СВЕДЕНИЯ О РОДСТВЕННЫХ ЗАЯВКАХ
Настоящая заявка претендует на приоритет австралийской заявки на патент №2013904595, поданной 28 ноября 2013 года и озаглавленной "Способ лечения нефропатии". Все содержание этой заявки включено в качестве ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка подается с перечнем последовательностей в электронном виде. Все содержание перечня последовательностей включено в данное описание в качестве ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящая заявка относится к способу лечения или профилактики нефропатии.
ВВЕДЕНИЕ
Нефропатия
Нефропатия представляет собой класс заболеваний, характеризующихся повреждением почки, и включает в себя нефрит (воспалительная болезнь почек) и нефроз (не воспалительное заболевание почек). Причины нефропатии включают хронические состояния (включая системную красную волчанку, сахарный диабет и высокое кровяное давление (гипертензию)), отложение IgA-антител в клубочках, введения анальгетиков, нехватку ксантиноксидазы, токсичность химиотерапевтических средств и долговременное воздействие свинца или его солей.
Нефропатия, связанная с диабетом (например, "диабетическая нефропатия") является наиболее частой причиной терминальной стадии почечной недостаточности в Соединенных Штатах и ряде других развитых стран. Например, диабетическая нефропатия составляет почти 35% терминальной стадии почечной недостаточности в США сегодня и обходится приблизительно 50000-65000$ на одного пациента в год, что превышает 2 млрд $ в год на всех американских пациентов. Примерно у 40% пациентов с сахарным диабетом 1 типа и 5-15 пациентов с диабетом 2-го типа в конечном итоге развивается почечная недостаточность в терминальной стадии.
Патофизиологические механизмы диабетической нефропатии выяснены не полностью. Самые ранние наглядные нарушения включают интраренальную гипертензию, гиперфильтрацию (увеличенная скорость клубочковой фильтрации) и микроальбуминурию. Клинически наиболее важным инструментом скрининга для выявления ранней нефропатии является обнаружение микроальбуминурии. Факторы риска развития диабетической нефропатии включают гипергликемию, гипертензию, наследственную предрасположенность к нефропатии и курение. Диабетическая нефропатия, как правило, рассматривается как результат гипертензии и гипергликемии при сахарном диабете, и многие исследователи рассматривают гипергликемию в качестве существенного фактора, способствующего развитию этого заболевания.
Медицинские вмешательства до сих пор не являются достаточно эффективными для лечения или профилактики прогрессирования диабетической нефропатии и развития терминальной стадии почечной недостаточности. В связи с этим, современные способы лечения направлены, в первую очередь, на уменьшение осложнений заболеваний следующим образом: 1) контроль артериального давления (ингибиторы АПФ-ингибиторы или блокаторы рецептора ангиотензина (БРА); 2) контроль гликемических значений; и 3) липопротеиновая диета, физические упражнения или другие изменения образа жизни. Тем не менее, современные способы лечения имеют ограниченное влияние на прогрессирующее снижение функции почек и пациенты все еще прогрессируют в направлении почечной заместительной терапии, либо диализа или трансплантации почки.
Другие стратегии лечения были сосредоточены на одном или нескольких факторах роста в качестве терапевтических мишеней, зачастую на тех, идентифицированных как активируемые в моделях нефропатии. Например, рассматривались способы лечения, направленные на ингибирование TGFβ, либо по отдельности, либо в комбинации с ингибиторами АПФ. Сосудистый эндотелиальный фактор роста А (VEGF-A) и другие факторы, вовлеченные в ангиогенез, также были изучены в качестве мишеней при лечении нефропатии.
VEGF и нефропатия
Семейство VEGF факторов роста включает в себя пять лигандов (VEGF-A, VEGF-B, VEGF-C, VEGF-D и фактор роста плаценты (PTGF)), которые могут связываться дифференцированно с тремя рецепторными тирозинкиназами (VEGFR-1, -2, и -3) и рецепторами семафорина (нейропилин 1 и 2). Семейство VEGP факторов роста вовлечено в нормальный и патологический ангиогенез и лимфангиогенез. VEGF-A связывается с VEGFR-1, VEGFR-2, NP-1 и NP-2; VEGF-B связывается с VEGFR-1 и NP-1. VEGF-C и VEGF-D в основном участвуют в лимфангиогенезе и связываются с VEGFR-2, VEGFR-3 и NP-2.
VEGF-A является наиболее хорошо изученным членом семейства VEGF, и роль VEGF-A и исследований ингибирования VEGF-A по отношению к почкам и нефропатии обеспечили смешанные результаты, что часто приводит к пагубным последствиям для почек. Некоторые из этих исследований включали введение растворимого рецептора VEGF 1 (также известного как sFlt-1), который может действовать в качестве антагониста VEGF-A, но также и VEGF-B и P1GF, и приводили к протеинурии и гипертензии (Nakagawa et al., Diabetes, 58, p1471-8, 2009). Kosugi et al. (Am J Physiol Renal Physiol 298: F609-F616, 2010) пришли к выводу, что лечение sFlt-1 вряд ли будет полезным при диабетической нефропатии и что необходимы альтернативные подходы к лечению диабетической почечной болезни. Недавнее исследование с участием антагониста VEGFR-1 (Yang et al., PLOS ONE, 9(4), e94540, 2014) у мышей db/db позволили заключить, что ингибирование VEGFR-1 усиливает диабетическую нефропатию, и действительно позволяет предположить, что активация VEGFR-1 может обеспечить терапевтическое средство при диабетической нефропатии 2 типа. Как обсуждалось выше, VEGF-B является одним из лигандов, который передает сигналы через VEGFR-1, и эти исследования с участием sFIt-1 или антагониста VEGFR-1 блокировали бы сигнальный путь VEGF-B, хотя и в неспецифической форме.
РЕЗЮМЕ
При разработке данного изобретения авторы изобретения действовали вопреки знаниям в данной области по отношению к сигнальному пути VEGF в нефропатии и изучали эффекты ингибирования сигнального пути VEGF-B в принятых мышиных моделях нефропатии, например мыши, получающие питание с высоким содержанием жиров и модели диабетической нефропатии, в том числе нефропатии, ассоциированной с сахарным диабетом 1 типа и диабетом 2 типа. Авторы настоящего изобретения исследовали влияние этого фактора роста путем предотвращения экспрессии VEGF-B (например, с использованием генетически модифицированных мышей, у которых экспрессия VEGF-B понижалась или исключалась) или путем введения антагониста VEGF-B (например, антагонистическое антитело). Авторы настоящего изобретения с удивлением обнаружили, что ингибирование сигнального пути VEGF-B оказывает благоприятное воздействие в различных животных моделях нефропатии. Например, авторы настоящего изобретения смогли предотвратить развитие нефропатии, например, диабетическую нефропатию и лечить (например, задержать прогрессирование) нефропатию, например, диабетическую нефропатию в зависимости от используемого режима лечения.
Авторы изобретения обнаружили, что антагонизм сигнального пути VEGF-B уменьшает или предотвращает, по меньшей мере, клубочковое мезангиальное расширение, клубочковый и тубулярный склероз, отложение мезангиального внеклеточного матрикса, аномальное утолщение клубочковой базальной мембраны и накопление липидов в почках, например, в почечных клубочках, потерю подоцитов, гипертензию, клубочковые сосудистые перестройки и сохраняет структуру подоцитов у пациентов с диабетом. В некоторых случаях (например, в эксперименте, описанном в настоящем документе) изменения в почках происходят, несмотря на умеренное воздействие на уровень глюкозы в крови у пациентов с диабетом, указывая, что ингибирование VEGP-B обеспечивает терапевтическое/профилактическое преимущество при нефропатии посредством пути, дополнительного или отличного от гликемического контроля.
Полученные авторами изобретения данные являются основой для способов лечения или профилактики нефропатии у индивидуума путем ингибирования сигнального пути VEGF-B. Например, настоящее изобретение относится к способу лечения или профилактики нефропатии у индивидуума, причем способ включает введение индивидууму соединения, которое ингибирует сигнальный путь VEGF-B.
В одном примере нефропатия представляет собой нефрит, т.е. воспалительное заболевание почек. Например, нефропатия представляет собой IgA-нефропатию или вызвана употреблением лекарственных препаратов (например, анальгетиков или химиотерапии), нехваткой ксантиноксидазы, поликистозным заболеванием почек или вызвана хроническим заболеванием, например, воспалительным или аутоиммунным заболеванием или диабетом.
Например, нефропатии представляет собой гломерулонефрит и /или гломерулосклероз. Так, например, гломерулонефрит и /или гломерулосклероз является пролиферативным гломерулонефритом и/или гломерулосклерозом.
В одном примере нефропатия или нефрит ассоциируется с или вызвана другим заболеванием. Например, нефропатия вызвана воспалительным или аутоиммунным заболеванием (например, системной красной волчанкой, синдромом Гудпасчера), васкулитом (например, гранулематозом Вегенера или микроскопическим полиангиитом) или диабетом.
В одном примере нефропатия или нефрит ассоциируется с или вызвана преддиабетом.
В одном примере нефропатия или нефрит ассоциируется с или вызвана диабетом типа 1.
В одном примере нефропатия или нефрит ассоциируется с или вызвана диабетом 2 типа.
Например, настоящее изобретение относится к способу лечения или профилактики диабетической нефропатии у индивидуума, страдающего от диабета, причем указанный способ включает введение индивидууму соединения, которое ингибирует сигнальный путь VEGF-B.
Как иллюстрируется в настоящем описании, авторы настоящего изобретения показали, что введение ингибитора сигнального пути VEGF-B индивидууму, страдающему от диабетической нефропатии, является эффективным при лечении этого состояния. Соответственно, настоящее изобретение дополнительно относится к способу лечения диабетической нефропатии, причем указанный способ включает введение индивидууму, страдающему от диабетической нефропатии, ингибитора сигнального пути VEGF-B.
В одном примере индивидуум находится под угрозой из развития нефропатии или у него развивается нефропатия (например, диабетическая нефропатия). Например, индивидуум страдает от микроальбуминурии или макроальбуминурии. В другом примере индивидуум страдает от гипертензии.
В одном из примеров настоящее изобретение предоставляет способ профилактики или замедления развития нефропатии (например, диабетической нефропатии), включающий введение индивидууму, страдающему от микроальбуминурии или макроальбуминурии (например, пациенты, страдающему от диабета и микроальбуминурии или микроальбуминурии), соединения, которое ингибирует сигнальный путь VEGF-B. В одном примере индивидуум страдает от микроальбуминурии.
В одном из примеров настоящее изобретение предоставляет способ профилактики или замедления развития нефропатии (например, диабетической нефропатии), причем способ включает введение индивидууму, страдающему от гипертензии (например, пациенту, страдающему от диабета и гипертензии) соединения, которое ингибирует сигнальный путь VEGF-B.
В одном примере соединение вводят в количестве, эффективном для достижения одного или нескольких из следующих эффектов:
уменьшение или предотвращение гипертензии;
уменьшение или предотвращение клубочкового и/или тубулярного склероза;
уменьшение или предотвращение отложения мезангиального внеклеточного матрикса и/или аномального утолщения клубочковой базальной мембраны;
уменьшение или предотвращение клубочкового мезангиального расширения;
уменьшение или предотвращение клубочковых сосудистых перестроек;
уменьшение или предотвращение накопления липидов в почках;
уменьшение или предотвращение клубочкового накопления липидов;
уменьшение или предотвращение клубочковых коллагеновых отложений и/или артериолярного гиалиноза; и/или
уменьшение или предотвращение макроальбуминурии.
В одном примере соединение, которое ингибирует сигнальный путь VEGF-B, специфически ингибирует сигнальный путь VEGF-B. Это вовсе не означает, что способ по настоящему изобретению не включает в себя ингибирование передачи сигналов многочисленными белками VEGF, а только то, что соединение (или его часть), которое ингибирует сигнальный путь VEGF-B, является специфическим для VEGF-B, например, не является общим ингибитором белков VEGF-B. Этот термин также не исключает, например, биспецифическое антитело или белок, содержащий связывающие домены этого, которые могут специфически ингибировать сигнальный путь VEGF-B с одним (или более) связывающими доменами и могут специфически ингибировать передачу сигналов другого белка с другим связывающим доменом.
В одном примере соединение, которое ингибирует сигнальный путь VEGF-B, связывается с VEGF-B. Например, соединение представляет собой белок, содержащий вариабельную область антитела, которая связывается с или специфически связывается с VEGF-B и нейтрализует сигнальный путь VEGF-B.
В одном примере соединение представляет собой миметик антитела. Например, соединение представляет собой белок, содержащий антигенсвязывающий домен иммуноглобулина, например, IgNAR, антитело верблюда или Т-клеточный рецептор.
В одном примере соединение представляет собой домен антитела, (например, включающий в себя только вариабельную область тяжелой цепи или вариабельную область легкой цепи, которая связывается с VEGF-B) или только тяжелоцепочечное антитело (например, антитело верблюда или IgNAR) или его вариабельную область.
В одном примере соединение представляет собой белок, содержащий Fv, например, белок выбирают из группы, состоящей из:
(I) одноцепочечный Fv-фрагмента (ScFv);
(II) димерного ScFv (ди-scPv); или
(IV) диатела;
(V) триатела;
(VI) тетратела;
(VII) Fab;
(VIII) F(аb')2;
(IX) Fv; или
(Х) один из (I)-(IX), связанный с константной областью антитела, Fc или константной областью тяжелой цепи (CH)2 и/или СН3.
В другом примере соединение представляет собой антитело. Типичные антитела представляют собой полноразмерные антитела и/или голые антитела.
В одном примере соединение представляет собой белок, который является рекомбинантным, химерным, CDR-привитым, гуманизированным, сингуманизированным, приматизированным, деиммунизированным или человеческим.
В одном примере соединение представляет собой белок, содержащий вариабельную область антитела, которая конкурентно ингибирует связывание антитела 2H10 с VEGF-B. В одном примере белок содержит вариабельную область тяжелой цепи (VН), содержащую последовательность, представленную в SEQ ID NO:3, и вариабельную область легкой цепи (VL), содержащую последовательность, представленную в SEQ ID NO:4.
В одном примере соединение представляет собой белок, содержащий гуманизированную вариабельную область антитела 2H10. Например, белок содержит вариабельную область, содержащую гипервариабельные участки (CDR) VH и VL антитела 2H10. Например, белок включает:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 25-34 из SEQ ID NO:3;
(b) CDR2, содержащий последовательность, представленную аминокислотами 49-65 из SEQ ID NO:3; а также
(c) CDR3, содержащий последовательность, представленную аминокислотами 98-108 из SEQ ID NO:3; и/или
(II) VL, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 23-33 из SEQ ID NO:4;
(b) CDR2, содержащий последовательность, представленную аминокислотами 49-5.5 из SEQ ID NO:4; а также
(c) CDR3, содержащий последовательность, представленную аминокислотами 88-96 из SEQ ID NO:4.
В одном примере соединение представляет собой белок, содержащий VH и VL, VH и VL являются гуманизированными вариабельными областями антитела 2H10. Например, белок включает:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 25-34 из SEQ ID NO:3;
(b) CDR2, содержащий последовательность, представленную аминокислотами 49-65 из SEQ ID NO:3; а также
(с) CDR3, содержащий последовательность, представленную аминокислотами 8-108 из SEQ ID NO:3; а также
(II) VL, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 23-33 из SEQ ID NO:4;
(b) CDR2, содержащий последовательность, представленную аминокислотами 49-55 из SEQ ID NO:4; а также
(с) CDR3, содержащий последовательность, представленную аминокислотами 88-96 из SEQ ID NO:4,
В одном примере вариабельная область или VH в любом из предыдущих пунктов содержит последовательность, представленную в SEQ ID NO:5.
В одном примере вариабельная область или VL в любом из предыдущих пунктов содержит последовательность, представленную в SEQ ID NO:6.
В одном примере соединение представляет собой антитело.
В одном примере соединение представляет собой антитело, содержащее VH, содержащую последовательность, представленную в SEQ ID NO:5 и VL, содержащую последовательность, представленную в SEQ ID NO:6.
В одном примере белок или антитело представляет собой любую форму белка или антитела, кодируемого нуклеиновой кислотой, кодирующей любой из вышеуказанных белков или антител.
В одном примере белок или антитело включает в себя:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:11, или содержащий аминокислотную последовательность SEQ. ID NO:17;
(b) CDR2, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:12, или содержащий аминокислотную последовательность SEQ ID NO:18; и
(с) CDR3, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:13, или содержащий аминокислотную последовательность SEQ ID NO:19; и/или
(II) VL, включающую:
(a) CDR1, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:14, или содержащий аминокислотную последовательность SEQ ID NO:20;
(b) CDR2, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:15, или содержащий аминокислотную последовательность SEQ ID NO:21; а также
(С) CDR3, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:16, или содержащий аминокислотную последовательность SEQ ID NO:22.
[1] В одном примере белок или антитело включает в себя:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:23, или содержащий аминокислотную последовательность SEQ ID NO:29;
(b) CDR2, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:24, или содержащий аминокислотную последовательность SEQ ID NO:30; а также
(с) CDR3, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:25, или содержащий аминокислотную последовательность SEQ ID NO:31; и /или
(II) VL, включающую:
(a) СDR1, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:26, или содержащий аминокислотную последовательность SEQ ID NO:32;
(b) CDR2, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:27, или содержащий аминокислотную последовательность SEQ ID NO:33; и
(с) CDR3, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:28, или содержащий аминокислотную последовательность SEQ ID NO:34.
В одном примере белок или антитело включает в себя:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:35, или содержащий аминокислотную последовательность SEQ ID NO:41;
(b) CDR2, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:36, или содержащий аминокислотную последовательность SEQ ID NO:42; а также
(с) CDR3, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:37, или содержащий аминокислотную последовательность SEQ ID NO:43; и/или
(II) VL включающую:
(a) CDR.1, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:38, или содержащий аминокислотную последовательность SEQ ID NO:44;
(b) CDR2, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:39, или содержащий аминокислотную последовательность SEQ ID NO:45; и
(с) CDR3, содержащий последовательность, кодируемую нуклеиновой кислотой, содержащей SEQ ID NO:40, или содержащий аминокислотную последовательность SEQ ID NO:46.
В одном примере соединение находится в пределах композиции. Например, композиция содержит белок, включающий вариабельную область VH или VL или антитело, как описано в настоящем документе. В одном примере композиция дополнительно содержит один или более вариантов белка или антитела. Например, она включает в себя вариант с отсутствием кодирования С-концевого остатка лизина, деамидированный вариант и/или гликозилированный вариант и/или вариант, содержащий пироглутамат, например, на N-конце белка и/или вариант с отсутствием N-концевого остатка, например, N-концевого глутамина антитела или V-области и/или вариант, включающий весь сигнал секреции или его часть. Дезамидированные варианты кодируемых остатков аспарагина могут приводить к образованию изоформ изоаспарагиновой и аспарагиновой кислоты или даже сукцинамида с участием соседнего аминокислотного остатка. Дезамидированные варианты кодируемых остатков глутамина могут привести к глутаминовой кислоте. Композиции, содержащие гетерогенную смесь таких последовательностей и вариантов, предназначены для включения, когда делается ссылка на определенную аминокислотную последовательность.
В одном примере соединение, которое ингибирует сигнальный путь VEGF-B, ингибирует или предотвращает экспрессию VEGF-B. Например, соединение выбирают из группы антисмысловой, siRNA, RNAi, рибозима и ДНКазима.
В одном примере VEGF-B является VEGF-B млекопитающего, например, VEGF-B человека. В одном примере индивидуум представляет собой млекопитающее, например, примата, такого как человек.
Способы лечения, описанные в настоящем документе, могут дополнительно включать в себя введение еще одного соединения для лечения или профилактики нефропатии.
Способы лечения диабетической нефропатии, описанные в настоящем документе, могут дополнительно включать введение еще одного соединения для лечения или профилактики (или задержки прогрессирования) диабета. Примеры таких соединений описаны в настоящем документе. Настоящее изобретение также относится к соединению, которое ингибирует сигнальный путь VEGF-B для применения при лечении или профилактики нефропатии.
Настоящее изобретение также относится к использованию соединения, которое ингибирует сигнальный путь VEGF-B, в производстве лекарственного средства для лечения или профилактики нефропатии.
Настоящее изобретение также предоставляет набор, содержащий соединение, которое ингибирует сигнальный путь VEGF-B в комплекте с инструкцией по применению при лечении или профилактике нефропатии.
Типичные примеры нефропатий и соединений описаны в настоящем документ, и должны применяться с соответствующими изменениями к примерам по раскрытию, изложенным в предыдущих трех пунктах.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1A содержит два графических представления, показывающих соотношение креатинин/альбулин в моче (ACR), измеренные способом ELISA для мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/- (n=5/группу, левый график) и тощих дикого типа (wt) и тощих Vegfb-/- мышей (n=4-6/группу, правый график). Значения представляют собой средние значения ± стандартная ошибка среднего**P<0,05, **P<0,01, ***P<0,001 по сравнению с контролями db/db.
Фигура 1B представляет собой графическое изображение, показывающее артериальное давление, определенное способом хвостовой манжеты (систолическое и диастолическое артериальное давление, как указано), у мышей db/db and db/db//Vegfb+/- (n=5/группа). Значения представляют собой средние значения ± стандартная ошибка среднего **P<0,05, **P<0,01 по сравнению с контролями db/db.
Фигура 2А представляет собой графическое изображение, показывающее количественное определение аномалий в компартментах канальцев, измеренных как утолщение базальной мембраны канальцев (BM) в срезах почек мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего **P<0,05, **P<0,01 по сравнению с контролями db/db. ##P<0,01 по сравнению с тощими 28 недельными старыми мышами дикого типа.
Фигура 2В представляет собой графическое изображение, показывающее количественное определение гломерулосклероза, измеренное как мезангиальное расширение в срезах почек мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего **P<0,05, **P<0,01, ***P<0,001 по сравнению с контролями db/db.
Фигура 2С представляет собой графическое изображение, показывающее количественное определение клубочкового апоптоза, измеренное как апоптотические клубочки на кадр (n=4/группу) в срезах почек мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего **P<0,05, **P<0,01, ***P<0,001 по сравнению с контролями db/db.
Фигура 3A является графическим представлением, показывающим количественное определение клубочковой толщины базальной мембраны, измеренной с помощью анализа с использованием просвечивающей электронной микроскопии (ПЭМ) срезов почек тощих wt, db/db, db/db//Vegfb+/- и db/db//Vegfb-/- животных (как показано), n=3-4/группу. Значения представляют собой средние значения ± стандартная ошибка среднего *###P<0,001 по сравнению с тощими db+ контролями и **P<0,01 по сравнению с контролями db/db.
Фигура 3В является графическим представлением, показывающим количественное определение числа щелей, измеренной с помощью анализа с использованием просвечивающей электронной микроскопии (ПЭМ) срезов почек тощих wt, db/db, db/db//Vegfb+/- и db/db//Vegfb-/- животных (как показано), n=3-4/группу. Значения представляют собой средние значения ± стандартная ошибка среднего *###P<0,001 по сравнению с тощими db+ контролями и **P<0,01 по сравнению с контролями db/db.
Фигура 4А является графическим представлением, показывающим количественное определение анализа масляным красный О (ORO) срезов почек db/db, db/db//Vegfb+/- и db/db//Vegfb-/- животных. Значения представляют собой средние значения ± стандартная ошибка среднего *P<0,05 по сравнению с контролями db/db.
Фигура 4B является графическим представлением, показывающим количественное определение окрашивание масляным красный О в клубочковых и канальцих компартментах (как указано). Животные db/db, db/db//Vegfb+/-, db/db//Vegfb-/- и животные дикие типа. Значения представляют собой средние значения ± стандартная ошибка среднего *P<0,05 и ###P<0,001 по сравнению с тощими контролями, ***P<0,001 по сравнению с контролями db/db.
Фигура 5A является графическим представлением, показывающим количественное определение окрашивания синаптоподином в клубочках из db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего, *P<0,05, **P<0,01, ***P<0,001 по сравнению с контролями db/db.
Фигура 5В является графическим представлением, показывающим количественное определение ПЕКАМ в клубочках из db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего,*P <0,05, **Р <0,01, ***р <0,001 по сравнению с контролями db/db.
Фигура 6А является графическим представлением, показывающим количественное определение окрашивания коллагеном IV в клубочках в срезах почек мышей db/db, db/db//Vegfb+/- db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего, *P<0,05, **P<0,01, ***P<0,001 по сравнению с контролями db/db.
Фигура 6B представляет собой графическое изображение, показывающее количественное определение окрашивания α-SMA в клубочках в срезах почек мышей db/db, db/db//Vegfb+/- and db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего, *P<0,05, **P<0,01, ***P<0,001 по сравнению с контролями db/db.
Фигура 6С представляет собой графическое изображение, показывающее толщину артериол в срезах почек мышей db/db, db/db//Vegfb+/- and db/db//Vegfb-/-. Значения представляют собой средние значения ± стандартная ошибка среднего,*P<0,05 по сравнению с контролями db/db.
Фигура 7 представляет собой графическое изображение, показывающее количественное определение окрашивания ORO как в клубочковом, так и в тубулярном компартменте у тощих дикого типа, db/db//BKS 6-weeks и db/db//BKS 21-недельных мышей, n=5-7. Значения представляют собой средние значения ± стандартная ошибка среднего, #P<0,05 and ##P<0,01, канальцев по сравнению с клубочками. *P<0,05 клубочки db/db//BKS 6-недель по сравнению с клубочками db/db//BKS 21-недельных.
Фигура 8 включает в себя два графических представления, показывающих, что клубочковое накопление липидов коррелирует с потерей подоцитов в прогрессировании DN у мышей db/db//BKS. График слева показывает количественное определение окрашивания адипофилином и график справа показывает количественное определение окрашивания синаптоподином. n=3-5/группу. Значения представляют собой средние значения ± стандартная ошибка среднего, #P<0,05, ##P<0,01, по сравнению с тощими db+-контролями. *P<0,05 db/db//BKS 6-недельных по сравнению с клубочками db/db//BKS 21-недельных.
Фигура 9 представляет собой графическое изображение, показывающее относительную экспрессию почечной мРНК Vegfb, Vegfr1, Fatp4 и Fatp3 у тощих db+, а также у 6-, 12- и 21-недельных db/db//BKS (как указано). Значения представляют собой средние значения ± стандартная ошибка среднего,*Р<0,05, **Р<0,01, по сравнению с контрольными животными худых. Vegfb, сосудистый эндотелиальный фактор роста B; Vegfr1, рецептор 1 сосудистого эндотелиального фактора роста; Fatp3, транспортер жирной кислоты 3; Fatp4, транспортер жирной кислоты 4.
Фигура 10А представляет собой графическое изображение, показывающее анализ соотношения альбумин/креатинин в моче у мышей, получающих питание с высоким содержанием жиров (HFD) WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных. Значения представляют собой средние значения ± стандартная ошибка среднего. #P<0,05, ##P<0,01, ###P<0,001 по сравнению с тощими контрольными животными. *Р<0,05, **Р<0,01 по сравнению wt HFD-питающимися.
Фигура 10В представляет собой графическое изображение, показывающее анализ альбумина в моче у HFD-питающийся WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных. Значения представляют собой средние значения ± стандартная ошибка среднего, #P<0,05, ##P<0,01, ###P<0,001 по сравнению с тощими контрольными животными. *Р<0,05, **Р<0,01 по сравнению wt HFD-питающимися.
Фигура 11А представляет собой графическое изображение, показывающее количественное определение клубочкового мезангиального расширения у HFD-питающийся WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных. n=5-10/группу. Значения представляют собой средние значения ± стандартная ошибка среднего, Масштабная линейка, 50 мм. #P<0,05, ###P<0,001 по сравнению с тощими контрольными животными. **P<0,01 по сравнению wt HFD-питающимися.
Фигура 11В представляет собой графическое изображение, показывающее количественное определение клубочковой зоны у HFD-питающийся Vegfb+/- и HFD-питающийся Vegfb-/- мышей. Аналил HFD-питающийся WT, Vegfb+/-, Vegfb-/- мышей и тощих контрольных животных. n=5-10/группу. Значения представляют собой средние значения ± стандартная ошибка среднего, Масштабная линейка, 50 мм. #P<0,05, ###P<0,001 по сравнению с тощими контрольными животными. **P<0,01 по сравнению с wt HFD-питающимися.
Фигура 12 представляет собой графическое изображение, показывающее количественное определение окрашивания ORO в срезах почек у HFD-питающийся WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных. n=5-10/группу. Значения представляют собой средние значения ± стандартная ошибка среднего, ##P<0,01 по сравнению с контрольными животными наклоняться. *Р<0,05 по сравнению с wt HFD-питающимися.
Фигура 13 представляет собой графическое изображение, показывающее количественное определение окрашивания адипофилином в клубочках срезов почек у HFD-питающийся WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных. Значения представляют собой средние значения ± стандартная ошибка среднего, #P<0,05 по сравнению с тощими контрольными животными. *Р<0,05 по сравнению с wt HFD-питающимися.
Фигура 14A представляет собой графическое изображение, показывающее количественное определение окрашивания коллагеном IV в срезах почек у HFD-питающийся WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных, n=5-10/группа. Значения представляют собой средние значения ± стандартная ошибка среднего, #P<0,05, ##P<0,01 по сравнению с тощими контрольными животными *P<0,05 **P<0,01 по сравнению с wt HFD-питающимися.
Фигура 14B представляет собой графическое изображение, показывающее количественное определение толщины артериол в срезах почек у HFD-питающийся WT, HFD-питающийся Vegfb+/-, HFD-питающийся Vegfb-/- мышей и тощих контрольных животных, n=5-10/группа. Значения представляют собой средние значения ± стандартная ошибка среднего, #P<0,05, ##P<0,01 по сравнению с тощими контрольными животными, *Р<0,05, **P<0,001 по сравнению с wt HFD-питающимися.
Фигуры 15A-F представляют собой графическое изображение, показывающее анализ мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело (обозначено как «С») в течение 8 недель. Фигура 15A показывает постпрандиальной уровень глюкозы в крови у не получавших лечение животных. Стрелки показывают уровни глюкозы на старте профилактического или терапевтического испытания. Фигура 15В показывает ACR у не получавших лечение db/db//BKS животных. Стрелки указывают ACR в начале профилактического или терапевтического испытания. Фигуры 15C и 15D показывают постпрандиальной уровень глюкозы в крови от профилактического лечения анти-VEGF-B у db/db//BKS, показано отдельно для самцов (6C, n=5/группа) и самок (6D, n=5/группа) мышей. Фигуры 15E и 15F показывают постпрандиальной уровень глюкозы в крови от терапевтического лечения анти-VEGF-B у db/db//BKS, показано отдельно для самцов (15E, n=5/группа) и самок (15F, n=5/группа) мышей. Период введения анти-VEGF-B (2H10) лечения указывается в соответствующих графах.
Фигура 16А является графическим изображением, показывающим количественное определение гломерулосклероза, измеренного по мезангиальному расширению в срезах почек у мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом, в течение 8 недель (как указано). Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05, **P<0,01 по сравнению с получавшими контрольное лечение мышами db/db//BKS.
Фигура 16В представляет собой графическое изображение, показывающее количественное определение тубулярного склероза, измеренная по размеру клубочков срезах почек мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом, в течение 8 недель (как показано) профилактическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *Р<0,05, **P<0,01 по сравнению с получавшими контрольное лечение мышами db/db//BKS.
Фигура 17А представляет собой графическое изображение, показывающее количественное определение клубочковое окрашивание подоцина срезах почек мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом профилактическим способом, или контрольных db+ мышей. Значения представляют собой средние значения ± стандартная ошибка среднего. ##P<0,01, по сравнению с тощими db+ контролями. **P<0,01, ***Р<0,001 по сравнению с получавшими контрольное лечение мышами db/db. n=3-7/группу.
Фигура 17В представляет собой графическое изображение, показывающее количественное определение клубочкового окрашивания пекама срезах почек мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом профилактическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05, **P<0,01 по сравнению с контрольной группой.
Фигура 18A представляет собой графическое изображение, показывающее количественное определение клубочкового окрашивания адипофилина в срезах почек мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом профилактическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. #P<0,05, по сравнению с тощими db+ контролями. (n=3-7/группа).
Фигура 18В представляет собой графическое изображение, показывающее количественное определение клубочкового окрашивания синаптоподина в срезах почек мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом профилактическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05. **P<0,01 по сравнению с контрольной группой. (n=3-5/группу).
Фигура 19 представляет собой графическое изображение, показывающее количественное определение клубочкового окрашивания коллагена IV в срезах почек мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом профилактическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *Р<0,05, **Р<0,01 по сравнению с контрольной группой. (n=3-5/группу).
Фигуры 20A-C представляют собой серию графических изображений, показывающих, что фармакологическое ингибирование VEGF-B при помощи 2H10 профилактическим способом улучшает липидный профиль плазмы у диабетических мышей db/db//BKS. Анализ мышей db/db//BKS, получавших лечение анти-VEGF-B (2H10) или контрольным антителом (n=8/группа) и тощих db/+ животных (n=3). Графики показывают плазменные уровни (Фигура 11 A) HDL-c и LDL-c, (Фигура 11В) неэтерифицированных FA (NEFA) и (Фигура 11В) кетонов. #/*Р<0,05, ##/**P<0,01 по сравнению с тощими db+ или по сравнению с получавшими контрольное лечение db/db. Значения представляют собой средние значения ± стандартная ошибка среднего.
Фигура 21А представляет собой графическое изображение, показывающее количественное определение толщины GBM в срезах почек тощих db+ и db/db//BKS мышей, профилактически получавших анти-VEGF-B (2H10) или контрольное антитело. n=3-4/группа. Значения представляют собой средние значения ± стандартная ошибка среднего, ##P<0,01 по сравнению с тощими db+ контролями и **P<0,01 по сравнению с получавшими контрольное лечение db/db.
Фигура 21В представляет собой графическое изображение, показывающее количественное определение числа щелей в срезах почек тощих db+ и db/db//BKS мышей, профилактически получавших анти-VEGF-B (2H10) или контрольное антитело. n=3-4/группа. Значения представляют собой средние значения ± стандартная ошибка среднего, ##P<0,01 по сравнению с тощими db+ контролями и **P<0,01 по сравнению с получавшими контрольное лечение db/db.
Фигура 22 представляет собой графическое изображение, показывающее количественное определение окрашивания масляным красным O как в клубочковых, так и в тубулярных компартментах в срезах почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело, и тощих db+ животных. n=3-7/группа. Значения представляют собой средние значения ± стандартная ошибка среднего. ###P<0,001, по сравнению с тощими db+ контролями. **P<0,01 по сравнению с получавшими контрольное лечение db/db.
Фигура 23А представляет собой графическое изображение, показывающее количественное определение гломерулосклероза, определенного по мезангиальному расширению в срезах почек самок мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель (как указано) в терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05, **P<0,01, ***Р<0,001 по сравнению с контрольной группой.
Фигура 23B представляет собой графическое изображение, показывающее количественное определение гломерулосклероза, определенного по мезангиальному расширению в срезах почек самцов мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель (как указано) терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего.
Фигура 24 представляет собой графическое изображение, показывающее количественное определение окрашивания масляным красным O срезов почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель (как указано) терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего.*P<0,05, **Р<0,01, ***P<0,001 по сравнению с контрольной группой.
На Фигуре 25А представлено графическое изображение, показывающее количественное определение клубочкового пекам-окрашивания в срезах почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего.*P<0,05, *Р<0,01 по сравнению с контрольной группой.
На Фигуре 25В представлено графическое изображение, показывающее количественное определение клубочкового окрашивания синаптоподина в срезах почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *Р<0,05, **P<0,01 по сравнению с контрольной группой (n= 4-5/группу).
Фигура 26A представляет собой графическое изображение, показывающее количественное определение клубочкового окрашивания коллагена IV в срезах почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *Р<0,05 по сравнению с контрольной группой.
Фигура 26B представляет собой графическое изображение, показывающее количественное определение клубочкового α-SMA окрашивания в срезах почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *Р<0,05, **Р<0,01 по сравнению с контрольной группой (n=4-5/группа).
Фигура 26C представляет собой графическое изображение, показывающее количественное определение клубочковой толщины артериол в срезах почек мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело терапевтическим способом. Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05, **Р<0,01 по сравнению с контрольной группой.(n=4-5/группа).
Фигура 27А представляет собой графическое изображение, показывающее ACR в моче, измеренный способом ELISA, у мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель или худых животных (n=3-7/группу). Значения представляют собой средние значения ± стандартная ошибка среднего. Начальная и конечная точка показывает ACR до и после периода лечения, соответственно; *Р<0,05, **Р<0,01 по сравнению с животными, получавшими контрольное лечение.
На Фигуре 27В представлено графическое изображение, показывающее скорость клубочковой фильтрации, измеренный как выведение креатинина у мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель или худых животных (n=3-7/группу). Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05, **P<0,01, ***P<0,001 по сравнению с db/db животными, получавшими контрольное лечение.
Фигура 27C представляет собой графическое представление, показывающее артериальное давление, определенное при помощи хвостовой манжеты, (систолическое и диастолическое артериальное давление) у мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель или худых животных (n=3-7/группу). Значения представляют собой средние значения ± стандартная ошибка среднего. *Р<0,05, **Р<0,01, ***P<0,001 по сравнению с db/db животными, получавшими контрольное лечение.
Фигура 27D представляет собой серию из трех графических представлений, показывающих массу тела (график слева), массу почек (центральный график) и отношение массы тела к массе почек (график справа) у мышей db/db//BKS, получавших анти-VEGF-B (2H10) или контрольное антитело в течение 8 недель или худых животных (n=4/группу). Значения представляют собой средние значения ± стандартная ошибка среднего. *P<0,05, *Р<0,01 ***P<0,001 по сравнению с db/db животными, получавшими контрольное лечение.
Фигура 28 представляет собой графическое представление, показывающее уровни глюкозы в крови у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых контрольных животных. Постпрандиальные уровни глюкозы в крови в конце периода лечения. Значения представляют собой средние значения ± стандартная ошибка среднего. ##P<0,01 по сравнению с худыми контрольными животными.
На Фигуре 29А представлен график, показывающий уровни альбумина в моче у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых контрольных животных (n=5-10 на группу). ACR, соотношение альбумин/креатинин. Значения представляют собой средние значения ± стандартная ошибка среднего. #P<0,05, ###P<0,001 по сравнению с худыми контрольными животными. ***P<0,001 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 29B представляет собой графическое изображение, показывающее соотношение альбумин/креатинин в моче, измеренное с помощью ELISA, у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых контрольных животных (n=5-10 на группу). Значения представляют собой средние значения ± стандартная ошибка среднего. #P<0,05, ###P<0,001 по сравнению с худыми контрольными животными. ***P<0,001 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 30А является графическим изображением, показывающим уровни триглицеридов в плазме HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых животных дикого типа (n=5-10 на группу). #P<0,05 »: ##P<0,01 по сравнению с худыми контрольными животными. Значения представляют собой средние значения ± стандартная ошибка среднего.*Р<0,05, по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 30B представляет собой графическое представление, показывающее плазменные уровни неэтерифицированных FA (NEFA) у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых животных дикого типа (n=5-10 на группу). #P<0,05, ##P<0,01, по сравнению с худыми контрольными животными. Значения представляют собой средние значения ± стандартная ошибка среднего.*Р<0,05, по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 30C представляет собой графическое изображение, показывающее плазменные уровни кетонов у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых животных дикого типа (n=5-10 на группу). #P<0,05, ##P<0,0I, по сравнению с контрольными животными наклоняться. Значения представляют собой средние значения ± стандартная ошибка среднего.*Р<0,05, по сравнению с HFD-питающимися, получавшими контрольное лечение.
На Фигуре 31А представлено графическое изображение, показывающее количественное определение клубочкового мезангиального расширения у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых животных дикого типа, n=5-10 на группу. Значения представляют собой средние значения ± стандартная ошибка среднего. #P<0,05, ###P<0,001 по сравнению с худыми контрольными животными.***P<0,001 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 31B представляет собой графическое изображение, показывающее количественное определение клубочковой области у HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых животных дикого типа (n=5-10 на группу). Значения представляют собой средние значения ± стандартная ошибка среднего. #P<0,05, ###P<0,001 по сравнению с худыми контрольными животными, ***P<0,001 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 32 представляет собой графическое изображение, показывающее количественное определение окрашивания масляным красным O клубочков срезов почек HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых контрольное животных. ##P<0,01 по сравнению с худым контролем и **P<0,01 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 33A представляет собой графическое изображение, показывающее количественное определение окрашивания коллагена IV в срезах почек HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых контрольное животных. #P<0,05 по сравнению с худыми контрольными животными. *P<0,05 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 33В представляет собой графическое изображение, показывающее количественное определение толщины артериол в срезах почек HFD-питающихся мышей, получавших терапевтически анти-VEGF-B (2H10) или контрольное антитело, и худых контрольное животных. #P<0,05 по сравнению с худыми контрольными животными. *Р<0,05 по сравнению с HFD-питающимися, получавшими контрольное лечение.
Фигура 34A представляет собой графическое изображение, показывающее уровни глюкозы в крови до и после инъекций стрептозотоцина (STZ). Худым контрольным животным STZ не вводили. После STZ-инъекций, животных с установленной гипергликемией (уровень глюкозы в крови более 12 ммоль) дозировали анти-VEGF-B (2H10) или контрольным антителом. (n=3-7/группа). Значения представляют собой средние значения ± стандартная ошибка среднего. ##P<0,01 ###P<0,001 по сравнению с худыми контрольными животными. STZ, стрептозотоцин.
Фигура 34В представляет собой графическое изображение, показывающее уровень глюкозы в крови у мышей, получавших STZ (отличных от худых контрольных животных) и получавших анти-VEGF-B (2H10) или контрольное антитело, и худых контрольных животных. Худым контрольным животным STZ не вводили (n=3-7/группа). Значения представляют собой средние значения ± стандартная ошибка среднего. ##P<0,01, ###P<0,001 по сравнению с худыми контрольными животными. STZ, стрептозотоцин.
Фигура 35А представляет собой графическое изображение, показывающее соотношение альбумин/креатинин (ACR) у мышей перед введением STZ и после введения STZ с дозированием анти-VEGF-B (2H10) или контрольного антитела в течение 1 недели. Худым контрольным животным и db/db животным STZ не вводили. Соотношение альбумин/креатинин в моче измеряли с помощью ELISA (n=3-6/группу). ACR, соотношение альбумин/креатинин. Значения представляют собой средние значения ± стандартная ошибка среднего. ##P<0,01 по сравнению с животными до введения STZ. *Р<0,05 по сравнению с контрольными STZ-получавшими животными. STZ, стрептозотоцин.
Фигура 35В представляет собой графическое изображение, показывающее уровень альбумина у мышей перед введением STZ и после введения STZ с дозированием анти-VEGF-B (2H10) или контрольного антитела в течение 1 недели. Худым контрольным животным STZ не вводили. Уровни альбумина в моче измеряли с помощью ELISA (n=3-6/группу). Значения представляют собой средние значения ± стандартная ошибка среднего. ##P<0,01 по сравнению с животными до введения STZ. *Р<0,05 по сравнению с контрольными STZ-получавшими животными. STZ, стрептозотоцин.
КЛЮЧ К ПЕРЕЧНЮ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 представляет собой аминокислотную последовательность VEGF-B186-изоформу человека, содержащую 21-аминокислотную N-концевую сигнальную последовательность
SEQ ID NO: 2 представляет собой аминокислотную последовательность VEGF-B167-изоформу человека, содержащую 21-аминокислотную N-концевую сигнальную последовательность
SEQ ID NO: 3 представляет собой аминокислотную последовательность из VH антитела 2H10.
SEQ ID NO: 4 представляет собой аминокислотную последовательность из VL 2H10 антител.
SEQ ID NO: 5 представляет собой аминокислотную последовательность из VH гуманизированной формы антитела 2H10.
SEQ ID NO: 6 представляет собой аминокислотную последовательность из VL гуманизированной формы антитела 2H10.
SBQ ID NO: 7 представляет собой аминокислотную последовательность из VH антитела 4E12,
SEQ ID NO: 8 представляет собой аминокислотную последовательность из VL антитела 4E12.
SEQ ID NO: 9 представляет собой аминокислотную последовательность из VH антитела 2F5.
SEQ ID NO: 10 представляет собой аминокислотную последовательность из VL антитела 2F5.
SEQ ID NO: 11 представляет собой нуклеотидную последовательность из VL CDR1 антитела 2H10.
SEQ ID NO: 12 представляет собой нуклеотидную последовательность из CDR2 антитела 2HIG.
SEQ ID NO: 13 представляет собой нуклеотидную последовательность из VL CDR3 антитела 2H10.
SEQ ID NO: 14 представляет собой нуклеотидную последовательность из VH CDR1 антитела 2R10.
SEQ ID NO: 15 представляет собой нуклеотидную последовательность из VH CDR2 антитела 2H10.
SEQ ID NO: 16 представляет собой нуклеотидную последовательность из VH CDR3 антитела 2H1Q.
SEQ ID NO: 17 представляет собой аминокислотную последовательность из VL CDR1 антитела 2H10.
SEQ ID NO: 18 представляет собой аминокислотную последовательность из VL CDR2 антитела 2H10.
SEQ ID NO: 19 представляет собой аминокислотную последовательность из VL CDR3 антитела 2H10.
SEQ ID NO: 20 представляет собой аминокислотную последовательность из VH CDR1 антитела 2H10.
SEQ ID NO: 21 представляет собой аминокислотную последовательность из VH CDR2 антитела 2H10.
SEQ ID NO: 22 представляет собой аминокислотную последовательность из VH CDR3 антитела 2HIQ.
SEQ ID NO: 23 представляет собой нуклеотидную последовательность из VL CDR1 антитела 2F5.
SEQ ID NO:; 24 представляет собой нуклеотидную последовательность из VL CDR2 антитела 2F5.
SEQ ID NO: 25 представляет собой нуклеотидную последовательность из VL CDR3 антитела 2F5.
SEQ ID NO: 26 представляет собой нуклеотидную последовательность из VH CDR1 антитела 2F5.
SEQ ID NO: 27 представляет собой нуклеотидную последовательность из VH CDR2 антитела 2F5.
SEQ ID NO: 28 представляет собой нуклеотидную последовательность из VH CDR3 антитела 2F5.
SEQ ID NO: 29 представляет собой аминокислотную последовательность из VL CDR1 антитела 2F5.
SEQ ID NO: 30 представляет собой аминокислотную последовательность из VL CDR2 антитела 2F5.
SEQ ID NO: 31 представляет собой аминокислотную последовательность из VL CDR3 антитела 2F5.
SEQ ID NO: 32 представляет собой аминокислотную последовательность из VH CDR1 антитела 2F5.
SEQ ID NO: 33 представляет собой аминокислотную последовательность из VH CDR2 антитела 2F5.
SEQ ID NO: 34 представляет собой аминокислотную последовательность из VH CDR3 антитела 2F5.
SEQ ID NO: 35 представляет собой нуклеотидную последовательность из VL CDR1 антитела 4E12.
SEQ ID NO: 36 представляет собой нуклеотидную последовательность из VL CDR2 антитела 4E12.
SEQ ID NO: 37 представляет собой нуклеотидную последовательность из VL CDR3 антитела 4E12.
SEQ ID NO: 38 представляет собой нуклеотидную последовательность из VH CDR1 антитела 4E12.
SEQ ID NO; 39 представляет собой нуклеотидную последовательность из VH CDR2 антитела 4E12.
SEQ ID NO: 40 представляет собой нуклеотидную последовательность из VH CDR3 антитела 4E12.
SEQ ID NO: 41 представляет собой аминокислотную последовательность из VL CDR1 антитела 4E12.
SEQ ID NO; 42 представляет собой аминокислотную последовательность из VL CDR2 антитела 4E12.
SEQ ID NO: 43 представляет собой аминокислотную последовательность из VL CDR3 антитела 4E12.
SEQ ID NO: 44 представляет собой аминокислотную последовательность из VH CDR1 антитела 4EI2.
SEQ ID NO: 45 представляет собой аминокислотную последовательность из VH CDR2 антитела 4EI2.
SEQ ID NO:; 46 представляет собой аминокислотную последовательность из VH CDR3 антитела 4EI2.
ПОДРОБНОЕ ОПИСАНИЕ
Общее
В данном описании, если конкретно не указано иное или иное требуется из контекста, ссылка на отдельную стадию, состав вещества, группу стадий или группу составов веществ следует принимать как охватываемые одним и множеством (т.е. одним или более) из этих стадий, составов веществ, групп стадий или групп составов веществ.
Специалистам в данной области техники должно быть понятно, что настоящее описание допускает изменения и модификации, отличные от тех, которые конкретно описаны. Следует понимать, что раскрытие включает в себя все такие изменения и модификации. Настоящее изобретение также включает все стадии, особенности, композиции и соединения, упомянутые или указанные в данном описании, в отдельности или в совокупности, а также любые и каждую из комбинаций или любые две или более из указанных стадий или особенностей.
Настоящее изобретение не должно быть ограничено в объеме конкретными примерами, описанными в настоящем документе, которые предназначены только для иллюстративных целей. Функционально эквивалентные продукты, композиции и способы, несомненно, находятся в пределах объема настоящего изобретения.
Любой из примеров осуществления настоящего изобретения в настоящем документе должен быть принят с внесением соответствующих изменений для применения к любому другому примеру по раскрытию, если конкретно не указано иное.
Любой из примеров по настоящему изобретению в отношении лечения или профилактики нефропатии должны быть приняты с соответствующими изменениями применительно к ингибированию или профилактике врожденного иммунного ответа (например, врожденного иммунного ответа в пищеварительной системе и/или системного врожденного иммунного ответа) у индивидуума, страдающего от нефропатии.
Если специально не указано иное, все технические и научные термины, используемые в настоящем документе, должны рассматриваться в том же значении, которое обычно подразумевается любым специалистом в данной области техники (например, в культуре клеток, молекулярной генетике, иммунологии, иммуногистохимии, химии белков и биохимии).
Если не указано иное, способы рекомбинантных белков, культур клеток и иммунологические способы, используемые в настоящем изобретении, представляют собой стандартные процедуры, хорошо известные специалистам в данной области техники. Такие способы описаны и объяснены по всей литературе, в источниках, таких как, J. Perbal, A Practical Guide to Molecular Cloning, John Wiley and Sons (1984), J. Sambrook et al. Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989), T.A. Brown (editor), Essential Molecular Biology: A Practical Approach, Volumes 1 and 2, IRL Press (1991), D.M. Glover and B.D. Hames (editors), DNA Cloning: A Practical Approach, Volumes 1-4, IRL Press (1995 and 1996), и F.M. Ausubel et al. (editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, including all updates until present), Ed Harlow and David Lane (editors) Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, (1988) и J.E. Coligan et al. (editors) Current Protocols in Immunology, John Wiley & Sons (включая все обновления до настоящего времени).
Описание и объяснения вариабельных областей и их частей, иммуноглобулинов, антител и их фрагментов в настоящем документе можно дополнительно прояснить в Kabat Sequences of Proteins of Immunological Interest, National Institutes of Health, Bethesda, Md., 1987 and 1991, Bork et al., J Mol. Biol. 242, 309-320, 1994, Chothia and Lesk J. Mol Biol. 196:901 -917, 1987, Chothia et al. Nature 342, 877-883, 1989 и/или или Al-Lazikani et al., J Mol Biol 273, 927-948, 1997.
Любое рассмотрение белка или антитела в данном описании следует понимать как включающее любые варианты белка или антитела, продуцируемые в процессе производства и/или хранения. Например, в процессе изготовления или хранения антитело может быть дезамидировано (например, по остатку аспарагина или глутамина) и/или обладать измененным гликозилированием и/или обладать остатком глутамина, преобразованного в пироглутамин и/или обладать удаленным или "отщепленным" N-концевым или C-концевым остатком и/или обладать частью или всей сигнальной последовательностью, не полностью процессированной и, как следствие, остающейся на конце антитела. Понятно, что композиция, содержащая определенную аминокислотную последовательность, может представлять собой гетерогенную смесь указанной или кодируемой последовательности и/или варианты этой указанной или кодируемой последовательности.
Термин "и/или", например, "X и/или Y" следует понимать либо как "X и Y", либо "X или Y", и он должны быть принят для обеспечения недвусмысленной поддержки для обоих значений или для любого из значений.
В данном описании слово "включать" или вариации, такие как "содержит" или "содержащий", следует понимать как подразумевающий включение указанного элемента, целого числа или стадии, или группы элементов, целых чисел или стадий, но не исключающий любого другого элемента, целого числа или стадии, или группы элементов, чисел или стадий.
Используемый в данном описании термин "полученный из" следует принимать для указания того, что заданное целое число может быть получено из определенного источника, хотя и не обязательно непосредственно из этого источника.
Выбранные определения
VEGF-B, как известно, существует в двух основных изоформах, именуемых как VEGF-B186 и VEGF-B167. Исключительно в целях терминологии, но не ограничения, типичные последовательности человеческого VEGF-B186 помещены в NCBI Reference Sequence: NP_003368.1, входящие номера NCBI для белков: NPJH 368, P49765 и AAL79001 и в SEQ ID NO: 1. В контексте настоящего раскрытия в последовательности VEGF-B186 может отсутствовать 21- аминокислотная N-концевая сигнальная последовательность (например, как приводится для аминокислот с 1 по 21 из SEQ ID NO: 1. Исключительно в целях терминологии, но не ограничения типичные последовательности человеческого VEGF-B167 приводится в NCBI Reference Sequence: NP_001230662.1, входящие номера NCBI для белков AAL7900Q и AAB06274 и в SEQ ID NO: 2. В контексте настоящего описания в последовательности VEGF-B167 может отсутствовать 21 аминокислотная N-концевая сигнальная последовательность (например, как установлено для аминокислот с 1 по 21 в SEQ ID NO: 2. Дополнительная последовательность VEGF-B может быть определена с использованием последовательностей, предоставленных в данном описании и/или в доступных для пользователей базах данных и/или определены с использованием стандартных способов (например, как описано в Ausubel et al., (editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, включая все обновления вплоть до настоящего времени) или Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989)). Ссылка на VEGF-B человека может быть сокращена до hVEGF-B. В одном примере ссылка в настоящем описании на VEGF-B представляет собой изоформу VEGF-B167.
Ссылка в настоящем документе на VEGF-B также включает пептид VEGF-B10-108, как описано в WO2006/012688.
Термин "нефропатия" следует понимать как означающий повреждение или заболевание почек. Этот термин охватывает все клинико-патологические изменения в почках, которые могут привести к фиброзу почек и/ или заболеваниям клубочков (например, гломерулосклероз, гломерулонефрит) и/или хронической почечной недостаточности, а также могут привести к конечной стадии почечной недостаточности и/или отказу почки. Типичные нефропатии включают гипертензивную нефропатию, диабетическую нефропатию, а также другие виды нефропатии, такие как анальгетическая нефропатия, иммунно-опосредованные гломерулопатии (например, IgA-нефропатия или болезнь Бергера, волчаночный нефрит), ишемическая нефропатия, ВИЧ-ассоциированная нефропатия, мембранозная нефропатия, гломерулонефрит, гломерулосклероз, радиоконтрастная медиа-индуцированная нефропатия, токсическая нефропатия, анальгетик-индуцированная нефротоксичность, цисплатиновая нефропатия, трансплантатная нефропатия и другие формы клубочковой патологии или повреждения; гломерулярное капиллярное повреждение (тубулярный фиброз). В некоторых примерах, термины "нефропатия" или "нефропатии" конкретно относится к расстройству или болезни, при которых наблюдается или наличие белков (т.е. протеинурии), таких как альбумин, в моче индивидуума и/или наличие почечной недостаточности. Нефропатия часто диагностируется на основании наличия альбумина в моче (микроальбуминурия или микроальбуминурия), повышения уровня мочевинного азота в крови, (например, уровни выше 20 мг/дл) и/или повышения уровня креатинина в сыворотке крови (например, уровни выше 1,3 мг/дл для мужчин и 1,1 мг/дл для женщин).
Термин "фиброз" относится к аномальному процессингу волокнистой ткани, или фиброидной или фиброзной дегенерации. Фиброз может возникнуть в результате различных травм или заболеваний. Фиброз, как правило, включает в себя аномальное продуцирование, накопление или отложение компонентов внеклеточного матрикса, в том числе сверхпродуцирование и увеличения отложения, например, коллагена и фибротектина. Используемые в настоящем описании термины "почечный фиброз" или "ренальный фиброз " или "фиброз почки" относятся к заболеваниям или расстройствам, связанным со сверхпродуцированием или аномальным отложением компонентов внеклеточного матрикса, в частности коллагена, что приводит к деградации или ухудшению функции почек.
Термин "нефрит" следует понимать в значении воспаления почки. В контексте настоящего описания, нефриты охватывает подмножество нефропатий, характеризуемых воспалением в почке. Воспаление может затрагивать клубочки, канальцы или интерстициальную ткань вокруг клубочков и канальцев. Как правило, нефрит представляет собой либо гломерулонефрит (т.е. воспаление клубочков), либо интерстициальный нефрит (т.е. воспаление пространства между почечными канальцами).
Термин "гломерулонефрит" охватывает класс заболеваний почек, которые могут быть разбиты на подклассы пролиферативных заболеваний и непролиферативных заболеваний. Как следует из названия, "пролиферативные" заболевания включают в себя формы гломерулонефрита, при которых наблюдается значительное увеличение числа клеток в клубочке, в то время как "непролиферативные" заболевания включают в себя формы гломерулонефрита, при которых такое увеличение числа клеток отсутствует. Примеры пролиферативных заболеваний включают IgA-нефропатию, постинфекционный гломерулонефрит, мембранопролиферативный гломерулонефрит и быстро прогрессирующий гломерулонефрит. Примеры непролиферирующих заболеваний включают болезнь минимальных изменений, очаговый сегментарный гломерулосклероз, полагают, заболевание базальной мембраны и перепончатый гломерулонефрит.
"Диабетическая нефропатия" представляет собой клинически четко определенную патологию, характеризующуюся протеинурией, гипертензией, отеками и почечной недостаточности. Характерные аспекты диабетической нефропатии включают гломерулосклероз, модификацию сосудистой структуры и тубулоинтерстициальную болезнь. Первым клиническим признаком диабетической нефропатии зачастую является наличие альбуминурии в моче, например, микроальбуминурии или макроальбуминурии. Диабетическая нефропатия, как правило, характеризуется следующим: 1) гломерулосклерозом, 2) модификацией сосудистой структуры, главным образом, в маленьких артериолах и 3) тубулоинтерстициальной болезнью. Наиболее характерным аспектом диабетической нефропатии является клубочковое повреждение, обнаруживаемое по расширению мезангиуми и по утолщению базальной мембраны, которая часто выглядит как диффузное рубцевание всего конгломерата. Первым клиническим признаком диабетической нефропатии является наличие альбуминурии или протеинурии.
Под "микроальбуминурией" подразумевается наличие 30-300 мг альбумина в моче, собранной за 24 часа, и/или 30-300 мг/л альбумина в одном образце. Как правило, каждое из вышесказанного следует определять, по меньшей мере, в двух из трех образцов в течение двух-трех месячного периода. Микроальбуминурия также может быть определена по отношению альбумина к креатинину (ACR) ≥3,5 мг/ммоль для женщин или ≥2,5 мг/ммоль для мужчин или от 30 до 300 мкг альбумина/мг креатинина. Уровни альбумина можно оценить с использованием, например, коммерчески доступных тест-полосок (например, включающих бромфеноловый синий в качестве индикатора).
Термин "макроальбуминурия" означает присутствие количеств альбумина, больших (или больших по ACR), чем наблюдается при микроальбуминурии.
Термин "протеинурия" означает, что количество общего белка в моче составляет около 30 мг/дл или соотношение белок/креатинин больше, чем 45 мг/ммоль.
"Гипертензия" относится к индивидууму (например, индивидууму человека), имеющему систолическое давление 140 мм рт.ст. или выше и/или диастолическое давление 90 мм рт.ст. или выше. В некоторых примерах способа или применения, описанного в настоящем документе, индивидуум является предгипертензивным, например, имеющим систолическое давление около 120-139 мм рт.ст. или выше и/или диастолическое давление 80-89 мм рт.ст. или выше.
Термин "рекомбинантный" следует понимать как означающий продукт искусственной генетической рекомбинации. Соответственно, в контексте рекомбинантного белка, содержащего вариабельную область антитела, этот термин не включает в себя антитело, природно присутствующее в организме индивидуума, которое является продуктом естественной рекомбинации, которая происходит во время созревания В-клеток. Тем не менее, если такое антитело выделяют, оно рассматривается как выделенный белок, содержащий вариабельную область антитела. Точно так же, если нуклеиновую кислоту, кодирующую белок, выделяют и экспрессируют с использованием рекомбинантных способов, то полученный белок представляет собой рекомбинантный белок, содержащий вариабельную область антитела. Рекомбинантный белок также охватывает белок, экспрессируемый искусственным рекомбинантным способом, когда он находится внутри клетки, ткани или индивидуума, в которых он экспрессирован.
Термин "белок" следует принимать как включающий одну полипептидную цепь, то есть ряд смежных аминокислот, соединенных пептидными связями или ряд полипептидных цепей, ковалентно или нековалентно связанных друг с другом (т.е. полипептидный комплекс). Например, ряд полипептидных цепей может быть ковалентно связан при помощи подходящей химической или дисульфидной связи. Примеры нековалентные связи включают водородные связи, ионные связи, ван-дер-ваальсовы силы и гидрофобные взаимодействия.
Термин "полипептид" или "полипептидная цепь" следует понимать, исходя из предыдущего абзаца, как означающий ряд последовательных аминокислот, соединенных пептидными связями.
Опытный специалист должен осознавать, что "антитело", как правило, рассматривается как белок, который содержит вариабельную область, состоящую из множества полипептидных цепей, например, полипептид, содержащий вариабельную область легкой цепи (VL) и полипептид, содержащий вариабельную область тяжелой цепи (VH). Также антитело, как правило, включает константные домены, некоторые из которых могут быть организованы в константную область, которая включает в себя: константный фрагмент или кристаллизующийся фрагмент (Fc), в случае тяжелой цепи. VH и VL взаимодействуют с образованием Fv, содержащего антигенсвязывающую область, которая способна специфически связываться с одним или несколькими близкородственными антигенами. Как правило, легкая цепь из млекопитающих является либо легкой цепью κ, либо легкой цепью λ и тяжелая цепь из млекопитающих представляет собой α, δ, ε, γ или μ. Антитела могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса. Термин "антитело" также включает гуманизированные антитела, приматизированные антитела, человеческие антитела, сингуманизированные антитела и химерные антитела.
Термины "полноразмерное антитело", "интактные антитела" или "целое антитело" используются взаимозаменяемо для обозначения антитела в его, по существу, интактной форме, в отличие от антигенсвязывающего фрагмента антитела. В частности, целые антитела включают антитела с тяжелой и легкой цепями, включая Fc-область. Константные домены могут представлять собой константные домены с последовательностью типа дикого (например, константные домены с последовательностью дикого типа человека) или варианты аминокислотной последовательности этого.
Используемый в данном описании термин "вариабельная область" относится к частям легких и/или тяжелой цепей антитела, как определено в настоящем документе, которая способна специфически связываться с антигеном и включает аминокислотные последовательности гипервариабельных участков (CDR); т.е. CDR1, CDR2 и С-DR3. и каркасные области (FR). Типичные вариабельные области включают три или четыре FR (например, FR1, FR2, FR3 и необязательно FR4) вместе с тремя CDR. В случае белка, полученного из IgNAR, в белке может отсутствовать CDR2. VH относится к вариабельной области тяжелой цепи. VL относится к вариабельной области легкой цепи.
Используемый в данном описании термин "гипервариабельные участки" (syn. CDR; т.е. CDR1, CDR2 и CDR3) относится к аминокислотным остаткам вариабельного домена антитела, наличие которого необходимо для связывания антигена. Каждый вариабельный домен содержит, как правило, три CDR-области, определенные как CDR1. CDR2 и CDR3. Положения аминокислот, присвоенные CDR и FR, могут быть определены в соответствии с Kabat Sequences of Proteins of Immunological Interest, National Institutes of Health, Bethesda, Md., 1987 и 1991 или других систем нумерации при выполнении данного описания, например, канонической системой нумерации Chothia and Lesk J. Mol Biol. 196: 901-917, 1987; Chothia et al. Nature 342, 877-883, 1989; и/или Al-Lazikani et al., J Mol Biol 273: 927-948, 1997; системой нумерации IMGT Lefranc et al., Devel. And Compar. Immunol., 27: 55-77, 2003; или системой нумерации AHO, Honnegher and Plükthun J. Mol. Biol., 309: 657-670, 2001.
"Каркасные области" (FR) представляют собой те остатки вариабельного домена, которые отличны от CDR-остатков.
Используемый в данном описании термин "Fv" был принят для обозначения любого белка, состоящего из нескольких полипептидов или одного полипептида, в котором VL и VH ассоциированы и образуют комплекс, имеющий сайт связывания с антигеном, т.е. способного специфически связываться с антигеном. VH. и VL, которые образуют участок связывания антигена, могут находиться в одной полипептидной цепи или в различных полипептидных цепях. Кроме того, Fv по раскрытию (а также как любой белок раскрытия) может иметь несколько сайтов связывания антигена, которые могут или не могут связывать один и тот же антиген. Этот термин следует понимать как охватывающий фрагменты, полученные непосредственно из антитела, а также белков, соответствующих такому фрагменту, полученных с использованием рекомбинантных способов. В некоторых примерах VH не связан с константным доменом тяжелой цепи (CH) 1 и/или VL не связан с константным доменом легкой цепи (CL). Типичные Fv-содержащие полипептиды или белки включают Fab-фрагмент, Fab'-фрагмент, F(ab')-фрагмент, scFv, диатело, триатело, тетратело или комплекс более высокого порядка, или любое из вышеперечисленного, связанного с константной областью или доменом этого, например, домен CH2 или CH3, например, минитело. "Fab-фрагмент" состоит из одновалентного антигенсвязывающего фрагмента антитела, и может быть получен расщеплением целого антитела ферментом папаином, чтобы получить фрагмент, состоящий из интактной легкой цепи и части тяжелой цепи, или может быть получен с использованием рекомбинантных способов. "Fab-фрагмент" антитела может быть получен путем обработки целого антитела пепсином, с последующим редуцированием, с получением молекулы, состоящей из интактной легкой цепи и части тяжелой цепи, содержащей VH и единственный константный домен. Получают два Fab'-фрагмента на антитело, обработанное таким образом. Fab'-фрагмент также может быть получен рекомбинантными способами. "F(ab')2 фрагмент" антитела состоит из димера двух Fab'-фрагментов, удерживаемых вместе двумя дисульфидными связями, и его получают обработкой целой молекулы антитела ферментом пепсином, без последующего восстановления. ʺFab2ʺ-фрагмент представляет собой рекомбинантный фрагмент, содержащий два Fab-фрагмента, связанных при помощи, например, лейциновой молнии или CH3-домена. «Одноцепочечный Fv» или ʺscFvʺ представляет собой рекомбинантную молекулу, содержащую фрагмент вариабельной области (Fv) антитела, в котором вариабельная область легкой цепи и вариабельная область тяжелой цепи ковалентно соединены подходящим гибким полипептидным линкером.
Используемый в данном описании термин "связывает" в связи с взаимодействием белка или его антигенсвязывающего участка с антигеном означает, что взаимодействие зависит от наличия определенной структуры (например, антигенной детерминанты или эпитопа) на антигене. Например, антитело распознает и связывается со специфической белковой структурой, а не с белками в целом. Если антитело связывается с эпитопом "А", присутствие молекулы, содержащей эпитоп "А" (или свободного, немаркированного "A"), в реакционной смеси, содержащей меченый "А" и белок, уменьшит количество меченого "A", связанного с антителом.
Используемый в данном описании термин "специфически связывается" или "связывается специфически" следует принимать в значении, что белок по раскрытию вступает в реакцию или ассоциируется чаще, быстрее, с большей продолжительностью и/или с более высокой аффинностью с определенным антигеном или клеткой, экспрессирующей таковой, чем с альтернативными антигенами или клетками. Например, белок связывается с VEGF-B с существенно большей аффинностью (например, от 20 раз или 40 раз или 60 раз или 80 раз до 100 раз или 150 раз или 200 раз выше), чем с другим фактором роста (например, VEGF-А) или антигенами, обычно распознаваемыми натуральными антителами (т.е. природными антителами, о которых известно, что они связываются с множеством антигенов, природно присутствующих в организме человека). Как правило, но не обязательно, ссылка на связывание означает специфическое связывание, и каждый термин следует понимать таким образом, чтобы обеспечивать недвусмысленное поддержание другого термина.
Используемый в данном описании термин "нейтрализовать" следует принимать в значении, что белок способен блокировать, уменьшать или препятствовать сигнальному пути VEGP-B в клетке через VEGF-R1. Способы определения нейтрализации известны в данной области техники и/или описаны в настоящем документе.
Используемый в данном описании термин "специфически ингибирует сигнальному пути VEGP-B" следует понимать как означающий, что соединение ингибирует сигнальный путь VEGP-B и не приводит к существенному или детектируемому ингибированию передачи сигналов с помощью одного или нескольких других белков VEGF, например, VEGF-A, VEGF-B, VEGF-C VEGF-D и/или PIGF.
Используемый в данном описании термин "несущественно ингибирует" следует понимать в значении, что уровень ингибирования передачи сигналов с помощью белка VEGF, за исключением VEGF-B (например, передача сигналов при помощи VEGF-A, VEGF-B, VEGF-C, VEGF-D и/или PIGF) в присутствии соединения, описанного в данном документе, не является статистически достоверно более низким, чем в отсутствие соединения, описанного в данном описании (например, в контрольном анализе, который можно проводить в присутствии изотипического контрольного антитела).
Используемый в данном описании термин "детектируемо не ингибирует" следует понимать как означающий, что соединение, как описано в настоящем документе, ингибирует передачу сигналов белками VEGF, за исключением VEGF-B (например, передача сигналов с помощью VEGF-A, VEGF-B, VEGF-С, VEGF-D и/или PIGF) не более чем на 10% или 8% или 6% или 5% или 4% или 3% или 2% или 1% от уровня передачи сигнала, детектируемого при отсутствии соединения, описанного в настоящем документе (например, в контрольном анализе, который можно проводить в присутствии изотипического контрольного антитела).
Используемые в настоящем описании термины "препятствующий", "предотвращать" или "предупреждение" включают введение соединения по раскрытию, чтобы таким образом остановить или препятствовать развитию, по меньшей мере, одного симптома состояния.
Используемые в настоящем описании термины "лечащий", "лечить" или "лечение" включают введение белка, описанного в настоящем документе, для ослабления или устранения тем самым, по меньшей мере, одного симптома определенного заболевания или состояния или для замедления прогрессирования заболевания или состояния.
Используемый в данном описании термин "индивидуум" следует понимать как означающий любое животное, в том числе человек, например млекопитающее. Типичные индивидуумы включают, в качестве неограничивающих примеров, людей и отличных от людей приматов. Например, индивидуум является человеком.
Лечение нефропатии
Раскрытие по настоящему описанию предоставляет, например, способ лечения или профилактики нефропатии у индивидуума, включающий введение индивидууму соединения, которое ингибирует сигнальный путь VEGF-B.
В одном примере индивидуум страдает сахарным диабетом. Например, у пациента, страдающего от диабета, имеется клинически принятый маркер диабета, такой как:
глюкоза в плазме натощак больше или равна 7 нмоль/л или 126 мг/дл;
повседневное содержание глюкозы в плазме (взятой в любое время дня) больше или равно 11,1 нмоль/л или 200 мг/дл с симптомами диабета.
Значение перорального теста на толерантность к глюкозе (ПТТГ) больше или равно 11,1 нмоль/л или 200 мг/дл, измеренное с двухчасовым интервалом. ПТТГ проводят с промежутком времени в два или три часа.
В одном примере индивидуум страдает от сахарного диабета 1 типа.
В одном примере индивидуум страдает от сахарного диабета 2 типа.
В одном примере индивидуум страдает от диабетической нефропатии. Например, индивидуум страдает от нефропатии, связанной с сахарным диабетом 1 типа. Например, индивидуум страдает от нефропатии, связанной с сахарным диабетом 2 типа.
В одном примере индивидуум подвержен риску развития диабетической нефропатии. Например, индивидуум подвержен риску развития нефропатии, связанной с диабетом 1 типа. Например, индивидуум подвержен риску развития нефропатии, связанной с сахарным диабетом 2 типа.
В одном примере индивидуум страдает от микроальбуминурии. В соответствии с этим примером, лечение в соответствии с настоящим изобретением может уменьшить выраженность микроальбуминурии (например, до менее чем приблизительно 30 мг альбумина за 24 часа сбора мочи и/или 30 мг/л альбумина в одном образце и/или ACR менее 3,5 мг/ммоль для женщин или менее 2,5 мг/ммоль для мужчин или меньше чем приблизительно 30 мкг альбумина/мг креатинина.
В другом примере лечение в соответствии с настоящим изобретением предотвращает или замедляет прогрессирование микроальбуминурии до макроальбуминурии.
В одном примере индивидуум страдает от пониженной скорости клубочковой фильтрации, например, как определяется по пониженной скорости выведения креатинина (например, ниже 90 мл/мм/1,73 м2). В одном примере способ по раскрытию повышает скорость клубочковой фильтрации.
В еще одном примере индивидуум страдает от гипертензии или прегипертензии. В одном примере способ по раскрытию эффективен в снижении систолического и/или диастолического давления крови у индивидуума, по меньшей мере, на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 мм ртутного столба или более.
В одном примере реализация способа, описанного в настоящем документе в соответствии с любым примером по раскрытию, приводит к повышению клинического ответа и/или замедлению прогрессирования заболевания.
Под термином "клинический ответ" понимают улучшение симптомов болезни. Клинический ответ может быть достигнут в течение определенного периода времени, например, в пределах, или около 8 недель с начала лечения, или от первоначального введения. Клинический ответ также может быть устойчивым в течение некоторого периода времени, например, в течение 24 недель или 48 недель.
Количественная оценка функции почек и показателей почечной дисфункции хорошо известны в данной области техники, и их можно найти, например, в Levey (Am J Kidney Dis. 22(1):207-214, 1993). Примерами анализов для определения функции/дисфункции почек являются: сывороточный уровень креатинина; скорость выведения креатинина; скорость выведения цистатина C, 24-часовое мочевое выведение креатинина, 24-часовой мочевая секреция белков; скорость клубочковой фильтрации (GFR); соотношение мочевого креатинина/альбумина (ACR); скорость экскреции альбумина (AER); и почечная биопсия.
Ингибиторы сигнального пути VEGF-B
Белки, содержащие вариабельные области антитела
Типичный ингибитор сигнального пути VEGF-B включает вариабельную область антитела, например, представляет собой антитело или фрагмент антитела, которое связывается с VEGF-B и нейтрализует сигнальный путь VEGF-B.
В одном примере вариабельная область антитела специфически связывается с VEGF-B.
Подходящие антитела и белки, включающие их вариабельные участки, известны в данной области техники.
Например, антитела против VEGF-B и их фрагменты описаны в WO2006/012688.
В одном из примеров антитело анти-VEGF-B или его фрагмент является антителом, которое конкурентно ингибирует связывание 2H10 или его антигенсвязывающего фрагмента с VEGF-B. В одном примере антитело анти-VEGF-B или его фрагмент представляет собой антитело 2H10 или его химерную, CDR-привитую или гуманизированную версии или его антигенсвязывающий фрагмент. В связи с этим, антитело 2H10 содержит VH, включающую последовательность, представленную в SEQ ID NO: 3, и VL включающую последовательность, представленную в SEQ ID NO: 4. Типичные химерные и гуманизированные варианты этого антитела описаны в WO2006/012688.
В одном примере антитело анти-VEGF-В или его фрагмент содержит VH, включающую последовательность, представленную в SEQ ID NO: 5, и VL, включающую последовательность, представленную в SEQ ID NO: 6.
В одном примере антитело анти-VEGF-B или его фрагмент представляет собой антитело, которое конкурентно ингибирует связывание 4E12 или его антигенсвязывающего фрагмента с VEGF-B. В одном примере антитело анти-VEGF-B или его фрагмент представляет собой антитело 4E12 или его химерную, CDR-привитую или гуманизированную версию или его антигенсвязывающий фрагмент. В связи с этим, антитело 4E12 содержит VH, включающую последовательность, представленную в SEQ ID NO: 7, и VL, включающую последовательность, представленную в SEQ ID NO: 8.
В одном примере соединение представляет собой белок, содержащий гуманизированную вариабельную область антитела 4Е12. Например, белок содержит вариабельную область, включающую гипервариабельные участки (CDR) VH и/или VL антитела 4E12. Например, белок включает:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 25-34 из SEQ ID NO: 7;
(b) CDR2, содержащий последовательность, представленную аминокислотами 49-65 из SEQ ID NO: 7; а также
(с) CDR3, содержащий последовательность, представленную аминокислотами 98-105 последовательности SEQ ID NO: 7; и/или
(II) VL, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 24-34 из SEQ ID NO: 8;
(b) CDR2, содержащий последовательность, представленную аминокислотами 50-56 из SEQ ID NO: 8; а также
(с) CDR3, содержащий последовательность, представленную аминокислотами 89-97 из SEQ ID NO: 8.
В одном примере антитело анти-VEGF-B или его фрагмент представляет собой антитело, которое конкурентно ингибирует связывание 2F5 или его антигенсвязывающего фрагмента с VEGP-B. В одном примере антитело анти-VEGF-B или его фрагмент представляет собой антитело 2F5 или его химерный, CDR-привитый или гуманизированный вариант или его антигенсвязывающий фрагмент. В связи с этим, антитело 2E5 содержит VH., включающую последовательность, представленную в SEQ ID NO: 9, и VL, включающую последовательность, представленную в SEQ ID NO: 10.
В одном примере соединение представляет собой белок, содержащий гуманизированную вариабельную область антитела 2F5. Например, белок включает вариабельную область, содержащую гипервариабельные участки (CDR) VH и/или VL антитела 2F5. Например, белок включает:
(I) VH, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 25-34 из SEQ ID NO: 9;
(b) CDR2, содержащий последовательность, представленную аминокислотами 49-65 из SEQ ID NO: 9; а также
(с) CDR3, содержащий последовательность, представленную аминокислотами 98-107 из SEQ ID NO: 9; и/или
(II) VL, включающую:
(a) CDR1, содержащий последовательность, представленную аминокислотами 24-34 из SEQ ID NO: 10;
(b) CDR2, содержащий последовательность, представленную аминокислотами 50-56 из SEQ ID NO: 10; а также
(с) CDR3, содержащий последовательность, представленную аминокислотами 89-96 из SEQ ID NO: 10.
В другом примере антитело или белок, содержащий его вариабельную область, продуцируют с использованием стандартного способа, например, известного в данной области техники или кратко описанного в настоящем документе.
Способы на основе иммунизации
Для получения антител VEGF-B или фрагмент несущий его эпитоп или часть, или его модифицированная форма или нуклеиновая кислота, его кодирующая ("иммуноген"), необязательно составленный с любым подходящим или желаемым адъювантом и/или фармацевтически приемлемым носителем, вводят индивидууму (например, не являющегося человеком индивидууму-животному, такому как мышь, крыса, курица и т.д.) в виде инъекционной композиции. Типичными отличными от человека животными являются млекопитающие, такие как мышевидные животные (например, крысы или мыши). Инъекция может быть интраназальной, внутримышечной, подкожной, внутривенной, внутрикожной, внутрибрюшинной, или при помощи другого известного способа. По желанию, иммуноген вводят множество раз. Средства для подготовки и характеристики антител известны в данной области (см области, например, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988). Способы получения антител анти-VEGF-B у мышей описаны в WO2006/012688.
Получения поликлональных антител можно контролировать путем отбора проб крови иммунизированных животных в различных точках после иммунизации. Второе, можно применять бустер-инъекцию, если это необходимо для достижения желаемого титра антитела. Процесс активизации и титрования повторяют до тех пор, пока соответствующий титр не будет достигнут. При получении желаемого уровня иммуногенности у иммунизированных животных отбирают кровь и сыворотку выделяют и хранят, и/или используют животных для генерации моноклональных антител (mAb).
Моноклональные антитела являются типичными антителами, рассматриваемыми настоящим изобретением. Как правило, продуцирование моноклональных антител включает в себя иммунизацию существа (например, грызуна, например, мыши или крысы) иммуногеном при условиях, достаточных для стимуляции антитело-продуцирующих клеток. В некоторых примерах мышь, генетически модифицированную для экспрессии человеческих антител и не для экспрессии белков мышиных антител, иммунизируют для получения антитела (например, как описано в PCT/US2007/008231 и/или Lonberg et al., Nature 368 (1994): 856-859). После иммунизации антитело-продуцирующие соматические клетки (например, B-лимфоциты) сливают с бессмертными клетками, например, бессмертными клетками миеломы. Различные способы получения таких слитых клеток (гибридом) известны в данной области техники и описаны, например, в Kohler and Milstein, Nature 256, 495-497, 1975. Гибридомные клетки затем можно культивировать в условиях, достаточных для выработки антител.
Настоящее изобретение рассматривает другие способы получения антител, например, технологию ABL-MYC (как описано, например, в Largaespada et al, Curr. Top. Microbiol. Immunol, 166, 91-96, 1990).
Способы на основе библиотек
Настоящее раскрытие также включает скрининг библиотек антител или белков, содержащих их антигенсвязывающие домены (например, включающие их вариабельные области), для идентификации VEGF-B-связывающего антитела или белка, содержащего его вариабельную область.
Примеры библиотек, предусмотренных настоящим раскрытием, включают наивные библиотеки (из не испытывающих воздействия существ), иммунизированные библиотеки (от существ, иммунизированных антигеном) или синтетические библиотеки. Нуклеиновые кислоты, кодирующие антитела и их области (например, вариабельные области), клонируют с помощью традиционных способов (например, как описано в Sambrook and Russell, eds, Molecular Cloning: A Laboratory Manual, 3rd Ed, vols. 1-3, Cold Spring Harbor Laboratory Press, 2001) и используют для кодирования и проявления белков с использованием способа, известного в данной области техники. Другие способы продуцирования библиотек белков описаны, например, в US6300064 (например, библиотека HuCAL MorphoSys AG); US5885793; US6204023; US6291158 или US6248516.
Белки по изобретению могут являться растворимыми секретируемыми белками или могут быть представлены в виде слитого белка на поверхности клетки или частицы (например, фага или другого вируса, рибосомы или споры). Различные форматы дисплейных библиотек известны в данной области техники. Например, библиотека представляет собой in vitro дисплейную библиотеку (например, рибосомную дисплейную библиотеку, ковалентную дисплейную библиотеку или дисплейную библиотеку мРНК, например, как описано в US7270969). В еще одном примере дисплейная библиотека является фаговой дисплейной библиотекой, в которой белки, содержащие антигенсвязывающие домены антител, экспрессируются с фага, например, как описано в US6300064; US5885793; US6204023; US6291158 или US6248516. Другие способы фагового дисплея известны в данной области техники и рассматриваются настоящим раскрытием. Аналогично, раскрытием рассматриваются способы клеточного дисплея, например, бактериальные дисплейные библиотеки, например, как описано в US5516637; дрожжевые дисплейные библиотеки, например, как описано в US6423538 или дисплейная библиотека млекопитающих.
Способы скрининга дисплейных библиотек известны в данной области техники. В одном примере дисплейную библиотеку по настоящему изобретению скринируют с использованием аффинной очистки, например, как описано в Scopes (В: Protein purification: principles and practice, Third Edition, Springer Verlag, 1994). Способы аффинной очистки, как правило, включают контактирование белков, содержащих антигенсвязывающие домены, отображенных в библиотеке, с антигеном-мишенью (например, VEGF-B), и, после промывки, элюирование тех доменов, которые остаются связанными с антигеном.
Любые вариабельные области или scFv, идентифицированные путем скрининга, легко модифицировать в полное антитело, при желании. Примеры способов модификации или переформатирования вариабельные области или scFvs в полное антитело описаны, например, в Jones et al., J Immunol Methods. 354:85-90, 2010; or Jostock et al., J Immunol Methods, 289: 65-80, 2004. В качестве альтернативы или дополнительно используются стандартные способы клонирования, например, как описано в Ausubel et al (В: Current Protocols in Molecular Biology. Wiley Interscience, ISBN 047 150338, 1987) и/или (Sambrook et al (В: Molecular Cloning: Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratories, New York, Third Edition 2001).
Деиммунизированные, химерные, гуманизированные, сингуманизированные, приматизированные и человеческие белки
Белки по настоящему изобретению могут представлять собой гуманизированный белок.
Термин "гуманизированный белок" следует понимать как относящийся к белку, содержащему вариабельную область, подобную таковой человека, которая включает CDR антитела из видов, отличных от человека (например, мышь или крыса или отличный от человека примат), пересаженный или встроенный в FR из антитела человека (этот тип антитела также называют "CDR-пересаженное антител"). Гуманизированные белки также включают белки, в которых один или несколько остатков белка человека модифицируют одной или более аминокислотных замен и/или один или несколько FR-остатков белка человека заменены соответствующими остатками, отличными от остатков у человека. Гуманизированные белки также могут содержать остатки, которые не встречаются ни в одном из антител человека или антител, отличных от антител человека. Любые дополнительные области белка (например, Fc-область), как правило, являются человеческими. Гуманизацию можно проводить с использованием способа, известного в данной области техники, например, US5225539, US6054297, US7566771 или 1185585089. Термин "гуманизированный белок" также охватывает супер-гуманизированный белок, например, как описано в US7732578.
Белки по настоящему изобретению могут представлять собой белки человека. Термин "белок человека", используемый в данном описании, относится к белкам, имеющим вариабельную и, необязательно, константную области антитела, обнаруживаемого в организме человека, например, в зародышевой линии человека или соматических клетках или в библиотеках, полученных с использованием таких областей. "Человеческие" антитела могут включать в себя аминокислотные остатки, не кодируемые последовательностями человека, например, мутации, введенные случайным образом, или сайт-направленные мутации in vitro (в частности, мутации, которые включают консервативные замены или мутации в небольшом количестве остатков белка, например, до 1, 2, 3, 4 или 5 остатков белка). Эти "человеческие антитела" не обязательно должны быть сгенерированы в результате иммунного ответа у человека, скорее, они могут быть получены с использованием рекомбинантных способов (например, скрининг библиотеки фагового дисплея) и/или с помощью трансгенного животного (например, мыши), содержащего нуклеиновую кислоту, кодирующую константную и/или вариабельную области антитела человека и/или с помощью направляемой селекции (например, как описано в или US5565332). Этот термин также включает формы таких антител со зрелой аффинностью. Для целей настоящего описания человеческий белок также будет рассматриваться как включающий белок, содержащий FR из антитела человека или FR, содержащие последовательности из консенсусной последовательности человеческих FR и в котором один или несколько CDR являются случайными или полу-случайными, например, как описано в US6300064 и/или US6248516.
Белки по настоящему изобретению могут быть сингуманизированными белками. Термин "сингуманизированный белок" относится к белку, полученному способом, описанным в WO2007/019620. Сингуманизированный белок включает вариабельную область антитела, причем вариабельная область содержит FR из вариабельной области антитела примата Нового Света и CDR из вариабельной области антитела примата, отличного от примата Нового Света. Например, сингуманизированный белок включает вариабельную область антитела, причем вариабельная область антитела включает FR из вариабельной области антитела примата Нового Света и CDR из антитела мыши или крысы.
Белки по настоящему описанию могут представлять собой приматизированные белки. "Приматизированный белок" включает вариабельную область(ти) антитела, генерируемого после иммунизации примата, не являющегося человеком (например, макаки-крабоеда). Необязательно, вариабельные области антитела примата, не являющегося человеком, связывают с константными областями человека для получения приматизированного антитела. Типичные способы получения приматизированных антител описаны в US6113898.
В одном примере белок по раскрытию представляет собой химерный белок. Термин "химерные белки" относится к белкам, в которых антигенсвязывающий домен происходит из определенных видов (например, мышевидных, таких как мышь или крыса) или принадлежит к конкретному классу или подклассу антител, в то время как остальная часть белка происходит из белка, полученного из других видов (таких как, например, человек или не являющийся человеком примат) или принадлежит к другому классу или подклассу антител. В одном примере химерный белок представляет собой химерное антитело, содержащее VL и/или VL. из антитела, отличного от антитела человека (например, мышиного антитела), а остальные участки антитела происходят из антитела человека. Продуцирование таких химерных белков известно в данной области техники и может быть достигнуто с помощью стандартных способов (как описано, например, в US6331415, US5807715, US4816567 и US4816397).
Настоящее изобретение также предусматривает деиммунизированный белок, например, как описано в WO2000/34317 и WO2004/108158. Деиммунизированные антитела и белки имеют один или несколько удаленных эпитопов, например, B-клеточные эпитопы или Т-клеточные эпитопы (т.е. мутантный), чтобы таким образом уменьшить вероятность того, что у индивидуума будет повышаться иммунный ответ против антитела или белка.
Другие белки, содержащие вариабельные области антител
Настоящее изобретение также охватывает другие белки, содержащие вариабельную область или антигенсвязывающий домен антитела, такие как:
(I) однодоменное антитело, которое представляет собой одну полипептидную цепь, содержащую все или часть VH или VL антитела (см., например, US624851);
(II) диатела, триатела и тетратела, например, как описано в US5844094 и/или US2008152586;
(III) scFv, например, как описано в IJS5260203;
(IV) минитела, например, как описано в US5837821;
(V) биспецифические белки "ключ и замок", как описано в US5731168;
(VI) гетероконьюгатные белки, например, как описано в US4676980;
(VII) гетероконьюгатные белки, полученные с использованием химического перекрестносшивающего средства, например, как описано в US4676980;
(VIII) фрагменты Fab'-SH, например, как описано в Shalaby et al, J. Exp. Med., 175: 217-225, 1992; или
(IX) Fab3 (например, как описано в EP19930302894).
Слияния с константным доменом
Настоящее изобретение включает белок, содержащий вариабельную область антитела и константную область или Fc или его домен, например, домен CH2 и/или CH3. Приемлемые константные области и/или домены будут очевидны для специалиста в данной области, и/или последовательности таких полипептидов легко доступны из общедоступных баз данных. Kabat et al. также предоставляют описание некоторых подходящих константных областей/доменов.
Константные области и/или домены этого могут быть использованы для обеспечения биологических активностей, таких как димеризация, увеличение времени жизни в сыворотке, например, путем связывания с FcRn (неонатальным Fc-рецептором), антиген-зависимая клеточная цитотоксичность (ADCC), комплемент-зависимая цитотоксичность (CDC), антиген-зависимый клеточной фагоцитоз (ADCP).
Настоящее изобретение также охватывает белки, содержащие мутантные константные области или домены, например, как описано в US7217797; US72I7798 или US20090041770 (с увеличенным временем полужизни) или US2005037000 (повышенный ADCC).
Стабилизированные белки
Нейтрализующие белки по настоящему изобретению могут содержать константную область IgG4 или стабилизирующую константную область IgG4. Термин "стабилизирующая константная область IgG4" следует понимать как означающий константную область IgG4, которая была модифицирована для уменьшения обмена Fab-плечами или склонности подвергаться обмену Fab-плечами или образования половины антитела или склонности образовывать половину антитела. "Обмен Fab-плечами" относится к типу белковой модификации IgG4 человека, в которой тяжелая цепь IgG4 и присоединенная легкая цепь (полумолекула) обмениваются парой тяжелой и легкой цепей из другой молекулы IgG4. Таким образом, молекулы IgG4 могут приобрести два различных Fab-плеча, распознающие два различных антигена (приводя к биспецифическим молекулам). Обмен Fab-плечами природно происходит in vivo и может индуцироваться in vitro очищенными клетками крови или восстанавливающими агентами, такими как восстановленный глутатион. «Половина антитела» образуется, когда антитело IgG4 диссоциирует с образованием двух молекул, каждое содержит одну тяжелую цепь и одну легкую цепь.
В одном примере стабилизирующая константная область IgG4 содержит пролин в положении 241 шарнирной области согласно системе Kabat (Kabat et al., Sequences of Proteins of Immunological Interest Washington DC United States Department of Health and Human Services, 1987 and/or 1991). Это положение соответствует положению 228 шарнирной области в соответствии с системой нумерации EU (Kabat et al., Sequences of Proteins of Immunological Interest Washington DC United States Department of Health and Human Services, 2001 и Edelman et al., Proc. Natl. Acad. USA, 63, 78-85, 1969). В IgG4 человека этим остатком, как правило, является серин. После замещения серина на пролин, шарнирная область IgG4 содержит последовательность CPPC. В связи с этим, специалист в данной области должен осознавать, что "шарнирная область" представляет собой богатую пролином часть константной области тяжелой цепи антитела, которая связывает области Fc и Fab, что придает подвижность двум Fab-плечам антитела. Шарнирная область включает остатки цистеина, которые участвуют в дисульфидных связях между тяжелыми цепями. Как правило, она определяется как простирающейся от Glu226 до Pro243 IgG1 человека в соответствии с системой нумерации Kabat. Шарнирные области других IgG-изотипов могут быть выровнены с последовательностью IgG1 путем размещения первого и последнего остатков цистеина, образующих дисульфидные (S-S) связи между тяжелыми цепями в одних и тех же положениях (смотри, например, WO2010/080538).
Дополнительные ингибиторы на основе белка сигнального пути VEGF-B
Другие белки, которые могут препятствовать продуктивному взаимодействию VEGF-B с его рецептором, включают мутантные белки VEGF-B.
В одном примере ингибитор представляет собой растворимый белок, содержащий один или несколько доменов VEGF-R1, которые связываются с VEGF-B (и, например, по существу не связываются с VEGF-A). В одном примере растворимый белок дополнительно содержит константную область антитела, такого как антитело IgGl. Например, растворимый белок дополнительно содержит Fc-область и, при необходимости, шарнирную область антитела, например антитела IgGl.
В одном примере ингибитор белка представляет собой миметик антитела, например, белковый каркас, включающий вариабельные области, которые связываются с белком-мишенью способом аналогично антителу. Описание типичных миметиков антител следует.
Иммуноглобулины и фрагменты иммуноглобулинов
Примером соединения по настоящему изобретению является белок, содержащий вариабельную область иммуноглобулина, такого как Т-клеточный рецептор или тяжелая цепь иммуноглобулина (например, IgNAR, антитело верблюдовых).
Тяжелоцепочечные иммуноглобулины
Тяжелоцепочечные иммуноглобулины структурно отличаются от многих других форм иммуноглобулинов (например, антител), поскольку они содержат тяжелую цепь, но не включают легкую цепь. Соответственно, эти иммуноглобулины также называют "антитела только с тяжелой цепью". Тяжелоцепочечные иммуноглобулины встречаются, например, у верблюдовых и хрящевых рыб (также называются IgNAR).
Вариабельные области, присутствующие в тяжелых цепях природных иммуноглобулинов, обычно называют "домены VHH» для верблюдовых Ig и V-НАР для IgNAR, для того, чтобы их можно было отличить от вариабельных областей тяжелых цепей, которые присутствуют в традиционных 4-цепочечных антителах (которые называются "VH-домены") и от вариабельных областей легкой цепи, которые присутствуют в обычных 4-цепочечных антителах (которые называют также "VL-доменами").
Тяжелоцепочечные иммуноглобулины не требуют наличия легких цепей для связывания с высокой аффинностью и с высокой специфичностью с соответствующим антигеном. Это означает, что отдельные фрагменты связывающего домена могут быть получены из тяжелоцепочечных иммуноглобулинов, которые легко экспрессировать и которые обычно стабильны и растворимы.
Общее описание тяжелоцепочечных иммуноглобулинов из верблюдовых и их вариабельных областей и способы их получения и/или выделения и/или применения находится inter alia (среди прочего) в следующих ссылках WO94/04678, WO97/49805 и WO 97/49805.
Общее описание тяжелоцепочечных иммуноглобулинов из хрящевых рыб и их вариабельных областей и способы их получения и/или выделения и/или применения находится inter alia (среди прочего) в WO2005/118629.
V-подобные белки
Примером соединения по раскрытию является Т-клеточный рецептор. Т-клеточные рецепторы имеют два V-домена, которые объединяются в структуру, похожую на Fv-модуль антитела. Novotny et al., Proc Natl Acad Sci USA 88: 8646-8650, 1991 описывает, как два V-домена Т-клеточного рецептора (называемые альфа и бета) могут быть слиты и экспрессированы в виде одной полипептидной цепи и, кроме того, как изменить поверхностные остатки для снижения гидрофобности непосредственно, по аналогии с антителом ScFv. Другие публикации, описывающие продуцирование одноцепочечных Т-клеточных рецепторов или мультимерных Т-клеточных рецепторов, содержащих два домена V-альфа и V-бета, включают WO1999/045110 or WO2011/107595.
Другие отличные от антитела белки, содержащие антигенсвязывающие домены, включают белки с V-подобными доменами, которые, как правило, являются мономерными. Примеры белков, содержащих такие V-подобные домены, включают CTLA-4, CD28 и ICOS. Дальнейшее раскрытие белков, содержащих такие V-подобные домены, включено в заявку WO1999/045110.
Аднектины
В одном примере соединением по раскрытию является аднектин. Аднектины основаны на десятом фибронектиновом домене типа III (10Fn3) фибронектина человека, в котором области петли изменены для придания связывания с антигеном. Например, можно сконструировать три петли на одном конце β-сэндвича 10Fn3-домена, чтобы дать возможность аднектину специфически распознавать антиген. Более подробную информацию см. в US20080139791 или WO2005/056764.
Антикалины
В другом примере соединением по раскрытию является антикалин. Антикалины получают из липокалинов, которые представляют собой семейство внеклеточных белков, которые транспортируют малые гидрофобные молекулы, такие как стероиды, билины, ретиноиды и липиды. Липокалины имеют жесткую β-складчатую вторичную структуру с множеством петель на открытом конце конической структуры, которая может быть сконструирована для связывания с антигеном. Такие сконструированные липокалины известны как антикалины. Для дополнительного описания антикалинов см. US7250297B1 или U520070224633.
Аффитела
В другом примере соединением по раскрытию является аффитело. Аффитело представляет собой каркас, полученный из Z-домена (антиген связывающего домена) белка А Staphylococcus aureus, который может быть сконструирован для связывания с антигеном. Z-домена состоит из трехспирального пучка приблизительно 58 аминокислот. Библиотеки были получены путем рандомизации поверхностных остатков. Более подробную информацию см. в EP1641818.
Авимеры
В другом примере соединением по раскрытию является авимер. Авимеры представляют собой полидоменными белки, полученные из каркасного семейства A-домена. Нативные домены F из примерно 35 аминокислот принимают определенную дисульфидную монолитную структуру. Разнообразие порождается перетасовкой природной изменчивости, проявляемой семейства А-доменов. Более подробную информацию см. в WO2002088171.
DARPины
В другом примере соединением по раскрытию является сконструированный белок анкириновых повторов (DARPин). DARPины получают из анкирина, который представляет собой семейство белков, которые опосредуют прикрепление интегральных мембранных белков цитоскелета. Один анкириновых повтор представляет собой 33-остаточный мотив, состоящий из двух α-спиралей и β-изгиба. Они могут быть сконструированы, чтобы связать различные антигены-мишени путем рандомизации остатков в первой α-спирали и β-изгибе каждого повтора. Их поверхность связывания может быть увеличен за счет увеличения количества модулей (способ созревания аффинности). Более подробную информацию см. в US20040132028.
Способы получения белков
Рекомбинантная экспрессия
В случае рекомбинантного белка кодирующая его нуклеиновая кислота может быть клонирована в векторы экспрессии, которые затем трансфицируют в клетки-хозяева, такие как клетки E. coli, дрожжевые клетки, клетки насекомых или клетки млекопитающих, таких как обезьяньи COS-клетки, клетки яичника китайского хомячка (СНО), клетки первичной почки человека (HEK) или клетки миеломы, которые не продуцируют антитела. Типичные клетки, используемые для экспрессии белка по настоящему раскрытию, представляют собой CHO-клетки, клетки миеломы или НЕК-клетки. Способы молекулярного клонирования для достижения этих целей известны в данной области и описаны, например, в Ausubel et al., (editors), Current Protocols in Molecular Biology, Greene Pub. Associates and Wiley-Interscience (1988, including all updates until present) или Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989). Широкое разнообразие способов клонирования и in vitro амплификации пригодны для конструирования рекомбинантных нуклеиновых кислот. Способы получения рекомбинантных антител также известны в данной области, см. US4816567 или US5530101.
После выделения нуклеиновую кислоту функционально связывают с промотором в экспрессионной конструкции или векторе экспрессии для дальнейшего клонирования (амплификация ДНК) или для экспрессии в бесклеточной системе или в клетках.
Используемый в настоящем описании термин «промотор» следует понимать в самом широком контексте, и он включает транскрипционные регуляторные последовательности геномного гена, включая TATA-бокс или инициирующий элемент, который требуется для точной инициации транскрипции, с дополнительными регуляторными элементами или без них (например, активирующие последовательности, сайты связывания транскрипционных факторов, энхансеров и сайленсеров), которые изменяют экспрессию нуклеиновой кислоты, например, в ответ на дифференцировочный и/или внешний стимул, или тканеспецифическим образом. В настоящем контексте термин «промотор» также используется для описания рекомбинантной, синтетической или слитой нуклеиновой кислоты, или производного, которое придает, активирует или усиливает экспрессию нуклеиновой кислоты, с которой она функционально связана. Типичные промоторы могут содержать дополнительные копии одного или нескольких специфических регуляторных элементов для дополнительного усиления экспрессии и/или изменения пространственной экспрессии и/или временной экспрессии указанной нуклеиновой кислоты.
Как используется в данном описании, термин "функционально связан с" означает позиционирование промотора по отношению к нуклеиновой кислоте таким образом, что экспрессия нуклеиновой кислоты находится под контролем промотора.
Доступны многие векторы для экспрессии в клетках. Компоненты вектора обычно включают, но не ограничиваются ими, один или несколько из следующих компонентов: сигнальную последовательность, последовательность, кодирующую антитело (например, на основе собранной информации, представленной в данном документе), энхансерный элемент, промотор и последовательность терминации транскрипции. Специалист должен быть осведомлен о подходящих последовательностях для экспрессии антитела. Типичные сигнальные последовательности включают прокариотические сигналы секреции (например, pelB, щелочной фосфатазы, пенициллиназы, IPP или термостабильного энтеротоксина 11), сигналы секреции дрожжей (например, лидер инвертазы, лидер α-фактора или лидер кислой фосфатазы) или сигналы секреции млекопитающих (например, gD-сигнал вируса простого герпеса). Типичные промоторы, активные в клетках млекопитающих, включают ранний промотор цитомегаловируса (CMV-IE), промотор фактор элонгации 1-α человека (EF1), промоторы малых ядерных РНК (U1а и U1b), (промотор тяжелой цепи α-миозина, промотор обезьяньего вируса 40 (SV40), промотор вируса саркомы Рауса (РСВ), основной поздний промотор аденовируса, промотор β-актина, гибридный регуляторный элемент, содержащий энхансер CMV/промотор β-актина или промотор иммуноглобулина или его активный фрагмент. Примеры подходящих линий клеток-хозяев млекопитающих представляют собой почечная линия CV1 обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); эмбриональная почечная линия человека (293 или 293 клеток, субклонированных для роста в суспензионной культуре, клетки почки детеныша хомячка (ВНК, АТСС CCL 10); или клетки яичника китайского хомячка (СНО).
Типичные промоторы, пригодные для экспрессии в клетках дрожжей, таких как, например, клетка дрожжей, выбранная из группы, включающей Pichia pastoris, Saccharomyces cerevisiae и S. pombe, включают в себя, но не ограничиваются ими, промотор ADH1, промотор GAL1 промотор GAL4, промотор CUP1, промотор PHO5, nmt-промотор, промотор RPR1 или промотор TEF1.
Средства для введения выделенной нуклеиновой кислоты или содержащей ее экспрессионной конструкции в клетку для экспрессии хорошо известны специалистам в данной области техники. Способ, используемый для данной клетки, зависит от известных успешных приемов. Средства для введения рекомбинантной ДНК в клетки включают микроинъекцию, трансфекцию, опосредованную DEAE-декстраном, трансфекцию, опосредованную липосомами, например, с использованием липофектамина (Gibco, MD, США) и/или целфектина (Gibep, MB, США), ПЭГ-опосредованное поглощение ДНК, электропорацию и бомбардировку микрочастицами, например, с использованием ДНК-покрытых частиц вольфрама или золота (Agracetus Inc., Висконсин, США) среди прочего.
Клетки-хозяева, используемые для получения антитела, можно культивировать в множестве сред, в зависимости от типа используемых клеток. Коммерчески доступные среды, такие как Ham's Fl0 (Sigma), Minimal Essential Medium ((MEM), (Sigma), RPMl-1640 (Sigma) и Dulbecco's Modified Eagle's Medium ((DMEM), Sigma) подходят для культивирования клеток млекопитающих. Среды для культивирования других типов клеток, рассмотренных выше, известны в данной области техники.
Очистка белков
После продуцирования/экспрессии белок по раскрытию очищают с помощью способа, известного в данной области техники. Такая очистка предоставляет белок по данному раскрытию, по существу свободный от неспецифического белка, кислот, липидов, углеводов и тому подобное. В одном примере белок находится в препарате, причем более чем приблизительно 90% (например, 95%, 98% или 99%) белка в препарате является белком по раскрытию. Стандартные способы очистки пептидов используют для получения выделенного белка по данному раскрытию, в том числе, но не ограничиваясь этим, различные протоколы жидкостной хроматографии высокого давления (или производительности) (ВЭЖХ) и выделения полипептидов, отличных от ВЭЖХ, такие как гель-проникающая хроматография, ионообменная хроматография, гидрофобная хроматография, смешанный режим хроматографии, способы разделения фаз, электрофоретических разделений, способы осаждения, способы всаливания/высаливания, иммунохроматографии и/или другие способы.
В одном примере аффинная очистка используется для выделения слитого белка, содержащего метку. Способы выделения белка с использованием аффинной хроматографии известны в данной области техники и описаны, например, в Scopes (In: Protein purification: principles and practice, Third Edition, Springer Verlag, 1994). Например, антитело или соединение, которое связывается с меткой (в случае полигистидинового тега это может быть, например, никель-NTA) иммобилизуют на твердом носителе. Образец, содержащий белок, затем вводят в контакт с иммобилизованным антителом или соединением, в течение времени и в условиях, достаточных для прохождения связывания. После промывания для удаления любого несвязанного или неспецифически связанного белка, белок элюируют.
В случае белка, содержащего Fc-область антитела, белок A или белок G или их модифицированные формы могут быть использованы для аффинной очистки. Белок А полезен для выделения очищенных белков, содержащих Fc область тяжелой цепи γ1, γ2 или γ4 человека. Белок G рекомендуется для всех мышиных Fc-изотипов и для γ3 человека.
Ингибиторы сигнального пути VEGF-B на основе нуклеиновых кислот
В одном примере осуществления раскрытия, терапевтические и/или профилактические способы, как описано в настоящем документе, в соответствии с любым примером раскрытия приводят к уменьшению экспрессии VEGF-B. Например, такой способ включает введение соединения, которое уменьшает транскрипцию и/или трансляцию нуклеиновой кислоты. В одном примере соединение представляет собой нуклеиновую кислоту, например, антисмысловой полинуклеотид, рибозим, PNA, интерферирующую РНК, малую интерферирующую РНК, микроРНК.
Антисмысловые нуклеиновые кислоты
Термин "антисмысловая нуклеиновая кислота» следует использовать для обозначения ДНК или РНК или производного этого (например, LNA или PNA), или комбинации этого, которая комплементарна, по меньшей мере, на части молекулы специфической мРНК, кодирующей полипептид, как описано в настоящем документе, в любом примере раскрытия и способной интерферировать с пост транскрипционными событиями, такими как трансляция мРНК. Использование антисмысловых способов известно в рассматриваемой области техники (см., например, Hartmann and Endres (editors), Manual of Antisense Methodology, Kluwer (1999)).
Антисмысловая нуклеиновая кислота по данному изобретению будет гибридизоваться с нуклеиновой кислотой-мишенью в физиологических условиях. Антисмысловые нуклеиновые кислоты включают последовательности, которые соответствуют структурным генам или кодирующим областям или последовательностям, которые осуществляют контроль за экспрессией генов или сплайсингом. Например, антисмысловая нуклеиновая кислота может соответствовать целевой кодирующей области нуклеиновой кислоты, кодирующей VEGF-B, или 5'-нетранслируемой области (UTR) или 3'-UTR или их комбинации. Она может быть частично комплементарна интронным последовательностям, которые могут быть сплайсированны во время или после транскрипции, например, только экзонным последовательностям гена-мишени. Длина антисмысловой последовательности должна составлять по меньшей мере 19 смежных нуклеотидов, например, по меньшей мере 50 нуклеотидов, например, по меньшей мере 100, 200, 500 или 1000 нуклеотидов нуклеиновой кислоты, кодирующей VEGF-B. Может быть использована полноразмерная последовательность, комплементарная всему транскрипту гена. Длина может составлять 100-2000 нуклеотидов. Степень идентичности антисмысловой последовательности с транскриптом-мишенью должна составлять не менее 90%, например, 95-100%.
Типичные антисмысловые нуклеиновые кислоты в отношении VEGF-B описаны, например, в WO2003/105754.
Каталитическая нуклеиновая кислота
Термин "каталитическая нуклеиновая кислота» относится к молекуле ДНК или ДНК-содержащей молекуле (также известный в данной области техники как "дезоксирибозим" или "ДНКзим") или РНК или РНК-содержащей молекуле (также известной как "рибозим" или "РНКзим"), который специфически распознает определенный субстрат и катализирует химическую модификацию этого субстрата. Основания нуклеиновых кислот в каталитической нуклеиновой кислоте могут представлять собой основания А, С, G, Т (и U для РНК).
Обычно каталитическая нуклеиновая кислота имеет антисмысловую последовательность для специфического распознавания нуклеиновой кислоты-мишени и ферментативную активность для расщепления нуклеиновой кислоты (также упоминаемой в настоящем документе как "каталитический домен"). Типы рибозимов, которые применяются в данном описании, представляют собой молотоглавый рибозим и шпилькообразный рибозим.
РНК-интерференция
РНК-интерференция (RNAi) полезна для специфически ингибирования продуцирования конкретного белка. Не ограничиваясь теорией, полагают, что эта технология основана на присутствии молекул дцРНК, которые содержат последовательность, которая по существу идентична мРНК представляющего интерес гена или его части, в данном случае мРНК, кодирующая VEGF-B. В целях удобства дцРНК может быть получена с одного промотора рекомбинантного вектора в клетке-хозяине, где смысловые и антисмысловые последовательности фланкированы неродственной последовательностью, что позволяет смысловым и антисмысловым последовательностям гибридизоваться с образованием молекул дцРНК с неродственной последовательностью, образующей структуру петли. Конструирование и продуцирование подходящих молекул дцРНК для данного изобретения является также в пределах возможностей специалиста в данной области техники, в частности, с учетом WO99/32619, WO99/53050, WO99/49029 и WO01/34815.
Длина смысловой и антисмысловой последовательностей, которые гибридизуются, должны составлять для каждой по меньшей мере 19 смежных нуклеотидов, например, по меньшей мере 30 или 50 нуклеотидов, например, по меньшей мере 100, 200, 500 или 1000 нуклеотидов. Можно использовать полноразмерную последовательность, соответствующую всему транскрипту гена. Длина может составлять 100-2000 нуклеотидов. Степень идентичности смысловой и антисмысловой последовательностей с транскриптом-мишенью должна составлять по меньшей мере 85%, например, по меньшей мере 90%, такой как 95-100%.
Примеры малых интерферирующих молекул РНК ("миРНК") содержат нуклеотидную последовательность, которая идентична примерно 19-21 смежных нуклеотидов мРНК мишени. Например, последовательность siRNA начинается с динуклеотида AA, имеет GC-содержание примерно 30-70% (например, 30-60%, например, 40-60%, например, приблизительно 45%-55%), и не имеет высокого процента идентичности с какой-либо нуклеотидной последовательностью, отличной от мишени в геноме млекопитающего, в который она должна быть введена, например, как определено с помощью стандартного поиска BLAST. Типичные миРНК, которые снижают экспрессию VEGF-B, коммерчески доступны от Santa Cruz Biotechnology или Novus Blologieals.
Короткие шпилечные РНК (shRNA), которые снижают экспрессию VEGF-B, также известны в данной области техники и коммерчески доступны от Santa Cruz Biotechnology.
Cкрининг-анализ
Соединения, которые ингибируют сигнальный путь VEGF-B, могут быть идентифицированы с использованием способов, известных в данной области, например, как описано ниже. Аналогичным образом, количества ингибиторов сигнального пути VEGF-B, пригодные для использования в способе, описанном в настоящем документе, можно определить или оценить с использованием способов, известных в данной области, например, как описано ниже.
Анализы нейтрализации
Для соединений, которые связываются с VEGF-B и ингибируют сигнальный путь, можно использовать анализ нейтрализации.
В одном примере анализ нейтрализации включает контактирование VEGF-B с соединением в присутствии или в отсутствие детектируемого меченого растворимого VEGF-R1 или контактирование детектируемо меченого VEGF-B с соединением в присутствии или в отсутствие клеток, экспрессирующих VEGF-Rl или растворимого VEGF-R1. Затем оценивали уровень VEGF-B, связанного с VEGF-Rl. Сниженный уровень связанного VEGF-B в присутствии соединения по сравнению по сравнению с таковым в отсутствие соединения указывает на то, что соединение ингибирует связывание VEGF-B с VEGF-R1 и, как следствие, сигнальный путь VEGF-B.
Другой анализ нейтрализации описан в WO2006/012688 и включает контактирование фрагмента VEGF-Rl, содержащего второй Ig-подобный домен, иммобилизованный на твердом носителе, с субнасыщающей концентрацией рекомбинантного VEGF-B, предварительно инкубированного с соединением. После промывания для удаления несвязанного белка, иммобилизованный белок контактируют с антителом анти-VEGF-B, и определяют количество связанного антитела (показатель иммобилизованного VEGF-B). Соединение, которое снижает уровень связанного антитела по сравнению с уровнем в отсутствие соединения, считается ингибитором сигнального пути VEGF-B.
В другом примере соединение, которое ингибирует сигнальный путь VEGF-B, идентифицируют с помощью клеток, пролиферация которых зависит от сигнального пути VEGF-B, например, клеток BaF3, модифицированных, как описано в WO2006/012688 для экспрессии химерного рецептора, включающего внутриклеточный домен рецептора эритропоэтина человека и внеклеточный домен VEGF-Rl. Клетки культивируют в присутствии VEGF-B, и в присутствии или в отсутствие соединения. Пролиферацию клеток затем оценивали с использованием стандартных способов, например, анализы формирования колоний, включение тимидина или поглощение другого подходящего маркера пролиферации клеток (например, анализ снижения MTS-окрашивания), Соединение, которое уменьшает уровень пролиферации в присутствии VEGF-B, считается ингибитором сигнального пути VEGF-B.
Соединения также можно оценивать по их способности связываться с VEGF-B с использованием стандартных способов. Способы оценки связывания с белком известны в рассматриваемой области техники, например, как описано в Scopes (В: Protein purification: principles and practice, Third Edition, Springer Verlag, 1994). Такой способ обычно включает мечение соединения и его контактирование с иммобилизованным VEGF-B. После промывки для удаления неспецифического связанного соединения, детектируют количество метки и, как следствие, связанного соединения. Конечно, соединение может быть иммобилизованным и VEGF-B маркированным. Анализы панорамирование типа также могут быть использованы. В качестве альтернативы или дополнительно, могут быть использованы анализы поверхностного плазмонного резонанса.
Анализ экспрессии
Соединение, которое уменьшает или предотвращает экспрессию VEGF-B, идентифицируют путем контактирования клетки с соединением и определение уровня экспрессии VEGF-B. Подходящие способы определения экспрессии гена на уровне нуклеиновых кислот известны в данной области техники и включают, например, количественную полимеразную цепную реакцию (кПЦР) или анализы на микрочипах. Подходящие способы определения экспрессии на уровне белка также известны в данной области техники и включают, например, твердофазный иммуноферментный анализ (ELISA), флуоресцентный иммуноферментный анализ (FLISA), иммунофлюоресценцию или Вестерн-блоттинг.
Анализы in vivo
Соединения, описанные в настоящем документе, могут быть протестированы на активность в моделях на животных. Примеры животных моделей диабета II типа и ожирения включают: мышиную Ob/Ob (моногенная модель ожирения, недостаточность по лептину), мышиную db/db (моногенная модель ожирения, лептин-устойчивость), Zucker (fa/fa) на крысах (моногенная модель ожирения, лептин-устойчивость), на крысах Гото-Какизаки, КК-мышиная, NSY-мышиная, на крысах OLETF, на израильских песчаных крысах, на получающих лечение стрептозотоцином крысах, питающихся обогащенной жиром пищей, на мышах CBA/Ca, на крысах Diabetic Torri, на новозеландских мышах с ожирением (см. Rees and Alcolado Diabet. Med. 22, 359-370, 2005).
Известные животные модели спонтанной 2-го типа диабетической нефропатии включают: спонтанно гипертензивная/NLH-тучная (SHR N-ср) крыса (модель ожирения, диабета типа 2 и нефропатии), тощая SHR/N-cp крыса и Вистар-Киото/NIH-тучная (WKY/N-cp) крыса. Каждая из этих моделей позволяет оценить роли гипертензии и ожирения в патогенезе диабетической нефропатии: крысы SHR/N-cp обладают аномальной толерантностью к глюкозе, гипертензией, и у них развивается заболевание почек, напоминающее диабетическую нефропатию человека, в то время как крысы WKY/N-cp также страдают ожирением и имеют гиперлипидемию, но у них контроль глюкозы менее контролируется, чем у крысы SHR/N-cp). Еще одной моделью является крыса LA/N-cp (также несет ген ожирения и проявляет гиперлипидемию) (см Kimmel et al. Acta Diabetologica, 29, 142-148, 1992). В качестве примера в данном контексте, мышь db/db также является эффективной моделью диабетической нефропатии.
Фармацевтические композиции и способы лечения
Соединение, которое ингибирует сигнальный путь VEGF-B (syn. активный ингредиент) пригодно для парентерального, местного, перорального или локального введение, аэрозольного введения, или чрескожного введения для профилактического или для терапевтического лечения. В одном примере соединение вводят парентерально, например, подкожно или внутривенно.
Состав соединения, подлежащего введению, будет варьироваться в зависимости от способа введения и выбранного состава (например, раствора, эмульсии, капсулы). Соответствующую фармацевтическую композицию, содержащую соединение, которое можно вводить, можно получить в физиологически приемлемом носителе. Для растворов или эмульсий подходящие носители включают, например, водные или спиртово/водные растворы, эмульсии или суспензии, в том числе солевые и забуференные среды. Парентеральные средства доставки могут включать раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия, лактат Рингера или фиксированные масла. Разнообразие соответствующих водных носителей известны специалисту в данной области техники, включая воду, забуференную воду, забуференный солевой раствор, полиолы (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль), раствор декстрозы и глицина. Внутривенные носители могут включать различные добавки, консерванты или жидкости, питательные вещества или электролитные наполнители (см., как правило, Remington's Pharmaceutical Science, 16th Edition, Mack, Ed. 1980). Композиции могут необязательно содержать фармацевтически приемлемые вспомогательные вещества, необходимые для соответствия физиологическим условиям, такие как pH-контролирующие и буферные средства и средства, регулирующие токсичность, например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция и лактат натрия. Соединение можно лиофилизировать для хранения и восстанавливать в соответствующем носителе непосредственно перед использованием в соответствии с известными способами лиофилизирования и восстановления.
Оптимальную концентрацию активного ингредиента(ов) в выбранной среде можно определить эмпирически, в соответствии со способами, известными специалистам в данной области техники, и она будет зависеть от желательного конечного фармацевтического состава.
Диапазоны доз для введения соединения по раскрытия являются достаточно большими, чтобы получить желаемый эффект. Например, композиция содержит терапевтически или профилактически эффективное количество соединения.
Используемый в данном описании термин "эффективное количество" следует понимать как означающий количество соединения, достаточное для ингибирования/уменьшения/предотвращения сигнального пути VEGF-B у индивидуума. Специалист должен осознавать, что такое количество будет варьировать в зависимости от, например, соединения и/или конкретного индивидуума и/или типа и/или тяжести нефропатии, подвергаемой лечению. Соответственно, этот термин не следует истолковывать как ограничивающий раскрытие определенным количеством, например, массой или числом соединений.
Как используется в данном описании, под термином "терапевтически эффективное количество" следует понимать достаточное количество соединения для снижения или подавления одного или более симптомов нефропатии.
Как используется в данном описании, под термином "профилактически эффективное количество" следует понимать количество соединения, достаточное для предотвращения или ингибирования или задержки начала одного или более детектируемых симптомов нефропатии.
В одном примере соединение вводят в количестве, эффективном, чтобы иметь один или несколько из следующих эффектов:
Снижение или предотвращение гипертензии;
снижение или предотвращение клубочкового и/или тубулярного склероза;
снижение или предотвращение мезангиального внеклеточного отложения матрикса и/или аномального утолщения клубочковой базальной мембраны;
снижение или предотвращение клубочкового мезангиального расширения;
снижение или предотвращение клубочковых сосудистых перестроек;
снижение или предотвращение накопления липидов в почках;
снижение или предотвращение клубочкового накопления липидов;
снижение или предотвращение клубочкового накопления коллагена и/или артериолярного гиатоза; и/или
снижение или предотвращение макроальбуминурии.
Дозировка не должна быть настолько большой, чтобы вызывать появление побочных эффектов, таких как синдром повышенной вязкости, отек легких, застойная сердечная недостаточность и тому подобное. Как правило, дозировка варьирует в зависимости от возраста, состояния, пола и степени заболевания у пациента и может быть определена специалистом в данной области техники. Дозировка может быть скорректирована лечащим врачом в случае каких-либо осложнений.
Дозировка может варьировать от примерно 0,1 мг/кг до 300 мг/кг, например, от примерно 0,2 мг/кг до примерно 200 мг/кг, например, от примерно 0,5 мг/кг до примерно 20 мг/кг, в одной или более дозах, вводимых ежедневно, в течение одного или нескольких дней.
В некоторых примерах, соединение вводят в начальный (или нагружающей) дозе, которая выше, чем последующие (поддерживающие дозы), например, соединение вводят при начальной дозе, составляющей от примерно 1 мг/кг до около 30 мг/кг. Соединение затем вводят в поддерживающей дозе от примерно 0,0001 мг/кг примерно до 1 мг/кг. Поддерживающие дозы могут вводиться через каждые 7-35 дней, например, каждые 14 или 21 или 28 дней.
В некоторых примерах используется режим повышения дозы, при котором соединение вначале вводят в более низкой дозе, чем используют в последующих дозах. Этот дозовый режим полезен в случае, если пациент изначально страдает от побочных реакций.
В случае индивидуума, который должным образом не реагирует на лечение, в течение недели могут вводиться многократные дозы. Альтернативно или в качестве дополнения можно вводить возрастающие дозы.
Индивидуума можно повторно лечить соединением, давая более чем одну экспозицию или набор доз, например, по меньшей мере, примерно две экспозиции соединения, например, от примерно 2 до 60 экспозиций, а более конкретно, примерно от 2 до 40 экспозиции, наиболее предпочтительно, примерно от 2 до 20 экспозиций.
В одном примере любое повторное лечение может предоставляться, если возвращаются признаки или симптомы болезни, например, когда микроальбуминурия прогрессирует.
В другом примере любое повторное лечение может предоставляться через определенные интервалы времени. Например, последующие экспозиции можно вводить через различные интервалы времени, такие как, например, примерно 24-28 недель или 48-56 недель или дольше. Например, такие экспозиции вводят с интервалом каждый примерно 24-26 недель или примерно 38-42 недель или примерно 30-54 недель.
Способ по настоящему изобретению может также включать в себя совместное введение, по меньшей мере, одного соединения по изобретению вместе с введением другого терапевтически эффективного средства для профилактики или лечения почечного заболевания или осложнения, нефропатии (например, диабетической нефропатии), диабета, дислипидемии, гипертензия и/или ожирения.
В одном примере соединение(я) по раскрытию используется в комбинации с по меньшей мере одним дополнительным известным соединением, которое в настоящее время используется или разрабатывается для профилактики или лечения диабета. Примеры таких известных соединений включают, в качестве неограничивающих примеров, общепринятые антидиабетические препараты, такие как сульфонилмочевины (например, гликазид, глипизид), метформин, глитазоны (например, розиглитазон, пиоглитазон), средства высвобождения прандиальной глюкозы (например, репаглинид, натеглинид), акарбоза и инсулин (в том числе все природные, синтетические и модифицированные формы инсулина, такие как инсулин человека, крупного рогатого скота или свиного происхождения, инсулин, суспендированный, например, в изофане или цинке, и его производные, такие как инсулин глулизин, инсулин лизпро, инсулин лизпро протамин, инсулин гларгин, инсулин детемир или инсулин аспарт).
В одном примере соединение(я) по раскрытию используется в комбинации с, по меньшей мере, одним дополнительным известным соединением, которое в настоящее время используется или разрабатывается для предотвращения или лечения почечного расстройства, такого как нефропатия, или ассоциированного расстройства или осложнения. Примеры таких известных соединений включают, но не ограничиваются ими: препараты ингибиторов АПФ (например, каптоприл. (Капотен™), эналаприл (Инновэйс™), фосихоприл (Старил™), лизиноприл (Зестрил™), периндоприл (Коверсил™), квинаприл (Аккупро™), транданалоприл (Гоптен™), лотенсин, моэксиприл, рамиприл); блокаторы ренин-ангиотензиновой системы (РАС); блокаторы рецепторов ангиотензина (БРА) (например, ольмесартан, ирбесартан, лозартан, валсартан, кандесартан, эпросартан, телмисартан и т.д.); ингибиторы протеинкиназы C (PKC) (например рубоксистаурин); ингибиторы AGE-зависимых путей (например, аминогуанидин, ALT-946, пиродоксамин (пирододорин), OPB-9295, алагебриум.); противовоспалительные средства (например, ингибиторы циклооксигеназы-2, микофенолата мофетил, мизорибин, пентоксифиллин), ГАГ (например сулодексид (патент США N5496807)); пиридоксамин (патент США №7030146); антагонисты эндотелина (например SPP 301), ингибиторы COX-2, антагонисты PPAR-гамма и другие соединения, такие как амифостин (используется при цисплатин-нефропатии), каптоприл (используется при диабетической нефропатии), циклофосфамид (используется при идиопатической мембранозной нефропатии), тиосульфат натрия (используется при цисплатин-нефропатии).
Кроме того, способы по раскрытию могут также включать совместное введение по меньшей мере одного другого терапевтического средства для лечения другого заболевания, прямо или косвенно связанного с диабетом и/или нефропатией, в том числе, в качестве неограничивающих примеров, дислипидемии, гипертензии, ожирения, нейропатии и/или ретинопатии и т.д. Дополнительными примерами средств, которые могут вводиться совместно с соединением (соединениями), в соответствии с изобретением, являются кортикостероиды; иммуносупрессивные лекарственные средства; антибиотики; гипотензивные и мочегонные препараты (такие как ингибиторы АПФ); гиполипидемические средства, такие как смола, усиливающая экскрецию желчи, холестирамин, холестипол, никотиновая кислота, и более конкретно, лекарственные средства и лекарственные препараты, используемые для снижения уровня холестерина и триглицеридов (например, фибраты (например, Гемфиброзил™) и ингибиторы ГМГ-КоА, такие как ловастатин™, аторвастатин™, рувастатин™, лескол™, лепитор™, мевакор™, правакор™, правастатин™, симвастатин™, зокор™, церивастатин™, и т.д.); соединения, которые ингибируют всасывание липидов в кишечнике (например, эзетиминд); никотиновая кислота; и витамин D.
Как очевидно из вышесказанного, настоящее изобретение относится к способам сопутствующего терапевтического лечения индивидуума, включающим введение индивидууму, нуждающемуся в этом, эффективного количества первого соединения и второго соединения, причем указанное средство представляет собой соединение по раскрытию (то есть, ингибитор сигнального пути VEGF-B), а второе средство предназначено для профилактики или лечения нефропатии, диабетической нефропатии, сахарного диабета, гипертензии, гиперлипидемии или ожирения.
Используемый в данном описании термин "сопутствующее", как в выражении «сопутствующее терапевтическое лечение" включает введение первого средства в присутствии второго средства. Способ сопутствующего терапевтического лечения включает способы, в которых первое, второе, третье или дополнительное средство вводятся совместно. Способ сопутствующего терапевтического лечения также включает в себя способы, в которых первое или дополнительное средства вводят в присутствии второго или дополнительного средств, причем второе или дополнительное средство, например, могут быть предварительно введены. Способ сопутствующего терапевтического лечения может быть выполнен пошагово различными действующими лицами. Например, одно действующее лицо может вводить индивидууму первое средство, в то время как другое действующее лицо может вводить индивидууму второе средство, и стадии введения могут выполняться одновременно, или почти одновременно, или разнесены по времени, при условии, что первое средство (и/или дополнительные средства) является после введения в присутствии второго средства (и/или дополнительных средств). Действующее лицо и индивидуум могут являться одним и тем же лицом (например, человеком).
В одном примере изобретение также относится к способу лечения или профилактики нефропатии у индивидуума, причем способ включает введение индивидууму первой фармацевтической композиции, содержащей по меньшей мере одно соединение по раскрытию и второй фармацевтической композиции, содержащей одно или более дополнительных соединений.
В одном примере способ раскрытия включает введение ингибитора сигнального пути VEGF-B индивидууму, страдающему от нефропатии (например, диабетической нефропатии) и получение другого лечения (например, при диабете).
Наборы
Другим примером изобретения предлагается набор, содержащий соединения, пригодные для лечения нефропатии, как описано выше.
В одном примере настоящего изобретения набор включает (a) контейнер, содержащий соединение, которое ингибирует сигнальный путь VEGF-B, как описано в настоящем документе, необязательно, в фармацевтически приемлемом носителе или разбавителе; и (b) вкладыш в упаковку с инструкциями для лечения нефропатии у индивидуума.
В соответствии с этим примером осуществления раскрытия, вкладыш в упаковку находится на контейнере или связан с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит или включает композицию, которая является эффективной для лечения нефропатии, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожной инъекции). По меньшей мере, одно активное средство в композиции представляет собой соединение, которое ингибирует сигнальный путь VEGF-B. На этикетке или вкладыше в упаковке указывается, что композиция используется для лечения пациента, подходящего для лечения, например, имеющего нефропатию или предрасположенного к ней, с предоставлением конкретных указаний относительно дозировочных количеств и интервалов для соединения и любого другого лекарственного средства. Набор может дополнительно содержать дополнительный контейнер, содержащий фармацевтически приемлемый разбавитель, буфер, такой как бактериостатическая вода для инъекций (BWFI), забуференнный фосфатом физиологический раствор, раствор Рингера и/или раствор декстрозы. Набор может дополнительно включать другие материалы, желательные с коммерческой и потребительской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Набор необязательно дополнительно содержит контейнер, содержащий второе лекарственное средство, причем соединение, которое ингибирует сигнальный путь VEGF-B, является первым лекарственным средством, и изделие, которое дополнительно содержит инструкции на вкладыше в упаковку для лечения индивидуума вторым лекарственным средством, в эффективном количестве. Второе лекарственное средство может быть любым из тех, которые указаны выше.
Настоящее изобретение включает следующие неограничивающие примеры.
Пример 1: Мыши c недостаточностью VEGF-B устойчивы к развитию диабетической нефропатии.
Диабетические мыши с недостаточностью VEGF-B обладают улучшенной функцией почек и сниженной гипертензией
Мыши C57BKS/Leprdb (db/db//BKS) получали из лаборатории Джексона (в качестве модели диабета и диабетической нефропатии) и скрещивали с мышами C57BL/6-Vegfb-/-. Мыши Db/db//Vegfb-/- были выведены путем скрещивания гетерозиготных мышей db/+//Vegfb+/- друг с другом, с получением db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. В исследовании были использованы 42-недельные самки db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Выработку мочи определяли путем фотографирования клеток, содержащих одно животное/генотип. Для определения глюкозурии и протеинурии анализом индикаторных полосок, мышей подвергали голоданию в течение 1 часа. Мочу затем собирали и концентрацию глюкозы и белка непосредственно измеряли с использованием индикаторных полосок в соответствии с инструкцией производителя (Uristix, Siemens). Для детекции ACR мочу объемом от 20 мкл до 200 мкл собирали от каждой мыши. Мочевой альбумин определяли с помощью набора Alhuwell M, и мочевой креатинин измеряли с использованием набора Креатинин Companion murine ELISA kit (Exocell, Филадельфия, штат Пенсильвания). Систолическое и диастолическое артериальное давление измеряли способом хвостовой манжеты с использованием установки CODA (Kent Scientific). Всех животных приучали к устройству для измерения артериального давления в течение 2-х недель. Все они прошли 1-2 цикла из 20 повторных измерений в день, в течение по меньшей мере 3-х дней.
Эти анализы показали, что делетирование Vegfb у диабетических мышей db/db BKS улучшает почечную физиологию и снижает повышенное артериальное давление. Например, животные db/db//Vegfb-/- вырабатывали меньше мочи, чем животные db/db и db/db//Vegfb+/-. Животные db/db//Vegfb-/- также имели меньше глюкозы и белка в моче, чем животные db/db и db/db//Vegfb+/-.
Фигура 1А показывает соотношение альбумин/креатинин в моче, измеренное способом ELISA для мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/- (n=5/группа, левый график) и тощих дикого типа и тощих мышей Vegfb-/-(n=4-6/группу, правый график). Эти данные показывают, что у животных db/db//Vegfb-/- было улучшенное соотношение креатинин/альбумин в моче по сравнению с животными db/db//Vegfb+/-. Подобный эффект наблюдали у тощих животных с недостаточностью Vegfb по сравнению с тощими животными дикого типа.
Фигура 1B показывает артериальное давление, определенное способом хвостовой манжеты (систолическое и диастолическое артериальное давление) у мышей db/db и db/db//Vegfb+/- (n=5/группу). Эти данные свидетельствуют о том, что животные db/db//Vegfb-/- обладали пониженным систолическим и диасистолическим артериальным давлением по сравнению с животными db/db.
Эти данные показывают, что диабетические мыши db/db и тощие мыши с пониженной экспрессией VEGF-B имеют улучшенную функцию почек с лучшей фильтрационной способностью, определенными по сниженной концентрации альбумина в моче и соотношению креатинин/альбумин в моче. Снижение экспрессии VEGF-B также понижало как систолическое, так и диастолическое артериальное давление у животных db/db.
Клубочковый и канальцевый склероз уменьшается после делеции Vegfb у мышей db/db BKS
42-недельных самок мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/- забивали и почки выделяли, фиксировали, заливали, разрезали и окрашивали Periodic acid-Schiff (PAS) для оценки клубочкового месангиального расширения и тубулярного склероза в соответствии с инструкциями изготовителя (Sigma). По меньшей мере, 10 клубочков или канальцев на животное, окрашенных PAS в каждом срезе, фотографировали с помощью светлопольная микроскопия (микроскоп Axio Vision, Carl Zeiss) при 20х увеличении. Клубочки количественно определяли при помощи программы Axio Vision Run wizard для I) PAS-окрашивания клубочков (пиксел2/мкм2) II) числа апоптотических клубочков на кадр. Клубочек обозначали как апоптический при интенсивном PAS-окрашивании и диаметром меньшим, чем 25 мкм. Канальцы количественно определяли с помощью программы Axio Vision Run wizard для I) толщины базальной мембраны канальцев (мкм2).
Как показано на Фигуре 2А, у мышей db/db развивался тубулярный склероз, в то время как этот дефект не наблюдался у мышей db/db//Vegfb+/- и db/db//Vegfb-/-. Кроме того, у мышей db/db развивался гломерулосклероз, а у мышей db/db//Vegfb+/- и db/db//Vegfb-/- нет (Фигура 2В). Эти изменения в мышей db/db были связаны с повышением уровня апоптоза клубочков по сравнению с мышами db/db//Vegfb+/- и db/db//Vegfb-/- (Фигура 2C).
Эти данные показывают, что клубочковый и канальцевый склероз уменьшается в почках диабетических db/db животных с пониженной экспрессии VEGF-B по сравнению с db/db животными.
Мезангиальные отложения внеклеточного матрикса и аномальное утолщение клубочковой базальной мембраны (GBM) уменьшаются у Vegfb -дефектных db/db BKS мышей
Биопсии почек с 42-недель самок мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/- были использованы для анализа при помощи просвечивающей электронной микроскопии (ПЭМ) по стандартным методикам. Кратко, ткани фиксировали в фиксирующем буферном растворе (2% глутаровый альдегид, 0,5% параформальдегид, 0,1 M какодилат, 0,1 М сахароза, 3 мМ СaСl2) и промывали в 0,1 М какодилатном буфере рН 7,4 перед окрашиванием в 2% OsO4 в какодилатном буфера в течение 1 ч при комнатной температуре. Образцы обезвоживали и en bloc окрашивание выполняли в 2% уранилацетате в абсолютном спирте в течение 1 ч при комнатной температуре. Затем образцы проводили через серию Эпон 812/ацетон и погружали при 60°C в чистый Эпон 812. Тонкие срезы толщиной 70 нм нарезали на ультратоме Leica EM UC6 и устанавливали на формвар-покрытых медных канавчатых сетках. Пост-окрашивание проводили в 2% водном ацетате рН 3,5 и цитрате, нагруженном Venable и Cogglesall. Сетки анализировали при помощи электронной микроскопии FEI TECNAI.
ТЭМ-анализ с высоким увеличением показал, что почечные аномалии, связанные с диабетической нефропатией, такие как мезангиальное расширение и утолщение GBM, уменьшаются у мышей db/db с пониженной экспрессией VEGF-В по сравнению с мышами db/db (Фигура 3А). Утолщение GBM характерно для диабетической нефропатии. Фигура 3В показывает, что образование щелей сохраняется у мышей db/db с пониженной экспрессией VEGF-В. Образование подоцитов, как обнаружено, также сохраняется у мышей db/db с пониженной экспрессией VEGF-В.
Делеция Vegfb уменьшает накопление липидов в почках мышей db/db BKS
Анализ с масляным красным О (ORO) проводили с использованием почек от 42-недельных самок db/db, db/db//Vegfb+/-, db/db//Vegfb-/- и тощих дикого типа мышей. Кратко, почки рассекали и быстро замораживали на сухом льду и погружали в Tissue-Tek® (Sakura) непосредственно на изложнице криостата. Криосрезы (12 мкм) погружали на 5-15 мин в рабочий раствор ORO (2,5 г масляного красного О (Sigma-Aldrich), растворенного в 400 мл 99% изопропилового спирта, дополнительно разбавленного 6:10 в Н2О, профильтрованного через фильтр 22 мкм (Corning)) и промывали 10 мин под проточной водой из крана, прежде чем они были установлены. Окрашенные срезы исследовали светлопольной микроскопией (микроскоп Axio Vision, Carl Zeiss) при 20-ти кратном увеличении, и делали не менее 10 кадров на срез. Для количественного определения липидных капель количество красных пикселей в каждом кадре количественно определяли, используя программу Axio Vision Run wizard для полного ORO-окрашивания (пиксель, a.u).
Почечные накопление липидных капель и структура улучшается у мышей db/db с пониженной экспрессии VEGF-В (Фигура 4А). В частности, липидных капель меньше, и они мельче в срезах почек Vegfb-дефектных мышей db/db. Фигура 4В показывает, что снижение уровней экспрессии VEGF-B у мышей db/db уменьшает накопление липидов в клубочках и обеспечивает меньше липидных капель и меньшего размера.
Предотвращение клубочковых перестроек у мышей db/db BKS, дефектных по Vegfb
Морфологию почечных клубочков оценивали в почках от 42-недельных самок мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Кратко, животных забивали и собрали почки, фиксировали в 4% ПФА в течение 48 часов, погружали и 3 мкм срезы подготавливали и впоследствии иммуноокрашивали на синаптоподин и/или пекам. Извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako) и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами кроличьими анти-синаптоподин (Santa Cruz), козьими анти-пекам (Abcam). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере, 10 клубочков на животное, окрашенных на синаптоподин и пекам в каждом срезе, были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на синаптоподин (пиксель2/мкм2) и клубочкового II) окрашивания на пекам (пиксель2/мкм2).
Фигуры 5А и 5B показывают повышенную экспрессию синаптоподина и пекама в клубочках мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Эти данные и анализ окрашенных клубочков указывают на то, что в целом морфология клубочков улучшилась в Vegfb дефектных мышах db/db. Экпрессия и структура подоцитов и эндотелиальных клеток, т.е. клеток, которые имеют решающее значение для процесса фильтрации, сохраняются у мышей db/db с пониженной экспрессией VEGF-В.
Отложения гломерулярного коллагена и гиалиноз артериол уменьшаются у Vegfb дефектных мышей db/db BKS
Отложения гломерулярного коллагена и гиалиноз артериол оценивали в почках от 42-недельных самок мышей db/db, db/db//Vegfb+/- и db/db//Vegfb-/-. Кратко, животных забивали и собрали почки, фиксировали в 4% ПФА в течение 48 часов, погружали и 3 мкм срезы подготавливали и впоследствии иммуноокрашивали на коллагена IV, пекам и/или α-ЅМА. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako) и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами, кроличьими анти-коллаген IV (abcam), козьими анти-пекам (abcam) или α-ЅМА (Sigma). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на коллаген IV, пекам или α-ЅМА в каждом срезе были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на коллагена IV (пиксель2/мкм2), клубочкового II) окрашивания α-ЅМА (пиксель2/мкм2) и III) толщины клубочковых артериол (мкм2).
Фигура 6 показывает, что уровень коллагена IV (Фигура 6А) и α-ЅМА (Фигура 6В) были снижены у мышей db/db//Vegfb+/- и db/db//Vegfb-/- по сравнению с мышами db/db. Кроме того, толщина артериол была уменьшена у мышей db/db//Vegfb+/- и db/db//Vegfb-/- по сравнению с мышами db/db. (Фигура 6В). Эти данные показывают, что патологические внутриклубочковые отложения коллагена (также известные как клубочковые отложения внеклеточного матрикса (ЕСМ)) и гиалиноз артериол уменьшаются у мышей db/db с пониженной экспрессией VEGF-В. Эти данные показывают, что общие гистологические патологии диабетической нефропатии, такие как клубочковые ЕСМ отложения и гиалиноз артериол, уменьшаются у мышей db/db с пониженной экспрессией VEGF-В.
Липиды накапливаются преимущественно в клубочках почек при прогрессировании диабетической нефропатии у мышей db/db BKS
Тощие db+, 6-недельные db/db и 21-недельные db/db BKS мыши были использованы для анализа с масляным красным О (ORO). Почки рассекали и быстро замораживали на сухом льду и погружали в Tissue-Tek® (Sakura) непосредственно на изложнице криостата. Криосрезы (12 мкм) погружали на 5-15 мин в рабочий раствор ORO (2,5 г масляного красного О (Sigma-Aldrich), растворенного в 400 мл 99% изопропилового спирта, дополнительно разбавленного 6:10 в Н2О, профильтрованного через фильтр 22 мкм (Corning)) и промывали 10 мин под проточной водой из крана, прежде чем они были установлены. Окрашенные срезы исследовали светлопольной микроскопией (микроскоп Axio Vision, Carl Zeiss) при 20-ти кратном увеличении, и делали не менее 10 кадров на срез. Для количественного определения липидных капель, количество красных пикселей в каждом кадре количественно определяли, используя программу Axio Vision Run wizard для полного ORO-окрашивания (пиксель, a.u).
Фигура 7 показывает, что поскольку макроальбуминурия развивается у мышей db/db//BKS, липиды накапливаются преимущественно в клубочках.
Клубочковое накопление липидов коррелирует с потерей подоцитов при прогрессировании диабетической нефропатии у мышей db/db//BKS
Тощие db+, 6-недельные db/db и 21-недельные db/db BKS мыши были использованы для анализа. Почки рассекали, фиксировали в 4% ПФА в течение 48 ч, а затем обрабатывали для погружения в парафин с помощью стандартных процедур. После погружения 3-мкм срезы подготавливали и затем иммуноокрашивали на синаптоподин и/или пекам. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: морской свинки против анти-адипофилина (Fitzgerald) и кроличьими анти-синаптоподин (Santa Cruz). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на адипофилин и синаптоподин в каждом срезе были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на адипофилин (пиксель2/мкм2) и клубочкового II) окрашивания на синаптоподин (пиксель2/мкм2).
На Фигуре 8 показано, что при развитии диабетической нефропатии липиды накапливаются в клубочках и подоцитов число снижается параллельно.
Сигнальный путь Vegfb активируется во время прогрессирования диабетической нефропатии
Почки из тощих db+, 6-недельных db/db и 21-недельных db/db BKS мышей препарировали и быстро замораживали в сухом льду. Тотальную РНК экстрагировали и очищали из почек, используя набор RNeasy Mini kit (Qiagen) в соответствии с инструкциями изготовителя. Первую цепь кДНК синтезировали из 0,5-1 мкг тотальной РНК с помощью набора iScript cDNA Synthesis Kit (Bio-Rad). Количественную ПЦР в режиме реального времени проводили с использованием KAPA SYBR FAST qPCR Kit Master Mix (2x) Universal (KAPA Biosystems) в амплификаторе Rotor-Gene Q (Qiagen) Real-Time PCR в соответствии с инструкциями производителя. Уровни экспрессии были нормированы на экспрессию L19 и микроглобулина β-2.
Как показано на Фигуре 9, при диабетической нефропатии почечная экспрессия Vegfb, нижележащие мишени VEGF-B, Fatp3 и Fatp4, и основной VEGF-B рецептор, Vegfr1 активируются. Эти данные указывают на то, что почечный сигнальный путь VEGF-В является подходящей мишенью для лечения диабетической нефропатии.
Функция почек улучшается у мышей Vegf-b+/ - и мышей Vegf-b-/ -, получающих питание с высоким содержанием жира
Самцов мышей дикого типа Vegf-b+/- и Vegf-b-/- кормили 60% питанием с высоким содержанием жира (HFD) (Research Diets, США) в течение 30 недель, начиная с возраста 5 недель. В исследование также были включены тощие контрольные животные дикого типа. Для детектирования АСR собирали мочу объемом от 20 мкл до 200 мкл от каждой мыши. Альбумин в моче детектировали, используя набор Albuwell M kit, креатинин определяли с помощью набора Creatinine Companion murine ELISA kit (Exocell, Филадельфия, Пенсильвания).
Фигура 10 показывает, что HFD-опосредованная утечка белка в моче снижалась у HFD-питающихся мышей с пониженной экспрессии VEGF-В. Таким образом, фильтрационная способность почек улучшалась, что определялось как по экскреции альбумина с мочой (Фигура 10В), так и по ACR (Фигура 10А).
Клубочковое мезангиальное расширение и гипертрофия уменьшаются у HFD ФРС VEGF-B+/- и VEGF-B-/- мышей
Самцов мышей дикого типа Vegf-b+/- и Vegf-b-/- кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также были включены тощие контрольные животные дикого типа. Животных умерщвляли при помощи анестезирующих средств и всю кровь удаляли путем пункции сердца. Почки рассекали, фиксировали в 4% ПФА в течение 48 ч, а затем обрабатывали для заливания парафином с помощью стандартных процедур. После заливания 3-мкм срезы готовили и окрашивали PAS (Sigma) в соответствии с инструкциями производителя. По меньшей мере 10 клубочков на животное, окрашенных PAS в каждом срезе, фотографировали при помощи светлопольной микроскопии (микроскоп Axio Vision, Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для PAS-окрашивания клубочков (пиксель2/мкм2).
Фигура 11 показывает, что HFD-индуцированное почечное клубочковое мезангиальное расширение (Фигура 11А) и гипертрофия (Фигура 11В) предотвращаются у HFD-питающихся мышей с пониженной экспрессией VEGF-В.
Клубочковое накопление липидов снижается у HFD-питающихся мышей Vegf-b+/ - и Vegf-b-/ -
Самцов мышей дикого типа Vegf-b+/- и Vegf-b-/- кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также были включены тощие контрольные животные дикого типа. Животных умерщвляли при помощи анестезирующих средств и всю кровь удаляли путем пункции сердца. Почки рассекали и быстро замораживали на сухом льду и погружали в Tissue-Tek® (Sakura) непосредственно на изложнице криостата. Криосрезы (12 мкм) погружали на 5-15 мин в рабочий раствор ORO (2,5 г масляного красного О (Sigma-Aldrich), растворенного в 400 мл 99% изопропилового спирта, дополнительно разбавленного 6:10 в Н2О, профильтрованного через фильтр 22 мкм (Corning)) и промывали 10 мин под проточной водой из крана. После этого срезы погружали в течение 3 секунд в раствор гематоксилина и промывали под проточной водой, прежде чем они были установлены. Окрашенные срезы исследовали с использованием светлопольной микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на ORO и гематоксилин в каждом срезе, фотографировали с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли с помощью программы Axio Vision Run wizard для клубочкового ORO-окрашивания (пиксель2/мкм2).
Фигура 12 и анализ окрашенных срезов почек показывает, что HFD увеличивает накопление липидов почек преимущественно в клубочках. Однако это накопление эктопических липидов снижается у HFD-питающихся мышей с пониженной экспрессии VEGF-B. Липидные капли уменьшались как по числу, так и по размеру.
Клубочковое накопление липидов уменьшается, а целостность подоцитов сохраняется у HFD-питающихся мышей Vegf-b+/ - и Vegf-b-/ -
Самцов мышей дикого типа Vegf-b+/- и Vegf-b-/- кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. Животных умерщвляли при помощи анестезирующих средств и всю кровь удаляли путем пункции сердца. Почки рассекали, фиксировали в 4% ПФА в течение 48 ч, а затем обрабатывали для погружения в парафин с помощью стандартных процедур. После погружения 3-мкм срезы подготавливали и затем иммуноокрашивали на синаптоподин и/или пекам. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: морской свинки против анти-адипофилина (Fitzgerald) и кроличьими анти-синаптоподин (Santa Cruz). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на адипофилин и синаптоподин в каждом срезе были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на адипофилин (пиксель2/мкм2) и клубочкового II) окрашивания на подоцин (пиксель2/мкм2).
Фигура 13 и анализ окрашенных срезов показывает, что HFD-направляемое эктопическое накопление липидов почек снижено у мышей с пониженной экспрессией VEGF-В. Параллельно возрастает структурная целостность и количество подоцитов.
Гломерулярные ECM-отложения и гиалиноз артериол уменьшаются у HFD-питающихся мышей Vegf-b+/ - и Vegf-b-/ -
Самцов мышей дикого типа, Vegf-b+/- и Vegf-b-/- кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также были включены тощие контрольные животные дикого типа. Животных умерщвляли при помощи анестезирующих средств и всю кровь удаляли путем пункции сердца. Почки рассекали, фиксировали в 4% ПФА в течение 48 ч, а затем обрабатывали для погружения в парафин с помощью стандартных процедур. После погружения 3-мкм срезы подготавливали и затем иммуноокрашивали на коллаген IV, пекам и/или α-SMA. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: кроличьими анти-коллаген IV (abcam), козьими анти-пекам (abcam) или α-SMA (Сигма). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на коллагена IV, пекам или α-SMA в каждом срезе были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на коллаген IV (пиксель2/мкм2) и II) толщина клубочковой артериолы (мкм2).
Фигура 14 и анализ окрашенных срезов показывают, что ключевые гистологические патологии при диабетической нефропатии, такие как увеличение клубочкового накопления ECM (Фигура 14А) и гиалиноз артериол (Фигура 14В) уменьшаются у HFD-питающихся мышей с пониженной экспрессией VEGF-В.
Пример 2: нейтрализующее антитело анти-VEGF-B излечивает или предотвращает прогрессирование диабетической нефропатии.
Антитело-опосредованное ингибирование VEGF-В умеренно влияет на уровни глюкозы в крови у диабетических мышей db/db//BKS
Мыши C57BKS/Leprdb (db/db/BKS) были приобретены в Jackson Laboratory, и им инъецировали внутрибрюшинно дважды в неделю, начиная с 6 (профилактическое испытание) или 12 (терапевтический испытание) недельного возраста и в продолжении 8 недель, 400 мкг либо 2H10 (нейтрализирующее антитело анти-VEGF-B) или изотипипически-соответствующее контрольное антитело. В профилактическом испытании, начальные значения ACR и глюкозы в крови составляли <50 мкг альбумина/мг креатинина и <15 мМ, соответственно. В терапевтическом испытании начальные значения ACR и глюкозы в крови составляли >150 мкг альбумина/мг креатинина и >15 мМ, соответственно. Уровни постпрандиальной глюкозы в крови мышей контролировали две недели после удаления пищи в течение 2 часов. Определение уровня глюкозы производили для крови, отобранной из хвостовой вены с помощью контурного глюкометра Байера. Для детектирования АСR мочу объемом от 20 мкл до 200 мкл собирали от каждой мыши. Альбумин определяли с помощью набора Alhuwell M, и мочевой креатинин измеряли с использованием набора Creatinine Companion murine ELISA kit (Exocell, Филадельфия, Пенсильвания).
Фигуры 15А и C-F показывают уровень глюкозы в крови мышей db/db/BKS, получавших терапевтическое или профилактическое лечение антителом 2H10. Как показано, введение антитела после того, как диабет прогрессировал в экспериментах, описанных в настоящем документе (как и в агрессивной модели сахарного диабета, используемой в настоящем документе) существенно не уменьшало уровни глюкозы в крови.
На Фигуре 15В показаны ACR-уровни у мышей db/db/BKS, получавших терапевтическое или профилактическое лечение антителом 2H10.
Эти данные показывают, что у мышей db/db//BKS развивается микроальбуминурия в возрасте 6 недель, и макроальбуминурия устанавливается в 12 недель. Ни профилактические, ни терапевтические введение антитела анти-VEGF-B мышам db/db//BKS в этих испытаниях не снижало детектируемо уровни глюкозы в крови.
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) уменьшает клубочковый и канальцевый склероз у мышей db/db//BK S
Почки от 6-8 недельных db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, как описано выше, собирали, фиксировали в 4% ПФА в течение 48 часов, заливали, и 3 мкм срезы готовили и окрашивали PAS (Sigma) в соответствии с инструкциями производителя. По меньшей мере 10 клубочков на животное, окрашенных PAS в каждом срезе фотографировали с использованием светлопольной микроскопии (микроскоп Axio Vision, Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли с помощью программы Axio Vision Run wizard для клубочкового PAS-окрашивания (пиксель2/мкм2).
Как показано на Фигурах 16А и 16В, мыши, получавшие лечение 2H10, показывают снижение уровня клубочкового склероза и канальцевого склероза. Эти данные показывают, что лечение с использованием анти-VEGF-B мышей db/db//BKS снижало как клубочковый склероз, так и канальцевый склероз, несмотря на заметные различия в уровнях глюкозы в крови.
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) препятствует сосудистым перестройкам у мышей db/db//BK S
Почки от 6-8 недельных db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, как описано выше, собирали, фиксировали в 4% ПФА в течение 48 часов, заливали, и 3 мкм срезы готовили и иммуноокрашивали на пекам или подоцин. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: козьими анти-пекам (abcam) и кроличьи анти-подоцин (sigma). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на пекам в каждом срезе, были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на пекам (пиксель2/мкм2) и II) окрашивания на подоцин (пиксель2/мкм2).
Как показано на Фигуре 17А, уровень окрашивания на подоцин сохранился у мышей, получавших 2H10. Уровень окрашивания на пекам был увеличен у мышей, получавших 2H10 (Фигура 17В). Эти данные и анализ окрашенных срезов показывают, что морфология кровеносных сосудов в клубочках улучшена у мышей db/db//BKS, получавших лечение 2H10, и что ингибирование VEGF-B сохраняет плотность и структуру сосудов. Эти данные также показывают, что ингибирование VEGF-B сохраняет плотность и структуру сосудов. Таким образом, как структура, так и целостность типов клеток, составляющих клубочковой фильтрационный барьер, подоцитов и эндотелиоцитов, сохраняются у мышей db/db//BKS, получавших лечение с использованием антитела анти-VEGF-B.
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) сохраняет структуру подоцитов у мышей db/db//BK S
Почки от 6-8 недельных db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, как описано выше, собирали, фиксировали в 4% ПФА в течение 48 часов, заливали, и 3 мкм срезы готовили и иммуноокрашивали на синаптоподин или пекам. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами кролика анти-синаптоподин (Santa Cruz). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на пекам в каждом срезе, фотографировали их с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на адипофилин (пиксель2/мкм2) и II) окрашивания на синаптоподин (пиксель2/мкм2).
Как показано на Фигуре 18А, уровень окрашивания на адипофилин был снижен у мышей, получавших 2H10. Фигура 18В показывает, что окрашивание на синаптоподин было увеличено у мышей, получавших лечение с использованием 2H10. Эти данные и анализ окрашенных срезов указывают на то, что структура и целостность типов клеток, составляющих клубочковый барьер фильтрации, подоцитов и эндотелиоцитов, сохраняются у мышей db/db//BKS, получавших лечение с использованием антитела анти-VEGF-B. Эти данные также показывают, что лечение с помощью антитела анти-VEGF-B резко снижается накопление эктопических клубочковых липидов и сохраняет экспрессию и морфологию подоцитов.
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) уменьшает клубочковое накопление коллагена (отложения внеклеточного матрикса) у мышей db/db//BK S
Почки от 6 недельных db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, как описано выше, собирали, фиксировали в 4% ПФА в течение 48 часов, заливали, и 3 мкм срезы готовили и иммуноокрашивали на клубочковый коллаген IV. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами кролика анти-синаптоподин (Santa Cruz). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на пекам в каждом срезе, фотографировали с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового окрашивания на коллаген IV (пиксель2/мкм2).
Как показано на фигуре 19, уровень окрашивание клубочкового коллагена IV был увеличено у мышей, получавших лечение с использованием 2H10. Эти данные и анализ окрашенных срезов показывают, что антитело-опосредованное ингибирование VEGF-В уменьшает интраклубочковое патологическое накопление коллагена (накопление внеклеточного матрикса) у мышей db/db//BKS, гистологической характеристике при диабетической нефропатии.
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) улучшает липидный профиль плазмы у мышей db/db//BK S
Кровь собирали от 6-8 недель самок мышей db/db//BKS и соответствующих им по возрасту и полу тощих db/+ путем пункции сердца, которые ранее получали лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель. Кровь центрифугировали при 14000 об/мин при 4°C в течение 10 минут, после чего отделяли плазму и замораживали в аликвотах при -80°C. Коммерчески доступные наборы использовали для ферментативного определения NEFA (Wako Chemicals, Нойс, Германия), бета-оксибутирата (Stanbio Laboratories, Берне, Техас, США) и HDL-c и LDL-c (BioVision, Mountain View, Калифорния, США).
Как показано на Фигуре 20, введение 2H10 защищало мышей db/db//BKS от повышенных уровней кетонов, ключевого признака сахарного диабета 2 типа. Лечение анти-VEGF-B повышало плазменные уровни HDL-c, также известного как носитель ʺхорошегоʺ холестерина. Ослабление сигнального пути VEGF-В при помощи 2H10 не оказывало никакого эффекта плазменные уровни LDL-с или NEFA.
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) предотвращает утолщение GBM и аномалии подоцитов
Почки собирали от 6-8 недельных самок db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, и использовали для анализа при помощи просвечивающей электронной микроскопии (ПЭМ). Препараты биопсий почек для ТЭМ проводили по стандартным методикам. Ткани фиксировали в фиксирующем буферном растворе (2% глутаровый альдегид, 0,5% параформальдегид, 0,1 M какодилат, 0,1 М сахароза, 3 мМ СaСl2) и промывали в 0,1 М какодилатном буфере рН 7,4 перед окрашиванием в 2% OsO4 в какодилатном буфера в течение 1 ч при комнатной температуре. Образцы обезвоживали и en bloc окрашивание выполняли в 2% уранилацетате в абсолютном спирте в течение 1 ч при комнатной температуре. Затем образцы проводили через серию Эпон 812/ацетон и погружали при 60°C в чистый Эпон 812. Тонкие срезы толщиной 70 нм нарезали на ультратоме Leica EM UC6 и устанавливали на формвар-покрытых медных канавчатых сетках. Пост-окрашивание проводили в 2% водном ацетате рН 3,5 и цитрате, нагруженном Venable и Cogglesall. Сетки анализировали при помощи электронной микроскопии FEI TECNAI.
ТЭМ-анализ с высоким увеличением показал, что при лечении 2H10 уменьшается утолщение GBM, первая почечная патология у мышей db/db (Фигура 21А). Кроме того, лечение анти-VEGF-B мышей db/db//BKS сохраняет морфологию подоцитов, что определяют по числу щелевых образований (Фигура 21В).
Профилактическое лечение с использованием анти-VEGF-B (с использованием 2H10) уменьшает клубочковое накопление липидов у мышей db/db//BKS
Кровь и почки собирали от 6-8 недельных мышей db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, или тощих мышей db/+. Почки рассекали и быстро замораживали на сухом льду и погружали в Tissue-Tek® (Sakura) непосредственно на изложнице криостата. Криосрезы (12 мкм) погружали на 5-15 мин в рабочий раствор ORO (2,5 г масляного красного О (Sigma-Aldrich), растворенного в 400 мл 99% изопропилового спирта, дополнительно разбавленного 6:10 в Н2О, профильтрованного через фильтр 22 мкм (Corning)) и промывали 10 мин под струей водопроводной воды. После этого срезы погружали в течение 3 секунд в раствор гематоксилина и промывали под проточной водой, прежде чем они были установлены. Окрашенные срезы исследовали с использованием светлопольной микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на ORO и гематоксилин в каждом срезе, фотографировали с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли с помощью программы Axio Vision Run wizard на клубочковое ORO-окрашивание (пиксель2/мкм2).
Специфическое увеличение накопления липидов было обнаружено в клубочках диабетических животных по сравнению с тощими недиабетическими животными (Фигура 22). Ингибирование VEGF-B в db/db//BKS резко снижало это эктопическое клубочковое накопление липидов. Количество и размер липидных капель было уменьшено. Анализ показал, что капли преимущественно накапливаются в эндотелиальных клетках и подоцитах.
Терапевтическое лечение с использованием анти-VEGF-B (с использованием) уменьшает клубочковый и канальцевый склероз у мышей с тяжелой степенью диабета.
Почки от 12-недельных самцов и самок db/db//BKS, получавших лечение 2H10 или изотипически подобранным контрольным антителом в течение 8 недель, собирали, фиксировали в 4% ПФА в течение 48 часов, заливали и 3 мкм срезы готовили и окрашивали PAS (Sigma) в соответствии с инструкциями производителя. По меньшей мере 10 клубочков на животное, окрашенных на PAS и каждом срезе, фотографировали с помощью светлопольной микроскопии (микроскоп Axio Vision, Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли с помощью программы Axio Vision Run wizard на клубочковое PAS-окрашивание (пиксель2/мкм2).
Результаты, показанные на Фигурах 23А и 23В, и анализ окрашенных срезов указывают на то, что терапевтическое ингибирование VEGF-B у мышей db/db//BKS снижает клубочковый и канальцевый склероз. Эффект детектируется только у особей женского пола, скорее всего потому, что болезнь более агрессивна у самцов мышей db/db//BKS (обычно наблюдаемый эффект).
Терапевтическое лечение с использованием анти-VEGF-B (2H10) уменьшает накопление липидов в почках мышей db/db//BKS с тяжелой степенью диабета
Почки 12-недельных самцов и самок db/db/BKS, получавших 2H10 или изотипически подобранное контрольное антитело в течение 8 недель, собирали и быстро замораживали на сухом льду и погружали в Tissue-Tek® (Sakura) непосредственно на изложнице криостата. Криосрезы (12 мкм) погружали на 5-15 мин в рабочий раствор ORO (2,5 г масляного красного О (Sigma-Aldrich), растворенного в 400 мл 99% изопропилового спирта, дополнительно разбавленного 6:10 в Н2О, профильтрованного через фильтр 22 мкм (Corning)) и промывали 10 мин под проточной водой из крана, прежде чем они были установлены. Окрашенные срезы исследовали светлопольной микроскопией (микроскоп Axio Vision, Carl Zeiss) при 20-ти кратном увеличении, и делали не менее 10 кадров на срез. Для количественного определения липидных капель количество красных пикселей в каждом кадре количественно определяли, используя программу Axio Vision Run wizard для полного ORO-окрашивания (пиксель, a.u).
Как показано на Фигуре 24, суммарное ORO-окрашивание уменьшалось у самок мышей db/db//BKS, получавших лечение антителом. Эти данные и анализ окрашенных срезов показывают, что как число, так и размер липидных капель было снижено у самок мышей db/db//BKS, получавших антитело анти-VEGF-B. Лечение антителом анти-VEGF-B не уменьшало накопление липидов в почках самцов мышей db/db//BKS, скорее всего, в результате более агрессивного развития заболевания у особей мужского пола.
Терапевтическое лечение с использованием анти-VEGF-B (с использованием 2H10) предотвращает перестройки клубочков у мышей db/db//BKS с тяжелой степенью диабета.
Почки от 12-недельных самок db/db//BKS получавших лечение 2H10 или изотипически подобранное контрольное антитело в течение 8 недель, собирали, фиксировали в 4% ПФА в течение 48 часов, погружали и 3 мкм срезы подготавливали и впоследствии иммуноокрашивали на синаптоподин или пекам. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367) и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: кроличьими анти-синаптоподин (Santa Cruz). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на пекам в каждом срезе, были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового окрашивания на синаптоподин или пекам (пиксель2/мкм2).
Фигуры 25А и 25В показывают повышение уровней пекама и уровней синаптоподина в клубочках мышей, получавших терапевтически антитело анти-VEGF. Эти данные и анализ окрашенных срезов показывают, что морфология клубочков улучшилась у мышей db/db//BKS, получавших антитело анти-VEGF-B. Экспрессия и структура подоцитов и эндотелиальных клеток, т.е. типов клеток, которые имеют решающее значение для процесса клубочковой фильтрации, сохраняются у получавших лечение мышей db/db//BKS. Таким образом, эти данные также указывают на то, что терапевтическое лечение с помощью антитела анти-VEGF также предотвращает сосудистую и подоцитарную потери у мышей db/db//BKS.
Терапевтическое лечение с использованием анти-VEGF-B (с использованием 2H10) уменьшает клубочковое накопление коллагена и гиалиноз артериол у мышей db/db//BKS с тяжелой степенью диабета.
Почки от 12-недельных самок db/db//BKS, получавших лечение 2H10 или изотипически подобранное контрольное антитело в течение 8 недель, собирали, фиксировали в 4% ПФА в течение 48 часов, погружали и 3 мкм срезы подготавливали и иммуноокрашивали на коллаген IV, пекам, и/или α-ЅМА. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367) и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: кроличьими анти-синаптоподин (Santa Cruz). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на пекам в каждом срезе, были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового окрашивания на коллаген IV, пекам и/или α-ЅМА (пиксель2/мкм2).
Фигуры 26А-C показывают снижение уровней коллагена IV, α-SMA и толщины артериол у мышей db/db//BKS, получавших 2H10. Эти данные и анализ окрашенных срезов показывают, что патологические отложения коллагена внутри клубочков (отложение внеклеточного матрикса) и гиалиноз артериол уменьшаются у мышей db/db//BKS, получавших антитело анти-VEGF-В.
Терапевтическое лечение с использованием анти-VEGF-B (2H10) предотвращает развитие макроальбуминурии у диабетических мышей db/db//BKS с установившейся микроальбуминурией
Мышам db/db/BKS вводили i.p. дважды в неделю, начиная с 6-8 недельного возраста, и в течение 8 недель 400 мкг либо 2H10, либо изотипически соответствующего контрольного антитела. Для детекции АСR мочу объемом от 20 мкл до 200 мкл собирали от каждой мыши. Альбумин в моче детектировали, используя комплект Albuwell M kit, креатинина в моче определяли с помощью набора Creatinine Companion murine ELISA kit (Exocell, Филадельфия, Пенсильвания). Систолическое и диастолическое артериальное давление измеряли способом хвостовой манжеты с использованием установки CODA (Kent Scientific). Всех животных приучали к устройству для измерения артериального давления в течение 2-х недель. Все они прошли 1-2 цикла из 20 повторных измерений в день, в течение по меньшей мере 3-х дней. Для почечных функциональных исследований на мышах, отдельных мышей содержали в метаболических клетках. Масса тела и объем мочи в течение 24 часов отслеживали в течение 2 дней подряд. Пробы мочи последних 24 часов были использованы для измерения креатинина в моче. После нахождения в метаболических клетках животных умерщвляли анестезирующими средствами, и всю кровь удаляли путем пункции сердца и использовали для измерения креатина плазмы. Почки рассекали и взвешивали на микроразмерных весах. Выведение креатинина рассчитывали по формуле ([Кр] в моче × объем мочи)/([Кр] в плазме × 24 ч).
Фигура 27А показывает лечение мышей db/db/BKS при помощи 2H10 предотвращает прогрессирование микроальбуминурии до макроальбуминурии, в частности, ACR существенно не меняется в течение исследования у проходящих лечение мышей. Фигура 27В показывает скорость клубочковой фильтрации (определялась как выведение креатинина) также была снижена у мышей db/db/BKS, получавших 2H10. Фигура 27С показывает, что развитие гипертензии предотвращалось путем ингибирования активности VEGF-B. Фигура 27D показывает, что снижение экспрессии VEGF-B не оказывало никакого эффекта на массу тела или почек. Эти данные указывают на то, что мыши db/db, получавшие антитело анти-VEGF-B, обладают улучшенной функцией почек с лучшей фильтрующей способностью, определенной по сниженной концентрации альбумина, соотношению альбумин/креатинин и выведению креатинина.
Терапевтическое лечение с использованием анти-VEGF-B (с использованием 2H10) HFD-питающихся мышей оказывает незначительное влияние на уровень глюкозы в крови
Мышей кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. Лечение антителом начинали на 11 неделе, и мышей лечили 2H10 или изотипически подобранным контрольным антителом в течение 20 недель. Уровни постпрандиального BG у мышей регистрировали в конце испытания, после удаления питания в течение 2 ч. Измерения уровня глюкозы производились для крови, отобранной из хвостовой вены с помощью контурного глюкометра Байера.
Фигура 28 показывает, что HFD повышает уровень глюкозы в крови, однако лечение анти-VEGF-B не предотвращает развитие гипергликемии.
Терапевтическое лечение с использованием анти-VEGF-B (2H10) HFD-питающихся мышей улучшает функцию почек без снижения уровня глюкозы в крови
Мышей кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также включали тощих контрольных животных дикого типа. Лечение антителом начинали на 11 неделе, и мышей лечили 2H10 или изотипически подобранным контрольным антителом в течение 20 недель. Для детектирования альбумина и ACR мочу объемом от 20 мкл до 200 мкл собирали от каждой мыши в конечной точке испытания. Альбумин детектировали, используя набор Albuwell M kit, креатинин определяли с помощью набора Creatinine Companion murine ELISA kit (Exocell, Филадельфия, Пенсильвания).
Несмотря на то, лечение при помощи анти-VEGF-B HFD-питающихся мышей не снижало уровни глюкозы в крови, лечение оказало существенное влияние на диабетическую почечную физиологию (Фигура 29). Фильтрационная способность почек была улучшена, что определяли по снижению экскреции альбумина (Фигура 29A), ACR (Фигура 29Б) и GFR. Лечение при помощи анти-VEGF-B HFD-питающихся мышей не оказывало никакого влияния на массу тела или почек, по сравнению с мышами, получавшими контрольное лечение.
Терапевтическое лечение с использованием анти-VEGF-B (2H10) HFD-питающихся мышей улучшает липидный профиль плазмы
Мышей кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также включали тощих контрольных животных дикого типа. Лечение антителом начинали на 11 неделе, и мышей лечили 2H10 или изотипически подобранным контрольным антителом в течение 20 недель. Животных умерщвляли с использованием изофлурановых анестетиков, и всю кровь удаляли путем пункции сердца. Кровь центрифугировали при 14000 об/мин, 4°C в течение 10 минут, после чего отделяли плазму и замораживали в аликвотах при -80°С. Использовали имеющиеся в продаже наборы для ферментативного определения NEFA (Wako Chemicals, Neuss, Германия), бета-гидроксибутирата (Stanbio Laboratories, Boerne, штат Техас, США) и триглицеридов (Sigma-Aldrich).
Фигура 30 показывает, что введение 2H10 HFD-питающимся мышам предохраняло от повышенных уровней видов липидов, связанных с T2D. Например, лечение с использованием антитела анти-VEGF-B снижало уровни триглицеридов в плазме (Фигура 30А), NEFA (Фигура 30В) и кетонов (Фигура 30С).
Терапевтическое лечение с использованием анти-VEGF-B (2H10) HFD-питающихся мышей предотвращает клубочковое мезангиальное расширение и склероз
Мышей кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также включали тощих контрольных животных дикого типа. Лечение антителом начинали на 11 неделе, и мышей лечили 2H10 или изотипически подобранным контрольным антителом в течение 20 недель. Животных умерщвляли с использованием изофлурановых анестетиков, и всю кровь удаляли путем пункции сердца.
Почки рассекали, фиксировали в 4% ПФА в течение 48 ч, а затем обрабатывали для заливания парафином с помощью стандартных процедур. После заливания 3-мкм срезы готовили и окрашивали PAS (Sigma) в соответствии с инструкциями производителя. По меньшей мере 10 клубочков на животное, окрашенных PAS в каждом срезе, фотографировали при помощи светлопольной микроскопии (микроскоп Axio Vision, Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для PAS-окрашивания клубочков (пиксель2/мкм2).
Данные, представленные на Фигуре 31, и анализ окрашенных срезов показывают, что введение 2H10 HFD-питающимся мышам защищало от клубочкового мезангиального расширения (Фигура 31А) и клубочковой гипертрофии (Фигура 31В).
Терапевтическое лечение с использованием анти-VEGF-B (2H10) HFD-питающихся мышей предотвращает клубочковое накопление липидов
Мышей кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также включали тощих контрольных животных дикого типа. Лечение антителом начинали на 11 неделе, и мышей лечили 2H10 или изотипически подобранным контрольным антителом в течение 20 недель. Животных умерщвляли с использованием изофлурановых анестетиков, и всю кровь удаляли путем пункции сердца.
Почки рассекали и быстро замораживали на сухом льду и погружали в Tissue-Tek® (Sakura) непосредственно на изложнице криостата. Криосрезы (12 мкм) погружали на 5-15 мин в рабочий раствор ORO (2,5 г масляного красного О (Sigma-Aldrich), растворенного в 400 мл 99% изопропилового спирта, дополнительно разбавленного 6:10 в Н2О, профильтрованного через фильтр 22 мкм (Corning)) и промывали 10 мин под проточной водой из крана. После этого срезы погружали в течение 3 секунд в раствор гематоксилина и промывали под проточной водой, прежде чем они были установлены. Окрашенные срезы исследовали при помощи светлопольной микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на ORO и гематоксилин в каждом срезе, фотографировали с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли с помощью программы Axio Vision Run wizard на клубочковое ORO-окрашивание (пиксель2/мкм2).
Данные, представленные на Фигуре 32, и анализ окрашенных срезов показывают, что введение 2H10 HFD-питающимся мышам защищало от эктопического накопления липидов в почках. Уменьшается количество и размер липидных капель.
Терапевтическое лечение с использованием анти-VEGF-B (2H10) HFD-питающихся мышей предотвращает клубочковое накопление липидов и сохраняет целостность подоцитов
Мышей кормили 60% HFD (Research Diets, США) вместо HFD в течение 30 недель, начиная с возраста 5 недель. В исследование также включали тощих контрольных животных дикого типа. Лечение антителом начинали на 11 неделе, и мышей лечили 2H10 или изотипически подобранным контрольным антителом в течение 20 недель. Животных умерщвляли с использованием анестетических средств, и всю кровь удаляли путем пункции сердца.
Почки рассекали, фиксировали в 4% ПФА в течение 48 ч, а затем обрабатывали для погружения в парафин с помощью стандартных процедур. После погружения 3-мкм срезы подготавливали и затем иммуноокрашивали на коллаген IV, пекам и/или α-SMA. Кратко, извлечение антигена проводили на 3 мкм срезе, используя раствор извлечения антигена Ph6 (Dako #S2367), и нагревали при 98°C в течение 10 минут. Срезы инкубировали при 4°C в течение 12 часов с первичными антителами: кроличьими анти-коллаген IV (abcam), козьими анти-пекам (abcam) или α-SMA (Сигма). Соответствующие флуоресцентно меченные вторичные антитела (Invitrogen, Alexa fluor) применяли, и срезы затем выдерживали в течение 1 ч при комнатной температуре, после чего они были подготовлены для микроскопии. По меньшей мере 10 клубочков на животное, окрашенных на коллагена IV, пекам или α-SMA в каждом срезе были сфотографированы с помощью микроскопа Axio Vision (Carl Zeiss) при 40-кратном увеличении. Клубочки количественно определяли, используя программу Axio Vision Run wizard для клубочкового I) окрашивания на коллаген IV (пиксель2/мкм2) и II) толщина клубочковой артериолы (мкм2).
Данные, представленные на Фигуре 33, и анализ окрашенных срезов показывают, что введение 2H10 HFD-питающимся мышам защищает от гистологических диабетических нефропатия-ассоциированных патологий, таких как увеличенное клубочковое ECM-накопление (Фигура 33А) и гиалиноз артериол (Фигура 33В).
Терапевтическое лечение с использованием анти-VEGF-B (2H10) STZ-инъецированных животных не оказывает влияния на уровень глюкозы в крови
Для индукции диабета I типа была использована стрептозотоциновая модель (STZ). Эксперименты проводились с самцами мышей Bl/6 от Charles River. Мыши голодали в течение 4 часов до инъекции STZ, и получали 50 мг стрептозотоцина на кг массы (Sigma-Aldrich) в 50 мМ цитрате натрия (рН 4,5) или натрийцитратном буфере внутрибрюшинно в день 1, 2, 3, 4 и 5. Уровни BG у мышей регистрировали до и через 1 неделю после введения STZ. Животные с уровнем глюкозы в крови выше 12 ммоль/л расценивались как гипергликемические. Животных разделяли случайным образом на две группы и лечили 2H10 или изотипически подобранным контрольным антителом два раза в неделю. Уровни постпрандиальной BG у мышей, получавших анти-VEGF-B (2H10) или контрольное антитело, и тощих контрольных животных регистрировали через две недели после удаления питания в течение 2 ч. Измерения уровня глюкозы производились для крови, отобранной из хвостовой вены, с помощью контурного глюкометра Байера.
Как показано на Фигуре 34А, STZ-инъекция самцам мышей индуцирует гипергликемию. Однако лечение с использованием анти-VEGF-B после инъекции STZ не оказывает большого влияния на уровень глюкозы в крови (Фигура 34В).
Терапевтическое лечение с использованием анти-VEGF-B STZ-инъецированных животных, используя 2H10, улучшает функцию почек
Для индукции сахарного диабета I типа была использована стрептозотоциновая модель (STZ). Эксперименты проводились с самцами мышей от Bl/6. Scanbur. Мыши голодали в течение 4 часов до инъекции STZ, и получали 50 мг стрептозотоцина на кг массы (Sigma-Aldrich) в 50 мМ натрийцитратном буфере (pH 4,5) внутрибрюшинно в день 1-5. Уровни BG у мышей регистрировали до и через 1 неделю после введения STZ. Животные с уровнем глюкозы в крови выше 12 ммоль/л расценивались как гипергликемические. Животных разделяли случайным образом на две группы и лечили 2H10 или изотипически подобранным контрольным антителом два раза в неделю, как указано выше.
Для детекции альбумина и ACR мочу объемом от 20 мкл до 200 мкл собирали от каждой мыши в конечной точке испытания (продолжающийся эксперимент, сообщается только об 1 неделе лечения. Альбумин определяли с помощью набора Alhuwell M, и мочевой креатинин измеряли с использованием набора Creatinine Companion murine ELISA kit (Exocell, Филадельфия, штат Пенсильвания).
Несмотря на незначительное воздействие на уровень глюкозы в крови у мышей, дозированных антителом анти-VEGF-B 2H10 после инъекции STZ, функция почек диабетических мышей значительно улучшалась при лечении антителом. Фильтрационная способность почек улучшалась, что определяли по уменьшенной экскреции альбумина (Фигура 35А) и ACR (Фигура 35В).
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CSL Limited
B Creative Sweden AB
<120> Способ лечения нефропатии
<130> 516049
<160> 46
<170> PatentIn version 3.5
<210> 1
<211> 207
<212> PRT
<213> Homo sapiens
<220>
<221> СИГНАЛ
<222> (1)..(21)
<223> Сигнальная последовательность
<400> 1
Met Ser Pro Leu Leu Arg Arg Leu Leu Leu Ala Ala Leu Leu Gln Leu
1 5 10 15
Ala Pro Ala Gln Ala Pro Val Ser Gln Pro Asp Ala Pro Gly His Gln
20 25 30
Arg Lys Val Val Ser Trp Ile Asp Val Tyr Thr Arg Ala Thr Cys Gln
35 40 45
Pro Arg Glu Val Val Val Pro Leu Thr Val Glu Leu Met Gly Thr Val
50 55 60
Ala Lys Gln Leu Val Pro Ser Cys Val Thr Val Gln Arg Cys Gly Gly
65 70 75 80
Cys Cys Pro Asp Asp Gly Leu Glu Cys Val Pro Thr Gly Gln His Gln
85 90 95
Val Arg Met Gln Ile Leu Met Ile Arg Tyr Pro Ser Ser Gln Leu Gly
100 105 110
Glu Met Ser Leu Glu Glu His Ser Gln Cys Glu Cys Arg Pro Lys Lys
115 120 125
Lys Asp Ser Ala Val Lys Pro Asp Arg Ala Ala Thr Pro His His Arg
130 135 140
Pro Gln Pro Arg Ser Val Pro Gly Trp Asp Ser Ala Pro Gly Ala Pro
145 150 155 160
Ser Pro Ala Asp Ile Thr His Pro Thr Pro Ala Pro Gly Pro Ser Ala
165 170 175
His Ala Ala Pro Ser Thr Thr Ser Ala Leu Thr Pro Gly Pro Ala Ala
180 185 190
Ala Ala Ala Asp Ala Ala Ala Ser Ser Val Ala Lys Gly Gly Ala
195 200 205
<210> 2
<211> 188
<212> PRT
<213> Homo sapiens
<220>
<221> СИГНАЛ
<222> (1)..(21)
<223> Сигнальная последовательность
<400> 2
Met Ser Pro Leu Leu Arg Arg Leu Leu Leu Ala Ala Leu Leu Gln Leu
1 5 10 15
Ala Pro Ala Gln Ala Pro Val Ser Gln Pro Asp Ala Pro Gly His Gln
20 25 30
Arg Lys Val Val Ser Trp Ile Asp Val Tyr Thr Arg Ala Thr Cys Gln
35 40 45
Pro Arg Glu Val Val Val Pro Leu Thr Val Glu Leu Met Gly Thr Val
50 55 60
Ala Lys Gln Leu Val Pro Ser Cys Val Thr Val Gln Arg Cys Gly Gly
65 70 75 80
Cys Cys Pro Asp Asp Gly Leu Glu Cys Val Pro Thr Gly Gln His Gln
85 90 95
Val Arg Met Gln Ile Leu Met Ile Arg Tyr Pro Ser Ser Gln Leu Gly
100 105 110
Glu Met Ser Leu Glu Glu His Ser Gln Cys Glu Cys Arg Pro Lys Lys
115 120 125
Lys Asp Ser Ala Val Lys Pro Asp Ser Pro Arg Pro Leu Cys Pro Arg
130 135 140
Cys Thr Gln His His Gln Arg Pro Asp Pro Arg Thr Cys Arg Arg Arg
145 150 155 160
Cys Arg Arg Arg Ser Phe Leu Arg Cys Gln Gly Arg Gly Leu Glu Leu
165 170 175
Asn Pro Asp Thr Cys Arg Cys Arg Lys Leu Arg Arg
180 185
<210> 3
<211> 119
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность из VH антитела 2H10
<400> 3
Val Gln Leu Gln Gln Pro Gly Thr Glu Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Phe Trp
20 25 30
Ile His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
35 40 45
His Ile Asn Pro Gly Asn Gly Gly Thr Asn Tyr Asn Glu Lys Phe Lys
50 55 60
Arg Met Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met
65 70 75 80
Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ser Tyr Ser Asn Tyr Val Arg Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser
115
<210> 4
<211> 106
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность из VL антитела 2H10
<400> 4
Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly Asp
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Phe Leu
20 25 30
Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile Tyr
35 40 45
Tyr Thr Ser Thr Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln Glu
65 70 75 80
Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Lys Thr Leu Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 5
<211> 119
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность VH гуманизированной формы антитела
2H10
<400> 5
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala Ser
1 5 10 15
Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Phe Trp
20 25 30
Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
35 40 45
His Ile Asn Pro Gly Asn Gly Gly Thr Asn Tyr Asn Glu Lys Phe Lys
50 55 60
Arg Arg Val Thr Met Thr Arg Asp Lys Ser Ile Ser Thr Ala Tyr Met
65 70 75 80
Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ser Tyr Ser Asn Tyr Val Arg Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 6
<211> 107
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность VL гуманизированной формы антитела 2H10
<400> 6
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Phe
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Thr Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Lys Thr Leu Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 7
<211> 116
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность VH антитела 4E12
<400> 7
Val Gln Pro Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Val Lys Met Ser Cys Lys Ala Ser Gly Asp Thr Phe Thr Asn Ser Trp
20 25 30
Ile Gly Trp Val Thr Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
35 40 45
Asp Ile Phe Pro Gly Ser Gly His Thr Asn Tyr Asn Glu Lys Phe Lys
50 55 60
Asn Arg Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr Met
65 70 75 80
Leu Leu Ser Ser Leu Thr Ser Asp Asp Ser Ala Val Tyr Tyr Cys Val
85 90 95
Ile Glu Asn Tyr Ala Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ala
115
<210> 8
<211> 107
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность VL антитела 4E12
<400> 8
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Ser Ser Val Gly
1 5 10 15
Asp Arg Val Ser Val Thr Cys Lys Ala Ser Gln Asn Val Asn Thr Asn
20 25 30
Val Ala Trp Tyr Gln Gln Lys Gln Gly Gln Ser Pro Arg Pro Leu Ile
35 40 45
Tyr Ser Ala Ser Ser Arg Cys Ser Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Phe Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His Ser Phe Pro Leu
85 90 95
Thr Phe Gly Ala Gly Ala Lys Leu Asp Leu Lys
100 105
<210> 9
<211> 118
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность VH антитела 2F5
<400> 9
Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Thr Ser
1 5 10 15
Val Arg Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Phe Tyr
20 25 30
Ile His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
35 40 45
Trp Phe Tyr Pro Gly Asn Val Asn Thr Asn Tyr Asn Glu Lys Leu Lys
50 55 60
Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Ala Ala Tyr Leu
65 70 75 80
Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys Thr
85 90 95
Arg Ser Pro Tyr Tyr Gly Tyr Val Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 10
<211> 107
<212> PRT
<213> искусственная
<220>
<223> аминокислотная последовательность VL антитела 2F5
<400> 10
Asp Ile Val Met Thr Gln Ser His Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Gly Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Ser Ser Ser Leu Thr
85 90 95
Phe Gly Ala Gly Ala Thr Lys Leu Glu Leu Lys
100 105
<210> 11
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2H10 VL CDR1
<400> 11
agggcaagtc aggacattag caatttttta aac 33
<210> 12
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2H10 VL CDR2
<400> 12
tacacatcaa cattacactc a 21
<210> 13
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2H10 VL CDR3
<400> 13
caacagggta aaacgcttcc tcccacg 27
<210> 14
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2H10 VH CDR1
<400> 14
ggctacactt tcactggctt ctggatacac 30
<210> 15
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2H10 VH CDR2
<400> 15
catattaatc ctggcaatgg tggcactaac tacaatgaga agttcaagag a 51
<210> 16
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2H10 VH CDR3
<400> 16
tcctatagta actacgtgcg ggctatggac tac 33
<210> 17
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2H10 VL CDR1
<400> 17
Arg Ala Ser Gln Asp Ile Ser Asn Phe Leu Asn
1 5 10
<210> 18
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2H10 VL CDR2
<400> 18
Tyr Thr Ser Thr Leu His Ser
1 5
<210> 19
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2H10 VL CDR3
<400> 19
Gln Gln Gly Lys Thr Leu Pro Pro Thr
1 5
<210> 20
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2H10 VH CDR1
<400> 20
Gly Tyr Thr Phe Thr Gly Phe Trp Ile His
1 5 10
<210> 21
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2H10 VH CDR2
<400> 21
His Ile Asn Pro Gly Asn Gly Gly Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Arg
<210> 22
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2H10 VH CDR3
<400> 22
Ser Tyr Ser Asn Tyr Val Arg Ala Met Asp Tyr
1 5 10
<210> 23
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2F5 VL CDR1
<400> 23
aaggccagtc aggatgtggg tactgctgta gcc 33
<210> 24
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2F5 VL CDR2
<400> 24
tgggcatcca cccggcacac t 21
<210> 25
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2F5 VL CDR3
<400> 25
caacaatata gcagctctct cacg 24
<210> 26
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2F5 VH CDR1
<400> 26
ggctacacct tcacaacctt ctatatacac 30
<210> 27
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2F5 VH CDR2
<400> 27
tggttttatc ctggaaatgt taataccaac tacaatgaga agctcaaggg c 51
<210> 28
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 2F5 VH CDR3
<400> 28
tccccttact acggctacgt ttttgactac 30
<210> 29
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2F5 VL CDR1
<400> 29
Lys Ala Ser Gln Asp Val Gly Thr Ala Val Ala
1 5 10
<210> 30
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2F5 VL CDR2
<400> 30
Trp Ala Ser Thr Arg His Thr
1 5
<210> 31
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2F5 VL CDR3
<400> 31
Gln Gln Tyr Ser Ser Ser Leu Thr
1 5
<210> 32
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2F5 VH CDR1
<400> 32
Gly Tyr Thr Phe Thr Thr Phe Tyr Ile His
1 5 10
<210> 33
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2F5 VH CDR2
<400> 33
Trp Phe Tyr Pro Gly Asn Val Asn Thr Asn Tyr Asn Glu Lys Leu Lys
1 5 10 15
Gly
<210> 34
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 2F5 VH CDR3
<400> 34
Ser Pro Tyr Tyr Gly Tyr Val Phe Asp Tyr
1 5 10
<210> 35
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 4E12 VL CDR1
<400> 35
aaggccagtc agaatgtgaa cactaatgta gcc 33
<210> 36
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 4E12 VL CDR2
<400> 36
tcggcatcct cccggtgcag t 21
<210> 37
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 4E12 VL CDR3
<400> 37
cagcaatatc acagctttcc gctcacg 27
<210> 38
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 4E12 VH CDR1
<400> 38
ggcgacacct tcaccaactc ctggataggc 30
<210> 39
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 4E12 VH CDR2
<400> 39
gatatttttc ctgggagtgg tcatactaac tacaatgaga agttcaagaa c 51
<210> 40
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность 4E12 VH CDR3
<400> 40
gagaattatg cctggtttgc ttat 24
<210> 41
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 4E12 VL CDR1
<400> 41
Lys Ala Ser Gln Asn Val Asn Thr Asn Val Ala
1 5 10
<210> 42
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 4E12 VL CDR2
<400> 42
Ser Ala Ser Ser Arg Cys Ser
1 5
<210> 43
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 4E12 VL CDR3
<400> 43
Gln Gln Tyr His Ser Phe Pro Leu Thr
1 5
<210> 44
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 4E12 VH CDR1
<400> 44
Gly Asp Thr Phe Thr Asn Ser Trp Ile Gly
1 5 10
<210> 45
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 4E12 VH CDR2
<400> 45
Asp Ile Phe Pro Gly Ser Gly His Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Asn
<210> 46
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность 4E12 VH CDR3
<400> 46
Glu Asn Tyr Ala Trp Phe Ala Tyr
1 5
<---
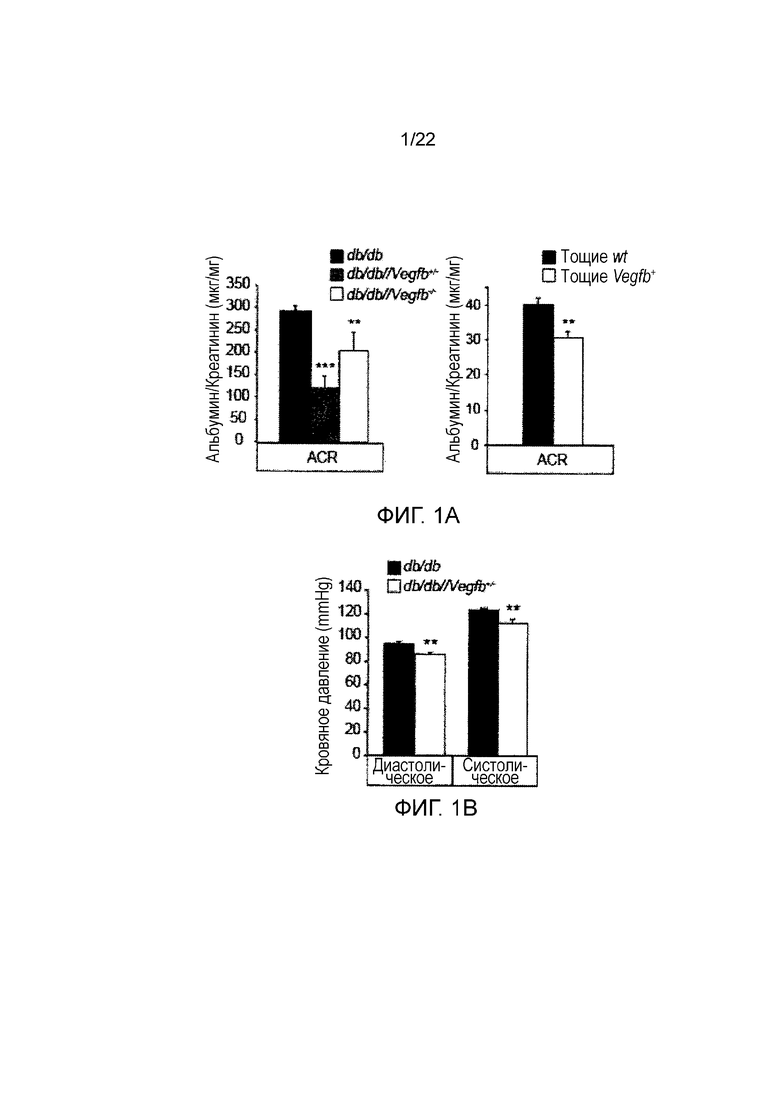
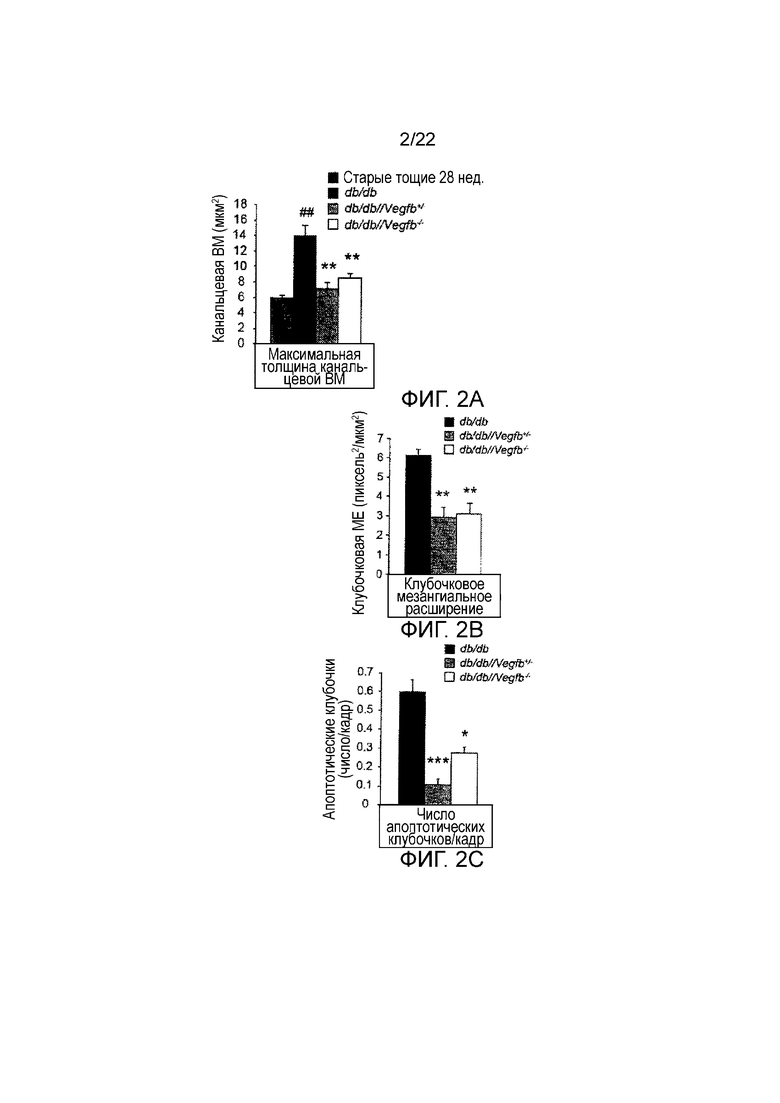
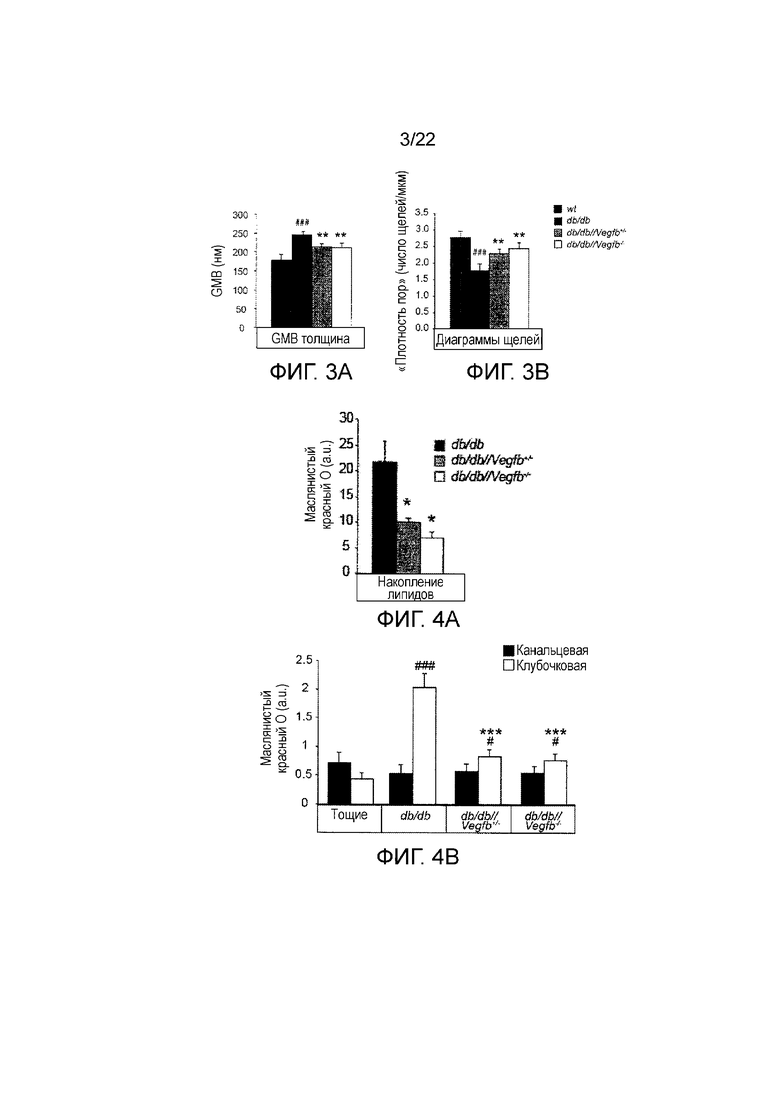
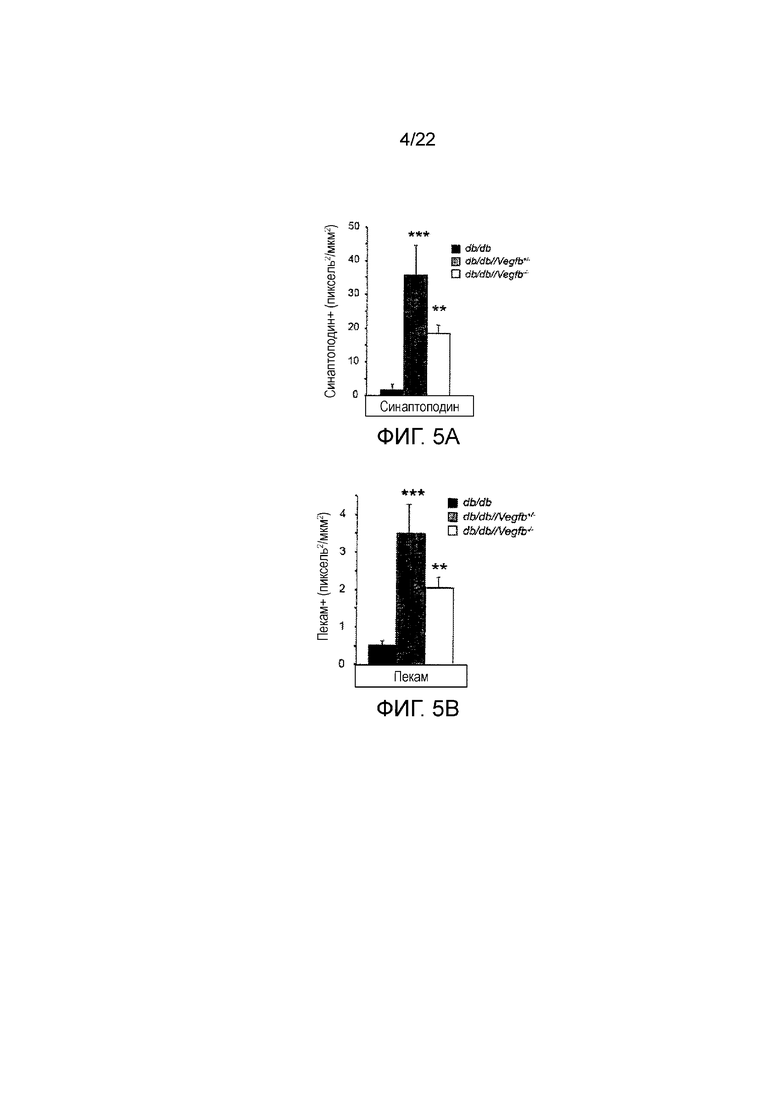
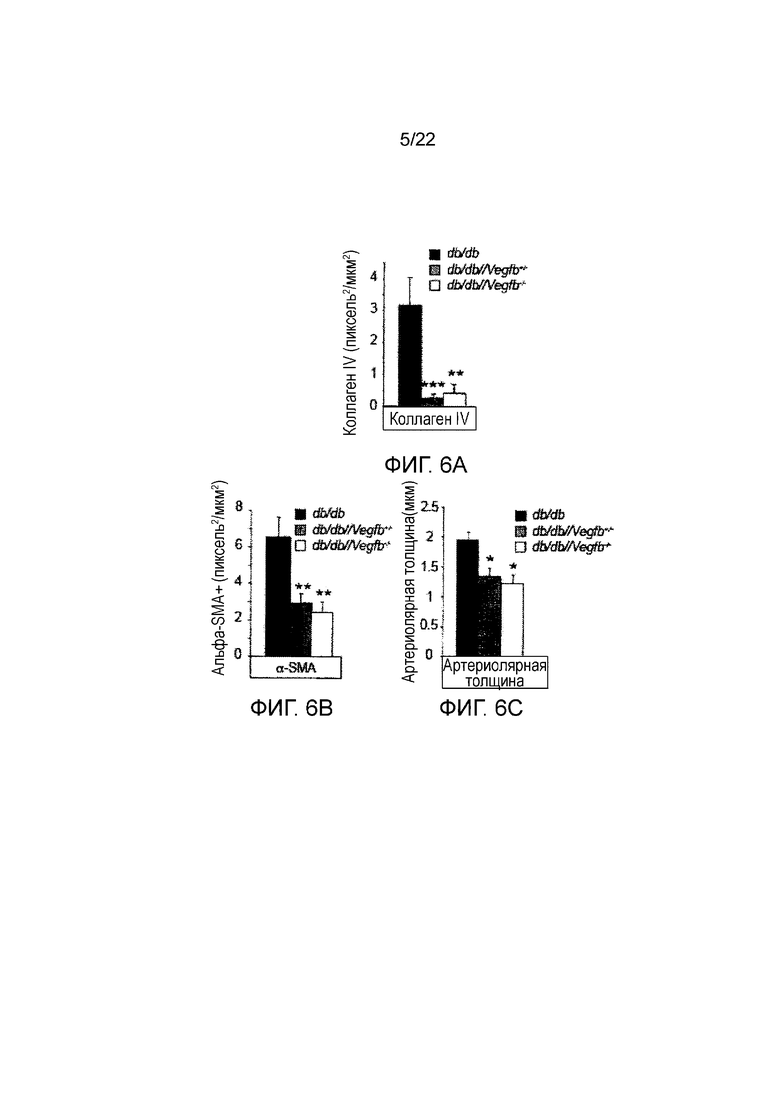
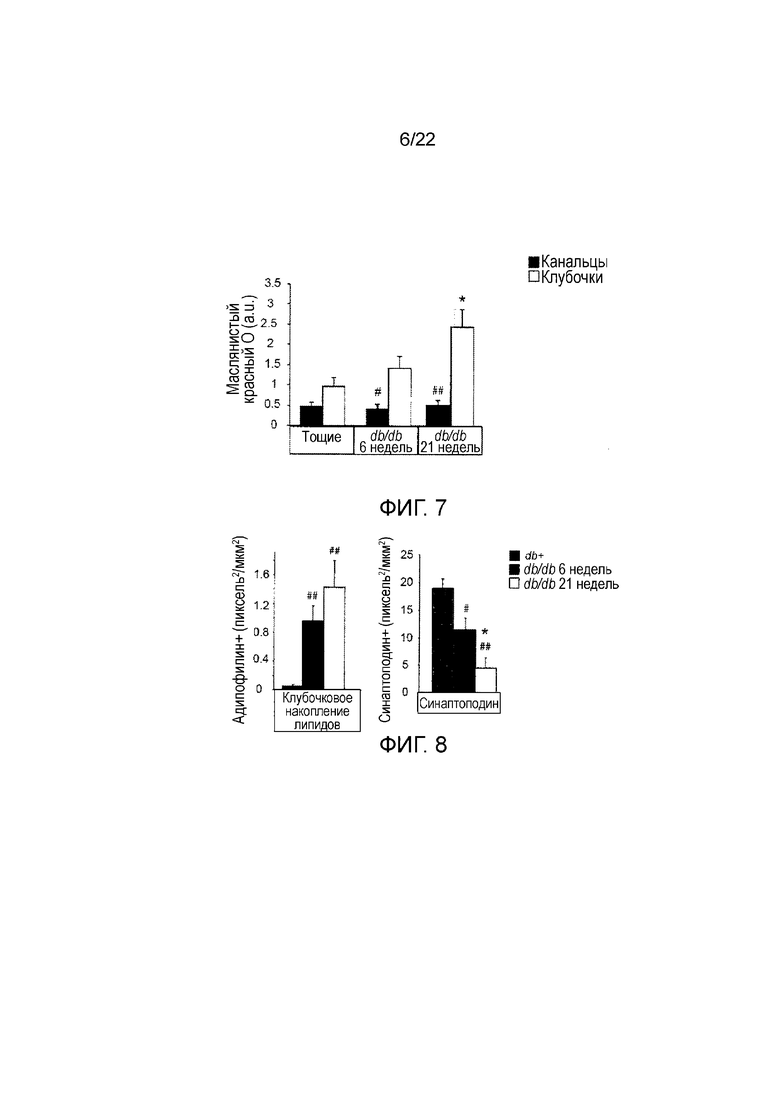
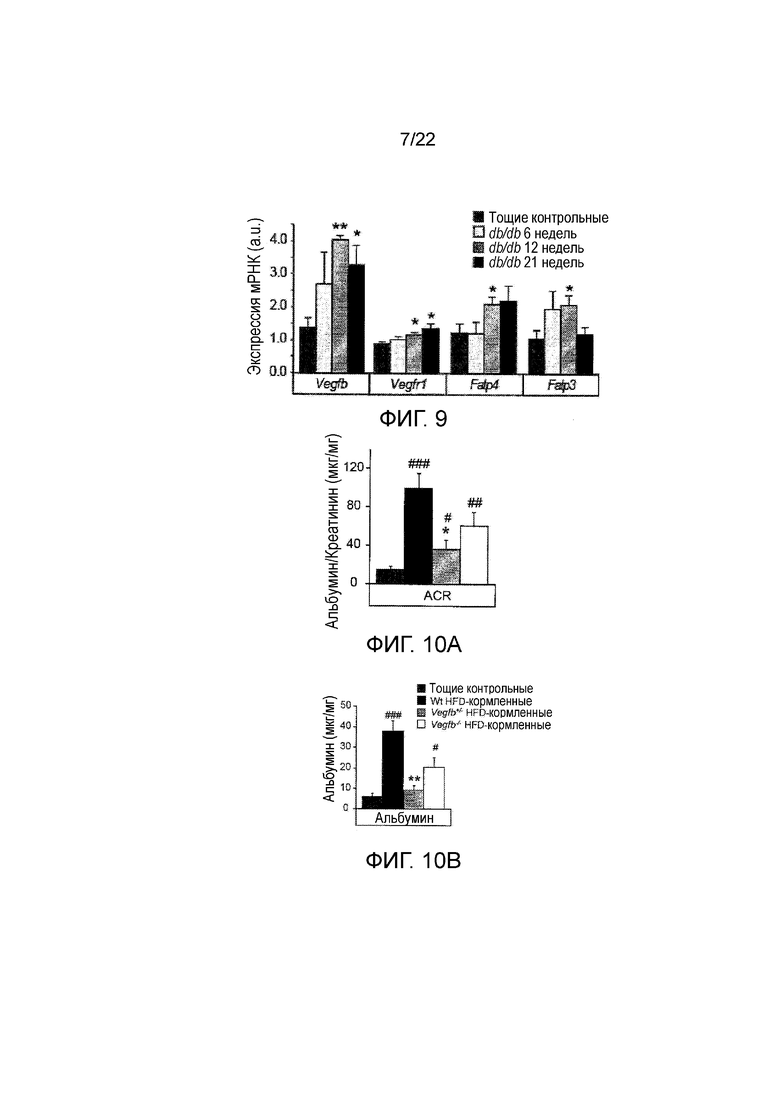
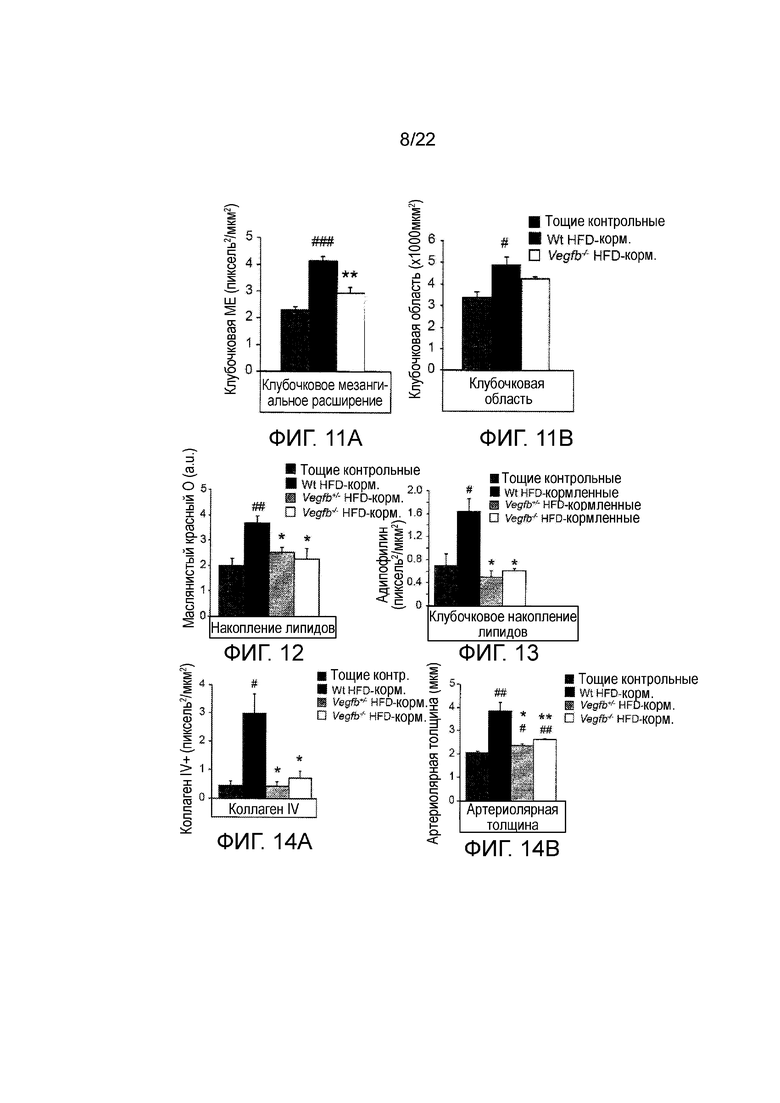
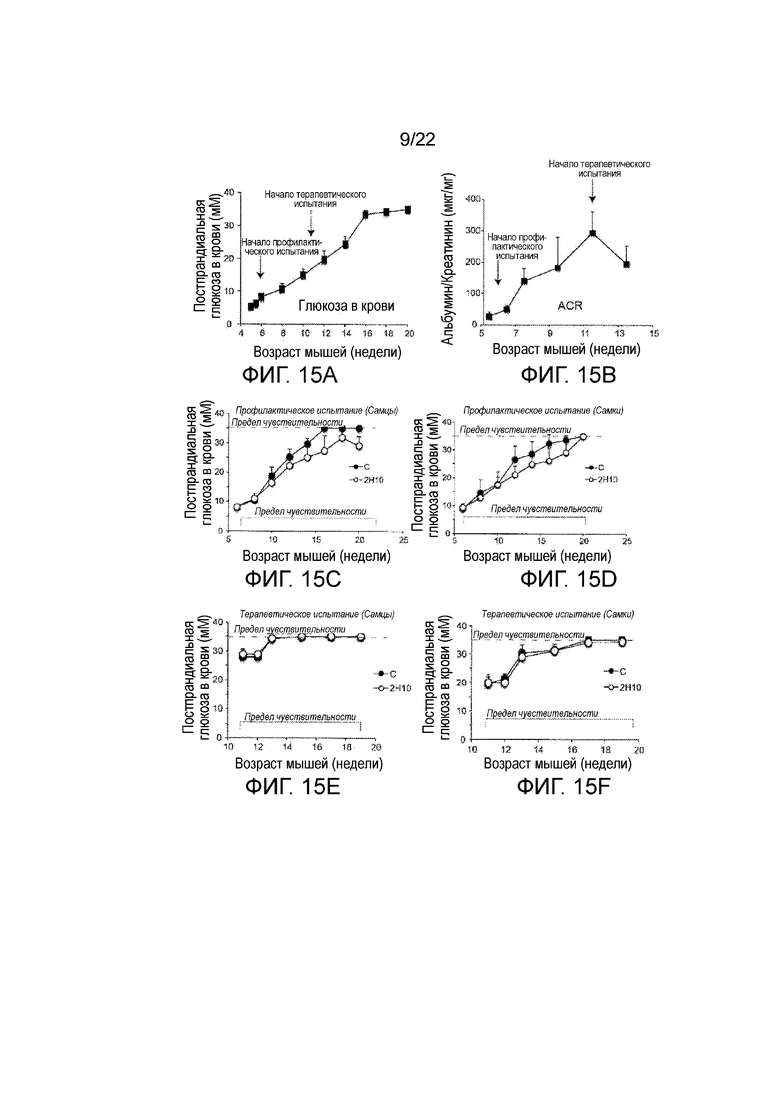
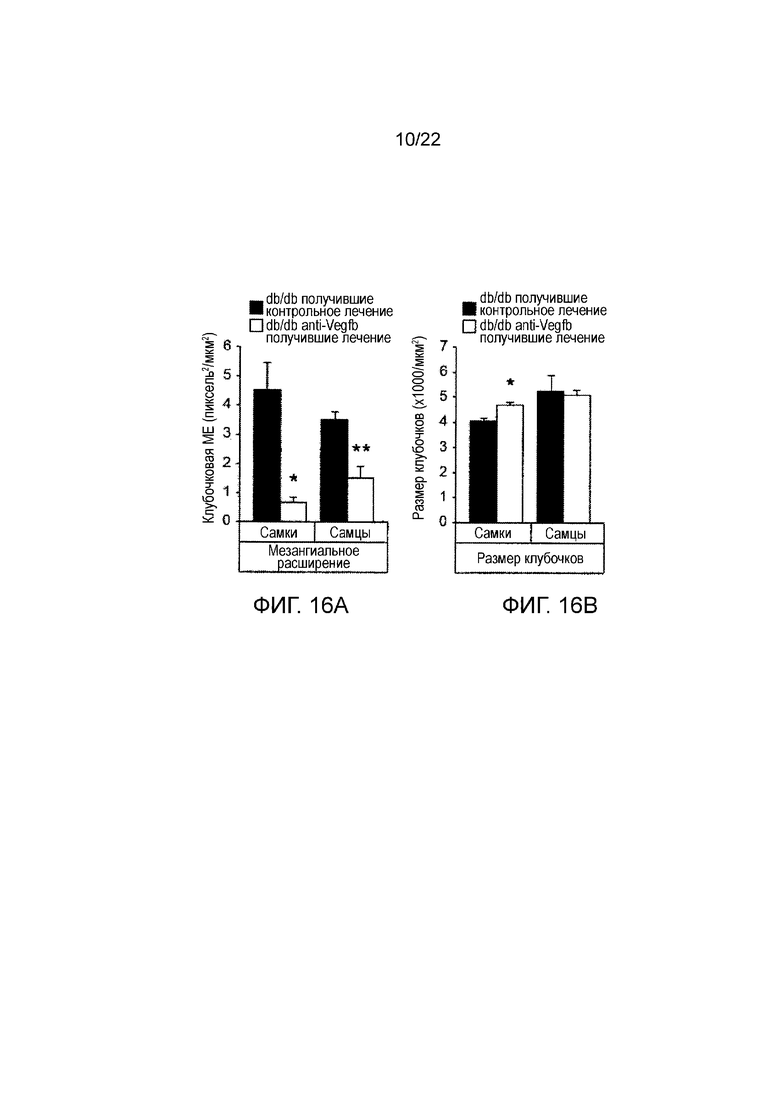
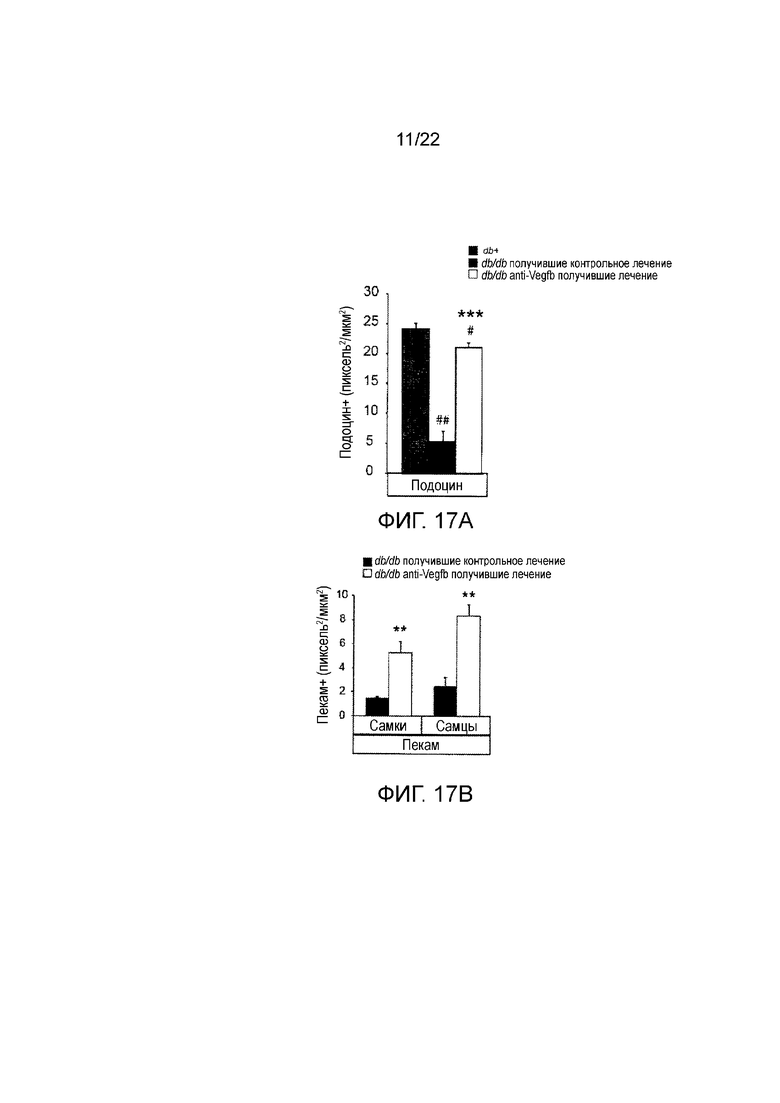
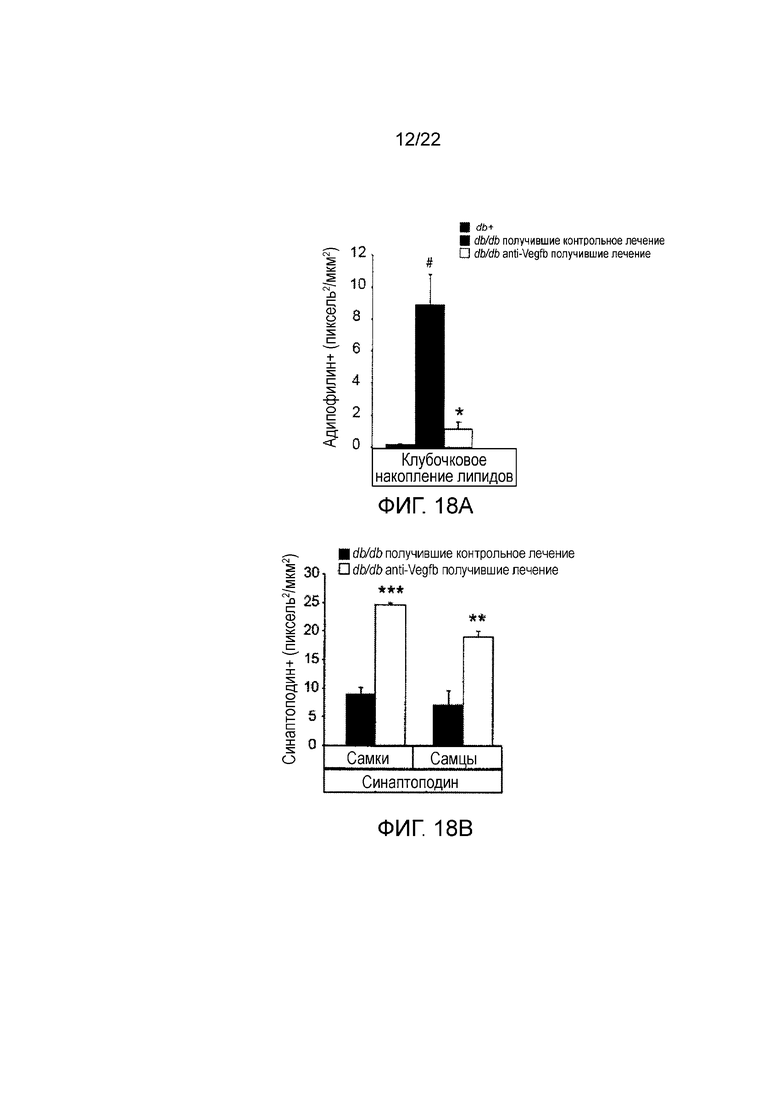
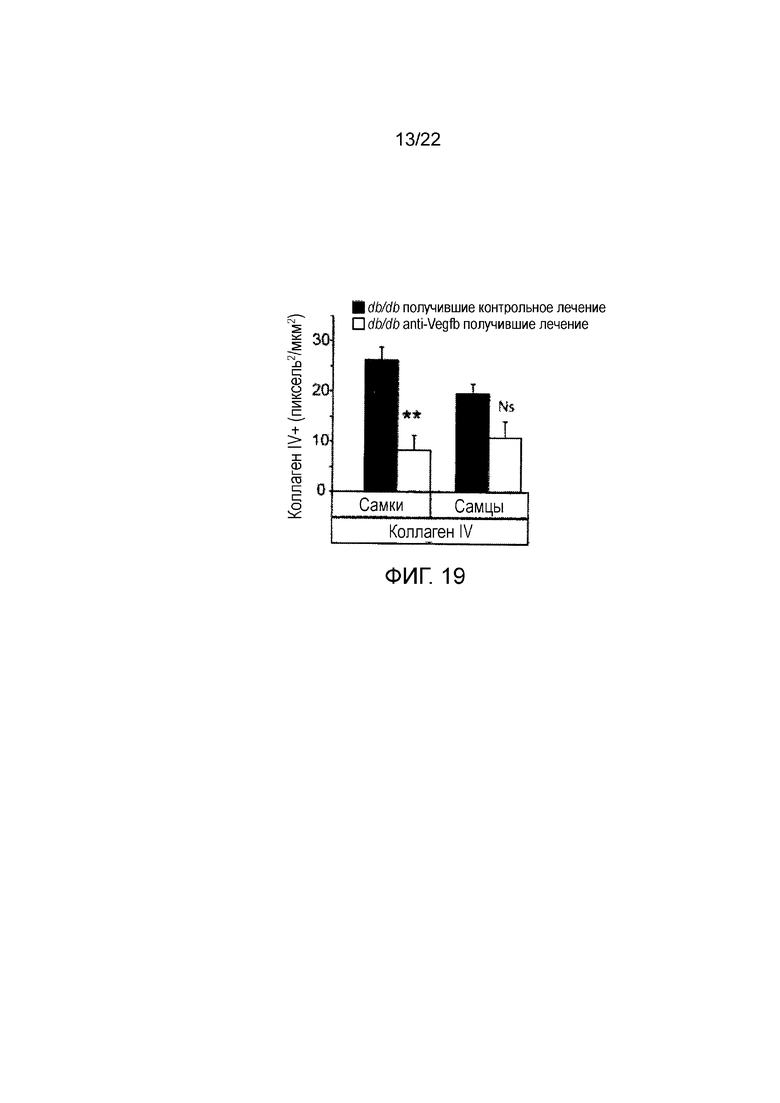
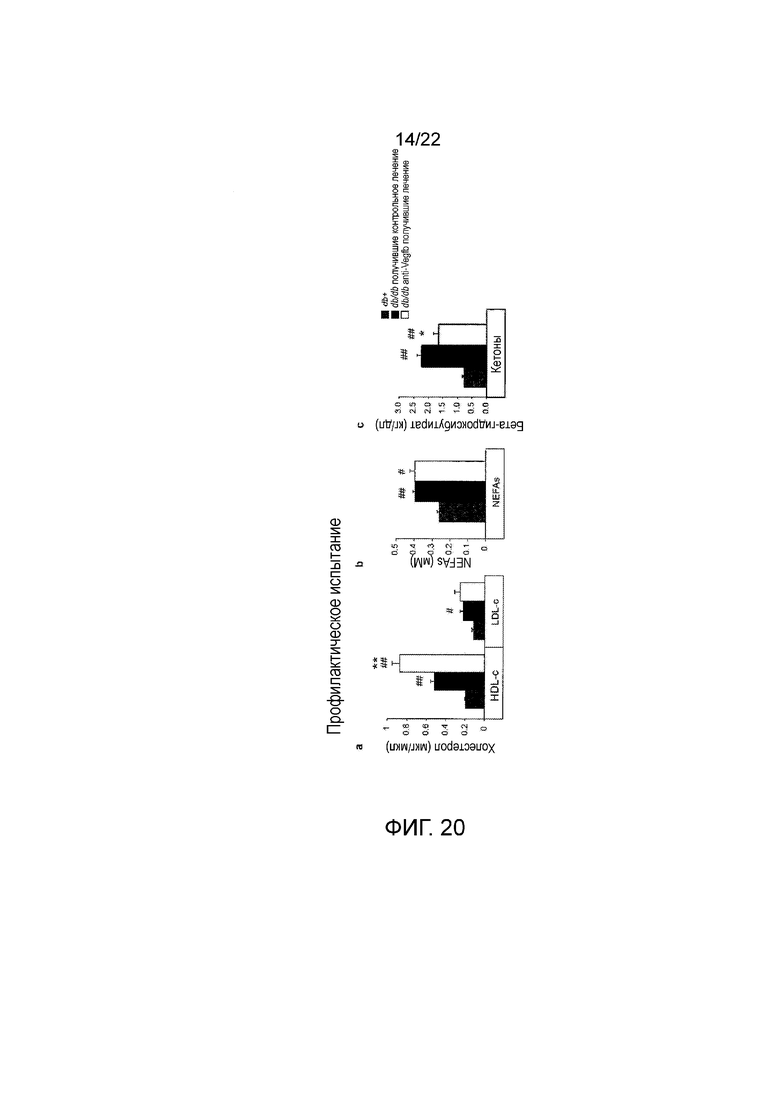
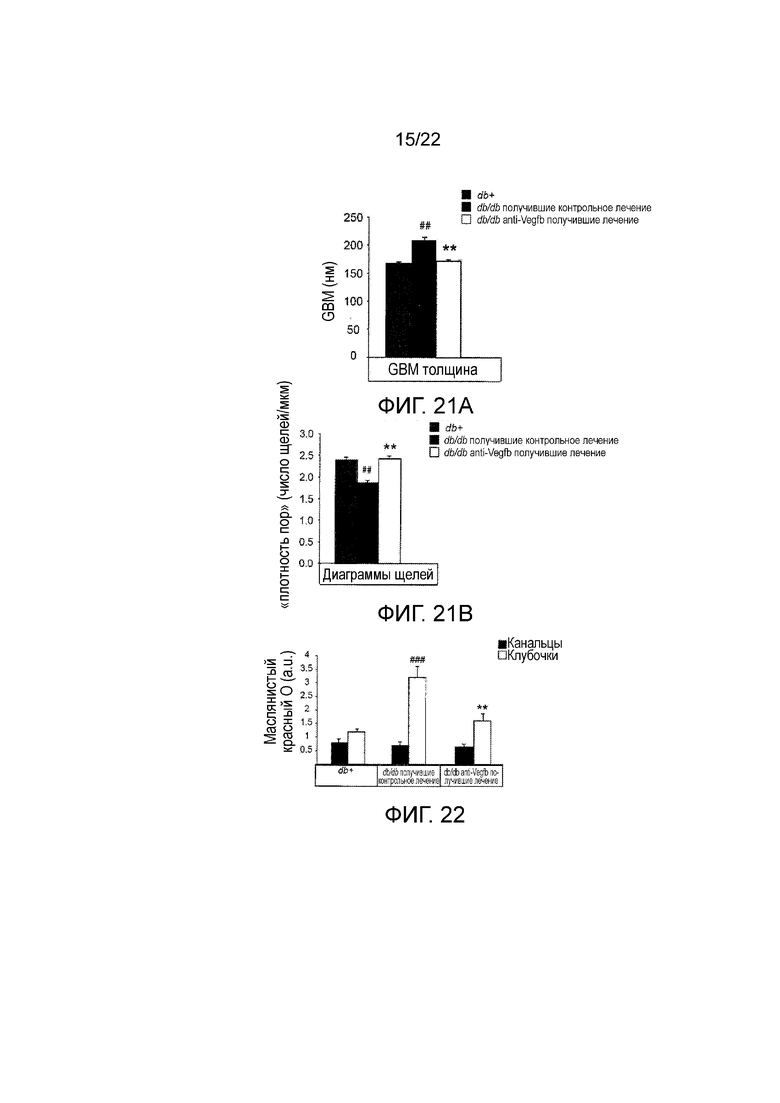
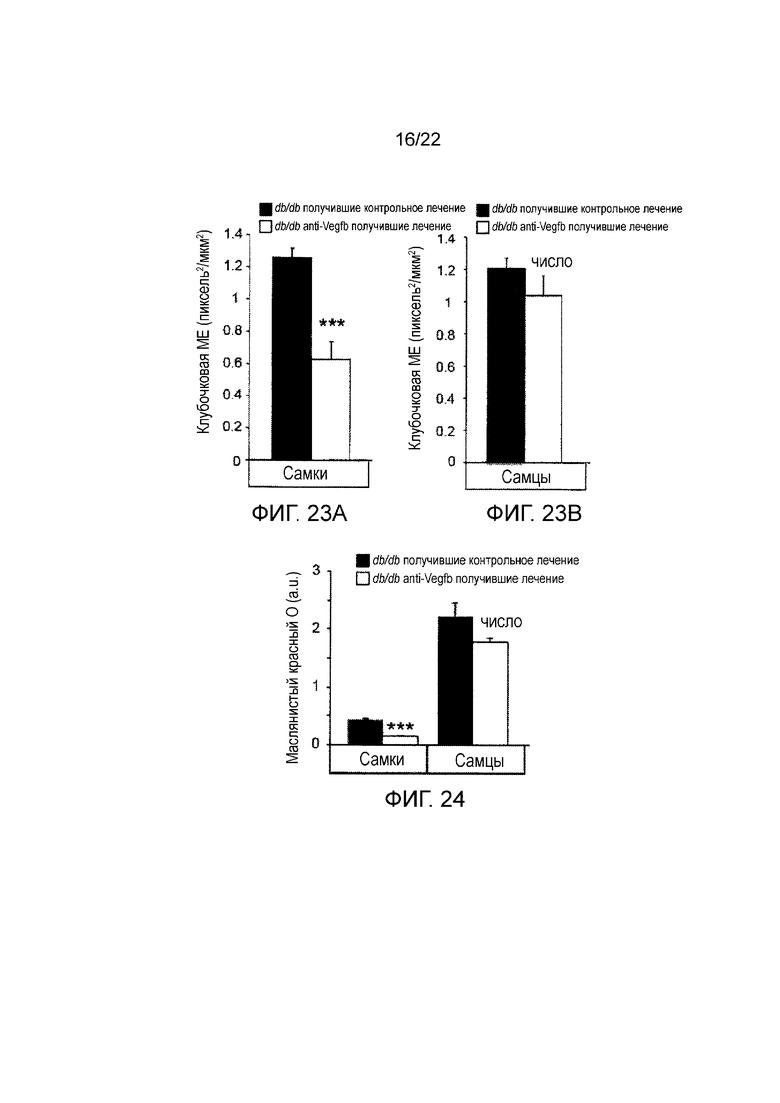
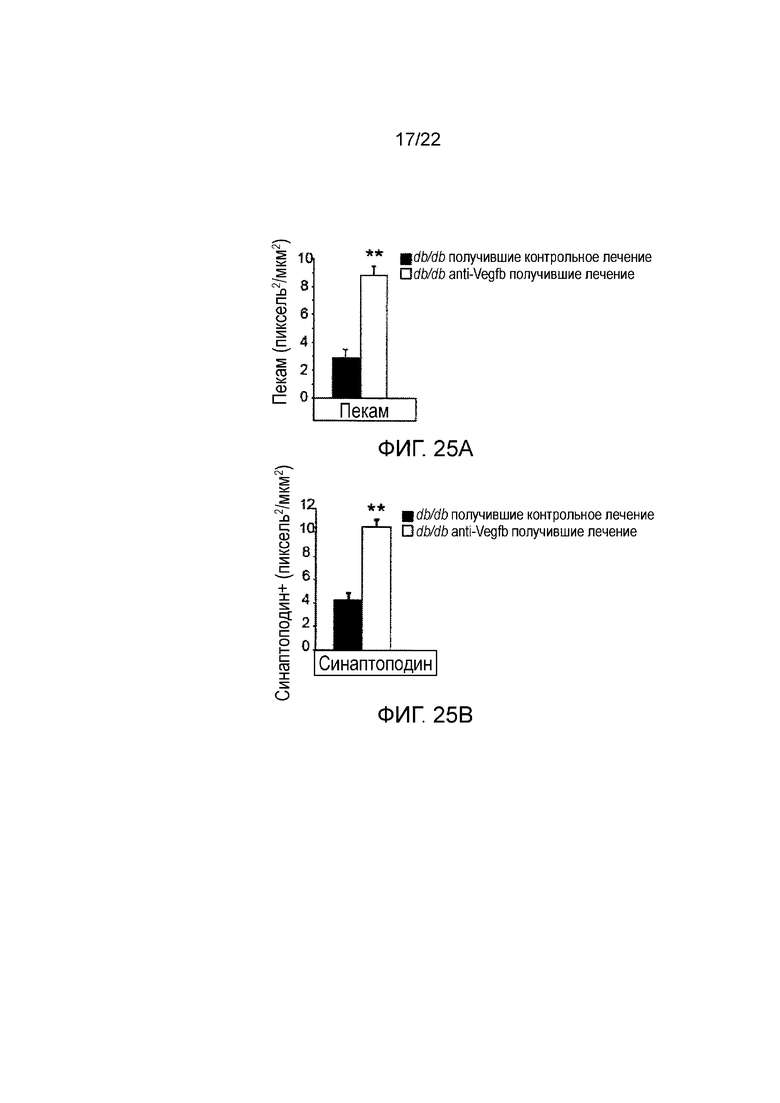
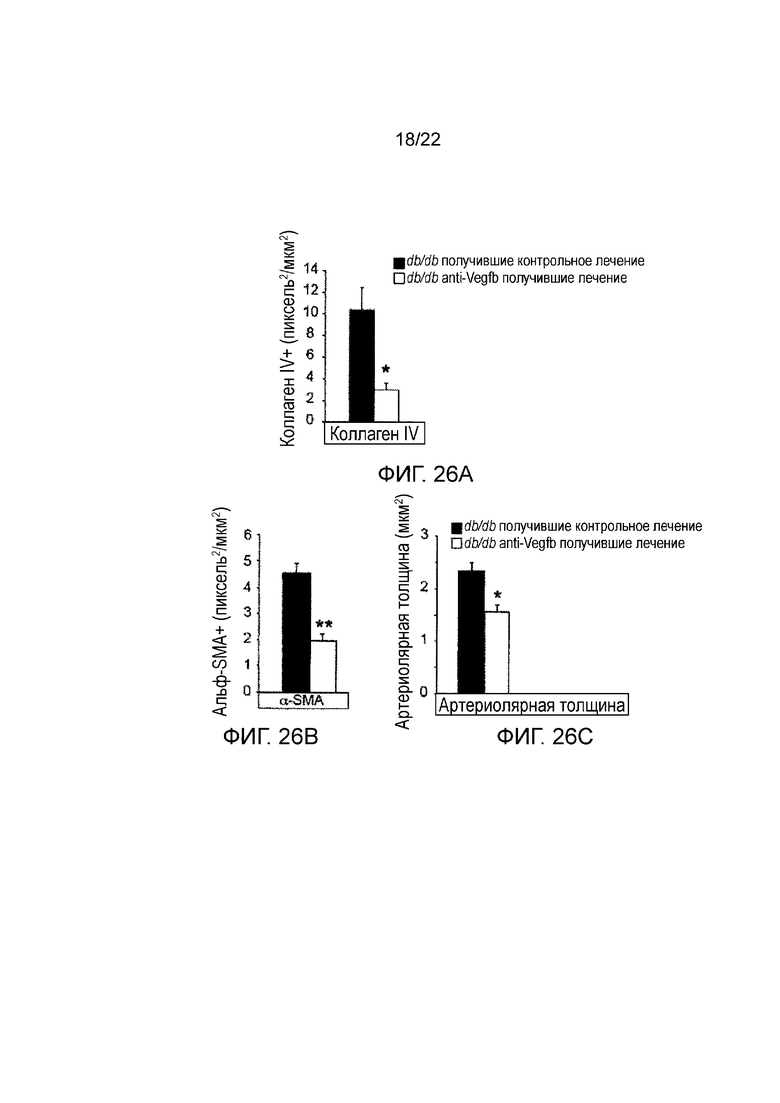
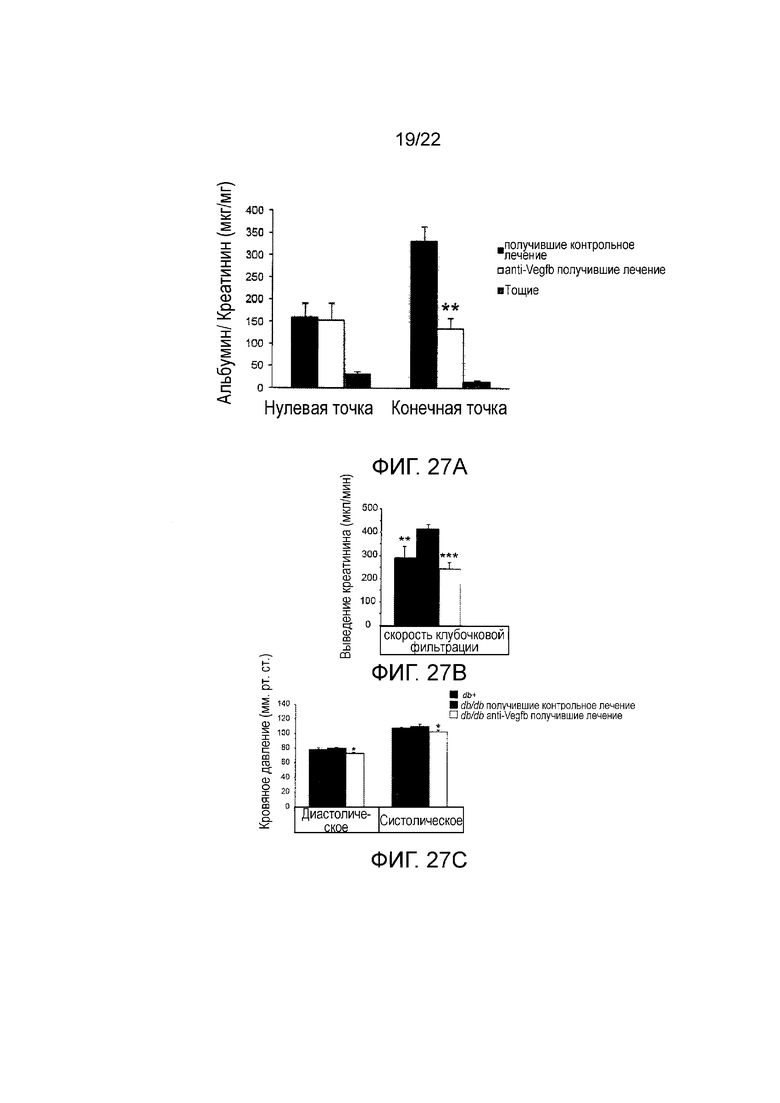
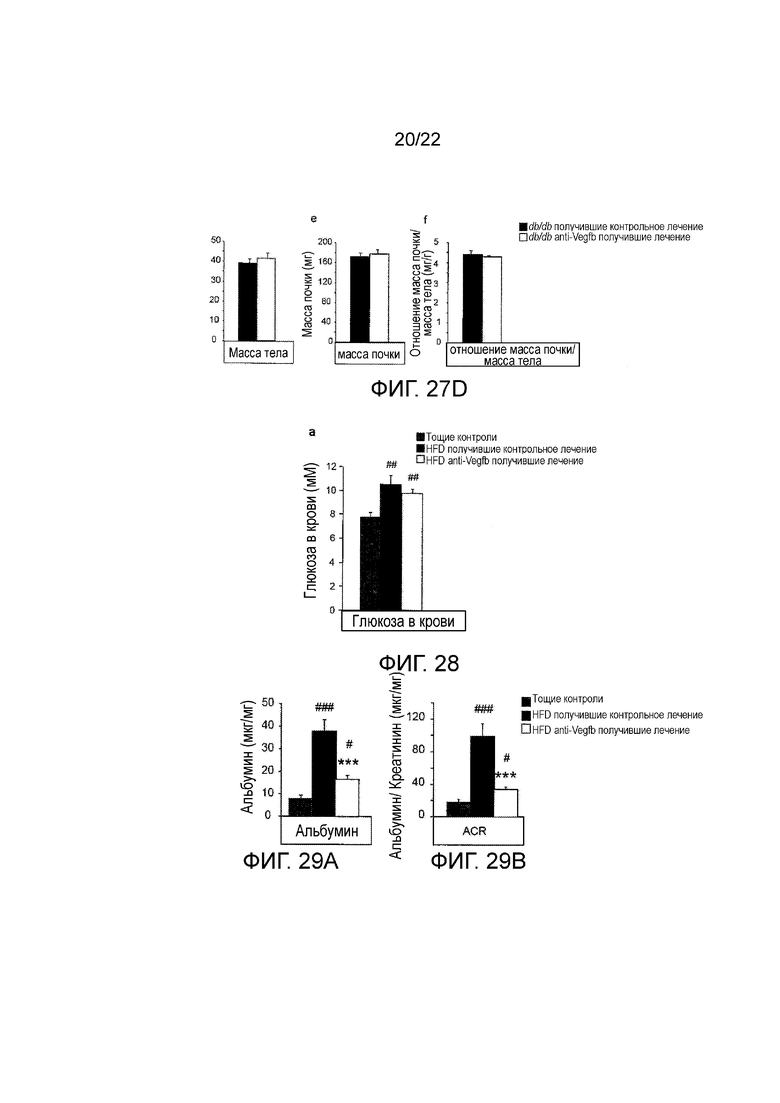
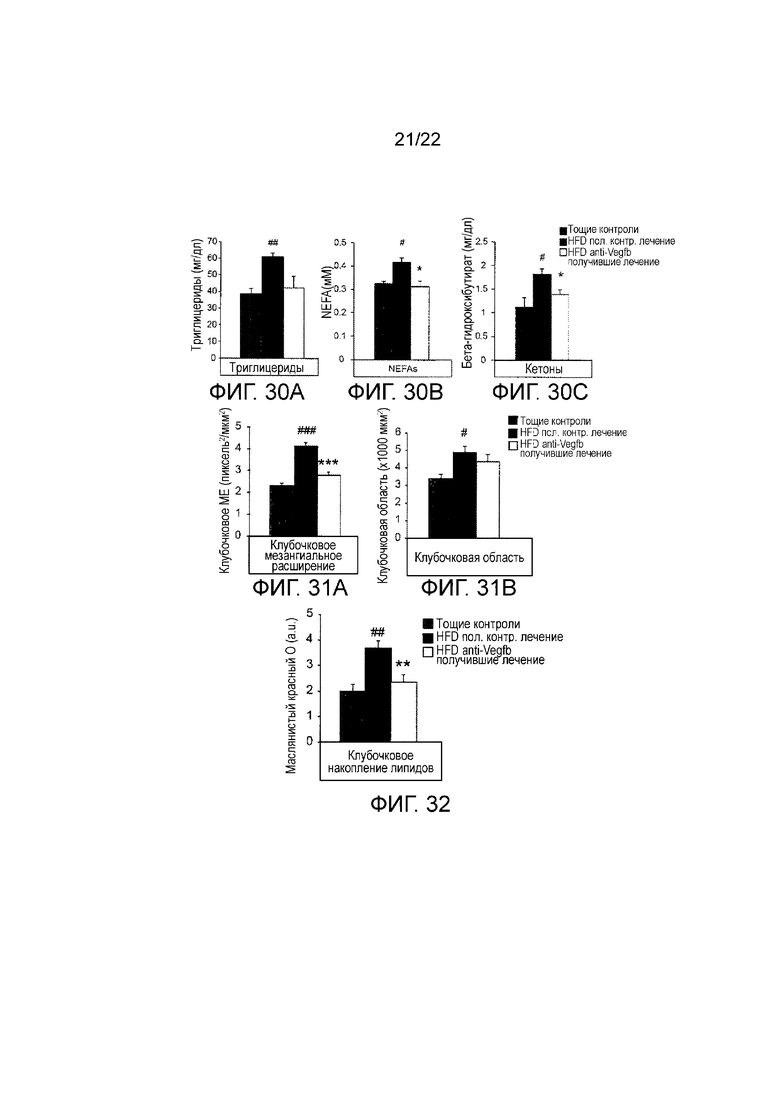
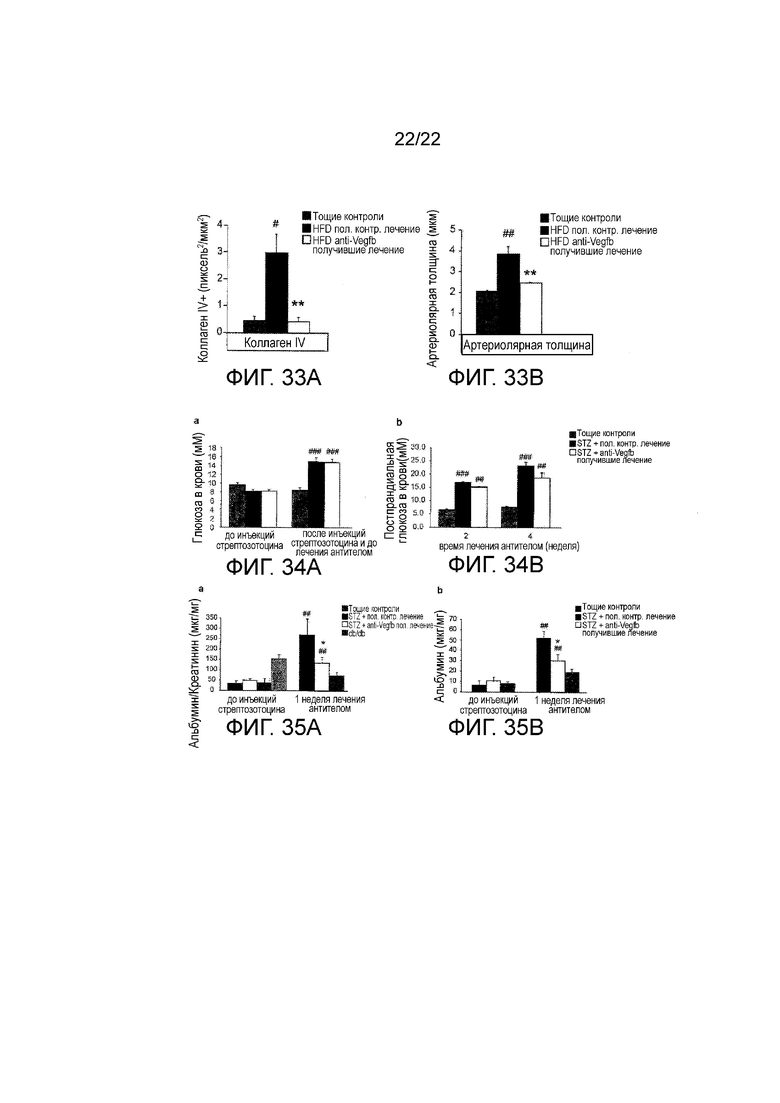
Настоящая группа изобретения относится к медицине, а именно к эндокринологии и нефрологии, и касается лечения диабетической нефропатии. Для этого вводят эффективное количество соединения, ингибирующего сигнальный путь VEGF-B. Это обеспечивает эффективную коррекцию функции почек за счет избирательного связывания ингибитора VEGF-B c рецептором VEGFR-2. 2 н. и 8 з.п. ф-лы, 35 ил., 2 пр.
1. Способ лечения диабетической нефропатии у индивидуума, страдающего от диабетической нефропатии, включающий введение указанному индивидууму соединения, которое ингибирует сигнальный путь VEGF-B, где соединение, которое ингибирует сигнальный путь VEGF-B, представляет собой:
(i) белок, содержащий вариабельную область антитела, которая связывается с или специфически связывается с VEGF-B и нейтрализует сигнальный путь VEGF-B; или
(ii) нуклеиновую кислоту, которая ингибирует или предотвращает экспрессию VEGF-B.
2. Способ предупреждения диабетической нефропатии у индивидуума, страдающего от диабета и микроальбуминурии или макроальбуминурии, включающий введение индивидууму соединения, которое ингибирует сигнальный путь VEGF-B, где соединение, которое ингибирует сигнальный путь VEGF-B, представляет собой:
(i) белок, содержащий вариабельную область антитела, которая связывается с или специфически связывается с VEGF-B и нейтрализует сигнальный путь VEGF-B; или
(ii) нуклеиновую кислоту, которая ингибирует или предотвращает экспрессию VEGF-B.
3. Способ по п. 1 или 2, в котором соединение вводят в количестве, которое эффективно, чтобы получить один или несколько из следующих эффектов:
уменьшение или предотвращение гипертензии;
уменьшение или предотвращение клубочкового и/или тубулярного склероза;
уменьшение или предотвращение отложения мезангиального внеклеточного матрикса и/или аномального утолщения клубочковой базальной мембраны;
уменьшение или предотвращение клубочкового мезангиального расширения;
уменьшение или предотвращение клубочковых сосудистых перестроек;
уменьшение или предотвращение накопления липидов в почках;
уменьшение или предотвращение клубочкового накопления липидов;
уменьшение или предотвращение клубочковых коллагеновых отложений и/или артериолярного гиалиноза; и/или
уменьшение или предотвращение макроальбуминурии.
4. Способ по п. 1 или 2, в котором соединение представляет собой белок, содержащий Fv.
5. Способ по п. 4, в котором белок выбран из группы, состоящей из:
(i) одноцепочечного Fv-фрагмента (ScFv);
(ii) димерного ScFv (ди-scPv);
(iii) диатела;
(iv) триатела;
(v) тетратела;
(vi) Fab;
(vii) F(аb')2;
(viii) Fv;
(iх) одного из (i)-(viii), связанного с константной областью антитела, Fc или константной областью тяжелой цепи (CH)2 или СН3 или полипептидом, содержащим домен CH2 и СН3; или
(x) антитела.
6. Способ по п. 1 или 2, в котором соединение представляет собой белок, содержащий вариабельную область антитела, которая конкурентно ингибирует связывание антитела, содержащего вариабельную область тяжелой цепи (VH), включающую последовательность, представленную в SEQ ID NO: 3, и вариабельную область легкой цепи (VL), включающую последовательность, представленную в SEQ ID NO: 4, с VEGF-B.
7. Способ по п. 6, в котором соединение представляет собой белок, содержащий гуманизированную форму вариабельной области указанного антитела, содержащего вариабельную область тяжелой цепи (VH), включающую последовательность, представленную в SEQ ID NO: 3, и вариабельную область легкой цепи (VL), включающую последовательность, представленную в SEQ ID NO: 4, или соединение представляет собой гуманизированную форму указанного антитела, содержащего вариабельную область тяжелой цепи (VH), включающую последовательность, представленную в SEQ ID NO: 3, и вариабельную область легкой цепи (VL), включающую последовательность, представленную в SEQ ID NO: 4.
8. Способ по п. 6, в котором соединение представляет собой антитело, содержащее VH, включающую последовательность, представленную в SEQ ID NO: 5 и VL, включающую последовательность, представленную в SEQ ID NO: 6.
9. Способ по п. 1 или 2, в котором соединение представляет собой нуклеиновую кислоту, выбранную из группы: антисмысловая нуклеиновая кислота, siRNA, RNAi, рибозим и ДНКазим.
10. Способ по п. 1 или 2, дополнительно включающий введение дополнительного соединения для лечения или предупреждения нефропатии или для лечения или предупреждения (или задержки прогрессирования) диабета.
| ПРОИЗВОДНЫЕ 2-АМИНОНИКОТИНАМИДА И ИХ ПРИМЕНЕНИЕ В КАЧЕСТВЕ ИНГИБИТОРОВ VEGF-РЕЦЕПТОРА ТИРОЗИНКИНАЗЫ | 2001 |
|
RU2296124C2 |
| HAGBERG C.E | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| ШИШКО О.Н | |||
| и др | |||
| "Роль эндотелиального фактора роста сосудов в патогенезе диабетической нефропатии", Оригинальные научные публикации, 29.10.2012, найдено 23.08.2018 из | |||

