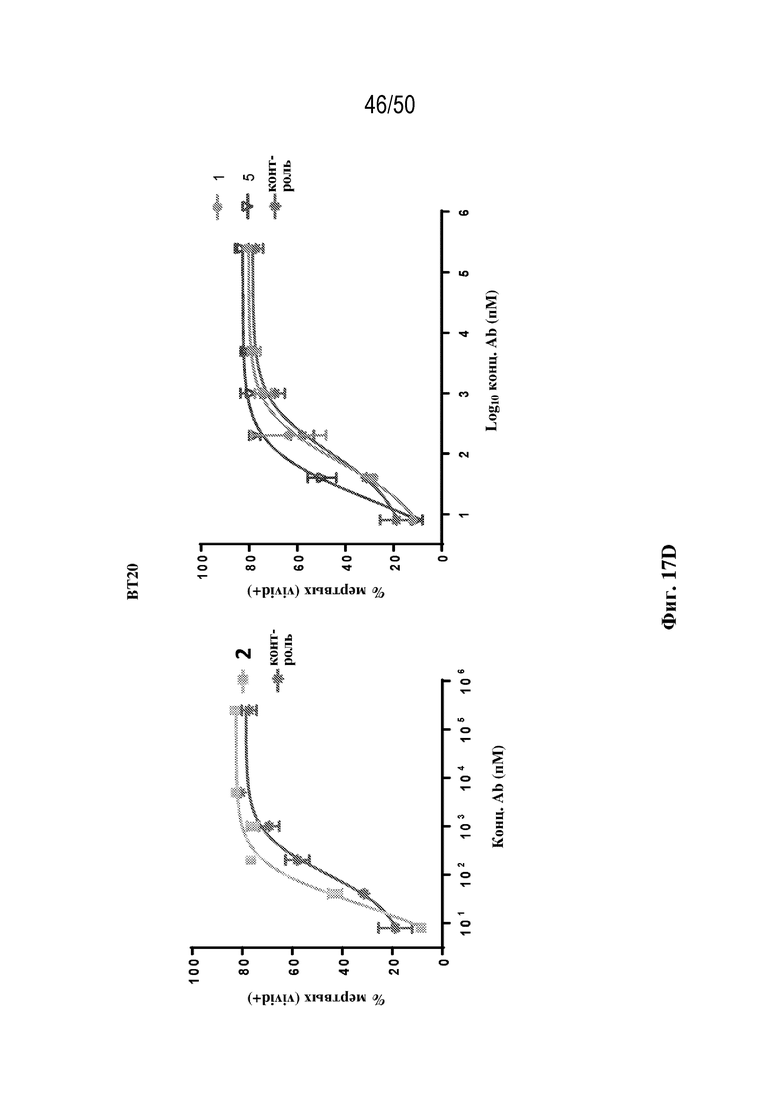
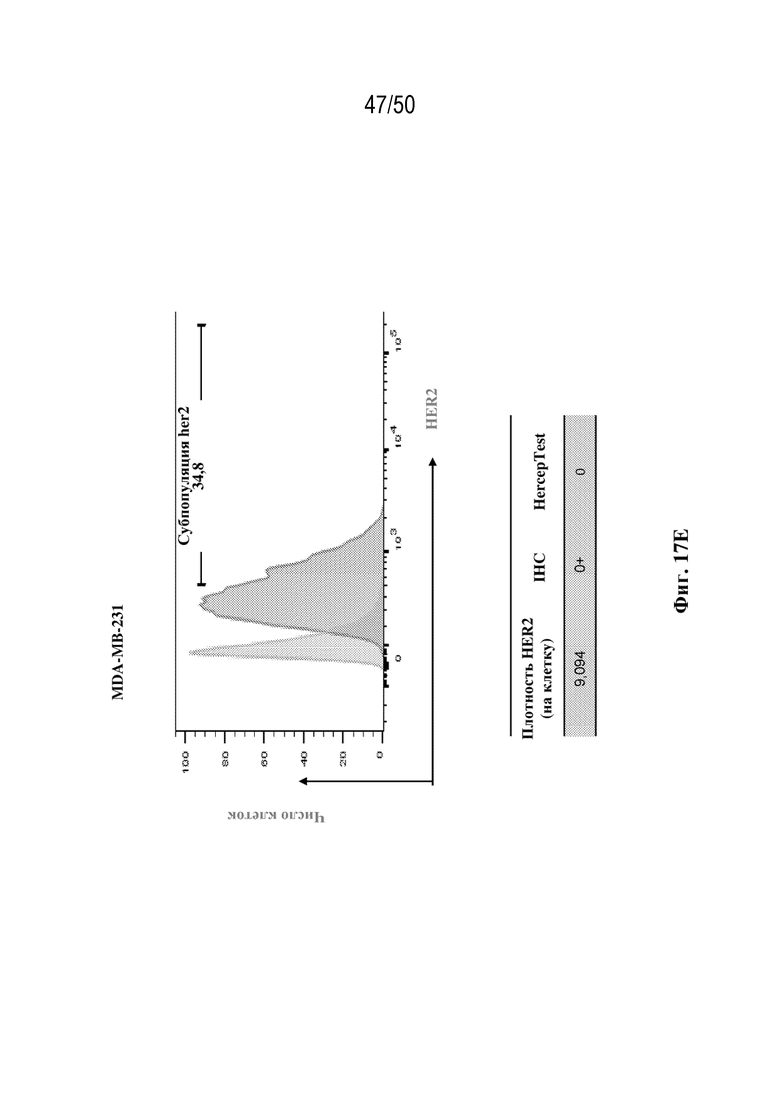
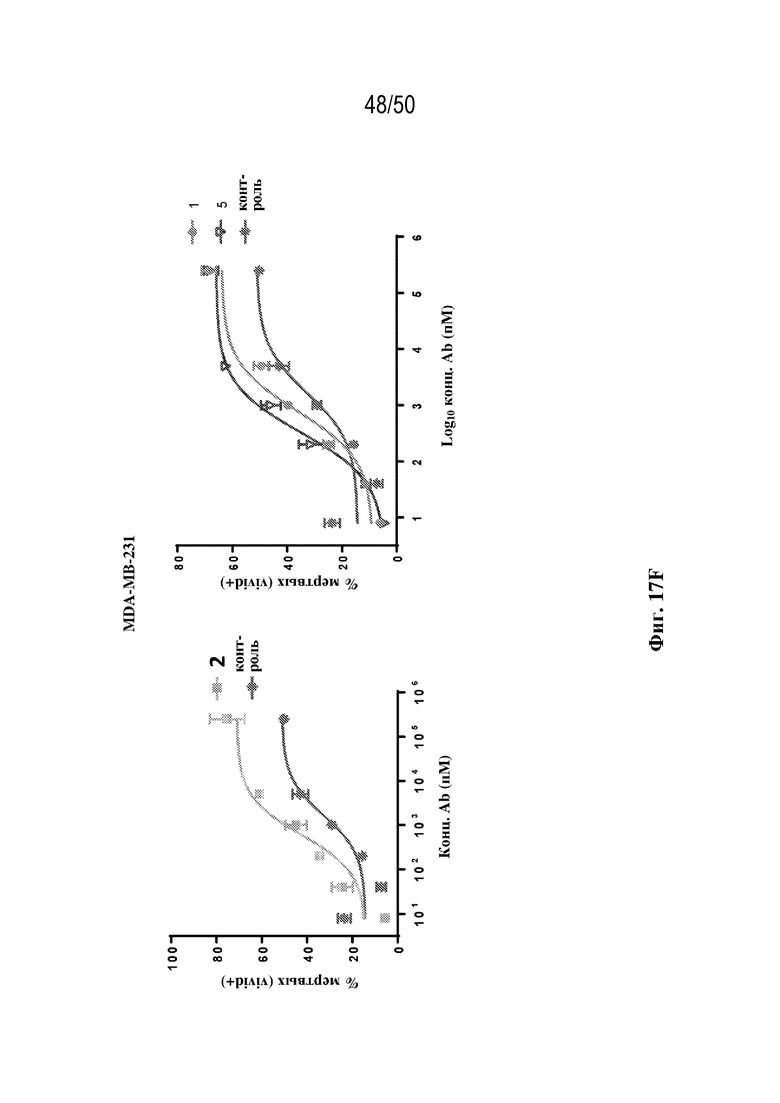

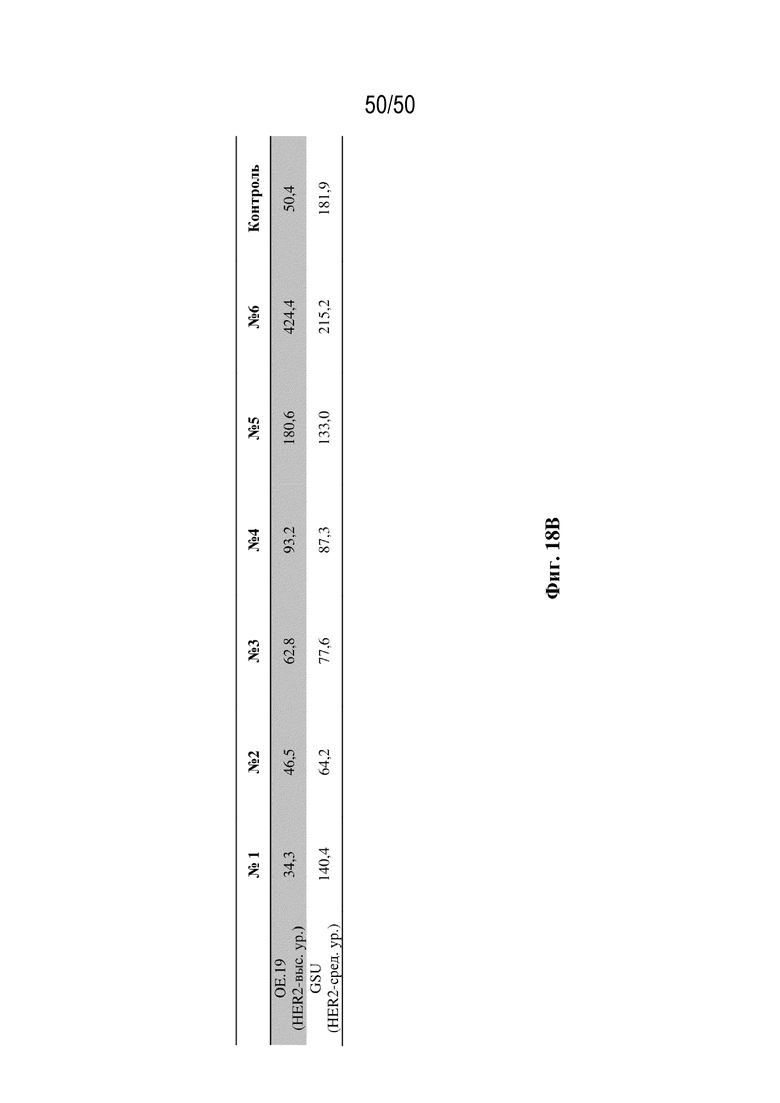
Изобретение относится к области биотехнологии, конкретно к получению триспецифических связывающих белков, и может быть использовано в медицине в терапии рака. Предложен триспецифический связывающий белок CD38/CD28xCD3, содержащий связывающий участок, который связывает полипептид CD28, связывающий участок, который связывает полипептид CD3, и связывающий участок, который связывает третий полипептид, такой как опухолевый белок-мишень HER2 человека. Изобретение обеспечивает активацию Т-клеток, посредством которой индуцируется потенциал иммунных клеток в отношении уничтожения близлежащих опухолевых клеток. 12 н. и 32 з.п. ф-лы, 18 ил., 12 табл., 7 пр.
1. Связывающий белок для профилактики или лечения рака, содержащий четыре полипептидные цепи, которые образуют три антигенсвязывающих участка, где первая полипептидная цепь содержит структуру, представленную формулой
VL2-L1-VL1-L2-CL [I];
и вторая полипептидная цепь содержит структуру, представленную формулой
VH1-L3-VH2-L4-CH1-шарнир-CH2-CH3 [II];
и третья полипептидная цепь содержит структуру, представленную формулой
VH3-CH1-шарнир-CH2-CH3 [III];
и четвертая полипептидная цепь содержит структуру, представленную формулой
VL3-CL [IV];
где
VL1 представляет собой первый вариабельный домен легкой цепи иммуноглобулина;
VL2 представляет собой второй вариабельный домен легкой цепи иммуноглобулина;
VL3 представляет собой третий вариабельный домен легкой цепи иммуноглобулина;
VH1 представляет собой первый вариабельный домен тяжелой цепи иммуноглобулина;
VH2 представляет собой второй вариабельный домен тяжелой цепи иммуноглобулина;
VH3 представляет собой третий вариабельный домен тяжелой цепи иммуноглобулина;
CL представляет собой константный домен легкой цепи иммуноглобулина;
CH1 представляет собой константный домен CH1 тяжелой цепи иммуноглобулина;
CH2 представляет собой константный домен CH2 тяжелой цепи иммуноглобулина;
CH3 представляет собой константный домен CH3 тяжелой цепи иммуноглобулина;
шарнир представляет собой шарнирную область иммуноглобулина, соединяющую домены CH1 и CH2; и
L1, L2, L3 и L4 представляют собой аминокислотные линкеры длиной от 0 до 100 аминокислот каждый,
где полипептид формулы I и полипептид формулы II образуют кроссоверную пару легкая цепь-тяжелая цепь и
где VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28;
где VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO: 55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO: 56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO: 57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHX1NX2X3TY, где X1 представляет собой E или Q, X2 представляет собой A или L и X3 представляет собой Q, R или F (SEQ ID NO: 180), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO: 64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO: 65); и
где VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид HER2 человека.
2. Связывающий белок по п. 1, где домен VH1 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYY (SEQ ID NO: 49), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGNVNT (SEQ ID NO: 50), и последовательность CDR-H3, содержащую аминокислотную последовательность TRSHYGLDWNFDV (SEQ ID NO: 51), и домен VL1 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QNIYVW (SEQ ID NO: 52), последовательность CDR-L2, содержащую аминокислотную последовательность KAS (SEQ ID NO: 53), и последовательность CDR-L3, содержащую аминокислотную последовательность QQGQTYPY (SEQ ID NO: 54).
3. Связывающий белок по п. 2, где домен VH1 содержит аминокислотную последовательность QVQLVQSGAEVVKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGSIYPGNVNTNYAQKFQGRATLTVDTSISTAYMELSRLRSDDTAVYYCTRSHYGLDWNFDVWGKGTTVTVSS (SEQ ID NO: 91) и/или домен VL1 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCQASQNIYVWLNWYQQKPGKAPKLLIYKASNLHTGVPSRFSGSGSGTDFTLTISSLQPEDIATYYCQQGQTYPYTFGQGTKLEIK (SEQ ID NO: 92).
4. Связывающий белок по п. 1, где последовательность CDR-L1 домена VL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из QSLVHQNAQTY (SEQ ID NO: 59), QSLVHENLQTY (SEQ ID NO: 60), QSLVHENLFTY (SEQ ID NO: 61) и QSLVHENLRTY (SEQ ID NO: 62).
5. Связывающий белок по п. 1, где последовательность CDR-L1 домена VL2 содержит аминокислотную последовательность QSLVHQNAQTY (SEQ ID NO: 59).
6. Связывающий белок по п. 4, где домен VH2 содержит аминокислотную последовательность QVQLVESGGGVVQPGRSLRLSCAASGFTFTKAWMHWVRQAPGKQLEWVAQIKDKSNSYATYYADSVKGRFTISRDDSKNTLYLQMNSLRAEDTAVYYCRGVYYALSPFDYWGQGTLVTVSS (SEQ ID NO: 93) или QVQLVESGGGVVQPGRSLRLSCAASGFTFTKAWMHWVRQAPGKQLEWVAQIKDKSNSYATYYASSVKGRFTISRDDSKNTLYLQMNSLRAEDTAVYYCRGVYYALSPFDYWGQGTLVTVSS (SEQ ID NO: 302), и/или домен VL2 содержит аминокислотную последовательность, выбранную из группы, состоящей из DIVMTQTPLSLSVTPGQPASISCKSSQSLVHQNAQTYLSWYLQKPGQSPQSLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCGQGTQYPFTFGSGTKVEIK (SEQ ID NO: 95), DIVMTQTPLSLSVTPGQPASISCKSSQSLVHENLQTYLSWYLQKPGQSPQSLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCGQGTQYPFTFGSGTKVEIK (SEQ ID NO: 96), DIVMTQTPLSLSVTPGQPASISCKSSQSLVHENLFTYLSWYLQKPGQSPQSLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCGQGTQYPFTFGSGTKVEIK (SEQ ID NO: 97) и DIVMTQTPLSLSVTPGQPASISCKSSQSLVHENLRTYLSWYLQKPGQSPQSLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCGQGTQYPFTFGSGTKVEIK (SEQ ID NO: 98).
7. Связывающий белок по п. 1, где
домен VH2 содержит аминокислотную последовательность QVQLVESGGGVVQPGRSLRLSCAASGFTFTKAWMHWVRQAPGKQLEWVAQIKDKSNSYATYYADSVKGRFTISRDDSSKNTLYLQMNSLRAEDTAVYYCRGVYYALSPFDYWGQGTLVTVSS (SEQ ID NO: 93); и
домен VL2 содержит аминокислотную последовательность DIVMTQTPLSLSVTPGQPASISCKSSQSLVHQNAQTYLSWYLQKPGQSPQSLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCGQGTQYPFTFGSGTKVEIK (SEQ ID NO: 95).
8. Связывающий белок по п. 1, где
(a) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO: 1) или GFNIRDTY (SEQ ID NO: 2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO: 3), IYPTQGYT (SEQ ID NO: 4) или IYPTNAYT (SEQ ID NO: 5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO: 6), SRWGGEGFYAMDY (SEQ ID NO: 7) или SRWGGSGFYAMDY (SEQ ID NO: 8), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9) или QDVQTA (SEQ ID NO: 10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12);
(b) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO: 1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO: 3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO: 6), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12);
(c) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO: 2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO: 4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO: 7), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12);
(d) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO: 2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO: 5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO: 8), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12);
(e) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO: 2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTQGYT (SEQ ID NO: 4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGSGFYAMDY (SEQ ID NO: 8), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12);
(f) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO: 2), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNAYT (SEQ ID NO: 5), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO: 7), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12); или
(g) домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIKDTY (SEQ ID NO: 1), последовательность CDR-H2, содержащую аминокислотную последовательность IYPTNGYT (SEQ ID NO: 3), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGDGFYAMDY (SEQ ID NO: 6), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVQTA (SEQ ID NO: 10), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12).
9. Связывающий белок по п. 8, где
(a) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYWGQGTLVTVSS (SEQ ID NO: 72), EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTQGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGEGFYAMDYWGQGTLVTVSS (SEQ ID NO: 73), EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTQGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGSGFYAMDYWGQGTLVTVSS (SEQ ID NO: 74), EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTNAYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGSGFYAMDYWGQGTLVTVSS (SEQ ID NO: 75) или EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTNAYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGEGFYAMDYWGQGTLVTVSS (SEQ ID NO: 76) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77) или DIQMTQSPSSLSASVGDRVTITCRASQDVQTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 78);
(b) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYWGQGTLVTVSS (SEQ ID NO: 72) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77);
(c) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTQGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGEGFYAMDYWGQGTLVTVSS (SEQ ID NO: 73) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77);
(d) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTNAYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGSGFYAMDYWGQGTLVTVSS (SEQ ID NO: 75) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77);
(e) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTQGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGSGFYAMDYWGQGTLVTVSS (SEQ ID NO: 74) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77);
(f) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTNAYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGEGFYAMDYWGQGTLVTVSS (SEQ ID NO: 76) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77); или
(g) домен VH3 содержит аминокислотную последовательность EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYWGQGTLVTVSS (SEQ ID NO: 72) и/или домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVQTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 78).
10. Связывающий белок по п. 1, где
(a) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 100 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 100; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 101 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 101; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 102 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 102; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 103 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 103;
(b) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 104 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 104; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 105 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 105; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 106 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 106; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 107 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 107;
(c) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 112 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 112; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 113 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 113; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 114 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 114; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 115 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 115;
(d) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 116 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 116; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 117 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 117; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 118 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 118; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 119 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 119;
(e) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 120 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 120; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 121 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 121; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 122 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 122; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 123 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 123;
(f) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 124 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 124; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 125 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 125; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 126 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 126; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 127 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 127;
(g) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 128 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 128; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 129 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 129; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 130 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 130; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 131 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 131;
(h) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 132 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 132; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 133 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 133; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 134 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 134; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 135 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 135;
(i) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 136 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 136; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 137 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 137; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 138 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 138; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 139 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 139;
(j) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 140 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 140; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 141 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 141; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 142 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 142; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 143 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 143;
(k) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 144 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 144; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 145 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 145; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 146 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 146; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 147 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 147;
(l) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 148 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 148; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 149 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 149; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 150 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 150; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 151 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 151;
(m) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 152 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 152; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 153 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 153; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 154 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 154; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 155 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 155;
(n) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 286 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 286; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 287 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 287; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 288 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 288; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 289 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 289;
(o) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 290 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 290; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 291 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 291; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 292 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 292; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 293 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 293;
(p) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 294 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 294; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 295 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 295; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 296 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 296; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 297 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 297; или
(q) первая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 298 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 298; вторая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 299 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 299; третья полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 300 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 300; и четвертая полипептидная цепь содержит аминокислотную последовательность под SEQ ID NO: 301 или аминокислотную последовательность, которая на по меньшей мере 95% идентична аминокислотной последовательности под SEQ ID NO: 301.
11. Связывающий белок по п. 1, где:
VH1 и VL1 образуют первый антигенсвязывающий участок, который связывает полипептид CD28, где домен VH1 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GYTFTSYY (SEQ ID NO: 49), последовательность CDR-H2, содержащую аминокислотную последовательность IYPGNVNT (SEQ ID NO: 50), и последовательность CDR-H3, содержащую аминокислотную последовательность TRSHYGLDWNFDV (SEQ ID NO: 51), и домен VL1 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QNIYVW (SEQ ID NO: 52), последовательность CDR-L2, содержащую аминокислотную последовательность KAS (SEQ ID NO: 53), и последовательность CDR-L3, содержащую аминокислотную последовательность QQGQTYPY (SEQ ID NO: 54);
VH2 и VL2 образуют второй антигенсвязывающий участок, который связывает полипептид CD3, где домен VH2 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFTFTKAW (SEQ ID NO: 55), последовательность CDR-H2, содержащую аминокислотную последовательность IKDKSNSYAT (SEQ ID NO: 56), и последовательность CDR-H3, содержащую аминокислотную последовательность RGVYYALSPFDY (SEQ ID NO: 57), и домен VL2 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QSLVHQNAQTY (SEQ ID NO: 59), последовательность CDR-L2, содержащую аминокислотную последовательность KVS (SEQ ID NO: 64), и последовательность CDR-L3, содержащую аминокислотную последовательность GQGTQYPFT (SEQ ID NO: 65); и
VH3 и VL3 образуют третий антигенсвязывающий участок, который связывает полипептид HER2, где домен VH3 содержит последовательность CDR-H1, содержащую аминокислотную последовательность GFNIRDTY (SEQ ID NO: 2), последовательность CDR-H2, содержащую аминокислоту кислотную последовательность IYPTQGYT (SEQ ID NO: 4), и последовательность CDR-H3, содержащую аминокислотную последовательность SRWGGEGFYAMDY (SEQ ID NO: 7), и домен VL3 содержит последовательность CDR-L1, содержащую аминокислотную последовательность QDVNTA (SEQ ID NO: 9), последовательность CDR-L2, содержащую аминокислотную последовательность SAS (SEQ ID NO: 11), и последовательность CDR-L3, содержащую аминокислотную последовательность QQHYTTP (SEQ ID NO: 12).
12. Связывающий белок по п. 11, где:
домен VH1 содержит аминокислотную последовательность QVQLVQSGAEVVKPGASVKVSCKASGYTFTSYYIHWVRQAPGQGLEWIGSIYPGNVNTNYAQKFQGRATLTVDTSISTAYMELSRLRSDDTAVYYCTRSHYGLDWNFDVWGKGTTVTVSS (SEQ ID NO: 91) и домен VL1 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCQASQNIYVWLNWYQQKPGKAPKLLIYKASNLHTGVPSRFSGSGSGTDFTLTISSLQPEDIATYYCQQGQTYPYTFGQGTKLEIK (SEQ ID NO: 92);
домен VH2 содержит аминокислотную последовательность QVQLVESGGGVVQPGRSLRLSCAASGFTFTKAWMHWVRQAPGKQLEWVAQIKDKSNSYATYYADSVKGRFTISRDDSSKNTLYLQMNSLRAEDTAVYYCRGVYYALSPFDYWGQGTLVTVSS (SEQ ID NO: 93) и домен VL2 содержит аминокислотную последовательность DIVMTQTPLSLSVTPGQPASISCKSSQSLVHQNAQTYLSWYLQKPGQSPQSLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCGQGTQYPFTFGSGTKVEIK (SEQ ID NO: 95); и
домен VH3 содержит аминокислотную последовательность EVQLVESGGLVQPGGSLRLSCAASGFNIRDTYIHWVRQAPGKGLEWVARIYPTQGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGEGFYAMDYWGQGTLVTVSS (SEQ ID NO: 73) и домен VL3 содержит аминокислотную последовательность DIQMTQSPSSLSASVGDRVTITCRASQDVNTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK (SEQ ID NO: 77).
13. Связывающий белок по п. 11, где домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-CH2-CH3 IgG4 человека и где домены шарнир-CH2-CH3 каждый содержит аминокислотные замены в положениях, соответствующих положениям 228, 234, 235 и 409 IgG4 человека в соответствии с индексом ЕС, где аминокислотные замены представляют собой S228P, F234A, L235A и R409K.
14. Связывающий белок по любому из пп. 1-13, где по меньшей мере один из L1, L2, L3 или L4 независимо имеет длину 0 аминокислот.
15. Связывающий белок по любому из пп. 1-13, где (a) каждый из L1, L2, L3 и L4 независимо имеет длину, составляющую ноль аминокислот, или содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO: 69), GGGGSGGGGSGGGGS (SEQ ID NO: 70), S, RT, TKGPS (SEQ ID NO: 68), GQPKAAP (SEQ ID NO: 67) и GGSGSSGSGG (SEQ ID NO: 71); или (b) каждый из L1, L2, L3 и L4 независимо содержит последовательность, выбранную из группы, состоящей из GGGGSGGGGS (SEQ ID NO: 69), GGGGSGGGGSGGGGS (SEQ ID NO: 70), S, RT, TKGPS (SEQ ID NO: 68), GQPKAAP (SEQ ID NO: 67) и GGSGSSGSGG (SEQ ID NO: 71).
16. Связывающий белок по любому из пп. 1-13, где L1 содержит последовательность GQPKAAP (SEQ ID NO: 67), L2 содержит последовательность TKGPS (SEQ ID NO: 68), L3 содержит последовательность S и L4 содержит последовательность RT.
17. Связывающий белок по любому из пп. 1-13, где по меньшей мере один из L1, L2, L3 или L4 содержит последовательность DKTHT (SEQ ID NO: 66).
18. Связывающий белок по п. 17, где L1, L2, L3 и L4 содержат, каждый индивидуально, последовательность DKTHT (SEQ ID NO: 66).
19. Связывающий белок по любому из пп. 1-13, где домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-CH2-CH3 IgG4 человека и где каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 234 и 235 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой F234A и L235A.
20. Связывающий белок по любому из пп. 1-13, где домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-CH2-CH3 IgG4 человека и где каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 233-236 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой E233P, F234V, L235A, и делецию в положении 236.
21. Связывающий белок по любому из пп. 1-13, где домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-CH2-CH3 IgG4 человека и где каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 228 и 409 IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S228P и R409K.
22. Связывающий белок по любому из пп. 1-13, где домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-CH2-CH3 IgG1 человека и где каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 234, 235 и 329 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой L234A, L235A и P329A.
23. Связывающий белок по любому из пп. 1-13, где домены шарнир-CH2-CH3 второй и третьей полипептидных цепей представляют собой домены шарнир-CH2-CH3 IgG1 человека и где каждый из доменов шарнир-CH2-CH3 содержит аминокислотные замены в положениях, соответствующих положениям 298, 299 и 300 IgG1 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S298N, T299A и Y300S.
24. Связывающий белок по любому из пп. 1-13, где домен шарнир-CH2-CH3 второй полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V; и где домен шарнир-CH2-CH3 третьей полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W.
25. Связывающий белок по любому из пп. 1-13, где домен шарнир-CH2-CH3 второй полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 354 и 366 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой S354C и T366W; и где домен шарнир-CH2-CH3 третьей полипептидной цепи содержит аминокислотные замены в положениях, соответствующих положениям 349, 366, 368 и 407 IgG1 или IgG4 человека в соответствии с EU-индексом, где аминокислотные замены представляют собой Y349C, T366S, L368A и Y407V.
26. Выделенная молекула нуклеиновой кислоты для экспрессии связывающего белка, содержащая нуклеотидную последовательность, кодирующую связывающий белок по любому из пп. 1-13.
27. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 26.
28. Выделенная клетка-хозяин для экспрессии связывающего белка, содержащая вектор экспрессии по п. 27, причем клетка-хозяин не является человеческой эмбриональной клеткой.
29. Выделенная клетка-хозяин по п. 28, где клетка-хозяин представляет собой клетку млекопитающего или клетку насекомого.
30. Фармацевтическая композиция для профилактики или лечения рака, содержащая эффективное количество связывающего белка по любому из пп. 1-13 и фармацевтически приемлемый носитель.
31. Способ профилактики или лечения рака у пациента, включающий введение пациенту терапевтически эффективного количества связывающего белка по любому из пп. 1-13 или фармацевтической композиции, содержащей эффективное количество связывающего белка по любому из пп. 1-13 и фармацевтически приемлемый носитель.
32. Способ профилактики или лечения рака у пациента, включающий введение пациенту терапевтически эффективного количества связывающего белка по любому из пп. 1-13 или фармацевтической композиции, содержащей эффективное количество связывающего белка по любому из пп. 1-13 и фармацевтически приемлемый носитель, где связывающий белок вводят совместно с химиотерапевтическим средством.
33. Способ по п. 31, где пациент представляет собой человека.
34. Способ по п. 31, где раковые клетки пациента экспрессируют HER2.
35. Способ по п. 34, где рак представляет собой рак молочной железы, рак толстой кишки, рак желудка или немелкоклеточный рак легкого (NSCLC).
36. Применение связывающего белка по любому из пп. 1-13 для профилактики или лечения рака у пациента.
37. Применение фармацевтической композиции по п. 30 для профилактики или лечения рака у пациента.
38. Применение связывающего белка по любому из пп. 1-13 для профилактики или лечения рака у пациента, где связывающий белок предназначен для совместного введения с химиотерапевтическим средством.
39. Применение по п. 37, где пациент представляет собой человека.
40. Применение по п. 36, где раковые клетки пациента экспрессируют HER2.
41. Применение по п. 40, где рак представляет собой рак молочной железы, рак толстой кишки, рак желудка или немелкоклеточный рак легкого (NSCLC).
42. Набор для получения связывающего белка, содержащий один или более векторов, кодирующих первую, вторую, третью и четвертую полипептидные цепи связывающего белка по любому из пп. 1-13.
43. Набор по п. 42, где набор содержит первый вектор, кодирующий первую полипептидную цепь связывающего белка, второй вектор, кодирующий вторую полипептидную цепь связывающего белка, третий вектор, кодирующий третью полипептидную цепь связывающего белка, и четвертый вектор, кодирующий четвертую полипептидную цепь связывающего белка.
44. Набор полинуклеотидов для получения связывающего белка, содержащий:
(a) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 189, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 190, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 191, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 192;
(b) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 193, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 194, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 195, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 196;
(c) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 197, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 198, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 199, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 200;
(d) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 201, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 202, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 203, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 204;
(e) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 205, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 206, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 207, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 208;
(f) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 209, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 210, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 211, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 212;
(g) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 213, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 214, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 215, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 216;
(h) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 217, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 218, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 219, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 220;
(i) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 221, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 222, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 223, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 224;
(j) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 225, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 226, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 227, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 228;
(k) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 229, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 230, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 231, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 232;
(l) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 233, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 234, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 235, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 236;
(m) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 237, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 238, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 239, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 240; или
(n) первый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 241, второй полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 242, третий полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 243, и четвертый полинуклеотид, содержащий полинуклеотидную последовательность под SEQ ID NO: 244.
| WO 2017180913 A2, 19.10.2017 | |||
| EA 201791666 A1, 30.11.2017 | |||
| WO 2018015340 A1, 25.01.2018 | |||
| WO 2012092612 A1, 05.07.2012 | |||
| US 2012076782 A1, 29.03.2012 | |||
| WO 2016205531 A2, 22.12.2016 | |||
| CHEN H.W | |||
| ET AL., Ex vivo expansion of dendritic-cell-activated antigen-specific CD4+ T cells with anti-CD3/CD28, interleukin-7, and interleukin-15: Potential for |

