Область техники, к которой относится изобретение
Настоящее раскрытие относится к антителу, ингибирующему функцию ангиопоэтина-2 (Ang-2) путем специфического связывания с Ang-2 и направлено на антитело анти-Ang2, кодирующую его нуклеиновую кислоту, вектор, содержащий нуклеиновую кислоту, клетку, трансформированную вектором, способ его получения, содержащий его ингибитор ангиогенеза, композицию для лечения заболевания, связанного с активацией и (или) избыточным производством ангиопоэтина-2, композицию для диагностики заболевания, связанного с активацией и (или) избыточным производством ангиопоэтина-2, композицию для лечения заболевания глаз или композицию для профилактики или лечения рака, а также композицию для объединения связывающегося с Ang2 антитела с другим лекарством, отличающимся от связывающегося с Ang2 антитела.
Уровень техники
Ангиогенез означает механизм, включающий рост новых кровеносных сосудов из уже существующих, и известно, что он играет жизненно важную роль в образовании органов, нормальном биологическом росте, заживлении ран и тому подобное. Известно также, что он играет важную роль в росте опухолей и метастазов, и известно, что нарушения ангиогенеза играют критическую роль в таких заболеваниях, как рост опухолей и метастазов, возрастная дегенерация макулы, диабетическая ретинопатия, псориаз, ревматоидный артрит и хроническое воспаление.
Таким образом, вовлеченные в развитие ангиогенеза факторы стали важной мишенью для развития новых терапевтических агентов для лечения таких заболеваний, как рак, и, поскольку количество больных диабетом быстро растет по причине старения населения и вестернизированной диеты, количество больных с неоваскулярными заболеваниями глаз быстро увеличивается. Примеры серьезных заболеваний глаз включают возрастную дегенерацию макулы (AMD), диабетическую ретинопатию (DR) и диабетический макулярный отек (DME). В частности, дегенерация макулы и диабетическая ретинопатия являются основными причинами слепоты в мире.
С прогрессированием возрастной дегенерации макулы связано множество факторов, но известно, что она ассоциирована с окислительным стрессом, воспалительным ответом и ангиогенезом. Однако, широко распространено мнение, что доминирующим фактором является фактор роста эндотелия сосудов (VEGF). Делались попытки разработать в качестве терапевтического агента ингибитор VEGF с использованием моноклонального антитела, фрагмента антитела или химерного белка, в качестве примера лекарственных препаратов можно назвать афлиберцепт и ранибизумаб. Как известно, механизм действия этих лекарств заключается в индуцировании ингибирования ангиогенеза путем передачи сигнала VEGF. Однако, известно, что 10-15% пациентов, получающих эти препараты, не отвечают на существующие виды лечения. Это связано с тем, что, как известно, существующие виды анти-VEGF лечения подавляют только патологический ангиогенез, в то время как на прогрессирование заболевания влияют другие механизмы с участием ангиогенных факторов. Известно, что ангиопоэтин-2 (ANG2) является цитокином, связывающимся с рецептором Tie2, присутствующим в энтотелиальных клетках стенки сосуда и стимулирующим ангиогенез. Благодаря экспериментам на животных и клиническим исследованиям известно, что, подавляя ингибирование передачи сигнала ANG2, ингибируется образование кровеносных сосудов в опухолях, и, таким образом, достигается противораковый эффект. Более того, известно, что экспрессия ANG является высокой в водостойкой жидкости глазных яблок пациентов с возрастной дегенерацией макулы. Таким образом, ожидается, что разработка анти-ANG2 терапевтического агента, а также анти-VEGF терапевтического агента и комплексной терапии будет полезно при лечении дегенерации макулы. Соответственно, авторы настоящего раскрытия сосредоточили свое внимание на Ang-2 для разработки терапевтического агента для лечения возрастной дегенерации макулы и диабетической ретинопатии.
Ангиопоэтин-2 (Ang2) является антагонистическим лигандом рецептора Tie2, присутствующего в клетках эндотелия сосудов, и ингибирует передачу сигнала Tie2, конкурируя с ангиопоэтином-1 (Ang1), являющимся агонистом Tie2, за связывание Tie2, а Ang1, являющийся лигандом для активации рецептора Tie2, представляет собой ключевой регулятор стабилизации кровеносных сосудов путем поддержания барьерной функции эндотелиальных клеток сосудов. В состоянии избыточной экспрессии VEGF или воспаления эндотелиальные клетки сосудов активируются и увеличивается проницаемость сосудов.
В этом отношении Ang1 стимулирует целостность контакта между эндотелиальными клетками кровеносных сосудов, тем самым индуцируя стабилизацию эндотелиальных клеток кровеносных сосудов и снижая проницаемость сосудов; в то же время Ang2 в активированных эндотелиальных клетках кровеносных сосудов связывается с Tie-2, тем самым принимая участие в миграции эндотелиальных клеток кровеносных сосудов и образовании неровностей. После этого стимулируется образование новых кровеносных сосудов.
В случае диабетической ретинопатии обнаружено, что передача сигнала PDGF важна для образования и созревания гематоретинального барьера путем регулирования периферических клеток сосудов, и было показано, что потеря периферических клеток сосудов в сосудах сетчатки взрослого человека усиливает ответ эндотелиальных клеток кровеносных сосудов на VEGF-A, тем самым активируя петлю FOXO1-Ang2, что приводит к обострению диабетической ретинопатии. Иными словами, понятно, что, индуцируя блокировку Ang2 и активацию Tie2, можно разработать новые виды лечения диабетической ретинопатии.
Кроме того, Ang-2 также вносит вклад в образование новых кровеносных сосудов пораженной раком ткани. Для формирования новых кровеносных сосудов в пораженной раком ткани возникает своего рода кооптация, при которой раковые клетки выбирают существующие кровеносные сосуды. Затем начинается дегенерация последних, при которой с участием Ang-2 нарушается их функция. По причине дегенерации имеющихся кровеносных сосудов окружение в раковой ткани становится гипоксическим, создавая условия для образования новых кровеносных сосудов. В описанных выше условиях индуцируется избыточная экспрессия фактора роста клеток сосудов эндотелия (VEGF), и, как упоминалось выше, ангиогенез. По этой причине Ang-2 был основной мишенью при разработке противораковых лекарств, действующих посредством ингибирования ангиогенеза.
С учетом описанного выше уровня техники авторы настоящего раскрытия попытались разработать антитела анти-Ang2. В результате авторы разработали анти-Ang2 антитело, демонстрирующее желаемую активность связывания с Ang2, и подтвердили, что такое анти-Ang2 антитело может использоваться в качестве направленного иммунного противоракового агента или терапевтического агента для лечения офтальмологических заболеваний, и, тем самым, завершили раскрытие настоящего изобретения.
Раскрытие
Техническая проблема
Таким образом, настоящее раскрытие сделано ввиду описанных выше проблем, и задачей настоящего раскрытия является предоставление нового антитела против Ang2 или его антигенсвязывающего фрагмента.
Еще одной задачей настоящего раскрытия является предоставление нуклеиновой кислоты, кодирующей антитело или его антигенсвязывающий фрагмент.
Дополнительной задачей настоящего раскрытия является предоставление вектора, включающего нуклеиновую кислоту, клетки, трансформированной вектором, и способа их конструирования.
Еще одной дополнительной задачей настоящего раскрытия является предоставление ингибитора ангиогенеза, включающего антитело или его антигенсвязывающий фрагмент, и композиции для лечения заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2.
Еще одной дополнительной задачей настоящего раскрытия является предоставление ингибитора ангиогенеза, включающего антитело или его антигенсвязывающий фрагмент, и композиции для диагностики заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2.
Еще одной дополнительной задачей настоящего раскрытия является предоставление композиции для профилактики или лечения заболеваний глаз, включающей антитело или его антигенсвязывающий фрагмент.
Еще одной дополнительной задачей настоящего раскрытия является предоставление композиции для профилактики или лечения опухолей или онкологических заболеваний, включающей антитело или его антигенсвязывающий фрагмент.
Еще одной дополнительной задачей настоящего раскрытия является предоставление композиции для совместного введения с анти-Ang2 антителом, включающей антитело или его антигенсвязывающий фрагмент.
Решение технической проблемы
В соответствии с аспектом настоящего раскрытия, описанные выше и другие задачи можно выполнить, получив антитело, связывающееся с ангиопоэтином-2 (Ang2), или его антигенсвязывающий фрагмент, включающие:
- вариабельный участок тяжелой цепи, включающий CDR1 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 1, 7, 13, 19 и 25, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 2, 8, 14, 20 и 26, а также CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 3, 9, 15, 21, 27, 51, 52 и 53; а также
- вариабельный участок легкой цепи, включающий CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 4, 10, 16, 22 и 28, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 5, 11, 17, 23 и 29, а также CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 6, 12, 18, 24 и 30.
В соответствии с другим аспектом настоящего раскрытия, предоставляется нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент.
В соответствии с дополнительным аспектом настоящего раскрытия, предоставляется вектор, включающий нуклеиновую кислоту.
В соответствии с дополнительным аспектом настоящего раскрытия, предоставляется клетка, трансформированная вектором.
В соответствии с дополнительным аспектом настоящего раскрытия, предоставляется способ получения антитела или его антигенсвязывающего фрагмента, включающий следующие процессы: (a) культивирование клеток и (b) восстановление антитела или его антигенсвязывающего фрагмента из культивируемых клеток.
Настоящее раскрытие также предоставляет ингибитор ангиогенеза, включающий антитело или его антигенсвязывающий фрагмент, и композицию для лечения заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2. Настоящее раскрытие также предоставляет ингибитор ангиогенеза, включающий антитело или его антигенсвязывающий фрагмент, и композицию для диагностики заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2. Настоящее раскрытие также предоставляет композицию для профилактики или лечения опухолей или онкологических заболеваний, включающую антитело или его антигенсвязывающий фрагмент. Настоящее раскрытие также предоставляет композицию для совместного введения с анти-Ang2 антителом, включающую антитело или его антигенсвязывающий фрагмент.
Описание чертежей
Описанные выше и другие задачи, особенности и прочие преимущества настоящего раскрытия будут понятнее из следующего далее подробного описания совместно с сопровождающими его фигурами, которые:
фиг. 1 - иллюстрирует результаты, подтверждающие, что выбранный моноклональный фаг scFv может ингибировать связывание Ang2/Tie2;
фиг. 2 - иллюстрирует результаты, полученные методом SDS-PAGE в восстанавливающих и невосстанавливающих условиях для продуктов, полученных посредством временной экспрессии и очистки отобранных антител анти-Ang2;
фиг. 3 - иллюстрирует результаты, полученные методом ELISA при оценке связывания антител анти-Ang2 с Ang2 и Ang1 человека и мыши, полученных посредством временной экспрессии и очистки;
фиг. 4 - иллюстрирует результаты, демонстрирующие способность выбранных антител анти-Ang2 нейтрализовывать связывание Ang2/Tie2 человека и мыши;
фиг. 5 - иллюстрирует результаты, демонстрирующие способность выбранных антител анти-Ang2 нейтрализовывать связывание Ang2/интегрин;
фиг. 6 - иллюстрирует результаты, демонстрирующие, что выбранные антитела анти-Ang2 могут ингибировать передачу сигнала Ang2/Tie2;
фиг. 7 - иллюстрирует результаты, подтверждающие чистоту выбранного антитела анти-Ang2 scFv, экспрессируемого в E. coli после процесса очистки;
фиг. 8 - иллюстрирует результаты, полученные методом ELISA при оценке связывания антитела анти-Ang2 scFv, экспрессируемого в E. coli, с белком Ang2 человека;
фиг. 9 - 11 - иллюстрируют результаты, подтверждающие in vivo эффективность выбранного антитела анти-Ang2 scFv на мышиной модели CNV; и
фиг. 12 - иллюстрирует результаты, подтверждающие противоопухолевый эффект антитела анти-Ang2 на модели трижды негативного рака молочной железы человека.
Подробное описание и примеры воплощений
Будут сделаны подробные ссылки на предпочтительные воплощения настоящего раскрытия, примеры которых проиллюстрированы на сопровождающих фигурах. Где это возможно, такие же номера ссылок будут использоваться на фигурах для обозначения таких же или подобных частей.
Если не утверждается иное, значение всех используемых здесь технических и научных терминов соответствует обычно понимаемому специалистом в той области техники, к которой относится настоящее раскрытие. В общем случае, используемая здесь номенклатура и способы постановки экспериментов, описанные ниже, хорошо известны и обычно используются в уровне техники.
Воплощение настоящего раскрытия относится к антителу, связывающемуся с ангиопоэтином-2 (Ang2) или его антигенсвязывающему фрагменту, включающему:
- вариабельный участок тяжелой цепи, включающий CDR1 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 1, 7, 13, 19 и 25, CDR2 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 2, 8, 14, 20 и 26, а также CDR3 тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 3, 9, 15, 21, 27, 51, 52 и 53; а также
- вариабельный участок легкой цепи, включающий CDR1 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 4, 10, 16, 22 и 28, CDR2 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 5, 11, 17, 23 и 29, а также CDR3 легкой цепи, выбранный из группы, состоящей из SEQ ID NO: 6, 12, 18, 24 и 30.
Здесь термин «антитело» означает анти-Ang2 антитело, специфически связывающееся с Ang2. В область действия настоящего раскрытия попадает не только полная форма антитела, специфически связывающаяся с Ang2, но также антигенсвязывающий фрагмент молекулы антитела.
Полное антитело представляет собой структуру, содержащую две легкие цепи полной длины и две тяжелые цепи полной длины, где каждая легкая цепь связывается с тяжелой цепью дисульфидными связями. Константная область тяжелой цепи может относиться к типам гамма (γ), мю (μ), альфа (α), дельта (δ) или эпсилон (ε), и эти типы далее делятся на категории гамма 1 (γ1), гамма 2 (γ2), гамма 3 (γ3), гамма 4 (γ4), альфа 1 (α1) или альфа 2 (α2). Константная область легкой цепи может относиться либо к типу каппа (κ), либо лямбда (λ)
Антигенсвязывающий фрагмент антитела, или фрагмент антитела, означает фрагмент с антигенсвязывающей функцией и включает Fab, Fab', F(ab')2 и Fv. Среди фрагментов антител Fab представляет собой структуру, содержащую вариабельные области легкой цепи и тяжелой цепи, константные области легкой цепи и первую константную область CH1 легкой цепи, и она содержит один антигенсвязывающий сайт. Fab' отличается от Fab тем, что Fab' содержит шарнирную область, включающую по меньшей мере один остаток цистеина у C-конца домена CH1 тяжелой цепи. Фрагмент F(ab')2 образуется, когда остатки цистеина Fab' соединяются дисульфидной связью в шарнирной области. Fv представляет собой минимальный фрагмент антитела, содержащий только вариабельные области тяжелой цепи и вариабельные области легкой цепи. Двухцепочечный Fv может иметь структуру, в которой вариабельные области тяжелой цепи связываются с вариабельными областями легкой цепи нековалентной связью, а одноцепочечный Fv (scFv) обычно имеет димерную структуру, как в случае двухцепочечного Fv, в котором вариабельные области тяжелой цепи ковалентно связаны с вариабельными областями легкой цепи посредством пептидного линкера, или вариабельные области тяжелой и легкой цепей непосредственно связаны друг с другом через свои С-концы. Антигенсвязывающий фрагмент можно получить с помощью протеазы (например, целое антитело подвергают рестриктазному расщеплению под действием папаина с получением Fab или расщеплению под действием пепсина с получением фрагмента F(ab')2) и можно приготовить методами генетической рекомбинации.
В соответствии с одним воплощением, антитело по настоящему раскрытию находится в форме Fv (например, scFv) или в форме полного антитела. Дополнительно, константная область тяжелой цепи может представлять любой изотип, выбранный из гамма (γ), мю (μ), альфа (α), дельта (δ), и эпсилон (ε). Например, константная область представляет собой гамма 1 (IgG1), гамма 3 (IgG3) или гамма 4 (IgG4). Константная область легкой цепи может быть каппа или дельта типа.
Используемый здесь термин «тяжелая цепь» включает тяжелые цепи полной длины, включающие вариабельную область VH, включая аминокислотные последовательности, содержащие последовательность вариабельной области, которая обеспечивает специфичность антигена, и три константных домена CH1, CH2 и CH3, а также их фрагменты. Используемый здесь термин «легкая цепь» включает легкие цепи полной длины, включающие вариабельную область VL, включая аминокислотные последовательности, содержащие последовательность вариабельной области, которая обеспечивает специфичность антигена, константную область CL и их фрагменты.
Антитело по настоящему раскрытию может представлять собой моноклональное антитело, биспецифическое антитело, антитело человека, гуманизированное антитело, химерное антитело, одноцепочечный фрагмент Fvs (scFV), одноцепочечное антитело, фрагмент Fab, фрагмент F(ab'), фрагмент дисульфидной связи Fvs (sdFV), антиидиотипное антитело (анти-Id) и связывающие эпитопы фрагменты этих антител, но настоящее раскрытие ими не ограничивается.
Моноклональное антитело означает антитело, полученное из фактически однородной популяции антител, то есть, идентичных антител, за исключением возможных природных мутаций, где индивидуальные антитела в популяции могут быть представлены в следовых количествах. Моноклональные антитела высокоспецифичны по отношению к одному антигенному сайту. В отличие от обычных (поликлональных) составов антител, включающих различные антитела для различных детерминант (эпитопов), каждое моноклональное антитело индуцируется против одного эпитопа антигена.
«Эпитоп» означает белковую детерминанту, с которой может специфически связываться антитело. Эпитопы обычно состоят из группы химически активных молекул поверхности, например, боковых цепей аминокислот или сахаров, и обычно имеют специфические характеристики трехмерной структуры и специфические свойства в отношении зарядов. Конформационные и неконформационные эпитопы различаются тем, что связь с первым разрушается в присутствии денатурирующего растворителя, но это не касается второго.
Нечеловеческое (например, мышиное) антитело в «гуманизированной форме» представляет собой химерное антитело, содержащее минимальную последовательность, полученную из нечеловеческого иммуноглобулина. В большинстве случаев гуманизированное антитело представляет собой нечеловеческий вид (антитело донора), который сохраняет желаемую активность, сродство и свойства остатков гипервариабельной области реципиента, например, иммуноглобулин человека (рецепторное антитело), замещенное остатком гипервариабельной области мыши, крысы, кролика или иных видов, отличающихся от человека.
«Антитело человека» представляет собой молекулу, производную от иммуноглобулина человека, это означает, что все аминокислотные последовательности, составляющие антитело, включая области, определяющие комплементарность и каркасные области, состоят из последовательностей иммуноглобулина человека.
Антитело человека включает не только «химерные» антитела, в которых часть тяжелой и (или) легкой цепи имеет последовательность, идентичную или гомологичную соответствующей последовательности антитела, полученного из специфического вида или принадлежащему к специфическому классу или подклассу антитела, в то время как остальные цепи идентичны или гомологичны антителам, полученным из других видов или антител другого класса или подкласса, но также и их фрагменты, демонстрирующие требуемую биологическую активность.
«Вариабельный домен антитела» здесь означает часть легкой или тяжелой цепи молекулы антитела, включающую аминокислотные последовательности определяющих комплементарность участков (CDR; CDR1, CDR2 и CDR3) или каркасных областей (FR). VH означает вариабельный домен тяжелой цепи. VL означает вариабельный домен легкой цепи.
«Определяющие комплементарность участки» (CDR; CDR1, CDR2 и CDR3) означает аминокислотные остатки вариабельного домена антитела, которые необходимы для связывания антигена. Каждый вариабельный домен, как правило, содержит три области CDR, обозначаемые CDR1, CDR2 и CDR3. Настоящее раскрытие включает вариабельную область тяжелой цепи, включающую CDR3 тяжелой цепи SEQ ID NO: 1, и вариабельную область легкой цепи, включающую CDR3 легкой цепи SEQ ID NO: 2.
В настоящем раскрытии антитело, связывающееся с Ang2 или его антигенсвязывающий фрагмент, может включать:
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 1, CDR2 тяжелой цепи SEQ ID NO: 2 и CDR3 тяжелой цепи SEQ ID NO: 3, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 4, CDR2 легкой цепи SEQ ID NO: 5 и CDR3 легкой цепи SEQ ID NO: 6;
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 7, CDR2 тяжелой цепи SEQ ID NO: 8 и CDR3 тяжелой цепи SEQ ID NO: 9, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 10, CDR2 легкой цепи SEQ ID NO: 11 и CDR3 легкой цепи SEQ ID NO: 12;
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 15, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18;
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 19, CDR2 тяжелой цепи SEQ ID NO: 20 и CDR3 тяжелой цепи SEQ ID NO: 21, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 22, CDR2 легкой цепи SEQ ID NO: 23 и CDR3 легкой цепи SEQ ID NO: 24;
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 25, CDR2 тяжелой цепи SEQ ID NO: 26 и CDR3 тяжелой цепи SEQ ID NO: 27, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 28, CDR2 легкой цепи SEQ ID NO: 29 и CDR3 легкой цепи SEQ ID NO: 30;
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 51, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18;
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 52, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18; или
вариабельную область тяжелой цепи, включающую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 53, а также вариабельную область легкой цепи, включающую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18.
«Каркасная область (FR)» представляет собой остаток вариабельного домена, отличающийся от остатков CDR. Каждый вариабельный домен как правило содержит четыре FR, обозначаемых как FR1, FR2, FR3 и FR4.
Фрагмент «Fv» представляет собой фрагмент антитела, содержащий сайты распознавания и связывания полноразмерного антитела. Эта область состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных прочной ковалентной связью, например, scFv.
Фрагмент «Fab» содержит вариабельный и константный домены легкой цепи и вариабельный и первый константный (CH1) домены тяжелой цепи. Фрагмент антитела «F(ab')2» обычно включает пару фрагментов Fab, ковалентно связанных около своих карбоксильных концов шарнирными цистеинами.
Фрагменты антитела «одноцепочечный Fv» или «scFv» включает домены VH и VL антитела, присутствующие в одной полипептидной цепи. Полипептид Fv может дополнительно включать полипептидный линкер между доменами VH и VL, которые позволяют scFv сформировать желаемую структуру для связывания антигена.
Анти-Ang2 антитела могут включать одинарную или двойную цепь. Функционально, сродство связывания антител анти-Ang2 находится в диапазоне 10-5 M - 10-12 M. Например, сродство связывания антител анти-Ang2 находится в диапазоне 10-6 M - 10-12 M, 10-7 M - 10-12 M, 10-8 M - 10-12 M, 10-9 M - 10-12 M, 10-5 M - 10-11 M, 10-6 M - 10-11 M, 10-7 M - 10-11 M, 10-8 M - 10-11 M, 10-9 M - 10-11 M, 10-10 M - 10-11 M, 10-5 M - 10-10 M, 10-6 M - 10-10 M, 10-7 M - 10-10 M, 10-8 M - 10-10 M, 10-9 M - 10-10 M, 10-5 M - 10-9 M, 10-6 M - 10-9 M, 10-7 M - 10-9 M, 10-8 M - 10-9 M, 10-5 M - 10-8 M, 10-6 M - 10-8 M, 10-7 M - 10-8 M, 10-5 M - 10-7 M, 10-6 M - 10-7 M или 10-5 M - 10-6 M.
Антитело, связывающееся с Ang2, или его антигенсвязывающий фрагмент может включать вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NOS: 32, 36, 40, 44, 48, 55, 57, 59 и 61. Дополнительно, антитело, связывающееся с Ang2, или его антигенсвязывающий фрагмент может включать вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NOS: 34, 38, 42, 46 и 50.
В частности, для разработки эффективного и высококонцентрированного состава антител анти-Ang2 в каркасной части вариабельной области тяжелой цепи была сделана мутация с целью усиления эффективности и растворимости. Для получения мутанта 12-й аминокислотный остаток вариабельной области тяжелой цепи валин заменили на серин. Соответственно, в эксперименте использовали антитело, включающее вариабельную область SEQ ID NO: 61 тяжелой цепи. В результате улучшенная эффективность и растворимость была подтверждена.
В соответствии с конкретным воплощением настоящего раскрытия, антитело, связывающееся с Ang2, или его антигенсвязывающий фрагмент могут включать:
вариабельную область тяжелой цепи SEQ ID NO: 32 и вариабельную область легкой цепи SEQ ID NO: 34;
вариабельную область тяжелой цепи SEQ ID NO: 36 и вариабельную область легкой цепи SEQ ID NO: 38;
вариабельную область тяжелой цепи SEQ ID NO: 40 и вариабельную область легкой цепи SEQ ID NO: 42;
вариабельную область тяжелой цепи SEQ ID NO: 44 и вариабельную область легкой цепи SEQ ID NO: 46;
вариабельную область тяжелой цепи SEQ ID NO: 48 и вариабельную область легкой цепи SEQ ID NO: 50;
вариабельную область тяжелой цепи SEQ ID NO: 55 и вариабельную область легкой цепи SEQ ID NO: 42;
вариабельную область тяжелой цепи SEQ ID NO: 57 и вариабельную область легкой цепи SEQ ID NO: 42;
вариабельную область тяжелой цепи SEQ ID NO: 59 и вариабельную область легкой цепи SEQ ID NO: 42; или
вариабельную область тяжелой цепи SEQ ID NO: 61 и вариабельную область легкой цепи SEQ ID NO: 42;
«Фаговый дисплей» представляет собой метод представления варианта полипептида как белка слияния по меньшей мере с фрагментом белка оболочки на поверхности фага, например, нитчатых фаговых частиц. Полезность фагового дисплея связана с тем, что с его помощью можно получать большие библиотеки рандомизированных вариантов белка, а также быстро и эффективно классифицировать последовательности, связывающиеся с целевым антигеном с высоким сродством. Дисплей пептидных и белковых библиотек с использованием фагов использовалось для скрининга миллионов полипептидов с целью выявления полипептида со специфическими свойствами связывания.
Технология фагового дисплея оказалась мощным инструментом для создания и скрининга новых белков, связывающихся со специфическими лигандами (например, антигенами). С помощью технологии фагового дисплея можно создать большую библиотеку вариантов белков и быстро найти последовательности, связывающиеся с целевым антигеном с высоким сродством. Нуклеиновую кислоту, кодирующую вариант полипептида, объединяют с белком оболочки вируса, например, это может быть нуклеотидная последовательность, кодирующая белок гена III или гена VIII. Была разработана одновалентная система фагового дисплея, при которой нуклеотидная последовательность, кодирующая белок или полипептид, объединяется с нуклеотидной последовательностью, кодирующей часть белка гена III. В одновалентной системе фагового дисплея химерный ген экспрессируется на низком уровне, и белок гена III дикого типа также экспрессируется, поддерживая инфицирующую способность частицы.
При разработке библиотек антител по технологии фагового дисплея важно показать экспрессию пептидов на поверхности нитчатого фага и экспрессию фрагментов функционального антитела в периплазме E. coli. Библиотеки антител или антигенсвязывающих полипептидов были разработаны различными способами, например, путем изменения отдельного гена введением в него случайной последовательность ДНК или клонированием родственного семейства генов. Можно провести скрининг библиотеки для определения экспрессии антител или антигенсвязывающих белков с требуемыми характеристиками.
При производстве антител с требуемыми характеристиками технология фазового дисплея характеризуется несколькими преимуществами перед традиционными гибридомными или рекомбинантными методами. Данная технология позволяет создавать за небольшой промежуток времени большие библиотеки антител с различными последовательностями, не используя животных. Получение гибридомных или гуманизированных антител может потребовать несколько месяцев. Дополнительно, так как иммунитет не требуется, библиотека фаговых антител позволяет создавать антитела против антигенов, характеризующихся токсичностью или низкой антигенностью. Библиотеки фаговых антител можно также использовать для создания и выявления новых терапевтических антител.
С помощью библиотек на основе фагового дисплея можно получить антитела человека от иммунизированных или неиммунизированных людей, из зародышевых последовательностей или из запасов иммуноглобулина несенситизированных В-клеток. Для подготовки несенситизированных или неиммунных антигенсвязывающих библиотек можно использовать различные лимфоидные ткани.
Для выделения новых терапевтических антител важны методы выявления и выделения высоко-аффинных антител из библиотек фагового дисплея. Выделение высоко-аффинных антител из библиотеки может зависеть от размера библиотеки, эффективности производства среди бактериальных клеток, а также от разнообразия библиотеки. Размер библиотеки уменьшается при неадекватном свертывании антитела или антигенсвязывающего белка и при неэффективном производстве по причине наличия стоп-кодонов. Экспрессия в бактериальных клетках может быть подавлена, если антитело или антигенсвязывающий домен свернут неправильно. Экспрессия может быть улучшена путем альтернативной мутации поверхности вариабельного/константного интерфейса или выбранных остатков CDR. Одним из элементов обеспечения правильного свертывания при создании фаговых библиотек антител в бактериальных клетках является последовательность каркасной области.
Для выделения высоко-аффинного антитела важно создавать различные библиотеки антител или антигенсвязывающих белков. Обнаружено, что области CDR3 часто участвуют в связывании антигенов. Области CDR3 тяжелой цепи значительно варьируют по размеру, последовательности и структурной конформации, и, таким образом, их можно использовать для получения различных библиотек.
Дополнительно, разнообразие можно повысить, рандомизируя области CDR вариабельных тяжелых и легких цепей, используя все 20 аминокислот в каждом положении. Использование всех 20 аминокислот может привести к получению большого разнообразия вариантов аминокислотных последовательностей антител и к повышению вероятности найти новые антитела.
Антитело или фрагмент антитела по настоящему раскрытию может включать описанную здесь последовательность анти-Ang2 антитела по настоящему раскрытию, а также ее биологические эквиваленты в диапазоне, когда они способны специфически распознавать Ang2. Например, в аминокислотную последовательность антитела могут быть внесены дополнительные изменения для улучшения сродства связывания и (или) других биологических свойств антитела. Такие изменения включают, например, делеции, вставки и (или) замены аминокислотных остатков антитела. Эти изменения в аминокислотах делают на основании относительного подобия боковых заместителей аминокислот, например, в отношении гидрофобности, гидрофильности, заряда, размера и т.д. Анализ размера, формы и типа боковых заместителей аминокислот показывает, что аргинин, лизин и гистидин являются положительно заряженными остатками, аланин, глицин и серин имеют похожие размеры, а фенилаланин, триптофан и тирозин похожую форму. Таким образом, на основании этих соображений, аргинин, лизин и гистидин; аланин, глицин и серин; а также фенилаланин, триптофан и тирозин можно считать биологическими функциональными эквивалентами.
С учетом описанных выше изменений в отношении биологически эквивалентной активности, антитела по настоящему раскрытию или кодирующие их молекулы нуклеиновых кислот интерпретируют так, чтобы они включали также последовательности, демонстрирующие значительную идентичность с последовательностями, охарактеризованными ID номерами. Значительная идентичность означает последовательность по меньшей мере с 90% гомологией и наиболее предпочтительно по меньшей мере с 95% гомологией, по меньшей мере с 96% гомологией, по меньшей мере с 97% гомологией, по меньшей мере с 98% гомологией, или по меньшей мере с 99% гомологией, когда описанную выше последовательность по настоящему раскрытию выравнивают с другой произвольной последовательностью для максимально возможного соответствия, и такие выровненные последовательности анализируют различными алгоритмами. Способы выравнивания последовательностей известны в уровне техники. В NCBI и подобных организациях можно получить инструмент Basic Local Alignment Search Tool (BLAST), и его можно использовать совместно с доступными в интернете программами по анализу последовательностей, такими как blastp, blasm, blastx, tblastn и tblastx. Инструмент BLAST можно получить по адресу www.ncbi.nlm.nih.gov/BLAST/. Информацию о том, как сравнивать гомологии последовательностей, используя эту программу, можно получить по адресу www.ncbi.nlm.nih.gov/BLAST/blast_help.html.
На основании этой информации, антитело или его антигенсвязывающий фрагмент по настоящему изобретению может иметь гомологию 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, в сравнении с последовательностью, указанной в описании. Эту гомологию можно определить сравнением и (или) выравниванием последовательностей способами, известными в уровне техники. Например, для определения процентной гомологии нуклеиновых кислот или белков по изобретению можно воспользоваться алгоритмами сравнения последовательностей (то есть, BLAST или BLAST 2.0), накладывать последовательности вручную или провести визуальное инспектирование.
Другое воплощение настоящего раскрытия относится к нуклеиновой кислоте, кодирующей антитело или его антигенсвязывающий фрагмент.
Антитело или его антигенсвязывающий фрагмент можно получить рекомбинантными способами, выделив нуклеиновую кислоту, кодирующую антитело или его антигенсвязывающий фрагмент по настоящему раскрытию. Нуклеиновую кислоту выделяют и вводят в реплицируемый вектор для дальнейшего клонирования (амплификации ДНК) или дальнейшей экспрессии. Таким образом, другое воплощение настоящего раскрытия относится к вектору, включающему нуклеиновую кислоту.
Термин «нуклеиновая кислота» в полном объеме включает молекулы ДНК (гДНК и кДНК) и РНК, а также нуклеотиды, являющиеся основными структурными единицами нуклеиновых кислот, включая не только природные нуклеотиды, но также сахара или их аналоги с модифицированными основаниями. Нуклеотидная последовательность, кодирующая вариабельные области тяжелой и легкой цепи по настоящему раскрытию, может быть модифицирована. Такие модификации включают вставки, делеции, а также неконсервативные или консервативные замены нуклеотидов.
Согласно специфическому воплощению настоящего раскрытия нуклеиновая кислота может включать нуклеиновую кислоту, кодирующую вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NOS: 31, 35, 39, 43, 47, 54, 56, 58 и 60. Дополнительно, нуклеиновая кислота может включать нуклеиновую кислоту, кодирующую вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NOS: 33, 37, 41, 45 и 49.
Конкретно, нуклеиновая кислота может включать:
нуклеиновую кислоту SEQ ID NO: 31, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 33, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 35, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 37, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 39, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 41, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 43, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 45, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 47, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 49, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 54, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 41, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 56, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 41, кодирующую вариабельную область легкой цепи;
нуклеиновую кислоту SEQ ID NO: 58, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 41, кодирующую вариабельную область легкой цепи; или
нуклеиновую кислоту SEQ ID NO: 60, кодирующую вариабельную область тяжелой цепи, и нуклеиновую кислоту SEQ ID NO: 41, кодирующую вариабельную область легкой цепи.
Кодирующее антитело ДНК легко выделить и синтезировать с использованием традиционных процедур (например, с использованием олигонуклеотидного зонда, способного специфически связывать ДНК, кодирующую тяжелые и легкие цепи антитела). Доступно множество векторов. Компоненты вектора обычно включают, но не ограничиваются, один или более следующих элементов: сигнальные последовательности, точки начала репликации, один или более маркерных генов, энхансерные элементы, промоторы и последовательности терминации транскрипции.
Термин «вектор», означающий здесь средство для экспрессии целевого гена в клетке-хозяине, включает плазмидные векторы, космидные векторы, бактериофаговые векторы, аденовирусные векторы, ретровирусные векторы, аденоассоциированные вирус-векторы и подобные им. В составе вектора нуклеиновая кислота, кодирующая антитело, функционально связана с промотором.
«Функционально связанный» означает функциональную связь между последовательностью, регулирующей экспрессию нуклеотидов (например, промотор, одиночную последовательность или набор сайтов связывания регуляторов транскрипции), и другой нуклеотидной последовательностью, и, таким образом, последовательность, регулирующая экспрессию нуклеотидов может регулировать транскрипцию и (или) трансляцию других нуклеотидных последовательностей.
Если прокариотическая клетка используется в качестве клетки-хозяина, вектор обычно включает сильный промотор, способный инициировать транскрипцию (например, промотор tac, промотор lac, промотор lacUV5, промотор lpp, промотор pLλ, промотор pRλ, промотор rac5, промотор amp, промотор recA, промотор SP6, промотор trp или промотор T7), связывающий рибосому сайт для инициирования трансляции и последовательность остановки транскрипции/трансляции. Дополнительно, например, если эукариотическая клетка используется в качестве клетки-хозяина, вектор может включать промотор, полученный из генома млекопитающих (например, промотор металлотионеина, промотор β-актина, промотор гемоглобина человека и промотор креатина мышц) или промотор, полученный из вирусов млекопитающих (например, поздний аденовирусный промотор, промотор 7,5К на основе вируса осповакцины, промотор SV40, цитомегаловирусный (CMV) промотор, tk промотор HSV, промотор на основе вируса опухоли молочной железы мыши (MMTV), промотор HIV LTR, промотор на основе вируса Молони, промотор на основе вируса Эпштейна-Барра (EBV) и промотор на основе вируса саркомы Рауса (RSV)), и обычно содержит последовательность полиаденилирования в качестве последовательности терминации транскрипции.
Иногда вектор может быть соединен с другими последовательностями для облегчения очистки экспрессируемого с его помощью антитела. Последовательности для присоединения включают, например, глутатион-S-трансферазу (Pharmacia, США), мальтозасвязывающий белок (NEB, США), FLAG (IBI, США), и 6x His (гексагистидин; Qiagen, США).
Вектор включает ген устойчивости к антибиотикам, обычно используемый в уровне техники в качестве селекционного маркера выбора, например, ген устойчивости к ампициллину, гентамицину, карбенициллину, хлорамфениколу, стрептомицину, канамицину, генетицину, неомицину и тетрациклину.
Другое воплощение настоящего раскрытия относится к клетке, трансформированной указанным выше вектором. Клетки, используемые для продуцирования антитела по настоящему раскрытию, могут быть прокариотическими, дрожжевыми или высшими эукариотическими клетками, но не ограничены ими.
Могут быть использованы такие прокариотические клетки-хозяева, как Escherichia coli, штаммы рода Bacillus, такие как Bacillus subtilis и Bacillus thuringiensis, Streptomyces, Pseudomonas (например, Pseudomonas putida), Proteus mirabilis и Staphylococcus (например, Staphylococcus carnosus)
Однако, наибольшее внимание уделяется животным клеткам, и примеры полезных линий клеток-хозяев включают COS-7, BHK, CHO, CHOK1, DXB-11, DG-44, CHO/-DHFR, CV1, COS-7, HEK293, BHK, TM4, VERO, HELA, MDCK, BRL 3A, W138, Hep G2, SK-Hep, MMT, TRI, MRC 5, FS4, 3T3, RIN, A549, PC12, K562, PER.C6, SP2/0, NS-0, U20S и HT1080, но настоящее раскрытие ими не ограничивается.
Другое воплощение настоящего раскрытия относится к способу производства антитела или его антигенсвязывающего фрагмента, включающему: (a) культивирование клеток; и (b) извлечение антитела или его антигенсвязывающего фрагмента из культивируемых клеток
Клетки можно культивировать в различных средах. Любая коммерчески доступная среда может использоваться в качестве культуральной среды без ограничений. Все другие важные добавки, известные обычному специалисту в данной области техники, также могут использоваться в подходящих концентрациях. Условия культивирования, такие как температура и pH, уже использовались с клетками-хозяевами, выбранными для экспрессии, что будет очевидно для обычного специалиста в данной области техники.
Извлечение антитела или его антигенсвязывающего фрагмента можно осуществить, удаляя примеси, например, центрифугированием или ультрафильтрацией, и очищая полученный продукт, например, методом аффинной хроматографии или подобными. Дополнительно можно использовать другие технологии очистки, такие как анионо- или катионобменная хроматография, хроматография гидрофобного взаимодействия, хроматография с гидроксиапатитом и подобные им.
Другое воплощение настоящего раскрытия относится к композиции для профилактики или лечения опухолей, включающей антитело в качестве активного ингредиента. Антитело может представлять собой фрагмент, включающий IgG или вариабельную область, а именно ScFv или Fab. Дополнительно, вариабельная область тяжелой цепи может представлять собой IgG1, IgG2, IgG3 или IgG4.
В настоящем раскрытии предлагается фармацевтическая композиция для профилактики или лечения заболеваний глаз, включающую (a) фармацевтически эффективное количество антитела против Ang2 или его антигенсвязывающего фрагмента по настоящему раскрытию; и (b) фармацевтически приемлемый носитель. В настоящем раскрытии также предлагается способ профилактики или лечения заболеваний глаз, включающий введение антитела или его антигенсвязывающего фрагмента пациенту с опухолями. В настоящем раскрытии предлагается применение антитела или его антигенсвязывающего фрагмента для ингибирования механизма Ang2 и для профилактики или лечения заболеваний глаз.
Что касается заболеваний глаз, роговица является лишенной сосудов тканью и должна всегда оставаться прозрачной, чтобы сохранять зрение. Однако, известно, что ангиогенез осуществляется также и в глазу, вызывая связанные с ангиогенезом заболевания глаза. Иными словами, образование новых кровеносных сосудов в роговице снижает прозрачность глаза, вызывая потерю зрения, и создание новых кровеносных сосудов в сетчатке приводит к образованию патологических кровеносных сосудов, вызывая экссудацию крови и затем индуцируя слепоту посредством дегенерации клеток сетчатки.
На основании этой информации настоящее раскрытие может быть использовано для профилактики или лечения заболеваний глаз, таких как незрелая ретинопатия, ангиогенез роговицы, диабетическая ретинопатия, хориоидальная неоваскулярная болезнь и дегенерация сетчатки (например, возрастная дегенерация сетчатки).
В настоящем раскрытии предлагается фармацевтическая композиция для профилактики или лечения опухолей, включающую (a) фармацевтически эффективное количество антитела против Ang2 или его антигенсвязывающего фрагмента по настоящему раскрытию; и (b) фармацевтически приемлемый носитель. В настоящем раскрытии также предлагается способ профилактики или лечения опухолей, включающий введение антитела или его антигенсвязывающего фрагмента пациенту с опухолями. В настоящем раскрытии предлагается применение антитела или его антигенсвязывающего фрагмента для ингибирования механизма Ang2 и его использования для профилактики или лечения опухолей.
Опухоли, к которым применимы композиция, как правило включают опухоли или раки, избыточно экспрессирующие Ang2, и опухоли или раки, не экспрессирующие избыточно Ang2. Неограничивающие примеры опухолей или раков, являющихся подходящими мишенями для лечения, включают меланому (например, метастатическую злокачественную меланому), рак почки (например, светлоклеточную карциному), рак простаты (например, гормонрефрактерную аденокарционму простаты), аденокарциному поджелудочной железы, рак молочной железы (например, трижды негативный рак молочной железы), рак толстой кишки, рак легкого (например, немелкоклеточный рак легкого), рак пищевода, карциному сквамозных клеток головы и шеи, рак печени, рак яичников, рак матки, рак щитовидной железы, глиобластому, глиому, лейкемию, лимфому и другие неопластические карциномы. Дополнительно, настоящее раскрытие включает рефрактерные или рецидивирующие раки, которые можно вылечить антителами по настоящему раскрытию.
Дополнительное воплощение настоящего раскрытия связано с фармацевтической композицией для ингибирования ангиогенеза, включающую анти-Ang2 антитело или его антигенсвязывающий фрагмент в качестве активного ингредиента. В другом воплощении предлагается фармацевтическая композиция для профилактики и (или) лечения заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2, включающую анти-Ang2 антитело или его антигенсвязывающий фрагмент в качестве активного ингредиента.
В настоящем раскрытии предлагается, например, способ ингибирования ангиогенеза, включающей введение терапевтически эффективного количества анти-Ang2 антитела или его антигенсвязывающий фрагмента пациенту, нуждающемуся в ингибировании ангиогенеза. Способ ингибирования ангиогенеза может дополнительно включать выявление пациента, нуждающегося в ингибировании, до введения. В другом воплощении предлагается способ профилактики и (или) лечения заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2, включая введение терапевтически эффективного количества анти-Ang2 антитела или его антигенсвязывающий фрагмента пациенту, нуждающемуся в профилактике и (или) лечении заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2. Способ может дополнительно включать до введения выявление пациента, нуждающегося в профилактике и (или) лечении заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2.
Фармацевтическая композиция может дополнительно включать фармацевтически приемлемый носитель, и носитель может быть одним из тех, которые обычно используются при составлении лекарств, и это может быть один или более носителей, выбранных из группы, состоящей из лактозы, декстрозы, сахарозы, сорбитола, маннитола, крахмала, гуммиарабика, кальция фосфата, альгината, желатина, кальция силиката, микрокристаллической целлюлозы, поливинилпирролидона, целлюлозы, воды, сиропа, метилцеллюлозы, метилгидроксибензоата, пропилгидроксибензоата, талька, магния стеарата и минерального масла, но настоящее раскрытие ими не ограничивается. Фармацевтическая композиция может дополнительно включать один или более объектов, выбранных из группы, состоящей из разбавителей, вспомогательных веществ, смазывающих компонентов, увлажняющих средств, подсластителей, вкусовых добавок, эмульсификаторов, суспендирующих агентов и консервантов, которые обычно используются при приготовлении фармацевтических композиций.
Эффективное количество фармацевтической композиции или антитела или его антигенсвязывающего фрагмента можно вводить перорально или парентерально. В случае парентерального введения могут использоваться внутривенная инъекция, подкожная инъекция, внутримышечная инъекция, внутрибрюшинная инъекция, эндотелиальное введение, топическое введение, интраназальное введение, внутрилегочное введение, ректальное введение и подобные им. В случае перорального введения, поскольку белок или пептид подвергается расщеплению, можно готовить такие оральные композиции, когда действующие вещества покрыты оболочкой или приготовлены так, чтобы они были защищены от разложения в желудке. Дополнительно, композицию можно вводить с помощью любого устройства, которое позволяет доставить действующее вещество к целевой клетке.
Содержание анти-Ang2 антитела или его антигенсвязывающего фрагмента в фармацевтической композиции можно определить различными способами, в зависимости таких факторов, как способ получения композиции, способ введения, возраст, масса тела и пол пациента, заболевания, режим питания, время введения, интервал введения, путь введения, скорость выделения и чувствительность отклика. Например, суточная доза анти-Ang2 антитела или его антигенсвязывающий фрагмента может находиться в диапазоне от 0,001 мг/кг до 1000 мг/кг, предпочтительно от 0,01 мг/кг до 100 мг/кг, еще более предпочтительно от 0,1 мг/кг до 50 мг/кг, и еще более предпочтительно от 0,1 мг/кг до 20 мг/кг, но настоящее раскрытие ими не ограничивается. Суточная доза может быть получена как одноразовая лекарственная форма в порционной упаковке, или может быть смешана в различных соотношениях, или может быть при приготовлении включена в многодозовый контейнер.
Фармацевтическую композицию можно вводить совместно с другими лекарствами, такими как ингибиторы ангиогенеза или терапевтические агенты для лечения заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2, вводимое количество и способ введения композиции, а также типы других лекарств можно назначить, в зависимости от состояния пациента.
Фармацевтическую композицию можно смешать в форме раствора в масле или водной среде, в форме суспензии, сиропа, эмульсии, экстракта, порошка, гранул, таблеток, капсул и т.д., и она может дополнительно включать диспергирующее вещество или стабилизатор препарата.
В частности, так как фармацевтическая композиция, включающая анти-Ang2 антитело или его антигенсвязывающий фрагмент, включает антитело или антигенсвязывающий фрагмент, ее можно готовить в составе иммунной липосомы. Содержащие антитела липосомы можно готовить способами, хорошо известными в уровне техники. Иммунная липосома представляет собой липидную композицию, включающую фосфатидилхолин, холестерин и фосфатидилэтаноламин, дериватизированный полиэтиленгликолем, и ее можно приготовить методом обращенно-фазового испарения (патентная публикация №10-2015-0089329). Например, фрагмент Fab' антитела можно конъюгировать с липосомой при помощи реакции дисульфидного замещения.
Следует отметить, что, поскольку анти-Ang2 антитело или его антигенсвязывающий фрагмент специфически связывается с ангиопоэтином-2, эту характеристику можно использовать для подтверждения того, происходит ли активация и (или) избыточное производство ангиопоэтина-2. Таким образом, в другом воплощении настоящего раскрытия предлагается фармацевтическая композиция, включающая анти-Ang2 антитело или его антигенсвязывающий фрагмент, для обнаружения активации и (или) избыточного производства ангиопоэтина-2 и (или) для диагностики заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2. В другом воплощении предлагается способ диагностики или способ предоставления информации о диагнозе, включающий: обработку взятого у пациента биологического образца анти-Ang2 антителом или его антигенсвязывающим фрагментом; подтверждение, происходит ли реакция антиген-антитело; и определение того, что, если реакция антиген-антитело обнаружена, у пациента наблюдаются симптомы активации и (или) избыточного производства ангиопоэтина-2, или имеется заболевание, связанное с активацией и (или) избыточным производством ангиопоэтина-2. Биологический образец может быть выбран из группы, состоящей из клеток, тканей и жидкостей организма, взятых у пациента.
Подтверждение реакции антиген-антитело можно сделать различными способами, известными в уровне техники. Например, реакцию можно подтвердить обычной ферментативной реакцией, детектированием флуоресценции, люминесценции и (или) радиации, и, в частности, можно измерить способом, выбранным из группы, состоящей из иммунохроматографии, иммуногистохимии, твердофазного иммуноферментного анализа (ELISA), радиоиммуноанализа (RIA), ферментативного иммуноанализа (EIA), флуоресцентного иммуноанализа (FIA), люминесцентного иммуноанализа (LIA) и вестерн-блоттинга, но настоящее раскрытие этими способами не ограничивается.
Пациент, которому вводят или проводят диагностику фармацевтической композицией, может быть млекопитающим, включая приматов, включая человека, обезьяну и т.д., а также грызунов, включая мышь, крысу и т.д.
Заболеванием, связанным с активацией и (или) избыточным производством ангиопоэтина-2, могут быть рак; метастатический рак; заболевания глаз, такие как ретинопатия недоношенных, ангиогенез роговицы, диабетическая ретинопатия, хориоидальная неоваскуляризация и макулярная дегенерация (например, возрастная макулярная дегенерация); астма, ревматоидный артрит, псориаз; воспалительные заболевания, такие как пневмония и хроническое воспаление; сердечно-сосудистые заболевания, такие как гипертензия или артериосклероз; или септицемия. При раке может иметь место избыточная экспрессия ангиопоэтина-2, это может быть солидная опухоль или рак крови, и это может быть без ограничений один или более раков, выбранных из группы, состоящей из сквамозной карциномы, мелкоклеточного рака легкого, немелкоклеточного рака легкого, аденокарциномы легкого, плоскоклеточный рак легкого, перитониальный рак, рак кожи, меланома кожи или глаз, рак прямой кишки, рак анального канала, рак пищевода, рак тонкой кишки, эндокринная аденокарцинома, рак паращитовидной железы, рак надпочечников, саркома мягких тканей, рак уретры, хронический или острый лейкоз, лимфоцитарная лимфома, гепатоцеллюлярная карцинома, рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластома, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, опухоль печени, рак молочной железы (в некоторых случаях трижды отрицательный рак молочной железы), рак толстой кишки, рак толстой кишки, карцинома эндометрия или матки, рак слюнной железы, рак почки, рак печени, рак простаты, рак вульвы, рак щитовидной железы, рак печени, рак головы и шеи, рак мозга и остеосаркома. Рак может быть первичным или метастатическим.
Примеры
Далее настоящее раскрытие будет описано более подробно со ссылкой на следующие примеры. Эти примеры предоставлены только для иллюстративных целей, и специалистам в данной области техники будет очевидно, что эти примеры не следует считать ограничивающими область настоящего раскрытия.
Пример 1. Выбор антител, связывающихся с Ang2
Для приготовления библиотек антител и библиотек для выбора антител, которые связываются с Ang2, используют библиотеку сенситизированных scFv человека (наивные ScFv человека), как в Корейской патентной заявке (патентная публикация №10-2008-0109417). 2 мкг/мл (100 мкл на лунку) антигена (hAng2-his: RND systems. Cat. No 623-AN/CF, hAng2-Fc: PharmAbcine) прибавляют в 96-луночный иммунопланшет и оставляют на ночь при 4°C. На следующий день покрытый антигеном планшет промывают 3 раза 5 мМ CaCl2 TBS, после чего прибавляют 200 мкл блокирующего буфера 2% BSA, после чего оставляют реакционную смесь при комнатной температуре на 2 часа. 50 мкл раствора банка XL1-Blue прибавляют к 2 мл среды роста 2x YT-TET (10 мкг/мл тетрациклиа), растят при 37°C и 200 об/мин около 2 часов, и затем прибавляют еще 13 мл смеси и культивируют банк клеток, пока ОП600 не достигнет 0,5. Через 2 часа блокирования промывают результат трижды 1X 5 мМ CaCl2 TBS. Группу фаговой библиотеки объединяют с каждой промытой лункой, после чего фаговую библиотеку и 4% BSA смешивают в одинаковых количествах, затем в каждую лунку прибавляют 200 мкл смеси, после чего покачивают при комнатной температуре в течение 30 минут и оставляют реакционную смесь на 2 часа. После завершения реакции фаговой библиотеки надосадочный раствор сливают, полученный раствор промывают 5 раз 0,1% TBST (5мМ CaCl2) и 5 раз TBS (5мМ CaCl2), затем в каждую лунку прибавляют 100 мкл 100 мМ триметиламина (TEA), после чего встряхивают при комнатной температуре 10 минут. Через 10 минут в каждую лунку прибавляют 50 мкл 1 M Tris (pH 7,5) и перемешивают. Надосадочный раствор прибавляют к 10 мл XL1-blue с ОП600 0,5 для инфицирования при 37°C и оставляют на 30 минут. После инфицирования в качестве титра на выходе используют 100 мкл, остаток центрифугируют при 6000 об/мин в течение 10 минут. Надосадочный раствор сливают, а осадок распределяют по большой квадратной пластине (34 мкг/мл CM + 1% глюкоза), после чего инкубируют в течение ночи при 30°C. Оставшиеся в качестве титра на выходе 100 мкл разбавляют в отношении 1/10, 1/100, или 1/1000, распределяют по пластине CM и оставляют на ночь при 37°C. На следующий день выросшие на квадратной пластине колонии переносят в 50 мл среды 2x YT, соскребают петлей и центрифугируют 10 минут при 6000 об/мин, чтобы слить надосадочный раствор, из осадка готовят первичный банк для пэннинга, 100 мл среды роста 2x YT (34 мкг/мл CM + 1% глюкоза) переносят в колбу Эрленмейера вместимостью 500 мл, после чего прибавляют туда клетки, так что ОП600 станет равным 0,2, затем дают возможность расти при 200 об/мин и 37°C, пока ОП600 не станет равной 0,5. После этого хелперный фаг (мутант M13KO7) прибавляют в количестве, в 20 раз превышающем ОП600 клеток. После прибавления хелперного фага и по завершении инфицирования в течение 30 минут при 37°C проводят центрифугирование при 6000 об/мин в течение 10 минут. Надосадочный раствор отбрасывают, и к клеткам прибавляют 100 мл среды 2xYT (34 мкг/мл CM + 70 мкг/мл Kan. + 1 мМ IPTG + 5 мМ MgCl2), затем их оставляют на ночь при 200 об/мин и 30°C. На следующий день выросшие клетки центрифугируют при 7000 об/мин в течение 10 минут, а потом еще раз тем же способом. Прибавляют 20% PEG/2,5 мМ NaCl к 1/5 (о/о) собранного надосадочного раствора и осаждают на льду 1 час. После осаждения проводят центрифугирование при 9000 об/мин в течение 1 часа. Надосадочный раствор сливают, а осадок взбалтывают в 3 мл TBS, фильтруют через фильтр с размером пор 0,45 мкм и хранят при 4°C для использования в последующем пэннинге. Эти процессы повторяют три или четыре раза, затем связавшиеся в антигеном антитела идентифицируют методом ELISA.
Пример 2. Скрининг моноклонального ScFv фага, который специфически связывается с Ang2 и нейтрализует связывание с Tie2 (ELISA связывания/конкурентный ELISA)
После завершения пэннига финальный круглый банк клеток разбавляют и высевают так, чтобы на агаровом планшете СМ сформировалось от 200 до 500 колоний, затем их оставляют на ночь при 37°C. На следующий день, когда колонии вырастут, в 96-луночный планшет вносят 200 мкл среды 2xYT (34 мкг/мл CM + 1% глюкоза), в каждую лунку по очереди прибавляют колонии и оставляют на ночь при 37°C и 3000 об/мин. На следующий день в новый 96-луночный планшет вносят 200 мкл среды 2xYT (34 мкг/мл CM + 1% глюкоза), и в каждую лунку прибавляют 20 мкл клеток, выращенных в предыдущий день, их выращивают при 37°C и 3000 об/мин в течение 1 часа и 10 минут. В каждую лунку добавляют 100 мкл 50% глицерина, оставшиеся клетки хранят при 70°C. Пока клетки растут, смешивают 1 мкл хэлперного фага и 19 мкл среды 2xYT, после чего в каждую лунку вносят по 20 мкл полученной смеси, затем инкубируют при 37°C в течение 30 минут. После завершения инкубации проводят центрифугирование при 3000 об/мин в течение 10 минут. Надосадочный раствор сливают и прибавляют 200 мкл среды 2xYT (34 мкг/мл CM + Kan. 70 мкг/мл + 1 мМ IPTG + 5 мМ MgCl2), после чего оставляют на одну ночь в Megagrow при 30°C и 3000 об/мин.
Чтобы отобрать фаги, специфически связывающиеся с Ang2, 1 мкг/мл (100 мкл/лунка) Ag (hAng2-Fc, hAng1-his: RND systems. Cat. No 923-AN/CF или mAng1-Fc, PharmAbcine) прибавляют на 96-луночный иммунопланшет и оставляют на ночь при 4°C. На следующий день клетки, выросшие накануне, центрифугируют при 3000 об/мин в течение 10 минут и хранят при 4°C. Ag промывают три раза средой 0,1% TBST (5 мМ CaCl2), затем прибавляют 200 мкл 2% блокирующего буфера BSA и инкубируют при 25°C в течение 2 часов. После завершения блокирования проводят промывание три раза 0,1% TBST (5 мМ CaCl2). 50 мкл 4% BSA и 50 мкл центрифугированного фага, сохранявшегося при 4°C, перемешивают в каждой лунке и встряхивают при комнатной температуре в течение 1 часа, чтобы началась реакция. После связывания фага трижды проводят промывание 0,1% TBST (5 мМ CaCl2), затем прибавляют 100 мкл конъюгированного с HRP мышиного анти-M13 Ab 1:3000 (Sino, 11973-MM05), и реакция продолжается 1 час при 25°C. После завершения реакции трижды проводят промывание 0,1% TBST (5 мМ CaCl2), прибавляют 100 мкл TMB (#BD TMB набор реактива субстрата 555214), что позволяет проявиться окрашиванию в течение 3-5 минут, затем в каждую лунку прибавляют 50 мкл стоп-раствора, после чего проводят анализ на считывающем устройстве ELISA.
Результаты измерения специфического связывания моноклонального scFv фага с антигеном Ang2 методом ELISA
Основные последовательности отобранных антител показаны в таблица 2 и 3 ниже.
Последовательности CDR антител, специфически связывающиеся с антигеном Ang2


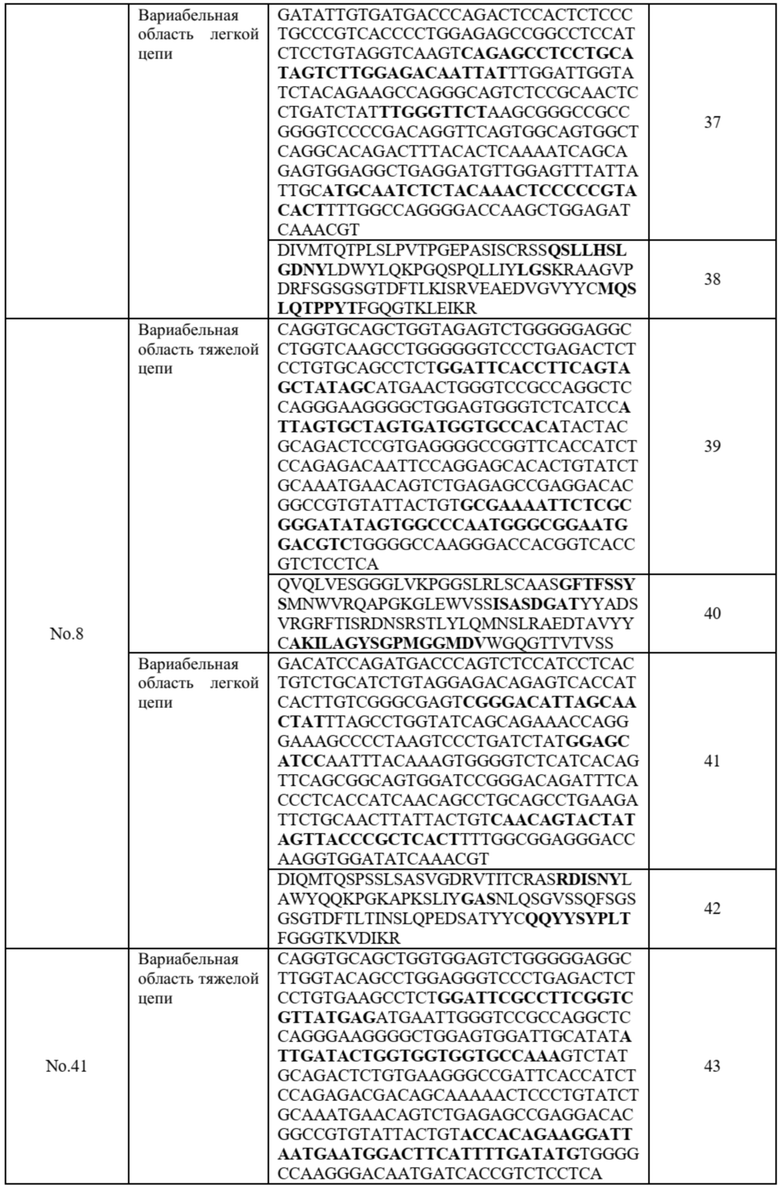

Чтобы выбрать фаги, которые нейтрализуют связывание Ang2/Tie2, методом конкурентного анализа ELISA, 1 мкг/мл (100 мкл на лунку) Ag (hTie2-Fc: PharmAbcine) прибавляют в 96-луночный иммунопланшет и оставляют на ночь при 4°C. Распределенный Ag дважды промывают 1X PBS, после чего прибавляют 200 мкл 3% блокирующего буфера BSA и инкубируют 2 часа при 25°C. После завершения блокировки дважды проводят промывание 0,1% PBST. В каждой лунке 20 мкл (5 мкг/мл) hAng2-his (RND, 623-AN/CF) смешивают с различными объемами (80 мкл, 40 мкл+1X PBS 40 мкл, 20 мкл+1X PBS 60 мкл) фага, который был охлажден и хранился при 4°C, и встряхивают при комнатной температуре в течение 1 часа, чтобы обеспечить протекание реакции. После связывания фага трижды проводят промывание 0,05% PBST, а затем реакцию при комнатной температуре в течение 1 часа с 0,5 мкг/мл мышиных антител анти-Ang2. После завершения связывания с антителами трижды проводят промывание 0,05% PBST, после чего прибавляют 100 мкл HRP-конъюгированного мышиного анти-IgG Ab 1:2000 (RND, HAF007) и продолжают реакцию при 25°C в течение 1 часа. После завершения трижды проводят промывание 0,05% PBST, после чего прибавляют 100 мкл TMB (#BD TMB набор реактива субстрата 555214)), что позволяет проявиться окрашиванию в течение 3-5 минут, затем в каждую лунку прибавляют 50 мкл стоп-раствора, после чего проводят анализ на считывающем устройстве ELISA. Результаты представлены на фиг. 1. Как видно из фиг. 1, способность выбранных фагов нейтрализовывать связывание Tie2/Ang2 была подтверждена.
Пример 3. Выбор антител с высоким сродством с Ang2 (скрининг скорости диссоциации)
Сродство связывания выбранных антител с антигеном измеряли с использованием Octet (Fortebio). С этой целью Ang2 иммобилизовали на биосенсоре, после чего прибавили кандидаты антител, экспрессированных в форме scFv, позволили им связаться, а затем провели измерение констант скорости диссоциации. Результаты представлены в таблице 4.
Константы скорости диссоциации антител, специфически связанных с антигеном Ang2
Пример 4. Экспрессия антитела анти-Ang2
Конверсию выбранных фагов scFv в форму IgG выполнили при помощи молекулярно-биологических технологий. Фагмиды экстрагировали из отобранных клонов E. coli, вариабельные области амплифицировали по технологии ПЦР. Амплифицированные вариабельные области тяжелой цепи ввели в вектор экспрессии (Invivogen, pfusess-hchg1), содержащий константные области тяжелой цепи, а амплифицированные вариабельные области легкой цепи ввели в вектор экспрессии (Invivogen, pfuse2ss-hclk), содержащий константные области легкой цепи, тем самым завершив клонирование ДНК IgG типа.
Временную экспрессию IgG выполнили с использованием системного набора для экспрессии Expi293F (Thermo Fisher Scientific, США). Включенные в набор клетки Expi293 суспендировали и инкубировали на орбитальном шейкере при 125 об/мин в условиях 37°C и 5% CO2, с использованием эксклюзивной среды. Каждые три дня клетки пересевали в концентрации 3×105 клеток/мл, и перед введением вектора экспрессии концентрацию клеток довели до 3×106 клеток/мл. Для введения гена использовали эксклюзивные реактив Expifectamine, получили комплекс липид-ДНК, содержащий 1 мкг вектора экспрессии ДНК и 2,7 мкл Expifectamine на 1 мл клеточной суспензии, его прибавили к клеточной суспензии и через 16-18 часов после введения прибавили энхансер 1/2, чтобы индуцировать экспрессию. Затем полученную суспензию культивировали 3-4 дня в тех же условиях, после чего центрифугировали, чтобы собрать содержащий IgG надосадочный раствор.
Пример 5. Очистка анти-Ang2 антитела
Собранный надосадочный раствор ввели в колонку с белком А (GE Healthcare) для очистки IgG методом аффинной хроматографии. Колонку уравновесили 20 мМ Tris-HCl, 50 мМ NaCl и 5 мМ EDTA (pH 7,0), после чего ввели надосадочный раствор, колонку промывали раствором 50 мМ Tris-HCl, 500 мМ NaCl, 5 мМ EDTA, 0,2% полисорбата 20 (pH 7,0), и после этого выполнили элюирование при помощи раствора 50 мМ NaCl, 0,1 M глицина-HCl (pH 3,5), а затем нейтрализацию 1 M Tris. Для элюирования белков растворитель заменили на PBS посредством диализа с использованием диализной мембраны MWCO 10000 spectra/por (Spectrum Labs, США). После этого белки сконцентрировали при помощи Vivaspin (Satorius, Германия) до требуемой концентрации, разлили и хранили при -80°C.
После очистки каждое антитело обработали в невосстанавливающем и восстанавливающем буфере для образца LDS (Thermo Fisher Scientific) и провели их электрофорез при помощи системы NuPAGE System (Thermo Fisher Scientific). В результате был получен IgG, включающий тяжелую цепь 50 кДа и легкую цепь 25 кДа с общей молекулярной массой около 150 кДа (фиг. 2).
Пример 6. Анализ специфичности связывания антитела Анти-Ang2
При выполнении анализа специфичности связывания измеряли константы связывания методом ELISA при помощи системы Biacore T200 (GE Healthcare Life Sciences).
1 мкг/мл (100 мкл на лунку) раствора меченого His Ang2 человека (R&D systems, 623-AN/CF) или мыши (sino, 50298-M07H) или меченого His Ang1 человека (R&D systems, 923-AN/CF) или мыши (sino, 50300-M07H) прибавили в 96-луночный иммунопланшет (Nunc, США) и оставили на одну ночь при 4°C для адсорбции. На следующий день раствор трижды промыли PBS, содержащим 0,05% Tween-20 (далее называется PBST), и в каждую лунку прибавили 200 мкл раствора 2% BSA/PBST, после чего оставили при комнатной температуре на 2 часа, чтобы осуществилась блокировка. После трехкратного промывания PBST по 100 мкл каждого испытуемого раствора антител добавили в каждую лунку в концентрации, вызывающей связывание при комнатной температуре в течение 1 часа, затем трижды выполнили промывание PBST, добавили 100 мкл HRP-конъюгированного античеловеческого IgG (каппа) козы (Bethyl lab #A80-115P), разбавленного в отношении 1:2000 и оставили реагировать при комнатной температуре на 1 час, тем самым индуцировав связывание, и после трехкратного промывания PBST провели окрашивание 100 мкл реактива субстрата TMB. Реакцию окрашивания остановили добавлением 50 мкл 2 н. H2SO4, и с помощью считывателя микропланшет Sunrise (TECAN, CH) определили специфическое оптическое поглощение ОП450-630 (фиг. 3). Как показано на фиг. 3, выбранные антитела специфически связываются с Ang2 человека и мыши и не связываются с Ang1 человека и мыши.
Для анализа сродства связывания выбранных антител анти-Ang2 выполнили анализ сродства Ang2 человека и мыши с использованием BIACORE® T200 (GE Healthcare). Использовали сенсорный чип на белок А, и эксперимент провели в соответствии с руководством производителя. Условия анализа следующие. Иммобилизовали 2000 единиц ответа (RU) белка А, для связывания использовали 25 RU предполагаемого антитела анти-Ang2, и для связывания использовали различные концентрации Ang2 человека и мыши. Исходная концентрация для анализа составляет 100 нМ и 150 нМ, соответственно. Анализ проходил при скорости потока 30 мкл/мин, связывание и время диссоциации Ang2 человека измеряли через 300 и 2000 секунд, соответственно, и связывание и время диссоциации Ang2 мыши измеряли через 300 и 1000 секунд, соответственно. В качестве аналитической модели использовали модель связывания 1:1. Результаты анализа представлены в таблице 5.
Пример 7. Подтверждение нейтрализующей способности антител Анти-Ang2 (Ang2/Tie2, Ang2/интегрин)
1 мкг/мл (100 мкл на лунку) раствора Ti32-Fc человека (R&D Systems, 313-TI) или мыши (R&D Systems, 762-T2-100), интегрина α3/β1 (R&D systems, 2840-A3-050) или интегрина α5/β1 (R&D systems, 3230-A5-050) внесли в 96-луночный иммунопланшет (Nunc, США), после чего оставили на ночь при 4°C для адсорбции. На следующий день раствор четыре раза промыли PBS, содержащим 0,05% Tween-20 (далее называется PBST), и в каждую лунку прибавили 200 мкл раствора 2% BSA/PBST, после чего оставили при комнатной температуре на 2 часа, чтобы осуществилась блокировка. Кандидаты антител анти-Ang2 последовательно разбавили в 4 раза с максимальной концентрацией 1000 нМ, биотинилированный белок Ang2 человека довели до конечной концентрации 100 нг/мл или 625 нг/мл с последующим связыванием при комнатной температуре в течение 1 часа, как описано ранее. Блокированный планшет четыре раза промыли PBST, после чего к планшету добавили образцы, в которых ранее была индуцирована реакция антиген-антитело, связывание проходило при комнатной температуре в течение 1 часа. Планшет трижды промыли PBST, прибавили 100 мкл HRP-конъюгированного стрептавидина (R&D Systems, DY998), разбавленного в отношении 1:200, затем реакция протекала при комнатной температуре 1 час, индуцируя при этом связывание, после чего планшет четыре раза промыли PBST, а затем осуществили окрашивание добавлением 100 мкл реактива субстрата TMB. Реакцию окрашивания остановили добавлением 50 мкл 2 н. H2SO4, и с помощью считывателя микропланшет Sunrise (TECAN, CH) определили специфическое оптическое поглощение ОП450-630 (фиг. 4 и 5). Как показано на фиг. 4, выбранные антитела нейтрализовывали Ang2/Tie2 человека и мыши. Эффективность нейтрализации оценили количественно, определив значения IC50, которые показаны в таблице 6. Дополнительно, как показано на фиг. 5, выбранные антитела также нейтрализовывали связывание интегрин/Ang2. Способность нейтрализовать integrin/Ang2 оценили количественно при помощи IC50, результаты показаны в таблице 7.
(конкурирование с Ang2 человека) (пМ)
(конкурирование с Ang2 мыши) (пМ)
(конкурирование с α5β1/hAng2 ) (пМ)
(конкурирование с α3β1/hAng2) (пМ)
Пример 8. Анализ влияния антител Анти-Ang2 на ингибирование передачи сигнала Ang2/Tie2 (анализ p-Tie2)
Клетки человека, избыточно экспрессирующие Tie2 (1×105) поместили в 96-луночный планшет и выращивали одну ночь при 37°C в инкубаторе с углекислым газом. Клетки выращивали одну ночь в бессывороточной среде, чтобы создать условия депривации сыворотки. Ang2 человека (5 мкг/мл) и анти-Ang2 человека в различных концентрациях взаимодействовали при комнатной температуре в течение 1 часа, как ранее описано, затем их поместили на содержащий клетки планшет, и реакция протекала 20 минут. В данном случае в планшете была предусмотрела лунка, не содержащая антитела и содержащая только белок Ang2, ее использовали для сравнения при анализе эффекта ингибирования передачи сигнала. Клетки лизировали лизирующим буфером и затем пересчитали. Для анализа реакции фосфорилирования использовали набор Human Phospho-Tie2 Duoset IC ELISA (R&D Systems, DYC2720-5), доступный в компании R&D Systems. В каждую лунку 96-луночного планшета прибавили 4 мкг/мл белка захвата Tie2 (Nunc, США) и оставили систему на ночь при 4°C для адсорбции. На следующий день в каждую лунку добавили по 200 мкл и оставили при комнатной температуре на 2 часа для блокировки. Добавили 50 мкг лизата клеток, и связывание протекало при комнатной температуре в течение 2 часов. После завершения реакции антитела против фосфотирозина разбавили в отношении 2700:1, и связывание проходило при комнатной температуре в течение 2 часов. По завершении реакции провели окрашивание с помощью реактива субстрата TMB. Реакцию окрашивания остановили добавлением 50 мкл 2 н. H2SO4, и с помощью считывателя микропланшетов Sunrise (TECAN, CH) определили специфическое оптическое поглощение ОП450-630 (фиг. 6). Как показано на фиг. 6, можно подтвердить, что при повышении концентрации антитела фосфорилирование уменьшается. Количественно степень фосфорилирования оценили, определив IC50, результаты показаны в таблице 8.
Пример 9. Конструирование и выбор вариантов для усиления сродства
Провели оптимизацию антител, чтобы усилить сродство антитела анти-Ang2, клон №8. Используя способ мягкой рандомизации для консервации и рандомизации 70% оригинальной последовательность ДНК №8, были приготовлены праймеры с рандомными мутациями в легкой цепи CDR3 и тяжелой цепи CDR3 этой последовательности. При помощи ПЦР с праймерами получили фрагменты ДНК, кодирующие затронутые мутацией вариабельные области легкой и тяжелой цепи. Фрагменты ДНК соответственно заместили на вариабельную область легкой цепи No. 8 scFv фагмида и вариабельную область тяжелой цепи No. 8 scFv фагмиды, тем самым завершив конструирование фаговой библиотеки ДНК scFv вариантов CDR3 легкой и тяжелой цепи.
Фаговую библиотеку ДНК scFv очистили системой фенол-хлороформ, после чего трансформировали в штамм E. coli XL-1 Blue методом электропорации. После подтверждения методом анализа эффективности трансформации и секвенирования ДНК того, что разнообразие достигнуто, 500 мл штамма культивировали, чтобы индуцировать экспрессию фага, и при помощи PEG-осаждения сконструировали фаговые библиотеки scFv вариантов CDR3 легкой и тяжелой цепи.
Биопэннинг провели способом, описанным в примере 1, с использованием всех мутантов фаговой библиотеки scFv. После этого в ходе скрининга измерили константу скорости диссоциации kdis как количественную оценку способности сохранять связывание. Далее показаны аминокислотные последовательности трех выбранных и оптимизированных клонов (таблица 9 и 10) и результаты измерения константы скорости диссоциации (таблица 11).

Пример 10. Производство оптимизированных антител ScFv Анти-Ang2
Оптимизированное антитело анти-Ang2 (No.O4) клонировали в вектор pET-22b (Novagen) для экспрессии в E. coli. Выделили колонии, полученные путем трансформации вектора в BL21 (De3), к среде LB (бульон Lysogeny) добавили 100 мкг/мл ампициллина, и 1% E. coli, предварительно выращиваемой в среде 37°C при 200 об/мин, инокулировали в среду LB, содержащую 100 мкг/мл ампициллина в качестве антибиотика. E. coli инкубировали при условиях 37°C и 200 об/мин, температуру инкубатора понизили до 20°C, после чего прибавили 0,5 мМ IPTG, а затем инкубировали в течение 16 часов.
Инкубированные E. coli собрали центрифугированием при 8000 об/мин в течение 10 минут. После удаления среды провели ресуспендирование в 10 мл (на массу (г) клеток) лизирующего буфера, содержащего 50 мМ Tris-HCl pH 7,4, 150 мМ NaCl и 1 мМ фенилметилсульфонилфторида (PMSF). Клетки разрушили генератором ультразвука в следующих условиях: мощность 20 Вт, пауза 3 сек, работа 3 сек, время 5 мин. Разрушенные клетки центрифугировали при 11000 об/мин в течение 1 часа для разделения надосадочного раствора и осадка.
ScFv экспрессировали в нерастворимой форме и промыли осадок для ренатурации. После гомогенизации в гомогенизаторе с использованием буфера 50 мМ Tris-HCl pH 7,4, 150 мМ NaCl осадок дважды промыли центрифугированием при 11000 об/мин в течение 1 часа. Остававшиеся в осадке извлеченные из E. coli вещества удалили с помощью буфера 50 мМ Tris-HCl pH 7,4, 150 мМ NaCl, 2 M мочевины, 0,5% Triton X-100, и трижды промыли буфером 50 мМ Tris-HCl pH 7,4, 150 мМ NaCl. Затем осадок ресуспендировали с помощью буфера 50 мМ Tris-HCl pH 7,4, 150 мМ NaCl, 8 M мочевины, 10 мМ DTT, после чего реакцию продолжали около 30 минут до получения scFv в несвернутой форме, а затем провели центрифугирование при 11000 об/мин в течение 1 часа для отделения осадка.
scFv антитело подвергли ступенчатому диализу для удаления мочевины, тем самым индуцируя ренатурацию. Диализ проводили с помощью диализного буфера, в основном, содержащего 50 мМ Tris-HCl pH 7,4 и 150 мМ NaCl, уменьшая концентрацию мочевины в 2 раза. Ренатурацию проводили путем подавления агрегации добавлением 0,1 М L-аргинина к фракциям, содержащим мочевину в концентрации 4-2-1 M, в которых в основном происходит правильное свертывание белка. Ренатурированное scFv антитело выделили и очистили с использованием колонок с HisTrap и Capo L.
Разделение и очистку выполнили в следующем порядке. Очистку проводили с использованием AKTA Prime (GE Healthcare) System, использовали упакованную колонку HisTrap на 5 мл. Буфер 50 мМ натрия фосфата, 400 мМ NaCl, 10 мМ имидазола с pH 7,4 в количестве 10 объемов колонки пропустили через колонку HisTrap для уравновешивания, и образец антитела scFv пропустили в колонку HisTrap со скоростью потока 5 мл/мин, чтобы образец антитела смог связаться со смолой внутри колонки. Для удаления присутствующих в смоле неспецифически связывающихся веществ пропустили около 10 объемов колонки буфера 50 мМ натрия фосфата, 400 мМ NaCl, 10 мМ имидазола с pH 7,4, после чего через колонку пропускали буфер 50 мМ натрия фосфата, 400 мМ NaCl, 300 мМ имидазола с pH 7,4 с градиентами концентрации, так что антитело scFv элюировало. Образец элюировавшего антитела подвергли следующей очистке с помощью 5 мл колонки Capto L. Колонку Capto L уравновесили физиологическим раствором с фосфатным буфером (PBS) pH 7,4, затем образец пропустили через колонку Capto L со скоростью 5 мл/мин, чтобы удалить неспецифически связанные вещества. Пропустили 10 объемов колонки буфера 0,1 M лимонной кислоты, 0,2 M Na2HPO4 pH 2,6, так чтобы элюировало антитело scFv. На фиг. 7 представлены результаты SDS-PAGE для белка, полученного в итоге после очистки. Видно, что полученное антитело scFv продемонстрировало высокую чистоту.
Пример 11. Анализ специфичности связывания оптимизированного анти-Ang2 антитела ScFv
С целью анализа специфичности связывания константы связывания измеряли с использованием метода ELISA и системы Octet (Pall ForteBio LLC, США).
1 мкг/мл (100 мкл на лунку) раствора меченного His белка Ang2 человека (R&D systems, 623-AN/CF) добавили в 96-луночный иммунопланшет (Nunc, США) и оставили на ночь при 4°C для адсорбции. На следующий день раствор трижды промыли PBS, содержащим 0,05% Tween-20 (далее называется PBST), и в каждую лунку прибавили 200 мкл раствора 2% BSA/PBST, после чего оставили при комнатной температуре на 2 часа, чтобы осуществилась блокировка. После трехкратного промывания PBST, по 100 мкл каждого испытуемого раствора антител добавили в каждую лунку в концентрации, вызывающей связывание при комнатной температуре в течение 1 часа, затем трижды выполнили промывание PBST, добавили 100 мкл HRP-конъюгированного анти-His меченного моноклонального антитела (MAB050H, R&D Systems, США), разбавленного в отношении 1:1000 и оставили реагировать при комнатной температуре на 1 час, тем самым индуцировав связывание, и после трехкратного промывания PBST провели окрашивание 100 мкл реактива субстрата TMB. Реакцию окрашивания остановили добавлением 50 мкл 2 н. H2SO4, и с помощью считывателя микропланшет Sunrise (TECAN, CH) определили специфическое оптическое поглощение ОП450 (фиг. 8). Как показано на фиг. 8, очищенное антитело связывается с Ang2.
Сродство связывания очищенного антитела scFv с Ang2 человека измеряли с использованием системы Octet (ForteBio Inc., США). С этой целью анти-Ang2 антитело иммобилизовали на биосенсоре и измерили кинетику связывания Ang2 человека в зависимости от концентрации. Это позволило рассчитать константу скорости связывания (ka), константу скорости диссоциации (kdis), и константу связывания (KD) (таблица 12).
Пример 12. Анализ in vivo эффективности выбранных антител Анти-Ang2 ScFv (модель CNV мыши)
Для подтверждения ингибирующего эффекта антитела анти-Ang2 scFv по отношению к ангиогенезу провели анализ эффективности препарата на модели индуцированной лазерным излучением хориоидальной неоваскуляризации мыши. При проверке эффективности в качестве контроля использовали афлиберцепт, являющийся коммерчески доступным лекарством. Мышам делали общую анестезию кетамином, после чего на глазные яблоки наносили анестезирующие глазные капли для дополнительной местной анестезии, затем на глаза наносили расширяющее зрачок лекарство, чтобы индуцировать мидриаз. Каждую мышь переносили на стенд и с помощью установки Micron-IV индуцировали лазерный ожог в условиях индукции CNV (длина волны 532 нм, диаметр 50 мкм, продолжительность 80 мс, уровень мощности 200 мВт), тем самым разрушая мембрану Бруха. Полученные в ходе индукции лазерного ожога поражения, при которых не наблюдались пузырьки, были классифицированы как неудачные лазерные ожоги и исключены из анализа результатов и статистической обработки на основании критерия исключения, полученного путем модификации критерия, предложенного Gong Y. и соавт.
Чтобы подтвердить ингибирующий ангиогенез эффект при индукции CNV и при действии лекарств, мышам сделали общую анестезию кетамином в день 10 после индукции CNV, после чего каждой мыши внутрибрюшинно ввели флуоресцентный контраст. Для дополнительной местной анестезии на глазные яблоки нанесли анестезирующие глазные капли, затем на глаза наносили расширяющее зрачок лекарство, чтобы индуцировать мидриаз. Каждую мышь переносили на стенд, выполняли настройку для визуализации глазного дна с использованием камеры от Micron-IV, после чего на соответствующий глаз наносили смазывающий гель и на роговицу глаза каждой мыши накладывали линзы оптической когерентной томографии (OCT). После получения FFA/OCT изображения на глаза мыши нанесли 1 каплю антибиотических глазных капель. Анализ изображений FFA и OCT провели с помощью программы Image-J. В этом отношении поражения CNV, соответствующие критерию исключения, предложенному Gong Y. и соавт., были исключены из итоговых результатов и статистического анализа.
Чтобы подтвердить восстанавливающий эффект на оптический нерв, выполнили испытание с помощью электроретинограммы (ERG). Перед оценкой ERG мышей оставили в темной комнате на 12 часов, чтобы индуцировать адаптацию к темноте. В день проведения оценки (день 11 после индукции CNV) мышам сделали общую анестезию реактивами Rompun® и Ketamine®, а затем на глаза нанесли Alcaine® для дополнительной местной анестезии, затем на глаза наносили расширяющее зрачок лекарство, чтобы индуцировать мидриаз. Каждую мышь поместили на стенд ERG и зонды ERG закрепили на хвосте, голове и роговице глаз, соответственно. ERG измерили как изменение потенциала сетчатки в ответ на стимул в виде одной вспышки (0,9 log cds/м2 (10 ответов/интенсивность)). После завершения оценки ERG на глаза мышей нанесли 1 каплю Tobrex. Анализ ERG выполнили с помощью программы LabScribeERG (iWorx Data Acquisition Software). В этом отношении глаза, соответствующие критерию исключения, предложенному Gong Y. и соавт., были исключены из итоговых результатов и статистического анализа.
Результаты, подтверждающие эффективность антитела анти-Ang2 scFv в отношении уменьшения ангиогенеза на мышиной модели CNV, показаны на фиг. 9. Статистически значимое уменьшение размера поражений CNV при FFA наблюдали в экспериментальной группе, которой вводили 10 мкг/мкл scFv (P<0,01), там наблюдался более мощный эффект, по сравнению с экспериментальной группой, которой давали афлиберцепт. Из фиг. 10 видно, что, даже при сравнении с контролем CNV при измерении OCT все экспериментальные группы, которым вводили scFv, демонстрировали зависящее от концентрации статистически значимое снижение объема поражения (P<0,01, P<0,0001, P<0,05, соответственно). Дополнительно, как показано на фиг. 11, когда потенциал сетчатки сравнили с контролем CNV методом скотопического ERG, наблюдали статистически значимое увеличение амплитуды В-волны (P<0,0001, соответственно).
Пример 13. Анализ противоопухолевой активности выбранного антитела Анти-Ang2 (модель TNBC)
Чтобы подтвердить наличие ингибирующего эффекта антитела анти-Ang2 против ангиогенеза, провели оценку противоопухолевой эффективности на модели трижды негативного рака молочной железы (TNBC) (MDA-MB-231). Чтобы определить противоопухолевую активность антитела анти-Ang2, оценили противораковую эффективность изотипического контроля, несвакумаба и антитела анти-Ang2 путем введения в хвостовую вену мыши NSG, которой была в левой бок трансплантирована линия клеток MDA-MB-231 рака молочной железы человека. Эксперимент провела компания Champions Oncology, Inc. (США)
Клеточную линию MDA-MB-231 рака молочной железы человека разморозили и культивировали в CO2-инкубаторе (Forma, США) при температуре 37°C и концентрации CO2 5%. В последний день культивирования все раковые клетки собрали и пересчитали, после чего довели концентрацию клеток до 1×108 клеток/мл при помощи бессывороточной среды. Такой скорректированный раствор культуры клеток ввели в левый бок каждой мыши в объеме 0,1 мл (1×107 клеток/мышь). Мышей случайным образом объединили в группы по 8 животных, пока объем опухолей не достигнет 100-150 мм3. Сразу после объединения начали введение лекарства. 10 мг/кг изотипического контроля в качестве отрицательного контроля, 3 мг/кг или 10 мг/кг антитела анти-Ang2 и 3 мг/кг или 10 мг/кг несвакумаба вводили мышам путем внутривенной инъекции дважды в неделю в течение 3 недель. После введения лекарства объем опухолей измеряли два раза в неделю в течение 20 дней. На протяжении времени измерения мыши не демонстрировали клинического токсического ответа. Группа, которой давали 10 мг/кг антитела анти-Ang2, продемонстрировала ингибирование роста опухоли приблизительно на 70%, что приблизительно в два раза превышает этот показатель для группы, получавшей 10 мг/кг несвакумаба. Объем опухоли рассчитали по следующему уравнению: Объем опухоли (TV) = ширина2 × длина × 0,52. Масса каждого животного одинаково увеличилась в среднем приблизительно на 5%, на основании чего подтвердили отсутствие особых токсических реакций (фиг. 12).
Ингибирование роста опухоли в ответ на введение антитела анти-Ang2 (мм3)
Изменение массы тела мышей (г) в зависимости от даты
Промышленная применимость
Антитело анти-Ang2 или его антигенсвязывающий фрагмент по настоящему раскрытию демонстрирует требуемую активность связывания с Ang2 и может эффективно использоваться в составе ингибиторов рака/опухоли или ангиогенеза и в ходе профилактики или лечения заболеваний, связанных с активацией и (или) избыточным производством ангиопоэтина-2. В соответствии с настоящим раскрытием, разработав терапевтические агенты, мишень которых отличается от мишеней существующих нацеленных на Ang2 терапевтических агентов, для лечения опухолей можно использовать как совместное лечение с существующими терапевтическими агентами, так и монотерапию.
Хотя специфические воплощения настоящего раскрытия были описаны подробно, специалисту в данной области техники будет очевидно, что эти подробные описания являются просто примерами и не ограничивают область настоящего раскрытия. Таким образом, основная область настоящего раскрытия будет определена в прилагаемой формуле изобретения и ее эквивалентах.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ФАРМАБЦИН ИНК.
<120> АНТИТЕЛО ПРОТИВ ANG2 И ЕГО ПРИМЕНЕНИЕ
<130> PP-B2372
<150> KR 19/021812
<151> 2019-02-25
<160> 61
<170> PatentIn version 3.5
<210> 1
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 1
Gly Phe Ser Phe Asp Asp Tyr Ala
1 5
<210> 2
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 2
Ile Lys Asp Asp Gly Ser Gln Thr
1 5
<210> 3
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 3
Thr Thr Glu Gly Leu Met Asn Gly Leu His Phe Asp Met
1 5 10
<210> 4
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 4
Ser Ser Asn Ile Gly Ala Gly Tyr Asp
1 5
<210> 5
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 5
Gly Asn Asn
1
<210> 6
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 6
Gln Ser Tyr Asp Ser Arg Leu Gly Val Val
1 5 10
<210> 7
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 7
Gly Tyr Ser Phe Thr Ser Tyr Trp
1 5
<210> 8
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 8
Ile Tyr Pro Gly Asn Ser Asp Thr
1 5
<210> 9
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 9
Thr Thr Glu Gly Leu Met Asn Gly Leu His Phe Asp Met
1 5 10
<210> 10
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 10
Gln Ser Leu Leu His Ser Leu Gly Asp Asn Tyr
1 5 10
<210> 11
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 11
Leu Gly Ser
1
<210> 12
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 12
Met Gln Ser Leu Gln Thr Pro Pro Tyr Thr
1 5 10
<210> 13
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 13
Gly Phe Thr Phe Ser Ser Tyr Ser
1 5
<210> 14
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 14
Ile Ser Ala Ser Asp Gly Ala Thr
1 5
<210> 15
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 15
Ala Lys Ile Leu Ala Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
1 5 10 15
<210> 16
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 16
Arg Asp Ile Ser Asn Tyr
1 5
<210> 17
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 17
Gly Ala Ser
1
<210> 18
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 18
Gln Gln Tyr Tyr Ser Tyr Pro Leu Thr
1 5
<210> 19
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 19
Gly Phe Ala Phe Gly Arg Tyr Glu
1 5
<210> 20
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 20
Ile Asp Thr Gly Gly Gly Ala Lys
1 5
<210> 21
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 21
Thr Thr Glu Gly Leu Met Asn Gly Leu His Phe Asp Met
1 5 10
<210> 22
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 22
Gln Ala Ile Ser Thr Trp
1 5
<210> 23
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 23
Thr Ala Ser
1
<210> 24
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 24
Gln Gln Leu Asn Ser Tyr Pro Tyr Thr
1 5
<210> 25
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 25
Gly Phe Thr Phe Asp Asp Cys Ala
1 5
<210> 26
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 26
Ile Ser Gly Asn Ser Lys Asn Val
1 5
<210> 27
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 27
Ala Arg Asp Pro Ala Tyr Ser Gln Phe Asp Tyr
1 5 10
<210> 28
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 28
Ser Ser Asn Val Gly Gly Tyr Pro
1 5
<210> 29
<211> 3
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 29
Thr Asp Tyr
1
<210> 30
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 30
Ala Thr Trp Asp Asp Asn Leu Asn Gly Tyr Val
1 5 10
<210> 31
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 31
cagatgcagc tggtgcagtc tgggggaggc ttggtacagc ctggcaggtc cctcaaactc 60
tcctgcgcag cctctggatt ctcctttgat gattatgcca tgcactgggt ccggcaagct 120
ccagggaagg ggctggagtg ggtggccacc ataaaggacg atggaagtca gacatactat 180
gtggactctg tgaagggccg attcaccatc tccagagaca acgccaagag ctcactgttt 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtac cacagaagga 300
ttaatgaatg gacttcattt tgatatgtgg ggccaaggga caatggtcac cgtctcctca 360
360
<210> 32
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 32
Gln Met Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Lys Asp Asp Gly Ser Gln Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Thr Glu Gly Leu Met Asn Gly Leu His Phe Asp Met Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 33
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 33
cagctcgtgc tgactcagcc gccctcagtg tctggggccc cagggcaggg ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtaccagcag 120
tttccaggaa cagcccccaa actcctcatc tctggtaaca ataatcggcc ctcagggctc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cactggactc 240
caggctgagg atgaggctga ttattactgc cagtcctatg acagcaggct gggtgtggtc 300
ttcggcggag ggaccaagct gaccgtccta ggt 333
<210> 34
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 34
Leu Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln Gly
1 5 10 15
Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr
20 25 30
Asp Val His Trp Tyr Gln Gln Phe Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Ser Gly Asn Asn Asn Arg Pro Ser Gly Leu Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Arg Leu
85 90 95
Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 35
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 35
caggtgcagc tggtggagtc tggagcagag gtgaaaagac ccggggagtc tctgaggatc 60
tcctgtaaga cttctggata cagctttacc agctactgga tccactgggt gcgccagatg 120
cccgggaaag aactggagtg gatggggagc atctatcctg ggaactctga taccagatac 180
agcccatcct tccaaggcca cgtcaccatc tcagccgaca gctccagcag caccgcctac 240
ctgcagtgga gcagcctgaa ggcctcggac accgccatgt attactgtac cacagaagga 300
ttaatgaatg gacttcattt tgatatgtgg ggccaaggga caatggtcac cgtctcctca 360
360
<210> 36
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 36
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Arg Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Thr Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile His Trp Val Arg Gln Met Pro Gly Lys Glu Leu Glu Trp Met
35 40 45
Gly Ser Ile Tyr Pro Gly Asn Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly His Val Thr Ile Ser Ala Asp Ser Ser Ser Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Thr Glu Gly Leu Met Asn Gly Leu His Phe Asp Met Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 37
<211> 342
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 37
gatattgtga tgacccagac tccactctcc ctgcccgtca cccctggaga gccggcctcc 60
atctcctgta ggtcaagtca gagcctcctg catagtcttg gagacaatta tttggattgg 120
tatctacaga agccagggca gtctccgcaa ctcctgatct atttgggttc taagcgggcc 180
gccggggtcc ccgacaggtt cagtggcagt ggctcaggca cagactttac actcaaaatc 240
agcagagtgg aggctgagga tgttggagtt tattattgca tgcaatctct acaaactccc 300
ccgtacactt ttggccaggg gaccaagctg gagatcaaac gt 342
<210> 38
<211> 114
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 38
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Leu Gly Asp Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Lys Arg Ala Ala Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ser
85 90 95
Leu Gln Thr Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg
<210> 39
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 39
caggtgcagc tggtagagtc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtgcta gtgatggtgc cacatactac 180
gcagactccg tgaggggccg gttcaccatc tccagagaca attccaggag cacactgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gaaaattctc 300
gcgggatata gtggcccaat gggcggaatg gacgtctggg gccaagggac cacggtcacc 360
gtctcctca 369
<210> 40
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 40
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ala Ser Asp Gly Ala Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ile Leu Ala Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 41
<211> 324
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 41
Gly Ala Cys Ala Thr Cys Cys Ala Gly Ala Thr Gly Ala Cys Cys Cys
1 5 10 15
Ala Gly Thr Cys Thr Cys Cys Ala Thr Cys Cys Thr Cys Ala Cys Thr
20 25 30
Gly Thr Cys Thr Gly Cys Ala Thr Cys Thr Gly Thr Ala Gly Gly Ala
35 40 45
Gly Ala Cys Ala Gly Ala Gly Thr Cys Ala Cys Cys Ala Thr Cys Ala
50 55 60
Cys Thr Thr Gly Thr Cys Gly Gly Gly Cys Gly Ala Gly Thr Cys Gly
65 70 75 80
Gly Gly Ala Cys Ala Thr Thr Ala Gly Cys Ala Ala Cys Thr Ala Thr
85 90 95
Thr Thr Ala Gly Cys Cys Thr Gly Gly Thr Ala Thr Cys Ala Gly Cys
100 105 110
Ala Gly Ala Ala Ala Cys Cys Ala Gly Gly Gly Ala Ala Ala Gly Cys
115 120 125
Cys Cys Cys Thr Ala Ala Gly Thr Cys Cys Cys Thr Gly Ala Thr Cys
130 135 140
Thr Ala Thr Gly Gly Ala Gly Cys Ala Thr Cys Cys Ala Ala Thr Thr
145 150 155 160
Thr Ala Cys Ala Ala Ala Gly Thr Gly Gly Gly Gly Thr Cys Thr Cys
165 170 175
Ala Thr Cys Ala Cys Ala Gly Thr Thr Cys Ala Gly Cys Gly Gly Cys
180 185 190
Ala Gly Thr Gly Gly Ala Thr Cys Cys Gly Gly Gly Ala Cys Ala Gly
195 200 205
Ala Thr Thr Thr Cys Ala Cys Cys Cys Thr Cys Ala Cys Cys Ala Thr
210 215 220
Cys Ala Ala Cys Ala Gly Cys Cys Thr Gly Cys Ala Gly Cys Cys Thr
225 230 235 240
Gly Ala Ala Gly Ala Thr Thr Cys Thr Gly Cys Ala Ala Cys Thr Thr
245 250 255
Ala Thr Thr Ala Cys Thr Gly Thr Cys Ala Ala Cys Ala Gly Thr Ala
260 265 270
Cys Thr Ala Thr Ala Gly Thr Thr Ala Cys Cys Cys Gly Cys Thr Cys
275 280 285
Ala Cys Thr Thr Thr Thr Gly Gly Cys Gly Gly Ala Gly Gly Gly Ala
290 295 300
Cys Cys Ala Ala Gly Gly Thr Gly Gly Ala Thr Ala Thr Cys Ala Ala
305 310 315 320
Ala Cys Gly Thr
<210> 42
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 42
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Arg Asp Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Gln Ser Gly Val Ser Ser Gln Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Ser Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 43
<211> 360
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 43
caggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggagggtc cctgagactc 60
tcctgtgaag cctctggatt cgccttcggt cgttatgaga tgaattgggt ccgccaggct 120
ccagggaagg ggctggagtg gattgcatat attgatactg gtggtggtgc caaagtctat 180
gcagactctg tgaagggccg attcaccatc tccagagacg acagcaaaaa ctccctgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtac cacagaagga 300
ttaatgaatg gacttcattt tgatatgtgg ggccaaggga caatgatcac cgtctcctca 360
360
<210> 44
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 44
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Glu Ala Ser Gly Phe Ala Phe Gly Arg Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Tyr Ile Asp Thr Gly Gly Gly Ala Lys Val Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Thr Glu Gly Leu Met Asn Gly Leu His Phe Asp Met Trp Gly Gln
100 105 110
Gly Thr Met Ile Thr Val Ser Ser
115 120
<210> 45
<211> 324
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 45
gacatccaga tgacccagtc tccttccacc ctgtctgcat ctgtaggaga cagagtcgcc 60
atcacttgcc gggccagtca ggctattagt acctggttgg cctggtatca gcagaaacct 120
ggtaaagccc ctaaactcct gatctatacg gcgtctactt tagaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag cctgcagcct 240
gatgattttg caacttatta ctgtcaacag cttaatagtt acccttacac tttcggcgga 300
gggaccaagg tggagatcaa acgt 324
<210> 46
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 46
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ala Ile Thr Cys Arg Ala Ser Gln Ala Ile Ser Thr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Thr Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105
<210> 47
<211> 354
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 47
caggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggcaggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttgat gattgtgcca tgcactgggt ccgacaagct 120
ccagggaagg gcctggagtg ggtctcaggt attagtggga atagtaaaaa cgtagcctat 180
gcggactctg tgaagggccg attcagcatc tccagagacg acgccaagaa ctccctgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gagagatccg 300
gcatacagcc agtttgacta ctggggccag ggaaccctga tcaccgtctc ctca 354
<210> 48
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 48
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Cys
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asn Ser Lys Asn Val Ala Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Ser Ile Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Pro Ala Tyr Ser Gln Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Ile Thr Val Ser Ser
115
<210> 49
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 49
cagtctgccc tgactcagcc tccctcactg tctgcgaccc ccgggcagag ggtcaccatc 60
tcttgctctg gaagcagctc caacgtcgga ggatatcctg tcaactggta ccagcaggtc 120
ccaggagcgg cccccaaact cctcatgtat actgattata agcggccctc aggtgtccct 180
gaccgattct ttggctccaa gtctggcact tcagcctccc tggccatcag tggcctccag 240
tctgaagatg aggctgatta ttactgtgct acatgggacg acaacctgaa tggctatgtc 300
ttcggaactg ggaccaaggt caccgtccta ggt 333
<210> 50
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 50
Gln Ser Ala Leu Thr Gln Pro Pro Ser Leu Ser Ala Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Val Gly Gly Tyr
20 25 30
Pro Val Asn Trp Tyr Gln Gln Val Pro Gly Ala Ala Pro Lys Leu Leu
35 40 45
Met Tyr Thr Asp Tyr Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Phe
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Asn Leu
85 90 95
Asn Gly Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 51
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 51
Ala Lys Thr Leu Ala Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
1 5 10 15
<210> 52
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 52
Ala Lys Ile Leu Val Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
1 5 10 15
<210> 53
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 53
Ala Lys Ser Leu Ala Ser Tyr Ser Gly Pro Met Gly Gly Met Asp Val
1 5 10 15
<210> 54
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 54
caggtgcagc tggtagagtc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtgcta gtgatggtgc cacatactac 180
gcagactccg tgaggggccg gttcaccatc tccagagaca attccaggag cacactgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gaaaactctc 300
gcgggatata gtggcccaat gggtggcatg gacgtctggg gccaagggac cacggtcacc 360
gtctcctca 369
<210> 55
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 55
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ala Ser Asp Gly Ala Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Thr Leu Ala Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 56
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 56
caggtgcagc tggtagagtc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtgcta gtgatggtgc cacatactac 180
gcagactccg tgaggggccg gttcaccatc tccagagaca attccaggag cacactgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gaaaatcctc 300
gtaggataca gtggcccaat gggcggaatg gacgtctggg gccaagggac cacggtcacc 360
gtctcctca 369
<210> 57
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 57
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ala Ser Asp Gly Ala Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ile Leu Val Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 58
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 58
caggtgcagc tggtagagtc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtgcta gtgatggtgc cacatactac 180
gcagactccg tgaggggccg gttcaccatc tccagagaca attccaggag cacactgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gaaaagcctt 300
gccagctata gtggcccaat gggcggcatg gacgtctggg gccaagggac cacggtcacc 360
gtctcctca 369
<210> 59
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 59
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ala Ser Asp Gly Ala Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ser Leu Ala Ser Tyr Ser Gly Pro Met Gly Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 60
<211> 369
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 60
caggtgcagc tggtagagtc tgggggaggc ctgttcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtgcta gtgatggtgc cacatactac 180
gcagactccg tgaggggccg gttcaccatc tccagagaca attccaggag cacactgtat 240
ctgcaaatga acagtctgag agccgaggac acggccgtgt attactgtgc gaaaactctc 300
gcgggatata gtggcccaat gggtggcatg gacgtctggg gccaagggac cacggtcacc 360
gtctcctca 369
<210> 61
<211> 123
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic Sequence
<400> 61
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ser Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ala Ser Asp Gly Ala Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Thr Leu Ala Gly Tyr Ser Gly Pro Met Gly Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ VISTA И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2750723C1 |
| АНТИТЕЛО ПРОТИВ TIE-2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2795455C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| АНТИТЕЛЬНЫЕ АГЕНТЫ, СПЕЦИФИЧНЫЕ К CD19 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773317C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| АНТИ-DR5-АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2735956C1 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
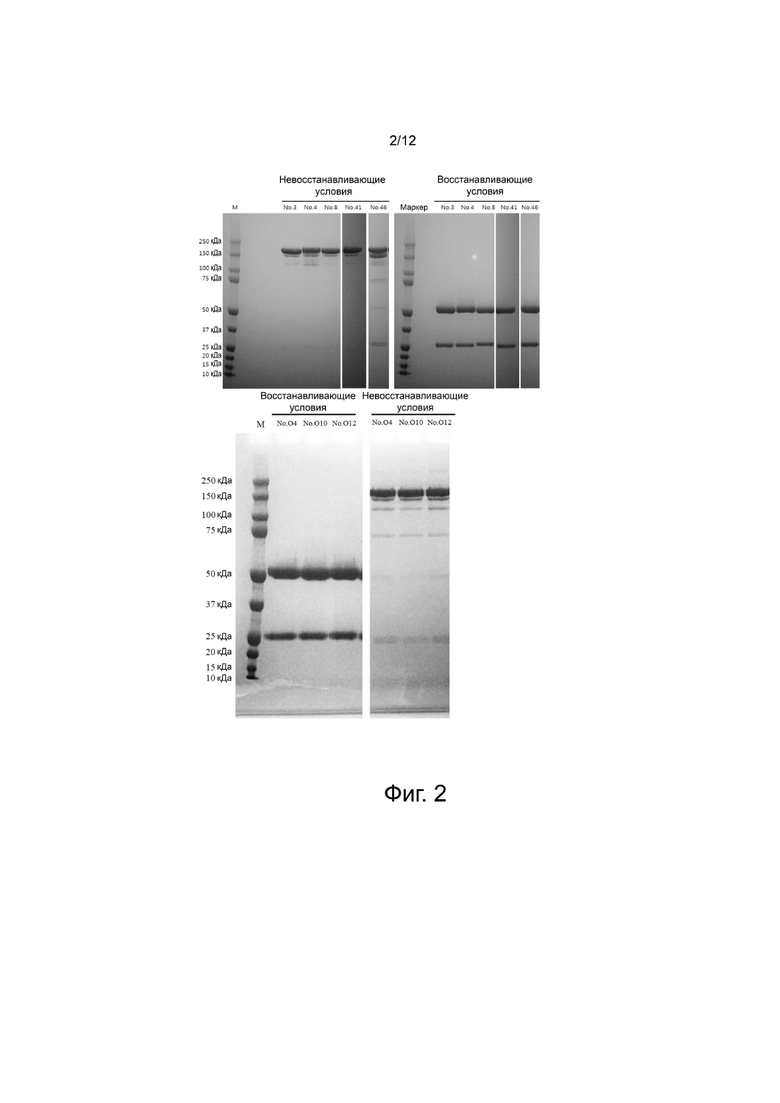

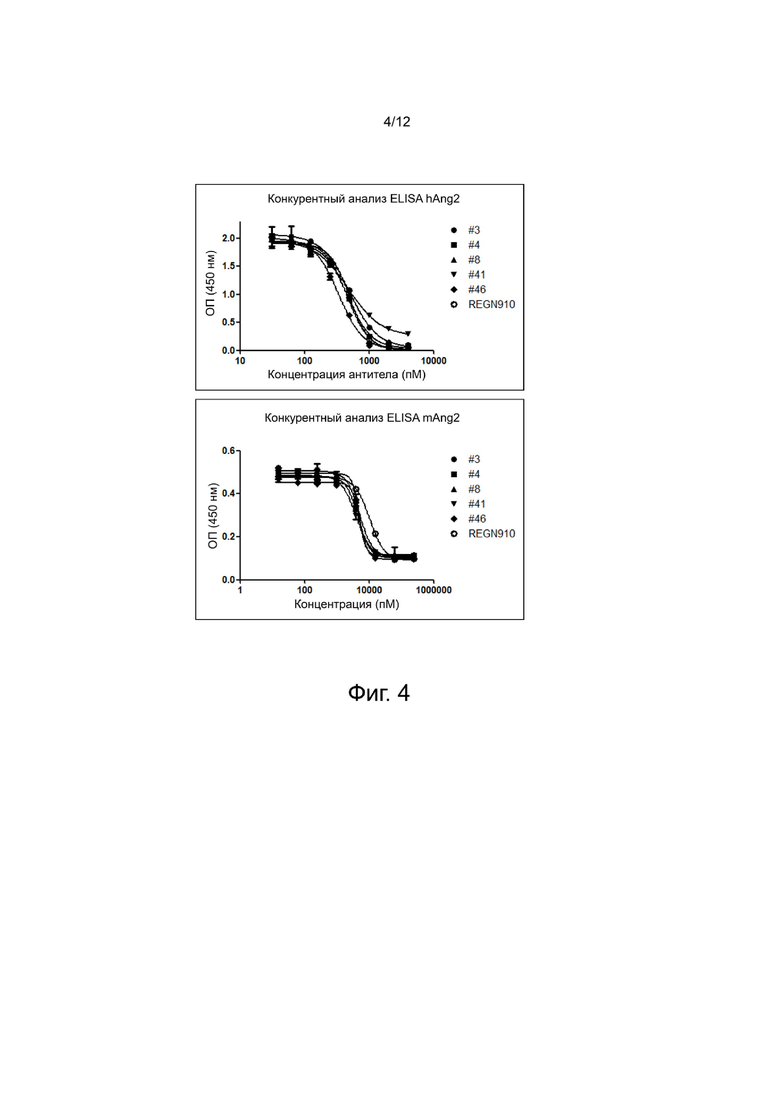
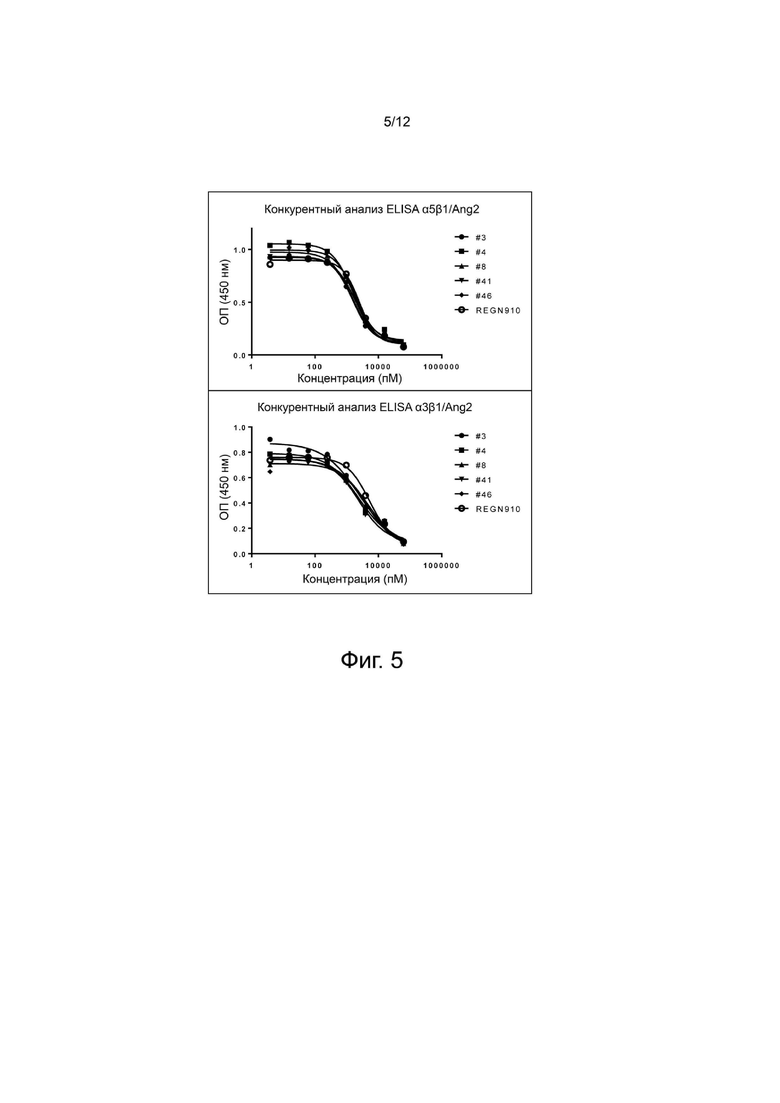
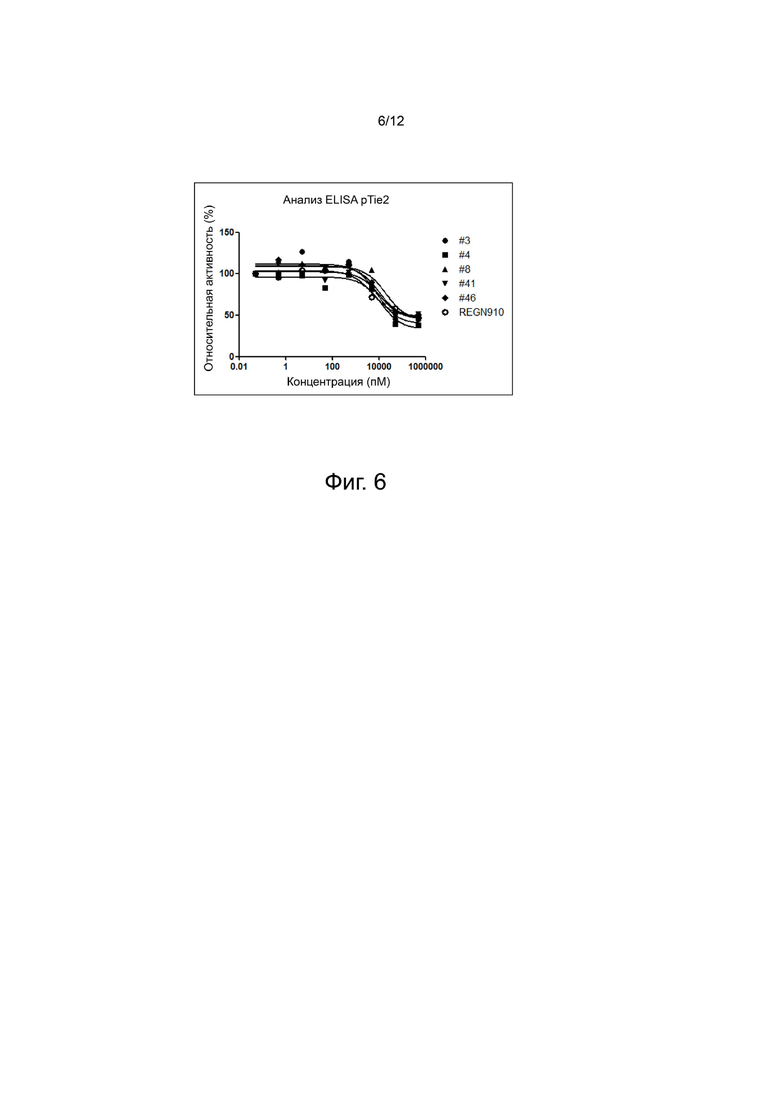
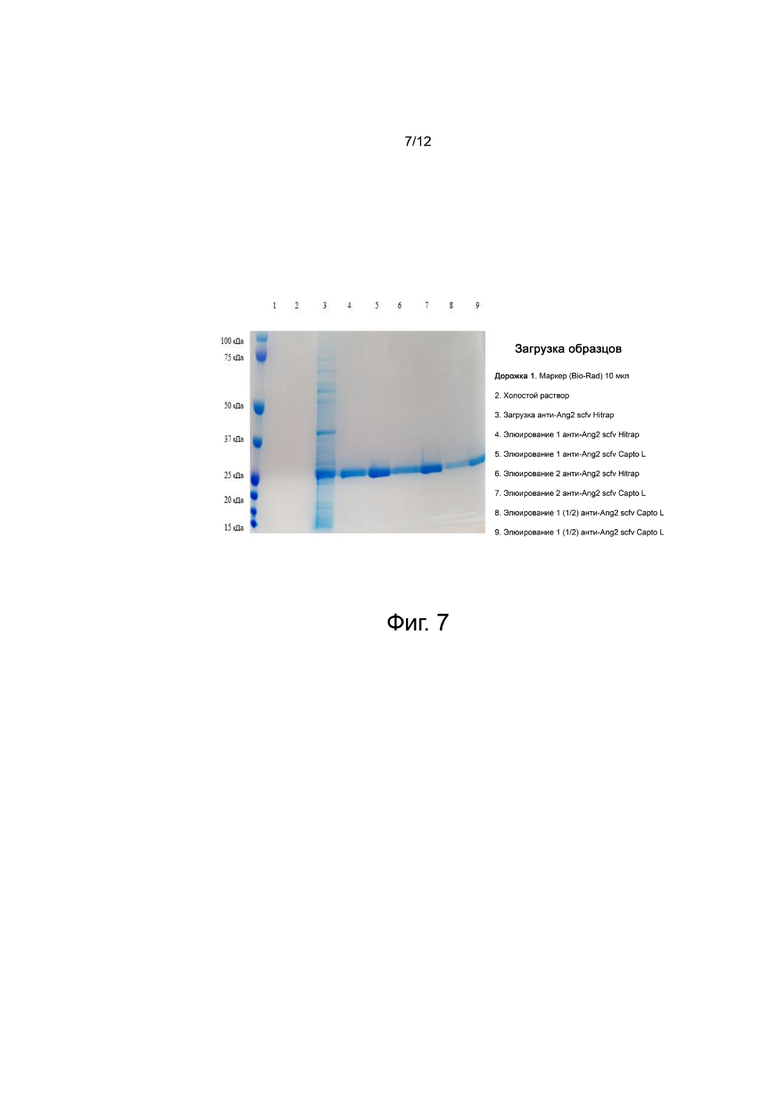

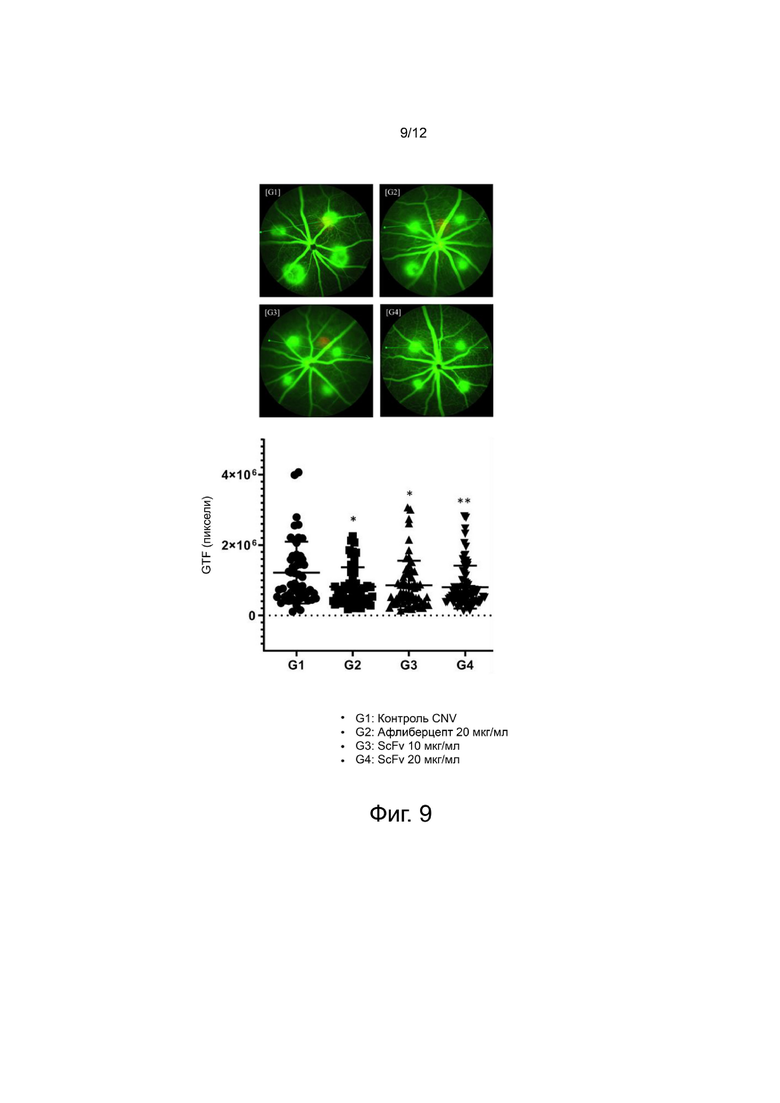
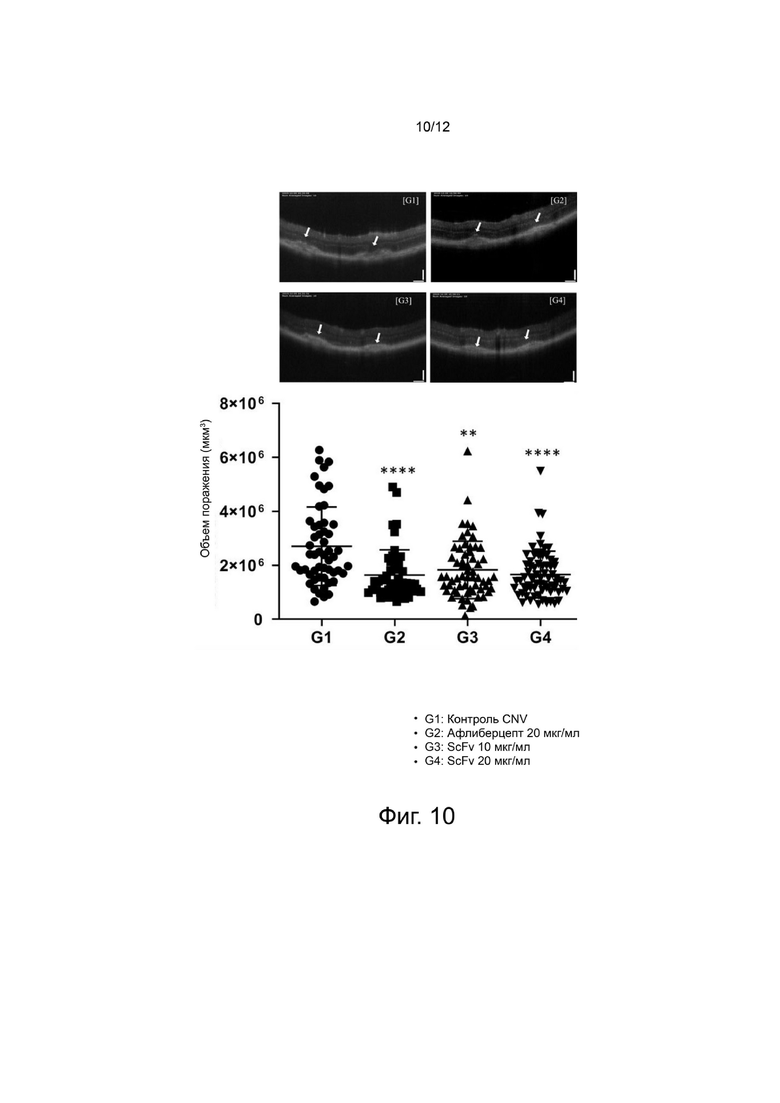
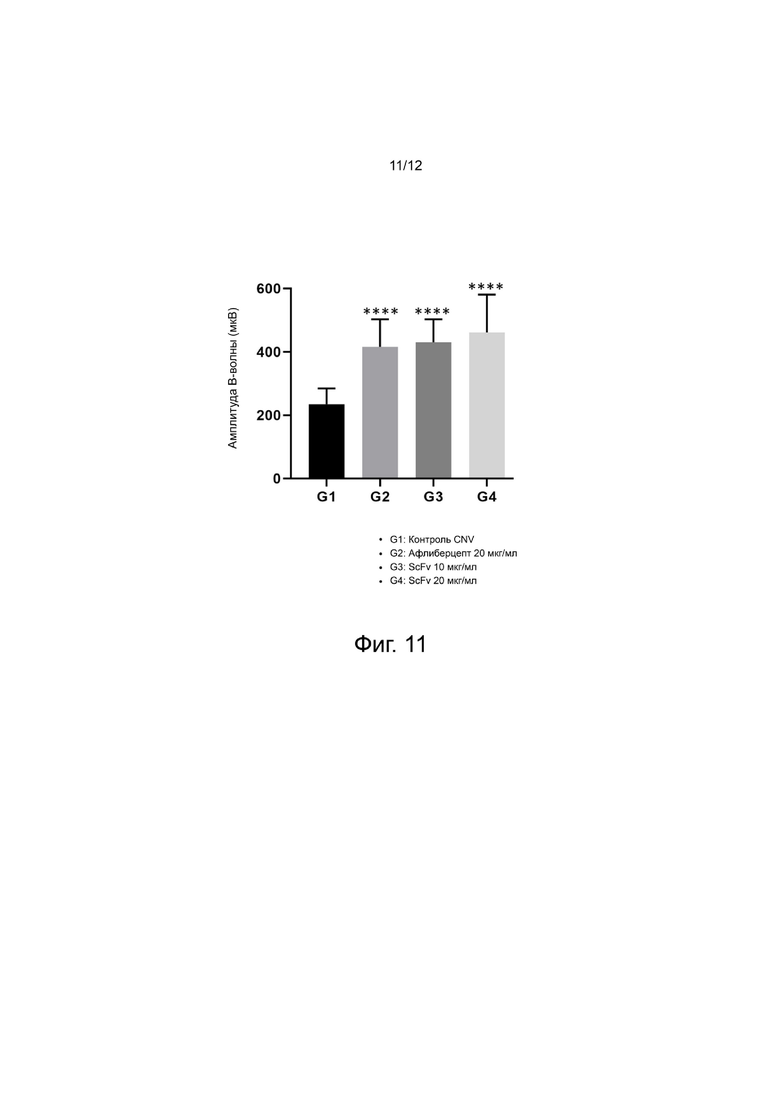
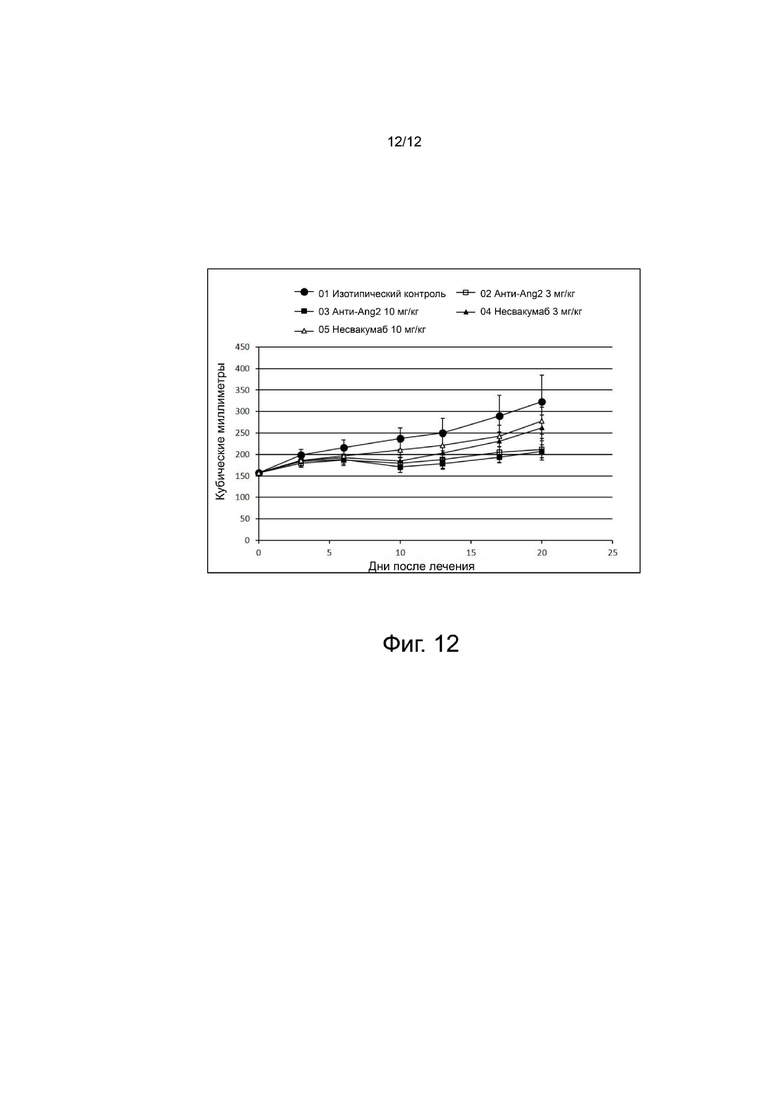
Изобретение относится к области биотехнологии. Предложено антитело, специфически связывающееся с ангиопоэтином-2 (Ang2), или его антигенсвязывающий фрагмент. Также предложены кодирующая это антитело нуклеиновая кислота, вектор экспрессии, содержащий нуклеиновую кислоту, клетка, трансформированная вектором, способ получения антитела. Кроме того, изобретение относится к вариантам композиции, включающей антитело по изобретению. Композиция может быть предназначена для профилактики или лечения заболевания, связанного с активацией и(или) избыточным производством Ang2, для диагностики указанного заболевания, для лечения заболеваний глаз, для профилактики или лечения онкологического заболевания, для объединения с другими терапевтическими препаратами. Изобретение обеспечивает ингибирование функции Ang2. 11 н. и 4 з.п. ф-лы, 12 ил., 14 табл., 13 пр.
1. Антитело, специфически связывающееся с ангиопоэтином-2 (Ang2), или его антигенсвязывающий фрагмент, содержащие:
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 1, CDR2 тяжелой цепи SEQ ID NO: 2 и CDR3 тяжелой цепи SEQ ID NO: 3, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 4, CDR2 легкой цепи SEQ ID NO: 5 и CDR3 легкой цепи SEQ ID NO: 6;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 7, CDR2 тяжелой цепи SEQ ID NO: 8 и CDR3 тяжелой цепи SEQ ID NO: 9, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 10, CDR2 легкой цепи SEQ ID NO: 11 и CDR3 легкой цепи SEQ ID NO: 12;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 15, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 19, CDR2 тяжелой цепи SEQ ID NO: 20 и CDR3 тяжелой цепи SEQ ID NO: 21, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 22, CDR2 легкой цепи SEQ ID NO: 23 и CDR3 легкой цепи SEQ ID NO: 24;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 25, CDR2 тяжелой цепи SEQ ID NO: 26 и CDR3 тяжелой цепи SEQ ID NO: 27, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 28, CDR2 легкой цепи SEQ ID NO: 29 и CDR3 легкой цепи SEQ ID NO: 30;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 51, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18;
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 52, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18; или
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи SEQ ID NO: 13, CDR2 тяжелой цепи SEQ ID NO: 14 и CDR3 тяжелой цепи SEQ ID NO: 53, а также вариабельную область легкой цепи, содержащую CDR1 легкой цепи SEQ ID NO: 16, CDR2 легкой цепи SEQ ID NO: 17 и CDR3 легкой цепи SEQ ID NO: 18.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 32, 36, 40, 44, 48, 55, 57, 59 и 61.
3. Антитело или его антигенсвязывающий фрагмент по п. 1, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 34, 38, 42, 46 и 50.
4. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3.
5. Нуклеиновая кислота по п. 4, где нуклеиновая кислота выбрана из группы, состоящей из SEQ ID NO: 31, 35, 39, 43, 47, 54, 56, 58 и 60, кодирующих вариабельную область тяжелой цепи.
6. Нуклеиновая кислота по п. 4, где нуклеиновая кислота выбрана из группы, состоящей из SEQ ID NO: 33, 37, 41, 45 и 49, кодирующих вариабельную область легкой цепи.
7. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 4.
8. Клетка для продуцирования антитела по любому из пп. 1-3, трансформированная вектором экспрессии по п. 7.
9. Способ производства антитела, связывающегося с Ang2, или его антигенсвязывающего фрагмента, причем способ содержит следующие процессы:
(а) культивирование клеток по п. 8, и
(b) восстановление антитела или его антигенсвязывающего фрагмента из культивируемых клеток.
10. Композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 в качестве действующего вещества для профилактики или лечения заболевания, связанного с активацией и(или) избыточным производством ангиопоэтина-2.
11. Композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 в качестве действующего вещества для ингибирования ангиогенеза.
12. Композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 в качестве действующего вещества для диагностики заболевания, связанного с активацией и(или) избыточным производством ангиопоэтина-2.
13. Композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 в качестве действующего вещества для профилактики или лечения заболеваний глаз.
14. Композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 в качестве действующего вещества для профилактики или лечения опухоли или рака.
15. Композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 для объединения с другими терапевтическими препаратами.
| WO 2013112438 A1, 01.08.2013 | |||
| WO 2015171747 A1, 12.11.2015 | |||
| СТАБИЛИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ АНГИОПОЭТИНА-2 И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2509085C2 |
| BROWN J.L | |||
| et al., A Human Monoclonal Anti-ANG2 Antibody Leads to Broad Antitumor Activity in Combination with VEGF Inhibitors and Chemotherapy Agents in Preclinical Models, Mol Cancer Ther, 2010, vol.9 (1), pp.145-156 | |||
| RENNEL E.S | |||
| et al., A Human Neutralizing | |||

