ПЕРЕКРЕСТНЫЕ ССЫЛКИ
[0001] По настоящей заявке испрашивается приоритет предварительной патентной заявки США № 62/445570, зарегистрированной 12 января 2017 года; предварительной патентной заявки США № 62/445557, зарегистрированной 12 января 2017 года; предварительной патентной заявки США № 62/447889, зарегистрированной 18 января 2017 года; предварительной патентной заявки США № 62/467032, зарегистрированной 3 марта 2017 года; предварительной патентной заявки США № 62/566199, зарегистрированной 29 сентября 2017 года и предварительной патентной заявки США № 62/577147, зарегистрированной 25 октября 2017 года, которые включены в настоящий документ в качестве ссылки.
ИНФОРМАЦИЯ ОТНОСИТЕЛЬНО СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит список последовательностей, который подан в электронном виде в формате ASCII и, таким образом, полностью включен в качестве ссылки. Указанная копия ASCII, созданная 3 января 2018 года, названа 47736-707_601_SL.txt, и ее размер составляет ≈599 кбайт.
ЗАЯВЛЕНИЕ О ФИНАНСИРУЕМОМ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА ИССЛЕДОВАНИИ
[0003] Настоящее изобретение осуществлено с поддержкой правительства Соединенных Штатов Америки в виде грана SBIR 1520545, выделенного национальным научным фондом (National Science Foundation). Правительство США обладает определенными правами на описываемый объект изобретения.
УРОВЕНЬ ТЕХНИКИ, ПРЕДШЕСТВУЮЩИЙ ИЗОБРЕТЕНИЮ
[0004] Растения связаны с микробиомом общим метаболомом. Многомерная зависимость между конкретным признаком сельскохозяйственной культуры и лежащим в его основе метаболоме характеризуется рельефом с множеством локальных максимумов. Оптимизация от меньшего локального максимума к другому, представляющему улучшенный признак, посредством изменения влияния микробиома на метаболом может являться желательным по целому ряду причин, например, для оптимизации сельскохозяйственных культур. Для удовлетворения потребностей растущего мирового населения необходимы экономически, экологически и социально устойчивые подходы к сельскому хозяйству и производству пищевых продуктов. Продовольственная и сельскохозяйственная организация ООН прогнозирует, что для удовлетворения потребностей растущего населения к 2050 году общее производство пищевых продуктов должно увеличиться на 70%, задача, которая осложняется многочисленными факторами, включая уменьшение ресурсов пресной воды, увеличивающуюся конкуренцию за пахотные земли, растущие цены на энергоносители, растущие затраты на производство и подобные требования для адаптации культур к давлению более засушливого, жаркого и более экстремального глобального климата.
[0005] Одной из представляющих интерес областей является улучшение фиксации азота. Газообразный азот (N2) является основным компонентом атмосферы Земли. Кроме того, элементарный азот (N) является важным компонентом множества химических соединений, составляющих живые организмы. Однако многие организмы не могут непосредственно использовать N2 для синтеза химических соединений, используемых в физиологических процессах, таких как рост и воспроизводство. Для использования N2 необходимо, чтобы N2 был комбинирован с водородом. Комбинирование водорода с N2 обозначают как фиксация азота. Фиксация азота, проводимая химически или биологически, требует затрат большого количества энергии. В биологических системах реакцию, которая приводит к фиксации азота, катализирует фермент, известный как нитрогеназа. Важной целью исследований фиксации азота является расширение этого фенотипа на небобовые растения, конкретно на важные агротехнические травы, такие как пшеница, рис и кукуруза. Несмотря на большой прогресс в понимании развития азотфиксирующего симбиоза между клубеньковыми бактериями и бобовыми, путь для использования этого знания для индукции азотфиксирующих клубеньков у небобовых культур все еще не ясен. Между тем, проблема обеспечения достаточных дополнительных источников азота, такого как в удобрениях, будет продолжать расти вместе с растущей необходимостью в увеличении производства пищевых продуктов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0006] По одному из аспектов изобретения предоставлена бактериальная композиция, содержащая по меньшей мере один генно-инженерный бактериальный штамм, который фиксирует атмосферный азот в сельскохозяйственной системе, в которую в качестве удобрения внесено более 9,1 кг азота на акр.
[0007] По другому аспекту изобретения предоставлена бактериальная композиция, которая содержит по меньшей мере один бактериальный штамм, выведенный для фиксации атмосферного азота в сельскохозяйственной системе, в которую в качестве удобрения внесено более 9,1 кг азота на акр.
[0008] По дополнительному аспекту изобретения предоставлена бактериальная композиция, которая содержит по меньшей мере один генно-инженерный бактериальный штамм, который фиксирует атмосферный азот, где этот по меньшей мере один генно-инженерный бактериальный штамм содержит экзогенно добавленную ДНК, где указанная экзогенно добавленная ДНК по меньшей мере на 80% идентична с соответствующим нативным бактериальным штаммом.
[0009] По дополнительному аспекту изобретения предоставлен семенной состав, содержащий семена растения, инокулированного бактериальной композицией.
[0010] Другой аспекту изобретения предоставлен способ выращивания сельскохозяйственной культуры с использованием множества семян с семенным составом, инокулированным бактериальной композицией.
[0011] По дополнительному аспекту изобретения предоставлен способ нанесения бактериальной композиции на поле.
[0012] По дополнительному аспекту изобретения предоставлена композиция удобрения, содержащая бактериальную композицию.
[0013] По другому аспекту изобретения предоставлен способ поддержания уровней азота в почве. Способ включает высаживание в почву поля сельскохозяйственной культуры, инокулированной генно-инженерными бактериями, которые фиксируют атмосферный азот. Способ также включает сбор урожая указанной сельскохозяйственной культуры, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 90% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
[0014] По дополнительному аспекту изобретения предоставлен способ доставки пробиотической добавки в сельскохозяйственную культуру. Способ включает покрытие семян сельскохозяйственной культуры покрытием для семян, пропиткой для семян или протравкой для семян. Указанные покрытие для семян, протравка для семян или пропитка для семян содержат живых представителей указанного пробиотика. Кроме того, способ включает внесение в почву поля указанных семян сельскохозяйственной культуры.
[0015] В дополнительном аспекте изобретения генно-инженерный бактериальный штамм представляет собой генно-инженерный штамм грамположительной бактерии. В определенных случаях генно-инженерный штамм грамположительной бактерии обладает измененным уровнем экспрессии регулятора кластера Nif. В определенных случаях генно-инженерный штамм грамположительной бактерии экспрессирует сниженное количество отрицательного регулятора кластера Nif. В определенных случаях генно-инженерный бактериальный штамм экспрессирует сниженное количество GlnR. В определенных случаях в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 75% идентичный с последовательностью из базы данных NifH лаборатории Дж. Зера (Zehr lab). В определенных случаях в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 85% идентичный с последовательностью из базы данных NifH лаборатории Дж. Зера. В определенных случаях в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 75% идентичный с последовательностью из базы данных NifH лаборатории Д. Бакли (Buckley lab). В определенных случаях в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 85% идентичный с последовательностью из базы данных NifH лаборатории Д. Бакли.
[0016] По другому аспекту изобретения предоставлен способ увеличения фиксации азота у небобового растения. Способ включает нанесение на растение множества не являющихся межродовыми бактерий, где указанное множество включает не являющихся межродовыми бактерий, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час. Кроме того, множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 1% или более фиксированного азота. Кроме того, каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
[0017] По дополнительному аспекту изобретения предоставлен способ увеличения фиксации азота у небобового растения. Способ включает нанесение на растение множества не являющихся межродовыми бактерий, где указанное множество включает не являющихся межродовыми бактерий, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и/или (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час. Кроме того, множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 1% или более фиксированного азота. Кроме того, каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
[0018] По дополнительному аспекту изобретения предоставлен способ выведения штаммов микроорганизмов для улучшения конкретных признаков агротехнического значения. Способ включает предоставление множества штаммов микроорганизмов, обладающих способностью к колонизации требуемой сельскохозяйственной культуры. Способ также включает улучшение регуляторных сетей, влияющих на признак, посредством внутригеномной перестановки. Кроме того, способ включает оценку штаммов микроорганизмов во множестве штаммов микроорганизмов для определения меры признака. Кроме того, способ включает отбор из множества штаммов микроорганизмов одного или нескольких штаммов микроорганизмов, которые обеспечивают улучшение признака в случае требуемой сельскохозяйственной культуры.
[0019] По другому аспекту изобретения предоставлен способ выведения штаммов микроорганизмов с улучшением конкретных признаков агротехнического значения. Способ включает предоставление множества штаммов микроорганизмов, обладающих способностью к колонизации требуемой сельскохозяйственной культуры. Способ также включает внесение генетического разнообразия во множество штаммов микроорганизмов. Кроме того, способ включает оценку штаммов микроорганизмов во множестве штаммов микроорганизмов для определения меры признака. Кроме того, способ включает отбор из множества штаммов микроорганизмов одного или нескольких штаммов микроорганизмов, которые обеспечивают улучшение признака в случае требуемой сельскохозяйственной культуры.
[0020] По другому аспекту изобретения предоставлен способ увеличения у небобового растения количества получаемого из атмосферы азота. Способ включает воздействие на указанное небобовое растение подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, или по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
[0021] По дополнительному аспекту изобретения предоставлен способ увеличения у растения кукурузы количества получаемого из атмосферы азота. Способ включает воздействие на указанное растение кукурузы подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, несущими генно-инженерные генетические вариации по меньшей мере в двух генах, выбранных из группы, состоящей из nifL, glnB, glnE и amtB.
[0022] По другому аспекту изобретения предоставлен способ увеличения у растения кукурузы количества получаемого из атмосферы азота. Способ включает воздействие на указанное растение кукурузы подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, несущими по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы, in planta, продуцируют у указанного растения кукурузы по меньшей мере 5% фиксированного азота, как измеряют посредством разведения 15N в сельскохозяйственных культурах, выращиваемых в полях, обрабатываемых удобрением, содержащим 1,2% 15N.
[0023] По дополнительному аспекту изобретения предоставлен способ увеличения фиксации азота у небобового растения. Способ включает нанесение на растение множества не являющихся межродовыми бактерий, где указанное множество включает не являющихся межродовыми бактерий, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час. Кроме того, произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,5 × 10-8 ммоль N на грамм сырой массы ткани корня растения в час. Кроме того, множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 1% или более фиксированного азота. Кроме того, каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
[0024] По другому аспекту изобретения предоставлен способ увеличения фиксации азота у небобового растения. Способ включает нанесение на растение множества бактерий, где указанное множество включает бактерии, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и/или (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час. Кроме того, множество бактерий, in planta, продуцирует в растении 1% или более фиксированного азота.
[0025] По дополнительному аспекту изобретения предоставлен не являющаяся межродовой популяция бактерий, способная к увеличению фиксации азота у небобового растения, содержащая множество не являющихся межродовыми бактерий, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и/или (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час. Кроме того, множество не являющихся межродовыми бактерий, in planta, продуцирует в растении, выращиваемого в присутствии множества не являющихся межродовыми бактерий, 1% или более фиксированного азота. Кроме того, каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
[0026] По дополнительному аспекту изобретения предоставлена популяция бактерий, способная к увеличению фиксации азота у небобового растения, где популяция бактерий содержит множество бактерий, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час. Кроме того, множество бактерий, in planta, продуцирует в растении 1% или более фиксированного азота.
[0027] В другом аспекте изобретения предоставлены бактерии, которые (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и/или (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
[0028] В дополнительном аспекте изобретения предоставлены не являющиеся межродовыми бактерии, которые несут по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии способны к фиксации атмосферного азота в присутствии экзогенного азота, и где указанные бактерии (i) обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и/или (ii) продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
[0029] В дополнительном аспекте изобретения предоставлен способ увеличения фиксации азота в растении, включающий введение в растение эффективного количества композиции, которая содержит очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 97% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269, 277-283; очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276, 284-295; и/или очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260, 296-303; и где растение, которому вводят эффективное количество композиции, демонстрирует увеличение фиксации азота по сравнению с растением без введения композиции.
[0030] По дополнительному аспекту изобретения предоставлены выделенные бактерии, содержащие последовательность нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 97% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269, 277-283; последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276, 284-295; и/или последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260, 296-303.
[0031] По другому аспекту изобретения предоставлен способ детекции последовательности с неприродным соединением, включающий: амплификацию нуклеотидной последовательности, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности с SEQ ID NO: 372-405.
[0032] По дополнительному аспекту изобретения предоставлен способ детекции последовательности с неприродным соединением, включающий: амплификацию нуклеотидной последовательности, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности фрагмента по меньшей мере с 10 последовательными парами оснований, содержащегося в SEQ ID NO: 372-405, где указанный фрагмент с последовательными парами оснований состоит из нуклеотидов на пересечении: расположенной выше последовательности, содержащей SEQ ID NO: 304-337, и расположенной ниже последовательности, содержащей SEQ ID NO: 338-371.
[0033] По дополнительному аспекту изобретения предоставлена последовательность с неприродным соединением, содержащая нуклеотидную последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности с SEQ ID NO: 372-405.
[0034] По дополнительному аспекту изобретения предоставлена последовательность с неприродным соединением, содержащая нуклеотидную последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности фрагмента по меньшей мере с 10 последовательными парами оснований, содержащегося в SEQ ID NO: 372-405, где указанный фрагмент с последовательными парами оснований состоит из нуклеотидов на пересечении: расположенной выше последовательности, содержащей SEQ ID NO: 304-337, и расположенной ниже последовательности, содержащей SEQ ID NO: 338-371.
[0035] По дополнительному аспекту изобретения предоставлена бактериальная композиция, содержащая по меньшей мере один реконструированный штамм бактерий который фиксирует атмосферный азот, по меньшей мере один реконструированный штамм бактерий, содержащий экзогенно добавленную ДНК, где указанная экзогенно добавленная ДНК, по меньшей мере на 80% идентична с соответствующим нативным штаммом бактерий.
[0036] По дополнительному аспекту изобретения предоставлен способ поддержания уровней азота в почве. Способ включает высаживание в почву поля сельскохозяйственной культуры, инокулированной реконструированными бактериями, которые фиксируют атмосферный азот. Способ также включает сбор урожая указанной сельскохозяйственной культуры, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 90% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
[0037] По другому аспекту изобретения предоставлен способ доставки пробиотической добавки в сельскохозяйственную культуру. Способ включает покрытие семян сельскохозяйственной культуры покрытием для семян, пропиткой для семян или протравкой для семян, где указанные покрытие для семян, протравка для семян или пропитка для семян содержат живых представителей указанного пробиотика. Способ также включает внесение указанных семян сельскохозяйственной культуры в почву поля.
[0038] По дополнительному аспекту изобретения предоставлен способ увеличения у небобового растения количества получаемого из атмосферы азота. Способ включает воздействие на указанное небобовое растение реконструированными не являющимися межродовыми микроорганизмами, где указанные реконструированные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, или по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
[0039] По дополнительному аспекту изобретения предоставлен способ увеличения у растения кукурузы количества получаемого из атмосферы азота. Способ включает воздействие на указанное растение кукурузы реконструированными не являющимися межродовыми микроорганизмами, несущими реконструированные генетические вариации по меньшей мере в двух генах, выбранных из группы, состоящей из nifL, glnB, glnE и amtB.
[0040] По другому аспекту изобретения предоставлен способ увеличения у растения кукурузы количества получаемого из атмосферы азота. Способ включает воздействие на указанное растение кукурузы реконструированными не являющимися межродовыми микроорганизмами, несущими по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота, где указанные реконструированные не являющиеся межродовыми микроорганизмы, in planta, продуцируют у указанного растения кукурузы по меньшей мере 5% фиксированного азота, как измеряют посредством разведения 15N в сельскохозяйственных культурах, выращиваем в полях, обрабатываемых удобрением, содержащим 1,2% 15N.
[0041] Дополнительные аспекты изобретения относятся к роду микроорганизмов, который подвергали эволюции и оптимизировали для фиксации азота у небобовых сельскохозяйственных культур in planta. В частности, описаны способы увеличения фиксации азота у небобового растения. Способы могут включать воздействие на растение множеством бактерий. Каждый представитель множества несет одну или несколько генетических вариаций, вносимых в один или несколько генов или некодирующих полинуклеотидов генетической регуляторной сети фиксации или ассимиляции азота бактерий так, что бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота. Бактерии не являются межродовыми микроорганизмами. Кроме того, бактерии, in planta, продуцируют в растении 1% или более фиксированного азота.
[0042] Дополнительные аспекты изобретения относятся к полезным выделенным микроорганизмам и композициям микроорганизмов. В частности, предоставлены выделенные и биологически чистые микроорганизмы, которые, в числе прочего, применимы при увеличении фиксации азота в сельскохозяйственной культуре. Описанные микроорганизмы можно использовать в их выделенном и биологически чистом состояниях, а также формулировать в композиции. Кроме того, в описании предоставлены композиции микроорганизмов, содержащие по меньшей мере два представителя описанных микроорганизмов, а также способы использования указанных композиций микроорганизмов.
ВКЛЮЧЕНИЕ В КАЧЕСТВЕ ССЫЛКИ
[0043] Все публикации, патенты и патентные заявки, указываемые в настоящем описании, включены в настоящий документ в качестве ссылки в той же степени, как если бы каждые конкретные публикация, патент или патентная заявка были конкретно и отдельно указаны для включения в качестве ссылки.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0044] Новые признаки изобретения подробно описаны в прилагаемой формуле изобретения. Лучшее понимание признаков и преимуществ настоящего изобретения можно получить при обращении к приводимому ниже подробному описанию, в котором приведены иллюстративные варианты осуществления, в которых использованы основы изобретения, и к сопровождающим фигурам, в которых:
[0045] На фигурах 1A-B представлено обогащение и выделение азотфиксирующих бактерий. (A) Для выделения единичных колоний азотфиксирующих бактерий использовали планшеты с агаром Nfb. (B) Полутвердый агар Nfb залитый в пробирку Balch. Стрелка указывает на пелликулу обогащенных азотфиксирующих бактерий.
[0046] На фигуре 2 представлен иллюстративный скрининг nifH посредством ПЦР. В этом скрининге положительные полосы наблюдали при ≈350 п.н. для двух колоний. Более низко расположенные полосы представляют собой димеры праймеров.
[0047] На фигуре 3 приведен пример скрининга посредством ПЦР колоний после мутагенеза с отбором CRISPR-Cas. Колонии CI006 подвергали скринингу с праймерами, специфичными для локуса nifL. Ожидаемый продукт ПЦР дикого типа составляет ≈2,2 т.п.н., тогда как ожидаемый мутант составляет ≈1,1 т.п.н. Семь из десяти подвергнутых скринингу колоний однозначно продемонстрировали требуемую делецию.
[0048] На фигурах 4A-D приведены фенотипы различных штаммов in vitro. Анализ восстановления ацетилена (ARA) активность мутантов штамма CI010 (фигура 4A) и мутантов штамма CI006 (фигура 4B), выращиваемых в среде для фиксации азота, дополненной глутамином в концентрации от 0 до 10 мМ. Активность дополнительных штаммов в ARA приведена на фигуре 4C, а профиль выделения двумя штаммами аммония с течением времени представлен на фигуре 4D.
[0049] На фигуре 5 приведен профиль экспрессии в культуре 9 различных генов у штаммов CI006, вовлеченных в диазатрофную фиксацию азота. Цифры представляют количества каждого транскрипта. Приведены различные условия (0, 1, 10 мМ глутамин и 0%, 10%, 20% атмосферный воздух в N2).
[0050] На фигуре 6 приведена колонизация CI006 корней кукурузы. Всходы кукурузы инокулировали CI006, несущими экспрессирующую RFP плазмиду. После двух недель роста и сохранения плазмиды при поливе соответствующим антибиотиком, корни собирали и проводили визуализацию посредством флуоресцентной микроскопии. Можно наблюдать колонизацию межклеточного пространства корней.
[0051] На фигуре 7 приведен азот, получаемый на уровне микроорганизмов у штамма дикого типа (CI050) и оптимизированного штамма (CM002).
[0052] На фигурах 8 представлена экспериментальная установка для анализа массы плодов Micro-Tom.
[0053] На фигуре 9 представлен скрининг 10 штаммов на увеличение массы плода растения в Micro-Tom. Представлены результаты шести повторений. Для столбца 3, p=0,07. Для столбца 7, p=0,05.
[0054] На фигурах 10A-C приведены дополнительные результаты для активности микроорганизмов-кандидатов и других мутантов-кандидатов, выращиваемых в средах для фиксации азота, дополненной глутамином в концентрации от 0 до 10 мМ, в ARA.
[0055] На фигуре 11 приведен двойной мутант, который демонстрирует более высокое выделение аммиака, чем одиночный мутант, из которого он получен.
[0056] На фигуре 12 приведен NDFA, получаемый в эксперименте по захвату газообразного 15N (с обратной экстраполяцией с использованием суток воздействия) для определения NDFA в растениях кукурузы в условиях добавления удобрения.
[0057] На фигуре 13 приведено значение NDFA, получаемое в эксперименте по захвату газообразного 15N (с обратной экстраполяцией с использованием суток воздействия) для определения NDFA в растениях Setaria в условиях добавления удобрения.
[0058] На фигуре 14A приведена степень включения газообразного 15N. Растения, инокулированные эволюционировавшим штаммом, по сравнению с неинокулированными растениями демонстрировали увеличение включения газообразного 15N.
[0059] На фигуре 14B приведены 4 недели после посадки, до 7% азота в растениях, инокулированных эволюционировавшим штаммом, происходит из фиксированного микроорганизмами азота.
[0060] На фигуре 14C показано увеличение площади листа (и других измерений биомассы, данные не показаны) у растений, инокулированных эволюционировавшим штаммом по сравнению с неинокулированными растениями или инокулированными растениями дикого типа.
[0061] На фигуре 15A приведены эволюционировавшие штаммы, которые демонстрируют значимо более высокую продукцию nifH в ткани корня, как определяют посредством транскриптомного исследования in planta.
[0062] На фигуре 15B показано, что уровень фиксированного азота, выявляемый в ткани растения, коррелирует с уровнем колонизации данного конкретного растения оптимизированным по HoME штаммом.
[0063] На фигуре 16A приведена карта структур почв различных полевых почв, тестируемых для колонизации. Почвы, которые исходно являлись источником незначительного количества микроорганизмов, указаны звездами.
[0064] На фигуре 16B приведен уровень колонизации штамма 1 и штамма 5, которые тестируют в четырех различных типах почв (окружности). Оба штамма продемонстрировали относительно устойчивый профиль колонизации в различных типах почв.
[0065] На фигуре 16C приведена колонизация штамма 1, протестированная в полевых испытаниях в течение вегетационного периода. Штамм 1 сохранялся в ткани кукурузы до 12 недели после посадки и начинал демонстрировать снижение колонизации после этого времени.
[0040] На фигуре 17A приведена схема выведения микроорганизмов по вариантам осуществления.
[0041] На фигуре 17B приведен расширенный вид определения состава микробиома, как приведено на фигуре 17A.
[0042] На фигуре 17C приведено получение образцов корней, используемых в примере 7.
[0043] На фигуре 18 приведена линия модифицированных штаммов, полученных из штамма CI006.
[0044] На фигуре 19 приведена линия модифицированных штаммов, полученных из штамма CI019.
[0045] На фигуре 20 приведена тепловая карта количества фунтов азота, доставляемых на акре/сезон микроорганизмами по настоящему изобретению, зарегистрированная как функция микроорганизмов на грамм сырой массы на ммоль азота/микроорганизм/час. Ниже тонкой линии, пересекающей более крупное изображение, находятся микроорганизмы, доставляющие менее одного фунта азота на акре/сезон, а выше линии находятся микроорганизмы, доставляющие более одного фунта азота на акре/сезон. В таблице ниже тепловой карты приведены точные значения ммоль N, продуцируемые микроорганизмом в час (ммоль N/микроорганизм/час) вместе с точными значениями КОЕ на грамм сырой массы (КОЕ/г св.масс.) для каждого микроорганизма, приведенного на тепловой карте. Микроорганизмы, используемые для тепловой карты, анализировали на продукцию N в кукурузе. Для штаммов дикого типа CI006 и CI019, данные по колонизации корня кукурузы получали с одного полевого участка. Для оставшихся штаммов, колонизацию полагали такой же, как уровень в поле у дикого типа. Активность фиксации N определяли с использованием анализа ARA in vitro с 5 мМ глутамином.
[0046] На фигуре 21 приведена урожайность растений под воздействием штамма CI006. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0047] На фигуре 22 приведена урожайность растений под воздействием штамма CM029. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0048] На фигуре 23 приведена урожайность растений под воздействием штамма CM038. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0049] На фигуре 24 приведена урожайность растений под воздействием штамма CI019. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0050] На фигуре 25 приведена урожайность растений под воздействием штамма CM081. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0051] На фигуре 26 приведена урожайность растений под воздействием штаммов CM029 и CM081. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0052] На фигуре 27 приведена урожайность растений в виде совокупных прироста/потери в бушелях. Область кругов соответствует относительной урожайности, тогда как затенение соответствует конкретному показателю MRTN. Ось x представляет собой значение p, а ось y представляет собой вероятность выигрыша.
[0053] На фигурах 28A-28E проиллюстрированы производные микроорганизмы, которые фиксируют и выделяют азот in vitro в условиях, сходных с высоконитратными сельскохозяйственными почвами. На фигуре 28A проиллюстрирована регуляторная сеть, контролирующая фиксацию и ассимиляцию азота у PBC6.1, включающая ключевые узлы NifL, NifA, GS, GlnE, изображенные в виде двухдоменого АТазного-AR фермента и AmtB. На фигуре 28B проиллюстрирован геном изолята PBC6.1 Kosakonia sacchari. Три траектории, описывающие геном, соответствуют данным транскрипции PBC6.1, PBC6.38 и дифференциальной экспрессии у штаммов, соответственно. На фигуре 28C проиллюстрирован кластер генов фиксации азота и данные по транскрипции увеличены для демонстрации более мелких подробностей. На фигуре 28D проиллюстрирована нитрогеназная активность при варьирующих концентрациях экзогенного азота, определенная анализом восстановления ацетилена. Штамм дикого типа демонстрирует подавление нитрогеназной активности по мере увеличения концентраций глутамина, тогда как производные штаммы демонстрируют различные степени устойчивости. Величины ошибок представляют собой стандартную ошибку среднего по меньшей мере трех биологических повторений. На фигуре 28E проиллюстрирован временное выделение аммиака производными штаммами, наблюдаемое при мМ концентрациях. По наблюдениям, штаммы дикого типа не выделяют фиксированный азот, и в средах накапливается незначительное количество аммиака. Величины ошибок представляют собой стандартную ошибку среднего.
[0054] На фигурах 29A-29C проиллюстрированы эксперименты в теплице, демонстрирующие фиксация азота микроорганизмами в кукурузе. На фигуре 29A проиллюстрирована колонизация микроорганизмами через шесть недель после инокуляция растений кукурузы штаммами, производными PBC6.1. Величины ошибок представляют собой стандартную ошибку среднего по меньшей мере восьми биологических повторений. На фигуре 29B проиллюстрирована транскрипция nifH in planta, определяемая посредством выделения тотальной РНК из корней и последующего анализа Nanostring. Только производные штаммы демонстрируют транскрипцию nifH в условиях корней. Величины ошибок представляют собой стандартную ошибку среднего по меньшей мере 3 биологических повторений. На фигуре 29C проиллюстрирована фиксация азота микроорганизмами, определяемая посредством разведения изотопного индикатора в тканях растений. Производные микроорганизмы демонстрируют значительный перенос фиксированного азота в растение. Величины ошибок представляют собой стандартную ошибку среднего по меньшей мере десяти биологических повторений.
[0055] На фигуре 30 проиллюстрирована колонизация PBC6.1 почти до 21% количества ассоциированной с корнями микробиоты в корнях кукурузы. Количественные данные основанные на секвенирование ампликонов 16S ризосферы и эндосферы растений кукурузы, инокулированных PBC6.1 и выращиваемых в тепличных условиях.
[0056] На фигуре 31 проиллюстрированы транскрипционные уровни nifA в штаммах, производных PBC6.1, коррелируемые с уровнями восстановления ацетилена. Анализ ARA проводили, как описано в разделе способов, после чего отбирали образец культуры и подвергали анализу кПЦР с определением уровней транскрипта nifA. Величины ошибок в каждом измерении представляют собой стандартную ошибку среднего по меньшей мере трех биологических повторений.
[0057] На фигуре 32 проиллюстрированы результаты тестового полевого эксперимента лета 2017 года. Полученные результаты урожайности демонстрируют, что микроорганизмы по изобретению могут служить в качестве потенциального заменителя удобрений. Например, использование микроорганизма по изобретению (т.е., 6-403) приводило к более высокому урожаю, чем у штамма дикого типа (WT) и к более высокому урожаю, чем у необработанного контроля (UTC). При обработке "-25 фунтов N" используют на 25 фунтов N на акр меньше, чем при стандартной сельскохозяйственной практике в этом регионе. Обработка UTC "100% N" предназначена для обозначения стандартной сельскохозяйственной практики в этом регионе, при которой 100% стандартного использования N применяет сельхозпроизводитель. Микроорганизм "6-403" депонирован под идентификатором NCMA 201708004, и его можно найти в таблице A. Он представляет собой мутанта Kosakonia sacchari (также называемого CM037) и представляет собой мутантный штамм-потомок CI006 дикого типа.
[0058] На фигуре 33 проиллюстрированы результаты тестового полевого эксперимента лета 2017 года. Полученные результаты урожайности демонстрируют, что микроорганизмы по изобретению в различных регионах действуют единообразно. Кроме того, результаты урожайности демонстрируют, что микроорганизмы по изобретению хорошо функционируют как в среде с недостаточным содержанием азота, так и в среде с достаточным содержанием азота. Микроорганизм "6-881" (также известный как CM094, PBC6.94) и являющийся мутантным потомком штамма Kosakonia sacchari CI006 дикого типа, депонирован под идентификатором NCMA 201708002, и его можно найти в таблице A. Микроорганизм "137-1034", который является мутантным потомком штамма Klebsiella variicola CI137 дикого типа, депонирован под идентификатором NCMA 201712001, и его можно найти в таблице A. Микроорганизм "137-1036", который является мутантным потомком штамма Klebsiella variicola CI137 дикого типа, депонирован под идентификатором NCMA 201712002, и его можно найти в таблице A. Микроорганизм "6-404" (также известный как CM38, PBC6.38), и который является мутантным потомком штамма Kosakonia sacchari CI006 дикого типа, депонирован под идентификатором NCMA 201708003, и его можно найти в таблице A. Условия "недостатка питания" соответствует условиям с 0% азота. Условия с "достаточным содержанием удобрения" соответствует условиям со 100% азота.
[0059] На фигуре 34 приведена линия модифицированных штаммов, полученных из штамма CI006 (также обозначаемого "6", дикий тип Kosakonia sacchari).
[0060] На фигуре 35 приведена линия модифицированных штаммов, полученных из штамма CI019 (также обозначаемого "19", дикий тип Rahnella aquatilis).
[0061] На фигуре 36 приведена линия модифицированных штаммов, полученных из штамма CI137 (также обозначаемого "137", дикий тип Klebsiella variicola).
[0062] На фигуре 37 приведена линия модифицированных штаммов, полученных из штамма 1021 (дикий тип Kosakonia pseudosacchari).
[0063] На фигуре 38 приведена линия модифицированных штаммов, полученных из штамма 910 (дикий тип Kluyvera intermedia).
[0064] На фигуре 39 приведена линия модифицированных штаммов, полученных из штамма 63 (дикий тип Rahnella aquatilis).
[0065] На фигуре 40 приведена тепловая карта количества фунтов азота, доставляемых на акре/сезон микроорганизмами по настоящему изобретению, зарегистрированная как функция микроорганизмов на грамм сырой массы на ммоль азота/микроорганизм/час. Ниже тонкой линии, пересекающей более крупное изображение, находятся микроорганизмы, доставляющие менее одного фунта азота на акре/сезон, а выше линии находятся микроорганизмы, доставляющие более одного фунта азота на акре/сезон. В таблица C в примере 12 приведены точные значения ммоль N, продуцируемых микроорганизмом в час (ммоль N/микроорганизм/час) вместе с точными значениями КОЕ на грамм сырой массы (КОЕ/г св.масс.) для каждого микроорганизма, приведенного на тепловой карте. Данные на фигуре 40 получают для штаммов микроорганизмов, оцениваемых на продукцию N в кукурузе в полевых условиях. Каждая точка представляет собой количество фунтов N/акр, продуцируемых микроорганизмом, с использованием данных по колонизации корня кукурузы с одного полевого участка. Активность фиксации N определяли с использованием анализа ARA in vitro с 5 мМ N в форме глутамина или фосфата аммония.
[0066] На фигуре 41 приведена тепловая карта количества фунтов азота, доставляемых на акре/сезон микроорганизмами по настоящему изобретению, зарегистрированная как функция микроорганизмов на грамм сырой массы на ммоль азота/микроорганизм/час. Ниже тонкой линии, пересекающей более крупное изображение, находятся микроорганизмы, доставляющие менее одного фунта азота на акре/сезон, а выше линии находятся микроорганизмы, доставляющие более одного фунта азота на акре/сезон. В таблица D в примере 12 приведены точные значения ммоль N, продуцируемых микроорганизмом в час (ммоль N/микроорганизм/час) вместе с точными значениями КОЕ на грамм сырой массы (КОЕ/г св.масс.) для каждого микроорганизма, приведенного на тепловой карте. Данные на фигуре 41 получают для штаммов микроорганизмов, оцениваемых на продукцию N в кукурузе в лабораторных и тепличных условиях. Каждая точка представляет собой количество фунтов N/акр, продуцируемое одним штаммом. Белые точки представляют штаммы, для которых данные по колонизации корня кукурузы получали в тепличных условиях. Черные точки представляют мутантные штаммы, для которых уровни колонизации корней кукурузы получают на основании средних полевых уровней колонизации корней кукурузы родительского штамма дикого типа. Заштрихованные точки представляют родительские штаммы дикого типа с их средними полевыми уровнями колонизации корней кукурузы. Во всех случаях активность фиксации N определяли посредством анализа ARA in vitro с 5 мМ N в форме глутамина или фосфата аммония.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0067] Хотя в настоящем документе представлены и описаны различные варианты осуществления изобретения, специалистам в данной области очевидно, что такие варианты осуществления предоставлены только в качестве примера. Специалисты в данной области могут проводить многочисленные вариации, изменения и замены без отклонения от изобретения. Следует понимать, что вместо вариантов осуществления изобретения, описываемых в настоящем документе, можно использовать различные альтернативы.
[0068] Повышенное использование удобрений сопряжено с экологическими проблемами и, вероятно, не представляется возможным для многих экономически напряженных регионов земного шара. Кроме того, многие промышленные предприятия в области микроорганизмов сфокусированы на создании межродовых микроорганизмов. Однако на подвергнутые инженерии микроорганизмы, которые характеризуют/классифицируют как межродовые, накладывается тяжелое регуляторное обременение. Эти межродовые микроорганизмы сталкиваются не только с более высоким регуляторным обременением, что затрудняет широкое распространение и внедрение, но также с большим вниманием общественности.
[0069] В настоящее время на рынке отсутствуют подвергнутые инженерии микроорганизмы, не являющиеся межродовыми и способные усиливать фиксацию азота у небобовых культур. Это отсутствие такого микроорганизма представляет собой недостающий элемент, помогающий создать действительно экологически чистую и более устойчивую сельскохозяйственную систему XXI века.
[0070] Настоящее изобретение решает указанные выше проблемы и предоставляет не являющийся межродовым микроорганизм, который был сконструирован для легкой фиксации азота в сельскохозяйственных культурах. Эти микроорганизмы не характеризуют/классифицируют как межродовые микроорганизмы и, следовательно, они не будут сталкиваться с их серьезным регулирующим обременением. Кроме того, исследованные не являющиеся межродовыми микроорганизмы могут поспособствовать сельхозпроизводителям XXI века стать менее зависимыми от использования постоянно растущего количества экзогенных азотных удобрений.
Определения
[0071] Термины "полинуклеотид", "нуклеотид", "нуклеотидная последовательность", "нуклеиновая кислота" и "олигонуклеотид" используют взаимозаменяемо. Они относятся к полимерной форме нуклеотидов любой длины, представляющих собой дезоксирибонуклеотиды, или рибонуклеотиды, или их аналоги. Полинуклеотиды могут обладать любой трехмерной структурой и могут выполнять любую функцию, известную или неизвестную. Неограничивающими примерами полинуклеотидов является следующее: кодирующие или некодирующие области гена или фрагмента гена, локусы (локус), определяемые на основе анализа сцепления, экзоны, интроны, информационная РНК (иРНК), транспортная РНК (тРНК), рибосомальная РНК (рРНК), малая интерферирующая РНК (миРНК), короткошпилечная РНК (кшРНК), микро-РНК (мкРНК), рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенная ДНК любой последовательности, выделенная РНК любой последовательности, зонды и праймеры нуклеиновой кислоты. Полинуклеотид может содержать один или несколько модифицированных нуклеотидов, таких как метилированные нуклеотиды и аналоги нуклеотидов. Если они присутствуют, модификации структуры нуклеотида можно обеспечивать до или после сборки полимера. Последовательности нуклеотидов могут прерываться ненуклеотидными компонентами. Полинуклеотид можно дополнительно модифицировать после полимеризации, например, посредством конъюгации с меченым компонентом.
[0072] "Гибридизация" относится к реакции, в которой реагируют один или несколько полинуклеотидов с формированием комплекса, который стабилизирован вследствие образования водородных связей между основаниями нуклеотидных остатков. Образование водородных связей может происходить при спаривании оснований по Уотсону-Крику, связывании по Хугстину или любым другим специфичным для последовательностей способом. Комплекс может содержать две цепи, формирующие дуплексную структуру, три или более цепи, формирующие мультицепочечный комплекс, одну самогибридизующуюся цепь или любую комбинацию из них. Реакция гибридизации может представлять собой этап в более сложном процессе, таком как инициация ПЦР или ферментативное расщепление полинуклеотида эндонуклеазой. Вторая последовательность, которая комплементарна первой последовательности обозначают как последовательность, "комплементарную" первой последовательности. Термин "гибридизующийся" в применении к полинуклеотиду относится к способности полинуклеотида к формированию комплекса, который стабилизирован вследствие образования водородных связей между основаниями нуклеотидных остатков в реакции гибридизации.
[0073] "Комплементарность" относится к способности нуклеиновой кислоты к формированию водородной связи(-ей) с другой последовательностью нуклеиновой кислоты посредством традиционного спаривания по Уотсону-Крику или других нетрадиционных типов спаривания. Процент комплементарности означает процент остатков в молекуле нуклеиновой кислоты, которые могут формировать водородные связи (например, посредством спаривания оснований по Уотсону-Крику) со второй последовательностью нуклеиновой кислоты (например, 5, 6, 7, 8, 9, 10 из 10 представляют собой комплементарность 50%, 60%, 70%, 80%, 90% и 100%, соответственно). "Идеальная комплементарность" означает, что се последовательные остатки последовательности нуклеиновой кислоты формируют водородные связи с таким же количеством последовательных остатков во второй последовательности нуклеиновой кислоты. Как используют в настоящем документе, "в значительной степени комплементарные" относится к степени комплементарности, которая составляет по меньшей мере 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% на области 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50 или более нуклеотидов, или относится к двум нуклеиновым кислотам, которые гибридизуются в жестких условиях. Идентичность последовательностей, например, для целей оценки процента комплементарности, можно определять любым подходящим алгоритмом выравнивания, включая в качестве неограничивающих примеров алгоритм Нидлмана-Вунша (см. например, средство выравнивания EMBOSS Needle, доступное на www.ebi.ac.uk/Tools/psa/emboss_needle/nucleotide.html, необязательно с установками по умолчанию), алгоритм BLAST (см. например, средство выравнивания BLAST, доступное на blast.ncbi.nlm.nih.gov/Blast.cgi, необязательно с настройками по умолчанию) или алгоритм Смита-Уотермана (см. например, средство выравнивания EMBOSS Water, доступное на www.ebi.ac.uk/Tools/psa/emboss_water/nucleotide.html, необязательно с настройками по умолчанию). Оптимальное выравнивание можно оценивать с использованием любых подходящих параметров выбранного алгоритма, включая параметры по умолчанию.
[0074] Как правило, "жесткие условия" гибридизации относятся к условиям, в которых нуклеиновая кислота, комплементарная последовательности-мишени, преимущественно гибридизуется с последовательностью-мишенью, и по существу не гибридизуется с не являющимися мишенями последовательностями. Как правило, жесткие условия зависят от последовательности и варьируют в зависимости от ряда факторов. Как правило, чем длиннее является последовательность, тем выше является температура при которой последовательность специфически гибридизуется с ее последовательностью-мишенью. Неограничивающие примеры жестких условий подробно описаны в Tijssen (1993), Laboratory Techniques In Biochemistry And Molecular Biology-Hybridization With Nucleic Acid Probes Part I, Second Chapter "Overview of principles of hybridization and the strategy of nucleic acid probe assay", Elsevier, N.Y.
[0075] Как используют в настоящем документе, "экспрессия" относится к процессу, посредством которого полинуклеотид транскрибируется с матрицы ДНК (например в иРНК или другой транскрипт РНК), и/или к процессу, посредством которого затем происходит трансляция транскрибируемой иРНК в пептиды, полипептиды или белки. Транскрипты и кодируемые полипептиды в совокупности можно обозначить как "продукт гена". Если полинуклеотид получен с геномной ДНК, экспрессия может включать сплайсинг иРНК в эукариотической клетке.
[0076] Термины "полипептид", "пептид" и "белок" в настоящем документе используют взаимозаменяемо для обозначения полимеров аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и он может прерываться не являющимися аминокислотами участками. Термины также включает модифицированный полимер из аминокислот; например, посредством формирования дисульфидной связи, гликозилирования, липидизации, ацетилирования, фосфорилирования или любой другой манипуляции, такой как конъюгация с меченым компонентом. Как используют в настоящем документе термин "аминокислота" включает природные и/или неприродные или синтетические аминокислоты, включая глицин и D или L оптические изомеры, и аналоги аминокислот и пептидомиметики.
[0077] Как используют в настоящем документе, термин "приблизительно" используют как синоним термина "примерно". Для иллюстрации, использование термина "приблизительно" в отношении количества означает, что значения немного выходят за указанные значения, например, плюс или минус от 0,1% до 10%.
[0078] Термин "биологически чистая культура" или "по существу чистая культура" относится к культуре вида бактерий, описываемого в настоящем документе, не содержащей других видов бактерий в количествах, достаточных для препятствия репликации культуры, или детектируемых стандартными бактериологическими методами.
[0079] "Продуктивность растений" в основном относится к любому показателю роста или развития растения, который является причиной, по которой это растение выращивают. Для продовольственных культур, таких как зерновые или овощи, "продуктивность растений" может относиться к урожаю зерновых или плодов, собираемых у конкретной сельскохозяйственной культуры. Как используют в настоящем документе, улучшенная продуктивность растений в широком смысле относится к улучшению урожая зерна, плодов, цветов или других частей растений, собираемых с различными целями, улучшению роста частей растений, включая стебли, листья и корни, к стимуляции роста растений, подержанию высокого содержания хлорофилла в листьях, увеличению количества плодов или семян, увеличению массы единичного плода или семени, уменьшению выброса NO2 вследствие сниженного использования азотных удобрений и подобным улучшениям роста и развития растений.
[0080] Микроорганизмы в продовольственных культурах и около них могут влиять на признаки этих культур. Признаки растений, на которые могут влиять микроорганизмы включают: урожайность (например, продукция зерна, производство биомассы, развитие плодов, строение цветов); питание (например, потребление азота, фосфора, калия, железа, питательных микроэлементов); способность к контролю абиотического стресса (например, засухоустойчивость, солеустойчивость, жароустойчивость) и способность к контролю биотического стресса (например, сельскохозяйственных вредителей, сорняков, насекомых, грибов и бактерий). Стратегии изменения признаков сельскохозяйственных культур включают: увеличение концентраций ключевых метаболитов; изменение временной динамики воздействия микроорганизмов на ключевые метаболиты; связывание продукции/разрушения метаболитов микроорганизмами с новыми параметрами окружающей среды; сокращение нежелательных метаболитов и улучшение баланса метаболитов или обуславливающих белков.
[0081] Как используют в настоящем документе, "регуляторная последовательность" относится к оператору, промотору, сайленсеру или терминатору.
[0082] Как используют в настоящем документе, "in planta" означает в растении, где растение дополнительно включает части, ткани, листья, корни, стебли, семена, семязачатки, пыльцу, цветы, плоды растения и т.д.
[0083] В определенных вариантах осуществления природные или эндогенные регуляторные последовательности генов по настоящему изобретению заменяют одной или несколькими внутриродовыми регуляторными последовательностями.
[0084] Как используют в настоящем документе, "вводимый" относится к введению посредством современной биотехнологии, а не к природному введению.
[0085] В определенных вариантах осуществления бактерии по настоящему изобретению модифицированы так, что они не являются природными бактериями.
[0086] В определенных вариантах осуществления бактерии по настоящему изобретению присутствуют в растении в количестве по меньшей мере 103 КОЕ, 104 КОЕ, 105 КОЕ, 106 КОЕ, 107 КОЕ, 108 КОЕ, 109 КОЕ, 1010 КОЕ, 1011 КОЕ или 1012 КОЕ на грамм сырой или сухой массы растения. В определенных вариантах осуществления бактерии по настоящему изобретению присутствуют в растении в количестве по меньшей мере приблизительно 103 КОЕ, приблизительно 104 КОЕ, приблизительно 105 КОЕ, приблизительно 106 КОЕ, приблизительно 107 КОЕ, приблизительно 108 КОЕ, приблизительно 109 КОЕ, приблизительно 1010 КОЕ, приблизительно 1011 КОЕ или приблизительно 1012 КОЕ на грамм сырой или сухой массы растения. В определенных вариантах осуществления бактерии по настоящему изобретению присутствуют в растении в количестве по меньшей мере от 103 до 109, от 103 до 107, от 103 до 105, от 105 до 109, от 105 до 107, от 106 до 1010, от 106 до 107 КОЕ на грамм сырой или сухой массы растения.
[0087] Удобрения и экзогенный азот по настоящему изобретению могут включать следующие азотсодержащие молекулы: аммоний, нитрат, нитрит, аммиак, глутамин и т.д. Источники азота по настоящему изобретению могут включать безводный аммиак, сульфат аммония, мочевину, фосфат диаммония, формальдегидмочевину, фосфат моноаммония, нитрат аммония, жидкие азотные удобрения, нитрат кальция, нитрат калия, нитрат натрия и т.д.
[0088] Как используют в настоящем документе, "экзогенный азот" относится к неатмосферному азоту, легкодоступному в почве, поле или среде для выращивания, который присутствует в условиях без ограничения азота, включая аммиак, аммоний, нитрат, нитрит, мочевина, мочевая кислота, аммонийные кислоты и т.д.
[0089] Как используют в настоящем документе, "условия без ограничения азота" относится к неатмосферному азоту, доступному в почве, поле, средах в концентрациях, больших чем приблизительно 4 мМ азот, как описано в Kant et al. (2010. J. Exp. Biol. 62(4):1499-1509), включенной в настоящий документ в качестве ссылки.
[0090] Как используют в настоящем документе, "межродовой микроорганизм" представляет собой микроорганизм, который сформирован посредством преднамеренной комбинации генетического материала, исходно выделенного из организмов различных таксономических родов. Взаимозаменяемо с "межродовым микроорганизмом" можно использовать "межродовой мутант". Иллюстративный "межродовой микроорганизм" включает микроорганизм, содержащий мобильный генетический элемент, который впервые идентифицирован в микроорганизме, принадлежащем роду, отличного от микроорганизма-реципиента. Дополнительное объяснение в числе прочего можно найти в 40 C.F.R. 725.3.
[0091] В различных аспектах микроорганизмы, исследованные в настоящем документе, представляют собой "не являющиеся межродовыми", что означает, что организмы микроорганизмы не являются межродовыми.
[0092] Как используют в настоящем документе, "внутриродовой микроорганизм" представляет собой микроорганизм, который сформирован посредством преднамеренной комбинации генетического материала, исходно выделенного из организмов одного и того же таксономического рода. Взаимозаменяемо с "внутриродовым микроорганизмом" можно использовать "внутриродовой мутант".
[0093] Как используют в настоящем документе, "вводимый генетический материал" означает генетический материал, который добавляют в геном реципиента и который остается в геноме реципиента в качестве компонента.
[0094] В определенных вариантах осуществления генетическая регуляторная сеть фиксации и ассимиляции азота содержит полинуклеотиды, кодирующие гены, и некодирующие последовательности, которые направляют, модулируют и/или регулируют фиксацию и/или ассимиляцию азота микроорганизмами и может содержать полинуклеотидные последовательности кластера nif (например, nifA, nifB, nifC,…nifZ), полинуклеотиды, кодирующие регулирующий метаболизм азота белок C, полинуклеотиды, кодирующие регулирующий метаболизм азота белок B, полинуклеотидные последовательности кластера gln (например, glnA и glnD), draT и переносчики/пермеазы аммония. В определенных случаях кластер Nif может содержать NifB, NifH, NifD, NifK, NifE, NifN, NifX, hesa и NifV. В определенных случаях кластер Nif может содержать подмножество NifB, NifH, NifD, NifK, NifE, NifN, NifX, hesa и NifV.
[0095] В определенных вариантах осуществления удобрение по настоящему изобретению содержит по меньшей мере 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 21%, 22%, 23%, 24%, 25%, 26%, 27%, 28%, 29%, 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% азота по массе.
[0096] В определенных вариантах осуществления удобрение по настоящему изобретению содержит по меньшей мере приблизительно 5%, приблизительно 6%, приблизительно 7%, приблизительно 8%, приблизительно 9%, приблизительно 10%, приблизительно 11%, приблизительно 12%, приблизительно 13%, приблизительно 14%, приблизительно 15%, приблизительно 16%, приблизительно 17%, приблизительно 18%, приблизительно 19%, приблизительно 20%, приблизительно 21%, приблизительно 22%, приблизительно 23%, приблизительно 24%, приблизительно 25%, приблизительно 26%, приблизительно 27%, приблизительно 28%, приблизительно 29%, приблизительно 30%, приблизительно 31%, приблизительно 32%, приблизительно 33%, приблизительно 34%, приблизительно 35%, приблизительно 36%, приблизительно 37%, приблизительно 38%, приблизительно 39%, приблизительно 40%, приблизительно 41%, приблизительно 42%, приблизительно 43%, приблизительно 44%, приблизительно 45%, приблизительно 46%, приблизительно 47%, приблизительно 48%, приблизительно 49%, приблизительно 50%, приблизительно 51%, приблизительно 52%, приблизительно 53%, приблизительно 54%, приблизительно 55%, приблизительно 56%, приблизительно 57%, приблизительно 58%, приблизительно 59%, приблизительно 60%, приблизительно 61%, приблизительно 62%, приблизительно 63%, приблизительно 64%, приблизительно 65%, приблизительно 66%, приблизительно 67%, приблизительно 68%, приблизительно 69%, приблизительно 70%, приблизительно 71%, приблизительно 72%, приблизительно 73%, приблизительно 74%, приблизительно 75%, приблизительно 76%, приблизительно 77%, приблизительно 78%, приблизительно 79%, приблизительно 80%, приблизительно 81%, приблизительно 82%, приблизительно 83%, приблизительно 84%, приблизительно 85%, приблизительно 86%, приблизительно 87%, приблизительно 88%, приблизительно 89%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98% или приблизительно 99% азота по массе.
[0097] В определенных вариантах осуществления удобрение по настоящему изобретению содержит приблизительно от 5% до 50%, приблизительно от 5% до 75%, приблизительно от 10% до 50%, приблизительно от 10% до 75%, приблизительно от 15% до 50%, приблизительно от 15% до 75%, приблизительно от 20% до 50%, приблизительно от 20% до 75%, приблизительно от 25% до 50%, приблизительно от 25% до 75%, приблизительно от 30% до 50%, приблизительно от 30% до 75%, приблизительно от 35% до 50%, приблизительно от 35% до 75%, приблизительно от 40% до 50%, приблизительно от 40% до 75%, приблизительно от 45% до 50%, приблизительно от 45% до 75% или приблизительно от 50% до 75% азота по массе.
[0098] В определенных вариантах осуществления увеличение фиксации азота и/или продукцию 1% или более азота в растении определяют относительно контрольных растений, которые не подвергали действию бактерий по настоящему изобретению. Все увеличения или уменьшения количеств бактерий определяют относительно контрольных растений. Все увеличения или уменьшения в растениях измеряют относительно контрольных растений.
[0099] Как используют в настоящем документе, "конститутивный промотор" представляет собой промотор, который является активным в большинстве условий и/или в течение большинства стадий развития. Существует несколько преимуществ использования в экспрессирующих векторах, используемых в биотехнологии, конститутивных промоторов, например: высокий уровень продукции белков, используемых для отбора трансгенных клеток или организмов; высокий уровень экспрессии репортерных белков или селективных маркеров, обеспечивая простые детекцию и количественный анализ; высокий уровень продукции фактора транскрипции, который является частью системы регуляции транскрипции; продукция соединений, у которых необходима повсеместная активность в организме; и продукция соединений, которые необходимы на всех стадиях развития. Неограничивающие иллюстративные конститутивные промоторы включают промотор 35S CaMV, промоторы опинов, промотор убиквитина, промотор алкогольдегидрогеназы и т.д.
[0100] Как используют в настоящем документе, "неконститутивный промотор" представляет собой промотор, который активен в определенных условиях, в определенных типах клеток и/или в течение определенных стадиях развития. Например, тканеспецифичные, тканепредпочтительные, специфичные для типов клеток, предпочтительные для типов клеток, индуцибельные промоторы и промоторы под контролем развития являются неконститутивными промоторы. Примеры промоторов под контролем развития включают промоторы, которые предпочтительно инициируют транскрипцию в определенных тканях.
[0101] Как используют в настоящем документе, "индуцибельный" или "реперссируемый" промотор представляет собой промотор, который находится под контролем химических веществ или факторов окружающей среды. Примеры условий окружающей среды, который могут влиять на транскрипцию под действием индуцибельных промоторов, включают анаэробные условия, определенные химические вещества, наличие света, кислые или основные условия и т.д.
[0102] Как используют в настоящем документе, "тканеспецифический" промотор представляет собой промотор, который инициирует транскрипцию только в определенных тканях. В отличие от конститутивной экспрессии генов, тканеспецифическая экспрессия является результатом нескольких взаимодействующих уровне регуляции генов. Таким образом, в данной области для достижения эффективной и надежной экспрессии трансгенов в конкретных тканях иногда предпочтительно использовать промоторы гомологичных или близкородственных видов. Это является одной из основных причин находимого как в научной, так и в патентной литературе большого количества тканеспецифичных промоторов, выделенных из конкретных тканей.
[0103] Как используют в настоящем документе, термин "функционально связанные" относится к ассоциации последовательностей нуклеиновых кислот в одном фрагменте нуклеиновой кислоты так, что функцию одной регулирует другая. Например, промотор функционально связан с кодирующей последовательностью, когда он может регулировать экспрессию этой кодирующей последовательности (т.е., эта кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации. В другом примере комплементарные области РНК по изобретению могут быть функционально связаны, непосредственно или опосредовано, со стороны 5'-конца от иРНК-мишени, или со стороны 3'-конца от иРНК-мишени, или с иРНК-мишенью, или первая комплементарная область находится со стороны 5'-конца, а ее комплементарная область находится со стороны 3'-конца от иРНК-мишени.
[0104] В определенных аспектах "нанесение на растение множества не являющихся межродовыми бактерий" включает любые средства, которыми растение (включая части растения, такие как семена, корень, стебель, ткань и т.д.) приводят в контакт (т.е., производят воздействие) с указанными бактериями на любом этапе жизненного цикла растения. Таким образом, "нанесение на растение множества не являющихся межродовыми бактерий" включает любые из приводимых ниже средств воздействия на растение (включая части растения, такие как семена, корень, стебель, ткань и т.д.) указанными бактериями: распыление на растение, капание на растение, нанесение в виде покрытия семян, внесение в поле, на которое затем высаживают семена, внесение в поле, на которое уже высажены семена, внесение в поле со взрослыми растениями и т.д.
[0105] Как используют в настоящем документе "MRTN" представляет собой сокращение для максимального возврата азота, и его используют в качестве экспериментального показателя в примерах. MRTN разработан в государственном университете штата Айова, а информацию можно найти на странице: http://cnrc.agron.iastate.edu/ MRTN представляет собой уровень азота, при котором чистая экономическая отдача при применении азота является максимальной. Подход к расчету MRTN является региональным подходом к разработке рекомендаций по уровням азота в кукурузе в конкретных штатах. Данные исследований уровня азота оценивали для Иллинойса, Айовы, Мичигана, Миннесоты, Огайо и Висконсина, где было доступно достаточное количество исследований для посадок кукурузы после сои и посадок кукурузы после кукурузы. Исследования проводили с использованием весеннего междурядного или разделенного предпосевного/междурядного внесения азота, и участки не подвергали ирригации за исключением участков, которые указаны для ирригируемых песков в Висконсине. MRTN разработан в государственном университете штата Айова ввиду явных различий способов определения предполагаемых уровней азота, необходимых для производства кукурузы, неправильных предположений относительно рекомендаций по уровню азота и опасений об уровнях внесения. Рассчитывая MRTN, практикующие специалисты могут определить следующее: (1) уровень азота, при котором чистая экономическая отдача при применении азота является максимальной, (2) экономически оптимальный уровень азота, который является точкой, где последнее увеличение уровня азота обеспечивает увеличение урожайности, достаточное для оплаты дополнительного азота, (3) значение увеличения зерен кукурузы, связанное с применением азота, и максимальный урожай, который является урожаем, при котором внесение большего количества азота не приводит к увеличению урожая кукурузы. Таким образом, расчеты MRTN предоставляют практикующим специалистам средства для максимизации посевов кукурузы в разных регионах при максимальной финансовой выгоде от применения азота.
[0106] Термин ммоль представляет собой сокращение для миллимоля, который представляет собой тысячную часть (10-3) моля, сокращенного в настоящем документе как моль.
[0107] Как используют в настоящем документе термины "микроорганизм" или "микроб" следует понимать широко. Эти термины, используемые взаимозаменяемо, в качестве неограничивающих примеров включают два домена прокариот, бактерии и археи. Термин также может включать эукариотические грибы и простейших.
[0108] Термин "консорциумы микроорганизмов" или "консорциум микроорганизмов" относится к подмножеству сообщества микроорганизмов конкретных видов микроорганизмов или штаммам видов, которое можно описать, как выполняющее общую функцию, или можно описать, как участвующее, или приводящее, или коррелирующее с требуемым параметром, таким как представляющий интерес фенотипический признак.
[0109] Термин "сообщество микроорганизмов" означает группу микроорганизмов, содержащую два или более видов или штаммов. В отличие от консорциумов микроорганизмов, сообщество микроорганизмов не должно выполнять общую функцию, или не должно участвовать, или приводить, или коррелировать с требуемым параметром, таким как представляющий интерес фенотипический признак.
[0110] Как используют в настоящем документе, "изолят", "выделенный", "выделенный микроорганизм" и подобные термины предназначены для обозначения того, что один или несколько микроорганизмов отделены по меньшей мере от одного из веществ, с которыми они ассоциированы в конкретной среде (например, почве, воде, ткани растения и т.д.). Таким образом, "выделенный микроорганизм" не находится в его природном окружении; точнее, что различными описанным в настоящем документе способами, микроорганизм удален из своего природного состояния и помещен в не встречающееся в природе состояние существования. Таким образом, выделенный штамм или выделенный микроорганизм может находиться, например, в виде биологически чистой культуры или в виде спор (или других форм штамма). В определенных аспектах выделенный микроорганизм может находиться в ассоциации с приемлемым носителем, который может являться приемлемым в сельском хозяйстве носителем.
[0111] В определенных аспектах изобретения выделенные микроорганизмы существуют в виде "выделенных и биологически чистых культур". Специалисту в данной области следует понимать, что выделенная и биологически чистая культура конкретного микроорганизма означает, что указанная культура по существу очищена от других живых организмов и содержит только конкретный рассматриваемый микроорганизм. Культура может содержать варьирующие концентрации указанного микроорганизма. В настоящем описании отмечено, что выделенные и биологически чистые микроорганизмы часто "необязательно отличаются от менее чистых или загрязненных материалов". См., например, In re Bergstrom, 427 F.2d 1394, (CCPA 1970) (где описаны очищенные простагландины), также см., In re Bergy, 596 F.2d 952 (CCPA 1979) (где описаны очищенные микроорганизмы), также см., Parke-Davis & Co. v. H.K. Mulford & Co., 189 F. 95 (S.D.N.Y. 1911) (Learned Hand, описывающий очищенный адреналин), частично подтверждено, частично рассмотрено, 196 F. 496 (2d Cir. 1912), каждый из которых включен в настоящий документ в качестве ссылки. Кроме того, в определенных аспектах, в описании предусмотрены определенные количественные меры для ограничений концентрации или чистоты, которые необходимо выявить в выделенной и биологически чистой культуре микроорганизмов. Наличие этих значений чистоты в определенных вариантах осуществления является дополнительным признаком, который отличает предоставляемые по настоящему описанию микроорганизмы от тех микроорганизмов, которые существуют в природном состоянии. См., например, Merck & Co. v. Olin Mathieson Chemical Corp., 253 F.2d 156 (4th Cir. 1958) (где описаны ограничения чистоты для витамина B12, продуцируемого микроорганизмами), включенную в настоящий документ в качестве ссылки.
[0112] Как используют в настоящем документе, "отдельные изоляты" следует понимать, как означающие композицию или культуру, содержащую преобладающие количества одного рода, вида или штамма микроорганизма после отделения от одного или нескольких других микроорганизмов.
[0113] Микроорганизмы по настоящему изобретению могут включать споры и/или вегетативные клетки. В определенных вариантах осуществления микроорганизмы по настоящему изобретению включают микроорганизмы в жизнеспособном, но не культивируемом (VBNC) состоянии. Как используют в настоящем документе, "спора" или "споры" относятся к структурам, продуцируемым бактериями и грибами, которые адаптированы для выживания и рассеивания. Как правило, споры характеризуют как покоящиеся структуры; однако споры способны к дифференцировке в процессе прорастания. Прорастание представляет собой дифференцировку спор в вегетативные клетки, которые способны к метаболической активности, росу и репродукции. Прорастание одной споры приводит к образованию одной грибковой или бактериальной вегетативной клетки. Споры грибов представляют собой единицы бесполой репродукция и в определенных случаях представляют собой необходимые структуры в жизненных циклах грибов. Бактериальные споры представляют собой структуры для условий выживания, которые, как правило, могут не способствовать выживанию или росту вегетативных клеток.
[0114] Как используют в настоящем документе, "композиция микроорганизмов" относится к композиции, содержащий один или несколько микроорганизмы по настоящему изобретению. В определенных вариантах осуществления композицию микроорганизмов вводят в растения (включая различные части растения) и/или на сельскохозяйственные поля.
[0115] Как используют в настоящем документе, "носитель", "приемлемый носитель" или "приемлемый в сельском хозяйстве носитель" относится к разбавителю, адъюванту, эксципиенту или носителю с которым микроорганизм можно вводить, который не оказывает вредного воздействия намикроорганизм.
Регуляция фиксации азота
[0116] В определенных случаях путь фиксации азота может действовать в качестве мишени генетической инженерии и оптимизации. Одним из признаков, которые могут являться мишенью для регуляции способами, описываемыми в настоящем документе, является фиксация азота. Азотные удобрения являются наибольшими эксплуатационными расходами на фермах и основным движущим фактором более высоких урожаев пропашных культур, таких как кукуруза и пшеница. В настоящем документе описаны продукты жизнедеятельности микроорганизмов, которые могут доставлять возобновляемые формы азота в небобовые сельскохозяйственные культуры. Хотя у некоторых эндофитов присутствуют генетические факторы, необходимые для фиксации азота в чистой культуре, основной технической проблемой является то, что эндофиты злаков и трав дикого типа прекращают фиксацию азота на удобряемых полях. Применение химических удобрений и остаточные уровни азота в полевых почвах подают микроорганизму сигнал выключения биохимического пути фиксации азота.
[0117] Для разработки микроорганизма, способного к фиксации и переноса азота в кукурузу в присутствии удобрения выгодными могут являться изменения на транскрипционном и посттрансляционном уровнях компонентов регуляторной сети фиксации азота. С этой целью в настоящем документе описана технология эволюции хозяина-микроорганизма (HoME), позволяющая обеспечить направленную эволюцию регуляторных сетей и получать новые фенотипы. Также в настоящем документе описаны уникальные, проприетарные библиотеки азотфиксирующих эндофитов, выделенных из кукурузы, совместно с обширными "омиксными" данными о взаимодействии микроорганизмов и растения-хозяина в различных условиях окружающей среды, таких как недостаток и избыток азота. В определенных вариантах осуществления эта технология обеспечивает направленную эволюцию генетической регуляторной сети эндофитов в направлении получения микроорганизмов, которые активно фиксируют азот даже в присутствии удобрения в полевых условиях. Также в настоящем документе описана оценка промышленного потенциала получаемых микроорганизмов, которые колонизирует ткани корня кукурузы и продуцируют азот у удобряемых растений, и оценка совместимости эндофитов со стандартными технологиями составления и различными почвами для определения возможности интеграции микроорганизмов в современные стратегии контроля азота
[0118] Для использования элементарного азота (N) для химического синтеза живые формы комбинируют газообразный азот (N2), доступный в атмосфере, с водородом в процессе, известном как фиксация азота. Вследствие энергоемкого характера биологической фиксации азота диазотрофы (бактерии и археи, которые фиксируют атмосферный газообразный азот) в процессе эволюции сформировали развитую и жесткую регуляцию кластера генов nif в ответ на кислород окружающей среды и доступный азот. Гены Nif кодируют ферменты, вовлеченные в фиксацию азота (такие как нитрогеназный комплекс), и белки, которые регулируют фиксацию азота. В Shamseldin (2013. Global J. Biotechnol. Biochem. 8(4):84-94) приведено подробное описание генов nif и их продуктов, и она включена в настоящий документ в качестве ссылки. В настоящем документе описаны способы получения растения с улучшенным признаком, включающие выделение бактерий из первого растения, внесение генетической вариации в ген выделенных бактерий для увеличения фиксации азота, воздействие вариантов бактерий на второе растение, выделение бактерий из второго растения с улучшенным признаком относительно первого растения и повторение этапов с бактериями, выделенными из второго растения.
[0119] У Proteobacteria регуляция фиксации азота сконцентрирована вокруг зависимого от фактора σ54 связывающего энхансеры белка NifA, положительного регулятора транскрипции кластера nif. Внутриклеточные уровни активного NifA контролируют два ключевых фактора: транскрипция оперона nifLA и ингибирование активности NifA белок-белковым взаимодействием с NifL. Оба эти процесса реагируют на внутриклеточные уровни глутамина посредством сигнального каскада белков PII. Этот каскад опосредован GlnD, который непосредственно определяет уровень глутамина и катализирует уридилирование или деуридилирование двух регуляторных белков PII, GlnB и GlnK, в ответ на отсутствие или наличие связанного глутамина, соответственно. В условиях избытка азота немодифицированный GlnB передает сигнал к деактивации промотора nifLA. Однако в условиях ограничения азота происходит посттрансляционная модификация GlnB, что ингибирует его активность и приводит к транскрипции оперона nifLA. Таким образом, существует жесткий контроль транскрипции nifLA в ответ на азот окружающей среды посредством сигнального каскада белков PII. На посттрансляционном уровне регуляции NifA GlnK ингибирует взаимодействие NifL/NifA в зависимости от общего уровня свободного GlnK в клетке.
[0120] Транскрипция NifA происходит с оперона nifLA, промотор которого активирует фосфорилированный NtrC, другой зависимый от фактора σ54 регулятор. Состояние фосфорилирования NtrC опосредовано гистидинкиназой NtrB, которая взаимодействует с деуридилированным GlnB, но не с уридилированным GlnB. В условиях избытка азота высокий внутриклеточный уровень глутамина приводит к деуридилированию GlnB, который затем взаимодействует с NtrB с деактивацией ее фосфорилазной активности и активацией ее фосфатазной активности, что приводит к дефосфорилированию NtrC и деактивации промотора nifLA. Однако в условиях ограничения азота низкий уровень внутриклеточного глутамина приводит к уридилированию GlnB, что ингибирует его взаимодействие с NtrB и обеспечивает фосфорилирование NtrC и транскрипцию оперона nifLA. Таким образом, существует жесткий контроль экспрессии nifLA в ответ на азот окружающей среды посредством сигнального каскада белков PII. Все из nifA, ntrB, ntrC и glnB представляют собой гены, которые можно подвергать мутированию в способах, описываемых в настоящем документе. Эти процессы также могут реагировать на внутриклеточные или внеклеточные уровни аммиака, мочевины или нитратов.
[0121] Также существует посттрансляционная регуляция активности NifA в ответ на азот окружающей среды, как правило, посредством опосредованного NifL ингибирования активности NifA. Как правило, взаимодействие на взаимодействие NifL и NifA влияет сигнальный каскад белков PII посредством GlnK, хотя характер взаимодействий между GlnK и NifL/NifA у диазотрофов значительно варьирует. У Klebsiella pneumoniae обе формы GlnK ингибируют взаимодействие NifL/NifA, и взаимодействие между GlnK и NifL/NifA определяют по общему уровню свободного GlnK в клетке. В условиях избытка азота деуридилированный GlnK взаимодействует с переносчиком аммиака AmtB, который служит для блокировки захвата аммиака посредством AmtB и изоляции GlnK на мембране, обеспечивая ингибирование NifA посредством NifL. С другой стороны, у Azotobacter vinelandii, для взаимодействия NifL/NifA и ингибирования NifA необходимо взаимодействие с деуридилированным GlnK, тогда как уридилирование GlnK ингибирует это взаимодействие с NifL. У диазотрофов с отсутствием гена nifL существуют данные о том, что ингибирование активности NifA происходит непосредственно при взаимодействии с деуридилированными формами GlnK и GlnB в условиях избытка азота. У некоторых бактерий кластер nif может находиться под регуляторным воздействием glnR, и дополнительно в определенных случаях это может включать отрицательную регуляцию. Независимо от механизма посттрансляционное ингибирование NifA является важным регулятором кластера nif у большинства известных диазотрофов. Дополнительно, гены, которые можно подвергать мутированию в способах, описываемых в настоящем документе, представляют собой nifL, amtB, glnK и glnR.
[0122] В дополнение к регуляции транскрипции кластера генов nif многие диазотрофы в процессе эволюции сформировали механизм прямой посттрансляционной модификации и ингибирования самого фермента нитрогеназы, известный как выключение нитрогеназы. Он опосредован АДФ-рибозилированием белка Fe (NifH) в условиях избытка азота, что нарушает его взаимодействие с белковым комплексом MoFe (NifDK) и устраняет нитрогеназную активность. DraT катализирует АДФ-рибозилирование белка Fe и выключение нитрогеназы, тогда как DraG катализирует удаление АДФ-рибозы и реактивацию нитрогеназы. Как и в случае транскрипции nifLA и ингибирования NifA, выключение нитрогеназы также регулирует сигнальный каскад белков PII. В условиях избытка азота деуридилированный GlnB взаимодействует с DraT и активирует его, тогда как деуридилированный GlnK взаимодействует с DraG и AmtB с формированием комплекса, изолируя DraG на мембране. В условиях ограничения азота уридилированные формы GlnB и GlnK не взаимодействуют с DraT и DraG, соответственно, что приводит к инактивации DraT и диффузии DraG к белку Fe, где он удаляет АДФ-рибозу и активирует нитрогеназу. В способах, описываемых в настоящем документе, также предусмотрено внесение генетических вариаций в гены nifH, nifD, nifK и draT.
[0123] Хотя некоторые эндофиты обладают способностью к фиксации азота in vitro, часто эти генетические факторы в полевых условиях подавлены высокими уровнями экзогенных химических удобрений. Для облегчения фиксации азота в полевых условиях можно разделить определение уровня экзогенного азота и экспрессию фермента нитрогеназы. Улучшение суммарной нитрогеназной активности по времени дополнительно способствует увеличению продукции азота для использования сельскохозяйственной культурой. Конкретные мишени генетической вариации для облегчения фиксации азота в полевых условиях способами, описываемыми в настоящем документе, включают один или несколько генов, выбранных из группы, состоящей из nifA, nifL, ntrB, ntrC, glnA, glnB, glnK, draT, amtB, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB и nifQ.
[0124] Дополнительной мишенью генетической вариации для облегчения фиксации азота в полевых условиях способами, описываемыми в настоящем документе, является белок NifA. Как правило, белок NifA является активатором экспрессии генов фиксации азота. Увеличение продукции NifA (конститутивно или в условиях высокого содержания аммиака) преодолевает природный путь определения уровня аммиака. Кроме того, снижение продукции белков NifL, известного ингибитора NifA, также приводит к увеличению уровня свободного активного NifA. Кроме того, к повышению общего уровня белков NifA также приводит увеличение уровня транскрипции оперона nifAL (конститутивно или в условиях высокого содержания аммиака). Повышенного уровня экспрессии nifAL достигают, изменяя сам промотор или снижая экспрессию NtrB (часть сигнального каскада ntrB и ntrC, который исходно может приводить к выключению оперона nifAL в условиях высокого содержания азота). Высокий уровень NifA, достигаемый этими или любыми другими способами, описываемыми в настоящем документе, увеличивает активность фиксации азота эндофитами.
[0125] Другой мишенью генетической вариации для облегчения фиксации азота в полевых условиях способами, описываемыми в настоящем документе, является сигнальный каскад PII GlnD/GlnB/GlnK. Внутриклеточный уровень глутамина определяет сигнальный каскад PII GlnD/GlnB/GlnK. Мутации активного центра GlnD, устраняющие активность удаления уридилата GlnD, разрывают каскад определения уровня азота. Кроме того, уменьшение концентрации GlnB замыкает каскад определения уровня глутамина. Эти мутации "обманом" приводят клетки к определению условий ограничения азота, таким образом, увеличивая уровень активности фиксации азота. Эти процессы также могут реагировать на внутриклеточные или внеклеточные уровни аммиака, мочевины или нитратов.
[0126] Также мишенью генетической вариации для облегчения фиксации азота в полевых условиях способами, описываемыми в настоящем документе, является белок amtB. Захват аммиака из окружающей среды можно уменьшать, снижая уровень экспрессии белка amtB. Без внутриклеточного аммиака эндофит неспособен к определению высокого уровня аммиака, что предотвращает отрицательную регуляцию генов фиксации азота. Любой аммиак, попадающий во внутриклеточный компартмент, преобразуется в глутамин. Внутриклеточный уровень глутамина является основным средством определения уровня азота. Снижение внутриклеточного уровня глутамина предотвращает определение клетками высоких уровней аммиака в окружающей среде. Это можно осуществлять, увеличивая уровень экспрессии глутаминазы, фермента, который преобразует глутамин в глутаминат. Кроме того, уровень внутриклеточного глутамина также можно уменьшать, снижая уровень глутаминсинтетазы (фермента, преобразующего аммиак в глутамин). Диазотрофы быстро ассимилируют фиксированный аммиак в глутамин и глутаминат для использования в клеточных процессах. Нарушение ассимиляции аммиака может обеспечить выведение фиксированного азота на экспорт из клетки в виде аммиака. Фиксированный аммиак преимущественно ассимилируется в глутамин глутаминсинтетазой (GS), кодируемой glnA, а затем в глутаминат глутаминоксоглутаратаминотрансферазой (GOGAT). В определенных примерах, glnS кодирует глутаминсинтетазу. GS посттрансляционно регулирует аденилилтрансфераза GS (GlnE), бифункциональный фермент, кодируемый glnE, который катализирует аденилирование и деаденилирование GS вследствие активности его аденилилтрансферазного (AT) и аденилилудаляющего (AR) доменов, соответственно. В условиях ограничения азота экспрессирован glnA и AR домен GlnE деаденилирует GS, обеспечивая ее активность. В условиях избытка азота экспрессия glnA выключена и глутамин аллостерически активирует AT домен GlnE, что вызывает аденилирование и деактивацию GS.
[0127] Кроме того, мишенью генетической вариации для облегчения фиксации азота в полевых условиях способами, описываемыми в настоящем документе, также может являться ген draT. После продукции клеткой ферментов фиксации азота другим уровнем, на котором клетка отрицательно регулирует активность фиксации в условиях высокого содержания азота, является выключение нитрогеназы. Это выключение можно устранить снижая уровень экспрессии DraT.
[0128] Способы придания микроорганизмам новых фенотипов можно проводить на транскрипционном, трансляционном и посттрансляционном уровнях. Транскрипционный уровень включает изменения в промоторе (такие как изменение аффинности к фактору сигма или участков связывания факторов транскрипции, включая делецию всего или части промотора) или изменение терминаторов и аттенюаторов транскрипции. Трансляционный уровень включает изменение в участках связывания рибосом и изменение сигналов деградации иРНК. Посттрансляционный уровень включает мутации активных центров ферментов и изменения белок-белковых взаимодействий. Эти изменения можно осуществлять множеством способов. Снижение уровня экспрессии (или полное устранение) можно осуществлять посредством замены природного участка связывания рибосомы (RBS) или промотора другими с меньшей силой/эффективностью. Участки старта ATG можно заменять на старт-кодоны GTG, TTG или CTG, что приводит к снижению трансляционной активности кодирующей области. Полного устранения экспрессии можно добиваться посредством нокаута (делеции) кодирующей области гена. Сдвиг открытой рамки считывания (ORF) с большой вероятностью приводит к преждевременному стоп-кодону в ORF, таким образом, приводя к получению нефункционального укороченного продукта. Подобным образом, вставка в рамку стоп-кодонов также приводит к получению нефункционального укороченного продукта. Для снижения эффективной концентрации конкретного гена также можно проводить добавление метки разрушения на N- или C-конец.
[0129] И наоборот, определенного уровня экспрессии генов, описываемых в настоящем документе, можно достигать с использованием более сильных промоторов. Для обеспечения высокой активности промотора в условиях высокого уровня азота (или в любых других условиях) можно получать профиль транскрипции всего генома в условиях высокого уровня азота, и из этого массива данных можно выбирать активные промоторы с желаемым уровнем транскрипции для замены слабого промотора. Для улучшения эффективности инициации трансляции слабые старт-кодоны можно заменять старт-кодоном ATG. Также можно заменять слабые участки связывания рибосом (RBS) другими RBS с улучшенной эффективностью инициации трансляции. Кроме того, для изменения активности фермента также можно проводить сайт-специфический мутагенез.
[0130] Увеличение уровня фиксации азота, которое происходит в растении, может приводить к уменьшению количества необходимых химических удобрений для получения сельскохозяйственных культур и уменьшению эмиссии парниковых газов (например, оксида одновалентного азота).
Получение популяций бактерий
Выделение бактерий
[0131] Микроорганизмы, пригодные в способах и композициях, описываемых в настоящем документе, можно получать посредством выделения микроорганизмов с поверхностей или из тканей природных растений. Микроорганизмы можно получать посредством размалывания семян с выделением микроорганизмов. Микроорганизмы можно получать посредством посадки семян в различные образцы почв и выделения микроорганизмов из тканей. Кроме того, микроорганизмы можно получать посредством инокуляции растений экзогенными микроорганизмами и определения того, какие микроорганизмы остаются в тканях растений. Неограничивающие примеры тканей растений могут включать семена, всходы, листья, срезы, растение, луковицы или клубни.
[0132] Способ получения микроорганизмов может представлять собой способ выделения бактерий из почв. Бактерии можно собирать из почв различного типа. В определенном примере почву можно характеризовать такими свойствами, как высокая или низкая плодородность, уровни влажности, уровни минералов и различные практические способы выращивания растений. Например, почва может быть включена в ротацию сельскохозяйственных культур, когда в одну и ту же почву в последовательные сезоны посадки высаживают различные сельскохозяйственные культуры. Последовательное выращивание различных сельскохозяйственных культур на одной и той же почве может предотвратить непропорциональное истощение определенных минералов. Бактерии можно выделять из растений, растущих в выбранных почвах. Проростки растений можно собирать на 2-6 неделях роста. Например, на один цикл сбора можно собрать по меньшей мере 400 изолятов. Типы почв и растений выявляют фенотипы растений, а также условия, которые обеспечивают последующее обогащение определенных фенотипов.
[0133] Микроорганизмы можно выделять из тканей растений для оценки признаков микроорганизмов. Для выделения различных типов ассоциативных микроорганизмов, таких как ризосферные бактерии, эпифиты или эндофиты, параметры обработки образцов тканей можно варьировать. Для обогащения бактериями, осуществляющими фиксацию азота, изоляты можно культивировать в не содержащих азота средах. Альтернативно, микроорганизмы можно получать из всемирных банков штаммов.
[0134] для оценки признаков микроорганизмов анализы проводят в полевых условиях. В определенных вариантах осуществления ткань растения можно обрабатывать для скрининга посредством высокопроизводительной обработки ДНК и РНК. Кроме того, для оценки таких характеристик растений, как колонизация можно использовать неинвазивные измерения. Измерения на микроорганизмах дикого типа можно проводить поочередно от растения к растению. Измерения на микроорганизмах дикого типа также можно проводить в полевых условиях с использованием способов средней производительности. Измерения можно проводить последовательно в течение времени. Можно использовать модельную растительную систему, включая в качестве неограничивающих примеров Setaria.
[0135] Микроорганизмы в растительной системе можно скринировать посредством транскрипционного профилирования микроорганизма в растительной системе. Примерами скрининга посредством транскрипционного профилирования являются использование способов количественной полимеразной цепной реакции (кПЦР), молекулярных штрих-кодов для детекции транскрипов, секвенирования нового поколения и мечения микроорганизмов флуоресцентными метками. Для оценки колонизации в теплице можно измерять факторы воздействия, включая в качестве неограничивающих примеров микробиом, абиотические факторы, почвенные условия, кислород, влажность, температуру, условия инокуляции и локализацию корней. Фиксацию азота у бактерий можно оценивать, определяя газообразный 15N/15N в удобрении (разведение) с использованием IRMS или NanoSIMS, как описано в настоящем документе. NanoSIMS представляет собой масс-спектрометрию вторичных ионов с высоким разрешением. Способ NanoSIMS представляет собой путь для исследования химической активности биологических образцов. Катализ восстановления реакций окисления, которые управляют метаболизмом микроорганизмов, может быть исследован на клеточном, субклеточном, молекулярном и элементном уровнях. NanoSIMS может обеспечивать высокое пространственное разрешение более 0,1 мкм. Посредством NanoSIMS можно детектировать использование изотопных индикаторов, таких как 13C, 15N и 18O. Таким образом, NanoSIMS можно использовать для определения химической активности азот в клетке.
[0136] Для анализа растений можно использовать автоматизированные теплицы. Параметры растений в ответ на воздействие микроорганизмов в качестве неограничивающих примеров включают определение биомассы, анализ хлоропластов, CCD-камеру, измерения волюметрической томографии.
[0137] Одним из способов обогащения популяции микроорганизмов является соответствие генотипу. Например, анализ полимеразной цепной реакцией (ПЦР) с направленным праймером или специфическим праймером. Для идентификации диазотрофов можно использовать праймеры, сконструированные для гена nifH, так как диазотрофы в процессе фиксации азота экспрессируют ген nifH. Популяцию микроорганизмов также можно обогащать посредством одноклеточных независимых от культивирования подходов и подходов к выделению на основе хемотаксиса. Альтернативно, направленное выделение микроорганизмов можно проводить посредством культивирования микроорганизмов на селективных средах. Для заранее спланированных подходов к обогащению популяций микроорганизмов требуемыми признаками можно руководствоваться биоинформатическими данными, и они описаны в настоящем документе.
Обогащение микроорганизмами со способностями к фиксации азота с использованием биоинформатики
[0138] Для идентификации и выделения способствующих росту растений ризобактерий (PGPR), которые выбирают на основе их способности к осуществлению фиксации азота можно использовать биоинформатические способы. Микроорганизмы с высокой способностью к фиксации азота могут способствовать благоприятным признакам в растениях. Биоинформатические способы анализа для идентификации PGPR в качестве неограничивающих примеров включают геномику, метагеномику, направленное выделение, секвенирование генов, секвенирование транскриптома и моделирование.
[0139] Геномный анализ можно использовать для идентификации PGPR и подтверждения присутствия мутаций способами секвенирования нового поколения, как описано в настоящем документе, и контроля версий микроорганизмов.
[0140] Метагеномику можно использовать для идентификации и выделения PGPR с использованием алгоритма прогнозирования для колонизации. Метаданные также можно использовать для идентификации присутствия подвергнутого генетической инженерии штамма в образцах из окружающей среды и теплицы.
[0141] Секвенирование транскриптома можно использовать для прогнозирования генотипов, приводящих к фенотипам PGPR. Кроме того, транскриптомные данные используют для идентификации промоторов для изменения экспрессии генов. Транскриптомные данные можно анализировать в сочетании с полногеномной последовательностью (WGS) с получением моделей метаболизма и генных регуляторных сетей.
Окультуривание микроорганизмов
[0142] Микроорганизмы, выделенные в природе, можно подвергать процессу окультуривания, где микроорганизмы преобразуют в форму, которая является генетически отслеживаемой и идентифицируемой. Одним из способов окультуривания микроорганизма является конструирование в нем свойства устойчивости к антибиотикам. Процесс конструирования устойчивости к антибиотикам может начинаться с определения устойчивости штамма микроорганизмов дикого типа к антибиотикам. Если бактерии чувствительны к антибиотику, то антибиотик может являться хорошим кандидатом для конструирования устойчивости к антибиотикам. Затем в геном микроорганизма рекомбинантными способами можно вводить ген устойчивости к антибиотику или противоселективный суицидный вектор. Противоселективный суицидный вектор может состоять из делеции представляющего интерес ген, селективного маркера и противоселективного маркера sacB. Противоселекцию можно использовать для замены нативных последовательностей ДНК микроорганизмов генами устойчивости к антибиотикам. Для одновременной оценки нескольких микроорганизмов, позволяя параллельное окультуривание, можно использовать способ средней производительности. Альтернативные способы окультуривания включают использования направленных нуклеаз для предотвращения выпетливания последовательностей суицидного вектора или получения промежуточных последовательностей векторов.
[0143] ДНК-содержащие векторы можно вводить в бактерии различными способами, включая электропорацию и химическую трансформацию. Для трансформации можно использовать стандартную библиотеку векторов. Примером способа редактирования генов является CRISPR с предшествующим тестированием Cas9 для гарантии активности Cas9 в микроорганизмах.
Нетрансгенная инженерия микроорганизмов
[0144] Популяцию микроорганизмов с благоприятными свойствами можно получать посредством направленной эволюции. Направленная эволюция представляет собой подход, где имитируют процесс природного отбора для эволюции белков или нуклеиновых кислот в направлении определенной пользователем цели. Примером направленной эволюции является способ, когда в популяцию микроорганизмов вносят случайные мутации, выбирают микроорганизмы с наиболее благоприятными свойствами и продолжают выращивание отобранных микроорганизмов. Наиболее благоприятные свойства у способствующих росту ризобактерий (PGPRs) могут присутствовать в фиксации азота. Способ направленной эволюции может являться повторяющимся и адаптивным на основе процесса отбора после каждого повторения.
[0145] Можно получать способствующие росту растений ризобактерии (PGPR) с высокой способностью к фиксации азота. Эволюцию PGPR можно проводить посредством внесения генетической вариации. Генетическую вариацию можно вносить посредством мутагенеза с использованием полимеразной цепной реакции, сайт-специфического мутагенеза, насыщающего мутагенеза, мутагенеза с перестановкой фрагментов, гомологичной рекомбинации, систем CRISPR/Cas9, химического мутагенеза и их сочетаний. Посредством этих подходов можно вносить случайные мутации в популяцию микроорганизмов. Например, мутантов можно получать с использованием синтетической ДНК или РНК посредством сайт-специфического мутагенеза. Мутантов можно получать с использованием средств, содержащихся в плазмидах, которые позже исправляют. Представляющие интерес гены можно идентифицировать с использованием библиотек других видов с улучшенными свойствами, включая в качестве неограничивающих примеров улучшенные свойства PGPR, улучшенную колонизацию злаковых, увеличенную чувствительность к кислороду, увеличенную фиксацию азота и увеличенное выведение аммиака. Внутриродовые гены можно конструировать на основе этих библиотек с использованием программного обеспечения, такого как программное обеспечение Geneious или Platypus design. Мутации можно конструировать с помощью машинного обучения. Мутации можно конструировать с помощью модели метаболизма. Автоматизированное конструирование мутаций можно проводить с использованием средств, подобных Platypus, и они будут формировать РНК для Cas-направленного мутагенеза.
[0146] Внутриродовые гены можно переносить в микроорганизм-хозяин. Кроме того, в микроорганизм также можно переносить репортерные системы. Репортерные системы характеризуют промоторы, определяют успешность трансформации, обеспечивают скрининг мутантов и действуют в качестве средств отрицательного отбора.
[0147] Микроорганизмы, несущие мутацию, можно культивировать посредством серийных пересевов. Колония микроорганизмов содержит один вариант микроорганизма. Колонии микроорганизмов подвергают скринингу с помощью автоматизированного сборщик колоний и жидкостного манипулятора. Мутанты с дупликацией гена и увеличенным количеством копий экспрессируют более высокий генотип требуемого признака.
Отбор способствующих росту растений микроорганизмов на основе фиксации азота
[0148] Колонии микроорганизмов можно скринировать с использованием различных анализов оценки фиксация азота. Одним из способов определения фиксации азота является анализ с применением одного фермента, в котором определяют выведение азота. Альтернативный способ представляют собой анализ восстановления ацетилена (ARA) с непрерывным получением образцов в течение времени. ARA можно проводить в высокопроизводительных планшетах из массивов микропробирок. ARA можно проводить для живых растений и тканей растений. Составы сред и концентрацию кислорода в средах при анализах ARA можно варьировать. Другим способом скрининга вариантов микроорганизмов является использование биосенсоров. Для исследования активности микроорганизмов можно использовать NanoSIMS и микроспектроскопию Raman. В определенных случаях бактерии также можно культивировать и наращивать способами ферментации в биореакторах. Биореакторы конструируют для увеличения устойчивости роста бактерий и снижения чувствительности бактерий к кислороду. Для оценки чувствительности к кислороду, потребности в питательных веществах, фиксации азота и выведения азота используют среду для микроферментеров на основе высокопроизводительных планшетов. Бактерии также можно совместно культивировать с конкурирующими или благоприятными микроорганизмами для определения скрытых путей. Проточную цитометрию можно использовать для скрининга бактерий, которые продуцируют высокие уровни азота с использованием химических, колориметрических или флуоресцентных индикаторов. Бактерии можно культивировать в присутствии или отсутствии источника азота. Например, бактерии можно культивировать с глутамином, аммиаком, мочевиной или нитратами.
Выведение микроорганизмов
[0149] Выведение микроорганизмов представляет собой способ систематической идентификации и улучшения роли видов в микробиоме сельскохозяйственной культуры. Способ включает три этапа: 1) отбор видов-кандидатов посредством картирования взаимодействий растение-микроорганизм и прогноза регуляторной сети, сцепленной с конкретным фенотипом, 2) практическое и прогнозируемое улучшение фенотипов микроорганизмов посредством внутривидового скрещивания регуляторных сетей и кластеров генов и 3) скрининг и отбор новых генотипов микроорганизмов, которые обеспечивают требуемые фенотипы сельскохозяйственных культур. Для систематического анализа улучшения штаммов получена модель, в которой сцеплена динамика колонизации сообщества микроорганизмов с генетической активностью ключевых видов. Модель используют для прогноза выведения генетических мишеней и увеличения частоты отбора улучшений кодируемых микробиомом свойств агротехнического значения. Для графического представления варианта осуществления процесса см., фигуру 17A. Конкретно, на фигуре 17A приведена схема выведения микроорганизмов по вариантам осуществления. Как проиллюстрировано на фигуре 17A, рациональное улучшение микробиома сельскохозяйственной культуры можно использовать для увеличения биоразнообразия почв, настройки воздействия ключевых видов и/или изменения сроков и экспрессии важных метаболических путей. С этой целью авторы изобретения разработали конвейер выведения микроорганизмов для идентификации и улучшения роли штаммов в микробиоме сельскохозяйственной культуры. Способ включает три этапы: 1) отбор видов-кандидатов посредством картирования взаимодействий растение-микроорганизм и прогноза регуляторной сети, сцепленной с конкретным фенотипом, 2) практическое и прогнозируемое улучшение фенотипов микроорганизмов посредством внутривидового скрещивания регуляторных сетей и кластеров генов и 3) скрининг и отбор новых генотипов микроорганизмов, которые обеспечивают требуемые фенотипы сельскохозяйственных культур. Для систематического анализа улучшения штаммов авторы изобретения используют модель, в которой сцеплена динамика колонизации сообщества микроорганизмов с генетической активностью ключевых видов. Этот процесс представляет собой методологию для выведения и отбора улучшений кодируемых микробиомом свойств агротехнического значения.
[0150] Получения бактерий для улучшения признаков растений (например, фиксации азота) можно достигать посредством серийного пересева. Получение этих бактерий в дополнение к идентификации бактерий и/или композиций, способных к обеспечению одного или нескольких улучшенных признаков в одном или нескольких растениях, можно проводить посредством отбора растений с конкретным улучшенным признаком, на который влияет микробная флора. Один из способов получения бактерий для улучшения признака растения включает этапы: (a) выделения бактерий из ткани или почвы первого растения; (b) внесение генетической вариации в одну или несколько бактерий с получением одного или нескольких вариантов бактерий; (c) воздействие вариантами бактерий на множество растений; (d) выделение бактерий из ткани или почвы одного из множества растений, где растение, из которого выделяют бактерии, обладает улучшенным признаком относительно других растений в множестве растений; и (e) повторение этапов (b)-(d) с бактериями, выделяемыми из растения с улучшенным признаком (этап (d)). Этапы (b)-(d) можно повторять любое количество раз (например, однократно, дважды, три раза, четыре раза, пять раз, десять раз или более) до достижения улучшенным признаком растения желаемого уровня. Кроме того, множество растений может представлять собой более двух растений, например, от 10 до 20 растений или 20 или более, 50 или более, 100 или более, 300 или более, 500 или более или 1000 или более растений.
[0151] В дополнение к получению растения с улучшенным признаком, получают популяцию бактерий, содержащую бактерии, несущие одну или несколько генетических вариаций, вносимых в один или несколько генов (например, генов, регулирующих фиксацию азота). Повторяя описанные выше этапы, можно получать популяцию бактерий, которая содержит наиболее подходящих представителей популяции, которая коррелирует с представляющим интерес признаком растения. Бактерии в этой популяции можно идентифицировать и определить их благоприятные свойства, например, посредством генетического и/или фенотипического анализа. Генетический анализ можно проводить у выделенных на этапе (a) бактерий. Фенотипическую и/или генотипическую информацию можно получать с использованием способов, включающих: высокопроизводительный скрининг химических компонентов растительного происхождения, способы секвенирования, включающие высокопроизводительное секвенирование генетического материала, способы дифференциального дисплея (включая ДДОТ-ПЦР и ДД-ПЦР), способы с микропанелями нуклеиновых кислот, РНК-сек (секвенирование методом дробовика всего генома) и кПЦР-РВ (количественную ПЦР с детекцией в реальном времени). Полученную информацию можно использовать для получения профилирующей популяцию информации об идентификационных характеристиках и активности присутствующих бактерий, например, филогенетического анализа или основанного на микропанелях скрининга нуклеиновых кислот, кодирующих компоненты оперонов рРНК или других таксономически информативных локусов. Примеры таксономически информативных локусов включают ген 16S рРНК, ген 23S рРНК, ген 5S рРНК, ген 5.8S рРНК, ген 12S рРНК, ген 18S рРНК, ген 28S рРНК, ген gyrB, ген rpoB, ген fusA, ген recA, ген coxl, ген nifD. Примеры способов таксономического профилирования для определения таксонов, присутствующих в популяции описаны в US 20140155283. Идентификация бактерий может включать характеризующую активность одного или нескольких генов или оного или нескольких путей передачи сигнала, например, генов, ассоциированных с путем фиксации азота. также в бактериальных популяциях могут присутствовать синергические взаимодействия (когда два компонента при их комбинации увеличивают требуемое действие более чем аддитивно) между различными видами бактерий.
Генетическая вариация - локализация и источники альтераций генома
[0152] Генетическая вариация может представлять собой ген, выбранный из группы, состоящей из: nifA, nifL, ntrB, ntrC, glnA, glnB, glnK, draT, amtB, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB и nifQ. Генетическая вариация может представлять собой вариацию в гене, кодирующем белок с функциональностью, выбранной из группы, состоящей из: глутаминсинтетазы, глутаминазы, аденилаттрансферазы глутаминсинтетазы, активатора транскрипции, антиактиватора транскрипции, пируват:флаводоксин-оксидоредуктазы, флаводоксина или АДФ-D-рибозилтрансферазы НАД+:динитрогеназоредуктазы. Генетическая вариация может представлять собой мутацию, которая приводит к одному или нескольким из: повышенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности NifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, AmtB; сниженной активности удаления аденилата GlnE или сниженной активности удаления уридилата GlnD. Внесение генетической вариации может включать вставку и/или делецию одного или нескольких нуклеотидов в намеченном участке, например, 1, 2, 3, 4, 5, 10, 25, 50, 100, 250, 500 или более нуклеотидов. Генетическая вариация, вносимая в одну или несколько бактерий способами, описываемыми в настоящем документе, может представлять собой нокаут-мутацию (например, делецию промотора, вставку или делецию с получением преждевременного стоп-кодона, делецию всего гена), или она может представлять собой удаление или устранение активности белкового домена (например, точечную мутацию влияющую на активный центр, или делецию части гена, кодирующей значимую часть белкового продукта), или она может представлять собой изменение или удаление регуляторной последовательности гена-мишени. Также можно вставлять одну или несколько регуляторных последовательностей, включая гетерологичные регуляторные последовательности и регуляторные последовательности, выявленные в геноме вида или рода бактерий, соответствующих бактериям, у которых проводят генетическую вариацию. Кроме того, регуляторные последовательности можно выбирать на основе уровня экспрессии гена в культуре бактерий или в ткани растения. Генетическая вариация может представлять собой предопределенную генетическую вариацию, которую конкретно вводят в намеченный участок. Генетическая вариация может представлять собой случайную мутацию в намеченном участке. Генетическая вариация может представлять собой вставку или делецию одного или нескольких нуклеотидов. В некоторых случаях в одну или несколько выделенных бактерий перед воздействием бактерий на растения для оценки улучшения признака вносят множество различных генетических вариаций (например, 2, 3, 4, 5, 10 или более). Множество генетических вариаций может являться любым из описанных выше типов, одним и тем же или различными типами и любой их комбинацией. В некоторых случаях последовательно проводят множество различных генетических вариаций, проводя первую генетическую вариацию после первого этапа выделения, вторую генетическую вариацию после второго этапа выделения, и т.д. так, чтобы накопить множество генетических вариаций в бактериях постепенно обеспечивая улучшенные признаки у ассоциированных растений.
Генетическая вариация - способы внесения альтераций генома
[0153] В основном термин "генетическая вариация" относится к любому изменению, вносимому в полинуклеотидную последовательность относительно эталонного полинуклеотида, такого как эталонный геном или его часть, или эталонный ген или его часть. Генетическую вариацию можно обозначать как "мутацию", а последовательность или организм, содержащие генетическую вариацию можно обозначать как "генетический вариант" или "мутант". Генетические вариации могут обладать рядом эффектов, таким как повышение или снижение определенного вида биологической активности, включение экспрессии гена, метаболизма и клеточной сигнализации. Генетические вариации можно конкретно вносить в намеченный участок или вносить случайным образом. Для внесения генетических вариаций доступен ряд средств и способов молекулярной биологии. Например, генетическую вариацию можно вносить посредством мутагенеза при полимеразной цепной реакции, сайт-специфического мутагенеза, насыщающего мутагенеза, мутагенеза с перестановкой фрагментов, гомологичной рекомбинации, рекомбинационной инженерии, опосредованной лямбда красным рекомбинация, систем CRISPR/Cas9, химического мутагенеза и их сочетаний. Химические способы внесения генетических вариаций включают воздействие на ДНК химическим мутагеном, например, этилметансульфонатом (EMS), метилметансульфонатом (MMS), N-нитрозомочевиной (EN U), N-метил-N-нитро-N'-нитрозогуанидином, 4-нитрохинолин-N-оксидом, диэтилсульфатом, бензопиреном, циклофосфамидом, блеомицином, триэтилмеламином, мономерным акриламидом, азотистым ипритом, винкристином, диэпоксиалканами (например, диэпоксибутаном), ICR-170, формальдегидом, гидрохлоридом прокарбазина, этиленоксидом, диметилнитрозамином, 7,12 диметилбенз(a)антраценом, хлорамбуцилом, гексаметилфосфамидом, бисульфаном и т.п. Индуцирующие мутации посредством облучения средства включают ультрафиолетовое облучение, γ-облучение, рентгеновские лучи и бомбардировку быстрыми нейтронами. Также генетические вариации можно вносить в нуклеиновые кислоты, например, с использованием триметилпсоралена с ультрафиолетовым облучением. Другим подходящим способом получения генетической вариации является случайная или направленная вставка мобильного элемента ДНК, например, транспозона. Генетические вариации можно вносить в нуклеиновую кислоту при амплификации в бесклеточной системе in vitro, например, с использованием способа полимеразной цепной реакции (ПЦР), такого как ПЦР со сниженной точностью. Генетические вариации можно вносить в нуклеиновые кислоты in vitro с использованием способы перестановки ДНК (например, перестановки экзонов, обмены доменов и т.п.). Генетические вариации также можно вносить в нуклеиновые кислоты как результат недостаточности в клетке ферментов репарации ДНК, например, полагают, что присутствие в клетке мутантного гена, кодирующего мутантный фермент репарации ДНК, приведет к получению высокой частоты мутаций (т.е., приблизительно от 1 мутации/100 генов до 1 мутации/10000 генов) в геноме клетки. Примеры генов, кодирующих ферменты репарации ДНК в качестве неограничивающих примеров включают Mut H, Mut S, Mut L, и Mut U и их гомологи у других видов (например, от MSH1 до MSH6, от PMS1 до PMS2, MLH1, GTBP, ERCC-1 и т.п.). Примеры описаний различных способов внесения генетических вариации приведены, например, в Stemple (2004) Nature 5:1-7; Chiang et al. (1993) PCR Methods Appl 2(3): 210-217; Stemmer (1994) Proc. Natl. Acad. Sci. USA 91:10747-10751 и в патентах США №№ 6033861 и 6773900.
[0154] Генетические вариации, вносимые в микроорганизмы, можно классифицировать как трансгенные, цисгенные, внутригеномные, внутриродовые, межродовые, синтетические, эволюционировавшие, полученные посредством перестановки или SNP.
[0155] Для обеспечения улучшений описанных выше свойств генетическую вариацию можно вносить во множество метаболических путей в микроорганизмах. Характерные пути включают пути захвата серы, биосинтез гликогена, путь регуляции глутамина, путь захвата молибдена, путь фиксации азота, ассимиляцию аммиака, экскрецию или секрецию аммиака, захват азота, биосинтез глутамина, анаэробное окисление аммония, растворение фосфата, транспорт органических кислот, продукцию органических кислот, продукцию агглютитинов, гены захвата свободных реакционноспособных радикалов кислорода, биосинтез индолуксусной кислоты, биосинтез трегалозы, ферменты или пути разрушения клеточной стенки растений, гены крепления корней, секрецию экзополисахаридов, путь синтеза глутаминатсинтазы, пути захвата железа, пути сидерофора, хитиназный путь, ACC-дезаминазу, биосинтез глутатиона, гены передачи сигнала посредством фосфатов, путь гашения, пути цитохромов, путь гемоглобина, путь бактериального гемоглобин-подобного белка, малых-РНК rsmZ, биосинтез ризобиотоксина, белок адгезии lapA, путь чувства кворума AHL, биосинтез феназина, биосинтез циклического липопептида и продукцию антибиотиков.
[0156] Для внесения требуемых мутаций можно использовать системы CRISPR/Cas9 (короткие палиндромные повторы, регулярно расположенные группами)/ассоциированные с CRISPR (Cas). CRISPR/Cas9 обеспечивают бактериям и археям приобретенный иммунитет против вирусов и плазмид посредством использования CRISPR РНК (кпРНК), направляя сайленсинг внедряющихся нуклеиновых кислот. Белок Cas9 (или его функциональный эквивалент и/или вариант, т.е., подобный Cas9 белок) в природных условиях обладает ДНК-эндонуклеазной активностью, которая зависит от ассоциации белка с двумя природными или синтетическими молекулами РНК, называемыми кпРНК и транскпРНК (также называемыми направляющими РНК). В некоторых случаях, две молекулы ковалентно связаны с формированием одной молекулы (также называемой одиночной направляющей РНК ("онРНК"). Таким образом, белок Cas9 или подобный Cas9 белок ассоциируют с направленной к ДНК РНК (где этот термин включает конфигурацию с двумя направляющими молекулами РНК и конфигурацию с одиночной направляющей молекулой РНК), которая активирует Cas9 или подобный Cas9 белок и направляет белок к последовательности нуклеиновой кислоты-мишени. Если Cas9 или подобный Cas9 белок сохраняют свою ферментативную функцию, они расщепляют ДНК-мишень с образованием двухцепочечного разрыва, что может приводить к изменению генома (т.е., редактированию: делеции, вставке (когда присутствует полинуклеотид-донор), замене и т.д.), таким образом, изменяя экспрессию генов. Определенные варианты Cas9 (которые включены в термин подобные Cas9) изменяют так, чтобы они обладали сниженной активностью расщепления ДНК (в некоторых случаях они расщепляют одну цепь вместо обеих цепей ДНК-мишени, тогда как в других случаях их активность сильно снижают до отсутствия активности расщепления ДНК). Дополнительные иллюстративные описания систем CRISPR для внесения генетических вариаций можно найти, например, в US 8795965.
[0157] В качестве способа циклической амплификации в мутагенезе при полимеразной цепной реакции (ПЦР) для внесения желаемых мутаций используют мутагенные праймеры. ПЦР проводят в виде циклов денатурации, отжига и достройки. После амплификации посредством ПЦР отбор мутантной ДНК и удаление исходной плазмидной ДНК можно проводить посредством: 1) замены dCTP в течение ПЦР гидроксиметилированным dCTP, с последующим расщепление рестрикционными ферментами с удалением только негидроксиметилированной исходной ДНК; 2) одновременного мутагенеза гена устойчивости к антибиотикам и исследуемого гена, изменяя у плазмиды устойчивость к антибиотикам на устойчивость к другим антибиотикам, где устойчивость к новым антибиотикам затем облегчает отбор требуемой мутации; 3) расщепление исходной метилированной матричной ДНК после внесения требуемой мутации рестрикционным ферментом DpnI, который расщепляет только метилированную ДНК, посредством чего происходит выделение мутантных неметилированных цепей; или 4) получения кольцевых молекул мутантных продуктов ПЦР в дополнительной реакции лигирования с увеличением эффективности трансформации мутантной ДНК. Дополнительное описание иллюстративных способов можно найти, например, в US 7132265, US 6713285, US 6673610, US 6391548, US 5789166, US 5780270, US 5354670, US 5071743 и US 20100267147.
[0158] В олигонуклеотид-специфическом мутагенезе, также называемом сайт-специфическим мутагенезом, как правило, используют синтетический праймер ДНК. Этот синтетический праймер содержит требуемую мутацию и комплементарен матричной ДНК вокруг участка мутации так, что он может гибридизоваться с ДНК в представляющем интерес гене. Мутация может представлять собой однонуклеотидную замену (точечную мутацию), замену нескольких нуклеотидов, делецию или вставку или их комбинацию. Затем одноцепочечный праймер достраивают с использованием ДНК-полимеразы, которая копирует остальную часть гена. Копированный таким образом ген содержит мутантный участок, и его затем можно вносить в клетку-хозяина в виде вектора и клонировать. В заключение мутантов можно отбирать посредством секвенирования ДНК для проверки того, что они содержат требуемую мутацию.
[0159] Генетические вариации можно вносить с использованием ПЦР со сниженной точностью. В этом способе представляющий интерес ген амплифицируют с использованием ДНК-полимеразы в условиях недостаточности точности репликации последовательности. Результатом является то, что продукты амплификации содержат по меньшей мере одну ошибку в последовательности. Когда ген амплифицируют и полученный продукт(ы) реакции содержит одно или несколько изменений последовательности по сравнению с матричной молекулой, полученные продукты являются мутантными по сравнению с матрицей. Другим способом внесения случайных мутаций является воздействие на клетки химическим мутагеном, таким как нитрозогуанидин или этилметансульфонат (Nestmann, Mutat Res 1975 June; 28(3):323-30), а затем у хозяина выделяют вектор, содержащий ген.
[0160] Другим способом случайного мутагенеза является насыщающий мутагенез, в котором стараются получить все или почти все возможные мутации в конкретном участке или небольшой области гена. В широком смысле насыщающий мутагенез состоит из мутагенеза полного набора мутагенных кассет (где длина каждой кассеты составляет, например, 1-500 оснований) в определенной полинуклеотидной последовательности для мутагенеза (где длина последовательности для мутагенеза составляет, например, от 15 до 100000 оснований). Таким образом, в каждую подвергаемую мутагенезу кассету вносят группу мутаций (например, в диапазоне от 1 до 100 мутаций). Группа мутаций, вносимых в одну кассету, может отличаться от второй группы мутаций, вносимых во вторую кассету в течение одного раунда насыщающего мутагенеза, или являться такой же. Примерами таких групп являются делеции, добавления, группы конкретных кодонов и группы конкретных нуклеотидных кассет.
[0161] Мутагенез с перестановкой фрагментов, также называемый перестановкой ДНК, является способом быстрой репродукции благоприятных мутаций. В примере способ перестановки, используют ДНКазу для фрагментации набора исходных генов на части длиной, например, приблизительно 50-100 п.н. После этого проводят полимеразную цепную реакцию (ПЦР) без праймеры - фрагменты ДНК с достаточным перекрыванием гомологичных последовательностей отжигаются друг с другом и затем их достраивает ДНК-полимераза. После достижения некоторыми молекулами ДНК размера исходных генов, проводят несколько циклов этой достройки посредством ПЦР. Затем эти гены можно амплифицировать посредством другой ПЦР, в этот раз с добавлением праймеров, которые сконструированы так, чтобы они были комплементарны концам цепей. Праймеры могут содержать дополнительные последовательности, добавляемые на их 5'-концы, такие как последовательности участков распознавания рестрикционных ферментов, необходимые для лигирования в клонирующий вектор. Дополнительные примеры способов перестановок приведены в US 20050266541.
[0162] Мутагенез посредством гомологичной рекомбинации включает рекомбинацию экзогенного фрагмента ДНК и полинуклеотидной последовательности-мишени. После образования двухцепочечных разрывов происходит удаление участков ДНК на 5'-концах разрывов в процессе, называемом резекцией. На последующем этапе внедрения цепи выступающий 3'-конец разорванной молекулы ДНК затем "внедряется" в сходную или идентичную молекулу ДНК, которая не подвергалась разрыву. Способ можно использовать для удаления генов, удаления экзонов, добавления генов и внесения точечных мутаций. Мутагенез посредством гомологичной рекомбинации может являться постоянным или условным. Как правило, также предоставляют матрицу рекомбинации. Матрица рекомбинации может представлять собой компонент другого вектора, содержащийся в отдельном векторе, или ее предоставляют в виде отдельного полинуклеотида. В определенных вариантах осуществления матрицу рекомбинации конструируют так, чтобы она служила в качестве матрицы в гомологичной рекомбинации, например, в пределах или рядом с последовательностью-мишенью, разрезаемой или расщепляемой сайт-специфической нуклеазой. Длина матричного полинуклеотида может являться любой подходящей длиной, например, длина может составлять приблизительно или более чем приблизительно 10, 15, 20, 25, 50, 75, 100, 150, 200, 500, 1000 или более нуклеотидов. В определенных вариантах осуществления матричный полинуклеотид комплементарен части полинуклеотида, содержащего последовательность-мишень. При оптимальном выравнивании матричныый полинуклеотид может перекрываться с одним или несколькими нуклеотидами последовательности-мишени (например, приблизительно или более чем приблизительно с 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100 или более нуклеотидами). В определенных вариантах осуществления, когда матричная последовательность и полинуклеотид, содержащий последовательность-мишень, оптимально выровнены, ближайший нуклеотид матричного полинуклеотида находится в пределах приблизительно 1, 5, 10, 15, 20, 25, 50, 75, 100, 200, 300, 400, 500, 1000, 5000, 10000 или более нуклеотидов от последовательности-мишени. Неограничивающие примеры сайт-специфических нуклеаз, пригодных в способах гомологичной рекомбинации, включают нуклеазы с цинковыми пальцами, нуклеазы CRISPR, нуклеазы TALE и мегануклеазу. Для дополнительного описания использования таких нуклеаз, см. например, US 8795965 и US 20140301990.
[0163] Для получения генетических вариаций можно использовать мутагены, посредством которых преимущественно получают точечные мутации и короткие делеции, вставки, трансверсии и/или транзиции, включая химические мутагены или облучение. В качестве неограничивающих примеров мутагены включают этилметансульфонат, метилметансульфонат, N-этил-N-нитрозомочевину, триэтилмеламин, N-метил-N-нитрозомочевину, прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, мономерный акриламид, мелфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитронитрозогуанидин, нитрозогуанидин, 2-аминопурин, 7,12-диметилбенз(a)антрацен, этиленоксид, гексаметилфосфамид, бисульфан, диэпоксиалканы (диэпоксиоктан, диэпоксибутан и т.п.), дигидрохлорид 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина и формальдегид.
[0164] Внесение генетической вариации может представлять собой незавершенный процесс так, что определенные бактерии в обрабатываемой популяции бактерий несут требуемую мутацию, тогда как другие нет. В определенных случаях для обогащения бактериями, несущими требуемую генетическую вариацию необходимо применять селекционное давление. Как правило, отбор успешных генетических вариантов включает отбор по или против определенной функциональности, придаваемой или устраняемой генетической вариацией, так как в случае вставки гена устойчивости к антибиотикам или устранения метаболической активности, обеспечивающей возможность преобразования нелетального соединения в летальный метаболит. Также селекционное давление можно применять на основе самой полинуклеотидной последовательности так, что необходимо внести только требуемую генетическую вариацию (например, также без необходимости селективного маркера). В этом случае, селекционное давление может включать расщепление геномов без генетической вариации, вносимой в участок-мишень так, что отбор эффективно направлен против эталонной последовательности в которую необходимо внести генетическую вариацию. Как правило, расщепление происходит в пределах 100 нуклеотидов от участка-мишени (например, в пределах 75, 50, 25, 10 или менее нуклеотидов от участка-мишени, включая расщепление в участке-мишени или в его пределах). Расщепление может направлять сайт-специфическая нуклеаза, выбранная из группы, состоящей из нуклеазы с цинковыми пальцами, нуклеазы CRISPR, нуклеазы TALE (TALEN) или мегануклеазы. Такой процесс аналогичен процессам увеличения гомологичной рекомбинации в участке-мишени, за исключением того, что не предоставлена матрица для гомологичной рекомбинации. В результате в бактериях без требуемой генетической вариации с большей вероятностью проходит расщепление, которое в отсутствии восстановления приводит к гибели клеток. Затем бактерии, выжившие в отборе, можно выделять для применения в воздействии на растения для оценки придаваемого улучшенного признака.
[0165] Для направления расщепления в участке-мишени можно использовать нуклеазу CRISPR. Используя Cas9 для уничтожения немутантных клеток, можно обеспечивать улучшенный отбор мутантных микроорганизмов. Затем растениям инокулируют мутантные микроорганизмы для подтверждения симбиоза и обеспечения эволюционного давления с отбором эффективных симбионтов. Затем микроорганизмы можно повторно выделять из тканей растений. В системах с нуклеазами CRISPR, применяемых для отбора относительно отсутствия вариантов, можно использовать элементы, сходные с описанными выше в отношении внесения генетических вариаций элементами, за исключением того, что не предоставляют матрицу для гомологичной рекомбинации. Таким образом, расщепление, направленное на участок-мишень увеличивает гибель подвергшихся ему клеток.
[0166] Доступны другие варианты специфически индуцируемого расщепления в участке-мишени, такие как нуклеазы с цинковыми пальцами, системы нуклеазы TALE (TALEN) и мегануклеаза. Нуклеазы с цинковыми пальцами (ZFN) представляют собой искусственные эндонуклеазы ДНК, получаемые посредством слияния связывающего ДНК домена цинковых пальцев с доменом расщепления ДНК. ZFN можно конструировать для расщепления требуемых последовательностей ДНК и это позволяет нуклеазам с цинковыми пальцами расщеплять уникальные последовательности-мишени. При введении в клетку ZFN можно использовать для редактирования ДНК-мишени в клетке (например, генома клетки) посредством индукции двухцепочечных разрывов. Подобные активаторам транскрипции эффекторные нуклеазы (TALEN) представляют собой искусственные эндонуклеазы ДНК, получаемые посредством слияния эффекторного ДНК-связывающего домена TAL (подобного активаторам транскрипции) с доменом расщепления ДНК. TALEN можно быстро конструировать для связывания практически любой требуемой последовательности ДНК и при введении в клетку, TALEN можно использовать для редактирования ДНК-мишени в клетке (например, генома клетки) посредством внесения двухцепочечных разрывов. Мегануклеазы (хоминг-эндонуклеаза) представляют собой эндодезоксирибонуклеазы, характеризууемые большим участком распознавания (двухцепочечных последовательностей ДНК длиной от 12 до 40 пар оснований). Мегануклеазы можно использовать для замещения, удаления или модификации последовательностей узконаправленным способом. Модифицируя узнаваемые ими последовательности посредством белковой инженерии, можно изменять последовательность-мишень. Мегануклеазы можно использовать для модификации всех типов геномов, бактерий, растений или животных и, как правило, их группируют в четыре семейства: семейство LAGLIDADG (SEQ ID NO:1), семейство GIY-YIG, семейство His-Cyst box и семейство HNH. Иллюстративные хоминг-эндонуклеазы включают I-SceI, I-CeuI, PI-PspI, PI-Sce, I-SceIV, I-CsmI, I-PanI, I-SceII, I-PpoI, I-SceIII, I-CreI, I-TevI, I-TevII и I-TevIII.
Генетическая вариация - Способы идентификации
[0167] Микроорганизмы по настоящему изобретению можно идентифицировать по одной или нескольким генетическим модификациям или альтерациям, которые внесены в указанный микроорганизм. Одним из способов, которым можно идентифицировать указанные генетические модификацию или альтерацию, является сравнение с SEQ ID NO, которая содержит часть геномной последовательности микроорганизма, достаточную для идентификации генетической модификации или альтерации.
[0168] Кроме того, в случае микроорганизмов, которые не должны нести генетических модификаций или альтераций (например, дикого типа), вносимых в их геномы, по описанию можно использовать последовательности нуклеиновых кислот 16S для идентификации указанных микроорганизмов. Последовательность нуклеиновой кислоты 16S представляет собой пример "молекулярного маркера" или "генетического маркера", который обозначает индикатор, который используют в способах визуализации различий характеристик последовательностей нуклеиновых кислот. Примерами других таких индикаторов являются метки полиморфизма длин рестрикционных фрагментов (ПДРФ), метки полиморфизма длин амплифицированных фрагментов (AFLP), однонуклеотидные полиморфизмы (SNP), мутации вставок, микросателлитные метки (SSR), характеризуемые последовательностями амплифицированные области (SCAR), метки рестрикционных полиморфизмов амплифицированных последовательностей (CAPS) или метки изоферментов или комбинации меток, описываемых в настоящем разделе, что определяет конкретное генетическое и хромосомное положение. Метки дополнительно включают полинуклеотидные последовательности, кодирующие 16S или 18S рРНК, и внутренние транскрибируемые спейсерные (ITS) последовательности, которые представляют собой последовательности, находящиеся между генами малой субъединицы и большой субъединицы рРНК, которые оказались особенно пригодными при определении сходства или различий при сравнении друг с другом. Кроме того, для идентификации микроорганизмов, описываемых в настоящем документе, по описанию использованы уникальные последовательности, выявленные в представляющих интерес генах (например, nif H,D,K,L,A, glnE, amtB и т.д.).
[0169] Первичная структура большой субъединицы рРНК 16S содержит конкретную комбинацию консервативных, вариабельных и гипервариабельных областей, которые эволюционируют с разными скоростями и позволяют различать как очень древние линии, такие как домены, так и более современные линии, такие как роды. Вторичная структура субъединицы 16S содержит приблизительно 50 спиралей, что приводит к спариванию оснований приблизительно 67% остатков. Эти высококонсервативные вторичные структурные признаки имеют большую функциональную важность, и их можно использовать для обеспечения позиционной гомологии при множественном выравнивании последовательностей и филогенетическом анализе. За последние несколько десятилетий ген 16S рРНК стал наиболее секвенированным таксономическим маркером, и он является краеугольным камнем современной систематической классификации бактерий и архей. (Yarza et al. 2014. Nature Rev. Micro. 12:635-45).
[0170] Таким образом, В определенных аспектах по описанию предусмотрена последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с любой последовательностью в таблицах E, F, G или H.
[0171] Таким образом, В определенных аспектах по описанию предусмотрен микроорганизм, который содержит последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с SEQ ID NO: 62-303. Эти последовательности и ассоциированные с ними описания можно найти в таблицах F, G и H.
[0172] В определенных аспектах по описанию предусмотрен микроорганизм, который содержит последовательность нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269, 277-283. Эти последовательности и ассоциированные с ними описания можно найти в таблицах G и H.
[0173] В определенных аспектах по описанию предусмотрен микроорганизм, который содержит последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276, 284-295. Эти последовательности и ассоциированные с ними описания можно найти в таблицах G и H.
[0174] В определенных аспектах по описанию предусмотрен микроорганизм, который содержит последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с SEQ ID NO: 177-260, 296-303. Эти последовательности и ассоциированные с ними описания можно найти в таблицах G и H.
[0175] В определенных аспектах по описанию предусмотрен микроорганизм, или праймер, или зонд, или последовательность с неприродным соединением, которые содержат, последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с SEQ ID NO: 304-424. Эти последовательности и ассоциированные с ними описания можно найти в таблице E.
[0176] В определенных аспектах по описанию предусмотрен микроорганизм, который содержит последовательность с неприродным соединением, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности с SEQ ID NO: 372-405. Эти последовательности и ассоциированные с ними описания можно найти в таблице E.
[0177] В определенных аспектах по описанию предусмотрен микроорганизм, который содержит аминокислотную последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична по последовательности с SEQ ID NO: 77, 78, 81, 82 или 83. Эти последовательности и ассоциированные с ними описания можно найти в таблицах F и H.
Генетическая вариация - Способы детекции: праймеры, зонды и анализы
[0178] В настоящем описании указаны праймеры, зонды и анализы, которые пригодны для детекции микроорганизмов, исследуемых в настоящем документе. В определенных аспектах по описанию предусмотрены способы детекции родительских штаммов дикого типа. В других аспектах по описанию предусмотрены способы детекции не являющихся межродовыми подвергнутых генетической инженерии микроорганизмов, полученных из штаммов дикого типа. В определенных аспектах настоящее изобретение относится к способам идентификации не являющихся межродовыми генетических альтераций в микроорганизме.
[0179] В определенных аспектах геноинженерные способы по настоящему изобретению приводят к получению в не являющихся межродовыми микроорганизмах последовательностей с неприродным "соединением" нуклеотидов. Эти неприродные соединения нуклеотидов можно использовать в качестве разновидности диагностики, что является показателем присутствия в микроорганизме, исследуемом в настоящем документе, конкретной генетической альтерации.
[0180] Способами по настоящему изобретению можно детектировать эти неприродные соединения нуклеотидов посредством использования специализированных способов количественной ПЦР, включая уникальным образом сконструированные праймеры и зонды. В определенных аспектах зонды по изобретению связываются с последовательностями с неприродным соединением нуклеотидов. В определенных аспектах используют традиционную ПЦР. В других аспектах используют ПЦР с детекцией в реальном времени. В определенных аспектах используют количественную ПЦР (кПЦР).
[0181] Таким образом, описание может включать использование двух общих способов детекции продуктов ПЦР с детекцией в реальном времени: (1) неспецифические флуоресцентные красители, которые интеркалируют любую двухцепочечную ДНК, и (2) специфичные для последовательности зонды ДНК, состоящие из олигонуклеотидов, меченных флуоресцентным репортером, который обеспечивает детекцию только после гибридизации зонда с его комплементарной последовательностью. В определенных аспектах посредством указанных праймеров можно амплифицировать и, таким образом, можно детектировать посредством неспецифического красителя или посредством использования специфического зонда для гибридизации, только неприродное соединение нуклеотидов. В других аспектах праймеры по изобретению выбирают так, что праймеры фланкируют любую из сторон последовательности с соединением так, что если реакция амплификации происходит, то присутствует указанная последовательность с соединением.
[0182] Аспекты по изобретению включают сами молекулы с последовательностями с неприродным соединением нуклеотидов, вместе с другими нуклеотидными молекулами, которые способны к связыванию с указанными последовательностями с неприродным соединением нуклеотидов в условиях гибридизации от умеренных до строгих. В определенных аспектах молекулы нуклеотидов, которые способны к связыванию с указанными последовательностями с неприродным соединением нуклеотидов в условиях от умеренных до строгих, обозначают как "нуклеотидные зонды".
[0183] В определенных аспектах геномную ДНК можно выделять из образцов и использовать для количественного определения присутствия микроорганизмов по изобретению с использованием кПЦР. Праймеры, используемые в реакции кПЦР, могут представлять собой праймеры, сконструированные посредством Primer Blast (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) для амплификации уникальных областей генома дикого типа или уникальных областей подвергнутых генетической инженерии не являющихся межродовыми мутантных штаммов. Реакцию кПЦР можно проводить с использованием набора SYBR GreenER qPCR SuperMix Universal (Thermo Fisher P/N 11762100) с использованием только прямого и обратного праймеров для амплификации; альтернативно, можно использовать Kapa Probe Force kit (Kapa Biosystems P/N KK4301) с праймерами для амплификации и зондом TaqMan, содержащим метку красителя FAM на 5'-конце, внутренний гаситель ZEN, и связывающим малую бороздку средством и флуоресцентным гасителем на 3'-конец (Integrated DNA Technologies).
[0184] Определенные последовательности праймеров, зондов и последовательности с неприродными соединениями приведены в таблице E. Эффективность реакции кПЦР можно определять с использованием стандартной кривой, получаемой на основе известного количества гДНК из генома-мишени. Данные можно нормализовать на количество копий генома на г сырой массы с использованием массы ткани и объема выделения.
[0185] Количественная полимеразная цепная реакция (кПЦР) представляет собой способ количественной, с детекцией в реальном времени, амплификации одной или нескольких последовательностей нуклеиновых кислот. Количественное определение в реальном времени в анализе ПЦР обеспечивает определение количества нуклеиновых кислот, получаемых на этапах амплификации ПЦР, сравнивая амплификацию представляющих интерес нуклеиновых кислот и соответствующей контрольной последовательности нуклеиновой кислоты, которая может действовать в качестве калибровочного стандарта.
[0186] Часто в анализах кПЦР используют зонды TaqMan, которые для количественного определения последовательностей-мишеней нуклеиновых кислот требуют увеличенной специфичности. Зонды TaqMan содержат олигонуклеотидный зонд с флуорофором, связанным с 5'-концом и гасителем связанным с 3'-концом зонда. Когда зонды TaqMan остаются как есть, с 5' и 3'-концами зонда в тесном контакте друг с другом, гаситель предотвращает передачу флуоресцентного сигнала с флуорофора. Зонды TaqMan сконструированы для отжига в области нуклеиновой кислоты, амплифицируемой посредством конкретного набора праймеров. Так как полимераза Taq продлевает праймер и синтезирует формируемую цепь, 5'-3' экзонуклеазная активность Taq полимеразы разрушает зонд, который отжегся с матрицей. Это разрушение зонда высвобождает флуорофор, таким образом, нарушая тесный контакт с гасителем и обеспечивая флуоресценцию флуорофора. Флуоресценция, детектируемая при анализе кПЦР, прямо пропорциональна высвобождаемому флуорофору и количеству матрицы ДНК присутствующей в реакции.
[0187] Свойства кПЦР позволяют практику исключать трудоемкий этап постамплификации препарата гель-электрофореза, который, как правило, необходим для наблюдения за амплифицированными продуктами традиционных анализов ПЦР. Преимущества кПЦР по сравнению с обычной ПЦР являются значительными и включают увеличенную скорость, простоту использования, воспроизводимость и количественные характеристики.
Улучшение свойств
[0188] Способы по настоящему изобретению можно использовать для придания или улучшения одного или нескольких из ряда требуемых признаков. Примеры признаков, которые можно придавать или улучшать, включают: биомассу корня, длину корня, рост, высоту побега, количество листьев, эффективность использования воды, общую биомассу, урожайность, размер плодов, размер семян, уровень фотосинтеза, засухостойкость, жароустойчивость, солеустойчивость, устойчивость к стрессу, вызванному нематодами, устойчивость к грибковому патогену, устойчивость к бактериальному патогену, устойчивость к вирусному патогену, уровень метаболита и экспрессия протеома. Требуемые признаки, включая рост, общую биомассу, биомассу корня и/или побега, пророст семян, жизнеспособность всходов, эффективность фотосинтеза, интенсивность транскрипции, количество или массу семян/плодов, урожай семян или плодов растения, содержание хлорофилла в листьях, скорость фотосинтеза, длина корня или любое их сочетание, можно использовать для определения роста и сравнения со скоростью роста эталонных сельскохозяйственных растений (например, растения без улучшенных признаков), выращиваемых в идентичных условиях.
[0189] Предпочтительный придаваемый или улучшаемый признак, как описано в настоящем документе, представляет собой фиксацию азота. В некоторых случаях, растения, получаемые способами, описываемыми в настоящем документе, демонстрируют отличия по признаку, которые представляют собой превышение по меньшей мере приблизительно на 5%, например, по меньшей мере приблизительно на 5%, по меньшей мере приблизительно на 8%, по меньшей мере приблизительно на 10%, по меньшей мере приблизительно на 15%, по меньшей мере приблизительно на 20%, по меньшей мере приблизительно на 25%, по меньшей мере приблизительно на 30%, по меньшей мере приблизительно на 40%, по меньшей мере приблизительно на 50%, по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 75%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, или по меньшей мере на 100%, по меньшей мере приблизительно на 200%, по меньшей мере приблизительно на 300%, по меньшей мере приблизительно на 400% или более по сравнению с эталонным сельскохозяйственным растением, выращиваемым в почве в тех же условиях. В дополнительных примерах растение, получаемое в способах, описываемых в настоящем документе, демонстрирует различия по признаку, которые по меньшей мере приблизительно на 5% больше, например, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, или по меньшей мере 100%, по меньшей мере приблизительно 200%, по меньшей мере приблизительно 300%, по меньшей мере приблизительно 400% или более чем у эталонного сельскохозяйственного растения, выращиваемого в сходных условиях в почве.
[0190] Улучшаемый признак можно оценивать в условиях, включающих воздействие одного или нескольких биотических или абиотических стрессовых факторов. Примеры стрессовых факторов включают абиотические стрессовые воздействия (такие как тепловой стресс, солевой стресс, вызываемый засухой стресс, холодовой стресс и стресс, вызванный недостаточностью питательных веществ) и биотические стрессовые воздействия (такие как стресс, вызываемый нематодами, стресс, вызываемый растительноядными насекомыми, стресс, вызываемый грибковыми патогенами, стресс, вызываемый бактериальными патогенами и стресс, вызываемый вирусными патогенами).
[0191] Признак, улучшаемый способами и композициями по настоящему изобретению, может представлять собой фиксацию азота, в том числе и в растении, ранее не способном к фиксации азота. В некоторых случаях бактерии, выделяемые способом, описываемым в настоящем документе, продуцируют 1% или более (например, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20% или более) азота растения, что может представлять собой увеличение способности к фиксации азота по меньшей мере в 2 раза (например, в 3 раза, в 4 раза, в 5 раз, в 6 раз, в 7 раз, в 8 раз, в 9 раз, в 10 раз, в 20 раз, в 50 раз, в 100 раз, в 1000 раз или более) по сравнению с бактериями, выделяемыми из первого растения до внесения какой-либо генетической вариации. В некоторых случаях бактерии продуцируют 5% или более азота растения. Желаемого уровня фиксации азота можно достигать после однократного или многократного повторения этапов внесения генетических вариаций, воздействия на множество растений и выделение бактерий из растений с улучшенным признаком (например, 1, 2, 3, 4, 5, 10, 15, 25 или более раз). В некоторых случаях, увеличенных уровней фиксации азота достигают в присутствии удобрения, дополненного глутамином, аммиаком или другим химическим источником азота. Известны способы оценки степени фиксации азота, примеры которых описаны в настоящем документе.
[0192] Выведение микроорганизмов представляет собой способ для систематической идентификации и улучшения роли видов в микробиоме сельскохозяйственной культуры. Способ включает три этапа: 1) отбор видов-кандидатов посредством картирования взаимодействий растение-микроорганизм и прогноза регуляторной сети, сцепленной с конкретным фенотипом, 2) практическое и прогнозируемое улучшение фенотипов микроорганизмов посредством внутривидового скрещивания регуляторных сетей и кластеров генов и 3) скрининг и отбор новых генотипов микроорганизмов, которые обеспечивают требуемые фенотипы сельскохозяйственных культур. Для систематического анализа улучшения штаммов получена модель, в которой сцеплена динамика колонизации сообщества микроорганизмов с генетической активностью ключевых видов. Модель используют для прогноза выведения генетических мишеней и увеличения частоты отбора улучшений кодируемых микробиомом свойств агротехнического значения.
Определение азота, доставляемого в значимых с сельскохозяйственной точки зрения полевых условиях
[0193] В полевых условиях доставляемое количество азота можно определять, как функцию колонизации умноженной на активность.

[0194] Приведенное выше уравнение требует (1) средней колонизации на единицу ткани растения и (2) активность в виде количества фиксированного азота или количества аммиака, экскретируемого каждой клеткой микроорганизма. Для преобразования в фунты азота на акр отслеживают физиологию роста кукурузы во времени, например, размер растения и ассоциированной корневой системы на всех стадиях созревания.
[0195] Фунты азота, доставляемые в сельскохозяйственную культуру на акр/сезон можно рассчитать по следующему уравнению:

[0196] Ткань растения(t) представляет собой сырую массу ткани растения кукурузы в течение времени роста (t). Значения для обоснованного проведения расчета подробно описаны в публикации под названием Roots, Growth and Nutrient Uptake (Mengel. Dept. of Agronomy Pub.# AGRY-95-08 (Rev. May-95. p. 1-8).
[0197] Колонизация (t) представляет собой количество представляющих интерес микроорганизмов, выявленное в ткани растения на грамм сырой массы ткани растения, в любое конкретное время, t, в течение сезона роста. В случае, когда доступна только одна временная точка, эту одну временную точку нормализуют как пиковый уровень колонизации за сезон, а уровень колонизации в оставшихся временных точках, таким образом, корректируют.
[0198] Активность(t) представляет собой скорость, с которой представляющие интерес микроорганизмы фиксируют N в единицу времени, в любое конкретное время, t, в течение сезона роста. В вариантах осуществления, описываемых в настоящем документе, этот уровень активности аппроксимируют посредством анализа восстановления ацетилена in vitro (ARA) в средах ARA в присутствии 5 мМ глутамина или анализа экскреции аммония в средах ARA media в присутствии 5 мМ ионов аммония.
[0199] Затем доставляемое количество азота рассчитывают посредством численного интегрирования указанной выше функции. В случаях, когда значения переменных, описанных выше, дискретно определяют в указанных временных точках, значения между этими временными точками аппроксимируют, проводя линейную интерполяцию.
Фиксация азота
[0200] В настоящем документе описаны способы увеличения фиксации азота в растении, включающие воздействие на растение бактериями, несущими одну или несколько генетических вариаций, вводимых в один или несколько генов, регулирующих фиксацию азота, где бактерии продуцируют в растении 1% или более азота (например, 2%, 5%, 10% или более), что может представлять собой по меньшей мере в 2 раза большую способность к фиксации азота по сравнению с растением в отсутствие бактерий. Бактерии могут продуцировать азот в присутствии удобрения, дополненного глутамином, мочевиной, нитратами или аммиаком. Генетические вариации могут представлять собой любые генетические вариации, описываемые в настоящем документе, включая примеры, приводимые выше, в любом количестве и в любой комбинации. Генетическую вариацию можно вносить в ген, выбранный из группы, состоящей из nifA, nifL, ntrB, ntrC, глутаминсинтетазы, glnA, glnB, glnK, draT, amtB, глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB и nifQ. Генетическая вариация может представлять собой мутацию, которая приводит к одному или нескольким из: повышенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности NifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, AmtB; сниженной активности удаления аденилата GlnE или сниженной активности удаления уридилата GlnD. Генетическая вариация, вносимая в одну или несколько бактерий способами, описываемыми в настоящем документе, может представлять собой нокаут-мутацию, или она может удалять регуляторную последовательность гена-мишени, или она может включать вставку гетерологичной регуляторной последовательности, например, вставку регуляторной последовательности, выявленной в геноме бактерий того же вида или рода. Регуляторную последовательность можно выбирать на основе уровня экспрессии гена в культуре бактерий или в ткани растения. Генетическую вариацию можно получать посредством химического мутагенеза. Растения, выращиваемые на этапе (c), можно подвергать воздействию биотических или абиотических стрессовых факторов.
[0201] Количественно фиксацию азота, происходящую в растениях, описываемых в настоящем документе, можно измерять различными способами, например, посредством анализа восстановления ацетилена (AR). Анализ восстановления ацетилена можно проводить in vitro или in vivo. Доказательство того, что конкретная бактерия предоставляет растению фиксированный азот, может включать: 1) общий N растения после инокуляции значимо увеличивается, предпочтительно с сопутствующим возрастанием концентрации N в растении; 2) симптомы недостаточности азота в условиях ограничения в N после инокуляции ослабляются (что должно включать возрастание сухой массы); 3) документацию фиксации N2 с использованием подхода с 15N (который может представлять собой эксперименты с разведением изотопа, анализы восстановления 15N2 или анализы распространенности 15N в природе); 4) встраивание фиксированного N в растение белок или метаболит и 5) отсутствие всех этих эффектов у растений без проведенной инокуляции или у растений, инокулированных мутантом инокулируемого штамма.
[0202] Регуляторный каскад фиксации азота дикого типа можно представить как цифровую логическую схему, где входящие O2 и NH4+ проходят через логический вентиль ИЛИ-НЕ, выход которого в дополнение к АТФ поступает в логический вентиль И. В определенных вариантах осуществления способы, описываемые в настоящем документе, нарушают действие NH4+ на эту схему в нескольких точках регуляторного каскада так, что микроорганизмы могут продуцировать азота даже на удобряемых полях. Однако способы, описываемые в настоящем документе, также предусматривают изменение влияния АТФ или O2 на схему или изменение схемы с использованием других регуляторных каскадов в клетке или изменение генетических схем, отличных от фиксации азота. Можно перестраивать генные кластеры с получением функциональных продуктов под контролем гетерологичной системы регуляции. Удаляя природные регуляторные элементы вне и в кодирующих последовательностях кластеров генов и заменяя их альтернативными системами регуляции, функциональные продукты сложных генетических оперонов и других кластеров генов можно контролировать и/или помещать в гетерологичные клетки, включая клетки других видов, отличных от видов, из которых получены природные гены. После перестройки синтетические кластеры генов можно контролировать посредством генетических схем или других индуцибельных систем регуляции, таким образом, контролируя экспрессию продуктов необходимым образом. Экспрессирующие кассеты можно конструировать так, чтобы они действовали в качестве логических вентилей, генераторов импульсов, осцилляторов, переключателей или устройств памяти. Контролирующую экспрессирующую кассету можно лигировать с промотором так, чтобы экспрессирующая кассета функционировала в качестве датчика состояний окружающей среды, такого как датчик кислорода, температуры, прикосновения, осмотического стресса, поверхностного напряжения или окислительно-восстановительного потенциала.
[0203] В качестве примера из кластера генов nif можно удалять гены nifL, nifA, nifT и nifX. Синтетические гены можно конструировать посредством рандомизации кодонов ДНК, кодирующей каждую аминокислотную последовательность. Отбор кодонов проводят, устанавливая, чтобы частота использования кодонов как можно больше отличалась от частоты использования кодонов в природном гене. Предлагаемые последовательности сканируют на наличие любых нежелательных свойств, таких как участки распознавания рестрикционных ферментов, участки распознавания транспозонов, повторяющиеся последовательности, промоторы сигма 54 и сигма 70, скрытые участки связывания рибосом и ро-независимые терминаторы. Синтетические участки связывания рибосом выбирают так, чтобы они соответствовали силе каждого соответствующего природного участка связывания рибосомы, например, конструируя плазмиду с флуоресцентным репортером, в которой 150 п.н., окружающих старт-кодон гена (от -60 до +90) слиты с геном флуоресцентного маркера. Этот химерный ген можно экспрессировать под контролем промотора Ptac, а флуоресценцию измерять посредством проточной цитометрии. Для получения синтетических участков связывания рибосом получают библиотеку репортерных плазмид с использованием 150 п.н. (от -60 до +90) синтетической экспрессирующей кассеты. В кратком изложении синтетическая экспрессирующая кассета может состоять из случайного ДНК-спейсера, вырожденной последовательности, кодирующей библиотеку RBS и кодирующей последовательности каждого синтетического гена. Подвергают скринингу множество клонов для идентификации синтетического участка связывания рибосомы, который наилучшим образом соответствует природному участку связывания рибосомы. Таким образом, конструируют синтетические опероны, которые состоят из таких же генов, что природные опероны, и тестируют на функциональную комплементацию. Дополнительное иллюстративное описание синтетических оперонов приведено в US20140329326.
Виды бактерий
[0204] Микроорганизмы пригодные в способах и композициях, описываемых в настоящем документе, можно получать из любого источника. В определенных случаях микроорганизмы могут представлять собой бактерии, археи, простейшие или грибы. Микроорганизмы по настоящему описанию могут представлять собой фиксирующие азот микроорганизмы, например, фиксирующие азот бактерии, фиксирующие азот археи, фиксирующие азот грибы, фиксирующие азот дрожжи или фиксирующие азот простейшие. Микроорганизмы, пригодные в способах и композициях, описываемых в настоящем документе, могут представлять формирующие споры микроорганизмы, например, формирующие споры бактерии. В определенных случаях бактерии, пригодные в способах и композициях, описываемых в настоящем документе, могут являться грамположительными бактериями или грамотрицательными бактериями. В определенных случаях бактерии могут представлять собой формирующие эндоспору бактерии Firmicute phylum. В определенных случаях бактерии могут представлять собой диазатроф. В определенных случаях бактерии могут не являться диазатрофом.
[0205] Способы и композиции по настоящему описанию можно использовать с использованием архей, таких как, например, Methanothermobacter thermoautotrophicus.
[0206] В определенных случаях бактерии, которые могут являться пригодными, в качестве неограничивающих примеров включают Agrobacterium radiobacter, Bacillus acidocaldarius, Bacillus acidoterrestris, Bacillus agri, Bacillus aizawai, Bacillus albolactis, Bacillus alcalophilus, Bacillus alvei, Bacillus aminoglucosidicus, Bacillus aminovorans, Bacillus amylolyticus (также известную как Paenibacillus amylolyticus) Bacillus amyloliquefaciens, Bacillus aneurinolyticus, Bacillus atrophaeus, Bacillus azotoformans, Bacillus badius, Bacillus cereus (synonyms: Bacillus endorhythmos, Bacillus medusa), Bacillus chitinosporus, Bacillus circulans, Bacillus coagulans, Bacillus endoparasiticus Bacillus fastidiosus, Bacillus firmus, Bacillus kurstaki, Bacillus lacticola, Bacillus lactimorbus, Bacillus lactis, Bacillus laterosporus (также известную как Brevibacillus laterosporus), Bacillus lautus, Bacillus lentimorbus, Bacillus lentus, Bacillus licheniformis, Bacillus maroccanus, Bacillus megaterium, Bacillus metiens, Bacillus mycoides, Bacillus natto, Bacillus nematocida, Bacillus nigrificans, Bacillus nigrum, Bacillus pantothenticus, Bacillus popillae, Bacillus psychrosaccharolyticus, Bacillus pumilus, Bacillus siamensis, Bacillus smithii, Bacillus sphaericus, Bacillus subtilis, Bacillus thuringiensis, Bacillus uniflagellatus, Bradyrhizobium japonicum, Brevibacillus brevis Brevibacillus laterosporus (ранее Bacillus laterosporus), Chromobacterium subtsugae, Delftia acidovorans, Lactobacillus acidophilus, Lysobacter antibioticus, Lysobacter enzymogenes, Paenibacillus alvei, Paenibacillus polymyxa, Paenibacillus popilliae (ранее Bacillus popilliae), Pantoea agglomerans, Pasteuria penetrans (ранее Bacillus penetrans), Pasteuria usgae, Pectobacterium carotovorum (ранее Erwinia carotovora), Pseudomonas aeruginosa, Pseudomonas aureofaciens, Pseudomonas cepacia (ранее known as Burkholderia cepacia), Pseudomonas chlororaphis, Pseudomonas fluorescens, Pseudomonas proradix, Pseudomonas putida, Pseudomonas syringae, Serratia entomophila, Serratia marcescens, Streptomyces colombiensis, Streptomyces galbus, Streptomyces goshikiensis, Streptomyces griseoviridis, Streptomyces lavendulae, Streptomyces prasinus, Streptomyces saraceticus, Streptomyces venezuelae, Xanthomonas campestris, Xenorhabdus luminescens, Xenorhabdus nematophila, Rhodococcus globerulus AQ719 (№ доступа NRRL B-21663), вид Bacillus AQ175 (№ доступа ATCC 55608), вид Bacillus AQ 177 (№ доступа ATCC 55609), вид Bacillus (№ доступа ATCC 53522) и штамм вида Streptomyces № доступа NRRL B-30145. В определенных случаях бактерия также может представлять собой Azotobacter chroococcum, Methanosarcina barkeri, Klesiella pneumoniae, Azotobacter vinelandii, Rhodobacter spharoides, Rhodobacter capsulatus, Rhodobcter palustris, Rhodosporillum rubrum, Rhizobium leguminosarum или Rhizobium etli.
[0207] В определенных случаях бактерия может представлять собой бактерию вида Clostridium, например, Clostridium pasteurianum, Clostridium beijerinckii, Clostridium perfringens, Clostridium tetani, Clostridium acetobutylicum.
[0208] В определенных случаях бактерии, используемые в способах и композициях по настоящему изобретению, могут представлять собой цианобактерии. Примеры родов цианобактерий включают Anabaena (например, вид Anagaena PCC7120), Nostoc (например, Nostoc punctiforme) или Synechocystis (например, вид Synechocystis PCC6803).
[0209] В определенных случаях бактерии, используемые в способах и композициях по настоящему изобретению, могут принадлежать к таксономической группе Chlorobi, например, Chlorobium tepidum.
[0210] В определенных случаях микроорганизмы, используемые в способах и композициях по настоящему изобретению, могут содержать ген, гомологичный известному гену NifH. Последовательности известных генов NifH можно найти, например, в базе данных лаборатории Дж. Зера NifH, (https://wwwzehr.pmc.ucsc.edu/nifH_Database_Public/, 4 апреля 2014 года), или базе данных лаборатории Д. Бакли NifH (http://www.css.cornell.edu/faculty/buckley/nifh.htm, и Gaby, John Christian, и Daniel H. Buckley. "A comprehensive aligned nifH gene database: a multipurpose tool for studies of nitrogen-fixing bacteria". Database 2014 (2014): bau001.). В определенных случаях микроорганизмы, используемые в способах и композициях по настоящему изобретению, могут содержать последовательность, которая кодирует полипептид по меньшей мере на 60%, 70%, 80%, 85%, 90%, 95%, 96%, 96%, 98%, 99% или более 99% идентичный по последовательности с последовательностью из базы данных NifH лаборатории Дж. Зера, (https://wwwzehr.pmc.ucsc.edu/nifH_Database_Public/, 4 апреля 2014 года). В определенных случаях микроорганизмы, используемые в способах и композициях по настоящему изобретению, могут содержать последовательность, которая кодирует полипептид по меньшей мере на 60%, 70%, 80%, 85%, 90%, 95%, 96%, 96%, 98%, 99% или более 99% идентичный по последовательности с последовательностью из базы данных NifH лаборатории Д. Бакли, (Gaby, John Christian, и Daniel H. Buckley. "A comprehensive aligned nifH gene database: a multipurpose tool for studies of nitrogen-fixing bacteria". Database 2014 (2014): bau001.).
[0211] Микроорганизмы, пригодные в способах и композициях, описываемых в настоящем документе, можно получать посредством выделения микроорганизмов с поверхностей или из тканей природных растений; измельчения семян с выделением микроорганизмов; высаживания семян в различные образцы почв и выделения микроорганизмов из тканей; или инокуляции растений экзогенными микроорганизмами и определения того, какие микроорганизмы обнаруживаются в тканях растений. Неограничивающие примеры тканей растений включают семена, всходы, листья, срезы, растения, луковицы или клубни. В некоторых случаях бактерии выделяют из семян. Параметры обработки образцов могут варьировать для выделения различных типов ассоциированных микроорганизмов, таких как ризосферные микроорганизмы, эпифиты или эндофиты. Вместо начального выделения из первого растения бактерии также можно получать из хранилищ, таких как коллекции природных штаммов. Микроорганизмы можно генотипировать и фенотипировать посредством секвенирование геномов выделенных микроорганизмов; профилирования составов сообществ in planta; характеристики функциональности транскриптомов сообществ или выделенных микроорганизмов; или скрининга свойств микроорганизмов с использованием селективных или фенотипических сред (например, фенотипы с фиксацией азота или растворением фосфатов). Выбранные штаммы или популяции кандидатов можно получать из данных секвенирования; данных фенотипирования; данных о растении (например, данных о геноме, фенотипе и/или урожае); данных о почве (например, pH, содержание N/P/K и/или биотическое сообщество основной почвы) или любой комбинации из них.
[0212] Бактерии и способы получения бактерий, описываемые в настоящем документе, можно применять для бактерий, способных к эффективному самостоятельному размножению на поверхностях листьев, поверхностях корней или внутри тканей растений, не вызывая разрушительной защитной реакции растений, или для бактерий, которые устойчивы к защитным реакциям растений. Бактерии, описываемые в настоящем документе, можно выделять посредством культивирования экстракта ткани растения или смыва поверхности листа в среде без добавления азота. Однако бактерии могут являться некультивируемыми, т.е. об их культивируемости может быть неизвестно, или их может быть трудно культивировать известными в данной области стандартными способами. Бактерии, описываемые в настоящем документе, могут являться эндофитными или эпифитными или бактериями, обитающими в ризосфере растения (ризосферные бактерии). Бактерии, получаемые после однократного или многократного повторения этапов внесения генетической вариации, воздействия на множество растений и выделения бактерий из растений с улучшенным признаком (например, 1, 2, 3, 4, 5, 10, 15, 25 или более раз) могут являться эндофитными, эпифитными или ризосферными. Эндофиты представляют собой организмы, которые проникают внутрь растений, не вызывая симптомов заболеваний или не вызывая формирования симбиотических структур, и представляют собой агротехнический интерес, так как они могут усиливать рост растений и улучшать питание растений (например, посредством фиксации азота). Бактерии могут представлять собой передающиеся с семенами эндофиты. Передающиеся с семенами эндофиты включают бактерии, ассоциированные с семенами трав или других растений или получаемые из них, такие как передающиеся с семенами бактериальные эндофиты, выявляемые в созревших, сухих, неповрежденных (например, без трещин, без видимой грибковой инфекции или без преждевременного пророста) семян. Передающиеся с семенами бактериальные эндофиты могут быть ассоциированы или их можно получать с поверхности семян; альтернативно или кроме того, они могут быть ассоциированы или их можно получать из внутреннего компартмента семян (например, семян со стерилизованной поверхностью). В определенных случаях передающиеся с семенами бактериальные эндофиты способны к репликации в тканях растений, например, внутри семян. В определенных случаях передающиеся с семенами бактериальные эндофиты также способны переживать обезвоживание.
[0213] Бактерии, выделяемые способами по изобретению или используемые в способах или композициях по изобретению, могут включать множество бактерий различных таксонов в комбинации. В качестве примера бактерии могут включать Proteobacteria (такие как Pseudomonas, Enterobacter, Stenotrophomonas, Burkholderia, Rhizobium, Herbaspirillum, Pantoea, Serratia, Rahnella, Azospirillum, Azorhizobium, Azotobacter, Duganella, Delftia, Bradyrhizobiun, Sinorhizobium и Halomonas), Firmicutes (такие как Bacillus, Paenibacillus, Lactobacillus, Mycoplasma и Acetabacterium) и Actinobacteria (такие как Streptomyces, Rhodacoccus, Microbacterium и Curtobacterium). Бактерии, используемые в способах и композиции по настоящему описанию, могут включать фиксирующие азот бактериальные консорциумы из двух или более видов. В определенных случаях один или несколько видов бактерий бактериальных консорциумов могут быть способными к фиксации азота. В определенных случаях один или несколько видов бактериальных консорциумов могут облегчать или увеличивать способность других бактерии фиксировать азот. Бактерии, которые фиксируют азот, и бактерии, которые увеличивают способность других бактерий фиксировать азот, могут являться одними и теми же или различными. В определенных примерах штамм бактерий может быть способным фиксировать азот при нахождении в комбинации с другим штаммом бактерий или в определенных бактериальных консорциумах, но могут являться неспособными к фиксации азота в монокультуре. Примеры родов бактерий, которые можно найти в фиксирующих азот бактериальных консорциумам в качестве неограничивающих примеров включают Herbaspirillum, Azospirillum, Enterobacter и Bacillus.
[0214] Бактерии, которые можно получать способами, описываемыми в настоящем документе, включают вид Azotobacter, вид Bradyrhizobium, вид Klebsiella и вид Sinorhizobium. В определенных случаях бактерии можно выбирать из группы, состоящей из: Azotobacter vinelandii, Bradyrhizobium japonicum, Klebsiella pneumoniae и Sinorhizobium meliloti. В определенных случаях бактерии могут представлять собой бактерии рода Enterobacter или Rahnella. В определенных случаях бактерии могут представлять собой бактерии рода Frankia или Clostridium. Примеры бактерий рода Clostridium в качестве неограничивающих примеров включают Clostridium acetobutilicum, Clostridium pasteurianum, Clostridium beijerinckii, Clostridium perfringens и Clostridium tetani. В определенных случаях бактерии могут представлять собой бактерии рода Paenibacillus, например, Paenibacillus azotofixans, Paenibacillus borealis, Paenibacillus durus, Paenibacillus macerans, Paenibacillus polymyxa, Paenibacillus alvei, Paenibacillus amylolyticus, Paenibacillus campinasensis, Paenibacillus chibensis, Paenibacillus glucanolyticus, Paenibacillus illinoisensis, Paenibacillus larvae subsp. Larvae, Paenibacillus larvae subsp. Pulvifaciens, Paenibacillus lautus, Paenibacillus macerans, Paenibacillus macquariensis, Paenibacillus macquariensis, Paenibacillus pabuli, Paenibacillus peoriae или Paenibacillus polymyxa.
[0215] В определенных примерах бактерии, выделенные способами по изобретению, могут являться представителями одного или нескольких из следующих таксонов: Achromobacter, Acidithiobacillus, Acidovorax, Acidovoraz, Acinetobacter, Actinoplanes, Adlercreutzia, Aerococcus, Aeromonas, Afipia, Agromyces, Ancylobacter, Arthrobacter, Atopostipes, Azospirillum, Bacillus, Bdellovibrio, Beijerinckia, Bosea, Bradyrhizobium, Brevibacillus, Brevundimonas, Burkholderia, Candidatus Haloredivivus, Caulobacter, Cellulomonas, Cellvibrio, Chryseobacterium, Citrobacter, Clostridium, Coraliomargarita, Corynebacterium, Cupriavidus, Curtobacterium, Curvibacter, Deinococcus, Delftia, Desemzia, Devosia, Dokdonella, Dyella, Enhydrobacter, Enterobacter, Enterococcus, Erwinia, Escherichia, Escherichia/Shigella, Exiguobacterium, Ferroglobus, Filimonas, Finegoldia, Flavisolibacter, Flavobacterium, Frigoribacterium, Gluconacetobacter, Hafnia, Halobaculum, Halomonas, Halosimplex, Herbaspirillum, Hymenobacter, Klebsiella, Kocuria, Kosakonia, Lactobacillus, Leclercia, Lentzea, Luteibacter, Luteimonas, Massilia, Mesorhizobium, Methylobacterium, Microbacterium, Micrococcus, Microvirga, Mycobacterium, Neisseria, Nocardia, Oceanibaculum, Ochrobactrum, Okibacterium, Oligotropha, Oryzihumus, Oxalophagus, Paenibacillus, Panteoa, Pantoea, Pelomonas, Perlucidibaca, Plantibacter, Polynucleobacter, Propionibacterium, Propioniciclava, Pseudoclavibacter, Pseudomonas, Pseudonocardia, Pseudoxanthomonas, Psychrobacter, Rahnella, Ralstonia, Rheinheimera, Rhizobium, Rhodococcus, Rhodopseudomonas, Roseateles, Ruminococcus, Sebaldella, Sediminibacillus, Sediminibacterium, Serratia, Shigella, Shinella, Sinorhizobium, Sinosporangium, Sphingobacterium, Sphingomonas, Sphingopyxis, Sphingosinicella, Staphylococcus, 25 Stenotrophomonas, Strenotrophomonas, Streptococcus, Streptomyces, Stygiolobus, Sulfurisphaera, Tatumella, Tepidimonas, Thermomonas, Thiobacillus, Variovorax, родов WPS-2 неопределенного положения, Xanthomonas и Zimmermannella.
[0216] В определенных случаях используют виды бактерий, выбранные по меньшей мере из одного из следующих родов: Enterobacter, Klebsiella, Kosakonia и Rahnella. В определенных случаях используют комбинацию видов бактерий следующих родов: Enterobacter, Klebsiella, Kosakonia и Rahnella. В определенных случаях используемые виды могут представлять собой один или несколько из: Enterobacter sacchari, Klebsiella variicola, Kosakonia sacchari и Rahnella aquatilis.
[0217] В определенных случаях грамположительный микроорганизм может содержать нитрогеназную систему с молибденом-железом, содержащую: nifH, nifD, nifK, nifB, nifE, nifN, nifX, hesA, nifV, nifW, nifU, nifS, nifI1 и nifI2. В определенных случаях грамположительный микроорганизм может содержать нитрогеназную систему с ванадием, содержащую: vnfDG, vnfK, vnfE, vnfN, vupC, vupB, vupA, vnfV, vnfR1, vnfH, vnfR2, vnfA (регулятор транскрипции). В определенных случаях грамположительный микроорганизм may have an iron-only нитрогеназа система comprising: anfK, anfG, anfD, anfH, anfA (регулятор транскрипции). В определенных случаях грамположительный микроорганизм может содержать нитрогеназную систему, содержащую glnB и glnK (белки, передающие сигнал от азота). Определенные примеры ферментов, вовлеченных в метаболизм азота в грамположительных микроорганизмах включают glnA (глутаминсинтетазу), gdh (глутаминатдегидрогеназу), bdh (3-гидроксибутиратдегидрогеназу), глутаминазу, gltAB/gltB/gltS (глутаминатсинтазу), asnA/asnB (аспартатаммиаклигазу/аспарагинсинтетазу) и ansA/ansZ (аспарагиназу). Определенные примеры белков, вовлеченных в транспорт азота у грамположительных микроорганизмов, включают amtB (транспортер аммония), glnK (регулятор транспорта аммония), glnPHQ/glnQHMP (АТФ-зависимые транспортеры глутамина/глутамината), glnT/alsT/yrbD/yflA (глутаминоподобные протонные совместные переносчики) и gltP/gltT/yhcl/nqt (глутаминоподобные протонные совместные переносчики).
[0218] Примеры грамположительных микроорганизмов, которые могут представлять особый интерес, включают Paenibacillus polymixa, Paenibacillus riograndensis, вид Paenibacillus, вид Frankia, вид Heliobacterium, Heliobacterium chlorum, вид Heliobacillus, вид Heliophilum, вид Heliorestis, Clostridium acetobutylicum, вид Clostridium, Mycobacterium flaum, вид Mycobacterium, вид Arthrobacter, вид Agromyces, Corynebacterium autitrophicum, вид Corynebacterium, вид Micromonspora, вид Propionibacteria, вид Streptomyces и вид Microbacterium.
[0219] Определенные примеры генетических альтераций, которые можно проводить в грамположительных микроорганизмах, включают: делецию glnR с удалением отрицательной регуляции BNF в присутствии азота окружающей среды, вставку различных промоторов непосредственно выше кластера nif с устранением регуляции посредством GlnR в ответ на азот окружающей среды, мутацию glnA со снижением уровня ассимиляции аммония по пути GS-GOGAT, делецию amtB с уменьшением потребления аммоний из среды, мутацию glnA так, что он конститутивно находится в состоянии ингибирования по типу обратной связи (FBI-GS), с уменьшением ассимиляции аммония по пути GS-GOGAT.
[0220] В определенных случаях основным регулятором метаболизма и фиксации N в видах Paenibacillus является glnR. В определенных случаях геном видов Paenibacillus может не содержать ген для продукции glnR. В определенных случаях геном вида Paenibacillus может не содержать ген для продукции glnE или glnD. В определенных случаях геном видов Paenibacillus может содержать ген для продукции glnB или glnK. Например, вид Paenibacillus WLY78 не содержит гена для glnB или его гомолога, выявленного у архебактерии Methanococcus maripaludis, nifI1 и nifI2. В определенных случаях геномы видов Paenibacillus могут варьировать. Например, у Paenibacillus polymixa E681 отсутствуют glnK и gdh, присутствует несколько переносчиков соединений азота, но по-видимому, только amtB находится под контролем GlnR. В другом примере вид Paenibacillus JDR2 содержит glnK, gdh и большинство других центральных генов метаболизма азота, содержит множество менее крупных переносчиков соединений азота, но не содержит glnPHQ под контролем GlnR. Paenibacillus riograndensis SBR5 содержит стандартный оперон glnRA, ген fdx, основной оперон nif, вторичный оперон nif и оперон anf (кодирующий содержащую только железо нитрогеназу). Выше каждого из этих оперонов выявлены предполагаемые участки glnR/tnrA. GlnR может регулировать все указанные выше опероны, за исключением оперона anf. GlnR может связываться с каждой из этих регуляторных последовательностей в виде димера.
[0221] Фиксирующие N штаммы Paenibacillus могут попадать в две подгруппы: подгруппу I, которая содержит только минимальный кластер генов nif, и подгруппу II, содержащую минимальный кластер и неохарактеризованный ген между nifX и hesA, и часто в других кластерах происходит дупликация определенных генов nif, таких как nifH, nifHDK, nifBEN, или кластеры, кодирующие гены нитрогеназы с ванадием (vnf) или содержащей только железо нитрогеназы).
[0222] В определенных случаях геном видов Paenibacillus может не содержать гена для продукции glnB или glnK. В определенных случаях геном видов Paenibacillus может содержать минимальный кластер nif с 9 генами, транскрибируемыми с промотора sigma-70. В определенных случаях кластер nif Paenibacillus могут отрицательно регулировать азот или кислород. В определенных случаях геном видов Paenibacillus может не содержать гена для продукции sigma-54. Например, вид Paenibacillus WLY78 не содержит гена sigma-54. В определенных случаях кластер nif могут регулировать glnR и/или TnrA. В определенных случаях активность кластера nif можно определять изменяя активность glnR и/или TnrA.
[0223] У Bacilli глутаминсинтетаза (GS) ингибирована по типу обратной связи высокими концентрациями внутриклеточного глутамина, вызывая сдвиг в подтверждении (обозначаемый как FBI-GS). Кластеры Nif содержат выраженные участки связывания для регуляторов GlnR и TnrA у различных видов Bacilli. GlnR связывает и подавляет экспрессию гена в присутствии избытка внутриклеточного глутамина и АМФ. Ролью GlnR может являться предотвращения притока и внутриклеточной продукции глутамина и аммония в условиях высокой доступности азота. TnrA может связывать и/или активировать (или подавлять) экспрессию генов в присутствии лимитирующего внутриклеточного глутамина и/или в присутствии FBI-GS. В определенных случаях активность кластера nif Bacilli можно изменять, изменяя активность GlnR.
[0224] Ингибированная по типу обратной связи глутаминсинтетаза (FBI-GS) может связывать GlnR и стабилизировать связывание GlnR с распознаваемыми последовательностями. Несколько видов бактерий содержат участок связывания GlnR/TnrA выше кластера nif. Изменение связывания FBI-GS и GlnR могут изменять активность пути nif.
Источники микроорганизмов
[0225] Бактерии (или любой микроорганизм по описанию) можно получать из любой обычной наземной окружающей среды, включая ее почвы, растения, грибы, животные (включая беспозвоночных) и другую биоту, включая осадки, воду и биоту озер и рек; из морской окружающей среды, ее биоты и осадков (например, морской воды, морского ила, морских растений, морских беспозвоночных (например, губок), морских позвоночных (например, рыб)); земной и морской геосферы (рыхлые поверхностные отложения и камни, например, раздробленные подпочвенные камни, песок и глины); криосферы и ее талых вод; атмосферы (например, отфильтрованной взвешенной пыли, облачных и дождевых капель); городской, промышленной и других техногенных окружающих сред (например, накопленные органический и минеральный материал на бетоне, желобах обочин, поверхностях крыш и дорожных покрытиях).
[0226] Растения, из которых получают бактерии (или любой микроорганизм по описанию), могут представлять собой растение с одним или несколькими требуемыми признаками, например, растения, которые в природе растут в конкретной окружающей среде или в определенных представляющих интерес условиях. В качестве примера, определенное растение может в природе расти в песчаном грунте или песке с высокой минерализацией, или при экстремальных температурах, или с малым количеством воды, или оно может быть устойчивым к определенным вредителям или заболеваниям, присутствующим в окружающей среда, и желательным может являться выращивание коммерческой сельскохозяйственной культуры в таких условиях, в частности, например, если они представляют собой единственные доступные в конкретном географическом положении условия. В качестве дополнительного примера бактерии можно выделять из коммерческой сельскохозяйственной культуры, выращиваемой в таких средах или более конкретно, из конкретной сельскохозяйственной культуры, в которой среди сельскохозяйственных культур, выращиваемых в любой конкретной окружающей среде, наилучшим образом представлен представляющий интерес признак: например, наиболее быстрорастущие растения среди сельскохозяйственных культур, выращиваемых в почвах с ограниченным уровнем солей, или наименее повреждаемые растения среди сельскохозяйственных культур, подвергшихся сильному повреждению насекомыми или эпидемическим заболеванием, или растения с требуемыми количествами определенных метаболитов и других соединений, включая содержание волокон, содержание масел и т.п., или растения, демонстрирующие требуемые цвета, вкус или запах. Бактерии можно выделять из представляющего интерес растения или любого материала, находящегося в исследуемой окружающей среде, включая грибы и другую животную и растительную биоту, почву, воду, осадки и друг составляющие окружающей среды, как указано ранее.
[0227] Бактерии (или любой микроорганизм по описанию) можно выделять из ткани растения. Это выделение можно проводить из любой подходящий ткани растения, включая например, корень, стебель и листья и репродуктивные ткани растения. В качестве примера, общепринятые способы выделения из растений, как правило, включают вырезание в стерильных условиях материала исследуемого растения (например, участки корня или стебля, листья), стерилизация поверхности подходящим раствором (например, 2% гипохлоритом натрия) с последующим помещением материала растения на питательную среду для роста микроорганизмов. Альтернативно, материал растения со стерилизованной поверхностью можно измельчать в стерильную жидкость (как правило, воду) и жидкую суспензию, содержащую небольшие части измельченного материала растения наносить на поверхность подходящей твердой агарозной среды или сред, которые могут являться селективными или нет (например, содержать только фитиновую кислоту в качестве источника фосфора). Этот подход особенно пригоден для бактерий, которые формируют изолированные колонии и которые можно индивидуально перенести на отдельные планшеты с питательной средой, и дополнительно очищать до одного вида хорошо известными способами. Альтернативно, образцы корней или листьев растение можно не подвергать поверхностной стерилизации, а только аккуратно промыть, таким образом, включая в процесс выделения обитающие на поверхности эпифитные микроорганизмы, или эпифитные микроорганизмы можно выделять отдельно, прикладывая и убирая части корней, стеблей или листьев растений на поверхности агарозной среда, а затем выделяя отдельные колонии, как описано выше. Этот подход особенно пригоден, например, для бактерий. Альтернативно, корни можно обрабатывать без отмывания небольших количеств почвы, налипших на корни, таким образом, включая микроорганизмы, которые колонизирует ризосферу растения. В других случаях можно отбирать почву, налипшую на корни, разбавлять и наносить на агар с подходящими селективными и неселективными средами с получением отдельных колоний ризосферных бактерий.
БУДАПЕШТСКИЙ ДОГОВОР О МЕЖДУНАРОДНОМ ПРИЗНАНИИ ДЕПОНИРОВАНИЯ МИКРООРГАНИЗМОВ ДЛЯ ЦЕЛЕЙ ПАТЕНТНОЙ ПРОЦЕДУРЫ
[0228] Депозиты микроорганизмов по настоящему изобретению сделаны в соответствии с положениями будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры (будапештского договора).
[0229] Авторы утверждают, что в соответствии с 37 C.F.R. 1.808(a) (2) "все ограничения, наложенные вкладчиком на доступность публике депонированного материала, будут безвозвратно сняты при выдаче патента". Это утверждение подпадает под действие пункта (b) данного раздела (т.е. 37 C.F.R. 1.808 (b)).
[0230] Биологически чистые культуры Rahnella aquatilis и Enterobacter sacchari депонированы 14 июля 2015 года в американскую коллекцию типовых культур (ATCC; International Depositary Authority), 10801 University Blvd., Manassas, VA 20110, USA, и им присвоены номера ATTC Patent Deposit Designation PTA-122293 и PTA-122294, соответственно. Соответствующая информация о депозите приведена ниже в таблице A.
[0231] Enterobacter sacchari в настоящее время переклассифицирована как Kosakonia sacchari, это название для этого организма можно использовать взаимозаменяемо на всем протяжении описания.
[0232] Многие микроорганизмы по настоящему изобретению получены из двух штаммов дикого типа, как представлено на фигуре 18 и фигуре 19. Штамм CI006 представляет собой вид бактерий, ранее классифицированный в роде Enterobacter (см. указанную выше переклассификацию в Kosakonia), и на фигуре 19 приведена линия мутантов, которые получены из CI006. Штамм CI019 представляет собой вид бактерий, классифицированный в роде Rahnella, и на фигуре 19 приведена линия мутантов, которые получены из CI019. В отношении фигуры 18 и фигуры 19, следует отметить, что штаммы, содержащие CM в названии, представляют собой мутанты штаммов, изображенные непосредственно слева от указанного штамма CM. Информация из депозита для CI006 Kosakonia дикого типа (WT) и CI019 Rahnella WT приведена ниже в таблице A.
[0233] Некоторые микроорганизмы, описанные в настоящей заявке, депонированы 06 января 2017 года или 11 августа 2017 года в Bigelow National Center for Marine Algae and Microbiota (NCMA), находящегося по адресу 60 Bigelow Drive, East Boothbay, Maine 04544, USA. Как указано выше, все депозиты сделаны в соответствии с условиями будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры. Номера доступа Bigelow National Center for Marine Algae and Microbiota и даты депозита для указанных выше депозитов по будапештскому договору приведены в таблице A.
[0234] Биологически чистые культуры Kosakonia sacchari (WT), Rahnella aquatilis (WT) и варианта штамма Kosakonia sacchari депонированы 06 января 2017 года в Bigelow National Center for Marine Algae and Microbiota (NCMA), находящийся по адресу 60 Bigelow Drive, East Boothbay, Maine 04544, USA, и им присвоены номера NCMA Patent Deposit Designation 201701001, 201701003 и 201701002, соответственно. Соответствующая информация о депозите приведена ниже в таблице A.
[0235] Биологически чистые культуры вариантов штаммов Kosakonia sacchari депонированы 11 августа 2017 года в Bigelow National Center for Marine Algae and Microbiota (NCMA), находящийся по адресу 60 Bigelow Drive, East Boothbay, Maine 04544, USA, и им присвоены номера NCMA Patent Deposit Designation 201708004, 201708003 и 201708002, соответственно. Соответствующая информация о депозите приведена ниже в таблице A.
[0236] Биологически чистая культура Klebsiella variicola (WT) депонирована 11 августа 2017 года в Bigelow National Center for Marine Algae and Microbiota (NCMA), находящийся по адресу 60 Bigelow Drive, East Boothbay, Maine 04544, USA, и ей присвоен номер NCMA Patent Deposit Designation 201708001. Биологически чистые культуры двух вариантов Klebsiella variicola депонированы 20 декабря 2017 года в Bigelow National Center for Marine Algae and Microbiota (NCMA), находящийся по адресу 60 Bigelow Drive, East Boothbay, Maine 04544, USA, и им присвоены номера NCMA Patent Deposit Designation 201712001 и 201712002, соответственно. Соответствующая информация о депозите приведена ниже в таблице A.
Таблица A: Микроорганизмы, депонированные в соответствии с будапештским договором
Выделенные и биологически чистые микроорганизмы
[0237] Настоящее изобретение в определенных вариантах осуществления относится к выделенным и биологически чистым микроорганизмам, которые в числе прочего имеют применения в сельском хозяйстве. Микроорганизмы по изобретению можно использовать в их выделенном и биологически чистом состояниях, а также сформулированными в композиции (для описания иллюстративных композиций см. раздел ниже). Кроме того, по изобретению предоставлены композиции микроорганизмов, содержащие по меньшей мере два представителя описанных выделенных и биологически чистых микроорганизмов, а также способы использования указанных композиций микроорганизмов. Кроме того, по изобретению предусмотрены способы модуляции фиксации азота в растениях посредством использование описанных выделенных и биологически чистых микроорганизмов.
[0238] В определенных аспектах выделенные и биологически чистые микроорганизмы по изобретению представляют собой микроорганизмы из таблицы A. В других аспектах выделенные и биологически чистые микроорганизмы по изобретению получают из микроорганизмов из таблицы A. Например, по настоящему документу предоставлены штамм, потомок, мутант или производное микроорганизма из таблицы A. По изобретению предусмотрены все возможные комбинации микроорганизмов, перечисленных в таблице A, где указанные комбинации иногда формируют консорциумы микроорганизмов. Микроорганизмы из таблицы A, индивидуально или в любой комбинации, можно комбинировать с любым растением, активным веществом (синтетическим, органическим и т.д.), адъювантом, носителем, добавков или биологическим компонентом, указанных в описании.
Композиции
[0239] Композиции, содержащие бактерии или популяции бактерий, полученных способами, описываемыми в настоящем документе, и/или обладающих характеристиками, как описано в настоящем документе, могут находиться в форме жидкости, пены или сухого продукта. Композиции, содержащие бактерии или популяции бактерий, полученных способами, описываемыми в настоящем документе, и/или обладающих характеристиками, как описано в настоящем документе, также можно использовать для улучшения свойств растений. В определенных примерах композиция, содержащая популяции бактерий, может находиться в форме a сухого порошка, взвеси порошка и воды или жидкой пропитки для семян. Композиции, содержащие популяции бактерий можно наносить на поверхность семян, и она может находиться в жидкой форме.
[0240] Композиция можно производить в биореакторах, таких как реакторы с непрерывно перемешиваемым резервуаром, реакторы периодического действия и на ферме. В определенных примерах композиции можно хранить в контейнере, таком как канистра или в миниобъеме. В определенных примерах композиции можно хранить в любом объекте, выбранном из группы, состоящей из бутылки, фляги, ампулы, упаковки, сосуда, мешка, коробки, ящика, обертки, картонной коробки, контейнера, накопитель, контейнер для перевозок, кузов грузового автомобиля и/или корпус.
[0241] Композиции также можно использовать для улучшения свойств растений. В определенных примерах одну или несколько композиций можно наносить на семена. В определенных примерах одну или несколько композиций можно наносить на проросток. В определенных примерах одну или несколько композиций можно наносить на поверхность семян. В определенных примерах одну или несколько композиций можно наносить в виде слоя на поверхности семян. В определенных примерах композиция, которую наносят на семена может находиться в жидкой форме, в форме сухого продукта, в форме пены, в форме взвеси порошка и воды, или в жидкой пропитки для семян. В определенных примерах одну или несколько композиций можно наносить на семена и/или проростки посредством распыления, погружения, покрытия, инкапсуляции и/или опыления семян и/или проростков одной или несколькими композициями. В определенных примерах на семена и/или проростки растений можно наносить несколько бактерий или популяции бактерий. В определенных примерах можно выбирать по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять, или более чем десять бактерии комбинации бактерий из одного из следующих родов: Acidovorax, Agrobacterium, Bacillus, Burkholderia, Chryseobacterium, Curtobacterium, Enterobacter, Escherichia, Methylobacterium, Paenibacillus, Pantoea, Pseudomonas, Ralstonia, Saccharibacillus, Sphingomonas и Stenotrophomonas.
[0242] В определенных примерах по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять или более чем десять бактерий и популяций бактерий эндофитной комбинации выбраны из одного из следующих семейств: Bacillaceae, Burkholderiaceae, Comamonadaceae, Enterobacteriaceae, Flavobacteriaceae, Methylobacteriaceae, Microbacteriaceae, Paenibacillileae, Pseudomonnaceae, Rhizobiaceae, Sphingomonadaceae, Xanthomonadaceae, Cladosporiaceae, Gnomoniaceae, Incertae sedis, Lasiosphaeriaceae, Netriaceae и Pleosporaceae.
[0243] В определенных примерах по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять или более десяти бактерий и популяций бактерий эндофитной комбинации выбраны из одного из следующих семейств: Bacillaceae, Burkholderiaceae, Comamonadaceae, Enterobacteriaceae, Flavobacteriaceae, Methylobacteriaceae, Microbacteriaceae, Paenibacillileae, Pseudomonnaceae, Rhizobiaceae, Sphingomonadaceae, Xanthomonadaceae, Cladosporiaceae, Gnomoniaceae, Incertae sedis, Lasiosphaeriaceae, Netriaceae, Pleosporaceae.
[0244] Примеры композиций могут включать покрытия семян для коммерчески важных сельскохозяйственных культур, например, сорго, канолы, томата, клубники, ячменя, риса, кукурузы и пшеницы. Примеры композиций также могут включать покрытия семян кукурузы, сои, канолы, сорго, картофеля, риса, овощей, злаковых и семена масличных культур. Семена, как предоставлено по настоящему документу, могут представлять собой генетически модифицированные организмы (GMO), не GMO, органические формы или общепринятые формы. В определенных примерах композиции можно распылять на надземные части растений или наносить на корни, внося их в борозды, в которые сажают семена растений, поливая почву или опуская корни в суспензию композиции. В определенных примерах композиции можно обезвоживать подходящим способом, который сохраняет жизнеспособность клеток и способность к искусственному заселению и колонизации растений-хозяев. Виды бактерий могут присутствовать в композициях в концентрации от 108 до 1010 КОЕ/мл. В определенных примерах композиции можно дополнять ионами металлов-микроэлементов, такими как ионы молибдена, ионы железа, ионы марганца или комбинации этих ионов. Концентрации ионов в примерах композиций, описываемых в настоящем документе, могут составлять приблизительно от 0,1 мМ до приблизительно 50 мМ. Определенные примеры композиций также можно формулировать c носителем, таким как бета-глюкан, карбоксилметилцеллюлоза (CMC), бактериальное внеклеточное полимерное вещество (EPS), сахар, молоко животного или другие подходящие носители. В определенных примерах в качестве носителя можно использовать торф или посадочные материалы или биополимеры, в которых композицию помещают в биополимер. Композиции, содержащие бактериальные популяции, описываемые в настоящем документе, могут улучшать признаки растений, например, стимулировать рост растений, поддерживать высокое содержание хлорофилла в листьях, увеличивать количества плодов или семян и увеличивать массу единичного плода или семени.
[0245] Композиции, содержащие бактериальные популяции, описываемые в настоящем документе, можно наносить на поверхность семян. Таким образом, предусмотрены композиции, содержащие семена, покрытые одним или несколькими видами бактерий, описанных в настоящем документе. Покрытие семян можно формулировать, смешивая популяцию бактерий с пористым, химически инертным гранулярным носителем. Альтернативно, композиции можно вносить непосредственно в борозды, в которые высаживают семена или распылять на листья растений или наносить посредством погружения корней в суспензию композиции. Можно использовать эффективное количество композиции для заполнения области грунта рядом с корнями растения с ростом жизнеспособных бактерий или заселения листьев растения с ростом жизнеспособных бактерий. Как правило, эффективное количество представляет собой количество, достаточное для получения растений с улучшенными признаками (например, с желаемым уровнем фиксации азота).
[0246] Бактериальные композиции, описываемые в настоящем документе, можно формулировать с использованием приемлемого в сельском хозяйстве носителя. Состав, пригодный для этих вариантов осуществления, может содержать по меньшей мере одного представителя, выбранного из группы, состоящей из придающего клейкость средства, стабилизатора микроорганизмов, фунгицида, антибактериального средства, консерванта, стабилизатора, поверхностно-активного вещества, противокомплексного средства, гербицида, нематоцида, инсектицида, регулятора роста растения, удобрения, родентицида, поглотителя влаги бактерицида, питательного вещества или любой их комбинации. В определенных примерах композиции могут быть пригодными для длительного хранения. Например, любая из композиций, описываемых в настоящем документе, может содержать приемлемый в сельском хозяйстве носитель (например, один или несколько из удобрений, таких как неприродное удобрение, обеспечивающих прилипание средств, таких как неприродное обеспечивающее прилипание средство, и пестицидов, таких как неприродный пестицид). Неприродное обеспечивающее прилипание средство может представлять собой, например, полимер, сополимер или синтетический воск. Например, любой из покрытых семян, всходов или растений, описываемых в настоящем документе, может содержать в покрытии семян такой приемлемый в сельском хозяйстве носитель. В любых из композиций или способов, описываемых в настоящем документе, приемлемый в сельском хозяйстве носитель может представлять собой или может содержать неприродное соединение (например, неприродное удобрение, неприродное обеспечивающее прилипание средство, такое как полимер, сополимер или синтетический воск, или неприродный пестицид). Неограничивающие примеры приемлемых в сельском хозяйстве носителей описаны ниже. В данной области известны дополнительные примеры приемлемых в сельском хозяйстве носителей.
[0247] В определенных случаях бактерии смешивают с приемлемым в сельском хозяйстве носителем. Носитель может представлять собой твердый носитель или жидкий носитель, и находиться в различных формах, включая микросферы, порошки, эмульсии и т.п. Носитель может представлять собой любой один или несколько из ряда носителей, которые обеспечивают ряд таких свойств, как увеличенная стабильность, смачиваемость или диспергируемость. В композиции можно включать средства для смачивания, такие как природные или синтетические поверхностно-активные вещества, которые могут быть неионными или ионными поверхностно-активными вещества, или их сочетание. Также для формулирования композиций, которые содержат выделенные бактерии, можно использовать эмульсии вода-в-масле (см., например, патент США № 7485451). Подходящие составы, которые можно получать, включают смачиваемые порошки, гранулы, гели, агарозные пластинки или шарики, загустители и т.п., микроинкапсулированные частицы и т.п., жидкости, такие как водные жидкости, водные суспензии, эмульсии вода-в-масле и т.д. Состав может содержать зерновые или бобовые продукты, например, молотые зерна или бобы, бульон или муку тонкого помола, получаемые из зерен или бобов, крахмал, сахар или масло.
[0248] В определенных вариантах осуществления сельскохозяйственный носитель может представлять собой почву или среду для выращивания растений. Другие сельскохозяйственные носители, которые можно использовать, включают воду, удобрения, растительные масла, увлажнители или их сочетания. Альтернативно, сельскохозяйственный носитель может быть твердым, таким как диатомовая земля, глина, диоксид кремния, альгинат, глинистый грунт, бентонит, вермикулит, семенные коробочки, другое растительные и животные продукты или их комбинации, включая гранулы, осадки или суспензии. В качестве носителей также предусмотрены смеси любых указанных выше ингредиентов, в качестве неограничивающих примеров такие как песта (мука тонкого помола и каолиновая глина), шарики на основе агара или муки тонкого помола в глине, песке или глинистый грунт и т.д. Составы могут содержать источники пищи для бактерий, такие как ячмень, рис или другие биологические материалы, такие как семена, части растений, выжимки сахарного тростника, шелуха или плодоножки после обработки зерна, измельченный растительный материал или древесину из отходов строительных участков, опилки или мелкие волокна после переработки бумаги, ткани или дерева.
[0249] Для помощи в стимуляции роста или обеспечения питательных веществ для семян, всходов или растений можно использовать, например, удобрение. Неограничивающие примеры удобрений включают азот, фосфор, калий, кальций, серу, магний, бор, хлорид, марганец, железо, цинк, медь, молибден и селен (или их соли). Дополнительные примеры удобрений, включают одну или несколько аминокислот, соли, углеводы, витамины, глюкозу, NaCl, дрожжевой экстракт, NH4H2PO4, (NH4)2SO4, глицерин, валин, L-лейцин, молочную кислоту, пропионовую кислоту, янтарную кислоту, яблочную кислоту, лимонную кислоту, тартрат KH, ксилозу, ликсозу и лецитин. В одном из вариантов осуществления состав может содержать придающее клейкость средство или клеящее средство (обозначаемое как адгезивное средство), помогающее связывать другие активные средства с веществом (например, с поверхностью семян). Такие средства пригодны для комбинации бактерий с носителями, которые могут содержать другие соединения (например, контрольные средства, которые являются небиологическими), с получением покрывающей композиции. Такие композиции помогают получать покрытия вокруг растения или семян с поддержанием контакта микроорганизмов и других средств с растением или частью растения. В одном из вариантов осуществления адгезивы выбраны из группы, состоящей из: альгината, камедей, различных видов крахмала, лецитинов, формононетина, поливинилового спирта, щелочного формононетината, гесперетина, поливинилацетата, цефалинов, гуммиарабика, ксантановой камеди, минерального масла, полиэтиленгликоля (ПЭГ), поливинилпирролидона (ПВП), арабино-галактана, метилцеллюлозы, ПЭГ 400, хитозана, полиакриламида, полиакрилата, аолиакрилонитрила, глицерина, триэтиленгликоля, винилацетата, геллановой камеди, полистирола, поливинила, карбоксиметилцеллюлозы, камеди гхатти и полиоксиэтилен-полиоксибутиленовых блок-сополимеров.
[0250] В определенных вариантах осуществления адгезивы могут представлять собой, например, воск, такой как карнаубский воск, пчелиный воск, китайский воск, шеллачный воск, спермацетовый воск, канделильский воск, гидрированное касторовое масло, орикурийский воск и воск рисовых отрубей, полисахарид (например, крахмал, декстрины, мальтодекстрины, альгинат и хитозаны), жир, масло, белок (например, желатин и зеины), гуммиарабик и шеллаки. Адгезивные средства могут представлять собой неприродные соединения, например, полимеры, сополимеры и воска. Например, неограничивающие примеры полимеров, которые можно использовать в качестве адгезивного средства, включают: поливинилацетаты, сополимеры поливинилацетатов, сополимеры этилена и винилацетата (EVA), поливиниловые спирты, сополимеры поливиниловых спиртов, соединения целлюлозы (например, различные виды этилцеллюлозы, метилцеллюлозы, гидроксиметилцеллюлозы, гидроксипропилцеллюлозы и карбоксиметилцеллюлозы), поливинилпирролидоны, винилхлорид, сополимеры винилиденхлорида, лигносульфонаты кальция, акриловые сополимеры, поливинилакрилаты, полиэтиленоксид, ациламидные полимеры и сополимеры, полигидроксиэтилакрилат, мономерные метилакриламиды и полихлоропрен.
[0251] В определенных примерах одно или несколько из адгезивных средств, противогрибковых средств, средств регуляции роста и пестицидов (например, инсектицидов) представляют собой неприродные соединения (например, в любой комбинации). Дополнительные примеры приемлемых в сельском хозяйстве носителей включают дисперсанты (например, поливинилпирролидон/винилацетат PVPIVA S-630), поверхностно-активные вещества, связывающие средства и наполнители.
[0252] Состав также может содержать поверхностно-активное вещество. Неограничивающие примеры поверхностно-активных веществ включают смеси азотных удобрений и поверхностно-активных веществ, такие как Prefer 28 (Cenex), Surf-N (US), Inhance (Brandt), P-28 (Wilfarm) и Patrol (Helena); этерифицированные масла семян включают Sun-It II (AmCy), MSO (UAP), Scoil (Agsco), Hasten (Wilfarm) и Mes-100 (Drexel); и органо-силиконовые поверхностно-активные вещества включают Silwet L77 (UAP), Silikin (Terra), Dyne-Amic (Helena), Kinetic (Helena), Sylgard 309 (Wilbur-Ellis) и Century (Precision). В одном из вариантов осуществления поверхностно-активное вещество присутствует в концентрации от 0,01% об./об. до 10% об./об. В другом варианте осуществления поверхностно-активное вещество присутствует в концентрации от 0,1% об./об. до 1% об./об.
[0253] В определенных случаях состав содержит стабилизатор микроорганизмов. Такое средство может включать поглотитель влаги, который может включать любое соединение или смесь соединений, которые можно классифицировать как поглотитель влаги вне зависимости от того используют ли соединение или соединения в таких концентрациях, что они фактически проявляют высушивающее действие на жидкое инокулируемое средство. Такие поглотители влаги в идеале совместимы с используемыми популяциями бактерий и должны поддерживать способность популяции микроорганизмов переживать нанесение на семена и переживать обезвоживание. Примеры подходящих поглотителей влаги включают одно или несколько из трегалозы, сахарозы, глицерина и метиленгликоля. Другие подходящие поглотители влаги в качестве неограничивающих примеров включают невосстанавливающие сахара и сахарные спирты (например, маннит или сорбит). Количество поглотителя влаги, вводимое в состав, может находиться в диапазоне приблизительно от 5% до приблизительно 50% по массе/объему, например, приблизительно от 10% до приблизительно 40%, приблизительно от 15% до приблизительно 35% или приблизительно от 20% до приблизительно 30%. В определенных случаях предпочтительно, чтобы состав содержал такие средства, как фунгицид, антибактериальное средство, гербицид, нематоцид, инсектицид, регулятор роста растения, родентицид, бактерицидное средство или питательное вещество. В определенных примерах средства могут включать защитные средства, которые обеспечивают защиту против патогенных микроорганизмов на поверхности семян. В определенных примерах защитные средства могут обеспечивать определенный уровень контроля почвенных патогенных микроорганизмов. В определенных примерах защитные средства могут быть эффективными преимущественно на поверхности семян.
[0254] В определенных примерах фунгицид может включать соединение или химическое или биологическое средство, которое может ингибировать рост гриба или уничтожать гриб. В определенных примерах фунгицид может включать соединения, которые могут являться фунгистатическими или фунгицидными. В определенных примерах фунгицид может представлять собой защитное средство или средства, которые эффективны преимущественно на поверхности семян, обеспечивая защиту против патогенных микроорганизмов на поверхности семян и обеспечивая определенный уровень контроля почвенных патогенных микроорганизмов. Неограничивающие примеры защитных фунгицидов включают каптан, манеб, тирам или флудиоксонил.
[0255] В определенных примерах фунгицид может представлять собой системный фунгицид, который может всасываться в растущий проросток и ингибировать или уничтожать гриб внутри тканей растений-хозяев. Системные фунгициды, используемые для пропитки семян в качестве неограничивающих примеров включают следующее: азоксистробин, карбоксин, мефеноксам, металаксил, тиабендазол, трифлоксистробин и различные триазоловые фунгициды, включающие дифеноконазол, ипконазол, тебуконазол и тритиконазол. Мефеноксам и металаксил преимущественно используют для воздействия на грибы водяной плесени Pythium и Phytophthora. Определенные фунгициды являются более предпочтительными, чем другие в зависимости от вида растения вследствие незначительных различий в чувствительности патогенных видов грибов или вследствие различий распределения фунгицидов или чувствительности растений. В определенных примерах фунгицид может представлять собой биологическое средство контроля, такое как бактерия или гриб. Такие организмы могут являться паразитами патогенных грибов или секретировать токсины или другие вещества, которые могут уничтожать или иным образом предотвращать рост грибов. В составе для семян в качестве контрольного средства можно использовать любой тип фунгицида, особенно фунгициды, которые широко используются у растений.
[0256] В определенных примерах композиция покрытия для семян содержит средство для контроля, которое обладает антибактериальными свойствами. В одном из вариантов осуществления средство для контроля с антибактериальными свойствами выбрано из соединений, описываемых в других разделах настоящего документа. В другом варианте осуществления соединение представляет собой стрептомицин, окситетрациклин, оксолиновая кислота или гентамицин. Другие примеры антибактериальных соединений, которые можно использовать в качестве части композиции покрытия для семян включают соединения на основе дихлорофена и гемиформаля бензилового спирта (Proxel®, производимый ICI, или Acticide® RS, производимый Thor Chemie, и Kathon® MK 25, производимый Rohm & Haas) и производные изотиазолинона, такие как алкилизотиазолиноны и бензизотиазолиноны (Acticide® MBS, производимые Thor Chemie).
[0257] В определенных примерах регулятор роста выбран из группы, состоящей из: абсцизовой кислотаы, амидохлора, анцимидола, 6-бензиламинопурина, брассинолида, бутралина, хлормеквата (хлорида хлормекват), холинхлорида, цикланилида, даминозида, дикегулака, диметипина, 2,6-диметилпуридина, этефона, флуметралина, флурпримидола, флутиацета, форхлорфенурона, гибберелловой кислоты, инабенфида, индол-3-уксусной кислоты, малеиновог гидразида, мефлуидида, мепиквата (хлорида мепиквата), нафталинуксусной кислоты, N-6-бензиладенина, паклобутразола, фосфотритиоата прогексадиона, 2,3,5-три-йодобензойной кислоты, тринексапакэтила и униконазола. Дополнительные неограничивающие примеры регуляторов роста включают брассиностероиды, цитокинины (например, кинетин и зеатин), ауксины (например, индолилуксусную кислоту и индолилацетиласпартат), флавоноиды и изофлавоноиды (например, формононетин и диосметин), фитоауксины (например, глицеоллин) и индуцируемые фитоалексинами олигосахариды (например, пектин, хитин, хитозан, полигалактуроновая кислота и олигогалактуроновая кислота) и гибеллерины. Такие средства в идеале совместимы с сельскохозяйственными семенами или всходами, на которые наносят состав (например, он не должен быть вреден для роста или здоровья растений). Кроме того, в идеале средство представляет собой средство, которое не представляет угрозу безопасности применения для человека, животного или промышленного (например, отсутствие угроз безопасности или соединение является в достаточной степени нестабильным так, что потребляемый растительный продукт, получаемый из растения, содержит незначительные количества этого соединения).
[0258] Определенные примеры антагонистичных нематодам средств биоконтроля, включают ARF18; 30 видов Arthrobotrys; виды Chaetomium; виды Cylindrocarpon; виды Exophilia; виды Fusarium; виды Gliocladium; виды Hirsutella; виды Lecanicillium; виды Monacrosporium; виды Myrothecium; виды Neocosmospora; виды Paecilomyces; виды Pochonia; виды Stagonospora; везикулярно-арбускулярные микоризные грибы, виды Burkholderia; виды Pasteuria, виды Brevibacillus; виды Pseudomonas и Rhizobacteria. Особенно предпочтительные антагонистичные нематодам средства биоконтроля включают ARF18, Arthrobotrys oligospora, Arthrobotrys dactyloides, Chaetomium globosum, Cylindrocarpon heteronema, Exophilia jeanselmei, Exophilia pisciphila, Fusarium aspergilus, Fusarium solani, Gliocladium catenulatum, Gliocladium roseum, Gliocladium vixens, Hirsutella rhossiliensis, Hirsutella minnesotensis, Lecanicillium lecanii, Monacrosporium drechsleri, Monacrosporium gephyropagum, Myrotehcium verrucaria, Neocosmospora vasinfecta, Paecilomyces lilacinus, Pochonia chlamydosporia, Stagonospora heteroderae, Stagonospora phaseoli, везикулярно-арбускулярные микоризные грибы, Burkholderia cepacia, Pasteuria penetrans, Pasteuria thornei, Pasteuria nishizawae, Pasteuria ramosa, Pastrueia usage, штамм G4 Brevibacillus laterosporus, Pseudomonas fluorescens и Rhizobacteria.
[0259] Определенные примеры питательных вещества можно выбрать из группы, состоящей из азотных удобрений, включающих в качестве неограничивающих примеров мочевину, нитрат аммония, сульфат аммония, безнапорные азотные растворы, водный аммиак, безводный аммиак, тиосульфат аммония, покрытую серой мочевину, мочевины-формальдегидов, IBDU, покрытую полимерами мочевину, нитрат кальция, уреаформ и метиленмочевину, фосфорные удобрения, такие как дифосфат аммония, монофосфат аммония, полифосфат аммония, концентрированный суперфосфат и тройной суперфосфат, и калийные удобрения, такие как хлорид калия, сульфат калия, сульфат калия-магния, нитрат калия. Такие композиции в композиции для покрытия семян могут находиться в виде свободных солей или ионов. Альтернативно, питательные вещества/удобрения можно формировать в виде комплексных или хелатированных соединений с обеспечением длительного высвобождения в течение продолжительного периода.
[0260] Определенные примеры родентицидов можно выбирать из группы веществ, состоящей из 2-изовалерилиндан-1,3-диона, 4-(хиноксалин-2-иламино)бензолсульфонамида, альфа-хлоргидрина, фосфида алюминия, альфа-нафтилтиомочевины, оксида мышьяка, карбоната бария, бистиосеми, бродифакума, бромадиолона, брометалина, цианида кальций, хлоралозы, хлорофацинона, холекальциферола, кумахлора, кумафурила, куматетралила, кримидина, дифенакума, дифетиалона, дифацинона, эргокальциферола, флокумафена, фторацетамида, флупропадина, гидрохлорида флупропадина, цианида водорода, йодометана, линдана, фосфида магния, метилбромида, норбормида, фосацетима, фосфина, фосфора, пиндона, арсенита калия, пиринурона, скиллирозида, арсенита натрия, цианида натрий, фторацетата натрия, стрихнина, сульфата талия, варфарина и фосфида цинка.
[0261] В жидкой форме, например, в растворе или суспензии, бактериальные популяции можно смешивать или суспендировать в воде или в водных растворах. Подходящие жидкие разбавители или носители включают воду, водные растворы, нефтяные дистилляты или другие жидкие носители.
[0262] Твердые композиции можно получать посредство диспергирования бактериальных популяций в соответствующим образом измельченном твердом носителе и на нем, таком как торф, пшеница, отруби, вермикулит, глина, тальк, бентонит, диатомовая земля, фуллерова глина, пастеризованная почва и т.п. Когда такие составы используют в качестве смачиваемых порошков, можно использовать биологически совместимые диспергирующие средства, такие как неионные, анионные, амфотерные или катионные диспергирующие средства и эмульгаторы.
[0263] Твердые носители, используемые в составе включают, например, минеральные носители, такие как каолиновая глина, пирофиллит, бентонит, монтмориллонит, диатомовая земля, кислая белая земля, вермикулит и перлит, и неорганические соли, такие как сульфат аммония, фосфат аммония, нитрат аммония, мочевина, хлорид аммония и карбонат кальция. Также можно использовать органические тонкодисперсные порошки, такие как пшеничная мука, пшеничные отруби и рисов отруби. Жидкие носители включают растительные масла, такие как соевое масло и хлопковое масло, глицерин, этиленгликоль, полиэтиленгликоль, пропиленгликоль, полипропиленгликоль и т.д.
Применение популяций бактерий для сельскохозяйственных культур
[0264] Композицию бактерий или популяцию бактерий, описываемые в настоящем документе, можно наносить в борозду, в форме талька или в виде пропитки для семян. Композицию можно наносить на упаковку семян в целом, миниобъеме, в мешке или в тальке.
[0265] Сельхозпроизводитель может высаживать обработанные семена и выращивать сельскохозяйственную культуру общепринятыми способами, двумя рядами или способами, не требующими обработки почвы. Семена можно распределять с использованием контрольного бункера или отдельного бункера. Семена также можно распределять с помощью сжатого воздуха или вручную. Посев семян можно проводить с использованием технологий с переменной скоростью. Кроме того, с использованием технологий с переменной скоростью может проводить нанесение бактерий или популяций бактерий, описываемых в настоящем документе. В определенных примерах бактерии можно наносить на семена кукурузы, сои, канолы, сорго, картофеля, риса, овощей, злаков, псевдозерновых и семена масличных культур. Примеры злаков могут включать ячмень, фонио, овес, траву пальмера, рожь, жемчужное просо, сорго, полбу, тэфф, тритикале и пшеницу. Примеры псевдозерновых могут включать грецкий орех, гречиху, полевой хвощ, чиа, лен, зерновой амарант, ганзу, киноа и кунжут. В определенных примерах семена могут являться генетически модифицированными организмами (GMO), не GMO, органическими или общепринятыми.
[0266] Кроме обработки сельскохозяйственных культур можно использовать добавки, такие как микроудобрение, PGR, гербицид, инсектицид и фунгицид. Примеры добавок включают защитные средства для сельскохозяйственных культур, такие как инсектициды, нематоциды, фунгициды, укрепляющие средства, такие как красители, полимеры, пеллетирование, примирование, и дезинфицирующие средства и другие средства, такие как инокулятор, PGR, смягчитель и питательные микроэлементы. PGR могут представлять собой природные или синтетические гормоны растений, которые влияют на рост корня, цветение или удлинение стебля. PGR могут включать ауксины, гиббереллины, цитокины, этилен и абсцизовую кислоту (ABA).
[0267] Композицию можно наносить в борозду в комбинации с жидким удобрением. В определенных примерах жидкое удобрение можно содержать в резервуарах. Удобрения NPK содержат макробиогенные вещества с натрием, фосфором и калием.
[0268] Композиция может улучшать свойства растений, например, способствуя росту растений, поддерживая высокое содержание хлорофилла в листьях, увеличивая количества плодов или семян и увеличивая массу единичных плодов или семян. Способы по настоящему изобретению можно использовать для добавления или улучшения одного или нескольких из ряда требуемых свойств. Примеры свойств которые можно добавлять или улучшать включают: биомассу корня, длина корня, высоту, высоту побега, количество листьев, эффективность использования воды, общую биомассу, урожайность, размер плода, размер зерен, интенсивность фотосинтеза, засухоустойчивость, жароустойчивость, солеустойчивость, устойчивость к стрессу с низким содержанием азота, эффективность использования азота, устойчивость к воздействию нематод, устойчивость к грибковым патогенным микроорганизмам, устойчивость к бактериальным патогенным микроорганизмам, устойчивость к вирусным патогенным микроорганизмам, уровень метаболитов, модуляцию уровня метаболитов, экспрессию протеома. Требуемые свойства, включая высоту, общую биомассу, биомассу корня и/или побега, прорастание семян, жизнеспособность проростков, эффективность фотосинтеза, интенсивность транспирации, количество или масса семян/плодов, урожайность зерна или плодов у растения, содержание хлорофилла в листьях, интенсивность фотосинтеза, длина корня или любое их сочетание можно использовать для определения роста и сравнивать со скоростью роста эталонных сельскохозяйственных растений (например, растений без добавления и/или улучшения свойств), выращиваемых в идентичных условиях. В определенных примерах требуемые свойства, включая высоту, общую биомассу, биомассу корней и/или побегов, прорастание семян, жизнеспособность проростков, эффективность фотосинтеза, интенсивность транспирации, количество или масса семян/плодов, урожайность зерна или плодов у растения, содержание хлорофилла в листьях, интенсивность фотосинтеза, длину корня или любое их сочетание можно использовать для определения роста и сравнения с скоростью роста эталонного сельскохозяйственного растения (например, растения без добавления и/или улучшения свойств), выращиваемых в сходных условиях.
[0269] Агротехнические признаки растения-хозяина в качестве неограничивающих примеров могут включать следующие: измененное содержание масла, измененное содержание белка, измененный состав углеводов семян, измененный состав масла семян и измененный состав белка семян, химическую устойчивость, устойчивость к холоду, замедленное старение, устойчивость к болезням, устойчивость к засухе, массу початка, улучшение роста, улучшение жизнеспособности, теплоустойчивость, устойчивость к гербицидам, устойчивость к травоядным животным, улучшенную фиксацию азота, улучшенное использование азота, улучшенную архитектуру корней, улучшенную эффективность использования воды, увеличенную биомассу, увеличенную длину корней, увеличенную массу семян, увеличенный рост побегов, увеличенную урожайность, увеличенный урожай при условиях с ограничением количества воды, массу зерен, содержание влаги в зернах, толерантность к металлам, количество початков, количество зерен в початке, количество стручков, улучшение питания, устойчивость к патогенным микроорганизмам, устойчивость к вредителям, улучшение способности к фотосинтезу, устойчивость к засолению, сохранение зеленого цвета, улучшение мощности, увеличенную сухую массу зрелых семян, увеличенную сырую массу зрелых семян, увеличенное количество зрелых семян на растение, увеличенное содержание хлорофилла, увеличенное количество стручков на растение, увеличенную длину стручков на растении, уменьшенное количество увядших листьев на растении, уменьшенное количество сильно увядших листьев на растении и увеличенное количество не увядших листьев на растение, детектируемаое изменение уровня метаболита, детектируемое изменение уровня транскрипта и детектируемое изменение протеома по сравнению с растением изолинии, выращенным из семян без указанного состава препарата для обработки семян.
[0270] В определенных случаях растения инокулируют бактериями или популяциями бактерий, которые выделены у того же вида растения в виде составляющей инокулируемого растения. Например, бактерия или популяция бактерий, которую в норме выявляют у одного сорта Zea mays (кукурузы), ассоциирована с составляющим растения другого сорта Zea mays, у которого в его природном состоянии существует недостаток указанных бактерий и популяций бактерий. В одном варианте осуществления бактерии и популяции бактерий получают у растения родственного вида в качестве составляющей инокулированного растения. Например, бактерии и популяции бактерий, которые обычно выявляют у Zea diploperennis, Iltis et al., (Diploperennial teosinte), используют для Zea mays (кукурузы) или наоборот. В определенных случаях растения инокулируют бактериями и популяциями бактерий, которые гетерологичны по отношению к элементу растения инокулируемого растения. В одном варианте осуществления бактерии и популяции бактерий получают у растения другого вида. Например, бактерии и популяции бактерий, которые обычно выявляют у двудольных растений, применяют для однодольных растений (например, инокулируют кукурузу бактериями и популяциями бактерий, полученными из бобов сои) или наоборот. В других случаях бактерии и популяции бактерий, которые для инокуляции в растение, получают у родственных видов растения, подвергаемого инокуляции. В одном варианте осуществления бактерии и популяции бактерий получают у родственного таксона, например, у родственных видов. Растение другого вида может являться сельскохозяйственным растением. В другом варианте осуществления бактерии и популяции бактерий являются частью разработанной композиции, инокулируемой в любой элемент растения-хозяина.
[0271] В определенных примерах бактерии или популяция бактерий являются экзогенными, где бактерии и популяция бактерий выделяют у растения, отличного от инокулируемого растения. Например, в одном из вариантов осуществления бактерии или популяция бактерий можно выделять из другого растения того же вида, что и инокулируемое растение. В определенных случаях бактерии или популяция бактерий можно выделять у видов, родственных инокулируемому растению.
[0272] В определенных примерах бактерии и популяции бактерий, описанные здесь, способны перемещаться от одного типа ткани к другому. Например, выявление и выделение бактерий и популяций бактерий по настоящему изобретению в зрелых тканях растений после нанесения покрытия на внешнюю поверхность семян демонстрирует их способность перемещаться из снаружи семян в вегетативные ткани созревающего растения. Таким ообразом, в одном из вариантов осуществления популяция бактерий и популяций бактерий способна перемещаться снаружи семян в вегетативные ткани растения. В одном из вариантов осуществления бактерии и популяции бактерий, которые наносят на семена растения, при прорастании семени в вегетативное состояние способны к локализации в других тканях растения. Например, бактерии и популяции бактерий могут быть способными к локализации в любой из тканей растения, в том числе: корне, добавочном корне, зародышевом корне, корневом волоске, побеге, листе, цветке, почке, метелке, меристеме, пыльце, пестике, завязи, тычинке, плоде, подземном побеге, корневом побеге, клубеньке, клубне, трихоме, защитных клетках, гидатоде, лепестке, чашелистике, оболочке зерна, оси колоса, сосудистом камбии, флоэме и ксилеме. В одном из вариантов осуществления бактерии и популяции бактерий способны к локализации в корне и/или корневых волосках растения. В другом варианте осуществления бактерии и популяции бактерий способны к локализации в фотосинтезирующих тканях, например листьях и побегах растения. В других случаях бактерии и популяции бактерий локализованы в сосудистых тканях растения, например, в ксилеме и флоэме. В еще одном из вариантов осуществления бактерии и популяции бактерий способны к локализации в репродуктивных тканях (цветке, пыльце, пестике, завязи, тычинке, плоде) растения. В другом варианте осуществления бактерии и популяции бактерий способны к локализации в корне, побегах, листьях и репродуктивных тканях растения. В еще одном из вариантов осуществления бактерии и популяции бактерий колонизируют плод или семенную ткань растения. В еще одном из вариантов осуществления бактерии и популяции бактерий способны колонизировать растение таким образом, что они находятся на поверхности растения (то есть их присутствие детектируют на внешней стороне растения или в эписфере растения). В других вариантах осуществления бактерии и популяции бактерий способны к локализации по существу во всем растении или во всех тканях растения. В определенных вариантах осуществления бактерии и популяции бактерий не локализованы в корне растения. В других случаях бактерии и популяции бактерий не локализованы в фотосинтезирующих тканях растения.
[0273] Эффективность композиций также можно оценивать определяя относительную зрелость сельскохозяйственной культуры или тепловые единицы сельскохозяйственной культуры (CHU). Например, популяцию бактерий можно наносить на кукурузу, и рост кукурузы можно оценивать в соответствии с относительной зрелость зерна кукурузы или по времени за которое зерно кукурузы достигает максимальной массы. Также для прогноза созревания кукурузы можно использовать тепловые единицы сельскохозяйственных культур (CHU). CHU определяют количество накопленной теплоты посредством определения суточного максимума температур при росте сельскохозяйственной культуры.
[0274] В примерах бактерии могут локализоваться в любой из тканей растения, в том числе: корне, добавочном корне, зародышевом корне, корневом волоске, побеге, листе, цветке, почке, метелке, меристеме, пыльце, пестике, завязи, тычинке, плоде, подземном побеге, корневом побеге, клубеньке, клубне, трихоме, защитных клетках, гидатоде, лепестке, чашелистике, оболочке зерна, оси колоса, сосудистом камбии, флоэме и ксилеме. В другом варианте осуществления бактерия или популяция бактерий способны к локализации в фотосинтезирующих тканях, например листьях и побегах растения. В других случаях бактерии и популяции бактерий локализуются в сосудистых тканях растения, например, в ксилеме и флоэме. В другом варианте осуществления бактерия или популяция бактерий способны к локализации в репродуктивных тканях (цветке, пыльце, пестике, завязи, тычинке или плоде) растения. В другом варианте осуществления бактерии и популяции бактерий способны к локализации в корне, побегах, листьях и репродуктивных тканях растения. В другом варианте осуществления бактерии или популяция бактерий колонизируют плод или семенную ткань растения. В еще одном варианте осуществления бактерии или популяция бактерий способны колонизировать растение так, что они находятся на поверхности растения. В другом варианте осуществления бактерии или популяция бактерий способны к локализации по существу во всем растении или во всех тканях растения. В определенных вариантах осуществления бактерии или популяция бактерий не локализованы в корне растения. В других случаях бактерии и популяции бактерий не локализованы в фотосинтезирующих тканях растения.
[0275] Эффективность бактериальных композиций, наносимых на сельскохозяйственные культуры можно оценивать, определяя различные свойства роста сельскохозяйственной культуры, включая в качестве неограничивающих примеров норму высева, мощность посева, силу корня, засухоустойчивость, высоту растения, сухую и тестовую массу.
Виды растений
[0276] Способы и бактерии, описываемые в настоящем документе, подходят для любого из множества растений, таких как растения родов Hordeum, Oryza, Zea и Triticeae. Другие неограничивающие примеры подходящих растений включают мхи, лишайники и водоросли. В некоторых случаях растения имеют экономическое, социальное и/или природоохранное значение, являясь продовольственными культурами, волокнистыми сельскохозяйственными культурами, масличными культурами, растениями, используемыми в лесной или целлюлозно-бумажной промышленности, сырьем для получения биотоплива и/или декоративными растениями. В определенных примерах растения можно использовать для продукции экономически значимых продуктов, таких как зерно, мука тонкого помола, крахмал, сироп, мука грубого помола, масло, пленка, упаковка, биологические добавки, волокнистая масса, корм для животных, корм для рыб, материал основы для промышленных химикатов, хлебопродукты, обработанный пищевой продукт для человека, сахар, спирт и/или белок. Неограничивающие примеры сельскохозяйственных культур включают кукурузу, рис, пшеницу, ячмень, сорго, просо, овес, рожь тритикале, гречиху, сахарную кукурузу, сахарный тростник, репчатый лук, томаты, клубнику и спаржу.
[0277] В определенных примерах растения, которые можно получать или улучшать способами и композициями, описываемыми в настоящем документе, может включать растения, которые являются важными или представляют интерес для сельского хозяйства, садоводства, биомассы для получения молекул биотоплива и других химических веществ, и/или для лесоводства. Определенные примеры этих растений могут включать ананас, банан, кокос, лилии, травяной горох и травы; и двудольные растения, такие как, например, горох, люцерна, физалис, дыня, нут, цикорий, клевер, кочанную капусту, чечевицу, сою, табак, картофель, батат, редис, огородную капусту, рапс, яблоневые деревья, виноград, хлопок, подсолнечник, арабидопсис, канолу, цитрусовые (включая апельсин, мандарин, кумкват, лимон, лайм, грейпфрут, тангерин, танжело, цитрон и помело), перец, фасоль и латук, Panicum virgatum (кисть), Sorghum bicolor (сорго, Судан), Miscanthus giganteus (мискантус), вид Saccharum (сахарный тростник), Populus balsamifera (тополь), Zea mays (кукуруза), Glycine max (соя), Brassica napus (канола), Triticum aestivum (пшеница), Gossypium hirsutum (хлопок), Oryza sativa (рис), Helianthus annuus (подсолнечник), Medicago sativa (люцерна), Beta vulgaris (сахарная свекла), Pennisetum glaucum (просо американское), виды Panicum виды Sorghum, виды Miscanthus, виды Saccharum, виды Erianthus, виды Populus, Secale cereale (рожь), виды Salix (верба), виды Eucalyptus (эвкалипт), виды Triticosecale (пшеница - 25 пшеница X рожь), Bamboo, Carthamus tinctorius (сафлор), Jatropha curcas (ятрофа), Ricinus communis (клещевина), Elaeis guineensis (масличная пальма), Phoenix dactylifera (финиковая пальма), Archontophoenix cunninghamiana (королевская пальма), Syagrus romanzoffiana (арекаструм), Linum usitatissimum (лен), Brassica juncea, Manihot esculenta (маниок), Lycopersicon esculentum (томат), Lactuca saliva (латук), Musa paradisiaca (банан), Solanum tuberosum (картофель), Brassica oleracea (брокколи, цветная капуста, брюссельская капуста), Camellia sinensis (чай), Fragaria ananassa (клубника), Theobroma cacao (какао), Coffea arabica (кофе), Vitis vinifera (виноград), Ananas comosus (ананас), Capsicum annum (острый и сладкий перец), Allium cepa (лук), Cucumis melo (дыня), Cucumis sativus (огурец), Cucurbita maxima (тыква), Cucurbita moschata (тыква), Spinacea oleracea (шпинат), Citrullus lanatus (арбуз), Abelmoschus esculentus (окра), Solanum melongena (баклажан), Papaver somniferum (опиумный мак), Papaver orientale, Taxus baccata, Taxus brevifolia, Artemisia annua, Cannabis saliva, Camptotheca acuminate, Catharanthus roseus, Vinca rosea, Cinchona officinalis, Coichicum autumnale, Veratrum californica, Digitalis lanata, Digitalis purpurea, виды Dioscorea 5, Andrographis paniculata, Atropa belladonna, Datura stomonium, виды Berberis, виды Cephalotaxus, Ephedra sinica, виды Ephedra, Erythroxylum coca, Galanthus wornorii, виды Scopolia, Lycopodium serratum (Huperzia serrata), виды Lycopodium, Rauwolfia serpentina, виды Rauwolfia, Sanguinaria canadensis, виды Hyoscyamus, Calendula officinalis, Chrysanthemum parthenium, Coleus forskohlii, Tanacetum parthenium, Parthenium argentatum (гваюла), виды Hevea (каучук), Mentha spicata (мята), Mentha piperita (мята), Bixa orellana, виды Alstroemeria, виды Rosa (роза), Dianthus caryophyllus (гвоздика), виды Petunia (петуния), Poinsettia pulcherrima (пуанзеция), Nicotiana tabacum (табак), Lupinus albus (люпин), Uniola paniculata (овес), Hordeum vulgare (ячмень) и виды Lolium (рожь).
[0278] В определенных примерах можно использовать однодольное растение. Однодольные растения принадлежат к порядкам Alismatales, Arales, Arecales, Bromeliales, Commelinales, Cyclanthales, Cyperales, Eriocaulales, Hydrocharitales, Juncales, Lilliales, Najadales, Orchidales, Pandanales, Poales, Restionales, Triuridales, Typhales, и Zingiberales. Растения, принадлежащие к классу Gymnospermae, представляют собой Cycadales, Ginkgoales, Gnetales и Pinales. В определенных примерах однодольное растение можно выбирать из группы, состоящей из кукурузы, риса, пшеницы, ячменя и сахарного тростника.
[0279] В определенных примерах можно использовать двудольное растение, включая растения, принадлежащие порядкам Aristochiales, Asterales, Batales, Campanulales, Capparales, Caryophyllales, Casuarinales, Celastrales, Cornales, Diapensales, Dilleniales, Dipsacales, Ebenales, Ericales, Eucomiales, Euphorbiales, Fabales, Fagales, Gentianales, Geraniales, Haloragales, Hamamelidales, Middles, Juglandales, Lamiales, Laurales, Lecythidales, Leitneriales, Magniolales, Malvales, Myricales, Myrtales, Nymphaeales, Papeverales, Piperales, Plantaginales, Plumb aginales, Podostemales, Polemoniales, Polygalales, Polygonales, Primulales, Proteales, Rafflesiales, Ranunculales, Rhamnales, Rosales, Rubiales, Salicales, Santales, Sapindales, Sarraceniaceae, Scrophulariales, Theales, Trochodendrales, Umbellales, Urticales и Violates. В определенных примерах двудольное растение можно выбирать из группы, состоящей из хлопка, сои, перца и томата.
[0280] В некоторых случаях улучшаемое растение трудно поддается экспериментальным условиям. Например, может пройти слишком много времени для достаточного роста сельскохозяйственной культуры, чтобы практически последовательно оценить улучшенный признак в нескольких повторениях. Таким образом, первое растение, из которого исходно выделяют бактерии, и/или множество растений, для которых применяют генетически модифицированные бактерии, могут представлять собой модельные растения, такие как растение, более поддающиеся анализу в требуемых условиях. Неограничивающие примеры модельных растений включают Setaria, Brachypodium и Arabidopsis. Способность бактерий, выделяемых способом по настоящему изобретению с использованием модельного растения, затем для подтверждения приобретения улучшенного признака можно применять для растений другого типа (например, сельскохозяйственной культуры).
[0281] Признаки, которые можно улучшать способами, описываемыми в настоящем документе, включают любые наблюдаемые характеристики растения, включая, например, скорость роста, рост, массу, цвет, вкус, запах, изменения продукции растением одного или нескольких соединений (включая например, метаболиты, белки, лекарственные средства, углеводы, масла и любые другие соединения). Также предусмотрен отбор растений на основе генотипической информации (например, включая профиль экспрессии генов растения в ответ на бактерии или идентификацию присутствия генетических маркеров, таких как генетические маркеры, ассоциированные с увеличенной фиксацией азота). Растения в противоположность присутствию определенного свойства или признака (таких как требуемые свойство или признак) также можно выбирать на основе отсутствия, подавления или ингибирования определенного свойства или признака (таких как нежелательные свойство или признак).
Концентрации и уровни применения сельскохозяйственных композиций
[0282] Как указано выше, сельскохозяйственные композиции по настоящему изобретению, которые содержат указанные микроорганизмы, можно наносить на растения множеством способов. В двух конкретных аспектах по изобретению предусмотрена обработка в бороздах или пропитка для семян.
[0283] Для варианты осуществления с пропиткой для семян микроорганизмы по изобретению могут находиться на семенах во множестве концентраций. Например, микроорганизмы в пропитке для семян могут присутствовать с концентрацией КОЕ на семя: 1×101, 1×102, 1×103, 1×104, 1×105, 1×106, 1×107, 1×108, 1×109, 1×1010 или более. В конкретных аспектах пропитка для семян композиции содержат приблизительно от 1×104 до приблизительно 1×108 КОЕ на семя. В других конкретных аспектах композиция пропитки для семян содержит приблизительно от 1×105 до приблизительно 1×107 КОЕ на семя. В других аспектах композиция пропитки для семян содержат приблизительно 1×106 КОЕ на семя.
[0284] В Соединенных Штатах Америки приблизительно 10% площади посева кукурузы засажено при плотности семян более чем приблизительно 36000 семян на акр; 1/3 площади посева кукурузы засажено при плотности семян приблизительно от 33000 до 36000 семян на акр; 1/3 площади посева кукурузы засажено при плотности семян приблизительно от 30000 до 33000 семян на акр, и оставшаяся часть площади является переменной. См., "Corn Seeding Rate Considerations", написанную Steve Butzen, доступную на https://www.pioneer.com/home/site/us/agronomy/library/corn-seeding-rate-considerations/
[0285] В таблице B ниже использованы различные концентрации КОЕ на семя в предусмотренном варианте осуществления пропитки для семян (поперечные ряды) и различные плотности посадки посевных площадей (1 колонка: 15K-41K) с расчетом общего количества КОЕ на акр, которое можно использовать в различных сельскохозяйственных условиях (т.е. концентрация пропитки для семян на семя × плотность посадки семян на акр). Таким образом, если использовать пропитка для семян с 1×106 КОЕ на семя и высаживать 30000 семян на акр, то общее содержание КОЕ на акр будет составлять 3×1010 (т.е. 30K × 1×106).
Таблица B: Расчет общего количества КОЕ на акр для вариантов осуществления пропитки для семян
[0286] Для вариантов осуществления обработки в борозде микроорганизмы по изобретению можно наносить с концентрацией КОЕ на акр: 1×106, 3,20×1010, 1,60×1011, 3,20×1011, 8,0×1011, 1,6×1012, 3,20×1012 или более. Таким образом, в определенных аспектах жидкие композиции для обработки в борозде можно наносить с концентрацией приблизительно от 1×106 до приблизительно 3×1012 КОЕ на акр.
[0287] В определенных аспектах композиции для обработки в борозде содержатся в жидком составе. В вариантах осуществления для обработке жидкостью в борозде микроорганизмы могут находиться в концентрации КОЕ в миллилитре: 1×101, 1×102, 1×103, 1×104, 1×105, 1×106, 1×107, 1×108, 1×109, 1×1010, 1×1011, 1×1012, 1×1013 или более. В определенных аспектах жидкие композиции для обработке в борозде содержат микроорганизмы в концентрации приблизительно от 1×106 до приблизительно 1×1011 КОЕ в миллилитре. В других аспектах жидкие композиции для обработке в борозде содержат микроорганизмы в концентрации приблизительно от 1×107 до приблизительно 1×1010 КОЕ в миллилитре. В других аспектах жидкие композиции для обработке в борозде содержат микроорганизмы в концентрации приблизительно от 1×108 до приблизительно 1×109 КОЕ в миллилитре. В других аспектах жидкие композиции для обработке в борозде содержат микроорганизмы в концентрации приблизительно до 1×1013 КОЕ в миллилитре.
ПРИМЕРЫ
[0288] В примерах, предоставляемых в настоящем документе, описаны способы выделения бактерий, анализа бактерий и растений и улучшения признаков растений. Примеры приведены исключительно с иллюстративными целями, и их не следует рассматривать, как каким-либо образом ограничивающие.
Пример 1: Выделение микроорганизмов из ткани растения
[0289] В различных сельскохозяйственных районах центральной Калифорнии получали верхние слои почвы. Получали двадцать почв с различными структурными характеристиками, включая плотную глину, торфяной суглинок, илистую глину и суглинок. Семена различных полевых кукурузы, сахарной кукурузы, старой негибридной кукурузу и томата высаживали в каждую почву, как представлено в таблице 1.
Таблица 1: Типы и сорта сельскохозяйственных культур, высаживаемых в почвы с различными характеристиками
[0290] Растения выкорчевывали через 2-4 недели роста и избыток почвы на поверхностях корней удаляли деионизированной водой. После удаления почвы у растений стерилизовали поверхность побелкой и интенсивно промывали в стерильной воде. Из растения вырезали 1 см участка очищенного корня и помещали в фосфатно-солевой буфер, содержащий 3 мм стальные гранулы. Посредством интенсивного перемешивания раствора на Qiagen TissueLyser II получали взвесь.
[0291] Взвесь корня и солевого раствора разбавляли и инокулировали на различные типы сред для выращивания для выделения ризосферных, эндофитных, эпифитных и других ассоциированных с растениями микроорганизмов. Для получения единичных колоний использовали агарозные среды R2A и Nfb, а для получения популяций фиксирующих азот бактерий использовали полутвердые скошенные среды Nfb. Через 2-4 недели инкубации в полутвердых скошенных средах Nfb популяции микроорганизмов собирали и штрихами наносили на агар R2A для получения единичных колоний, как показано на фигуре 1A-B. Единичные колонии ресуспендировали в смеси R2A и глицерина, подвергали анализу ПЦР и замораживали при -80°C для дальнейшего анализа. Получали приблизительно 1000 единичных колоний и обозначали как "выделенные микроорганизмы".
[0292] Затем изоляты подвергали скринингу колоний посредством ПЦР с детекцией присутствия генов nifH для идентификации диазотрофов. Для анализа присутствия кластера nif в каждом изоляте использовали ранее описанный набор праймеров Ueda 19F/388R, для которого показано, что посредством него при скринингах детектируют более 90% диазотрофов (Ueda et al. 1995; J. Bacteriol. 177: 1414-1417). Накалывали единичные колонии выделенных изолятов, ресуспендировали в PBS и использовали в качестве матрицы для ПЦР колоний, как представлено на фигуре 2. Колонии изолятов, для которых получали положительные полосы при ПЦР повторно высевали штрихом, и дважды повторяли ПЦР колоний и повторное высевание штрихом для предотвращения ложно-положительной идентификации диазотрофов. Затем выделенные изоляты обозначали как "микроорганизмы-кандидаты".
Пример 2: Характеристика выделенных микроорганизмов
Секвенирование, анализ и филогенетическая характеристика
[0293] Для получения предварительных филогенетических идентификационных характеристик выделенных микроорганизмов и микроорганизмов-кандидатов использовали секвенирование 16S рДНК с набором праймеров 515f-806r (см. например, Vernon et al.; BMC Microbiol. 2002 Dec 23;2:39.). Микроорганизмы содержали различные рода, включая: Enterobacter, Burkholderia, Klebsiella, Bradyrhizobium, Rahnella, Xanthomonas, Raoultella, Pantoea, Pseudomonas, Brevundimonas, Agrobacterium и Paenibacillus, как представлено в таблице 2.
Таблица 2: Разнообразие микроорганизмов, выделяемых из растений томата, как определено посредством глубокого секвенирования 16S рДНК.
[0294] Затем геномы 39 микроорганизмов-кандидатов секвенировали с использованием платформы Illumina Miseq. Геномную ДНК из чистых культур выделяли с использованием мини-набора для ДНК QIAmp (QIAGEN), а библиотеки тотальной ДНК для секвенирования получали у независимого поставщика (SeqMatic, Hayward). Затем проводили сборку генома посредством конвейера A5 (Tritt et al., 2012; PLoS One 7(9):e42304). Проводили идентификацию и аннотацию генов, и гены, связанные с регуляцией и проявлением фиксации азота отмечали в качестве мишеней мутагенеза.
Транскриптомное профилирование микроорганизмов-кандидатов
[0295] Для идентификации промоторов, которые активны в присутствии азота окружающей среды, проводили транскриптомное профилирование штамма CI010. Штамм CI010 культивировали в определенной среде без азота, дополненной 10 мМ глутамином. Из этих культур выделяли тотальную РНК (набор QIAGEN RNeasy) и подвергали секвенированию RNAseq посредством HiSeq Illumina (SeqMatic, Fremont CA). Прочтения секвенирования картировали на данные генома CI010 с использованием Geneious и идентифицировали высокоэкспрессированные гены под контролем проксимальных промоторов транскрипции.
[0296] В таблицах 3A-C перечислены гены и их относительные уровни экспрессии, определяемые посредством секвенирования RNAseq тотальной РНК. Последовательности проксимальных промоторов регистрировали для применения в мутагенезе метаболических путей nif, метаболических путей, связанных с утилизацией азота, или других генов с требуемым уровнем экспрессии.
Таблица 3A
Таблица 3B
Таблица 3С





Таблица штаммов
Таблица последовательностей штаммов
Оценка удобства манипуляций с генами
[0297] Микроорганизмы-кандидаты характеризовали на основе способности к трансформации и удобства манипуляций с генами. Во-первых, определяли оптимальное использование источника углерода посредством выращивания на небольшой панели соответствующей среды, а также кривую роста в средах без азота и в богатых средах. Во-вторых, определяли природную устойчивость к антибиотикам каждого штамма посредством метода реплик и выращивания в жидкой культуре, содержащей панель антибиотиков, используемых в качестве селективных маркеров мутагенеза. В-третьих, каждый штамм тестировали на его способность к трансформации посредством электропорации коллекции плазмид. Коллекция плазмид содержит комбинаторное расширение семи участки начала репликации, т.е. p15a, pSC101, CloDF, colA, RK2, pBBR1 и pRO1600 и четыре маркера устойчивости к антибиотикам, т.е. CmR, KmR, SpecR и TetR. Эту системную оценку совместимости участков начала репликации и маркеров устойчивости использовали для идентификации векторов для основанного на плазмидах мутагенеза микроорганизмов-кандидатов.
Пример 3: Мутагенез микроорганизмов-кандидатов
Опосредованный Red лямбда нокаут
[0298] Получали несколько мутантных микроорганизмов-кандидатов с использованием плазмиды pKD46 или ее производное, содержащие маркер устойчивости к канамицину (Datsenko et al. 2000; PNAS 97(12): 6640-6645). Нокаут-кассеты конструировали с гомологией 250 п.н., фланкирующей ген-мишень, и получали посредством ПЦР с достройкой перекрывающихся участков. Микроорганизмы-кандидаты трансформировали pKD46, культивировали в присутствии арабинозы для индукции механизм экспрессии Red лямбда, подготавливали для электропорация и трансформировали нокаут-кассетами с получением мутантных штаммов-кандидатов. Для получения тринадцати кандидатов, мутантных по регуляторным генам фиксации азота nifL, glnB и amtB, использовали четыре микроорганизма-кандидата и один лабораторный штамм, Klebsiella oxytoca M5A1, как представлено в таблице 4.
Таблица 4: Список одиночных нокаут-мутантов, полученных с использованием опосредованного Red лямбда мутагенеза
Олигонуклеотид-специфический мутагенез с отбором Cas9
[0299] Для локализации изменений генома в гене rpoB DH10B E. coli использовали олигонуклеотид-специфический мутагенез, а мутантов отбирали с использованием системы CRISPR-Cas. Проведено конструирование мутагенного олигонуклеотида (ss1283: "G*T*T*G*ATCAGACCGATGTTCGGACCTTCcaagGTTTCGATCGGACATACGCGACCGTAGTGGGTCGGGTGTACGTCTCGAACTTCAAAGCC" (SEQ ID NO:2), где * означает тиофосфатную связь) для придания устойчивости к рифампицину посредством мутации 4 п.н. в гене rpoB. В клетках, содержащих плазмиду, кодирующую Cas9, проводили индукцию экспрессии Cas9, подготавливали их для электропорации, а затем проводили электропорацию мутагенным олигонуклеотидом и плазмидой, кодирующей конститутивную экспрессию направляющей РНК (нРНК), которая направляет расщепление Cas9 последовательности rpoB WT. Подвергнутые электропорации клетки восстанавливали в неселективных средах в течение ночи для обеспечения достаточной сегрегации полученных мутантных хромосом. После высевания на селекционную среду для отбора кодирующей нРНК плазмиды, показано, что две из десяти подвергаемых скринингу колоний содержат требуемую мутацию, тогда как остальные, как показано, являются "ускользнувшими" мутантами, полученными вследствие протоспейсерной мутации в плазмиде с нРНК или потери плазмиды с Cas9.
Опосредованный Red лямбда мутагенез с отбором посредством Cas9
[0300] Получали мутанты микроорганизмов-кандидатов CI006 и CI010 с использованием опосредованного Red лямбда мутагенеза с отбором посредством CRISPR-Cas. Нокаут-кассеты содержали эндогенный промотор, идентифицированный посредством транскриптомного профилирования (как описано в примере 2 и приведено в таблицах 3A-C) и области гомологии ≈250 п.н., фланкирующие мишень делеции. CI006 и CI010 трансформировали плазмидами, кодирующими систему рекомбинации Red лямбда (гены exo, beta, gam) под контролем промотора, индуцируемого арабинозой, и Cas9 под контролем промотора, индуцируемого IPTG. У полученных трансформантов индуцировали системы рекомбинации Red и Cas9 и получали штаммы для электропорации. Затем в компетентные клетки трансформировали нокаут-кассеты и плазмиду с кодируемой селектирующей нРНК. После высевания на селективные среды с антибиотиками для плазмиды Cas9 и плазмиды с нРНК, 7 из 10 подвергнутых скринингу колоний продемонстрировали требуемую нокаут-мутация, как представлено на фигуре 3.
Пример 4: Фенотипирование молекул-кандидатов in vitro
[0301] У различных мутантов определяли влияние экзогенного азота на биосинтез и активность нитрогеназы. Для определения нитрогеназной активности в условиях чистой культуры использовали анализ восстановления ацетилена (ARA) (Temme et. al. 2012; 109(18): 7085-7090). Штаммы выращивали в герметически закрытых тестовых пробирках и восстановление ацетилена в этилен количественно определяли с использованием газового хроматографа Agilent 6890. Активность микроорганизмов-кандидатов и других мутантов-кандидатов, выращиваемых в средах для фиксации азота, дополненных глутамином в концентрации от 0 до 10 мМ, в ARA представлена на фигурах 4A-B и фигурах 10A-C.
[0302] В анаэробных условиях культивирования определяли диапазон концентраций глутамина и аммиака с количественным определением влияния на активность фиксации азота. В клетках дикого типа по мере роста концентрации глутамина активность быстро снижалась. Однако в ряду исходных нокаут-мутаций был подтвержден класс мутаций, обеспечивающих экспрессию генов фиксации азота в условиях концентрации глутамина, которые в ином случае подавляют эту активность у дикого типа. Как видно на фигуре 4C, этот профиль получали у четырех разных видов диазотрофов. Кроме того, перенастраивая регуляторную сеть с использованием идентифицированных генетических компонентов, уровень активности фиксации азота регулировали предсказуемым образом. Это можно наблюдать на фигуре 4B, на который проиллюстрированы штаммы CM023, CM021, CM015 и CI006. Штамм CM023 представляет собой слабо эволюционировавший штамм; штамм CM021 представляет собой сильно эволюционировавший штамм; штамм CM015 представляет собой средне эволюционировавший штамм; штамм CI006 представляет собой штамм дикого типа (штамм 2). Аммиак, выделяемый в супернатанты культур, тестировали с использованием ферментативного анализа (MEGAZYME). В анализе определяли количество расходуемого NADPH при оптической плотности 340 нм. Анализ проводили в бактериальных культурах, выращиваемых в не содержащей азот, анаэробной среде с исходной плотностью 109 КОЕ/мл. Как видно на фигуре 4D, в панели из шести эволюционировавших штаммов, один из штаммов выделял до 100 мкМ аммиака в течение периода 48 часов. Кроме того, как видно на фигуре 11 двойной мутант демонстрировал большее выделение аммиака, чем одиночный мутант, из которого он получен. Это демонстрирует способность микроорганизмов продуцировать аммиак в количествах, превышающих их физиологические потребности.
Транскриптомное профилирование чистых культур
[0303] Транскрипционная активность CI006 определяли с использованием платформы Nanostring Elements. Клетки выращивали в не содержащей азот среде и через 4 часа инкубации собирали 108 клеток. Выделяли тотальную РНК с использованием набора Qiagen RNeasy. Очищенную РНК отправляли в Core Diagnostics в Palo Alto, CA, для гибридизации с зондами и анализа Digital Analyzer, как представлено на фигуре 5.
Пример 5: Фенотипирование микроорганизмов-кандидатов in planta
Колонизация растений микроорганизмами-кандидатами
[0304] Колонизацию желательных растения-хозяев микроорганизмами-кандидатами количественно определяли посредством экспериментов с кратковременным ростом растений. Растения кукурузы инокулировали штаммами, экспрессирующими RFP с плазмиды или с интегрированной посредством Tn5 экспрессирующей RFP кассеты. Растения выращивали в стерилизованном песке и нестерильной торфяной среде, а инокуляцию проводили посредством нанесения 1 мл культуры клеток пипеткой непосредственно на появившийся колеоптиль растения через трое суток после пророста. Плазмиды сохраняли посредством полива растений раствором, содержащим соответствующий антибиотик. Через три недели, собирали корни растений, три раза промывали в стерильной воде для удаления видимой почвы и разделяли на два образца. Один образец корней анализировали посредством флуоресцентной микроскопии для определения профилей локализации микроорганизмов-кандидатов. Микроскопию проводили на 10 мм частях лучших интактных корней растений, как представлено на фигуре 6.
[0305] Разработан второй количественный способ оценки колонизации. Проводили количественной анализ ПЦР на препаратах тотальной ДНК из корней растений, инокулированных эндофитами. Семена кукурузы (Dekalb DKC-66-40) проращивали в предварительно автоклавированном песке в горшке 6,35 см на 6,35 см на 25,4 см. Через сутки после посадки проводили пропитку 1 мл ночной культуры эндофитов (среда SOB) непосредственно в участке расположения семян. 1 мл этой ночной культуры приблизительно эквивалентен приблизительно 109 КОЕ, с разбросом в пределах 3 раз друг от друга в зависимости от используемого штамма. Каждый проросток удобряли 3 раза в неделю 50 мл модифицированного раствора Хогланда, дополненного 2,5 мМ или 0,25 мМ нитратом аммония. Через четыре недели после посадки собирали образцы корней для выделения ДНК. Остатки почвы смывали с использованием распыления воды под давлением. Затем эти образцы ткани гомогенизировали с использованием QIAGEN Tissuelyzer, и затем выделяли ДНК с использованием мини-набор для выделения ДНК QIAmp (QIAGEN) по рекомендованному протоколу. С этими выделенными образцами ДНК проводили анализ кПЦР с использованием устройства для ОТ-ПЦР Stratagene Mx3005P с использованием праймеров, которые сконструированы (с использованием Primer BLAST NCBI) специфичными к локусам, в каждом геноме эндофитов. Количественно определяли присутствие копий геномов эндофитов. Для дальнейшего подтверждения идентификации эндофитов проводили секвенирование продуктов амплификации ПЦР и подтверждали наличие правильной последовательности. Обобщенная информация о профилях колонизации штаммов CI006 и CI008 из числа микроорганизмов-кандидатов представлена в таблице 5. У штамма CI008 продемонстрирован уровень колонизации до 107× КОЕ/г сырой массы корня.
Таблица 5: Колонизация кукурузы, определяемая посредством кПЦР
Профилирование РНК in planta
[0306] Биосинтез компонентов пути nif in planta оценивали, определяя транскрипцию генов nif. Тотальную РНК получали из тканей корней растений, инокулированных CI006 (способы посадки описаны ранее). Выделение РНК проводили с использованием мини-набора RNEasy по рекомендуемому протоколу (QIAGEN). Затем тотальную РНК этих тканей растений анализировали с использованием наборов Nanostring Elements (NanoString Technologies, Inc.) с использованием зондов, специфичных к генам nif в геноме штамма CI006. Данные по экспрессии генов nif in planta обобщены в таблице 6. Экспрессию генов nifH детектировали в инокулированных штаммом CM013 растениях, в то время как у инокулированных CI006 растений экспрессию nifH не детектировали. Штамм CM013 представляет собой производное штамма CI006, у которого ген nifL подвергнут нокауту.
[0307] Высокоэкспрессированные гены CM011, ранжированные по транскриптам в т.н. на миллион (TPM), определяли in planta в условиях добавления удобрений. Промоторы, контролирующие экспрессию некоторых из этих высокоэкспрессированных генов, использовали в качестве матриц для гомологичной рекомбинации в являющихся мишенями локусах фиксации и ассимиляции азота. Проводили выделение образцов РНК выращиваемых в теплице инокулированных CM011 растений, удаляли рРНК с использованием набора Ribo-Zero, секвенировали с использованием платформы Truseq Illumina и обратно картировали на геном CM011. Высокоэкспрессированные гены CM011 приведены в таблице 7.
Таблица 6: Экспрессия nifH in planta
Таблица 7
Анализ 15N
[0308] В основном способе демонстрации фиксации используют изотоп азота 15N, который находится в атмосфере с установленным уровнем относительно 14N. Добавляя в каждое удобрение или атмосферу повышенные уровни 15N, можно наблюдать фиксацию или непосредственно в увеличенных количествах 15N, фиксируемого из атмосферы, дополненной газообразным 15N2 (Yoshida 1980), или опосредовано посредством разведения обогащенного удобрения атмосферным газообразным N2 в тканях растений (Iniguez 2004). Способ разведения позволяет наблюдать накопление фиксированного азота в течение роста растения, тогда как способ с газообразным 15N2 ограничен определением фиксации, которая происходит в течение короткого интервала, в течение которого растение можно выращивать в ограниченной атмосфере (измерения скорости). Таким образом, способ с использованием газа лучше подходит для определения специфичности (так как любые повышенные уровни 15N2 в растении, превышающие атмосферный уровень, можно однозначно соотнести с фиксацией), но им нельзя демонстрировать суммарную активность.
[0309] Для определения активности фиксация улучшенных штаммов относительно штаммов дикого типа и не подвергаемых инокуляции растений кукурузы проводили оба типа анализа, и у нескольких из улучшенных штаммов in planta наблюдали увеличенные уровни фиксация (фигура 12, фигура 14A и фигура 14B). Эти анализы являются важными для демонстрации того, что активность исследуемых штаммов in vitro соответствует результатам in vivo. Кроме того, эти анализы позволяют определять влияние удобрения на активность штамма, позволяя предполагать соответствующую функциональность в сельскохозяйственных условиях. Подобные результаты наблюдали, когда штаммами дикого типа и улучшенными штаммами инокулировали растения Setaria (фигура 13). Активность фиксации in planta, приведенная на фигурах 14A-14C, дополнительно подтверждена транскриптомными данными. Эволюционировавшие штаммы демонстрируют увеличенный уровень транскриптов nifH относительно вариантов дикого типа. Кроме того, обеспечиваемый микроорганизмами уровень азота in planta также коррелировал с уровнем колонизации растения в расчете на растение. Эти результаты (фигура 12, фигура 13, фигуры 14A-14C, фигура 15A и фигура 15B) поддерживают гипотезу о том, что вероятной причиной увеличения получаемого из атмосферы азота, наблюдаемого в тканях растений, является микроорганизм, вследствие улучшенной регуляции кластера генов nif. В дополнение к прямому определению фиксация определяли эффект инокуляции растений улучшенными штаммами в анализе биомассы растений в условиях недостатка азота. Хотя биомасса растения может быть связана со множеством возможных взаимодействий микроорганизмов с растением, можно ожидать, что добавление фиксированного азота может влиять на фенотип растения в условиях недостатка азота. Инокулированные растения выращивали в условиях полного отсутствия азота, наблюдали и значимое увеличение площади листьев, сырой и сухой массы побегов и сырой и сухой массы корней у инокулированных растений относительно необработанных контрольных растений (фигура 14C). Хотя эти различия нельзя приписать исключительно фиксации азота, они поддерживают вывод, что улучшенные штаммы активно предоставляют азот растениям. Выращивали растения кукурузы и Setaria и инокулировали как описано выше. Растениям посредством полива регулярно добавляли удобрение, содержавшее 1,2% 15N. Фиксацию азота микроорганизмами количественно определяли, определяя уровень 15N в ткани растения. На 4 неделе после посадки получали ткань четвертого листа и высушивали. Высушенные образцы листьев гомогенизировали с использованием гранул (QIAGEN Tissuelyzer) и разделяли на аликвоты в жестяные капсулы для IRMS (MBL Stable Isotope Laboratory at The Ecosystems Center, Woods Hole, MA). Рассчитывали азот, получаемый из атмосферы (NDFA), и продукция азота CI050 и CM002 приведена на фигуре 7.
Анализ продукции фитогормонов
[0310] Ранее для исследования влияния индол-3-уксусной кислоты на созревание плодов посредством анализа in vitro использовали сорт карликовых томатов (Solanum lycopersicum) "Micro-Tom" (Cohen 1996; J Am Soc Hortic Sci 121: 520-524). Для оценки продукции и секреции фитогормонов микроорганизмами-кандидатами разработан анализ планшетного скрининга с использованием незрелых плодов Micro-Tom. Получали двенадцатилуночные тестовые планшеты для тканевых культур, заполняя лунки агарозной средой, позволяя ей отвердевать и нанося на поверхность агара 10 мкл ночной культуры микроорганизмов, как представлено на фигуре 8. Лунки с агаром содержат увеличенные количества гибберелловой кислоты (GA), но в качестве положительного контроля и стандартнов не использовали ни одной культуры бактерий. В агар в участок нанесения бактериальной культуры стеблем вперед помещали цветы, срезанные с растущих растений Micro-Tom через сутки после начала цветения. За этими цветами наблюдали в течение 2-3 недель, после чего собирали и взвешивали плоды. Увеличение массы плода растения в нескольких повторениях свидетельствовал о продукции гормонов растений инокулируемыми микроорганизмами, как представлено на фигуре 9.
Пример 6: Циклическaя эволюция хозяин-микроорганизм
[0311] Растения кукурузы инокулировали CM013 и выращивали 4 недели приблизительно до стадии роста V5. Растения, демонстрирующие улучшенное накопление азота из микробных источников по результатам анализа 15N, выкорчевывали, и корни отмывали с использованием воды под давлением для удаления основной почвы. Производили срез корня массой 0,25 г и промывали в растворе PBS для удаления мелких частичек почвы и неприкрепившихся микроорганизмов. Образцы ткани гомогенизировали с использованием 3 мм стальных гранул в QIAGEN TissueLyser II. Гомогенат разбавляли и наносили в планшеты на агарозные среды SOB. Единичные колонии ресуспендировали в жидких средах и подвергали анализу ПЦР 16s рДНК и мутаций, уникальных для инокулируемого штамма. Процесс выделения микроорганизма, мутагенеза, инокуляции и повторного выделения можно повторять несколько раз с улучшением признаков микроорганизма, признаков растений и способности микроорганизмов к колонизации.
Пример 7: Совместимость в различных географических условиях
[0312] Способность улучшенных микроорганизмов к колонизации инокулированного растения является критичной для успешного существования растения в полевых условиях. Хотя описанные способы выделения разработаны для выбора из почвенных микроорганизмов, которые могут иметь тесную связь с сельскохозяйственными культурами, такими как кукуруза, многие штаммы не могут эффективно колонизировать в целому ряду генотипов растений, типов окружающей среды, типов почв или условий инокуляции. Так как колонизация представляет собой сложный процесс, требующий целого ряда взаимодействий между штаммом микроорганизмов и растением-хозяином, основным способом для выбора приоритетных штаммов для дальнейшей разработки стал скрининг на способность к колонизации. В ранних попытках оценки колонизации использовали флуоресцентно меченые штаммы, которая являлась эффективной, но отнимала много времени и не являлась масштабируемой относительно штаммов. Так как активность колонизации не поддается непосредственному улучшению, крайне важно отбирать потенциальных продуцирующих кандидатов из штаммов, которые являются природными колонизаторами.
[0313] Разработан анализ для тестирования на стабильную колонизацию штаммами дикого типа любого данного растения-хозяина с использованием кПЦР и праймеров, сконструированных так, чтобы они являлись специфичными для штамма в образце сообщества. Этот анализ предназначен для быстрого определения скорости колонизации микроорганизмов из образцов ткани кукурузы. Исходные тесты с использованием штаммов, оцениваемых как вероятные колонизаторы, с использованием флуоресцентной микроскопии и планшетных способов, продемонстрировали, что подход кПЦР может являться количественным и масштабируемым.
[0314] Типичный анализ проводят следующим образом: растения, в основном сорта кукурузы и пшеницы, выращивают в торфяной горшковой смеси в теплице в шести повторениях на штамм. Через четыре или пять суток после посадки 1 мл пропитки культур бактерий в ранней стационарной фазе, разбавленной до OD590 0,6-1,0 (приблизительно 5×108 КОЕ/мл), пипетируют на восходящий колеоптиль. Растения поливают только водопроводной водой и оставляют рати в течение четырех недели с последующим забором образцов, во время которого растения выкорчевывают и корни тщательно промывают с удалением большинства остатков торфа. Вырезают образцы чистых корней и гомогенизируют с получением взвеси клеток дебриса растения и ассоциированных бактериальных клеток. Авторы разработали протокол высокопроизводительного выделения ДНК, который позволяет эффективно получать смесь ДНК растения и бактерий для использования в качестве матрицы для кПЦР. На основе экспериментом во резкому возрастанию количества бактериальных клеток, этим способом выделения ДНК получают количественный образец бактериальной ДНК относительно сырой массы корней. Каждый штамм оценивают с использованием штаммоспецифичных праймеров, сконструированных с использованием Primer BLAST (Ye 2012) и сравнивают с фоновой амплификацией у не подвергаемых инокуляции растений. Так как у некоторых праймеров у не подвергаемых инокуляции растений выявлена побочная амплификация, колонизацию определяют по наличию амплификации или повышенной амплификации соответствующего продукта по сравнению с фоновым уровнем.
[0315] Этот анализ использовали для определения совместимости продукта микроорганизмов в различных почвенных географических условиях. Качества полевых почв и полевые условия могут оказывать очень большое влияние на действие продукта микроорганизмов. pH почвы, способность к удержанию воды и конкурирующие микроорганизмы являются только малым количеством примеров факторов почвы, которые могут влиять на жизнеспособность инокулята и его способность к колонизации. Анализ колонизации проводили с использованием трех различных типов почв, образцы которых в качестве среды для выращивания растений получены с сельскохозяйственных полей в Калифорнии (фигура 16A). Для аппроксимации реальных сельскохозяйственных условий использовали промежуточную плотность инокуляции. В течение 3 недель штамм 5 колонизировал все растения на уровне от 106 до 107 КОЕ/г сырой массы. Через 7 недель роста растений, эволюционировавшая версия штамма 1 демонстрировала высокие уровни колонизации (106 КОЕ/г сырой массы) во всех типах почв. (фигура 16B).
[0316] Кроме того, для оценки колонизации в сложных полевых условиях на поле площадью 1 акр в Сан-Луис-Обиспо в июне 2015 года начато испытание с оценкой влияния и колонизации семи штаммов дикого типа на два сорта полевой кукурузы. Агротехническое проектирование и проведение испытания выполняла по контракту полевая исследовательская организация, Pacific Ag Research. Для инокуляции, использовали тот же способ покрытия семян торфяной культуры, который был испытан в экспериментах по способам инокуляции. В течение вегетационного периода собирали образцы растений для оценки колонизации корня и внутренности стебля. Образцы собирали из трех повторных участков после каждой обработки через четыре и восемь недель после посадки, и из всех шести повторений после каждой обработки незадолго перед сбором через 16 недель. Дополнительные образцы собирали из всех шести повторных участков после обработок путем инокуляции штамма 1 и штамма 2, а также необработанных контролей, через 12 недель. Число клеток на грамм сырой масса отмытых корней оценивали также, как и в других анализах колонизации с использованием кПЦР и штаммоспецифичных праймеров. Два штамма, штамм 1 и штамм 2, продемонстрировали стабильную и повсеместную колонизацию корней, которая достигала максимума через 12 недель, а затем резко снижалась (фигура 16C). Хотя штамм 2, по видимому, присутствовал в количествах, на порядок меньших, чем штамм 1, выявлено, что он его количества от растению к растению являются более постоянными. По видимому, ни один из штаммов внутренность стебля эффективно не колонизирует. В поддержку данных кПЦР о колонизации, оба штамма успешно повторно выделяли из образцов корней с использованием планшетов и секвенирование 16S для идентификации изолятов соответствующей последовательности.
Пример 8: Выведение микроорганизмов
[0317] Примеры выведения микроорганизмов можно обобщить на схеме фигуры 17A. На фигуре 17A приведено выведение микроорганизмов, где сначала можно определить состав микробиома и идентифицировать представляющий интерес вид. Метаболизм микробиома можно картировать и связать с генетикой. Затем можно провести направленную генетическую вариацию способами, включающими в качестве неограничивающих примеров конъюгацию и рекомбинацию, химический мутагенез, адаптивную эволюцию и редактирование генов. Производные микроорганизмы используют для инокуляции сельскохозяйственных культур. В определенных примерах выбраны сельскохозяйственные культуры с наилучшими фенотипами.
[0318] Как предоставлено на фигуре 17A сначала можно определить состав микробиома и идентифицировать представляющий интерес вид. На фигуре 17B приведен расширенный вид этапа определения микробиома. Метаболизм микробиома можно картировать и связать с генетикой. Метаболизм азота может включать вход аммиака (NH4+) из ризосферы в цитозоль бактерий посредством транспортера AmtB. Глутаминсинтетаза и АТФ осуществляют катализ преобразования аммиака и L-глутамината (L-Glu) в глутамин. Глутамин может приводить к формированию биомассы (росту растений), а также он может ингибировать экспрессию оперона nif. Затем, можно провести направленную генетическую вариацию способами, включающими в качестве неограничивающих примеров конъюгацию и рекомбинацию, химический мутагенез, адаптивную эволюцию и редактирование генов. Производные микроорганизмы используют для инокуляции сельскохозяйственных культур. Выбраны сельскохозяйственные культуры с наилучшими фенотипами.
Пример 9: Полевые испытания с микроорганизмами по изобретению - лето 2016 года
[0319] Для оценки воздействия штаммов по настоящему изобретению на рост и продуктивность кукурузы при различных режимах азота проводили полевые испытания.
[0320] Испытания проводили с (1) обработкой семи участков шестью штаммами и контролем - у четырех основных участков устанавливали 0, 15, 85 и 100% максимального возврата азота (MRTN) с региональным подтверждением. Контроль (только UTC) проводили при 10 100% MRTN плюс 5, 10 или 15 фунтов. Обработки проводили в четырех повторениях.
[0321] Участки кукурузы (минимум) составляли 4 ряда по 9,144 метра длиной с 124 участки на регион. Все наблюдения проводили в центральных двух рядах участков, а все получения образца с разрушением проводили во внешних рядах. Образцы семян перед использованием охлаждали до 1,5-2 часов.
[0322] Региональная сельскохозяйственная практика: Семена представляли собой коммерческую кукурузу без общепринятой обработки фунгицидами и инсектицидами. Все пропитки семян проводили одним специалистом по пропитке семян для обеспечения однородности. Дату посадки, норму высева, контроль сорняков/насекомых и т.д. оставляли как при региональной сельскохозяйственной практике. За исключением применения фунгицидов, использовали стандартные способы регуляции.
[0323] Характеристики почв: Оценивали текстуру почв и плодородность почв. Образцы почв для каждого повторения засеивали предварительно для гарантии остаточных уровней нитратов ниже 22,7 кг/акр. Получали образцы грунта от 0 см до 30 см. Почву дополнительно характеризовали по pH, CEC, общим K и P.
[0324] Оценки: Начальную популяцию растений оценивали через 14 суток после посадки (DAP)/акр и дополнительно оценивали на: (1) мощность (по шкале от 1 до 10, w/10=отлично) 14 DAP & V10; (2) ведение протокола рейтингов заболеваний в любой момент, когда на участках проявляются симптомы; (3) запись любых различий при полегании, если на участке происходит полегание; (4) урожайность (Bu/акр), скорректированная на стандартный процент влажности; (5) массу теста и (6) процент влажности зерен.
[0325] Требования к получению образцов: Образец почва получали в трех временных точках (до начала испытания, V10-VT, 1 неделя после сбора урожая). Во всех шести регионах и на всех участках отбирали образец с 10 граммами на образец (124 участка × 3 временные точки × 6 регионов).
[0326] Получение образца колонизации: Образцы колонизация собирали в двух временных точках (V10 и VT) для пяти регионов и шести временных точках (V4, V8, V10, VT, R5 и после сбора урожая). Образцы собирали следующим образом: (1) от 0% и 100% MRTN, 60 участков на регион; (2) 4 растения на участок, случайно выбранных из внешних рядов; (3) 5 граммов корня, 20,3 см стебля и вернхие три листа, каждый раздельно упакованный в мешок и с присвоенным ID - 12/мешков на участок; (4) пять регионов (60 участков × 2 временные точки × 12 мешков/участок); и один регион (60 участков × 6 временных точек × 12 мешков/участок. См., фигуру 17C, иллюстрирующую получение образцов колонизации.
[0327] Определение нормализованного разностного вегетационного индекса (NDVI) проводили с использованием инструмента Greenseeker в двух временных точках (V4 - V6 и VT). Анализировали каждый участок во всех шести регионах (124 участка × 2 временных точки × 6 регионов).
[0328] Анализ корней проводили с использованием Win Rhizo из оного региона, который лучше всего иллюстрировал различия обработки. Случайным образом проводили отбор десяти растений на участок (по 5 рядом с каждым внешним рядом; предпочтительно отбирали растения на этапе V3-V4) и осторожно промывали для удаления как можно большего количества загрязнений. Десять корни помещали в пластиковый пакет и помечали. Анализ корней проводили с использованием анализа корней WinRhizo.
[0329] Характеристики стеблей проводили для всех шести регионов между R2 и R5. Регистрировали диаметр стебля десяти растений на участок при высоте 15 см, а также длину первого междоузлия выше отметки 15 см. Проводили мониторинг десяти растений; пять последовательных растений от центра двух внутренних рядов. Проводили оценку в шести регионах (124 участка × 2 измерения × 6 регионов).
[0330] Нитраты тканей анализировали на всех участках и во всех регионах. Участок стебля длиной 20,3 см начинающийся от 15 см выше почва когда кукуруза находилась между первой и третей неделями после формирования черной точки; влагалища листьев удаляли. Оценивали все регионы и участки (6 регионов × 124 участка).
[0331] Для всех регионов от начала посадки до сбора урожая регистрировали следующие погодные данные: суточные максимальные и минимальные температуры, температура почвы при посеве, суточные осадки плюс полив (если применимо) и любые необычные погодные явления, такие как сильный дождь, ветер, холод или жара.
[0332] В таблице 8 приведены данные по урожайности во всех шести регионах. Уровень азота значимо влиял на урожайность, но штаммы по уровням азота влияния не оказывали. Однако при наименьшем уровне азота штаммы CI006, CM029 и CI019 численно превзошли UTC на 4-6 bu/акр. Урожайность также численно увеличилась на 2-4 bu/акр под действием штаммов CM029, CI019 и CM081 при 15% MRTN.
Таблица 8: Данные по урожайности для всех шести регионов
[0333] В таблице 9 представлен другой подход к анализу. В таблице представлены четыре региона, где реакция на азот была наибольшей, что позволяет предполагать, что доступный остаточный азот был наименьшим. Этот подход не меняет оценку того, что уровень азота значимо влияет на урожайность, чего не делали штаммы при усреднении по всем уровням азота. Однако числовое преимущество урожайности при наименьшем уровне N является более выраженным для всех штаммов, особенно для CI006, CM029 и CM029/CM081, где урожайность вырастала от 8 до 10 bu/акр. При 15% MRTN штамм CM081 превосходил UTC на 5 bu.
Таблица 9: Данные по урожайности в четырех регионах
[0334] Результаты полевых испытаний также проиллюстрированы на фигурах 21-27. Результаты свидетельствуют, что микроорганизмы по изобретению способны увеличивать урожайность растений, что указывает на способность исследуемых микроорганизмов увеличивать фиксацию азота у важной сельскохозяйственной культуры, т.е. кукурузы.
[0335] Полевые результаты дополнительно подтверждают описанные способы не являющейся межродовой модификации генома отобранных штаммов микроорганизмов для получения сельскохозяйственно значимых результатов в полевых условиях при применении указанных инженерных штаммов к сельскохозяйственной культуре.
[0336] На фигуре 18 приведена линия модифицированных штаммов, полученных из штамма CI006 (дикий тип Kosakonia sacchari). Полевые данные демонстрируют, что подвергнутое генетической инженерии производное штаммов CI006 дикого типа, т.е. CM029, может приводить к численно релевантным результатам в полевых условиях. Например, таблица 8 демонстрирует, что при 0% MRTN CM029 приводил к урожайности 147,0 bu/акр по сравнению с необработанным контролем при 141,2 bu/акр (увеличение на 5,8 bu/акр). Таблица 8 также демонстрирует, что при 15% MRTN CM029 приводил к урожайности 167,3 bu/акр по сравнению с необработанным контролем при 165,1 bu/акр (увеличение на 2,2 bu/акр). Таблица 9 поддерживает эти заключения и демонстрирует, что при 0% MRTN CM029 приводил к урожайности 140,7 bu/акр по сравнению с необработанным контролем при 131,9 bu/акр (увеличение на 8,8 bu/акр). Таблица 9 также демонстрирует, что при 15% MRTN CM029 приводил к урожайности 164,1 bu/акр по сравнению с необработанным контролем при 161,3 bu/акр (увеличение на 2,8 bu/акр).
[0337] На фигуре 19 приведена линия модифицированных штаммов, полученных из штамма CI019 (дикий тип Rahnella aquatilis). Полевые данные демонстрируют, что подвергнутое генетической инженерии производное штаммов CI019 дикого типа, т.е. CM081, может приводить к численно релевантным результатам в полевых условиях. Например, таблица 8 демонстрирует, что при 15% MRTN CM081 приводил к урожайности 169,3 bu/акр по сравнению с необработанным контролем при 165,1 bu/акр (увеличение на 4,2 bu/акр). Таблица 9 поддерживает эти заключения и демонстрирует, что при 0% MRTN CM081 приводил к урожайности 136,3 bu/акр по сравнению с необработанным контролем при 131,9 bu/акр (увеличение на 4,4 bu/акр). Таблица 9 также демонстрирует, что при 15% MRTN CM081 приводил к урожайности 166,8 bu/акр по сравнению с необработанным контролем при 161,3 bu/акр (увеличение на 5,5 bu/акр).
[0338] Кроме того, в таблице 9 можно наблюдать, что комбинация CM029/см081 при 0% MRTN приводила к урожайности 141,4 bu/акр по сравнению с необработанным контролем при 131,9 bu/акр (увеличение на 9,5 bu/акр).
Пример 10: Полевые испытания с микроорганизмами по изобретению
[0339] В природе, включая сельскохозяйственные почвы, можно найти большое разнообразие фиксирующих азот бактерий можно. Однако потенциал микроорганизма для предоставления достаточного количество азота сельскохозяйственным культурам, чтобы обеспечить снижение использования удобрений, может быть лимитирован подавлением нитрогеназных генов в удобряемых почвах, а также низким количеством в тесной ассоциации с корнями сельскохозяйственных культур. Идентификация, выделение и выведение микроорганизмов, которые тесно ассоциированы с ключевыми коммерческими сельскохозяйственными культурами, может разрушить и улучшить регуляторные сети, связывающие чувствительность к азоту и фиксацию азота, и разблокировать значительный вклад ассоциированных с сельскохозяйственными культурами микроорганизмов в уровень азота. С этой целью идентифицированы фиксирующие азот микроорганизмы, ассоциированные и колонизирующие корневую систему кукурузы.
[0340] Собирали образцы корней растений кукурузы, выращиваемых в агротехнически значимых почвах, и из ризосферы и эндосферы выделяли популяции микроорганизмов. Выделяли геномную ДНК этих образцов с последующим секвенированием ампликонов 16S для профилирования состава сообщества. Выделен микроорганизм Kosakonia sacchari (штамм PBC6.1) и классифицирован посредством 16S рРНК и полногеномного секвенирования. Он представляет собой особенно интересный фиксатор азота, способный к колонизации почти 21% численности ассоциированной с корнями микробиоты (фигура 30). Для оценки чувствительности штамма к экзогенному азоту определяли уровни фиксации азота в чистой культуре с использованием классического анализа восстановления ацетилена (ARA) и различных уровней добавления глутамина. Вид демонстрировал высокий уровень активности фиксации азота в не содержащих азота средах, но экзогенный фиксированный азот подавлял экспрессию гена nif и нитрогеназную активность (штамм PBC6.1, фигура 28C и 28D). Кроме того, когда определяли аммиак, выделенный в супернатант PBC6.1, выращиваемого в азотфиксирующих условиях, можно было выявить очень небольшое выделение фиксированного азота (фигура 28E).
[0341] Авторы выдвинули гипотезу, что PBC6.1 мог бы стать значительным источником фиксированного азота на удобряемых полях, если бы регуляторные сети, контролирующие метаболизм азот были переконструированы с обеспечением оптимальной экспрессии нитрогеназы и выделения аммиака в присутствии фиксированного азота. Для обеспечения широкой фенотипической реконструкции без вставок трансгенов или синтетических регуляторных элементов в геноме PBC6.1 должно существовать достаточное генетическое разнообразие. Геном выделенного штамма составляет по меньшей мере 5,4 т.п.н. и содержит канонический кластер генов фиксации азота. Связанные пути метаболизма азота в PBC6.1 являются сходными с путями метаболизма азота модельного организма для фиксации азота, Klebsiella oxytoca m5al.
[0342] Идентифицировано несколько узлов генной регуляторной сети, которые могут усиливать фиксацию азота и последующую его передачу растению-хозяину, особенно в условиях высоких экзогенных концентраций фиксированного азота (рис. 28А). Оперон nifLA напрямую регулирует остальную часть кластера nif посредством активации транскрипции с помощью NifA и азот- и кислород-зависимой репрессии NifA посредством NifL. Разрушение nifL может устранить ингибирование NifA и улучшить экспрессию nif в присутствии кислорода и экзогенного фиксированного азота. Кроме того, экспрессия nifA под контролем азот-независимого промотора может отделить биосинтез нитрогеназы от регуляции азотчувствительным комплексом NtrB/NtrC. Ассимиляцию фиксированного азота микроорганизмом в глютамин посредством глутаминсинтетазы (GS) обратимо регулирует двухдоменный фермент аденилаттрансфераза (АТаза) GlnE посредством аденилирования и деаденилирования GS с ослаблением и восстановлением активности, соответственно. Укорочение белка GlnE с удалением его удаляющего аденилат (AR) домена может приводить к конститутивно аденилированной глутаминсинтетазе, ограничивающей ассимиляцию аммиака микроорганизмом и увеличивающей содержание внутри- и внеклеточного аммиака. Наконец, снижение экспрессии AmtB, переносчика, ответственного за захват аммиака, может приводить к увеличению содержания внеклеточного аммиака. Для получения рационально сконструированных фенотипов микроорганизмов без использования трансгенов использовали два подхода: получение безмаркерных делеций геномных последовательностей, кодирующих домены белка или целые гены, и перестройка регуляторных сетей посредством внутригеномной перегруппировки промоторов. Посредством повторяющегося процесса мутагенеза получено несколько нетрансгенных производных штаммов PBC6.1 (таблица 10).
Таблица 10. Список выделенных и производных штаммов K. sacchari используемых в настоящей работе. Prm, промоторная последовательность, полученная из генома PBC6.1; ΔglnEAR1 и ΔglnEAR2, различные укороченные варианты последовательности гена glnE, удаляющего удаляющий аденилат домен.
[0343] Проведено несколько анализов in vitro для характеристики конкретных фенотипов производных штаммов. Для оценки чувствительности штамма к экзогенному азоту использовали ARA, в котором PBC6.1 демонстрировал подавление активности нитрогеназы при высоких концентрациях глютамина (фиг. 28D). В отличие от этого у большинства производных штаммов выявлен дерепрессированный фенотип с варьирующими уровнями восстановления ацетилена, наблюдаемыми при высоких концентрациях глютамина. Скорости транскрипции nifA в образцах, анализируемых посредством кПЦР, хорошо коррелировали со скоростями восстановления ацетилена (рис. 31), свидетельствуя в пользу гипотезы о том, что разрушение nifL и вставка азот-независимого промотора для контроля nifA может приводить к дерепрессии кластера nif. Штаммы с измененной активностью GlnE или AmtB продемонстрировали заметно повышенные уровни экскреции аммония по сравнению со штаммами дикого типа или производными без этих мутаций (фиг. 28E), иллюстрируя влияние этих генотипов на ассимиляцию и обратный захват аммиака.
[0344] Проведено два эксперимента для изучения взаимодействия производных PBC6.1 с растениями кукурузы и количественной оценки введения фиксированного азота в ткани растений. Во-первых, в исследовани в теплице количественно определяли уровни фиксации азота микроорганизмами с использованием изотопных индикаторов. В кратком изложении, растения выращивают с меченным 15N удобрением, а разбавленные концентрации 15N в тканях растений указывают на вклад фиксированного азота из микроорганизмов. Саженцы кукурузы инокулировали отобранными штаммами микроорганизмов, и растения выращивали до стадии роста V6. Затем растения разрушали для обеспечения возможности определения колонизации микроорганизмами и экспрессии генов, а также измерения соотношений 15N/14N в тканях растений посредством масс-спектрометрии с соотношением изотопов (IRMS). Анализ находящейся на воздухе ткани продемонстрировал небольшой незначимый вклад PBC6.38 и значимый вклад PBC6.94 (p=0,011) в уровень азота в растениях. Приблизительно 20% азота, выявленного в надземных листьях кукурузы, продуцировано PBC6.94, а остальное поступило из семян, почвенной смеси или "фоновой" фиксации другими почвенными микроорганизмами (рис. 29C). Это демонстрирует, что конвейер размножения микроорганизмов авторов изобретения может обеспечивать получение штаммов, способных вносить существенный вклад азота в растения в присутствии азотных удобрений. Определена транскрипция микроорганизмов в тканях растений, и экспрессию кластера генов nif наблюдали у производных штаммов, но не у штамма дикого типа (рис. 29B), что демонстрирует важность дерепрессии nif для вклада BNF в сельскохозяйственные культуры в условиях добавки удобрений. Колонизация корней, определяемая посредством кПЦР, демонстрировала, что для каждого из протестированных штаммов плотность колонизации различна (фигура 29А). У PBC6.38 и PBC6.94 наблюдали 50-кратное различие колонизации. Это различие может указывать на то, что PBC6.94 снизил приспособленность к ризосфере по сравнению с PBC6.38 в результате высокого уровня фиксации и выведения.
Способы
Среды
[0345] Минимальная среда содержит (на литр) 25 г Na2HPO4, 0,1 г CaCL2⋅2H2O, 3 г KH2PO4, 0,25 г MgSO4⋅7H2O, 1 г NaC1, 2,9 мг FeCl3, 0,25 мг Na2MoO4⋅2H2O и 20 г сахарозу. Среда для выращивания определена как минимальная среда, дополненная 50 мл 200 мМ глутамин на литр.
Выделение диазотрофов
[0346] Саженцы кукурузы выращивали из семян (DKC 66-40, DeKalb, IL) в течение двух недель в условиях теплицы, с контролем от 22°C (ночью) до 26°C (днем), и обеспечивали 16-часовые световые циклы в почве, собранной в San Joaquin County, CA. Корни собирали и промывали стерильной деионизированной водой с удалением основной почвы. Ткани корня гомогенизировали с помощью 2-миллиметровых шариков из нержавеющей стали в лизаторе тканей (TissueLyser II, Qiagen P/N 85300) в течение трех минут при установке 30, и образцы центрифугировали в течение 1 минуты при 13000 об/мин для отделения ткани от бактерий, ассоциированных с корнем. Супернатанты разделяли на две фракции, и одну использовали для характеристики микробиома посредством секвенирования ампликона 16S рРНК, а оставшуюся фракцию разбавляли и высевали на среду, не содержащую азота (NfB), с добавлением 1,5% агара. Планшеты инкубировали при 30°С в течение 5-7 суток. Образовавшиеся колонии тестировали на присутствие гена nifH посредством ПЦР колоний с праймерами Ueda19f и Ueda406r. Геномную ДНК штаммов с положительной на nifH ПЦР колонии (QIAamp DNA Mini Kit, Cat No. 51306, QIAGEN, Germany) выделяли и секвенировали (Illumina MiSeq v3, SeqMatic, Fremont, CA). После сборки и аннотации последовательности изоляты, содержащие кластеры генов фиксации азота, использовали в последующих исследованиях.
Профилирование микробиома из саженцев после выделения
[0347] Из ассоциированных с корнями бактерий выделяли геномную ДНК с использованием набора ZR-96 Genomic DNA I (Zymo Research P/N D3011), и получали ампликоны 16S рРНК с использованием праймеров со штрихкодом Nextera, направленных к 799f и 1114r. Библиотеки ампликонов очищали и секвенировали с использованием платформы Illumina MiSeq v3 (SeqMatic, Fremont, CA). Считывания классифицировали по таксонам с использованием Kraken с использованием базы данных minikraken (фигура 30).
Анализ восстановления ацетилена (ARA)
[0348] Использовали модифицированную версию анализа восстановления ацетилена с определением нитрогеназной активности в условия культивирования чистых культур. Штаммы выращивали из одной колонии в SOB (RPI, P/N S25040-1000) при 30°C с перемешиванием при 200 об/мин в течение 24 часов, а затем субкультивировали 1:25 в среду для выращивания и аэробно выращивали в течение 24 часов (30°C, 200 об/мин). Затем 1 мл культуры из минимальной среды добавляли к 4 мл минимальной среды, дополненной от 0 до 10 мМ глутамином, в воздухонепроницаемых пробирках Hungate и выращивали анаэробно в течение 4 часов (30°C, 200 об/мин). Удаляли 10% свободного пространства, затем посредством инъекции проводили замещение равным объемом ацетилена, и инкубацию продолжали в течение 1 часа. Затем удаляли 2 мл свободного пространства посредством газонепроницаемого шприца для количественного определения продукции для количественного определения продукции этилена с использованием газового хроматографа Agilent 6850, оборудованного пламенно-ионизационного детектором (FID).
Анализ выделения аммония
[0349] Выведение фиксированного азота в форме аммиака определяли с использованием периодической ферментации в анаэробных биореакторах. Штаммы выращивали из одной колонии в 1 мл/лунку SOB в 96-луночных планшетах DeepWell. Планшеты инкубировали при 30°C с перемешиванием при 200 об/мин в течение 24 часов, а затем разбавляли 1:25 в свежем планшете, содержащем 1 мл/лунку среды для выращивания. Клетки инкубировали в течение 24 часов (30°C, 200 об/мин), а затем разбавляли 1:10 в свежем планшете, содержащем минимальную среду. Планшет переносили в анаэробную камеру с газовой смесью >98,5% азота, 1,2-1,5% водорода и < 30 м.д. кислорода и инкубировали с 1350 об/мин, комнатной температуре в течение 66-70 час. Биомассу исходной культуры сравнивали с конечной биомассой посредством определения оптической плотности при 590 нм. Затем клетки разделяли посредством центрифугирования, и супернатант из реакционного бульона анализировали на свободный аммиак с использованием набора для анализа аммиака Megazyme (P/N K-AMIAR) с нормализацией на биомассу в каждой временной точке.
Выделение ассоциированного с корнями микробиома
[0350] Корни осторожно встряхивали для удаления осыпающихся частиц и корневые системы разделяли и вымачивали в стабилизирующем РНК растворе (Thermo Fisher P/N AM7021) в течение 30 минут. Затем корни недолго промывали стерильной деионизированной водой. Образцы гомогенизировали с использованием перемешивания с шариками с использованием шарикоподшипников из нержавеющей стали размером 1,27 см в тканевом лизаторе (TissueLyser II, Qiagen P/N 85300) в 2 мл буфера для лизиса (Qiagen P/N 79216). Выделение геномной ДНК проводили с использованием набора ZR-96 Quick-gDNA (Zymo Research P/N D3010), а выделение РНК проводили с использованием набор RNeasy (Qiagen P/N 74104).
Анализ колонизации корня
[0351] Через четверо суток после посадки на почву выше посаженных семян наносили 1 мл бактериальной ночной культуры (приблизительно 109 КОЕ). Саженцы удобряли три раза в неделю 25 мл модифицированного раствора Хогланда, дополненного 0,5 мМ нитратрм аммония. Через четыре недели после посадки собирали образцы корней и выделяли тотальную геномную ДНК (гДНК). Колонизацию корня количественно определяли с использованием кПЦР с праймерами, сконструированными для амплификации уникальных областей геномов дикого типа или производного штамма. Эффективность реакции кПЦР определяли с использованием стандартной кривой, получаемой на основе известных количеств гДНК из генома-мишени. Данные нормализовали на число копий генома на грамм сырой массы с использованием массы ткани и объема выделения. Для каждого эксперимента количество колонизаций сравнивали с саженцами необработанного контроля.
Транскриптомика in planta
[0352] Транскрипционное профилирование ассоциированных с корнями микроорганизмов проводили у саженцев, выращиваемы и обрабатываемых, как описано в разделе "Анализ колонизации корня". Очищенную РНК секвенировали с использованием платформы Illumina NextSeq (SeqMatic, Fremont, CA). Считывания картировали на геном инокулированного штамма с использованием bowtie2 с использованием параметров '-very-sensitive-local' и минимального показателя выравнивания 30. Покрытие генома рассчитывали с использованием samtools. Дифференциальное покрытие нормализовали на экспрессию генов домашнего хозяйства и визуализировали по геному с использованием Circos и по генному кластеру nif с использованием DNAplotlib. Кроме того, транскрипционный профиль in planta количественно определяли посредством направленного анализ Nanostring. Очищенные РНК обрабатывали на nCounter Sprint (Core Diagnostics, Hayward, CA).
Тепличное исследование разведения 15N
[0353] Эксперимент по разведению удобрения с 15N проводили для оценки активности оптимизированного штамма in planta. Среду для посадки, содержащую минимальный фоновый N получали с использованием смеси вермикулита и промытого песка (5 промываний в DI H2O). Песчаную смесь автоклавировали в течение 1 часа при 122°C и отмеряли приблизительно 600 г в 656 мл горшки, которые пропитывали стерильной DI H2O и за 24 часа перед посадкой оставляли стекать. Поверхность семян кукурузы (DKC 66-40) стерилизовали в 0,625% гипохлорите натрия в течение 10 минут, затем пять раз промывали в стерильной дистиллированной воде и высаживали на глубину 1 см. Растения содержали под флуоресцентными лампами в течение четырех недель с 16 часовой длиной суток при комнатных температурах в среднем от 22°C (ночью) до 26°C (днем).
[0354] Через пять суток после посадки саженцы инокулировали 1 мл суспензии клеток вымачивая непосредственно над растущим колеоптилем. Инокулят получали из 5 мл ночной культуры в SOB, которую дважды центрифугировали и ресуспендировали в 5 мл PBS для удаления остаточного SOB с последующим конечным разведением до OD 1,0 (приблизительно 109 КОЕ/мл). Контрольные растения обрабатывали стерильным PBS, и каждую обработку проводили для десяти повторов растений.
[0355] Растения удобряли 25 мл раствора удобрения, содержащего 2% обогащенного 15N 2 мМ KNO3, на 5, 9, 14 и 19 сутки после посадки, и тем же раствором без KNO3 на 7, 12, 16 и 18 сутки после посадки. Раствор удобрения содержал (на литр) 3 ммоль CaCl2, 0,5 ммоль KH2PO4, 2 ммоль MgSO4, 17,9 мкмоль FeSO4, 2,86 мг H3BO3, 1,81 мг MnCl2⋅4H2O, 0,22 мг ZnSO4⋅7H2O, 51 мкг CuSO4⋅5H2O, 0,12 мг Na2MoO4⋅2H2O и 0,14 нмоль NiCl2. Все горшки по мере необходимости смачивали стерильной DI H2O для поддержания соответствующей влажности почвы без стекания.
[0356] Через четыре недели растения собирали и разделяли на самом нижнем узле на образцы для выделения гДНК и РНК корня и находящиеся на воздухе ткани для IRMS. Находящиеся на воздухе ткани по мере необходимости протирали для удаления песка, целиком помещали в бумажные пакеты и высушивали по меньшей мере в течение 72 часов при 60°C. После полного высыхания тотальную находящуюся на воздухе ткань гомогенизировали посредством перемешивания с гранулами и 5-7 мг образцы анализировали посредством масс-спектрометрии с соотношением изотопов (IRMS) на δ15N в MBL Stable Isotope Laboratory (The Ecosystems Center, Woods Hole, MA). Процент NDFA рассчитывали с использованием следующей формулы: %NDFA=(δ15N среднего UTC - δ15N образца)/(δ15N среднего UTC) × 100.
Пример 11: Полевые испытания с микроорганизмами по изобретению - лето 2017 года
[0357] Для оценки воздействия штаммов по настоящему изобретению рост и продуктивность кукурузы при различных режимах азота проводили полевые испытания. Приводимые ниже полевые данные демонстрируют, что не являющиеся межродовыми микроорганизмы по изобретению способны к фиксации атмосферного азота и доставке указанного азота в растение, что приводит к увеличенной урожайности в среде с ограниченным содержанием азота, а также в среде без ограничения азота.
[0358] Испытания проводили в семи регионы Соединенных Штатов в шести географически различных среднезападных регионах. Для обработки удобрениями использовали пять режимов азота: 100% от стандартной сельскохозяйственной практики участка/региона, 100% минус 25 фунтов, 100% минус 50 фунтов, 100% синус 75 фунтов и 0%; все данные приведены на акр. Фунты азота на акр для режима 100% зависели от стандартной сельскохозяйственной практики участка/региона. Указанные выше режимы азота находились в диапазоне приблизительно от 153 фунтов на акр до приблизительно 180 фунтов на акр со средним приблизительно 164 фунтов азота на акр.
[0359] Для каждого режима удобрения проводили 14 обработок. Для каждого режима проводили шесть повторений, и использовали схему с расщепленными делянками. 14 обработок включали: 12 различных микроорганизмов, 1 UTC c тем же уровнем удобрения как и основной участок и 1 UTC со 100% азота. В режиме со 100% азота 2-й UTC представлял собой 100 плюс 25 фунтов.
[0360] Участки кукурузы, минимально, состояли из 4 рядов по 9,144 м в длину (76,2 см между рядами) с 420 участками на регион. Если не указано иначе, все наблюдения проводили в центральных двух рядах участков, а все получения образца с разрушением проводили во внешних рядах. Образцы семян перед использованием охлаждали до 1,5-2 часов.
[0361] Региональная сельскохозяйственная практика: Семена представляли собой коммерческую кукурузу, используемую с коммерческой пропиткой для семян без биологического совместного применения. Норма высева, дата посадки, контроль сорняков/насекомых, времена сбора урожая и другие стандартные практики ведения хозяйства соответствовали нормам местных сельскохозяйственных практик для регионов за исключением применения фунгицидов (если требовалось).
[0362] Нанесение микроорганизмов: Микроорганизмы наносили на семена в пропитке для семян на семена, которым уже проводили нормальную химическую обработку. Семена покрывали ферментационным бульоном, содержащим микроорганизмы.
[0363] Характеристики почв: Оценивали текстуру почв и плодородность почв. Использовали стандартные способы получения образцов почв, которые включали образцы грунта на глубинах 0-30 см и 30-60 см. Стандарт получения образца почвы включал определение азота нитратов, азота аммония, общего азота, органических веществ и CEC. Стандарт получения образца почвы дополнительно включал определение pH, общего калия и общего фосфора. Для определения уровней азотных удобрений получали предпосадочные образцы почв из каждого региона для гарантии, что 0-12" и потенциально 12" до 24" почвенных регионов для азота нитратов.
[0364] Перед посадкой и внесением удобрений собирали по 2 мл образцов почв от 0 до 6-12" из UTC. Собирали по одному образцу на повтор на азотную область с использованием середины ряда. (5 режимов удобрения × 6 повторений=тридцать образцов почв).
[0365] После посадки (V4-V6) собирали по 2 мл образцов почв от 0 до 6-12" из UTC. Собирали по одному образцу на повтор на каждую азотную область с использованием середины ряда. (5 режимов удобрения × 6 повторений=тридцать образцов почв).
[0366] После сбора урожая (V4-V6) собирали по 2 мл образцы почв от 0 до 6-12" из UTC. Собирали по одному образцу на повтор на каждую азотную область с использованием середины ряда. Дополнительно после сбора урожая собирали образцы почвы на 0-12" из UTC и потенциально 12-24" из UTC (5 режимов удобрения × 6 повторений=тридцать образцов почв).
[0367] Образец почвы для V6-V10 для каждого режима удобрения (исключая обработку 100% и 100%+25 фунтов [в блоке 100%] для всех режимов удобрения при 0-12" и 12-24". (5 режимов удобрения × 2 глубины=10 образцов на регион).
[0368] После сбора урожая образец почва для каждого режима удобрения (исключая обработку 100% и 100%+25 фунтов [в блоке 100%] для всех режимов удобрения при 0-12" и 12-24". (5 режимов удобрения × 2 глубины=10 образцов на регион).
[0369] Анализ: Исходную популяцию растений оценивали при ≈50% UTC, а конечную популяцию растений оценивали перед сбором. Анализ включал (1) потенциально температуру (датчик температуры); (2) мощность (по шкале 1-10 с 10=отлично) при V4 и V8-V10; (3) высота растения при V8-V10 и V14; (4) урожайность (бушелей/акр), скорректированная на стандартный процент влажности; (5) тестовая масса; (6) процент влажности зерен; (7) тесты нитратов стебля при черной точке (420 участков × 7 регионов); (8) колонизацию с 1 растением на участок в пакете с застежкой на молнии при 0% и 100% удобрения при V4-V6 (1 растение × 14 обработок × 6 повторений × 2 режима удобрения=168 растений); (9) транскриптомика с 1 растением на участок в пакете с застежкой на молнии при 0% и 100% удобрения при V4-V6 (1 растение × 14 обработок × 6 повторений × 2 режима удобрения=168 растения); (10) определение нормализованного разностного вегетационного индекса (NDVI) или нормализованной разности красного края (NDRE) с использованием инструмента Greenseeker в два момента времени (V4-V6 и VT) для оценки каждого участка во всех 7 регионах (420 участки × 2 момента времени × 7 регионов=5880 точек данных); (11) характеристики стебля, определяемые во всех 7 регионах между R2 и R5 посредством регистрации диаметра стебля 10 растений/участок при высоте 15 см, регистрация длины первого междоузлия выше отметки 15 см, проводили мониторинг 10 растений (5 последовательных растений от центра двух внутренних рядов) (420 участки × 10 растений × 7 регионов=29400 точек данных).
[0370] Схема мониторинга: Практические исполнители посещали все испытания на стадии V3-V4 для оценки ранней сезонной реакции на обработки и в течение стадии репродуктивного роста для мониторинга созревания. Местный сотрудник непрерывно производил исследовательские испытания.
[0371] Информация о погоде: Собирали данные о погоде, охватывающие период от посадки до сбора урожая, и они состояли из суточных минимальных и максимальных температур, температуры почвы при посеве, суточных осадков и ирригации (если применяли) и необычных погодных явлений, таких как сильный ветер, дождь, холод, жара.
[0372] Представление данных: Включая данные, указанные выше, в полевых испытаниях получили точки данных, включая текстуры почвы; расстояние между рядами; размеры участка; ирригацию; обработку почвы; предыдущий урожай; норму высева; популяцию растений; сезонные поступления удобрений, включая источник, норму, сроки и место размещения; размеры площади уборки, способ сбора урожая, например, ручным или машинным способом и используемые измерительные инструменты (весы, монитор урожайности и т. д.)
[0373] Результаты: Избранные результаты указанного выше полевого испытания приведены на фигуре 32 и фигуре 33.
[0374] На фигуре 32, можно видеть, что использование микроорганизма по изобретению (т.е., 6-403) приводило к более высокому урожаю, чем у штамма дикого типа (дикий тип) и к более высокому урожаю, чем у необработанного контроля (UTC). При обработке "-25 lbs N" использовали на 25 фунтов N на акр менее, чем при стандартной сельскохозяйственной практике в регионе. Обработка UTC "100% N" предназначена для обозначения стандартной сельскохозяйственной практики региона, при которой 100% стандартного использования N применяет сельхозпроизводитель. Микроорганизм "6-403" депонирован под идентификатором NCMA 201708004, и его можно найти в таблице A. Он представляет собой мутант Kosakonia sacchari (также называемый CM037) и является мутантным потомком штамма CI006 дикого типа.
[0375] На фигуре 33 полученные результаты урожайности демонстрируют, что микроорганизмы по настоящему изобретению действуют в различных регионах действуют единообразно. Кроме того, результаты урожайности демонстрируют, что микроорганизмы по настоящему изобретению хорошо функционируют как в среде с недостаточным содержанием азота (то есть в среде с ограничением азота), так и в среде, которая с достаточным содержанием азота (то есть среда без ограничения азота). Микроорганизм "6-881" (также известный как CM094, PBC6.94), и который является мутантным потомком штамма Kosakonia sacchari CI006 дикого типа, депонирован под идентификатором NCMA 201708002, и его можно найти в таблице A. Микроорганизм "137-1034", который является мутантным потомком штамма Klebsiella variicola CI137 дикого типа, депонирован под идентификатором NCMA 201712001, и его можно найти в таблице A. Микроорганизм "137-1036", который является мутантным потомком штамма Klebsiella variicola CI137 дикого типа, депонирован под идентификатором NCMA 201712002, и его можно найти в таблице A. Микроорганизм "6-404" (также известный как CM38, PBC6.38), и который является мутантным потомком штамма Kosakonia sacchari CI006 дикого типа, депонирован под идентификатором NCMA 201708003, и его можно найти в таблице A.
Пример 12: Род не являющихся межродовыми микроорганизмов, эффективный для сельскохозяйственных систем
[0376] Микроорганизмы по настоящему изобретению оценивали и сравнивали друг с другом по продукции азота, происходящей на акре в течение сезона. См. фигуру 20, фигуру 40 и фигуру 41
[0377] Авторы изобретения выдвинули гипотезу, что для того, чтобы популяция подвергнутых генетической инженерии не являющихся межродовыми микроорганизмов была эффективна в современной сельскохозяйственной системе пропашных культур, популяция микроорганизмов должна продуцировать по меньшей мере один фунт или более азота на акр за сезон.
[0378] Исходя из этого, авторы изобретения неожиданно выявили функциональный род микроорганизмов, которые в числе прочего способны вносить вклад в: повышение урожайности у небобовых сельскохозяйственных культур; и/или снижение зависимости сельхозпроизводителя от применения экзогенного азота; и/или способность производить по меньшей мере один фунт азота на акр за сезон, даже в неограничивающих азот средах, где указанный род определен произведением колонизирующей способность × ммоль N, продуцируемых на микроорганизмом в час (т.е. линии, разделяющая фигуры 20, 40 и 41).
[0379] В отношении фигур 20, 40 и 41, определенные данные использования микроорганизмов по изобретению объединены для изображения тепловой карты количества фунтов азота, доставляемых на акре/сезон микроорганизмами по изобретению, которая зарегистрирована как функция микроорганизмов на грамм сырой массы на ммоль азота/микроорганизм/час. Ниже тонкой линии, пересекающей более крупное изображение, находятся микроорганизмы, доставляющие менее одного фунта азота на акре/сезон, а выше линии находятся микроорганизмы, доставляющие более одного фунта азота на акре/сезон.
[0380] Полевые данные и тепловая карта колонизация дикого типа: Микроорганизмы, используемые в тепловой карте фигуры 20, анализировали на продукцию N в кукурузе. Для штаммов дикого типа CI006 и CI019 данные по колонизации корня кукурузы получали с одного полевого участка. Для оставшихся штаммов колонизацию полагали такой же, как уровень в поле у дикого типа. Активность фиксации N определяли с использованием анализа ARA in vitro с 5 мМ глутамином. В таблице ниже тепловая карта на фигуре 20 дает точное значение ммоль N, продуцируемых микроорганизмом в час (ммоль N/микроорганизм/час), вместе с точными значениями КОЕ на грамм сырой массы (КОЕ/г св.масс.) для каждого микроорганизма, приведенного на тепловой карте.
[0381] Тепловая карта полевых данных: Данные, используемые в тепловой карте фигуры 40, получали у штаммов микроорганизмов, оцениваемых на продукцию N в кукурузе в полевых условиях. Каждая точка представляет собой фунты N/акр, продуцируемые микроорганизмом, с использованием данных по колонизации корня кукурузы с одного полевого участка. Активность фиксации N определяли с использованием анализа ARA in vitro с 5 мМ N в форме глутамина или фосфата аммония. В приводимой ниже таблице C даны точные значения ммоль N, продуцируемых микроорганизмом в час (ммоль N/микроорганизм/час) вместе с точными значениями КОЕ на грамм сырой массы (КОЕ/г св.масс.) для каждого микроорганизма, приведенного на тепловой карте фигуры 40.
[0382] Тепловая карта на основе тепличных и лабораторных данных: Данные, используемые в тепловой карте фигуры 41, получали у штаммов микроорганизмов, оцениваемых на продукцию N в кукурузе в лабораторных и тепличных условиях. Каждая точка представляет собой фунты N/акр, продуцируемые одним штаммом. Белые точки представляют штаммы, у которых данные по колонизации корня кукурузы собраны в тепличных условиях. Черные точки представляют мутантные штаммы, для которых уровни колонизации корней кукурузы получают на основе средних полевых уровней колонизации корней кукурузы родительского штамма дикого типа. Заштрихованные точки представляют родительские штаммы дикого типа с их средними полевыми уровнями колонизации корней кукурузы. Во всех случаях, активность фиксации N определяли посредством анализа ARA in vitro с 5 мМ N в форме глутамина или фосфата аммония. В приводимой ниже таблице D даны точные значения ммоль N, продуцируемых микроорганизмом в час (ммоль N/микроорганизм/час) вместе с точными значениями КОЕ на грамм сырой массы (КОЕ/г св.масс.) для каждого микроорганизма, приведенного на тепловой карте of фигура 41.
Таблица C: фигура 40 - Тепловая карта для полевых данных
Таблица D: фигура 41 - Тепловая карта для тепличных и лабораторных данных
[0383] Выводы: Данные на фигурах 20, 40, 41 и в таблицах C и D иллюстрируют большое количество представителей описанного рода (т.е. микроорганизмов справа от линии на фигурах). Кроме того, эти многочисленные представители происходят из разнообразного набора таксономических родов, которые можно найти в приводимых выше таблицах C и D. Кроме того, авторы изобретения выявили множество генетических свойств, которые отображают зависимость структура/функция, которая выявлено у многих микроорганизмов. Эту генетическую зависимость можно найти во множестве таблиц по изобретению, где приведены генетические модификации, проводимые авторами изобретения, которые включают внесение по меньшей мере одной генетической вариации по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота.
[0384] Таким образом, недавно выявленный род подтверждается: (1) надежным набором данных, (2) большим количеством представителей, (3) представителями из разных таксономических родов и (4) классами генетических модификаций, которые определяют зависимость структура/функция в лежащей в основе генетической архитектуре представителей рода.
Пример 13: Способы и анализы для детекции не являющихся межродовыми подвергнутых генетической инженерии микроорганизмов
[0385] В настоящем описании указаны праймеры, зонды и анализы, которые пригодны для детекции микроорганизмов, используемых в различных указанных выше примерах. Посредством анализов можно детектировать последовательности с неприродным "соединением" нуклеотидов в полученных/мутантных не являющихся межродовыми микроорганизмах. Эти неприродные соединения нуклеотидов можно использовать в качестве типа диагностики, которая является показателем присутствия конкретной генетической альтерация в микроорганизме.
[0386] Способами по настоящему изобретению можно детектировать эти неприродные соединения нуклеотидов с использованием специализированных способов количественной ПЦР, включая уникальным образом сконструированные праймеры и зонды. Зонды могут связываться с последовательностями с неприродным соединением нуклеотидов. Т.е. можно использовать специфичные для последовательностей ДНК-содержащие зонды, состоящие из олигонуклеотидов, которые мечены флуоресцентным репортером, обеспечивающем детекцию только после гибридизации зонда с комплементарной ему последовательностью. Количественные способы могут гарантировать, что амплификацию посредством указанных праймеров пройдет только неприродное соединение нуклеотидов, и, таким образом, его можно детектировать посредством неспецифического красителя или посредством использования специфического зонда для гибридизации. Другим аспектом способа является выбор праймеров так, чтобы праймеры фланкировали какой-либо конец последовательности с соединением так, что если происходит реакция амплификации, то указанная последовательность с соединением присутствует.
[0387] Таким образом, из образцов можно выделять геномную ДНК и использовать для количественного определения присутствия микроорганизмов по изобретению с применением кПЦР. Праймеры, используемые в реакции кПЦР, могут представлять собой праймеры, сконструированные посредством Primer Blast (https://www.ncbi.nlm.nih.gov/tools/праймер-blast/) с амплификацией уникальных областей генома дикого типа или уникальных областей подвергнутых генетической инженерии не являющихся межродовыми мутантных штаммов. Реакцию кПЦР можно проводить с использованием набор SYBR GreenER qPCR SuperMix Universal (Thermo Fisher P/N 11762100), с использованием только прямого и обратного праймеров для амплификации; альтернативно, можно использовать набор Kapa Probe Force (Kapa Biosystems P/N KK4301) с праймерами для амплификации и зондом TaqMan, содержащем красящую метку FAM dye на 5'-конец, внутренний гаситель ZEN, и связывающее малую бороздку средство и флуоресцентный гаситель на 3'-конце (Integrated DNA Technologies).
[0388] Определенные последовательности праймеров, зондов и последовательности с неприродным соединением, которые можно использовать в способах кПЦР, перечислены в приводимой ниже таблице E. Конкретно, последовательности с неприродным соединением можно найти в SEQ ID NO: 372-405.
Таблица E: Детекция микроорганизмов
GAACGTCGCCAG
GTGGCACAAATT
GTCAGAACTACG
ACACGACTAACC
GACCGCAGGAG
TGTGCGATGACC
CTGAATATGATG
ATGGA/TTCTTGG
TTCTCTGGAGCG
CTTTATCGGCAT
CCTGACTGAAGA
ATTTGCAGGCTT
CTTCCCAACCTG
GCTTGCACCCGT
GCAGGTAGTTGT
GATGAACAT-3'
CGGAAAACGAG
TTCAAACGGCGC
GTCCCAATCGTATTAATGGCGAGATTCGCGCCACGGAAGTTCGCTTAACAGGTCTGGAAGGCGAGCAGCTTGGTATT/GCGATAGAACTCACTTCACGCCCCGAAGGGGGAAGCTGCCTGACCCTACGATTCCCGCTATTTCATTCACTGACCGGAGGTTCAAAATGACCCAGCGAAC
-3'
GAAATCGAACAT
TTCCGTAATACC
GCCATTACCCAG
GAGCCGTTCTGG
TTGCACAGCGGA
AAACGTTAACGA
AAGGATATTTCG
CATG/TCCCTGTG
CGCCGCGTCGCC
GATGGTGGCCAG
CCAACTGGCGCG
CTACCCGATCCT
GCTCGATGAACT
GCTCGACCCGAA
CACGCTCTATCA
ACCGACGG-3'
GCCAGGTGGCAC
AAATTGTCAGAA
CTACGACACGAC
TAACCGACCGCA
GGAGTGTGCGAT
GACCCTGAATAT
GATGATGGATGC
CAGC/CGTTCTGT
AATAATAACCGG
ACAATTCGGACT
GATTAAAAAAG
CGCCCTCGCGGC
GCTTTTTTTATAT
TCTCGACTCCAT
TTAAAATAAAAA
ATCCAATC-3'
AGTTTGTGTAAT
ACTTGTAACGCT
ACATGGAGATTA
ACTCAATCTAGA
GGGTATTAATAA
TGAATCGTACTA
AACTGGTACTGG
GCGC/AACTCAC
TTCACGCCCCGA
AGGGGGAAGCT
GCCTGACCCTAC
GATTCCCGCTAT
TTCATTCACTGA
CCGGAGGTTCAA
AATGACCCAGCG
AACCGAGTCG-3'
GCCAGGTGGCAC
AAATTGTCAGAA
CTACGACACGAC
TAACCGACCGCA
GGAGTGTGCGAT
GACCCTGAATAT
GATGATGGATGC
CAGC/CGCGTCA
GGTTGAACGTAA
AAAAGTCGGTCT
GCGCAAAGCAC
GTCGTCGTCCGC
AGTTCTCCAAAC
GTTAATTGGTTT
CTGCTTCGGCAG
AACGATTGGC-3'
AAATGGCTGGG
ATTGTTCATTTTT
TGTTTGCCTTAC
AACGAGAGTGA
CAGTACGCGCGG
GTAGTTAACTCA
ACATCTGACCGG
TCGAT/AACTCA
CTTCACGCCCCG
AAGGGGGAAGC
TGCCTGACCCTA
CGATTCCCGCTA
TTTCATTCACTG
ACCGGAGGTTCA
AAATGACCCAGC
GAACCGAGTCG-
3'
GGCCACCACGCC
AGACCACACGAT
AGTGATGGCAAC
ACTTTCCAGCTG
CACCAGCACCTG
ATGGCCCATGGT
CACACCTTCAGC
GAAA/CCGATCC
CCATCACTGTGT
GTCTTGTATTAC
AGTGCCGCTTCG
TCGGCTTCGCCG
GTACGAATACGA
ATGACGCGTTGC
AGCTCAGCAACG
AAAATTTTG-3'
TCTGAATGAAGC
TCTTGAAAAAGC
TGAGGAAGCGG
GCGTCGATTTAG
TAGAAATCAGTC
CGAATGCCGAGC
CGCCAGTTTGTC
GAATC/CCGTCTC
TGAAGCTCTCGG
TGAACATTGTTG
CGAGGCAGGAT
GCGAGCTGGTTG
TGTTTTGACATT
ACCGATAATGTG
CCGCGTGAACGG
GTGCGTTATG-3'
GTTCGC
CTCACA
GG
TCGC
AACA
ATGT
TCAC
GCGCGCATTTGT
TATGCCAATCCG
GCATTCTGCCGC
CAGACGGGTTTT
GCACTTGAGACA
CTTTTGGGCGAG
AACCACCGTCTG
CTGG/TGAACAT
CACTGATGCACA
AGCTACCTATGT
CGAAGAATTAAC
TAAAAAACTGCA
AGATGCAGGCAT
TCGCGTTAAAGC
CGACTTGAGAAA
TGAGAAGAT-3'
GTTATAATGCCG
CGCCCTCATATT
GTGGGGATTTCT
TAATGACCTATC
CTGGGTCCTAAA
GTTGTAGTTGAC
ATTAGCGGAGCA
CTAAC/CCGTCTC
TGAAGCTCTCGG
TGAACATTGTTG
CGAGGCAGGAT
GCGAGCTGGTTG
TGTTTTGACATT
ACCGATAATGTG
CCGCGTGAACGG
GTGCGTTATG-3'
CATATT
GTGGGG
AT
TAAC
GCAC
CCGT
TCA
M/T
A
AC
C
CG
T
C/Z
EN/
T
CT
G
AAG
CTC
TC
G
GT/
3IA
BkF
Q/
GCGCGCATTTGT
TATGCCAATCCG
GCATTCTGCCGC
CAGACGGGTTTT
GCACTTGAGACA
CTTTTGGGCGAG
AACCACCGTCTG
CTGG/TACAGTA
GCGCCTCTCAAA
AATAGATAAAC
GGCTCATGTACG
TGGGCCGTTTAT
TTTTTCTACCCA
TAATCGGGAACC
GGTGTTATAATG
CCGCGCCCTC-3'
AGTTTGTGTAAT
ACTTGTAACGCT
ACATGGAGATTA
ACTCAATCTAGA
GGGTATTAATAA
TGAATCGTACTA
AACTGGTACTGG
GCGC/AACTCAC
TTCACACCCCGA
AGGGGGAAGTT
GCCTGACCCTAC
GATTCCCGCTAT
TTCATTCACTGA
CCGGAGGTTCAA
AATGACCCAGCG
AACCGAGTCG-3'
GGTACT
GGGCGC
AACT
TCGA
AGCG
CCAG
ACGG
TAT
FA
M/A
AG
TTG
CCT
/ZE
N/G
AC
CCT
AC
GA
TTC
CC/
3IA
BkF
Q/
GGCCTGGTGGCA
CAAATTGTCAGA
ACTACGACACGA
CTAACTGACCGC
AGGAGTGTGCG
ATGACCCTGAAT
ATGATGATGGAT
GCCGGC/CGTCC
TGTAATAATAAC
CGGACAATTCGG
ACTGATTAAAAA
AGCGCCCTTGTG
GCGCTTTTTTTA
TATTCCCGCCTC
CATTTAAAATAA
AAAATCCAATC-
3'
GGCCTGGTGGCA
CAAATTGTCAGA
ACTACGACACGA
CTAACTGACCGC
AGGAGTGTGCG
ATGACCCTGAAT
ATGATGATGGAT
GCCGGC/GGACA
TCATCGCGACAA
ACAATATTAATA
CCGGCAACCACA
CCGGCAATTTAC
GAGACTGCGCA
GGCATCCTTTCT
CCCGTCAATTTC
TGTCAAATAAAG
-3'
CTCTTT
GCATGG
TT
AGTC
TCGT
AAAT
TGCC
FA
M/C
A
GG
A
GT
G
T/Z
EN/G
CG
A
TG
A
CC
C
TG
A
AT/
3IA
BkF
Q
GGATGAATGTGC
CATTAAATGCGC
AGCATAATGGTG
CGTTGTGCGGGA
AAACTGCTTTTT
TTTGAAAGGGTT
GGTCAGTAGCGG
AAAC/AACTCAC
TTCACACCCCGA
AGGGGGAAGTT
GCCTGACCCTAC
GATTCCCGCTAT
TTCATTCACTGA
CCGGAGGTTCAA
AATGACCCAGCG
AACCGAGTCG-3'
GAAATCGAACAT
TTCCGTAATACC
GCGATTACCCAG
GAGCCGTTCTGG
TTGCACAGCGGA
AAACGTTAACGA
AAGGATATTTCG
CATG/TTTAACG
ATCTGATTGGCG
ATGATGAAACG
GATTCGCCGGAA
GATGCGCTTTCT
GAGAGCTGGCG
CGAATTGTGGCA
GGATGCGTTGCA
GGAGGAGGATT-
3'
GAAATCGAACAT
TTCCGTAATACC
GCGATTACCCAG
GAGCCGTTCTGG
TTGCACAGCGGA
AAACGTTAACGA
AAGGATATTTCG
CATG/GCACTGA
AACACCTCATTT
CCCTGTGTGCCG
CGTCGCCGATGG
TTGCCAGTCAGC
TGGCGCGCTACC
CGATCCTGCTTG
ATGAATTGCTCG
ACCCGAATA-3'
TTTCTGGTTAAA
CGGGCAACCTCG
TTAACTGACTGA
CTAGCCTGGGCA
AACTGCCCGGGC
TTTTTTTTGCAA
GGAATCTGATTT
CATG/GCGCTCA
AACAGTTAATCC
GTCTGTGTGCCG
CCTCGCCGATGG
TCGCGACACAAC
TTGCACGTCATC
CTTTATTGCTCG
ATGAACTGCTCG
ACCCGCGCA-3'
GCGCGCATTTGT
TATGCCAATCCG
GCATTCTGCCGC
CAGACGGGTTTT
GCACTTGAGACA
CTTTTGGGCGAG
AACCACCGTCTG
CTGG/AGTCTGA
ACTCATCCTGCG
GCAGTCGGTGAG
ACGTATTTTTGA
CCAAAGAGTGAT
CTACATCACGGA
ATTTTGTGGTTG
TTGCTGCTTAAA
AGGGCAAAT-3'
CCAGCTTACAAA
TTGCCTGATTGC
GGCCCCGATGGC
CGGTATCACTGA
CCGACCATTTCG
TGCCTTATGTCA
TGCGATGGGGGC
TGGG/CCGTCTCT
GAAGCTCTCGGT
GAACATTGTTGC
GAGGCAGGATG
CGAGCTGGTTGT
GTTTTGACATTA
CCGATAATGTGC
CGCGTGAACGG
GTGCGTTATG-3'
GGAGGAATAAG
GTATTAAAGGCG
GAAAACGAGTTC
AAACGGCACGTC
CGAATCGTATCA
ATGGCGAGATTC
GCGCCCTGGAAG
TTCGC/GCCATTG
AGCTGGCTTCCC
GACCGCAGGGC
GGCACCTGCCTG
ACCCTGCGTTTC
CCGCTGTTTAAC
ACCCTGACCGGA
GGTGAAGCATG
ATCCCTGAATC-
3'
AGCATG
GACGTA
A
GTGT
TAAA
CAGC
GGG
AAA
FA
M/A
A
CG
G
CA
C
G/Z
EN/
T
CC
G
AA
T
CG
T
AT
C
AA/
3IA
BkF
Q/
TTACCCCGCCGC
GTTTTGCGCACG
GTGTCGGACAAT
TTGTCATAACTG
CGACACAGGAG
TTTGCGATGACC
CTGAATATGATG
CTCGA/AGCGTC
AGGTACCGGTCA
TGATTCACCGTG
CGATTCTCGGTT
CCCTGGAGCGCT
TCATTGGCATCC
TGACCGAAGAGT
TCGCTGGCTTCT
TCCCAACCTG-3'
TTGAATATTTCC
GTCGCCAGGCGC
TGGCTGCCGAGC
CGTTCTGGCTGC
ATAGTGGAAAA
CGATAATTTCAG
GCCAGGGAGCC
CTTATG/GCGCTG
AAGCACCTGATC
ACGCTCTGCGCG
GCGTCGCCGATG
GTCGCCAGCCAG
CTGGCGCGCCAC
CCGCTGCTGCTG
GATGAGCTGCTG
GATCCCAACA-3'
TTCTGG
CTGCAT
AG
TCGG
CTGA
TAGA
GG
FA
M/T
TAT
GG
CG
C/Z
EN/
TG
AA
GC
AC
CT
GA
TC
A/3I
ABk
FQ/
TTACCCCGCCGC
GTTTTGCGCACG
GTGTCGGACAAT
TTGTCATAACTG
CGACACAGGAG
TTTGCGATGACC
CTGAATATGATG
CTCGA/GCCCGC
TGACCGACCAGA
ACTTCCACCTTG
GACTCGGCTATA
CCCTTGGCGTGA
CGGCGCGCGATA
ACTGGGACTACA
TCCCCATTCCGG
TGATCTTACC-3'
AAAGTTGCACTG
GACAAAAGGTA
CCACAATTGGTG
TACTGATACTCG
ACACAGCATTAG
TGTCGATTTTTC
ATATAAAGGTAA
TTTTG/GCCATTG
AGCTGGCTTCCC
GACCGCAGGGC
GGCACCTGCCTG
ACCCTGCGTTTC
CCGCTGTTTAAC
ACCCTGACCGGA
GGTGAAGCATG
ATCCCTGAATC-
3'
TTACCCCGCCGC
GTTTTGCGCACG
GTGTCGGACAAT
TTGTCATAACTG
CGACACAGGAG
TTTGCGATGACC
CTGAATATGATG
CTCGA/GCTAAA
GTTCTCGGCTAA
TCGCTGATAACA
TTTGACGCAATG
CGCAATAAAAG
GGCATCATTTGA
TGCCCTTTTTGC
ACGCTTTCATAC
CAGAACCTGGC-
3'
AATAGCAGGGA
AGAGGCGCCAG
AACCGCCAGCGT
TGAAGCAGTTTG
AACGCGTTCAGT
GTATAATCCGAA
ACTTAATTTCGG
TTTGGA/GCCATT
GAGCTGGCTTCC
CGACCGCAGGG
CGGCACCTGCCT
GACCCTGCGTTT
CCCGCTGTTTAA
CACCCTGACCGG
AGGTGAAGCAT
GATCCCTGAATC
-3'
TTACCCCGCCGC
GTTTTGCGCACG
GTGTCGGACAAT
TTGTCATAACTG
CGACACAGGAG
TTTGCGATGACC
CTGAATATGATG
CTCGA/CGCCGT
CCTCGCAGTACC
ATTGCAACCGAC
TTTACAGCAAGA
AGTGATTCTGGC
ACGCATGGAAC
AAATTCTTGCCA
GTCGGGCTTTAT
CCGATGACGAA-
3'
GTATTCAATTAC
TTAGGCATTTAC
TTAACGCAGGCA
GGCAATTTTGAT
GCTGCCTATGAA
GCGTTTGATTCT
GTACTTGAGCTT
GATC/GCCATTG
AGCTGGCTTCCC
GACCGCAGGGC
GGCACCTGCCTG
ACCCTGCGTTTC
CCGCTGTTTAAC
ACCCTGACCGGA
GGTGAAGCATG
ATCCCTGAATC-
3'
TCTGAATGAAGC
TCTTGAAAAAGC
TGAGGAAGCGG
GCGTCGATTTAG
TAGAAATCAGTC
CGAATGCCGAGC
CGCCAGTTTGTC
GAATC/TCTTTAG
ATCTCTCGGTCC
GCCCTGATGGCG
GCACCTTGCTGA
CGTTACGCCTGC
CGGTACAGCAG
GTTATCACCGGA
GGCTTAAAATGA
CCCAGTTACC-3'
CGAGTT
CAACCG
GC
GGAC
CGAG
AGAT
CTAA
GGTTATTCCGGG
TGAGTATATGTG
TGATTTGGGTTC
CGGCATTGCGCA
ATAAAGGGGAG
AAAGACATGAG
CATCACGGCGTT
ATCAGC/TGAAT
ATCACTGACTCA
CAAGCTACCTAT
GTCGAAGAATTA
ACTAAAAAACTG
CAAGATGCAGG
CATTCGCGTTAA
AGCCGACTTGAG
AAATGAGAAGAT-3'
ATGTCGGGCGTT
TCACAACACAAA
ATGTTGTAAATG
CGACACAGCCG
GGCCTGAAACCA
GGAGCGTGTGAT
GACCTTTAATAT
GATGC/CTGGGG
TCACTGGAGCGC
TTTATCGGCATC
CTGACCGAAGA
ATTTGCCGGTTT
CTTCCCGACCTG
GCTGGCCCCTGT
TCAGGTTGTGGT
GATGAATATCA-
3'
TTCAAACGGCAC
GTCCGAATCGTA
TCAATGGCGAGA
TTCGCGCCCAGG
AAGTTCGCTTAA
CTGGTCTGGAAG
GTGAGCAGCTGG
GTATT/GCAATA
GAACTAACTACC
CGCCCTGAAGGC
GGTACCTGCCTG
ACCCTGCGATTC
CCGTTATTTCAT
TCACTGACCGGA
GGCCCACGATGA
CCCAGCGACC-3'
Таблица F: Подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы
Таблица G: Подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы
Таблица H
[0389] Использование терминов в единственном числе в рамках описания изобретения (особенно в рамках приводимой ниже формулы изобретения) следует рассматривать как относящиеся к единственному и множественному числу, если в настоящем документе не указано иначе или явно не продиктовано контекстом. Если не указано иначе, термины "содержащий", "обладающий", "включающий" и "вмещающий" следует рассматривать как открытые термины (т.е., означающие "включающие в качестве неограничивающих примеров"). Если в настоящем документе не указано иначе указание диапазонов значений в настоящем документе служит исключительно в качестве способа сокращения индивидуального указания каждого отдельного значения, находящегося в указанном диапазоне и каждое отдельное значение включено в описание также, как если бы оно было индивидуально указано в настоящем документе. Например, если указан диапазон 10-15, то также включены 11, 12, 13 и 14. Если в настоящем документе не указано иначе или иначе явно не продиктовано контекстом, все способы, описываемые в настоящем документе, можно проводить в любом подходящем порядке. Если не указано иначе, использование любого и всех примеров или иллюстративных формулировок (например, "такой как"), приводимое в настоящем документе, предназначено исключительно для лучшего описания изобретения и не предназначено для ограничения объема изобретения. Ни одну из формулировок в описании не следует рассматривать как указание того, что какой-либо незаявляемый элемент существенен для практического осуществления изобретения.
[0390] Хотя в настоящем документы приведены и описаны предпочтительные варианты осуществления настоящего изобретения, специалистам в данной области очевидно, что такие варианты осуществления приведены исключительно в качестве примера. Специалисты в данной области могут проводить множество вариаций, изменений и изменений без отклонения от изобретения. Следует понимать, что в практическом осуществлении изобретения можно использовать различные альтернативы вариантам осуществления изобретения, описываемым в настоящем документе. Следует понимать, что объем изобретения определен приводимой ниже формулой изобретения и что таким образом охвачены способы и структуры в объеме этой формулы изобретения и их эквиваленты.
ПУНКТЫ
1. Способ увеличения фиксации азота у небобового растения, включающий:
a. нанесение на растение множества не являющихся межродовыми бактерий, где указанное множество включает не являющихся межродовыми бактерий, которые:
i. обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или
ii. продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и
где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 1% или более фиксированного азота, и
где каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
2. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
3. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит бактерии, которые: продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
4. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
5. Способ по п. 1, где по меньшей мере одна генетическая вариация содержит введенную контрольную последовательность, функционально связанную по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
6. Способ по п. 1, где по меньшей мере одна генетическая вариация содержит промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
7. Способ по п. 1, где по меньшей мере одна генетическая вариация содержит индуцибельный промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
8. Способ по п. 1, где множество не являющихся межродовыми бактерий не содержит конститутивный промотор, функционально связанный с геном генетической регуляторной сети фиксации или ассимиляции азота.
9. Способ по п. 1, где множество не являющихся межродовыми бактерий не содержит конститутивный промотор, функционально связанный с геном в генном кластере nif.
10. Способ по п. 1, где множество не являющихся межродовыми бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
11. Способ по п. 1, где множество не являющихся межродовыми бактерий, наносимых на растение, не стимулирует увеличения захвата экзогенного неатмосферного азота.
12. Способ по п. 1, где растение выращивают в почве поля, куда добавили по меньшей мере приблизительно 22,7 кг азот-содержащего удобрения на акр, и где азот-содержащее удобрение содержит по меньшей мере приблизительно 5% азота по массе.
13. Способ по п. 1, где растение выращивают в почве поля, куда добавили по меньшей мере приблизительно 22,7 кг азот-содержащего удобрения на акр, и где азот-содержащее удобрение содержит по меньшей мере приблизительно 5% азота по массе, и где азот-содержащее удобрение содержит аммоний или содержащую аммоний молекулу.
14. Способ по п. 1, где экзогенный азот выбран из удобрения, содержащего один или несколько из: глутамина, аммиака, аммония, мочевины, нитрата, нитрита, содержащих аммоний молекул, содержащих нитраты молекул и содержащих нитриты молекул.
15. Способ по п. 1, где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
16. Способ по п. 1, где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
17. Способ по п. 1, где множество не являющихся межродовыми бактерий, in planta, фиксирует атмосферный азот в нелимитирующих азот условиях.
18. Способ по п. 1, где множество не являющихся межродовыми бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
19. Способ по п. 1, где фиксированный азот, продуцируемый множеством не являющихся межродовыми бактерий, определяют посредством разведения обогащенного удобрения атмосферным газообразным N2 в ткани растения.
20. Способ по п. 1, где по меньшей мере один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота выбран из группы, состоящей из: nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
21. Способ по п. 1, где по меньшей мере одна генетическая вариация представляет собой мутацию, которая приводит к одному или нескольким из: увеличенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности nifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, AmtB; сниженной аденилил-удаляющей активности GlnE или сниженной уридилат-удаляющей активности GlnD.
22. Способ по п. 1, где по меньшей мере одна генетическая вариация выбрана из: (A) нокаут-мутации; (B) удаления или устранения регуляторной последовательность гена-мишени; (C) наличия вставки гетерологичной регуляторной последовательности или (D) делеции домена.
23. Способ по п. 1, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL.
24. Способ по п. 1, где по меньшей мере одна генетическая вариация представляет собой мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
25. Способ по п. 1, где по меньшей мере одна генетическая вариация представляет собой мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
26. Способ по п. 1, где по меньшей мере одна генетическая вариация выбрана из: мутантного гена nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL; мутантного гена glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена; мутантного гена amtB, который приводит к отсутствию экспрессии указанного гена amtB; и их сочетаний.
27. Способ по п. 1, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL, и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
28. Способ по п. 1, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL, и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена, и мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
29. Способ по п. 1, где растение содержит семена, стебель, цветок, плод, листья или ризому.
30. Способ по п. 1, где множество не являющихся межродовыми бактерий формулируют в композицию.
31. Способ по п. 1, где множество не являющихся межродовыми бактерий формулируют в композицию, содержащую приемлемый в сельском хозяйстве носитель.
32. Способ по п. 1, где множество не являющихся межродовыми бактерий вносят в борозды, в которые высажены семена указанного растения.
33. Способ по п. 1, где множество не являющихся межродовыми бактерий формулируют в жидкую композицию для внесения в борозды, содержащую бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
34. Способ по п. 1, где множество не являющихся межродовыми бактерий наносят на семена указанного растения.
35. Способ по п. 1, где множество не являющихся межродовыми бактерий формулируют в покрытие для семян и наносят на семена указанного растения.
36. Способ по п. 1, где множество не являющихся межродовыми бактерий формулируют в покрытие для семян и наносят на семена указанного растения в концентрации приблизительно от 1 × 105 до приблизительно 1 × 107 КОЕ на семя.
37. Способ по п. 1, где растение представляет собой злаковую сельскохозяйственную культуру.
38. Способ по п. 1, где растение выбрано из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
39. Способ по п. 1, где растение представляет собой кукурузу.
40. Способ по п. 1, где растение представляет собой сельскохозяйственную культуру.
41. Способ по п. 1, где растение представляет собой генетически модифицированный организм.
42. Способ по п. 1, где растение не является генетически модифицированным организмом.
43. Способ по п. 1, где растение подвергали генной инженерии или выводили для эффективного использования азота
44. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит по меньшей мере два различных вида бактерий.
45. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит по меньшей мере два различных штамма одного и того же вида бактерий.
46. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит бактерии, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
47. Способ по п. 1, где множество не являющихся межродовыми бактерий являются эндофитными, эпифитными или ризосферными.
48. Способ по п. 1, где множество не являющихся межродовыми бактерий содержит бактерии, выбранные из: бактерий, депонированных как PTA-122293, бактерий, депонированных как PTA-122294, бактерий, депонированных как NCMA 201701002, бактерий, депонированных как NCMA 201708004, бактерий, депонированных как NCMA 201708003, бактерий, депонированных как NCMA 201708002, бактерий, депонированных как NCMA 201712001, бактерий, депонированных как NCMA 201712002, и их сочетаний.
49. Бактериальная композиция, содержащая:
по меньшей мере один генно-инженерный бактериальный штамм который фиксирует атмосферный азот в сельскохозяйственной системе, в которую в качестве удобрения внесено более 9,1 кг азота на акр.
50. Бактериальная композиция, содержащая:
по меньшей мере один штамм бактерий, который выведен для фиксации атмосферного азота в сельскохозяйственной системе, в которую в качестве удобрения внесено более 9,1 кг азота на акр.
51. Бактериальн композиция по п. 49 или п. 50, где указанное удобрение представляет собой химическое удобрение, выбранное из группы, состоящей из безводного аммиака, сульфата аммония, мочевины, дифосфата аммония, уреаформа, UAN (мочевино-аммониевый нитрат), монофосфата аммония, нитрата аммония, растворов азота, нитрата кальция, нитрата калия и нитрата натрия.
52. Бактериальная композиция, содержащая:
по меньшей мере один генно-инженерный бактериальный штамм который фиксирует атмосферный азот, по меньшей мере один генно-инженерный бактериальный штамм, содержащий экзогенно добавленную ДНК, где указанная экзогенно добавленная ДНК, по меньшей мере на 80% идентична с соответствующим нативным штаммом бактерий.
53. Бактериальная композиция по п. 52, где указанная экзогенно добавленная ДНК, по меньшей мере на 85% идентична с соответствующим нативным штаммом бактерий.
54. Бактериальная композиция по п. 52, где указанная экзогенно добавленная ДНК, по меньшей мере на 90% идентична с соответствующим нативным штаммом бактерий.
55. Бактериальная композиция по п. 52, где указанная экзогенно добавленная ДНК, по меньшей мере на 95% идентична с соответствующим нативным штаммом бактерий.
56. Бактериальная композиция по п. 52, где указанная экзогенно добавленная ДНК, по меньшей мере на 99% идентична с соответствующим нативным штаммом бактерий.
57. Бактериальная композиция по п. 52, где указанная экзогенно добавленная ДНК происходит из того же штамма бактерий, что и указанный соответствующий нативный штамм бактерий.
58. Бактериальная композиция по любому из предшествующих пп., где указанная бактериальная композиция представляет собой композицию удобрения.
59. Бактериальная композиция по любому из предшествующих пп., где указанный по меньшей мере один генно-инженерный бактериальный штамм содержит по меньшей мере одну вариацию в гене или межгенной области в пределах 10000 п.н. от гена, выбранного из группы, состоящей из: nifA, nifL, ntrB, ntrC, glnA, glnB, glnK, draT, amtB, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB и nifQ.
60. Бактериальная композиция по любому из предшествующих пп., дополнительно содержащая по меньшей мере один дополнительный компонент, выбранный из группы, состоящей из придающего клейкость средства, стабилизатора микроорганизмов, фунгицида, антибактериального средства, консерванта, стабилизатора, поверхностно-активного вещества, антикомплексого средства, гербицида, нематоцида, инсектицида, регулятора роста растения, удобрения, родентицида, влагопоглотителя, бактерицидного средства и питательного вещества.
61. Бактериальная композиция по любому из предшествующих пп., дополнительно содержащая по меньшей мере один дополнительный компонент, выбранный из группы, состоящей из красителя, праймера, гранулы, дезинфицирующего средства, регулятора роста растений, предохранителя и нематоцида.
62. Бактериальная композиция по любому из предшествующих пп., где указанную бактериальную композицию формулируют для внесения в поле.
63. Бактериальная композиция по п. 62, где указанную бактериальную композицию формулируют для внесения в борозду.
64. Бактериальная композиция по п. 62, где указанную бактериальную композицию формулируют для нанесения в качестве покрытия для семян, протравки для семян или пропитки для семян.
65. Бактериальная композиция по п. 61 или п. 62, где указанную бактериальную композицию формулируют для нанесения во время, до или после посадки семян.
66. Семенной состав, содержащий семена растения, которые инокулированы бактериальной композицией по любому из предшествующих пп.
67. Способ выращивания сельскохозяйственной культуры с использованием множества семян с семенным составом по п. 66.
68. Способ по п. 67, дополнительно включающий сбор урожая указанной сельскохозяйственной культуры.
69. Способ внесения бактериальной композиции по любому из предшествующих пп. в поле.
70. Способ по п. 69, где указанную бактериальную композицию вносят в указанное поле в форме, выбранной из группы, состоящей из жидкой формы, сухой формы, гранул, порошка и осадка.
71. Способ по п. 69, где указанную бактериальную композицию вносят в указанное поле в виде покрытия для семян, протравки для семян или пропитки для семян.
72. Способ по п. 69, где указанную бактериальную композицию вносят в указанное поле в качестве обработки борозды.
73. Способ по п. 69, где указанную бактериальную композицию вносят в указанное поле во время, до или после посадки семян.
74. Композиция удобрения, содержащая бактериальную композицию по любому из предшествующих пп.
75. Композиция удобрения по п. 74, где указанная композиция удобрения представляет собой композиция покрытия для семян, протравки для семян или пропитки для семян.
76. Композиция удобрения по п. 74, где указанная композиция удобрения представляет собой композицию для внесения в борозду.
77. Композиция удобрения по п. 74, где указанную композицию удобрения предоставляют для сельскохозяйственной культуры во время, до или после посадки.
78. Композиция удобрения по п. 74, дополнительно содержащая пористый носитель.
79. Композиция удобрения по п. 74, дополнительно содержащая дополнительный синергический компонент, который при комбинации с указанной бактериальной композицией увеличивает положительное удобряющее действие указанной композиции удобрения на сельскохозяйственную культуру, которое превышает суммарное положительное действие ее индивидуальных компонентов.
80. Способ поддержания уровней азота в почве, включающий:
высаживание в почву поля сельскохозяйственной культуры, инокулированной генно-инженерными бактериями, которые фиксируют атмосферный азот; и
сбор урожая указанной сельскохозяйственной культуры, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 90% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
81. Способ по п. 80, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 80% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
82. Способ по п. 80, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 70% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
83. Способ по п. 80, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 60% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
84. Способ по п. 80, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 50% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
85. Способ по п. 80, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 40% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
86. Способ по п. 80, где указанная генно-инженерная бактерия содержит бактериальную композицию по любому из предшествующих пп.
87. Способ по п. 80, где указанная генно-инженерная бактерия состоит из бактериальной композиции по любому из предшествующих пп.
88. Способ доставки пробиотической добавки в сельскохозяйственную культуру, включающий:
покрытие семян сельскохозяйственной культуры покрытием для семян, пропиткой для семян или протравкой для семян, где указанные покрытие для семян, протравка для семян или пропитка для семян содержат живых представителей указанного пробиотика; и
внесение в почву поля указанных семян сельскохозяйственной культуры.
89. Способ по п. 88, где указанные покрытие для семян, протравку для семян или пропитку для семян наносят на указанные семена сельскохозяйственной культуры одним слоем.
90. Способ по п. 88, где указанное покрытие для семян наносят на указанные семена сельскохозяйственной культуры несколькими слоями.
91. Способ по п. 88, где указанное покрытие для семян наносят на указанные семена сельскохозяйственной культуры в смеси.
92. Способ по п. 88, где указанные семена сельскохозяйственной культуры являются неклубеньковыми.
93. Способ по п. 88, где указанное покрытие для семян содержит бактериальную композицию по любому из предшествующих пп.
94. Способ по любому из предшествующих пп., где генно-инженерный бактериальный штамм представляет собой генно-инженерный штамм грамположительной бактерии.
95. Способ по п. 94, где генно-инженерный штамм грамположительной бактерии обладает измененным уровнем экспрессии регулятора кластера Nif.
96. Способ по п. 94, где генно-инженерный штамм грамположительной бактерии экспрессирует сниженное количество отрицательного регулятора кластера Nif.
97. Способ по п. 94, где генно-инженерный бактериальный штамм экспрессирует сниженное количество of GlnR.
98. Способ по любому из предшествующих пп., где в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 75% идентичный с последовательностью из базы данных NifH лаборатории Дж. Зера.
99. Способ по любому из предшествующих пп., где в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 85% идентичный с последовательностью из базы данных NifH лаборатории Дж. Зера.
100. Способ по любому из предшествующих пп., где в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 75% идентичный с последовательностью из базы данных NifH лаборатории Д. Бакли.
101. Способ по любому из предшествующих пп., где в геноме генно-инженерного бактериального штамма закодирован полипептид по меньшей мере на 85% идентичный с последовательностью из базы данных NifH лаборатории Д. Бакли.
102. Способ выведения штаммов микроорганизмов для улучшения конкретных признаков агротехнического значения, включающий:
предоставление множества штаммов микроорганизмов, обладающих способностью к колонизации требуемой сельскохозяйственной культуры;
улучшение регуляторных сетей, влияющих на признак посредством внутригеномной перестановки;
оценку штаммов микроорганизмов во множестве штаммов микроорганизмов для определения меры признака; и
отбор из множества штаммов микроорганизмов одного или нескольких штаммов микроорганизмов, которые обеспечивают улучшение признака в случае требуемой сельскохозяйственной культуры.
103. Способ по п. 102, где конкретный признак, подвергаемый улучшению, представляет собой способность штамма микроорганизмов фиксировать азот.
104. Способ по п. 103, где конкретный признак, подвергаемый улучшению, представляет собой способность штамма микроорганизмов фиксировать атмосферный азот в присутствии N-удобряем условия роста.
105. Способ выведения штаммов микроорганизмов для улучшения конкретных признаков агротехнического значения, включающий:
предоставление множества штаммов микроорганизмов, обладающих способностью к колонизации требуемой сельскохозяйственной культуры;
внесение генетического разнообразия во множество штаммов микроорганизмов;
оценку штаммов микроорганизмов во множестве штаммов микроорганизмов для определения меры признака; и
отбор из множества штаммов микроорганизмов одного или нескольких штаммов микроорганизмов, которые обеспечивают улучшение признака в случае требуемой сельскохозяйственной культуры.
106. Способ по п. 105, где конкретный признак, подвергаемый улучшению, представляет собой способность штамма микроорганизмов фиксировать азот.
107. Способ по п. 106, где конкретный признак, подвергаемый улучшению, представляет собой способность штамма микроорганизмов фиксировать атмосферный азот в присутствии N-удобряем условия роста.
108. Способ увеличения у небобового растения количества получаемого из атмосферы азота, включающий:
воздействие на указанное небобовое растение подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы несут по меньшей мере одна генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, или по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
109. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть фиксации азота.
110. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть ассимиляции азота.
111. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть ассимиляции азота.
112. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы вносят в борозды, в которые высаживают семена указанного небобового растения.
113. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы наносят на семена указанного небобового растения.
114. Способ по п. 108, где указанное небобовое растение представляет собой небобовую сельскохозяйственную культуру, выбранную из группы, состоящей из сорго, канолы, томата, клубники, ячменя, риса, кукурузы, пшеницы, картофеля, просо, злаковых, зерновых и кукурузы.
115. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы колонизируют по меньшей мере корень указанного небобового растения так, что указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы присутствуют в указанном небобовом растении в количестве по меньшей мере 105 колониеобразующих единиц на грамм сырой массы ткани.
116. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы способны к фиксации атмосферного азота в нелимитирующих азот условиях.
117. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы, in planta, экскретирует азот-содержащие продукты фиксации азота.
118. Способ по п. 108, где указанную по меньшей мере одну генетическую вариацию вносят в ген, выбранный из группы, состоящей из nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
119. Способ по п. 108, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы, in planta, продуцируют в указанном небобовом растении по меньшей мере 1% фиксированного азота.
120. Способ по п. 119, где указанный фиксированный азот в указанном небобовом растении, продуцируемый указанными подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, определяют посредством разведения 15N в сельскохозяйственных культурах, выращиваемых в полях, обрабатываемых удобрением, содержащим 1,2% 15N.
121. Способ по п. 119, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы, in planta, продуцируют в указанном небобовом растении 5% фиксированного азота или более.
122. Способ по п. 108, где указанные не являющиеся межродовыми микроорганизмы подвергают генетической инженерии с использованием по меньшей мере инженерии одного типа, выбранной из группы, состоящей из направленного мутагенеза, случайного мутагенеза и направленной эволюции.
123. Способ увеличения количества получаемого из атмосферы азота у растения кукурузы, включающий
воздействие на указанное растение кукурузы подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, несущими генно-инженерные генетические вариации по меньшей мере в двух генах, выбранных из группы, состоящей из nifL, glnB, glnE и amtB.
124. Способ по п. 123, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы, in planta, экскретирует азот-содержащие продукты фиксации азота.
125. Способ по п. 123, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы вносят в борозды, в которые высажены семена указанного растения кукурузы.
126. Способ по п. 123, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы наносят на семена указанного растения кукурузы.
127. Способ увеличения количества получаемого из атмосферы азота у растения кукурузы, включающий:
воздействие на указанное растение кукурузы подвергнутыми генетической инженерии не являющимися межродовыми микроорганизмами, несущими по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота, где указанные подвергнутые генетической инженерии не являющиеся межродовыми микроорганизмы, in planta, продуцируют у указанного растения кукурузы по меньшей мере 5% фиксированного азота, как измеряют посредством разведения 15N в сельскохозяйственных культурах, выращиваемых в полях, обрабатываемых удобрением, содержащим 1,2% 15N.
128. Способ увеличения фиксации азота у небобового растения, включающий:
a. нанесение на растение множества не являющихся межродовыми бактерий, где указанное множество включает не являющихся межродовыми бактерий, которые:
i. обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и
ii. продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и
где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,5 × 10-8 ммоль N на грамм сырой массы ткани корня растения в час, и
где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 1% фиксированного азота или более, и
где каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
129. Способ по п. 128, где по меньшей мере одна генетическая вариация содержит введенную контрольную последовательность, функционально связанную по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
130. Способ по п. 128, где по меньшей мере одна генетическая вариация содержит промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
131. Способ по п. 128, где по меньшей мере одна генетическая вариация содержит индуцибельный промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
132. Способ по п. 128, где множество не являющихся межродовыми бактерий не содержит конститутивный промотор, функционально связанный с геном генетической регуляторной сети фиксации или ассимиляции азота.
133. Способ по п. 128, где множество не являющихся межродовыми бактерий не содержит конститутивный промотор, функционально связанный с геном в генном кластере nif.
134. Способ по п. 128, где множество не являющихся межродовыми бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
135. Способ по п. 128, где множество не являющихся межродовыми бактерий, наносимых на растение, не стимулирует увеличения захвата экзогенного неатмосферного азота.
136. Способ по п. 128, где растение выращивают в почве поля, куда добавили по меньшей мере приблизительно 22,7 кг азот-содержащего удобрения на акр, и где азот-содержащее удобрение содержит по меньшей мере приблизительно 5% азота по массе.
137. Способ по п. 128, где растение выращивают в почве поля, куда добавили по меньшей мере приблизительно 22,7 кг азот-содержащего удобрения на акр, и где азот-содержащее удобрение содержит по меньшей мере приблизительно 5% азота по массе, и где азот-содержащее удобрение содержит аммоний или содержащую аммоний молекулу.
138. Способ по п. 128, где экзогенный азот выбран из удобрения, содержащего один или несколько из: глутамина, аммиака, аммония, мочевины, нитрата, нитрита, содержащих аммоний молекул, содержащих нитраты молекул и содержащих нитриты молекул.
139. Способ по п. 128, где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
140. Способ по п. 128, где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
141. Способ по п. 128, где множество не являющихся межродовыми бактерий, in planta, фиксирует атмосферный азот в нелимитирующих азот условиях.
142. Способ по п. 128, где множество не являющихся межродовыми бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
143. Способ по п. 128, где фиксированный азот, продуцируемый множеством не являющихся межродовыми бактерий, определяют посредством разведения обогащенного удобрения атмосферным газообразным N2 в ткани растения.
144. Способ по п. 128, где по меньшей мере один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота выбраны из группы, состоящей из: nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
145. Способ по п. 128, где по меньшей мере одна генетическая вариация представляет собой мутацию, которая приводит к одному или нескольким из: увеличенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности nifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, AmtB; сниженной аденилил-удаляющей активности GlnE или сниженной уридилат-удаляющей активности GlnD.
146. Способ по п. 128, где по меньшей мере одна генетическая вариация выбрана из: (A) нокаут-мутации; (B) удаления или устранения регуляторной последовательность гена-мишени; (C) наличия вставки гетерологичной регуляторной последовательности или (D) делеции домена.
147. Способ по п. 128, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL.
148. Способ по п. 128, где по меньшей мере одна генетическая вариация представляет собой мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
149. Способ по п. 128, где по меньшей мере одна генетическая вариация представляет собой мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
150. Способ по п. 128, где по меньшей мере одна генетическая вариация выбрана из: мутантного гена nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL; мутантного гена glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена; мутантного гена amtB, который приводит к отсутствию экспрессии указанного гена amtB; и их сочетаний.
151. Способ по п. 128, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL, и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
152. Способ по п. 128, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL, и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена и мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
153. Способ по п. 128, где растение содержит семена, стебель, цветок, плод, листья или ризому.
154. Способ по п. 128, где множество не являющихся межродовыми бактерий формулируют в композицию.
155. Способ по п. 128, где множество не являющихся межродовыми бактерий формулируют в композицию, содержащую приемлемый в сельском хозяйстве носитель.
156. Способ по п. 128, где множество не являющихся межродовыми бактерий вносят в борозды, в которые высажены семена указанного растения.
157. Способ по п. 128, где множество не являющихся межродовыми бактерий формулируют в жидкую композицию для внесения в борозду, содержащую бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
158. Способ по п. 128, где множество не являющихся межродовыми бактерий наносят на семена указанного растения.
159. Способ по п. 128, где множество не являющихся межродовыми бактерий формулируют в покрытие для семян и наносят на семена указанного растения.
160. Способ по п. 128, где множество не являющихся межродовыми бактерий формулируют в покрытие для семян и наносят на семена указанного растения в концентрации приблизительно от 1 × 105 до приблизительно 1 × 107 КОЕ на семя.
161. Способ по п. 128, где растение представляет собой злаковую сельскохозяйственную культуру.
162. Способ по п. 128, где растение выбрано из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
163. Способ по п. 128, где растение представляет собой кукурузу.
164. Способ по п. 128, где растение представляет собой сельскохозяйственную культуру.
165. Способ по п. 128, где растение представляет собой генетически модифицированный организм.
166. Способ по п. 128, где растение не является генетически модифицированным организмом.
167. Способ по п. 128, где растение подвергали генной инженерии или выводили для эффективного использования азота
168. Способ по п. 128, где множество не являющихся межродовыми бактерий содержит по меньшей мере два различных вида бактерий.
169. Способ по п. 128, где множество не являющихся межродовыми бактерий содержит по меньшей мере два различных штамма одного и того же вида бактерий.
170. Способ по п. 128, где множество не являющихся межродовыми бактерий содержит бактерии, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
171. Способ по п. 128, где множество не являющихся межродовыми бактерий являются эндофитными, эпифитными или ризосферными.
172. Способ по п. 128, где множество не являющихся межродовыми бактерий содержит бактерии, выбранные из: бактерий, депонированных как PTA-122293, бактерий, депонированных как PTA-122294, бактерий, депонированных как NCMA 201701002, бактерий, депонированных как NCMA 201708004, бактерий, депонированных как NCMA 201708003, бактерий, депонированных как NCMA 201708002, бактерий, депонированных как NCMA 201712001, бактерий, депонированных как NCMA 201712002, и их сочетаний.
173. Способ увеличения фиксации азота у небобового растения, включающий:
a. нанесение на растение множества бактерий, где указанное множество включает бактерии, которые:
i. обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или
ii. продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и
где множество бактерий, in planta, продуцирует в растении 1% фиксированного азота или более.
174. Способ по п. 173, где множество бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
175. Способ по п. 173, где множество бактерий содержит бактерии, которые: продуцируют фиксированный N в количестве по меньшей мере приблизительно 173 × 10-17 ммоль N на бактериальную клетку в час.
176. Способ по п. 173, где множество бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
177. Способ по п. 173, где множество бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,5 × 10-8 ммоль N на грамм сырой массы ткани корня растения в час.
178. Способ по п. 173, где множество бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
179. Способ по п. 173, где множество бактерий, наносимых на растение, не стимулирует увеличения захвата экзогенного неатмосферного азота.
180. Способ по п. 173, где растение содержит семена, стебель, цветок, плод, листья или ризому.
181. Способ по п. 173, где множество бактерий формулируют в композицию.
182. Способ по п. 173, где множество бактерий формулируют в композицию, содержащую приемлемый в сельском хозяйстве носитель.
183. Способ по п. 173, где множество бактерий вносят в борозды, в которые высажены семена указанного растения.
184. Способ по п. 173, где множество бактерий формулируют в жидкую композицию для внесения в борозду, содержащую бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
185. Способ по п. 173, где множество бактерий наносят на семена указанного растения.
186. Способ по п. 173, где множество бактерий формулируют в покрытие для семян и наносят на семена указанного растения.
187. Способ по п. 173, где множество бактерий формулируют в покрытие для семян и наносят на семена указанного растения в концентрации приблизительно от 1 × 105 до приблизительно 1 × 107 КОЕ на семя.
188. Способ по п. 173, где растение представляет собой злаковую сельскохозяйственную культуру.
189. Способ по п. 173, где растение выбрано из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
190. Способ по п. 173, где растение представляет собой кукурузу.
191. Способ по п. 173, где растение представляет собой сельскохозяйственную культуру.
192. Способ по п. 173, где растение представляет собой генетически модифицированный организм.
193. Способ по п. 173, где растение не является генетически модифицированным организмом.
194. Способ по п. 173, где растение подвергали генной инженерии или выводили для эффективного использования азота
195. Способ по п. 173, где множество бактерий содержит по меньшей мере два различных вида бактерий.
196. Способ по п. 173, где множество бактерий содержит по меньшей мере два различных штамма одного и того же вида бактерий.
197. Способ по п. 173, где множество бактерий содержит бактерии, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
198. Способ по п. 173, где множество бактерий являются эндофитными, эпифитными или ризосферными.
199. Способ по п. 173, где множество бактерий выбраны из: бактерий, депонированных как NCMA 201701003, бактерий, депонированных как NCMA 201701001, и бактерий, депонированных как NCMA 201708001.
200. Не являющаяся межродовой популяция бактерий, способная к увеличению фиксации азота у небобового растения, содержащая:
a. множество не являющихся межродовыми бактерий, которые:
i. обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или
ii. продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и
где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 1% фиксированного азота или более, выращиваем в присутствии множества не являющихся межродовыми бактерий, и
где каждый представитель множества не являющихся межродовыми бактерий несет по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющиеся межродовыми бактерии становятся способны к фиксации атмосферного азота в присутствии экзогенного азота.
201. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
202. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит бактерии, которые: продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
203. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
204. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация содержит введенную контрольную последовательность, функционально связанную по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
205. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация содержит промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
206. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация содержит индуцибельный промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
207. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий не содержит конститутивный промотор, функционально связанный с геном генетической регуляторной сети фиксации или ассимиляции азота.
208. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий не содержит конститутивный промотор, функционально связанный с геном в генном кластере nif.
209. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
210. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий, наносимых на растение, не стимулирует увеличения захвата экзогенного неатмосферного азота.
211. Не являющаяся межродовой популяция бактерий по п. 200, где растение выращивают в почве поля, куда добавили по меньшей мере приблизительно 22,7 кг азот-содержащего удобрения на акр, и где азот-содержащее удобрение содержит по меньшей мере приблизительно 5% азота по массе.
212. Не являющаяся межродовой популяция бактерий по п. 200, где растение выращивают в почве поля, куда добавили по меньшей мере приблизительно 22,7 кг азот-содержащего удобрения на акр, и где азот-содержащее удобрение содержит по меньшей мере приблизительно 5% азота по массе, и где азот-содержащее удобрение содержит аммоний или содержащую аммоний молекулу.
213. Не являющаяся межродовой популяция бактерий по п. 200, где экзогенный азот выбран из удобрения, содержащего один или несколько из: глутамина, аммиака, аммония, мочевины, нитрата, нитрита, содержащих аммоний молекул, содержащих нитраты молекул и содержащих нитриты молекул.
214. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
215. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
216. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий, in planta, фиксирует атмосферный азот в нелимитирующих азот условиях.
217. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
218. Не являющаяся межродовой популяция бактерий по п. 200, где фиксированный азот, продуцируемый множеством не являющихся межродовыми бактерий, определяют посредством разведения обогащенного удобрения атмосферным газообразным N2 в ткани растения.
219. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота выбраны из группы, состоящей из: nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
220. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация представляет собой мутацию, которая приводит к одному или нескольким из: увеличенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности nifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, AmtB; сниженной аденилил-удаляющей активности GlnE или сниженной уридилат-удаляющей активности GlnD.
221. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация выбрана из: (A) нокаут-мутации; (B) удаления или устранения регуляторной последовательность гена-мишени; (C) наличия вставки гетерологичной регуляторной последовательности или (D) делеции домена.
222. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL.
223. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация представляет собой мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
224. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация представляет собой мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
225. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация выбрана из: мутантного гена nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL; мутантного гена glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена; мутантного гена amtB, который приводит к отсутствию экспрессии указанного гена amtB; и их сочетаний.
226. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
227. Не являющаяся межродовой популяция бактерий по п. 200, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена и мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
228. Не являющаяся межродовой популяция бактерий по п. 200, где растение содержит семена, стебель, цветок, плод, листья или ризому.
229. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий формулируют в композицию.
230. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий формулируют в композицию, содержащую приемлемый в сельском хозяйстве носитель.
231. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий формулируют в жидкую композицию для внесения в борозду, содержащую бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
232. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий формулируют в покрытие для семян.
233. Не являющаяся межродовой популяция бактерий по п. 200, где растение представляет собой злаковую сельскохозяйственную культуру.
234. Не являющаяся межродовой популяция бактерий по п. 200, где растение выбрано из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
235. Не являющаяся межродовой популяция бактерий по п. 200, где растение представляет собой кукурузу.
236. Не являющаяся межродовой популяция бактерий по п. 200, где растение представляет собой сельскохозяйственную культуру.
237. Не являющаяся межродовой популяция бактерий по п. 200, где растение представляет собой генетически модифицированный организм.
238. Не являющаяся межродовой популяция бактерий по п. 200, где растение не является генетически модифицированным организмом.
239. Не являющаяся межродовой популяция бактерий по п. 200, где растение подвергали генной инженерии или выводили для эффективного использования азота
240. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит по меньшей мере два различных вида бактерий.
241. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит по меньшей мере два различных штамма одного и того же вида бактерий.
242. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит бактерии, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
243. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий являются эндофитными, эпифитными или ризосферными.
244. Не являющаяся межродовой популяция бактерий по п. 200, где множество не являющихся межродовыми бактерий содержит бактерии, выбранные из: бактерий, депонированных как PTA-122293, бактерий, депонированных как PTA-122294, бактерий, депонированных как NCMA 201701002, бактерий, депонированных как NCMA 201708004, бактерий, депонированных как NCMA 201708003, бактерий, депонированных как NCMA 201708002, бактерий, депонированных как NCMA 201712001, бактерий, депонированных как NCMA 201712002, и их сочетаний.
245. Популяция бактерий, способная к увеличению фиксации азота у небобового растения, содержащая:
a. множество бактерий, которые:
i. обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или
ii. продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и
где множество бактерий, in planta, продуцирует в растении 1% фиксированного азота или более.
246. Популяция бактерий по п. 245, где множество бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
247. Популяция бактерий по п. 245, где множество бактерий содержит бактерии, которые: продуцируют фиксированный N в количестве по меньшей мере приблизительно 245 × 10-17 ммоль N на бактериальную клетку в час.
248. Популяция бактерий по п. 245, где множество бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
249. Популяция бактерий по п. 245, где множество бактерий содержит бактерии, которые: обладают средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцируют фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,5 × 10-8 ммоль N на грамм сырой массы ткани корня растения в час.
250. Популяция бактерий по п. 245, где множество бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
251. Популяция бактерий по п. 245, где множество бактерий, наносимых на растение, не стимулирует увеличения захвата экзогенного неатмосферного азота.
252. Популяция бактерий по п. 245, где растение содержит семена, стебель, цветок, плод, листья или ризому.
253. Популяция бактерий по п. 245, где множество бактерий формулируют в композицию.
254. Популяция бактерий по п. 245, где множество бактерий формулируют в композицию, содержащую приемлемый в сельском хозяйстве носитель.
255. Популяция бактерий по п. 245, где множество бактерий вносят в борозды, в которые высажены семена указанного растения.
256. Популяция бактерий по п. 245, где множество бактерий формулируют в жидкую композицию для внесения в борозду, содержащую бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
257. Популяция бактерий по п. 245, где множество бактерий наносят на семена указанного растения.
258. Популяция бактерий по п. 245, где множество бактерий формулируют в покрытие для семян и наносят на семена указанного растения.
259. Популяция бактерий по п. 245, где множество бактерий формулируют в покрытие для семян и наносят на семена указанного растения в концентрации приблизительно от 1 × 105 до приблизительно 1 × 107 КОЕ на семя.
260. Популяция бактерий по п. 245, где растение представляет собой злаковую сельскохозяйственную культуру.
261. Популяция бактерий по п. 245, где растение выбрано из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
262. Популяция бактерий по п. 245, где растение представляет собой кукурузу.
263. Популяция бактерий по п. 245, где растение представляет собой сельскохозяйственную культуру.
264. Популяция бактерий по п. 245, где растение представляет собой генетически модифицированный организм.
265. Популяция бактерий по п. 245, где растение не является генетически модифицированным организмом.
266. Популяция бактерий по п. 245, где растение подвергали генной инженерии или выводили для эффективного использования азота
267. Популяция бактерий по п. 245, где множество бактерий содержит по меньшей мере два различных вида бактерий.
268. Популяция бактерий по п. 245, где множество бактерий содержит по меньшей мере два различных штамма одного и того же вида бактерий.
269. Популяция бактерий по п. 245, где множество бактерий содержит бактерии, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
270. Популяция бактерий по п. 245, где множество бактерий являются эндофитными, эпифитными или ризосферными.
271. Популяция бактерий по п. 245, где множество бактерий выбраны из: бактерий, депонированных как NCMA 201701003, бактерий, депонированных как NCMA 201701001, и бактерий, депонированных как NCMA 201708001.
272. Бактерия, которая:
i. обладает средней колонизирующей способность на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или
ii. продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
273. Не являющаяся межродовой бактерия, несущая: по меньшей мере одну генетическую вариацию, внесенную по меньшей мере в один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота так, что не являющаяся межродовой бактерия способна к фиксации атмосферного азота в присутствии экзогенного азота, и где указанная бактерия:
i. обладает средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения; и/или
ii. продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
274. Не являющаяся межродовой бактерия по п. 273, где бактерия обладает средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
275. Не являющаяся межродовой бактерия по п. 273, где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
276. Не являющаяся межродовой бактерия по п. 273, где бактерия обладает средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час.
277. Не являющаяся межродовой бактерия по п. 273, где бактерия обладает средней колонизирующей способностью на единицу ткани корня растения по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения и продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час, и где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,5 × 10-8 ммоль N на грамм сырой массы ткани корня растения в час.
278. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация содержит введенную контрольную последовательность, функционально связанную по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
279. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация содержит промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
280. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация содержит индуцибельный промотор, функционально связанный по меньшей мере с одним геном генетической регуляторной сети фиксации или ассимиляции азота.
281. Не являющаяся межродовой бактерия по п. 273, где бактерия не содержит конститутивный промотор, функционально связанный с геном генетической регуляторной сети фиксации или ассимиляции азота.
282. Не являющаяся межродовой бактерия по п. 273, где бактерия не содержит конститутивный промотор, функционально связанный с геном в генном кластере nif.
283. Не являющаяся межродовой бактерия по п. 273, где бактерия, in planta, экскретирует азот-содержащие продукты фиксации азота.
284. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере один ген или некодирующий полинуклеотид генетической регуляторной сети фиксации или ассимиляции азота выбраны из группы, состоящей из: nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
285. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация представляет собой мутацию, которая приводит к одному или нескольким из: увеличенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности nifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, AmtB; сниженной аденилил-удаляющей активности GlnE или сниженной уридилат-удаляющей активности GlnD.
286. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация выбрана из: (A) нокаут-мутации; (B) удаления или устранения регуляторной последовательность гена-мишени; (C) наличия вставки гетерологичной регуляторной последовательности или (D) делеции домена.
287. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL.
288. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация представляет собой мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
289. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация представляет собой мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
290. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация выбрана из: мутантного гена nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL; мутантного гена glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена; мутантного гена amtB, который приводит к отсутствию экспрессии указанного гена amtB; и их сочетаний.
291. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
292. Не являющаяся межродовой бактерия по п. 273, где по меньшей мере одна генетическая вариация представляет собой мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена и мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
293. Не являющаяся межродовой бактерия по п. 273, формулируемая в композицию.
294. Не являющаяся межродовой бактерия по п. 273, формулируемая в композицию, содержащую приемлемый в сельском хозяйстве носитель.
295. Не являющаяся межродовой бактерия по п. 273, формулируемая в жидкую композицию для внесения в борозду.
296. Не являющаяся межродовой бактерия по п. 273, формулируемая в покрытие для семян.
297. Не являющаяся межродовой бактерия по п. 273, где указанная бактерия выбрана из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
298. Не являющаяся межродовой бактерия по п. 273, где указанная бактерия является эндофитной, эпифитной или ризосферной.
299. Не являющаяся межродовой бактерия по п. 273, где указанная бактерия выбрана из: бактерий, депонированных как PTA-122293, бактерий, депонированных как PTA-122294, бактерий, депонированных как NCMA 201701002, бактерий, депонированных как NCMA 201708004, бактерий, депонированных как NCMA 201708003, бактерий, депонированных как NCMA 201708002, бактерий, депонированных как NCMA 201712001, бактерий, депонированных как NCMA 201712002, и их сочетаний.
300. Способ увеличения фиксации азота в растении, включающий введение в растение эффективного количество композиции, содержащей:
i. очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 97% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269, 277-283;
ii. очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276, 284-295; и/или
iii. очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260, 296-303; и
где растение, которому вводят эффективное количество композиции, демонстрирует увеличение фиксации азота по сравнению с растением без введения композиции.
301. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 97% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269 и 277-283.
302. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 99% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269 и 277-283.
303. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты 16S, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269 и 277-283.
304. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
305. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 95% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
306. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 99% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
307. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
308. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
309. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 95% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
310. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, которая по меньшей мере приблизительно на 99% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
311. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с последовательностью нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
312. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии по меньшей мере с одной генетической вариацией, внесенной в ген, выбранный из группы, состоящей из nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
313. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии по меньшей мере с одной генетической вариацией, внесенной в ген, выбранный из группы, состоящей из nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента, где генетическая вариация (A) представляет собой нокаут-мутацию; (B) удаляет или устраняет регуляторную последовательность гена-мишени; (C) содержит вставку гетерологичной регуляторной последовательности или (D) является делецией домена.
314. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии по меньшей мере с одной мутацией, которая приводит к одному или нескольким из: увеличенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности nifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, или AmtB; сниженной аденилил-удаляющей активности GlnE или сниженной уридилат-удаляющей активности GlnD.
315. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с мутантным геном nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL.
316. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с мутантным геном glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
317. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с мутантным геном amtB, который приводит к отсутствию экспрессии указанного гена amtB.
318. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии по меньшей мере с одной из следующих генетических альтераций: мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL; мутантного гена glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена; мутантного гена amtB, который приводит к отсутствию экспрессии указанного гена amtB; и их сочетаний.
319. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с мутантным геном nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
320. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с мутантным геном nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена и мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
321. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL.
322. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL, где модификация представляет собой разрушение, нокаут или укорочение.
323. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL, и где бактерии дополнительно содержат промотор, функционально связанный с последовательностью nifA.
324. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL, и где у бактерий отсутствует гомолог nifL.
325. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE.
326. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE, где модификация представляет собой разрушение, нокаут или укорочение.
327. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE, и где в последовательности белка glnE отсутствует аденилил-удаляющий (AR) домен.
328. Способ по п. 300, где композиция содержит: очищенную популяцию бактерий, которая содержит бактерии с модификацией glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE, и где в последовательности белка glnE отсутствует аденилил-удаляющая (AR) активность.
329. Способ по п. 300, где очищенная популяция бактерий, in planta, обеспечивает растению по меньшей мере 1% фиксированного азота.
330. Способ по п. 300, где очищенная популяция бактерий, in planta, обеспечивает растению по меньшей мере 5% фиксированного азота.
331. Способ по п. 300, где очищенная популяция бактерий, in planta, обеспечивает растению по меньшей мере 10% фиксированного азота.
332. Способ по п. 300, где очищенная популяция бактерий способна к фиксации атмосферного азота в нелимитирующих азот условиях.
333. Способ по п. 300, где очищенная популяция бактерий, in planta, экскретирует азот-содержащие продукты фиксации азота.
334. Способ по п. 300, где очищенная популяция бактерий, in planta, обеспечивает растению по меньшей мере 1% фиксированного азота, и где указанный фиксированный азот измеряют посредством разведения обогащенного удобрения атмосферным газообразным N2 в ткани растения.
335. Способ по п. 300, где очищенная популяция бактерий колонизирует корень указанного растения и присутствует в количестве по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
336. Способ по п. 300, где растение содержит семена, стебель, цветок, плод, листья или ризому.
337. Способ по п. 300, где композиция содержит: приемлемый в сельском хозяйстве носитель.
338. Способ по п. 300, где композицию, содержащую очищенную популяцию бактерий, вносят в борозды, в которые высажены семена указанного растения.
339. Способ по п. 300, где композицию, содержащую очищенную популяцию бактерий, формулируют в виде жидкой композиции для внесения в борозду, содержащей бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
340. Способ по п. 300, где композицию, содержащую очищенную популяцию бактерий, наносят на семена указанного растения.
341. Способ по п. 300, где композицию, содержащую очищенную популяцию бактерий, формулируют в виде покрытия для семян и наносят на семена указанного растения.
342. Способ по п. 300, где композицию, содержащую очищенную популяцию бактерий, формулируют в виде покрытия для семян и наносят на семена указанного растения в концентрации приблизительно от 1 × 105 до приблизительно 1 × 107 КОЕ на семя.
343. Способ по п. 300, где растение является небобовым.
344. Способ по п. 300, где растение представляет собой злаковую сельскохозяйственную культуру.
345. Способ по п. 300, где растение выбрано из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
346. Способ по п. 300, где растение представляет собой кукурузу.
347. Способ по п. 300, где растение представляет собой бобовое растение.
348. Способ по п. 300, где растение представляет собой зерновую сельскохозяйственную культуру.
349. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
350. Способ по п. 300, где очищенная популяция бактерий содержит бактерии рода Rahnella.
351. Способ по п. 300, где очищенная популяция бактерий содержит бактерии вида Rahnella aquatilis.
352. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как PTA-122293.
353. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201701003.
354. Способ по п. 300, где очищенная популяция бактерий содержит бактерии рода Kosakonia.
355. Способ по п. 300, где очищенная популяция бактерий содержит бактерии вида Kosakonia sacchari.
356. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как PTA-122294.
357. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201701001.
358. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201701002.
359. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201708004.
360. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201708003.
361. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201708002.
362. Способ по п. 300, где очищенная популяция бактерий содержит бактерии рода Klebsiella.
363. Способ по п. 300, где очищенная популяция бактерий содержит бактерии вида Klebsiella variicola.
364. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201708001.
365. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201712001.
366. Способ по п. 300, где очищенная популяция бактерий содержит бактерии, депонированные как NCMA 201712002.
367. Выделенные бактерии, содержащие:
i. последовательность нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 97% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269, 277-283;
ii. последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276, 284-295; и/или
iii. последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260, 296-303.
368. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 97% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269 и 277-283.
369. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты 16S, которая по меньшей мере приблизительно на 99% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269 и 277-283.
370. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты 16S, выбранную из SEQ ID NO: 85, 96, 111, 121, 122, 123, 124, 136, 149, 157, 167, 261, 262, 269 и 277-283.
371. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
372. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 95% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
373. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 99% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
374. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, выбранную из SEQ ID NO: 86-95, 97-110, 112-120, 125-135, 137-148, 150-156, 158-166, 168-176, 263-268, 270-274, 275, 276 и 284-295.
375. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 90% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
376. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 95% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
377. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, которая по меньшей мере приблизительно на 99% идентична последовательности нуклеиновой кислоты, выбранной из SEQ ID NO: 177-260 и 296-303.
378. Выделенные бактерии по п. 367, содержащие: последовательность нуклеиновой кислоты, выбранную из SEQ ID NO: 177-260 и 296-303.
379. Выделенные бактерии по п. 367, содержащие: по меньшей мере одну генетическую вариацию, внесенную в ген, выбранный из группы, состоящей из nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента.
380. Выделенные бактерии по п. 367, содержащие: по меньшей мере одну генетическую вариацию, внесенную в ген, выбранный из группы, состоящей из nifA, nifL, ntrB, ntrC, полинуклеотида, кодирующего глутаминсинтетазу, glnA, glnB, glnK, drat, amtB, полинуклеотида, кодирующего глутаминазу, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB, nifQ и гена, ассоциированного с биосинтезом нитрогеназного фермента, где генетическая вариация (A) представляет собой нокаут-мутацию; (B) удаляет или устраняет регуляторную последовательность гена-мишени; (C) содержит вставку гетерологичной регуляторной последовательности или (D) является делецией домена.
381. Выделенные бактерии по п. 367, содержащие: по меньшей мере одну мутацию, которая приводит к одному или нескольким из: увеличенной экспрессии или активности NifA или глутаминазы; сниженной экспрессии или активности nifL, NtrB, глутаминсинтетазы, GlnB, GlnK, DraT, или AmtB; сниженной аденилил-удаляющей активности GlnE или сниженной уридилат-удаляющей активности GlnD.
382. Выделенные бактерии по п. 367, содержащие: мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL.
383. Выделенные бактерии по п. 367, содержащие: мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
384. Выделенные бактерии по п. 367, содержащие: мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
385. Выделенные бактерии по п. 367, содержащие: по меньшей мере одну из следующих генетических альтераций: мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL; мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена; мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB; и их сочетания.
386. Выделенные бактерии по п. 367, содержащие: мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена.
387. Выделенные бактерии по п. 367, содержащие: мутантный ген nifL, который изменен так, чтобы содержать гетерологичный промотор, встроенный в указанный ген nifL и мутантный ген glnE, который приводит к образованию укороченного белка glnE с отсутствием аденилил-удаляющего (AR) домена и мутантный ген amtB, который приводит к отсутствию экспрессии указанного гена amtB.
388. Выделенные бактерии по п. 367, содержащие: модификацию nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL.
389. Выделенные бактерии по п. 367, содержащие: модификацию nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL, где модификация представляет собой разрушение, нокаут или укорочение.
390. Выделенные бактерии по п. 367, содержащие: модификацию nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL, и где бактерии дополнительно содержат промотор, функционально связанный с последовательностью nifA.
391. Выделенные бактерии по п. 367, содержащие: модификацию nifL, которые экспрессируют белок NifL менее чем приблизительно 50% бактерий с отсутствием модификации nifL, и где у бактерий отсутствует гомолог nifL.
392. Выделенные бактерии по п. 367, содержащие: модификацию glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE.
393. Выделенные бактерии по п. 367, содержащие: модификацию glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE, где модификация представляет собой разрушение, нокаут или укорочение.
394. Выделенные бактерии по п. 367, содержащие: модификацию glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE, и где в последовательности белка glnE отсутствует аденилил-удаляющий (AR) домен.
395. Выделенные бактерии по п. 367, содержащие: модификацию glnE, которые экспрессируют белок glnE менее чем приблизительно 50% бактерий с отсутствием модификации glnE, и где в последовательности белка glnE отсутствует аденилил-удаляющий (AR) активность.
396. Выделенные бактерии по п. 367, где бактерии, in planta, обеспечивают растению по меньшей мере 1% фиксированного азота, воздействующего на указанные бактерии.
397. Выделенные бактерии по п. 367, где бактерии, in planta, обеспечивают растению по меньшей мере 5% фиксированного азота, воздействующего на указанные бактерии.
398. Выделенные бактерии по п. 367, где бактерии, in planta, обеспечивают растению по меньшей мере 10% фиксированного азота, воздействующего на указанные бактерии.
399. Выделенные бактерии по п. 367, где бактерии способны к фиксации атмосферного азота в нелимитирующих азот условиях.
400. Выделенные бактерии по п. 367, где бактерии, in planta, экскретирует азот-содержащие продукты фиксации азота.
401. Выделенные бактерии по п. 367, где бактерии, in planta, обеспечивают растению по меньшей мере 1% фиксированного азота, воздействующего на указанные бактерии, и где указанный фиксированный азот измеряют посредством разведения обогащенного удобрения атмосферным газообразным N2 в ткани растения.
402. Выделенные бактерии по п. 367, где бактерии колонизируют корень растения, воздействующего на указанные бактерии в концентрации по меньшей мере приблизительно 1,0 × 104 бактериальных клеток на грамм сырой массы ткани корня растения.
403. Выделенные бактерии по п. 367, формулируемые в сельскохозяйственную композицию.
404. Выделенные бактерии по п. 367, формулируемые во вносимую в борозду композицию.
405. Выделенные бактерии по п. 367, формулируемые в виде жидкой композиции для внесения в борозду, которая содержит бактерии в концентрации приблизительно от 1 × 107 до приблизительно 1 × 1010 КОЕ в миллилитре.
406. Выделенные бактерии по п. 367, формулируемые в виде пропитки для семян или покрытия для семян.
407. Выделенные бактерии по п. 367, формулируемые в виде пропитки для семян или покрытия для семян, которые содержит бактерии в концентрации приблизительно от 1 × 105 до приблизительно 1 × 107 КОЕ на семя.
408. Выделенные бактерии по п. 367, где бактерии контактируют с растением и увеличивают фиксацию азота в растении.
409. Выделенные бактерии по п. 367, наносимые на небобовое растение.
410. Выделенные бактерии по п. 367, наносимые на злаковую сельскохозяйственную культуру.
411. Выделенные бактерии по п. 367, наносимые на растение, выбранное из группы, состоящей из: кукурузы, риса, пшеницы, ячменя, сорго, просо, овса, ржи и тритикале.
412. Выделенные бактерии по п. 367, наносимые на кукурузу.
413. Выделенные бактерии по п. 367, наносимые на бобовое растение.
414. Выделенные бактерии по п. 367, наносимые на зерновую сельскохозяйственную культуру.
415. Выделенные бактерии по п. 367, выбранные из: Rahnella aquatilis, Klebsiella variicola, Achromobacter spiritinus, Achromobacter marplatensis, Microbacterium murale, Kluyvera intermedia, Kosakonia pseudosacchari, вида Enterobacter, Azospirillum lipoferum, Kosakonia sacchari и их сочетаний.
416. Выделенные бактерии по п. 367, где бактерии принадлежат роду Rahnella.
417. Выделенные бактерии по п. 367, где бактерии принадлежат виду Rahnella aquatilis.
418. Выделенные бактерии по п. 367, депонированы как PTA-122293.
419. Выделенные бактерии по п. 367, депонированы как NCMA 201701003.
420. Выделенные бактерии по п. 367, где бактерии принадлежат роду Kosakonia.
421. Выделенные бактерии по п. 367, где бактерии принадлежат виду Kosakonia sacchari.
422. Выделенные бактерии по п. 367, депонированы как PTA-122294.
423. Выделенные бактерии по п. 367, депонированы как NCMA 201701001.
424. Выделенные бактерии по п. 367, депонированы как NCMA 201701002.
425. Выделенные бактерии по п. 367, депонированы как NCMA 201708004.
426. Выделенные бактерии по п. 367, депонированы как NCMA 201708003.
427. Выделенные бактерии по п. 367, депонированы как NCMA 201708002.
428. Выделенные бактерии по п. 367, где бактерии принадлежат роду Klebsiella.
429. Выделенные бактерии по п. 367, где бактерии принадлежат виду Klebsiella variicola.
430. Выделенные бактерии по п. 367, депонированы как NCMA 201708001.
431. Выделенные бактерии по п. 367, депонированы как NCMA 201712001.
432. Выделенные бактерии по п. 367, депонированы как NCMA 201712002.
433. Композиция, содержащая любую одну или несколько бактерий по пп. 415-432.
434. Способ детекции последовательности с неприродным соединением, включающий: амплификацию нуклеотидной последовательности, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности с SEQ ID NO: 372-405.
435. Способ по п. 434, где указанную амплификацию проводят посредством полимеразной цепной реакции.
436. Способ по п. 434, где указанную амплификацию проводят посредством количественной полимеразной цепной реакции.
437. Способ детекции последовательности с неприродным соединением, включающий: амплификацию нуклеотидной последовательности, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности фрагмента по меньшей мере с 10 последовательными парами оснований, содержащегося в SEQ ID NO: 372-405, где указанный фрагмент с последовательными парами оснований состоит из нуклеотидов на пересечении: расположенной выше последовательности, содержащей SEQ ID NO: 304-337, и расположенной ниже последовательности, содержащей SEQ ID NO: 338-371.
438. Способ по п. 437, где указанную амплификацию проводят посредством полимеразной цепной реакции.
439. Способ по п. 437, где указанную амплификацию проводят посредством количественной полимеразной цепной реакции.
440. Последовательность с неприродным соединением, содержащая: нуклеотидную последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности с SEQ ID NO: 372-405.
441. Последовательность с неприродным соединением, содержащая: нуклеотидную последовательность, которая по меньшей мере приблизительно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности фрагмента по меньшей мере с 10 последовательными парами оснований, содержащегося в SEQ ID NO: 372-405, где указанный фрагмент с последовательными парами оснований состоит из нуклеотидов на пересечении: расположенной выше последовательности, содержащей SEQ ID NO: 304-337, и расположенной ниже последовательности, содержащей SEQ ID NO: 338-371.
442. Бактериальная композиция, содержащая:
по меньшей мере один реконструированный штамм бактерий который фиксирует атмосферный азот, по меньшей мере один реконструированный штамм бактерий, содержащий экзогенно добавленную ДНК, где указанная экзогенно добавленная ДНК, по меньшей мере на 80% идентична с соответствующим нативным штаммом бактерий.
443. Способ поддержания уровней азота в почве, включающий:
высаживание в почву поля сельскохозяйственной культуры, инокулированной реконструированными бактериями, которые фиксируют атмосферный азот; и
сбор урожая указанной сельскохозяйственной культуры, где между посадкой и сбором урожая в указанную почву указанного поля вносят не более 90% дозы азота, необходимой для получения указанной сельскохозяйственной культуры.
444. Способ доставки пробиотической добавки в сельскохозяйственную культуру, включающий:
покрытие семян сельскохозяйственной культуры покрытием для семян, пропиткой для семян или протравкой для семян, где указанные покрытие для семян, протравка для семян или пропитка для семян содержат живых представителей указанного пробиотика; и
внесение указанных семян сельскохозяйственной культуры в почву поля.
445. Способ увеличения у небобового растения количества получаемого из атмосферы азота, включающий:
воздействие на указанное небобовое растение реконструированными не являющимися межродовыми микроорганизмами, где указанные реконструированные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, или по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
446. Способ увеличения у растения кукурузы количества получаемого из атмосферы азота, включающий:
воздействие на указанное растение кукурузы реконструированными не являющимися межродовыми микроорганизмами, несущими реконструированные генетические вариации по меньшей мере в двух генах, выбранных из группы, состоящей из nifL, glnB, glnE и amtB.
447. Способ увеличения у растения кукурузы количества получаемого из атмосферы азота, включающий:
воздействие на указанное растение кукурузы реконструированными не являющимися межродовыми микроорганизмами, несущими по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота, где указанные реконструированные не являющиеся межродовыми микроорганизмы, in planta, продуцируют у указанного растения кукурузы по меньшей мере 5% фиксированного азота, как измеряют посредством разведения 15N в сельскохозяйственных культурах, выращиваемых в полях, обрабатываемых удобрением, содержащим 1,2% 15N.
448. Способ по любому из предыдущих пп., где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час в присутствии экзогенного азота.
449. Бактерия по любому из предыдущих пп., где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час в присутствии экзогенного азота.
450. Популяция бактерий по любому из предыдущих пп., где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час в присутствии экзогенного азота.
451. Выделенные бактерии по любому из предыдущих пп., где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час в присутствии экзогенного азота.
452. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час в присутствии экзогенного азота.
453. Не являющаяся межродовой бактерия по любому из предыдущих пп., где бактерия продуцирует фиксированный N в количестве по меньшей мере приблизительно 1 × 10-17 ммоль N на бактериальную клетку в час в присутствии экзогенного азота.
454. Способ по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
455. Способ по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
456. Бактерия по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
457. Популяция бактерий по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
458. Выделенные бактерии по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
459. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
460. Не являющаяся межродовой бактерия по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 5% фиксированного азота или более.
461. Способ по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
462. Способ по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
463. Бактерия по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
464. Популяция бактерий по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
465. Выделенные бактерии по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
466. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
467. Не являющаяся межродовой бактерия по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 10% фиксированного азота или более.
468. Способ по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
469. Способ по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
470. Бактерия по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
471. Популяция бактерий по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
472. Выделенные бактерии по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
473. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
474. Не являющаяся межродовой бактерия по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 15% фиксированного азота или более.
475. Способ по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
476. Способ по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
477. Бактерия по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
478. Популяция бактерий по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
479. Выделенные бактерии по любому из предыдущих пп., где множество бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
480. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
481. Не являющаяся межродовой бактерия по любому из предыдущих пп., где множество не являющихся межродовыми бактерий, in planta, продуцирует в растении 20% фиксированного азота или более.
482. Способ по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-7 ммоль N на грамм сырой массы ткани корня растения в час.
483. Бактерия по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-7 ммоль N на грамм сырой массы ткани корня растения в час.
484. Популяция бактерий по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-7 ммоль N на грамм сырой массы ткани корня растения в час.
485. Выделенные бактерии по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-7 ммоль N на грамм сырой массы ткани корня растения в час.
486. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-7 ммоль N на грамм сырой массы ткани корня растения в час.
487. Не являющаяся межродовой бактерия по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-7 ммоль N на грамм сырой массы ткани корня растения в час.
488. Способ по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-6 ммоль N на грамм сырой массы ткани корня растения в час.
489. Бактерия по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-6 ммоль N на грамм сырой массы ткани корня растения в час.
490. Популяция бактерий по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-6 ммоль N на грамм сырой массы ткани корня растения в час.
491. Выделенные бактерии по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-6 ммоль N на грамм сырой массы ткани корня растения в час.
492. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-6 ммоль N на грамм сырой массы ткани корня растения в час.
493. Не являющаяся межродовой бактерия по любому из предыдущих пп., где произведение (i) средней колонизирующей способности на единицу ткани корня растения и (ii) продуцируемого фиксированного N на бактериальную клетку в час составляет по меньшей мере приблизительно 2,0 × 10-6 ммоль N на грамм сырой массы ткани корня растения в час.
494. Способ по любому из предыдущих пп., где растение реконструировано или выведено для эффективного использования азота.
495. Бактериальная композиция по любому из предыдущих пп., где указанный по меньшей мере один реконструированный штамм бактерий содержит по меньшей мере одну вариацию в гене или межгенной области в пределах 10000 п.н. от гена, выбранного из группы, состоящей из: nifA, nifL, ntrB, ntrC, glnA, glnB, glnK, draT, amtB, glnD, glnE, nifJ, nifH, nifD, nifK, nifY, nifE, nifN, nifU, nifS, nifV, nifW, nifZ, nifM, nifF, nifB и nifQ.
496. Способ по любому из предыдущих пп., где указанная реконструированная бактерия содержит бактериальную композицию по любому из предшествующих пп.
497. Способ по любому из предыдущих пп., где указанная реконструированная бактерия состоит из бактериальной композиции по любому из предшествующих пп.
498. Способ по любому из предыдущих пп., где реконструированный штамм бактерий представляет собой реконструированный грамположительный штамм бактерий.
499. Способ по любому из предыдущих пп., где реконструированный грамположительный штамм бактерий обладает измененным уровнем экспрессии регулятора кластера Nif.
500. Способ по любому из предыдущих пп., где реконструированный грамположительный штамм бактерий экспрессирует сниженное количество отрицательного регулятора кластера Nif.
501. Способ по любому из предыдущих пп., где реконструированный штамм бактерий экспрессирует сниженное количество GlnR.
502. Способ по любому из предыдущих пп., где геном реконструированного штамма бактерий кодирует полипептид по меньшей мере на 75% идентичный с последовательностью из базы данных NifH лаборатории Дж. Зера.
503. Способ по любому из предыдущих пп., где геном реконструированного штамма бактерий кодирует полипептид по меньшей мере на 85% идентичный с последовательностью из базы данных NifH лаборатории Дж. Зера.
504. Способ по любому из предыдущих пп., где геном реконструированного штамма бактерий кодирует полипептид по меньшей мере на 75% идентичный с последовательностью из базы данных NifH лаборатории Д. Бакли.
505. Способ по любому из предыдущих пп., где геном реконструированного штамма бактерий кодирует полипептид по меньшей мере на 85% идентичный с последовательностью из базы данных NifH лаборатории Д. Бакли.
506. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть фиксации азота.
507. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть ассимиляции азота.
508. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в указанную генетическую регуляторную сеть ассимиляции азота.
509. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы вносят в борозды, в которые высаживают семена указанного небобового растения.
510. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы наносят на семена указанного небобового растения.
511. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы колонизируют по меньшей мере корень указанного небобового растения так, что указанные реконструированные не являющиеся межродовыми микроорганизмы присутствует в указанном небобовом растении в количестве по меньшей мере 105 колониеобразующих единиц на грамм сырой массы ткани.
512. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы способны к фиксации атмосферного азота в нелимитирующих азот условиях.
513. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы, in planta, экскретирует азот-содержащие продукты фиксации азота.
514. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы, in planta, продуцируют в указанном небобовом растении по меньшей мере 1% фиксированного азота.
515. Способ по любому из предыдущих пп., где указанный фиксированный азот в указанном небобовом растении, продуцируемый указанными реконструированными не являющимися межродовыми микроорганизмы определяют посредством разведения 15N в сельскохозяйственных культурах, выращиваемых в полях, обрабатываемых удобрением, содержащим 1,2% 15N.
516. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы, in planta, продуцируют в указанном небобовом растении 5% фиксированного азота или более.
517. Способ по любому из предыдущих пп., где указанные не являющиеся межродовыми микроорганизмы реконструированы с использованием по меньшей мере инженерии одного типа, выбранной из группы, состоящей из направленного мутагенеза, случайного мутагенеза и направленной эволюции.
518. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы, in planta, экскретирует азот-содержащие продукты фиксации азота.
519. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы вносят в борозды, в которые высажены семена указанного растения кукурузы.
520. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы наносят на семена указанного растения кукурузы.
521. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
522. Способ по любому из предыдущих пп., где указанные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
523. Способ по любому из предыдущих пп., где указанные генно-инженерные не являющиеся межродовыми микроорганизмы несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
524. Способ по любому из предыдущих пп., где указанные реконструированные не являющиеся межродовыми бактерии несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
525. Способ по любому из предыдущих пп., где указанные не являющиеся межродовыми бактерии несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
526. Способ по любому из предыдущих пп., где указанные генно-инженерные не являющиеся межродовыми бактерии несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
527. Бактерия по любому из предыдущих пп., где указанная бактерия несет по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
528. Популяция бактерий по любому из предыдущих пп., где бактерии в указанной популяции бактерий несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
529. Выделенные бактерии по любому из предыдущих пп., где указанные выделенные бактерии несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
530. Не являющаяся межродовой популяция бактерий по любому из предыдущих пп., где не являющиеся межродовыми бактерии в указанной не являющейся межродовой популяции бактерий несут по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
531. Не являющаяся межродовой бактерия по любому из предыдущих пп., где указанная не являющаяся межродовой бактерия несет по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть фиксации азота, и по меньшей мере одну генетическую вариацию, внесенную в генетическую регуляторную сеть ассимиляции азота.
532. Композиция, содержащая любую одну или несколько бактерий по любому из предыдущих пп.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> PIVOT BIO, INC.
<120> СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УЛУЧШЕНИЯ ПРИЗНАКОВ РАСТЕНИЙ
<130> 47736-707.601
<140>
<141>
<150> 62/577,147
<151> 2017-10-25
<150> 62/566,199
<151> 2017-09-29
<150> 62/467,032
<151> 2017-03-03
<150> 62/447,889
<151> 2017-01-18
<150> 62/445,557
<151> 2017-01-12
<150> 62/445,570
<151> 2017-01-12
<160> 424
<170> PatentIn version 3.5
<210> 1
<211> 9
<212> Белок
<213> Неизвестно
<220>
<223> Описание неизвестного:
последовательность пептидного мотива семейства "LAGLIDADG"
<400> 1
Leu Ala Gly Leu Ile Asp Ala Asp Gly
1 5
<210> 2
<211> 90
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический олигонуклеотид
<400> 2
gttgatcaga ccgatgttcg gaccttccaa ggtttcgatc ggacatacgc gaccgtagtg 60
ggtcgggtgt acgtctcgaa cttcaaagcc 90
<210> 3
<211> 257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 3
gcctctcggg gcgctttttt ttattccggc actagccgct attaataaaa atgcaaatcg 60
gaatttacta tttaacgcga gattatctaa gatgaatccg atggaagcgc gctgttttca 120
ctcgcctttt taaagttacg tgatgatttc gatgcttctt tgagcgaacg atcaaaaata 180
agcgtattca ggtaaaaaaa tattctcatc acaaaaaagt ttgtgtaata cttgtaacgc 240
tacatggaga ttaactc 257
<210> 4
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 4
ggttcacata aacataatta tcgccacggc gatagccgta cgctttttgc gtcacaacat 60
ccatggtgaa gccggctttt tcaagaacac gcgccacctc atcgggtctt aaatacatac 120
tcattcctca ttatctttta ccgcacgtta accttacctt attcattaaa ggcaacgctt 180
tcggaatatt ccataaaggg ctatttacag cataattcaa aatcttgtcc tacacttata 240
gactcaatgg aattaaggga 260
<210> 5
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 5
gcgcggaaaa tcgacgcata gcgcattctc agaagccggc ctggtctcgg tggaaaagcg 60
aatctttccc acgaccgccg ggcctttaac aaaagaatca atgacctgat taatgtcgct 120
atccattctc tctccgcgta atgcgatctt ttttcatcat acctaacaaa ctggcagagg 180
gaaaagccgc gcggtttttc tgcgaagtgt attgtaagat ttgtttgata tgttatatcg 240
taacatatta ttgcaaacat 260
<210> 6
<211> 259
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 6
ctgacgaagc gagttacatc accggtgaaa ctctgcacgt caacggcgga atgtatatgg 60
tctgaccgag atttgcgcaa aacgctcagg aaccgcgcag tctgtgcggt tcactgtaat 120
gttttgtaca aaatgatttg cgttatgagg gcaaacagcc gcaaaatagc gtaaaatcgt 180
ggtaagacct gccgggattt agttgcaaat ttttcaacat tttatacact acgaaaacca 240
tcgcgaaagc gagttttga 259
<210> 7
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 7
acgcctgggg cgccgaccag cgggaagagt gatttggcca acgaggcgcc gctctgaatg 60
gaaatcatgg cgattaaaat aaccagtatc ggcaaccatg ccggtacctt acgagacgag 120
ccgggcatcc tttctcctgt caattttgtc aaatgcggta aaggttccag tgtaattgaa 180
ttaccccgcg ccggttgagc taatgttgaa aaaaagggtc ttaaaagcag tacaataggg 240
cgggtctgaa gataatttca 260
<210> 8
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 8
tctgattcct gatgaaaata aacgcgacct tgaagaaatt ccggataacg ttatcgccga 60
tttagatatc catccggtga aacgaatcga ggaagttctg gcacttgcgc tacagaacga 120
accgtttgga atggaagtcg tcacggcaaa atagtgattt cgcgcaaata gcgctaagaa 180
aaatagggct ggtaagtaaa ttcgtacttg ccagcctttt tttgtgtagc taacttagat 240
cgctggcagg ggggtcaatt 260
<210> 9
<211> 259
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 9
gtaagaaagt cggcctgcgt aaagcacgtc gtcgtcctca gttctccaaa cgttaattgt 60
tttctgctca cgcagaacaa tttgcgaaaa aacccgcttc ggcgggtttt tttatggata 120
aatttgccat tttccctcta caaacgcccc attgttacca ctttttcagc atttccagaa 180
tcccctcacc acaacgtctt caaaatctgg taaactatca tccaattttc tgcccaaatg 240
caggtgattg ttcattttt 259
<210> 10
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 10
gtcaaagccg tattatcgac cccttaggga caacgcttgc cggggcggga gagcggccgc 60
agttgatttt tgccgaactt tcagctgatt atattcagca ggtacgcgag cgcctgccgg 120
tgttgcgcaa tcgccgcttt gcgccaccgc aattattatg acgttttttt aaacaaggct 180
tgattcacct tgttacagat tgctattgtg tccgcgcgtc aaatagccgt taattgtatg 240
cgtgtatgat ggcgtattcg 260
<210> 11
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 11
gaggcggtgg ttgaccgtat cggtcccgag catcatgagc tttcggggcg agcgaaagat 60
atgggatcgg cggcggtact gctggcgatt atcatcgcgc tgatcgcgtg gggaacgctg 120
ctgtgggcga actaccgcta agtcttgtcg tagctgctcg caaaacggaa agaaactcct 180
gatttttgtg tgaaatgtgg ttccaaaatc accgttagct gtatatactc acagcataac 240
tgtatataca cccagggggc 260
<210> 12
<211> 260
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 12
taagaaaagc ggcctgtacg aagacggcgt acgtaaagac aggctggata acgacgatat 60
gatcgatcag ctggaagcgc gtattcgcgc taaagcatcg atgctggatg aggcgcgtcg 120
tatcgatatc cagcaggttg aagcgaaata acgtgttggg aagcgatacg cttcccgtgt 180
atgattgaac ctgcgggcgc gaggcgccgg ggttcatttt tgtatatata aagagaataa 240
acgtggcaaa gaacattcaa 260
<210> 13
<211> 237
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 13
atgaatcgta ctaaactggt actgggcgcg gtaatcctgg gttctactct gctggctggt 60
tgctccagca atgctaaaat cgatcagctg tcttctgacg ttcagactct gaacgctaaa 120
gttgaccagc tgagcaacga cgtgaacgca atgcgttccg acgttcaggc tgctaaagat 180
gacgcagctc gcgctaacca gcgtctggac aacgcagcta ctaaataccg taagtaa 237
<210> 14
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 14
atggccaacc gagcaaaccg caacaacgta gaagagagcg ctgaagatat ccataacgat 60
gtcagccaat tagcggatac gctggaagag gtgctgaaat cgtggggcag cgacgccaaa 120
gacgaagcgg aggccgcgcg caaaaaagcg caggcgctgc tgaaagagac ccgcgcccgg 180
cttaacggca acaaccgcgt ccagcaggcg gcgtgcgacg ccatgggctg cgctgacagc 240
tacgtgcgcg acaaaccgtg gcaaagcgtc ggcgccgcag cagccgttgg ggtatttatt 300
ggcgtattac tgaatttacg tcgataa 327
<210> 15
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 15
atgaccaaaa agatttccgc cctagcgttt ggcattggca tggtaatggc gagcagccag 60
gcttttgccc acggtcacca tagtcatggc ccggcgctga ccgaagcgga acaaaaggcg 120
agtgaaggca tttttgctga ccaggacgta aaggacaggg cgctgagcga ctgggagggg 180
atctggcagt cggttaaccc ctatctgctg aacggggatt tagatccggt tctggagcag 240
aaggccaaaa aggccggtaa aagcgtggcg gaatatcggg aatattataa gaagggctac 300
gctaccgatg tcgaccagat tggtatcgag gataacgtca tggagtttca cgtcgggaaa 360
accgtcaacg cctgtaagta cagctattcc ggttacaaaa ttctgaccta cgcatccggt 420
aaaaaaggcg tgcgctacct gttcgaatgc cagcaggcgg attcaaaagc gccgaagttt 480
gttcagttta gcgatcacac catcgcgcca cgcaagtccc agcatttcca catctttatg 540
ggcaatgagt cccaggaagc gctgctgaaa gagatggata actggccaac ctactatcct 600
tatgcgctgc ataaagagca gattgtcgac gaaatgctgc accactaa 648
<210> 16
<211> 237
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 16
atgagcacta tcgaagaacg cgttaagaaa attatcggcg aacagctggg cgttaagcag 60
gaagaagtta ccaacaatgc ttccttcgtt gaagacctgg gcgctgattc tcttgacacc 120
gttgagctgg taatggctct ggaagaagag tttgatactg agattccgga cgaagaagct 180
gagaaaatca ctactgttca ggctgccatt gattacatca acggccacca ggcgtaa 237
<210> 17
<211> 513
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 17
atgaataaaa ttgcacgttt ttcagcactg gccgttgttc tggctgcatc cgtaggtacc 60
actgctttcg ctgcgacttc taccgttacc ggtggctacg cgcagagcga catgcagggt 120
gaagcgaaca aagctggcgg tttcaacctg aagtaccgct acgagcaaga caacaacccg 180
ctgggtgtta tcggttcttt cacctacacc gaaaaagatc gttctgaatc tggcgtttac 240
aaaaaaggcc agtactacgg catcaccgca ggtccggctt accgtctgaa cgactgggct 300
agcatctacg gcgtagtggg tgttggttac ggtaaattcc aggacaacag ctacccgaac 360
aaatctgata tgagcgacta cggtttctct tacggcgctg gtctgcagtt caacccgatc 420
gaaaacgttg ccctggactt ctcctacgag cagtctcgca ttcgtaacgt tgacgttggc 480
acctggattg ctggcgtagg ttaccgcttc taa 513
<210> 18
<211> 273
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 18
gtgaataaat ctcaactgat tgacaaaatt gctgccggtg cggacatttc taaagccgca 60
gctggacgtg cgttagatgc tttaatcgct tctgttactg aatctctgca ggctggagat 120
gacgttgcgc tggtagggtt tggtactttt gctgttaaag agcgcgctgc ccgtactggt 180
cgcaatccgc aaacaggcaa agaaatcacc attgctgctg ctaaagttcc gggtttccgc 240
gcaggtaaag cgctgaaaga cgcggtaaac tga 273
<210> 19
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 19
atggctgtcg ctgccaacaa acgttcggta atgacgctgt tttctggtcc tactgacatc 60
tatagccatc aggtccgcat cgtgctggcc gaaaaaggtg ttagttttga gatagagcac 120
gtggagaagg acaacccgcc tcaggatctg attgacctca acccgaatca aagcgtaccg 180
acgcttgtgg atcgtgagct cactctgtgg gaatctcgca tcattatgga atatctggat 240
gagcgtttcc cgcatccgcc gctcatgccg gtttacccgg tggcgcgtgg ggaaagccgt 300
ctgtatatgc agcgtatcga aaaggactgg tattcgttga tgaataccat tcagaccggt 360
accgctgcgc aggctgatac tgcgcgtaag cagctgcgtg aagaactaca ggcgattgcg 420
ccagttttca cccagaagcc ctacttcctg agcgatgagt tcagcctggt ggactgctac 480
ctggcaccac tgctgtggcg tctgccggtt ctcggcgtag agctggtcgg cgctggcgcg 540
aaagagctta aaggctatat gactcgcgta tttgagcgcg actctttcct cgcttcttta 600
actgaagccg aacgtgaaat gcgtctcggt cggggctaa 639
<210> 20
<211> 204
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 20
atgggtgaga ttagtattac caaactgctg gtagtcgcag cgctgattat cctggtgttt 60
ggtaccaaaa agttacgcac gctgggtgga gacctgggct cggctatcaa aggctttaaa 120
aaagccatga gcgatgacga tgacagtgcg aagaagacca gtgctgaaga agcgccggca 180
cagaagctct ctcataaaga gtaa 204
<210> 21
<211> 609
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 21
atgaaagcgt taacgaccag gcagcaagag gtgtttgatc tcattcggga tcatatcagc 60
cagacgggca tgccgccgac gcgtgcggag attgctcagc gcttggggtt tcgctcccca 120
aacgcggcgg aagagcatct gaaagcgctg gcgcgtaaag gcgcaatcga gatcgtttcc 180
ggcgcctccc gcggtattcg tctgctgacg gaagaagaaa ccggtctgcc gcttattggc 240
cgcgtcgcgg caggtgagcc gctgctagcg cagcagcaca ttgaaggcca ctaccaggtg 300
gacccggcca tgtttaagcc gaacgccgat tttctgctgc gtgttagcgg tatgtcgatg 360
aaggatatcg gtattctcga tggcgacctg ctggctgtcc ataaaacgca ggatgtgcgc 420
aatggtcagg tggttgtggc gcgtatcgac gaagaagtga ccgtgaagcg tctgaaaaaa 480
cagggtaacg tcgtggaatt gctgccggaa aacagcgaat tctcgccgat cgtggtcgac 540
cttcgcgaac aaagctttac tattgaaggc ctggccgtcg gcgttatccg caacggcaac 600
tggcaataa 609
<210> 22
<211> 1245
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 22
atgaacgatt atctgccggg cgaaaccgct ctctggcagc gcattgaagg ctcactgaag 60
caggtgcttg gtagctacgg ttacagcgaa atccgtttgc cgattgtaga gcagaccccg 120
ttattcaaac gcgctatcgg cgaagtgacc gacgtggttg aaaaagagat gtacaccttt 180
gaggaccgta acggcgatag cctgactcta cgtccggaag gcacggctgg ctgcgtacgc 240
gccggtatcg aacatggtct cctgtacaat caagaacagc gcctgtggta cattgggccg 300
atgttccgcc acgaacgtcc gcaaaaaggc cgctaccgtc agttccacca gattggcgcc 360
gaagcgtttg gcctgcaggg gccggatatc gatgccgagc tgattatgct gaccgcccgc 420
tggtggcgcg agctgggcat ctccggccac gttgcgctgg agctgaactc tatcggttcg 480
ctggaggctc gcgctaacta tcgcgacgcg ctggtggcct atcttgagca gtttaaagat 540
aagctggacg aagactgcaa acgccgcatg tacaccaacc cgctgcgcgt gctggattct 600
aaaaacccgg acgtccaggc gctgctgaac gacgccccga cgctgggcga ctatcttgat 660
gaagagtcca aaacgcattt tgccgggctg tgcgcgctgc tggatgatgc cggtattcgc 720
tataccgtga atcagcgtct ggtacgcggt ctcgactact acaaccgcac cgtgtttgag 780
tgggtcacca ccagcctcgg ttcccagggc accgtctgcg ccggaggccg ttacgatggt 840
ctggttgagc agcttggcgg tcgcgctacc cctggcgtcg gctttgcgat ggggctggaa 900
cgtcttgttt tactggttca ggcagtgaat ccggaattta aagccgatcc tgttgtcgat 960
atatacctgg tagcctccgg aactgacacc cagtccgcag caatgcgtct ggctgaacag 1020
gtacgcgatg cgttacccgg cgttaagctg atgaccaacc atggcggcgg caactttaag 1080
aagcagtttg cgcgcgctga taaatggggc gctcgcgttg cgctggtgct gggcgaatca 1140
gaaatcgccg acggaaacgt ggtagtgaaa gatttacgct caggtgagca aactaccgta 1200
acgcaggata gcgttgctgc gcatttgcgc acacttctgg gttaa 1245
<210> 23
<211> 1413
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 23
atgaaaaaga ccaaaattgt ttgcaccatc ggtccgaaaa ccgaatccga agagatgttg 60
accaaaatgc tggacgcggg catgaacgtt atgcgtctga acttctctca cggtgactat 120
gcggaacacg gtcagcgcat ccagaatctg cgcaatgtga tgagtaaaac cggtaagaaa 180
gcggcaatcc tgctggacac caaaggtccg gaaatccgta ccattaagct ggaaggcggc 240
aacgacgtct ccctgaaagc gggccagacc ttcaccttca ccaccgataa atccgttgtc 300
ggtaataacg aaatcgttgc ggtgacctat gaaggcttca ccagcgacct gagcgttggc 360
aacacggtac tggttgacga tggtctgatc ggtatggaag tgaccgctat cgaaggcaac 420
aaagttgttt gtaaagtgct gaacaacggc gacctcggcg agaacaaagg cgttaacctg 480
ccgggcgtat ctatcgcgct gccggcgctg gctgaaaaag acaaacagga tctgatcttc 540
ggttgcgaac agggcgttga ctttgttgcg gcatccttta tccgtaagcg ttctgacgtt 600
gttgaaatcc gtgagcacct gaaagcccac ggcggcgaga agatccagat catctccaaa 660
atcgaaaacc aggaaggcct gaacaacttc gacgaaatcc tcgaagcctc tgacggcatc 720
atggtagccc gtggcgacct gggcgttgaa atcccggttg aagaagttat cttcgcgcag 780
aagatgatga tcgagaaatg tatccgcgcg cgtaaagtcg ttatcaccgc gacccagatg 840
ctggattcca tgatcaaaaa cccgcgtccg acccgtgcgg aagcaggcga cgtggccaac 900
gccatcctcg acggcaccga cgcagttatg ctgtccggcg aatccgcgaa aggtaaatac 960
ccgctggaag cggtcaccat catggcgacc atctgcgaac gtaccgaccg cgtcatgacc 1020
agccgtcttg agtacaacaa cgacaaccgt aagctgcgca tcaccgaagc ggtgtgccgc 1080
ggtgcggtag aaacggctga aaaactggaa gcgccgctga tcgttgtggc aacccagggc 1140
ggtaaatccg cgcgcgccgt acgtaaatac ttcccggatg ccactatcct ggcgctgacc 1200
accaacgaaa ccaccgcgcg tcagctggtg ctgagcaaag gcgttgtggc acagctggtt 1260
gaagatatct cctctaccga tgcgttctac atccagggta aagaactggc gctgcagagc 1320
ggtctggcgc gtaaaggcga cgtggttgtt atggtttccg gcgcgttagt cccgagcgga 1380
accaccaata ccgcttccgt gcacgtgctg taa 1413
<210> 24
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 24
atgtatttaa gacccgatga ggtggcgcgt gttcttgaaa aagccggctt caccatggat 60
gttgtgacgc aaaaagcgta cggctatcgc cgtggcgata attatgttta tgtgaaccgt 120
gaagctcgta tggggcgtac cgcgttaatt attcatccgg ctttaaaaga gcgcagcaca 180
acgcttgcgg agcccgcgtc ggatatcaaa acctgcgatc attatgagca gttcccgctc 240
tatttagcgg gggatgctca acagcattat ggtattccac acgggttcag ttcgcgaatg 300
gcgcttgagc gttttctgag tggcctgttt ggcgaaacgc agtatagctg a 351
<210> 25
<211> 864
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 25
atggatagcg acattaatca ggtcattgat tcttttgtta aaggcccggc ggtcgtggga 60
aagattcgct tttccaccga gaccaggccg gcttctgaga atgcgctatg cgtcgatttt 120
ccgcgcctcg aaatcatgct tgcgggtcag cttcacgatc cggcgattaa agccgatcgc 180
gcccagctca tgccgcacga tgtgctgtat attcccgctg gcggatggaa tgacccgcaa 240
tggctggcgc cctccactct gctcactatc ttatttggta aacagcagct ggaattcgtc 300
ctgcgccact gggacggcag cgcgcttaac gtgctggata aacagcaggt tccgcgccgc 360
ggtccccggg tcggctcttt tctgctgcag gcgctgaatg aaatgcagat gcagccgcgg 420
gagcagcaca cggcccgctt tattgtcacc agcctgctca gccactgtgc cgatctgctg 480
ggcagccagg tacaaacctc atcgcgcagc caggcgcttt ttgaagcgat tcgtaagcat 540
attgacgccc actttgccga cccgttaacc cgggagtcgg tggcgcaggc gttttacctc 600
tcgccaaact atctatccca cctgttccag aaatgcgggc caatgggctt taacgagtat 660
ctgaatcaca tccgcctgga gcaggccaga atgctgttaa aaggccacga tatgaaagtg 720
aaagatatcg cccacgcctg cggtttcgcc gacagcaact acttctgccg cctgtttcgc 780
aaaaacaccg aacgctcgcc gtcggagtat cgccgtcaat atcacagcca gctgacggaa 840
aaaacagccc cggcaaaaaa ctag 864
<210> 26
<211> 735
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 26
atgagttttg aaggaaaaat cgcgctggtt accggtgcaa gtcgcgggat tggccgcgca 60
atcgctgaaa cgctcgttgc ccgtggcgcg aaagttatcg ggactgcgac cagcgaaagc 120
ggcgcgcagg cgatcagcga ttatttaggt gctaacggta aaggtctgct gctgaatgtg 180
accgatcctg catctattga atctgttctg ggaaatattc gcgcagaatt tggtgaagtt 240
gatatcctgg tgaacaatgc cgggatcact cgtgataacc tgttaatgcg catgaaagat 300
gatgagtgga acgatattat cgaaaccaac ctgtcatctg ttttccgtct gtcaaaagcg 360
gtaatgcgcg ctatgatgaa aaagcgtcat ggacgtatta tcactatcgg ttctgtggtt 420
ggtaccatgg gaaatgcggg tcaggccaac tacgctgcgg cgaaagcggg tctgattggc 480
ttcagtaaat cactggctcg cgaagttgcg tcccgcggta ttactgtaaa cgttgttgct 540
ccgggcttta ttgaaacgga catgacgcgt gcgctgaccg atgagcagcg tgcgggtacg 600
ctggcggcag ttcctgcggg gcgcctcggc tctccaaatg aaatcgccag tgcggtggca 660
tttttagcct ctgacgaagc gagttacatc accggtgaaa ctctgcacgt caacggcgga 720
atgtatatgg tctga 735
<210> 27
<211> 71
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический олигонуклеотид
<400> 27
atgcccggct cgtctcgtaa ggtaccggca tggttgccga tactggttat tttaatcgcc 60
atgatttcca t 71
<210> 28
<211> 2355
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 28
atgaatcctg agcgttctga acgcattgaa atccccgtat tgccgttgcg cgatgtggtg 60
gtttatccgc acatggtcat acccctgttt gtagggcggg aaaaatctat ccgttgtctc 120
gaagcagcca tggaccatga taaaaaaatc atgctggttg cgcagaaaga agcctcgacg 180
gatgagccgg gtgtaaacga tcttttcacc gtcgggaccg tggcgtctat tttgcaaatg 240
ctgaagctac cggacggtac tgttaaagtg ctggtcgaag gtttgcagcg cgcgcgcatc 300
tctgcgctgt ctgataatgg cgaacatttt tcggcgaagg cggaatacct tgaatcgccg 360
gcgattgacg aacgcgagca ggaagtgctg gttcgtaccg ctatcagcca gtttgaaggc 420
tacatcaagc tgaacaaaaa aatccctccg gaagtgctga cgtcgctgaa tagcatcgac 480
gatccggcgc gtctggcgga taccatcgct gcgcatatgc cgctgaagct ggcggacaaa 540
cagtccgtgc tggagatgtc cgacgttaac gagcgtctgg aatatctgat ggcgatgatg 600
gagtcggaaa tcgatctgct gcaggtggag aagcgtattc gcaaccgcgt gaaaaagcag 660
atggagaaat ctcagcgcga gtactatctg aatgagcaaa tgaaagccat tcaaaaagag 720
ctcggcgaga tggacgacgc cccggacgag aacgaagcgc tgaagcgtaa gatcgacgcg 780
gcgaaaatgc cgaaagaggc aaaagagaaa accgaagcgg aactgcaaaa actgaaaatg 840
atgtccccga tgtcggcgga agcgaccgtc gttcgcggct acatcgactg gatggtgcag 900
gtaccgtgga acgctcgcag caaggttaaa aaagacctgc gtcaggctca ggagatcctc 960
gataccgatc actacggcct tgagcgcgtg aaggatcgca ttcttgagta cctcgcggtg 1020
cagagccgtg ttaacaagct caaagggccg atcctgtgcc tggttgggcc tccgggggta 1080
ggtaaaacct ctctcggcca atccatcgcc aaagcaactg gacgcaaata tgtgcgtatg 1140
gcgctgggcg gcgtgcgtga tgaagcggaa atccgcggtc accgccgtac ctatattggc 1200
tcaatgccgg gcaaactgat ccagaaaatg gctaaagtgg gcgttaaaaa cccgctgttc 1260
ttgctggatg agatcgacaa gatgtcttct gacatgcgcg gcgatccggc ctcggcgctg 1320
ctggaggtgt tggatccgga acagaacgtg gcctttaacg accactatct ggaagtggat 1380
tacgatctca gcgacgtgat gttcgttgcg acctctaact ccatgaacat cccggcgccg 1440
ctgctggatc gtatggaagt gatccgcctc tccggctata ccgaagatga gaagctaaac 1500
atcgccaaac gccatctgct gtcaaaacag attgagcgta acgcgctcaa gaaaggcgag 1560
ctgacggtgg atgacagcgc gattatcggc atcattcgct actacacccg tgaagcaggc 1620
gtgcgtggtc tggagcgtga aatctcgaaa ctgtgccgca aagcggtgaa acagctgctg 1680
ctggataagt cgctgaaaca catcgagatt aacggcgaca acctgcacga tttccttggc 1740
gtgcagcgct acgactatgg tcgtgcggat agcgaaaacc gcgtaggtca ggtgaccgga 1800
ctggcgtgga cggaagtggg cggcgatctg ctgaccattg aaaccgcctg cgttccgggt 1860
aaaggcaaac tgacctacac cggttcactg ggtgaagtca tgcaggaatc catccaggcg 1920
gcgctgacgg tggttcgttc acgtgcggat aagctgggta ttaactcaga cttttacgaa 1980
aaacgtgata ttcacgttca cgtgccggaa ggcgcgacgc cgaaggatgg tccaagcgcc 2040
ggtatcgcga tgtgcaccgc gctggtttcc tgtctgacgg gtaatccggt acgcgccgac 2100
gtggcgatga ccggtgagat taccctccgt ggccaggtat tgccgattgg tggtctgaag 2160
gaaaaactgt tggccgcgca tcgcggcggc attaagactg ttctgattcc tgatgaaaat 2220
aaacgcgacc ttgaagaaat tccggataac gttatcgccg atttagatat ccatccggtg 2280
aaacgaatcg aggaagttct ggcacttgcg ctacagaacg aaccgtttgg aatggaagtc 2340
gtcacggcaa aatag 2355
<210> 29
<211> 393
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 29
atggctgaaa atcaatacta cggcaccggt cgccgcaaaa gttccgcagc tcgcgttttc 60
atcaaaccgg gcaacggtaa aatcgttatc aaccagcgtt ctctggaaca gtacttcggt 120
cgtgaaactg cccgcatggt agttcgtcag ccgctggaac tggtcgacat ggttgagaaa 180
ttagatctgt acatcaccgt taaaggtggt ggtatctctg gtcaggctgg tgcgatccgt 240
cacggtatca cccgcgctct gatggagtac gacgagtccc tgcgtggcga actgcgtaaa 300
gctggtttcg ttactcgtga tgctcgtcag gttgaacgta agaaagtcgg cctgcgtaaa 360
gcacgtcgtc gtcctcagtt ctccaaacgt taa 393
<210> 30
<211> 789
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 30
atgtttgttg ctgccggaca atttgccgta acgccggact ggacgggaaa cgcgcagacc 60
tgcgtcagca tgatgcgcca ggccgcggag cggggggcgt cgcttctggt tctgcctgag 120
gcgttgctgg cgcgagacga taacgatgcg gatttatcgg ttaaatccgc ccagcagctg 180
gatggcggct tcttacagct cttgctggcg gagagcgaaa acagcgcttt gacgacggtg 240
ctgaccctgc atatcccttc cggcgaaggt cgagcgacga atacgctggt ggccctgcgt 300
caggggaaga ttgtggcgca atatcagaaa ctgcatctct atgatgcgtt caatatccag 360
gaatccaggc tggtcgatgc cgggcggcaa attccgccgc tgatcgaagt cgacgggatg 420
cgcgtcgggc tgatgacctg ctacgattta cgtttccctg agctggcgct gtcgttagcg 480
ctcagcggcg cgcagctcat agtgttgcct gccgcgtggg taaaagggcc gctgaaggaa 540
catcactggg cgacgctgct ggcggcgcgg gcgctggata caacctgcta tattgtcgcc 600
gcaggagagt gcgggacgcg taatatcggt caaagccgta ttatcgaccc cttagggaca 660
acgcttgccg gggcgggaga gcggccgcag ttgatttttg ccgaactttc agctgattat 720
attcagcagg tacgcgagcg cctgccggtg ttgcgcaatc gccgctttgc gccaccgcaa 780
ttattatga 789
<210> 31
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 31
atggccaata ataccactgg gttaacccga attattaaag cggccgggta ttcctggaaa 60
ggattccgtg cggcgtgggt caatgaggcc gcatttcgtc aggaaggcat cgcggccgtt 120
attgccgtgg cgatcgcctg ctggttggac gtcgatgcca tcacgcgggt gctgctcatt 180
agctcggtcc tgttagtgat gatagttgaa attatcaata gcgcgattga ggcggtggtt 240
gaccgtatcg gtcccgagca tcatgagctt tcggggcgag cgaaagatat gggatcggcg 300
gcggtactgc tggcgattat catcgcgctg atcgcgtggg gaacgctgct gtgggcgaac 360
taccgctaa 369
<210> 32
<211> 1122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 32
atgcataacc aggctccgat tcaacgtaga aaatcaaaac gaatttacgt tgggaatgtg 60
ccgattggcg atggcgcccc catcgccgta cagtcgatga caaacacgcg caccaccgat 120
gtggcggcga cggtaaatca aattaaagcc ctcgagcgcg ttggcgcgga tatcgtgcgc 180
gtttcggtgc cgacgatgga tgcggcggaa gcgttcaaac ttatcaaaca gcaggttaac 240
gtcccgctgg ttgccgatat ccacttcgat taccgcattg cgctgaaggt agcggaatac 300
ggcgttgatt gcctgcgtat taacccgggc aatatcggca acgaagagcg tatccgcatg 360
gtggtggact gcgctcgcga taaaaatatt cctatccgta tcggggtaaa cgccggttct 420
ctggaaaaag atctccagga aaaatacggc gaaccgactc cgcaggcgct gctggaatcg 480
gcaatgcgcc atgttgatca tctcgatcgt ctcaacttcg atcagtttaa agtcagcgta 540
aaagcctccg atgtgttcct cgcggttgaa tcctatcgcc tgttggcgaa acagatcgat 600
cagcctctgc acctcgggat caccgaagcg ggcggcgcgc gcagcggcgc ggtgaagtcc 660
gcgatcggcc tcggcctgct gctgtctgaa gggattggcg atacgctgcg cgtctctctg 720
gcggcggatc ccgttgaaga gatcaaagtg ggcttcgata ttctcaagtc gctgcgtatt 780
cgctctcgcg ggatcaactt tattgcctgc ccgacctgtt cacgtcagga gtttgacgtt 840
atcggtaccg ttaacgcgct ggagcagcgc ctggaagata tcattacgcc gatggatatt 900
tcgatcattg gctgcgtggt aaacggtccc ggcgaggcgc tggtttccac cctcggcgta 960
accggcggca ataagaaaag cggcctgtac gaagacggcg tacgtaaaga caggctggat 1020
aacgacgata tgatcgatca gctggaagcg cgtattcgcg ctaaagcatc gatgctggat 1080
gaggcgcgtc gtatcgatat ccagcaggtt gaagcgaaat aa 1122
<210> 33
<211> 876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 33
atgagccata ttcaacggga aacgtcttgc tccaggccgc gattaaattc caacatggat 60
gctgatttat atgggtataa atgggctcgc gataatgtcg ggcaatcagg tgcgacaatc 120
tatcgattgt atgggaagcc cgatgcgcca gagttgtttc tgaaacatgg caaaggtagc 180
gttgccaatg atgttacaga tgagatggtc agactaaact ggctgacgga atttatgcct 240
cttccgacca tcaagcattt tatccgtact cctgatgatg catggttact caccactgcg 300
atccccggga aaacagcatt ccaggtatta gaagaatatc ctgattcagg tgaaaatatt 360
gttgatgcgc tggcagtgtt cctgcgccgg ttgcattcga ttcctgtttg taattgtcct 420
tttaacagcg atcgcgtatt tcgtctcgct caggcgcaat cacgaatgaa taacggtttg 480
gttgatgcga gtgattttga tgacgagcgt aatggctggc ctgttgaaca agtctggaaa 540
gaaatgcata agcttttgcc attctcaccg gattcagtcg tcactcatgg tgatttctca 600
cttgataacc ttatttttga cgaggggaaa ttaataggtt gtattgatgt tggacgagtc 660
ggaatcgcag accgatacca ggatcttgcc atcctatgga actgcctcgg tgagttttct 720
ccttcattac agaaacggct ttttcaaaaa tatggtattg ataatcctga tatgaataaa 780
ttgcagtttc atttgatgct cgatgagttt ttctaataag cctgcctggt tctgcgtttc 840
ccgctcttta ataccctgac cggaggtgag caatga 876
<210> 34
<211> 1491
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 34
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gagcgtgttg 120
agtcatcctg actagctgag atgagggctc gccccctcgt cccgacactt ccagatcgcc 180
atagcgcaca gcgcctcgag cggtggtaac ggcgcagtgg cggttttcat ggcttgttat 240
gactgttttt ttggggtaca gtctatgcct cgggcatcca agcagcaagc gcgttacgcc 300
gtgggtcgat gtttgatgtt atggagcagc aacgatgtta cgcagcaggg cagtcgccct 360
aaaacaaagt taaacatcat gagggaagcg gtgatcgccg aagtatcgac tcaactatca 420
gaggtagttg gcgtcatcga gcgccatctc gaaccgacgt tgctggccgt acatttgtac 480
ggctccgcag tggatggcgg cctgaagcca cacagtgata ttgatttgct ggttacggtg 540
accgtaaggc ttgatgaaac aacgcggcga gctttgatca acgacctttt ggaaacttcg 600
gcttcccctg gagagagcga gattctccgc gctgtagaag tcaccattgt tgtgcacgac 660
gacatcattc cgtggcgtta tccagctaag cgcgaactgc aatttggaga atggcagcgc 720
aatgacattc ttgcaggtat cttcgagcca gccacgatcg acattgatct ggctatcttg 780
ctgacaaaag caagagaaca tagcgttgcc ttggtaggtc cagcggcgga ggaactcttt 840
gatccggttc ctgaacagga tctatttgag gcgctaaatg aaaccttaac gctatggaac 900
tcgccgcccg actgggctgg cgatgagcga aatgtagtgc ttacgttgtc ccgcatttgg 960
tacagcgcag taaccggcaa aatcgcgccg aaggatgtcg ctgccgactg ggcaatggag 1020
cgcctgccgg cccagtatca gcccgtcata cttgaagcta gacaggctta tcttggacaa 1080
gaagaagatc gcttggcctc gcgcgcagat cagttggaag aatttgtcca ctacgtgaaa 1140
ggcgagatca ccaaggtagt cggcaaataa tgtctaacaa ttcgttcaag ccgacgccgc 1200
ttcgcggcgc ggcttaactc aagcgttaga tgcactaagc acataattgc tcacagccaa 1260
actatcaggt caagtctgct tttattattt ttaagcgtgc ataataagcc ctacacaaat 1320
ggtacccgac cggtggtgaa tttaatctcg ctgacgtgta gacattccct tatccagacg 1380
ctgatcgccc atcatcgcgg ttctttagat ctctcggtcc gccctgatgg cggcaccttg 1440
ctgacgttac gcctgccggt acagcaggtt atcaccggag gcttaaaatg a 1491
<210> 35
<211> 1021
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 35
ctgatccttc aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa 60
tctctgatgt tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc 120
ttacataaac agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc 180
caggccgcga ttaaattcca acatggatgc tgatttatat gggtataaat gggctcgcga 240
taatgtcggg caatcaggtg cgacaatcta tcgattgtat gggaagcccg atgcgccaga 300
gttgtttctg aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtcag 360
actaaactgg ctgacggaat ttatgcctct tccgaccatc aagcatttta tccgtactcc 420
tgatgatgca tggttactca ccactgcgat ccccgggaaa acagcattcc aggtattaga 480
agaatatcct gattcaggtg aaaatattgt tgatgcgctg gcagtgttcc tgcgccggtt 540
gcattcgatt cctgtttgta attgtccttt taacagcgat cgcgtatttc gtctcgctca 600
ggcgcaatca cgaatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa 660
tggctggcct gttgaacaag tctggaaaga aatgcataag cttttgccat tctcaccgga 720
ttcagtcgtc actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt 780
aataggttgt attgatgttg gacgagtcgg aatcgcagac cgataccagg atcttgccat 840
cctatggaac tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata 900
tggtattgat aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt 960
ctaataagcc ttgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 1020
a 1021
<210> 36
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 36
atgaagatag caacaatgaa aacaggtctg ggagcgttgg ctcttcttcc ctgatccttc 60
aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa tctctgatgt 120
tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc ttacataaac 180
agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc ccgtccgcgc 240
ttaaactcca acatggacgc tgatttatat gggtataaat gggctcgcga taatgtcggg 300
caatcaggtg cgacaatcta tcgcttgtat gggaagcccg atgcgccaga gttgtttctg 360
aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtccg tctcaactgg 420
ctgacggagt ttatgcctct cccgaccatc aagcatttta tccgtactcc tgatgatgcg 480
tggttactca ccaccgcgat tcctgggaaa acagccttcc aggtattaga agaatatcct 540
gattcaggtg aaaatattgt tgatgcgctg gccgtgttcc tgcgccggtt acattcgatt 600
cctgtttgta attgtccttt taacagcgat cgtgtatttc gtcttgctca ggcgcaatca 660
cgcatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa tggctggcct 720
gttgaacaag tctggaaaga aatgcacaag ctcttgccat tctcaccgga ttcagtcgtc 780
actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt aataggttgt 840
attgatgttg gacgggtcgg aatcgcagac cgttaccagg accttgccat tctttggaac 900
tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata tggtattgat 960
aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt ctaataagcc 1020
tgtgaagggc tggacgtaaa cagccacggc gaaaacgcct acaacgcctg a 1071
<210> 37
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 37
atgaccctga atatgatgct cgataacgcc gtacccgagg cgattgccgg ctgatccttc 60
aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa tctctgatgt 120
tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc ttacataaac 180
agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc ccgtccgcgc 240
ttaaactcca acatggacgc tgatttatat gggtataaat gggctcgcga taatgtcggg 300
caatcaggtg cgacaatcta tcgcttgtat gggaagcccg atgcgccaga gttgtttctg 360
aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtccg tctcaactgg 420
ctgacggagt ttatgcctct cccgaccatc aagcatttta tccgtactcc tgatgatgcg 480
tggttactca ccaccgcgat tcctgggaaa acagccttcc aggtattaga agaatatcct 540
gattcaggtg aaaatattgt tgatgcgctg gccgtgttcc tgcgccggtt acattcgatt 600
cctgtttgta attgtccttt taacagcgat cgtgtatttc gtcttgctca ggcgcaatca 660
cgcatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa tggctggcct 720
gttgaacaag tctggaaaga aatgcacaag ctcttgccat tctcaccgga ttcagtcgtc 780
actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt aataggttgt 840
attgatgttg gacgggtcgg aatcgcagac cgttaccagg accttgccat tctttggaac 900
tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata tggtattgat 960
aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt ctaataagcc 1020
ttggttctgc gtttcccgct ctttaatacc ctgaccggag gtgagcaatg a 1071
<210> 38
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 38
atgaccctga atatgatgat ggatgccggc ggacatcatc gcgacaaaca atattaatac 60
cggcaaccac accggcaatt tacgagactg cgcaggcatc ctttctcccg tcaatttctg 120
tcaaataaag taaaagaggc agtctacttg aattaccccc ggctggttga gcgtttgttg 180
aaaaaaagta actgaaaaat ccgtagaata gcgccactct gatggttaat taacctattc 240
aattaagaat tatctggatg aatgtgccat taaatgcgca gcataatggt gcgttgtgcg 300
ggaaaactgc ttttttttga aagggttggt cagtagcgga aacaactcac ttcacacccc 360
gaagggggaa gttgcctgac cctacgattc ccgctatttc attcactgac cggaggttca 420
aaatga 426
<210> 39
<211> 446
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 39
atgaccctga atatgatgat ggatgccggc tcaccacggc gataaccata ggttttcggc 60
gtggccacat ccatggtgaa tcccactttt tccagcacgc gcgccacttc atcgggtctt 120
aaatacatag attttcctcg tcatctttcc aaagcctcgc caccttacat gactgagcat 180
ggaccgtgac tcagaaaatt ccacaaacga acctgaaagg cgtgattgcc gtctggcctt 240
aaaaattatg gtctaaacta aaatttacat cgaaaacgag ggaggatcct atgtttaaca 300
aaccgaatcg ccgtgacgta gatgaaggtg ttgaggatat taaccacgat gttaaccagc 360
tcgaactcac ttcacacccc gaagggggaa gttgcctgac cctacgattc ccgctatttc 420
attcactgac cggaggttca aaatga 446
<210> 40
<211> 452
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 40
atgaccctga atatgatgat ggatgccggc tgacgaggca ggttacatca ctggtgaaac 60
cctgcacgtc aatggcggaa tgtatatggt ttaaccacga tgaaaattat ttgcgttatt 120
agggcgaaag gcctcaaaat agcgtaaaat cgtggtaaga actgccggga tttagttgca 180
aatttttcaa cattttatac actacgaaaa ccatcgcgaa agcgagtttt gataggaaat 240
ttaagagtat gagcactatc gaagaacgcg ttaagaaaat tatcggcgaa cagctgggcg 300
ttaagcagga agaagttacc aacaatgctt ccttcgttga agacctgggc gctgattctc 360
ttgacaccga actcacttca caccccgaag ggggaagttg cctgacccta cgattcccgc 420
tatttcattc actgaccgga ggttcaaaat ga 452
<210> 41
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 41
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 42
<211> 463
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 42
atgaccctga atatgatgat ggatgccggc atattgacac catgacgcgc gtaatgctga 60
ttggttctgt gacgctggta atgattgtcg aaattctgaa cagtgccatc gaagccgtag 120
tagaccgtat tggtgcagaa ttccatgaac tttccgggcg ggcgaaggat atggggtcgg 180
cggcggtgct gatgtccatc ctgctggcga tgtttacctg gatcgcatta ctctggtcac 240
attttcgata acgcttccag aattcgataa cgccctggtt ttttgcttaa atttggttcc 300
aaaatcgcct ttagctgtat atactcacag cataactgta tatacaccca gggggcggga 360
tgaaagcatt aacggccagg aactcacttc acaccccgaa gggggaagtt gcctgaccct 420
acgattcccg ctatttcatt cactgaccgg aggttcaaaa tga 463
<210> 43
<211> 428
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 43
atgaccctga atatgatgat ggatgccggc atcatattgc gctccctggt tatcatttgt 60
tactaaatga aatgttataa tataacaatt ataaatacca catcgctttc aattcaccag 120
ccaaatgaga ggagcgccgt ctgacatagc cagcgctata aaacatagca ttatctatat 180
gtttatgatt aataactgat ttttgcgttt tggatttggc tgtggcatcc ttgccgctct 240
tttcgcagcg tctgcgtttt tgccctccgg tcagggcatt taagggtcag caatgagttt 300
ttacgcaatt acgattcttg ccttcggcat gtcgatggat gctttaactc acttcacacc 360
ccgaaggggg aagttgcctg accctacgat tcccgctatt tcattcactg accggaggtt 420
caaaatga 428
<210> 44
<211> 452
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 44
atgaccctga atatgatgat ggatgccggc cgcgtcaggt tgaacgtaaa aaagtcggtc 60
tgcgcaaagc acgtcgtcgt ccgcagttct ccaaacgtta attggtttct gcttcggcag 120
aacgattggc gaaaaaaccc ggtgcgaacc gggttttttt atggataaag atcgtgttat 180
ccacagcaat ccattgatta tctcttcttt ttcagcattt ccagaatccc ctcaccacaa 240
agcccgcaaa atctggtaaa ctatcatcca attttctgcc caaatggctg ggattgttca 300
ttttttgttt gccttacaac gagagtgaca gtacgcgcgg gtagttaact caacatctga 360
ccggtcgata actcacttca caccccgaag ggggaagttg cctgacccta cgattcccgc 420
tatttcattc actgaccgga ggttcaaaat ga 452
<210> 45
<211> 410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 45
atgaccctga atatgatgat ggatgccggc cctgtatgaa gatggcgtgc gcaaagatcg 60
cctggataac agcgatatga ttagccagct tgaagcccgc attcgcgcga aagcgtcaat 120
gctggacgaa gcgcgtcgta tcgatgtgca acaggtagaa aaataaggtt gctgggaagc 180
ggcaggcttc ccgtgtatga tgaacccgcc cggcgcgacc cgttgttcgt cgcggccccg 240
agggttcatt ttttgtatta ataaagagaa taaacgtggc aaaaaatatt caagccattc 300
gcggcatgaa cgattatctg cctggcgaac tcacttcaca ccccgaaggg ggaagttgcc 360
tgaccctacg attcccgcta tttcattcac tgaccggagg ttcaaaatga 410
<210> 46
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 46
atgaaaaaga ttgatgcgat tattaaacct ttcaaactgg atgacgtgcg ctgatccttc 60
aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa tctctgatgt 120
tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc ttacataaac 180
agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc ccgtccgcgc 240
ttaaactcca acatggacgc tgatttatat gggtataaat gggctcgcga taatgtcggg 300
caatcaggtg cgacaatcta tcgcttgtat gggaagcccg atgcgccaga gttgtttctg 360
aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtccg tctcaactgg 420
ctgacggagt ttatgcctct cccgaccatc aagcatttta tccgtactcc tgatgatgcg 480
tggttactca ccaccgcgat tcctgggaaa acagccttcc aggtattaga agaatatcct 540
gattcaggtg aaaatattgt tgatgcgctg gccgtgttcc tgcgccggtt acattcgatt 600
cctgtttgta attgtccttt taacagcgat cgtgtatttc gtcttgctca ggcgcaatca 660
cgcatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa tggctggcct 720
gttgaacaag tctggaaaga aatgcacaag ctcttgccat tctcaccgga ttcagtcgtc 780
actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt aataggttgt 840
attgatgttg gacgggtcgg aatcgcagac cgttaccagg accttgccat tctttggaac 900
tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata tggtattgat 960
aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt ctaataagcc 1020
tcgcgcgtga ttcgtatccg caccggcgaa gaagacgacg cggcgattta a 1071
<210> 47
<211> 1295
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 47
atgaccatga acctgatgac ggatgtcgtc tcagccaccg ggatcgccgg gttgctttca 60
cgacaacacc cgacgctgtt ttttacacta attgaacagg cccccgtggc gatcacgctg 120
acggataccg ctgcccgcat tgtctatgcc aacccgggcg tgttgagtca tcctgactag 180
ctgagatgag ggctcgcctg atccttcaac tcagcaaaag ttcgatttat tcaacaaagc 240
cacgttgtgt ctcaaaatct ctgatgttac attgcacaag ataaaaatat atcatcatga 300
acaataaaac tgtctgctta cataaacagt aatacaaggg gtgttatgag ccatattcaa 360
cgggaaacgt cttgctccag gccgcgatta aattccaaca tggatgctga tttatatggg 420
tataaatggg ctcgcgataa tgtcgggcaa tcaggtgcga caatctatcg attgtatggg 480
aagcccgatg cgccagagtt gtttctgaaa catggcaaag gtagcgttgc caatgatgtt 540
acagatgaga tggtcagact aaactggctg acggaattta tgcctcttcc gaccatcaag 600
cattttatcc gtactcctga tgatgcatgg ttactcacca ctgcgatccc cgggaaaaca 660
gcattccagg tattagaaga atatcctgat tcaggtgaaa atattgttga tgcgctggca 720
gtgttcctgc gccggttgca ttcgattcct gtttgtaatt gtccttttaa cagcgatcgc 780
gtatttcgtc tcgctcaggc gcaatcacga atgaataacg gtttggttga tgcgagtgat 840
tttgatgacg agcgtaatgg ctggcctgtt gaacaagtct ggaaagaaat gcataagctt 900
ttgccattct caccggattc agtcgtcact catggtgatt tctcacttga taaccttatt 960
tttgacgagg ggaaattaat aggttgtatt gatgttggac gagtcggaat cgcagaccga 1020
taccaggatc ttgccatcct atggaactgc ctcggtgagt tttctccttc attacagaaa 1080
cggctttttc aaaaatatgg tattgataat cctgatatga ataaattgca gtttcatttg 1140
atgctcgatg agtttttcta ataagcctga ccggtggtga atttaatctc gctgacgtgt 1200
agacattcat cgatctgcat ccacggtccg gcggcggtac ctgcctgacg ctacgtttac 1260
cgctctttta tgaactgacc ggaggcccaa gatga 1295
<210> 48
<211> 1491
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 48
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gagcgtgttg 120
agtcatcctg actagctgag atgagggctc gccccctcgt cccgacactt ccagatcgcc 180
atagcgcaca gcgcctcgag cggtggtaac ggcgcagtgg cggttttcat ggcttgttat 240
gactgttttt ttggggtaca gtctatgcct cgggcatcca agcagcaagc gcgttacgcc 300
gtgggtcgat gtttgatgtt atggagcagc aacgatgtta cgcagcaggg cagtcgccct 360
aaaacaaagt taaacatcat gagggaagcg gtgatcgccg aagtatcgac tcaactatca 420
gaggtagttg gcgtcatcga gcgccatctc gaaccgacgt tgctggccgt acatttgtac 480
ggctccgcag tggatggcgg cctgaagcca cacagtgata ttgatttgct ggttacggtg 540
accgtaaggc ttgatgaaac aacgcggcga gctttgatca acgacctttt ggaaacttcg 600
gcttcccctg gagagagcga gattctccgc gctgtagaag tcaccattgt tgtgcacgac 660
gacatcattc cgtggcgtta tccagctaag cgcgaactgc aatttggaga atggcagcgc 720
aatgacattc ttgcaggtat cttcgagcca gccacgatcg acattgatct ggctatcttg 780
ctgacaaaag caagagaaca tagcgttgcc ttggtaggtc cagcggcgga ggaactcttt 840
gatccggttc ctgaacagga tctatttgag gcgctaaatg aaaccttaac gctatggaac 900
tcgccgcccg actgggctgg cgatgagcga aatgtagtgc ttacgttgtc ccgcatttgg 960
tacagcgcag taaccggcaa aatcgcgccg aaggatgtcg ctgccgactg ggcaatggag 1020
cgcctgccgg cccagtatca gcccgtcata cttgaagcta gacaggctta tcttggacaa 1080
gaagaagatc gcttggcctc gcgcgcagat cagttggaag aatttgtcca ctacgtgaaa 1140
ggcgagatca ccaaggtagt cggcaaataa tgtctaacaa ttcgttcaag ccgacgccgc 1200
ttcgcggcgc ggcttaactc aagcgttaga tgcactaagc acataattgc tcacagccaa 1260
actatcaggt caagtctgct tttattattt ttaagcgtgc ataataagcc ctacacaaat 1320
ggtacccgac cggtggtgaa tttaatctcg ctgacgtgta gacattccct tatccagacg 1380
ctgatcgccc atcatcgcgg ttctttagat ctctcggtcc gccctgatgg cggcaccttg 1440
ctgacgttac gcctgccggt acagcaggtt atcaccggag gcttaaaatg a 1491
<210> 49
<211> 1021
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 49
ctgatccttc aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa 60
tctctgatgt tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc 120
ttacataaac agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc 180
caggccgcga ttaaattcca acatggatgc tgatttatat gggtataaat gggctcgcga 240
taatgtcggg caatcaggtg cgacaatcta tcgattgtat gggaagcccg atgcgccaga 300
gttgtttctg aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtcag 360
actaaactgg ctgacggaat ttatgcctct tccgaccatc aagcatttta tccgtactcc 420
tgatgatgca tggttactca ccactgcgat ccccgggaaa acagcattcc aggtattaga 480
agaatatcct gattcaggtg aaaatattgt tgatgcgctg gcagtgttcc tgcgccggtt 540
gcattcgatt cctgtttgta attgtccttt taacagcgat cgcgtatttc gtctcgctca 600
ggcgcaatca cgaatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa 660
tggctggcct gttgaacaag tctggaaaga aatgcataag cttttgccat tctcaccgga 720
ttcagtcgtc actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt 780
aataggttgt attgatgttg gacgagtcgg aatcgcagac cgataccagg atcttgccat 840
cctatggaac tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata 900
tggtattgat aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt 960
ctaataagcc ttgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 1020
a 1021
<210> 50
<211> 1071
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 50
atgaccctga atatgatgct cgataacgcc gtacccgagg cgattgccgg ctgatccttc 60
aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa tctctgatgt 120
tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc ttacataaac 180
agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc ccgtccgcgc 240
ttaaactcca acatggacgc tgatttatat gggtataaat gggctcgcga taatgtcggg 300
caatcaggtg cgacaatcta tcgcttgtat gggaagcccg atgcgccaga gttgtttctg 360
aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtccg tctcaactgg 420
ctgacggagt ttatgcctct cccgaccatc aagcatttta tccgtactcc tgatgatgcg 480
tggttactca ccaccgcgat tcctgggaaa acagccttcc aggtattaga agaatatcct 540
gattcaggtg aaaatattgt tgatgcgctg gccgtgttcc tgcgccggtt acattcgatt 600
cctgtttgta attgtccttt taacagcgat cgtgtatttc gtcttgctca ggcgcaatca 660
cgcatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa tggctggcct 720
gttgaacaag tctggaaaga aatgcacaag ctcttgccat tctcaccgga ttcagtcgtc 780
actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt aataggttgt 840
attgatgttg gacgggtcgg aatcgcagac cgttaccagg accttgccat tctttggaac 900
tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata tggtattgat 960
aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt ctaataagcc 1020
ttggttctgc gtttcccgct ctttaatacc ctgaccggag gtgagcaatg a 1071
<210> 51
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 51
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 52
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 52
atgaccctga atatgatgat ggatgccggc ggacatcatc gcgacaaaca atattaatac 60
cggcaaccac accggcaatt tacgagactg cgcaggcatc ctttctcccg tcaatttctg 120
tcaaataaag taaaagaggc agtctacttg aattaccccc ggctggttga gcgtttgttg 180
aaaaaaagta actgaaaaat ccgtagaata gcgccactct gatggttaat taacctattc 240
aattaagaat tatctggatg aatgtgccat taaatgcgca gcataatggt gcgttgtgcg 300
ggaaaactgc ttttttttga aagggttggt cagtagcgga aacaactcac ttcacacccc 360
gaagggggaa gttgcctgac cctacgattc ccgctatttc attcactgac cggaggttca 420
aaatga 426
<210> 53
<211> 452
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 53
atgaccctga atatgatgat ggatgccggc tgacgaggca ggttacatca ctggtgaaac 60
cctgcacgtc aatggcggaa tgtatatggt ttaaccacga tgaaaattat ttgcgttatt 120
agggcgaaag gcctcaaaat agcgtaaaat cgtggtaaga actgccggga tttagttgca 180
aatttttcaa cattttatac actacgaaaa ccatcgcgaa agcgagtttt gataggaaat 240
ttaagagtat gagcactatc gaagaacgcg ttaagaaaat tatcggcgaa cagctgggcg 300
ttaagcagga agaagttacc aacaatgctt ccttcgttga agacctgggc gctgattctc 360
ttgacaccga actcacttca caccccgaag ggggaagttg cctgacccta cgattcccgc 420
tatttcattc actgaccgga ggttcaaaat ga 452
<210> 54
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 54
atgaccctga atatgatgat ggatgccggc ggacatcatc gcgacaaaca atattaatac 60
cggcaaccac accggcaatt tacgagactg cgcaggcatc ctttctcccg tcaatttctg 120
tcaaataaag taaaagaggc agtctacttg aattaccccc ggctggttga gcgtttgttg 180
aaaaaaagta actgaaaaat ccgtagaata gcgccactct gatggttaat taacctattc 240
aattaagaat tatctggatg aatgtgccat taaatgcgca gcataatggt gcgttgtgcg 300
ggaaaactgc ttttttttga aagggttggt cagtagcgga aacaactcac ttcacacccc 360
gaagggggaa gttgcctgac cctacgattc ccgctatttc attcactgac cggaggttca 420
aaatga 426
<210> 55
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 55
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 56
<211> 1491
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 56
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gagcgtgttg 120
agtcatcctg actagctgag atgagggctc gccccctcgt cccgacactt ccagatcgcc 180
atagcgcaca gcgcctcgag cggtggtaac ggcgcagtgg cggttttcat ggcttgttat 240
gactgttttt ttggggtaca gtctatgcct cgggcatcca agcagcaagc gcgttacgcc 300
gtgggtcgat gtttgatgtt atggagcagc aacgatgtta cgcagcaggg cagtcgccct 360
aaaacaaagt taaacatcat gagggaagcg gtgatcgccg aagtatcgac tcaactatca 420
gaggtagttg gcgtcatcga gcgccatctc gaaccgacgt tgctggccgt acatttgtac 480
ggctccgcag tggatggcgg cctgaagcca cacagtgata ttgatttgct ggttacggtg 540
accgtaaggc ttgatgaaac aacgcggcga gctttgatca acgacctttt ggaaacttcg 600
gcttcccctg gagagagcga gattctccgc gctgtagaag tcaccattgt tgtgcacgac 660
gacatcattc cgtggcgtta tccagctaag cgcgaactgc aatttggaga atggcagcgc 720
aatgacattc ttgcaggtat cttcgagcca gccacgatcg acattgatct ggctatcttg 780
ctgacaaaag caagagaaca tagcgttgcc ttggtaggtc cagcggcgga ggaactcttt 840
gatccggttc ctgaacagga tctatttgag gcgctaaatg aaaccttaac gctatggaac 900
tcgccgcccg actgggctgg cgatgagcga aatgtagtgc ttacgttgtc ccgcatttgg 960
tacagcgcag taaccggcaa aatcgcgccg aaggatgtcg ctgccgactg ggcaatggag 1020
cgcctgccgg cccagtatca gcccgtcata cttgaagcta gacaggctta tcttggacaa 1080
gaagaagatc gcttggcctc gcgcgcagat cagttggaag aatttgtcca ctacgtgaaa 1140
ggcgagatca ccaaggtagt cggcaaataa tgtctaacaa ttcgttcaag ccgacgccgc 1200
ttcgcggcgc ggcttaactc aagcgttaga tgcactaagc acataattgc tcacagccaa 1260
actatcaggt caagtctgct tttattattt ttaagcgtgc ataataagcc ctacacaaat 1320
ggtacccgac cggtggtgaa tttaatctcg ctgacgtgta gacattccct tatccagacg 1380
ctgatcgccc atcatcgcgg ttctttagat ctctcggtcc gccctgatgg cggcaccttg 1440
ctgacgttac gcctgccggt acagcaggtt atcaccggag gcttaaaatg a 1491
<210> 57
<211> 1491
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 57
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gagcgtgttg 120
agtcatcctg actagctgag atgagggctc gccccctcgt cccgacactt ccagatcgcc 180
atagcgcaca gcgcctcgag cggtggtaac ggcgcagtgg cggttttcat ggcttgttat 240
gactgttttt ttggggtaca gtctatgcct cgggcatcca agcagcaagc gcgttacgcc 300
gtgggtcgat gtttgatgtt atggagcagc aacgatgtta cgcagcaggg cagtcgccct 360
aaaacaaagt taaacatcat gagggaagcg gtgatcgccg aagtatcgac tcaactatca 420
gaggtagttg gcgtcatcga gcgccatctc gaaccgacgt tgctggccgt acatttgtac 480
ggctccgcag tggatggcgg cctgaagcca cacagtgata ttgatttgct ggttacggtg 540
accgtaaggc ttgatgaaac aacgcggcga gctttgatca acgacctttt ggaaacttcg 600
gcttcccctg gagagagcga gattctccgc gctgtagaag tcaccattgt tgtgcacgac 660
gacatcattc cgtggcgtta tccagctaag cgcgaactgc aatttggaga atggcagcgc 720
aatgacattc ttgcaggtat cttcgagcca gccacgatcg acattgatct ggctatcttg 780
ctgacaaaag caagagaaca tagcgttgcc ttggtaggtc cagcggcgga ggaactcttt 840
gatccggttc ctgaacagga tctatttgag gcgctaaatg aaaccttaac gctatggaac 900
tcgccgcccg actgggctgg cgatgagcga aatgtagtgc ttacgttgtc ccgcatttgg 960
tacagcgcag taaccggcaa aatcgcgccg aaggatgtcg ctgccgactg ggcaatggag 1020
cgcctgccgg cccagtatca gcccgtcata cttgaagcta gacaggctta tcttggacaa 1080
gaagaagatc gcttggcctc gcgcgcagat cagttggaag aatttgtcca ctacgtgaaa 1140
ggcgagatca ccaaggtagt cggcaaataa tgtctaacaa ttcgttcaag ccgacgccgc 1200
ttcgcggcgc ggcttaactc aagcgttaga tgcactaagc acataattgc tcacagccaa 1260
actatcaggt caagtctgct tttattattt ttaagcgtgc ataataagcc ctacacaaat 1320
ggtacccgac cggtggtgaa tttaatctcg ctgacgtgta gacattccct tatccagacg 1380
ctgatcgccc atcatcgcgg ttctttagat ctctcggtcc gccctgatgg cggcaccttg 1440
ctgacgttac gcctgccggt acagcaggtt atcaccggag gcttaaaatg a 1491
<210> 58
<211> 1021
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 58
ctgatccttc aactcagcaa aagttcgatt tattcaacaa agccacgttg tgtctcaaaa 60
tctctgatgt tacattgcac aagataaaaa tatatcatca tgaacaataa aactgtctgc 120
ttacataaac agtaatacaa ggggtgttat gagccatatt caacgggaaa cgtcttgctc 180
caggccgcga ttaaattcca acatggatgc tgatttatat gggtataaat gggctcgcga 240
taatgtcggg caatcaggtg cgacaatcta tcgattgtat gggaagcccg atgcgccaga 300
gttgtttctg aaacatggca aaggtagcgt tgccaatgat gttacagatg agatggtcag 360
actaaactgg ctgacggaat ttatgcctct tccgaccatc aagcatttta tccgtactcc 420
tgatgatgca tggttactca ccactgcgat ccccgggaaa acagcattcc aggtattaga 480
agaatatcct gattcaggtg aaaatattgt tgatgcgctg gcagtgttcc tgcgccggtt 540
gcattcgatt cctgtttgta attgtccttt taacagcgat cgcgtatttc gtctcgctca 600
ggcgcaatca cgaatgaata acggtttggt tgatgcgagt gattttgatg acgagcgtaa 660
tggctggcct gttgaacaag tctggaaaga aatgcataag cttttgccat tctcaccgga 720
ttcagtcgtc actcatggtg atttctcact tgataacctt atttttgacg aggggaaatt 780
aataggttgt attgatgttg gacgagtcgg aatcgcagac cgataccagg atcttgccat 840
cctatggaac tgcctcggtg agttttctcc ttcattacag aaacggcttt ttcaaaaata 900
tggtattgat aatcctgata tgaataaatt gcagtttcat ttgatgctcg atgagttttt 960
ctaataagcc ttgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 1020
a 1021
<210> 59
<211> 1491
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 59
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gagcgtgttg 120
agtcatcctg actagctgag atgagggctc gccccctcgt cccgacactt ccagatcgcc 180
atagcgcaca gcgcctcgag cggtggtaac ggcgcagtgg cggttttcat ggcttgttat 240
gactgttttt ttggggtaca gtctatgcct cgggcatcca agcagcaagc gcgttacgcc 300
gtgggtcgat gtttgatgtt atggagcagc aacgatgtta cgcagcaggg cagtcgccct 360
aaaacaaagt taaacatcat gagggaagcg gtgatcgccg aagtatcgac tcaactatca 420
gaggtagttg gcgtcatcga gcgccatctc gaaccgacgt tgctggccgt acatttgtac 480
ggctccgcag tggatggcgg cctgaagcca cacagtgata ttgatttgct ggttacggtg 540
accgtaaggc ttgatgaaac aacgcggcga gctttgatca acgacctttt ggaaacttcg 600
gcttcccctg gagagagcga gattctccgc gctgtagaag tcaccattgt tgtgcacgac 660
gacatcattc cgtggcgtta tccagctaag cgcgaactgc aatttggaga atggcagcgc 720
aatgacattc ttgcaggtat cttcgagcca gccacgatcg acattgatct ggctatcttg 780
ctgacaaaag caagagaaca tagcgttgcc ttggtaggtc cagcggcgga ggaactcttt 840
gatccggttc ctgaacagga tctatttgag gcgctaaatg aaaccttaac gctatggaac 900
tcgccgcccg actgggctgg cgatgagcga aatgtagtgc ttacgttgtc ccgcatttgg 960
tacagcgcag taaccggcaa aatcgcgccg aaggatgtcg ctgccgactg ggcaatggag 1020
cgcctgccgg cccagtatca gcccgtcata cttgaagcta gacaggctta tcttggacaa 1080
gaagaagatc gcttggcctc gcgcgcagat cagttggaag aatttgtcca ctacgtgaaa 1140
ggcgagatca ccaaggtagt cggcaaataa tgtctaacaa ttcgttcaag ccgacgccgc 1200
ttcgcggcgc ggcttaactc aagcgttaga tgcactaagc acataattgc tcacagccaa 1260
actatcaggt caagtctgct tttattattt ttaagcgtgc ataataagcc ctacacaaat 1320
ggtacccgac cggtggtgaa tttaatctcg ctgacgtgta gacattccct tatccagacg 1380
ctgatcgccc atcatcgcgg ttctttagat ctctcggtcc gccctgatgg cggcaccttg 1440
ctgacgttac gcctgccggt acagcaggtt atcaccggag gcttaaaatg a 1491
<210> 60
<211> 1491
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 60
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gagcgtgttg 120
agtcatcctg actagctgag atgagggctc gccccctcgt cccgacactt ccagatcgcc 180
atagcgcaca gcgcctcgag cggtggtaac ggcgcagtgg cggttttcat ggcttgttat 240
gactgttttt ttggggtaca gtctatgcct cgggcatcca agcagcaagc gcgttacgcc 300
gtgggtcgat gtttgatgtt atggagcagc aacgatgtta cgcagcaggg cagtcgccct 360
aaaacaaagt taaacatcat gagggaagcg gtgatcgccg aagtatcgac tcaactatca 420
gaggtagttg gcgtcatcga gcgccatctc gaaccgacgt tgctggccgt acatttgtac 480
ggctccgcag tggatggcgg cctgaagcca cacagtgata ttgatttgct ggttacggtg 540
accgtaaggc ttgatgaaac aacgcggcga gctttgatca acgacctttt ggaaacttcg 600
gcttcccctg gagagagcga gattctccgc gctgtagaag tcaccattgt tgtgcacgac 660
gacatcattc cgtggcgtta tccagctaag cgcgaactgc aatttggaga atggcagcgc 720
aatgacattc ttgcaggtat cttcgagcca gccacgatcg acattgatct ggctatcttg 780
ctgacaaaag caagagaaca tagcgttgcc ttggtaggtc cagcggcgga ggaactcttt 840
gatccggttc ctgaacagga tctatttgag gcgctaaatg aaaccttaac gctatggaac 900
tcgccgcccg actgggctgg cgatgagcga aatgtagtgc ttacgttgtc ccgcatttgg 960
tacagcgcag taaccggcaa aatcgcgccg aaggatgtcg ctgccgactg ggcaatggag 1020
cgcctgccgg cccagtatca gcccgtcata cttgaagcta gacaggctta tcttggacaa 1080
gaagaagatc gcttggcctc gcgcgcagat cagttggaag aatttgtcca ctacgtgaaa 1140
ggcgagatca ccaaggtagt cggcaaataa tgtctaacaa ttcgttcaag ccgacgccgc 1200
ttcgcggcgc ggcttaactc aagcgttaga tgcactaagc acataattgc tcacagccaa 1260
actatcaggt caagtctgct tttattattt ttaagcgtgc ataataagcc ctacacaaat 1320
ggtacccgac cggtggtgaa tttaatctcg ctgacgtgta gacattccct tatccagacg 1380
ctgatcgccc atcatcgcgg ttctttagat ctctcggtcc gccctgatgg cggcaccttg 1440
ctgacgttac gcctgccggt acagcaggtt atcaccggag gcttaaaatg a 1491
<210> 61
<211> 1563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 61
atgtttaacg atctgattgg cgatgatgaa acggattcgc cggaagatgc gctttctgag 60
agctggcgcg aattgtggca ggatgcgttg caggaggagg attccacgcc cgtgctggcg 120
catctctcag aggacgatcg ccgccgcgtg gtggcgctga ttgccgattt tcgcaaagag 180
ttggataaac gcaccattgg cccgcgaggg cggcaggtac tcgatcactt aatgccgcat 240
ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac cgctgtcacg cctgacgccg 300
ctgctcaccg gaattattac ccgcaccact taccttgagc tgctaagtga atttcccggc 360
gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg 420
cgctacccga tcctgcttga tgaattgctc gacccgaata cgctctatca accgacggcg 480
atgaatgcct atcgcgatga gctgcgccaa tacctgctgc gcgtgccgga agatgatgaa 540
gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc agttgctgcg cgtggcggcg 600
gcggatattg ccggtacgtt gccagtaatg aaagtgagcg atcacttaac ctggctggcg 660
gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc agatggtggc gcgttatggc 720
cagccaacgc atctgcacga tcgcgaaggg cgcggttttg cggtggtcgg ttatggcaag 780
ctgggcggct gggagctggg ttacagctcc gatctggatc tggtattcct gcacgactgc 840
ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg gtcgccagtt ctatttgcgt 900
ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt cgtccggcat cctttatgaa 960
gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc tggtcactac tacggaatcg 1020
ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac atcaggcgct ggcccgtgcg 1080
cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg acgccattcg ccgcgatatt 1140
ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg tgcgagaaat gcgcgagaaa 1200
atgcgtgccc atcttggcaa caagcataaa gaccgcttcg atctgaaagc cgatgaaggc 1260
ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc tgcgctttgc ccatgacaag 1320
ccgaaactga cgcgctggtc ggataatgtg cgcattctcg aagggctggc gcaaaacggc 1380
atcatggagg agcaggaagc gcaggcattg acgctggcgt acaccacatt gcgtgatgag 1440
ctgcaccacc tggcgctgca agagttgccg ggacatgtgg cgctctcctg ttttgtcgcc 1500
gagcgtgcgc ttattaaaac cagctgggac aagtggctgg tggaaccgtg cgccccggcg 1560
taa 1563
<210> 62
<211> 1536
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> рДНК 16S - контиг 5, штамм CI006
<400> 62
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gaacggtagc acagagagct tgctctcggg tgacgagtgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagaggggga ccttcgggcc tcttgccatc agatgtgccc agatgggatt 240
agctagtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggg agtaaggtta ataaccttat tcattgacgt 480
tacccgcaga agaagcaccg gctaactccg tgccagcagc cgcggtaata cggagggtgc 540
aagcgttaat cggaattact gggcgtaaag cgcacgcagg cggtctgtca agtcggatgt 600
gaaatccccg ggctcaacct gggaactgca tccgaaactg gcaggcttga gtctcgtaga 660
gggaggtaga attccaggtg tagcggtgaa atgcgtagag atctggagga ataccggtgg 720
cgaaggcggc ctcctggacg aagactgacg ctcaggtgcg aaagcgtggg gagcaaacag 780
gattagatac cctggtagtc cacgccgtaa acgatgtcta tttggaggtt gtgcccttga 840
ggcgtggctt ccggagctaa cgcgttaaat agaccgcctg gggagtacgg ccgcaaggtt 900
aaaactcaaa tgaattgacg ggggcccgca caagcggtgg agcatgtggt ttaattcgat 960
gcaacgcgaa gaaccttacc tggtcttgac atccacagaa ctttccagag atggattggt 1020
gccttcggga actgtgagac aggtgctgca tggctgtcgt cagctcgtgt tgtgaaatgt 1080
tgggttaagt cccgcaacga gcgcaaccct tatcctttgt tgccagcggt ccggccggga 1140
actcaaagga gactgccagt gataaactgg aggaaggtgg ggatgacgtc aagtcatcat 1200
ggcccttacg accagggcta cacacgtgct acaatggcgc atacaaagag aagcgacctc 1260
gcgagagtaa gcggacctca taaagtgcgt cgtagtccgg attggagtct gcaactcgac 1320
tccatgaagt cggaatcgct agtaatcgtg gatcagaatg ccacggtgaa tacgttcccg 1380
ggccttgtac acaccgcccg tcacaccatg ggagtgggtt gcaaaagaag taggtagctt 1440
aaccttcggg agggcgctta ccactttgtg attcatgact ggggtgaagt cgtaacaagg 1500
taaccgtagg ggaacctgcg gttggatcac ctcctt 1536
<210> 63
<211> 1537
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> рДНК 16S - контиг 8, штамм CI006
<220>
<221> модифицированное основание
<222> (450)..(450)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (452)..(452)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (455)..(455)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (473)..(473)
<223> a, c, g или t
<400> 63
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gaacggtagc acagagagct tgctctcggg tgacgagtgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagaggggga ccttcgggcc tcttgccatc agatgtgccc agatgggatt 240
agctagtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggn antanggtta ataacctgtg ttnattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc atccgaaact ggcaggcttg agtctcgtag 660
agggaggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg cctcctggac gaagactgac gctcaggtgc gaaagcgtgg ggagcaaaca 780
ggattagata ccctggtagt ccacgccgta aacgatgtct atttggaggt tgtgcccttg 840
aggcgtggct tccggagcta acgcgttaaa tagaccgcct ggggagtacg gccgcaaggt 900
taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggtcttga catccacaga acttagcaga gatgctttgg 1020
tgccttcggg aactgtgaga caggtgctgc atggctgtcg tcagctcgtg ttgtgaaatg 1080
ttgggttaag tcccgcaacg agcgcaaccc ttatcctttg ttgccagcgg ttaggccggg 1140
aactcaaagg agactgccag tgataaactg gaggaaggtg gggatgacgt caagtcatca 1200
tggcccttac gaccagggct acacacgtgc tacaatggcg catacaaaga gaagcgacct 1260
cgcgagagta agcggacctc ataaagtgcg tcgtagtccg gattggagtc tgcaactcga 1320
ctccatgaag tcggaatcgc tagtaatcgt ggatcagaat gccacggtga atacgttccc 1380
gggccttgta cacaccgccc gtcacaccat gggagtgggt tgcaaaagaa gtaggtagct 1440
taaccttcgg gagggcgctt accactttgt gattcatgac tggggtgaag tcgtaacaag 1500
gtaaccgtag gggaacctgc ggttggatca cctcctt 1537
<210> 64
<211> 1540
<212> ДНК
<213> вид Rahnella
<220>
<223> рДНК 16S, штамм CI019
<220>
<221> модифицированное основание
<222> (70)..(70)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (267)..(267)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (455)..(455)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (458)..(458)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (473)..(473)
<223> a, c, g или t
<220>
<221> модифицированное основание
<222> (1135)..(1135)
<223> a, c, g или t
<400> 64
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcan cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgaaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggctnacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcancanac ttaatacgtg tgntgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgagcttaa cttgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccaga gaatttgcca gagatggcga 1020
agtgccttcg ggaactctga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc acgtnatggt 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgaga gcaagcggac ctcataaagt atgtcgtagt ccggattgga gtctgcaact 1320
cgactccatg aagtcggaat cgctagtaat cgtagatcag aatgctacgg tgaatacgtt 1380
cccgggcctt gtacacaccg cccgtcacac catgggagtg ggttgcaaaa gaagtaggta 1440
gcttaacctt cgggagggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac 1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt 1540
<210> 65
<211> 882
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifH, штамм CI006
<400> 65
atgaccatgc gtcaatgcgc catttacggc aaaggtggga tcggcaaatc gaccaccaca 60
cagaacctgg tcgccgcgct ggcggagatg ggtaaaaaag tcatgattgt cggctgtgac 120
ccgaaagccg attccacgcg tttgatcctg catgcgaaag cgcagaacac cattatggag 180
atggctgctg aagtcggctc cgtggaagac ctggagttag aagacgtgct gcaaatcggt 240
tacggcggcg tgcgctgcgc agagtccggc ggcccggagc caggcgtggg ctgtgccggt 300
cgcggggtga tcaccgcgat taacttcctc gaagaagaag gcgcttacgt gccggatctc 360
gattttgttt tctacgacgt gctgggcgac gtggtatgcg gtggtttcgc catgccgatt 420
cgtgaaaaca aagcgcagga gatctacatc gtttgctctg gcgaaatgat ggcgatgtac 480
gccgccaaca acatctccaa aggcatcgtg aaatacgcca aatccggtaa agtgcgcctc 540
ggcgggctga tttgtaactc gcgccagacc gaccgtgaag atgaactgat cattgcgctg 600
gcagaaaaac tcggcacgca gatgatccac tttgttcccc gcgacaacat tgtgcagcgt 660
gcggaaatcc gccgtatgac ggttatcgaa tatgacccga cctgcaatca ggcgaacgaa 720
tatcgcagcc ttgccagcaa aatcgtcaac aacaccaaaa tggtggtgcc caccccctgc 780
accatggatg aactggaaga actgctgatg gagttcggca ttatggatgt ggaagacacc 840
agcatcattg gtaaaaccgc cgccgaagaa aacgccgtct ga 882
<210> 66
<211> 1449
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> NifD, штамм CI006
<400> 66
atgagcaatg caacaggcga acgcaacctg gagataatcg agcaggtgct cgaggttttc 60
ccggagaaga cgcgcaaaga acgcagaaaa cacatgatgg tgacggaccc ggagcaggaa 120
agcgtcggta agtgcatcat ctctaaccgc aaatcgcagc caggcgtgat gaccgtgcgc 180
ggctgctcgt atgccggttc gaaaggggtg gtatttgggc caatcaagga tatggcgcat 240
atctcgcatg gcccaatcgg ctgcggccaa tactcccgcg ccgggcggcg gaactactac 300
accggcgtca gcggcgtgga cagcttcggc acgctcaact tcacctccga ttttcaggag 360
cgcgacatcg tgtttggcgg cgataaaaag ctcgccaaac tgattgaaga gctggaagag 420
ctgttcccgc tgaccaaagg catttcgatt cagtcggaat gcccggtcgg cctgattggc 480
gatgacattg aggccgtcgc gaacgccagc cgcaaagcca tcaacaaacc ggttattccg 540
gtgcgttgcg aaggctttcg cggcgtgtcg caatccctcg gtcaccatat tgccaacgat 600
gtgatccgcg actgggtgct ggataaccgc gaaggcaaac cgttcgaatc caccccttac 660
gatgtggcga tcatcggcga ttacaacatc ggcggcgatg cctgggcttc gcgcattttg 720
ctcgaagaga tgggcttgcg ggtggtggca cagtggtctg gcgacggtac gctggtggag 780
atggaaaaca cgccgttcgt caaactgaac ctggtgcatt gttaccgctc aatgaactac 840
atctcgcgcc atatggagga gaagcacggt attccgtgga tggaatacaa cttctttggt 900
ccgacgaaaa tcgcggaatc gctgcgcaaa atcgccgacc agtttgacga caccattcgc 960
gccaacgccg aagcggtgat cgccagatac caggcgcaaa acgacgccat tatcgccaaa 1020
tatcgcccgc gtctggaggg gcgcaaagtg ctgctttata tgggcgggct gcgtccgcgc 1080
catgtgattg gcgcctatga agacctggga atggagatca tcgctgccgg ttatgagttc 1140
ggtcataacg atgattacga ccgcaccttg ccggatctga aagagggcac gctgctgttt 1200
gatgatgcca gcagttatga gctggaggcg ttcgtcaacg cgctgaaacc ggatctcatc 1260
ggttccggca tcaaagagaa gtacatcttt cagaaaatgg gcgtgccgtt tcgccagatg 1320
cactcctggg attactccgg cccgtaccac ggctatgacg gcttcgccat cttcgcccgc 1380
gatatggata tgacgctcaa caaccccgcg tggggccagt tgaccgcgcc gtggctgaaa 1440
tccgcctga 1449
<210> 67
<211> 1563
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifK, штамм CI006
<400> 67
atgagccaga ctgctgagaa aatacagaat tgccatcccc tgtttgaaca ggatgcttac 60
cagacgctgt ttgccggtaa acgggcactc gaagaggcgc actcgccgga gcgggtgcag 120
gaagtgtttc aatggaccac taccccggaa tatgaagcgc tgaactttaa acgcgaagcg 180
ctgactatcg acccggcaaa agcctgccag ccgctgggcg cggtgctctg ttcgctgggg 240
tttgccaata ccctaccgta tgtgcacggt tcacagggtt gcgtggccta tttccgcacg 300
tactttaacc gccactttaa agaaccggtg gcctgcgtgt cggattcaat gacggaagac 360
gcggcggtgt tcggcgggaa taacaacctc aacaccggct tacaaaacgc cagcgcgctg 420
tataaaccgg agattatcgc cgtctctacc acctgtatgg cggaagtgat cggtgatgat 480
ttgcaggcct ttatcgccaa cgccaaaaaa gatggttttc tcgatgccgc catccccgtg 540
ccctacgcgc acacccccag ttttatcggc agccatatca ccggctggga taacatgttt 600
gaaggttttg cccggacctt tacggcagac catgaagctc agcccggcaa actttcacgc 660
atcaacctgg tgaccgggtt tgaaacctat ctcggcaatt tccgcgtgct gaaacgcatg 720
atggaacaaa tggaggtgcc ggcgagtgtg ctctccgatc cgtcggaagt gctggatact 780
cccgccaacg ggcattacca gatgtacgcg ggcgggacga cgcagcaaga gatgcgcgag 840
gcgccggatg ctatcgacac cctgttgctg cagccctggc aactggtgaa aagcaaaaaa 900
gtggtgcagg agatgtggaa tcagcccgcc accgaggttt ctgttcccgt tgggctggca 960
ggaacagacg aactgttgat ggcgattagc cagttaaccg gcaaggccat tcccgattca 1020
ctggcgctgg agcgcgggcg gctggtcgat atgatgctcg attcccacac ctggttgcac 1080
ggtaaaaaat tcggcctgtt tggcgatccg gattttgtca tgggattgac ccgtttcctg 1140
ctggagctgg gctgcgaacc gaccgttatc ctctgccaca acggtaacaa acgctggcag 1200
aaagcaatga agaaaatgct tgacgcctcg ccgtacggcc aggagagcga agtgtttatc 1260
aactgcgatt tgtggcattt ccgctcgctg atgtttaccc gccagccgga ttttatgatt 1320
ggcaactcgt acggcaagtt cattcagcgc gacaccttag ccaaaggcga gcagtttgaa 1380
gttccgctga tccgcctcgg ttttcccctg ttcgaccgcc accatctgca ccgccagacc 1440
acctggggct acgagggcgc catgagcatt ctcactaccc ttgtgaatgc ggtactggag 1500
aaagtggaca aagagaccat caagctcggc aaaaccgact acagcttcga tcttatccgt 1560
taa 1563
<210> 68
<211> 1488
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifL, штамм CI006
<400> 68
atgaccctga atatgatgat ggatgccggc gcgcccgagg caatcgccgg tgcgctttcg 60
cgacaccatc ctgggctgtt ttttaccatc gttgaagaag cgcccgtcgc catttcgctg 120
actgatgccg acgcacgcat tgtctatgcc aacccggctt tctgccgcca gaccggctat 180
gaactagaag cgttgttgca gcaaaatccc cgcctgcttg caagtcgcca aaccccacgg 240
gaaatctatc aggatatgtg gcacaccttg ttacaacgcc gaccgtggcg cgggcaattg 300
attaaccgcc accgcgacgg cagcctgtat ctggtcgaga tcgatatcac cccggtgatt 360
aacccgtttg gcgaactgga acactacctg gcaatgcagc gcgatatcag cgccagttat 420
gcgctggagc agcggttgcg caatcacatg acgctgaccg aagcggtgct gaataacatt 480
ccggcggcgg tggttgtagt ggatgaacgc gatcatgtgg ttatggataa ccttgcctac 540
aaaacgttct gtgccgactg cggcggaaaa gagctcctga gcgaactcaa tttttcagcc 600
cgaaaagcgg agctggcaaa cggccaggtc ttaccggtgg tgctgcgcgg tgaggtgcgc 660
tggttgtcgg tgacctgctg ggcgctgccg ggcgtcagcg aagaagccag tcgctacttt 720
attgataaca ggctgacgcg cacgctggtg gtgatcaccg acgacaccca acaacgccag 780
cagcaggaac agggccgact tgaccgcctt aaacagcaga tgaccaacgg caaactactg 840
gcagcgatcc gcgaagcgct tgacgccgcg ctgatccagc ttaactgccc catcaatatg 900
ctggcggcgg cgcgacgttt aaacggcagt gataacaaca atgtggcgct cgacgccgcg 960
tggcgcgaag gtgaagaggc gatggcgcgg ctgaaacgtt gccgcccgtc gctggaactg 1020
gaaagtgcgg ccgtctggcc gctgcaaccc ttttttgacg atctgcgcgc gctttatcac 1080
acccgctacg agcaggggaa aaatttgcag gtcacgctgg attcccatca tctggtggga 1140
tttggtcagc gtacgcaact gttagcctgc ctgagtctgt ggctcgatcg cacgctggat 1200
attgccgccg ggctgggtga tttcaccgcg caaacgcaga tttacgcccg cgaagaagag 1260
ggctggctct ctttgtatat cactgacaat gtgccgctga tcccgctgcg ccacacccac 1320
tcgccggatg cgcttaacgc tccgggaaaa ggcatggagc tgcgcctgat ccagacgctg 1380
gtggcacacc accacggcgc aatagaactc acttcacacc ccgaaggggg aagttgcctg 1440
accctacgat tcccgctatt tcattcactg accggaggtt caaaatga 1488
<210> 69
<211> 1575
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifA, штамм CI006
<400> 69
atgacccagc gaaccgagtc gggtaatacc gtctggcgct tcgatttgtc ccagcagttc 60
actgcgatgc agcgcataag cgtggtactc agccgggcga ccgaggtcga tcagacgctc 120
cagcaagtgc tgtgcgtatt gcacaatgac gcctttttgc agcacggcat gatctgtctg 180
tacgacagcc agcaggcgat tttgaatatt gaagcgttgc aggaagccga tcagcagtta 240
atccccggca gctcgcaaat ccgctatcgt ccgggcgaag ggctggtcgg gacggtgctt 300
tcgcagggcc aatcattagt gctggcgcgc gttgctgacg atcagcgctt tcttgaccgg 360
ctcgggttgt atgattacaa cctgccgttt atcgccgtgc cgctgatagg gccagatgcg 420
cagactttcg gtgtgctgac ggcacaaccc atggcgcgtt acgaagagcg attacccgcc 480
tgcacccgct ttctggaaac ggtcgctaac ctggtcgcgc aaaccgtgcg tttgatggca 540
ccaccggcag tgcgcccttc cccgcgcgcc gccataacac aggccgccag cccgaaatcc 600
tgcacggcct cacgcgcatt tggttttgaa aatatggtcg gtaacagtcc ggcgatgcgc 660
cagaccatgg agattatccg tcaggtttcg cgctgggaca ccaccgttct ggtacgcggc 720
gagagtggca ccggcaagga gctgattgcc aacgccatcc accaccattc gccgcgtgcc 780
ggtgcgccat ttgtgaaatt caactgtgcg gcgctgccgg acacactgct ggaaagcgaa 840
ttgttcggtc acgagaaagg ggcatttacc ggcgcggtac gccagcgtaa aggccgtttt 900
gagctggccg atggcggcac gctgtttctt gacgagatcg gcgagagtag cgcctcgttt 960
caggctaagc tgctgcgcat tttgcaggaa ggcgaaatgg aacgcgtcgg cggcgacgag 1020
acattgcaag tgaatgtgcg cattattgcc gcgacgaacc gcaatcttga agatgaagtc 1080
cggctggggc actttcgcga agatctctat tatcgcctga atgtgatgcc catcgccctg 1140
ccgccactac gcgaacgcca ggaggacatt gccgagctgg cgcactttct ggtgcgtaaa 1200
atcgcccata accagagccg tacgctgcgc attagcgagg gcgctatccg cctgctgatg 1260
agctacaact ggcccggtaa tgtgcgcgaa ctggaaaact gccttgagcg ctcagcggtg 1320
atgtcggaga acggtctgat cgatcgggat gtgattttgt ttaatcatcg cgaccagcca 1380
gccaaaccgc cagttatcag cgtctcgcat gatgataact ggctcgataa caaccttgac 1440
gagcgccagc ggctgattgc ggcgctggaa aaagcgggat gggtacaagc caaagccgcg 1500
cgcttgctgg ggatgacgcc gcgccaggtc gcctatcgta ttcagacgat ggatataacc 1560
ctgccaaggc tataa 1575
<210> 70
<211> 876
<212> ДНК
<213> вид Rahnella
<220>
<223> nifH, штамм CI019
<400> 70
atggcaatgc gtcaatgtgc aatctacggg aaagggggta ttggtaaatc caccactacc 60
caaaaccttg tagcggctct ggccgaaatg aataagaagg tcatgatcgt cggctgtgac 120
cctaaggctg attcaacccg cctcattctg catgcgaaag cacagaacac catcatggaa 180
atggccgctg aagtgggctc cgtggaagat ctggagctgg aagatgtgat gcaaatcggc 240
tatggcggcg tgcgctgtgc ggaatcaggc ggccctgagc ctggtgtggg ttgtgccgga 300
cgcggggtga tcaccgccat caacttcctc gaagaagaag gcgcgtatgt gccggatctg 360
gatttcgtgt tttacgacgt attgggcgat gtggtctgtg gcggtttcgc gatgccaatt 420
cgcgaaaaca aagcgcagga aatctacatc gtatgctccg gtgaaatgat ggcgatgtat 480
gccgccaaca acatttccaa aggcatcgtg aaatacgcga aatcgggcaa agttcgcctg 540
gccgggctga tctgtaactc ccgccagacg gatcgcgaag atgaactgat catcgcgctg 600
gctgaaaaac ttggcacgca aatgatccac ttcgtgccgc gtgacaacat tgtgcaacgc 660
gctgaaatcc gccgcatgac ggtcatcgaa tacgacccga cttgtgcgca ggcagatcag 720
tatcgtgcac tggcgaacaa aatcgtcaac aacaccaaaa tggtggtgcc gacaccggtc 780
accatggatg agctggaagc cctgttaatg gaatttggca ttatggaaga agaagacctg 840
accatcgtcg gtcgtaccgc cgccgaagag gcgtga 876
<210> 71
<211> 1449
<212> ДНК
<213> вид Rahnella
<220>
<223> nifD, штамм CI019
<400> 71
atgaccagtg aaacacgcga acgtaacgag gcattgatcc aggaagtgct ggagatcttc 60
cccgagaagg cgcttaaaga tcgtaagaaa cacatgatga ccaccgaccc ggcgatggaa 120
tctgtcggca agtgtattgt ctcaaaccgc aaatcacagc cgggcgtgat gaccgtgcga 180
ggctgcgctt acgccggttc caaaggcgtg gtctttggcc cgatcaaaga catggcgcat 240
atctcccacg gcccggttgg ttgcggccag tattctcgtg ccggacgccg taactattac 300
accggctgga gcggcgtgaa cagctttggc accctcaact tcaccagtga ttttcaggaa 360
cgggacatcg tatttggcgg cgataaaaag ctcgacaaac tgatcgacga actggagatg 420
ttgttcccgc tgaccaaagg catttcggta cagtcggaat gtccggtcgg tctgatcggc 480
gatgacattt ctgccgtcgc caaagccagc agcgccaaaa tcggtaagcc ggtcgtgccg 540
gtacgctgcg aggggttccg cggtgtgtcg caatcgctcg gccatcacat tgctaacgat 600
gtcatccgcg actgggtgct ggataaccgc gaaggcaatg aatttgaaac cacgccttac 660
gacgtggcga ttatcggcga ctacaacatc ggcggtgacg cctgggcctc acgtattctg 720
ctcgaagaaa tggggctgcg tgtggtggcg cagtggtccg gcgacggcac gctggtggag 780
atggaaaaca ccccgaaagt cgcactcaat ctggtgcact gctaccgctc gatgaactac 840
atctcccgtc atatggaaga aaaacacggc attccgtgga tggaatacaa cttctttggc 900
ccgaccaaaa ttgcggaatc tctgcgcgaa atcgcggcgc gttttgacga taccatccgg 960
aaaaacgccg aagcggtgat tgaaaaatat caggcgcaaa cgcaggcggt gatcgacaaa 1020
taccgtccgc gtctggaagg caaaaaggtg ctgttgtatc tcggcggttt acgtccgcgc 1080
cacatcatcg gggcgtatga agatctggga atggaaatca tcggtaccgg ctatgaattc 1140
ggtcataacg atgattacga ccgcacctta ccgatgctca aagaaggcac gttgctgttc 1200
gatgacctga gcagttatga gctggaagcg ttcgttaaag cgctgaaacc ggatcttgtc 1260
gggtcaggca tcaaagaaaa atacattttc cagaaaatgg gcgtgccgtt ccgccagatg 1320
cactcctggg attattccgg cccttatcac ggctacgacg gtttcggcat ttttgcccgt 1380
gacatggaca tgacgctgaa caatccgggc tggagtcagc tgaccgcccc ctggttgaaa 1440
tcggcctga 1449
<210> 72
<211> 1569
<212> ДНК
<213> вид Rahnella
<220>
<223> nifK, штамм CI019
<400> 72
atgagtcaag atcttggcac cccaaaatcc tgtttcccgc tgttcgagca ggatgaatac 60
cagagtatgt ttacccacaa acgcgcgctg gaagaagcac acggcgaggc gaaagtgcgg 120
gaagtgtttg aatggaccac cacgcaggaa tatcaggatc tgaacttctc gcgtgaagcg 180
ctgaccgtcg acccggcgaa agcctgccag ccgttaggcg cggtactttg cgcgctgggt 240
tttgccaaca cgttgccgta tgtccacggt tcacaaggct gtgtggcgta tttccgtacc 300
tattttaatc gtcatttcaa agagccggtg gcctgtgttt ccgactcaat gaccgaagat 360
gccgccgttt ttggcggaaa taacaacatg aatgtcggtc tggaaaacgc cagcgcgctg 420
tacaagccgg aaattattgc tgtctccacc acctgtatgg cggaagtgat cggtgatgac 480
ctgcaggctt ttatcgccaa cgccaaaaaa gacggatttg tggatgccgg tatgccaatc 540
ccgtatgccc atacaccgag ttttctgggc agtcatgtca ccggctggga caacatgttt 600
gaaggcttcg cccgtacctt taccaccgac gccacgcggg aatatcagcc gggcaaactt 660
gccaaactga acgtggtgac cggttttgaa acttatctcg gcaactaccg ggttattcac 720
cgcatgatga gccagatggg ggtcgaatgc agcgtcttgt ccgatccgtc tgaagtgctc 780
gacaccccgg ctgacggcca ataccgcatg tatgccggcg gcaccacgca aaccgaaatg 840
cgtgatgcac cggatgccat cgacaccttg ctgctgcaac cgtggcaatt gcagaaaacc 900
aaaaaagtgg tgcagggcga ctggaatcag ccgggcaccg aagtcagtgt accgattggc 960
ctggcggcga ccgatgcctt gctgatgacg gtaagcgaac tgaccggcaa accgatagct 1020
gacacgctgg cgactgaacg tggccgtctg gtggacatga tgctcgattc ccacacctgg 1080
ctgcatggca agcgtttcgg tctctacggt gacccggatt ttgtgatggg catgaccgca 1140
ttcctgctgg aactgggctg tgaaccgacc accattctca gccataacgg caacaaacgc 1200
tggcagaaag ccatgaagaa aatgctggct gattcgcctt acgggcagga cagcgaagtg 1260
tatgtgaact gcgatctgtg gcatttccgc tcgctgatgt ttacccgtaa accggacttt 1320
atgatcggca actcttacgg aaaattcatt cagcgtgaca cgctggccaa aggcgaacag 1380
ttcgaagtgc cgctgatccg catcggtttt ccgatttttg accggcacca tttgcaccgt 1440
cagaccacct ggggatacga aggggcgatg agcatactga cgcaactggt gaatgcggta 1500
cttgaacaac tggatcgcga aaccatgaag ctcggcaaaa ccgactacaa cttcgacctg 1560
atccgctaa 1569
<210> 73
<211> 1488
<212> ДНК
<213> вид Rahnella
<220>
<223> nifL, штамм CI019
<400> 73
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg ccagccagca gacgccgaaa 240
catatctatg acgaaatgtg gcgcactttg ttgcagggca aatcctggaa cggccaactg 300
atcaaccggc gtaataaccg ttcgctttat ctggcggatg tcactatcac gcctgtttta 360
ggcgcggacg ggcaggtgga gcattacctc ggcatgcaca aagatatcag cgagaaatac 420
gcgctggaac agcggttgcg caaccacatc accttgttca cggaggtgct gaacaatatt 480
cccgccgccg tggtggtggt ggatgagcag gacaatgtgg tgatggacaa tctggcctac 540
aaaacccttt gcgcggactg cggcggcaaa gagctgctgg ctgaaatggg ctatccgcaa 600
ctcaaagaga tgctcaacag tggcgaaccg gtgccggttt ccatgcgcgg caacgtacgc 660
tggttttctt tcggtcaatg gttattgcag ggcgttaatg aagaggccag ccgctttttt 720
accggcatta ccgcgccggg aaaactgatt gttctgaccg actgcaccga tcagcatcac 780
cggcagcagc agggttatct tgaccggctt aagcaaaaac tcaccaacgg caaattattg 840
gcggccatcc gtgagtcgct cgatgccgcg cttatccagc tcaacgggcc aatcaatatg 900
ctggcggctg cgcgtcgtct taacggcgaa gaaggcaaca acatggcgct ggaattcgcc 960
tggcgcgaag gcgagcaggc ggtgagtcgc ttacaggcct gccgtccgtc gctggatttt 1020
gagccgcagg cagaatggcc ggtcagtgaa ttctttgacg atctgagcgc gctgtacgac 1080
agccattttc tcagtgacgg tgaattgcgt tacgtggtca tgccatctga tctgcacgct 1140
gtcgggcaac gaacgcaaat ccttaccgcg ctgagcttat ggattgatca cacgctgtca 1200
caggcgcagg ccatgccgtc tctgaagctc tcggtgaaca ttgttgcgag gcaggatgcg 1260
agctggttgt gttttgacat taccgataat gtgccgcgtg aacgggtgcg ttatgcccgc 1320
ccggaagcgg cgttttcccg tccggggaat ggcatggagc tgcgccttat ccagacgctg 1380
atcgcccatc atcgcggttc tttagatctc tcggtccgcc ctgatggcgg caccttgctg 1440
acgttacgcc tgccggtaca gcaggttatc accggaggct taaaatga 1488
<210> 74
<211> 1557
<212> ДНК
<213> вид Rahnella
<220>
<223> nifA, штамм CI019
<400> 74
atgacccagt tacctaccgc gggcccggtt atccggcgct ttgatatgtc tgcccagttt 60
acggcgcttt atcgcatcag cgtggcgctg agtcaggaaa gcaacaccgg gcgcgcactg 120
gcggcgatcc tcgaagtgct tcacgatcat gcatttatgc aatacggcat ggtgtgtctg 180
tttgataaag aacgcaatgc actctttgtg gaatccctgc atggcatcga cggcgaaagg 240
aaaaaagaga cccgccatgt ccgttaccgc atgggggaag gcgtgatcgg cgcggtgatg 300
agccagcgtc aggcgctggt gttaccgcgc atttcagacg atcagcgttt tctcgaccgc 360
ctgaatattt acgattacag cctgccgttg attggcgtgc cgatccccgg tgcggataat 420
cagccatcgg gcgtgctggt ggcacagccg atggcgttgc acgaagaccg gctgactgcc 480
agtacgcggt ttttagaaat ggtcgccaat ctcatcagcc agccactgcg ttctgccacg 540
cccccggaat cattgcctgc tcaaacgccg gtccggtgca gtgttccgcg ccagtttggt 600
ttcgagcaga tggtcgggaa aagtcaggcg atgcgccaga cgatggacat tttacggcag 660
gtttccaaat gggataccac ggttctggtg cgtggtgaaa gcggcaccgg caaggaactt 720
atcgccaatg ccattcatta caactcaccc cgtgcggccg cgccatttgt gaaattcaac 780
tgcgccgcgc tgccggataa cctgctggaa agcgaactgt tcggtcatga aaaaggggcc 840
ttcaccggcg ctatccgtac ccgtaaaggc cgctttgaac tggcggacgg gggcacgtta 900
ttcctcgatg aaatcggcga atcgagcgcg tcgtttcagg ccaaattgct gcgcattttg 960
caggaaggtg aaatggaacg ggtcggcggc gataccacgc tgaaagttga tgtgcgcatt 1020
attgctgcca ccaaccgtaa tcttgaagag gaagtgcgtg ccgggaattt tcgcgaagac 1080
ctgtattatc gcctgaacgt gatgccggtt tcgctgcctg cactgcgtga aaggctggat 1140
gatatcgccg atctggcgcc gtttctggtc aaaaagattg cgctgcgtca ggggcgggaa 1200
ctgcgcatca gcgacggtgc ggtgcgtctg ctgatgacct acagctggcc aggcaacgtg 1260
cgtgaactgg aaaactgtct cgaacgggcg tcggtaatga ccgatgaagg gctgatcgac 1320
cgcgacgtga tcctgttcaa tcaccatgaa tccccggcgc tgtccgtcaa acccggcctg 1380
ccgctcgcga cagatgaaag ctggctggat caggaactcg acgaacgcca gcgggtgatt 1440
gccgcactgg agaaaaccgg ctgggtgcag gccaaagcgg cccgactgct gggcatgaca 1500
ccgcgccaga ttgcctaccg tatccagatt atggacatca acatgcaccg tatctga 1557
<210> 75
<211> 1314
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> Prm5 с фланкирующими областями длиной 500 п.н., штамм CI006
<400> 75
aaaactaccg ccgcaattaa tgaacccaac gctactgttg ccgggccatg ctcttccccg 60
gcgcgctgcc cggaaaggat atagattgcc cagcacgcgc cagcaccaag cgcgaacgcc 120
gcgccagtga gatcaacatg tgaaacattt tcgcccagcg gcagcagata caagaggcca 180
agtaccgcca ggatcaccca gatgaaatcc accgggcggc gtgaggcaaa aagcgccacc 240
gccagcgggc cggtaaattc cagcgccacc gcaacgccga gcggtatcgt ctggatcgat 300
aaatagaaca tatagttcat ggcgccgagc gacaggccat aaaacagcag tggcaggcgt 360
tgttcacggg taaaatgtaa acgccagggc ttgaacacta cgaccaaaat aagggtgcca 420
agtgcgagac gcagcgcggt gacgccgggt gcgccaacaa tcggaaacag tgatttcgcc 480
agcgacgcgc ctccctgaat ggacatcatc gcgacaaaca atattaatac cggcaaccac 540
accggcaatt tacgagactg cgcaggcatc ctttctcccg tcaatttctg tcaaataaag 600
taaaagaggc agtctacttg aattaccccc ggctggttga gcgtttgttg aaaaaaagta 660
actgaaaaat ccgtagaata gcgccactct gatggttaat taacctattc aattaagaat 720
tatctggatg aatgtgccat taaatgcgca gcataatggt gcgttgtgcg ggaaaactgc 780
ttttttttga aagggttggt cagtagcgga aactttctgt tacatcaaat ggcgctttag 840
accccaattc ccgcaaagag tttcttaact aattttgata tatttaaacg cgtaggacgt 900
aggatttact tgaagcacat ttgaggtgga ttatgaaaaa aattgcatgt ctttcagcac 960
tggccgcact tctggcggtt tctgcaggtt ccgcagtagc agcaacttca accgtaactg 1020
gcggctacgc tcagagcgac gctcagggta ttgctaacaa aactaacggt ttcaacctga 1080
aatatcgcta cgagcaggac aacaacccgc tgggtgttat cggttccttt acttacactg 1140
aaaaagatcg caccgaaagc agcgtttata acaaagcgca gtactacggc atcaccgcag 1200
gcccggctta ccgcatcaac gactgggcga gcatctacgg tgttgtgggt gtaggttacg 1260
gtaaattcca gcagactgta gacaccgcta aagtgtctga caccagcgac tacg 1314
<210> 76
<211> 3413
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> оперон nifLA - выше межгенной области и CDS nifL и nifA, штамм CI006
<400> 76
aacacacgct cctgttgaaa aagagatccc gccgggaaat gcggtgaacg tgtctgatat 60
tgcgaagagt gtgccagttt tggtcgcggg caaaacctgc accagtttgg ttattaatgc 120
accagtctgg cgcttttttt cgccgagttt ctcctcgcta atgcccgcca ggcgcggctt 180
tggcgctgat agcgcgctga ataccgatct ggatcaaggt tttgtcgggt tatcagccaa 240
aaggtgcact ctttgcatgg ttatacgtgc ctgacatgtt gtccgggcga caaacggcct 300
ggtggcacaa attgtcagaa ctacgacacg actaactgac cgcaggagtg tgcgatgacc 360
ctgaatatga tgatggatgc cggcgcgccc gaggcaatcg ccggtgcgct ttcgcgacac 420
catcctgggc tgttttttac catcgttgaa gaagcgcccg tcgccatttc gctgactgat 480
gccgacgcac gcattgtcta tgccaacccg gctttctgcc gccagaccgg ctatgaacta 540
gaagcgttgt tgcagcaaaa tccccgcctg cttgcaagtc gccaaacccc acgggaaatc 600
tatcaggata tgtggcacac cttgttacaa cgccgaccgt ggcgcgggca attgattaac 660
cgccaccgcg acggcagcct gtatctggtc gagatcgata tcaccccggt gattaacccg 720
tttggcgaac tggaacacta cctggcaatg cagcgcgata tcagcgccag ttatgcgctg 780
gagcagcggt tgcgcaatca catgacgctg accgaagcgg tgctgaataa cattccggcg 840
gcggtggttg tagtggatga acgcgatcat gtggttatgg ataaccttgc ctacaaaacg 900
ttctgtgccg actgcggcgg aaaagagctc ctgagcgaac tcaatttttc agcccgaaaa 960
gcggagctgg caaacggcca ggtcttaccg gtggtgctgc gcggtgaggt gcgctggttg 1020
tcggtgacct gctgggcgct gccgggcgtc agcgaagaag ccagtcgcta ctttattgat 1080
aacaggctga cgcgcacgct ggtggtgatc accgacgaca cccaacaacg ccagcagcag 1140
gaacagggcc gacttgaccg ccttaaacag cagatgacca acggcaaact actggcagcg 1200
atccgcgaag cgcttgacgc cgcgctgatc cagcttaact gccccatcaa tatgctggcg 1260
gcggcgcgac gtttaaacgg cagtgataac aacaatgtgg cgctcgacgc cgcgtggcgc 1320
gaaggtgaag aggcgatggc gcggctgaaa cgttgccgcc cgtcgctgga actggaaagt 1380
gcggccgtct ggccgctgca accctttttt gacgatctgc gcgcgcttta tcacacccgc 1440
tacgagcagg ggaaaaattt gcaggtcacg ctggattccc atcatctggt gggatttggt 1500
cagcgtacgc aactgttagc ctgcctgagt ctgtggctcg atcgcacgct ggatattgcc 1560
gccgggctgg gtgatttcac cgcgcaaacg cagatttacg cccgcgaaga agagggctgg 1620
ctctctttgt atatcactga caatgtgccg ctgatcccgc tgcgccacac ccactcgccg 1680
gatgcgctta acgctccggg aaaaggcatg gagctgcgcc tgatccagac gctggtggca 1740
caccaccacg gcgcaataga actcacttca caccccgaag ggggaagttg cctgacccta 1800
cgattcccgc tatttcattc actgaccgga ggttcaaaat gacccagcga accgagtcgg 1860
gtaataccgt ctggcgcttc gatttgtccc agcagttcac tgcgatgcag cgcataagcg 1920
tggtactcag ccgggcgacc gaggtcgatc agacgctcca gcaagtgctg tgcgtattgc 1980
acaatgacgc ctttttgcag cacggcatga tctgtctgta cgacagccag caggcgattt 2040
tgaatattga agcgttgcag gaagccgatc agcagttaat ccccggcagc tcgcaaatcc 2100
gctatcgtcc gggcgaaggg ctggtcggga cggtgctttc gcagggccaa tcattagtgc 2160
tggcgcgcgt tgctgacgat cagcgctttc ttgaccggct cgggttgtat gattacaacc 2220
tgccgtttat cgccgtgccg ctgatagggc cagatgcgca gactttcggt gtgctgacgg 2280
cacaacccat ggcgcgttac gaagagcgat tacccgcctg cacccgcttt ctggaaacgg 2340
tcgctaacct ggtcgcgcaa accgtgcgtt tgatggcacc accggcagtg cgcccttccc 2400
cgcgcgccgc cataacacag gccgccagcc cgaaatcctg cacggcctca cgcgcatttg 2460
gttttgaaaa tatggtcggt aacagtccgg cgatgcgcca gaccatggag attatccgtc 2520
aggtttcgcg ctgggacacc accgttctgg tacgcggcga gagtggcacc ggcaaggagc 2580
tgattgccaa cgccatccac caccattcgc cgcgtgccgg tgcgccattt gtgaaattca 2640
actgtgcggc gctgccggac acactgctgg aaagcgaatt gttcggtcac gagaaagggg 2700
catttaccgg cgcggtacgc cagcgtaaag gccgttttga gctggccgat ggcggcacgc 2760
tgtttcttga cgagatcggc gagagtagcg cctcgtttca ggctaagctg ctgcgcattt 2820
tgcaggaagg cgaaatggaa cgcgtcggcg gcgacgagac attgcaagtg aatgtgcgca 2880
ttattgccgc gacgaaccgc aatcttgaag atgaagtccg gctggggcac tttcgcgaag 2940
atctctatta tcgcctgaat gtgatgccca tcgccctgcc gccactacgc gaacgccagg 3000
aggacattgc cgagctggcg cactttctgg tgcgtaaaat cgcccataac cagagccgta 3060
cgctgcgcat tagcgagggc gctatccgcc tgctgatgag ctacaactgg cccggtaatg 3120
tgcgcgaact ggaaaactgc cttgagcgct cagcggtgat gtcggagaac ggtctgatcg 3180
atcgggatgt gattttgttt aatcatcgcg accagccagc caaaccgcca gttatcagcg 3240
tctcgcatga tgataactgg ctcgataaca accttgacga gcgccagcgg ctgattgcgg 3300
cgctggaaaa agcgggatgg gtacaagcca aagccgcgcg cttgctgggg atgacgccgc 3360
gccaggtcgc ctatcgtatt cagacgatgg atataaccct gccaaggcta taa 3413
<210> 77
<211> 495
<212> Белок
<213> Kosakonia sacchari
<220>
<223> NifL, штамм CI006
<400> 77
Met Thr Leu Asn Met Met Met Asp Ala Gly Ala Pro Glu Ala Ile Ala
1 5 10 15
Gly Ala Leu Ser Arg His His Pro Gly Leu Phe Phe Thr Ile Val Glu
20 25 30
Glu Ala Pro Val Ala Ile Ser Leu Thr Asp Ala Asp Ala Arg Ile Val
35 40 45
Tyr Ala Asn Pro Ala Phe Cys Arg Gln Thr Gly Tyr Glu Leu Glu Ala
50 55 60
Leu Leu Gln Gln Asn Pro Arg Leu Leu Ala Ser Arg Gln Thr Pro Arg
65 70 75 80
Glu Ile Tyr Gln Asp Met Trp His Thr Leu Leu Gln Arg Arg Pro Trp
85 90 95
Arg Gly Gln Leu Ile Asn Arg His Arg Asp Gly Ser Leu Tyr Leu Val
100 105 110
Glu Ile Asp Ile Thr Pro Val Ile Asn Pro Phe Gly Glu Leu Glu His
115 120 125
Tyr Leu Ala Met Gln Arg Asp Ile Ser Ala Ser Tyr Ala Leu Glu Gln
130 135 140
Arg Leu Arg Asn His Met Thr Leu Thr Glu Ala Val Leu Asn Asn Ile
145 150 155 160
Pro Ala Ala Val Val Val Val Asp Glu Arg Asp His Val Val Met Asp
165 170 175
Asn Leu Ala Tyr Lys Thr Phe Cys Ala Asp Cys Gly Gly Lys Glu Leu
180 185 190
Leu Ser Glu Leu Asn Phe Ser Ala Arg Lys Ala Glu Leu Ala Asn Gly
195 200 205
Gln Val Leu Pro Val Val Leu Arg Gly Glu Val Arg Trp Leu Ser Val
210 215 220
Thr Cys Trp Ala Leu Pro Gly Val Ser Glu Glu Ala Ser Arg Tyr Phe
225 230 235 240
Ile Asp Asn Arg Leu Thr Arg Thr Leu Val Val Ile Thr Asp Asp Thr
245 250 255
Gln Gln Arg Gln Gln Gln Glu Gln Gly Arg Leu Asp Arg Leu Lys Gln
260 265 270
Gln Met Thr Asn Gly Lys Leu Leu Ala Ala Ile Arg Glu Ala Leu Asp
275 280 285
Ala Ala Leu Ile Gln Leu Asn Cys Pro Ile Asn Met Leu Ala Ala Ala
290 295 300
Arg Arg Leu Asn Gly Ser Asp Asn Asn Asn Val Ala Leu Asp Ala Ala
305 310 315 320
Trp Arg Glu Gly Glu Glu Ala Met Ala Arg Leu Lys Arg Cys Arg Pro
325 330 335
Ser Leu Glu Leu Glu Ser Ala Ala Val Trp Pro Leu Gln Pro Phe Phe
340 345 350
Asp Asp Leu Arg Ala Leu Tyr His Thr Arg Tyr Glu Gln Gly Lys Asn
355 360 365
Leu Gln Val Thr Leu Asp Ser His His Leu Val Gly Phe Gly Gln Arg
370 375 380
Thr Gln Leu Leu Ala Cys Leu Ser Leu Trp Leu Asp Arg Thr Leu Asp
385 390 395 400
Ile Ala Ala Gly Leu Gly Asp Phe Thr Ala Gln Thr Gln Ile Tyr Ala
405 410 415
Arg Glu Glu Glu Gly Trp Leu Ser Leu Tyr Ile Thr Asp Asn Val Pro
420 425 430
Leu Ile Pro Leu Arg His Thr His Ser Pro Asp Ala Leu Asn Ala Pro
435 440 445
Gly Lys Gly Met Glu Leu Arg Leu Ile Gln Thr Leu Val Ala His His
450 455 460
His Gly Ala Ile Glu Leu Thr Ser His Pro Glu Gly Gly Ser Cys Leu
465 470 475 480
Thr Leu Arg Phe Pro Leu Phe His Ser Leu Thr Gly Gly Ser Lys
485 490 495
<210> 78
<211> 524
<212> Белок
<213> Kosakonia sacchari
<220>
<223> NifA, штамм CI006
<400> 78
Met Thr Gln Arg Thr Glu Ser Gly Asn Thr Val Trp Arg Phe Asp Leu
1 5 10 15
Ser Gln Gln Phe Thr Ala Met Gln Arg Ile Ser Val Val Leu Ser Arg
20 25 30
Ala Thr Glu Val Asp Gln Thr Leu Gln Gln Val Leu Cys Val Leu His
35 40 45
Asn Asp Ala Phe Leu Gln His Gly Met Ile Cys Leu Tyr Asp Ser Gln
50 55 60
Gln Ala Ile Leu Asn Ile Glu Ala Leu Gln Glu Ala Asp Gln Gln Leu
65 70 75 80
Ile Pro Gly Ser Ser Gln Ile Arg Tyr Arg Pro Gly Glu Gly Leu Val
85 90 95
Gly Thr Val Leu Ser Gln Gly Gln Ser Leu Val Leu Ala Arg Val Ala
100 105 110
Asp Asp Gln Arg Phe Leu Asp Arg Leu Gly Leu Tyr Asp Tyr Asn Leu
115 120 125
Pro Phe Ile Ala Val Pro Leu Ile Gly Pro Asp Ala Gln Thr Phe Gly
130 135 140
Val Leu Thr Ala Gln Pro Met Ala Arg Tyr Glu Glu Arg Leu Pro Ala
145 150 155 160
Cys Thr Arg Phe Leu Glu Thr Val Ala Asn Leu Val Ala Gln Thr Val
165 170 175
Arg Leu Met Ala Pro Pro Ala Val Arg Pro Ser Pro Arg Ala Ala Ile
180 185 190
Thr Gln Ala Ala Ser Pro Lys Ser Cys Thr Ala Ser Arg Ala Phe Gly
195 200 205
Phe Glu Asn Met Val Gly Asn Ser Pro Ala Met Arg Gln Thr Met Glu
210 215 220
Ile Ile Arg Gln Val Ser Arg Trp Asp Thr Thr Val Leu Val Arg Gly
225 230 235 240
Glu Ser Gly Thr Gly Lys Glu Leu Ile Ala Asn Ala Ile His His His
245 250 255
Ser Pro Arg Ala Gly Ala Pro Phe Val Lys Phe Asn Cys Ala Ala Leu
260 265 270
Pro Asp Thr Leu Leu Glu Ser Glu Leu Phe Gly His Glu Lys Gly Ala
275 280 285
Phe Thr Gly Ala Val Arg Gln Arg Lys Gly Arg Phe Glu Leu Ala Asp
290 295 300
Gly Gly Thr Leu Phe Leu Asp Glu Ile Gly Glu Ser Ser Ala Ser Phe
305 310 315 320
Gln Ala Lys Leu Leu Arg Ile Leu Gln Glu Gly Glu Met Glu Arg Val
325 330 335
Gly Gly Asp Glu Thr Leu Gln Val Asn Val Arg Ile Ile Ala Ala Thr
340 345 350
Asn Arg Asn Leu Glu Asp Glu Val Arg Leu Gly His Phe Arg Glu Asp
355 360 365
Leu Tyr Tyr Arg Leu Asn Val Met Pro Ile Ala Leu Pro Pro Leu Arg
370 375 380
Glu Arg Gln Glu Asp Ile Ala Glu Leu Ala His Phe Leu Val Arg Lys
385 390 395 400
Ile Ala His Asn Gln Ser Arg Thr Leu Arg Ile Ser Glu Gly Ala Ile
405 410 415
Arg Leu Leu Met Ser Tyr Asn Trp Pro Gly Asn Val Arg Glu Leu Glu
420 425 430
Asn Cys Leu Glu Arg Ser Ala Val Met Ser Glu Asn Gly Leu Ile Asp
435 440 445
Arg Asp Val Ile Leu Phe Asn His Arg Asp Gln Pro Ala Lys Pro Pro
450 455 460
Val Ile Ser Val Ser His Asp Asp Asn Trp Leu Asp Asn Asn Leu Asp
465 470 475 480
Glu Arg Gln Arg Leu Ile Ala Ala Leu Glu Lys Ala Gly Trp Val Gln
485 490 495
Ala Lys Ala Ala Arg Leu Leu Gly Met Thr Pro Arg Gln Val Ala Tyr
500 505 510
Arg Ile Gln Thr Met Asp Ile Thr Leu Pro Arg Leu
515 520
<210> 79
<211> 2850
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> glnE, штамм CI006
<400> 79
atgccgcacc acgcaggatt gtcgcagcac tggcaaacgg tattttctcg tctgccggaa 60
tcgctcaccg cgcagccatt gagcgcgcag gcgcagtcag tgctcacttt tagtgatttt 120
gttcaggaca gcatcatcgc gcatcctgag tggctggcag agcttgaaag cgcgccgccg 180
cctgcgaacg aatggcaaca ctatgcgcaa tggctgcaag cggcgctgga tggcgtcacc 240
gatgaagcct cgctgatgcg cgcgctgcgg ctgtttcgcc gtcgcatcat ggtgcgcatc 300
gcctggagcc aggcgttaca gttggtggcg gaagaagata tcctgcaaca gcttagcgtg 360
ctggcggaaa ccctgatcgt cgccgcgcgc gactggcttt atgaggcctg ctgccgtgag 420
tggggaacgc cgagcaatcc acaaggcgtg gcgcagccga tgctggtact cggcatgggc 480
aaactgggtg gcggcgaact caatttctca tccgatatcg atttgatttt cgcctggccg 540
gaaaatggcg caacgcgcgg tggacgccgt gagctggata acgcgcaatt tttcactcgc 600
cttggtcaac ggctgattaa agtcctcgac cagccaacgc aggatggctt tgtctaccgc 660
gtcgatatgc gcttgcgccc gtttggcgac agcggcccgc tggtgctgag ctttgccgcg 720
ctggaagatt actaccagga gcaggggcgc gattgggaac gctacgcgat ggtgaaagcg 780
cgcattatgg gcgataacga cggcgaccat gcgcgggagt tgcgcgcaat gctgcgcccg 840
tttgttttcc gccgttatat cgacttcagc gtgattcagt ccctgcgtaa catgaaaggc 900
atgattgccc gcgaagtgcg tcgccgtggc ctgaaggaca acattaagct cggcgcgggc 960
gggatccgcg aaatagaatt tatcgtccag gttttccagc tgattcgcgg cggtcgcgag 1020
cctgcactgc aatcgcgttc actgttgccg acgcttgctg ccatagatca actgcatctg 1080
ctgccggatg gcgacgcaac ccggctgcgc gaggcgtatt tgtggctgcg acggctggag 1140
aacctgctgc aaagcatcaa tgacgaacag acacagacgc tgccgggcga tgaactgaat 1200
cgcgcgcgcc tcgcctgggg aatgggcaaa gatagctggg aagcgctctg cgaaacgctg 1260
gaagcgcata tgtcggcggt gcgtcagata tttaacgatc tgattggcga tgatgaaacg 1320
gattcgccgg aagatgcgct ttctgagagc tggcgcgaat tgtggcagga tgcgttgcag 1380
gaggaggatt ccacgcccgt gctggcgcat ctctcagagg acgatcgccg ccgcgtggtg 1440
gcgctgattg ccgattttcg caaagagttg gataaacgca ccattggccc gcgagggcgg 1500
caggtactcg atcacttaat gccgcatctg ctcagcgatg tatgctcgcg cgacgatgcg 1560
ccagtaccgc tgtcacgcct gacgccgctg ctcaccggaa ttattacccg caccacttac 1620
cttgagctgc taagtgaatt tcccggcgca ctgaaacacc tcatttccct gtgtgccgcg 1680
tcgccgatgg ttgccagtca gctggcgcgc tacccgatcc tgcttgatga attgctcgac 1740
ccgaatacgc tctatcaacc gacggcgatg aatgcctatc gcgatgagct gcgccaatac 1800
ctgctgcgcg tgccggaaga tgatgaagag caacagcttg aggcgctgcg gcagtttaag 1860
caggcgcagt tgctgcgcgt ggcggcggcg gatattgccg gtacgttgcc agtaatgaaa 1920
gtgagcgatc acttaacctg gctggcggaa gcgattattg atgcggtggt gcagcaagcc 1980
tgggggcaga tggtggcgcg ttatggccag ccaacgcatc tgcacgatcg cgaagggcgc 2040
ggttttgcgg tggtcggtta tggcaagctg ggcggctggg agctgggtta cagctccgat 2100
ctggatctgg tattcctgca cgactgcccg atggatgtga tgaccgatgg cgagcgtgaa 2160
atcgatggtc gccagttcta tttgcgtctc gcgcagcgcg tgatgcacct gtttagcacg 2220
cgcacgtcgt ccggcatcct ttatgaagtt gatgcgcgtc tgcgtccatc tggcgctgcg 2280
gggatgctgg tcactactac ggaatcgttc gccgattacc agcaaaacga agcctggacg 2340
tgggaacatc aggcgctggc ccgtgcgcgc gtggtgtacg gcgatccgca actgaccgcc 2400
gaatttgacg ccattcgccg cgatattctg atgacgcctc gcgacggcgc aacgctgcaa 2460
accgacgtgc gagaaatgcg cgagaaaatg cgtgcccatc ttggcaacaa gcataaagac 2520
cgcttcgatc tgaaagccga tgaaggcggt atcaccgaca tcgagtttat cgcccaatat 2580
ctggtgctgc gctttgccca tgacaagccg aaactgacgc gctggtcgga taatgtgcgc 2640
attctcgaag ggctggcgca aaacggcatc atggaggagc aggaagcgca ggcattgacg 2700
ctggcgtaca ccacattgcg tgatgagctg caccacctgg cgctgcaaga gttgccggga 2760
catgtggcgc tctcctgttt tgtcgccgag cgtgcgctta ttaaaaccag ctgggacaag 2820
tggctggtgg aaccgtgcgc cccggcgtaa 2850
<210> 80
<211> 1563
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> glnE_KO1, штамм CI006
<400> 80
atgtttaacg atctgattgg cgatgatgaa acggattcgc cggaagatgc gctttctgag 60
agctggcgcg aattgtggca ggatgcgttg caggaggagg attccacgcc cgtgctggcg 120
catctctcag aggacgatcg ccgccgcgtg gtggcgctga ttgccgattt tcgcaaagag 180
ttggataaac gcaccattgg cccgcgaggg cggcaggtac tcgatcactt aatgccgcat 240
ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac cgctgtcacg cctgacgccg 300
ctgctcaccg gaattattac ccgcaccact taccttgagc tgctaagtga atttcccggc 360
gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg 420
cgctacccga tcctgcttga tgaattgctc gacccgaata cgctctatca accgacggcg 480
atgaatgcct atcgcgatga gctgcgccaa tacctgctgc gcgtgccgga agatgatgaa 540
gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc agttgctgcg cgtggcggcg 600
gcggatattg ccggtacgtt gccagtaatg aaagtgagcg atcacttaac ctggctggcg 660
gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc agatggtggc gcgttatggc 720
cagccaacgc atctgcacga tcgcgaaggg cgcggttttg cggtggtcgg ttatggcaag 780
ctgggcggct gggagctggg ttacagctcc gatctggatc tggtattcct gcacgactgc 840
ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg gtcgccagtt ctatttgcgt 900
ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt cgtccggcat cctttatgaa 960
gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc tggtcactac tacggaatcg 1020
ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac atcaggcgct ggcccgtgcg 1080
cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg acgccattcg ccgcgatatt 1140
ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg tgcgagaaat gcgcgagaaa 1200
atgcgtgccc atcttggcaa caagcataaa gaccgcttcg atctgaaagc cgatgaaggc 1260
ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc tgcgctttgc ccatgacaag 1320
ccgaaactga cgcgctggtc ggataatgtg cgcattctcg aagggctggc gcaaaacggc 1380
atcatggagg agcaggaagc gcaggcattg acgctggcgt acaccacatt gcgtgatgag 1440
ctgcaccacc tggcgctgca agagttgccg ggacatgtgg cgctctcctg ttttgtcgcc 1500
gagcgtgcgc ttattaaaac cagctgggac aagtggctgg tggaaccgtg cgccccggcg 1560
taa 1563
<210> 81
<211> 949
<212> Белок
<213> Kosakonia sacchari
<220>
<223> glnE, штамм CI006
<400> 81
Met Pro His His Ala Gly Leu Ser Gln His Trp Gln Thr Val Phe Ser
1 5 10 15
Arg Leu Pro Glu Ser Leu Thr Ala Gln Pro Leu Ser Ala Gln Ala Gln
20 25 30
Ser Val Leu Thr Phe Ser Asp Phe Val Gln Asp Ser Ile Ile Ala His
35 40 45
Pro Glu Trp Leu Ala Glu Leu Glu Ser Ala Pro Pro Pro Ala Asn Glu
50 55 60
Trp Gln His Tyr Ala Gln Trp Leu Gln Ala Ala Leu Asp Gly Val Thr
65 70 75 80
Asp Glu Ala Ser Leu Met Arg Ala Leu Arg Leu Phe Arg Arg Arg Ile
85 90 95
Met Val Arg Ile Ala Trp Ser Gln Ala Leu Gln Leu Val Ala Glu Glu
100 105 110
Asp Ile Leu Gln Gln Leu Ser Val Leu Ala Glu Thr Leu Ile Val Ala
115 120 125
Ala Arg Asp Trp Leu Tyr Glu Ala Cys Cys Arg Glu Trp Gly Thr Pro
130 135 140
Ser Asn Pro Gln Gly Val Ala Gln Pro Met Leu Val Leu Gly Met Gly
145 150 155 160
Lys Leu Gly Gly Gly Glu Leu Asn Phe Ser Ser Asp Ile Asp Leu Ile
165 170 175
Phe Ala Trp Pro Glu Asn Gly Ala Thr Arg Gly Gly Arg Arg Glu Leu
180 185 190
Asp Asn Ala Gln Phe Phe Thr Arg Leu Gly Gln Arg Leu Ile Lys Val
195 200 205
Leu Asp Gln Pro Thr Gln Asp Gly Phe Val Tyr Arg Val Asp Met Arg
210 215 220
Leu Arg Pro Phe Gly Asp Ser Gly Pro Leu Val Leu Ser Phe Ala Ala
225 230 235 240
Leu Glu Asp Tyr Tyr Gln Glu Gln Gly Arg Asp Trp Glu Arg Tyr Ala
245 250 255
Met Val Lys Ala Arg Ile Met Gly Asp Asn Asp Gly Asp His Ala Arg
260 265 270
Glu Leu Arg Ala Met Leu Arg Pro Phe Val Phe Arg Arg Tyr Ile Asp
275 280 285
Phe Ser Val Ile Gln Ser Leu Arg Asn Met Lys Gly Met Ile Ala Arg
290 295 300
Glu Val Arg Arg Arg Gly Leu Lys Asp Asn Ile Lys Leu Gly Ala Gly
305 310 315 320
Gly Ile Arg Glu Ile Glu Phe Ile Val Gln Val Phe Gln Leu Ile Arg
325 330 335
Gly Gly Arg Glu Pro Ala Leu Gln Ser Arg Ser Leu Leu Pro Thr Leu
340 345 350
Ala Ala Ile Asp Gln Leu His Leu Leu Pro Asp Gly Asp Ala Thr Arg
355 360 365
Leu Arg Glu Ala Tyr Leu Trp Leu Arg Arg Leu Glu Asn Leu Leu Gln
370 375 380
Ser Ile Asn Asp Glu Gln Thr Gln Thr Leu Pro Gly Asp Glu Leu Asn
385 390 395 400
Arg Ala Arg Leu Ala Trp Gly Met Gly Lys Asp Ser Trp Glu Ala Leu
405 410 415
Cys Glu Thr Leu Glu Ala His Met Ser Ala Val Arg Gln Ile Phe Asn
420 425 430
Asp Leu Ile Gly Asp Asp Glu Thr Asp Ser Pro Glu Asp Ala Leu Ser
435 440 445
Glu Ser Trp Arg Glu Leu Trp Gln Asp Ala Leu Gln Glu Glu Asp Ser
450 455 460
Thr Pro Val Leu Ala His Leu Ser Glu Asp Asp Arg Arg Arg Val Val
465 470 475 480
Ala Leu Ile Ala Asp Phe Arg Lys Glu Leu Asp Lys Arg Thr Ile Gly
485 490 495
Pro Arg Gly Arg Gln Val Leu Asp His Leu Met Pro His Leu Leu Ser
500 505 510
Asp Val Cys Ser Arg Asp Asp Ala Pro Val Pro Leu Ser Arg Leu Thr
515 520 525
Pro Leu Leu Thr Gly Ile Ile Thr Arg Thr Thr Tyr Leu Glu Leu Leu
530 535 540
Ser Glu Phe Pro Gly Ala Leu Lys His Leu Ile Ser Leu Cys Ala Ala
545 550 555 560
Ser Pro Met Val Ala Ser Gln Leu Ala Arg Tyr Pro Ile Leu Leu Asp
565 570 575
Glu Leu Leu Asp Pro Asn Thr Leu Tyr Gln Pro Thr Ala Met Asn Ala
580 585 590
Tyr Arg Asp Glu Leu Arg Gln Tyr Leu Leu Arg Val Pro Glu Asp Asp
595 600 605
Glu Glu Gln Gln Leu Glu Ala Leu Arg Gln Phe Lys Gln Ala Gln Leu
610 615 620
Leu Arg Val Ala Ala Ala Asp Ile Ala Gly Thr Leu Pro Val Met Lys
625 630 635 640
Val Ser Asp His Leu Thr Trp Leu Ala Glu Ala Ile Ile Asp Ala Val
645 650 655
Val Gln Gln Ala Trp Gly Gln Met Val Ala Arg Tyr Gly Gln Pro Thr
660 665 670
His Leu His Asp Arg Glu Gly Arg Gly Phe Ala Val Val Gly Tyr Gly
675 680 685
Lys Leu Gly Gly Trp Glu Leu Gly Tyr Ser Ser Asp Leu Asp Leu Val
690 695 700
Phe Leu His Asp Cys Pro Met Asp Val Met Thr Asp Gly Glu Arg Glu
705 710 715 720
Ile Asp Gly Arg Gln Phe Tyr Leu Arg Leu Ala Gln Arg Val Met His
725 730 735
Leu Phe Ser Thr Arg Thr Ser Ser Gly Ile Leu Tyr Glu Val Asp Ala
740 745 750
Arg Leu Arg Pro Ser Gly Ala Ala Gly Met Leu Val Thr Thr Thr Glu
755 760 765
Ser Phe Ala Asp Tyr Gln Gln Asn Glu Ala Trp Thr Trp Glu His Gln
770 775 780
Ala Leu Ala Arg Ala Arg Val Val Tyr Gly Asp Pro Gln Leu Thr Ala
785 790 795 800
Glu Phe Asp Ala Ile Arg Arg Asp Ile Leu Met Thr Pro Arg Asp Gly
805 810 815
Ala Thr Leu Gln Thr Asp Val Arg Glu Met Arg Glu Lys Met Arg Ala
820 825 830
His Leu Gly Asn Lys His Lys Asp Arg Phe Asp Leu Lys Ala Asp Glu
835 840 845
Gly Gly Ile Thr Asp Ile Glu Phe Ile Ala Gln Tyr Leu Val Leu Arg
850 855 860
Phe Ala His Asp Lys Pro Lys Leu Thr Arg Trp Ser Asp Asn Val Arg
865 870 875 880
Ile Leu Glu Gly Leu Ala Gln Asn Gly Ile Met Glu Glu Gln Glu Ala
885 890 895
Gln Ala Leu Thr Leu Ala Tyr Thr Thr Leu Arg Asp Glu Leu His His
900 905 910
Leu Ala Leu Gln Glu Leu Pro Gly His Val Ala Leu Ser Cys Phe Val
915 920 925
Ala Glu Arg Ala Leu Ile Lys Thr Ser Trp Asp Lys Trp Leu Val Glu
930 935 940
Pro Cys Ala Pro Ala
945
<210> 82
<211> 520
<212> Белок
<213> Kosakonia sacchari
<220>
<223> GlnE_KO1, штамм CI006
<400> 82
Met Phe Asn Asp Leu Ile Gly Asp Asp Glu Thr Asp Ser Pro Glu Asp
1 5 10 15
Ala Leu Ser Glu Ser Trp Arg Glu Leu Trp Gln Asp Ala Leu Gln Glu
20 25 30
Glu Asp Ser Thr Pro Val Leu Ala His Leu Ser Glu Asp Asp Arg Arg
35 40 45
Arg Val Val Ala Leu Ile Ala Asp Phe Arg Lys Glu Leu Asp Lys Arg
50 55 60
Thr Ile Gly Pro Arg Gly Arg Gln Val Leu Asp His Leu Met Pro His
65 70 75 80
Leu Leu Ser Asp Val Cys Ser Arg Asp Asp Ala Pro Val Pro Leu Ser
85 90 95
Arg Leu Thr Pro Leu Leu Thr Gly Ile Ile Thr Arg Thr Thr Tyr Leu
100 105 110
Glu Leu Leu Ser Glu Phe Pro Gly Ala Leu Lys His Leu Ile Ser Leu
115 120 125
Cys Ala Ala Ser Pro Met Val Ala Ser Gln Leu Ala Arg Tyr Pro Ile
130 135 140
Leu Leu Asp Glu Leu Leu Asp Pro Asn Thr Leu Tyr Gln Pro Thr Ala
145 150 155 160
Met Asn Ala Tyr Arg Asp Glu Leu Arg Gln Tyr Leu Leu Arg Val Pro
165 170 175
Glu Asp Asp Glu Glu Gln Gln Leu Glu Ala Leu Arg Gln Phe Lys Gln
180 185 190
Ala Gln Leu Leu Arg Val Ala Ala Ala Asp Ile Ala Gly Thr Leu Pro
195 200 205
Val Met Lys Val Ser Asp His Leu Thr Trp Leu Ala Glu Ala Ile Ile
210 215 220
Asp Ala Val Val Gln Gln Ala Trp Gly Gln Met Val Ala Arg Tyr Gly
225 230 235 240
Gln Pro Thr His Leu His Asp Arg Glu Gly Arg Gly Phe Ala Val Val
245 250 255
Gly Tyr Gly Lys Leu Gly Gly Trp Glu Leu Gly Tyr Ser Ser Asp Leu
260 265 270
Asp Leu Val Phe Leu His Asp Cys Pro Met Asp Val Met Thr Asp Gly
275 280 285
Glu Arg Glu Ile Asp Gly Arg Gln Phe Tyr Leu Arg Leu Ala Gln Arg
290 295 300
Val Met His Leu Phe Ser Thr Arg Thr Ser Ser Gly Ile Leu Tyr Glu
305 310 315 320
Val Asp Ala Arg Leu Arg Pro Ser Gly Ala Ala Gly Met Leu Val Thr
325 330 335
Thr Thr Glu Ser Phe Ala Asp Tyr Gln Gln Asn Glu Ala Trp Thr Trp
340 345 350
Glu His Gln Ala Leu Ala Arg Ala Arg Val Val Tyr Gly Asp Pro Gln
355 360 365
Leu Thr Ala Glu Phe Asp Ala Ile Arg Arg Asp Ile Leu Met Thr Pro
370 375 380
Arg Asp Gly Ala Thr Leu Gln Thr Asp Val Arg Glu Met Arg Glu Lys
385 390 395 400
Met Arg Ala His Leu Gly Asn Lys His Lys Asp Arg Phe Asp Leu Lys
405 410 415
Ala Asp Glu Gly Gly Ile Thr Asp Ile Glu Phe Ile Ala Gln Tyr Leu
420 425 430
Val Leu Arg Phe Ala His Asp Lys Pro Lys Leu Thr Arg Trp Ser Asp
435 440 445
Asn Val Arg Ile Leu Glu Gly Leu Ala Gln Asn Gly Ile Met Glu Glu
450 455 460
Gln Glu Ala Gln Ala Leu Thr Leu Ala Tyr Thr Thr Leu Arg Asp Glu
465 470 475 480
Leu His His Leu Ala Leu Gln Glu Leu Pro Gly His Val Ala Leu Ser
485 490 495
Cys Phe Val Ala Glu Arg Ala Leu Ile Lys Thr Ser Trp Asp Lys Trp
500 505 510
Leu Val Glu Pro Cys Ala Pro Ala
515 520
<210> 83
<211> 341
<212> Белок
<213> Kosakonia sacchari
<220>
<223> 3АТ-азный домен, штамм CI006
<400> 83
Glu Glu Gln Gln Leu Glu Ala Leu Arg Gln Phe Lys Gln Ala Gln Leu
1 5 10 15
Leu Arg Val Ala Ala Ala Asp Ile Ala Gly Thr Leu Pro Val Met Lys
20 25 30
Val Ser Asp His Leu Thr Trp Leu Ala Glu Ala Ile Ile Asp Ala Val
35 40 45
Val Gln Gln Ala Trp Gly Gln Met Val Ala Arg Tyr Gly Gln Pro Thr
50 55 60
His Leu His Asp Arg Glu Gly Arg Gly Phe Ala Val Val Gly Tyr Gly
65 70 75 80
Lys Leu Gly Gly Trp Glu Leu Gly Tyr Ser Ser Asp Leu Asp Leu Val
85 90 95
Phe Leu His Asp Cys Pro Met Asp Val Met Thr Asp Gly Glu Arg Glu
100 105 110
Ile Asp Gly Arg Gln Phe Tyr Leu Arg Leu Ala Gln Arg Val Met His
115 120 125
Leu Phe Ser Thr Arg Thr Ser Ser Gly Ile Leu Tyr Glu Val Asp Ala
130 135 140
Arg Leu Arg Pro Ser Gly Ala Ala Gly Met Leu Val Thr Thr Thr Glu
145 150 155 160
Ser Phe Ala Asp Tyr Gln Gln Asn Glu Ala Trp Thr Trp Glu His Gln
165 170 175
Ala Leu Ala Arg Ala Arg Val Val Tyr Gly Asp Pro Gln Leu Thr Ala
180 185 190
Glu Phe Asp Ala Ile Arg Arg Asp Ile Leu Met Thr Pro Arg Asp Gly
195 200 205
Ala Thr Leu Gln Thr Asp Val Arg Glu Met Arg Glu Lys Met Arg Ala
210 215 220
His Leu Gly Asn Lys His Lys Asp Arg Phe Asp Leu Lys Ala Asp Glu
225 230 235 240
Gly Gly Ile Thr Asp Ile Glu Phe Ile Ala Gln Tyr Leu Val Leu Arg
245 250 255
Phe Ala His Asp Lys Pro Lys Leu Thr Arg Trp Ser Asp Asn Val Arg
260 265 270
Ile Leu Glu Gly Leu Ala Gln Asn Gly Ile Met Glu Glu Gln Glu Ala
275 280 285
Gln Ala Leu Thr Leu Ala Tyr Thr Thr Leu Arg Asp Glu Leu His His
290 295 300
Leu Ala Leu Gln Glu Leu Pro Gly His Val Ala Leu Ser Cys Phe Val
305 310 315 320
Ala Glu Arg Ala Leu Ile Lys Thr Ser Trp Asp Lys Trp Leu Val Glu
325 330 335
Pro Cys Ala Pro Ala
340
<210> 84
<211> 1342
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> Prm5 встроенный в область nifL, штамм CI006, в настоящее время штамм CM029
<400> 84
ccgagcgtcg gggtgcctaa tatcagcacc ggatacgaga gaaaagtgtc tacatcggtt 60
cggttgatat tgaccggcgc atccgccagc ccgcccagtt tctggtggat ctgtttggcg 120
attttgcggg tcttgccggt gtcggtgccg aaaaaaatac caatatttgc cataacacac 180
gctcctgttg aaaaagagat cccgccggga aatgcggtga acgtgtctga tattgcgaag 240
agtgtgccag ttttggtcgc gggcaaaacc tgcaccagtt tggttattaa tgcaccagtc 300
tggcgctttt tttcgccgag tttctcctcg ctaatgcccg ccaggcgcgg ctttggcgct 360
gatagcgcgc tgaataccga tctggatcaa ggttttgtcg ggttatcagc caaaaggtgc 420
actctttgca tggttatacg tgcctgacat gttgtccggg cgacaaacgg cctggtggca 480
caaattgtca gaactacgac acgactaact gaccgcagga gtgtgcgatg accctgaata 540
tgatgatgga tgccggcgga catcatcgcg acaaacaata ttaataccgg caaccacacc 600
ggcaatttac gagactgcgc aggcatcctt tctcccgtca atttctgtca aataaagtaa 660
aagaggcagt ctacttgaat tacccccggc tggttgagcg tttgttgaaa aaaagtaact 720
gaaaaatccg tagaatagcg ccactctgat ggttaattaa cctattcaat taagaattat 780
ctggatgaat gtgccattaa atgcgcagca taatggtgcg ttgtgcggga aaactgcttt 840
tttttgaaag ggttggtcag tagcggaaac aactcacttc acaccccgaa gggggaagtt 900
gcctgaccct acgattcccg ctatttcatt cactgaccgg aggttcaaaa tgacccagcg 960
aaccgagtcg ggtaataccg tctggcgctt cgatttgtcc cagcagttca ctgcgatgca 1020
gcgcataagc gtggtactca gccgggcgac cgaggtcgat cagacgctcc agcaagtgct 1080
gtgcgtattg cacaatgacg cctttttgca gcacggcatg atctgtctgt acgacagcca 1140
gcaggcgatt ttgaatattg aagcgttgca ggaagccgat cagcagttaa tccccggcag 1200
ctcgcaaatc cgctatcgtc cgggcgaagg gctggtcggg acggtgcttt cgcagggcca 1260
atcattagtg ctggcgcgcg ttgctgacga tcagcgcttt cttgaccggc tcgggttgta 1320
tgattacaac ctgccgttta tc 1342
<210> 85
<211> 1270
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (186)..(186)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (458)..(458)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1041)..(1041)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1131)..(1132)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1135)..(1135)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1139)..(1140)
<223> a, c, t, g, неизвестное или другое
<400> 85
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcag cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgnaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggctcacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcatcanac ttaatacgtg tggtgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgcgcttaa cgtgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccacg gaattcgcca gagatggctt 1020
agtgccttcg ggaaccgtga nacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc nngtnatgnn 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgagg 1270
<210> 86
<211> 876
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifH
<400> 86
atggcaatgc gtcaatgcgc aatctacggg aaagggggta ttgggaaatc caccactacc 60
caaaaccttg tagcggctct ggccgaaatg aataagaagg tcatgatcgt cggctgtgac 120
cctaaggctg attcaacccg cctcattctg catgcgaaag cacagaacac catcatggaa 180
atggccgctg aagtgggctc cgtggaagat ctggagctgg aagatgtgat gcaaatcggc 240
tatggcggcg tgcgctgtgc ggaatcaggc ggccctgagc ctggtgtggg ttgtgccgga 300
cgcggggtga tcaccgccat caacttcctc gaagaagaag gcgcgtatgt gccggatctg 360
gattttgtgt tttacgacgt attgggcgat gtggtctgtg gcggtttcgc gatgccaatt 420
cgcgaaaaca aagcgcagga aatctacatc gtgtgctccg gtgaaatgat ggcgatgtat 480
gccgccaaca acatttccaa aggcatcgtg aaatacgcga aatcgggcaa agttcgcctg 540
gccgggctga tctgtaactc ccgccagacg gatcgcgaag atgaactgat catcgcgctg 600
gctgaaaaac ttggcacgca aatgatccac ttcgtgccgc gtgacaacat tgtgcaacgc 660
gctgaaatcc gccgcatgac ggtcatcgaa tacgacccga cttgtgcgca ggcagatcag 720
tatcgtgcac tggcgaacaa aatcgtcaac aacaccaaaa tggtggtgcc gacaccggtc 780
accatggatg agctggaagc cctgttaatg gaatttggca ttatggaaga agaagacctg 840
gccatcgtcg gtcgtaccgc cgccgaagag gcgtga 876
<210> 87
<211> 1374
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifD1
<400> 87
atgaaggcaa aagagattct ggcgctgatt gatgagccag cctgtgagca taaccacaag 60
cagaagtcgg gttgcagcct gccgaaaccg ggcgcgacgg caggcggttg tgcgtttgat 120
ggcgcgcaga ttgcgctgct gccggtcgcg gacgtcgcgc atctggtgca cggcccgatt 180
ggctgtaccg gcagttcatg ggacaaccgt ggcagccgca gttccgggcc ttccatcaac 240
cgcatgggct tcaccaccga catgagcgag caggatgtga ttatggggcg cggcgagcga 300
cgcttatttc acgccgtgca gcacatcgtc agccattacc atccggtggc ggtctttatt 360
tacaacacct gcgtacccgc gatggaaggg gatgacgttg aagccgtgtg tcgcgccgca 420
tcggccgctg ccggtgtgcc ggttatttca gtcgatgccg ccggtttcta cggcagcaaa 480
aatctcggta accgcattgc cggggacgtg atggtcaaaa aggtgatcgg ccagcgcgaa 540
cccgcgccgt ggccggaaaa ctcaccgatc cccgccggac accgccacag catcagcctg 600
attggcgaat tcaatattgc cggcgagttc tggcacgttc tgccgctgct cgatgagctc 660
gggatccgcg tgctgtgcag cctttccggg gattcccgtt ttgctgaaat ccagactatg 720
caccgtggcg aagccaacat gctggtgtgc tcgcgggcgc tgatcaacgt cgcccgaaaa 780
atggaagagc gttaccagat cccatggttt gaaggcagtt tttatggcct gcgttccatg 840
gctgattccc tgcgcacgat cgccgtgctg ctcaaagacc cggatttaca ggcgcgcaca 900
gaacgtctga ttgagcgcga ggaggcggcg acacatcttg cgcttgcgcc ttaccgtgcg 960
cggctcagcg ggcgcaaggc gctgctgtat accggtggcg tgaaatcctg gtcggtggtc 1020
tcggcgttac aggatttagg catcacggtg gtggcgaccg gcacccgaaa atcaaccgaa 1080
gaagacaagc agcgtattcg cgaactgatg ggtgaagacg tgctgatgct cgacgaaggc 1140
aatgccagaa ccttgctcga caccctctat cgtttcggcg gcgacatcat gatcgccggg 1200
ggccgcaaca tgtataccgc gtacaaagcc cgcctgccgt tcctggatat caatcaggag 1260
cgcgagcatg cgtttgccgg atatcacggg ctggtaaatc tggccgaaca gttgtgtatc 1320
accctggaaa gcccggtctg ggcgcaggtc aaccgtctgg cgccgtggcg ctaa 1374
<210> 88
<211> 1449
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifD2
<400> 88
atgaccagtg aaacacgcga acgtaacgag gcattgatcc aggaagtgct ggagatcttc 60
cccgagaagg cgcttaaaga tcgtaagaaa cacatgatga ccaccgaccc ggcgatggaa 120
tctgtcggca agtgtattgt ctcaaaccgc aaatcacagc cgggcgtgat gaccgtgcga 180
ggctgcgctt acgccggttc caaaggcgtg gtctttggcc cgatcaaaga catggcgcat 240
atctcccacg gcccggttgg ttgcggccag tattcccgtg ccggacgccg taactattac 300
accggctgga gcggcgtgaa cagctttggc accctcaact tcaccagtga ttttcaggaa 360
cgggacatcg tatttggcgg cgataaaaag ctcgacaaat tgatcgatga actggagatg 420
ttgttcccgc tgagcaaagg catttcggtg cagtcggaat gtccggtcgg tctgatcggc 480
gatgacattt ctgccgtcgc caaagccagc agcgccaaaa tcggtaagcc ggtcgtgccg 540
gtacgctgcg aggggttccg cggtgtgtcg caatcgctcg gccatcacat tgctaacgat 600
gtcatccgcg actgggtgct ggataaccgc gaaggcaatg aatttgaaac cacgccttac 660
gacgtggcga ttatcggcga ctacaacatc ggcggtgacg cctgggcctc acgtattctg 720
ctcgaagaaa tggggctgcg cgtggtggcg cagtggtccg gcgacggcac gctggtggag 780
atggaaaaca ccccgaaagt cgcgctcaat ctggtgcact gctaccgctc gatgaactac 840
atctcccgtc atatggaaga aaaacacggc attccgtgga tggaatacaa cttctttggc 900
ccgaccaaaa ttgcggaatc tctgcgcgaa atcgcggcgc gttttgacga taccatccgg 960
aaaaacgccg aagcggtgat tgaaaaatat caggcgcaaa cgcaggcggt gatcgacaaa 1020
taccgtccgc gtctggaagg caaaaaggtg ctgttgtatc tcggcggttt acgtccgcgc 1080
cacatcatcg gggcgtatga agatctggga atggaaatca tcggtaccgg ctatgaattc 1140
ggtcataacg atgattacga ccgcacctta ccgatgctca aagaaggcac gttgctgttc 1200
gatgacctga gcagttatga gctggaagcg ttcgttaaag cgctgaaacc ggatcttgtc 1260
gggtcaggta tcaaagaaaa atacattttc cagaaaatgg gcgtgccgtt ccgccagatg 1320
cactcctggg attattccgg cccttatcac ggctacgacg gtttcggcat ttttgcccgt 1380
gacatggaca tgacgctgaa caatccgggc tggagtcagc tgaccgcccc ctggttgaaa 1440
acggcctga 1449
<210> 89
<211> 1386
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifK1
<400> 89
atggctcaaa ttctgcgtaa tgccaagccg cttgccacca cgcctgtcaa aagcgggcaa 60
ccgctcgggg cgatcctggc cagtcagggg ctggaaaatt gcatcccgct ggttcacggc 120
gcgcaaggtt gtagcgcgtt cgccaaagtt ttcttcatcc agcattttca cgatccgatc 180
ccgttgcagt ccacggcgat ggaatcgacc acgactatca tgggctcgga tggcaacgtc 240
agtactgcgt tgaccacgtt gtgtcagcgc agtaatccaa aagccattgt gattttgagc 300
accggactgt cagaagcgca gggcagtgat ttgtcgatgg cgctgcgtga gtttcgcgac 360
aaagaaccgc gctttaatgc catcgctatt ctgaccgtta acacgccgga tttttacggc 420
tcgctggaaa acggctacag cgcgctgatg gaaagcgtga tcactcagtg ggtgccggaa 480
aagccgccga ccggcatgcg taacaagcgc gtgaacctgc tggtgagcca tctgctgacg 540
ccgggggatc tggaattact gcgcagctat gtcgaagcct ttggcctgca accggtgatc 600
ctgccggatt tatcacagtc gctggacgga catctggcga atggcgattt caatccggtc 660
acgcagggcg gcacgtcgca acgccagatt gaacaaatgg ggcagagcct gaccaccatt 720
accattggca gttcgctcaa ctgcgccgcc agtctgatgg cgatgcgcag ccgtggcatg 780
gcgctgaacc tgccgcacct gatgacgctg gaaaacatgg acagtctgat ccgccatctg 840
catcaggtgt caggccgcga ggtaccggca tggattgagc gccagcgcgg gcaactgctg 900
gacgccatga tcgactgcca tacctggctg cagtcacagc gtattgcgct ggcggcagaa 960
gcggatttgc tggtggcgtg gtgcgatttc gctcagagcc agggaatgcg cgtcgggccg 1020
gtgattgcgc cggttaatca gcagtcactg gccgggctgc cggtcgaaca ggtggtgatc 1080
ggcgatctgg aagatttaca aacccggctc gacagctacc cggtttcact gctggtggcg 1140
aactcccacg ctgcaccact ggcggaaaaa aacggtatcg cgctggtacg tgccggtttc 1200
ccgctttacg accgtctcgg ggaatttcgc cgcgtgcggc agggctatgc gggtattcgc 1260
gacaccttgt tcgaactcgc gaacctgatg caggcgcgcc atcacatgct gacggcgtat 1320
cactcaccgc ttaggcaggt gttcggcctg agcccggtac cggaggccag tcatgaggcg 1380
cgctaa 1386
<210> 90
<211> 1569
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifK2
<400> 90
atgagtcaag atcttggcac cccaaaatcc tgtttcccgc tgttcgagca ggatgaatac 60
cagaatatgt ttacccacaa acgcgcgctg gaagaagcac acggcgaggc gaaagtgcgg 120
gaagtgtttg aatggaccac cacgcaggaa tatcaggatc tgaacttctc gcgtgaagcg 180
ctgaccgtcg acccggcgaa agcctgccag ccgttaggcg cggtactttg cgcgctgggt 240
tttaccaaca cgttgccgta tgtccatggt tcacaaggct gtgtggcgta tttccgtacc 300
tattttaatc gtcatttcaa agagccggtg gcctgtgttt ccgactcaat gaccgaagat 360
gccgccgttt ttggcggaaa taacaacatg aatgtcggtc tggaaaacgc cagcgcgctg 420
tacaagccgg aaattattgc ggtctccacc acctgtatgg cggaagtgat cggtgatgac 480
ctgcaggctt ttatcgccaa cgccaaaaaa gacggatttg tggatgccgg tatgccaatc 540
ccgtatgccc atacaccgag ttttctgggc agtcatgtca ccggctggga caacatgttt 600
gaaggcttcg cccgtacctt taccaccgac gccacgcggg aatatcagcc gggcaaactt 660
gccaaactga acgtggtgac cggttttgaa acttatctcg gcaactaccg ggttattcac 720
cgcatgatga gccagatggg ggtcgaatgc agcgtcttgt ccgatccgtc tgaagtgctc 780
gacaccccgg ctgacggcca ataccgcatg tatgccggcg gcaccacgca aaccgaaatg 840
cgtgatgcac cggatgccat cgacaccttg ctgctgcaac cgtggcaatt acagaaaacc 900
aaaaaggtgg tgcagggcga ctggaatcag ccgggcaccg aagtcagtgt accgattggc 960
ctggcggcga ccgatgcctt gctgatgacg gtaagcgaac tgaccggcaa accgatagct 1020
gacgcgctgg cgactgaacg tggccgtctg gtggacatga tgctcgattc tcacacctgg 1080
ctgcacggca agcgtttcgg tctctacggt gacccggatt ttgtgatggg catgaccgca 1140
ttcctgctgg aactgggctg tgaaccgacc accattctca gccataacgg caacaaacgc 1200
tggcagaaag ccatgaagaa aatgctggct gattcgcctt acggacagga cagcgaagtg 1260
tatgtgaact gcgatctgtg gcatttccgc tcgctgatgt ttacccgtaa accggacttt 1320
atgatcggca actcttacgg aaaattcatt cagcgtgaca cgctggccaa aggcgaacag 1380
ttcgaagtgc cgctgatccg tatcggattc ccgatttttg accggcacca tttgcaccgt 1440
cagaccacct ggggatacga gggcgcgatg agcatcctga cgcaactggt gaatgcggtg 1500
ctcgaacagc tggatcgcga aaccatgaag ctcggcaaaa ccgactacaa cttcgatctg 1560
atccgctaa 1569
<210> 91
<211> 1488
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifL
<400> 91
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat catcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg ccagccagca gacgccgaaa 240
catatctatg acgaaatgtg gcgcactttg ttgcagggca aatcctggaa cggccaactg 300
atcaaccggc gtaataaccg ttcgctttat ctggcggatg tcactatcac gcctgtttta 360
ggcgcggacg ggcaggtgga gcattacctc ggcatgcaca aagatatcag cgagaaatac 420
gcgctggagc agcggttgcg caaccacatc accttgttca cggaggtgct gaacaatatt 480
cccgccgccg tggtggtggt ggatgagcag gacaatgtgg tgatggacaa tctggcctac 540
aaaaccctgt gcgctgactg cggcggaaaa gagctgttgg ccgaaatggg ctatccgcaa 600
ctcaaagaga tgctcaacag tggcgaaccg gtgccggttt ccatgcgcgg caacgtacgc 660
tggttttctt tcggtcagtg gtcattgcag ggcgttaatg aagaggccag ccgctttttt 720
accggcatta ccgcgccggg aaaactgatt gttctcaccg actgcaccga tcagcatcac 780
cggcagcagc agggttatct tgaccggctc aagcaaaaac ttaccaacgg caaattgctg 840
gcagccatcc gcgagtcgct tgatgccgcg ctgattcagc tcaacgggcc aattaatatg 900
ctggcggctg cgcgtcgtct taacggcgaa gaaggcaaca acatggcgct ggaattcgcc 960
tggcgcgaag gcgagcaggc ggtgagtcgc ttacaggcct gccgtccgtc gctggatttt 1020
gagccgcagg cagaatggcc ggtcagtgaa ttcttcgacg atctgagcgc gctgtacgac 1080
agccattttc tcagtgacgg tgaattgcgt tacgtggtca tgccatctga tctgcacgct 1140
gtcgggcaac gaacgcaaat ccttaccgcg ctgagcttat ggattgatca cacgctgtca 1200
caggcgcagg ccatgccgtc tctgaagctc tcggtgaaca ttgttgcgaa gcaggatgcg 1260
agctggttgt gttttgacat taccgataat gtgccgcgtg aacgggtgcg ttatgcccgc 1320
ccggaagcgg cgttttcccg tccggggaat ggcatggagc tgcgccttat ccagacgctg 1380
atcgcccatc atcgcggttc tttagatctc tcggtccgcc ctgatggcgg caccttgctg 1440
acgttacgcc tgccggtaca gcaggttatc accggaggct taaaatga 1488
<210> 92
<211> 1557
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifA
<400> 92
atgacccagt tacctaccgc gggcccggtt atccggcgct ttgatatgtc tgcccagttt 60
acggcgcttt atcgcatcag cgtggcgctg agtcaggaaa gcaataccgc gcgcgcactg 120
gcggcgatcc tcgaagtgct tcacgatcat gcatttatgc aatacggcat ggtgtgtctg 180
ttcgataaag aacgcaatgc actgtttgtg gaatccctgc atggcatcga cggcgaaagg 240
aaaaaagaga cccgccatgt ccgttaccgc atgggggaag gcgtgatcgg cgcggtgatg 300
agccagcgtc aggcgctggt gttaccgcgc atttcagacg atcagcgttt tctcgaccgc 360
ctgaatattt acgattacag cctgccgctg attggtgtgc cgatccccgg tgcggataat 420
cagcctgcgg gtgtgctggt ggcacagccg atggcgttgc acgaagaccg gctggctgcc 480
agtacgcggt ttttagaaat ggtcgccaat ctcatcagcc agccactgcg ttctgccacg 540
cccccggaat cattgcctgc tcaaacgccg gtccggtgca gtgttccgcg ccagtttggt 600
tttgagcaga tggtcgggaa aagtcaggcg atgcgccaga cgatggacat tttacggcag 660
gtttccaaat gggataccac ggttctggtg cgtggtgaaa gcggcaccgg caaggaactt 720
atcgccaatg ccattcatta caactcaccc cgtgcggccg cgccatttgt gaaattcaac 780
tgcgccgcgc tgccggataa cctgctggaa agcgaactgt tcggtcatga aaaaggggcc 840
ttcaccggcg ctatacgcac ccgaaaaggc cgctttgaac tggcggacgg gggcacgtta 900
ttcctcgatg aaatcggcga atcgagcgcg tcgtttcagg ccaaattgct gcgcattttg 960
caggaaggtg aaatggaacg ggtcggcggc gataccacgc tgaaagttga tgtgcgcatt 1020
attgctgcca ccaaccgtaa tcttgaggag gaagtgcgtg ccgggaattt tcgcgaagac 1080
ctgtattatc gcctgaacgt gatgccggtt tcgctgcctg cactgcgtga aaggctggat 1140
gatatcgccg atctggcgcc gtttctggtc aaaaagattg cgctgcgtca ggggcgggaa 1200
ctgcgcatca gtgatggtgc ggtgcgtctg ctgatgacct acagctggcc aggcaacgtg 1260
cgtgaactgg aaaactgcct cgaacgggcg tcggtaatga ccgatgaagg gctgatcgac 1320
cgcgacgtga tcctgttcaa tcaccatgag tccccggcgc tgtccgtcaa acccggtctg 1380
ccgctggcga cagatgaaag ctggctggat caggaactcg acgaacgcca gcgggtgatt 1440
gctgcactgg agaaaaccgg ctgggtgcag gccaaagcgg cccgactgct gggcatgaca 1500
ccgcgccaga ttgcctaccg tatccagatt atggacatca acatgcaccg tatctga 1557
<210> 93
<211> 2838
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> glnE
<400> 93
atgttgccac tttcttctgt tttgcaaagc cacgcgcaga gtttgcctga acgctggcat 60
gaacatcctg aaaacctgcc cctccccgat gatgaacagc tggctgtgct gagcagcagt 120
gaattcatga cggacagttt gctggctttt ccgcagtggt ggcatgaaat tgtccaaaat 180
ccccctcagg cgcaggagtg gcaactttac cgtcagtggc tggatgaatc gctgacgcag 240
gtgactgacg aagccgggtt aatgaaagct ttgcgtctgt tccgccgccg tattctgacc 300
cgcattgcgt ggtcacagtc cgcgcaaacc agcgaagcaa aagatacgct tcaccagctg 360
agtgaactgg cggaattatt gattgtcagc gcccgtgact ggctgtatgc cgcttgctgt 420
cgcgagttcg gtacgccggt caatgccgca ggggaaccgc agagaatgct gatcctcggg 480
atgggcaaac tcggcggtgg cgagctgaat ttctcatcgg acatcgacct gatttttgct 540
tatccggaaa atggccagac acgcggcggt cggcgtgaac tggataacgc acaatttttc 600
acccggctcg gccagcgtct gatcaaagcg ctggatcagc ccactatcga cggttttgtc 660
tatcgcgtgg acatgcgttt gcgtccgttc ggcgacagtg gcccgctggt gatgagcttc 720
ccggcactgg aagattatta tcaggaacag gggcgcgact gggaacgcta cgcaatggtg 780
aaagcgcgtc tgatgggcgg cgcggaggac atcagcagtc aggaattgcg taaaatgctg 840
atgccttttg tcttccgccg ttatatcgat ttcagtgtga tccagtccct gcgtaacatg 900
aaaggcatga tcgcccgcga agtacgccgc cgtggtctga aagacaacat caaactcggc 960
gcaggcggta ttcgtgaaat tgaatttatc gtgcaggtat ttcagctgat ccgtggcggt 1020
cgtgaaccgg cattgcagca gcgtgcgttg ttgccaacgc ttcaggcgct ggaaaatctg 1080
gggctgctgc cggtagagca ggtgttgcag ttgcgtaaca gctatctgtt cctgcgacgt 1140
ctggaaaacc tgttgcaggc cattgctgac gagcaaacgc aaaccttacc gtccgatgag 1200
ctgaatcagg cgcgtctggc gtgggggatg aattacgctg gctggccgca gcttctggat 1260
gcagtgaatg ctcacatgca ggccgtacgc gcggtattta acgatctgat tggcgatgac 1320
acgccagatg ccgaagatga cgtgcaactc tcccggttca gcagtttatg gattgatacg 1380
cttgagcctg acgagctggc tccgctggtg ccgcaacttg acgaaaatgc gcaacggcat 1440
gttttacatc agattgctga ttttcgccgt gacgtggata aacgcacgat agggccacgt 1500
gggcgtgatc agttggattt gctgatgccg cgtttactgg cccaggtctg cacctataaa 1560
aatgcggatg tgacgttaca gcgcctgatg cagttgctgc tcaatatcgt cacgcgcacg 1620
acgtatatcg agctgctggt ggaatatccc ggtgcgctca aacagttaat acgtctgtgc 1680
gctgcctcgc cgatggtggc gacgcaactt gcgcgtcatc ctttattgct cgacgaactg 1740
ctcgacccgc gcacgcttta ccagccgatt gagccgggcg cgtaccgtga tgaactgcgg 1800
caatatctga tgcgggtgcc aaccgaagac gaagaacaac agcttgaagc cgtgcgccag 1860
ttcaaacagg cacagcattt acgtattgcg gccggggata tttccggtgc gttgccggtg 1920
atgaaagtca gtgaccattt aacctacctt gcggaggcca ttctcgacgt tgtggtgcaa 1980
caggcgtggg aacaaatggt cgtaaaatac ggtcagccaa cccatcttca gcaccgtaaa 2040
gggcgcggtt ttgccgtggt gggttacgga aaactcggtg gctgggagct gggttacagc 2100
tcggatctgg atctggtctt cctgctcgat tgcgcgccgg aagtcatgac cgacggcgaa 2160
cgcagcattg acgggcgtca gttttatctg cggctggcgc agcgcatcat gcatttattc 2220
agcacccgta cgtcgtcagg cattctttat gaggttgacc cgcgtctgcg gccttccggt 2280
gcttccggca tgctggtcag caccatcgaa gcttttgcgg attatcaggc caacgaagcc 2340
tggacatggg agcatcaggc gctggttcgc gcgcgtgtgg tttatggtga tccgcaactg 2400
acgcagcaat ttaatgccac gcgtcgcgac attctttgcc gccagcgcga tgccgacggc 2460
ttgcgtaagg aagtccgtga aatgcgcgag aaaatgtatg cccatctggg cagcaaaaga 2520
gccgacgagt ttgatctgaa agccgatccg ggtggcataa cggatattga attcatcgca 2580
caatatctgg ttctgcgttt cgcgcatgat gagccgaagc tgacccgctg gtctgataac 2640
gtgcggattt tcgaactgat ggcgcgacat gacatcatgc cggaagagga agcacgccat 2700
ctgacgcagg cttacgtgac attgcgcgat gaaattcatc atctggcgtt gcaggaacac 2760
agcgggaaag tggccgcaga cagctttgcc actgagcgcg cgcaaatccg cgccagctgg 2820
gcaaactggc ttggctga 2838
<210> 94
<211> 1290
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> amtB
<400> 94
atgaaaaaac ttttatccat gatggggctt ggtgcagtgg ctttgctacc ttcgcttgcc 60
atggcagcag caccagcagc ggcaaacggt gctgataacg cctttatgat gatttgtacc 120
gcgctggtat tgttcatgac cgtacccggt gtggcgttgt tctacggcgg cttactgcgt 180
tctaaaaacg ttttgtccat gctgactcag gttattgtta cctttgctct ggtctgcgtc 240
ctgtggatcc tctacggtta cagccttgcc ttcagtgaag gtaacgcgtt cttcggtggt 300
ttcagcaacg taatgatgaa aggcattggc ctggattctg tgactggcac cttctcgcag 360
atgatccacg ttgcattcca gtgttcattt gcctgcatca ctgtagcgct gatcgtaggt 420
ggtattgctg aacgtgtgcg tttctcagca gttctgattt tcactgtgat ctggctgact 480
ttctcttata ttccgatggc tcacatggta tgggcaggcg gtttcctggc tgctgacggt 540
gcgctggact ttgccggtgg taccgttgtt catatcaatg ccgcaattgc tggcctggta 600
ggggcttatc tgctgggtaa acgcgccggt tttggcaaag aagctttcaa accacacaac 660
ctgccaatgg tcttcactgg cgcctcaatc ctgtatgtgg gctggttcgg cttcaatgcg 720
ggttcagcaa gtgccgcaag ctctgttgcc gcgctggctt tcctgaacac tgtcattgct 780
actgctggcg caatcctgtc ctggacgctg gttgagtgga tggtgcgcgg taagccctca 840
ctgctgggcg caagctccgg tgctatcgca ggtctggtgg ctatcacgcc tgcatgtggt 900
acggtcggcg taggtggtgc tctgattatc ggtctggtag gcggtatcac tggtctgtgg 960
ggggttgtta ccctgaaaaa atggctgcgt gttgatgaca cctgtgatgt gttcggtgtt 1020
catggcgtgt gcggtatcgt aggttgtctg ctgacgggtg tattcacgtc cagttcactt 1080
ggcggcgtgg gctacgcaga aggcgtgacc atgggccatc aggtttgggt gcagttcttc 1140
agcgtgtgcg taacattggt ctggtcaggc gttgttgcct tcatcggtta caaagtggct 1200
gacatgatcg taggtctgcg tgttcctgaa gaacaagaac gcgaaggtct ggacgttaac 1260
agccacggcg aaaacgctta caaccaataa 1290
<210> 95
<211> 500
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> PinfC
<400> 95
tgaatatcac tgactcacaa gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg 60
caggcattcg cgttaaagcc gacttgagaa atgagaagat tggctttaaa attcgcgaac 120
acacgctacg ccgtgttcct tatatgttag tttgtggcga taaagaggtc gaagcaggca 180
aagttgctgt tcgtactcgt cgcggcaaag acttaggaag catggatgtt agcgaagtcg 240
ttgacaaact gctggcggaa atccgcagca gaagtcatca tcaactggag gaataaagta 300
ttaaaggcgg aaaacgagtt caaccggcgc gtcctaatcg cattaacaaa gagattcgcg 360
cgcaagaagt tcgcctcacc ggcgtcgatg gcgagcagat tggtattgtc agtctgaatg 420
aagctcttga aaaagctgag gaagcgggcg tcgatttagt agaaatcagt ccgaatgccg 480
agccgccagt ttgtcgaatc 500
<210> 96
<211> 1085
<212> ДНК
<213> Klebsiella variicola
<220>
<223> 16S
<400> 96
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggtag cacagagagc ttgctctcgg gtgacgagcg gcggacgggt gagtaatgtc 120
tgggaaactg cctgatggag ggggataact actggaaacg gtagctaata ccgcataacg 180
tcgcaagacc aaagtggggg accttcgggc ctcatgccat cagatgtgcc cagatgggat 240
tagctagtag gtggggtaac ggctcaccta ggcgacgatc cctagctggt ctgagaggat 300
gaccagccac actggaactg agacacggtc cagactccta cgggaggcag cagtggggaa 360
tattgcacaa tgggcgcaag cctgatgcag ccatgccgcg tgtgtgaaga aggccttcgg 420
gttgtaaagc actttcagcg gggaggaagg cggtgaggtt aataacctca ccgattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc attcgaaact ggcaggctag agtcttgtag 660
aggggggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg ccccctggac aaagactgac gctcaggtgc gaaagcgtgg ggagcaaaca 780
ggattagata ccctggtagt ccacgctgta aacgatgtcg atttggaggt tgtgcccttg 840
aggcgtggct tccggagcta acgcgttaaa tcgaccgcct ggggagtacg gccgcaaggt 900
taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggtcttga catccacaga actttccaga gatggattgg 1020
tgccttcggg aactgtgaga caggtgctgc atggctgtcg tcagctcgtg ttgtgaaatg 1080
ttggg 1085
<210> 97
<211> 882
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifH1
<400> 97
atgaccatgc gtcaatgcgc tatctacggt aaaggcggta tcggtaaatc caccaccacc 60
cagaatctcg tcgcggccct cgccgagatg ggtaagaaag tgatgatcgt cggctgcgat 120
ccgaaagcgg attccacccg tctgatcctc cacgctaaag cccagaacac catcatggag 180
atggcggcgg aagtgggctc ggtcgaggat ctggagctcg aagacgttct gcaaatcggc 240
tatggcgatg tccgttgcgc cgaatccggc ggcccggagc caggcgtcgg ctgcgccgga 300
cgcggggtga tcaccgccat caacttcctc gaggaagaag gcgcctatga agaagatttg 360
gatttcgtct tctatgacgt cctcggcgac gtggtctgcg gcggcttcgc tatgccgatc 420
cgcgaaaaca aagcccagga gatctacatc gtctgctccg gcgagatgat ggcgatgtat 480
gccgccaaca atatctccaa agggatcgtg aagtacgcca aatccggcaa ggtgcgcctc 540
ggcggcctga tctgtaactc gcgcaaaacc gaccgggaag acgaactgat catcgccctg 600
gcggagaagc ttggcacgca gatgatccac ttcgttcccc gcgacaacat tgtgcagcgc 660
gcggagatcc gccggatgac ggtgatcgag tacgacccga cctgtcagca ggcgaatgaa 720
tatcgtcaac tggcgcagaa gatcgtcaat aacaccaaaa aagtggtgcc gacgccgtgc 780
accatggacg agctggaatc gctgctgatg gagttcggca tcatggaaga agaagacacc 840
agcatcattg gtaaaaccgc cgctgaagaa aacgcggcct ga 882
<210> 98
<211> 1113
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifH2
<400> 98
atggttagga aaagtagaag taaaaataca aatatagaac taactgaaca tgaccattta 60
ttaataagtc aaataaaaaa gcttaaaaca caaaccactt gcttttttaa taataaagga 120
ggggttggga agactacatt agtagcaaat ttaggagcag agctatcaat aaactttagt 180
gcaaaagttc ttattgtgga tgccgaccct caatgtaatc tcacgcagta tgtattaagt 240
gatgaagaaa ctcaggactt atatgggcaa gaaaatccag atagtattta tacagtaata 300
agaccactat cctttggtaa aggatatgaa agtgacctcc ctataaggca tgtagagaat 360
ttcggttttg acataattgt cggtgaccct agacttgctt tacaggaaga ccttttagct 420
ggagactggc gagatgccaa aggcggtggg atgcgaggaa ttaggacaac ttttgtattt 480
gcagagttaa ttaagaaagc tcgtgagcta aattatgatt ttgttttctt tgacatggga 540
ccatcattag gcgcaatcaa cagggcagta ttactggcaa tggaattctt tgtcgtccca 600
atgtcaatcg atgtattttc actatgggct attaaaaata ttggctccac ggtttcaata 660
tggaaaaaag aattagacac agggattcgg ctctcagagg aacctagcga attatcacaa 720
ttatcacctc aaggaaaact aaagtttctc ggttacgtca cccaacaaca taaagaacgc 780
tctggatacg atacaattca gcttgagaat actgaggaag aaataaaatc gaaacgtcgg 840
gtaaaggcgt atgaagacat tggagaggtg tttccttcta aaattactga gcatctttct 900
aaactttatg catcaaaaga tatgaaccca caccttggag atatacgtca tttaggtagt 960
ttagctccga aatcacaatc acaacacgtt ccgatgatat cagtgtctgg tacaggaaat 1020
tacaccagac ttagaaaaag cgcgcgtgaa ctttatcgag atattgcaag aagatactta 1080
gagaacattc agactgctaa tggcgagaaa tag 1113
<210> 99
<211> 1374
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifD1
<400> 99
atgaagggaa aggaaattct ggcgctgctg gacgaacccg cctgcgagca caaccagaag 60
caaaaatccg gctgcagcgc ccctaagccc ggcgctaccg ccggcggttg cgccttcgac 120
ggcgcgcaga taacgctcct gcccatcgcc gacgtcgcgc acctggtgca cggccccatc 180
ggctgcgcgg gcagctcgtg ggataaccgc ggcagcgtca gcgccggccc ggccctcaac 240
cggctcggct ttaccaccga tcttaacgaa caggatgtga ttatgggccg cggcgaacgc 300
cgcctgttcc acgccgtgcg tcacatcgtc gaccgctatc atccggcggc ggtctttatc 360
tacaacacct gcgtaccggc gatggagggc gatgacatcg aggcggtctg ccaggccgca 420
cagaccgcca ccggcgtccc ggtcatcgct attgacgccg ccggtttcta cggcagtaaa 480
aatcttggca accgaatggc gggcgacgtg atgctcaggc aggtgattgg ccagcgcgaa 540
ccggccccgt ggccagacaa cacgcccttt gccccggccc agcgccacga tatcggcctg 600
attggcgaat tcaatatcgc cggcgagttc tggcaggtcc agccgctgct cgacgagctg 660
gggatccgcg tcctcggcag cctctccggc gacggccgct ttgccgagat ccagaccctg 720
caccgggcgc aggccaatat gctggtgtgc tcgcgcgcgc tgatcaacgt cgcccggggg 780
ctggagctgc gctacggcac gccgtggttt gaaggcagct tctacgggat ccgcgccacc 840
tccgacgcct tgcgccagct ggcgacgctg ctgggggatg acgacctgcg ccgccgcacc 900
gaggcgctga tcgcccgcga agagcaggcg gcggagcagg ctcttgcgcc gtggcgtgag 960
cagctccgcg ggcgcaaagt gctgctctat accggcggcg tgaaatcctg gtcggtggta 1020
tcggccctgc aggatctcgg catgaccgtg gtggccaccg gcacgcgcaa atccaccgag 1080
gaggacaaac agcggatccg tgagctgatg ggcgacgagg cggtgatgct tgaggagggc 1140
aatgcccgca ccctgctcga cgtggtgtac cgctatcagg ccgacctgat gatcgccggc 1200
ggacgcaata tgtacaccgc ctggaaagcc cggctgccgt ttctcgatat caatcaggag 1260
cgcgagcacg cctacgccgg ctatcagggc atcatcaccc tcgcccgcca gctctgtctg 1320
accctcgcca gccccgtctg gccgcaaacg catacccgcg ccccgtggcg ctag 1374
<210> 100
<211> 1449
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifD2
<400> 100
atgaccaacg caacaggcga acgtaacctt gcgctcatcc aggaagtcct ggaggtgttt 60
cccgaaaccg cgcgcaaaga gcgcagaaag cacatgatga tcagcgatcc gcagatggag 120
agcgtcggca agtgcattat ctcgaaccgt aaatcgcagc ccggggtgat gaccgtgcgc 180
ggctgcgcct atgcgggctc gaaaggggtg gtgtttgggc caatcaaaga catggcccat 240
atctcgcacg gccccatcgg ctgcggccag tattcccgcg ccggacggcg caactactat 300
accggcgtca gcggtgtcga cagcttcggc accctgaact tcacctctga ttttcaggag 360
cgcgatattg ttttcggcgg cgataaaaag ctgaccaaac tgatcgaaga gatggagctg 420
ctgttcccgc tgaccaaagg gatcaccatc cagtcggagt gcccggtggg cctgatcggc 480
gatgacatca gcgccgtagc caacgccagc agcaaggcgc tggataaacc ggtgatcccg 540
gtgcgctgcg aaggctttcg cggcgtatcg caatcgctgg gccaccatat cgccaacgac 600
gtggtgcgcg actgggtgct gaacaatcgc gaagggcagc cgtttgccag caccccgtac 660
gatgttgcca tcattggcga ttacaacatc ggcggcgacg cctgggcctc gcgcattctg 720
ctggaagaga tggggctgcg cgtagtggcg cagtggtccg gcgacggcac cctggtggag 780
atggagaaca ccccattcgt taagcttaac ctcgtccact gctaccgttc gatgaactat 840
atcgcccgcc atatggagga gaaacatcag atcccatgga tggaatataa cttcttcggc 900
ccgaccaaaa tcgccgaatc gctgcgcaag atcgccgatc aatttgatga caccattcgc 960
gccaatgcgg aagcggtgat cgccaaatat gaggggcaga tggcggccat catcgccaaa 1020
tatcgcccgc ggctggaggg gcgcaaagtg ctgctgtaca tgggggggct gcggccgcgc 1080
cacgtcatcg gcgcctatga ggatctcggg atggagatca tcgccgccgg ctacgagttt 1140
gcccataacg atgattacga ccgcaccctg ccggacctga aagagggcac cctgctgttt 1200
gacgatgcca gcagctatga gctggaggcc ttcgtcaaag cgctgaaacc tgacctcatc 1260
ggctccggga tcaaagagaa atatatcttc cagaaaatgg gggtgccgtt ccgccagatg 1320
cactcctggg actattccgg cccctatcac ggctatgacg gcttcgccat ctttgcccgc 1380
gatatggata tgaccctgaa caatccggcg tggaacgaac tgactgcccc gtggctgaag 1440
tctgcgtga 1449
<210> 101
<211> 1386
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifK1
<400> 101
atggcagata ttatccgcag tgaaaaaccg ctggcggtga gcccgattaa aaccgggcaa 60
ccgctcgggg cgatcctcgc cagcctcggg ctggcccagg ccatcccgct ggtccacggc 120
gcccagggct gcagcgcctt cgccaaagtt ttctttattc agcatttcca tgacccggtg 180
ccgctgcagt cgacggccat ggatccgacc gccacgatca tgggggccga cggcaatatc 240
ttcaccgcgc tcgacaccct ctgccagcgc cacagcccgc aggccatcgt gctgctcagc 300
accggtctgg cggaagcgca gggcagcgat atcgcccggg tggtgcgcca gtttcgcgag 360
gcgcatccgc gccataacgg cgtggcgatc ctcaccgtca ataccccgga tttttttggc 420
tctatggaaa acggctacag cgcggtgatc gagagcgtga tcgagcagtg ggtcgcgccg 480
acgccgcgtc cggggcagcg gccccggcgg gtcaacctgc tggtcagcca cctctgttcg 540
ccaggggata tcgaatggct gggccgctgc gtggaggcct ttggcctgca gccggtgatc 600
ctgccggacc tctcgcagtc aatggatggc cacctcggtg aaggggattt tacgcccctg 660
acccagggcg gcgcctcgct gcgccagatt gcccagatgg gccagagtct gggcagcttc 720
gccattggcg tgtcgctcca gcgggcggca tcgctcctga cccaacgcag ccgcggcgac 780
gtgatcgccc tgccgcatct gatgaccctc gaccattgcg atacctttat ccatcagctg 840
gcgaagatgt ccggacgccg cgtaccggcc tggattgagc gccagcgtgg ccagctgcag 900
gatgcgatga tcgactgcca tatgtggctt cagggccagc gcatggcgat ggcggcggag 960
ggcgacctgc tggcggcgtg gtgtgatttc gcccgcagcc aggggatgca gcccggcccg 1020
ctggtcgccc ccaccagcca ccccagcctg cgccagctgc cggtcgagca agtcgtgccg 1080
ggggatcttg aggatctgca gcagctgctg agccaccaac ccgccgatct gctggtggct 1140
aactctcacg cccgcgatct ggcggagcag tttgccctgc cgctgatccg cgtcggtttt 1200
cccctcttcg accggctcgg tgagtttcgt cgcgtccgcc aggggtacgc cggtatgcga 1260
gatacgctgt ttgaactggc caatctgctg cgcgaccgcc atcaccacac cgccctctac 1320
cgctcgccgc ttcgccaggg cgccgacccc cagccggctt caggagacgc ttatgccgcc 1380
cattaa 1386
<210> 102
<211> 1563
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifK2
<400> 102
atgagccaaa cgatcgataa aattcacagc tgttatccgc tgtttgaaca ggatgaatac 60
cagaccctgt tccagaataa aaagaccctt gaagaggcgc acgacgcgca gcgtgtgcag 120
gaggtttttg cctggaccac caccgccgag tatgaagcgc tgaacttcca gcgcgaggcg 180
ctgaccgtcg acccggccaa agcctgccag ccgctcggcg ccgtactctg cgcgctgggg 240
ttcgccggca ccctgcccta cgtgcacggc tcccagggct gcgtcgccta ttttcgcacc 300
tactttaacc gccattttaa agagccggtc gcctgcgtct ccgactccat gaccgaggac 360
gcggcggtgt tcggcggcaa caacaacatg aatctgggcc tgcagaatgc cagcgcgctg 420
tataaacccg agattatcgc cgtctccacc acctgtatgg ccgaggtgat cggcgacgat 480
ctgcaggcgt ttatcgccaa cgccaaaaaa gagggatttg ttgacgaccg catcgccatt 540
ccttacgccc atacccccag ctttatcggc agccatgtca ccggctggga caatatgttc 600
gaagggttcg cgaagacctt taccgctgac tacgccgggc agccgggcaa acagcaaaag 660
ctcaatctgg tgaccggatt tgagacctat ctcggcaact tccgcgtgct gaagcggatg 720
atggcgcaga tggatgtccc gtgcagcctg ctctccgacc catcagaggt gctcgacacc 780
cccgccgacg gccattaccg gatgtacgcc ggcggcacca gccagcagga gatcaaaacc 840
gcgccggacg ccattgacac cctgctgctg cagccgtggc agctggtgaa aagcaaaaag 900
gtggttcagg agatgtggaa ccagcccgcc accgaggtgg ccgttccgct gggcctggcc 960
gccaccgacg cgctgctgat gaccgtcagt cagctgaccg gcaaaccgat cgccgacgct 1020
ctgaccctgg agcgcggccg gctggtcgac atgatgctgg attcccacac ctggctgcat 1080
ggcaaaaaat tcggcctcta cggcgatccg gatttcgtga tggggctgac gcgcttcctg 1140
ctggagctgg gctgcgagcc gacggtgatc ctcagtcata acgccaataa acgctggcaa 1200
aaagcgatga agaaaatgct cgatgcctcg ccgtacggtc aggaaagcga agtgttcatc 1260
aactgcgacc tgtggcactt ccggtcgctg atgttcaccc gtcagccgga ctttatgatc 1320
ggtaactcct acggcaagtt tatccagcgc gataccctgg caaagggcaa agccttcgaa 1380
gtgccgctga tccgtctggg ctttccgctg ttcgaccgcc atcatctgca ccgccagacc 1440
acctggggct atgaaggcgc aatgaacatc gtcacgacgc tggtgaacgc cgtgctggaa 1500
aaactggacc acgacaccag ccagttgggc aaaaccgatt acagcttcga cctcgttcgt 1560
taa 1563
<210> 103
<211> 1485
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifL
<400> 103
atgaccctga atatgatgct cgataacgcc gcgccggagg ccatcgccgg cgcgctgact 60
caacaacatc cggggctgtt ttttaccatg gtggaacagg cctcggtggc catctccctc 120
accgatgcca gcgccaggat catttacgcc aacccggcgt tttgccgcca gaccggctat 180
tcgctggcgc aattgttaaa ccagaacccg cgcctgctgg ccagcagcca gacgccgcgc 240
gagatctatc aggagatgtg gcataccctg ctccagcgtc agccctggcg cggtcagctg 300
attaatcagc gtcgggacgg cggcctgtac ctggtggaga ttgacatcac cccggtgctt 360
agcccgcaag gggaactgga gcattatctg gcgatgcagc gggatatcag cgtcagctac 420
accctcgaac agcggctgcg caaccatatg accctgatgg aggcggtgct gaataatatc 480
cccgccgccg tggtagtggt ggacgagcag gatcgggtgg tgatggacaa cctcgcctac 540
aaaaccttct gcgctgactg cggcggccgg gagctgctca ccgagctgca ggtctcccct 600
ggccggatga cgcccggcgt ggaggcgatc ctgccggtgg cgctgcgcgg ggccgcgcgc 660
tggctgtcgg taacctgctg gccgttgccc ggcgtcagtg aagaggccag ccgctacttt 720
atcgacagcg cgctggcgcg gaccctggtg gtgatcgccg actgtaccca gcagcgtcag 780
cagcaggagc aagggcgcct tgaccggctg aagcagcaaa tgaccgccgg caagctgctg 840
gcggcgatcc gcgagtcgct ggacgccgcg ctgatccagc tgaactgccc gattaatatg 900
ctggcggcag cccgtcggct gaacggcgag ggaagcggga atgtggcgct ggaggccgcc 960
tggcgtgaag gggaagaggc gatggcgcgg ctccagcgct gtcgcccatc gctggaactc 1020
gaaaaccccg ccgtctggcc gctgcagccc tttttcgacg atctgtgcgc cctctaccgt 1080
acacgcttcg atcccgacgg gctgcaggtc gacatggcct caccgcatct gatcggcttt 1140
ggccagcgca ccccactgct ggcgtgctta agcctgtggc tcgatcgcac cctggccctc 1200
gccgccgaac tcccctccgt gccgctggcg atgcagctct acgccgagga gaacgacggc 1260
tggctgtcgc tgtatctgac tgacaacgta ccgctgctgc aggtgcgcta cgctcactcc 1320
cccgacgcgc tgaactcgcc gggcaaaggc atggagctgc ggctgatcca gaccctggtg 1380
gcgcaccatc gcggggccat tgagctggct tcccgaccgc agggcggcac ctgcctgacc 1440
ctgcgtttcc cgctgtttaa caccctgacc ggaggtgaag catga 1485
<210> 104
<211> 1575
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifA
<400> 104
atgatccctg aatccgaccc ggacaccacc gtcagacgct tcgacctctc tcagcagttc 60
accgccatgc agcggataag cgtggtgctg agccgggcca ccgaggccag caaaacgctg 120
caggaggtgc tcagcgtatt acacaacgat gcctttatgc agcacgggat gatctgcctg 180
tacgacagcg agcaggagat cctcagtatc gaagcgctgc agcaaaccgg ccagcagccc 240
ctccccggca gcacgcagat ccgctatcgc cccggcgagg gactggtggg gaccgtgctg 300
gcccaggggc agtcgctggt gctgccccgg gtcgccgacg atcagcgttt tctcgaccgc 360
ctgagcctct acgattacga tctgccgttt atcgccgtac cgttgatggg gcccaacgcc 420
cggccaatag gggtgctggc ggcccagccg atggcgcgcc aggaagagcg gctgccggcc 480
tgcacccgtt ttctcgaaac cgtcgccaac ctcgtcgccc agaccatccg gctgatgatc 540
cttccggcct cacccgccct gtcgagccgc cagccgccga aggtggaacg gccgccggcc 600
tgctcgtcgt cgcgcggcgt gggccttgac aatatggtcg gcaagagccc ggcgatgcgc 660
cagatcgtgg aggtgatccg tcaggtttcg cgctgggaca ccaccgtgct ggtacgcggc 720
gaaagcggca ccgggaaaga gctgatcgcc aacgccatcc atcaccattc gccacgggct 780
ggcgccgcct tcgtcaaatt taactgcgcg gcgctgccgg acaccctgct ggaaagcgaa 840
ctgttcggcc atgagaaagg cgcctttacc ggggcggtgc gtcagcgtaa aggacgtttt 900
gagctggcgg atggcggcac cctgttcctc gatgagattg gtgaaagcag cgcctcgttc 960
caggccaagc tgctgcgtat cctccaggag ggggagatgg agcgggtcgg cggcgatgag 1020
accctgcggg tgaatgtccg catcatcgcc gccaccaacc gtcacctgga ggaggaggtc 1080
cggctgggcc atttccgcga ggatctctac tatcgtctga acgtgatgcc catcgccctg 1140
cccccgctgc gcgagcgtca ggaggacatc gccgagctgg cgcacttcct ggtgcgcaaa 1200
atcggccagc atcaggggcg cacgctgcgg atcagcgagg gcgcgatccg cctgctgatg 1260
gagtacagct ggccgggtaa cgttcgcgaa ctggagaact gcctcgaacg atcggcggtg 1320
atgtcggaga gtggcctgat cgatcgcgac gtgatcctct tcactcacca ggatcgtccc 1380
gccaaagccc tgcctgccag cgggccagcg gaagacagct ggctggacaa cagcctggac 1440
gaacgtcagc gactgatcgc cgcgctggaa aaagccggct gggtgcaggc caaggcggca 1500
cggctgctgg ggatgacgcc gcgccaggtc gcttatcgga tccagatcat ggatatcacc 1560
ctgccgcgtc tgtag 1575
<210> 105
<211> 2838
<212> ДНК
<213> Klebsiella variicola
<220>
<223> glnE
<400> 105
atgatgccgc tttctccgca attacagcag cactggcaga cggtcgctga ccgtctgcca 60
gcggattttc ccattgccga actgagccca caggccaggt cggtcatggc gttcagcgat 120
tttgtcgaac agagtgtgat cgcccagccg ggctggctga atgagcttgc ggactcctcg 180
ccggaggcgg aagagtggcg gcattacgag gcctggctgc aggatcgcct gcaggccgtc 240
actgacgaag cggggttgat gcgagagctg cgtctcttcc gccgccagat gatggtccgc 300
atcgcctggg cgcaggcgct gtcgctggtg agcgaagaag agactctgca gcagctgagc 360
gtcctggcgg agaccctgat tgtcgccgcc cgcgactggc tgtacgccgc ctgctgtaag 420
gagtggggaa cgccatgcaa tgccgagggc cagccgcagc cgctgctgat cctcgggatg 480
ggaaagctgg gcggcggcga gctgaacttc tcttccgata tcgatctgat ctttgcctgg 540
cctgagcatg gcgccacccg cggcggccgc cgcgagctgg ataacgccca gttctttacc 600
cgtctggggc agcggctgat caaggccctt gaccagccga cgcaggacgg ctttgtctat 660
cgggttgaca tgcgcctgcg gccgtttggc gacagtgggc cgctggtact cagttttgcg 720
gcgctggaag attattacca ggagcagggt cgggactggg aacgctatgc gatggtgaaa 780
gcgcggatca tgggcgataa cgacggcgtg tacgccagcg agttgcgcgc gatgctccgt 840
cctttcgtct tccgccgtta tatcgacttc agcgtgatcc agtcgctgcg taacatgaaa 900
ggcatgatcg cccgcgaagt gcggcgtcgc gggctgaaag acaacatcaa gctcggcgcc 960
ggcgggatcc gtgaaattga gtttatcgtt caggtctttc aactgatccg cggtggtcgc 1020
gaacctgcac tgcagcagcg cgccctgctg ccgacgctgg cggcgattga tgagctacat 1080
ctgctgccgg aaggcgacgc ggcgctgctg cgcgaggcct atctgttcct gcgccggctg 1140
gaaaacctgc tgcaaagcat caacgatgag cagacccaga ccctgccgca ggatgaactt 1200
aaccgcgcca ggctggcgtg ggggatgcat accgaagact gggagacgct gagcgcgcag 1260
ctggcgagcc agatggccaa cgtgcggcga gtgtttaatg aactgatcgg cgatgatgag 1320
gatcagtccc cggatgagca actggccgag tactggcgcg agctgtggca ggatgcgctg 1380
gaagaagatg acgccagccc ggcgctggcg catttaaacg ataccgaccg ccgtagcgtg 1440
ctggcgctga ttgccgattt tcgtaaagag ctggatcggc gcaccatcgg cccgcgcggc 1500
cgccaggtgc tggatcagct gatgccgcat ctgctgagcg aaatctgctc gcgcgccgat 1560
gcgccgctgc ctctggcgcg gatcacgccg ctgttgaccg ggatcgtcac ccgtaccacc 1620
tatcttgagc tgctgagcga attccccggc gcgctgaagc acctgatcac gctctgcgcg 1680
gcgtcgccga tggtcgccag ccagctggcg cgccacccgc tgctgctgga tgagctgctg 1740
gatcccaaca ccctctatca gccgacggcg accgatgcct atcgcgacga gctgcgccag 1800
tacctgctgc gcgtgccgga agaggatgaa gagcagcagc tggaggcgtt gcgccagttt 1860
aagcaggcgc agcagctgca tatcgcggcg gcggatatcg ctggtaccct gccggtgatg 1920
aaggtcagcg atcacttaac ctggcttgcc gaagcgatcc tcgacgcggt ggtgcagcag 1980
gcatgggggc agatggtcgc tcgctacggc cagccgaccc acctgcacga tcgccagggt 2040
cgcggcttcg ccgtcgtcgg ctacggtaag cttggcggct gggagctggg ctacagctcc 2100
gatctcgatc tggtgttcct ccatgactgc ccggcggagg tgatgaccga cggcgagcgg 2160
gagattgacg gccgtcagtt ctacctgcgg ctggcccagc ggatcatgca cctgttcagc 2220
acccgcacct cgtccggtat tctctacgaa gtggacgccc ggctgcgtcc ttctggcgcg 2280
gcggggatgc tggtcaccac cgccgacgcg tttgctgact atcagcagaa cgaagcctgg 2340
acgtgggaac atcaggcgct ggtgcgcgcc cgcgtggtct atggcgaccc ggcgctgcag 2400
gcgcgctttg acgccattcg tcgcgatatc ctgaccaccc cgcgggaggg gatgaccctg 2460
cagaccgagg ttcgcgagat gcgcgagaag atgcgcgccc accttggcaa caaacatccc 2520
gatcgttttg atatcaaagc cgatgccggc gggatcaccg atattgaatt tattactcag 2580
tatctggtcc tacgctatgc cagtgacaag ccgaagctga cccgctggtc tgacaacgtg 2640
cgtattcttg agctgctggc gcagaacgac atcatggacg aggaggaggc gcgcgcctta 2700
acgcatgcgt acaccacctt gcgtgatgcg ctccatcacc tggccctgca ggagcagccg 2760
ggacacgtgg cgccagaggc cttcagccgg gagcgtcagc aggtcagcgc cagctggcag 2820
aagtggctga tggcttaa 2838
<210> 106
<211> 500
<212> ДНК
<213> Klebsiella variicola
<220>
<223> PinfC
<400> 106
agcgtcaggt accggtcatg attcaccgtg cgattctcgg ttccctggag cgcttcattg 60
gcatcctgac cgaagagttc gctggcttct tcccaacctg gattgcacca gtgcaggtag 120
tggtcatgaa tattaccgat tctcaggctg aatacgttaa cgaattgacg cgtaaactac 180
aaaatgcggg cattcgtgta aaagcagact tgagaaatga gaagattggc tttaaaatcc 240
gcgagcacac tttacgtcgt gtcccgtata tgttggtctg tggcgacaaa gaagtcgaag 300
ccggcaaagt ggccgtgcgc acccgtcgcg ggaaagacct cggcagcatg gacgtaagtg 360
aagtgattga gaagctgcaa caagagattc gcagccgcag tcttcaacaa ctggaggaat 420
aaggtattaa aggcggaaaa cgagttcaaa cggcacgtcc gaatcgtatc aatggcgaga 480
ttcgcgccct ggaagttcgc 500
<210> 107
<211> 1287
<212> ДНК
<213> Klebsiella variicola
<220>
<223> amtB
<400> 107
atgaaaatgg caacaatgaa atcgggtctg ggggcattag cccttcttcc gggactggca 60
atggccgcgc ccgcagtggc ggacaaagcc gataacgcgt ttatgatgat ttgcaccgcg 120
ctggttctgt ttatgaccat cccggggatc gcgctgtttt acggcggcct gatccgcggc 180
aaaaacgtcc tttccatgct gactcaggtg attgtgacct ttggcctggt ctgcgtactg 240
tgggtgattt atggctatac cctggccttc ggcaccggcg gcagcttctt cggtagtttt 300
gactgggtga tgctgaaaaa tattgaactg aaagcgctga tgggcacctt ctatcagtac 360
atccacgtgg ccttccaggg ctcgttcgcc tgtatcaccg tcgggctgat cgtgggggcg 420
ctggctgagc gtattcgttt ctccgccgtg ctgatttttg tggtggtgtg gatgacgctc 480
tcttatgttc cgattgcgca catggtctgg ggcggcggtc tgctggcgac ccacggcgcg 540
ctggacttcg cgggcggcac cgttgtacac atcaacgccg cggttgccgg gctggtgggt 600
gcgtacatga tgggcaaacg tgtgggcttc ggcaaagaag cgttcaaacc gcacaatctg 660
ccgatggtgt tcaccggaac cgccatcctc tacgtgggct ggttcggctt caacgccggc 720
tccgccagcg cagcgaacga aattgccgca ttggctttcg tcaacaccgt cgtcgccaca 780
gcggctgcca tcctggcgtg gacctttggc gaatgggccc tgcgcggtaa accttcactg 840
ctgggcgcct gctccggggc gattgccggt ctggttggcg tcacaccagc ctgtgggtat 900
atcggtgtcg gtggggcgtt gattgtgggt atcgcatctg gtctggcggg catctggggc 960
gtaacggcgc tgaaacgctg gctgcgggtt gatgaccctt gcgacgtctt cggcgtccac 1020
ggcgtctgcg gcatcgtcgg ctgtatcctg accggtatct tcgcggccac ctctctgggc 1080
ggcgtgggtt atgcagaagg cgtcaccatg ggccatcagc tgctggtgca actcgagagt 1140
atcgcgatta ccatcgtctg gtcgggcgtt gtcgctttca ttggctacaa agtggcggac 1200
atgaccgtgg ggctgcgcgt accagaagag caggagcgcg aaggactgga cgtcaacagc 1260
catggcgaaa acgcctacaa cgcctga 1287
<210> 108
<211> 299
<212> ДНК
<213> Klebsiella variicola
<220>
<223> Prm8.2
<400> 108
cgccgtcctc gcagtaccat tgcaaccgac tttacagcaa gaagtgattc tggcacgcat 60
ggaacaaatt cttgccagtc gggctttatc cgatgacgaa cgcgcacagc ttttatatga 120
gcgcggagtg ttgtatgata gtctcggtct gagggcatta gcgcgaaatg atttttcaca 180
agcgctggca atccgacccg atatgcctga agtattcaat tacttaggca tttacttaac 240
gcaggcaggc aattttgatg ctgcctatga agcgtttgat tctgtacttg agcttgatc 299
<210> 109
<211> 300
<212> ДНК
<213> Klebsiella variicola
<220>
<223> Prm6.2
<400> 109
gctaaagttc tcggctaatc gctgataaca tttgacgcaa tgcgcaataa aagggcatca 60
tttgatgccc tttttgcacg ctttcatacc agaacctggc tcatcagtga ttttttttgt 120
cataatcatt gctgagacag gctctgaaga gggcgtttat acaccaaacc attcgagcgg 180
tagcgcgacg gcaagtcagc gttctccttt gcaatagcag ggaagaggcg ccagaaccgc 240
cagcgttgaa gcagtttgaa cgcgttcagt gtataatccg aaacttaatt tcggtttgga 300
<210> 110
<211> 400
<212> ДНК
<213> Klebsiella variicola
<220>
<223> Prm1.2
<400> 110
gcccgctgac cgaccagaac ttccaccttg gactcggcta tacccttggc gtgacggcgc 60
gcgataactg ggactacatc cccattccgg tgatcttacc attggcgtca ataggttacg 120
gtccggcgac tttccagatg acctatattc ccggcaccta caataacggt aacgtttact 180
tcgcctgggc tcgtatacag ttttaattcg ctaagtctta gcaataaatg agataagcgg 240
tgtgtcttgt ggaaaaacaa ggactaaagc gttacccact aaaaaagata gcgactttta 300
tcacttttta gcaaagttgc actggacaaa aggtaccaca attggtgtac tgatactcga 360
cacagcatta gtgtcgattt ttcatataaa ggtaattttg 400
<210> 111
<211> 1536
<212> ДНК
<213> Klebsiella variicola
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (245)..(245)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (452)..(452)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (454)..(454)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1131)..(1132)
<223> a, c, t, g, неизвестное или другое
<400> 111
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gagcggtagc acagagagct tgctctcggg tgacgagcgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagtggggga ccttcgggcc tcatgccatc agatgtgccc agatgggatt 240
agctngtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggc gntnaggtta ataaccttgt cgattgacgt 480
tacccgcaga agaagcaccg gctaactccg tgccagcagc cgcggtaata cggagggtgc 540
aagcgttaat cggaattact gggcgtaaag cgcacgcagg cggtctgtca agtcggatgt 600
gaaatccccg ggctcaacct gggaactgca ttcgaaactg gcaggctaga gtcttgtaga 660
ggggggtaga attccaggtg tagcggtgaa atgcgtagag atctggagga ataccggtgg 720
cgaaggcggc cccctggaca aagactgacg ctcaggtgcg aaagcgtggg gagcaaacag 780
gattagatac cctggtagtc cacgctgtaa acgatgtcga tttggaggtt gtgcccttga 840
ggcgtggctt ccggagctaa cgcgttaaat cgaccgcctg gggagtacgg ccgcaaggtt 900
aaaactcaaa tgaattgacg ggggcccgca caagcggtgg agcatgtggt ttaattcgat 960
gcaacgcgaa gaaccttacc tggtcttgac atccacagaa ctttccagag atggattggt 1020
gccttcggga actgtgagac aggtgctgca tggctgtcgt cagctcgtgt tgtgaaatgt 1080
tgggttaagt cccgcaacga gcgcaaccct tatcctttgt tgccagcggt nnggccggga 1140
actcaaagga gactgccagt gataaactgg aggaaggtgg ggatgacgtc aagtcatcat 1200
ggcccttacg accagggcta cacacgtgct acaatggcat atacaaagag aagcgacctc 1260
gcgagagcaa gcggacctca taaagtatgt cgtagtccgg attggagtct gcaactcgac 1320
tccatgaagt cggaatcgct agtaatcgta gatcagaatg ctacggtgaa tacgttcccg 1380
ggccttgtac acaccgcccg tcacaccatg ggagtgggtt gcaaaagaag taggtagctt 1440
aaccttcggg agggcgctta ccactttgtg attcatgact ggggtgaagt cgtaacaagg 1500
taaccgtagg ggaacctgcg gttggatcac ctcctt 1536
<210> 112
<211> 882
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifH
<400> 112
atgaccatgc gtcaatgcgc tatctacggt aaaggcggta tcggtaaatc caccaccacc 60
cagaatctcg tcgcggccct cgccgagatg ggtaagaaag tgatgatcgt cggctgcgat 120
ccgaaagcgg actccacccg tctgatcctt cacgctaaag cccagaacac catcatggag 180
atggcggcgg aagtgggctc ggtcgaggat ctggagctcg aagacgttct gcaaatcggc 240
tatggcgatg tccgttgcgc cgaatccggc ggcccggagc caggcgtcgg ctgcgccgga 300
cgcggggtga tcaccgccat caacttcctc gaggaagaag gcgcctatga ggaagatttg 360
gatttcgtct tctatgacgt cctcggcgac gtagtctgcg gcggcttcgc catgccgatc 420
cgcgaaaaca aagcccagga gatctacatc gtctgctccg gcgagatgat ggcgatgtat 480
gccgccaaca atatctccaa ggggatcgtg aagtacgcga aatctggcaa ggtgcgcctc 540
ggcggcctga tctgtaactc gcgcaaaacc gaccgggaag acgaactgat catcgccctg 600
gcggagaagc ttggcacgca gatgatccac ttcgttcccc gcgacaacat tgtgcagcgc 660
gcggagatcc gccggatgac ggtgatcgag tacgacccga cctgtcagca ggcgaatgaa 720
tatcgtcaac tggcgcagaa gatcgtcaat aacaccaaaa aagtggtgcc aacgccgtgc 780
accatggacg agctggaatc gctgctgatg gagttcggca tcatggaaga agaagacacc 840
agcatcattg gtaaaaccgc cgctgaagaa aacgcggcct ga 882
<210> 113
<211> 1449
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifD1
<400> 113
atgaccaacg caacaggcga acgtaacctt gcgctcatcc aggaagtcct ggaggtgttt 60
cccgaaaccg cgcgcaaaga gcgcagaaag cacatgatga tcagcgatcc gcagatggag 120
agcgtcggca agtgcattat ctcgaaccgt aaatcgcagc ccggggtgat gaccgtgcgt 180
ggctgcgcct atgcgggctc gaaaggggtg gtgtttgggc caatcaaaga catggcccat 240
atctcgcacg gccccatcgg ctgcggccag tactcgcgcg ccggacggcg caactactat 300
accggcgtca gcggtgtcga cagcttcggc accctgaact tcacctctga ttttcaggag 360
cgcgatattg ttttcggcgg cgataaaaag ctgaccaaac tgatcgaaga gatggagctg 420
ctgttcccgc tgaccaaagg gatcaccatc cagtcggagt gcccggtggg cctgatcggc 480
gatgacatca gcgccgtggc caacgccagc agcaaggcgc tggataaacc ggtgatcccg 540
gtgcgctgcg aaggctttcg cggcgtatcg caatcgctgg gccaccatat cgccaacgac 600
gtggtgcgcg actgggtgct gaacaatcgc gaagggcagc cgtttgccag caccccgtat 660
gatgttgcca tcattggcga ttacaacatc ggcggcgacg cctgggcctc gcgcattctg 720
ctggaagaga tggggctgcg cgtagtggcg cagtggtccg gcgacggcac cctggtggag 780
atggagaaca ccccattcgt taagcttaac ctcgtccact gctaccgttc gatgaactat 840
atcgcccgcc atatggagga gaaacatcag atcccgtgga tggaatataa cttcttcggc 900
ccgaccaaaa tcgccgaatc gctgcgcaag atcgccgatc aatttgatga caccattcgc 960
gccaatgcgg aagcggtgat cgccaaatat gaggggcaga tggcggccat catcgccaaa 1020
tatcgcccgc ggctggaggg gcgcaaagtg ctgctgtaca tgggggggct gcggccgcgc 1080
cacgtcatcg gcgcctatga ggatctcggg atggagatca tcgccgccgg ctacgagttt 1140
gcccataacg atgattacga ccgcaccctg ccggacctga aagagggcac cctgctgttt 1200
gacgatgcca gcagctatga gctggaggcc ttcgtcaaag cgctgaaacc tgacctcatc 1260
ggctccggga tcaaagagaa atatatcttc cagaaaatgg gggtgccgtt ccgccagatg 1320
cactcctggg actattccgg cccctatcac ggctatgacg gcttcgccat ctttgcccgc 1380
gatatggata tgaccctgaa caatccggcg tggaacgaac tgactgcccc gtggctgaag 1440
tctgcgtga 1449
<210> 114
<211> 1374
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifD2
<400> 114
atgaagggaa aggaaattct ggcgctgctg gacgaacccg cctgcgagca caaccagaag 60
caaaaatccg gctgcagcgc tcctaagccc ggcgcaaccg ccggcggctg cgccttcgac 120
ggcgcgcaga taacgctcct gcccatcgcc gacgtcgcgc acctggtgca cggccccatc 180
ggctgcgcgg gcagctcgtg ggataaccgc ggcagcgtca gcgccggccc ggccctcaac 240
cggctcggct ttaccaccga tcttaacgaa caggatgtga ttatgggccg cggcgaacgc 300
cgcctgttcc acgccgtccg tcacatcgtc gaccgctatc atccggcggc ggtctttatc 360
tacaacacct gcgtaccggc gatggagggg gatgacctgg aggccgtctg ccaggccgca 420
cagaccgcca ccggcgtccc ggtcatcgcc attgacgccg ccggtttcta cggcagtaaa 480
aatcttggca accgaatggc gggcgacgtg atgctcaggc aggtgattgg ccagcgcgaa 540
ccggccccgt ggccagacaa cacgcccttt gccccggccc agcgccacga tatcggcctg 600
attggcgaat tcaatatcgc cggcgagttc tggcaggtcc agccgctgct cgacgagctg 660
gggatccgcg tcctcggcag cctctccggc gacggccgct ttgccgagat ccagaccctg 720
caccgggcgc aggccaatat gctggtgtgc tcgcgcgcgc tgatcaacgt cgcccggggg 780
ctggagctgc gctacggcac gccgtggttt gaaggcagct tctacgggat ccgcgccacc 840
tccgacgcct tgcgccagct ggcggcgctg ctgggggatg acgacctgtg ccgccgcacc 900
gaggcgctga tcgcccgcga agagcaggcg gcggagcagg cgctggcgcc gtggcgcgag 960
cagctccgtg ggcgcaaagt gttgctctac accggcggcg tgaaatcctg gtcggtggta 1020
tcagccctgc aggatctcgg catgaccgtg gtggccaccg gcacgcggaa atccaccgag 1080
gaggacaaac agcggatccg tgagctgatg ggcgacgagg cggtgatgct tgaggagggc 1140
aatgcccgca ccctgctcga cgtggtgtac cgctatcagg ccgacctgat gatcgccggc 1200
ggacgcaata tgtacaccgc ctggaaagcc cggctgccgt ttctcgatat caatcaggag 1260
cgcgagcacg cctacgccgg ctatcagggc atcatcaccc tcgcccgcca gctctgtctg 1320
accctcgcca gtcccgtctg gccgcaaacg catacccgcg ccccgtggcg ctag 1374
<210> 115
<211> 1386
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifK1
<400> 115
atggcagaca ttatccgcag tgaaaaaccg ctggcggtga gcccgattaa aaccgggcaa 60
ccgctcgggg cgatcctcgc cagcctcggg ctggcccagg ccatcccgct ggtccacggc 120
gcccagggct gcagcgcctt cgccaaagtt ttctttattc agcatttcca tgacccggtg 180
ccgctgcagt cgacggccat ggatccgacc gccacgatca tgggggccga cggcaatatc 240
ttcaccgcgc tcgacaccct ctgccagcgc cacagcccgc aggccatcgt gctgctcagc 300
accggtctgg cggaagcgca gggcagcgat atcgcccggg tggtgcgcca gtttcgtgag 360
gcgcatccgc gccataacgg cgtggcgatc ctcaccgtca ataccccgga tttttttggc 420
tcgatggaaa acggctacag cgcggtgatc gagagcgtga tcgagcagtg ggtcgcgccg 480
acgccgcgtc cggggcagcg gccccggcgg gtcaacctgc tggtcagcca cctctgttcg 540
ccaggggata tcgaatggct gggccgctgc gtggaggcct ttggcctgca gccggtgatc 600
ctgccggacc tctcgcagtc aatggatggc cacctcggtg aaggggattt tacgcccctg 660
acccagggcg gcgcctcgct gcgccagatt gcccagatgg gccagagtct gggcagcttc 720
gccattggcg tgtcgctcca gcgggcggca tcgctcctga cccaacgcag ccgcggcgac 780
gtgatcgccc tgccgcatct gatgaccctc gaccattgcg atacctttat ccatcagctg 840
gcgaagatgt ccggacgccg cgtaccggcc tggattgagc gccagcgcgg ccagctgcag 900
gatgcgatga tcgactgcca tatgtggctt cagggccagc gcatggcgat ggcggcggag 960
ggcgacctgc tggcggcgtg gtgtgatttc gcccgcagcc aggggatgca gcccggcccg 1020
ctggtcgccc ccaccagcca ccccagcctg cgccagctgc cggtcgatca ggtcgtgccg 1080
ggggatcttg aggatctgca gcagctgctg agccaccaac ccgccgatct gctggtggct 1140
aactctcacg cccgcgatct ggcggagcag tttgccctgc cgctgatccg cgtcggtttt 1200
cccctcttcg accggctcgg tgagtttcgt cgcgtccgcc aggggtacgc cggtatgcga 1260
gatacgctgt ttgagctggc caatctgctg cgcgaccgcc atcaccacac cgccctctac 1320
cgctcgccgc ttcgccaggg cgccgacccc ctgccggctt caggagacgc ttatgccgcc 1380
cattaa 1386
<210> 116
<211> 183
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifK2
<400> 116
gtgccgctga tccgtctggg ctttccgctg ttcgaccgcc atcatctgca ccgccagacc 60
acctggggct atgaaggcgc aatgaacatc gtcacgacgc tggtgaacgc cgtgctggaa 120
aaactggacc acgacaccag ccagttgggc aaaaccgatt acagcttcga cctcgttcgt 180
taa 183
<210> 117
<211> 1485
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifL
<400> 117
atgaccctga atatgatgct cgataacgcc gcaccggagg ccatcgccgg cgcgctgact 60
caacaacatc cggggctgtt ttttaccatg gtggaacagg cctcggtggc catatccctc 120
accgatgcca gcgccaggat catttacgcc aacccagcgt tttgccgcca gaccggctat 180
tcgctggcgc aattgttaaa ccagaacccg cgcctgctgg ccagcagcca gacgccgcgc 240
gcgatctatc aggagatgtg gcataccctg ctccagcgtc agccctggcg cggtcagctg 300
attaatcagc gtcgggacgg cggcctgtgc ctggtggaga ttgacatcac cccggtgctt 360
agcccgcaag gggaactgga gcattatctg gcgatgcagc gggatatcag cgtcagctac 420
accctcgaac aacggctgcg caaccatatg accctgatgg aggcggtgct gaataatatc 480
cccgccgccg tggtggtggt ggacgagcag gatcgggtgg tgatggacaa cctcgcctac 540
aaaaccttct gcgctgactg cggcggccgg gagctgctca ccgagctgca ggtctcccct 600
ggccggatga cgcccggcgt ggaggcgatc ctgccggtag cgctgcgcgg ggccgcgcgc 660
tggctgtcgg taacctgctg gccgttgccc ggcgtcagtg aagaggccag ccgctacttt 720
atcgacagcg cgctggcgcg gaccctggtg gtgatcgccg actgtaccca gcagcgtcag 780
cagcaggagc aaggacgcct tgaccggctg aagcagcaaa tgaccgccgg caagctgctg 840
gcggcgatcc gcgagtcgct ggacgccgcg ctgatccagc tgaactgccc gattaatatg 900
ctggcggcag cccgtcggct gaacggcgag ggaagcggga atgtggcgct ggaggccgcc 960
tggcgtgaag gggaagaggc gatggcgcgg ctccagcgct gtcgcccatc gctggaactc 1020
gaaaaccccg ccgtctggcc gctgcagccc tttttcgacg atctgtgcgc cctctaccgt 1080
acccgcttcg atcccgacgg gctgcaggtc gacatggcct caccgcatct gatcggcttt 1140
ggccagcgca ccccgctgct ggcgtgctta agcctgtggc tcgaccgcac cctggccctc 1200
gccgccgaat tgccctccgt gccgctggcg atgcagctct atgccgagga gaacgacggc 1260
tggctgtcgc tgtacctgac tgataacgta ccgctgttgc aggtgcgcta cgcccactcc 1320
cccgacgcgc tgaactcgcc gggtaaaggc atggagctgc ggctgatcca gaccctggtg 1380
gcgcaccatc gcggggccat tgagctggct tcccgaccgc agggcggcac ctgcctgacc 1440
ctgcgtttcc cgctgtttaa caccctgacc ggaggtgaag catga 1485
<210> 118
<211> 1575
<212> ДНК
<213> Klebsiella variicola
<220>
<223> nifA
<400> 118
atgatccctg aatccgaccc ggacaccacc gtcagacgct tcgacctctc tcagcagttc 60
accgccatgc agcggataag cgtggtgctg agccgggcca ccgaggccag caaaacgctg 120
caggaggtac tcactgtatt gcacaacgat gcctttatgc agcacgggat gatctgcctg 180
tacgacagcg agcaggagat cctcagtatc gaagcgctgc agcaaaccgg ccagcagccc 240
ctccccggca gcacgcagat ccgctatcgc cccggcgagg gactggtggg gaccgtgctg 300
gcccaggggc agtcgctggt gctgccccgg gtcgccgacg atcagcgttt tctcgaccgc 360
ctgagcctct acgattacga tctgccgttt atcgccgtac cgttgatggg gcccaacgcc 420
cggccaatag gggtgctggc ggcccagccg atggcgcgcc aggaagagcg gctgccggcc 480
tgcacccgtt ttctcgaaac cgtcgccaac ctcgtcgccc agaccatccg gctgatgatc 540
cttccggcct cacccgccct gtcgagccgc cagccgccga aggtggaacg gccgccggcc 600
tgctcgtcgt cgcgcggcgt gggccttgac aatatggtcg gcaagagccc ggcgatgcgc 660
cagatcgtgg aggtgatccg tcaggtttcg cgctgggaca ccaccgtgct ggtgcgcggt 720
gaaagcggca ccgggaaaga gctgatcgcc aacgccatcc atcaccattc gccacgggct 780
ggcgccgcct tcgtcaaatt taactgcgcg gcgctgccgg acaccctgct ggaaagcgaa 840
ctgttcggcc atgagaaagg cgcctttacc ggggcggtgc gtcagcgtaa aggacgtttt 900
gagctggcgg atggcggcac cctgttcctc gatgagattg gtgaaagcag cgcctcgttc 960
caggccaagc tgctgcgtat cctccaggag ggggagatgg agcgggtcgg cggcgatgag 1020
accctgcggg tgaatgtccg catcatcgcc gccaccaacc gtcacctgga ggaggaggtc 1080
cggctgggcc atttccgcga ggatctctat tatcgtctga acgtgatgcc catcgccctg 1140
cccccgctgc gcgagcgtca ggaggacatc gccgagctgg cgcacttcct ggtgcgcaaa 1200
atcggccagc atcaggggcg cacgctgcgg atcagcgagg gcgcgatccg cctgctgatg 1260
gagtacagct ggccgggtaa cgttcgcgaa ctggagaact gcctcgaacg atcggcggtg 1320
atgtcggaga gtggcctgat cgatcgcgac gtgatcctct tcactcacca ggatcgtccc 1380
gccaaagccc tgcctgccag cgggccagcg gaagacagct ggctggacaa cagcctggac 1440
gaacgtcagc gactgatcgc cgcgctggaa aaagccggct gggtgcaggc caaggcggca 1500
cggctgctgg ggatgacgcc gcgccaggtc gcttaccgga tccagatcat ggatatcacc 1560
ctgccgcgtc tgtag 1575
<210> 119
<211> 2838
<212> ДНК
<213> Klebsiella variicola
<220>
<223> glnE
<400> 119
atgatgccgc tttctccgca attacagcag cactggcaga cggtcgctga ccgtctgcca 60
gcggattttc ccattgcaga actgagccca caggccaggt cggtcatggc gttcagcgat 120
tttgtcgaac agagtgtgat cgcccagccg ggctggctga atgagcttgc ggactcctcg 180
ccggaggcgg aagagtggcg gcattacgag gcctggctgc aggatcgcct gcaggccgtc 240
actgacgaag cggggttgat gcgagagctg cgtctcttcc gccgccagat gatggtccgc 300
atcgcctggg cgcaggcgct gtcgctggtg agcgaagaag agaccctgca gcagctgagc 360
gccctggcgg agaccctgat tgtcgccgcc cgcgactggc tctacgccgc ctgctgtaag 420
gagtggggaa cgccatgcaa tgccgagggc cagccgcagc cgctgctgat cctcgggatg 480
ggaaagctgg gcggcggcga gctgaacttc tcttccgata tcgatctgat ctttgcctgg 540
cctgagcatg gcgccacccg cggcggccgc cgcgagctgg ataacgccca gttctttacc 600
cgtctggggc agcggctgat caaggccctt gaccagccga cgcaggacgg ctttgtctat 660
cgggttgaca tgcgcctgcg gccgtttggc gacagtgggc cgctggtact cagctttgcg 720
gcactggaag attattacca ggagcagggt cgggactggg aacgctatgc gatggtgaaa 780
gcgcggatca tgggcgataa cgacggcgtg tacgccagcg agttgcgcgc gatgctccgt 840
cctttcgtct tccgccgtta tatcgacttc agcgtgatcc agtcgctgcg taacatgaaa 900
ggcatgatcg cccgcgaagt gcggcgtcgc gggctgaaag acaacatcaa gctcggcgcc 960
ggcgggatcc gtgaaattga gtttatcgtt caggtctttc agctgatccg cggtggtcgc 1020
gaacctgcac tgcagcagcg cgccctgctg ccgacgctgg cggcgattga tgagctacat 1080
ctgctgccgg aaggcgacgc ggcgctgctg cgcgaggcct atctgttcct gcgccggctg 1140
gaaaacctgc tgcaaagcat caacgatgaa cagacccaga ccctgccgca ggatgaactt 1200
aaccgcgcca ggctggcgtg ggggatgcat accgaagact gggagacgct gagcgcgcag 1260
ctggcgagcc agatggccaa cgtgcggcga gtgtttaatg aactgatcgg cgatgatgag 1320
gatcagtccc cggatgagca actggccgag tactggcgcg agctgtggca ggatgcgctg 1380
gaagaagatg acgccagccc ggcgctggcg catttaaacg ataccgaccg ccgtagcgtg 1440
ctggcgctga ttgccgattt tcgtaaagag ctggatcggc gcaccatcgg cccgcgcggc 1500
cgccaggtgc tggatcagct gatgccgcat ctgctgagcg aaatctgctc gcgtgccgat 1560
gcgccgctgc ctctggcgcg gatcacgccg ctgttgaccg ggatcgtcac ccgtaccacc 1620
tatcttgagc tgctgagcga attccccggc gcgctgaagc acctgatcac gctctgcgcg 1680
gcgtcgccga tggtcgccag ccagctggcg cgccacccgc tgctgctgga tgagctgctg 1740
gatcccaaca ccctctatca gccgacggcg accgatgcct atcgcgacga gctgcgccag 1800
tacctgctgc gcgtgccgga agaggatgaa gagcagcagc tggaggcgtt gcgccagttt 1860
aagcaggcgc agcagctgca tatcgcggcg gcggatatcg ctggtaccct gccggtgatg 1920
aaggtcagcg atcacttaac ctggcttgcc gaagcgatcc tcgacgcggt ggtgcagcag 1980
gcatgggggc agatggtcgc tcgctacggt cagccgaccc acctgcacga tcgccagggt 2040
cgcggcttcg ccgttgtcgg ctacggtaag ctcggcggct gggagctggg ctacagctcc 2100
gatctcgatc tggtgttcct ccatgactgc ccggcggagg tgatgaccga cggcgagcgg 2160
gagattgacg gccgtcagtt ctacctgcgg ctggcccagc ggatcatgca cctgttcagc 2220
acccgcacct cgtccggtat tctctacgaa gtggacgccc ggctgcgtcc ttctggcgcg 2280
gcggggatgc tggtcaccac cgccgacgcg tttgctgact atcagcagaa cgaagcctgg 2340
acgtgggaac atcaggcgct ggtgcgcgcc cgcgtggtct atggcgaccc ggcgctgcag 2400
gcgcgctttg acgccattcg tcgcgatatc ctgaccaccc cgcgggaggg gacgaccctg 2460
cagaccgagg ttcgcgagat gcgcgagaag atgcgcgccc accttggcaa caaacatccc 2520
gatcgttttg atatcaaagc cgatgccggc gggatcaccg atattgaatt tattactcag 2580
tatctggtcc tacgctatgc cagtgacaag ccgaagctga cccgctggtc tgacaacgtg 2640
cgtattcttg agctgctggc gcagaacgac atcatggacg aggaggaggc gcgcgcctta 2700
acgcatgcat acaccacctt gcgtgatgcg ctccatcacc tggccctgca ggagcagccg 2760
ggacacgtgg cgccagaggc cttcagccgg gagcgtcagc aggtcagcgc cagctggcag 2820
aagtggctga tggcttaa 2838
<210> 120
<211> 1287
<212> ДНК
<213> Klebsiella variicola
<220>
<223> amtB
<400> 120
atgaaaatgg caacaatgaa atcgggtctg ggggcattag cccttcttcc gggactggca 60
atggccgcgc ccgcagtggc ggacaaagcc gataacgcgt ttatgatgat ttgcaccgcg 120
ctggttctgt ttatgaccat cccggggatc gcgctgtttt acggcggcct gatccgcggc 180
aaaaacgtcc tttccatgct gactcaggtg attgtgacct ttggcctggt ctgcgtactg 240
tgggtgattt atggctatac cctggccttc ggcaccggcg gcagcttctt cggtagcttt 300
gactgggtga tgctgaaaaa tattgaactg aaagcgctga tgggcacctt ctatcagtac 360
atccacgtgg ccttccaggg ctcgttcgcc tgtatcaccg tcgggctgat cgtgggggcg 420
ctggctgagc gtattcgttt ctccgccgtg ctgattttcg tggtggtgtg gatgacgctc 480
tcttatgttc cgattgcgca catggtctgg ggcggcggtc tgctggcgac ccacggcgcg 540
ctggacttcg cgggcggcac cgttgtacac atcaacgccg cggttgccgg gctggtgggt 600
gcgtatatga tgggcaaacg tgtgggcttc ggcaaagaag cgttcaaacc gcacaatctg 660
ccgatggtgt tcaccggaac cgccatcctc tacgtgggct ggttcggctt caacgccggc 720
tccgccagcg cagcgaacga aattgccgca ctggctttcg tcaacaccgt cgtcgccaca 780
gcggcagcca tcctggcctg gacctttggc gaatgggctc tgcgcggcaa accttcactg 840
ctgggcgcct gctccggggc gattgccggt ctggttggcg tcacaccagc ctgtgggtat 900
atcggtgtcg gtggggcgtt gattgtgggt atcgcatctg gtctggcggg catctggggc 960
gtaacggcgc tgaaacgctg gctgcgggtt gatgaccctt gcgacgtctt cggcgtccac 1020
ggcgtctgcg gcatcgtcgg ctgtatcctg accggtatct tcgcggccac ctctctgggc 1080
ggcgtgggtt atgcagaagg cgtcaccatg ggccatcagc tgctggtgca actcgagagt 1140
atcgcgatta ccatcgtctg gtcgggcgtt gtcgctttca ttggctacaa agtggcggac 1200
atgaccgtgg ggctgcgcgt accagaagag caggagcgcg aaggactgga cgtcaacagc 1260
catggcgaaa acgcctacaa cgcctga 1287
<210> 121
<211> 1435
<212> ДНК
<213> Achromobacter spiritinus
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (255)..(255)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (454)..(454)
<223> a, c, t, g, неизвестное или другое
<400> 121
ctgaagagtt tgatcctggc tcagattgaa cgctagcggg atgccttaca catgcaagtc 60
gaacggcagc acggacttcg gtctggtggc gagtggcgaa cgggtgagta atgtatcgga 120
acgtgcctag tagcggggga taactacgcg aaagcgtagc taataccgca tacgccctac 180
gggggaaagc aggggatcgc aagaccttgc actattagag cggccgatat cggattagct 240
agttggtggg gtaanggctc accaaggcga cgatccgtag ctggtttgag aggacgacca 300
gccacactgg gactgagaca cggcccagac tcctacggga ggcagcagtg gggaattttg 360
gacaatgggg gaaaccctga tccagccatc ccgcgtgtgc gatgaaggcc ttcgggttgt 420
aaagcacttt tggcaggaaa gaaacgtcat gggntaatac cccgtgaaac tgacggtacc 480
tgcagaataa gcaccggcta actacgtgcc agcagccgcg gtaatacgta gggtgcaagc 540
gttaatcgga attactgggc gtaaagcgtg cgcaggcggt tcggaaagaa agatgtgaaa 600
tcccagagct taactttgga actgcatttt taactaccgg gctagagtgt gtcagaggga 660
ggtggaattc cgcgtgtagc agtgaaatgc gtagatatgc ggaggaacac cgatggcgaa 720
ggcagcctcc tgggataaca ctgacgctca tgcacgaaag cgtggggagc aaacaggatt 780
agataccctg gtagtccacg ccctaaacga tgtcaactag ctgttggggc cttcgggcct 840
tagtagcgca gctaacgcgt gaagttgacc gcctggggag tacggtcgca agattaaaac 900
tcaaaggaat tgacggggac ccgcacaagc ggtggatgat gtggattaat tcgatgcaac 960
gcgaaaaacc ttacctaccc ttgacatgtc tggaattctg aagagattcg gaagtgctcg 1020
caagagaacc ggaacacagg tgctgcatgg ctgtcgtcag ctcgtgtcgt gagatgttgg 1080
gttaagtccc gcaacgagcg caacccttgt cattagttgc tacgaaaggg cactctaatg 1140
agactgccgg tgacaaaccg gaggaaggtg gggatgacgt caagtcctca tggcccttat 1200
gggtagggct tcacacgtca tacaatggtc gggacagagg gtcgccaacc cgcgaggggg 1260
agccaatccc agaaacccga tcgtagtccg gatcgcagtc tgcaactcga ctgcgtgaag 1320
tcggaatcgc tagtaatcgc ggatcagcat gtcgcggtga atacgttccc gggtcttgta 1380
cacaccgccc gtcacaccat gggagtgggt tttaccagaa gtagttagcc taacc 1435
<210> 122
<211> 1528
<212> ДНК
<213> Achromobacter marplatensis
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (255)..(255)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (999)..(999)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1009)..(1009)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1437)..(1437)
<223> a, c, t, g, неизвестное или другое
<400> 122
ctgaagagtt tgatcctggc tcagattgaa cgctagcggg atgccttaca catgcaagtc 60
gaacggcagc acggacttcg gtctggtggc gagtggcgaa cgggtgagta atgtatcgga 120
acgtgcctag tagcggggga taactacgcg aaagcgtagc taataccgca tacgccctac 180
gggggaaagc aggggatcgc aagaccttgc actattagag cggccgatat cggattagct 240
agttggtggg gtaanggctc accaaggcga cgatccgtag ctggtttgag aggacgacca 300
gccacactgg gactgagaca cggcccagac tcctacggga ggcagcagtg gggaattttg 360
gacaatgggg gaaaccctga tccagccatc ccgcgtgtgc gatgaaggcc ttcgggttgt 420
aaagcacttt tggcaggaaa gaaacgtcat gggttaatac cccgtgaaac tgacggtacc 480
tgcagaataa gcaccggcta actacgtgcc agcagccgcg gtaatacgta gggtgcaagc 540
gttaatcgga attactgggc gtaaagcgtg cgcaggcggt tcggaaagaa agatgtgaaa 600
tcccagagct taactttgga actgcatttt taactaccgg gctagagtgt gtcagaggga 660
ggtggaattc cgcgtgtagc agtgaaatgc gtagatatgc ggaggaacac cgatggcgaa 720
ggcagcctcc tgggataaca ctgacgctca tgcacgaaag cgtggggagc aaacaggatt 780
agataccctg gtagtccacg ccctaaacga tgtcaactag ctgttggggc cttcgggcct 840
tagtagcgca gctaacgcgt gaagttgacc gcctggggag tacggtcgca agattaaaac 900
tcaaaggaat tgacggggac ccgcacaagc ggtggatgat gtggattaat tcgatgcaac 960
gcgaaaaacc ttacctaccc ttgacatgtc tggaattcng aagagattng gaagtgctcg 1020
caagagaacc ggaacacagg tgctgcatgg ctgtcgtcag ctcgtgtcgt gagatgttgg 1080
gttaagtccc gcaacgagcg caacccttgt cattagttgc tacgaaaggg cactctaatg 1140
agactgccgg tgacaaaccg gaggaaggtg gggatgacgt caagtcctca tggcccttat 1200
gggtagggct tcacacgtca tacaatggtc gggacagagg gtcgccaacc cgcgaggggg 1260
agccaatccc agaaacccga tcgtagtccg gatcgcagtc tgcaactcga ctgcgtgaag 1320
tcggaatcgc tagtaatcgc ggatcagcat gtcgcggtga atacgttccc gggtcttgta 1380
cacaccgccc gtcacaccat gggagtgggt tttaccagaa gtagttagcc taaccgnaag 1440
gggggcgatt accacggtag gattcatgac tggggtgaag tcgtaacaag gtagccgtat 1500
cggaaggtgc ggctggatca cctccttt 1528
<210> 123
<211> 1522
<212> ДНК
<213> Microbacterium murale
<220>
<223> 16S
<400> 123
tacggagagt ttgatcctgg ctcaggatga acgctggcgg cgtgcttaac acatgcaagt 60
cgaacggtga acacggagct tgctctgtgg gatcagtggc gaacgggtga gtaacacgtg 120
agcaacctgc ccctgactct gggataagcg ctggaaacgg cgtctaatac tggatatgtg 180
acgtggccgc atggtctgcg tctggaaaga atttcggttg gggatgggct cgcggcctat 240
cagcttgttg gtgaggtaat ggctcaccaa ggcgtcgacg ggtagccggc ctgagagggt 300
gaccggccac actgggactg agacacggcc cagactccta cgggaggcag cagtggggaa 360
tattgcacaa tgggcgcaag cctgatgcag caacgccgcg tgagggatga cggccttcgg 420
gttgtaaacc tcttttagca gggaagaagc gaaagtgacg gtacctgcag aaaaagcgcc 480
ggctaactac gtgccagcag ccgcggtaat acgtagggcg caagcgttat ccggaattat 540
tgggcgtaaa gagctcgtag gcggtttgtc gcgtctgctg tgaaatccgg aggctcaacc 600
tccggcctgc agtgggtacg ggcagactag agtgcggtag gggagattgg aattcctggt 660
gtagcggtgg aatgcgcaga tatcaggagg aacaccgatg gcgaaggcag atctctgggc 720
cgtaactgac gctgaggagc gaaagggtgg ggagcaaaca ggcttagata ccctggtagt 780
ccaccccgta aacgttggga actagttgtg gggtccattc cacggattcc gtgacgcagc 840
taacgcatta agttccccgc ctggggagta cggccgcaag gctaaaactc aaaggaattg 900
acggggaccc gcacaagcgg cggagcatgc ggattaattc gatgcaacgc gaagaacctt 960
accaaggctt gacatatacg agaacgggcc agaaatggtc aactctttgg acactcgtaa 1020
acaggtggtg catggttgtc gtcagctcgt gtcgtgagat gttgggttaa gtcccgcaac 1080
gagcgcaacc ctcgttctat gttgccagca cgtaatggtg ggaactcatg ggatactgcc 1140
ggggtcaact cggaggaagg tggggatgac gtcaaatcat catgcccctt atgtcttggg 1200
cttcacgcat gctacaatgg ccggtacaaa gggctgcaat accgcgaggt ggagcgaatc 1260
ccaaaaagcc ggtcccagtt cggattgagg tctgcaactc gacctcatga agtcggagtc 1320
gctagtaatc gcagatcagc aacgctgcgg tgaatacgtt cccgggtctt gtacacaccg 1380
cccgtcaagt catgaaagtc ggtaacacct gaagccggtg gcctaaccct tgtggaggga 1440
gccgtcgaag gtgggatcgg taattaggac taagtcgtaa caaggtagcc gtaccggaag 1500
gtgcggctgg atcacctcct tt 1522
<210> 124
<211> 1537
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (455)..(455)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1004)..(1004)
<223> a, c, t, g, неизвестное или другое
<400> 124
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgaacggtag cacagagagc ttgctctcgg gtgacgagtg gcggacgggt gagtaatgtc 120
tgggaaactg cccgatggag ggggataact actggaaacg gtagctaata ccgcataatg 180
tcgcaagacc aaagaggggg accttcgggc ctcttgccat cggatgtgcc cagatgggat 240
tagcttgttg gtgaggtaat ggctcaccaa ggcgacgatc cctagctggt ctgagaggat 300
gaccagccac actggaactg agacacggtc cagactccta cgggaggcag cagtggggaa 360
tattgcacaa tgggcgcaag cctgatgcag ccatgccgcg tgtgtgaaga aggccttcgg 420
gttgtaaagc actttcagcg gggaggaagg cgatncggtt aataaccgtg ttgattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc attcgaaact ggcaggcttg agtcttgtag 660
aggggggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg ccccctggac aaagactgac gctcaggtgc gaaagcgtgg ggagcaaaca 780
ggattagata ccctggtagt ccacgccgta aacgatgtcg acttggaggt tgtgcccttg 840
aggcgtggct tccggagcta acgcgttaag tcgaccgcct ggggagtacg gccgcaaggt 900
taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggtcttga catccacgga attnggcaga gatgccttag 1020
tgccttcggg aaccgtgaga caggtgctgc atggctgtcg tcagctcgtg ttgtgaaatg 1080
ttgggttaag tcccgcaacg agcgcaaccc ttatcctttg ttgccagcgg tccggccggg 1140
aactcaaagg agactgccag tgataaactg gaggaaggtg gggatgacgt caagtcatca 1200
tggcccttac gaccagggct acacacgtgc tacaatggca tatacaaaga gaagcgacct 1260
cgcgagagca agcggacctc ataaagtatg tcgtagtccg gattggagtc tgcaactcga 1320
ctccatgaag tcggaatcgc tagtaatcgt ggatcagaat gccacggtga atacgttccc 1380
gggccttgta cacaccgccc gtcacaccat gggagtgggt tgcaaaagaa gtaggtagct 1440
taaccttcgg gagggcgctt accactttgt gattcatgac tggggtgaag tcgtaacaag 1500
gtaaccgtag gggaacctgc ggttggatca cctcctt 1537
<210> 125
<211> 882
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifH
<400> 125
atgaccatgc gtcaatgcgc catttatggc aaaggtggga tcggcaaatc caccaccacg 60
caaaacctcg tcgccgctct cgcggaaatg ggtaaaaaag tgatgatcgt cggctgcgac 120
ccgaaagcgg actccacccg tctgatcctg catgcgaaag cacagaacac cattatggag 180
atggccgccg aagtgggttc agtggaagac cttgaactgg aagatgtgct gcaaatcggt 240
tacggcggcg tgcgttgtgc agaatccggc ggcccggagc caggcgtggg ttgtgcaggc 300
cgcggcgtta ttaccgccat taacttcctt gaagaagaag gcgcctatgt cagcgacctc 360
gactttgtct tctatgacgt cctcggtgac gtggtctgcg gcgggttcgc catgccgatt 420
cgtgaaaaca aagcgcaaga gatctatatc gtctgctccg gggaaatgat ggcgatgtat 480
gccgctaaca acatctccaa aggcatcgtg aaatacgcta aatccggcaa ggtgcgcctg 540
ggcgggctga tttgtaactc ccgtcagacc gaccgcgaag atgaactgat catcgcgctg 600
gcagaaaaac tgggcaccca gatgattcac tttgtgccac gcgacaacat cgtccagcgc 660
gcggaaattc gccgtatgac ggttatcgaa tatgacccga aatgcaacca ggccgacgaa 720
taccgcgcgc tggcgaacaa gatcgtcaac aacaccctga tggtcgtccc gaccccttgc 780
accatggatg aactggaaga gctgctgatg gaattcggca ttatggatgt ggaagacgcc 840
agcatcatcg gtaaaaccgc cgccgaagaa aacgcggcct ga 882
<210> 126
<211> 660
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> CDS железо-молибденового кофактора динитрогеназы
<400> 126
atgaacgata acgatgtcct tttctggcgc atgctggcgc tatttcagtg tctgccggaa 60
ctgcaacccg cgcagatcct ggcctggctg acaggagaac gcgacgacgc cttaaccccg 120
gcgtacctcg ataagcttaa cgtccgcgaa ctggaagcga ccttcccgtc tgaaacggcg 180
atgatgtcgc ccgcacgctg gagccgcgtt aacgcgtgcc ttcacggtac gctgcccgca 240
cacctgcagg taaaaagcac cactcgtcag gggcaattac gggtagcctt ttgttcacag 300
gatggattgc tgatcaatgg tcattttggt caggggcggc tgttttttat ctacgccttt 360
gatgaacagg gcggatggct acacgcgtta cgccgtcttc cctcggcccc gcaaacccag 420
gagccgaatg aagttcgcgc gcagctcctg agtgattgcc acctgctgtt ttgtgaagcc 480
attggcggcc ctgcggcggc ccggctgatt cgtcacaata tccacccgat gaaagtgtcg 540
ccagggatgt ccattgccgc ccagtgtgat gccattaccg cactgctgag cggacgtctg 600
ccaccgtggc tggcaaaacg tcttgagaaa gccaacccgc tggaagagcg ggtgttttaa 660
<210> 127
<211> 1374
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifD1
<400> 127
atgaagggaa atgacattct cgcgctgctg gatgaacccg cctgcgaaca caatcacaaa 60
cagaaatccg gctgtagcgc ccctaaaccc ggtgccacgg cgggcggttg cgcgttcgac 120
ggcgcgcaaa tcaccctgtt gccgctgtcg gatgtggcgc acctggtcca cggaccgatt 180
ggctgcacgg gaagctcctg ggataaccgg ggcagtatga gctccggccc cagtctcaac 240
cggctcggct ttaccaccga cctgaacgag caggatgtca ttatggggcg cggcgaacgg 300
cggcttttcc acgcggtgcg tcatatcgtc aaccgttatc accctgccgc cgtgtttatc 360
tataacacct gcgttccggc gatggagggt gatgatattg acgccgtctg tcaggcggcg 420
gaaaccgcca ccggcgtgcc agtgattgcc gttgatgccg ccgggttcta tggcagcaaa 480
aaccttggca accgtctcgc gggtgaagtg atggttaaca aggtcattgg acggcgcccg 540
cccgccccct ggccggacga tacccccttc gcgccggaac accgccacga tatcggcctg 600
attggcgaat ttaatatcgc cggggagttc tggcacgttc agccgctgct cgatgagctg 660
ggtattcgcg tgctgggcag cctttccggg gatggccgtt ttagtgaaat ccagaccctg 720
caccacgcgc aggtcaatat gctggtctgc tcaagagcgc tgatcaatgt tgcccgcacc 780
ctggaacagc gctatggcac cccctggttt gagggcagtt tttacggcgt gcgcgctacc 840
tccgatgccc tgcgtcaact ggcatccctg cttggcgaca gcgatctgat tgcccgcacc 900
gaagccgtta ttgcccgcga agaagccacg gcaaatcagg cgctcgcccc gtggcgcgaa 960
cggctacagg gtcgcaaagt gctgctctat accggtgggg tgaaatcctg gtcggtggtc 1020
tccgcattgc aggatttagg gatgaccgtc gtggcgactg gcacccgcaa atctaccgaa 1080
gaagataagc agcgtattcg cgaattaatg ggcgatgacg cgctaatgct ggaagaaggc 1140
aacgcccgca ccctgctgga tgtggtgtac cgctatcagg cggatttgat gatcgctggg 1200
gggcgtaaca tgtataccgc gtacaaagcg cggctgccgt ttctggatat caaccaggag 1260
cgtgaacacg cctttgcggg ttatcgcggc atcgtcaccc tcgcccaaca gctttgccag 1320
actattgaaa gccccgtctg gccgcaaaca cacgcccgcg cgccgtggca ataa 1374
<210> 128
<211> 1449
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifD2
<400> 128
atgagcaatg caacaggcga acgtaatctg gaaattatcc aggaagtgct ggagatcttt 60
cccgaaaaaa cgcgcaaaga acgcagaaag cacatgatgg tgaccgaccc ggagatggaa 120
agcgtcggga aatgcatcat ctctaaccgc aaatcgcagc cgggtgtgat gactgtccgc 180
ggctgctcct acgccgggtc gaaaggcgtg gtttttgggc cgattaaaga tatggcccac 240
atctcccacg gcccgatcgg ctgtgggcag tactcccgtg ccgggcggcg caactactac 300
accggggtca gcggcgttga ttccttcggg acgctgaact ttacctctga ttttcaggag 360
cgcgatatcg tcttcggcgg cgataaaaag ctcaccaaac tgattgagga gatggaggaa 420
ctgttcccgc tgaccaaagg catctccatt cagtcggagt gcccggtagg tttaatcggt 480
gacgatatcg aagcggtggc gaatgccagt aaaaaagcgc tcaacaagcc ggtgatcccg 540
gtgcgttgcg aaggctttcg cggcgtgtcg cagtcgctcg gtcaccatat cgccaacgac 600
gttatccgcg actgggtgct ggataaccgc gaagggaagc ccttcgaatc taccccctat 660
gacgtggcca tcatcggcga ttacaacatc gggggggatg cctgggcgtc gcgcattctg 720
cttgaagaga tggggttacg cgtggtggcg cagtggtccg gtgacggcac gctggtagag 780
atggaaaaca ccccgttcgt caagctgaac ctggtgcact gctaccgctc tatgaactac 840
atctctcgcc atatggaaga gaaacacggt atcccgtgga tggagtacaa cttcttcggc 900
ccgaccaaaa tcgccgaatc gctgcgtaag atcgccgatc aatttgacga caccatccgc 960
gccaatgcgg aagcggtgat cgccaaatat caggcgcaaa acgatgcgat tatcgccaaa 1020
taccgcccgc gtctcgaagg ccgcaaggtg ctgctctata tgggtggcct gcgtcctcgc 1080
cacgtgattg gcgcgtatga ggatttgggc atggagattg tcgccgccgg gtatgaattt 1140
gcccataacg acgattacga ccgcaccctg ccggacctca aagagggcac gctgttgttc 1200
gacgatgcca gcagttatga actggaagcc ttcgtgaagg cgattaagcc ggacctcatt 1260
ggctcaggca tcaaggaaaa atacattttc cagaaaatgg gggtaccgtt tcgccagatg 1320
cactcctggg attactccgg cccgtatcac ggctatgacg gctttgccat ctttgcccgc 1380
gatatggaca tgacgctcaa caatcccgcc tggggcgagt tgaccgcacc ctggctgaaa 1440
tcagcctga 1449
<210> 129
<211> 1383
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifK1
<400> 129
atggcagata tcatccgtaa tcagaaaccg ctggcggtaa gcccggtaaa aagcggccag 60
ccgttaggcg ccattctggc gagcctcggc tttgagcaca gtattccact ggtgcacggt 120
gcgcagggat gcagcgcgtt cgccaaagtg ttttttatcc aacattttca tgaccctatt 180
ccgctgcaat ccacggcgat ggaccccacc tcaacggtca tgggggcgga cggcaatatc 240
cttgccgcgc tcaatacgct gtgccagcgc aacaccccga aagctatcgt cctgttgagt 300
accggcctgt ctgaggcgca gggcagcgat atcagccgcg tggtacgtca gtttcgtgag 360
gattttcccc gccacaaaaa tatcgccctc ctgacggtca acaccccgga tttttacggc 420
acgctggaga acggctttag tgcggtggtg gaaagcgtca tcgaacagtg ggtgccggaa 480
aagcctcagc atggcctgcg taaccggcgg gtcaacttgt tgttaagtca cctgctgacg 540
cccggtgatg ttgagttgct gcgcagctac gtggaggctt ttggcctgca accggtgatc 600
gtgccggatc tttcacagtc gctggatggt cacctggcaa gcggtgattt ttcgccggtc 660
actcaggggg gaacgcccct gcgcattatc gaacagatgg gacagagcct gtgcacgttt 720
gctattggcg tgtcgctgtc ccgtgcggca tcgctgctgg cacagcgtag ccgtggcgag 780
gtgatcgtgc ttccccatct gatgaccatg gaacattgcg accgttttat tcatcaactg 840
aagatcattt ccgggcgcga ggttcccgcc tggattgagc gccagcgcgg acaattgcag 900
gatgcgatga tcgattgtca tatgtggttg caggataccc ggctcgcgct ggccgccgag 960
ggcgatctgc tggcgggctg gtgtgatttc gcccgtagcc agggcatgct ccccggcccc 1020
gttgtggcgc cggtcagcca gccgggcctg caacagcttc ccgtggagaa agtggtcatt 1080
ggcgatctgg aagatatgca ggatttactc tgcgctatgc ctgctgacct gctggtcgcc 1140
aactcccatg ccgcagacct ggccgaacaa ttctccatcc cgctgatccg cgccgggttc 1200
cctatcttcg acaggcttgg cgaatttcgt cgcgtgcgtc agggataccc cggcattcgc 1260
gacacgctgt ttgagctggc gaacctgatg cgcgaacgtc atcaccacct gcccgtctac 1320
cgctcccccc tgcgccagca atttgcccag gacgctgacg gaggccgcta tgcaacatgt 1380
taa 1383
<210> 130
<211> 1563
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifK2
<400> 130
atgagccaaa ctgctgagaa aattgtcacc tgtcatccgc tgtttgaaca ggacgaatac 60
cagacgctgt ttcgcaataa gcgcggtctg gaagaggcgc acgacccgca gcgcgtgcaa 120
gaggtttttg aatggaccac cacggcggag tatgaagcgc tgaactttaa gcgtgaagcg 180
ttaaccgtcg atccggcaaa ggcctgccag cctttaggat cggtactctg ctcgctgggt 240
tttgccaata cgctgcctta tgtgcacggt tcccagggct gtgtggccta tttccgcacc 300
tattttaacc gtcatttcaa agagccgatc gcttgcgttt ccgactctat gacagaggat 360
gcggcggtct tcggcggcaa caacaacctt aacaccgggt tgcaaaatgc cagcgccctg 420
tacaaaccgg aaattgtcgc tgtctccact acctgtatgg cggaggtcat cggcgatgac 480
ctgcaggcct ttatcgccaa cgccaaaaag gacgggttta ttgatgccgc cattccggtg 540
ccctacgccc atacgccaag ttttatcggt agccacatca ccggctggga caacatgttt 600
gaaggtttcg cccgggcatt taccgccgat cacgtggcgc aaccgggcaa actggcgaag 660
ctaaacctgg tgaccggttt tgaaacctat cttggcaatt accgcgtgct caaacgcatg 720
atggcccaga tggaggtgcc ctgtagcctg ctgtctgacc cgtctgaggt gttagatacg 780
ccagccgacg gccactatcg catgtatgcg ggcggcacaa cgcaacaaga gatgcgcgac 840
gcccccgatg ctatcgacac cctgctgctg caaccctggc atctggtgaa gagtaaaaaa 900
gtggtgcagg agtcctgggg ccagcccgcc acagaagtgt ccatcccaat gggactgacc 960
gggaccgacg aactgctgat ggcagtcagt cagttaaccg gcaaaccggt ggccgatgaa 1020
ctgacgctgg agcgtgggcg cctggtggat atgattctcg attcacacac ctggctgcac 1080
ggtaagaaat tcggtctcta cggcgatccg gattttgtga tggggctgac gcgtttcctg 1140
ctggaactgg gctgcgagcc gacggttatc ctctgtcata acggtagcaa gcgctggcag 1200
aaagcgatga agaaaatgct tgaggcatcg ccctacggtc aggagagcga agtgttcatc 1260
aactgcgatc tgtggcattt ccgctcgctg atgtttaccc gcaaaccgga ctttatgatc 1320
ggcaactcgt acgccaaatt catccagcgt gacacgctgg cgaaaggcga acagtttgaa 1380
gttccgctga tccgtcttgg cttcccgttg ttcgaccgcc accacctgca tcgccagacc 1440
acatggggtt atgaaggggc gatgaatatc gtcaccaccc tggtcaacgc cgtgctggaa 1500
aaagtcgacc gcgataccat caaactgggc aaaacggact acagcttcga ccttgtccgc 1560
taa 1563
<210> 131
<211> 1488
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifL
<400> 131
atgaccttta atatgatgct ggagaccagc gcaccgcagc acattgcggg caacctctca 60
cttcaacatc ccggactgtt ttccacgatg gttgaacagg ctccgatcgc gatttcgctg 120
accgacccgg acgcgaggat tctgtacgct aatccggcct tttgtcgcca gaccggttat 180
agcctggaag agctgctcaa ccagaaccat cgcatactgg caagccaaca gacgccgcgc 240
agcatttatc aggaactgtg gcaaacgctg ctgcaacaga tgccctggcg cggtcagctc 300
atcaatcgcc gtcgggatgg cagcctttat ctggctgagg tcgatatcac cccggtcgtc 360
aacaaacagg gcgaactgga acactacctc gccatgcaac gtgatatcag cgccagctat 420
gcgctcgaac agcgattgcg caatcacacc accatgagcg aggcggtgct gaacaacatt 480
cctgccgccg tggtggtggt caacgagcag gaccaggtag tcatggacaa cctcgcctac 540
aaaaccttct gtgccgactg cggtggcaag gagctgctca ccgaactgga tttctcccgg 600
cgcaaaagcg atctctatgc cgggcaaata ctgcctgtgg tgctgcgcgg cgccgtgcgc 660
tggctctctg tcacctgctg gaccttgccg ggggtgagcg aagaagccag ccgctacttt 720
attgataccg cgctgccccg caccctggtg gtgatcaccg actgcaccca gcaacaacaa 780
caggccgaac agggccgtct cgatcgtctc aaacaggaga tgaccaccgg gaagctgctg 840
gccgcgatcc gtgaatcgtt ggatgccgcg ctggttcagc taaactgccc catcaatatg 900
ctggcggcgg cgcgacgtct caacggtgaa gataaccata acgtggcgct ggatgccgcg 960
tggcgcgagg gggaagaggc gctggcccgc ctgcaacgct gccgcccttc tctcgatctg 1020
gaagagagcg cgctgtggcc tctgcaaccg ctgtttgacg acctgcgcgc cctttaccat 1080
acccgctata acaatggcga aaatctgcac gttgaaatgg cctctccgca tctggcgggg 1140
tttggtcagc gcacgcagat ccttgcctgt ctcagtttgt ggctcgaccg tacgctggcc 1200
ctcgccgccg cgctaccgga cagaacgctg catacccagc tttacgcccg tgaagaagat 1260
ggctggctgt ccatttggct gacagataat gtgccgctca tccatgtgcg atacgcccac 1320
tcccccgatg ccctgaacgc ccccggcaaa gggatggagc tgcgattgat tcaaaccctg 1380
gttgcccatc atcgcggcgc aatagaacta actacccgcc ctgaaggcgg tacctgcctg 1440
accctgcgat tcccgttatt tcattcactg accggaggcc cacgatga 1488
<210> 132
<211> 1572
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> nifA
<400> 132
atgacccagc gacccgagtc gggcaccacc gtctggcgtt ttgatctctc acagcaattt 60
accgccatgc agcgcatcag cgtggtgttg agtcgcgcaa ccgagataag ccagacgctg 120
caggaggtgc tgtgtgttct gcataatgac gcatttatgc aacacggcat gctgtgtctg 180
tatgacaacc agcaggaaat tctgagtatt gaagccttgc aggaggcaga ccaacatctg 240
atccccggca gctcgcaaat tcgctatcgc cctggcgaag ggctggtagg agccgtactg 300
tcccagggac aatctcttgt gctgccgcgt gtcgccgacg atcaacgctt tctcgacagg 360
cttggcatct atgattacaa cctgccgttt atcgccgtcc ccttaatggg gccaggcgcg 420
cagacgattg gcgtgctcgc cgcgcagccg atggcgcgtc tggaggagcg gcttccttcc 480
tgtacgcgct ttctggaaac cgtcgccaat ctggtcgcac agacagtccg gctgatgacc 540
ccgcctgccg ccgccacacc gcgcgccgcg attgcccaga ccgaacgcca gcgcaactgt 600
ggcactcctc gccccttcgg ctttgagaat atggtgggca aaagcccggc catgcagcag 660
acaatggaca ttatccgcca ggtttcgcgc tgggatacca cggtactggt gcgcggcgaa 720
agcggcaccg gtaaagaact tatcgccaat gctattcatc acaactcccc tcgcgccgcc 780
gcgccctttg tgaaatttaa ctgcgcggcg ctaccggata cgctactgga gagcgaattg 840
ttcggccatg aaaaaggggc gttcaccggc gcggttcgcc agcgtaaagg acgttttgaa 900
ctggccgatg gcggcacact gtttcttgat gaaattggcg aaagcagcgc ctcgttccag 960
gccaaactgc tgcgtatttt gcaggagggt gaaatggagc gcgttggcgg cgacgaaacc 1020
ctgcgcgtca atgtgcgtat catcgccgcc accaaccgga atctggaaga agaggtgcgg 1080
atgggcaatt tccgcgagga tctctattat cgcctcaacg taatgcccat ctccctgccc 1140
ccgctgcgtg aacgtcagga ggacattgcc gagctggcgc actttctggt gcgcaaaatc 1200
gcccataacc aggggcgtac gctgcgcatc agtgatggcg ccatccgtct gctgatgggt 1260
tacaactggc ccggtaacgt gcgtgagctg gaaaattgcc tggaacgttc ggcagtgatg 1320
tcagaaaacg gcctgatcga ccgcgatgtg gtgctcttta accaccgtga gaacacgcca 1380
aaactcgcta tcgccgccgc gccaaaagag gatagctggc ttgatcaaac gctggatgaa 1440
cgtcaacggc tgattgccgc gctggaaaaa gccgggtggg tgcaggccaa agcggcgcgt 1500
ctgctgggta tgacgccccg tcaggtcgcc tatcggatac aaattatgga tatcagcatg 1560
cccaggatgt ga 1572
<210> 133
<211> 2853
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> glnE
<400> 133
atgatgccgc actctccaca gctacagcag cactggcaaa ctgtactggc ccgcttgcct 60
gagtcattca gtgaaacacc gcttagtgaa caagcgcagt tagtgcttac tttcagtgat 120
tttgtgcagg atagccttgc cgcgcatcct gactggctgg ctgagctgga aagcgcaccg 180
ccacaggcgg acgagtggaa gcagtatgcg caaacccttc gcgaatcgct ggaaggtgtg 240
ggagatgagg catcattaat gcgtgcgctg cgcctgttcc gtcgccatat gatggtgcgc 300
attgcctggg cgcagtcgct ggcgctggtg gcagaagatg agacgttgca gcagttgagc 360
gtactggcgg agaccctgat cgtcgctgca cgcgactggc tttacgatgc ctgctgtcgc 420
gagtggggaa cgccgtgcaa tcagcagggg gaaccgcagc cgttgctgat cctgggcatg 480
ggcaagctgg gtggcgggga gcttaacttt tcgtccgata tcgatctgat ttttgcctgg 540
ccggaaaacg gttcaacgcg cggtgggcga cgcgaacttg ataacgccca gttttttact 600
cgcttgggac agcgcctgat caaagtgctc gaccagccga cgcaggatgg ctttgtctat 660
cgcgtggata tgcggctgcg cccgtttggc gacagcggtc cgctggtgct gagttttgcc 720
gcgctggaag attattatca ggagcagggg cgcgactggg aacgttatgc gatggtgaaa 780
gcccgcatta tgggcgataa ggacgatgtt tacgctggcg aattacgggc catgctgcgg 840
ccgttcgtct tccgtcgcta tatcgatttc agcgttattc agtctctgcg taacatgaaa 900
gggatgattg cccgcgaagt gcgccgccgt ggtctgaaag ataacattaa gctgggcgcg 960
ggcggcatcc gtgagattga gtttatcgtt caggtgttcc agttgatacg cggtgggcgc 1020
gagccgtcgt tgcagtcccg ttcactgtta ccgacgctgg acgctatcga taagctgggt 1080
ttgctgccgc ctggcgatgc accggcgtta cgccaggcct atttgtatct gcgccgtctg 1140
gaaaacctgc tgcaaagcat taacgacgaa caaacgcaga cgctgccgac agatgaactc 1200
aatcgcgcgc gtctggcctg ggggatgcgg gtcgcagact gggaaaccct gaccgctgag 1260
cttgaaaagc agatgtctgc cgtacgaggg atattcaaca ccctgattgg cgatgacgaa 1320
gccgaagagc agggggatgc gctctgcggg caatggagtg agttgtggca ggatgcgttt 1380
caggaagatg acagcacgcc tgtgctggcg cacctttctg acgatgatcg ccgccgcgtg 1440
gtcgcgatga ttgctgattt tcgcaaagag ctggataaac gcaccattgg cccacgcggc 1500
cgccaggtgc tcgaccatct gatgccgcat ctgttgagtg atgtctgctc ccgtgaggat 1560
gcccctgtac cgttgtctcg cgtgacgccg ctgttaacgg gaattgtcac gcgtacgacg 1620
tatcttgagc tgctcagcga gtttcctggt gcgcgtaagc atctgatttc actctgtgcc 1680
gcctcgccga tggtggccag taagctggcg cgctatccgt tattgctgga tgagttgctc 1740
gatccgaata ccctttatca gcccacggcg atgaatgcct accgggatga gctacgtcag 1800
tatctgctgc gtgtgccgga tgacgatgaa gagcagcaac tggaggcgtt acgccagttt 1860
aaacaggctc aattgttgcg tgtggcggca gcagatctgg caggcacact ccccgtgatg 1920
aaagtgagcg atcacttaac atggcttgcc gaagccatca ttgaagccgt ggtacaacag 1980
gcgtggagcc tgatggtatc gcgttatggg cagccgaaac acttacgcga ccgtgaaggc 2040
cgtgggtttg cagtggtcgg ttacggcaaa ctgggcggtt gggagctggg ctatagttcc 2100
gatctggatt tgattttcct tcatgactgt ccggtggacg tgatgactga cggcgagcgg 2160
gaaatcgatg gccgccaatt ttatctgcgc cttgcccagc gcgtgatgca cctgttcagt 2220
acgcgcacct catccgggat cctgtatgag gtagacgcgc gcttgcgccc gtccggtgcg 2280
gcgggaatgc tggtgacctc aaccgaatcc tttgccgact accagcgcac cgaagcctgg 2340
acctgggaac atcaggcgct ggttcgcgcc cgcgttgtct atggcgatcc acaattaaac 2400
gcgcaatttg atgccatccg ccgcgatatc accatgaccg tgcgtaatgg tgcaacgtta 2460
caaaccgagg tgcgcgagat gcgcgaaaaa atgcgcgccc acttgagcaa taagcacaag 2520
gatcgctttg atattaaagc cgatgagggt ggaattaccg atatcgaatt tatcacccag 2580
tatctggtgc tgcgttatgc ccatgccaaa ccgaaactga cgcgctggtc ggacaatgtc 2640
cgcattctgg aagggctggc gcaaaacggc attatggaag agcaggaagc gcaggcactt 2700
accaccgcct atacaacgtt gcgtgatgag ctgcatcacc tggcgctaca ggagctgcca 2760
ggacatgttc cggaggcatg ttttgtcgct gaacgcgcga tggtgcgagc ctgctggaac 2820
aagtggttgg tggagccgtg cgaggacgcg taa 2853
<210> 134
<211> 1290
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> amtB
<400> 134
atgaagaaag cactattaaa agcgggtctg gcctcgctgg cattactgcc gtgtctggct 60
atggcagccg atccggttgt cgtcgataaa gccgacaatg cctttatgat gatttgcacc 120
gcgctggtgc tgtttatgtc aattccgggc atcgccctgt tctatggtgg tttaatccgc 180
ggtaaaaacg tcctttctat gctgacacag gttgcggtta cgttcgcact ggtgtgcgtg 240
ctgtgggtgg tttacggcta ctctctggcc tttggcactg gcggcagctt cttcggtagc 300
ttcgactggg tgatgctgaa aaatattgag ctgaaagcgc tgatgggcac catctatcag 360
tacattcacg ttgcgttcca gggctcgttt gcctgtatta ccgtcggcct gattgtcggt 420
gcgctggcag aacgtatccg tttctccgca gtactgattt tcgtcgtggt atggctgacg 480
ctgtcctacg tgccgatcgc acacatggtc tggggcggcg gtctgctggc aacccatggc 540
gccatggatt ttgcgggcgg tacagtcgtt cacatcaacg cagccgttgc aggcctggtg 600
ggtgcttacc tgattggcaa acgtgtcggt ttcggtaaag aagcgtttaa accgcacaac 660
ctgccgatgg tgtttaccgg tacggcaatc ctctactttg gctggttcgg attcaacgcg 720
ggttctgcaa gcgcggcgaa cgaaattgcg ggtctggctt ttgttaacac cgtcgtggca 780
acagcgggtg caatcctctc ctgggtcttc ggtgagtggg cgctgcgcgg caaaccgtct 840
ctgttgggtg cctgttctgg tgcgattgct ggcctcgtgg gtatcacccc ggcgtgtggt 900
tacgttggtg tgggtggcgc gctgatcgtg ggcatcgttg caggcctggc gggtctgtgg 960
ggcgttaccg cgctgaaacg ctggctgcgt gttgacgacc cgtgtgatgt cttcggtgtt 1020
cacggcgtgt gcggtatcgt aggttgtatc atgacaggta tcttcgcagc cacttcactg 1080
ggcggcgtgg gttatgccga aggcgtgacc atgggccatc aggttctggt acaactggaa 1140
agtatcgcca ttactatcgt atggtctggt atcgtcgcct ttatcggtta caaactggct 1200
gatatgacag tgggtctgcg tgttccggaa gatcaggaac gcgaagggct ggacgtcaac 1260
agccacggcg agaacgccta caacgcctga 1290
<210> 135
<211> 498
<212> ДНК
<213> Kluyvera intermedia
<220>
<223> PinfC
<400> 135
ctggggtcac tggagcgctt tatcggcatc ctgaccgaag aatttgccgg tttcttcccg 60
acctggctgg cccctgttca ggttgtggtg atgaatatca ctgattctca agctgaatat 120
gtcaacgaat tgacccgtaa attgcaaaat gcgggcattc gtgtaaaagc ggacttgaga 180
aacgagaaga ttggctttaa aatccgcgag cacactttac gtcgtgtccc ttatatgttg 240
gtctgtggtg ataaagaggt ggaagcaggc aaagtggccg ttcgcacccg ccgcggtaaa 300
gacctgggca gcctggacgt aagtgaagtg attgagaagc tgcaacaaga gattcgcagc 360
cgcagtcttc aacaactgga ggaataaggt attaaaggcg gaaaacgagt tcaaacggca 420
cgtccgaatc gtatcaatgg cgagattcgc gcccaggaag ttcgcttaac tggtctggaa 480
ggtgagcagc tgggtatt 498
<210> 136
<211> 1537
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (454)..(454)
<223> a, c, t, g, неизвестное или другое
<400> 136
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgaacggtag cacagagagc ttgctctcgg gtgacgagtg gcggacgggt gagtaatgtc 120
tgggaaactg cctgatggag ggggataact actggaaacg gtagctaata ccgcataacg 180
tcgcaagacc aaagaggggg accttcgggc ctcttgccat cagatgtgcc cagatgggat 240
tagctagtag gtggggtaac ggctcaccta ggcgacgatc cctagctggt ctgagaggat 300
gaccagccac actggaactg agacacggtc cagactccta cgggaggcag cagtggggaa 360
tattgcacaa tgggcgcaag cctgatgcag ccatgccgcg tgtatgaaga aggccttcgg 420
gttgtaaagt actttcagcg gggaggaagg cganacggtt aataaccgtg ttgattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc atccgaaact ggcaggcttg agtctcgtag 660
agggaggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg cctcctggac gaagactgac gctcaggtgc gaaagcgtgg ggagcaaaca 780
ggattagata ccctggtagt ccacgccgta aacgatgtct atttggaggt tgtgcccttg 840
aggcgtggct tccggagcta acgcgttaaa tagaccgcct ggggagtacg gccgcaaggt 900
taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggtcttga catccacaga acttgccaga gatggcttgg 1020
tgccttcggg aactgtgaga caggtgctgc atggctgtcg tcagctcgtg ttgtgaaatg 1080
ttgggttaag tcccgcaacg agcgcaaccc ttatcctttg ttgccagcgg tccggccggg 1140
aactcaaagg agactgccag tgataaactg gaggaaggtg gggatgacgt caagtcatca 1200
tggcccttac gaccagggct acacacgtgc tacaatggcg catacaaaga gaagcgacct 1260
cgcgagagca agcggacctc ataaagtgcg tcgtagtccg gattggagtc tgcaactcga 1320
ctccatgaag tcggaatcgc tagtaatcgt ggatcagaat gccacggtga atacgttccc 1380
gggccttgta cacaccgccc gtcacaccat gggagtgggt tgcaaaagaa gtaggtagct 1440
taaccttcgg gagggcgctt accactttgt gattcatgac tggggtgaag tcgtaacaag 1500
gtaaccgtag gggaacctgc ggttggatca cctcctt 1537
<210> 137
<211> 882
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifH
<400> 137
atgaccatgc gtcaatgcgc catttacggc aaaggtggga tcggtaaatc gaccaccaca 60
cagaacctgg tcgccgcgct ggcggagatg ggtaagaaag tcatgatcgt cggctgcgat 120
ccgaaagccg actccacgcg tttgatcctg catgcgaaag cgcagaacac cattatggag 180
atggccgccg aagtcggctc cgtcgaagac ctggaattag aagacgtgct gcaaatcggt 240
tacggcggcg tgcgctgcgc ggaatccggt ggcccggagc caggtgtggg ttgtgccggt 300
cgtggcgtga tcaccgcgat taacttcctc gaagaagaag gcgcttacgt gccggatctg 360
gattttgttt tctacgacgt gctgggcgac gtggtatgcg gtggtttcgc catgccgatt 420
cgtgaaaaca aagcgcagga gatctacatc gtttgctctg gcgaaatgat ggcgatgtac 480
gccgccaata acatctccaa aggcatcgtg aaatatgcca aatccggtaa agtgcgcctc 540
ggcgggctga tttgtaactc gcgccagacc gaccgcgaag atgaactcat cattgcgctg 600
gcggaaaaac tcggcacgca aatgatccac tttgttcccc gcgacaacat tgtgcagcgt 660
gcggaaatcc gccgtatgac ggttatcgaa tatgacccga cctgcaatca ggccaacgaa 720
tatcgcagcc ttgccagcaa aatcgtcaac aacaccaaaa tggtggtacc aaccccctgc 780
accatggatg aactggaaga actgctgatg gagttcggca ttatggatgt ggaagacgcc 840
agcatcattg gtaaaaccgc cgccgaagaa aacgccgtct ga 882
<210> 138
<211> 1449
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifD1
<400> 138
atgagcaatg caacaggcga acgtaacctg gaaatcatcg agcaggtgct ggaggttttc 60
ccggaaaaga cgcgcaaaga gcgcagaaaa cacatgatgg tgacggaccc ggagcaggag 120
agcgtcggca agtgcatcat ctctaaccgc aaatcgcagc cgggcgtgat gaccgtgcgt 180
ggctgctcgt atgccggatc aaaaggggtg gtatttgggc caatcaaaga tatggcgcat 240
atctcccacg gcccgatcgg ctgcgggcag tactcccgcg ccgggcggcg taactactat 300
accggcgtca gcggcgtgga cagtttcggc acgctcaact tcacctccga tttccaggag 360
cgcgacatcg tgtttggcgg cgacaaaaag ctcgccaaac tgattgaaga gctggaagaa 420
ctgtttccgc tgaccaaagg catttcgatt cagtcggaat gcccggtcgg cctgattggc 480
gatgatattg aagccgtggc gaacgccagc cgcaaagcga tcaacaaacc ggttattccg 540
gtgcgttgcg aaggctttcg cggcgtgtcg caatccctcg gtcaccatat tgccaacgat 600
gtgatccgcg actgggtact ggataaccgc gaaggcaaac cgtttgaatc caccccttac 660
gatgtggcga tcatcggcga ttacaacatc ggtggcgacg cctgggcctc gcgcattttg 720
ctcgaagaga tggggttgcg ggtggtcgcg cagtggtccg gcgacggtac gctggtggag 780
atggaaaaca cgccgttcgt caaactgaac ctggtgcact gctaccgctc gatgaactac 840
atctcgcgcc atatggagga gaagcacggt attccgtgga tggaatacaa cttctttggc 900
ccgacgaaaa tcgcggaatc gctgcgcaaa atcgccgacc tgttcgacga caccattcgc 960
gccaacgccg aagcggtgat cgcccgatac caggcgcaga acgacgccat tatcgccaaa 1020
tatcgcccac gtctggaggg tcgcaaagtg ttgctctata tgggcgggct gcgtccgcgc 1080
catgtgattg gcgcctatga agatctggga atggagatca tcgccgccgg ttatgagttt 1140
ggtcataacg acgattacga ccgcaccctg ccggatctga aagagggcac gctgctgttt 1200
gatgacgcca gcagctatga gctggaggcg tttgtcaacg cgctgaaacc ggatctcatc 1260
ggttccggca tcaaagagaa gtacatcttt cagaaaatgg gcgtgccgtt tcgccagatg 1320
cactcctggg attactccgg cccgtaccac ggctatgacg gcttcgccat cttcgcccgc 1380
gatatggata tgacgctcaa caaccccgcc tggggtcagt tgaccgcgcc gtggcttaaa 1440
tccgcctga 1449
<210> 139
<211> 1374
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifD2
<400> 139
atgaagggga acgacatcct ggctctgctc gatgaaccag cctgcgagca taaccataaa 60
cagaaaaccg gctgtagcgc gccaaaaccc ggcgccaccg ccggaggctg cgccttcgac 120
ggcgcacaga tcaccctgct gccactttcc gatgtggcgc atctggtaca tggcccgatt 180
ggctgcgccg gcagctcatg ggataaccgt ggcagcctga gttctggccc gctgattaac 240
cgactcggat tcaccactga tttgaacgaa caggatgtca tcatggggcg cggcgagcgg 300
cggttgtttc acgcggtgcg ccatattgtc gagcgctatc acccggcggc ggtatttatt 360
tacaacacct gcgttccggc tatggaaggc gatgacattg acgcggtctg ccaggccgcc 420
gcgaccgcca ccggtgtgcc cgtgattgcc gtagatgtgg ccggttttta cggtagcaaa 480
aacctgggta accgcctcgc gggcgaggtg atggtgaaaa aagttatcgg cgggcgcgaa 540
cccgcgccgt ggccggacaa tacacctttt gccccggcgc accgccatga cataggcctg 600
attggcgaat ttaacatcgc cggcgagttc tggcatatcc agccgctgct tgatgagctg 660
ggtattcgcg tccttggctc cctttccggc gacgggcgct ttgccgagat ccagacgttg 720
caccgcgcgc aggtcaatat gctggtgtgc tccagggcgc tgattaatgt cgccagatcg 780
cttgaacaac gttatggcac accctggttt gaaggcagtt tttatggcgt tcgcgccacc 840
tccgatgccc tgcgccagct ggcaacactc accggcgata gcgatttaat ggcgcgaacc 900
gaacggctga tcgcacgtga agagcaagcc acagaacagg cgctagcacc gctgcgtgaa 960
cggttacacg gccggaaagt gctgctctat accggtggcg tgaaatcctg gtcggtggtt 1020
tcggcgctgc aggatctcgg catgacggtc gttgctaccg gaacgcgcaa atccaccgaa 1080
gaggataaac aacgcatccg tgaactgatg ggcgatgacg ccatcatgct ggatgaaggc 1140
aatgcccgcg ccttgctgga tgtggtctat cgctacaaag ccgacatgat gatcgcgggc 1200
gggcgcaaca tgtacaccgc ctataaagcg cgtctgccct ttctggatat caaccaggag 1260
cgtgaacacg cgtttgccgg ttatcgcggc atcatcacgc ttgccgaaca actttgtcag 1320
acgctggaaa gcccggtctg gccgcaaaca catgcccgcg ccccgtggca ataa 1374
<210> 140
<211> 1563
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifK1
<400> 140
atgagccaga ctgctgagaa aatacagaat tgccatcccc tgtttgaaca ggacgcctac 60
cagacactat ttgccggtaa acgggcactc gaagaggctc actcgccgga gcgggtgcag 120
gaagtgtttc aatggaccac caccccggaa tacgaagcgc tgaacttcaa acgcgaagcg 180
ctgactatcg acccggcaaa agcctgccag ccgctggggg cggtgctctg ttcgctgggg 240
tttgccaaca ccctgccgta tgtgcacggt tcacagggtt gtgtggccta tttccgtacg 300
tactttaacc gccacttcaa agaaccggtg gcctgcgtgt cggattcgat gacggaagac 360
gcggccgtgt tcggcgggaa taacaacctc aacaccgggt tacaaaacgc cagcgcactg 420
tataaaccgg agattatcgc cgtctctacc acctgtatgg cggaagtgat cggtgatgat 480
ttacaggcgt ttatcgccaa cgccaaaaaa gatggttttc tcgatgccgc catccccgtg 540
ccctacgccc acacccccag ttttatcggt agccatatca ccggctggga caacatgttt 600
gaaggttttg cccgtacctt taccgcaaac catcagccac agcccggtaa actttcacgc 660
ctgaacctgg tgaccgggtt tgaaacctat ctcggcaatt tccgcgtgct gaaacgcatg 720
atggaacaaa tggaggtgca ggcgagtgtg ctctccgatc cgtcggaggt gctggacacc 780
cccgccaatg gccattacca gatgtacgcg ggcggtacga cgcagcaaga gatgcgcgag 840
gcaccggatg ccatcgacac cctgctgctg caaccgtggc agctggtgaa aagcaaaaaa 900
gtggtgcagg agatgtggaa tcagcccgcc accgaggttg ccattcccgt cgggctggca 960
ggcacagacg aactgttgat ggcgattagc cagttaaccg gcaaagccat tcccgattcg 1020
ctggcgctgg agcgcgggcg gctggtcgat atgatgctcg actcccacac ctggttacac 1080
ggtaaaaaat tcggtctgtt tggcgatccg gattttgtca tgggattgac ccgcttcctg 1140
ctggaactgg gctgtgaacc tgccgtcatc ctctgccata acggtaacaa acgctggcaa 1200
aaagcgatga agaaaatgct cgatgcttca ccgtacggcc aggagagcga agtgtttatc 1260
aactgcgact tgtggcattt ccgctcgctg atgttcaccc gccagccgga ttttatgatt 1320
ggcaactcgt acgccaagtt tattcagcgc gacaccttag ccaagggcga acagtttgaa 1380
gtcccgctga tccgcctcgg ttttccgctg ttcgaccgtc accatctgca ccgccagacc 1440
acctggggct acgagggcgc gatgagcatt ctcacgacgc tggtgaatgc ggtactggag 1500
aaagtggaca aagagaccat caagctcggc aaaaccgact acagcttcga tcttatccgt 1560
taa 1563
<210> 141
<211> 1386
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifK2
<400> 141
atggctgata ttgttcgtag taaaaaaccg ctggcggtga gcccgataaa aagcggccag 60
ccgctggggg cgatcctggc aagcctgggt ttcgaacagt gcataccgct ggtacacggc 120
gctcaggggt gcagcgcgtt cgcgaaagtg ttctttattc aacattttca cgacccgatc 180
ccgctgcaat cgacggcgat ggacccgact tccaccatta tgggcgccga tgaaaacatt 240
tttaccgcgc tcaatgttct ctgccagcgc aacgccgcga aagccatcgt gctgctcagc 300
accgggctgt cagaagccca gggcagcgat atttcacgag tggtgcgcca gtttcgtgat 360
gactttccgc ggcataaaaa cgtggcgctg ctcaccgtca acaccccgga tttctacggc 420
tcgctggaaa acggctacag cgccgtgctg gaaagcatga ttgaacagtg ggtgcccgcg 480
cagcccgccg ccagcctgcg caaccgtcgc gtcaacctgc tggtcagcca tttactgacg 540
ccgggcgata tcgaactgtt acgcagttat gtggaagcat tcggtctgca accggtgatt 600
gtgccggatc tatcgcagtc gctggacgga catctggcca acggtgattt ttcgcccgtc 660
acccaggggg gaacaccgct gcgcatgatt gaacagatgg ggcaaaacct ggccactttt 720
gtgattggcc actcgctggg gcgggcggcg gcgttactgg cgcagcgcag ccgtggcgag 780
gtgatcgccc tgccgcatct gatgacgctt gatgcgtgcg acacctttat ccatcgcctg 840
aaaaccctct ccgggcgcga cgtgcccgcg tggattgagc gccagcgcgg gcaagtgcag 900
gatgcgatga tcgattgcca tatgtggttg cagggcgcgg ctatcgccat ggccgcagaa 960
ggcgatcacc tggcggcatg gtgcgatttc gcccgcagcc agggcatgat ccccggcccg 1020
gttgtcgcgc cggtcagcca gccggggttg caaaatctgc cggttgaaat ggtggtcatc 1080
ggcgatctgg aagatatgca ggatcggctt tgcgcgacgc ccgccgcgtt actggtggcc 1140
aattctcatg ccgccgatct cgccacgcag tttgatatgt cgcttatccg cgccgggttt 1200
ccggtgtatg accggctggg ggaatttcgt cggctgcgcc aggggtatag cggcattcgt 1260
gacacgctgt ttgagctggc gaatgtgatg cgcgaacgcc attgcccgct tgcaacctac 1320
cgctcgccgc tgcgtcagcg cttcggcgac aacgttacgc caggagatcg gtatgccgca 1380
tgttaa 1386
<210> 142
<211> 1488
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifL
<400> 142
atgaccctga atatgatgat ggatgccagc gcgcccgagg ccatcgccgg tgcgctttcg 60
caacaacatc ctgggctgtt ttttaccatc gttgaagaag cccccgtcgc tatttcacta 120
accgatgccg aggcacgtat tgtctatgcc aacccggcat tctgccgcca gaccggctat 180
gagcttgagg agttgttgca gcaaaatccc cgcctgcttg ccagtcagca gaccccacgg 240
gaaatctacc aggatatgtg gcacaccctg ttacaacgtc gaccatggcg cgggcaattg 300
atcaaccgcc accgtgacgg cagccttttt ctggttgaga tcgatatcac cccggtgatt 360
aacccgtttg gcgaactgga acactacctg gccatgcagc gcgatatcag cgccggttat 420
gcgctggagc agcggttgcg taatcacatg gcgctgaccg aagcggtgct gaataacatt 480
ccggcggcgg tggtcgtggt cgatgaacgc gatcgtgtgg ttatggataa cctcgcctat 540
aaaactttct gtgctgattg cggcggaaaa gagctactga gcgaactcca tttttcagcc 600
cgtaaagcgg agctggcaaa cggccaggtc ttaccggtgg tgctgcgcgg cgcggtgcgc 660
tggttgtcgg tcacctgctg ggcgctgcca ggcgtcagcg aagaagccag tcgctacttt 720
attgataata ccttgacgcg cacgctggtg gtcatcaccg acgacaccca gcagcgccag 780
cagcaagagc aaggacggct tgaccgcctt aaacagcaga tgaccagcgg caaactgctg 840
gcggcgatcc gcgaagcgct tgacgccgcg ctgatccagc ttaactgccc catcaatatg 900
ctggcggcgg cgcggcgttt aaacggcagt gataacagca acgtagcgct ggacgccgcg 960
tggcgcgaag gtgaagaagc gatggcgcgg ctgaaacggt gccgcccgtc gctggagctg 1020
gaaagtgccg ccgtctggcc gctgcaaccc ttttttgacg acttgcgcgc gctttatcac 1080
acccgctacg agcagggtaa aaatttgcag gtcacgctgg attcgacgca tctggtggga 1140
tttggtcagc gaacccaact gctggcctgc ctgagtctgt ggctcgatcg cacgctggat 1200
attgccgtcg ggctgcgtga tttcaccgcc caaacgcaga tttacgcccg ggaagaagcg 1260
ggctggctct cgttgtatat cactgacaat gtgccgttga ttccgctgcg ccatacccat 1320
tcgccggatg cgcttaacgc accgggaaaa ggtatggagt tgcggctgat ccagacgctg 1380
gtagcgcatc acaacggcgc gatagaactc acttcacgcc ccgaaggggg aagctgcctg 1440
accctacgat tcccgctatt tcattcactg accggaggtt caaaatga 1488
<210> 143
<211> 1575
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> nifA
<400> 143
atgacccagc gaaccgagtc gggtaatacc gtctggcgct tcgatttatc ccagcagttc 60
accgcgatgc agcggataag cgtggttctc agccgggcga ccgaggttga acagacactc 120
cagcaggtgc tgtgcgtatt gcacaatgac gcctttttgc agcacggcat gatctgtctg 180
tacgacagcc agcaggcgat tttgactatt gaagcgttgc aggaagccga tcagcagttg 240
atccccggca gctcgcaaat tcgctaccgt ccgggtgaag ggctggtcgg gacggtgctt 300
tcgcaggggc aatcgttagt gctggcgcgt gtggctgacg atcagcgctt tcttgaccgc 360
ctgggactgt atgattacaa cctgccgttt atcgccgtgc cgctgatagg gccggatgcg 420
cagacttttg gcgtgctgac ggcgcaaccg atggcgcgtt acgaagagcg gttacccgcc 480
tgcacccgct ttctggaaac ggtcgcgaat ctggtggcgc agaccgtgcg tttgatgacg 540
ccgccggctg cacgcccttc cccacgcgct gccatcacgc caaccgccag cccgaaatcg 600
tgcagtactt cacgcgcgtt cggcttcgaa aatatggtcg gcaacagccc ggcaatgcgc 660
cagaccatgg agattatccg tcaggtttcg cgctgggata ccaccgttct ggtgcgcggc 720
gagagcggca ccggcaagga actgattgcc aacgccatcc atcacaattc gccgcgcgcc 780
agtgcgccat ttgtgaaatt caactgtgcg gcgctgccgg acacattgct tgaaagcgaa 840
ttatttggtc atgaaaaagg cgcctttacc ggcgcggtac gccagcgtaa aggccgtttt 900
gagctggccg atggcggcac gctgtttctt gacgaaattg gggaaagcag cgcctcgttt 960
caggctaagc tgctgcgtat tttgcaggag ggcgaaatgg aacgcgtcgg tggtgacgag 1020
acattgcaag tgaatgtgcg catcattgcc gcgacgaacc gcaaccttga agatgaagta 1080
cgcctgggac attttcgcga agatctctat taccgcctga atgtgatgcc catcgccctg 1140
ccgccgctgc gcgaacgcca ggacgacatc gccgaactgg cacattttct ggtgcgtaaa 1200
atcgcccaca accagaaccg cacgctgcgc attagcgagg gcgctatccg cctgctgatg 1260
agctacagct ggcccggcaa tgtgcgcgaa ctggaaaact gccttgagcg ctctgcggtg 1320
atgtcggaaa acggtctgat cgatcgggac gtgattttat ttaatcatcg cgaccagcca 1380
gccaaaccgc cggttatcag cgtcacgccc gacgataact ggctcgataa cacccttgac 1440
gagcgccagc ggctgattgc cgcgctggaa aaagcgggat gggtacaagc caaagccgcc 1500
cgcttgctgg ggatgacgcc gcgccaggtc gcttatcgta ttcagaccat ggatatcacc 1560
ctgccaaggc tataa 1575
<210> 144
<211> 2850
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> glnE
<400> 144
atgccgcacc acgcaggatt gtcgcagcac tggcaaacgg ttttttctcg tctgccggaa 60
gcgctcaccg cgcaaccatt gagcgcgcag gcgcagtcag tgctcacttt tagtgatttt 120
gttcaggaca gcatcatcgt gcatcctgag tggctggcag agcttgaaag cgcaccgccg 180
ccagcgaacg agtggcaaca ctacgcgcaa tggctgcaag cggcgctgga gggcgtcacc 240
gatgaaacct cgctgatgcg cacgctgcgg ctgtttcgcc gtcgcattat ggtgcgcatc 300
gcctggagtc aggcgctaca gttggtggcg gaagaggata tcctgcaaca gctcagcgtg 360
ctggcggaaa ctctgatcgt cgccgcgcgc gactggctct atgacgcctg ctgccgtgag 420
tggggaacgc cgtgcaatcc gcaaggcgtc gcgcagccga tgctggtgct cggcatgggc 480
aaacttggcg gcggcgaact caatttctca tccgatatcg atttgatttt tgcctggccg 540
gaaaatggca ccacgcgcgg cggacgccgt gaactggata acgcgcagtt ttttacccgc 600
cttggtcaac ggctaattaa agtcctcgac cagcccacgc aggatggctt tgtctaccgc 660
gtcgatatgc gcttgcgtcc ctttggcgac agcggcccgc tggtgctgag ttttgccgcg 720
ctggaagatt actaccagga gcaggggcgc gactgggaac gatacgcgat ggtgaaagcg 780
cgcattatgg gggacaacga cggcgaccat gcgcgagagt tgcgcgccat gctgcgcccg 840
ttcgttttcc gccgctatat cgacttcagc gtgatccagt ctctgcgcaa catgaaaggc 900
atgattgccc gcgaagtgcg gcgtcgcggc ctgaaggaca acataaaact cggcgcgggc 960
ggtattcgcg aaatagagtt tatcgtgcag gttttccagt tgattcgcgg cggtcgcgag 1020
cctgcgctgc aatcgcgttc gctgttgccg acgcttgctg ccattgatca actacatctg 1080
ctgccggatg gtgatgcacc ccggctgcgc gaggcgtatt tgtggctgcg acggctggaa 1140
aacttgctgc aaagcattaa tgacgaacag acacagacgc tgccggccga tgatttgaat 1200
cgcgcgcgcc tcgcctgggg aatgggcaaa gagagctggg aagcgctctg cgaaacgctg 1260
gaagcgcata tgtcggcggt gcggcagatt ttcaacgatc tgattggcga tgatgaaacg 1320
gattcgccgg aagatgcgct ttctgagggc tggcgcgaat tgtggcagga tgcgttgcag 1380
gaagaggact ctacgcccgt gctggcgcat ctttccgagg acgatcgccg ccgcgtggtg 1440
gcgctgattg ctgattttcg caaagagctg gataaacgca ccattggccc gcgcgggcga 1500
caggtactcg atcacttaat gccgcatctg ctcagcgatg tatgctcgcg tgacgatgcg 1560
ccagtgccgc tgtcgcgtct gacgccgctg ctcaccggta ttattacgcg caccacttac 1620
cttgagctgc tgagtgaatt ccccggtgcg ctgaaacacc tcatttccct gtgcgccgcg 1680
tcgccgatgg tggccagcca actggcgcgc tacccgatcc tgctcgatga actgctcgac 1740
ccgaacacgc tctatcaacc gacggcgatg aacgcctatc gcgatgaact gcgacaatac 1800
ctgttgcgcg tgccggaaga ggatgaagag cagcaactgg aggcgctacg gcagtttaag 1860
caggcgcagt tgttgcgcgt agcggcggcg gatatcgccg gtacgttacc cgtcatgaaa 1920
gtgagcgatc acttaacctg gctggcggaa gcgattatcg atgcggtggt gcagcaagcc 1980
tggaaccaga tggtggcgcg ttacggccag ccgacgcatc tgcacgatcg cgaagggcgc 2040
ggtttcgccg tggtcggtta cggcaaactt ggcggctggg aattaggtta cagctccgat 2100
ctggatctgg tgttcctgca cgactgcccc atggatgtga tgaccgatgg cgagcgtgaa 2160
atcgatggcc gccagttcta tttgcgcctc gcgcagcgcg tgatgcacct gttcagcacg 2220
cgcacgtcgt ccggcattct ttatgaagtc gatgcgcgtt tgcgcccgtc cggcgcggcc 2280
ggaatgctgg tgaccactgc ggaagcgttc gccgattatc aaaaaaatga agcctggaca 2340
tgggagcatc aggcgctggc gcgtgcgcgc gtggtgtacg gcgatccgca actgaccgcc 2400
gaatttgacg ccattcgccg cgatatcctg atgacctccc gcgatgccgc taccctgcaa 2460
accgaagtgc gggaaatgcg tgagaaaatg cgcgcccatc ttggtaacaa gcacaaagac 2520
cgtttcgatc tgaaagccga tgaaggcggt atcaccgata ttgagtttat cgctcagtat 2580
ctggtgctgc gctttgccca tgagaagccg aaactgacgc gctggtcgga taatgtgcgc 2640
atcctcgaag ggctggcgca aaacggcatc atggatgagc aggaagcgca ggcattgacg 2700
ctggcgtaca ccacgttgcg tgatgagctg caccacctgg cgctgcaaga gctgccagga 2760
catgtggcgc tctcctgttt tgtcgccgag cgtgcgctta tcaaaaccag ctgggacaag 2820
tggctggtgg aaccgtgcgc cccggcgtaa 2850
<210> 145
<211> 1287
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> amtB
<400> 145
atgaaaaaca caacattaaa aacggctctt gcttcgctgg cgttgctgcc aggcctggcg 60
atggcggctc ccgctgtggc ggataaagcc gacaacggct ttatgatgat ttgcaccgcg 120
ctggtgctgt ttatgaccat tccgggcatt gcgctgttct acggcggttt gatccgcggt 180
aaaaacgtgc tgtcgatgct gacgcaggtt gccgtcacct tcgctctggt gtgcatcctg 240
tgggtggttt acggctactc tctggcattt ggcgagggca acagcttctt cggcagtttc 300
aactgggcga tgttgaaaaa catcgaattg aaagccgtga tgggcagcat ttatcagtac 360
atccacgtgg cgttccaggg ctcctttgct tgtatcaccg ttggcctgat tgtcggtgcg 420
ctggctgagc gtattcgctt ctctgcggtg ctgatttttg tggtggtatg gctgacgctt 480
tcttatgtgc cgattgcgca catggtctgg ggtggcggtc tgctggcaac ccacggcgcg 540
ctggatttcg cgggcggtac ggttgttcac atcaacgccg cgatcgcagg tctggtgggg 600
gcttacctga ttggcaaacg cgtgggcttt ggcaaagaag cgttcaaacc gcataacctg 660
ccgatggtct tcaccggcac cgcgatcctc tatgttggct ggtttggctt caacgccggc 720
tctgcaagct cggcgaacga aatcgctgcg ctggctttcg tgaacacggt tgttgccact 780
gcggccgcta ttctggcgtg ggtatttggc gagtgggcaa tgcgcggtaa gccgtctctg 840
ctcggtgcct gttctggtgc catcgcgggt ctggttggta tcaccccggc gtgcggttat 900
gtgggtgtcg gcggcgcgct gattgtgggt ctgattgccg gtctggcagg gctgtggggc 960
gttactgcac tgaaacgtat gttgcgtgtt gatgacccat gcgatgtctt cggtgtgcac 1020
ggcgtgtgcg gcatcgtggg ttgtatcctg accggtatct tcgcgtctac gtcgctgggc 1080
ggtgtcggtt tcgctgaagg ggtgaccatg ggccatcagg tactggtaca gctggaaagc 1140
gttgccatca ctatcgtgtg gtctggcgtg gtggccttta tcggttacaa actggcggat 1200
atgacggtag gcctgcgcgt accggaagag caagagcgtg aagggctgga tgtgaacagc 1260
cacggcgaaa atgcgtataa cgcctga 1287
<210> 146
<211> 500
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> PinfC
<400> 146
ttcttggttc tctggagcgc tttatcggca tcctgactga agaatttgca ggcttcttcc 60
caacctggct tgcacccgtg caggtagttg tgatgaacat cactgattcg caggctgaat 120
acgttaacga attgacccgt aaactgcaaa atgcgggcat tcgtgtaaaa gcagacttga 180
gaaacgagaa gattggcttt aaaatccgcg agcacacttt acgtcgtgtc ccttatatgc 240
tggtttgtgg tgacaaagag gtcgaagccg gcaaagttgc tgtgcgtacc cgtcgcggta 300
aagacctggg tagcctggac gtaaatgatg ttatcgagaa gctgcaacaa gagattcgca 360
gccgcagtct tcaacaactg gaggaataag gtattaaagg cggaaaacga gttcaaacgg 420
cgcgtcccaa tcgtattaat ggcgagattc gcgccacgga agttcgctta acaggtctgg 480
aaggcgagca gcttggtatt 500
<210> 147
<211> 348
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> Prm1
<400> 147
cgttctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctc gcggcgcttt 60
ttttatattc tcgactccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcgc 348
<210> 148
<211> 339
<212> ДНК
<213> Kosakonia pseudosacchari
<220>
<223> Prm7
<400> 148
cgcgtcaggt tgaacgtaaa aaagtcggtc tgcgcaaagc acgtcgtcgt ccgcagttct 60
ccaaacgtta attggtttct gcttcggcag aacgattggc gaaaaaaccc ggtgcgaacc 120
gggttttttt atggataaag atcgtgttat ccacagcaat ccattgatta tctcttcttt 180
ttcagcattt ccagaatccc ctcaccacaa agcccgcaaa atctggtaaa ctatcatcca 240
attttctgcc caaatggctg ggattgttca ttttttgttt gccttacaac gagagtgaca 300
gtacgcgcgg gtagttaact caacatctga ccggtcgat 339
<210> 149
<211> 1538
<212> ДНК
<213> вид Enterobacter
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (62)..(62)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (265)..(265)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (457)..(457)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (469)..(469)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (539)..(539)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (551)..(552)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (559)..(559)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (561)..(561)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (576)..(576)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (587)..(587)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (591)..(591)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (655)..(657)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (665)..(665)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (678)..(678)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (706)..(706)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (710)..(710)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (714)..(714)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (718)..(718)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (735)..(735)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (755)..(755)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (759)..(759)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (796)..(796)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1006)..(1007)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1016)..(1017)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1133)..(1133)
<223> a, c, t, g, неизвестное или другое
<400> 149
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gngcggaagc acaggagagc ttgctctctg ggtgacgagc ggcggacggg tgagtaatgt 120
ctgggaaact gcctgatgga gggggataac tactggaaac ggtagctaat accgcataac 180
gtcgcaagac caaagagggg gaccttcggg cctcttgcca tcagatgtgc ccagatggga 240
ttagctagta ggtggggtaa cggcncacct aggcgacgat ccctagctgg tctgagagga 300
tgaccagcca cactggaact gagacacggt ccagactcct acgggaggca gcagtgggga 360
atattgcaca atgggcgcaa gcctgatgca gccatgccgc gtgtatgaag aaggccttcg 420
ggttgtaaag tactttcagc ggggaggaag gtgttgnggt taataaccnc agcaattgac 480
gttacccgca gaagaagcac cggctaactc cgtgccagca gccgcggtaa tacggaggnt 540
gcaagcgtta nncggaatna ntgggcgtaa agcgtncgca ggcggtntgt naagtcggat 600
gtgaaatccc cgggctcaac ctgggaactg cattcgaaac tggcaggcta gagtnnngta 660
gaggngggta gaattccngg tgtagcggtg aaatgcgtag agatcnggan gaanaccngt 720
ggcgaaggcg gcccnctgga caaagactga cgctnaggng cgaaagcgtg gggagcaaac 780
aggattagat accctngtag tccacgccgt aaacgatgtc gacttggagg ttgtgccctt 840
gaggcgtggc ttccggagct aacgcgttaa gtcgaccgcc tggggagtac ggccgcaagg 900
ttaaaactca aatgaattga cgggggcccg cacaagcggt ggagcatgtg gtttaattcg 960
atgcaacgcg aagaacctta cctactcttg acatccagag aacttnncag agatgnnttg 1020
gtgccttcgg gaactctgag acaggtgctg catggctgtc gtcagctcgt gttgtgaaat 1080
gttgggttaa gtcccgcaac gagcgcaacc cttatccttt gttgccagcg gtncggccgg 1140
gaactcaaag gagactgcca gtgataaact ggaggaaggt ggggatgacg tcaagtcatc 1200
atggccctta cgagtagggc tacacacgtg ctacaatggc gcatacaaag agaagcgacc 1260
tcgcgagagc aagcggacct cataaagtgc gtcgtagtcc ggattggagt ctgcaactcg 1320
actccatgaa gtcggaatcg ctagtaatcg tagatcagaa tgctacggtg aatacgttcc 1380
cgggccttgt acacaccgcc cgtcacacca tgggagtggg ttgcaaaaga agtaggtagc 1440
ttaaccttcg ggagggcgct taccactttg tgattcatga ctggggtgaa gtcgtaacaa 1500
ggtaaccgta ggggaacctg cggttggatc acctcctt 1538
<210> 150
<211> 882
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifH
<400> 150
atgaccatgc gtcaatgtgc catttacggc aaaggtggta tcggtaaatc cactaccacg 60
caaaacctgg tcgccgcgct ggcggagatg ggcaagaaag taatgatcgt cggctgcgac 120
ccgaaagcag actccactcg tctgatcctg catgcgaaag cgcagaacac cattatggag 180
atggcggctg aagtcggctc cgtggaagac cttgaactgg aagatgtgct gcaaatcggt 240
tacggcgacg tacgctgcgc agaatccggc ggcccggaac caggcgttgg ctgtgctggt 300
cgcggggtaa ttaccgccat caacttcctg gaagaagaag gcgcctatgt tcccgacctc 360
gatttcgtct tttacgacgt gttgggcgac gtggtgtgcg gggggttcgc catgccgatt 420
cgcgaaaaca aagcgcagga gatctacatc gtctgctccg gcgaaatgat ggcgatgtac 480
gccgccaaca acatctctaa aggcatcgtg aaatacgcca aatccggcaa agtgcgcctt 540
ggcgggctga tctgtaactc ccgtcagacc gaccgcgaag atgagctgat catagcgctg 600
gcggaaaaac tcggcaccca gatgatccac ttcgtgccgc gcgacaacat cgtgcaacgc 660
gctgaaatcc gccgtatgac ggtgattgag tacgatccga aatgcaacca ggccaatgaa 720
taccgcacgc tggcgaacaa gatcgtcaac aacaccaaaa tggtcgtgcc aacgcccatc 780
accatggacg aactggaaga gctgttgatg gaattcggca ttatggatgt ggaagacacc 840
agcattatcg gtaaaaccgc cgcagaagaa aacgcggttt ga 882
<210> 151
<211> 1449
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifD1
<400> 151
atgagcaatg caacaggcga acgtaatctg gagatcatcc aggaagtgct ggagatcttt 60
ccggaaaaaa cgcgcaaaga acgcagaaag cacatgatgg tgagcgaccc ggagatggaa 120
agcgtcggga aatgcatcat ctccaaccgt aagtcgcagc ccggcgtaat gaccgtgcgc 180
ggttgctctt acgccggttc taaaggggtg gtattcgggc cgatcaaaga tatggcccat 240
atttcccacg gcccggtcgg ctgcggtcag tactcccgcg ccgggcggcg taactactac 300
accggcgtca gcggtgtgga tagcttcggt acgctcaact ttacctccga ttttcaggag 360
cgcgatatcg tgtttggcgg cgataaaaag ctgaccaaac tgattgaaga gatggagacg 420
ctgttcccgc tgaccaaagg gatctccatt cagtccgaat gcccggtcgg cctgattggc 480
gacgacattg aagccgttgc caacgccagc cgcaaagcca tcaataaacc ggtcattccg 540
gtgcgctgcg aaggttttcg cggcgtttcc cagtcactcg gtcaccacat tgccaacgac 600
gtgatccgcg actgggtact ggataaccgc gaaggcaagc cgtttgaggc cggtccttat 660
gacgtggcga tcatcggcga ttacaacatc ggcggcgatg cctgggcgtc gcgcattttg 720
ctcgaagaga tgggcctgcg cgtggtggcg cagtggtccg gcgacggcac gctggttgag 780
atggagaaca cgccgttcgt caaactcaac cttgtgcact gctaccgctc aatgaactat 840
atctcccgcc atatggagga gaaacacggt attccgtgga tggagtacaa cttcttcggt 900
ccgaccaaag tcgccgaatc gttgcgcaaa atcgccgata tgtttgatga caccattcgc 960
gccaacgccg aagcggtgat cgccaaatat caggcgcaga acgacgccat catcgccaaa 1020
taccgtccgc gtctggaagg ccgcaaagtg ctgctgtata tgggcggttt acgtcctcgc 1080
catgtgattg gcgcttatga agatctgggg atggaaatta tcgctgcggg ttatgaattc 1140
gcccacaacg atgactacga ccgcaccctg ccggatctga aagaaggcac cttgctgttc 1200
gacgatgcca gcagttatga actggaagcc tttgtcaaag cgctgaagcc ggatctgatc 1260
ggctccggca ttaaagagaa gtacatcttc cagaaaatgg gcgtgccgtt tcgccagatg 1320
cactcctggg attactccgg cccctatcac ggttatgacg gctttgccat cttcgcccgc 1380
gatatggata tgacgatcaa caaccccgcg tggggccagt tgaccgcgcc gtggctgaaa 1440
tccgcctga 1449
<210> 152
<211> 1092
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifD2
<220>
<221> модифицированное основание
<222> (709)..(709)
<223> a, c, t, g, неизвестное или другое
<400> 152
atgggacgcg gcgagcgccg cctgttccat gccgtgcgcc acatcgtcaa ccgctaccac 60
ccggccgccg tctttatcta taacacctgc gttcccgcga tggagggcga cgatatcgaa 120
gccgtctgcc aggcggcaga aaccgccatc ggcgtaccgg tgattgccgt tgatgtcgcc 180
gggttttacg gcagcaaaaa tctcggcaac cggttggccg gtgaagtgat ggtgaaaaag 240
gtgattggcg ggcgtgaacc cgcgccgtgg ccggaagata ccccttttgc cccggcgcac 300
cgccacgata tcgggctgat tggcgaattc aatattgccg gagagttctg gcatattcag 360
ccgctgctcg atgagctggg tattcgcgtg ctcggcagcc tctccggcga cgggcgcttc 420
agtgaaatcc agacgctgca ccgggcgcag gtcaatatgc tggtctgctc cagggcgctg 480
atcaacgtcg cccgctcgct ggagcagcgc tacggcacgc cgtggtttga aggcagtttt 540
tatggtgttc gcgccacctc tgacgccctg cgccaactgg cggcgctgac cggagaccgc 600
gatctgatgc agcgcaccga acagctcatt gcccgcgaag agcagcaaac agagcaggcg 660
ctggccccgc tgcgcgagcg cctgcgcggg cgcaaagcgc tgctctatnc cggcggcgtg 720
aaatcctggt cggtggtttc ggcgcttcag gatctgggca tggaagtggt ggcgaccggc 780
acgcgcaaat ccaccgaaga ggataaacag cgcatccgcg aactgatggg cgccgacgcg 840
ctgatgcttg atgaaggtaa cgcccgctcg ctgctggacg tggtttaccg ctacaaggcg 900
gacatgatga tcgccggggg acgcaatatg tacaccgcct acaaagcgcg gctgccgttc 960
ctcgatatca atcaggagcg cgagcacgcc tttgccggct accgcggcat tgtcaccctg 1020
gccgaacagc tctgcctgac catggaaagc ccggtctggc cgcaaaccca ttcccgcgca 1080
ccgtggcaat aa 1092
<210> 153
<211> 846
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifK
<400> 153
atgatggagc aaatggacgt gccgtgcagc ctgctttccg atccctccga agtgctggat 60
accccggctg acgggcatta ccacatgtat gcgggcggta cgacccagca ggagatgcgc 120
gaagcgcctg acgctatcga caccctgctg ctgcaaccct ggcaactggt gaaaaccaaa 180
aaagtggtgc aggaaagctg gaaccagccc gctaccgagg tgcaaatccc aatggggctg 240
gccggaaccg acgagctgct gatgacggta agccagttaa ccggcaaagc cattccggat 300
agcttagcgc tggaacgcgg tcggctggtg gatatgatgc tcgactccca cacctggctg 360
cacggcaaga aattcggcct gttcggtgac ccggattttg tcatggggct gacccgcttc 420
ctgctggaac tgggctgcga accgacggtg attctgtgcc ataacggcag caagcgctgg 480
cagaaagcga tgaagaaaat gcttgaagcc tcgccgtacg ggaaagagag cgaagtcttt 540
atcaactgcg atttgtggca tttccgctcg ctgatgttta cccgtcagcc ggactttatg 600
atcggcaact cctacgccaa gtttatccag cgcgatacgc tggcgaaggg tgagcagttt 660
gaagtgccgc tgatccgcct ggggttcccg ctgttcgatc gccaccatct gcaccgccag 720
accacctggg gttacgaagg ggccatgagt atcctcacca cgctggttaa tgcggtgctg 780
gagaaagtcg acagagagac catcaagctc ggcaaaaccg actacagctt cgatcttatc 840
cgttaa 846
<210> 154
<211> 1095
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifL
<220>
<221> модифицированное основание
<222> (942)..(942)
<223> a, c, t, g, неизвестное или другое
<400> 154
atgcagcgcg acatcagcac cagctacgcg ctggaacaac ggctgcgcaa tcatatgacg 60
ctgaccgaag ccgtcttgaa taacattccg gcggcggttg tagtggtgga tgaacgcgat 120
cgggtggtga tggataacct cgcctacaaa accttttgcg ccgattgcgg cggtaaagaa 180
ctactcaccg aaatcaactt ttccgcccat aaggcggagc tggcgcaggg cctggtactg 240
ccggtagtgc tgcgcggcac cgtgcgctgg ttgtccgtta cctgttgggc gctgccgggc 300
gtcagcgaag aagcaggccg ctactttatt gatagcgccg tgccgcgcac gctggtggtg 360
atcaccgata atactcagca gcagcaacaa caggagcagg ggcgtcttga tcgtctgaag 420
cagcagataa ccagcggtaa attgctggcg gcgatccgcg aatcgctgga cgccgcgctg 480
gtacaactca attgcccaat taatatgctg gccgccgcac gccgcttaaa tggcgacgag 540
catagcaatc tggcgctgga tgccgcatgg cgtgaaggcg aagaagcgat ggcgcggttg 600
cagcgctgcc gcccgtcgct ggaactggaa agcccggcag tctggccgct ccagccgttc 660
cttgacgatc tgcgtgccct gtatcacacc cgatataacc agggcgaaaa cctgcaaatt 720
gagctggaat cccccgacct ggtgggcttt ggccagcgaa cacaactgct tgcctgcctg 780
agcctgtggc tcgacagaac cctggatatt gccgcggagc tacgtgattt cacggtacag 840
actcaacttt acgcccgcga agagagcggc tggctgtcgt tctatttaaa cgacaatgtg 900
ccgctgattc aggtgcgcta cacccattca cccgatgcac tnaatgcgcc cggtaaaggc 960
atggagctgc ggctgatcca gacgctggtc gcccaccatc gaggcgcaat agaactgacc 1020
tcacgccctc agggaggcac ctgtctgatc ctgcgtttcc cattatttta ctcgctgaca 1080
ggaggctcac tatga 1095
<210> 155
<211> 219
<212> ДНК
<213> вид Enterobacter
<220>
<223> частичный ген nifA
<400> 155
atgactcagc gaaccgagtc gggtacaacc gtctggcgct ttgacctctc ccaacagttt 60
acagccatgc agcgtatcag tgtggtgtta agccgcgcga cggagatcgg gcagacgcta 120
caggaagtgc tgtgcgtgct gcacaacgat gcctttatgc agcacgggat gatctgtccg 180
tacgcgcggg tgcgcgtctt cgcgagcgta tggctttga 219
<210> 156
<211> 1635
<212> ДНК
<213> вид Enterobacter
<220>
<223> glnE
<220>
<221> модифицированное основание
<222> (234)..(234)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (274)..(274)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (1535)..(1535)
<223> a, c, t, g, неизвестное или другое
<400> 156
atgcgcgtgg aagactggtc aacgctgacc gaacggctcg atgcccatat ggcaggcgtg 60
cgccgaatct ttaacgaact gatcggtgat gacgaaagtg agtcgcagga cgatgcgctc 120
tccgagcact ggcgcgagct gtggcaggac gcgcttcagg aagatgacac cacgccggtg 180
ctgacgcact taaccgacga cgcgcgccat cgcgtggtgg cgctgatcgc tganttccgt 240
cttgagctga acaaacgcgc catcggcccg cgtngtcgcc aggtgctgga tcacctgatg 300
ccgcacctgc tgagcgaagt ctgctcgcgt gccgatgcgc cggtgccgct gtcgcggatg 360
atgcccctgc tgagcgggat tatcacccgt actacctacc ttgaactcct gagcgagttc 420
cctggcgcgc ttaagcacct gatttcactc tgcgccgcgt cgccgatggt ggccaacaag 480
ctggcgcgtt acccgctgct gctggatgag ctgctcgatc cgaataccct ttatcaaccg 540
acggcgaccg acgcctaccg ggacgaactg cgtcagtatc tgctgcgcgt gccggaagaa 600
gacgaagagc aacagctgga ggcgctgcgt cagtttaagc aggcccagat gctgcgcgtg 660
gcggccgcag atattgccgg aacgctgccg gtgatgaaag tgagcgatca cttaacctgg 720
cttgcggaag cgattatcga cgcggtggtg catcaggcct gggtgcagat ggtggcgcgc 780
tatggccagc cgaaacatct ggctgaccgt gatggtcgcg gcttcgcggt ggtgggttac 840
ggtaagctcg gcggttggga gctgggctat agctccgatc tggatttaat cttcctccac 900
gactgcccgg ttgatgtgat gaccgacggc gagcgcgaga ttgacgggcg tcagttctac 960
ctgcgcctgg cgcagcgcat catgcacctg ttcagcaccc gcacctcgtc gggcattttg 1020
tatgaagtgg atgcccgtct gcgcccgtcc ggcgcggcgg gcatgctggt cacctcgacg 1080
gagtccttcg ctgattacca gaagaatgaa gcctggacgt gggagcatca ggcgctggtg 1140
cgcgcccgtg tggtgtatgg cgatccgctg ctgaaaacgc agtttgacgt gattcgtaag 1200
gaagtcatga ccaccgtgcg cgatggcagc acgctgcaaa cggaagtgcg cgaaatgcgc 1260
gagaaaatgc gcgcgcactt aggcaataaa catcgcgatc gctttgatat taaagccgat 1320
gagggcggta ttaccgatat tgagtttatt acccagtatc tggtgttgct gcacgcgcat 1380
gacaagccga agctgacgcg ctggtcggat aacgtgcgca ttctggaact gctggcgcaa 1440
aacgacatta tggacgagca ggaggcgcag gccttaaccc gtgcctatac aacgcttcgc 1500
gatgagctcc atcatctggc gttgcaggag cagcngggac acgtggcgct ggactgtttc 1560
accgctgaac gcgctcaggt aacggccagc tggcagaagt ggctggtgga accgtgcgta 1620
acaaatcaag tgtga 1635
<210> 157
<211> 1316
<212> ДНК
<213> вид Enterobacter
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (43)..(43)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (247)..(247)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (784)..(785)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (794)..(795)
<223> a, c, t, g, неизвестное или другое
<400> 157
agatgtgccc agatgggatt agctagtagg tggggtaacg gcncacctag gcgacgatcc 60
ctagctggtc tgagaggatg accagccaca ctggaactga gacacggtcc agactcctac 120
gggaggcagc agtggggaat attgcacaat gggcgcaagc ctgatgcagc catgccgcgt 180
gtatgaagaa ggccttcggg ttgtaaagta ctttcagcgg ggaggaaggt gttgtggtta 240
ataaccncag caattgacgt tacccgcaga agaagcaccg gctaactccg tgccagcagc 300
cgcggtaata cggagggtgc aagcgttaat cggaattact gggcgtaaag cgcacgcagg 360
cggtctgtca agtcggatgt gaaatccccg ggctcaacct gggaactgca ttcgaaactg 420
gcaggctaga gtcttgtaga ggggggtaga attccaggtg tagcggtgaa atgcgtagag 480
atctggagga ataccggtgg cgaaggcggc cccctggaca aagactgacg ctcaggtgcg 540
aaagcgtggg gagcaaacag gattagatac cctggtagtc cacgccgtaa acgatgtcga 600
cttggaggtt gtgcccttga ggcgtggctt ccggagctaa cgcgttaagt cgaccgcctg 660
gggagtacgg ccgcaaggtt aaaactcaaa tgaattgacg ggggcccgca caagcggtgg 720
agcatgtggt ttaattcgat gcaacgcgaa gaaccttacc tactcttgac atccagagaa 780
cttnncagag atgnnttggt gccttcggga actctgagac aggtgctgca tggctgtcgt 840
cagctcgtgt tgtgaaatgt tgggttaagt cccgcaacga gcgcaaccct tatcctttgt 900
tgccagcggt ccggccggga actcaaagga gactgccagt gataaactgg aggaaggtgg 960
ggatgacgtc aagtcatcat ggcccttacg agtagggcta cacacgtgct acaatggcgc 1020
atacaaagag aagcgacctc gcgagagcaa gcggacctca taaagtgcgt cgtagtccgg 1080
attggagtct gcaactcgac tccatgaagt cggaatcgct agtaatcgta gatcagaatg 1140
ctacggtgaa tacgttcccg ggccttgtac acaccgcccg tcacaccatg ggagtgggtt 1200
gcaaaagaag taggtagctt aaccttcggg agggcgctta ccactttgtg attcatgact 1260
ggggtgaagt cgtaacaagg taaccgtagg ggaacctgcg gttggatcac ctcctt 1316
<210> 158
<211> 882
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifH
<400> 158
atgaccatgc gtcaatgtgc catttacggc aaaggtggta tcggtaaatc cactaccacg 60
caaaacctgg tcgccgcgct ggcggagatg ggcaagaaag taatgatcgt cggctgcgac 120
ccgaaagcag actccactcg tctgatcctg catgcgaaag cgcagaacac cattatggag 180
atggcggctg aagtcggctc cgtggaagac cttgaactgg aagatgtgct gcaaatcggt 240
tacggcgacg tacgctgcgc agaatccggc ggcccggaac caggcgttgg ctgtgctggt 300
cgcggggtaa ttaccgccat caacttcctg gaagaagaag gcgcctatgt tcccgacctc 360
gatttcgtct tttacgacgt gttgggcgac gtggtgtgcg gggggttcgc catgccgatt 420
cgcgaaaaca aagcgcagga gatctacatc gtctgctccg gcgaaatgat ggcgatgtac 480
gccgccaaca acatctctaa aggcatcgtg aaatacgcca aatccggcaa agtgcgcctt 540
ggcgggctga tctgtaactc ccgtcagacc gaccgcgaag atgagctgat catagcgctg 600
gcggaaaaac tcggcaccca gatgatccac ttcgtgccgc gcgacaacat cgtgcaacgc 660
gctgaaatcc gccgtatgac ggtgattgag tacgatccga aatgcaacca ggccaatgaa 720
taccgcacgc tggcgaacaa gatcgtcaac aacaccaaaa tggtcgtgcc aacgcccatc 780
accatggacg aactggaaga gctgttgatg gaattcggca ttatggatgt ggaagacacc 840
agcattatcg gtaaaaccgc cgcagaagaa aacgcggttt ga 882
<210> 159
<211> 1449
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifD1
<400> 159
atgagcaatg caacaggcga acgtaatctg gagatcatcc aggaagtgct ggagatcttt 60
ccggaaaaaa cgcgcaaaga acgcagaaag cacatgatgg tgagcgaccc ggagatggaa 120
agcgtcggga aatgcatcat ctccaaccgt aagtcgcagc ccggcgtaat gaccgtgcgc 180
ggttgctctt acgccggttc taaaggggtg gtattcgggc cgatcaaaga tatggcccat 240
atttcccacg gcccggtcgg ctgcggtcag tactcccgcg ccgggcggcg taactactac 300
accggcgtca gcggtgtgga tagcttcggt acgctcaact ttacctccga ttttcaggag 360
cgcgatatcg tgtttggcgg cgataaaaag ctgaccaaac tgattgaaga gatggagacg 420
ctgttcccgc tgaccaaagg gatctccatt cagtccgaat gcccggtcgg cctgattggc 480
gacgacattg aagccgttgc caacgccagc cgcaaagcca tcaataaacc ggtcattccg 540
gtgcgctgcg aaggttttcg cggcgtttcc cagtcactcg gtcaccacat tgccaacgac 600
gtgatccgcg actgggtact ggataaccgc gaaggcaagc cgtttgaggc cggtccttat 660
gacgtggcga tcatcggcga ttacaacatc ggcggcgatg cctgggcgtc gcgcattttg 720
ctcgaagaga tgggcctgcg cgtggtggcg cagtggtccg gcgacggcac gctggttgag 780
atggagaaca cgccgttcgt caaactcaac cttgtgcact gctaccgctc aatgaactat 840
atctcccgcc atatggagga gaaacacggt attccgtgga tggagtacaa cttcttcggt 900
ccgaccaaag tcgccgaatc gttgcgcaaa atcgccgata tgtttgatga caccattcgc 960
gccaacgccg aagcggtgat cgccaaatat caggcgcaga acgacgccat catcgccaaa 1020
taccgtccgc gtctggaagg ccgcaaagtg ctgctgtata tgggcggttt acgtcctcgc 1080
catgtgattg gcgcttatga agatctgggg atggaaatta tcgctgcggg ttatgaattc 1140
gcccacaacg atgactacga ccgcaccctg ccggatctga aagaaggcac cttgctgttc 1200
gacgatgcca gcagttatga actggaagcc tttgtcaaag cgctgaagcc ggatctgatc 1260
ggctccggca ttaaagagaa gtacatcttc cagaaaatgg gcgtgccgtt tcgccagatg 1320
cactcctggg attactccgg cccctatcac ggttatgacg gctttgccat cttcgcccgc 1380
gatatggata tgacgatcaa caaccccgcg tggggccagt tgaccgcgcc gtggctgaaa 1440
tccgcctga 1449
<210> 160
<211> 1374
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifD2
<400> 160
atgaagggga acgagatcct ggctttgctc gatgaacctg cctgcgagca caaccataaa 60
cagaaatccg gctgcagcgc gccgaaaccc ggcgcgacag cgggcggctg cgcctttgac 120
ggtgcgcaga tcaccctgct gccactctcc gatgttgccc acctggtaca cggccccatt 180
ggttgtaccg gtagctcatg ggataaccgt ggcagcttca gttccggccc gacgatcaac 240
cggctgggtt ttaccaccga tctgagcgaa caggatgtga tcatgggacg cggcgagcgc 300
cgcctgttcc atgccgtgcg ccacatcgtc aaccgctacc acccggccgc cgtctttatc 360
tataacacct gcgttcccgc gatggagggc gacgatatcg aagccgtctg ccaggcggca 420
gaaaccgcca tcggcgtacc ggtgattgcc gttgatgtcg ccgggtttta cggcagcaaa 480
aatctcggca accggttggc cggtgaagtg atggtgaaaa aggtgattgg cgggcgtgaa 540
cccgcgccgt ggccggaaga tacccctttt gccccggcgc accgccacga tatcgggctg 600
attggcgaat tcaatattgc cggagagttc tggcatattc agccgctgct cgatgagctg 660
ggtattcgcg tgctcggcag cctctccggc gacgggcgct tcagtgaaat ccagacgctg 720
caccgggcgc aggtcaatat gctggtctgc tccagggcgc tgatcaacgt cgcccgctcg 780
ctggagcagc gctacggcac gccgtggttt gaaggcagtt tttatggtgt tcgcgccacc 840
tctgacgccc tgcgccaact ggcggcgctg accggagacc gcgatctgat gcagcgcacc 900
gaacagctca ttgcccgcga agagcagcaa acagagcagg cgctggcccc gctgcgcgag 960
cgcctgcgcg ggcgcaaagc gctgctctat accggcggcg tgaaatcctg gtcggtggtt 1020
tcggcgcttc aggatctggg catggaagtg gtggcgaccg gcacgcgcaa atccaccgaa 1080
gaggataaac agcgcatccg cgaactgatg ggcgccgacg cgctgatgct tgatgaaggt 1140
aacgcccgct cgctgctgga cgtggtttac cgctacaagg cggacatgat gatcgccggg 1200
ggacgcaata tgtacaccgc ctacaaagcg cggctgccgt tcctcgatat caatcaggag 1260
cgcgagcacg cctttgccgg ctaccgcggc attgtcaccc tggccgaaca gctctgcctg 1320
accatggaaa gcccggtctg gccgcaaacc cattcccgcg caccgtggca ataa 1374
<210> 161
<211> 1563
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifK1
<400> 161
atgagccaaa gtgctgagaa aattcaaaac tgtcatccgc tgtttgaaca ggatgcgtac 60
cagatgctgt ttaaagataa acggcaactg gaagaggccc acgatccggc gcgcgtgcag 120
gaggtctttc aatggaccac caccgccgag tatgaagcgc ttaactttca acgcgaagcg 180
ctgactatcg atccggccaa agcctgccag ccgctgggtg cggtactgtg ctcgctgggc 240
tttgccaata ccctgcccta tgttcacggc tcccaggggt gcgtggccta tttccgcacc 300
tattttaacc gtcactttaa agagccgatt gcctgtgttt ctgactcgat gacggaagat 360
gcggcagtat tcggcggcaa caacaacctg aacaccgggt tgcagaacgc cagcgccctc 420
tacaagccgg aaatcattgc cgtctccacc acctgtatgg cggaggtcat cggcgacgac 480
ctgcaggcgt ttattgctaa cgccaaaaaa gacggcttta tcgacgcggc gatcccggtg 540
ccttacgcgc acacgccaag ctttatcggc agccatatca ccggctggga caatatgttt 600
gagggcttcg cccgtacctt taccgccgat tacagcggac aaccgggcaa attaccgcgt 660
atcaatctgg tcagcggatt tgaaacctat ctcggtaatt tccgcgtgct gaaacgcatg 720
atggagcaaa tggacgtgcc gtgcagcctg ctttccgatc cctccgaagt gctggatacc 780
ccggctgacg ggcattacca catgtatgcg ggcggtacga cccagcagga gatgcgcgaa 840
gcgcctgacg ctatcgacac cctgctgctg caaccctggc aactggtgaa aaccaaaaaa 900
gtggtgcagg aaagctggaa ccagcccgct accgaggtgc aaatcccaat ggggctggcc 960
ggaaccgacg agctgctgat gacggtaagc cagttaaccg gcaaagccat tccggatagc 1020
ttagcgctgg aacgcggtcg gctggtggat atgatgctcg actcccacac ctggctgcac 1080
ggcaagaaat tcggcctgtt cggtgacccg gattttgtca tggggctgac ccgcttcctg 1140
ctggaactgg gctgcgaacc gacggtgatt ctgtgccata acggcagcaa gcgctggcag 1200
aaagcgatga agaaaatgct tgaagcctcg ccgtacggga aagagagcga agtctttatc 1260
aactgcgatt tgtggcattt ccgctcgctg atgtttaccc gtcagccgga ctttatgatc 1320
ggcaactcct acgccaagtt tatccagcgc gatacgctgg cgaagggtga gcagtttgaa 1380
gtgccgctga tccgcctggg gttcccgctg ttcgatcgcc accatctgca ccgccagacc 1440
acctggggtt acgaaggggc catgagtatc ctcaccacgc tggttaatgc ggtgctggag 1500
aaagtcgaca gagagaccat caagctcggc aaaaccgact acagcttcga tcttatccgt 1560
taa 1563
<210> 162
<211> 1386
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifK2
<400> 162
atggcagaaa ttatccgtag taaaaagccg ctggccgtca gcccggtaaa aagtggccag 60
ccgctgggcg cgattctggc gagcatgggc tttgaacaga gcattccgct ggttcatggc 120
gctcaggggt gcagcgcctt cgcgaaggtc ttttttatcc agcattttca cgatccgatc 180
ccgctgcaat cgacggcaat ggacccgaca tcgaccatta tgggtgccga tgagaacatc 240
tttaccgcgc tgaatgtgct gtgttcacgc aacaacccga aagcgattgt tctgctgagc 300
actggccttt ccgaggcgca gggcagcgat atttcgcgcg tggtgcgcca gttccgcgat 360
gaatatccgc gccataaagg ggtggcgctg ctgaccgtca acacgccgga tttttacggc 420
agcctggaaa acggctacag cgcggtgctg gagagcatgg ttgaacagtg ggtgccggaa 480
aaaccgcagc cgggcgtgcg caatcgccgc gtgaacctgc tgctcagcca tttgcttacg 540
ccgggcgaca ttgagctgct gcgaagttat gtcgaggcat ttggcctgca gccggtgatg 600
gtgccggatc tttcccagtc gctggatggc catctcgcca gcggggattt ctcgccaatt 660
acccagggcg gcagcagcct gcggctgatt gaacagatgg gacagagtct tggcacgttc 720
gccattggcg tatccctctc ccgcgccgcg caattgctgg cgcagcgcag ccatgcggaa 780
gtggtcaccc tgccgcatct gatgaccatg agccagtgcg atacgtttat tcatcaactg 840
aagcgcctct ccgggcgcga tgttccggcg tggatcgaac gccagcgcgg gcaactgcag 900
gatgcgatga tcgattgtca tatgtggttg cagggcgcgc ctgtcgcgct ggccgccgag 960
ggcgatctgc tcgccgcctg gtgcgatttc gcctgcgata tgggcatggt gcccggcccg 1020
gtggtggcgc cggtgagcca gaaagggttg caggatctgc cggtcgaaaa agtcattatc 1080
ggcgatctgg aggatatgca ggatctgttg tgtgaaacgc ctgcatcgct gctcgtctct 1140
aattctcacg ccgctgattt ggccgggcag ttcgacattc cgctggtgcg cgccggtttc 1200
cccctgttcg accgtctggg cgagtttcgc cgcgtgcgcc agggttacgc cgggatgcgc 1260
gacaccttgt ttgagctggc gaatgcgctg cgcgatcgcc atcatcatct tgccgcttat 1320
cactcgccgc tgcgccagcg tttttacgaa cccgcatctt cgggaggtga ctatgcaaca 1380
tgttaa 1386
<210> 163
<211> 1488
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifL
<400> 163
atgaccctga atatgatgat ggacgccacc gcgcccgccg agatcgccgg agcgctctca 60
caacagcatc ccgggttgtt tttcaccatg gttgaacagg cgcccgtcgc gatttcactg 120
accgatgccg atgcccacat tctctacgcc aaccccgcgt tttgtcgcca gtcggggtat 180
gaactggaag agttgttgca gcaaaacccg cgcctgcttg ccagtaagca gacgccgcgt 240
gaaatctacc aggaaatgtg gcacaccctg ctgcaacacc gtccgtggcg cggacaactg 300
atcaaccgtc gccgcgacgg cagcctgttt ctggtggaaa tcgacatcac cccactgttt 360
gatgcgttcg gcaaactcga acattacctg gccatgcagc gcgacatcag caccagctac 420
gcgctggaac aacggctgcg caatcatatg acgctgaccg aagccgtctt gaataacatt 480
ccggcggcgg ttgtagtggt ggatgaacgc gatcgggtgg tgatggataa cctcgcctac 540
aaaacctttt gcgccgattg cggcggtaaa gaactactca ccgaaatcaa cttttccgcc 600
cataaggcgg agctggcgca gggcctggta ctgccggtag tgctgcgcgg caccgtgcgc 660
tggttgtccg ttacctgttg ggcgctgccg ggcgtcagcg aagaagcagg ccgctacttt 720
attgatagcg ccgtgccgcg cacgctggtg gtgatcaccg ataatactca gcagcagcaa 780
caacaggagc aggggcgtct tgatcgtctg aagcagcaga taaccagcgg taaattgctg 840
gcggcgatcc gcgaatcgct ggacgccgcg ctggtacaac tcaattgccc aattaatatg 900
ctggccgccg cacgccgctt aaatggcgac gagcatagca atctggcgct ggatgccgca 960
tggcgtgaag gcgaagaagc gatggcgcgg ttgcagcgct gccgcccgtc gctggaactg 1020
gaaagcccgg cagtctggcc gctccagccg ttccttgacg atctgcgtgc cctgtatcac 1080
acccgatata accagggcga aaacctgcaa attgagctgg aatcccccga cctggtgggc 1140
tttggccagc gaacacaact gcttgcctgc ctgagcctgt ggctcgacag aaccctggat 1200
attgccgcgg agctacgtga tttcacggta cagactcaac tttacgcccg cgaagagagc 1260
ggctggctgt cgttctattt aaacgacaat gtgccgctga ttcaggtgcg ctacacccat 1320
tcacccgatg cactcaatgc gcccggtaaa ggcatggagc tgcggctgat ccagacgctg 1380
gtcgcccacc atcgaggcgc aatagaactg acctcacgcc ctcagggagg cacctgtctg 1440
atcctgcgtt tcccattatt ttactcgctg acaggaggct cactatga 1488
<210> 164
<211> 1575
<212> ДНК
<213> вид Enterobacter
<220>
<223> nifA
<400> 164
atgactcagc gaaccgagtc gggtacaacc gtctggcgct ttgacctctc ccaacagttt 60
acagccatgc agcgtatcag tgtggtgtta agccgcgcga cggagatcgg gcagacgcta 120
caggaagtgc tgtgcgtgct gcacaacgat gcctttatgc agcacgggat gatctgtctg 180
tacgacagta agcaagcgat cctttccatt gaagccttgc atgaggccga tcagcagtta 240
attcccggca gttcacagat tcgctaccgt ccgggcgaag ggctggtagg cacggtgctt 300
tcacagggac agtcgctggt actgccctgt gtctccgacg atcggcgttt tctcgatcgc 360
ctgggattgt atgattacag cttgccgttt atcgccgtgc cgctgatggg gccaaactcg 420
cagcctatcg gcgtgctggc cgcccagcct atggcgcgtt acgaggagcg gctgcccgcc 480
tgcacgcgtt ttcttgaaac cgtcgccaat ctggtggcgc aaaccgttcg cctgatgaca 540
ccgcccagcg tcgcgtctcc accccgtgct gctgccgcgc agattgccag ccagcgcggg 600
tgcgcgtctt cgcgagcgta tggctttgaa aacatggtcg gtaaaagcgc ggctatgcgt 660
cagacgctgg aaattattcg ccaggtatca cgctgggaca ccaccgtgct ggtgcgtggc 720
gaaagcggaa ccggtaaaga gttgatagcc aacgctatcc accacaattc accgcgcgcc 780
gccgcgccgt ttgtcaaatt caactgcgcg gcgctgcccg atacgctgct ggagagtgaa 840
ctcttcggtc atgaaaaagg cgcgtttacc ggcgcggtgc gccagcgcaa aggccgtttc 900
gaactggcgg atggcggtac gctgtttctt gatgagatcg gcgaaagtag cgcctcgttt 960
caggcgaaat tgctgcgtat cttgcaggaa ggcgaaatgg aacgcgtcgg cggcgacgaa 1020
acgctgcggg tgaatgtacg gatcattgcc gccaccaacc gcaatctgga agaggaagtg 1080
cggctgggta attttcgcga agatctctac tatcgcctta atgtgatgcc gatctccctg 1140
cccccgctcc gcgagcgtca ggaggacatc gtcgagctgg cgcattttct ggtgcgcaaa 1200
atcgcgcaaa accagaaccg cacgctgcgc atcagcgatg gcgcgatccg tttgttgatg 1260
agctatagct ggcctggaaa cgtgcgtgag ctggaaaact gccttgagcg atcggcggtg 1320
atgtcggaaa acgggctgat cgatcgcgac gtgattttgt ttcaccacag ggaaaatctg 1380
ccaaaaacgc cacagaccag tgcgccgcgc gaagagagct ggctcgatca gaacctcgat 1440
gagcgacaaa gattgatcgc cgcgctggag aaagccggtt gggtacaggc aaaagccgcg 1500
cgcctgctgg gaatgacccc gcgccaggtg gcctatcgta ttcagacgat ggacattgcc 1560
atgccgagat tgtag 1575
<210> 165
<211> 2856
<212> ДНК
<213> вид Enterobacter
<220>
<223> glnE
<400> 165
atgccgcttt cttcgcagtt acagcagcag tggcagaccg tttgcgaacg tctgcctgag 60
tcattaccgg cgtcatcgtt aagcgagcag gcaaagagcg tgctcgtctt cagtgatttt 120
gtgcaggaaa gtatcaccgc caacccgaac tggctggcgg aacttgagaa cgcaccaccg 180
caggcagaag agtggcggca ctatgctggc tggctgcaaa ctgtactcga agacgttacg 240
gatgaggcca cgctgatgcg cgtcctgcgc cagttccgtc gtcgggtgat ggtgcgcatt 300
gcctgggctc aggcgctgga actggtgagc gaagagagta cgctgcagca gttaagcgag 360
ctggcgcaaa cgttgattgt cgccgcgcga gactggctct atgccgcctg ctgtaaagag 420
tggggcacgc cgtgcagcga ggaaggggtt cctcagccgc tgttgattct ggggatggga 480
aagctgggcg gctgcgagct gaacttctcc tctgatatcg acctgatttt tgcctggccg 540
gagaacggct ccacgcgcgg aggccgccgc gagctggaca acgcgcagtt ctttacccgt 600
ctcggccagc gcctgattaa agcgctggat cagcccacgc aggacggttt tgtttaccgc 660
gtggacatgc gcctgcgtcc gtttggcgac agcgggccgc tggtgctgag ctttgcggcg 720
ctggaagatt attaccagga gcaaggtcgc gactgggagc gttacgcgat ggtcaaagcg 780
cggatcatgg gcgacagcga cgacgcttat gccaacgagc tgcgcgccat gctgcgtccg 840
ttcgtgttcc gtcgctatat cgacttcagc gtcatccagt ccctgcgaaa tatgaaaggg 900
atgattgccc gcgaggtgcg ccgccgtggg ctgaaagaca atatcaagct cggtgcgggc 960
ggcatccgcg aaatcgaatt tatcgtccag gtcttccagc ttattcgcgg cggacgcgag 1020
ccgtcgctgc agtcccgttc cttattaccg acgctgagcg ccattgcgca gctgcatctc 1080
ctgccggacg gcgacgcgca aaccctgcgc gaggcctatc ttttcctgcg tcgtctggaa 1140
aacctgctgc aaagcattaa tgacgaacag acccaaaccc tgccgggcga cgaccttaac 1200
cgggcgcgtc tggcctgggg aatgcgcgtg gaagactggt caacgctgac cgaacggctc 1260
gatgcccata tggcaggcgt gcgccgaatc tttaacgaac tgatcggtga tgacgaaagt 1320
gagtcgcagg acgatgcgct ctccgagcac tggcgcgagc tgtggcagga cgcgcttcag 1380
gaagatgaca ccacgccggt gctgacgcac ttaaccgacg acgcgcgcca tcgcgtggtg 1440
gcgctgatcg ctgatttccg tcttgagctg aacaaacgcg ccatcggccc gcgtggtcgc 1500
caggtgctgg atcacctgat gccgcacctg ctgagcgaag tctgctcgcg tgccgatgcg 1560
ccggtgccgc tgtcgcggat gatgcccctg ctgagcggga ttatcacccg tactacctac 1620
cttgaactcc tgagcgagtt ccctggcgcg cttaagcacc tgatttcact ctgcgccgcg 1680
tcgccgatgg tggccaacaa gctggcgcgt tacccgctgc tgctggatga gctgctcgat 1740
ccgaataccc tttatcaacc gacggcgacc gacgcctacc gggacgaact gcgtcagtat 1800
ctgctgcgcg tgccggaaga agacgaagag caacagctgg aggcgctgcg tcagtttaag 1860
caggcccaga tgctgcgcgt ggcggccgca gatattgccg gaacgctgcc ggtgatgaaa 1920
gtgagcgatc acttaacctg gcttgcggaa gcgattatcg acgcggtggt gcatcaggcc 1980
tgggtgcaga tggtggcgcg ctatggccag ccgaaacatc tggctgaccg tgatggtcgc 2040
ggcttcgcgg tggtgggtta cggtaagctc ggcggttggg agctgggcta tagctccgat 2100
ctggatttaa tcttcctcca cgactgcccg gttgatgtga tgaccgacgg cgagcgcgag 2160
attgacgggc gtcagttcta cctgcgcctg gcgcagcgca tcatgcacct gttcagcacc 2220
cgcacctcgt cgggcatttt gtatgaagtg gatgcccgtc tgcgcccgtc cggcgcggcg 2280
ggcatgctgg tcacctcgac ggagtccttc gctgattacc agaagaatga agcctggacg 2340
tgggagcatc aggcgctggt gcgcgcccgt gtggtgtatg gcgatccgct gctgaaaacg 2400
cagtttgacg tgattcgtaa ggaagtcatg accaccgtgc gcgatggcag cacgctgcaa 2460
acggaagtgc gcgaaatgcg cgagaaaatg cgcgcgcact taggcaataa acatcgcgat 2520
cgctttgata ttaaagccga tgagggcggt attaccgata ttgagtttat tacccagtat 2580
ctggtgttgc tgcacgcgca tgacaagccg aagctgacgc gctggtcgga taacgtgcgc 2640
attctggaac tgctggcgca aaacgacatt atggacgagc aggaggcgca ggccttaacc 2700
cgtgcctata caacgcttcg cgatgagctc catcatctgg cgttgcagga gcagccggga 2760
cacgtggcgc tggactgttt caccgctgaa cgcgctcagg taacggccag ctggcagaag 2820
tggctggtgg aaccgtgcgt aacaaatcaa gtgtga 2856
<210> 166
<211> 1290
<212> ДНК
<213> вид Enterobacter
<220>
<223> amtB
<400> 166
atgaagatag caacacttaa aacgggtctg ggttcgctgg cactgctgcc gggcctggcg 60
ctggctgctg cacctgcggt ggcagacaaa gccgataacg cctttatgat gatcagcacc 120
gcgctggtgc tgttcatgtc cattccgggc attgcgctgt tctatggcgg cctgatccgt 180
ggcaaaaacg ttctctccat gctgacgcag gttgccgtaa cgttcgcgct ggtctgcgta 240
ctgtgggtgg tttacggtta ctcgctggct ttcggcacgg gcaacgcgtt ctttggtaac 300
ttcgactggg tgatgctgaa aaatattgaa ctgaccgcgc tgatgggcag tttctaccag 360
tatattcacg ttgctttcca gggctcgttc gcctgcatta ccgtcgggct gattgtaggc 420
gcgcttgccg agcgtattcg tttctctgcg gtcctgatct tcgtggtggt ctggctgacg 480
ctctcctatg tgccgattgc gcacatggtc tggggtggcg gtctgctggc gacgcatggc 540
gcgctggact tcgcgggcgg taccgttgtg cacattaacg ccgcggtagc gggtctggtt 600
ggcgcatacc tgattggcaa acgcgtgggc ttcggtaaag aagcgttcaa accgcacaac 660
ctgccgatgg tcttcaccgg taccgcgatc ctctactttg gctggtttgg tttcaacgcc 720
ggctcagcaa gtgccgcgaa cgaaatcgcc gcgctggcct tcgtgaatac cgttgtggcc 780
acggcaggtg caatcctctc ctgggtcttt ggcgagtggg ctgtgcgcgg taaaccttct 840
ctgctgggtg cctgttcggg ggcgattgct ggtctggtcg gtatcacccc agcatgtggt 900
tatgtcggtg tgggtggcgc gctgctggtc ggcctggtgt caggtctggc gggtctgtgg 960
ggcgtgacgg cgctgaaacg tattctgcgc gttgatgacc cttgcgatgt gtttggcgtg 1020
cacggcgtgt gcggcatcgt cggctgtatc atgaccggta tctttgcagc gaaatcgctg 1080
ggtggcgtgg gctacgcaga aggcgtcacc atggcccatc aggtgctggt gcagctggaa 1140
agtattgctg tcaccgtggt gtggtctgcc gttgtcgctt tcattggcta caaactggcg 1200
gacatgacgg ttggtctgcg cgtgccggaa gagcaggaac gcgaaggtct ggacgtcaac 1260
agccacggcg agaatgcgta taacgcatga 1290
<210> 167
<211> 1299
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> 16S
<220>
<221> модифицированное основание
<222> (294)..(294)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (299)..(299)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (302)..(302)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (309)..(310)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (332)..(332)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (346)..(346)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (351)..(351)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (438)..(438)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (445)..(445)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (454)..(454)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (469)..(469)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (477)..(477)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (492)..(492)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (504)..(504)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (555)..(555)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (775)..(775)
<223> a, c, t, g, неизвестное или другое
<400> 167
gccgagagag gggcccgcgt cggattaggt agttggtgag gtaatggctc accaagcctt 60
cgatccgtag ctggtctgag aggatgatca gccacactgg gactgagaca cggcccagac 120
tcctacggga ggcagcagtg gggaatattg gacaatgggc gcaagcctga tccagcaatg 180
ccgcgtgagt gatgaaggcc ttagggttgt aaagctcttt cgcacgcgac gatgatgacg 240
gtagcgtgag aagaagcccc ggctaacttc gtgccagcag ccgcggtaat acgnagggng 300
cnagcgttnn tcggaattac tgggcgtaaa gngcgcgtag gcggcntgtt nagtcagaag 360
tgaaagcccc gggctcaacc tgggaatagc ttttgatact ggcaggcttg agttccggag 420
aggatggtgg aattcccngt gtagnggtga aatncgtaga tattgggang aacaccngtg 480
gcgaaggcgg cnatctggac gganactgac gctgaggcgc gaaagcgtgg ggagcaaaca 540
ggattagata ccctngtagt ccacgccgta aacgatgaat gctagacgtc ggggtgcatg 600
cacttcggtg tcgccgctaa cgcattaagc attccgcctg gggagtacgg ccgcaaggtt 660
aaaactcaaa ggaattgacg ggggcccgca caagcggtgg agcatgtggt ttaattcgaa 720
gcaacgcgca gaaccttacc aacccttgac atgtccactt tgggctcgag agatngggtc 780
cttcagttcg gctgggtgga acacaggtgc tgcatggctg tcgtcagctc gtgtcgtgag 840
atgttgggtt aagtcccgca acgagcgcaa cccctaccgt cagttgccat cattcagttg 900
ggcactctgg tggaaccgcc ggtgacaagc cggaggaagg cggggatgac gtcaagtcct 960
catggccctt atgggttggg ctacacacgt gctacaatgg cggtgacagt gggaagcgaa 1020
gtcgcgagat ggagcaaatc cccaaaagcc gtctcagttc ggatcgcact ctgcaactcg 1080
agtgcgtgaa gttggaatcg ctagtaatcg cggatcagca cgccgcggtg aatacgttcc 1140
cgggccttgt acacaccgcc cgtcacacca tgggagttgg ttttacccga aggtggtgcg 1200
ctaaccgcaa ggaggcagcc aaccacggta aggtcagcga ctggggtgaa gtcgtaacaa 1260
ggtagccgta ggggaacctg cggctggatc acctccttt 1299
<210> 168
<211> 897
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> nifH
<400> 168
atggccaaag cgcctctgcg tcagatcgcc ttttacggca agggcggtat cggcaagtcc 60
accacctctc agaacacgct ggccgcgctg gtcgagctgg atcagaggat cctgatcgtc 120
ggctgcgacc cgaaggccga ctcgacccgc ctgatcctgc acgcaaaggc ccaggacacc 180
gtcctgcatc tggccgccga ggccggctcg gtcgaggatc tggagctcga ggacgttctc 240
aagatcggct acaagaacat caagtgcgtc gagtccggcg gtccggagcc gggggtcggc 300
tgcgccggcc gcggcgtcat cacctcgatc aacttcctgg aagagaacgg cgcctacgac 360
gacgtggact atgtgtccta cgacgtgctg ggcgacgtgg tctgcggcgg cttcgccatg 420
ccgatccgcg agaacaaggc ccaggaaatc tacatcgtca tgtccggcga gatgatggcg 480
ctgtacgccg ccaacaacat cgccaagggc atcctgaagt acgcgcacag cggcggcgtc 540
cgtctcggcg gcctgatctg caacgagcgc cagaccgaca aggaatggga tctggccgac 600
gcgctggcca agcgcctggg ctccaagctg atccacttcg tgccgcgcga caacatcgtc 660
cagcacgccg agctgcgccg catgacggtc atcgagtacg ccccggacag caagcaggcc 720
ggcgaatacc gcgcgctcgc caacaagatc catgcgaact ccggccaggg ttgcatcccg 780
accccgatca ccatggaaga gctggaagag atgctgatgg acttcggcat catgaagacc 840
gaggagcagc agctcgccga gctcgccgcc aaggaagcgg cgaaggccgg cgcctga 897
<210> 169
<211> 1440
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> nifD1
<400> 169
atgagcctgt ccgagaacac cacggtcgac gtcaagaacc tcgtcaacga agtcctcgaa 60
gcctatcccg aaaaatcccg caagcgccgc gccaagcacc tgaacgtgct ggaggccgag 120
gccaaggact gcggcgtcaa gtcgaacgtc aagtccatcc ccggcgtcat gaccatccgc 180
ggctgcgcct atgccggctc caagggcgtg gtgtggggtc cgatcaagga catgatccac 240
atctcccacg gtccggtcgg ctgcggctac tactcctggt ccggccgccg caactactac 300
atcggcgaca ccggtgtgga cagctggggc acgatgcact tcacctccga cttccaggag 360
aaggacatcg tcttcggcgg cgacaagaag ctgcacaagg tcatcgagga aatcaacgag 420
ctgttcccgc tggtgaacgg catctcgatc cagtcggaat gcccgatcgg cctgatcggc 480
gacgacatcg aggctgtcgc ccgcgccaag tcggcggaaa tcggcaagcc ggtcatcccc 540
gtgcgctgcg aaggcttccg cggcgtgtcc cagtcgctgg gccaccacat cgccaacgac 600
gccatccgag actgggtgtt cgagaagacg gaacccaagg ccggcttcgt ctccaccccc 660
tatgacgtca ccatcatcgg cgactacaac atcggcggcg acgcctggtc gtcccgcatc 720
ctgctggagg agatcggcct gcgcgtgatc gcccagtggt cgggcgacgg cacgctcgcc 780
gaactggaga acacgccgaa ggccaaggtc aacctgatcc actgctaccg ctcgatgaac 840
tacatcgcgc gccacatgga agagaagttc aacattcctt ggatggaata caacttcttc 900
ggcccgagcc agatcgccga atccctgcgc aagatcgccg ctctcttcga cgacaagatc 960
aaggagaacg ccgagaaggt catcgcccgc taccagccga tggtcgatgc ggtcatcgcc 1020
aagtacaagc cgcggctcga aggcaagaag gtcatgatct acgtcggcgg cctgcgtccc 1080
cgccacgtcg tcgatgccta ccatgacctc ggcatggaga tcaccggcac cggctacgag 1140
ttcgcccaca acgacgacta tcagcgcacg cagcactacg tgaaggaagg cacgctgatc 1200
tacgacgacg tcaccgcgtt cgaactggag aagttcgtcg aggcgatgcg tcccgacctc 1260
gtcgcgtcgg gcatcaagga aaagtacgtg ttccagaaga tgggcctgcc gttccgccag 1320
atgcacagct gggactactc cggcccgtac cacggctatg acggcttcgc gatcttcgcc 1380
cgcgacatgg acctggccat caacaacccc gtctggggcg tgatgaaggc cccgttctga 1440
<210> 170
<211> 1419
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> nifD2
<400> 170
atgctccagg acaagatcca ggatgtcttc aacgaaccgg gctgcgcgac caaccaagcc 60
aaatcggcca aggagaagaa gaagggctgc accaagtcgc tgaaaccggg ggcggcagcc 120
ggcggctgcg cctatgacgg ggcgatgatc gtgctccagc cgatcgccga cgccgcccat 180
ctggtccatg gccccatcgc ctgcctcgga aacagttggg acaaccgcgg ctccaaatcc 240
tccggctcgc agctctaccg caccggcttc accaccgatc tgtcggaact ggacgtcatc 300
ggcggcggcg agaagaagct ctaccgcgcc atcaaggaga tcgttcagca atacgacccg 360
ccggccgtct tcgtctatca gacctgcgtg cccgccatga ccggcgacga catcgccgcg 420
gtctgcaagt tcgccacgca gaagctgggc aagccggtga tcccggtgga ctcgccgggc 480
ttcgtcgggt cgaagaatct cggcaacaag ctggccggcg aagccctgct ggagcatgtc 540
atcggcacgg tcgaaccgga ctacaccacc ccgaccgacg tctgcatcat cggcgaatac 600
aaccttgccg gcgagctgtg gctggtcaag ccgctgctgg acgagatcgg catccgcctc 660
ctgtcctgca tttccggcga cggccgctac cgggaggtgg cgcaggccca ccgcgcccgc 720
gtcaccatga tggtgtgcag ccaggcgctg gtgaatgtcg ggcgcaagat ggaggagcgc 780
tacggcatcc cctatttcga ggggtccttc tacggcgtgt ccgacatgtc ggacaccctg 840
cgcaccatga cccgcatgct ggtggagcgc ggcgccgaca agggcctgat cgaccgggcg 900
gagggcgtga tcgcgcggga ggaaagccgg gtctggcgcc ggctggaacc ctacaagccg 960
cgcttcgacg gcaagcgcgt ccttctcttc accggcggcg tcaagagctg gtcgatggtc 1020
agcgcgctgg agggtgcggg gctgaccatc ctcggcacct ccaccaagaa atcgaccagg 1080
gaggacaagg agcgcatcaa gaagatgaag ggcgaagagt tccaccagtg ggacgatttg 1140
aagccgcgcg acatctacag gatgctggcc gacgatcagg ccgacatcat gatgtccggc 1200
ggccgctcgc agttcatctc gctgaaggcc aaggttccct ggctcgacat caaccaggag 1260
cgccaccacg cctatgccgg ctatgacggc atcgtcaatc tctgcgagga gatcgacaaa 1320
acgctgtcga atccgatctg gcgtcaggtg cgtcagccgg caccgtggga gtccggcgcg 1380
tcctccaccc ttctggcttc ctcgatggcg gcggagtga 1419
<210> 171
<211> 1383
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> nifK
<400> 171
atgtcccaca tccagcgctt cccctccgcc gccaaggccg cctccaccaa cccgctgaag 60
atgagccagc cgctgggtgc ggctctggcc tatctcggcg tcgaccgctg cctgccgctg 120
ttccatggct cgcagggctg caccgccttc gggctggtcc tgctggtgcg ccatttccgc 180
gaggcgatcc cgctccagac cacggcgatg gatcaggtcg ccaccatcct cggcggctac 240
gacaatctgg agcaggcgat ccgcaccatc gtcgagcgca accagcccgc catgatcggc 300
gtcgccacca ccggcgtcac cgagaccaag ggcgaggata tggccggaca gtacacgctg 360
ttccgccagc gcaaccccga cttggccgac acggccctgg tcttcgccaa cacccccgac 420
ttcgccggcg gcttcgagga cggcttcgcc gccgcggtca ccgcgatggt cgagcggttg 480
gtcgaaccgt cgccggtgcg catcccgacc caggtcaacg tgctggccgg ctgccatctg 540
tcccccggcg acgtggagga actgcgcgac atcatcgaag gcttcggcct gtcgccgatc 600
ttcctgcccg acctgtcgct gtcgatggcg ggccgccagc cggccgactt caccgccacc 660
tcgctgggcg gcgtgaccgt cgatcagatc cgcgccatgg gcgcttcggc cctcaccatc 720
gtggtcggtg agcatatgcg ggtggccggt aacgcgctgg agctgaagac cgacgtgccc 780
agccatttct tcaaccgcct gaccgggctg gaggcgacgg acaagctggt ccggctgctg 840
atggagttgt cgggcaagcc ggcgcccgcc cggctgcggc gccagcgcga aagcctggtc 900
gatgccatgc tcgacgggca tttcttctac agccgcaagc gcatcgccgt cgcgctggag 960
cccgacctgc tctatgccgt caccggcttc ctcgccgaca tgggggccga ggtgatcgcc 1020
gcggtgtccc cgacgcagag cccggtgctg gagcggttga aggccgccac catcatggtc 1080
ggcgatcatt ccgacgtgga gacgctggcc cgcgacgccg acctgatcgt ctccaactcg 1140
cacgggcggc agggagccgc gcggatcggc gtggctctgc accgcatggg cctgccgctg 1200
ttcgaccggc tgggggccgg cctgcgcgtc caggtcggct accgcggcac gcgggaactg 1260
ctgtgcgaca tcggcaacct gttcctcgcc cgcgagatgg accacgagca cgggcacgag 1320
agccacgacc acggggaatc ccacggctgc ggaggcggat catgcggatg caacgccgtc 1380
tga 1383
<210> 172
<211> 1560
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> nifK1
<400> 172
atgaccgaca agctttcgca gagcgccgac aaggtcctcg accactacac cctcttccgg 60
cagcccgaat acgcggcgat gttcgagaag aagaagaccg agttcgagta cggccattcg 120
gacgaggaag tcgcccgcgt gtccgaatgg accaagtccg aggactacaa ggcgaagaac 180
ttcgcccgtg aagcggtcgt catcaacccg accaaggcct gccagccgat cggcgcaatg 240
ttcgccgccc agggcttcga aggcaccctg cccttcgtcc acggctccca gggctgcgtc 300
gcctattacc gcacccacct gacccgtcac ttcaaggagc cgaacagcgc ggtctcctcg 360
tcgatgacgg aggacgcggc ggtgttcggc ggcctgaaca acatgatcga cggcctggcg 420
aacgcctatg cgctctacaa gccgaagatg atcgcggtga tgaccacctg catggccgaa 480
gtcatcggcg acgatttgca gggcttcatc gccaatgcga agaccaagga cagcgtcccg 540
gccgacttcc cggtccccta cgcccacacc ccggccttcg tcggcagcca catcgtcggc 600
tacgacaaca tgatcaaggg gatcctgacc aacttctggg gtacgtcgga gaatttcgac 660
acacccaaga ccgagcagat caacctgatc ccgggcttcg acggcttcgc cgtcggcaac 720
aaccgcgaac tgaagcgcat cgccggcgaa ttcggcgtga agctgcaaat cctgtccgac 780
gtgtccgaca atttcgacac gccgatggat ggcgagtacc gcatgtatga cggcggcacc 840
accatcgagg agaccaagga ggccctgcac gccaaggcca ccatctccat gcaggagtac 900
aacacgaccc agaccctgca attctgcaag gagaagggtc aggaagtcgc caagttcaac 960
tacccgatgg gcgtcaccgg caccgacgag ctgctgctga agctcgccga actgtcgggc 1020
aagccggtcc cggccagcct gaagctggag cgcggccgtc tggtcgacgc catcgccgac 1080
agccacaccc acatgcacgg caagcgcttc gccgtctatg gcgacccgga cttctgcctg 1140
ggcatgtcca agttcctgct ggagctgggt gcggagccgg tgcacatcct gtcgacgtcg 1200
ggctccaaga agtgggagaa gcaggtccag aaggtgctgg acggctcgcc cttcggcgcc 1260
tcgggcaagg cccatggcgg caaggatctg tggcacctgc gttcgctgat cttcaccgac 1320
aaggtggact acatcatcgg caacagctac ggcaagtatc tggagcgcga caccaaggtt 1380
ccgctgatcc gcctgaccta cccgatcttc gaccgccacc accaccaccg ctacccgacc 1440
tggggctacc agggcgcgct gaacgtgctg gtacggatcc tggaccggat cttcgaggac 1500
atcgacgcca acaccaacat cgtcggccag accgactact cgttcgacct gatccgctga 1560
<210> 173
<211> 1530
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> nifA
<400> 173
atgttgacct ctgatattgt tggcaaattg cgctgcatcg cagcagaccc caaagcgggc 60
atcgcaaggg gcctcgacac cgggacgacg aagatcggtc ccgtttggga gggtgacgtg 120
ggcgacaccg tggatttcga agcgctgcgc cagcgggcgg tccactccct gttcgaacat 180
ctggaatcca tgtgcgtcgg cgccgtcgcc gtcgaccaca ccggccgcat cgcctggatg 240
gacgagaagt acaaggctct gctgggcgtt cccgacgacc cgcgcggccg gcaggtggag 300
gacgtcatcc ccaacagcca gctgcgccgg gtgatcgaca gcggccagcc gcagccgctg 360
gacatcatgg agttcgacga ccggtccttc gtggtgacgc ggatgccgct gttcggcacc 420
gacggttcga tcatcggcgc catcggcttc gtgctgttcg accgcgccga atatctccgc 480
ccgctggtcc gcaaatacga gaagatgcag gaggagctgg cccgcaccca gcaggagctg 540
gcgcatgagc gccgcgccaa atactccttc tcgcagttcc tgggcgccag cgaatcgatc 600
cgcgagatca agcggctggg gcgccgcgcc gcccagatgg attcgaccgt cctgctgctg 660
ggcgaaaccg ggaccggcaa ggagctgctg gcccaggcca tccattccgc cagcccgcgg 720
gcgtccaagc ccttcgtcgg cgtcaatgtc gccgccattc cggaaaccct gctggaggcg 780
gagttcttcg gcgtcgcccc cggcgccttc accggcgccg accgccgcca ccgcgacggc 840
aagttccagc tcgccaacgg cggcaccctg ttcctcgacg agatcggcga catgccgctg 900
ccggtgcagg ccaagcttct gcgcgtgctg caggagcggg agatcgagcc gctcggctcc 960
aacaaggtgg tgcgggtcga tgtccgcatc atcgccgcca ccagccgtga cctgcacgcc 1020
ctggtgcgtg agaagcagtt ccgcgccgac ctctattacc ggctgaatgt ggtgccgatc 1080
accctgccgc cgctgcgcga ccggccggag gacatcgaga gcatcgccga ccgcatcctg 1140
gaacagctgg cgatccagca gggcacgccg ccgcgcgagc tgctggaatc ggcggtgcag 1200
gtgctgcgcg actatgactg gcccggcaat gtgcgcgagc tttacaacac gctggaacgg 1260
gtggtggcgc tgaccgatgc gccgatcctg accgcgccgc acatccgcag cgtgctgccc 1320
ggccagcatc cggccggcgc gtcggccctg ccgctggcgg ccggcgcgcg gccgttgcag 1380
gaggtgctgc acgccgccga gcgccacgcc atcgccgcgg cgcttgagga ggcgaacggc 1440
gtcaaggcgc gggcggcgaa gctgctgggc atttcgcgcg cgtcgctgta cgaacgcatg 1500
gtgacgctgg ggttgggggc gacgcagtag 1530
<210> 174
<211> 3030
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> glnE
<400> 174
atgccgagtc ccatcgcgtt ctcaagcccc ttgccgaagc ctttcgacag cgcgcaggcg 60
gcgctgggga tggagcgctg gcgccagcag gccgccgcgg cggagccgga gacccgcgcc 120
tgggcggaag ccttcgccga ttcggagacc ggccgggcgc tgatcggggc ggtgtgcggc 180
aacagcccgt atctcggcca cagcctgacg cgggagttgc ccttcgtcgc ccgtacagtg 240
caggacggct tcgacgacac cttcgccgcg ctgatcgccg ctctccatgc cgagcatggc 300
gaggagaaat cgatggaccg gctgatggcc ggcctgcggg tggcgaagcg gcgggcggcg 360
ctgctgatcg cgctggccga catcgccggc gcgtggccgc tgttccgcgt caccggcgcc 420
ctgtcggagc tggcggagac gggggtgcag ctggccgcga atttcctgct gcgccgcgcc 480
agggaggcgg ggacgctgac gctgccggat ccgcagcgac cgtgggtcgg ttcgggcctg 540
atcgttttgg gcatgggtaa gcttggcggg cgcgaactca actattccag cgacatcgac 600
ctgatcgtcc tgtatgacga cgctgttgtg cagacgcccc agccggacaa cctcgcgcga 660
accttcatca ggctcgcacg cgatcttgtc cgcattatgg atgaacggac caaggacggc 720
tacgtcttcc gcaccgacct tcggcttagg cccgatcccg gcgccacgcc gctggcggtt 780
tccgtctccg cagccgaaat ttattacggc agcgtcggtc agaactggga acgcgcggcg 840
atgatcaagg cccgtcccat cgccggcgat ctggaggcgg gcgcctcctt tgtccgcttc 900
ctggagccct tcgtctggcg ccgcaacctg gatttcgccg ccatccagga catccattcg 960
atcaaacgcc agatcaacgc ccacaagggc caccgcgagg tgacggtcaa cggccacgac 1020
atcaaggtcg gccgcggcgg catccgcgag atcgagttct tcgcccagac ccagcagctg 1080
atcttcggcg ggcgcgaccc gcgcgtgcga atcgctccga ccctgatggc gaacgaggcg 1140
ctgcgcgacg tcggccgcgt gccgccgcag acggtggaag agcttgccgg ggcctatcat 1200
ttcctgcgcc gtgtcgaaca tcgcatccag atgatcgacg accagcagac ccatcgtatt 1260
cccgccgacg atgccggggt ggcgcatttg gccaccttcc tcggctatga cgaccccgcc 1320
gccttccggg cggaactgct ggcgacgctg gggcaggtgg aggaccgcta tgccgagctg 1380
ttcgaggagg cgccgtcgct ttccggcccc ggcaatctgg tcttcaccgg caccgacccc 1440
gatccgggca cgatggagac gctgaagggc atgggcttcg ccgatccggc ccgcgtcatc 1500
agcgtggtgt cggcctggca tcgcggccgc taccgcgcca cccggtcggg ccgggcgcgg 1560
gagctgctga cggagctgac gccggccctg ctgagtgcgc tgaccaagac cccggccccc 1620
gattcggcgc tgatgaactt cgacgatttc ctcggcaagc tgccggccgg cgtcggtctg 1680
ttctcgctgt tcgtcgccaa tccctggctc ctggagctgg tggcggagat catgggcatc 1740
gcgccgcaga tggcgcagac gctgtcgcgc aacccgtcgc tgctcgacgc cgtgctgtcg 1800
ccggacttct tcgacccgct gccgggcaag gaggacgggc tggccgacga ccacgcccgc 1860
gtgatggcgc cggcccgcga tttcgaggat gcgctgaccc tgtcgcggcg ctggaccaac 1920
gaccagcgct tccgcgccgg ggtgcatatc ctgcgcggca tcaccgatgg cgaccgctgc 1980
ggcgccttcc tggccgatct ggccgacatc gtcgtccccg accttgcccg ccgggtggag 2040
gaggagttcg cccagcgcca cggccatatt cccggcggcg cctgggtggt ggtggcgatg 2100
ggcaagctcg gcagccggca gctgaccatc acgtccgaca tcgacctgat cgtcatctac 2160
gatgtggcgc cgggccaagg gggcgggggc ggtccccgct tgtcggatgg tgccaagccg 2220
ctgtcgccca acgagtatta catcaagctg actcagcgtc tgaccaacgc cattaccgcg 2280
ccgatgggcg acggccggct ctacgaggtc gacatgcggc tgcgcccgtc gggcaacgcc 2340
gggccgctcg ccacctcgct ggacgctttc ctgaaatatc aggcgaccga tgcctggacc 2400
tgggagcata tggccctgac ccgcgcccgg gtgatcggcg gtgatgcgga gctggccggg 2460
cgggtgtcgg cagcgatccg ctcggtgctg acggcgccgc gcgatgccga ccggctgctg 2520
tgggacgtgg ccgacatgcg gcggcggatc gagaaggagt tcgggacgac caatgtctgg 2580
aacgtcaaat acgcccgcgg cggcctgatc gacatcgagt tcatcgccca gtacctgcaa 2640
ctgcgccatg gtcacgagcg gccggacatc ctgcacatcg gcaccgccaa ggcgctgggc 2700
tgcgccgccc ggacgggcgc gctggcgccg gaggtggcgg aggatctgga gacgacgctg 2760
cggctgtggc ggcgggtgca gggctttctg cggttgacca ccgccggggt gctcgatccc 2820
aatcaggtgt cgcccagcct gctggccggg ctggtccgcg ccgcctttcc tgctgacttt 2880
cagggcgagc gtgagcctgg cactgttgac ttccccgaac tggaccacaa aatccgtgcc 2940
gtcgccgccc gcgcccatgg tcatttcaag accctggtcg aggaaccggc gggccgtctg 3000
gccccacccg ccaccacgcc tccagcctga 3030
<210> 175
<211> 1440
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> amtB1
<400> 175
atgaaccgtc tgttccttat ggccgcaccg atgatggcgg ttgctctggg cgcggtcggc 60
atgccggccg cagcccttgc ccaggatccg gcggctgccg ccgctgccgc ggctgcggct 120
gcggctgccg ccgctgctgc cgcaccggcg gctccggcgc tgaatggcgg cgacaccgcc 180
tggatgctca tctccaccgc gctggtgctg atgatgacca tccccggcct ggcgctgttc 240
tacggcggca tggtccgcaa gatgaacgtg ctgtcgacgg tgatgcagag cttcgccatc 300
acctgcctga tcagcgtcct gtggtacgtc atcggctaca gcctggcctt caccggcacc 360
ggtgcctatg tcggcggtct cgaccggctg ttcctcaacg ggctcgactt cacgaaggcc 420
ttcgtgctgg gcgaggcgac cgggtcgggc gtcccgacga ccatccccga gccggtcttc 480
atgatgttcc agatgacctt tgcgatcatc accccggccc tgatcaccgg cgccttcgcc 540
gaccgcatga agttctcctc cctgctggtc ttcaccgcgc tgtggtcgat cgtggtctat 600
gcgccgatcg cccactgggt ctggtacccg tcgggcttcc tgttcggcct gggcgtgctg 660
gacttcgccg gcggcacggt cgtgcacatc aacgccggcg tcgccggcct ggtcgccgcg 720
ctggtgatcg gcaagcgcaa gggctacccg aaggaagcct tcatgccgca caacctggtg 780
ctgtcgctga tcggcgcctc gctgctgtgg gtcggctggt tcggcttcaa cgccggttcg 840
gccctgaccg ccggtccgcg tgccggcatg gcgctggccg ccacgcacat cgccaccgcc 900
ggtgccgcca tgggctggct gttcgcggag tggatcgtca agggcaagcc gtcgatcctc 960
ggcatcatct ccggcgccgt cgccggcctg gtcgcggtga ccccggccgc cggcttcgtc 1020
gacccgacgg gcgccatcgt catcggcatc gtcgccggcg tggtctgctt ctggtcggcc 1080
accagcctca agcacatgct gggctatgac gacagcctgg acgccttcgg cgtgcacggc 1140
gtcggcggcc tgatcggcgc catcctgacc ggcgtcttcg ccaagatgtc ggtgtccaac 1200
agcgaaggcg gcttcgcctc cgtcctgcag gccgacccga aggccacgct gggcctgctg 1260
gaaggcaacg ccgccgccgt ctggatccag gtccagggcg tcctctacac catggtctgg 1320
tgcgccatcg ccaccttcgt cctgctgaag atcgtcgatg tggtcatggg cctgcgcgtc 1380
gaagaggatg tggagcgcga cggtctcgac ctcgccctgc atggcgagag catccactaa 1440
<210> 176
<211> 1227
<212> ДНК
<213> Azospirillum lipoferum
<220>
<223> amtB2
<400> 176
atggatgcgg caaagacggg tggcgacgtc cttttcgtgc tgatgggcgc ggtgatggtg 60
ctggcgatgc attgcggctt cgccctgctg gaggtcggga cggtccggcg caagaatcag 120
gtcaacgcgc tggtgaagat cctgtcggac ttcgccatgt cgaccatcgc ctattttttc 180
gtcggttatg ccgtggccta cggcatcgac ttcttcgccg acgcccacac gctggtcggc 240
aagggaagcg gcgggttcgc ggcctatggc tacgatctgg tgaagttctt cttcctggcg 300
accttcgccg ccgcggtgcc ggccatcgtc tcgggcggca tcgccgagcg tgctaggttc 360
tggccgcagg ccgccgccac gctggcgctg atcgcgctgt tctatccatt gctggaaggc 420
acggtctggg gcacccgctt cggcctgcaa agctggatgg ccgcgacctt cggccagcct 480
ttccacgact tcgccggatc tgtggtggtg catgccttcg gcggctgggt ggcgctgggt 540
gccgtgctga acctcggcaa ccgccgcggc cgctaccgtc cgaacggctc gctgatcgcc 600
attccgccgt cgaacatccc cttcctggcg ctgggcgcct gggtgctgtg cgtggggtgg 660
ttcggcttca acgtgatgag cgcccaggtg ctggatggcg tgacgggtct ggtggcgctg 720
aactcgctga tggcgatggt cggcggcatc gtcacctcgc tggtgatcag ccgcaccgat 780
cccggcttcg tccacaacgg cgcgctggcc ggtctggtgg cggtctgcgc cgggtccgac 840
gtgatgcacc cgctgggcgc gctggtcacc ggcggcatcg ccgggctgct gttcgtctgg 900
gccttcaaca aatgccagat cgactggaag atcgacgacg tgctgggcgt ctggccgctg 960
cacggtctgt gcggcctgac cggcggcctg ctggccggcg tcttcgggca ggaggcactg 1020
ggcggccttg gcggcgtgtc gatcctcagc cagatcgtcg gcacggcaag cggcgccagc 1080
ttcggattcg tctcgggtct ggcggtctac ggcctgctgc gcgtcaccgt cggcatccgc 1140
ctcgatcccg agcaggagta caagggcgcc gacttgtcgt tgcaccatat caccgcgtac 1200
ccggaagagg acgcgccgac cctgtaa 1227
<210> 177
<211> 613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC
<400> 177
atgaccctga atatgatgat ggattcttgg ttctctggag cgctttatcg gcatcctgac 60
tgaagaattt gcaggcttct tcccaacctg gcttgcaccc gtgcaggtag ttgtgatgaa 120
catcactgat tcgcaggctg aatacgttaa cgaattgacc cgtaaactgc aaaatgcggg 180
cattcgtgta aaagcagact tgagaaacga gaagattggc tttaaaatcc gcgagcacac 240
tttacgtcgt gtcccttata tgctggtttg tggtgacaaa gaggtcgaag ccggcaaagt 300
tgctgtgcgt acccgtcgcg gtaaagacct gggtagcctg gacgtaaatg atgttatcga 360
gaagctgcaa caagagattc gcagccgcag tcttcaacaa ctggaggaat aaggtattaa 420
aggcggaaaa cgagttcaaa cggcgcgtcc caatcgtatt aatggcgaga ttcgcgccac 480
ggaagttcgc ttaacaggtc tggaaggcga gcagcttggt attgcgatag aactcacttc 540
acgccccgaa gggggaagct gcctgaccct acgattcccg ctatttcatt cactgaccgg 600
aggttcaaaa tga 613
<210> 178
<211> 1613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC с фланкирующей областью длиной 500 п.н.
<400> 178
accggataag agagaaaagt gtcgacgtcg gtccggttga tattgaccgg cgcatccgcc 60
agctcgccca gtttttggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgcgg 180
ggaaaatgcg gtgaacatgt cagctattgc gaagagtgtg ccagttttgc tcacgggcaa 240
aagctgcacc agaatgggta ttaatgcacc agcctggcgc tttttttcgc ggcacgtccc 300
ctcgctaatg cccgtctggc gcggctttga cgctgataag gcgctgaata ccgatctgga 360
tcaaggtttt gtcgggttat cgtccaaaag gtgcactctt tgcatggtta taagtgcctg 420
acatggtgtc cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta 480
accgaccgca ggagtgtgcg atgaccctga atatgatgat ggattcttgg ttctctggag 540
cgctttatcg gcatcctgac tgaagaattt gcaggcttct tcccaacctg gcttgcaccc 600
gtgcaggtag ttgtgatgaa catcactgat tcgcaggctg aatacgttaa cgaattgacc 660
cgtaaactgc aaaatgcggg cattcgtgta aaagcagact tgagaaacga gaagattggc 720
tttaaaatcc gcgagcacac tttacgtcgt gtcccttata tgctggtttg tggtgacaaa 780
gaggtcgaag ccggcaaagt tgctgtgcgt acccgtcgcg gtaaagacct gggtagcctg 840
gacgtaaatg atgttatcga gaagctgcaa caagagattc gcagccgcag tcttcaacaa 900
ctggaggaat aaggtattaa aggcggaaaa cgagttcaaa cggcgcgtcc caatcgtatt 960
aatggcgaga ttcgcgccac ggaagttcgc ttaacaggtc tggaaggcga gcagcttggt 1020
attgcgatag aactcacttc acgccccgaa gggggaagct gcctgaccct acgattcccg 1080
ctatttcatt cactgaccgg aggttcaaaa tgacccagcg aaccgagtcg ggtaataccg 1140
tctggcgctt cgatttatcc cagcagttca ccgcgatgca gcggataagc gtggttctca 1200
gccgggcgac cgaggttgaa cagacactcc agcaggtgct gtgcgtattg cacaatgacg 1260
cctttttgca gcacggcatg atctgtctgt acgacagcca gcaggcgatt ttgactattg 1320
aagcgttgca ggaagccgat cagcagttga tccccggcag ctcgcaaatt cgctaccgtc 1380
cgggtgaagg gctggtcggg acggtgcttt cgcaggggca atcgttagtg ctggcgcgtg 1440
tggctgacga tcagcgcttt cttgaccgcc tgggactgta tgattacaac ctgccgttta 1500
tcgccgtgcc gctgataggg ccggatgcgc agacttttgg cgtgctgacg gcgcaaccga 1560
tggcgcgtta cgaagagcgg ttacccgcct gcacccgctt tctggaaacg gtc 1613
<210> 179
<211> 1185
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 179
tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg cgcgctaccc gatcctgctc 60
gatgaactgc tcgacccgaa cacgctctat caaccgacgg cgatgaacgc ctatcgcgat 120
gaactgcgac aatacctgtt gcgcgtgccg gaagaggatg aagagcagca actggaggcg 180
ctacggcagt ttaagcaggc gcagttgttg cgcgtagcgg cggcggatat cgccggtacg 240
ttacccgtca tgaaagtgag cgatcactta acctggctgg cggaagcgat tatcgatgcg 300
gtggtgcagc aagcctggaa ccagatggtg gcgcgttacg gccagccgac gcatctgcac 360
gatcgcgaag ggcgcggttt cgccgtggtc ggttacggca aacttggcgg ctgggaatta 420
ggttacagct ccgatctgga tctggtgttc ctgcacgact gccccatgga tgtgatgacc 480
gatggcgagc gtgaaatcga tggccgccag ttctatttgc gcctcgcgca gcgcgtgatg 540
cacctgttca gcacgcgcac gtcgtccggc attctttatg aagtcgatgc gcgtttgcgc 600
ccgtccggcg cggccggaat gctggtgacc actgcggaag cgttcgccga ttatcaaaaa 660
aatgaagcct ggacatggga gcatcaggcg ctggcgcgtg cgcgcgtggt gtacggcgat 720
ccgcaactga ccgccgaatt tgacgccatt cgccgcgata tcctgatgac ctcccgcgat 780
gccgctaccc tgcaaaccga agtgcgggaa atgcgtgaga aaatgcgcgc ccatcttggt 840
aacaagcaca aagaccgttt cgatctgaaa gccgatgaag gcggtatcac cgatattgag 900
tttatcgctc agtatctggt gctgcgcttt gcccatgaga agccgaaact gacgcgctgg 960
tcggataatg tgcgcatcct cgaagggctg gcgcaaaacg gcatcatgga tgagcaggaa 1020
gcgcaggcat tgacgctggc gtacaccacg ttgcgtgatg agctgcacca cctggcgctg 1080
caagagctgc caggacatgt ggcgctctcc tgttttgtcg ccgagcgtgc gcttatcaaa 1140
accagctggg acaagtggct ggtggaaccg tgcgccccgg cgtaa 1185
<210> 180
<211> 2085
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 180
taaagcgagc gctcacttac gtgatctgtt gacgcagtcc gaagcgacca ttacttcagc 60
cgtttcagca gatacggcgg tgtggagtgc gcaatcagcc ctggcgaaac tggtgctcac 120
cgagtggtta gtgacgcagg gctggcgaac cttccttgat gaaaaagcgc aggctaagtt 180
tgccgactcc tttaaacgct ttgctgacgt tcatctgtca cgcagcgccg ccgagctgaa 240
aaaagccttt gcccagccgc tgggcgacag ctatcgcgac cagttaccgc ggctggcgcg 300
tgatatcgac agcgcgttat tgctggccgg acattacgat cgcgcgcgcg ccgtggagtg 360
gctggaaaac tggcaggggc ttcagcacgc tattgaaacg cgccagagag ttgaaatcga 420
acatttccgt aataccgcca ttacccagga gccgttctgg ttgcacagcg gaaaacgtta 480
acgaaaggat atttcgcatg tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg 540
cgcgctaccc gatcctgctc gatgaactgc tcgacccgaa cacgctctat caaccgacgg 600
cgatgaacgc ctatcgcgat gaactgcgac aatacctgtt gcgcgtgccg gaagaggatg 660
aagagcagca actggaggcg ctacggcagt ttaagcaggc gcagttgttg cgcgtagcgg 720
cggcggatat cgccggtacg ttacccgtca tgaaagtgag cgatcactta acctggctgg 780
cggaagcgat tatcgatgcg gtggtgcagc aagcctggaa ccagatggtg gcgcgttacg 840
gccagccgac gcatctgcac gatcgcgaag ggcgcggttt cgccgtggtc ggttacggca 900
aacttggcgg ctgggaatta ggttacagct ccgatctgga tctggtgttc ctgcacgact 960
gccccatgga tgtgatgacc gatggcgagc gtgaaatcga tggccgccag ttctatttgc 1020
gcctcgcgca gcgcgtgatg cacctgttca gcacgcgcac gtcgtccggc attctttatg 1080
aagtcgatgc gcgtttgcgc ccgtccggcg cggccggaat gctggtgacc actgcggaag 1140
cgttcgccga ttatcaaaaa aatgaagcct ggacatggga gcatcaggcg ctggcgcgtg 1200
cgcgcgtggt gtacggcgat ccgcaactga ccgccgaatt tgacgccatt cgccgcgata 1260
tcctgatgac ctcccgcgat gccgctaccc tgcaaaccga agtgcgggaa atgcgtgaga 1320
aaatgcgcgc ccatcttggt aacaagcaca aagaccgttt cgatctgaaa gccgatgaag 1380
gcggtatcac cgatattgag tttatcgctc agtatctggt gctgcgcttt gcccatgaga 1440
agccgaaact gacgcgctgg tcggataatg tgcgcatcct cgaagggctg gcgcaaaacg 1500
gcatcatgga tgagcaggaa gcgcaggcat tgacgctggc gtacaccacg ttgcgtgatg 1560
agctgcacca cctggcgctg caagagctgc caggacatgt ggcgctctcc tgttttgtcg 1620
ccgagcgtgc gcttatcaaa accagctggg acaagtggct ggtggaaccg tgcgccccgg 1680
cgtaagtgtg gtatcatcgc gcgcaaattt tgtatctctc aggagacagg aatgaaagtt 1740
acgctgccag agttcaatca agccggtgtc atggtggtgg gtgatgtgat gctggatcgc 1800
tactggtacg gcccaaccag ccgcatttct ccggaagcgc cagttccggt tgttaaagtc 1860
gatactattg aagagcgacc gggcggtgcg gcaaacgtgg cgatgaacat tgcctcgctg 1920
ggcgcaacgg cgcgtctggt tggcctgact ggcattgatg atgcggcgcg cgcgctgagc 1980
aaagcgctgg cggatgttaa tgttaaatgt gacttcgttt ctgttccgac tcaccccacc 2040
atcactaagc tgcgcgtgct gtcgcgtaac cagcaactga ttcgc 2085
<210> 181
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 181
atgaccctga atatgatgat ggatgccagc cgttctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctc gcggcgcttt ttttatattc tcgactccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac gccccgaagg gggaagctgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 182
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 182
accggataag agagaaaagt gtcgacgtcg gtccggttga tattgaccgg cgcatccgcc 60
agctcgccca gtttttggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgcgg 180
ggaaaatgcg gtgaacatgt cagctattgc gaagagtgtg ccagttttgc tcacgggcaa 240
aagctgcacc agaatgggta ttaatgcacc agcctggcgc tttttttcgc ggcacgtccc 300
ctcgctaatg cccgtctggc gcggctttga cgctgataag gcgctgaata ccgatctgga 360
tcaaggtttt gtcgggttat cgtccaaaag gtgcactctt tgcatggtta taagtgcctg 420
acatggtgtc cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta 480
accgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccagc cgttctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctc gcggcgcttt ttttatattc 600
tcgactccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac gccccgaagg 900
gggaagctgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttatccca gcagttcacc 1020
gcgatgcagc ggataagcgt ggttctcagc cgggcgaccg aggttgaaca gacactccag 1080
caggtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gactattgaa gcgttgcagg aagccgatca gcagttgatc 1200
cccggcagct cgcaaattcg ctaccgtccg ggtgaagggc tggtcgggac ggtgctttcg 1260
caggggcaat cgttagtgct ggcgcgtgtg gctgacgatc agcgctttct tgaccgcctg 1320
ggactgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc ggatgcgcag 1380
acttttggcg tgctgacggc gcaaccgatg gcgcgttacg aagagcggtt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 183
<211> 1185
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 183
tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg cgcgctaccc gatcctgctc 60
gatgaactgc tcgacccgaa cacgctctat caaccgacgg cgatgaacgc ctatcgcgat 120
gaactgcgac aatacctgtt gcgcgtgccg gaagaggatg aagagcagca actggaggcg 180
ctacggcagt ttaagcaggc gcagttgttg cgcgtagcgg cggcggatat cgccggtacg 240
ttacccgtca tgaaagtgag cgatcactta acctggctgg cggaagcgat tatcgatgcg 300
gtggtgcagc aagcctggaa ccagatggtg gcgcgttacg gccagccgac gcatctgcac 360
gatcgcgaag ggcgcggttt cgccgtggtc ggttacggca aacttggcgg ctgggaatta 420
ggttacagct ccgatctgga tctggtgttc ctgcacgact gccccatgga tgtgatgacc 480
gatggcgagc gtgaaatcga tggccgccag ttctatttgc gcctcgcgca gcgcgtgatg 540
cacctgttca gcacgcgcac gtcgtccggc attctttatg aagtcgatgc gcgtttgcgc 600
ccgtccggcg cggccggaat gctggtgacc actgcggaag cgttcgccga ttatcaaaaa 660
aatgaagcct ggacatggga gcatcaggcg ctggcgcgtg cgcgcgtggt gtacggcgat 720
ccgcaactga ccgccgaatt tgacgccatt cgccgcgata tcctgatgac ctcccgcgat 780
gccgctaccc tgcaaaccga agtgcgggaa atgcgtgaga aaatgcgcgc ccatcttggt 840
aacaagcaca aagaccgttt cgatctgaaa gccgatgaag gcggtatcac cgatattgag 900
tttatcgctc agtatctggt gctgcgcttt gcccatgaga agccgaaact gacgcgctgg 960
tcggataatg tgcgcatcct cgaagggctg gcgcaaaacg gcatcatgga tgagcaggaa 1020
gcgcaggcat tgacgctggc gtacaccacg ttgcgtgatg agctgcacca cctggcgctg 1080
caagagctgc caggacatgt ggcgctctcc tgttttgtcg ccgagcgtgc gcttatcaaa 1140
accagctggg acaagtggct ggtggaaccg tgcgccccgg cgtaa 1185
<210> 184
<211> 2085
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 184
taaagcgagc gctcacttac gtgatctgtt gacgcagtcc gaagcgacca ttacttcagc 60
cgtttcagca gatacggcgg tgtggagtgc gcaatcagcc ctggcgaaac tggtgctcac 120
cgagtggtta gtgacgcagg gctggcgaac cttccttgat gaaaaagcgc aggctaagtt 180
tgccgactcc tttaaacgct ttgctgacgt tcatctgtca cgcagcgccg ccgagctgaa 240
aaaagccttt gcccagccgc tgggcgacag ctatcgcgac cagttaccgc ggctggcgcg 300
tgatatcgac agcgcgttat tgctggccgg acattacgat cgcgcgcgcg ccgtggagtg 360
gctggaaaac tggcaggggc ttcagcacgc tattgaaacg cgccagagag ttgaaatcga 420
acatttccgt aataccgcca ttacccagga gccgttctgg ttgcacagcg gaaaacgtta 480
acgaaaggat atttcgcatg tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg 540
cgcgctaccc gatcctgctc gatgaactgc tcgacccgaa cacgctctat caaccgacgg 600
cgatgaacgc ctatcgcgat gaactgcgac aatacctgtt gcgcgtgccg gaagaggatg 660
aagagcagca actggaggcg ctacggcagt ttaagcaggc gcagttgttg cgcgtagcgg 720
cggcggatat cgccggtacg ttacccgtca tgaaagtgag cgatcactta acctggctgg 780
cggaagcgat tatcgatgcg gtggtgcagc aagcctggaa ccagatggtg gcgcgttacg 840
gccagccgac gcatctgcac gatcgcgaag ggcgcggttt cgccgtggtc ggttacggca 900
aacttggcgg ctgggaatta ggttacagct ccgatctgga tctggtgttc ctgcacgact 960
gccccatgga tgtgatgacc gatggcgagc gtgaaatcga tggccgccag ttctatttgc 1020
gcctcgcgca gcgcgtgatg cacctgttca gcacgcgcac gtcgtccggc attctttatg 1080
aagtcgatgc gcgtttgcgc ccgtccggcg cggccggaat gctggtgacc actgcggaag 1140
cgttcgccga ttatcaaaaa aatgaagcct ggacatggga gcatcaggcg ctggcgcgtg 1200
cgcgcgtggt gtacggcgat ccgcaactga ccgccgaatt tgacgccatt cgccgcgata 1260
tcctgatgac ctcccgcgat gccgctaccc tgcaaaccga agtgcgggaa atgcgtgaga 1320
aaatgcgcgc ccatcttggt aacaagcaca aagaccgttt cgatctgaaa gccgatgaag 1380
gcggtatcac cgatattgag tttatcgctc agtatctggt gctgcgcttt gcccatgaga 1440
agccgaaact gacgcgctgg tcggataatg tgcgcatcct cgaagggctg gcgcaaaacg 1500
gcatcatgga tgagcaggaa gcgcaggcat tgacgctggc gtacaccacg ttgcgtgatg 1560
agctgcacca cctggcgctg caagagctgc caggacatgt ggcgctctcc tgttttgtcg 1620
ccgagcgtgc gcttatcaaa accagctggg acaagtggct ggtggaaccg tgcgccccgg 1680
cgtaagtgtg gtatcatcgc gcgcaaattt tgtatctctc aggagacagg aatgaaagtt 1740
acgctgccag agttcaatca agccggtgtc atggtggtgg gtgatgtgat gctggatcgc 1800
tactggtacg gcccaaccag ccgcatttct ccggaagcgc cagttccggt tgttaaagtc 1860
gatactattg aagagcgacc gggcggtgcg gcaaacgtgg cgatgaacat tgcctcgctg 1920
ggcgcaacgg cgcgtctggt tggcctgact ggcattgatg atgcggcgcg cgcgctgagc 1980
aaagcgctgg cggatgttaa tgttaaatgt gacttcgttt ctgttccgac tcaccccacc 2040
atcactaagc tgcgcgtgct gtcgcgtaac cagcaactga ttcgc 2085
<210> 185
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 185
atgaccctga atatgatgat ggatgccagc cgttctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctc gcggcgcttt ttttatattc tcgactccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac gccccgaagg gggaagctgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 186
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 186
accggataag agagaaaagt gtcgacgtcg gtccggttga tattgaccgg cgcatccgcc 60
agctcgccca gtttttggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgcgg 180
ggaaaatgcg gtgaacatgt cagctattgc gaagagtgtg ccagttttgc tcacgggcaa 240
aagctgcacc agaatgggta ttaatgcacc agcctggcgc tttttttcgc ggcacgtccc 300
ctcgctaatg cccgtctggc gcggctttga cgctgataag gcgctgaata ccgatctgga 360
tcaaggtttt gtcgggttat cgtccaaaag gtgcactctt tgcatggtta taagtgcctg 420
acatggtgtc cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta 480
accgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccagc cgttctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctc gcggcgcttt ttttatattc 600
tcgactccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac gccccgaagg 900
gggaagctgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttatccca gcagttcacc 1020
gcgatgcagc ggataagcgt ggttctcagc cgggcgaccg aggttgaaca gacactccag 1080
caggtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gactattgaa gcgttgcagg aagccgatca gcagttgatc 1200
cccggcagct cgcaaattcg ctaccgtccg ggtgaagggc tggtcgggac ggtgctttcg 1260
caggggcaat cgttagtgct ggcgcgtgtg gctgacgatc agcgctttct tgaccgcctg 1320
ggactgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc ggatgcgcag 1380
acttttggcg tgctgacggc gcaaccgatg gcgcgttacg aagagcggtt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 187
<211> 1185
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 187
tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg cgcgctaccc gatcctgctc 60
gatgaactgc tcgacccgaa cacgctctat caaccgacgg cgatgaacgc ctatcgcgat 120
gaactgcgac aatacctgtt gcgcgtgccg gaagaggatg aagagcagca actggaggcg 180
ctacggcagt ttaagcaggc gcagttgttg cgcgtagcgg cggcggatat cgccggtacg 240
ttacccgtca tgaaagtgag cgatcactta acctggctgg cggaagcgat tatcgatgcg 300
gtggtgcagc aagcctggaa ccagatggtg gcgcgttacg gccagccgac gcatctgcac 360
gatcgcgaag ggcgcggttt cgccgtggtc ggttacggca aacttggcgg ctgggaatta 420
ggttacagct ccgatctgga tctggtgttc ctgcacgact gccccatgga tgtgatgacc 480
gatggcgagc gtgaaatcga tggccgccag ttctatttgc gcctcgcgca gcgcgtgatg 540
cacctgttca gcacgcgcac gtcgtccggc attctttatg aagtcgatgc gcgtttgcgc 600
ccgtccggcg cggccggaat gctggtgacc actgcggaag cgttcgccga ttatcaaaaa 660
aatgaagcct ggacatggga gcatcaggcg ctggcgcgtg cgcgcgtggt gtacggcgat 720
ccgcaactga ccgccgaatt tgacgccatt cgccgcgata tcctgatgac ctcccgcgat 780
gccgctaccc tgcaaaccga agtgcgggaa atgcgtgaga aaatgcgcgc ccatcttggt 840
aacaagcaca aagaccgttt cgatctgaaa gccgatgaag gcggtatcac cgatattgag 900
tttatcgctc agtatctggt gctgcgcttt gcccatgaga agccgaaact gacgcgctgg 960
tcggataatg tgcgcatcct cgaagggctg gcgcaaaacg gcatcatgga tgagcaggaa 1020
gcgcaggcat tgacgctggc gtacaccacg ttgcgtgatg agctgcacca cctggcgctg 1080
caagagctgc caggacatgt ggcgctctcc tgttttgtcg ccgagcgtgc gcttatcaaa 1140
accagctggg acaagtggct ggtggaaccg tgcgccccgg cgtaa 1185
<210> 188
<211> 2085
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 188
taaagcgagc gctcacttac gtgatctgtt gacgcagtcc gaagcgacca ttacttcagc 60
cgtttcagca gatacggcgg tgtggagtgc gcaatcagcc ctggcgaaac tggtgctcac 120
cgagtggtta gtgacgcagg gctggcgaac cttccttgat gaaaaagcgc aggctaagtt 180
tgccgactcc tttaaacgct ttgctgacgt tcatctgtca cgcagcgccg ccgagctgaa 240
aaaagccttt gcccagccgc tgggcgacag ctatcgcgac cagttaccgc ggctggcgcg 300
tgatatcgac agcgcgttat tgctggccgg acattacgat cgcgcgcgcg ccgtggagtg 360
gctggaaaac tggcaggggc ttcagcacgc tattgaaacg cgccagagag ttgaaatcga 420
acatttccgt aataccgcca ttacccagga gccgttctgg ttgcacagcg gaaaacgtta 480
acgaaaggat atttcgcatg tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg 540
cgcgctaccc gatcctgctc gatgaactgc tcgacccgaa cacgctctat caaccgacgg 600
cgatgaacgc ctatcgcgat gaactgcgac aatacctgtt gcgcgtgccg gaagaggatg 660
aagagcagca actggaggcg ctacggcagt ttaagcaggc gcagttgttg cgcgtagcgg 720
cggcggatat cgccggtacg ttacccgtca tgaaagtgag cgatcactta acctggctgg 780
cggaagcgat tatcgatgcg gtggtgcagc aagcctggaa ccagatggtg gcgcgttacg 840
gccagccgac gcatctgcac gatcgcgaag ggcgcggttt cgccgtggtc ggttacggca 900
aacttggcgg ctgggaatta ggttacagct ccgatctgga tctggtgttc ctgcacgact 960
gccccatgga tgtgatgacc gatggcgagc gtgaaatcga tggccgccag ttctatttgc 1020
gcctcgcgca gcgcgtgatg cacctgttca gcacgcgcac gtcgtccggc attctttatg 1080
aagtcgatgc gcgtttgcgc ccgtccggcg cggccggaat gctggtgacc actgcggaag 1140
cgttcgccga ttatcaaaaa aatgaagcct ggacatggga gcatcaggcg ctggcgcgtg 1200
cgcgcgtggt gtacggcgat ccgcaactga ccgccgaatt tgacgccatt cgccgcgata 1260
tcctgatgac ctcccgcgat gccgctaccc tgcaaaccga agtgcgggaa atgcgtgaga 1320
aaatgcgcgc ccatcttggt aacaagcaca aagaccgttt cgatctgaaa gccgatgaag 1380
gcggtatcac cgatattgag tttatcgctc agtatctggt gctgcgcttt gcccatgaga 1440
agccgaaact gacgcgctgg tcggataatg tgcgcatcct cgaagggctg gcgcaaaacg 1500
gcatcatgga tgagcaggaa gcgcaggcat tgacgctggc gtacaccacg ttgcgtgatg 1560
agctgcacca cctggcgctg caagagctgc caggacatgt ggcgctctcc tgttttgtcg 1620
ccgagcgtgc gcttatcaaa accagctggg acaagtggct ggtggaaccg tgcgccccgg 1680
cgtaagtgtg gtatcatcgc gcgcaaattt tgtatctctc aggagacagg aatgaaagtt 1740
acgctgccag agttcaatca agccggtgtc atggtggtgg gtgatgtgat gctggatcgc 1800
tactggtacg gcccaaccag ccgcatttct ccggaagcgc cagttccggt tgttaaagtc 1860
gatactattg aagagcgacc gggcggtgcg gcaaacgtgg cgatgaacat tgcctcgctg 1920
ggcgcaacgg cgcgtctggt tggcctgact ggcattgatg atgcggcgcg cgcgctgagc 1980
aaagcgctgg cggatgttaa tgttaaatgt gacttcgttt ctgttccgac tcaccccacc 2040
atcactaagc tgcgcgtgct gtcgcgtaac cagcaactga ttcgc 2085
<210> 189
<211> 452
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm7
<400> 189
atgaccctga atatgatgat ggatgccagc cgcgtcaggt tgaacgtaaa aaagtcggtc 60
tgcgcaaagc acgtcgtcgt ccgcagttct ccaaacgtta attggtttct gcttcggcag 120
aacgattggc gaaaaaaccc ggtgcgaacc gggttttttt atggataaag atcgtgttat 180
ccacagcaat ccattgatta tctcttcttt ttcagcattt ccagaatccc ctcaccacaa 240
agcccgcaaa atctggtaaa ctatcatcca attttctgcc caaatggctg ggattgttca 300
ttttttgttt gccttacaac gagagtgaca gtacgcgcgg gtagttaact caacatctga 360
ccggtcgata actcacttca cgccccgaag ggggaagctg cctgacccta cgattcccgc 420
tatttcattc actgaccgga ggttcaaaat ga 452
<210> 190
<211> 1451
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm7 с фланкирующей областью длиной 500 п.н.
<400> 190
accggataag agagaaaagt gtcgacgtcg gtccggttga tattgaccgg cgcatccgcc 60
agctcgccca gtttttggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgcgg 180
ggaaaatgcg gtgaacatgt cagctattgc gaagagtgtg ccagttttgc tcacgggcaa 240
aagctgcacc agaatgggta ttaatgcacc agcctggcgc tttttttcgc ggcacgtccc 300
ctcgctaatg cccgtctggc gcggctttga cgctgataag gcgctgaata ccgatctgga 360
tcaaggtttt gtcgggttat cgtccaaaag gtgcactctt tgcatggtta taagtgcctg 420
acatggtgtc cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta 480
accgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccagc cgcgtcaggt 540
tgaacgtaaa aaagtcggtc tgcgcaaagc acgtcgtcgt ccgcagttct ccaaacgtta 600
attggtttct gcttcggcag aacgattggc gaaaaaaccc ggtgcgaacc gggttttttt 660
atggataaag atcgtgttat ccacagcaat ccattgatta tctcttcttt ttcagcattt 720
ccagaatccc ctcaccacaa agcccgcaaa atctggtaaa ctatcatcca attttctgcc 780
caaatggctg ggattgttca ttttttgttt gccttacaac gagagtgaca gtacgcgcgg 840
gtagttaact caacatctga ccggtcgata actcacttca cgccccgaag ggggaagctg 900
cctgacccta cgattcccgc tatttcattc actgaccgga ggttcaaaat gacccagcga 960
accgagtcgg gtaataccgt ctggcgcttc gatttatccc agcagttcac cgcgatgcag 1020
cggataagcg tggttctcag ccgggcgacc gaggttgaac agacactcca gcaggtgctg 1080
tgcgtattgc acaatgacgc ctttttgcag cacggcatga tctgtctgta cgacagccag 1140
caggcgattt tgactattga agcgttgcag gaagccgatc agcagttgat ccccggcagc 1200
tcgcaaattc gctaccgtcc gggtgaaggg ctggtcggga cggtgctttc gcaggggcaa 1260
tcgttagtgc tggcgcgtgt ggctgacgat cagcgctttc ttgaccgcct gggactgtat 1320
gattacaacc tgccgtttat cgccgtgccg ctgatagggc cggatgcgca gacttttggc 1380
gtgctgacgg cgcaaccgat ggcgcgttac gaagagcggt tacccgcctg cacccgcttt 1440
ctggaaacgg t 1451
<210> 191
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 191
atggcgctga agcacctgat cacgctctgc gcggcgtcgc cgatggtcgc cagccagctg 60
gcgcgccacc cgctgctgct ggatgagctg ctggatccca acaccctcta tcagccgacg 120
gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc tgcgcgtgcc ggaagaggat 180
gaagagcagc agctggaggc gttgcgccag tttaagcagg cgcagcagct gcatatcgcg 240
gcggcggata tcgctggtac cctgccggtg atgaaggtca gcgatcactt aacctggctt 300
gccgaagcga tcctcgacgc ggtggtgcag caggcatggg ggcagatggt cgctcgctac 360
ggccagccga cccacctgca cgatcgccag ggtcgcggct tcgccgtcgt cggctacggt 420
aagcttggcg gctgggagct gggctacagc tccgatctcg atctggtgtt cctccatgac 480
tgcccggcgg aggtgatgac cgacggcgag cgggagattg acggccgtca gttctacctg 540
cggctggccc agcggatcat gcacctgttc agcacccgca cctcgtccgg tattctctac 600
gaagtggacg cccggctgcg tccttctggc gcggcgggga tgctggtcac caccgccgac 660
gcgtttgctg actatcagca gaacgaagcc tggacgtggg aacatcaggc gctggtgcgc 720
gcccgcgtgg tctatggcga cccggcgctg caggcgcgct ttgacgccat tcgtcgcgat 780
atcctgacca ccccgcggga ggggatgacc ctgcagaccg aggttcgcga gatgcgcgag 840
aagatgcgcg cccaccttgg caacaaacat cccgatcgtt ttgatatcaa agccgatgcc 900
ggcgggatca ccgatattga atttattact cagtatctgg tcctacgcta tgccagtgac 960
aagccgaagc tgacccgctg gtctgacaac gtgcgtattc ttgagctgct ggcgcagaac 1020
gacatcatgg acgaggagga ggcgcgcgcc ttaacgcatg cgtacaccac cttgcgtgat 1080
gcgctccatc acctggccct gcaggagcag ccgggacacg tggcgccaga ggccttcagc 1140
cgggagcgtc agcaggtcag cgccagctgg cagaagtggc tgatggctta a 1191
<210> 192
<211> 2191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 192
cgtaaggcga ccacccagct ccgcgcgttg ctgaacgacg ctgaagccgt tctgctggcc 60
gcggacaccg ccgacgaggc gttattccgc accgaggtcg tcggcgccaa actggccctg 120
actgaatggc tggtccagcg cggctggcgt ccgttcctca acgaggcagg agagaaaaaa 180
atagccggat cgttcaaacg gtttgccgat attaacctct cgcgggtggc ggccgagctg 240
cgcagcgccg tgcagcatct ggcggttgaa gatgccgccg accagttgcc gaagctgtcc 300
cgcgacatcg acagcgtcca gctgctggcg ggcgcctatg gcgacgccgt cgcgccgtgg 360
ctggagaact ggcaggagct tcaccgtgca atagcacatg acgatcgcag cgtctttgaa 420
tatttccgtc gccaggcgct ggctgccgag ccgttctggc tgcatagtgg aaaacgataa 480
tttcaggcca gggagccctt atggcgctga agcacctgat cacgctctgc gcggcgtcgc 540
cgatggtcgc cagccagctg gcgcgccacc cgctgctgct ggatgagctg ctggatccca 600
acaccctcta tcagccgacg gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc 660
tgcgcgtgcc ggaagaggat gaagagcagc agctggaggc gttgcgccag tttaagcagg 720
cgcagcagct gcatatcgcg gcggcggata tcgctggtac cctgccggtg atgaaggtca 780
gcgatcactt aacctggctt gccgaagcga tcctcgacgc ggtggtgcag caggcatggg 840
ggcagatggt cgctcgctac ggccagccga cccacctgca cgatcgccag ggtcgcggct 900
tcgccgtcgt cggctacggt aagcttggcg gctgggagct gggctacagc tccgatctcg 960
atctggtgtt cctccatgac tgcccggcgg aggtgatgac cgacggcgag cgggagattg 1020
acggccgtca gttctacctg cggctggccc agcggatcat gcacctgttc agcacccgca 1080
cctcgtccgg tattctctac gaagtggacg cccggctgcg tccttctggc gcggcgggga 1140
tgctggtcac caccgccgac gcgtttgctg actatcagca gaacgaagcc tggacgtggg 1200
aacatcaggc gctggtgcgc gcccgcgtgg tctatggcga cccggcgctg caggcgcgct 1260
ttgacgccat tcgtcgcgat atcctgacca ccccgcggga ggggatgacc ctgcagaccg 1320
aggttcgcga gatgcgcgag aagatgcgcg cccaccttgg caacaaacat cccgatcgtt 1380
ttgatatcaa agccgatgcc ggcgggatca ccgatattga atttattact cagtatctgg 1440
tcctacgcta tgccagtgac aagccgaagc tgacccgctg gtctgacaac gtgcgtattc 1500
ttgagctgct ggcgcagaac gacatcatgg acgaggagga ggcgcgcgcc ttaacgcatg 1560
cgtacaccac cttgcgtgat gcgctccatc acctggccct gcaggagcag ccgggacacg 1620
tggcgccaga ggccttcagc cgggagcgtc agcaggtcag cgccagctgg cagaagtggc 1680
tgatggctta actataaaat cgggtgtgct attatcgcgc gcaaagtttg cgtctcgcag 1740
gagagagtca tgaaagtaac gctgccggag tttgaacgtg caggagtgtt ggtggtgggt 1800
gatgtgatgc tggaccgcta ctggtacggc cccaccagtc gtatttcccc ggaagccccg 1860
gtgccggtgg tgaaggtgga aaatatcgaa gaacgtcctg gcggcgcggc aaacgtagcg 1920
atgaacatcg cctccctggg ggcaacgtcg cgcctggtgg gattgaccgg gattgatgac 1980
gctgcccgcg cgctgagcca ggcgctggcc aatgtgaatg tgaagtgcga cttcgtctcc 2040
gtcccgactc acccgaccat caccaagctg cgggtgctgt cgcgcaatca gcagctgatc 2100
cgcctcgact ttgaagaggg cttctccggc gtggatccgc agccgatgca tgagcgcatt 2160
cagcaggcgc tgggagccat tggcgcactg g 2191
<210> 193
<211> 613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC
<400> 193
atgaccctga atatgatgct agaagcgtca ggtaccggtc atgattcacc gtgcgattct 60
cggttccctg gagcgcttca ttggcatcct gaccgaagag ttcgctggct tcttcccaac 120
ctggattgca ccagtgcagg tagtggtcat gaatattacc gattctcagg ctgaatacgt 180
taacgaattg acgcgtaaac tacaaaatgc gggcattcgt gtaaaagcag acttgagaaa 240
tgagaagatt ggctttaaaa tccgcgagca cactttacgt cgtgtcccgt atatgttggt 300
ctgtggcgac aaagaagtcg aagccggcaa agtggccgtg cgcacccgtc gcgggaaaga 360
cctcggcagc atggacgtaa gtgaagtgat tgagaagctg caacaagaga ttcgcagccg 420
cagtcttcaa caactggagg aataaggtat taaaggcgga aaacgagttc aaacggcacg 480
tccgaatcgt atcaatggcg agattcgcgc cctggaagtt cgcgccattg agctggcttc 540
ccgaccgcag ggcggcacct gcctgaccct gcgtttcccg ctgtttaaca ccctgaccgg 600
aggtgaagca tga 613
<210> 194
<211> 1613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC с фланкирующей областью длиной 500 п.н.
<400> 194
ggccgtcgcc cagcgtcggc gtccccaaca gcagggccgg gtaggccagc aggtccgcca 60
gcgtggcgcg gttaatattg accggggcgg cggcggcctc ccccagctgc ttgtggatca 120
ttttcgcgat cttgcgggtt ttaccggtat cggtaccaaa gaaaatgcca atgttcgcca 180
tagtacgctc ctgtcggaat ggtgttgaaa aaaggaatga cgacagaggt attgcgaagg 240
ctgtgccagg ttgccctgca ccgcgacggc ccatccctgc cccatcagga tcgcttcgca 300
tcacgatgcc gcgcgccaaa ggcgcacccg gcggggcgaa aggtaaaaat ccgtgaattt 360
tccccctgtc ggatcaatgt ttcgcgtggt cgttccgata agggcgcaca ctttgcatgg 420
ttatccgggt tcggcttacc ccgccgcgtt ttgcgcacgg tgtcggacaa tttgtcataa 480
ctgcgacaca ggagtttgcg atgaccctga atatgatgct agaagcgtca ggtaccggtc 540
atgattcacc gtgcgattct cggttccctg gagcgcttca ttggcatcct gaccgaagag 600
ttcgctggct tcttcccaac ctggattgca ccagtgcagg tagtggtcat gaatattacc 660
gattctcagg ctgaatacgt taacgaattg acgcgtaaac tacaaaatgc gggcattcgt 720
gtaaaagcag acttgagaaa tgagaagatt ggctttaaaa tccgcgagca cactttacgt 780
cgtgtcccgt atatgttggt ctgtggcgac aaagaagtcg aagccggcaa agtggccgtg 840
cgcacccgtc gcgggaaaga cctcggcagc atggacgtaa gtgaagtgat tgagaagctg 900
caacaagaga ttcgcagccg cagtcttcaa caactggagg aataaggtat taaaggcgga 960
aaacgagttc aaacggcacg tccgaatcgt atcaatggcg agattcgcgc cctggaagtt 1020
cgcgccattg agctggcttc ccgaccgcag ggcggcacct gcctgaccct gcgtttcccg 1080
ctgtttaaca ccctgaccgg aggtgaagca tgatccctga atccgacccg gacaccaccg 1140
tcagacgctt cgacctctct cagcagttca ccgccatgca gcggataagc gtggtgctga 1200
gccgggccac cgaggccagc aaaacgctgc aggaggtgct cagcgtatta cacaacgatg 1260
cctttatgca gcacgggatg atctgcctgt acgacagcga gcaggagatc ctcagtatcg 1320
aagcgctgca gcaaaccggc cagcagcccc tccccggcag cacgcagatc cgctatcgcc 1380
ccggcgaggg actggtgggg accgtgctgg cccaggggca gtcgctggtg ctgccccggg 1440
tcgccgacga tcagcgtttt ctcgaccgcc tgagcctcta cgattacgat ctgccgttta 1500
tcgccgtacc gttgatgggg cccaacgccc ggccaatagg ggtgctggcg gcccagccga 1560
tggcgcgcca ggaagagcgg ctgccggcct gcacccgttt tctcgaaacc gtc 1613
<210> 195
<211> 1155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> делеция 36 п.н. glnE-delta-AR-2
<400> 195
atggcgctga agcacctgat cacgctctgc gcggcgtcgc cgatggtcgc cagccagctg 60
gcgcgccacc cgctgctgct ggatgagctg ctggatccca acaccctcta tcagccgacg 120
gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc tgcgcgtgcc ggaagaggat 180
gaagagcagc agctgcatat cgcggcggcg gatatcgctg gtaccctgcc ggtgatgaag 240
gtcagcgatc acttaacctg gcttgccgaa gcgatcctcg acgcggtggt gcagcaggca 300
tgggggcaga tggtcgctcg ctacggccag ccgacccacc tgcacgatcg ccagggtcgc 360
ggcttcgccg tcgtcggcta cggtaagctt ggcggctggg agctgggcta cagctccgat 420
ctcgatctgg tgttcctcca tgactgcccg gcggaggtga tgaccgacgg cgagcgggag 480
attgacggcc gtcagttcta cctgcggctg gcccagcgga tcatgcacct gttcagcacc 540
cgcacctcgt ccggtattct ctacgaagtg gacgcccggc tgcgtccttc tggcgcggcg 600
gggatgctgg tcaccaccgc cgacgcgttt gctgactatc agcagaacga agcctggacg 660
tgggaacatc aggcgctggt gcgcgcccgc gtggtctatg gcgacccggc gctgcaggcg 720
cgctttgacg ccattcgtcg cgatatcctg accaccccgc gggaggggat gaccctgcag 780
accgaggttc gcgagatgcg cgagaagatg cgcgcccacc ttggcaacaa acatcccgat 840
cgttttgata tcaaagccga tgccggcggg atcaccgata ttgaatttat tactcagtat 900
ctggtcctac gctatgccag tgacaagccg aagctgaccc gctggtctga caacgtgcgt 960
attcttgagc tgctggcgca gaacgacatc atggacgagg aggaggcgcg cgccttaacg 1020
catgcgtaca ccaccttgcg tgatgcgctc catcacctgg ccctgcagga gcagccggga 1080
cacgtggcgc cagaggcctt cagccgggag cgtcagcagg tcagcgccag ctggcagaag 1140
tggctgatgg cttaa 1155
<210> 196
<211> 2155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> делеция 36 п.н. glnE-delta-AR-2
<400> 196
cgtaaggcga ccacccagct ccgcgcgttg ctgaacgacg ctgaagccgt tctgctggcc 60
gcggacaccg ccgacgaggc gttattccgc accgaggtcg tcggcgccaa actggccctg 120
actgaatggc tggtccagcg cggctggcgt ccgttcctca acgaggcagg agagaaaaaa 180
atagccggat cgttcaaacg gtttgccgat attaacctct cgcgggtggc ggccgagctg 240
cgcagcgccg tgcagcatct ggcggttgaa gatgccgccg accagttgcc gaagctgtcc 300
cgcgacatcg acagcgtcca gctgctggcg ggcgcctatg gcgacgccgt cgcgccgtgg 360
ctggagaact ggcaggagct tcaccgtgca atagcacatg acgatcgcag cgtctttgaa 420
tatttccgtc gccaggcgct ggctgccgag ccgttctggc tgcatagtgg aaaacgataa 480
tttcaggcca gggagccctt atggcgctga agcacctgat cacgctctgc gcggcgtcgc 540
cgatggtcgc cagccagctg gcgcgccacc cgctgctgct ggatgagctg ctggatccca 600
acaccctcta tcagccgacg gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc 660
tgcgcgtgcc ggaagaggat gaagagcagc agctgcatat cgcggcggcg gatatcgctg 720
gtaccctgcc ggtgatgaag gtcagcgatc acttaacctg gcttgccgaa gcgatcctcg 780
acgcggtggt gcagcaggca tgggggcaga tggtcgctcg ctacggccag ccgacccacc 840
tgcacgatcg ccagggtcgc ggcttcgccg tcgtcggcta cggtaagctt ggcggctggg 900
agctgggcta cagctccgat ctcgatctgg tgttcctcca tgactgcccg gcggaggtga 960
tgaccgacgg cgagcgggag attgacggcc gtcagttcta cctgcggctg gcccagcgga 1020
tcatgcacct gttcagcacc cgcacctcgt ccggtattct ctacgaagtg gacgcccggc 1080
tgcgtccttc tggcgcggcg gggatgctgg tcaccaccgc cgacgcgttt gctgactatc 1140
agcagaacga agcctggacg tgggaacatc aggcgctggt gcgcgcccgc gtggtctatg 1200
gcgacccggc gctgcaggcg cgctttgacg ccattcgtcg cgatatcctg accaccccgc 1260
gggaggggat gaccctgcag accgaggttc gcgagatgcg cgagaagatg cgcgcccacc 1320
ttggcaacaa acatcccgat cgttttgata tcaaagccga tgccggcggg atcaccgata 1380
ttgaatttat tactcagtat ctggtcctac gctatgccag tgacaagccg aagctgaccc 1440
gctggtctga caacgtgcgt attcttgagc tgctggcgca gaacgacatc atggacgagg 1500
aggaggcgcg cgccttaacg catgcgtaca ccaccttgcg tgatgcgctc catcacctgg 1560
ccctgcagga gcagccggga cacgtggcgc cagaggcctt cagccgggag cgtcagcagg 1620
tcagcgccag ctggcagaag tggctgatgg cttaactata aaatcgggtg tgctattatc 1680
gcgcgcaaag tttgcgtctc gcaggagaga gtcatgaaag taacgctgcc ggagtttgaa 1740
cgtgcaggag tgttggtggt gggtgatgtg atgctggacc gctactggta cggccccacc 1800
agtcgtattt ccccggaagc cccggtgccg gtggtgaagg tggaaaatat cgaagaacgt 1860
cctggcggcg cggcaaacgt agcgatgaac atcgcctccc tgggggcaac gtcgcgcctg 1920
gtgggattga ccgggattga tgacgctgcc cgcgcgctga gccaggcgct ggccaatgtg 1980
aatgtgaagt gcgacttcgt ctccgtcccg actcacccga ccatcaccaa gctgcgggtg 2040
ctgtcgcgca atcagcagct gatccgcctc gactttgaag agggcttctc cggcgtggat 2100
ccgcagccga tgcatgagcg cattcagcag gcgctgggag ccattggcgc actgg 2155
<210> 197
<211> 412
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm8.2
<400> 197
atgaccctga atatgatgct cgacgccgtc ctcgcagtac cattgcaacc gactttacag 60
caagaagtga ttctggcacg catggaacaa attcttgcca gtcgggcttt atccgatgac 120
gaacgcgcac agcttttata tgagcgcgga gtgttgtatg atagtctcgg tctgagggca 180
ttagcgcgaa atgatttttc acaagcgctg gcaatccgac ccgatatgcc tgaagtattc 240
aattacttag gcatttactt aacgcaggca ggcaattttg atgctgccta tgaagcgttt 300
gattctgtac ttgagcttga tcgccattga gctggcttcc cgaccgcagg gcggcacctg 360
cctgaccctg cgtttcccgc tgtttaacac cctgaccgga ggtgaagcat ga 412
<210> 198
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm8.2 с фланкирующей областью длиной 500 п.н.
<400> 198
cccaacagca gggccgggta ggccagcagg tccgccagcg tggcgcggtt aatattgacc 60
ggggcggcgg cggcctcccc cagctgcttg tggatcattt tcgcgatctt gcgggtttta 120
ccggtatcgg taccaaagaa aatgccaatg ttcgccatag tacgctcctg tcggaatggt 180
gttgaaaaaa ggaatgacga cagaggtatt gcgaaggctg tgccaggttg ccctgcaccg 240
cgacggccca tccctgcccc atcaggatcg cttcgcatca cgatgccgcg cgccaaaggc 300
gcacccggcg gggcgaaagg taaaaatccg tgaattttcc ccctgtcgga tcaatgtttc 360
gcgtggtcgt tccgataagg gcgcacactt tgcatggtta tccgggttcg gcttaccccg 420
ccgcgttttg cgcacggtgt cggacaattt gtcataactg cgacacagga gtttgcgatg 480
accctgaata tgatgctcga cgccgtcctc gcagtaccat tgcaaccgac tttacagcaa 540
gaagtgattc tggcacgcat ggaacaaatt cttgccagtc gggctttatc cgatgacgaa 600
cgcgcacagc ttttatatga gcgcggagtg ttgtatgata gtctcggtct gagggcatta 660
gcgcgaaatg atttttcaca agcgctggca atccgacccg atatgcctga agtattcaat 720
tacttaggca tttacttaac gcaggcaggc aattttgatg ctgcctatga agcgtttgat 780
tctgtacttg agcttgatcg ccattgagct ggcttcccga ccgcagggcg gcacctgcct 840
gaccctgcgt ttcccgctgt ttaacaccct gaccggaggt gaagcatgat ccctgaatcc 900
gacccggaca ccaccgtcag acgcttcgac ctctctcagc agttcaccgc catgcagcgg 960
ataagcgtgg tgctgagccg ggccaccgag gccagcaaaa cgctgcagga ggtgctcagc 1020
gtattacaca acgatgcctt tatgcagcac gggatgatct gcctgtacga cagcgagcag 1080
gagatcctca gtatcgaagc gctgcagcaa accggccagc agcccctccc cggcagcacg 1140
cagatccgct atcgccccgg cgagggactg gtggggaccg tgctggccca ggggcagtcg 1200
ctggtgctgc cccgggtcgc cgacgatcag cgttttctcg accgcctgag cctctacgat 1260
tacgatctgc cgtttatcgc cgtaccgttg atggggccca acgcccggcc aataggggtg 1320
ctggcggccc agccgatggc gcgccaggaa gagcggctgc cggcctgcac ccgttttctc 1380
gaaaccgtc 1389
<210> 199
<211> 1155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> делеция 36 п.н. glnE-delta-AR-2
<400> 199
atggcgctga agcacctgat cacgctctgc gcggcgtcgc cgatggtcgc cagccagctg 60
gcgcgccacc cgctgctgct ggatgagctg ctggatccca acaccctcta tcagccgacg 120
gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc tgcgcgtgcc ggaagaggat 180
gaagagcagc agctgcatat cgcggcggcg gatatcgctg gtaccctgcc ggtgatgaag 240
gtcagcgatc acttaacctg gcttgccgaa gcgatcctcg acgcggtggt gcagcaggca 300
tgggggcaga tggtcgctcg ctacggccag ccgacccacc tgcacgatcg ccagggtcgc 360
ggcttcgccg tcgtcggcta cggtaagctt ggcggctggg agctgggcta cagctccgat 420
ctcgatctgg tgttcctcca tgactgcccg gcggaggtga tgaccgacgg cgagcgggag 480
attgacggcc gtcagttcta cctgcggctg gcccagcgga tcatgcacct gttcagcacc 540
cgcacctcgt ccggtattct ctacgaagtg gacgcccggc tgcgtccttc tggcgcggcg 600
gggatgctgg tcaccaccgc cgacgcgttt gctgactatc agcagaacga agcctggacg 660
tgggaacatc aggcgctggt gcgcgcccgc gtggtctatg gcgacccggc gctgcaggcg 720
cgctttgacg ccattcgtcg cgatatcctg accaccccgc gggaggggat gaccctgcag 780
accgaggttc gcgagatgcg cgagaagatg cgcgcccacc ttggcaacaa acatcccgat 840
cgttttgata tcaaagccga tgccggcggg atcaccgata ttgaatttat tactcagtat 900
ctggtcctac gctatgccag tgacaagccg aagctgaccc gctggtctga caacgtgcgt 960
attcttgagc tgctggcgca gaacgacatc atggacgagg aggaggcgcg cgccttaacg 1020
catgcgtaca ccaccttgcg tgatgcgctc catcacctgg ccctgcagga gcagccggga 1080
cacgtggcgc cagaggcctt cagccgggag cgtcagcagg tcagcgccag ctggcagaag 1140
tggctgatgg cttaa 1155
<210> 200
<211> 2155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> делеция 36 п.н. glnE-delta-AR-2
<400> 200
cgtaaggcga ccacccagct ccgcgcgttg ctgaacgacg ctgaagccgt tctgctggcc 60
gcggacaccg ccgacgaggc gttattccgc accgaggtcg tcggcgccaa actggccctg 120
actgaatggc tggtccagcg cggctggcgt ccgttcctca acgaggcagg agagaaaaaa 180
atagccggat cgttcaaacg gtttgccgat attaacctct cgcgggtggc ggccgagctg 240
cgcagcgccg tgcagcatct ggcggttgaa gatgccgccg accagttgcc gaagctgtcc 300
cgcgacatcg acagcgtcca gctgctggcg ggcgcctatg gcgacgccgt cgcgccgtgg 360
ctggagaact ggcaggagct tcaccgtgca atagcacatg acgatcgcag cgtctttgaa 420
tatttccgtc gccaggcgct ggctgccgag ccgttctggc tgcatagtgg aaaacgataa 480
tttcaggcca gggagccctt atggcgctga agcacctgat cacgctctgc gcggcgtcgc 540
cgatggtcgc cagccagctg gcgcgccacc cgctgctgct ggatgagctg ctggatccca 600
acaccctcta tcagccgacg gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc 660
tgcgcgtgcc ggaagaggat gaagagcagc agctgcatat cgcggcggcg gatatcgctg 720
gtaccctgcc ggtgatgaag gtcagcgatc acttaacctg gcttgccgaa gcgatcctcg 780
acgcggtggt gcagcaggca tgggggcaga tggtcgctcg ctacggccag ccgacccacc 840
tgcacgatcg ccagggtcgc ggcttcgccg tcgtcggcta cggtaagctt ggcggctggg 900
agctgggcta cagctccgat ctcgatctgg tgttcctcca tgactgcccg gcggaggtga 960
tgaccgacgg cgagcgggag attgacggcc gtcagttcta cctgcggctg gcccagcgga 1020
tcatgcacct gttcagcacc cgcacctcgt ccggtattct ctacgaagtg gacgcccggc 1080
tgcgtccttc tggcgcggcg gggatgctgg tcaccaccgc cgacgcgttt gctgactatc 1140
agcagaacga agcctggacg tgggaacatc aggcgctggt gcgcgcccgc gtggtctatg 1200
gcgacccggc gctgcaggcg cgctttgacg ccattcgtcg cgatatcctg accaccccgc 1260
gggaggggat gaccctgcag accgaggttc gcgagatgcg cgagaagatg cgcgcccacc 1320
ttggcaacaa acatcccgat cgttttgata tcaaagccga tgccggcggg atcaccgata 1380
ttgaatttat tactcagtat ctggtcctac gctatgccag tgacaagccg aagctgaccc 1440
gctggtctga caacgtgcgt attcttgagc tgctggcgca gaacgacatc atggacgagg 1500
aggaggcgcg cgccttaacg catgcgtaca ccaccttgcg tgatgcgctc catcacctgg 1560
ccctgcagga gcagccggga cacgtggcgc cagaggcctt cagccgggag cgtcagcagg 1620
tcagcgccag ctggcagaag tggctgatgg cttaactata aaatcgggtg tgctattatc 1680
gcgcgcaaag tttgcgtctc gcaggagaga gtcatgaaag taacgctgcc ggagtttgaa 1740
cgtgcaggag tgttggtggt gggtgatgtg atgctggacc gctactggta cggccccacc 1800
agtcgtattt ccccggaagc cccggtgccg gtggtgaagg tggaaaatat cgaagaacgt 1860
cctggcggcg cggcaaacgt agcgatgaac atcgcctccc tgggggcaac gtcgcgcctg 1920
gtgggattga ccgggattga tgacgctgcc cgcgcgctga gccaggcgct ggccaatgtg 1980
aatgtgaagt gcgacttcgt ctccgtcccg actcacccga ccatcaccaa gctgcgggtg 2040
ctgtcgcgca atcagcagct gatccgcctc gactttgaag agggcttctc cggcgtggat 2100
ccgcagccga tgcatgagcg cattcagcag gcgctgggag ccattggcgc actgg 2155
<210> 201
<211> 413
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm6.2
<400> 201
atgaccctga atatgatgct cgagctaaag ttctcggcta atcgctgata acatttgacg 60
caatgcgcaa taaaagggca tcatttgatg ccctttttgc acgctttcat accagaacct 120
ggctcatcag tgattttttt tgtcataatc attgctgaga caggctctga agagggcgtt 180
tatacaccaa accattcgag cggtagcgcg acggcaagtc agcgttctcc tttgcaatag 240
cagggaagag gcgccagaac cgccagcgtt gaagcagttt gaacgcgttc agtgtataat 300
ccgaaactta atttcggttt ggagccattg agctggcttc ccgaccgcag ggcggcacct 360
gcctgaccct gcgtttcccg ctgtttaaca ccctgaccgg aggtgaagca tga 413
<210> 202
<211> 1413
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm6.2 с фланкирующей областью длиной 500 п.н.
<400> 202
ggccgtcgcc cagcgtcggc gtccccaaca gcagggccgg gtaggccagc aggtccgcca 60
gcgtggcgcg gttaatattg accggggcgg cggcggcctc ccccagctgc ttgtggatca 120
ttttcgcgat cttgcgggtt ttaccggtat cggtaccaaa gaaaatgcca atgttcgcca 180
tagtacgctc ctgtcggaat ggtgttgaaa aaaggaatga cgacagaggt attgcgaagg 240
ctgtgccagg ttgccctgca ccgcgacggc ccatccctgc cccatcagga tcgcttcgca 300
tcacgatgcc gcgcgccaaa ggcgcacccg gcggggcgaa aggtaaaaat ccgtgaattt 360
tccccctgtc ggatcaatgt ttcgcgtggt cgttccgata agggcgcaca ctttgcatgg 420
ttatccgggt tcggcttacc ccgccgcgtt ttgcgcacgg tgtcggacaa tttgtcataa 480
ctgcgacaca ggagtttgcg atgaccctga atatgatgct cgagctaaag ttctcggcta 540
atcgctgata acatttgacg caatgcgcaa taaaagggca tcatttgatg ccctttttgc 600
acgctttcat accagaacct ggctcatcag tgattttttt tgtcataatc attgctgaga 660
caggctctga agagggcgtt tatacaccaa accattcgag cggtagcgcg acggcaagtc 720
agcgttctcc tttgcaatag cagggaagag gcgccagaac cgccagcgtt gaagcagttt 780
gaacgcgttc agtgtataat ccgaaactta atttcggttt ggagccattg agctggcttc 840
ccgaccgcag ggcggcacct gcctgaccct gcgtttcccg ctgtttaaca ccctgaccgg 900
aggtgaagca tgatccctga atccgacccg gacaccaccg tcagacgctt cgacctctct 960
cagcagttca ccgccatgca gcggataagc gtggtgctga gccgggccac cgaggccagc 1020
aaaacgctgc aggaggtgct cagcgtatta cacaacgatg cctttatgca gcacgggatg 1080
atctgcctgt acgacagcga gcaggagatc ctcagtatcg aagcgctgca gcaaaccggc 1140
cagcagcccc tccccggcag cacgcagatc cgctatcgcc ccggcgaggg actggtgggg 1200
accgtgctgg cccaggggca gtcgctggtg ctgccccggg tcgccgacga tcagcgtttt 1260
ctcgaccgcc tgagcctcta cgattacgat ctgccgttta tcgccgtacc gttgatgggg 1320
cccaacgccc ggccaatagg ggtgctggcg gcccagccga tggcgcgcca ggaagagcgg 1380
ctgccggcct gcacccgttt tctcgaaacc gtc 1413
<210> 203
<211> 513
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1.2
<400> 203
atgaccctga atatgatgct cgagcccgct gaccgaccag aacttccacc ttggactcgg 60
ctataccctt ggcgtgacgg cgcgcgataa ctgggactac atccccattc cggtgatctt 120
accattggcg tcaataggtt acggtccggc gactttccag atgacctata ttcccggcac 180
ctacaataac ggtaacgttt acttcgcctg ggctcgtata cagttttaat tcgctaagtc 240
ttagcaataa atgagataag cggtgtgtct tgtggaaaaa caaggactaa agcgttaccc 300
actaaaaaag atagcgactt ttatcacttt ttagcaaagt tgcactggac aaaaggtacc 360
acaattggtg tactgatact cgacacagca ttagtgtcga tttttcatat aaaggtaatt 420
ttggccattg agctggcttc ccgaccgcag ggcggcacct gcctgaccct gcgtttcccg 480
ctgtttaaca ccctgaccgg aggtgaagca tga 513
<210> 204
<211> 1513
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1.2 с фланкирующей областью длиной 500 п.н.
<400> 204
ggccgtcgcc cagcgtcggc gtccccaaca gcagggccgg gtaggccagc aggtccgcca 60
gcgtggcgcg gttaatattg accggggcgg cggcggcctc ccccagctgc ttgtggatca 120
ttttcgcgat cttgcgggtt ttaccggtat cggtaccaaa gaaaatgcca atgttcgcca 180
tagtacgctc ctgtcggaat ggtgttgaaa aaaggaatga cgacagaggt attgcgaagg 240
ctgtgccagg ttgccctgca ccgcgacggc ccatccctgc cccatcagga tcgcttcgca 300
tcacgatgcc gcgcgccaaa ggcgcacccg gcggggcgaa aggtaaaaat ccgtgaattt 360
tccccctgtc ggatcaatgt ttcgcgtggt cgttccgata agggcgcaca ctttgcatgg 420
ttatccgggt tcggcttacc ccgccgcgtt ttgcgcacgg tgtcggacaa tttgtcataa 480
ctgcgacaca ggagtttgcg atgaccctga atatgatgct cgagcccgct gaccgaccag 540
aacttccacc ttggactcgg ctataccctt ggcgtgacgg cgcgcgataa ctgggactac 600
atccccattc cggtgatctt accattggcg tcaataggtt acggtccggc gactttccag 660
atgacctata ttcccggcac ctacaataac ggtaacgttt acttcgcctg ggctcgtata 720
cagttttaat tcgctaagtc ttagcaataa atgagataag cggtgtgtct tgtggaaaaa 780
caaggactaa agcgttaccc actaaaaaag atagcgactt ttatcacttt ttagcaaagt 840
tgcactggac aaaaggtacc acaattggtg tactgatact cgacacagca ttagtgtcga 900
tttttcatat aaaggtaatt ttggccattg agctggcttc ccgaccgcag ggcggcacct 960
gcctgaccct gcgtttcccg ctgtttaaca ccctgaccgg aggtgaagca tgatccctga 1020
atccgacccg gacaccaccg tcagacgctt cgacctctct cagcagttca ccgccatgca 1080
gcggataagc gtggtgctga gccgggccac cgaggccagc aaaacgctgc aggaggtgct 1140
cagcgtatta cacaacgatg cctttatgca gcacgggatg atctgcctgt acgacagcga 1200
gcaggagatc ctcagtatcg aagcgctgca gcaaaccggc cagcagcccc tccccggcag 1260
cacgcagatc cgctatcgcc ccggcgaggg actggtgggg accgtgctgg cccaggggca 1320
gtcgctggtg ctgccccggg tcgccgacga tcagcgtttt ctcgaccgcc tgagcctcta 1380
cgattacgat ctgccgttta tcgccgtacc gttgatgggg cccaacgccc ggccaatagg 1440
ggtgctggcg gcccagccga tggcgcgcca ggaagagcgg ctgccggcct gcacccgttt 1500
tctcgaaacc gtc 1513
<210> 205
<211> 1155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> делеция 36 п.н. glnE-delta-AR-2
<400> 205
atggcgctga agcacctgat cacgctctgc gcggcgtcgc cgatggtcgc cagccagctg 60
gcgcgccacc cgctgctgct ggatgagctg ctggatccca acaccctcta tcagccgacg 120
gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc tgcgcgtgcc ggaagaggat 180
gaagagcagc agctgcatat cgcggcggcg gatatcgctg gtaccctgcc ggtgatgaag 240
gtcagcgatc acttaacctg gcttgccgaa gcgatcctcg acgcggtggt gcagcaggca 300
tgggggcaga tggtcgctcg ctacggccag ccgacccacc tgcacgatcg ccagggtcgc 360
ggcttcgccg tcgtcggcta cggtaagctt ggcggctggg agctgggcta cagctccgat 420
ctcgatctgg tgttcctcca tgactgcccg gcggaggtga tgaccgacgg cgagcgggag 480
attgacggcc gtcagttcta cctgcggctg gcccagcgga tcatgcacct gttcagcacc 540
cgcacctcgt ccggtattct ctacgaagtg gacgcccggc tgcgtccttc tggcgcggcg 600
gggatgctgg tcaccaccgc cgacgcgttt gctgactatc agcagaacga agcctggacg 660
tgggaacatc aggcgctggt gcgcgcccgc gtggtctatg gcgacccggc gctgcaggcg 720
cgctttgacg ccattcgtcg cgatatcctg accaccccgc gggaggggat gaccctgcag 780
accgaggttc gcgagatgcg cgagaagatg cgcgcccacc ttggcaacaa acatcccgat 840
cgttttgata tcaaagccga tgccggcggg atcaccgata ttgaatttat tactcagtat 900
ctggtcctac gctatgccag tgacaagccg aagctgaccc gctggtctga caacgtgcgt 960
attcttgagc tgctggcgca gaacgacatc atggacgagg aggaggcgcg cgccttaacg 1020
catgcgtaca ccaccttgcg tgatgcgctc catcacctgg ccctgcagga gcagccggga 1080
cacgtggcgc cagaggcctt cagccgggag cgtcagcagg tcagcgccag ctggcagaag 1140
tggctgatgg cttaa 1155
<210> 206
<211> 2155
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> делеция 36 п.н. glnE-delta-AR-2
<400> 206
cgtaaggcga ccacccagct ccgcgcgttg ctgaacgacg ctgaagccgt tctgctggcc 60
gcggacaccg ccgacgaggc gttattccgc accgaggtcg tcggcgccaa actggccctg 120
actgaatggc tggtccagcg cggctggcgt ccgttcctca acgaggcagg agagaaaaaa 180
atagccggat cgttcaaacg gtttgccgat attaacctct cgcgggtggc ggccgagctg 240
cgcagcgccg tgcagcatct ggcggttgaa gatgccgccg accagttgcc gaagctgtcc 300
cgcgacatcg acagcgtcca gctgctggcg ggcgcctatg gcgacgccgt cgcgccgtgg 360
ctggagaact ggcaggagct tcaccgtgca atagcacatg acgatcgcag cgtctttgaa 420
tatttccgtc gccaggcgct ggctgccgag ccgttctggc tgcatagtgg aaaacgataa 480
tttcaggcca gggagccctt atggcgctga agcacctgat cacgctctgc gcggcgtcgc 540
cgatggtcgc cagccagctg gcgcgccacc cgctgctgct ggatgagctg ctggatccca 600
acaccctcta tcagccgacg gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc 660
tgcgcgtgcc ggaagaggat gaagagcagc agctgcatat cgcggcggcg gatatcgctg 720
gtaccctgcc ggtgatgaag gtcagcgatc acttaacctg gcttgccgaa gcgatcctcg 780
acgcggtggt gcagcaggca tgggggcaga tggtcgctcg ctacggccag ccgacccacc 840
tgcacgatcg ccagggtcgc ggcttcgccg tcgtcggcta cggtaagctt ggcggctggg 900
agctgggcta cagctccgat ctcgatctgg tgttcctcca tgactgcccg gcggaggtga 960
tgaccgacgg cgagcgggag attgacggcc gtcagttcta cctgcggctg gcccagcgga 1020
tcatgcacct gttcagcacc cgcacctcgt ccggtattct ctacgaagtg gacgcccggc 1080
tgcgtccttc tggcgcggcg gggatgctgg tcaccaccgc cgacgcgttt gctgactatc 1140
agcagaacga agcctggacg tgggaacatc aggcgctggt gcgcgcccgc gtggtctatg 1200
gcgacccggc gctgcaggcg cgctttgacg ccattcgtcg cgatatcctg accaccccgc 1260
gggaggggat gaccctgcag accgaggttc gcgagatgcg cgagaagatg cgcgcccacc 1320
ttggcaacaa acatcccgat cgttttgata tcaaagccga tgccggcggg atcaccgata 1380
ttgaatttat tactcagtat ctggtcctac gctatgccag tgacaagccg aagctgaccc 1440
gctggtctga caacgtgcgt attcttgagc tgctggcgca gaacgacatc atggacgagg 1500
aggaggcgcg cgccttaacg catgcgtaca ccaccttgcg tgatgcgctc catcacctgg 1560
ccctgcagga gcagccggga cacgtggcgc cagaggcctt cagccgggag cgtcagcagg 1620
tcagcgccag ctggcagaag tggctgatgg cttaactata aaatcgggtg tgctattatc 1680
gcgcgcaaag tttgcgtctc gcaggagaga gtcatgaaag taacgctgcc ggagtttgaa 1740
cgtgcaggag tgttggtggt gggtgatgtg atgctggacc gctactggta cggccccacc 1800
agtcgtattt ccccggaagc cccggtgccg gtggtgaagg tggaaaatat cgaagaacgt 1860
cctggcggcg cggcaaacgt agcgatgaac atcgcctccc tgggggcaac gtcgcgcctg 1920
gtgggattga ccgggattga tgacgctgcc cgcgcgctga gccaggcgct ggccaatgtg 1980
aatgtgaagt gcgacttcgt ctccgtcccg actcacccga ccatcaccaa gctgcgggtg 2040
ctgtcgcgca atcagcagct gatccgcctc gactttgaag agggcttctc cggcgtggat 2100
ccgcagccga tgcatgagcg cattcagcag gcgctgggag ccattggcgc actgg 2155
<210> 207
<211> 613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC
<400> 207
atgaccctga atatgatgct agaagcgtca ggtaccggtc atgattcacc gtgcgattct 60
cggttccctg gagcgcttca ttggcatcct gaccgaagag ttcgctggct tcttcccaac 120
ctggattgca ccagtgcagg tagtggtcat gaatattacc gattctcagg ctgaatacgt 180
taacgaattg acgcgtaaac tacaaaatgc gggcattcgt gtaaaagcag acttgagaaa 240
tgagaagatt ggctttaaaa tccgcgagca cactttacgt cgtgtcccgt atatgttggt 300
ctgtggcgac aaagaagtcg aagccggcaa agtggccgtg cgcacccgtc gcgggaaaga 360
cctcggcagc atggacgtaa gtgaagtgat tgagaagctg caacaagaga ttcgcagccg 420
cagtcttcaa caactggagg aataaggtat taaaggcgga aaacgagttc aaacggcacg 480
tccgaatcgt atcaatggcg agattcgcgc cctggaagtt cgcgccattg agctggcttc 540
ccgaccgcag ggcggcacct gcctgaccct gcgtttcccg ctgtttaaca ccctgaccgg 600
aggtgaagca tga 613
<210> 208
<211> 1613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC с фланкирующей областью длиной 500 п.н.
<400> 208
ggccgtcgcc cagcgtcggc gtccccaaca gcagggccgg gtaggccagc aggtccgcca 60
gcgtggcgcg gttaatattg accggggcgg cggcggcctc ccccagctgc ttgtggatca 120
ttttcgcgat cttgcgggtt ttaccggtat cggtaccaaa gaaaatgcca atgttcgcca 180
tagtacgctc ctgtcggaat ggtgttgaaa aaaggaatga cgacagaggt attgcgaagg 240
ctgtgccagg ttgccctgca ccgcgacggc ccatccctgc cccatcagga tcgcttcgca 300
tcacgatgcc gcgcgccaaa ggcgcacccg gcggggcgaa aggtaaaaat ccgtgaattt 360
tccccctgtc ggatcaatgt ttcgcgtggt cgttccgata agggcgcaca ctttgcatgg 420
ttatccgggt tcggcttacc ccgccgcgtt ttgcgcacgg tgtcggacaa tttgtcataa 480
ctgcgacaca ggagtttgcg atgaccctga atatgatgct agaagcgtca ggtaccggtc 540
atgattcacc gtgcgattct cggttccctg gagcgcttca ttggcatcct gaccgaagag 600
ttcgctggct tcttcccaac ctggattgca ccagtgcagg tagtggtcat gaatattacc 660
gattctcagg ctgaatacgt taacgaattg acgcgtaaac tacaaaatgc gggcattcgt 720
gtaaaagcag acttgagaaa tgagaagatt ggctttaaaa tccgcgagca cactttacgt 780
cgtgtcccgt atatgttggt ctgtggcgac aaagaagtcg aagccggcaa agtggccgtg 840
cgcacccgtc gcgggaaaga cctcggcagc atggacgtaa gtgaagtgat tgagaagctg 900
caacaagaga ttcgcagccg cagtcttcaa caactggagg aataaggtat taaaggcgga 960
aaacgagttc aaacggcacg tccgaatcgt atcaatggcg agattcgcgc cctggaagtt 1020
cgcgccattg agctggcttc ccgaccgcag ggcggcacct gcctgaccct gcgtttcccg 1080
ctgtttaaca ccctgaccgg aggtgaagca tgatccctga atccgacccg gacaccaccg 1140
tcagacgctt cgacctctct cagcagttca ccgccatgca gcggataagc gtggtgctga 1200
gccgggccac cgaggccagc aaaacgctgc aggaggtgct cagcgtatta cacaacgatg 1260
cctttatgca gcacgggatg atctgcctgt acgacagcga gcaggagatc ctcagtatcg 1320
aagcgctgca gcaaaccggc cagcagcccc tccccggcag cacgcagatc cgctatcgcc 1380
ccggcgaggg actggtgggg accgtgctgg cccaggggca gtcgctggtg ctgccccggg 1440
tcgccgacga tcagcgtttt ctcgaccgcc tgagcctcta cgattacgat ctgccgttta 1500
tcgccgtacc gttgatgggg cccaacgccc ggccaatagg ggtgctggcg gcccagccga 1560
tggcgcgcca ggaagagcgg ctgccggcct gcacccgttt tctcgaaacc gtc 1613
<210> 209
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 209
atggcgctga agcacctgat cacgctctgc gcggcgtcgc cgatggtcgc cagccagctg 60
gcgcgccacc cgctgctgct ggatgagctg ctggatccca acaccctcta tcagccgacg 120
gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc tgcgcgtgcc ggaagaggat 180
gaagagcagc agctggaggc gttgcgccag tttaagcagg cgcagcagct gcatatcgcg 240
gcggcggata tcgctggtac cctgccggtg atgaaggtca gcgatcactt aacctggctt 300
gccgaagcga tcctcgacgc ggtggtgcag caggcatggg ggcagatggt cgctcgctac 360
ggccagccga cccacctgca cgatcgccag ggtcgcggct tcgccgtcgt cggctacggt 420
aagcttggcg gctgggagct gggctacagc tccgatctcg atctggtgtt cctccatgac 480
tgcccggcgg aggtgatgac cgacggcgag cgggagattg acggccgtca gttctacctg 540
cggctggccc agcggatcat gcacctgttc agcacccgca cctcgtccgg tattctctac 600
gaagtggacg cccggctgcg tccttctggc gcggcgggga tgctggtcac caccgccgac 660
gcgtttgctg actatcagca gaacgaagcc tggacgtggg aacatcaggc gctggtgcgc 720
gcccgcgtgg tctatggcga cccggcgctg caggcgcgct ttgacgccat tcgtcgcgat 780
atcctgacca ccccgcggga ggggatgacc ctgcagaccg aggttcgcga gatgcgcgag 840
aagatgcgcg cccaccttgg caacaaacat cccgatcgtt ttgatatcaa agccgatgcc 900
ggcgggatca ccgatattga atttattact cagtatctgg tcctacgcta tgccagtgac 960
aagccgaagc tgacccgctg gtctgacaac gtgcgtattc ttgagctgct ggcgcagaac 1020
gacatcatgg acgaggagga ggcgcgcgcc ttaacgcatg cgtacaccac cttgcgtgat 1080
gcgctccatc acctggccct gcaggagcag ccgggacacg tggcgccaga ggccttcagc 1140
cgggagcgtc agcaggtcag cgccagctgg cagaagtggc tgatggctta a 1191
<210> 210
<211> 2191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 210
cgtaaggcga ccacccagct ccgcgcgttg ctgaacgacg ctgaagccgt tctgctggcc 60
gcggacaccg ccgacgaggc gttattccgc accgaggtcg tcggcgccaa actggccctg 120
actgaatggc tggtccagcg cggctggcgt ccgttcctca acgaggcagg agagaaaaaa 180
atagccggat cgttcaaacg gtttgccgat attaacctct cgcgggtggc ggccgagctg 240
cgcagcgccg tgcagcatct ggcggttgaa gatgccgccg accagttgcc gaagctgtcc 300
cgcgacatcg acagcgtcca gctgctggcg ggcgcctatg gcgacgccgt cgcgccgtgg 360
ctggagaact ggcaggagct tcaccgtgca atagcacatg acgatcgcag cgtctttgaa 420
tatttccgtc gccaggcgct ggctgccgag ccgttctggc tgcatagtgg aaaacgataa 480
tttcaggcca gggagccctt atggcgctga agcacctgat cacgctctgc gcggcgtcgc 540
cgatggtcgc cagccagctg gcgcgccacc cgctgctgct ggatgagctg ctggatccca 600
acaccctcta tcagccgacg gcgaccgatg cctatcgcga cgagctgcgc cagtacctgc 660
tgcgcgtgcc ggaagaggat gaagagcagc agctggaggc gttgcgccag tttaagcagg 720
cgcagcagct gcatatcgcg gcggcggata tcgctggtac cctgccggtg atgaaggtca 780
gcgatcactt aacctggctt gccgaagcga tcctcgacgc ggtggtgcag caggcatggg 840
ggcagatggt cgctcgctac ggccagccga cccacctgca cgatcgccag ggtcgcggct 900
tcgccgtcgt cggctacggt aagcttggcg gctgggagct gggctacagc tccgatctcg 960
atctggtgtt cctccatgac tgcccggcgg aggtgatgac cgacggcgag cgggagattg 1020
acggccgtca gttctacctg cggctggccc agcggatcat gcacctgttc agcacccgca 1080
cctcgtccgg tattctctac gaagtggacg cccggctgcg tccttctggc gcggcgggga 1140
tgctggtcac caccgccgac gcgtttgctg actatcagca gaacgaagcc tggacgtggg 1200
aacatcaggc gctggtgcgc gcccgcgtgg tctatggcga cccggcgctg caggcgcgct 1260
ttgacgccat tcgtcgcgat atcctgacca ccccgcggga ggggatgacc ctgcagaccg 1320
aggttcgcga gatgcgcgag aagatgcgcg cccaccttgg caacaaacat cccgatcgtt 1380
ttgatatcaa agccgatgcc ggcgggatca ccgatattga atttattact cagtatctgg 1440
tcctacgcta tgccagtgac aagccgaagc tgacccgctg gtctgacaac gtgcgtattc 1500
ttgagctgct ggcgcagaac gacatcatgg acgaggagga ggcgcgcgcc ttaacgcatg 1560
cgtacaccac cttgcgtgat gcgctccatc acctggccct gcaggagcag ccgggacacg 1620
tggcgccaga ggccttcagc cgggagcgtc agcaggtcag cgccagctgg cagaagtggc 1680
tgatggctta actataaaat cgggtgtgct attatcgcgc gcaaagtttg cgtctcgcag 1740
gagagagtca tgaaagtaac gctgccggag tttgaacgtg caggagtgtt ggtggtgggt 1800
gatgtgatgc tggaccgcta ctggtacggc cccaccagtc gtatttcccc ggaagccccg 1860
gtgccggtgg tgaaggtgga aaatatcgaa gaacgtcctg gcggcgcggc aaacgtagcg 1920
atgaacatcg cctccctggg ggcaacgtcg cgcctggtgg gattgaccgg gattgatgac 1980
gctgcccgcg cgctgagcca ggcgctggcc aatgtgaatg tgaagtgcga cttcgtctcc 2040
gtcccgactc acccgaccat caccaagctg cgggtgctgt cgcgcaatca gcagctgatc 2100
cgcctcgact ttgaagaggg cttctccggc gtggatccgc agccgatgca tgagcgcatt 2160
cagcaggcgc tgggagccat tggcgcactg g 2191
<210> 211
<211> 1188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 211
atggcgctca aacagttaat ccgtctgtgt gccgcctcgc cgatggtcgc gacacaactt 60
gcacgtcatc ctttattgct cgatgaactg ctcgacccgc gcacgcttta ccagccgatt 120
gagccgggcg cttaccgcga cgaactgcgt cagtatctga tgcgggtgcc aacagaagac 180
gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg cccagcattt gcgtatcgca 240
gccggggata tttccggggc attgccggtg atgaaagtca gtgaccattt aacctacctt 300
gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg aacaaatggt cgtaaaatac 360
gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt ttgccgtggt cggttacggg 420
aaactcggtg gctgggagct gggttatagc tcagatctgg atctggtctt cctgctcgat 480
tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg acggacgtca gttttatctt 540
cggctggcgc agcgcattat gcacttattc agcacccgga catcgtcagg cattctttac 600
gaggttgatc cgcgtctgcg accttccggc gcatccggca tgctggtcag taccattgaa 660
gcgtttgcag attatcaggc caatgaagcc tggacgtggg agcatcaggc gctggttcgc 720
gcgcgcgtgg tttacgggga tccgcaactg acacagcaat ttaacgccac gcgtcgcgac 780
attctttgcc gccagcgcga tggcgacggc ctgcgtaagg aggtccgtga aatgcgcgag 840
aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt ttgatctgaa agccgatccg 900
ggtggcatca cggatattga attcattgca caatacctgg ttctgcgttt cgcgcatgat 960
gagccgaagc tgacgcgctg gtctgataac gtgcggattt ttgaactgat ggcacgatat 1020
gacatcatgc cggaagagga agcgcgccat ctgacgcagg cttatgtgac gctgcgcgat 1080
gaaattcatc atctggcgtt gcaggaacac agcgggaaag tggccgcgga cagctttgct 1140
actgagcgcg cgcagatccg tgccagctgg gcaaagtggc tcggctga 1188
<210> 212
<211> 2188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 212
cggtactgga acagaaatcg gcggatgcgc aggaaatttg ttatgacacg gcctgtctga 60
agtgcaagtt agtgcttact tcctggctgg caacctcagg ctggacgccg tttattgatg 120
ataaatctgc gaagaaactg gacgcttcct tcaaacgttt tgctgacatc atgctcggtc 180
gtaccgcagc ggatctgaaa gaagcctttg cgcagccact gacggaagaa ggttatcgcg 240
atcagctggc gcgcctgaaa cgccagatca ttaccttcca tttgcttgcc ggtgcttacc 300
ctgaaaaaga cgtcgatgcg tatattgccg gctgggtgga cctgcaacag gccatcgttc 360
agcagcaaca cgcctgggag gattcggccc gttctcacgc ggtgatgatg gatgctttct 420
ggttaaacgg gcaacctcgt taactgactg actagcctgg gcaaactgcc cgggcttttt 480
tttgcaagga atctgatttc atggcgctca aacagttaat ccgtctgtgt gccgcctcgc 540
cgatggtcgc gacacaactt gcacgtcatc ctttattgct cgatgaactg ctcgacccgc 600
gcacgcttta ccagccgatt gagccgggcg cttaccgcga cgaactgcgt cagtatctga 660
tgcgggtgcc aacagaagac gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg 720
cccagcattt gcgtatcgca gccggggata tttccggggc attgccggtg atgaaagtca 780
gtgaccattt aacctacctt gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg 840
aacaaatggt cgtaaaatac gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt 900
ttgccgtggt cggttacggg aaactcggtg gctgggagct gggttatagc tcagatctgg 960
atctggtctt cctgctcgat tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg 1020
acggacgtca gttttatctt cggctggcgc agcgcattat gcacttattc agcacccgga 1080
catcgtcagg cattctttac gaggttgatc cgcgtctgcg accttccggc gcatccggca 1140
tgctggtcag taccattgaa gcgtttgcag attatcaggc caatgaagcc tggacgtggg 1200
agcatcaggc gctggttcgc gcgcgcgtgg tttacgggga tccgcaactg acacagcaat 1260
ttaacgccac gcgtcgcgac attctttgcc gccagcgcga tggcgacggc ctgcgtaagg 1320
aggtccgtga aatgcgcgag aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt 1380
ttgatctgaa agccgatccg ggtggcatca cggatattga attcattgca caatacctgg 1440
ttctgcgttt cgcgcatgat gagccgaagc tgacgcgctg gtctgataac gtgcggattt 1500
ttgaactgat ggcacgatat gacatcatgc cggaagagga agcgcgccat ctgacgcagg 1560
cttatgtgac gctgcgcgat gaaattcatc atctggcgtt gcaggaacac agcgggaaag 1620
tggccgcgga cagctttgct actgagcgcg cgcagatccg tgccagctgg gcaaagtggc 1680
tcggctgagg gtttttattc ggctaacagg cgcttgtgat attatccggc gcattgtatt 1740
tacccgattt gatttatctg ttttggagtc ttgggatgaa agtgactttg cctgattttc 1800
accgcgcagg tgtgctggtt gtcggtgacg taatgttaga ccgttactgg tatggcccga 1860
ccaatcgtat ttctccggaa gctccggtgc cggtggtgaa ggtcagtacc attgaagagc 1920
ggcctggcgg tgcagctaac gtggcgatga acatttcatc tctgggcgcc tcttcctgtc 1980
tgatcggcct gaccggcgta gacgacgctg cgcgtgccct cagtgagcgt ctggcagaag 2040
tgaaagttaa ctgcgatttc gtcgcactat ccacacatcc taccatcacc aaactgcgaa 2100
ttttgtcccg taaccagcaa ctgatccgcc tcgactttga ggaaggtttt gaaggcgttg 2160
atctcgagcc gatgctgacc aaaataga 2188
<210> 213
<211> 635
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm6.1
<400> 213
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg aatttttttt cacaaagcgt 240
agcgttattg aatcgcacat tttaaactgt tggccgctgt ggaagcgaat attggtgaaa 300
ggtgcggttt taaggccttt ttctttgact ctctgtcgtt acaaagttaa tatgcgcgcc 360
ctccgtctct gaagctctcg gtgaacattg ttgcgaggca ggatgcgagc tggttgtgtt 420
ttgacattac cgataatgtg ccgcgtgaac gggtgcgtta tgcccgcccg gaagcggcgt 480
tttcccgtcc ggggaatggc atggagctgc gccttatcca gacgctgatc gcccatcatc 540
gcggttcttt agatctctcg gtccgccctg atggcggcac cttgctgacg ttacgcctgc 600
cggtacagca ggttatcacc ggaggcttaa aatga 635
<210> 214
<211> 1635
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm6.1 с фланкирующей областью длиной 500 п.н.
<400> 214
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
aatttttttt cacaaagcgt agcgttattg aatcgcacat tttaaactgt tggccgctgt 780
ggaagcgaat attggtgaaa ggtgcggttt taaggccttt ttctttgact ctctgtcgtt 840
acaaagttaa tatgcgcgcc ctccgtctct gaagctctcg gtgaacattg ttgcgaggca 900
ggatgcgagc tggttgtgtt ttgacattac cgataatgtg ccgcgtgaac gggtgcgtta 960
tgcccgcccg gaagcggcgt tttcccgtcc ggggaatggc atggagctgc gccttatcca 1020
gacgctgatc gcccatcatc gcggttcttt agatctctcg gtccgccctg atggcggcac 1080
cttgctgacg ttacgcctgc cggtacagca ggttatcacc ggaggcttaa aatgacccag 1140
ttacctaccg cgggcccggt tatccggcgc tttgatatgt ctgcccagtt tacggcgctt 1200
tatcgcatca gcgtggcgct gagtcaggaa agcaacaccg ggcgcgcact ggcggcgatc 1260
ctcgaagtgc ttcacgatca tgcatttatg caatacggca tggtgtgtct gtttgataaa 1320
gaacgcaatg cactctttgt ggaatccctg catggcatcg acggcgaaag gaaaaaagag 1380
acccgccatg tccgttaccg catgggggaa ggcgtgatcg gcgcggtgat gagccagcgt 1440
caggcgctgg tgttaccgcg catttcagac gatcagcgtt ttctcgaccg cctgaatatt 1500
tacgattaca gcctgccgtt gattggcgtg ccgatccccg gtgcggataa tcagccatcg 1560
ggcgtgctgg tggcacagcc gatggcgttg cacgaagacc ggctgactgc cagtacgcgg 1620
tttttagaaa tggtc 1635
<210> 215
<211> 635
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm6.1
<400> 215
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg aatttttttt cacaaagcgt 240
agcgttattg aatcgcacat tttaaactgt tggccgctgt ggaagcgaat attggtgaaa 300
ggtgcggttt taaggccttt ttctttgact ctctgtcgtt acaaagttaa tatgcgcgcc 360
ctccgtctct gaagctctcg gtgaacattg ttgcgaggca ggatgcgagc tggttgtgtt 420
ttgacattac cgataatgtg ccgcgtgaac gggtgcgtta tgcccgcccg gaagcggcgt 480
tttcccgtcc ggggaatggc atggagctgc gccttatcca gacgctgatc gcccatcatc 540
gcggttcttt agatctctcg gtccgccctg atggcggcac cttgctgacg ttacgcctgc 600
cggtacagca ggttatcacc ggaggcttaa aatga 635
<210> 216
<211> 1635
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm6.1 с фланкирующей областью длиной 500 п.н.
<400> 216
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
aatttttttt cacaaagcgt agcgttattg aatcgcacat tttaaactgt tggccgctgt 780
ggaagcgaat attggtgaaa ggtgcggttt taaggccttt ttctttgact ctctgtcgtt 840
acaaagttaa tatgcgcgcc ctccgtctct gaagctctcg gtgaacattg ttgcgaggca 900
ggatgcgagc tggttgtgtt ttgacattac cgataatgtg ccgcgtgaac gggtgcgtta 960
tgcccgcccg gaagcggcgt tttcccgtcc ggggaatggc atggagctgc gccttatcca 1020
gacgctgatc gcccatcatc gcggttcttt agatctctcg gtccgccctg atggcggcac 1080
cttgctgacg ttacgcctgc cggtacagca ggttatcacc ggaggcttaa aatgacccag 1140
ttacctaccg cgggcccggt tatccggcgc tttgatatgt ctgcccagtt tacggcgctt 1200
tatcgcatca gcgtggcgct gagtcaggaa agcaacaccg ggcgcgcact ggcggcgatc 1260
ctcgaagtgc ttcacgatca tgcatttatg caatacggca tggtgtgtct gtttgataaa 1320
gaacgcaatg cactctttgt ggaatccctg catggcatcg acggcgaaag gaaaaaagag 1380
acccgccatg tccgttaccg catgggggaa ggcgtgatcg gcgcggtgat gagccagcgt 1440
caggcgctgg tgttaccgcg catttcagac gatcagcgtt ttctcgaccg cctgaatatt 1500
tacgattaca gcctgccgtt gattggcgtg ccgatccccg gtgcggataa tcagccatcg 1560
ggcgtgctgg tggcacagcc gatggcgttg cacgaagacc ggctgactgc cagtacgcgg 1620
tttttagaaa tggtc 1635
<210> 217
<211> 786
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm7.1
<400> 217
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg ttaaaaacgt gaccacgagc 240
attaataaac gccacgaaat gtggcgttta tttattcaaa aagtatcttc tttcataaaa 300
agtgctaaat gcagtagcag caaaattggg ataagtccca tggaatacgg ctgttttcgc 360
tgcaattttt aactttttcg taaaaaaaga tgtttctttg agcgaacgat caaaatatag 420
cgttaaccgg caaaaaatta ttctcattag aaaatagttt gtgtaatact tgtaacgcta 480
catggagatt aacttaatct agagggtttt ataccgtctc tgaagctctc ggtgaacatt 540
gttgcgaggc aggatgcgag ctggttgtgt tttgacatta ccgataatgt gccgcgtgaa 600
cgggtgcgtt atgcccgccc ggaagcggcg ttttcccgtc cggggaatgg catggagctg 660
cgccttatcc agacgctgat cgcccatcat cgcggttctt tagatctctc ggtccgccct 720
gatggcggca ccttgctgac gttacgcctg ccggtacagc aggttatcac cggaggctta 780
aaatga 786
<210> 218
<211> 1786
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm7.1 с фланкирующей областью длиной 500 п.н.
<400> 218
gtttcgtctc gaggccgggc aactgagcgg ccccgttgaa accgacctgg gctggcatct 60
gttgttgtgc gaacaaattc gcctgccgca acccttgccg aaagccgaag ccttaacgcg 120
ggtgcgtcag caactgattg cccggcaaca gaaacattat cagcgccagt ggctgcaaca 180
actgatcaac gcctgagcct gttctccttc ttgttgatgc agacgggtta atgcccgttt 240
tgcacgaaaa atgcacataa attgcctgcg ttgccttata acagcgcagg gaaatcctgc 300
ctccggcctt gtgccacacc gcgctttgcc tggtttgtgg taaaaactgg cccgctttgc 360
atcctgatgc ttaaaacacc ccgttcagat caacctttgg gcagataagc ccgcgaaagg 420
cctgcaaatt gcacggttat tccgggtgag tatatgtgtg atttgggttc cggcattgcg 480
caataaaggg gagaaagaca tgagcatcac ggcgttatca gcatcatttc ctgaggggaa 540
tatcgccagc cgcttgtcgc tgcaacatcc ttcactgttt tataccgtgg ttgaacaatc 600
ttcggtggcg atttcgctga ccgatccgca ggcgcgcatt tgttatgcca atccggcatt 660
ctgccgccag acgggttttg cacttgagac acttttgggc gagaaccacc gtctgctggt 720
taaaaacgtg accacgagca ttaataaacg ccacgaaatg tggcgtttat ttattcaaaa 780
agtatcttct ttcataaaaa gtgctaaatg cagtagcagc aaaattggga taagtcccat 840
ggaatacggc tgttttcgct gcaattttta actttttcgt aaaaaaagat gtttctttga 900
gcgaacgatc aaaatatagc gttaaccggc aaaaaattat tctcattaga aaatagtttg 960
tgtaatactt gtaacgctac atggagatta acttaatcta gagggtttta taccgtctct 1020
gaagctctcg gtgaacattg ttgcgaggca ggatgcgagc tggttgtgtt ttgacattac 1080
cgataatgtg ccgcgtgaac gggtgcgtta tgcccgcccg gaagcggcgt tttcccgtcc 1140
ggggaatggc atggagctgc gccttatcca gacgctgatc gcccatcatc gcggttcttt 1200
agatctctcg gtccgccctg atggcggcac cttgctgacg ttacgcctgc cggtacagca 1260
ggttatcacc ggaggcttaa aatgacccag ttacctaccg cgggcccggt tatccggcgc 1320
tttgatatgt ctgcccagtt tacggcgctt tatcgcatca gcgtggcgct gagtcaggaa 1380
agcaacaccg ggcgcgcact ggcggcgatc ctcgaagtgc ttcacgatca tgcatttatg 1440
caatacggca tggtgtgtct gtttgataaa gaacgcaatg cactctttgt ggaatccctg 1500
catggcatcg acggcgaaag gaaaaaagag acccgccatg tccgttaccg catgggggaa 1560
ggcgtgatcg gcgcggtgat gagccagcgt caggcgctgg tgttaccgcg catttcagac 1620
gatcagcgtt ttctcgaccg cctgaatatt tacgattaca gcctgccgtt gattggcgtg 1680
ccgatccccg gtgcggataa tcagccatcg ggcgtgctgg tggcacagcc gatggcgttg 1740
cacgaagacc ggctgactgc cagtacgcgg tttttagaaa tggtcg 1786
<210> 219
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1.2
<400> 219
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg tgaacatcac tgatgcacaa 240
gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg caggcattcg cgttaaagcc 300
gacttgagaa atgagaagat tggctttaaa attcgcgaac acacgctacg ccgtgttcct 360
tatatgttag tttgtggcga taaagaggtc gaagcaggca aagttgctgt tcgtacccgc 420
cgcggcaaag acttaggaag catggatgtt agcgaagtcg ttgacaaact gctggcggaa 480
atccgcagca gaagtcttca tcaactggag gaataaagta ttaaaggcgg aaaacgagtt 540
caaccggcgc gtcctaatcg cattaacaaa gagattcgcg cgcaagaagt tcgcctcaca 600
ggcgtcgatg gcgagcagat tggtattgtc agtctgaatg aagctcttga aaaagctgag 660
gaagcgggcg tcgatttagt agaaatcagt ccgaatgccg agccgccagt ttgtcgaatc 720
ccgtctctga agctctcggt gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt 780
gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg cccgcccgga agcggcgttt 840
tcccgtccgg ggaatggcat ggagctgcgc cttatccaga cgctgatcgc ccatcatcgc 900
ggttctttag atctctcggt ccgccctgat ggcggcacct tgctgacgtt acgcctgccg 960
gtacagcagg ttatcaccgg aggcttaaaa tga 993
<210> 220
<211> 1993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1.2 с фланкирующей областью длиной 500 п.н.
<400> 220
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
tgaacatcac tgatgcacaa gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg 780
caggcattcg cgttaaagcc gacttgagaa atgagaagat tggctttaaa attcgcgaac 840
acacgctacg ccgtgttcct tatatgttag tttgtggcga taaagaggtc gaagcaggca 900
aagttgctgt tcgtacccgc cgcggcaaag acttaggaag catggatgtt agcgaagtcg 960
ttgacaaact gctggcggaa atccgcagca gaagtcttca tcaactggag gaataaagta 1020
ttaaaggcgg aaaacgagtt caaccggcgc gtcctaatcg cattaacaaa gagattcgcg 1080
cgcaagaagt tcgcctcaca ggcgtcgatg gcgagcagat tggtattgtc agtctgaatg 1140
aagctcttga aaaagctgag gaagcgggcg tcgatttagt agaaatcagt ccgaatgccg 1200
agccgccagt ttgtcgaatc ccgtctctga agctctcggt gaacattgtt gcgaggcagg 1260
atgcgagctg gttgtgtttt gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg 1320
cccgcccgga agcggcgttt tcccgtccgg ggaatggcat ggagctgcgc cttatccaga 1380
cgctgatcgc ccatcatcgc ggttctttag atctctcggt ccgccctgat ggcggcacct 1440
tgctgacgtt acgcctgccg gtacagcagg ttatcaccgg aggcttaaaa tgacccagtt 1500
acctaccgcg ggcccggtta tccggcgctt tgatatgtct gcccagttta cggcgcttta 1560
tcgcatcagc gtggcgctga gtcaggaaag caacaccggg cgcgcactgg cggcgatcct 1620
cgaagtgctt cacgatcatg catttatgca atacggcatg gtgtgtctgt ttgataaaga 1680
acgcaatgca ctctttgtgg aatccctgca tggcatcgac ggcgaaagga aaaaagagac 1740
ccgccatgtc cgttaccgca tgggggaagg cgtgatcggc gcggtgatga gccagcgtca 1800
ggcgctggtg ttaccgcgca tttcagacga tcagcgtttt ctcgaccgcc tgaatattta 1860
cgattacagc ctgccgttga ttggcgtgcc gatccccggt gcggataatc agccatcggg 1920
cgtgctggtg gcacagccga tggcgttgca cgaagaccgg ctgactgcca gtacgcggtt 1980
tttagaaatg gtc 1993
<210> 221
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1.2
<400> 221
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg tgaacatcac tgatgcacaa 240
gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg caggcattcg cgttaaagcc 300
gacttgagaa atgagaagat tggctttaaa attcgcgaac acacgctacg ccgtgttcct 360
tatatgttag tttgtggcga taaagaggtc gaagcaggca aagttgctgt tcgtacccgc 420
cgcggcaaag acttaggaag catggatgtt agcgaagtcg ttgacaaact gctggcggaa 480
atccgcagca gaagtcttca tcaactggag gaataaagta ttaaaggcgg aaaacgagtt 540
caaccggcgc gtcctaatcg cattaacaaa gagattcgcg cgcaagaagt tcgcctcaca 600
ggcgtcgatg gcgagcagat tggtattgtc agtctgaatg aagctcttga aaaagctgag 660
gaagcgggcg tcgatttagt agaaatcagt ccgaatgccg agccgccagt ttgtcgaatc 720
ccgtctctga agctctcggt gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt 780
gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg cccgcccgga agcggcgttt 840
tcccgtccgg ggaatggcat ggagctgcgc cttatccaga cgctgatcgc ccatcatcgc 900
ggttctttag atctctcggt ccgccctgat ggcggcacct tgctgacgtt acgcctgccg 960
gtacagcagg ttatcaccgg aggcttaaaa tga 993
<210> 222
<211> 1993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1.2 с фланкирующей областью длиной 500 п.н.
<400> 222
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
tgaacatcac tgatgcacaa gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg 780
caggcattcg cgttaaagcc gacttgagaa atgagaagat tggctttaaa attcgcgaac 840
acacgctacg ccgtgttcct tatatgttag tttgtggcga taaagaggtc gaagcaggca 900
aagttgctgt tcgtacccgc cgcggcaaag acttaggaag catggatgtt agcgaagtcg 960
ttgacaaact gctggcggaa atccgcagca gaagtcttca tcaactggag gaataaagta 1020
ttaaaggcgg aaaacgagtt caaccggcgc gtcctaatcg cattaacaaa gagattcgcg 1080
cgcaagaagt tcgcctcaca ggcgtcgatg gcgagcagat tggtattgtc agtctgaatg 1140
aagctcttga aaaagctgag gaagcgggcg tcgatttagt agaaatcagt ccgaatgccg 1200
agccgccagt ttgtcgaatc ccgtctctga agctctcggt gaacattgtt gcgaggcagg 1260
atgcgagctg gttgtgtttt gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg 1320
cccgcccgga agcggcgttt tcccgtccgg ggaatggcat ggagctgcgc cttatccaga 1380
cgctgatcgc ccatcatcgc ggttctttag atctctcggt ccgccctgat ggcggcacct 1440
tgctgacgtt acgcctgccg gtacagcagg ttatcaccgg aggcttaaaa tgacccagtt 1500
acctaccgcg ggcccggtta tccggcgctt tgatatgtct gcccagttta cggcgcttta 1560
tcgcatcagc gtggcgctga gtcaggaaag caacaccggg cgcgcactgg cggcgatcct 1620
cgaagtgctt cacgatcatg catttatgca atacggcatg gtgtgtctgt ttgataaaga 1680
acgcaatgca ctctttgtgg aatccctgca tggcatcgac ggcgaaagga aaaaagagac 1740
ccgccatgtc cgttaccgca tgggggaagg cgtgatcggc gcggtgatga gccagcgtca 1800
ggcgctggtg ttaccgcgca tttcagacga tcagcgtttt ctcgaccgcc tgaatattta 1860
cgattacagc ctgccgttga ttggcgtgcc gatccccggt gcggataatc agccatcggg 1920
cgtgctggtg gcacagccga tggcgttgca cgaagaccgg ctgactgcca gtacgcggtt 1980
tttagaaatg gtc 1993
<210> 223
<211> 1188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 223
atggcgctca aacagttaat ccgtctgtgt gccgcctcgc cgatggtcgc gacacaactt 60
gcacgtcatc ctttattgct cgatgaactg ctcgacccgc gcacgcttta ccagccgatt 120
gagccgggcg cttaccgcga cgaactgcgt cagtatctga tgcgggtgcc aacagaagac 180
gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg cccagcattt gcgtatcgca 240
gccggggata tttccggggc attgccggtg atgaaagtca gtgaccattt aacctacctt 300
gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg aacaaatggt cgtaaaatac 360
gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt ttgccgtggt cggttacggg 420
aaactcggtg gctgggagct gggttatagc tcagatctgg atctggtctt cctgctcgat 480
tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg acggacgtca gttttatctt 540
cggctggcgc agcgcattat gcacttattc agcacccgga catcgtcagg cattctttac 600
gaggttgatc cgcgtctgcg accttccggc gcatccggca tgctggtcag taccattgaa 660
gcgtttgcag attatcaggc caatgaagcc tggacgtggg agcatcaggc gctggttcgc 720
gcgcgcgtgg tttacgggga tccgcaactg acacagcaat ttaacgccac gcgtcgcgac 780
attctttgcc gccagcgcga tggcgacggc ctgcgtaagg aggtccgtga aatgcgcgag 840
aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt ttgatctgaa agccgatccg 900
ggtggcatca cggatattga attcattgca caatacctgg ttctgcgttt cgcgcatgat 960
gagccgaagc tgacgcgctg gtctgataac gtgcggattt ttgaactgat ggcacgatat 1020
gacatcatgc cggaagagga agcgcgccat ctgacgcagg cttatgtgac gctgcgcgat 1080
gaaattcatc atctggcgtt gcaggaacac agcgggaaag tggccgcgga cagctttgct 1140
actgagcgcg cgcagatccg tgccagctgg gcaaagtggc tcggctga 1188
<210> 224
<211> 2188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 224
cggtactgga acagaaatcg gcggatgcgc aggaaatttg ttatgacacg gcctgtctga 60
agtgcaagtt agtgcttact tcctggctgg caacctcagg ctggacgccg tttattgatg 120
ataaatctgc gaagaaactg gacgcttcct tcaaacgttt tgctgacatc atgctcggtc 180
gtaccgcagc ggatctgaaa gaagcctttg cgcagccact gacggaagaa ggttatcgcg 240
atcagctggc gcgcctgaaa cgccagatca ttaccttcca tttgcttgcc ggtgcttacc 300
ctgaaaaaga cgtcgatgcg tatattgccg gctgggtgga cctgcaacag gccatcgttc 360
agcagcaaca cgcctgggag gattcggccc gttctcacgc ggtgatgatg gatgctttct 420
ggttaaacgg gcaacctcgt taactgactg actagcctgg gcaaactgcc cgggcttttt 480
tttgcaagga atctgatttc atggcgctca aacagttaat ccgtctgtgt gccgcctcgc 540
cgatggtcgc gacacaactt gcacgtcatc ctttattgct cgatgaactg ctcgacccgc 600
gcacgcttta ccagccgatt gagccgggcg cttaccgcga cgaactgcgt cagtatctga 660
tgcgggtgcc aacagaagac gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg 720
cccagcattt gcgtatcgca gccggggata tttccggggc attgccggtg atgaaagtca 780
gtgaccattt aacctacctt gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg 840
aacaaatggt cgtaaaatac gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt 900
ttgccgtggt cggttacggg aaactcggtg gctgggagct gggttatagc tcagatctgg 960
atctggtctt cctgctcgat tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg 1020
acggacgtca gttttatctt cggctggcgc agcgcattat gcacttattc agcacccgga 1080
catcgtcagg cattctttac gaggttgatc cgcgtctgcg accttccggc gcatccggca 1140
tgctggtcag taccattgaa gcgtttgcag attatcaggc caatgaagcc tggacgtggg 1200
agcatcaggc gctggttcgc gcgcgcgtgg tttacgggga tccgcaactg acacagcaat 1260
ttaacgccac gcgtcgcgac attctttgcc gccagcgcga tggcgacggc ctgcgtaagg 1320
aggtccgtga aatgcgcgag aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt 1380
ttgatctgaa agccgatccg ggtggcatca cggatattga attcattgca caatacctgg 1440
ttctgcgttt cgcgcatgat gagccgaagc tgacgcgctg gtctgataac gtgcggattt 1500
ttgaactgat ggcacgatat gacatcatgc cggaagagga agcgcgccat ctgacgcagg 1560
cttatgtgac gctgcgcgat gaaattcatc atctggcgtt gcaggaacac agcgggaaag 1620
tggccgcgga cagctttgct actgagcgcg cgcagatccg tgccagctgg gcaaagtggc 1680
tcggctgagg gtttttattc ggctaacagg cgcttgtgat attatccggc gcattgtatt 1740
tacccgattt gatttatctg ttttggagtc ttgggatgaa agtgactttg cctgattttc 1800
accgcgcagg tgtgctggtt gtcggtgacg taatgttaga ccgttactgg tatggcccga 1860
ccaatcgtat ttctccggaa gctccggtgc cggtggtgaa ggtcagtacc attgaagagc 1920
ggcctggcgg tgcagctaac gtggcgatga acatttcatc tctgggcgcc tcttcctgtc 1980
tgatcggcct gaccggcgta gacgacgctg cgcgtgccct cagtgagcgt ctggcagaag 2040
tgaaagttaa ctgcgatttc gtcgcactat ccacacatcc taccatcacc aaactgcgaa 2100
ttttgtcccg taaccagcaa ctgatccgcc tcgactttga ggaaggtttt gaaggcgttg 2160
atctcgagcc gatgctgacc aaaataga 2188
<210> 225
<211> 663
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm3.1
<400> 225
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg tacagtagcg cctctcaaaa 240
atagataaac ggctcatgta cgtgggccgt ttattttttc tacccataat cgggaaccgg 300
tgttataatg ccgcgccctc atattgtggg gatttcttaa tgacctatcc tgggtcctaa 360
agttgtagtt gacattagcg gagcactaac ccgtctctga agctctcggt gaacattgtt 420
gcgaggcagg atgcgagctg gttgtgtttt gacattaccg ataatgtgcc gcgtgaacgg 480
gtgcgttatg cccgcccgga agcggcgttt tcccgtccgg ggaatggcat ggagctgcgc 540
cttatccaga cgctgatcgc ccatcatcgc ggttctttag atctctcggt ccgccctgat 600
ggcggcacct tgctgacgtt acgcctgccg gtacagcagg ttatcaccgg aggcttaaaa 660
tga 663
<210> 226
<211> 1663
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm3.1 с фланкирующей областью длиной 500 п.н.
<400> 226
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
tacagtagcg cctctcaaaa atagataaac ggctcatgta cgtgggccgt ttattttttc 780
tacccataat cgggaaccgg tgttataatg ccgcgccctc atattgtggg gatttcttaa 840
tgacctatcc tgggtcctaa agttgtagtt gacattagcg gagcactaac ccgtctctga 900
agctctcggt gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt gacattaccg 960
ataatgtgcc gcgtgaacgg gtgcgttatg cccgcccgga agcggcgttt tcccgtccgg 1020
ggaatggcat ggagctgcgc cttatccaga cgctgatcgc ccatcatcgc ggttctttag 1080
atctctcggt ccgccctgat ggcggcacct tgctgacgtt acgcctgccg gtacagcagg 1140
ttatcaccgg aggcttaaaa tgacccagtt acctaccgcg ggcccggtta tccggcgctt 1200
tgatatgtct gcccagttta cggcgcttta tcgcatcagc gtggcgctga gtcaggaaag 1260
caacaccggg cgcgcactgg cggcgatcct cgaagtgctt cacgatcatg catttatgca 1320
atacggcatg gtgtgtctgt ttgataaaga acgcaatgca ctctttgtgg aatccctgca 1380
tggcatcgac ggcgaaagga aaaaagagac ccgccatgtc cgttaccgca tgggggaagg 1440
cgtgatcggc gcggtgatga gccagcgtca ggcgctggtg ttaccgcgca tttcagacga 1500
tcagcgtttt ctcgaccgcc tgaatattta cgattacagc ctgccgttga ttggcgtgcc 1560
gatccccggt gcggataatc agccatcggg cgtgctggtg gcacagccga tggcgttgca 1620
cgaagaccgg ctgactgcca gtacgcggtt tttagaaatg gtc 1663
<210> 227
<211> 1188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 227
atggcgctca aacagttaat ccgtctgtgt gccgcctcgc cgatggtcgc gacacaactt 60
gcacgtcatc ctttattgct cgatgaactg ctcgacccgc gcacgcttta ccagccgatt 120
gagccgggcg cttaccgcga cgaactgcgt cagtatctga tgcgggtgcc aacagaagac 180
gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg cccagcattt gcgtatcgca 240
gccggggata tttccggggc attgccggtg atgaaagtca gtgaccattt aacctacctt 300
gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg aacaaatggt cgtaaaatac 360
gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt ttgccgtggt cggttacggg 420
aaactcggtg gctgggagct gggttatagc tcagatctgg atctggtctt cctgctcgat 480
tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg acggacgtca gttttatctt 540
cggctggcgc agcgcattat gcacttattc agcacccgga catcgtcagg cattctttac 600
gaggttgatc cgcgtctgcg accttccggc gcatccggca tgctggtcag taccattgaa 660
gcgtttgcag attatcaggc caatgaagcc tggacgtggg agcatcaggc gctggttcgc 720
gcgcgcgtgg tttacgggga tccgcaactg acacagcaat ttaacgccac gcgtcgcgac 780
attctttgcc gccagcgcga tggcgacggc ctgcgtaagg aggtccgtga aatgcgcgag 840
aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt ttgatctgaa agccgatccg 900
ggtggcatca cggatattga attcattgca caatacctgg ttctgcgttt cgcgcatgat 960
gagccgaagc tgacgcgctg gtctgataac gtgcggattt ttgaactgat ggcacgatat 1020
gacatcatgc cggaagagga agcgcgccat ctgacgcagg cttatgtgac gctgcgcgat 1080
gaaattcatc atctggcgtt gcaggaacac agcgggaaag tggccgcgga cagctttgct 1140
actgagcgcg cgcagatccg tgccagctgg gcaaagtggc tcggctga 1188
<210> 228
<211> 2188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 228
cggtactgga acagaaatcg gcggatgcgc aggaaatttg ttatgacacg gcctgtctga 60
agtgcaagtt agtgcttact tcctggctgg caacctcagg ctggacgccg tttattgatg 120
ataaatctgc gaagaaactg gacgcttcct tcaaacgttt tgctgacatc atgctcggtc 180
gtaccgcagc ggatctgaaa gaagcctttg cgcagccact gacggaagaa ggttatcgcg 240
atcagctggc gcgcctgaaa cgccagatca ttaccttcca tttgcttgcc ggtgcttacc 300
ctgaaaaaga cgtcgatgcg tatattgccg gctgggtgga cctgcaacag gccatcgttc 360
agcagcaaca cgcctgggag gattcggccc gttctcacgc ggtgatgatg gatgctttct 420
ggttaaacgg gcaacctcgt taactgactg actagcctgg gcaaactgcc cgggcttttt 480
tttgcaagga atctgatttc atggcgctca aacagttaat ccgtctgtgt gccgcctcgc 540
cgatggtcgc gacacaactt gcacgtcatc ctttattgct cgatgaactg ctcgacccgc 600
gcacgcttta ccagccgatt gagccgggcg cttaccgcga cgaactgcgt cagtatctga 660
tgcgggtgcc aacagaagac gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg 720
cccagcattt gcgtatcgca gccggggata tttccggggc attgccggtg atgaaagtca 780
gtgaccattt aacctacctt gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg 840
aacaaatggt cgtaaaatac gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt 900
ttgccgtggt cggttacggg aaactcggtg gctgggagct gggttatagc tcagatctgg 960
atctggtctt cctgctcgat tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg 1020
acggacgtca gttttatctt cggctggcgc agcgcattat gcacttattc agcacccgga 1080
catcgtcagg cattctttac gaggttgatc cgcgtctgcg accttccggc gcatccggca 1140
tgctggtcag taccattgaa gcgtttgcag attatcaggc caatgaagcc tggacgtggg 1200
agcatcaggc gctggttcgc gcgcgcgtgg tttacgggga tccgcaactg acacagcaat 1260
ttaacgccac gcgtcgcgac attctttgcc gccagcgcga tggcgacggc ctgcgtaagg 1320
aggtccgtga aatgcgcgag aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt 1380
ttgatctgaa agccgatccg ggtggcatca cggatattga attcattgca caatacctgg 1440
ttctgcgttt cgcgcatgat gagccgaagc tgacgcgctg gtctgataac gtgcggattt 1500
ttgaactgat ggcacgatat gacatcatgc cggaagagga agcgcgccat ctgacgcagg 1560
cttatgtgac gctgcgcgat gaaattcatc atctggcgtt gcaggaacac agcgggaaag 1620
tggccgcgga cagctttgct actgagcgcg cgcagatccg tgccagctgg gcaaagtggc 1680
tcggctgagg gtttttattc ggctaacagg cgcttgtgat attatccggc gcattgtatt 1740
tacccgattt gatttatctg ttttggagtc ttgggatgaa agtgactttg cctgattttc 1800
accgcgcagg tgtgctggtt gtcggtgacg taatgttaga ccgttactgg tatggcccga 1860
ccaatcgtat ttctccggaa gctccggtgc cggtggtgaa ggtcagtacc attgaagagc 1920
ggcctggcgg tgcagctaac gtggcgatga acatttcatc tctgggcgcc tcttcctgtc 1980
tgatcggcct gaccggcgta gacgacgctg cgcgtgccct cagtgagcgt ctggcagaag 2040
tgaaagttaa ctgcgatttc gtcgcactat ccacacatcc taccatcacc aaactgcgaa 2100
ttttgtcccg taaccagcaa ctgatccgcc tcgactttga ggaaggtttt gaaggcgttg 2160
atctcgagcc gatgctgacc aaaataga 2188
<210> 229
<211> 613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC
<400> 229
atgagcatca cggcgttatc agctgaatat cactgactca caagctacct atgtcgaaga 60
attaactaaa aaactgcaag atgcaggcat tcgcgttaaa gccgacttga gaaatgagaa 120
gattggcttt aaaattcgcg aacacacgct acgccgtgtt ccttatatgt tagtttgtgg 180
cgataaagag gtcgaagcag gcaaagttgc tgttcgtact cgtcgcggca aagacttagg 240
aagcatggat gttagcgaag tcgttgacaa actgctggcg gaaatccgca gcagaagtca 300
tcatcaactg gaggaataaa gtattaaagg cggaaaacga gttcaaccgg cgcgtcctaa 360
tcgcattaac aaagagattc gcgcgcaaga agttcgcctc accggcgtcg atggcgagca 420
gattggtatt gtcagtctga atgaagctct tgaaaaagct gaggaagcgg gcgtcgattt 480
agtagaaatc agtccgaatg ccgagccgcc agtttgtcga atctctttag atctctcggt 540
ccgccctgat ggcggcacct tgctgacgtt acgcctgccg gtacagcagg ttatcaccgg 600
aggcttaaaa tga 613
<210> 230
<211> 1613
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC с фланкирующей областью длиной 500 п.н.
<400> 230
tgtttcgtct cgaagccggg caactgagca gccccgttga aaccgaactg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgag gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaatcatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttggtg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aactgccttc gctgccttat aacagcgcat ggaaatcctg 300
cctcctgcct tgtgccacgc cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg tttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agctgaatat cactgactca 540
caagctacct atgtcgaaga attaactaaa aaactgcaag atgcaggcat tcgcgttaaa 600
gccgacttga gaaatgagaa gattggcttt aaaattcgcg aacacacgct acgccgtgtt 660
ccttatatgt tagtttgtgg cgataaagag gtcgaagcag gcaaagttgc tgttcgtact 720
cgtcgcggca aagacttagg aagcatggat gttagcgaag tcgttgacaa actgctggcg 780
gaaatccgca gcagaagtca tcatcaactg gaggaataaa gtattaaagg cggaaaacga 840
gttcaaccgg cgcgtcctaa tcgcattaac aaagagattc gcgcgcaaga agttcgcctc 900
accggcgtcg atggcgagca gattggtatt gtcagtctga atgaagctct tgaaaaagct 960
gaggaagcgg gcgtcgattt agtagaaatc agtccgaatg ccgagccgcc agtttgtcga 1020
atctctttag atctctcggt ccgccctgat ggcggcacct tgctgacgtt acgcctgccg 1080
gtacagcagg ttatcaccgg aggcttaaaa tgacccagtt acctaccgcg ggcccggtta 1140
tccggcgctt tgatatgtct gcccagttta cggcgcttta tcgcatcagc gtggcgctga 1200
gtcaggaaag caataccgcg cgcgcactgg cggcgatcct cgaagtgctt cacgatcatg 1260
catttatgca atacggcatg gtgtgtctgt tcgataaaga acgcaatgca ctgtttgtgg 1320
aatccctgca tggcatcgac ggcgaaagga aaaaagaaac ccgccatgtc cgttaccgca 1380
tgggggaagg cgtgatcggc gcggtgatga gccagcgtca ggcgctggtg ttaccgcgca 1440
tttcagacga tcagcgtttt ctcgaccgcc tgaatattta cgattacagc ctgccgctga 1500
ttggtgtgcc gatccccggt gcggataatc agcctgcggg tgtgctggtg gcacagccga 1560
tggcgttgca cgaagaccgg ctggctgcca gtacgcggtt tttagaaatg gtc 1613
<210> 231
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm5
<400> 231
atgaccctga atatgatgat ggatgccggc ggacatcatc gcgacaaaca atattaatac 60
cggcaaccac accggcaatt tacgagactg cgcaggcatc ctttctcccg tcaatttctg 120
tcaaataaag taaaagaggc agtctacttg aattaccccc ggctggttga gcgtttgttg 180
aaaaaaagta actgaaaaat ccgtagaata gcgccactct gatggttaat taacctattc 240
aattaagaat tatctggatg aatgtgccat taaatgcgca gcataatggt gcgttgtgcg 300
ggaaaactgc ttttttttga aagggttggt cagtagcgga aacaactcac ttcacacccc 360
gaagggggaa gttgcctgac cctacgattc ccgctatttc attcactgac cggaggttca 420
aaatga 426
<210> 232
<211> 1426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm5 с фланкирующей областью длиной 500 п.н.
<400> 232
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc ggacatcatc 540
gcgacaaaca atattaatac cggcaaccac accggcaatt tacgagactg cgcaggcatc 600
ctttctcccg tcaatttctg tcaaataaag taaaagaggc agtctacttg aattaccccc 660
ggctggttga gcgtttgttg aaaaaaagta actgaaaaat ccgtagaata gcgccactct 720
gatggttaat taacctattc aattaagaat tatctggatg aatgtgccat taaatgcgca 780
gcataatggt gcgttgtgcg ggaaaactgc ttttttttga aagggttggt cagtagcgga 840
aacaactcac ttcacacccc gaagggggaa gttgcctgac cctacgattc ccgctatttc 900
attcactgac cggaggttca aaatgaccca gcgaaccgag tcgggtaata ccgtctggcg 960
cttcgatttg tcccagcagt tcactgcgat gcagcgcata agcgtggtac tcagccgggc 1020
gaccgaggtc gatcagacgc tccagcaagt gctgtgcgta ttgcacaatg acgccttttt 1080
gcagcacggc atgatctgtc tgtacgacag ccagcaggcg attttgaata ttgaagcgtt 1140
gcaggaagcc gatcagcagt taatccccgg cagctcgcaa atccgctatc gtccgggcga 1200
agggctggtc gggacggtgc tttcgcaggg ccaatcatta gtgctggcgc gcgttgctga 1260
cgatcagcgc tttcttgacc ggctcgggtt gtatgattac aacctgccgt ttatcgccgt 1320
gccgctgata gggccagatg cgcagacttt cggtgtgctg acggcacaac ccatggcgcg 1380
ttacgaagag cgattacccg cctgcacccg ctttctggaa acggtc 1426
<210> 233
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 233
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 234
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 234
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc cgtcctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt ttttatattc 600
ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac accccgaagg 900
gggaagttgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttgtccca gcagttcact 1020
gcgatgcagc gcataagcgt ggtactcagc cgggcgaccg aggtcgatca gacgctccag 1080
caagtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gaatattgaa gcgttgcagg aagccgatca gcagttaatc 1200
cccggcagct cgcaaatccg ctatcgtccg ggcgaagggc tggtcgggac ggtgctttcg 1260
cagggccaat cattagtgct ggcgcgcgtt gctgacgatc agcgctttct tgaccggctc 1320
gggttgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc agatgcgcag 1380
actttcggtg tgctgacggc acaacccatg gcgcgttacg aagagcgatt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 235
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 235
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 236
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 236
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc cgtcctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt ttttatattc 600
ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac accccgaagg 900
gggaagttgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttgtccca gcagttcact 1020
gcgatgcagc gcataagcgt ggtactcagc cgggcgaccg aggtcgatca gacgctccag 1080
caagtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gaatattgaa gcgttgcagg aagccgatca gcagttaatc 1200
cccggcagct cgcaaatccg ctatcgtccg ggcgaagggc tggtcgggac ggtgctttcg 1260
cagggccaat cattagtgct ggcgcgcgtt gctgacgatc agcgctttct tgaccggctc 1320
gggttgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc agatgcgcag 1380
actttcggtg tgctgacggc acaacccatg gcgcgttacg aagagcgatt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 237
<211> 1188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 237
atggcactga aacacctcat ttccctgtgt gccgcgtcgc cgatggttgc cagtcagctg 60
gcgcgctacc cgatcctgct tgatgaattg ctcgacccga atacgctcta tcaaccgacg 120
gcgatgaatg cctatcgcga tgagctgcgc caatacctgc tgcgcgtgcc ggaagatgat 180
gaagagcaac agcttgaggc gctgcggcag tttaagcagg cgcagttgct gcgcgtggcg 240
gcggcggata ttgccggtac gttgccagta atgaaagtga gcgatcactt aacctggctg 300
gcggaagcga ttattgatgc ggtggtgcag caagcctggg ggcagatggt ggcgcgttat 360
ggccagccaa cgcatctgca cgatcgcgaa gggcgcggtt ttgcggtggt cggttatggc 420
aagctgggcg gctgggagct gggttacagc tccgatctgg atctggtatt cctgcacgac 480
tgcccgatgg atgtgatgac cgatggcgag cgtgaaatcg atggtcgcca gttctatttg 540
cgtctcgcgc agcgcgtgat gcacctgttt agcacgcgca cgtcgtccgg catcctttat 600
gaagttgatg cgcgtctgcg tccatctggc gctgcgggga tgctggtcac tactacggaa 660
tcgttcgccg attaccagca aaacgaagcc tggacgtggg aacatcaggc gctggcccgt 720
gcgcgcgtgg tgtacggcga tccgcaactg accgccgaat ttgacgccat tcgccgcgat 780
attctgatga cgcctcgcga cggcgcaacg ctgcaaaccg acgtgcgaga aatgcgcgag 840
aaaatgcgtg cccatcttgg caacaagcat aaagaccgct tcgatctgaa agccgatgaa 900
ggcggtatca ccgacatcga gtttatcgcc caatatctgg tgctgcgctt tgcccatgac 960
aagccgaaac tgacgcgctg gtcggataat gtgcgcattc tcgaagggct ggcgcaaaac 1020
ggcatcatgg aggagcagga agcgcaggca ttgacgctgg cgtacaccac attgcgtgat 1080
gagctgcacc acctggcgct gcaagagttg ccgggacatg tggcgctctc ctgttttgtc 1140
gccgagcgtg cgcttattaa aaccagctgg gacaagtggc tggtggaa 1188
<210> 238
<211> 2206
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 238
gcgcaaagcg agtgctcact tacgtgatct gttgacacaa tctgaagcga ccataacttc 60
tgccgtttca gcgaatacgg cggtgtggag cgcacaatca gccctggcga agctggtgct 120
caccgagtgg ctagtgacgc agggctggcg aaccttcctt gatgaaaaag cgcaggccaa 180
attcgccgac tcctttaaac gctttgctga catccatctg tcacgcagcg ccgccgagct 240
gaaaaaagcc tttgcccaac cgctgggcga cagctatcgc gaccagttgc cgcgcctggc 300
gcgtgatatc gactgcgcgt tactgctggc cgggcattac gatcgcgcgc gcgccgtgga 360
atggctggaa aactggcagg ggcttcagca cgccattgaa acgcgccaga gagtcgaaat 420
cgaacatttc cgtaataccg cgattaccca ggagccgttc tggttgcaca gcggaaaacg 480
ttaacgaaag gatatttcgc atggcactga aacacctcat ttccctgtgt gccgcgtcgc 540
cgatggttgc cagtcagctg gcgcgctacc cgatcctgct tgatgaattg ctcgacccga 600
atacgctcta tcaaccgacg gcgatgaatg cctatcgcga tgagctgcgc caatacctgc 660
tgcgcgtgcc ggaagatgat gaagagcaac agcttgaggc gctgcggcag tttaagcagg 720
cgcagttgct gcgcgtggcg gcggcggata ttgccggtac gttgccagta atgaaagtga 780
gcgatcactt aacctggctg gcggaagcga ttattgatgc ggtggtgcag caagcctggg 840
ggcagatggt ggcgcgttat ggccagccaa cgcatctgca cgatcgcgaa gggcgcggtt 900
ttgcggtggt cggttatggc aagctgggcg gctgggagct gggttacagc tccgatctgg 960
atctggtatt cctgcacgac tgcccgatgg atgtgatgac cgatggcgag cgtgaaatcg 1020
atggtcgcca gttctatttg cgtctcgcgc agcgcgtgat gcacctgttt agcacgcgca 1080
cgtcgtccgg catcctttat gaagttgatg cgcgtctgcg tccatctggc gctgcgggga 1140
tgctggtcac tactacggaa tcgttcgccg attaccagca aaacgaagcc tggacgtggg 1200
aacatcaggc gctggcccgt gcgcgcgtgg tgtacggcga tccgcaactg accgccgaat 1260
ttgacgccat tcgccgcgat attctgatga cgcctcgcga cggcgcaacg ctgcaaaccg 1320
acgtgcgaga aatgcgcgag aaaatgcgtg cccatcttgg caacaagcat aaagaccgct 1380
tcgatctgaa agccgatgaa ggcggtatca ccgacatcga gtttatcgcc caatatctgg 1440
tgctgcgctt tgcccatgac aagccgaaac tgacgcgctg gtcggataat gtgcgcattc 1500
tcgaagggct ggcgcaaaac ggcatcatgg aggagcagga agcgcaggca ttgacgctgg 1560
cgtacaccac attgcgtgat gagctgcacc acctggcgct gcaagagttg ccgggacatg 1620
tggcgctctc ctgttttgtc gccgagcgtg cgcttattaa aaccagctgg gacaagtggc 1680
tggtggaacc gtgcgccccg gcgtaagtgt ggtatcatcg cgcgcaaatt ttgtatctct 1740
caggagacag gaatgaaagt gacgctgcca gagtttaagc aagccggtgt aatggtggtg 1800
ggtgatgtga tgctggatcg ttactggtat ggcccaacca gccgtatctc tccggaagcg 1860
ccagtcccgg ttgttaaagt cgataccatt gaagagcgtc ctggcggcgc ggcaaacgtg 1920
gcgatgaata tcgcctcact gggcgccacg gcgcgtctgg ttggcctgac tggcattgac 1980
gatgcggcgc gcgcgctgag caaagcgctg gccgatgtta acgttaaatg tgacttcgtt 2040
tctgttccga cgcatcccac catcactaag ctgcgcgtgc tgtcgcgtaa ccagcagctg 2100
attcgcctgg actttgaaga gggttttgaa ggagtcgatc cgcaaccgat gcatgaacgc 2160
atcagccagg cgcttggtaa tattggcgcg ctggtgctgt cggatt 2206
<210> 239
<211> 1563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-1
<400> 239
atgtttaacg atctgattgg cgatgatgaa acggattcgc cggaagatgc gctttctgag 60
agctggcgcg aattgtggca ggatgcgttg caggaggagg attccacgcc cgtgctggcg 120
catctctcag aggacgatcg ccgccgcgtg gtggcgctga ttgccgattt tcgcaaagag 180
ttggataaac gcaccattgg cccgcgaggg cggcaggtac tcgatcactt aatgccgcat 240
ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac cgctgtcacg cctgacgccg 300
ctgctcaccg gaattattac ccgcaccact taccttgagc tgctaagtga atttcccggc 360
gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg 420
cgctacccga tcctgcttga tgaattgctc gacccgaata cgctctatca accgacggcg 480
atgaatgcct atcgcgatga gctgcgccaa tacctgctgc gcgtgccgga agatgatgaa 540
gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc agttgctgcg cgtggcggcg 600
gcggatattg ccggtacgtt gccagtaatg aaagtgagcg atcacttaac ctggctggcg 660
gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc agatggtggc gcgttatggc 720
cagccaacgc atctgcacga tcgcgaaggg cgcggttttg cggtggtcgg ttatggcaag 780
ctgggcggct gggagctggg ttacagctcc gatctggatc tggtattcct gcacgactgc 840
ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg gtcgccagtt ctatttgcgt 900
ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt cgtccggcat cctttatgaa 960
gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc tggtcactac tacggaatcg 1020
ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac atcaggcgct ggcccgtgcg 1080
cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg acgccattcg ccgcgatatt 1140
ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg tgcgagaaat gcgcgagaaa 1200
atgcgtgccc atcttggcaa caagcataaa gaccgcttcg atctgaaagc cgatgaaggc 1260
ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc tgcgctttgc ccatgacaag 1320
ccgaaactga cgcgctggtc ggataatgtg cgcattctcg aagggctggc gcaaaacggc 1380
atcatggagg agcaggaagc gcaggcattg acgctggcgt acaccacatt gcgtgatgag 1440
ctgcaccacc tggcgctgca agagttgccg ggacatgtgg cgctctcctg ttttgtcgcc 1500
gagcgtgcgc ttattaaaac cagctgggac aagtggctgg tggaaccgtg cgccccggcg 1560
taa 1563
<210> 240
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 240
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 241
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 241
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc cgtcctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt ttttatattc 600
ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac accccgaagg 900
gggaagttgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttgtccca gcagttcact 1020
gcgatgcagc gcataagcgt ggtactcagc cgggcgaccg aggtcgatca gacgctccag 1080
caagtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gaatattgaa gcgttgcagg aagccgatca gcagttaatc 1200
cccggcagct cgcaaatccg ctatcgtccg ggcgaagggc tggtcgggac ggtgctttcg 1260
cagggccaat cattagtgct ggcgcgcgtt gctgacgatc agcgctttct tgaccggctc 1320
gggttgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc agatgcgcag 1380
actttcggtg tgctgacggc acaacccatg gcgcgttacg aagagcgatt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 242
<211> 2563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-1 с фланкирующей областью длиной 500 п.н.
<400> 242
gcgcaaagcg agtgctcact tacgtgatct gttgacacaa tctgaagcga ccataacttc 60
tgccgtttca gcgaatacgg cggtgtggag cgcacaatca gccctggcga agctggtgct 120
caccgagtgg ctagtgacgc agggctggcg aaccttcctt gatgaaaaag cgcaggccaa 180
attcgccgac tcctttaaac gctttgctga catccatctg tcacgcagcg ccgccgagct 240
gaaaaaagcc tttgcccaac cgctgggcga cagctatcgc gaccagttgc cgcgcctggc 300
gcgtgatatc gactgcgcgt tactgctggc cgggcattac gatcgcgcgc gcgccgtgga 360
atggctggaa aactggcagg ggcttcagca cgccattgaa acgcgccaga gagtcgaaat 420
cgaacatttc cgtaataccg cgattaccca ggagccgttc tggttgcaca gcggaaaacg 480
ttaacgaaag gatatttcgc atgtttaacg atctgattgg cgatgatgaa acggattcgc 540
cggaagatgc gctttctgag agctggcgcg aattgtggca ggatgcgttg caggaggagg 600
attccacgcc cgtgctggcg catctctcag aggacgatcg ccgccgcgtg gtggcgctga 660
ttgccgattt tcgcaaagag ttggataaac gcaccattgg cccgcgaggg cggcaggtac 720
tcgatcactt aatgccgcat ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac 780
cgctgtcacg cctgacgccg ctgctcaccg gaattattac ccgcaccact taccttgagc 840
tgctaagtga atttcccggc gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga 900
tggttgccag tcagctggcg cgctacccga tcctgcttga tgaattgctc gacccgaata 960
cgctctatca accgacggcg atgaatgcct atcgcgatga gctgcgccaa tacctgctgc 1020
gcgtgccgga agatgatgaa gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc 1080
agttgctgcg cgtggcggcg gcggatattg ccggtacgtt gccagtaatg aaagtgagcg 1140
atcacttaac ctggctggcg gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc 1200
agatggtggc gcgttatggc cagccaacgc atctgcacga tcgcgaaggg cgcggttttg 1260
cggtggtcgg ttatggcaag ctgggcggct gggagctggg ttacagctcc gatctggatc 1320
tggtattcct gcacgactgc ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg 1380
gtcgccagtt ctatttgcgt ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt 1440
cgtccggcat cctttatgaa gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc 1500
tggtcactac tacggaatcg ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac 1560
atcaggcgct ggcccgtgcg cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg 1620
acgccattcg ccgcgatatt ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg 1680
tgcgagaaat gcgcgagaaa atgcgtgccc atcttggcaa caagcataaa gaccgcttcg 1740
atctgaaagc cgatgaaggc ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc 1800
tgcgctttgc ccatgacaag ccgaaactga cgcgctggtc ggataatgtg cgcattctcg 1860
aagggctggc gcaaaacggc atcatggagg agcaggaagc gcaggcattg acgctggcgt 1920
acaccacatt gcgtgatgag ctgcaccacc tggcgctgca agagttgccg ggacatgtgg 1980
cgctctcctg ttttgtcgcc gagcgtgcgc ttattaaaac cagctgggac aagtggctgg 2040
tggaaccgtg cgccccggcg taagtgtggt atcatcgcgc gcaaattttg tatctctcag 2100
gagacaggaa tgaaagtgac gctgccagag tttaagcaag ccggtgtaat ggtggtgggt 2160
gatgtgatgc tggatcgtta ctggtatggc ccaaccagcc gtatctctcc ggaagcgcca 2220
gtcccggttg ttaaagtcga taccattgaa gagcgtcctg gcggcgcggc aaacgtggcg 2280
atgaatatcg cctcactggg cgccacggcg cgtctggttg gcctgactgg cattgacgat 2340
gcggcgcgcg cgctgagcaa agcgctggcc gatgttaacg ttaaatgtga cttcgtttct 2400
gttccgacgc atcccaccat cactaagctg cgcgtgctgt cgcgtaacca gcagctgatt 2460
cgcctggact ttgaagaggg ttttgaagga gtcgatccgc aaccgatgca tgaacgcatc 2520
agccaggcgc ttggtaatat tggcgcgctg gtgctgtcgg att 2563
<210> 243
<211> 1563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-1
<400> 243
atgtttaacg atctgattgg cgatgatgaa acggattcgc cggaagatgc gctttctgag 60
agctggcgcg aattgtggca ggatgcgttg caggaggagg attccacgcc cgtgctggcg 120
catctctcag aggacgatcg ccgccgcgtg gtggcgctga ttgccgattt tcgcaaagag 180
ttggataaac gcaccattgg cccgcgaggg cggcaggtac tcgatcactt aatgccgcat 240
ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac cgctgtcacg cctgacgccg 300
ctgctcaccg gaattattac ccgcaccact taccttgagc tgctaagtga atttcccggc 360
gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg 420
cgctacccga tcctgcttga tgaattgctc gacccgaata cgctctatca accgacggcg 480
atgaatgcct atcgcgatga gctgcgccaa tacctgctgc gcgtgccgga agatgatgaa 540
gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc agttgctgcg cgtggcggcg 600
gcggatattg ccggtacgtt gccagtaatg aaagtgagcg atcacttaac ctggctggcg 660
gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc agatggtggc gcgttatggc 720
cagccaacgc atctgcacga tcgcgaaggg cgcggttttg cggtggtcgg ttatggcaag 780
ctgggcggct gggagctggg ttacagctcc gatctggatc tggtattcct gcacgactgc 840
ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg gtcgccagtt ctatttgcgt 900
ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt cgtccggcat cctttatgaa 960
gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc tggtcactac tacggaatcg 1020
ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac atcaggcgct ggcccgtgcg 1080
cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg acgccattcg ccgcgatatt 1140
ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg tgcgagaaat gcgcgagaaa 1200
atgcgtgccc atcttggcaa caagcataaa gaccgcttcg atctgaaagc cgatgaaggc 1260
ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc tgcgctttgc ccatgacaag 1320
ccgaaactga cgcgctggtc ggataatgtg cgcattctcg aagggctggc gcaaaacggc 1380
atcatggagg agcaggaagc gcaggcattg acgctggcgt acaccacatt gcgtgatgag 1440
ctgcaccacc tggcgctgca agagttgccg ggacatgtgg cgctctcctg ttttgtcgcc 1500
gagcgtgcgc ttattaaaac cagctgggac aagtggctgg tggaaccgtg cgccccggcg 1560
taa 1563
<210> 244
<211> 2563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-1 с фланкирующей областью длиной 500 п.н.
<400> 244
gcgcaaagcg agtgctcact tacgtgatct gttgacacaa tctgaagcga ccataacttc 60
tgccgtttca gcgaatacgg cggtgtggag cgcacaatca gccctggcga agctggtgct 120
caccgagtgg ctagtgacgc agggctggcg aaccttcctt gatgaaaaag cgcaggccaa 180
attcgccgac tcctttaaac gctttgctga catccatctg tcacgcagcg ccgccgagct 240
gaaaaaagcc tttgcccaac cgctgggcga cagctatcgc gaccagttgc cgcgcctggc 300
gcgtgatatc gactgcgcgt tactgctggc cgggcattac gatcgcgcgc gcgccgtgga 360
atggctggaa aactggcagg ggcttcagca cgccattgaa acgcgccaga gagtcgaaat 420
cgaacatttc cgtaataccg cgattaccca ggagccgttc tggttgcaca gcggaaaacg 480
ttaacgaaag gatatttcgc atgtttaacg atctgattgg cgatgatgaa acggattcgc 540
cggaagatgc gctttctgag agctggcgcg aattgtggca ggatgcgttg caggaggagg 600
attccacgcc cgtgctggcg catctctcag aggacgatcg ccgccgcgtg gtggcgctga 660
ttgccgattt tcgcaaagag ttggataaac gcaccattgg cccgcgaggg cggcaggtac 720
tcgatcactt aatgccgcat ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac 780
cgctgtcacg cctgacgccg ctgctcaccg gaattattac ccgcaccact taccttgagc 840
tgctaagtga atttcccggc gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga 900
tggttgccag tcagctggcg cgctacccga tcctgcttga tgaattgctc gacccgaata 960
cgctctatca accgacggcg atgaatgcct atcgcgatga gctgcgccaa tacctgctgc 1020
gcgtgccgga agatgatgaa gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc 1080
agttgctgcg cgtggcggcg gcggatattg ccggtacgtt gccagtaatg aaagtgagcg 1140
atcacttaac ctggctggcg gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc 1200
agatggtggc gcgttatggc cagccaacgc atctgcacga tcgcgaaggg cgcggttttg 1260
cggtggtcgg ttatggcaag ctgggcggct gggagctggg ttacagctcc gatctggatc 1320
tggtattcct gcacgactgc ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg 1380
gtcgccagtt ctatttgcgt ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt 1440
cgtccggcat cctttatgaa gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc 1500
tggtcactac tacggaatcg ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac 1560
atcaggcgct ggcccgtgcg cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg 1620
acgccattcg ccgcgatatt ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg 1680
tgcgagaaat gcgcgagaaa atgcgtgccc atcttggcaa caagcataaa gaccgcttcg 1740
atctgaaagc cgatgaaggc ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc 1800
tgcgctttgc ccatgacaag ccgaaactga cgcgctggtc ggataatgtg cgcattctcg 1860
aagggctggc gcaaaacggc atcatggagg agcaggaagc gcaggcattg acgctggcgt 1920
acaccacatt gcgtgatgag ctgcaccacc tggcgctgca agagttgccg ggacatgtgg 1980
cgctctcctg ttttgtcgcc gagcgtgcgc ttattaaaac cagctgggac aagtggctgg 2040
tggaaccgtg cgccccggcg taagtgtggt atcatcgcgc gcaaattttg tatctctcag 2100
gagacaggaa tgaaagtgac gctgccagag tttaagcaag ccggtgtaat ggtggtgggt 2160
gatgtgatgc tggatcgtta ctggtatggc ccaaccagcc gtatctctcc ggaagcgcca 2220
gtcccggttg ttaaagtcga taccattgaa gagcgtcctg gcggcgcggc aaacgtggcg 2280
atgaatatcg cctcactggg cgccacggcg cgtctggttg gcctgactgg cattgacgat 2340
gcggcgcgcg cgctgagcaa agcgctggcc gatgttaacg ttaaatgtga cttcgtttct 2400
gttccgacgc atcccaccat cactaagctg cgcgtgctgt cgcgtaacca gcagctgatt 2460
cgcctggact ttgaagaggg ttttgaagga gtcgatccgc aaccgatgca tgaacgcatc 2520
agccaggcgc ttggtaatat tggcgcgctg gtgctgtcgg att 2563
<210> 245
<211> 426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm5
<400> 245
atgaccctga atatgatgat ggatgccggc ggacatcatc gcgacaaaca atattaatac 60
cggcaaccac accggcaatt tacgagactg cgcaggcatc ctttctcccg tcaatttctg 120
tcaaataaag taaaagaggc agtctacttg aattaccccc ggctggttga gcgtttgttg 180
aaaaaaagta actgaaaaat ccgtagaata gcgccactct gatggttaat taacctattc 240
aattaagaat tatctggatg aatgtgccat taaatgcgca gcataatggt gcgttgtgcg 300
ggaaaactgc ttttttttga aagggttggt cagtagcgga aacaactcac ttcacacccc 360
gaagggggaa gttgcctgac cctacgattc ccgctatttc attcactgac cggaggttca 420
aaatga 426
<210> 246
<211> 1426
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm5 с фланкирующей областью длиной 500 п.н.
<400> 246
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc ggacatcatc 540
gcgacaaaca atattaatac cggcaaccac accggcaatt tacgagactg cgcaggcatc 600
ctttctcccg tcaatttctg tcaaataaag taaaagaggc agtctacttg aattaccccc 660
ggctggttga gcgtttgttg aaaaaaagta actgaaaaat ccgtagaata gcgccactct 720
gatggttaat taacctattc aattaagaat tatctggatg aatgtgccat taaatgcgca 780
gcataatggt gcgttgtgcg ggaaaactgc ttttttttga aagggttggt cagtagcgga 840
aacaactcac ttcacacccc gaagggggaa gttgcctgac cctacgattc ccgctatttc 900
attcactgac cggaggttca aaatgaccca gcgaaccgag tcgggtaata ccgtctggcg 960
cttcgatttg tcccagcagt tcactgcgat gcagcgcata agcgtggtac tcagccgggc 1020
gaccgaggtc gatcagacgc tccagcaagt gctgtgcgta ttgcacaatg acgccttttt 1080
gcagcacggc atgatctgtc tgtacgacag ccagcaggcg attttgaata ttgaagcgtt 1140
gcaggaagcc gatcagcagt taatccccgg cagctcgcaa atccgctatc gtccgggcga 1200
agggctggtc gggacggtgc tttcgcaggg ccaatcatta gtgctggcgc gcgttgctga 1260
cgatcagcgc tttcttgacc ggctcgggtt gtatgattac aacctgccgt ttatcgccgt 1320
gccgctgata gggccagatg cgcagacttt cggtgtgctg acggcacaac ccatggcgcg 1380
ttacgaagag cgattacccg cctgcacccg ctttctggaa acggtc 1426
<210> 247
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 247
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 248
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 248
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc cgtcctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt ttttatattc 600
ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac accccgaagg 900
gggaagttgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttgtccca gcagttcact 1020
gcgatgcagc gcataagcgt ggtactcagc cgggcgaccg aggtcgatca gacgctccag 1080
caagtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gaatattgaa gcgttgcagg aagccgatca gcagttaatc 1200
cccggcagct cgcaaatccg ctatcgtccg ggcgaagggc tggtcgggac ggtgctttcg 1260
cagggccaat cattagtgct ggcgcgcgtt gctgacgatc agcgctttct tgaccggctc 1320
gggttgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc agatgcgcag 1380
actttcggtg tgctgacggc acaacccatg gcgcgttacg aagagcgatt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 249
<211> 1188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 249
atggcactga aacacctcat ttccctgtgt gccgcgtcgc cgatggttgc cagtcagctg 60
gcgcgctacc cgatcctgct tgatgaattg ctcgacccga atacgctcta tcaaccgacg 120
gcgatgaatg cctatcgcga tgagctgcgc caatacctgc tgcgcgtgcc ggaagatgat 180
gaagagcaac agcttgaggc gctgcggcag tttaagcagg cgcagttgct gcgcgtggcg 240
gcggcggata ttgccggtac gttgccagta atgaaagtga gcgatcactt aacctggctg 300
gcggaagcga ttattgatgc ggtggtgcag caagcctggg ggcagatggt ggcgcgttat 360
ggccagccaa cgcatctgca cgatcgcgaa gggcgcggtt ttgcggtggt cggttatggc 420
aagctgggcg gctgggagct gggttacagc tccgatctgg atctggtatt cctgcacgac 480
tgcccgatgg atgtgatgac cgatggcgag cgtgaaatcg atggtcgcca gttctatttg 540
cgtctcgcgc agcgcgtgat gcacctgttt agcacgcgca cgtcgtccgg catcctttat 600
gaagttgatg cgcgtctgcg tccatctggc gctgcgggga tgctggtcac tactacggaa 660
tcgttcgccg attaccagca aaacgaagcc tggacgtggg aacatcaggc gctggcccgt 720
gcgcgcgtgg tgtacggcga tccgcaactg accgccgaat ttgacgccat tcgccgcgat 780
attctgatga cgcctcgcga cggcgcaacg ctgcaaaccg acgtgcgaga aatgcgcgag 840
aaaatgcgtg cccatcttgg caacaagcat aaagaccgct tcgatctgaa agccgatgaa 900
ggcggtatca ccgacatcga gtttatcgcc caatatctgg tgctgcgctt tgcccatgac 960
aagccgaaac tgacgcgctg gtcggataat gtgcgcattc tcgaagggct ggcgcaaaac 1020
ggcatcatgg aggagcagga agcgcaggca ttgacgctgg cgtacaccac attgcgtgat 1080
gagctgcacc acctggcgct gcaagagttg ccgggacatg tggcgctctc ctgttttgtc 1140
gccgagcgtg cgcttattaa aaccagctgg gacaagtggc tggtggaa 1188
<210> 250
<211> 2206
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 250
gcgcaaagcg agtgctcact tacgtgatct gttgacacaa tctgaagcga ccataacttc 60
tgccgtttca gcgaatacgg cggtgtggag cgcacaatca gccctggcga agctggtgct 120
caccgagtgg ctagtgacgc agggctggcg aaccttcctt gatgaaaaag cgcaggccaa 180
attcgccgac tcctttaaac gctttgctga catccatctg tcacgcagcg ccgccgagct 240
gaaaaaagcc tttgcccaac cgctgggcga cagctatcgc gaccagttgc cgcgcctggc 300
gcgtgatatc gactgcgcgt tactgctggc cgggcattac gatcgcgcgc gcgccgtgga 360
atggctggaa aactggcagg ggcttcagca cgccattgaa acgcgccaga gagtcgaaat 420
cgaacatttc cgtaataccg cgattaccca ggagccgttc tggttgcaca gcggaaaacg 480
ttaacgaaag gatatttcgc atggcactga aacacctcat ttccctgtgt gccgcgtcgc 540
cgatggttgc cagtcagctg gcgcgctacc cgatcctgct tgatgaattg ctcgacccga 600
atacgctcta tcaaccgacg gcgatgaatg cctatcgcga tgagctgcgc caatacctgc 660
tgcgcgtgcc ggaagatgat gaagagcaac agcttgaggc gctgcggcag tttaagcagg 720
cgcagttgct gcgcgtggcg gcggcggata ttgccggtac gttgccagta atgaaagtga 780
gcgatcactt aacctggctg gcggaagcga ttattgatgc ggtggtgcag caagcctggg 840
ggcagatggt ggcgcgttat ggccagccaa cgcatctgca cgatcgcgaa gggcgcggtt 900
ttgcggtggt cggttatggc aagctgggcg gctgggagct gggttacagc tccgatctgg 960
atctggtatt cctgcacgac tgcccgatgg atgtgatgac cgatggcgag cgtgaaatcg 1020
atggtcgcca gttctatttg cgtctcgcgc agcgcgtgat gcacctgttt agcacgcgca 1080
cgtcgtccgg catcctttat gaagttgatg cgcgtctgcg tccatctggc gctgcgggga 1140
tgctggtcac tactacggaa tcgttcgccg attaccagca aaacgaagcc tggacgtggg 1200
aacatcaggc gctggcccgt gcgcgcgtgg tgtacggcga tccgcaactg accgccgaat 1260
ttgacgccat tcgccgcgat attctgatga cgcctcgcga cggcgcaacg ctgcaaaccg 1320
acgtgcgaga aatgcgcgag aaaatgcgtg cccatcttgg caacaagcat aaagaccgct 1380
tcgatctgaa agccgatgaa ggcggtatca ccgacatcga gtttatcgcc caatatctgg 1440
tgctgcgctt tgcccatgac aagccgaaac tgacgcgctg gtcggataat gtgcgcattc 1500
tcgaagggct ggcgcaaaac ggcatcatgg aggagcagga agcgcaggca ttgacgctgg 1560
cgtacaccac attgcgtgat gagctgcacc acctggcgct gcaagagttg ccgggacatg 1620
tggcgctctc ctgttttgtc gccgagcgtg cgcttattaa aaccagctgg gacaagtggc 1680
tggtggaacc gtgcgccccg gcgtaagtgt ggtatcatcg cgcgcaaatt ttgtatctct 1740
caggagacag gaatgaaagt gacgctgcca gagtttaagc aagccggtgt aatggtggtg 1800
ggtgatgtga tgctggatcg ttactggtat ggcccaacca gccgtatctc tccggaagcg 1860
ccagtcccgg ttgttaaagt cgataccatt gaagagcgtc ctggcggcgc ggcaaacgtg 1920
gcgatgaata tcgcctcact gggcgccacg gcgcgtctgg ttggcctgac tggcattgac 1980
gatgcggcgc gcgcgctgag caaagcgctg gccgatgtta acgttaaatg tgacttcgtt 2040
tctgttccga cgcatcccac catcactaag ctgcgcgtgc tgtcgcgtaa ccagcagctg 2100
attcgcctgg actttgaaga gggttttgaa ggagtcgatc cgcaaccgat gcatgaacgc 2160
atcagccagg cgcttggtaa tattggcgcg ctggtgctgt cggatt 2206
<210> 251
<211> 199
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-amtB
<400> 251
tttcgctgaa ggtgtgacca tgggccatca ggtgctggtg cagctggaaa gtgttgccat 60
cactatcgtg tggtctggcg tggtggcctt tattggttac aaactggcgg acatgacggt 120
aggcctgcgc gtaccggaag aacaagaacg tgaagggctg gatgtaaaca gccacggcga 180
aaacgcctat aacgcctga 199
<210> 252
<211> 1199
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-amtB с фланкирующей областью длиной 500 п.н.
<400> 252
tttcctttct gactctgccc gtccgggcgc actaacggcc tgaaatactc cctcttttca 60
ttcctggcac aacgattgca atgtctgttg cgtgttagct gcggccatta tcgaattcga 120
ctggaggggg atctatgaag ctggttaccg tggtgattaa gccattcaaa cttgaagacg 180
tgcgtgaagc gctttcttct attggtattc aagggttgac cgtaactgaa gtgaaaggct 240
ttggccgtca gaagggtcac gctgagctgt accgcggtgc ggaatatagc gttaatttcc 300
tgccgaaagt gaaaattgat gtggcgatcg ctgacgatca actcgatgaa gtaatcgatg 360
tgatcagcaa agcggcctac accggaaaaa ttggcgacgg caaaattttc gttgctgagc 420
tgcaacgcgt cattcgtatt cgtaccggcg aagccgacga agcggcactg taatacaaga 480
cacacagtga tggggatcgg tttcgctgaa ggtgtgacca tgggccatca ggtgctggtg 540
cagctggaaa gtgttgccat cactatcgtg tggtctggcg tggtggcctt tattggttac 600
aaactggcgg acatgacggt aggcctgcgc gtaccggaag aacaagaacg tgaagggctg 660
gatgtaaaca gccacggcga aaacgcctat aacgcctgat tgcgttgagt tatctcctga 720
gcataaaaaa gcctccattc ggaggctttt ctttttttaa gtttaaagcg cggttagttg 780
cgattgcgca tgacgccttc ctgcacgctg gacgcgacca gcacaccctc ttgcgtatag 840
aactcgccgc gcacaaaacc gcgagcgctg gaggctgacg tgctttccac actgtagagc 900
agccattcgt tcatattaaa cgggcgatgg aaccacatgg agtggtcaat ggtggcaacc 960
tgcataccgc gctcaaggaa gcccacgccg tgcggctgaa gtgcaaccgg caggaagtta 1020
aagtctgagg catatccaag cagatattga tgtacgcgaa aatcgtccgg caccgtgccg 1080
tttgcgcgga tccatacctg gcgggtggga tcggcaacgt ggcctttcag cgggttatga 1140
aactcaaccg ggcggatctc cagtggttta tcactaagaa acttctcttt ggcctgcgg 1199
<210> 253
<211> 1563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-1
<400> 253
atgtttaacg atctgattgg cgatgatgaa acggattcgc cggaagatgc gctttctgag 60
agctggcgcg aattgtggca ggatgcgttg caggaggagg attccacgcc cgtgctggcg 120
catctctcag aggacgatcg ccgccgcgtg gtggcgctga ttgccgattt tcgcaaagag 180
ttggataaac gcaccattgg cccgcgaggg cggcaggtac tcgatcactt aatgccgcat 240
ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac cgctgtcacg cctgacgccg 300
ctgctcaccg gaattattac ccgcaccact taccttgagc tgctaagtga atttcccggc 360
gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg 420
cgctacccga tcctgcttga tgaattgctc gacccgaata cgctctatca accgacggcg 480
atgaatgcct atcgcgatga gctgcgccaa tacctgctgc gcgtgccgga agatgatgaa 540
gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc agttgctgcg cgtggcggcg 600
gcggatattg ccggtacgtt gccagtaatg aaagtgagcg atcacttaac ctggctggcg 660
gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc agatggtggc gcgttatggc 720
cagccaacgc atctgcacga tcgcgaaggg cgcggttttg cggtggtcgg ttatggcaag 780
ctgggcggct gggagctggg ttacagctcc gatctggatc tggtattcct gcacgactgc 840
ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg gtcgccagtt ctatttgcgt 900
ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt cgtccggcat cctttatgaa 960
gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc tggtcactac tacggaatcg 1020
ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac atcaggcgct ggcccgtgcg 1080
cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg acgccattcg ccgcgatatt 1140
ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg tgcgagaaat gcgcgagaaa 1200
atgcgtgccc atcttggcaa caagcataaa gaccgcttcg atctgaaagc cgatgaaggc 1260
ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc tgcgctttgc ccatgacaag 1320
ccgaaactga cgcgctggtc ggataatgtg cgcattctcg aagggctggc gcaaaacggc 1380
atcatggagg agcaggaagc gcaggcattg acgctggcgt acaccacatt gcgtgatgag 1440
ctgcaccacc tggcgctgca agagttgccg ggacatgtgg cgctctcctg ttttgtcgcc 1500
gagcgtgcgc ttattaaaac cagctgggac aagtggctgg tggaaccgtg cgccccggcg 1560
taa 1563
<210> 254
<211> 2563
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-1 с фланкирующей областью длиной 500 п.н.
<400> 254
gcgcaaagcg agtgctcact tacgtgatct gttgacacaa tctgaagcga ccataacttc 60
tgccgtttca gcgaatacgg cggtgtggag cgcacaatca gccctggcga agctggtgct 120
caccgagtgg ctagtgacgc agggctggcg aaccttcctt gatgaaaaag cgcaggccaa 180
attcgccgac tcctttaaac gctttgctga catccatctg tcacgcagcg ccgccgagct 240
gaaaaaagcc tttgcccaac cgctgggcga cagctatcgc gaccagttgc cgcgcctggc 300
gcgtgatatc gactgcgcgt tactgctggc cgggcattac gatcgcgcgc gcgccgtgga 360
atggctggaa aactggcagg ggcttcagca cgccattgaa acgcgccaga gagtcgaaat 420
cgaacatttc cgtaataccg cgattaccca ggagccgttc tggttgcaca gcggaaaacg 480
ttaacgaaag gatatttcgc atgtttaacg atctgattgg cgatgatgaa acggattcgc 540
cggaagatgc gctttctgag agctggcgcg aattgtggca ggatgcgttg caggaggagg 600
attccacgcc cgtgctggcg catctctcag aggacgatcg ccgccgcgtg gtggcgctga 660
ttgccgattt tcgcaaagag ttggataaac gcaccattgg cccgcgaggg cggcaggtac 720
tcgatcactt aatgccgcat ctgctcagcg atgtatgctc gcgcgacgat gcgccagtac 780
cgctgtcacg cctgacgccg ctgctcaccg gaattattac ccgcaccact taccttgagc 840
tgctaagtga atttcccggc gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga 900
tggttgccag tcagctggcg cgctacccga tcctgcttga tgaattgctc gacccgaata 960
cgctctatca accgacggcg atgaatgcct atcgcgatga gctgcgccaa tacctgctgc 1020
gcgtgccgga agatgatgaa gagcaacagc ttgaggcgct gcggcagttt aagcaggcgc 1080
agttgctgcg cgtggcggcg gcggatattg ccggtacgtt gccagtaatg aaagtgagcg 1140
atcacttaac ctggctggcg gaagcgatta ttgatgcggt ggtgcagcaa gcctgggggc 1200
agatggtggc gcgttatggc cagccaacgc atctgcacga tcgcgaaggg cgcggttttg 1260
cggtggtcgg ttatggcaag ctgggcggct gggagctggg ttacagctcc gatctggatc 1320
tggtattcct gcacgactgc ccgatggatg tgatgaccga tggcgagcgt gaaatcgatg 1380
gtcgccagtt ctatttgcgt ctcgcgcagc gcgtgatgca cctgtttagc acgcgcacgt 1440
cgtccggcat cctttatgaa gttgatgcgc gtctgcgtcc atctggcgct gcggggatgc 1500
tggtcactac tacggaatcg ttcgccgatt accagcaaaa cgaagcctgg acgtgggaac 1560
atcaggcgct ggcccgtgcg cgcgtggtgt acggcgatcc gcaactgacc gccgaatttg 1620
acgccattcg ccgcgatatt ctgatgacgc ctcgcgacgg cgcaacgctg caaaccgacg 1680
tgcgagaaat gcgcgagaaa atgcgtgccc atcttggcaa caagcataaa gaccgcttcg 1740
atctgaaagc cgatgaaggc ggtatcaccg acatcgagtt tatcgcccaa tatctggtgc 1800
tgcgctttgc ccatgacaag ccgaaactga cgcgctggtc ggataatgtg cgcattctcg 1860
aagggctggc gcaaaacggc atcatggagg agcaggaagc gcaggcattg acgctggcgt 1920
acaccacatt gcgtgatgag ctgcaccacc tggcgctgca agagttgccg ggacatgtgg 1980
cgctctcctg ttttgtcgcc gagcgtgcgc ttattaaaac cagctgggac aagtggctgg 2040
tggaaccgtg cgccccggcg taagtgtggt atcatcgcgc gcaaattttg tatctctcag 2100
gagacaggaa tgaaagtgac gctgccagag tttaagcaag ccggtgtaat ggtggtgggt 2160
gatgtgatgc tggatcgtta ctggtatggc ccaaccagcc gtatctctcc ggaagcgcca 2220
gtcccggttg ttaaagtcga taccattgaa gagcgtcctg gcggcgcggc aaacgtggcg 2280
atgaatatcg cctcactggg cgccacggcg cgtctggttg gcctgactgg cattgacgat 2340
gcggcgcgcg cgctgagcaa agcgctggcc gatgttaacg ttaaatgtga cttcgtttct 2400
gttccgacgc atcccaccat cactaagctg cgcgtgctgt cgcgtaacca gcagctgatt 2460
cgcctggact ttgaagaggg ttttgaagga gtcgatccgc aaccgatgca tgaacgcatc 2520
agccaggcgc ttggtaatat tggcgcgctg gtgctgtcgg att 2563
<210> 255
<211> 461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1
<400> 255
atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg acaattcgga 60
ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat ttaaaataaa 120
aaatccaatc ggatttcact atttaaactg gccattatct aagatgaatc cgatggaagc 180
tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag tcggtgcttc tttgagcgaa 240
cgatcaaatt taagtggatt cccatcaaaa aaatattctc aacctaaaaa agtttgtgta 300
atacttgtaa cgctacatgg agattaactc aatctagagg gtattaataa tgaatcgtac 360
taaactggta ctgggcgcaa ctcacttcac accccgaagg gggaagttgc ctgaccctac 420
gattcccgct atttcattca ctgaccggag gttcaaaatg a 461
<210> 256
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm1 с фланкирующей областью длиной 500 п.н.
<400> 256
accggatacg agagaaaagt gtctacatcg gttcggttga tattgaccgg cgcatccgcc 60
agcccgccca gtttctggtg gatctgtttg gcgattttgc gggtcttgcc ggtgtcggtg 120
ccgaaaaaaa taccaatatt tgccataaca cacgctcctg ttgaaaaaga gatcccgccg 180
ggaaatgcgg tgaacgtgtc tgatattgcg aagagtgtgc cagttttggt cgcgggcaaa 240
acctgcacca gtttggttat taatgcacca gtctggcgct ttttttcgcc gagtttctcc 300
tcgctaatgc ccgccaggcg cggctttggc gctgatagcg cgctgaatac cgatctggat 360
caaggttttg tcgggttatc agccaaaagg tgcactcttt gcatggttat acgtgcctga 420
catgttgtcc gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta 480
actgaccgca ggagtgtgcg atgaccctga atatgatgat ggatgccggc cgtcctgtaa 540
taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt ttttatattc 600
ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg gccattatct 660
aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt ttattgaaag 720
tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa aaatattctc 780
aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc aatctagagg 840
gtattaataa tgaatcgtac taaactggta ctgggcgcaa ctcacttcac accccgaagg 900
gggaagttgc ctgaccctac gattcccgct atttcattca ctgaccggag gttcaaaatg 960
acccagcgaa ccgagtcggg taataccgtc tggcgcttcg atttgtccca gcagttcact 1020
gcgatgcagc gcataagcgt ggtactcagc cgggcgaccg aggtcgatca gacgctccag 1080
caagtgctgt gcgtattgca caatgacgcc tttttgcagc acggcatgat ctgtctgtac 1140
gacagccagc aggcgatttt gaatattgaa gcgttgcagg aagccgatca gcagttaatc 1200
cccggcagct cgcaaatccg ctatcgtccg ggcgaagggc tggtcgggac ggtgctttcg 1260
cagggccaat cattagtgct ggcgcgcgtt gctgacgatc agcgctttct tgaccggctc 1320
gggttgtatg attacaacct gccgtttatc gccgtgccgc tgatagggcc agatgcgcag 1380
actttcggtg tgctgacggc acaacccatg gcgcgttacg aagagcgatt acccgcctgc 1440
acccgctttc tggaaacggt c 1461
<210> 257
<211> 199
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-amtB
<400> 257
tttcgctgaa ggtgtgacca tgggccatca ggtgctggtg cagctggaaa gtgttgccat 60
cactatcgtg tggtctggcg tggtggcctt tattggttac aaactggcgg acatgacggt 120
aggcctgcgc gtaccggaag aacaagaacg tgaagggctg gatgtaaaca gccacggcga 180
aaacgcctat aacgcctga 199
<210> 258
<211> 1199
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-amtB с фланкирующей областью длиной 500 п.н.
<400> 258
tttcctttct gactctgccc gtccgggcgc actaacggcc tgaaatactc cctcttttca 60
ttcctggcac aacgattgca atgtctgttg cgtgttagct gcggccatta tcgaattcga 120
ctggaggggg atctatgaag ctggttaccg tggtgattaa gccattcaaa cttgaagacg 180
tgcgtgaagc gctttcttct attggtattc aagggttgac cgtaactgaa gtgaaaggct 240
ttggccgtca gaagggtcac gctgagctgt accgcggtgc ggaatatagc gttaatttcc 300
tgccgaaagt gaaaattgat gtggcgatcg ctgacgatca actcgatgaa gtaatcgatg 360
tgatcagcaa agcggcctac accggaaaaa ttggcgacgg caaaattttc gttgctgagc 420
tgcaacgcgt cattcgtatt cgtaccggcg aagccgacga agcggcactg taatacaaga 480
cacacagtga tggggatcgg tttcgctgaa ggtgtgacca tgggccatca ggtgctggtg 540
cagctggaaa gtgttgccat cactatcgtg tggtctggcg tggtggcctt tattggttac 600
aaactggcgg acatgacggt aggcctgcgc gtaccggaag aacaagaacg tgaagggctg 660
gatgtaaaca gccacggcga aaacgcctat aacgcctgat tgcgttgagt tatctcctga 720
gcataaaaaa gcctccattc ggaggctttt ctttttttaa gtttaaagcg cggttagttg 780
cgattgcgca tgacgccttc ctgcacgctg gacgcgacca gcacaccctc ttgcgtatag 840
aactcgccgc gcacaaaacc gcgagcgctg gaggctgacg tgctttccac actgtagagc 900
agccattcgt tcatattaaa cgggcgatgg aaccacatgg agtggtcaat ggtggcaacc 960
tgcataccgc gctcaaggaa gcccacgccg tgcggctgaa gtgcaaccgg caggaagtta 1020
aagtctgagg catatccaag cagatattga tgtacgcgaa aatcgtccgg caccgtgccg 1080
tttgcgcgga tccatacctg gcgggtggga tcggcaacgt ggcctttcag cgggttatga 1140
aactcaaccg ggcggatctc cagtggttta tcactaagaa acttctcttt ggcctgcgg 1199
<210> 259
<211> 607
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC
<400> 259
atgaccttta atatgatgcc tggggtcact ggagcgcttt atcggcatcc tgaccgaaga 60
atttgccggt ttcttcccga cctggctggc ccctgttcag gttgtggtga tgaatatcac 120
tgattctcaa gctgaatatg tcaacgaatt gacccgtaaa ttgcaaaatg cgggcattcg 180
tgtaaaagcg gacttgagaa acgagaagat tggctttaaa atccgcgagc acactttacg 240
tcgtgtccct tatatgttgg tctgtggtga taaagaggtg gaagcaggca aagtggccgt 300
tcgcacccgc cgcggtaaag acctgggcag cctggacgta agtgaagtga ttgagaagct 360
gcaacaagag attcgcagcc gcagtcttca acaactggag gaataaggta ttaaaggcgg 420
aaaacgagtt caaacggcac gtccgaatcg tatcaatggc gagattcgcg cccaggaagt 480
tcgcttaact ggtctggaag gtgagcagct gggtattgca atagaactaa ctacccgccc 540
tgaaggcggt acctgcctga ccctgcgatt cccgttattt cattcactga ccggaggccc 600
acgatga 607
<210> 260
<211> 1607
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::PinfC с фланкирующей областью длиной 500 п.н.
<400> 260
ggtacgacaa aaacgtctcc agcgacgtgc ggttaatatt gactggcgca tccgccacat 60
cccccagttt ttgctggatc agtttggcga ttttgcgggt ttttcccgtg tcactgccaa 120
aaaaaatacc aatgttagcc atgtcgcgct cctgttgaga aagaataagg ccgcctgcaa 180
acggcggata tcccttctcc tgttgcgaag gctgtgccag gtttttttaa ggccttctgt 240
gcactgaaat gggtgaaaaa atgactcttt tttgtgcagg caccgtcctc tctccgctat 300
ccagacctgc tttgaaggcc tctgagggcc aaatcagggc caaaacacga atcaggatca 360
atgtttcggc gcgttacctg ttcgaaaggt gcactctttg catggttaat cacacccaat 420
cagggctgcg gatgtcgggc gtttcacaac acaaaatgtt gtaaatgcga cacagccggg 480
cctgaaacca ggagcgtgtg atgaccttta atatgatgcc tggggtcact ggagcgcttt 540
atcggcatcc tgaccgaaga atttgccggt ttcttcccga cctggctggc ccctgttcag 600
gttgtggtga tgaatatcac tgattctcaa gctgaatatg tcaacgaatt gacccgtaaa 660
ttgcaaaatg cgggcattcg tgtaaaagcg gacttgagaa acgagaagat tggctttaaa 720
atccgcgagc acactttacg tcgtgtccct tatatgttgg tctgtggtga taaagaggtg 780
gaagcaggca aagtggccgt tcgcacccgc cgcggtaaag acctgggcag cctggacgta 840
agtgaagtga ttgagaagct gcaacaagag attcgcagcc gcagtcttca acaactggag 900
gaataaggta ttaaaggcgg aaaacgagtt caaacggcac gtccgaatcg tatcaatggc 960
gagattcgcg cccaggaagt tcgcttaact ggtctggaag gtgagcagct gggtattgca 1020
atagaactaa ctacccgccc tgaaggcggt acctgcctga ccctgcgatt cccgttattt 1080
cattcactga ccggaggccc acgatgaccc agcgacccga gtcgggcacc accgtctggc 1140
gttttgatct ctcacagcaa tttaccgcca tgcagcgcat cagcgtggtg ttgagtcgcg 1200
caaccgagat aagccagacg ctgcaggagg tgctgtgtgt tctgcataat gacgcattta 1260
tgcaacacgg catgctgtgt ctgtatgaca accagcagga aattctgagt attgaagcct 1320
tgcaggaggc agaccaacat ctgatccccg gcagctcgca aattcgctat cgccctggcg 1380
aagggctggt aggagccgta ctgtcccagg gacaatctct tgtgctgccg cgtgtcgccg 1440
acgatcaacg ctttctcgac aggcttggca tctatgatta caacctgccg tttatcgccg 1500
tccccttaat ggggccaggc gcgcagacga ttggcgtgct cgccgcgcag ccgatggcgc 1560
gtctggagga gcggcttcct tcctgtacgc gctttctgga aaccgtc 1607
<210> 261
<211> 1536
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> 16S-1
<400> 261
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gaacggtagc acagagagct tgctctcggg tgacgagtgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagaggggga ccttcgggcc tcttgccatc agatgtgccc agatgggatt 240
agctagtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggg agtaaggtta ataaccttat tcattgacgt 480
tacccgcaga agaagcaccg gctaactccg tgccagcagc cgcggtaata cggagggtgc 540
aagcgttaat cggaattact gggcgtaaag cgcacgcagg cggtctgtca agtcggatgt 600
gaaatccccg ggctcaacct gggaactgca tccgaaactg gcaggcttga gtctcgtaga 660
gggaggtaga attccaggtg tagcggtgaa atgcgtagag atctggagga ataccggtgg 720
cgaaggcggc ctcctggacg aagactgacg ctcaggtgcg aaagcgtggg gagcaaacag 780
gattagatac cctggtagtc cacgccgtaa acgatgtcta tttggaggtt gtgcccttga 840
ggcgtggctt ccggagctaa cgcgttaaat agaccgcctg gggagtacgg ccgcaaggtt 900
aaaactcaaa tgaattgacg ggggcccgca caagcggtgg agcatgtggt ttaattcgat 960
gcaacgcgaa gaaccttacc tggtcttgac atccacagaa ctttccagag atggattggt 1020
gccttcggga actgtgagac aggtgctgca tggctgtcgt cagctcgtgt tgtgaaatgt 1080
tgggttaagt cccgcaacga gcgcaaccct tatcctttgt tgccagcggt ccggccggga 1140
actcaaagga gactgccagt gataaactgg aggaaggtgg ggatgacgtc aagtcatcat 1200
ggcccttacg accagggcta cacacgtgct acaatggcgc atacaaagag aagcgacctc 1260
gcgagagtaa gcggacctca taaagtgcgt cgtagtccgg attggagtct gcaactcgac 1320
tccatgaagt cggaatcgct agtaatcgtg gatcagaatg ccacggtgaa tacgttcccg 1380
ggccttgtac acaccgcccg tcacaccatg ggagtgggtt gcaaaagaag taggtagctt 1440
aaccttcggg agggcgctta ccactttgtg attcatgact ggggtgaagt cgtaacaagg 1500
taaccgtagg ggaacctgcg gttggatcac ctcctt 1536
<210> 262
<211> 1537
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> 16S-2
<220>
<221> модифицированное основание
<222> (450)..(450)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (452)..(452)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (455)..(455)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (473)..(473)
<223> a, c, t, g, неизвестное или другое
<400> 262
ttgaagagtt tgatcatggc tcagattgaa cgctggcggc aggcctaaca catgcaagtc 60
gaacggtagc acagagagct tgctctcggg tgacgagtgg cggacgggtg agtaatgtct 120
gggaaactgc ctgatggagg gggataacta ctggaaacgg tagctaatac cgcataacgt 180
cgcaagacca aagaggggga ccttcgggcc tcttgccatc agatgtgccc agatgggatt 240
agctagtagg tggggtaacg gctcacctag gcgacgatcc ctagctggtc tgagaggatg 300
accagccaca ctggaactga gacacggtcc agactcctac gggaggcagc agtggggaat 360
attgcacaat gggcgcaagc ctgatgcagc catgccgcgt gtgtgaagaa ggccttcggg 420
ttgtaaagca ctttcagcgg ggaggaaggn antanggtta ataacctgtg ttnattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc atccgaaact ggcaggcttg agtctcgtag 660
agggaggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg cctcctggac gaagactgac gctcaggtgc gaaagcgtgg ggagcaaaca 780
ggattagata ccctggtagt ccacgccgta aacgatgtct atttggaggt tgtgcccttg 840
aggcgtggct tccggagcta acgcgttaaa tagaccgcct ggggagtacg gccgcaaggt 900
taaaactcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggtcttga catccacaga acttagcaga gatgctttgg 1020
tgccttcggg aactgtgaga caggtgctgc atggctgtcg tcagctcgtg ttgtgaaatg 1080
ttgggttaag tcccgcaacg agcgcaaccc ttatcctttg ttgccagcgg ttaggccggg 1140
aactcaaagg agactgccag tgataaactg gaggaaggtg gggatgacgt caagtcatca 1200
tggcccttac gaccagggct acacacgtgc tacaatggcg catacaaaga gaagcgacct 1260
cgcgagagta agcggacctc ataaagtgcg tcgtagtccg gattggagtc tgcaactcga 1320
ctccatgaag tcggaatcgc tagtaatcgt ggatcagaat gccacggtga atacgttccc 1380
gggccttgta cacaccgccc gtcacaccat gggagtgggt tgcaaaagaa gtaggtagct 1440
taaccttcgg gagggcgctt accactttgt gattcatgac tggggtgaag tcgtaacaag 1500
gtaaccgtag gggaacctgc ggttggatca cctcctt 1537
<210> 263
<211> 882
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifH
<400> 263
atgaccatgc gtcaatgcgc catttacggc aaaggtggga tcggcaaatc gaccaccaca 60
cagaacctgg tcgccgcgct ggcggagatg ggtaaaaaag tcatgattgt cggctgtgac 120
ccgaaagccg attccacgcg tttgatcctg catgcgaaag cgcagaacac cattatggag 180
atggctgctg aagtcggctc cgtggaagac ctggagttag aagacgtgct gcaaatcggt 240
tacggcggcg tgcgctgcgc agagtccggc ggcccggagc caggcgtggg ctgtgccggt 300
cgcggggtga tcaccgcgat taacttcctc gaagaagaag gcgcttacgt gccggatctc 360
gattttgttt tctacgacgt gctgggcgac gtggtatgcg gtggtttcgc catgccgatt 420
cgtgaaaaca aagcgcagga gatctacatc gtttgctctg gcgaaatgat ggcgatgtac 480
gccgccaaca acatctccaa aggcatcgtg aaatacgcca aatccggtaa agtgcgcctc 540
ggcgggctga tttgtaactc gcgccagacc gaccgtgaag atgaactgat cattgcgctg 600
gcagaaaaac tcggcacgca gatgatccac tttgttcccc gcgacaacat tgtgcagcgt 660
gcggaaatcc gccgtatgac ggttatcgaa tatgacccga cctgcaatca ggcgaacgaa 720
tatcgcagcc ttgccagcaa aatcgtcaac aacaccaaaa tggtggtgcc caccccctgc 780
accatggatg aactggaaga actgctgatg gagttcggca ttatggatgt ggaagacacc 840
agcatcattg gtaaaaccgc cgccgaagaa aacgccgtct ga 882
<210> 264
<211> 1449
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifD2
<400> 264
atgagcaatg caacaggcga acgcaacctg gagataatcg agcaggtgct cgaggttttc 60
ccggagaaga cgcgcaaaga acgcagaaaa cacatgatgg tgacggaccc ggagcaggaa 120
agcgtcggta agtgcatcat ctctaaccgc aaatcgcagc caggcgtgat gaccgtgcgc 180
ggctgctcgt atgccggttc gaaaggggtg gtatttgggc caatcaagga tatggcgcat 240
atctcgcatg gcccaatcgg ctgcggccaa tactcccgcg ccgggcggcg gaactactac 300
accggcgtca gcggcgtgga cagcttcggc acgctcaact tcacctccga ttttcaggag 360
cgcgacatcg tgtttggcgg cgataaaaag ctcgccaaac tgattgaaga gctggaagag 420
ctgttcccgc tgaccaaagg catttcgatt cagtcggaat gcccggtcgg cctgattggc 480
gatgacattg aggccgtcgc gaacgccagc cgcaaagcca tcaacaaacc ggttattccg 540
gtgcgttgcg aaggctttcg cggcgtgtcg caatccctcg gtcaccatat tgccaacgat 600
gtgatccgcg actgggtgct ggataaccgc gaaggcaaac cgttcgaatc caccccttac 660
gatgtggcga tcatcggcga ttacaacatc ggcggcgatg cctgggcttc gcgcattttg 720
ctcgaagaga tgggcttgcg ggtggtggca cagtggtctg gcgacggtac gctggtggag 780
atggaaaaca cgccgttcgt caaactgaac ctggtgcatt gttaccgctc aatgaactac 840
atctcgcgcc atatggagga gaagcacggt attccgtgga tggaatacaa cttctttggt 900
ccgacgaaaa tcgcggaatc gctgcgcaaa atcgccgacc agtttgacga caccattcgc 960
gccaacgccg aagcggtgat cgccagatac caggcgcaaa acgacgccat tatcgccaaa 1020
tatcgcccgc gtctggaggg gcgcaaagtg ctgctttata tgggcgggct gcgtccgcgc 1080
catgtgattg gcgcctatga agacctggga atggagatca tcgctgccgg ttatgagttc 1140
ggtcataacg atgattacga ccgcaccttg ccggatctga aagagggcac gctgctgttt 1200
gatgatgcca gcagttatga gctggaggcg ttcgtcaacg cgctgaaacc ggatctcatc 1260
ggttccggca tcaaagagaa gtacatcttt cagaaaatgg gcgtgccgtt tcgccagatg 1320
cactcctggg attactccgg cccgtaccac ggctatgacg gcttcgccat cttcgcccgc 1380
gatatggata tgacgctcaa caaccccgcg tggggccagt tgaccgcgcc gtggctgaaa 1440
tccgcctga 1449
<210> 265
<211> 1563
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifK2
<400> 265
atgagccaga ctgctgagaa aatacagaat tgccatcccc tgtttgaaca ggatgcttac 60
cagacgctgt ttgccggtaa acgggcactc gaagaggcgc actcgccgga gcgggtgcag 120
gaagtgtttc aatggaccac taccccggaa tatgaagcgc tgaactttaa acgcgaagcg 180
ctgactatcg acccggcaaa agcctgccag ccgctgggcg cggtgctctg ttcgctgggg 240
tttgccaata ccctaccgta tgtgcacggt tcacagggtt gcgtggccta tttccgcacg 300
tactttaacc gccactttaa agaaccggtg gcctgcgtgt cggattcaat gacggaagac 360
gcggcggtgt tcggcgggaa taacaacctc aacaccggct tacaaaacgc cagcgcgctg 420
tataaaccgg agattatcgc cgtctctacc acctgtatgg cggaagtgat cggtgatgat 480
ttgcaggcct ttatcgccaa cgccaaaaaa gatggttttc tcgatgccgc catccccgtg 540
ccctacgcgc acacccccag ttttatcggc agccatatca ccggctggga taacatgttt 600
gaaggttttg cccggacctt tacggcagac catgaagctc agcccggcaa actttcacgc 660
atcaacctgg tgaccgggtt tgaaacctat ctcggcaatt tccgcgtgct gaaacgcatg 720
atggaacaaa tggaggtgcc ggcgagtgtg ctctccgatc cgtcggaagt gctggatact 780
cccgccaacg ggcattacca gatgtacgcg ggcgggacga cgcagcaaga gatgcgcgag 840
gcgccggatg ctatcgacac cctgttgctg cagccctggc aactggtgaa aagcaaaaaa 900
gtggtgcagg agatgtggaa tcagcccgcc accgaggttt ctgttcccgt tgggctggca 960
ggaacagacg aactgttgat ggcgattagc cagttaaccg gcaaggccat tcccgattca 1020
ctggcgctgg agcgcgggcg gctggtcgat atgatgctcg attcccacac ctggttgcac 1080
ggtaaaaaat tcggcctgtt tggcgatccg gattttgtca tgggattgac ccgtttcctg 1140
ctggagctgg gctgcgaacc gaccgttatc ctctgccaca acggtaacaa acgctggcag 1200
aaagcaatga agaaaatgct tgacgcctcg ccgtacggcc aggagagcga agtgtttatc 1260
aactgcgatt tgtggcattt ccgctcgctg atgtttaccc gccagccgga ttttatgatt 1320
ggcaactcgt acggcaagtt cattcagcgc gacaccttag ccaaaggcga gcagtttgaa 1380
gttccgctga tccgcctcgg ttttcccctg ttcgaccgcc accatctgca ccgccagacc 1440
acctggggct acgagggcgc catgagcatt ctcactaccc ttgtgaatgc ggtactggag 1500
aaagtggaca aagagaccat caagctcggc aaaaccgact acagcttcga tcttatccgt 1560
taa 1563
<210> 266
<211> 1488
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifL
<400> 266
atgaccctga atatgatgat ggatgccggc gcgcccgagg caatcgccgg tgcgctttcg 60
cgacaccatc ctgggctgtt ttttaccatc gttgaagaag cgcccgtcgc catttcgctg 120
actgatgccg acgcacgcat tgtctatgcc aacccggctt tctgccgcca gaccggctat 180
gaactagaag cgttgttgca gcaaaatccc cgcctgcttg caagtcgcca aaccccacgg 240
gaaatctatc aggatatgtg gcacaccttg ttacaacgcc gaccgtggcg cgggcaattg 300
attaaccgcc accgcgacgg cagcctgtat ctggtcgaga tcgatatcac cccggtgatt 360
aacccgtttg gcgaactgga acactacctg gcaatgcagc gcgatatcag cgccagttat 420
gcgctggagc agcggttgcg caatcacatg acgctgaccg aagcggtgct gaataacatt 480
ccggcggcgg tggttgtagt ggatgaacgc gatcatgtgg ttatggataa ccttgcctac 540
aaaacgttct gtgccgactg cggcggaaaa gagctcctga gcgaactcaa tttttcagcc 600
cgaaaagcgg agctggcaaa cggccaggtc ttaccggtgg tgctgcgcgg tgaggtgcgc 660
tggttgtcgg tgacctgctg ggcgctgccg ggcgtcagcg aagaagccag tcgctacttt 720
attgataaca ggctgacgcg cacgctggtg gtgatcaccg acgacaccca acaacgccag 780
cagcaggaac agggccgact tgaccgcctt aaacagcaga tgaccaacgg caaactactg 840
gcagcgatcc gcgaagcgct tgacgccgcg ctgatccagc ttaactgccc catcaatatg 900
ctggcggcgg cgcgacgttt aaacggcagt gataacaaca atgtggcgct cgacgccgcg 960
tggcgcgaag gtgaagaggc gatggcgcgg ctgaaacgtt gccgcccgtc gctggaactg 1020
gaaagtgcgg ccgtctggcc gctgcaaccc ttttttgacg atctgcgcgc gctttatcac 1080
acccgctacg agcaggggaa aaatttgcag gtcacgctgg attcccatca tctggtggga 1140
tttggtcagc gtacgcaact gttagcctgc ctgagtctgt ggctcgatcg cacgctggat 1200
attgccgccg ggctgggtga tttcaccgcg caaacgcaga tttacgcccg cgaagaagag 1260
ggctggctct ctttgtatat cactgacaat gtgccgctga tcccgctgcg ccacacccac 1320
tcgccggatg cgcttaacgc tccgggaaaa ggcatggagc tgcgcctgat ccagacgctg 1380
gtggcacacc accacggcgc aatagaactc acttcacacc ccgaaggggg aagttgcctg 1440
accctacgat tcccgctatt tcattcactg accggaggtt caaaatga 1488
<210> 267
<211> 1575
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifA
<400> 267
atgacccagc gaaccgagtc gggtaatacc gtctggcgct tcgatttgtc ccagcagttc 60
actgcgatgc agcgcataag cgtggtactc agccgggcga ccgaggtcga tcagacgctc 120
cagcaagtgc tgtgcgtatt gcacaatgac gcctttttgc agcacggcat gatctgtctg 180
tacgacagcc agcaggcgat tttgaatatt gaagcgttgc aggaagccga tcagcagtta 240
atccccggca gctcgcaaat ccgctatcgt ccgggcgaag ggctggtcgg gacggtgctt 300
tcgcagggcc aatcattagt gctggcgcgc gttgctgacg atcagcgctt tcttgaccgg 360
ctcgggttgt atgattacaa cctgccgttt atcgccgtgc cgctgatagg gccagatgcg 420
cagactttcg gtgtgctgac ggcacaaccc atggcgcgtt acgaagagcg attacccgcc 480
tgcacccgct ttctggaaac ggtcgctaac ctggtcgcgc aaaccgtgcg tttgatggca 540
ccaccggcag tgcgcccttc cccgcgcgcc gccataacac aggccgccag cccgaaatcc 600
tgcacggcct cacgcgcatt tggttttgaa aatatggtcg gtaacagtcc ggcgatgcgc 660
cagaccatgg agattatccg tcaggtttcg cgctgggaca ccaccgttct ggtacgcggc 720
gagagtggca ccggcaagga gctgattgcc aacgccatcc accaccattc gccgcgtgcc 780
ggtgcgccat ttgtgaaatt caactgtgcg gcgctgccgg acacactgct ggaaagcgaa 840
ttgttcggtc acgagaaagg ggcatttacc ggcgcggtac gccagcgtaa aggccgtttt 900
gagctggccg atggcggcac gctgtttctt gacgagatcg gcgagagtag cgcctcgttt 960
caggctaagc tgctgcgcat tttgcaggaa ggcgaaatgg aacgcgtcgg cggcgacgag 1020
acattgcaag tgaatgtgcg cattattgcc gcgacgaacc gcaatcttga agatgaagtc 1080
cggctggggc actttcgcga agatctctat tatcgcctga atgtgatgcc catcgccctg 1140
ccgccactac gcgaacgcca ggaggacatt gccgagctgg cgcactttct ggtgcgtaaa 1200
atcgcccata accagagccg tacgctgcgc attagcgagg gcgctatccg cctgctgatg 1260
agctacaact ggcccggtaa tgtgcgcgaa ctggaaaact gccttgagcg ctcagcggtg 1320
atgtcggaga acggtctgat cgatcgggat gtgattttgt ttaatcatcg cgaccagcca 1380
gccaaaccgc cagttatcag cgtctcgcat gatgataact ggctcgataa caaccttgac 1440
gagcgccagc ggctgattgc ggcgctggaa aaagcgggat gggtacaagc caaagccgcg 1500
cgcttgctgg ggatgacgcc gcgccaggtc gcctatcgta ttcagacgat ggatataacc 1560
ctgccaaggc tataa 1575
<210> 268
<211> 2850
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> glnE
<400> 268
atgccgcacc acgcaggatt gtcgcagcac tggcaaacgg tattttctcg tctgccggaa 60
tcgctcaccg cgcagccatt gagcgcgcag gcgcagtcag tgctcacttt tagtgatttt 120
gttcaggaca gcatcatcgc gcatcctgag tggctggcag agcttgaaag cgcgccgccg 180
cctgcgaacg aatggcaaca ctatgcgcaa tggctgcaag cggcgctgga tggcgtcacc 240
gatgaagcct cgctgatgcg cgcgctgcgg ctgtttcgcc gtcgcatcat ggtgcgcatc 300
gcctggagcc aggcgttaca gttggtggcg gaagaagata tcctgcaaca gcttagcgtg 360
ctggcggaaa ccctgatcgt cgccgcgcgc gactggcttt atgaggcctg ctgccgtgag 420
tggggaacgc cgagcaatcc acaaggcgtg gcgcagccga tgctggtact cggcatgggc 480
aaactgggtg gcggcgaact caatttctca tccgatatcg atttgatttt cgcctggccg 540
gaaaatggcg caacgcgcgg tggacgccgt gagctggata acgcgcaatt tttcactcgc 600
cttggtcaac ggctgattaa agtcctcgac cagccaacgc aggatggctt tgtctaccgc 660
gtcgatatgc gcttgcgccc gtttggcgac agcggcccgc tggtgctgag ctttgccgcg 720
ctggaagatt actaccagga gcaggggcgc gattgggaac gctacgcgat ggtgaaagcg 780
cgcattatgg gcgataacga cggcgaccat gcgcgggagt tgcgcgcaat gctgcgcccg 840
tttgttttcc gccgttatat cgacttcagc gtgattcagt ccctgcgtaa catgaaaggc 900
atgattgccc gcgaagtgcg tcgccgtggc ctgaaggaca acattaagct cggcgcgggc 960
gggatccgcg aaatagaatt tatcgtccag gttttccagc tgattcgcgg cggtcgcgag 1020
cctgcactgc aatcgcgttc actgttgccg acgcttgctg ccatagatca actgcatctg 1080
ctgccggatg gcgacgcaac ccggctgcgc gaggcgtatt tgtggctgcg acggctggag 1140
aacctgctgc aaagcatcaa tgacgaacag acacagacgc tgccgggcga tgaactgaat 1200
cgcgcgcgcc tcgcctgggg aatgggcaaa gatagctggg aagcgctctg cgaaacgctg 1260
gaagcgcata tgtcggcggt gcgtcagata tttaacgatc tgattggcga tgatgaaacg 1320
gattcgccgg aagatgcgct ttctgagagc tggcgcgaat tgtggcagga tgcgttgcag 1380
gaggaggatt ccacgcccgt gctggcgcat ctctcagagg acgatcgccg ccgcgtggtg 1440
gcgctgattg ccgattttcg caaagagttg gataaacgca ccattggccc gcgagggcgg 1500
caggtactcg atcacttaat gccgcatctg ctcagcgatg tatgctcgcg cgacgatgcg 1560
ccagtaccgc tgtcacgcct gacgccgctg ctcaccggaa ttattacccg caccacttac 1620
cttgagctgc taagtgaatt tcccggcgca ctgaaacacc tcatttccct gtgtgccgcg 1680
tcgccgatgg ttgccagtca gctggcgcgc tacccgatcc tgcttgatga attgctcgac 1740
ccgaatacgc tctatcaacc gacggcgatg aatgcctatc gcgatgagct gcgccaatac 1800
ctgctgcgcg tgccggaaga tgatgaagag caacagcttg aggcgctgcg gcagtttaag 1860
caggcgcagt tgctgcgcgt ggcggcggcg gatattgccg gtacgttgcc agtaatgaaa 1920
gtgagcgatc acttaacctg gctggcggaa gcgattattg atgcggtggt gcagcaagcc 1980
tgggggcaga tggtggcgcg ttatggccag ccaacgcatc tgcacgatcg cgaagggcgc 2040
ggttttgcgg tggtcggtta tggcaagctg ggcggctggg agctgggtta cagctccgat 2100
ctggatctgg tattcctgca cgactgcccg atggatgtga tgaccgatgg cgagcgtgaa 2160
atcgatggtc gccagttcta tttgcgtctc gcgcagcgcg tgatgcacct gtttagcacg 2220
cgcacgtcgt ccggcatcct ttatgaagtt gatgcgcgtc tgcgtccatc tggcgctgcg 2280
gggatgctgg tcactactac ggaatcgttc gccgattacc agcaaaacga agcctggacg 2340
tgggaacatc aggcgctggc ccgtgcgcgc gtggtgtacg gcgatccgca actgaccgcc 2400
gaatttgacg ccattcgccg cgatattctg atgacgcctc gcgacggcgc aacgctgcaa 2460
accgacgtgc gagaaatgcg cgagaaaatg cgtgcccatc ttggcaacaa gcataaagac 2520
cgcttcgatc tgaaagccga tgaaggcggt atcaccgaca tcgagtttat cgcccaatat 2580
ctggtgctgc gctttgccca tgacaagccg aaactgacgc gctggtcgga taatgtgcgc 2640
attctcgaag ggctggcgca aaacggcatc atggaggagc aggaagcgca ggcattgacg 2700
ctggcgtaca ccacattgcg tgatgagctg caccacctgg cgctgcaaga gttgccggga 2760
catgtggcgc tctcctgttt tgtcgccgag cgtgcgctta ttaaaaccag ctgggacaag 2820
tggctggtgg aaccgtgcgc cccggcgtaa 2850
<210> 269
<211> 1536
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> 16S-3
<220>
<221> модифицированное основание
<222> (766)..(776)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (905)..(905)
<223> a, c, t, g, неизвестное или другое
<400> 269
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgaacggtag cacagagagc ttgctctcgg gtgacgagtg gcggacgggt gagtaatgtc 120
tgggaaactg cctgatggag ggggataact actggaaacg gtagctaata ccgcataacg 180
tcgcaagacc aaagaggggg accttcgggc ctcttgccat cagatgtgcc cagatgggat 240
tagctagtag gtggggtaac ggctcaccta ggcgacgatc cctagctggt ctgagaggat 300
gaccagccac actggaactg agacacggtc cagactccta cgggaggcag cagtggggaa 360
tattgcacaa tgggcgcaag cctgatgcag ccatgccgcg tgtgtgaaga aggccttcgg 420
gttgtaaagc actttcagcg gggaggaagg gagtaaggtt aataaccttg ctcattgacg 480
ttacccgcag aagaagcacc ggctaactcc gtgccagcag ccgcggtaat acggagggtg 540
caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag gcggtctgtc aagtcggatg 600
tgaaatcccc gggctcaacc tgggaactgc atccgaaact ggcaggcttg agtctcgtag 660
agggaggtag aattccaggt gtagcggtga aatgcgtaga gatctggagg aataccggtg 720
gcgaaggcgg cctcctggac gaagactgac gctcaggtgc gaaagnnnnn nnnnnnaaca 780
ggattagata ccctggtagt ccatgccgta aacgatgtct actagccgtt ggggcctttg 840
aggctttagt ggcgcagcta acgcgataag tagaccgcct ggggagtacg gtcgcaagac 900
taaanctcaa atgaattgac gggggcccgc acaagcggtg gagcatgtgg tttaattcga 960
tgcaacgcga agaaccttac ctggccttga catagtaaga attttccaga gatggattgg 1020
tgccttcggg aacttacata caggtgctgc atggctgtcg tcagctcgtg tcgtgagatg 1080
ttgggttaag tcccgcaacg agcgcaaccc ttgtcattag ttgctacatt tagttgggca 1140
ctctaatgag actgccggtg acaaaccgga ggaaggtggg gatgacgtca agtcctcatg 1200
gcccttatag gtggggctac acacgtcata caatggctgg tacaaagggt tgccaacccg 1260
cgagggggag ctaatcccat aaaaccagtc gtagtccgga tcgcagtctg caactcgact 1320
gcgtgaagtc ggaatcgcta gtaatcgtgg atcagaatgt cacggtgaat acgttcccgg 1380
gtcttgtaca caccgcccgt cacaccatgg gagcgggttc tgccagaagt agttagctta 1440
accgcaagga gggcgattac cacggcaggg ttcgtgactg gggtgaagtc gtaacaaggt 1500
agccgtatcg gaaggtgcgg ctggatcacc tccttt 1536
<210> 270
<211> 882
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifD1
<400> 270
atgaccatgc gtcaatgcgc catttacggc aaaggtggga tcggcaaatc gaccaccaca 60
cagaacctgg tcgccgcgct ggcggagatg ggtaaaaaag tcatgattgt cggctgtgac 120
ccgaaagccg attccacgcg tttgatcctg catgcgaaag cgcagaacac cattatggag 180
atggctgctg aagtcggctc cgtggaagac ctggagttag aagacgtgct gcaaatcggt 240
tacggcggcg tgcgctgcgc agagtccggc ggcccggagc caggcgtggg ctgtgccggt 300
cgcggggtga tcaccgcgat taacttcctc gaagaagaag gcgcttacgt gccggatctc 360
gattttgttt tctacgacgt gctgggcgac gtggtatgcg gtggtttcgc catgccgatt 420
cgtgaaaaca aagcgcagga gatctacatc gtttgctctg gcgaaatgat ggcgatgtac 480
gccgccaaca acatctccaa aggcatcgtg aaatacgcca aatccggtaa agtgcgcctc 540
ggcgggctga tttgtaactc gcgccagacc gaccgtgaag atgaactgat cattgcgctg 600
gcagaaaaac tcggcacgca gatgatccac tttgttcccc gcgacaacat tgtgcagcgt 660
gcggaaatcc gccgtatgac ggttatcgaa tatgacccga cctgcaatca ggcgaacgaa 720
tatcgcagcc ttgccagcaa aatcgtcaac aacaccaaaa tggtggtgcc caccccctgc 780
accatggatg aactggaaga actgctgatg gagttcggca ttatggatgt ggaagacacc 840
agcatcattg gtaaaaccgc cgccgaagaa aacgccgtct ga 882
<210> 271
<211> 1386
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> nifK1
<400> 271
atggccgaaa ttctgcgcag taaaaaaccg ctggcggtca gcccgataaa aagcggccag 60
ccgctggggg cgatcctcgc aagcctgggt gtcgaacagt gcataccgct ggtacacggc 120
gcacagggat gtagcgcgtt cgcgaaggtg ttctttattc aacattttca cgatccgatc 180
ccgctgcaat cgacggcgat ggatccgact tccaccatta tgggcgccga tgaaaacatt 240
tttaccgcgc tcaatgtgct ctgccagcgc aacgccgcga aagccattgt gctgctcagc 300
accgggcttt cagaagccca gggcagcgac atttcgcggg tggtgcgcca gtttcgtgat 360
gattttcccc ggcataaagg cgttgcgctg ctcaccgtca acacacccga tttctacggc 420
tcgctggaaa acggctacag cgccgtgctg gaaagcatga ttgaacagtg ggtacccgca 480
cagcccgccg ccagcctgcg caaccgccgt gtcaacctgc tggtcagcca tttactgaca 540
ccaggcgata tcgaactgtt gcgcagttat gttgaagcct tcggcctgca accggtgatt 600
gtgccggatc tgtcgctgtc gctggacggg catctggcag acggtgattt ttcgcctgtt 660
acccaagggg gaacatcgct gcgcatgatt gaacagatgg ggcaaaacct ggccaccttt 720
gtgattggcg cctcgctggg ccgtgcggcg gcgttactgg cgcagcgcag ccgtggcgag 780
gtgatcgccc tgccgcatct gatgacgctt gcagcctgcg acacgtttat tcatcgactg 840
aaaaccctct ccgggcgcga tgtccccgcg tggattgagc gccagcgcgg ccaagttcag 900
gatgcgatga tcgattgcca tatgtggctg cagggtgcgg ctatcgccat ggcagcagaa 960
ggcgatcacc tggcggcatg gtgcgatttc gcccgcagcc agggcatgat ccccggcccg 1020
attgtcgcac cggtcagcca gccggggttg caaaatctgc cggttgaaac cgtggttatc 1080
ggcgatctgg aagatatgca ggatcggctt tgcgcgacgc ccgccgcgtt actggtggcc 1140
aattctcatg ccgccgatct cgccacgcag tttgatttgt cacttatccg cgccgggttc 1200
ccggtgtatg accggctggg ggaatttcgt cgcctgcgcc aggggtacag cggcattcgt 1260
gacacgctgt ttgagctggc gaatgtgatg cgcgagcgcc atcacccgct tgcaacctac 1320
cgctcgccgc tgcgccagca cgccgacgac aacgttacgc ctggagatct gtatgccgca 1380
tgttaa 1386
<210> 272
<211> 1287
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> amtB
<400> 272
atgaaaaaca caacattaaa aacagcgctt gcttcgctgg cgttactgcc tggcctggcg 60
atggcggctc ccgctgtggc ggataaagcc gacaacggct ttatgatgat ttgcaccgcg 120
ctggtgctgt ttatgaccat tccgggcatt gcgctgttct acggcggttt gatccgcggt 180
aaaaacgtgc tgtcgatgct gacgcaggtt gccgtcacct tcgcactggt gtgcattctg 240
tgggtggtgt atggctactc gctggcattt ggcgagggca acagcttctt cgggagtttt 300
aactgggcga tgttgaaaaa catcgaactg aaagccgtga tgggcagcat ttatcagtat 360
atccacgtgg cgttccaggg ttccttcgcc tgtatcaccg ttggcctgat tgtcggtgca 420
ctggctgagc gtattcgctt ctctgcggtg ctgatttttg tggtggtatg gctgacgctt 480
tcttacgtgc cgattgcaca catggtgtgg ggcggcggtc tgctggcaac ccacggtgcg 540
ctggatttcg caggcggtac ggttgttcac atcaacgctg cgattgcagg tctggtgggg 600
gcttacctga ttggcaaacg cgtgggcttt ggcaaagaag cattcaaacc gcataacctg 660
ccgatggtct tcactggcac cgctatcctg tatgttggct ggtttggttt caacgccggc 720
tccgcaagct cggcgaacga aattgctgcg ctggccttcg tgaacactgt cgttgccact 780
gctgccgcta ttctggcgtg ggtatttggc gaatgggcaa tgcgcggcaa gccgtctctg 840
ctcggtgcct gttctggtgc catcgcgggt ctggttggta tcacccccgc ctgtggttat 900
gtgggtgtcg gcggtgcgct gattgtgggt ctgattgccg gtctggctgg gctgtggggc 960
gttactgcgc tgaaacgtat gttgcgtgtc gatgacccgt gtgacgtatt cggtgtgcac 1020
ggcgtgtgcg gcatcgtggg ctgtatcctg acgggtatct tcgcctctac gtcgctgggt 1080
ggtgtcggtt tcgctgaagg tgtgaccatg ggccatcagg tgctggtgca gctggaaagt 1140
gttgccatca ctatcgtgtg gtctggcgtg gtggccttta ttggttacaa actggcggac 1200
atgacggtag gcctgcgcgt accggaagaa caagaacgtg aagggctgga tgtaaacagc 1260
cacggcgaaa acgcctataa cgcctga 1287
<210> 273
<211> 348
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> Prm1
<400> 273
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc ggatttcact atttaaactg 120
gccattatct aagatgaatc cgatggaagc tcgctgtttt aacacgcgtt ttttaacctt 180
ttattgaaag tcggtgcttc tttgagcgaa cgatcaaatt taagtggatt cccatcaaaa 240
aaatattctc aacctaaaaa agtttgtgta atacttgtaa cgctacatgg agattaactc 300
aatctagagg gtattaataa tgaatcgtac taaactggta ctgggcgc 348
<210> 274
<211> 313
<212> ДНК
<213> Kosakonia sacchari
<220>
<223> Prm5
<400> 274
ggacatcatc gcgacaaaca atattaatac cggcaaccac accggcaatt tacgagactg 60
cgcaggcatc ctttctcccg tcaatttctg tcaaataaag taaaagaggc agtctacttg 120
aattaccccc ggctggttga gcgtttgttg aaaaaaagta actgaaaaat ccgtagaata 180
gcgccactct gatggttaat taacctattc aattaagaat tatctggatg aatgtgccat 240
taaatgcgca gcataatggt gcgttgtgcg ggaaaactgc ttttttttga aagggttggt 300
cagtagcgga aac 313
<210> 275
<211> 1485
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifL
<400> 275
atgaccctga atatgatgct cgataacgcc gcgccggagg ccatcgccgg cgcgctgact 60
caacaacatc cggggctgtt ttttaccatg gtggaacagg cctcggtggc catctccctc 120
accgatgcca gcgccaggat catttacgcc aacccggcgt tttgccgcca gaccggctat 180
tcgctggcgc aattgttaaa ccagaacccg cgcctgctgg ccagcagcca gacgccgcgc 240
gagatctatc aggagatgtg gcataccctg ctccagcgtc agccctggcg cggtcagctg 300
attaatcagc gtcgggacgg cggcctgtac ctggtggaga ttgacatcac cccggtgctt 360
agcccgcaag gggaactgga gcattatctg gcgatgcagc gggatatcag cgtcagctac 420
accctcgaac agcggctgcg caaccatatg accctgatgg aggcggtgct gaataatatc 480
cccgccgccg tggtagtggt ggacgagcag gatcgggtgg tgatggacaa cctcgcctac 540
aaaaccttct gcgctgactg cggcggccgg gagctgctca ccgagctgca ggtctcccct 600
ggccggatga cgcccggcgt ggaggcgatc ctgccggtgg cgctgcgcgg ggccgcgcgc 660
tggctgtcgg taacctgctg gccgttgccc ggcgtcagtg aagaggccag ccgctacttt 720
atcgacagcg cgctggcgcg gaccctggtg gtgatcgccg actgtaccca gcagcgtcag 780
cagcaggagc aagggcgcct tgaccggctg aagcagcaaa tgaccgccgg caagctgctg 840
gcggcgatcc gcgagtcgct ggacgccgcg ctgatccagc tgaactgccc gattaatatg 900
ctggcggcag cccgtcggct gaacggcgag ggaagcggga atgtggcgct ggaggccgcc 960
tggcgtgaag gggaagaggc gatggcgcgg ctccagcgct gtcgcccatc gctggaactc 1020
gaaaaccccg ccgtctggcc gctgcagccc tttttcgacg atctgtgcgc cctctaccgt 1080
acacgcttcg atcccgacgg gctgcaggtc gacatggcct caccgcatct gatcggcttt 1140
ggccagcgca ccccactgct ggcgtgctta agcctgtggc tcgatcgcac cctggccctc 1200
gccgccgaac tcccctccgt gccgctggcg atgcagctct acgccgagga gaacgacggc 1260
tggctgtcgc tgtatctgac tgacaacgta ccgctgctgc aggtgcgcta cgctcactcc 1320
cccgacgcgc tgaactcgcc gggcaaaggc atggagctgc ggctgatcca gaccctggtg 1380
gcgcaccatc gcggggccat tgagctggct tcccgaccgc agggcggcac ctgcctgacc 1440
ctgcgtttcc cgctgtttaa caccctgacc ggaggtgaag catga 1485
<210> 276
<211> 1575
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifA
<400> 276
atgatccctg aatccgaccc ggacaccacc gtcagacgct tcgacctctc tcagcagttc 60
accgccatgc agcggataag cgtggtgctg agccgggcca ccgaggccag caaaacgctg 120
caggaggtgc tcagcgtatt acacaacgat gcctttatgc agcacgggat gatctgcctg 180
tacgacagcg agcaggagat cctcagtatc gaagcgctgc agcaaaccgg ccagcagccc 240
ctccccggca gcacgcagat ccgctatcgc cccggcgagg gactggtggg gaccgtgctg 300
gcccaggggc agtcgctggt gctgccccgg gtcgccgacg atcagcgttt tctcgaccgc 360
ctgagcctct acgattacga tctgccgttt atcgccgtac cgttgatggg gcccaacgcc 420
cggccaatag gggtgctggc ggcccagccg atggcgcgcc aggaagagcg gctgccggcc 480
tgcacccgtt ttctcgaaac cgtcgccaac ctcgtcgccc agaccatccg gctgatgatc 540
cttccggcct cacccgccct gtcgagccgc cagccgccga aggtggaacg gccgccggcc 600
tgctcgtcgt cgcgcggcgt gggccttgac aatatggtcg gcaagagccc ggcgatgcgc 660
cagatcgtgg aggtgatccg tcaggtttcg cgctgggaca ccaccgtgct ggtacgcggc 720
gaaagcggca ccgggaaaga gctgatcgcc aacgccatcc atcaccattc gccacgggct 780
ggcgccgcct tcgtcaaatt taactgcgcg gcgctgccgg acaccctgct ggaaagcgaa 840
ctgttcggcc atgagaaagg cgcctttacc ggggcggtgc gtcagcgtaa aggacgtttt 900
gagctggcgg atggcggcac cctgttcctc gatgagattg gtgaaagcag cgcctcgttc 960
caggccaagc tgctgcgtat cctccaggag ggggagatgg agcgggtcgg cggcgatgag 1020
accctgcggg tgaatgtccg catcatcgcc gccaccaacc gtcacctgga ggaggaggtc 1080
cggctgggcc atttccgcga ggatctctac tatcgtctga acgtgatgcc catcgccctg 1140
cccccgctgc gcgagcgtca ggaggacatc gccgagctgg cgcacttcct ggtgcgcaaa 1200
atcggccagc atcaggggcg cacgctgcgg atcagcgagg gcgcgatccg cctgctgatg 1260
gagtacagct ggccgggtaa cgttcgcgaa ctggagaact gcctcgaacg atcggcggtg 1320
atgtcggaga gtggcctgat cgatcgcgac gtgatcctct tcactcacca ggatcgtccc 1380
gccaaagccc tgcctgccag cgggccagcg gaagacagct ggctggacaa cagcctggac 1440
gaacgtcagc gactgatcgc cgcgctggaa aaagccggct gggtgcaggc caaggcggca 1500
cggctgctgg ggatgacgcc gcgccaggtc gcttatcgga tccagatcat ggatatcacc 1560
ctgccgcgtc tgtag 1575
<210> 277
<211> 1540
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-1
<400> 277
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcag cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgaaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggctcacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcatcatac ttaatacgtg tggtgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgagcttaa cttgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccaga gaatttgcca gagatggcga 1020
agtgccttcg ggaactctga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc acgtaatggt 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgaga gcaagcggac ctcataaagt atgtcgtagt ccggattgga gtctgcaact 1320
cgactccatg aagtcggaat cgctagtaat cgtagatcag aatgctacgg tgaatacgtt 1380
cccgggcctt gtacacaccg cccgtcacac catgggagtg ggttgcaaaa gaagtaggta 1440
gcttaacctt cgggagggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac 1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt 1540
<210> 278
<211> 1540
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-2
<220>
<221> модифицированное основание
<222> (267)..(267)
<223> a, c, t, g, неизвестное или другое
<400> 278
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcat cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgaaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggctnacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcatcatac ttaatacgtg tggtgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgagcttaa cttgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccaga gaatttgcca gagatggcga 1020
agtgccttcg ggaactctga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc acgtgatggt 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgaga gcaagcggac ctcataaagt atgtcgtagt ccggattgga gtctgcaact 1320
cgactccatg aagtcggaat cgctagtaat cgtagatcag aatgctacgg tgaatacgtt 1380
cccgggcctt gtacacaccg cccgtcacac catgggagtg ggttgcaaaa gaagtaggta 1440
gcttaacctt cgggagggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac 1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt 1540
<210> 279
<211> 1540
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-3
<220>
<221> модифицированное основание
<222> (455)..(455)
<223> a, c, t, g, неизвестное или другое
<400> 279
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcag cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgaaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggctcacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcancatac ttaatacgtg tggtgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgagcttaa cttgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccaga gaatttgcca gagatggcga 1020
agtgccttcg ggaactctga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc acgtgatggt 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgaga gcaagcggac ctcataaagt atgtcgtagt ccggattgga gtctgcaact 1320
cgactccatg aagtcggaat cgctagtaat cgtagatcag aatgctacgg tgaatacgtt 1380
cccgggcctt gtacacaccg cccgtcacac catgggagtg ggttgcaaaa gaagtaggta 1440
gcttaacctt cgggagggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac 1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt 1540
<210> 280
<211> 1540
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-4
<220>
<221> модифицированное основание
<222> (70)..(70)
<223> a, c, t, g, неизвестное или другое
<400> 280
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcan cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgaaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggcttacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcatcacac ttaatacgtg tggtgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgagcttaa cttgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccaga gaatttgcca gagatggcga 1020
agtgccttcg ggaactctga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc acgtgatggt 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgaga gcaagcggac ctcataaagt atgtcgtagt ccggattgga gtctgcaact 1320
cgactccatg aagtcggaat cgctagtaat cgtagatcag aatgctacgg tgaatacgtt 1380
cccgggcctt gtacacaccg cccgtcacac catgggagtg ggttgcaaaa gaagtaggta 1440
gcttaacctt cgggagggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac 1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt 1540
<210> 281
<211> 1540
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-5
<400> 281
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcag cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tctgggaaac tgcctgatgg agggggataa ctactggaaa cggtagctaa taccgcatga 180
cctcgaaaga gcaaagtggg ggatcttcgg acctcacgcc atcggatgtg cccagatggg 240
attagctagt aggtgaggta atggctcacc taggcgacga tccctagctg gtctgagagg 300
atgaccagcc acactggaac tgagacacgg tccagactcc tacgggaggc agcagtgggg 360
aatattgcac aatgggcgca agcctgatgc agccatgccg cgtgtgtgaa gaaggcctta 420
gggttgtaaa gcactttcag cgaggaggaa ggcatcacac ttaatacgtg tgttgattga 480
cgttactcgc agaagaagca ccggctaact ccgtgccagc agccgcggta atacggaggg 540
tgcaagcgtt aatcggaatt actgggcgta aagcgcacgc aggcggtttg ttaagtcaga 600
tgtgaaatcc ccgagcttaa cttgggaact gcatttgaaa ctggcaagct agagtcttgt 660
agaggggggt agaattccag gtgtagcggt gaaatgcgta gagatctgga ggaataccgg 720
tggcgaaggc ggccccctgg acaaagactg acgctcaggt gcgaaagcgt ggggagcaaa 780
caggattaga taccctggta gtccacgctg taaacgatgt cgacttggag gttgtgccct 840
tgaggcgtgg cttccggagc taacgcgtta agtcgaccgc ctggggagta cggccgcaag 900
gttaaaactc aaatgaattg acgggggccc gcacaagcgg tggagcatgt ggtttaattc 960
gatgcaacgc gaagaacctt acctactctt gacatccaga gaatttgcca gagatggcga 1020
agtgccttcg ggaactctga gacaggtgct gcatggctgt cgtcagctcg tgttgtgaaa 1080
tgttgggtta agtcccgcaa cgagcgcaac ccttatcctt tgttgccagc acgtgatggt 1140
gggaactcaa aggagactgc cggtgataaa ccggaggaag gtggggatga cgtcaagtca 1200
tcatggccct tacgagtagg gctacacacg tgctacaatg gcatatacaa agagaagcga 1260
actcgcgaga gcaagcggac ctcataaagt atgtcgtagt ccggattgga gtctgcaact 1320
cgactccatg aagtcggaat cgctagtaat cgtagatcag aatgctacgg tgaatacgtt 1380
cccgggcctt gtacacaccg cccgtcacac catgggagtg ggttgcaaaa gaagtaggta 1440
gcttaacctt cgggagggcg cttaccactt tgtgattcat gactggggtg aagtcgtaac 1500
aaggtaaccg taggggaacc tgcggttgga tcacctcctt 1540
<210> 282
<211> 1009
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-6
<220>
<221> модифицированное основание
<222> (296)..(296)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (311)..(311)
<223> a, c, t, g, неизвестное или другое
<220>
<221> модифицированное основание
<222> (973)..(973)
<223> a, c, t, g, неизвестное или другое
<400> 282
gtagctaata ccgcatgacc tcgaaagagc aaagtggggg atcttcggac ctcacgccat 60
cggatgtgcc cagatgggat tagctagtag gtgaggtaat ggctcaccta ggcgacgatc 120
cctagctggt ctgagaggat gaccagccac actggaactg agacacggtc cagactccta 180
cgggaggcag cagtggggaa tattgcacaa tgggcgcaag cctgatgcag ccatgccgcg 240
tgtgtgaaga aggccttagg gttgtaaagc actttcagcg aggaggaagg catcanactt 300
aatacgtgtg ntgattgacg ttactcgcag aagaagcacc ggctaactcc gtgccagcag 360
ccgcggtaat acggagggtg caagcgttaa tcggaattac tgggcgtaaa gcgcacgcag 420
gcggtttgtt aagtcagatg tgaaatcccc gagcttaact tgggaactgc atttgaaact 480
ggcaagctag agtcttgtag aggggggtag aattccaggt gtagcggtga aatgcgtaga 540
gatctggagg aataccggtg gcgaaggcgg ccccctggac aaagactgac gctcaggtgc 600
gaaagcgtgg ggagcaaaca ggattagata ccctggtagt ccacgctgta aacgatgtcg 660
acttggaggt tgtgcccttg aggcgtggct tccggagcta acgcgttaag tcgaccgcct 720
ggggagtacg gccgcaaggt taaaactcaa atgaattgac gggggcccgc acaagcggtg 780
gagcatgtgg tttaattcga tgcaacgcga agaaccttac ctactcttga catccagaga 840
atttgccaga gatggcgaag tgccttcggg aactctgaga caggtgctgc atggctgtcg 900
tcagctcgtg ttgtgaaatg ttgggttaag tcccgcaacg agcgcaaccc ttatcctttg 960
ttgccagcac gtnatggtgg gaactcaaag gagactgccg gtgataaac 1009
<210> 283
<211> 1519
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> 16S-7
<400> 283
attgaagagt ttgatcatgg ctcagattga acgctggcgg caggcctaac acatgcaagt 60
cgagcggcag cgggaagtag cttgctactt tgccggcgag cggcggacgg gtgagtaatg 120
tcctgatgga ggggataact actggaacgg tagctaatac cgcacctcga aagagcaaag 180
tgggggatct tcggacctca cgccatcgga tgtgcccaga tgggattagc tagtaggtga 240
ggtaatggct cacctaggcg acgatcccta gctggtctga gaggatgacc agccacactg 300
gaactgagac acggtccaga ctcctacggg aggcagcagt ggggaatatt gcacaatggg 360
cgcaagcctg atgcagccat gccgcgtgtg tgaagaaggc cttagggttg taaagcactt 420
tcagcgagga ggaaggcatc atacttaata cgtgtggtga ttgacgttac tcgcagaaga 480
agcaccggct aactccgtgc cagcagccgc ggtaatacgg agggtgcaag cttaatcgga 540
attactgggc gtaaagcgca cgcaggcggt tgttaagtca gatgtgaaat ccccgagctt 600
aacttgggaa ctgcatttga aactggcaag ctagagtctt gtagaggggg gtagaattcc 660
aggtgtagcg gtgaaatgcg tagagatctg gaggaatacc ggtggcgaag gcggccccct 720
ggacaaagac tgacgctcag gtgcgaaagc gtggggagca aacaggatta ataccctggt 780
agtccacgct gtaacgatgt cgacttggag gttgtgccct gaggcgtggc ttccggagct 840
aacgcgttaa gtcgaccgcc tggggagtac ggccgcaagg ttaaaactca aatgaattga 900
cgggggcccg cacaagcggt ggagcatgtg gtttaattcg atgcaacgcg aagaacctta 960
cctactcttg acatccagag aatttgccag agatggcgaa gtgccttcgg gaactctgag 1020
acaggtgctg catggctgtc gtcagctcgt gttgtgaaat gttgggttaa gtcccgcaac 1080
gagcgcaacc cttatccttt gttgccagca cgtaatggtg ggaactcaaa ggagactgcc 1140
ggtgataaac cggaggaagg tggggatgac gtcaagtcat catggccctt acgagtaggg 1200
ctacacacgt gctacaatgg catatacaaa gagaagcgaa ctcgcgagag caagcggacc 1260
tcataaagta tgtcgtagtc cggattggag tctgcaactc gactccatga agtcggaatc 1320
gctagtaatc gtagatcaga atgctacggt gaatacgttc ccgggccttg tacacaccgc 1380
ccgtcacacc atgggagtgg gttgcaaaag aagtaggtag cttaaccttc gggagggcgc 1440
ttaccacttt gtgattcatg actggggtga agtcgtaaca aggtaaccgt aggggaacct 1500
gcggttggat cacctcctt 1519
<210> 284
<211> 882
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifH1
<400> 284
atgaccatgc gtcaatgcgc tatctacggt aaaggcggta tcggtaaatc caccaccacc 60
cagaatctcg tcgcggccct cgccgagatg ggtaagaaag tgatgatcgt cggctgcgat 120
ccgaaagcgg attccacccg tctgatcctc cacgctaaag cccagaacac catcatggag 180
atggcggcgg aagtgggctc ggtcgaggat ctggagctcg aagacgttct gcaaatcggc 240
tatggcgatg tccgttgcgc cgaatccggc ggcccggagc caggcgtcgg ctgcgccgga 300
cgcggggtga tcaccgccat caacttcctc gaggaagaag gcgcctatga agaagatttg 360
gatttcgtct tctatgacgt cctcggcgac gtggtctgcg gcggcttcgc tatgccgatc 420
cgcgaaaaca aagcccagga gatctacatc gtctgctccg gcgagatgat ggcgatgtat 480
gccgccaaca atatctccaa agggatcgtg aagtacgcca aatccggcaa ggtgcgcctc 540
ggcggcctga tctgtaactc gcgcaaaacc gaccgggaag acgaactgat catcgccctg 600
gcggagaagc ttggcacgca gatgatccac ttcgttcccc gcgacaacat tgtgcagcgc 660
gcggagatcc gccggatgac ggtgatcgag tacgacccga cctgtcagca ggcgaatgaa 720
tatcgtcaac tggcgcagaa gatcgtcaat aacaccaaaa aagtggtgcc gacgccgtgc 780
accatggacg agctggaatc gctgctgatg gagttcggca tcatggaaga agaagacacc 840
agcatcattg gtaaaaccgc cgctgaagaa aacgcggcct ga 882
<210> 285
<211> 1113
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifH2
<400> 285
atggttagga aaagtagaag taaaaataca aatatagaac taactgaaca tgaccattta 60
ttaataagtc aaataaaaaa gcttaaaaca caaaccactt gcttttttaa taataaagga 120
ggggttggga agactacatt agtagcaaat ttaggagcag agctatcaat aaactttagt 180
gcaaaagttc ttattgtgga tgccgaccct caatgtaatc tcacgcagta tgtattaagt 240
gatgaagaaa ctcaggactt atatgggcaa gaaaatccag atagtattta tacagtaata 300
agaccactat cctttggtaa aggatatgaa agtgacctcc ctataaggca tgtagagaat 360
ttcggttttg acataattgt cggtgaccct agacttgctt tacaggaaga ccttttagct 420
ggagactggc gagatgccaa aggcggtggg atgcgaggaa ttaggacaac ttttgtattt 480
gcagagttaa ttaagaaagc tcgtgagcta aattatgatt ttgttttctt tgacatggga 540
ccatcattag gcgcaatcaa cagggcagta ttactggcaa tggaattctt tgtcgtccca 600
atgtcaatcg atgtattttc actatgggct attaaaaata ttggctccac ggtttcaata 660
tggaaaaaag aattagacac agggattcgg ctctcagagg aacctagcga attatcacaa 720
ttatcacctc aaggaaaact aaagtttctc ggttacgtca cccaacaaca taaagaacgc 780
tctggatacg atacaattca gcttgagaat actgaggaag aaataaaatc gaaacgtcgg 840
gtaaaggcgt atgaagacat tggagaggtg tttccttcta aaattactga gcatctttct 900
aaactttatg catcaaaaga tatgaaccca caccttggag atatacgtca tttaggtagt 960
ttagctccga aatcacaatc acaacacgtt ccgatgatat cagtgtctgg tacaggaaat 1020
tacaccagac ttagaaaaag cgcgcgtgaa ctttatcgag atattgcaag aagatactta 1080
gagaacattc agactgctaa tggcgagaaa tag 1113
<210> 286
<211> 1374
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifD1
<400> 286
atgaagggaa aggaaattct ggcgctgctg gacgaacccg cctgcgagca caaccagaag 60
caaaaatccg gctgcagcgc ccctaagccc ggcgctaccg ccggcggttg cgccttcgac 120
ggcgcgcaga taacgctcct gcccatcgcc gacgtcgcgc acctggtgca cggccccatc 180
ggctgcgcgg gcagctcgtg ggataaccgc ggcagcgtca gcgccggccc ggccctcaac 240
cggctcggct ttaccaccga tcttaacgaa caggatgtga ttatgggccg cggcgaacgc 300
cgcctgttcc acgccgtgcg tcacatcgtc gaccgctatc atccggcggc ggtctttatc 360
tacaacacct gcgtaccggc gatggagggc gatgacatcg aggcggtctg ccaggccgca 420
cagaccgcca ccggcgtccc ggtcatcgct attgacgccg ccggtttcta cggcagtaaa 480
aatcttggca accgaatggc gggcgacgtg atgctcaggc aggtgattgg ccagcgcgaa 540
ccggccccgt ggccagacaa cacgcccttt gccccggccc agcgccacga tatcggcctg 600
attggcgaat tcaatatcgc cggcgagttc tggcaggtcc agccgctgct cgacgagctg 660
gggatccgcg tcctcggcag cctctccggc gacggccgct ttgccgagat ccagaccctg 720
caccgggcgc aggccaatat gctggtgtgc tcgcgcgcgc tgatcaacgt cgcccggggg 780
ctggagctgc gctacggcac gccgtggttt gaaggcagct tctacgggat ccgcgccacc 840
tccgacgcct tgcgccagct ggcgacgctg ctgggggatg acgacctgcg ccgccgcacc 900
gaggcgctga tcgcccgcga agagcaggcg gcggagcagg ctcttgcgcc gtggcgtgag 960
cagctccgcg ggcgcaaagt gctgctctat accggcggcg tgaaatcctg gtcggtggta 1020
tcggccctgc aggatctcgg catgaccgtg gtggccaccg gcacgcgcaa atccaccgag 1080
gaggacaaac agcggatccg tgagctgatg ggcgacgagg cggtgatgct tgaggagggc 1140
aatgcccgca ccctgctcga cgtggtgtac cgctatcagg ccgacctgat gatcgccggc 1200
ggacgcaata tgtacaccgc ctggaaagcc cggctgccgt ttctcgatat caatcaggag 1260
cgcgagcacg cctacgccgg ctatcagggc atcatcaccc tcgcccgcca gctctgtctg 1320
accctcgcca gccccgtctg gccgcaaacg catacccgcg ccccgtggcg ctag 1374
<210> 287
<211> 1449
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifD2
<400> 287
atgaccaacg caacaggcga acgtaacctt gcgctcatcc aggaagtcct ggaggtgttt 60
cccgaaaccg cgcgcaaaga gcgcagaaag cacatgatga tcagcgatcc gcagatggag 120
agcgtcggca agtgcattat ctcgaaccgt aaatcgcagc ccggggtgat gaccgtgcgc 180
ggctgcgcct atgcgggctc gaaaggggtg gtgtttgggc caatcaaaga catggcccat 240
atctcgcacg gccccatcgg ctgcggccag tattcccgcg ccggacggcg caactactat 300
accggcgtca gcggtgtcga cagcttcggc accctgaact tcacctctga ttttcaggag 360
cgcgatattg ttttcggcgg cgataaaaag ctgaccaaac tgatcgaaga gatggagctg 420
ctgttcccgc tgaccaaagg gatcaccatc cagtcggagt gcccggtggg cctgatcggc 480
gatgacatca gcgccgtagc caacgccagc agcaaggcgc tggataaacc ggtgatcccg 540
gtgcgctgcg aaggctttcg cggcgtatcg caatcgctgg gccaccatat cgccaacgac 600
gtggtgcgcg actgggtgct gaacaatcgc gaagggcagc cgtttgccag caccccgtac 660
gatgttgcca tcattggcga ttacaacatc ggcggcgacg cctgggcctc gcgcattctg 720
ctggaagaga tggggctgcg cgtagtggcg cagtggtccg gcgacggcac cctggtggag 780
atggagaaca ccccattcgt taagcttaac ctcgtccact gctaccgttc gatgaactat 840
atcgcccgcc atatggagga gaaacatcag atcccatgga tggaatataa cttcttcggc 900
ccgaccaaaa tcgccgaatc gctgcgcaag atcgccgatc aatttgatga caccattcgc 960
gccaatgcgg aagcggtgat cgccaaatat gaggggcaga tggcggccat catcgccaaa 1020
tatcgcccgc ggctggaggg gcgcaaagtg ctgctgtaca tgggggggct gcggccgcgc 1080
cacgtcatcg gcgcctatga ggatctcggg atggagatca tcgccgccgg ctacgagttt 1140
gcccataacg atgattacga ccgcaccctg ccggacctga aagagggcac cctgctgttt 1200
gacgatgcca gcagctatga gctggaggcc ttcgtcaaag cgctgaaacc tgacctcatc 1260
ggctccggga tcaaagagaa atatatcttc cagaaaatgg gggtgccgtt ccgccagatg 1320
cactcctggg actattccgg cccctatcac ggctatgacg gcttcgccat ctttgcccgc 1380
gatatggata tgaccctgaa caatccggcg tggaacgaac tgactgcccc gtggctgaag 1440
tctgcgtga 1449
<210> 288
<211> 1386
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifK1
<400> 288
atggcagata ttatccgcag tgaaaaaccg ctggcggtga gcccgattaa aaccgggcaa 60
ccgctcgggg cgatcctcgc cagcctcggg ctggcccagg ccatcccgct ggtccacggc 120
gcccagggct gcagcgcctt cgccaaagtt ttctttattc agcatttcca tgacccggtg 180
ccgctgcagt cgacggccat ggatccgacc gccacgatca tgggggccga cggcaatatc 240
ttcaccgcgc tcgacaccct ctgccagcgc cacagcccgc aggccatcgt gctgctcagc 300
accggtctgg cggaagcgca gggcagcgat atcgcccggg tggtgcgcca gtttcgcgag 360
gcgcatccgc gccataacgg cgtggcgatc ctcaccgtca ataccccgga tttttttggc 420
tctatggaaa acggctacag cgcggtgatc gagagcgtga tcgagcagtg ggtcgcgccg 480
acgccgcgtc cggggcagcg gccccggcgg gtcaacctgc tggtcagcca cctctgttcg 540
ccaggggata tcgaatggct gggccgctgc gtggaggcct ttggcctgca gccggtgatc 600
ctgccggacc tctcgcagtc aatggatggc cacctcggtg aaggggattt tacgcccctg 660
acccagggcg gcgcctcgct gcgccagatt gcccagatgg gccagagtct gggcagcttc 720
gccattggcg tgtcgctcca gcgggcggca tcgctcctga cccaacgcag ccgcggcgac 780
gtgatcgccc tgccgcatct gatgaccctc gaccattgcg atacctttat ccatcagctg 840
gcgaagatgt ccggacgccg cgtaccggcc tggattgagc gccagcgtgg ccagctgcag 900
gatgcgatga tcgactgcca tatgtggctt cagggccagc gcatggcgat ggcggcggag 960
ggcgacctgc tggcggcgtg gtgtgatttc gcccgcagcc aggggatgca gcccggcccg 1020
ctggtcgccc ccaccagcca ccccagcctg cgccagctgc cggtcgagca agtcgtgccg 1080
ggggatcttg aggatctgca gcagctgctg agccaccaac ccgccgatct gctggtggct 1140
aactctcacg cccgcgatct ggcggagcag tttgccctgc cgctgatccg cgtcggtttt 1200
cccctcttcg accggctcgg tgagtttcgt cgcgtccgcc aggggtacgc cggtatgcga 1260
gatacgctgt ttgaactggc caatctgctg cgcgaccgcc atcaccacac cgccctctac 1320
cgctcgccgc ttcgccaggg cgccgacccc cagccggctt caggagacgc ttatgccgcc 1380
cattaa 1386
<210> 289
<211> 1563
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> nifK2
<400> 289
atgagccaaa cgatcgataa aattcacagc tgttatccgc tgtttgaaca ggatgaatac 60
cagaccctgt tccagaataa aaagaccctt gaagaggcgc acgacgcgca gcgtgtgcag 120
gaggtttttg cctggaccac caccgccgag tatgaagcgc tgaacttcca gcgcgaggcg 180
ctgaccgtcg acccggccaa agcctgccag ccgctcggcg ccgtactctg cgcgctgggg 240
ttcgccggca ccctgcccta cgtgcacggc tcccagggct gcgtcgccta ttttcgcacc 300
tactttaacc gccattttaa agagccggtc gcctgcgtct ccgactccat gaccgaggac 360
gcggcggtgt tcggcggcaa caacaacatg aatctgggcc tgcagaatgc cagcgcgctg 420
tataaacccg agattatcgc cgtctccacc acctgtatgg ccgaggtgat cggcgacgat 480
ctgcaggcgt ttatcgccaa cgccaaaaaa gagggatttg ttgacgaccg catcgccatt 540
ccttacgccc atacccccag ctttatcggc agccatgtca ccggctggga caatatgttc 600
gaagggttcg cgaagacctt taccgctgac tacgccgggc agccgggcaa acagcaaaag 660
ctcaatctgg tgaccggatt tgagacctat ctcggcaact tccgcgtgct gaagcggatg 720
atggcgcaga tggatgtccc gtgcagcctg ctctccgacc catcagaggt gctcgacacc 780
cccgccgacg gccattaccg gatgtacgcc ggcggcacca gccagcagga gatcaaaacc 840
gcgccggacg ccattgacac cctgctgctg cagccgtggc agctggtgaa aagcaaaaag 900
gtggttcagg agatgtggaa ccagcccgcc accgaggtgg ccgttccgct gggcctggcc 960
gccaccgacg cgctgctgat gaccgtcagt cagctgaccg gcaaaccgat cgccgacgct 1020
ctgaccctgg agcgcggccg gctggtcgac atgatgctgg attcccacac ctggctgcat 1080
ggcaaaaaat tcggcctcta cggcgatccg gatttcgtga tggggctgac gcgcttcctg 1140
ctggagctgg gctgcgagcc gacggtgatc ctcagtcata acgccaataa acgctggcaa 1200
aaagcgatga agaaaatgct cgatgcctcg ccgtacggtc aggaaagcga agtgttcatc 1260
aactgcgacc tgtggcactt ccggtcgctg atgttcaccc gtcagccgga ctttatgatc 1320
ggtaactcct acggcaagtt tatccagcgc gataccctgg caaagggcaa agccttcgaa 1380
gtgccgctga tccgtctggg ctttccgctg ttcgaccgcc atcatctgca ccgccagacc 1440
acctggggct atgaaggcgc aatgaacatc gtcacgacgc tggtgaacgc cgtgctggaa 1500
aaactggacc acgacaccag ccagttgggc aaaaccgatt acagcttcga cctcgttcgt 1560
taa 1563
<210> 290
<211> 2838
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> glnE
<400> 290
atgatgccgc tttctccgca attacagcag cactggcaga cggtcgctga ccgtctgcca 60
gcggattttc ccattgccga actgagccca caggccaggt cggtcatggc gttcagcgat 120
tttgtcgaac agagtgtgat cgcccagccg ggctggctga atgagcttgc ggactcctcg 180
ccggaggcgg aagagtggcg gcattacgag gcctggctgc aggatcgcct gcaggccgtc 240
actgacgaag cggggttgat gcgagagctg cgtctcttcc gccgccagat gatggtccgc 300
atcgcctggg cgcaggcgct gtcgctggtg agcgaagaag agactctgca gcagctgagc 360
gtcctggcgg agaccctgat tgtcgccgcc cgcgactggc tgtacgccgc ctgctgtaag 420
gagtggggaa cgccatgcaa tgccgagggc cagccgcagc cgctgctgat cctcgggatg 480
ggaaagctgg gcggcggcga gctgaacttc tcttccgata tcgatctgat ctttgcctgg 540
cctgagcatg gcgccacccg cggcggccgc cgcgagctgg ataacgccca gttctttacc 600
cgtctggggc agcggctgat caaggccctt gaccagccga cgcaggacgg ctttgtctat 660
cgggttgaca tgcgcctgcg gccgtttggc gacagtgggc cgctggtact cagttttgcg 720
gcgctggaag attattacca ggagcagggt cgggactggg aacgctatgc gatggtgaaa 780
gcgcggatca tgggcgataa cgacggcgtg tacgccagcg agttgcgcgc gatgctccgt 840
cctttcgtct tccgccgtta tatcgacttc agcgtgatcc agtcgctgcg taacatgaaa 900
ggcatgatcg cccgcgaagt gcggcgtcgc gggctgaaag acaacatcaa gctcggcgcc 960
ggcgggatcc gtgaaattga gtttatcgtt caggtctttc aactgatccg cggtggtcgc 1020
gaacctgcac tgcagcagcg cgccctgctg ccgacgctgg cggcgattga tgagctacat 1080
ctgctgccgg aaggcgacgc ggcgctgctg cgcgaggcct atctgttcct gcgccggctg 1140
gaaaacctgc tgcaaagcat caacgatgag cagacccaga ccctgccgca ggatgaactt 1200
aaccgcgcca ggctggcgtg ggggatgcat accgaagact gggagacgct gagcgcgcag 1260
ctggcgagcc agatggccaa cgtgcggcga gtgtttaatg aactgatcgg cgatgatgag 1320
gatcagtccc cggatgagca actggccgag tactggcgcg agctgtggca ggatgcgctg 1380
gaagaagatg acgccagccc ggcgctggcg catttaaacg ataccgaccg ccgtagcgtg 1440
ctggcgctga ttgccgattt tcgtaaagag ctggatcggc gcaccatcgg cccgcgcggc 1500
cgccaggtgc tggatcagct gatgccgcat ctgctgagcg aaatctgctc gcgcgccgat 1560
gcgccgctgc ctctggcgcg gatcacgccg ctgttgaccg ggatcgtcac ccgtaccacc 1620
tatcttgagc tgctgagcga attccccggc gcgctgaagc acctgatcac gctctgcgcg 1680
gcgtcgccga tggtcgccag ccagctggcg cgccacccgc tgctgctgga tgagctgctg 1740
gatcccaaca ccctctatca gccgacggcg accgatgcct atcgcgacga gctgcgccag 1800
tacctgctgc gcgtgccgga agaggatgaa gagcagcagc tggaggcgtt gcgccagttt 1860
aagcaggcgc agcagctgca tatcgcggcg gcggatatcg ctggtaccct gccggtgatg 1920
aaggtcagcg atcacttaac ctggcttgcc gaagcgatcc tcgacgcggt ggtgcagcag 1980
gcatgggggc agatggtcgc tcgctacggc cagccgaccc acctgcacga tcgccagggt 2040
cgcggcttcg ccgtcgtcgg ctacggtaag cttggcggct gggagctggg ctacagctcc 2100
gatctcgatc tggtgttcct ccatgactgc ccggcggagg tgatgaccga cggcgagcgg 2160
gagattgacg gccgtcagtt ctacctgcgg ctggcccagc ggatcatgca cctgttcagc 2220
acccgcacct cgtccggtat tctctacgaa gtggacgccc ggctgcgtcc ttctggcgcg 2280
gcggggatgc tggtcaccac cgccgacgcg tttgctgact atcagcagaa cgaagcctgg 2340
acgtgggaac atcaggcgct ggtgcgcgcc cgcgtggtct atggcgaccc ggcgctgcag 2400
gcgcgctttg acgccattcg tcgcgatatc ctgaccaccc cgcgggaggg gatgaccctg 2460
cagaccgagg ttcgcgagat gcgcgagaag atgcgcgccc accttggcaa caaacatccc 2520
gatcgttttg atatcaaagc cgatgccggc gggatcaccg atattgaatt tattactcag 2580
tatctggtcc tacgctatgc cagtgacaag ccgaagctga cccgctggtc tgacaacgtg 2640
cgtattcttg agctgctggc gcagaacgac atcatggacg aggaggaggc gcgcgcctta 2700
acgcatgcgt acaccacctt gcgtgatgcg ctccatcacc tggccctgca ggagcagccg 2760
ggacacgtgg cgccagaggc cttcagccgg gagcgtcagc aggtcagcgc cagctggcag 2820
aagtggctga tggcttaa 2838
<210> 291
<211> 449
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> Prm4
<400> 291
agtctgaact catcctgcgg cagtcggtga gacgtatttt tgaccaaaga gtgatctaca 60
tcacggaatt ttgtggttgt tgctgcttaa aagggcaaat ctacccttag aatcaactgt 120
tatatcaggg ggattcagag agatattagg aatttgcaca agcgcacaat ttaaccacat 180
catgataacg ccatgtaaaa caaagataaa aaaacaaaat gcagtgactt acatcgcaag 240
caaggcattt tcttatccaa ttgctcaaag tttggccttt catatcgcaa cgaaaatgcg 300
taatatacgc gcccttgcgg acatcagtat ggtcattcct agttcatgcg catcggacac 360
caccagctta caaattgcct gattgcggcc ccgatggccg gtatcactga ccgaccattt 420
cgtgccttat gtcatgcgat gggggctgg 449
<210> 292
<211> 500
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> Prm1.2
<400> 292
tgaacatcac tgatgcacaa gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg 60
caggcattcg cgttaaagcc gacttgagaa atgagaagat tggctttaaa attcgcgaac 120
acacgctacg ccgtgttcct tatatgttag tttgtggcga taaagaggtc gaagcaggca 180
aagttgctgt tcgtacccgc cgcggcaaag acttaggaag catggatgtt agcgaagtcg 240
ttgacaaact gctggcggaa atccgcagca gaagtcttca tcaactggag gaataaagta 300
ttaaaggcgg aaaacgagtt caaccggcgc gtcctaatcg cattaacaaa gagattcgcg 360
cgcaagaagt tcgcctcaca ggcgtcgatg gcgagcagat tggtattgtc agtctgaatg 420
aagctcttga aaaagctgag gaagcgggcg tcgatttagt agaaatcagt ccgaatgccg 480
agccgccagt ttgtcgaatc 500
<210> 293
<211> 170
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> Prm3.1
<400> 293
tacagtagcg cctctcaaaa atagataaac ggctcatgta cgtgggccgt ttattttttc 60
tacccataat cgggaaccgg tgttataatg ccgcgccctc atattgtggg gatttcttaa 120
tgacctatcc tgggtcctaa agttgtagtt gacattagcg gagcactaac 170
<210> 294
<211> 142
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> Prm6.1
<400> 294
aatttttttt cacaaagcgt agcgttattg aatcgcacat tttaaactgt tggccgctgt 60
ggaagcgaat attggtgaaa ggtgcggttt taaggccttt ttctttgact ctctgtcgtt 120
acaaagttaa tatgcgcgcc ct 142
<210> 295
<211> 293
<212> ДНК
<213> Rahnella aquatilis
<220>
<223> Prm7.1
<400> 295
ttaaaaacgt gaccacgagc attaataaac gccacgaaat gtggcgttta tttattcaaa 60
aagtatcttc tttcataaaa agtgctaaat gcagtagcag caaaattggg ataagtccca 120
tggaatacgg ctgttttcgc tgcaattttt aactttttcg taaaaaaaga tgtttctttg 180
agcgaacgat caaaatatag cgttaaccgg caaaaaatta ttctcattag aaaatagttt 240
gtgtaatact tgtaacgcta catggagatt aacttaatct agagggtttt ata 293
<210> 296
<211> 1188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2
<400> 296
atggcgctca aacagttaat ccgtctgtgt gccgcctcgc cgatggtcgc gacacaactt 60
gcacgtcatc ctttattgct cgatgaactg ctcgacccgc gcacgcttta ccagccgatt 120
gagccgggcg cttaccgcga cgaactgcgt cagtatctga tgcgggtgcc aacagaagac 180
gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg cccagcattt gcgtatcgca 240
gccggggata tttccggggc attgccggtg atgaaagtca gtgaccattt aacctacctt 300
gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg aacaaatggt cgtaaaatac 360
gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt ttgccgtggt cggttacggg 420
aaactcggtg gctgggagct gggttatagc tcagatctgg atctggtctt cctgctcgat 480
tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg acggacgtca gttttatctt 540
cggctggcgc agcgcattat gcacttattc agcacccgga catcgtcagg cattctttac 600
gaggttgatc cgcgtctgcg accttccggc gcatccggca tgctggtcag taccattgaa 660
gcgtttgcag attatcaggc caatgaagcc tggacgtggg agcatcaggc gctggttcgc 720
gcgcgcgtgg tttacgggga tccgcaactg acacagcaat ttaacgccac gcgtcgcgac 780
attctttgcc gccagcgcga tggcgacggc ctgcgtaagg aggtccgtga aatgcgcgag 840
aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt ttgatctgaa agccgatccg 900
ggtggcatca cggatattga attcattgca caatacctgg ttctgcgttt cgcgcatgat 960
gagccgaagc tgacgcgctg gtctgataac gtgcggattt ttgaactgat ggcacgatat 1020
gacatcatgc cggaagagga agcgcgccat ctgacgcagg cttatgtgac gctgcgcgat 1080
gaaattcatc atctggcgtt gcaggaacac agcgggaaag tggccgcgga cagctttgct 1140
actgagcgcg cgcagatccg tgccagctgg gcaaagtggc tcggctga 1188
<210> 297
<211> 2188
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> glnE-delta-AR-2 с фланкирующей областью длиной 500 п.н.
<400> 297
cggtactgga acagaaatcg gcggatgcgc aggaaatttg ttatgacacg gcctgtctga 60
agtgcaagtt agtgcttact tcctggctgg caacctcagg ctggacgccg tttattgatg 120
ataaatctgc gaagaaactg gacgcttcct tcaaacgttt tgctgacatc atgctcggtc 180
gtaccgcagc ggatctgaaa gaagcctttg cgcagccact gacggaagaa ggttatcgcg 240
atcagctggc gcgcctgaaa cgccagatca ttaccttcca tttgcttgcc ggtgcttacc 300
ctgaaaaaga cgtcgatgcg tatattgccg gctgggtgga cctgcaacag gccatcgttc 360
agcagcaaca cgcctgggag gattcggccc gttctcacgc ggtgatgatg gatgctttct 420
ggttaaacgg gcaacctcgt taactgactg actagcctgg gcaaactgcc cgggcttttt 480
tttgcaagga atctgatttc atggcgctca aacagttaat ccgtctgtgt gccgcctcgc 540
cgatggtcgc gacacaactt gcacgtcatc ctttattgct cgatgaactg ctcgacccgc 600
gcacgcttta ccagccgatt gagccgggcg cttaccgcga cgaactgcgt cagtatctga 660
tgcgggtgcc aacagaagac gaagaacagc agcttgaagc cgtgcgccag ttcaaacagg 720
cccagcattt gcgtatcgca gccggggata tttccggggc attgccggtg atgaaagtca 780
gtgaccattt aacctacctt gccgaggcca ttctcgatgt cgtggtgcag catgcgtggg 840
aacaaatggt cgtaaaatac gggcagcccg cgcatcttca gcaccgtgag gggcgcggtt 900
ttgccgtggt cggttacggg aaactcggtg gctgggagct gggttatagc tcagatctgg 960
atctggtctt cctgctcgat tgcgcgccgg aggtgatgac ggacggcgaa cgcagcatcg 1020
acggacgtca gttttatctt cggctggcgc agcgcattat gcacttattc agcacccgga 1080
catcgtcagg cattctttac gaggttgatc cgcgtctgcg accttccggc gcatccggca 1140
tgctggtcag taccattgaa gcgtttgcag attatcaggc caatgaagcc tggacgtggg 1200
agcatcaggc gctggttcgc gcgcgcgtgg tttacgggga tccgcaactg acacagcaat 1260
ttaacgccac gcgtcgcgac attctttgcc gccagcgcga tggcgacggc ctgcgtaagg 1320
aggtccgtga aatgcgcgag aaaatgtatg cccatctggg gagtaaaaaa gcccacgagt 1380
ttgatctgaa agccgatccg ggtggcatca cggatattga attcattgca caatacctgg 1440
ttctgcgttt cgcgcatgat gagccgaagc tgacgcgctg gtctgataac gtgcggattt 1500
ttgaactgat ggcacgatat gacatcatgc cggaagagga agcgcgccat ctgacgcagg 1560
cttatgtgac gctgcgcgat gaaattcatc atctggcgtt gcaggaacac agcgggaaag 1620
tggccgcgga cagctttgct actgagcgcg cgcagatccg tgccagctgg gcaaagtggc 1680
tcggctgagg gtttttattc ggctaacagg cgcttgtgat attatccggc gcattgtatt 1740
tacccgattt gatttatctg ttttggagtc ttgggatgaa agtgactttg cctgattttc 1800
accgcgcagg tgtgctggtt gtcggtgacg taatgttaga ccgttactgg tatggcccga 1860
ccaatcgtat ttctccggaa gctccggtgc cggtggtgaa ggtcagtacc attgaagagc 1920
ggcctggcgg tgcagctaac gtggcgatga acatttcatc tctgggcgcc tcttcctgtc 1980
tgatcggcct gaccggcgta gacgacgctg cgcgtgccct cagtgagcgt ctggcagaag 2040
tgaaagttaa ctgcgatttc gtcgcactat ccacacatcc taccatcacc aaactgcgaa 2100
ttttgtcccg taaccagcaa ctgatccgcc tcgactttga ggaaggtttt gaaggcgttg 2160
atctcgagcc gatgctgacc aaaataga 2188
<210> 298
<211> 524
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::null-v1
<400> 298
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg agtctgaact catcctgcga 240
tgggggctgg gccgtctctg aagctctcgg tgaacattgt tgcgaggcag gatgcgagct 300
ggttgtgttt tgacattacc gataatgtgc cgcgtgaacg ggtgcgttat gcccgcccgg 360
aagcggcgtt ttcccgtccg gggaatggca tggagctgcg ccttatccag acgctgatcg 420
cccatcatcg cggttcttta gatctctcgg tccgccctga tggcggcacc ttgctgacgt 480
tacgcctgcc ggtacagcag gttatcaccg gaggcttaaa atga 524
<210> 299
<211> 1524
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::null-v1 с фланкирующей областью длиной 500 п.н.
<400> 299
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
agtctgaact catcctgcga tgggggctgg gccgtctctg aagctctcgg tgaacattgt 780
tgcgaggcag gatgcgagct ggttgtgttt tgacattacc gataatgtgc cgcgtgaacg 840
ggtgcgttat gcccgcccgg aagcggcgtt ttcccgtccg gggaatggca tggagctgcg 900
ccttatccag acgctgatcg cccatcatcg cggttcttta gatctctcgg tccgccctga 960
tggcggcacc ttgctgacgt tacgcctgcc ggtacagcag gttatcaccg gaggcttaaa 1020
atgacccagt tacctaccgc gggcccggtt atccggcgct ttgatatgtc tgcccagttt 1080
acggcgcttt atcgcatcag cgtggcgctg agtcaggaaa gcaacaccgg gcgcgcactg 1140
gcggcgatcc tcgaagtgct tcacgatcat gcatttatgc aatacggcat ggtgtgtctg 1200
tttgataaag aacgcaatgc actctttgtg gaatccctgc atggcatcga cggcgaaagg 1260
aaaaaagaga cccgccatgt ccgttaccgc atgggggaag gcgtgatcgg cgcggtgatg 1320
agccagcgtc aggcgctggt gttaccgcgc atttcagacg atcagcgttt tctcgaccgc 1380
ctgaatattt acgattacag cctgccgttg attggcgtgc cgatccccgg tgcggataat 1440
cagccatcgg gcgtgctggt ggcacagccg atggcgttgc acgaagaccg gctgactgcc 1500
agtacgcggt ttttagaaat ggtc 1524
<210> 300
<211> 266
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::null-v2
<400> 300
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg ttaaagcctg ccggtacagc 240
aggttatcac cggaggctta aaatga 266
<210> 301
<211> 1266
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::null-v2 с фланкирующей областью длиной 500 п.н.
<400> 301
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
ttaaagcctg ccggtacagc aggttatcac cggaggctta aaatgaccca gttacctacc 780
gcgggcccgg ttatccggcg ctttgatatg tctgcccagt ttacggcgct ttatcgcatc 840
agcgtggcgc tgagtcagga aagcaacacc gggcgcgcac tggcggcgat cctcgaagtg 900
cttcacgatc atgcatttat gcaatacggc atggtgtgtc tgtttgataa agaacgcaat 960
gcactctttg tggaatccct gcatggcatc gacggcgaaa ggaaaaaaga gacccgccat 1020
gtccgttacc gcatggggga aggcgtgatc ggcgcggtga tgagccagcg tcaggcgctg 1080
gtgttaccgc gcatttcaga cgatcagcgt tttctcgacc gcctgaatat ttacgattac 1140
agcctgccgt tgattggcgt gccgatcccc ggtgcggata atcagccatc gggcgtgctg 1200
gtggcacagc cgatggcgtt gcacgaagac cggctgactg ccagtacgcg gtttttagaa 1260
atggtc 1266
<210> 302
<211> 943
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm4
<400> 302
atgagcatca cggcgttatc agcatcattt cctgagggga atatcgccag ccgcttgtcg 60
ctgcaacatc cttcactgtt ttataccgtg gttgaacaat cttcggtggc gatttcgctg 120
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 180
gcacttgaga cacttttggg cgagaaccac cgtctgctgg agtctgaact catcctgcgg 240
cagtcggtga gacgtatttt tgaccaaaga gtgatctaca tcacggaatt ttgtggttgt 300
tgctgcttaa aagggcaaat ctacccttag aatcaactgt tatatcaggg ggattcagag 360
agatattagg aatttgcaca agcgcacaat ttaaccacat catgataacg ccatgtaaaa 420
caaagataaa aaaacaaaat gcagtgactt acatcgcaag caaggcattt tcttatccaa 480
ttgctcaaag tttggccttt catatcgcaa cgaaaatgcg taatatacgc gcccttgcgg 540
acatcagtat ggtcattcct agttcatgcg catcggacac caccagctta caaattgcct 600
gattgcggcc ccgatggccg gtatcactga ccgaccattt cgtgccttat gtcatgcgat 660
gggggctggg ccgtctctga agctctcggt gaacattgtt gcgaggcagg atgcgagctg 720
gttgtgtttt gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg cccgcccgga 780
agcggcgttt tcccgtccgg ggaatggcat ggagctgcgc cttatccaga cgctgatcgc 840
ccatcatcgc ggttctttag atctctcggt ccgccctgat ggcggcacct tgctgacgtt 900
acgcctgccg gtacagcagg ttatcaccgg aggcttaaaa tga 943
<210> 303
<211> 1943
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> delta-nifL::Prm4 с фланкирующей областью длиной 500 п.н.
<400> 303
tgtttcgtct cgaggccggg caactgagcg gccccgttga aaccgacctg ggctggcatc 60
tgttgttgtg cgaacaaatt cgcctgccgc aacccttgcc gaaagccgaa gccttaacgc 120
gggtgcgtca gcaactgatt gcccggcaac agaaacatta tcagcgccag tggctgcaac 180
aactgatcaa cgcctgagcc tgttctcctt cttgttgatg cagacgggtt aatgcccgtt 240
ttgcacgaaa aatgcacata aattgcctgc gttgccttat aacagcgcag ggaaatcctg 300
cctccggcct tgtgccacac cgcgctttgc ctggtttgtg gtaaaaactg gcccgctttg 360
catcctgatg cttaaaacac cccgttcaga tcaacctttg ggcagataag cccgcgaaag 420
gcctgcaaat tgcacggtta ttccgggtga gtatatgtgt gatttgggtt ccggcattgc 480
gcaataaagg ggagaaagac atgagcatca cggcgttatc agcatcattt cctgagggga 540
atatcgccag ccgcttgtcg ctgcaacatc cttcactgtt ttataccgtg gttgaacaat 600
cttcggtggc gatttcgctg accgatccgc aggcgcgcat ttgttatgcc aatccggcat 660
tctgccgcca gacgggtttt gcacttgaga cacttttggg cgagaaccac cgtctgctgg 720
agtctgaact catcctgcgg cagtcggtga gacgtatttt tgaccaaaga gtgatctaca 780
tcacggaatt ttgtggttgt tgctgcttaa aagggcaaat ctacccttag aatcaactgt 840
tatatcaggg ggattcagag agatattagg aatttgcaca agcgcacaat ttaaccacat 900
catgataacg ccatgtaaaa caaagataaa aaaacaaaat gcagtgactt acatcgcaag 960
caaggcattt tcttatccaa ttgctcaaag tttggccttt catatcgcaa cgaaaatgcg 1020
taatatacgc gcccttgcgg acatcagtat ggtcattcct agttcatgcg catcggacac 1080
caccagctta caaattgcct gattgcggcc ccgatggccg gtatcactga ccgaccattt 1140
cgtgccttat gtcatgcgat gggggctggg ccgtctctga agctctcggt gaacattgtt 1200
gcgaggcagg atgcgagctg gttgtgtttt gacattaccg ataatgtgcc gcgtgaacgg 1260
gtgcgttatg cccgcccgga agcggcgttt tcccgtccgg ggaatggcat ggagctgcgc 1320
cttatccaga cgctgatcgc ccatcatcgc ggttctttag atctctcggt ccgccctgat 1380
ggcggcacct tgctgacgtt acgcctgccg gtacagcagg ttatcaccgg aggcttaaaa 1440
tgacccagtt acctaccgcg ggcccggtta tccggcgctt tgatatgtct gcccagttta 1500
cggcgcttta tcgcatcagc gtggcgctga gtcaggaaag caacaccggg cgcgcactgg 1560
cggcgatcct cgaagtgctt cacgatcatg catttatgca atacggcatg gtgtgtctgt 1620
ttgataaaga acgcaatgca ctctttgtgg aatccctgca tggcatcgac ggcgaaagga 1680
aaaaagagac ccgccatgtc cgttaccgca tgggggaagg cgtgatcggc gcggtgatga 1740
gccagcgtca ggcgctggtg ttaccgcgca tttcagacga tcagcgtttt ctcgaccgcc 1800
tgaatattta cgattacagc ctgccgttga ttggcgtgcc gatccccggt gcggataatc 1860
agccatcggg cgtgctggtg gcacagccga tggcgttgca cgaagaccgg ctgactgcca 1920
gtacgcggtt tttagaaatg gtc 1943
<210> 304
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 304
tggtgtccgg gcgaacgtcg ccaggtggca caaattgtca gaactacgac acgactaacc 60
gaccgcagga gtgtgcgatg accctgaata tgatgatgga 100
<210> 305
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 305
cggaaaacga gttcaaacgg cgcgtcccaa tcgtattaat ggcgagattc gcgccacgga 60
agttcgctta acaggtctgg aaggcgagca gcttggtatt 100
<210> 306
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 306
cgccagagag ttgaaatcga acatttccgt aataccgcca ttacccagga gccgttctgg 60
ttgcacagcg gaaaacgtta acgaaaggat atttcgcatg 100
<210> 307
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 307
cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta accgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccagc 100
<210> 308
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 308
tcaacctaaa aaagtttgtg taatacttgt aacgctacat ggagattaac tcaatctaga 60
gggtattaat aatgaatcgt actaaactgg tactgggcgc 100
<210> 309
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 309
cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta accgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccagc 100
<210> 310
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 310
aattttctgc ccaaatggct gggattgttc attttttgtt tgccttacaa cgagagtgac 60
agtacgcgcg ggtagttaac tcaacatctg accggtcgat 100
<210> 311
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 311
gtaaccaata aaggccacca cgccagacca cacgatagtg atggcaacac tttccagctg 60
caccagcacc tgatggccca tggtcacacc ttcagcgaaa 100
<210> 312
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 312
tggtattgtc agtctgaatg aagctcttga aaaagctgag gaagcgggcg tcgatttagt 60
agaaatcagt ccgaatgccg agccgccagt ttgtcgaatc 100
<210> 313
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 313
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 60
gcacttgaga cacttttggg cgagaaccac cgtctgctgg 100
<210> 314
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 314
cgggaaccgg tgttataatg ccgcgccctc atattgtggg gatttcttaa tgacctatcc 60
tgggtcctaa agttgtagtt gacattagcg gagcactaac 100
<210> 315
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 315
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 60
gcacttgaga cacttttggg cgagaaccac cgtctgctgg 100
<210> 316
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 316
tcaacctaaa aaagtttgtg taatacttgt aacgctacat ggagattaac tcaatctaga 60
gggtattaat aatgaatcgt actaaactgg tactgggcgc 100
<210> 317
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 317
gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta actgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccggc 100
<210> 318
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 318
gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta actgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccggc 100
<210> 319
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 319
taagaattat ctggatgaat gtgccattaa atgcgcagca taatggtgcg ttgtgcggga 60
aaactgcttt tttttgaaag ggttggtcag tagcggaaac 100
<210> 320
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 320
cgccagagag tcgaaatcga acatttccgt aataccgcga ttacccagga gccgttctgg 60
ttgcacagcg gaaaacgtta acgaaaggat atttcgcatg 100
<210> 321
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 321
cgccagagag tcgaaatcga acatttccgt aataccgcga ttacccagga gccgttctgg 60
ttgcacagcg gaaaacgtta acgaaaggat atttcgcatg 100
<210> 322
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 322
gatgatggat gctttctggt taaacgggca acctcgttaa ctgactgact agcctgggca 60
aactgcccgg gctttttttt gcaaggaatc tgatttcatg 100
<210> 323
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 323
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 60
gcacttgaga cacttttggg cgagaaccac cgtctgctgg 100
<210> 324
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 324
catcggacac caccagctta caaattgcct gattgcggcc ccgatggccg gtatcactga 60
ccgaccattt cgtgccttat gtcatgcgat gggggctggg 100
<210> 325
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 325
tcttcaacaa ctggaggaat aaggtattaa aggcggaaaa cgagttcaaa cggcacgtcc 60
gaatcgtatc aatggcgaga ttcgcgccct ggaagttcgc 100
<210> 326
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 326
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga 100
<210> 327
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 327
atcgcagcgt ctttgaatat ttccgtcgcc aggcgctggc tgccgagccg ttctggctgc 60
atagtggaaa acgataattt caggccaggg agcccttatg 100
<210> 328
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 328
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga 100
<210> 329
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 329
tcacttttta gcaaagttgc actggacaaa aggtaccaca attggtgtac tgatactcga 60
cacagcatta gtgtcgattt ttcatataaa ggtaattttg 100
<210> 330
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 330
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga 100
<210> 331
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 331
gttctccttt gcaatagcag ggaagaggcg ccagaaccgc cagcgttgaa gcagtttgaa 60
cgcgttcagt gtataatccg aaacttaatt tcggtttgga 100
<210> 332
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 332
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga 100
<210> 333
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 333
gatatgcctg aagtattcaa ttacttaggc atttacttaa cgcaggcagg caattttgat 60
gctgcctatg aagcgtttga ttctgtactt gagcttgatc 100
<210> 334
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 334
tggtattgtc agtctgaatg aagctcttga aaaagctgag gaagcgggcg tcgatttagt 60
agaaatcagt ccgaatgccg agccgccagt ttgtcgaatc 100
<210> 335
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 335
tgcaaattgc acggttattc cgggtgagta tatgtgtgat ttgggttccg gcattgcgca 60
ataaagggga gaaagacatg agcatcacgg cgttatcagc 100
<210> 336
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 336
tcagggctgc ggatgtcggg cgtttcacaa cacaaaatgt tgtaaatgcg acacagccgg 60
gcctgaaacc aggagcgtgt gatgaccttt aatatgatgc 100
<210> 337
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. выше соединения
<400> 337
cggaaaacga gttcaaacgg cacgtccgaa tcgtatcaat ggcgagattc gcgcccagga 60
agttcgctta actggtctgg aaggtgagca gctgggtatt 100
<210> 338
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 338
ttcttggttc tctggagcgc tttatcggca tcctgactga agaatttgca ggcttcttcc 60
caacctggct tgcacccgtg caggtagttg tgatgaacat 100
<210> 339
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 339
gcgatagaac tcacttcacg ccccgaaggg ggaagctgcc tgaccctacg attcccgcta 60
tttcattcac tgaccggagg ttcaaaatga cccagcgaac 100
<210> 340
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 340
tccctgtgcg ccgcgtcgcc gatggtggcc agccaactgg cgcgctaccc gatcctgctc 60
gatgaactgc tcgacccgaa cacgctctat caaccgacgg 100
<210> 341
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 341
cgttctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctc gcggcgcttt 60
ttttatattc tcgactccat ttaaaataaa aaatccaatc 100
<210> 342
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 342
aactcacttc acgccccgaa gggggaagct gcctgaccct acgattcccg ctatttcatt 60
cactgaccgg aggttcaaaa tgacccagcg aaccgagtcg 100
<210> 343
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 343
cgcgtcaggt tgaacgtaaa aaagtcggtc tgcgcaaagc acgtcgtcgt ccgcagttct 60
ccaaacgtta attggtttct gcttcggcag aacgattggc 100
<210> 344
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 344
aactcacttc acgccccgaa gggggaagct gcctgaccct acgattcccg ctatttcatt 60
cactgaccgg aggttcaaaa tgacccagcg aaccgagtcg 100
<210> 345
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 345
ccgatcccca tcactgtgtg tcttgtatta cagtgccgct tcgtcggctt cgccggtacg 60
aatacgaatg acgcgttgca gctcagcaac gaaaattttg 100
<210> 346
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 346
ccgtctctga agctctcggt gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt 60
gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg 100
<210> 347
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 347
tgaacatcac tgatgcacaa gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg 60
caggcattcg cgttaaagcc gacttgagaa atgagaagat 100
<210> 348
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 348
ccgtctctga agctctcggt gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt 60
gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg 100
<210> 349
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 349
tacagtagcg cctctcaaaa atagataaac ggctcatgta cgtgggccgt ttattttttc 60
tacccataat cgggaaccgg tgttataatg ccgcgccctc 100
<210> 350
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 350
aactcacttc acaccccgaa gggggaagtt gcctgaccct acgattcccg ctatttcatt 60
cactgaccgg aggttcaaaa tgacccagcg aaccgagtcg 100
<210> 351
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 351
cgtcctgtaa taataaccgg acaattcgga ctgattaaaa aagcgccctt gtggcgcttt 60
ttttatattc ccgcctccat ttaaaataaa aaatccaatc 100
<210> 352
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 352
ggacatcatc gcgacaaaca atattaatac cggcaaccac accggcaatt tacgagactg 60
cgcaggcatc ctttctcccg tcaatttctg tcaaataaag 100
<210> 353
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 353
aactcacttc acaccccgaa gggggaagtt gcctgaccct acgattcccg ctatttcatt 60
cactgaccgg aggttcaaaa tgacccagcg aaccgagtcg 100
<210> 354
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 354
tttaacgatc tgattggcga tgatgaaacg gattcgccgg aagatgcgct ttctgagagc 60
tggcgcgaat tgtggcagga tgcgttgcag gaggaggatt 100
<210> 355
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 355
gcactgaaac acctcatttc cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg 60
cgctacccga tcctgcttga tgaattgctc gacccgaata 100
<210> 356
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 356
gcgctcaaac agttaatccg tctgtgtgcc gcctcgccga tggtcgcgac acaacttgca 60
cgtcatcctt tattgctcga tgaactgctc gacccgcgca 100
<210> 357
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 357
agtctgaact catcctgcgg cagtcggtga gacgtatttt tgaccaaaga gtgatctaca 60
tcacggaatt ttgtggttgt tgctgcttaa aagggcaaat 100
<210> 358
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 358
ccgtctctga agctctcggt gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt 60
gacattaccg ataatgtgcc gcgtgaacgg gtgcgttatg 100
<210> 359
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 359
gccattgagc tggcttcccg accgcagggc ggcacctgcc tgaccctgcg tttcccgctg 60
tttaacaccc tgaccggagg tgaagcatga tccctgaatc 100
<210> 360
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 360
agcgtcaggt accggtcatg attcaccgtg cgattctcgg ttccctggag cgcttcattg 60
gcatcctgac cgaagagttc gctggcttct tcccaacctg 100
<210> 361
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 361
gcgctgaagc acctgatcac gctctgcgcg gcgtcgccga tggtcgccag ccagctggcg 60
cgccacccgc tgctgctgga tgagctgctg gatcccaaca 100
<210> 362
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 362
gcccgctgac cgaccagaac ttccaccttg gactcggcta tacccttggc gtgacggcgc 60
gcgataactg ggactacatc cccattccgg tgatcttacc 100
<210> 363
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 363
gccattgagc tggcttcccg accgcagggc ggcacctgcc tgaccctgcg tttcccgctg 60
tttaacaccc tgaccggagg tgaagcatga tccctgaatc 100
<210> 364
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 364
gctaaagttc tcggctaatc gctgataaca tttgacgcaa tgcgcaataa aagggcatca 60
tttgatgccc tttttgcacg ctttcatacc agaacctggc 100
<210> 365
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 365
gccattgagc tggcttcccg accgcagggc ggcacctgcc tgaccctgcg tttcccgctg 60
tttaacaccc tgaccggagg tgaagcatga tccctgaatc 100
<210> 366
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 366
cgccgtcctc gcagtaccat tgcaaccgac tttacagcaa gaagtgattc tggcacgcat 60
ggaacaaatt cttgccagtc gggctttatc cgatgacgaa 100
<210> 367
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 367
gccattgagc tggcttcccg accgcagggc ggcacctgcc tgaccctgcg tttcccgctg 60
tttaacaccc tgaccggagg tgaagcatga tccctgaatc 100
<210> 368
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 368
tctttagatc tctcggtccg ccctgatggc ggcaccttgc tgacgttacg cctgccggta 60
cagcaggtta tcaccggagg cttaaaatga cccagttacc 100
<210> 369
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 369
tgaatatcac tgactcacaa gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg 60
caggcattcg cgttaaagcc gacttgagaa atgagaagat 100
<210> 370
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 370
ctggggtcac tggagcgctt tatcggcatc ctgaccgaag aatttgccgg tttcttcccg 60
acctggctgg cccctgttca ggttgtggtg atgaatatca 100
<210> 371
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 100 п.н. ниже соединения
<400> 371
gcaatagaac taactacccg ccctgaaggc ggtacctgcc tgaccctgcg attcccgtta 60
tttcattcac tgaccggagg cccacgatga cccagcgacc 100
<210> 372
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / PinfC
<400> 372
tggtgtccgg gcgaacgtcg ccaggtggca caaattgtca gaactacgac acgactaacc 60
gaccgcagga gtgtgcgatg accctgaata tgatgatgga ttcttggttc tctggagcgc 120
tttatcggca tcctgactga agaatttgca ggcttcttcc caacctggct tgcacccgtg 180
caggtagttg tgatgaacat 200
<210> 373
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> PinfC / разрушенный ген nifL
<400> 373
cggaaaacga gttcaaacgg cgcgtcccaa tcgtattaat ggcgagattc gcgccacgga 60
agttcgctta acaggtctgg aaggcgagca gcttggtatt gcgatagaac tcacttcacg 120
ccccgaaggg ggaagctgcc tgaccctacg attcccgcta tttcattcac tgaccggagg 180
ttcaaaatga cccagcgaac 200
<210> 374
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 5'-UTR и ATG / укороченный ген glnE
<400> 374
cgccagagag ttgaaatcga acatttccgt aataccgcca ttacccagga gccgttctgg 60
ttgcacagcg gaaaacgtta acgaaaggat atttcgcatg tccctgtgcg ccgcgtcgcc 120
gatggtggcc agccaactgg cgcgctaccc gatcctgctc gatgaactgc tcgacccgaa 180
cacgctctat caaccgacgg 200
<210> 375
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm1
<400> 375
cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta accgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccagc cgttctgtaa taataaccgg 120
acaattcgga ctgattaaaa aagcgccctc gcggcgcttt ttttatattc tcgactccat 180
ttaaaataaa aaatccaatc 200
<210> 376
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm1 / разрушенный ген nifL
<400> 376
tcaacctaaa aaagtttgtg taatacttgt aacgctacat ggagattaac tcaatctaga 60
gggtattaat aatgaatcgt actaaactgg tactgggcgc aactcacttc acgccccgaa 120
gggggaagct gcctgaccct acgattcccg ctatttcatt cactgaccgg aggttcaaaa 180
tgacccagcg aaccgagtcg 200
<210> 377
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm7
<400> 377
cgggcgaacg tcgccaggtg gcacaaattg tcagaactac gacacgacta accgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccagc cgcgtcaggt tgaacgtaaa 120
aaagtcggtc tgcgcaaagc acgtcgtcgt ccgcagttct ccaaacgtta attggtttct 180
gcttcggcag aacgattggc 200
<210> 378
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm4 / разрушенный ген nifL
<400> 378
aattttctgc ccaaatggct gggattgttc attttttgtt tgccttacaa cgagagtgac 60
agtacgcgcg ggtagttaac tcaacatctg accggtcgat aactcacttc acgccccgaa 120
gggggaagct gcctgaccct acgattcccg ctatttcatt cactgaccgg aggttcaaaa 180
tgacccagcg aaccgagtcg 200
<210> 379
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> от 5'-UTR до ATG - 4 п.н. гена amtB / разрушенный ген amtB
<400> 379
gtaaccaata aaggccacca cgccagacca cacgatagtg atggcaacac tttccagctg 60
caccagcacc tgatggccca tggtcacacc ttcagcgaaa ccgatcccca tcactgtgtg 120
tcttgtatta cagtgccgct tcgtcggctt cgccggtacg aatacgaatg acgcgttgca 180
gctcagcaac gaaaattttg 200
<210> 380
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm1.2 / разрушенный ген nifL
<400> 380
tggtattgtc agtctgaatg aagctcttga aaaagctgag gaagcgggcg tcgatttagt 60
agaaatcagt ccgaatgccg agccgccagt ttgtcgaatc ccgtctctga agctctcggt 120
gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt gacattaccg ataatgtgcc 180
gcgtgaacgg gtgcgttatg 200
<210> 381
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm1.2
<400> 381
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 60
gcacttgaga cacttttggg cgagaaccac cgtctgctgg tgaacatcac tgatgcacaa 120
gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg caggcattcg cgttaaagcc 180
gacttgagaa atgagaagat 200
<210> 382
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm3.1 / разрушенный ген nifL
<400> 382
cgggaaccgg tgttataatg ccgcgccctc atattgtggg gatttcttaa tgacctatcc 60
tgggtcctaa agttgtagtt gacattagcg gagcactaac ccgtctctga agctctcggt 120
gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt gacattaccg ataatgtgcc 180
gcgtgaacgg gtgcgttatg 200
<210> 383
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm3.1
<400> 383
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 60
gcacttgaga cacttttggg cgagaaccac cgtctgctgg tacagtagcg cctctcaaaa 120
atagataaac ggctcatgta cgtgggccgt ttattttttc tacccataat cgggaaccgg 180
tgttataatg ccgcgccctc 200
<210> 384
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm1 / разрушенный ген nifL
<400> 384
tcaacctaaa aaagtttgtg taatacttgt aacgctacat ggagattaac tcaatctaga 60
gggtattaat aatgaatcgt actaaactgg tactgggcgc aactcacttc acaccccgaa 120
gggggaagtt gcctgaccct acgattcccg ctatttcatt cactgaccgg aggttcaaaa 180
tgacccagcg aaccgagtcg 200
<210> 385
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm1
<400> 385
gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta actgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccggc cgtcctgtaa taataaccgg 120
acaattcgga ctgattaaaa aagcgccctt gtggcgcttt ttttatattc ccgcctccat 180
ttaaaataaa aaatccaatc 200
<210> 386
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm5
<400> 386
gggcgacaaa cggcctggtg gcacaaattg tcagaactac gacacgacta actgaccgca 60
ggagtgtgcg atgaccctga atatgatgat ggatgccggc ggacatcatc gcgacaaaca 120
atattaatac cggcaaccac accggcaatt tacgagactg cgcaggcatc ctttctcccg 180
tcaatttctg tcaaataaag 200
<210> 387
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm5 / разрушенный ген nifL
<400> 387
taagaattat ctggatgaat gtgccattaa atgcgcagca taatggtgcg ttgtgcggga 60
aaactgcttt tttttgaaag ggttggtcag tagcggaaac aactcacttc acaccccgaa 120
gggggaagtt gcctgaccct acgattcccg ctatttcatt cactgaccgg aggttcaaaa 180
tgacccagcg aaccgagtcg 200
<210> 388
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 5'-UTR и ATG / укороченный ген glnE
<400> 388
cgccagagag tcgaaatcga acatttccgt aataccgcga ttacccagga gccgttctgg 60
ttgcacagcg gaaaacgtta acgaaaggat atttcgcatg tttaacgatc tgattggcga 120
tgatgaaacg gattcgccgg aagatgcgct ttctgagagc tggcgcgaat tgtggcagga 180
tgcgttgcag gaggaggatt 200
<210> 389
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 5'-UTR и ATG / укороченный ген glnE
<400> 389
cgccagagag tcgaaatcga acatttccgt aataccgcga ttacccagga gccgttctgg 60
ttgcacagcg gaaaacgtta acgaaaggat atttcgcatg gcactgaaac acctcatttc 120
cctgtgtgcc gcgtcgccga tggttgccag tcagctggcg cgctacccga tcctgcttga 180
tgaattgctc gacccgaata 200
<210> 390
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 5'-UTR и ATG / укороченный ген glnE
<400> 390
gatgatggat gctttctggt taaacgggca acctcgttaa ctgactgact agcctgggca 60
aactgcccgg gctttttttt gcaaggaatc tgatttcatg gcgctcaaac agttaatccg 120
tctgtgtgcc gcctcgccga tggtcgcgac acaacttgca cgtcatcctt tattgctcga 180
tgaactgctc gacccgcgca 200
<210> 391
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm4
<400> 391
accgatccgc aggcgcgcat ttgttatgcc aatccggcat tctgccgcca gacgggtttt 60
gcacttgaga cacttttggg cgagaaccac cgtctgctgg agtctgaact catcctgcgg 120
cagtcggtga gacgtatttt tgaccaaaga gtgatctaca tcacggaatt ttgtggttgt 180
tgctgcttaa aagggcaaat 200
<210> 392
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm4 / разрушенный ген nifL
<400> 392
catcggacac caccagctta caaattgcct gattgcggcc ccgatggccg gtatcactga 60
ccgaccattt cgtgccttat gtcatgcgat gggggctggg ccgtctctga agctctcggt 120
gaacattgtt gcgaggcagg atgcgagctg gttgtgtttt gacattaccg ataatgtgcc 180
gcgtgaacgg gtgcgttatg 200
<210> 393
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> PinfC / разрушенный ген nifL
<400> 393
tcttcaacaa ctggaggaat aaggtattaa aggcggaaaa cgagttcaaa cggcacgtcc 60
gaatcgtatc aatggcgaga ttcgcgccct ggaagttcgc gccattgagc tggcttcccg 120
accgcagggc ggcacctgcc tgaccctgcg tttcccgctg tttaacaccc tgaccggagg 180
tgaagcatga tccctgaatc 200
<210> 394
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / PinfC
<400> 394
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga agcgtcaggt accggtcatg 120
attcaccgtg cgattctcgg ttccctggag cgcttcattg gcatcctgac cgaagagttc 180
gctggcttct tcccaacctg 200
<210> 395
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> 5'-UTR и ATG / укороченный ген glnE
<400> 395
atcgcagcgt ctttgaatat ttccgtcgcc aggcgctggc tgccgagccg ttctggctgc 60
atagtggaaa acgataattt caggccaggg agcccttatg gcgctgaagc acctgatcac 120
gctctgcgcg gcgtcgccga tggtcgccag ccagctggcg cgccacccgc tgctgctgga 180
tgagctgctg gatcccaaca 200
<210> 396
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm1.2
<400> 396
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga gcccgctgac cgaccagaac 120
ttccaccttg gactcggcta tacccttggc gtgacggcgc gcgataactg ggactacatc 180
cccattccgg tgatcttacc 200
<210> 397
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm1.2 / разрушенный ген nifL
<400> 397
tcacttttta gcaaagttgc actggacaaa aggtaccaca attggtgtac tgatactcga 60
cacagcatta gtgtcgattt ttcatataaa ggtaattttg gccattgagc tggcttcccg 120
accgcagggc ggcacctgcc tgaccctgcg tttcccgctg tttaacaccc tgaccggagg 180
tgaagcatga tccctgaatc 200
<210> 398
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm6.2
<400> 398
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga gctaaagttc tcggctaatc 120
gctgataaca tttgacgcaa tgcgcaataa aagggcatca tttgatgccc tttttgcacg 180
ctttcatacc agaacctggc 200
<210> 399
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm6.2 / разрушенный ген nifL
<400> 399
gttctccttt gcaatagcag ggaagaggcg ccagaaccgc cagcgttgaa gcagtttgaa 60
cgcgttcagt gtataatccg aaacttaatt tcggtttgga gccattgagc tggcttcccg 120
accgcagggc ggcacctgcc tgaccctgcg tttcccgctg tttaacaccc tgaccggagg 180
tgaagcatga tccctgaatc 200
<210> 400
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / Prm8.2
<400> 400
tccgggttcg gcttaccccg ccgcgttttg cgcacggtgt cggacaattt gtcataactg 60
cgacacagga gtttgcgatg accctgaata tgatgctcga cgccgtcctc gcagtaccat 120
tgcaaccgac tttacagcaa gaagtgattc tggcacgcat ggaacaaatt cttgccagtc 180
gggctttatc cgatgacgaa 200
<210> 401
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> Prm8.2 / разрушенный ген nifL
<400> 401
gatatgcctg aagtattcaa ttacttaggc atttacttaa cgcaggcagg caattttgat 60
gctgcctatg aagcgtttga ttctgtactt gagcttgatc gccattgagc tggcttcccg 120
accgcagggc ggcacctgcc tgaccctgcg tttcccgctg tttaacaccc tgaccggagg 180
tgaagcatga tccctgaatc 200
<210> 402
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> PinfC / разрушенный ген nifL
<400> 402
tggtattgtc agtctgaatg aagctcttga aaaagctgag gaagcgggcg tcgatttagt 60
agaaatcagt ccgaatgccg agccgccagt ttgtcgaatc tctttagatc tctcggtccg 120
ccctgatggc ggcaccttgc tgacgttacg cctgccggta cagcaggtta tcaccggagg 180
cttaaaatga cccagttacc 200
<210> 403
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / PinfC
<400> 403
tgcaaattgc acggttattc cgggtgagta tatgtgtgat ttgggttccg gcattgcgca 60
ataaagggga gaaagacatg agcatcacgg cgttatcagc tgaatatcac tgactcacaa 120
gctacctatg tcgaagaatt aactaaaaaa ctgcaagatg caggcattcg cgttaaagcc 180
gacttgagaa atgagaagat 200
<210> 404
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> разрушенный ген nifL / PinfC
<400> 404
tcagggctgc ggatgtcggg cgtttcacaa cacaaaatgt tgtaaatgcg acacagccgg 60
gcctgaaacc aggagcgtgt gatgaccttt aatatgatgc ctggggtcac tggagcgctt 120
tatcggcatc ctgaccgaag aatttgccgg tttcttcccg acctggctgg cccctgttca 180
ggttgtggtg atgaatatca 200
<210> 405
<211> 200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<220>
<223> PinfC / разрушенный ген nifL
<400> 405
cggaaaacga gttcaaacgg cacgtccgaa tcgtatcaat ggcgagattc gcgcccagga 60
agttcgctta actggtctgg aaggtgagca gctgggtatt gcaatagaac taactacccg 120
ccctgaaggc ggtacctgcc tgaccctgcg attcccgtta tttcattcac tgaccggagg 180
cccacgatga cccagcgacc 200
<210> 406
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 406
caagaagttc gcctcacagg 20
<210> 407
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 407
tgcctcgcaa caatgttcac 20
<210> 408
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 408
cgccctcata ttgtggggat 20
<210> 409
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 409
ggcataacgc acccgttca 19
<210> 410
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический зонд
<400> 410
tctgaagctc tcggt 15
<210> 411
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 411
taaactggta ctgggcgcaa ct 22
<210> 412
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 412
caaatcgaag cgccagacgg tat 23
<210> 413
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический зонд
<400> 413
gaccctacga ttccc 15
<210> 414
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 414
ggtgcactct ttgcatggtt 20
<210> 415
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 415
gcgcagtctc gtaaattgcc 20
<210> 416
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический зонд
<400> 416
gcgatgaccc tgaat 15
<210> 417
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 417
ctcggcagca tggacgtaa 19
<210> 418
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 418
agggtgttaa acagcgggaa a 21
<210> 419
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический зонд
<400> 419
tccgaatcgt atcaa 15
<210> 420
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 420
gagccgttct ggctgcatag 20
<210> 421
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 421
gccgtcggct gatagagg 18
<210> 422
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический зонд
<400> 422
tgaagcacct gatca 15
<210> 423
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 423
ggaaaacgag ttcaaccggc 20
<210> 424
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический праймер
<400> 424
gggcggaccg agagatctaa 20
<---
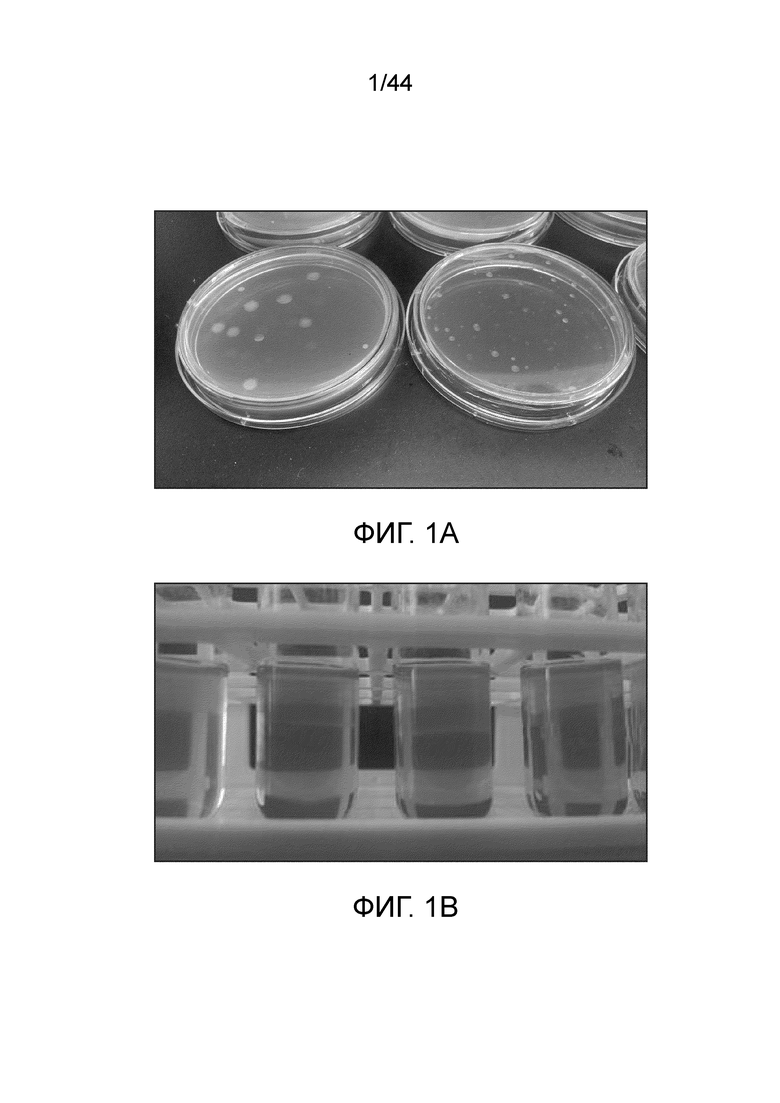
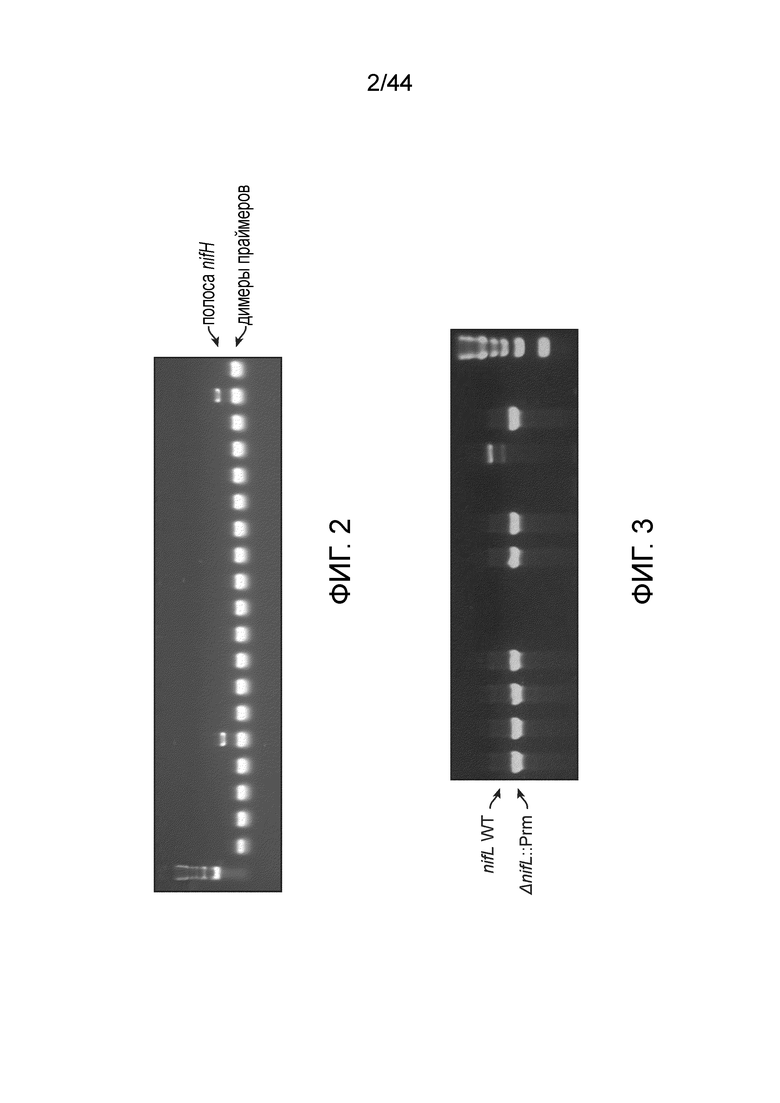
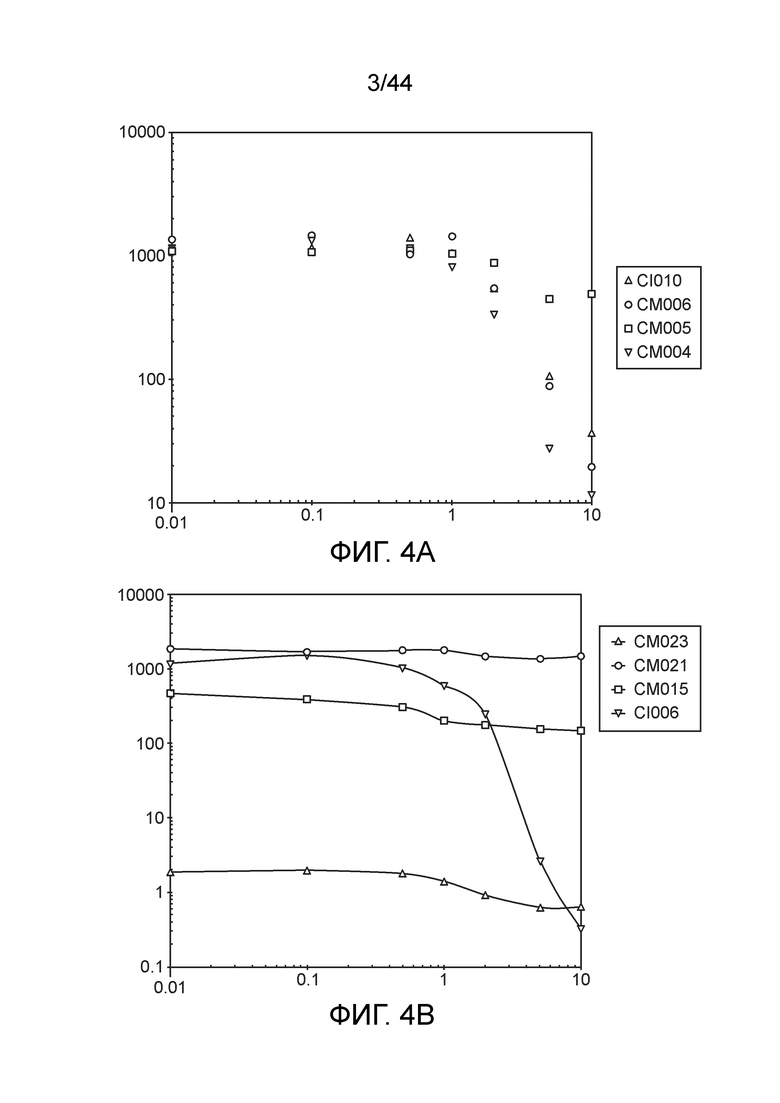
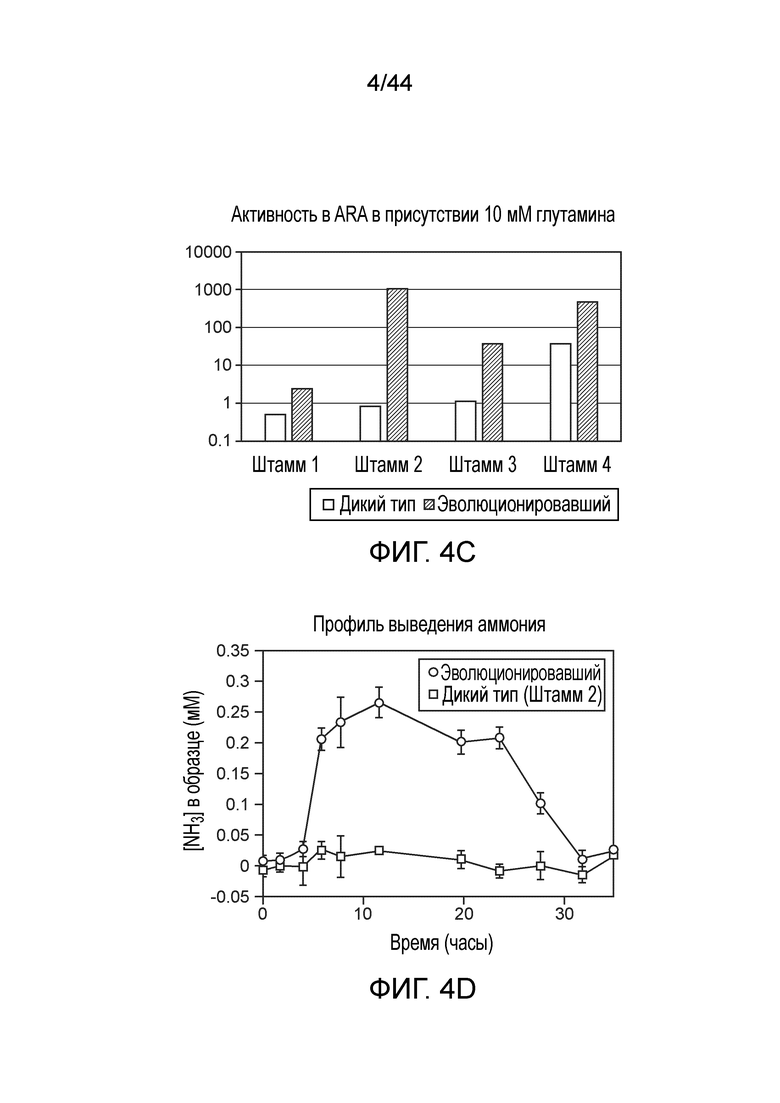
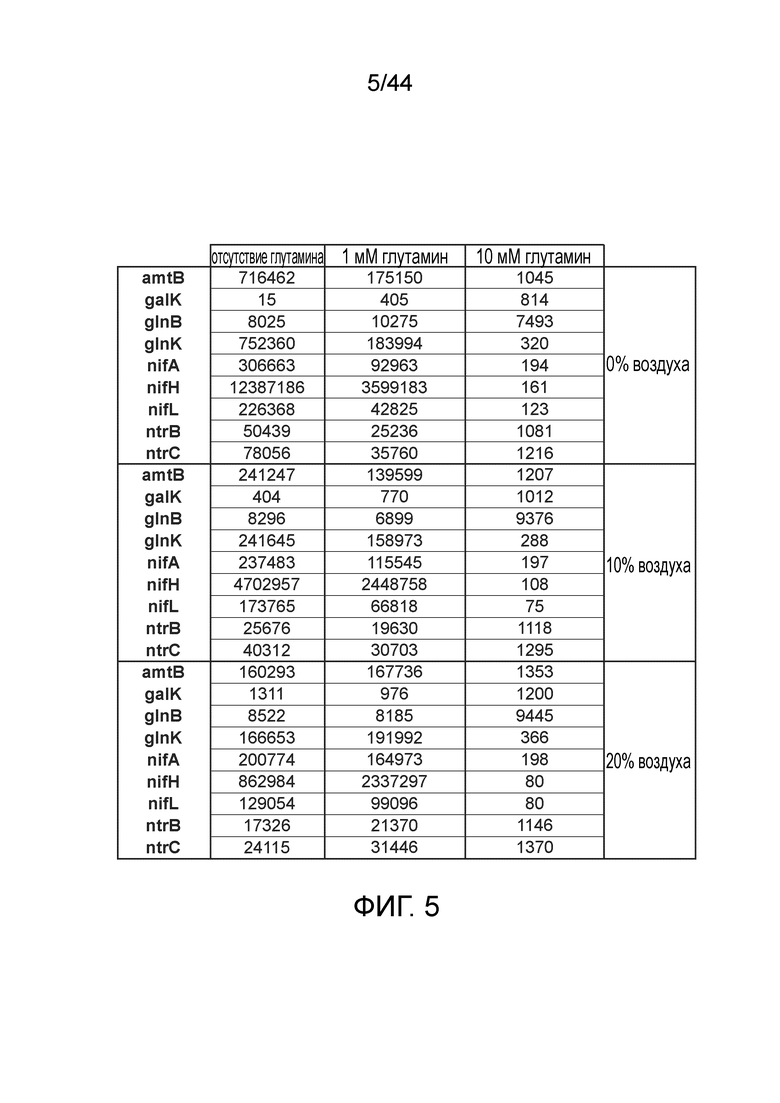
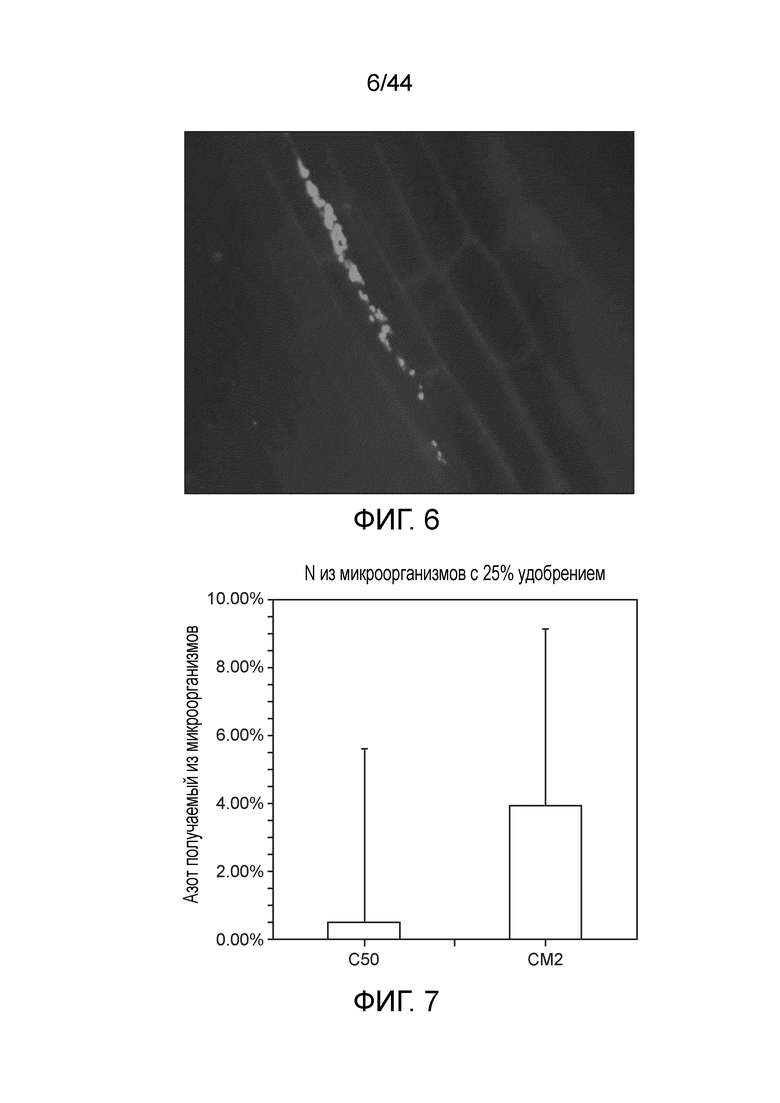
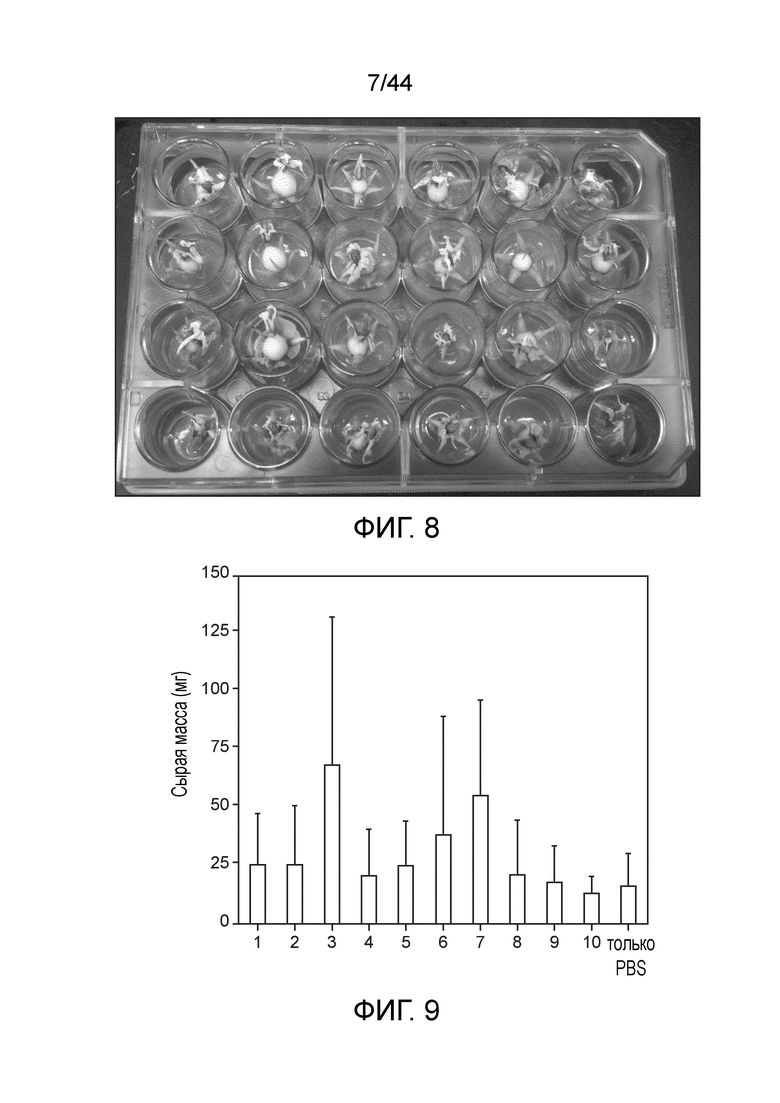
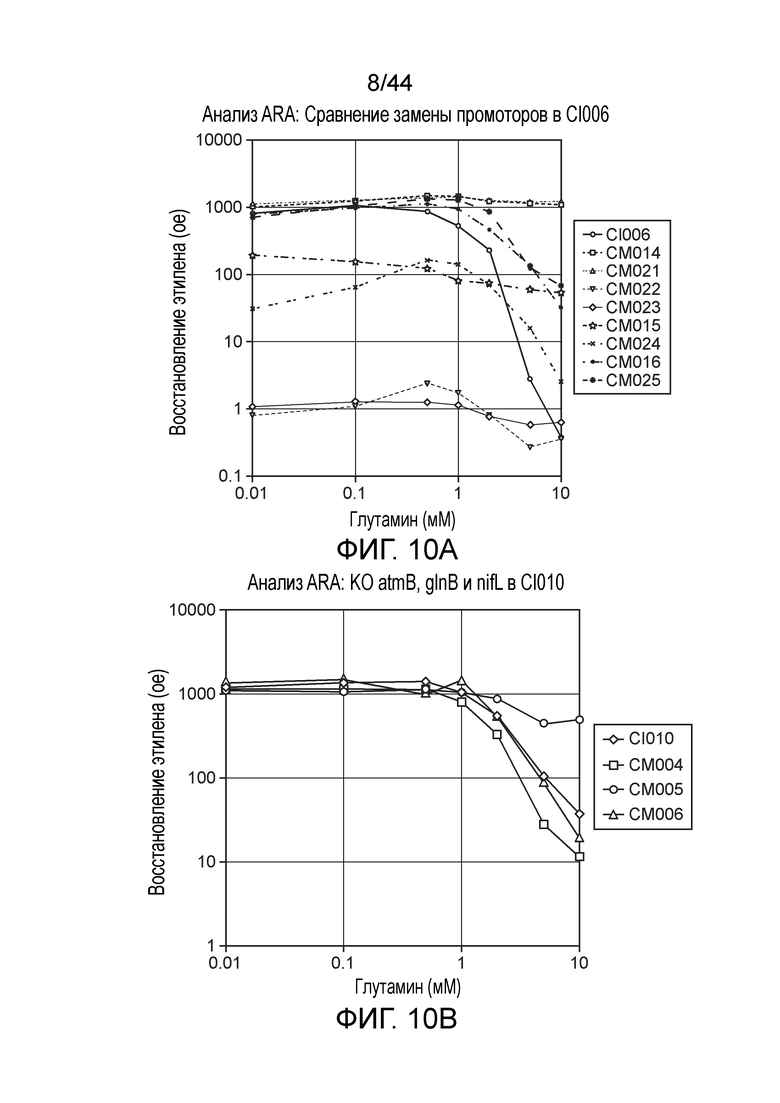
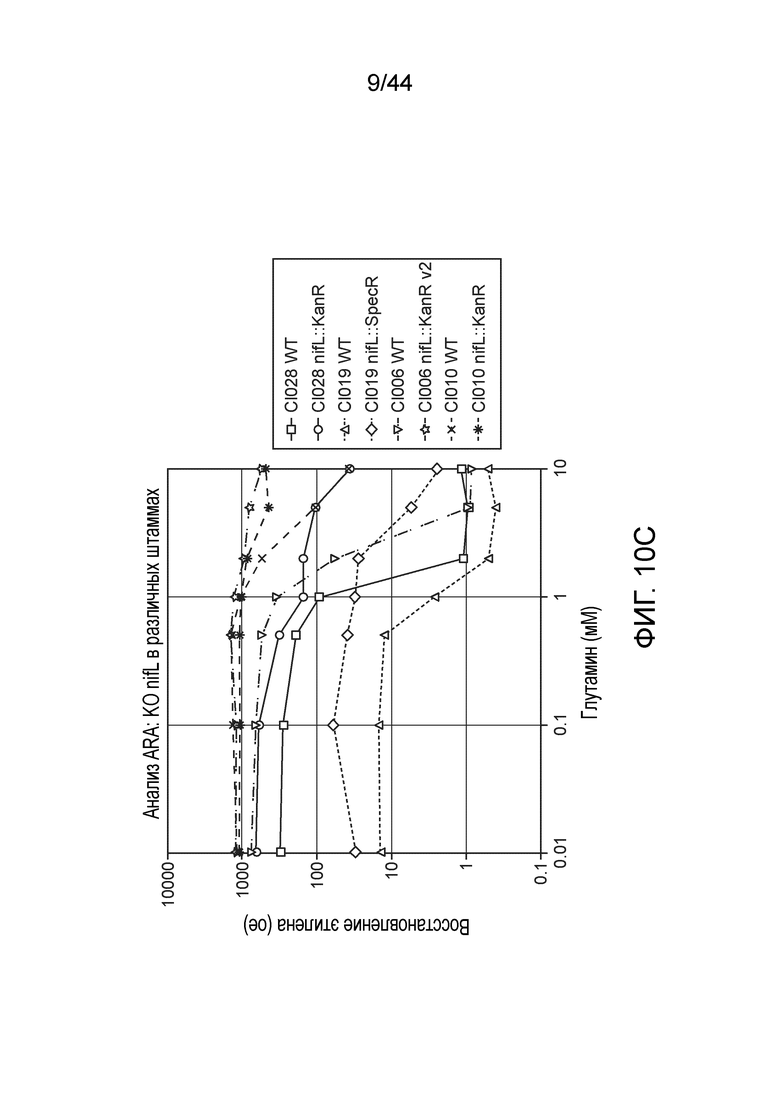
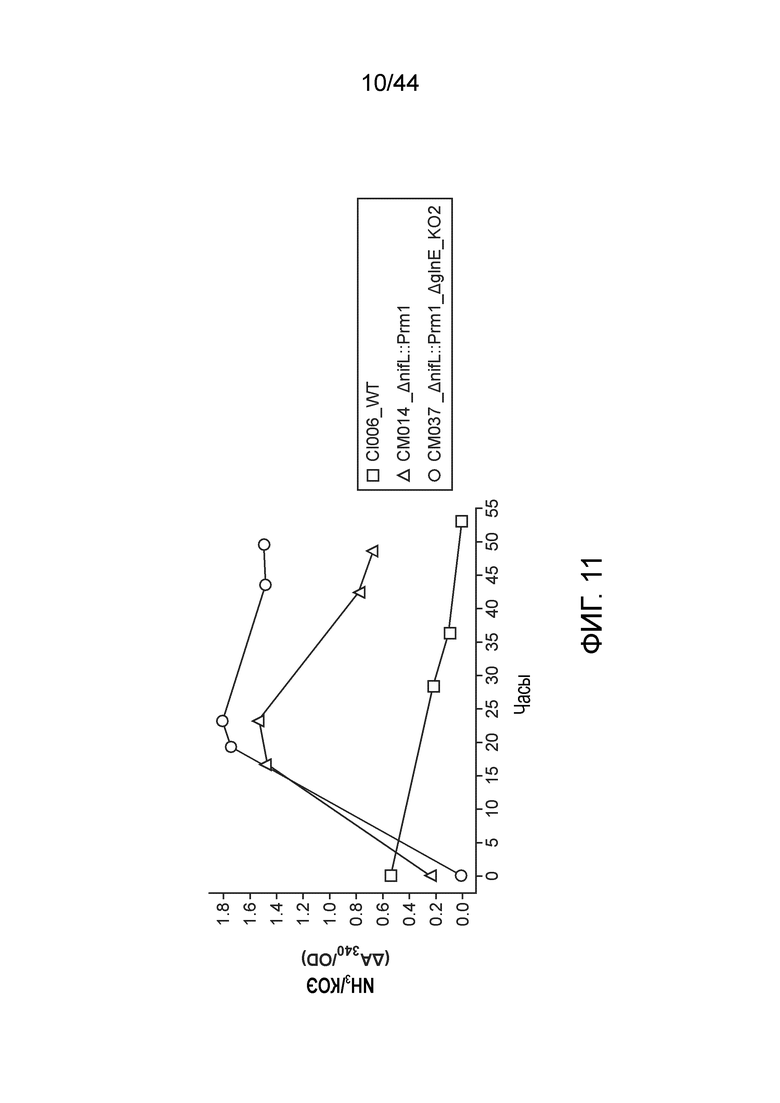
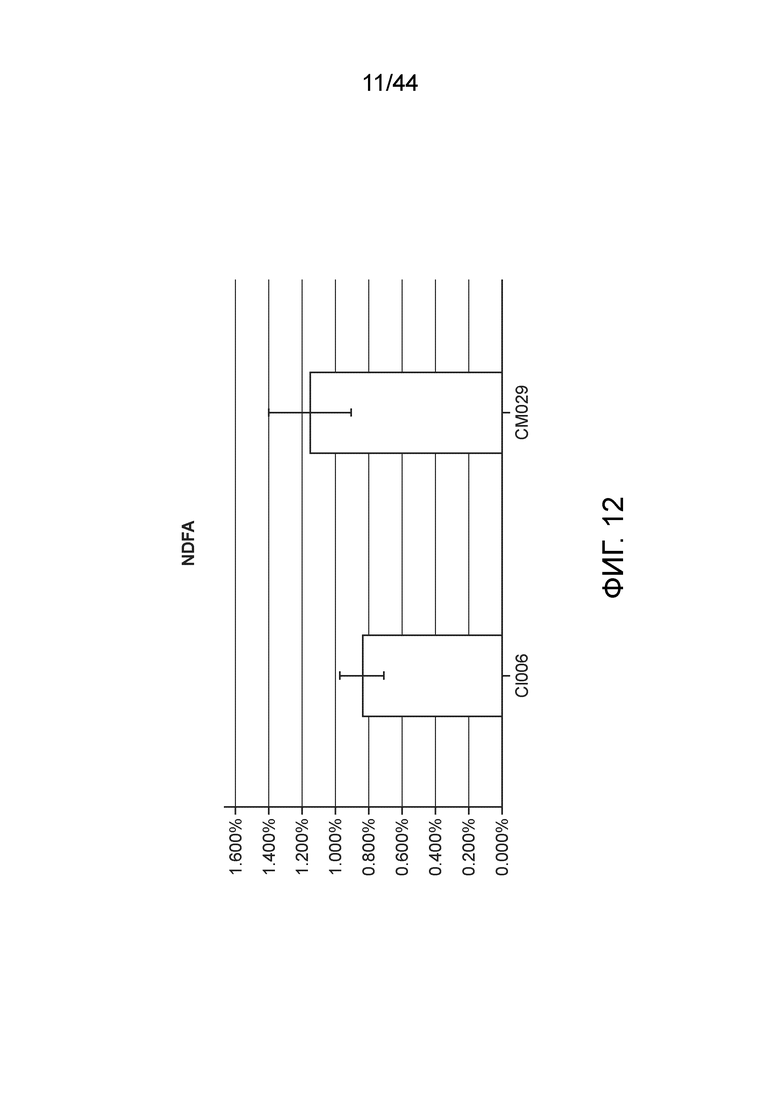
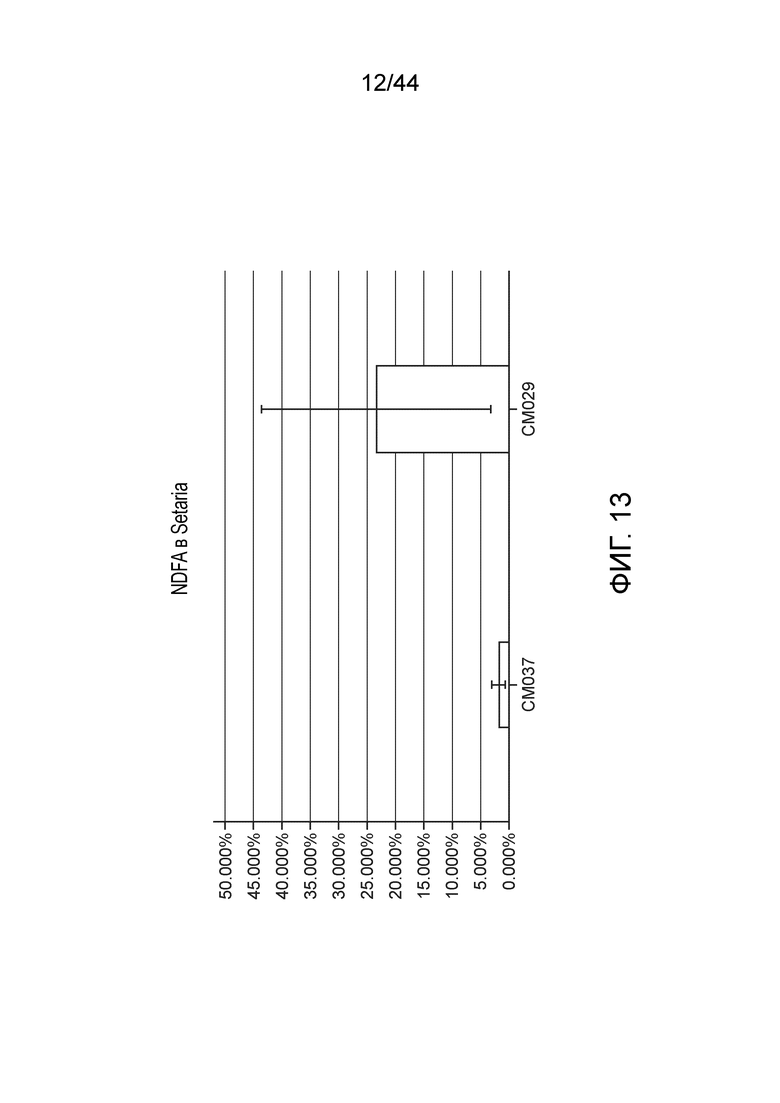
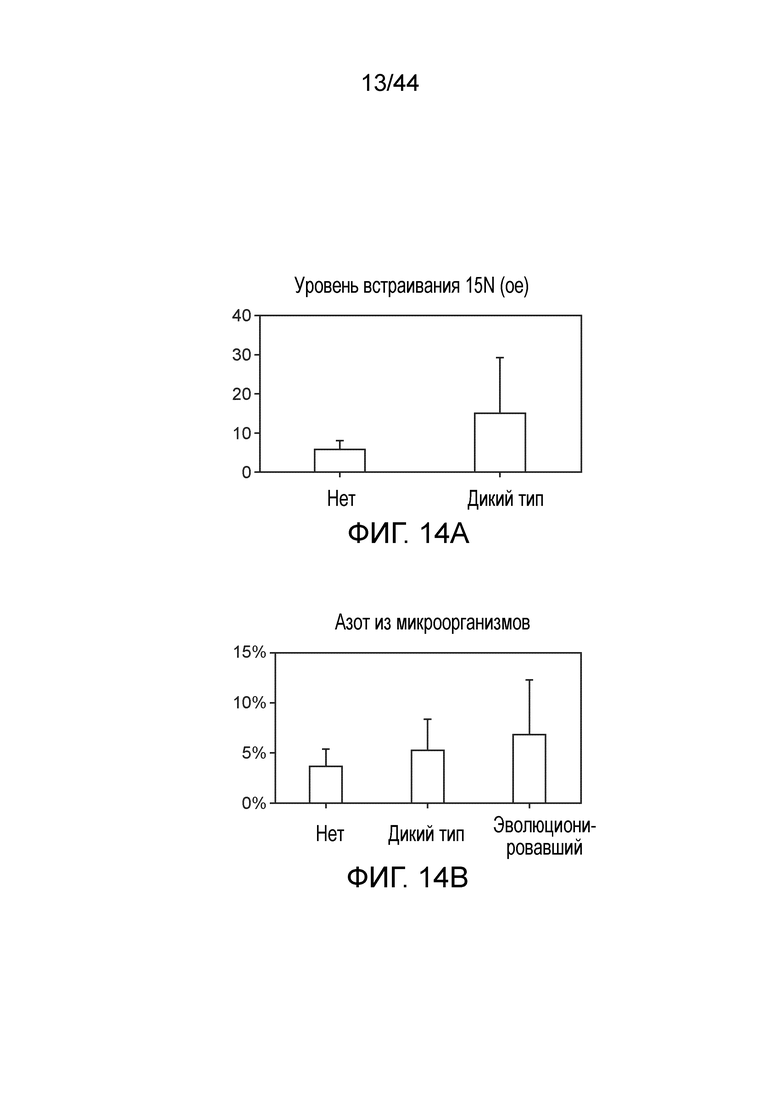
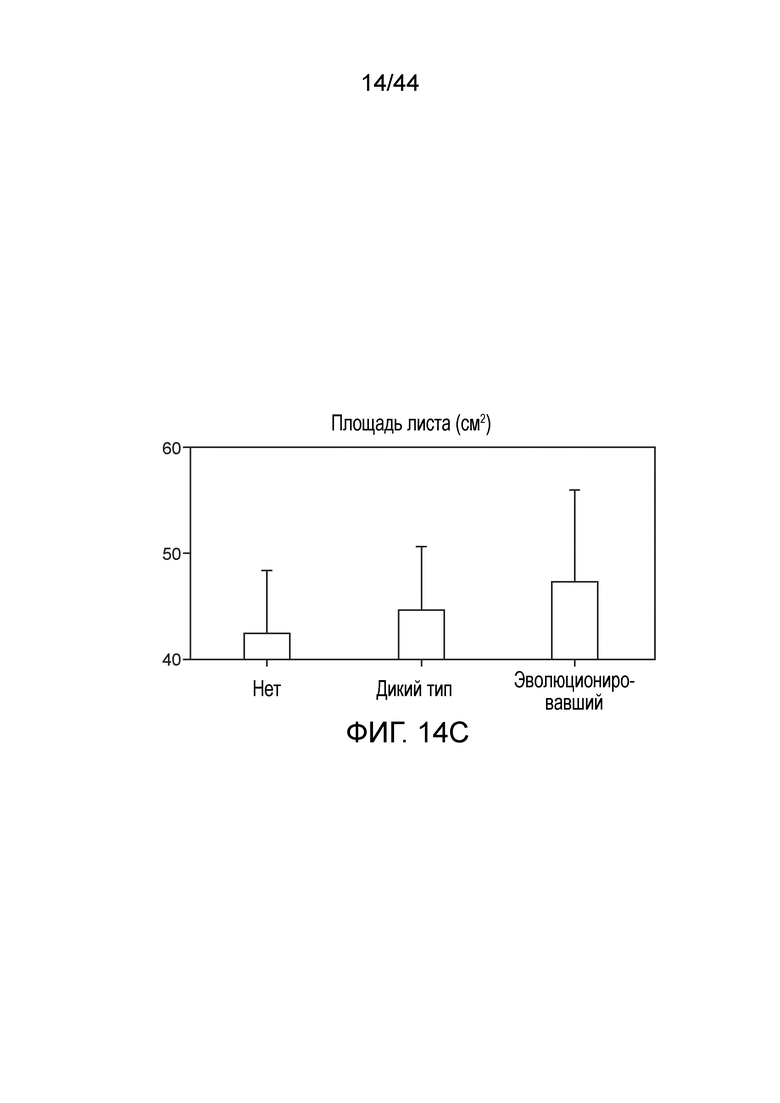
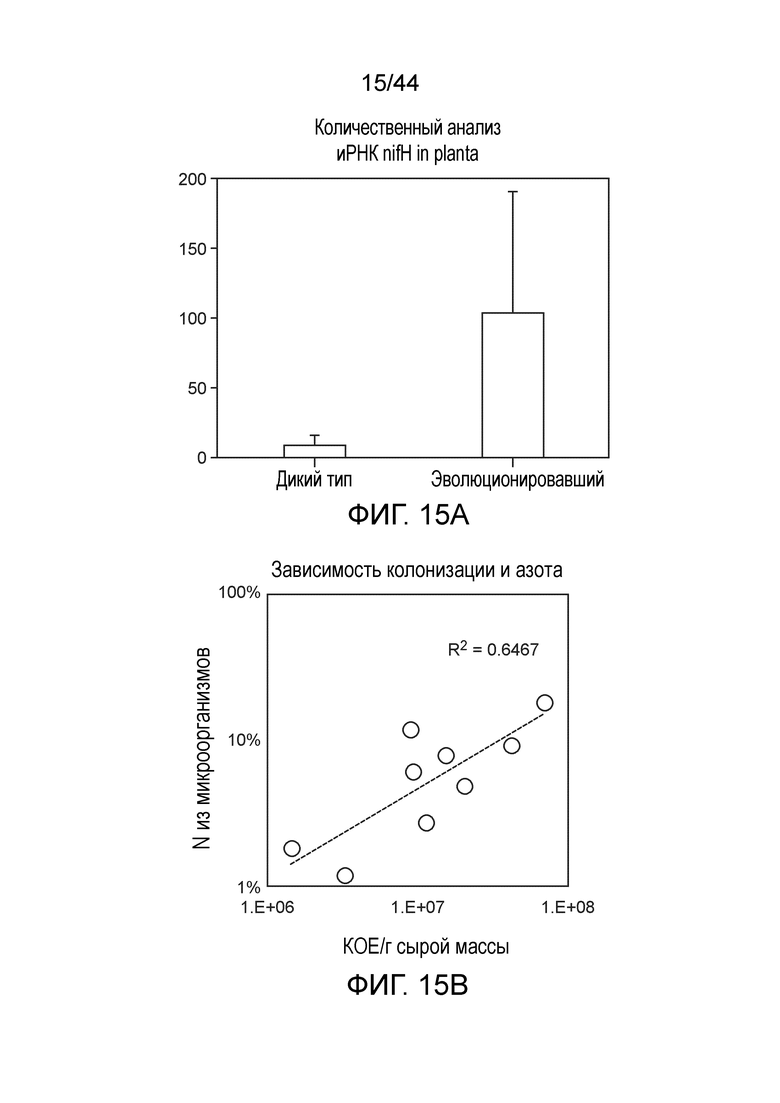
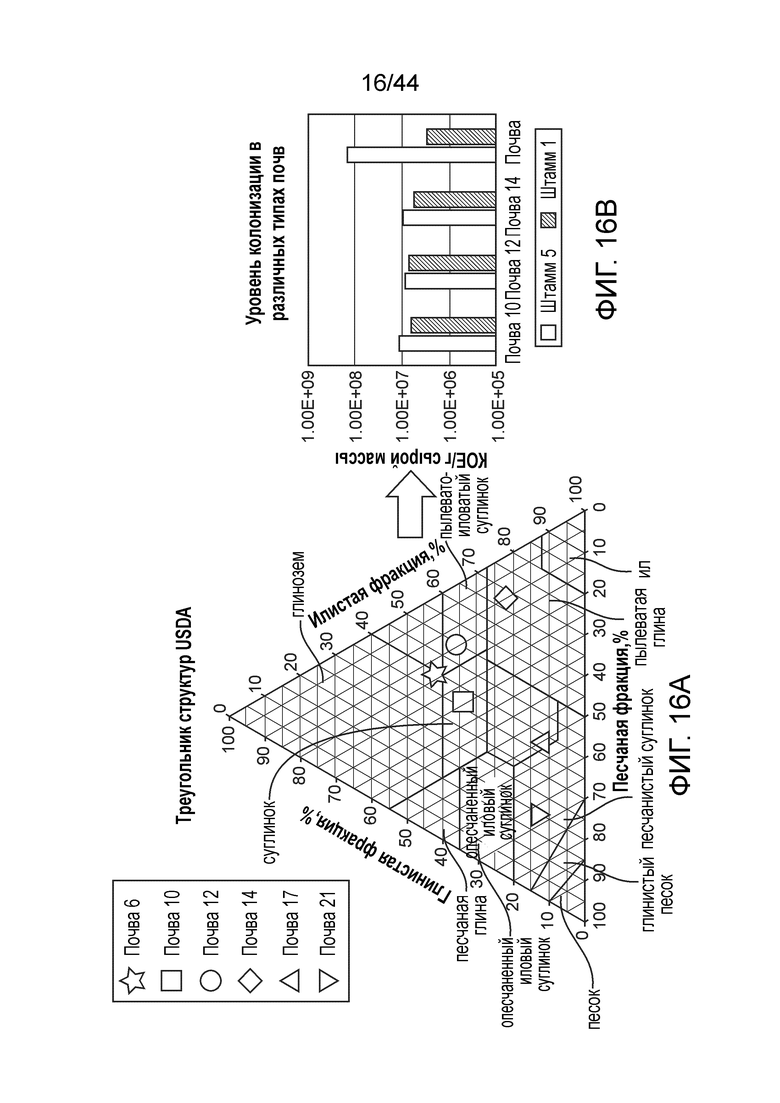
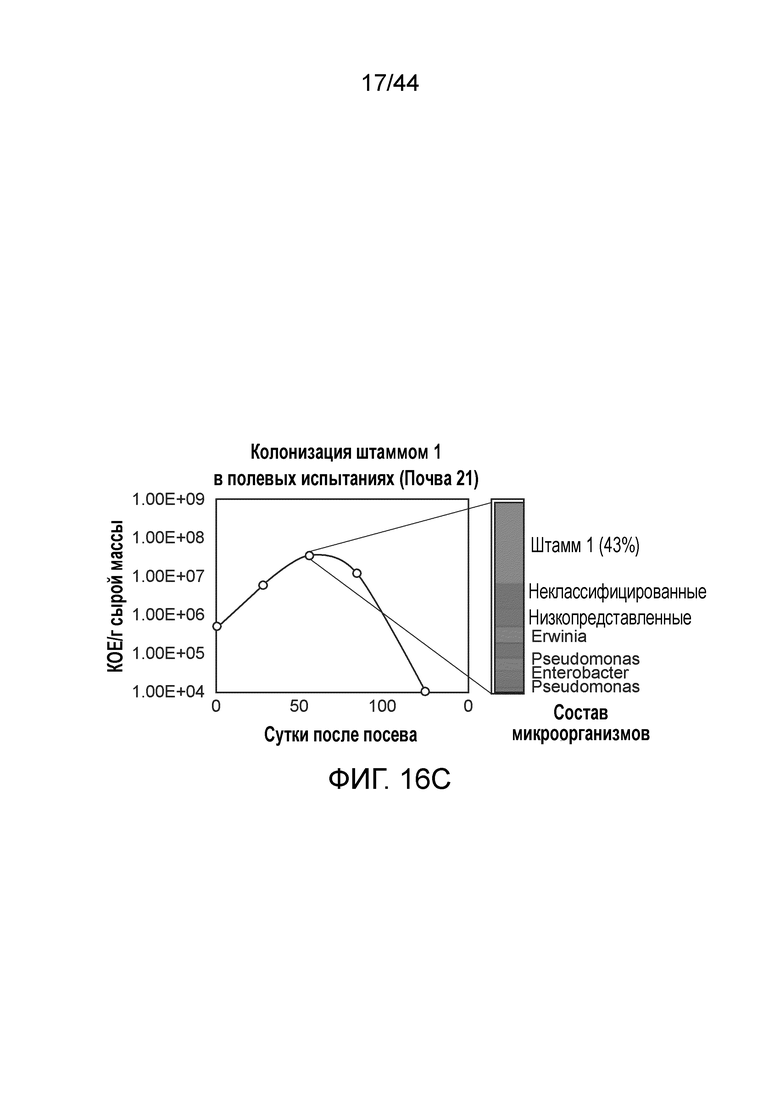
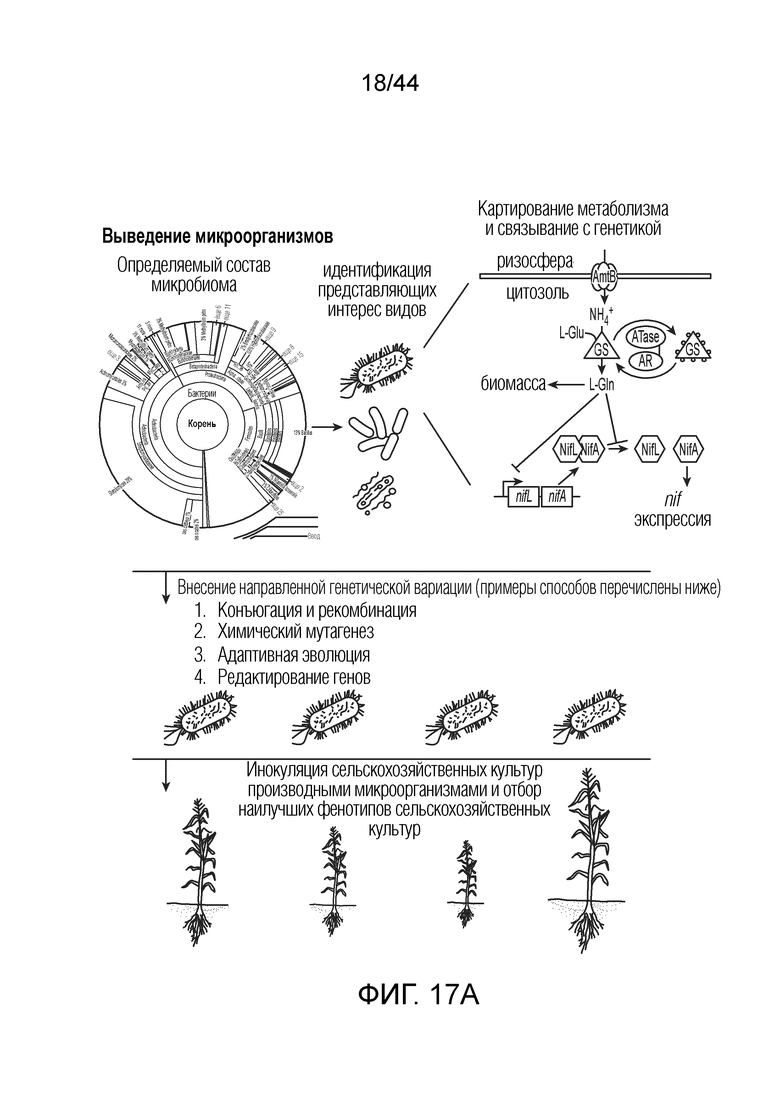
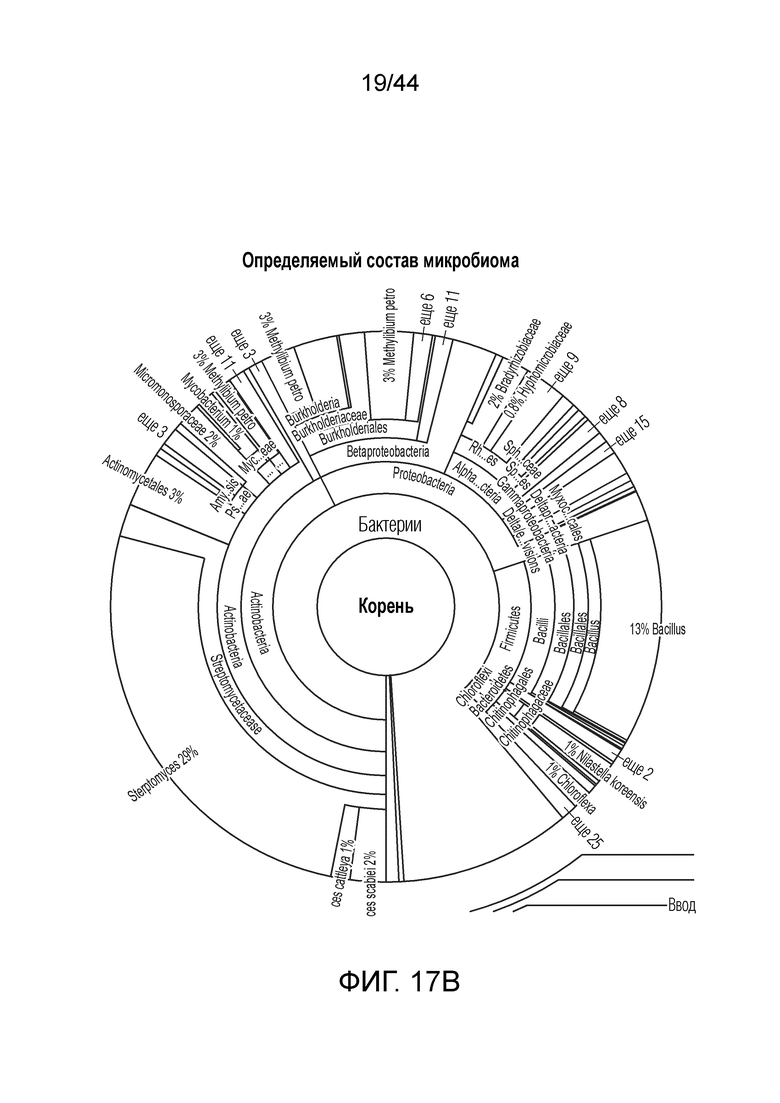
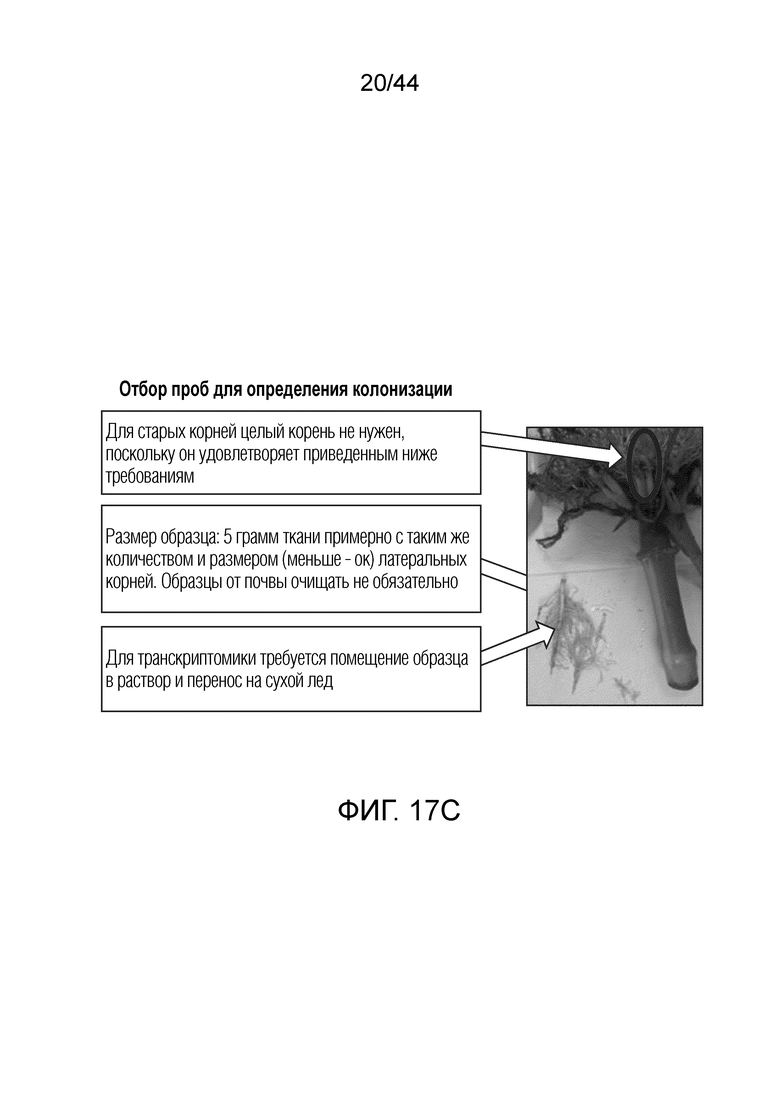
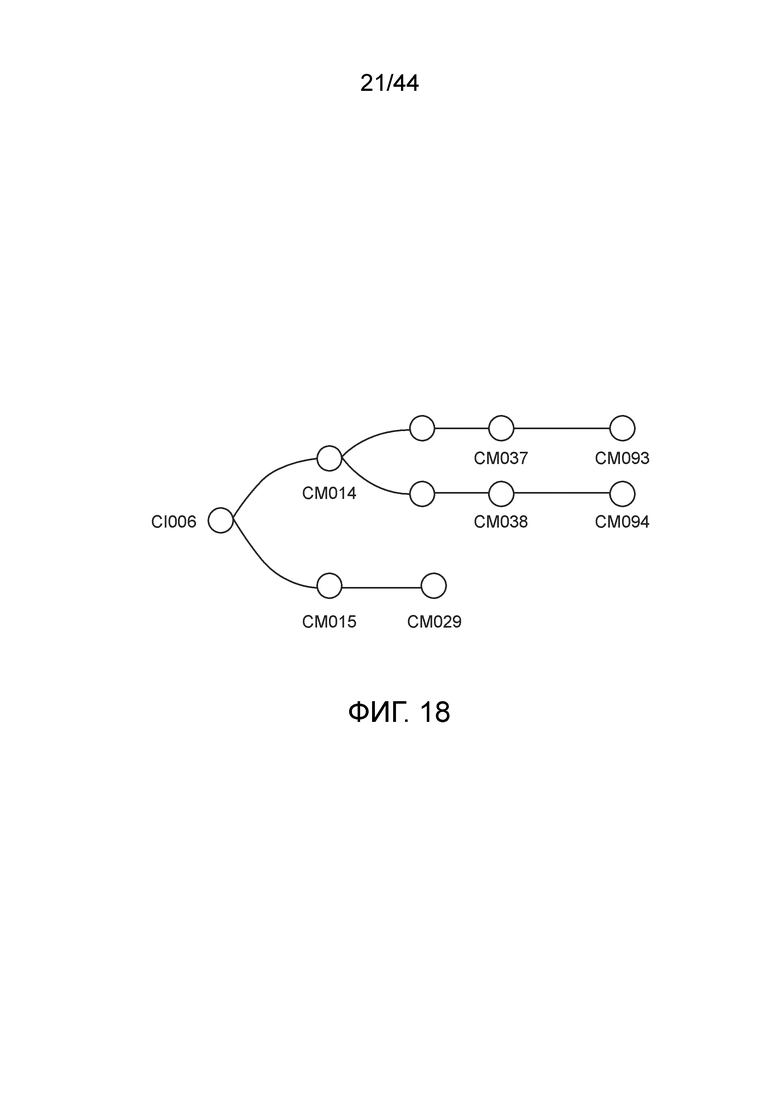
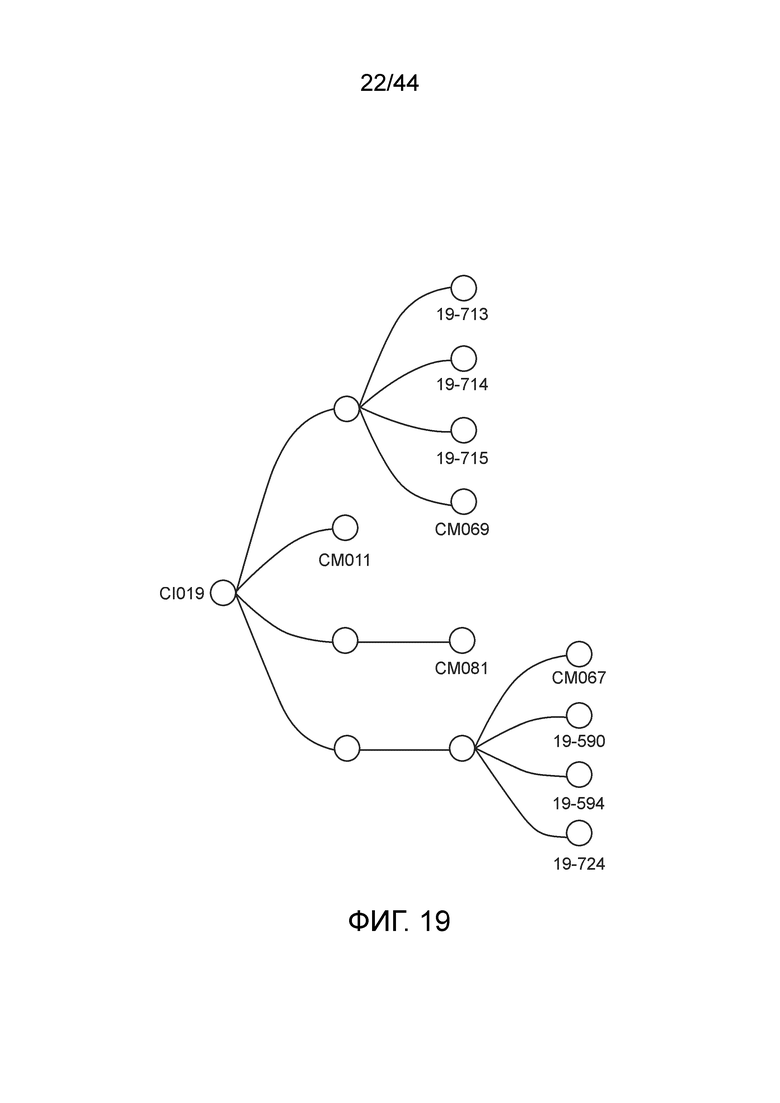
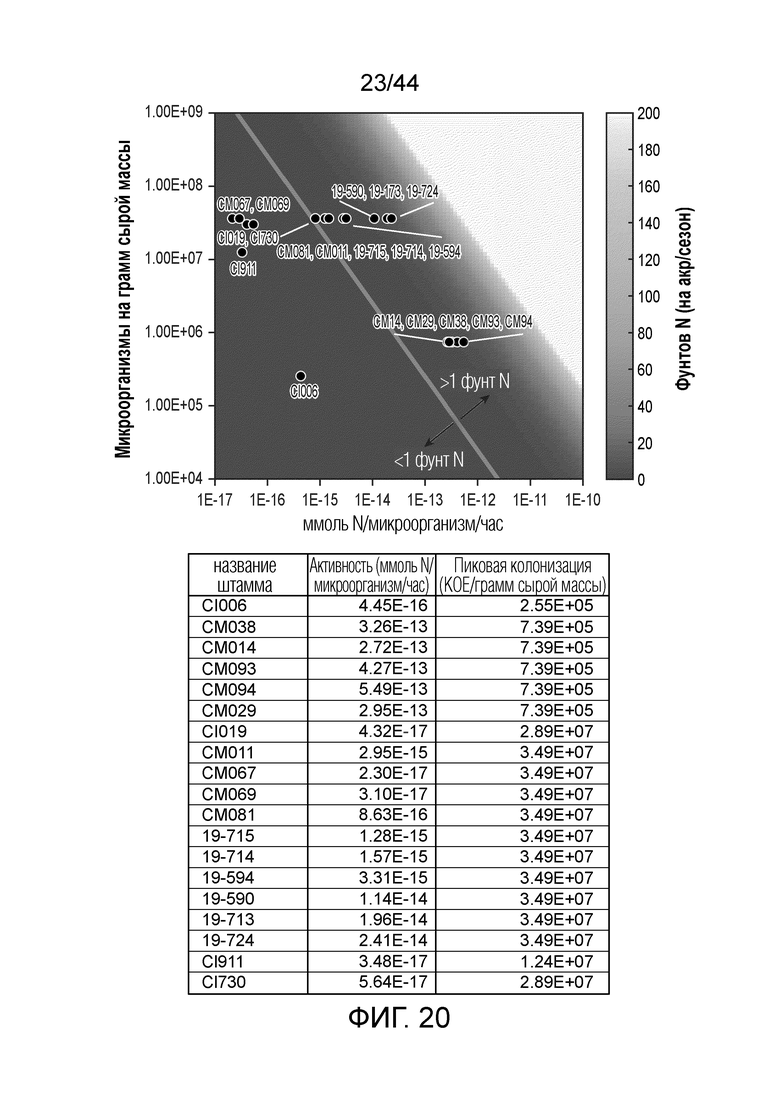
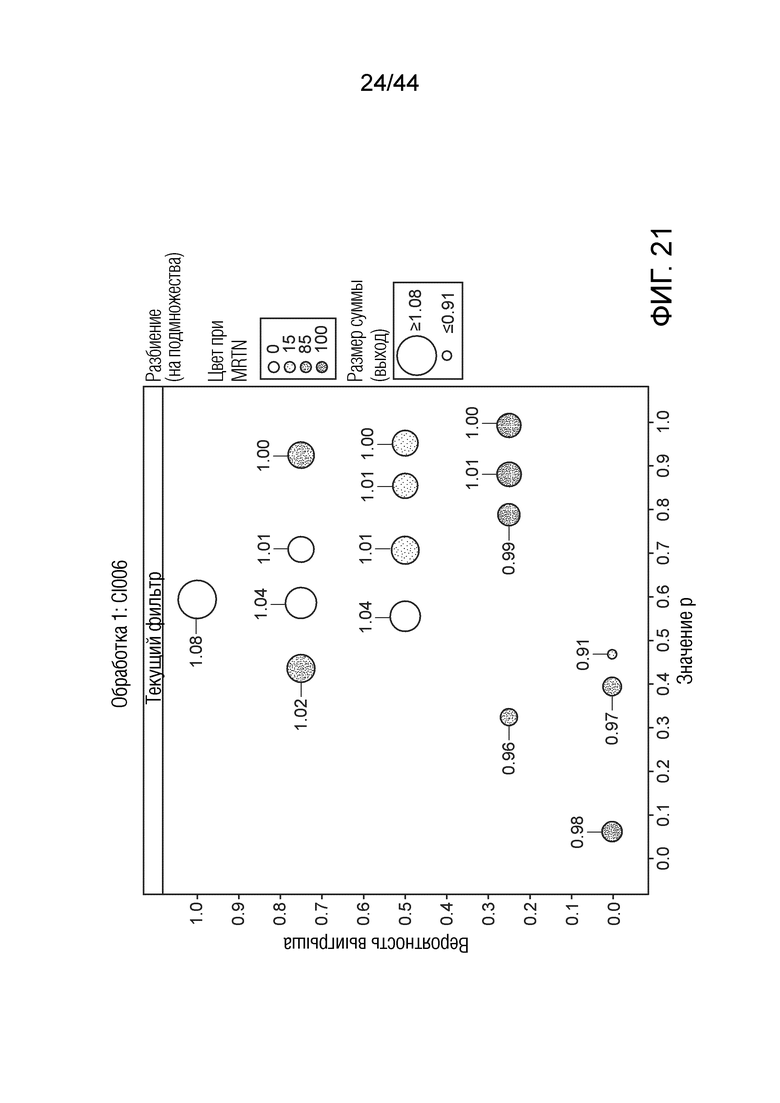
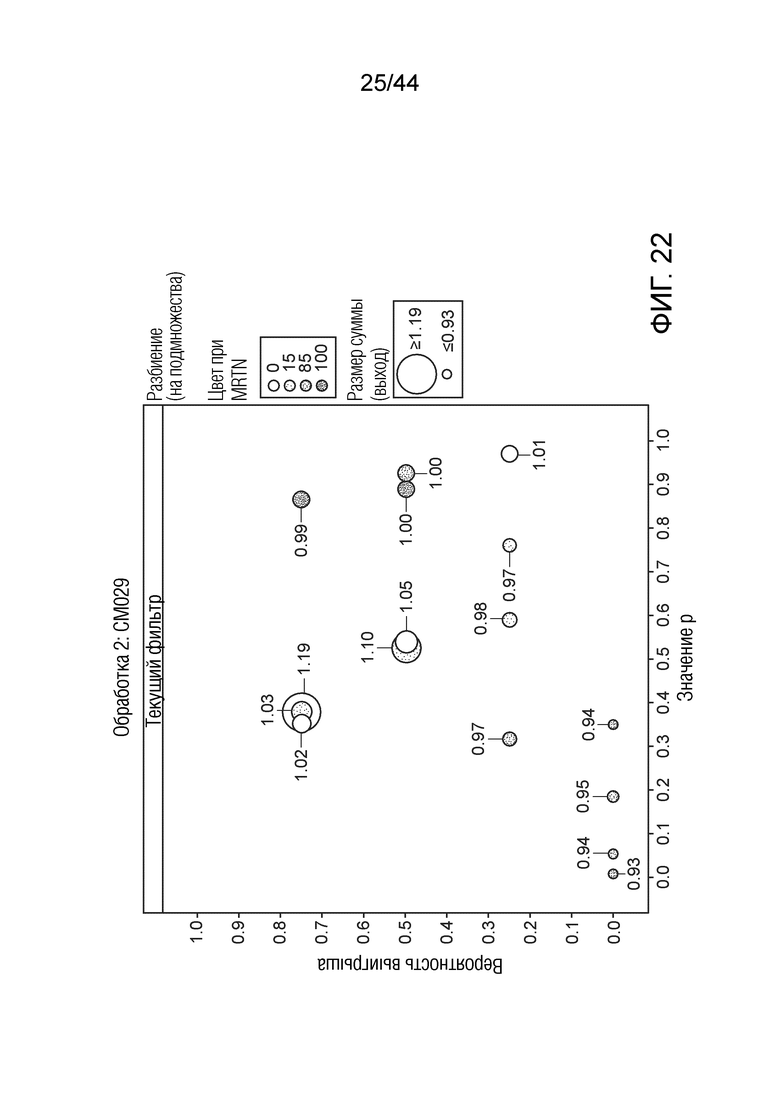
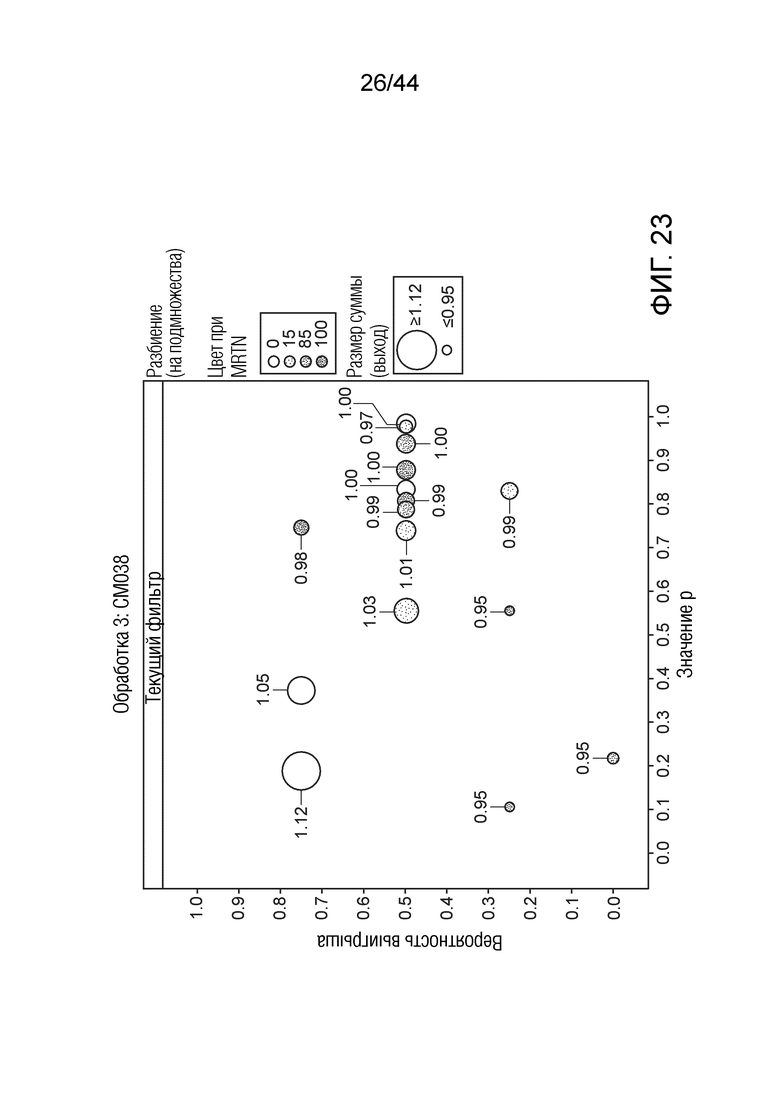
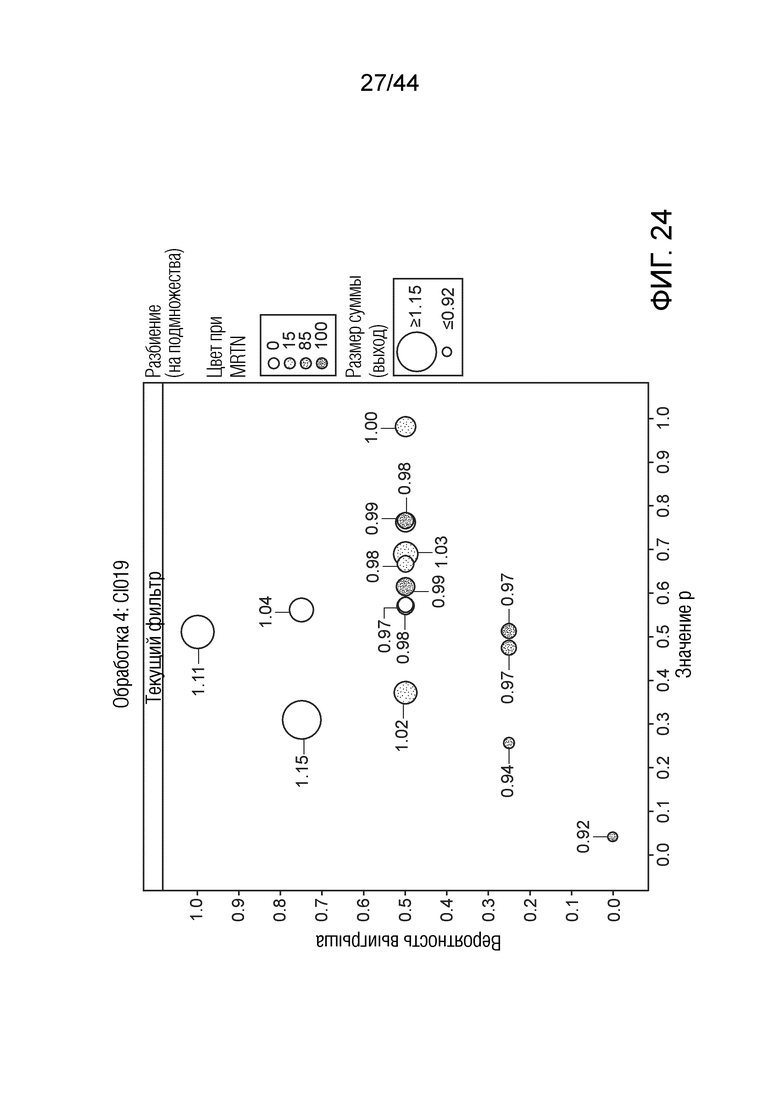
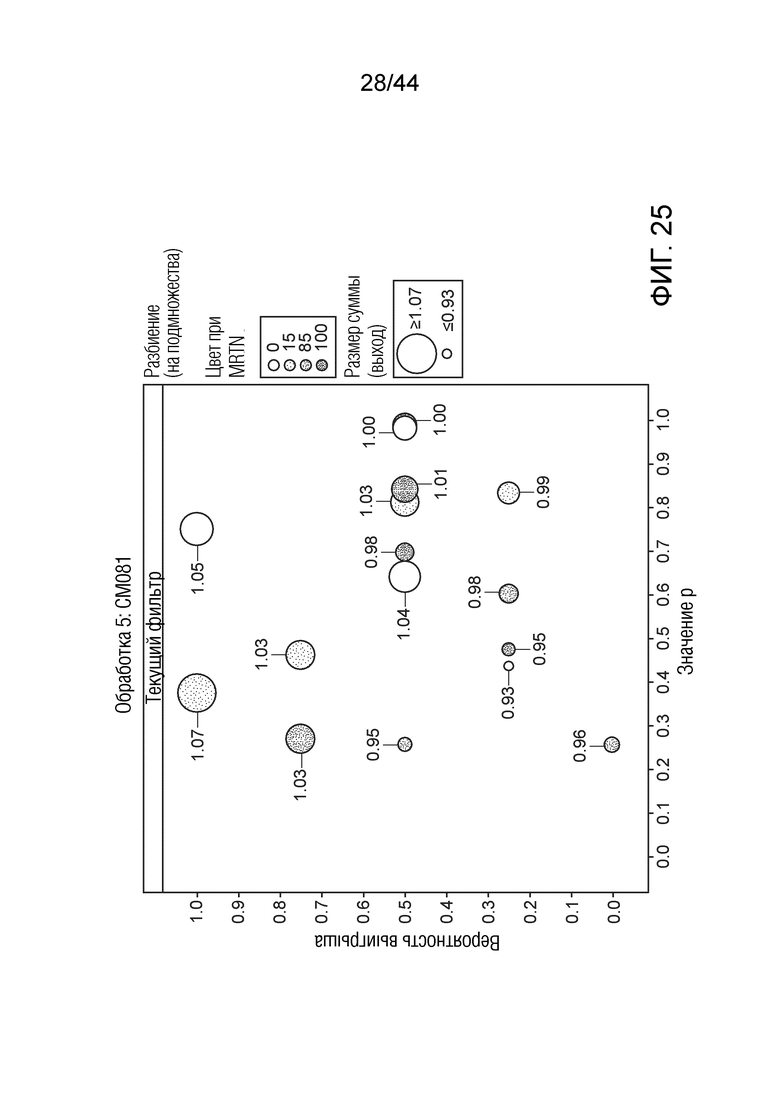
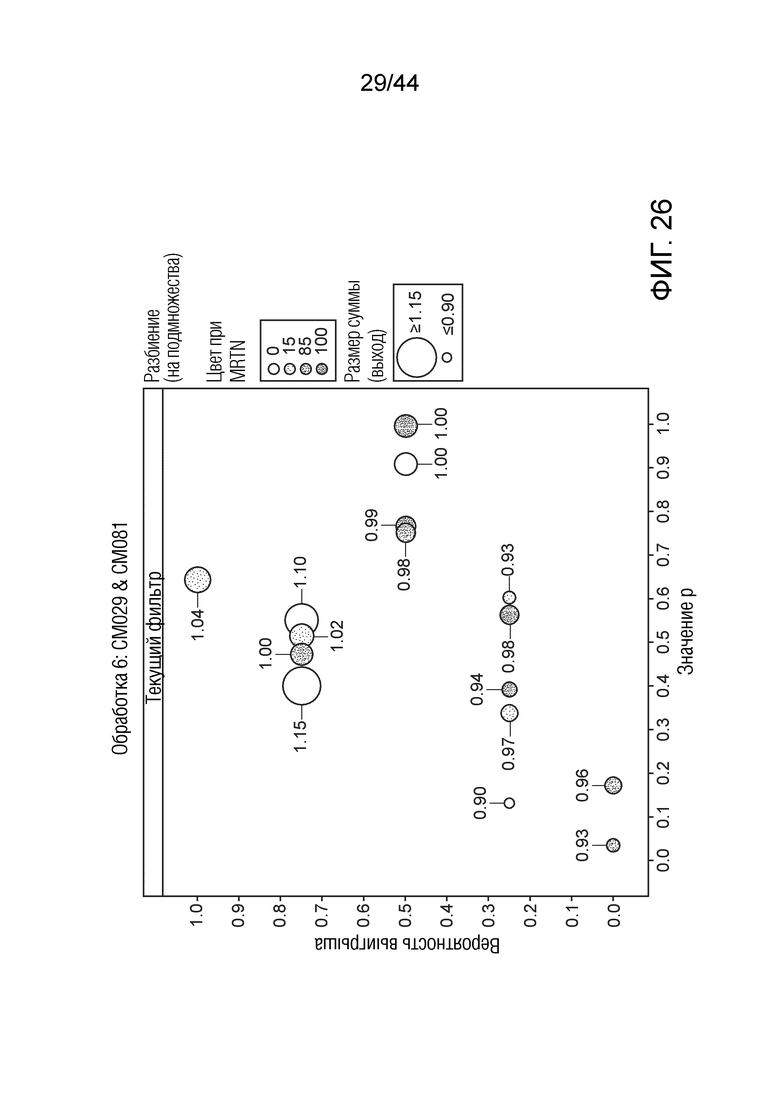
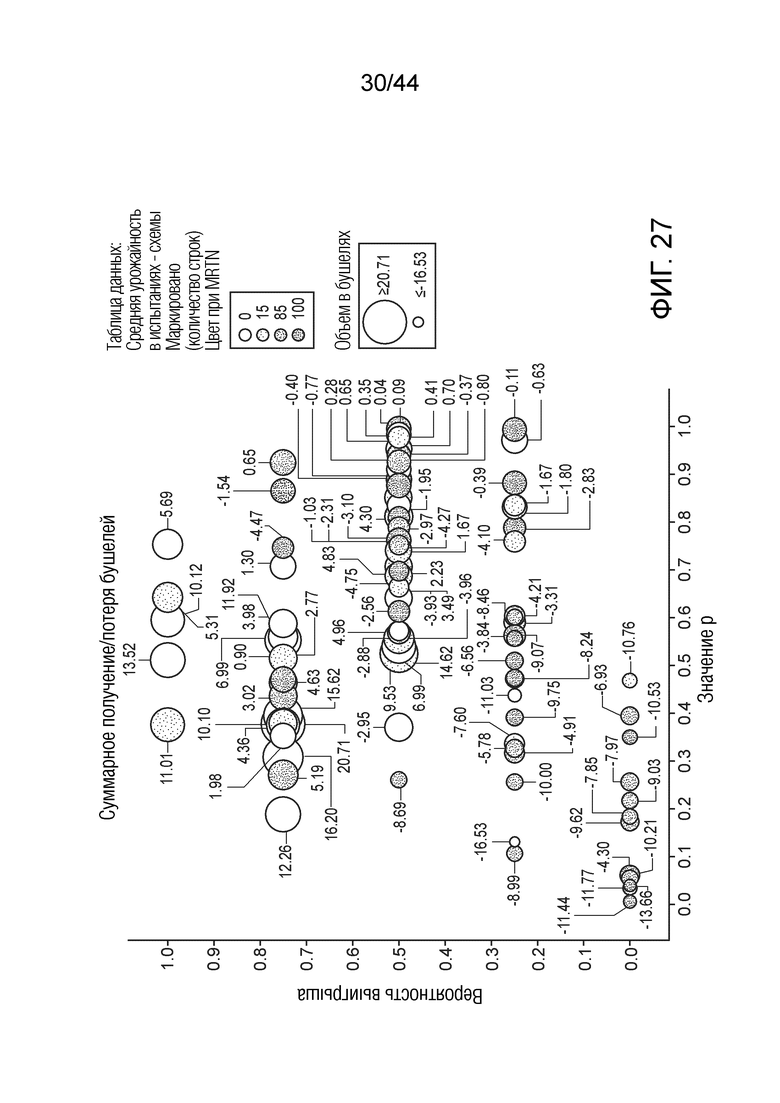
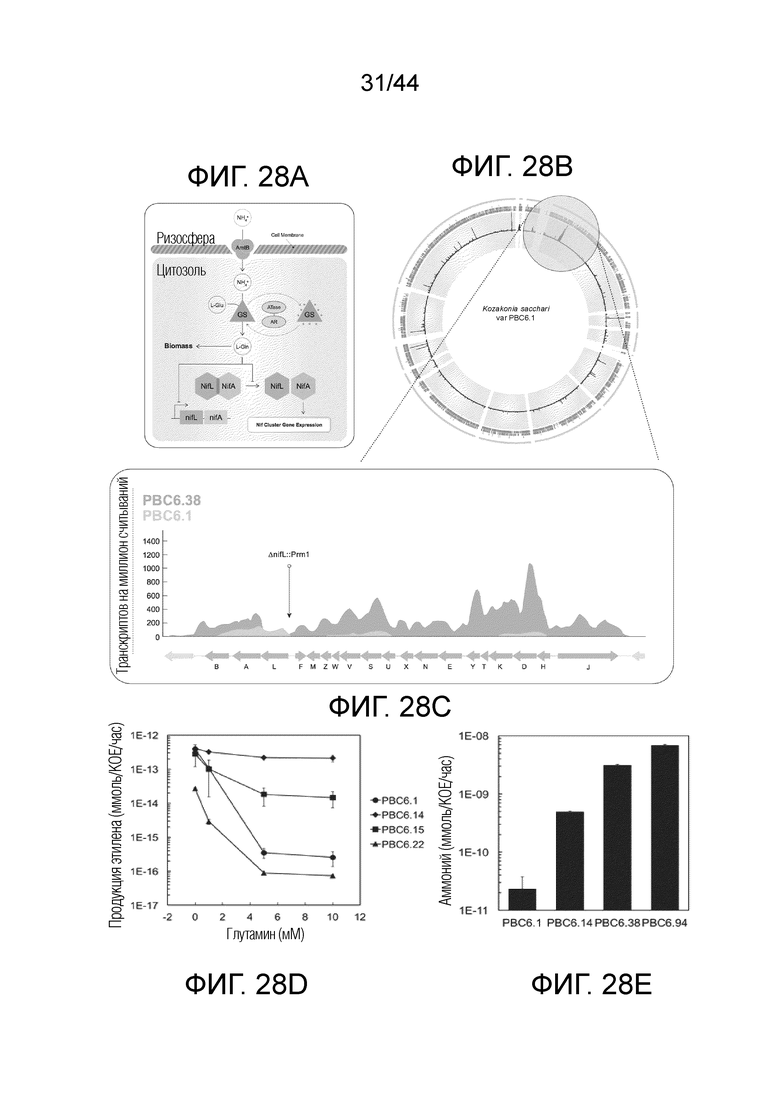

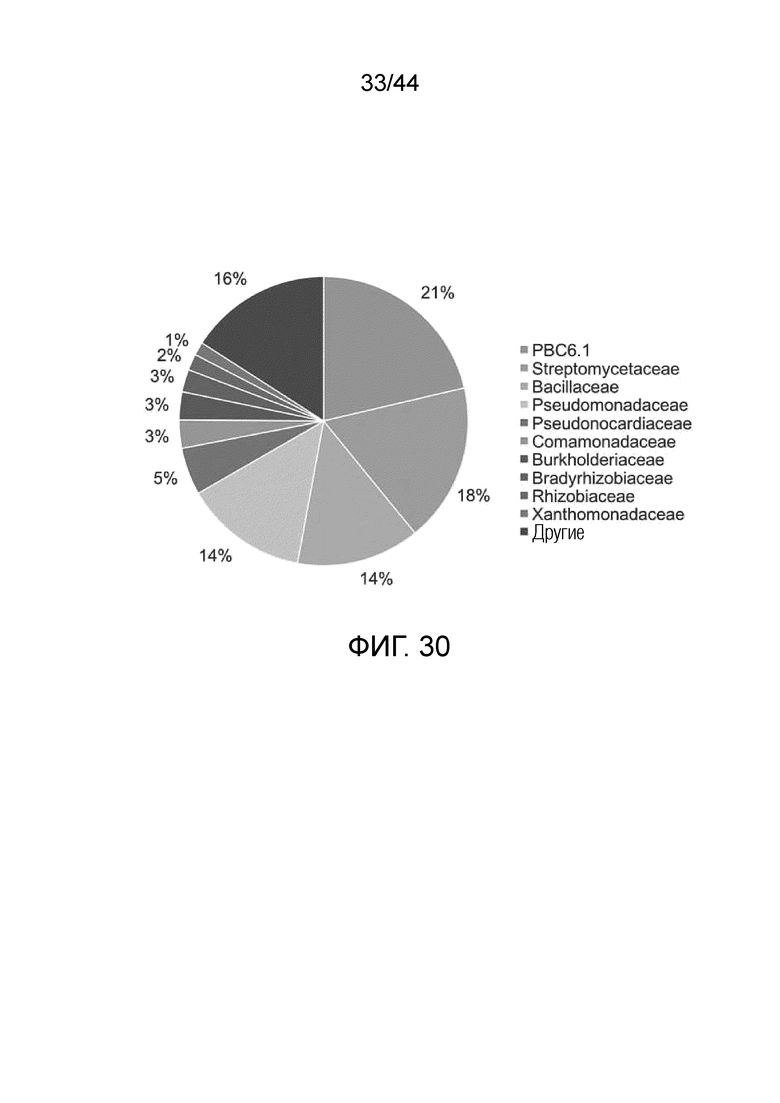
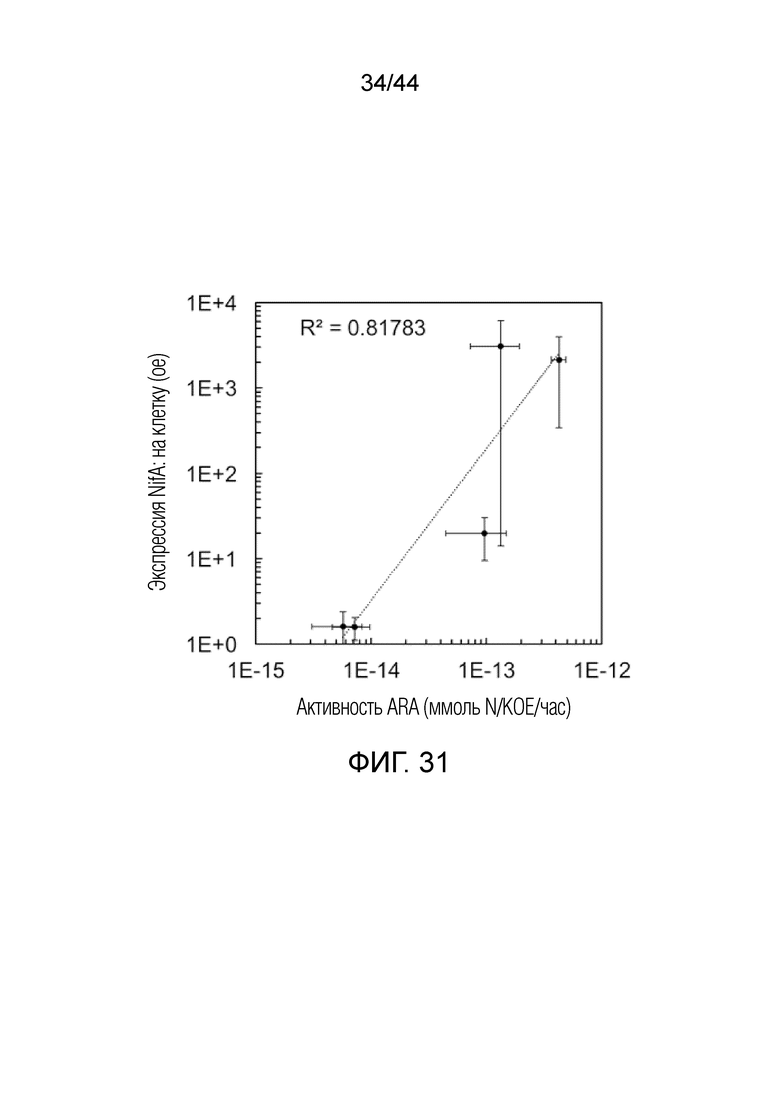
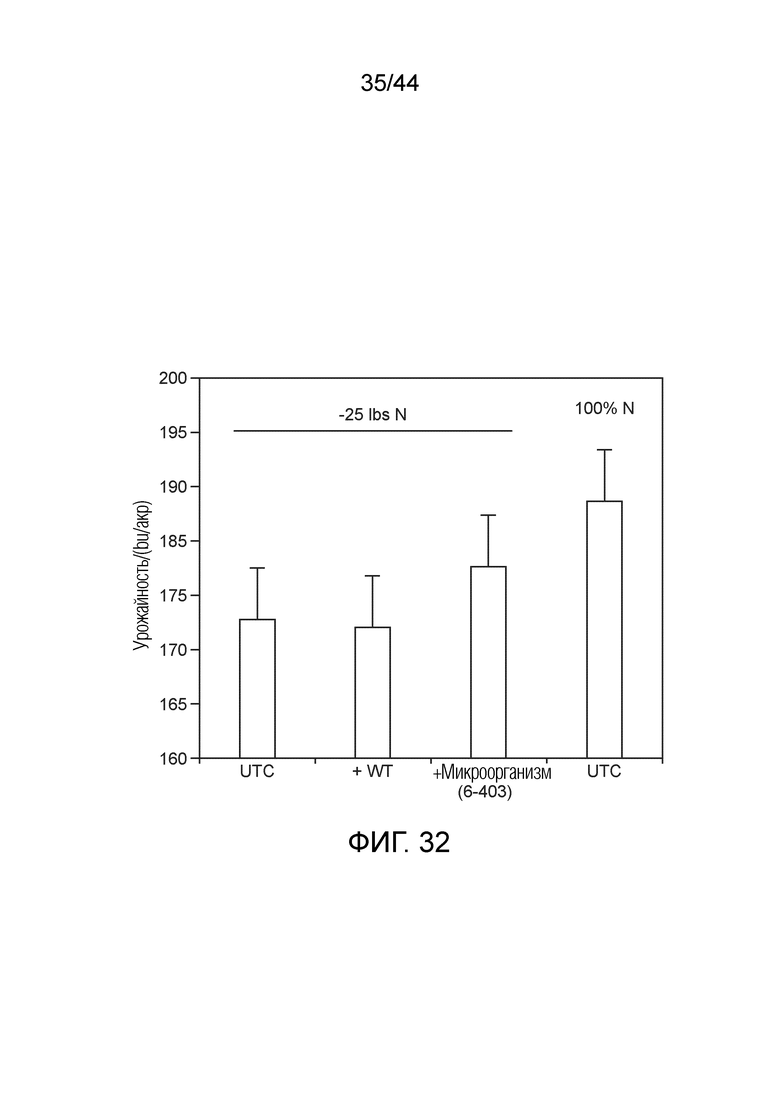

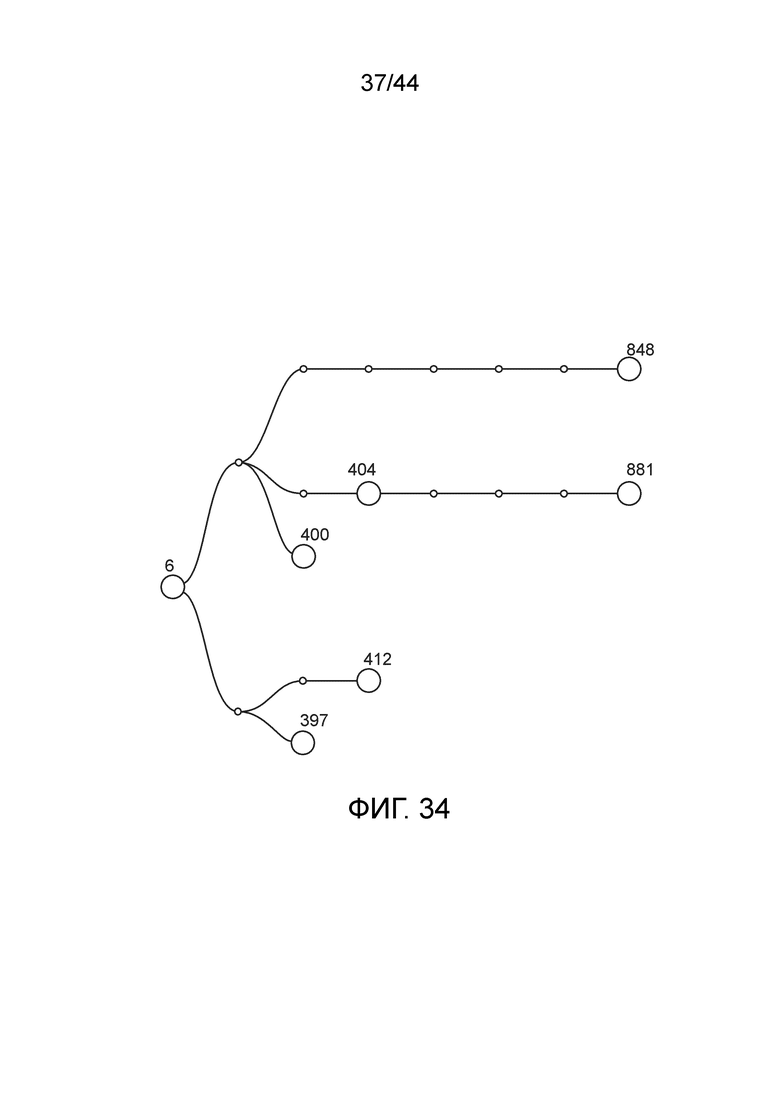
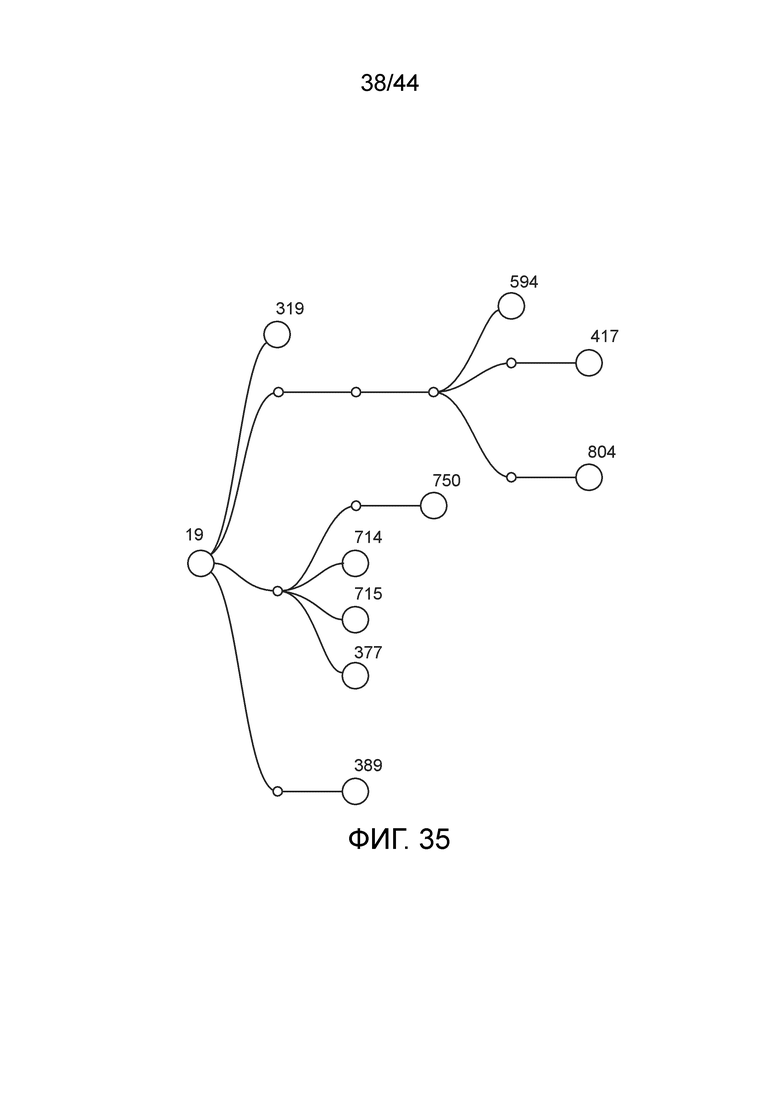
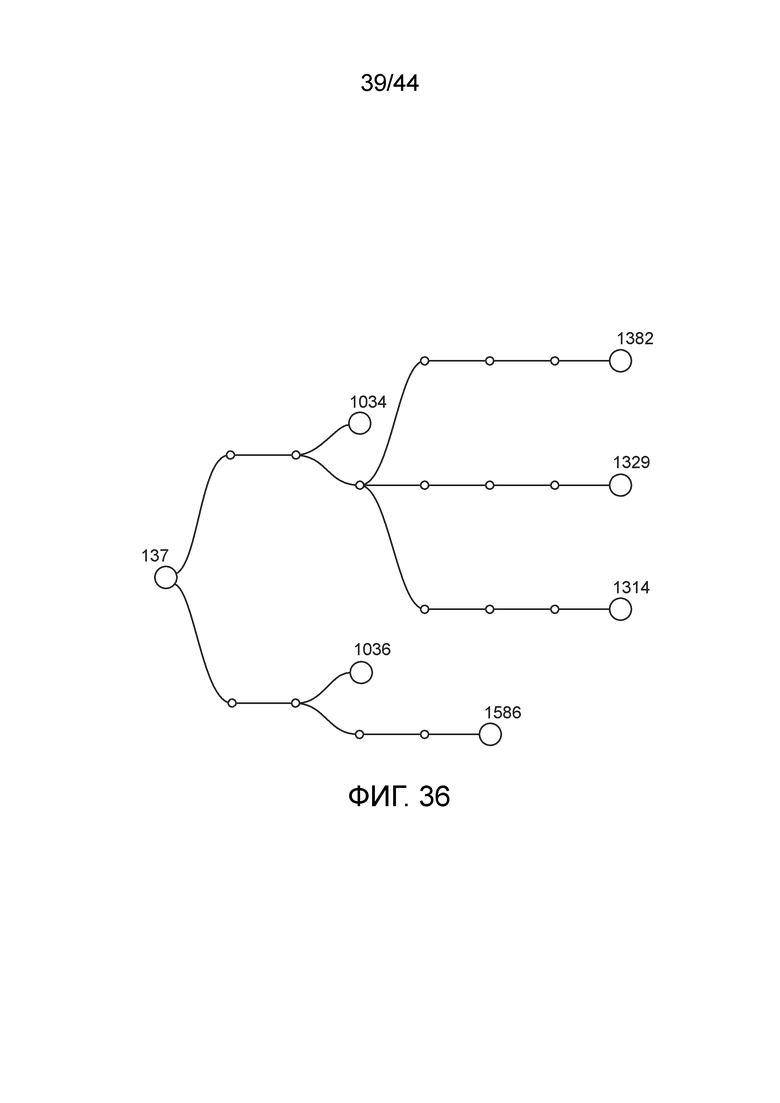
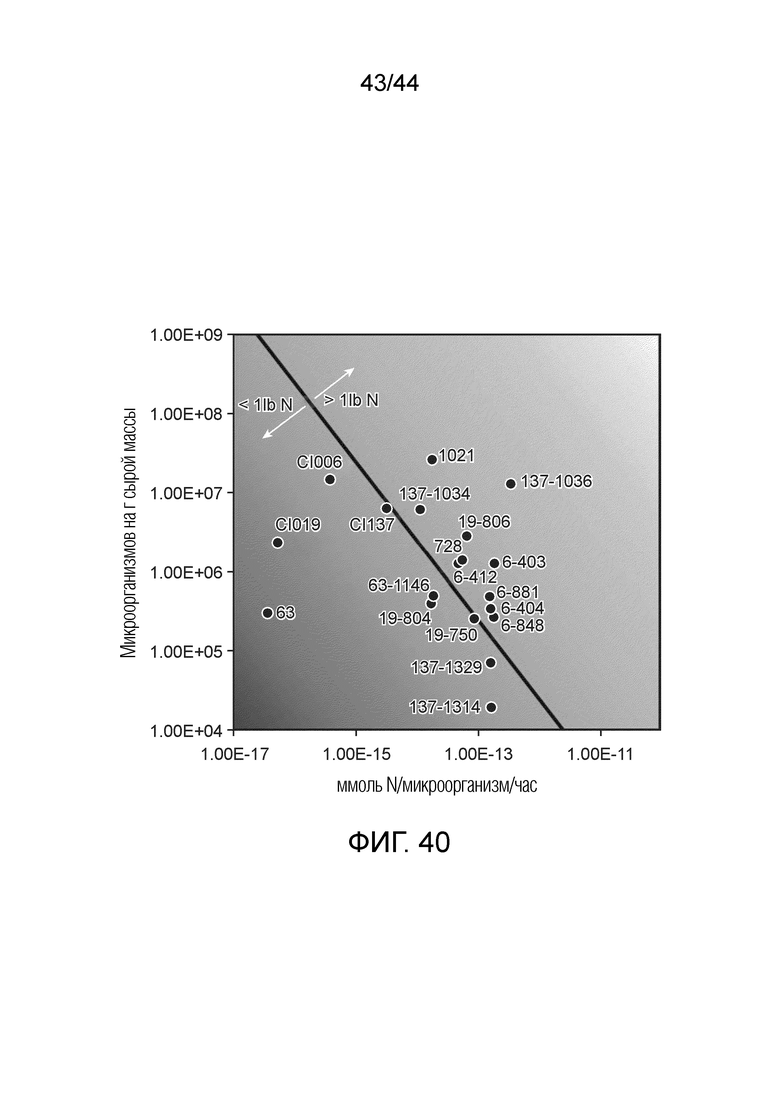
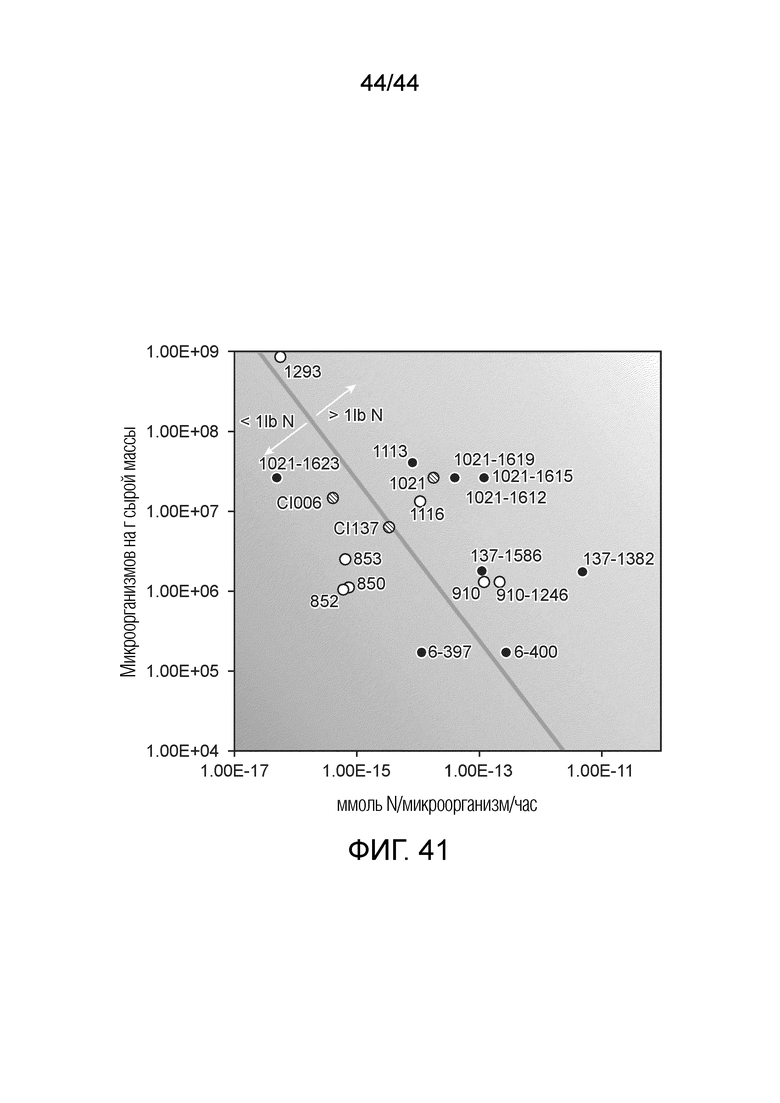
Изобретение относится к биотехнологии. Предложен способ идентификации штамма микроорганизмов, который производит по меньшей мере один фунт азота на акр за сезон. Указанный способ включает анализ двух или более штаммов микроорганизмов-кандидатов на колонизирующую способность и продуцирование азота на микроорганизм в час. При этом идентифицируют по меньшей мере один штамм микроорганизмов из указанных двух или более штаммов микроорганизмов-кандидатов, который имеет произведение (i) колонизирующей способности и (ii) N, продуцируемого на микроорганизм в час по меньшей мере примерно 2,5 × 10-8 ммоль N на грамм сырой массы ткани корня растения в час. При этом по меньшей мере один штамм микроорганизмов из указанных двух или более штаммов микроорганизмов-кандидатов имеет среднюю колонизирующую способность на единицу ткани корня растения по меньшей мере 1,79× 105 бактериальных клеток на грамм сырой массы ткани корня растения. Идентифицированный штамм с указанными характеристиками (i) колонизирующей способности и (ii) N, продуцируемого на микроорганизм в час используют в способе увеличения фиксации азота у небобовых растений в поле, для колонизации небобовой сельскохозяйственной культуры, для получения композиция для повышения урожайности небобовой сельскохозяйственной культуры. Изобретение обеспечивает увеличение фиксации азота у небобовых растений. 4 н. и 25 з.п. ф-лы, 41 ил., 22 табл., 13 пр.
1. Способ идентификации штамма микроорганизмов, который производит по меньшей мере один фунт азота на акр за сезон, указанный способ включает:
a) анализ двух или более штаммов микроорганизмов-кандидатов на колонизирующую способность и продуцирование азота на микроорганизм в час; и
b) идентификацию по меньшей мере одного штамма микроорганизмов из указанных двух или более штаммов микроорганизмов-кандидатов, который имеет произведение (i) колонизирующей способности и (ii) N, продуцируемого на микроорганизм в час по меньшей мере примерно 2,5×10-8 ммоль N на грамм сырой массы ткани корня растения в час,
при этом по меньшей мере один штамм микроорганизмов из указанных двух или более штаммов микроорганизмов-кандидатов имеет среднюю колонизирующую способность на единицу ткани корня растения по меньшей мере 1,79×105 бактериальных клеток на грамм сырой массы ткани корня растения.
2. Способ по п. 1, отличающийся тем, что указанный способ включает анализ указанных двух или более штаммов микроорганизмов-кандидатов в полевых условиях.
3. Способ по п. 2, отличающийся тем, что указанный способ включает идентификацию по меньшей мере одного штамма микроорганизмов, выбранного из группы, состоящей из Kosakonia sacchari, Rahnella aquatilis, Klebsiella varicola и Kosakonia pseudosacchari.
4. Способ по п. 3, отличающийся тем, что указанный штамм Kosakonia sacchari содержит бактерии, депонированные под обозначением патентного депозитария № 201701002, 201708004, 201708003 или 201708002.
5. Способ по п. 37, отличающийся тем, что указанный штамм Rahnella aquatilis содержит бактерии, депонированные под номером доступа PTA-122293.
6. Способ по п. 3, отличающийся тем, что указанный штамм Klebsiella varicola содержит бактерии, депонированные под обозначением патентного депозитария № 201712001 или 201712002.
7. Способ по п. 1, отличающийся тем, что указанный способ включает анализ указанных двух или более штаммов микроогранизмов-кандидатов в лабораторных и/или тепличных условиях.
8. Способ по п. 7, отличающийся тем, что указанный способ включает идентификацию по меньшей мере одного штамма микроорганизмов, выбранного из группы, состоящей из Kosakonia sacchari, Klebsiella Varicola, Kosakonia pseudosacchari, Azospirillum lipoferum, Enterobacter sp. и Kluyvera intermedia.
9. Способ по п.1, отличающийся тем, что указанный способ включает анализ указанных двух или более штаммов микроорганизмов-кандидатов, когда штаммы микроорганизмов связаны с растением кукурузы.
10. Способ по п.1, отличающийся тем, что указанный способ включает анализ продуцируемого азота с использованием анализа восстановления ацетилена.
11. Способ по п.1, отличающийся тем, что указанный способ включает анализ указанных двух или более штаммов микроорганизмов-кандидатов в присутствии экзогенного азота.
12. Способ по п.1, в котором идентификация по меньшей мере одного штамма микроорганизмов включает идентификацию генно-инженерного штамма микроорганизмов.
13. Способ увеличения фиксации азота у небобовых растений в поле, указанный способ включает применение к указанным небобовым растениям на указанном поле двух или более штаммов микроорганизмов, причем указанные два или более штаммов микроорганизмов включают штаммы микроорганизмов, имеющие произведение (i) способности к колонизации и (ii) N, продуцируемого на микроорганизм в час по меньшей мере примерно 2,5×10-8 ммоль N на грамм сырой массы ткани корня растения в час, при этом по меньшей мере один штамм микроорганизмов из указанных двух или более штаммов микроорганизмов-кандидатов имеет среднюю колонизирующую способность на единицу ткани корня растения по меньшей мере 1,79×105 бактериальных клеток на грамм сырой массы ткани корня растения.
14. Способ по п.13, в котором указанные два или более штаммов микроорганизмов in planta продуцирует 5% или более фиксированного азота в указанном небобовом растении.
15. Способ по п.13, отличающийся тем, что указанный способ включает применение по меньшей мере одного штамма микроорганизмов, выбранного из группы, состоящей из Kosakonia sacchari, Rahnella aquatilis, Klebsiella varicola и Kosakonia pseudosacchari.
16. Способ по п.15, в котором указанный штамм Kosakonia sacchari содержит бактерии, депонированные обозначением патентного депозитария № 201701002, 201708004, 201708003 или 201708002.
17. Способ по п.15, в котором указанный штамм Rahnella aquatilis содержит бактерии, депонированные под номером доступа PTA-122293.
18. Способ по п.15, в котором указанный штамм Klebsiella varicola содержит бактерии, депонированные обозначением патентного депозитария № 201712001 или 201712002.
19. Небобовая сельскохозяйственная культура, колонизированная штаммом микроорганизмов, для повышения урожайности небобовой сельскохозяйственной культуры, причем указанный штамм микроорганизмов имеет произведение (i) способности к колонизации и (ii) N, продуцируемого на микроорганизм в час по меньшей мере примерно 2,5×10-8 ммоль N на грамм сырой массы ткани корня растения в час,
где указанный штамм микроорганизмов производит не менее одного фунта азота на акр за сезон, и при этом указанный штамм микроорганизмов имеет среднюю колонизирующую способность на единицу ткани корня растения по меньшей мере 1,79×105 бактериальных клеток на грамм сырой массы ткани корня растения.
20. Небобовое растение по п.19, где указанное растение выбрано из группы, состоящей из кукурузы, риса, пшеницы, ячменя, сорго, проса, овса, ржи и тритикале.
21. Композиция для повышения урожайности небобовой сельскохозяйственной культуры, где указанная композиция содержит:
a) два или более штаммов микроорганизмов, которые имеют произведение (i) способности к колонизации и (ii) N, продуцируемого на микроорганизм в час по меньшей мере примерно 2,5×10-8 ммоль N на грамм сырой массы ткани корня растения в час, при этом указанные два или более штаммов микроорганизмов имеют среднюю колонизирующую способность на единицу ткани корня растения по меньшей мере 1,79×105 бактериальных клеток на грамм сырой массы ткани корня растения; и
b) семена небобовой сельскохозяйственной культуры, где указанные два или более штаммов микроорганизмов обеспечивают по меньшей мере один фунт азота на акр за сезон в поле небобовой сельскохозяйственной культуры, полученных из семян указанной сельскохозяйственной культуры.
22. Композиция по п.21, в которой указанная композиция содержит два или более штамма микроорганизмов.
23. Композиция по п.21, в которой указанные два или более штаммов микроорганизмов включает по меньшей мере один штамм микроорганизмов, выбранный из группы, состоящей из Kosakonia sacchari, Rahnella aquatilis, Klebsiella varicola и Kosakonia pseudosacchari.
24. Композиция по п.23, в которой указанный штамм Kosakonia sacchari содержит бактерии, депонированные под обозначением патентного депозитария № 201701002, 201708004, 201708003 или 201708002.
25. Композиция по п.23, где указанный штамм Rahnella aquatilis содержит бактерии, депонированные под номером доступа PTA-122293.
26. Композиция по п.23, отличающаяся тем, что указанный штамм Klebsiella varicola содержит бактерии, депонированные под обозначением патентного депозитария № 201712001 или 201712002.
27. Способ по п.1 или 13, в котором два или более штаммов микроорганизмов являются внутриродовыми.
28. Небобовое растение, колонизированное штаммом микроорганизмов, по п.19, в котором два или более штаммов микроорганизмов являются внутриродовыми.
29. Композиция по п. 21, в которой два или более штаммов микроорганизмов являются внутриродовыми.
| US 2009137390 A, 28.05.2009 | |||
| WO 2013132518 A1, 12.09.2013 | |||
| Q | |||
| AN et al | |||
| Способ получения на волокне оливково-зеленой окраски путем образования никелевого лака азокрасителя | 1920 |
|
SU57A1 |
| JOURNAL OF APPLIED MICROBIOLOGY, vol | |||
| Клапанный регулятор для паровозов | 1919 |
|
SU103A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
