Область техники
[0001]
Настоящее изобретение относится к технологиям, способам, средствам и т.п. для лечения или предупреждения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы.
Уровень техники
[0002]
Зрительная информация распознается, когда свет попадает на роговицу, которая представляет собой прозрачную ткань передней части глазного яблока, достигает сетчатки и возбуждает нервные клетки сетчатки, и сгенерированный электрический сигнал передается через оптический нерв в зрительную кору головного мозга. Для достижения хорошего зрения необходимо, чтобы роговица была прозрачной. Прозрачность роговицы поддерживается путем поддержания постоянного содержания воды насосной и барьерной функциями эндотелиальных клеток роговицы.
[0003]
Эндотелиальные клетки роговицы человека при рождении присутствуют с плотностью приблизительно 3000 на 1 мм2. Однако в результате повреждения способность клеток к регенерации высоко ограничивается. Эндотелиальная дистрофия роговицы Фукса представляет собой заболевание, при котором возникает аномалия в эндотелиальных клетках внутри роговицы, вызывая отек роговицы и т.п. Ее причина неизвестна. При эндотелиальной дистрофии роговицы Фукса внеклеточный матрикс, такой как коллаген и т.п., депонируется на одной части задней поверхности десцеметовой оболочки на задней стороне роговицы, что приводит к "каплям" на роговице и утолщению десцеметовой оболочки. "Капли" на роговице и утолщение десцеметовой оболочки являются причинами фотофобии и неясного зрения у пациентов с эндотелиальной дистрофией роговицы Фукса, и они значительно ухудшают QOL пациентов. Для эндотелиальной дистрофии роговицы Фукса не рассматривается эффективного способа терапии кроме кератопластики. Однако донорство роговицы в Японии является недостаточным. Хотя ежегодно приблизительно 2600 пациентов ожидают кератопластики, количество случаев кератопластики, проводимой внутри страны, составляет приблизительно 1700.
[0004]
Что касается эндотелиальной дистрофии роговицы Фукса, описана культура эндотелиальных клеток роговицы, полученных от пациентов с эндотелиальной дистрофией роговицы Фукса (непатентные документы 1 и 3), а также их иммобилизация (непатентный документ 2), в то время как пригодные клетки для скрининга терапевтического средства и препятствующего прогрессированию лекарственного средства, которые сохраняют характеристики заболевания, такие как сопутствующая сверхпродукция внеклеточного матрикса, не описаны. Таким образом, для разработки терапевтического средства существуют ограничения. В настоящее время отсутствует клинически применяемое терапевтическое средство и необходимо полагаться на кератопластику.
[0005]
Кроме того, в патентном документе 1 описан ингибиторный пептид TGF-бета 1 для проведения терапии против фиброза и/или помутнения роговицы. В патентном документе 2 описаны антитела, которые связываются с TGF-бета 1, 2 или 3. В патентном документе 3 описано, что агонист или активатор Nrf2 можно использовать для терапии эндотелиального нарушения роговицы. В патентном документе 4 описан пептид, который может связываться с трансформирующим фактором роста TGF-бета 1 и который может стать мощным ингибирующим средством против биологической активности TGF-бета 1 посредством прямого связывания с этим цитокином. В патентном документе 5 описано средство, подавляющее образование рубцов, включающее полипептид BMP-7. В патентном документе 6 в общем описано нарушение роговицы в качестве заболевания, против которого эффект ингибирования TGF-бета является терапевтически или профилактически эффективным.
Список ссылок
Патентные документы
[0006]
[PTL 1] Выложенная публикация PCT национальной фазы Японии № 2013-520405
[PTL 2] Международная публикация № WO 2012/167143, брошюра
[PTL 3] Международная публикация № WO 2012/009171, брошюра
[PTL 4] Выложенная публикация PCT национальной фазы Японии № 2007-525204
[PTL 5] Выложенная публикация PCT национальной фазы Японии № 2006-508169
[PTL 6] Международная публикация № WO 2004/018430, брошюра
[Непатентные документы]
[0007]
[NPL 1] Zaniolo K, et al., Exp. Eye Res. Vol. 94(1):22-31, 2012.
[NPL 2]Azizi B, et al., Invest Ophthalmol Vis. Sci. 2;52(13):9291-9297, 2011.
[NPL 3] Kelliher C. et al., Exp. Eye Res. Vol. 93(6), 880-888, 2011.
Сущность изобретения
Решение проблемы
[0008]
Авторы изобретения открыли, что ингибирование каскада трансформирующего фактора роста-бета (TGF-бета) может подавлять отложение внеклеточного матрикса (ECM), такого как коллаген и т.п., как наблюдают при эндотелиальной дистрофии роговицы Фукса и т.п.; нашли способы, которые могут лечить или предупреждать обусловленное ECM нарушение; и осуществили настоящее изобретение. Таким образом, изобретение настоящей заявки относится к следующим изобретениям:
[0009]
(1) Лекарственное средство для лечения или предупреждения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы, где лекарственное средство содержит средство, ингибирующее сигнал TGF-бета.
[0010]
(2) Лекарственное средство для лечения или предупреждения согласно 1, где заболевание, нарушение или состояние представляет собой нарушение, обусловленное эндотелиальной дистрофией роговицы Фукса.
[0011]
(3) Лекарственное средство для лечения или предупреждения согласно 1 или 2, где заболевание, нарушение или состояние включает по меньшей мере одно заболевание, нарушение или состояние, выбранное из группы, состоящей из фотофобии, неясного зрения, нарушения зрения, боли в глазу, слезоотделения, гиперемии, боли, буллезной кератопатии, неприятных ощущений в глазу, снижения контрастности, бликов, отека стромы роговицы, буллезной кератопатии и помутнения роговицы при эндотелиальной дистрофии роговицы Фукса.
[0012]
(4) Лекарственное средство для лечения или предупреждения согласно любому из 1-3, где средство, ингибирующее сигнал TGF-бета, включает по меньшей мере одно из 4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)-1H-имидазол-2-ил]бензамида, BMP-7, антитела против TGF-бета, антитела против рецептора TGF-бета, миРНК к TGF-бета, миРНК к рецептору TGF-бета, кшРНК к TGF-бета, кшРНК к рецептору TGF-бета, аптамера к TGF-бета, аптамера к рецептору TGF-бета и антисмыслового олигонуклеотида к TGF-бета, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-ил-проп-2-еноил))-1,2,3,4-тетрагидроизохинолона, 3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1H-пиразол-1-карботиоамида, 2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридина, 6-(4-(пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил)пиразолo[1,5-a]пиримидина, 2-(5-хлор-2-фторфенил)-4-[(4-пиридинил)амино]птеридина, 4-[3-(2-пиридинил)-1H-пиразол-4-ил]хинолина, их фармацевтически приемлемых солей или сольватов, или сольватов фармацевтически приемлемых солей.
[0013]
(5) Лекарственное средство для лечения или предупреждения согласно любому из 1-4, где средство, ингибирующее сигнал TGF-бета, включает 4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)-1H-имидазол-2-ил]бензамид или его фармацевтически приемлемую соль.
[0014]
(6) Лекарственное средство для лечения или предупреждения по любому из 1-5, где эндотелий роговицы представляет собой эндотелий роговицы примата.
[0015]
(7) Лекарственное средство для лечения или предупреждения согласно любому из 1-6, где эндотелий роговицы представляет собой эндотелий роговицы человека.
[0016]
(8) Лекарственное средство для лечения или предупреждения согласно любому из 1-7, содержащее дополнительный медицинский компонент.
[0017]
(9) Лекарственное средство для лечения или предупреждения согласно любому из 1-8, представляющее собой глазные капли.
[0018]
(10) Вещество, ингибирующее сигнал TGF-бета, для лечения или предупреждения нарушения, обусловленного с аномалией внеклеточного матрикса (ECM) в эндотелии роговицы.
[0019]
(10A) Вещество, ингибирующее сигнал TGF-бета, согласно (10), где вещество, ингибирующее сигнал TGF-бета, имеет характеристику ингибирующего средства согласно любому из (1)-(9).
[0020]
(11) Способ лечения или предупреждения нарушения, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы у индивидуума, где способ включает стадию введения эффективного количества средства, ингибирующего сигнал TGF-бета, индивидууму.
[0021]
В рамках настоящего изобретения в дополнение к разъясненным комбинациям, предполагается дальнейшее комбинирование и предоставление упомянутых выше одной или нескольких характеристик. Следующие варианты осуществления и преимущества в соответствии с настоящим изобретением при необходимости станут понятными специалистам в данной области при прочтении и осмыслении следующего подробного описания изобретения.
Полезные эффекты изобретения
[0022]
Настоящее изобретение относится к лекарственному средству, которое может лечить или предупреждать заболевание, обусловленное аномалией внеклеточного матрикса (ECM), такое как эндотелиальная дистрофия роговицы Фукса, для которой традиционно единственным способом лечения является кератопластика, а также к способу, который может реализовать глазные капли и т.п. против этого заболевания.
Краткое описание чертежей
[0023]
[Фиг.1] На фиг.1 демонстрируется положение, что экспрессия Snail1 и ZEB1 в эндотелиальных клетках, полученных от пациента с эндотелиальной дистрофией роговицы Фукса, стимулируется. На фиг.1 представлен результат анализа способом ПЦР в реальном времени уровня экспрессии гена, связанного с эпителиально-мезенхимальным переходом (EMT), связанным с продукцией внеклеточного матрикса. Использовали iFCED, которые были получены путем получения и культивирования эндотелиальных клеток роговицы от трех пациентов, у которых возникла буллезная кератопатия при клиническом диагнозе эндотелиальной дистрофии роговицы Фукса и которые подверглись эндотелиальной кератопластике десцеметовой оболочки (DMEK), при информированном согласии и одобрении Этического комитета. В культивируемые эндотелиальные клетки роговицы, полученные от пациента с эндотелиальной дистрофией роговицы Фукса вносили гены SV40 и hTERT с помощью лентивируса с получением иммобилизованного штамма. Эндотелиальные клетки роговицы, культивируемые из роговицы для научных целей, импортированной в качестве контроля из банка роговичных трансплантатов Сиэтла, иммобилизовывали сходным образом для получения штамма иммобилизованных клеток (iHCEC). Для iHCEC и iFECD, проводили поддерживающее культивирование в DMEM+10% FBS. На каждом графике слева представлены iHCEC и справа представлены iFECD. На A представлена относительная экспрессия Snail1, на B представлена относительная экспрессия Snail2, и на C представлена относительная экспрессия ZEB1. С помощью ПЦР в реальном времени было установлено значительное усиление экспрессии Snail1 и ZEB1 в iFECD по сравнению с iHCEC.
[0024]
[Фиг.2] На фиг.2 демонстрируется положение, что TGF-бета увеличивает экспрессию Snail1, ZEB1 и белков, образующих матрикс in vitro. На фиг.2 представлен результат стимуляции посредством TGF-бета, который, как известно, ускоряет экспрессию Snail1 и ZEB1, для подтверждения того, что стимуляция экспрессия Snail1 и ZEB1 связана с продукцией внеклеточного матрикса. Белым цветом показаны iHCEC и черным цветом показаны iFECD. На A представлена относительная экспрессия Snail1, на B представлена относительная экспрессия ZEB1, на C представлена относительная экспрессия коллагена типа I, на D представлена относительная экспрессия коллагена типа IV, на E представлена относительная экспрессия коллагена типа VIII, и на F представлена относительная экспрессия фибронектина. Было подтверждено, что TGF-бета значительно ускоряет экспрессию Snail1 и ZEB1 в iFECD (A, B). Далее, когда величину экспрессии генов образующего внеклеточный матрикс белка анализировали с помощью ПЦР в реальном времени, экспрессия коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина была значительно ускоренной.
[0025]
[Фиг.3] На фиг.3 демонстрируется положение, что TGF-бета ускоряет продукцию белка в модели оценки белка, образующего матрикс in vitro. На фиг.3 представлен результат исследования того, ускоряется ли продукция внеклеточного матрикса в iFECD посредством TGF-бета. iHCEC и iFECD культивировали в DMEM на Transwell без сыворотки, через одну неделю после этого их фиксировали в состоянии монослоя и окрашивали HE (на левой панели представлены микрофотографии окрашивания HE, в верхнем ряду представлены iHCEC, и в нижнем ряду представлены iFECD. На левой стороне представлены контроли и на правой стороне представлен результат стимуляции TGF-бета). На правом графике представлены измеренные значения толщины. На графике с левой стороны представлен контроль, и с правой стороны представлен результат стимуляции посредством TGF-бета, белым цветом показаны iHCEC, и черным цветом показаны iFECD. Каждый из * и # показывает статистическую значимость при p<0,05. Что касается iHCEC и iFECD, было установлено, что стимуляция посредством TGF-бета обеспечивала значительно утолщенный внеклеточный матрикс. Кроме того, было установлено, что в присутствии TGF-бета, iFECD продуцировали значительно утолщенный внеклеточный матрикс по сравнению с iHCEC. Представленное выше описание показывает, что в эндотелиальных клетках роговицы пациента с эндотелиальной дистрофией роговицы Фукса уровень экспрессии Snail1 и ZEB1 является высоким и что продуцируемое количество внеклеточного матрикса в ответ на стимуляцию TGF-бета является значительно более высоким, чем в эндотелиальных клетках роговицы здорового индивидуума.
[0026]
[Фиг.4] На фиг.4 показано, что ZEB1 и Snail1 подавляют экспрессию генов белка, образующего матрикс in vitro. На фиг.4 представлен результат исследования эффекта подавления Snail1 и ZEB1 на продукцию внеклеточного матрикса с использованием миРНК, чтобы продемонстрировать, что стимуляция экспрессии Snail1 и ZEB1 вызывает продукцию внеклеточного матрикса. Белым цветом показаны iHCEC и черным цветом показаны iFECD. На A-E показан результат для миРНК к Snail1 и на F-J показан результат для миРНК к Snail1. На A представлен результат для ZEB1, на F показан результат для SNAIL1, на B и G показан результат для коллагена типа I, на C и H показан результат для коллагена типа IV, на D и I показан результат для коллагена типа VIII, и на E и J показан результат для фибронектина. * Указывает на статистическую значимость при p<0,01. Было подтверждено, что миРНК подавляет экспрессию Snail1 и ZEB1 (A, F). Подавление экспрессии Snail1 или ZEB1 посредством миРНК значительно подавляет экспрессию коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина.
[0027]
[Фиг.5] На фиг.5 демонстрируется положение, что ZEB1 или Snail1 подавляют экспрессию белков, образующих матрикс in vitro. На фиг.5 показан результат исследования экспрессии коллагена типа I, коллагена типа IV и фибронектина путем иммунного окрашивания аналогичным образом. С левой стороны показан результат для контрольной миРНК, в середине показан результат для миРНК к ZEB, и с правой стороны показан результат для миРНК для SNAIL1. На верхних двух рядах показан коллаген типа I, на двух средних рядах показан коллаген типа IV, на двух нижних рядах показан фибронектин. Каждый верхний ряд пары из двух рядов демонстрирует iHCEC и каждый ее нижний ряд демонстрирует iFECD. Было подтверждено, что подавление экспрессии Snail1 или ZEB1 посредством миРНК также подавляет экспрессию коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина на уровне белка.
[0028]
[Фиг.6] На фиг.6 показано, что подавление ZEB1 или Snail1 может подавлять сверхпродукцию матрикса in vitro в клетках эндотелиальной дистрофии роговицы Фукса. На фиг.6, кроме того, показан результат культивирования iHCEC и iFECD в DMEM на Transwell без сыворотки, и фиксации их через одну неделю в состоянии монослоя, и окрашивания HE (на левой панели показана микрофотография окрашивания HE, в верхнем ряду показаны iHCEC и в нижнем ряду показаны iFECD. С левой стороны показан результат для контрольной миРНК, в середине показан результат для миРНК ZEB1, и с правой стороны показан результат для миРНК SNAI). На правом графике показаны измеренные величины толщины. На графике с левой стороны показана контрольная миРНК, в середине показана миРНК к ZEB1, с правой стороны показан результат стимуляции посредством миРНК к SNAIL, белым цветом показаны iHCEC и черным цветом показаны iFECD. * Указывает на статистическую значимость при p < 0,01. Подавление экспрессии Snail1 или ZEB1 посредством миРНК подавляло сверхпродукцию внеклеточного матрикса в iFECD до нормального уровня.
[0029]
[Фиг.7] На фиг.7 демонстрируется положение, что ингибирование сигнала TGF-бета подавляет экспрессию Snail1, ZEB1 и белка, образующего матрикс in vitro. На фиг.7 показан результат ингибирования сигнала TGF-бета с помощью средства, ингибирующего сигнал TGF-бета, SB431542 (0 мкМ, 1 мкМ, 3 мкМ и 10 мкМ). Белым цветом показаны iHCEC и черным цветом показаны iFECD. На A показан результат для Snail1, на B показан результат для ZEB1, на D показан результат для коллагена типа I, на E показан результат для коллагена типа IV, на F показан результат для коллагена типа VIII, на G показан фибронектин. * Указывает на статистическую значимость при p<0,01. Посредством ПЦР в реальном времени было установлено, что в результате ингибирования сигнала TGF-бета уровень экспрессии Snail1 и ZEB1 значительно снижался. Кроме того, когда с помощью ПЦР в реальном времени анализировали уровень экспрессии гена белка, образующего внеклеточный матрикс, в iFCED при использовании SB431542, экспрессия коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина значительно подавлялась.
[0030]
[Фиг.8] На фиг.8 демонстрируется положение, что ингибирование сигнала TGF-бета может контролировать экспрессию белка, образующего матрикс in vitro. На фиг.8 показан результат исследования экспрессии коллагена типа I, коллагена типа IV и фибронектина посредством иммунного окрашивания аналогично тому, как на фиг.7. С левой стороны показан результат для контроля и с правой стороны показан результат стимуляции посредством SB431542. Два верхних ряда демонстрируют коллаген типа I, два средних ряда демонстрируют коллаген типа IV, и два нижних ряда демонстрируют фибронектин. Каждый верхний ряд пар из двух рядов демонстрирует iHCEC и каждый их нижний ряд демонстрирует iFECD. Было подтверждено, что ингибирование сигнала TGF-бета с использованием SB431542 также подавляет экспрессию коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина на уровне белка.
[0031]
[Фиг.9] На фиг.9 демонстрируется положение, что ингибирование сигнала TGF-бета может подавлять сверхпродукцию матрикса in vitro в клетках эндотелиальной дистрофии роговицы Фукса. Кроме того, на фиг.9 показан результат культивирования iHCEC и iFECD в DMEM на Transwell без сыворотки, и фиксации их через одну неделю в состоянии монослоя, и окрашивания HE (на левой панели верхний ряд демонстрирует iHCEC и нижний ряд демонстрирует iFECD. С левой стороны показан контроль и с правой стороны показа результат стимуляции посредством SB431542). На правом графике показаны измеренные величины толщины. С левой стороны показан контроль, с правой стороны показан результат стимуляции посредством SB431542, белым цветом показаны iHCEC и черным цветом показаны iFECD. * Представляет собой статистическую значимость при p<0,01. Ингибирование сигнала TGF-бета с использованием SB431542 подавляло продукцию внеклеточного матрикса iFECD до нормального уровня.
Описание вариантов осуществления
[0032]
Настоящее изобретение описано далее. На протяжении всего описания выражение в форме единственного числа следует понимать как охватывающее это понятие в множественной форме, если нет иных конкретных указаний. Кроме того, термины, используемые в настоящем описании, следует понимать как используемые в значении, которое обычно используется в данной области, если нет иных конкретных указаний. Таким образом, если не определено иначе, вся терминология и научные технические термины, которые используют в настоящем описании, имеют то же значение, что и термины, обычно подразумеваемые специалистами в области, к которой относится настоящее изобретение. В случае противоречия преимуществом обладает настоящее описание (включая определения).
Определения
[0033]
Как используют в рамках изобретения, "iFECD" (иммобилизованная или иммортализованная клетка эндотелиальной дистрофии роговицы Фукса) представляет собой сокращенное обозначение для иммобилизованной клетки, полученной от пациента с эндотелиальной дистрофией роговицы Фукса.
[0034]
Как используют в рамках изобретения, "HCEC" (эндотелиальные клетки роговицы человека) является сокращенным обозначением для эндотелиальных клеток роговицы человека. "iHCEC" является сокращенным обозначением для иммобилизованных (или иммортализованных) эндотелиальных клеток роговицы человека.
[0035]
Как используют в рамках изобретения, "трансформирующий фактор роста-бета (трансформирующий фактор роста-бета; также упоминаемый под сокращенным названием TGF-бета)" используют в значении, сходном со значением, используемым в данной области; и трансформирующий фактор роста-бета представляет собой гомодимерный многофункциональный цитокин с молекулярной массой 25 кДа, который проявляет различные типы биологической активности. TGF-бета участвует в патогенезе различных вызывающих склероз заболеваний, ревматоидного артрита и пролиферативной витреоретинопатии, и он в значительной степени вовлечен в выпадение волос, подавление действия иммунокомпетентных клеток, подавление сверхпродукции протеаз для предупреждения деградации легочных тканей и предотвращения эмфиземы, и подавление роста злокачественных клеток и т.п. У человека существует три изоформы TGF-бета, а именно, с TGF-бета 1 по TGF-бета 3. TGF-бета продуцируется в качестве неактивного латентного типа с молекулярной массой приблизительно 300 кДа, который не способен связываться с рецептором. TGF-бета активируется на поверхности клетки-мишени или на ее периферии, становясь активным типом, способным связываться с рецептором, таким образом, оказывая свое действие.
[0036]
Не желая ограничиваться теорией, полагают, что действие TGF-бета в клетке-мишени передается каскадом фосфорилирования набора белков, осуществляющего передачу информации, упоминаемого как Smad. Сначала, когда активный TGF-бета связывается с рецептором TGF-бета типа II, присутствующим на поверхности клетки-мишени, образуется рецепторный комплекс, который состоит из двух молекул рецептора типа II и двух молекул рецептора TGF типа I, и рецептор типа II фосфорилирует рецептор типа I. Далее фосфорилированный рецептор типа I фосфорилирует Smad2 или Smad3, и фосфорилированный Smad2 или Smad3 образует комплекс с Smad4, и комплекс переносится в ядро, связывается с последовательностью-мишенью, упоминаемой как CAGA-бокс, которая присутствует в промоторной области гена-мишени, и индуцирует транскрипционную экспрессию гена-мишени вместе с коактиватором.
[0037]
Каскад передачи сигнала трансформирующего (трансформационного) фактора роста-бета (TGF-бета) способен регулировать множество типов клеточной активности, таких как рост и дифференцировка клеток, остановка роста, апоптоз и эпителиально-мезенхимальный переход (EMT), путем регуляции его генов-мишеней. Представители семейства TGF-бета, включая сам TGF-бета (такой как TGF-бета 1, TGF-бета 2 и TGF-бета 3), активин и костный морфогенетический белок (BMP), являются мощными агентами, регулирующими рост, дифференцировку, миграцию и апоптоз клеток.
[0038]
TGF-бета представляет собой белок массой приблизительно 24 кДа, который продуцируется многими клетками, включая B-лимфоциты, T-лимфоциты и активированные макрофаги, и многими другими типами клеток. Эффекты TGF-бета на иммунную систему включают индукцию рецептора IL-2, ингибирование индуцируемого IL-1 роста тимусных клеток и блокирование индуцируемой IFN-гамма активации макрофагов. Полагают, что TGF-бета вовлечен в разнообразные патологические состояния (Border et al., (1992) J. Clin. Invest. 90:1) и в достаточной степени подтверждается, что он функционирует в качестве либо ингибирующего опухоль вещества, либо стимулятора опухоли.
[0039]
TGF-бета опосредует его передачу сигнала через два серин/треонин-киназных рецептора клеточной поверхности: TGF-бета RII и ALK5. Передача сигнала TGF-бета инициируется лиганд-индуцируемой димеризацией рецептора, которая позволяет TGF-бета RII фосфорилировать рецептор ALK5. Его фосфорилирование является таким, что киназа ALK5 активируется, а затем активированная ALK5 фосфорилирует нижеследующий эффекторный белок Smad (гомолог MAD у позвоночных или белок "Mothers against DPP (decapentaplegic)"), Smad2 или 3. Комплекс p-Smad2/3 с Smad4 входит в ядро, активируя транскрипцию гена-мишени.
[0040]
Smad3 является представителем подгруппы R-Smad (рецептор-активируемый Smad) среди Smad и он является прямым медиатором активации транскрипции рецептором TGF-бета. Стимуляция TGF-бета вызывает фосфорилирование и активацию Smad2 и Smad3, которые образуют комплекс с Smad4 ("общий Smad" или "co-Smad" у позвоночных), который накапливается в ядре, регулируя транскрипцию гена-мишени. R-Smad локализуется в цитоплазме и образует комплекс с co-Smad через лиганд-индуцируемое фосфорилирование рецептором TGF-бета; и комплекс перемещается в ядро, а затем регулирует экспрессию генов, которые ассоциированы с хроматином и кооперативным фактором транскрипции. Каждый из Smad6 и Smad7 является ингибиторным Smad ("I-Smad"), т.е. они транскрипционно индуцируются посредством TGF-бета и функционируют в качестве ингибирующего средства для передачи сигнала TGF-бета (Feng et al., (2005) Annu. Rev. Cell. Dev. Biol. 21:659). Smad6/7 ингибируют рецептор-опосредуемую активацию R-Smad, проявляя их ингибиторный эффект; и они ассоциированы с рецептором типа I, который конкурентно препятствует мобилизации и фосфорилированию R-Smad. Известно, что Smad6 и Smad7 восполняют E3-убиквитинлигазу, которая вызывает убиквитинилирование и деградацию, белка, взаимодействующего с Smad6/7.
[0041]
Что касается каскада передачи сигнала TGF-бета, кроме того, существует другой каскад, передачу в котором осуществляет BMP-7 и т.п., который, как считают, проявляет функции через ALK-1/2/3/6, а затем через Smad1/5/8. Что касается каскада передачи сигнала TGF-бета, также см. J. Massagu’e, Annu. Rev. Biochem. 1998. 67: 753-91; Vilar JMG, Jansen R, Sander C (2006) PLoS Comput Biol 2(1):e3; Leask, A., Abraham, D. J. FASEB J.18,816-827 (2004); Coert Margadant & Arnoud Sonnenberg EMBO reports (2010) 11, 97-105; Joel Rosenbloom et al., Ann Intern Med. 2010; 152: 159-166 и т.п.
[0042]
Как используют в рамках изобретения, "средство, ингибирующее сигнал трансформирующего фактора роста (TGF)-бета" относится к любому фактору, который ингибирует передачу сигнала TGF. Когда осуществляется противодействие TGF-бета, средство, ответственное за это, может указываться как антагонист. Однако в случае настоящего изобретения антагонист TGF-бета охватывается средством, ингибирующим сигнал TGF-бета. Поскольку этот ингибирующий агент обычно представляет собой вещество, "вещество, ингибирующее сигнал TGF-бета," можно использовать взаимозаменяемо со "средством, ингибирующим сигнал TGF-бета".
[0043]
Таким образом, средство, ингибирующее сигнал TGF-бета. используемое в рамках настоящего изобретения, как правило, включает, но не ограничивается ими, антагонист TGF-бета, антагонист рецептора TGF-бета и средство, ингибирующее Smad3, ловушку лиганда (антитело против лиганда, рецептор-приманка), антисмысловой олигонуклеотид, средство, ингибирующее киназу рецептора TGF-бета, пептидный аптамер, миРНК, кшРНК и т.п. (см. Connolly E., et al., Int. J. Biol. Sci. 2012; 8(7): 964-978, фиг.3 и т.п).
[0044]
Иллюстративное средство, ингибирующее сигнал TGF-бета, которое можно использовать в рамках настоящего изобретения, может включать, но не ограничиваться ими, SB431542 (4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)]-1H-имидазол-2-ил]бензамид), BMP-7, антитело против TGF-бета, антитело против рецептора TGF-бета, миРНК к TGF-бета, миРНК к рецептору TGF-бета, антисмысловой олигонуклеотид к TGF-бета, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-илпроп-2-еноил))-1,2,3,4-тетрагидроизохинолон, A83-01 (3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1H-пиразол-1-карботиоамид), ингибитор TLK StemoleculeTM (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин), ингибитор BMP StemoleculeTM LDN-193189 (6-(4-(пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил)пиразолo[1,5-a]пиримидин), SD-208 (2-(5-хлор-2-фторфенил)-4-[(4-пиридинил)амино]птеридин), LY364947 (4-[3-(2-пиридинил)-1H-пиразол-4-ил]-хинолин), их фармацевтически приемлемые соли и сольваты, и сольваты фармацевтически приемлемых солей, и т.п.
[0045]
Другие средства, ингибирующие сигнал TGF-бета, включают, но не ограничиваются ими, моноклональное антитело и поликлональное антитело к одной или нескольким изоформам TGF-бета (патент США № 5571714; также см. международную публикацию № WO 97/13844 и международную публикацию № WO 00/66631), рецептор TGF-бета, растворимую форму такого рецептора (например, растворимый рецептор TGF-бета типа III) или антитело, направленное на рецептор TGF-бета (патент США № 5693607, патент США № 6001969, патент США № 6010872, патент США № 6086867, патент США № 6201108; международная публикация № WO 98/48024; международная публикация № WO 95/10610; международная публикация № WO 93/09228; международная публикация № WO 92/00330), латентный и ассоциированный пептид (международная публикация № WO 91/08291), большой латентный TGF-бета (международная публикация № WO 94/09812), фетуин (патент США № 5821227), другой протеогликан, такой как декорин и бигликан, фибромодулин, люмикан и эндоглин, и т.п. (международная публикация № WO 91/10727; патент США № 5654270, патент США № 5705609, патент США № 5726149; патент США № 5824655; международная публикация № WO 91/04748; патент США № 5830847, патент США № 6015693; международная публикация № WO 91/10727; международная публикация № WO 93/09800 и международная публикация № WO 94/10187), соматостатин (международная публикация № WO 98/08529), маннозо-6-фосфорную кислоту или маннозо-1-фосфорную кислоту (патент США № 5520926), пролактин (международная публикация № WO 97/40848), инсулиноподобный фактор роста II (международная публикация № WO 98/17304), IP-10 (международная публикация № WO 97/00691), Arg-Gly-Asp-содержащий пептид (Pfeffer, патент США № 5958411; международная публикация № WO 93/10808), экстракты растений, грибов и бактерий (EP-A-813875; выложенная японская публикация № 8-119984; и Matsunaga et al., патент США № 5693610), антисмысловой олигонуклеотид (патент США № 5683988; патент США № 5772995; патент США № 5821234, патент США № 5869462 и международная публикация № WO 94/25588), белок, ассоциированный с передачей сигнала TGF-бета, включая Smad и MAD (EP-A-874046; международная публикация № WO 97/31020; международная публикация № WO 97/38729; международная публикация № WO 98/03663; международная публикация № WO 98/07735; международная публикация № WO 98/07849; международная публикация № WO 98/45467; международная публикация № WO 98/53068; международная публикация № WO 98/55512; международная публикация № WO 98/56913; международная публикация № WO 98/53830; международная публикация № WO 99/50296; патент США № 5834248; патент США № 5807708 и патент США № 5948639), Ski и Sno (Vogel, 1999, Science, 286:665; и Stroschein et al.,1999, Science, 286:771-774), один или несколько одноцепочечных олигонуклеотидных аптамеров или экспрессирующие плазмиды, кодирующие их, пригодные для ингибирования или препятствования связыванию TGF-бета с рецептором того же происхождения и любым мутантом, фрагментом или производным молекулы, указанной выше, которые сохраняют способность ингибировать активность TGF-бета. Средство, ингибирующее TGF-бета, может представлять собой антагонист TGF-бета, и может представлять собой моноклональное антитело человека или гуманизированное моноклональное антитело (или F(ab)2-фрагмент, Fv-фрагмент, одноцепочечное антитело, и другие формы или фрагменты антитела, сохраняющие способность связываться с TGF-бета, их фрагмент и т.п.), которые блокируют связывание TGF-бета с рецептором. Рецептор TGF-бета и связывающий TGF-бета фрагмент и, в частности, растворимый фрагмент рецептора TGF-бета представляют собой антагонисты TGF-бета, которые являются пригодными в способе в соответствии с настоящим изобретением. В определенном варианте осуществления ингибирующее средство, предпочтительное для функционирования TGF-бета, представляет собой растворимый рецептор TGF-бета и, в частности, рецептор TGF-бета типа II (TGFBIIR) или рецептор TGF-бета типа III (TGFBIIIR или бета-гликан), включая, например, внеклеточный домен TGFBIIR или TGFBIIIR, предпочтительно рекомбинантный растворимый рецептор TGF-бета (rsTGFBIIR или rsTGFBIIIR). Рецептор TGF-бета и связывающий TGF-бета фрагмент рецептор TGF-бета, в частности, растворимый фрагмент, представляют собой антагонисты TGF-бета, пригодные в способе в соответствии с настоящим изобретением. Рецепторы TGF-бета и нуклеиновые кислоты, кодирующие их, достаточно известны в данной области. Последовательность нуклеиновой кислоты, кодирующая рецептор TGF-бета 1 типа, описана в GenBank под номером доступа L15436 и в патенте США № 5538892 (Donahoe et al.). Последовательность нуклеиновой кислоты для рецептора TGF-бета 2 типа является общедоступной под номером доступа GenBank AW236001, AI35790, AI279872, AI074706 и AA808255. Последовательность нуклеиновой кислоты рецептора TGF-бета 3 типа также является общедоступной под номером доступа GenBank NM003243, AI887852, AI817295 и AI681599.
[0046]
Кроме того, другие средства, ингибирующие сигнал TGF-бета, или его антагонисты и способы их получения достаточно известны в данной области, в дополнение к многим другим, которые в настоящее время находятся на стадии разработки. В способе в соответствии с настоящим изобретением могут быть пригодными любые эффективные антагонисты TGF-бета, и, таким образом, конкретные используемые агенты, ингибирующие сигнал TGF-бета, или его антагонисты не являются средствами с ограниченными характеристиками. Примеры таких антагонистов включают моноклональные и поликлональные антитела к TGF-бета одного или нескольких изотипов (патент США № 5571714 и международная публикация № WO 97/13844), рецептор TGF-бета, его фрагмент, его производное и антитело к рецептору TGF-бета (патент США № 5693607, патент США № 6008011, патент США № 6001969 и патент США № 6010872, и международная публикация № WO 92/00330, международная публикация № WO 93/09228, международная публикация № WO 95/10610 и международная публикация № WO 98/48024); ассоциированный с латентностью пептид (ассоциированный с латентностью пептид; международная публикация № WO 91/08291), большой латентный TGF-бета (международная публикация № WO 94/09812), фетуин (патент США № 5821227), другой протеогликан, такой как декорин и бигликан, фибромодулин, люмикан, эндоклин и т.п. (патент США № 5583103, патент США № 5654270, патент США № 5705609, патент США № 5726149, патент США № 5824655, патент США № 5830847, патент США № 6015693 и международная публикация № WO 91/04748, международная публикация № WO 91/10727, международная публикация № WO 93/09800 и международная публикация № WO 94/10187).
[0047]
Следующие примеры такого антагониста включают множество других белков, ассоциированных с передачей сигнала TGF-бета, включая соматостатин (международная публикация № WO 98/08529), маннозо-6-фосфорную кислоту или маннозо-1-фосфорную кислоту (патент США № 5520926), пролактин (международная публикация № WO 97/40848), инсулиноподобный фактор роста II (международная публикация № WO 98/17304), IP-10 (международная публикация № WO 97/00691), аргинин (arg)-глицин (gly)-аспарагиновая кислота (asp)-содержащий пептид (патент США № 5958411 и международная публикация № WO 93/10808), экстракты растений, грибов и бактерий (публикация патентной заявки Европы № 813875, выложенную публикацию Японии № 8-119984 и патент США № 5693610), антисмысловой олигонуклеотид (патент США № 5683988, патент США № 5772995, патент США № 5821234 и Патент США № 5869462, и международная публикация № WO 94/25588), и Smad и MAD (патентная заявка Европы № EP874046, международная публикация № WO 97/31020, международная публикация № WO 97/38729, международная публикация № WO 98/03663, международная публикация № WO 98/07735, международная публикация № WO 98/07849, международная публикация № WO 98/45467, международная публикация № WO 98/53068, международная публикация № WO 98/55512, международная публикация № WO 98/56913, международная публикация № WO 98/53830 и международная публикация № WO 99/50296 и патент США № 5834248, патент США № 5807708 и патент США № 5948639), и Ski и Sno (G.Vogel, Science, 286:665(1999) и Stroschein et al., Science, 286:771-74(1999)), и любой фрагмент и производное упомянутой выше молекулы, сохраняющий способность ингибировать активность TGF-бета.
[0048]
Антагонисты TGF-бета, пригодные для применения в рамках настоящего изобретения, также включают функциональный мутант, мутант, производное и аналог упомянутого выше антагониста TGF-бета при условии, что их способность ингибирования уровня или активности TGF-бета сохраняется. "Мутант", "производное" и "аналог", как используют в рамках изобретения, относится к молекуле, имеющей форму или структуру, сходную с формой или структурой ее исходного соединения, и сохраняющей способность действовать в качестве антагониста TGF-бета. Например, любой из антагонистов TGF-бета в рамках настоящего изобретения может быть кристаллизованным, и пригодные аналоги можно получать рациональным конструированием на основе участков, которые играют роль в формировании (одного или нескольких) активных центров. Вместо этого специалисты в данной области могут изменять функциональную группу известных антагонистов или могут проводить скрининг такой измененной молекулы в отношении увеличения активности, времени полужизни, биодоступности или других желаемых характеристик без ненужных экспериментов. Когда антагонист TGF-бета представляет собой полипептид, можно получать фрагмент и вариант полипептида для обеспечения простоты доставки, увеличения активности, времени полужизни и т.п. (например, гуманизированные антитела или функциональные фрагменты антител, обсуждаемые выше). Исходя из уровня техники в области получения синтетических и рекомбинантных полипептидов, такой вариант можно получать без ненужных экспериментов. Специалисты в данной области также могут конструировать новое ингибирующее средство, исходя из знания кристаллической структуры и/или активного центра средства, ингибирующего TGF-бета, как описано в настоящем описании. Ингибирующее полипептид средство, такое как растворимый рецептор TGF-бета, можно эффективно вводить посредством генного переноса. Таким образом, определенный вариант осуществления способа в соответствии с настоящим изобретением включает применение вектора, пригодного для экспрессии рецептора TGF-бета или связывающегося партнера, предпочтительно растворимого рецептора или растворимого связывающегося партнера. В предпочтительном варианте осуществления введение растворимого антагониста TGF-бета можно осуществлять посредством генного переноса, в котором используется вектор, содержащий кДНК, кодирующую растворимый антагонист, или кДНК, кодирующую внеклеточный домен рецептора TGF-бета типа II (rsTGFBIIR) или рецептор TGF-бета типа III (rsTGFBIIIR). Этот вектор обеспечивает экспрессию in situ растворимого антагониста TGF-бета в клетке, которая трансфицирована с использованием вектора, ингибирует активность TGF-бета и подавляет опосредуемый TGF-бета фиброгенез. Можно использовать любой пригодный вектор. Предпочтительные векторы включают аденовирусный вектор, лентивирусный вектор, вектор на основе вируса Эпштейна-Барр (EBV), вектор на основе аденоассоциированного вируса (AAV) и ретровирусный вектор, разработанный для генного переноса. Также можно использовать другие невекторые способы, пригодные для генного переноса, такие как способы переноса комплекса липид/ДНК, конъюгата белок/ДНК и голой ДНК. Следующие пригодные антагонисты TGF-бета, разработанные для доставки посредством генного переноса с использованием аденовируса, включают, но не ограничиваются ими, химерную кДНК, кодирующую внеклеточный домен рецептора TGF-бета II типа, слитую с Fc-доменом Ig (Isaka et al.,1999, Kidney Int., 55:pp.465-475), аденовирусный вектор для генного переноса доминантно-негативного мутанта рецептора TGF-бета типа II (Zhao et al.,1998, Mech. Dev., 72:pp.89-100) и аденовирусный вектор для генного переноса декорина, который представляет собой связывающий TGF-бета протеогликан (Zhao et al., 1999, Am. J. Physiol., 277: pp. L412-L422). Опосредуемый аденовирусом генный перенос имеет чрезвычайно высокую эффективность по сравнению с другими способами доставки генов.
[0049]
Рецептор TGF-бета и связывающий TGF-бета фрагмент, растворимый фрагмент и т.п. рецептора TGF-бета являются антагонистами TGF-бета, пригодными в рамках настоящего изобретения. Рецепторы TGF-бета и нуклеиновые кислоты, кодирующие их, достаточно известны в данной области. Последовательность нуклеиновой кислоты, кодирующая рецептор TGF-бета 1 типа, описана в GenBank под номером доступа L15436 и в патенте США № 5538892, выданном Donahoe et al. Последовательность нуклеиновой кислоты рецептора TGF-бета 2 типа также является общедоступной под номером доступа GenBank AW236001; AI35790; AI279872; AI074706 и AA808255. Последовательность нуклеиновой кислоты рецептора TGF-бета 3 типа также является общедоступной под номером доступа GenBank NM003243; AI887852; AI817295 и AI681599. В одном иллюстративном варианте осуществления антагонист TGF-бета представляет собой антитело, которое блокирует связывание TGF-бета с его рецептором или с его фрагментами, такими как F(ab)2-фрагмент, Fv-фрагмент, одноцепочечное антитела и другие типы "антител", сохраняющие способность связываться с TGF-бета. Антитело может быть химеризованным или гуманизированным. В рамках настоящего изобретения химеризованное антитело включает константную область антитела человека и вариабельную область не являющихся человеческими антител, таких как антитела мыши. Гуманизированное антитело включает константную область и каркасную вариабельную область (т.е. вариабельные области, отличные от гипервариабельных областей) антитела человека и гипервариабельную область не являющихся человеческими антител, таких как антитела мыши. Безусловно, антитело может представлять собой любые другие типы производных антител, такие как антитела человека, отобранные из системы фагового дисплея или продуцированные в XenoMouse.
[0050]
Накапливаются данные, касающиеся Smad. Каскад передачи сигнала TGF-бета инициируется, когда эта молекула связывается с гетеродимерным комплексом клеточной поверхности, состоящим из серин/треонин-киназного рецептора типа I (TbRI) и типа II (TbRII) и индуцирует этот гетеродимерный комплекс клеточной поверхности. Затем, гетеродимерный рецептор далее передает сигнал через фосфорилирование белка-мишени Smad. Как описано выше, существует три функциональных класса белка Smad, и они представляют собой, например, Smad (R-Smad), ограниченные рецепторами, такие как Smad2 и Smad3, комедиатор (Co-Smad), который также обозначается как Smad4, и ингибиторный Smad (I-Smad). После фосфорилирования гетеродимерного рецепторного комплекса этот R-Smad образует комплекс с Co-Smad, перемещается в ядро и, действуя вместе с другими соответствующими белками, регулирует транскрипцию гена-мишени (Derynck, R., et al. (1998) Cell 95: 737-740); Massague, J. и Wotton, D. (2000) EMBO J. 19:1745). Нуклеотидная последовательность и аминокислотная последовательность Smad3 человека описаны, например, в GenBank под номером доступа gi:42476202. Нуклеотидная последовательность и аминокислотная последовательность Smad3 мыши описана, например, в GenBank под номером доступа gi: 31543221. Как описано выше, стимуляция TGF-бета обеспечивает фосфорилирование и активацию Smad2 и Smad3, которые образуют комплекс с Smad4 (также называемый "общим Smad" или "co-Smad"), и комплекс накапливается в ядре, регулируя транскрипцию гена-мишени. Таким образом, ингибирования сигнала TGF-бета также можно достигать путем ингибирования Smad2, 3 или co-Smad (Smad4). R-Smad локализуется в цитоплазме и образует комплекс с co-Smad посредством лиганд-индуцируемого фосфорилирования рецептором TGF-бета, перемещаясь в ядро, в котором они регулируют экспрессию генов, ассоциированную с хроматином и кооперативным фактором транскрипции. Таким образом, ингибирования сигнала TGF-бета также можно достигать путем ингибирования R-Smad либо прямо, либо непрямо. Smad6 и Smad7 представляют собой ингибиторные Smad (I-Smad), и, таким образом, TGF-бета транскрипционно индуцирует их функционирование в качестве средства, ингибирующего передачу сигнала TGF-бета (Feng et al., (2005) Annu. Rev. Cell. Dev. Biol. 21: 659). Smad6/7 препятствуют рецептор-опосредуемой активации R-Smad, тем самым, проявляя их ингибиторный эффект. Они ассоциированы с рецептором типа I, который конкурентно ингибирует мобилизацию и фосфорилирование R-Smad. Известно, что Smad6 и Smad7 дополняют E3-убиквитинлигазу, которая обеспечивает убиквитинилирование и деградацию белка, взаимодействующего со Smad6/7. Таким образом, в рамках настоящего изобретения Smad6 и 7 могут функционировать в качестве средства, ингибирующего сигнал TGF-бета.
[0051]
Средства, ингибирующие Smad3, которые можно использовать в рамках настоящего изобретения, могут включать, но не ограничиваться ими, антисмысловой нуклеотид, миРНК, антитело и т.п., и, кроме того, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-ил-проп-2-еноил))-1,2,3,4-тетрагидроизохинолон и т.п., коммерчески доступный от Calbiochem, в качестве низкомолекулярного соединения.
[0052]
Как используют в рамках изобретения, "вещество (например, нуклеиновая кислота) для подавления экспрессии (TGF-бета и т.п.)" конкретно не ограничивается при условии, что такое вещество представляет собой вещество, которое подавляет транскрипцию гена-мишени мРНК, вещество, которое деградирует транскрибированную мРНК (например, нуклеиновая кислота), или вещество (например, нуклеиновая кислота), которое подавляет трансляцию белка с мРНК. Что касается веществ, их примерами являются миРНК, антисмысловой олигонуклеотид и рибозим, а также нуклеиновые кислоты экспрессирующих векторов. Среди них предпочтительными являются миРНК и экспрессирующие векторы для них, и особенно предпочтительными являются миРНК. "Вещество, которое подавляет экспрессию гена" включает, в дополнение к веществам, описанным выше, белок, пептид и другие низкомолекулярные соединения. Следует отметить, что ген-мишень в рамках настоящего изобретения означает любой ген, который ассоциирован с каскадом передачи сигнала TGF-бета.
[0053]
Что касается способа ингибирования экспрессии конкретного эндогенного гена, такого как TGF-бета, на который осуществляется нацеливание в рамках настоящего изобретения, специалистам в данной области хорошо известен способ с использованием антисмысловой технологии. Что касается действий антисмысловой нуклеиновой кислоты, посредством которых она ингибирует экспрессию гена-мишени, существует множество факторов, которые представлены ниже. В частности, такие факторы представляют собой: ингибирование инициации транскрипта вследствие образования триплекса; ингибирование транскрипции вследствие образования гибрида с участком, в котором локально образуется структура открытой петли под действием РНК-полимеразы; ингибирование транскрипции вследствие образования гибрида с РНК, синтез которой осуществляется; ингибирование сплайсинга вследствие образования гибрида в точке соединения интрона и экзона; ингибирование сплайсинга вследствие образования гибрида с участком образования сплайсеосомы; ингибирование переноса из ядра в цитоплазму вследствие образования гибрида с мРНК; ингибирование сплайсинга вследствие образования гибрида с участком кэппирования или участком добавления поли (A); ингибирование инициации трансляции вследствие образования гибрида с участком связывания фактора инициации трансляции; ингибирование трансляции вследствие образования гибрида с участком связывания рибосомы вблизи кодона инициации; ингибирование удлинения пептидной цепи вследствие образования гибрида с участком связывания полисомы или областью трансляции мРНК; и ингибирование экспрессиии гена вследствие образования гибрида с участком взаимодействия нуклеиновой кислоты и белка и т.п. По существу, антисмысловая нуклеиновая кислота ингибирует множество процессов, таких как транскрипция, сплайсинг или трансляция, ингибируя экспрессию гена-мишени (Hirashima and Inoue, Shinsei Kagaku Jikken Kouza [New Chemical Experiment Course] 2, Nucleic Acid, IV Idenshi no Fukusei to Hatsugen [Duplication and Expression of Gene], Edited by the Japanese Biochemical Society, Tokyo Kagaku Dozin, 1993, 319-347).
[0054]
Антисмысловая нуклеиновая кислота, используемая в рамках настоящего изобретения, может ингибировать экспрессию и/или функцию гена (нуклеиновой кислоты), кодирующего представителя и т.п. каскада передачи сигнала упомянутого выше TGF-бета посредством любого из упомянутых выше действий. В одном варианте осуществления считается, что для ингибирования трансляции гена является эффективным, когда конструируют антисмысловую последовательность, комплементарную нетранслируемой области вблизи 5’-конца мРНК гена, кодирующего упомянутый выше TGF-бета и т.п. Кроме того, является возможным использование последовательности, комплементарной кодирующей области или 3’-нетранслируемой области. По существу, область трансляции гена, кодирующего упомянутый выше TGF-бета и т.п., а также нуклеиновая кислота, включающая антисмысловую последовательность для последовательности нетранслируемой области, включены в антисмысловую нуклеиновую кислоту, которую используют в рамках настоящего изобретения. Используемую антисмысловую нуклеиновую кислоту связывают с нижеследующим подходящим промотором и предпочтительно связывают с последовательностью, включающей сигнал терминации транскрипции со стороны ближе к 3’-концу. Нуклеиновой кислотой, полученной таким образом, можно трансформировать желаемое животное (клетку) с использованием общеизвестного способа. Хотя последовательность антисмысловой нуклеиновой кислоты предпочтительно представляет собой последовательность, комплементарную гену или части гена, кодирующего TGF-бета и т.п. животного (клетки), подлежащего трансформации, она не должна быть полностью комплементарной при условии, что последовательность может эффективно подавлять экспрессию генов. Транскрибированная РНК предпочтительно имеет 90% или более, и наиболее предпочтительно 95% или более, комплементарность продукту транскрипции гена-мишени. Для эффективного ингибирования экспрессии гена-мишени с использованием антисмысловой нуклеиновой кислоты длина антисмысловой нуклеиновой кислоты предпочтительно составляет по меньшей мере 12 оснований или более, но менее 25 оснований. Однако антисмысловая нуклеиновая кислота в соответствии с настоящим изобретением не обязательно ограничивается этой длиной и антисмысловая нуклеиновая кислота может составлять, например, 11 оснований или менее, 100 оснований или более или 500 оснований или более. Хотя антисмысловая нуклеиновая кислота может состоять только из ДНК она также может включать нуклеиновые кислоты, отличные от ДНК, такие как замкнутая нуклеиновая кислота (LNA). В одном варианте осуществления антисмысловая нуклеиновая кислота, используемая в рамках настоящего изобретения, может представлять собой LNA-содержащую антисмысловую нуклеиновую кислоту, включающую LNA на 5’-конце и LNA на 3’-конце. Более того, в одном варианте осуществления, когда в рамках настоящего изобретения используют антисмысловую нуклеиновую кислоту, антисмысловую последовательность можно конструировать на основе последовательности нуклеиновой кислоты, такой как TGF-бета, с использованием способа, описанного, например, в Hirashima and Inoue, Shinsei Kagaku Jikken Kouza [New Chemical Experiment Course] 2, Nucleic Acid, IV Idenshi no Fukusei to Hatsugen [Duplication и Expression of Gene], Edited by the Japanese Biochemical Society, Tokyo Kagaku Dozin, 1993, 319-347.
[0055]
Ингибирование экспрессии TGF-бета и т.п. также можно осуществлять с использованием рибозима или ДНК, кодирующей рибозим. Рибозим относится к молекуле РНК, обладающей каталитической активностью. Существуют различные типы рибозимов, имеющих различные типы активности, и исследования, фокусирующиеся, в частности, на рибозиме в качестве фермента для расщепления РНК, обеспечили возможность конструирования рибозима для расщепления РНК сайт-специфическим образом. Хотя рибозимы включают рибозимы размером 400 нуклеотидов или более, такие как рибозимы интронного типа группы I и РНК M1, включенная в РНК-азу P, также существуют рибозимы, имеющие домен активности не более чем из 40 нуклеотидов, такие как рибозимы, называемые рибозимами типа головки молотка или типа шпильки (Makoto Koizumi and Eiko Ohtsuka, Tanpakushitu Kakusan Kouso [Protein Nucleic Acid Enzyme], 1990, 35, 2191).
[0056]
Например, саморасщепляюющийся домен рибозима типа головки молотка расщепляет участок вблизи 3’-стороны C15 в последовательности, обозначаемой как G13U14C15, и образование пар U14 и A9 считается важным для его активности; и показано, что расщепление может осуществляться по A15 или U15 вместо C15 (Koizumi, M. et al., FEBS Lett, 1988, 228, 228). Если конструируют рибозим, в котором участок связывания вещества комплементарен последовательности РНК вблизи участка-мишени, можно создавать рибозим с расщеплением РНК ферментом рестрикции, который распознает последовательность, такую как UC, UU или UA в РНК-мишени (Koizumi, M. et al., FEBS Lett, 1988, 239, 285., Makoto Koizumi and Eiko Ohtsuka, Tanpakushitu Kakusan Kouso [Protein Nucleic Acid Enzyme], 1990, 35, 2191., Koizumi, M. et al., Nucl. Acids Res., 1989, 17, 7059).
[0057]
Кроме того, для целей настоящего изобретения также является пригодным рибозим типа шпильки. Такой рибозим встречается, например, на отрицательной цепи сателлитной РНК вируса кольцевой пятнистости табака (Buzayan, JM., Nature, 1986, 323, 349.). Показано, что из рибозима типа шпильки можно конструировать рибозим со специфическим к мишени расщеплением РНК (Kikuchi, Y. & Sasaki, N., Nucl. Acids Res, 1991, 19, 6751., Kikuchi, Yo, Kagaku to Seibutu [Chemistry and Living Organism], 1992, 30,112.). По существу, продукт транскрипции гена, кодирующего TGF-бета и т.п., специфически расщепляют с использованием рибозима, так что экспрессия гена может ингибироваться.
[0058]
Подавление экспрессии эндогенного гена TGF-бета и т.п. также можно проводить посредством РНК-интерференции (далее сокращенно обозначаемой как "РНК-i") с использованием двухцепочечной РНК, имеющей последовательность, идентичную или сходную с последовательностью гена-мишени. Что касается РНК-i, когда двухцепочечная РНК (дцРНК) захватывается непосредственно в клетку, экспрессия гена, имеющего последовательность, гомологичную дцРНК, подавляется, и этот способ в настоящее время привлекает внимание. В клетках млекопитающих чтобы индуцировать РНК-i, используют дцРНК с короткой цепью (миРНК). По сравнению с мышами с нокаутом РНК-i имеет множество преимуществ, таких как высокая стабильность эффекта, простота экспериментирования и низкая стоимость. миРНК подробно описаны в другой части настоящего описания.
[0059]
Как используют в рамках изобретения, "миРНК" относится к молекуле РНК, имеющей часть в виде двухцепочечной РНК, состоящую из 15-40 оснований, и миРНК имеет функцию расщепления мРНК гена-мишени, имеющего последовательность, комплементарную антисмысловой цепи указанной миРНК, и подавления экспрессии гена-мишени. Более конкретно, миРНК в соответствии с настоящим изобретением представляет собой РНК, включающую часть в виде двухцепочечной РНК, состоящую из смысловой цепи РНК, состоящей из последовательности, гомологичной непрерывной последовательности РНК в мРНК TGF-бета и т.п., и антисмысловой цепи РНК, состоящей из последовательности, комплементарной смысловой последовательности РНК. Производство и конструирование миРНК и мутантных миРНК, описанных ниже, входят в пределы способностей специалистов в данной области. Идея выбора какой-либо непрерывной области РНК в мРНК, которая является продуктом транскрипции последовательности TGF-бета и т.п., и создания двухцепочечной РНК, соответствующей этой области, является действием, которое специалисты в данной области могут осуществлять в рамках их обычной творческой способности. Более того, идея выбора последовательности миРНК с более мощным эффектом РНК-i из последовательности мРНК, которая является продуктом транскрипции рассматриваемой последовательности, может быть соответствующим образом осуществлена специалистами в данной области с использованием общеизвестного способа. Более того, Если идентифицирована одна из цепей, специалист в данной области без труда определит последовательность оснований другой цепи (комплементарной цепи). Специалисты в данной области могут соответствующим образом создавать миРНК с использованием коммерчески доступного устройства для синтеза нуклеиновых кислот. Кроме того, для синтеза желаемой РНК обычно можно использовать организацию, осуществляющую синтез.
[0060]
Длина части в виде двухцепочечной РНК составляет, в основном, 15-40 оснований, предпочтительно 15-30 оснований, более предпочтительно 15-25 оснований, еще более предпочтительно 18-23 оснований и наиболее предпочтительно 19-21 оснований. Понятно, что ее верхние и нижние пределы не ограничиваются указанными пределами, а пределы могут представлять собой любые комбинации из приведенных пределов. Что касается концевой структуры смысловой цепи или антисмысловой цепи миРНК, отсутствуют конкретные ограничения и ее можно соответствующим образом выбирать в зависимости от назначения. Например, концевая структура может представлять собой структуру, имеющую тупой конец, или структуру, имеющую выступающий конец ("липкий" конец), и предпочтительным является тип с выступающим 3’-концом. миРНК, имеющая выступающий конец, состоящий из нескольких оснований, предпочтительно 1-3 оснований и еще более предпочтительно 2 оснований на 3’-конце смысловой цепи РНК и антисмысловой цепи РНК часто имеет значительный эффект ингибирования экспрессии гена-мишени, что является предпочтительным. Тип оснований выступающего конца конкретно не ограничен, и этот тип может представлять собой либо основания, составляющие РНК, либо основания, составляющие ДНК. Предпочтительные последовательности выступающих концов могут включать dTdT (2 п.н. из дезокси T) на 3’-конце и т.п. Например, предпочтительные миРНК включают, но не ограничиваются ими, миРНК, в которых dTdT (2 п.н. из дезокси T) добавлен на 3’-конце смысловой и антисмысловой цепей всех миРНК.
[0061]
Более того, также является возможным использование миРНК, в которой от одного до нескольких нуклеотидов удалены, заменены, встроены и/или добавлены на любой или обеих из смысловой цепи и антисмысловой цепи упомянутой выше миРНК. В этом отношении принцип от одного до нескольких оснований конкретно не ограничен, но предпочтительно их число составляет от 1 до 4 оснований, еще более предпочтительно от 1 до 3 оснований, наиболее предпочтительно от 1 до 2 оснований. Конкретные примеры рассматриваемой мутации включают, но не ограничиваются ими, мутации, в которых количество оснований на 3’-выступающей части составляет от 0 до 3, мутации, в которых последовательность оснований 3’-выступающей части заменена на другую последовательность оснований, мутации, в которых длина упомянутой выше смысловой цепи РНК и антисмысловой цепи РНК отличается на 1-3 основания вследствие встраивания, добавления или делеции оснований, мутации, в которых основание в смысловой цепи и/или антисмысловой цепи заменены другим основанием и т.п. Однако для смысловой цепи и антисмысловой цепи необходимо, чтобы она была способна гибридизоваться с этими мутантными миРНК, и для этих мутантных миРНК необходимо, чтобы они обладали способностью ингибировать экспрессию генов эквивалентно миРНК, которые не имеют мутации.
[0062]
Более того, миРНК может представлять собой миРНК (короткая шпилечная РНК; кшРНК), в которой один из концов имеет молекулу замкнутой структуры, такой как шпилечная структура. кшРНК представляет собой смысловую цепь РНК с конкретной последовательностью гена-мишени, антисмысловую цепь РНК, состоящую из последовательности, комплементарной последовательности смысловой цепи, и РНК, включающую линкерную последовательность для связывания обеих этих цепей, где часть в виде смысловой цепи и часть в виде антисмысловой гибридизуются с образованием части в виде двухцепочечной РНК.
[0063]
миРНК желательно не проявляет так называемого эффекта вне мишени при клиническом применении. Эффект вне мишени относится к эффекту подавления экспрессии другого гена с частичной гомологией с используемой миРНК, отличного от гена-мишени. Чтобы избежать эффекта вне мишени, можно заранее подтвердить, что миРНК-кандидат не обладает перекрестной реактивностью, с использованием ДНК-микрочипа и т.п. Более того, эффекта вне мишени можно избежать путем проверки того, существует ли ген, включающий часть, обладающую высокой гомологией с последовательностью миРНК-кандидата, отличный от гена-мишени, с использованием общеизвестной базы данных, предоставленной NCBI (National Center for Biotechnology Information) и т.п.
[0064]
Для получения миРНК в соответствии с настоящим изобретением можно соответствующим образом использовать общеизвестный способ, такой как способ с использованием химического синтеза и способ с использованием технологии рекомбинации генов. В случае способа с использованием синтеза двухцепочечную РНК можно синтезировать на основе информации о последовательности с использованием обычного способа. Кроме того, в способе с использованием технологии рекомбинации генов также возможно создавать такую миРНК путем конструирования экспрессирующего вектора, кодирующего последовательность смысловой цепи и последовательность антисмысловой цепи, и введения вектора в клетку-хозяина, а затем получения РНК смысловой цепи и РНК антисмысловой цепи, каждую из которых получают путем транскрипции. Более того, также можно получать желаемую двухцепочечную РНК путем экспрессии кшРНК, которая включает смысловую цепь конкретной последовательности гена-мишени, антисмысловую цепь, состоящую из последовательности, комплементарной последовательности смысловой цепи, и линкерную последовательность для связывания обеих цепей, и которая формирует шпилечную структуру.
[0065]
Что касается миРНК, все нуклеиновые кислоты, составляющие миРНК, или их часть, могут представлять собой природные нуклеиновые кислоты или модифицированные нуклеиновые кислоты при условии, что такие нуклеиновые кислоты обладают активностью подавления экспрессии гена-мишени.
[0066]
миРНК в соответствии с настоящим изобретением необязательно должна представлять собой пару двухцепочечных РНК к последовательности-мишени, и она может представлять собой смесь множества ("множество" конкретно не ограничено, а предпочтительно относится к небольшому количеству приблизительно из 2-5) двухцепочечных РНК к области, которая включает последовательность-мишень. В этом отношении специалисты в данной области могут соответствующим образом создавать миРНК в качестве смеси нуклеиновых кислот, соответствующей последовательности-мишени, с использованием коммерчески доступного устройства для синтеза нуклеиновых кислот и фермента DICER; и, что касается синтеза желаемой РНК, обычно можно использовать организацию, осуществляющую синтез. Следует отметить, что миРНК в соответствии с настоящим изобретением включает так называемый "коктейль миРНК". Более того, следует отметить, что миРНК в соответствии с настоящим изобретением является такой, что не все нуклеотиды должны быть рибонуклеотидами (РНК). В частности, в рамках настоящего изобретения один или множество рибонуклеотидов, составляющих миРНК, могут представлять собой соответствующий дезоксирибонуклеотид. Термин "соответствующий" относится к одному и тому же типу оснований (аденин, гуанин, цитозин, тимин (урацил)), хотя структура сахарной части отличается. Например, дезоксирибонуклеотид, соответствующий рибонуклеотиду, имеющему аденин, относится к дезоксирибонуклеотиду, имеющему аденин.
[0067]
Более того, ДНК (вектор), которая может экспрессировать упомянутую выше РНК в соответствии с настоящим изобретением, также включена в предпочтительный вариант осуществления нуклеиновой кислоты, которая может подавлять экспрессию TGF-бета и т.п. Например, ДНК (вектор), которая может экспрессировать упомянутую выше двухцепочечную РНК в соответствии с настоящим изобретением, является такой ДНК, которая имеет структуру, в которой ДНК, кодирующая одну из цепей двухцепочечной РНК, и ДНК, кодирующая другую из цепей двухцепочечной РНК, связаны с промотором, так что каждая из ДНК может экспрессироваться. Упомянутую выше ДНК в соответствии с настоящим изобретением могут соответствующим образом получать специалисты в данной области с использованием обычных способов генной инженерии. Более конкретно, экспрессирующий вектор в соответствии с настоящим изобретением можно получать путем встраивания соответствующим образом ДНК, кодирующей РНК в соответствии с настоящим изобретением, в различные общеизвестные экспрессирующие векторы.
[0068]
В рамках настоящего изобретения в качестве нуклеиновой кислоты для подавления экспрессии гена-мишени можно использовать модифицированную нуклеиновую кислоту. Модифицированная нуклеиновая кислота означает нуклеиновую кислоту, в которой модификация находится в нуклеозиде (часть основания, часть сахара) и/или межнуклеозидном участке связывания, и которая имеет структуру, отличающуюся от структуры природной нуклеиновой кислоты. "Модифицированный нуклеозид", который составляет модифицированную нуклеиновую кислоту, включает, например, абазический нуклеозид; арабинонуклеозид, 2’-дезоксиуридин, альфа-дезоксирибонуклеозид, бета-L-дезоксирабонуклеозид, нуклеозид, имеющий другую модификацию сахара; пептидно-нуклеиновую кислоту (PNA), пептидно-нуклеиновую кислоту, связывающую фосфатную группу (PHONA), замкнутую нуклеиновую кислоту (LNA), морфолино-нуклеиновую кислоту и т.п. Упомянутый выше нуклеозид, имеющий модификацию, включает нуклеозиды, имеющие замещенную пентозу, такую как 2’-O-метилрибоза, 2’-дезокси-2’-фторрибоза, 3’-O-метилрибоза и т.п.; 1’,2’-дезоксирибозу; арабинозу; замещенный сахар арабинозу; и гексозу, и модификацию сахара альфа-аномера. Эти нуклеозиды могут представлять собой модифицированное основание, в котором часть основания является модифицированной. Такие модифицированные основания включают, например, пиримидин, такой как 5-гидроксицитозин, 5-фторурацил, 4-тиоурацил и т.п.; пурин, такой как 6-метиладенин, 6-тиогуанозин и т.п.; и другие гетероциклические основания.
[0069]
"Модифицированное межнуклеозидное связывание", которое обеспечивает модифицированную нуклеиновую кислоту, включает неприродное межнуклеозидное связывание, такое как алкильный линкер, глицерильный линкер, аминолинкер, связывание через полиэтиленгликоль, связывание между метилфосфонатными нуклеозидами; метилфосфонотиоат, сложный фосфотриэфир, сложный фосфотиотриэфир, фосфоротиоат, фосфородитиоат, пролекарство в виде сложного триэфира, сульфон, сульфонамид, сульфамат, формацеталь, N-метилгидроксиламин, карбонат, карбамат, морфолино, боранофосфонат, фосфорамидат и т.п.
[0070]
Последовательность нуклеиновой кислоты, включенная в двухцепочечную миРНК в соответствии с настоящим изобретением, может включать миРНК, направленную на представителя TGF-бета или других представителей передачи сигнала TGF-бета и т.п.
[0071]
Также является возможным введение нуклеиновой кислоты или средства в соответствии с настоящим изобретением в фосфолипидную эндоплазматическую сеть, такую как липосомы и т.п., и введение эндоплазматической сети. Эндоплазматическую сеть, в которой находятся миРНК или кшРНК, можно вводить в заданную клетку с использованием способа липофекции. Затем полученную клетку вводят системно, например внутривенно, внутриартериально и т.п. Эндоплазматическую сеть также можно вводить локально в требуемую область глаза и т.п. Хотя миРНК проявляет в высокой степени превосходный эффект посттранскрипционного подавления in vitro, она быстро деградируется in vivo вследствие нуклеазной активности в сыворотке крови. Таким образом, длительность действия является ограниченной и вследствие этого существует необходимость в разработке лучшей и более эффективной системы доставки. В качестве одного примера в Ochiya, T et al., Nature Med., 5:707-710, 1999, Curr. Gene Ther., 1: 31-52, 2001 описано следующее: биосовместимый материал ателоколлаген смешивают с нуклеиновой кислотой с получением комплекса, действие которого состоит в защите нуклеиновой кислоты от деградирующего фермента в живом организме и который является носителем, являющимся чрезвычайно пригодным в качестве носителя для миРНК. Хотя можно использовать такую форму, способ введения нуклеиновой кислоты или лекарственного средства в соответствии с настоящим изобретением не ограничивается этим способом. По существу, вследствие быстрой деградации под действием фермента, деградирующего нуклеиновую кислоту, в сыворотке крови живого организма, становится возможным достижение длительного эффекта. Например, в Takeshita F. PNAS. (2003) 102(34) 12177-82, Minakuchi Y Nucleic Acid Research (2004) 32 (13) e109 описано следующее: ателоколлаген, происходящий из кожи животных семейства бычьих, образует комплекс с нуклеиновой кислотой, который обладает действием защиты нуклеиновой кислоты от деградирующего фермента в живом организме и который является чрезвычайно пригодным в качестве носителя для миРНК. Такой способ можно использовать.
[0072]
Как используют в рамках изобретения, "средство" используют в широком значении и оно может представлять собой любое вещество или другие элементы (например, энергию, такую как свет, радиационное излучение, тепло и электричество) при условии, что намеченная цель может быть достигнута. Примеры такого вещества включают, но не ограничиваются ими, белки, полипептиды, олигопептиды, пептиды, полинуклеотиды, олигонуклеотиды, нуклеотиды, нуклеиновые кислоты (например, включая ДНК, такую как кДНК и геномную НК, и РНК, такую как мРНК), полисахариды, олигосахариды, жиры, органические низкомолекулярные соединения (например, гормоны, лиганды, передающие информацию вещества, органические низкомолекулярные соединения, молекулы, синтезированные комбинаторной химией, низкомолекулярные соединения, которые можно использовать в качестве фармацевтического продукта (например, низкомолекулярный лиганд) и т.п.) и их составные молекулы. Репрезентативные примеры средства, специфичного к полинуклеотиду, включают, но не ограничиваются ими, полинуклеотид, обладающий комплементарностью с определенной гомологией последовательности (например, 70% или более идентичность последовательности) относительно последовательности полинуклеотида, полипептид, такой как фактор транскрипции, связывающийся с промоторной областью. Репрезентативные примеры средства, специфичного к полипептиду, включают, но не ограничиваются ими, антитело, специфически направленное на полипептид или его производное или аналог (например, одноцепочечное антитело), специфический лиганд или рецептор, когда полипептид представляет собой рецептор или лиганд, и субстрат, когда полипептид представляет собой фермент.
[0073]
Как используют в рамках изобретения, "заболевание, нарушение или состояние, ассоциированное с аномалией внеклеточного матрикса (ECM) в эндотелии роговицы" относится к заболеваниям, нарушениям или состояниям, ассоциированным с аномалией внеклеточного матрикса (ECM) при заболеваниях, нарушениях или состояниях эндотелия роговицы. По существу, примеры могут включать нарушения, связанные с эндотелиальной дистрофией роговицы Фукса, птеригий, аллергические заболевания, кератит, язву роговицы и т.п.
[0074]
Как используют в рамках изобретения, "нарушение, обусловленное с эндотелиальной дистрофией роговицы Фукса," относится к любым нарушениям, связанным с эндотелиальной дистрофией роговицы Фукса. Среди них настоящее изобретение, в частности, направлено на нарушения, обусловленные аномалией внеклеточного матрикса (ECM), однако оно не ограничивается этими нарушениями. Примеры нарушений, обусловленных эндотелиальной дистрофией роговицы Фукса, которые ассоциированы с такой аномалией внеклеточного матрикса (ECM), включают, но не ограничиваясь ими, фотофобию, неясное зрение, нарушение зрения, боль в глазу, слезоотделение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, снижение контрастности, блики, отек стромы роговицы, буллезную кератопатию, помутнение роговицы и т.п.
(Основные способы)
[0075]
Молекулярно-биологические способы, биохимические способы и микробиологические способы, используемые в настоящем описании, хорошо известны и широко используются в данной области, и описаны, например, в Sambrook J. et al., (1989). Molecular Cloning: A Laboratory Manual, Cold Spring Harbor и the 3rd Ed. (2001); Ausubel, F. M. (1987). Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience; Ausubel, F. M. (1989). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience; Innis, M. A. (1990). PCR Protocols: A Guide to Methods and Applications, Academic Press; Ausubel, F. M. (1992). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Greene Pub. Associates; Ausubel, F. M. (1995). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Greene Pub. Associates; Innis, M. A. et al. (1995). PCR Strategies, Academic Press; Ausubel, F. M. (1999). Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Wiley, and annual updates; Sninsky, J. J. et al., (1999). PCR Applications: Protocols for Functional Genomics, Academic Press, Gait, M. J. (1985). Oligonucleotide Synthesis: A Practical Approach, IRLPress; Gait, M. J. (1990). Oligonucleotide Synthesis: A Practical Approach, IRL Press; Eckstein, F. (1991). Oligonucleotides and Analogues: A Practical Approach, IRL Press; Adams, R. L. et al. (1992). The Biochemistry of the Nucleic Acids, Chapman & Hall; Shabarova, Z. et al. (1994). Advanced Organic Chemistry of Nucleic Acid, Weinheim; Blackburn, G. M. et al. (1996). Nucleic Acids in Chemistry и Biology, Oxford University Press; Hermanson, G. T. (1996). Bioconjugate Techniques, Academic Press, Experimental Medicine, separate volume, "Gene Introduction & Expression Analysis Experimental Method" Yodosha Co., Ltd., 1997, и т.п. Что касается эндотелиальных клеток роговицы, хорошо известен отчет Nancy Joyce et al., {Joyce, 2004 #161} {Joyce, 2003 #7}, хотя исследователи в настоящее время осуществляют эффективные способы культивирования путем проведения трансформации по принципу культивирования фибробластов через длительное культивирование и субкультивирование, как описано выше. Что касается этого, их соответствующие части (могут представлять собой целый документ) включены в настоящее описание в качестве ссылок.
Описание предпочтительных вариантов осуществления
[0076]
Далее описаны предпочтительные варианты осуществления, однако следует понимать, что варианты осуществления являются иллюстрацией настоящего изобретения, и объем настоящего изобретения не ограничивается такими предпочтительными вариантами осуществления. Также следует понимать, что специалисты в данной области могут без труда осуществить изменение, замену и т.п. в рамках объема настоящего изобретения относительно представленных ниже предпочтительных примеров.
(Лекарственное средство для лечения или предупреждения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы, содержащее средство, ингибирующее сигнал TGF-бета)
[0077]
В одном аспекте настоящее изобретение относится лекарственным средствам для лечения или предупреждения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM), в эндотелии роговицы, причем лекарственные средства содержат средство, ингибирующее сигнал TGF-бета. В рамках настоящего изобретения было обнаружено, что заболевание, нарушение или состояние, ассоциированное с ECM в эндотелии роговицы, и, неожиданно, с аномалией ECM, можно уменьшать или устранять путем введения средства, ингибирующего сигнал TGF-бета. Таким образом, может быть понятно, что, исходя из предшествующей информации, нельзя ожидать применения такого средства, ингибирующего сигнал TGF-бета, для лечения или предупреждения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы.
[0078]
В предпочтительном варианте осуществления заболевание, нарушение или состояние в рамках настоящего изобретения представляют собой нарушение, обусловленное эндотелиальной дистрофией роговицы Фукса. Для эндотелиальной дистрофии роговицы Фукса в настоящее время отсутствует радикальный способ или технология лечения, и терапия эндотелиальной дистрофии роговицы Фукса основана на кератопластике. Поскольку настоящее изобретение может лечить аномалию внеклеточного матрикса (ECM), которая вызывает одну важную аномалию или нарушение при эндотелиальной дистрофии роговицы Фукса, следует понимать, что оно является полезным для лечения или предупреждения эндотелиальной дистрофии роговицы Фукса.
[0079]
В одном определенном варианте осуществления заболевания, нарушения или состояния в рамках настоящего изобретения включают фотофобию, неясное зрение, нарушение зрения, боль в глазу, слезоотделение, гиперемию, боль, буллезную кератопатию, неприятные ощущения в глазу, снижение контрастности, блики, отек стромы роговицы, буллезную кератопатию, помутнение роговицы при эндотелиальной дистрофии роговицы Фукса.
[0080]
Субъект введения (трансплантации) лекарственного средства или способа в соответствии с настоящим изобретением включает млекопитающих (например, человека, мышей, крыс, хомячков, кроликов, кошек, собак, коров, лошадей, овец, обезьян и т.п.), и субъектом предпочтительно являются приматы, и особенно предпочтительно люди. Терапия эндотелия роговицы у приматов не достигала существенных результатов, и с этой точки зрения настоящее изобретение относится инновационному терапевтическому способу и лекарственному средству.
[0081]
Каскады передачи сигнала TGF-бета ориентировочно классифицируют на систему Smad2/3 через ALK4, 5 или 7 и систему Smad1/5/8 через ALK1, 2, 3 или 6. Хорошо известно, что обе из них связаны с фиброзом (J. Massagu’e, Annu. Rev. Biochem. 1998,67:753-91; Vilar JMG, Jansen R, Sander C (2006) PLoS Comput Biol 2(1):e3; Leask, A., Abraham, D. J. FASEB J. 18, 816-827 (2004); Coert Margadant & Arnoud Sonnenberg EMBO reports (2010) 11, 97-105; Joel Rosenbloom et al., Ann Intern Med. 2010; 152:159-166.). Также известно, что BMP-7 может подавлять сигнал TGF-бета, подавляя фиброз (помимо указанной выше литературы, Ralf Weiskirchen, et al., Frontiers in Bioscience 14, 4992-5012, June 1, 2009; Elisabeth M Zeisberg et al., Nature Medicine 13, 952-961 (2007); Michael Zeisberg et al., Nature Medicine 9, 964-968 (2003)). Однако в этих источниках литературы описано вовлечение TGF-бета в сифилитический интерстициальный кератит, который является особенным заболеванием, или подобное состояние, сопровождающее ткань в форме мембраны, фактически состоящую из внеклеточного субстрата, такого как коллаген и т.п., вследствие тяжелого нарушения, созданного искусственно. Однако трудно ожидать терапевтического эффекта из этого описания. Кроме того, в этих источниках литературы показано, что фиброз в момент тяжелого нарушения роговицы вызывается IL-1-бета или активацией каскада p38 MAPK, хотя в них показано, что при использовании кролика фиброз, наблюдаемый, когда тяжелое воспаление в живом организме происходит вследствие чрезмерного обморожения, сопровождается активацией p38 MAPK, и ингибирующее средство может частично подавлять фиброз. Эта информация показывает, что активация p38 MAPK сопровождает такое состояние, при котором происходит очень мощное воспаление в живом организме и ткани в виде мембраны, состоящей из внеклеточного субстрата. В них не упомянуто, что средство, ингибирующее сигнал TGF-бета, является эффективным для лечения или предупреждения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы при эндотелиальной дистрофии роговицы Фукса и т.п. В них не указано ничего о поддержании нормального состояния. Как описано выше, ранее считали трудным культивирование клеток эндотелия роговицы при сохранении нормальных функций. Наконец в предшествующих сообщениях, заболевание, нарушение или состояние, обусловленное аномалией внеклеточного матрикса (ECM) в эндотелии при эндотелиальной дистрофии роговицы Фукса и т.п., нельзя лечить или предупреждать, не говоря о том, что не считалось возможным лечить или предупреждать заболевание, нарушение или состояние, обусловленное аномалией внеклеточного матрикса (ECM) в эндотелии роговицы при эндотелиальной дистрофии роговицы Фукса и т.п. путем подавления каскадов передачи сигнала TGF-бета.
[0082]
Средство, ингибирующее сигнал TGF-бета, используемое в рамках настоящего изобретения, может представлять собой любое средство при условии, что средство ингибирует каскад передачи сигнала TGF-бета. Кроме того, каскад передачи сигнала TGF-бета, подлежащий ингибированию, может быть ассоциирован с фактором, ассоциированным с любым сигналом при условии, что такой каскад передачи сигнала в конечном итоге оказывает эффект, сходный (противоположный в случае ингибирующего средства, антагониста и т.п.) с каскадом передачи сигнала TGF-бета, таким как BMP-7, в дополнение к каскадам передачи сигнала, с которыми TGF-бета и рецептор TGF-бета прямо ассоциированы, как хорошо известно.
[0083]
В рамках настоящего изобретения является возможным включение только средства, ингибирующего сигнал TGF-бета, и также при необходимости является возможным включение нескольких его типов в комбинации с друг с другом.
[0084]
В одном варианте осуществления средство, ингибирующее сигнал TGF-бета, включает по меньшей мере один из антагониста TGF-бета, антагониста рецептора TGF-бета или ингибирующего Smad3 средства, ингредиентов, проиллюстрированных в других частях настоящего описания, их фармацевтически приемлемых солей или сольватов, или сольватов фармацевтически приемлемых солей. Что касается антагониста TGF-бета, антагониста рецептора TGF-бета, и ингибирующего Smad3 средства, можно использовать любое из этих средств, описанных в других частях настоящего описания.
[0085]
В одном варианте осуществления средства, ингибирующие сигнал TGF-бета, которые можно использовать в рамках настоящего изобретения, включают по меньшей мере одно из SB431542 (4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)-1H-имидазол-2-ил]бензамид), BMP-7, антитела против TGF-бета, антитела против рецептора TGF-бета, миРНК к TGF-бета, миРНК к рецептору TGF-бета, антисмыслового олигонуклеотида к TGF-бета, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1H-пирроло[2,3-b]пиридин-3-ил-проп-2-еноил))-1,2,3,4-тетрагидроизохинолона, A83-01 (3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1H-пиразол-1-карботиоамида), ингибитора TLK StemoleculeTM (2-(3-(6-метилпиридин-2-ил)-1H-пиразол-4-ил)-1,5-нафтиридин), ингибитора BMP StemoleculeTM LDN-193189 (6-(4-(пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил)пиразолo[1,5-a]пиримидин), SD-208 (2-(5-хлор-2-фторфенил)-4-[(4-пиридинил)амино]птеридин), LY364947 (4-[3-(2-пиридинил)-1H-пиразол-4-ил]-хинолин), компонентов, проиллюстрированных в других частях настоящего описания, их фармацевтически приемлемых солей или сольватов, или сольватов фармацевтически приемлемых солей. Следует отметить, что упомянутые выше антитела могут представлять собой, но не ограничиваться ими, нейтрализующие антитела. Без связи с какой-либо теорией, эффект лечения или предупреждения заболевания, нарушения или состояния, ассоциированных с аномалией внеклеточного матрикса (ECM) в эндотелии роговицы при эндотелиальной дистрофии роговицы Фукса и т.п. наблюдают как для SB431542, который достигает эффекта через Smad2/3 (родственные для ALK4, 5 и 7), так и для BMP-7, который достигает эффекта через Smad1/5/8 (родственные для ALK1, 2, 3 и 6). Таким образом, понятно, что даже если они являются средствами, ингибирующими сигнал TGF-бета, в каком-либо из этих каскадов, может быть достигнут эффект настоящего изобретения.
[0086]
В предпочтительном варианте осуществления средства, ингибирующие сигнал TGF-бета, используемые в рамках настоящего изобретения, включают SB431542 (4-[4-(1, 3-бензодиоксол-5-ил)2-пиридинил)-1H-имидазол-2-ил]бензамид). Причина этого состоит в том, что достигалось улучшение при заболевании, нарушении или состоянии, обусловленном аномалией внеклеточного матрикса (ECM) в эндотелии роговицы при эндотелиальной дистрофии роговицы Фукса и т.п. В предпочтительном варианте осуществления SB431542 включают в концентрации от приблизительно 0,1 мкм до приблизительно 10 мкМ при применении, предпочтительно включают в концентрации от приблизительно 1 мкМ до приблизительно 10 мкМ при применении, и, кроме того, предпочтительно включают в концентрации приблизительно 1 мкМ при применении.
[0087]
Концентрация средства, ингибирующего сигнал TGF-бета, используемого в рамках настоящего изобретения, обычно составляет приблизительно от 0,1 до 100 мкмоль/л, предпочтительно приблизительно от 0,1 до 30 мкмоль/л, и более предпочтительно приблизительно 1 мкмоль/л; когда используют несколько их типов, концентрацию можно менять соответствующим образом, и примеры других диапазонов концентраций обычно представляют собой приблизительно от 0,001 до 100 мкмоль/л, предпочтительно приблизительно от 0,01 до 75 мкмоль/л, приблизительно от 0,05 до 50 мкмоль/л, приблизительно от 1 до 10 мкмоль/л, приблизительно от 0,01 до 10 мкмоль/л, приблизительно от 0,05 до 10 мкмоль/л, приблизительно от 0,075 до 10 мкмоль/л, приблизительно от 0,1 до 10 мкмоль/л, приблизительно от 0,5 до 10 мкмоль/л, приблизительно от 0,75 до 10 мкмоль/л, приблизительно от 1,0 до 10 мкмоль/л, приблизительно от 1,25 до 10 мкмоль/л, приблизительно от 1,5 до 10 мкмоль/л, приблизительно от 1,75 до 10 мкмоль/л, приблизительно от 2,0 до 10 мкмоль/л, приблизительно от 2,5 до 10 мкмоль/л, приблизительно от 3,0 до 10 мкмоль/л, приблизительно от 4,0 до 10 мкмоль/л, приблизительно от 5,0 до 10 мкмоль/л, приблизительно от 6,0 до 10 мкмоль/л, приблизительно от 7,0 до 10 мкмоль/л, приблизительно от 8,0 до 10 мкмоль/л, приблизительно от 9,0 до 10 мкмоль/л, приблизительно от 0,01 до 50 мкмоль/л, приблизительно от 0,05 до 5,0 мкмоль/л, приблизительно от 0,075 до 5,0 мкмоль/л, приблизительно от 0,1 до 5,0 мкмоль/л, приблизительно от 0,5 до 5,0 мкмоль/л, приблизительно от 0,75 до 5,0 мкмоль/л, приблизительно от 1,0 до 5,0 мкмоль/л, приблизительно от 1,25 до 5,0 мкмоль/л, приблизительно от 1,5 до 5,0 мкмоль/л, приблизительно от 1,75 до 5,0 мкмоль/л, приблизительно от 2,0 до 5,0 мкмоль/л, приблизительно от 2,5 до 5,0 мкмоль/л, приблизительно от 3,0 до 5,0 мкмоль/л, приблизительно от 4,0 до 5,0 мкмоль/л, приблизительно от 0,01 до 3,0 мкмоль/л, приблизительно от 0,05 до 3,0 мкмоль/л, приблизительно от 0,075 до 3,0 мкмоль/л, приблизительно от 0,1 до 3,0 мкмоль/л, приблизительно от 0,5 до 3,0 мкмоль/л, приблизительно от 0,75 до 3,0 мкмоль/л, приблизительно от 1,0 до 3,0 мкмоль/л, приблизительно от 1,25 до 3,0 мкмоль/л, приблизительно от 1,5 до 3,0 мкмоль/л, приблизительно от 1,75 до 3,0 мкмоль/л, приблизительно от 2,0 до 3,0 мкмоль/л, приблизительно от 0,01 до 1,0 мкмоль/л, приблизительно от 0,05 до 1,0 мкмоль/л, приблизительно от 0,075 до 1,0 мкмоль/л, приблизительно от 0,1 до 1,0 мкмоль/л, приблизительно от 0,5 до 1,0 мкмоль/л, приблизительно от 0,75 до 1,0 мкмоль/л, приблизительно от 0,09 до 35 мкмоль/л и приблизительно от 0,09 до 3,2 мкмоль/л, и более предпочтительно может включать, но не ограничиваться ими, приблизительно от 0,05 до 1,0 мкмоль/л, приблизительно от 0,075 до 1,0 мкмоль/л, приблизительно от 0,1 до 1,0 мкмоль/л, приблизительно от 0,5 до 1,0 мкмоль/л и приблизительно от 0,75 до 1,0 мкмоль/л.
[0088]
В предпочтительном варианте осуществления используемое средство, ингибирующее сигнал TGF-бета, включает 4-[4-(1,3-бензодиоксол-5-ил)2-пиридинил)-1H-имидазол-2-ил]бензамид или его фармацевтически приемлемую соль.
[0089]
В другом предпочтительном варианте осуществления средство, ингибирующее сигнал TGF-бета, используемое в рамках настоящего изобретения, включает BMP-7. Это является следствием того, что фиброз подавлялся, и, более того, было показано, что белок, отвечающий за нормальные функции, сохранялся, и трансплантат приматам был переносимым. В предпочтительном варианте осуществления BMP-7 включают в концентрации от приблизительно 10 нг/мл до приблизительно 1000 нг/мл при применении, и более предпочтительно, включают в концентрации от приблизительно 100 нг/мл до приблизительно 1000 нг/мл при применении. BMP-7 может быть включен в концентрации приблизительно 100 нг/мл при применении, или он может быть включен в концентрации приблизительно 1000 нг/мл.
[0090]
Лекарственное средство для лечения или предупреждения в соответствии с настоящим изобретением может содержать дополнительный медицинский компонент. Репрезентативные примеры таких медицинских продуктов включают средства, ингибирующие Rho-киназу, и стероиды. Без связи с какой-либо теорией, из-за того, что включение средства, ингибирующего Rho-киназау, стимулирует адгезию клеток эндотелия роговицы, предупреждая слущивание клеток, оно обеспечивает образование слоя эндотелиальных клеток роговицы, имеющего хорошую морфологию и высокую клеточную плотность и, тем самым, эффект средства, ингибирующего сигнал TGF-β, может быть усилен. В рамках настоящего изобретения один тип средства, ингибирующего Rho-киназу, может быть включен отдельно, или несколько их типов можно использовать в комбинации и включать при необходимости.
[0091]
Примеры средств, ингибирующих Rho-киназу, которые можно использовать в рамках настоящего изобретения, включают соединения, описанные в следующих документах: патент США № 4678783, патент Японии № 3421217, международная публикация № WO 95/28387, международная публикация № WO 99/20620, международная публикация № WO 99/61403, международная публикация № WO 02/076976, международная публикация № WO 02/076977, международная публикация № WO 2002/083175, международная публикация № WO 02/100833, международная публикация № WO 03/059913, международная публикация № WO 03/062227, международная публикация № WO 2004/009555, международная публикация № WO 2004/022541, международная публикация № WO 2004/108724, международная публикация № WO 2005/003101, международная публикация № WO 2005/039564, международная публикация № WO 2005/034866, международная публикация № WO 2005/037197, международная публикация № WO 2005/037198, международная публикация № WO 2005/035501, международная публикация № WO 2005/035503, международная публикация № WO 2005/035506, международная публикация № WO 2005/080394, международная публикация № WO 2005/103050, международная публикация № WO 2006/057270, международная публикация № WO 2007/026664 и т.п. Каждое из таких соединений можно получать способами, описанными в документах, в которых описаны соответствующие соединения. Конкретные примеры могут включать 1-(5-изохинолинсульфонил)гомопиперазин или его соль (например, фасудил (1-(5-изохинолинсульфонил)гомопиперазин)), (R)-(+)-транс-(4-пиридил)-4-(1-аминоэтил)циклогексанкарбоксамид или его соль (например, Y-27632 ((R)-(+)-транс-(4-пиридил)-4-(1-аминоэтил)циклогексанкарбоксамид дигидрохлорид моногидрат) и т.п.) и т.п.
[0092]
Концентрация средства, ингибирующего Rho-киназу, в рамках настоящего изобретения обычно составляет от приблизительно 1 до 100 мкмоль/л, предпочтительно, от приблизительно 5 до 20 мкмоль/л, и более предпочтительно от приблизительно 10 мкмоль/л; когда используют нескольких типов, концентрацию можно изменять соответствующим образом, и примеры других диапазонов концентраций обычно представляют собой от приблизительно 0,001 до 100 мкмоль/л, предпочтительно, от приблизительно 0,01 до 75 мкмоль/л, от приблизительно 0,05 до 50 мкмоль/л, от приблизительно 1 до 10 мкмоль/л, от приблизительно 0,01 до 10 мкмоль/л, от приблизительно 0,05 до 10 мкмоль/л, от приблизительно 0,075 до 10 мкмоль/л, от приблизительно 0,1 до 10 мкмоль/л, от приблизительно 0,5 до 10 мкмоль/л, от приблизительно 0,75 до 10 мкмоль/л, от приблизительно 1,0 до 10 мкмоль/л, от приблизительно 1,25 до 10 мкмоль/л, от приблизительно 1,5 до 10 мкмоль/л, от приблизительно 1,75 до 10 мкмоль/л, от приблизительно 2,0 до 10 мкмоль/л, от приблизительно 2,5 до 10 мкмоль/л, от приблизительно 3,0 до 10 мкмоль/л, от приблизительно 4,0 до 10 мкмоль/л, от приблизительно 5,0 до 10 мкмоль/л, от приблизительно 6,0 до 10 мкмоль/л, от приблизительно 7,0 до 10 мкмоль/л, от приблизительно 8,0 до 10 мкмоль/л, от приблизительно 9,0 до 10 мкмоль/л, от приблизительно 0,01 до 50 мкмоль/л, от приблизительно 0,05 до 5,0 мкмоль/л, от приблизительно 0,075 до 5,0 мкмоль/л, от приблизительно 0,1 до 5,0 мкмоль/л, от приблизительно 0,5 до 5,0 мкмоль/л, от приблизительно 0,75 до 5,0 мкмоль/л, от приблизительно 1,0 до 5,0 мкмоль/л, от приблизительно 1,25 до 5,0 мкмоль/л, от приблизительно 1,5 до 5,0 мкмоль/л, от приблизительно 1,75 до 5,0 мкмоль/л, от приблизительно 2,0 до 5,0 мкмоль/л, от приблизительно 2,5 до 5,0 мкмоль/л, от приблизительно 3,0 до 5,0 мкмоль/л, от приблизительно 4,0 до 5,0 мкмоль/л, от приблизительно 0,01 до 3,0 мкмоль/л, от приблизительно 0,05 до 3,0 мкмоль/л, от приблизительно 0,075 до 3,0 мкмоль/л, от приблизительно 0,1 до 3,0 мкмоль/л, от приблизительно 0,5 до 3,0 мкмоль/л, от приблизительно 0,75 до 3,0 мкмоль/л, от приблизительно 1,0 до 3,0 мкмоль/л, от приблизительно 1,25 до 3,0 мкмоль/л, от приблизительно 1,5 до 3,0 мкмоль/л, от приблизительно 1,75 до 3,0 мкмоль/л, от приблизительно 2,0 до 3,0 мкмоль/л, от приблизительно 0,01 до 1,0 мкмоль/л, от приблизительно 0,05 до 1,0 мкмоль/л, от приблизительно 0,075 до 1,0 мкмоль/л, от приблизительно 0,1 до 1,0 мкмоль/л, от приблизительно 0,5 до 1,0 мкмоль/л, от приблизительно 0,75 до 1,0 мкмоль/л, от приблизительно 0,09 до 35 мкмоль/л, и от приблизительно 0,09 до 3,2 мкмоль/л, и более предпочтительно могут включать, но не ограничиваться ими, от приблизительно 0,05 до 1,0 мкмоль/л, от приблизительно 0,075 до 1,0 мкмоль/л, от приблизительно 0,1 до 1,0 мкмоль/л, от приблизительно 0,5 до 1,0 мкмоль/л и от приблизительно 0,75 до 1,0 мкмоль/л.
[0093]
Настоящее изобретение можно вводить в качестве глазных капель.
[0094]
Дозировка и частота введения варьируются в зависимости от симптомов, возраста, массы тела и форм введения. В случае применения в качестве глазного лосьона, например, для здоровых взрослых, состав, содержащий эффективный ингредиент в количестве приблизительно от 0,0001 до 0,1 масс./об.%, и предпочтительно приблизительно от 0,003 до 0,03 масс./об.%, можно вводить от 1до 10 раз в сутки, предпочтительно от 1 до 6 раз в сутки и более предпочтительно от 1 до 3 раз в сутки и в количестве в диапазоне приблизительно от 0,01 до 0,1 мл за один раз. Когда лекарственное средство в соответствии с настоящим изобретением вводят в переднюю камеру, можно использовать лекарственное средство в концентрации, составляющей от одной десятой до одной тысячной от упомянутых выше концентраций. Специалисты в данной области могут соответствующим образом выбирать тип и концентрацию средства, ингибирующего сигнал TGF-бета, средства, ингибирующего Rho-киназу и т.п. в зависимости от болезненного состояния.
[0095]
В другом аспекте настоящее изобретение относится к веществу, ингибирующему сигнал TGF-бета, для лечения или предупреждения нарушения, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы. Вещество, ингибирующее сигнал TGF-бета, может использоваться взаимозаменяемо со средством, ингибирующим сигнал TGF-бета. В этом применении, что касается аномалии внеклеточного матрикса (ECM) в эндотелии роговицы и средства, ингибирующего сигнал TGF-бета, можно использовать любые варианты осуществления, описанные в настоящем описании.
[0096]
В другом аспекте настоящее изобретение относится к способу лечения или предупреждения нарушения, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы у индивидуума, где способ включает стадию введения индивидууму эффективного количества средства, ингибирующего сигнал TGF-бета. В этом способе, что касается аномалии внеклеточного матрикса (ECM) в эндотелии роговицы и средства, ингибирующего сигнал TGF-бета, можно использовать любые варианты осуществления, описанные в настоящем описании.
[0097]
Субъект введения (трансплантации) лекарственного средства или способа в соответствии с настоящим изобретением включает млекопитающих (например, человека, мышей, крыс, хомячков, кроликов, кошек, собак, коров, лошадей, овец, обезьян и т.п.), и субъектом предпочтительно являются приматы, и особенно предпочтительно люди. Терапия эндотелия роговицы у приматов не достигала существенных результатов, и с этой точки зрения настоящее изобретение относится инновационному терапевтическому способу и лекарственному средству.
[0098]
Справочная литература, такая как научная литература, патенты и патентные заявки, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок в той же степени, как если бы весь документ был конкретно описан.
[0099]
Как описано выше, настоящее изобретение проиллюстрировано путем демонстрации предпочтительных вариантов осуществления для облегчения понимания. Настоящее изобретение проиллюстрировано ниже с помощью примеров. Упомянутая выше иллюстрация и представленные ниже примеры предоставлены не для цели ограничения настоящего изобретения, а только для цели иллюстрации. Таким образом, объем настоящего изобретения не ограничивается вариантами осуществления и примерами, конкретно описанными в настоящем описании, и ограничивается только объемом формулы изобретения.
Примеры
[0100]
Далее описаны примеры обычного культивирования клетки эндотелия роговицы в соответствии с настоящим изобретением. Если применимо, стандарты, предусмотренные Ministry of Health, Labour and Welfare, Ministry of Education, Culture, Sports, Science and Technology и т.п. использовали для обработки биологических образцов и т.п.; и если применимо, обработку проводили на основе Хельсинской декларации или этических кодов в соответствии с декларацией. Для донирования глаз, использованных для исследования, было получено согласие близких родственников всех умерших доноров. Настоящее исследование было одобрено экспертным советом организации Erlangen university (Doyle), глазного банка SightLifeTM (Seattle, WA) и соответствующих организаций.
[0101]
При эндотелиальной дистрофии роговицы Фукса эндотелиальные клетки роговицы приводят к гибели клеток и, когда оставшиеся эндотелиальные клетки роговицы не могут компенсировать насосную функцию и барьерную функцию, прозрачность роговицы не может сохраняться, что приводит к потере зрения вследствие помутнения роговицы. Кроме того, известно, что эндотелиальные клетки роговицы у пациентов с эндотелиальной дистрофией роговицы Фукса сверхпродуцируют внеклеточный матрикс, вызывая образование "капель" и утолщение десцеметовой оболочки. Поскольку образование "капель" и утолщение десцеметовой оболочки вызывают рассеяние света и т.п., они вызывают снижение остроты зрения, фотофобию, неясность зрения, что значительно ухудшает QOL пациентов. В качестве модели использовали иммобилизованную линию эндотелия роговицы (iFECD), происходящую из клеток пациентов с эндотелиальной дистрофией роговицы Фукса, и сравнивали с иммобилизованной линией клеток эндотелия роговицы (iHCEC), происходящей из клеток здоровых доноров, для выяснения причины продуцирования внеклеточного матрикса и для идентификации мишени терапии.
(Пример получения: создание модели линии иммобилизованных клеток эндотелия роговицы (iFECD), полученных от пациента с эндотелиальной дистрофией роговицы Фукса)
[0102]
В настоящем примере получали линии иммобилизованных клеток эндодтелия роговицы (iFECD) из клеток эндотелия роговицы, полученных от пациента с эндотелиальной дистрофией роговицы Фукса.
Способ культивирования
[0103]
Клетки эндотелия роговицы для исследования механически счищали с базальной мембраны роговицы, приобретенной в Глазном банке Сиэтла. После использования коллагеназы для открепления и сбора эндотелиальных клеток с базальной мембраны, клетки подвергали первичному культивированию. Что касается среды, в качестве базальной среды использовали среду Opti-MEM I Reduced-Serum Medium, Liquid (INVITROGEN, каталожный номер № 31985-070), в которую были добавлены 8% FBS (BIOWEST, каталожный номер № S1820-500), 200 мг/мл CaCl2⋅2H2O (SIGMA, каталожный номер № C7902-500G), 0,08% хондроитинсульфат (SIGMA, каталожный номер № C9819-5G), 20 мкг/мл аскорбиновой кислоты (SIGMA, каталожный номер № A4544-25G), 50 мкг/мл гентамицина (INVITROGEN, каталожный номер № 15710-064) и 5 нг/мл EGF (INVITROGEN, каталожный номер № PHG0311), и которая была акклиматизирована для фидерных клеток 3T3. Кроме того, клетки культивировали в базальной среде, в которую были добавлены SB431542 (1 мкмоль/л) и SB203580 (4−(4−фторфенил)−2−(4−метилсульфонилфенил)−5(4−пиридил)имидазол <4−[4−(4−фторфенил)−2−(4−метилсульфинилфенил)−1H−имидазол−5−ил]пиридин) (1 мкмоль/л) (обозначаемая как "акклиматизированная среда SB203580+SB431542+3T3").
Способ получения
[0104]
Эндотелиальные клетки роговицы от трех пациентов, у которых возникла буллезная керптопатия при клиническом диагнозе эндотелиальной дистрофии роговицы Фукса и которые подверглись трансплантации эндотелия роговицы (керптопластика десцеметовой оболочки=DMEK), получали при письменном согласии и одобрении Этического комитета. При DMEK, механически патологические эндотелиальные клетки роговицы и десцеметову оболочку, которая является базальной мембраной, счищали вместе и погружали в Optisol-GS (Bausch & Lomb Incorporated), который представляет собой раствор для хранения роговицы. После этого проводили обработку коллагеназой для ферментативного сбора эндотелиальных клеток роговицы, которые затем культивировали в среде, кондиционируемой SB203580+SB431542+3T3. Что касается культивируемых эндотелиальных клеток роговицы, полученных от пациентов с эндотелиальной дистрофией роговицы Фукса, гены большого T-антигена SV40 и hTERT амплифицировали способами ПЦР и вносили в лентивирусный вектор (pLenti6,3_V5−TOPO; Life Technologies Inc.). Затем лентивирусный вектор использовали для инфицирования клеток 293T (RCB2202; Riken Bioresource Center, Ibaraki, Japan) с помощью реагента для трансфекции (Fugene HD; Promega Corp., Madison, WI), вместе с тремя типами хелперных плазмид (pLP1, pLP2, pLP/VSVG; Life Technologies Inc.). Культуральный супернатант, содержащий вирусы, собирали через 48 часов после инфицирования. Его добавляли с использованием 5 мкг/мл полибрена в культуральные растворы культивируемых эндотелиальных клеток роговицы, полученных от пациентов с эндотелиальной дистрофией роговицы Фукса, для внесения генов большого T-антигена SV40 и hTERT. Проводили подтверждение с помощью изображений фазово-контрастной микроскопии иммобилизованной линии эндотелиальных клеток (iFECD), полученных от пациентов с эндотелиальной дистрофией роговицы Фукса. Эндотетиальные клетки роговицы, культивированные из роговицы для исследований, импортированной в качестве контроля из Глазного банка Сиэтла, иммобилизовывали сходным способом для получения иммобилизованной клеточной линии (iHCEC) нормальных эндотелиальных клеток роговицы. При изучении изображений, полученных с помощью фазово-контрастного микроскопа, линии иммобилизованных эндотелиальных клеток роговицы (iHCEC), полученной от здоровых доноров, и линии иммобилизованных клеток эндотелия роговицы (iFECD), как iHCEC, так и iFECD имеют однослойную многоугольную форму, как и нормальные эндотелиальные клетки роговицы. В случаях iHCEC и iFECD, SB431542, для которых поддерживающее культивирование осуществляли в DMEM+10%FBS, получали от TOCRIS Corporation (каталожный номер: 1614). SB203580 получали от CALBIOCHEM (каталожный номер: 559389).
(Пример получения 2: подтверждение нормальных функций линии иммобилизованных клеток роговицы (iFECD))
[0105]
В настоящем примере была подтверждена нормальная функция линии иммобилизованных клеток эндотелия (iFECD).
(Иммунное окрашивание Na+/K+-ATP-азой и ZO-1)
[0106]
Сначала для подтверждения нормальных функций линии иммобилизованных клеток эндотелия роговицы (iFECD) проводили иммунное окрашивание Na+/K+-ATP-азы и ZO-1. Это проводили для подтверждения насосной функции и барьерной функции, которые являются функциями эндотелиальных клеток роговицы. Каждый из Na+/K+-ATP-азы и ZO-1 проявляет нормальную насосную функцию и барьерную функцию, которые являются функциями эндотелиальных клеток роговицы. Способ является следующим.
(Способ наблюдения клеток, включающий окрашивание и т.п. (гистологический тест))
[0107]
Клетки наблюдали под фазово-контрастным микроскопом. Кроме того, после фиксации клеток ZO-1 и Na+/K+-ATP-азу использовали в качестве связанных с функцией маркеров для проведения иммунного окрашивания и наблюдения под флуоресцентным микроскопом. Для испытания с окрашиванием тканей культивируемые клетки помещают в Lab-TekTM Chamber SlidesTM (NUNC A/S, Roskilde, Дания), а затем фиксируют при комнатной температуре (RT) в течение 10 минут в 4% формальдегиде и инкубируют с 1% бычьим сывороточным альбумином (BSA) в течение 30 минут. В частности, культивируемые клетки на Lab-TekTM Chamber SlidesTM (NUNC A/S, Roskilde, Denmark) фиксируют при комнатной температуре в течение 10 минут в 4% формальдегиде, а затем инкубируют с 1% бычьим сывороточным альбумином (BSA) в течение 30 минут. Для исследования типа экспрессии клеток, ZO-1 (Zymed Laboratories, Inc., South San Francisco, CA), который представляет собой белок плотных контактов, и Na+/K+-ATP-азу (Upstate Biotec, Inc., Lake Placid, NY), которая представляет собой белок, связанный с насосной функцией, подвергали иммуногистохимическому анализу. ZO-1 и Na+/K+-ATP-азу использовали в качестве маркеров, связанных с функциями клеток. Окрашивание ZO-1 и Na+/K+-ATP-азы проводили с использованием разведений 1:200 поликлонального антитела против ZO-1 и моноклонального антитела против Na+/K+-ATP-азы, соответственно. Для вторичного антитела использовали разведение 1:2000 меченного Alexa Fluor (зарегистрированный торговый знак) 488 или меченного Alexa Fluor (зарегистрированный торговый знак) 594 антитела козы против IgG мыши (Life Technologies). Затем ядра клеток окрашивали DAPI (Vector Laboratories, Inc., Burlingame, CA) или PI (Sigma-Aldrich). Затем предметное стекло наблюдали под флуоресцентным микроскопом (TCS SP2 AOBS; Leica Microsystems, Welzlar, Германия).
[0108]
При наблюдении результата как iHCEC, так и iFECD экспрессировали Na+/K+-ATP-азу и ZO-1 во всех клетках, и было показано, что полученная иммобилизованная линия сохраняла нормальные функции.
[0109]
Кроме того, показаны изображения морфологического исследования iHCEC и iFECD под трансмиссионным электронным микроскопом. iHCEC и iFECD культивировали в DMEM на Transwell без сыворотки. Через одну неделю их фиксировали в состоянии монослоя и, когда морфологию наблюдали под трансмиссионным электронным микроскопом, было показано, что они представляют собой однослойные клетки, которые не показывали наличия морофологически заметной аномалии.
[0110]
Кроме того, известно, что клетки эндотелия роговицы у пациента с эндотелиальной дистрофией роговицы Фукса сверхпродуцирует матрикс, вызывая образование "капель" и утолщение десцеметовой оболочки. Таким образом, что касается экспрессии коллагена типа I, коллагена типа IV и фибронектина, которые представляют собой белки, составляющие внеклеточный матрикс, iHCEC и iFECD культивировали на чашках для культивирования и подвергали иммунному окрашиванию. Было показано, что в iFECD экспрессия коллагена типа I, коллагена типа IV и фибронектина была увеличена по сравнению с iHCEC. Кроме того, когда уровень экспрессии генов культивируемых iHCEC и iFECD исследовали способом ПЦР в реальном времени, было показано, что коллаген типа I и фибронектин имели значительно повышенный уровень экспрессии, и что имелась тенденция к стимуляции экспрессии коллагена типа IV.
[0111]
Проводили исследование того, осуществляют ли iFECD сверхпродукцию внеклеточного матрикса, аналогично эндотелию роговицы у пациентов с эндотелиальной дистрофией роговицы Фукса. iHCEC и iFECD культивировали в DMEM на Transwell без сыворотки, после чего их фиксировали в состоянии монослоя и окрашивали HE. Для iFECD было показано, что продуцировался значительно утолщенный внеклеточный матрикс по сравнению с iHCEC. Как описано выше, получали модельные клетки для заболевания, которые имеют характеристику сверхпродукции внеклеточного матрикса у пациентов с эндотелиальной дистрофией роговицы Фукса. Поскольку ожидается, что анализ с использованием модельных клеток для заболевания внесет вклад в понимание патологического состояния эндотелиальной дистрофии роговицы Фукса, в отношении которого существует множество неясных моментов, эти клетки использовали, чтобы попытаться разработать терапевтическое лекарственное средство от эндотелиальной дистрофии роговицы Фукса следующим образом.
(Пример 1: анализ с использованием ПЦР в реальном времени уровня экспрессии гена, связанного с эпителиально-мезенхимальным переходом (EMT), связанным с продукцией внеклеточного матрикса)
[0112]
Что касается iHCEC и iFECD, в настоящем примере показан результат анализа способом ПЦР в реальном времени уровня экспрессии гена, связанного с эпителиально-мезенхимальным переходом (EMT), связанным с продукцией внеклеточного матрикса.
(ПЦР в реальном времени)
[0113]
Способ ПЦР в реальном времени: кроме того, ПЦР проводили способом Taqman для Snail1, Snail2 или ZEB1 в соответствии со следующим способом. Зонд Taqman приобретали от INVITROGEN. Уровень мРНК коллагена типа I, коллагена типа IV и фибронектина исследовали способом ПЦР в реальном времени. RNEasy (QIAGEN, каталожный номер: 74106) использовали для экстракции тотальной РНК из клеток. Экстрагированную РНК подвергали реакции обратной транскрипции (42°C, 60 минут) с ReverTra Ace (TOYOBO Co., Ltd., каталожный номер: TRT-101), и коллаген типа I, коллаген типа IV и фибронектин амплифицировали с помощью реагента TaqMan Fast Advanced mastermix (Applied Biosystems) с использованием GAPDH в качестве внутреннего стандарта. В реакции ПЦР, которую проводили в системе для ПЦР в реальном времени StepOneTM (Applied Biosystems) использовали зонды (набор меченых праймеров, доступный от Applied Biosystems), описанные ниже.
Snail1, Hs00195591_ml SNAI1
Snail2, Hs00950344_ml SNAI2
ZEB1, Hs00232783_ml ZEB1
GAPDH TaqMan®pre, разработанный Assay Reagents Human GADPH (каталожный номер №: 4333764F).
Результат
[0114]
Результат показан на фиг.1. Как показано на фиг.1, когда уровень экспрессии гена, связанного с эпителиально-мезенхимальным переходом (EMT), связанным с продукцией внеклеточного матрикса, анализировали с использованием ПЦР в реальном времени, для Snail1 и ZEB1 было выявлено, что в iFECD их экспрессия была значительно увеличена по сравнению с iHCEC.
(Стимуляция экспрессии Snail1 и ZEB1 посредством TGF-бета)
[0115]
Для подтверждения того, что стимуляция экспрессии Snail1 и ZEB1 связана со сверхпродукцией внеклеточного матрикса, проводили стимуляцию посредством TGF-бета, о котором известно, что он ускоряет экспрессию Snail1 и ZEB1. Способ является таким, как описано ниже. iFECD и iHCEC культивировали в DMEM, содержавшей 10% эмбриональную телячью сыворотку, и культивировали в течение ночи в DMEM без 10% эмбриональной телячьей сыворотки. После этого экспрессию Snail1, ZEB1, коллагена типа I, коллагена типа IV, коллагена типа VIII, фибронектина исследовали способом ПЦР в реальном времени. Зонды, описанные ниже, использовали в реакции ПЦР, которую проводили в системе для ПЦР в реальном времени StepOneTM (Applied Biosystems).
Snail1: Hs00195591_ml SNAI1
ZEB1: Hs00232783_ml ZEB1
коллаген типа I: Hs00164004_ml COL1A1
коллаген типа IV: Hs00266327_ml COL4A1
коллаген типа VIII: Hs00697025_ml COL8A2
фибронектин: Hs01549976_ml FN1
GAPDH TaqMan®pre, разработанный Assay Reagents Human GADPH (cat №: 4333764F).
[0116]
Результат показан на фиг.2. Было подтверждено, что TGF-бета значительно повышает экспрессию Snail1 и ZEB1 в iFECD (A, B). Таким образом, когда уровень экспрессии гена белка, составляющего, внеклеточный матрикс, анализировали с использованием ПЦР в реальном времени, экспрессия коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина значительно ускорялась.
(Пример 2: ускорение продукции внеклеточного матрикса посредством iFECD с помощью TGF-бета)
[0117]
В данном примере проводили исследование того, ускоряется ли продукция внеклеточного матрикса в iFECD посредством TGF-бета.
[0118]
iHCEC и iFECD культивировали в DMEM на Transwell без сыворотки, и через одну неделю после этого их фиксировали в состоянии монослоя и окрашивали HE. Методика является такой, как описано ниже. Если требовалось, проводили депарафинизацию (например, чистым этанолом) и промывание водой, и образец погружали в гематоксилин из Omni на 10 минут. После этого, проводили промывание проточной водой и использовали аммиачную воду в течение 30 секунд для развития окраски. После этого, проводили промывание проточной водой в течение 5 минут, окрашивание в течение 2 минут разбавленным в десять раз раствором эозина гидрохлорида, дегидратацию, осветление и заливку. Было выявлено, что стимуляция iHCEC и iFECD посредством TGF-бета вызывала значительное утолщение внеклеточного матрикса. Кроме того, было выявлено, что в присутствии TGF-бета, iFECD продуцировали значительно утолщенный внеклеточный матрикс по сравнению с iHCEC.
[0119]
Это показывает, что в эндотелиальных клетках роговицы у пациентов с эндотелиальной дистрофией роговицы Фукса уровень экспрессии Snail1 и ZEB1 является высоким и что уровень продукции внеклеточного матрикса в ответ на стимуляцию TGF-бета является значительно более высоким, чем в эндотелиальных клетках роговицы здорового индивидуума.
(Пример 3: эффект подавления Snail1 и ZEB1 с использованием миРНК на продукцию внеклеточного матрикса)
[0120]
В настоящем примере эффект подавления Snail1 и ZEB1 с использованием миРНК на продукцию внеклеточного матрикса исследовали, чтобы продемонстрировать, что стимуляция экспрессии Snail1 и ZEB1 вызывает продукцию внеклеточного матрикса. Методика эксперимента описана ниже.
Способ
[0121]
iFECD и iHCEC высевали и инкубировали с Snail1 Stealth RNAi™ (Life Technologies Corp., Carlsbad, CA) или ZEB1 Stealth RNAi™ (Life Technologies Corp., Carlsbad, CA) и Lipofectamine™ RNAiMAX (Life Technologies Corp., Carlsbad, CA) при 37°C в течение 12 часов. В качестве контроля использовали случайную последовательность РНК-i. После этого клетки пассировали и использовали в эксперименте. Три типа каждой из Snail1 Stealth RNAi™ и ZEB1 Stealth RNAi™ использовали для проведения эксперимента, и репрезентативные примеры показаны в качестве результатов. Клетки, в которых был осуществлен нокдаун Snail1 или ZEB1 посредством миРНК, высевали и экспрессию Snail1, ZEB1, коллагена типа I, коллагена типа IV, коллагена типа VIII, и фибронектина исследовали способом ПЦР в реальном времени. В реакции ПЦР, которую проводили в системе для ПЦР в реальном времени StepOneTM (Applied Biosystems), использовали зонды, описанные ниже.
Материалы
миРНК
миРНК к Snail1 (SNAI1 HSS143995*, SNAI1 HSS143996, SNAI1 HSS143997)
миРНК к ZEB1 (ZEB1 HSS110548*, ZEB1 HSS110549, ZEB1 HSS186235)
Следует отметить, что миРНК, показанная в результате, была описана как *.
[0122]
Зонды в способе ПЦР в реальном времени
Snail1 Hs00195591_ml SNAI1
ZEB1, Hs00232783_ml ZEB1
Коллаген типа I, Hs00164004_ml COL1A1
Коллаген типа IV, Hs00266327_ml COL4A1
Коллаген типа VIII, Hs00697025_ml COL8A2
Фибронектин, Hs01549976_ml FN1
GAPDH TaqMan®pre, разработанный Assay Reagents Human GADPH (каталожный номер №: 4333764F).
Результат
[0123]
Результат показан на фиг.4. Было подтверждено, что миРНК подавляет экспрессию Snail1 и ZEB1 (A, F). Подавление экспрессии Snail1 или ZEB1 посредством миРНК значительно подавляло экспрессию коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина. Из этого результата было обнаружено, что ZEB1 или Snail1 подавляют экспрессию составляющего внеклеточный матрикс белка.
(Коррекция экспрессии коллагена типа I, коллагена типа IV и фибронектина посредством иммунного окрашивания)
[0124]
Затем с помощью иммунного окрашивания было подтверждено, что экспрессия коллагена типа I, коллагена типа IV и фибронектина подавлялась. Способ иммунного окрашивания является сходным с описанным выше примером получения 2. Что касается антител, для проведения эксперимента вместо этого использовали антитела против коллагена типа I, коллагена типа IV и фибронектина.
Антитело против коллагена типа I: антитело против коллагена типа I (кролика, поликлональное)
(антитела и способы анализа ROCKLLANDTM, каталожный номер №: 600-401-103S)
Антитело против коллагена типа IV: коллаген типа IV (кролика, поликлональное) (Abcam, каталожный номер №: ab6586)
Антитело против фибронектина: антитело против фибронектина (мыши, моноклональное) (BD Biosciences, каталожный номер №: 610077)
[0125]
Для испытания с окрашиванием тканей культивируемые клетки помещали в Lab-TekTM Chamber SlidesTM (NUNC A/S, Roskilde, Дания), фиксировали 4% формальдегидом в течение 10 минут при комнатной температуре (RT), и инкубировали с 1% бычьим сывороточным альбумином (BSA) в течение 30 минут. В частности, культивируемые клетки на Lab-TekTM Chamber SlidesTM (NUNC A/S, Roskilde, Дания) фиксировали при комнатной температуре в течение 10 минут в 4% формальдегиде, а затем инкубировали с 1% бычьим сывороточным альбумином (BSA) в течение 30 минут. Для исследования экспрессии внеклеточного матрикса, продуцируемого клетками, использовали разведение 1:200 каждого из антител против коллагена типа I, коллагена типа IV и фибронектина. Для вторичного антитела использовали разведение 1:2000 меченного Alexa Fluor (зарегистрированный торговый знак) 488, или меченного Alexa Fluor (зарегистрированный торговый знак) 594 антитела козы против IgG мыши (Life Technologies). Затем ядра клеток окрашивали DAPI (Vector Laboratories, Inc., Burlingame, CA) или PI (Sigma-Aldrich). Затем предметное стекло исследовали под флуоресцентным микроскопом (TCS SP2 AOBS; Leica Microsystems, Welzlar, Германия).
Результат
[0126]
Результат представлен на фиг.5. Как показано на фиг.5, было подтверждено, что подавление экспрессии Snail1 или ZEB1 посредством миРНК также подавляло экспрессию коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина на уровне белка.
(Пример 4: подавление сверхпродукции внеклеточного матрикса в iFECD посредством подавления экспрессии Snail1 или ZEB1)
[0127]
В настоящем примере было подтверждено, что подавление экспрессии Snail1 или ZEB1 подавляло сверхпродукцию внеклеточного матрикса в iFECD.
[0128]
Далее, iHCEC и iFECD культивировали в DMEM на Transwell без сыворотки, через одну неделю после чего их фиксировали в состоянии монослоя и окрашивали HE. Окрашивание HE проводили в соответствии с методикой описанных выше примеров.
Результат
[0129]
Результат показан на фиг. 6. Как показано на фиг.6, подавление экспрессии Snail1 или ZEB1 посредством миРНК подавляло сверхпродукцию в iFECD внеклеточного матрикса до нормального уровня. Таким образом, было обнаружено, что подавление ZEB1 или Snail может подавлять сверхпродукцию внеклеточного матрикса в клетке при эндотелиальной дистрофии роговицы Фукса.
(Пример 5: коррекция аномалии внеклеточного матрикса в эндотелии роговицы средством, ингибирующим сигнал TGF-бета)
[0130]
Затем проводили исследование того, можно ли средство, ингибирующее сигнал TGF-бета, SB431542, использовать для ингибирования сигнала TGF-бета и коррекции аномалии внеклеточного матрикса в эндотелии роговицы. SB431542 получали от TOCRIS Corporation (каталожный номер: 1614).
(Исследование с помощью ПЦР в реальном времени)
[0131]
Уровень экспрессии подтверждали с помощью ПЦР в реальном времени. ПЦР в реальном времени проводили в соответствии с описанными выше примерами. Следующие зонды использовали для коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина.
Коллаген типа I, Hs00164004_ml COL1A1
Коллаген типа IV, Hs00266327_ml COL4A1
Коллаген типа VIII, Hs00697025_ml COL8A2
Фибронектин, Hs01549976_ml FN1
GAPDH TaqMan®pre, разработанный Assay Reagents Human GADPH (каталожный номер №: 4333764F)
(Результат)
[0132]
Результат показан на фиг.7. Как показано на фиг.7, с помощью ПЦР в реальном времени было выявлено значительное снижение экспрессии Snail1 и ZEB1. Кроме того, когда уровень экспрессии гена белка, составляющего внеклеточный матрикс, в iFCED анализировали с использованием SB431542 способом ПЦР в реальном времени, экспрессия коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина значительно подавлялась.
(Исследование экспрессии коллагена типа I, коллагена типа IV и фибронектина посредством иммунного окрашивания)
[0133]
Затем аналогичным образом исследовали экспрессию коллагена типа I, коллагена типа IV и фибронектина посредством иммунного окрашивания. Иммунное окрашивание проводили в соответствии с описанными выше примерами при условии, что следующие антитела использовали в качестве антител к коллагену типа I, коллагену типа IV и фибронектину.
Антитело против коллагена типа I: антитело против коллагена типа I (кролика, поликлональное)
(антитела и способы анализа ROCKLLANDTM, каталожный номер №: 600-401-103S)
Антитело против коллагена типа IV: коллаген типа IV (кролика, поликлональное) (Abcam, каталожный номер №: ab6586)
Антитело против фибронектина: антитело против фибронектина (мыши, моноклональное) (BD Biosciences, каталожный номер №: 610077)
Результат
[0134]
Результат показан на фиг.8. Как показано на фиг.8, было подтверждено, что ингибирование сигнала TGF-бета с использованием SB431542 также подавляло экспрессию коллагена типа I, коллагена типа IV, коллагена типа VIII и фибронектина на белковом уровне.
(Пример 6: эффект средства, ингибирующего сигнал TGF-бета, в случае фиксации состояния монослоя через одну неделю)
[0135]
В настоящем примере был подтвержден эффект средства, ингибирующего сигнал TGF-бета, в случае фиксации в состоянии монослоя через одну неделю.
[0136]
Кроме того, iHCEC и iFECD культивировали на проницаемых подложках Transwell: 0,4-мкм, шестиячеечные планшеты (Costar, Cat №: 3450) без сыворотки, и через одну неделю после этого их фиксировали в состоянии монослоя и окрашивали HE. Окрашивание HE проводили в соответствии с методикой описанных выше примеров.
Результат
[0137]
Результат показан на фиг.9. Как показано на фиг.9, ингибирование сигнала TGF-бета с использованием SB431542 также подавляло сверхпродукцию внеклеточного матрикса в iFECD до нормального уровня.
[0138]
Представленное выше описание показывает, что у пациента с эндотелиальной дистрофией роговицы Фукса продуцируется избыточное количество внеклеточного матрикса под действием сигнала TGF-бета путем стимуляции Snail1 или ZEB1 по сравнению со здоровым индивидуумом. Кроме того, оно показывает, что подавление посредством миРНК и т.п. генов, связанных с EMT, таких как Snail1, ZEB1 и т.п., и генов, связанных с продукцией белка, может подавлять продукцию внеклеточного матрикса. Кроме того, оно показывает, что ингибирование сигнала TGF-бета также может подавлять продукцию внеклеточного матрикса. В нем показана возможность того, что подавление ингибирования сигнала TGF-бета, связанных с EMT генов, таких как Snail1, ZEB1 и т.п., или их сигналов может подавлять сверхпродукцию внеклеточного матрикса эндотелиальных клеток роговицы у пациентов с эндотелиальной дистрофией роговицы Фукса и подавлять образование "капель" и утолщение десцеметовой оболочки.
[0139]
Как описано выше, настоящее изобретение иллюстрируется предпочтительными вариантами осуществления настоящего изобретения. Однако будет понятно, что объем настоящего изобретения следует интерпретировать только в рамках формулы изобретения. Будет понятно, что содержание патентов, патентных заявок и источников литературы, цитированных в настоящем описании, включено в настоящее описание в качестве ссылки, как если бы их содержание само по себе конкретно было описано в настоящем описании.
Промышленная применимость
[0140]
Предусматриваются способы, доступные для промышленности (промышленность культивирования клеток, фармацевтическая промышленность и т.п.), связанные с терапевтическими или профилактическими средствами от заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ECM) в эндотелии роговицы, в частности, аномалией внеклеточного матрикса (ECM) при эндотелиальной дистрофии роговицы Фукса, и фотофобии, где лекарственные средства содержат средство, ингибирующее сигнал TGF-бета.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО ПРОТИВ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ГИБЕЛЬЮ КЛЕТОК ЭНДОТЕЛИЯ РОГОВИЦЫ, ОБУСЛОВЛЕННОЙ СОСТОЯНИЕМ ЭНДОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА | 2014 |
|
RU2712967C2 |
| СОДЕРЖАЩЕЕ ИНГИБИТОР mTOR ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ ГЛАЗНЫХ СИМПТОМОВ, НАРУШЕНИЙ ИЛИ ЗАБОЛЕВАНИЙ И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2782613C2 |
| КОМПОЗИЦИЯ ИЛИ СПОСОБ, ВКЛЮЧАЮЩИЕ (T)EW-7197, ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ ЗАБОЛЕВАНИЙ ЭНДОТЕЛИЯ РОГОВИЦЫ | 2018 |
|
RU2798396C2 |
| ПРИМЕНЕНИЕ ЛАМИНИНА В КУЛЬТУРЕ КЛЕТОК ЭНДОТЕЛИЯ РОГОВИЦЫ | 2014 |
|
RU2704984C1 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ФИБРОЗА ПОЧЕК | 2011 |
|
RU2635460C2 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ФИБРОЗА ПОЧЕК | 2011 |
|
RU2711531C2 |
| УЛУЧШЕННЫЙ СПОСОБ ДИФФЕРЕНЦИРОВКИ | 2017 |
|
RU2772435C2 |
| АНТАГОНИСТЫ РЕЦЕПТОРА И ЛИГАНДОВ ALK1 И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2559532C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ЛЕГОЧНОЙ ГИПЕРТЕНЗИИ | 2017 |
|
RU2748278C2 |
| АНТАГОНИСТЫ ALK1 И ИХ ПРИМЕНЕНИЕ В ЛЕЧЕНИИ ПОЧЕЧНО-КЛЕТОЧНОЙ КАРЦИНОМЫ | 2013 |
|
RU2633638C2 |





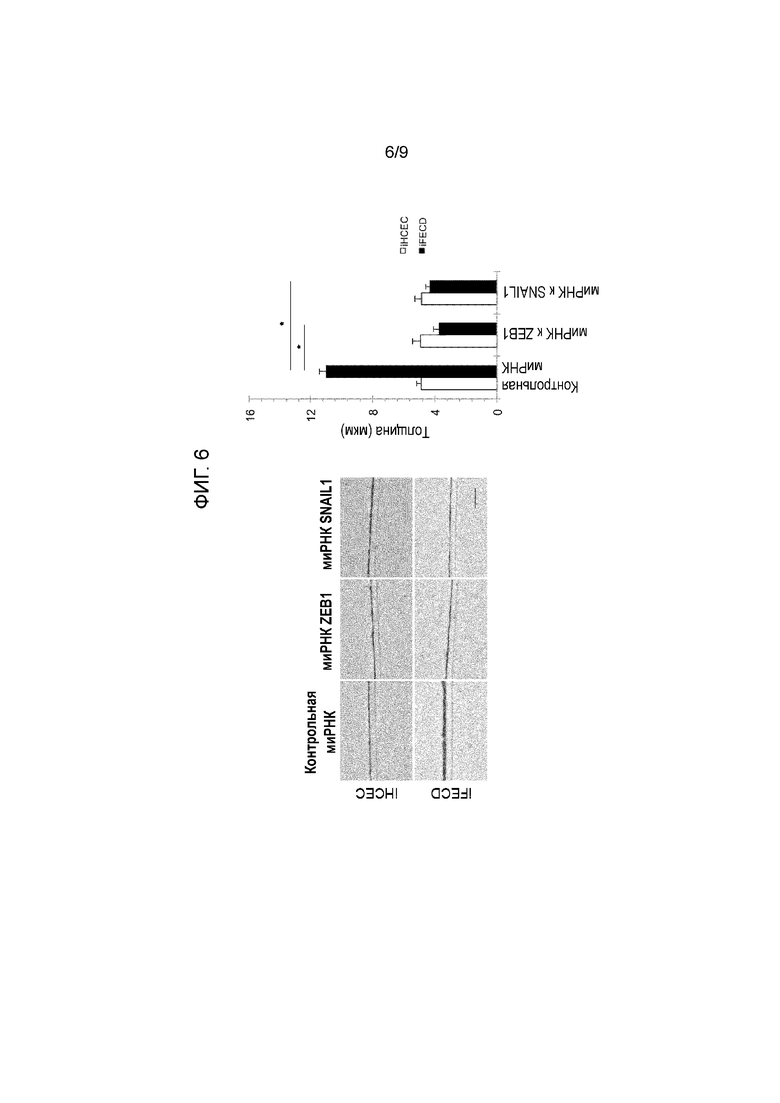
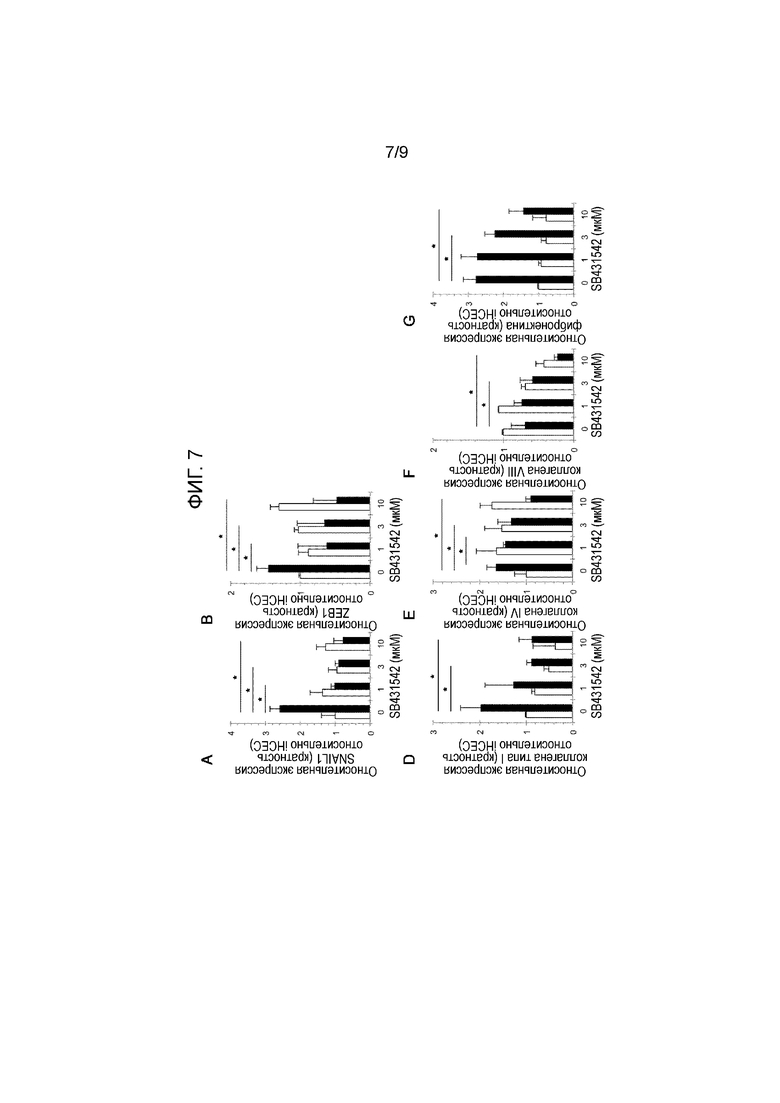
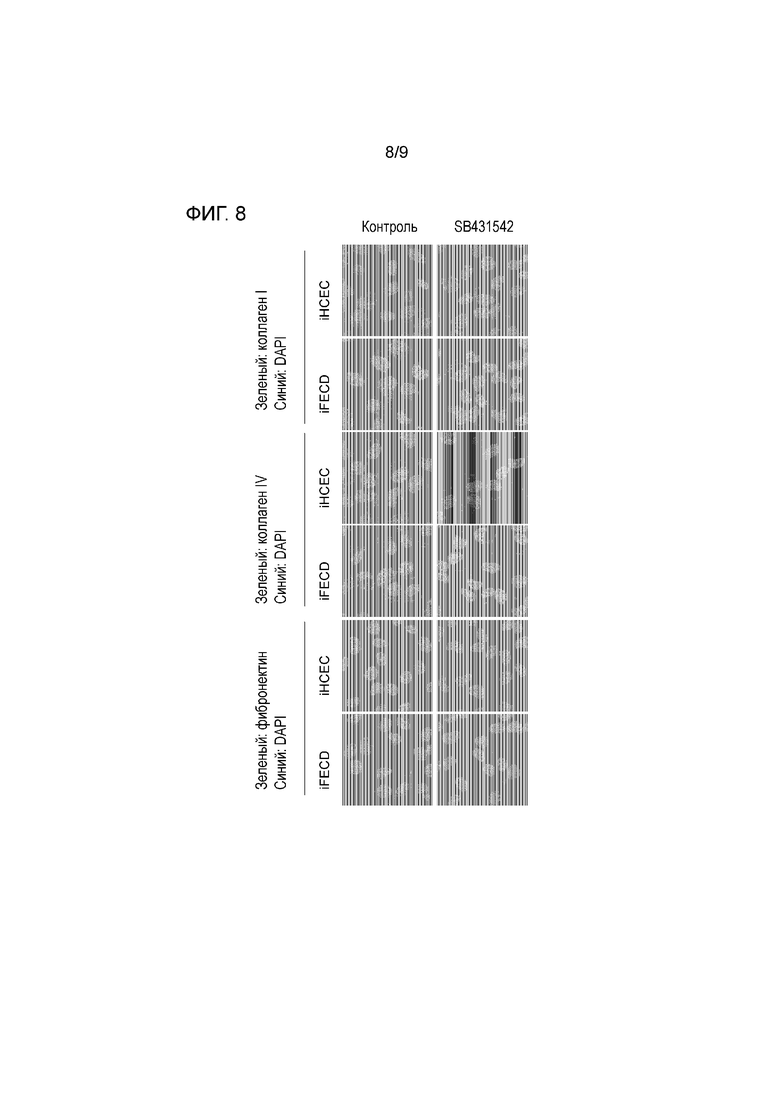

Изобретение относится к области медицины, а именно к офтальмологии, и предназначено для лечения заболевания, нарушения или состояния, обусловленного эндотелиальной дистрофией роговицы Фукса. Применяют лекарственное средство, содержащее средство, ингибирующее сигнал TGF-бета, для лечения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ЕСМ) в эндотелии роговицы у субъекта, где заболевание, нарушение или состояние обусловлено эндотелиальной дистрофией роговицы Фукса. Лекарственное средство представляет собой глазные капли. Использование изобретения обеспечивает эффективное лечение заболевания, нарушения или состояния, обусловленного эндотелиальной дистрофией роговицы Фукса, за счет ингибирования сверхэкспрессии ЕСМ. 6 з.п. ф-лы, 9 ил., 6 пр.
1. Применение лекарственного средства для лечения заболевания, нарушения или состояния, обусловленного аномалией внеклеточного матрикса (ЕСМ) в эндотелии роговицы у субъекта, имеющего заболевание, нарушение или состояние, где лекарственное средство содержит средство, ингибирующее сигнал TGF-бета, где лекарственное средство представляет собой глазные капли, где заболевание, нарушение или состояние обусловлено эндотелиальной дистрофией роговицы Фукса.
2. Применение по п.1, где заболевание, нарушение или состояние включает по меньшей мере одно заболевание, нарушение или состояние, выбранное из группы, состоящей из фотофобии, неясного зрения, нарушения зрения, боли в глазу, слезоотделения, гиперемии, боли, буллезной кератопатии, неприятных ощущений в глазу, снижения контрастности, бликов, отека стромы роговицы, буллезной кератопатии и помутнения роговицы при эндотелиальной дистрофии роговицы Фукса.
3. Применение по п.1, где средство, ингибирующее сигнал TGF-бета, включает по меньшей мере одно из 4-[4-(1,3-бензодиоксол-5-ил)-5-(2-пиридинил)-1Н-имидазол-2-ил]бензамида, ВМР-7, антитела против TGF-бета, антитела против рецептора TGF-бета, миРНК к TGF-бета, миРНК к рецептору TGF-бета, кшРНК к TGF-бета, кшРНК к рецептору TGF-бета, аптамера к TGF-бета, аптамера к рецептору TGF-бета, антисмыслового олигонуклеотида к TGF-бета, 6,7-диметокси-2-((2E)-3-(1-метил-2-фенил-1Н-пирроло[2,3-b]пиридин-3-ил-проп-2-еноил))-1,2,3,4-тетрагидроизохинолона, 3-(6-метил-2-пиридинил)-N-фенил-4-(4-хинолинил)-1Н-пиразол-1-карботиоамида, 2-(3-(6-метилпиридин-2-ил)-1Н-пиразол-4-ил)-1,5-нафтиридина, 6-(4-{пиперидин-1-ил)этокси)фенил)-3-(пиридин-4-ил)пиразоло[1,5-а]пиримидина, 2-(5-хлор-2-фторфенил)-4- [ (4-пиридинил)амино]птеридина, 4- [3- (2-пиридинил)-1Н-пиразол-4-ил]хинолина, их фармацевтически приемлемых солей или сольватов, или сольватов фармацевтически приемлемых солей.
4. Применение по п.1, где средство, ингибирующее сигнал TGF-бета, включает 4-[4-(1,3-бензодиоксол-5- ил)-5-(2-пиридинил)-1Н-имидазол-2-ил]бензамид или его фармацевтически приемлемую соль.
5. Применение по п.1, где эндотелий роговицы представляет собой эндотелий роговицы примата.
6. Применение по п.1, где эндотелий роговицы представляет собой эндотелий роговицы человека.
7. Применение по п.1, содержащее дополнительный лекарственный компонент.
| US 2010222280 A1, 02.09.2010 | |||
| WO 2009146408 A1, 03.12.2009 | |||
| OKUMURA N et al | |||
| Inhibition of TGF-beta signaling enables human corneal endothelial cell expansion in vitro for use in regenerative medicine | |||
| PLoS One, 25.02.2013, 8(2), p.e58000 | |||
| WO 2013086236 A2, 13.06.2013 | |||
| WO 2012073238 A1, 07.06.2012. |

