ОБЛАСТЬ ТЕХНИКИ
[0001]
Настоящее изобретение относится к методу, способу, средству и т.п. для лечения или профилактики заболевания, расстройства или состояния, связанного со стрессом эндоплазматического ретикулума (ER) и гибелью клеток.
УРОВЕНЬ ТЕХНИКИ
[0002]
Визуальная информация распознается после того, как свет попадает в роговую оболочку, которая представляет собой прозрачную ткань в передней части глазного яблока, достигает сетчатки и возбуждает нервные клетки сетчатки, после чего сгенерированный электрический сигнал передается через оптический нерв в зрительную зону коры головного мозга. Для хорошего зрения нужно, чтобы роговица была прозрачной. Прозрачность роговой оболочки достигается путем поддержания постоянного содержания воды посредством насосных и барьерных функций эндотелиальных клеток роговицы.
[0003]
Плотность эндотелиальных клеток роговицы человека при рождении составляет примерно 3000 клеток на 1 мм2. Однако эндотелиальные клетки роговицы человека обладают крайне ограниченной способностью к регененрации после повреждения.
[0004]
В настоящее время проводятся исследования по болезненным состояниям эндотелиальных клеток роговицы. В непатентном документе 1 описано исследование по взаимосвязи человеческих эндотелиальных клеток роговицы и окислительного стресса. В непатентном документе 2 описано основное исследование по взаимосвязи человеческих эндотелиальных клеток роговицы и стресса эндоплазматического ретикулума. В непатентном документе 3 описано основное исследование по взаимосвязи человеческих эндотелиальных клеток роговицы и окислительного стресса.
СПИСОК ЛИТЕРАТУРЫ
[Непатентные документы]
[0005]
[NPL 1] Onouchi H. et al., Biomedical Gerontology: Vol.34, No.2, 51 (2010)
[NPL 2] William L. Corwin et al., Cryobiology: Vol.63, No.1, 46−55 (2011)
[NPL 3] Ula V. Jurkunas et al., Am J Pathol: Vol.177, No.5, 2278−2289 (2010)
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[Решение задачи]
[0006]
Для поддержания эндотелия роговой оболочки в хорошем состоянии требуются разные факторы. Авторы настоящего изобретения сконцентрировали свое внимание, в числе прочих факторов, на взаимосвязи стресса эндоплазматического ретикулума (ER) и состояния эндотелиальных клеток роговицы, и обнаружили, что стрессовое состояние можно улучшить путем ингибирования пути трансформирующего фактора роста-β (TGF-β), кроме того, они разработали способ, позволяющий лечить или предотвращать расстройства, связанные со стрессом ER, и тем самым завершили настоящее изобретение. Таким образом, настоящее изобретение относится к следующим изобретениям:
(1) Терапевтическое или профилактическое средство против заболевания, расстройства или состояния, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки, включающее в себя ингибитор сигнального пути TGFβ.
(2) Терапевтическое или профилактическое средство по пункту (1), где заболевание, расстройство или состояние представляет собой расстройство, связанное с дистрофией эндотелия роговицы Фукса.
(3) Терапевтическое или профилактическое средство по пункту (1) или (2), где терапевтическое или профилактическое средство подавляет заболевание, расстройство или состояние, включающее в себя расстройство эндотелиальных клеток роговицы при дистрофии эндотелия роговицы Фукса.
(4) Терапевтическое или профилактическое средство по любому из пунктов (1)-(3), где заболевание, расстройство или состояние представляет собой по меньшей мере одно заболевание, расстройство или состояние, выбранное из группы, включающей уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету и отек стромы роговой оболочки.
(5) Терапевтическое или профилактическое средство по любому из пунктов (1)-(4), где ингибитор сигнального пути TGFβ включает по меньшей мере одно средство, выбранное из группы, включающей 4−[4−(1,3−бензодиоксол−5-ил)−5−(2−пиридинил)−1H−имидазол−2-ил]бензамид, BMP−7, антитело к TGF−β, антитело к рецептору TGF−β, миРНК TGF−β, миРНК рецептора TGF−β, кшРНК TGF−β, кшРНК рецептора TGF−β, аптамер TGF−β, аптамер рецептора TGF−β, антисмысловой олигонуклеотид TGF−β, 6,7−диметокси−2−((2E)−3−(1−метил−2−фенил−1H−пирроло[2,3−b]пиридин−3-илпроп-2-еноил))−1,2,3,4−тетрагидроизохинолон, 3−(6−метил−2−пиридинил)−N−фенил−4−(4−хинолинил)−1H−пиразол−1−карботиоамид, 2−(3−(6−метилпиридин−2-ил)−1H−пиразол−4-ил)−1,5−нафтиридин, 6−(4−(пиперидин−1-ил)этокси)фенил)−3−(пиридин−4-ил)пиразоло[1,5−a]пиримидин, 2−(5−хлор−2−фторфенил)−4−[(4−пиридинил)амино]птеридин, 4−[3−(2−пиридинил)−1H−пиразол−4-ил]−хинолин, A−83−01 (3−(6−метил−2−пиридинил)−N−фенил−4−(4−хинолинил)−1H−пиразол−1−карботиоамид), фармацевтически приемлемые соли или сольваты указанных соединений, или сольваты их фармацевтически приемлемых солей.
(6) Терапевтическое или профилактическое средство по любому из пунктов (1)-(5), где ингибитор сигнального пути TGF-β представляет собой 4−[4−(1,3−бензодиоксол−5-ил)−5−(2−пиридинил)−1H−имидазол−2-ил]бензамид или его фармацевтически приемлемую соль.
(7) Терапевтическое или профилактическое средство по любому из пунктов (1)-(6), где эндотелий роговой оболочки представляет собой эндотелий примата.
(8) Терапевтическое или профилактическое средство по любому из пунктов (1)-(7), где эндотелий роговой оболочки представляет собой эндотелий человека.
(9) Терапевтическое или профилактическое средство по любому из пунктов (1)-(8), содержащее дополнительный фармацевтический ингредиент.
(10) Терапевтическое или профилактическое средство по любому из пунктов (1)-(9), которое представляет собой глазные капли.
(11) Средство, ингибирующее сигнальный путь TGF-β, предназначенное для лечения или профилактики расстройства, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки.
(11A) Средство, ингибирующее сигнальный путь TGFβ, по пункту (11), которое обладает признаком ингибитора, описанным в одном из пунктов (1)-(10).
(12) Способ лечения или профилактики расстройства, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки у индивидуума, где способ включает стадию введения индивидууму эффективного количества ингибитора сигнального пути TGFβ.
(12A) Способ по пункту (12), характеризующийся признаком, описанным в одном из пунктов (1)-(10).
[0007]
В другом аспекте настоящее изобретение относится к следующим изобретениям:
(A1) Терапевтическое или профилактическое средство против заболевания, расстройства или состояния, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки, включающее в себя ингибитор сигнального пути TGF-β.
(A2) Терапевтическое или профилактическое средство по пункту (A1), где заболевание, расстройство или состояние связано с митохондриальной недостаточностью.
(A3) Терапевтическое или профилактическое средство по пункту (A1) или (A2), где заболевание, расстройство или состояние связано с апоптозом вследствие митохондриальной недостаточности.
(A4) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A3), где заболевание, расстройство или состояние связано с дистрофией эндотелия роговицы Фукса.
(A5) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A4), где терапевтическое или профилактическое средство подавляет заболевание, расстройство или состояние, включающее в себя расстройство функционирования клеток эндотелия роговицы при дистрофии эндотелия роговицы Фукса.
(A6) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A5), где терапевтическое или профилактическое средство подавляет по меньшей мере одно заболевание, расстройство или состояние, выбранное из группы, включающей уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету и отек стромы роговой оболочки.
(A7) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A6), где ингибитор сигнального пути TGF-β включает по меньшей мере одно средство, выбранное из группы, включающей 4−[4−(1,3−бензодиоксол−5-ил)−5−(2−пиридинил)−1H−имидазол−2-ил]бензамид, BMP−7, антитело к TGF−β, антитело к рецептору TGF−β, миРНК TGF−β, миРНК рецептора TGF−β, кшРНК TGF−β, кшРНК рецептора TGF−β, аптамер TGF−β, аптамер рецептора TGF−β, антисмысловой олигонуклеотид TGF−β, 6,7−диметокси−2−((2E)−3−(1−метил−2−фенил−1H−пирроло[2,3−b]пиридин−3-ил−проп-2-еноил))−1,2,3,4−тетрагидроизохинолон, 3−(6−метил−2−пиридинил)−N−фенил−4−(4−хинолинил)−1H−пиразол−1−карботиоамид, 2−(3−(6−метилпиридин−2-ил)−1H−пиразол−4-ил)−1,5−нафтиридин, 6−(4−(пиперидин−1-ил)этокси)фенил)−3−(пиридин−4-ил)пиразоло[1,5−a]пиримидин, 2−(5−хлор−2−фторфенил)−4−[(4−пиридинил)амино]птеридин, 4−[3−(2−пиридинил)−1H−пиразол−4-ил]−хинолин, A−83−01 (3−(6−метил−2−пиридинил)−N−фенил−4−(4−хинолинил)−1H−пиразол−1−карботиоамид), фармацевтически приемлемые соли или сольваты указанных соединений, или сольваты их фармацевтически приемлемых солей.
(A8) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A7), где ингибитор сигнального пути TGF-β представляет собой 4−[4−(1,3−бензодиоксол−5-ил)−5−(2−пиридинил)−1H−имидазол−2-ил]бензамид или его фармацевтически приемлемую соль.
(A9) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A8), дополнительно включающее в себя терапевтическое средство против митохондриальной недостаточности, вызванной стрессом ER.
(A10) Терапевтическое или профилактическое средство по пункту (A9), где терапевтическое средство против митохондриальной недостаточности, вызванной стрессом ER, выбрано из группы, включающей BiP индуктор X (BIX), 4−фенилмасляную кислоту (PBA), триметиламин N−оксид (TMAO), тауроурсодезоксихолевую кислоту (TUDCA) и терпенон (также продаваемый под названием селбекс).
(A11) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A10), где эндотелий роговой оболочки представляет собой эндотелий примата.
(A12) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A11), где эндотелий роговой оболочки представляет собой эндотелий человека.
(A13) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A12), включающее в себя дополнительный фармацевтический ингредиент.
(A14) Терапевтическое или профилактическое средство по любому из пунктов (A1)-(A13), которое представляет собой глазные капли.
(A14A) Терапевтическое или профилактическое средство по любому из вышеуказанных пунктов, где заболевание, расстройство или состояние сопровождается экспрессией агресомы.
(A14B) Терапевтическое или профилактическое средство по любому из вышеуказанных пунктов, где заболевание, расстройство или состояние представляет собой заболевание, расстройство или состояние, связанное с агресомой.
(A14C) Терапевтическое или профилактическое средство по любому из вышеуказанных пунктов, где заболевание, расстройство или состояние сопровождается неправильной укладкой белка.
(A14D) Терапевтическое или профилактическое средство по любому из вышеуказанных пунктов, где заболевание, расстройство или состояние обусловлено неправильной укладкой белка.
(A15) Средство, ингибирующее сигнальный путь TGF-β, предназначенное для лечения или профилактики расстройства, связанного со стрессом эндоплазматического ретикулума (ER).
(A15A) Средство, ингибирующее сигнальный путь TGFβ, по пункту (A15), которое обладает признаком ингибитора, описанным в одном из пунктов (A1)-(A14) и (A14A)-(A14D).
(A16) Способ лечения или профилактики расстройства, связанного со стрессом эндоплазматического ретикулума (ER), у индивидуума, где способ включает стадию введения индивидууму эффективного количества ингибитора сигнального пути TGFβ.
(A16A) Способ по пункту (A16), характеризующийся признаком, описанным в одном из пунктов (A1)-(A14) и (A14A)-(A14D).
[0008]
Стресс эндоплазматического ретикулума представляет собой стрессовое состояние клеток, обусловленное накоплением в клетках белков, укладка которых не завершается нормальной конфигурацией. Стресс эндоплазматического ретикулума не индуцируется в нормальном внеклеточном матриксе. В данной связи невозможно предсказать, исходя из сведений, касающихся данного факта, таких как информация по внеклеточному матриксу, что облегчения клеточного расстройства, обусловленного стрессом эндоплазматического ретикулума при дистрофии эндотелия роговицы Фукса, в соответствии с настоящим изобретением можно достичь путем подавления сигнального пути TGFβ.
[0009]
Предполагается, что один или несколько из вышеуказанных признаков, описанных в настоящем изобретении, могут существовать в других комбинациях, помимо явно указанных комбинаций. Другие варианты осуществления и преимущества настоящего изобретения станут ясны для специалистов в данной области после прочтения и осознания нижеследующего Подробного описания в необходимом объеме.
[Полезные эффекты изобретения]
[0010]
Настоящее изобретение относится к лекарственному средству, позволяющему лечить или предотвращать заболевание, связанное со стрессом эндоплазматического ретикулума (ER), для лечения которого в настоящее время отсутствуют другие способы, кроме роговичной трансплантации, причем указанное средство также может существовать в виде глазных капель и т.п.
[Краткое описание чертежей]
[0011]
[Фигура 1] На фигуре 1 приведена гипотетическая схема патологии заболевания, связанного со стрессом эндоплазматического ретикулума (такого как дистрофия эндотелия роговицы Фукса), где основное внимание уделяется взаимосвязи стресса эндоплазматического ретикулума и апоптоза, в соответствии с настоящим изобретением.
[Фигура 2] На фигуре 2 показаны морфологические аномалии эндоплазматического ретикулума и митохондрий при дистрофии эндотелия роговицы Фукса. Слева показаны иммобилизованные эндотелиальные клетки роговицы человека (iHCEC), а справа показаны иммобилизованные клетки эндотелия, пораженные дистрофией роговицы Фукса (iFECD). Как в верхнем, так и в нижнем ряду показаны изображения, полученные с помощью просвечивающего электронного микроскопа. * обозначает митохондрии, ** обозначает эндоплазматический ретикулум, а *** обозначает внеклеточный матрикс.
[Фигура 3] На фигуре 3 показаны результаты иммуноокрашивания, демонстрирующие, что стресс эндоплазматического ретикулума в эндотелиальных клетках роговицы усиливается при дистрофии эндотелия роговицы Фукса. Показано, что в iFECD наблюдается выраженная экспрессия белков, ассоциированных со стрессом эндоплазматического ретикулума, таких как GRP78 и GADD153. В верхнем ряду в iHCEC, показанных в левой панели, наблюдается слабая зеленая флуоресценция GRP78, тогда как в iFECD, показанных в правой панели, зеленая флуоресценция является значительной. С другой стороны, в нижнем ряду зеленая флуоресценция GADD153 в левой панели еле заметна, а в правой панели она является выраженной.
[Фигура 4] На фигуре 4 показана диаграмма, демонстрирующая, что TGF-β стимулирует стресс ER в эндотелиальных клетках роговицы. В верхней панели показаны iHCEC, а в нижней панели показаны iFECD. В каждой панели, в направлении слева направо, показаны контрольные клетки (отсутствие стимуляции) и клетки, стимулированные TGF-β или TG. В каждой панели приведены, в направлении сверху вниз, GRP78, IRE1 и GAPDH. Показано, что экспрессия шаперона GRP78 и индикатора стресса IRE1 индуцируется под действием TGFβ, причем их экспрессия происходит на повышенном уровне, особенно в iFECD. TG обозначает тапсигаргин, который используют в качестве положительного контроля для стресса ER.
[Фигура 5] На фигуре 5 показаны результаты, демонстрирующие, что в эндотелиальных клетках роговицы при дистрофии эндотелия роговицы Фукса нблюдается более высокий уровень стресса эндоплазматического ретикулума, индуцированного TGF-бета, чем в контрольных клетках (эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса, обладают повышенной чувствительностью к стрессу эндоплазматического ретикулума). Слева показаны iHCEC, а справа показаны iFECD. С каждой стороны, в верхней левой части показаны результаты, полученные для контрольных клеток, а в верхней правой части показаны результаты добавления SB431542 к контрольным клеткам, в средней части слева показаны результаты, полученные для клеток, стимулированных TGF-β2, а в средней части справа показаны результаты добавления SB431542 к клеткам, стимулированным TGF-β2. Внизу слева показаны результаты стимуляции TG, а внизу справа показаны результаты добавления SB431542 к клеткам, стимулированным TG.
[Фигура 6] На фигуре 6 показаны результаты, демонстрирующие, что в эндотелиальных клетках роговицы при дистрофии эндотелия роговицы Фукса нблюдается более высокий уровень стресса эндоплазматического ретикулума, индуцированного TGFβ, чем в контрольных клетках (эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса, обладают повышенной чувствительностью к стрессу эндоплазматического ретикулума). Это является типичным примером оценки апоптоза в клетках, приведенных на фигуре 5, методом проточной цитометрии. Слева показаны iHCEC, а справа показаны iFECD. С каждой стороны, в верхней левой части показаны результаты, полученные для контрольных клеток, а в верхней правой части показаны результаты добавления SB431542 к контрольным клеткам. Внизу слева показаны результаты стимуляции TGF-β2, а внизу справа показаны результаты добавления SB431542 к клеткам, стимулированным TGF-β2.
[Фигура 7] На фигуре 7 показан график, демонстрирующий, что SB431542 подавляет апоптоз при дистрофии эндотелия роговицы Фукса. На оси y показан процент аннексин V-положительных клеток. На оси x показаны, в направлении слева направо, контрольные клетки, клетки, обработанные SB431542, TGFβ2 и TGFβ+SB431542. Белым цветом обозначены iHCEC, а черным цветом обозначены iFECD. * указывает статистическую значимость (p<0,01). Вертикальные черточки соответствуют стандартному отклонению.
[Фигура 8] На фигуре 8 показан результат эксперимента, подобного эксперименту, приведенному на фигуре 7, в котором используют A-83-01 и ингибитор ALK5. На оси y показан процент аннексин V-положительных клеток. На оси x показаны, в направлении слева направо, контрольные клетки, клетки, обработанные TGFβ2, TGFβ2+SB431542, TGFβ2+A-83-01 и TGFβ2+ингибитор ALK5. Белым цветом обозначены iHCEC, а черным цветом обозначены iFECD. * указывает статистическую значимость (p<0,01). Вертикальные черточки соответствуют стандартному отклонению.
[Фигура 9] На фигуре 9 показана диаграмма, демонстрирующая, что TGF-β стимулирует стресс ER в эндотелиальных клетках роговицы. Показано, что стресс ER индуцируется под действием TGF-β в iFECD. Результаты вестерн-блоттинга для разных моментов времени после обработки TGF-β2 представлены в виде 2 колонок для каждого момента времени (в каждой паре из 2 колонок левая колонка соответствует iHCEC, а правая колонка соответствует iFECD), включающего в себя, в направлении слева направо, 0 часов, 3 часа и 6 часов после обработки. Начиная с верхнего ряда, показаны PERK, фосфорилированный PERK, ATF6, GRP78, CHOP и GAPDH. PERK и ATF6 являются индикаторами стресса ER, GRP78 представляет собой молекулярный шаперон, ответственный за целостность ER, а CHOP представляет собой фактор, индуцирующий апоптоз, опосредованный стрессом ER. GAPDH является контролем. Показано, что TGF-β стимулирует фосфорилирование индикатора стресса ER, PERK, и экспрессию ATF6. Кроме того, повышается экспрессия шаперона GRP78. Также показано, что в результате стресса ER повышается экспрессия CHOP и индуцируется апоптоз.
[Фигура 10] На фигуре 10 показано изображение окрашивания флуоресцентным красителем JC-1 (5,5',6,6'−тетрахлор−1,1',3,3'−тетраэтилбензимидазолилкарбоцианина иодид). В iFECD наблюдается деполяризация митохондрий. Левая колонка соответствует iHCEC, а правая колонка соответствует iFECD. В верхнем ряду показано окрашивание JC1 (зеленым цветом), в нижнем ряду показано окрашивание JC1 (красным цветом). Зеленый указывает на митохондрии, а красный указывает на мембранный потенциал митохондрий. Синий соответствует DAPI (окрашивает ядро). В iFECD наблюдается уменьшение мембранного потенциала митохондрий по сравнению с iHCEC. В случае зеленого окрашивания, указывающего на митохондрии (верхняя секция), зеленая флуоресценция, указывающая на митохондрии, наблюдается как в левой, так и в правой панелях. В случае красного окрашивания, указывающего на мембранный потенциал митохондрий (нижняя секция), красная флуоресценция преимущественно наблюдается в левой панели, однако в незначительной степени ее можно видеть и в правой.
[Фигура 11] На фигуре 11 показаны мембранные потенциалы митохондрий, измеренные с использованием MitoTracker® методом проточной цитометрии. Деполяризация митохондрий наблюдается в iFECD. На левой панели показана интенсивность флуоресценции митохондриальной мембраны. На правой панели показаны iHCEC в левом графике и iFECD в правом графике. На левой панели показан процент деполяризованных клеток. Левая колонка соответствует iHCEC, а правая колонка соответствует iFECD. В iFECD мембранный потенциал митохондрий значительно ниже, чем в iHCEC. ** указывает на статистическую значимость (p<0,05). Цвет левой панели (зеленый и красный) указывает на наличие деполяризованных клеток с пониженным мембранным потенциалом (зеленый) и недеполяризованных клеток с отсутствием снижения мембранного потенциала (красный).
[Фигура 12] На фигуре 12 показано просачивание цитохрома C из митохондрий в цитоплазму. Просачивание цитохрома C из митохондрий измеряют методом вестерн-блоттинга. В iFECD просачивание цитохрома C наблюдается в большей степени, чем в iHCEC. Стауроспорин используют в качестве контроля. Две левые колонки соответствуют iHCEC (левая сторона указывает на отсутствие стауроспорина, а правая сторона указывает на присутствие стауроспорина), а две правые колонки соответствуют iFECD (левая сторона указывает на отсутствие стауроспорина, а правая сторона указывает на присутствие стауроспорина). Начиная с верхнего ряда, показано окрашивание цитохрома C, GAPDH и VDAC. GAPDH представляет собой цитозольный маркер, а VDAC представляет собой митохондриальный маркер.
[Фигура 13] На фигуре 13 показана роль митохондриальной недостаточности в развитии апоптоза. В данной схеме стауроспосрином индуцируют митохондриальное расстройство и методом вестерн-блоттинга измеряют ассоциированные с апоптозом белки, чтобы подтвердить, что митохондриальное расстройство индуцирует апоптоз. Начиная от левой колонки показаны контрольные результаты и результаты, полученные через 15 минут, 30 минут, 1 час, 3 часа и 4 часа после стимуляции стауроспорином. Начиная с верхнего ряда, показаны результаты по каспазе 9, каспазе 3, PARP и GADPH (контроль). Наблюдается активация каспазы 9, которая обычно происходит в результате митохондриального расстройства, а также активация каспазы 3 и PARP, свидетельствующая о том, что митохондриальное расстройство вызывает апоптоз.
[Фигура 14] На фигуре 14 показана диаграмма, демонстрирующая, что дистрофия эндотелия роговицы Фукса характеризуется высоким уровнем денатурированных белков и усиливается в результате стимуляции под действием TGF-β. На левой панели приведены изображения контрольных клеток (отсутствие стимуляции TGFβ: верхний ряд) и клеток, стимулированных TGF-β (10 нг/мл; нижний ряд) (40-кратное увеличение). Каждый из графиков в верхней правой части демонстрирует результат, полученный помощью проточного цитометра. В направлении слева направо показано сравнение iHCEC (черный) с iFECD (красный), сравнение iHCEC (черный) с iHCEC+TGFβ (красный) и сравнение iFECD (черный) с iFECD+TGFβ (красный). По оси y откладывают число клеток, а по оси x откладывают интенсивность флуоресценции агресом. На левом графике наблюдается сдвиг всего пика iHCEC (черный) влево относительно iFECD (красный). На среднем графике пик iHCEC (черный) почти полностью перекрывается с пиком iHCEC+TGFβ (красный). На правом графике пик iFECD (черный) сдвинут вверх влево по сравнению с iFECD+TGFβ (красный). В нижней правой секции сравнивается интенсивность флуоресценции агресом в качестве контроля с TGFβ-стимулированными iHCEC и iFECD. Белый столбик соответствует iHCEC, а черный столбик соответствует iFECD. * указывает, что p<0,05.
[Описание вариантов осуществления]
[0012]
Далее приводится описание настоящего изобретения. Если не указано иное, используемые на протяжении всего описания выражения, связанные с единственным числом, следует понимать как охватывающие соответствующее множественное число. Таким образом, если не указано иначе, грамматические формы, обозначающие единственное число, следует понимать как охватывающие соответствующие грамматические формы, относящиеся к множественному числу. Кроме того, если не указано иначе, термины, присутствующие в настоящем описании, имеют значения, традиционно используемые в данной области. Так, если не указано иначе, все терминологии и научно-технические термины используются в традиционном значении, известном специалистам в области, к которой принадлежит настоящее изобретение. В случае противоречия настоящее описание (включая определения) имеет преимущество.
[0013]
(Определения)
В рамках изобретения "iFECD" (иммобилизованные клетки эндотелия, пораженного дистрофией роговицы Фукса) представляет собой сокращенное название иммобилизованных клеток эндотелия, пораженного дистрофией роговицы Фукса.
[0014]
В рамках изобретения "HCEC" представляет собой сокращенное название эндотелиальных клеток роговицы человека. А "iHCEC" представляет собой сокращенное название иммобилизованных эндотелиальных клеток роговицы человека.
[0015]
В рамках изобретения термин "трансформирующий фактор роста-β (также сокращенно обозначаемый TGF-β)" имеет значение, используемое в данной области. Он представляет собой гомодимерный полифункциональный цитокин с молекулярной массой 25 кДа, выполняющий ряд биологических функций, например, он участвует в патогенезе разных склеротических заболеваний, ревматоидного артрита и пролиферативной витреоретинопатии, играет важную роль в развитии алопеции, подавляет функционирование иммунокомпетентных клеток и одновременно чрезмерную продукцию протеаз, предотвращая деградацию легочной ткани, приводящую к эмфиземе легких, и подавляет рост раковых клеток. У людей существуют три изоформы, от TGF-β1 до β3. TGF-β продуцируется в виде неактивной латентной формы с молекулярной массой примерно 300 кДа, которая не способна связываться с рецептором. Ее действие осуществляется путем активации на поверхности мишени или в окружающей ее среде с превращением в активную форму, способную связываться с рецептором.
[0016]
Не желая быть связанными с какой-либо теорией, авторы полагают, что действие TGF-β в клетке-мишени осуществляется через канал фосфорилирования ряда белков, отвечающих за передачу информации, называемых Smad. Вначале активированный TGF-β связывается с рецептором TGF-β типа II на поверхности клетки-мишени, образуя комплекс с рецептором, состоящий из двух молекул рецепторов типа II и двух молекул рецепторов TGF-β типа I, причем рецепторы типа II фосфорилируют рецепторы типа I. Известно, что фосфорилированные рецепторы типа I затем фосфорилируют Smad2 или Smad3, а Smad2 или Smad3 образуют комплекс со Smad4, который мигрирует в ядро и связывается с последовательностью-мишенью, называемой блок CAGA, присутствующей в участке промотора гена-мишени, индуцируя транскрипцию и экспрессию целевого гена с соактиватором.
[0017]
Сигнальный путь трансформирующего фактора роста-β (TGF-β) участвует в осуществлении многих клеточных функций, таких как рост и дифференциация клеток, прекращение роста, апоптоз и эпителиально-мезенхимальный переход (EMT), путем модуляции целевого гена. Члены семейства TGF-β, включающие в себя сам TGF-β (например, TGF-β1, TGF-β2 и TGF-β3), активин и костные морфогенетические белки (BMP), являются активными модуляторами роста, дифференциации, миграции и апоптоза клеток.
[0018]
TGF-β представляет собой белок размером примерно 24 кДа, продуцируемый многими клетками, такими как B-клетки, T-клетки и активированные макрофаги, а также клетками разных других типов. Эффекты TGF-β, оказываемые на иммунную систему, включают в себя индукцию рецептора IL-2, ингибирование IL-1-индуцированного роста тимоцитов и блокирование IFN-γ-индуцированной активации макрофагов. Известно, что TGF-β участвует в развитии разных патологических состояний (Border et al. (1992) J. Clin. Invest. 90:1), причем доказано, что он функционирует либо как вещество, подавляющее развитие опухоли, либо как вещество, стимулирующее развитие опухоли.
[0019]
Сигнальный путь TGF-β опосредуется двумя клеточными поверхностными рецепторами с серин/треонин-киназной активностью, TGF-βRII и ALK5. Сигнальный путь TGF-β инициируется лиганд-индуцированной димеризацией рецептора, после чего TGF-βRII может фосфорилировать рецептор ALK5. Фосорилирование активирует киназную активность ALK5 и затем активированный ALK5 фосфорилирует нижестоящий эфекторный белок Smad (позвоночный гомолог MAD или "Mothers against DPP (декапентаплегический фактор)", Smad2 или Smad3. Комплекс p-Smad2/3 со Smad4 поступает в ядро и активирует транскрипцию целевого гена.
[0020]
Smad3 является членом подгруппы R-Smad (рецептор-активируемый Smad) и непосредственным медиатором активации транскрипции под действием рецептора TGF-β. Стимуляция TGF-β приводит к фосфорилированию и активации Smad2 и Smad3, которые образуют комплекс со Smad4 ("сомедиаторный Smad" или "co-Smad" у позвоночных). Данный комплекс накапливается в ядре и модулирует транскрипцию целевого гена. R-Smad локализуется в цитоплазме и образует комплекс с co-Smad после лиганд-индуцированного фосфорилирования под действием рецептора TGF-β и мигрирует в ядро, где он модулирует экспрессию гена, ассоциированную с кооперативным фактором транскрипции и хроматином. Smad6 и Smad7 представляют собой ингибиторные Smad ("I-Smad"), то есть, они индуцируются на уровне транскрипции под действием TGF-β и функционируют как ингибиторы сигнального пути TGF-β (Feng et al. (2005) Annu. Rev. Cell. Dev. Biol. 21:659). Smad6/7 препятствуют рецептор-опосредованной активации R-Smad, осуществляя свой ингибиторный эффект; они связываются с рецептором типа I, конкурентно препятствуя мобилизации и фосфорилированию R-Smad. Известно, что Smad6 и Smad7 пополняют запасы убихитинлигазы E3, которая индуцирует убихитинирование и деградацию белков, взаимодействующих со Smad6/7.
В одном варианте осуществления заболевание, расстройство или состояние, на которое направлено настоящее изобретение, сопровождается повышенной экспрессией агресом. Альтернативно заболевание, расстройство или состояние на которое направлено настоящее изобретение, представляет собой заболевание, расстройство или состояние, ассоциированное с агресомами.
В другом варианте осуществления заболевание, расстройство или состояние, на которое направлено настоящее изобретение, сопровождается аномальной укладкой белка. Альтернативно, заболевание, расстройство или состояние, на которое направлено настоящее изобретение, обусловлено аномальной укладкой белка.
Известно, что в клетках млекопитающих белки, агрегированные вследствие отсутствия укладки, неправильной укладки, аномалий в деградации белков и т.п. (также называемые неполностью уложенные белки или денатурированные белки (неуложенные белки)), подвергаются убихитинированию и накапливаются вблизи центросомы посредством перемещения в микротрубочках, опосредованного динеином, образуя тельца включения, называемые агресомами. Как правило, агресомы образуются в результате термического шока, вирусной инфекции, окислительного стресса и т.п. У людей известны некоторые заболевания, в которых участвуют тельца включения, такие как тельца Льюиса, присутствующие в нервных клетках при болезни Паркинсона, тельца Мэллори, присутствующие в клетках печени при алкогольных болезнях печени, и стеклоподобные тельца в астроцитах при боковом амиотрофическом склерозе.
[0021]
Сигнальный путь TGF-β также может проходить по другим маршрутам, с использованием передачи BMP-7 и т.п., и осуществлением функции посредством ALK-1/2/3/6 и Smad1/5/8. Описание сигнальных путей TGF-β можно найти в J. Massagu'e, Annu. Rev. Biochem. 1998. 67: 753-91; Vilar JMG, Jansen R, Sander C (2006) PLoS Comput Biol 2 (1):e3; Leask, A., Abraham, D. J. FASEB J. 18, 816-827 (2004); Coert Margadant & Arnoud Sonnenberg EMBO reports (2010) 11, 97-105; Joel Rosenbloom et al., Ann Intern Med. 2010; 152: 159-166 и т.п.
[0022]
В рамках изобретения термин "ингибитор сигнального пути трансформирующего фактора роста (TGF)-β " относится к любому фактору, ингибирующему сигнальный путь TGF. TGF-β, выполняющий антагонистическую функцию, можно назвать антагонистом. Однако, в соответствии с настоящим изобретением, антагонисты TGF-β охватываются термином ингибиторы сигнального пути TGF-β. Поскольку такой ингибитор, как правило, представляет собой вещество, термины "вещество, ингибирующее сигнальный путь TGF-β " и "ингибитор сигнального пути TGF-β " используются как взаимозаменяемые. "TGF-β " также можно обозначать "TGFβ", который имеет такое же значение.
[0023]
Таким образом, типичные ингибиторы сигнального пути TGF-β, используемые в настоящем изобретении, включают в себя, без ограничения, антагонисты TGF-β, антагонисты рецептора TGF-β, ингибиторы Smad3, средства, улавливающие лиганды (рецепторы-ловушки, антитела против лигандов), антисмысловые олигонуклеотиды, ингибиторы TGF-β -рецепторной киназы, пептидные аптамеры, миРНК, кшРНК и т.п. (см. Connolly E., et al. Int. J. Biol. Sci. 2012; 8(7): 964−978, фиг. 3 и т.п.).
[0024]
Примеры ингибиторов сигнального пути TGF-β, которые можно использовать в настоящем изобретении, включают в себя, без ограничения, SB431542 (4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)]-1H-имидазол-2-ил]бензамид), BMP-7, антитела против TGF-β, антитела против рецептора TGF-β, миРНК TGF-β, миРНК рецептора TGF-β, антисмысловые олигонуклеотиды TGF-β, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-илпроп-2-еноил))-1,2,3,4-тетрагидроизохинолон, A83-01 (3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1H-пиразол-1-карботиоамид), ингибиторы Stemolecule™ TLK (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин), ингибитор Stemolecule™ BMP LDN-193189 (6-(4-(пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил)пиразоло[1,5-a]пиримидин), SD-208 (2-(5-хлор-2-фторфенил)-4-[(4-пиридинил)амино]птеридин), LY364947 (4-[3-(2-пиридинил)-1H-пиразол-4-ил]хинолин), их фармацевтически приемлемые соли или сольваты, сольваты фармацевтически приемлемых солей и т.п. Ингибиторы сигнального пути TGF-β, композиции, лекарственные средства, терапевтические средства и профилактические средства настоящего изобретения могут находиться в виде нейтральной формы, в виде соли, или в смеси с другим пролекарством (таким как сложный эфир и т.п.). В рамках изобретения термин "соль" включает, например, анионные соли, образованные любой кислой (например, карбоксильной) группой, и катионные соли, образованные любой основной (например, амино) группой. Соли включают в себя неорганические и органические соли, такие, как, например, соли, описанные в Berge et al., J.Pharm.Sci., 1977, 66, 1−19. Кроме того, соли включают в себя, например, соли металлов, соли аммония, соли, образованные органическими основаниями, соли, образованные неорганическими кислотами, соли, образованные органическими кислотами и т.п. В одном варианте осуществления настоящего изобретения "сольват" представляет собой соединение, образованное растворенным веществом и растворителем. Например, сольваты описаны в J. Honiget al., The Van Nostrand Chemist's Dictionary P650 (1953). Если растворитель представляет собой воду, полученный сольват называют гидратом. Предпочтительно растворитель не препятствует биологической активности растворенного вещества. Примеры таких предпочтительных растворителей конкретно не ограничиваются, но включают в себя воду и разные буферы. В одном варианте осуществления настоящего изобретения примеры "химической модификации" включают в себя модификацию PEG или его производными, модификацию флуоресцеином, модификацию биотином и т.п. Фармацевтически приемлемые соли включают в себя соли, образованные свободной карбоксильной группой хлористоводородной кислоты, фосфорной кислоты, уксусной кислоты, щавелевой кислоты, винной кислоты и т.п., соли, образованные свободной аминогруппой, например, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и т.п., а также соли, образованные гидроксидом натрия, калия, аммония, кальция, железа и т.п.
[0025]
Другие ингибиторы сигнального пути TGF-β включают в себя, без ограничения, моноклональные и поликлональные антитела против одной или нескольких изоформ TGF-β (патент США № 5571714; а также международная публикация № WO 97/13844 и международная публикация № WO 00/66631), рецепторы TGF-β, растворимые формы таких рецепторов (например, растворимый рецептор TGF-β типа III), или антитела против рецептора TGF-β (патент США № 5693607, патент США № 6001969, патент США № 6010872, патент США № 6086867, патент США № 6201108; международная публикация № WO 98/48024; международная публикация № WO 95/10610; международная публикация № WO 93/09228; международная публикация № WO 92/00330), латентно ассоциированные пептиды (международная публикация № WO 91/08291), большой латентный комплекс TGF-β (международная публикация № WO 94/09812), фетуин (патент США № 5821227), декорин и другие протеогликаны, такие как бигликан, фибромодулин, лумикан и эндоглин (международная публикация № WO 91/10727; патент США № 5654270, патент США № 5705609, патент США № 5726149; патент США № 5824655; международная публикация № WO 91/04748; патент США № 5830847, патент США № 6015693; международная публикация № WO 91/10727; международная публикация № WO 93/09800; и международная публикация № WO 94/10187), соматостатин (международная публикация № WO 98/08529), маннозо-6-фосфорную кислоту или маннозо-1-фосфорную кислоту (патент США № 5520926), пролактин (международная публикация № WO 97/40848), инсулиноподобный фактор роста II (международная публикация № WO 98/17304), IP-10 (международная публикация № WO 97/00691), Arg-Gly-Asp-содержащие пептиды (Pfeffer, патент США № 5958411; международная публикация № WO 93/10808), растительные, грибковые и бактериальные экстракты (EP-A-813875; выложенная публикация Японии № 8-119984; и Matsunaga et al., патент США № 5693610), антисмысловые олигонуклеотиды (патент США № 5683988; патент США № 5772995; патент США № 5821234, патент США № 5869462; и международная публикация № WO 94/25588), белки, ассоциированные с сигнальным путем TGF-β, такие как Smad и MAD (EP-A-874046; международная публикация № WO 97/31020; международная публикация № WO 97/38729; международная публикация № WO 98/03663; международная публикация № WO 98/07735; международная публикация № WO 98/07849; международная публикация № WO 98/45467; международная публикация № WO 98/53068; международная публикация № WO 98/55512; международная публикация № WO 98/56913; международная публикация № WO 98/53830; международная публикация № WO 99/50296; патент США № 5834248; патент США № 5807708; и патент США № 5948639), Ski и Sno (Vogel, 1999, Science, 286:665; и Stroschein et al., 1999, Science, 286:771 to 774), один или несколько одноцепочечных олигонуклеотидных аптамеров или кодирующие их экспрессионные плазмиды, способные ингибировать связывание, или препятствовать связыванию TGF-β с рецептором того же происхождения, а также любые мутанты, фрагменты или производные указанных выше молекул, которые сохраняют способность ингибировать активность TGF-β. Ингибитор TGF-β может представлять собой антагонист TGF-β, а также человеческое моноклональное антитело, или гуманизированное моноклональное антитело (или фрагмент F(ab)2, фрагмент Fv, одноцепочечное антитело, а также другие формы или фрагменты антител, сохраняющие способность связываться с TGF-β, его фрагментом и т.п.), которые блокируют связывание TGF-β с рецептором. Рецептор TGF-β, фрагмент рецептора, связывающий TGF-β, особенно растворимый фрагмент рецептора TGF-β, являются антагонистами TGF-β, подходящими для применения в способе настоящего изобретения. В отдельном варианте осуществления предпочтительный ингибитор TGF-β представляет собой растворимый рецептор TGF-β, особенно рецептор TGF-β типа II (TGFBIIR), или рецептор TGF-β типа III (TGFBIIIR или бета-гликан), включающий в себя, например, внеклеточный домен TGFBIIIR или TGFBIIR, предпочтительно рекомбинантный растворимый рецептор TGF-β (rsTGFBIIR или rsTGFBIIIR). Рецептор TGF-β, фрагмент рецептора TGF-β, связывающий TGF-β, особенно растворимый фрагмент, являются антагонистами TGF-β, подходящими для применения в способе настоящего изобретения. Рецепторы TGF-β и кодирующие их нуклеиновые кислоты хорошо известны в данной области. Нуклеотидная последовательность, кодирующая рецептор TGF-β типа 1, описана в GenBank под номером доступа L15436 и в патенте США № 5538892 (Donahoe et al.). Нуклеотидная последовательность рецептора TGF-β типа 2 находится в GenBank в открытом доступе под номерами AW236001, AI35790, AI279872, AI074706 и AA808255. Нуклеотидная последовательность рецептора TGF-β типа 3 находится в GenBank в открытом доступе под номерами NM003243, AI887852, AI817295 и AI681599.
[0026]
Кроме того, другие ингибиторы или антагонисты сигнального пути TGF-β, или способы их промышленного получения хорошо известны в данной области, помимо многих, находящихся в настоящий момент на стадии разработки. Поскольку в способе настоящего изобретения можно использовать любой эффективный антагонист TGF-β, конкретные используемые ингибиторы или антагонисты сигнального пути TGF-β не имеют ограничивающих характеристик. Примеры таких антагонистов включают в себя моноклональные и поликлональные антитела против TGF-β одного или нескольких изотипов (патент США № 5571714 и международная публикация № WO 97/13844), рецепторы TGF-β, их фрагменты, их производные, антитела против рецептора TGF-β (патент США № 5693607, патент США № 6008011, патент США № 6001969 и патент США № 6010872, международная публикация № WO 92/00330, международная публикация № WO 93/09228, международная публикация № WO 95/10610 и международная публикация № WO 98/48024); латентно ассоциированные пептиды (международная публикация № WO 91/08291), большой латентный комплекс TGF-β (международная публикация № WO 94/09812), фетуин (патент США № 5821227), декорин и другие протеогликаны, такие как бигликан, фибромодулин, лумикан, эндоглин (патент США № 5583103, патент США № 5654270, патент США № 5705609, патент США № 5726149, патент США № 5824655, патент США № 5830847, патент США № 6015693, международная публикация № WO 91/04748, международная публикация № WO 91/10727, международная публикация № WO 93/09800 и международная публикация № WO 94/10187).
[0027]
Другие примеры таких антагонистов включают в себя огромное количество других белков, ассоциированных с сигнальным путем TGF-β, таких как соматостатин (международная публикация № WO 98/08529), маннозо-6-фосфорная кислота или маннозо-1-фосфорная кислота (патент США № 5520926), пролактин (международная публикация № WO 97/40848), инсулиноподобный фактор роста II (международная публикация № WO 98/17304), IP-10 (международная публикация № WO 97/00691), аргинин (arg)-глицин (gly)-аспарагиновая кислота (asp)-содержащие пептиды (патент США № 5958411 и международная публикация № WO 93/10808), растительные, грибковые и бактериальные экстракты (публикация европейской патентной заявки № 813875, выложенная публикация Японии № 8-119984 и патент США № 5693610), антисмысловые олигонуклеотиды (патент США № 5683988, патент США № 5772995, патент США № 5821234, патент США № 5869462 и международная публикация № WO 94/25588), Smad и MAD (европейская патентная заявка № EP 874046, международная публикация № WO 97/31020, международная публикация № WO 97/38729, международная публикация № WO 98/03663, международная публикация № WO 98/07735, международная публикация № WO 98/07849, международная публикация № WO 98/45467, международная публикация № WO 98/53068, международная публикация № WO 98/55512, международная публикация № WO 98/56913, международная публикация № WO 98/53830 и международная публикация № WO 99/50296, а также патент США № 5834248, патент США № 5807708 и патент США № 5948639), Ski и Sno (G. Vogel, Science, 286: 665 (1999) и Stroschein et al., Science, 286:771-74 (1999)), а также любые фрагменты и производные вышеуказанных молекул, сохраняющие способность ингибировать активность TGF-β.
[0028]
Антагонисты TGF-β, подходящие для применения в настоящем изобретении, также включают в себя функциональные мутанты, мутанты, производные и аналоги вышеупомянутых антагонистов TGF-β, при условии, что они сохраняют способность уменьшать количество или ингибировать активность TGF-β. Термины "мутант", "производное" и "аналог" в соответствии с данным описанием относятся к молекуле, подобной исходной молекуле по форме или структуре и сохраняющей способность функционировать как антагонист TGF-β. Например, любой из раскрытых здесь антагонистов TGF-β может находиться в кристаллической форме и подходящие аналоги можно рационально сконструировать на основе координат, описывающих форму (одного или нескольких) активных участков. Кроме того, специалисты в данной области могут изменить функциональную группу известного антагониста, или провести скрининг с целью поиска такой измененной молекулы, обладающей улучшенными характеристиками, включающими в себя активность, период полужизни, биодоступность или другие желательные характеристики, без излишнего экспериментирования. Если антагонист TGF-β представляет собой полипептид, можно получить фрагмент или вариант полипептида (например, гуманизированное антитело или описанный выше функциональный фрагмент антитела), характеризующийся более простой доставкой, повышенной активностью, увеличенным периодом полужизни и т.п. С учетом технического уровня получения синтетических и рекомбинантных полипептидов, присутствующего в данной области, такой вариант можно получить без излишнего экспериментирования. Специалисты в данной области также могут сконструировать новый ингибитор на основе информации по кристаллической структуре и/или активному участку описанного здесь ингибитора TGF-β. Полипептидный ингибитор, такой как растворимый рецептор TGF-β, можно эффективно вводить посредством переноса генов. Таким образом, конкретный вариант осуществления способа настоящего изобретения включает применение вектора, обеспечивающего экспрессию рецептора или партнера TGF-β по связыванию, предпочтительно растворимого рецептора или партнера по связыванию. В предпочтительном варианте осуществления растворимый антагонист TGF-β можно ввести путем переноса генов с использованием вектора, содержащего кДНК, кодирующую растворимый антагонист, или кДНК, кодирующую внеклеточный домен рецептора TGF-β типа II (rsTGFBIIR) или рецептора TGF-β типа III (rsTGFBIIIR). В качестве такого вектора можно использовать любой подходящий вектор, способный обеспечивать экспрессию растворимого антагониста TGF-β in situ в клетке, трансфицированной таким вектором, ингибировать активность TGF-β и подавлять TGF-β -опосредованный фиброгенез. Предпочтительные векторы включают в себя аденовирусный вектор, лентивирусный вектор, вектор на основе вируса Эпштейна-Барра (EBV), вектор на основе адено-ассоциированного вируса (AAV) и ретровирусный вектор, разработанные для переноса генов. Также можно использовать способы переноса генов, в которых вместо вектора используют, например, комплекс липид/ДНК, конъюгат белок/ДНК и оголенную ДНК. Другие подходящие антагонисты TGF-β, разработанные для доставки посредством аденовирусного переноса генов, включают в себя, без ограничения, химерную кДНК, кодирующую внеклеточный домен рецептора TGF-β типа II, гибридизованный с Fc-доменом Ig (Isaka et al., 1999, Kidney Int., 55: pp. 465 to 475), аденовирусный вектор для переноса генов, кодирующий доминантно-негативный мутант рецептора TGF-β типа II (Zhao et al., 1998, Mech. Dev., 72: pp. 89 to 100), и аденовирусный вектор для переноса генов, кодирующий декорин, который представляет собой TGF-β -связывающий протеогликан (Zhao et al., 1999, Am. J. Physiol., 277: pp. L412 to L422). Опосредованный аденовирусами перенос генов характеризуется крайне высокой эффективностью по сравнению с другими способами доставки.
[0029]
Рецепторы TGF-β, TGF-β -связывающие фрагменты и растворимые фрагменты рецепторов TGF-β и т.п. являются антагонистами TGF-β, подходями для применения в настоящем изобретении. Рецепторы TGF-β и кодирующие их нуклеиновые кислоты хорошо известны в данной области. Нуклеотидные последовательности, кодирующие рецептор TGF-β типа 1, описаны в GenBank под номером доступа L15436 и патенте США № 5538892, Donahoe et al. Нуклеотидные последовательности рецептора TGF-β типа 2 находятся в свободном доступе в GenBank под номерами доступа AW236001; AI35790; AI279872; AI074706; и AA808255. Нуклеотидные последовательности рецептора TGF-β типа 3 находятся в свободном доступе в GenBank под номерами доступа NM003243; AI887852; AI817295; и AI681599. В одном иллюстративном варианте осуществления антагонист TGF-β представляет собой антитело, которое блокирует связывание TGF-β с его рецептором, или фрагмент F(ab)2, фрагмент Fv, одноцепочечное антитело или фрагмент других форм "антитела", сохраняющий способность связываться с TGF-β. Такое антитело может быть химерным или гуманизированным. В соответствии с данным описанием, химерное антитело содержит константный домен человеческого антитела и вариабельный домен нечеловеческого антитела, такого как мышиное антитело. Гуманизированное антитело содержит константный домен и каркасные участки вариабельного домена (т.е., участки вариабельного домена, отличные от гипервариабельных участков) человеческого антитела и гипервариабельные участки нечеловеческого антитела, такого как мышиное антитело. Само собой разумеется, что такое антитело может представлять собой производное антитела любого другого типа, такого как человеческое антитело, выбранное или отобранное из системы фагового дисплея, или полученное с использованием технологии XenoMouse.
[0030]
Число данных, касающихся Smad, непрерывно растет. Сигнальный путь TGF-β инициируется, когда участвующая в нем молекула связывается с гетеродимерным комплексом клеточной поверхности, содержащим серин/треонин-киназные рецепторы типа I (TbRI) и типа II (TbRII), активируя указанный гетеродимерный комплекс клеточной поверхности. Затем гетеродимерный рецептор передает сигнал посредством фосфорилирования на нижестоящий белок-мишень Smad. Как указано выше, существует три функциональных класса белков Smad, включающих в себя Smad (R-Smad), регулируемые рецептором, такие как Smad2 и Smad3, сомедиаторные Smad (Co-Smad), которые также обозначают Smad4, и ингибиторные Smad (I-Smad). После фосфорилирования гетеродимерным рецепторным комплексом R-Smad образует комплекс с Co-Smad, перемещается в ядро и модулирует транскрипцию целевого гена вместе с другими белками (Derynck, R., et al. (1998) Cell 95: 737-740); Massague, J. and Wotton, D. (2000) EMBO J. 19:1745). Нуклеотидная и аминокислотная последовательности человеческого Smad3 описаны, например, в GenBank под номером доступа gi: 42476202. Нуклеотидная и аминокислотная последовательности мышиного Smad3 описаны, например, в GenBank под номером доступа gi: 31543221. Как указано выше, стимуляция TGF-β приводит к фосфорилированию и активации Smad2 и Smad3, которые образуют комплекс со Smad4 (также называемым "сомедиаторный Smad" или "co-Smad"), после чего комплекс накапливается в ядре и модулирует транскрипцию целевого гена. Следовательно, блокирования сигнального пути TGF-β также можно достичь путем ингибирования Smad2, 3 или co-Smad (Smad4). R-Smad локализуется в цитоплазме и образует комплекс с co-Smad после лиганд-индуцированного фосфорилирования под действием рецептора TGF-β, после чего комплекс перемещается в ядро, где он модулирует экспрессию генов в ассоциации с хроматином и кооперативным фактором транскрипции. Следовательно, блокирования сигнального пути TGF-β также можно достичь путем непосредственного или косвенного ингибирования R-Smad. Smad6 и Smad7 представляют собой ингибиторные Smad (I-Smad), то есть, они индуцируются на уровне транскрипции под действием TGF-β и функционируют как ингибиторы сигнального пути TGF-β (Feng et al., (2005) Annu. Rev. Cell. Dev. Biol. 21: 659). Smad6/7 препятствуют рецептор-опосредованной активации R-Smad, осуществляя свой ингибиторный эффект. Они связываются с рецептором типа I, конкурентно препятствуя мобилизации и фосфорилированию R-Smad. Известно, что Smad6 и Smad7 пополняют запасы убихитинлигазы E3, которая индуцирует убихитинирование и деградацию белков, взаимодействующих со Smad6/7. Таким образом, Smad6 и 7 можно использовать в настоящем изобретении в качестве ингибитора сигнального пути TGF-β.
[0031]
Ингибиторы Smad3, которые можно использовать в настоящем изобретении, включают в себя, без ограничения, антисмысловые нуклеотиды, миРНК, антитела и т.п., а также низкомолекулярные соединения, такие как 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-илпроп-2-еноил))-1,2,3,4-тетрагидроизохинолон и т.п., поставляемые Calbiochem.
[0032]
В рамках изобретения "вещество (например, нуклеиновая кислота), подавляющее экспрессию (TGF-β и т.п.)" специально не ограничивается и включает, например, вещество, которое подавляет транскрипцию мРНК целевого гена, вещество (такое как нуклеиновая кислота), которое разрушает транскрибированную мРНК, или вещество (такое как нуклеиновая кислота), которое подавляет трансляцию белка с мРНК. Примеры таких веществ включают в себя миРНК, антисмысловые олигонуклеотиды, рибозимы, векторы экспрессии и другие нуклеиновые кислоты, среди которых предпочтительными являются миРНК и вектор экспрессии, а особенно предпочтительной является миРНК. "Вещество, подавляющее экспрессию гена" может включать в себя белки, пептиды, а также другие низкомолекулярные вещества, помимо описанных выше. Следует отметить, что целевой ген в соответствии с настоящим изобретением может представлять собой любой ген, связанный с сигнальным путем TGF-β.
[0033]
Способ с использованием антисмысловых технологий хорошо известен специалистам в данной области как способ ингибирования экспрессии конкретного эндогенного гена, такого как TGF-β, на который направлено настоящее изобретение. Антисмысловая нуклеиновая кислота может ингибировать экспрессию целевого гена посредством ряда механизмов, включающих в себя, например, ингибирование инициации транскрипции вследствие образования триплекса; ингибирование транскрипции вследствие образования гибрида, содержащего участок, в котором под действием РНК-полимеразы локально образуется открытая петлеобразная структура; ингибирование транскрипции вследствие образования гибрида с РНК, которая находится в процессе синтеза; ингибирование сплайсинга вследствие образования гибрида в участке соединения интрона и экзона; ингибирование сплайсинга вследствие образования гибрида в участке образования сплайсингосомы; ингибирование миграции из ядра в цитоплазму вследствие образования гибрида с мРНК; ингибирование сплайсинга вследствие образования гибрида в участке кэпирования или в участке добавления поли(A); ингибирование инициации трансляции вследствие образования гибрида с участком, связывающим фактор инициации трансляции; ингибирование трансляции вследствие образования гибрида с участком связывания рибосомы вблизи кодона инициации; ингибирование элонгации пептидной цепи вследствие образования гибрида с участком связывания полисомы или участком трансляции мРНК; ингибирование экспрессии гена вследствие образования гибрида с участком взаимодействия нуклеиновой кислоты и белка, и т.п. Таким образом, антисмысловая нуклеиновая кислота может ингибировать разные процессы, такие как транскрипция, сплайсинг и трансляция, подавляя экспрессию целевого гена (Hirashima and Inoue, Shinsei Kagaku Jikken Kouza [New Biochemical Experiment Course] 2, Nucleic Acid, IV Idenshi no Fukusei to Hatsugen [Duplication and Expression of Gene], Edited by the Japanese Biochemical Society, Tokyo Kagaku Dojin, 1993, 319-347).
[0034]
Антисмысловая нуклеиновая кислота, используемая в настоящем изобретении, может ингибировать экспрессию и/или функцию гена (нуклеиновой кислоты), кодирующего член сигнального пути TGF-β, описанного выше, и т.п., посредством любого из вышеуказанных механизмов. В одном варианте осуществления эффективного ингибирования трансляции гена достигают путем конструирования антисмысловой последовательности, комплементарной нетранслируемому участку вблизи 5′-конца мРНК гена, кодирующего TGF-β, описанного выше, и т.п. Кроме того, можно использовать последовательность, комплементарную кодирующему участку или 3′-нетранслируемому участку. Таким образом, нуклеиновые кислоты, содержащие антисмысловые последовательности, комплементарные не только транслируемому участку, но и нетранслируемому участку гена, кодирующего TGF-β, описанного выше, и т.п., охватываются термином "антисмысловые нуклеиновые кислоты, подходящие для применения в настоящем изобретении". Используемую антисмысловую нуклеиновую кислоту присоединяют ниже соответствующего промотора, и предпочтительно ее присоединяют к последоватлеьности, включающей сигнал терминации транскрипции на 3′-конце. Полученную таким образом нуклеиновую кислоту можно использовать для трансформации целевого животного (клетки) с помощью известного способа. Хотя последовательность антисмысловой нуклеиновой кислоты предпочтительно представляет собой последовательность, комплементарную гену, кодирующему TGF-β, и т.п. животного (клетки), подлежащего трансформации, или его части, она не обязательно должна быть полностью комплементарной, при условии, что последовательность может эффективно подавлять экспрессию гена. Предпочтительно транскрибированная РНК комплементарна продукту транскрипции целевого гена на 90% или более, наиболее предпочтительно на 95% или более. Антисмысловая нуклеиновая кислота, способная эффективно ингибировать экспрессию целевого гена, предпочтительно содержит в длину, по меньшей мере, 12 оснований или более, но менее 25 оснований. Однако длина антисмысловой нуклеиновой кислоты настоящего изобретения не ограничивается указанными значениями. Ее длина может составлять, например, 11 оснований или менее, 100 оснований или более, или 500 оснований или более. Антисмысловая нуклеиновая кислота может состоять не только из ДНК, она также может содержать нуклеиновые кислоты, отличные от ДНК, такие как запертые нуклеиновые кислоты (LNA). В одном варианте осуществления антисмысловая нуклеиновая кислота, используемая в настоящем изобретении, может представлять собой антисмысловую нуклеиновую кислоту, содержащую LNA на 5′-конце или на 3′-конце. В одном варианте осуществления антисмысловую нуклеиновую кислоту, используемую в настоящем изобретении, конструируют на основе нуклеотидной последовательности TGF-β и т.п., с помощью способа, описанного, например, в Hirashima and Inoue, Shinsei Kagaku Jikken Kouza [New Biochemical Experiment Course] 2, Nucleic Acid, IV Idenshino Fukuseito Hatsugen [Duplication and Expression of Gene], Edited by the Japanese Biochemical Society, Tokyo Kagaku Dojin, 1993, 319-347.
[0035]
Экспрессию TGF-β и т.п. также можно ингибировать с ипользованием рибозима или ДНК, кодирующей рибозим. Рибозим представляет собой молекулу РНК, обладающую каталитической активностью. Существуют рибозимы с разными типами активности, однако исследователи, уделяя основное внимание рибозимам, функционирующим как ферменты, расщепляющие РНК, сделали возможным конструирование рибозим, расщепляющих РНК в сайт-специфичной манере. Хотя существуют рибозимы размером 400 нуклеотидов или более, такие как рибозимы интрона группы I и РНК Ml, входящей в состав РНКазы P, также существуют рибозимы, содержащие активный домен размером примерно 40 нуклеотидов, такие как рибозимы типа головки молотка и рибозимы типа шпильки (Makoto Koizumi and Eiko Ohtsuka, Protein Nucleic Acid And Enzyme, 1990, 35, 2191).
[0036]
Например, саморасщепляющий домен рибозимов типа головки молотка расщепляет 3'-конец C15 в последовательности, называемой G13U14C15. Считается, что образование пары оснований U14 и A9 играет важную роль в проявлении активности рибозима. Кроме того, показано, что расщепление может осуществляться по A15 или U15 вместо C15 (Koizumi, M. et al., FEBS Lett, 1988, 228, 228). Можно сконструировать рибозим, субстрат-связывающий участок которого будет комплементарен последовательности РНК вблизи целевого участка, с получением рибозима, расщепляющего РНК, как фермент рестрикции, и распознающего такую последовательность, как UC, UU или UA в молекуле РНК-мишени (Koizumi, M. et al., FEBS Lett, 1988, 239, 285., Makoto Koizumi and Eiko Ohtsuka, Protein Nucleic Acid And Enzyme, 1990, 35, 2191., Koizumi, M. et al., Nucl. Acids Res., 1989, 17, 7059).
[0037]
Кроме того, в целях настоящего изобретения можно использовать рибозимы типа шпильки. Такой рибозим можно найти, например, в отрицательной цепи сателлитной РНК вируса кольцевой пятнистости табака (Buzayan, J M., Nature, 1986, 323, 349). Также показано, что специфичный к мишени РНК-расщепляющий рибозим можно получить из рибозима типа шпильки (Kikuchi, Y. & Sasaki, N., Nucl. Acids Res, 1991, 19, 6751., Kikuchi, Yo, Kagaku to Seibutsu [Chemistry and Organism], 1992, 30, 112). Таким образом, продукт транскрипции гена, кодирующего TGF-β, и т.п. можно специфически расщеплять с помощью рибозима, ингибируя экспрессию гена.
[0038]
Экспрессию эндогенного гена TGF-β и т.п. также можно подавить путем РНК-интерференции (далее сокращенно обозначаемой "РНКи") с использованием двухцепочечной РНК, имеющей последовательность, идентичную или подобную последовательности целевого гена. РНКи представляет собой технологию, которой в настоящее время уделяется много внимания, и которая заключается в том, что после непосредственного введения в клетку двухцепочечной РНК (дцРНК) экспрессия гена, имеющего последовательность, гомологичную дцРНК, подавляется. Для индукции РНКи в клетках млекопитающих используют короткоцепочечную дцРНК (миРНК). По сравнению с нокаутными мышами РНКи имеет много преимуществ, включающих в себя стабильный эффект, простоту экспериментирования и низкую стоимость. миРНК подробно описаны в других разделах настоящего документа.
[0039]
В рамках изобретения термин "миРНК" относится к молекуле РНК, содержащей фрагмент двухцепочечной РНК, состоящий из 15-40 оснований. миРНК функционирует путем расщепления мРНК целевого гена, имеющего последовательность, комплементарную антисмысловой цепи указанной миРНК, подавляя экспрессию целевого гена. Более конкретно, миРНК в соответствии с настоящим изобретением представляет собой РНК, содержащую фрагмент двухцепочечной РНК, состоящий из смысловой цепи РНК, последовательность которой гомологична непрерывной последовательности мРНК TGF-β и т.п., и антисмысловой цепи РНК, последовательность которой комплементарна смысловой последовательности РНК. Способы промышленного получения и конструирования миРНК и мутантной миРНК, описанные ниже, относятся к сфере компетенции специалистов в данной области. Методология выбора любого непрерывного участка мРНК, которая представляет собой продукт транскрипции последовательности TGF-β и т.п., и получения двухцепочечной РНК, соответствующей данному участку, является исключительно предметом обычной творческой способности специалистов в данной области. Более того, последовательность миРНК, обеспечивающая более интенсивный эффект РНКи от последовательности мРНК, которая представляет собой продукт транскрипции последовательности, может быть выбрана соответственно специалистами в данной области с помощью известного способа. Кроме того, если одна из цепей идентифицирована, специалисты в данной области могут легко определить последовательность оснований другой цепи (комплементарной цепи). Специалисты в данной области могут получить соответствующую миРНК с помощью коммерчески доступного синтезатора нуклеиновых кислот. Кроме того, для синтеза целевой РНК можно использовать единую службу синтеза.
[0040]
Длина фрагмента двухцепочечной РНК составляет от 15 до 40 оснований, предпочтительно от 15 до 30 оснований, более предпочтительно от 15 до 25 оснований, еще более предпочтительно от 18 до 23 оснований, и наиболее предпочтительно от 19 до 21 оснований. Следует понимать, что ее верхние и нижние пределы не ограничиваются указанными значениями и могут представлять собой любые сочетания данных значений. Конечная структура смысловой или антисмысловой цепи миРНК конкретно не ограничивается и может быть соответственно выбрана в зависимости от цели. Например, конечная структура может иметь тупые концы или выступающие концы (липкие), хотя предпочтительным является выступающий 3′-конец. миРНК, содержащая липкий конец, состоящий из нескольких оснований, предпочтительно 1-3 оснований, еще более предпочтительно, 2 оснований на 3′-конце смысловой цепи РНК и антисмысловой цепи РНК, зачастую является предпочтительной, поскольку она оказывает интенсивный эффект при подавлении экспрессии целевого гена. Тип основания на липком конце специально не ограничивается и может представлять собой либо основание, входящее в состав РНК, либо основание, входящее в состав ДНК. Предпочтительные последовательности липкого конца включают в себя dTdT (2 п.о. дезокси T) на 3′-конце и т.п. Примеры предпочтительных миРНК включают в себя, без ограничения, молекулы миРНК, содержащие dTdT (2 п.о. дезокси T), присоединенные к 3′-концу смысловой и антисмысловой цепей.
[0041]
Кроме того, можно использовать миРНК, содержащую делеции, замены, вставки и/или добавления одного или нескольких нуклеотидов в одной из смысловой и антисмысловой цепей, или в обоих цепях вышеописанной миРНК. В данной связи число оснований специально не ограничивается, однако предпочтительно оно варьирует от 1 до 4 оснований, более предпочтительно от 1 до 3 оснований и наиболее предпочтительно от 1 до 2 оснований. Конкретные примеры таких мутаций включают в себя, без ограничения, мутации, в результате которых число оснований в 3′-концевом липком фрагменте составляет от 0 до 3, мутации, в результате которых последовательность оснований 3′-концевого липкого фрагмента изменяется на другую последовательность оснований, мутации, в результате которых длина вышеописанной смысловой цепи РНК и антисмысловой цепи РНК изменяется на 1-3 основания вследствие вставки, добавления или делеции оснований, мутации, в результате которых основание в смысловой цепи и/или в антисмысловой цепи заменяется на другое основание, и т.п. Однако смысловая цепь и антисмысловая цепь в таких мутантных миРНК должны обладать способностью к гибридизации, а указанные мутантные миРНК должны обладать способностью подавлять экспрессию генов, как и миРНК, не содержащие мутаций.
[0042]
Кроме того, миРНК может представлять собой молекулу, в которой один конец имеет закрытую структуру, например, миРНК со шпилечной структурой (короткая шпилечная РНК; кшРНК). кшРНК представляет собой РНК, содержащую смысловую цепь РНК конкретной последовательности целевого гена, антисмысловую цепь РНК, содержащую последовательность, комплементарную последовательности смысловой цепи, и линкерную последовательность, соединяющую две цепи, где фрагмент смысловой цепи и фрагмент антисмысловой цепи гибридизуются, образуя фрагмент двухцепочечной РНК.
[0043]
Желательно, чтобы миРНК не оказывала так называемый нецелевой эффект при клиническом применении. Нецелевой эффект представляет собой эффект подавления экспрессии другого гена, частично гомологичного используемой миРНК, отличного от целевого гена. Чтобы избежать нецелевого эффекта, можно предварительно показать, используя микрочипы ДНК и т.п., что миРНК-кандидат не обладает перекрестной реакционноспособностью. Кроме того, чтобы избежать нецелевого эффекта, можно определить, используя известную базу данных, предоставляемую NCBI (Национальный центр биотехнологической информации), и т.п., существует ли ген, содержащий фрагмент, в высокой степени гомологичный последовательности миРНК-кандидата, отличный от целевого гена.
[0044]
Для получения миРНК настоящего изобретения можно соответственно использовать известный способ, такой как способ химического синтеза или способ рекомбинации генов. На основании информации о последовательности можно синтезировать двухцепочечную РНК, используя традиционный способ синтеза. В случае использования рекомбинантного способа, миРНК можно получить путем конструирования вектора экспрессии, кодирующего последовательность смысловой цепи или антисмысловой цепи, и введения вектора в клетку-хозяина, с последующим получением каждой из смысловой цепи РНК и антисмысловой цепи РНК, продуцированных в результате транскрипции. Кроме того, целевую двухцепочечную РНК можно получить путем экспрессии кшРНК, образующей шпилечную структуру, которая содержит смысловую цепь конкретной последовательности целевого гена, антисмысловую цепь, содержащую последовательность, комплементарную последовательности смысловой цепи, линкерную последовательность, соединяющую две цепи.
[0045]
В случае миРНК, вся нуклеиновая кислота, составляющая миРНК, или ее часть, может представлять собой природную или модифицированную нуклеиновую кислоту, при условии, что такая нуклеиновая кислота обладает способностью подавлять экспрессию целевого гена.
[0046]
миРНК настоящего изобретения не обязательно образует пары двухцепочечных РНК с целевой последовательностью. Она может представлять собой смесь нескольких пар (термин "несколько" конкретно не ограничивается, однако предпочтительно он относится к небольшому числу, составляющему примерно от 2 до 5) двухцепочечных РНК с участком, содержащим целевую последовательность. С учетом вышесказанного, специалисты в данной области могут соответственно получить миРНК в виде смеси нуклеиновых кислот, соответствующих целевой последовательности, с использованием коммерчески доступного синтезатора нуклеиновых кислот и фермента DICER. Кроме того, для синтеза целевой РНК можно использовать единую службу синтеза. Следует отметить, что миРНК настоящего изобретения включает так называемый "коктейль миРНК". В случае миРНК настоящего изобретения не все нуклеотиды должны представлять собой рибонуклеотиды (РНК). А именно, в соответствии с настоящим изобретением, один или несколько рибонуклеотидов, составляющих миРНК, могут представлять собой соответствующие дезоксирибонуклеотиды. Термин "соответствующий" относится к молекуле, содержащей основания такого же типа (аденин, гуанин, цитозин, тимин (урацил)), но другую структуру сахарного фрагмента. Например, дезоксирибонуклеотид, соответствующий рибонуклеотиду, содержащему аденин, представляет собой дезоксирибонуклеотид, содержащий аденин.
[0047]
Кроме того, ДНК (вектор), способная экспрессировать вышеописанную РНК настоящего изобретения, также составляет предпочтительный вариант осуществления нуклеиновой кислоты, способный подавлять экспрессию TGF-β и т.п. Например, ДНК (вектор), способная экспрессировать вышеописанную двухцепочечную РНК настоящего изобретения, представляет собой ДНК, в которой фрагмент ДНК, кодирующий одну из цепей двухцепочечной РНК, и фрагмент ДНК, кодирующий другую цепь двухцепочечной РНК, соединены с промотором так, чтобы обеспечить экспрессию каждой ДНК. Специалисты в данной области могут соответственно получить вышеописанную ДНК настоящего изобретения с помощью традиционного метода генной инженерии. Более конкретно, вектор экспрессии настоящего изобретения можно получить путем соответствующей вставки ДНК, кодирующей РНК настоящего изобретения, в разные известные векторы экспрессии.
[0048]
В настоящем изобретении в качестве нуклеиновой кислоты, подавляющей экспрессию целевого гена, можно использовать модифицированную нуклеиновую кислоту. Модифицированная нуклеиновая кислота представляет собой нуклеиновую кислоту, которая содержит модификацию в нуклеозиде (во фрагменте основания, или во фрагменте сахара) и/или межнуклеозидном участке связывания, и имеет структуру, отличную от структуры природной нуклеиновой кислоты. Примеры "модифицированного нуклеозида", входящего в состав модифицированной нуклеиновой кислоты, включают в себя: нуклеозид с удаленным азотистым основанием; арабинонуклеозид, 2′-дезоксиуридин, α-дезоксирибонуклеозид, β-L-дезоксирибонуклеозид и нуклеозид, содержащий другую модификацию сахара; пептидо-нуклеиновую кислоту (PNA), связанную по фосфатной группе пептидо-нуклеиновую кислоту (PHONA), запертую нуклеиновую кислоту (LNA), морфолинонуклеиновую кислоту и т.п. Описанные выше нуклеозиды, содержащие модификацию сахара, включают в себя 2′-O-метилрибозу, 2′-дезокси-2′-фторрибозу, 3′-O-метилрибозу и другие замещенные пентозы; 1′,2′-дезоксирибозу; арабинозу; сахарный остаток, содержащий замещенную арабинозу; и нуклеозид, содержащий в качестве модифицированного сахара альфа-аномер и гексозу. Указанные нуклеозиды могут содержать модифицированное основание, с модификацией по фрагменту основания. Примеры таких модифицированных оснований включают в себя 5-гидроксицитозин, 5-фторурацил, 4-тиоурацил и другие пиримидины; 6-метиладенин, 6-тиогуанозин и другие пурины; а также другие гетероциклические основания.
[0049]
Примеры "модифицированной межнуклеозидной связи", входящей в состав модифицированной нуклеиновой кислоты, включают в себя неприродные межнуклеозидные связи, такие как алкильный линкер, глицерильный линкер, аминолинкер, поли(этиленгликолевая) связь, метилфосфонатная межнуклеозидная связь; метилфосфонотиоат, фосфотриэфир, фосфотиотриэфир, фосфоротиоат, фосфородитиоат, триэфирные пролекарства, сульфон, сульфонамид, сульфамат, формацеталь, N-метилгидроксиламин, карбонат, карбамат, морфолино, боранофосфонат, фосфорамидат и т.п.
[0050]
Двухцепочечная миРНК настоящего изобретения может содержать нуклеотидную последовательность, направленную на TGF-β, других членов сигнального пути TGF-β и т.п.
[0051]
Нуклеиновую кислоту или средство настоящего изобретения можно вводить в фосфолипиды эндоплазматического ретикулума (везикулы), такие как липосомы, с последующим введением в эндоплазматический ретикулум. Эндоплазматический ретикулум, содержащий миРНК или кшРНК, можно ввести в выбранные клетки методом липофекции. Затем осуществляют системное введение полученной клетки, например, внутривенное, внутриартериальное и т.п. Эндоплазматический ретикулум также можно вводить местно в желательный участок глаза и т.п. Хотя in vitro миРНК проявляет очень хороший, специфичный пост-транскрипционный подавляющий эффект, in vivo миРНК быстро разлагается в сыворотке под действием нуклеаз. Поскольку продолжительность ее действия является ограниченной, существует потребность в разработке более совершенной и более эффективной системы доставки. В качестве примера Ochiya, T et al., Nature Med., 5:707-710, 1999, Curr. Gene Ther., 1: 31-52, 2001 описывают, что биосовместимое вещество ателоколлаген, образующее комплекс с нуклеиновой кислотой при смешивании с ней, представляет собой носитель, защищающий нуклеиновую кислоту от разрушающих ферментов в живом организме, и может весьма успешно использоваться в качестве носителя миРНК. Хотя можно использовать такую форму, способ введения нуклеиновой кислоты или лекарственного средства настоящего изобретения не ограничивается указанным методом. Таким образом, хотя в сыворотке живого организма нуклеиновая кислота быстро разрушается под действием ферментов, расщепляющих нуклеиновые кислоты, данный метод позволяет продлить действие нуклеиновой кислоты на продолжительный период времени. Например, Takeshita F. PNAS, (2003) 102 (34) 12177-82, Minakuchi Y Nucleic Acids Research (2004) 32 (13) e109 описывают, что ателоколлаген, полученый из кожи крупного рогатого скота, образует комплекс с нуклеиновой кислотой, защищающий ее от ферментативного разрушения в живом организме, и может весьма успешно использоваться в качестве носителя миРНК. Такой метод подходит для применения в настоящем изобретении.
[0052]
В рамках изобретения термин "средство" используется в широком смысле и может относиться к любым веществам или другим факторам (например, к энергии, такой как свет, излучение, тепло и электричество) при условии, что они позволяют достичь намеченной цели. Примеры таких веществ включают в себя, без ограничения, белки, полипептиды, олигопептиды, пептиды, полинуклеотиды, олигонуклеотиды, нуклеотиды, нуклеиновые кислоты (например, включающие в себя ДНК, такие как кДНК и геномная ДНК, и РНК, такие как мРНК), полисахариды, олигосахариды, жиры, органические низкомолекулярные вещества (такие как гормоны, лиганды, вещества, передающие информацию, органические маленькие молекулы, молекулы, синтезированные методами комбинаторной химии, маленькие молекулы, которые можно использовать в качестве лекарственных средств (например, низкомолекулярные лиганды) и т.п.), и молекулы, содержащие их сочетания. Типичные примеры средств, специфичных к полинуклеотидам, включают в себя, без ограничения, полинуклеотид, комплементарный последовательности, обладающей определенной степенью гомологии (например, идентичной на 70% или более) по отношению к последовательности полинуклеотида, полипептид, такой как фактор транскрипции, связывающийся с участком промотора, и т.п. Типичные примеры средств, специфичных к полипептидам, включают в себя, без ограничения, антитело, специфичное к полипептиду, или его производному или аналогу (например, одноцепочечное антитело), специфический лиганд или рецептор, если полипептид представляет собой рецептор или лиганд, субстрат, если полипептид представляет собой фермент, и т.п.
[0053]
В рамках изобретения термин "заболевание, расстройство или состояние, связанное со стрессом эндоплазматического ретикулума (ER)" в "эндотелии роговой оболочки" относится к заболеванию, расстройству или состоянию эндотелия роговой оболочки, связанному со стрессом эндоплазматического ретикулума (ER).
[0054]
В рамках изобретения "заболевание, расстройство или состояние, связанное со стрессом эндоплазматического ретикулума (ER)" в эндотелии роговой оболочки включает в себя любое заболевание, расстройство или состояние, связанное со стрессом ER. Его примеры включают в себя, без ограничения, уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету, отек стромы роговой оболочки и т.п. Стресс ER и митохондриальное расстройство обычно встречаются одновременно, и при этом все симптомы, как правило, обусловлены митохондриальным расстройством. Так, связь между ними можно детектировать или диагностировать путем подтверждения митохондриальной недостаточности. Кроме того, многие заболевания, расстройства или состояния связаны с апоптозом, обусловленным митохондриальной недостаточностью, и их можно детектирвать или диагностировать путем подтверждения апоптоза.
[0055]
В рамках изобретения термин "терапевтическое средство против митохондриального расстройства (обусловленного стрессом ER)" относится к терапевтическому средству, по существу аналогичному терапевтическому средству против стресса ER. Примеры такого средства включают в себя, без ограничения, BiP индуктор X (BIX), 4−фенилмасляную кислоту (PBA), триметиламина N−оксид (TMAO), тауроурсодезоксихолевую кислоту (TUDCA), терпенон (также продаваемый под названием селбекс) и т.п. (например, см. Journal of the Japanese Society of Psychiatry and Neurology (2012) Vol 114 No 2 p. 115).
[0056]
В рамках изобретения термин "заболевание, расстройство или состояние, связанное с дистрофией эндотелия роговицы Фукса" относится к любому заболеванию, расстройству или состоянию, связанному с расстройством эндотелия роговицы Фукса, среди которых особое внимание в соответствии с настоящим изобретением уделяется, без ограничения, заболеванию, расстройству или состоянию, связанному со стрессом эндоплазматического ретикулума (ER). Возможные примеры такого заболевания, расстройства или состояния, связанного с дистрофией эндотелия роговицы Фукса, обусловленной стрессом эндоплазматического ретикулума (ER), включают в себя заболевания, расстройства или состояния, ассоциированные с нарушением функционирования клеток эндотелия роговицы. Альтернативно примеры других форм заболевания, расстройства или состояния, связанного с дистрофией эндотелия роговицы Фукса, включают в себя, без ограничения, уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, pain, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету, отек стромы роговой оболочки и т.п. Дистрофия эндотелия роговицы Фукса представляет собой заболевание, вызываемое аномалией в эндотелиальных клетках внутри роговой оболочки, приводящей к отеку стромы роговой оболочки. Ее причина неизвестна. При дистрофии эндотелия роговицы Фукса внеклеточный матрикс, такой как коллаген, откладывается на части черной поверхности десцеметовой мембраны с обратной стороны роговой оболочки, приводя к гипертрофии гутты роговой оболочки и десцеметовой мембраны. Гипертрофия гутты роговой оболочки и десцеметовой мембраны является причиной светобоязни или нечеткости зрения, ставя под угрозу QOL пациентов с дистрофией эндотелия роговицы Фукса. Как известно, в настоящее время отсутствуют эффективные способы терапевтического лечения дистрофии эндотелия роговицы Фукса, помимо трансплантации роговицы. Однако в Японии сущесутвует недостаток доноров роговицы, так, число пациентов, ожидающих трансплантации роговицы, составляет примерно 2600, а число операций по трансплантации роговицы в Японии составляет примерно 1700 в год.
[0057]
Существуют описания иммобилизации (Azizi B, et al. Invest Ophthalmol Vis Sci.2; 52(13): 9291−9297.2011) и культивирования клеток эндотелия роговицы, полученных от пациентов с дистрофией эндотелия роговицы Фукса (Zaniolo K, et al. Exp Eye Res.; 94(1): 22−31.2012 and Kelliher C. et al. ExpEye Res Vol.93 (6), 880−888, 2011). Однако отсутствуют описания клеток, подходящих для скрининга терапевтических средств или средств, предотвращающих развитие, которые сохраняют признаки заболевания, такие как избыточная продукция внеклеточного матрикса. Таким образом, разработка терапевтических средств является недостаточной и в настоящее время в клинике отсутствуют средства для лечения данного заболевания. Следовательно, выбор отсутствует и единственным способом лечения является трансплантация роговицы.
[0058]
(Общие методы)
Методы молекулярной биологии, биохимические методы и микробиологические методы, подходящие для применения в настоящем изобретении, хорошо известны и широко используются в данной области. Они описаны, например, в Sambrook J. et al. (1989). Molecular Cloning: A Laboratory Manual, Cold Spring Harbor and 3rd Ed. thereof (2001); Ausubel, F. M. (1987). Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience; Ausubel, F. M. (1989). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience; Innis, M. A. (1990). PCR Protocols: A Guide to Methods and Applications, Academic Press; Ausubel, F. M. (1992). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Greene Pub. Associates; Ausubel, F. M. (1995). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Greene Pub. Associates; Innis, M. A. et al. (1995). PCR Strategies, Academic Press; Ausubel, F. M. (1999). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Wiley, and annual updates; Sninsky, J. J. et al. (1999). PCR Applications: Protocols for Functional Genomics, Academic Press, Gait, M. J. (1985). Oligonucleotide Synthesis: A Practical Approach, IRL Press; Gait, M. J. (1990). Oligonucleotide Synthesis: A Practical Approach, IRL Press; Eckstein, F. (1991). Oligonucleotides and Analogues: A Practical Approach, IRL Press; Adams, R. L. et al. (1992). The Biochemistry of the Nucleic Acids, Chapman & Hall; Shabarova, Z. et al. (1994). Advanced Organic Chemistry of Nucleic Acids, Weinheim; Blackburn, G. M. et al. (1996). Nucleic Acids in Chemistry and Biology, Oxford University Press; Hermanson, G. T. (1996). Bioconjugate Techniques, Academic Press, Jikken Igaku Bessatsu [Experimental Medicine, Separate Volume], "Idenshi Dounyu & Hatsugen Kaiseki Jikkenho [Method of Gene Introduction & Expression Analysis Experimental Technique" Yodosha Co., Ltd., 1997, и т.п. Хорошо известно описание эндотелиальных клеток роговицы от Nancy Joyce et al., {Joyce, 2004 #161} {Joyce, 2003 #7}, тогда как в настоящее время исследователи с целью эффективного культивирования проводят трансформацию в фибробласт-подобной манере посредством длительного культивирования и субкультивирования, как описано выше. Соответствующие фрагменты (которые могут включать в себя все фрагменты) указанных работ включены в настоящее описание в качестве ссылки.
[0059]
(Описание предпочтительного варианта осуществления)
Ниже описаны предпочтительные варианты осуществления. Однако следует понимать, что варианты осуществления приведены для иллюстрации настоящего изобретения, и что объем настоящего изобретения не ограничивается такими предпочтительными вариантами осуществления. Следует также понимать, что специалисты в данной области могут легко осуществить модификации, изменения и т.п. в пределах объема настоящего изобретения со ссылкойна нижеследующие предпочтительные примеры. Варианты осуществления могут соответственно включать в себя любые сочетания вариантов осуществления.
[0060]
(Лечение или профилактика заболевания, расстройства или состояния, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки, с использованием ингибитора сигнального пути TGFβ)
В одном аспекте настоящее изобретение предлагает терапевтическое или профилактическое средство против заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки, содержащее ингибитор сигнального пути TGFβ. При осуществлении настоящего изобретения неожиданно было обнаружено, что стресс ER можно уменьшить или устранить, либо поддержать на нормальном уровне, или возвратить к нормальному уровню путем введения ингибитора сигнального пути TGFβ с целью лечения заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки. Таким образом, такое применение ингибитора сигнального пути TGFβ для лечения или профилактики заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки, является неожиданным с точки зрения совокупности традиционных знаний.
[0061]
В предпочтительном варианте осуществления целевое заболевание, расстройство или состояние настоящего изобретения представляет собой расстройство, связанное с дистрофией эндотелия роговицы Фукса. В настоящее время не существует фундаментального терапевтического метода лечения дистрофии эндотелия роговицы Фукса, поэтому лечение дистрофии эндотелия роговицы Фукса зависит только от трансплантации роговицы. Поскольку настоящее изобретение предлагает способ лечения стресса ER, который является одной из важных причин аномалий или нарушений, возникающих при дистрофии эндотелия роговицы Фукса, понятно, что настоящее изобретение можно использовать для лечения или профилактики дистрофии эндотелия роговицы Фукса. В частности, обнаружено, что стресс ER тесно связан апоптозом. Таким образом, можно предположить, что настоящее изобретение предлагает фундаментальный способ лечения дистрофии эндотелия роговицы Фукса. Настоящее изобретение предлагает способ лечения или профилактики нарушений в эндотелиальных клетках роговицы при дистрофии эндотелия роговицы Фукса, а также при состояниях, включающих в себя уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету, отек стромы роговой оболочки и т.п.
[0062]
Ниже приводится подтверждение связи дистрофии эндотелия роговицы Фукса с TGF-β -индуцированным стрессом эндоплазматического ретикулума (ER) в эндотелиальных клетках роговицы. А именно, с использованием роговицы нормальных доноров HCEC (iHCEC) и иммобилизованной клеточной модели (iFECD) эндотелиальных клеток роговицы пациента-человека (HCEC) с дистрофией эндотелия роговицы Фукса (FECD), полученной авторами настоящего изобретения, показано, что стресс ER участвует в развитии FECD, индуцированной сигнальным путем TGFβ. Также показано, что стресс ER может участвовать в гибели клеток при FECD. Таким образом, очевидно, что ингибирование сигнального пути TGFβ является вероятным эффективным способом терапевтического лечения FECD.
[0063]
В соответствии с настоящим изобретением, при анализе iFECD и iHCEC с помощью просвечивающего электронного микроскопа (TEM) с целью выявления морфологических изменений в ER при дистрофии эндотелия роговицы Фукса, рост клеток наблюдается в iFECD, но не в iHCEC. Экспрессию индикаторов стресса ER (PERK и ATF6) стимулируют в присутствии или в отсутствии TGFβ и анализируют методом вестерн-блоттинга, кроме того, продукцию белков внеклеточного матрикса и их солокализацию с ER тестируют методом иммуноокрашивания коллагена типа I и IV, фибронектина и белкового маркера ER PDI, и в результате демонстрируют, что коллаген типа I и IV и фибронектин в iFECD экспрессируются на более высоком уровне, чем в iHCEC, однако обнаружено, что они солокализуются с ER. Кроме того, авторам настоящего изобретения известно, что фосфорилирование PERK и расщепление ATF6 увеличивается под действием TGFβ, причем в iFECD сильнее, чем в iHCEC. Чтобы исследовать участие сигнального пути TGFβ в апоптозе, клетки обрабатывают TGFβ и методом проточной цитометрии определяют апоптотические клетки, положительные по аннексину V. Процент аннексин V-положительных апоптотических клеток среди iHCEC не повышается по сравнению с уровнем, наблюдающимся до стимуляции TGFβ (например, процент апоптотических клеток может составлять 11,1±0,6% до стимуляции и 12,3±0,5% после стимуляции). В то же время, как известно, TGFβ повышает число аннексин V-положительных клеток после стимуляции iFECD по сравнению с числом, наблюдающимся до стимуляции (например, процент апоптотических клеток может составлять 19,4±1,4% до стимуляции и 29,9±1,5% после стимуляции, p<0,01). Помимо вышеуказанного, показано, что повышается экспрессия шаперона GRP78. Кроме того, показано, что в результате стресса ER повышается экспрессия CHOP и индуцируется апоптоз. Такая стимуляция фосфорилирования и экспрессии в iFECD является более заметной, чем в iHCEC. Показано, что повышение можно наблюдать через 3 часа, а через 6 часов данный эффект усиливается. Таким образом, показано, что он увеличивается со временем.
[0064]
Кроме того, в одном конкретном варианте осуществления настоящего изобретения показано, что митохондриальная недостаточность участвует в развитии дистрофии эндотелия роговицы Фукса на уровне функционирования эндотелиальных клеток роговицы. Как показано в Примерах, участие митохондриальной недостаточности в FECD демонстрируетсяс использованием клеточной модели роговицы нормальных доноров HCEC (iHCEC) и иммобилизованной клеточной модели человеческих эндотелиальных клеток роговицы (HCEC) пациентов с FECD (iFECD).
[0065]
Анализ iFECD и iHCEC с помощью просвечивающего электронного микроскопа (TEM) демонстрирует, что в iFECD наблюдается рост митохондрий, а в iHCEC митохондрии являются морфологически нормальными. Таким образом, показано, что при дистрофии эндотелия роговицы Фукса в митоходриях появляются морфологические нарушения. Кроме того, с использованием красителя JC-1, MitoTracker®, и т.п. показано, что мембранный потенциал митохондрий (MMP) в iFECD выше, чем в iHCEC. Также показано, что истечение цитохрома C из митохондрий в цитоплазму в iFECD выше, чем в iHCEC, относительно уровня цитохрома C в митохондриальной фракции, определенного методом Вестерн-блоттинга. Как показано методом Вестерн-блоттинга, каспаза 9, каспаза 3 и поли(АДФ-рибоза)-полимераза (PARP) расщепляются под действием стимулятора митохондриального стресса (такого как стауроспорин). Таким образом, показано, что митохондриальная недостаточность участвует в апоптозе. Известно, что стресс ER индуцирует митохондриальное расстройство во многих клетках. Таким образом, можно легко сделать вывод, что митохондриальное расстройство можно облегчить путем контролирования стресса ER. Таким образом, специалисты в данной области на основании результатов, полученных при осуществлении настоящего изобретения, могут сделать вывод, что митохондриальная недостаточность участвует в патологии FECD и может использоваться в качестве мишени потенциального терапевтического средства.
[0066]
В качестве мишеней для введения (трансплантации) лекарственного средства, или в способе настоящего изобретения можно использовать млекопитающих (таких как люди, мыши, крысы, хомяки, кролики, кошки, собаки, коровы, лошади, овцы, обезьяны и т.п.). Однако предпочтительными являются приматы и особенно предпочтительными - люди. До настоящего момента не были достигнуты удовлетворительные результаты по разработке способа терапевтического лечения эндотелия роговицы. С учетом вышесказанного, настоящее изобретение предлагает инновационные терапевтические способы и лекарственные средства.
[0067]
Сигнальные пути TGF-β примерно подразделяют на системы Smad2/3, опосредуемые ALK4, 5 или 7, и системы Smad1/5/8, опосредуемые ALK1, 2, 3 или 6, которые, как хорошо известно, связаны с фиброзом (J. Massagu'e, Annu. Rev. Biochem. 1998.67: 753−91; Vilar JMG, Jansen R, Sander C(2006) PLoS Comput Biol 2(1): e3; Leask, A., Abraham, D.J. FASEB J.18, 816−827 (2004); Coert Margadant & Arnoud Sonnenberg EMBO reports (2010) 11, 97−105; Joel Rosenbloom et al., Ann Intern Med.2010; 152:159−166). Известно, что BMP-7 может ингибировать сигнальные пути TGF-β, подавляя фиброз (помимо вышеуказанных документов, Ralf Weiskirchen, et al., Frontiers in Bioscience 14, 4992−5012, June 1, 2009; Elisabeth M Zeisberg et al., Nature Medicine 13, 952−961 (2007); Michael Zeisberg et al., Nature Medicine 9, 964−968(2003)). Хотя в указанных документах описано участие TGF-β в состояниях, реально сопровождающихся образованием мембранозной ткани, состоящей из внеклеточного матрикса, такого как коллаген, вследствие искусственно вызванного тяжелого нарушения или редкого заболевания, сифилитического интерстициального кератита, трудно предстказать терапевтический эффект. Кроме того, показано, что фиброз при тяжелом расстройстве роговичной оболочки обусловлен активацией p38 MAPK в процессе фиброза, индуцированного IL-1β. При этом показано с использованием кроликов, что фиброз, возникающий в результате тяжелого воспаления в живом организме вследствие холодового повреждения, сопровождается активацией p38 MAPK и может частично подавляться ингибитором. Полученные данные демонстрируют, что активация p38 MAPK происходит в ситуации, когда в живом организме наблюдается сильное воспаление и образование мембранозной ткани, состояшей из внеклеточного матрикса. Такие данные не демонстрируют, что с помощью ингибитора сигнального пути TGF-β можно лечить или предотвращать заболевание, расстройство или состояние, связанное со стрессом ER эпителия роговицы (например, такое расстройство, как дистрофии эндотелия роговицы Фукса), или поддерживать нормальное состояние. В данной связи считается трудным культивировать эндотелиальные клетки роговицы, поддерживая их нормальное функционирование. В конечном счете, ранее полученные данные не позволяют разработать способ лечения или профилактики заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки, такого как дистрофия эндотелия роговицы Фукса. Подавление сигнального пути TGF-β с целью лечения или профилактики заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки (например, такого расстройства, как дистрофия эндотелия роговицы Фукса) даже не считалось возможным.
[0068]
Любое средство можно использовать в качестве подходящего для настоящего изобретения ингибитора сигнального пути TGF-β, при условии, что оно может ингибировать сигнальный путь TGF-β. Кроме того, хорошо известно, что сигнальный путь TGF-β, подлежащий ингибированию, может представлять собой фактор, связанный с любым сигнальным путем, при условии, что в конечном счете он оказывает эффект, подобный эффекту (в отличие от ингибиторов, антагонистов и т.п.) сигнального пути TGF-β, как в случае BMP-7, помимо факторов, с которыми ассоциированы TGF-β и рецепторы TGF-β.
[0069]
В настоящем изобретении можно использовать один ингибитор сигнального пути TGF-β, или несколько типов такого ингибитора, по необходимости.
[0070]
В одном варианте осуществления средство, ингибирующее сигнальный путь TGF-β, включает по меньшей мере одно из таких средств, как антагонист TGF-β, антагонист рецептора TGF-β или ингибитор Smad3, другие ингредиенты, указанные в настоящем описании, их фармацевтически приемлемые соли или сольваты, или сольваты их фармацевтически приемлемых солей. Любой ингибитор, описанный в других разделах настоящего документа, можно использовать в качестве ингибитора антагониста TGF-β, антагониста рецептора TGF-β и ингибитора Smad3.
[0071]
В одном варианте осуществления ингибитор сигнального пути TGF-β, который можно использовать в настоящем изобретении, включает по меньшей мере одно из таких средств как SB431542 (4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)-1H-имидазол-2-ил]бензамид), BMP-7, антитело к TGF-β, антитело к рецептору TGF-β, миРНК TGF-β, миРНК рецептора TGF-β, антисмысловой олигонуклеотид TGF-β, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-илпроп-2-еноил))-1,2,3,4-тетрагидроизохинолон, A83-01 (3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1H-пиразол-1-карботиоамид), ингибитор Stemolecule™ TLK (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин), ингибитор Stemolecule™ BMP LDN-193189 (6-(4-(пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил) пиразоло[1,5-a]пиримидин), SD-208 (2-(5-хлор-2-фторфенил)-4-[(4-пиридинил)амино]птеридин), LY364947 (4-[3-(2-пиридинил)-1H-пиразол-4-ил]хинолин), другие ингредиенты, описанные в данном документе, их фармацевтически приемлемые соли или сольваты, или сольваты их фармацевтически приемлемых солей. Указанные антитела могут представлять собой нейтрализующие антитела, но не ограничиваются ими. Не желая быть связанными с какой-либо теорией, авторы настоящего изобретения понимают, что эффект настоящего изобретения можно достичь с помощью ингибитора сигнального пути TGF-β, или любого из указанных путей. В разделе Примеры также демонстрируется эффект ингибитора ALK-4, 5 и 7 A83-01 (3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1H-пиразол-1-карботиоамида). Так, следует понимать, что в конкретных примерах настоящего изобретения также можно использовать ингибитор ALK-4, 5, 7. Кроме того, в разделе Примеры также демонстрируется эффект ингибитора ALK-5 2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридина. Так, следует понимать, что в конкретных примерах настоящего изобретения также можно использовать ингибитор ALK-5.
[0072]
В предпочтительном варианте осуществления средство, ингибирующее сигнальный путь TGF-β, подходящее для применения в настоящем изобретении, включает SB431542 (4-[4-(1,3-бензодиоксол-5-ил)2-пиридинил)-1H-имидазол-2-ил]бензамид). Показано, что данное средство улучшает заболевание, расстройство или состояние, связанное со стрессом ER в эндотелии роговой оболочки (например, расстройство, связанное с дистрофией эндотелия роговицы Фукса и т.п.). В предпочтительном варианте осуществления SB431542 используют в концентрации, составляющей примерно от 0,1 мкМ до 10 мкМ, предпочтительно в концентрации, составляющей примерно от 1 мкМ до 10 мкМ, и еще более предпочтительно в концентрации, составляющей примерно 1 мкМ.
[0073]
Концентрация ингибитора сигнального пути TGF-β, используемого в настоящем изобретении, обычно составляет примерно от 0,1 до 100 мкмоль/л, предпочтительно примерно от 0,1 до 30 мкмоль/л, и более предпочтительно примерно 1 мкмоль/л. Если используют несколько типов ингибиторов, концентрацию можно изменить соответствующим образом. Примеры других диапазонов концентраций, как правило, включают в себя, без ограничения, примерно от 0,001 до 100 мкмоль/л, предпочтительно примерно от 0,01 до 75 мкмоль/л, примерно от 0,05 до 50 мкмоль/л, примерно от 1 до 10 мкмоль/л, примерно от 0,01 до 10 мкмоль/л, примерно от 0,05 до 10 мкмоль/л, примерно от 0,075 до 10 мкмоль/л, примерно от 0,1 до 10 мкмоль/л, примерно от 0,5 до 10 мкмоль/л, примерно от 0,75 до 10 мкмоль/л, примерно от 1,0 до 10 мкмоль/л, примерно от 1,25 до 10 мкмоль/л, примерно от 1,5 до 10 мкмоль/л, примерно от 1,75 до 10 мкмоль/л, примерно от 2,0 до 10 мкмоль/л, примерно от 2,5 до 10 мкмоль/л, примерно от 3,0 до 10 мкмоль/л, примерно от 4,0 до 10 мкмоль/л, примерно от 5,0 до 10 мкмоль/л, примерно от 6,0 до 10 мкмоль/л, примерно от 7,0 до 10 мкмоль/л, примерно от 8,0 до 10 мкмоль/л, примерно от 9,0 до 10 мкмоль/л, примерно от 0,01 до 50 мкмоль/л, примерно от 0,05 до 5,0 мкмоль/л, примерно от 0,075 до 5,0 мкмоль/л, примерно от 0,1 до 5,0 мкмоль/л, примерно от 0,5 до 5,0 мкмоль/л, примерно от 0,75 до 5,0 мкмоль/л, примерно от 1,0 до 5,0 мкмоль/л, примерно от 1,25 до 5,0 мкмоль/л, примерно от 1,5 до 5,0 мкмоль/л, примерно от 1,75 до 5,0 мкмоль/л, примерно от 2,0 до 5,0 мкмоль/л, примерно от 2,5 до 5,0 мкмоль/л, примерно от 3,0 до 5,0 мкмоль/л, примерно от 4,0 до 5,0 мкмоль/л, примерно от 0,01 до 3,0 мкмоль/л, примерно от 0,05 до 3,0 мкмоль/л, примерно от 0,075 до 3,0 мкмоль/л, примерно от 0,1 до 3,0 мкмоль/л, примерно от 0,5 до 3,0 мкмоль/л, примерно от 0,75 до 3,0 мкмоль/л, примерно от 1,0 до 3,0 мкмоль/л, примерно от 1,25 до 3,0 мкмоль/л, примерно от 1,5 до 3,0 мкмоль/л, примерно от 1,75 до 3,0 мкмоль/л, примерно от 2,0 до 3,0 мкмоль/л, примерно от 0,01 до 1,0 мкмоль/л, примерно от 0,05 до 1,0 мкмоль/л, примерно от 0,075 до 1,0 мкмоль/л, примерно от 0,1 до 1,0 мкмоль/л, примерно от 0,5 до 1,0 мкмоль/л, примерно от 0,75 до 1,0 мкмоль/л, примерно от 0,09 до 35 мкмоль/л, примерно от 0,09 до 3,2 мкмоль/л, и более предпочтительно примерно от 0,05 до 1,0 мкмоль/л, примерно от 0,075 до 1,0 мкмоль/л, примерно от 0,1 до 1,0 мкмоль/л, примерно от 0,5 до 1,0 мкмоль/л, и примерно от 0,75 до 1,0 мкмоль/л.
[0074]
В предпочтительном варианте осуществления используемый ингибитор сигнального пути TGF-β представляет собой 4-[4-(1,3-бензодиоксол-5-ил)2-пиридинил)-1H-имидазол-2-ил]бензамид или его фармацевтически приемлемую соль.
[0075]
В другом предпочтительном варианте осуществления используемый в настоящем изобретении ингибитор сигнального пути TGF-β представляет собой BMP-7. Причины использования данного ингибитора: он подавляет фиброз; показано, что он способствует сохранению нормальных функций, и его применение может заменить трансплантацию у приматов. В предпочтительном варианте осуществления BMP-7 используют в концентрации, составляющей примерно от 10 нг/мл до 1000 нг/мл, более предпочтительно в концентрации, составляющей примерно от 100 нг/мл до 1000 нг/мл. BMP-7 можно использовать в концентрации, составляющей примерно 100 нг/мл или в концентрации, составляющей примерно 1000 нг/мл.
[0076]
В одном варианте осуществления с помощью ингибитора сигнального пути TGF-β можно лечить митохондриальную недостаточность. Подлежащая лечению митохондриальная недостаточность может представлять собой, например, дистрофию эндотелия роговицы Фукса, но не ограничивается ею.
[0077]
Альтернативно средство для лечения митохондриальной недостаточности можно использовать при лечении болезней глаза, особенно болезней эндотелия роговицы, отдельно или в сочетании с другим средством, таким как ингибитор сигнального пути TGF-β. Примеры средств, используемых для лечения заболевания, расстройства или состояния, связанного с митохондриальной недостаточностью, включают в себя, без ограничения, витамины, используемые в качестве специфических элементов митохондриальной дыхательной цепи (такие как витамин E или его производные), кофакторы, кофермент Q (убихитин), никотинамид, рибофлавин, карнитин, биотин, липоевую кислоту и т.п.
[0078]
Примеры заболеваний, обусловленных митохондриальной недостаточностью, могут включать в себя набухание митохондрий, вызываемое латентной дисфункцией митохондрий, дисфункцию, вызываемую окислительным стрессом (например, обусловленным свободными радикалами или активными формами кислорода), дисфункцию, вызываемую генетическим фактором, а также заболевания, обусловленные недостаточностью механизма окислительного фосфорилирования, участвующего в продукции энергии в митохондриях. Существует много конкретных примеров заболеваний, развитие которых обусловлено вышеуказанными патологическими факторами, при этом заболевания, относящиеся к области офтальмологии, включают в себя, без ограничения, атрофию зрительного нерва, невропатию зрительного нерва, пигментный ретинит и катаракту. Кроме того, в качестве другой формы вышеуказанного заболевания, расстройства или состояния, связанного с дистрофией эндотелия роговицы Фукса, примеры заболевания, расстройства или состояния, связанного с митохондриальной недостаточностью эндотелия роговой оболочки, включают в себя уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету, отек стромы роговой оболочки и т.п.
[0079]
Терапевтическое или профилактическое средство настоящего изобретения может содержать дополнительный фармацевтический ингредиент. Типичные примеры такого фармацевтического продукта включают в себя ингибиторы киназы Rho, стероиды и т.п. Не желая быть связанными с какой-либо теорией, авторы полагают, что путем включения ингибитора киназы Rho, позволяющего предотвартить потерю клеток в результате стимуляции адгезии эндотелиальных клеток роговицы и образования слоя клеток эндотелия роговицы, характеризующегося превосходной морфологией клеток и высокой клеточной плотностью, можно повысить эффект ингибитора сигнального пути TGF-β. В настоящем изобретении можно использовать один тип ингибитора киназы Rho, или сочетание нескольких типов ингибиторов киназы Rho, по необходимости.
[0080]
Примеры ингибиторов киназы Rho, подходящих для применения в настоящем изобретении, включают в себя соединения, раскрытые в следующих документах: патент США № 4678783, патент Японии № 3421217, международная публикация № WO 95/28387, международная публикация № WO 99/20620, международная публикация № WO 99/61403, международная публикация № WO 02/076976, международная публикация № WO 02/076977, международная публикация № WO 2002/083175, международная публикация № WO 02/100833, международная публикация № WO 03/059913, международная публикация № WO 03/062227, международная публикация № WO 2004/009555, международная публикация № WO 2004/022541, международная публикация № WO 2004/108724, международная публикация № WO 2005/003101, международная публикация № WO 2005/039564, международная публикация № WO 2005/034866, международная публикация № WO 2005/037197, международная публикация № WO 2005/037198, международная публикация № WO 2005/035501, международная публикация № WO 2005/035503, международная публикация № WO 2005/035506, международная публикация № WO 2005/080394, международная публикация № WO 2005/103050, международная публикация № WO 2006/057270, международная публикация № WO 2007/026664 и т.п. Каждое из таких соединений можно получить в промышленности с помощью способов, описанных в документах, раскрывающих соответствующие соединения. Примеры таких соединений включают в себя 1-(5-ихохинолинсульфонил)гомопиперазин или его соль (например, фасудил (1-(5-изохинолинсульфонил)гомопиперазин)), (R)-(+)-транс-(4-пиридил)-4-(1-аминоэтил)-циклогексанкарбоксамид), или его соль (например, Y-27632 ((R)-(+)-транс-(4-пиридил)-4-(1-аминоэтила) дигидрохлорид циклогексанкарбоксамида моногидрат) и т.п.) и т.п.
[0081]
Концентрация ингибитора киназы Rho в настоящем изобретении, как правило, составляет примерно от 1 до 100 мкмоль/л, предпочтительно примерно от 5 до 20 мкмоль/л, и более предпочтительно примерно 10 мкмоль/л. При использовании нескольких типов указанного ингибитора, концентрацию можно изменить соответствующим образом. Примеры других диапазонной концентраций, как правило, включают в себя, без ограничения, примерно от 0,001 до 100 мкмоль/л, предпочтительно примерно от 0,01 до 75 мкмоль/л, примерно от 0,05 до 50 мкмоль/л, примерно от 1 до 10 мкмоль/л, примерно от 0,01 до 10 мкмоль/л, примерно от 0,05 до 10 мкмоль/л, примерно от 0,075 до 10 мкмоль/л, примерно от 0,1 до 10 мкмоль/л, примерно от 0,5 до 10 мкмоль/л, примерно от 0,75 до 10 мкмоль/л, примерно от 1,0 до 10 мкмоль/л, примерно от 1,25 до 10 мкмоль/л, примерно от 1,5 до 10 мкмоль/л, примерно от 1,75 до 10 мкмоль/л, примерно от 2,0 до 10 мкмоль/л, примерно от 2,5 до 10 мкмоль/л, примерно от 3,0 до 10 мкмоль/л, примерно от 4,0 до 10 мкмоль/л, примерно от 5,0 до 10 мкмоль/л, примерно от 6,0 до 10 мкмоль/л, примерно от 7,0 до 10 мкмоль/л, примерно от 8,0 до 10 мкмоль/л, примерно от 9,0 до 10 мкмоль/л, примерно от 0,01 to 50 мкмоль/л, примерно от 0,05 до 5,0 мкмоль/л, примерно от 0,075 до 5,0 мкмоль/л, примерно от 0,1 до 5,0 мкмоль/л, примерно от 0,5 до 5,0 мкмоль/л, примерно от 0,75 до 5,0 мкмоль/л, примерно от 1,0 до 5,0 мкмоль/л, примерно от 1,25 до 5,0 мкмоль/л, примерно от 1,5 до 5,0 мкмоль/л, примерно от 1,75 до 5,0 мкмоль/л, примерно от 2,0 до 5,0 мкмоль/л, примерно от 2,5 до 5,0 мкмоль/л, примерно от 3,0 до 5,0 мкмоль/л, примерно от 4,0 до 5,0 мкмоль/л, примерно от 0,01 до 3,0 мкмоль/л, примерно от 0,05 до 3,0 мкмоль/л, примерно от 0,075 до 3,0 мкмоль/л, примерно от 0,1 до 3,0 мкмоль/л, примерно от 0,5 до 3,0 мкмоль/л, примерно от 0,75 до 3,0 мкмоль/л, примерно от 1,0 до 3,0 мкмоль/л, примерно от 1,25 до 3,0 мкмоль/л, примерно от 1,5 до 3,0 мкмоль/л, примерно от 1,75 до 3,0 мкмоль/л, примерно от 2,0 до 3,0 мкмоль/л, примерно от 0,01 до 1,0 мкмоль/л, примерно от 0,05 до 1,0 мкмоль/л, примерно от 0,075 до 1,0 мкмоль/л, примерно от 0,1 до 1,0 мкмоль/л, примерно от 0,5 до 1,0 мкмоль/л, примерно от 0,75 до 1,0 мкмоль/л, примерно от 0,09 до 35 мкмоль/л, примерно от 0,09 до 3,2 мкмоль/л, и более предпочтительно примерно от 0,05 до 1,0 мкмоль/л, примерно от 0,075 до 1,0 мкмоль/л, примерно от 0,1 до 1,0 мкмоль/л, примерно от 0,5 до 1,0 мкмоль/л, и примерно от 0,75 до 1,0 мкмоль/л.
[0082]
Композицию настоящего изобретения можно вводить в виде глазных капель.
[0083]
Величина дозы и частота ее введения варьируют в зависимости от симптомов, возраста, массы или формата введения доз. Например, композицию в виде глазных капель, содержащую примерно 0,0001-0,1 масс/об% эффективного ингредиента, предпочтительно примерно 0,003-0,03 масс/об%, как правило, можно вводить 1-10 раз в день, предпочтительно 1-6 раз, более предпочтительно 1-3 раза, в объеме, составляющем для взрослых примерно 0,01-0,1 мл на дозу. Если лекарственное средство настоящего изобретения вводят в переднюю камеру, его концентрация в композиции может составлять от 1/10 до 1/1000 от указанной концентрации. Специалисты в данной области могут соответствующим образом выбрать тип и концентрацию ингибитора сигнального пути TGFβ, ингибитора киназы Rho и т.п., в зависимости от состояния заболевания.
[0084]
В другом аспекте настоящее изобретение предлагает вещество, ингибирующее сигнальный путь TGFβ, для лечения или профилактики расстройства, связанного со стрессом ER в эндотелии роговой оболочки. Термин "вещество, ингибирующее сигнальный путь TGFβ", можно использовать как взаимозаменяемый с термином "ингибитор сигнального пути TGFβ". Любой описанный здесь вариант осуществления можно использовать в применении к стрессу ER в эндотелии роговицы и к ингибитору сигнального пути TGF-β.
[0085]
В другом аспекте настоящее изобретение предлагает способ лечения или профилактики заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки у индивидуума, где способ включает стадию введения индивидууму эффективного количества ингибитора сигнального пути TGF-β. Любой описанный здесь вариант осуществления данного способа можно использовать в применении к ингибитору сигнального пути TGF-β и к заболеванию, расстройству или состоянию, связанному со стрессом ER в эндотелии роговой оболочки.
[0086]
Мишени для введения (трансплантации) лекарственного средства настоящего изобретения, или применения способа настоящего изобретения включают в себя млекопитающих (таких как люди, мыши, крысы, хомяки, кролики, кошки, собаки, коровы, лошади, овцы, обезьяны и т.п.). Однако предпочтительными являются приматы и особенно предпочтительными - люди. До настоящего момента не были достигнуты удовлетворительные результаты по разработке способа терапевтического лечения эндотелия роговицы у приматов. С учетом вышесказанного, настоящее изобретение предлагает инновационные терапевтические способы и лекарственные средства. Количество терапевтического средства настоящего изобретения, эффективное для лечения конкретного заболевания, расстройства или состояния, может варьировать в зависимости от характеристик расстройства или состояния. Однако специалисты в данной области могут определить эффективное количество с помощью стандартного клинического метода на основании приведенного здесь описания. Кроме того, в некоторых случаях оптимальный диапазон можно определить с помощью анализа in vitro. Поскольку точная доза, используемая в смеси, может варьировать в зависимости от пути введения или тяжести заболевания или расстройства, дозу следует определять с учетом мнения врача или состояния каждого пациента. Однако доза, не ограничиваясь конкретными значениями, может составлять, например, 0,001, 1, 5, 10, 15, 100 или 1000 мг/кг массы тела, или она может находиться в диапазоне между любыми двумя из указанных значений. Интервал между введением доз конкретно не ограничивается, однако его примеры могут включать в себя введение 1 или 2 доз в течение 1, 7, 14, 21 или 28 дней, или 1 или 2 доз в течение диапазона между любыми 2 из указанных значений. Доза, интервал между введением доз и способ введения дозы могут быть выбраны соответствующим образом в зависимости от возраста пациента, массы пациента, наблюдающихся у пациента симптомов, органа-мишени и т.п. Кроме того, терапевтическое средство предпочтительно содержит активный ингредиент в терапевтически эффективном количестве, или в количестве, которое эффективно обеспечивает желательный результат. Вывод о наличии терапевтического эффекта можно сделать на основании значимого снижения уровня терапевтического маркера после введения лекарственного средства. Эффективную дозу можно определить с помощью кривой доза-ответ, полученной in vitro или в результате тестирования животной модели.
[0087]
Цитирующиеся здесь литературные источники, такие как научные статьи, патенты и патентные заявки, включены в данный документ в качестве ссылки в такой степени, как если бы было конкретно приведено полное описание каждого документа.
[0088]
Как указано выше, настоящее изобретение раскрывается путем демонстрации предпочтительных вариантов осуществления, облегчающих понимание. Далее настоящее изобретение описывается посредством примеров. Приведенное выше описание и нижеследующие примеры предназначаются не для ограничения настоящего изобретения, а лишь для его иллюстрации. Таким образом, объем настоящего изобретения ограничивается не вариантами осуществления и примерами, конкретно описанными в данном документе, а только формулой изобретения.
[ПРИМЕРЫ]
[0089]
Ниже приведены примеры настоящего изобретения. Биологические примеры и т.п., когда это применимо, проводятся в соответствии со стандартами, установленными Министерством здравоохранения, труда и социального обеспечения, Министерством образования, культуры, спорта, науки и техники, и т.п., и, если это применимо, в соответствии с Хельсинкской декларацией или этическими нормами и правилами, утвержденными на ее основе. Согласие на донорство глаз, используемых в исследовании, получают от близких родственников всех умерших доноров. Настоящее исследование одобрено Комитетом по этике или соответствующим органом Банка роговиц университета Эрлангена и SightLife™ (Seattle, WA).
[0090]
Дистрофия эндотелия роговицы Фукса приводит к гибели эндотелиальных клеток роговицы. Если оставшиеся эндотелиальные клетки роговицы не могут компенсировать утрату насосной или барьерной функции, прозрачность роговицы перестает поддерживаться, приводя к слепоте вследствие помутнения роговицы. Кроме того, известно, что эндотелиальные клетки роговицы пациентов с дистрофией эндотелия роговицы Фукса продуцируют избыточное количество внеклеточного матрикса, приводя к гипертрофии десцеметовой мембраны и образованию гутты. Образование гутты и гипертрофия десцеметовой мембраны обуславливают светорассеяние и т.п., вызывая снижение остроты зрения, светобоязнь или нечеткость зрения, подвергая значительному риску QOL пациента. Чтобы определить причину продукции внеклеточного матрикса и идентифицировать терапевтическую мишень, в качестве модели используют иммобилизованные линии клеток эндотелия роговицы (iFECD), полученных от пациента с дистрофией эндотелия роговицы Фукса, сравнивая их с иммобилизованными линиями клеток эндотелия роговицы (iHCEC), полученных от здорового донора.
[0091]
(Пример способа получения 1: Получение модели, включающей иммобилизованные линии клеток эндотелия роговицы (iFECD), полученных от пациента с дистрофией эндотелия роговицы Фукса)
В настоящем примере иммобилизованную линию клеток эндотелия роговицы (iFECD) получают из эндотелиальных клеток роговицы, полученных от пациентов с дистрофией эндотелия роговицы Фукса.
[0092]
(Способ культивирования)
Эндотелиальные клетки роговицы механически удаляют с базальной мембраны роговицы, полученной из Банка роговицы Сиэтла. Клетки эндотелия роговицы отделяют от базальной мембраны с помощью коллагеназы, собирают и затем подвергают первичному культивированию. В качестве основной среды используют среду с пониженным содержанием сыворотки Opti-MEM I в жидком виде (INVITROGEN, № по каталогу: 31985-070), к которой добавляют 8% FBS (BIOWEST, № по каталогу: S1820-500), 200 мг/мл CaCl2•2H2O (SIGMA, № по каталогу: C7902-500G), 0,08% хондроитинсульфата (SIGMA, № по каталогу: C9819-5G), 20 мкг/мл аскорбиновой кислоты (SIGMA, № по каталогу: A4544-25G), 50 мкг/мл гентамицина (INVITROGEN, № по каталогу: 15710-064) и 5 нг/мл EGF (INVITROGEN, № по каталогу: PHG0311), и которую затем акклиматизируют с использованием питающих клеток 3T3. Затем клетки культивируют в основной среде, к которой добавляют SB431524 (1 мкмоль/л) и SB203580 (4−(4−фторфенил)−2−(4−метилсульфонилфенил)−5(4−пиридил)имидазол<4−[4−(4−фторфенил)−2−(4−метилсульфинилфенил)−1H−имидазол−5-ил]пиридин) (1 мкмоль/л) (такую среду в рамках изобретения также называют "SB203580+SB431542+3T3-акклиматизированная среда").
[0093]
(Способ приобретения)
Эндотелиальные клетки роговицы приобретают, после согласования с Комитетом по этике и получения письменного согласия, от 3 пациентов-людей, страдающих от буллезной кератопатии, в соответствии с клиническим диагнозом Дистрофия эндотелия роговицы Фукса, и подвергшихся трансплантации эндотелия роговицы (эндотелиальная кератопластика десцеметовой мембраны=DMEK). В процессе DMEK патологические эндотелиальные клетки роговицы механически удаляют с десцеметовой мембраны, базальной мембраны, и погружают в растовр для хранения роговицы Optisol-GS (Bauch & Lomb). После ферментативной обработки коллагеназой эндотелиальные клетки роговицы собирают и культивируют в SB203580+SB431542+3T3-акклиматизированной среде. Для культивированных эндотелиальных клеток роговицы, полученных от пациента с дистрофией эндотелия роговицы Фукса, большой T-антиген и ген hTERT SV40 амплифицируют методом ПЦР и вводят в лентивирусный вектор (pLenti6.3_V5−TOPO; Life Technologies Inc). Затем лентивирусным вектором инфицируют клетки 293T (RCB2202; Riken Bioresource Center, Ibaraki, Japan), используя реагент для трансфекции (Fugene HD; Promega Corp., Madison, WI) и три типа хелперных плазмид (pLP1, pLP2, pLP/VSVG; Life Technologies Inc.). Культуральный супернатант, содержащий вирусы, собирают через 48 после инфицирования. К культуральному раствору культивируемых эндотелиальных клеток роговицы, полученных от пациента с дистрофией эндотелия роговицы Фукса, добавляют 5 мкг/мл полибрена и затем вводят большой T-антиген и ген hTERT SV40. Анализируют изображения иммобилизованной линии клеток эндотелия роговицы (iFECD) от пациента с дистрофией эндотелия роговицы Фукса, полученные с помощью фазоразностного микроскопа. Эндотелиальные клетки, полученные из предназначенной для исследовательских целей роговицы, импортированной из Банка роговицы Сиэтла, иммобилизуют таким же способом с получением иммобилизованной линии нормальных эндотелиальных клеток роговицы (iHCEC), которую используют в качестве контроля. Анализ изображений иммобилизованной линии клеток эндотелия роговицы (iFECD) и иммобилизованной линии клеток эндотелия роговицы здорового донора (iHCEC), полученных с помощью фазоразностного микроскопа, показывает, что и iHCEC, и iFECD образуют слой клеток полигональной формы, как и нормальные эндотелиальные клетки роговицы. IHCEC и iFECD поддерживают и культивируют в среде DMEM+10% FBS. SB431542 получают от TOCRIS (номер по каталогу: 1614). SB203580 получают от CALBIOCHEM (номер по каталогу: 559389).
[0094]
(Пример способа получения 2: Подтверждение нормального функционирования иммобилизованной линии клеток эндотелия роговицы (iFECD))
В настоящем примере подтверждают нормальное функционирование иммобилизованной линии клеток эндотелия роговицы (iFECD).
[0095]
(Иммуноокрашивание Na+/K+-АТФазой и ZO-1)
Вначале проводят иммуноокрашивание по Na+/K+-АТФазе и ZO-1, чтобы подтвердить нормальное функционирование иммобилизованной линии клеток эндотелия роговицы (iFECD). Данное иммуноокрашивание проводят для анализа функций эндотелиальных клеток роговицы, таких как насосная и барьерная функции. По Na+/K+-АТФае и ZO-1 можно определить нормальные насосную и барьерную функции эндотелиальных клеток роговицы, соответственно. Технология иммуноокрашивания описана ниже.
[0096]
(Способ наблюдения клеток с помощью окрашивания и т.п. (Гистологический тест))
Клетки наблюдают с помощью фазоразностного микроскопа. После иммобилизации клеток проводят иммуноокрашивание, используя ZO-1 и Na+/K+-АТФазу в качестве маркеров функционирования, и наблюдают клетки с помощью флуоресцентного микроскопа. Для тестирования путем окрашивания ткани культивированные клетки помешают в Lab−TekTM Chamber SlidesTM (NUNC A/S, Roskilde, Denmark), иммобилизуют в 4% формальдегиде в течение 10 минут при комнатной температуре (КТ) и инкубируют в течение 30 минут в присутствии 1% бычьего сывороточного альбумина (BSA). А именно, клетки, культивированные на Lab−TekTM Chamber SlidesTM (NUNC A/S, Roskilde, Denmark), иммобилизуют в 4% формальдегиде в течение 10 минут при комнатной температуре и инкубируют в течение 30 минут в присутствии 1% бычьего сывороточного альбумина (BSA). Чтобы исследовать фенотип клеток, проводят иммуногистохимический анализ по ZO-1, который представляет собой белок, отвечающий за плотные контакты, и по Na+/K+-АТФазе, которая представляет собой белок, отвечающий за насосную функцию. ZO-1 и Na+/K+-АТФазу используют в качестве маркеров, ассоциированных с клеточными функциями. ZO-1 и Na+/K+-АТФазу окрашивают поликлональным антителом против ZO-1 (Zymed Laboratories, Inc., South San Francisco, CA) и моноклональным антителом против Na+/K+-АТФазы (Upstate Biotec, Inc., Lake Placid, NY), соответственно, в разведении 1:200. В качестве вторичных антител используют козлиные антитела против мышиных IgG, меченные Alexa Fluor® 488 или Alexa Fluor® 594 (Life Technologies), в разведении 1:2000. Затем клеточные ядра окрашивают DAPI (Vector Laboratories, Inc., Burlingame, CA) или PI (Sigma-Aldrich). И наконец, предметные стекла наблюдают с помощью флуоресцентного микроскопа (TCS SP2 AOBS; Leica Microsystems, Welzlar, Germany).
[0097]
Полученные результаты показывают, что Na+/K+-АТФаза и ZO-1 экспрессируются во всех клетках iHCEC и iFECD, свидетельствуя о том, что в полученной иммобилизованной клеточной линии сохраняются нормальные функции.
[0098]
Кроме того, изображения, полученные с помощью просвечивающего электронного микроскопа (данные не показаны), анализируют на морфологические изменения в iHCEC и iFECD. IHCEC и iFECD культивируют на Transwell в бессывороточной среде DMEM и иммобилизуют через неделю, когда клетки находятся в состоянии слияния, после чего изучают морфологию с помощью просвечивающего электронного микроскопа. Результаты демонстрируют, что iHCEC и iFECD образуют слой клеток, в котором отсутствуют видимые морфологические аномалии.
[0099]
Кроме того, показано, что эндотелиальные клетки роговицы пациентов с дистрофией эндотелия роговицы Фукса продуцируют избыточное количество внеклеточного матрикса, приводя к образованию гутты и гипертрофии десцеметовой мембраны. В данной связи, чтобы определить экспрессию коллагена типа I, коллагена типа IV и фибронектина, которые являются белками внеклеточного матрикса, iHCEC и iFECD культивируют в чашках для культивирования и подвергают иммуноокрашиванию. Показано, что экспрессия коллагена типа I, коллагена типа IV и фибронектина в iFECD выше, чем в iHCEC. Кроме того, анализ уровней экспрессии генов в iHCEC и iFECD методом ПЦР в режиме реального времени показывает, что наблюдается значительное повышение уровней экспрессии коллагена типа I и фибронектина, и существует тенденция к повышению экспрессии коллагена типа IV. Определяют, продуцируют ли iFECD избыточное количество внеклеточного матрикса, как клетки эндотелия роговицы пациентов с дистрофией эндотелия роговицы Фукса. IHCEC и iFECD культивируют на Transwell в бессывороточной среде DMEM и иммобилизуют через неделю, когда клетки находятся в состоянии слияния, после чего проводят окрашивание HE. В iFECD наблюдается более значительное превышение продукции внеклеточного матрикса, чем в iHCEC. С учетом вышесказанного, можно сделать вывод, что получена клеточная модель заболевания, характеризующаяся избыточной продукцией внеклеточного матрикса, свойственной пациентам с дистрофией эндотелия роговицы Фукса. Поскольку ожидается, что анализ с использованием клеточной модели заболевания может вносить вклад в исследование патологии дистрофии эндотелия роговицы Фукса, в которой существует много неясных моментов, клетки далее используют для разработки терапевтического средства против дистрофии эндотелия роговицы Фукса, как типичного примера заболевания, связанного со стрессом эндоплазматического ретикулума.
[0100]
(Пример 1: Наблюдение морфологических особенностей эндоплазматического ретикулума и митохондрий при дистрофии эндотелия роговицы Фукса)
В настоящем примере, как типичном примере исследования стресса эндоплазматического ретикулума, определяют присутствие морфологических аномалий в эндоплазматическом ретикулуме и митохондриях клеток модели дистрофии эндотелия роговицы Фукса, полученной в вышеописанном Примере получения.
[0101]
(Наблюдение органелл с помощью электронного микроскопа)
Затем клетки наблюдают с помощью электронного микроскопа, чтобы подтвердить повышение продукции ECM и стресса ER. Предварительную иммобилизацию проводят путем высевания клеток в трансвел и иммобилизации их в течение трех часов при комнатной температуре с использованием 2,5% глутаральдегида, pH 7,2, разбавленного 0,1 M раствором какодилата натрия, после чего промывают три раза 0,1 M раствором какодилата натрия. Затем проводят пост-иммобилизацию в течение 1 часа при комнатной температуре с использованием 1% осмиевой кислоты, разбавленной 0,1 M раствором какодилата натрия, и промывают три раза дистиллированной водой. Чтобы исследовать локализацию и морфологические изменения органелл, клетки погружают на 1 час в 0,5% раствор уранилацетата при комнатной температуре. Чтобы провести обезвоживание, клетки погружают на 10 минут в 70% этанол и в 90% этанол, и на 20 минут в 100% этанол. Затем клетки погружают в пропиленксид на 30 минут и на 1 час при комнатной температуре в смесь, состоящую из 1 части пропиленоксида и 1 части аралдитовой смолы. Трансвел погружают в смесь, содержащую аралдитовую смолу, и выдерживают в течение 2 часов при комнатной температуре. Смесь, содержащую аралдитовую смолу, заменяют три раза через каждые 2 часа. Через 12 часов аралдитовую смолу снова заменяют три раза через каждые 2 часа. Затем клетки погружают в смесь, содержащую аралдитовую смолу, инкубируют 24 часа при 60°C и отверждают. С помощью микротома (Leica EMUC7, Leica Microsystems, Welzlar, Germany) получают срез толщиной 300 нм и помещают его на решетку. После окрашивания решетки срез наблюдают с помощью электронного микроскопа.
[0102]
(Результаты)
Результаты показаны на фигуре 2. Обнаружено, что при дистрофии эндотелия роговицы Фукса в эндоплазматическом ретикулуме и митохондриях наблюдаются морфологические аномалии. Более конкретно, в iHCEC эндоплазматический ретикулум и митохондрии имеют обычную форму, а внеклеточный матрикс отсутствует. С другой стороны, в iFECD наблюдается морфологическая аномалия, заключающаяся в увеличении объема эндоплазматического ретикулума и митохондрий. Кроме того, между клетками присутствует внеклеточный матрикс.
[0103]
Полученные результаты демонстрируют, что в клетках модели дистрофии эндотелия роговицы Фукса наблюдаются морфологические аномалии эндоплазматического ретикулума и митохондрий.
[0104]
(Пример 2: Повышение стресса эндоплазматического ретикулума в эндотелиальных клетках роговицы при дистрофии эндотелия роговицы Фукса)
В настоящем примере наблюдают экспрессию молекулярных шаперонов GRP78 и GADD153 в ответ на стресс эндоплазматического ретикулума, чтобы определить, действительно ли в клетках модели дистрофии эндотелия роговицы Фукса усиливается стресс эндоплазматического ретикулума. GADD153 известен как белок, который экспрессируется/индуцируется в ответ на ряд стрессов, в особенности, в ответ на стресс эндоплазматического ретикулума, прерывает клеточный цикл и участвует в апоптозе (в качестве ссылки можно привести Verfaillie T, et al. Cancer Lett. 2013 May 28; 332(2): 249−64).
[0105]
(Материалы и методы)
(Наблюдение экспрессии с использованием иммуноокрашивания)
Повышение уровня экспрессии GRP78 и GADD153, связанных со стрессом ER, подтверждают методом иммуноокрашивания. Метод иммуноокрашивания проводят по способу, описанному в приведенном выше Примере способа получения 2. В данном эксперименте антитела заменяют на антитела против GRP78 и GADD153.
*Антитела против GRP78: (sc−376768, SANTA CRUZ BIOTECHNOLOGY)
*Антитела против GADD153: (sc−575, SANTA CRUZ BIOTECHNOLOGY)
Коротко говоря, эксперимент проводят по описанному ниже способу.
[0106]
Для проведения анализа путем окрашивания ткани культивированные клетки иммобилизуют в течение 10 минут при комнатной температуре (КТ) с использованием 4% формальдегида и инкубируют в течение 30 минут в присутствии 1% раствора бычьего сывороточного альбумина (BSA). Антитела против GRP78 и GADD153 используют в разведении 1:200. В качестве вторичных антител используют козлиные антитела против мышиных IgG, меченные Alexa Fluor® 488 (Life Technologies), в разведении 1:2000. Затем клеточные ядра окрашивают DAPI (Vector Laboratories, Inc., Burlingame, CA) или PI (Sigma-Aldrich). И наконец, предметные стекла наблюдают с помощью флуоресцентного микроскопа (TCS SP2 AOBS; Leica Microsystems, Welzlar, Germany).
*Вестерн-блоттинг: Для электрофореза используют белок, экстрагированный и полученный с использованием буфера RIPA, содержащего 7,5% полиакриламида. Выделенный белок переносят на мембрану из PVDF (PALL LIFE SCIENCE (номер по каталогу: EH-2222)). К забуференному трисом физиологическому раствору (10 мМ Tris-HCl, pH 7,4; 100 мM NaCl) (TBS-T), содержащему 0,1% (об/об) полиэтиленсорбитанмонолаурата (Nacalai Tesque, номер по каталогу: 28353-85), добавляют 5% обезжиренное сухое молоко (CELL SIGNALING, номер по каталогу: 9999) и проводят блокирование путем инкубации пропитанной мембраны в течение 1 часа. Мембрану погружают в раствор, содержащий антитела против GRP78, разбавленные в 1000 раз TBS-T, содержащим 5% обезжиренное сухое молоко, и подвергают взаимодействию в течение 1 часа при комнатной температуре. После трехкратного промывания TBS-T, инкубации с комплексом антитела против мышиных IgG с HRP (CELL SIGNALING (номер по каталогу: 7074P2)) и промывания, детектируют светящуюся полосу с использованием набора для детекции результатов вестерн-блоттинга (GE Healthcare Japan (номер по каталогу: RPN2135V)).
[0107]
В настоящем способе используют перечисленные ниже первичные антитела.
*Антитело к GRP78 (SANTA CRUZ BIOTECHNOLOGY, SC−376768)
*Антитело к IRE1 (Cell Signaling Technology, 14C10)
*Антитело к GAPDH (MEDICAL & BIOLOGICAL LABORATORIES, M171−3)
Далее описаны вторичные антитела.
*HRP-меченное антитело к кроличьих IgG или антитело к кроличьих IgG (Cell Signaling Technology) (разведение 1:5000)
Проводят инкубацию с первичными и вторичными антителами. Мембрану подвергают воздействию света с использованием набора для детекции результатов вестерн-блоттинга ECL Advance (GE Healthcare, Piscataway, NJ) и затем исследуют с помощью системы визуализации LAS4000S (Fuji Film, Tokyo)
[0108]
(Стимуляция клеток)
Клетки стимулируют в течение 24 часов TGFβ2, 10 нг/мл (R&D SYSTEMS). Клетки, стимулированные в течение 24 часов 10 мкМ тапсигартином (TG; Alomone Laboratories Ltd., Jerusalem, Israel), используют в качестве положительного контроля стресса ER.
[0109]
(Результаты)
Результаты показаны на фигуре 3. В клетках модели дистрофии эндотелия роговицы Фукса наблюдают повышение экспрессии GRP78 и GADD153, свидетельствующее о более сильном стрессе эндоплазматического ретикулума, чем в эндотелиальных клетках нормальной роговицы.
[0110]
(Пример 3: TGF-β стимулирует стресс ER в эндотелиальных клеткахк роговицы)
Чтобы подтвердить, что TGF-β стимулирует стресс ER в эндотелиальных клетках роговицы, в настоящем примере проводят вестерн-блоттинг по способу примера 2 и определяют тенденцию GRP78 помимо IRE1, который представляет собой мембранный белок, участвующий в сигнальном пути, опосредующем стресс, на мембране эндоплазматического ретикулума. Клетки стимулируют TGFβ2, 10 нг/мл (R&D SYSTEMS). Клетки, стимулированные тапсигартином (TG; Alomone Laboratories Ltd., Jerusalem, Israel), используют в качестве положительного контроля стресса ER.
[0111]
(Материалы и методы)
Вестерн-блоттинг проводят по способу, описанному в примере 2. Помимо антител, описанных в примере 2, используют перечисленные ниже антитела.
*Антитела против IRE1: (14C10, Cell Signaling Technology)
*TGF-β2: (302−B2−002, R&D SYSTEMS)
[0112]
(Результаты)
Результаты показаны на фигуре 4. Экспрессию шаперона GRP78 и индикатора стресса IRE1 индуцируют с помощью TGFβ в клеточной модели дистрофии эндотелия роговицы Фукса и нормальных клетках. Уровень экспрессии является высоким, особенно в клетках модели дистрофии эндотелия роговицы Фукса. TG обозначает тапсигаргин, который используют для получения положительного контроля стресса ER.
[0113]
(Пример 4: Эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса, являются в высокой степени чувствительными к стрессу эндоплазматического ретикулума)
В настоящем примере ингибитор сигнального пути TGFβ SB431542 вводят, чтобы определить, устраняется ли усиление стресса ER, и продемонстрировать, что эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса, являются высокочувствительными к стрессу эндоплазматического ретикулума.
[0114]
(Материалы и методы)
SB431542 получают от TOCRIS (номер по каталогу: 1614).
[0115]
(Способ)
Человеческие эндотелиальные клетки роговицы (iHCEC) или эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса (iFECD), полученные в Примерах способов получения, высевают в чашки для культивирования, покрытые с использованием смеси для нанесения покрытия FNC, и культивируют в течение примерно 3 дней до достижения субслияния в условиях 5% CO2 при 37°C. Затем добавляют TGFβ2, SB431542 и TG, после чего клетки инкубируют в течение 24 часов в условиях 5% CO2 при 37°C. С помощью фазоразностного микроскопа наблюдают морфологию и апоптоз клеток.
[0116]
(Результаты)
Результаты показаны на фиг. 5. С помощью TGFβ2 разрушают только клетки iFECD, причем наблюдается большое число плавающих клеток по сравнению с iHCEC. Плавающие клетки уничтожают с использованием SB431542. Тем не менее, большое число плавающих клеток наблюдается, в основсном, в линии iFECD вследствие обработки TG. Однако число плавающих клеток не уменьшается под действием SB431542. Это означает, что SB431542 не является неспецифическим супрессором повреждения клеток, нго является специфичным к повреждению клеток, индуцированному сигнальным путем TGFβ. На фигуре 6 показан типичный пример результатов анализа апоптоза в клетках, представленных на фигуре 5, методом проточной цитометрии, а на фигуре 7 показан график, изображающий аннексин V-положительные апоптотические клетки на вертикальной оси.
[0117]
(Материалы и методы)
*Проточная цитометрия:
Человеческие эндотелиальные клетки роговицы (iHCEC) или эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса (iFECD), полученные в Примерах способов получения, высевают в чашки для культивирования, покрытые смесью FNC для нанесения покрытия, и культивируют в течение примерно 3 дней до достижения субслияния в условиях 5% CO2 при 37°C. Затем добавляют TGFβ2 и SB431542, после чего клетки инкубируют в течение 24 часов в условиях 5% CO2 при 37°C. Клетки отделяют с помощью ACCUMAX. Аннексин V- или PI-положительные клетки измеряют в присутствии аннексина V и PI с помощью проточного цитометра (FACS Aria II (BD Biosciences, Franklin Lakes, NJ)), используя набор для определения апоптоза MEBCYTO (Аннексин V−FITC Kit) 100 test (4700, MEDICAL & BIOLOGICAL LABORATORIES).
[0118]
(Результаты)
Результаты приведены на фигуре 7. Как показано, при дистрофии эндотелия роговицы Фукса число апоптотических клеток увеличивается под действием TGFβ2, причем указанное увеличение устраняется после обработки SB431542. Полученные результаты позволяют предположить, что апоптоз, спонтанно развивающийся при дистрофии эндотелия роговицы Фукса, можно лечить, или предотвращать его развитие, путем подавления стресса, ассоциированного с эндоплазматическим ретикулумом (ER), вследствие ингибирования сигнального пути TGFβ.
[0119]
(Пример 7: ингибитор сигнального пути TGFβ подавляет апоптоз при дистрофии эндотелия роговицы Фукса)
В настоящем изобретении определяют, действительно ли ингибитор сигнального пути TGFβs, отличный от SB431542 (A-83-01, ингибиторы ALK5), может подавлять апоптоз при дистрофии эндотелия роговицы Фукса. Для этого считают число аннексин V-положительных клеток, используемых в данном примере в качестве индикатора апоптоза.
[0120]
(Способ)
Человеческие эндотелиальные клетки роговицы (iHCEC) или эндотелиальные клетки роговицы, пораженной дистрофией эндотелия роговицы Фукса (iFECD), полученные в Примерах способов получения, высевают в чашки для культивирования, покрытые смесью FNC для нанесения покрытия, и культивируют в течение примерно 3 дней до достижения субслияния в условиях 5% CO2 при 37°C. Затем добавляют TGFβ2, SB431542 и ингибитор сигнального пути TGFβ A-83-01 (Wako, Osaka; № по каталогу: 018−22521), ингибитор ALK5 (Wako, № по каталогу: 012−23021; CAS No. 446859−33−2; 2−(3−(6−метилпиридин−2-ил)−1H−пиразол−4-ил)−1,5−нафтиридин) и клетки инкубируют в течение 24 часов в условиях 5% CO2 при 37°C. Клетки отделяют с помощью ACCUMAX. Аннексин V- или PI-положительные клетки измеряют в присутствии аннексина V и иодида пропидия с помощью проточного цитометра (FACS Aria II (BD Biosciences, Franklin Lakes, NJ)), используя набор для определения апоптоза MEBCYTO (Аннексин V−FITC Kit) 100 test (4700, MEDICAL & BIOLOGICAL LABORATORIES).
[0121]
(Результаты)
Результаты приведены на фигуре 8. Как и в примере 6, при дистрофии эндотелия роговицы Фукса число апоптотических клеток увеличивается под действием TGFβ2, причем указанное увеличение устраняется после обработки SB431542. Кроме того, обнаружено, что A-83-01 и ингибиторы ALK5, которые являются ингибиторами сигнального пути TGFβ, отличными от SB431542, уменьшают число апоптотических клеток.
[0122]
Приведенные выше данные демонстрируют, что заболевание, расстройство или состояние, ассоциированное со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки, в частности апоптоз, можно лечить или предотвращатьс использованием не только SB431542, но и ингибитора сигнального пути TGFβ.
[0123]
(Пример 9: Эксперимент с использованием антитела против IGF-β2 и ингибитора Smad)
Эксперимент проводят по способу, аналогичному способу эксперимента 8, используя антитело к TGF-β2 (R&D SYSTEMS; № по каталогу: AB−112−NA) и ингибитор Smad (CALBIOCHEM; № по каталогу: 556405).
[0124]
(Пример 10: Стимуляция стресса ER под действием TGF-β при дистрофии эндотелия роговицы Фукса)
В настоящем примере вестерн-блоттинг проводят по способу примера 2 с целью исследования экспрессии молекулярного шаперона GRP78 и CHOP, участвующих в апоптозе, развивающемся в ответ на стресс ER, чтобы подтвердить, что при дистрофии эндотелия роговицы Фукса TGF-β индуцирует более сильный стресс. Кроме того, анализируют экспрессию фосфорилированного PERK и ATF6, участвующих в ассоциированном со стрессом сигнальном пути на мембране эндоплазматического ретикулума. Стимуляцию осуществляют с помощью TGFβ2, 10 нг/мл (R&D SYSTEMS). Также анализируют хронологические изменения.
[0125]
(Материалы и методы)
Вестерн-блоттинг проводят по способу, описанному в примере 2. Помимо антител, описанных в примере 2, используют перечисленные ниже антитела.
*Антитела против PERK: (D11A8, Cell Signaling Technology)
*Антитела против фосфорилированного PERK: (sc−32577, SANTA CRUZ BIOTECHNOLOGY)
*Антитела против ATF6: (73−505, Bio Academia)
*Антитела против GRP78: (SC−376768, SANTA CRUZ BIOTECHNOLOGY)
*Антитела против CHOP: (L63F7, Cell Signaling Technology)
*TGF-β2: (302−B2−002, R&D SYSTEMS)
[0126]
(Результаты)
Результаты приведены на фигуре 9. Показано, что TGF-β стимулирует фосорилирование PERK, котоый представляет собой индикатор стресса ER, и экспрессию ATF6. Также показано, что повышается экспрессия шаперона GRP78. В iFECD наблюдается более значительная стимуляция фосорилирования и экспрессии, чем в iHCEC. Таким образом, показано, что TGF-β повышает уровень маркера стресса ER. Кроме того, показано, что экспрессия CHOP вызывает апоптоз, обусловленный стрессом ER, и усиливает его. Также показано, что усиление стресса наблюдается через 3 часа, а эффект такого усиления увеличивается через 6 часов.
[0127]
(Пример 11: Митохондриальная недостаточность в эндотелиальных клетках роговицы, пораженной дистрофией эндотелия роговицы Фукса)
В настоящем примере проводят эксперимент, подтверждающий присутствие митохондриальной недостаточности в эндотелиальных клетках роговицы, пораженной дистрофией эндотелия роговицы Фукса.
[0128]
(Способ)
Как описано в приведенных выше примерах способов получения, авторы настоящего изобретения получили линию клеток роговицы нормальных доноров HCEC (iHCEC) и иммобилизованную клеточную модель (iFECD) человеческих эндотелиальных клеток роговицы (HCEC), полученной от пациентов с FECD, которые используют в настоящем примере. Целью примера является применение клеточной модели для исследования митохондриальной недостаточности при FECD.
[0129]
Чтобы исследовать морфологические изменения в митохондриях, определяют мембранный потенциал митохондрий в клетках iFECD и iHCEC, используя краситель JC-1 (BD Bioscience, 551302) и MitoTracker® Red CMXRos (Life Technologies, M−7512).
Высвобождение цитохрома C из митохондрий в цитоплазму определяют путем выделения митохондрий с помощью набора для выделения митохондрий (Thermo SCIENTIFIC, 89874) и проведения вестерн-блоттинга (антитело к цитохрома C, abcam, ab13575).
[0130]
Уровни каспазы 9, каспазы 3 и поли(АДФ-рибоза)полимеразы (PARP) анализируют методом вестерн-блоттинга в клетках iHCEC, стимулированных стимулятором митохондриального стресса стауроспорином (abcam, ab120056), чтобы определить участие митохондриальной недостаточности в апоптозе.
[0131]
(Результаты и обсуждение)
На фигуре 10 приведены результаты анализа мембранного потенциала митохондрий с использованием в качестве флуоресцентного зонда красителя JC-1. Зеленая флуоресценция указывает на митохондрии, а красная флуоресценция указывает на мембранный потенциал митохондрий. Уменьшение красной флуоресценции свидетельствует о деполяризации мембранного потенциала ed. Красная флуоресценция в клетках iFECD слабее, чем в клетках iHCEC. На фигуре 11 показаны значения мембранного потенциала митохондрий, измеренные методом проточной цитометрии. Результаты анализа с использованием JC-1 и MitoTracker® демонстрируют, что мембранный потенциал митохондрий в клетках iFECD ниже, чем в клетках iHCEC (данные результаты приведены на фигурах 10 и 11).
[0132]
Анализ методом вестерн-блоттинга демонстиррует, что уровень цитохрома C в митохондриальной фракции клеток iFECD выше, чем в митохондриальной фракции клеток iHCEC (фигура 12). Результаты анализа истечения приведенные на фигуре 12, демонстирируют, что в эндотелиальных клетках роговицы, пораженной дистрофией эндотелия роговицы Фукса, присутствует митохондриальное расстройство.
[0133]
(Пример 12: Взаимозависимость митохондриального расстройства и апоптоза)
В настоящем примере методом вестерн-блоттинга, проводимого по способу, описанному в примере 2, исследуют экспрессию каспазы 9, каспазы 3 и PARP, которые представляют собой белки, ассоциированные с апоптозом, чтобы определить, приводит ли митохондриальное расстройство в эндотелиальных клетках роговицы к повреждению клеток. Путем стимуляции стауроспорином индуцируют митохондриальное расстройство, чтобы исследовать хронологические изменения.
[0134]
(Материалы и методы)
Вестерн-блоттинг проводят по способу, описанному в примере 2. Помимо антител, используемых в примере 2, используют перечисленные ниже антитела.
*Антитела против каспазы 9: (9508S, Cell Signaling Technology)
*Антитела против каспазы 3: (9662S, Cell Signaling Technology)
*Антитела против PARP: (9542S, Cell Signaling Technology)
*Стауроспорин: (ab120056, abcam)
[0135]
(Результаты)
Результаты показаны на фигуре 13. На фигуре 13 показаны результаты анализа нижестоящих сигнальных путей методом вестерн-блоттинга в клетках iHCEC, стимулированных фактором, вызывающим митохондриальный стресс, стауроспорином. Митохондриальное расстройство индуцируют стауроспорином, активируя каспазу 9, каспазу 3 и PARP. Морфология митохондрий в клетках iHCEC является нормальной, однако в клетках iFECD с помощью электронного микроскопа наблюдают увеличение митохондрий в объеме. Клетки, характеризующиеся пониженным мембранным потенциалом митохондрий, составляют 11,7±1,7% в линии iHCEC, а в линии iFECD их число значительно увеличивается до 41,9±10,2% (p<0,05). Кроме того, в клетках iFECD, в отличие от клеток iHCEC, обнаружено сильное истечение цитохрома C в цитоплазму. Полученные результаты демонстрируют, что митохондриальный стресс индуцирует истечение цитохрома C, при этом также наблюдается активация каспазы 9, которая, как известно, активируется в результате митохондриального расстройства, а также активация каспазы 3 и PARP. Таким образом, очевидно, что митохондриальное расстройство индуцирует апоптоз. Приведенные выше результаты также свидетельствуют о том, что митохондриальное расстройство участвует в патологии FECD.
[0136]
Приведенные выше результаты, полученные в настоящем примере, свидетельствуют о том, что митохондриальная недостаточность участвует в патологии FECD и может использоваться в качестве мишени потенциального терапевтического средства. Поскольку известно, что стресс ER приводит к митохондриальному расстройству в клетках разных видов, лекарственное средство, улучшающее стресс ER, может приводить к подавлению митохондриального расстройства и использоваться для лечения дистрофии эндотелия роговицы Фукса.
[0137]
(Пример 13: Количество денатурированных белков увеличивается при дистрофии эндотелия роговицы Фукса)
С помощью настоящего примера демонстрируют, что количество денатурированных белков увеличивается при дистрофии эндотелия роговицы Фукса.
[0138]
Чтобы исследовать накопление денатурированных белков (также упоминающихся как неуложенные белки или неполностью уложенные белки) в клетках модели дистрофии эндотелия роговицы Фукса, измеряют экспрессию агресом, которые бразуются в ответ на накопление денатурированных белков. Известно, что белки, агрегированные вследствие денатурации (или неправильной укладки) или нарушения деградации белков, подвергаются убихитинированию и накоплению вблизи центросомы посредством перемещения в микротрубочках, опосредованного динеином, с образованием телец включения, называемых агресомами.
[0139]
Как правило, агресомы образуются в результате термического шока, вирусной инфекции, окислительного стресса и т.п. У людей известны заболевания, в которых участвуют тельца включения, такие как тельца Льюиса, присутствующие в нервных клетках при болезни Паркинсона, тельца Мэллори, присутствующие в клетках печени при алкогольных болезнях печени, и стеклоподобные тельца в астроцитах при боковом амиотрофическом склерозе.
[0140]
(Материалы и методы)
Накопление агресом при дистрофии эндотелия роговицы Фукса подтверждают методом проточной цитометрии.
(Используемое оборудование)
*Набор для детекции агресом ProteoStat®: (EZN−51035−K100, Enabling Discovery in Life Science®)
Ниже приведено краткое описание испоьзуемой методики.
1x105 человеческих эндотелиальных клеток роговицы (iHCEC) или эндотелиальных клеток роговицы, пораженной дистрофией эндотелия роговицы Фукса (iFECD), высевают на 6-луночный планшет, покрытый с использованием смеси для нанесения покрытия FNC, и культивируют в течение примерно 2 дней до достижения субслияния в условиях 5% CO2 при 37°C. Затем добавляют TGFβ2 (10 нг/мл; R&D SYSTEMS), после чего клетки инкубируют в течение 24 часов в условиях 5% CO2 при 37°C. Клетки отделяют с использованием ACCUMAXTM (Funakoshi) и измеряют агресомы с помощью проточного цитометра (FACS Aria II (BD Biosciences, Franklin Lakes, NJ), испоьзуя набор для детекции агресом ProteoStat®: (EZN−51035−K100, Enabling Discovery in Life Science®).
[0141]
(Результаты)
Результаты приведены на фигуре 14. Интенсивность флуоресценции в агресомах в 1,22 раза выше, причем интенсивность в клетках, пораженных дистрофией эндотелия роговицы Фукса (iFECD) значительно превышает интенсивность, наблюдающуюся в человеческих эндотелиальных клетках роговицы (iHCEC). Кроме того, при дистрофии эндотелия роговицы Фукса стимуляция под действием TGF-β2 приводит к увеличению количества агресом в 1,11 раз и заметному уровню гибели клеток.
Приведенные выше результаты демонстрируют, что при дистрофии эндотелия роговицы Фукса в клетках присутствует много денатурированных белков и их количество увеличивается после стимуляции сигнального пути TGF-β. Можно предположить, что стресс эндоплазматического ретикулума, обусловленный денатурированными белками, связан с патологией дистрофии эндотелия роговицы Фукса (FECD).
[0142]
(Пример 13: Денатурированные белки (неуложенные белки) накапливаются при дистрофии эндотелия роговицы Фукса)
С помощью настоящего примера демонстрируют, что денатурированные белки (неуложенные белки) накапливаются при дистрофии эндотелия роговицы Фукса.
[0143]
Чтобы исследовать накопление денатурированных белков в клетках модели дистрофии эндотелия роговицы Фукса, измеряют экспрессию агресом, которые образуются в ответ на накопление денатурированных белков. Известно, что белки, агрегированные вследствие денатурации (или неправильной укладки) или нарушения деградации белков, подвергаются убихитинированию и накоплению вблизи центросомы посредством перемещения в микротрубочках, опосредованного динеином, с образованием телец включения, называемых агресомами.
[0144]
Как правило, агресомы образуются в результате термического шока, вирусной инфекции, окислительного стресса и т.п. У людей известны заболевания, в которых участвуют тельца включения, такие как тельца Льюиса, присутствующие в нервных клетках при болезни Паркинсона, тельца Мэллори, присутствующие в клетках печени при алкогольных болезнях печени, и стеклоподобные тельца в астроцитах при боковом амиотрофическом склерозе.
[0145]
(Материалы и методы)
Культуры человеческих эндотелиальных клеток роговицы (iHCEC) или эндотелиальных клеток роговицы, пораженной дистрофией эндотелия роговицы Фукса (iFECD), высевают в чашки для культивирования, покрытые с использованием смеси для нанесения покрытия FNC, и культивируют в течение примерно 3 дней до достижения субслияния в условиях 5% CO2 при 37°C. Клетки отделяют с использованием ACCUMAXTM (Funakoshi), агресомы окрашивают, используя набор для детекции агресом ProteoStat®: (EZN−51035−K100, Enabling Discovery in Life Science®), и измеряют с помощью проточного цитометра (FACS Aria II (BD Biosciences, Franklin Lakes, NJ).
[0146]
(Результаты)
В клетках iFECD уровень экспрессии агресом выше, чем в клетках iHCEC. Это может свидетельствовать о том, что денатурированные белки накапливаются в клетках модели дистрофии эндотелия роговицы Фукса.
Результаты, полученные по агресомам и денатурированным белкам, демонстрируют, что стресс эндоплазматического ретикулума, обусловленный накоплением агресом и денатурированных белков, участвует в развитии дистрофии эндотелия роговицы Фукса в тесной связи с сигнальными путями TGF-β.
[0147]
(Пример 15: Диагноз с использованием агресом в качестве индикатора)
С помощью настоящего примера демонстрируют, что агресомы можно использовать для диагностики. Агресомы можно измерять с помощью способов, описанных в примерах 13-14 и т.п.
Путем введения красителя, способного окрашивать агресомы, в переднюю камеру и т.п. можно определить отложение агресом в эндотелиальных клетках роговицы и затем использовать полученные результаты для постановки диагноза.
[0148]
(Пример 16: Пример получения глазных капель)
Пример получения глазных капель
Ниже приведена композиция, содержащая тестируемое вещество в соответствующей концентрации.
[0149]
SB431542 (доступный от TOCRIS и др.)
0,1-30 мМ, предпочтительно 1-10 мМ
Хлорид натрия
0,85 г
Дигидрат дигидрофосфата натрия
0,1 г
Бензалкония хлорид
0,005 г
Гидроксид натрия
подходящее количество
Очищенная вода
подходящее количество
Общее количество
100 мг (pH 7,0).
[0150]
Глазные капли также можно разбавить основным реагентом.
[0151]
Ниже приведен состав основного реагента
[0152]
Хлорид натрия
0,85 г
Дигидрат дигидрофосфата натрия
0,1 г
Бензалкония хлорид
0,005 г
Гидроксид натрия
подходящее количество
Очищенная вода
подходящее количество
Общее количество
100 мг (pH 7,0).
[0153]
Как указано выше, настоящее изобретение иллюстрируется посредством предпочтительных вариантов осуществления. Однако следует понимать, что объем настоящего изобретения определяется исключительно Формулой изобретения. Также следует понимать, что любой патент, любая патентная заявка и любые ссылки, цитирующиеся в настоящем описании, включены в данный документ в такой степени, как если бы их содержание было специально описано здесь. Настоящая заявка испрашивает приоритет на основании патентных заявок Японии №№ 2013-227048 и 2014-184172, содержание которых включено в данный документ в качестве ссылки.
[Промышленное применение]
[0154]
Настоящее изобретение предлагает подходящий для применения в промышленности (такой как микробиологическая промышленность, фармацевтическая промышленность и т.п.) способ получения терапевтического профилактического средства против заболевания, расстройства или состояния, связанного со стрессом ER в эндотелии роговой оболочки, в особенности, заболевания, расстройства или состояния, связанного со стрессом ER или апоптозом при дистрофии эндотелия роговицы Фукса, включающего в себя ингибитор сигнального пути TGFβ.

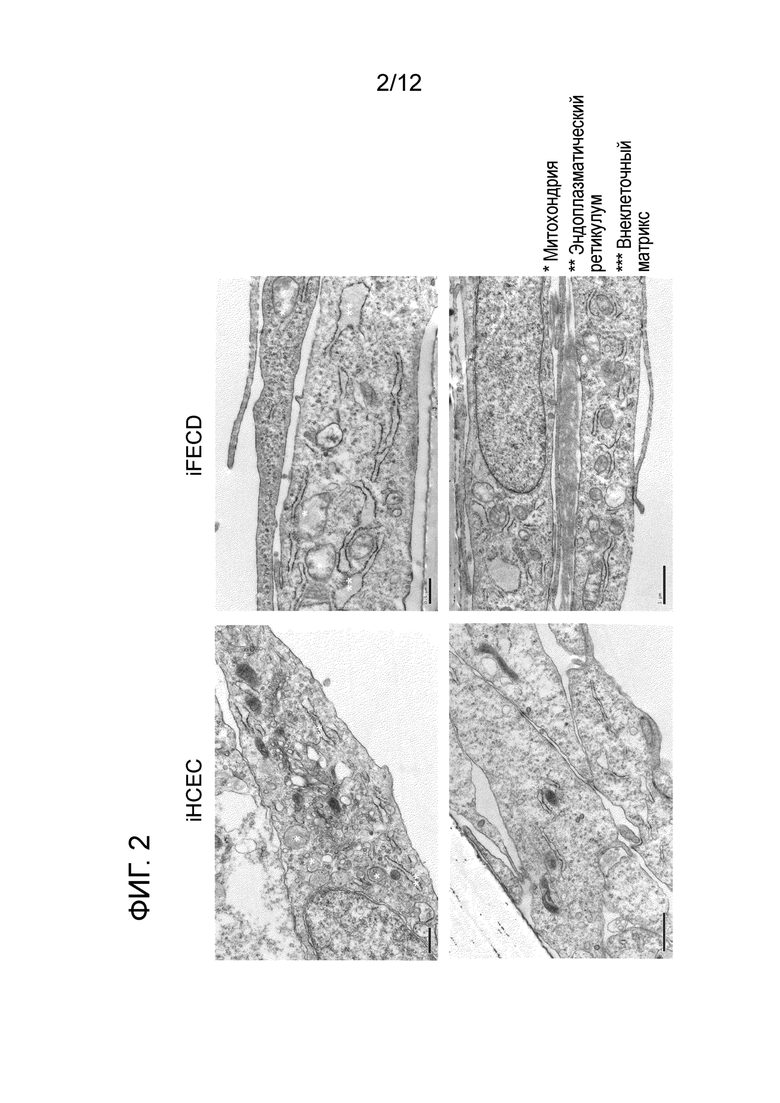
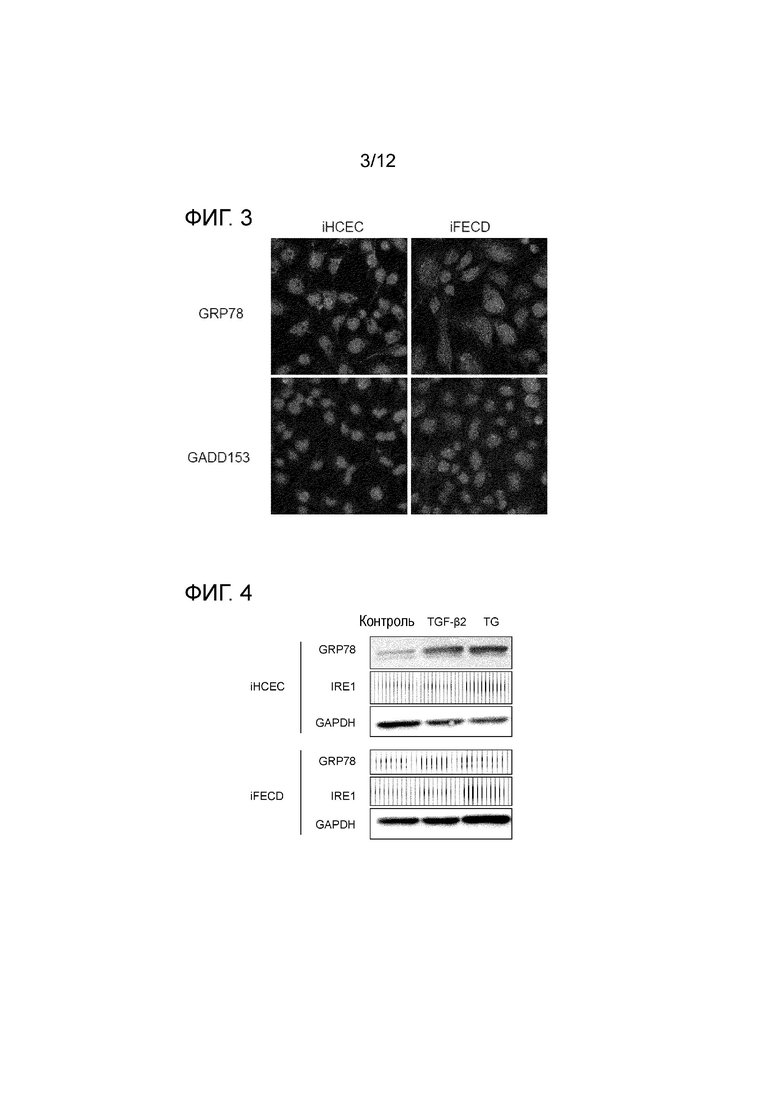

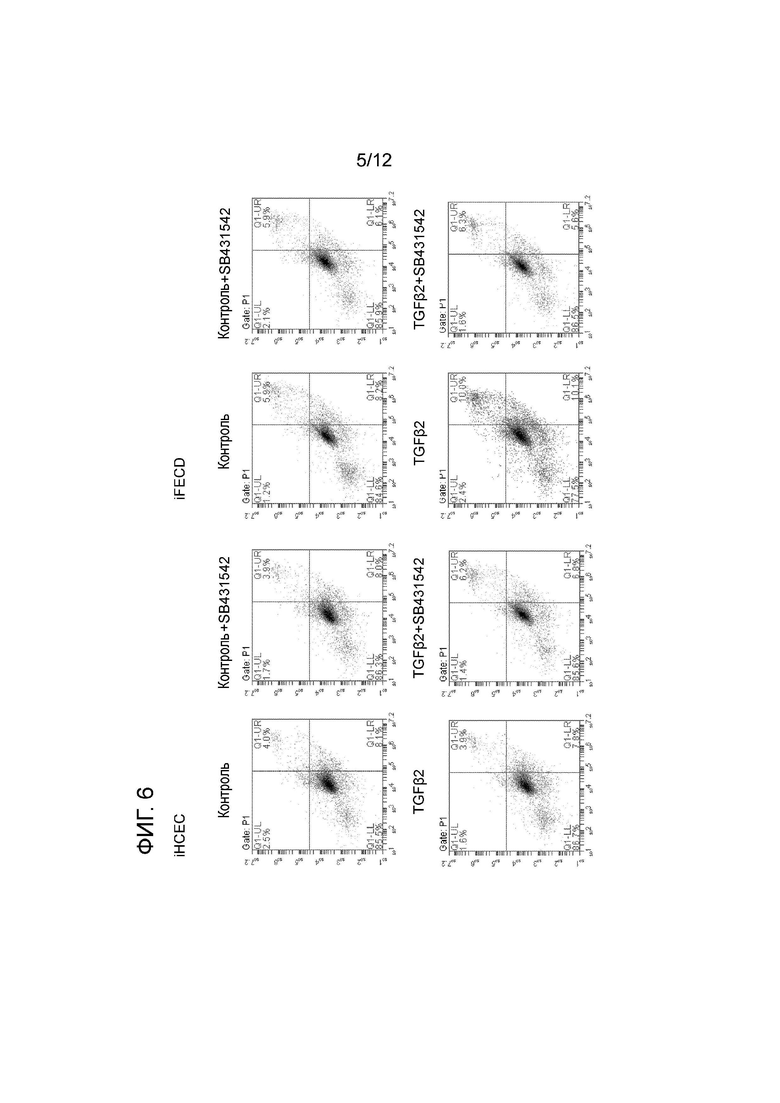
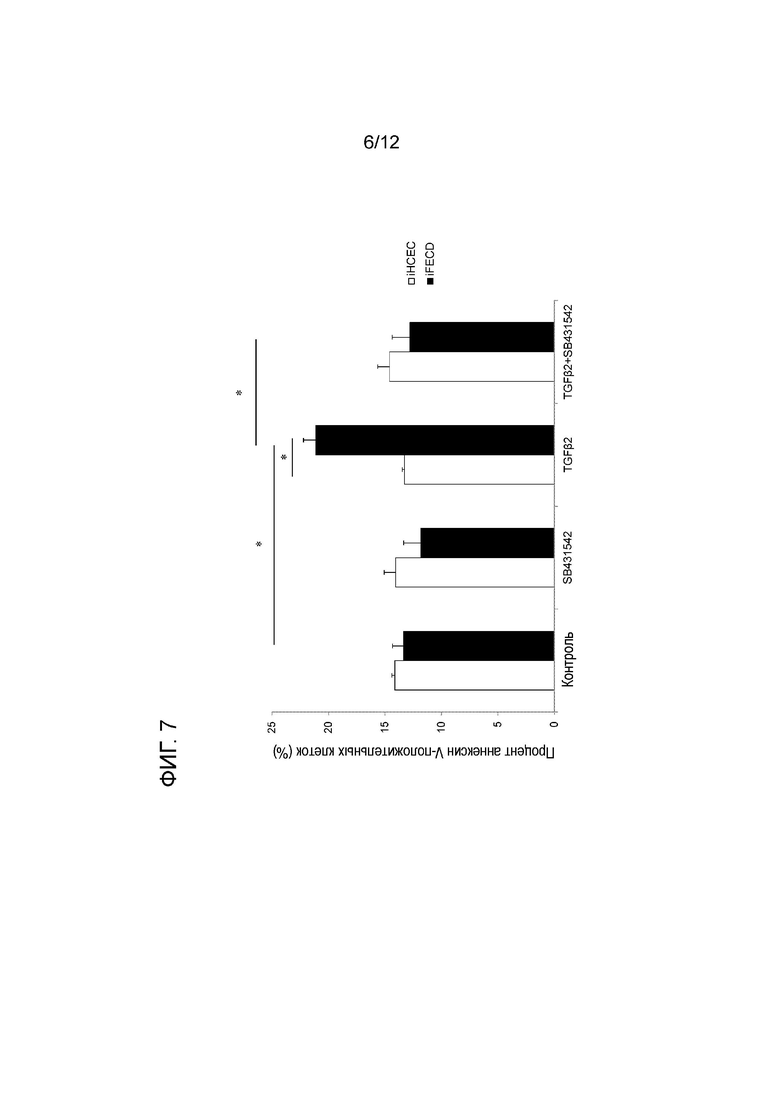
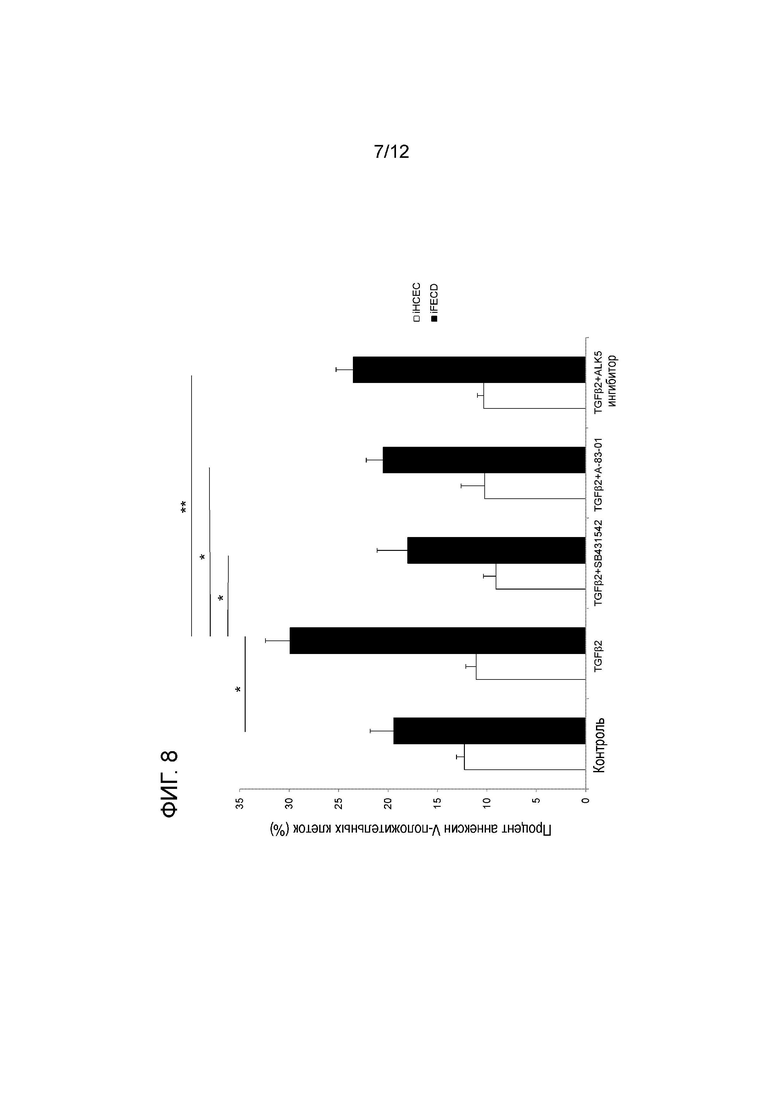


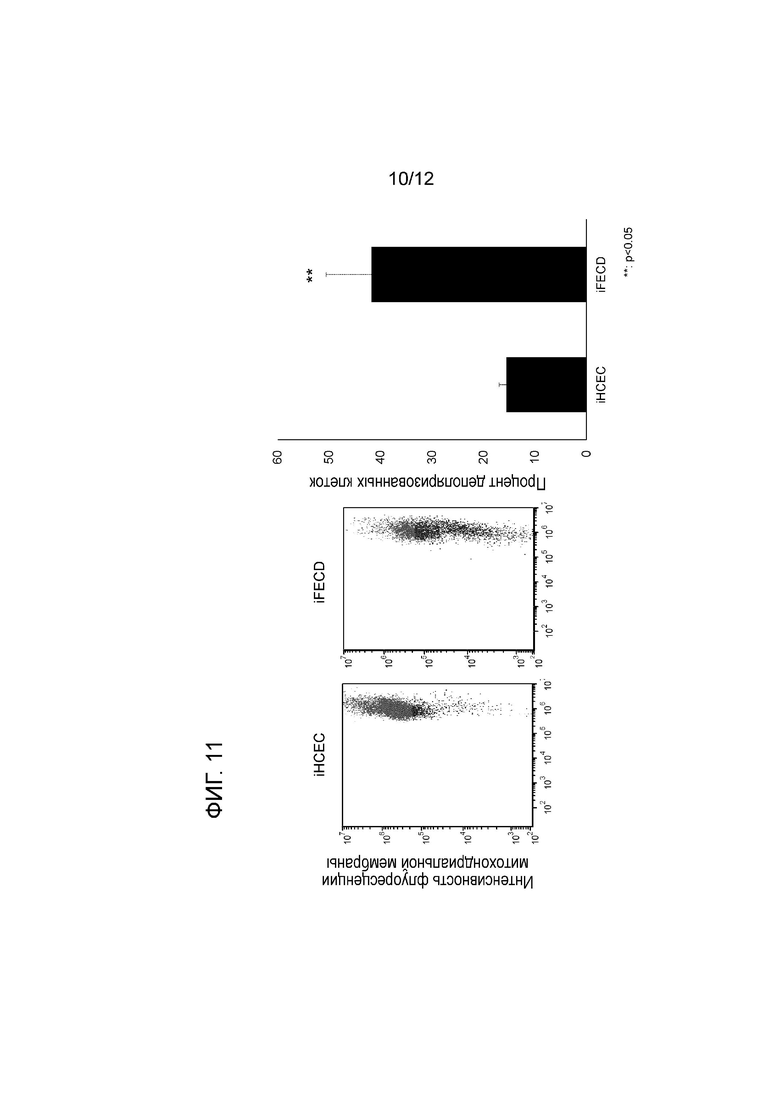
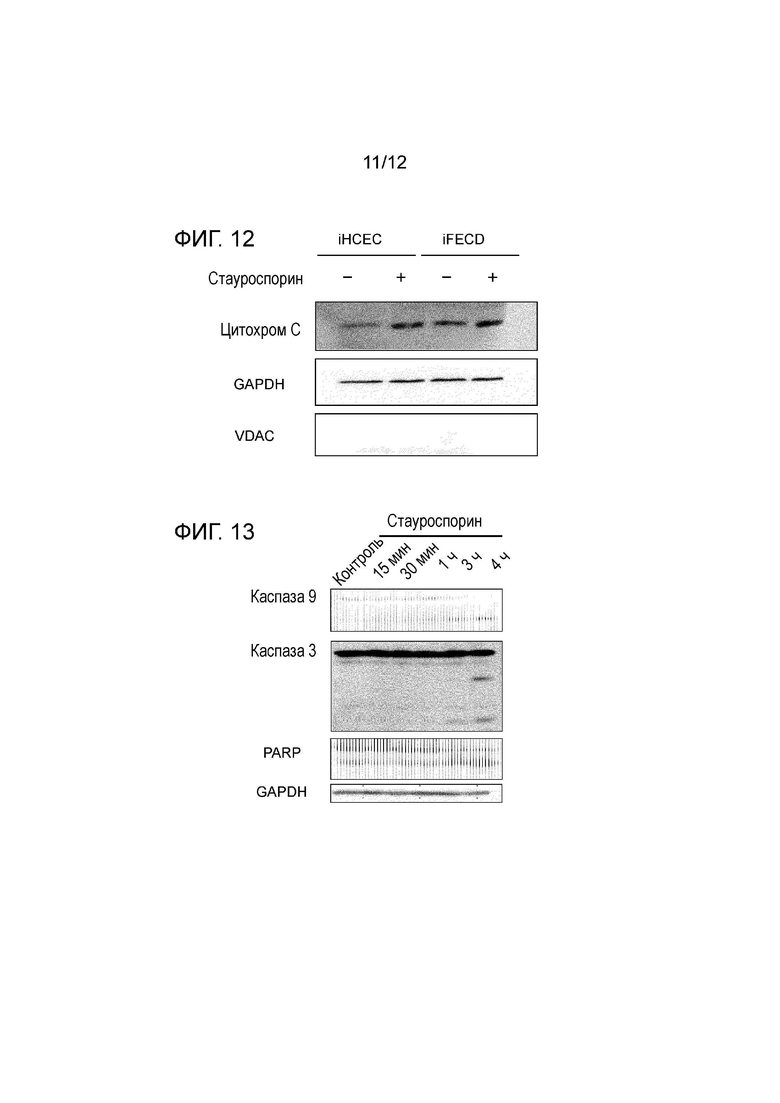
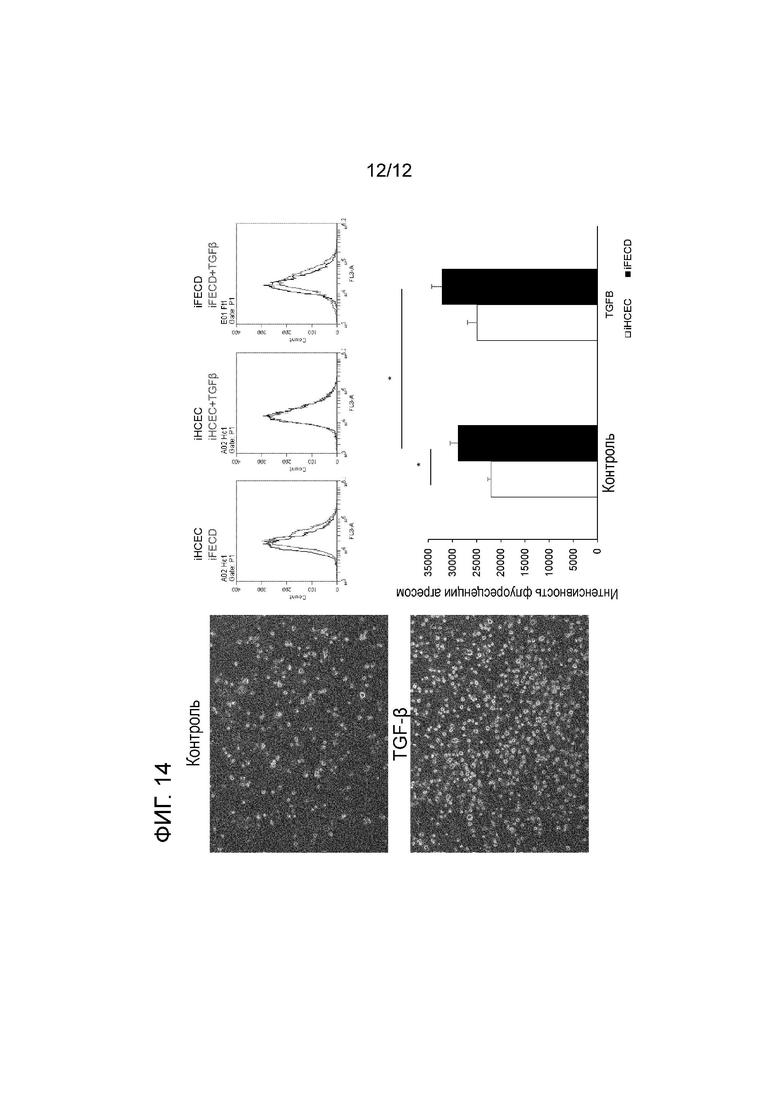
Группа изобретений относится к медицине, а именно к офтальмологии, и может быть использована для лечения или профилактики заболевания, расстройств или состояний, вызванных стрессом эндоплазматического ретикулума в эндотелии роговой оболочки глаза. Для этого используют терапевтическое или профилактическое средство в виде глазных капель, содержащее ингибитор сигнального пути NGF-beta. Предпочтительно ингибитор сигнального пути содержит 4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)-1H-имидазол-2-ил]бензамид, а также дополнительный фармацевтический ингредиент, выбранный из ингибиторов киназы Rho или стероидов. Группа изобретений обеспечивает способ получения терапевтического профилактического средства против заболеваний, расстройств или состояний, вызванных стрессом эндоплазматического ретикулума в эндотелии роговой оболочки глаза. 3 н. и 12 з.п. ф-лы, 14 ил., 15 пр.
1. Терапевтическое или профилактическое средство для лечения или профилактики заболевания, расстройства или состояния, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки, содержащее ингибитор сигнального пути TGF-β, причем терапевтическое или профилактическое средство представляет собой глазные капли.
2. Терапевтическое или профилактическое средство по п.1, где заболевание, расстройство или состояние связано с митохондриальной недостаточностью.
3. Терапевтическое или профилактическое средство по п.1, где заболевание, расстройство или состояние связано с апоптозом, обусловленным митохондриальной недостаточностью.
4. Терапевтическое или профилактическое средство по п.1, где заболевание, расстройство или состояние связано с дистрофией эндотелия роговицы Фукса.
5. Терапевтическое или профилактическое средство по п.1, где терапевтическое или профилактическое средство подавляет заболевание, расстройство или состояние, включающее в себя нарушение функционирования эндотелиальных клеток роговицы при дистрофии эндотелия роговицы Фукса.
6. Терапевтическое или профилактическое средство по п.1, где терапевтическое или профилактическое средство подавляет по меньшей мере одно заболевание, расстройство или состояние, выбранное из группы, включающей уменьшение плотности эндотелия роговицы, образование гутты, гипертрофию десцеметовой мембраны, гипертрофию роговой оболочки, нарушение функционирования эндотелия роговицы, помутнение стромы роговицы, светобоязнь, нечеткость зрения, нарушение зрения, офтальмалгию, слезотечение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, пониженную контрастность, чувствительность к яркому свету и отек стромы роговой оболочки.
7. Терапевтическое или профилактическое средство по п.1, где ингибитор сигнального пути TGF-β включает по меньшей мере одно из следующих средств: 4−[4−(1,3−бензодиоксол−5-ил)−5−(2−пиридинил)−1H−имидазол−2-ил]бензамид, BMP−7, антитело к TGF−β, антитело к рецептору TGF−β, миРНК TGF−β, миРНК рецептора TGF−β, кшРНК TGF−β, кшРНК рецептора TGF−β, аптамер TGF−β, аптамер рецептора TGF−β, антисмысловой олигонуклеотид TGF−β, 6,7−диметокси−2−((2E)−3−(1−метил−2−фенил−1H−пирроло[2,3−b]пиридин−3-илпроп-2-еноил))−1,2,3,4−тетрагидроизохинолон, 3−(6−метил−2−пиридинил)−N−фенил−4−(4−хинолинил)−1H−пиразол−1−карботиоамид, 2−(3−(6−метилпиридин−2-ил)−1H−пиразол−4-ил)−1,5−нафтиридин, 6−(4−(пиперидин−1-ил)этокси)фенил)−3−(пиридин−4-ил)пиразоло[1,5−a]пиримидин, 2−(5−хлор−2−фторфенил)−4−[(4−пиридинил)амино]птеридин, 4−[3−(2−пиридинил)−1H−пиразол−4-ил]−хинолин, A−83−01 (3−(6−метил−2−пиридинил)−N−фенил−4−(4−хинолинил)−1H−пиразол−1−карботиоамид), фармацевтически приемлемая соль, или фармацевтически приемлемый сольват одного из указанных соединений, или сольват фармацевтически приемлемой соли одного из указанных соединений.
8. Терапевтическое или профилактическое средство по п.1, где ингибитор сигнального пути TGF-β включает 4−[4−(1,3−бензодиоксол−5-ил)−5−(2−пиридинил)−1H−имидазол−2-ил]бензамид или его фармацевтически приемлемую соль.
9. Терапевтическое или профилактическое средство по п.1, дополнительно включающее в себя терапевтическое средство против митохондриальной недостаточности, вызванной стрессом ER.
10. Терапевтическое или профилактическое средство по п.9, где терапевтическое средство против митохондриальной недостаточности, вызванной стрессом ER, выбрано из группы, включающей BiP индуктор X (BIX), 4−фенилмасляную кислоту (PBA), N−оксид триметиламина (TMAO), тауроурсодезоксихолевую кислоту (TUDCA) и тепренон.
11. Терапевтическое или профилактическое средство по п.1, где эндотелий роговой оболочки принадлежит примату.
12. Терапевтическое или профилактическое средство по п.1, где эндотелий роговой оболочки принадлежит человеку.
13. Терапевтическое или профилактическое средство по п.1, содержащее дополнительный фармацевтический ингредиент, причем дополнительный фармацевтический ингредиент выбран из ингибиторов киназы Rho или стероидов.
14. Применение средства, ингибирующего сигнальный путь TGF-β, для лечения или профилактики расстройства, связанного со стрессом эндоплазматического ретикулума (ER), в эндотелии роговой оболочки, причем средство, ингибирующее сигнальный путь TGF-β, вводят в форме глазных капель.
15. Способ лечения или профилактики расстройства, связанного со стрессом эндоплазматического ретикулума (ER) в эндотелии роговой оболочки у индивидуума, где способ включает стадию введения индивидууму эффективного количества ингибитора сигнального пути TGF-β, причем ингибитор сигнального пути TGF-β вводят в форме глазных капель.
| JPWO 2013100208 A1, 04.07.2013 | |||
| US 2004038856 A1, 26.02.2004 | |||
| ПЕПТИДЫ, ИНГИБИРУЮЩИЕ ТРАНСФОРМИРУЮЩИЙ ФАКТОР РОСТА TGFБЕТА1 | 1999 |
|
RU2232771C2 |
| OKUMURA N | |||
| et al, Inhibition of TGF-b Signaling Enables Human Corneal Endothelial Cell Expansion In Vitro for Use in Regenerative Medicine, PLOS ONE, 2013, 8, pp | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

