Настоящее изобретение относится к рекомбинантной сигнальной последовательности и к слитому белку, содержащему сигнальную последовательность и рекомбинантный белок. Описаны способы продуцирования рекомбинантного белка с использованием рекомбинантной сигнальной последовательности. В частности, рекомбинантная сигнальная последовательность обеспечивает способ контролирования вязкости ферментации и/или контролирования базовой (доиндукционной) экспрессии рекомбинантного белка.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Крупномасштабное и рентабельное изготовление, выделение и очистка рекомбинантных белков являются важными задачами для биотехнологической промышленности.
Микробные клетки-хозяева являются широко используемыми организмами для продуцирования рекомбинантных белков. Факторы, которые необходимо учитывать во время продуцирования, включают рост клетки-хозяина и экспрессию рекомбинантного белка, титр белка, локализацию белка (например внутриклеточная, внеклеточная, периплазматическая и т.д.) и селективное выделение и очистку рекомбинантного белка из конечной локализации. Баланс и оптимизация этих разных факторов не являются простыми.
Грамотрицательная эубактерия Escherichia coli (Е. coli) является надежной экспрессионной системой для применения в промышленных биопроцессах благодаря ее хорошо охарактеризованной генетике; ее способности быстро накапливать биомассу с использованием недорогого субстрата; легкости масштабирования процесса и большого числа доступных штаммов-хозяев и экспрессионных векторов. Е. coli является обычным хозяином для продукции терапевтических белков, не требующих сложных посттрансляционных модификаций, таких как гликозилирование. Для образования дисульфидной связи экспрессия может быть направлена в окислительную среду периплазмы для облегчения правильного сворачивания. Это может быть достигнуто путем использования N-концевых сигнальных последовательностей, которые распознаются компонентами клеточной секреторной системы, для инициации их нацеливания и прохождения через специализированные транспортные системы.
Соответственно, существует потребность в улучшенных способах продуцирования, выделения и очистки рекомбинантных белков.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложена рекомбинантная сигнальная последовательность, содержащая (а) последовательность MGRISSGG (SEQ ID NO: 1) или ее вариант, последовательность которого отличается 1, 2 или 3 аминокислотными заменами, делениями или вставками, и (б) гетерологичную сигнальную последовательность, расположенную в направлении С-конца относительно (а).
В изобретении также предложен рекомбинантный слитый белок, содержащий (а) сигнальную последовательность, как определено; и (б) гетерологичный рекомбинантный белок, расположенный в направлении С-конца относительно (а).
В изобретении также предложена рекомбинантная нуклеиновокислотная последовательность, которая кодирует сигнальную последовательность или слитый белок, как определено.
В изобретении также предложена рекомбинантная нуклеиновокислотная последовательность, которая кодирует сигнальную последовательность, содержащую последовательность MGRISSGG (SEQ ID NO: 1), где в нуклеиновокислотной последовательности кодоны были оптимизированы для экспрессии в клетке-хозяине.
В изобретении также предложен рекомбинантный экспрессионный вектор, содержащий нуклеиновокислотная последовательность, как определено.
В изобретении также предложен способ продуцирования рекомбинантного белка в клетке-хозяине, включающий:
(а) культивирование клетки-хозяина в условиях, которые обеспечивают экспрессию рекомбинантного белка с рекомбинантного экспрессионного вектора, как определено, присутствующего в клетке-хозяине; и
(б) выделение рекомбинантного белка.
В изобретении также предложен способ контролирования вязкости бульона клеток-хозяев, экспрессирующих рекомбинантный белок, включающий:
культивирование клетки-хозяина в условиях, которые обеспечивают экспрессию рекомбинантного белка, где клетка-хозяин содержит рекомбинантную нуклеиновокислотную последовательность, которая кодирует (1) рекомбинантную сигнальную последовательность, содержащую последовательность MGRISSGG (SEQ ID NO: 1) или ее вариант, последовательность которого отличается 1, 2 или 3 аминокислотными заменами, делециями или вставками, и (2) рекомбинантный белок.
В изобретении также предложен способ контролирования базовой экспрессии рекомбинантного белка, экспрессируемого бульоном клеток-хозяев, включающий: культивирование клетки-хозяина в доиндукционных условиях,
где клетка-хозяин содержит рекомбинантную нуклеиновокислотную последовательность, которая кодирует (1) рекомбинантную сигнальную последовательность, содержащую последовательность MGRISSGG (SEQ ID NO: 1) или ее вариант, последовательность которого отличается 1, 2 или 3 аминокислотными заменами, делениями или вставками, и (2) рекомбинантный белок.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Влияние разных N-концевых сигнальных последовательностей на экспрессию dAb (доменное антитело) из Е. coli W3110 в культурах в 24 dwp (планшет с 24 глубокими лунками). Обозначения: В - dAb, локализованные внеклеточно (супернатант); С - dAb, локализованные внутриклеточно (цельноклеточный лизат: периплазматические плюс цитозольные). В+С - общее количество dAb, В/(В+С) - высвобожденная доля. Сложение В и С обеспечивает визуальную оценку общего количества образовавшегося dAb. Образцы отбирали через 24 часа после индукции, восстанавливали и визуализировали полосы посредством SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) с окрашиванием кумасси. «Std» обозначает стандарт dAb с концентрацией 0,5 мг⋅мл-1.
Фиг. 2. Послеиндукционные профили ферментации для культур с подпиткой Е. coli W3110 с высокой плотностью клеток. N-концевая сигнальная последовательность и базовый (доиндукционный) уровень экспрессии: (Δ), OmpASS, 0,30 г. л-1, ( ) YceISS, 0,64 г. л-1 или (◊), IvySS, 0,061 г. л-1. Все результаты получены после индукции образования dAb при помощи IPTG (изопропилтиогалактопиранозид). (А) Биомассу оценивали по OD600нм (оптическая плотность при длине волны 600 нм); (Б) концентрацию внеклеточной растворимой дцДНК; выход dAb определяли (В) внутриклеточно (цельноклеточный лизат), (Г) внеклеточно (супернатант), в виде (Д) общего количества образовавшегося dAb (внутриклеточное плюс внеклеточное) и в виде (Е) процентной доли общего количества dAb во внеклеточном пространстве. Вязкость бульона μ описывают в показателях степенной зависимости, μ=K γn-1 где γ представляет собой скорость сдвига, (Ж) K представляет собой показатель консистенции потока и (З) n представляет собой показатель поведения потока. Результаты были получены от усреднения данных для параллельных ферментеров (n равен 2-4) и были добавлены планки погрешностей, соответствующие одному стандартному отклонению.
) YceISS, 0,64 г. л-1 или (◊), IvySS, 0,061 г. л-1. Все результаты получены после индукции образования dAb при помощи IPTG (изопропилтиогалактопиранозид). (А) Биомассу оценивали по OD600нм (оптическая плотность при длине волны 600 нм); (Б) концентрацию внеклеточной растворимой дцДНК; выход dAb определяли (В) внутриклеточно (цельноклеточный лизат), (Г) внеклеточно (супернатант), в виде (Д) общего количества образовавшегося dAb (внутриклеточное плюс внеклеточное) и в виде (Е) процентной доли общего количества dAb во внеклеточном пространстве. Вязкость бульона μ описывают в показателях степенной зависимости, μ=K γn-1 где γ представляет собой скорость сдвига, (Ж) K представляет собой показатель консистенции потока и (З) n представляет собой показатель поведения потока. Результаты были получены от усреднения данных для параллельных ферментеров (n равен 2-4) и были добавлены планки погрешностей, соответствующие одному стандартному отклонению.
Фиг. 3. Выравнивания аминокислотных последовательностей сигнальных последовательностей и оценка их индексов адаптации кодонов. (А) Выравнивания аминокислотных последовательностей для YceISS (SEQ ID NO: 35) и IvySS (SEQ ID NO: 34) соотнесены с OmpASS (SEQ ID NO: 33). (Б) Адаптивность каждого положения кодона рассчитывали как отношение частоты данного кодона относительно наиболее многочисленного синонимичного кодона с отсылкой к таблице использования кодонов Е. coli, полученной из базы данных использования кодонов Е. coli в Kazusa. Аминокислотные последовательности в «Б» выравнивали от первого метионина после кассеты Ivy'SS.
Фиг. 4. Профили послеиндукционной ферментации для культур с подпиткой Е. coli W3110 с высокой плотностью клеток. N-концевая сигнальная последовательность и базовый (доиндукционный) уровень экспрессии: (Δ), IvyTruncSS, 0,90 г. л-1, (), Ivy'OmpASS, 0,086 г. л-1, (◊), IvyOpt1SS, 0,16 г. л-1 и (♦), IvyOpt2SS, 0,089 г. л-1. Подробности см. на Фиг. 2. Результаты были получены от образцов одной ферментации (m равно 3).
Фиг. 5. (А) Общая продуктивность, рассчитанная как масса внеклеточного dAb на литр бульона на общее послеиндукционное время. (Б) Эффективность центрифугирования собранных культур Е. coli, экспрессирующих dAb с разными N-концевыми сигнальными последовательностями; OmpASS (48 ч после индукции), YceISS (48 ч), IvyTruncSS (55 ч), Ivy'OmpA (153 ч) и Ivy кодон-оптимизированными сигнальными последовательностями IvyOpt1 (48 ч) и IvyOpt2 (55 ч). Клеточные бульоны подвергали воздействию следующих условий: ( ) нет сдвига; (
) нет сдвига; ( ) высокое напряжение сдвига (максимальная интенсивность рассеивания энергии (ε) 0,53×106 Вт.кг-1 за 20 с), и затем обработка посредством USD центрифугирования (V равен 2,0 мл, t равно 420 с, Σ равна 0,164 м2, (см. текст), V/tΣ=2,9×10-8 м.с-1). Результаты были получены от ферментеров в повторности (n равно 3) для OmpASS и YceISS или образцов из одного ферментера для остальных сигнальных последовательностей. Характеристику образца для (Б) была проведена в трех повторностях (m равно 3). Результаты показаны как среднее значение плюс/минус SD (стандартное отклонение).
) высокое напряжение сдвига (максимальная интенсивность рассеивания энергии (ε) 0,53×106 Вт.кг-1 за 20 с), и затем обработка посредством USD центрифугирования (V равен 2,0 мл, t равно 420 с, Σ равна 0,164 м2, (см. текст), V/tΣ=2,9×10-8 м.с-1). Результаты были получены от ферментеров в повторности (n равно 3) для OmpASS и YceISS или образцов из одного ферментера для остальных сигнальных последовательностей. Характеристику образца для (Б) была проведена в трех повторностях (m равно 3). Результаты показаны как среднее значение плюс/минус SD (стандартное отклонение).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее описание изобретения включает осознание того, что использование специфических сигнальных последовательностей обеспечивает преимущества в показателях ферментации, экспрессии рекомбинантного белка и извлекаемости белка.
Сигнальные последовательности исследовали для периплазматической секреции рекомбинантного белка. Данные сигнальные последовательности обеспечивают преимущество маловязкого бульона, возникающее из-за низкой базовой экспрессии. Последовательность из восьми аминокислот, содержащаяся в сигнальной последовательности, идентифицирована как ключевая для того, чтобы привести к такой низкой базовой экспрессии. Оптимизация кодонов сигнальных последовательностей была успешной в сохранении низкой базовой экспрессии и, следовательно, маловязких бульонов при увеличении продуктивности ферментации рекомбинантного белка. Следовательно, достигается комбинация высокой продуктивности и практически осуществимого осветления посредством полномасштабного центрифугирования.
Последовательность «MGRISSGG» (SEQ ID NO: 1) можно использовать N-терминально (5') по отношению к любой сигнальной последовательности. Например, «MGRISSGG» (SEQ ID NO: 1) можно использовать N-терминально по отношению к гетерологичной сигнальной последовательности. Гетерологичная сигнальная последовательность может представлять собой последовательность, которая не является эндогенной сигнальной последовательностью IVY. Например, гетерологичная сигнальная последовательность представляет собой периплазматическую сигнальную последовательность из Е. coli. Например, периплазматическая сигнальная последовательность Е. coli представляет собой OmpA, MalE, PelB, ОтрТ или LamB. Например, "MGRISSGG" (SEQ ID NO: 1) может быть N-терминальной к MKKTAIAIAVALAGFATVAQA (SEQ ID NO: 33) или к любой из сигнальных последовательностей в Таблице 3, или к периплазматической сигнальной последовательности Е. coli.
Рекомбинантная сигнальная последовательность содержит (а) последовательность MGRISSGG (SEQ ID NO: 1) или ее вариант, последовательность которого отличается 1, 2 или 3 аминокислотными заменами, делениями или вставками, и (б) гетерологичную сигнальную последовательность С-терминально по отношению к (а). Гетерологичная сигнальная последовательность может находиться непосредственно С-терминально к MGRISSGG (SEQ ID NO: 1). Например, рекомбинантная сигнальная последовательность может содержать последовательность MGRISSGG (SEQ ID NO: 1) непосредственно N-терминально к гетерологичной периплазматической сигнальной последовательности Е. coli. Например, рекомбинантная сигнальная последовательность может содержать последовательность MGRISSGG (SEQ ID NO: 1) непосредственно N-терминально к гетерологичной сигнальной последовательности MKKTAIAIAVALAGFATVAQA (SEQ ID NO: 33) с образованием рекомбинантной сигнальной последовательности MGRISSGGMKKTAIAIAVALAGFATVAQA (SEQ ID NO: 55).
Последовательность "MGRISSGG" (SEQ ID NO: 1) может отличаться 1, 2 или 3 аминокислотными заменами, делениями или вставками. Последовательность "MGRISSGG" (SEQ ID NO: 1) плюс гетерологичная сигнальная последовательность могут отличаться 1-10 аминокислотными заменами, делениями или вставками. Вариант может сохранять активность "MGRISSGG" (SEQ ID NO: 1).
Аминокислотная замена может включать замену по меньшей мере одной аминокислоты на аминокислоту того же самого класса, как определено ниже в Таблице 1.
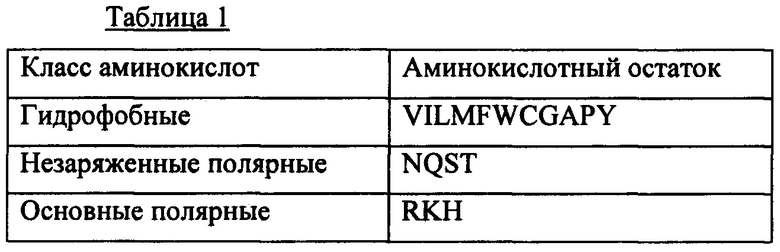
Такая по меньшей мере одна аминокислотная замена, делеция или вставка может сохранять первичную структуру последовательности "MGRISSGG" (SEQ ID NO: 1) путем поддержания консенсусной последовательности с: (1) одной или более гидрофобными аминокислотами, (2) одной или более основными полярными аминокислотами, (3) одной или более гидрофобными аминокислотами и (4) одной или более незаряженными полярными аминокислотами, как определено в Таблице 1.
Активность "MGRISSGG" (SEQ ID NO: 1), при нахождении ее N-терминально (5') к сигнальной последовательности, может быть определена как контроль базовой экспрессии (доиндукционной экспрессии) рекомбинантного белка. Базовая экспрессия происходит во время доиндукционной экспоненциальной фазы роста клетки-хозяина. Во время этой фазы роста рекомбинантный белок не должен экспрессироваться или его экспрессия минимизирована, и, вместо этого, метаболическая нагрузка клеток сфокусирована на росте. Однако многие сигнальные последовательности допускают некоторую базовую экспрессию рекомбинантного белка. Активностью "MGRISSGG" (SEQ ID NO: 1) или ее варианта может быть регулирование базовой экспрессии рекомбинантного белка до уровня 0,5 г/л или менее на момент индукции, т.е. доиндукционого уровня экспрессии рекомбинантного белка величиной 0,5 г/л или менее. Использование сигнальной последовательности, содержащей "MGRISSGG" (SEQ ID NO: 1), может приводить к низкому доиндукционному уровню экспрессии рекомбинантного белка. Например, доиндукционный уровень экспрессии рекомбинантного белка может составлять 0,5 г/л или менее, 0,4 г/л или менее, 0,3 г/л или менее, 0,2 г/л или менее, 0,1 г/л или менее, или 0,05 г/л или менее.
Возможно, кроме того, что активность "MGRISSGG" (SEQ ID NO: 1), при нахождении ее N-терминально (5') к сигнальной последовательности, может быть определена как регулирование вязкости бульона клеток-хозяев, экспрессирующего рекомбинантный белок. Бульон клеток-хозяев состоит из клеток-хозяев и супернатанта, содержащего культуральные среды. Вязкость бульона может меняться во время ферментации в следующие моменты времени: до индукции, в момент индукции и после индукции. Использование сигнальной последовательности, содержащей "MGRISSGG" (SEQ ID NO: 1), может приводить к низкой вязкости бульона клеток-хозяев во время ферментации. Вязкость можно измерять, используя обычные в данной области техники способы. Например, вязкость можно измерять, используя показатель консистенции потока (K) или кажущуюся вязкость (μ).
Например, бульон клеток-хозяев имеет вязкость, эквивалентную показателю консистенции потока K 0,1 Нсnм-2 или менее. Например, бульон клеток-хозяев имеет вязкость K 0,05 Нсnм-2 или менее, K 0,04 Нсnм-2 или менее, K 0,03 Нсnм-2 или менее, K 0,02 Нсnм-2 или менее, или K 0,01 Нсnм-2 или менее.
Например, «низкая» вязкость может быть эквивалентна значению К примерно от 0,001 Нсnм-2 до 0,01 Нсnм-2, «средняя» вязкость может быть эквивалентна значению К примерно от 0,01 Нсnм-2 до 0,1 Нсnм-2 и «высокая» вязкость может быть эквивалентна значению К примерно более чем 0,1 Нnм-2.
Альтернативно, например, бульон клеток-хозяев имеет кажущуюся вязкость μ (определенную при скорости сдвига 1 с-1) 0,1 Нсм-2 или менее. Например, бульон клеток-хозяев имеет кажущуюся вязкость μ 0,05 Нсм-2 или менее, μ 0,04 Нсм-2 или менее, μ 0,03 Нсм-2 или менее, μ 0,02 Нсм-2 или менее или μ 0,01 Нсм-2 или менее. Кажущаяся вязкость будет меняться в зависимости от применяемой скорости сдвига, значения могут быть рассчитаны из значений n показателя поведения потока и выбранной скорости сдвига.
В одном воплощении это вязкость бульона в точке индукции.
Нуклеиновокислотная последовательность, которая кодирует рекомбинантную сигнальную последовательность, содержащую "MGRISSGG" (SEQ ID NO: 1), может быть кодон-оптимизированной для экспрессии в клетке-хозяине. Способы оптимизации кодонов (также известной как адаптация кодонов) известны в данной области. Например, индекс адаптации кодонов (CAI) может быть соотнесен с использованием кодонов для всех природных генов конкретной линии клеток или только для высокоэкспрессируемых генов. Могут быть соотнесены все гены Е. coli или могут быть соотнесены все гены Е. coli K12 или гены Е. coli K12 класса II (высокоэкспрессируемые).
Нуклеиновокислотная последовательность, которая кодирует сигнальную последовательность, содержащую "MGRISSGG" (SEQ ID NO: 1), может иметь балл CAI 0,40 или выше, 0,45 или выше, 0,50 или выше, 0,55 или выше, 0,60 или выше, 0,65 или выше, 0,70 или выше, 0,75 или выше, 0,80 или выше, 0,85 или выше, или 0,90 или выше.
Например, нуклеиновокислотная последовательность, которая кодирует сигнальную последовательность, содержащую "MGRISSGG" (SEQ ID NO: 1), может содержать кодон-оптимизированную последовательность:
 (SEQ ID NO: 49) или
(SEQ ID NO: 49) или
 (SEQ ID NO: 50).
(SEQ ID NO: 50).
Возможно, кроме того, что активность "MGRISSGG" (SEQ ID NO: 1) или ее варианта, или кодон-оптимизированной последовательности может заключаться в регулировании экспрессии рекомбинантного белка после индукции до уровня 5 г/л или более. Использование сигнальной последовательность, содержащей "MGRISSGG" (SEQ ID NO: 1), может приводить к высокому послеиндукционному уровню экспрессии рекомбинантного белка. Например, послеиндукционный уровень экспрессии рекомбинантного белка может составлять 5 г/л и более, 6 г/л и более, 7 г/л и более, 8 г/л и более, 9 г/л и более, или 10 г/л и более. Уровень экспрессии рекомбинантного белка может относиться к общему продуцированному белку или внеклеточному белку.
Возможно, кроме того, что активность "MGRISSGG" (SEQ ID NO: 1) или ее варианта, или кодон-оптимизированной последовательности может представлять собой регулирование продуктивности по рекомбинантному белку после индукции до уровня 0,1 г/л/ч или более. Использование сигнальной последовательности, содержащей "MGRISSGG" (SEQ ID NO: 1), может приводить к высокой продуктивности по рекомбинантному белку после индукции. Например, послеиндукционная продуктивность по рекомбинантному белку может составлять 0,10 г/л/ч или более, 0,11 г/л/ч или более, 0,12 г/л/ч или более, 0,13 г/л/ч или более, 0,14 г/л/ч или более, или 0,15 г/л/ч или более. Продуктивность по рекомбинантному белку может относиться к общему продуцированному или внеклеточному белку. Время (часы) может быть рассчитано с момента индукции.
Возможно, кроме того, что активность "MGRISSGG" (SEQ ID NO: 1) или ее варианта, или кодон-оптимизированной последовательности может представлять собой регулирование % оставшихся твердых веществ (при определении путем измерения оптической плотности при 600 нм) после центрифугирования сбора (т.е. осветления) до уровня 20% или менее. Использование сигнальной последовательности, содержащей "MGRISSGG" (SEQ ID NO: 1), может приводить к низкому % твердых веществ, остающихся после центрифугирования сбора. Например, % твердых веществ, остающихся после центрифугирования сбора, может составлять 20% или менее, 15% или менее, 10% или менее, 9% или менее, 8% или менее, 7% или менее, 6% или менее, или 5% или менее.
Термин «культура клеток» при использовании в настоящем описании изобретения приводится в самом широком значении, а именно относится к массовому росту клеток в среде для роста клеток. Термины «ферментирование» и «культивирование» при использовании в данном описании изобретения означает массовое выращивание клеток в среде для роста клеток. Термины «ферментирование» и «культивирование» используются здесь взаимозаменяемо. Экспоненциальная фаза представляет собой период, характеризующийся удвоением клеток, следовательно массовым ростом. Число новых клеток, появляющихся в единицу времени, значительно возрастает и пропорционально существующей популяции. Клетки могут находиться в экспоненциальной фазе роста до индукции. Например, базовая экспрессия рекомбинантного белка происходит во время доиндукционной экспоненциальной фазы.
Стационарной фазой является фаза, в которой скорость роста и скорость гибели клеток являются равными, что приводит к фазе линейного роста. Например, экспрессия рекомбинантного белка индуцируется во время стационарной фазы. Клетки часто достигают стационарной фазы из-за фактора, лимитирующего рост, такого как исчерпание незаменимого питательного вещества и/или образование ингибирующего продукта, такого как органическая кислота. Для поддержания постоянной биомассы в стационарной фазе и, следовательно, массового роста, клетки продолжают расти.
Биомассу можно оценивать по OD600нм. Биомасса «с высокой плотностью клеток» может иметь OD600нм по меньшей мере 30. Например, высокая плотность клеток может относиться к OD600нм по меньшей мере 30, по меньшей мере 40, по меньшей мере 50 или по меньшей мере 60. Альтернативно, биомассу можно измерять по сухой клеточной массе. Биомасса «с высокой плотностью клеток» может иметь сухую клеточную массу по меньшей мере 10 гл-1. Например, высокая плотность клеток может относиться к сухой клеточной массе по меньшей мере 10 гл-1, по меньшей мере 15 гл-1, по меньшей мере 20 гл-1 или по меньшей мере 25 гл-1. Альтернативно, биомассу можно измерять по сырой клеточной массе. Биомасса «с высокой плотностью клеток» может иметь сырую клеточную массу по меньшей мере 50 гл-1. Например, высокая плотность клеток может относиться к сырой клеточной массе по меньшей мере 50 гл-1, по меньшей мере 70 гл-1, по меньшей мере 85 гл-1 или по меньшей мере 100 гл-1.
Все термины - внеклеточная среда, супернатант, внеклеточное окружение, внеклеточное пространство, культуральная среда и ферментационная среда -используются здесь для описания внешней среды клеток во время культивирования клеток, в момент индукции и в момент сбора клеток.
Термин «индукция» или «индуцирование экспрессии» при использовании в данном описании изобретения относится к моменту, в который инициируют индукцию экспрессии рекомбинантного белка. Например, экспрессию рекомбинантного белка можно индуцировать путем добавления индуктора или изменения температуры, когда индукция является температурозависимой. Термин «после индукции» используют здесь для описания времени, прошедшего после момента, в который инициируют индукцию.
Термин «сбор» используется здесь для обозначения окончания ферментации. Сбор может происходить в любой момент времени во время ферментации, который считается достаточным для завершения процесса ферментации и выделения экспрессируемого рекомбинантного белка. Например, выделение белка начинают в момент сбора. Время сбора может зависеть от оптимальной концентрации рекомбинантного белка в супернатанте. Рекомбинантный белок может быть выделен и очищен непосредственно из внеклеточной среды культуры клеток в момент сбора. Альтернативно, рекомбинантный белок может быть выделен и очищен из периплазмы клетки-хозяина.
Подразумевается, что «примерно» при использовании в данном описании изобретения в отношении измеряемого значения, такого как количество, молекулярная масса, продолжительность времени и тому подобное, включает отклонения плюс/минус 1%, плюс/минус 0,75%, плюс/минус 0,5%, плюс/минус 0,25%, плюс/минус 0,2% и плюс/минус 0,1% от указанного значения, поскольку такие отклонения подходят для осуществления описанных способов.
Рекомбинантный белок
Рекомбинантный белок может включать антигенсвязывающий белок, например моноклональное антитело, фрагмент антитела или доменное антитело (dAb).
Рекомбинантный белок может включать вирусный белок, бактериальный токсин, бактериальный анатоксин или раковый антиген.
Термин «рекомбинантный белок» при использовании в данном описании изобретения относится к любому белку и/или полипептиду, который можно вводить млекопитающему, чтобы вызвать биологический или медицинский ответа ткани, системы, животного или человека. Рекомбинантный белок может вызывать более одного биологического или медицинского ответа. Кроме того, термин «терапевтически эффективное количество» означает любое количество, которое, по сравнению с соответствующим субъектом, который не получал такое количество, приводит к излечению, предупреждению или уменьшению интенсивности заболевания, расстройства или побочного эффекта, или к уменьшению скорости развития заболевания или расстройства, но не ограничивается ими. В объем данного термина также включены количества, эффективные для улучшения нормальной физиологической функции, а также количества, эффективные, чтобы вызвать у пациента физиологическую функцию, которая усиливает или способствует терапевтическому эффекту второго фармацевтического агента.
Термин «антигенсвязывающий белок» при использовании в данном описании изобретения, относится к антителам, фрагментам антител и другим белковым конструкциям, таким как домены, которые способны связываться с антигеном.
Термин «антитело» используется здесь в самом широком смысле для обозначения молекул с иммуноглобулин-подобным доменом. Термин «иммуноглобулин-подобный домен» при использовании в данном описании изобретения относится к семейству полипептидов, которые сохраняют сворачивание иммуноглобулина, характерное для молекул антител, которые содержат два бета-листа и, обычно, консервативную дисульфидную связь. Это семейство включает моноклональные (например IgG, IgM, IgA, IgD или IgE), рекомбинантные, поликлональные, химерные, гуманизированные, биспецифичные и гетероконъюгатные антитела; единичный вариабельный домен, доменное антитело, антигенсвязывающие фрагменты, иммунологически эффективные фрагменты, Fab, F(ab')2, Fv, Fv, связанный дисульфидной связью, одноцепочечный Fv, диатела, TANDABS™ и т.д.
Выражение «единичный вариабельный домен» относится к вариабельному домену антигенсвязывающего белка (например VH, VHH, VL), который специфично связывается с антигеном или эпитопом, независимо от другой вариабельной области или домена. «Доменное антитело» или «dAb» может рассматривать как то же самое, что и «единичный вариабельный домен».
Термин «домен» при использовании в данном описании изобретения относится к свернутой белковой структуре, которая сохраняет свою третичную структуру независимо от остальной части белка. В общем случае, домены отвечают за дискретные функциональные свойства белков и во многих случаях могут быть добавлены, удалены или перенесены на другие белки без потери функции остальной части белка и/или домена. Под единичным вариабельным доменом антитела или единичным вариабельным доменом иммуноглобулина подразумевают свернутый полипептидный домен, содержащий характерные последовательности вариабельного домена антитела. Он, следовательно, включает полные вариабельные домены антитела и модифицированные вариабельные домены, например те, в которых одна или более петель заменены последовательностями, которые не являются характерными для вариабельных доменов антитела, или вариабельные домены антитела, которые были укорочены или содержат N- или С-концевые удлинения, а также свернутые фрагменты вариабельных доменов, которые сохраняют, по меньшей мере частично, активность и специфичность связывания полноразмерного домена.
Доменное антитело может присутствовать в формате (например гомо- или гетеромультимера) с другими вариабельными областями или вариабельными доменами, где другие области или домены не требуются для связывания антигена единичным вариабельным доменом иммуноглобулина (т.е. где единичный вариабельный домен иммуноглобулина связывается с антигеном независимо от дополнительных вариабельных доменов).
Доменное антитело (dAb™) может представлять собой вариабельный домен человеческого антитела. dAb™ может иметь человеческое происхождение. Другими словами dAb™ может быть основано на каркасной последовательности человеческого Ig.
Термин «антигенсвязывающий сайт» при использовании в данном описании изобретения относится к сайту на антигенсвязывающем белке, который способен специфично связываться с антигеном, он может представлять собой единичный домен, или он может представлять собой парные домены VH/VL, которые могут находиться на стандартном антителе. Одноцепочечные Fv (ScFv)-домены также могут обеспечивать антигенсвязывающие сайты.
Антигенсвязывающий белок может содержать дополнительные антигенсвязывающие сайты для других антигенов. Например, антигенсвязывающий белок может обладать специфичностью в отношении более чем одного антигена, например двух антигенов или в отношении трех антигенов или в отношении четырех антигенов.
Антигенсвязывающий белок может состоять, или по существу состоит, из Fc-области антитела или ее части, связанной на каждом конце, прямо или опосредованно (например через линкерную последовательность), со связывающим доменом. Такой антигенсвязывающий белок может содержать два связывающих домена, разделенных Fc-областью или ее частью. Под разделенными подразумевается, что связывающие домены не связаны друг с другом непосредственно и могут быть расположены на противоположных концах (С- и N-конце) Fc-области или любой другой каркасной области.
Антигенсвязывающий белок может содержать две каркасные области, каждая из которых связана с двумя связывающими доменами, например на N- и С-концах каждой каркасной области, либо прямо, либо опосредованно через линкер. Каждый связывающий домен может связываться с отличным антигеном.
Антигенсвязывающий белок может принимать формат белкового каркаса mAbdAb. Аббревиатуры «mAbdAb» и «dAbmAb» используются взаимозаменяемо, и подразумевается, что они имеют одинаковое значение при использовании здесь. Такие антигенсвязывающие белки содержат белковый каркас, например Ig-каркас, такой как IgG, например моноклональное антитело, который связан с дополнительным связывающим доменом, например доменным антителом. mAbdAb имеет по меньшей мере два антигенсвязывающих сайта, по меньшей мере один из которых происходит из доменного антитела, и меньшей мере один из парного домена VH/VL.
Доменные антитела могут существовать и связываться с мишенью в мономерной или мультимерной (например димерной) формах, и могут быть использованы в комбинации с другими молекулами для подходов форматирования и направленной доставки. Например, может быть получен антигенсвязывающий белок, имеющий многочисленные домены, в котором один из данных доменов связывается с сывороточными белками, такими как альбумин. Известны доменные антитела, которые связываются с сывороточным альбумином (AlbudAb), и они могут сами по себе предоставлять домену-партнеру слияния пролонгированный период полувыведения из сыворотки.
dAb также могут быть конъюгированными с другими молекулами, например в виде конъюгата dAb или слияния dAb с другими молекулами, например лекарственным средством, другим белком, молекулой антитела или фрагментом антитела. Например, dAb™ может быть представлено в виде форматированного dAb™, например, dAb™ может быть представлено в виде слияния dAb-Fc или конъюгата. Альтернативно, форматированное dAb™ может быть представлено в виде mAbdAb. dAb™ может быть представлено в виде слияния или конъюгата с белками или полипептидами, пролонгирующими период полувыведения, например с другим dAb™, которое связывается с сывороточным альбумином (AlbudAb), или с химической группировкой, пролонгирующей период полувыведения, такой как полиэтиленгликоль (ПЭГ). dAb™ может быть представлено в виде слияния или конъюгата с другими терапевтическими или активными молекулами.
При использовании здесь термин «лекарственное средство» относится к любому соединению (например к малой органической молекуле, нуклеиновой кислоте, полипептиду), которое может быть введено субъекту с получением полезного терапевтического или диагностического эффекта посредством связывания с биологической молекулой-мишенью у субъекта и/или изменения ее функции. Молекула-мишень может представлять собой эндогенную молекулу-мишень, кодируемую геномом субъекта (например фермент, рецептор, фактор роста, цитокин, кодируемый геномом субъекта), или экзогенную молекулу-мишень, кодируемую геномом патогена. Лекарственное средство может представлять собой dAb™ или mAb.
Термин «конъюгат dAb» относится к композиции, содержащей dAb™, с которым химически конъюгировано лекарственное средство посредством ковалентной или нековалентной связи. Предпочтительно dAb™ и лекарственное средство связаны ковалентно. Такая ковалентная связь может быть реализована через пептидную связь или другими средствами, например посредством модифицированной боковой цепи. Нековалентное связывание может быть прямым (например электростатическое взаимодействие, гидрофобное взаимодействие) или опосредованным (например посредством нековалентного связывания партнеров по комплементарному связыванию (например биотина и авидина), где один партнер ковалентно связан с лекарственным средством, а партнер по комплементарному связыванию ковалентно связан с dAb™). При использовании партнеров по комплементарному связыванию один из партнеров связывания может быть ковалентно связан с лекарственным средством прямо или через подходящую линкерную группировку, и партнер по комплементарному связыванию может быть ковалентно связан с dAb™ прямо или через подходящую линкерную группировку.
При использовании здесь термин «слияние dAb» относится к слитому белку, который содержит dAb™ и полипептидное лекарственное средство (которое может представлять собой dAb™ или mAb). dAb™ и полипептидное лекарственное средство присутствуют в виде дискретных частей (группировок) одной непрерывной полипептидной цепи.
Таким образом, способы по данному раскрытию могут применяться к одному или более из следующего: терапевтический белок, моноклональное антитело (mAb), доменное антитело (dAb™), конъюгат dAb™, слияние dAb™, mAbdAb или любой другой описанный здесь антигенсвязывающий белок.
В одном воплощении антигенсвязывающий белок представляет собой dAb™, которое препятствует передаче сигналов посредством TNFα (фактор некроза опухолей-альфа). В одном воплощении dAb™ нейтрализует TNFα. В одном воплощении dAb™ специфично связывается с TNFα или с рецептором TNFα. В одном воплощении dAb™ специфично связывается с TNFR1 (рецептор 1 фактор некроза опухолей). В одном воплощении антигенсвязывающий белок представляет собой VH dAb™ (анти-TNFR1) / DOM0101) с SEQ ID NO: 53 (DOM0101).
Экспрессия белка: клетка-хозяин
Подходящие клетки-хозяева включают микробные клетки, такие как грамотрицательные бактерии, например Escherichia coli (например W3110 и BL21), Pseudomonas и Salmonella. В конкретном воплощении клетка-хозяин представляет собой Escherichia coli. В одном воплощении штаммом Е. coli является W3110.
В данном описании изобретения также описан вектор, содержащий молекулу рекомбинантной нуклеиновой кислоты, кодирующую сигнальную последовательность и рекомбинантный белок. Такие векторы используют для генетической инженерии клеток с целью экспрессии нужного белкового продукта. Данный вектор может представлять собой экспрессионный вектор, содержащий один или более элементов или последовательностей контроля экспрессии, которые связаны функциональным образом с сигнальной последовательностью и рекомбинантной нуклеиновой кислотой. Примеры векторов включают плазмиды и фагмиды.
Подходящие экспрессионные векторы могут содержать целый ряд компонентов, например точку начала репликации, селектируемый маркерный ген, один или более элементов контроля экспрессии, такие как элемент контроля транскрипции (например промотор, энхансер, терминатор) и/или один или более сигналов трансляции, сигнальную последовательность или лидерную последовательность. Элементы контроля экспрессии и сигнальная последовательность, при их наличии, могут быть предоставлены вектором или другим источником. Например, для управления экспрессией могут быть использованы последовательности контроля транскрипции и/или трансляции клонированной нуклеиновой кислоты, кодирующей цепь антитела.
Для экспрессии в желательной клетке может быть обеспечен промотор. Промоторы могут быть конститутивными или индуцируемыми. Например, промотор может быть связан функциональным образом с нуклеиновой кислотой, кодирующей антитело, цепь антитела или ее часть, так что он управляет транскрипцией этой нуклеиновой кислоты. Можно использовать целый ряд подходящих промоторов для грамотрицательных бактерий (например промоторы lac, tac, trp, phoA, lambdapL, Т3, T7 (T7A1, T7A2, T7A3) для E. coli). Последовательности оператора, которые можно использовать, включают lac, gal, deo и gin. Можно использовать одну или более чем одну совершенную палиндромную последовательность оператора. В одном воплощении экспрессия рекомбинантного белка по изобретению находится под контролем индуцируемого промотора. Например, промоторы lac, tac и trc Е. coli индуцируются лактозой или негидролизуемым аналогом лактозы - изопропил-β-D-1-тиогалактопиранозидом (IPTG), а промоторы phoA, trp и araBAD индуцируются фосфатом, триптофаном и L-арабинозой, соответственно.
Кроме того, экспрессионные векторы обычно содержат селектируемый маркер для селекции клеток, несущих вектор, и, в случае реплицируемого экспрессионного вектора, точку начала репликации. Гены, кодирующие продукты, которые придают резистентность к антибиотикам или лекарственным средствам, являются обычными селектируемыми маркерами и могут быть использованы (например ген лактамазы (резистентность к ампициллину), ген Tet для резистентности к тетрациклину). Маркерные гены дигидрофолатредуктазы обеспечивают отбор с метотрексатом в целом ряде клеток.
Экспрессионный вектор, описанный в WO2007/088371 (например pAVE037, pAVE007 или pAVE011) можно использовать для экспрессии белка. В одном воплощении вектор представляет собой pAVE011. Альтернативно, для экспрессии белка можно использовать такой вектор, как pJExpress401.
Также известны типичные альтернативные экспрессионные векторы и методики (например для использования с СНО (клетки яичника китайского хомяка), PerC6 и т.д.).
Клетка-хозяин содержит молекулу рекомбинантной нуклеиновой кислоты или вектор, описанный выше.
Культуры клеток-хозяев по настоящему изобретению можно культивировать в любой среде, которая поддерживает рост клетки-хозяина и экспрессию рекомбинантного белка. Такие среды хорошо известны специалистам в данной области техники.
Направленность белка
Хотя экспрессия рекомбинантного белка происходит в цитоплазме, конечная локализация рекомбинантного белка может быть цитоплазматической, периплазматической или внеклеточной, в зависимости от природы рекомбинантного белка, используемой клетки-хозяина и используемых условий ферментации.
Использование сигнальных последовательностей в настоящем изобретении может быть реализовано в отношении рекомбинантного белка, который нацелен в периплазму клетки (например у грамотрицательной бактерии).
У грамотрицательных бактерий некоторые секретируемые белки экспортируются через внутреннюю и наружную мембраны за одну стадию посредством путей секреции I типа, III типа, IV типа или VI типа, тогда как другие белки сначала экспортируются в периплазму посредством универсальных Sec- или Tat-путей и затем перемещаются через наружную мембрану главным образом посредством механизмов II типа или V типа. Система II типа включает двухстадийный процесс, в котором незрелый белок, содержащий лидерную последовательность Sec, экспортируется в периплазму с использованием Sec-пути. Сигнальная последовательность удаляется посредством протеолиза, приводя к зрелому, процессированному белку, присутствующему в периплазме, и секретируется или нет данный белок в культуральную среду в значительной степени зависит от характеристик лидерной последовательности, белка, клетки и условий культивирования. Также в случае лизиса клетки (автолиза) можно предположить то, что большая часть белка в культуральной среде происходит из периплазмы и, следовательно, подвергается процессингу. Рекомбинантный белок может активно секретироваться в культуральную среду при помощи сигнальной последовательности; или пассивно из периплазмы в культуральную среду посредством других клеточных путей, известных в данной области.
Процессинг сигнальной последовательности включает отщепление и удаление сигнальной последовательности от белка. Однако некоторые аминокислоты данного сигнала могут оставаться на N-конце белка, так что сигнальная последовательность не подвергается правильному процессингу. Сигнальная последовательность может быть процессированной на 90% или более, так что 10% или менее данной последовательности остается на N-конце белка. Сигнальная последовательность может быть процессированной по меньшей мере на 91, 92, 93, 94, 95, 96, 97, 98 или 99%. Сигнальная последовательность может быть процессированной примерно на 100%, так что на N-конце белка после прохождения через секреторный путь клетки ничего не остается.
Сигнальная последовательность может представлять собой сигнальную последовательность с периплазматической направленностью, например N-концевую последовательность с периплазматической направленностью. Сигнальные последовательности для направления белков в периплазму или внеклеточную среду известны в данной области. Сигнальная последовательность может включать гетерологичную сигнальную последовательность.
Сбор
Сбор представляет собой конец ферментации. Сбор может происходить в любой момент времени во время ферментации, который считается достаточным для завершения процесса ферментации и выделения экспрессируемого рекомбинантного белка. Сбор может происходить через 10-160 часов после индукции. Например, сбор может происходить через 15-60 часов после индукции. Сбор может происходить через 24, 48 или 55 часов после индукции. При сборе содержание твердых веществ популяции микробных клеток может составлять 5-30% от сырой массы клеток (WCW).
Объем ферментера может составлять:
(1) примерно 10000 литров; примерно 5000 литров; примерно 2000 литров; примерно 1000 литров; примерно 500 литров; примерно 125 литров; примерно 50 литров; примерно 20 литров; примерно 10 литров; примерно 5 литров; или
(2) от 5 до 10000 литров; от 10 до 5000 литров; от 20 до 2000 литров; от 50 до 1000 литров.
Сбор может содержать клетки, которые лизировались естественным образом, также известным как автолиз. Например, 1-50% клеток в сборе могут претерпевать автолиз. Альтернативно, 20-50%, или 30-50%, или 40-50% клеток в сборе подверглись автолизу. Альтернативно, 10% или более; 20% или более; 30% или более; 40% или более; или 50% или более клеток в сборе подверглись автолизу. Автолиз может быть опосредованно определен по концентрации ДНК в осветленном сборе или по электрической емкости.
Сбор может включать возможную стадию опустошения ферментера от клеток и внеклеточных сред (т.е. от клеточной культуры или бульона).
Возможная предобработка сбора
В зависимости от клетки-хозяина и рекомбинантного белка, предобработка сбора представляет собой способ кондиционирования сбора. Данную стадию можно проводить в ферментере или после того, как сбор удалили из ферментера. Предобработка включает: тепловое, механическое или химическое лизирование сбора (например посредством гомогенизации, замораживания-оттаивания, лизиса) и экстракцию периплазмы. По меньшей мере один периплазматический экстракт может быть экстрагирован с использованием способов, известных в данной области. Альтернативно, если во внеклеточной среде уже присутствует достаточное количество продукта, тогда такая предобработка может не потребоваться.
Осветление
Осветление представляет собой процесс удаления твердых частиц. Осветление может уменьшать нагрузку на последующие хроматографические стадии во время очистки. Типичные стадии осветления включают стадию отстаивания, также известную как осаждение (например посредством силы тяжести), и/или стадию центрифугирования, и/или стадию фильтрования.
Стадия центрифугирования может представлять собой непрерывное центрифугирование (например с непрерывной зоной подпитки). Сама центрифуга может работать «периодически» или «с перерывами» или «непрерывно» в отношении разгрузки твердых веществ. Например, в качестве стадии непрерывного центрифугирования можно использовать трубчатую центрифугу.
Очистка рекомбинантного белка
Рекомбинантный белок может быть выделен непосредственно из культуральной среды. После выделения рекомбинантного белка следует очистка, чтобы обеспечить надлежащую чистоту рекомбинантного белка. При очистке можно использовать одну или более хроматографических стадий, например одну или более хроматографических смол; и/или одну или более стадий фильтрования. Например, для очистки рекомбинантного белка можно использовать аффинную хроматографию с использованием смол, таких как белок А или L. Альтернативно или дополнительно, для очистки рекомбинантного белка можно использовать ионообменную смолу, такую как катионообменная смола.
ПРИМЕРЫ
Для экспрессии доменного антитела (dAb) в штамме-хозяине Е. coli W3110 исследовали целый ряд нативных эндогенных N-концевых сигнальных последовательностей (SS) Е. coli. Две последовательности с многообещающими характеристиками - YceISS и IvySS - были отобраны для биореакторных исследований при сравнении с используемой в настоящее время сигнальной последовательностью OmpASS. Для исследованных биореакторных условий использование OmpASS обеспечивало высокий титр продукта за короткое время ферментации, но с высоковязким бульоном, который, как было предсказано исследованиями в ультра-уменьшенном масштабе, представляет собой сложную проблему при осветлении посредством полномасшабного центрифугирования. Использование YceISS приводило к эффективности, сопоставимой с эффективностью для OmpASS в показателях титра, вязкости и сложности центрифугирования, тогда как использование IvySS приводило к маловязкому бульону, который было относительно легко центрифугировать, и к высокому титру продукта, но достигаемому за более длительное время ферментации. Оптимизация кодонов IvySS приводила к аналогично маловязкому бульону, подходящему для центрифугирования, и к высокому титру, но теперь за более короткое время ферментации, сравнимое со временем контроля с использованием OmpASS. Улучшения, достигаемые при использовании IvySS или его оптимизированных версий, по-видимому, связаны с пониженной базовой экспрессией dAb, т.е. до индукции. Считается, что эта пониженная экспрессия обусловлена присутствием кассеты из восьми кодонов с N-конца сигнальной последовательности, которая при удалении приводила к высоковязким бульонам, аналогичным бульону, полученному с использованием OmpASS. Исследуется связь между базовой экспрессией и вязкостью бульона.
Материалы и методы
Молекулярное клонирование кассет секреции. Штаммы Е. coli One Shot™ ТОР10 (Life Technologies, Калифорния, США) и W3110 (АТСС 27325) использовали в качестве штаммов-хозяев для клонирования и экспрессии рекомбинантного белка соответственно. Олигонуклеотиды (Таблица 2), разработанные для получения кассет секреции dAb с разными расположенными выше сигнальными последовательностями, конструировали согласно спецификации (Invitrogen, Paisley, Великобритания). Использованными реагентами были рестрикционные ферменты (Ndel и Xhol), ТА ДНК-лигаза (New England Biolabs UK Ltd., Hertfordshire, Великобритания) и ДНК-полимераза (мастер-микс Phusion™ для высокоточной флэш-ПЦР (полимеразная цепная реакция), Finnzymes Оу, Эспоо, Финляндия). Экспрессионный вектор pAVEway™ - pAVE011 (ниже обозначенный как p11), содержащий ген резистентности к тетрациклину - tetA, и промотор Т7А3, индуцируемый изопропил-β-D-1-тиогалактопиранозидом (IPTG) (WO2007088371; Fujifilm Diosynth Biotechnologies UK Ltd., Биллингем, Великобритания), использовали в качестве каркаса для кассет секреции dAb.
Таблица 2. Олигонуклеотиды, использованные для конструирования конструкций гена dAb с разными сигнальными последовательностями. Рестрикционные сайты Ndel (CATATG) и XhoI (CTCGAG) подчеркнуты.

Выбранные семь эндогенных сигнальных последовательностей Е. coli (Таблица 3) генетически сливали выше гена, кодирующего домен Vн антитела против TNFR1 (рецептор 1 фактора некроза опухолей) с молекулярной массой примерно 13,1 кДа (ниже обозначенного как dAb, SEQ ID NO: 53). Вплоть до четырех последовательных ПЦР-реакций использовали для удлинения вышерасположенного конца dAb с помощью ДНК для желательной сигнальной последовательности. Для этого требовались 3 или 4 прямых олигонуклеотидов (один для каждой последовательной ПЦР-реакции), обратный олигонуклеотид SEQ ID NO: 32 (см. Таблицу 2) и экспрессионная плазмида pi 1 в качестве исходной матрицы (pDAB01, см. Таблицу 3). ПЦР-реакционные смеси содержали подходящий прямой (fwd) олигонуклеотид и обратный (rev) олигонуклеотид SEQ ID NO: 32 (Таблица 2), ДНК-матрицу и мастер-микс для высокоточной флэш-ПЦР Phusion™. Осуществляли термические циклы со следующими условиями: 98°С в течение 10 с, затем 30 циклов при 98°С в течение 1 с, 55°С в течение 5 с и 72°С в течение 15 с, и конечная инкубация при 72°С в течение 1 мин. После каждого цикла ПЦР продукт очищали с использованием колоночной хроматографии (набор для очистки ПЦР QIAquick™, QIAGEN Ltd., Манчестер, Великобритания) и использовали в качестве ДНК-матрицы для следующей последовательной ПЦР-реакции, пока не была получена кассета слияния сигнальная последовательность-dAb, фланкированная рестрикционными сайтами NdeI и XhoI. Кассеты секреции отщепляли посредством NdeI и XhoI и лигировали в экспрессионный вектор p11, расщепленный NdeI-XhoI, с использованием ДНК-лигазы Т4, затем трансформировали в клетки Е. coli One Shot™ ТОР10 посредством теплового шока и подвергали селекции на чашках с vLB агаром и тетрациклином. Плазмиды pDAB02 - pDAB08 (Таблица 3) подтверждали путем ДНК-секвенирования, затем трансформировали в клетки Е. coli W3110 для исследований экспрессии белка. Клеточные банки (глицериновые концентраты, 20% об./об.) каждой полученной конструкции хранили при -80°С.
Таблица 3. Экспрессионные плазмиды и сигнальные последовательности Е. coli, использованные для исследований экспрессии dAb.
a Название сигнальной последовательности и [число кодонов]; b Аминокислотная последовательность сигнальных последовательностей с С-концевым сайтом расщепления на конце последовательности. с Индекс адаптации кодонов (CAI) для каждой сигнальной последовательности анализировали, используя пакет EMBOSS, относительно использования кодонов в генах класса II (высокоэкспрессируемые) Е. coli (Eecol_high.cut).

* Аминокислотная последовательность такая же как для Ivyss. Последовательность кодона в положении 23 была изменена с gcc на gct.
Таблица 4. Экспрессионные плазмиды, содержащие модифицированные сигнальные последовательности Ivy Е. coli для исследования влияния индекса адаптации кодонов (CAI) на скорость экспрессии модельного dAb и последующих эффектов на вязкость бульона во время ферментации с подпиткой клеток в высокой плотности. a Название сигнальной последовательности и [число кодонов]. b Индекс использования кодонов для каждой сигнальной последовательности был проанализирован с использованием пакета EMBOSS относительно использования кодонов у генов класса II (высокоэкспрессируемых) Е. coli (Eecoli_high.cut).

Таблица 5. Влияние N-концевой сигнальной пептидной последовательности на уровень базовой экспрессии dAb, вязкость бульона и исходную продуктивность во время ферментации Е. coli с высокой плотностью клеток. a Исходная продуктивность представляет собой общее количество dAb, экспрессированного за первые 24 ч после индукции (Фиг. 2Д и 4Д). b N-концевая кассета содержит кодоны для первых восьми аминокислот из IvySS.

Кассеты секреции для dAb-экспресионных плазмид pDAB021 и pDAB022 (Таблица 4), содержащие кодон-оптимизированные сигнальные последовательности Ivy - IvyOpt1SS и IvyOpt2SS - получали посредством ПЦР-амплификации гена, кодирующего dAb, из pDAB01 (как описано выше) с использованием fwd олигонуклеотидов 23-25 и 26-28, соответственно, и обратного олигонуклеотида SEQ ID NO: 32 (Таблица 2). Кассету секреции для pDAB023 (Таблица 4), содержащую укороченную сигнальную последовательность Ivy (IvyTruncSS) выше dAb, получали посредством ПЦР-амплификации с использованием pDAB02 в качестве матрицы (Таблица 3) и fwd олигонуклеотида 29 и rev олигонуклеотида 31. Кассета секреции для pDAB011 кодировала dAb с расположенной выше сигнальной последовательностью Ivy'OmpASS, состоящей из первых 8 кодонов IvySS, слитых с OmpASS. Ее получали посредством амплификации гена, кодирующего OmpASS-dAb, из pDAB01 (Таблица 3) с использованием fwd олигонуклеотида 30 и rev олигонуклеотида SEQ ID NO: 32, Все кассеты секреции затем лигировали в вектор p11 в NdeI/XhoI и трансформировали в штамм Е. coli W3110.
Индекс адаптации кодонов (CAI) (Sharp and Li 1987) для каждой сигнальной последовательности определяли с использованием пакета EMBOSS (Rice et al. 2000) относительно использования кодонов только у генов класса II (высокоэкспрессируемых) (Eecoli_high.cut). IvySS была кодон-оптимизирована для экспрессии в Е. coli с использованием алгоритма адаптации кодонов (программа Leto, версия 1.0.26, Entelechon GmbH, Регенсбург, Германия). Порог для кодонов (Leto) был установлен на уровне 70% относительно всех генов Е. coli K12, а также только для класса II генов Е. coli K12 (высокоэкспрессируемых) с получением IvyOpt1SS и IvyOpt2SS соответственно (Таблица 4).
Ферментация. Все среды и реактивы были приобретены у Sigma-Aldrich (Дорсет, Великобритания), если не определено иное. Инокуляцию и культивирование в планшете с 24 глубокими лунками (24 dwp) проводили в среде Луриа Бертрани (vLB) (рН 7,0): 10 г. л-1 Difco™ Select Soytone (Becton Dickinson and Co. (BD), Нью-Джерси, США), 5 г. л-1 дрожжевого экстракта Bacto™ (BD) и 5 г. л-1 NaCl. 10-кратно концентрированный глицериновый стимулятор (рН 7,0) для культур в 24 dwp содержал: 70 г. л-1 глицерина, 50 г. л-1 дрожжевого экстракта (BD), 25 г. л-1 (NH4)2SO4 и 1,0 мкг.мл-1 тиамина. Ферментацию при высокой плотности клеток проводили в комплексной среде, как описано в других источниках (Voulgaris et al. 2015). Во все ферментационные среды (исключащие подпитку ТВ/SB) добавляли 15 мкг.мл-1 тетрациклина (Tet) для селективности клона Е. coli, содержащего экспрессионную плазмиду p11.
Для экспрессии в планшете с глубокими лунками культуры инокулята в среде vLB (20 мл) были приготовлены в 125 мл встряхиваемых колбах Эрленмейера с перегородками с вентилируемыми крышками (Corning Life Sciences, Амстердам, Нидерланды) с 0,5% об./об. Е. coli W3110, трансформированного экспрессионной плазмидой pDAB. Культуры инкубировали в течение ночи при 23°С, 230 об./мин на орбитальном шейкере с 25 мм смещением (Climo-shaker ISF1-X, Kühner Shaker Ltd., Дербишир, Великобритания) до OD600 нм 1-2. Их использовали для инокулирования свежей среды vLB до OD600 нм 0.05, распределяли в лунки квадратной формы с пирамидальным дном (рабочий объем 2 мл) планшетов для микротитрования с 24 глубокими лунками (24 dwp; Porvair Sciences Ltd., Врексхем, Великобритания) и покрывали газопроницаемой системой крышки для высокой плотности клеток (Applikon Biotechnology, Tewkesbury, Великобритания). Планшеты инкубировали при 37°С, 230 об./мин до OD600 нм 0,7-1,0, и затем индуцировали образование dAb при помощи 0,1 мМ IPTG с одновременным добавлением 10× глицеринового стимулятора (состав см. выше). Культуры инкубировали в течение еще 24 ч при 23°С, 230 об./мин. Собранные культуры (2 мл) осаждали центрифугированием (16100 x g в течение 10 мин) и бульонный супернатант экстрагировали. Осадки ресуспендировали в 1 мл 0,1 М PBS (фосфатно-солевой буферный раствор) и снова центрифугировали, как описано выше. Бульонный супернатант и осадки хранили при -20°С.
Для экспрессии в биореакторе культуры инокулята в среде vLB (200 мл) были приготовлены с 0,5% об./об. глицериновыми маточными растворами в 500 мл встряхиваемых колбах Эрленмейера с перегородками с вентилируемыми крышками (Corning Life Sciences). Культуры инкубировали при 37°С, 230 об./мин, на орбитальном шейкере в течение 4-5 ч до OD600 нм 1-2 и использовали в концентрации 0,05% об./об. для инокулирования реактора с рабочим объемом 1 л (биореактор SR1000DLL, диаметр сосуда 100 мм, соотношение измерений 2,4:1, с верхнеприводными тройными 6-лопастными мешалками Раштона диаметром 46 мм, DASGIP AG, Юлих, Германия), содержащего комплексную среду, указанную выше. Данный реактор поддерживали при: 30°С, рН 7,0 плюс/минус 0,05 (с использованием 25% об./об. H3PO4 и 30% об./об. NH4OH); уровне растворенного кислорода (DO) 30 плюс/минус 5% (посредством каскадного контроля с использованием: (1) скорости лопастной мешалки (400-1200 об./мин); (2) скорости потока газа (1-2 об./об./мин) и (3) содержания кислорода (21-100%)). Подпиточную среду автоматически добавляли со скоростью 6,0 мл.л-1⋅ч-1, начиная через 10 мин после резкого скачка DO, указывающего на полное израсходование глицерина. Продуцирование dAb индуцировали добавлением 0,2 мМ IPTG, когда культура достигала OD600нм 75 плюс/минус 3 (что соответствует биомассе примерно 25 г dcw (сухая масса клеток).л-1), и скорость подпитки снижали до 3,6 мл.л-1⋅ч-1 на остальное время ферментации. Записывали значения рН в реальном времени, уровень растворенного кислорода, скорость встряхивания, температуру, скорость потока воздуха, процентное содержание кислорода, скорость поглощения кислорода, скорость выделения диоксида углерода (DASGIP Control, Юлих, Германия). Реологические характеристики свежего клеточного бульона измеряли (программируемый вискозиметр DV-II+, оснащенный либо ротором ULA, 1,233 Н, либо SSA, 1,32 Н, Brookfield Engineering Laboratories Inc., Массачусетс, США) во время прогонок с возрастающим и снижающимся напряжением сдвига (от 1,3 до 110 с-1 за пять приращений с выдерживанием 30 с при каждом приращении) при 25°С. Реологию используют в качестве показателя обрабатываемости бульона, например посредством центрифугирования.
Обработка клеточного бульона. Чтобы подвернуть образцы клеточного бульона воздействию максимальной интенсивности рассеивания энергии (ε) 0,53×106 Вт.кг-1 в течение 20 с (Chatel et al. 2014а), использовали устройство для создания усилия сдвига в ультрамалом масштабе (20 мл камера из нержавеющей стали, диаметр 50 мм × высота 10 мм, оснащенная вращающимся диском из нержавеющей стали, диаметр 40 мм × 0,1 мм толщина 12000 об./мин, сконструированная в механической мастерской UCL (Университетский колледж Лондона)). Такое значение 8 типично имитирует то, что происходит в подпиточной зоне центрифуги промышленного масштаба с высоким напряжением сдвига и непрерывным током. Затем образцы центрифугировали (2 мл бульона, подвергнутого или неподвергнутого напряжению сдвига) (3400xg в течение 7 мин при 21°С, центрифуга 5415R, Эппендорф, Кембридж, Великобритания), и отделяли супернатанты (0,8 мл). Их характеризовали в показателях оставшихся твердых веществ путем измерения OD600нм с поправкой на исходное значения OD600нм для хорошо осветленного образца (16100xg в течение 30 мин). Условия центрифугирования были охарактеризованы в показателях группы V/tΣ, где V представляет собой центрифугированный объем (2,0 мл), t представляет собой время центрифугирования (420 с), и Σ представляет собой эквивалентную площадь осаждения центрифужной пробирки для конкретной скорости вращения и геометрии ротора центрифуги (составляет 0,164 м2 -детали расчета см. в Chatel et al. 2014).
Аналитические способы. Клеточные осадки оттаивали и ресуспендировали в 1 мл (образцы из 24 dwp) или 4 мл (образцы из 1 л биореактора) 50 мМ Tris-HCl, рН 7,5. 1 мл каждого ресуспендированного осадка лизировали (ультразвуковой измельчитель Soniprep 150 Plus, оснащенный экспоненциальным микрозондом, №партии 38121-114А, MSE (Великобритания) Ltd., Лондон, Великобритания) с использованием двух импульсов 30 с для включения и 10 с для выключения при амплитуде 8,0. Лизированный цельноклеточный образец затем центрифугировали при 16100xg в течение 20 мин, и отделяли супернатант.Образцы ферментационного бульона и цельноклеточные лизаты затем фильтровали с использованием 0,22 мкм фильтровальной пластинки, центрифугируя при 3000 g в течение 30 мин (мембрана GHP, фильтровальная пластинка AcroPrep™ 96, Pall Life Sciences, Портсмут, Великобритания). Экспрессию dAb из 24 dwp культур Е. coli анализировали посредством SDS-PAGE. Образцы восстанавливали, денатурировали и загружали на гель SDS-PAGE (NuPAGE® 4-12% Bis-Tris, Invitrogen) с выполнением гель-электрофореза в подвижном буфере 1× MES (морфолиноэтансульфоновая кислота) (Invitrogen) согласно инструкциям изготовителя. Белок обнаруживали посредством окрашивания кумасси, и продукт количественно измеряли с использованием денситометрической программы (Image Lab, версия 3.0, Bio-Rad Laboratories). Для ферментации Е. coli при высокой плотности клеток титр продукта dAb в супернатанте бульона и в цельноклеточном лизате определяли посредством ВЭЖХ (высокоэффективная жидкостная хроматография) на белке A (HP1100 с охлаждаемым автосэмплером, Agilent Technologies UK Limited, Чешир, Великобритания), оснащенном 1 мл колонкой HiTrap MabSelect Xtra (GE Healthcare Life Science, Бёкингемшир, Великобритания). Колонку уравновешивали и промывали 0,1 М PBS, рН 7,3, а для элюирования продукта (концентрацию белка отслеживали при 280 нм) использовали 20 мМ HCl, рН 1,8. Концентрацию dAb определяли с использованием стандартной кривой. Молекулярную массу очищенного dAb определяли, используя жидкостную хроматографию-времяпролетную масс-спектрометрию) (способ GSK (ГлаксоСмитКлайн) собственной разработки с использованием Agilent HP1100, Agilent Technologies UK Ltd., соединенного с масс-спектрометром Micromass LCT, Waters, Массачусетс, США, оснащенным зондом электрораспылительной ионизации (ESI), и контролировали с использованием программы MassLynx, версия 4.1, Waters). Уровень двухцепочечной ДНК измеряли с использованием флюорометрии (набор для анализа Qubit™ dsDNA BR, Life Technologies) согласно инструкциям изготовителя.
Пример 1
На Фиг. 1 показаны конечные уровни биомассы и гели SDS-PAGE исходного скрининга N-концевых сигнальных последовательностей, подробно показывающие экспрессию dAb в показателях внеклеточной и внутриклеточной локализации.
Влияние разных N-концевых сигнальных последовательностей на экспрессию dAb из Е. coli W3110 в 24 dwp ферментациях показано на Фиг. 1. Обозначения: В - dAb, локализованное внеклеточно (супернатант); С - dAb, локализованное внутриклеточно (цельноклеточный лизат: периплазматическое плюс цитозольное). В + С - общее количество dAb, В/(В + С) - высвобожденная доля. Сложение В и С обеспечивает визуальную оценку общего количества образовавшегося dAb. Образцы отбирали через 24 часа после индукции, восстанавливали и полосы визуализировали посредством SDS-PAGE с окрашиванием кумасси. «Std» обозначает стандарт dAb с концентрацией 0,5 мг⋅мл-1.
Использование сигнальной последовательности OmpA (OmpASS), IvySS и YcelSS приводит к высоким уровням экспрессии dAb на единицу объема бульона (бульон это клетки плюс супернатант, т.е. внеклеточная среда), тогда как использование сигнальных последовательностей YgiWSS, YncESS, FimASS, YehZSS и YcdOSS обеспечивает умеренные уровни экспрессии. OmpASS, IvySS, YceISS и FimASS приводят к высоким уровням внутриклеточного продукта на единицу OD600нм, тогда как YgiWSS, YncESS, YehZSS и YcdOSS приводят к умеренным уровням. Имеется некоторая положительная корреляция между внутриклеточным уровнем продукта на единицу OD600нм и высвобождением продукта в супернатант для OmpASS, YceISS, YgiWSS, FimASS и YehZSS. Использование YceISS, YncESS и YcdOSS, по-видимому, приводит к высоким уровням высвобождения, и для YceISS, это плюс получаемый высокий титр, делает ее одним кандидатом для дальнейшего исследования. IvySS демонстрирует относительно низкое высвобождение в супернатант и высокую OD600 нм что может означать дополнительный потенциал для образования dAb. Это плюс образующийся высокий уровень общего продукта и относительно низкое пропорциональное высвобождение dAb в бульон идентифицирует ее как второго кандидата для дальнейшего исследования. Эти два штамма переносят для более подробного исследования роста в биореакторах по сравнению с использованием OmpASS в качестве контроля.
Пример 2
Влияние использования OmpASS, YceISS и IvySS на ферментацию с подпиткой Е. coli W3110 показано на Фиг. 2. Ферментацию отслеживали в послеиндукционной фазе по OD600нм в качестве показателя роста клеток, высвобождению дцДНК в качестве показателя лизиса, внеклеточному и внутриклеточному dAb в качестве показателя продуктивности и локализации продукта, реологии в качестве показателя обратываемости бульона, например посредством центрифугирования.
На Фиг. 2 показаны профили послеиндукционной ферментации для культур с подпиткой Е. coli W3110 с высокой плотностью клеток. N-концевая сигнальная последовательность и базовый (доиндукционный) уровень экспрессии: (Δ), OmpASS, 0,30 г. л-1, () YceISS, 0,64 г. л-1 или (◊), IvySS, 0,061 г. л-1. Все результаты получены после индукции образования dAb с использованием IPTG. (А) Биомассу оценивали по OD600нм; (Б) концентрация внеклеточной растворимой дцДНК; выход dAb определяли (В) внутриклеточно (цельноклеточный лизат), (Г) внеклеточно (супернатант), в виде (Д) общего количества образовавшегося dAb (внутриклеточный плюс внеклеточный) и в виде (Е) процента от общего количества dAb во внеклеточном пространстве. Вязкость бульона μ описывают в показателях степенной зависимости, μ=K γn-1, где γ представляет собой скорость сдвига, (Ж) K представляет собой показатель консистенции потока и (З) n представляет собой показатель поведения потока. Результаты были получены от ферментеров в повторности (n равен 2-4), усреднены, и были добавлены планки погрешностей, соответствующие одному стандартному отклонению.

Штамм с использованием OmpASS (Δ) демонстрировал типичную фазу быстрого роста, с последующим автолизом клеток, о чем свидетельствует одновременное уменьшение OD600нм (Фиг. 2А) и увеличение уровня внеклеточной дцДНК (Фиг. 2Б). Это совпадало с быстрым образованием dAb с высвобождением во внеклеточную среду/супернатант (Фиг. 2В и Г). Общее количество образовавшегося dAb обобщено на Фиг. 2Д, причем почти все находятся внеклеточно к концу ферментации (Фиг. 2Е). В момент индукции значительное количество (0,30 г. л-1) dAb уже экспрессировалось в клетках (Фиг. 2В и легенда). Это сопровождалось образованием высоковязкого, подвергающегося сдвиговому разжижению бульона к началу фазы индукции (Фиг. 2Ж и З). Вязкость бульона продолжала возрастать в течение 24 ч во время послеиндукционной фазы (Фиг. 2Ж) с параллельным увеличением уровней внеклеточной дцДНК.
Тенденции, аналогичные использованию культуры OmpASS, наблюдали для использования YceISS (). Здесь уровень экспрессии продукта в момент индукции (0,64 г. л-1) выше, чем для штаммов, использующих OmpASS. Это сопровождается большей вязкостью бульона в момент индукции и, затем, пониженной скоростью роста и немного более низкими уровнями продукта либо в виде общего, либо в виде внеклеточного dAb.
В отличие от этого, использование IvySS (◊) приводило к низким уровням экспрессии dAb 0,061 г. л-1 во время индукции и к маловязкому бульону. За этим следовало увеличение вязкости на протяжении всего развития ферментации, параллельно с повышенным высвобождением дцДНК (Фиг. 2Б). Рост клеток продолжается в течение более длительного периода перед началом лизиса примерно в 72 ч, т.е. по сравнению с примерно 12 ч, наблюдаемыми для использования OmpASS и YceISS. Полученным эффектом является меньшая скорость экспрессии dAb и отсроченное высвобождение dAb во внеклеточное пространство (Фиг. 2Г и Е). В целом представляется, что использование IvySS по сравнению с OmpASS и YceISS предлагает более жесткий контроль образования dAb до индукции.
Пример 3
В результате выравнивание аминокислотных последовательностей OmpASS, YceISS и IvySS (Фиг. 3А) и оценки их индекса адаптации кодонов (CAI) (Фиг. 3Б и Таблица 3) идентифицировали кассету из восьми аминокислот (Ivy'SS) на N-конце IvySS, которая значительно отличалась от OmpASS и YceISS с точки зрения аминокислотной последовательности, но также содержала большую долю редких кодонов (Таблица 3).
На Фиг. 3 показаны выравнивания аминокислотных последовательностей сигнальных последовательностей и оценка их индексов адаптации кодонов. (А) Выравнивания аминокислотных последовательностей YceISS (SEQ ID NO: 35) и IvySS (SEQ ID NO: 34) приведены в сравнении с OmpASS (SEQ ID NO: 33). Консервативные аминокислоты подчеркнуты. (Б) Адаптивность каждого положения кодона рассчитывали как отношение частоты данного кодона относительно наиболее многочисленного синонимичного кодона со ссылкой на таблицу использования кодонов Е. coli, полученную из базы данных использования кодонов Е. coli Kazusa. Аминокислотные последовательности в «Б» выравниваются от первого метионина после кассеты Ivy'SS.
Предполагалось, что присутствие данной восьмикодоновой кассеты или образующихся в результате аминокислот IvySS может влиять на уровень и скорость экспрессии dAb до индукции (т.е. на базовый уровень). Для исследования влияния идентифицированной кассеты Ivy'SS в облегчении образования dAb во время ферментации с подпиткой при высокой плотности клеток, были разработаны две модифицированные сигнальные последовательности: IvyTruncSS - сигнальная последовательность, основанная на последовательности IvySS с удаленным Ivy'SS; и также Ivy'OmpASS, которая представляет собой продукт слияния кодонов Ivy'SS с N-концом OmpASS. Кроме того, были созданы две кодон-оптимизированные сигнальные последовательности IvySS -IvyOpt1SS и IvyOpt2SS (см. подробности на Фиг. 3Б и в разделе Материалы и методы) -для исследования, можно ли увеличить скорость экспрессии dAb без изменения базового уровня экспрессии dAb и вязкости бульона. Полученные в результате значения CAI для четырех модифицированных сигнальных последовательностей, родственных Ivy, приведены в Таблице 4.
Пример 4
Рост культур Е. coli W3110, экспрессирующих dAb с модифицированной сигнальной последовательностью, описан на Фиг. 4. Все модифицированные сигнальные последовательности, представленные на Фиг. 4, эффективно отщеплялись, приводя к правильно свернутой молекуле dAb.
На Фиг. 4 показаны профили послеиндукционной ферментации для культур с подпиткой Е. coli W3110 с высокой плотностью клеток. N-концевая сигнальная последовательность и базовый (до индукции) уровень экспрессии: (Δ), IvyTruncSS, 0,90 г. л-1(), Ivy'OmpASS, 0,086 г. л-1, (◊), IvyOpt1SS, 0,16 г. л-1 и ( ), IvyOpt2SS, 0,089 г. л-1. Подробности см. на Фиг. 2. Результаты были получены от образцов одиночных ферментаций (m равно 3).
), IvyOpt2SS, 0,089 г. л-1. Подробности см. на Фиг. 2. Результаты были получены от образцов одиночных ферментаций (m равно 3).

Использование IvyTruncSS приводило к более раннему прекращению роста, большему высвобождению дцДНК и меньшему образованию dAb, чем наблюдалось для использования IvySS (Фиг. 2). Уровень базовой экспрессии dAb во время индукции выше, чем для использования IvySS, и вязкость бульона непосредственно до и после индукции значительно выше. Следовательно, присутствие Ivy'SS, по-видимому, является критически важным для уменьшения ранней экспрессии до индукции и, следовательно, результирующих высоких уровней вязкости (Фиг. 4Ж). Повышенная наблюдаемая скорость экспрессии (Фиг. 4Д) согласуется с удалением последовательности, содержащей большую долю редких кодонов. Использование Ivy'OmpASS приводило к росту, высвобождению дцДНК, образованию и локализации dAb, и профилям вязкости, аналогичным наблюдаемым при использовании IvySS (Фиг. 2), подтверждая важность Ivy'SS в уменьшении базовой экспрессии dAb. Опять, низкая вязкость наблюдается для Ivy'OmpASS в начале фазы индукции, и последующее увеличение вязкости объясняют присутствием внеклеточной дцДНК. Аналогично, как и в случае с IvySS, увеличенное время ферментации было необходимым для получения значительных уровней экспрессии dAb и высвобождения во внеклеточную среду. Использование IvyOpt1SS и IvyOpt2SS приводило к низкой базовой экспрессии dAb в момент индукции (обозначения на Фиг. 4). Результатом является маловязкий бульон (Фиг. 4Ж). Как и ожидалось, сниженная частота редких кодонов в оптимизированных сигнальных последовательностях приводит к повышенной скорости образования dAb (Фиг. 4Д). Опять, как и при использовании IvySS, увеличение вязкости бульона после индукции объясняют высвобождением дцДНК. IvyOpt2SS, по-видимому, дает меньшую базовую экспрессию, более высокий титр dAb и менее вязкий бульон, чем IvyOpt1SS.
Пример 5
Продуктивность в ферментере, измеренная для общего титра после индукции, сравнивается на Фиг. 5А для ряда исследованных сигнальных последовательностей. Низкие значения dAb для штаммов, использующих IvySS и Ivy'OmpASS, очевидно обусловлены увеличенным временем (примерно 150 ч), необходимым для образования и высвобождения продукта. Все штаммы, использующие IvyOpt1SS, IvyOpt2SS и IvyTruncSS, обеспечивают максимальную концентрацию dAb в пределах 24 ч после индукции и высокие уровни продуктивности в ферментере. Масс-спектрометрический анализ (здесь не показан) свидетельствовал о том, что сигнальная последовательность была полностью удалена из dAb во всех исследованных конструкций, показанных на Фиг. 5. Осветление конечных бульонов с использованием USD центрифугирования обобщено на Фиг. 5Б. Использованные условия являются типичными для центрифуг промышленного масштаба с непрерывным потоком с тарельчатым сепаратором или сплошным ротором, работающих на нижней границе этой производительности по скорости потока и оснащенных зоной подпитки с высоким напряжением сдвига. Подпитка без усилия сдвига, как для настольной центрифуги, имитируется в качестве сравнительного контроля.
На Фиг. 5 (А) показана общая продуктивность, рассчитанная как масса внеклеточного dAb на литр бульона на общее время после индукции; и на (Б) показана эффективность центрифугирования собранных культур Е. coli, экспрессирующих dAb с разными N-концевыми сигнальными последовательностями: OmpASS (48 ч после индукции), YceISS (48 ч), IvyTruncSS (55 ч), Ivy'OmpA (153 ч) и кодон-оптимизированными сигнальными последовательностями Ivy - IvyOpt1 (48 ч) и IvyOpt2 (55 ч). Клеточные бульоны подвергали воздействию следующих условий: () нет усилия сдвига; () высокое напряжение сдвига (максимальная интенсивность рассеивания энергии (ε) 0,53×106 Вт.кг-1 за 20 с) и затем обработка посредством USD центрифугирования (V равен 2,0 мл, t равно 420 с, Σ равна 0,164 м2, (см. текст), V/tΣ=2,9×10-8 м.с-1). Результаты были получены от ферментеров в повторности (n равно 3) для OmpASS и YceISS или одиночных образцов из ферментера для остальных сигнальных последовательностей. Характеристика образца для (Б) была произведена в тройной повторности (m равно 3). Результаты показаны как среднее значение плюс/минус SD.
Высокие вязкости бульонов, полученных с использованием OmpASS и YceISS, приводили к плохо осветленному супернатанту с примерно 50% и примерно 80% твердых веществ, все еще остающихся в супернатанте соответственно. Все остальные бульоны осветляют до более чем в 10 раз большей степени (примерно 1-4% остающихся твердых веществ), так что различия в эффективности являются менее очевидными. Бульон средней вязкости, полученный при использовании IvyTruncSS (Фиг. 4Ж), привел к осветленному бульону на верхней границе данного интервала (примерно 4%). Все остальные бульоны осветляют в большей степени, отражая даже меньшие вязкости конечных бульонов. Почти во всех случаях влияние высокого напряжения сдвига приводило к 5-30%-ному улучшению осветления, указывая на вероятное уменьшение вязкости, например из-за распада жидкостных структур (таких как полимерные мостики). Несколько штаммов приводили как к высокой продуктивности в ферментере, так и к хорошему центрифужному осветлению. Штаммы с использованием IvyOpt2SS и затем IvyOpt1SS обеспечивают самые многообещающие варианты на основе данного исследования. Использование IvyTruncSS также хорошо работает, но указано, что осветление центрифугированием является более затруднительным.
Краткие выводы из примеров
Были идентифицированы две сигнальные последовательности - IvySS и YceISS, которые обеспечивают секрецию гетерологичного белка в периплазматическое пространство Е. coli. Было показано то, что штамм с использованием YceISS работает в 1 л ферментере с подпиткой с емкостью с перемешиванием сравнимым образом с использованием OmpASS (Фиг. 2). Использование IvySS приводило к значительно меньшим вязкостям бульона по сравнению с вязкостями, полученными с использованием OmpASS и YceISS, хотя и с меньшими скоростями продукции (Фиг. 2 и Таблица 3) и меньшими общими продуктивностями (Фиг. 5А). Авторы изобретения, во-первых, обсуждают причину образования высоковязких бульонов с использованными здесь средами и стратегией ферментации и, во-вторых, разработку сигнальных последовательностей на основе IvySS.
Исходные исследования с использованием OmpASS, YceISS и IvySS свидетельствуют о том, что высокая базовая (т.е. доиндукционная) экспрессия dAb приводит к образованию высоковязкого ферментационного бульона в начале индукции (Фиг. 2). Такие высокие вязкости сохранялись на протяжении всей остальной части ферментации с любым дополнительным увеличением, вероятно объясняемым высвобождением ДНК при повышенном автолизе клеток. Эта связь между базовой экспрессией и вязкостью поддерживается наблюдениями с использованием ряда других сигнальных последовательностей (Фиг. 4) и обобщена в Таблице 5. Предварительные исследования, не показанные здесь, продемонстрировали высоковязкий бульон (K примерно 1,1 Нсnм-2) для IvySS при проведении индукции во время, а не после окончания, экспоненциальной фазы роста, имитируя высокую базовую экспрессию.
Высокие вязкости, наблюдаемые ко времени индукции, вероятно могут быть объяснены образованием внеклеточных полимерных веществ (EPS) во время ферментации, которые являются главным компонентом микробных биопленок. Было показано то, что биосинтез EPS контролируют такие факторы, как фаза роста, состав среды, температура, ограничение количества кислорода, недостаточность азота и катионов. Что касается других микробных систем, сам экспоненциальный рост был связан с образованием EPS и биопленки. С образованием EPS также был связан ацетат. Было показано то, что образование биопленки и EPS в общем случае приводит к уменьшению скорости роста. Наблюдения в данном исследовании свидетельствуют о возможности того, что экспрессия dAb в сочетании с эффектом условий среды до индукции может стимулировать образование EPS.
Для исследованных здесь ферментаций стратегии контроля в отношении DO являются такими, что ограничение кислорода (DO примерно 0%), вызывающее условия гипоксии, имеет место в фазе экспоненциального роста до индукции. И гипоксия, и экспоненциальный рост связаны с образованием побочных продуктов ферментативного метаболизма и сливных продуктов метаболизма соответственно, таких как ацетат. Такие условия, в сочетании с присутствием высоких базовых уровней dAb, возможно приводят к образованию EPS. После индукции условия являются таковыми, что любой образующийся ацетат вероятно будет израсходован до начала подпитки питательными веществами. С этого момента контролируемая постоянная скорость подпитки питательными веществами будет означать, что клетки находятся в линейной или стационарной фазе роста, и контроль DO эффективно поддерживается на уровне 30%; т.е. маловероятны какие-либо значимые уровни присутствующего образования ацетата и, следовательно, отсутствует дальнейшее образование EPS, несмотря на высокие уровни dAb. Следовательно, контроль условий ферментации до индукции можно использовать, чтобы избежать образования ацетата. Альтернативно, корректировка сред может уменьшить образование биопленки, например меньшее присутствие сои или использование глюкозы, а не глицерина (как в данной работе) в качестве источника углерода. В данном исследовании ферментации проводили с использованием среды, содержащей сойтон; недостатки высоковязкого бульона при использовании OmpACC для получения того же самого dAb были преодолены путем использования бессоевых ферментационных сред. Базовая экспрессия (и, следовательно, для исследованных здесь систем - образование EPS) может быть снижена посредством использования более строго контролируемой системы экспрессии, но это может создать другие технологические проблемы. Другим возможным путем ликвидации образования EPS является использование штамма Е. coli, дефицитного по образованию биопленки. Целью будущей работы будет подтверждение влияния EPS, приводящего к высоким вязкостям, наблюдаемым в данном исследовании. Внимание здесь сосредоточено на том, как можно улучшить выбранные сигнальные последовательности для достижения маловязкой ферментации с использованием существующих сред и рабочих условий ферментации.
Два аспекта структуры IvySS, по-видимому, относятся к его эффективности в определении базовой экспрессии и послеиндукционной продуктивности dAb. Во-первых, присутствие восьмикодоновой кассеты (Ivy'SS) в начале IvySS приводит к строгому контролю экспрессии белка перед индукцией, т.е. к низкой базовой экспрессии. Во-вторых, низкий индекс адаптации кодонов (CAI), по-видимому, приводит к низкой скорости экспрессии белка. Влияние выбора сигнальной последовательности, включая модифицированные сигнальные последовательности Ivy, для сохранения низкой базовой экспрессии при увеличении продуктивности обобщен в Таблице 5. Когда Ivy'SS присутствовал на N-конце сигнальной последовательности, уровень базовой экспрессии до индукции экспрессии dAb был низким, и был получен бульон низкой вязкости. Низкие базовые уровни, достигаемые с IvyOpt1SS и IvyOpt2SS (Фиг. 4), свидетельствуют о том, что именно аминокислотная последовательность, а не присутствие редких кодонов в последовательности гена, определяет уровень базовой экспрессии.
Исходная продуктивность возможно связана с CAI, который, в свою очередь, связан со скоростью элонгации трансляции и образования dAb. Предполагали, что присутствие редких кодонов на 5'-конце мРНК приводит к медленной элонгации образующегося полипептида во время трансляции, к уменьшению заторов рибосомального транспорта, что приводит к более низкой исходной продуктивности клеток. Согласно Таблице 5, низкое значение отношения CAI к длине последовательности приводит к низкой исходной продуктивности, например, как видно для IvySS и Ivy'OmpSS, тогда как для всех других сигнальных последовательностей наблюдается высокое значение CAI на число кодонов, приводящее к высокой исходной продуктивности. В идеале данный анализ должен быть взвешенным в отношении N-терминального конца конструкции. Это особенно необходимо, когда для двух штаммов присутствуют редкие кодоны, дающие низкую исходную продуктивность, как и для штаммов с использованием IvySS и Ivy'OmpSS. Увеличение исходной продуктивности dAb для IvyOpt1SS и IvyOpt2SS по сравнению с продуктивностью, полученной с использованием IvySS, может быть обусловлено либо модифицированной последовательностью кодонов, либо удалением редких кодонов.
IvyOpt2SS сконструирована посредством оптимизации на основе эталонного набора оптимальных кодонов, часто представленных в генах, которые сильно и непрерывно экспрессируются во время экспоненциального роста, тогда как IvyOpt1SS сконструирована на основе экспрессии всех генов Е. coli. Первая приводит к высокому CAI на число кодонов (Таблица 5), и это может привести к отмеченной большей общей продуктивности (Фиг. 5А). В будущей работе будет дополнительно исследован потенциал такой оптимизации для дополнительного улучшения общего уровня экспрессии при одновременном предотвращении значительной базовой экспрессии, приводящей к высоковязким бульонам.
Журнальные ссылки
Rice Р, Longden I, Bleasby А. 2000. EMBOSS: The European Molecular Biology Open Software Suite. Trend Genet 16(6): 276-277.
Voulgaris I, Finka G, Uden M, Hoare M. 2015. Enhancing the selective extracellular location of a recombinant E.coli domain antibody by management of fermentation conditions. Appl Microbiol Biotechnol 99(20): 8441-8453.
Chatel A, Kumpalume P, Hoare M. 2014a. Ultra scale-down characterization of the impact of conditioning methods for harvested cell broths on clarification by continuous centrifugation-Recovery of domain antibodies from rec E. coli. Biotechnol Bioeng 111(5): 913-924.
Sharp PM, Li W-H. 1987. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res 15(3): 1281-1295.
Другие последовательности
SEQ ID NO: 53: аминокислотная последовательность DOM0101 EVQLLESGGGLVQPGGSLRLSCAASGFTFAHETMVWVRQAPGKGLEWVSHIPPDGQDP FYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYHCALLPKRGPWFDYWGQGTLV TVSS
SEQ ID NO: 54: последовательность ДНК DOM0101 - (нет сигнальной последовательности)
GAAGTACAACTGCTGGAGAGCGGTGGCGGCCTGGTTCAACCGGGTGGTTCCCTGCG
CCTGTCCTGTGCGGCATCTGGTTTCACCTTCGCACACGAAACCATGGTGTGGGTTCG
CCAAGCTCCGGGCAAAGGCCTGGAATGGGTAAGCCACATTCCTCCAGATGGCCAGG
ACCCATTCTATGCGGATTCCGTTAAGGGTCGCTTTACCATTTCTCGTGATAACTCCAA
AAACACCCTGTACCTGCAGATGAACTCCCTGCGCGCCGAGGATACTGCGGTGTACC
ATTGTGCGCTGCTGCCTAAACGTGGCCCGTGGTTCGATTACTGGGGTCAGGGTACTC
TGGTCACCGTAAGCAGC
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GlaxoSmithKline Intellectual Property Development Limited
<120> Method of Producing a Recombinant Protein
<130> PB66104
<150> US62/332636
<151> 2016-05-06
<150> US62/338045
<151> 2016-05-18
<160> 55
<170> PatentIn version 3.5
<210> 1
<211> 8
<212> PRT
<213> Escherichia coli
<400> 1
Met Gly Arg Ile Ser Ser Gly Gly
1 5
<210> 2
<211> 49
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 2
ctctggtcat cgctaccagt gcaatggcgg aagtacaact gctggagag 49
<210> 3
<211> 51
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 3
tgatgtttaa ggcaataacg acagtcgccg ctctggtcat cgctaccagt g 51
<210> 4
<211> 51
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 4
gaccatatgg gcaggataag ctcgggagga atgatgttta aggcaataac g 51
<210> 5
<211> 44
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 5
ttctctgccg gttcagcggt tgccgaagta caactgctgg agag 44
<210> 6
<211> 41
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 6
ggtttaacct tcgcgtccct gatgttctct gccggttcag c 41
<210> 7
<211> 45
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 7
gaccatatga aaaaaagcct gcttggttta accttcgcgt ccctg 45
<210> 8
<211> 42
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 8
gtgcagcgca ccggtgatgg cagaagtaca actgctggag ag 42
<210> 9
<211> 42
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 9
cagtaatcgc agtaatggcc ctgtgcagcg caccggtgat gg 42
<210> 10
<211> 47
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 10
gaccatatga aaaaattcgc agcagtaatc gcagtaatgg ccctgtg 47
<210> 11
<211> 44
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 11
gtttcatcat tcagtacgca ggccgaagta caactgctgg agag 44
<210> 12
<211> 47
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 12
ttactgttag gttcattgct tgttgtttca tcattcagta cgcaggc 47
<210> 13
<211> 49
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 13
ttttcatcgc gcctgcgtgg ttcattactg ttaggttcat tgcttgttg 49
<210> 14
<211> 44
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 14
gaccatatgc atttacgtca tctgttttca tcgcgcctgc gtgg 44
<210> 15
<211> 45
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 15
cctcagttct acagcggctc tggccgaagt acaactgctg gagag 45
<210> 16
<211> 48
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 16
caatcgttgt tctgtcggct ctgtccctca gttctacagc ggctctgg 48
<210> 17
<211> 48
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 17
gaccatatga aaattaaaac tctggcaatc gttgttctgt cggctctg 48
<210> 18
<211> 48
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 18
ggcagccgtg agcctgccgc tacaagcaga agtacaactg ctggagag 48
<210> 19
<211> 50
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 19
ctgggcaggt tcactggtta tgttggcagc cgtgagcctg ccgctacaag 50
<210> 20
<211> 45
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 20
gaccatatgc cactcttaaa gctctgggca ggttcactgg ttatg 45
<210> 21
<211> 48
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 21
gttttcttct gcttttatgg ctaacgccga agtacaactg ctggagag 48
<210> 22
<211> 53
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 22
acgcattgca gttgagcgtg gctgcgctgt tttcttctgc ttttatggct aac 53
<210> 23
<211> 48
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 23
gaccatatga ccattaactt ccgccgtaac gcattgcagt tgagcgtg 48
<210> 24
<211> 51
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 24
gctctggtta ttgctactag cgctatggct gaagtacaac tgctggagag c 51
<210> 25
<211> 52
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 25
atgatgttca aagcaatcac cacagtggca gctctggtta ttgctactag cg 52
<210> 26
<211> 55
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 26
gaccatatgg gacgtatttc ttcgggcgga atgatgttca aagcaatcac cacag 55
<210> 27
<211> 52
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 27
gctctggtta ttgcgacctc tgctatggca gaagtacaac tgctggagag cg 52
<210> 28
<211> 50
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 28
atgatgttca aagcaatcac taccgttgca gctctggtta ttgcgacctc 50
<210> 29
<211> 53
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 29
gaccatatgg gtcgtatctc ttccggcggt atgatgttca aagcaatcac tac 53
<210> 30
<211> 42
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 30
gaccatatga tgtttaaggc aataacgaca gtcgccgctc tg 42
<210> 31
<211> 50
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 31
gaccatatgg gcaggataag ctcgggagga atgaagaaaa ctgctatcgc 50
<210> 32
<211> 36
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 32
cttactcgag tcattagctg cttacggtga ccagag 36
<210> 33
<211> 21
<212> PRT
<213> Escherichia coli
<400> 33
Met Lys Lys Thr Ala Ile Ala Ile Ala Val Ala Leu Ala Gly Phe Ala
1 5 10 15
Thr Val Ala Gln Ala
20
<210> 34
<211> 28
<212> PRT
<213> Escherichia coli
<400> 34
Met Gly Arg Ile Ser Ser Gly Gly Met Met Phe Lys Ala Ile Thr Thr
1 5 10 15
Val Ala Ala Leu Val Ile Ala Thr Ser Ala Met Ala
20 25
<210> 35
<211> 22
<212> PRT
<213> Escherichia coli
<400> 35
Met Lys Lys Ser Leu Leu Gly Leu Thr Phe Ala Ser Leu Met Phe Ser
1 5 10 15
Ala Gly Ser Ala Val Ala
20
<210> 36
<211> 20
<212> PRT
<213> Escherichia coli
<400> 36
Met Lys Lys Phe Ala Ala Val Ile Ala Val Met Ala Leu Cys Ser Ala
1 5 10 15
Pro Val Met Ala
20
<210> 37
<211> 30
<212> PRT
<213> Escherichia coli
<400> 37
Met His Leu Arg His Leu Phe Ser Ser Arg Leu Arg Gly Ser Leu Leu
1 5 10 15
Leu Gly Ser Leu Leu Val Val Ser Ser Phe Ser Thr Gln Ala
20 25 30
<210> 38
<211> 23
<212> PRT
<213> Escherichia coli
<400> 38
Met Lys Ile Lys Thr Leu Ala Ile Val Val Leu Ser Ala Leu Ser Leu
1 5 10 15
Ser Ser Thr Ala Ala Leu Ala
20
<210> 39
<211> 23
<212> PRT
<213> Escherichia coli
<400> 39
Met Pro Leu Leu Lys Leu Trp Ala Gly Ser Leu Val Met Leu Ala Ala
1 5 10 15
Val Ser Leu Pro Leu Gln Ala
20
<210> 40
<211> 26
<212> PRT
<213> Escherichia coli
<400> 40
Met Thr Ile Asn Phe Arg Arg Asn Ala Leu Gln Leu Ser Val Ala Ala
1 5 10 15
Leu Phe Ser Ser Ala Phe Met Ala Asn Ala
20 25
<210> 41
<211> 63
<212> DNA
<213> Escherichia coli
<400> 41
atgaagaaaa ctgctatcgc tattgcggtt gctctggcag gttttgccac ggttgcgcag 60
gcc 63
<210> 42
<211> 84
<212> DNA
<213> Escherichia coli
<400> 42
atgggcagga taagctcggg aggaatgatg tttaaggcaa taacgacagt cgccgctctg 60
gtcatcgcta ccagtgcaat ggcg 84
<210> 43
<211> 66
<212> DNA
<213> Escherichia coli
<400> 43
atgaaaaaaa gcctgcttgg tttaaccttc gcgtccctga tgttctctgc cggttcagcg 60
gttgcc 66
<210> 44
<211> 60
<212> DNA
<213> Escherichia coli
<400> 44
atgaaaaaat tcgcagcagt aatcgcagta atggccctgt gcagcgcacc ggtgatggca 60
<210> 45
<211> 90
<212> DNA
<213> Escherichia coli
<400> 45
atgcatttac gtcatctgtt ttcatcgcgc ctgcgtggtt cattactgtt aggttcattg 60
cttgttgttt catcattcag tacgcaggcc 90
<210> 46
<211> 69
<212> DNA
<213> Escherichia coli
<400> 46
atgaaaatta aaactctggc aatcgttgtt ctgtcggctc tgtccctcag ttctacagcg 60
gctctggcc 69
<210> 47
<211> 69
<212> DNA
<213> Escherichia coli
<400> 47
atgccactct taaagctctg ggcaggttca ctggttatgt tggcagccgt gagcctgccg 60
ctacaggcg 69
<210> 48
<211> 78
<212> DNA
<213> Escherichia coli
<400> 48
atgaccatta acttccgccg taacgcattg cagttgagcg tggctgcgct gttttcttct 60
gcttttatgg ctaacgcc 78
<210> 49
<211> 84
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 49
atgggacgta tttcttcggg cggaatgatg ttcaaagcaa tcaccacagt ggcagctctg 60
gttattgcta ctagcgctat ggct 84
<210> 50
<211> 84
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 50
atgggtcgta tctcttccgg cggtatgatg ttcaaagcaa tcactaccgt tgcagctctg 60
gttattgcga cctctgctat ggca 84
<210> 51
<211> 60
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 51
atgatgttta aggcaataac gacagtcgcc gctctggtca tcgctaccag tgcaatggcg 60
<210> 52
<211> 87
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 52
atgggcagga taagctcggg aggaatgaag aaaactgcta tcgctattgc ggttgctctg 60
gcaggttttg ccacggttgc gcaggcc 87
<210> 53
<211> 119
<212> PRT
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 53
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ala His Glu
20 25 30
Thr Met Val Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser His Ile Pro Pro Asp Gly Gln Asp Pro Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr His Cys
85 90 95
Ala Leu Leu Pro Lys Arg Gly Pro Trp Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 54
<211> 357
<212> DNA
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 54
gaagtacaac tgctggagag cggtggcggc ctggttcaac cgggtggttc cctgcgcctg 60
tcctgtgcgg catctggttt caccttcgca cacgaaacca tggtgtgggt tcgccaagct 120
ccgggcaaag gcctggaatg ggtaagccac attcctccag atggccagga cccattctat 180
gcggattccg ttaagggtcg ctttaccatt tctcgtgata actccaaaaa caccctgtac 240
ctgcagatga actccctgcg cgccgaggat actgcggtgt accattgtgc gctgctgcct 300
aaacgtggcc cgtggttcga ttactggggt cagggtactc tggtcaccgt aagcagc 357
<210> 55
<211> 29
<212> PRT
<213> Artificial
<220>
<223> Синтетическая последовательность
<400> 55
Met Gly Arg Ile Ser Ser Gly Gly Met Lys Lys Thr Ala Ile Ala Ile
1 5 10 15
Ala Val Ala Leu Ala Gly Phe Ala Thr Val Ala Gln Ala
20 25
<---
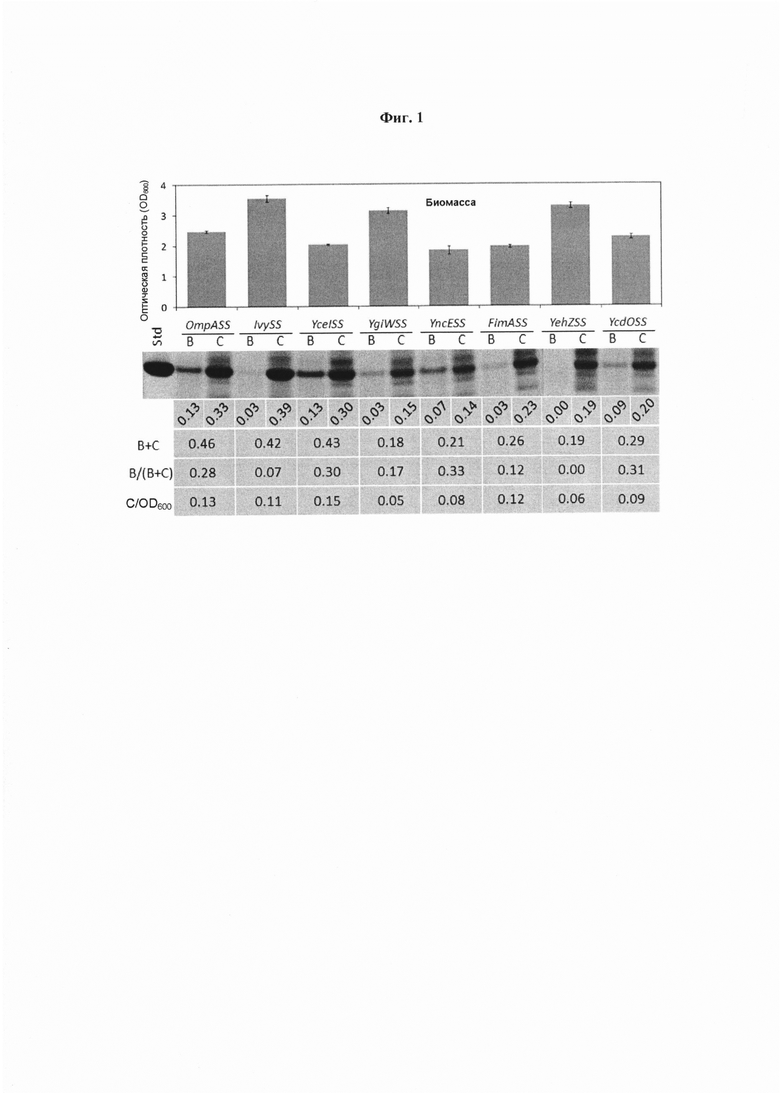
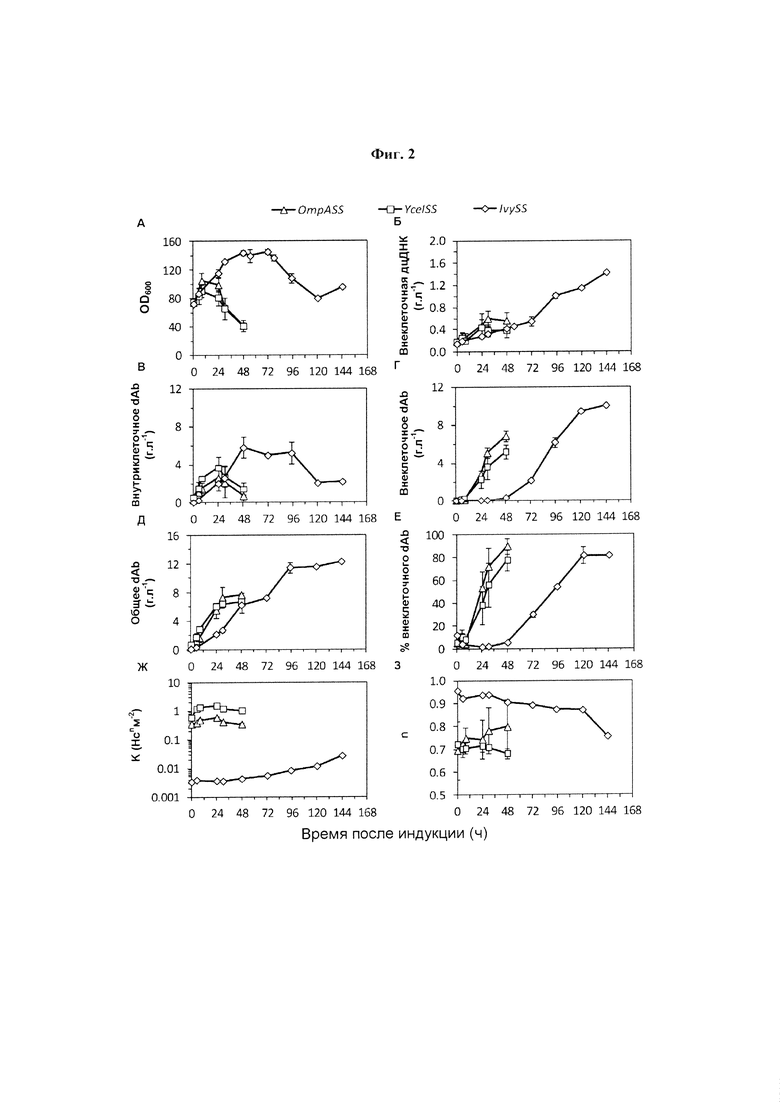
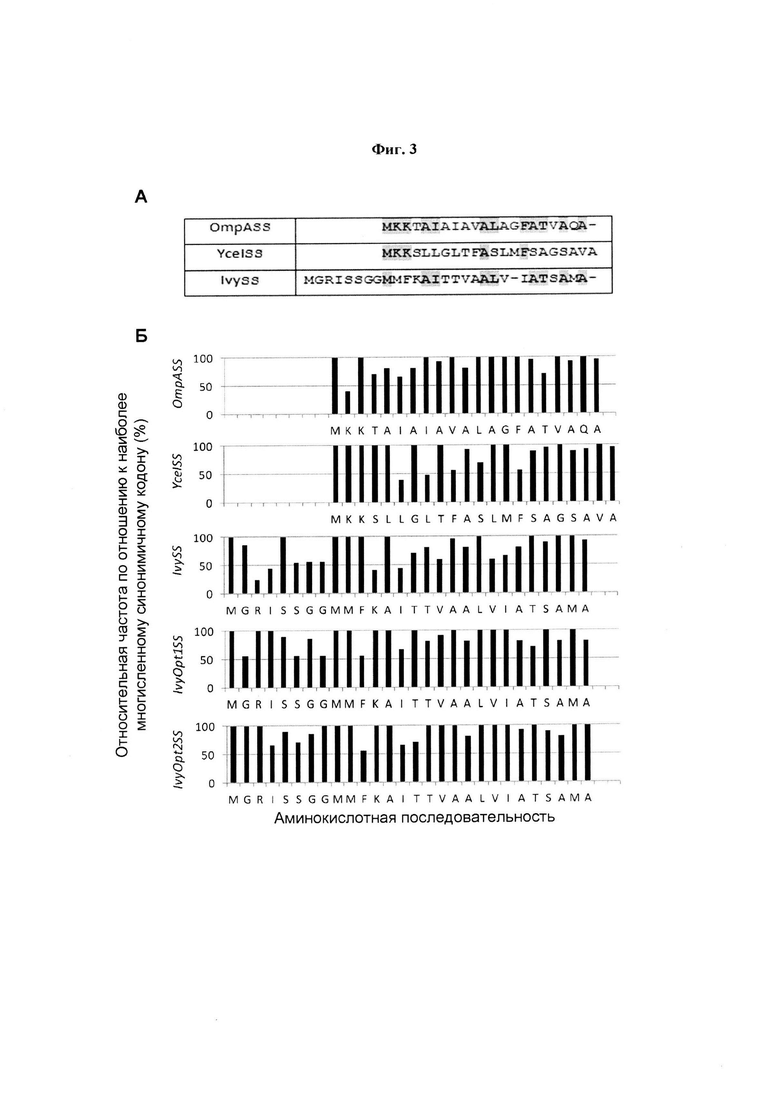
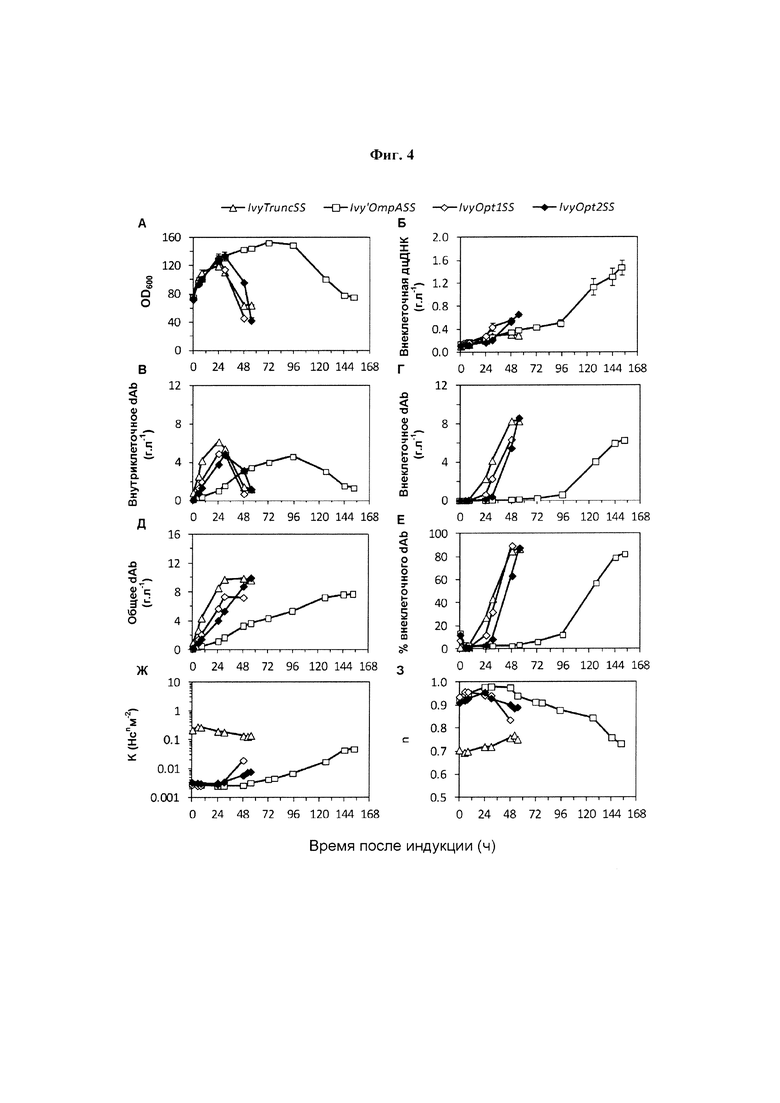
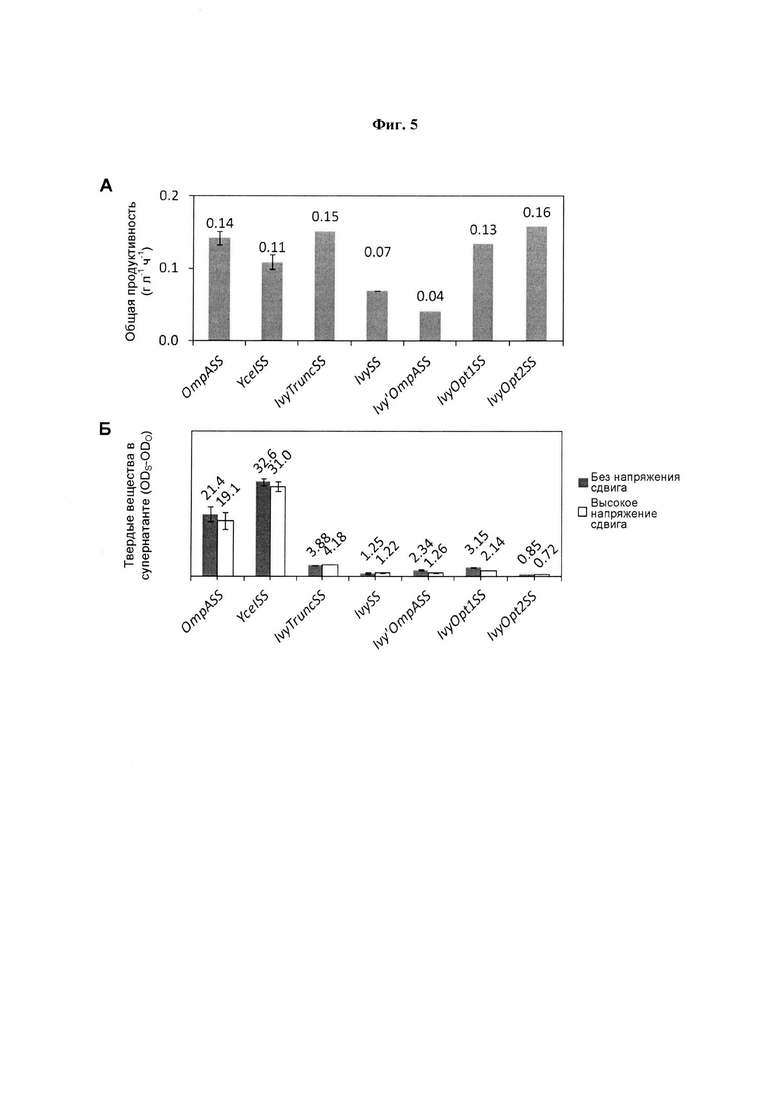
Настоящее изобретение относится к области биотехнологии, конкретно к рекомбинантным сигнальным последовательностям, происходящим из Е. coli, и может быть использовано для рекомбинантной продукции белков в Е. coli. Изобретение также относится к слитому белку, содержащему данную сигнальную последовательность, к рекомбинантному белку и к способам продуцирования данного рекомбинантного белка. Изобретение обеспечивает снижение вязкости бульона клеток-хозяев E. coli, где экспрессируется рекомбинантный антигенсвязывающий белок, а также снижение базовой экспрессии рекомбинантного антигенсвязывающего белка, экспрессируемого бульоном клеток-хозяев E. coli, при культивировании клетки-хозяина в доиндукционных условиях. 9 н. и 10 з.п. ф-лы, 5 ил., 7 табл., 5 пр.
1. Рекомбинантная сигнальная последовательность для периплазматичской секреции рекомбинантного белка, состоящая из (а) последовательности MGRISSGG (SEQ ID NO: 1) или ее варианта, последовательность которого отличается 1 аминокислотной заменой, делецией или вставкой, и (б) гетерологичной сигнальной последовательности в направлении С-конца относительно (а), где указанная гетерологичная сигнальная последовательность представляет собой периплазматическую сигнальную последовательность из E. coli.
2. Рекомбинантный слитый белок, вызывающий биологический или медицинский ответ ткани, системы, животного или человека, содержащий (а) рекомбинантную сигнальную последовательность по п. 1, которая функциональным образом связана с (б) гетерологичным рекомбинантным белком в направлении С-конца относительно (а), где указанный гетерологичный рекомбинантный белок представляет собой антигенсвязывающий белок.
3. Нуклеиновая кислота, которая кодирует рекомбинантную сигнальную последовательность по п. 1.
4. Нуклеиновая кислота по п. 3, где нуклеиновая кислота кодон-оптимизирована для экспрессии в клетке-хозяине.
5. Нуклеиновая кислота по п. 4, где кодон-оптимизированная нуклеиновая кислота кодирует сигнальную последовательность, содержащую последовательность MGRISSGGMKKTAIAIAVALAGFATVAQA (SEQ ID NO: 55).
6. Нуклеиновая кислота по любому из пп. 3-5, где нуклеиновая кислота была кодон-оптимизирована для экспрессии в клетке-хозяине и имеет индекс адаптации кодонов (CAI) 0,40 или выше.
7. Нуклеиновая кислота, которая кодирует рекомбинантный слитый белок по п. 2.
8. Нуклеиновая кислота по п. 7, где нуклеиновая кислота была кодон-оптимизирована для экспрессии в клетке-хозяине и имеет индекс адаптации кодонов (CAI) 0,40 или выше.
9. Рекомбинантный экспрессионный вектор, содержащий нуклеиновую кислоту по любому из пп. 3-6.
10. Рекомбинантный экспрессионный вектор, содержащий нуклеиновую кислоту по любому из пп. 7, 8.
11. Способ продуцирования рекомбинантного антигенсвязывающего белка в клетке-хозяине E. coli, включающий:
(а) культивирование клетки-хозяина в условиях, которые обеспечивают экспрессию рекомбинантного белка с рекомбинантного экспрессионного вектора по п. 10, присутствующего в клетке-хозяине; и
(б) выделение рекомбинантного белка.
12. Способ по п. 11, где рекомбинантный белок выделяют из периплазмы или внеклеточной среды.
13. Способ по любому из пп. 11, 12, где уровень доиндукционной экспрессии рекомбинантного белка составляет 0,5 г/л или менее; и/или уровень послеиндукционной экспрессии рекомбинантного белка составляет 5 г/л или более.
14. Способ по любому из пп. 11-13, где сигнальная последовательность отщепляется от рекомбинантного белка.
15. Применение рекомбинантной сигнальной последовательности по п. 1 для снижения вязкости бульона клеток-хозяев E. coli, где экспрессируется рекомбинантный антигенсвязывающий белок.
16. Применение рекомбинантной сигнальной последовательности по п. 1 для снижения базовой экспрессии рекомбинантного антигенсвязывающего белка, экспрессируемого бульоном клеток-хозяев E. coli, при культивировании клетки-хозяина в доиндукционных условиях.
17. Применение по любому из пп. 15 или 16, где рекомбинантный белок выделяют из периплазмы или внеклеточной среды.
18. Применение по любому из пп. 15-17, где уровень доиндукционной экспрессии рекомбинантного белка составляет 0,5 г/л или менее; и/или уровень послеиндукционной экспрессии рекомбинантного белка составляет 5 г/л или более.
19. Применение по любому из пп. 15-18, где сигнальная последовательность отщепляется от рекомбинантного белка.
| US 2014244228 А1, 28.08.2014 | |||
| WO 2009021548 A1, 19.02.2009 | |||
| US 2009011995 A1, 08.01.2009 | |||
| YIZHOU ZHOU ET AL., Enhancing full-length antibody production by signal peptide engineering, MICROBIAL CELL FACTORIES, 2016, v.15, n.1, p.1-11 | |||
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИАНГИОГЕННОГО ПОЛИПЕПТИДА | 2012 |
|
RU2488635C1 |
| ORLANDO M., Modification of proteins and low molecular weight substances with | |||

