Настоящее изобретение касается способа добавления одного или нескольких L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты, способа амплификации целевой L-нуклеиновой кислоты, белка, содержащего фрагмент, проявляющий ферментативную активность, полимераз, содержащих аминокислотную последовательность, состоящую из D-аминокислот, варианта полимеразы, являющейся полимеразой дикого типа, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 15, варианта полимеразы, являющейся полимеразой дикого типа, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 1, использования белка, содержащего фрагмент, проявляющий ферментативную активность, в способе добавления одного или нескольких L-нуклеотидов, использования белка, содержащего фрагмент, проявляющий ферментативную активность, в способе амплификации целевой L-нуклеиновой кислоты, способа идентификации целевой молекулы, связывающей молекулу L-нуклеиновой кислоты, способа продуцирования белка и полимеразы, соответственно, и способа лигирования первого D-пептида или первого D-белка и второго D-пептида или второго D-белка друг с другом.
Доступность генной технологии в широком смысле внесла большой вклад в достигнутый за последние десятилетия прогресс в области медицины и диагностики, а также в фундаментальных исследованиях. Возможности синтеза, обеспечиваемые генной технологией, выходят за пределы химического синтеза. Генная технология и особенно генная инженерия позволяют производить практически неограниченные количества L-пептидов и L-белков, используя ферментативные механизмы прокариотических и эукариотических клеток. Ферменты и особенно полимеразы, в виде форм дикого типа или вариантов таких форм дикого типа, позволяют проводить синтез D-нуклеиновых кислот, связывая «строительные кирпичики» таких D-нуклеиновых кислот, т.е. D-нуклеотиды, в последовательность, которая, хотя и может быть получена методами химического синтеза, но, во всяком случае, не с приемлемым выходом.
Вследствие хиральной специфичности, ферменты, используемые в генной технологии, могут использовать только строительные блоки и субстраты, соответственно, хиральность которых совпадает с их собственной хиральностью. Строительные блоки и субстраты, соответственно, противоположной хиральности не могут быть объектами воздействия ферментной активности. Вследствие принципа взаимности хиральных взаимодействий переработка строительных блоков и субстратов, соответственно, противоположной хиральности требует использования ферментов, также имеющих противоположную хиральность.
Этот принцип взаимности хиральных взаимодействий, например, интенсивно используется для обеспечения целенаправленного связывания L-нуклеиновых кислот, которые также известны как шпигельмеры (spiegelmers). В настоящее время, шпигельмеры идентифицируют с помощью способа, использующего, на первой стадии, библиотеку D-нуклеиновых кислот для in vitro отбора против энантиомерной формы целевой молекулы или целевой структуры, такой как D-пептиды или D-белки. На второй стадии идентифицированные таким образом D-нуклеиновые кислоты, связывающиеся с энантиомерной формой целевой молекулы или целевой структуры, получают в виде соответствующих L-нуклеиновых кислот. В результате принципа взаимности хиральных взаимодействий такие L-нуклеиновые кислоты, т.е. шпигельмеры, способны связываться с истинной или действительной целевой молекулой, такой как L-пептиды или L-белки, а не с ее энантиомерной формой, такой как D-пептиды или D-белки, используемые в процессе отбора. Предпочтительно, такая истинная или действительная целевая молекула или целевая структура представляет собой целевую молекулу или целевую структуру, присутствующую в биологической системе, такой как организм человека или животного. Способы получения таких шпигельмеров описаны, например, в книге "The Aptamer Handbook" (eds. Klussmann, 2006).
Одним из путей облегчения процесса идентификации шпигельмеров может быть переработка процесса таким образом, чтобы L-нуклеиновые кислоты выбирали непосредственно из библиотеки L-нуклеиновых кислот с использованием целевой молекулы или целевой структуры в энантиомерной форме, демонстрируемой истинной или действительной целевой молекулой или целевой структурой. Поскольку частью процесса является амплификация таких L-нуклеиновых кислот, первоначально связывающихся с целевой молекулой и целевой структурой, соответственно, потребуется полимераза, добавляющая по меньшей мере один нуклеотид к L-праймеру. На сегодняшний день не известно ни одной полимеразы, состоящей из L-аминокислот, способной на это. В связи с этим существует потребность в полимеразе и подобных ферментах, состоящих из D-аминокислот. Поскольку генная технология не может обеспечить такие функционально активные полимеразы, состоящие из D-аминокислот, необходимо использовать химический синтез. Однако синтез D-белков или D-полипептидов ограничен сравнительно малыми молекулами. Наибольшим синтезированным до настоящего времени D-белком является D-белковая форма ангиогенного белка фактора роста сосудистого эндотелия (сокращенно, VEGF-A), состоящего из 102 D-аминокислот (Mandai et al., 2012). Полимеразы, однако, типично состоят из более чем 300 аминокислот.
Таким образом, проблемой, лежащей в основе настоящего изобретения, является создание способа, позволяющего добавлять по меньшей мере один нуклеотид к L-нуклеиновой кислоте, такой как праймер. Дополнительной проблемой, лежащей в основе настоящего изобретения, является создание способа амплификации целевой L-нуклеиновой кислоты с использованием L-нуклеотидов. Еще одной дополнительной проблемой, лежащей в основе настоящего изобретения, является создание средств, позволяющих реализовать такие способы.
Эти и другие проблемы, лежащие в основе настоящего изобретения, решаются с помощью объектов независимых пунктов прилагаемой формулы изобретения. Предпочтительные варианты реализации могут быть взяты из зависимых пунктов приложенной формулы изобретения.
Проблема, лежащая в основе данной заявки, также решается в первом аспекте, который также является первым вариантом реализации первого аспекта, с помощью способа добавления одного или нескольких L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты, который включает стадию проведения реакции одного или нескольких L-нуклеотидов с первой L-нуклеиновой кислотой в присутствии белка, содержащего фрагмент, проявляющий ферментативную активность, которая способна добавлять один или несколько L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты.
Во втором варианте реализации первого аспекта, который также является вариантом реализации первого варианта реализации первого аспекта, фрагмент, проявляющий ферментативную активность, состоит из аминокислотной последовательности, состоящей из D-аминокислот.
В третьем варианте реализации первого аспекта, который также является вариантом реализации первого и второго вариантов реализации первого аспекта, фрагмент, проявляющий ферментативную активность, проявляет полимеразную активность.
В четвертом варианте реализации первого аспекта, который также является вариантом реализации первого, второго и третьего вариантов реализации первого аспекта, ферментативная активность представляет собой полимеразную активность.
В пятом варианте реализации первого аспекта, который также является вариантом реализации четвертого варианта реализации первого аспекта, полимеразная активность представляет собой активность термостабильной полимеразы.
В шестом варианте реализации первого аспекта, который также является вариантом реализации третьего, четвертого и пятого вариантов реализации первого аспекта, аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает по меньшей мере 300 аминокислот.
В седьмом варианте реализации первого аспекта, который также является вариантом реализации шестого варианта реализации первого аспекта, аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает от 300 до 900 аминокислот, предпочтительно, от 300 до 600 аминокислот, более предпочтительно, от 300 до 360 аминокислот, наиболее предпочтительно, от 340 до 360 аминокислот.
В восьмом варианте реализации первого аспекта, который также является вариантом реализации четвертого, пятого, шестого и седьмого вариантов реализации первого аспекта, полимеразная активность представляет собой ДНК-полимеразную активность.
В девятом варианте реализации первого аспекта, который также является вариантом реализации восьмого варианта реализации первого аспекта, ДНК-полимеразная активность представляет собой ДНК-зависимую ДНК-полимеразную активность.
В десятом варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого и девятого вариантов реализации первого аспекта, фрагмент, проявляющий ферментативную активность, является ферментом.
В одиннадцатом варианте реализации первого аспекта, который также является вариантом реализации третьего, четвертого, пятого, шестого, седьмого, восьмого и девятого вариантов реализации первого аспекта, фрагмент, проявляющий ферментативную активность, является полимеразой.
В двенадцатом варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, восьмого, девятого, десятого и одиннадцатого вариантов реализации первого аспекта, фрагмент, проявляющий полимеразную активность, выбирают из группы полимеразы X вируса африканской чумы свиней, корового домена полимеразы X Thermus thermophilus, крысиной полимеразы-бета, эукариотической полимеразы-бета, фрагмента Кленова, экзополимеразы Кленова, ДНК-полимеразы Т4, ДНК-полимеразы Phi29, секвеназы, ДНК-полимеразы Т7, полимеразы SP6, ДНК-полимеразы I, полимеразы-лямбда, и вариантов всех и любой из них, в которых, предпочтительно, фрагмент, проявляющий полимеразную активность, представляет собой полимеразу X вируса африканской чумы свиней или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 1, аминокислотной последовательности, соответствующей SEQ ID NO: 2, аминокислотной последовательности, соответствующей SEQ ID NO: 3, и аминокислотной последовательности, соответствующей SEQ ID NO: 4.
В 13-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого и одиннадцатого вариантов реализации первого аспекта, фрагмент, проявляющий полимеразную активность, выбирают из группы полимеразы DPO4, ДНК-полимеразы Thermococcus litoralis, ДНК-полимеразы Pyrococcus sp., ДНК-полимеразы Pyrococcus furiosus, полимеразы Pfuturbo, ДНК-полимеразы Sulfolobus solfataricus, ДНК-полимеразы Thermococcus gorgonarius, полимеразы KOD, полимеразы Taq, полимеразы Tth, полимеразы Pyrobest, полимеразы Pwo, полимеразы Sac, полимеразы Bst, полимеразы Рос, полимеразы Pab, полимеразы Mth, полимеразы Pho, полимеразы ES4, полимеразы EX-Taq, полимеразы LA-Taq, полимераз Expand, полимераз Platinum Taq, высокоточной (Hi-Fi) полимеразы, полимеразы Tbr, полимеразы Tfl, полимеразы Tru, полимеразы Тас, полимеразы Tne, полимеразы Tma, полимеразы Tih, полимеразы Tfi, AmpliTaq, фрагмента Стоффеля, ДНК-полимеразы 9°Nm, Therminator, Therminator II, высокоточной (High Fidelity) полимеразы Phusion, Paq5000, Pfx-50, Proofstart, FideliTaq, элонгазы, и их вариантов, в которых, предпочтительно, фрагмент, проявляющий полимеразную активность, представляет собой полимеразу Dpo4 или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 15, аминокислотной последовательности, соответствующей SEQ ID NO: 16, аминокислотной последовательности, соответствующей SEQ ID NO: 17, аминокислотной последовательности, соответствующей SEQ ID NO: 18, аминокислотной последовательности, соответствующей SEQ ID NO: 19, аминокислотной последовательности, соответствующей SEQ ID NO: 20, аминокислотной последовательности, соответствующей SEQ ID NO: 21, и аминокислотной последовательности, соответствующей SEQ ID NO: 22.
В 14-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого и 13-ого вариантов реализации первого аспекта, стадия проведения реакции осуществляется в условиях, позволяющих добавлять по меньшей мере один или нескольких L-нуклеотидов к первой L-нуклеиновой кислоте, предпочтительно, позволяющих добавлять от 5 до 20000 L-нуклеотидов, предпочтительно, от 10 до 2000 L-нуклеотидов, более предпочтительно, от 50 до 500 L-нуклеотидов, наиболее предпочтительно, от 50 до 100 L-нуклеотидов.
В 15-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го и 14-го вариантов реализации первого аспекта, добавление по меньшей мере одного или нескольких L-нуклеотидов к первой L-нуклеиновой кислоте представляет собой ковалентное связывание по меньшей мере одного или нескольких L-нуклеотидов с первой L-нуклеиновой кислотой, предпочтительно, путем образования 3'-5' фосфодиэфирной связи между 3'ОН первой L-нуклеиновой кислоты и 5'-фосфатом одного из по меньшей мере одного или нескольких L-нуклеотидов.
В 16-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го и 15-го вариантов реализации первого аспекта, первая L-нуклеиновая кислота представляет собой праймер, состоящий из ДНК, РНК, модифицированной ДНК, модифицированной РНК или их комбинаций.
В 17-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го, 15-го и 16-го вариантов реализации первого аспекта, первая L-нуклеиновая кислота состоит из L-нуклеотидов и, необязательно, модификации.
В 18-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го, 15-го и 16-го и 17-го вариантов реализации первого аспекта, первая L-нуклеиновая кислота состоит из L-нуклеотидов.
В 19-м варианте реализации первого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го, 15-го и 16-го, 17-ого и 18-го вариантов реализации первого аспекта, реакция дополнительно включает вторую L-нуклеиновую кислоту, при том что одна молекула первой L-нуклеиновой кислоты гибридизуется с одной молекулой второй L-нуклеиновой кислоты, предпочтительно, путем спаривания оснований по Уотсону-Крику.
В 20-м варианте реализации первого аспекта, который также является вариантом реализации 19-го варианта реализации первого аспекта, фрагмент, проявляющий полимеразную активность, синтезирует третью L-нуклеиновую кислоту, которая является комплементарной ко второй L-нуклеиновой кислоте, при том что третья L-нуклеиновая кислота содержит первую L-нуклеиновую кислоту и L-нуклеотиды, добавленные к 3'-концу первой L-нуклеиновой кислоты.
Проблема, лежащая в основе данной заявки, также решается во втором аспекте, который также является первым вариантом реализации второго аспекта, с помощью способа амплификации целевой L-нуклеиновой кислоты в присутствии L-нуклеотидов и белка, содержащего фрагмент, проявляющий ферментативную активность, способную амплифицировать целевую L-нуклеиновую кислоту.
Во втором варианте реализации второго аспекта, который также является вариантом реализации первого варианта реализации второго аспекта, фрагмент, проявляющий ферментативную активность, состоит из аминокислотной последовательности, состоящей из D-аминокислот.
В третьем варианте реализации второго аспекта, который также является вариантом реализации первого и второго вариантов реализации второго аспекта, фрагмент, проявляющий ферментативную активность, проявляет полимеразную активность.
В четвертом варианте реализации второго аспекта, который также является вариантом реализации первого, второго и третьего вариантов реализации второго аспекта, ферментативная активность представляет собой полимеразную активность.
В пятом варианте реализации второго аспекта, который также является вариантом реализации четвертого варианта реализации второго аспекта, полимеразная активность представляет собой активность термостабильной полимеразы.
В шестом варианте реализации второго аспекта, который также является вариантом реализации третьего, четвертого и пятого вариантов реализации второго аспекта, аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает по меньшей мере 300 аминокислот.
В седьмом варианте реализации второго аспекта, который также является вариантом реализации шестого варианта реализации второго аспекта, аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает от 300 до 900 аминокислот, предпочтительно, от 300 до 600 аминокислот, более предпочтительно, от 300 до 360 аминокислот, наиболее предпочтительно, от 340 до 360 аминокислот.
В восьмом варианте реализации второго аспекта, который также является вариантом реализации четвертого, пятого и шестого вариантов реализации второго аспекта, полимеразная активность представляет собой ДНК-полимеразную активность.
В девятом варианте реализации второго аспекта, который также является вариантом реализации восьмого варианта реализации второго аспекта, ДНК-полимеразная активность представляет собой ДНК-зависимую ДНК-полимеразную активность.
В десятом варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого и девятого вариантов реализации второго аспекта, фрагмент, проявляющий ферментативную активность, является ферментом.
В одиннадцатом варианте реализации второго аспекта, который также является вариантом реализации третьего, четвертого, пятого, шестого, седьмого, восьмого и девятого вариантов реализации второго аспекта, фрагмент, проявляющий полимеразную активность, является полимеразой.
В двенадцатом варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, восьмого, девятого, десятого и одиннадцатого вариантов реализации второго аспекта, фрагмент, проявляющий полимеразную активность, выбирают из группы, состоящей из полимеразы X вируса африканской чумы свиней, корового домена полимеразы X Thermus thermophilus, крысиной полимеразы-бета, эукариотической полимеразы-бета, фрагмента Кленова, экзополимеразы Кленова, ДНК-полимеразы Т4, ДНК-полимеразы РЫ29, секвеназы, ДНК-полимеразы Т7, полимеразы SP6, ДНК-полимеразы I, полимеразы-лямбда и вариантов всех и любой из них, в которых, предпочтительно, фрагмент, проявляющий полимеразную активность, представляет собой полимеразу X вируса африканской чумы свиней или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 1, аминокислотной последовательности, соответствующей SEQ ID NO: 2, аминокислотной последовательности, соответствующей SEQ ID NO: 3, и аминокислотной последовательности, соответствующей SEQ ID NO: 4.
В 13-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого и одиннадцатого вариантов реализации второго аспекта, фрагмент, проявляющий полимеразную активность, выбирают из группы, состоящей из полимеразы DPO4, ДНК-полимеразы Thermococcus litoralis, ДНК-полимеразы Pyrococcus sp., ДНК-полимеразы Pyrococcus furiosus, полимеразы Pfuturbo, ДНК-полимеразы Sulfolobus solfataricus, ДНК-полимеразы Thermococcus gorgonarius, полимеразы KOD, полимеразы Taq, полимеразы Tth, полимеразы Pyrobest, полимеразы Pwo, полимеразы Sac, полимеразы Bst, полимеразы Рос, полимеразы Pab, полимеразы Mth, полимеразы Pho, полимеразы ES4, полимеразы EX-Taq, полимеразы LA-Taq, полимераз Expand, полимераз Platinum Taq, высокоточной (Hi-Fi) полимеразы, полимеразы Tbr, полимеразы Tfl, полимеразы Tru, полимеразы Тас, полимеразы The, полимеразы Tma, полимеразы Tih, полимеразы Tfi, AmpliTaq, фрагмента Стоффеля, ДНК-полимеразы 9°Nm, Therminator, Therminator II, высокоточной полимеразы Phusion, Paq5000, Pfx-50, Proofstart, FideliTaq, элонгазы и их вариантов, где, предпочтительно, фрагмент, проявляющий полимеразную активность, представляет собой полимеразу Dpo4 или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 15, аминокислотной последовательности, соответствующей SEQ ID NO: 16, аминокислотной последовательности, соответствующей SEQ ID NO: 17, аминокислотной последовательности, соответствующей SEQ ID NO: 18, аминокислотной последовательности, соответствующей SEQ ID NO: 19, аминокислотной последовательности, соответствующей SEQ ID NO: 20, аминокислотной последовательности, соответствующей SEQ ID NO: 21, и аминокислотной последовательности, соответствующей SEQ ID NO: 22.
В 14-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого и 13-го вариантов реализации второго аспекта, стадия проведения реакции осуществляется в условиях, обеспечивающих возможность амплификации целевой L-нуклеиновой кислоты.
В 15-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го и 14-го вариантов реализации второго аспекта, способ использует по меньшей мере один праймер, предпочтительно, два праймера, и меньшей мере один праймер состоит из L-нуклеотидов и, необязательно, модификации.
В 16-м варианте реализации второго аспекта, который также является вариантом реализации 15-го варианта реализации второго аспекта, праймеры состоят из L-нуклеотидов.
В 17-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го, 15-го и 16-го вариантов реализации второго аспекта, целевая L-нуклеиновая кислота состоит из L-нуклеотидов.
В 18-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-ого, 14-ого, 15-ого, 16-го и 17-го вариантов реализации второго аспекта, способ представляет собой полимеразную цепную реакцию.
В 19-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го, 15-го, 16-го, 17-го и 18-го вариантов реализации второго аспекта, целевая L-нуклеиновая кислота состоит из L-ДНК.
В 10-м варианте реализации второго аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого, 13-го, 14-го, 15-го, 16-го, 17-го, 18-го и 19-го вариантов реализации второго аспекта, целевая L-нуклеиновая кислота состоит из от 20 до 20000 L-нуклеотидов, предпочтительно, 30-2000 L-нуклеотидов, более предпочтительно, 40-500 L-нуклеотидов, наиболее предпочтительно, от 50 до 100 L-нуклеотидов.
Проблема, лежащая в основе данной заявки, также решается в третьем аспекте, который также является первым вариантом реализации третьего аспекта, с помощью белка, содержащего проявляющий ферментативную активность фрагмент, который состоит из аминокислотной последовательности, состоящей из D-аминокислот, причем ферментативная активность способна добавлять один или несколько L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты.
Во втором варианте реализации третьего аспекта, который также является вариантом реализации первого варианта реализации третьего аспекта, фрагмент, проявляющий ферментативную активность, является фрагментом, проявляющим полимеразную активность.
В третьем варианте реализации третьего аспекта, который также является вариантом реализации первого варианта реализации третьего аспекта, ферментативная активность представляет собой полимеразную активность.
В четвертом варианте реализации третьего аспекта, который также является вариантом реализации третьего вариант реализации третьего аспекта, полимеразная активность представляет собой активность термостабильной полимеразы.
В пятом варианте реализации третьего аспекта, который также является вариантом реализации второго, третьего и четвертого вариантов реализации третьего аспекта, аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает по меньшей мере 300 аминокислот.
В шестом варианте реализации третьего аспекта, который также является вариантом реализации пятого варианта реализации третьего аспекта, аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает от 300 до 900 аминокислот, предпочтительно, от 300 до 600 аминокислот, более предпочтительно, от 300 до 360 аминокислот, наиболее предпочтительно, от 340 до 360 аминокислот.
В седьмом варианте реализации третьего аспекта, который также является вариантом реализации третьего, четвертого, пятого и шестого вариантов реализации третьего аспекта, полимеразная активность представляет собой ДНК-полимеразную активность.
В восьмом варианте реализации третьего аспекта, который также является вариантом реализации седьмого варианта реализации третьего аспекта, ДНК-полимеразная активность представляет собой ДНК-зависимую ДНК-полимеразную активность.
В девятом варианте реализации третьего аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого и восьмого вариантов реализации третьего аспекта, фрагмент, проявляющий ферментативную активность, является ферментом.
В десятом варианте реализации третьего аспекта, который также является вариантом реализации второго, третьего, четвертого, пятого, шестого, седьмого и восьмого вариантов реализации третьего аспекта, фрагмент, проявляющий ферментативную активность, является полимеразой.
В одиннадцатом варианте реализации третьего аспекта, который также является вариантом реализации первого, второго, третьего, седьмого, восьмого, девятого и десятого вариантов реализации третьего аспекта, фрагмент, проявляющий полимеразную активность, выбирают из группы, состоящей из полимеразы X вируса африканской чумы свиней, корового домена полимеразы X Thermus thermophilus, крысиной полимеразы-бета, эукариотической полимеразы-бета, фрагмента Кленова, экзополимеразы Кленова, ДНК-полимеразы Т4, ДНК-полимеразы РЫ29, секвеназы, ДНК-полимеразы Т7, полимеразы SP6, ДНК-полимеразы I, полимеразы-лямбда, и вариантов всех и любой из них, в которых, предпочтительно, фрагмент, проявляющий полимеразную активность, представляет собой полимеразу X вируса африканской чумы свиней или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 1, аминокислотной последовательности, соответствующей SEQ ID NO: 2, аминокислотной последовательности, соответствующей SEQ ID NO: 3, и аминокислотной последовательности, соответствующей SEQ ID NO: 4.
В двенадцатом варианте реализации третьего аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого, шестого, седьмого, восьмого, девятого и десятого вариантов реализации третьего аспекта, фрагмент, проявляющий полимеразную активность, выбирают из группы, состоящей из полимеразы DPO4, ДНК-полимеразы Thermococcus litoralis, ДНК-полимеразы Pyrococcus sp., ДНК-полимеразы Pyrococcus furiosus, полимеразы Pfuturbo, ДНК-полимеразы Sulfolobus solfataricus, ДНК-полимеразы Thermococcus gorgonarius, полимеразы KOD, полимеразы Taq, полимеразы Tth, полимеразы Pyrobest, полимеразы Pwo, полимеразы Sac, полимеразы Bst, полимеразы Рос, полимеразы Pab, полимеразы Mth, полимеразы Pho, полимеразы ES4, полимеразы EX-Taq, полимеразы LA-Taq, полимераз Expand, полимераз Platinum Taq, высокоточной (Hi-Fi) полимеразы, полимеразы Tbr, полимеразы Tfl, полимеразы Tru, полимеразы Тас, полимеразы The, полимеразы Tma, полимеразы Tih, полимеразы Tfi, AmpliTaq, фрагмента Стоффеля, ДНК-полимеразы 9°Nm, Therminator, Therminator И, высокоточной полимеразы Phusion, Paq5000, Pfx-50, Proofstart, FideliTaq, элонгазы и их вариантов, в которых, предпочтительно, фрагмент, проявляющий полимеразную активность, представляет собой полимеразу Dpo4 или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 15, аминокислотной последовательности, соответствующей SEQ ID NO: 16, аминокислотной последовательности, соответствующей SEQ ID NO: 17, аминокислотной последовательности, соответствующей SEQ ID NO: 18, аминокислотной последовательности, соответствующей SEQ ID NO: 19, аминокислотной последовательности, соответствующей SEQ ID NO: 20, аминокислотной последовательности, соответствующей SEQ ID NO: 21, и аминокислотной последовательности, соответствующей SEQ ID NO: 22.
Проблема, лежащая в основе данной заявки, также решается в четвертом аспекте, который также является первым вариантом реализации четвертого аспекта, с помощью полимеразы, содержащей аминокислотную последовательность, соответствующую SEQ ID NO: 15, где аминокислоты аминокислотной последовательности являются D-аминокислотами.
Проблема, лежащая в основе данной заявки, также решается в пятом аспекте, который также является первым вариантом реализации пятого аспекта, с помощью полимеразы, содержащей аминокислотную последовательность, соответствующую SEQ ID NO: 1, где аминокислоты аминокислотной последовательности являются D-аминокислотами.
Проблема, лежащая в основе данной заявки, также решается в шестом аспекте, который также является первым вариантом реализации шестого аспекта, с помощью варианта полимеразы, являющейся полимеразой дикого типа, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 15, и где вариант полимеразы обладает полимеразной активностью, предпочтительно, активностью термостабильной полимеразы.
Во втором варианте реализации шестого аспекта, который также является вариантом реализации первого варианта реализации шестого аспекта, вариант полимеразы включает аминокислотную последовательность, которая отличается от аминокислотной последовательности полимеразы дикого типа по меньшей мере в одной аминокислотной позиции.
В третьем варианте реализации шестого аспекта, который также является вариантом реализации первого и второго вариантов реализации шестого аспекта, аминокислотная последовательность варианта полимеразы отличается от аминокислотной последовательности полимеразы дикого типа в одной или двух аминокислотных позициях.
В четвертом варианте реализации шестого аспекта, который также является вариантом реализации первого, второго и третьего вариантов реализации шестого аспекта, аминокислотная последовательность варианта полимеразы отличается от аминокислотной последовательности полимеразы дикого типа в аминокислотной позиции 155 и/или 203 аминокислотной последовательности, соответствующей SEQ ID NO: 15 или в соответствующих им аминокислотных позициях, при том, что, предпочтительно, аминокислота (аминокислоты) в позиции 155 и/или 203 замещена/замещены цистеином.
В пятом варианте реализации шестого аспекта, который также является вариантом реализации первого, второго, третьего и четвертого вариантов реализации шестого аспекта, вариант полимеразы состоит из аминокислотной последовательности, выбранной из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 16, аминокислотной последовательности, соответствующей SEQ ID NO: 17, и аминокислотной последовательности, соответствующей SEQ ID NO: 18.
В шестом варианте реализации шестого аспекта, который также является вариантом реализации первого, второго и третьего вариантов реализации шестого аспекта, вариант полимеразы состоит из аминокислотной последовательности, выбранной из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 19, аминокислотной последовательности, соответствующей SEQ ID NO: 20, аминокислотной последовательности, соответствующей SEQ ID NO: 21, и аминокислотной последовательности, соответствующей SEQ ID NO: 22.
В седьмом варианте реализации шестого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого и шестого вариантов реализации шестого аспекта, аминокислоты аминокислотной последовательности варианта полимеразы являются D-аминокислотами.
В восьмом варианте реализации шестого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого и шестого вариантов реализации шестого аспекта, аминокислоты аминокислотной последовательности варианта полимеразы являются L-аминокислотами.
Проблема, лежащая в основе данной заявки, также решается в седьмом аспекте, который также является первым вариантом реализации седьмого аспекта, с помощью варианта полимеразы, являющейся полимеразой дикого типа, при том, что полимераза дикого типа состоит из аминокислотной последовательности, соответствующей SEQ ID NO: 1, и при том, что вариант полимеразы обладает полимеразной активностью.
Во втором варианте реализации седьмого аспекта, который также является вариантом реализации первого варианта реализации седьмого аспекта, вариант полимеразы включает аминокислотную последовательность, где аминокислотная последовательность варианта полимеразы отличается от аминокислотной последовательности полимеразы дикого типа по меньшей мере в одной аминокислотной позиции.
В третьем варианте реализации седьмого аспекта, который также является вариантом реализации первого и второго вариантов реализации седьмого аспекта, аминокислотная последовательность варианта полимеразы отличается от аминокислотной последовательности полимеразы дикого типа в одной аминокислотной позиции.
В четвертом варианте реализации седьмого аспекта, который также является вариантом реализации первого, второго и третьего вариантов реализации седьмого аспекта, аминокислотная последовательность варианта полимеразы отличается от аминокислотной последовательности полимеразы дикого типа по меньшей мере в одной аминокислотной позиции, где по меньшей мере аминокислотную позицию выбирают из аминокислотной позиции 80, аминокислотной позиции 86 и аминокислотной позиции 124, которые все принадлежат аминокислотной последовательности, соответствующей SEQ ID NO: 1, или в соответствующей ей аминокислотной позиции.
В пятом варианте реализации седьмого аспекта, который также является вариантом реализации первого, второго, третьего и четвертого вариантов реализации седьмого аспекта, вариант полимеразы состоит из аминокислотной последовательности, выбранной из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 2, аминокислотной последовательности, соответствующей SEQ ID NO: 3, и аминокислотной последовательности, соответствующей SEQ ID NO: 4.
В шестом варианте реализации седьмого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого и пятого вариантов реализации седьмого аспекта, аминокислоты аминокислотной последовательности варианта полимеразы являются D-аминокислотами.
В седьмом варианте реализации седьмого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого, пятого и шестого вариантов реализации седьмого аспекта, аминокислоты аминокислотной последовательности варианта полимеразы являются L-аминокислотами.
Проблема, лежащая в основе данной заявки, также решается в восьмом аспекте, который также является первым вариантом реализации восьмого аспекта, путем использования белка, содержащего фрагмент, проявляющий ферментативную активность, в способе добавления одного или нескольких L-нуклеотидов к 3'-концу L-нуклеиновой кислоты.
Проблема, лежащая в основе данной заявки, также решается в девятом аспекте, который также является первым вариантом реализации девятого аспекта, путем использования белка, содержащего фрагмент, проявляющий ферментативную активность, в способе амплификации целевой L-нуклеиновой кислоты в присутствии L-нуклеотидов.
Во втором варианте реализации девятого аспекта, который также является вариантом реализации первого варианта реализации девятого аспекта, способ амплификации целевой L-нуклеиновой кислоты представляет собой полимеразную цепную реакцию.
Во втором варианте реализации восьмого аспекта, который также является вариантом реализации первого варианта реализации восьмого аспекта, и в третьем варианте реализации девятого аспекта, белок представляет собой белок в соответствии с любым вариантом реализации третьего, четвертого, пятого, шестого и седьмого аспектов, где белок состоит из аминокислотной последовательности, причем аминокислоты аминокислотной последовательности являются D-аминокислотами.
Проблема, лежащая в основе данной заявки, также решается в десятом аспекте, который также является первым вариантом реализации десятого аспекта, с помощью способа идентификации целевой молекулы, связывающей молекулу L-нуклеиновой кислоты, включающего следующие стадии:
(a) генерирование гетерогенной популяции молекул L-нуклеиновой кислоты;
(b) введение в контакт гетерогенной популяции молекул L-нуклеиновой кислоты стадии (а) с целевой молекулой;
(c) отделение молекул L-нуклеиновой кислоты, не связанных целевой молекулой; и
(d) амплификация молекул L-нуклеиновой кислоты, связанных целевой молекулой, на которой используется белок в соответствии с любым вариантом реализации третьего, четвертого, пятого, шестого и седьмого аспектов,
при том, что белок состоит из аминокислотной последовательности, состоящей из D-аминокислот.
Во втором варианте реализации десятого аспекта, который также является вариантом реализации первого варианта реализации десятого аспекта, способ дополнительно включает стадию:
(e) секвенирования молекул L-нуклеиновой кислоты, связанных целевой молекулой; и
(f) синтеза молекул нуклеиновой кислоты, нуклеотидная последовательность которых идентична нуклеотидной последовательности молекул L-нуклеиновой кислоты, секвенированной на стадии (е).
В третьем варианте реализации десятого аспекта, который также является вариантом реализации первого и второго вариантов реализации десятого аспекта, молекулы нуклеиновой кислоты гетерогенной популяции молекул L-нуклеиновой кислоты стадии (а) содержат на их 5'-конце и их 3'-конце сайт связывания праймера и, соответственно, последовательность, комплементарную сайту связывания праймера, что обеспечивает возможность амплификации молекул L-нуклеиновой кислоты, полученных на стадии (d), с помощью полимеразной цепной реакции, при том, что полимераза, используемая в полимеразной цепной реакции, представляет собой белок в соответствии с любым вариантом реализации третьего, четвертого, пятого, шестого и седьмого аспектов; праймеры, используемые в полимеразной цепной реакции, состоят из L-нуклеотидов; и нуклеотиды, используемые в полимеразной цепной реакции, являются L-нуклеотидами.
В четвертом варианте реализации десятого аспекта, который также является вариантом реализации первого, второго и третьего вариантов реализации десятого аспекта, после стадии (d) вводят следующую стадию:
(da) введение в контакт амплифицированных молекул нуклеиновой кислоты с целевой молекулой, при том, что стадия (b) и, необязательно, стадии (с) и/или (d) проводятся перед стадией (е), а стадии (da), (b), (с) и, необязательно, (d) проводятся в таком порядке один или несколько раз.
В пятом варианте реализации десятого аспекта, который также является вариантом реализации первого, второго, третьего и четвертого вариантов реализации десятого аспекта, целевая молекула, связывающая L-нуклеиновую кислоту, представляет собой ДНК.
В шестом варианте реализации десятого аспекта, который также является вариантом реализации первого, второго, третьего, четвертого и пятого вариантов реализации десятого аспекта, целевая молекула, связывающая молекулу L-нуклеиновой кислоты, состоит из L-нуклеотидов.
Проблема, лежащая в основе данной заявки, также решается в одиннадцатом аспекте, который также является первым вариантом реализации одиннадцатого аспекта, с помощью способа продуцирования белка в соответствии с любым вариантом реализации третьего, четвертого, пятого, шестого и седьмого аспектов, где
a) два или больше фрагментов белка в соответствии с любым вариантом реализации третьего, четвертого, пятого, шестого и седьмого аспектов, являются химически синтезированными, посредством чего фрагменты целиком образуют аминокислотную последовательность белка, предпочтительно, фрагменты синтезированы методом твердофазного пептидного синтеза, и
b) фрагменты стадии а) лигируют друг с другом путем сегментной конденсации, нативного химического лигирования, ферментативного лигирования или их комбинаций,
при том, что белок состоит из аминокислотной последовательности, состоящей из D-аминокислот.
Во втором варианте реализации одиннадцатого аспекта, который также является вариантом реализации первого варианта реализации одиннадцатого аспекта, ферментом, используемым в ферментативном лигировании, является Clostripain.
Проблема, лежащая в основе данной заявки, также решается в двенадцатом аспекте, который также является первым вариантом реализации двенадцатого аспекта, с помощью способа лигирования первого D-пептида или первого D-белка и второго D-пептида или второго D-белка друг с другом путем ферментативного лигирования, при котором
первый D-пептид или первый D-белок защищен на своем N-конце защитной группой, и защищен на его С-конце 4-гуанидинофенилэфирной группой, и
второй D-пептид или второй D-белок имеет свободный N-конец и содержит тиоалкилэфирную или тиоарилэфирную группу на своем С-конце.
Во втором варианте реализации двенадцатого аспекта, который также является вариантом реализации первого варианта реализации двенадцатого аспекта, ферментом, используемым при ферментативном лигировании, является Clostripain.
Авторы данного изобретения неожиданно обнаружили, что можно химически синтезировать белки, состоящие из D-аминокислот, являющиеся функционально активными, в результате чего такие белки имеют размер, типично демонстрируемый полимеразой. Более конкретно, авторы данного изобретения нашли способ, обеспечивающий возможность синтеза таких D-белков и D-полимераз, т.е. полимераз, состоящих из D-аминокислот, которые проявляют активность полимераз. На основании этого неожиданного результата становятся доступными белки и ферментативные активности, необходимые для ферментативного синтеза L-нуклеиновых кислот и молекул L-нуклеиновой кислоты. Такой ферментативный синтез L-нуклеиновых кислот и молекул L-нуклеиновой кислоты включает, без ограничений, способ добавления одного или нескольких L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты и способ амплификации целевой L-нуклеиновой кислоты в присутствии L-нуклеотидов как L-нуклеиновой кислоты, т.е. продукт амплификации является L-нуклеиновой кислотой.
Поскольку такие способы и ферментативные активности являются частью альтернативного способа идентификации шпигельмеров с их использованием, такой альтернативный способ идентификации шпигельмеров может быть теперь реализован на практике.
Авторы данного изобретения разработали способ добавления одного или нескольких L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты, который включает стадию проведения реакции одного или нескольких L-нуклеотидов с первой L-нуклеиновой кислотой в присутствии белка, содержащего фрагмент, проявляющий ферментативную активность, способную добавлять один или несколько L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты.
В предпочтительном варианте реализации ферментативная активность способна добавлять от 5 до 20000 L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты, предпочтительно, от 10 до 2000 L-нуклеотидов, более предпочтительно, от 50 до 500 L-нуклеотидов, наиболее предпочтительно, от 50 до 100 L-нуклеотидов.
Термин «добавление», в контексте данного изобретения, обозначает ковалентное связывание между молекулами, в соответствии с настоящим изобретением, ковалентное связывание L-нуклеиновой кислоты и по меньшей мере одного или нескольких L-нуклеотидов с L-нуклеиновой кислотой, предпочтительно, путем образования 3'-5' фосфодиэфирной связи между 3'ОН первой L-нуклеиновой кислоты и 5'-фосфатом одного из по меньшей мере одного или нескольких L-нуклеотидов. В соответствии с настоящим изобретением L-нуклеотид, добавленный к L-нуклеиновой кислоте, образует 3'-конец L-нуклеиновой кислоты, удлиненной указанным L-нуклеотид ом.
В предпочтительном варианте реализации способ добавления одного или нескольких L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты включает вторую L-нуклеиновую кислоту, при том, что одна молекула первой L-нуклеиновой кислоты гибридизуется с одной молекулой второй L-нуклеиновой кислоты, предпочтительно, путем спаривания оснований по Уотсону-Крику. В более предпочтительном варианте реализации способ позволяет синтезировать третью L-нуклеиновую кислоту, которая является комплементарной ко второй L-нуклеиновой кислоте, при том, что третья L-нуклеиновая кислота содержит первую L-нуклеиновую кислоту и L-нуклеотиды, добавленные к 3'-концу первой L-нуклеиновой кислоты, т.е. к первой L-нуклеиновой кислоте прибавляют один или несколько L-нуклеотидов к 3'-концу первой L-нуклеиновой кислоты с получением третьей L-нуклеиновой кислоты.
Белок, содержащий фрагмент, проявляющий ферментативную активность, в соответствии с настоящим изобретением включает белки, состоящие исключительно из фрагмента, проявляющего ферментативную активность, и белки, содержащие фрагмент, проявляющий ферментативную активность, и другие остатки или части, не обладающие ферментативной активностью. В соответствии с настоящим изобретением аминокислотная последовательность фрагмента, проявляющего ферментативную активность, включает от 300 до 900 аминокислот, предпочтительно, от 300 до 600 аминокислот, более предпочтительно, от 300 до 360 аминокислот, наиболее предпочтительно, от 340 до 360 аминокислот.
Белок, содержащий фрагмент, проявляющий ферментативную активность, в соответствии с настоящим изобретением, предпочтительно представляет собой фрагмент, проявляющий полимеразную активность. Белок, содержащий фрагмент, проявляющий полимеразную активность, в соответствии с настоящим изобретением, включает полимеразы, состоящие исключительно из фрагмента, проявляющего полимеразную активность, и полимеразы, содержащие фрагмент, проявляющий полимеразную активность, и другие остатки или части, не обладающие ферментативной активностью. В соответствии с настоящим изобретением аминокислотная последовательность фрагмента, проявляющего полимеразную активность, включает от 300 до 900 аминокислот, предпочтительно, от 300 до 600 аминокислот, более предпочтительно, от 300 до 360 аминокислот, наиболее предпочтительно, от 340 до 360 аминокислот.
Фрагмент, проявляющий полимеразную активность, в соответствии с настоящим изобретением предпочтительно является термостабильным фрагментом, проявляющим полимеразную активность, более предпочтительно, термостабильным ДНК-фрагментом, проявляющим полимеразную активность, и наиболее предпочтительно, термостабильны ДНК-зависимым ДНК-фрагментом, проявляющим полимеразную активность.
Фрагмент, проявляющий полимеразную активность, в соответствии с настоящим изобретением предпочтительно является ДНК-фрагментом, проявляющим полимеразную активность, более предпочтительно, ДНК-зависимым ДНК-фрагментом, проявляющим полимеразную активность, или термостабильным ДНК-фрагментом, проявляющим полимеразную активность, наиболее предпочтительно, термостабильным ДНК-зависимым ДНК-фрагментом, проявляющим полимеразную активность.
Термин «ферментативная активность», используемый в данном документе, обозначает катализ специфической реакции, предпочтительно, прибавления одного или нескольких нуклеотидов к 3'-концу нуклеиновой кислоты, амплификации нуклеиновой кислоты и/или полимеразной активности, более предпочтительно, прибавления одного или нескольких L-нуклеотидов к 3'-концу L-нуклеиновой кислоты и амплификации L-нуклеиновой кислоты.
Термин «полимеразная активность», в соответствии с настоящим изобретением, обозначает способность фермента к полимеризации L-нуклеотидов и/или полимеризации L-нуклеотидов с L-нуклеиновой кислотой, притом что, предпочтительно, L-нуклеотиды представляют собой L-нуклеозидтрифосфаты.
Полимеразная активность в соответствии с настоящим изобретением предпочтительно является активностью термостабильной полимеразы, более предпочтительно, термостабильной ДНК-полимеразной активностью, и наиболее предпочтительно, термостабильной ДНК-зависимой ДНК-полимеразной активностью.
Полимеразная активность в соответствии с настоящим изобретением предпочтительно является ДНК-полимеразной активностью, более предпочтительно, ДНК-зависимой ДНК-полимеразной или термостабильной ДНК-полимеразной активностью, наиболее предпочтительно, термостабильной ДНК-зависимой ДНК-полимеразной активностью.
Известные полимеразы получены из природных источников или представляют собой оптимизированные или мутированные варианты полимераз из природных источников. Полимеразы состоят из хиральных строительных блоков, т.е. L-аминокислот. Следовательно, структура полимераз по своему существу также является хиральной, что приводит к стереоспецифическому распознаванию субстратов. Поэтому, эти ферменты признают только молекулы субстратов в соответствующей хиральной конфигурации. Вследствие этого известные полимеразы полимеризуют D-нуклеотиды или D-нуклеозидтрифосфаты, с использованием в качестве матрицы нити D-нуклеиновой кислоты, состоящей из D-нуклеотидов, для синтеза нити комплементарной D-нуклеиновой кислоты, состоящей из D-нуклеотидов. Дополнительно к матричной нити, полимераза необязательно использует праймер, который гибридизуется с матричной нитью и состоит из D-нуклеотидов. Поскольку природные нуклеиновые кислоты состоят из D-нуклеотидов и могут быть процессированы, например, амплифицированы, белками и ферментами, в частности, состоящими из L-аминокислот, L-нуклеиновая кислота не распознается такими белками и ферментами, соответственно, состоящими из L-аминокислот. Соответственно, L-нуклеиновые кислоты, которые связываются с целевой молекулой или целевой структурой, также называемые шпигельмерами, не могут быть получены непосредственно с помощью in vitro процесса селекции с использованием природных вариантов такой целевой молекулы или целевой структуры.
Авторы данного изобретения неожиданно обнаружили, что возможно получить полимеразу, которая может прибавлять нуклеотид L-нуклеиновой кислоты к праймеру, состоящему из L-нуклеотидов, который гибридизуется с матричной нитью L-нуклеиновой кислоты. Более того, авторы данного изобретения неожиданно обнаружили, что можно получить полимеразу, которая может быть использована для амплификации L-нуклеиновой кислоты, предпочтительно, с помощью способа, известного как полимеразная цепная реакция (сокр. ПЦР).
Полимераза представляет собой фермент, который полимеризует нуклеозидтрифосфаты. Полимеразы используют нить матричной нуклеиновой кислоты для синтеза нити нуклеиновой кислоты, которая является комплементарной к нити матричной нуклеиновой кислоты. В дополнение к нити матричной нуклеиновой кислоты, полимераза необязательно использует праймер, который гибридизуется на основе комплементарности оснований с нитью матричной нуклеиновой кислоты. Нить матричной нуклеиновой кислоты, праймер и нить нуклеиновой кислоты, синтезированная полимеразой, могут независимо представлять собой ДНК или РНК. Полимераза, в значении, предпочтительно используемом в данном документе, включает ДНК-полимеразу и РНК-полимеразу, предпочтительно, ДНК-зависимую ДНК-полимеразу, РНК-зависимую ДНК-полимеразу, такую как обратная транскриптаза, РНК-зависимую РНК-полимеразу и РНК-зависимую ДНК-полимеразу. Более предпочтительно, полимераза представляет собой термостабильную полимеразу. Полимераза не должна обязательно содержать все аминокислоты, присутствующие в соответствующем ферменте нативного или дикого типа, а только те, которые являются достаточными для того, чтобы позволить полимеразе проявлять желательную каталитическую активность. В одном варианте реализации полимеразная активность представляет собой каталитическую активность, которую выбирают из группы, содержащей каталитические активности, включающие, например, 5-3'-полимеризацию, 5'-3'-экзонуклеазную и 3'-5'-экзонуклеазную активности.
Полимеразы в соответствии с настоящим изобретением состоят из D-аминокислот и полимеризуют L-нуклеотиды или L-нуклеозидтрифосфаты, притом что полимеразы в соответствии с настоящим изобретением используют в качестве матричной нити L-нуклеиновую кислоту, состоящую из L-нуклеотидов, для синтеза комплементарной L-нити нуклеиновой кислоты, состоящей из L-нуклеотидов. Дополнительно к матричной нити, полимеразы в соответствии с настоящим изобретением необязательно используют праймер, который гибридизуется с матричной нитью и состоит из L-нуклеотидов. Матричная нить, праймер и синтезированная нить нуклеиновой кислоты могут независимо быть L-ДНК или L-PHK. Полимеразы в соответствии с настоящим изобретением включают ДНК-полимеразы, состоящие из D-аминокислот, и РНК-полимеразы, состоящие из D-аминокислот, предпочтительно, ДНК-зависимые ДНК-полимеразы, состоящие из D-аминокислот, РНК-зависимые ДНК-полимеразы, такие как обратные транскриптазы, состоящие из D-аминокислот, РНК-зависимые РНК-полимеразы, состоящие из D-аминокислот, и РНК-зависимые ДНК-полимеразы, состоящие из D-аминокислот. Более предпочтительно, полимераза в соответствии с настоящим изобретением представляет собой термостабильную полимеразу, состоящую из D-аминокислот. Полимераза в соответствии с настоящим изобретением не должна обязательно содержать все аминокислоты, присутствующие в нативном ферменте, а только те, которые являются достаточными для того, чтобы позволить полимеразам в соответствии с настоящим изобретением проявлять желательную каталитическую активность. Каталитические активности включают, например, 5-3'-полимеризацию, 5'-3'-экзонуклеазную и 3-5'-экзонуклеазную активности.
Полимераза, состоящая исключительно из L-аминокислот, предпочтительно называется в данном документе «алло-L-полимераза».
Полимераза, состоящая исключительно из D-аминокислот, предпочтительно называется в данном документе «алло-D-полимераза».
В предпочтительном варианте реализации полимеразу в соответствии с настоящим изобретением выбирают из группы, состоящей из полимеразы X вируса африканской чумы свиней, корового домена полимеразы X Thermus thermophilus, крысиной полимеразы-бета, эукариотической полимеразы-бета, фрагмента Кленова, экзополимеразы Кленова, ДНК-полимеразы Т4, ДНК-полимеразы Phi29, секвеназы, ДНК-полимеразы Т7, полимеразы SP6, ДНК-полимеразы I, полимеразы-лямбда, полимеразы DPO4, ДНК-полимеразы Thermococcus litoralis, ДНК-полимеразы Pyrococcus sp., ДНК-полимеразы Pyrococcus furiosus, полимеразы Pfuturbo™, ДНК-полимеразы Sulfolobus solfataricus, ДНК-полимеразы Thermococcus gorgonarius, полимеразы KOD, полимеразы Taq, полимеразы Tth, полимеразы Pyrobest, полимеразы Pwo, полимеразы Sac, полимеразы Bst, полимеразы Рос, полимеразы Pab, полимеразы Mth, полимеразы Pho, полимеразы ES4, полимеразы EX-Taq™, полимеразы LA-Taq™, полимеразы Expand™, полимеразы Platinum™ Taq, полимеразы Hi-Fi™, полимеразы Tbr, полимеразы Til, полимеразы Тш, полимеразы Tac, полимеразы The, полимеразы Tma, полимеразы Tih, полимеразы Tfi, AmpliTaq™, фрагмента Стоффеля, ДНК-полимеразы 9°Nm™, Therminator™, Therminator II™, полимеразы Phusion High Fidelity™, Paq5000™, Pfx-50™, Proofstart™, FideliTaq™, Elongase™, и вариантов всех и любой из них.
В более предпочтительном варианте реализации, полимераза в соответствии с настоящим изобретением представляет собой полимеразу X вируса африканской чумы свиней, состоящую из аминокислотной последовательности, соответствующей SEQ ID NO: 1. В другом более предпочтительном варианте реализации, полимераза в соответствии с настоящим изобретением является вариантом полимеразы X вируса африканской чумы свиней, наиболее предпочтительно, вариантом полимеразы X вируса африканской чумы свиней с аминокислотной последовательностью, выбранной из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 2, аминокислотной последовательности, соответствующей SEQ ID NO: 3, и аминокислотной последовательности, соответствующей SEQ ID NO: 4.
В более предпочтительном варианте реализации, полимераза в соответствии с настоящим изобретением представляет собой полимеразу Dpo4, состоящую из аминокислотной последовательности, соответствующей SEQ ID NO: 15. В другом более предпочтительном варианте реализации, полимераза в соответствии с настоящим изобретением является вариантом полимеразы Dpo4, наиболее предпочтительно, вариантом полимеразы Dpo4, состоящим из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 16, аминокислотной последовательности, соответствующей SEQ ID NO: 17, аминокислотной последовательности, соответствующей SEQ ID NO: 18, аминокислотной последовательности, соответствующей SEQ ID NO: 19, аминокислотной последовательности, соответствующей SEQ ID NO: 20, аминокислотной последовательности, соответствующей SEQ ID NO: 21, и аминокислотной последовательности, соответствующей SEQ ID NO: 22.
Вариант полимеразы представляет собой полимеразу, которая отличается от аминокислотной последовательности полимеразы в одной или нескольких аминокислотных позициях. Позиция аминокислоты в аминокислотной последовательности предпочтительно определяется ее позицией по отношению к N-концу и С-концу полимеразы и/или ее позицией по отношению к аминокислотам, окружающим аминокислоту, так чтобы:
a) если полимераза укорочена на N-конце, то позиция аминокислоты определяется ее позицией по отношению к С-концу полимеразы и по отношению к аминокислотам, окружающим аминокислоту,
b) если полимераза укорочена на С-конце, то позиция аминокислоты определяется ее позицией по отношению к N-концу полимеразы и по отношению к аминокислотам, окружающим аминокислоту, и
b) если полимераза укорочена на N-конце и С-конце, то позиция аминокислоты определяется ее позицией по отношению к аминокислотам, окружающим аминокислоту.
Полимеразы в соответствии с настоящим изобретением, которые являются термостабильными, являются относительно устойчивыми к воздействию повышенных температур. В одном конкретном неограничивающем примере, полимераза со свойствами термостабильности является устойчивой к воздействию температуры, равной по меньшей мере 50°С, например, 50°С, 60°С, 75°С, 80°С, 82°С, 85°С, 88°С, 90°С, 92°С, 95°С, или еще более высоких температур.
В процессе полимеризации L-нуклеозидтрифосфатов полимераза прибавляет один нуклеозидтрифосфат к другому нуклеозидтрифосфату, предпочтительно, с образованием олигонуклеотида, также называемого нуклеиновой кислотой. В предпочтительном варианте реализации, полимераза прибавляет только один нуклеозидтрифосфат к одному нуклеозидтрифосфату или к терминальному нуклеотиду нуклеиновой кислоты, например, если нуклеотид представляет собой терминирующие нуклеотиды цепи, такие как дидезоксинуклеотид. Такие терминирующие нуклеотиды цепи используются для секвенирования нуклеиновых кислот и известны квалифицированным специалистам в данной области техники.
Процесс полимеризации L-нуклеозидтрифосфатов может быть использован для амплификации L-нуклеиновой кислоты, предпочтительно, целевой L-нуклеиновой кислоты.
Амплификация представляет собой любой процесс, увеличивающий число копий нуклеиновой кислоты, предпочтительно, целевой L-нуклеиновой кислоты.
В предпочтительном варианте реализации, целевая L-нуклеиновая кислота состоит из от 20 до 20000 L-нуклеотидов, предпочтительно, от 30 до 2000 L-нуклеотидов, более предпочтительно, от 40 до 500 L-нуклеотидов, наиболее предпочтительно, от 50 до 100 L-нуклеотидов.
Примером амплификации является процесс, в котором нуклеиновая кислота вводится в контакт с парой праймеров, в условиях, обеспечивающих возможность для гибридизации праймеров с матричной нуклеиновой кислотой. Праймеры удлиняются полимеразой путем прибавления одного или нескольких нуклеозидтрифосфатов к праймеру в пригодных условиях, диссоциации от матричной нуклеиновой кислоты, и затем повторного отжига, удлинения и диссоциации для амплификации числа копий молекул нуклеиновой кислоты. Продукт in vitro амплификации может быть охарактеризован путем электрофореза, с помощью методов гидролиза рестриктазами, гибридизации или лигирования олигонуклеотидов и/или секвенированием нуклеиновой кислоты, с использованием стандартных методик.
Альтернативно, квалифицированному специалисту в данной области техники известны методики in vitro амплификации, включая бестранскрипционную изотермическую амплификацию, амплификацию с перемещением цепи и бестранскрипционную амплификацию РНК NASBA™.
Некоторые методы амплификации основаны на термоциклировании, состоящем из циклов повторяющегося нагревания и охлаждения реакции для плавления нуклеиновой кислоты, предпочтительно, двухцепочечной нуклеиновой кислоты, и ферментативной репликации нуклеиновой кислоты. Такие стадии термоциклирования необходимы, во-первых, для физического разделения двух цепей двухцепочечной нуклеиновой кислоты при высокой температуре в результате процесса, называемого плавлением нуклеиновой кислоты. При более низкой температуре, каждая цепь затем используется в качестве матрицы при синтезе нуклеиновой кислоты полимеразой для селективной амплификации целевой нуклеиновой кислоты. Праймеры, содержащие последовательности, комплементарные к целевому участку наряду с полимеразой (которая дала название методу), являются ключевыми компонентами, обеспечивающими возможность селективной и повторной амплификации. Поскольку метод амплификации основан на процессах термоциклирования, образующаяся нуклеиновая кислота сама используется в качестве матрицы для репликации, запуская цепную реакцию, в которой матричная нуклеиновая кислота экспоненциально амплифицируется.
Наиболее известным методом амплификации, использующим термическую амплификацию, является полимеразная цепная реакция (сокр. ПЦР).
Праймеры представляют собой короткие молекулы нуклеиновой кислоты, состоящие из ДНК или РНК или их комбинаций, предпочтительно, ДНК-олигонуклеотиды длиной 10 нуклеотидов или больше. Более предпочтительно, более длинные праймеры могут иметь длину около 15, 20, или 25 нуклеотидов. Праймеры могут быть подвергнуты отжигу с комплементарной целевой нитью нуклеиновой кислоты путем гибридизации нуклеиновой кислоты для образования гибрида между праймером и целевой нитью нуклеиновой кислоты, и затем праймер удлиняется полимеразой вдоль нити нуклеиновой кислоты. Для амплификации нуклеиновой кислоты могут быть использованы пары праймеров, например, методом ПЦР или другими способами амплификации нуклеиновых кислот, известными специалистам.
Использование полимеразы по настоящему изобретению, состоящей из D-аминокислот, делает необходимым, чтобы праймер и комплементарная целевая нить нуклеиновой кислоты состояли из L-нуклеотидов. Предпочтительно, по меньшей мере один праймер состоит из L-нуклеотидов и, необязательно, модификации.
Способы получения и использования праймеров и зондов нуклеиновых кислот описаны, например, в Sambrook et al. (Sambrock et al., 1989). Пары ПЦР-праймеров могут быть получены из известных последовательностей, например, путем использования компьютерных программ, предназначенных для этой цели, таких как Primer. Рядовому специалисту в данной области техники будет понятно, что специфичность конкретного зонда или праймера увеличивается с его длиной.
Полимеразы в соответствии с настоящим изобретением состоят из D-аминокислот. Вследствие этого полимеразы в соответствии с настоящим изобретением, состоящие из D-аминокислот, не могут быть выделены из природных источников и не могут быть получены путем рекомбинантной экспрессии с использованием бактерий, дрожжей, грибов, вирусов или животных клеток, и их приходится получать химическим способом, предпочтительно таким, как твердофазный пептидный синтез (сокр. ТФПС) в комбинации со способами лигирования.
Твердофазный пептидный синтез является современной технологией синтеза пептидов или фрагментов белков: мелкие твердые бусины, нерастворимые, но пористые, обрабатывают функциональными фрагментами («линкерами»), на которых могут быть построены пептидные цепи. Пептид будет оставаться ковалентно присоединенным к бусине до отщепления от нее с помощью реагента, такого как безводный фтористый водород или трифторуксусная кислота. Пептид, таким образом, «иммобилизован» на твердой фазе и может удерживаться в процессе фильтрации, в то время как реагенты, находящиеся в жидкой фазе, и побочные продукты синтеза смываются. Общий принцип ТФПС заключается в повторяющихся циклах присоединение - промывка - снятие защиты - промывка. Свободный N-концевой амин присоединенного к твердой фазе пептида связывается (см. ниже) с единичным N-защищенным аминокислотным звеном. У этого звена затем удаляется защитная группа, высвобождая новый N-концевой амин, к которому может быть присоединена дополнительная аминокислота. Превосходство этой методики частично заключается в возможности проведения циклов промывки после каждой реакции, с удалением избытка реагента, в то время как весь растущий пептид, представляющий интерес, остается ковалентно присоединенным к нерастворимой смоле. Существуют две преимущественно используемые формы ТФПС - Fmoc и Boc. N-Концы аминокислотных мономеров защищают какой-либо из этих двух групп и присоединяют к аминокислотной цепи со снятой защитой. ТФПС имеет ограничения по выходу, и обычно пептиды и белки в пределах 70 аминокислот находятся на пределе возможностей синтеза. Сложность синтеза также зависит от последовательности. Синтетические олигопептиды и белки большего размера могут быть получены с использованием способов лигирования, таких как конденсация фрагментов, нативное химическое лигирование или ферментативное лигирование, для связывания двух пептидов друг с другом. Однако самым большим D-белком, синтезированным до настоящего времени, является D-белковая форма ангиогенного белка фактора роста сосудистого эндотелия (сокр. VEGF-A), состоящая из 102 D-аминокислот (Mandai et al., 2012),
Конденсация фрагментов использует пептиды, причем боковые цепи аминокислот пептида являются полностью защищенными химическими группами, и присоединение пептидов происходит в растворе.
Нативное химическое лигирование проводится в водном растворе. Проблемой является получение необходимых незащищенных пептид-тиоэфирных строительных блоков. При нативном химическом лигировании, тиолатная группа N-концевого цистеинового остатка незащищенного пептида 2 атакует С-концевой тиоэфир второго незащищенного пептида 1 в водном буфере при рН 7,0, 20°С<Т<37°С. Эта обратимая стадия переэтерификации является хемоселективной и региоселективной и приводит к образованию тиоэфирного промежуточного соединения 3. Это промежуточное соединение подвергается перегруппировке в результате внутримолекулярного S,N-ацильного переноса, что приводит к образованию нативной амидной («пептидной») связи 4 в сайте лигирования.
Как показано в примерах, авторы изобретения смогли неожиданно продемонстрировать, что С-концевой тиоэфир, необходимый для нативного химического лигирования, является стабильным в условиях ферментативного лигирования, так что нативное химическое лигирование и ферментативное лигирование могут быть использованы в комбинации.
Ферментативное лигирование D-пептидов работает за счет использования протеаз, и включает следующие стадии: (а) приготовление аминного компонента, в котором указанный аминный компонент является уникальным D-пептидом, (b) приготовление карбоксильного компонента, в котором указанный карбоксильный компонент включает отходящую группу и является уникальным D-пептидом, и (с) реакция аминного компонента и карбоксильного компонента в присутствии протеазы для образования пептидной связи между аминовым компонентом и карбоксильным компонентом с отщеплением отходящей группы с образованием уникального D-полипептида (см. WO 2003047743). Предпочтительно, протеаза представляет собой Clostripain.
Полимераза по настоящему изобретению также должна включать полимеразы, которая является по существу гомологичные полимеразе по настоящему изобретению и, в частности, конкретной последовательности (последовательностям), раскрытой (раскрытым) в данном документе. Термин «по существу гомологичный» следует понимать как гомологию, составляющую по меньшей мере 75%, предпочтительно, 85%, более предпочтительно, 90%, и наиболее предпочтительно, более 95%, 96%, 97%, 98% или 99%.
Фрагмент, проявляющий полимеразную активность, по настоящему изобретению должен также включать фрагменты, проявляющие полимеразную активность, по существу гомологичные фрагменту, проявляющему полимеразную активность, по настоящему изобретению и, в частности, конкретной последовательности (последовательностям), раскрытой (раскрытым) в данном документе. Термин «по существу гомологичный» следует понимать как гомологию, составляющую по меньшей мере 75%, предпочтительно, 85%, более предпочтительно, 90% и наиболее предпочтительно, более 95%, 96%, 97%, 98% или 99%.
Фактический процент гомологии аминокислот, присутствующих в полимеразе по настоящему изобретению или фрагменте, проявляющем полимеразную активность, по настоящему изобретению, будет зависеть от общего числа аминокислот, присутствующих в полимеразе или фрагменте, проявляющем полимеразную активность. Процент модификации может определяться по отношению к общему числу аминокислот, присутствующих в полимеразе или фрагменте, проявляющем полимеразную активность.
Гомология между двумя полимеразами или двумя фрагментами, проявляющими полимеразную активность, может быть определена, как известно квалифицированному специалисту в данной области техники. Более конкретно, алгоритм сравнения последовательностей может быть использован для расчета процента гомологии последовательностей для тестируемой последовательности (последовательностей) по сравнению с референсной последовательностью, на основании заданных параметров программы. Тестируемая последовательность, предпочтительно, является полимеразой или фрагментом, проявляющим полимеразную активность, которые считаются гомологичными или должны быть проверены на гомологичность, и в случае ее наличия, должна быть определена ее степень по отношению к другой полимеразе или фрагменту, проявляющему полимеразную активность, вследствие чего такая другая полимераза или фрагмент, проявляющий полимеразную активность, также называются референсной гомологической последовательностью. Оптимальное выравнивание аминокислотных последовательностей полимеразы для сравнения может быть проведено, например, по алгоритму локальной гомологии Смита-Уотермана (Smith & Waterman, 1981), по алгоритму выравнивания гомологии Нидлмена-Вунша (Needleman & Wunsch, 1970), по методу поиска подобия Пирсона-Липмана (Pearson & Lipman, 1988), с помощью компьютеризованных вариантов реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете прикладного программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis.), или путем визуального сравнения.
Одним из примеров алгоритма, пригодного для определения процента идентичности последовательностей, является алгоритм, используемый в средстве поиска основного локального выравнивания (Basic Local Alignment Search Tool, здесь и далее «BLAST»), см., например, Altschul et al. (Altschul et al. 1990, и Altschul et al., 1997). Программное обеспечение для проведения анализа BLAST является общедоступным через Национальный центр биотехнологической информации (National Center for Biotechnology Information, здесь и далее «NCBI»). Параметры по умолчанию, используемые для определения идентичности последовательностей, с использованием программного обеспечения, доступного от NCBI, например, BLASTN (для нуклеотидных последовательностей) и BLASTP (для аминокислотных последовательностей), описаны в McGinnis et al. (McGinnis et al., 2004).
Полимераза по настоящему изобретению должна также включать полимеразу, имеющую определенную степень идентичности по сравнению с полимеразой по настоящему изобретению и, в частности, с конкретной полимеразой по настоящему изобретению, раскрытую в данном документе и определенную с помощью ее аминокислотной последовательности. Более предпочтительно, настоящее изобретение также включает полимеразы, имеющие идентичность, равную по меньшей мере 75%, предпочтительно, 85%, более предпочтительно, 90%, и наиболее предпочтительно, более чем 95%, 96%, 97%, 98% или 99% по сравнению с полимеразой по настоящему изобретению, и в частности, с конкретной полимеразой по настоящему изобретению, раскрытые в данном документе и определенные с помощью их аминокислотных последовательностей или их части.
Фрагмент, проявляющий полимеразную активность, по настоящему изобретению, должен также включать фрагмент, проявляющий полимеразную активность, который имеет определенную степень идентичности по сравнению с фрагментом, проявляющим полимеразную активность, по настоящему изобретению, и в частности, с конкретным фрагментом, проявляющим полимеразную активность, по настоящему изобретению, раскрытый в данном документе и определенный с помощью его аминокислотной последовательности. Более предпочтительно, настоящее изобретение также включает такие фрагменты, проявляющие полимеразную активность, которые имеют идентичность, равную по меньшей мере 75%, предпочтительно, 85%, более предпочтительно, 90%, и наиболее предпочтительно, более чем 95%, 96%, 97%, 98% или 99%, по сравнению с фрагментом, проявляющим полимеразную активность, по настоящему изобретению, и в частности, с конкретным фрагментом, проявляющим полимеразную активность, по настоящему изобретению, раскрытые в данном документе и определенные с помощью их аминокислотных последовательностей или их части.
В связи с данной заявкой, термины «молекула нуклеиновой кислоты» и «нуклеиновая кислота» используются взаимозаменяемо, если явным образом не будет указано иное.
В предпочтительно используемом в данном документе значении, термины «нуклеиновая кислота» и «нуклеиновая кислота» относятся к полинуклеотидам или олигонуклеотидам, таким как дезоксирибонуклеиновая кислота (сокр. ДНК) и рибонуклеиновая кислота (сокр. РНК). Более того, термин «нуклеиновая кислота» включает множество нуклеиновых кислот. Термины «нуклеиновая кислота» и «нуклеиновые кислоты» следует также понимать как включающие, в качестве эквивалентов, варианты и аналоги РНК или ДНК, полученные из нуклеотидных аналогов, одно- (смысловых или антисмысловых) и двухцепочечных полинуклеотидов или олигонуклеотидов. Дезоксирибонуклеотиды включают дезоксиаденозин, дезоксицитидин, дезоксигуанозин и дезокситимидин. Рибонуклеотиды включают аденозин, цитидин, гуанозин и уридин. Указание на молекулу нуклеиновой кислоты как «полинуклеотид» используется в самом широком смысле для обозначения двух или больше нуклеотидов или нуклеотидных аналогов, соединенных ковалентной связью, включая одноцепочечные или двухцепочечные молекулы. Термин «олигонуклеотид» также используется в данном документе для обозначения двух или больше нуклеотидов или нуклеотидных аналогов, соединенных ковалентной связью, хотя, как определено в данном документе, олигонуклеотиды содержат менее ста нуклеотидов.
Нуклеиновая кислота характеризуется тем, что все последовательные нуклеотиды, образующие нуклеиновую кислоту, связаны или соединены друг с другом с помощью одной или более чем одной ковалентной связи. Более конкретно, каждый из таких нуклеотидов связан или соединен с двумя другими нуклеотидами, предпочтительно, с помощью фосфодиэфирных связей или других связей, образуя цепочку последовательно расположенных нуклеотидов. В такой конструкции, однако, два терминальных нуклеотида, т.е., предпочтительно, нуклеотиды на 5'-конце и на 3'-конце, связаны каждый только с одним нуклеотидом, при условии, что такая конструкция является линейной, а не кольцевой и, таким образом, образует линейную, а не кольцевую, молекулу.
В другом варианте реализации данной заявки нуклеиновая кислота включает по меньшей мере две группы последовательно расположенных нуклеотидов, таким образом, что в каждой группе последовательно расположенных нуклеотидов каждый нуклеотид связан или соединен с двумя другими нуклеотидами, предпочтительно, с помощью фосфодиэфирных связей или других связей, образуя цепочку последовательно расположенных нуклеотидов. В такой конструкции, однако, два терминальных нуклеотида, т.е., предпочтительно, нуклеотиды на 5'-конце и на 3'-конце, связаны каждый только с одним нуклеотидом. В таком варианте реализации две группы последовательно расположенных нуклеотидов, однако, не связаны или не соединены друг с другом с помощью ковалентной связи, которая соединяет один нуклеотид одной группы и один нуклеотид другой или иной группы, предпочтительно, ковалентной связи, образованной между сахарным фрагментом одного из указанных двух нуклеотидов и фосфорным фрагментом другого из указанных двух нуклеотидов или нуклеозидов. В альтернативном варианте реализации две группы последовательно соединенных нуклеотидов, однако, связаны или соединены друг с другом с помощью ковалентной связи, которая связывает один нуклеотид одной группы и один нуклеотид другой или иной группы, предпочтительно, ковалентной связи, образованной между сахарным фрагментом одного из указанных двух нуклеотидов и фосфорным фрагментом другого из указанных двух нуклеотидов или нуклеозидов. Предпочтительно, по меньшей мере две группы последовательно соединенных нуклеотидов не связаны с помощью каких-либо ковалентных связей. В другом предпочтительном варианте реализации по меньшей мере две группы соединены с помощью ковалентной связи, которая отличается от фосфодиэфирной связи.
Термин «нуклеиновая кислота», предпочтительно, также охватывает как D-нуклеиновую кислоту, так и L-нуклеиновую кислоту. Предпочтительно, нуклеиновая кислота является L-нуклеиновой кислотой. Дополнительно возможно, чтобы одна или несколько частей нуклеиновой кислоты присутствовали в виде D-нуклеиновой кислоты, и по меньшей мере одна или несколько частей нуклеиновой кислоты представляли собой L-нуклеиновую кислоту. Термин «часть» нуклеиновой кислоты может обозначать всего лишь один нуклеотид. Такая нуклеиновая кислота в общем называется в данном документе D- и L-нуклеиновой кислотой, соответственно. Таким образом, в предпочтительном варианте реализации, нуклеиновая кислота в соответствии с настоящим изобретением состоит из L-нуклеотидов и содержит по меньшей мере один D-нуклеотид. Предпочтительно, такой D-нуклеотид присоединен на конце любой из цепей и любой нуклеиновой кислоты.
L-Нуклеиновая кислота, используемая в данном документе, представляет собой нуклеиновую кислоту, состоящую из L-нуклеотидов, предпочтительно, состоящую исключительно из L-нуклеотидов.
D-Нуклеиновая кислота, используемая в данном документе, представляет собой нуклеиновую кислоту, состоящую из D-нуклеотидов, предпочтительно, состоящую исключительно из D-нуклеотидов.
Также, если не указано иное, любая нуклеотидная последовательность описывается в данном документе в направлении 5'→3'.
Независимо от того, состоит или нет нуклеиновая кислота из D-нуклеотидов, L-нуклеотидов или комбинации обоих, где комбинация является, например, статистической комбинацией или упорядоченной последовательностью отрезков, состоящих из по меньшей мере одного L-нуклеотида и по меньшей мере одной D-нуклеиновой кислоты, молекула нуклеиновой кислоты может состоять из дезоксирибонуклеотида (дезоксирибонуклеотидов), рибонуклеотида (рибонуклеотидов) или их комбинаций.
Независимо от того, является ли нуклеиновая кислота D-нуклеиновой кислотой, L-нуклеиновой кислотой, их смесью, ДНК или РНК, или всеми без исключения их комбинациями, термин «нуклеиновая кислота», в значении, предпочтительно используемом в данном документе, должен также охватывать одноцепочечную нуклеиновую кислоту и двухцепочечную нуклеиновую кислоту, причем, предпочтительно, молекула нуклеиновой кислоты, используемая в способе в соответствии с настоящим изобретением, представляет собой одноцепочечную нуклеиновую кислоту.
Термин «нуклеиновая кислота» в значении, предпочтительно используемом в данном документе, должен также охватывать модифицированную нуклеиновую кислоту. Модифицированная нуклеиновая кислота может быть молекулой нуклеотид-модифицированной РНК или нуклеотид-модифицированной ДНК, причем молекулы РНК или ДНК являются в значительной степени модифицированными по индивидуальным нуклеотидам для повышения стабильности путем модификации нуклеаза-резистентными группами, например, 2'-амино, 2'-С-аллил, 2'-фтор, 2'-O-метил, 2'-Н (см. обзор Usman & Cedergren, 1992).
Термин «нуклеиновая кислота», в значении, предпочтительно используемом в данном документе, должен также охватывать полностью закрытую нуклеиновую кислоту. Полностью закрытая, т.е. кольцевая структура нуклеиновой кислоты реализуется в случае, когда нуклеиновая кислота, нуклеотидная последовательность которой должна быть определена в соответствии с настоящим изобретением, замыкается, предпочтительно, посредством ковалентной связи, вследствие чего, более предпочтительно, такая ковалентная связь образуется между 5'-концом и 3'-концом последовательностей молекул нуклеиновой кислоты, раскрытых в данном документе.
Термин «нуклеиновая кислота», в предпочтительно используемом значении, должен также охватывать любые молекулы нуклеиновой кислоты, которые включают фрагмент, не являющийся молекулой нуклеиновой кислоты. Такой фрагмент, не являющийся молекулой нуклеиновой кислоты, может быть выбран из группы, включающей пептиды, олигопептиды, полипептиды, белки, углеводы, различные группы, как будет описано более подробно далее. Термин «нуклеиновая кислота», таким образом, должен также охватывать конъюгаты и/или комплексы, содержащие по меньшей мере один фрагмент нуклеиновой кислоты и по меньшей мере один дополнительный фрагмент, который может быть использован для содействия доставке молекул нуклеиновой кислоты в биологическую систему, такую как клетка. Предусматриваемые конъюгаты и комплексы могут придавать терапевтическую активность путем перемещения терапевтических соединений через клеточные мембраны, изменения фармакокинетических характеристик и/или модулирования локализации нуклеиновой кислоты по изобретению. Такие виды конъюгатов и комплексов, предпочтительно, пригодные для доставки молекул через клеточные мембраны, включают, без ограничений, малые молекулы, липиды, фосфолипиды, нуклеозиды, нуклеотиды, нуклеиновые кислоты, антитела, токсины, отрицательно заряженные полимеры и другие полимеры, например, белки, пептиды, гормоны, углеводы, полиэтиленгликоли или полиамины. В общем, описанные переносчики предназначены для использования индивидуально или в составе многокомпонентной системы, со способными к разложению линкерами или без них. Ожидается, что такие соединения будут улучшать доставку и/или локализацию молекул нуклеиновой кислоты в ряде клеточных типов, полученных из разных тканей, в присутствии или отсутствии сыворотки (см. патент США US 5854038). Конъюгаты молекул, описанные в данном документе, могут быть присоединены к биологически активным молекулам через биоразлагаемые линкеры, такие как линкерные молекулы биоразлагаемой нуклеиновой кислоты.
Как будет детальнее описано ниже в связи с нуклеиновой кислотой, последовательность которой должна быть определена, фрагмент, не являющийся нуклеиновой кислотой, может быть ПЭГ-фрагментом, т.е. полиэтиленгликольным фрагментом, или ГЭК-фрагментом, т.е. гидроксиэтилкрахмальным фрагментом.
Фрагмент, не являющийся нуклеиновой кислотой, и, предпочтительно, ПЭГ- и/или ГЭК-фрагмент могут быть присоединены к молекуле нуклеиновой кислоты непосредственно или через линкер. Настоящее изобретение также предусматривает, что молекула нуклеиновой кислоты включает одну или несколько модификаций, предпочтительно, один или несколько ПЭГ- и/или ГЭК-фрагментов. В варианте реализации индивидуальная линкерная молекула присоединяет несколько ПЭГ-фрагментов или ГЭК-фрагментов к молекуле нуклеиновой кислоты. Линкер, используемый в связи с настоящим изобретением, сам по себе может быть линейным или разветвленным. Такие виды линкеров известны квалифицированным специалистам в данной области техники и дополнительно описаны в патентных заявках WO 2005/074993 и WO 2003/035665.
В предпочтительном варианте реализации линкер представляет собой биоразлагаемый линкер. Биоразлагаемый линкер позволяет модифицировать характеристики молекул нуклеиновой кислоты по показателям, в числе прочего, времени пребывания в организме животного, предпочтительно, в организме человека, вследствие удаления модификации из молекул нуклеиновой кислоты. Использование биоразлагаемого линкера может обеспечить возможность лучшего контроля времени пребывания молекул нуклеиновой кислоты. Предпочтительным вариантом реализации таких биоразлагаемых линкеров являются такие биоразлагаемые линкеры, как описанные в международных патентных заявках WO 2006/052790, WO 2008/034122, WO 2004/092191 и WO 2005/099768, без ограничения ими, причем в международных патентных заявках WO 2004/092191 и WO 2005/099768 линкер является частью полимерного олигонуклеотидного пролекарства, которое состоит из одной или двух модификаций, описанных в данном документе, молекулы нуклеиновой кислоты и биоразлагаемого линкера между ними.
В предпочтительно используемом в данном документе значении, «нуклеотиды» включают, без ограничений, природные моно-, ди- и трифосфаты ДНК-нуклеозидов: моно-, ди- и трифосфат дезоксиаденозина; моно-, ди- и трифосфат дезоксигуанозина; моно-, ди- и трифосфат дезокситимидина; и моно-, ди- и трифосфат дезоксицитидина (обозначаемые в данном документе dA, dG, dT и dC, или A, G, Τ и С, соответственно). Термин «нуклеотиды» также включает природные моно-, ди- и трифосфаты РНК-нуклеозидов: моно-, ди- и трифосфат аденозина; моно-, ди- и трифосфат гуанина; моно-, ди- и трифосфат уридина; и моно-, ди- и трифосфат цитидина (обозначаемые в данном документе A, G, U и С, соответственно), относящиеся к комбинации основание-сахар-фосфат, являющейся мономерным звеном молекулы нуклеиновой кислоты, т.е. молекулы ДНК и молекулы РНК. Однако, другими словами, термин «нуклеотиды» относится к любому соединению, содержащему циклический сахар фуранозидного типа (p-D/L-рибоза в РНК и Р-D/L-2'-дезоксирибоза в ДНК), который фосфорилирован в 5'-позиции и имеет пуриновое или пиримидиновое основание, присоединенное в С-1' позиции сахара посредством - гликозил C1'-N связи. Нуклеотиды могут быть природными или синтетическими, включая нуклеотид, подвергнутый массовой модификации, включая, в числе прочих, нуклеотиды, содержащие модифицированные нуклеозиды с модифицированными основаниями (например, 5-метилцитозин) и модифицированными группами сахара (например, 2'-O-метилрибозил, 2'-O-метоксиэтилрибозил, 2'-фторрибозил, 2'-аминорибозил и т.п.).
Термин «гетероциклическое основание нуклеиновых кислот» (nucleobase) охватывает природные гетероциклические основания нуклеиновых кислот аденин (А), гуанин (G), цитозин (С), тимин (Т) и урацил (U), а также неприродные гетероциклические основания нуклеиновых кислот, такие как ксантин, диаминопурин, 8-оксо-М6-метиладенин, 7-деазаксантин, 7-деазагуанин, N4,N4-этаноцитозин, N6,N6-этано-2,6-диаминопурин, 5-метилцитозин, 5~(С3-С6)-алкинилцитозин, 5-фторурацил, 5-бромурацил, псевдоизоцитозин, 2-гидрокси-5-метил-4-триазолопиридин, изоцитозин, изогуанин, инозин и неприродные гетероциклические основания нуклеиновых кислот, описанные в патенте США US 5432272, в публикации Freier & Altmann (Freier & Altmann, 1997). Термин «гетероциклическое основание нуклеиновых кислот», таким образом, включает не только известные пуриновые и пиримидиновые гетероциклы, но также их гетероциклические аналоги и таутомеры.
Настоящее изобретение предусматривает, что одноцепочечная нуклеиновая кислота может образовывать четко определенные и стабильные трехмерные структуры и специфически связываться с целевой молекулой, такой как антитела.
Такие молекулы нуклеиновой кислоты, состоящие из D-нуклеотидов, называются аптамерами. Аптамеры могут быть идентифицированы по отношению к нескольким целевым молекулам, например, малым молекулам, белкам, нуклеиновым кислотам и даже клеткам, тканям и организмам, и могут ингибировать in vitro и/или in vivo функцию конкретной целевой молекулы. Аптамеры обычно идентифицируют путем мишень-направленного процесса селекции, называемого in vitro селекцией или систематической эволюцией лигандов экспоненциальным обогащением (сокр. SELEX) (Bock et al, 1992; Ellington & Szostak, 1990; Tuerk & Gold, 1990). Немодифицированные аптамеры быстро выводятся из кровотока, со временем полувыведения от минут до часов, преимущественно, вследствие нуклеазной деградации и клиренса из организма почками, в результате характерного для аптамеров низкого молекулярного веса. Таким образом, для терапевтического применения аптамеров, они должны быть модифицированы в позиции 2' сахарного (например, рибозного) скелета (Burmester et al., 2006).
Повсеместно распространенные нуклеазы, ответственные за нестабильность аптамеров, состоят из хиральных строительных блоков, т.е. L-аминокислот. Следовательно, структура нуклеаз также по своему существу является хиральной, приводя к стереоспецифическому распознаванию субстратов. Вследствие этого, такие ферменты принимают только молекулы субстратов в соответствующей хиральной конфигурации. Поскольку аптамеры и природные молекулы нуклеиновой кислоты состоят из D-нуклеотидов, L-олигонуклеотид должен избегать ферментативного узнавания и последующей деградации. По этому же принципу, к сожалению, в данном случае, природа не предусмотрела ферментативной активности для амплификации таких зеркально отраженных нуклеиновых кислот. Соответственно, аптамеры L-нуклеиновой кислоты не могут быть непосредственно получены с помощью процесса SELEX. Принципы стереохимии, однако, указывают на существование обходного пути, который, в конечном счете, приводит к желательным функциональным аптамерам L-нуклеиновой кислоты.
Если in vitro выделенный (О-)аптамер связывается со своей природной мишенью, структурное зеркальное отображение этого аптамера связывается с имеющим такие же характеристики зеркальным отображением природной мишени. При этом оба партнера взаимодействия имеют одинаковую (неприродную) хиральность. Вследствие гомохиральности жизни и большинства биохимических соединений, такие энантио-РНК лиганды, конечно, будут находить ограниченное практическое применение. Если, с другой стороны, процесс SELEX проводить с использованием (неприродной) зеркально отображенной мишени, то будет получен аптамер, распознающий такую (неприродную) мишень. Соответствующее зеркальное отображение конфигурации указанного аптамера - желательный L-аптамер - в свою очередь распознает природную мишень. Этот процесс селекции зеркального отображения для получения биостабильной молекулы нуклеиновой кислоты был опубликован впервые в 1996 г. (Klussmann et al., 1996; Nolte et al., 1996) и приводит к получению функциональных лигандов зеркального отображения молекулы нуклеиновой кислоты, которые проявляют не только высокую аффинность и специфичность к данной молекуле-мишени, но и, в то же время, также биологическую стабильность. Настоящее изобретение предусматривает, что одноцепочечная молекула нуклеиновой кислоты является такой лиганд-связывающей L-нуклеиновой кислотой, которая называется «шпигельмером» (от немецкого слова «Spiegel», т.е. зеркало) (см. 'The Aptamer Handbook'; eds. Klussmann, 2006).
В числе прочего, нуклеиновые кислоты в соответствии с настоящим изобретением могут содержать модификацию, которая предпочтительно обеспечивает возможность детектирования нуклеиновых кислот в соответствии с настоящим изобретением. Такая модификация, предпочтительно, выбрана из группы, включающей радиоактивные, ферментативные и флуоресцентные метки. Такая модификация также выбрана из D-нуклеотидов, которые сами по себе могут быть модифицированы посредством модификации, выбранной из группы, включающей радиоактивные, ферментативные и флуоресцентные метки.
Различные SEQ ID Nos., химическая природа нуклеиновых кислот, пептидов, олигопептидов и белков, используемых в данном документе, их фактические последовательности и внутренние ссылочные номера, приведены в следующей таблице.
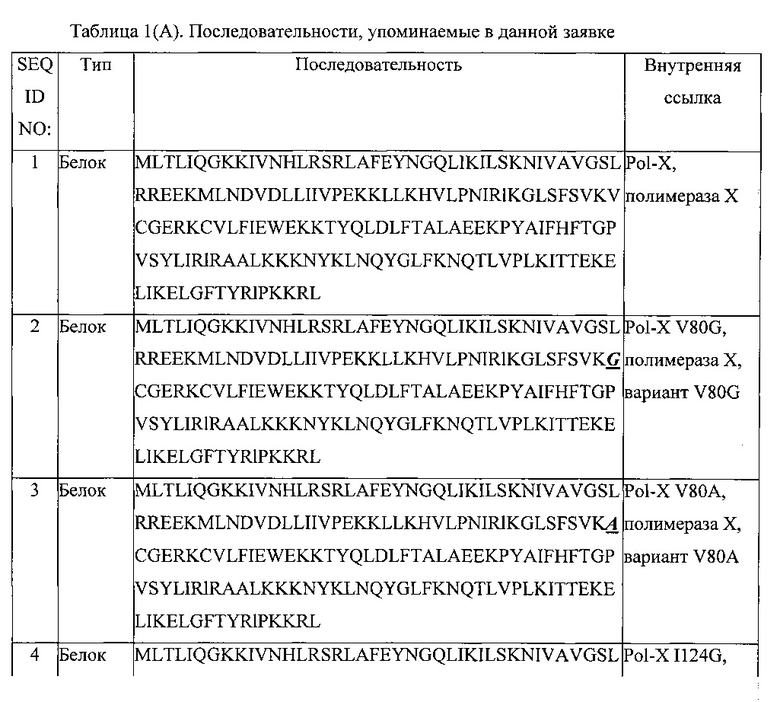
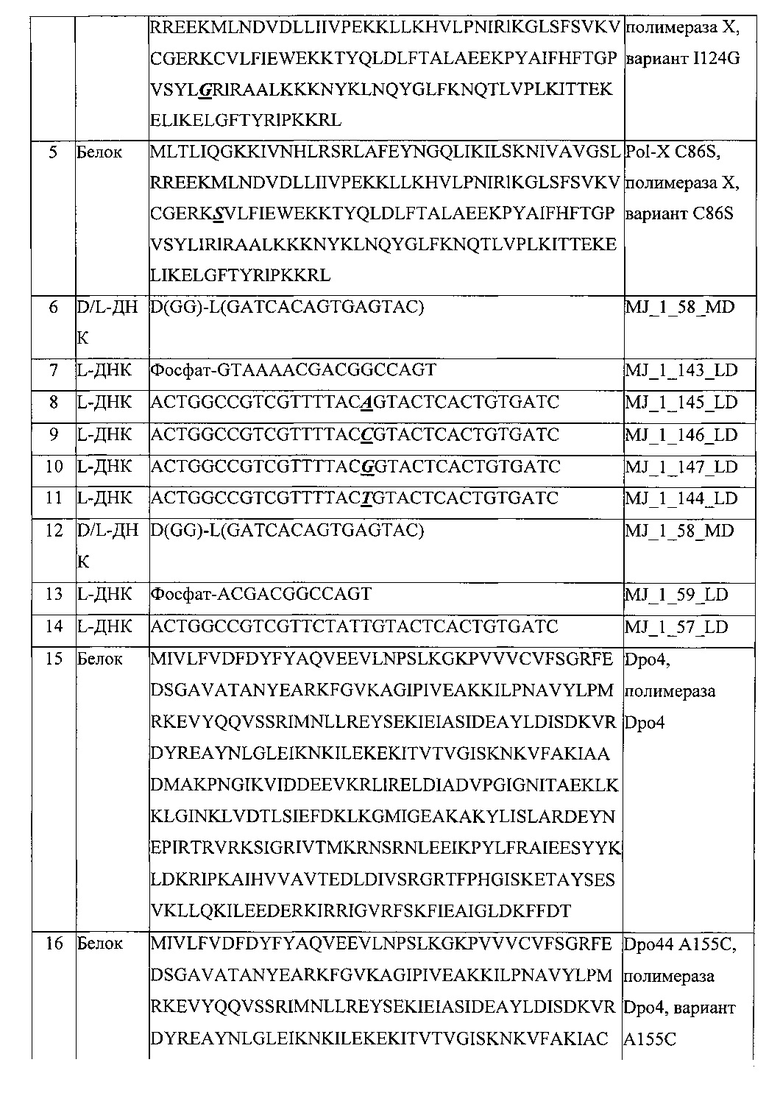





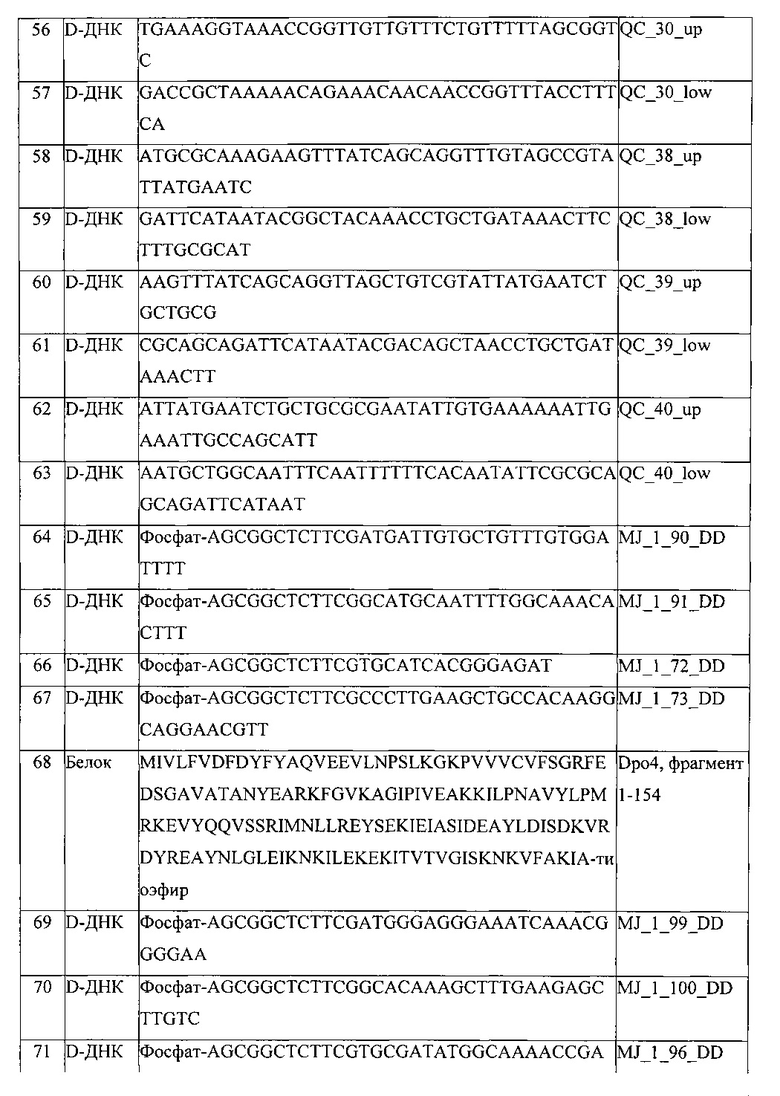


Следует понимать, что выше представлены молекулы, как они были использованы в связи с настоящим изобретением. Приложенные перечни последовательностей отображают только их простые аминокислотные или нуклеотидные последовательности без каких-либо дополнительных признаков указанных молекул, указанных в вышеприведенной таблице.
Настоящее изобретение дополнительно проиллюстрировано фигурами, примерами и перечнями последовательностей, из которых можно уяснить дополнительные признаки, варианты реализации и преимущества, притом что
Фиг. 1А иллюстрирует состав D-ДНК матрицы с 1 пробелом для испытаний на активность L-полимеразы X;
Фиг. 1 В иллюстрирует состав D-ДНК матрицы с 6 пробелами для испытаний на активность L-полимеразы X;
Фиг. 2А-В иллюстрирует результаты анализов синтезированного D-полипептидного продукта Ac-MLTLIQGKKIVNHLRSRLAFEYNGQLIKILSKNIVAVGSL-OGp (1) методами ультраэффективной жидкостной хроматографии (УЭЖХ) (А) и масс-спектрометрии (В);
Фиг. 3А-В иллюстрирует результаты анализов синтезированного D-полипептидного продукта H-RREEKLNDVDLLIIVPEKKLLKHVLPNIRIKGLSFSVKA-SMe (2) методами УЭЖХ (А) и масс-спектрометрии (В);
Фиг. 4А-В иллюстрирует результаты анализов синтезированного D-полипептидного продукта H-CGERKCVLFIEWEKKTYQLDLFTALAEEKPYAIFHFTGP VSYLIRIRAALKKKNYKLNQYGLFKNQTLVPLKITTEKELIKELGFTYRIPKKRL-OH (3) методами УЭЖХ (А) и масс-спектрометрии (В);
Фиг. 5А-В иллюстрирует результаты анализов синтезированного D-полипептидного продукта Ac-MLTLIQGKKIVNHLRSRLAFEYNGQLIKILSKNIVAVGS LRREEKMLNDVDLLIIVPEKKLLKHVLPNIRIKGLSFSVKA-SMe (4) методами УЭЖХ (А) и масс-спектрометрии (В);
Фиг. 6А-В иллюстрирует результаты анализов нативного химического лигирования D-полипептидного продукта Ac-MLTLIQGKKIVNHLRSRLAFEYNGQLIKILSKNIVAVG
SLRREEKMLNDVDLLIIVPEKKLLKHVLPNIRIKGLSFSVKACGERKCVLFIEW
EKKTYQLDLFTALAEEKPYAIFHFTGPVSYLIRIRAALKKKNYKLNQYGLF
KNQTLVPLKITTEKELIKELGFTYRIPKKRL-OH (5) методами ДСН-ПААГ (А) и масс-спектрометрии (В);
Фиг. 7 иллюстрирует состав L-ДНК матриц с 1 пробелом для испытаний на активность D-полимеразы X;
Фиг. 8 иллюстрирует гель-электрофорез анализа активности удлинения L-ДНК D-полимеразой X на субстратах с 1 пробелом;
Фиг. 9А иллюстрирует состав L-ДНК матрицы с 6 пробелами для испытаний на активность D-полимеразы X;
Фиг. 9В иллюстрирует гель-электрофорез анализа активности удлинения L-ДНК D-полимеразой X на субстрате с 6 пробелами;
Фиг. 10А иллюстрирует субстрат D-ДНК комплекса праймер-матрица для анализа активности L-полимеразы X;
Фиг. 10В иллюстрирует гель-электрофорез анализа активности удлинения D-ДНК L-полимеразой X, проводившегося при постоянной температуре;
Фиг. 10С иллюстрирует гель-электрофорез анализа активности удлинения D-ДНК L-полимеразой X, проводившегося с использованием термоциклирования;
Фиг. 11А-В иллюстрирует результаты анализов синтетической алло-L-полимеразы dpo4, вариант A155C, методами ДСН-ПААГ (А) и жидкостной хроматомасс-спектрометрии с электрораспылительной ионизацией ЖК/МС (ЭРИ) (В);
Фиг. 12А иллюстрирует гель-электрофорез D-ДНК анализов ПЦР-активности L-полимеразы dpo4, варианты A155C, V203C, C31S и A155C/V203C;
Фиг. 12 В иллюстрирует гель-электрофорез D-ДНК анализов ПЦР-активности рекомбинантной и синтетической L-полимеразы dpo4;
Фиг. 13 иллюстрирует результаты анализов синтезированного D-полипептидного продукта
H-RTFPHGISKETA YSES VKLLQKILEEDERKIRRIGVRFSKFIEAIGLDKFFDT-NH2 (1) методом масс-спектрометрии;
Фиг. 14 иллюстрирует результаты анализов синтезированного D-полипептидного продукта
Boc-VDTLSIEFDKLKGMIGEAKAKYLISLARDEYNEPIRTRVRKSIGRIVTMKRNSRNLEE IKPYLFRAIEESYYKLDKRIPKAIHVVAVTEDLDIVSRG-OH (2) методом масс-спектрометрии;
Фиг. 15 иллюстрирует результаты анализов конденсации фрагментов D-полипептидного продукта H-VDTLSIEFDKLKGMIGEAKAKYLISLARDEYNEPIRTRVRKSIGRIVTM
KRNSRNLEEIKPYLFRAIEESYYKLDKRIPKAIHVVAVTEDLDIVSRGRTFPHG
ISKETAYSESVKLLQKILEEDERKIRRIGVRFSKFIEAIGLDKFFDT-NH2 (3) методом масс-спектрометрии;
Фиг. 16А-В иллюстрирует результаты анализов синтезированного D-полипептидного продукта Z-CDMAKPNGIKVIDDEEVKRLlRELDIADVPGIGNITAEKLKKLGINKL-бензил-тиоэфир (4) методами ОФ-ВЭЖХ (А) и масс-спектрометрии (В);
Фиг. 17А-В иллюстрирует результаты анализов синтезированного D-полипептидного продукта H-RKEVYQQVSSRIMNLLREYSEKIEIASIDEAYLDISDKVRDYREAYNLGLEIKNKILEKEKITVTVGISKNKVFAKIA-SMe (7) методами УЭЖХ (А) и масс-спектрометрии (В);
Фиг. 18А-В показывает результаты анализов синтезированного D-полипептидного продукта
Ac-MIVLFVDFDYFYAQVEEVLNPSLKGKPVVVCVFSGRFEDSGAVATANY
EARKFGVKAGIPIVEAKKILPNAVYLPM-OGp (6) методами УЭЖХ (А) и масс-спектрометрии (В);
Фиг. 19А-В иллюстрирует результаты анализов Clostripain-медиированного D-полипептидного продукта лигирования Ac-MIVLFVDFDYFYAQVEEVLNPSLKGKPVVVCVFSGRFEDSGAVATANYEARKFGVK AGIPIVEAKKILPNAVYLPMRKEVYQQVSSRIMNLLREYSEKIEIASIDEAYLDISDKVRD YREAYNLGLEIKNKILEKEKITVTVGISKNKVFAKIA-SMe (8) методами ДСН-ПААГ (А) и масс-спектрометрии с электрораспылительной ионизацией (В);
Фиг. 20А-В иллюстрирует результаты анализов продукта нативного химического лигирования алло-L-полимеразы dpo4, фрагмент 155-352 (V203C), методами ДСН-ПААГ (А) и жидкостной хроматомасс-спектрометрии с электрораспылительной ионизацией ЖК/МС (ЭРИ) (В).
Примеры
Аббревиатуры, используемые в примерах.
ACN - ацетонитрил (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
DCM - дихлорметан (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
DIPEA - Ν,Ν-диизопропиламин (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
EDT (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
Fmoc - 9-флуоренилметоксикарбонил
HATU - (2-(7-аза-1Н-бензотриазол-1-ил)-1,1,3,3-тетраметилурония гексафторфосфат) (CreoSalus, Louisville KY, USA)
HFIP -1,1,1,3,3,3-гексафторфосфат (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
HPLC - высокоэффективная жидкостная хроматография (ВЭЖХ) (иногда называется жидкостной хроматографией высокого давления)
MeIm - метилимидазол (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
MeOH - метанол (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
MSNT - 1-(мезитилен-2-сульфонил)-3-нитро-1,2,4-триазол (Merck KGaA, Darmstadt, Germany)
NMP - N-метилпирролидон (Iris Biotech GmbH, Marktredwitz, Deutschland)
PyBOP - (бензотриазол-1-илокси)трипирролидинофосфония гексафторфосфат (MERCK KGAA, DARMSTADT, GERMANY)
SDS - додецилсульфат натрия (ДСН) (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
TBTU O-(бензотриазол-1-ил)-N,N,N',N'-тетраметилурония тетрафторборат (Merck KGaA, Darmstadt, Germany)
tBu - (трет-бутил-)
TFA - трифторуксусная кислота (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
TFE - 1,1,1-трифторэтанол (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
THF (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
TIS (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
TLC - тонкослойная хроматография (TCX)
Tris - трис(гидроксиметил)аминометан (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland)
UPLC - ультраэффективная жидкостная хроматография (УЭЖХ).
Пример 1 - Рекомбинантная экспрессия и очистка дикого типа и вариантов полимеразы X
Полимераза X вируса африканской чумы свиней (сокр. ASFV) была описана и охарактеризована Oliveros et al. в 1997. Ген дикого типа полимеразы X имеет открытую рамку считывания (сокр. ORF) всего 525 пар оснований, включая инициирующий кодон и терминирующий кодон (Oliveros et al., 1997). Кодированный белок имеет длину всего 174 аминокислоты. Этот пример описывает, как полимераза X, а также ее варианты, была экспрессирована в E.coli и очищена с использованием His6-метки.
1.1 Экспрессионные конструкты
Поскольку частота использования кодонов ASFV отличается от E.coli, синтетический ген полимеразы X с оптимизированной для E.coli частотой использования кодонов был приобретен у фирмы Gene Art AG (Regensburg, Germany). Синтетическая генная последовательность была введена в вектор pCR4-Blunt-TOPO (компания-разработчик: Invitrogen, Karlsruhe, Germany). Кодон-оптимизированная открытая рамка считывания, включающая инициирующий кодон и два терминирующих кодона, имела следующую последовательность:

Для получения экспрессионного конструкта полимеразы X, также называемого алло-L-полимераза X, ген полимеразы X вырезали из pCR4-Blunt-TOPO с помощью BamHI и PstI и субклонировали в вектор pRSET-A (Invitrogen, Karlsruhe, Germany). Субклонирование добавляло His6-метку на N-конце, и ставило ген под контроль промотора Т7. Конструкт был назван pMJ14 и использовался для экспрессии алло-L-полимеразы X в E.coli. Белок полимеразы X, экспрессированный из pMJI4, имел следующую последовательность из 210 аминокислот:
MRGSHHHHHHGMASMTGGQQMGRDLYDDDDKDRWGSMLTLIQGKKIVNHLRSRLAFEYNGQLIKILSKNIVAVGSLRREEKMLNDVDLLIIVPEKKLLKHVLPNIRIKGLSFSVKVCGERKCVLFIEWEKKTYQLDLFTALAEEKPYAIFHFTGPVSYLIR1RAALKKKNYKLNQYGLFKNQTLVPLKITTEKELIKELGFTYRIPKKRL.
Начальные 36 аминокислот представляли His6-метку, включая несколько спейсерных аминокислот и некоторые другие метки последовательности (лидерный пептид Т7 гена 10, анти-Express-эпитоп). Концевой фрагмент из 174 аминокислот был идентичным белковой последовательности полимеразы X, присутствующей в ASFV.
Экспрессионные конструкты для вариантов алло-L-полимеразы X были приготовлены с использованием коммерчески доступного набора QuikChange (Stratagene GmbH, Waldbronn, Germany) в соответствии с протоколом производителя. Плазмида pMJ14 была использована в качестве матрицы. Олигонуклеотиды, необходимые для QuikChange, были или синтезированы на оборудовании NOXXON (QC10_up, QC10_low), или приобретены у фирмы Purimex (Grebenstein, Germany) (QC26_up, QC26_low, QC27_up, QC27_low, QC31_up, QC31_low). Следующие варианты экспрессионных конструктов были приготовлены и использованы для экспрессии вариантов алло-L-полимеразы в E.coli:

1.2 Экспрессия белка в E.coli
Алло-L-полимераза X была экспрессирована в E.coli с использованием экспрессионного конструкта pMJ14. Варианты алло-L-полимеразы X были экспрессированы из pMJ130, pMJ356, pMJ357 или pMJ412. Для экспрессии соответствующий экспрессионный конструкт трансформировали в компетентный штамм E.coli «BL-21 (DE3) pLysS» (Novagen/VWR, Dresden, Germany) и поддерживали с антибиотиком ампициллином. Культуру выращивали при 37°С в среде 2YT до достижения значения оптической плотности при 600 нм ок. 0,6. Затем индуцировали экспрессию белка прибавлением изопропил-бета-D-1- тиогалактопиранозида (сокр. IPTG) до конечной концентрации 0,4 мМ. Экспрессия проводилась в течение 4 часов при 30°С. Клетки собирали центрифугированием и либо хранили при -80°С, либо немедленно обрабатывали.
1.3 Очистка белка
Свежие или замороженные клетки E.coli ресуспендировали на льду в буфере для лизирования и связывания (50 мМ Na-фосфата, рН 7,5, 500 мМ NaCl, 40 мМ имидазола) и лизировали с использованием клеточного дезинтегратора «French Press» (G. Heinemann,; Schwabisch Gmund, Germany). Очистка проводилась при 4°C с использованием материала «Ni-NTA Superflow» (Qiagen, Hilden, Germany). Постадийное элюирование проводили с использованием буфера для элюирования (50 мМ Na-фосфат, рН 7,5, 500 мМ NaCl, 200 мМ имидазола). Фракции анализировали методом ДСН-ПААГ (Invitrogen, Karlsruhe, Germany), объединяли в пул и, при необходимости, дополнительно очищали анионообменной хроматографией на системе «AKTA purifier» с использованием материала «Q Sepharose fast flow» (GE healthcare, Freiburg, Germany). Идентичность белка подтверждали методом масс-спектрометрии с матрично-активированной лазерной десорбцией/ионизацией (MALDI) и требуемые фракции были объединены в пул, сконцентрированы и повторно буферизованы. Очищенный белок хранили при -20°С в буфере, состоящем из 25 мМ Na-фосфата, рН 7,5,250 мМ NaCl, 50% глицерина. Концентрации белка оценивали методом анализа белка с помощью бицинхониновой кислоты (ВСА) (Pierce/Perbio Science, Bonn, Germany) с использованием в качестве стандарта бычий сывороточный альбумин (сокр. BSA).
Пример 2 - Подтверждение активности полимеразы X и вариантов полимеразы X
Анализы активности алло-L-полимеразы X и вариантов алло-L-полимеразы X (см. Пример 1) были проведены с разными типами комплексов субстратов, образуемых олигонуклеотидами, где субстраты и олигонуклеотиды состоят из D-ДНК-нуклеотидов.
2.1 Анализы активности на субстратах с 1-нуклеотидным пробелом
Перечень олигонуклеотидов для субстратов с 1 пробелом:

Субстратные комплексы были получены путем отжига матричной нити ДНК-олигонуклеотида, состоящего из 33 нуклеотидов (также называемого нижней нитью) с двумя разными ДНК-олигонуклеотидами, состоящими из 15 и 17 нуклеотидов, соответственно, которые гибридизуются с матричной нитью на ее 5'-конце и 3'-конце, соответственно, приводя к образованию пробела в один нуклеотид в верхней нити. Комплексы содержали А, С, G или Т в позиции матрицы в пробеле. Перед отжигом, олигонуклеотид SP-1, состоящий из 15 нуклеотидов, метили радиоактивной меткой 32Р на его 5'-конце путем стандартной киназной реакции с использованием гамма-32Р-аденозинтрифосфата (АТФ) и полинуклеотидкиназы Т4. Отжиг проводили в 10 мМ Tris-HCl, 5 мМ MgCl2, рН 8,0, путем нагревания в течение 10 мин. при 65°С и медленного охлаждения. Невключенный гамма-32Р-АТФ удаляли очисткой на NAP-колонках (GE healthcare).
В анализе активности, алло-L-полимеразу X и ее варианты объединяли с: субстратными комплексами с 1 пробелом в D-конфигурации (см. Фиг. 1А). В качестве отрицательного контроля, каждый субстрат также инкубировали без алло-L-полимеразы X и ее вариантов и D-дезоксинуклеотидтрифосфатов (dNTP). В зависимости от основания матрицы в комплексе с 1 пробелом, при проведении анализа прибавляли только соответствующий D-dNTP. Типичный анализ на 6 мкл содержал 50 нМ субстратного комплекса, 1,7 нг/мкл L-алло-L-полимеразы X или ее вариантов, 8 мкМ одного D-dNTP и буфер (50 мМ Tris-HCl, 10 мМ MgCl2, 4% глицерина, 0,1 мг/мл бычьего сывороточного альбумина (BSA), рН 7,5). D-dNTP были приобретены у фирмы Rovalab (Teltow, Germany). Время инкубирования составляло 30 минут при 37°С. Весь объем анализа смешивали с буфером для образца/красителем, загружали на денатурирующий гель для секвенирования и разделяли в течение 4 часов. Гель экспонировали на экране Kodak K в течение ночи при -80°С и считывали с использованием системы BioRad Fx Phosphoimager.
Алло-L-полимераза X и варианты I124G, V80A и V80G были активными в таких условиях и заполняли 1-нуклеотидный пробел между двумя верхними нитями ДНК-олигонуклеотида.
2.2 Анализ активности на субстрате с 6-нуклеотидным пробелом
Перечень олигонуклеотидов для субстрата с 6-нуклеотидным пробелом:
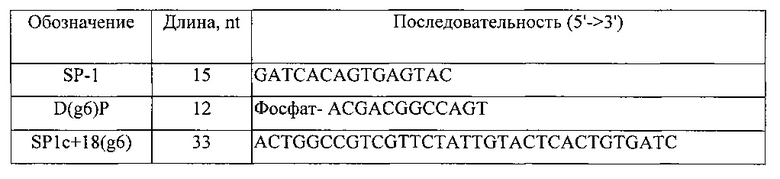
Субстратные комплексы были получены путем отжига матричной нити ДНК-олигонуклеотида, состоящего из 33 нуклеотидов (называемого нижней нитью) с двумя разными ДНК-олигонуклеотидами, состоящими из 15 и 12 нуклеотидов, соответственно, которые гибридизуются с матричной нитью на ее 5'-конце и 3'-конце, соответственно, приводя к образованию пробела в шесть нуклеотидов в верхней нити. Перед отжигом, олигонуклеотид SP-1, состоящий из 15 нуклеотидов, метили радиоактивной меткой 32Р на его 5'-конце путем стандартной киназной реакции с: использованием гамма-32Р-аденозинтрифосфата (АТФ) и полинуклеотидкиназы Т4. Отжиг проводили в 10 мМ Tris-HCl, 5 мМ MgCl2, рН 8,0, путем нагревания в течение 10 мин при 65°С и медленного охлаждения. Невключенный гамма-32Р-АТФ удаляли очисткой на NAP-колонках (GE healthcare, Freiburg, Germany).
В анализе активности, алло-Ь-полимеразу X и ее варианты объединяли с субстратным комплексом в D-конфигурации с 6 пробелами (Фиг. 1 В). В качестве отрицательного контроля, субстрат также инкубировали без алло-L-полимеразы X или ее вариантов и дезоксинуклеотидтрифосфатов (D-dNTP). Типичный анализ на 6 мкл содержал 50 нМ субстратного комплекса, до 1,3 нг/мкл алло-L-полимеразы X или ее вариантов, 8 мкМ каждого из D-dNTP и буфер (50 мМ Tris-HCl, 10 мМ MgCl2, 4% глицерина, 0,1 мг/мл бычьего сывороточного альбумина (BSA), рН 7,5). D-dNTP были приобретены у фирмы Rovalab (Teltow, Germany). Типичное время инкубирования составляло 30 минут при 37°С. Весь объем анализа смешивали с буфером для образца/красителем, загружали на денатурирующий гель для секвенирования и разделяли в течение 4 часов. Гель экспонировали на экране Kodak K в течение ночи при -80°С и считывали с использованием системы BioRad Fx Phosphoimager.
Алло-L-полимераза X и варианты (за исключением C86S) были активными в таких условиях и заполняли пробел в 6 нуклеотидов между двумя верхними нитями ДНК-олигонуклеотида.
Пример 3 - Синтез варианта полимеразы Pol X, состоящего из D-аминокислот
В примере описан синтез алло-О-полимеразы X, вариант V80A. Аминокислотная последовательность алло-О-полимеразы X, вариант V80A, имеет вид:

Все используемые аминокислоты были защищены в соответствии с требованиями стратегии твердофазного пептидного синтеза Fmoc/tBu (Eric Atherton et al., 1981). Все используемые аминокислоты являются D-аминокислотами (Bachem, Bubendorf, Switzerland).
3.1 Синтез HO-Gp(Boc)2
трет-Бутилоксикарбонил-защищенный 4-гуанидинофенол был синтезирован по аналогии с Sekizaki et al. (Sekizaki et al., 1996). В соответствии с этой методикой, 40 ммоль N,N'-бис-(трет-бутилоксикарбонил)-S-метилизотиомочевины (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland) и 60 ммоль 4-аминофенола (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland) растворяли в 250 мл ТГФ в круглодонной колбе на 500 мл. После этого раствор продували аргоном в течение 10 мин и выдерживали при перемешивании в течение 120 часов, закрытым трубкой с CaCl2.
После выпаривания растворителя остаток осаждали охлажденным на льду метанолом. Осадок высушивали под вакуумом над Р4О10. Наконец, продукт очищали с использованием флеш-хроматографии с DCM. Фракции, содержащие продукт, объединяли, и растворитель выпаривали при пониженном давлении. Для проведения анализов использовали ТСХ, обращенно-фазовую ВЭЖХ, масс-спектрометрию и ЯМР. Экспериментально определенная масса соответствует расчетной массе, равной 351 Да.
3.2 Синтез H-D-Leu-OGp(Boc)2
1 ммоль Z-D-Leu-OH (Bachem, Bubendorf, Switzerland), 0,9 экв. TBTU и 0,9 экв. HO-Gp(Boc)2 растворяли в 10 мл ДМФ. После прибавления 2 экв. DIPEA раствор перемешивали в течение 2 часов. После выпаривания растворителя сырой продукт очищали флеш-хроматографией с использованием DCM. Чистые фракции Z-D-Leu-OGp(Boc)2 объединяли и растворитель выпаривали.
Z-D-Leu-OGp(Boc)2 растворяли в 10 мл МеОН и продували аргоном. Гидролитическое расщепление N-концевой Z-группы проводили путем прибавления катализатора Pd/C и Н2 в течение 2 часов. После фильтрации H-D-Leu-OGp(Boc)2, МеОН выпаривали при пониженном давлении. Анализы проводили с использованием обращенно-фазовой ВЭЖХ и масс-спектрометрии. Точная масса продукта была найдена равной 465 Да и соответствует расчетной массе.
3.3 Синтез алло-D-пептида AcMLTLIOGKKIVNHLRSRLAFEYNGOLIKILSKNIVAVGSL-OGp (1)
0,10 ммоль смолы TentaGel-R-Trityl (Rapp Polymere, Tubingen, Deutschland) загружали Fmoc-D-Ser(tBu)-OH (Bachem, Bubendorf, Switzerland), как описано Barios et al. (Barios et al., 1989). После этого 0,10 ммоль смолы инкубировали два раза по 30 мин с 0,6 ммоль тионилхлорида и затем промывали DCM. После этого, смолу инкубировали 90 мин с 0,6 ммоль Fmoc-D-Ser(tBu)-OH, 2,4 ммоль DIPEA в 6 мл DCM. Затем смолу блокировали три раза по 10 мин. с использованием раствора 10% МеОН (об./об.), 10% DIPEA (об./об.) в DCM и промывали DCM. Автоматизированный синтез проводили с использованием ABI 433 (Applied Biosystems, Foster City, USA) по протоколу FASTmoc. 10 экв. аминокислоты активировали с использованием 9 экв. HATU и 20 экв. DIPEA в NMP. Время реакции сочетания составляло 45 мин. и снятие Fmoc-защиты проводили три раза по 7 мин. с помощью 20% (об./об.) пиперидина (Sigma-Aldrich Chemie GmbH, Schnelldorf, Deutschland) в NMP.
Расщепление полностью защищенного пептида в кислотной форме проводили путем инкубирования пептидильной смолы дважды в 10 мл 30% (об./об.) HFIP в DCM в течение 2 часов. После фильтрации пептида растворитель выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Осажденный пептид выделяли и сушили.
Растворяли 0,01 ммоль полностью защищенного пептида, 4 экв. РуВОР и 5 экв. H-D-Leu-OGp(Boc)2 в 6 мл NMP. После прибавления 10 экв. DIPEA смесь перемешивали в течение 4 часов. После этого, растворитель удаляли выпариванием и остаток осаждали охлажденным на льду диэтиловым эфиром. Осажденный сложный эфир пептида высушивали и затем защитные группы отщепляли с использованием 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA пептид осаждали охлажденным на льду диэтиловым эфиром. Очистку методом обращенно-фазовой ВЭЖХ сложного эфира пептида проводили на колонке C18 (Phenomenex, Aschaffenburg, Germany) с использованием градиента ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ВЭЖХ/УЭЖХ (Фиг. 2А) и масс-спектрометрии (Фиг. 2В). Измеренная масса продукта, равная 4654,7 Да, соответствует теоретической массе 4652,7 Да.
3.4 Синтез алло-D-пептида H-RREEKLNDVDLLIIVPEKKLLKHVLPNIRIKGLSFSVKA-SMe (2)
0,10 ммоль смолы TentaGel-R-NH2 (Rapp Polymère, Tübingen, Deutschland) загружали Fmoc-D-Ala-OH с использованием 5 экв. аминокислоты, 4,9 экв. HATU и 10 экв. DIPEA в течение 45 мин. в 6 мл NMP. После этого смолу промывали ТГФ. Конверсию в Fmoc-d-Ala-ψ[CS-NH]-R-TentaGel проводили путем инкубирования с 4 экв. реагента Лоусона в ТГФ при 80°С в течение 2 часов. После этого смолу промывали NMP. Затем приготовленную таким образом смолу использовали для автоматизированного пептидного синтеза, как описано выше (см. Пример 1.3). После этого соответствующий тиоэфир получали путем инкубирования с метилйодидом в ДМФ в течение ночи в соответствии с Sharma et al. (Sharma et al., 2011). После фильтрации смолы, растворитель, содержащий тиоэфир пептида, выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Отщепление защитных групп боковых цепей проводили с помощью 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA, пептид осаждали охлажденным на льду диэтиловым эфиром. Очистку методом обращенно-фазовой ВЭЖХ тиоэфира пептида проводили на колонке C18 (Phenomenex, Aschaffenburg, Germany) с использованием градиента ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ВЭЖХ/УЭЖХ (Фиг. 3А) и масс-спектрометрии (Фиг. 3В). Экспериментально определенная масса продукта (4683,8 Да) соответствует теоретическому значению, равному 4681,7 Да.
3.5 Синтез алло-Р-пептида H-CGERKCVLFIEWEKKTYOLDLFTALAEEKPYAIFHFTGPVSYLIRIRAALKKKNYKLNOYGLFKNOTLVPLKITTEKELIKELGFTYRIPKKRL-OH (3)
0,10 ммоль смолы TentaGel-R-PHB (Rapp Polymère, Tübingen, Deutschland) загружали Fmoc-D-Leu-OH с использованием 6 экв. аминокислоты, 6 экв. MSNT и 4,5 экв. Melm в DCM. Автоматизированный синтез проводили с использованием ABI 433 по протоколу FASTmoc. Активировали 10 экв. аминокислоты с использованием 9 экв. HATU и 20 экв. DIPEA в NMP. Время реакции сочетания составляло 45 мин., и снятие Fmoc-защиты проводили три раза по 7 мин. с помощью 20% (об./об.) пиперидина в NMP. Стадии двойного сопряжения проводили после 42 аминокислот. Отщепление N-терминального Fmoc-защищенного пептида проводили с использованием 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA, пептид осаждали охлажденным на льду диэтиловым эфиром. Очистку методом обращенно-фазовой ВЭЖХ сырого пептида с защищенным N-концом проводили на колонке C18 с использованием градиента ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали. Fmoc-защищенный пептид затем растворяли и перемешивали в 20% пиперидине (piperine) в ДМФ для отщепления N-концевой Fmoc-группы. Спустя 20 мин. растворитель выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. После этого осажденный сырой пептид очищали с использованием обращенно-фазовой ВЭЖХ на колонке C18 с градиентом ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ВЭЖХ/УЭЖХ (Фиг. 4А) и масс-спектрометрии (Фиг. 4В). Определенная масса продукта (11184,3 Да) соответствует теоретической массе, равной 11178,2 Да.
3.6 Синтез алло-D-пептида Ac-MLTLIOGKKIVNHLRSRLAFEYNGOLIKILSKNIVAVGSLRREEKMLNDVDLLIIVPEKKLLKHVLPNIRIKGLSFSVKA-SMe (4) путем протеаза-катализованного лигирования пептида 1 с пептидом 2
Пептид 1 растворяли в концентрации 0,2 мМ и пептид 2 растворяли в концентрации 0,6 мМ в натрий-фосфатном буфере (100 мМ, рН 8,5, с 100 мМ NaCl), содержащем 2% Triton X100 (Sigma Aldrich Chemie GmbH, Schnelldorf, Germany). После прибавления 20 мкМ Clostripain (эндопротеаза Arg-C, Worthington Biochemical Corporation, Lakewood, N.J., USA) реакционную смесь встряхивали в течение ночи при 37°С. Осажденные пептиды центрифугировали, растворяли в Н2О/ACN/муравьиной кислоте 60/40/0,5 и очищали обращенно-фазовой ВЭЖХ с использованием колонки RP-18 (Phenomenex, Aschaffenburg, Germany) с градиентом ACN в воде от 30% до 60% в течение 30 мин. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный пептид анализировали методами обращенно-фазовой УЭЖХ (Фиг. 5А) и ESI-масс-спектрометрии (Фиг. 5В). Теоретический молекулярный вес (Мтеор = 9199,3 Да) соответствует наблюдаемому молекулярному весу (Мнабл. = 9199,4 Да).
3.7 Синтез алло-D-полимеразы X, вариант V80A, путем нативного химического лигирования пептида 4 с пептидом 3
Оба пептида 3 и 4 растворяли с концентрацией 0,2 M в TRIS-буфере (рН 8,6), содержащем 6М гуанидин-HCl (Sigma Aldrich Chemie GmbH, Schnelldorf, Germany), 200 мМ меркаптофенилуксусной кислоты (Sigma Aldrich Chemie GmbH, Schnelldorf, Germany) и 5 мМ (mm) трис(2-карбоксиэтил)фосфина гидрохлорида (Sigma Aldrich Chemie GmbH, Schnelldorf, Germany). Реакционную смесь встряхивали 72 ч при комнатной температуре. Затем смесь очищали обращенно-фазовой ВЭЖХ с использованием колонки RP-8 (Phenomenex, Aschaffenburg, Germany) с градиентом ACN в воде от 30% до 60% в течение 30 мин. Фракции, содержащие продукт лигирования, объединяли в пул и сушили. Сухой порошок растворяли в воде и очищали эксклюзионной хроматографией с использованием колонки SEC3000 (Phenomenex, Aschaffenburg, Germany) с натрий-фосфатным буфером (Sigma Aldrich Chemie GmbH, Schnelldorf, Germany) (50 мМ, pH 6,8, 0,5% ДСН) в качестве элюента. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ДСН-ПААГ (Фиг. 6А) и ESI-масс-спектрометрии (Фиг. 6В). Четкая полоса была обнаружена на дорожке 7 между 14,4 кДа и 21,5 кДа, указывающая на чистую полноразмерную полимеразу. Теоретический молекулярный вес (Мтеор. = 20342 Да) соответствует наблюдаемому молекулярному весу (Мнабл. = 20361 Да), что подтверждается методом ESI-MS.
Пример 4 - Подтверждение активности варианта синтетической полимеразы X, состоящего из D-аминокислот
Сухую алло-D-полимеразу X, вариант V80A, в соответствии с Примером 3, растворяли в 6М гуанидиния гидрохлориде и подвергали повторной укладке при 4°С путем постадийного диализа в коммерчески доступных диализаторах (Pierce/PerBio, Bonn, Germany) с отсекающим значением молекулярного веса, равным 3500. Конечный буфер содержал 50 мМ фосфата натрия, 500 мМ хлорида натрия, рН 7,5. Концентрацию белка оценивали методом электрофореза на полиакриламидном геле (ПААГ) в присутствии додецилсульфата натрия (ДСН) на предварительно отлитых гелях (Invitrogen, Karlsruhe, Germany) с использованием серии стандартов с известными концентрациями белка с последующим окрашиванием SYPRO-RED (Invitrogen, Karlsruhe, Germany) и денситометрическим анализом полос на сканере BioRad Fx.
Анализ активности алло-D-полимеразы X, вариант V80A, проводили с двумя разными типами субстрата:
4.1 Анализы активности на субстратах с 1-нуклеотидным пробелом
Субстраты были получены путем отжига 33-мерной нижней нити ДНК-олигонуклеотида с двумя 17-мерными верхними нитями ДНК-олигонуклеотидов, что приводило к образованию пробела в 1 нуклеотид в верхней нити. Олигонуклеотиды были синтезированы в L-конфигурации. Перед отжигом, олигонуклеотид MJ_1_58_MD 17-мерной верхней нити метили радиоактивной меткой 32Р на его 5'-конце путем стандартной киназной реакции с использованием гамма-32Р-аденозинтрифосфата (гамма-32Р-АТФ) и полинуклеотидкиназы Т4. Для облегчения киназной реакции L-олигонуклеотида MJ_1_58_MD, во время синтеза олигонуклеотида на 5'-конце добавляли два гуанозиновых основания в D-конфигурации. Отжиг проводили в 10 мМ Tris-HCl, 5 мМ MgCb, рН 8,0, путем нагревания в течение 10 мин. при 65°С и медленного охлаждения. Невключенный гамма-32Р-АТФ удаляли очисткой на колонках NAP (GE healthcare, Freiburg, Germany). Комплексы содержали А, С, G или Τ в позиции пробела матрицы. Структура субстратных комплексов приведена на Фиг. 7.
В анализе активности, синтетическую алло-D-полимеразу X, вариант V80A, объединяли с субстратными комплексами в L-конфигурации с 1 пробелом. В качестве отрицательного контроля, каждый субстрат также инкубировали без алло-D-полимеразы X, вариант V80A, и L-дезоксинуклеотидтрифосфатов (dNTP). В зависимости от основания матрицы в комплексе с 1 пробелом, при проведении анализа прибавляли только соответствующий L-dNTP. Типичный анализ на 6 мкл содержал 50 нМ субстратного комплекса, 1,7 нг/мкл алло-D-полимеразы X, вариант V80A, 8 мкМ одного из L-dNTP и буфер (50 мМ Tris-HCl, 10 мМ MgCb, 4% глицерина, 0,1 мг/мл бычьего сывороточного альбумина (BSA), рН 7,5). L-dNTP были синтезированы на заказ фирмой Rasayan, Inc. (Encinitas, СА, USA). Время инкубирования составляло 30 минут при 37°С. Весь объем анализа смешивали с буфером для образца/красителем, загружали на денатурирующий гель для секвенирования и разделяли в течение 4 часов. Гель экспонировали на экране Kodak K в течение ночи при -80°С и считывали с использованием системы BioRad Fx Phosphoimager.
Как видно на Фиг. 8, алло-D-полимераза X, вариант V80A, дает продукты удлинения на субстратах L-ДНК с 1 пробелом, тем самым подтверждая активность синтетического белка. Примечательно, что только алло-D-полимераза X, вариант V80A, в комбинации с L-субстратом и L-dNTP, давала какие-либо продукты удлинения. Также, только образцы, содержащие L-dNTP, соответствующие основанию их матрицы, давали продукт удлинения. Это означает, что для получения каких-либо продуктов удлинения на A-комплексе должен присутствовать нуклеотид dTTP, на С-комплексе - нуклеотид dGTP, на G-комплексе - нуклеотид dCTP, и на Т-комплексе - нуклеотид dATP.
4.2 Анализ активности на субстратах с 6-нуклеотидным пробелом
Субстраты были получены путем отжига 33-мерной нижней нити ДНК-олигонуклеотида с двумя 17-мерной и 12-мерной верхними нитями ДНК-олигонуклеотида, приводя к образованию пробела в 6 нуклеотидов в верхней нити. Олигонуклеотиды были синтезированы в L-конфигурации. Перед отжигом 17-мерный олигонуклеотид MJ_1_58_MD верхней нити (L-конфигурация) метили радиоактивной меткой 32Р на его 5'-конце путем стандартной киназной реакции с использованием гамма- Р-аденозинтрифосфата (АТФ) и полинуклеотидкиназы Т4. Для облегчения киназной реакции L-олигонуклеотида MJ_1_58_MD, два гуанозиновых основания в D-конфигурации добавляли на 5'-конце во время синтеза олигонуклеотида. Отжиг проводили в 10 мМ Tris-HCl, 5 мМ MgCl2, рН 8,0, путем нагревания в течение 10 мин. при 65°С и медленного охлаждения. Невключенный гамма-32Р-АТФ удаляли очисткой на колонках NAP (GE healthcare, Freiburg, Germany). Структура субстратных комплексов приведена на Фиг. 9А.
В анализе активности синтетическую алло-D-полимеразу X, вариант V80A, объединяли с субстратным комплексом с 6 пробелами в L-конфигурации. В качестве отрицательного контроля, субстрат также инкубировали без алло-D-полимеразы X, вариант V80A, и дезоксинуклеотидтрифосфатов (L-dNTP). Типичный анализ на 6 мкл содержал 50 нМ субстратного комплекса, до 1,3 нг/мкл алло-D-полимеразы X, вариант V80A, 8 мкМ каждого из L-dNTP и буфер (50 мМ Tris-HCl, 10 мМ MgCl2, 4% глицерина, 0,1 мг/мл бычьего сывороточного альбумина (BSA), рН 7,5). L-dNTP были синтезированы на заказ фирмой Rasayan, Inc. (Encinitas, СА, USA). Типичное время инкубирования составляло 30 минут при 37°С, но, в зависимости от активности партии, использовали более продолжительное инкубирование. Весь объем анализа смешивали с буфером для образца/красителем, загружали на денатурирующий гель для секвенирования и разделяли в течение 4 часов. Гель экспонировали на экране Kodak K в течение ночи при -80°С и считывали с использованием системы BioRad Fx Phosphoimager.
Как видно на Фиг. 9В, синтетическая алло-D-полимераза X, вариант V80A, давала N+6 продукты удлинения на субстратах L-ДНК с 6 пробелами, тем самым подтверждая активность синтетического белка. Однако синтез продукта удлинения N+6 был менее наглядным, чем продукта удлинения N+1 на том же самом комплексе с 6 пробелами. Также для заполнения 6 пробелов требовалось большее время инкубирования. Примечательно, что только алло-D-полимераза X, вариант V80A, в комбинации с L-субстратом плюс L-dNTP, давала какие-либо продукты удлинения.
Пример 5 - Синтез ДНК полимеразой X и вариантами полимеразы X Полимераза X вируса африканской чумы свиней (сокр. ASFV) описана в литературе (Oliveros, 1997) как высоко дистрибутивный фермент с функцией заполнения пробелов. В Примере 2 было продемонстрировано, что алло-L-полимераза X и ее варианты катализируют включение только очень ограниченного числа нуклеотидов после каждого инициирования на субстратах с пробелами. В данном документе мы раскрываем способ, который позволяет использовать алло-L-полимеразу X и ее варианты для синтеза более длинных ДНК, и продемонстрировал полную полимеризацию 83-мерной нити.
5.1 Субстрат праймер-матрица
Комплекс праймер-матрица был использован для тестирования активности алло-L-полимеразы X и ее вариантов. Этот же комплекс был также использован для тестирования вариантов V80G и V80A алло-D-полимеразы X.
Перечень D-олигонуклеотидов для комплекса праймер-матрица без пробелов:

Субстрат получали путем отжига матричной нити ДНК-олигонуклеотида, состоящего из 83 нуклеотидов (MJ_1_1_DD) с ДНК-олигонуклеотидом, состоящим из 19 нуклеотидов. Олигонуклеотиды были синтезированы NOXXON. Олигонуклеотид MJ_1_33_DD несет флуоресцентный краситель Atto-532 (AttoTec, Siegen, Germany). Отжиг проводили в 10 мМ Tris-HCl, 5 мМ MgCb, рН 8,0, путем нагревания в течение 10 мин. при 65°С и медленного охлаждения. Комплекс праймер-матрица изображен на Фигуре 10А.
5.2 Реакция при постоянной температуре
В анализе активности, алло-L-полимеразу X или варианты V80G или V80A алло-L-полимеразы X объединяли с комплексом праймер-матрица в D-конфигурации. Типичный анализ на 6 мкл содержал 50 нМ субстратного комплекса, до 1,3 нг/мкл алло-L-полимеразы X или вариантов V80G или V80A алло-L-полимеразы X, 8 мкМ каждого из D-dNTP и буфер (50 мМ Tris-HCl, 10 мМ MgCl2, 4% глицерина, 0,1 мг/мл бычьего сывороточного альбумина (BSA), рН 7,5). D-dNTP были приобретены у фирмы Rovalab (Teltow, Germany). Время инкубирования составляло 30 минут при 37°С для образцов Ро1-Х. В качестве отрицательного контроля, субстрат также инкубировали без какой-либо алло-L-полимеразы X или вариантов V80G или V80A алло-L-полимеразы X и без дезоксинуклеотидтрифосфатов (D-dNTP). Положительный контроль проводили с использованием полимеразы Taq (Invitrogen, Karlsruhe, Germany) в конечной концентрации 0,083 U/мкл в буфере Taq, поставленном производителем. Образцы Taq инкубировали 30 минут при 60°С. Весь объем анализа смешивали с буфером для образца/красителем, и разделяли на денатурирующем геле. Гель считывали с использованием системы BioRad Fx Phosphoimager.
Алло-L-полимераза X или варианты V80G или V80A алло-L-полимеразы X были активными, но были неспособны завершить полимеризацию полной 83-мерной матрицы. Положительный контроль с полимеразами Taq продемонстрировал полную полимеризацию 83-мерной матрицы, см. Фигуру 10В.
5.3 Реакция в условиях термоциклирования
Основываясь на предположении, что алло-L-полимераза X после инициирования катализирует включение только одного нуклеотида и затем останавливается, оставаясь на ДНК-субстрате, мы использовали повторные тепловые импульсы (50°С, 2 минуты) для того, чтобы позволить алло-L-полимеразе X диссоциировать и повторно ассоциировать с матрицей. Используя такую процедуру многократного термоциклирования, мы смогли осуществить полную полимеризацию 83-мера с помощью алло-L-полимеразы X. Реакции и контроли были организованы, как описано выше для постоянной температуры, за исключением того, что температурный профиль образцов для алло-L-полимеразы X осуществляли следующим образом:
от 5 до 25 циклов (30 минут при 20°С // 2 минуты при 50°С)
затем конечная стадия 30 минут при 20°С.
Было обнаружено, что начиная с 15-го цикла, алло-L-полимераза X была способна полимеризовать полную 83-мерную матричную нить, аналогично положительному контролю, см. Фигуру 10С.
Пример 6 - Удлинение праймера вариантом синтетической полимеразы X, состоящим из D-аминокислот
Способ, раскрытый в этом примере, использует субстраты в L-конфигурации для тестирования полимеразы в алло-D-конфигурации.
6.1 Субстрат праймер-матрица
Перечень L-олигонуклеотидов для комплекса праймер-матрица без пробелов:

Субстрат получают путем отжига 83-мерной нижней нити ДНК-олигонуклеотида с 19-мерной верхней нитью ДНК-олигонуклеотида. Олигонуклеотиды синтезировали на собственных производственных мощностях NOXXON в L-конфигурации. Перед отжигом, 21-мерную верхнюю нить олигонуклеотида MJ_1_109_MD метили радиоактивным 32Р на его 5'-конце путем стандартной киназной реакции с использованием гамма-32Р-аденозинтрифосфата (АТФ) и полинуклеотидкиназы Т4. Для облегчения киназной реакции L-олигонуклеотида MJ_1_109_MD, два гуанозиновых основания в D-конфигурации добавляли на 5'-конце во время синтеза олигонуклеотида. Отжиг проводили в 10 мМ Tris-HCl, 5 мМ MgCl2, рН 8,0 путем нагревания в течение 10 мин. при 65°С и медленного охлаждения. Невключенный гамма-32Р-АТФ удаляли очисткой на колонках NAP (GE healthcare, Freiburg, Germany).
6.2 Реакция при постоянной температуре
В анализе активности синтетическую алло-D-полимеразу X, вариант V80A, объединяли с комплексом праймер-матрица в L-конфигурации. Типичный анализ на 6 мкл содержит 50 нМ субстратного комплекса, до 1,3 нг/мкл варианта алло-L-полимеразы X, 8 мкМ каждого из L-dNTP и буфер (50 мМ Tris-HCl, 10 мМ MgCb, 4% глицерина, 0,1 мг/мл бычьего сывороточного альбумина (BSA), рН 7,5). L-dNTP были приобретены у фирмы Rasayan, Inc. (Encinitas, СА, USA). Время инкубирования составляло по меньшей мере 30 минут при 37°С. В качестве отрицательного контроля, субстрат также инкубировали без какой-либо полимеразы и без дезоксинуклеотидтрифосфатов (L-dNTP). Весь объем анализа смешивали с буфером для образца/красителем, и разделяли на денатурирующем геле. Гель считывали с использованием системы BioRad Fx Phosphoimager.
Синтетическая алло-D-полимераза X, вариант V80A, является активной в таких условиях, но - аналогично ее двойнику алло-L-полимеразе X – не способна полимеризовать полную 83-нуклеотидную матричную нить.
6.3 Реакция в условиях термоциклирования
Аналогично Примеру 5, используется процедура многократного термоциклирования для обеспечения возможности полной полимеризации 83-мерного L-субстрата с помощью алло-L-полимеразы X, вариант V80A. Реакции и контроли были организованы, как описано выше для постоянной температуры, за исключением того, что температурный профиль осуществляли следующим образом: от 5 до 25 циклов (30 минут при 20°С // 2 минуты при 50°С), затем конечная стадия 30 минут при 20°С.
Было обнаружено, что алло-D-полимераза X, вариант V80A - аналогично ее двойнику алло-L-полимеразе X - способна полимеризовать полную 83-мерную матричную нить при использовании удлинения с термоциклированием.
Пример 7 - Рекомбинантная экспрессия и очистка полимеразы Dpo4 и вариантов полимеразы Dpo4, полностью состоящих из L-аминокислот
Полимераза Dpo4 была первоначально обнаружена у Sulfolobus Solfataricus (сокр. Sso) (Boudsocq, 2001). Ген дикого типа имеет открытую рамку считывания (сокр. ORF) из 1059 пар оснований, включая инициирующий кодон и терминирующий кодон. Кодированный белок имеет длину 352 аминокислоты. Этот пример описывает, как полимераза Dpo4 и варианты полимеразы Dpo4 были экспрессированы в E.coli и очищены с использованием Strep-Tag.
7.1 Экспрессионные конструкты
Поскольку частота использования кодонов Sso отличается от E.coli, синтетический ген с оптимизированной для E.coli частотой использования кодонов для полимеразы Dpo4 Sso дикого типа был приобретен у фирмы GeneArt AG (Regensburg, Germany). Последовательность синтетического гена была вставлена в вектор pENTRY-IBA10 (компания-разработчик: IBA GmbH, Gottingen, Germany). Кодон-оптимизированная открытая рамка считывания, включающая инициирующий кодон, но не включающая терминирующий кодон, имела следующую последовательность:


Для получения экспрессионного конструкта полимеразы Dpo4, также называемого алло-L-полимеразой Dpo4, ген субклонировали из pENTRY-IBA10 в вектор pASG-IBA5 (IBA GmbH, Göttingen, Germany), с использованием коммерчески доступного набора для клонирования StarGate (IBA GmbH). Субклонирование добавляло метку Strep-Tag II на N-конце и терминирующий кодон на С-конце, и ставило ген под контроль промотора tet. Конструкт был назван pMJ343 и использовался для экспрессии алло-L-полимеразы Dpo4 в E.coli. Алло-L-полимераза Dpo4, экспрессированная из pMJ343, имела следующую последовательность из 368 аминокислот:

Начальные 14 аминокислот представляют Strep-Tag II, включая несколько спейсерных аминокислот, последние 2 аминокислоты представляют собой спейсерные аминокислоты, и средняя часть из 352 аминокислот была идентична последовательности полимеразы Dpo4, присутствующей в Sso.
Экспрессионные конструкты вариантов алло-L-полимеразы Dpo4 были получены с использованием коммерчески доступного набора QuikChange (Stratagene GmbH, Waldbronn, Germany) в соответствии с протоколом производителя. Плазмида pMJ343 служила матрицей. Олигонуклеотиды, необходимые для QuikChange, были или синтезированы NOXXON (QC_38_up, QC_38_low, QC_39_up, QC_39_low, QC_40_up, QC_40_low), или приобретены у фирмы Purimex (Grebenstein, Germany) (QC_28_up, QC_28_low, QC_29_up, QC_29_low, QC_30_up, QC_30_low).
Следующие варианты экспрессионных конструктов были получены и использованы для экспрессии вариантов алло-L-полимеразы Dpo4 в E.coli.
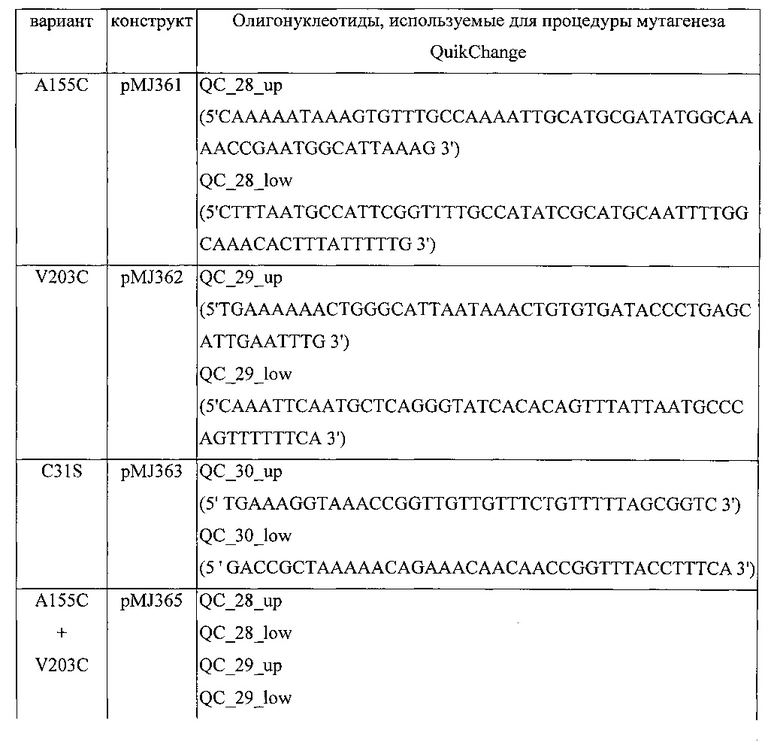

7.2 Экспрессия белка в E.coli
Алло-L-полимераза Dpo4 была экспрессирована в E.coli с использованием! экспрессионного конструкта pMJ343. Мутантные варианты алло-L-полимеразы Dpo4 были' экспрессированы из pMJ361, pMJ362, pMJ363, pMJ365, pMJ502, pMJ503 или pMJ504. Для экспрессии, соответствующий экспрессионный конструкт трансформировали в штамм E.coli «NEB express» (New England Biolabs, Frankfurt am Main, Germany) с использованием раствора для трансформации и хранения transformation and Storage Solutions (Epicentre/Biozym, Hessisch Oldendorf, Germany) и поддерживали с антибиотиком ампициллином. Экспрессию проводили в среде «EnBase Flo» или «EnPresso» (BioSilta, Oulu, Finland) в течение 48 ч при 30°С с использованием 200 нг/мл ангидротетрациклина; (IBA GmbH, Gottingen, Germany) в качестве индуктора. Клетки собирали центрифугированием и или хранили при -80°С, или немедленно обрабатывали.
7.3 Очистка белка
Свежие или замороженные клетки E.coli ресуспендировали на льду в «буфере W» (100 мМ Tris-HCl, рН 8,0, 150 мМ NaCl, 1 мМ EDTA) и лизировали с использованием клеточного дезинтегратора «French Press» (G. Heinemann, Schwâbisch Gmünd, Germany). Очистку проводили при 4°С на системе "АКТА Express", оснащенной колонками 5 мл StrepTrap HP (GE healthcare, Freiburg, Germany). Постадийное элюирование проводили с буферов W, содержащим 2,5 мМ дестиобиотина (IBA GmbH, Gôttingen, Germany). Фракции анализировали методом ДСН-ПААГ (Invitrogen, Karlsruhe, Germany), объединяли в пул и, при необходимости, дополнительно очищали анионообменной хроматографией на системе «AKTA purifier», оснащенной колонками «Q HP» (GE healthcare, Freiburg, Germany). Идентичность белка подтверждали методом жидкостной хроматомасс-спектрометрии (LC-MS) и точные фракции были объединены в пул, сконцентрированы и повторно буферизованы с использованием концентрирующих устройств VivaSpin 15R с отсечкой по молекулярному весу 10000 (MWCO) (VivaSciences/Sartorius Stedim Biotech, Göttingen, Germany). Очищенный белок хранили при -20°С в буфере, состоящем из 100 мМ КС1, 10 мМ Tris-HCl рН 7,4, 0,1 мМ EDTA, 1 мМ DTT, 50% глицерина. Концентрации белка оценивали методом гель-денситометрии с использованием стандарта бычьего сывороточного альбумина (сокр. BSA) на ДСН-ПААГ и окрашивания SYPRO Red (Invitrogen, Karlsruhe, Germany).
Пример 8 - Продуцирование синтетической полимеразы Dpo4, состоящей из двух фрагментов
Алло-L-полимераза Dpo4 имела длину 352 аминокислоты. Для получения алло-L-полимеразы Dpo4 химическими методами, такую синтетическую алло-L-полимеразу Dpo4 нужно было собирать из более коротких фрагментов, которые могут быть синтезированы методами твердофазного пептидного синтеза, причем указанные более короткие фрагменты должны быть лигированы методом лигирования пептидов, таким как нативное химическое лигирование. Этот пример описывает, как фрагменты лигирования 1-154, 155-352, 155-202 и 203-352 Dpo4 были экспрессированы в E.coli, очищены и лигированы друг с другом путем нативного химического лигирования для получения синтетической алло-L-полимеразы Dpo4.
8.1 Экспрессионные конструкты
Как раскрыто в Примере 7, ген алло-L-полимеразы Dpo4 был получен как синтетический генный конструкт из коммерческого источника (GeneArt, Regensburg, Germany). Все фрагменты этого примера были клонированы на основании данной кодон-оптимизированной последовательности. Были приготовлены следующие экспрессионные конструкты фрагментов 1-154, 155-352, 155-202 и 203-352 алло-L-полимеразы Dpo4, вариант А155С:

8.1.1 Фрагмент 1-154-тиоэфир алло-L-полимеразы Dpo4, вариант А155С - Экспрессионный конструкт pMJ370
Этот конструкт содержит фрагмент 1-154 алло-L-полимеразы Dpo4, вариант А155С, за которым следует интеин Мхе GyrA, используемый для получения тиоэфира, и хитин-связывающий домен (CBD). Конструкт был собран из двух ПЦР-продуктов. ПЦР-продукт 1 был получен с использованием pMJ343 в качестве матрицы и праймеров MJ_1_90_DD (5'-фосфат-AGCGGCTCTTCGATGATTGTGCTGTTTGTGGATTTT-3') и MJ_1_91_DD (5'-фосфат-AGCGGCTCTTCGGCATGCAATTTTGGCAAACACTTT-3') для амплификации фрагмента 1-154 алло-L-полимеразы Dpo4, вариант А155С. ПЦР-продукт 2 был получен с использованием pTWIN1 (New England Biolabs, Frankfurt am Main, Germany) в качестве матрицы и праймеров MJ_1_72JDD (5'-фосфат-AGCGGCTCTTCGTGCATCACGGGAGAT-3') и MJ_1_73_DD (5'-фосфат-AGCGGCTCTTCGCCCTTGAAGCTGCCACAAGGCAGGAACGTT-3') для амплификации интеина Мхе Gyr А и CBD. Праймеры MJ_1_90_DD и MJ_1_91_DD были получены от фирмы Purimex (Grebenstein, Germany), а праймеры MJ_1_72_DD и MJ_1_73_DD были получены от фирмы IBA GmbH (Gottingen, Germany). Два продукта ПЦР очищали на геле на системе flash-gel (LONZA, Basel, Switzerland) и клонировали вместе в pENTRY-IBA20 с использованием набора для клонирования StarGate Mutagenesis ENTRY (IBA GmbH, Gottingen, Germany), получая pMJ366. Субклонирование из pMJ366 в pASG-IBAwtl (IBA GmbH, Gottingen, Germany) с использованием набора для клонирования StarGate Transfer (IBA GmbH, Gottingen, Germany) давало pMJ370. Конструкт pMJ370 кодирует фрагмент 1-154-тиоэфир алло-L-полимеразы Dpo4, вариант А155С, со следующей белковой последовательностью длиной 154 аминокислоты (после отщепления интеина/продуцирования тиоэфира):

EAYNLGLEIKNKILEKEKITVTVGISKNKVFA-тиоэфир.
8.1.2 Фрагмент 155-352 алло-L-полимеразы Dpo4, вариант А155С - Экспрессионный конструкт pMJ384
Этот конструкт содержит метку «Profinity eXact» со следующим за ней фрагментом 155-352 алло-L-полимеразы Dpo4, вариант А155С. Метка «Profinity eXact» использовалась для очистки и протеолитического расщепления. Конструкт собирали из двух ПЦР-продуктов. ПЦР-продукт 1 был получен с использованием pPAL7 (Bio-Rad, München, Germany) в качестве матрицы и праймеров MJ_1_99_DD (5'-фосфат-AGCGGCTCTTCGATGGGAGGGAAATCAAACGGGGAA-3') и MJ_1_100_DD (5'-фосфат-AGCGGCTCTTCGGCACAAAGCTTTGAAGAGCTTGTC-3') для амплификации метки «Profinity eXact». ПЦР-продукт 2 был получен с использованием pMJ361 в качестве матрицы и праймеров MJ_1_96_DD (5'-фосфат-AGCGGCTCTTCGTGCGATATGGCAAAACCGAATGGCATTAAA-3') и MJ_1_97_DD (5'-фосфат-AGCGGCTCTTCGTGCGATATGGCAAAACCGAATGGCATTAAA-3') для амплификации фрагмента 155-352 dpo4, содержащего мутацию А155С. Все праймеры MJ_1_96_DD, MJ_1_97_DD, MJ_1_99_DD и MJ_1_100_DD были получены от фирмы Purimex (Grebenstein, Germany). 2 ПЦР-продукта очищали на геле на системе flash-gel (LONZA, Basel, Switzerland) и клонировали вместе в pENTRY-IBA20 с использованием набора для клонирования StarGate Mutagenesis ENTRY (IBA GmbH, Gottingen, Germany), получая pMJ382. Субклонирование из pMJ382 в pASG-IBA5 (IBA GmbH, Gottingen, Germany) с использованием набора для клонирования StarGate Transfer (IBA GmbH, Gottingen, Germany) давало pMJ384. Конструкт pMJ384 кодирует фрагмент 155-352 алло-L-полимеразы Dpo4, содержащий мутацию А155С со следующей белковой последовательностью длиной 198 аминокислот (после протеолитического расщепления):

8.1.3 Фрагмент 203-352 алло-L-полимеразы Рро4, вариант А155С - Экспрессионный конструкт pMJ385
Этот конструкт содержит метку «Profinity eXact» со следующим за ней фрагментом 203-352 dpo4, содержащим мутацию V203C. Метка «Profinity eXact» использовалась для очистки и протеолитического расщепления. Конструкт собирали из двух ПЦР-продуктов. ПЦР-продукт 1 был получен с использованием pPAL7 (Bio-Rad, Munchen, Germany) в качестве матрицы и праймеров MJ_1_99_DD (5'-фосфат-AGCGGCTCTTCGATGGGAGGGAAATCAAACGGGGAA-3') и MJ_1_100_DD (5'-фосфат-AGCGGCTCTTCGGCACAAAGCTTTGAAGAGCTTGTC-3') для амплификации метки «Profinity eXact». ПЦР-продукт 2 был получен с использованием pMJ362 в качестве матрицы и праймеров MJ_1_98_DD (5'-фосфат-AGCGGCTCTTCGTGTGATACCCTGAGCATTGAATTT-3') и MJ_1_97_DD (5'-фосфат-AGCGGCTCTTCGCCCTTAGGTATCAAAAAATTTATCCAGG-3') для амплификации фрагмента 203-352 dpo4, содержащего мутацию V203C. Все праймеры MJ_1_97_DD, MJ_1_98_DD, MJ_1_99_DD и MJ_1_100_DD были получены от фирмы Purimex (Grebenstein, Germany). 2 ПЦР-продукта очищали на геле на системе flash-gel (LONZA, Basel, Switzerland) и клонировали вместе в pENTRY-IBA20 с использованием набора для клонирования StarGate Mutagenesis ENTRY (IBA GmbH, Gottingen, Germany), получая pMJ383. Субклонирование из pMJ383 в pASG-IBA5 (IBA GmbH, Gottingen, Germany) с использованием набора для клонирования StarGate Transfer (IBA GmbH, Gottingen, Germany) давало pMJ385. Конструкт pMJ385 кодирует фрагмент 203-352 V203C dpo4 со следующей белковой последовательностью длиной 150 аминокислот (после протеолитического расщепления):
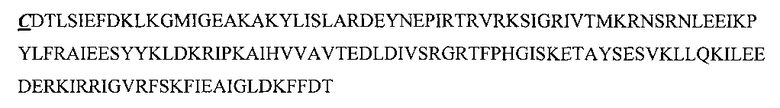
8.1.4 Фрагмент 155-202 алло-L-полимеразы Рро4 - Экспрессионный конструкт pMJ388
Этот конструкт содержит фрагмент 155-202 dpo4 со следующим за ним интеином Мхе GyrA, который использовался для получения тиоэфира, и хитин-связывающим доменом (CBD). Конструкт собирали из двух ПЦР-продуктов. ПЦР-продукт 1 был получен с использованием pMJ343 в качестве матрицы и праймеров MJ_1_101_DD (5'-фосфат-AGCGGCTCTTCGATGGCAGATATGGCAAAACCGAAT-3') и MJ l_102_DD (5'-фосфат-AGCGGCTCTTCGGCACAGTTTATTAATGCCCAGTTT-3') для амплификации фрагмента 155-202 dpo4. ПЦР-продукт 2 был получен с использованием pTWINl (New England Biolabs, Frankfurt am Main, Germany) в качестве матрицы и праймеров MJ_1_72_DD (5'-фосфат-AGCGGCTCTTCGTGCATCACGGGAGAT-3') и MJ_1_73_DD (5'-фосфат-AGCGGCTCTTCGCCCTTGAAGCTGCCACAAGGCAGGAACGTT-3') для амплификации интеина Мхе Gyr А и CBD. Праймеры MJ_1_101_DD и MJ_1_102_DD были получены от фирмы Purimex (Grebenstein, Germany), а праймеры MJ_1_72_DD и MJ_1_73_DD были получены от фирмы IBA GmbH (Gôttingen, Germany). 2 ПЦР-продукта очищали на геле на системе flash-gel (LONZA, Basel, Switzerland) и клонировали вместе в pENTRY-IBA20 с использованием набора для клонирования StarGate Mutagenesis ENTRY (IBA GmbH, Gôttingen, Germany), получая pMJ386. Субклонирование из pMJ386 в pASG-IBAwtl (IBA GmbH, Gôttingen, Germany) с использованием набора для клонирования StarGate Transfer (IBA GmbH, Gôttingen, Germany) давало pMJ388. Конструкт pMJ388 кодирует фрагмент 155-202 dpo4 со следующей белковой последовательностью длиной 48 аминокислот (после E.coli-медиированного отщепления начального метионина и после отщепления интеина/продуцирования тиоэфира):
ADMAKPNGIKVIDDEEVKRLIRELDIADVPGIGNITAEKLKKLGINKL-тиоэфир.
8.2 Экспрессия белка в E.coli
Для экспрессии соответствующий экспрессионный конструкт трансформировали в штамм E.coli «NEB express» (New England Biolabs, Frankfurt am Main, Germany) с использованием раствора для трансформации и хранения «Transformation and Storage Solution» (Epicentre/Biozym, Hessisch Oldendorf, Germany) и поддерживали с антибиотиком ампициллином. Экспрессию проводили в среде «EnBase Fio» или «EnPresso» (BioSilta, Oulu, Finland) при температуре окружающей среды с использованием 200 нг/мл ангидротетрациклина (IBA GmbH, Gôttingen, Germany) в качестве индуктора в течение ночи. Клетки собирали центрифугированием и либо хранили при -80°С, либо немедленно обрабатывали.
8.3 Очистка и получение тиоэфира из конструктов pMJ370 и pMJ388 с интеином Мхе Gyr А
Свежие или замороженные клетки E.coli ресуспендировали на льду в «буфере для колонки» (20 мМ HEPES, рН 8,5, 500 мМ NaCl) и лизировали с использованием клеточного дезинтегратора «French Press» (G. Heinemann, Schwabisch Gmund, Germany). Очистку проводили при 4°С на системе «АКТА Express» (GE healthcare, Freiburg, Germany), оснащенной колонками, содержащими хитин-связывающие бусины (New Englands Biolabs, Frankfurt am Main, Germany). После нанесения клеточного лизата и промывания буфером для колонок до базовой линии колонки инкубировали 20 часов при 4°С с 50 мМ 2-меркаптоэтансульфоната (сокр. MESNA) в буфере для колонок для индуцирования интеин-медиированного отщепления белка и образования тиоэфира. Отщепленный белок, несущий тиоэфир, вымывали из колонки с помощью буфера для колонок, концентрировали и подвергали гель-фильтрации с использованием материала среды BioGel Р60 (BioRad, München, Germany) в буфере, состоящем из 5 мМ Bis-Tris, рН 6,5, 250 мМ NaCl. Концентрации белка оценивали методами гель-денситометрии с использованием стандарта бычьего сывороточного альбумина (BSA) на ДСН-ПААГ и окрашивания SYPRO Red (Invitrogen, Karlsruhe, Germany). Идентичность белка и присутствие тиоэфира были подтверждены методом жидкостной хроматомасс-спектрометрии (LC-MS).
8.4 Очистка и протеолитическое отщепление от конструкта pMJ384 с меткой «Profínitv eXact»
Очистку фрагмента 155-352 алло-L-полимеразы Dpo4, вариант А155С, из pMJ384 проводили следующим образом: Свежие или замороженные клетки E.coli ресуспендировали на льду в «буфере W» (100 мМ Tris-HCl, рН 8,0, 150 мМ NaCl, 1 мМ EDTA) и лизировали с использованием клеточного дезинтегратора «French Press» (G. Heinemann, Schwâbisch Gmünd, Germany). Очистку проводили при 4°С на системе «АКТА Express», оснащенной колонками 5 мл StrepTrap HP (GE healthcare, Freiburg, Germany). Постадийное элюирование проводили с использованием буфера W, содержащего 2,5 мМ дестиобиотина (IBA GmbH, Gottingen, Germany). Элюированный белок подвергали замене буфера в буфере для элюирования «Profinity eXact» (0,1 M Na-фосфат, рН 7,2, 0,1 M NaF) с использованием обессоливающей колонки HiPrep 26/10 (GE healthcare, Freiburg, Germany) и затем медленно прокачивали насосом через колонку Profinity eXact. Образец концентрировали, дополняли трис(2-карбоксиэтил)фосфином) (ТСЕР) до конечной концентрации 1 мМ и наносили на гель для фильтрации с использованием препаративной (prep grade) колонки HiLoad 16/60 Superdex 75 (GE healthcare, Freiburg, Germany), проявляли в буфере, состоящем из 5 мМ Bis-Tris, рН 6,5, 250 мМ NaCl. Белок концентрировали и хранили при -80°С. Концентрации белка оценивали методами гель-денситометрии с использованием стандарта бычьего сывороточного альбумина (BSA) на ДСН-ПААГ и окрашивания SYPRO Red (Invitrogen, Karlsruhe, Germany). Идентичность белка подтверждали методом жидкостной хроматомасс-спектрометрии (LC-MS).
8.5 Очистка и протеолитическое отщепление от конструкта pMJ385 с меткой «Profinity eXact»
Очистку фрагмента 203-352 V203C dpo4 от pMJ385 проводили следующим образом: Тельца включения готовили и денатурировали, как описано в руководстве к системе повторной укладки белка «i-FOLD protein refolding System» (Novagen/Merck-Millipore, Darmstadt, Germany). Солюбилизированный белок подвергали замене буфера на «буфер W» (100 мМ Tris-HCl, рН 8,0, 150 мМ NaCl, 1 мМ EDTA) с использованием мелкодисперсного материала Sephadex G-25 (GE healthcare, Freiburg, Germany) и очищали при 4°С на системе «AKTA Express», оснащенной колонками StrepTrap HP (GE healthcare, Freiburg, Germany). Постадийное элюирование проводили с использованием буфера W, содержащего 2,5 мМ дестиобиотина (IBA GmbH, Gôttingen, Germany). Элюированный белковый раствор дополняли NaF до конечной концентрации 0,1 M и трис(2-карбоксиэтил)фосфином) (ТСЕР) до конечной концентрации 1 мМ и затем медленно прокачивали насосом через колонку Profinity eXact. Выходящий поток разбавляли 1:3 деионизированной водой и доводили рН до 7,2 с помощью НС1. Образец дополнительно очищали катионообменной хроматографией с использованием колонок HiTrap SP HP (GE healthcare, Freiburg, Germany), уравновешенных в «буфере А» (50 мМ Na-фосфата, рН 7,2, 1 мМ 2-меркаптоэтанола). Постадийное элюирование проводили с использованием 17%, 25% и 100% «буфера В» (50 мМ Na-фосфата, рН 7,2, 1 M NaCl, 1 мМ 2-меркаптоэтанола). Фракции были объединены в пул, сконцентрированы и нанесены на гель для фильтрации с использованием препаративной колонки HiLoad 16/60 Superdex 75 (GE healthcare, Freiburg, Germany), проявленной деионизированной водой. Белок резко замораживали в жидком азоте и лиофилизировали. Концентрации белка оценивали методами гель-денситометрии с использованием стандарта бычьего сывороточного альбумина (BSA) на ДСН-ПААГ и окрашивания SYPRO Red (Invitrogen, Karlsruhe, Germany). Идентичность белка подтверждали методом жидкостной хроматомасс-спектрометрии (LC-MS).
8.6 Синтез синтетической алло-L-полимеразы Рро4, вариант А155С, путем нативного химического лигирования фрагментов 1-154-тиоэфир и 155-352
Фрагменты 1-154-тиоэфир и 155-352 алло-L-полимеразы Dpo4, вариант А155С, растворяли по 50 мкМ в TRIS-буфере (рН 8,6), содержащем 2% Triton Х100, 1% тиофенола и 5 мМ трис(2-карбоксиэтил)фосфина гидрохлорида. Реакционную смесь встряхивали в течение 72 ч при комнатной температуре. Затем анализировали успешность лигирования методами ДСН-ПААГ (Фиг. 11А) и LC-ESI-масс-спектрометрии (колонка RP18, градиент ACN с 0,1% TFA в воде с 0,1% TFA 5-95% за 20 мин, Фиг. 11В). Была обнаружена четкая полоса на дорожке около 41 кДа, указывающая на полноразмерную полимеразу. Теоретический молекулярный вес (Мтеор. = 40223 Да) соответствует наблюдаемому молекулярному весу (Мнабл. = 40265 Да), определенному методом ESI-MS.
8.7 Синтез фрагмента 155-352 путем нативного химического лигирования фрагментов 155-202-тиоэфир и 203-352
Фрагменты 155-202-тиоэфир и 203-352 V203C алло-L-полимеразы Dpo4 растворяли с концентрацией 0,2 M в TRIS-буфере (рН 8,6), содержащем 2% ДСН, 1% тиофенола и 5 мМ трис(2-карбоксиэтил)фосфина гидрохлорида. Реакционную смесь встряхивали в течение 72 ч при комнатной температуре. Анализ успешности лигирования проводили методами ДСН-ПААГ (Фиг. 20А) и LC-ESI-масс-спектрометрии (колонка RP18, градиент ACN с 0,1% TFA в воде с 0,1% TFA 5-95% за 20 мин., Фиг. 20В). Была обнаружена четкая полоса на дорожке 7 около 21,5 кДа, указывающая на продукт лигирования. Теоретический молекулярный вес (Мтеор. = 22749 Да) соответствует наблюдаемому молекулярному весу (Мнабл. = 22769 Да), определенному методом ESI-MS.
Пример 9 - Подтверждение активности полимеразы Dpo4 и вариантов полимеразы Dpo4, состоящих из L-аминокислот
Этот пример описывает испытания на активность методом ГЩР для алло-Ь-полимеразы Dpo4 и вариантов алло-L-полимеразы Dpo4 в соответствии с Примером 7 и для синтетической алло-L-полимеразы Dpo4 в соответствии с Примером 8.
9.1 Матрицы для анализа ПЦР-активности
Матрица для ПЦР-реакции представляет собой 83-мерный одноцепочечный D-ДНК-олигонуклеотид (MJ_1_1_DD)? из которого, в первом термическом цикле, получают противоположную нить. После этого обе нити служат матрицей для экспоненциальной амплификации. Матрица ДНК-олигонуклеотид и ДНК праймеры синтезированы NOXXON в D-конфигурации.
Перечень олигонуклеотидов для анализа ПЦР-активности:
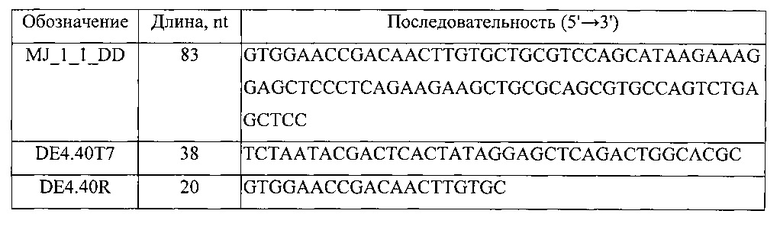
9.2 ПЦР-реакции
15 мкл ПЦР-реакций содержали 0,2 мМ каждого из четырех D-dNTP, 10 нМ матрицы 83-мерной однонитевой ДНК (ssDNA) (MJ_1_1_DD), 1 мкМ прямого и обратного праймеров, буфер 1× ThermoPol (Invitrogen, 20 мМ Tris-HCl, 10 мМ КС1, 10 мМ (NH4)2SO4, 2 мМ MgSO4, 0,1% Triton Х-100, рН 8,8 при 25°С) и по меньшей мере 0,67 нг/мкл алло-L-полимеразы Dpo4 или варианта алло-L-полимеразы Dpo4 или синтетической алло-L-полимеразы Dpo4. Прямой праймер представлял собой DE4.40T7, обратным праймером был DE4.40R, которые давали ПЦР-продукт длиной 102 пары оснований. D-dNTP были приобретены у фирмы Rovalab (Teltow, Germany). Эксперименты с отрицательными контролями проводили без использования алло-L-полимеразы Dpo4 или варианта алло-L-полимеразы Dpo4 или синтетической алло-L-полимеразы Dpo4. Эксперименты с положительными контролями проводили с использованием коммерчески доступной алло-L-о!ро4 (New England Biolabs, Frankfurt am Main, Germany).
Программа термоциклирования состояла из 1 цикла (85°С, 3 мин), затем по меньшей мере 7 циклов (85°С, 30 с / 56°С, 1 мин. / 60°С, 4 мин.), затем выдерживания при 4°С. Аликвоты 4 мкл ПЦР-реакций смешивали с буфером для загрузки образца и анализировали на ТВЕ-ПААГ (полиакриламидный гель с буфером трис-борат-ЭДТА) или на агарозных гелях (LONZA, Cologne, Germany). На гель наносили линейку стандартных ДНК-маркеров молекулярного веса, включающую, в том числе, полосу 100 п.о.
9.3 Подтверждение активности
Были проведены испытания алло-L-полимеразы Dpo4, вариантов А155С, V203C, C31S, A155C/V203C, S85C, S86C, S96C алло-L-полимеразы Dpo4 и синтетической алло-L-полимеразы Dpo4. Все исследуемые полимеразы были способны амплифицировать матричную нить в ПЦР-реакции. Фиг. 12А показывает анализ ПЦР-реакций, проведенных с вариантами А155С, V203C, C31S, A155C/V203C алло-L-полимеразы Dpo4, которые все продемонстрировали полосу в диапазоне значений около 100 п.о. при сравнении с линейкой стандартных ДНК-маркеров молекулярного веса. Ожидаемый размер ПЦР-продукта составлял 102 пары оснований. Указанная полоса не появляется у отрицательных контролей, не содержащих полимеразу. Также указанная полоса 102 п.о. мигрирует выше чем 83-мерная матрица. Фиг. 12В показывает анализ ПЦР-реакций, проведенных с рекомбинантной алло-L-полимеразой dpo4 и с синтетической алло-L-полимеразой dpo4, которая была получена путем лигирования фрагментов. Гель показывает полосы в диапазоне значений около 100 п.о. при сравнении с линейкой стандартных ДНК-маркеров молекулярного веса. Ожидаемый размер ПЦР-продукта составлял 102 пар оснований. Указанная полоса является очень слабой в отрицательных контролях, не содержащих полимеразу. Рекомбинантная алло-L-полимераза dpo4 и синтетическая алло-L-полимераза dpo4 демонстрируют идентичную активность, что можно увидеть при сравнении дорожек 2 и 3 на Фиг. 12В.
Пример 10 - Синтез варианта полимеразы Dpo4, состоящего из D-аминокислот
В Примере описан синтез алло-D-полимеразы Dpo, вариант A155C/V203C. Аминокислотная последовательность алло-D-полимеразы Dpo, вариант A155C/V203C, представляет собой

Все используемые аминокислоты являются защищенными в соответствии с требованиями стратегии твердофазного пептидного синтеза Fmoc/tBu (Eric Atherton et al., 1981). Все используемые аминокислоты являются D-аминокислотами (Bachem, Bubendorf, Switzerland).
10.1 Синтез H-D-Met-OGp(Boc)2
Растворяли 1 ммоль Z-D-Met-OH, 0,9 экв. TBTU и 0,9 ммоль HO-Gp(Boc)2 в 10 мл ДМФ. После прибавления 2 экв. DIPEA раствор перемешивали в течение 2 часов. После выпаривания растворителя сырой продукт очищали флеш-хроматографией с использованием DCM. Чистые фракции Z-D-Met-OGp(Boc)2 объединяли и растворитель выпаривали.
Z-D-Met-OGp(Boc)2 растворяли в 10 мл МеОН и продували аргоном. Гидролитическое отщепление N-концевой Z-группы проводили путем прибавления катализатора Pd/C и Н2 в течение 2 часов. После фильтрации H-D-Met-OGp(Boc)2 МеОН выпаривали при пониженном давлении. Анализы проводили с использованием обращенно-фазовой ВЭЖХ и масс-спектрометрии. Расчетная масса 482 Да соответствует определенной массе, равной 483 Да.
10.2 Синтез полностью защищенного алло-D-пептида H-RTFPHGISKETAYSESVKLLQKILEEDERKIRRIGVRFSKFIEAIGLDKFFDT-NH2 (1)
0,1 ммоль смолы Fmoc-Sieber rink amide NovaSynTG загружали после снятия Fmoc-защиты с помощью Fmoc-D-Thr(tBu)-OH с использованием 5 экв. аминокислоты, 4,9 экв. HATU и 10 экв. DIPEA в течение 45 мин в 6 мл NMP.
Автоматизированный синтез проводили с использованием ABI 433 по протоколу FASTmoc. Активировали 10 экв. аминокислоты с использованием 9 экв. HATU и 20 экв. DIPEA в NMP. Время реакции сочетания составляло 45 мин., и снятие Fmoc-защиты проводили три раза по 7 мин с помощью 20% (об./об.) пиперидина в NMP. Двойное сочетание проводили после присоединения 42 аминокислот.
Отщепление полностью защищенного пептида в кислотной форме проводили путем инкубирования пептидильной смолы дважды в 10 мл 1% (об./об.) TFA в DCM в течение 2 часов. После фильтрации пептида растворитель выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Осажденный пептид выделяли и сушили.
Конечный продукт анализировали методами ВЭЖХ и масс-спектрометрии (Фиг. 13). Расчетная масса для продукта (6244 Да) соответствует измеренной массе (6249 Да).
10.3 Синтез полностью защищенного алло-D-пептида
Boc-VDTLSIEFPKLKGMIGEAKAKYLISLARPEYNEPIRTRVRKSIGRIVT
MKRNSRNLEEIKPYLFRAIEESYYKLPKRIPKAIHVVAVTEDLDIVSRG-OH (2)
Загружали 0,10 ммоль смолы TentaGel-R-Trityl с помощью Fmoc-P-Gly-OH, как описано Barios et al. (Barios et al., 1989). После этого 0,10 ммоль смолы инкубировали два раза по 30 мин. с 0,6 ммоль тионилхлорида и затем промывали РСМ. После этого, смолу инкубировали 90 мин. с 0,6 ммоль Fmoc-Gly-OH, 2,4 ммоль РГРЕА в 6 мл РСМ. Затем смолу блокировали три раза по 10 мин. с использованием раствора 10% МеОН (об./об.), 10% РГРЕА (об./об.) в РСМ и промывали РСМ. Автоматизированный синтез проводили с использованием ABI 433 по протоколу FASTmoc. Активировали 10 экв. аминокислоты с использованием 9 экв. HATU и 20 экв. DIPEA в NMP. Время реакции сочетания составляло 45 мин. и снятие Fmoc-защиты проводили три раза по 7 мин. с помощью 20% (об./об.) пиперидина в NMP. Двойное сочетание проводили после присоединения 39 аминокислот.
Отщепление полностью защищенного пептида в кислотной форме проводили путем инкубирования пептидильной смолы дважды в 10 мл 30% (об./об.) HFIP в РСМ в течение 2 часов. После фильтрации пептида растворитель выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Осажденный пептид выделяли и сушили. Конечный продукт анализировали методами ВЭЖХ и масс-спектрометрии (Фиг. 14). Экспериментально определенная масса продукта (11289 Да) соответствовала теоретическому значению (11286 Да).
10.4 Синтез алло-D-пептида
H-VPTLSIEFPKLKGMIGEAKAKYLISLARPEYNEPIRTRVRKSIGRIVTMKRNSRNLEEI
KPYLFRAIEESYYKLPKRIPKAIHVVAVTEPLPIVSRGRTFPHGISKETAYSESVKLLOK
ILEEPERKIRRIGVRFSKFIEAIGLPKFFPT-NH2 (3) путем конденсации фрагментов пептида 1 с пептидом 2.
Растворяли 5 мкмоль (2) и 1 экв. (I) в 25% (об./об.) TFE в РСМ. После прибавления 5 экв. РуВОР и 10 экв. DIPEA смесь перемешивали в течение ночи. После выпаривания растворителя пептид осаждали с использованием охлажденного на льду диэтилового эфира и фильтровали.
Отщепление защитных групп боковой цепи пептида проводили с помощью 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA пептид осаждали охлажденным на льду диэтиловым эфиром. Очистку пептида с Fmoc-защищенной N-концевой группой методом обращенно-фазовой ВЭЖХ проводили на колонке C18 с использованием градиента ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ВЭЖХ и масс-спектрометрии (Фиг. 15). Экспериментально определенная масса (17531 Да) соответствует теоретической молекулярной массе (17512 Да).
10.5 Синтез алло-D-пептида Z-CDMAKPNGIKVIDDEEVKRLIRELDIADVPGIGNITAEKLKKLGINKL-бензил-тиоэфир (4)
0,10 ммоль смолы TentaGel-R-Trityl загружали Fmoc-D-Leu-OH, как описано Barios et al. (Barios et al., 1989). После этого 0,10 ммоль смолы инкубировали два раза по 30 мин с 0,6 ммоль тионилхлорида и затем промывали DCM. После этого, смолу инкубировали 90 мин с 0,6 ммоль Fmoc-D-Leu-OH, 2,4 ммоль DIPEA в 6 мл DCM. Затем смолу блокировали три раза по 10 мин с использованием раствора 10% МеОН (об./об.), 10% DIPEA (об./об.) в DCM и промывали DCM. Автоматизированный синтез проводили с использованием ABI 433 по протоколу FASTmoc. Активировали 10 экв. аминокислоты с использованием 9 экв. HATU и 20 экв. DIPEA в NMP. Время реакции сочетания составляло 45 мин, и снятие Fmoc-защиты проводили три раза по 7 мин. с помощью 20% (об./об.) пиперидина в NMP.
Отщепление полностью защищенного пептида в кислотной форме проводили путем инкубирования пептидильной смолы дважды в 10 мл 30% (об./об.) HFIP в DCM в течение 2 часов. После фильтрации пептида растворитель выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Осажденный пептид выделяли и сушили.
Пептид с защищенной N-концевой Z- и полностью защищенными боковыми цепями (4) растворяли с концентрацией 1 мМ в ДМФ. После прибавления 5 экв. РуВОР, 10 экв. DIPEA и 30 экв. бензилмеркаптана смесь перемешивали в течение 4 ч. Затем ДМФ выпаривали, пептид осаждали и промывали охлажденным на льду диэтиловым эфиром. Защитные группы боковой цепи удаляли путем обработки 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA, пептид осаждали и промывали охлажденным на льду диэтиловым эфиром. Пептид-бензил-тиоэфир затем очищали методом обращенно-фазовой ВЭЖХ и анализировали методами обращенно-фазовой ВЭЖХ (Фиг. 16А) и ESI-MS (Фиг. 16В). Теоретический молекулярный вес (Мтеор. = 5527 Да) соответствует наблюдаемому молекулярному весу (Мнабл. = 5533 Да), определенному методом ESI-MS.
10.6 Синтез алло-D-пептида
H-RKEVYOOVSSRIMNLLREYSEKIEIASIDEAYLDISDKVRDYREAYNLGLEIKNKIL
EKEKITVTVGISKNKVFAKIA-SMe (7)
0,10 ммоль смолы TentaGel-R-NH2 загружали Fmoc-D-Ala-OH с использованием 5 экв. аминокислоты, 4,9 экв. HATU и 10 экв. DIPEA в течение 45 мин. в 6 мл NMP. После этого смолу промывали ТГФ. Конверсию в Fmoc-D-Ala-ψ[CS-NH]-R-TentaGel проводили путем инкубирования с 4 экв. реагента Лоусона в ТГФ при 80°С в течение 2 часов. После этого смолу промывали NMP. Затем приготовленную таким образом смолу использовали для автоматизированного пептидного синтеза, как описано выше (ABI 433, протокол FASTmoc, активировали 10 экв. аминокислоты с использованием 9 экв. HATU и 20 экв. DIPEA в NMP; время реакции сочетания составляло 45 мин., и снятие Fmoc-защиты проводили три раза по 7 мин. с помощью 20% (об./об.) пиперидина в NMP). Стадии двойного сопряжения проводили после присоединения 44 аминокислот.
После этого получали соответствующий тиоэфир путем инкубирования с метилйодидом в ДМФ в течение ночи в соответствии с Sharma et al. (Sharma et al., 2011). После фильтрации смолы, растворитель, содержащий пептид-тиоэфир, выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Отщепление защитных групп боковых цепей проводили с помощью 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA пептид осаждали охлажденным на льду диэтиловым эфиром. Очистку пептида-тиоэфира методом обращенно-фазовой ВЭЖХ проводили на колонке С18 с использованием градиента ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ВЭЖХ (Фиг. 17 А) и масс-спектрометрии (Фиг. 17 В). Молекулярная масса продукта, определенная методом масс-спектрометрии (9155 Да), соответствовала расчетной массе (9150 Да).
10.7 Синтез
Ac-MIVLFVDFDYFYAQVEEVLNPSLKGKPVVVCVFSGRFEDSGAVATAN
YEARKFGVKAGIPIVEAKKILPNAVYLPM-OGp (6)
0,10 ммоль смолы TentaGel-R-Trityl загружали Fmoc-D-Pro-OH, как описано Barios et al. (Barios et al., 1989). После этого 0,10 ммоль смолы инкубировали два раза по 30 мин с 0,6 ммоль тионилхлорида и затем промывали DCM. После этого смолу инкубировали 90 мин. с 0,6 ммоль Fmoc-D-Pro-OH, 2,4 ммоль DIPEA в 6 мл DCM. Затем смолу блокировали три раза по 10 мин. с использованием раствора 10% МеОН (об./об.), 10% DIPEA (об./об.) в DCM и промывали DCM. Автоматизированный синтез проводили с использованием ABI 433 по протоколу FASTmoc. Активировали 10 экв. аминокислоты с использованием 9 экв. HATU и 20 экв. DIPEA в NMP. Время реакции сочетания составляло 45 мин, и снятие Fmoc-защиты проводили три раза по 7 мин с помощью 20% (об./об.) пиперидина в NMP. Двойное сочетание проводили после присоединения 46 аминокислот. Ацетилирование N-конца проводили с помощью 10% (об./об.) уксусного ангидрида и 10% (об./об.) DIPEA в ДМФ три раза по 10 мин.
Отщепление полностью защищенного пептида в кислотной форме проводили путем инкубирования пептидильной смолы дважды в 10 мл 30% (об./об.) HFIP в DCM в течение 2 часов. После фильтрации пептида, растворитель выпаривали и остаток осаждали с использованием охлажденного на льду диэтилового эфира. Осажденный пептид выделяли и сушили.
Растворяли 0,01 ммоль полностью защищенного пептида, 4 экв. РуВОР и 5 экв. H-D-Met-OGp(Boc)2 в 6 мл NMP. После прибавления 10 экв. DIPEA смесь перемешивали в течение 4 часов. После этого, растворитель удаляли выпариванием и остаток осаждали охлажденным на льду диэтиловым эфиром. Осажденный сложный эфир пептида высушивали и затем защитные группы отщепляли с использованием 2,5% EDT, 2,5% воды, 2,5% TIS в TFA в течение 2 часов. После выпаривания TFA пептид осаждали охлажденным на льду диэтиловым эфиром. Очистку методом обращенно-фазовой ВЭЖХ сложного эфира пептида проводили на колонке С18 с использованием градиента ACN/вода. Фракции, содержащие продукт, объединяли и лиофилизировали.
Конечный продукт анализировали методами ВЭЖХ (Фиг. 18 А) и масс-спектрометрии (Фиг. 18 В). Экспериментально определенная масса (8547 Да) соответствовала теоретической молекулярной массе (8541 Да).
10.8 Синтез алло-D-пептида
H-CDMAKPNGIKVIDDEEVKRLIRELDIADVPGIGNITAEKLKKLGINKLCD
TLSIEFDKLKGMIGEAKAKYLISLARPEYNEPIRTRVRKSIGRIVTMKRNSR
NLEEIKPYLFRAIEESYYKLDKRIPKAIHVVAVTEPLPIVSRGRTFPHGISK
ETAYSESVKLLQKILEEPERKIRRIGVRFSKFIEAIGLPKFFPT-OH (5) путем нативного химического лигирования пептида 4 с пептидом 3
Оба пептида 3 и 4 растворяли до 0,2 M в TRIS-буфере (рН 8,6), содержащем 2% ДСН, 1% тиофенола и 5 мМ трис(2-карбоксиэтил)фосфина гидрохлорида. Реакционную смесь встряхивали 72 ч при комнатной температуре. Затем смесь очищали методом обращенно-фазовой ВЭЖХ. Для удаления защитной группы N-концевой Z-группы пептид растворяли в 270 экв. TFA и 50 экв. тиоанизола и встряхивали в течение 6 ч при комнатной температуре (Yoshiaki Kisoet al., 1980). После выпаривания TFA, пептид осаждали и промывали охлажденным на льду диэтиловым эфиром и снова очищали методом обращенно-фазовой ВЭЖХ (Phenomenex, Aschaffenburg, Germany). Анализ свободного пептида 5 проводили методами ДСН-ПААГ, обращенно-фазовой УЭЖХ и ESI-масс-спектрометрии. Определена точная масса продукта.
10.9 Синтез алло-D-пептида Ac-MIVLFVDFDYFYAQVEEVLNPSLKGKPVVVCVFSG
RFEDSGAVATANYEARKFGVKAGIPIVEAKKILPNAVYLPMRKEVYQQVSSRIMNL
LREYSEKIEIASIDEAYLDISDKVRDYREAYNLGLEIKNKILEKEKITVTVGISKNKVF
AKIA-SMe (8) путем протеаза-катализированного лигирования пептида 6 с пептидом 7
Пептид 6 растворяли в концентрации 0,2 мМ и пептид 7 растворяли в концентрации 0,6 мМ в натрий-фосфатном буфере (100 мМ, рН 8,5, со 100 мМ NaCl), содержащем 4 M мочевины. После прибавления 20 мкМ Clostripain (эндопротеаза Arg-C, Worthington Biochemical Corporation, Lakewood, N.J., USA), реакционную смесь встряхивали в течение ночи при 37°С.Осажденные пептиды центрифугировали, растворяли в Н2О/муравьиной кислоте 80/20 и очищали обращенно-фазовой ВЭЖХ с использованием колонки RP-18 (Phenomenex, Aschaffenburg, Germany) с градиентом ACN в воде от 5% до 95% в течение 30 мин. Конечный пептид анализировали методами ДСН-ПААГ (Фиг. 19А) и ESI-масс-спектрометрии (Фиг. 19В). Полоса, указывающая на продукт лигирования, была обнаружена на дорожке 1 между 14,4 кДа и 21,5 кДа. Теоретический молекулярный вес продукта лигирования (Мтеор. = 17476 Да) соответствует наблюдаемому молекулярному весу (Мнабл. = 17486 Да), определенному методом ESI-MS.
10.10 Синтез алло-D-полимеразы Dpo, вариант A115C/V203C, путем нативного химического лигирования пептида 8 с пептидом 5
Оба пептида 5 и 8 растворяли в концентрации 0,2 M в TRIS-буфере (рН 8,6), содержащем 2% Triton XI00, 1% тиофенола и 5 мМ трис(2-карбоксиэтил)фосфина гидрохлорида. Реакционную смесь встряхивали 72 ч при комнатной температуре. Затем смесь очищали методом обращенно-фазовой ВЭЖХ и анализировали методами ДСН-ПААГ, обращенно-фазовой УЭЖХ и ESI-масс-спектрометрии. Была определена точная масса продукта лигирования.
Пример 11 - Подтверждение активности синтетической полимеразы dpo4, состоящей из D-аминокислот
Этот пример описывает испытания на активность методом ПЦР алло-D-полимеразы Dpo4, вариант A155C/V203C, в соответствии с Примером 10.
Неожиданно, синтетическая алло-D-полимераза Dpo4, вариант A155C/V203C, проявляет активность без дополнительных операций повторной укладки цепи и, таким образом, используется без процедуры дополнительной повторной укладки. Концентрацию белка оценивали методом электрофореза на полиакриламидном геле (сокр. ПААГ) в присутствии додецилсульфата натрия (сокр. ДСН) на предварительно отлитых гелях (Invitrogen, Karlsruhe, Germany) с использованием серии стандартов с известными концентрациями белка с последующим окрашиванием SYPRO-RED (Invitrogen, Karlsruhe, Germany) и денситометрическим анализом полос на сканере BioRad Fx.
11.1 Матрицы для анализа ПЦР-активности
Матрица для ПЦР-реакции представляет собой 83-мерный одноцепочечный L-ДНК-олигонуклеотид (MJ_1_105_LD), из которого, на первом термическом цикле, получают противоположную нить. После этого обе нити служат матрицами для экспоненциальной амплификации. Матрица ДНК-олигонуклеотид и ДНК праймеры были синтезированы NOXXON в L-конфигурации.
Перечень олигонуклеотидов для анализа ПЦР-активности:

11.2 ПЦР-реакции
15 мкл ПЦР-реакции содержат 0,2 мМ каждого из четырех L-dNTP, 10 нМ матрицы 83-мерной одноцепочечной ДНК (MJ_1_105_LD), 1 мкМ прямого и обратного праймеров, буфер 1× ThermoPol (Invitrogen, 20 мМ Tris-HCl, 10 мМ KCl, 10 мМ (NH4)2SO4, 2 мМ MgSO4, 0,1% Triton Х-100, рН 8,8 при 25°С) и по меньшей мере 0,67 нг/мкл алло-D-полимеразы Dpo4, вариант A155C/V203C. Прямой праймер представляет собой MJ_oligo_187_LD, дающий ПЦР-продукт 102 п.о., отличающийся от 83-мерной матрицы. Обратный праймер представляет собой MJ_oligo_189_LD. L-dNTP были синтезированы на заказ фирмой Rasayan, Inc. (Encinitas, СА, USA).
Негативные контроли проводят без использования алло-D-полимеразы Dpo4, вариант A155C/V203C.
Программа термоциклирования состоит из 1 цикла (85°С, 3 мин), затем по меньшей мере 7 циклов (85°С, 30 с / 56°С, 1 мин/60°С, 4 мин), затем выдерживания при 4°С. Аликвоты 4 мкл ПЦР-реакций смешивали с буфером для загрузки образца и анализировали на ТВЕ-гелях. На гель наносили линейку стандартных ДНК-маркеров молекулярного веса, содержащих, в том числе полосу 100 п.о.
11.3 Подтверждение активности
ПЦР-реакция с алло-D-полимеразой Dpo4, вариант A155C/V203C, и L-ДНК субстратом и L-нуклеотидами дает полосу в диапазоне значений около 100 п. о. при сравнении с линейкой стандартных ДНК-маркеров молекулярного веса. Указанная полоса не появляется у отрицательных контролей, не содержащих полимеразу. Также указанная полоса 102 п.о. мигрирует выше чем 83-мерная матрица. Зависящее от алло-D-полимеразы Dpo4, вариант A155C/V203C, появление продукта амплификации L-ДНК, таким образом, подтверждает активность синтетической алло-D-полимеразы Dpo4, вариант A155C/V203C, в процессе термической амплификации.
Пример 12 - Синтез D- или L-нуклеиновых кислот
L-ДНК нуклеиновые кислоты или D-ДНК нуклеиновые кислоты были получены методами твердофазного синтеза с помощью синтезатора ABI 394 (Applied Biosystems, Foster City, С A, USA) с использованием 2'TBDMS ДНК фосфорамидитной химии со стандартными защитными группами экзоциклических аминов (Damha and Ogilvie, 1993). Для синтеза ДНК были использованы dA(N-Bz)-, dC(N-Ac)-, dG(N-ibu)-, и dT в D- и L-конфигурациях. Все фосфорамидиты были приобретены у фирмы ChemGenes, Wilmington, MA. После синтеза и снятия защиты L-ДНК нуклеиновые кислоты или D-ДНК нуклеиновые кислоты очищали методом гель-электрофореза.
Использованная литература
Полные библиографические данные документов, упоминаемых в данном описании, если не указано иное, приведены ниже, посредством чего содержание указанных документов включено в данное описание с помощью ссылок.
Altschul S.F., Gish W., et al. (1990) Basic local alignment search tool. J Mol Biol. 215(3):403-10.
Altschul S.F., Madden T.L., et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25(17):3389-402.
Atherton E., Logan C.J. and Sheppard R.C. (1981) Peptide synthesis. Part 2. Procedures for solid-phase synthesis using Na-fluorenylmethoxycarbonylamino-acids on polyamide supports. Synthesis of substance Ρ and of acyl carrier protein 65-74 decapeptide J. Chem. Soc, Perkin Trans. 1, 538-546.
Barios, K., Gatos, D., Kallitsis, J., Papaphotiu, G., Sotiriu, P., Yao, W.Q. and Schafer, W. (1989) Darstellung geschiitzter Peptid-Fragmente unter Einsatz substituierter Triphenylmethyl-Harze. Tetrahedron Letters. 30(30):3943-3946.
Bock L.C., Griffin L.C., Latham J.A., Vermaas E.H. and Toole J.J. (1992) Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature. 355(6360):564-6.
Boudsocq, F., S. Iwai, et al. (2001). "Sulfolobus solfataricus P2 DNA polymerase IV (Dpo4): an archaeal DinB-like DNA polymerase with lesion-bypass properties akin to eukaryotic poleta." Nucleic Acids Res 29(22): 4607-16. Burmeister P.E. et al (2006) 2'-Deoxy purine, 2'-O-methyl pyrimidine (dRmY) aptamers as candidate therapeutics. Oligonucleotides. 16(4):337-51.
Damha MJ, Ogilvie KK. (1993) Oligoribonucleotide synthesis. The silyl-phosphoramidite method. Methods Mol Biol. 20:81-114.
Ellington A.D. and Szostak J.W. (1990) In vitro selection of RNA molecules that bind specific ligands. Nature. 346(6287):818-22.
Freier S.M. and Altmann K.H. (1997) The ups and downs of nucleic acid duplex stability: structure-stability studies on chemically-modified DNA:RNA duplexes. Nucleic Acids Res. 25(22):4429-43.
Klussmann S. (2006). The Aptarner Handbook - Functional Oligonucleotides and their Applications. Edited by S. Klussmann. WILEY-VCH, Weinheim, Germany, ISBN 3-527-31059-2.
Klussmann S., Nolte Α., Bald R., Erdmann V.A. and Fürste J.P. (1996) Mirror-image RNA that binds D-adenosine. Nat Biotechnol. 14(9): 1112-5.
Kusser W. (2000) Chemically modified nucleic acid aptamers for in vitro selections: evolving evolution. J Biotechnol 74(1): 27-38.
Mairal T., Ozalp V.C., Lozano Sanchez P., et al. (2008) Aptamers: molecular tools for analytical applications. Anal Bioanal Chem. 390(4):989-1007.
MandaI K., Uppalapati M., Ault-Riché D., Kenney J., Lowitz J., Sidhu S.S. and Kent S.B.H. (2012) Chemical synthesis and X-ray structure of a heterochiral {D-protein antagonist plus vascular endothelial growth factor} protein complex by racemic crystallography. PNAS. 109(37).
McGinnis S., Madden T.L. et al. (2004) BLAST: at the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 32 (Web Server issue): W20-5.
Needleman and Wunsch (1970) A general method applicable to the search for similarities in the amino acid sequence of two proteins. J Mol Biol. 48(3):443-53.
Nolte Α., Klussmann S., Bald R., Erdmann V.A. and Fürste J.P. (1996) Mirror-design of L-oligonucleotide ligands binding to L-arginine. Nat Biotechnol. 14(9): 1116-9.
Oliveros, M., R.J. Yanez, et al. (1997). "Characterization of an African swine fever virus 20-kDa DNA polymerase involved in DNA repair." J Biol Chem 272(49): 30899-910.
Pearson and Lipman (1988) Improved tools for biological sequence comparison. Proc. Nat'l. Acad. Sci. USA 85: 2444.
Sambrook et al. (ed.), (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Sekizaki, H., Itoh, K., Toyota, E. and Tanizawa, K. (1996) Synthesis and tryptic hydrolysis of p-guanidinophenyl esters derived from amino acids and peptides. Chemical & Pharmaceutical Bulletin. 44(8): 1577-9.
Sekizaki, H., Itoh, K., Toyota, E. and Tanizawa, K. (1996) Trypsin-catalyzed peptide synthesis and various p-guanidinophenyl esters as acyl donors. Chemical & Pharmaceutical Bulletin. 44(8): 1585-7.
Sharma, I. and Crich, D. (2011) Direct Fmoc-Chemistry-Based Solid-Phase Synthesis of Peptidyl Thioesters. Journal of Organic Chemistry. 76(16):6518-24.
Smith and Waterman (1981), Adv. Appl. Math. 2: 482.
Tuerk C. and Gold L. (1990) Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science. 249(4968):505-10.
Usman N. and Cedergren R. (1992) Exploiting the chemical synthesis of RNA. Trends Biochem Sci. 17(9):334-9.
Venkatesan N., Kim S.J., et al. (2003) Novel phosphoramidite building blocks in synthesis and applications toward modified oligonucleotides. Curr Med Chem 10(19): 1973-91.
Wincott F, DiRenzo A, et al. (1995). Synthesis, deprotection, analysis and purification of RNA and ribozymes. Nucleic Acids Res.;23(14):2677-84.
Признаки настоящего изобретения, раскрытые в описании, формуле изобретения и/или чертежах, могут по отдельности и в любой их комбинации служить материалом для реализации изобретения в его различных формах.
| название | год | авторы | номер документа |
|---|---|---|---|
| НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ, УЧАСТВУЮЩИЕ В УВЕЛИЧЕНИИ ИЛИ УМЕНЬШЕНИИ СКОРОСТИ ОВУЛЯЦИИ У МЛЕКОПИТАЮЩИХ | 2001 |
|
RU2283866C2 |
| Мутантные полимеразы вируса гепатита В | 2012 |
|
RU2625021C2 |
| ГЕНЫ CORYNEBACTERIUM GLUTAMICUM, КОДИРУЮЩИЕ БЕЛКИ, УЧАСТВУЮЩИЕ В СИНТЕЗЕ МЕМБРАН И МЕМБРАННОМ ТРАНСПОРТЕ | 2000 |
|
RU2312145C2 |
| ПРОНИКАЮЩИЕ В КЛЕТКУ ПЕПТИДЫ И ПОЛИПЕПТИДЫ ДЛЯ КЛЕТОК МИКРООРГАНИЗМОВ | 2008 |
|
RU2526511C2 |
| СПОСОБ ВЫЯВЛЕНИЯ ПАРВОВИРУСА B19 ЧЕЛОВЕКА В БИОЛОГИЧЕСКОМ ОБРАЗЦЕ (ВАРИАНТЫ) И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2002 |
|
RU2301263C2 |
| МОДИФИЦИРОВАННАЯ ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЁ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 1997 |
|
RU2235773C2 |
| РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 1998 |
|
RU2238978C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УСИЛИВАЮЩЕЙ ЦЕЛЛЮЛОЛИТИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДА | 2008 |
|
RU2510417C2 |
| БЫСТРЫЙ СПОСОБ КЛОНИРОВАНИЯ И ЭКСПРЕССИИ СЕГМЕНТОВ ГЕНА РОДСТВЕННОЙ ВАРИАБЕЛЬНОЙ ОБЛАСТИ АНТИТЕЛА | 2012 |
|
RU2612910C2 |
| ГЕНЫ CORYNEBACTERIUM GLUTAMICUM, КОДИРУЮЩИЕ БЕЛКИ РЕЗИСТЕНТНОСТИ И ТОЛЕРАНТНОСТИ К СТРЕССАМ | 2000 |
|
RU2303635C2 |

















































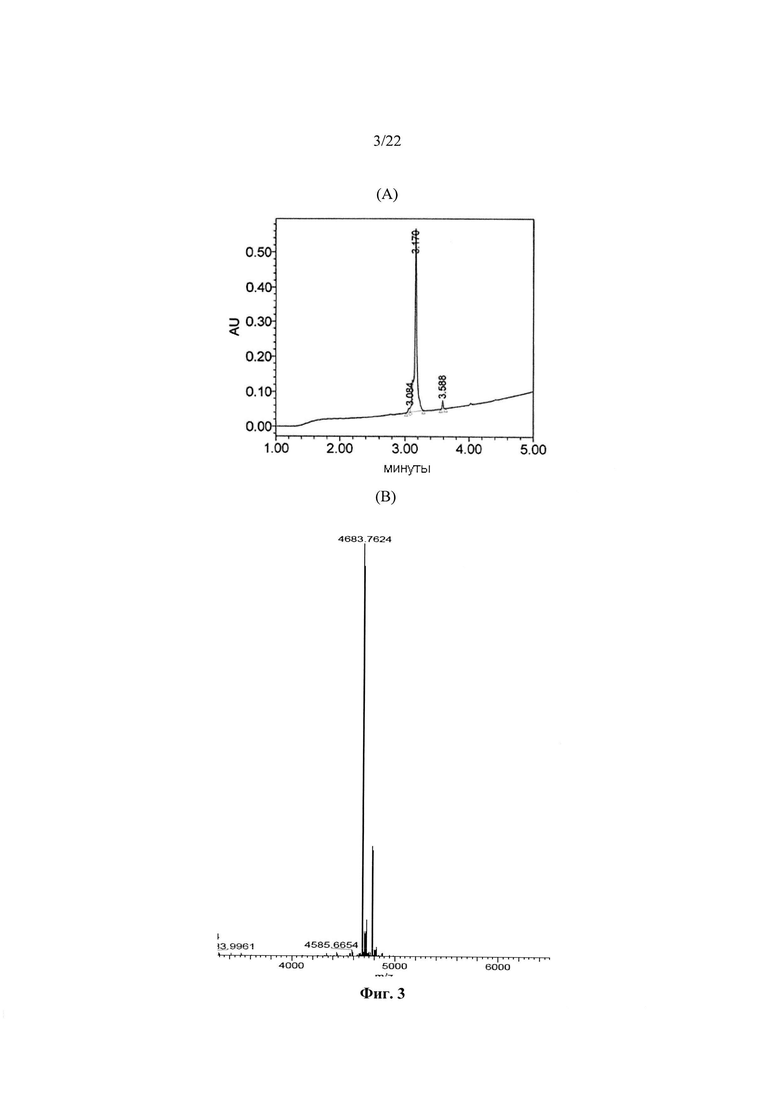
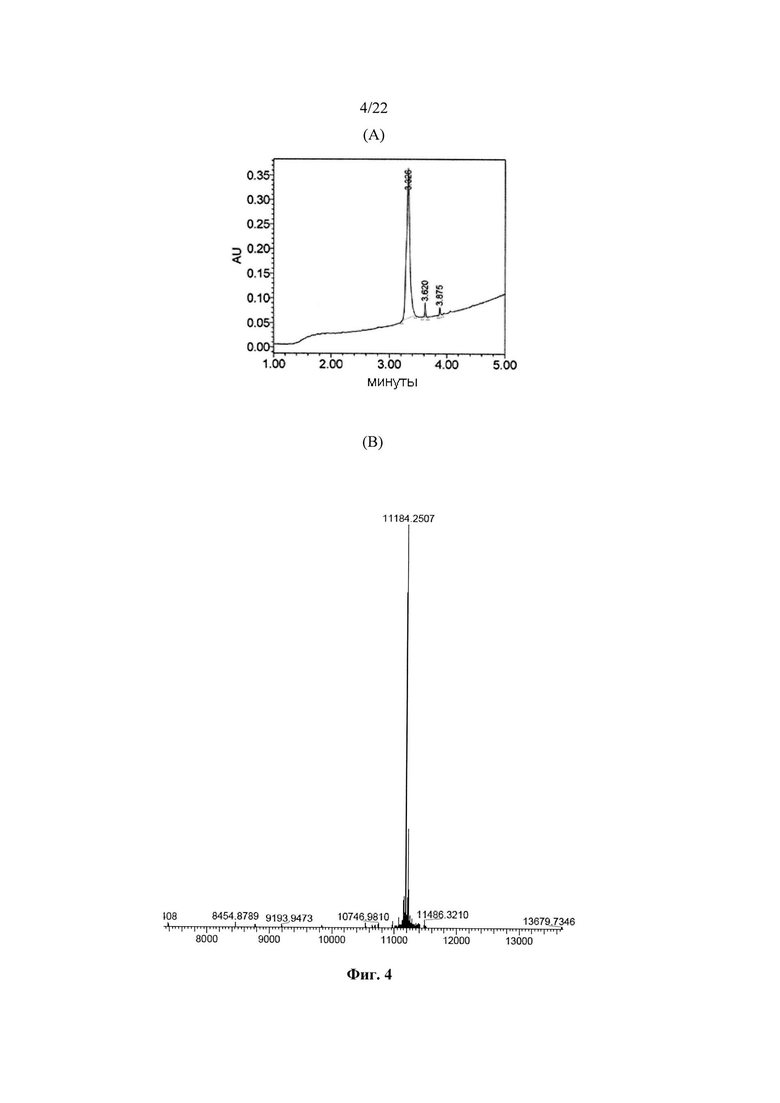
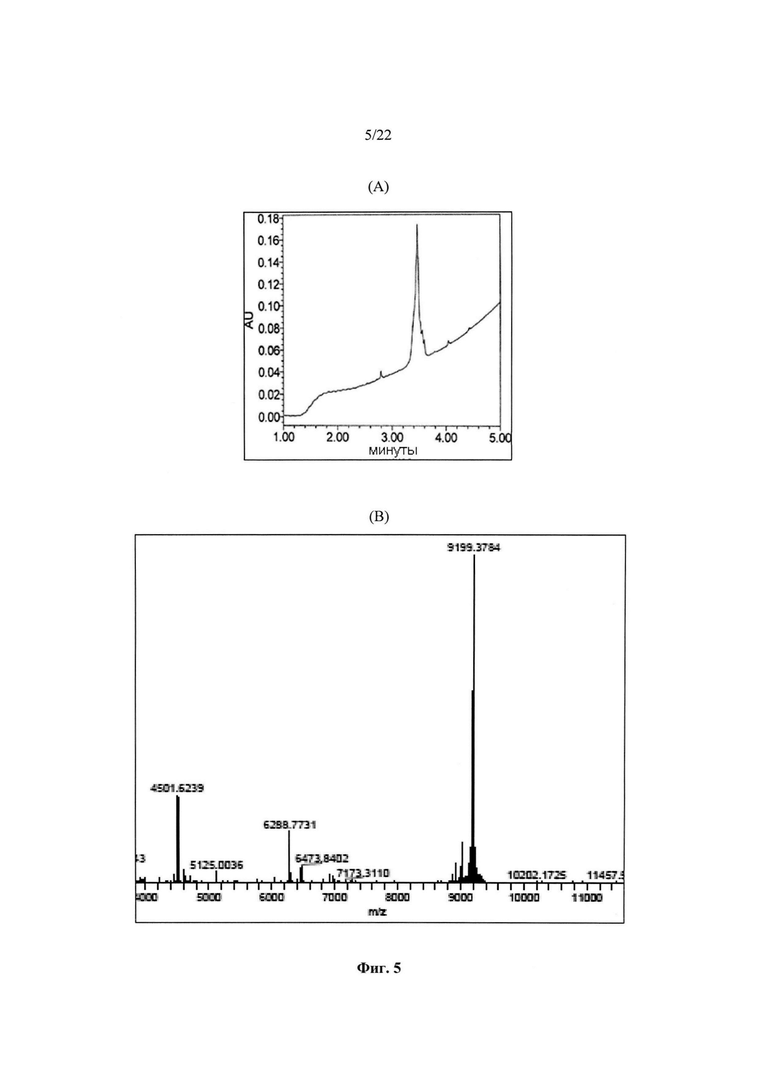




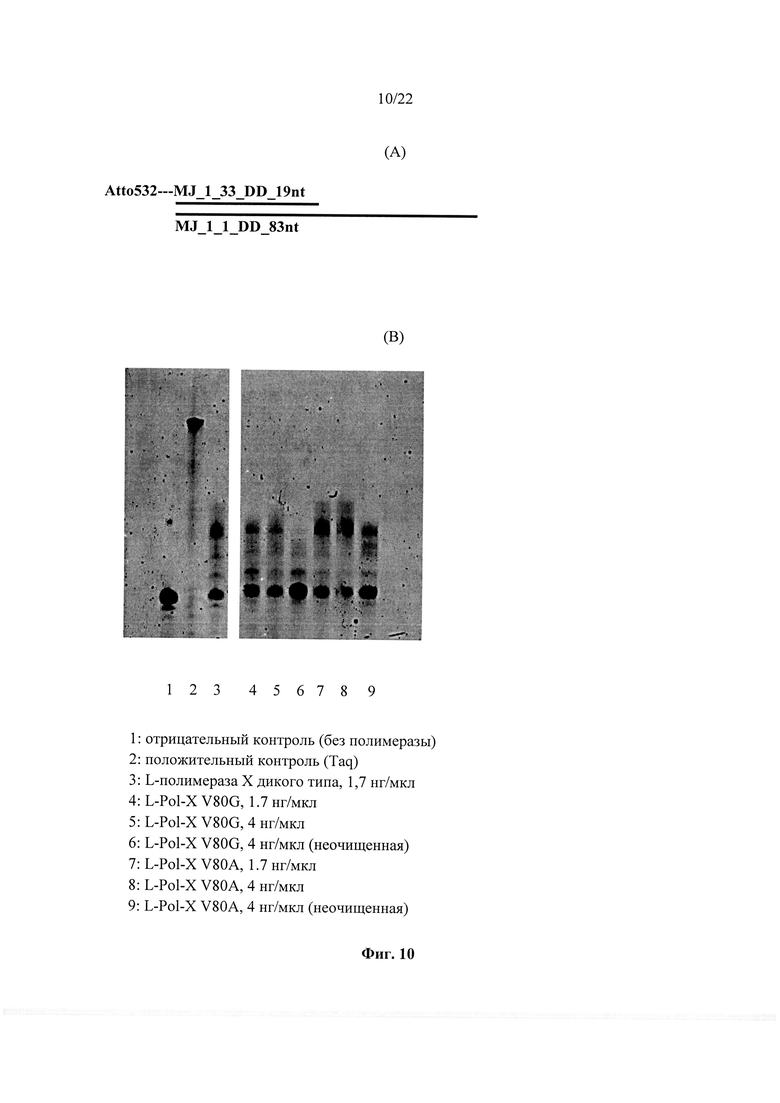
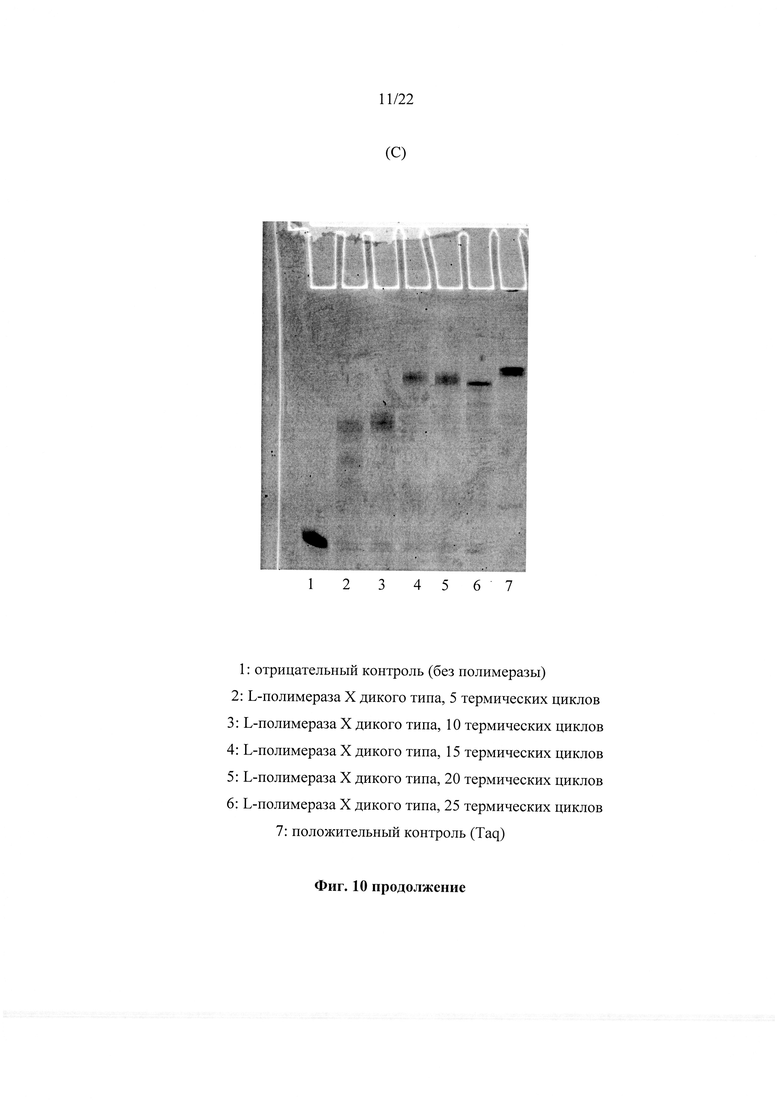
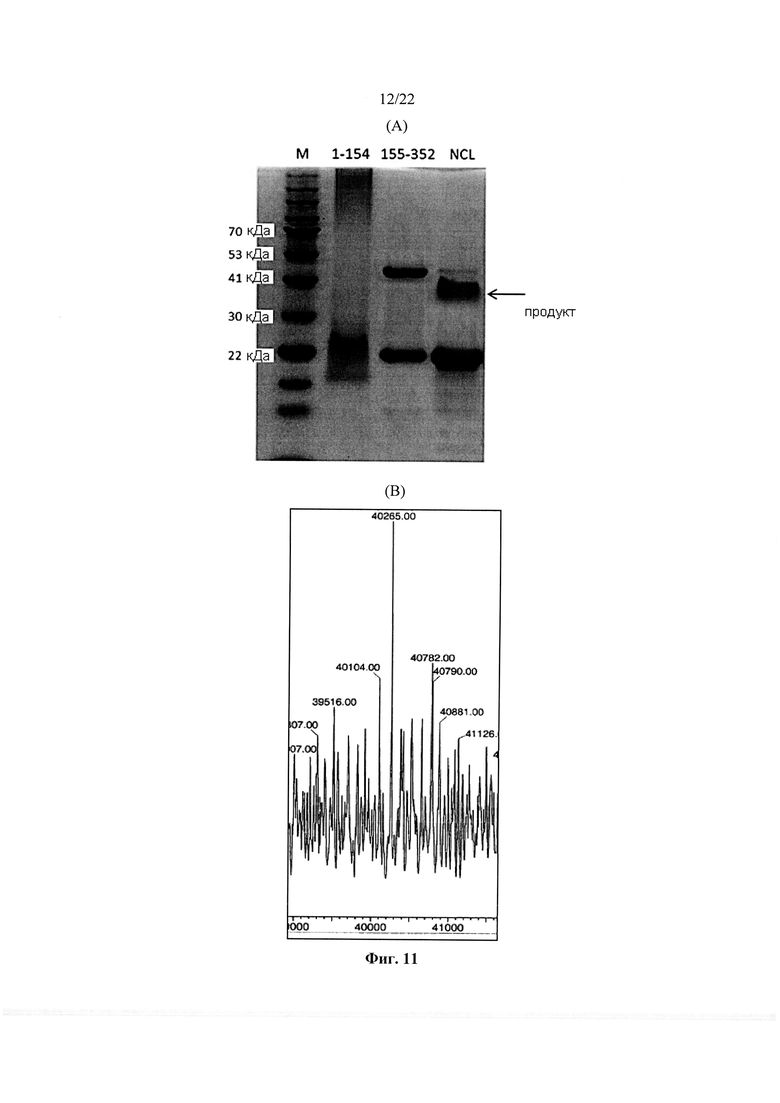

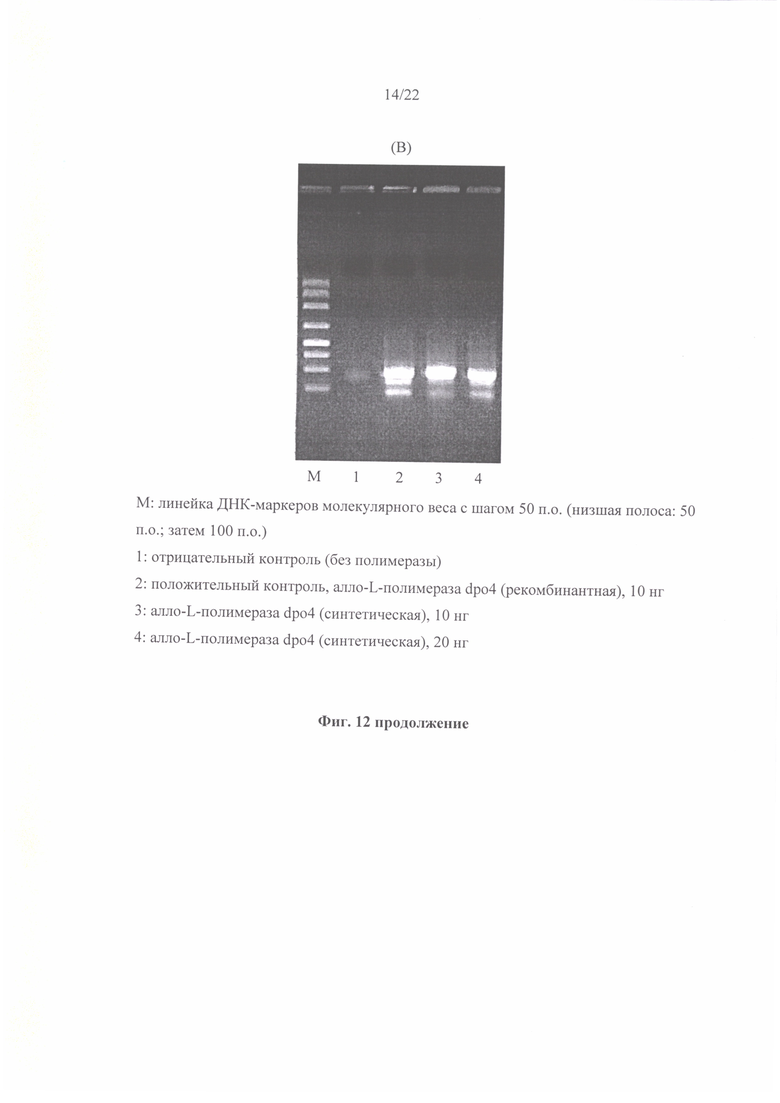
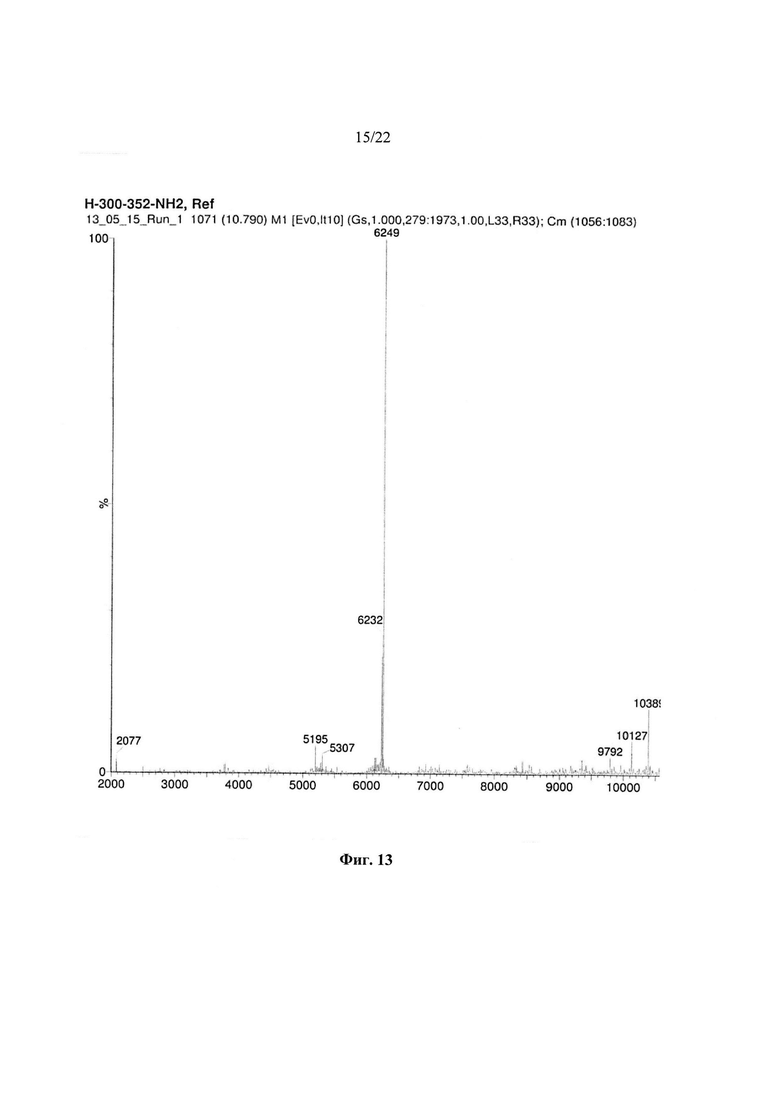


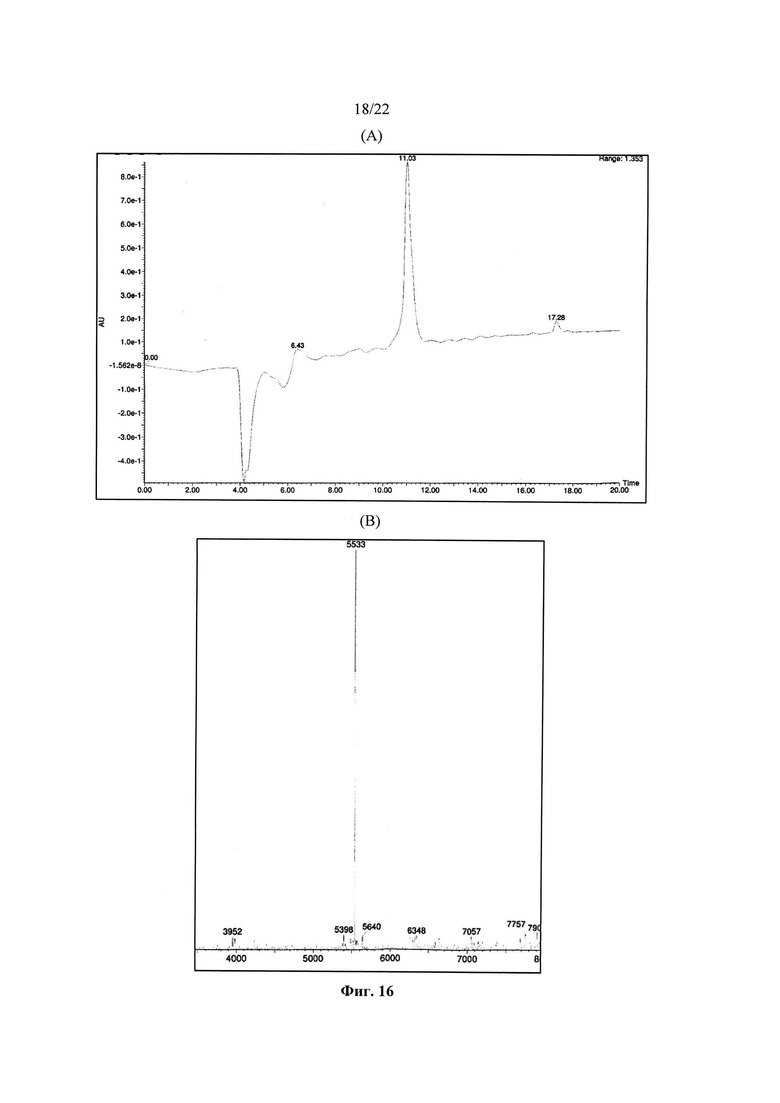

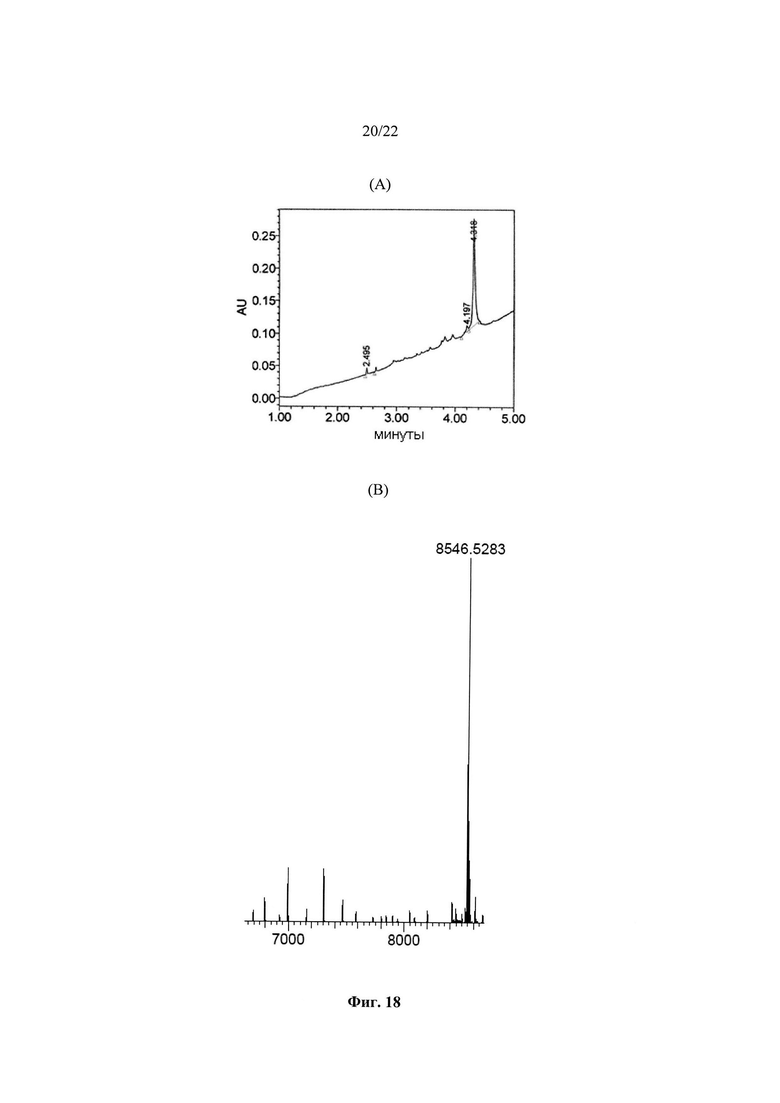
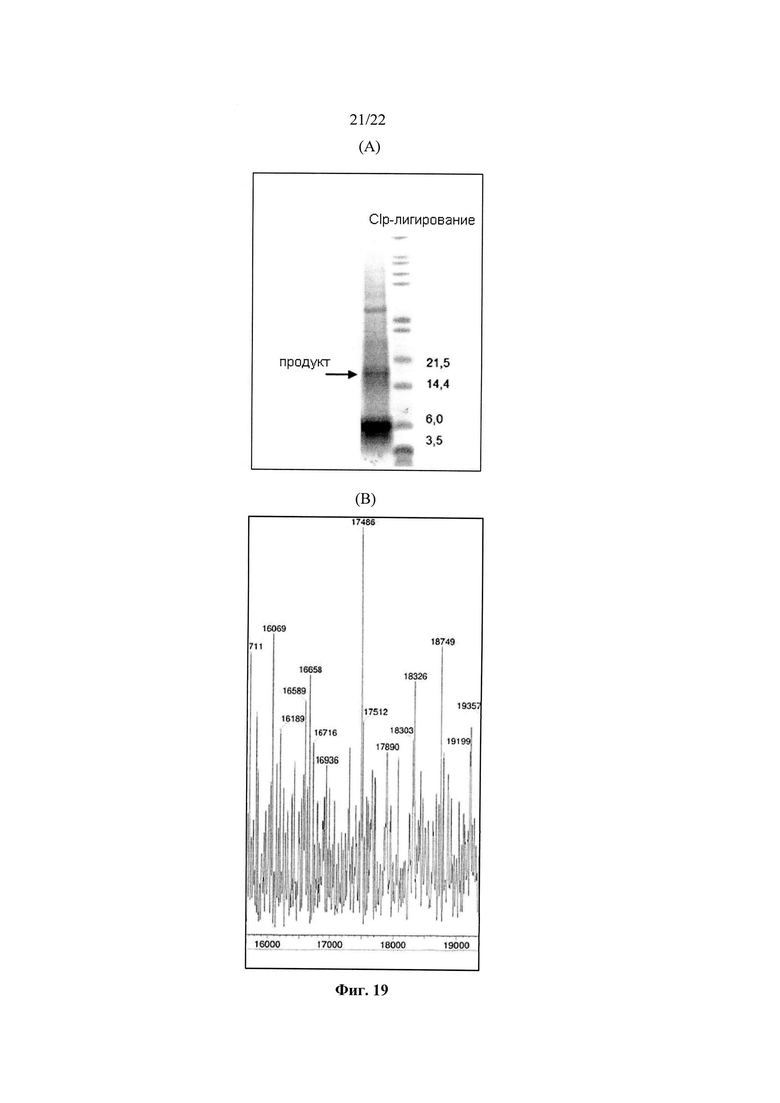
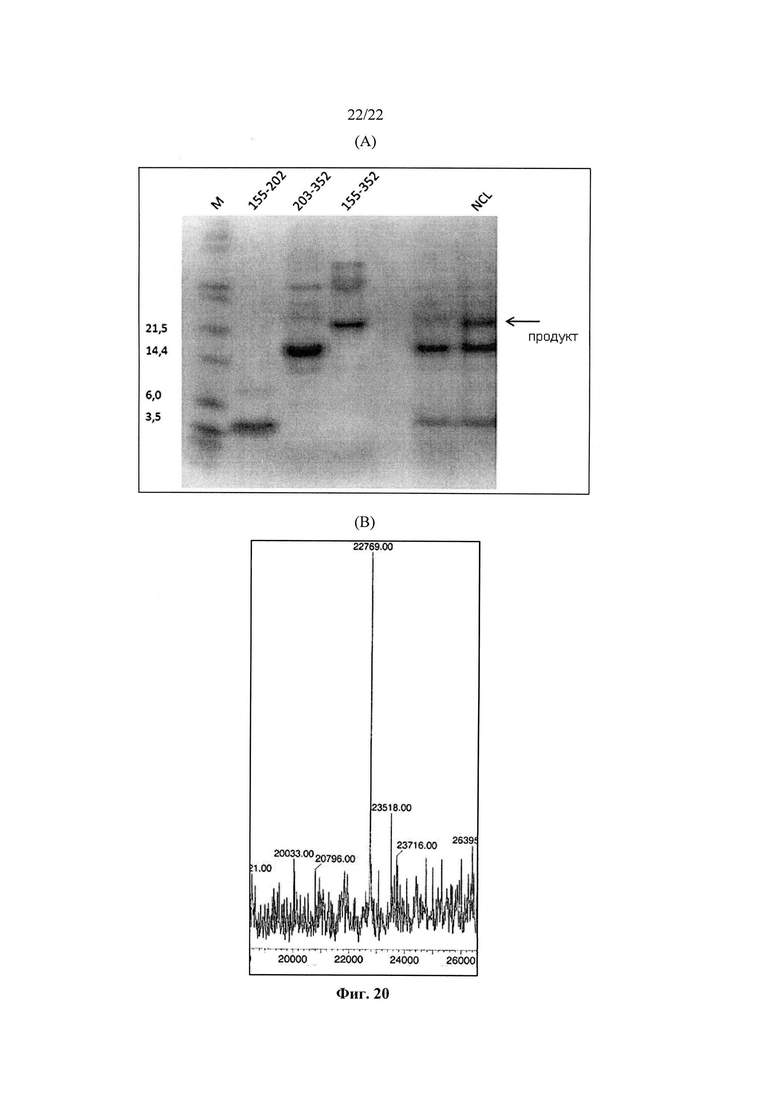
Изобретение относится к ферментативному синтезу L-нуклеиновых кислот в присутствии полимеразы. Предложен способ добавления одного или нескольких L-нуклеотидов к 3’-концу L-нуклеиновой кислоты, включающий стадию проведения реакции одного или нескольких L-нуклеотидов с L-нуклеиновой кислотой в присутствии полимеразы, где указанная полимераза способна добавлять один или несколько L-нуклеотидов к 3’-концу указанной L-нуклеиновой кислоты, при этом указанная полимераза состоит из аминокислотной последовательности, причём аминокислоты указанной аминокислотной последовательности представляют собой D-аминокислоты. Предложено также применение полимеразы, которая состоит из последовательности аминокислот, причём аминокислоты указанной последовательности аминокислот представляют собой D-аминокислоты, в способе добавления одного или нескольких L-нуклеотидов к 3’-концу L-нуклеиновой кислоты. Изобретение обеспечивает эффективное добавление одного или нескольких L-нуклеотидов к 3’-концу L-нуклеиновой кислоты в присутствии полимеразы, состоящей из D-аминокислот. 2 н. и 5 з.п. ф-лы, 20 ил., 10 табл., 12 пр.
1. Способ добавления одного или нескольких L-нуклеотидов к 3’-концу L-нуклеиновой кислоты, включающий стадию проведения реакции одного или нескольких L-нуклеотидов с L-нуклеиновой кислотой в присутствии полимеразы, где указанная полимераза способна добавлять один или несколько L-нуклеотидов к 3’-концу указанной L-нуклеиновой кислоты, при этом указанная полимераза состоит из аминокислотной последовательности, причём аминокислоты указанной аминокислотной последовательности представляют собой D-аминокислоты.
2. Способ по п.1, отличающийся тем, что указанная полимераза представляет собой термостабильную полимеразу.
3. Способ по п.1, отличающийся тем, что аминокислотная последовательность указанной полимеразы включает по меньшей мере 300 аминокислот.
4. Способ по п.1, отличающийся тем, что указанная полимераза выбрана из группы, состоящей из полимеразы X вируса африканской чумы свиней, корового домена полимеразы X Thermus thermophilus, крысиной полимеразы-бета, эукариотической полимеразы-бета, фрагмента Кленова, экзополимеразы Кленова, ДНК-полимеразы T4, ДНК-полимеразы Phi29, секвеназы, ДНК-полимеразы T7, полимеразы SP6, ДНК-полимеразы I, полимеразы-лямбда, и вариантов всех и любой из них, притом что предпочтительно указанная полимераза представляет собой полимеразу X вируса африканской чумы свиней или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 1, аминокислотной последовательности, соответствующей SEQ ID NO: 2, аминокислотной последовательности, соответствующей SEQ ID NO: 3, и аминокислотной последовательности, соответствующей SEQ ID NO: 4.
5. Способ по п.1, отличающийся тем, что указанная полимераза выбрана из группы полимеразы DPO4, ДНК-полимеразы Thermococcus litoralis, ДНК-полимеразы Pyrococcus sp., ДНК-полимеразы Pyrococcus furiosus, полимеразы Pfuturbo, ДНК-полимеразы Sulfolobus solfataricus, ДНК-полимеразы Thermococcus gorgonarius, полимеразы KOD, полимеразы Taq, полимеразы Tth, полимеразы Pyrobest, полимеразы Pwo, полимеразы Sac, полимеразы Bst, полимеразы Poc, полимеразы Pab, полимеразы Mth, полимеразы Pho, полимеразы ES4, полимеразы EX-Taq, полимеразы LA-Taq, полимераз Expand, полимераз Platinum Taq, высокоточной (Hi-Fi) полимеразы, полимеразы Tbr, полимеразы Tfl, полимеразы Tru, полимеразы Tac, полимеразы Tne, полимеразы Tma, полимеразы Tih, полимеразы Tfi, AmpliTaq, фрагмента Стоффеля, ДНК-полимеразы 9°Nm, Therminator, Therminator II, высокоточной полимеразы Phusion, Paq5000, Pfx-50, Proofstart, FideliTaq, элонгазы, и их вариантов, притом что предпочтительно указанная полимераза представляет собой полимеразу Dpo4 или ее вариант, состоящие из аминокислотных последовательностей, выбранных из группы, состоящей из аминокислотной последовательности, соответствующей SEQ ID NO: 15, аминокислотной последовательности, соответствующей SEQ ID NO: 16, аминокислотной последовательности, соответствующей SEQ ID NO: 17, аминокислотной последовательности, соответствующей SEQ ID NO: 18, аминокислотной последовательности, соответствующей SEQ ID NO: 19, аминокислотной последовательности, соответствующей SEQ ID NO: 20, аминокислотной последовательности, соответствующей SEQ ID NO: 21, и аминокислотной последовательности, соответствующей SEQ ID NO: 22.
6. Способ по любому из пп.1-5, отличающийся тем, что реакция включает дополнительную L-нуклеиновую кислоту, причём одна молекула указанной L-нуклеиновой кислоты гибридизуется с одной молекулой указанной дополнительной L-нуклеиновой кислоты, предпочтительно, путем спаривания оснований по Уотсону-Крику.
7. Применение полимеразы, которая состоит из последовательности аминокислот, причём аминокислоты указанной последовательности аминокислот представляют собой D-аминокислоты, в способе добавления одного или нескольких L-нуклеотидов к 3’-концу L-нуклеиновой кислоты.
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| KEEFE A.D | |||
| ET AL | |||
| Aptamers as therapeutics // NATURE REVIEWS, Drug Discovery, JULY 2010, volume 9, 537-550 | |||
| Биохимия: Учебник / Под ред | |||
| Е.С | |||
| Северина | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| - М.: ГЭОТАР-МЕД, 2004, с | |||
| Двухколейная подвесная дорога | 1919 |
|
SU151A1 |

