Область изобретения
Описание настоящего изобретения относится к микроорганизму, продуцирующему L-валин, и способу получения L-валина с использованием этого микроорганизма.
Предшествующий уровень техники
L-Валин, представляющий собой аминокислоту с разветвленной цепью, биосинтезируется в микроорганизмах, начиная с пировиноградной кислоты через ацетомолочную кислоту, дигидроксиизовалериановую кислоту и кетоизовалериановую кислоту. Эти промежуточные метаболиты образуются в реакции, катализируемой синтазой ацетогидроксикислоты, изомероредуктазой ацетогидроксикислоты, дегидратазой ацетогидроксикислоты и трансаминазой В. Тем не менее, эти ферменты также вовлечены в биосинтез L-изолейцина, начинающийся с кетомасляной кислоты и пировиноградной кислота, и L-лейцин также биосинтезируется с промежуточного метаболита, представляющего собой кетоизовалериановую кислоту, через 2-изопропиляблочную кислоту, 3-изопропиляблочную кислоту и кетоизокапроновую кислоту. Таким образом, поскольку ферменты, используемые в путях биосинтеза аминокислот с разветвленной цепью, т.е. L-валин, L-изолейцин и L-лейцин, идентичны, известно, что промышленное получение одного типа аминокислоты с разветвленной цепью посредством ферментации затруднительно. Кроме того, ингибирование по механизму обратной связи осуществляется со стороны конечного продукта L-валина или его производных, что затрудняет промышленное массовое производство L-валина.
Тем не менее, до настоящего времени проводились исследования способа получения валина путем ингибирования по механизму обратной связи (Патент США №10457919), но не было никаких исследований увеличения способности продуцировать валин путем комбинации ферментов, обладающих усиленной или уменьшенной активностью, в соответствии с описанием настоящего изобретения.
Описание изобретения
Техническая проблема
Соответственно, авторы настоящего изобретения провели непрерывное исследование эффективного способа получения L-валина, и в качестве результата подтвердили то, что микроорганизм, обладающий усиленной активностью дегидратазы дигидроксикислоты; уменьшенной активностью трансаминазы С; ослабленной активностью пируватдегидрогеназы; уменьшенной активностью цитратсинтазы; или их комбинацией, обладает превосходной способностью продуцировать L-валин по сравнению с микроорганизмом дикого типа, таким образом, завершая настоящее изобретение.
Техническое решение
Одна из задач описания настоящего изобретения заключается в том, чтобы предложить микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты; и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы С;
(2) ослабленной активностью пируватдегидрогеназы;
(3) уменьшенной активностью цитратсинтазы.
Еще одна задача описания настоящего изобретения заключается в том, чтобы предложить способ получения L-валина, включающий выращивание микроорганизма.
Благоприятные действия
При выращивании микроорганизма, продуцирующего L-валин, в соответствии с описанием настоящего изобретения, L-валин может продуцироваться с высоким выходом. Соответственно, можно ожидать эффектов удобства получения и уменьшения стоимости получения с точки зрения промышленного аспекта.
Подробное описание предпочтительных воплощений
Далее, описание настоящего изобретения будет описано подробно.
В то же самое время, каждое описание и раскрытые здесь воплощения могут применяться здесь в различных описаниях и воплощениях в отношении общих свойств. То есть, все комбинации различных компонентов, раскрытых здесь, включены в объем описания настоящего изобретения. Кроме того, объем описания настоящего изобретения не должен ограничиваться приведенным ниже конкретным описанием.
Дополнительно, специалисты в данной области техники могут быть способны понять или подтвердить с использованием исключительно обычного экспериментального пути множество эквивалентов конкретных аспектов описанного здесь изобретения. Кроме того, также предполагается, что эти эквиваленты включены в описание настоящего изобретения.
В одном из аспектов описания настоящего изобретения предложен микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты; и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы С;
(2) ослабленной активностью пируватдегидрогеназы; и
(3) уменьшенной активностью цитратсинтазы.
В одном из воплощений микроорганизм, продуцирующий валин, может представлять собой микроорганизм, обладающий усиленной активностью дегидратазы дигидроксикислоты и уменьшенной активностью трансаминазы С.
В еще одном воплощении микроорганизм, продуцирующий валин, может представлять собой микроорганизм, обладающий усиленной активностью дегидратазы дигидроксикислоты и ослабленной активностью пируватдегидрогеназы.
В еще одном воплощении микроорганизм, продуцирующий валин, может представлять собой микроорганизм, обладающий усиленной активностью дегидратазы дигидроксикислоты и уменьшенной активностью цитратсинтазы.
В еще одном воплощении микроорганизм, продуцирующий валин, может представлять собой микроорганизм, обладающий усиленной активностью дегидратазы дигидроксикислоты и уменьшенной активностью цитратсинтазы, и дополнительно обладающий уменьшенной активностью трансаминазы С или ослабленной активностью пируватдегидрогеназы, но не ограничивается им.
Используемый здесь термин «дегидратаза дигидроксикислоты» представляет собой фермент, вовлеченный в синтез кетоизовалерата, представляющего собой предшественник валина, в пути биосинтеза L-валина, начиная с пирувата через ацетолактат, дигидроксиизовалерат в кетоизовалерат. В описании настоящего изобретения можно способствовать получению L-валина путем усиления активности дегидратазы дигидроксикислоты, и, таким образом, увеличения синтеза кетоизовалерата.
Используемый здесь термин «трансаминаза С» представляет собой фермент, вовлеченный в путь синтеза L-аланина из пирувата.
Используемый здесь термин «пируватдегидрогеназа» представляет собой фермент, вовлеченный в синтез ацетил-соА из пировиноградной кислоты.
Используемый здесь термин «цитратсинтаза» представляет собой фермент, синтезирующий цитрат из ацетил-соА.
Использованный здесь термин «усиление» означает то, что активность белка увеличивается по сравнению с его эндогенной активностью. Усиление может быть использовано взаимозаменяемо с терминами, такими как активация, повышающая регуляция, сверхэкспрессия, увеличение и т.п. В частности, активация, усиление, повышающая регуляция, сверхэкспрессия и увеличение могут включать оба случая, при которых демонстрируется активность, которая исходно не обнаруживалась, или активность усиливается по сравнению с эндогенной активностью или активностью перед модификацией. «Эндогенная активность» относится к активности конкретного полипептида, исходно присутствующей в родительском штамме перед трансформацией, или не модифицированном микроорганизме, когда свойство изменяют путем генетической модификации, вызванной природными или искусственными факторами, и может быть использована взаимозаменяемо с «активностью перед модификацией». «Усиление», «повышающая регуляция», «сверхэкспрессия» или «увеличение» активности полипептида по сравнению с его эндогенной активностью означает то, что активность и/или концентрация (уровень экспрессии) полипептида усиливается по сравнению с конкретным полипептидом, исходно присутствующим в родительском штамме перед трансформацией или в немодифицированном микроорганизме.
Усиление может быть достигнуто путем введения чужеродного полипептида или путем усиления активности и/или увеличения концентрации (уровня экспрессии) эндогенного полипептида. Усилена ли активность дегидратазы дигидроксикислоты или не усилена можно подтвердить путем увеличения уровня полипептида, уровня экспрессии или количества продукта, экскретируемого из полипептида.
Усиление активности регулятора дегидратазы дигидроксикислоты может быть достигнуто путем применения различных способов, хорошо известных в области техники, и эти способы не ограничены при условии, что они усиливают активность полипептида-мишени по равнению с активностью микроорганизма перед модификацией. В частности, может быть использована генетическая инженерия и/или белковая инженерия, хорошо известные специалистам в данной области техники, которые представляют собой обычные для молекулярной биологии способы, но способ не ограничен ими (например, Sitnicka et al. Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol. 2. 1-16, Sambrook et al. Molecular Cloning 2012 и т.п.).
В частности, усиление активности дегидратазы дигидроксикислоты в соответствии с описанием настоящего изобретения может быть достигнуто путем:
1) увеличения числа копий полинуклеотида, кодирующего полипептид, в клетках;
2) замены последовательности, регулирующей экспрессию гена, кодирующего полипептид, в хромосоме на последовательность, обладающую более сильной активностью;
3) модификации нуклеотидной последовательности, кодирующей стартовый кодон или 5'-UTR генного транскрипта, кодирующего полипептид;
4) модификации аминокислотной последовательности полипептида, таким образом, что активность полипептида усиливается;
5) модификации полинуклеотидной последовательности, кодирующей полипептид, таким образом, что активность полипептида усиливается (например, путем модификации полинуклеотидной последовательности гена полипептида для кодирования полипептида, модифицированного таким образом, чтобы усилить активность полипептида);
6) введения чужеродного полипептида, демонстрирующего активность полипептида, или кодирующего его чужеродного полинуклеотида;
7) оптимизации кодона полинуклеотида, кодирующего полипептид;
8) анализа третичной структуры полипептида и, таким образом, выбора и модификации экспонируемого сайта или его химической модификации; или
9) комбинации двух или более чем двух, выбранных из (1)-(8) выше, но не ограничиваясь этим.
Конкретнее,
1) Способ увеличения числа копий полинуклеотида в клетке, кодирующего полипептид, может быть достигнуто путем введения вектора, который функционально связан с полинуклеотидом, кодирующим полипептид, и способен реплицироваться и функционировать независимо от клетки-хозяина в клетке-хозяине. В качестве альтернативы, этот способ может быть достигнут путем введения одной копии или двух копий полинуклеотидов, кодирующих полипептид, в хромосому клетки-хозяина. Введение в хромосому может быть осуществлено путем введения вектора, способного встраивать полинуклеотид в хромосому клетки-хозяина, в клетку-хозяина, но не ограничиваясь этим. Вектор является таким, как описано выше.
2) Способ замены области, регулирующей экспрессию (или последовательности, регулирующей экспрессию) гена, кодирующего полипептид, в хромосоме, на последовательность, обладающую сильной активностью, может быть достигнуто, например, путем индукции модификации в последовательности путем делеции, вставки, не консервативной или консервативной замены, или путем любой их комбинации для дополнительного усиления активности области, регулирующей экспрессию, или путем замены последовательности на последовательность, обладающую более сильной активностью. Область, регулирующая экспрессию, может включать без ограничения промотор, операторную последовательность, последовательность, кодирующую сайт связывания с рибосомой, последовательность, регулирующую терминацию транскрипции или трансляции и т.п. В одном из примеров способ может включать замену исходного промотора на сильный промотор, но не ограничиваться этим.
Примеры известного сильного промотора могут включать промоторы CJ1-CJ7 (патент США № US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13(sm3) (патент США № US 10584338 В2), промотор 02 (патент США №US 10273491 В2), промотор tkt и промотор уссА, но не ограничиваются этим.
3) Способ модификации нуклеотидной последовательности, кодирующей стартовый кодон или 5'-UTR генного транскрипта, кодирующего полипептид, может быть достигнут, например, путем замены нуклеотидной последовательности на нуклеотидную последовательность, кодирующую другой стартовый кодон, обладающий более высоким уровнем экспрессии полипептида, чем эндогенный стартовый кодон, но не ограничивается этим.
4) и 5) Способы модификации аминокислотной последовательности или полинуклеотидной последовательности могут быть достигнуты путем индукции модификации в последовательности путем делеции, вставки, не консервативной или консервативной замены аминокислотной последовательности полипептида или полинуклеотидной последовательности, кодирующей полипептид, или путем любой их комбинации для усиления активности полипептида или путем замены последовательности на аминокислотную последовательность или полинуклеотидную последовательность, модифицированную таким образом, чтобы усилить активность, но не ограничиваются этим. Используемый здесь вектор может дополнительно включать маркер для отбора для подтверждения встраивания в хромосому. Маркер для отбора является таким как описано выше.
6) Способ введения чужеродного полинуклеотида, демонстрирующего активностью полипептида может быть достигнут путем введения в клетку чужеродного полинуклеотида, кодирующего полипептид, который демонстрирует активность, идентичную/похожую на активность полипептида. Чужеродный полинуклеотид может быть использован без ограничения независимо от его происхождения или последовательности при условии, что он демонстрирует активность, идентичную/похожую на активность полипептида. Введение может быть осуществлено специалистами в данной области техники путем подходящего выбора способа трансформации, известного в области техники, и экспрессия введенного полинуклеотида в клетке-хозяине обеспечивает продукцию полипептида, таким образом, увеличивая его активность.
7) Способ оптимизации кодона полинуклеотида, кодирующего полипептид, может быть достигнут путем оптимизации кодона эндогенного полинуклеотида для увеличения транскрипции или трансляции в клетке-хозяине, или путем оптимизации его кодонов таким образом, чтобы дать возможность для достижения оптимизированной транскрипции и трансляции чужеродного полинуклеотида в клетке-хозяине.
Кроме того, 8) способ анализа третичной структуры полипептида и, таким образом, отбора и модификации экспонируемого сайта или его химической модификации может быть достигнут, например, путем сравнения информации о последовательности анализируемого полипептида с базой данных, в которой хранится информация о последовательностях известных белков, для определения кандидатов белков-матриц в соответствии со степенью сходства последовательности, и, таким образом, подтверждения структуры, основанной на информации, таким образом, отбора и трансформации или модификации экспонируемого сайту, который предполагается модифицировать или химически модифицировать.
Вектор в соответствии с описанием настоящего изобретения представляет собой молекулу ДНК, используемую в качестве посредника для искусственного переноса чужеродного генетического материала в другие клетки, и может включать конструкцию ДНК, содержащую нуклеотидную последовательность полинуклеотида, кодирующего полипептид-мишень, функционально связанный с подходящей областью, регулирующей экспрессию (последовательностью, регулирующей экспрессию), таким образом, что она способна экспрессировать полипептид-мишень в подходящей клетке-хозяине. Область, регулирующая экспрессию, может включать промотор, способный инициировать транскрипцию, любую операторную последовательность для регулирования транскрипции, последовательность, кодирующую подходящий сайт связывания мРНК с рибосомой, и последовательность, регулирующую терминацию транскрипции и трансляцию. После трансформации в подходящую клетку-хозяин вектор может реплицироваться или функционировать независимо от генома хозяина или может быть встроен в ее геном.
Вектор, используемый в описании настоящего изобретения, не ограничен конкретным образом, и может быть использован любой вектор, известный в области техники. Примеры обычно используемого вектора могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A и т.п., и в качестве плазмидного вектора могут быть использованы векторы на основе pDZ, pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и т.п. В частности, могут быть использованы векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.п.
В одном из примеров полинуклеотид, кодирующий желаемый полипептид, может быть встроен в хромосому с использованием вектора для внутриклеточного встраивания хромосом. Встраивание полинуклеотида в хромосому может быть произведено без ограничения при помощи любого способа, известного в области техники, например, при помощи гомологичной рекомбинации. Вектор может дополнительно включать селективный маркер для подтверждения встраивания в хромосому. Селективный маркер предназначен для отбора клеток, трансформированных вектором, то есть для подтверждения того, что встроена желаемая молекула нуклеиновой кислоты, и могут быть использованы маркеры, обеспечивающие селективные фенотипы, такие как резистентность к лекарственным средствам, потребность в питательных веществах, резистентность к цитотоксическим агентам и экспрессия поверхностных полипептидов. При обработке селективным агентом только клетки, экспрессирующие селективные маркеры, могут выжить или демонстрировать другие фенотипы в среде, и, таким образом, могут быть отобраны трансформированные клетки.
Использованный здесь термин «трансформация» относится к введению вектора, содержащего полинуклеотид, кодирующий полипептид-мишень, в клетку-хозяина или микроорганизм таким образом, чтобы полипептид, кодируемый полинуклеотидом, мог экспрессироваться в клетке-хозяине. Трансформированный полинуклеотид может быть представлен в форме, встроенной в хромосому клетки-хозяина, или в форме, расположенной за пределами хромосомы, при условии, что полипептид экспрессируется в клетке-хозяине. При условии, что трансформируемый полинуклеотид может экспрессироваться в клетке-хозяине, не важно, интегрируется ли трансформированный полинуклеотид в хромосому клетки-хозяина или располагается экстрахромосомно, и могут быть включены оба случая. Кроме того, полинуклеотид может включать ДНК и/или РНК, кодирующую полипептид-мишень. Полинуклеотид может быть введен в любой форме при условии, что он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессирующейся кассеты, которая представляет собой генетическую конструкцию, включающую все элементы, необходимые для ее автономной экспрессии. Как правило, экспрессирующаяся кассета может включать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания с рибосомой и сигнал терминации трансляции. Экспрессирующаяся кассета может находиться в форме самореплицирующегося экспрессирующегося вектора. Кроме того, полинуклеотид может быть введен в клетку-хозяина в своей исходной форме и функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине, но не ограничиваться этим.
Кроме того, использованный здесь термин «функционально связанный» означает то, что последовательность гена функционально связана с промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего белок-мишень в соответствии с описанием настоящего изобретения.
Такое усиление белковой активности может означать то, что усиливается активность соответствующего белка, которой он исходно не обладает, или его активность или концентрация в общем увеличена на 1%, 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500%, и максимально на 1000% или 2000% или более, основываясь на активности или концентрации белка дикого типа или исходного микробного штамма, но не ограничена ими.
В частности, усиление активности дегидратазы дигидроксикислоты может означать то, что активность дегидратазы дигидроксикислоты у микроорганизма усилена по сравнению с активностью у микроорганизма дикого типа, микроорганизма до модификации или микроорганизма, имеющего не модифицированный белок, таким образом, увеличивая синтез кетоизовалерата, который является предшественником L-валина, из дигидроксиизовалерата, приводя в результате к увеличению продукции L-валина.
Используемый здесь термин «уменьшение» представляет собой всеобъемлющую концепцию, включающую случай, при котором активность белка, представленного у микроорганизма в его естественном состоянии или до модификации, ослаблена или устранена (удалена), т.е. по сравнению с эндогенной активностью или одной копией гена, кодирующего белок в клетке, и может означать то, что активность составляет 0% или более до 100% или менее.
Такое «уменьшение активности» белка означает то, что активность самого белка устранена или реализуется действие меньше чем его исходная функция, но конкретно не ограничена этим. То есть, уменьшение активности в частности включает как «устранение активности», так и «ослабление активности».
«Устранение активности» может означать то, что фермент или белок не экспрессируется совсем по сравнению с природным штаммом дикого типа, представляющего собой родительский штамм или штамм, имеющий не модифицированный белок, или его активность не обнаруживается даже тогда, когда фермент или белок экспрессируется.
В описании настоящего изобретения устранение активности может быть достигнуто путем применения различных способов, хорошо известных в области техники. Примеры этих способов включают: 1) устранение фрагмента или всего гена, кодирующего белок; 2) модификацию последовательности гена, кодирующего белок, таким образом то, что активность белка устранена или ослаблена; 3) введение антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующего белок; 4) добавление последовательности, комплементарной последовательности Шайна-Дальгарно (SD) относительно переднего конца последовательности SD гена, кодирующего белок, с образованием вторичной структуры, таким образом, ингибируя рибосомальное прикрепление; 5) инженерия путем обратной транкрипции (RTE), при которой добавляют промотор, который должен обратимо транскрибироваться, по 3' концу открытой рамки считывания (ORF) последовательности гена, кодирующей белок; или их комбинацию, но конкретно не ограничены этим.
Дополнительно, «ослабление активности» может означать то, что реализуется действие, меньшее чем исходная функция, и может быть достигнуто при помощи способов, включающих: устранение части гена, кодирующего белок, на хромосоме; замена гена, кодирующего белок на хромосоме, на мутантный ген, таким образом, что активность белка уменьшается; введение мутации в регулирующую экспрессию последовательность гена, кодирующего белок на хромосоме; замена регулирующей экспрессию последовательности гена, кодирующего белок, на последовательность, обладающую слабой активностью (например, замена промотора гена на промотор, который слабее чем эндогенный промотор) и т.п., но не ограничивающихся ими, и без ограничения могут быть использованы известные способы ослабления активности.
Такое уменьшение активности белка может означать то, что активность соответствующего белка устранена или его активность или концентрация в общем уменьшена на 0%, 1%, 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% относительно активности или концентрации в белке дикого типа или исходном штамме микроба, но не ограничиваться этим.
В частности, активность трансаминазы С у микроорганизма может быть уменьшена по сравнению с активностью природного штамма дикого типа, родительского штамма до мутации или штамма, имеющего не модифицированный белок. В одном из воплощений микроорганизм может не обладать активностью белка трансаминазы С вследствие делеции гена трансаминазы С или может обладать ослабленной активностью, поскольку последовательность, кодирующая стартовый кодон гена трансаминазы С, модифицирована до GTG, таким образом, уменьшая экспрессию белка трансаминазы С.
Дополнительно, в частности, активность пируватдегидрогеназы у микроорганизма может быть уменьшена по сравнению с активностью природного штамма дикого типа, родительского штамма до мутации или штамма, имеющего не модифицированный белок. В одном из воплощений микроорганизм может не обладать активностью белка пируватдегидрогеназы вследствие делеции гена пируватдегидрогеназы или может обладать ослабленной активностью, поскольку последовательность, кодирующая стартовый кодон гена пируватдегидрогеназы, модифицирована до GTG, таким образом, уменьшая экспрессию белка пируватдегидрогеназы. В еще одном воплощении, поскольку последовательность гена пируватдегидрогеназы мутирована у микроорганизма, микроорганизм может экспрессировать мутант пируватдегидрогеназы, обладающий ослабленной активностью по сравнению с активностью белка дикого типа, при котором аминокислота, соответствующая 432ому или 435ому положению от N-конца аминокислотной последовательности SEQ ID NO: 3 заменена на другую аминокислоту. В частности, мутант пируватдегидрогеназы, обладающий ослабленной активностью по сравнению с активностью белка дикого типа, может включать последовательность SEQ ID NO: 5 или SEQ ID NO: 6, но не ограничен ими.
Кроме того, в частности, активность цитратсинтазы у микроорганизма может быть уменьшена по сравнению с активностью природного штамма дикого типа, родительского штамма до мутации или штамма, имеющего не модифицированный белок. В одном из воплощений микроорганизм может не обладать активностью белка цитратсинтазы вследствие делеции гена цитратсинтазы или может обладать ослабленной активностью, поскольку последовательность, кодирующая стартовый кодон гена цитратсинтазы, модифицирована до GTG, таким образом, уменьшая экспрессию белка цитратсинтазы. В еще одном воплощении, поскольку последовательность гена цитратсинтазы мутирована у микроорганизма, микроорганизм может экспрессировать мутант цитратсинтазы, обладающий уменьшенной активностью по сравнению с активностью белка дикого типа, при котором аминокислота, соответствующая 241ому или 312ому положению от N-конца аминокислотной последовательности SEQ ID NO: 4 заменена на другую аминокислоту. В частности, мутант цитратсинтазы, обладающий уменьшенной активностью по сравнению с активностью белка дикого типа, может включать последовательность SEQ ID NO: 7 или SEQ ID NO: 8, но не ограничен ими.
Выражение «замена на другую аминокислоту» не ограничено при условии, что аминокислота до замены заменяется на другую аминокислоту. В частности, она может быть заменена на любую аминокислоту, выбранную из лизина, гистидина, глутаминовой кислоты, аспарагиновой кислоты, глицина, аланина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, цистеина, тирозина, аспарагина, аргинина и глутамина.
Конкретнее, пируватдегидрогеназа, обладающая ослабленной активностью, может представлять собой пируватдегидрогеназу, в которой аминокислота, соответствующая 432ому или 435ому положению от N-конца аминокислотной последовательности SEQ ID NO: 3 заменена на не полярную аминокислоту.
Дополнительно, конкретнее, цитратсинтаза, обладающая уменьшенной активностью, может представлять собой цитратеинтазу, в которой аминокислота, соответствующая 241ому или 312ому положению от N-конца аминокислотной последовательности SEQ ID NO: 4 заменена на полярную или не полярную аминокислоту.
Использованный здесь термин «соответствующий» относится к аминокислотному остатку в положении, указанном в белке или пептиде, или аминокислотному остатку, который похож на, идентичен или гомологичен остатку, указанному в белке или пептиде. Использованная здесь «соответствующая область» как правило относится к похожей или соответствующей позиции в родственном белке или референсном белке.
В описании настоящего изобретения может быть использована специфическая нумерация положений аминокислотных остатков в используемом здесь белке. Например, возможно перенумеровать положения аминокислотных остатков в белке в соответствии с описанием настоящего изобретения в соответствующие положения путем выравнивания последовательности белка в соответствии с описанием настоящего изобретения с желаемым белком, с которым производится сравнение.
Использованный здесь термин «гомология» или «идентичность» относится к степени сходства между двумя данными аминокислотными последовательностями или нуклеотидными последовательностями, и она может быть выражена в процентах. Часто термины «гомология» и «идентичность» можно использовать взаимозаменяемо друг с другом.
Гомологию или идентичность последовательности консервативных полинуклеотидов или полипептидов можно определить путем стандартного алгоритма выравнивания, и вместе с ним можно использовать штраф за пропуск в последовательности, установленный в используемой программе. По существу, гомологичные или идентичные последовательности как правило могут гибридизоваться с полными последовательностями или частью последовательностей в умеренно строгих или очень строгих условиях. Понятно то, что также включена гибридизация с полинуклеотидами, содержащими общий кодон или вырожденные кодоны в гибридизируемых полинуклеотидах.
Имеют ли любые две полинуклеотидные или полипептидные последовательности гомологию, сходство или идентичность может быть, например, определено с использованием известного компьютерного алгоритма, такого как программа «FASTA» (Pearson et al., (1988) [Proc. Natl. Acad. Sci. USA 85]: 2444) с использованием параметров по умолчанию. Альтернативно, они могут быть определены с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), который осуществляют с использованием программы Needleman пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16:276 277) (предпочтительно, версия 5.0.0 или более поздняя) (программный пакет GCG (Devereux, J. et al., Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F. et al., J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, и CARILLO et al. (1988) SIAM J Applied Math 48:1073). Например, гомология, сходство или идентичность могут быть определены с использованием BLAST или ClustalW Национального Центра Биотехнологической Информации (NCBI).
Гомологию, сходство или идентичность между полинуклеотидами или полипептидами можно определить, например, путем сравнения информации о последовательностях с помощью, например, компьютерной программы GAP, такой как Needleman et al., (1970), J Mol Biol. 48:443, как раскрыто в Smith and Waterman, Adv. Appl. Math (1981) 2:482. В общем, программа GAP определяет гомологию, сходство или идентичность путем деления количества выровненных одинаковых символов (т.е. нуклеотидов или аминокислот) на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) матрицу двоичного сравнения (содержащую величину 1 в случае идентичности и 0 в случае не идентичности) и взвешенную матрицу сравнения в соответствии с Gribskov, et al. (1986), Nucl. Acids Res. 14:6745, как описано в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence and Structure, National Biomedical Research Foundation, pp. 353-358 (1979) (или матрица замен EDNAFULL (версия EMBOSS NCBI NUC4.4); (2) штраф 3,0 за каждый пропуск и дополнительно штраф 0,10 за каждый символ в каждом пропуске (или штраф за внесение пропуска 10 и штраф за удлинение пропуска 0,5) и (3) отсутствие штрафа за концевые пропуски.
Используемый здесь термин «вариант» относится к белку, имеющему одну или более чем одну аминокислоту, отличающуюся от описанной последовательности путем консервативных замен и/или модификаций, таким образом, что функции или свойства белка сохраняются. Варианты отличаются от последовательностей, идентифицированных путем замены, делеции или вставки нескольких аминокислот. Такие варианты как правило могут быть идентифицированы путем модификации одной или более чем одной из вышеприведенных аминокислотных последовательностей белка и путем оценки свойств модифицированного белка. То есть, способность вариантов может быть усилена, не изменена или уменьшена по сравнению с нативным белком. Другие варианты могут включать варианты, в которых фрагмент удален из N-и/или С-конца зрелого белка. Термин «вариант» может быть использован взаимозаменяемо с терминами, такими как модификация, модифицированный белок, модифицированный полипептид, мутант, мутеин, дивергент, вариант и т.п., при условии, что термины используют для обозначения вариации, но термины не ограничены ими. Для задачи описания настоящего изобретения вариант может представлять собой варианты, в которых активность белка уменьшена или ослаблена по сравнению с активностью природного белка дикого типа или не модифицированного белка, но не ограничен ими.
Используемый здесь термин «консервативная замена» относится к замене аминокислоты на другую аминокислоту, обладающую похожими структурными и/или химическими свойствами. Такая аминокислотная замена как правило может происходить на основе сходства полярности, заряда (основной, кислотный), растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатка.
Кроме того, вариант также может включать делецию или добавление аминокислот, которые обладают минимальным влиянием на свойства и вторичную структуру полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью по N-концу, вовлеченной в перенос белков котрансляционно или посттрансляционно. Кроме того, полипептид также может быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
Используемый здесь термин «микроорганизм, продуцирующий L-валин», относится к микроорганизму, способному продуцировать избыточное количество L-валина из источника углерода в среде по сравнению с микроорганизмом дикого типа или не модифицированным микроорганизмом. Кроме того, микроорганизм, продуцирующий L-валин, может представлять собой рекомбинантный микроорганизм. В частности, микроорганизм не ограничен конкретно его типом при условии, что он может продуцировать L-валин, и может представлять собой микроорганизм рода Enterobacter, рода Escherichia, рода Erwinia, рода Serratia, рода Providencia, рода Corynebacterium и рода Brevibacterium. Конкретнее, он может представлять собой микроорганизм рода Corynebacterium или рода Escherichia.
Еще конкретнее, микроорганизм рода Escherichia может представлять собой Escherichia coli, и микроорганизм рода Corynebacterium может представлять собой Corynebacterium glutamicum, и может быть включен любой микроорганизм рода Corynebacterium или рода Escherichia, способный увеличивать продукцию L-валина, в который введена усиленная активность дегидратазы дигидроксикислоты; и любая одна или более чем одна из комбинаций, выбранных из уменьшенной активности трансаминазы С; уменьшенной активности пируватдегидрогеназы; или уменьшенной активности цитратсинтазы.
Родительский штамм микроорганизма, продуцирующего L-валин, который модифицирован таким образом, что обладает усиленной активностью дегидратазы дигидроксикислоты; и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы С;
(2) ослабленной активностью пируватдегидрогеназы; и
(3) уменьшенной активностью цитратсинтазы, не ограничен конкретным образом при условии, что он представляет собой микроорганизм, продуцирующий L-валин.
Микроорганизм, продуцирующий L-валин, сам может представлять природный микроорганизм или микроорганизм, обладающий улучшенной способностью продуцировать L-валин путем встраивания гена, связанного с механизмом внешней продукции L-валина, или усилением или уменьшением (ослаблением или подавлением) активности эндогенного гена.
В одном из воплощений микроорганизм может представлять собой микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты по сравнению со штаммом дикого типа или родительским штаммом до мутации, и уменьшенной активностью трансаминазы С по сравнению со штаммом дикого типа или родительским штаммом до мутации. В частности, дегидратаза дигидроксикислоты может кодироваться геном ilvD, и трансаминаза С может кодироваться геном avtA. Гены могут быть получены из Corynebacterium glutamicum, но не ограничены ими.
В еще одном воплощении микроорганизм может представлять собой микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты по сравнению со штаммом дикого типа или родительским штаммом до мутации, и ослабленной активностью пируватдегидрогеназы по сравнению со штаммом дикого типа или родительским штаммом до мутации. В частности, дегидратаза дигидроксикислоты может кодироваться геном ilvD, и пируватдегидрогеназа может кодироваться геном асеЕ. Гены могут быть получены из Corynebacterium glutamicum, но не ограничены ими.
В еще одном воплощении микроорганизм может представлять собой микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты по сравнению со штаммом дикого типа или родительским штаммом до мутации и уменьшенной активностью цитратсинтазы по сравнению со штаммом дикого типа или родительским штаммом до мутации. В частности, дегидратаза дигидроксикислоты может кодироваться геном ilvD, и цитратсинтаза может кодироваться геном gltA. Гены могут быть получены из Corynebacterium glutamicum, но не ограничены ими.
Дополнительно к вышеприведенному микроорганизму он может представлять собой микроорганизм, продуцирующий L-валин, у которого активность трансаминазы С уменьшена или активность пируватдегидрогеназы уменьшена по сравнению со штаммом дикого типа или родительским штаммом до мутации, или может представлять собой микроорганизм, продуцирующий L-валин, у которого активность трансаминазы С уменьшена и активность пируватдегидрогеназы уменьшена по сравнению со штаммом дикого типа или родительским штаммом до мутации.
В еще одном аспекте описания настоящего изобретения предложен способ получения L-валина, включающий: культивирование микроорганизма, продуцирующего L-валин, который отличается усиленной активностью дегидратазы дигидроксикислоты; и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы С;
(2) ослабленной активностью пируватдегидрогеназы; и
(3) уменьшенной активностью цитратсинтазы.
«L-валин» в соответствии с описанием настоящего изобретения может включать не только сам L-валин, а также и его соль.
Использованный здесь термин «культивирование» означает, что микроорганизм выращивают в надлежащим образом контролируемых условиях окружающей среды. Процесс культивирования в соответствии с описанием настоящего изобретения может быть осуществлен в подходящей культуральной среде и условиях выращивания, известных в области техники. Такой процесс культивирования может быть легко скорректирован для применения специалистом в данной области техники в соответствии с выбранным штаммом. В частности, выращивание может представлять собой периодическое выращивание, непрерывное выращивание и выращивание подпитываемой культуры, но не ограничиваться этим.
Использованный здесь термин «среда» относится к смеси веществ, которая содержит в качестве основного ингредиента питательные вещества, требующиеся для культивирования микроорганизма, и она обеспечивает питательные вещества и факторы роста, а также воду, которые необходимы для жизнедеятельности и роста. В частности, среда и другие условия культивирования, используемые для культивирования микроорганизма в соответствии с описанием настоящего изобретения, могут представлять собой любую среду, используемую для обычного культивирования микроорганизмов без какого-либо конкретного ограничения. Тем не менее, микроорганизм в соответствии с описанием настоящего изобретения может быть выращен в аэробных условиях в обычной среде, содержащей подходящий источник углерода, источник азота, источник фосфора, неорганическое соединение, аминокислоту и/или витамин, при корректировании температуры, рН и т.п.
В описании настоящего изобретения источник углерода может включать углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза и т.п.; сахарные спирты, такие как маннит, сорбит и т.п.; органические кислоты, такие как пировиноградная кислота, молочная кислота, лимонная кислота и т.п.; аминокислоты, такие как глутаминовая кислота, метионин, лизин и т.п. Кроме того, источник углерода может включать природные органические питательных вещества, такие как гидролизат крахмала, мелассы, сырые мелассы, рисовые отруби, маниок, выжимки сахарного тростника и жидкий кукурузный экстракт и т.п. В частности, могут быть использованы углеводы, такие как глюкоза и стерилизованные предварительно обработанные мелассы (т.е. мелассы, преобразованные в редуцированные сахара), и, кроме того, различные другие источники углерода в подходящем количестве могут быть использованы без ограничения. Эти источники углерода могут быть использованы сами по себе или в комбинации двух или более чем двух из них, но не ограничиваться этим.
Источник азота может включать источники неорганического азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония, нитрат аммония и т.п.; аминокислоты, такие как глутаминовая кислота, метионин, глютамин и т.п.; и источники органического азота, такие как пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат казеина, рыба или продукты ее деградации, обезжиренный соевый жмых или продукт его деградации и т.п. Эти источники азота могут быть использованы сами по себе или в комбинации двух или более чем двух из них, но не ограничиваться этим.
Источник фосфора может включать первичный кислый фосфат калия, гидрофосфат калия или соответствующие содержащие натрий соли. Примеры неорганических соединений могут включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и т.д. Кроме того, могут быть включены аминокислоты, витамины и/или соответствующие предшественники. Эти компоненты или предшественники могут быть добавлены к среде партиями или непрерывным образом, но эти источники фосфора не ограничиваются этим.
В описании настоящего изобретения рН среды может быть скорректирован путем добавления соединения, такого как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота, серная кислота и т.п., подходящим образом во время культивирования микроорганизма. Кроме того, во время культивирования образование пены может быть предотвращено путем использования пеногасителя, такого как сложный эфир полигликоля и жирной кислоты. Кроме того, кислород или содержащий кислород газ могут быть введены в среду для поддержания в среде аэробных условий; или азот, водород или углекислый газ могут быть введены для поддержания анаэробных и микроаэробных условий без нагнетания в нее газа, но газ не ограничен ими.
Температура среды может находиться в диапазоне от 20°С до 45°С, в частности, от 30°С до 37°С, но не ограничиваться этим. Культивирование может осуществляться до получения желательного продуцируемого количества полезных веществ, в частности, в течение приблизительно от 10 до 100 часов, но не ограничиваться этим.
L-валин, продуцирумый путем выращивания в соответствии с описанием настоящего изобретения, может быть высвобожден в среду или оставаться в клетках.
Способ получения L-валина в соответствии с описанием настоящего изобретения дополнительно может включать стадию приготовления микроорганизма в соответствии с описанием настоящего изобретения, стадию приготовления среды для культивирования штамма или их комбинацию (независимо от последовательности, в любой последовательности), например, до стадии выращивания.
Способ получения L-валина в соответствии с описанием настоящего изобретения дополнительно может включать стадию выделения L-валина из культуральной среды (среды, в которой осуществлялось выращивание) или микроорганизма в соответствии с описанием настоящего изобретения. Стадия выделения может быть дополнительно включена после стадии культивирования.
На стадии выделения желаемый L-валин может быть собран с использованием способа культивирования микроорганизма в соответствии с описанием настоящего изобретения, например, с использованием подходящего способа, известного в области техники, в соответствии с периодическим способом культивирования, непрерывным способом культивирования или культивированием с подпиткой. Например, могут быть использованы способы, такие как центрифугирование, фильтрование, обработка агентом, осаждающим белок (способ высаливания), экстракция, ультразвуковое разрушение, ультрафильтрация, диализ, различные виды хроматографий, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография и т.п., HPLC (высокоэффективная жидкостная хроматография) или их комбинация, и желаемый L-валин может быть выделен из культурной среды или микроорганизма с использованием подходящих способов, известных в области техники.
Кроме того, способ получения L-валина в соответствии с описанием настоящего изобретения дополнительно может включать стадию очистки, которая может быть осуществлена с использованием подходящего способа, известного в области техники. В одном из примеров, когда способ получения L-валина в соответствии с описанием настоящего изобретения включает как стадию выделения, так и стадию очистки, тогда стадия выделения и стадия очистки могут быть осуществлены непрерывно или с перерывами независимо от последовательности, или могут быть осуществлены одновременно, или могут быть объединены в одну стадию, но способ не ограничивается этим.
В способе в соответствии с описанием настоящего изобретения полинуклеотид, вектор, микроорганизм, L-валин и т.п. являются такими, как описано в других аспектах выше.
В еще одном аспекте описания настоящего изобретения предложен способ увеличения способности продуцировать L-валин, включающий введение модификации в микроорганизм, которая характеризуется усиленной активностью дегидратазы дигидроксикислоты; и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы С;
(2) ослабленной активностью пируватдегидрогеназы; и
(3) уменьшенной активностью цитратсинтазы.
В еще одном аспекте описания настоящего изобретения предложено применение микроорганизма, обладающего усиленной активностью дегидратазы дигидроксикислоты; и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы С;
(2) ослабленной активностью пируватдегидрогеназы; и
(3) уменьшенной активностью цитратсинтазы, при получении L-валина.
Способ осуществления изобретения
Далее описание настоящего изобретения будет описано более подробно со ссылкой на следующие примеры. Тем не менее, эти примеры представляют собой всего лишь предпочтительные примеры, приведенные для иллюстративных задач, и, таким образом, не предполагается, что объем описания настоящего изобретения ограничен этими примерами.
Пример 1: Конструирование штамма, основанного на получении валила, и его оценка
Один из типов мутаций [ilvN(A42V); Biotechnology and Bioprocess Engineering, June 2014, Volume 19, Issue 3, pp 456-467] вводили в штаммы дикого типа Corynebacterium glutamicum АТСС14067 и АТСС13869 для создания штамма, обладающего усиленной способностью продуцировать L-валин.
В частности, геномную ДНК штамма дикого типа Corynebacterium glutamicum АТСС 14067 экстрагировали с использованием мининабора для экстракции всей ДНК G-spin Total DNA (intron, № по каталогу 17045) в соответствии с протоколом, приведенным в наборе. ПЦР (полимеразную цепную реакцию) осуществляли с использованием пары праймеров в соответствии с SEQ ID NO: 9 и 10 и пары праймеров в соответствии с SEQ ID NO: 11 и 12, основанных на геномной ДНК в качестве матрицы с получением фрагмента гена 537 п.о., соответственно. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут.
ПЦР с перекрывающимися праймерами осуществляли на основе двух фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 9 и 12 с получением продукта ПЦР длиной 1044 п. о. (далее названного как «фрагмент 2 с введенной мутацией»).
Полученный таким образом фрагмент 2 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования вектора, содержащего фрагмент 2 с введенной мутацией. Вектор для введения мутации A42V в ген ilvN назван pDZ-ilvN(A42V).

Затем pDZ-ilvN(A42V) трансформировали в каждый из штаммов дикого типа Corynebacterium glutamicum АТСС14067 и АТСС13869 для индукции гомологичной рекомбинации в хромосоме (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Штаммы, в который был введен вектор в хромосому, путем рекомбинации гомологичных последовательностей отбирали в среде, содержащей 25 мг/л канамицина.
Затем фрагменты гена амплифицировали на основании трансформантов Corynebacterium glutamicum, отобранных выше при помощи ПЦР с использованием пары праймеров в соответствии с SEQ ID NO: 18 и 21, и введение мутации подтверждали при помощи анализа путем секвенирования. Рекомбинантные штаммы были названы Corynebacterium glutamicum CJ7V и CJ8V, соответственно.
Эксперимент по определению ферментационного титра осуществляли на основе штаммов Corynebacterium glutamicum АТСС14067 и АТСС13869 дикого типа, и штаммов CJ7V и CJ8V, сконструированных выше. Каждый штамм подвергали субкультивированию в питательной среде и затем инокулировали в колбу с угловыми перегородками объемом 250 мл, содержащей 25 мл продуцирующей среды, и выращивали при перемешивании при 30°С в течение 72 часов при 200 об./мин. Затем концентрации L-валина анализировали с использованием HPLC (высокоэффективной жидкостной хроматографии), и проанализированные концентрации L-валина представлены в таблице 2 ниже.
Питательная среда (рН 7,2)
10 г глюкозы, 5 г мясного бульона, 10 г полипептона, 2,5 г хлорида натрия, 5 г дрожжевого экстракта, 20 г агара, 2 г мочевины (на основе 1 л дистиллированной воды).
Продуцирующая среда (рН 7,0)
100 г глюкозы, 40 г (NH4)2SO4, 2,5 г соевого белка, 5 г кукурузного экстракта, 3 г мочевины, 1 г KH2PO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1 мг тиамина-HCl, 2 мг пантотената кальция, 3 мг никотинамида и 30 г СаСО3 (на основе 1 л дистиллированной воды).
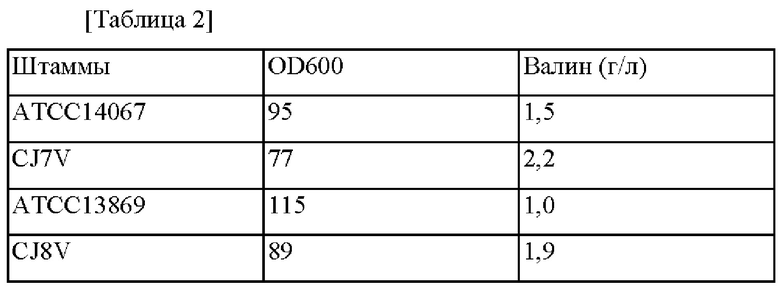
В соответствии с представленным в вышеприведенных результатах подтвердили то, что способность продуцировать L-валин увеличивалась в штаммах CJ7V и CJ8V, в которые была введена мутация гена ilvN(A42V), по сравнению с штаммами Corynebacterium glutamicum АТСС 14067 и АТСС 13 869 дикого типа.
Пример 2: Конструирование штаммов, обладающих высокой валинпродуцирующей способностью и их оценка
Пример 2-1. Конструирование штаммов с усиленным геном биосинтеза валина ilvD и их оценка
Вектор pDZ-Pcj7-ilvD для замены промотора гена ilvD конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 13 и 14, SEQ ID NO: 15 и 16, и SEQ ID NO: 17 и 18 для конструирования штамма, обладающего высокой валинпродуцирующей способностью, при которой усилена экспрессия гена биосинтеза валина ilvD.

В частности, для конструирования pDZ-Pcj7-ilvD ПЦР осуществляли с использованием пар праймеров в соответствии с SEQ ID NO: 13 и 14, SEQ ID NO: 15 и 16, и SEQ ID NO: 17 и 18, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС 14067 в качестве матрицы с получением, соответственно, фрагмента гена. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут.
ПЦР осуществляли с перекрывающимися праймерами, основанными на фрагментах, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 13 и 18 с получением фрагмента 3 с введенной мутацией. Полученный таким образом фрагмент 3 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования вектора, который был назван pDZ-Pcj7-ilvD.
Затем сконструированный выше pDZ-Pcj7-ilvD трансформировали в каждый из CJ7V, CJ8V и KCCM11201P, которые представляют собой штаммы, основанные на получении валина, для индукции гомологичной рекомбинации в хромосоме (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Штаммы, в который был введен вектор в хромосому путем рекомбинации гомологичных последовательностей, отбирали в среде, содержащей 25 мг/л канамицина. Затем фрагменты гена амплифицировали на основании трансформантов Corynebacterium glutamicum, отобранных выше, при помощи ПЦР с использованием пары праймеров в соответствии с SEQ ID NO: 13 и 18, и введение мутации подтверждали при помощи анализа путем секвенирования гена. Отобранные рекомбинантные штаммы были названы Corynebacterium glutamicum CJ7V:ilvD, CJ8V:ilvD, KCCM11201P:ilvD, соответственно. Ферментационный титр отобранных штаммов с усиленным геном ilvD определяли тем же самым образом, как в примере 1, и результаты представлены ниже.
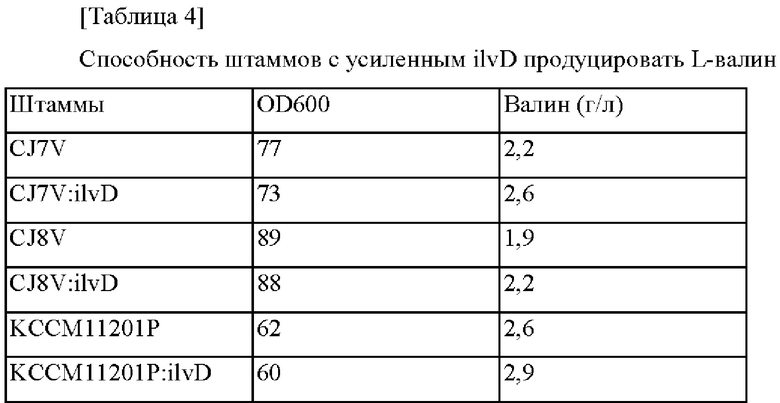
В соответствии с представленными выше результатами, когда усиливали ilvD, представляющий собой один из генов биосинтеза валина, тогда подтверждали то, что способность продуцировать валин у всех продуцирующих валин штаммов CJ7V, CJ8V и KCCM11201P увеличивалась.
Пример 2-2. Конструирование штаммов с делецией avtA и с ослабленным alg, и их оценка
Для конструирования штаммов, обладающих высокой способностью продуцировать валин, была сделана попытка ослабить (модифицируя стартовый кодон гена avtA при помощи GTG) или удалить avtA. Соответственно, вектор для конструирования штамма с ослабленным avtA (путем модификации стартового кодона гена avtA при помощи GTG) конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 19 и 20, и SEQ ID NO: 21 и 22 ниже, и вектор для конструирования устранения активности avtA с использованием пар праймеров в соответствии с SEQ ID NO: 23 и 24 и SEQ ID NO: 25 и 26 ниже, которые были названы, соответственно, pDZ-avtA(Alg) и pDZ-avtA(del).
В частности, для конструирования вектора с ослабленным avtA (pDZ-avtA(Alg)) ПЦР осуществляли с использованием пар праймеров в соответствии с SEQ ID NO: 19 и 20 и SEQ ID NO: 21 и 22, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС 14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 19 и 22 с получением фрагмента 4 с введенной мутацией. Полученный таким образом фрагмент 4 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-avtA(Alg).
Для конструирования вектора с делецией avtA (pDZ-avtA(del)) осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 23 и 24 и SEQ ID NO: 25 и 26, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 23 и 26 с получением фрагмента 5 с введенной мутацией. Полученный таким образом фрагмент 5 с введенной мутацией обрабатывали ферментом рестрикции Xbal (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования вектора pDZ-avtA(del).
Затем pDZ-avtA(del) и pDZ-avtA(Alg) трансформировали в каждый из CJ7V:ilvD, CJ8V:ilvD и KCCM11201P:ilvD, которые представляют собой штаммы, продуцирующие валин, для индукции гомологичной рекомбинации в хромосоме (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Штаммы, в которые были введены векторы в хромосому путем рекомбинации гомологичных последовательностей, отбирали в среде, содержащей 25 мг/л канамицина.
Затем фрагменты гена амплифицировали на основании трансформантов Corynebacterium glutamicum, отобранных выше при помощи ПЦР, с использованием пар праймеров в соответствии с SEQ ID NO: 19 и 20, SEQ ID NO: 21 и 22, SEQ ID NO: 23 и 24 и SEQ ID NO: 25 и 26, и введение мутации подтверждали при помощи анализа путем секвенирования гена тем же самым образом, как в примере 1. Рекомбинантные штаммы были названы Corynebacterium glutamicum CJ7V:ilvD-avtA(del), CJ7V:ilvD-avtA (alg), CJ8V:ilvD-avtA(del), CJ8V:ilvD-avtA (alg), KCCM11201P:ilvD-avtA(del) и KCCM11201P:ilvD-avtA (alg), как представлено ниже, и оценку титра осуществляли тем же самым образом, как в примере 1.
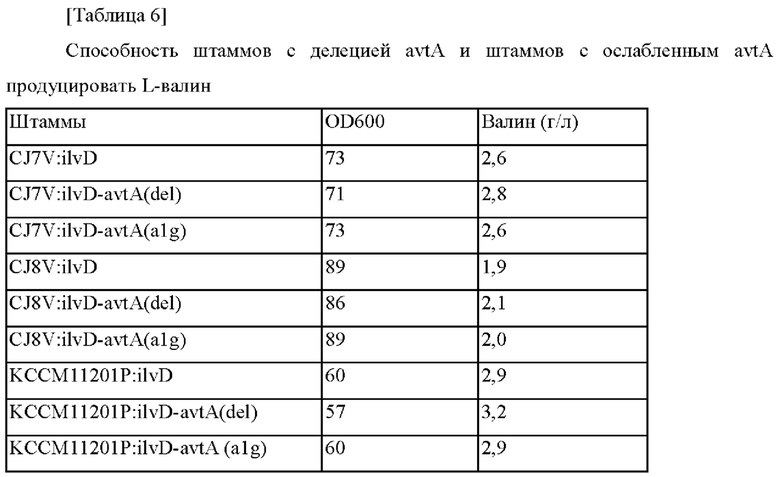
В соответствии с представленными выше результатами, когда avtA подвергали делеции, тогда способность продуцировать валин усиливалась, и когда экспрессию avtA ослабляли, тогда способность продуцировать валин увеличивалась в штамме CJ8V:ilvD-avtA (alg) по сравнению с штаммом CJ8V:ilvD, демонстрируя равный или более высокий уровень способности продуцировать валин по сравнению с контрольными штаммами с усиленным ilvD. Подтвердили то, что способность продуцировать валин усиливалась во всех случаях штаммов с усиленным ilvD, и штаммов с делецией avtA или штаммов с ослабленным avtA по сравнению с штаммами CJ7V, CJ8V и KCCM11201P, как представлено в таблице 4.
Пример 2-3. Конструирование штаммов с делецией асеЕ и штаммов с ослабленным асеЕ (alg, Q432A, K435A) и их оценка
Для конструирования штаммов, обладающих высокой способностью продуцировать валин, была сделана попытка ослабить или удалить асеЕ. Соответственно, для конструирования штаммов с ослабленным асеЕ, вектор для конструирования штамма, в котором стартовый кодон гена асеЕ был модифицирован до GTG, конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 27 и 28 и SEQ ID NO: 29 и 30, вектор для конструирования штамма aceE(Q432A) конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 27 и 32 и SEQ ID NO: 33 и 36, и вектор для конструирования штамма асеЕ(K435A) конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 27 и 34 и SEQ ID NO: 35 и 36. Дополнительно, вектор для конструирования штамма с делецией асеЕ (aceE(del)) конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 37 и 38 и SEQ ID NO: 39 и 40. Сконструированные таким образом векторы были названы, соответственно, pDZ-aceE(Alg), pDZ-aceE(Q432A), pDZ-aceE(K435A) и pDZ-aceE(del).
В частности, для конструирования вектора для ослабления стартового кодона асеЕ осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 27 и 28 и SEQ ID NO: 29 и 30, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС 14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 27 и 30 с получением фрагмента 6 с введенной мутацией. Полученный таким образом фрагмент 6 с введенной мутацией обрабатывали ферментом рестрикции Xbal (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-aceE(Alg).
Для конструирования вектора для введения мутации aceE(Q432A) осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 27 и 28 и SEQ ID NO: 33 и 36, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 27 и 36 с получением фрагмента 7 с введенной мутацией. Полученный таким образом фрагмент 7 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-aceE(Q432A).
Для конструирования вектора для введения мутации асеЕ(K435A) осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 27 и 34 и SEQ ID NO: 35 и 36, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 27 и 36 с получением фрагмента 8 с введенной мутацией. Полученный таким образом фрагмент 8 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-aceE(K435A).
Для введения вектора, используемого для введения мутации aceE(del) осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 37 и 38 и SEQ ID NO: 39 и 40, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС 14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов, полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 37 и 40 с получением фрагмента 9 с введенной мутацией. Полученный таким образом фрагмент 9 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-aceE(del).

Затем pDZ-aceE(del), pDZ-aceE(Alg), pDZ-aceE(Q432A) и pDZ-aceE(K435A) трансформировали в каждый из CJ7V:ilvD, CJ8V:ilvD и KCCM11201P:ilvD, которые представляют собой штаммы, продуцирующие валин, для индукции гомологичной рекомбинации в хромосоме (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Штаммы, в которые были введены векторы в хромосому путем рекомбинации гомологичных последовательностей, отбирали в среде, содержащей 25 мг/л канамицина.
Затем фрагменты гена амплифицировали на основании трансформантов Corynebacterium glutamicum, отобранных выше при помощи ПЦР, с использованием пары праймеров в соответствии с SEQ ID NO: 27 и 36, и введение мутации подтверждали при помощи анализа путем секвенирования гена. Рекомбинантные штаммы были названы Corynebacterium glutamicum CJ7V:ilvD-aceE(del), CJ7V:ilvD-aceE (alg), CJ7V:ilvD-aceE(Q432A), CJ7V:ilvD-aceE(K435A), CJ8V:ilvD-aceE(del), CJ8V:ilvD-aceE (alg), CJ8V:ilvD-aceE(Q432A), CJ8V:ilvD-aceE(K435A), KCCM11201P:ilvD-aceE(del), KCCM11201P:ilvD-aceE(alg), KCCM11201P:ilvD-aceE(Q432A), и KCCM11201P:ilvD-aceE(K435A), как представлено ниже, и оценку титра осуществляли тем же самым образом, как в примере 1.
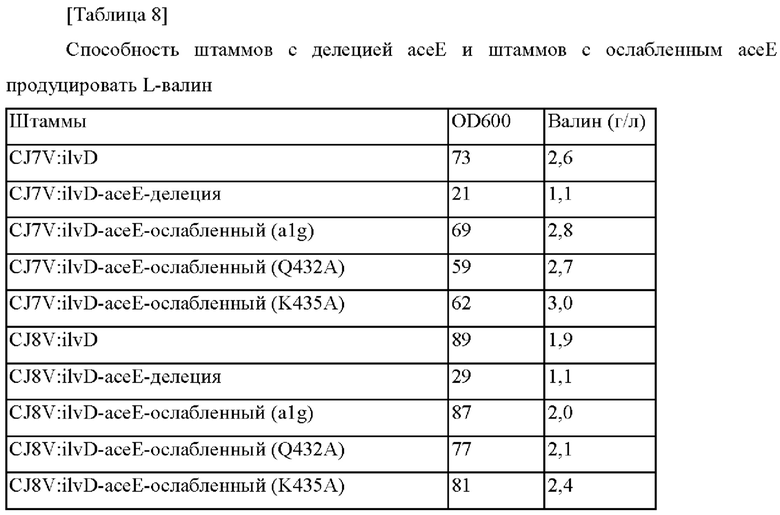
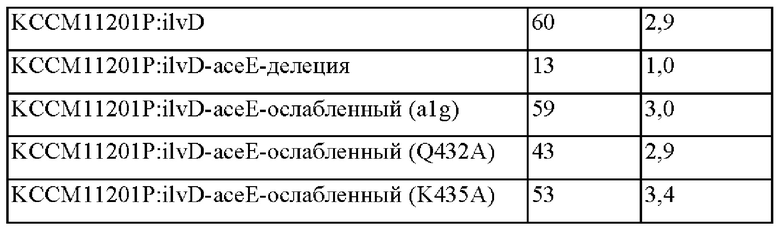
В соответствии с представленными выше результатами, когда асеЕ дополнительно подвергали делеции в штаммах, обладающих усиленной активностью дегидратазы дигидроксикислоты (ilvD), тогда скорости роста и потребления сахара быстро уменьшались, приводя в результате к уменьшенной способности продуцировать валин. В то же самое время, в случае штаммов, обладающих ослабленным асеЕ, подтвердили то, что скорости роста и потребления сахара были не значимыми, и способность продуцировать валин увеличивалась, хотя имелись различия в зависимости от уровня ослабления.
Пример 2-4. Конструирование штаммов с ослабленным gltA (alg, N241T, M312I) и их оценка
Для конструирования штаммов, обладающих высокой способностью продуцировать валин, была сделана попытка ослабить gltA. Соответственно, вектор для конструирования штамма с ослабленным gltA, в котором стартовый кодон гена gltA модифицирован до GTG, конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 41 и 42 и SEQ ID NO: 43 и 44, вектор для конструирования штамма gltA(N241T) конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 45 и 46 и SEQ ID NO: 47 и 50, и вектор для конструирования штамма gltA(M312I) конструировали с использованием пар праймеров в соответствии с SEQ ID NO: 45 и 48 и SEQ ID NO: 49 и 50. Сконструированные таким образом векторы были названы, соответственно, pDZ-gltA(Alg), pDZ-gltA(N241T) и pDZ-gltA(M312I).
Для введения вектора для введения мутации, ослабляющей стартовый кодон gltA, осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 41 и 42 и SEQ ID NO: 43 и 44, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС 14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов (А, В), полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 41 и 44 с получением фрагмента 10 с введенной мутацией. Полученный таким образом фрагмент 10 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-gltA(Alg).
Для введения вектора для введения мутации gltA(N241T) осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 45 и 46 и SEQ ID NO: 47 и 50, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов (А, В), полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 45 и 50 с получением фрагмента 11 с введенной мутацией. Полученный таким образом фрагмент 11 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-gltA(N241T).
Для введения вектора для введения мутации gltA(M312I) осуществляли ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 45 и 48 и SEQ ID NO: 49 и 50, основанных на геномной ДНК штамма дикого типа Corynebacterium glutamicum АТСС14067 в качестве матрицы. Условия ПЦР были следующими: денатурация при 94°С в течение 5 минут и затем 25 циклов денатурации при 94°С в течение 30 секунд; отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд, а затем полимеризация при 72°С в течение 7 минут. Осуществляли ПЦР с перекрывающимися праймерами, основанными на каждом из фрагментов (А, В), полученных выше, в качестве матрицы с использованием пары праймеров в соответствии с SEQ ID NO: 45 и 50 с получением фрагмента 12 с введенной мутацией. Полученный таким образом фрагмент 12 с введенной мутацией обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MaA) и затем лигировали в вектор pDZ, который был обработан тем же самым ферментом рестрикции, с использованием лигазы Т4 (New England Biolabs, Beverly, MA) для конструирования ДНК вектора pDZ-gltA(M312I).
Затем pDZ-gltA(Alg), pDZ-gltA(N241T) и pDZ-gltA(M312I) трансформировали в каждый из CJ7V:ilvD, CJ8V:ilvD и KCCM11201P:ilvD, которые представляют собой штаммы, продуцирующие валин, для индукции гомологичной рекомбинации в хромосоме (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Штаммы, в которые были введены векторы в хромосому путем рекомбинации гомологичных последовательностей, отбирали в среде, содержащей 25 мг/л канамицина.
Затем фрагменты гена амплифицировали на основании трансформантов Corynebacterium glutamicum, отобранных выше, при помощи ПЦР с использованием пары праймеров в соответствии с SEQ ID NO: 45 и 50, и введение мутации подтверждали при помощи анализа путем секвенирования гена. Рекомбинантные штаммы были названы Corynebacterium glutamicum CJ7V:ilvD-gltA (alg), CJ7V:ilvD-gltA(N241T), CJ7V:ilvD-gltA(M312I), CJ8V:ilvD-gltA(alg), CJ8V:ilvD-gltA(N241T), CJ8V:ilvD-gltA(M312I), KCCM11201P:ilvD-gltA(alg), KCCM11201P:ilvD-gitA(N241T), и KCCM11201P:ilvD-gltA(M312I), и оценку титра осуществляли тем же самым образом, как в примере 1.
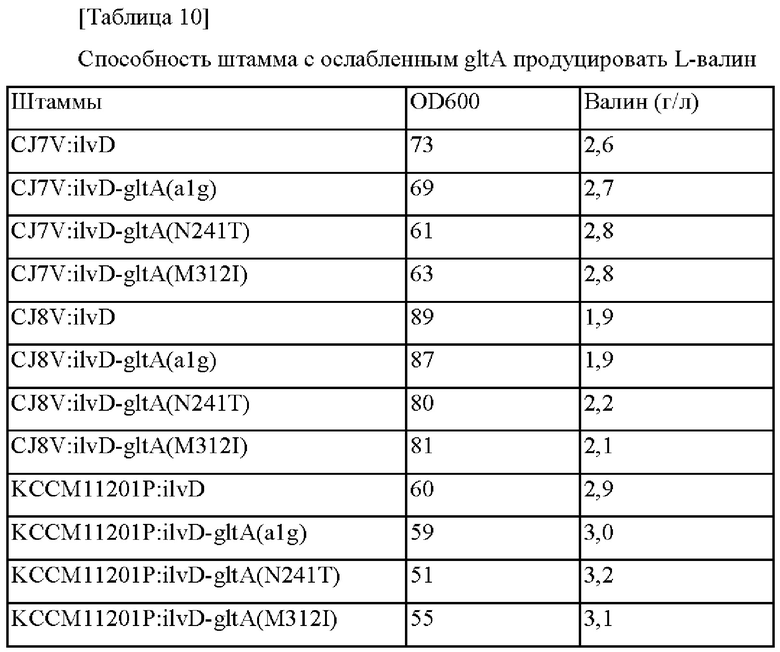
В соответствии с представленными выше результатами, когда gltA был ослаблен в штаммах, обладающих усиленной активностью дегидратазы дигидроксикислоты (ilvD), тогда подтвердили то, что скорости роста и потребления сахара были не значимыми и способность продуцировать валин увеличивалась, хотя существовали различия в зависимости от уровня ослабления.
Пример 2-5. Конструирование комбинации штаммов для показателей эффективных мутантов и их оценки
На основании результатов, подтвержденных в примерах 2-2 - 2-4, сделана попытка определить то, существовало ли синергетическое действие в отношении способности продуцировать валин, когда комбинировали различные мутации. Векторы pDZ-avtA(del) и pDZ-aceE(K435A) трансформировали в каждый из CJ7V:ilvD-gltA-ослабленный(N241T), CJ8V:ilvD-gltA-ослабленный(N241T) и KCCM11201P:ilvD-gltA-ослабленный(М241Т), которые представляют собой штаммы, продуцирующие валин, сконструированные в примере 2-4 (van der Rest et al., Appl Microbiol Biotechnol 52:541-545, 1999). Штаммы, в которых векторы были введены в хромосому путем рекомбинации гомологичных последовательностей, отбирали в среде, содержащей 25 мг/л канамицина.
Затем фрагменты гена амплифицировали на основании трансформантов Corynebacterium glutamicum, в которых была завершена вторичная рекомбинация, при помощи ПЦР с использованием пар праймеров в соответствии с SEQ ID NO: 23 и 26 и SEQ ID NO: 27 и 36, и затем штаммы с введенной мутацией подтверждали путем анализа секвенирования гена. Рекомбинантные штаммы были названы, как представлено ниже, и оценку титра осуществляли тем же самым образом, как в примере 1.
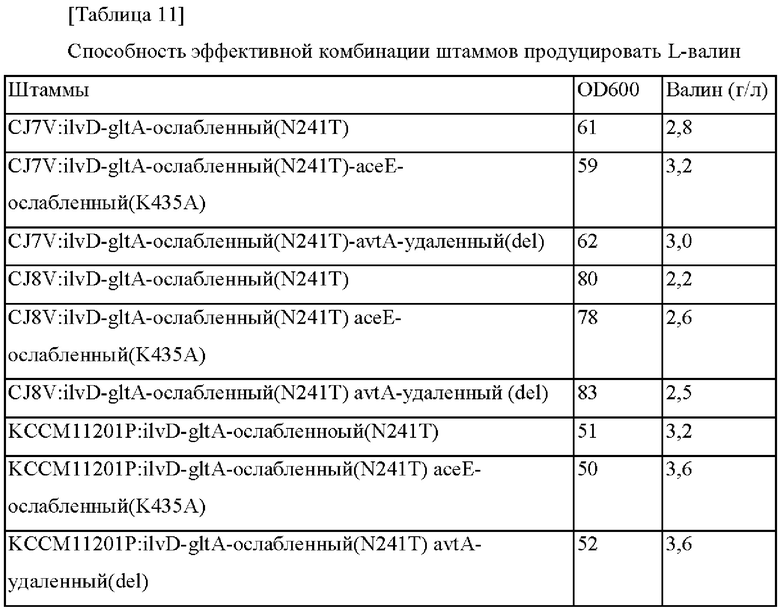
В соответствии с представленными выше результатами подтвердили то, что когда штаммы с ослабленным асеЕ и штаммы с делецией avtA вводили в штаммы с усиленным ilvD и ослабленным gltA, тогда скорости роста и потребления сахара были на том же самом уровне, и способность продуцировать валин дополнительно увеличивалась.
Вектор pDZ-aceE(K435A) трансформировали в KCCM11201P:ilvD-gltA-ослабленный (N241T) штамм, и штамм, в который была введена мутация асе(K435A) в хромосоме путем рекомбинации гомологичных последовательностей, был назван СА08-1592. СА08-1592 был депонирован в Корейский центр культур микроорганизмов (KCCM) в соответствии с Будапештским договором 3 июля 2020 года под номером KCCM12761P.
В соответствии с вышеизложенным специалист в данной области техники, в отношении которого направлено описание настоящего изобретения, будет в состоянии понять, что описание настоящего изобретения может быть воплощено в других конкретных формах без модификации технических понятий или существенных характеристик описания настоящего изобретения. В этой связи примеры воплощений, раскрытые здесь, приведены исключительно в иллюстративных целях, и их не следует истолковывать как ограничивающие объем описания настоящего изобретения. Наоборот, описание настоящего изобретения предназначено для того, чтобы охватить не только примеры воплощений, а также и различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем описания настоящего изобретения, определенные в формуле изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ L-ВАЛИН, И СПОСОБ ПОЛУЧЕНИЯ L-
ВАЛИНА С ЕГО ИСПОЛЬЗОВАНИЕМ
<130> OPA21084-PCT
<150> KR 10-2020-0111084
<151> 2020-09-01
<160> 50
<170> KoPatentIn 3.0
<210> 1
<211> 613
<212> PRT
<213> Unknown
<220>
<223> Дегидратаза дигидроксикислоты Corynebacterium sp
<400> 1
Met Ile Pro Leu Arg Ser Lys Val Thr Thr Val Gly Arg Asn Ala Ala
1 5 10 15
Gly Ala Arg Ala Leu Trp Arg Ala Thr Gly Thr Lys Glu Asn Glu Phe
20 25 30
Gly Lys Pro Ile Val Ala Ile Val Asn Ser Tyr Thr Gln Phe Val Pro
35 40 45
Gly His Val His Leu Lys Asn Val Gly Asp Ile Val Ala Asp Ala Val
50 55 60
Arg Lys Ala Gly Gly Val Pro Lys Glu Phe Asn Thr Ile Ala Val Asp
65 70 75 80
Asp Gly Ile Ala Met Gly His Gly Gly Met Leu Tyr Ser Leu Pro Ser
85 90 95
Arg Glu Ile Ile Ala Asp Ser Val Glu Tyr Met Val Asn Ala His Thr
100 105 110
Ala Asp Ala Met Val Cys Ile Ser Asn Cys Asp Lys Ile Thr Pro Gly
115 120 125
Met Leu Asn Ala Ala Met Arg Leu Asn Ile Pro Val Val Phe Val Ser
130 135 140
Gly Gly Pro Met Glu Ala Gly Lys Ala Val Val Val Asp Gly Val Ala
145 150 155 160
His Ala Pro Thr Asp Leu Ile Thr Ala Ile Ser Ala Ser Ala Ser Asp
165 170 175
Ala Val Asp Asp Ala Gly Leu Ala Ala Val Glu Ala Ser Ala Cys Pro
180 185 190
Thr Cys Gly Ser Cys Ser Gly Met Phe Thr Ala Asn Ser Met Asn Cys
195 200 205
Leu Thr Glu Ala Leu Gly Leu Ser Leu Pro Gly Asn Gly Ser Thr Leu
210 215 220
Ala Thr His Ala Ala Arg Arg Ala Leu Phe Glu Lys Ala Gly Glu Thr
225 230 235 240
Val Val Glu Leu Cys Arg Arg Tyr Tyr Gly Glu Glu Asp Glu Ser Val
245 250 255
Leu Pro Arg Gly Ile Ala Thr Lys Lys Ala Phe Glu Asn Ala Met Ala
260 265 270
Leu Asp Met Ala Met Gly Gly Ser Thr Asn Thr Ile Leu His Ile Leu
275 280 285
Ala Ala Ala Gln Glu Gly Glu Val Asp Phe Asp Leu Ala Asp Ile Asp
290 295 300
Glu Leu Ser Lys Asn Val Pro Cys Leu Ser Lys Val Ala Pro Asn Ser
305 310 315 320
Asp Tyr His Met Glu Asp Val His Arg Ala Gly Gly Ile Pro Ala Leu
325 330 335
Leu Gly Glu Leu Asn Arg Gly Gly Leu Leu Asn Lys Asp Val His Ser
340 345 350
Val His Ser Asn Asp Leu Glu Gly Trp Leu Asp Asp Trp Asp Ile Arg
355 360 365
Ser Gly Lys Thr Thr Glu Val Ala Thr Glu Leu Phe His Ala Ala Pro
370 375 380
Gly Gly Ile Arg Thr Thr Glu Ala Phe Ser Thr Glu Asn Arg Trp Asp
385 390 395 400
Glu Leu Asp Thr Asp Ala Ala Lys Gly Cys Ile Arg Asp Val Glu His
405 410 415
Ala Tyr Thr Ala Asp Gly Gly Leu Val Val Leu Arg Gly Asn Ile Ser
420 425 430
Pro Asp Gly Ala Val Ile Lys Ser Ala Gly Ile Glu Glu Glu Leu Trp
435 440 445
Asn Phe Thr Gly Pro Ala Arg Val Val Glu Ser Gln Glu Glu Ala Val
450 455 460
Ser Val Ile Leu Thr Lys Thr Ile Gln Ala Gly Glu Val Leu Val Val
465 470 475 480
Arg Tyr Glu Gly Pro Ser Gly Gly Pro Gly Met Gln Glu Met Leu His
485 490 495
Pro Thr Ala Phe Leu Lys Gly Ser Gly Leu Gly Lys Lys Cys Ala Leu
500 505 510
Ile Thr Asp Gly Arg Phe Ser Gly Gly Ser Ser Gly Leu Ser Ile Gly
515 520 525
His Val Ser Pro Glu Ala Ala His Gly Gly Val Ile Gly Leu Ile Glu
530 535 540
Asn Gly Asp Ile Val Ser Ile Asp Val His Asn Arg Lys Leu Glu Val
545 550 555 560
Gln Val Ser Asp Glu Glu Leu Gln Arg Arg Arg Asp Ala Met Asn Ala
565 570 575
Ser Glu Lys Pro Trp Gln Pro Val Asn Arg Asn Arg Val Val Thr Lys
580 585 590
Ala Leu Arg Ala Tyr Ala Lys Met Ala Thr Ser Ala Asp Lys Gly Ala
595 600 605
Val Arg Gln Val Asp
610
<210> 2
<211> 383
<212> PRT
<213> Unknown
<220>
<223> Трансаминаза C Corynebacterium sp
<400> 2
Met Lys Pro Ser Thr Arg Ser Asn Val Gln Pro Phe Arg Val Met Gln
1 5 10 15
Met Leu Asp Arg Val His Arg Arg Arg Arg Glu Gly Lys Asp Thr Ile
20 25 30
Met Phe Cys Ala Gly Gln Pro Ser Thr Gly Ala Pro Glu Ala Val Ile
35 40 45
Glu Glu Ala Glu Ile Ala Leu Arg Ser Gly Pro Leu Gly Tyr Thr Glu
50 55 60
Val Ile Gly Asp Arg Glu Phe Arg Glu Arg Ile Ala Asp Trp His Ser
65 70 75 80
Ala Thr Tyr Asp Val Asp Thr Asn Pro Asp Asn Val Ile Val Thr Thr
85 90 95
Gly Ser Ser Gly Gly Phe Val Ala Ser Phe Ile Ala Thr Leu Asp His
100 105 110
Gly Asp Tyr Val Ala Met Pro Thr Pro Gly Tyr Pro Ala Tyr Arg Asn
115 120 125
Ile Leu Glu Ser Leu Gly Ala Lys Val Leu Asn Leu Arg Cys Thr Ala
130 135 140
Glu Thr Arg Phe Gln Pro Thr Ala Gln Met Leu Glu Glu Leu Pro His
145 150 155 160
Lys Pro Lys Ala Val Ile Val Thr Ser Pro Gly Asn Pro Thr Gly Thr
165 170 175
Ile Ile Asp Pro Glu Glu Leu Glu Arg Ile Ala Lys Trp Cys Asp Asp
180 185 190
Asn Asp Ala Val Leu Ile Ser Asp Glu Asp Tyr His Gly Met Ser Phe
195 200 205
Gly Arg Pro Leu Ala Thr Ala His Gln Phe Ser Lys Asn Ala Ile Val
210 215 220
Val Gly Thr Leu Ser Lys Tyr Phe Ser Met Thr Gly Trp Arg Val Gly
225 230 235 240
Trp Ile Ile Val Pro Asp Glu Leu Val Thr Ser Ile Glu Asn Leu Gln
245 250 255
Ala Ser Leu Ser Leu Cys Ala Pro Ala Ile Gly Gln Ala Ala Gly Arg
260 265 270
Ala Ala Phe Thr Leu Glu Ala Gly Ala Glu Leu Asp Ala His Val Glu
275 280 285
Ala Tyr Arg Glu Ala Arg Glu Val Phe Val Asp Lys Leu Pro Glu Ile
290 295 300
Gly Leu Gly Thr Phe Ala Asp Pro Asp Gly Gly Leu Tyr Leu Trp Val
305 310 315 320
Asp Val Ser Ala Tyr Thr Asp Asp Ser Glu Glu Trp Val Leu Arg Leu
325 330 335
Leu Asp Glu Ala Gly Val Ala Val Ala Pro Gly Val Asp Phe Asp Pro
340 345 350
Glu Glu Gly His Lys Trp Ile Arg Leu Ser Leu Cys Ala Ser Lys Glu
355 360 365
Asp Thr Ile Glu Gly Val Arg Lys Ile Gly Glu Phe Ile Lys Lys
370 375 380
<210> 3
<211> 922
<212> PRT
<213> Unknown
<220>
<223> Пируватдегидрогеназа Corynebacterium sp
<400> 3
Met Ala Asp Gln Ala Lys Leu Gly Gly Lys Pro Ser Asp Asp Ser Asn
1 5 10 15
Phe Ala Met Ile Arg Asp Gly Val Ala Ser Tyr Leu Asn Asp Ser Asp
20 25 30
Pro Glu Glu Thr Asn Glu Trp Met Asp Ser Leu Asp Gly Leu Leu Gln
35 40 45
Glu Ser Ser Pro Glu Arg Ala Arg Tyr Leu Met Leu Arg Leu Leu Glu
50 55 60
Arg Ala Ser Ala Lys Arg Val Ser Leu Pro Pro Met Thr Ser Thr Asp
65 70 75 80
Tyr Val Asn Thr Ile Pro Thr Ser Met Glu Pro Glu Phe Pro Gly Asp
85 90 95
Glu Glu Met Glu Lys Arg Tyr Arg Arg Trp Ile Arg Trp Asn Ala Ala
100 105 110
Ile Met Val His Arg Ala Gln Arg Pro Gly Ile Gly Val Gly Gly His
115 120 125
Ile Ser Thr Tyr Ala Gly Ala Ala Pro Leu Tyr Glu Val Gly Phe Asn
130 135 140
His Phe Phe Arg Gly Lys Asp His Pro Gly Gly Gly Asp Gln Ile Phe
145 150 155 160
Phe Gln Gly His Ala Ser Pro Gly Met Tyr Ala Arg Ala Phe Met Glu
165 170 175
Gly Arg Leu Ser Glu Asp Asp Leu Asp Gly Phe Arg Gln Glu Val Ser
180 185 190
Arg Glu Gln Gly Gly Ile Pro Ser Tyr Pro His Pro His Gly Met Lys
195 200 205
Asp Phe Trp Glu Phe Pro Thr Val Ser Met Gly Leu Gly Pro Met Asp
210 215 220
Ala Ile Tyr Gln Ala Arg Phe Asn Arg Tyr Leu Glu Asn Arg Gly Ile
225 230 235 240
Lys Asp Thr Ser Asp Gln His Val Trp Ala Phe Leu Gly Asp Gly Glu
245 250 255
Met Asp Glu Pro Glu Ser Arg Gly Leu Ile Gln Gln Ala Ala Leu Asn
260 265 270
Asn Leu Asp Asn Leu Thr Phe Val Val Asn Cys Asn Leu Gln Arg Leu
275 280 285
Asp Gly Pro Val Arg Gly Asn Thr Lys Ile Ile Gln Glu Leu Glu Ser
290 295 300
Phe Phe Arg Gly Ala Gly Trp Ser Val Ile Lys Val Val Trp Gly Arg
305 310 315 320
Glu Trp Asp Glu Leu Leu Glu Lys Asp Gln Asp Gly Ala Leu Val Glu
325 330 335
Ile Met Asn Asn Thr Ser Asp Gly Asp Tyr Gln Thr Phe Lys Ala Asn
340 345 350
Asp Gly Ala Tyr Val Arg Glu His Phe Phe Gly Arg Asp Pro Arg Thr
355 360 365
Ala Lys Leu Val Glu Asn Met Thr Asp Glu Glu Ile Trp Lys Leu Pro
370 375 380
Arg Gly Gly His Asp Tyr Arg Lys Val Tyr Ala Ala Tyr Lys Arg Ala
385 390 395 400
Leu Glu Thr Lys Asp Arg Pro Thr Val Ile Leu Ala His Thr Ile Lys
405 410 415
Gly Tyr Gly Leu Gly His Asn Phe Glu Gly Arg Asn Ala Thr His Gln
420 425 430
Met Lys Lys Leu Thr Leu Asp Asp Leu Lys Leu Phe Arg Asp Lys Gln
435 440 445
Gly Ile Pro Ile Thr Asp Glu Gln Leu Glu Lys Asp Pro Tyr Leu Pro
450 455 460
Pro Tyr Tyr His Pro Gly Glu Asp Ala Pro Glu Ile Lys Tyr Met Lys
465 470 475 480
Glu Arg Arg Ala Ala Leu Gly Gly Tyr Leu Pro Glu Arg Arg Glu Asn
485 490 495
Tyr Asp Pro Ile Gln Val Pro Pro Leu Asp Lys Leu Arg Ser Val Arg
500 505 510
Lys Gly Ser Gly Lys Gln Gln Ile Ala Thr Thr Met Ala Thr Val Arg
515 520 525
Thr Phe Lys Glu Leu Met Arg Asp Lys Gly Leu Ala Asp Arg Leu Val
530 535 540
Pro Ile Ile Pro Asp Glu Ala Arg Thr Phe Gly Leu Asp Ser Trp Phe
545 550 555 560
Pro Thr Leu Lys Ile Tyr Asn Pro His Gly Gln Asn Tyr Val Pro Val
565 570 575
Asp His Asp Leu Met Leu Ser Tyr Arg Glu Ala Pro Glu Gly Gln Ile
580 585 590
Leu His Glu Gly Ile Asn Glu Ala Gly Ser Met Ala Ser Phe Ile Ala
595 600 605
Ala Gly Thr Ser Tyr Ala Thr His Gly Lys Ala Met Ile Pro Leu Tyr
610 615 620
Ile Phe Tyr Ser Met Phe Gly Phe Gln Arg Thr Gly Asp Ser Ile Trp
625 630 635 640
Ala Ala Ala Asp Gln Met Ala Arg Gly Phe Leu Leu Gly Ala Thr Ala
645 650 655
Gly Arg Thr Thr Leu Thr Gly Glu Gly Leu Gln His Met Asp Gly His
660 665 670
Ser Pro Val Leu Ala Ser Thr Asn Glu Gly Val Glu Thr Tyr Asp Pro
675 680 685
Ser Phe Ala Tyr Glu Ile Ala His Leu Val His Arg Gly Ile Asp Arg
690 695 700
Met Tyr Gly Pro Gly Lys Gly Glu Asp Val Ile Tyr Tyr Ile Thr Ile
705 710 715 720
Tyr Asn Glu Pro Thr Pro Gln Pro Ala Glu Pro Glu Gly Leu Asp Val
725 730 735
Glu Gly Leu His Lys Gly Ile Tyr Leu Tyr Ser Arg Gly Glu Gly Thr
740 745 750
Gly His Glu Ala Asn Ile Leu Ala Ser Gly Val Gly Met Gln Trp Ala
755 760 765
Leu Lys Ala Ala Ser Ile Leu Glu Ala Asp Tyr Gly Val Arg Ala Asn
770 775 780
Ile Tyr Ser Ala Thr Ser Trp Val Asn Leu Ala Arg Asp Gly Ala Ala
785 790 795 800
Arg Asn Lys Ala Gln Leu Arg Asn Pro Gly Ala Asp Ala Gly Glu Ala
805 810 815
Phe Val Thr Thr Gln Leu Lys Gln Thr Ser Gly Pro Tyr Val Ala Val
820 825 830
Ser Asp Phe Ser Thr Asp Leu Pro Asn Gln Ile Arg Glu Trp Val Pro
835 840 845
Gly Asp Tyr Thr Val Leu Gly Ala Asp Gly Phe Gly Phe Ser Asp Thr
850 855 860
Arg Pro Ala Ala Arg Arg Phe Phe Asn Ile Asp Ala Glu Ser Ile Val
865 870 875 880
Val Ala Val Leu Asn Ser Leu Ala Arg Glu Gly Lys Ile Asp Val Ser
885 890 895
Val Ala Ala Gln Ala Ala Glu Lys Phe Lys Leu Asp Asp Pro Thr Ser
900 905 910
Val Ser Val Asp Pro Asn Ala Pro Glu Glu
915 920
<210> 4
<211> 437
<212> PRT
<213> Unknown
<220>
<223> Цитратсинтаза Corynebacterium sp
<400> 4
Met Phe Glu Arg Asp Ile Val Ala Thr Asp Asn Asn Lys Ala Val Leu
1 5 10 15
His Tyr Pro Gly Gly Glu Phe Glu Met Asp Ile Ile Glu Ala Ser Glu
20 25 30
Gly Asn Asn Gly Val Val Leu Gly Lys Met Leu Ser Glu Thr Gly Leu
35 40 45
Ile Thr Phe Asp Pro Gly Tyr Val Ser Thr Gly Ser Thr Glu Ser Lys
50 55 60
Ile Thr Tyr Ile Asp Gly Asp Ala Gly Ile Leu Arg Tyr Arg Gly Tyr
65 70 75 80
Asp Ile Ala Asp Leu Ala Glu Asn Ala Thr Phe Asn Glu Val Ser Tyr
85 90 95
Leu Leu Ile Asn Gly Glu Leu Pro Thr Pro Asp Glu Leu His Lys Phe
100 105 110
Asn Asp Glu Ile Arg His His Thr Leu Leu Asp Glu Asp Phe Lys Ser
115 120 125
Gln Phe Asn Val Phe Pro Arg Asp Ala His Pro Met Ala Thr Leu Ala
130 135 140
Ser Ser Val Asn Ile Leu Ser Thr Tyr Tyr Gln Asp Gln Leu Asn Pro
145 150 155 160
Leu Asp Glu Ala Gln Leu Asp Lys Ala Thr Val Arg Leu Met Ala Lys
165 170 175
Val Pro Met Leu Ala Ala Tyr Ala His Arg Ala Arg Lys Gly Ala Pro
180 185 190
Tyr Met Tyr Pro Asp Asn Ser Leu Asn Ala Arg Glu Asn Phe Leu Arg
195 200 205
Met Met Phe Gly Tyr Pro Thr Glu Pro Tyr Glu Ile Asp Pro Ile Met
210 215 220
Val Lys Ala Leu Asp Lys Leu Leu Ile Leu His Ala Asp His Glu Gln
225 230 235 240
Asn Cys Ser Thr Ser Thr Val Arg Met Ile Gly Ser Ala Gln Ala Asn
245 250 255
Met Phe Val Ser Ile Ala Gly Gly Ile Asn Ala Leu Ser Gly Pro Leu
260 265 270
His Gly Gly Ala Asn Gln Ala Val Leu Glu Met Leu Glu Asp Ile Lys
275 280 285
Asn Asn His Gly Gly Asp Ala Thr Ala Phe Met Asn Lys Val Lys Asn
290 295 300
Lys Glu Asp Gly Val Arg Leu Met Gly Phe Gly His Arg Val Tyr Lys
305 310 315 320
Asn Tyr Asp Pro Arg Ala Ala Ile Val Lys Glu Thr Ala His Glu Ile
325 330 335
Leu Glu His Leu Gly Gly Asp Asp Leu Leu Asp Leu Ala Ile Lys Leu
340 345 350
Glu Glu Ile Ala Leu Ala Asp Asp Tyr Phe Ile Ser Arg Lys Leu Tyr
355 360 365
Pro Asn Val Asp Phe Tyr Thr Gly Leu Ile Tyr Arg Ala Met Gly Phe
370 375 380
Pro Thr Asp Phe Phe Thr Val Leu Phe Ala Ile Gly Arg Leu Pro Gly
385 390 395 400
Trp Ile Ala His Tyr Arg Glu Gln Leu Gly Ala Ala Asp Asn Lys Ile
405 410 415
Asn Arg Pro Arg Gln Val Tyr Thr Gly Asn Glu Ser Arg Lys Leu Val
420 425 430
Pro Arg Glu Glu Arg
435
<210> 5
<211> 922
<212> PRT
<213> Unknown
<220>
<223> Вариант пируватдегидрогеназы Corynebacterium sp
<400> 5
Met Ala Asp Gln Ala Lys Leu Gly Gly Lys Pro Ser Asp Asp Ser Asn
1 5 10 15
Phe Ala Met Ile Arg Asp Gly Val Ala Ser Tyr Leu Asn Asp Ser Asp
20 25 30
Pro Glu Glu Thr Asn Glu Trp Met Asp Ser Leu Asp Gly Leu Leu Gln
35 40 45
Glu Ser Ser Pro Glu Arg Ala Arg Tyr Leu Met Leu Arg Leu Leu Glu
50 55 60
Arg Ala Ser Ala Lys Arg Val Ser Leu Pro Pro Met Thr Ser Thr Asp
65 70 75 80
Tyr Val Asn Thr Ile Pro Thr Ser Met Glu Pro Glu Phe Pro Gly Asp
85 90 95
Glu Glu Met Glu Lys Arg Tyr Arg Arg Trp Ile Arg Trp Asn Ala Ala
100 105 110
Ile Met Val His Arg Ala Gln Arg Pro Gly Ile Gly Val Gly Gly His
115 120 125
Ile Ser Thr Tyr Ala Gly Ala Ala Pro Leu Tyr Glu Val Gly Phe Asn
130 135 140
His Phe Phe Arg Gly Lys Asp His Pro Gly Gly Gly Asp Gln Ile Phe
145 150 155 160
Phe Gln Gly His Ala Ser Pro Gly Met Tyr Ala Arg Ala Phe Met Glu
165 170 175
Gly Arg Leu Ser Glu Asp Asp Leu Asp Gly Phe Arg Gln Glu Val Ser
180 185 190
Arg Glu Gln Gly Gly Ile Pro Ser Tyr Pro His Pro His Gly Met Lys
195 200 205
Asp Phe Trp Glu Phe Pro Thr Val Ser Met Gly Leu Gly Pro Met Asp
210 215 220
Ala Ile Tyr Gln Ala Arg Phe Asn Arg Tyr Leu Glu Asn Arg Gly Ile
225 230 235 240
Lys Asp Thr Ser Asp Gln His Val Trp Ala Phe Leu Gly Asp Gly Glu
245 250 255
Met Asp Glu Pro Glu Ser Arg Gly Leu Ile Gln Gln Ala Ala Leu Asn
260 265 270
Asn Leu Asp Asn Leu Thr Phe Val Val Asn Cys Asn Leu Gln Arg Leu
275 280 285
Asp Gly Pro Val Arg Gly Asn Thr Lys Ile Ile Gln Glu Leu Glu Ser
290 295 300
Phe Phe Arg Gly Ala Gly Trp Ser Val Ile Lys Val Val Trp Gly Arg
305 310 315 320
Glu Trp Asp Glu Leu Leu Glu Lys Asp Gln Asp Gly Ala Leu Val Glu
325 330 335
Ile Met Asn Asn Thr Ser Asp Gly Asp Tyr Gln Thr Phe Lys Ala Asn
340 345 350
Asp Gly Ala Tyr Val Arg Glu His Phe Phe Gly Arg Asp Pro Arg Thr
355 360 365
Ala Lys Leu Val Glu Asn Met Thr Asp Glu Glu Ile Trp Lys Leu Pro
370 375 380
Arg Gly Gly His Asp Tyr Arg Lys Val Tyr Ala Ala Tyr Lys Arg Ala
385 390 395 400
Leu Glu Thr Lys Asp Arg Pro Thr Val Ile Leu Ala His Thr Ile Lys
405 410 415
Gly Tyr Gly Leu Gly His Asn Phe Glu Gly Arg Asn Ala Thr His Ala
420 425 430
Met Lys Lys Leu Thr Leu Asp Asp Leu Lys Leu Phe Arg Asp Lys Gln
435 440 445
Gly Ile Pro Ile Thr Asp Glu Gln Leu Glu Lys Asp Pro Tyr Leu Pro
450 455 460
Pro Tyr Tyr His Pro Gly Glu Asp Ala Pro Glu Ile Lys Tyr Met Lys
465 470 475 480
Glu Arg Arg Ala Ala Leu Gly Gly Tyr Leu Pro Glu Arg Arg Glu Asn
485 490 495
Tyr Asp Pro Ile Gln Val Pro Pro Leu Asp Lys Leu Arg Ser Val Arg
500 505 510
Lys Gly Ser Gly Lys Gln Gln Ile Ala Thr Thr Met Ala Thr Val Arg
515 520 525
Thr Phe Lys Glu Leu Met Arg Asp Lys Gly Leu Ala Asp Arg Leu Val
530 535 540
Pro Ile Ile Pro Asp Glu Ala Arg Thr Phe Gly Leu Asp Ser Trp Phe
545 550 555 560
Pro Thr Leu Lys Ile Tyr Asn Pro His Gly Gln Asn Tyr Val Pro Val
565 570 575
Asp His Asp Leu Met Leu Ser Tyr Arg Glu Ala Pro Glu Gly Gln Ile
580 585 590
Leu His Glu Gly Ile Asn Glu Ala Gly Ser Met Ala Ser Phe Ile Ala
595 600 605
Ala Gly Thr Ser Tyr Ala Thr His Gly Lys Ala Met Ile Pro Leu Tyr
610 615 620
Ile Phe Tyr Ser Met Phe Gly Phe Gln Arg Thr Gly Asp Ser Ile Trp
625 630 635 640
Ala Ala Ala Asp Gln Met Ala Arg Gly Phe Leu Leu Gly Ala Thr Ala
645 650 655
Gly Arg Thr Thr Leu Thr Gly Glu Gly Leu Gln His Met Asp Gly His
660 665 670
Ser Pro Val Leu Ala Ser Thr Asn Glu Gly Val Glu Thr Tyr Asp Pro
675 680 685
Ser Phe Ala Tyr Glu Ile Ala His Leu Val His Arg Gly Ile Asp Arg
690 695 700
Met Tyr Gly Pro Gly Lys Gly Glu Asp Val Ile Tyr Tyr Ile Thr Ile
705 710 715 720
Tyr Asn Glu Pro Thr Pro Gln Pro Ala Glu Pro Glu Gly Leu Asp Val
725 730 735
Glu Gly Leu His Lys Gly Ile Tyr Leu Tyr Ser Arg Gly Glu Gly Thr
740 745 750
Gly His Glu Ala Asn Ile Leu Ala Ser Gly Val Gly Met Gln Trp Ala
755 760 765
Leu Lys Ala Ala Ser Ile Leu Glu Ala Asp Tyr Gly Val Arg Ala Asn
770 775 780
Ile Tyr Ser Ala Thr Ser Trp Val Asn Leu Ala Arg Asp Gly Ala Ala
785 790 795 800
Arg Asn Lys Ala Gln Leu Arg Asn Pro Gly Ala Asp Ala Gly Glu Ala
805 810 815
Phe Val Thr Thr Gln Leu Lys Gln Thr Ser Gly Pro Tyr Val Ala Val
820 825 830
Ser Asp Phe Ser Thr Asp Leu Pro Asn Gln Ile Arg Glu Trp Val Pro
835 840 845
Gly Asp Tyr Thr Val Leu Gly Ala Asp Gly Phe Gly Phe Ser Asp Thr
850 855 860
Arg Pro Ala Ala Arg Arg Phe Phe Asn Ile Asp Ala Glu Ser Ile Val
865 870 875 880
Val Ala Val Leu Asn Ser Leu Ala Arg Glu Gly Lys Ile Asp Val Ser
885 890 895
Val Ala Ala Gln Ala Ala Glu Lys Phe Lys Leu Asp Asp Pro Thr Ser
900 905 910
Val Ser Val Asp Pro Asn Ala Pro Glu Glu
915 920
<210> 6
<211> 922
<212> PRT
<213> Unknown
<220>
<223> Вариант пируватдегидрогеназы Corynebacterium sp
<400> 6
Met Ala Asp Gln Ala Lys Leu Gly Gly Lys Pro Ser Asp Asp Ser Asn
1 5 10 15
Phe Ala Met Ile Arg Asp Gly Val Ala Ser Tyr Leu Asn Asp Ser Asp
20 25 30
Pro Glu Glu Thr Asn Glu Trp Met Asp Ser Leu Asp Gly Leu Leu Gln
35 40 45
Glu Ser Ser Pro Glu Arg Ala Arg Tyr Leu Met Leu Arg Leu Leu Glu
50 55 60
Arg Ala Ser Ala Lys Arg Val Ser Leu Pro Pro Met Thr Ser Thr Asp
65 70 75 80
Tyr Val Asn Thr Ile Pro Thr Ser Met Glu Pro Glu Phe Pro Gly Asp
85 90 95
Glu Glu Met Glu Lys Arg Tyr Arg Arg Trp Ile Arg Trp Asn Ala Ala
100 105 110
Ile Met Val His Arg Ala Gln Arg Pro Gly Ile Gly Val Gly Gly His
115 120 125
Ile Ser Thr Tyr Ala Gly Ala Ala Pro Leu Tyr Glu Val Gly Phe Asn
130 135 140
His Phe Phe Arg Gly Lys Asp His Pro Gly Gly Gly Asp Gln Ile Phe
145 150 155 160
Phe Gln Gly His Ala Ser Pro Gly Met Tyr Ala Arg Ala Phe Met Glu
165 170 175
Gly Arg Leu Ser Glu Asp Asp Leu Asp Gly Phe Arg Gln Glu Val Ser
180 185 190
Arg Glu Gln Gly Gly Ile Pro Ser Tyr Pro His Pro His Gly Met Lys
195 200 205
Asp Phe Trp Glu Phe Pro Thr Val Ser Met Gly Leu Gly Pro Met Asp
210 215 220
Ala Ile Tyr Gln Ala Arg Phe Asn Arg Tyr Leu Glu Asn Arg Gly Ile
225 230 235 240
Lys Asp Thr Ser Asp Gln His Val Trp Ala Phe Leu Gly Asp Gly Glu
245 250 255
Met Asp Glu Pro Glu Ser Arg Gly Leu Ile Gln Gln Ala Ala Leu Asn
260 265 270
Asn Leu Asp Asn Leu Thr Phe Val Val Asn Cys Asn Leu Gln Arg Leu
275 280 285
Asp Gly Pro Val Arg Gly Asn Thr Lys Ile Ile Gln Glu Leu Glu Ser
290 295 300
Phe Phe Arg Gly Ala Gly Trp Ser Val Ile Lys Val Val Trp Gly Arg
305 310 315 320
Glu Trp Asp Glu Leu Leu Glu Lys Asp Gln Asp Gly Ala Leu Val Glu
325 330 335
Ile Met Asn Asn Thr Ser Asp Gly Asp Tyr Gln Thr Phe Lys Ala Asn
340 345 350
Asp Gly Ala Tyr Val Arg Glu His Phe Phe Gly Arg Asp Pro Arg Thr
355 360 365
Ala Lys Leu Val Glu Asn Met Thr Asp Glu Glu Ile Trp Lys Leu Pro
370 375 380
Arg Gly Gly His Asp Tyr Arg Lys Val Tyr Ala Ala Tyr Lys Arg Ala
385 390 395 400
Leu Glu Thr Lys Asp Arg Pro Thr Val Ile Leu Ala His Thr Ile Lys
405 410 415
Gly Tyr Gly Leu Gly His Asn Phe Glu Gly Arg Asn Ala Thr His Gln
420 425 430
Met Lys Ala Leu Thr Leu Asp Asp Leu Lys Leu Phe Arg Asp Lys Gln
435 440 445
Gly Ile Pro Ile Thr Asp Glu Gln Leu Glu Lys Asp Pro Tyr Leu Pro
450 455 460
Pro Tyr Tyr His Pro Gly Glu Asp Ala Pro Glu Ile Lys Tyr Met Lys
465 470 475 480
Glu Arg Arg Ala Ala Leu Gly Gly Tyr Leu Pro Glu Arg Arg Glu Asn
485 490 495
Tyr Asp Pro Ile Gln Val Pro Pro Leu Asp Lys Leu Arg Ser Val Arg
500 505 510
Lys Gly Ser Gly Lys Gln Gln Ile Ala Thr Thr Met Ala Thr Val Arg
515 520 525
Thr Phe Lys Glu Leu Met Arg Asp Lys Gly Leu Ala Asp Arg Leu Val
530 535 540
Pro Ile Ile Pro Asp Glu Ala Arg Thr Phe Gly Leu Asp Ser Trp Phe
545 550 555 560
Pro Thr Leu Lys Ile Tyr Asn Pro His Gly Gln Asn Tyr Val Pro Val
565 570 575
Asp His Asp Leu Met Leu Ser Tyr Arg Glu Ala Pro Glu Gly Gln Ile
580 585 590
Leu His Glu Gly Ile Asn Glu Ala Gly Ser Met Ala Ser Phe Ile Ala
595 600 605
Ala Gly Thr Ser Tyr Ala Thr His Gly Lys Ala Met Ile Pro Leu Tyr
610 615 620
Ile Phe Tyr Ser Met Phe Gly Phe Gln Arg Thr Gly Asp Ser Ile Trp
625 630 635 640
Ala Ala Ala Asp Gln Met Ala Arg Gly Phe Leu Leu Gly Ala Thr Ala
645 650 655
Gly Arg Thr Thr Leu Thr Gly Glu Gly Leu Gln His Met Asp Gly His
660 665 670
Ser Pro Val Leu Ala Ser Thr Asn Glu Gly Val Glu Thr Tyr Asp Pro
675 680 685
Ser Phe Ala Tyr Glu Ile Ala His Leu Val His Arg Gly Ile Asp Arg
690 695 700
Met Tyr Gly Pro Gly Lys Gly Glu Asp Val Ile Tyr Tyr Ile Thr Ile
705 710 715 720
Tyr Asn Glu Pro Thr Pro Gln Pro Ala Glu Pro Glu Gly Leu Asp Val
725 730 735
Glu Gly Leu His Lys Gly Ile Tyr Leu Tyr Ser Arg Gly Glu Gly Thr
740 745 750
Gly His Glu Ala Asn Ile Leu Ala Ser Gly Val Gly Met Gln Trp Ala
755 760 765
Leu Lys Ala Ala Ser Ile Leu Glu Ala Asp Tyr Gly Val Arg Ala Asn
770 775 780
Ile Tyr Ser Ala Thr Ser Trp Val Asn Leu Ala Arg Asp Gly Ala Ala
785 790 795 800
Arg Asn Lys Ala Gln Leu Arg Asn Pro Gly Ala Asp Ala Gly Glu Ala
805 810 815
Phe Val Thr Thr Gln Leu Lys Gln Thr Ser Gly Pro Tyr Val Ala Val
820 825 830
Ser Asp Phe Ser Thr Asp Leu Pro Asn Gln Ile Arg Glu Trp Val Pro
835 840 845
Gly Asp Tyr Thr Val Leu Gly Ala Asp Gly Phe Gly Phe Ser Asp Thr
850 855 860
Arg Pro Ala Ala Arg Arg Phe Phe Asn Ile Asp Ala Glu Ser Ile Val
865 870 875 880
Val Ala Val Leu Asn Ser Leu Ala Arg Glu Gly Lys Ile Asp Val Ser
885 890 895
Val Ala Ala Gln Ala Ala Glu Lys Phe Lys Leu Asp Asp Pro Thr Ser
900 905 910
Val Ser Val Asp Pro Asn Ala Pro Glu Glu
915 920
<210> 7
<211> 922
<212> PRT
<213> Unknown
<220>
<223> Вариант цитратсинтазы Corynebacterium sp
<400> 7
Met Ala Asp Gln Ala Lys Leu Gly Gly Lys Pro Ser Asp Asp Ser Asn
1 5 10 15
Phe Ala Met Ile Arg Asp Gly Val Ala Ser Tyr Leu Asn Asp Ser Asp
20 25 30
Pro Glu Glu Thr Asn Glu Trp Met Asp Ser Leu Asp Gly Leu Leu Gln
35 40 45
Glu Ser Ser Pro Glu Arg Ala Arg Tyr Leu Met Leu Arg Leu Leu Glu
50 55 60
Arg Ala Ser Ala Lys Arg Val Ser Leu Pro Pro Met Thr Ser Thr Asp
65 70 75 80
Tyr Val Asn Thr Ile Pro Thr Ser Met Glu Pro Glu Phe Pro Gly Asp
85 90 95
Glu Glu Met Glu Lys Arg Tyr Arg Arg Trp Ile Arg Trp Asn Ala Ala
100 105 110
Ile Met Val His Arg Ala Gln Arg Pro Gly Ile Gly Val Gly Gly His
115 120 125
Ile Ser Thr Tyr Ala Gly Ala Ala Pro Leu Tyr Glu Val Gly Phe Asn
130 135 140
His Phe Phe Arg Gly Lys Asp His Pro Gly Gly Gly Asp Gln Ile Phe
145 150 155 160
Phe Gln Gly His Ala Ser Pro Gly Met Tyr Ala Arg Ala Phe Met Glu
165 170 175
Gly Arg Leu Ser Glu Asp Asp Leu Asp Gly Phe Arg Gln Glu Val Ser
180 185 190
Arg Glu Gln Gly Gly Ile Pro Ser Tyr Pro His Pro His Gly Met Lys
195 200 205
Asp Phe Trp Glu Phe Pro Thr Val Ser Met Gly Leu Gly Pro Met Asp
210 215 220
Ala Ile Tyr Gln Ala Arg Phe Asn Arg Tyr Leu Glu Asn Arg Gly Ile
225 230 235 240
Lys Asp Thr Ser Asp Gln His Val Trp Ala Phe Leu Gly Asp Gly Glu
245 250 255
Met Asp Glu Pro Glu Ser Arg Gly Leu Ile Gln Gln Ala Ala Leu Asn
260 265 270
Asn Leu Asp Asn Leu Thr Phe Val Val Asn Cys Asn Leu Gln Arg Leu
275 280 285
Asp Gly Pro Val Arg Gly Asn Thr Lys Ile Ile Gln Glu Leu Glu Ser
290 295 300
Phe Phe Arg Gly Ala Gly Trp Ser Val Ile Lys Val Val Trp Gly Arg
305 310 315 320
Glu Trp Asp Glu Leu Leu Glu Lys Asp Gln Asp Gly Ala Leu Val Glu
325 330 335
Ile Met Asn Asn Thr Ser Asp Gly Asp Tyr Gln Thr Phe Lys Ala Asn
340 345 350
Asp Gly Ala Tyr Val Arg Glu His Phe Phe Gly Arg Asp Pro Arg Thr
355 360 365
Ala Lys Leu Val Glu Asn Met Thr Asp Glu Glu Ile Trp Lys Leu Pro
370 375 380
Arg Gly Gly His Asp Tyr Arg Lys Val Tyr Ala Ala Tyr Lys Arg Ala
385 390 395 400
Leu Glu Thr Lys Asp Arg Pro Thr Val Ile Leu Ala His Thr Ile Lys
405 410 415
Gly Tyr Gly Leu Gly His Asn Phe Glu Gly Arg Asn Ala Thr His Gln
420 425 430
Met Lys Ala Leu Thr Leu Asp Asp Leu Lys Leu Phe Arg Asp Lys Gln
435 440 445
Gly Ile Pro Ile Thr Asp Glu Gln Leu Glu Lys Asp Pro Tyr Leu Pro
450 455 460
Pro Tyr Tyr His Pro Gly Glu Asp Ala Pro Glu Ile Lys Tyr Met Lys
465 470 475 480
Glu Arg Arg Ala Ala Leu Gly Gly Tyr Leu Pro Glu Arg Arg Glu Asn
485 490 495
Tyr Asp Pro Ile Gln Val Pro Pro Leu Asp Lys Leu Arg Ser Val Arg
500 505 510
Lys Gly Ser Gly Lys Gln Gln Ile Ala Thr Thr Met Ala Thr Val Arg
515 520 525
Thr Phe Lys Glu Leu Met Arg Asp Lys Gly Leu Ala Asp Arg Leu Val
530 535 540
Pro Ile Ile Pro Asp Glu Ala Arg Thr Phe Gly Leu Asp Ser Trp Phe
545 550 555 560
Pro Thr Leu Lys Ile Tyr Asn Pro His Gly Gln Asn Tyr Val Pro Val
565 570 575
Asp His Asp Leu Met Leu Ser Tyr Arg Glu Ala Pro Glu Gly Gln Ile
580 585 590
Leu His Glu Gly Ile Asn Glu Ala Gly Ser Met Ala Ser Phe Ile Ala
595 600 605
Ala Gly Thr Ser Tyr Ala Thr His Gly Lys Ala Met Ile Pro Leu Tyr
610 615 620
Ile Phe Tyr Ser Met Phe Gly Phe Gln Arg Thr Gly Asp Ser Ile Trp
625 630 635 640
Ala Ala Ala Asp Gln Met Ala Arg Gly Phe Leu Leu Gly Ala Thr Ala
645 650 655
Gly Arg Thr Thr Leu Thr Gly Glu Gly Leu Gln His Met Asp Gly His
660 665 670
Ser Pro Val Leu Ala Ser Thr Asn Glu Gly Val Glu Thr Tyr Asp Pro
675 680 685
Ser Phe Ala Tyr Glu Ile Ala His Leu Val His Arg Gly Ile Asp Arg
690 695 700
Met Tyr Gly Pro Gly Lys Gly Glu Asp Val Ile Tyr Tyr Ile Thr Ile
705 710 715 720
Tyr Asn Glu Pro Thr Pro Gln Pro Ala Glu Pro Glu Gly Leu Asp Val
725 730 735
Glu Gly Leu His Lys Gly Ile Tyr Leu Tyr Ser Arg Gly Glu Gly Thr
740 745 750
Gly His Glu Ala Asn Ile Leu Ala Ser Gly Val Gly Met Gln Trp Ala
755 760 765
Leu Lys Ala Ala Ser Ile Leu Glu Ala Asp Tyr Gly Val Arg Ala Asn
770 775 780
Ile Tyr Ser Ala Thr Ser Trp Val Asn Leu Ala Arg Asp Gly Ala Ala
785 790 795 800
Arg Asn Lys Ala Gln Leu Arg Asn Pro Gly Ala Asp Ala Gly Glu Ala
805 810 815
Phe Val Thr Thr Gln Leu Lys Gln Thr Ser Gly Pro Tyr Val Ala Val
820 825 830
Ser Asp Phe Ser Thr Asp Leu Pro Asn Gln Ile Arg Glu Trp Val Pro
835 840 845
Gly Asp Tyr Thr Val Leu Gly Ala Asp Gly Phe Gly Phe Ser Asp Thr
850 855 860
Arg Pro Ala Ala Arg Arg Phe Phe Asn Ile Asp Ala Glu Ser Ile Val
865 870 875 880
Val Ala Val Leu Asn Ser Leu Ala Arg Glu Gly Lys Ile Asp Val Ser
885 890 895
Val Ala Ala Gln Ala Ala Glu Lys Phe Lys Leu Asp Asp Pro Thr Ser
900 905 910
Val Ser Val Asp Pro Asn Ala Pro Glu Glu
915 920
<210> 8
<211> 437
<212> PRT
<213> Unknown
<220>
<223> Вариант цитратсинтазы Corynebacterium sp
<400> 8
Met Phe Glu Arg Asp Ile Val Ala Thr Asp Asn Asn Lys Ala Val Leu
1 5 10 15
His Tyr Pro Gly Gly Glu Phe Glu Met Asp Ile Ile Glu Ala Ser Glu
20 25 30
Gly Asn Asn Gly Val Val Leu Gly Lys Met Leu Ser Glu Thr Gly Leu
35 40 45
Ile Thr Phe Asp Pro Gly Tyr Val Ser Thr Gly Ser Thr Glu Ser Lys
50 55 60
Ile Thr Tyr Ile Asp Gly Asp Ala Gly Ile Leu Arg Tyr Arg Gly Tyr
65 70 75 80
Asp Ile Ala Asp Leu Ala Glu Asn Ala Thr Phe Asn Glu Val Ser Tyr
85 90 95
Leu Leu Ile Asn Gly Glu Leu Pro Thr Pro Asp Glu Leu His Lys Phe
100 105 110
Asn Asp Glu Ile Arg His His Thr Leu Leu Asp Glu Asp Phe Lys Ser
115 120 125
Gln Phe Asn Val Phe Pro Arg Asp Ala His Pro Met Ala Thr Leu Ala
130 135 140
Ser Ser Val Asn Ile Leu Ser Thr Tyr Tyr Gln Asp Gln Leu Asn Pro
145 150 155 160
Leu Asp Glu Ala Gln Leu Asp Lys Ala Thr Val Arg Leu Met Ala Lys
165 170 175
Val Pro Met Leu Ala Ala Tyr Ala His Arg Ala Arg Lys Gly Ala Pro
180 185 190
Tyr Met Tyr Pro Asp Asn Ser Leu Asn Ala Arg Glu Asn Phe Leu Arg
195 200 205
Met Met Phe Gly Tyr Pro Thr Glu Pro Tyr Glu Ile Asp Pro Ile Met
210 215 220
Val Lys Ala Leu Asp Lys Leu Leu Ile Leu His Ala Asp His Glu Gln
225 230 235 240
Asn Cys Ser Thr Ser Thr Val Arg Met Ile Gly Ser Ala Gln Ala Asn
245 250 255
Met Phe Val Ser Ile Ala Gly Gly Ile Asn Ala Leu Ser Gly Pro Leu
260 265 270
His Gly Gly Ala Asn Gln Ala Val Leu Glu Met Leu Glu Asp Ile Lys
275 280 285
Asn Asn His Gly Gly Asp Ala Thr Ala Phe Met Asn Lys Val Lys Asn
290 295 300
Lys Glu Asp Gly Val Arg Leu Ile Gly Phe Gly His Arg Val Tyr Lys
305 310 315 320
Asn Tyr Asp Pro Arg Ala Ala Ile Val Lys Glu Thr Ala His Glu Ile
325 330 335
Leu Glu His Leu Gly Gly Asp Asp Leu Leu Asp Leu Ala Ile Lys Leu
340 345 350
Glu Glu Ile Ala Leu Ala Asp Asp Tyr Phe Ile Ser Arg Lys Leu Tyr
355 360 365
Pro Asn Val Asp Phe Tyr Thr Gly Leu Ile Tyr Arg Ala Met Gly Phe
370 375 380
Pro Thr Asp Phe Phe Thr Val Leu Phe Ala Ile Gly Arg Leu Pro Gly
385 390 395 400
Trp Ile Ala His Tyr Arg Glu Gln Leu Gly Ala Ala Asp Asn Lys Ile
405 410 415
Asn Arg Pro Arg Gln Val Tyr Thr Gly Asn Glu Ser Arg Lys Leu Val
420 425 430
Pro Arg Glu Glu Arg
435
<210> 9
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 9
aatttctaga ggcagaccct attctatgaa gg 32
<210> 10
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 10
agtgtttcgg tctttacaga cacgagggac 30
<210> 11
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 11
gtccctcgtg tctgtaaaga ccgaaacact 30
<210> 12
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 12
aatttctaga cgtgggagtg tcactcgctt gg 32
<210> 13
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 13
ttcgagctcg gtacccggtc tagagcactt tcgctcgcac c 41
<210> 14
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 14
gatttgaaaa gcgcatcaga aacatcccag cgctac 36
<210> 15
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 15
gtagcgctgg gatgtttctg atgcgctttt caaatc 36
<210> 16
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 16
gaaacactat gatcccactt cgttcaaaag tcaccaccgt c 41
<210> 17
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 17
gacggtggtg acttttgaac gaagtgggat catagtgttt c 41
<210> 18
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 18
gcatgcctgc aggtcgactc tagagcgtgt gcaacgccgt c 41
<210> 19
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 19
gctcggtacc cggggatcct ctagaccgct tccttggctg cctgaagatg 50
<210> 20
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 20
tgcttggctt cacaagagac aagcct 26
<210> 21
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 21
aggcttgtct cttgtgaagc caagca 26
<210> 22
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 22
gcctgcaggt cgacctagat ctagacctca tcagagataa gaacagcatc 50
<210> 23
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 23
gctcggtacc cggggatcct ctagatactc cggtctgctt tatgcaggta 50
<210> 24
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 24
aactaaccta gtcgcttaag agacaagcct atctgc 36
<210> 25
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 25
gcagataggc ttgtctctta agcgactagg ttagtt 36
<210> 26
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 26
gcctgcaggt cgacctagat ctagaaagtg ccacgagcat ttcatcagct 50
<210> 27
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 27
gctcggtacc cggggatcct ctagataccg tccaaccggt actttgaacc 50
<210> 28
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 28
ttgatcggcc acttccacac 20
<210> 29
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 29
ggtgtggaag tggccgatca a 21
<210> 30
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 30
gcctgcaggt cgacctagat ctagagatcg tcttcagaaa ggcgaccctc 50
<210> 31
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 31
gctcggtacc cggggatcct ctagaaccgt ggcatcaagg acacctctga 50
<210> 32
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 32
agcttcttca ttgcgtgggt tg 22
<210> 33
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 33
caacccacgc aatgaagaag ct 22
<210> 34
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 34
aagcgtcagt gccttcatct g 21
<210> 35
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 35
ccagatgaag gcactgacgc tt 22
<210> 36
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 36
gcctgcaggt cgacctagat ctagaagatg gagtcaccgg tgcgctggaa 50
<210> 37
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 37
gctcggtacc cggggatcct ctagaacctt tccctggaat tttttccttt 50
<210> 38
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 38
tgtcccttga ggtgatttcc acacctcctg ttggaatg 38
<210> 39
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 39
tccaacagga ggtgtggaaa tcacctcaag ggacaga 37
<210> 40
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 40
gcctgcaggt cgacctagat ctagatgctg cgcggcaagc gccgtggatt 50
<210> 41
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 41
gctcggtacc cggggatcct ctagaaccct gaagctgtca gttcctagca 50
<210> 42
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 42
ccctttcaaa cacatttgtt c 21
<210> 43
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 43
cgaacaaatg tgtttgaaag gg 22
<210> 44
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 44
gcctgcaggt cgacctagat ctagatgcgc ggtgtgcgta cgcagccagc 50
<210> 45
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 45
gctcggtacc cggggatcct ctagatatgt gagcactggc tctaccgagt 50
<210> 46
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 46
aggtggagca ggtctgctcg tg 22
<210> 47
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 47
cacgagcaga cctgctccac ct 22
<210> 48
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 48
tgtccgaagc cgatgaggcg gac 23
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 49
cgtccgcctc atcggcttcg gaca 24
<210> 50
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 50
gcctgcaggt cgacctagat ctagagtcag agattacgag atcttccggt 50
30
<---
Изобретение относится к биотехнологии. Предложен микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты по сравнению с эндогенной активностью. Также микроорганизм обладает любой одной или более чем одной из комбинаций, выбранных из уменьшенной активности трансаминазы C по сравнению с эндогенной активностью, ослабленной активности пируватдегидрогеназы по сравнению с эндогенной активностью и ослабленной активности цитратсинтазы по сравнению с эндогенной активностью. Предложен также способ получения L-валина с использованием указанного микроорганизма. Изобретение обеспечивает получение L-валина с высоким выходом. 2 н. и 13 з.п. ф-лы, 11 табл., 2 пр.
1. Микроорганизм, продуцирующий L-валин, обладающий усиленной активностью дегидратазы дигидроксикислоты по сравнению с эндогенной активностью и любой одной или более чем одной из комбинаций, выбранных из (1)-(3) ниже:
(1) уменьшенной активностью трансаминазы C по сравнению с эндогенной активностью;
(2) ослабленной активностью пируватдегидрогеназы по сравнению с эндогенной активностью; и
(3) ослабленной активностью цитратсинтазы по сравнению с эндогенной активностью.
2. Микроорганизм по п. 1, обладающий усиленной активностью дегидратазы дигидроксикислоты и уменьшенной активностью трансаминазы C.
3. Микроорганизм по п. 1, обладающий усиленной активностью дегидратазы дигидроксикислоты и ослабленной активностью пируватдегидрогеназы.
4. Микроорганизм по п. 1, обладающий усиленной активностью дегидратазы дигидроксикислоты и ослабленной активностью цитратсинтазы.
5. Микроорганизм по п. 4, дополнительно обладающий уменьшенной активностью трансаминазы C или ослабленной активностью пируватдегидрогеназы.
6. Микроорганизм по п. 1, где дегидратаза дигидроксикислоты кодируется геном ilvD.
7. Микроорганизм по п. 1, где трансаминаза C кодируется геном avtA.
8. Микроорганизм по п. 1, где пируватдегидрогеназа кодируется геном aceE.
9. Микроорганизм по п. 1, где цитратсинтаза кодируется геном gltA.
10. Микроорганизм по п. 1, где ослабленная пируватдегидрогеназа отличается тем, что аминокислота, соответствующая положению 432 или 435 относительно N-конца аминокислотной последовательности SEQ ID NO: 3, соответственно заменена на аланин.
11. Микроорганизм по п. 1, где цитратсинтаза, обладающая ослабленной активностью по сравнению с эндогенной активностью, отличается тем, что аминокислота, соответствующая положению 241 или 312 относительно N-конца аминокислотной последовательности SEQ ID NO: 4, заменена на треонин или изолейцин соответственно.
12. Микроорганизм по п. 1, где микроорганизм, продуцирующий L-валин, представляет собой микроорганизм рода Corynebacterium.
13. Микроорганизм по п. 12, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
14. Способ получения L-валина, включающий культивирование микроорганизма по любому из пп. 1-13.
15. Способ по п. 14, дополнительно включающий выделение L-валина из культивируемого микроорганизма или среды.
| US 2010151449 A1, 17.06.2010 | |||
| US 9879289 B2, 30.01.2018 | |||
| Мутант пируватдегидрогеназы, микроорганизм, содержащий мутант, и способ получения L-аминокислоты с использованием микроорганизма | 2016 |
|
RU2683208C1 |
| US 20170051323 A1, 23.02.2017 | |||
| EP 3561055 A1, 30.10.2019. | |||

