Релевантная информация
По настоящей заявке испрашивается приоритет US 61/133212, поданной 25 июня 2008 года, US 61/075697, поданной 25 июня 2008 года, US 61/155041, поданной 24 февраля 2009 года и US 61/075692, поданной 25 июня 2008 года.
Содержания любых патентов, заявок на патент и ссылок, цитируемых в этом описании, включены в настоящее описание в качестве ссылки в полном объеме.
Уровень техники
Считается, что ангиогенез участвует в патогенезе различных нарушений, включая солидные опухоли, внутриглазные неоваскулярные синдромы, такие как пролиферативные ретинопатии или возрастная дегенерация желтого пятна (AMD), ревматоидный артрит и псориаз (Folkman et al. J. Biol. Chem. 267:10931-10934 (1992); Klagsbrun et al. Annu. Rev. Physiol. 53:217-239 (1991); и Garner A, Vascular diseases. In: Pathobiology of ocular disease. A dynamic approach. Garner A, Klinrworth G K, Eds. 2nd Edition Marcel Dekker, NY, pp 1625-1710 (1994)). При солидных опухолях, ангиогенез и рост новой сосудистой сети обеспечивает выживание опухоли, и между плотностью микрососудов в срезах опухолей и выживанием пациентов в случае рака молочной железы и других раковых опухолей была продемонстрирована четкая корреляция (Weidner et al. N Engl J Med 324:1-6 (1991); Horak et al. Lancet 340:1120-1124 (1992); и Macchiarini et al. Lancet 340: 145-146 (1992)).
Эндотелиальный фактор роста сосудов (VEGF) является известным регуляторным фактором ангиогенеза и неоваскуляризации, и было показано, что он является ключевым медиатором неоваскуляризации, связанной с опухолями и внутриглазными нарушениями (Ferrara et al. Endocr. Rev. 18:4-25 (1997)). мРНК VEGF сверхэкспрессируется во многих опухолях человека, и концентрация VEGF в глазных жидкостях в высокой степени коррелируют с наличием активной пролиферации кровеносных сосудов у пациентов с диабетической ретинопатией и другими связанными с ишемией ретинопатиями. (Berkman et al, J Clin Invest 91:153-159 (1993); Brown et al. Human Pathol. 26:86-91 (1995); Brown et al. Cancer Res. 53:4727-4735 (1993); Mattern et al. Brit. J. Cancer. 73:931-934 (1996); и Dvorak et al. Am J. Pathol. 146:1029-1039 (1995); Aiello et al. N. Engl. J. Med. 331:1480-1487 (1994)). Кроме того, недавние исследования показали наличие локализованного VEGF в хороидальных неоваскулярных мембранах у пациентов, пораженных AMD (Lopez et al. Invest. Ophtalmo. Vis. Sci. 37:855-868 (1996)). Нейтрализующие антитела против VEGF могут быть использованы для подавления роста различных линий опухолевых клеток человека у «голых» мышей, а также ингибирования внутриглазного ангиогенеза на моделях ишемических ретинальных нарушений (Kim et al. Nature 362:841-844 (1993); Warren et al. J. Clin. Invest 95:1789-1797 (1995); Borgstrom et al. Cancer Res. 56:4032-4039 (1996); и Melnyk et al. Cancer Res. 56:921-924 (1996)) (Adamis et al. Arch. Opthalmol. 114:66-71 (1996)).
Таким образом, существует потребность в моноклональных антителах против VEGF, которые могут быть использованы для лечения солидных опухолей и различных неоваскулярных внутриглазных заболеваний.
Сущность изобретения
Настоящее изобретение относится к растворимым и стабильным анти-VEGF-иммуносвязывающим агентам, содержащим CDR моноклональных антител кролика. Указанные антитела предназначены для диагностики и/или лечения VEGF-опосредованных нарушений. Также описаны гибридомы, нуклеиновые кислоты, векторы и клетки-хозяева для экспрессии рекомбинантных антител по настоящему изобретению, способы их выделения и использование указанных антител в медицине.
Краткое описание фигур
Фиг.1 иллюстрирует кинетику связывания выбранных scFv с hVEGF165 с использованием Biacore (hVEGF165). Фиг.1а показывает данные, полученные для 511max: Ka (1/Ms): 6,59E+05; SE (ka): 1,10E+03; kd(1/s):4,40E-05; SE(kd):6,30E-07; KD(M):6,67E-11. Фиг.1b показывает данные, полученные для 578max: Ka (1/Ms): 7,00E+05; SE (ka): 1,40E+03; kd(1/s): 3,07E-04; SE(kd): 8,50E-07; KD(M): 4,39E-10.
Фиг.2 иллюстрирует видоспецифичность, показывая кинетику связывания 578max с VEGF человека, мыши и крысы. Фиг.2а показывает данные, полученные для VEGF165 человека: Ka (1/Ms): 7,00E+05; SE (ka): 1,40E+03; kd(1/s): 3,07E-04; SE(kd): 8,50E-07; KD(M): 4,39E-10. Фиг.2b показывает данные, полученные для VEGF164 мыши: Ka (1/Ms): 1,03E+06; SE (ka): 2,30E+03; kd(1/s): 4,40E-04; SE(kd): 9,40E-07; KD(M): 4,29E-10. Фиг.2c показывает данные, полученные для VEGF164 крысы: Ka (1/Ms): 8,83E+05; SE (ka): 2,50E+03; kd(1/s): 5,28E-04; SE(kd): 1,20E-06; KD(M): 5,98E-10.
Фиг.3 иллюстрирует кинетику связывания 578max с VEGF-изоформами (hVEGF121 и hVEGF110). Фиг.3a показывает данные, полученные для VEGF165 человека: Ka (1/Ms): 7,00E+05; SE (ka): 1,4E+03; kd(1/s): 3,07E-04; SE(kd): 8,50E-07; KD(M): 4,39E-10. Фиг.3b показывает данные, полученные для VEGF121 человека: Ka (1/Ms): 5,87E+05; SE (ka): 1,20E+03; kd(1/s): 5,58E-04; SE(kd): 9,60E-07; KD(M): 9,50E-11. Фиг. 3c показывает данные, полученные для VEGF110 человека: Ka (1/Ms): 5,23E+05; SE (ka): 1,30E+03; kd(1/s): 7,22E-04; SE(kd): 8,10E-07; KD(M): 1,38E-09.
Фиг.4 изображает кинетику связывания 578max, 578minmax и 578wt с hVEGF165. Фиг.4а показывает данные, полученные для 578max: Ka (1/Ms): 7,00E+05; SE (ka): 1,40E+03; kd(1/s): 3,07E-04; SE(kd): 8,50E-07; KD(M): 4,39E-10. Фиг.4b показывает данные, полученные для 578minmax: Ka (1/Ms): 8,06E+05; SE (ka): 2,10E+03; kd(1/s): 5,04E-04; SE(kd): 1,10E-06; KD(M): 6,25E-10. Фиг.4c показывает данные, полученные для 578wt-His: Ka (1/Ms): 8,45E+05; SE (ka): 1,60E+03; kd(1/s): 1,69E-04; SE(kd): 7,60E-07; KD(M): 2,00E-10.
Фиг.5 иллюстрирует термическую стабильность 578max, 578minmax и 578minmax_DHP (разворачивание, измеренное посредством FT-IR). Фиг.5а: 578minmax (ESBA903): Tm=71,1°C; Фиг.5b: 578minmax_DHP (#961): Tm=70,2°C; Фиг.5c: 578max (#821): Tm=70,4°C.
Фиг.6 иллюстрирует денатурацию и осаждение производных 578 после теплового стресса (фиг.6а: 50°C, фиг.6b: 60°C, фиг.6c: 70°C) в течение 30 минут.
Фиг.7 иллюстрирует растворимость 578max, 578minmax и 578minmax_DHP (определяемую по осаждению сульфатом аммония). Фиг.7a: 578max (#821). V50 был равен 27,24%. Фиг.7b: 578minmax (ESBA903). V50 был равен 28,13. Фиг.7c: 578minmax_DHP (#961). V50 был равен 32,36%.
Фиг.8 иллюстрирует конкурентный ELISA VEGFR2 относительно HUVEC-анализа в качестве способов для измерения эффективности. Фиг.8a: Сравнение Lucentis и 511max (#802) в конкурентном ELISA VEGFR2. R2 Lucentis: 0,9417; R2 ESBA802: 0,9700. EC50 Lucentis: 7,137 нМ; EC50 #802: 0,8221 нМ. Фиг.8b: Сравнение Lucentis и 578max (#821) в конкурентном ELISA VEGFR2. Фиг.8c: Сравнение Lucentis, 511maxC-his и 534max в HUVEC-анализе. R2 Lucentis 0,9399; R2 EP511maxC-his: 0,9313, R2 EP534max: 0,7391. EC50 Lucentis: 0,08825 нМ, EC50 511maxC-his: 0,7646 нМ, EC50 534max: 63,49 нМ. Фиг.8d: Сравнение Lucentis, 578min и 578max в HUVEC-анализе. R2 Lucentis: 0,9419, R2 EP578min: 0,8886, R2 EP578max: 0,9274. EC50 Lucentis: 0,1529 нМ, EC50 578min: 1,528 нМ, EC50 578max: 0,1031 нМ.
Фиг.9 иллюстрирует действия 578minmax на пролиферацию HUVEC, индуцируемую hVEGF165. Параметры этого анализа были следующими: 0,08 нМ (3 нг/мл); инкубирование с VEGF и тест-изделием: 96 часов. EC50 была равна 0,08959 нМ для Lucentis и 0,05516 нМ для 578minmax, в то время как R2 был равен 0,9066 для Lucentis и 0,9622 для 578minmax.
Фиг.10 иллюстрирует действия 578minmax на пролиферации HUVEC, индуцируемые VEGF164 мыши и VEGF164 крысы. Параметры этого анализа были следующими: концентрация VEGF164 мыши: 0,08 нМ (3 нг/мл); концентрация VEGF164 крысы: 0,3 нМ (11,3 нг/мл). Обе концентрации выбирали при EC90 для VEGF-индуцированной пролиферации HUVEC; инкубирование с VEGF и тест-изделием: 96 часов. Фиг.10a иллюстрирует данные, полученные для VEGF мыши. EC50 была равна 0,1196 нМ для V1253 и 0,06309 нМ для 578minmax, в то время как R2 был равен 0,02744 для Lucentis, 0,9348 для V1253 и 0,9767 для EP578minmax. Lucentis не ингибировал пролиферацию HUVEC, индуцированную VEGF мыши. Фиг.10b иллюстрирует данные, полученные для VEGF крысы. EC50 была равна 1,597 нМ для V1253 и 0,06974 нМ для 578minmax, в то время как R2 был равен 0,7664 для V1253 и 0,6635 для 578minmax.
Фиг.11 иллюстрирует исследования эффективности с использованием анализа Miles в голых морских свинках (часть I). Голым морским свинкам вводили внутривенно краситель альмаровый синий. Спустя один час после инъекции красителя премикс 2 hVEGF (2,61 нМ) и Lucentis, ESBA903 или #802, соответственно, инъецировали в кожу животного 3. Спустя один час после инъекции этих растворов животных 3 эвтаназировали и шкурки собирали, очищали и фотографировали цифровым способом с использованием падающего света и рассеянного света. Площадь красителя Evans Blue, который просачивался в места инъекции, оценивали с использованием изображения J и строили график зависимости площади удерживания от дозы.
Фиг.12 иллюстрирует исследования эффективности с использованием анализа Miles в голых морских свинках (часть II). Фиг.12a показывает результаты, полученные для #803 (511max). EC50 была равна 5,990 нМ и имела статистический разброс между 2,060 и 17,41 нМ, тогда как R2 был равен 0,5800. Фиг.12b показывает результаты, полученные для ESBA903 (578minmax). EC50 был равен 3,989 и имел статистический разброс между 1,456 и 10,93 нМ, тогда как R2 был равен 0,3920. Фиг.12c показывает зону просачивания красителя для Lucentis. EC50 для Lucentis не могла быть рассчитана вследствие плохой подгонки кривой.
Фиг.13 иллюстрирует исследования эффективности с использованием модифицированного анализа Miles в крысах (предварительно смешанные hVEGF165 и 578minmax (ESBA903)). Фиг.13а иллюстрирует понижающую проницаемость эффективность Авастина на VEGF-индуцированном ретинальном васкулярном просачивании в крысах – доза-ответ. Авастин ингибирует hVEGF-индуцируемую ретинальную васкулярную проницаемость. Предварительное смешивание перед инъекцией. Приблизительно эквимолярные, 3-кратный или 10-кратный избыток. *p<0,05 (VEGF vs. BSA), **p<0,05 (Авастин-обработанные vs. VEGF). Фиг.13b показывает понижающую проницаемость эффективность ESBA903 после VEGF-индуцируемого ретинального васкулярного просачивания в крысах. Доза-ответ (предварительно смешанные, ivt). Полное ингибирование hVEGF-индуцируемой ретинальной васкулярной проницаемости ESBA903. Предварительное смешивание перед инъекцией. Приблизительно эквимолярные, 3-кратный или 10-кратный избыток. *p<0,05 (VEGF s. BSA), **p<0,05 (ESBA903-обработанные vs. VEGF).
Фиг.14 иллюстрирует исследования эффективности с использованием модифицированного анализа Miles в крысах (местное введение 578minmax (ESBA903)). Понижающую проницаемость AL-51287 (ESBA903) после VEGF-индуцированного ретинального васкулярного просачивания в крысах испытывали после местного введения. За пять дней перед обработкой, 4 капли/день с 10 нг/мл готовой формы ESBA903. *p<0,05 (VEGF vs. BSA), **p<0,05 (VEGF vs. AL-51287), ***p=0,060 (AL-51287 vs. AL-52667), ****(VEGF vs. AL-39324); p<0,05 (AL-39324 vs. носителя-ссылочного контроля). AL-51287: ESBA903; AL-52657: местный носитель-ссылочный контроль; AL-39324: ингибитор-малая молекула RTK.
Фиг.15 иллюстрирует определение CDR1 VH, используемое в настоящем описании.
Подробное описание
Изобретение относится к растворимым и стабильным анти-VEGF-связывающим агентам, содержащим CDR из моноклональных антител кролика. Указанные иммуносвязывающие агенты предназначены для диагностики и/или лечения VEGF-опосредованных нарушений. Также описаны гибридомы, нуклеиновые кислоты, векторы и клетки-хозяева для экспрессии рекомбинантных антител по настоящему изобретению, способы их выделения и использование указанных антител в медицине.
Определения
Для облегчения понимания настоящего изобретения некоторые термины будут определены следующим образом. Дополнительные определения представлены во всем подробном описании.
Термин "VEGF" относится к состоящему из 165 аминокислот эндотелиальному фактору роста сосудов и соответствующим состоящим из 121 аминокислоты, 189 аминокислот и 206 аминокислот эндотелиальным факторам роста сосудов, описанным Leung et al, Science 246:1306 (1989) и Houck et al, Mol. Endocrin. 5:1806 (1991), а также к природным аллельным и процессированным формам этих факторов роста.
Термин "VEGF-рецептор" или "VEGFr" относится к клеточному рецептору VEGF, обычно рецептору клеточной поверхности, обнаруживаемому на эндотелиальных клетках сосудов, а также его вариантам, которые сохраняют способность связываться с hVEGF. Одним из примеров VEGF-рецептора является fms-подобная тирозинкиназа (flt), трансмембранный рецептор в семействе тирозинкиназ. DeVries et al, Science 255:989 (1992); Shibuya et al, Oncogene 5:519 (1990). Рецептор flt содержит внеклеточный домен, трансмембранный домен и внутриклеточный домен с тирозинкиназной активностью. Внеклеточный домен участвует в связывании с VEGF, тогда как внутриклеточный домен участвует в трансдукции сигнала. Другим примером VEGF-рецептора является рецептор flk-1 (также называемый KDR). Matthews et al, Proc. Nat. Acad. Sci. 88:9026 (1991); Terman et al, Oncogene 6:1677 (1991); Terman et al, Biochem. Biophys. Res. Commun. 187:1579 (1992). Связывание VEGF с рецептором flt приводит к образованию по меньшей мере двух высокомолекулярных комплексов со средней молекулярной массой 205000 и 300000 Дальтон. Считается, что комплекс 300000 Дальтон является димером, содержащим две молекулы рецептора, связанные с единственной молекулой VEGF.
Термин "кролик” относится в контексте настоящей заявки к животному, принадлежащему семейству заячьих.
Термин "антитело" является в контексте настоящей заявки синонимом для "иммуноглобулина". Антитела по настоящему изобретению могут быть полноразмерными иммуноглобулинами или их фрагментами, содержащими по меньшей мере один вариабельный домен иммуноглобулина, такими как отдельные вариабельные домены, Fv (Skerra A. and Pluckthun, A. (1988) Science 240:1038-41), scFv (Bird, R.E. et al (1988) Science 242:423-26; Huston, J.S. et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-83), Fab, F(ab’)2 или другие фрагменты, хорошо известные специалисту в данной области.
Термин "CDR" относится к одному из шести гипервариабельных районов в вариабельных доменах антитела, которые в основном способствуют связыванию антитела с антигеном. Одно из наиболее часто используемых определений для этих шести CDR представлено в статье Kabat E.A. et al, (1991) Sequences of proteins of immunological interest. NIH Publication 91-3242). В контексте настоящей заявки, определение CDR Кабата относятся только к CDR1, CDR2 и CDR3 вариабельного домена легкой цепи (CDR L1, CDR L2, CDR L3 или L1, L2, L3), а также к CDR2 и CDR3 вариабельного домена тяжелой цепи (CDR H2, CDR H3 или H2, H3). Однако CDR1 тяжелой цепи (CDR H1 или H1) определяется в данном случае следующими остатками (согласно нумерации Кабата): он начинается с положения 26 и заканчивается перед положением 36. Он является в основном слиянием CDR H1, по-разному определяемым Кабатом и Хотиа (см. также фиг.15 для иллюстрации).
Термин "каркас антитела", или иногда только "каркас", относится в данном случае к части вариабельного домена, либо VL, либо VH, которая служит в качестве каркаса для антигенсвязывающих петель (CDR) вариабельного домена. В сущности, он является вариабельным доменом без CDR.
Термин "одноцепочечное антитело", "одноцепочечный Fv" или "scFv" относится к молекуле, содержащей вариабельный домен тяжелой цепи антитела (или район; VН) и вариабельный домен легкой цепи (или район; VL), связанные линкером. Такие молекулы scFv могут иметь общие структуры: NH2-VL-линкер-VH-COOH или NH2-VH-линкер-VL-COOH.
В контексте настоящей заявки, "идентичность" обозначает совпадение последовательности двух полипептидов, молекул или двух нуклеиновых кислот. Если какое-либо положение в обеих из этих двух сравниваемых последовательностей занято одним и тем же основанием или мономерной аминокислотной субъединицей (например, если какое-либо положение в каждой из двух молекул ДНК занято аденином или какое-либо положение в каждом из двух полипептидов занято лизином), то соответствующие молекулы являются идентичными в этом положении. "Процентная идентичность" двух последовательностей является функцией числа совпадающих положений, являющихся общими для этих двух последовательностей, деленного на число сравниваемых положений х 100. Например, если совпадают 6 из 10 положений в двух последовательностях, то эти последовательности имеют 60% идентичность. В качестве примера, DNA-последовательности CTGACT и CAGGTT имеют 50% идентичность (совпадают 3 из 6 общих положений). Обычно сравнение делается при сопоставлении двух последовательностей для получения максимальной идентичности. Такое сопоставление может быть обеспечено с использованием, например, способа Needleman et al. (1970) J. Mol. Biol. 48: 443-453, осуществляемого предпочтительно компьютерными программами, такими как программа Align (DNAstar, Inc.). Процентная идентичность двух аминокислотных последовательностей может быть также определена с использованием алгоритма E. Meyers и W. Miller (Comput. Appl. Biosci., 4:1 1-17 (1988)), который включен в программу ALIGN (версию 2.0), с использованием таблицы PAM120 взвешенных остатков, штрафа за удлинение 12 и штрафа за пропуск 4. Кроме того, процентная идентичность между двумя аминокислотными последовательностями может быть определена с использованием алгоритма Needleman и Wunsch (J. Mol. Biol. 48:444-453 (1970)), который был включен в программу GAP в пакете программ GCG.software (доступном в www.gcg.com), с использованием либо матрицы Blossum 62, либо матрицы PAM250 и величины пропуска 16, 14, 12, 10, 8, 6 или 4 и величины удлинения 1, 2, 3, 4, 5 или 6.
"Сходными" последовательностями являются последовательности, которые при сравнении имеют идентичные и сходные аминокислотные остатки, где сходные остатки являются консервативными заменами соответствующих аминокислотных остатков при выравнивании ссылочной последовательности. В этом отношении, "консервативная замена" остатка в эталонной последовательности является заменой остатком, который является физически или функционально сходным с соответствующим ссылочным остатком, например, который имеет сходные размер, форму, электрический заряд, химические свойства, в том числе способность образовывать ковалентные или водородные связи, или т.п. Таким образом, “модифицированная консервативной заменой” последовательность является последовательностью, которая отличается от эталонной последовательности или последовательности дикого типа тем, что присутствуют одна или несколько консервативных замен. “Процентное сходство” двух последовательностей является функцией числа положений, которые содержат совпадающие остатки или консервативные замены, общие у этих двух последовательностей, разделенного на число сравниваемых положений, х 100. Например, если 6 из 10 положений в двух последовательностях совпадают и 2 из 10 положений содержат консервативные замены, то эти две последовательности имеют 80% положительное сходство.
В контексте настоящей заявки, термин "модификации консервативными последовательностями" обозначает аминокислотные модификации, которые не оказывают отрицательного влияния на свойства связывания или не изменяют свойства связывания антитела, содержащего эту аминокислотную последовательность. Такие модификации с консервативными заменами включают нуклеотидные и аминокислотные замены, добавления и делеции. Например, модификации могут быть введены стандартными способами, известными в данной области, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены включают в себя замены, в которых аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в данной области. Эти семейства включают в себя аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновую кислоту, глутаминовую кислоту), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, прогнозируемый не являющийся незаменимым аминокислотный остаток в антителе против VEGF человека, предпочтительно заменяют другим аминокислотным остатком того же самого семейства боковых цепей. Способы идентификации нуклеотидных и аминокислотных консервативных замен, которые не влияют на связывание с антигеном, хорошо известны в данной области (см., например, Brummell et al, Biochem. 32:1180-1187 (1993); Kobayashi et al. Protein Eng. 12(10):879-884 (1999) и Burks et al. Proc. Natl. Acad. Sci. USA 94:412-417 (1997)).
"Аминокислотной консенсусной последовательностью" называют в контексте настоящей заявки аминокислотную последовательность, которая может быть получена с использованием матрицы из по меньшей мере двух, и предпочтительно, более, выравненных аминокислотных последовательностей и допускает пропуски в этом сопоставлении, так чтобы можно было определить наиболее частый аминокислотный остаток в каждом положении. Консенсусной последовательностью является последовательность, которая содержит аминокислоты, которые наиболее часто присутствуют в каждом положении. В случае, когда две или более аминокислоты представлены одинаково в одном положении, эта консенсусная последовательность включает в себя обе или все эти аминокислоты.
Аминокислотную последовательность белка можно анализировать на различных уровнях. Например, консервативность или вариабельность может проявляться на уровне единственного остатка, на уровне множественных остатков, множественных остатков с пропусками и тому подобное. Остатки могут проявлять консервативность идентичного остатка или могут быть консервативными на уровне класса. Примеры классов аминокислот включают полярные, но не заряженные R-группы (серин, треонин, аспарагин и глутамин); положительно заряженные R-группы (лизин, аргинин и гистидин); отрицательно заряженные R-группы (глутаминовая кислота и аспарагиновая кислота); гидрофобные R-группы (аланин, изолейцин, лейцин, метионин, фенилаланин, триптофан, валин и тирозин) и особые аминокислоты (цистеин, глицин и пролин). Специалисту в данной области известны другие классы, и они могут быть определены с использованием структурных определений или других данных для оценки заменяемости. В этом смысле, заменяемой аминокислотой может быть названа любая аминокислота, которая может быть заменена и может сохранять функциональную консервативность в этом положении.
Однако следует учитывать, что аминокислоты в одном и том же классе могут применяться по своим биофизическим свойствам. Например, понятно, что некоторые гидрофобные R-группы (например, аланин, серин или треонин) являются более гидрофильными (т.е. имеют более высокую гидрофильность или более низкую гидрофобность), чем другие гидрофобные R-группы (например, валин или лейцин). Относительная гидрофильность или гидрофобность может быть определена общепринятыми в данной области способами (см., например, Rose et al, Science, 229: 834-838 (1985) и Cornette et al., J. Mol Biol, 195: 659-685 (1987)).
В контексте настоящей заявки, при сравнении одной аминокислотной последовательности (например, первой последовательности VH или VL) с одной или несколькими дополнительными аминокислотными последовательностями (например, одной или несколькими VH или VL в базе данных), положение аминокислоты в одной последовательности (например, первой последовательности VH или VL) может сравниваться с “соответствующим положением” в одной или нескольких дополнительных аминокислотных последовательностях. В контексте настоящей заявки, "соответствующее положение" представляет эквивалентное положение в последовательности(ях), сравниваемых при оптимальном сопоставлении этих последовательностей, т.е. при сравнении этих последовательностей с достижением максимальной процентной идентичности или высочайшего процентного сходства.
В контексте настоящей заявки, термин "база данных антител" относится к коллекции двух или более аминокислотных последовательностей антител ("множеству" последовательностей) и обычно относится к коллекции десятков, сотен или даже тысяч аминокислотных последовательностей. База данных последовательностей может хранить аминокислотные последовательности, например, коллекции VH-районов антител, VL-районов антител или обоих или может хранить коллекцию scFv-последовательностей, содержащих VH- и VL-районы. Предпочтительно, эта база данных хранит в доступной для поиска, фиксированной среде, например, в компьютере в доступной для поиска компьютерной программе. В одном из вариантов осуществления, база данных антител является базой данных, содержащей последовательности антител эмбрионального типа или состоящей из последовательностей антител эмбрионального типа. В другом варианте осуществления, база данных антител является базой данных, содержащей зрелые (т.е. экспрессируемые) последовательности антител или состоящей из зрелых (т.е. экспрессируемых) последовательностей антител (например, базой данных Кабата зрелых последовательностей антител, например, базой данных KBD). Еще в одном из вариантов осуществления, эта база данных содержит функционально выбранные последовательности или состоит из функционально выбранных последовательностей (например, последовательностей, выбранных на основе QC-анализа).
Термин "иммуносвязывающий агент" относится к молекуле, которая содержит весь антигенсвязывающий сайт или часть антигенсвязывающего сайта антитела, например, весь вариабельный домен тяжелой цепи и/или легкой цепи или часть вариабельного домена тяжелой цепи и/или легкой цепи, так что этот иммуносвязывающий агент связывает антиген-мишень. Не ограничивающие примеры иммуносвязывающих агентов включают полноразмерные молекулы иммуноглобулинов и одноцепочечные scFv, а также фрагменты антител, включающие, но не ограничивающиеся ими, (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CHI; (ii) F(ab’)2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанные дисульфидным мостиком в шарнирной области; (iii) Fab’-фрагмент, который по существу является Fab с частью шарнирного района (см. Fundamental Immunology (Paul ed., 3.sup.rd ed. 1993); (iv) Fd-фрагмент, состоящий из доменов VH и CН1; (v) Fv-фрагмент, состоящий из доменов VL и VH единственного плеча антитела, (vi) однодоменное антитело, такое как Dab-фрагмент (Ward et al, (1989) Nature 341:544-546), который состоит из домена VH или VL, верблюда (см. Hamers-Casterman, et al, Nature 363:446-448 (1993) и Dumoulin, et al, Protein Science 11:500-515 (2002)) или антитело акулы (например, нанотела Ig-NAR акулы, Nanobodies®); и (vii) нанотело, район тяжелой цепи, содержащий вариабельный домен и два константных домена.
В контексте настоящей заявки, термин "функциональное свойство" является свойством полипептида (например, иммуносвязывающего агента), в отношении которого является желательным и/или выгодным для специалиста в данной области улучшение (например, относительно общепринятого полипептида), например, для улучшения свойств приготовления или терапевтической эффективности этого полипептида. В одном из вариантов осуществления, этим функциональным свойством является стабильность (например, термостабильность). В другом варианте осуществления, этим функциональным свойством является растворимость (например, при клеточных условиях). Еще в одном из вариантов осуществления, этим функциональным свойством является отсутствие агрегации. Еще в одном из вариантов осуществления, этим функциональным свойством является экспрессия белка (например, в прокариотической клетке). Еще в одном из вариантов осуществления, этим функциональным свойством является эффективность рефолдинга после солюбилизации тельца включения в соответствующем процессе очистки. В некоторых вариантах осуществления, антигенсвязывающая активность не является функциональным свойством, которое желательно улучшить.
Термин "эпитоп" или "антигенная детерминанта" относится к сайту на антигене (например, на VEGF), с которым специфически связывается иммуноглобулин или антитело. Эпитоп обычно включает по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 последовательных или непоследовательных аминокислот в уникальной пространственной конформации. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, G. E. Morris, Ed. (1996).
Термины "специфическое связывание", "селективное связывание", "селективно связывается" и "специфически связывается" относятся к связыванию антитела с эпитопом на предварительно определенном антигене. Обычно, это антитело связывается с аффинностью (KD), приблизительно меньшей, чем 10-7 M, например, меньшей, чем приблизительно 10-8M, 10-9M или 10-10M или даже более низкой.
Термин "KD" или "Kd" относится к константе диссоциации в равновесном состоянии конкретного взаимодействия антитело-антиген. Обычно антитела по настоящему изобретению связываются с VEGF с константой диссоциации равновесного состояния (KD), меньшей, чем приблизительно 10-7M, например, меньшей, чем приблизительно 10-8M, 10-9M или 10-10M, или даже более низкой, например, определенной технологией резонанса поверхностных плазмонов (SPR) в приборе BIACORE.
Термины "нейтрализует VEGF", "ингибирует VEGF" и "блокирует VEGF" используются взаимозаменяемо для обозначения способности антитела по настоящему изобретению препятствовать взаимодействию антитела по настоящему изобретению с одним или несколькими VEGF-рецепторами, такими как VEGFR-1 и/или VEGFR-2, и, например, запуску трансдукции сигнала.
"Рекомбинантный иммуносвязывающий агент" относится в контексте настоящей заявки к иммуносвязывающему агенту, продуцируемому экспрессией из рекомбинантной ДНК.
"Химерный" иммуносвязывающий агент в контексте настоящей заявки имеет часть тяжелой цепи и/или легкой цепи, идентичную соответствующей последовательности или гомологичную соответствующим последовательностям антитела, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, тогда как остальная часть цепи (цепей) является идентичной с соответствующими последовательностями или гомологичной соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител, а также фрагменты таких антител. Используемое в данном случае гуманизированное антитело является субпопуляцией химерных антител.
"Гуманизированные антитела" обозначают в контексте настоящей заявки иммуносвязывающие агенты, которые были синтезированы с использованием технологии рекомбинантных ДНК для исключения иммунной реакции на чужеродные антигены. Гуманизация является хорошо известным способом уменьшения иммуногенности моноклональных антител, полученных из ксеногенных источников. Она включает выбор акцепторного каркаса, предпочтительно, акцепторного каркаса человека, степени CDR из донорского иммуносвязывающего агента, подлежащей встраиванию в этот акцепторный каркас и замены остатков из донорского каркаса в акцепторный каркас. Общий способ трансплантации CDR в акцепторные каркасы человека был описан Winter в патенте США 5225539, содержание которого включено в настоящее описание в качестве ссылки в полном объеме. В US 6407213, содержание которого включено в настоящее описание в качестве ссылки в полном объеме, раскрыт ряд аминокислотных положений этого каркаса, где замена из донорского иммуносвязывающего агента является предпочтительной.
Термин "молекулы нуклеиновой кислоты" относится к молекулам ДНК и молекулам РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но, предпочтительно, она является двухцепочечной ДНК. Нуклеиновая кислота является "функционально связанной", если она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности.
Термин "вектор" относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. Одним типом вектора является "плазмида", которая является кольцевой двухцепочечной ДНК-петлей, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации и эписомальные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть введены в геном клетки-хозяина и посредством этого реплицируются вместе с геномом хозяина.
Термин "клетка-хозяин" относится к клетке, в которую был введен экспрессирующий вектор. Клетки-хозяева могут включать в себя бактериальные, микробные клетки, клетки растений или клетки животных. Бактерии, которые восприимчивы к трансформации, включают в себя члены семейства Enterobacteriaceae, такие как штаммы Escherichia coli или Salmonella; Bacillaceae, такие как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Подходящие микроорганизмы включают в себя Saccharomyces cerevisiae и Pichia pastoris. Подходящие линии клеток-хозяев животных включают в себя CHO (линии яичника китайского хомячка) и клетки NS0.
Термины "лечить", "лечение" или "терапия" относятся к терапевтическим или профилактическим мерам, описанным в настоящем описании. Способы "терапии" предусматривают введение больному антитела по настоящему изобретению, например, больному с VEGF-опосредованным нарушением, или больному, который в конечном счете может приобрести такое нарушение, для профилактики, лечения, задержки, уменьшения тяжести или ослабления одного или нескольких симптомов этого нарушения или рецидива нарушения или для продления выживания индивида за пределы выживания, которые ожидались в отсутствие такого лечения.
Термин "VEGF-опосредованное нарушение" относится к любому нарушению, возникновению, прогрессированию или хроническим симптомам или состояниям заболевания, которое требует участия VEGF. Примеры VEGF-опосредованных нарушений включают, но не ограничиваются ими, возрастную дегенерацию желтого пятна, неоваскулярную глаукому, диабетическую ретинопатию, ретинопатию недоношенных, ретролентальную фиброплазию, карциномы молочной железы, карциномы легких, желудочные карциномы, эзофагеальные карциномы, колоректальные карциномы, карциномы печени, карциномы яичника, комы, арренобластомы, карциномы шейки матки, эндометриальную карциному, эндометриальную гиперплазию, эндометриоз, фибросаркомы, хориокарциному, рак головы и шеи, назофарингеальную карциному, карциномы гортани, гепатобластому, саркому Капоши, меланому, кожные карциномы, гемангиомы, кавернозную гемангиому, гемангиобластому, карциномы поджелудочной железы, ретинобластому, астроцитому, глиобластому, Шванному, олигодендроглиому, медуллобластому, нейробластомы, рабдомиосаркому, остеогенную саркому, лейомиосаркомы, карциномы мочеполовых путей, карциномы щитовидной железы, опухоль Вильмса, почечно-клеточную карциному, рак предстательной железы, аномальную пролиферацию сосудов, ассоциированную с факоматозом, эдему (например, ассоциированную с опухолями головного мозга), синдром Мейжа, ревматоидный артрит, псориаз и атеросклероз.
Термин "эффективная доза" или "эффективная дозировка" относится к количеству, достаточному для достижения или по меньшей мере для частичного достижения желаемого эффекта. Термин "терапевтически эффективная доза" определяется как количество, достаточное для излечения или по меньшей мере задержки этого заболевания и его осложнений в пациенте, уже страдающем от этого заболевания. Количества, эффективные для этого применения, будут зависеть от тяжести подлежащего лечению нарушения и общего состояния собственной иммунной системы пациента.
Термин "индивид" относится к любому человеку или животному (не человеку). Например, способы и композиции по настоящему изобретению могут быть использованы для лечения индивида с VEGF-опосредованным нарушением.
Термин "Min-трансплантат" или "min" относится в контексте настоящей заявки к гуманизированному вариабельному домену, который был получен трансплантацией кроличьих CDR из кроличьего вариабельного домена в природный акцепторный каркас человека (FW 1.4, SEQ ID NO: 172). В этих каркасных районах не проводят никаких изменений. Сам каркас предварительно выбран с точки зрения желаемых функциональных свойств (растворимости и стабильности).
Термин "Max-трансплантат" или "max" относится в контексте настоящей заявки к гуманизированному вариабельному домену, который был получен трансплантацией кроличьих CDRs из кроличьего вариабельного домена в "модифицированный консервативными кроличьими остатками” акцепторный каркас человека "RabTor" (rFW1.4, SEQ ID NO: 173), или в его производное, называемое rFW1.4(v2) (SEQ ID NO: 174). Этот каркас "RabTor" получали включением консервативных кроличьих остатков (или же остатков, которые являются довольно вариабельными в других видах) в положениях каркаса, обычно участвующих в структуре и стабильности вариабельного домена кролика, с целью генерирования универсально применимого каркаса, который принимает фактически любой набор кроличьих CDR без необходимости трансплантации остатков донорского каркаса, других, чем остатки в положениях, которые являются различными в их предполагаемой последовательности-предшественнике, например, которые были изменены во время соматической гипермутации и, следовательно, возможно, влияют на связывание антигена. Определено, что эта предполагаемая последовательность-предшественник является самой близкой копией зародышевой линии кролика, и в случае, когда эта самая близкая копия зародышевой линии не может быть установлена, консенсусом кроличьей подгруппы или консенсусом последовательностей кролика с высоким процентным сходством.
Термин "Min-Max" или "minmax" относится в контексте настоящей заявки к гуманизированному вариабельному домену, содержащему вариабельную легкую цепь, соединенную с вариабельной тяжелой цепью "Max-graft".
Термин "Max-Min" или "maxmin" относится в контексте настоящей заявки к гуманизированному вариабельному домену, содержащему вариабельную легкую цепь "Max-graft", соединенную с вариабельной тяжелой цепью "Min-graft".
Для этих полученных иммуносвязывающих агентов использовали различные номенклатуры. Они обычно идентифицируются по номеру (например, №578). В случаях, когда использовался префикс, такой как ЕР или Epi (например, ЕР №578, который идентичен Epi 578), посредством этого указывался тот же самый иммуносвязывающий агент. Иногда иммуносвязывающий агент получал второе обозначение, которое идентифицируется префиксом "ESBA". Например, ESBA903 обозначает тот же самый иммуносвязывающий агент, что и 578minmax или EP578minmax или Epi578minmax.
Если нет другого определения, все технические и научные термины, используемые в настоящем описании, имеют значение, обычно понимаемое специалистом в данной области, к которой относится это изобретение. Хотя в практике или испытании настоящего изобретения могут использоваться способы и материалы, сходные с описанными или эквивалентные описанным в настоящем описании, ниже описаны подходящие способы и материалы. В случае противоречия, эталоном будет являться настоящее описание, включающее в себя определения. Кроме того, эти материалы, способы и примеры являются только иллюстративными и не предназначены для ограничения.
Различные аспекты по настоящему изобретению описаны более подробно в следующих подразделах. Понятно, что различные варианты, преференции и диапазоны могут произвольно комбинироваться. Кроме того, в зависимости от конкретного варианта осуществления, выбранные определения, варианты или диапазоны могут не применяться.
Анти-VEGF-иммуносвязывающие агенты
В одном из аспектов настоящее изобретение относится к иммуносвязывающим агентам, которые связывают VEGF и, следовательно, блокируют функцию VEGF in vivo. CDR этих иммуносвязывающих агентов происходят из кроличьих моноклональных антител против VEGF, которые получены у кроликов, иммунизированых VEGF человека и/или его фрагментом (SEQ ID NO: 1). Насколько известно авторам по настоящему изобретению, впервые моноклональные антитела против VEGF получены у кроликов и подробно охарактеризованы. Неожиданно, было обнаружено, что аффинность (Kd) была чрезвычайно высокой.
В некоторых вариантах осуществления, изобретение относится к иммуносвязывающему агенту, который специфически связывается с VEGF, содержащему последовательность по меньшей мере одного из CDRH1, CDRH2, CDRH3, CDRL1, CDRL2 или CDRL3. Примеры аминокислотных последовательностей CDR, которые можно использовать в иммуносвязывающих агентах по настоящему изобретению, представлены в SEQ ID NО: 2-72 (таблицы 1-6).
Таблица 1
Аминокислотные последовательности CDR Н1 анти-VEGF-иммуносвязывающих агентов по настоящему изобретению
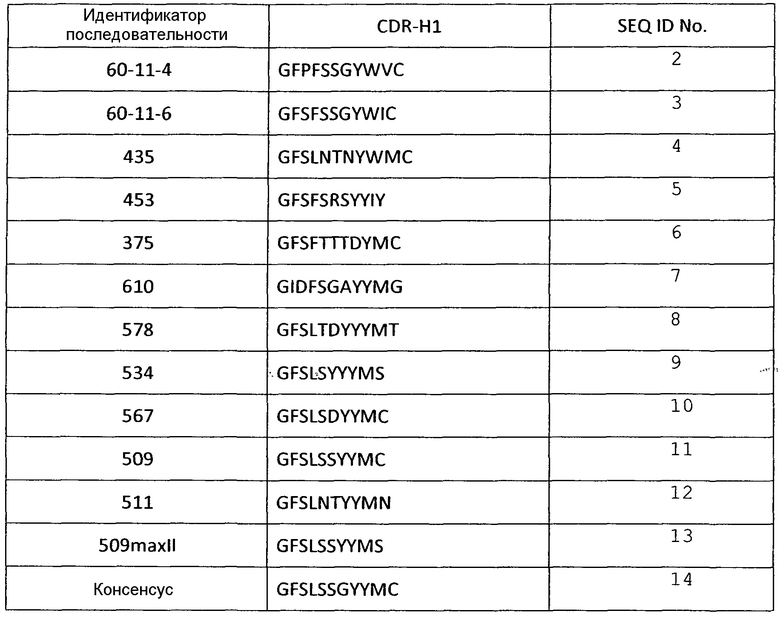
Таблица 2
Аминокислотные последовательности CDR Н2 анти-VEGF-иммуносвязывающих агентов по настоящему изобретению
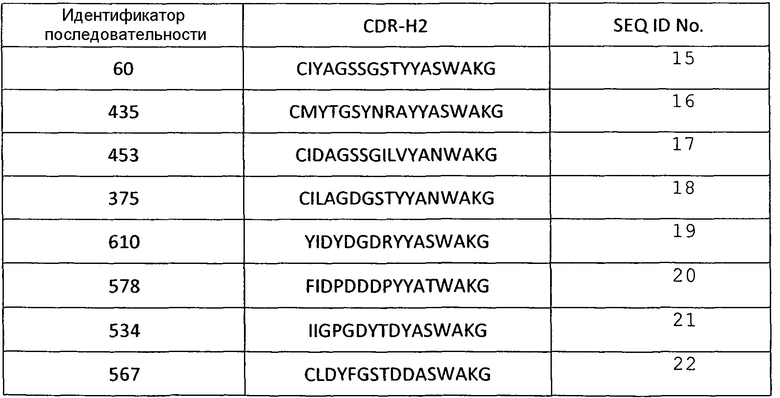

Таблица 3
Аминокислотные последовательности CDR Н3 анти-VEGF-иммуносвязывающих агентов по настоящему изобретению
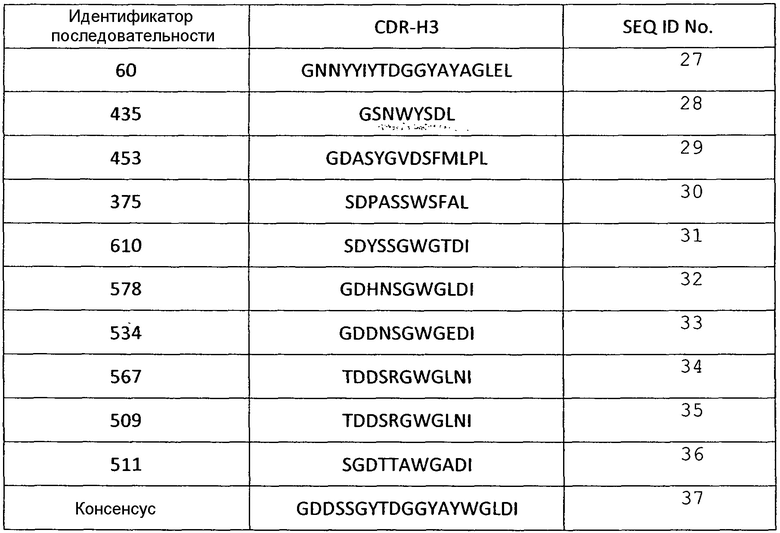
Таблица 4
Аминокислотные последовательности CDR L1 анти-VEGF-иммуносвязывающих агентов по настоящему изобретению
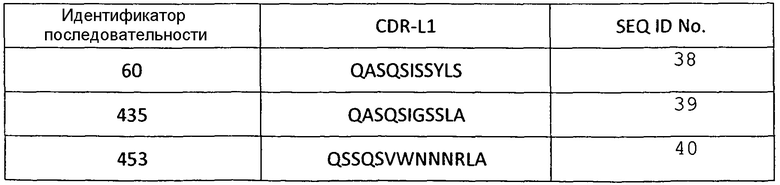
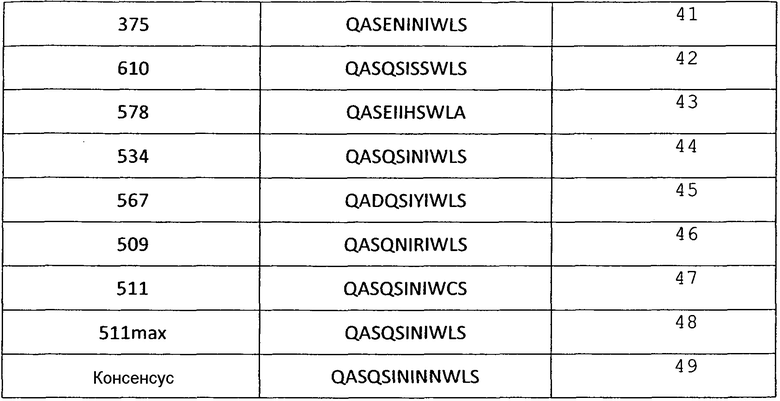
Таблица 5
Аминокислотные последовательности CDR L2 анти-VEGF-иммуносвязывающих агентов по настоящему изобретению
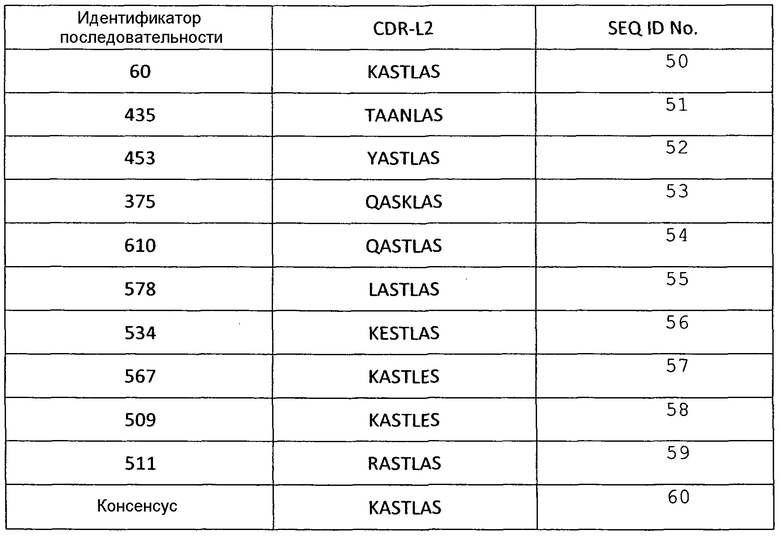
Таблица 6
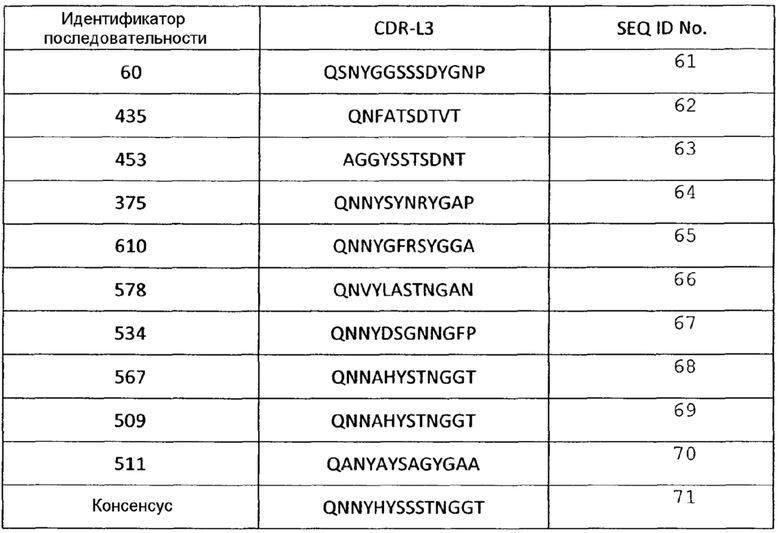
Аминокислотные последовательности CDR L3 анти-VEGF-иммуносвязывающих агентов по настоящему изобретению
В одном из вариантов осуществления изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющий по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с консенсусной последовательностью из группы, состоящей из SEQ ID NO: 14, SEQ ID NO: 26, SEQ ID NO: 37, SEQ ID NO: 49, SEQ ID NO: 60 и SEQ ID NO: 71. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 14, SEQ ID NO: 26 и SEQ ID NO: 37, и/или CDR VL указанного иммуносвязывающего агента содержит CDR из группы, состоящей из SEQ ID NO: 49, SEQ ID NO: 60 и SEQ ID NO: 71. Предпочтительно, CDR выбран из группы, состоящей из SEQ ID NO: 2 - SEQ ID NO: 13, SEQ ID NO: 15 - SEQ ID NO: 25, SEQ ID NO: 27 - SEQ ID NO: 36, SEQ ID NO: 38 - SEQ ID NO: 48, SEQ ID NO: 50 - SEQ ID NO: 59 и SEQ ID NO: 61 - SEQ ID NO: 70.
В другом варианте осуществления, настоящее изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с консенсусной последовательностью из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 15, SEQ ID NO: 27, SEQ ID NO: 38, SEQ ID NO: 50 и SEQ ID NO: 61. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 15 и SEQ ID NO: 27 и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 50 и SEQ ID NO: 61. В другом предпочтительном варианте осуществления, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 15 и SEQ ID NO: 27, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 38, SEQ ID NO: 50 и SEQ ID NO: 61.
В другом варианте осуществления, настоящее изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 16, SEQ ID NO: 28, SEQ ID NO: 39, SEQ ID NO: 51 и SEQ ID NO: 62. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 4, SEQ ID NO: 16, SEQ ID NO: 28, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 39, SEQ ID NO: 51 и SEQ ID NO: 62.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 5, SEQ ID NO: 17, SEQ ID NO: 29, SEQ ID NO: 40, SEQ ID NO: 52 и SEQ ID NO: 63. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 5, SEQ ID NO: 17, SEQ ID NO: 29, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 40, SEQ ID NO: 52 и SEQ ID NO: 63.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичноcть с последовательностью из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 18, SEQ ID NO: 30, SEQ ID NO: 41, SEQ ID NO: 53 и SEQ ID NO: 64. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 6, SEQ ID NO: 18 и SEQ ID NO: 30, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 41, SEQ ID NO: 53 и SEQ ID NO: 64.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 19, SEQ ID NO: 31 , SEQ ID NO: 42, SEQ ID NO: 54 и SEQ ID NO: 65. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 19 и SEQ ID NO: 31, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 42, SEQ ID NO: 54 и SEQ ID NO: 65.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 8, SEQ ID NO: 20, SEQ ID NO: 32, SEQ ID NO: 43, SEQ ID NO: 55 и SEQ ID NO: 66. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 8, SEQ ID NO: 20 и SEQ ID NO: 32, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 55 и SEQ ID NO: 66.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 9, SEQ ID NO: 21, SEQ ID NO: 33, SEQ ID NO: 44, SEQ ID NO: 56 и SEQ ID NO: 67. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 9, SEQ ID NO: 21 и SEQ ID NO: 33, и/или CDR VL иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 44, SEQ ID NO: 56 и SEQ ID NO: 67.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 10, SEQ ID NO: 22, SEQ ID NO: 34, SEQ ID NO: 45, SEQ ID NO: 57 и SEQ ID NO: 68. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 10, SEQ ID NO: 22 и SEQ ID NO: 34, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 45, SEQ ID NO: 57 и SEQ ID NO: 68.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 11, SEQ ID NO: 13, SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 35, SEQ ID NO: 46, SEQ ID NO: 58 и SEQ ID NO: 69. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 11, SEQ ID NO: 23 и SEQ ID NO: 35, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 46, SEQ ID NO: 58 и SEQ ID NO: 69. Альтернативно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 13, SEQ ID NO: 25 и SEQ ID NO: 35, и/или CDR VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 46, SEQ ID NO: 58 и SEQ ID NO: 69.
В другом варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему по меньшей мере один CDR, имеющему по меньшей мере 75% сходство, предпочтительно, по меньшей мере 75% идентичность, более предпочтительно, по меньшей мере 80%, 85%, 90% 95%, еще более предпочтительно, 100% идентичность с последовательностью из группы, состоящей из SEQ ID NO: 12, SEQ ID NO: 24, SEQ ID NO: 36, SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 59, и SEQ ID NO: 70. Предпочтительно, VH указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 12, SEQ ID NO: 24 и SEQ ID NO: 36. Дополнительно или альтернативно, VL указанного иммуносвязывающего агента содержат CDR из группы, состоящей из SEQ ID NO: 47, SEQ ID NO: 48, SEQ ID NO: 59 и SEQ ID NO: 70, например, SEQ ID NO: 47, SEQ ID NO: 59 и SEQ ID NO: 70; или SEQ ID NO: 48, SEQ ID NO: 59 и SEQ ID NO: 70.
В еще более предпочтительном варианте осуществления, описанный в настоящем описании иммуносвязывающий агент нейтрализует VEGF человека и перекрестно реагирует с крысиным/мышиным VEGF или его частью.
Иммуносвязывающий агент может содержать антитело или любой альтернативный связывающий каркас, способный вмещать CDR. CDR, представленные как SEQ ID NО: 2-72, могут быть трансплантированы в любой подходящий связывающий каркас известными в данной области способами (см., например, Riechmann, L. et al. (1998) Nature 332:323-327; Jones, P. et al. (1986) Nature 321:522-525; Queen, C. et al. (1989) Proc. Natl. Acad. See. U.S.A. 86:10029-10033; патент США 5225539 Winter и патенты США 5530101; 5585089; 5693762 и 6180370, Queen et al.). Однако предпочтительно описанные в настоящем описании иммуносвязывающие агенты являются гуманизированными и, следовательно, подходят для терапевтического использования.
В случае антител, кроличьи CDR, представленные как SEQ ID NО: 2-72, могут быть трансплантированы в каркасные районы любого антитела любого вида. Однако ранее было показано, что антитела или производные антител, содержащие каркасы, идентифицированные в так называемом скрининге "контроля качества" (WO 0148017), характеризуются обычно высокой стабильностью и/или растворимостью и, следовательно, могут также использоваться в контексте внеклеточных применений, таких как нейтрализация VEGF человека. Кроме того, дополнительно было обнаружено, что одна конкретная комбинация этих растворимых и стабильных каркасов VL (вариабельной легкой цепи) и VH (вариабельной тяжелой цепи) является особенно подходящей для вмещения кроличьих CDR. Таким образом, в одном из вариантов осуществления, CDR, представленные как SEQ ID NО: 2-72, трансплантируют в каркасы антител человека, полученные скринингом "контроля качества", описанным в EP 1479694. Аминокислотные последовательности примерных каркасов для применения в настоящем изобретении, представлены в SEQ ID NО: 172-174. Неожиданно было обнаружено, что после трансплантации в указанный каркас или его производные, может полностью сохраняться петлевая конформация большого разнообразия кроличьих CDR независимо от последовательности донорского каркаса. Кроме того, указанный каркас или его производные, содержащие различные кроличьи CDR, хорошо экспрессируются и продуцируются в отличие от одиночных цепей дикого типа кролика и все еще сохраняют почти полностью аффинность исходных донорских кроличьих антител.
Таким образом, в одном из предпочтительных вариантов осуществления, CDR и/или CDR-мотивы, описанные в настоящем описании, присутствуют в каркасной последовательности вариабельного района тяжелой цепи, имеющей по меньшей мере 80% идентичность последовательности, еще более предпочтительно, по меньшей мере 85%, 90% 95%, даже более предпочтительно, 100% идентичность последовательности SEQ ID NO: 169. В предпочтительном варианте осуществления, каркасная последовательность вариабельного района тяжелой цепи содержит SEQ ID NO: 170 или SEQ ID NO: 171.
В одном из предпочтительных вариантов осуществления, CDR и/или CDR-мотивы, описанные в данном случае, присутствуют в каркасной последовательности вариабельного района легкой цепи, которая по меньшей мере на 85% идентична, еще более предпочтительно, по меньшей мере на 90% 95%, еще более предпочтительно, на 100% идентична последовательности SEQ ID NO: 167, более предпочтительно, содержащей SEQ ID NO: 167 или SEQ ID NO: 168.
В кроличьих антителах, CDR могут содержать остатки цистеина, которые связываются дисульфидной связью с остатками цистеина в каркасе антитела. Таким образом, может быть необходимым, при трансплантации кроличьих CDR, содержащих остатки цистеина, в не-кроличий каркас, вводить остатки цистеина в не-кроличий каркас, например, мутагенезом, для облегчения стабилизации кроличьего CDR посредством дисульфидной связи.
В других вариантах осуществления, изобретение относится к иммуносвязывающему агенту, который специфически связывается с VEGF, содержащему по меньшей мере одну из аминокислотных последовательностей VL или VH. Примеры аминокислотных последовательностей VH или VL для применения в иммуносвязывающих агентах по настоящему изобретению представлены в SEQ ID NO: 72-106 и 107-166, соответственно.
В одном из предпочтительных вариантов осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 107, SEQ ID NO: 108, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 130 и SEQ ID NO: 131 (VH 60-11-4, VH 60-11-6, VH 60-11-4min, VH 60-11-6min, VH 60-11-4max и VH 60-11-6max, соответственно);
и/или VL, которая по меньшей мере 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 72, SEQ ID NO:82 и SEQ ID NO: 93 (VL 60, VL 60min, VL 60max, соответственно).
В другом предпочтительном варианте осуществления, изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 109, SEQ ID NO: 120 и SEQ ID NO: 132 (VH 435, VH 435min и VH 435max, соответственно;
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 73, SEQ ID NO: 83 и SEQ ID NO:94 (VL 435, VL 435min и VL 435max, соответственно).
Предпочтительно, указанный иммуносвязывающий агент имеет по меньшей мере 80%, более предпочтительно, по меньшей мере 85%, 90%, 95%, наиболее предпочтительно, 100% идентичность SEQ ID NO: 175 (435max).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 110, SEQ ID NO: 121 и SEQ ID NO: 133 (VH 453, VH 453min и VH 453max, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 74, SEQ ID NO: 84 и SEQ ID NO: 95 (VL 453, VL 453min и VL 453max, соответственно).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 111, SEQ ID NO: 122 и SEQ ID NO: 134 (VH 375, VH 375min и VH 375max, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 75, SEQ ID NO: 85 и SEQ ID NO: 96 (VL 375, VL 375min и VL 375max, соответственно).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 112, SEQ ID NO: 123 и 135 (VH 610, VH 610min и VH 610max, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 76, SEQ ID NO: 86 и SEQ ID NO: 97 (VL 610, VL 610min и VL 610max, соответственно).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 113, SEQ ID NO: 124, SEQ ID NO: 129, SEQ ID NO: 136, SEQ ID NO: 142, SEQ ID NO: 144, SEQ ID NO: 146, SEQ ID NO: 147, SEQ ID NO: 148, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151, SEQ ID NO: 152, SEQ ID NO: 153, SEQ ID NO: 154, SEQ ID NO: 155, SEQ ID NO: 156, SEQ ID NO: 157, SEQ ID NO: 158, SEQ ID NO: 159, SEQ ID NO: 160, SEQ ID NO: 161, SEQ ID NO: 162, SEQ ID NO: 163, SEQ ID NO: 164, SEQ ID NO: 165 и SEQ ID NO: 166 (VH 578 и ее варианты);
и/или VL, которая по меньшей мере 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 77, SEQ ID NO: 87, SEQ ID NO: 92, SEQ ID NO: 98, SEQ ID NO: 103, SEQ ID NO: 104 и SEQ ID NO: 105 (VL 578 и ее варианты).
Предпочтительно, указанный иммуносвязывающий агент имеет по меньшей мере 80%, более предпочтительно, по меньшей мере 85%, 90%, 95%, наиболее предпочтительно, 100% идентичность с SEQ ID NO: 178 (578min), SEQ ID NO: 179 (578max) или SEQ ID NO: 180 (578minmax).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 114, SEQ ID NO: 125 и SEQ ID NO: 137 (VH 534, VH 534min и VH 534max, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 78, SEQ ID NO: 88 и SEQ ID NO: 99 (VL 534, VL 534min и VL 534max, соответственно).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 115, SEQ ID NO: 126, SEQ ID NO: 138 и SEQ ID NO: 143 (VH 567, VH 567min и VH 567max, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 79, SEQ ID NO: 89 и SEQ ID NO: 100 (VL 567, VL 567min и VL 567max, соответственно).
Предпочтительно, указанный иммуносвязывающий агент имеет по меньшей мере 80%, более предпочтительно, по меньшей мере 85%, 90%, 95%, наиболее предпочтительно, 100% идентичность с SEQ ID NO: 177 (567min).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 116, SEQ ID NO: 127, SEQ ID NO: 139 и SEQ ID NO: 140 (VH 509, VH 509min, VH 509max и VH 509maxII, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 80, SEQ ID NO: 90 и SEQ ID NO: 101 (VL 509, VL 509min и VL 509max, соответственно).
В другом предпочтительном варианте осуществления изобретение относится к иммуносвязывающему агенту, содержащему VH, который по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентичен последовательности, выбранной из группы, состоящей из SEQ ID NO: 117, SEQ ID NO: 128, SEQ ID NO: 141 и SEQ ID NO: 145 (VH 511, VH 511min, VH 511max и VH 511maxDHP, соответственно);
и/или VL, которая по меньшей мере на 80%, более предпочтительно, по меньшей мере на 85%, 90%, 95%, наиболее предпочтительно, на 100% идентична последовательности, выбранной из группы, состоящей из SEQ ID NO: 81, SEQ ID NO: 91, SEQ ID NO: 102 и SEQ ID NO: 106 (VL 511, VL 511min, VL 511max и VL 511minC41L, соответственно).
Предпочтительно, указанный иммуносвязывающий агент имеет по меньшей мере 80%, более предпочтительно, по меньшей мере 85%, 90%, 95%, наиболее предпочтительно, 100% идентичность с SEQ ID NO: 176 (511max).
В некоторых вариантах осуществления, это изобретение дополнительно относится к иммуносвязывающему агенту, который специфически связывается с VEGF, содержащему аминокислотную последовательность с существенным сходством с аминокислотной последовательностью, представленной в SEQ ID NO: 2-166 и в SEQ ID NО: 175-180, где этот иммуносвязывающий агент по существу сохраняет или улучшает желаемые функциональные свойства анти-VEGF-иммуносвязывающего агента по настоящему изобретению. Предпочтительные процентные сходства включают в себя, но не ограничиваются ими, по меньшей мере 50%, 60%, 70%, 75%, 80%, 85%, 90% или 95% идентичность.
В некоторых вариантах осуществления, это изобретение дополнительно относится к иммуносвязывающему агенту, который специфически связывается с VEGF, содержащему аминокислотную последовательность с существенной идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO: 2-166 и в SEQ ID NO: 175-180, где этот иммуносвязывающий агент по существу сохраняет или улучшает желаемые функциональные свойства анти-VEGF-иммуносвязывающего агента по настоящему изобретению. Предпочтительные процентные идентичности включают в себя, но не ограничиваются ими, по меньшей мере 50%, 60%, 70%, 75%, 80%, 85%, 90% или 95% идентичность.
В некоторых вариантах осуществления, это изобретение дополнительно относится к иммуносвязывающему агенту, который специфически связывается с VEGF, содержащему аминокислотную последовательность с консервативными заменами относительно аминокислотной последовательности, представленной в SEQ ID NО: 2-166 и в SEQ ID NО: 175-180, где этот иммуносвязывающий агент по существу сохраняет или улучшает желаемые функциональные свойства анти-VEGF-иммуносвязывающего агента по настоящему изобретению.
В некоторых вариантах осуществления, это изобретение относится к иммуносвязывающим агентам, которые связываются специфически с VEGF человека и перекрестно реагируют с молекулами VEGF других видов, например, VEGF мыши, VEGF крысы, VEGF кролика или VEGF морской свинки. В одном конкретном варианте осуществления, анти-VEGF-иммуносвязывающий агент может связываться специфически с VEGF человека и VEGF крысы/мыши.
В некоторых вариантах осуществления, это изобретение относится к иммуносвязывающим агентам, которые связываются специфически с VEGF человека и не реагируют перекрестно с молекулами VEGF других видов, например, VEGF мыши, VEGF крысы, VEGF кролика или VEGF морской свинки.
В некоторых вариантах осуществления, это изобретение относится к иммуносвязывающим агентам, которые связываются специфически с VEGF человека, причем эти иммуносвязывающие агенты являются аффинно зрелыми.
В одном из вариантов осуществления, антитела и фрагменты антител по настоящему изобретению являются одноцепочечными антителами (scFv) или Fab-фрагментами. В случае антител scFv, выбранный VL-домен может быть связан с выбранным VH-доменом в любой ориентации посредством гибкого линкера. Подходящее состояние типичного линкера состоит из повторяемых аминокислотных последовательностей GGGGS или их вариантов. В предпочтительном варианте по настоящему изобретению линкер с аминокислотной последовательностью (GGGGS)4 представлен в SEQ ID NO: 181, но возможны также варианты из 1-3 повторов (Holliger et al. (1993), Proc. Natl. Acad. Sci. USA 90:6444-6448). Другие линкеры, которые могут быть использованы для по настоящему изобретению, описаны Alfthan et al. (1995), Protein Eng. 8:725-731, Choi et al. (2001), Eur. J. Immunol. 31:94-106, Hu et al. (1996), Cancer Res. 56:3055-3061, Kipriyanov et al. (1999), J. Mol. Biol. 293:41-56 и Roovers et al. (2001), Cancer Immunol. Immunother. 50:51-59. Расположение может быть либо VL-линкер-VH, либо VH-линкер-VL, причем предпочтительной является первая ориентация. Однако обсуждаются также антитела с единственными доменами VH или VL. В случае, Fab-фрагментов, выбранные вариабельные домены VL легкой цепи сливают с константным районом каппа-цепи Ig человека, тогда как подходящие вариабельные домены VH сливают с первым (N-концевым) константным доменом CH1 IgG человека. На С-конце константного домена или в других сайтах вариабельного или константного домена может образовываться межцепочечный дисульфидный мостик. Альтернативно, эти две цепи могут быть связаны гибким линкером, что приводит к одноцепочечному Fab-антителу.
Антитела или производные антител по изобретению могут иметь аффинность в отношении VEGF человека с константами диссоциации Kd в диапазоне 10-14М – 10-5M. В одном из предпочтительных вариантов осуществления по настоящему изобретению Kd равна ≤1 нМ. Аффинность антитела в отношении антигена может быть определена экспериментально с использованием подходящего способа (Berzofsky et al. "Antibody-Antigen Interactions", in Fundamental Immunology, Paul, W.E., Ed, Raven Press: New York, NY (1992); Kuby, J. Immunology, W.H. Freeman and Company: New York, NY) и описанных в данном случае способов.
От компании Epitomics доступно антитело против VEGF, которое является кроличьим моноклональным антителом (VEGF (C-term) Rabbit Antibody, Cat.№ 1909-1). Указанное антитело направлено против остатков на С-конце VEGF человека и, следовательно, не способно нейтрализовать VEGF. Таким образом, указанное антитело не пригодно для терапевтических применений. Кроме того, указанный моноклональный IgG не является гуманизированным антителом, а является природным кроличьим полноразмерным иммуноглобулином. Кроме того, было показано, что это антитело не узнает нативную форму VEGF.
Иммуносвязывающие агенты, которые связывают одни и те же эпитопы на VEGF
В другом аспекте, это изобретение относится к антителам, которые связываются с эпитопом на VEGF, распознаваемым антителом, содержащим любую из аминокислотных последовательностей SEQ ID NO 2-211. Такие антитела могут быть идентифицированы на основании их способности перекрестно взаимодействовать с антителом, содержащим любые одну или несколько из аминокислотных последовательностей, SEQ ID NО: 2-211, в стандартном анализе связывания VEGF, включающем, но не ограничивающемся им, ELISA. Способность тестируемого антитела ингибировать связывание с VEGF человека антитела, содержащего любые одну или несколько аминокислотных последовательностей, SEQ ID NО: 2-211, демонстрирует, что это тестируемое антитело может перекрестно конкурировать, следовательно, взаимодействовать с перекрывающимся (совпадающим частично) эпитопом на VEGF человека, в качестве антитела, содержащего одну или несколько из аминокислотных последовательностей SEQ ID NО: 2-211.
Дополнительно или альтернативно, такие антитела могут быть также идентифицированы стандартными способами картирования эпитопов для определения, связываются ли они с одними и теми же пептидными иммуногенами. Для дополнительного определения точных молекулярных детерминант для взаимодействия антитело/VEGF могут быть также использованы способы структурного моделирования, включающие в себя, но не ограничивающиеся ими, ЯМР, рентгеновскую кристаллографию, моделирование на основе компьютера или томографию белков (Banyay et al, 2004 ASSAY and Drug Development Technologies (2), 5, Page 516-567). Действительно, была установлена кристаллическая структура VEGF и известны поверхностные аминокислотные остатки, участвующие в связывании VEGFr (Fuh, et al, 2006, J. Biol. Chem., 281, 6625-6631). Таким образом, при условии, что в данной области доступна аминокислотная последовательность этого пептидного иммуногена и структура VEGF, специалист в данной области вполне может идентифицировать антитела, которые связываются с эпитопом на VEGF, узнаваемым антителами, содержащими любые одну или несколько аминокислотных последовательностей, SEQ ID NO: 2-211.
В некоторых вариантах осуществления, антитела, которые связываются с эпитопом на VEGF, распознаваемым антителами, содержащими любые одну или несколько аминокислотных последовательностей, SEQ ID NO: 2-211, связываются с VEGF с аффинностью 107M-1, например, по меньшей мере 107M-1, по меньшей мере 108M-1, по меньшей мере 109M-1, по меньшей мере 1010M-1, по меньшей мере 1011M-1, по меньшей мере 1012M-1 или по меньшей мере 1013M-1.
В некоторых вариантах осуществления, антитела, которые связываются с эпитопом на VEGF, распознаваемым антителом, содержащим любые одну или несколько аминокислотных последовательностей, SEQ ID NO: 2-211, связываются специфически с VEGF человека и не реагируют перекрестно с молекулами VEGF других видов, например, VEGF мыши, VEGF крысы, VEGF кролика или VEGF морской свинки.
В некоторых вариантах осуществления, антитела, которые связываются с эпитопом на VEGF, распознаваемым антителом, содержащим любые одну или несколько аминокислотные последовательности, SEQ ID NO: 2-211, перекрестно реагируют с молекулами VEGF других видов, например, VEGF мыши, VEGF крысы или VEGF кролика.
Оптимизированные варианты
Антитела по настоящему изобретению могут быть дополнительно оптимизированы в отношении увеличенных функциональных свойств, например, в отношении увеличенной растворимости и/или стабильности.
В некоторых вариантах осуществления, антитела по настоящему изобретению оптимизируют в соответствии с методологией "функционального консенсуса", описанной в заявке PCT с порядковым номером PCT/EP2008/001958, озаглавленной "Конструирование на основе последовательности и оптимизация одноцепочечных антител", поданной 12 марта 2008 года, которая включена в настоящее описание в качестве ссылки. Например, иммуносвязывающие агенты VEGF по настоящему изобретению могут сравниваться с базой данных функционально-выбранных scFv для идентификации положений аминокислотных остатков, которые являются либо более, либо менее толерантными в отношении вариабельности, чем соответствующее положение(я), в иммуносвязывающем агенте VEGF, показывая посредством этого, что такое идентифицированное положение(я) остатков может быть пригодно для конструирования для улучшения функциональности, такой как стабильность и/или растворимость.
Примеры положений каркаса для замены описаны в заявке РСТ РСТ/СН2008/000284 "Способы модификации антител и модифицированные антитела с улучшенными функциональными свойствами”, поданной 25 июня 2008 года, и заявки РСТ PCT/CH2008/000284 "Конструирование на основе инженерии и оптимизация одноцепочечных антител", поданной 25 июня 2008 года. Например, одна или несколько из следующих замен могут быть введены в положение аминокислот (Нумерация AHo приводится для каждого положения аминокислоты, в перечне ниже) в вариабельном районе тяжелой цепи иммуносвязывающего агента по настоящему изобретению:
(a) Q или E в положении аминокислоты 1;
(b) Q или E в положении аминокислоты 6;
(c) T, S или A в положении аминокислоты 7, более предпочтительно, T или A, еще более предпочтительно, T;
(d) A, T, P, V или D, более предпочтительно, T, P, V или D, в положении аминокислоты 10,
(e) L или V, более предпочтительно, L, в положении аминокислоты 12,
(f) V, R, Q, M или K, более предпочтительно, V, R, Q или M в положении аминокислоты 13;
(g) R, M, E, Q или K, более предпочтительно, R, M, E или Q, еще более предпочтительно, R или E, в положении аминокислоты 14;
(h) L или V, более предпочтительно, L, в положении аминокислоты 19;
(i) R, T, K или N, более предпочтительно, R, T или N, еще более предпочтительно, N, в положении аминокислоты 20;
(j) I, F, L или V, более предпочтительно, I, F или L, еще более предпочтительно, I или L, в положении аминокислоты 21;
(k) R или K, более предпочтительно, K, в положении аминокислоты 45;
(l) T, P, V, A или R, более предпочтительно, T, P, V или R, еще более предпочтительно, R, в положении аминокислоты 47;
(m) K, Q, H или E, более предпочтительно, K, H или E, еще более предпочтительно, K, в положении аминокислоты 50;
(n) M или I, более предпочтительно, I, в положении аминокислоты 55;
(o) K или R, более предпочтительно, K, в положении аминокислоты 77;
(p) A, V, L или I, более предпочтительно, A, L или I, еще более предпочтительно, A, в положении аминокислоты 78;
(q) E, R, T или A, более предпочтительно, E, T или A, еще более предпочтительно, E, в положении аминокислоты 82;
(r) T, S, I или L, более предпочтительно, T, S или L, еще более предпочтительно, T, в положении аминокислоты 86;
(s) D, S, N или G, более предпочтительно, D, N или G, еще более предпочтительно, N, в положении аминокислоты 87;
(t) A, V, L или F, более предпочтительно, A, V или F, еще более предпочтительно, V, в положении аминокислоты 89;
(u) F, S, H, D или Y, более предпочтительно, F, S, H или D, в положении аминокислоты 90;
(v) D, Q или E, более предпочтительно, D или Q, еще более предпочтительно, D, в положении аминокислоты 92;
(w) G, N, T или S, более предпочтительно, G, N или T, еще более предпочтительно, G, в положении аминокислоты 95;
(x) T, A, P, F или S, более предпочтительно, T, A, P или F, еще более предпочтительно, F, в положении аминокислоты 98;
(y) R, Q, V, I, M, F или L, более предпочтительно, R, Q, I, M, F или L, еще более предпочтительно, Y, даже более предпочтительно, L, в положении аминокислоты 103; и
(z) N, S или A, более предпочтительно, N или S, еще более предпочтительно, N, в положении аминокислоты 107.
Дополнительно или альтернативно, одна или несколько замен могут быть введены в вариабельный район легкой цепи иммуносвязывающего агента по настоящему изобретению:
(aa) Q, D, L, E, S, или I, более предпочтительно, L, E, S или I, еще более предпочтительно, L или E, в положении аминокислоты 1;
(bb) S, A, Y, I, P или T, более предпочтительно, A, Y, I, P или T, еще более предпочтительно, P или T в положении аминокислоты 2;
(cc) Q, V, T или I, более предпочтительно, V, T или I, еще более предпочтительно, V или T, в положении аминокислоты 3;
(dd) V, L, I или M, более предпочтительно, V или L, в положении аминокислоты 4;
(ee) S, E или P, более предпочтительно, S или E, еще более предпочтительно, S, в положении аминокислоты 7;
(ff) T или I, более предпочтительно, I, в положении аминокислоты 10;
(gg) A или V, более предпочтительно, A, в положении аминокислоты 11;
(hh) S или Y, более предпочтительно, Y, в положении аминокислоты 12;
(ii) T, S или A, более предпочтительно, T или S, еще более предпочтительно, T, в положении аминокислоты 14;
(jj) S или R, более предпочтительно, S, в положении аминокислоты 18;
(kk) T или R, более предпочтительно, R, в положении аминокислоты 20;
(11) R или Q, более предпочтительно, Q, в положении аминокислоты 24;
(mm) H или Q, более предпочтительно, H, в положении аминокислоты 46;
(nn) K, R или I, более предпочтительно, R или I, еще более предпочтительно, R, в положении аминокислоты 47;
(oo) R, Q, K, E, T, или M, более предпочтительно, Q, K, E, T или M, в положении аминокислоты 50;
(pp) K, T, S, N, Q или P, более предпочтительно, T, S, N, Q или P, в положении аминокислоты 53;
(qq) I или M, более предпочтительно, M, в положении аминокислоты 56;
(rr) H, S, F или Y, более предпочтительно, H, S или F, в положении аминокислоты 57;
(ss) I, V или T, более предпочтительно, V или T, R, еще более предпочтительно, T, в положении аминокислоты 74;
(tt) R, Q или K, более предпочтительно, R или Q, еще более предпочтительно, R, в положении аминокислоты 82;
(uu) L или F, более предпочтительно, F, в положении аминокислоты 91;
(vv) G, D, T или A, более предпочтительно, G, D или T, еще более предпочтительно, T, в положении аминокислоты 92;
(xx) S или N, более предпочтительно, N, в положении аминокислоты 94;
(yy) F, Y или S, более предпочтительно, Y или S, еще более предпочтительно, S, в положении аминокислоты 101; и
(zz) D, F, H, E, L, A, T, V, S, G или I, более предпочтительно, H, E, L, A, T, V, S, G или I, еще более предпочтительно, A или V, в положении аминокислоты 103.
Система нумерации AHo описана дополнительно в Honegger, A. and Pluckthun, A. (2001) J. Mol. Biol. 309:657-670. Альтернативно, может быть использована система нумерации Кабата, описанная дополнительно в Kabat et al. (Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication № 91-3242). Таблицы переведения для этих двух систем нумерации, используемых для идентификации положений аминокислотных остатков в вариабельных районах тяжелой и легкой цепи обеспечены в A. Honegger, J.Mol.Biol. 309 (2001) 657-670.
В других вариантах осуществления, иммуносвязывающие агенты по настоящему изобретению содержат одну или несколько усиливающих растворимость и/или стабильность мутаций, описанных в предварительной заявке на патент США с порядковым номером 61/075692 "Оптимизация растворимости иммуносвязывающих агентов", поданной 25 июня 2008 года. В некоторых предпочтительных вариантах осуществления, этот иммуносвязывающий агент содержит увеличивающую растворимость мутацию в положении аминокислоты, выбранном из группы положений аминокислот тяжелой цепи, состоящей из 12, 103 и 144 (AHo Numbering convention). В одном из предпочтительных вариантов осуществления, этот иммуносвязывающий агент содержит одну или несколько замен, выбранных из группы, состоящей из: (a) серина (S) в положении аминокислоты 12 тяжелой цепи; (b) серина (S) или треонина (T) в положении аминокислоты 103 тяжелой цепи и (c) серина (S) или треонина (T) в положении аминокислоты 144 тяжелой цепи. В другом варианте осуществления иммуносвязывающий агент содержит следующие замены: (a) серин (S) в положении 12 аминокислоты тяжелой цепи; (b) серин (S) или треонин (T) в положении 103 аминокислоты тяжелой цепи и (c) серин (S) или треонин (T) в положении 144 аминокислоты тяжелой цепи
Гибридомы, экспрессирующие кроличьи антитела против VEGF
В другом аспекте изобретение относится к гибридоме, экспрессирующей моноклональное антитело, содержащее любые одну или несколько аминокислотных последовательностей, представленные SEQ ID NO 72-81 и SEQ ID NO 107-117. Способы генерирования гибридом из кроличьих В-клеток хорошо известны в данной области и описаны, например, в заявке на патент США 2005/0033031.
Получение анти-VEGF-иммуносвязывающих агентов
Антитела или производные антител по изобретению могут быть получены обычными способами в области рекомбинантной генетики. На основании информации об этих последовательностях этих полипептидов, кДНК, кодирующие их, могут быть получены посредством синтеза генов (www.genscript.com). Эти кДНК могут быть клонированы в подходящие векторные плазмиды. После получения ДНК, кодирующей VL- и/или VH-домен, может выполняться сайт-направленный мутагенез, например, при помощи ПЦР с использованием мутагенных праймеров, с получением различных производных. Наилучшая "исходная последовательность" может быть выбрана в зависимости от изменений, желаемых в этих VL- и/или VH- последовательностях.
Способы введения или трансплантации CDR в каркасные районы включают в себя способы, приведенные, например, в Riechmann, L. et al. (1998) Nature 332:323-327; Jones, P. et al. (1986) Nature 321:522-525; Queen, C. et al. (1989) Proc. Natl. Acad. See. U.S.A. 86:10029-10033; U.S. Patent № 5225539 to Winter, и патентах США с номерами 5530101; 5585089; 5693762 и 6180370 to Queen et al, а также описанные в предварительной заявке США 61/075697, "Гуманизация кроличьих антител с использованием универсальных каркасов антител", поданной 25 июня 2008 года.
Стандартные способы клонирования и мутагенеза, хорошо известные специалисту в данной области, могут быть использованы для присоединения линкеров, перетасовки доменов или создания конструкции для получения Fab-фрагментов. Основные протоколы, описывающие общие способы по изобретению, описаны в Molecular Cloning, A Laboratory Manual (Sambrook & Russell, 3rd ed. 2001) и в Current Protocols in Molecular Biology (Ausubel et al, 1999).
ДНК-последовательность, несущую ген, кодирующий полипептид scFv, или в случае Fab-фрагментов, кодирующую либо два отдельных гена, либо бицистронный оперон, содержащий два гена для слияний VL-Cκ и VH-CH1, клонируют в подходящий экспрессирующий вектор, предпочтительно вектор с индуцируемым промотором. Следует обратить внимание на то, присутствует ли перед каждым геном подходящий сайт связывания рибосом, который гарантирует трансляцию. Должно быть понятно, что антитела по настоящему изобретению содержат описанные последовательности, а не состоят из них. Например, стратегии клонирования могут требовать создания конструкции, из которой может быть получено антитело с одним или несколькими дополнительными остатками на N-концевой стороне. Конкретно, метионин, происходящий из стартового кодона, может присутствовать в конечном белке в случаях, когда он не был отщеплен посттрансляционно. Большинство конструкций для scFv-антител имеют дополнительный аланин на N-концевой стороне. В предпочтительном варианте осуществления по настоящему изобретению, выбран экспрессирующий вектор для периплазматической экспрессии в E. coli (Krebber, 1997). Указанный вектор содержит промотор перед отщепляемой сигнальной последовательностью. Затем кодирующую последовательность для этого пептида антитела сливают в рамке считывания с отщепляемой сигнальной последовательность. Это позволяет нацеливать экспрессируемый полипептид на бактериальную периплазму, в которой отщепляется эта сигнальная последовательность. Затем антитело укладывается. В случае Fab-фрагментов, слитые пептиды как VL-Cκ, так и VH-CH1 должны быть связаны с сигналом экспорта. После достижения этими пептидами периплазмы, образуется ковалентная связь S-S при С-концевых цистеинах. Если предпочтительной является цитоплазматическая экспрессия антител, указанные антитела обычно могут быть получены при высоких выходах из телец включения, которые могут быть легко отделены от других клеточных фрагментов и клеточного белка. В этом случае, эти тельца включения солюбилизируют в денатурирующем агенте, таком как, например, гидрохлорид гуанидина (GndHCl), и затем повторно укладывают при помощи процедур ренатурации, хорошо известных специалистам в данной области.
Плазмиды, экспрессирующие scFv или Fab-полипептиды вводят в подходящего хозяина, предпочтительно, бактериальную, дрожжевую клетку или клетку млекопитающего, наиболее предпочтительно, в подходящий штамм E. coli, например, JM83, для периплазматической экспрессии или в BL21 для экспрессии в тельцах включения. Этот полипептид может быть собран либо из периплазмы, либо из телец включения и очищен с использованием стандартных способов, таких как ионообменная хроматография, хроматография обращенной фазой, аффинная хроматография и/или гель-фильтрация, известных специалисту в данной области.
Антитела или производные антител по настоящему изобретению могут быть охарактеризованы в отношении выхода, растворимости и стабильности in vitro. Связывающие способности в отношении VEGF, предпочтительно против VEGF человека, могут быть тестированы in vitro при помощи ELISA или резонанса поверхностных плазмонов (BIACore), с использованием рекомбинантного VEGF человека, как описано в WO9729131, причем последний способ позволяет также определять константу скорости диссоциации koff, которая должна быть предпочтительно менее 10-3с-1. Предпочтительными являются величины Kd≤10 нМ.
Кроме антител с сильной связывающей аффинностью в отношении VEGF человека, желательно также отобрать антитела против VEGF, которые имеют и другие благоприятные свойства с точки зрения терапевтической перспективы. Например, это антитело может быть антителом, которое ингибирует рост клеток HUVEC в ответ на VEGF (см. пример 3). В одном из вариантов осуществления, это антитело может быть способным ингибировать пролиферацию клеток HUVEC в ответ на почти максимальную эффективную концентрацию VEGF (0,08 нМ). Предпочтительно, это антитело имеет величину эффективной дозы 50 (ED50), не большую, чем приблизительно 5 нМ, предпочтительно не большую, чем приблизительно 1 нМ, предпочтительно не большую, чем приблизительно 1 нМ, предпочтительно не большую, чем приблизительно 0,5 нМ и наиболее предпочтительно, не большую, чем приблизительно 0,06 нМ, для ингибирования VEGF-индуцированной пролиферации эндотелиальных клеток в этом "анализе роста эндотелиальных клеток", т.е. при этих концентрациях это антитело способно ингибировать VEGF-индуцированный рост эндотелиальных клеток in vitro, например, на 50% или более.
Биспецифические молекулы
В другом аспекте, изобретение относится к биспецифическим молекулам, содержащим антитело против VEGF, или его фрагмент, по изобретению. Антитело по изобретению, или его антигенсвязывающие части, могут быть преобразованы путем введения функциональных групп или связаны с функциональной молекулой, например, другим пептидом или белком (например, другим антителом или лигандом для рецептора) с получением биспецифической молекулы, которая связывается по меньшей мере с двумя различными сайтами связывания или молекулами-мишенями. Антитело по изобретению может быть преобразовано путем введения функциональных групп или связано с функциональной молекулой с получением биспецифической молекулы, которая связывается по меньшей мере с двумя различными сайтами связывания и/или молекулами-мишенями; такие мультиспецифические молекулы также предназначены для включения в термин "биспецифическая молекула" в контексте настоящей заявки. Для создания биспецифической молекулы по изобретению, антитело по изобретению может быть функционально связано (например, химическим связыванием, генетическим слиянием, нековалентной ассоциацией или иным образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, фрагмент антитела, опухолеспецифические или патогенспецифические антигены, пептид или связывающий миметик, так что возникает биспецифическая молекула. Таким образом, изобретение включает биспецифические молекулы, содержащие по меньшей мере одну первую связывающую молекулу, имеющую специфичность в отношении VEGF, и вторую связывающую молекулу, имеющую специфичность в отношении одного или нескольких дополнительных эпитопов-мишеней.
В одном из вариантов осуществления, биспецифические молекулы по настоящему изобретению содержат специфичность связывания по меньшей мере одного антитела, или фрагмента этого антитела, в том числе, например, Fab, Fab’, F(ab’)2, Fv или одноцепочечного Fv. Это антитело может быть димером легкой цепи или тяжелой цепи или любым его минимальным фрагментом, таким как Fv или одноцепочечная конструкция, как описано в работе Ladner et al. патент США 4946778, содержание которого включено в настоящее описание в качестве ссылки в полном объеме.
Хотя предпочтительными являются моноклональные антитела, другими антителами, которые могут быть использованы в биспецифических молекулах по настоящему изобретению, являются мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы по изобретению могут быть получены конъюгацией связывающих компонент специфичностей с использованием известных в данной области способов. Например, каждая связывающая специфичность биспецифической молекулы может быть генерирована отдельно и затем они могут быть конъюгированы одна с другой. Когда этими связывающими специфичностями являются белки или пептиды, для ковалентной конъюгации могут быть использованы связывающие или сшивающие агенты. Примеры сшивающих агентов включают в себя белок А, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5’-дитиобис-(2-нитробензойную кислоту) (DTNB), o-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (см., например, Karpovsky et al. (1984) J. Exp. Med. 160:1686; Liu, MA et al. (1985) Proc. Natl. Acad. Sci USA 82:8648). Другие способы включают в себя способы, описанные в Paulus (1985) Behring Ins. Mitt. № 78, 118-132; Brennan et al. (1985) Science 229:81-83), и Glennie et al. (1987) J. Immunol. 139: 2367-2375). Предпочтительными конъюгирующими агентами являются SATA и сульфо-SMCC, оба из которых доступны из Pierce Chemical Co. (Rockford, IL).
Если эти связывающие специфичности являются антителами, то могут быть конъюгированы через образование сульфгидрильных связей, например, через шарнирные области С-концов двух тяжелых цепей или другие сайты, независимо от того, являются ли они природно-встречающимися или вводятся искусственно. В особенно предпочтительном варианте осуществления, шарнирную область модифицируют таким образом, что она содержит нечетное число сульфгидрильных остатков, предпочтительно один остаток, перед конъюгацией.
Альтернативно, обе связывающие специфичные части могут кодироваться в одном и том же векторе и экспрессироваться и собираться в одной и той же клетке-хозяине. Этот способ особенно эффективен, когда биспецифическая молекула представляет собой слитый белок mAb x mAb, mAb x Fab, Fab x F(ab’)2 или лиганд x Fab. Биспецифическая молекула по настоящему изобретению может быть одноцепочечной молекулой, содержащей одноцепочечное антитело и детерминанту связывания, или одноцепочечной биспецифической молекулой, содержащей две детерминанты связывания. Биспецифические молекулы могут содержать по меньшей мере две одноцепочечные молекулы. Далее, биспецифической молекулой может быть scFv, который специфически связывается с первой мишенью, причем VH и VL указанного scFv связаны с гибким линкером, содержащим домен, обеспечивающий специфическое связывание со второй мишенью. Подходящие линкеры описаны в предварительной заявке США № 60/937820. Способы получения биспецифических молекул описаны, например, в патенте США № 5260203; патенте США № 5455030; патенте США № 4881175; патенте США № 5132405; патенте США № 5091513; патенте США № 5476786; патенте США № 5013653; патенте США № 5258498 и патенте США № 5482858.
Связывание биспецифических молекул с их специфическими мишенями может быть подтверждено, например, твердофазным иммуноферментным анализом (ELISA), радиоиммуноанализом (RIA), FACS-анализом, биоанализом (например, ингибирования роста) или иммуноблот-анализом. Каждый из этих анализов обычно детектирует присутствие представляющих особый интерес комплексов белок-антитело с использованием меченого реагента (например, антитела), специфического для этого представляющего интерес комплекса. Например, комплексы антитела против VEGF могут быть детектированы с использованием, например, фермент-связанного антитела или фрагмента антитела, которое узнает комплексы антитело-VEGF и специфически связывается с комплексами антитело-VEGF. Альтернативно, эти комплексы могут быть радиоактивно мечеными и использоваться в радиоиммуноанализе (RIA) (см., например, статью Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986, которая включена в настоящее описание в качестве ссылки). Радиоактивный изотоп может быть детектирован такими способами, как применение γ-счетчика или сцинтилляционного счетчика, или авторадиографией.
Иммуноконъюгат
В другом аспекте, настоящее изобретение относится к антителу против VEGF, или его фрагменту, конъюгированному с терапевтической частью молекулы, такой как цитотоксин, лекарственное средство (например, иммуносупрессор) или радиотоксин. Такие конъюгаты называют в данном случае "иммуноконъюгатами". Иммуноконъюгаты, которые включают в себя один или несколько цитотоксинов, называют "иммунотоксинами". Цитотоксин или цитотоксический агент включает любой агент, который является вредным для клеток (например, убивает клетки). Примеры включают в себя таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропанолол и пуромицин и их аналоги или гомологи. Терапевтические агенты включают в себя также, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацилдекарбазин), алкилирующие агенты (например, меклоретамин, тиоепа хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклотосфамид, бусульфан, дибромоманнит, стрептозоцин, митомицин С и цис-дихлордиамин-платину (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)), и антимитотические агенты (например, винкристин и винбластин).
Другие предпочтительные примеры терапевтических цитотоксинов, которые могут быть конъюгированы с антителом по настоящему изобретению, включают в себя дуокармицины, калихеамицины, майтансины и ауристатины и их производные. Примером конъюгата калихеамицин-антитело является коммерчески доступный Mylotarg™ (Wyeth-Ayerst).
Цитотоксины могут быть конъюгированы с антителами по настоящему изобретению с использованием линкерной технологии, доступной в данной области. Примеры типов линкеров, которые использовали для конъюгирования цитотоксина с антителом, включают в себя, но не ограничиваются ими, гидразоны, тиоэфиры, эфиры, дисульфиды и пептидсодержащие линкеры. Может быть выбран линкер, который, например, является чувствительным к расщеплению низким рН в лизосомном компартменте, или чувствительным к расщеплению протеазами, таким как протеазы, преимущественно экспрессируемые в опухолевой ткани, такие как катепсины (например, катепсины В, С, D).
В отношении дополнительного обсуждения типов цитотоксинов, линкеров и способов конъюгирования терапевтических агентов с антителами, см. также Saito, G. et al. (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail, P.A. et al. (2003) Cancer Immunol. Immunother. 52:328-337; Payne, G. (2003) Cancer Cell 3:207-212; Allen, T.M. (2002) Nat. Rev. Cancer 2:750-763; Pastan, I. and Kreitman, R. J. (2002) Curr. Opin. Investig. Drugs 3:1089-1091; Senter, P.D. and Springer, CJ. (2001) Adv. Drug Deliv. Rev. 53:247-264.
Антитела по настоящему изобретению могут быть также конъюгированы с радиоактивным изотопом для получения цитотоксических радиофармацевтических веществ, также называемых радиоиммуноконъюгатами. Примеры радиоактивных изотопов, которые могут быть конъюгированы с антителами для диагностического или терапевтического использования, включают в себя, но не ограничиваются ими, иод131, индий111, иттрий90 и лютеций177. Способы получения радиоиммуноконъюгатов установлены в данной области. Примеры радиоиммуноконъюгатов являются коммерчески доступными, в том числе Zevalin™ (IDEC Pharmaceuticals) и Bexxar™ (Corixa Pharmaceuticals), и сходные способы могут быть использованы для получения радиоиммуноконъюгатов с использованием антител по настоящему изобретению.
Конъюгаты антител по настоящему изобретению могут быть использованы для модификации конкретной биологической реакции, и эта лекарственная часть молекулы не должна рассматриваться как ограничиваемая классическими химическими терапевтическими агентами. Например, эта лекарственная часть может быть белком или полипептидом, обладающим желаемой биологической активностью. Такие белки могут включать, например, ферментативно активный токсин, или его активные фрагменты, такой как абрин, рицин А, экзотоксин псевдомонаса или дифтерийный токсин; белок, такой как фактор некроза опухолей или интерферон-γ; или модификаторы биологической реакции, такие как, например, лимфокины, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарный макрофагальный колониестимулирующий фактор ("GM-CSF"), гранулоцитарный колониестимулирующий фактор ("G-CSF") или другие факторы роста.
Способы конъюгации такой терапевтической части с антителами хорошо известны, см., например, Arnon et al, "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al, "Antibodies For Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", in Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", in Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985) и Thorpe et al, "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982).
Применения антител против VEGF
Для терапевтического использования, антитела против VEGF по изобретению вводят млекопитающему, предпочтительно, человеку, в фармацевтически приемлемой лекарственной форме, такой как формы, обсуждаемые в данном случае, в том числе в форме, которая может вводиться человеку внутривенно, в виде болюса или непрерывной инфузией на протяжении некоторого периода времени, местным, внутриглазным, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным, внутрисуставным, внутрисиновиальным, интратекальным, пероральным или ингаляционным способами. Эти антитела могут также удобным образом вводиться внутриопухолевым, околоопухолевым способами введения, введением в повреждение или введением около повреждения, для вызывания местных, а также системных терапевтических эффектов. Ожидается, что внутрибрюшинный способ особенно эффективен, например, для лечения опухолей яичников.
Для профилактики или лечения заболевания, подходящая доза антитела будет зависеть от типа подлежащего лечению заболевания, определенного выше, тяжести и течения этого заболевания от того, вводится ли это антитело для профилактической или терапевтической целей, предыдущей терапии, клинической истории пациента и реакции на это антитело и от решения лечащего врача. Это антитело вводят удобным образом пациенту один раз или на протяжении ряда обработок.
Антитела против VEGF могут использоваться для лечения VEGF-опосредованных заболеваний, описанных в настоящем описании. Например, возрастная дегенерация желтого пятна (AMD) является главной причиной тяжелой потери зрения в пожилой популяции. Экссудативная форма AMD характеризуется хороидальной неоваскуляризацией и отслойкой ретинальных пигментных эпителиальных клеток. Поскольку хороидальная неоваскуляризация ассоциируется с разительным ухудшением прогноза, антитела против VEGF по настоящему изобретению особенно полезны в уменьшении тяжести AMD. Прогресс этой терапии может быть легко подвергнут мониторингу общепринятыми способами, включающими в себя офтальмоскопию, микроскопию глазного дна и глазную компьютерную томографию.
Рассматриваются все одобренные FDA (Департаментом по контролю за качеством пищевых продуктов, медикаментов и косметических средств (США)) дозы и схемы лечения, подходящие для применения с Lucentis. Другие дозы и схемы лечения описаны в предварительной заявке США с порядковым номером 61/075641, "Улучшенные препараты иммуносвязывающего агента и способы введения", поданной 25 июня 2008 года, и предварительной заявке США № 61/058504, которые включены в настоящее описание в качестве ссылки.
Согласно другому варианту по настоящему изобретению, эффективность этого антитела для профилактики или лечении заболевания может быть улучшена введением этого антитела последовательно или в комбинации с другим агентом, который является эффективным для этих целей, такого как фактор некроза опухолей (TNF), антитело, способное ингибировать или нейтрализовать ангиогенную активность кислого или основного фактора роста фибробластов (FGF) или фактора роста гепатоцитов (HGF), антитело, способное ингибировать или нейтрализовать коагулянтные активности тканевого фактора, белка С или белка S (см. Esmon et al, патентная публикация PCT № WO 91/01753, опубликованная 21 февраля 1991), антитело, способное связываться с рецептором HER2 (см. Hudziak et al, патентная публикация PCT № WO 89/06692, опубликованная 27 июля 1989), или один или несколько общепринятых терапевтических средств, таких как, например, алкилирующие агенты, фотокоагулянты (такие как вертепорфин), антагонисты фолиевой кислоты, антиметаболиты метаболизма нуклеиновых кислот, антибиотики, пиримидиновые аналоги, 5-фторурацил, цисплатин, пуриновые нуклеозиды, амины, аминокислоты, триазолнуклеозиды или кортикостероиды. Такие другие агенты могут присутствовать во вводимой композиции или могут вводиться отдельно. Кроме того, это антитело предпочтительно вводят последовательно или в комбинации с радиологическими обработками, независимо от того, включают ли они облучение или введение радиоактивных веществ.
Антитела по изобретению могут быть использованы в качестве агента аффинной очистки. В этом способе, эти антитела иммобилизуют на твердой фазе, такой как смола Сефадекс, или на фильтровальной бумаге, с использованием способов, хорошо известных в данной области. Это иммобилизованное антитело контактирует с пробой, содержащей подлежащий очистке белок VEGF (или его фрагмент), и после этого этот носитель промывают подходящим растворителем, который будет удалять по существу весь материал в этой пробе, за исключением белка VEGF, который связан с иммобилизованным антителом. Наконец, этот носитель промывают другим подходящим растворителем, таким как глициновый буфер, рН 5,0, который будет высвобождать белок VEGF из этого антитела.
Антитела против VEGF могут быть также эффективны в диагностических анализах на белок VEGF, например, детектировании его экспрессии в конкретных клетках, тканях или сыворотке. Такие диагностические анализы могут быть использованы в диагностике рака.
Для диагностических применений, это антитело обычно метят детектируемой частью молекулы. Доступны многочисленные метки, которые могут быть обычно сгруппированы в следующие категории:
(a) Радиоизотопы, такие как 111In, 99Tc, 14С, 131I, 125I, 3H, 32P или 35S. Это антитело может быть помечено радиоизотопом с использованием способов, описанных, например, в Current Protocols in Immunology, Volumes 1 and 2, Coligen et al, Ed. Wiley-Interscience, New York, N.Y., Pubs. (1991), и радиоактивность может быть измерена с использованием сцинтилляционного счета.
(b) Доступными являются флуоресцентные метки, такие как хелаты редкоземельных металлов (хелаты европия), или флуоресцеин и его производные, родамин и его производные, дансил, Лиссамин, фикоэритрин и Техасский Красный. Эти флуоресцентные метки могут быть конъюгированы с этим антителом с использованием способов, описанных, например, в Current Protocols in Immunology, выше. Флуоресценция может быть определена количественно с использованием флуориметра.
(c) Доступными являются различные фермент-субстратные метки и патент США № 4275149 обеспечивает обзор некоторых из них. Этот фермент обычно катализирует химическое изменение хромогенного субстрата, которое может быть измерено при помощи разных способов. Например, этот фермент может катализировать изменение окраски в субстрате, которое может быть измерено спектрофотометрически. Альтернативно, этот фермент может изменять флуоресценцию или хемилюминесценцию этого субстрата. Способы количественного измерения изменения флуоресценции описаны выше. Хемилюминесцентный субстрат становится электронно возбужденным посредством химической реакции и может затем испускать свет, который может быть измерен (например, при помощи хемилюминометра) или передает энергию флуоресцентному акцептору. Примеры ферментативных меток включают в себя люциферазы (например, люциферазу светляка и бактериальную люциферазу; патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, уреазу, пероксидазу, например, пероксидазу хрена (HRPO), щелочную фосфатазу, бета-галактозидазу, глюкоамилазу, лизоцим, сахаридоксидазы (например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и т.п. Способы конъюгирования ферментов с антителами описаны в O'Sullivan et ai, Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (ed J. Langone & H. Van Vunakis), Academic press, New York, 73:147-166 (1981). Примеры фермент-субстратных комбинаций включают в себя, например:
(i) Пероксидазу хрена (HRPO) с пероксидом водорода в качестве субстрата, где эта водородпероксидаза окисляет предшественник красителя (например, гидрохлорид ортофенилендиамина (OPD) или гидрохлорид 3,3’,5,5’-тетраметилбензидина (TMB));
(ii) щелочную фосфатазу (AP) с пара-нитрофенилфосфатом в качестве хромогенного субстрата; и; и
(iii) бета-D-галактозидазу (бета-D-Gal) с хромогенным субстратом (например, П-нитрофенил-бета-D-галактозидазой) или флуорогенным субстратом 4-метилумбеллиферил-бета-D-галактозидазой.
В другом варианте осуществления по настоящему изобретению, антитело против VEGF не должно быть меченым, и его присутствие может быть детектировано с использованием меченого антитела, которое связывается с антителом против VEGF
Антитела по изобретению могут быть использованы в любом известном способе анализа, таком как анализы конкурентного связывания, прямые и опосредованные сэндвич-анализы и анализы иммунопреципитации. Zola, Monoclonal Antibodies: A Manual of Techniques, pp.147-158 (CRC Press, Inc. 1987).
Анализы конкурентного связывания основаны на способности меченого стандарта конкурировать с аналитом тест-пробы за связывание с ограниченным количеством антитела. Количество белка VEGF в этой тест-пробе обратно пропорционально количеству стандарта, который становится связанным с этими антителами. Для облегчения определения количества стандарта, который становится связанным, эти антитела обычно являются несолюбилизированными перед конкуренцией или после конкуренции, так что стандарт и аналит, которые связаны с этими антителами, могут быть удобным образом отделены от стандарта и аналита, которые остаются несвязанными.
Сэндвич-анализы включают использование двух антител, каждое из которых способно связываться с отличающейся иммуногенной частью, или эпитопом детектируемого белка. В сэндвич-анализе, тест-проба аналита связывается первым антителом, который иммобилизован на твердом носителе, и после этого второе антитело связывается с этим аналитом, образуя нерастворимый состоящий из трех частей комплекс. См., например, патент США № 4376110. Это второе антитело само может быть меченым детектируемой частью молекулы (прямой сэндвич-анализ) или может быть измерено с использованием анти-иммуноглобулин-антитела, которое помечено детектируемой частью молекулы (непрямой сэндвич-анализ). Например, одним типом сэндвич-анализа является анализ ELISA, в котором детектируемой частью молекулы является фермент.
Для иммуногистохимии, проба опухоли может быть свежей или замороженной или может быть залита в парафин и фиксирована консервантом, таким как, например, формалин.
Эти антитела могут быть также использованы для диагностических анализов in vivo. Обычно, антитело метят радионуклидом (таким как 111In, 99Tc, 14С, 131I, 125I, 3H, 32P или 35S), так что опухоль может быть локализована с использованием иммуносцинтиографии.
Антитело по изобретению может быть обеспечено в наборе, упакованной комбинации реагентов в заранее заданных количествах с инструкциями в отношении выполнения этого диагностического анализа. Если это антитело помечено ферментом, набор будет включать в себя субстраты и кофакторы, требуемые для этого фермента (например, предшественник субстрата, который обеспечивает детектируемый хромофор или флуорофор). Кроме того, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или лизисный буфер) и т.п. Относительные количества различных реагентов могут широко варьироваться для обеспечения концентраций в растворе этих реагентов, которые существенно оптимизируют чувствительность этого анализа. В частности, эти реагенты могут быть обеспечены в виде сухих порошков, обычно лиофилизированных, включающих в себя эксципиенты, которые при растворении будут обеспечивать раствор реагентов, имеющий подходящие концентрации.
Фармацевтические препараты
В одном из аспектов изобретение относится к фармацевтическим готовым формам, содержащим антитела против VEGF для лечения VEGF-опосредованных заболеваний. Термин "фармацевтическая композиция" относится к препаратам, которые находятся в такой форме, которая позволяет биологической активности этого антитела или производного антитела быть явно эффективной, и которые не содержат дополнительных компонентов, которые являются токсичными в отношении индивидов, которым должна быть введена эта готовая форма. "Фармацевтически приемлемыми" эксципиентами (носителями, добавками) являются эксципиенты, которые могут быть обоснованно введены индивиду-млекопитающему для обеспечения эффективной дозы используемого активного ингредиента.
"Стабильной" готовой формой является форма, в которой это антитело или производное антитела по существу сохраняет его физическую стабильность и/или химическую стабильность и/или биологическую активность при хранении. Различные аналитические способы измерения стабильности белка доступны в данной области и обсуждаются, например, в обзоре Peptide and Protein Drug Delivery, 247-301, Vincent Lee Ed., Marcel Dekker, Inc., New York, N.Y., Pubs. (1991) и Jones, A. Adv. Drug Delivery Rev. 10: 29-90 (1993). Стабильность может быть измерена при выбранной температуре в течение выбранного периода времени. Предпочтительно, эта готовая форма является стабильной при комнатной температуре (приблизительно 30°C) или при 40°C в течение по меньшей мере 1 недели и/или стабильной при приблизительно 2-8°C в течение по меньшей мере 3 месяцев - 2 лет. Кроме того, эта готовая форма является предпочтительно стабильной после замораживания (например, до -70°С) и оттаивания этой готовой формы.
Антитело или производное антитела "сохраняет его физическую стабильность" в фармацевтической готовой форме, если оно удовлетворяет определенным документальным указаниям в отношении агрегации, деградации, осаждения и/или денатурации после визуального обследования окраски и/или прозрачности или после измерения рассеивания УФ-света или посредством эксклюзионной хроматографии на основе размера или другими подходящими общепринятыми в данной области способами.
Антитело или производное антитела "сохраняет его химическую стабильность" в фармацевтической готовой форме, если химическая стабильность в конкретное время является такой, что этот белок может все еще сохранять его биологическую активность, как определено ниже. Химическая стабильность может быть оценена детектированием и количественным измерением химически измененных форм этого белка. Химическое изменение может включать в себя модификацию размера (например, клиппирование), которая может оцениваться с использованием, например, электрофореза в ДСН-ПААГ и/или масс-спектрометрии MALDI/TOF MS (использующей матрикс малых органических молекул для лазерной десорбционной-ионизационной/определяющей время полета ионов масс-спектрометрии (matrix-assisted laser desorption ionization/time-of-flight mass spectrometry)). Другие типы химического изменения включают в себя изменение заряда (например, встречающееся как следствие деамидирования), которое может оцениваться, например, ионообменной хроматографией.
Антитело или производное антитела "сохраняет его биологическую активность" в фармацевтической готовой форме, если биологическая активность этого антитела в конкретное время находится в пределах приблизительно 10% (в пределах ошибок этого анализа) биологической активности, проявляемой в момент приготовления этой фармацевтической готовой формы, например, при определении в анализе связывания антигена. Другие анализы "биологической активности” для антител объяснены в данном случае ниже.
Термин "изотоническая” обозначает, что представляющая интерес готовая форма имеет по существу такое же осмотическое давление, что и кровь человека. Изотонические готовые формы будут обычно иметь осмотическое давление от приблизительно 250 до 350 мОсм. Изотоничность может быть измерена с использованием, например, давления (упругости) пара или с использованием осмометра типа обледенения (ice-freezing).
"Полиол" является веществом со множественными гидроксильными группами и включает в себя сахара (редуцирующие и не редуцирующие сахара), сахароспирты и сахарокислоты. Предпочтительные в данном случае полиолы имеют молекулярную массу, которая меньше приблизительно 600 кД (например, в диапазоне от приблизительно 120 до приблизительно 400 кД). "Редуцирующий сахар" является сахаром, который содержит гемиацетальную группу, которая восстанавливает ионы металла или реагирует ковалентно с лизином и другими аминогруппами в белках, а "нередуцирующий сахар" является сахаром, который не имеет свойств редуцирующего сахара. Примерами редуцирующих сахаров являются фруктоза, манноза, мальтоза, лактоза, арабиноза, ксилоза, рибоза, рамноза, галактоза и глюкоза. Нередуцирующие сахара включают в себя сахарозу, трегалозу, сорбозу, мелецитозу и раффинозу. Примерами сахароспиртов являются маннит, ксилит, эритрит, треитол, сорбит и глицерин. Что касается сахарокислот, они включают в себя глюконат и его соли металлов. Если желательно, чтобы эта готовая форма была стабильной при замораживании-оттаивании, этим полиолом предпочтительно является полиол, который не кристаллизуется при температурах замерзания (например, -20°С), так что он дестабилизирует антитело в этой готовой форме. Нередуцирующие сахара, такие как сахароза и трегалоза, являются предпочтительно полиолами в этом изобретении, причем наиболее предпочтительной является трегалоза вследствие превосходящей стабильности раствора трегалозы.
В контексте настоящей заявки, "буфер" относится к забуференному раствору, который выдерживает изменения рН посредством действия его кислотно-основных конъюгированных компонентов. Буфер по настоящему изобретению имеет рН в диапазоне от приблизительно 4,5 до приблизительно 8,0; предпочтительно от приблизительно 5,5 до приблизительно 7. Примеры буферов, которые будут контролировать этот рН в данном диапазоне, включают в себя ацетатный (например, ацетат натрия), сукцинатный (такой как сукцинат натрия), глюконатный, гистидиновый, цитратный буферы и буферы других органических кислот. Когда желательной является стабильная к замораживанию-оттаиванию готовая форма, этот буфер предпочтительно не является фосфатным.
В фармакологическом смысле, в контексте настоящей заявки, "терапевтически эффективным количеством" антитела или производного антитела называют количество, эффективное для профилактики или лечения нарушения, для лечения которого является эффективным это антитело или производное этого антитела. "Заболевание/нарушение" является любым состоянием, которое могло бы выиграть от лечения этим антителом или производным этого антитела. Это состояние включает в себя хронические и острые нарушения или заболевания, в том числе те патологические состояния, которые предрасполагают это млекопитающее к рассматриваемому нарушению.
"Консервант" является соединением, которое может быть включено в эту готовую форму для существенного уменьшения бактериального действия в ней с облегчением посредством этого, например, приготовления готовой формы многоцелевого применения. Примеры потенциальных консервантов включают в себя хлорид октадецилдиметилбензиламмония, хлорид гексаметония, хлорид бензалкония (смесь хлоридов алкилбензилдиметиламмония, в которых алкильные группы являются соединениями с длинной цепью), и хлорид бензетония. Другие типы консервантов включают в себя ароматические спирты, такие как фенол, бутиловый и бензиловый спирт, алкилпарабены, такие как метил- или пропилпарабен, катехин, резорцин, циклогексанол, 3-пентанол и м-крезол. Наиболее предпочтительным консервантом является в данном случае бензиловый спирт.
Настоящее изобретение обеспечивает также фармацевтические композиции, содержащие одно или несколько соединений антител или производных антител, вместе по меньшей мере с одним физиологически приемлемым носителем или эксципиентом. Фармацевтические композиции могут содержать, например, один или несколько из воды, буферов (например, нейтральный забуференный солевой раствор или забуференный фосфатом солевой раствор), этанол, минеральное масло, растительное масло, диметилсульфоксид, углеводы (например, глюкозу, маннозу, сахарозу или декстраны), маннит, белки, адъюванты, полипептиды или аминокислоты, такие как глицин, антиоксиданты, хелатообразующие агенты, такие как ЭДТА, или глутатион и/или консерванты. Как отмечалось выше, в обеспеченные в данном случае фармацевтические композиции могут быть включены (но необязательно) и другие активные ингредиенты.
Носитель является веществом, которое может быть ассоциировано с антителом или производным антитела перед введением его пациенту, часто с целью контролирования стабильности или биодоступности этого соединения. Носители для применения в таких готовых формах обычно являются биосовместимыми и могут также быть биодеградируемыми. Носители включают в себя, например, одновалентные или многовалентные молекулы, такие как сывороточный альбумин (например, человека или коровы), яичный альбумин, пептиды, полилизин и полисахариды, такие как аминодекстран и полиамидоамины. Носители включают в себя также материалы твердого носителя, такие как гранулы и микрочастицы, содержащие, например, полилактат-полигликолат, поли(сополимер лактида с гликолидом), полиакрилат, латекс, крахмал, целлюлозу или декстран. Носитель может нести эти соединения различными путями, в том числе образованием ковалентной связи (непосредственно или через линкерную группу), нековалентным взаимодействием или в виде смеси.
Фармацевтические композиции могут быть приготовлены для любого подходящего способа введения, в том числе, например, местного, перорального, назального, ректального или парентерального введения. В некоторых вариантах осуществления, предпочтительными являются композиции в форме, подходящей для местного применения, например, в виде глазных капель. Другие формы включают в себя, например, пилюли, таблетки, пастилки, лепешки, водные или масляные суспензии, диспергируемые порошки или гранулы, эмульсию, жесткие или мягкие капсулы или сиропы или эликсиры. В других вариантах осуществления, обеспеченные в данном случае композиции могут быть приготовлены в виде лиофилизата. Термин парентеральные, в контексте настоящей заявки, включает в себя подкожную, внутрикожную, внутрисосудистую (например, внутривенную), внутримышечную, спинальную, внутричерепную, внутриоболочечную и внутрибрюшинную инъекцию, а также любой подобный способ инъекции или инфузии.
Фармацевтическая композиция может быть приготовлена в виде стерильной инъецируемой водной или масляной суспензии, в которой модулятор, в зависимости от используемых носителя и концентрации, либо суспендирован, либо растворен в этом носителе. Такая композиция может быть приготовлена в соответствии с известной техникой с использованием подходящих диспергирующих, увлажняющих агентов и/или суспендирующих агентов, таких как вышеупомянутые агенты. Среди приемлемых носителей и растворителей, которые могут быть использованы, находятся вода, 1,3-бутандиол, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве растворителя или суспендирующей среды могут быть использованы стерильные нелетучие масла. Для этой цели может быть использовано любое легкое нелетучее масло, включающее в себя синтетические моно- или диглицериды. Кроме того, в приготовлении инъецируемых композиций могут быть использованы жирные кислоты, такие как олеиновая кислота, и в этом носителе могут быть растворены адъюванты, такие как местные анестетики, консерванты и/или буферящие агенты.
Фармацевтические композиции могут быть приготовлены в виде готовых форм пролонгированного высвобождения (т.е. в виде такой готовой формы, как капсула, которая осуществляет медленное высвобождение модулятора после введения). Такие готовые формы могут быть обычно приготовлены с использованием хорошо известной технологии и введены, например, перорально, ректально или подкожной имлантацией или имплантацией в желаемом участке-мишени. Носители для применения в таких готовых формах являются биосовместимыми и могут быть также биодеградируемыми; предпочтительно эта готовая форма обеспечивает относительно постоянный уровень высвобождения модулятора. Количество антитела или производного антитела, содержащееся в форме пролонгированного высвобождения, зависит, например, от участка имплантации, скорости и ожидаемой продолжительности высвобождения и природы подлежащего лечению или предупреждению заболевания/нарушения.
Антитело или производные антитела, обеспеченные в данном случае, обычно вводят в количестве, которое дает концентрацию в общей воде организма (например, крови, плазме, сыворотке, CSF, синовиальной жидкости, лимфе, клеточной интерстициальной жидкости, слезах или моче), которая является достаточной для детектируемого связывания с VEGF и предотвращения или подавления VEGF-опосредованных заболеваний/нарушений. Доза считается эффективной, если она приводит к явной пользе пациента, как описано в данном случае. Предпочтительные системные дозы находятся в диапазоне от приблизительно 0,1 мг до приблизительно 140 мг на килограмм массы тела в день (приблизительно 0,5 мг - приблизительно 7 г на пациента в день), причем пероральные дозы обычно являются приблизительно в 5-20 раз более высокими, чем внутривенные дозы. Количество антитела или производного антитела, которое может быть объединено с материалами носителя для получения единой лекарственной формы, будет варьироваться в зависимости от хозяина, получающего лечение, и конкретного способа введения. Унифицированные (стандартные) лекарственные формы будут обычно содержать приблизительно 1 мг - приблизительно 500 мг активного ингредиента.
Фармацевтические композиции могут быть упакованы для лечения состояний, чувствительных к антителу или производному антитела, нацеленному на VEGF. Упакованные фармацевтические композиции могут включать в себя контейнер, содержащий эффективное количество по меньшей мере одного антитела или производного антитела, описанного в данном случае, и инструкции (например, этикетку), указывающие, что содержащаяся в контейнере композиция предназначена для применения для лечения заболевания/нарушения, отвечающего на антитело или производное антитела после введения пациенту.
Антитела или производные антител по настоящему изобретению могут быть также химически модифицированы. Предпочтительными модифицирующими группами являются полимеры, например, необязательно замещенный имеющий прямую или разветвленную цепь полиалкеновый, полиалкениленовый, или полиоксиалкиленовый полимер или разветвленный или неразветвленный полисахарид. Такая эффекторная группа может увеличивать период полужизни этого антитела in vivo. Конкретные примеры синтетических полимеров включают в себя необязательно замещенный имеющий прямую или разветвленную цепь поли(этиленгликоль) (ПЭГ), поли(пропиленгликоль), поли(виниловый спирт) или их производные. Конкретные природно-встречающиеся полимеры включают в себя лактозу, амилозу, декстран, гликоген или их производные. Размер этого полимера может варьироваться по желанию, но обычно будет находиться в диапазоне средней молекулярной массы 500 Да – 50000 Да. Для местного применения, в котором это антитело предназначено для проникновения в ткань, предпочтительная молекулярная масса этого полимера равна приблизительно 5000 Да. Эта молекула полимера может быть присоединена к антителу, в частности, к С-концевой стороне тяжелой цепи Fab-фрагмента, через ковалентно связанный пептид шарнирной области, как описано в WO 0194585. Что касается присоединения ПЭГ-частей, делается ссылка на "Poly(ethyleneglycol) Chemistry, Biotechnological and Biomedical Applications", 1992, J. Milton Harris (ed), Plenum Press, New York и "Bioconjugation Protein Coupling Techniques for the Biomedical Sciences", 1998, M. Aslam and A. Dent, Grove Publishers, New York.
После приготовления представляющего интерес антитела или производного антитела, как описано в данном случае, готовят содержащую его фармацевтическую композицию. Антитело, которое должно быть приготовлено в виде этой композиции, не подвергали предшествующей лиофилизации, и представляющая интерес готовая форма является в данном случае водной готовой формой. Предпочтительно, антителом или производным антитела в этой готовой форме является фрагмент антитела, такой как scFv. Терапевтически эффективное количество антитела, присутствующее в этой готовой форме, определяют, например, с учетом желаемых объемов дозы и способа(ов) введения. Примерная концентрация антитела в этой готовой форме равна приблизительно 0,1 мг/мл - приблизительно 50 мг/мл, предпочтительно, приблизительно 0,5 мг/мл - приблизительно 40 мг/мл и наиболее предпочтительно, приблизительно 10 мг/мл - приблизительно 20 мг/мл.
Готовят водную готовую форму, содержащую антитело или производное антитела в рН-забуференном растворе. Буфер по настоящему изобретению имеет рН в диапазоне от приблизительно 4,5 до приблизительно 8,0, предпочтительно, от приблизительно 5,5 до приблизительно 7. Примеры буферов, которые будут контролировать рН в этом диапазоне, включают в себя ацетатный (например, ацетат натрия), сукцинатный (такой как сукцинат натрия), глюконатный, гистидиновый, цитратный буферы и буферы других органических кислот. Концентрация буфера может быть от приблизительно 1 мМ до приблизительно 50 мМ, предпочтительно от приблизительно 5 мМ до приблизительно 30 мМ, в зависимости, например, от этого буфера и желаемой изотоничности готовой формы.
В эту готовую форму включают полиол, который действует в качестве агента тоничности и может стабилизировать это антитело. В предпочтительных вариантах осуществления, эта готовая форма не содержит влияющего на тоничность количества соли, такой как хлорид натрия, так как оно может вызывать осаждение антитела или его производного и/или приводить к окислению при низком рН. В предпочтительных вариантах осуществления, полиол является нередуцирующим сахаром, таким как сахароза или трегалоза. Полиол добавляют к готовой форме в количестве, которое может варьироваться в зависимости от желаемой изотоничности этой готовой формы. Предпочтительно, эта водная готовая форма является изотонической, и в этом случае подходящие концентрации полиола в этой готовой форме находятся, например, в диапазоне от приблизительно 1% до приблизительно 15% м/о, предпочтительно в диапазоне от приблизительно 2% до приблизительно 10% м/о. Однако могут быть также подходящими гипертонические или гипотонические готовые формы. Количество добавляемого полиола может также изменяться в зависимости от молекулярной массы полиола. Например, может добавляться более низкое количество моносахарида (например, маннита) в сравнении с дисахаридом (таким как трегалоза).
К этой готовой форме антитела и/или производного антитела добавляют также поверхностно-активный агент. Примеры поверхностно-активных агентов включают в себя неионогенные поверхностно-активные агенты, такие как полисорбаты (например, полисорбаты 20, 80 и т.д.) или полоксамеры (например, полоксамер 188). Количество добавляемого поверхностно-активного агента является таким, что он уменьшает агрегацию приготовленного антитела/производного антитела и/или минимизирует образование частиц в этой готовой форме и/или уменьшает адсорбцию. Например, поверхностно-активный агент может присутствовать в этой готовой форме в количестве от приблизительно 0,001% до приблизительно 0,5%, предпочтительно, от приблизительно 0,005% до приблизительно 0,2% и наиболее предпочтительно, от приблизительно 0,01% до приблизительно 0,1%.
В одном из вариантов осуществления, эта готовая форма содержит вышеуказанные агенты (т.е. антитело или производное антитела, буфер, полиол и поверхностно-активный агент) и является по существу свободной от одного или нескольких консервантов, таких как бензиловый спирт, фенол, м-крезол, хлорбутанол и хлорид бензетония. В другом варианте осуществления, консервант может быть включен в эту готовую форму, особенно, когда эта готовая форма является многодозовой готовой формой. Концентрация консерванта может находиться в диапазоне от приблизительно 0,1% до приблизительно 2%, наиболее предпочтительно, от приблизительно 0,5% до приблизительно 1%. Один или несколько других фармацевтически приемлемых носителей, эксципиентов или стабилизаторов, таких как описанные в Remington's Pharmaceutical Sciences 21st edition, Osol, A. Ed. (2006), могут быть включены в готовую форму, при условии, что они не оказывают вредного действия на желаемые характеристики этой готовой формы. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов при используемых дозах и концентрациях и включают в себя: дополнительные буферящие агенты, сорастворители, антиоксиданты, в том числе аскорбиновую кислоту и метионин, хелатообразующие агенты, такие как ЭДТА, комплексы с металлами (например, комплексы Zn-белок), биодеградируемые полимеры, такие как полиэфиры, и/или солеобразующие противоионы, такие как натрий.
Готовые формы, которые должны использоваться для введения in vivo, должны быть стерильными. Это легко осуществляется фильтрованием через мембраны стерильного фильтрования до или после приготовления этой готовой формы.
Готовую форму вводят млекопитающему, нуждающемуся в лечении этим антителом, предпочтительно человеку, в соответствии с известными способами, такими как внутривенное введение в виде болюса или непрерывной инфузии на протяжении некоторого периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным, внутрисуставным, внутрисиновиальным, интратекальным, пероральным, местным или ингаляционным способами. В предпочтительных вариантах осуществления эту готовую форму вводят млекопитающему местным внесением глазных капель на поверхность глаза. Для этих целей, эта готовая форма может вноситься при помощи, например, аппликатора для глазных капель.
Подходящая доза (“терапевтически эффективное количество”) этого антитела будет зависеть, например, от подлежащего лечению состояния, тяжести и течения этого состояния, от того, вводится ли это антитело для превентивной или терапевтической целей, предыдущей терапии, клинической истории пациента и реакции на это антитело, типа используемого антитела и от решения лечащего врача. Это антитело или производное антитела вводят подходящим образом этому пациенту один раз или на протяжении ряда обработок, и оно может вводиться этому пациенту в любое время от диагностики и далее. Это антитело или производное антитела может вводиться в виде единственного лекарственного средства обработки или вместе с другими лекарственными средствами или терапиями, применимыми в лечении рассматриваемого состояния.
В качестве обычного предложения, терапевтически эффективное количество антитела или производного антитела будет вводиться в диапазоне приблизительно 0,1 - приблизительно 50 мг/кг массы тела пациента посредством одного или нескольких введений, причем обычный диапазон используемого антитела равен приблизительно 0,3 - приблизительно 20 мг/кг, более предпочтительно, приблизительно 0,3 - приблизительно 15 мг/кг, вводимых, например, один раз в день. Однако могут быть применимы и другие схемы введения доз. Прогресс этой терапии может быть легко подвергнут мониторингу с использованием общепринятых способов.
Рассматриваются одобренные FDA (Департаментом по контролю за качеством пищевых продуктов, медикаментов и косметических средств (США)) дозы и схемы лечения, подходящие для применения с Lucentis.
Другие дозы и схемы лечения описаны в предварительной заявке США с порядковым номером 61/075641 "Улучшенные препараты иммуносвязывающего агента и способы введения", поданной 25 июня 2008 года, которая особо включена в настоящее описание в качестве ссылки.
Изделия
В другом варианте осуществления настоящее изобретение, относится к изделию, содержащему контейнер, в котором находится водная фармацевтическая готовая форма по настоящему изобретению, и обеспечены инструкции для его использования. Подходящие контейнеры включают в себя, например, склянки, флаконы, аппликаторы глазных капель и шприцы. Этот контейнер может быть выполнен из различных материалов, таких как стекло или пластик. Примерным контейнером является одноразовый стеклянный или пластиковый флакон на 3-20 кубических сантиметров. Альтернативно, для многодозовой готовой формы этот контейнер может быть стеклянным флаконом на 3-100 см3. Этот контейнер содержит готовую форму, и этикетка на этом контейнере или приложенная к этому контейнеру может содержать инструкции для применения. Это изделие может дополнительно включать в себя другие материалы, желательные с коммерческой точки зрения и с точки зрения пользователя, включающие в себя другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши упаковки с инструкциями для применения.
Пояснение примерами
Данное описание дополнительно иллюстрируется следующими примерами, которые не должны рассматриваться в качестве дополнительного ограничения. Содержания всех фигур и всех ссылок, патентов и опубликованных заявок на патенты, цитируемых по всей этой заявке, специально включены в настоящее описание в качестве ссылки в их полном объеме.
Во всех этих примерах использовали следующие материалы и способы, если нет других указаний.
Общие материалы и способы
В общем, практика по настоящему изобретению использует, если нет других указаний, общепринятые способы химии, молекулярной биологии, технологии рекомбинантных ДНК, иммунологии (в частности, технологии антител) и стандартные способы получения полипептидов. См., например, Sambrook, Fritsch and Maniatis, Molecular Cloning: Cold Spring Harbor Laboratory Press (1989); Antibody Engineering Protocols (Methods in Molecular Biology), 510, Paul, S., Humana Pr (1996); Antibody Engineering: A Practical Approach (Practical Approach Series, 169), McCafferty, Ed., Irl Pr (1996); Antibodies: A Laboratory Manual, Harlow et al, C.S.H.L. Press, Pub. (1999); и Current Protocols in Molecular Biology, eds. Ausubel et al, John Wiley & Sons (1992).
Измерения термостабильности
Спектры, полученные с использованием спектроскопии ослабленного полного отражения/инфракрасной спектроскопии с Фурье-преобразованием (ATR-FTIR), получали для различных отдельных цепей и модифицированных молекул с использованием ячейки FT-IR Bio-ATR в тензоре (Bruker). Эти молекулы концентрировали до 3 мг/мл и диализовали в течение ночи при 4оС против PBS, рН 6,5, и протекающий поток буфера собирали в качестве «слепого» опыта. Профили денатурации получали термообработкой этих молекул широким диапазоном температур в 5°С-стадиях (25-95°С). Все манипуляции со спектрами выполняли с использованием компьютерной программы OPUS. Фон основного буфера и транзиторный атмосферный фон (CO2 и H2O) вычитали из спектра белка. Затем полученный спектр белка корректировали относительно фона и определяли спектры амида I белка из ширины самого широкого разделяемого пика в ожидаемой области. Спектры вторых производных получали для спектров полосы амида I с использованием полиномиальной (многочленной) функции третьей степени со сглаживающей функцией. Изменения в структуре белков оценивали анализом вторых производных амида I с использованием линейной калибровочной кривой для начальных расчетов подгонки кривой с предположением 0% денатурации для 3 более низких измерений и 100% денатурации для 3 более высоких измерений. Профили денатурации использовали для аппроксимирования средних точек термических развертывающих переходов (ТМ) для каждого варианта с применением сигмоидной модели Болтцмана.
Измерения растворимости
Относительную растворимость различных молекул scFv измеряли после усиления агрегации белка и осаждения в присутствии сульфата аммония. Сульфат аммония добавляли к этому белку в водном растворе с получением приращений 5% насыщения в конечной смеси соль-белок. Осаждение в динамическом диапазоне определяли эмпирически и интервалы насыщения уменьшали в этом диапазоне до насыщения с 2,5% интервалами в конечной смеси. После добавления сульфата аммония, пробы осторожно перемешивали и центрифугировали 30 минут при 6000 об/мин. Остающийся белок в супернатантах извлекали для каждого процента насыщения сульфата аммония. Кривые растворимости определяли измерением концентрации белка в супернатанте при помощи измерения UV-VIS (УФ-видимый свет) с использованием спектрофотометра NanoDropTM-1000. Измерения остающегося растворимого белка в супернатантах нормализовали и использовали для оценивания средних точек относительной растворимости для каждого варианта с применением сигмоидной модели Болтцмана.
Краткосрочный тест стабильности
Молекулы scFv испытывали после двухнедельной инкубации при 40°С на присутствие растворимых агрегатов и продукты деградации. Белки с концентрацией 10 мг/мл диализовали в течение ночи при 4°С против PBS с широким диапазоном рН (3,5, 4,5, 5,5, 6,5, 7,0, 7,5 и 8,5). Контрольные молекулы с той же самой концентрацией в стандартном буфере PBS (рН 6,5) хранили при -80°С на протяжении периода 2 недель. Определение полос деградации электрофорезом в ДСН/ПААГ выполняли при временных точках t=0 и t=14 дней и растворимые агрегаты оценивали в SEC-HPLC. Определение остающейся активности после 2 недель при 40°С выполняли при помощи Biacore.
ПРИМЕР 1
СТРАТЕГИЯ ИММУНИЗАЦИИ ДЛЯ ГЕНЕРИРОВАНИЯ АНТИТЕЛ ПРОТИВ VEGF
В этом примере описывается стратегия иммунизации, которая использует новый антигенный VEGF-произведенный пептид, для генерирования антител, способных узнавать VEGFА человека, мыши и кролика.
Из исследований с использованием аланин-сканирующего мутагенеза, выполняемых в Genentech, известны остатки VEGFA, которые являются решающими для высокоаффинного взаимодействия с VEGFr (Fuh, G. et al, (2006) J. Biol Chem. 281, 6625-6631). Хотя этот рецепторсвязывающий сайт, возможно, представляет собой конформационный эпитоп, большинство этих решающих остатков лежат на альфа-спирали, на первых 10 аминокислотах зрелого VEGFА.
Кроличий VEGFA содержит три изменения аминокислот в этой альфа-спирали при сравнении с последовательностью человека; в отличие от этого, мышиный VEGFA является идентичным VEGFА человека в этом районе. Таким образом, для генерирования мышь-человек перекрестно реагирующих антител, кролик представляет подходящий вид для иммунизации. Кроме того, иммунизация кролика может привести к Ab с более высокой аффинностью, чем иммунизация мыши.
Как описано выше, взаимодействие с остатками на N-концевой альфа-спирали VEGFA, по-видимому, является наиболее важной для связывания с VEGFR1. Таким образом, этот состоящий из 10 аминокислот сегмент может быть использован в качестве эпитопа для иммунизации. Альтернативно, может быть инъецирован полноразмерный VEGFА, однако, другие пептидные сегменты на VEGFA являются более иммуногенными, понижая таким образом шанс индуцирования нейтрализующих антител. Эта гипотеза подтверждается тем фактом, что два разных пептида, оба из которых лежат вблизи С-конца VEGFA, являются потенциально иммуногенными, как спрогнозировано с помощью способа Johnson и Wolf. Этот способ прогнозирует только минорный иммуногенный потенциал для N-концевой альфа-спирали. Таким образом, иммунизация пептидом, составляющим только альфа-спираль, может быть более простой, чем иммунизация полноразмерным VEGFA. Вероятность индуцирования сильной иммунной реакции может быть дополнительно увеличена слиянием или химическим связыванием пептида гемоцианина морского моллюска (KLH).
Четыре стратегии иммунизации выполняли следующим образом
A. Предварительная иммунизация кроликов полноразмерным VEGFA165 человека для усиления возможности получения конформационных связывающих агентов. Второй бустинг пептидом из сегмента аминокислот 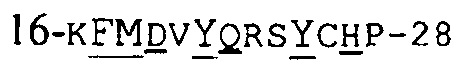 (подчеркнутые: взаимодействие с рецептором; дважды подчеркнутые, дивергентные в кролике, Cys участвует в дисульфидной связи согласно структуре кристалла). Cys, содержащийся в этой пептидной последовательности, мог бы быть использован для связывания KLH и, следовательно, мог не быть экспонирован в виде свободного Cys. Конечный пептид мог бы выглядеть следующим образом: KFMDVYQRSY-Cys-KLH.
(подчеркнутые: взаимодействие с рецептором; дважды подчеркнутые, дивергентные в кролике, Cys участвует в дисульфидной связи согласно структуре кристалла). Cys, содержащийся в этой пептидной последовательности, мог бы быть использован для связывания KLH и, следовательно, мог не быть экспонирован в виде свободного Cys. Конечный пептид мог бы выглядеть следующим образом: KFMDVYQRSY-Cys-KLH.
B. Предварительная иммунизация мышей полноразмерным VEGFA165 для усиления возможности получения конформационных связывающих агентов. Второй бустинг пептидом из сегмента аминокислот 16- 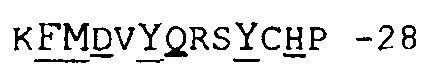 (Cys участвует в дисульфидной связи согласно структуре кристалла). Cys, содержащийся в этой пептидной последовательности, мог бы быть использован для связывания KLH и, следовательно, мог не быть экспонирован в виде свободного Cys. Конечный пептид мог бы выглядеть следующим образом: KFMDVYQRSY-Cys-KLH.
(Cys участвует в дисульфидной связи согласно структуре кристалла). Cys, содержащийся в этой пептидной последовательности, мог бы быть использован для связывания KLH и, следовательно, мог не быть экспонирован в виде свободного Cys. Конечный пептид мог бы выглядеть следующим образом: KFMDVYQRSY-Cys-KLH.
C. Предварительная иммунизация кроликов/мышей пептидом из сегмента 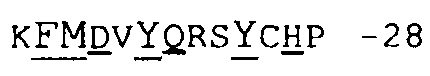 (конечный пептид: KFMDVYQRSY-Cys-KLH). Второй бустинг полноразмерным VEGFA16S для усиления вероятности получения конформационных связывающих агентов.
(конечный пептид: KFMDVYQRSY-Cys-KLH). Второй бустинг полноразмерным VEGFA16S для усиления вероятности получения конформационных связывающих агентов.
D. Иммунизация полноразмерным VEGFA165 в кроликах.
ПРИМЕР 2
Трансплантация CDR и функциональная гуманизация моноклональных антител против VEGF
Трансплантация кроличьих CDR
В отличие от традиционных способов гуманизации, которые используют акцепторный каркас антитела человека, который имеет наибольшую гомологию последовательности с донорским антителом не человека, кроличьи CDR трансплантировали либо в каркас FW1.4 (SEQ ID NO: 172) для генерирования Min-graft, либо в модифицированный частями кроличьего антитела "rabbitized" каркас rFW1.4 (SEQ ID NO: 173) или его вариант rFW1.4(v2) (SEQ ID NO: 174) для генерирования Max-graft. Оба каркаса отбирали сначала на желаемые функциональные свойства (растворимость и стабильность), структурную пригодность для вмещения большого разнообразия кроличьих CDR и приемлемую гомологию с кроличьей консенсусной последовательностью вариабельного домена. Каркас rFW1.4 является производным FW1.4, которое было дополнительно сконструировано с той целью, чтобы служить в качестве универсального акцепторного каркаса для фактически любого набора кроличьих CDR. Хотя стабильная и растворимая каркасная последовательность FW1.4 проявляет высокую гомологию с кроличьими антителами, она не является доступной наиболее гомологичной последовательностью.
Идентификация остатков, потенциально участвующих в связывании
Для каждой последовательности вариабельного домена кролика, идентифицировали ближайшую копию зародышевой линии кролика. Если эта ближайшая зародышевая линия не могла быть установлена, эту последовательность сравнивали против консенсуса этой подгруппы или консенсуса кроличьих последовательностей с высоким процентным сходством. Редкие каркасные последовательности считали возможным результатом соматической гипермутации и, следовательно, играющими роль в связывании антигена. Таким образом, такие остатки обсуждали для трансплантации на акцепторный каркас rFW1.4 или rFW1.4(v2) для генерирования Max-graft. В частности, остатки, потенциально участвующие в прямом контакте антигена или влияющие на размещение VL и VH, трансплантировали. Дополнительные остатки, описанные, как влияющие на структуру CDR, заменяли, если требовалось. При трансплантации CDR на FW1.4 (Min-grafts) не производили каркасных замен. Например, для генерирования остатка 578minmax VH 94 (H94), rFW1.4 мутировали в соответствующий остаток в донорской последовательности. Кроличье антитело 578 содержит Gly в H94, тогда как обе, наиболее гомологичная зародышевая линия, и кроличья консенсусная последовательность, содержат Arg в положении H94. Gly имеет исключительную гибкость (положительные углы phi), которая не обнаруживается в отношении других аминокислот. Это предполагает роль в угле закручивания основной цепи и возможное сильное влияние петлевой конформации с последствиями в отношении активности. Дополнительные примеры положений в каркасе, которые были трансплантированы для получения Max-graft, как описано в данном случае, могут быть идентифицированы сопоставлением последовательностей каркасных районов rFW1.4, rFW1.4(v2) и представляющих интерес обеспеченных в данном случае последовательностей scFv. Для указанной цели может быть использован инструментарий Web, известный в данной области (например, ClustalW, доступный с 23 июня 2009 года в http://www.ebi.ac.uk/Tools/clustalw2/index.html или MultiAlin, доступный с 23 июня 2009 года в http://bioinfo.genotoul.fr/multalin). Все положения каркаса, в которых rFW1.4 и rFW1.4(v2) содержат один и тот же остаток и в которых представляющий интерес scFv обнаруживает другой остаток, являются положениями каркаса, которые были трансплантированы для получения Max-graft.
Перетасовка доменов
Вариабельные легкие цепи Min-graft комбинировали с вариабельной тяжелой цепью Max-graft для идентификации оптимальных комбинаций в отношении биофизических свойств (растворимости и стабильности) и активности.
Клонирование и экспрессия scFv
Описанные и охарактеризованные в данном случае scFvs получали следующим образом. Гуманизированные последовательности VL (SEQ ID NO: 82-106) соединяли с гуманизированными последовательностями VH (SEQ ID NO: 118-166) через линкер SEQ ID NO: 181 с получением scFv следующей ориентации: NH2-VL-linker-VH-COOH. Во многих случаях ДНК-последовательности, кодирующие различные scFv, синтезировали de novo в предоставляющей услуги компании (SP-компании) Entelechon GmbH (www.entelechon.com). Полученные ДНК-инсерты клонировали в бактериальный экспрессирующий вектор pGMP002 через сайты рестрикции NcoI и HindIII, введенные в 5’- и 3’-конец ДНК-последовательности scFv, соответственно. Между ДНК-последовательностью VL-домена и VH-домена расположен сайт рестрикции BamHI. В некоторых случаях кодирующую scFv ДНК не синтезировали de novo, и экспрессирующие scFv конструкции клонировали перетасовкой доменов. Таким образом, VL-домены вырезали и вводили в новые конструкции через сайты рестрикции NcoI и BamHI, VH-домены через сайты рестрикции BamHI и HindIII. В других случаях точковые мутации вводили в VH- и/или VL-домен с использованием известных в данной области ПЦР-способов сборки. Клонирование GMP002 описано в примере 1 WO2008006235. Получение scFv выполняли аналогично получению ESBA 105, как описано в примере 1 WO2008006235.
ПРИМЕР 3
АНАЛИЗ BIACORE СВЯЗЫВАНИЯ анти-VEGF-scFv
В этом примере тестировали Biacore-связывающую способность scFvs и аффинность связывания измеряли с использованием примерного способа резонанса поверхностных плазмонов при помощи BIAcore™-T100. VEGF-белки, тестируемые на связывание этими кандидатными scFv, в этом примере и последующих примерах, включают в себя очищенный экспрессируемый Escherichia coli рекомбинантный VEGF165 человека (PeproTech EC Ltd.), рекомбинантный VEGF121 человека (PeproTech EC Ltd.), рекомбинантный VEGF110 человека (ESBATech AG), рекомбинантный мышиный VEGF164 (PeproTech EC Ltd.), рекомбинантный крысиный VEGF164 (Biovision), рекомбинантный кроличий VEGF110 (ESBATech AG) и рекомбинантный PLGF человека (PeproTech EC Ltd.). Для эксперимента с использованием резонанса поверхностных плазмонов, биосенсорные чипы (микропроцессоры) из карбоксиметилированного декстрана (CM4, GE Healthcare) активировали гидрохлоридом N-этил-N’-(3-диметиламинопропил)карбодиимида и N-гидроксисукцинимидом в соответствии с инструкциями поставщика. Каждую из 6 различных форм VEGF, представленных выше, связывали с 1 из 4 разных проточных ячеек на сенсорном чипе СМ4 с использованием стандартной процедуры связывания амина. Диапазоны реакций, полученных с этими иммобилизованными молекулами VEGF после связывания и блокирования, были равны ~250-500 единицам реакции (RU) для hVEGF165, ~200 RU для hVEGF110, hVEGF121, мышиного VEGF164, крысиного VEGF164 и кроличьего VEGF110 и ~400 RU для PLGF. 4-ую проточную ячейку каждого чипа обрабатывали подобным образом, за исключением того, что перед блокированием в них не иммобилизовали белки, и эту проточную ячейку использовали в качестве включенного в этот эксперимент эталона. Различные концентрации анти-VEGF-scFv (например, 90 нМ, 30 нМ, 10 нМ, 3,33 нМ, 1,11 нМ, 0,37 нМ, 0,12 нМ и 0,04 нМ) в буфере HBS-EP (0,01M HEPES, pH 7,4 или 5, 0,15M NaCl, 3 мМ ЭДТА, 0,005% поверхностно-активный агент P20) инъецировали в эти проточные ячейки при скорости потока 30 мкл/мин в течение 5 минут. Диссоциации анти-VEGF-scFv из VEGF на чипе CM4 давали протекать в течение 10 мин при 25°C. Генерировали сенсограммы для каждой пробы анти-VEGF-scFv после коррекции с использованием включенного в измерения эталона с последующим вычитанием пробы буфера. Среднюю (кажущуюся) константу скорости диссоциации (kd), среднюю константу скорости ассоциации (ka) и среднюю константу диссоциации в равновесном состоянии (KD) рассчитывали с использованием взаимно однозначной модели связывания Langmuir с применением версии 1.1 программы BIAcore T100 evaluation.
В качестве примерного результата, некоторые основные кандидатные анти-VEGF-scFv приведены в списке таблицы 7, показывающей их аффинность связывания с hVEGF165. Их эффективность в качестве ингибиторов VEGF, которая измеряется с использованием конкурентного ELISA VEGFR и/или HUVEC-анализа и описывается в последних примерах, показана также в таблице 7. Кривые кинетики некоторых примерных главных кандидатов, например, 51lmax и 578max, в отношении их связывания с hVEGF165 иллюстрированы на фиг.1. Определяли также их константы аффинности (kd, ka и KD). Некоторые главные кандидаты проявляют также видоспецифичность в их связывании с различными белками VEGF из различных источников. Например, некоторые данные аффинности, измеренные при рН 5 с использованием VEGF164 мыши и крысы в качестве партера связывания, показаны в таблицах 8а и b. Один примерный главный кандидат, 578minmax, имеет KD of 5,76Е-10M и 7,48Е-10M в его связывании с VEGF164, соответственно, при рН 5 (таблицы 8а и 8b) и 2,73Е-11 и 2,19Е-11 при рН 7,4 (данные не показаны). Эта видоспецифичность дополнительно иллюстрируется на фиг.4 в кинетических кривых и данных аффинности в отношении связывания между белками 578minmax и VEGF человека, мыши или крысы.
Наряду с видоспецифичностью в их связывании с VEGF из различных организмов, многие ведущие кандидатные scFv обнаруживают также различные аффинности связывания в отношении различных изоформ VEGF. Например, данные аффинности, измеренные при рН 5,0 для связывания нескольких scFv-кандидатов с VEGF165, VEGF121 и VEGF110 человека, сравниваются в таблице 9. В тех же самых экспериментах использовали белок PIGF в качестве отрицательного контроля без способности связывания с этими кандидатными scFv. Различные кинетические кривые и данные аффинности, например, для связывания между 578Max и изоформами VEGF, иллюстрируются также на фиг.3.
Данное изобретение описывает также производные, происходящие из вышеупомянутых главных кандидатных анти-VEGF-scFv. Некоторые главные производные кандидатов 578 и 511, перечисленные в таблице 10, иллюстрируются в отношении их аффинности и эффективности (измеренных при рН 5,0). В этом эксперименте использовали Biacore-измерение для аффинности этих производных в отношении hVEGF165, тогда как конкурентный ELISA hVEGFR2 и/или HUVEC-анализ использовали для определения их эффективности ингибирования VEGF (таблица 10). Три производных, 578max, 578minmax и 578 wt-His, дополнительно иллюстрированы в их кинетических кривых и данных аффинности в отношении связывания с hVEGF165 на фиг.4.
Для производных главных кандидатов определяли их биофизические характеристики, и они представлены на фиг.5-7 и в таблице 11. Эти характеристики включают в себя, как показано в таблице 11, Tm, определенную при помощи FTIR, процент потери β-складки или белка после инкубирования при 60°C в течение 30 мин, растворимость, определенную осаждением сульфатом аммония, выход рефолдинга во время процесса получения и уровни экспрессии в E. coli. Три производных, 578max, 578minmax и 578minmax_DHP, характеризовали в отношении их термостабильности в их кривых разворачивания в зависимости от разных температур, посредством FT-IR (фиг.5).
Таблица 7
Обзор аффинности и эффективности главных кандидатов
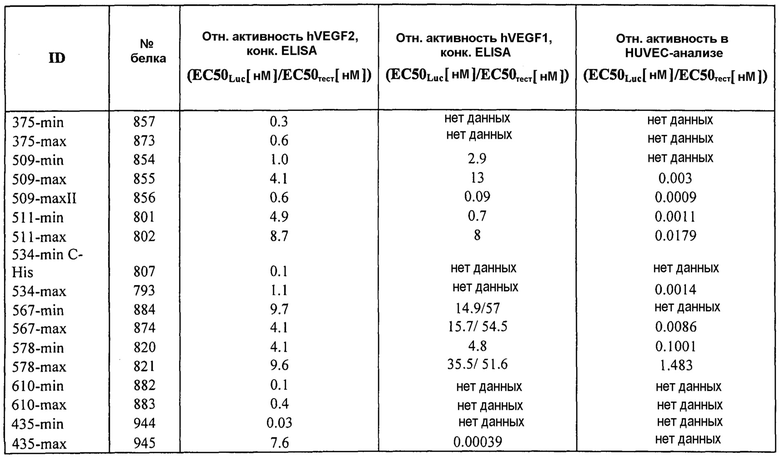
(таблица 7, продолжение)
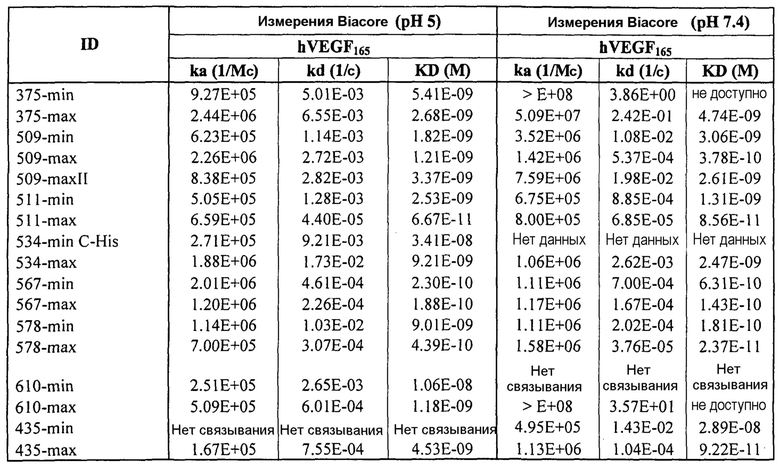
Таблица 8а
Видоспецифичность отобранных главных кандидатов (VEGF 164 мыши и крысы)
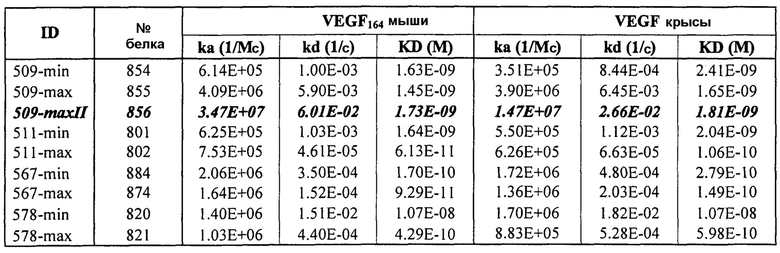
Таблица 8b
Видоспецифичность отобранных разработанных кандидатов
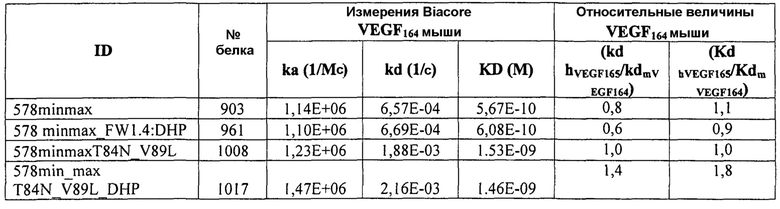
(продолжение)
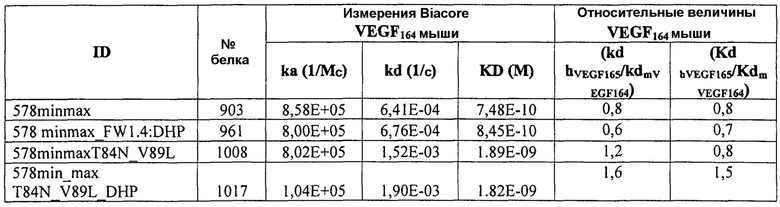
Таблица 9
Связывание отобранных главных кандидатов изоформ VEGF (VEGF121 человека и hVEGF110)
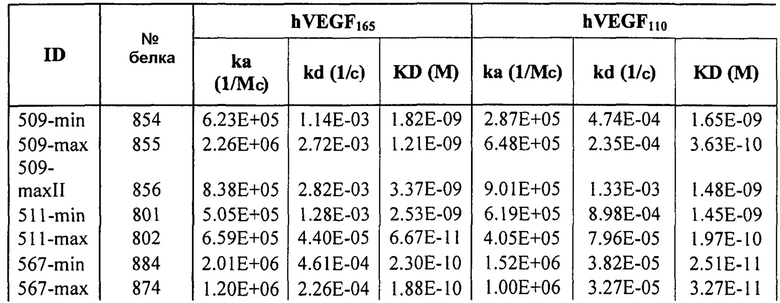

(продолжение)
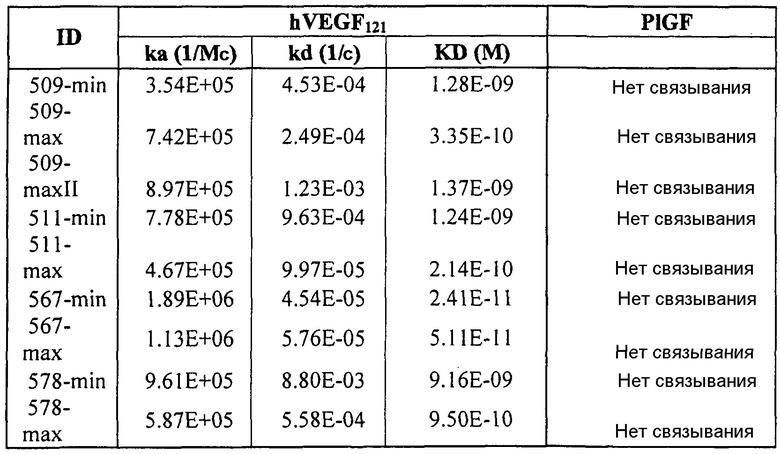
Таблица 10
Обзор аффинности и эффективности главных производных (578 и 511)
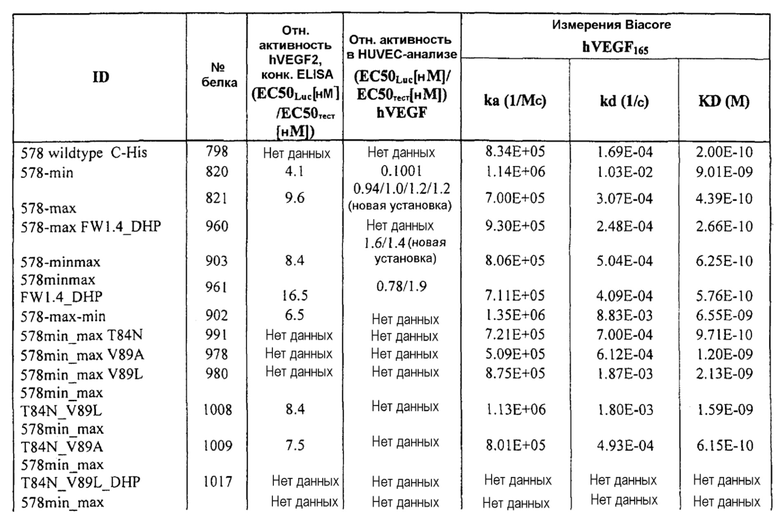
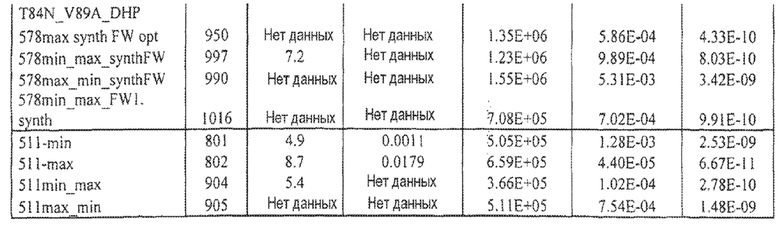
Таблица 11
Обзор биофизических характеристик главных производных (578 и 511)
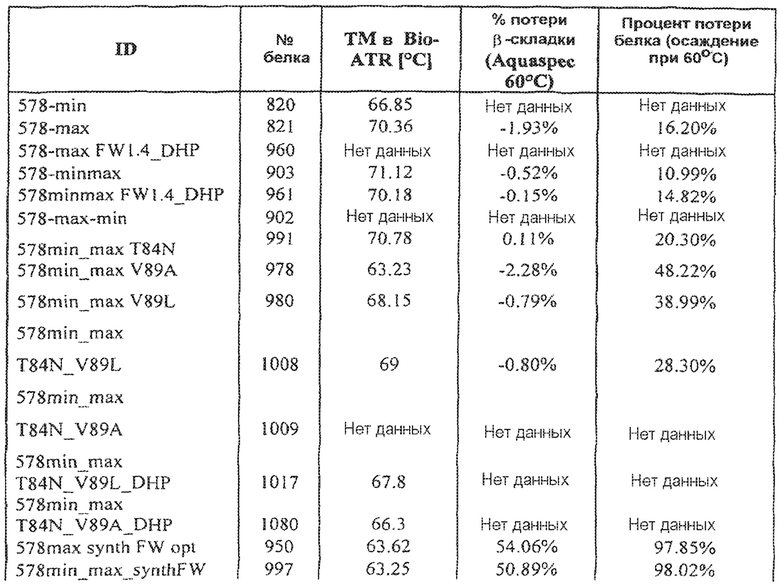
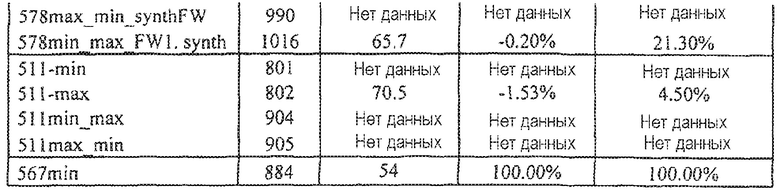
(продолжение)


Некоторые производные, перечисленные на фиг.6, сравнивали в отношении их денатурации и осаждения после теплового стресса (например, при 50оC, 60оC или 70оC) в течение 30 минут. 578max, 578minmax и 578minmax_DHP дополнительно иллюстрировали в отношении их растворимости, которую определяли посредством осаждения сульфатом аммония. Как показано на фиг.7, сравнивали процент растворимых белков этих производных при различных концентрациях сульфата аммония.
ПРИМЕР 4
АНАЛИЗЫ БЛОКИРОВАНИЯ РЕЦЕПТОРА VEGF
Для кандидатов анти-VEGF-scFv или их производных, описанных в данном изобретении, измеряли также их эффективность в качестве ингибиторов VEGF, наряду с их аффинностью связывания с VEGF в примере 3. Способы измерения их эффективности включают в себя, например, конкурентный ELISA VEGFR, иллюстрируемый в этом примере, и HUVEC-анализы (фиг.8).
Конкурентные анализы ELISA VEGFR включают в себя, например, анализы блокирования рецептора VEGFR2 и анализы блокирования VEGFR1. Для анализа блокирования рецептора VEGFR2, VEGF165 человека наносили в виде покрытия на 96-луночный планшет Maxisorp ELISA (Nunc) при 0,05 мкг/мл в PBS и блокировали с использованием PBS с 0,1% BSA и 0,2% Твином 20 (PBST). 500 нг/мл рекомбинантного химерного антитела VEGFR2 человека/Fc (R&D Systems Inc.), состоящего из аминокислотных остатков 1-764 внеклеточного домена VEGFR2 человека, слитого с меченым 6х гистидинами Fc IgG1 человека, сначала инкубировали с 3-кратно серийно разведенными анти-VEGF-scFv в PBST. После 30-60 минут инкубирования при комнатной температуре, эти смеси переносили в планшет с иммобилизованным VEGF165 человека и инкубировали в течение 90 минут. Связывание этой химеры VEGFR2/Fc с иммобилизованным VEGF165 детектировали козьим (Fab2) против Fcγ IgG человека, связанным с пероксидазой хрена (Jackson ImmunoResearch) с последующим добавлением субстрата (BM Blue POD substrate, Roche Diagnostics). Оптическую плотность при 450 нм (OD 450 нм) измеряли при помощи микропланшет-ридера Sunrise (Tecan). Данные анализировали с использованием подгонки четырехпараметрической логистической кривой и величины EC50 рассчитывали из кривых доза-ответ этих scFv. Примерная эффективность главных кандидатов или их производных, измеренная анализом блокирования рецептора VEGFR2, приведена в списке в таблице 7 и 9.
Для анализа блокировании рецептора VEGFR1, VEGF165 человека наносили в виде покрытия на 96-луночный планшет Maxisorp ELISA (Nunc) при 0,0125 мкг/мл в PBS и блокировали с использованием PBS с 0,4% BSA и 0,1% Твином 20 (PBST). 100 нг/мл рекомбинантного химерного антитела VEGFR1 человека/Fc (R&D Systems Inc.), состоящего из аминокислотных остатков 1-687 внеклеточного домена VEGFR1 человека, слитого с меченым 6х гистидинами Fc IgG1 человека, сначала инкубировали с 3-кратно серийно разведенными анти-VEGF-scFv в PBST. После 30-60 минут инкубирования при комнатной температуре, эти смеси переносили в планшет с иммобилизованным VEGF165 человека и инкубировали в течение 90 минут. Связывание этой химеры VEGFR1/Fc с иммобилизованным VEGF165 детектировали козьим (Fab2) против Fcγ IgG человека, связанным с пероксидазой хрена (Jackson ImmunoResearch) с последующим добавлением субстрата (BM Blue POD substrate, Roche Diagnostics). Оптическую плотность при 450 нм (OD 450 нм) измеряли при помощи микропланшет-ридера Sunrise (Tecan). Данные анализировали, как описано выше, и величины EC50 рассчитывали из кривых доза-ответ этих scFv. Примерная эффективность главных кандидатов, измеренная анализом блокирования рецептора VEGFR1, приведена в списке в таблице 7.
ПРИМЕР 5
HUVEC-АНАЛИЗ ИНГИБИРОВАНИЯ VEGF
Этот пример иллюстрирует HUVEC-анализы в качестве другого способа измерения эффективности описанных кандидатных анти-VEGF-scFv или их производных в качестве ингибиторов VEGF.
Эндотелиальные клетки пупочной вены человека (HUVEC) (Promocell), объединенные из нескольких доноров, использовали при пассаже 2 – пассаже 14. Клетки высевали при 1000 клетках на лунку в 50 мкл полной среды для выращивания эндотелиальных клеток (ECGM) (Promocell), которая содержала 0,4% ECGS/H, 2% фетальную телячью сыворотку, 0,1 нг/мл эпидермального фактора роста, 1 мкг/мл гидрокортизона, 1 нг/мл основного фактора фибробластов и 1% пенициллин/стрептомицин (Gibco). Спустя 7-8 часов, добавляли к клеткам 50 мкл истощенной среды (ECGM без добавок, содержащая 0,5% инактивированную нагреванием FCS и 1% пенициллин-стрептомицин), и эти клетки лишали питательной среды в течение 15-16 часов. Готовили 3-кратные серийные разведения анти-VEGF-scFv (0,023-150 нМ) и получали один из следующих VEGF: рекомбинантный VEGF165 человека (0,08 нМ), рекомбинантный VEGF165 мыши (0,08 нМ) или рекомбинантный VEGF165 крысы (0,3 нМ) в истощенной среде и предынкубировали в течение 30-60 мин при комнатной температуре. Различные концентрации VEGF использовали для компенсации в отношении их различных относительных биологических активностей. Использовали концентрации, которые стимулируют субмаксимальную VEGF-индуцированную пролиферацию (EC90). 100 мкл этих смесей добавляли в 96-луночные планшеты для культуры ткани, содержащие суспензию HUVEC, и инкубировали в течение 4 дней в увлажненном термостате 37°C/5% CO2. Пролиферацию HUVEC оценивали измерением оптической плотности при 450 нм (в качестве ссылочной длины волны использовали 620 нм) после добавления 20 мкл на лунку реагента пролиферации WST-1 (Roche), с использованием микропланшетридера Sunrise (Tecan). Данные анализировали с использованием подгонки четырехпараметрической логистической кривой и концентрацию анти-VEGF-scFv, необходимую для ингибирования пролиферации HUVEC на 50% (EC50), получали из кривых ингибирования.
Примерная эффективность главных кандидатов или их производных, измеренная анализами HUVEC, приведена в списке в таблице 7. Кроме того, ингибирование hVEGF165-индуцированной пролиферации HUVEC одним производным главных кандидатов, 578minmax, иллюстрировано на фиг.9. Было определено, что EC50 578minmax для ингибирования hVEGF165-индуцированной пролиферации равна 0,06 нМ (фиг.9). Эффективность 578minmax в качестве ингибитора VEGF приблизительно в 1,6 раз лучше в сравнении Lucentis. Ингибирование hVEGF165-индуцированной пролиферации HUVEC с использованием 578minmax также иллюстрируется на фиг.10. EC50 578minmax для ингибирования индуцированной VEGF164 мыши и крысы пролиферации клеток равна 0,06 нМ и 0,07 нМ, соответственно (фиг.10). Таким образом, мышиный и крысиный VEGF являются равнозначными VEGF человека в отношении их ингибирования примерным производным (578minmax). В этом документе Lucentis также не ингибирует пролиферацию, индуцируемую VEGF грызунов.
ПРИМЕР 6
ДЕЙСТВИЯ анти-VEGF-scFv НА ИНДУЦИРОВАННУЮ HVEGF165 ПРОНИЦАЕМОСТЬ СОСУДОВ В БЕСШЕРСТНЫХ МОРСКИХ СВИНКАХ
В этом примере, действие анти-VEGF-scFv на индуцированную VEGF165 человека проницаемость сосудов оценивали на морской свинке с использованием анализа Miles. Тридцать участков нанесения на одно животное отмечали на спине бесшерстных самцов морских свинок с использованием постоянного маркера. В день обработки каждому животному вводили внутривенно 1 мл 1% раствора красителя Evans blue под общей анестезией. Спустя один час после инъекции красителя, инъецировали 0,1 мл тест-раствора, содержащего 2,61 нМ рекомбинантного VEGF165 человека (PeproTech EC Ltd.), и различные концентрации анти-VEGF-scFv (0 нМ, 0,085 нМ, 0,256 нМ, 0,767 нМ, 2,3 нМ, 6,9 нМ, 20,7 нМ, 62,1 нМ; n=7 животных на тестируемый компонент) в трех повторностях в метки на спине (3 инъекции на каждую концентрацию тестируемого компонента). В качестве отрицательного контроля во всех животных служил PBS. В качестве дополнительного контроля, во всех животных инъецировали 6,9 нМ Lucentis (Nоvartis).
Спустя один час после инъекции тест-раствора, животных эвтанизировали и шкурки собирали, очищали и фотографировали цифровым фотоаппаратом с использованием падающего света и проходящего света. Площадь красителя Evans blue, который просачивался в виде транссудации в участках инъекции, оценивали с использованием ImageJ. Для каждого животного, концентрацию анти-VEGF-scFv в зависимости от площади просачивания анализировали с использованием подгонки 4-параметрической логистической кривой. Концентрацию анти-VEGF-scFv, требующуюся для ингибирования просачивания из сосудов на 50% (EC50) получали из кривых ингибирования.
Протокол этого эксперимента иллюстрируется на фиг.11. Кроме того, эффективность кандидатных scFv, ESBA903 (578minmax) и 802 (511max), в ингибировании hVEGF иллюстрировалась на фиг.11, в виде различных размеров площадей, содержащих краситель Evans blue, просачивавшийся из сосудистой системы в кожу. Данные эффективности для 903 и 802 показаны на фиг.12. При 6,9 нМ, 903 и 802 показали более сильное ингибирование индуцированного VEGF просачивания сосудов в кожу в сравнении с Lucentis во всх тестируемых животных (фиг.12).
ПРИМЕР 7
ДЕЙСТВИЯ МЕСТНОЙ ОБРАБОТКИ анти-VEGF-scFv НА VEGF165-ИНДУЦИРОВАННОЕ ПРОСАЧИВАНИЕ СОСУДОВ СЕТЧАТКИ В КРЫСАХ
В этом примере, локальная эффективность 578minmax демонстрируется с использованием модифицированного анализа Miles. Эти модификации включают в себя, например, исследования с предварительно смешанными интравитреальными инъекциями и локальным нанесением scFv.
Предварительно смешанные различные концентрации анти-VEGF-scFv (10-, 3- и 1-кратный молярный избыток в сравнении с VEGF) и VEGF (500 нг) наносили посредством единой интравитреальной инъекции. Авастин (Roche) (10-, 3- и 1-кратный молярный избыток в сравнении с VEGF) использовали в качестве положительного контроля. Носитель для 578minmax (цитратный буфер, 20 мМ Na-цитрат, 125 мМ NaCl, pH 7) использовали в качестве отрицательного контроля. Как показано на фиг.13, предварительное смешивание с VEGF165 облегчало 578minmax (ESBA903) выполнять полное ингибирование hVEGF-индуцированной проницаемости сосудов сетчатки. В этом эксперименте, ингибирующее действие 578minmax (ESBA903) было более существенным в сравнении с Авастином.
Для местного нанесения, за пять дней перед стимуляцией VEGF, взрослые крысы Sprague-Dawley получали 578minmax (1% = 10 мг/мл) через билатеральное местное q.i.d введение (4 капли/день) до дня перфузии (День 6). Носитель для 578minmax (местное введение доз) и Alcon RTKi (10 мг/кг/день, пероральное зондовое введение) использовали в качестве отрицательного и положительного контролей. В день 5, крыс анестезируют и их зрачки расширяются. Все животные получают интравитреальные инъекции 500 нг hrVEGF (10 мкл) в оба глаза. Спустя 24 часа после инъекции VEGF, выполняют внутривенную инфузию красителя Evans blue на всех животных во время общей анестезии. После циркуляции этого красителя в течение 90 минут крыс эвтанизируют. Берут пробы крови, затем крыс перфузируют стерильным солевым раствором, затем оба глаза каждой крысы немедленно энуклеируют и сетчатки собирают с использованием хирургического микроскопа. Как для проб сетчатки, так и для проб плазмы, используют 60 мкл супернатанта для измерения абсорбции красителя Evans blue при помощи спектрофотометра при 620/740 нМ. Разрушение гематоретинального барьера и последующую проницаемость сосудов сетчатки, измеренные по абсорбции красителя, рассчитывают в виде среднего ± s.e.m. нетто ABS/сырая масса/плазма ABS. Используют однофакторный ANOVA для определения общего различия между способами обработки, где P≤0,05 является значимым. Как иллюстрировано на фиг.14, локальное введение (5 дней предобработки, 4 капли в день) 578minmax (903) значимо ингибировало hVEGF-индуцированную проницаемость сосудов. Это является первой демонстрацией локально эффективного антитела, применимого для лечения внутриглазного заболевания.
ЭКВИВАЛЕНТЫ
Многочисленные модификации и альтернативные варианты по настоящему изобретению будут очевидны специалистам в данной области в связи с предыдущим описанием. Таким образом, это описание должно пониматься только как иллюстративное и предназначено для разъяснения специалистам в данной области наилучшего способа проведения по настоящему изобретению. Детали этой структуры могут варьироваться по существу без отклонения от идеи по настоящему изобретению, и резервируется эксклюзивное применение всех модификаций, которые находятся в объеме прилагаемой формулы изобретения. Предполагается, что данное изобретение ограничивается только до степени, требуемой прилагаемой формулой изобретения и применимыми нормами права.
Весь литературный и подобный материал, цитируемый в этой заявке, включающий в себя патенты, заявки на патент, статьи, книги, учебники, диссертации, веб-страницы, фигуры и/или приложения, независимо от формата таких литературных и подобных материалов, особо включены в качестве ссылки в их полном объеме. В случае, когда один или несколько из этих включенных литературных и подобных материалов отличается от этого описания или противоречит этому описанию в том числе определенным терминам, применению терминов, описанным способам или т.п., контролем является это описание.
Заголовки разделов, используемые в данном случае, предназначены только для организационных целей и не должны пониматься как ограничивающие каким-либо образом описанный предмет изобретения.
Хотя настоящее изобретение было описано вместе с различными вариантами и примерами, не предполагается, что данные описания ограничены такими вариантами или примерами. Напротив, данные изобретения включают в себя различные альтернативы, модификации и эквиваленты, как будет понятно специалистам в данной области.
Данная формула изобретения не должна пониматься как ограничивающая порядок или элементы изобретения, если это не указано. Должно быть понятно, что могут быть произведены различные изменения в форме или деталях без отклонения от объема прилагаемой формулы изобретения. Таким образом, заявляются все варианты, которые находятся в пределах объема и идеи следующей формулы изобретения, и их эквиваленты.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1
Пептидный иммуноген
KFMDVYQRSYC
VL-последовательности:
SEQ ID NO. 72: 60
EVVMAQTPASVEAAVGGTVTIKCQASQSISSYLSWYQQKPGQPPKLLIYKASTLASGVPSRFKGSRS GTEYTLTISDLECADAATYYCQSNYGGSSSDYGNPFGGGTEAVVK
SEQ ID NO. 73: 435
AFELTQTPSSVEAAVGGTVTIKCQASQSIGSSLAWYQQKPGQRPKLLIYTAANLASGVPSRFRGSRSG AAFTLTISDLECADAATYYCQNFATSDTVTFGGGTEVWT
SEQ ID NO. 74: 453
AVVLTQTPSPVSAAVGGTVSISCQSSQSVWNNNRLAWFQQKSGQPPKLLIYYASTLASGVPSRFKG SGSGTEFTLTISDVQCDDAATYYCAGGYSSTSDNTFGGGTEVVVK
SEQ ID NO. 75: 375
DIVMTQTPASVEATVGGTITINCQASENINIWLSWYQQKPGQPPKLLIYQASKLASGVPSRFKGSGSGTQFTLTISDLECADAATΎYCQNNYSYNRYGAPFGGGTEVVVK
SEQ ID NO. 76: 610
DVVMTQTPASVSEPVGGTVTIKCQASQSISSWLSWYQQKPGQPPKLLIYQASTLASGVPPRSSGSGSGTEYTLTISDLECADAATYFCQNNYGFRSYGGAFGGGTEVVVK
SEQ ID NO.77: 578
DVVMTQTPSSVSAAVGDTVTINCQASEIIHSWLAWYQQKPGQPPKLLIYLASTLASGVPSRFKGSGSGTQFTLTISDLECADAAIYYCQNVYLASTNGANFGGGTEVVVK
SEQ ID NO. 78: 534
DVVMTQTPSSVSAAVGDTVTIKCQASQSINIWLSWYQQKSGQPPKLLVYKESTLASGVPSRFRGSGSGTQFTLTISDLECADAATYYCQNNYDSGNNGFPFGGGTEVVVK
SEQ ID NO. 79: 567
DVVMTQTPSSVSAAVGDTVTINCQADQSIYIWLSWYQQKPGQPPKLLIYKASTLESGVPSRFKGSGSGTQFTLTISDLECADAATYYCQNNAHYSTNGGTFGGGTEVVVK
SEQ ID NO. 80: 509
DVVMTQTPSSVSAAVGDTVTIKCQASQNIRIWLSWYQQKPGQPPKLLIYKASTLESGVPSRFKGSGSGTEFTLTISDLECADAATYYCQNNAHYSTNGGTFGGGTEVVVK
SEQ ID NO. 81: 511
EVVMTQTPASVEAAVGGTVTIKCQASQSINIWCSWYQQKPGHPPKLLIYRASTLASGVSSRFKGSGSGTEFTLTISDLECADAATYYCQANYA YSAGYGAAFGGGTEVVVK
SEQ ID NO. 82: 60min
EIVMTQSPSTLSASVGDRVIITCQASQSISSYLSWYQQKPGKAPKLLIYKASTLASGVPSRFSGSGSGA EFTLTISSLQPDDFATYYCQSNYGGSSSDYGNPFGQGTKLTVLG
SEQ ID NO. 83: 435min
EIVMTQSPSTLSASVGDRVIITCQASQSIGSSLAWYQQKPGKAPKLLIYTAANLASGVPSRFSGSGSG AEFTLTISSLQPDDFATYYCQNFATSDTVTFGQGTKLTVLG
SEQ ID NO. 84: 453min
EIVMTQSPSTLSASVGDRVIITCQSSQSVWNNNRLAWYQQKPGKAPKLLIYYASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCAGGYSSTSDNTFGQGTKLTVLG
SEQ ID NO. 85: 375min
EIVMTQSPSTLSASVGDRVIITCQASENINIWLSWYQQKPGKAPKLLIYQASKLASGVPSRFSGSGSG AEFTLTISSLQPDDFATYYCQNNYSYNRYGAPFGQGTKLTVLG
SEQ ID NO. 86: 610min
EIVMTQSPSTLSASVGDRVIITCQASQSISSWLSWYQQKPGKAPKLLIYQASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNNYGFRSYGGAFGQGTKLTVLG
SEQ ID NO. 87: 578min
EIVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLG
SEQ ID NO. 88: 534min
EIVMTQSPSTLSASVGDRVIITCQASQSINIWLSWYQQKPGKAPKLLIYKESTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNNYDSGNNGFPFGQGTKLTVLG
SEQ ID NO. 89: 567min
EIVMTQSPSTLSASVGDRVIITCQADQSIYIWLSWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNNAHYSTNGGTFGQGTKLTVLG
SEQ ID NO. 90: 509min
EIVMTQSPSTLSASVGDRVIITCQASQNIRIWLSWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNNAHYSTNGGTFGQGTKLTVLG
SEQ ID NO. 91: 511min
EIVMTQSPSTLSASVGDRVIITCQASQSINIWCSWYQQKPGKAPKLLIYRASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQANYAYSAGYGAAFGQGTKLTVLG
SEQ ID NO. 92: 578min_Pref subst
EIVLTQSPSSLSASVGDRVTITCQASEIIHSWLAWYQQRPGKAPKLLISLASTLASGVPSRFSGSGSGTDFTFTISSLQPEDFAVYYCQNVYLASTNGANFGQGTKVEIKR
SEQ ID NO. 93: 60max
EIVMTQSPSTLSASVGDRVIITCQASQSISSYLSWYQQKPGKAPKLLIYKASTLASGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQSNYGGSSSDYGNPFGQGTKLTVLG
SEQ ID NO. 94: 435max
EIVMTQSPSTLSASVGDRVIIKCQASQSIGSSLAWYQQKPGKAPKLLIYTAANLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNFATSDTVTFGQGTKLTVLG
SEQ ID NO. 95: 453max
EIVMTQSPSTLSASVGDRVIITCQSSQSVWNNNRLAWYQQKPGKAPKLLIYYASTLASGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCAGGYSSTSDNTFGQGTKLTVLG
SEQ ID NO. 96: 375max
EIVMTQSPSTLSASVGDRVIITCQASENINIWLSWYQQKPGKAPKLLIYQASKLASGVPSRFSGSGSGTQFTLTISSLQPDDFATYYCQNNYSYNRYGAPFGQGTKLTVLG
SEQ ID NO. 97: 610max
EIVMTQSPSTLSASVGDRVIITCQASQSISSWLSWYQQKPGKAPKLLIYQASTLASGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQNNYGFRSYGGAFGQGTKLTVLG
SEQ ID NO. 98: 578max
EIVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGTQFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLG
SEQ ID NO. 99: 534max
EIVMTQSPSTLSASVGDRVIITCQASQSINIWLSWYQQKPGKAPKLLIYKESTLASGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQNNYDSGNNGFPFGQGTKLTVLG
SEQ ID NO. 100: 567max
EIVMTQSPSTLSASVGDRVIITCQADQSIYIWLSWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGT QFTLTISSLQPDDFATYYCQNNAHYSTNGGTFGQGTKLTVLG
SEQ ID NO. 101: 509max
EIVMTQSPSTLSASVGDRVIITCQASQNIRIWLSWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQNNAHYSTNGGTFGQGTKLTVLG
SEQ ID NO. 102: 511max
EIVMTQSPSTLSASVGDRVIITCQASQSINIWLSWYQQKPGKAPKLLIYRASTLASGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQANYAYSAGYGAAFGQGTKLTVLG
SEQ ID NO. 103: 578max_Pref subst
EIVMTQSPSSLSASVGDRVTITCQASEIIHSWLAWYQQRPGKAPKLLISLASTLASGVPSRFSGSGSGTQFTFTISSLQPEDFAVYYCQNVYLASTNGANFGQGTKVEIKR
SEQ ID NO. 104: 578min VL: E1D
DIVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLG
SEQ ID NO. 105: 578min VL: 12V
EVVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLG
SEQ ID NO. 106: 511min VL: C41L
EIVMTQSPSTLSASVGDRVIITCQASQSINIWLSWYQQKPGKAPKLLIYRASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQANYAYSAGYGAAFGQGTKLTVLG
VH-последовательности:
SEQ ID NO. 107: 60-11-4
QSLEESGGDLVKPGASLTLTCTASGFPFSSGYWVCWVRQAPGKGLEWIACIYAGSSGSTYYASWAKGRFTISKTSSTTVTLQMTSLTAADTATYFCARGNNYYIYTDGGYAYAGLELWGPGILVTVSS
SEQ ID NO. 108: 60-11-6
QSLEESGGDLVKPGASLTLTCTASGFSFSSGYWICWVRQAPGKGLEWIACIYAGSSGSTYYASWAKGRFTISKTSSTTVTLQMTSLTAADTATYFCARGNNYYIYTDGGYAYAGLELWGPGILVTVSS
SEQ ID NO. 109: 435
QSLEESGGDLVQPGASLTLTCKVSGFSLNTNYWMCWVRQAPGKGLEWIGCMYTGSYNRAYYASW AKGRFTSSKTSSTTVTLEMTSLTAADTATYFCAKGSNWYSDLWGPGTLVTVSS
SEQ ID NO. 110: 453
QERLVESGGGLVQPEGSLTLTCKASGFSFSRSYYIYWVRQAPGKGLEWIACIDAGSSGILVYANWAK GRFTISKTSSTTVTLQMTSLTAADTATYFCARGDASYGVDSFMLPLWGPGTLVTVSS
SEQ ID NO. 111: 375
QSLEESGGGLVQPEGSLTLTCKASGFSFTTTDYMCWVRQAPGKGLEWIGCILAGDGSTYYANWAKGRFTGSKTSSTTVDLKMTGLTAADTATYFCARSDPASSWSFALWGPGTLVTVSS
SEQ ID NO. 112: 610
QSLEESGGRLVTPGTPLTLTCTASGIDFSGAYYMGWVRQAPGKGLEWIGYIDYDGDRYYASWAKGRFTISKTSTTVDLKITSPTTEDTATYFCARSDYSSGWGTDIWGPGTLVTVSL
SEQ ID NO. 113: 578
QSVEESGGRLVTPGTPLTLTCTASGFSLTDYYYMTWVRLAPGKGLEYIGFIDPDDDPYYATWAKGRFTISRTSTTVNLKMTSPTTEDTATYFCAGGDHNSGWGLDIWGPGTLVTVSL
SEQ ID NO. 114: 534
QSLEESGGRLVTPGTPLTLTCTASGFSLSYYYMSWVRQAPGKGLEWIGIIGPGDYTDYASWAKGRFTISKTSTTVDLKITSPTTEDTATYFCGRGDDNSGWGEDIWGPGTLVTVSL
SEQ ID NO. 115: 567
QSVEESGGRLVTPGAPLTLTCSVSGFSLSDYYMCWVRQAPGKGLQWIGCLDYFGSTDDASWAKGRFTISKTSTAVDLKITSPTTEDTATYFCARTDDSRGWGLNIWGPGTLVTVSL
SEQ ID NO. 116: 509
QSLEESGGRLVTPGTPLTLTCTASGFSLSSYYMCWVRQAPGKGLEWIGCLDYVGDTDYASWAKGRFTISKASTTVDLKITSLTTEDTATYFCARTDDSRGWGLNIWGPGTLVTVSL
SEQ ID NO. 117: 511
QSVEESGGRLVTPGTPLTLTCTVSGFSLNTYYMNWVRQAPGKGLEWIGIIAPDDTTYYASWAKSRSTITRDTNENTVTLKMTSLTTEDTATYFCARSGDTTAWGADIWGPGTLVTVSL
SEQ ID NO. 118: 60-11-4min
EVQLVESGGGLVQPGGSLRLSCAASGFPFSSGYWVCWVRQAPGKGLEWVSCIYAGSSGSTYYASWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGNNYYIYTDGGYAYAGLELWGQGTLVTVSS
SEQ ID NO. 119: 60-11-6min
EVQLVESGGGLVQPGGSLRLSCAASGFSFSSGYWICWVRQAPGKGLEWVSCIYAGSSGSTYYAS
WAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGNNYYIYTDGGYAYAGLELWGQGTLVTVSS
SEQ ID NO. 120: 435min
EVQLVESGGGLVQPGGSLRLSCAASGFSLNTNYWMCWVRQAPGKGLEWVSCMYTGSYNRAYYA SWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGSNWYSDLWGQGTLVTVSS
SEQ ID NO. 121: 453min
EVQLVESGGGLVQPGGSLRLSCAASGFSFSRSYYIYWVRQAPGKGLEWVSCIDAGSSGILVYANWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGDASYGVDSFMLPLWGQGTLVTVSS
SEQ ID NO. 122: 375min
EVQLVESGGGLVQPGGSLRLSCAASGFSFTTTDYMCWVRQAPGKGLEWVSCILAGDGSTYYANW
AKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSDPASSWSFALWGQGTLVTVSS
SEQ ID NO. 123: 610min
EVQLVESGGGLVQPGGSLRLSCAASGIDFSGAYYMGWVRQAPGKGLEWVSYIDYDGDRYYASWA KGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSDYSSGWGTDIWGQGTLVTVSS
SEQ ID NO. 124: 578min
EVQLVESGGGLVQPGGSLRLSCAASGFSLTDYYYMTWVRQAPGKGLEWVSFIDPDDDPYYATWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 125: 534min
EVQLVESGGGLVQPGGSLRLSCAASGFSLSYYYMSWVRQAPGKGLEWVSIIGPGDYTDYASWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGDDNSGWGEDIWGQGTLVTVSS
SEQ ID NO. 126: 567min
EVQLVESGGGLVQPGGSLRLSCAASGFSLSDYYMCWVRQAPGKGLEWVSCLDYFGSTDDASWAK GRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKTDDSRGWGLNIWGQGTLVTVSS
SEQ ID NO. 127: 509min
EVQLVESGGGLVQPGGSLRLSCAASGFSLSSYYMCWVRQAPGKGLEWVSCLDYVGDTDYASWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKTDDSRGWGLNIWGQGTLVTVSS
SEQ ID NO. 128: 511min
EVQLVESGGGLVQPGGSLRLSCAASGFSLNTYYMNWVRQAPGKGLEWVSIIAPDDTTYYASWAKSRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSGDTTAWGADIWGQGTLVTVSS
SEQ ID NO. 129: 578min_Pref subst
substQVQLVQTGGGLVQPGGSLRLSCAASGFSLTDYYYMTWVRQAPGKGLEWVSFIDPDDDPYYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTALYYCAKGDH NSGWGLDIWGQGTLVTVSS
SEQ ID NO. 130: 60-11-4max
EVQLVESGGGLVQPGGSLRLSCTASGFPFSSGYWVCWVRQAPGKGLEWVGCIYAGSSGSTYYASWAKGRFTISKDTSKNTVYLQMNSLRAEDTAVYYCARGNNYYIYTDGGYAYAGLELWGQGTLVTVSS
SEQ ID NO. 131: 60-11-6max
EVQLVESGGGLVQPGGSLRLSCTASGFSFSSGYWICWVRQAPGKGLEWVGCIYAGSSGSTYYASWAKGRFTISKDTSKNTVYLQMNSLRAEDTAVYYCARGNNYYIYTDGGYAYAGLELWGQGTLVTVSS
SEQ ID NO. 132: 435max
EVQLVESGGGLVQPGGSLRLSCKVSGFSLNTNYWMCWVRQAPGKGLEWVGCMYTGSYNRAYYASWAKGRFTSSKDTSKNTVYLQMNSLRAEDTAVYYCAKGSNWYSDLWGQGTLVTVSS
SEQ ID NO. 133: 453max
EVQLVESGGGLVQPGGSLRLSCKASGFSFSRSYYIYWVRQAPGKGLEWVGCIDAGSSGILVYANWAKGRFTISKDTSKNTVYLQMNSLRAEDTAVYYCARGDASYGVDSFMLPLWGQGTLVTVSS
SEQ ID NO. 134: 375max
EVQLVESGGGLVQPGGSLRLSCKASGFSFTTTDYMCWVRQAPGKGLEWVGCILAGDGSTYYANWAKGRFTGSKDTSKNTVYLQMNSLRAEDTAVYYCARSDPASSWSFALWGQGTLVTVSS
SEQ ID NO. 135: 610max
EVQLVESGGGLVQPGGSLRLSCTASGIDFSGAYYMGWVRQAPGKGLEWVGYIDYDGDRYYASWAKGRFTISKDTSKNTVYLQMNSLRAEDTAVYYCARSDYSSGWGTDIWGQGTLVTVSS
SEQ ID NO. 136: 578max
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 137: 534max
EVQLVESGGGLVQPGGSLRLSCTASGFSLSYYYMSWVRQAPGKGLEWVGIIGPGDYTDYASWAKGRFTISKDTSKNTVYLQMNSLRAEDTAVYYCARGDDNSGWGEDIWGQGTLVTVSS
SEQ ID NO. 138: 567max
EVQLVESGGGLVQPGGSLRLSCSVSGFSLSDYYMCWVRQAPGKGLEWVGCLDYFGSTDDASWAKGRFTISKDTSKNTVYLQMNSLRAEDTAVYYCARTDDSRGWGLNIWGQGTLVTVSS
SEQ ID NO. 139: 509max
EVQLVESGGGLVQPGGSLRLSCTASGFSLSSYYMCWVRQAPGKGLEWVGCLDYVGDTDYASWAK
GRFTISKDASKNTVYLQMNSLRAEDTAVYYCARTDDSRGWGLNIWGQGTLVTVSS
SEQ ID NO. 140: 509maxll
EVQLVESGGGLVQPGGSLRLSCTASGFSLSSYYMSWVRQAPGKGLEWVGILDYVGDTDYASWAKGRFTISKDASKNTVYLQMNSLRAEDTAVYYCARTDDSRGWGLNIWGQGTLVTVSS
SEQ ID NO. 141: 511max
EVQLVESGGGLVQPGGSLRLSCTVSGFSLNTYYMNWVRQAPGKGLEWVGIIAPDDTTYYASWAKSRSTISRDTSKNTVYLQMNSLRAEDTAVYYCARSGDTTAWGADIWGQGTLVTVSS
SEQ ID NO. 142: 578max_Pref subst
substQVQLVQTGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTALYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 143: 567minDHP
EVQLVESGGGSVQPGGSLRLSCAASGFSLSDYYMCWVRQAPGKGLEWVSCLDYFGSTDDASWAK GRFTISRDNSKNTLYLQMNSLRAEDTATYYCAKTDDSRGWGLNIWGQGTTVTVSS
SEQ ID NO. 144: 578maxDHP
EVQLVESGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWA
KGRFTISRDTSKNTVYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
SEQ ID NO. 145: 511maxDHP
EVQLVESGGGSVQPGGSLRLSCTVSGFSLNTYYMNWVRQAPGKGLEWVGIIAPDDTTYYASWAKSRSTISRDTSKNTVYLQMNSLRAEDTATYYCARSGDTTAWGADIWGQGTTVTVSS
SEQ ID NO. 146: 578max_Pref subst_DHP
QVQLVQTGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
SEQ ID NO. 147: 578max VH: El_
VQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 148: 578max VH: V2Q
EQQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 149: 578max VH: Q46L
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRLAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 150: 578max VH: W54Y
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEYVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 151: 578max VH: V55I
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWIGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 152: 578max VH: D83A
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRATSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 153: 578max VH: N87A
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKATVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 154: 578max VH: Y105F
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYFCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 155: 578max VH: D83_
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 156: 578max VH: N87_
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 157: 578max VH: T84N
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 158: 578max VH: V89L
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTLYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 159: 578max VH: V89A
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTAYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 160: 578maxDHP VH: T84N
EVQLVESGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDNSKNTVYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
SEQ ID NO. 161: 578maxDHP VH: V89L
EVQLVESGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYΫATWAKGRFTISRDTSKNTLYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
SEQ ID NO. 162: 578maxDHP VH: V89A
EVQLVESGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTAYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
SEQ ID NO. 163: 578max VH:T84N, V89A
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDNSKNTAYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 164: 578max VH:T84N, V89L
EVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 165: 578maxDHP VH: T84N, V89A
EVQLVESGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDNSKNTAYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
SEQ ID NO. 166: 578maxDHP VH: T84N, V89L
EVQLVESGGGSVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDNSKNTLYLQMNSLRAEDTATYYCAGGDHNSGWGLDIWGQGTTVTVSS
Каркасные последовательности (X остатки являются сайтами инсертирования CDR и могут быть любой природно-встречающейся аминокислотой. Могут присутствовать по меньшей мере 3 и до 50 аминокислот):
SEQ ID NO. 167: Вариабельные легкие цепи FW1.4 и rFW1.4
EIVMTQSPSTLSASVGDRVIITC(X)n=3-50 WYQQKPGKAPKLLIY(X)n=3-50
GVPSRFSGSGSGAEFTLTISSLQPDDFATYC(X)n=3-50 FGQGTKLT VLG
SEQ ID NO. 168: rFW1.4 вариант 2 (v2)
EIVMTQSPSTLSASVGDRVIITC(X)n=3-50 WYQQKPG KAPKLLIY(X)n=3-50
GVPSRFSGSGSGAEFTLTISSLQPDDFATYC(X)n=3-50 FGQGTKLT VLG
SEQ ID NO. 169: Вариабельная тяжелая цепь FW1.4
EVQLVESGGGLVQPGGSLRLSCAAS(X)n=3-50 WVRQAPGKGLEWVS (X)n=3-50
RFTISRDNSKNTLYLQMNSLRAEDTA VYYCAK(X)n=3-50 WGQGTL VTVSS
SEQ ID NO. 170: Вариабельная тяжелая цепь rFW1.4
EVQLVESGGGLVQPGGSLRLSCTAS(X)n=3-50 WVRQAPGKGLEWVG(X)n=3-50
RFTISRDTSKNTVYLQMNSLRAEDTA VYYCAR(X)n=3-50 WGQGTLV TVS
SEQ ID NO. 171: Вариабельная тяжелая цепь rFW1.4 вариант 2 (v2)
EVQLVESGGGLVQPGGSLRLSCTVS(X)n=3-50 WVRQAPGKG LEWVG(X)n=3-5O
RFTISKDTSKNTVYLQMNSLRAEDTAVYYCAR(X)n=3-50 WGQGTLVTVSS
Каркасные последовательности ScFv:
SEQ ID NO. 172: FW1.4
EIVMTQSPSTLSASVGDRVIITC(X)n=3-50 WYQQKPGKAPKLLIY(X)n-3-50
GVPSRFSGSGSGAEFTLTISSLQPDDFAT^YC(X)n=3-50 FGQGTKLTVLG
GGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAAS(X)n=3-50 WVRQAPGKG
LEWVS(X)n=3-S0 RFTISRDNSKNTLYLQMNSLRAEDTA VYYCAK(X)n=3-50
WGQGTLVTVSS
SEQ ID NO. 173: rFW1.4
EIVMTQSPSTLSASVGDRVIITC(X)n=3-50 WYQQKPGKAPKLLIY(X)n=3-50
GVPSRFSGSGSGTEFTLTISSLQPDDFATYYC(X)n=3-50 FGQGTKLTVLG
GGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCTAS(X)n=3-50 WVRQAPGKGLEWVG(X)n-3-50 RFTISRDTSKNTVYLQMNS LRAEDTA VYYCAR(X)n=3-50
WGQGTLVTVSS
SEQ ID NO. 174: rFW1.4 вариант 2 (v2)
EIVMTQSPSTLSASVGDRVIITC(X)n=3-50 WYQQKPGKAPKLLIY(X)n=3-50
GVPSRFSGSGSGTEFTLTISSLQPDDFATYYC(X)n=3-50 FGQGTKLTVLG
GGGGSGGGGSGGGGSGGGGS EVQLVESGGGLVQPGGSLRLSCTVS(X)n=3-50
WVRQAPGKGLEWVG(X)n=3-50 RFTISKDTSKNTVYLQMNSLR AEDTA VYYCAR(X)n=3-50
WGQGTLVTVSS
Последовательности анти-VEGF-scFv:
SEQ ID NO. 175: 435_max
EIVMTQSPSTLSASVGDRVIITCQASQSIGSSLAWYQQKPGKAPKLLIYTAANLASGVPSRFSGSRSGAEFTLTISSLQPDDFATYYCQNFATSDTVTFGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCKASGFSLNTNYWMCWVRQAPGKGLEWVGCMYTGSYNRAYYASWAKGRFTSSKDTSKNTVYLQMNSLRAEDTAVYYCAKGSNWYSDLWGQGTLVTVSS
SEQ ID NO. 176: 511_max
EIVMTQSPSTLSASVGDRVIITCQASQSINIWLSWYQQKPGKAPKLLIYRASTLASGVPSRFSGSGSGTEFTLTISSLQPDDFATYYCQANYAYSAGYGAAFGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCTVSGFSLNTYYMNWVRQAPGKGLEWVGIIAPDDTTYYASWAKSRSTISRDTSKISITVYLQMNSLRAEDTAVYYCARSGDTTAWGADIWGQGTLVTVSS
SEQ ID NO. 177: 567_min
EIVMTQSPSTLSASVGDRVIITCQADQSIYIWLSWYQQKPGKAPKLLIYKASTLESGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNNAHYSTNGGTFGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAASGFSLSDYYMCWVRQAPGKGLEWVSCLDYFGSTDDASWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKTDDSRGWGLNIWGQGTLVTVSS
SEQ ID NO. 178: 578min
EIVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAASGFSLTDYYYMTWVRQAPGKGLEWVSFIDPDDDPYYATWAKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 179: 578max
EIVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGTQFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 180: 578minmax (ESBA903)
EIVMTQSPSTLSASVGDRVIITCQASEIIHSWLAWYQQKPGKAPKLLIYLASTLASGVPSRFSGSGSGAEFTLTISSLQPDDFATYYCQNVYLASTNGANFGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCTASGFSLTDYYYMTWVRQAPGKGLEWVGFIDPDDDPYYATWAKGRFTISRDTSKNTVYLQMNSLRAEDTAVYYCAGGDHNSGWGLDIWGQGTLVTVSS
SEQ ID NO. 181 линкер
GGGGSGGGGSGGGGSGGGGS
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ESBATEch AG
<120> Стабильные и растворимые антитела, ингибирующие VEGF
<130> VEGF binders
<140> Us61/133,212
<141> 2008-06-25
<160> 180
<170> PatentIn version 3.5
<210> 1
<211> 11
<212> Белок
<213> Homo sapiens
<400> 1
Lys Phe Met Asp Val Tyr Gln Arg Ser Tyr Cys
1 5 10
<210> 2
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 2
Gly Phe Pro Phe Ser Ser Gly Tyr Trp Val Cys
1 5 10
<210> 3
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 3
Gly Phe Ser Phe Ser Ser Gly Tyr Trp Ile Cys
1 5 10
<210> 4
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 4
Gly Phe Ser Leu Asn Thr Asn Tyr Trp Met Cys
1 5 10
<210> 5
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 5
Gly Phe Ser Phe Ser Arg Ser Tyr Tyr Ile Tyr
1 5 10
<210> 6
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 6
Gly Phe Ser Phe Thr Thr Thr Asp Tyr Met Cys
1 5 10
<210> 7
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 7
Gly Ile Asp Phe Ser Gly Ala Tyr Tyr Met Gly
1 5 10
<210> 8
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 8
Gly Phe Ser Leu Thr Asp Tyr Tyr Tyr Met Thr
1 5 10
<210> 9
<211> 10
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 9
Gly Phe Ser Leu Ser Tyr Tyr Tyr Met Ser
1 5 10
<210> 10
<211> 10
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 10
Gly Phe Ser Leu Ser Asp Tyr Tyr Met Cys
1 5 10
<210> 11
<211> 10
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 11
Gly Phe Ser Leu Ser Ser Tyr Tyr Met Cys
1 5 10
<210> 12
<211> 10
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 12
Gly Phe Ser Leu Asn Thr Tyr Tyr Met Asn
1 5 10
<210> 13
<211> 10
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 13
Gly Phe Ser Leu Ser Ser Tyr Tyr Met Ser
1 5 10
<210> 14
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 14
Gly Phe Ser Leu Ser Ser Gly Tyr Tyr Met Cys
1 5 10
<210> 15
<211> 18
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 15
Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 16
<211> 18
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 16
Cys Met Tyr Thr Gly Ser Tyr Asn Arg Ala Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 17
<211> 18
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 17
Cys Ile Asp Ala Gly Ser Ser Gly Ile Leu Val Tyr Ala Asn Trp Ala
1 5 10 15
Lys Gly
<210> 18
<211> 17
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 18
Cys Ile Leu Ala Gly Asp Gly Ser Thr Tyr Tyr Ala Asn Trp Ala Lys
1 5 10 15
Gly
<210> 19
<211> 16
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 19
Tyr Ile Asp Tyr Asp Gly Asp Arg Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 20
<211> 16
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 20
Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala Lys Gly
1 5 10 15
<210> 21
<211> 16
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 21
Ile Ile Gly Pro Gly Asp Tyr Thr Asp Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 22
<211> 16
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 22
Cys Leu Asp Tyr Phe Gly Ser Thr Asp Asp Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 23
<211> 16
<212> Белок
<213> Искусственная последовательность
<400> 23
Cys Leu Asp Tyr Val Gly Asp Thr Asp Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 24
<211> 16
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 24
Ile Ile Ala Pro Asp Asp Thr Thr Tyr Tyr Ala Ser Trp Ala Lys Ser
1 5 10 15
<210> 25
<211> 16
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 25
Ile Leu Asp Tyr Val Gly Asp Thr Asp Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 26
<211> 18
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 26
Cys Ile Asp Ala Gly Ser Asp Gly Asp Thr Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 27
<211> 19
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 27
Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr Ala Tyr Ala Gly
1 5 10 15
Leu Glu Leu
<210> 28
<211> 8
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 28
Gly Ser Asn Trp Tyr Ser Asp Leu
1 5
<210> 29
<211> 14
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 29
Gly Asp Ala Ser Tyr Gly Val Asp Ser Phe Met Leu Pro Leu
1 5 10
<210> 30
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 30
Ser Asp Pro Ala Ser Ser Trp Ser Phe Ala Leu
1 5 10
<210> 31
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 31
Ser Asp Tyr Ser Ser Gly Trp Gly Thr Asp Ile
1 5 10
<210> 32
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 32
Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile
1 5 10
<210> 33
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 33
Gly Asp Asp Asn Ser Gly Trp Gly Glu Asp Ile
1 5 10
<210> 34
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 34
Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile
1 5 10
<210> 35
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 35
Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile
1 5 10
<210> 36
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 36
Ser Gly Asp Thr Thr Ala Trp Gly Ala Asp Ile
1 5 10
<210> 37
<211> 19
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 37
Gly Asp Asp Ser Ser Gly Tyr Thr Asp Gly Gly Tyr Ala Tyr Trp Gly
1 5 10 15
Leu Asp Ile
<210> 38
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 38
Gln Ala Ser Gln Ser Ile Ser Ser Tyr Leu Ser
1 5 10
<210> 39
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 39
Gln Ala Ser Gln Ser Ile Gly Ser Ser Leu Ala
1 5 10
<210> 40
<211> 13
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 40
Gln Ser Ser Gln Ser Val Trp Asn Asn Asn Arg Leu Ala
1 5 10
<210> 41
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 41
Gln Ala Ser Glu Asn Ile Asn Ile Trp Leu Ser
1 5 10
<210> 42
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 42
Gln Ala Ser Gln Ser Ile Ser Ser Trp Leu Ser
1 5 10
<210> 43
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 43
Gln Ala Ser Glu Ile Ile His Ser Trp Leu Ala
1 5 10
<210> 44
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 44
Gln Ala Ser Gln Ser Ile Asn Ile Trp Leu Ser
1 5 10
<210> 45
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 45
Gln Ala Asp Gln Ser Ile Tyr Ile Trp Leu Ser
1 5 10
<210> 46
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 46
Gln Ala Ser Gln Asn Ile Arg Ile Trp Leu Ser
1 5 10
<210> 47
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 47
Gln Ala Ser Gln Ser Ile Asn Ile Trp Cys Ser
1 5 10
<210> 48
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 48
Gln Ala Ser Gln Ser Ile Asn Ile Trp Leu Ser
1 5 10
<210> 49
<211> 13
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 49
Gln Ala Ser Gln Ser Ile Asn Ile Asn Asn Trp Leu Ser
1 5 10
<210> 50
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 50
Lys Ala Ser Thr Leu Ala Ser
1 5
<210> 51
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 51
Thr Ala Ala Asn Leu Ala Ser
1 5
<210> 52
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 52
Tyr Ala Ser Thr Leu Ala Ser
1 5
<210> 53
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 53
Gln Ala Ser Lys Leu Ala Ser
1 5
<210> 54
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 54
Gln Ala Ser Thr Leu Ala Ser
1 5
<210> 55
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 55
Leu Ala Ser Thr Leu Ala Ser
1 5
<210> 56
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 56
Lys Glu Ser Thr Leu Ala Ser
1 5
<210> 57
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 57
Lys Ala Ser Thr Leu Glu Ser
1 5
<210> 58
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 58
Lys Ala Ser Thr Leu Glu Ser
1 5
<210> 59
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 59
Arg Ala Ser Thr Leu Ala Ser
1 5
<210> 60
<211> 7
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 60
Lys Ala Ser Thr Leu Ala Ser
1 5
<210> 61
<211> 14
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 61
Gln Ser Asn Tyr Gly Gly Ser Ser Ser Asp Tyr Gly Asn Pro
1 5 10
<210> 62
<211> 10
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 62
Gln Asn Phe Ala Thr Ser Asp Thr Val Thr
1 5 10
<210> 63
<211> 11
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 63
Ala Gly Gly Tyr Ser Ser Thr Ser Asp Asn Thr
1 5 10
<210> 64
<211> 12
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 64
Gln Asn Asn Tyr Ser Tyr Asn Arg Tyr Gly Ala Pro
1 5 10
<210> 65
<211> 12
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 65
Gln Asn Asn Tyr Gly Phe Arg Ser Tyr Gly Gly Ala
1 5 10
<210> 66
<211> 12
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 66
Gln Asn Val Tyr Leu Ala Ser Thr Asn Gly Ala Asn
1 5 10
<210> 67
<211> 12
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 67
Gln Asn Asn Tyr Asp Ser Gly Asn Asn Gly Phe Pro
1 5 10
<210> 68
<211> 12
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 68
Gln Asn Asn Ala His Tyr Ser Thr Asn Gly Gly Thr
1 5 10
<210> 69
<211> 12
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 69
Gln Asn Asn Ala His Tyr Ser Thr Asn Gly Gly Thr
1 5 10
<210> 70
<211> 13
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 70
Gln Ala Asn Tyr Ala Tyr Ser Ala Gly Tyr Gly Ala Ala
1 5 10
<210> 71
<211> 14
<212> Белок
<213> Неизвестный
<220>
<223> CDR, происходящий из кроличьего антитела
<400> 71
Gln Asn Asn Tyr His Tyr Ser Ser Ser Thr Asn Gly Gly Thr
1 5 10
<210> 72
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 72
Glu Val Val Met Ala Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Arg Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Asn Tyr Gly Gly Ser Ser
85 90 95
Ser Asp Tyr Gly Asn Pro Phe Gly Gly Gly Thr Glu Ala Val Val Lys
100 105 110
<210> 73
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 73
Ala Phe Glu Leu Thr Gln Thr Pro Ser Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Gly Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Arg Pro Lys Leu Leu Ile
35 40 45
Tyr Thr Ala Ala Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Arg Gly
50 55 60
Ser Arg Ser Gly Ala Ala Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Asn Phe Ala Thr Ser Asp Thr
85 90 95
Val Thr Phe Gly Gly Gly Thr Glu Val Val Val Thr
100 105
<210> 74
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 74
Ala Val Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Ser Ile Ser Cys Gln Ser Ser Gln Ser Val Trp Asn Asn
20 25 30
Asn Arg Leu Ala Trp Phe Gln Gln Lys Ser Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Tyr Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Thr Ser Asp Asn Thr Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 75
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 75
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Thr Val Gly
1 5 10 15
Gly Thr Ile Thr Ile Asn Cys Gln Ala Ser Glu Asn Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Ser Tyr Asn Arg
85 90 95
Tyr Gly Ala Pro Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 76
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 76
Asp Val Val Met Thr Gln Thr Pro Ala Ser Val Ser Glu Pro Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Thr Leu Ala Ser Gly Val Pro Pro Arg Ser Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Phe Cys Gln Asn Asn Tyr Gly Phe Arg Ser
85 90 95
Tyr Gly Gly Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 77
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 77
Asp Val Val Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Asp Thr Val Thr Ile Asn Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Ile Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 78
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 78
Asp Val Val Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Asp Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Ser Gly Gln Pro Pro Lys Leu Leu Val
35 40 45
Tyr Lys Glu Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Arg Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Asp Ser Gly Asn
85 90 95
Asn Gly Phe Pro Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 79
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 79
Asp Val Val Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Asp Thr Val Thr Ile Asn Cys Gln Ala Asp Gln Ser Ile Tyr Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 80
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 80
Asp Val Val Met Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Asp Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Asn Ile Arg Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 81
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 81
Glu Val Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Cys Ser Trp Tyr Gln Gln Lys Pro Gly His Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Arg Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ala Asn Tyr Ala Tyr Ser Ala
85 90 95
Gly Tyr Gly Ala Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 82
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 82
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Asn Tyr Gly Gly Ser Ser
85 90 95
Ser Asp Tyr Gly Asn Pro Phe Gly Gln Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 83
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 83
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Gly Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Thr Ala Ala Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Phe Ala Thr Ser Asp Thr
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 84
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 84
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ser Ser Gln Ser Val Trp Asn Asn
20 25 30
Asn Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Tyr Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Thr Ser Asp Asn Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 85
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 85
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Asn Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Ser Tyr Asn Arg
85 90 95
Tyr Gly Ala Pro Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 86
<211> 111
<212> Белок
<213> Искусственная последовательность
<400> 86
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Gly Phe Arg Ser
85 90 95
Tyr Gly Gly Ala Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 87
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 87
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 88
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 88
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Glu Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Asp Ser Gly Asn
85 90 95
Asn Gly Phe Pro Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 89
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 89
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Asp Gln Ser Ile Tyr Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 90
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 90
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Asn Ile Arg Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 91
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 91
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Cys Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Arg Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ala Asn Tyr Ala Tyr Ser Ala
85 90 95
Gly Tyr Gly Ala Ala Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 92
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 92
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Arg Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Ser Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
<210> 93
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 93
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ser Asn Tyr Gly Gly Ser Ser
85 90 95
Ser Asp Tyr Gly Asn Pro Phe Gly Gln Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 94
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 94
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Lys Cys Gln Ala Ser Gln Ser Ile Gly Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Thr Ala Ala Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Phe Ala Thr Ser Asp Thr
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105
<210> 95
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 95
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ser Ser Gln Ser Val Trp Asn Asn
20 25 30
Asn Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Tyr Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Ser
85 90 95
Thr Ser Asp Asn Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 96
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 96
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Asn Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Ser Tyr Asn Arg
85 90 95
Tyr Gly Ala Pro Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 97
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 97
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Gly Phe Arg Ser
85 90 95
Tyr Gly Gly Ala Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 98
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 98
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 99
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 99
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Glu Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Tyr Asp Ser Gly Asn
85 90 95
Asn Gly Phe Pro Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 100
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 100
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Asp Gln Ser Ile Tyr Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 101
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 101
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Asn Ile Arg Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 102
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 102
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Arg Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ala Asn Tyr Ala Tyr Ser Ala
85 90 95
Gly Tyr Gly Ala Ala Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 103
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 103
Glu Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Arg Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Ser Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
<210> 104
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 104
Asp Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 105
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 105
Glu Val Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 106
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-последовательность
<400> 106
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Arg Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ala Asn Tyr Ala Tyr Ser Ala
85 90 95
Gly Tyr Gly Ala Ala Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 107
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 107
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Pro Phe Ser Ser Gly Tyr
20 25 30
Trp Val Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr Ala Tyr
100 105 110
Ala Gly Leu Glu Leu Trp Gly Pro Gly Ile Leu Val Thr Val Ser Ser
115 120 125
<210> 108
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 108
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Ser Gly Tyr
20 25 30
Trp Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr Ala Tyr
100 105 110
Ala Gly Leu Glu Leu Trp Gly Pro Gly Ile Leu Val Thr Val Ser Ser
115 120 125
<210> 109
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 109
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Lys Val Ser Gly Phe Ser Leu Asn Thr Asn Tyr
20 25 30
Trp Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Cys Met Tyr Thr Gly Ser Tyr Asn Arg Ala Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ser Ser Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Glu Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Lys Gly Ser Asn Trp Tyr Ser Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 110
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 110
Gln Glu Arg Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Glu Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Lys Ala Ser Gly Phe Ser Phe Ser Arg Ser
20 25 30
Tyr Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Ala Cys Ile Asp Ala Gly Ser Ser Gly Ile Leu Val Tyr Ala Asn
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val
65 70 75 80
Thr Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe
85 90 95
Cys Ala Arg Gly Asp Ala Ser Tyr Gly Val Asp Ser Phe Met Leu Pro
100 105 110
Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 111
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 111
Gln Ser Leu Glu Glu Ser Gly Gly Gly Leu Val Gln Pro Glu Gly Ser
1 5 10 15
Leu Thr Leu Thr Cys Lys Ala Ser Gly Phe Ser Phe Thr Thr Thr Asp
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Cys Ile Leu Ala Gly Asp Gly Ser Thr Tyr Tyr Ala Asn Trp Ala
50 55 60
Lys Gly Arg Phe Thr Gly Ser Lys Thr Ser Ser Thr Thr Val Asp Leu
65 70 75 80
Lys Met Thr Gly Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Ser Asp Pro Ala Ser Ser Trp Ser Phe Ala Leu Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 112
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 112
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Ile Asp Phe Ser Gly Ala Tyr
20 25 30
Tyr Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asp Tyr Asp Gly Asp Arg Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile
65 70 75 80
Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Ser
85 90 95
Asp Tyr Ser Ser Gly Trp Gly Thr Asp Ile Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Leu
115
<210> 113
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 113
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr Tyr
20 25 30
Tyr Met Thr Trp Val Arg Leu Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asn Leu Lys Met
65 70 75 80
Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Gly Gly
85 90 95
Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Leu
115
<210> 114
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 114
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Tyr Tyr Tyr
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Gly Pro Gly Asp Tyr Thr Asp Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Gly Arg Gly Asp
85 90 95
Asp Asn Ser Gly Trp Gly Glu Asp Ile Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Val Ser Leu
115
<210> 115
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 115
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Ala Pro
1 5 10 15
Leu Thr Leu Thr Cys Ser Val Ser Gly Phe Ser Leu Ser Asp Tyr Tyr
20 25 30
Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Ile Gly
35 40 45
Cys Leu Asp Tyr Phe Gly Ser Thr Asp Asp Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Ala Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Thr Asp
85 90 95
Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Val Ser Leu
115
<210> 116
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 116
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr Tyr
20 25 30
Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Cys Leu Asp Tyr Val Gly Asp Thr Asp Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Ala Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Thr Asp
85 90 95
Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Val Ser Leu
115
<210> 117
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 117
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Asn Thr Tyr Tyr
20 25 30
Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Ala Pro Asp Asp Thr Thr Tyr Tyr Ala Ser Trp Ala Lys Ser
50 55 60
Arg Ser Thr Ile Thr Arg Asp Thr Asn Glu Asn Thr Val Thr Leu Lys
65 70 75 80
Met Thr Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg
85 90 95
Ser Gly Asp Thr Thr Ala Trp Gly Ala Asp Ile Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Leu
115
<210> 118
<211> 130
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 118
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Pro Phe Ser Ser Gly
20 25 30
Tyr Trp Val Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Lys Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr
100 105 110
Ala Tyr Ala Gly Leu Glu Leu Trp Gly Gln Gly Thr Leu Val Thr Val
115 120 125
Ser Ser
130
<210> 119
<211> 130
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 119
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Lys Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr
100 105 110
Ala Tyr Ala Gly Leu Glu Leu Trp Gly Gln Gly Thr Leu Val Thr Val
115 120 125
Ser Ser
130
<210> 120
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 120
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Asn Thr Asn
20 25 30
Tyr Trp Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Cys Met Tyr Thr Gly Ser Tyr Asn Arg Ala Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Lys Gly Ser Asn Trp Tyr Ser Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 121
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 121
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Arg Ser
20 25 30
Tyr Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Cys Ile Asp Ala Gly Ser Ser Gly Ile Leu Val Tyr Ala Asn
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Lys Gly Asp Ala Ser Tyr Gly Val Asp Ser Phe Met Leu
100 105 110
Pro Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 122
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 122
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Thr Thr Thr
20 25 30
Asp Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Cys Ile Leu Ala Gly Asp Gly Ser Thr Tyr Tyr Ala Asn Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
65 70 75 80
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Lys Ser Asp Pro Ala Ser Ser Trp Ser Phe Ala Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 123
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 123
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ile Asp Phe Ser Gly Ala
20 25 30
Tyr Tyr Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Tyr Ile Asp Tyr Asp Gly Asp Arg Tyr Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ser Asp Tyr Ser Ser Gly Trp Gly Thr Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 124
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 125
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 125
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Tyr Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Gly Pro Gly Asp Tyr Thr Asp Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Gly Asp Asp Asn Ser Gly Trp Gly Glu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 126
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 126
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Asp Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Cys Leu Asp Tyr Phe Gly Ser Thr Asp Asp Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 127
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 127
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Cys Leu Asp Tyr Val Gly Asp Thr Asp Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 128
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 128
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Asn Thr Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ile Ile Ala Pro Asp Asp Thr Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Ser Gly Asp Thr Thr Ala Trp Gly Ala Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 129
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 129
Gln Val Gln Leu Val Gln Thr Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ser Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 130
<211> 130
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 130
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Pro Phe Ser Ser Gly
20 25 30
Tyr Trp Val Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr
100 105 110
Ala Tyr Ala Gly Leu Glu Leu Trp Gly Gln Gly Thr Leu Val Thr Val
115 120 125
Ser Ser
130
<210> 131
<211> 130
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 131
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Gly Asn Asn Tyr Tyr Ile Tyr Thr Asp Gly Gly Tyr
100 105 110
Ala Tyr Ala Gly Leu Glu Leu Trp Gly Gln Gly Thr Leu Val Thr Val
115 120 125
Ser Ser
130
<210> 132
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 132
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Val Ser Gly Phe Ser Leu Asn Thr Asn
20 25 30
Tyr Trp Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Cys Met Tyr Thr Gly Ser Tyr Asn Arg Ala Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ser Ser Lys Asp Thr Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Lys Gly Ser Asn Trp Tyr Ser Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 133
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 133
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Phe Ser Phe Ser Arg Ser
20 25 30
Tyr Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Cys Ile Asp Ala Gly Ser Ser Gly Ile Leu Val Tyr Ala Asn
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr
65 70 75 80
Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Gly Asp Ala Ser Tyr Gly Val Asp Ser Phe Met Leu
100 105 110
Pro Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 134
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 134
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Phe Ser Phe Thr Thr Thr
20 25 30
Asp Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Cys Ile Leu Ala Gly Asp Gly Ser Thr Tyr Tyr Ala Asn Trp
50 55 60
Ala Lys Gly Arg Phe Thr Gly Ser Lys Asp Thr Ser Lys Asn Thr Val
65 70 75 80
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Ser Asp Pro Ala Ser Ser Trp Ser Phe Ala Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 135
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 135
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Ile Asp Phe Ser Gly Ala
20 25 30
Tyr Tyr Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Tyr Ile Asp Tyr Asp Gly Asp Arg Tyr Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Asp Tyr Ser Ser Gly Trp Gly Thr Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 136
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 136
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 137
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 137
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Ser Tyr Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Gly Pro Gly Asp Tyr Thr Asp Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Asp Asp Asn Ser Gly Trp Gly Glu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 138
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 138
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Val Ser Gly Phe Ser Leu Ser Asp Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Cys Leu Asp Tyr Phe Gly Ser Thr Asp Asp Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 139
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 139
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Cys Leu Asp Tyr Val Gly Asp Thr Asp Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ala Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 140
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 140
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Leu Asp Tyr Val Gly Asp Thr Asp Tyr Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Asp Ala Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 141
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 141
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Val Ser Gly Phe Ser Leu Asn Thr Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ala Pro Asp Asp Thr Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Ser Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ser Gly Asp Thr Thr Ala Trp Gly Ala Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 142
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 142
Gln Val Gln Leu Val Gln Thr Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 143
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 143
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Asp Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Cys Leu Asp Tyr Phe Gly Ser Thr Asp Asp Ala Ser Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Lys Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 144
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 144
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 145
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 145
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Val Ser Gly Phe Ser Leu Asn Thr Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ile Ile Ala Pro Asp Asp Thr Thr Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Ser Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Ser Gly Asp Thr Thr Ala Trp Gly Ala Asp Ile Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 146
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 146
Gln Val Gln Leu Val Gln Thr Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 147
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 147
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
1 5 10 15
Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr Tyr
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 148
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 148
Glu Gln Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 149
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 149
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Leu Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 150
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 150
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 151
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 151
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 152
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 152
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Ala Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 153
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 153
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Ala Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 154
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 154
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 155
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 155
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Thr Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 156
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 156
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 157
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 157
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 158
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 158
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 159
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 159
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 160
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 160
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 161
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 161
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 162
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 162
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 163
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 163
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 164
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 164
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 165
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 165
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 166
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-последовательность
<400> 166
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu Thr Asp Tyr
20 25 30
Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala Thr Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 167
<211> 381
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-акцепторная последовательность
<220>
<221> CDR
<222> (24)..(123)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (139)..(238)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (271)..(370)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<400> 167
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Tyr Gln Gln Lys
115 120 125
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Xaa Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Gly Val
225 230 235 240
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr
245 250 255
Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Xaa Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
370 375 380
<210> 168
<211> 381
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VL-акцепторная последовательность
<220>
<221> CDR
<222> (24)..(123)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (139)..(238)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (271)..(370)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<400> 168
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Tyr Gln Gln Lys
115 120 125
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Xaa Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Gly Val
225 230 235 240
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr
245 250 255
Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Xaa Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
370 375 380
<210> 169
<211> 382
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-акцепторная последовательность
<220>
<221> CDR
<222> (26)..(125)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (140)..(239)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (272)..(371)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<400> 169
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Val Arg
115 120 125
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Arg
225 230 235 240
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
245 250 255
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Xaa Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
370 375 380
<210> 170
<211> 381
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-акцепторная последовательность
<220>
<221> CDR
<222> (26)..(125)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (140)..(239)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (272)..(371)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<400> 170
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Val Arg
115 120 125
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Arg
225 230 235 240
Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr Leu Gln Met
245 250 255
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Xaa Trp Gly Gln Gly Thr Leu Val Thr Val Ser
370 375 380
<210> 171
<211> 382
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - VH-акцепторная последовательность
<220>
<221> CDR
<222> (26)..(125)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (140)..(239)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (272)..(371)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<400> 171
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Val Ser Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Val Arg
115 120 125
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Arg
225 230 235 240
Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr Val Tyr Leu Gln Met
245 250 255
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Xaa Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
370 375 380
<210> 172
<211> 783
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - акцепторная последовательность
<220>
<221> CDR
<222> (24)..(123)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (139)..(238)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (271)..(370)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (427)..(526)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (541)..(640)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (673)..(772)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<400> 172
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Tyr Gln Gln Lys
115 120 125
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Xaa Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Gly Val
225 230 235 240
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr
245 250 255
Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Xaa Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly
370 375 380
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
385 390 395 400
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
405 410 415
Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Xaa Xaa Xaa Xaa Xaa Xaa
420 425 430
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
435 440 445
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
450 455 460
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
465 470 475 480
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
485 490 495
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
500 505 510
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Val
515 520 525
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Xaa Xaa Xaa Xaa
530 535 540
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
545 550 555 560
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
565 570 575
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
580 585 590
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
595 600 605
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
610 615 620
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
625 630 635 640
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
645 650 655
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
660 665 670
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
675 680 685
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
690 695 700
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
705 710 715 720
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
725 730 735
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
740 745 750
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
755 760 765
Xaa Xaa Xaa Xaa Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
770 775 780
<210> 173
<211> 783
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - акцепторная последовательность
<220>
<221> CDR
<222> (24)..(123)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (139)..(238)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (271)..(370)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (427)..(526)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (541)..(640)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<220>
<221> CDR
<222> (673)..(772)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот.
<400> 173
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Tyr Gln Gln Lys
115 120 125
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Xaa Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Gly Val
225 230 235 240
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr
245 250 255
Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Xaa Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly
370 375 380
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
385 390 395 400
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
405 410 415
Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Xaa Xaa Xaa Xaa Xaa Xaa
420 425 430
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
435 440 445
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
450 455 460
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
465 470 475 480
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
485 490 495
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
500 505 510
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Val
515 520 525
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly Xaa Xaa Xaa Xaa
530 535 540
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
545 550 555 560
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
565 570 575
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
580 585 590
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
595 600 605
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
610 615 620
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
625 630 635 640
Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Val Tyr Leu Gln
645 650 655
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
660 665 670
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
675 680 685
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
690 695 700
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
705 710 715 720
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
725 730 735
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
740 745 750
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
755 760 765
Xaa Xaa Xaa Xaa Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
770 775 780
<210> 174
<211> 783
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv - акцепторная последовательность
<220>
<221> CDR
<222> (24)..(123)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (139)..(238)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (271)..(370)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (427)..(526)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (541)..(640)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<220>
<221> CDR
<222> (673)..(772)
<223> Могут присутствовать по меньшей мере 3 и до 50 аминокислот
<400> 174
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
20 25 30
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
35 40 45
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
50 55 60
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
85 90 95
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
100 105 110
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Tyr Gln Gln Lys
115 120 125
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Xaa Xaa Xaa Xaa Xaa Xaa
130 135 140
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
145 150 155 160
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
165 170 175
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
180 185 190
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
195 200 205
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
210 215 220
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Gly Val
225 230 235 240
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr
245 250 255
Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Xaa Xaa
260 265 270
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
275 280 285
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
290 295 300
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
305 310 315 320
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
325 330 335
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
340 345 350
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
355 360 365
Xaa Xaa Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly
370 375 380
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
385 390 395 400
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
405 410 415
Gly Ser Leu Arg Leu Ser Cys Thr Val Ser Xaa Xaa Xaa Xaa Xaa Xaa
420 425 430
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
435 440 445
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
450 455 460
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
465 470 475 480
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
485 490 495
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
500 505 510
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Trp Val
515 520 525
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly Xaa Xaa Xaa Xaa
530 535 540
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
545 550 555 560
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
565 570 575
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
580 585 590
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
595 600 605
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
610 615 620
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
625 630 635 640
Arg Phe Thr Ile Ser Lys Asp Thr Ser Lys Asn Thr Val Tyr Leu Gln
645 650 655
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
660 665 670
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
675 680 685
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
690 695 700
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
705 710 715 720
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
725 730 735
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
740 745 750
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
755 760 765
Xaa Xaa Xaa Xaa Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
770 775 780
<210> 175
<211> 248
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv
<400> 175
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Gly Ser Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Thr Ala Ala Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Phe Ala Thr Ser Asp Thr
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
130 135 140
Gly Ser Leu Arg Leu Ser Cys Lys Ala Ser Gly Phe Ser Leu Asn Thr
145 150 155 160
Asn Tyr Trp Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
165 170 175
Trp Val Gly Cys Met Tyr Thr Gly Ser Tyr Asn Arg Ala Tyr Tyr Ala
180 185 190
Ser Trp Ala Lys Gly Arg Phe Thr Ser Ser Lys Asp Thr Ser Lys Asn
195 200 205
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Lys Gly Ser Asn Trp Tyr Ser Asp Leu Trp Gly Gln
225 230 235 240
Gly Thr Leu Val Thr Val Ser Ser
245
<210> 176
<211> 251
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv
<400> 176
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Gln Ser Ile Asn Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Arg Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Ala Asn Tyr Ala Tyr Ser Ala
85 90 95
Gly Tyr Gly Ala Ala Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
130 135 140
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Val Ser Gly Phe Ser
145 150 155 160
Leu Asn Thr Tyr Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly
165 170 175
Leu Glu Trp Val Gly Ile Ile Ala Pro Asp Asp Thr Thr Tyr Tyr Ala
180 185 190
Ser Trp Ala Lys Ser Arg Ser Thr Ile Ser Arg Asp Thr Ser Lys Asn
195 200 205
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Arg Ser Gly Asp Thr Thr Ala Trp Gly Ala Asp Ile
225 230 235 240
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 177
<211> 250
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv
<400> 177
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Asp Gln Ser Ile Tyr Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Asn Ala His Tyr Ser Thr
85 90 95
Asn Gly Gly Thr Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu
145 150 155 160
Ser Asp Tyr Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
165 170 175
Glu Trp Val Ser Cys Leu Asp Tyr Phe Gly Ser Thr Asp Asp Ala Ser
180 185 190
Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
195 200 205
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
210 215 220
Tyr Cys Ala Lys Thr Asp Asp Ser Arg Gly Trp Gly Leu Asn Ile Trp
225 230 235 240
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 178
<211> 251
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv
<400> 178
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu
145 150 155 160
Thr Asp Tyr Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly
165 170 175
Leu Glu Trp Val Ser Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala
180 185 190
Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
195 200 205
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Lys Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile
225 230 235 240
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 179
<211> 251
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv
<400> 179
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu
145 150 155 160
Thr Asp Tyr Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly
165 170 175
Leu Glu Trp Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala
180 185 190
Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn
195 200 205
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile
225 230 235 240
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<210> 180
<211> 251
<212> Белок
<213> Искусственная последовательность
<220>
<223> Рекомбинантный scFv
<400> 180
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Gln Ala Ser Glu Ile Ile His Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ala Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Asn Val Tyr Leu Ala Ser Thr
85 90 95
Asn Gly Ala Asn Phe Gly Gln Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
130 135 140
Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Ser Leu
145 150 155 160
Thr Asp Tyr Tyr Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly
165 170 175
Leu Glu Trp Val Gly Phe Ile Asp Pro Asp Asp Asp Pro Tyr Tyr Ala
180 185 190
Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn
195 200 205
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
210 215 220
Tyr Tyr Cys Ala Gly Gly Asp His Asn Ser Gly Trp Gly Leu Asp Ile
225 230 235 240
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
245 250
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭЛЕМЕНТЫ, СВЯЗЫВАЮЩИЕСЯ С ИЛ-1 БЕТА | 2013 |
|
RU2711118C2 |
| CD131-СВЯЗЫВАЮЩИЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773927C2 |
| СТАБИЛЬНЫЕ СОСТАВЫ АНТИТЕЛ ПРОТИВ TIGIT, ОТДЕЛЬНО И В КОМБИНАЦИИ С АНТИТЕЛАМИ ПРОТИВ РЕЦЕПТОРА 1 ПРОГРАММИРУЕМОЙ СМЕРТИ (PD-1), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2820576C2 |
| АНТИТЕЛО К TIGIT И ЕГО ИСПОЛЬЗОВАНИЕ | 2019 |
|
RU2786434C2 |
| МОНОКЛОНАЛЬНЫЕ АГЕНТЫ, СВЯЗЫВАЮЩИЕ сМЕТ, ИХ КОНЪЮГАТЫ С ЛЕКАРСТВЕННЫМИ СРЕДСТВАМИ И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2744914C2 |
| АНТИТЕЛО К PD-1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2807484C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| АНТИТЕЛО К PD-L1, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2778085C2 |
| СПОСОБ ОЧИСТКИ ДЛЯ УДАЛЕНИЯ ВАРИАНТОВ АНТИТЕЛ С СУЛЬФАТИРОВАНИЕМ ТИРОЗИНА; ОЧИЩЕННЫЕ КОМПОЗИЦИИ | 2017 |
|
RU2793400C2 |
| БИСПЕЦИФИЧЕСКИЙ БЕЛОК | 2019 |
|
RU2784486C1 |
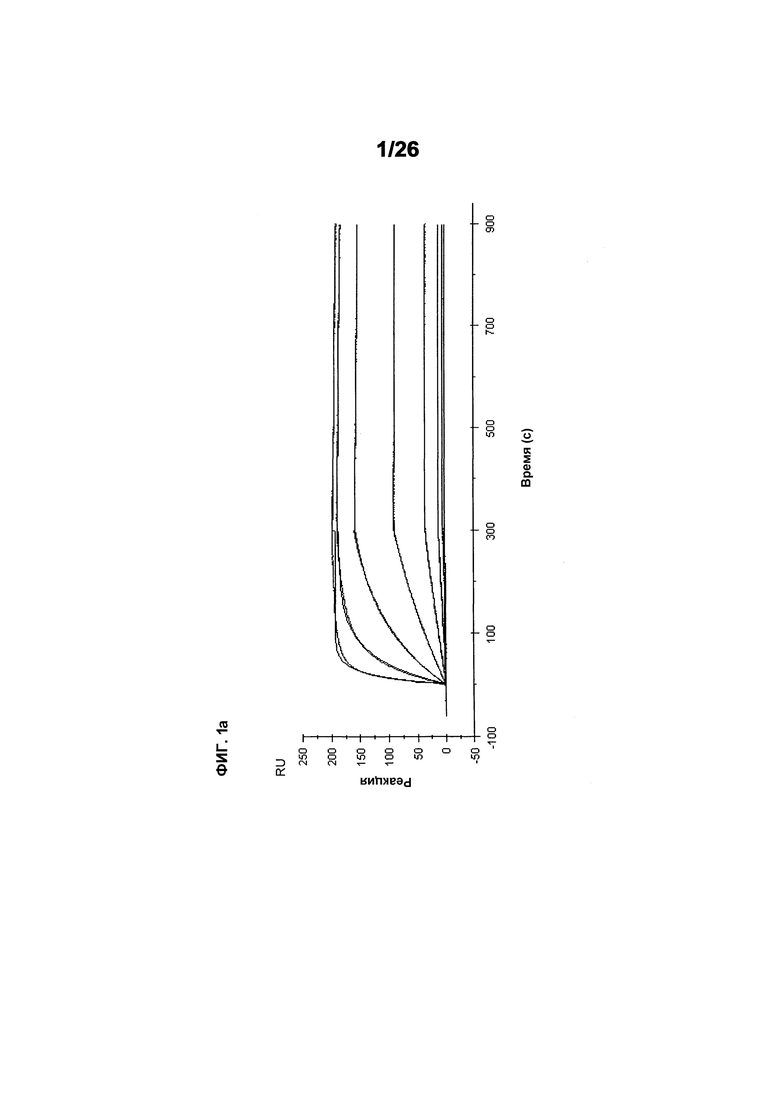
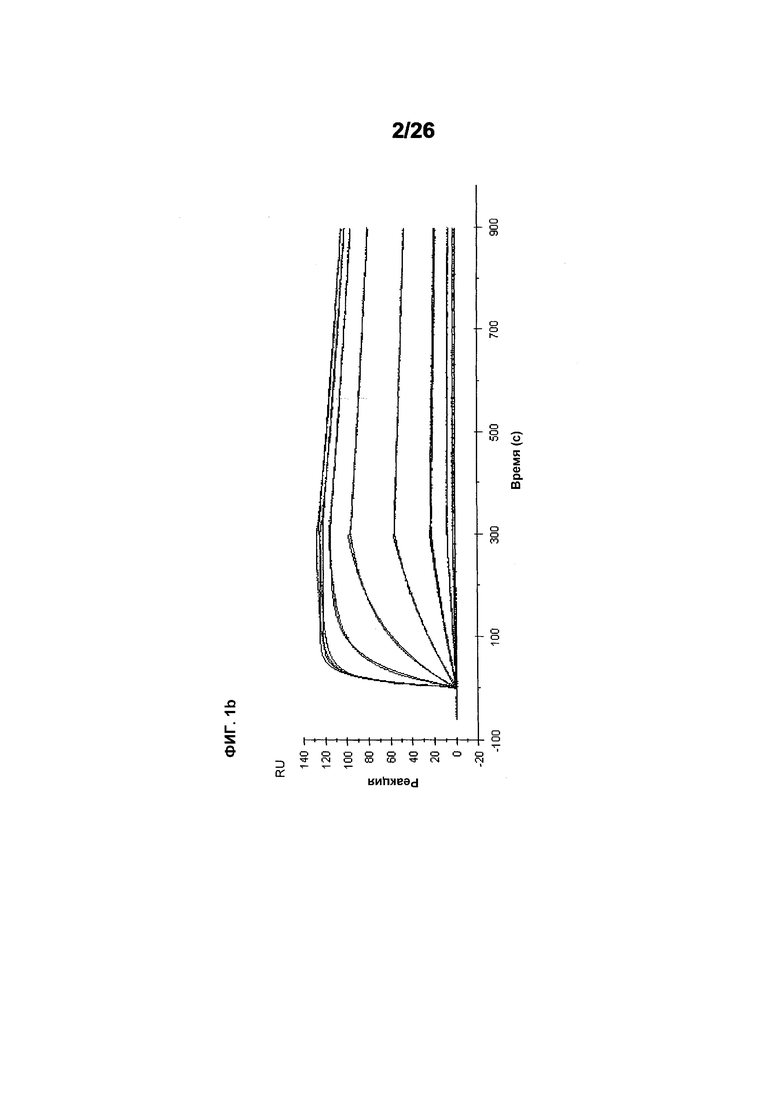
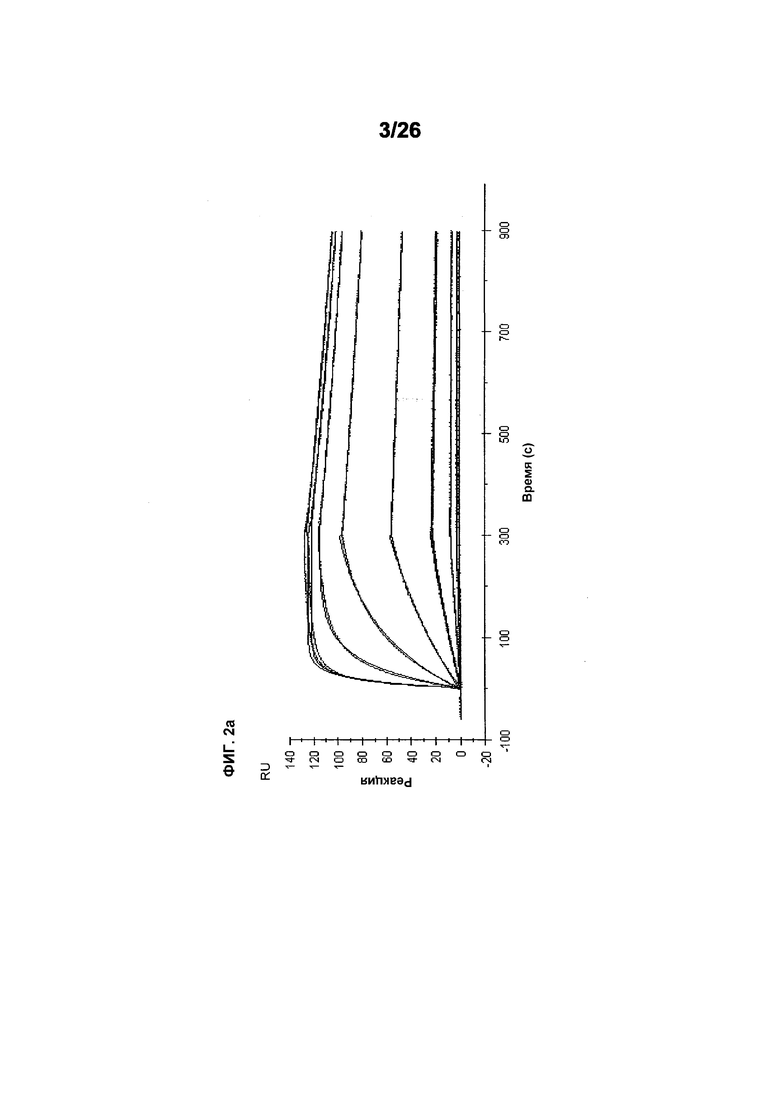
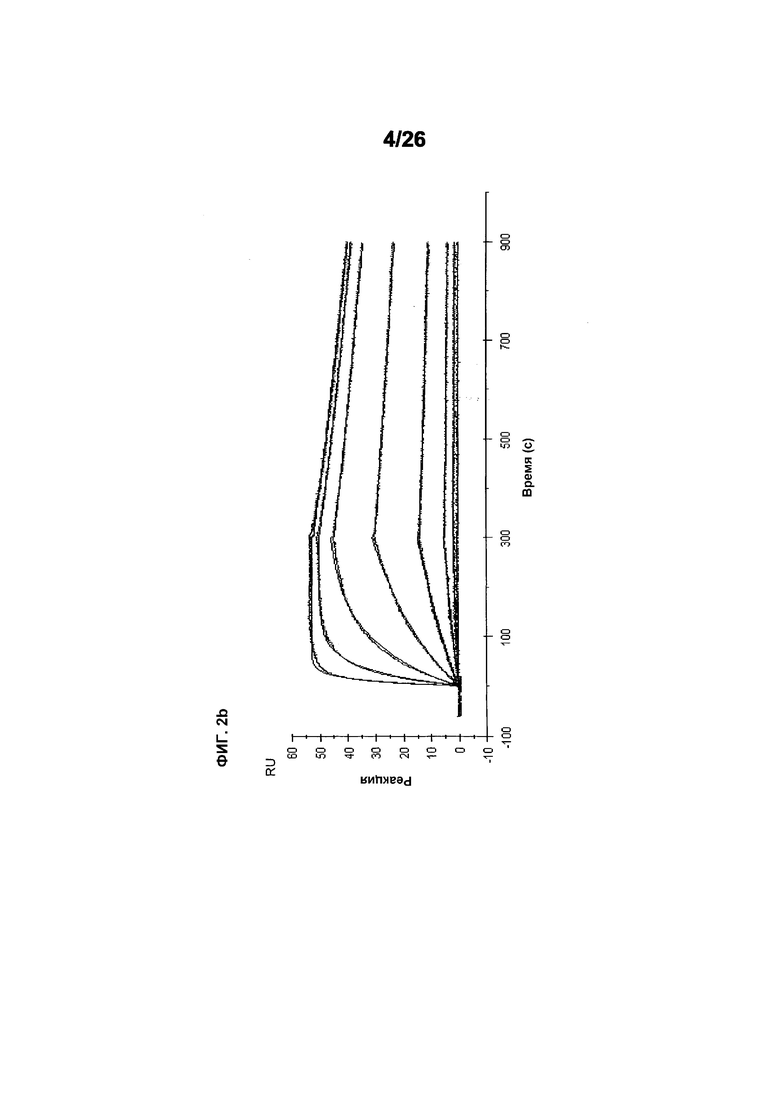
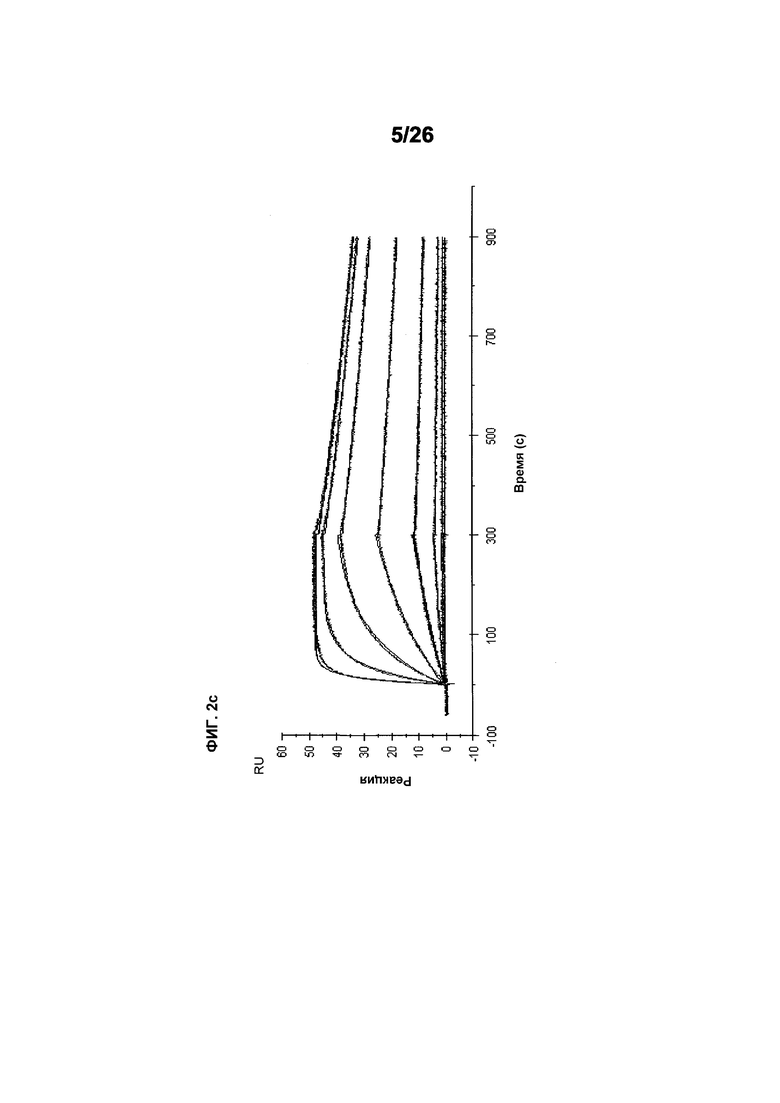
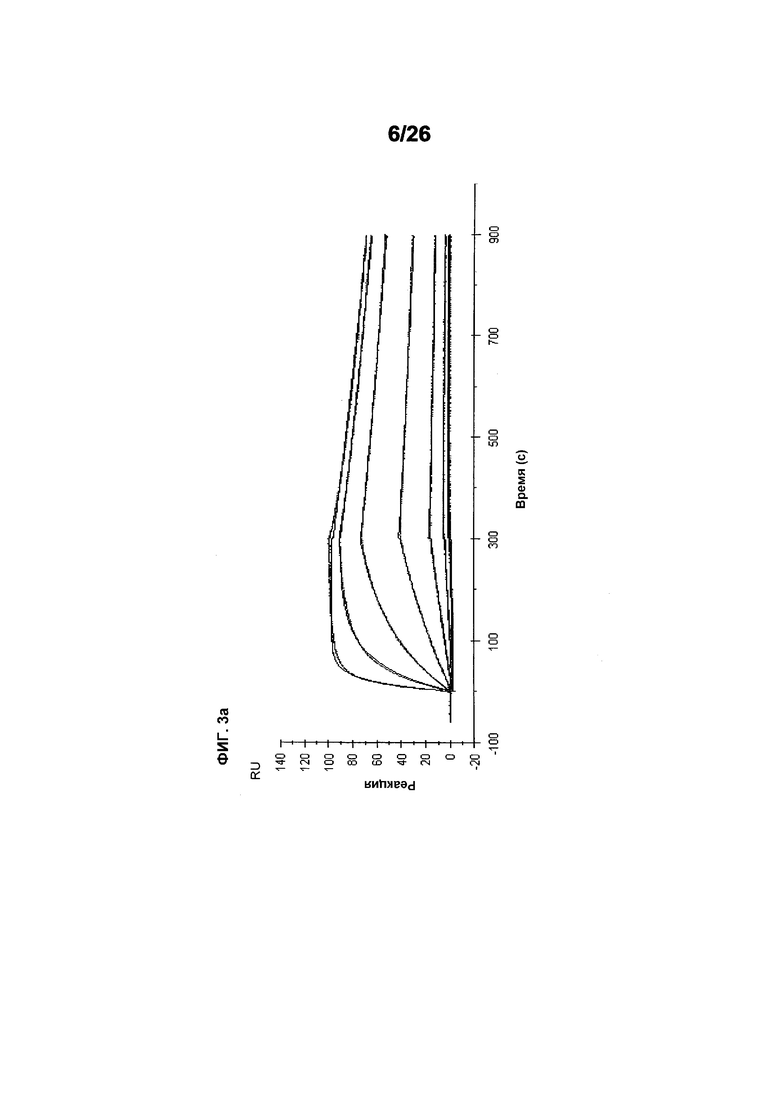
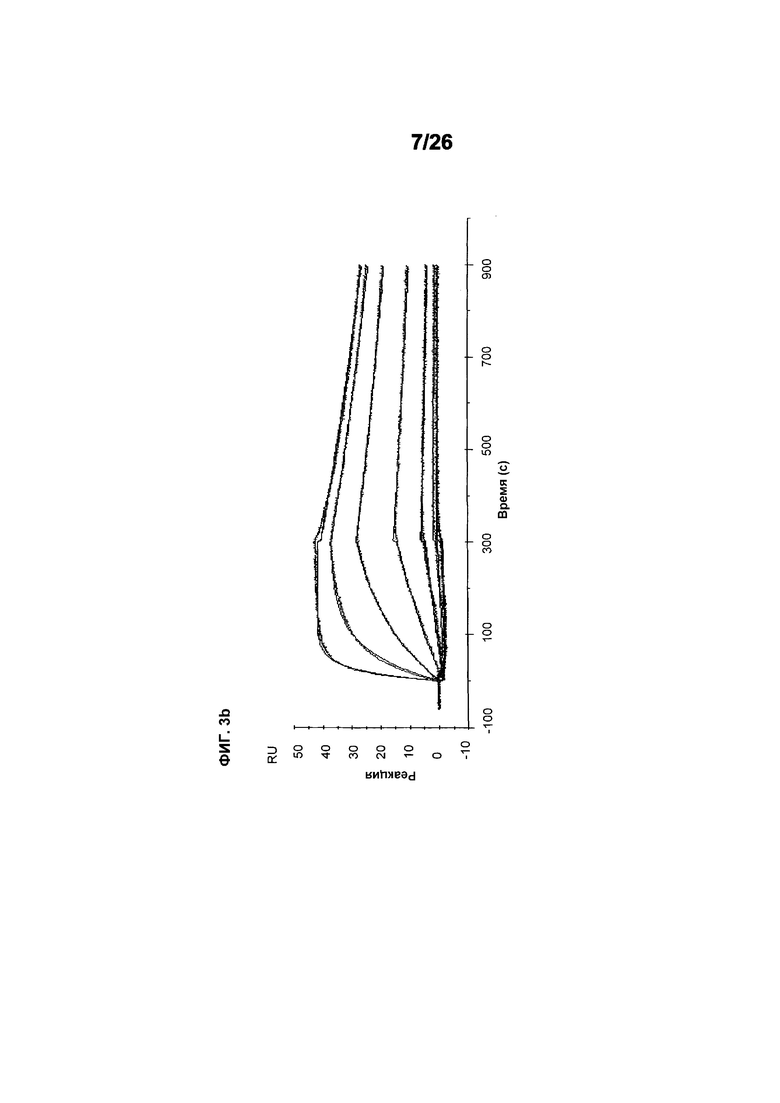
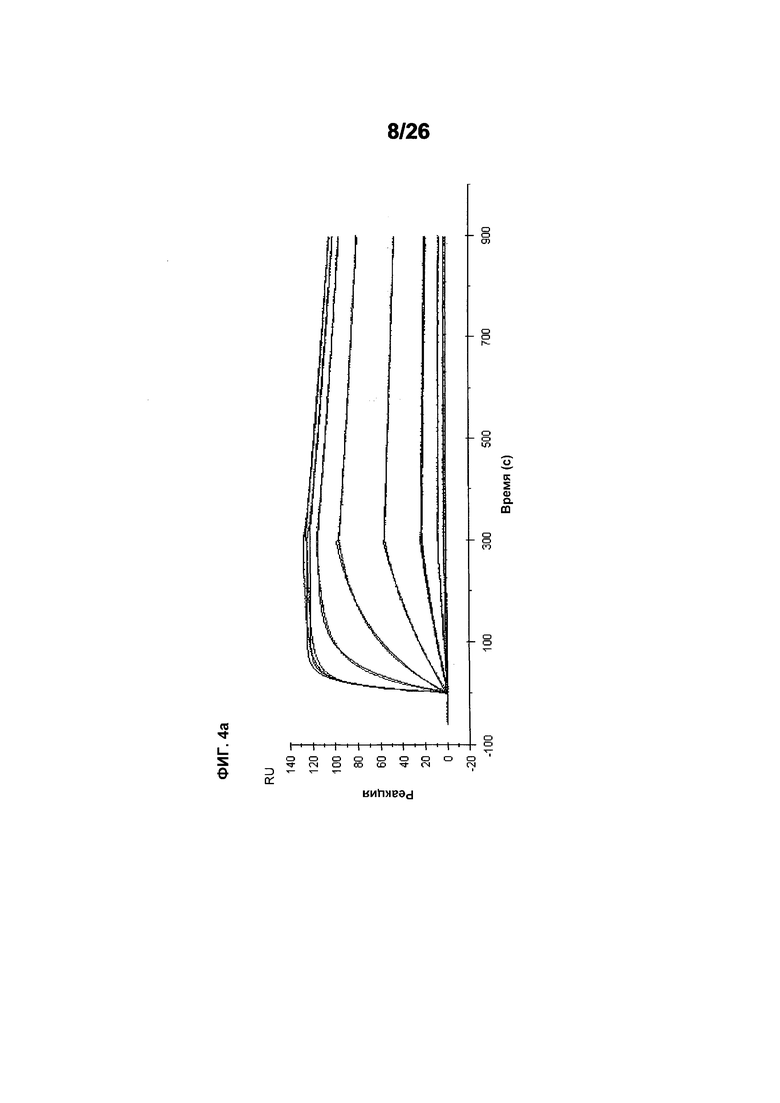
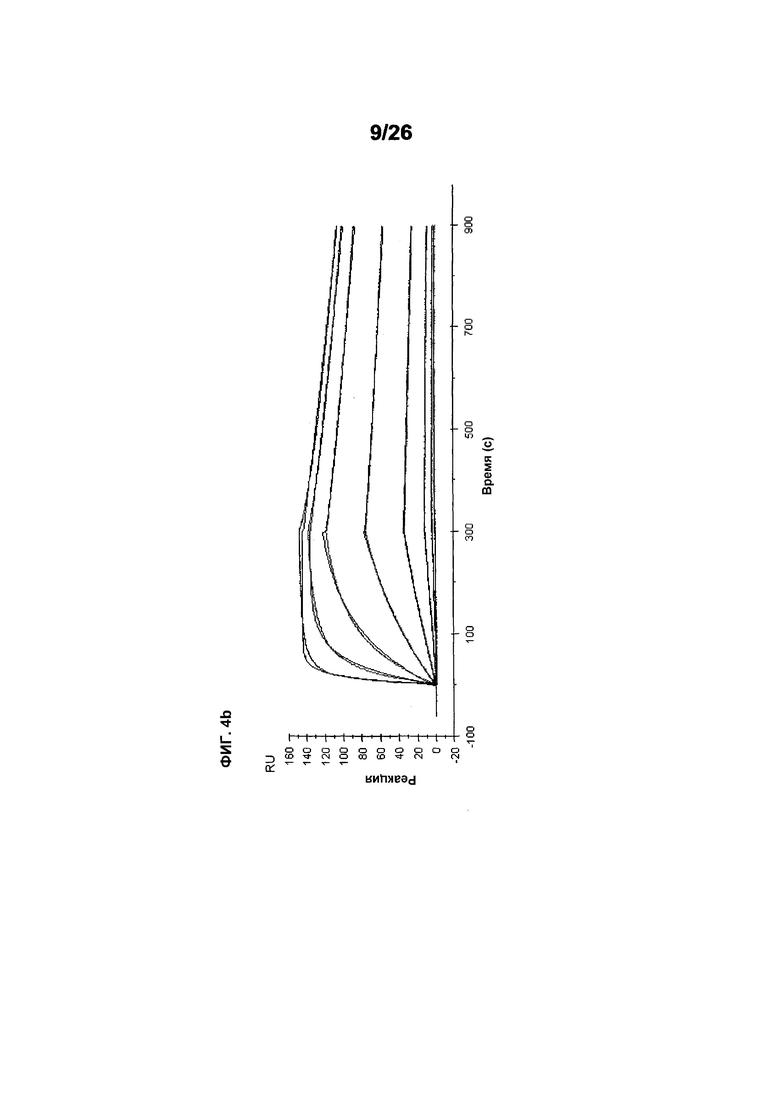
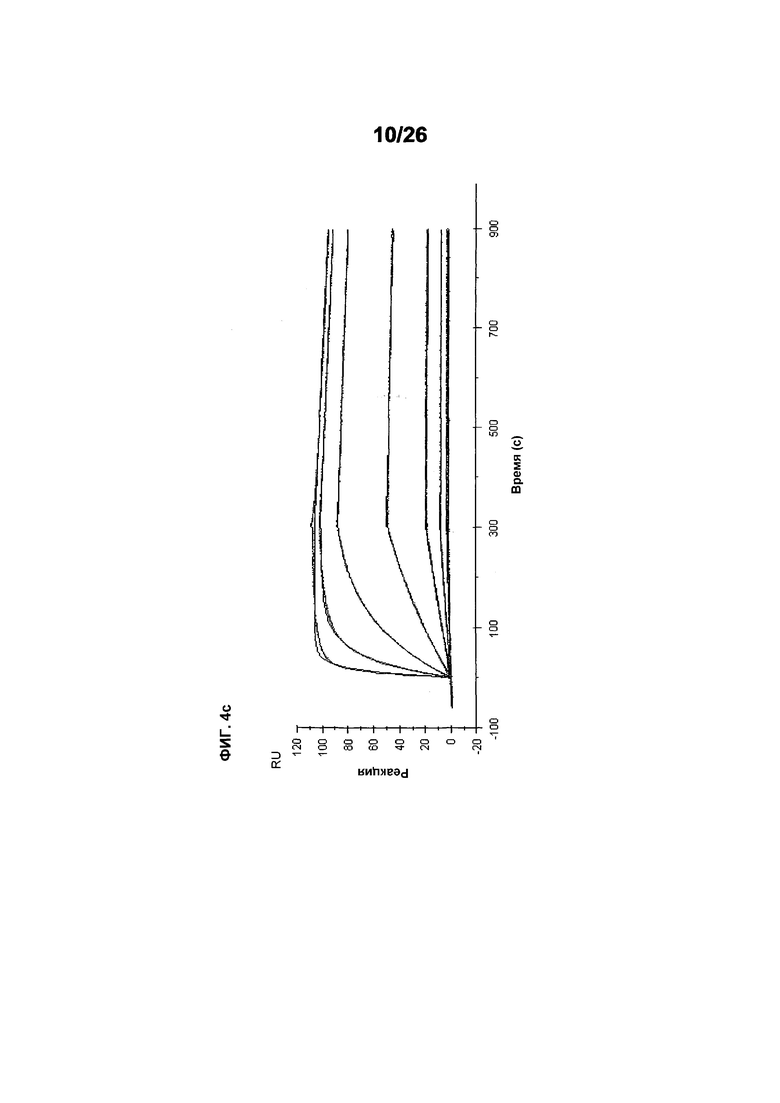
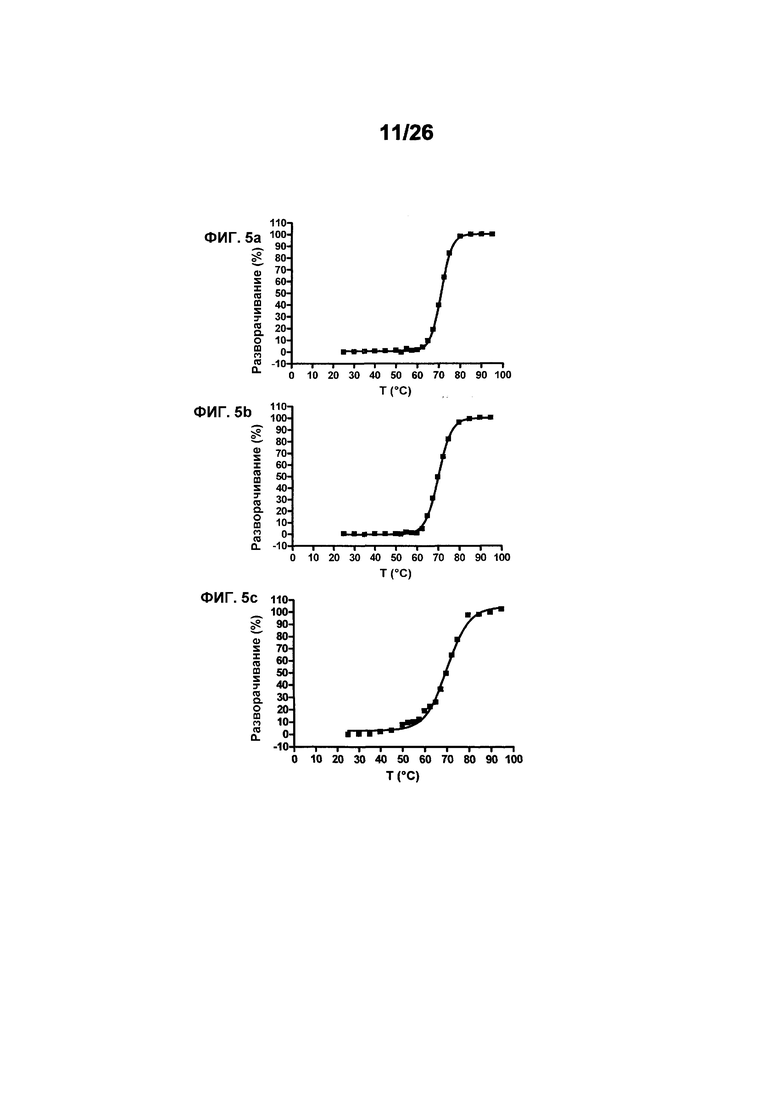
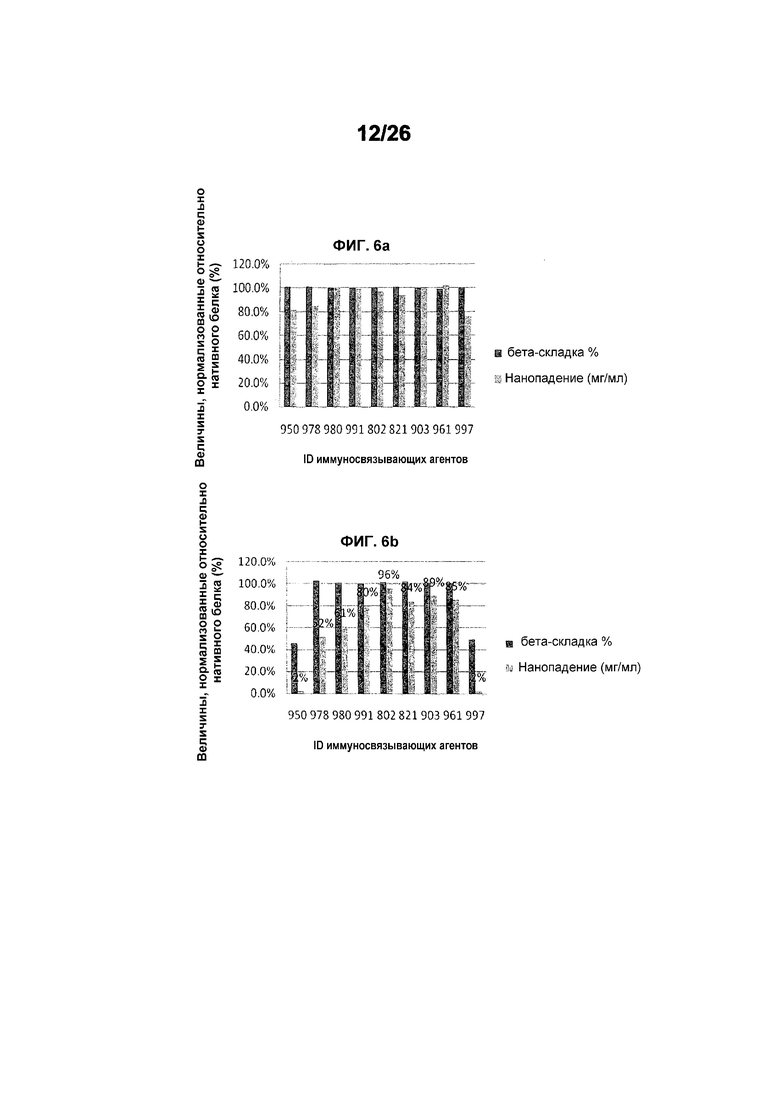
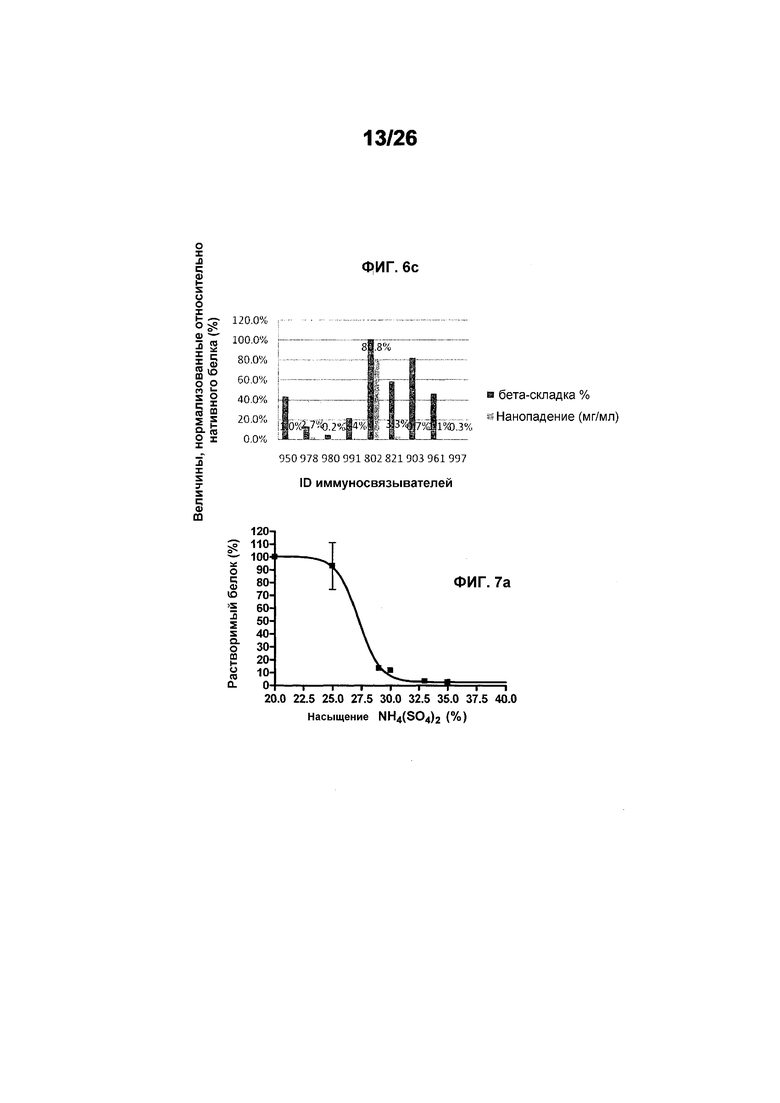
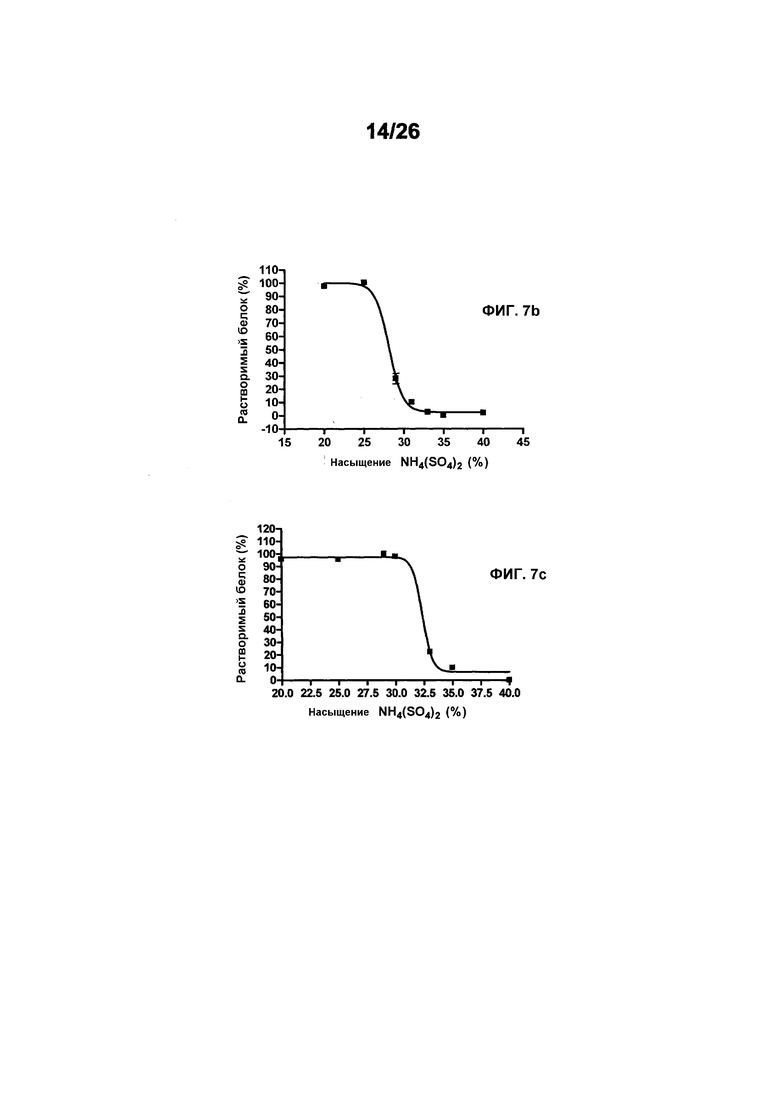
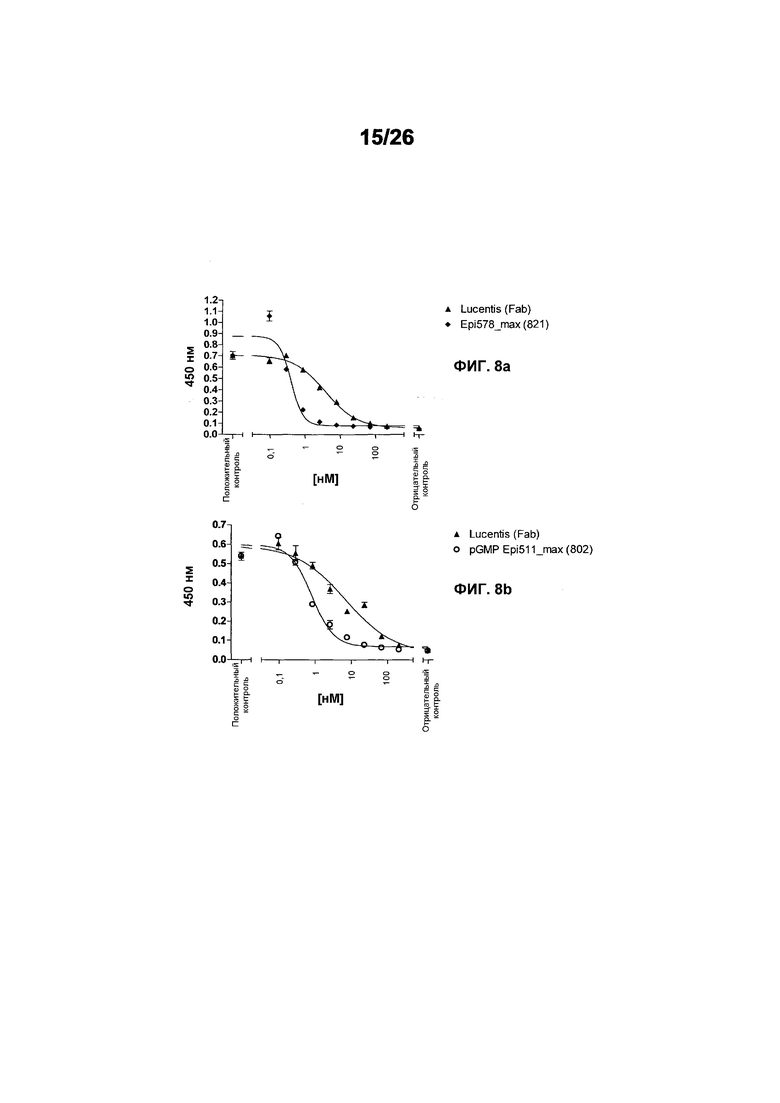
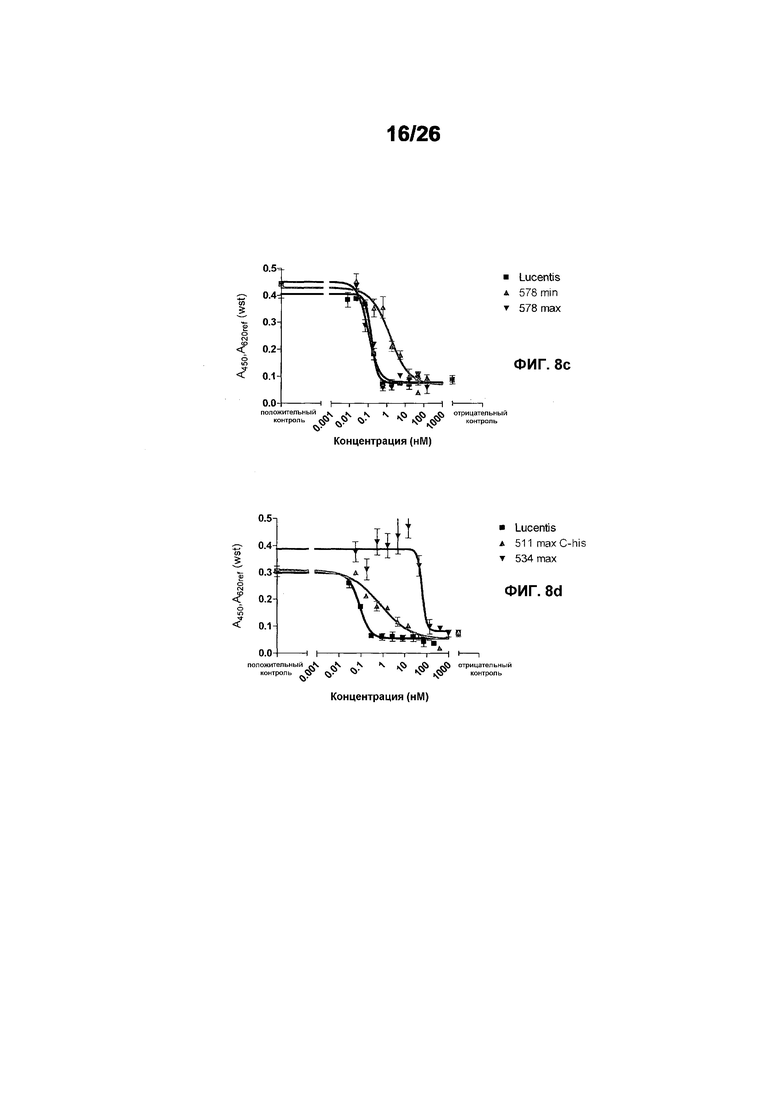
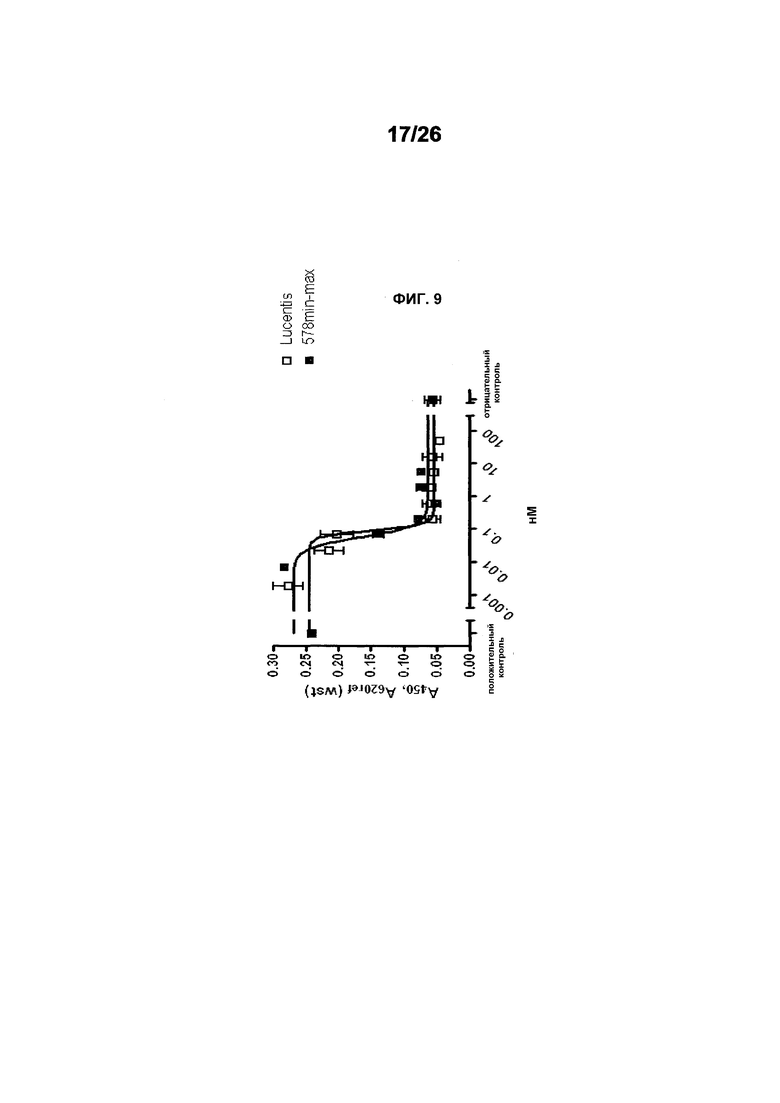
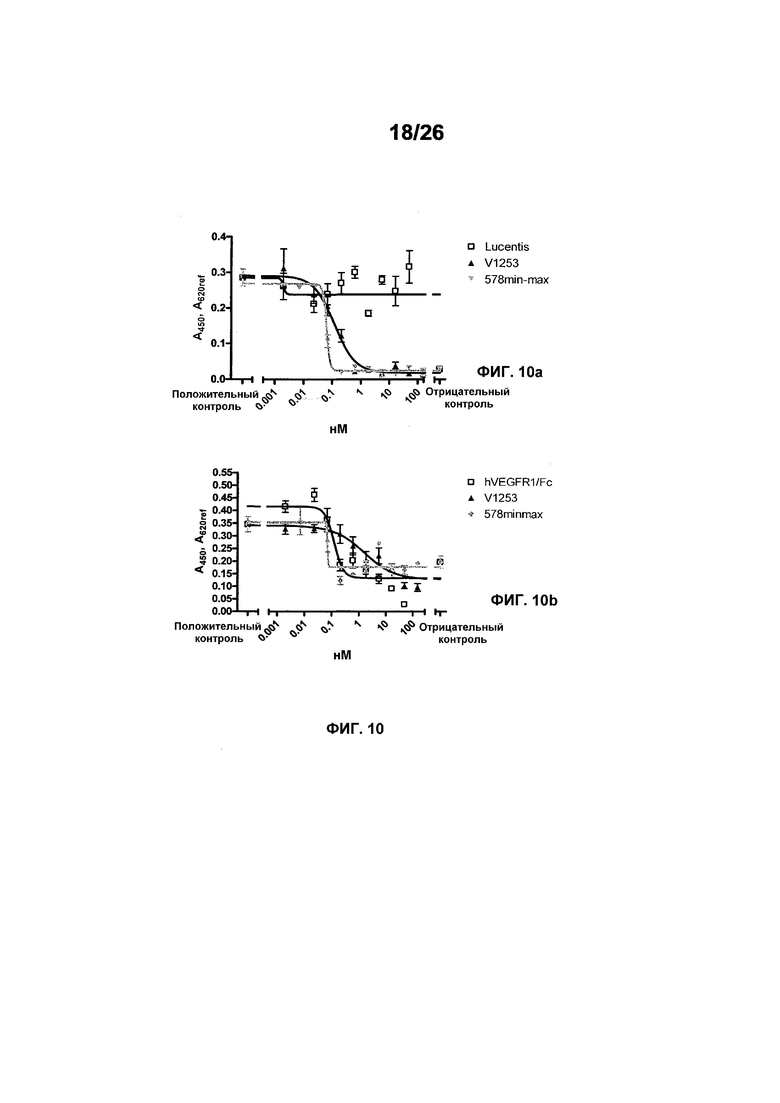
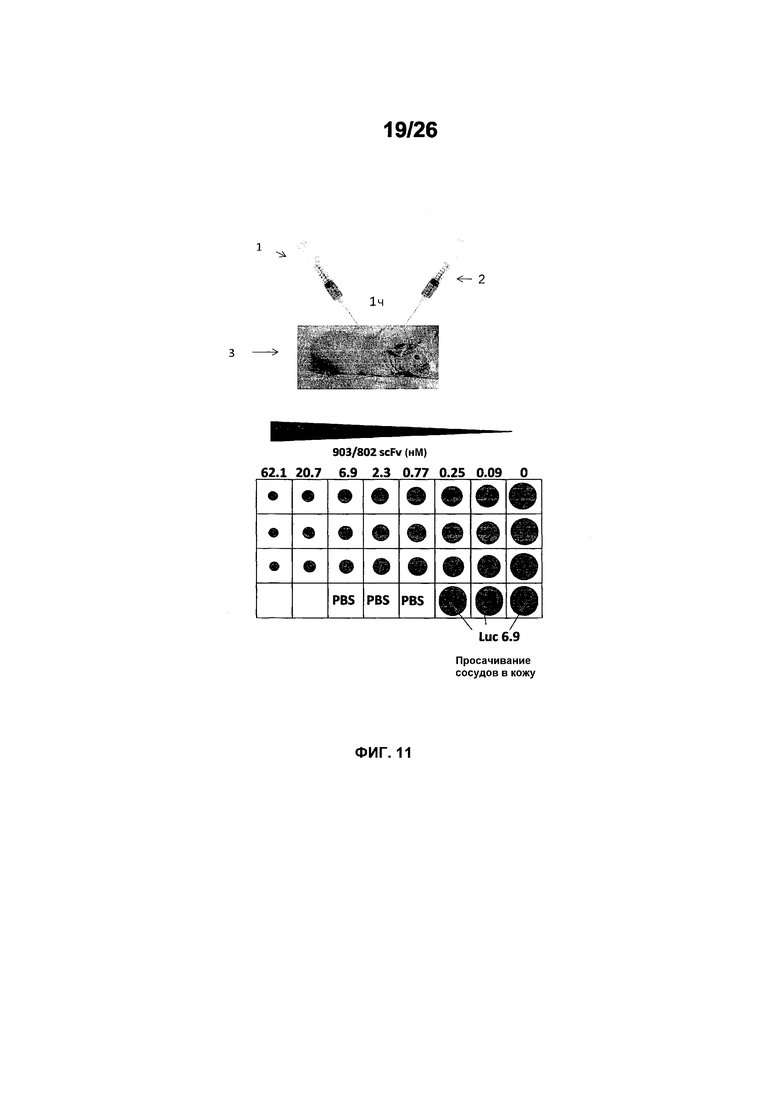
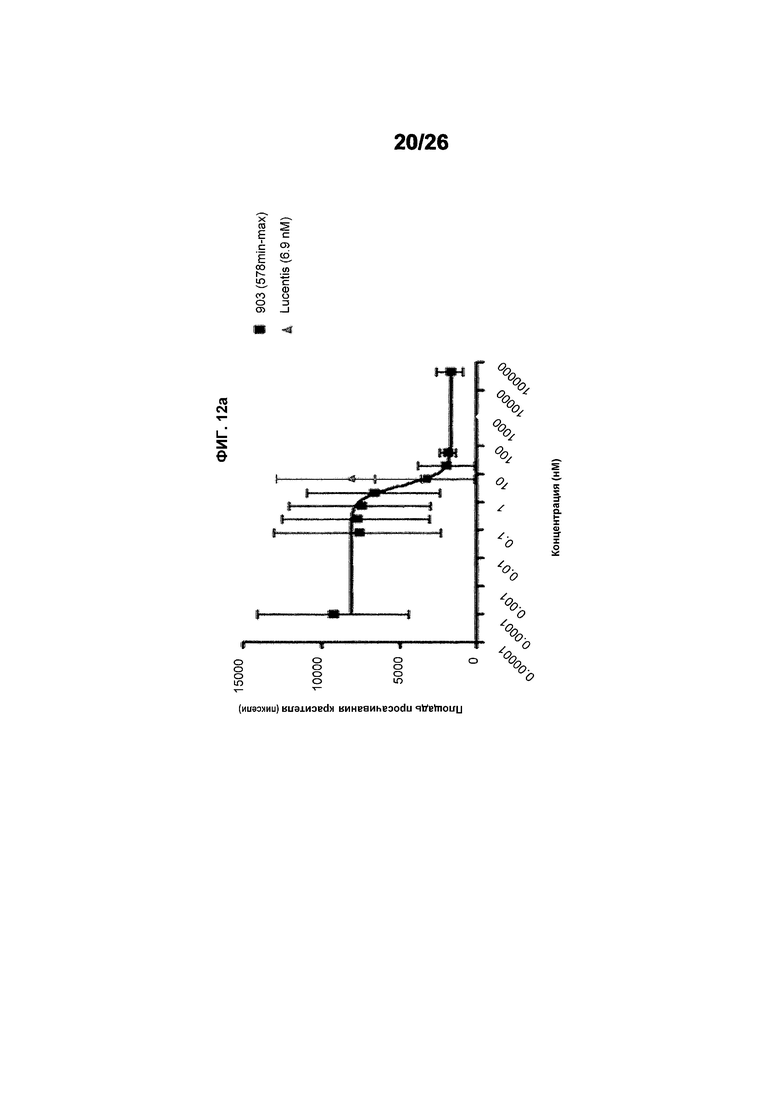
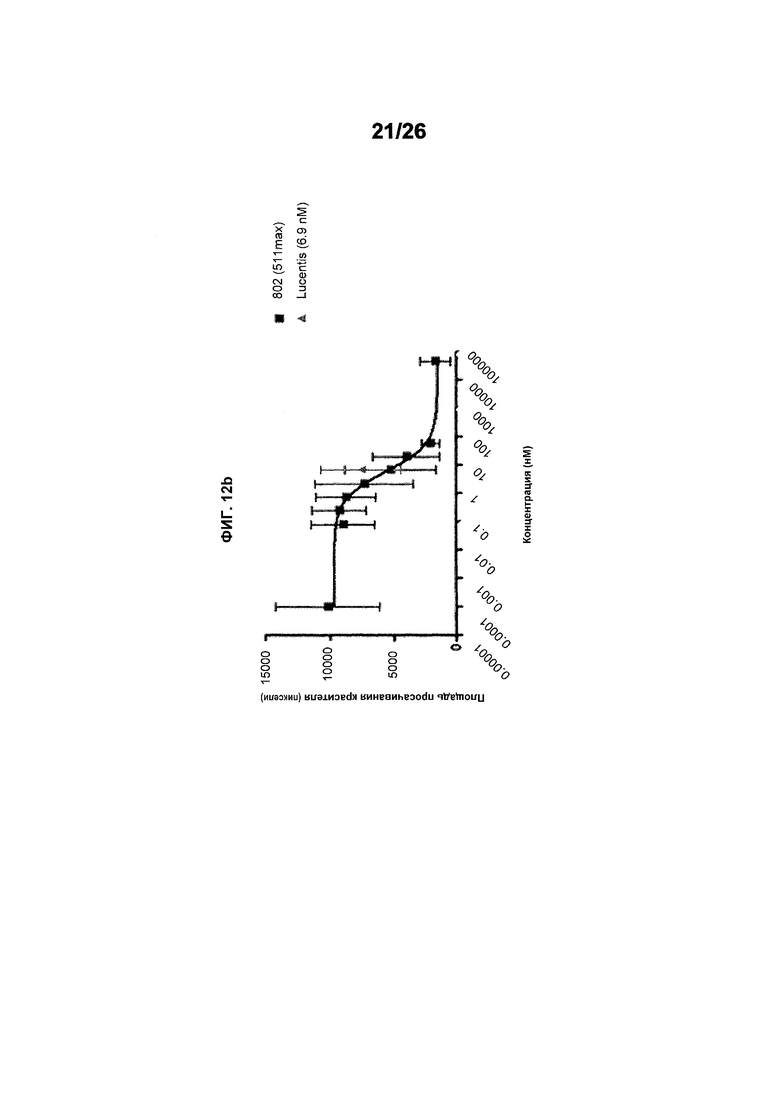
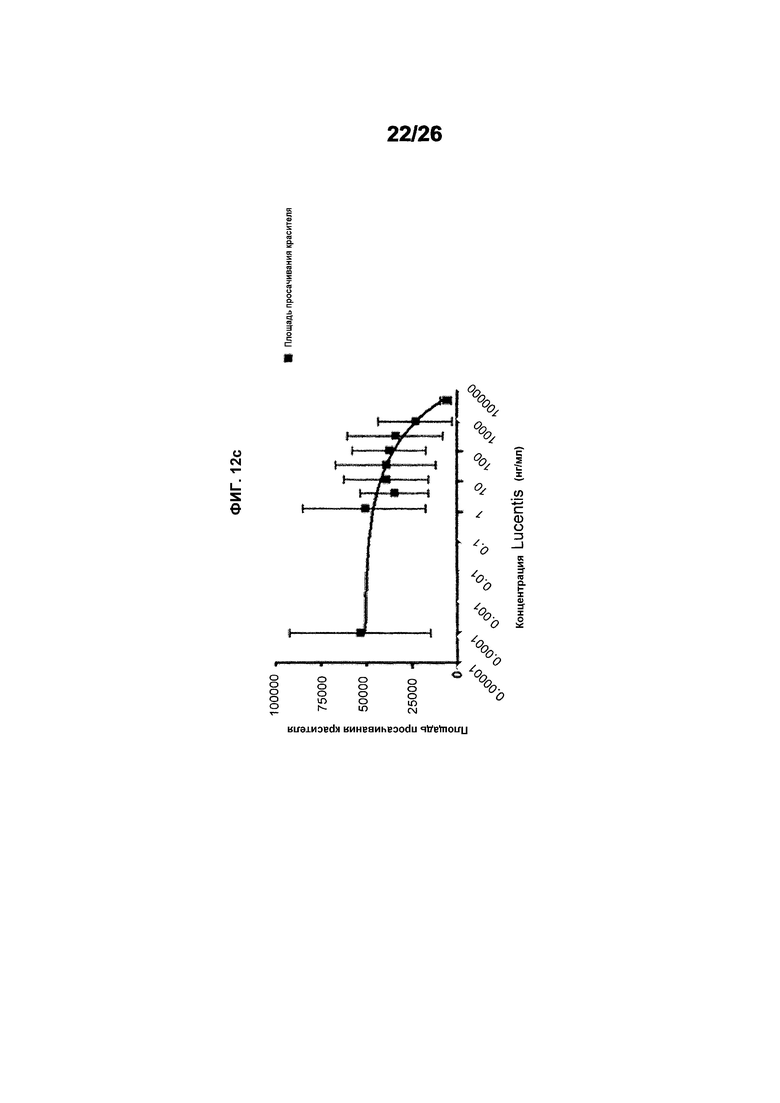

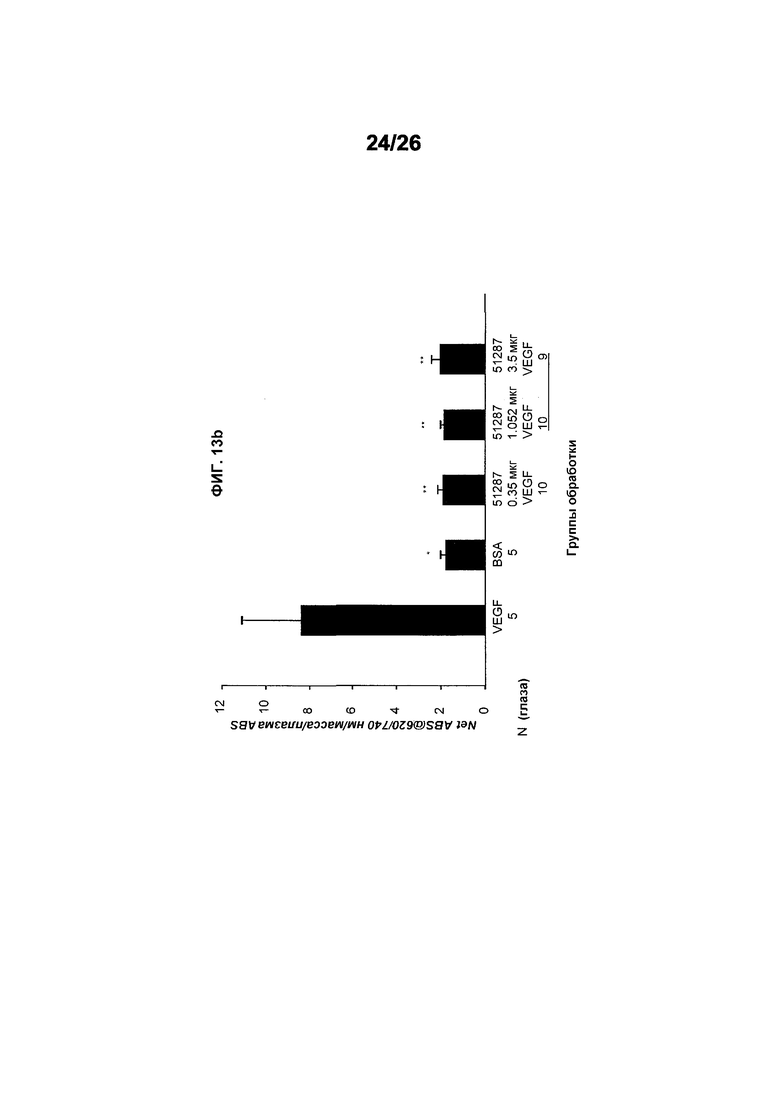
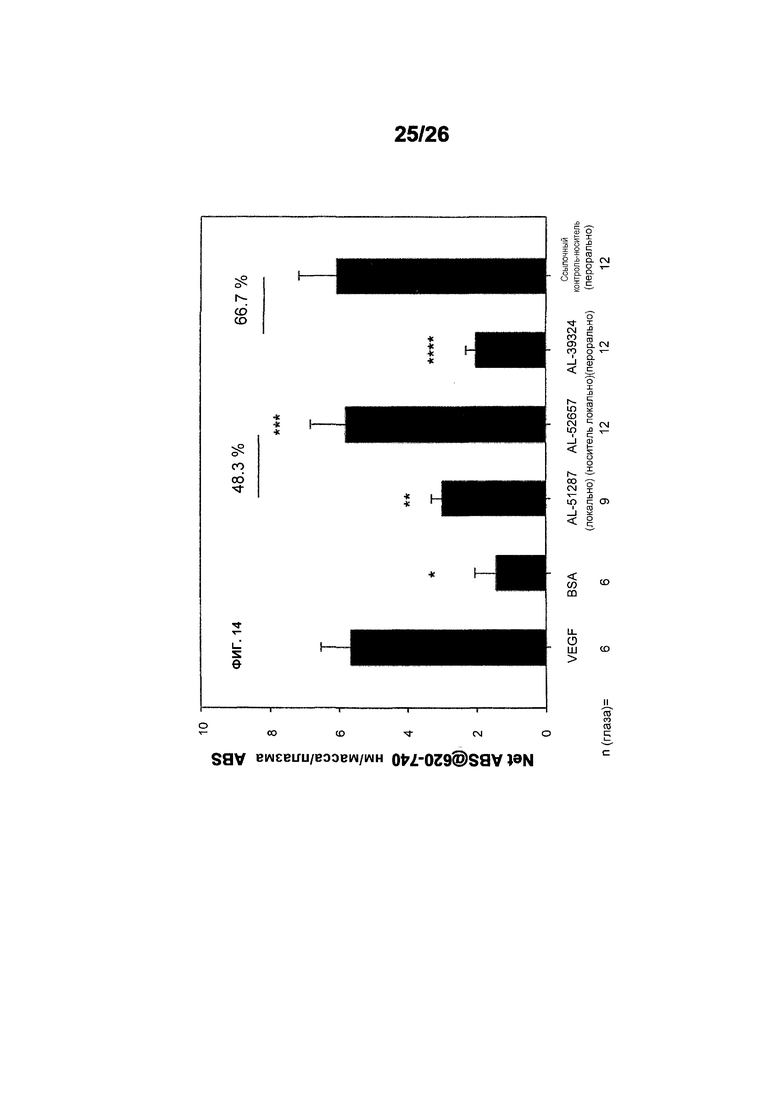
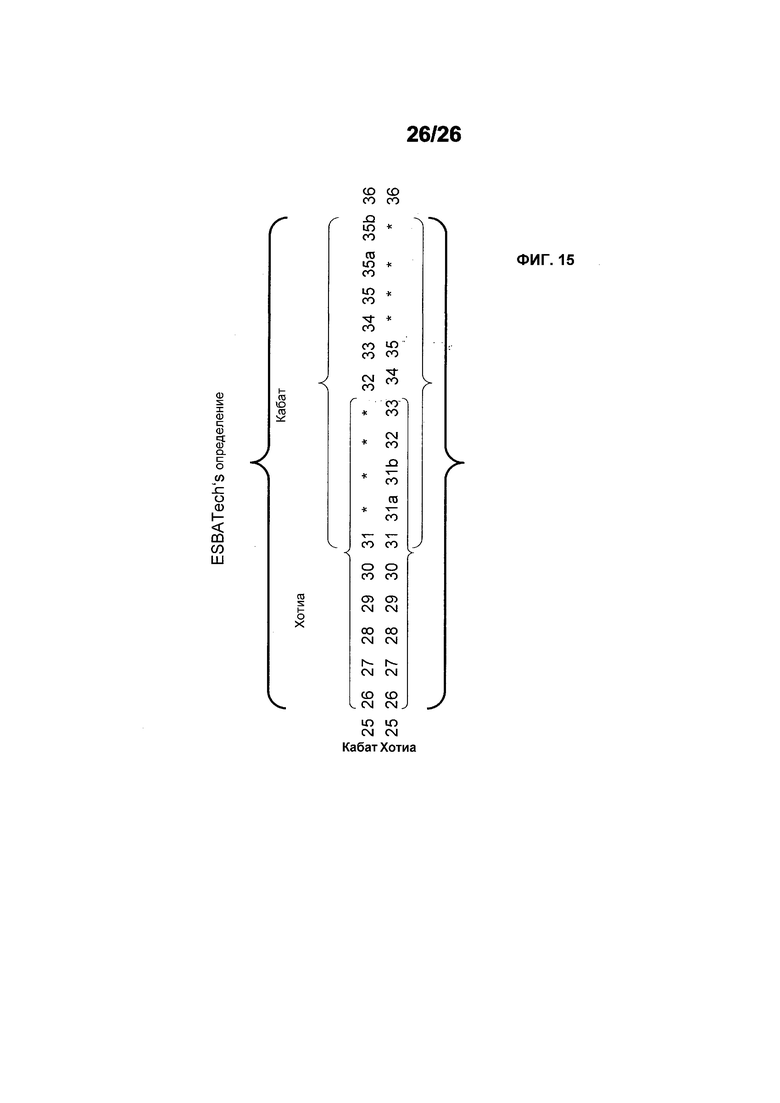
Изобретение относится к области биохимии, в частности к гуманизированному антителу или его антигенсвязывающему фрагменту, которое нейтрализует VEGF человека, а также к содержащим его композиции и набору. Также раскрыта выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент, а также содержащие ее вектор и клетка-хозяин. Изобретение также относится к способу ингибирования пролиферации эндотелиальных клеток и проницаемости сосудов, опосредованных VEGF человека, у больного с заболеванием, опосредованным VEGF человека, с использованием вышеуказанного антитела. Изобретение может быть эффективно использовано для лечения заболевания, опосредованного VEGF человека. 15 н. и 17 з.п. ф-лы, 15 ил., 12 табл., 7 пр.
1. Гуманизированное антитело или его антигенсвязывающий фрагмент, которое(ый) нейтрализует VEGF человека, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где
VH содержит CDRH1, CDRH2 и CDRH3 с последовательностями SEQ ID NO: 8, SEQ ID NO: 20 и SEQ ID NO: 32 соответственно и VL содержит CDRL1, CDRL2 и CDRL3 c последовательностями SEQ ID NO: 43, SEQ ID NO: 55 и SEQ ID NO: 66,
где вариабельная область тяжелой цепи имеет последовательность SEQ ID NO: 164;
где вариабельная область легкой цепи имеет последовательность SEQ ID NO: 87;
где вариабельная область тяжелой цепи и вариабельная область легкой цепи связаны линкером с последовательностью SEQ ID NO: 181; и
где гуманизированное антитело содержит N-концевой метионин.
2. Гуманизированное антитело или его антигенсвязывающий фрагмент, которое(ый) нейтрализует VEGF человека, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где
VH содержит CDRH1, CDRH2 и CDRH3 с последовательностями SEQ ID NO: 8, SEQ ID NO: 20 и SEQ ID NO: 32 соответственно и последовательность VH по крайней мере на 95% идентична последовательности SEQ ID NO: 164; и
VL содержит CDRL1, CDRL2 и CDRL3 c последовательностями SEQ ID NO: 43, SEQ ID NO: 55 и SEQ ID NO: 66 и последовательность VL по крайней мере на 95% идентична последовательности SEQ ID NO: 87;
и где антитело или его антигенсвязывающий фрагмент связывается с VEGF165 человека с аффинностью (Kd) ≤ 1x10 нМ.
3. Гуманизированное антитело или антиген-связывающий фрагмент по п.2, где фрагмент представляет собой scFv, Fab-фрагмент, Fab’-фрагмент или F(ab’)2-фрагмент.
4. Гуманизированное антитело или его антиген-связывающий фрагмент по п.2, где вариабельная область тяжелой цепи содержит последовательность, которая на 100% идентична последовательности SEQ ID NO: 164.
5. Гуманизированное антитело или его антиген-связывающий фрагмент по п.2, где вариабельная область легкой цепи содержит последовательность, которая на 100% идентична последовательности SEQ ID NO: 87.
6. Гуманизированное антитело или его антигенсвязывающий фрагмент по п.2, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO: 164, и вариабельную область легкой цепи, содержащую последовательность SEQ ID NO: 87.
7. Гуманизированное антитело или его антигенсвязывающий фрагмент по п.6, содержащее(ий) линкерную последовательность с последовательностью SEQ ID NO: 181, связывающую вариабельную область тяжелой цепи и вариабельную область легкой цепи.
8. Гуманизированное антитело или его антигенсвязывающий фрагмент по п.7, где антигенсвязывающий фрагмент представляет собой одноцепочечное антитело (scFv).
9. Гуманизированное антитело или его антигенсвязывающий фрагмент по п.8, где антигенсвязывающий фрагмент содержит N-концевой метионин, полученный из старт-кодона, и присутствует в векторе экспрессии, который экспрессирует антигенсвязывающий фрагмент.
10. Гуманизированное антитело или его антигенсвязывающий фрагмент по п.2, где антитело или его антигенсвязывающий фрагмент связывается с VEGF165 человека с аффинностью (Kd) ≤ 1 нM.
11. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по п.1 или 2 и содержащая вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где
VH содержит CDR группы, состоящей из SEQ ID NO: 8, SEQ ID NO: 20 и SEQ ID NO: 32, и VL содержит CDR группы, состоящей из SEQ ID NO: 43, SEQ ID NO: 55 и SEQ ID NO: 66.
12. Выделенная молекула нуклеиновой кислоты по п.11, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи с аминокислотной последовательностью, которая по крайней мере на 95% идентична последовательности SEQ ID NO: 164, и вариабельную область легкой цепи с аминокислотной последовательностью, которая по крайней мере на 95% идентична последовательности SEQ ID NO: 87.
13. Выделенная молекула нуклеиновой кислоты по п.12, где вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 164 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 87.
14. Выделенная молекула нуклеиновой кислоты по п.13, где вариабельная область тяжелой цепи и вариабельная область легкой цепи связаны с помощью последовательности SEQ ID NO: 181.
15. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п.11.
16. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п.12.
17. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п.13.
18. Клетка-хозяин для репликации вектора экспрессии по п.15, где клетка-хозяин содержит указанный вектор экспрессии.
19. Клетка-хозяин для репликации вектора экспрессии по п.16, где клетка-хозяин содержит указанный вектор экспрессии.
20. Клетка-хозяин для репликации вектора экспрессии по п.17, где клетка-хозяин содержит указанный вектор экспрессии.
21. Композиция для ингибирования пролиферации эндотелиальных клеток, опосредованной VEGF человека, содержащая эффективное количество гуманизированного антитела или его антигенсвязывающего фрагмента по п.1 и фармацевтически приемлемый носитель.
22. Композиция по п.21, составленная для местного, внутриглазного, перорального, назального, ректального или парентерального введения.
23. Композиция для ингибирования пролиферации эндотелиальных клеток, опосредованной VEGF человека, содержащая эффективное количество гуманизированного антитела или его антигенсвязывающего фрагмента по п.10 и фармацевтически приемлемый носитель.
24. Фармацевтическая композиция для ингибирования пролиферации эндотелиальных клеток и проницаемости сосудов, опосредованной VEGF человека, у больного с заболеванием, опосредованным VEGF человека, где фармацевтическая композиция содержит рекомбинантное анти-VEGF антитело или его антигенсвязывающий фрагмент в концентрации от около 0,1 до около 50 мг/мл, где антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 164 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 87.
25. Фармацевтическая композиция по п.24, где антигенсвязывающим фрагментом является Fab, F(ab')2 или Fab’.
26. Фармацевтическая композиция по п.24, где антиген-связывающим фрагментом является scFv.
27. Фармацевтическая композиция по п.26, где рекомбинантное анти-VEGF антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, которые связаны с помощью последовательности SEQ ID NO: 181.
28. Фармацевтическая композиция по п.24, где рекомбинантное антитело или его антигенсвязывающий фрагмент находится в концентрации от около 0,5 до около 40 мг/мл.
29. Фармацевтическая композиция по любому из пп.24-28, составленная для системного, внутриопухолевого, околоопухолевого, для введения в повреждение или около повреждения, местного, внутриглазного, перорального, назального, ректального или парентерального введения.
30. Набор для ингибирования пролиферации эндотелиальных клеток и проницаемости сосудов, опосредованных VEGF человека, отличающийся тем, что содержит фармацевтическую композицию по п.24.
31. Способ ингибирования пролиферации эндотелиальных клеток и проницаемости сосудов, опосредованных VEGF человека, у больного с заболеванием, опосредованным VEGF человека, включающий введение больному гуманизированного антитела или антигенсвязывающего фрагмента по п.1 или 2, обеспечивая лечение VEGF-опосредованного заболевания, которое выбрано из группы, состоящей из возрастной дегенерации желтого пятна, неоваскулярной глаукомы, диабетической ретинопатии, ретинопатии недоношенных, ретролентальной фиброплазии, карциномы молочной железы, карциномы легких, карциномы желудка, эзофагеальной карциномы, колоректальной карциномы, карциномы печени, карциномы яичника, комы, арренобластомы, карциномы шейки матки, карциномы эндометрия, гиперплазии эндометрия, эндометриоза, фибросаркомы, хориокарциномы, рака головы и шеи, назофарингеальной карциномы, карциномы гортани, гепатобластомы, саркомы Капоши, меланомы, карциномы кожи, гемангиомы, кавернозной гемангиомы, гемангиобластомы, карциномы поджелудочной железы, ретинобластомы, астроцитомы, глиобластомы, Шванномы, олигодендроглиомы, медуллобластомы, нейробластомы, рабдомиосаркомы, остеогенный саркомы, лейомиосаркомы, карциномы мочеполовых путей, карциномы щитовидной железы, опухоли Вильмса, почечно-клеточной карциномы, рака предстательной железы, патологической пролиферации сосудов, связанной с факоматозом, водянки, синдрома Мейжа, ревматоидного артрита, псориаза и атеросклероза.
32. Способ лечения заболевания, опосредованного VEGF человека, у больного, включающий введение больному фармацевтической композиции по любому из пп.24-29, обеспечивая, таким образом, лечение у пациента VEGF-опосредованного заболевания, которое выбрано из группы, состоящей из возрастной дегенерации желтого пятна, неоваскулярной глаукомы, диабетической ретинопатии, ретинопатии недоношенных, ретролентальной фиброплазии, карциномы молочной железы, карциномы легких, карциномы желудка, эзофагеальной карциномы, колоректальной карциномы, карциномы печени, карциномы яичника, комы, арренобластомы, карциномы шейки матки, карциномы эндометрия, гиперплазии эндометрия, эндометриоза, фибросаркомы, хориокарциномы, рака головы и шеи, назофарингеальной карциномы, карциномы гортани, гепатобластомы, саркомы Капоши, меланомы, карциномы кожи, гемангиомы, кавернозной гемангиомы, гемангиобластомы, карциномы поджелудочной железы, ретинобластомы, астроцитомы, глиобластомы, Шванномы, олигодендроглиомы, медуллобластомы, нейробластомы, рабдомиосаркомы, остеогенный саркомы, лейомиосаркомы, карциномы мочеполовых путей, карциномы щитовидной железы, опухоли Вильмса, почечно-клеточной карциномы, рака предстательной железы, патологической пролиферации сосудов, связанной с факоматозом, водянки, синдрома Мейжа, ревматоидного артрита, псориаза и атеросклероза.
| SOPHIA RAN and CARLA MOUTA-BELLUM, Generation of new rabbit monoclonal antibody RAM-1 against human VEGF-C, Proc Amer Assoc Cancer Res, 2005, Volume 46 | |||
| US 2003023046 A1, 30.01.2003 | |||
| WO 2007019620 A1, 22.02.2007 | |||
| MIKMIKHAIL POPKOV et al., Human/mouse cross-reactive anti-VEGF receptor 2 recombinant antibodies selected from an immune b9 allotype |

