Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам, которые связываются специфически с хелатными радионуклидами, включая биспецифические антитела. Оно относится также к применению указанных биспецифических антител в таких областях, как радиоиммуновизуализация и радиоиммунотерапия. Кроме того, оно относится к очищающим агентам и композициям, которые можно применять в указанных способах.
Предпосылки создания изобретения
Для направленного переноса лекарственных средств к раковым клеткам созданы моноклональные антитела. Путем конъюгации токсического агента с антителом, которое связывается с опухолеассоциированным антигеном, можно обеспечивать более специфический цитолиз опухолей с меньшей степенью повреждения окружающих тканей.
При осуществлении претаргетной радиоиммунотерапии (PRIT) используют конструкцию антитела, которая обладает высокой аффинностью к опухолеассоциированому антигену, с одной стороны, и радиоактивно меченное соединение, с другой стороны. На первой стадии вводят антитело, и оно локализуется внутри опухоли. После этого вводят радиоактивно меченное соединение. Поскольку радиоактивно меченное соединение является небольшим, оно может быстро достигать опухоли и быстро выводиться, что снижает радиационную нагрузку вне опухоли (Goldenberg и др., Theranostics 2(5), 2012, сс. 523-540). Помимо непосредственного уничтожающего клетку действия PRIT может действовать также в качестве индуктора гибели иммуногенных клеток и в качестве возможного партнера для комбинации, применяемой для противораковой иммунотерапии и в подходах эндогенной вакцинации. Аналогичную процедуру можно применять также для визуализации. Для предварительного таргетинга можно применять биспецифическое антитело или системы на основе авидина-биотина, хотя последние обладают недостатком, связанным с тем, что система авидин/стрептавидин является иммуногенной.
Радионуклиды, предназначенные для применения в PRIT, как правило, находятся в форме хелата, нагруженного представляющим интерес радионуклидом.
Su с соавторами (Nucl Med Biol 32, 2005, сс. 741-747) изучали систему для предварительного таргетинга, включающую антитело, на основе МАт-стрептавидина и DOTA. Было установлено, что система радиоактивно меченный 212Pb-DOTA-биотин была нестабильной, при этом более 30% свободного 212Bi (продукт распада 212Pb) высвобождалось из 212Pb-DOTA.
В WO 2010/099536 описано биспецифическое антитело, которое обладает способностью связываться с комплексами, включающими DOTA и иттрий, лютений и гадолиний. Однако DOTA не обладает способностью стабильно связываться со всеми радионуклидами, и может характеризоваться медленной скоростью комплексообразования (Yong и Brechbiel, Dalton Trans. 40(23), 21 июня 2001 г., с. 6068-6076). Отсутствие у хелатора способности стабильно связываться с радионуклидом создает риск снижения уровня доставки радиации к опухоли с возрастанием при этом токсичности.
Краткое изложение сущности изобретения
В настоящем изобретении предложены антитела, которые связываются с хелатом металла, содержащим DOTAM и свинец (Pb). DOT AM обладает способностью стабильно хелатировать Pb с образованием комплекса Pb[DOTAM].
Антитела, предлагаемые в настоящем изобретении, связываются с хелатом, содержащим DOTAM и Pb, в котором Pb может представлять собой либо стабильный (нерадиоактивный) изотоп или радиоактивный изотоп. Радиоизотопы свинца могут найти применение для радиоиммуновизуализации и радиоиммунотерапии.
Предпочтительно антитела, предлагаемые в настоящем изобретении, обладают чрезвычайно высокой аффинностью к хелату Pb-DOTAM, находящейся в диапазоне концентраций от пикомолярных (пМ) до фентомолярных (фМ).
Антитела дополнительно связываются с висмутом (Bi), хелатированным DOTAM. 212Pb представляет собой родительский радионуклид 212Bi и может служить в качестве генератора in vivo 212Bi. Способность антител связываться с хелатным Bi, а также с хелатным Pb повышает возможность их применения в области радиоиммунотерапии, при этом изотоп Bi образуется как продукт распада изотопа Ph. В некоторых вариантах осуществления изобретения антитела могут связываться как с хелатом Bi-DOTAM, так и с хелатом Pb-DOTAM с очень высокой аффинностью в диапазоне концентраций от пМ до фМ.
Кроме того, антитела, предлагаемые в настоящем изобретении, необязательно или предпочтительно обладают селективностью в отношении хелата Bi-DOTAM и хелата Pb-DOTAM по сравнению с другими хелатными комплексами металлов, такими как хелат Cu-DOTAM.
Одним из вариантов осуществления настоящего изобретения является антитело, содержащее антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, в котором антигенсвязывающий сайт содержит тяжелую цепь, которая содержит по меньшей мере одну, две или три последовательности CDR тяжелой цепи: в котором:
а) CDR1 тяжелой цепи содержит аминокислотную последовательность GFSLSTYSMS (SEQ ID NO: 1);
б) CDR2 тяжелой цепи содержит аминокислотную последовательность FIGSRGDTYYASWAKG (SEQ ID NO: 2);
в) CDR3 тяжелой цепи содержит аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3);
и/или в котором антигенсвязывающий сайт содержит легкую цепь, которая содержит по меньшей мере одну, две или три последовательности CDR легкой цепи: в котором:
г) CDR1 легкой цепи содержит аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4);
д) CDR2 легкой цепи содержит аминокислотную последовательность QASKLAS (SEQ ID NO: 5);
е) CDR3 легкой цепи содержит аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6).
В некоторых вариантах осуществления изобретения антигенсвязывающий сайт содержит и легкую цепь, и тяжелую цепь, указанные выше.
В другом варианте осуществления настоящего изобретения предложено антитело, которое содержит антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, в котором указанный антигенсвязывающий сайт содержит по меньшей мере:
а) CDR2 тяжелой цепи, содержащий аминокислотную последовательность FIGSRGDTYYASWAKG (SEQ ID NO: 2) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 2, где указанные замены не включают Phe50, Asp56 и/или Tyr58 и необязательно не включают также Gly52 и/или Arg54;
б) CDR3 тяжелой цепи, содержащий аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 3, где указанные замены не включают Glu95, Arg96, Asp97, Pro98 и необязательно не включают также Ala100C, Tyr100D и/или Pro100E, и/или необязательно не включают также Tyr99;
в) CDR1 легкой цепи, содержащий аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 4, где указанные замены не включают Tyr28 и Asp32;
г) CDR3 легкой цепи, содержащий аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 6, где указанные замены не включают Gly91, Tyr92, Asp93, Thr95c и Tyr96.
Нумерация остатков соответствует нумерации по Кэботу.
В некоторых вариантах осуществления изобретения антитело включает также CDR1 тяжелой цепи и CDR2 легкой цепи, которые необязательно представляют собой:
I) CDR1 тяжелой цепи, содержащий аминокислотную последовательность GFSLSTYSMS (SEQ ID NO: 1) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 1;
II) CDR2 легкой цепи, содержащий аминокислотную последовательность QASKLAS (SEQ ID NO: 5) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 5, которые необязательно не включают Gln50.
В любых вариантах осуществления настоящего изобретения, которые относятся к вариантам последовательности, содержащей указанные выше CDR, белок может быть инвариантным касательно одного или нескольких указанных выше остатков.
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, связывается с тем же эпитопом или с перекрывающимся эпитопом хелатного радионуклида, с которым связывается представленное в настоящем описании антитело.
В некоторых вариантах осуществления изобретения антитело связывается с тем же эпитопом или с перекрывающимся эпитопом, что и эпитоп, который связывается Fab PRIT-0213 или PRIT-0214. Например, антитело может связываться с тем же эпитопом или с перекрывающимся эпитопом, что и:
I) антитело, которое имеет вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 7, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 8; или
II) антитело, которое имеет вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 9, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 10.
В одном из вариантов осуществления изобретения антигенсвязывающий сайт содержит по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из:
а) CDR1 тяжелой цепи, который содержит аминокислотную последовательность GFSLSTYSMS (SEQ ID NO: 1);
б) CDR2 тяжелой цепи, который содержит аминокислотную последовательность FIGSRGDTYYASWAKG (SEQ ID NO: 2);
в) CDR3 тяжелой цепи, который содержит аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3);
г) CDR1 легкой цепи, который содержит аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4);
д) CDR2 легкой цепи, который содержит аминокислотную последовательность QASKLAS (SEQ ID NO: 5);
е) CDR3 легкой цепи, который содержит аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6).
Необязательно антитело, указанное в любом из описанных выше объектов изобретения, является человеческим, химерным или гуманизированным.
Необязательно антигенсвязывающий сайт может содержать вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 7 и SEQ ID NO: 9, или ее вариант, содержащий аминокислотную последовательность, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% SEQ ID NO: 7 или SEQ ID NO: 9.
Необязательно антигенсвязывающий сайт может содержать вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 8 и SEQ ID NO: 10, или ее вариант, содержащий аминокислотную последовательность, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% SEQ ID NO: 8 или SEQ ID NO: 10.
Необязательно антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, может содержать вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 7 и SEQ ID NO: 9, или ее вариант, указанный выше, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 8 и SEQ ID NO: 10, или ее вариант, указанный выше. Например, антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, может содержать вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 7 или ее вариант, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 8 или ее вариант. В другом варианте осуществления изобретения он может содержать вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 9 или ее вариант, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 10 или ее вариант.
Антитело может находиться в любом формате, включая цельные антитела и фрагменты антител. Антитело может быть моноспецифическим. В указанной форме антитело находит применение, например, в схемах сортировки и очистки, например, для успешного разделения радиоактивно меченных фрагментов.
В некоторых объектах изобретения антитело, которое специфически связывается с хелатом Pb-DOTAM, сшивают со связывающимся с клеткой агентом/таргетирующим фрагментом с образованием таргетного агента. Указанный агент можно применять, например, для претаргетной радиоиммунотерапии или претаргетной радиоиммуновизуализации.
Сшивание предпочтительно можно осуществлять путем экспрессии слитого полипептида или белка. Слияние может быть непосредственным или через линкер. Слитый полипептид или белок можно получать рекомбинантно, для чего не требуется химическая конъюгация.
В некоторых вариантах осуществления изобретения таргетирующий фрагмент (содержащий антигенсвязывающий сайт для мишени) представляет собой антитело или его фрагмент. Это означает, что в некоторых вариантах осуществления изобретения описанное выше антитело может находиться в форме мультиспецифического (например, биспецифического) антитела, которое будет дополнительно описано ниже.
Другим объектом настоящего изобретения является также мультиспецифическое антитело/содержащий антитело комплекс, пригодное/пригодный для направленного переноса хелата Pb-DOTAM к клетке-мишени.
Таким образом, другим объектом настоящего изобретения является биспецифическое или мультиспецифическое антитело, которое специфически связывается как с хелатом Pb-DOTAM, так и с антигеном-мишенью, например, антигеном, который экспрессируется на поверхности клетки-мишени. Биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический в отношении хелатированного DOTAM свинца, и по меньшей мере один антигенсвязывающий сайт для антигена-мишени. В некоторых вариантах осуществления изобретения антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, может представлять собой сайт, указанный в любых описанных выше вариантах осуществления изобретения.
Антиген-мишень может представлять собой любой антиген, дополнительно указанный в настоящем описании, например, опухолеспецифический антиген. В некоторых вариантах осуществления изобретения он может представлять собой белок или полипептид, экспрессируемый патогеном, таким как прокариотический организм или вирус.
В некоторых вариантах осуществления изобретения опухолеассоциированный антиген может представлять собой СЕА (карциноэмбриональный антиген). Так, в некоторых вариантах осуществления изобретения биспецифическое антитело может содержать по меньшей мере один антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический в отношении СЕА. СЕА является предпочтительным в контексте настоящего изобретения, поскольку он относительно медленно интернализируется, и поэтому высокий процент биспецифического антитела должен оставаться доступным для связывания с радионуклидом на поверхности клетки после начальной обработки. Могут оказаться предпочтительными другие слабо интернализирующиеся антигены-мишени/ассоциированные с опухолью антигены, и они указаны в настоящем описании. Другими примерами опухолеассоциированного антигена, который можно применять в настоящем изобретении, является CD20 или HER2.
В некоторых вариантах осуществления изобретения, в которых антиген-мишень представляет собой СЕА, антигенсвязывающий сайт, специфический в отношении СЕА, может содержать тяжелую цепь, которая содержит по меньшей мере один, два или три CDR тяжелой цепи, в которой:
г) CDR1 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 11;
д) CDR2 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 12;
е) CDR3 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 13;
и/или антигенсвязывающий сайт, специфический в отношении СЕА, может содержать легкую цепь, которая содержит по меньшей мере один, два или три CDR тяжелой цепи, в которой:
а) CDR1 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 14;
б) CDR2 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 15;
в) CDR3 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 16.
В некоторых вариантах осуществления изобретения антигенсвязывающий сайт, специфический в отношении СЕА, может содержать по меньшей мере один, два, три, четыре, пять или шесть (т.е. все) CDR, выбранные из:
а) CDR1 тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 11;
б) CDR2 тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 12;
в) CDR3 тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 13;
г) CDR1 легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 14;
д) CDR2 легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 15;
е) CDR3 легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 16.
В некоторых вариантах осуществления изобретения антигенсвязывающий сайт для СЕА может содержать вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 17, или его вариант, содержащий аминокислотную последовательность, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% SEQ ID NO: 17.
Необязательно антигенсвязывающий сайт может содержать вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 18, или его вариант, содержащий аминокислотную последовательность, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% SEQ ID NO: 18.
Необязательно антигенсвязывающий сайт, специфический в отношении СЕА, может содержать вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 17, или его вариант, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 18, или его вариант.
Различные возможные форматы биспецифических антител или мультиспецифических антител известны в данной области, в том числе дополнительно указанные в настоящем описании. Антитела, предлагаемые в изобретении, могут иметь любой из указанных форматов. Например, в некоторых вариантах осуществления изобретения биспецифическое антитело может быть двухвалентным, трехвалентным или четырехвалентным.
Предпочтительно предлагаемые в настоящем изобретении антитела могут включать Fc-область. Присутствие Fc-области обладает преимуществами в контексте радиоиммунотерапии и радиовизуализации, например, пролонгируя время полужизни белка в кровотоке и/или обеспечивая более высокое поглощение опухолью, по сравнению с использованием фрагментов меньшего размера.
В некоторых вариантах осуществления изобретения, в которых присутствует Fc-область, может оказаться предпочтительным создавать Fc-область со сниженной эффекторной функцией. Это может включать замену одного или нескольких остатков Fc-области в положениях 234, 235, 238, 265, 269, 270, 297, 327 и/или 329, например, одного или нескольких остатков в положениях 234, 235 и/или 329. В некоторых вариантах осуществления изобретения Fc-область можно создавать так, чтобы она включала замену Pro 329 на Gly, Leu 234 на Ala и/или Leu 235 на Ala (нумерация согласно EU-индексу).
Известны различные форматы мультиспецифических антител, которые включают Fc-домен.
В одном из вариантов осуществления изобретения биспецифическое или мультиспецифическое антитело может содержать I) Fc-домен, II) по меньшей мере один Fab-, cross-Fab-, Fv-, scFab- или scFv-фрагмент или однодоменное антитело (VHH), который/которое содержит антигенсвязывающий сайт, специфический в отношении хелата Pb-DOTAM, и III) и по меньшей мере один Fab-, cross-Fab-, Fv-, scFab- или scFv-фрагмент или однодоменное антитело (VHH), который/которое содержит антигенсвязывающий сайт, специфический в отношении антигена-мишени.
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если биспецифическое или мультиспецифическое антитело является многовалентным, например, двухвалентным в отношении антигена-мишени (например, опухолеассоциированного антигена). Это обладает преимуществом с позиции увеличения авидности.
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если биспецифическое или мультиспецифическое антитело является одновалентным в отношении Pb-DOTAM. Это снижает риск образования высокомолекулярного комплекса при применении очищающего агента (см. дополнительно представленное ниже обсуждение).
Таким образом, в некоторых вариантах осуществления изобретения антитело может быть трехвалентным: т.е. двухвалентным в отношении антигена-мишени и одновалентным в отношении Pb-DOTAM.
В одном приведенном в качестве примера формате биспецифическое или мультиспецифическое антитело может представлять собой полноразмерное антитело (например, IgG), содержащее первую и вторую тяжелые цепи антитела и первую и вторую легкие цепи антитела, в котором первая тяжелая цепь и первая легкая цепь объединены с образованием антигенсвязывающего сайта для первого антигена, и в котором вторая тяжелая цепь и вторая легкая цепь объединены с образованием антигенсвязывающего сайта для второго антигена. Необязательно дополнительные антигенсвязывающие фрагменты могут быть слиты, например, через полипептидный линкер, с N- или С-концом первой и/или второй тяжелой цепи для увеличения валентности в отношении одного или обоих антигенов. Например, дополнительный антигенсвязывающий фрагмент для первого антигена можно сливать с N-концом одной или обеих молекул тяжелой цепи.
В другом приведенном в качестве примера формате биспецифическое или мультиспецифическое антитело может представлять собой полноразмерное антитело (например, IgG), содержащее антигенсвязывающий сайт для первого антигена (например, которое может быть двухвалентным в отношении первого антигена), и оно дополнительно содержит по меньшей мере один антигенсвязывающий фрагмент, специфический в отношении второго антигена. В различных вариантах осуществления изобретения антигенсвязывающий фрагмент может представлять собой Fab-фрагмент, молекулу кроссовер-Fab, scFab, Fv-молекулу, scFv или однодоменное антитело (VHH) или может представлять собой часть второго полноразмерного антитела. Например, антитело может представлять собой полноразмерное антитело, содержащее антигенсвязывающий сайт для первого антигена, и оно дополнительно может содержать по меньшей мере вариабельный домен второй тяжелой цепи и вариабельный домен второй легкой цепи, которые вместе образуют антигенсвязывающий сайт для второго антигена. Либо первый, либо второй антиген представляет собой хелат Pb-DOTAM, а другой антиген представляет собой антиген-мишень.
В некоторых вариантах представленных в настоящем описании форматов второй антиген представляет собой хелат Pb-DOTAM, а первый антиген представляет собой мишень, например, опухолеассоциированный антиген (СЕА, CD20 или ERBB2 в некоторых вариантах осуществления изобретения).
В другом приведенном в качестве примера формате биспецифическое или мультиспецифическое антитело может представлять собой полноразмерное антитело, содержащее антигенсвязывающий сайт для первого антигена (например, которое может быть двухвалентным в отношении первого антигена), в котором N- или С-конец одной из тяжелых цепей связан через полипептидный линкер с первым полипептидом, и в котором первый полипептид объединен со вторым полипептидом с образованием Fab или cross-Fab, содержащего сайт связывания для второго антигена. Например, указанный формат может содержать:
I) первый полипептид, состоящий из VH-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VL- и CL-домена; или
II) первый полипептид, состоящий из VL-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VH- и CL-доменов; или
III) третий полипептид, состоящий из VH-домена и CL-домена, который объединен со вторым полипептидом, состоящим из VL- и СН1-доменов;
в результате первый и второй полипептиды вместе образуют антигенсвязывающий сайт для второго антигена.
В некоторых вариантах осуществления изобретения слияние может находиться на N-конце одной из тяжелых цепей полноразмерного антитела.
В другом конкретном варианте осуществления изобретения антитело может представлять собой биспецифическое антитело, которое содержит:
а) полноразмерное антитело, специфически связывающее первый антиген и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела;
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH); или
II) вариабельного домена тяжелой цепи антитела (VH) и константного домена антитела (СН1); или
III) вариабельного домена тяжелой цепи антитела (VH) и константного домена легкой цепи антитела (CL);
где указанный полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL); или
II) вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL), или
III) вариабельного домена легкой цепи антитела (VL) и константного домена тяжелой цепи антитела антитело (СН1);
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
и где вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для второго антигена.
В указанном формате либо первый, либо второй антиген может представлять собой хелат Pb-DOTAM. Другой должен представлять собой антиген-мишень, например, опухолеассоциированный антиген.
В некоторых вариантах осуществления изобретения второй антиген представляет собой хелат Pb-DOTAM, а первый антиген представляет собой мишень, например, опухолеассоциированный антиген (СЕА, CD20 или ERBB2 в некоторых вариантах осуществления изобретения).
Описанное выше антитело может быть трехвалентным. В другом возможном варианте осуществления изобретения дополнительные антигенсвязывающие фрагменты можно сливать для повышения валентности в отношении одного или обоих антигенов, что будет дополнительно обсуждаться в настоящем описании.
Необязательно указанный линкер (и любой линкер указанный в контексте настоящего описания) может представлять собой пептид, состоящий по меньшей мере из 5 аминокислот, предпочтительно из 25-50 аминокислот. Линкер может представлять собой жесткий линкер или гибкий линкер. В некоторых вариантах осуществления изобретения он представляет собой гибкий линкер, содержащий или состоящий из остатков Thr, Ser, Gly и/или Ala. Например, он может содержать или состоять из остатков Gly и Ser. В некоторых вариантах осуществления изобретения он может включать повторяющийся мотив, такой как (Gly-Gly-Gly-Gly-Ser)n, в котором n обозначает, например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. В некоторых вариантах осуществления изобретения линкер может представлять собой или может содержать последовательность GGGGSGGGGSGGGGSGGGGS (SEQ ID NO: 26). Можно применять другие линкеры и их может идентифицировать специалист в данной области.
Дополнительные подробности, касающиеся указанного формата антител представлены в WO 2010/115589 А1 (фирма Roche Glycart AG), содержание которой полностью включено в настоящее описание в качестве ссылки.
Необязательно,
I) в константном домене CL первой легкой цепи полноразмерного антитела, указанного в подпункте (а), аминокислота в положении 124 заменена независимо на лизин (К), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном из предпочтительных вариантов осуществления изобретения независимо на лизин (K) или аргинин (R)), а в константном домене СН1 первой тяжелой цепи антитела, указанного в подпункте (а), аминокислота в положении 147 или аминокислота в положении 213 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота); или
II) в константном домене CL второй легкой цепи, указанном в подпункте (б), аминокислота в положении 124 заменена независимо на лизин (K), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном из предпочтительных вариантов осуществления изобретения независимо на лизин (K) или аргинин (R)), а в константном домене СН1 второй тяжелой цепи, указанном в подпункте (б), аминокислота в положении 147 или аминокислота в положении 213 заменена независимо на глутаминовую кислоту (нумерация согласно EU-индексу Кэбота).
В одном из вариантов осуществления изобретения биспецифическое антитело, предлагаемое в настоящем изобретении, может иметь описанную выше трехвалентную структуру и может содержать:
а) полноразмерное антитело, которое специфически связывается с СЕА и состоит из двух тяжелых цепей антитела и двух легких цепей антитела;
в котором тяжелые цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-450 (включительно, при использовании последовательной нумерации) SEQ ID NO: 22 или 23 (т.е. части последовательности, простирающейся до линкера);
и в котором легкие цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO 21; и/или
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7; или
II) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7, и константного домена тяжелой цепи антитела (СН1); или
III) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7, и константного домена легкой цепи антитела;
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
и/или
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8; или
II) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8, и константного домена легкой цепи антитела (CL); или
III) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8, и константного домена тяжелой цепи антитела (СН1);
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
где вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте в), вместе образуют антигенсвязывающий сайт для хелата Pb-DOTAM.
В одном из примеров одна из тяжелых цепей полноразмерного антитела содержит так называемые «knob-мутации» (мутации, приводящие к образованию «выступа») (T366W и необязательно одну из S354C или Y349C, предпочтительно S354C), а другая содержит так называемые «hole-мутации» (мутации, приводящие к образованию «впадины») (T366S, L368A и Y407V и необязательно Y349C или S354C, предпочтительно Y349C) (см., например, Carter Р. и др., Immunotechnol. 2, 1996, с. 73) (нумерация согласно EU-индексу). В некоторых вариантах осуществления изобретения две тяжелые цепи антитела, указанного в подпункте (а), содержат I) первую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-450 (включительно, при использовании последовательной нумерации) SEQ ID NO: 23 (т.е. последовательности, простирающейся до линкера), и имеющую С в положении 349, S в положении 366, А в положении 368 и V в положении 407 (EU-нумерация); и II) вторую тяжелую цепь антитела, идентичную по меньше мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-450 (включительно, при использовании последовательной нумерации) SEQ ID NO: 22, и имеющую С в положении 354 и W в положении 366 (EU-нумерация).
Упоминаемые в контексте настоящего изобретения «фиксированные» остатки относятся к мутациям типа «knob-into-hole» (типа «выступ-во-впадину») в СН3 или представляют собой другие остатки, такие как образующие дисульфидные мостики остатки, которые спариваются с соответствующими остатками на СН3 другой тяжелой цепи, благоприятствуя образованию требуемой молекулы. Возможные остатки, которые могут присутствовать на другой тяжелой цепи, можно получать из последовательностей, представленных в таблице 2 (например, последовательности 19 и 20 или 22 и 23). Например, в одном из вариантов осуществления изобретения, если первая тяжелая цепь антитела имеет С в положении 349, то вторая тяжелая цепь антитела имеет С в положении 354.
Необязательно линкер представляет собой описанный выше линкер.
В другом варианте осуществления изобретения биспецифическое антитело, предлагаемое в настоящем изобретении, может иметь описанную выше трехвалентную структуру и может содержать:
а) полноразмерное антитело, которое специфически связывается с СЕА и состоит из двух тяжелых цепей антитела и двух легких цепей антитела;
в котором тяжелые цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-450 (включительно, при использовании последовательной нумерации) SEQ ID NO: 19 или 20 (т.е. части последовательности, простирающейся до линкера);
и в котором легкие цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO: 21;
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 9; или
II) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 9, и константного домена тяжелой цепи антитела (СН1); или
III) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 9, и константного домена легкой цепи антитела;
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 10; или
II) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 10, и константного домена легкой цепи антитела (CL); или
III) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 10, и константного домена тяжелой цепи антитела (СН1);
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
где вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для хелата Pb-DOTAM.
Как описано выше, в одном из указанных выше вариантов осуществления изобретения одна из тяжелых цепей полноразмерного антитела может содержать так называемые «knob-мутации» (T366W и необязательно одну из S354C или Y349C, предпочтительно S354C), а другая может содержать так называемые «hole-мутации» (T366S, L368A и Y407V и необязательно Y349C или S354C, предпочтительно Y349C) (см., например, Carter Р. и др., Immunotechnol. 2, 1996, с. 73) (нумерация согласно EU-индексу).
В одном из вариантов осуществления изобретения две тяжелые цепи антитела, указанного в подпункте (а), могут содержать I) первую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-450 SEQ ID NO: 22, которая имеет С в положении 354 и W в положении 366 (EU-нумерация); и II) вторую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-450 SEQ ID NO: 23, которая имеет С в положении 349, S в положении 366, А в положении 368 и V в положении 407 (EU-нумерация).
Необязательно линкер представляет собой описанный выше линкер.
В другом варианте осуществления изобретения биспецифическое антитело содержит:
I) первую тяжелую цепь, имеющую аминокислотную последовательность, которая идентична по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 22,
II) вторую тяжелую цепь, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 23,
III) две легкие цепи антитела, идентичные по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO: 21.
В следующем варианте осуществления изобретения биспецифическое антитело представляет собой молекулу, обозначенную в настоящем описании как PRIT-0213, которая содержит
I) первую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 22;
II) вторую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 23; и
III) две легкие цепи антитела, имеющие аминокислотную последовательность SEQ ID NO: 21.
В другом варианте осуществления изобретения биспецифическое антитело содержит:
I) первую тяжелую цепь, имеющую аминокислотную последовательность, которая идентична по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 19,
II) вторую тяжелую цепь, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 20,
III) две легкие цепи антитела, идентичные по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO: 21.
В следующем варианте осуществления изобретения биспецифическое антитело представляет собой молекулу, обозначенную в настоящем описании как PRIT-0214, которая содержит
I) первую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 19;
II) вторую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 20; и
III) две легкие цепи антитела, имеющие аминокислотную последовательность SEQ ID NO: 21.
В одном из вариантов осуществления изобретения биспецифическое антитело, предлагаемое в настоящем изобретении, может иметь описанную выше трехвалентную структуру и может содержать:
а) полноразмерное антитело, которое специфически связывается с ERBB2 и состоит из двух тяжелых цепей антитела и двух легких цепей антитела;
в котором тяжелые цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-449 (включительно, при использовании последовательной нумерации) SEQ ID NO: 36 или 37 (т.е. части последовательности, простирающейся до линкера);
и в котором легкие цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO 38; и/или
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7 или 9 (в некоторых вариантах осуществления изобретения предпочтительно 7); или
II) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7 или 9 (в некоторых вариантах осуществления изобретения предпочтительно 7), и константного домена тяжелой цепи антитела (СН1); или
III) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7 или 9 (в некоторых вариантах осуществления изобретения предпочтительно 7), и константного домена легкой цепи антитела (CL);
где указанный полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела; и/или
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8 или 10 (в некоторых вариантах осуществления изобретения предпочтительно 8); или
II) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8 или 10 (в некоторых вариантах осуществления изобретения предпочтительно 8), и константного домена легкой цепи антитела (CL); или
III) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8 или 10 (в некоторых вариантах осуществления изобретения предпочтительно 8), и константного домена тяжелой цепи антитела (СН1);
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
где вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для хелата Pb-DOTAM.
В одном из примеров одна из тяжелых цепей полноразмерного антитела содержит так называемые «knob-мутации» (T366W и необязательно одну из S354C или Y349C, предпочтительно S354C), а другая содержит так называемые «hole-мутации» (T366S, L368A и Y407V и необязательно S354C или Y349C, предпочтительно Y349C) (см., например, Carter Р. и др., Immunotechnol. 2, 1996, с. 73) (нумерация согласно EU-индексу).
В одном из вариантов осуществления изобретения две тяжелые цепи антитела, указанного в подпункте (а), содержат I) первую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-449 SEQ ID NO: 37 (т.е. последовательности, простирающейся до линкера), и которая имеет С в положении 349, S в положении 366, А в положении 368 и V в положении 407 (EU-нумерация); и II) вторую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-449 (включительно, при использовании последовательной нумерации) SEQ ID NO: 36, которая имеет С в положении 354 и W в положении 366 (EU-нумерация).
Необязательно линкер представляет собой описанный выше линкер.
В другом варианте осуществления изобретения биспецифическое антитело содержит:
I) первую тяжелую цепь, имеющую аминокислотную последовательность, которая идентична по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 36,
II) вторую тяжелую цепь, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 37,
III) две легкие цепи антитела, идентичные по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO: 38.
В следующем варианте осуществления изобретения биспецифическое антитело представляет собой молекулу, обозначенную в настоящем описании как P1AD9827, которая содержит
I) первую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 36;
II) вторую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 37; и
III) две легкие цепи антитела, имеющие аминокислотную последовательность SEQ ID NO: 38.
В другом варианте осуществления изобретения биспецифическое антитело, предлагаемое в настоящем изобретении, может иметь описанную выше трехвалентную структуру и может содержать:
а) полноразмерное антитело, которое специфически связывается с CD20 и состоит из двух тяжелых цепей антитела и двух легких цепей антитела;
в котором тяжелые цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-448 (включительно, при использовании последовательной нумерации) SEQ ID NO: 47 или 48 (т.е. части последовательности, простирающейся до линкера);
и в котором легкие цепи идентичны по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO 49; и/или
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7 или 9 (в некоторых вариантах осуществления изобретения предпочтительно 7); или
II) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7 или 9 (в некоторых вариантах осуществления изобретения предпочтительно 7), и константного домена тяжелой цепи антитела (СН1); или
III) указанного вариабельного домена тяжелой цепи антитела (VH), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7 или 9 (в некоторых вариантах осуществления изобретения предпочтительно 7), и константного домена легкой цепи антитела (CL);
где указанный полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела; и/или
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8 или 10 (в некоторых вариантах осуществления изобретения предпочтительно 8); или
II) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8 или 10 (в некоторых вариантах осуществления изобретения предпочтительно 8), и константного домена легкой цепи антитела; или
III) указанного вариабельного домена легкой цепи антитела (VL), идентичного по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% вариабельному домену легкой цепи, имеющему SEQ ID NO: 8 или 10 (в некоторых вариантах осуществления изобретения предпочтительно 8), и константного домена тяжелой цепи антитела;
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
где вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для хелата Pb-DOTAM.
В одном из примеров одна из тяжелых цепей полноразмерного антитела содержит так называемые «knob-мутации» (T366W и необязательно одну из S354C или Y349C, предпочтительно S354C), а другая содержит так называемые «hole-мутации» (T366S, L368A и Y407V и необязательно Y349C или S354C предпочтительно Y349C) (см., например, Carter Р. и др., Immunotechnol. 2, 1996, с. 73) (нумерация согласно EU-индексу).
В некоторых вариантах осуществления изобретения две тяжелые цепи антитела, указанного в подпункте (а), содержат I) первую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-448 (включительно, при использовании последовательной нумерации) SEQ ID NO: 48 (т.е. последовательности, простирающейся до линкера), и которая имеет С в положении 349, S в положении 366, А в положении 368 и V в положении 407 (EU-нумерация); и II) вторую тяжелую цепь антитела, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% аминокислотам тяжелой цепи 1-449 (включительно, при использовании последовательной нумерации) SEQ ID NO: 47, которая имеет С в положении 354 и W в положении 366 (EU-нумерация).
Необязательно линкер представляет собой описанный выше линкер.
В другом варианте осуществления изобретения биспецифическое антитело содержит:
I) первую тяжелую цепь, имеющую аминокислотную последовательность, которая идентична по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 47,
II) вторую тяжелую цепь, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% тяжелой цепи, имеющей SEQ ID NO: 48,
III) две легкие цепи антитело, идентичные по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% легкой цепи, имеющей SEQ ID NO: 49.
В следующем варианте осуществления изобретения биспецифическое антитело представляет собой молекулу, обозначенную как P1AD9826, которая содержит
I) первую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 47;
II) вторую тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 48; и
III) две легкие цепи антитела, имеющие аминокислотную последовательность SEQ ID NO: 49.
Следующим объектом настоящего изобретения является полинуклеотид или набор полинуклеотидов, кодирующих любое из антител, указанных в настоящем описании. В других вариантах осуществления настоящего изобретения предложен вектор или набор экспрессионных векторов, содержащих указанный полинуклеотид или указанные полинуклеотиды, необязательно экспрессионный вектор. Следующими объектами настоящего изобретения является прокариотическая или эукариотическая клетка-хозяин, содержащая вектор, предлагаемый в настоящем изобретении. Кроме того, предложен способ получения антитела, включающий культивирование клетки-хозяина таким образом, чтобы получать антитело.
Биспецифические или мультиспецифические антитела, указанные в настоящем описании, могут найти применение в различных сферах, включая применения в области терапии и диагностики, такие как претаргетная радиоиммунотерапии и претаргетная радиоиммуновизуализация.
Таким образом, следующим объектом настоящего изобретения является любое биспецифическое или мультиспецифическое антитело, указанное в настоящем описании, предназначенное для применения в претаргетной радиовизуализации. В указанных вариантах осуществления изобретения хелатный Pb предпочтительно представляет собой 203Pb.
Способ направленного переноса радиоизотопа в ткань или орган для визуализации может включать:
I) введение индивидууму мультиспецифического или биспецифического антитела, указанного в настоящем описании, где антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень; и
II) последующее введение индивидууму радионуклида Pb, хелатированного DOTAM или его функциональным вариантом, где радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, связывается с антителом, локализованным на поверхности клетки-мишени.
Необязательно между стадиями (I) и (II) вводят очищающий агент, где очищающий агент связывается с антигенсвязывающим сайтом, специфическим в отношении хелата Pb-DOTAM. Очищающий агент блокирует антигенсвязывающий сайт, специфический для Pb-DOTAM, предупреждая связывание циркулирующего антитела с хелатным радионуклидом Pb. В альтернативном или дополнительном варианте очищающий агент может повышать скорость очищения (клиренса) организма от антитела. Альтернативно этому, «очищающий агент» можно обозначать как «блокирующий агент»: ниже в описании указанные понятия могут использоваться взаимозаменяемо.
Очищающий агент может содержать комплекс иона металла с DOTAM или его функциональным вариантом, где указанный комплекс распознается антигенсвязывающим сайтом, специфическим для Pb-DOTAM. Предпочтительно ион металла представляет собой стабильный изотоп или практически стабильный изотоп. Под «стабильным изотопом» подразумевается изотоп, который не подвергается радиоактивному распаду. Под «практически стабильным изотопом» подразумевается изотоп, который подвергается радиоактивному распаду с очень длинным периодом полураспада, что делает его безопасным для применения. Предпочтительно ион металла выбирают из ионов Pb, Са и Bi. Например, очищающий агент может содержать стабильный изотоп Pb в комплексе с DOTAM или его функциональным вариантом, Ca в комплексе с DOTAM или его функциональным вариантом или 209Bi (практически стабильный изотоп с периодом полураспада 1,9×1019 лет) в комплексе с DOTAM или его функциональным вариантом. Pb может представлять собой встречающийся в естественных условиях свинец, который представляет собой смесь стабильных (нерадиоактивных) изотопов 204Pb, 206Pb, 207Pb и 208Pb.
DOTAM или его функциональный вариант конъюгируют с очищающим фрагментом. Указанный фрагмент придает очищающему агенту низкий уровень поглощения опухолью, например, благодаря его размеру и/или большому гидродинамическому радиусу. Приемлемые очищающие фрагменты будут дополнительно обсуждены ниже. В некоторых вариантах осуществления изобретения очищающий агент может содержать DOTAM или его функциональный вариант, конъюгированный с декстраном или его производным.
В другом варианте способов визуализации мультиспецифическое или биспецифическое антитело в момент введения может быть связано с хелатным радионуклидом Pb.
Необязательно в одном из вариантов осуществления изобретения способ включает также:
III) визуализацию ткани или органа, в которых локализован радионуклид Pb, хелатированный DOTAM или его функциональным вариантом.
В некоторых вариантах осуществления изобретения антиген-мишень может представлять собой опухолеспецифический антиген, а визуализация может представлять собой способ визуализации опухоли или опухолей.
Следующим объектом настоящего изобретения является антитело, указанное в настоящем описании, предназначенное для применения в способе претаргетной радиоиммунотерапии. В указанных вариантах осуществления изобретения хелатный Pb предпочтительно представляет собой 212Pb.
Способ направленного переноса радиоизотопа в ткань или орган для терапии может включать:
I) введение индивидууму мультиспецифического или биспецифического антитела, указанного в настоящем описании, где антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень; и
II) последующее введение индивидууму радионуклида Pb, хелатированного DOTAM или его функциональным вариантом, где радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, связывается с антителом, локализованным на поверхности клетки.
Необязательно между стадиями (I) и (II) вводят описанный выше очищающий агент.
В некоторых вариантах осуществления изобретения антиген-мишень представляет собой опухолеассоциированный антиген, и способ представляет собой способ лечения рака. В других вариантах осуществления изобретения антиген-мишень может представлять собой антиген, ассоциированный с инфекцией, например, белок, который экспрессируется прокариотической или инфицированной вирусом клеткой.
В некоторых вариантах осуществления изобретения антитела, указанные в настоящем описании, можно вводить в качестве компонента комбинированной терапии. Например, их можно вводить в комбинации с одним или несколькими радиосенсибилизаторами и/или химиотерапевтическими агентами: радиосенсибилизатор или химиотерапевтический агент и антитело можно вводить одновременно или последовательно в любом порядке.
Способы радиовизуализации и радиоиммунотерапии, указанные в настоящем описании, можно объединять, например, путем введения антитела и как 203Pb-DOTAM, так и 212Pb-DOTAM, например, в виде смеси.
В настоящем изобретении предложены также фармацевтические композиции, содержащие антитело, предлагаемое в настоящем изобретении, и фармацевтически приемлемый эксципиент.
Следующими вариантами осуществления настоящего изобретения является набор, который содержит антитело, предлагаемое в настоящем изобретении, и один, два, три, четыре или все следующие компоненты:
I) фармацевтически приемлемый эксципиент;
II) радионуклид Pb, хелатированный DOTAM или его функциональным вариантом;
III) очищающий агент, указанный в настоящем описании;
IV) один или несколько дополнительных химиотерапевтических агентов; и/или
V) один или несколько радиосенсибилизаторов.
Еще одним объектом изобретения являются разработанный при создании настоящего изобретения новый очищающий агент. Указанный очищающий агент можно применять в любом из способов диагностирования, визуализации или лечения, которые указаны в настоящем описании.
Одним из объектов настоящего изобретения является очищающий агент, содержащий декстран или его производное, конъюгированный/конъюгированное с хелатором, выбранным из DOTAM и функционального варианта DOTAM, где указанный хелатор образует комплекс с Pb, Zn, Са или Bi. Очищающий агент, как правило, включает хелатный ион металла, такой как ион Pb, Zn, Са или Bi.
Очищающий агент может содержать аминодекстран, сшитый с DOTAM или функциональным вариантом DOTAM. Например, очищающий агент может содержать DOTAM, сшитый с аминодекстраном посредством сочетания с изотиоцианатом (например, соединение, которое можно получать посредством взаимодействия аминодекстрана с пара-SCN-Bn-TCMC).
Одним из потенциальных недостатков, связанных с применением очищающих агентов, является возможность того, что они могут проникать в опухоли, отрицательно влияя на последующее связывание радиолигандов.
При создании настоящего изобретения было установлено также, что можно достигать высокого уровня клиренса крови в сочетании с низкой скоростью проникновения очищающего агента в опухоли, если применяют очищающий агент на основе декстрана, который имеет I) высокую молекулярную массу и II) который был подвергнут такому отсечению по молекулярной массе, которое обеспечивает удаление фрагментов ниже определенного размера.
Таким образом, предпочтительный очищающий агент может представлять собой агент, в котором I) средняя молекулярная масса декстрана или его производного в очищающем агента составляет 200-800 кДа, необязательно выше чем 300, 350, 400 или 450 кДа и необязательно ниже чем 700, 650, 600 или 550 кДа, необязательно составляет примерно 500 кДа, и II) из декстрана, производных декстрана или очищающих агентов удалены компоненты с молекулярной массой ниже, чем конкретная отсекаемая молекулярная масса, где отсекаемая молекулярная масса составляет 50 кДа или выше, 100 кДа или выше или 200 кДа или выше, необязательно находится в диапазоне 50-250 кДа или 50-200 кДа, необязательно 100-200 кДа и необязательно составляет примерно 100, 150 или 200 кДа.
Другим объектом настоящего изобретения являются способы получения очищающего агента и применения очищающих агентов в способах радиоиммунотерапии или радиоиммуновизуализации.
Указанные и другие объекты изобретения будут дополнительно обсуждены ниже.
Чертежи
На чертежах показано:
на фиг. 1 схематическое изображение возможного формата биспецифических антител. Указанный формат включает два антигенсвязывающих сайта для одной мишени (А) и один антигенсвязывающий сайт для второй мишени (Б) (формат 2:1);
на фиг. 2 - структура PRIT-0213 в комплексе с Pb-DOTAM;
на фиг. 3 - схематическое изображение сайта взаимодействия PRIT-0213 в комплексе с Pb-DOTAM (нумерация согласно Кэботу);
на фиг. 4 - данные о распределении 212Pb через 24 ч после инъекции радиоактивно меченного DOTAM (%ID/г±SD, n=3). Мишенью для PRIT-0206, -0207, -0208 и -0165 является Т84.66, а мишенью для PRIT-0186, -0187 и -0156 является СН1А1А. PRIT-0175 представляет собой не связывающийся с СЕА контроль;
на фиг. 5 данные о распределении 203Pb, выраженные в виде количеств импульсов в минуту (СРМ), через 96 ч после инъекции антител PRIT, предварительно связанных с радиоактивно меченным DOTAM (СРМ±SD, n=3). PRIT-0205, -0206, -0207, -0208 и -0209 представляют собой полностью гуманизированные конструкции, a PRIT-0165 и -0175 представляют собой положительный и отрицательный контроли соответственно;
на фиг. 6 - данные о накоплении 203Pb-DOTAM-BsAb в опухолях ВхРС3/клиренсе 203Pb-DOTAM-BsAb из крови, выраженные в виде СРМ±SD (n=3), в различные моменты времени после инъекции PRIT-антител, предварительно связанных с радиоактивно меченным DOTAM. PRIT-0206 представляет собой полностью гуманизированную версию PRIT-0165; PRIT-0175 представляет собой не связывающийся с СЕА контроль;
на фиг. 7 - данные о распределении радиоактивности в отобранных тканях через 2 ч после инъекции 212Pb-меченных очищающих агентов в несущих MKN45-опухоли мышах (%ID/г±SD, n=3);
на фиг. 8 - данные о распределении радиоактивности в отобранных тканях и моче через 24 ч после инъекции 212Pb-меченных очищающих агентов в несущих MKN45-опухоли мышах (%ID/г±SD, n=3);
на фиг. 9 - данные о распределении радиоактивности по органам в отобранных тканях и моче через 24 ч после инъекции 212Pb-меченных очищающих агентов в несущих MKN45-опухоли мышах (%ID/г±SD, n=3);
на фиг. 10 - данные о распределении радиоактивности в отобранных тканях через 1 неделю после инъекции 203Pb-меченных очищающих агентов в свободных от опухолей мышах (%ID/г±SD, n=3).
на фиг. 11 - данные о распределении радиоактивности по органам в отобранных тканях через 1 недели после инъекции 203Pb-меченных очищающих агентов в свободных от опухолей мышах (%ID/г±SD, n=3);
на фиг. 12 данные об уровне радиоактивности в крови через 4 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3). Заштрихованный прямоугольник соответствует контролю без СА (без очищающего агента), с которым сравнивали все реагенты-кандидаты. Звездочками обозначен уровень статистической значимости от низкого (*) до высокого (***);
на фиг. 13 данные о среднем уровне радиоактивности в крови через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3). Заштрихованный прямоугольник соответствует контролю без СА (без очищающего агента), с которым сравнивали все реагенты-кандидаты;
на фиг. 14 - данные об уровне радиоактивности в крови через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3);
на фиг. 15 - данные о распределении 212Pb через 24 ч после инъекции радиоактивно меченного DOTAM (%ID/г±SD, n=3) при использовании 30 или 100 мкг биспецифического антитела и 10 100 мкг очищающих агентов после фильтрации с отсечением по молекулярной массе 100 или 30 кДа, или вообще без использования очищающего агента (ЗФР);
на фиг. 16 - данные о воздействии на активность концентрации 212Pb в крови и опухоли при применении возрастающих количеств очищающего агента (0-100 мкг). Опухоли предварительно таргетировали, используя 100 мкг PRIT-0165, через 4 дня после этого добавляли Dex500, который подвергали диафильтрации с отсечением по молекулярной массе 100 кДа, или ЗФР. 212Pb-DOTAM вводили через 2 ч после очищающего агента. Символами обозначена величина %ID/г через 24 ч после инъекции радиоактивного вещества, а линией обозначены обработанные методом линейной регрессии данные для опухолей;
на фиг. 17 - данные о распределении радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3). Темно-серыми и черными прямоугольниками обозначены положительные контроли без СА (без очищающего агента), с которыми сравнивали реагенты-кандидаты;
на фиг. 18 данные о содержании 212Pb в крови и опухолях через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3) и соответствующие соотношения опухоль/кровь. Темно-серыми и черными прямоугольниками обозначены положительные контроли без СА (без очищающего агента), с которыми сравнивали реагенты-кандидаты;
на фиг. 19 данные о соотношении опухоль/кровь через 24 ч после инъекции 212Pb-DOTAM в зависимости от количества очищающего агента (СА) (PJRD08-46) и ТСМС-насыщения (9-, 20-, 39- или 84 к 1). Пунктирными линиями обозначены результаты аппроксимации соответствующих данных методом линейной регрессии (R2=0,82) и результаты, полученные путем подгонки с помощью нелинейной кривой (R2=0,74);
на фиг. 20 данные о распределении 212Pb через 24 ч после инъекции радиоактивно меченного DOTAM (%ID/г±SD, n=3). Прямоугольниками с белым и серым фоном обозначены данные для вариантов, таргетирующих Т84.66 и СН1А1А соответственно, черными прямоугольниками обозначены данные для не связывающегося с СЕА контроля;
на фиг. 21 данные о распределении радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SEM, n=3) в циклах 1 и 2 полученные на модели ВхРС3;
на фиг. 22 - данные о среднем весе тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на модели ВхРС3. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 23 данные об изменении среднего веса тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на модели ВхРС3, выраженном в виде процента от начального веса тела. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 24 данные о росте опухолей, выраженные в виде средних значений со стандартной ошибкой в группах А-Ж, полученные на модели ВхРС3 (n=8). Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 25 - кривые индивидуального роста опухолей в группах А-Ж, полученные на модели ВхРС3. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM;
на фиг. 26 кривые Каплана-Мейера, демонстрирующие выживаемость в группах А-Ж, полученные на модели ВхРС3 (n=8). Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM;
на фиг. 27 - данные о распределении радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3) в циклах 1 и 2 обработки, полученные на модели LS174T;
на фиг. 28 данные о среднем весе тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на модели LS174T3. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 29 - данные об изменении среднего веса тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на модели LS174T3, выраженное в виде процента от начального веса тела. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 30 - данные о росте опухолей в группах А-Ж, выраженные в виде средних значений со стандартной ошибкой в группах, полученные на модели LS174T3 (n=8). Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 31 - кривые индивидуального роста опухолей в группах А-Ж, полученные на модели LS174T3. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM;
на фиг. 32 - кривые Каплана-Мейера, демонстрирующие выживаемость в группах А-Ж, полученные на модели LS174T3 (n=8). Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM;
на фиг. 33 - данные о распределении радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3). Серыми прямоугольниками обозначено накопление в ткани после инъекции в различных количествах очищающего агента (СА) Dex500-(50%); черный прямоугольник соответствует контролю без СА (без очищающего агента);
на фиг. 34 - данные о содержании 212Pb в крови и опухолях через 24 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3) и соответствующие соотношения опухоль/кровь. Серыми прямоугольниками обозначено накопление в ткани после инъекции в различных количествах очищающего агента (СА) Dex500-(50%); черный прямоугольник соответствует контролю без СА (без очищающего агента);
на фиг. - 35 данные об уровне радиоактивности в крови через 4 ч после инъекции 212Pb-DOTAM (%ID/г±SD, n=3);
на фиг. 36 - данные о связывании антитела (PRIT-0165) с MKN-45-клетками, для оценки которого применяли либо вторичное идентифицирующее антитело (правая панель, Alexa 488), либо DOTAM-ФИТЦ (левая панель, ФИТЦ-А);
на фиг. 37 - возможные форматы биспецифических антител, антигеном-мишенью которых является, представленный в качестве примера СЕА. Можно применять также другие антигены-мишени;
на фиг. 38 - данные о связывании PIAD8927 с KPL-4-клетками, демонстрирующие способность связываться с Her2: детекция антител с использованием человеческих IgG-специфических вторичных антител;
на фиг. 39 - данные о связывании P1AD8927 с KPL-4-клетками, демонстрирующие способность связываться с DOTAM: скорректированная для изотипа детекция с использованием Pb-DOTAM-ФИТЦ;
на фиг. 40 - данные о связывании P1AD8926 с Raji-клетками, демонстрирующие способность связываться с CD20: детекция антител с использованием человеческих IgG-специфических вторичных антител;
на фиг. 41 - данные о связывании P1AD8926 с Raji-клетками, демонстрирующие способность связываться с DOTAM: скорректированная для изотипа детекция с использованием Pb-DOTAM-ФИТЦ;
на фиг. 42 - план исследования согласно протоколу 103, в котором изучали CEA-PRIT на несущих s.c. ВхРС3-опухоли SCID-мышах (ч = часы, d = дни, w = недели);
на фиг. 43 - на панели А - средний уровень накопления 212Pb в собранных тканях после обоих циклов обработки, выраженный в виде % ID/г±SD (n=3). На панели Б- поглощение индивидуальной опухолью 212Pb для каждой мыши наряду с объемами соответствующих опухолей (мм3) в момент эвтаназии;
на фиг. 44 - кривые среднего роста опухолей в группах А-Ж со стандартной ошибкой, полученные на модели ВхРС3 (n=10). Кривые обрывали при n<5. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM (30 или 10 мкКи) некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 45 - кривые роста индивидуальных опухолей в группах А-Ж, полученные на ВхРС3-модели (n=10). Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM (30 или 10 мкКи);
на фиг. 46 - кривые Каплана-Мейера, демонстрирующие выживаемость в группах А-Ж, полученные на модели ВхРС3 (n=10). Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM (30 или 10 мкКи);
на фиг. 47 - данные об уменьшении среднего веса тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на модели ВхРС3. Кривые обрывали в каждой группе при n<5. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана эксперимента;
на фиг. 48 - данные о распределении 203Pb-BsAb (20 мкКи, 100 мкг) у SCID-мышей, несущих s.c. ВхРС3-опухоли. Мышам инъецировали имеющий радиоактивность 20 мкКи предварительно связанный 203Pb-DOTAM-CEA-DOTAM или 203Pb-DOTAM-DIG-DOTAM (отрицательный контроль), после чего органы изымали в день 1, 4, 7 или 10 после инъекции для оценки накопленной радиоактивности в собранных тканях (% ID/г±SD, n=5);
на фиг. 49 - данные о накоплении 203Pb-BsAb в s.c. ВхРС3-опухолях в дни 1-10 после инъекции 20 мкКи/100 мкг предварительно связанного 203Pb-DOTAM-CEA-DOTAM или 203Pb-DOTAM-DIG-DOTAM (отрицательный контроль) (% ID/г±SD, n=5);
на фиг. 50 - данные о распределении 212Pb у SCID-мышей, несущих s.c. ВхРС3-опухоли. Мышам инъецировали CEA-DOTAM BsAb и СА до введения 212Pb-DOTAM через период времени от 5 мин до 48 ч после инъекции радиоактивного соединения (% ID/г±SD, n=5). *Кумулятивное содержание 212Pb в моче и экскрементах в течение времени, т.е. в каждый момент времени, включая предыдущее значение. Установленная величина % ID/г в моче основывалась на данных, полученных с использованием 1/5 (10 мл) собранной мочи/промывочного раствора для 5 мышей (50 мл);
на фиг. 51 - план исследования согласно протоколу 131, в котором изучали распределение in vivo 212Pb после осуществления PRIT с использованием СЕА-DOTAM BsAb, Pb-DOTAM-декстран-500 СА и 212Pb-DOTAM, погашенного с использованием любого из 5 различных металлов (Zn, Gd, Cu, Са или Pb), у несущих s.c. ВхРС3-опухоли SCID-мышей (d = дни, ч = часы);
на фиг. 52 данные о распределении 212Pb у несущих опухоли SCID-мышей через 2 ч после инъекции предварительно таргетированного CEA-DOTAM-212Pb-DOTAM (% ID/г±SD, n=4);
на фиг. 53 - данные о распределении 212Pb в отобранных здоровых тканях несущих опухоли SCID-мышей через 2 ч после инъекции 212Pb-DOTAM, погашенного различными металлами (% ID/г);
на фиг. 54 - данные о распределении 212Pb у несущих опухоли SCID-мышей через 24 ч после инъекции 212Pb-DOTAM, предварительно таргетированного CD20-DOTAM BsAb или отрицательным контролем DIG-DOTAM (% ID/г±SD, n=3);
на фиг. 55 - данные о распределении 212Pb у несущих опухоли SCID-мышей через 24 ч после инъекции 212Pb-DOTAM, предварительно таргетированного HER2-DOTAM BsAb или отрицательным контролем DIG-DOTAM (% ID/г±SD, n=3);
на фиг. 56 - план исследования согласно протоколу 154, в котором изучали биораспределение in vivo 212Pb-DOTAM после обработки CEA-PRIT с использованием различных конструкций BsAb, у SCID-мышей, несущих s.c. опухоли HPAF-II (ч = часы, d = дни);
на фиг. 57 - данные о распределении 212Pb у несущих опухоли SCID-мышей через 6 ч после инъекции 212Pb-DOTAM, предварительно таргетированного либо отрицательным контролем DIG-DOTAM, либо стандартным DOTAM-BsAb, либо одной из альтернативных конструкций BsAb (% ID/г±SD, n=3);
на фиг. 58 - данные о содержании в крови и накоплении в опухолях 212Pb через 6 ч после инъекции 212Pb-DOTAM, предварительно таргетированного либо отрицательным контролем DIG-DOTAM, либо стандартным CEA-DOTAM BsAb, либо одной из альтернативных конструкций BsAb (% ID/г±SD, n=3);
на фиг. 59 - схема эксперимента согласно протоколу 162. Для осуществления CD20-PRIT для обработки SCID-мышей, несущих s.c.-опухоли WSU-DLCL2, использовали CD20-DOTAM BsAb, Са-ООТАМ-декстран-500 СА и 212Pb-DOTAM; 1-ую стадию RIT осуществляли с использованием CD20-DOTAM BsAb, предварительно связанного с 212Pb-DOTAM (212Pb-DOTAM-CD20-DOTAM), на SCID-мышах, несущих s.с.-опухоли WSU-DLCL2;
на фиг. 60 данные о распределении 212Pb у несущих опухоли SCID-мышей после инъекции 212Pb-DOTAM, предварительно таргетированного CD20-DOTAM, или предварительно связанного 212Pb-DOTAM-CD20-DOTAM. Уровень радиоактивности в органах и тканях выражали в виде среднего значения % ID/г и стандартного отклонения (SD; n=3);
на фиг. 61 - данные о среднем росте s.с.-опухолей WSU-DLCL2 в группах А-Ж, выраженные в виде мм3 ± SEM (n=10);
на фиг. 62 - данные о изменении среднего веса тела мышей после различных обработок, выраженные в виде % от начального веса тела ± SEM. Пунктирными линиями обозначена инъекция 212Pb или антитела в зависимости от схемы обработки.
Определения
Для целей настоящего описания «акцепторный человеческий каркасный участок» означает каркасный участок, который содержит аминокислотную последовательность каркасного участка вариабельного домена легкой цепи (VL) или каркасный участок вариабельного домена тяжелой цепи (VH), имеющий происхождение из каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, указанного ниже. Акцепторный человеческий каркасный участок, «имеющий происхождение из» каркасного участка человеческого иммуноглобулина или человеческого консенсусного каркасного участка, может иметь такую же аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых вариантах осуществления изобретения количество аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых вариантах осуществления изобретения последовательность человеческого акцепторного каркасного участка VL идентична последовательности каркасного участка VL человеческого иммуноглобулина или последовательности человеческого консенсусного каркасного участка.
Понятие «аффинность» относится к суммарной силе всех нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). В контексте настоящего описания, если не указано иное, «аффинность связывания» относится к присущей молекуле аффинности связывания, отражающей взаимодействие по типу 1:1 между компонентами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X к ее партнеру Y, как правило, можно характеризовать с помощью константы диссоциации (Kd). Аффинность можно оценивать с помощью общепринятых методов, известных в данной области, включая те, которые представлены в настоящем описании. Ниже в качестве иллюстрации представлены конкретные варианты измерения аффинности связывания.
Антитело «с созревшей аффинностью» означает антитело с одним или несколькими изменениями в одном или нескольких гипервариабельных участках (HVR) по сравнению с родительским антителом, которое не содержит указанных изменений, указанные изменения приводят к повышению аффинности антитела к антигену.
Понятия «антитело к Pb-DOTAM», «антитело, которое связывается с Pb-DOTAM», «антитело, которое связывается с хелатом Pb-DOTAM» и эквивалентные понятия относятся к антителу, которое обладает способностью связываться с хелатом Pb-DOTAM, с аффинностью, достаточной для того, чтобы антитело можно было применять в схеме сортировки/очистки для разделения меченных Pb-DOTAM-фрагментов, и/или такой, чтобы антитело обладало способностью локализовать Pb-DOTAM в сайте антитела, например, для направленного переноса Pb-DOTAM в клетку. Понятия «антитело к мишени» и «антитело, которое связывается с мишенью» относятся к антителу, которое обладает способностью связываться с мишенью с аффинностью, достаточной для того, чтобы антитело можно было применять в терапевтических и/или диагностических целях, включая локализацию антитела в мишени, например, экспрессируемой на поверхности клетки. В одном из вариантов осуществления изобретения уровень связывания антитела с неродственным фрагментом и/или белком, неродственным мишени, составляет менее чем примерно 10% от связывания антитела к Pb-DOTAM или мишени по данным измерений, полученным, например, с помощью радиоиммуноанализа (РИА). В некоторых вариантах осуществления изобретения антитело имеет константу диссоциации (Kd), характеризующую связывание с Pb-DOTAM или с мишенью ≤1 мкМ, ≤100 нM, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
В контексте настоящего описания понятие «антитело» используется в его наиболее широком смысле, и оно относится к различным структурам антител, включая (но, не ограничиваясь только ими) моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они обладают требуемой антигенсвязывающей активностью.
Понятие «фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, связывающуюся с антигеном, с которым связывается интактное антитело. Примеры фрагментов антитела включают (но, не ограничиваясь только ими) молекулы Fv, Fab, Fab', Fab'-SH, F(ab')2; димерных антител (диабоди); линейных антител; одноцепочечных антител (например, scFv) и мультиспецифических антител, образованных из фрагментов антител.
Понятие «антитело, которое связывается с тем же эпитопом», что и референс-антитело, относится к антителу, которое блокирует связывание референс-антитела с его антигеном при оценке с помощью анализа в условиях конкуренции (конкурентный анализ) на 50%, и наоборот, референс-антитело блокирует связывание антитела с его антигеном при оценке с помощью конкурентного анализа на 50%. Пример конкурентного анализа представлен в настоящем описании.
Понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи имеет происхождение из конкретного источника или вида, а остальная часть тяжелой и/или легкой цепи имеет происхождение из другого источника или вида.
Понятие «класс» антитела относится к типу константного домена или константной области, который/которая входит в его тяжелую цепь. Известно пять основных классов антител: IgA, IgD, IgE, IgG и IgM и некоторые из них можно подразделять дополнительно на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, обозначают как α, δ, ε, γ и μ, соответственно.
Понятие «цитотоксический агент» в контексте настоящего описания относится к субстанции, которая ингибирует клеточную функцию или препятствует ей и/или вызывает клеточную гибель или деструкцию. Цитотоксические агенты включают (но, не ограничиваясь только ими) радиоактивные изотопы (например, 225Ac, 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 213Bi, 32P, 212Pb и радиоактивные изотопы Lu); химиотерапевтические агенты или лекарственные средства (например, метотрексат, адриамицин, алкалоиды барвинка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты); ингибирующие рост агенты; ферменты и их фрагменты, такие как нуклеолитические ферменты; антибиотики; токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты; и различные противоопухолевые или противораковые агенты, указанные ниже.
Понятие «эффекторные функции» относится к тем видам биологической активности, которые связаны с Fc-областью антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и комплементзависимую цитотоксичность (CDC); связывание Fc-рецептора; антитело-обусловленную клеточнозависимую цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора) и активацию В-клеток.
Понятие «эффективное количество» агента, например, фармацевтической композиции, означает количество, эффективное в дозах и в течение периода времени, необходимых для достижения требуемого терапевтического или профилактического результата.
Понятие «Fc-область» в контексте настоящего описания относится к C-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Понятие включает нативную последовательность Fc-областей и вариант Fc-областей. В одном из вариантов осуществления изобретения Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако С-концевой лизин (Lys447) Fc-области может присутствовать или отсутствовать. Если специально не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, которую обозначают также как EU-индекс, описанной у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
«Каркасный участок» или «FR» означает остатки вариабельного домена, отличные от остатков гипервариабельного участка (HVR). FR вариабельного домена, как правило, состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Таким образом, последовательности HVR и FR, как правило, имеют следующее расположение в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Понятия «полноразмерное антитело», «интактное антитело» и «цельное антитело» в контексте настоящего описания используют взаимозаменяемо для обозначения антитела, имеющего структуру, практически сходную с нативной структурой антитела, или имеющего тяжелые цепи, которые содержат представленную в настоящем описании Fc-область. Полноразмерное антитело может представлять собой, например, IgG.
Понятия «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используют взаимозаменяемо, и они относятся к клеткам, в которые интродуцирована экзогенная нуклеиновая кислота, включая потомство указанных клеток. К клеткам-хозяевам относятся «трансформанты» и «трансформированные клетки», которые включают первично трансформированную клетку и полученное из нее потомство безотносительно к количеству пересевов. Потомство может не быть полностью идентичным по составу нуклеиновых кислот родительской клетке, но может содержать мутации. Под объем изобретения подпадает мутантное потомство, которое обладает такой же функцией или биологической активностью, которая обнаружена в результате скрининга или отбора у исходной трансформированной клетки.
«Человеческое антитело» представляет собой антитело, имеющее аминокислотную последовательность, которая соответствует последовательности антитела, продуцируемого в организме человека или человеческими клетками, или имеющее происхождение из нечеловеческого источника, в котором использован спектр человеческих антител или другие кодирующие человеческое антитело последовательности. Из указанного определения человеческого антитела специально исключено гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки.
«Человеческий консенсусный каркасный участок» представляет собой каркасный участок, в который входят наиболее часто встречающиеся аминокислотные остатки в выбранных последовательностях каркасных участков VL или VH человеческого иммуноглобулина. Как правило, выбор последовательностей VL или VH человеческого иммуноглобулина осуществляют из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу, описанную у Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., публикация NIH 91-3242, Bethesda MD, т.т .1-3, 1991. В одном из вариантов осуществления изобретения касательно VL подгруппа представляет собой подгруппу каппа I согласно Kabat и др., выше. В одном из вариантов осуществления изобретения касательно VH подгруппа представляет собой подгруппу III согласно Kabat и др., выше.
Понятие «гуманизированное» антитело относится к химерному антителу, которое содержит аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых вариантах осуществления изобретения гуманизированное антитело может содержать практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют участкам нечеловеческого антитела, а все или практически все FR соответствуют участкам человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, имеющей происхождение из человеческого антитела. Понятие «гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подвергли гуманизации.
Понятие «гипервариабельный участок» или «HVR» в контексте настоящего описания относится к каждому из участков вариабельного домена антитела, последовательности которых являются гипервариабельными («определяющие комплементарность участки» или «CDR») и/или формируют петли определенной структуры («гипервариабельные петли»), и/или содержат контактирующие с антигеном остатки («контакты с антигеном»). Как правило, антитела содержат шесть HVR; три в VH (H1, Н2, Н3) и три в VL (L1, L2, L3). Примеры HVR включают
(а) гипервариабельные петли, включающие аминокислотные остатки 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (Н3) (Chothia и Lesk, J. Mol. Biol. 196, 1987, cc. 901-917);
(б) CDR, включающие аминокислотные остатки 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD,1991);
(в) области контакта с антигеном, включающие аминокислотные остатки 27с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (Н3) (MacCallum и др., J. Mol. Biol. 262, 1996, cc. 732-745); и
(г) комбинации остатков, указанных в подпунктах (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Вместо вышеуказанного, последовательность CDR-H1, указанная в настоящем описании, может простираться от положение по Кэботу 26 до положения по Кэботу 35 (от Kabat26 до Kabat35).
В одном из вариантов осуществления изобретения остатки HVR или CDR включают остатки, указанные конкретно в таблице 2 или в ином месте в настоящей заявке.
Если специально не указано иное, то остатки в HVR/CDR и другие остатки в вариабельном домене (например, остатки в FR) нумеруют согласно Kabat и др., выше.
«Иммуноконъюгат» представляет собой антитело, конъюгированное с одной или несколькими гетерологичной(ыми) молекулой(ами), включая (но, не ограничиваясь только им) цитотоксический агент.
«Индивидуум» или «субъект» представляет собой млекопитающее. К млекопитающим относятся (но, не ограничиваясь только ими) одомашненные животные (например, коровы, овцы, кошки, собаки и лошади), приматы (например, человек и приматы кроме человека, такие как обезьяны), кролики и грызуны (например, мыши и крысы). В некоторых вариантах осуществления изобретения индивидуум или субъект представляет собой человека.
Указанные в настоящем описании молекулы могут быть «выделены». «Выделенное» антитело представляет собой антитело, которое отделено от компонента его естественного окружения. В некоторых вариантах осуществления изобретения антитело очищают до чистоты, превышающей 95% или 99% по данным, например, электрофоретических анализов (таких, например, как ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ), капиллярный электрофорез) или хроматографических анализов (таких, например, как ионообменная хроматография или ЖХВР с обращенной фазой). Обзор методов оценки чистоты антител см., например, у Flatman и др., J. Chromatogr. В 848, 2007, сс. 79-87.
Понятие «молекула нуклеиновой кислоты» или «полинуклеотид» включает любое соединение и/или любую субстанцию, которое/которая содержит полимер нуклеотидов. Каждый нуклеотид состоит из основания, в частности пуринового или пиримидинового основания (т.е. цитозина (С), гуанина (G), аденина (А), тимина (Т) или урацила (U)), сахара (т.е. дезоксирибозы или рибозы) и фосфатной группы. Часто молекула нуклеиновой кислоты описывается последовательностью оснований, при этом указанные основания образуют первичную структуру (линейную структуру) молекулы нуклеиновой кислоты. Последовательность оснований, как правило, представляют в направлении 5’→3’. В настоящем описании под понятие молекула нуклеиновой кислоты подпадают дезоксирибонуклеиновая кислота (ДНК), включая, например, комплементарную ДНК (кДНК) и геномную ДНК, рибонуклеиновая кислота (РНК), в частности, матричная РНК (мРНК), синтетические формы ДНК или РНК и смешанные полимеры, содержащие две или большее количество указанных молекул. Нуклеиновая кислота может быть линейной или кольцевой. Кроме того, понятие «нуклеиновая кислота» включает как смысловые, так и антисмысловые цепи, а также одноцепоченые и двухцепочечные формы. Кроме того, указанная в настоящем описании молекула нуклеиновой кислоты может содержать встречающиеся в естественных условиях или не встречающиеся в естественных условиях нуклеотиды. Примеры не встречающихся в естественных условиях нуклеотидов включают модифицированные нуклеотидные основания с дериватизированными сахарами или варианты фосфатного каркаса, или химически модифицированные остатки. Молекулы нуклеиновых кислот включают также молекулы ДНК и РНК, пригодные в качестве вектора для непосредственной экспрессии антитела, предлагаемого в изобретении, in vitro и/или in vivo, например, в хозяине или пациенте. Указанные векторы на основе ДНК (например, кДНК) или РНК (например, мРНК) могут представлять собой немодифицированный или модифицированный вектор. Например, мРНК можно химически модифицировать для повышения стабильности РНК-вектора и/или экспрессии кодируемой молекулы таким образом, чтобы мРНК можно было бы инъецировать индивидууму с целью образования антитела in vivo (см., например, Stadler и др., Nature Medicine 2017, публикация онлайн 12 июня 2017 г., doi:10.1038/nm.4356 или ЕР 2101823 В1).
«Выделенная» нуклеиновая кислота представляет собой молекулу нуклеиновой кислоты, которая отделена от компонента ее естественного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетке, которая в норме включает молекулу нуклеиновой кислоты, но в которой молекула нуклеиновой кислоты присутствует вне хромосомы или в которой ее локализация на хромосоме отличается от ее встречающейся в естественных условиях локализации на хромосоме.
Понятие «выделенная нуклеиновая кислота, кодирующая антитело» относится к одной или нескольким молекулам нуклеиновых кислот, кодирующих тяжелые и легкие цепи антитела (или их фрагменты), включая указанную(ые) молекулу(ы) нуклеиновой(ых) кислоты(т) в одном векторе или в различных векторах, и указанную(ые) молекулу(ы) нуклеиновой(ых) кислоты(т), присутствующую(ие) в одном или нескольких положениях в клетке-хозяине.
Понятие «моноклональное антитело» в контексте настоящего описания относится к антителу, полученному из популяции практически гомогенных антител, т.е. индивидуальные антитела, входящие в популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител, например, содержащих мутации, встречающиеся в естественных условиях или возникающие в процессе производства препарата моноклонального антитела, указанные варианты, как правило, присутствуют в минорных количествах. В отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты на антигене. Таким образом, прилагательное «моноклональный» определяет особенность антитела, характеризуя его как полученное из практически гомогенной популяции антител, а не сконструированное в соответствии с требованиями к получению антитела с помощью какого-либо конкретного метода. Например, моноклональные антитела, которые можно применять согласно настоящему изобретению, можно создавать с помощью различных технологий, включая (но, не ограничиваясь только ими) метод гибридом, методы рекомбинантной ДНК, методы фагового дисплея и методы, основанные на использовании трансгенных животных, которые содержат все или часть локусов человеческого иммуноглобулина, указанные методы и другие, приведенные в качестве примера методы получения моноклональных антител, представлены в настоящем описании.
Понятие «голое антитело» относится к антителу, неконъюгированному с гетерологичным фрагментом (например, цитотоксическим фрагментом) или радиоактивной меткой. Голое антитело может присутствовать в фармацевтической композиции.
Понятие «нативные антитела» относится к встречающимся в естественных условиях молекулам иммуноглобулинов с различными структурами. Например, нативные антитела в виде IgG представляют собой гетеротетрамерные гликопротеины с молекулярной массой примерно 150000 Да, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными мостиками. В направлении от N-конца к С-концу, каждая тяжелая цепь имеет вариабельную область (VH), которую называют также вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следует три константных домена (CH1, СН2 и СН3). Аналогично этому, в направлении от N-конца к С-концу, каждая легкая цепь имеет вариабельную область (VL), которую называют также вариабельным легким доменом или вариабельным доменом легкой цепи, за которой следует константный домен легкой цепи (CL). Легкая цепь антитела в зависимости от аминокислотной последовательности ее константного домена может принадлежать к одному из двух типов, которые обозначают каппа (κ) и лямбда (λ).
Понятие «листовка-вкладыш в упаковку» относится к инструкциям, которые обычно входят в поступающие в продажу упаковки терапевтических продуктов, содержащим информацию о показаниях, применении, дозе, пути введения, комбинированной терапии, противопоказаниях и/или мерах предосторожности, которые связаны с применением указанных терапевтических продуктов.
«Процент (%) идентичности аминокислотной последовательности» относительно полипептидной референс-последовательности определяют как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в полипептидной референс-последовательности, после выравнивая последовательностей и при необходимости интродукции брешей для достижения максимального процента идентичности последовательностей, и не рассматривая какие-либо консервативные замены в качестве компонента при оценке идентичности последовательностей. Сравнительный анализ для целей определения процента идентичности аминокислотных последовательностей можно осуществлять различными путями, известными в данной области, используя, например, публично доступные компьютерные программы, такие как программа BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определять соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Однако для целей настоящего изобретения величину % идентичности аминокислотных последовательностей получают с использованием предназначенной для сравнения последовательностей компьютерной программы ALIGN-2. Предназначенная для сравнения последовательностей компьютерная программа ALIGN-2 разработана фирмой Genentech, Inc., и исходный код помещен на хранение вместе с документацией для пользователя в US Copyright Office, Washington D.C., 20559, где он зарегистрирован под регистрационным номером US Copyright Registration, № TXU510087. Программа ALIGN-2 представляет собой публично доступную программу фирмы Genentech, Inc., Южный Сан-Франциско, шт. Калифорния, или ее можно компилировать из исходного кода. Программу ALIGN-2 можно компилировать для применения в операционной системе UNIX, включая цифровую версию UNIX V4.0D. В программе ALIGN-2 все параметры для сравнения последовательностей являются заданными и не должны изменяться.
В ситуациях, когда ALIGN-2 применяют для сравнения аминокислотных последовательностей, % идентичности аминокислотных последовательностей данной аминокислотной последовательности А относительно или по сравнению с данной аминокислотной последовательностью Б (которую другими словами можно обозначать как данная аминокислотная последовательность А, которая имеет или отличается определенным % идентичности аминокислотной последовательности относительно или по сравнению с данной аминокислотной последовательностью Б), рассчитывают следующим образом:
100 × частное X/Y,
где X обозначает количество аминокислотных остатков, оцененных программой сравнительного анализа последовательностей ALIGN-2 как идентичные совпадения при сравнительном анализе последовательностей А и Б с помощью указанной программы, и где Y обозначает общее количество аминокислотных остатков в Б. Должно быть очевидно, что, когда длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, то % идентичности аминокислотной последовательности А относительно аминокислотной последовательности Б не должен быть равен % идентичности аминокислотной последовательности Б относительно аминокислотной последовательности А. Если специально не указано иное, то в контексте настоящего описания все величины % идентичности аминокислотных последовательностей получают согласно процедуре, описанной в последнем из предшествующих параграфов, с помощью компьютерной программы ALIGN-2.
Понятие «фармацевтическая композиция» относится к препарату, который находится в такой форме, которая обеспечивает биологическую активность входящего в ее состав действующего вещества, которое должно обладать эффективностью, и которая не содержит дополнительных компонентов, обладающих неприемлемой токсичностью для индивидуума, которому следует вводить композицию.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтической композиции, отличному от действующего вещества, который является нетоксичным для индивидуума. Фармацевтически приемлемые носители включают (но, не ограничиваясь только ими) буфер, эксципиент, стабилизатор или консервант.
В контексте настоящего описания понятие «лечение» (и его грамматические вариации, такие как «лечить» или «процесс лечения») относится к клиническому вмешательству с целью изменения естественного течения болезни у индивидуума, подлежащего лечению, и его можно осуществлять либо для профилактики, либо в процессе развития клинической патологии. Требуемыми действиями лечения являются (но, не ограничиваясь только ими) предупреждение возникновения или рецидива болезни, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий болезни, предупреждение метастазов, снижение скорости развития болезни, облегчение или временное ослабление болезненного состояния и ремиссия или улучшение прогноза. В некоторых вариантах осуществления изобретения антитела, предлагаемые в изобретении, применяют для задержки развития болезни или замедления прогрессирования болезни.
Понятие «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL соответственно) нативного антитела, как правило, имеют сходные структуры, при этом каждый домен содержит четыре консервативных каркасных участка (FR) и три гипервариабельных участка (HVR) (см., например, Kindt и др., Kuby Immunology, 6-ое изд., изд-во W.H. Freeman and Co., 2007, с. 91). Одного VH- или VL-домена может быть достаточно для обеспечения специфичности связывания антигена. Кроме того, антитела, которые связываются с конкретным антигеном, можно выделять, используя VH- или VL-домен из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов соответственно (см., например, Portolano и др., J. Immunol. 150, 1993, сс. 880-887; Clarkson и др., Nature 352, 1991, сс. 624-628).
Понятие «вектор» в контексте настоящего описания относится к молекуле нуклеиновой кислоты, которая обладает способностью к размножению другой нуклеиновой кислоты, с которой она связана. Понятие включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он интродуцирован. Некоторые векторы обладают способностью контролировать экспрессию нуклеиновых кислот, с которыми они функционально связаны. Указанные векторы в контексте настоящего описания обозначают как «экспрессионные векторы».
Понятия «Pb» или «свинец» в контексте настоящего описания включает его ионы, например, Pb(II). Таким образом, специалисту в данной области должно быть очевидно, что, например, понятия свинец, Pb, 212Pb или 203Pb включают ионные формы элемента, в частности, Pb(II). В различных объектах изобретения Pb может представлять собой радиоизотоп (например, при применении в способе радиоиммунотерапии или радиоиммуновизуализации) или может представлять собой стабильный не радиоактивный изотоп (например, который может являться предпочтительным в контексте очищающего агента). «DOTAM» имеет химическое название:
1,4,7,10-тетракис(карбамоилметил)-1,4,7,10-тетраазациклододекан, который представляет собой соединение следующей формулы:
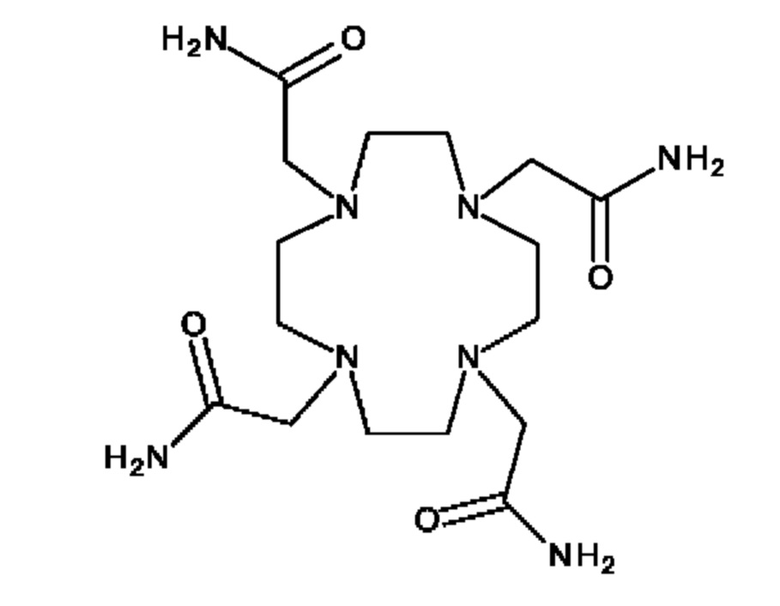
212Pb-DOTAM имеет следующую структуру:
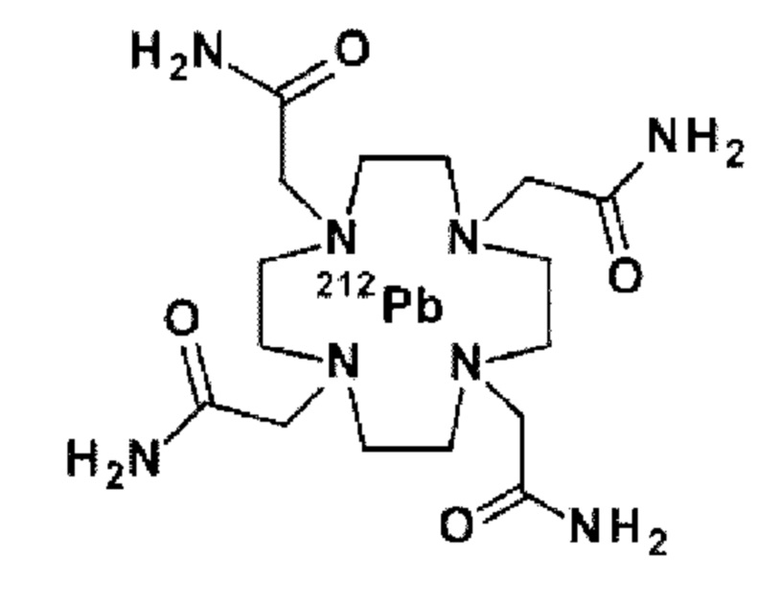
Согласно некоторым объектам и вариантам осуществления настоящего изобретения можно применять также функциональные варианты или производные DOTAM, включающие ион металла. Приемлемые варианты/производные DOTAM имеют структуру, которая в некоторой степени отличается от структуры DOTAM, но сохраняет способность функционировать (т.е. сохраняет достаточную активность, позволяющую использовать их для одной или нескольких целей, указанных в настоящем описании). В указанных объектах и вариантах осуществления изобретения DOTAM или функциональный вариант/функциональное производное DOTAM может представлять собой один из активных вариантов, описанных в WO 2010/099536.
Приемлемые функциональные варианты/производные могут представлять собой соединение следующей формулы:
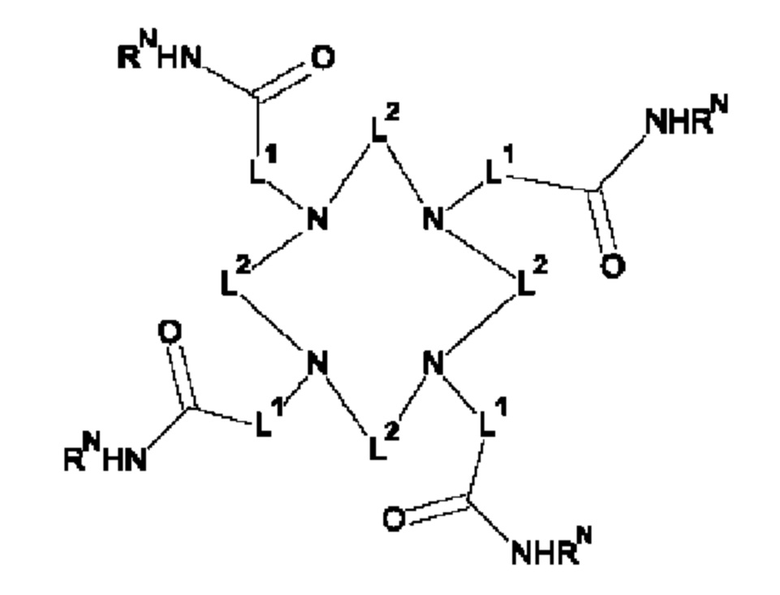
или его фармацевтически приемлемую соль; в которой
RN обозначает H, C1-С6алкил, C1-С6галоалкил, С2-С6алкенил, С2-С6алкинил, С3-С7циклоалкил, С3-С7циклоалкил-C1-С4алкил, С2-С7гетероциклоалкил, С2-С7гетероциклоалкил-C1-С4алкил, фенил, фенил-C1-С4алкил, C1-С7гетероарил и C1-С7гетероарил-C1-С4алкил; где C1-С6алкил, C1-С6галоалкил, С2-С6алкенил и С2-С6алкинил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными Rw-группами; и где указанный С3-С7циклоалкил, С3-С7циклоалкил-C1-С4алкил, С2-С7гетероциклоалкил, С2-С7гетероциклоалкил-C1-С4алкил, фенил, фенил-C1-С4алкил, C1-С7гетероарил и C1-С7гетероарил-C1-С4алкил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными Rx-группами;
L1 независимо обозначает C1-С6алкилен, C1-С6алкенилен или C1-С6алкиниленен, каждый из которых необязательно замещен 1, 2 или 3 группами, независимо выбранными из R1-групп;
L2 обозначает С2-С4алкилен с прямой цепью, который необязательно замещен независимо выбранной R1-группой; и который необязательно замещен 1, 2, 3 или 4 группами, независимо выбранными из C1-С4алкила и/или C1-С4 галоалкила;
R1 независимо выбран из D1-D2-D3, галогена, цианогруппы, нитрогруппы, гидроксила, C1-С6алкоксигруппы, C1-C6 галоалкоксигруппы, C1-С6алкилтиогруппы, C1-С6алкилсульфинила, C1-С6алкилсульфонила, аминогруппы, C1-С6алкиламиногруппы, ди-C1-С6алкиламиногруппы, C1-С4алкилкарбонила, карбоксигруппы, C1-С6алкоксикарбонила, C1-С6алкилкарбониламиногруппы, ди-C1-С6алкилкарбониламиногруппы, C1-С6алкоксикарбониламиногруппы, C1-С6алкоксикарбонил-(C1-С6алкил)аминогруппы, карбамила, C1-С6алкилкарбамила и ди-C1-С6алкилкарбамила;
D1 каждый независимо друг от друга выбран из C1-С6арил-C1-С4алкила, C1-С9гетероарил-C1-С4алкила, С3-С10циклоалкил-C1-С4алкила, С2-С9гетероциклоалкил-C1-С4алкила, C1-С8алкилена, C1-С8алкенилена и C1-С8алкиниленена; где указанный C1-С8алкилен, C1-С8алкенилен и C1-С8алкиниленен необязательно замещены 1, 2, 3 или 4 независимо выбранными R4-группами; и где указанный C1-С6арил-C1-С4алкил, C1-С9гетероарил-C1-С4алкил, С3-С10циклоалкил-C1-С4алкил, С2-С9гетероциклоалкил-C1-С4алкил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными R5-группами;
D2 каждый независимо друг от друга отсутствует или обозначает C1-С20алкилен с прямой цепью, где от 1 до 6 несмежных метиленовых групп в указанном C1-С20алкилене с прямой цепью каждая необязательно замещена независимо выбранным -D4-фрагментом, при условии, что по меньшей мере одно метиленовое звено в указанном C1-С20алкилене с прямой цепью обязательно замещено -D4-фрагментом; где указанный C1-С20алкилен с прямой цепью необязательно замещен одной или несколькими группами, независимо выбранными из галогена, цианогруппы, гидроксила, C1-С4алкила, C1-С4 галоалкила, C1-С4алкоксигруппы, C1-С4 галоалкоксигруппы, аминогруппы, C1-С4алкиламиногруппы, ди-C1-С4алкиламиногруппы, C1-С4алкилкарбонила, карбоксигруппы, C1-С4алкоксикарбонила, C1-С4алкилкарбониламиногруппы, ди-C1-С4алкилкарбониламиногруппы, C1-С4алкоксикарбониламиногруппы, C1-С4алкоксикарбонил-(C1-С4алкил)аминогруппы, карбамила, C1-С4алкилкарбамила и ди-C1-С4алкилкарбамила;
D3 каждый независимо друг от друга выбран из Н, галогена, цианогруппы, нитрогруппы, гидроксила, C1-С6алкила, C1-C6 галоалкила, С2-С6алкенила, С2-С6алкинила, С3-С14циклоалкила, С3-С14циклоалкил-C1-С4алкила, С2-С14гетероциклоалкила, С2-С14гетероциклоалкил-C1-С4алкила, C6-С14арила, C6-С14арил-C1-С4алкила, C1-С13 гетероарила, C1-С13гетероарил-C1-С4алкила; где указанный C1-С6алкил, C1-С6галоалкил, С2-С6алкенил, С2-С6алкинил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными R6 -группами; и где указанный С3-С14циклоалкил, С3-С14циклоалкил-C1-С4алкил, С2-С14гетероциклоалкил, С2-С14гетероциклоалкил-C1-С4алкил, C6-С14арил, C6-С14арил- C1-С4алкил, C1-С13гетероарил, C1-С13гетероарил-C1-С4алкил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными R7-группами;
D4 каждый независимо друг от друга выбран из -О-, -S-, -NRaC(=O)-, -NRaC(=S)-, -NRbC(=O)NRc-, -NRbC(=S)NRc-, -S(=O)-, -S(=O)2-, -S(=O)NRa-, -C(=O)-, -C(=S)-, -C(=O)O-, -OC(=O)NRa-, -OC(=S)NRa-, -NRa-, -NRbS(=O)NRc- и NRbS(=O)2NRO-;
R4 и R6 каждый независимо друг от друга выбран из галогена, цианогруппы, нитрогруппы, гидроксила, C1-С4алкоксигруппы, C1-С4галоалкоксигруппы, C1-С4алкилтиогруппы, C1-С4алкилсульфинила, C1-С4алкилсульфонила, аминогруппы, C1-С4алкиламиногруппы, ди-C1-С4алкиламиногруппы, C1-С4алкилкарбонила, карбоксигруппы, C1-С4алкоксикарбонила, C1-С4алкилкарбониламиногруппы, ди-C1-С4алкилкарбониламиногруппы, C1-С4алкоксикарбониламиногруппы, C1-С4алкоксикарбонил-(C1-С4алкил)аминогруппы, карбамила, C1-С4алкилкарбамила и ди-C1-С4алкилкарбамила;
R5 каждый независимо друг от друга выбран из галогена, цианогруппы, нитрогруппы, гидроксила, C1-С4алкила, С2-С4алкенила, С2-С4алкинила, C1-С4алкоксигруппы, C1-С4галоалкоксигруппы, C1-С4алкилтиогруппы, C1-С4алкилсульфинилв, C1-С4алкилсульфонила, аминогруппы, C1-С4алкиламиногруппы, ди-C1-С4алкиламиногруппы, C1-С4алкилкарбонила, карбоксигруппы, C1-С4алкоксикарбонила, C1-С4алкилкарбониламиногруппы, ди-C1-С4алкилкарбониламиногруппы, C1-С4алкоксикарбониламино группы, C1-С4алкоксикарбонил-(C1-С4алкил)аминогруппы, карбамила, C1-С4алкилкарбамила и ди-C1-С4алкилкарбамила;
R7 каждый независимо друг от друга выбран из галогена, цианогруппы, нитрогруппы, гидроксила, C1-С6алкила, С2-С6алкенила, С2-С6алкинила, С3-С7циклоалкила, С3-С7циклоалкил-C1-С4алкила, С2-С7гетероциклоалкила, С2-С7гетероциклоалкил-C1-С4алкила, фенила, фенил-C1-С4алкила, C1-С7гетероарила, C1-С7гетероарил-C1-С4алкила, -ORO, -SRO, -S(=O)RP, -S(=O)2RP, -S(=O)NRsRt, -C(=O)Rp, -C(=O)ORp, -C(=O)NRsRt, -OC(=O)Rp, -OC(=O)NRsRt, -NRsRt, -NRqC(=O)Rr, -NRqC(=O)ORr, -NRqC(=O)NRr, -NRqS(=O)2Rr и -NRpS(=O)2NRsRt; где указанный C1-С6алкил, C2-С6алкенил, С2-С6алкинил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными R’-группами; и где указанный С3-С7циклоалкил, С3-С7циклоалкил-C1-С4алкил, С2-С7гетероциклоалкил, С2-С7гетероциклоалкил-C1-С4алкил, фенил, фенил-C1-С4алкил, C1-С7гетероарил, C1-С7гетероарил-C1-С4алкил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными R’’-группами;
Ra, Rb и Rc каждый независимо друг от друга выбран из Н, C1-С6алкила, C1-С6галоалкила, С2-С6алкенила, С2-С6алкинила, С3-С7циклоалкила, С3-С7циклоалкил-C1-С4алкила, С2-С7гетероциклоалкила, С2-С7гетероциклоалкил-C1-С4алкила, фенила, фенил-C1-С4алкила, C1-С7 гетероарила, C1-С7 гетероарил-C1-С4алкила; где указанный C1-С6алкил, C1-С6галоалкил, С2-С6алкенил, С2-С6алкинил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными Rw-группами; и где указанный С3-С7циклоалкил-C1-С4алкил, С2-С7гетероциклоалкил, С2-С7гетероциклоалкил-C1-С4алкил, фенил, фенил-1-С4алкил, C1-С7гетероарил, C1-С7гетероарил-C1-С4алкил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными Rx-группами;
Ro, Rp, Rq, Rr, Rs и Rt каждый независимо выбран из Н, C1-С6алкила, C1-С6галоалкила, С2-С6алкенила, С2-С6алкинила, С3-С7циклоалкила, С3-С7циклоалкил-C1-С4алкила, С2-С7гетероциклоалкила, С2-С7гетероциклоалкил-C1-С4алкила, фенила, фенил-C1-С4алкила, C1-С7 гетероарила, C1-С7гетероарил-C1-С4алкила; где указанный C1-С6алкил, C1-С6галоалкил, С2-С6алкенил, С2-С6алкинил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными Ry-группами; и где указанный С3-С7циклоалкил-C1-С4алкил, С2-С7гетероциклоалкил, С2-С7гетероциклоалкил-C1-С4алкил, фенил, фенил-C1-С4алкил, C1-С7гетероарил, C1-С7гетероарил-C1-С4алкил каждый необязательно замещен 1, 2, 3 или 4 независимо выбранными Rz-группами;
R’, Rw и Ry каждый независимо друг от друга выбран из гидроксила, цианогруппы, нитрогруппы, C1-С4алкоксигруппы, C1-С4галоалкоксигруппы, аминогруппы, C1-С4алкиламиногруппы и ди-C1-С4алкиламиногруппы; и
R’’, Rx и Rz каждый независимо друг от друга выбран из гидроксила, галогена, цианогруппы, нитрогруппы, C1-С4алкила, C1-С4галоалкила, C1-С4алкоксигруппы, C1-С4галоалкоксигруппы, аминогруппы, C1-С4алкиламиногруппы и ди-C1-С4алкиламиногруппы;
при условии, что валентность каждого атома в необязательно замещенных фрагментах не превышена.
Предпочтительно функциональные варианты/производные указанной выше формулы имеют аффинность в отношении антитела, предлагаемого в настоящем изобретении, сопоставимую или более высокую по сравнению с аффинностью DOTAM, и имеют силу связывания с Pb, сопоставимую или более высокую по сравнению с DOTAM (при измерении «аффинности» по константе диссоциации, как описано выше). Например, константа диссоциации функционального варианта/производного в отношении антитела, предлагаемого в настоящем изобретении или Pb может быть выше в 1,1 раза или менее, в 1,2 раза или менее, в 1,3 раза или менее, в 1,4 раза или менее, в 1,5 раза или менее или в 2 раза или менее, чем константа диссоциации DOTAM, характеризующая связывание с этим же антителом/Pb.
RN каждый может обозначать Н, C1-С6алкил или C1-С6галоалкил; предпочтительно Н, C1-С4алкил или C1-С4 галоалкил. Наиболее предпочтительно RN каждый обозначает Н.
Предпочтительно, если в вариантах DOTAM 1, 2, 3 или наиболее предпочтительно все L2 обозначают С2алкилен. Важно отметить, что содержащие С2алкилен варианты DOTAM могут обладать наиболее высокой аффинностью к Pb. Необязательными заместителями L2 могут быть R1, C1-С4алкил или C1-С4галоалкил. Приемлемо, когда необязательными заместителями L2 являются C1-С4алкил или C1-С4 галоалкил.
Необязательно L2 каждый может быть не замещен С2алкиленом СН2СН2-.
L1 каждый предпочтительно обозначает C1-С4алкилен, более предпочтительно С1алкилен, такой как -СН2-.
Функциональные варианты/производные могут включать также DOTAM или описанное выше соединение, конъюгированный/конъюгированное с одним или несколькими дополнительными фрагментами, например, с низкомолекулярным соединением, полипептидом или углеводом. Указанное присоединение может происходить через один из углеродов каркаса макроциклического кольца. Низкомолекулярное соединение может представлять собой, например, краситель (такой как Alexa 647 или Alexa 488), биотин или фрагмент биотина. Полипептид может представлять собой, например, олигопептид, например, терапевтический пептид или полипептид, такой как антитело. Примеры углеводов включают декстран, линейные или разветвленные полимеры или сополимеры (например, полиалкилен, поли(этиленлизин), полиметакрилат, полиаминокислоты, поли- или олигосахариды, дендримеры).
Функциональный вариант/функциональное производное DOTAM может представлять собой соединение следующей формулы:
в которой Z каждый независимо обозначает R1, указанный выше; р, q, r и s обозначают 0, 1 или 2; и сумма p+q+r+s составляет 1 или более. Предпочтительно р, q, r и s обозначают 0 или 1 и/или сумма p+q+r+s составляет 1. Например, в соединении сумма p+q+r+s может быть равна 1, и Z обозначает пара-SCN-бензильный фрагмент такое соединение поступает в продажу от фирмы Macrocyclics, Inc. (Плано, шт. Техас).
Подробное описание изобретения
Композиции и способы
Антитела
Один из объектов изобретения базируется, в том числе, на создании антител, которые связываются специфически с Pb-DOTAM (т.е. хелатом, который содержит DOTAM в комплексе с Pb, который обозначают также в настоящем описании как «хелат Pb-DOTAM»).
В некоторых вариантах осуществления изобретения антитело, которое связывается специфически с Pb-DOTAM, может обладать одним или несколькими следующими свойствами:
• связывается специфически с Pb-DOTAM и Bi-DOTAM;
• обладает избирательным действием в отношении Pb-DOTAM по сравнению с другими хелатами металлов, такими как Cu-DOTAM;
• связывается с Pb-DOTAM с очень высокой аффинностью;
• связывается с тем же эпитопом на Pb-DOTAM, что и антитела, указанные в настоящем описании, например, PRIT-0213 или PRIT-0214, и/или имеет такие же участвующие к контакте остатки, что и указанные антитела.
Радиоизотопы Pb применяют в методах диагностирования и терапии. Конкретные радиоизотопы свинца, которые можно применять в настоящем изобретении, включают 212Pb и 203Pb. Стабильные изотопы свинца можно применять также в очищающих агентах, например, 204Pb, 206Pb, 207Pb или 208Pb. Pb может представлять собой встречающийся в естественных условиях свинец, который является смесью стабильных (нерадиоактивных) изотопов 204Pb, 206Pb, 207Pb и 208Pb.
Радионуклиды, которые испускают α-частицы, потенциально являются более специфическими в отношении уничтожения опухолевых клеток, повреждая в меньшей степени окружающую ткань, чем β-излучатели, благодаря сочетанию короткой длины пути и высокой линейной передачи энергии. 212Bi является излучателем α-частиц, но его непосредственному применению мешает его короткий период полураспада. 212Pb является родительским радионуклидом 212Bi и может служить в качестве генератора in vivo 212Bi, что позволяет эффективно преодолевать недостаток, связанный с коротким периодом полураспада 212Bi (Yong и Brechbiel, Dalton Trans. 40(23), 21 июня 2001 г., сс. 6068-6076).
203Pb можно применять в качестве визуализирующего изотопа. Таким образом, антитело, связанное с 203Pb-DOTAM, может найти применение для радиоиммуновизуализации (RII).
Как правило, радиоактивные металлы применяют в форме хелатов. В некоторых объектах настоящего изобретения DOTAM применяют в качестве хелатирующего агента. DOTAM представляет собой стабильный хелатор Pb(II) (Yong и Brechbiel, Dalton Trans. 40(23), 21 июня 2001 г., сс. 6068-6076; Chappell и др., Nuclear Medicine and Biology, т. 27, 2000, сс. 93-100). Таким образом, DOTAM является наиболее ценным для применения в сочетании с изотопами свинца, указанными выше, такими как 212Pb и 203Pb.
Как указано выше, антитела, предлагаемые в настоящем изобретении, связываются с Pb-DOTAM. В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если аффинность связывания антител с Pb-DOTAM характеризуется величиной Kd, составляющей 100пМ, 50пМ, 20пМ, 10пМ, 5пМ, 1пМ или менее, например, 0,9пМ или менее, 0,8пМ или менее, 0,7пМ или менее, 0,6пМ или менее или 0,5пМ или менее.
Антитела связываются также с Bi, образующим хелат с DOTAM. В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если антитела связываются с Bi-DOTAM (т.е. хелатом, содержащим DOTAM в комплексе с висмутом, который обозначают также в настоящем описании как «хелат Bi-DOTAM»), при этом аффинность связывания характеризуется величиной Kd, составляющей 1нМ, 500пМ, 200пМ, 100пМ, 50пМ, 10пМ или менее, например, 9пМ, 8пМ, 7пМ, 6пМ, 5пМ или менее.
В некоторых вариантах осуществления изобретения антитела могут связываться с Bi-DOTAM и с Pb-DOTAM со сходной аффинностью. Например, может оказаться предпочтительным, если соотношение аффинности, например, соотношение величин Kd для Bi-DOTAM/Pb-DOTAM, находится в диапазоне 0,1-10, например 1-10.
Примеры величин аффинности для приведенного в качестве примера антитела, предлагаемого в изобретении (PRIT-0213), представлены ниже:

Аффинность определяли с помощью кинетического анализа исключения (KinExA) в условиях равновесия.
Кроме того, антитела, предлагаемые в изобретении, предпочтительно обладают избирательным действием в отношении Bi-DOTAM и/или Pb-DOTAM по сравнению с другими хелатными металлами, такими как Cu-DOTAM. Например, соотношение аффинностей, например, соотношение величин Kd, для Pb-DOTAM /Cu-DOTAM, может составлять по меньшей мере 100000.
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если антитела связываются с Pb-DOTAM и/или Bi-DOTAM с аффинностью (например, с характеризующей аффинность величиной Kd), равной или более высокой по сравнению с аффинностью биспецифического антитела (которое обозначено в настоящем описании как PRIT-0213), имеющего:
I) первую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 22;
II) вторую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 23; и
III) две легкие цепи антитела, которые имеют аминокислотную последовательность SEQ ID NO: 21.
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если антитела связываются с хелатом Pb-DOTAM и/или с хелатом Bi-DOTAM с аффинностью (например, с характеризующей аффинность величиной Kd), равной или более высокой по сравнению с аффинностью биспецифического антитела (которое обозначено в настоящем описании как PRIT-0214), имеющего:
I) первую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 19;
II) вторую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 20; и
III) две легкие цепи антитела, которые имеют аминокислотную последовательность SEQ ID NO: 21.
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, связывается с тем же эпитопом или с перекрывающимся эпитопом хелатного радионуклида, что и антитело, представленное в настоящем описании.
В некоторых вариантах осуществления изобретения антитело связывается с тем же эпитопом или с перекрывающимся эпитопом хелата Pb-DOTAM (Pb-DOTAM), что и Fab PRIT-0213, имеющий
I) первую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 22;
II) вторую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 23; и
III) две легкие цепи антитела, которые имеют аминокислотную последовательность SEQ ID NO: 21.
Эпитоп хелатного радионуклида (например, Pb-DOTAM), который связывается данным антителом, можно определять, и его можно сравнивать с эпитопом хелатного радионуклида, который связывается антителом, представленным в настоящем описании (например, Fab PRIT-0213).
В примере 14 настоящего описания дана характеризация взаимодействия связывания между Fab PRIT-0213 и Pb-DOTAM на основе определения кристаллической структуры Fab PRIT-0213 в комплексе с Pb-DOTAM при разрешении 1,40  и анализа указанной структуры с использованием программы для анализа поверхности раздела и сборки белков (PISA) (Krissinel и Henrick, J Mol Biol 372(3), 2007, cc. 774-797).
и анализа указанной структуры с использованием программы для анализа поверхности раздела и сборки белков (PISA) (Krissinel и Henrick, J Mol Biol 372(3), 2007, cc. 774-797).
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, может характеризоваться взаимодействием с одним или несколькими из следующих сайтов с Pb-DOTAM, например, определенными с помощью PISA-анализа структуры антитела в комплексе с Pb-DOTAM: взаимодействием по типу «ребро-к-грани» с областью азациклододеканового кольца ниже азациклододеканового кольца (например, тетрациклододеканового кольца), с N6, N7, N8, N5 и/или С12. В некоторых вариантах осуществления изобретения антитело может характеризоваться взаимодействием с одним или несколькими следующими сайтами Pb-DOTAM: N7, N8, взаимодействием по типу «ребро-к-грани» с азациклододекановым кольцом, тетрациклододекановым кольцом и/или N6.
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, может характеризоваться одним или несколькими из следующих взаимодействий с Pb-DOTAM, например, определенных с помощью PISA-анализа структуры антитела в комплексе с Pb-DOTAM: неполярным взаимодействием по типу «ребро-к-грани» с азациклододекановым кольцом, полярным взаимодействием с N8, водородной связью с N7, водородной связью с N8, полярным взаимодействием с N5, неполярным взаимодействием с С12, полярным взаимодействием с N7, полярным взаимодействием (водородная связь) с N6 и/или полярным взаимодействием с тетрациклододекановым кольцом.
В некоторых вариантах осуществления изобретения антитело, предлагаемое в настоящем изобретении, может характеризоваться одним или несколькими из следующих взаимодействий с Pb-DOTAM, например, определенных с помощью PISA-анализа структуры антитела в комплексе с Pb-DOTAM: водородной связью между одним или несколькими остатками CDR3 тяжелой цепи антитела и N7, водородная связь между одним или несколькими остатками CDR3 тяжелой цепи антитела и N8, неполярным взаимодействием между одним или несколькими остатками CDR2 тяжелой цепи антитела по типу «ребро-к-грани» относительно азациклододеканового кольца, неполярным взаимодействием между CDR3 легкой цепи антитело и тетрациклододекановым кольцом, и/или неполярным взаимодействием между CDR1 легкой цепи антитела и N6.
В других вариантах осуществления изобретения антитела могут иметь такие же участвующие в контакте остатки, что и указанные в настоящем описании: например, эти остатки могут быть инвариантными. Указанные остатки могут включать следующие:
а) в CDR2 тяжелой цепи: Phe50, Asp56 и/или Tyr58 и необязательно также Gly52 и/или Arg 54;
б) в CDR3 тяжелой цепи: Glu95, Arg96, Asp97, Pro98, Tyr99, Ala100C и/или Tyr100D и необязательно также Pro100E;
в) в CDR1 легкой цепи: Tyr28 и/или Asp32;
г) в CDR3 легкой цепи: Gly91, Tyr92, Asp93, Thr95c и/или Tyr96;
д) в CDR2 легкой цепи: необязательно Gln50.
Описанные выше антитела, предлагаемые в настоящем изобретении, являются некоторыми объектами и вариантами осуществления изобретения. Другие важные объекты и варианты осуществления изобретения обсуждены ниже. Во всех вариантах осуществления изобретения антитела сохраняют способность связываться с Pb-DOTAM и предпочтительно также с Bi-DOTAM, еще более предпочтительно с описанной выше аффинностью и/или селективностью.
Одним из вариантов осуществления изобретения может являться антитело к Pb-DOTAM, содержащее по меньшей мере один, два, три, четыре, пять или шесть CDR, которые выбраны из (a) CDR-H1, содержащего аминокислотную последовательность SEQ ID NO: 1; (б) CDR-H2, содержащего аминокислотную последовательность SEQ ID NO: 2; (в) CDR-H3, содержащего аминокислотную последовательность SEQ ID NO: 3; (г) CDR-L1, содержащего аминокислотную последовательность SEQ ID NO: 4; (д) CDR-L2, содержащего аминокислотную последовательность SEQ ID NO: 5; и (е) CDR-L3, содержащего аминокислотную последовательность SEQ ID NO: 6.
Одним из вариантов осуществления изобретения является антитело, содержащее по меньшей мере одну, по меньшей мере две или все три последовательности CDR VH, которые выбраны из (a) CDR-H1, содержащего аминокислотную последовательность SEQ ID NO: 1; (б) CDR-H2, содержащего аминокислотную последовательность SEQ ID NO: 2; и (в) CDR-H3, содержащего аминокислотную последовательность SEQ ID NO: 3. В одном из вариантов осуществления изобретения антитело содержит CDR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3. В другом варианте осуществления изобретения антитело содержит CDR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3, и CDR-L3, содержащий аминокислотную последовательность SEQ ID NO: 6. В другом варианте осуществления изобретения антитело содержит CDR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3, CDR-L3, содержащий аминокислотную последовательность SEQ ID NO: 6, и CDR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2. В другом варианте осуществления изобретения антитело содержит CDR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3, CDR-L3, содержащий аминокислотную последовательность SEQ ID NO: 6, CDR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2, и CDR-L1, содержащий аминокислотную последовательность SEQ ID NO: 4. В следующем варианте осуществления изобретения антитело содержит (a) CDR-H1, содержащий аминокислотную последовательность SEQ ID NO: 1; (б) CDR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2; и (в) CDR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3.
Другим объектом изобретения является антитело, которое содержит по меньшей мере одну, по меньшей мере две или все три последовательности CDR VL, которые выбраны из (a) CDR-L1, содержащего аминокислотную последовательность SEQ ID NO: 4; (б) CDR-L2, содержащего аминокислотную последовательность SEQ ID NO: 5; и (в) CDR-L3, содержащего аминокислотную последовательность SEQ ID NO: 6. В одном из вариантов осуществления изобретения антитело содержит (a) CDR-L1, содержащий аминокислотную последовательность SEQ ID NO: 4; (б) CDR-L2, содержащий аминокислотную последовательность SEQ ID NO: 5; и (в) CDR-L3, содержащий аминокислотную последовательность SEQ ID NO: 6.
В другом объекте изобретения антитело, предлагаемое в изобретении, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности CDR VH, которые выбраны из (I) CDR-H1, содержащего аминокислотную последовательность SEQ ID NO: 1, (II) CDR-H2, содержащего аминокислотную последовательность SEQ ID NO: 2, и (III) CDR-Н3, содержащего аминокислотную последовательность SEQ ID NO: 3; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три последовательности CDR VL, которые выбраны из (I) CDR-L1, содержащего аминокислотную последовательность SEQ ID NO: 4, (II) CDR-L2, содержащего аминокислотную последовательность SEQ ID NO: 5, и (в) CDR-L3, содержащего аминокислотную последовательность SEQ ID NO: 6.
Другим объектом изобретения является антитело, которое содержит (а) CDR-H1, содержащий аминокислотную последовательность SEQ ID NO: 1; (б) CDR-H2, содержащий аминокислотную последовательность SEQ ID NO: 2; (в) CDR-H3, содержащий аминокислотную последовательность SEQ ID NO: 3; (г) CDR-L1, содержащий аминокислотную последовательность SEQ ID NO: 4; (д) CDR-L2, содержащий аминокислотную последовательность SEQ ID NO: 5; и (е) CDR-L3, содержащий аминокислотную последовательность SEQ ID NO: 6.
В некоторых вариантах осуществления изобретения антитела могут содержать один или несколько CDR-H1, CDR-H2, CDR-H3, CDR-L1, CDR-L2 и/или CDR-L3, которые имеют замены по сравнению с аминокислотными последовательностями SEQ ID NO: 1-6 соответственно, например, 1, 2 или 3 замены. Может оказаться предпочтительным, если указанные замены не находятся в инвариантных положениях, указанных выше.
Например, в некоторых вариантах осуществления изобретения CDR-H2 может содержать аминокислотную последовательность FIGSRGDTYYASWAKG (SEQ ID NO: 2) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 2, где указанные замены не включают Phe50, Asp56 и/или Tyr58 и необязательно не включают также Gly52 и/или Arg 54 (во всех случаях нумерация по Кэботу).
В некоторых вариантах осуществления изобретения CDR-H2 может иметь замены в одном или нескольких положениях, которые указаны ниже. В настоящем описании и приведенных ниже таблицах замены представляют собой замены относительно остатков зародышевой линии (подчеркнуты) или аминокислот, которые теоретически стерически соответствуют или встречаются также в кристаллической структуре в сайте. В некоторых вариантах осуществления изобретения указанные выше остатки могут оставаться фиксированными, а другие остатки можно заменять в соответствии с представленной ниже таблицей: в других вариантах осуществления изобретения замены любого остатка можно осуществлять согласно указанной ниже таблице.
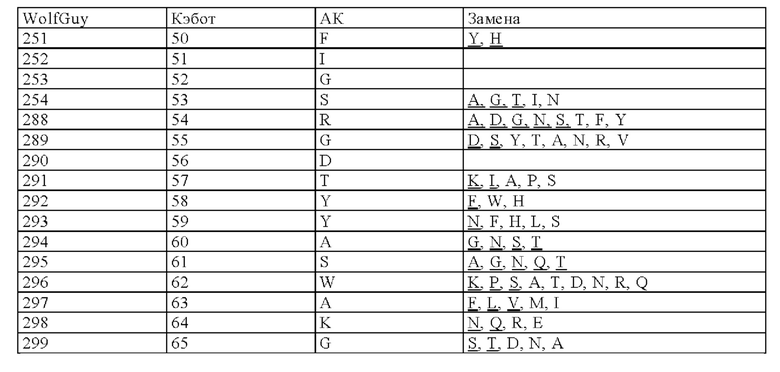
Необязательно CDR-H3 может содержать аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 3, где указанные замены не включают Glu95, Arg96, Asp97, Pro98 и необязательно не включают также Ala100C, Tyr100D и/или Pro100E, и/или необязательно не включают также Tyr99. Например, в некоторых вариантах осуществления изобретения замены не включают Glu95, Arg96, Asp97, Pro98, Tyr99 Ala100C и Tyr100D.
В некоторых вариантах осуществления изобретения CDR-H3 может иметь замены в одном или нескольких положениях, которые указаны ниже. В некоторых вариантах осуществления изобретения указанные выше остатки могут оставаться фиксированными, а другие остатки можно заменять в соответствии с представленной ниже таблицей: в других вариантах осуществления изобретения замены любого остатка можно осуществлять согласно указанной ниже таблице.
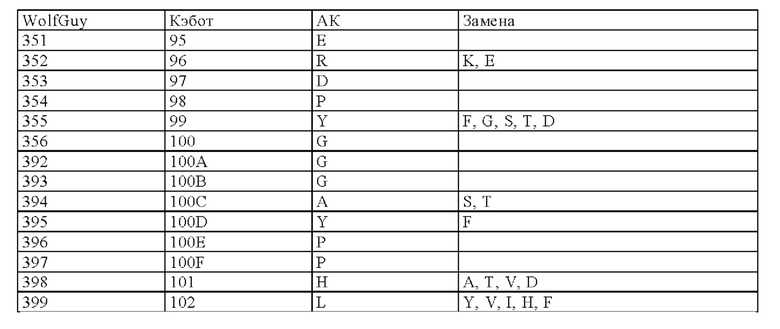
Необязательно CDR-L1 может содержать аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 4, где указанные замены не включают Tyr28 и/или Asp32 (нумерация по Кэботу).
В некоторых вариантах осуществления изобретения CDR-L1 может иметь замены в одном или нескольких положениях, которые указаны ниже. И в этом случае в некоторых вариантах осуществления изобретения указанные выше остатки могут оставаться фиксированными, а другие остатки можно заменять в соответствии с представленной ниже таблицей: в других вариантах осуществления изобретения замены любого остатка можно осуществлять согласно указанной ниже таблице.
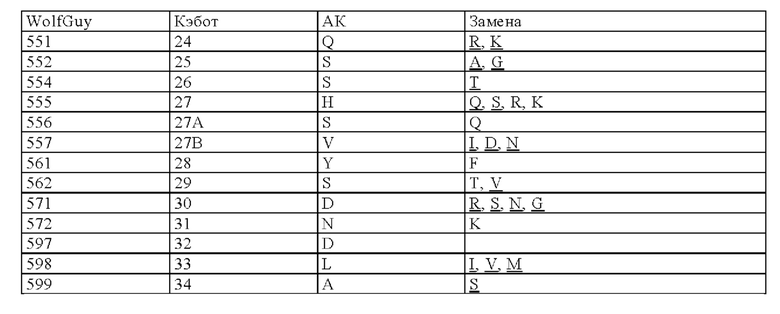
Необязательно CDR-L3 может содержать аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 6, где указанные замены не включают Gly91, Tyr92, Asp93, Thr95c и/или Tyr96 (по Кэботу).
В некоторых вариантах осуществления изобретения CDR-L3 может иметь замены в следующих положениях, которые указаны ниже. (Поскольку большинство остатков доступны для растворителей и не имеют контактов с антигеном, то возможно наличие целого ряда замен). И в этом случае в некоторых вариантах осуществления изобретения указанные выше остатки могут оставаться фиксированными, а другие остатки можно заменять в соответствии с представленной ниже таблицей: в других вариантах осуществления изобретения замены любого остатка можно осуществлять согласно указанной ниже таблице.
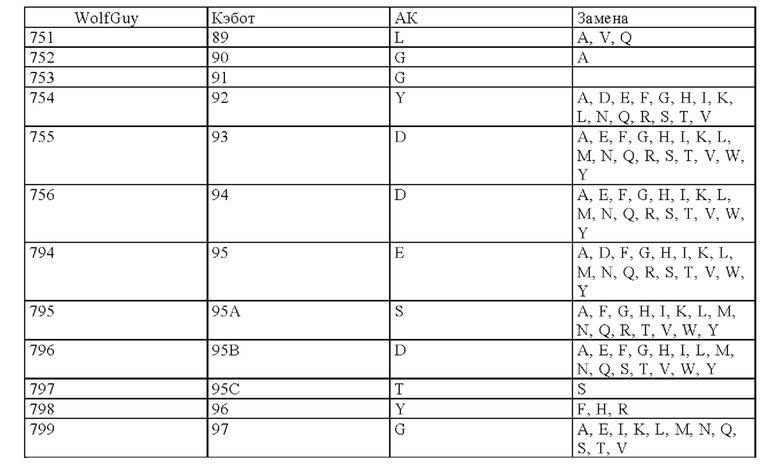
Антитело может содержать также CDR-H1 и CDR-L2, которые необязательно имеют последовательность SEQ ID NO: 1 или SEQ ID NO: 5 соответственно или ее вариант, имеющий по меньшей мере 1, 2 или 3 замены относительно указанных последовательностей, необязательно консервативные замены.
В любом из указанных выше вариантов осуществления изобретения антитело к Pb-DOTAM может быть гуманизировано. В одном из вариантов осуществления изобретения антитело к Pb-DOTAM содержит CDR, представленные в любом из указанных выше вариантов осуществления изобретения, а также содержит акцепторный человеческий каркасный участок, например, каркасный участок человеческого иммуноглобулина или человеческий консенсусный каркасный участок. В другом варианте осуществления изобретения антитело к Pb-DOTAM содержит CDR, представленные в любом из указанных выше вариантов осуществления изобретения, а также содержит каркасные участки, имеющие происхождение из vk 1 39 и/или vh 2 26. В случае vk 1 39 в некоторых вариантах осуществления изобретения они могут не иметь обратных мутаций. В случае vh 2 26 остаток А1а49 зародышевой линии может в результате обратной мутации быть заменен на Gly49.
Необязательно антигенсвязывающий сайт может содержать вариабельный домен тяжелой цепи (VH), который содержит аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 7 или SEQ ID NO 9, или ее вариант, который содержит аминокислотную последовательность, идентичную по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% SEQ ID NO: 7 или SEQ ID NO: 9. В некоторых вариантах осуществления изобретения VH-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело, которое содержит указанную последовательность, сохраняет способность связываться с Pb-DOTAM, предпочтительно с указанной в настоящем описании аффинностью. VH-последовательность может сохранять инвариантные остатки, которые указаны выше. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислотных остатков замены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 7 или SEQ ID NO: 9. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне CDR (т.е. FR). Необязательно антитело содержит VH-последовательность SEQ ID NO: 7 или SEQ ID NO: 9, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три CDR, которые выбраны из: (a) CDR-Н1, содержащего аминокислотную последовательность SEQ ID NO: 1, (б) CDR-Н2, содержащего аминокислотную последовательность SEQ ID NO: 2, и (в) CDR-H3, содержащего аминокислотную последовательность SEQ ID NO: 3.
Другим объектом изобретения является антитело к Pb-DOTAM, где антитело содержит вариабельный домен легкой цепи (VL), последовательность которого идентична по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 8 или SEQ ID NO: 10. В некоторых вариантах осуществления изобретения VL-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антитело, которое содержит указанную последовательность, сохраняет способность связываться с Pb-DOTAM, предпочтительно с указанной в настоящем описании аффинностью. VL-последовательность может сохранять инвариантные остатки, которые указаны выше. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислотных остатков заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 8 или SEQ ID NO: 10. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне CDR (т.е. FR). Необязательно антитело содержит VL-последовательность SEQ ID NO: 8 или SEQ ID NO: 10, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три CDR, которые выбраны из: (a) CDR-L1, содержащего аминокислотную последовательность SEQ ID NO: 4, (б) CDR-L2, содержащего аминокислотную последовательность SEQ ID NO: 5, и (в) CDR-L3, содержащего аминокислотную последовательность SEQ ID NO: 6.
Другим объектом изобретения является антитело к Pb-DOTAM, где антитело содержит VH-домен, указанный в любом из представленных выше вариантов осуществления изобретения, и VL-домен, указанный в любом из представленных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит VH- и VL последовательности SEQ ID NO: 7 и SEQ ID NO: 8 соответственно, включая пост-трансляционные модификации указанных последовательностей. В другом варианте осуществления изобретения антитело содержит VH- и VL-последовательности SEQ ID NO: 9 и SEQ ID NO: 10 соответственно, включая пост-трансляционные модификации указанных последовательностей.
В другом объекте изобретения антитело, представленное в любом из указанных выше вариантов осуществления изобретения, представляет собой моноклональное антитело, включая химереное, гуманизированное или человеческое антитело. В одном из вариантов осуществления изобретения антитело представляет собой фрагмент антитела, например, Fv-, Fab-, Fab’-scFab-, scFv-, димерные антитела (диабоди) или F(ab’)2-фрагмент. Фрагменты антител включают (но, не ограничиваясь только ими) Fab-, Fab’-, Fab’-SH-, F(ab’)2-, Fv- и scFv- фрагменты и другие фрагменты, описанные ниже. Обзор некоторых фрагментов антител см., например, у Hudson и др., Nat Med 9, 2003, сс. 129-134. Обзор scFv-фрагментов см., например, у  в: The Pharmacology of Monoclonal Antibodies, т.113, под ред. Rosenburg и Moore, изд-во Springer-Verlag, New York, 1994, сс. 269-315; см. также WO 93/16185; и US №№5571894 и 5587458. Обсуждение Fab- и Р(ab’)2-фрагментов, содержащих остатки эпитопа, связывающегося с рецептором спасения, и обладающих удлиненным временем полужизни in vivo, см. в US №5869046.
в: The Pharmacology of Monoclonal Antibodies, т.113, под ред. Rosenburg и Moore, изд-во Springer-Verlag, New York, 1994, сс. 269-315; см. также WO 93/16185; и US №№5571894 и 5587458. Обсуждение Fab- и Р(ab’)2-фрагментов, содержащих остатки эпитопа, связывающегося с рецептором спасения, и обладающих удлиненным временем полужизни in vivo, см. в US №5869046.
Димерные антитела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими (см., например, ЕР 404097; WO 1993/01161; Hudson и др., Nat Med 9, 2003, сс. 129-134; и Hollinger и др., Proc Natl Acad Sci USA 90, 1993, сс. 6444-6448). Тримерные и тетрамерные антитела описаны также у Hudson и др., Nat Med 9, 2003, сс. 129-134.
Однодоменные антитела представляют собой фрагменты антител, содержащие весь вариабельный домен тяжелой цепи или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В некоторых вариантах осуществления изобретения однодоменное антитело представляет собой человеческое однодоменное антитело (фирма Domantis, Inc.,, Валтам, шт. Массачусетс; см., например, US №6248516 В1).
Фрагменты антител можно создавать с помощью различных методик, включая (но, не ограничиваясь только ими) протеолитическое расщепление интактного антитела, а также получение с использованием рекомбинантных клеток-хозяев (например, Е. coli или фага), как указано в настоящем описании.
В другом варианте осуществления изобретения антитело представляет собой полноразмерное антитело, например, интактное антитело IgG-класса или антитело другого класса или изотипа, указанного в настоящем описании
Таргетные агенты
В некоторых объектах изобретения антитело, которое специфически связывается с хелатированным DOTAM Pb, сшивают со связывающимся с клеткой агентом/таргетирующим фрагментом с получением таргетного агента. Необязательно антитело, которое специфически связывается с хелатированным DOTAM РЬ, может представлять собой антитело, указанное в любом из представленных выше вариантов осуществления изобретения.
Связывание можно предпочтительно осуществлять путем экспрессии в виде слитого полипептида или белка. Слияние можно осуществлять непосредственно или через линкер. Слитый полипептид или белок можно получать рекомбинантно, в результате чего исчезает необходимость в применении химических методов конъюгации. Необязательно указанный линкер может представлять собой пептид, состоящий по меньшей мере из 5 аминокислот, предпочтительно из 25-50 аминокислот. Линкер может представлять собой жесткий линкер или гибкий линкер. В некоторых вариантах осуществления изобретения он представляет собой гибкий линкер, содержащий или состоящий из остатков Thr, Ser, Gly и/или Ala. Например, он может содержать или состоять из остатков Gly и Ser. В некоторых вариантах осуществления изобретения он может включать повторяющийся мотив, такой как (Gly-Gly-Gly-Gly-Ser)n, в котором n обозначает, например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. В некоторых вариантах осуществления изобретения линкер может представлять собой или может содержать последовательность GGGGSGGGGSGGGGSGGGGS (SEQ ID NO: 26). Можно применять другие линкеры и их может идентифицировать специалист в данной области.
Таким образом, предложен содержащий мультиспецифическое (например, биспецифическое) антитело комплекс, который специфически связывается как с хелатом Pb-DOTAM, так и с другим антигеном-мишенью, например, антигеном, который присутствует на поверхности клетки-мишени.
Поскольку в изобретении предложены способы лечения и продукты, предназначенные для применения в способах лечения, то его можно применять для любого состояния, поддающегося лечению с использованием цитотоксической активности, целенаправленно воздействующей на пораженные заболеванием клетки пациента. Лечение предпочтительно относится к лечению опухоли или рака (например, рака поджелудочной, молочной или предстательной железы). Однако область применения изобретения не ограничена опухолями и раками. Например, лечение может представлять собой также лечение вирусной инфекции. Иммунотоксины против вирусных антигенов, экспрессируемых на поверхности инфицированных клеток, изучены в отношении целого ряда вирусных инфекций, таких как ВИЧ, бешенство и EBV. Cai и Berger (Antiviral Research 90(3), 2011, cc. 143-50) применяли иммунотоксин, содержащий РЕ38, для таргетного цитолиза клеток, инфицированных ассоциированной с вирусом герпеса саркомой Капоши. Кроме того, Resimmune® (A-dmDT390-bisFv(UCHT1)) избирательно уничтожает человеческие злокачественные Т-клетки и кратковременно истощает здоровые Т-клетки и, как считается, обладает потенциалом в отношении лечения обусловленных Т-клетками аутоиммунных заболеваний, таких как рассеянный склероз и реакция «трансплантат-против-хозяина», а также Т-клеточных раков крови, в отношении которых проходят клинические испытания.
Таким образом, приемлемые антигены-мишени могут включать антигены раковых клеток, прежде всего антигены человеческих раковых клеток, вирусные антигены или микробные антигены.
Таргетные антитела, представленные в настоящем описании, создают для связывания с пораженными заболеванием клетками, такими как опухолевые клетки, через антигены их клеточной поверхности. Антигены, как правило, представляют собой поверхностные антигены здоровых клеток, для которых характерна либо сверхэкспрессия, либо экспрессия в аномальное время. В идеальном варианте антиген-мишень экспрессируется только на пораженных заболеванием клетках (таких как опухолевые клетки), хотя это редко встречается на практике. В результате антигены-мишени, как правило, выбирают на основе различной экспрессии в пораженной заболеванием и здоровой ткани.
Таким образом, таргетное антитело может специфически связываться с любым приемлемым маркером клеточной поверхности. Выбор конкретного таргетирующего фрагмента и/или маркера клеточной поверхности можно осуществлять в зависимости от конкретной клеточной популяции, подлежащей таргетингу. Маркеры клеточной поверхности известны в данной области (см., например, Mufson и др., Front. Biosci., 11, 2006, сс. 337-43; Frankel и др., Clin. Cancer Res., 6, 2000, сс. 326-334 и Kreitman и др., AAPS Journal, 8(3), 2006, Е532-Е551) и могут представлять собой, например, белок или углевод. В одном из вариантов осуществления изобретения таргетирующий фрагмент (связывающийся с клеткой агент) представляет собой лиганд, который специфически связывается с рецептором на клеточной поверхности. Примеры лигандов включают (но, не ограничиваясь только ими) сосудистый эндотелиальный фактор роста (VEGF), Fas, связанный с TNF апоптоз-индуцирующий лиганд (TRAIL), цитокин (например, IL-2, IL-15, IL-4, IL-13), лимфокин, гормон и фактор роста (например, трансформирующий фактор роста (TGFa), фактор роста нервов, эпидермальный фактор роста).
Маркер клеточной поверхности может представлять собой, например, опухолеассоциированный антиген.
В контексте настоящего описания понятие «опухолеассоциированный антиген» или «опухолеспецифический антиген» относится к любой молекуле (например, белку, пептиду, липиду, углеводу и т.д.), которая исключительно или преимущественно экспрессируется или сверхэкспрессируется опухолевыми клетками и/или раковыми клетками, в результате чего антиген является ассоциированным с опухолью(ями) и/или раком(ами). Опухолеассоциированный антиген может экспрессироваться также здоровыми, неопухолевыми, нераковыми клетками. Однако в этих случаях экспрессия опухолеассоциированного антигена здоровыми, неопухолевыми, нераковыми клетками не является столь же сильной, как экспрессия опухолевыми или раковыми клетками. В этом контексте опухолевые или раковые клетки могут сверхэкспрессировать существенно более высокие уровни антигена по сравнению с экспрессией антигена здоровыми, неопухолевыми, нераковыми клетками. Кроме того, опухолеассоциированный антиген может экспрессироваться также клетками на другой стадии развития или созревания. Например, опухолеассоциированный антиген может экспрессироваться также клетками на эмбриональной или фетальной стадии, т.е. клетками, которые в норме не встречаются у взрослых хозяев. Альтернативно этому, опухолеассоциированный антиген может экспрессироваться также стволовыми клетками или клетками-предшественниками, которые в норме не встречаются у взрослых хозяев.
Опухолеассоциированный антиген может представлять собой антиген, экспрессируемый любой клеткой любого рака или опухоли, включая виды раков и опухолей, которые указаны в настоящем описании. Опухолеассоциированный антиген может представлять собой опухолеассоциированный антиген только одного типа рака или опухоли, такой как опухолеассоциированный антиген, который ассоциирован или характерен только для одного типа рака или опухоли. Альтернативно этому, опухолеассоциированный антиген может представлять собой опухолеассоциированный антиген (например, может быть характерным) для нескольких типов раков или опухолей. Например, опухолеассоциированный антиген может экспрессироваться раковыми клетками, как молочной железы, так и предстательной железы и не экспрессироваться вообще здоровыми, неопухолевыми или нераковыми клетками.
Примеры опухолеассоциированных антигенов, с которыми связывающийся с клетками агент может специфически связываться, включают (но, не ограничиваясь только ими) муцин 1 (MUC1; опухолеассоциированный эпителиальный муцин), преимущественно экспрессируемый антиген меланомы (PRAME), карциноэмбриональный антиген (СЕА), простатспецифический мембранный антиген (PSMA), PSCA, ЕрСАМ, Trop2, колониестимулирующий фактор гранулоцитов-макрофагов (GM-CSFR), CD56, рецептор человеческого эпидермального фактора роста 2 (HER2/neu) (который обозначают также как erbB-2), CDS, CD7, родственный тирозиназе белок (TRP) I и TRP2. В предпочтительном варианте осуществления изобретения маркер клеточной поверхности, с которым таргетирующий фрагмент (связывающийся с клеткой агент) специфически связывается, выбирают из группы, состоящей из кластера дифференцировки (CD) 19, CD20, CD21, CD22, CD25, CD30, CD33 (связывающий сиаловую кислоту Ig-подобный лектин 3, поверхностный антиген миелоидных клеток), CD79b, CD123 (рецептор интерлейкина 3 альфа), рецептора трансферрина, EGF-рецептора, мезотелина, кадхерина, антигена Льюиса Y, глипикана-3, FAP (фибробласт-активирующий белок альфа), PSMA (простатспецифический мембранный антиген), СА9=CAIX (угольная ангидраза IX), L1 САМ (молекула адгезии нервной клетки L1), эндосиалина, HER3 (активированная конформация члена семейства рецепторов эпидермального фактора роста 3), комплекса Alkl/BMP9 (киназа анапластической лимфомы 1/участвующий в остеогенезе белок 9), TPBG=5Т4 (трофобластический гликопротеин), ROR1 (поверхностный антиген типа рецептороной тирозинкиназы), HER1 (активированная конформация рецептора эпидермального фактора роста) и CLL1 (семейство лектиновых доменов С-типа, семейство 12, член А). Мезотелин экспрессируется, например, при раке яичников, мезотелиоме, немелкоклеточном раке легких, аденокарциноме легких, раке фаллопиевых труб, раке головы и шеи, раке шейки матки и раке поджелудочной железы. CD22 экспрессируется, например, при волосатоклеточном лейкозе, хроническом лимфоцитарном лейкозе (CLL), пролимфоцитарном лейкозе (PLL), неходжкинской лимфоме, лимфоме из малых лимфоцитов (SLL) и остром лимфатическом лейкозе (ALL). CD25 экспрессируется, например, при лейкозах и лимфомах, включая волосатоклеточный лейкоз и лимфому Ходжкина. Антиген Льюиса Y экспрессируется, например, при раке мочевого пузыря, раке молочной железы, раке яичника, колоректальном раке, раке пищевода, раке легких, раке желудка и раке поджелудочной железы. CD33 экспрессируется, например, при остром миелоидном лейкозе (AML), хроническом миеломоноцитарном лейкозе (CML) и миелопролиферативных нарушениях.
В одном из вариантов осуществления изобретения таргетирующий фрагмент представляет собой антитело (включая фрагмент антитела), которое специфически связывается с мишенью, например, опухолеассоциированным антигеном. В указанных вариантах осуществления изобретения агент может представлять собой биспецифическое или мультиспецифическое антитело.
Примеры антител, которые специфически связываются с опухолеассоциированными антигенами, включают (но, не ограничиваясь только ими) антитела против рецептора трансферрина (например, НВ21 и его варианты), антитела против CD22 (например, RFB4 и его варианты), антитела против CD25 (например, анти-Тас и его варианты), антитела против мезотелина (например, SS 1, MORAb-009, SS, HN1, HN2, MN, MB, и их варианты) и антитела против Льюиса Y (например, ВЗ и его варианты). В этом контексте таргетирующий фрагмент (связывающийся с клетками агент) может представлять собой антитело, выбранное из группы, которая состоит из В3, RFB4, SS, SS1, MN, MB, HN1, HN2, НВ21 и MORAb-009 и их антигенсвязывающих участков. Дополнительные примеры таргетирующих фрагментов, пригодных для применения в заявляемых химерных молекулах, описаны, например, в US №№5242824 (антитело к рецептору трансферрина); 5846535 (антитело к CD25); 5889157 (антитело к антигену Льюиса Y); 5981726 (антитело к антигену Льюиса Y); 5990296 (антитело к антигену Льюиса Y); 7081518 (антитело к мезотелину); 7355012 (антитело к CD22 и к CD25); 7368110 (антитело к мезотелину); 7470775 (антитело к CD30); 7521054 (антитело к CD25) и 7541034 (антитело к CD22); в публикации заявки на патент США 2007/0189962 (антитело в CD22); у Frankel и др., Clin. Cancer Res., 6, 2000, cc. 326-334 и Kreitman и др., AAPS Journal, 8(3), 2006, E532-E551), содержание каждого документа включено в настоящее описание в качестве ссылки. Антитела могут образовываться против специфических для опухоли родственных антигенов-мишеней, включая: Cripto, CD30, CD19, CD33, гликопротеин NMB, CanAg, Her2 (ErbB2/Neu), CD56 (NCAM), CD22 (Siglec2), CD33 (Siglec3), CD79, CD138, PSCA, PSMA (простатспецифический мембранный антиген), ВСМА, CD20, CD70, Е-селектин, EphB2, меланотрансферрин, Мис16 и TMEFF2.
В некоторых вариантах осуществления настоящего изобретения может оказаться предпочтительным, если опухолеассоциированный антиген представляет собой карциноэмбриональный антиген (СЕА). СЕА может иметь аминокислотную последовательность человеческого СЕА, в частности карциноэмбриональному антигену родственная молекула адгезии 5 (СЕАСАМ5), которая зарегистрирована в UniProt (www.uniprot.org), регистрационный номер Р06731 (версия 119) или в NCBI (www.ncbi.nlm.nih.gov/) RefSeq NP 004354.2. Антитела, образовавшиеся против СЕА, включают Т84.66 и его гуманизированные и химерные версии, такие как T84.66-LCHA, которые описаны в WO 2016/075278 А1 и/или WO 2017/055389, CH1A1a, антитело к СЕА, описанное в WO 2011/034660, и СЕА hMN-14, указанное ниже в таблице 2 (см. также US №6676924 и US №5874540).
СЕА является предпочтительным в контексте настоящего изобретения, поскольку он относительно медленно интернализуется, и поэтому высокий процент антитела должен оставаться доступным на поверхности клетки после начальной обработки, связываясь с радионуклидом. Предпочтительными могут являться также другие отличающиеся низким уровнем интернализации антигены-мишени/опухолеассоциированные антигены. Например, в некоторых вариантах осуществления изобретения опухолеассоциированный антиген может представлять собой CD20 или HER2. В GenBank под регистрационными номерами NP 001005862, NP 004439, ХР 005257196 и ХР 005257197 представлены белковые последовательности Her2, по данным регистрации в GenBank на 4 октября 2013 г., и в базе данных SwissProt, код доступа Р11836 представлена последовательность CD20. В следующих вариантах осуществления изобретения мишень может представлять собой EGP-1 (эпителиальный гликопротеин-1, известный также как трофобласт-2), специфический для ободочной кишки антиген-р (CSAp) или панкреатический муцин MUC1 (см., например, у Goldenberg и др., Theranostics 2(5), 2012, публикация включена в настоящее описание в качестве ссылки). В указанной ссылке описаны также антитела, такие как Mu-9, связывающееся с CSAp (см. также Sharkey и др., Cancer Res. 63, 2003, сс. 354-363), hPAM4, связывающееся с MUC1 (см. также Gold и др., Cancer Res. 68, 2008, сс. 4819-4826), валтузумаб, связывающийся с CD20 (см. также Sharkey и др., Cancer Res. 68, 2008, сс. 5282-5290), и hRS7, связывающееся с EGP-1 (см. также Cubas и др., Biochim Biophys Acta 1796, 2009, сс. 309-14). Любое из этих антител или их антигенсвязывающих участков можно применять согласно настоящему изобретению, т.е. можно включать в антитела, указанные в настоящем описании.
Мультиспецифические антитела
Как обсуждалось выше, в некоторых вариантах осуществления изобретения антитело, указанное в настоящем описании, представляет собой мультиспецифическое антитело, например, биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, имеющие связывающие специфичности в отношении по меньшей мере двух различных сайтов. Биспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
В последние годы разработано широкое разнообразие форматов рекомбинантных антител, например, четырехвалентные биспецифические антитела, полученные путем слияния, например, антитела IgG-формата и одноцепочечных доменов (см., например, Coloma M.J. и др., Nature Biotech 15, 1997, сс. 159-163; WO 2001/077342 и Morrison S.L., Nature Biotech 25, 2007, с.с. 1233-1234).
Кроме того, разработано несколько других новых форматов антител, в которых уже не сохраняется коровая структура (IgA, IgD, IgE, IgG или IgM), таких как диа-, триа- или тетрабоди, минибоди, несколько одноцепочечных форматов (scFv, Bis-scFv), которые обладают способностью связываться с двумя или большим количеством антигенов (Holliger Р. и др., Nature Biotech 23, 2005, сс. 1126-1136; Fischer N., Leger О., Pathobiology 74, 2007, сс. 3-14; Shen J. и др., Journal of Immunological Methods 318,2007, cc. 65-74; Wu С.и др., Nature Biotech. 25, 2007, cc. 1290-1297).
Во всех указанных форматах используют линкеры либо для слияния корового антитела (IgA, IgD, IgE, IgG или IgM) с дополнительным связывающим белком (например, scFv), либо для слияния, например, двух Fab-фрагментов или scFv (Fischer N., Leger О., Pathobiology 74, 2007, cc. 3-14). Следует иметь в виду, что может требоваться сохранение эффекторных функций, таких, например, как комплементзависимая цитотоксичность (CDC) или антитело-обусловленная клеточнозависимая цитотоксичность (ADCC), которое опосредуются связыванием Fc-рецептора, для сохранения высокой степени сходства со встречающимися в естественных условиях антителами.
В WO 2007/024715 описаны иммуноглобулины с двойными вариабельными доменами в качестве сконструированных многовалентных и мультиспецифических связывающих белков. Процесс получения биологически активных димеров антител описан в US №6897044. Многовалентная конструкция Fv антитела по меньшей мере с четырьмя вариабельными доменами, которые связаны друг с другом через пептидные линкеры, описана в US №7129330. Димерные и мультимерные антигенсвязывающие структуры описаны в US 2005/0079170. Трех- или четырехвалентные моноспецифические антигенсвязывающие белки, содержащие три или четыре Fab-фрагмента, которые ковалентно связаны друг с другом с помощью соединяющей структуры, в которых белок не представляет собой встречающийся в естественных условиях иммуноглобулин, описаны в US №6511663. В WO 2006/020258 описаны четырехвалентные биспецифические антитела, которые могут эффективно экспрессироваться в прокариотических и эукариотических клетках, и которые можно применять в терапевтических и диагностических методах. Метод разделения или предпочтительно синтеза димеров, которые связаны по меньшей мере через один межцепочечный дисульфидный мостик, из димеров, которые не связаны по меньшей мере через один межцепочечный дисульфидный мостик, из смеси, содержащей два типа полипептидных димеров, описан в US 2005/0163782. Биспецифические четырехвалентные рецепторы описаны в US №5959083. Сконструированные антитела с тремя или большим количеством функциональных антигенсвязывающих сайтов описаны в WO 2001/077342.
Мультиспецифические и многовалентные антигенсвязывающие полипептиды описаны в WO 1997/001580. В WO 1992/004053 описаны гомоконъюгаты, как правило, полученные из моноклональных антител IgG-класса, которые связываются с одной и той же антигенной детерминантой, ковалентно связанные с помощью синтетического перекретносшивающего агента. Олигомерные моноклональные антитела с высокой авидностью к антигену описаны в WO 1991/06305, при этом секретируются олигомеры, как правило, IgG-класса, имеющие два или большее количество иммуноглобулиновых мономеров, ассоциированных друг с другом, с образованием четырехвалентных или шестивалентных молекул IgG. В US №6350860 описаны овечьи антитела и созданные конструкции антител, которые можно применять для лечения заболеваний, при которых активность интерферона гамма является патогенной. В US 2005/0100543 описаны обладающие способностью к таргетингу конструкции, которые представляют собой многовалентные носители биспецифических антител, т.е. каждая молекула обладающей способностью к таргетингу конструкции может служить в качестве носителя двух или большего количества биспецифических антител. Генетически сконструированные биспецифические четырехвалентные антитела описаны в WO 1995/009917. В WO 2007/109254 описаны стабилизирующие связывающие молекулы, которые состоят из стабилизированного scFv или содержат его.
Мультиспецифические антитела могут находиться также в асимметричной форме с кроссовером доменов в одном или нескольких связывающих плечах одной и той же антигенной специфичности, т.е. путем обмена VH/VL-доменов (см., например, WO 2009/080252 и WO 2015/150447), CH1/CL-доменов (см., например, WO 2009/080253) или полных Fab-плечей (см., например, WO 2009/080251, WO 2016/016299, см. также Schaefer и др., PNAS, 108, 2011, сс.1187-1191 и Klein и др., MAbs 8, 2016, сс. 1010-1020). В одном из объектов изобретения мультиспецифическое антитело содержит кросс-Fab-фрагмент. Понятие «кросс-Fab-фрагмент» или «xFab-фрагмент» или «кроссовер-Fab-фрагмент» относится к Fab-фрагменту, в котором либо вариабельные области, либо константные области тяжелой и легкой цепей обменены. Kpocc-Fab-фрагмент содержит полипептидную цепь, состоящую из вариабельной области легкой цепи (VL) и константной области 1 тяжелой цепи (СН1), и полипептидную цепь, состоящую из вариабельной области тяжелой цепи (VH) и константной области легкой цепи (CL). Асимметричные Fab-плечи можно создавать также путем интродукции мутаций заряженных или незаряженных аминокислот в области поверхности раздела для непосредственной коррекции спаривания Fab (см., например, WO 2016/172485).
Любой из указанных выше форматов можно применять для мультиспецифических антител, предлагаемых в настоящем изобретении.
В одном из приведенных в качестве примера форматов биспецифическое антитело представляет собой «тримерайзер» (trimerizer), например, описанный в WO 214/180754. Это обозначение относится к тримерной антигенсвязывающей молекуле, содержащей три слитых полипептида, каждый из которых содержит по меньшей мере один антигенсвязывающий фрагмент, слитый с доменом тримеризации, который имеет происхождение из человеческого матриксного белка хряща (СМР), где указанный домен тримеризации обладает способностью опосредовать стабильную ассоциацию тримерной антигенсвязывающей молекулы. Антигенсвязывающие фрагменты могут представлять собой, например, молекулу Fab, кроссовер-молекулу Fab, scFab, молекулу Fv, scFv или однодоменное антитело (VHH). В некоторых вариантах осуществления изобретения каждый из слитых белков содержит два (первый и второй) антигенсвязывающих фрагмента, например, когда первый антигенсвязывающий фрагмент слит с N-концевой аминокислотой указанного домена тримеризации, а второй антигенсвязывающий фрагмент слит с С-концевой аминокислотой указанного домена тримеризации, в обоих случаях необязательно через пептидный линкер. В таком формате либо первый, либо второй антигенсвязывающий фрагмент может связываться с хелатом Pb-DOTAM. Другой должен связываться с антигеном-мишенью, например, опухолеассоциированным антигеном. Каждая из трех антигенсвязывающих молекул, слитых с С-концом, может специфически связываться с одним и тем же антигеном; каждая из трех антигенсвязывающих молекул, слитых N-концом, может специфически связываться с другим антигеном.
СМР-домен тримеризации, который можно применять в настоящем изобретении, получали из белка хряща человека, который представлен ниже, и в одном из вариантов осуществления изобретения он содержит последовательность, идентичную по меньшей мере на 95%, и наиболее предпочтительно идентичную по меньшей мере на 98%, последовательности домена тримеризации, представленной ниже. В одном из вариантов осуществления изобретения указанный домен тримеризации содержит последовательность указанного домена тримеризации.
Последовательность белка хряща человека (496 ак)

Примером последовательность домена тримеризации (39 ак) является последовательность:
Другой приведенный в качестве примера формат содержит полноразмерное антитело (например, IgG), которое содержит первую и вторую тяжелые цепи и первую и вторую легкие цепи антитела, в котором первая тяжелая цепь и первая легкая цепь объединены с образованием антигенсвязывающего сайта для первого антигена и в котором вторая тяжелая цепь и вторая легкая цепь объединены с образованием антигенсвязывающего сайта для второго антигена.
Правильной сборке гетеродимерных тяжелых цепей может способствовать, например, применение мутаций «knob into hole» и/или других модификаций, которые будут описаны ниже.
Правильной сборке легких цепей с соответствующей тяжелой цепью может способствовать применение технологии «Cross-mab». При применении указанного подхода либо первую тяжелую цепь и первую легкую цепь, либо вторую тяжелую цепь и вторую легкую цепь можно объединять с получением cross-Fab-фрагмента (в то время как сборка других цепей соответствует каноническому Fab). Таким образом, в одном из вариантов осуществления изобретения первая тяжелая цепь может содержать VL-домен вместо VH-домена (например, VL-СН1-шарнир-СН2-CH3), а первая легкая цепь может содержать VH-домен вместо VL-домена (например, VH-CL), или первая тяжелая цепь может содержать CL-домен вместо CH1-домена (например, VH-CL-шарнир-СН2-CH3), а первая легкая цепь может содержать CH1-домен вместо CL-домена (например, VL-CH1). В указанном варианте осуществления изобретения вторая тяжелая цепь и вторая легкая цепь имеют каноническую структуру домена (например, VH-СН1-шарнир-СН2-CH3 и VL-CL соответственно). В альтернативном варианте осуществления изобретения вторая тяжелая цепь может содержать VL-домен вместо VH-домена (например, VL-CH1-шарнир-СН2-CH3), а вторая легкая цепь может содержать VH-домен вместо VL-домена (например, VH-CL), или вторая тяжелая цепь может содержать CL-домен вместо CH1-домена (например, VH-CL-шарнир-CH2-CH3), а вторая легкая цепь может содержать CH1-домен вместо CL-домена (например, VL-CH1). В указанном варианте осуществления изобретения первая тяжелая цепь и первая легкая цепь имеют каноническую структуру домена.
В некоторых вариантах осуществления изобретения правильной сборке легких цепей с соответствующей тяжелой цепью может в дополнительном или альтернативном варианте способствовать применение модификации зарядов, что будет дополнительно описано ниже.
Одно из таких антител представлено на фиг. 37 в виде Р1АЕ1768. В нем вторая тяжелая цепь содержит CL-домен вместо НС1-домена (например, VH-CL-шарнир-СН2-CH3), а вторая легкая цепь содержит CH1-домен вместо CL-домена (например, VL-CH1); первая тяжелая цепь и первая легкая цепь имеют каноническую структуру домена. Fab с канонической структурой содержит модификацию заряда. Так, в одном из вариантов осуществления изобретения антитело, предлагаемое в изобретении, содержит первую и вторую тяжелые цепи, имеющие SEQ ID NO: 59 и 58 соответственно, и первую и вторую легкие цепи, имеющие SEQ ID NO: 57 и 60 соответственно.
В некоторых вариантах указанного выше формата формат может быть двухвалентным. В другом возможном варианте осуществления изобретения дополнительные антигенсвязывающие фрагменты можно сливать, например, с первой и/или второй тяжелой цепью для повышения валентности в отношении одного или обоих антигенов. Например, дополнительный антигенсвязывающий фрагмент для первого антигена можно сливать с N-концом одной или обеих молекул тяжелых цепей. Антитело может быть многовалентным, например, двухвалентным в отношении первого антигена (например, опухолеассоциированного антигена) и одновалентным в отношении второго антигена (например, хелата Pb-DOTAM).
Дополнительный антигенсвязывающий фрагмент может представлять собой, например, scFab, например, содержащий антигенсвязывающий сайт для первого антигена (например, опухолеассоциированного антигена). scFab содержит VH- и СН1-домены, сцепленные через полипептидный линкер с VL- и CL-доменам, так, чтобы они экспрессировались в виде одной цепи. Иными словами, scFab содержит полипептидный линкер между Fd и легкой цепью.
В другом варианте осуществления изобретения дополнительный антигенсвязывающий фрагмент представляет собой Fab или cross-Fab. Например, N- или С-конец одной из тяжелых цепей может быть сцеплен через полипептидный линкер с первым полипептидом, состоящим из VH-домена и CH1-домена, который связан со вторым полипептидом, состоящим из VL- и CL-доменов, с образованием Fab. В другом варианте осуществления изобретения N-или С-конец одной из тяжелых цепей может быть сцеплен через полипептидный линкер с первым полипептидом, состоящим из VL-домена и СН1-домена, который связан со вторым полипептидом, состоящим из VH- и CL- доменов. В другом варианте осуществления изобретения N- или С-конец одной из тяжелых цепей может быть сцеплен через полипептидный линкер с первым полипептидом, состоящим из VH-домена и CL-домена, который связан со вторым полипептидом, состоящим из VL- и СН1-домена.
В указанном формате может оказаться предпочтительным, чтобы связывающиеся плечи, специфические в отношении одного и того же антигена, образовывались путем ассоциации с одной и той же легкой цепью. Таким образом, антигенсвязывающие фрагменты/плечи для первого антигена могут представлять собой cross-Fab, а антигенсвязывающий(ие) фрагмент(ы) /плечо(и) для второго антигена могут представлять собой канонические Fab. Альтернативно этому, антигенсвязывающие фрагменты/плечи для первого антигена могут представлять собой канонические Fab, а антигенсвязывающий(ие) фрагмент(ы)/плечо(и) для второго антигена могут представлять собой cross-Fab.
Формат может включать также модификацию заряда, что будет дополнительно описано ниже.
Одним из вариантов указанного формата является многовалентное антитело, содержащее полноразмерное антитело, которое содержит первую и вторую тяжелые цепи антитела и первую и вторую легкие цепи антитела, в котором первая тяжелая цепь и первая легкая цепь объединены с образованием Fab, который содержит антигенсвязывающий сайт для первого антигена (например, опухолеспецифического антигена, например, СЕА), и в котором вторая тяжелая цепь и вторая легкая цепь объединены с образованием cross-Fab, который содержит антигенсвязывающий сайт для второго антигена (например, хелата Pb-DOTAM) (например, вторая тяжелая цепь имеет VL-домен вместо VH-домена, а вторая легкая цепь имеет VH-домен вместо VL-домена); и в котором либо первая, либо вторая тяжелая цепь антитела слита через линкер с полипептидом, содержащим СН1- и VH-домены, и указанный первый полипептид объединен со вторым полипептидом, содержащим CL и VL, в результате первый и второй полипептиды объединены с образованием Fab, который содержит антигенсвязывающий сайт для первого антигена.
Слияние может иметь место на N-конце одной из тяжелых цепей полноразмерного антитела, необязательно второй тяжелой цепи.
Необязательно можно применять также модификацию заряда. Например, Fab, содержащие антигенсвязывающий сайт для первого антигена, могут содержать модифицирующие заряд замены, описанные ниже.
Например, указанный формат представляет собой Р1АЕ1769, указанные на фиг. 37. Таким образом, в одном из вариантов осуществления изобретения антитело, предлагаемое в изобретении, содержит первую и вторую тяжелые цепи, имеющие SEQ ID NO: 64 и 63 соответственно, и первую и вторую легкие цепь, имеющие SEQ ID NO: 62 и 61 соответственно.
Другой приведенный в качестве примера формат содержит полноразмерное антитело, такое как IgG, содержащее антигенсвязывающий сайт для первого антигена (например, которое может быть двухвалентным в отношении первого антигена), сцепленный с антигенсвязывающим фрагментом для второго антигена.
Например, антигенсвязывающий фрагмент для второго антигена может представлять собой scFab, содержащий антигенсвязывающий сайт для второго антигена (например, хелата Pb-DOTAM). В некоторых вариантах осуществления изобретения scFab может быть слит с С-концом одной из двух тяжелых цепей полноразмерного антитела, например, на С-конце его СН3-домена. Правильной сборке гетеродимерных тяжелых цепей может способствовать, например, применение мутаций «knob into hole» и/или других модификаций, которые будут описаны ниже.
Одно из таких антител представлено в качестве примера на фиг. 37 в виде Р1АЕ1770. Таким образом, в одном из вариантов осуществления изобретения антитело, предлагаемое в изобретении, содержит тяжелые цепи, имеющие SEQ ID NO: 66 и 67, и легкую цепь, имеющую SEQ ID NO: 65.
Другой приведенный в качестве примера формат содержит полноразмерное антитело, содержащее антигенсвязывающий сайт для первого антигена (например, которое может быть двухвалентным в отношении первого антигена), в котором N- или С-конец одной из тяжелых цепей сцеплен через полипептидный линкер с первым полипептидом, и в котором первый полипептид объединен со вторым полипептидом с образованием Fab или cross-Fab, содержащего сайт связывания для второго антигена. Например, указанный формат может содержать:
I) первый полипептид, состоящий из VH-домена и CH1-домена, который находится в ассоциации со вторым полипептидом, состоящим из VL- и CL-доменов; или
II) первый полипептид, состоящий из VL-домена и CH1-домена, который находится в ассоциации со вторым полипептидом, состоящим из VH- и CL-доменов; или
III) первый полипептид, состоящий из VH-домена и CL-домена, который находится в ассоциации со вторым полипептидом, состоящим из VL- и СН1-доменов;
в результате чего первый и второй полипептид вместе образуют антигенсвязывающий сайт для второго антигена.
Правильной сборке гетеродимерных тяжелых цепей может способствовать, например, применение мутаций «knob into hole» и/или других модификаций, которые будут описаны ниже, включая модификации зарядов. Например, Fab-домены полноразмерного антитела могут включать модификации зарядов.
В одном из вариантов осуществления изобретения первый полипептид сцеплен через полипептидный линкер с С-концом одной из тяжелых цепей, например, на С-конце ее СН3-домена. Первый полипептид может содержать N-концевой VL-домен и С-концевой CH1-домен. Таким образом, тяжелая цепь, имеющая слияние, может содержать в направлении от N-конца к С-концу VH-СН1-шарнир-СН2-СН3-линкер-VL-СН1. Легкая цепь может содержать VH-CL. Fab-фрагменты полноразмерного антитела могут включать модифицирующие заряды замены. Один из таких вариантов представлен на фиг. 37 в виде Р1АЕ1767. Таким образом, в одном из вариантов осуществления изобретения антитело, предлагаемое в изобретении, содержит тяжелые цепи, имеющие SEQ ID NO: 63 и 64, и легкие цепи, имеющие SEQ ID NO: 61 и 62.
В другом варианте осуществления изобретения первый полипептид сцеплен через полипептидный линкер с N-концом VH-домена тяжелой цепи. Первый полипептид может содержать N-концевой VL-домен и С-концевой СН1- домен. Таким образом, тяжелая цепь имеющая слияние, может содержать в направлении от N-конца к С-концу VL-CH1-линкер-VH-CH1-шарнир-CH2-CH3. Легкая цепь может содержать VH-CL.
В другом приведенном в качестве примера формате антитело может содержать полноразмерное антитело, специфически связывающее первый антиген и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, в котором С-конец каждой из тяжелых цепей слит с антигенсвязывающим фрагментом, который специфически связывается со вторым антигеном.
В одном из вариантов осуществления изобретения первый антиген представляет собой мишень, например, опухолеспецифический антиген, а второй представляет собой хелат Pb-DOTAM, но может также иметь место обратная ситуация.
В другом приведенном в качестве примера формате антитело может представлять собой биспецифическое антитело, которое содержит:
а) полноразмерное антитело, специфически связывающее первый антиген и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела;
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH); или
II) вариабельного домена тяжелой цепи антитела (VH) и константного домена тяжелой цепи антитела (СН1); или
III) вариабельного домена тяжелой цепи антитела (VH) и константного домена легкой цепи антитела (CL);
в котором указанный полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL); или
II) вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL); или
III) вариабельного домена легкой цепи антитела (VL) и константного домена тяжелой цепи антитела (СН1);
в котором указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
и в котором вариабельный домен тяжелой цепи антитела (VH) пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для второго антигена.
В указанном формате, если первый полипептид, представляет собой указанный в подпункте 6(I), то второй полипептид представляет собой указанный в подпункте в(I); если первый полипептид, представляет собой указанный в подпункте б(II), то второй полипептид представляет собой указанный в подпункте в(II); и, если первый полипептид, представляет собой указанный в подпункте б(III), то второй полипептид представляет собой указанный в подпункте в(III). Можно применять также модифицирующие заряды замены, например, в Fab полноразмерного антитела.
В указанном формате либо первый, либо второй антиген может представлять собой хелат Pb-DOTAM. Другой может представлять собой мишень, например, опухолеассоциированный антиген, например, СЕА, CD20 или ERBB2. В некоторых вариантах осуществления изобретения второй антиген представляет собой хелатированный DOTAM Pb, а первый антиген представляет собой мишень.
Антитело, описанное выше, может быть трехвалентным. В другом возможном варианте осуществления изобретения можно сливать дополнительные антигенсвязывающие фрагменты для повышения валентности в отношении одного или обоих антигенов. Например, дополнительный антигенсвязывающий фрагмент для первого антигена можно сливать с карбоксильным концом любой или обеих тяжелых цепей полноразмерного антитела (например, для опухолеассоциированного антигена), например, так, чтобы антитело имело валентность 4 в отношении первого антигена (когда он слит с карбоксильным концом обеих тяжелых цепей) и валентность 1 в отношении второго антигена.
Примерами указанных выше форматов, в которых антитело, указанное в подпункте (б), состоит из VH-домена, а антитело, указанное в подпункте (в), состоит из VL-домена, являются PRIT-213 и PRIT214. Примером указанного выше формата, в котором полипептид, указанный в подпункте (б), состоит из VH-домена и CL-домена, а указанный в подпункте (в) состоит из VL-домена и CH1-домена, является антитело Р1АЕ1766, указанное на фиг. 37. Таким образом, в одном из вариантов осуществления изобретения антитело, предлагаемое в изобретении, содержит первую и вторую тяжелые цепи, имеющие SEQ ID NO: 51 и 52 соответственно, и легкую цепь, имеющую SEQ ID NO: 50.
Необязательно применяемый согласно настоящему изобретению формат мультиспецифических антител может представлять собой трехвалентный формат, описанный в WO 2010/115589 А1 (фирма Roche Glycart AG), содержание которой полностью включено в настоящее описание в качестве ссылки.
В WO 2010/115589 описана необязательная стабилизация структуры, в которой полипептид вариабельной области тяжелой цепи антитела (VH), указанный в подпункте (б), и полипептид вариабельной области легкой цепи антитела (VL), указанный в подпункте (в), сцеплены и стабилизированы через межцепочечный дисульфидный мостик, например, путем интродукции дисульфидной связи между следующими положениями:
I) положение 44 в вариабельном домене тяжелой цепи и положение 100 в вариабельном домене легкой цепи,
II) положение 105 в вариабельном домене тяжелой цепи и положение 43 в вариабельном домене легкой цепи или
III) положение 101 в вариабельном домене тяжелой цепи и положение 100 в вариабельном домене легкой цепи (во всех случаях нумерация согласно EU-индексу Кэбота).
В WO 2010/115589 описано также, что СН3-домены указанного полноразмерного антитела, предлагаемого в изобретении, можно изменять с использованием технологии «knob-into-holes» (взаимодействие по типу «выступ-во-впадину»), которая описана подробно с помощью нескольких примеров, например, в WO 96/027011, у Ridgway J.B. и др., Protein Eng 9, 1996, сс. 617-621 и Merchant A.M. и др., Nat Biotechnol 16, 1998, сс. 677-681.
Таким образом, в некоторых вариантах осуществления изобретения указанное трехвалентное биспецифическое антитело характеризуется также следующим: СН3-домен одной из тяжелых цепей полноразмерного антитела и СН3-домен другой тяжелой цепи полноразмерного антитела, которые оба встречаются на поверхности раздела, которая содержит исходную поверхность раздела между СН3-доменами антитела; где указанная поверхность раздела изменена для того, чтобы способствовать образованию трехвалентного биспецифического антитела, где изменение отличается тем, что:
а) СН3-домен одной тяжелой цепи изменен таким образом, что в исходной поверхности раздела СН3-домен одной тяжелой цепи, который встречается в исходной поверхности раздела СН3-домена другой тяжелой цепи в трехвалентном биспецифическом антителе,
аминокислотный остаток заменен аминокислотным остатком, который имеет больший объем боковой цепи, что приводит к образованию выпуклости на поверхности раздела СН3-домена первой тяжелой цепи, которая может помещаться в полость в поверхности раздела СН3-домена другой тяжелой цепи и
б) СН3-домен другой тяжелой цепи изменен таким образом, что в исходной поверхности раздела второго СН3-домена, который встречается в исходной поверхностью раздела первого СН3-домена в трехвалентном биспецифическом антителе, аминокислотный остаток заменен аминокислотным остатком, который имеет меньший объем боковой цепи, что приводит к образованию полости в поверхности раздела второго СН3-домена, в которую может помещаться выпуклость на поверхности раздела первого СН3-домена.
Указанный аминокислотный остаток, имеющий больший объем боковой цепи, необязательно можно выбирать из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W). Указанный аминокислотный остаток, имеющий меньший объем боковой цепи, можно необязательно выбирать из группы, состоящей из аланина (А), серина (S), треонина (Т), валина (V).
Необязательно в некоторых вариантах осуществления изобретения оба СН3-домена дополнительно изменены путем интродукции цистеина (С) в качестве аминокислоты в соответствующих положениях каждого СН3-домена, так, чтобы мог образоваться дисульфидный мостик между обоими СН3-доменами.
Вышеизложенные и другие детали, касающиеся формата биспецифического трехвалентного антитела, описанные в WO 2010/115589 А1, можно применять в настоящем изобретении.
В контексте настоящего описании понятие «полноразмерное антитело» означает антитело, состоящее из двух «тяжелых цепей полноразмерного антитела» и двух «легких цепей полноразмерного антитела». «Тяжелая цепь полноразмерного антитела» может представлять собой полипептид, состоящий в направлении от N-конца к С-концу из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 тяжелой цепи антитела (СН1), шарнирной области антитела (HR), константного домена 2 тяжелой цепи антитела (СН2) и константного домена 3 тяжелой цепи антитела (CH3), что сокращенно обозначают как VH-CH1-HR-CH2-CH3; и необязательно константного домена 4 тяжелой цепи антитела (СН4) в случае антитела подкласса IgE. Предпочтительно «тяжелая цепь полноразмерного антитела» представляет собой полипептид, состоящий в направлении от N-конца к С-концу из VH, CH1, HR, СН2 и CH3. Не подразумевается, что образование cross-Mab исключено из определения «полноразмерный» - поэтому тяжелая цепь может иметь обмен VH-домена на VL-домен или обмен CH1-домена на CL-домен. «Легкая цепь полноразмерного антитела» может представлять собой полипептид, состоящий в направлении от N-конца к С-концу из вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL), что сокращенного обозначают как VL-CL. Альтернативно этому, в случае cross-Mab, может иметь место обмен VL-домена на VH-домен или обмен CL-домена на CH1-домен. Константный домен легкой цепи антитела (CL) может относиться к κ- (каппа) или γ-(лямбда)-типу. Две цепи полноразмерного антитела сцеплены друг с другом с помощью межполипептидных дисульфидных связей между CL-доменом и CH1-доменом и между шарнирными областями тяжелых цепей полноразмерного антитела. Примерами типичных полноразмерных антител являются встречающиеся в естественных условиях антитела типа IgG (например, IgGl и IgG2), IgM, IgA, IgD и IgE.) Полноразмерные антитела, предлагаемые в изобретении, могут иметь происхождение из одного вида, например, человека, или они могут представлять собой химерные или гуманизированные антитела. Полноразмерные антитела, представленные в настоящем описании, содержат два антигенсвязывающих сайта, каждый из которых образован парой VH и VL. С-конец тяжелой или легкой цепи указанного полноразмерного антитела означает последнюю аминокислоту на С-конце указанной тяжелой или легкой цепи.
N-конец вариабельного домена тяжелой цепи антитела (VH) в полипептиде, указанном в подпункте б), и вариабельного домена легкой цепи антитела (VL) в полипептиде, указанном в подпункте в), означает последнюю аминокислоту на N-конце VH- или VL-домена.
В любых описанных выше форматах первый антиген может представлять собой опухолеассоциированный антиген, а второй антиген может представлять собой Pb-DOTAM, (но в некоторых вариантах осуществления изобретения может иметь место обратная ситуация).
В любых описанных выше форматах правильной сборке гетеродимеров тяжелых цепей может способствовать применение модификаций последовательности тяжелой цепи. В одном из вариантов осуществления изобретения применяют технологию «knob-into-hole». Поверхности раздела двух СН3-доменов можно изменять для повышения гетеродимеризации обеих тяжелых цепей, которые содержат два указанных СН3-домена. Каждый из двух СН3-доменов (двух тяжелых цепей) может иметь «выступ», а второй -«впадину». Например, один содержит так называемые «приводящие к образованию выступа мутации» (T366W и необязательно одну из S354C или Y349C, предпочтительно S354C), а другой содержит так называемые «приводящие к образованию впадины мутации» (T366S, L368A и Y407V и необязательно Y349C или S354C, предпочтительно Y349C) (см., например, Carter Р. и др., Immunotechnol. 2, 1996, с. 73) согласно нумерации на основе EU-индекса.
В дополнительном или альтернативном варианте можно использовать интродукцию дисульфидного мостика для стабилизации гетеродимеров (Merchant A.M. и др., Nature Biotech 16, 1998, сс. 677-681; Atwell S. и др., J. Mol. Biol. 270, 1997, cc. 26-35) и повышения выхода. Примеры включают интродукцию дисульфид ной связи между следующими положениями:
I) положение 44 в вариабельном домене тяжелой цепи и положение 100 в вариабельном домене легкой цепи,
II) положение 105 в вариабельном домене тяжелой цепи и положение 43 в вариабельном домене легкой цепи или
III) положение 101 в вариабельном домене тяжелой цепи и положение 100 в вариабельном домене легкой цепи (во всех случаях нумерация согласно EU-индексу Кэбота).
Модификации зарядов
Мультиспецифические антитела, предлагаемые в изобретении, могут содержать аминокислотные замены в Fab-молекулах, входящих в их состав, которые являются особенно эффективными в отношении снижения ошибочного спаривания легких цепей с несоответствующими им тяжелыми цепями (побочные продукты бенс-джонсовского типа), которое может иметь место при получении на основе Fab би-/мультиспецифических антигенсвязывающих молекул с VH/VL-обменом в одном (или в нескольких, в случае молекул, содержащих более двух антигенсвязывающих Fab-молекул) из их связывающих плечей (см. также публикацию РСТ WO 2015/150447, прежде всего представленные в ней примеры, содержание которой полностью включено в настоящее описание в качестве ссылки). Относительное содержание требуемых мультиспецифических антител по сравнению с нежелательными побочными продуктами, прежде всего побочными продуктами бенс-джонсовского типа, встречающимися в одном из их связывающих плечей, можно повышать путем интродукции заряженных аминокислот с противоположными зарядами в специфические аминокислотные положения в СН1- и CL-доменах Fab-молекулы (иногда в контексте настоящего описания обозначают как «модификации зарядов»).
Таким образом, в некоторых вариантах осуществления изобретения антитела, предлагаемые в настоящем изобретении, которые содержат Fab-молекулы, содержат по меньшей мере один Fab с константным доменом тяжелой цепи СН1, который содержит модификации зарядов, указанные в настоящем описании, и константный домен легкой цепи CL, который содержит модификации зарядов, указанные в настоящем описании.
Модификации зарядов осуществляют либо в канонической(их) Fab-молекуле(ах), которые содержатся в антителах, предлагаемых в настоящем изобретении (например, которые представлены на фиг 37: Р1АЕ1766, Р1АЕ1767 Р1АЕ1768, Р1АЕ1769), либо в кроссовер-ЕаЬ-молекуле(ах), которая(ые) содержится(атся) в антителах, предлагаемых в настоящем изобретении (но не в обеих молекулах). В конкретных вариантах осуществления изобретения модификации зарядов осуществляют либо в канонической(их) Fab- молекуле(ах), которая(ые) содержится(атся) в антителах, предлагаемых в настоящем изобретении (которая(ые) в некоторых вариантах осуществления изобретения специфически связывается(ются) с антигеном клетки-мишени).
Таким образом, в некоторых вариантах осуществления изобретения антитела, предлагаемые в настоящем изобретении, содержат а) первый антигенсвязывающий фрагмент, связывающийся с первым антигеном (например, опухолеассоциированным антигеном), и б) второй связывающий фрагмент, который связывается со вторым антигеном (например, DOTAM-Pb), где первый и второй антигенсвязывающие фрагменты биспецифической антигенсвязывающей молекулы оба представляют собой Fab-молекулы, и один из антигенсвязывающих фрагментов (в некоторых вариантах осуществления изобретения предпочтительно второй антигенсвязывающий фрагмент) представляет собой cross-Fab-фрагмент, одна из Fab-молекул содержит СН1-домен, который содержит указанные в настоящем описании модификации зарядов, и CL-домен, который содержит указанные в настоящем описании модификации зарядов. Может оказаться предпочтительным, если Fab, содержащий модификации зарядов, представляет собой канонический (без кроссовера доменов) Fab, например, в некоторых вариантах осуществления изобретения представляет собой антигенсвязывающий фрагмент, который связывается с первым антигеном.
Антитела, предлагаемые в изобретении, могут дополнительно содержать третью Fab-молекулу, которая специфически связывается с первым антигеном. В конкретных вариантах осуществления изобретения указанная третья Fab-молекула идентична первой Fab-молекуле, указанной в подпункте а). В этих вариантах осуществления изобретения аминокислотные замены, которые описаны в представленных ниже вариантах осуществления изобретения (модификации зарядов), можно осуществлять в константном домене CL и константном домене СН1 и в первой Fab-молекуле, и в третьей Fab-молекуле. Альтернативно этому, аминокислотные замены, которые описаны в представленных ниже вариантах осуществления изобретения, можно осуществлять в константном домене CL и константном домене СН1 второй Fab-молекулы, указанной в подпункте б), но не в константном домене CL и константном домене СН1 первой Fab-молекулы и третьей Fab-молекулы.
В некоторых вариантах осуществления изобретения в Fab-молекуле, содержащей константный домен легкой цепи CL, который содержит модификации зарядов, и константный домен тяжелой цепи СН1, который содержит модификации зарядов, модификации зарядов в константном домене легкой цепи CL находятся в положении 124 и необязательно в положении 123 (нумерация согласно Кэботу), а модификации зарядов в константном домене тяжелой цепи СН1 находятся в положении 147 и/или 213 (нумерация согласно Кэботу).
В некоторых вариантах осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена независимо на лизин (K), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном предпочтительном варианте осуществления изобретения независимо на лизин (K)), и в константном домене тяжелой цепи СН1 аминокислота в положении 147 и/или аминокислота в положении 213 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В другом варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена независимо на лизин (K), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном предпочтительном варианте осуществления изобретения независимо на лизин (K) или аргинин (R)), и в константном домене тяжелой цепи СН1 аминокислота в положении 147 или аминокислота в положении 213 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В другом варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена независимо на лизин (K) или аргинин (R) (нумерация согласно Кэботу) (в одном предпочтительном варианте осуществления изобретения независимо на лизин (K) или аргинин (R)), и в константном домене тяжелой цепи СН1 аминокислота в положении 213 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В следующем варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена независимо на лизин (K), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном предпочтительном варианте осуществления изобретения независимо на лизин (K) или аргинин (R)) (нумерация согласно Кэботу), и в константном домене тяжелой цепи СН1 аминокислота в положении 147 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В следующем варианте осуществления изобретения в константном домене легкой цепи аминокислота в положении 124 заменена независимо на лизин (K), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном предпочтительном варианте осуществления изобретения независимо на лизин (K) или аргинин (R)), и аминокислота в положении 123 заменена независимо на лизин (K), аргинин (R) или гистидин (Н) (нумерация согласно Кэботу) (в одном предпочтительном варианте осуществления изобретения независимо на лизин (K) или аргинин (R)),
и в константном домене тяжелой цепи СН1 аминокислота в положении 147 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота) и аминокислота в положении 213 заменена независимо на глутаминовую кислоту (Е) или аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В другом варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена на лизин (K) (нумерация согласно Кэботу) и аминокислота в положении 123 заменена на аргинин (R) (нумерация согласно Кэботу), и в константном домене тяжелой цепи СН1 аминокислота в положении 147 заменена на глутаминовую кислоту (Е) (нумерация согласно EU-индексу Кэбота) и аминокислота в положении 213 заменена на глутаминовую кислоту (Е) (нумерация согласно EU-индексу Кэбота).
В следующем варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена на лизин (K) (нумерация согласно Кэботу) и аминокислота в положении 123 заменена на лизин (K) (нумерация согласно Кэботу), и в константном домене тяжелой цепи СН1 аминокислота в положении 147 заменена на глутаминовую кислоту (Е) (нумерация согласно EU-индексу Кэбота) и аминокислота в положении 213 заменена на глутаминовую кислоту (Е) (нумерация согласно EU-индексу Кэбота).
В следующем варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена на лизин (K) (нумерация согласно Кэботу) и аминокислота в положении 123 заменена на аргинин (R) (нумерация согласно Кэботу),
и в константном домене тяжелой цепи СН1 аминокислота в положении 147 заменена на глутаминовую кислоту (Е) (нумерация согласно EU-индексу Кэбота) и аминокислота в положении 213 заменена на аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В следующем варианте осуществления изобретения в константном домене легкой цепи CL аминокислота в положении 124 заменена на лизин (K) (нумерация согласно Кэботу) и аминокислота в положении 123 заменена на лизин (K) (нумерация согласно Кэботу),
и в константном домене тяжелой цепи СН1 аминокислота в положении 147 заменена на глутаминовую кислоту (Е) (нумерация согласно EU-индексу Кэбота) и аминокислота в положении 213 заменена на аспарагиновую кислоту (D) (нумерация согласно EU-индексу Кэбота).
В одном из вариантов осуществления изобретения антитело содержит первую тяжелую цепь и первую легкую цепь, которые специфически связываются с первым антигеном, и вторую тяжелую цепь и вторую легкую цепь, которые специфически связываются со вторым антигеном, в котором а) константный домен CL первой легкой цепи и константный домен СН1 первой тяжелой цепи содержат варианты с заменами зарядов, указанные в настоящем описании; и
б) константный домен легкой цепи CL и константный домен СН1 тяжелой цепи второй легкой цепи и второй тяжелой цепи заменены друг на друга (образуя тем самым cross-Fab). Пример такого расположения показан для Р1АЕ1768.
В другом варианте осуществления изобретения антитело содержит
полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей и двух легких цепей, где константные домены СН1 двух тяжелых цепей и константные домены CL двух легких цепей полноразмерного антитела содержат модификации зарядов, указанные в настоящем описании; и
scFab, содержащий VH- и CH1-домены, сцепленные через полипептидный линкер с VL- и CL-доменами (VH-СН1-линкер-VL-СЬ), где scFab слит с N-концом одной из тяжелых цепей, и где scFab образует антигенсвязывающий сайт для второго антигена. Примером такого расположения является Р1АЕ1770.
В другом варианте осуществления изобретения мультиспецифическое антитело содержит полноразмерное антитело, которое содержит антигенсвязывающий сайт для первого антигена (например, который может быть двухвалентным в отношении первого антигена), где константные домены СН1 двух тяжелых цепей и константные домены CL двух легких цепей полноразмерного антитело содержат модификации зарядов, указанные в настоящем описании; и
в котором С-конец одной из тяжелых цепей (например, С-конец ее СН3-домена) сцеплен через полипептидный линкер с первым полипептидом и в котором первый полипептид связан со вторым полипептидом с образованием cross-Fab, который содержит связывающий сайт для второго антигена.
Первый полипептид может содержать N-концевой VL-домен и С-концевой СН1-домен. Таким образом, тяжелая цепь, имеющая слияние, может содержать в направлении от N-конца к С-концу VH-СН1-шарнир-СН2-СН3-линкер-VL-СН1. Легкая цепь может содержать VH-CL. Примером такого расположения является Р1АЕ1767.
В другом варианте осуществления изобретения антитело может представлять собой биспецифическое антитело, которое содержит:
а) полноразмерное антитело, специфически связывающее первый антиген и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, в котором СН1-домены тяжелых цепей и CL-домены легких цепей содержат модификации зарядов, указанные в настоящем описании;
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH); или
II) вариабельного домена тяжелой цепи антитела (VH) и константного домена тяжелой цепи антитела (СН1); или
III) вариабельного домена тяжелой цепи антитела (VH) и константного домена легкой цепи антитела (CL),
где указаны полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL); или
II) вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL); или
III) вариабельного домена легкой цепи антитела (VL) и константного домена тяжелой цепи антитела (СН1),
где указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
и где вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для второго антигена. Примерами такого расположения являются PRIT-213 и р1АЕ1766.
В некоторых вариантах осуществления изобретения антитело, предлагаемое в изобретении, содержит а) первый антигенсвязывающий фрагмент, который связывается с первым антигеном, б) второй антигенсвязывающий фрагмент, который связывается со вторым антигеном, и в) третий антигенсвязывающий фрагмент, который связывается с первым антигеном, где первый, второй и
третий антигенсвязывающие фрагменты антитела все представляют собой Fab-молекулы, и в одном из антигенсвязывающих фрагментов (прежде всего во втором антигенсвязывающем фрагменте) вариабельные домен VL и VH легкой цепи Fab и тяжелой цепи Fab соответственно заменены друг на друга, где
I) аминокислотные замены, указанные в представленных выше вариантах осуществления изобретения, осуществлены в константном домене CL и константном домене СН1 и первой Fab-молекулы, и третьей Fab-молекулы, но не в константном домене CL и константном домене СН1 второй Fab-молекулы, указанной в подпункте б); или
II) аминокислотные замены, указанные в представленных выше вариантах осуществления изобретения, осуществлены в константном домене CL и константном домене СН1 второй Fab-молекулы, указанной в подпункте б), но не в константном домене CL и константном домене СН1 первой Fab-молекулы и третьей Fab-молекулы. В некоторых вариантах осуществления изобретения модификации зарядов присутствуют в каноническом (не имеющем обменов) Fab: так, например, если второй антигенсвязывающий фрагмент представляет собой cross-Fab, то предпочтительным является вариант (I).
В одном из конкретных вариантов осуществления изобретения многовалентное антитело, предлагаемое в изобретении, содержит
полноразмерное антитело, содержащее первая и вторую тяжелые цепи антитела и первую и вторую легкие цепи антитела, в котором первая тяжелая цепь и первая легкая цепь объединены с образованием Fab, который содержит антигенсвязывающий сайт для первого антигена (например,
опухолеспецифического антигена, например, CEA), и в котором вторая тяжелая цепь и вторая легкая цепь объединены с образованием cross-Fab, который содержит антигенсвязывающий сайт для второго антигена (например, хелата Pb-DOTAM) (например, вторая тяжелая цепь имеет VL-домен вместо VH-домена, а вторая легкая цепь имеет VH-домен вместо VL-домена);
и в котором либо первая, либо вторая тяжелая цепь антитела слита через линкер с полипептидом, содержащим СН1- и VH-домены, и указанный первый полипептид объединен со вторым полипептидом, содержащим CL и VL, в результате первый и второй полипептиды объединены с образованием Fab, который содержит антигенсвязывающий сайт для первого антигена, в котором
CH1-домен первой тяжелой цепи и CL-домен первой легкой цепи содержат модификации зарядов, указанные в настоящем описании.
CH1-домен первого полипептида и CL-домен второго полипептида также могут содержать модификации зарядов, указанные в настоящем описании.
Слияние может происходить на N-конце одной из тяжелых цепей полноразмерного антитела, необязательно второй тяжелой цепи. Примером такого расположения является Р1АЕ1769.
Мультиспецифические антитела, связывающиеся с Pb-DOTAM и CEA
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если антитела, предлагаемые в настоящем изобретении, представляют собой мультиспецифические, например, биспецифические антитела, которые связываются и с Pb-DOTAM, и с СЕА. Так, они содержат антигенсвязывающий сайт для хелата Pb-DOTAM и антигенсвязывающий сайт для СЕА. В указанных вариантах осуществления изобретения антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, может соответствовать любому из вариантов осуществления изобретения, указанных в настоящем описании. Формат может представлять собой любой из форматов, указанных в настоящем описании.
Необязательно связывание антигенсвязывающего сайта с СЕА может характеризоваться величиной Kd, составляющей 1 нМ или менее, 500 пМ или менее, 200 пМ или менее или 100 пМ или менее, при одновалентном связывании.
Необязательно антигенсвязывающий сайт, который связывается с СЕА, может содержать по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 11; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 14; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 15; и (е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 16.
Необязательно антигенсвязывающий сайт, который связывается с СЕА, может содержать по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VH, выбранные из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 11; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12; и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13. В одном из вариантов осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13. В другом варианте осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13, и CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 16. В следующем варианте осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13, CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 16, и CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12. В другом варианте осуществления изобретения антитело содержит (a) CDR-Н1, который содержит аминокислотную последовательность SEQ ID NO: 11; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12; и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13.
Необязательно антигенсвязывающий сайт, который связывается с СЕА, содержит по меньшей мере одну, по меньшей мере две или все CDR-последовательности VL, выбранные из (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 14; (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 15; и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 16. В одном из вариантов осуществления изобретения антитело содержит (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 13, (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 14; и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 15.
Необязательно антигенсвязывающий сайт, который связывается с СЕА, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VH, выбранные из (I) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 11, (II) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12, и (III) CDR-H3, который содержит аминокислотную последовательность, выбранную из SEQ ID NO: 13; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VL, выбранные из (I) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 14, (II) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 15, и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 16.
В другом объекте изобретения антигенсвязывающий сайт, который связывается с СЕА, содержит (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 11; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 14; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 15; и (е) CDR-L3, который содержит аминокислотную последовательность, выбранную из SEQ ID NO: 16.
В одном из указанных выше вариантов осуществления изобретения мультиспецифическое антитело может быть гуманизировано. В одном из вариантов осуществления изобретения анти-СЕА антигенсвязывающий сайт содержит CDR, представленные в любом из вышеуказанных вариантов осуществления изобретения, и дополнительно содержит человеческий акцепторный каркасный участок, например, каркасный участок человеческого иммуноглобулина или человеческий консенсусный каркасный участок.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с СЕА, содержит последовательность вариабельного домена тяжелой цепи (VH), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 17. В некоторых вариантах осуществления изобретения VH-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антигенсвязывающий сайт, содержащий указанную последовательность, сохраняет способность связываться с СЕА, предпочтительно с указанной выше аффинностью. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 17. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антигенсвязывающий сайт, который связывается с СЕА, содержит VH- последовательность SEQ ID NO: 17, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три CDR, выбранных из: (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 11, (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 12, и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 13.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с СЕА, содержит последовательность вариабельного домена легкой цепи (VL), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 18. В некоторых вариантах осуществления изобретения VL-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антигенсвязывающий сайт, содержащий указанную последовательность, сохраняет способность связываться с СЕА, предпочтительно с указанной выше аффинностью. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 18. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антигенсвязывающий сайт, который связывается с СЕА, содержит VL-последовательность SEQ ID NO: 18, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три CDR, выбранных из: (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 14, (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 15, и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 16.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с СЕА, содержит VH, указанный в любом из представленных выше вариантов осуществления изобретения, и VL, указанный в любом из представленных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит VH- и VL-последовательности SEQ ID NO: 17 и SEQ ID NO: 18 соответственно, включая пост-трансляционные модификации указанных последовательностей.
В некоторых вариантах осуществления изобретения мультиспецифическое антитело может связываться с тем же эпитопом СЕА, что и антитело PRIT-0213 или PRIT-0214, представленное в настоящем описании.
Мультиспецифические антитела, связывающиеся с Pb-DOTAM и ERBB2
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если антитела, предлагаемые в настоящем изобретении, представляют собой мультиспецифические, например, биспецифические антитела, которые связываются и с Pb-DOTAM, и с ERBB2. Так, они содержат антигенсвязывающий сайт для хелата Pb-DOTAM и антигенсвязывающий сайт для ERBB2. В указанных вариантах осуществления изобретения антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, может соответствовать любому из вариантов осуществления изобретения, указанных в настоящем описании. Формат может представлять собой любой из форматов, указанных в настоящем описании.
Необязательно связывание антигенсвязывающего сайта с ERBB2 может характеризоваться величиной Kd, составляющей 1 нМ или менее, 500 пМ или менее, 200 пМ или менее или 100 пМ или менее, при одновалентном связывании.
Необязательно антигенсвязывающий сайт, который связывается с ERBB2, может содержать по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32; и (е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33.
Необязательно антигенсвязывающий сайт, который связывается с ERBB2, может содержать по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VH, выбранные из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29; и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30. В одном из вариантов осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30. В другом варианте осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30, и CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33. В следующем варианте осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30, CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33, и CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29. В другом варианте осуществления изобретения антитело содержит (a) CDR-Н1, который содержит аминокислотную последовательность SEQ ID NO: 28; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29; и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30.
Необязательно антигенсвязывающий сайт, который связывается с ERBB2, содержит по меньшей мере одну, по меньшей мере две или все CDR-последовательности VL, выбранные из (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31; (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32; и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33. В одном из вариантов осуществления изобретения антитело содержит (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31, (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32; и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33.
Необязательно антигенсвязывающий сайт, который связывается с ERBB2, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VH, выбранные из (I) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28, (II) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29, и (III) CDR-H3, который содержит аминокислотную последовательность, выбранную из SEQ ID NO: 30; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VL, выбранные из (I) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31, (II) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32, и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33.
В другом объекте изобретения антигенсвязывающий сайт, который связывается с ERBB2 содержит (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32; и (е) CDR-L3, который содержит аминокислотную последовательность, выбранную из SEQ ID NO: 33.
В одном из указанных выше вариантов осуществления изобретения мультиспецифическое антитело может быть гуманизировано. В одном из вариантов осуществления изобретения анти-ERBB2 антигенсвязывающий сайт содержит CDR, представленные в любом из вышеуказанных вариантов осуществления изобретения, и дополнительно содержит человеческий акцепторный каркасный участок, например, каркасный участок человеческого иммуноглобулина или человеческий консенсусный каркасный участок.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с ERBB2, содержит последовательность вариабельного домена тяжелой цепи (VH), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 34. В некоторых вариантах осуществления изобретения VH-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антигенсвязывающий сайт, содержащий указанную последовательность, сохраняет способность связываться с ERBB2, предпочтительно с указанной выше аффинностью. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 34. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антигенсвязывающий сайт, который связывается с ERBB2, содержит VH-последовательность SEQ ID NO: 34, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три CDR, выбранных из: (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28, (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29, и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с ERBB2, содержит последовательность вариабельного домена легкой цепи (VL), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 35. В некоторых вариантах осуществления изобретения VL-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антигенсвязывающий сайт, содержащий указанную последовательность, сохраняет способность связываться с ERBB2, предпочтительно с указанной выше аффинностью. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 35. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR).
Необязательно антигенсвязывающий сайт, который связывается с ERBB2, содержит VL-последовательность SEQ ID NO: 35, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три CDR, выбранных из: (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31, (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32, и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с ERBB2, содержит VH, указанный в любом из представленных выше вариантов осуществления изобретения, и VL, указанный в любом из представленных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит VH- и VL-последовательности SEQ ID NO: 34 и SEQ ID NO: 35 соответственно, включая пост-трансляционные модификации указанных последовательностей.
В некоторых вариантах осуществления изобретения мультиспецифическое антитело может связываться с тем же эпитопом ERBB2, что и антитело P1AD9827, представленное в настоящем описании.
Мультиспецифические антитела, связывающиеся с Pb-DOTAM и CD20
В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если антитела, предлагаемые в настоящем изобретении, представляют собой мультиспецифические, например, биспецифические антитела, которые связываются и с Pb-DOTAM, и с CD20. Так, они содержат антигенсвязывающий сайт для хелата Pb-DOTAM и антигенсвязывающий сайт для CD20. В указанных вариантах осуществления изобретения антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, может соответствовать любому из вариантов осуществления изобретения, указанных в настоящем описании. Формат может представлять собой любой из форматов, указанных в настоящем описании.
Необязательно связывание антигенсвязывающего сайта с CD20 может характеризоваться величиной Kd, составляющей 1 нМ или менее, 500 пМ или менее, 200 пМ или менее или 100 пМ или менее, при одновалентном связывании.
Необязательно антигенсвязывающий сайт, который связывается с CD20, может содержать по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43; и (е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44.
Необязательно антигенсвязывающий сайт, который связывается с CD20, может содержать по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VH, выбранные из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40; и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41. В одном из вариантов осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41. В другом варианте осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41, и CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44. В следующем варианте осуществления изобретения антитело содержит CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41, CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44, и CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40. В другом варианте осуществления изобретения антитело содержит (a) CDR-Н1, который содержит аминокислотную последовательность SEQ ID NO: 39; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40; и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41.
Необязательно антигенсвязывающий сайт, который связывается с CD20, содержит по меньшей мере одну, по меньшей мере две или все CDR-последовательности VL, выбранные из (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42; (6) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43; и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44. В одном из вариантов осуществления изобретения антитело содержит (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42, (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43; и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44.
Необязательно антигенсвязывающий сайт, который связывается с CD20, содержит (а) VH-домен, содержащий по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VH, выбранные из (I) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39, (II) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40, и (III) CDR-H3, который содержит аминокислотную последовательность, выбранную из SEQ ID NO: 41; и (б) VL-домен, содержащий по меньшей мере одну, по меньшей мере две или все три CDR-последовательности VL, выбранные из (I) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42, (II) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43, и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44.
В другом объекте изобретения антигенсвязывающий сайт, который связывается с CD20, содержит (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43; и (е) CDR-L3, который содержит аминокислотную последовательность, выбранную из SEQ ID NO: 44.
В одном из указанных выше вариантов осуществления изобретения мультиспецифическое антитело может быть гуманизировано. В одном из вариантов осуществления изобретения анти- CD20 антигенсвязывающий сайт содержит CDR, представленные в любом из вышеуказанных вариантов осуществления изобретения, и дополнительно содержит человеческий акцепторный каркасный участок, например, каркасный участок человеческого иммуноглобулина или человеческий консенсусный каркасный участок.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с CD20, содержит последовательность вариабельного домена тяжелой цепи (VH), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 45. В некоторых вариантах осуществления изобретения VH-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антигенсвязывающий сайт, содержащий указанную последовательность, сохраняет способность связываться с CD20, предпочтительно с указанной выше аффинностью. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 45. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антигенсвязывающий сайт, который связывается с CD20, содержит VH- последовательность SEQ ID NO: 45, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VH содержит один, два или три CDR, выбранных из: (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39, (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40, и (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с CD20, содержит последовательность вариабельного домена легкой цепи (VL), идентичную по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% аминокислотной последовательности SEQ ID NO: 46. В некоторых вариантах осуществления изобретения VL-последовательность, идентичная по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, содержит замены (например, консервативные замены), инсерции или делеции относительно референс-последовательности, но антигенсвязывающий сайт, содержащий указанную последовательность, сохраняет способность связываться с CD20, предпочтительно с указанной выше аффинностью. В некоторых вариантах осуществления изобретения в целом от 1 до 10 аминокислот заменены, встроены путем инсерции или удалены путем делеции в SEQ ID NO: 46. В некоторых вариантах осуществления изобретения замены, инсерции или делеции затрагивают области вне HVR (т.е. FR). Необязательно антигенсвязывающий сайт, который связывается с CD20, содержит VL-последовательность SEQ ID NO: 46, включая пост-трансляционные модификации указанной последовательности. В конкретном варианте осуществления изобретения VL содержит один, два или три CDR, выбранных из: (a) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42, (б) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43, и (в) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44.
В другом варианте осуществления изобретения антигенсвязывающий сайт, который связывается с CD20, содержит VH, указанный в любом из представленных выше вариантов осуществления изобретения, и VL, указанный в любом из представленных выше вариантов осуществления изобретения. В одном из вариантов осуществления изобретения антитело содержит VH- и VL-последовательности SEQ ID NO: 45 и SEQ ID NO: 46 соответственно, включая пост-трансляционные модификации указанных последовательностей.
В некоторых вариантах осуществления изобретения мультиспецифическое антитело может связываться с тем же эпитопом CD20, что и антитело P1AD9826, представленное в настоящем описании.
Варианты антител
Некоторые варианты осуществления изобретения относятся к вариантам аминокислотных последовательностей антител, представленных в настоящем описании. Например, может требоваться повышать аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела можно получать путем интродукции соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем синтеза пептидов. Указанные модификации включают, например, делеции и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечной конструкции можно использовать любую комбинацию делеций, инсерций и замен, при условии, что конечная конструкция обладает требуемыми характеристиками, например, способностью связываться с антигеном. Полученные путем замены, инсерций и делеций варианты Некоторыми вариантами осуществления изобретения являются варианты антител, которые имеют одну или несколько аминокислотных замен. Сайты, представляющие интерес для замещающего мутагенеза, включают HVR (CDR) и FR. Консервативные замены представлены ниже в таблице 1 под заголовком «предпочтительные замены». Более важные замены представлены ниже в таблице 1 под заголовком «приведенные в качестве примера замены», и они описаны ниже со ссылкой на классы боковых цепей аминокислот. Аминокислотные замены можно интродуцировать в представляющее интерес антитело и продукты подвергать скринингу в отношении требуемой активности, например, сохранения/повышения способности к связыванию антигена, пониженной иммуногенности или повышенной ADCC или CDC.
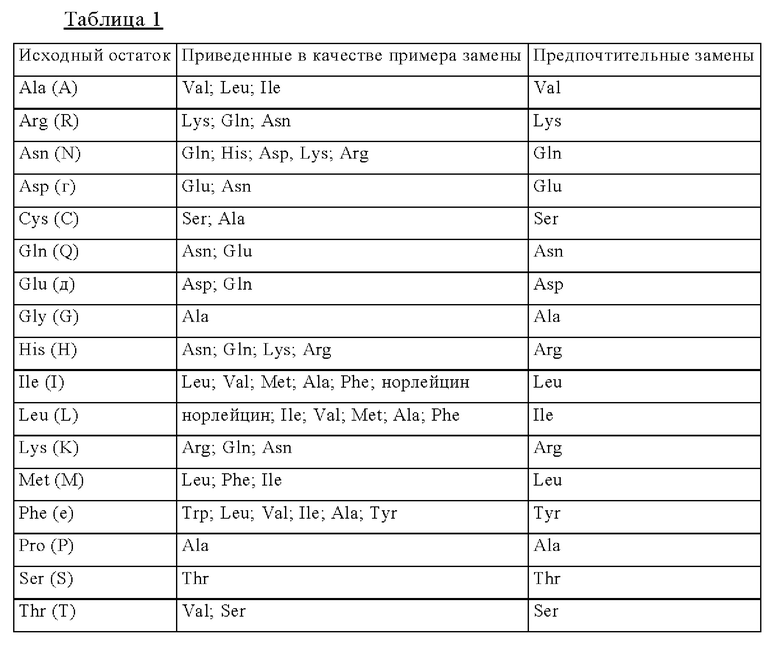

Аминокислоты можно группировать на основе общих свойств боковых цепей следующим образом:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, He;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gin;
(3) кислотные: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Тгр, Tyr, Phe. Под неконсервативными заменами подразумевают замену представителя одного из указанных классов на представителя из другого класса.
Один из типов полученного в результате замены варианта включает замену одного или нескольких остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный(ые) в результате вариант(ы), отобранный(ые) для дополнительного исследования, должен(ы) иметь модификации (например, улучшения) некоторых биологических свойств (например, повышенную аффинность, пониженную иммуногенность) относительно родительского антитела, и/или должен(ы) иметь практически сохраненные определенные биологические свойства родительского антитела. Примером полученного в результате замены варианта является антитело с созревшей аффинностью, которое можно создавать, например, с использованием технологий созревания аффинности на основе фагового дисплея, например, указанных в настоящем описании. В целом, метод состоит в следующем: один или несколько остатков HVR подвергают мутации и варианты антител экспонируют на фаге и подвергают скринингу в отношении конкретной биологической активности (например, аффинности связывания).
Изменения (например, замены) можно осуществлять в HVR, например, для повышения аффинности антитела. Указанные изменения можно делать в «горячих (активных) точках» в HVR, т.е. остатках, кодируемых кодонами, которые с высокой частотой подвергаются мутации в процессе соматического созревания (см., например, Chowdhury, Methods Mol. Biol. 207, 2008, cc. 179-196), и/или в остатках, которые контактируют с антигеном, с последующим тестированием варианта VH или VL в отношении аффинности связывания. Созревание аффинности путем создания и повторной селекции из вторичных библиотек описано, например, у Hoogenboom и др. в: Methods in Molecular Biology 178, 2002, cc. 1-37 (под ред. O'Brien и др., изд-во Human Press, Totowa, NJ, 2001). В некоторых вариантах осуществления процесса созревания аффинности интродуцируют разнообразие в гены вариабельной области, выбранные для созревания, с помощью любого из широкого разнообразия методов (например, ПЦР пониженной точности, перестановка цепи или сайтнаправленный мутагенез с использованием олигонуклеотидов). Затем создают вторичную библиотеку. Далее библиотеку подвергают скринингу для идентификации любых вариантов антител с требуемой аффинностью. Другой метод, применяемый для интродукции разнообразия, включает подходы, мишенью которых является HVR, при которых рандомизируют несколько остатков HVR (например, одновременно 4-6 остатков). Остатки HVR, участвующие в связывании антигена, можно специфически идентифицировать, например, используя аланин-сканирующий мутагенез или моделирование. Часто мишенями являются, прежде всего CDR-H3 и CDR-L3.
В некоторых вариантах осуществления изобретения замены, инсерций или делеций могут затрагивать один или несколько HVR, если указанные изменения не снижают существенно способность антитела связываться с антигеном. Например, в HVR могут быть сделаны консервативные изменения (например, консервативные замены, указанные в настоящем описании), которые не снижают существенно аффинность связывания. Указанные изменения, например, могут находиться вне принимающих участие в контактировании с антигеном остатков HVR. В некоторых вариантах осуществления изобретения в последовательностях вариантов VH и VL, указанных выше, каждый HVR либо является неизмененным, либо содержит не более одной, двух или трех аминокислотных замен.
Ценным методом идентификации остатков или областей антитела, которые можно подвергать мутагенезу, является так называемый «аланин-сканирующий мутагенез», описанный у Cunningham и Wells, Science 244, 1989, сс. 1081-1085. При осуществлении этого метода остаток или группу остатков-мишеней (например, заряженные остатки, такие как arg, asp, his, lys и glu) идентифицируют и заменяют на нейтральную или отрицательно заряженную аминокислоту (например, аланин или полиаланин) для решения вопроса о том, будет ли это влиять на взаимодействие антитела с антигеном. Дополнительные замены можно интродуцировать в те положения аминокислот, для которых продемонстрирована функциональная чувствительность к начальным заменам. В альтернативном или дополнительном варианте можно анализировать кристаллическую структуру комплекса антиген-антитело для идентификации точек контакта между антителом и антигеном. Указанные контактирующие остатки и соседние остатки можно рассматривать в качестве мишеней или исключать из рассмотрения в качестве кандидатов для замены. Варианты можно подвергать скринингу для решения вопроса о том, обладают ли они требуемыми свойствами.
Инсерции в аминокислотные последовательности включают амино- и/или карбоксиконцевые слияния, варьирующие по длине от одного остатка до полипептидов, содержащих сто или большее количество остатков, а также инсерции одного или нескольких аминокислотных остатков внутрь последовательности. Примером результата осуществления концевых инсерции является антитело с N-концевым метионильным остатком. Другие инсерционные варианты молекул антител включают слияние N- или С-концов антитела с ферментом (например, для ADEPT (антитело-направляемый фермент пролекарственной терапии) или полипептидом, который удлиняет время полужизни антитела в сыворотке.
Варианты гликозилирования
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, изменяют с целью повышения или снижения степени гликозилирования антитела. Добавление или делецию сайтов гликозилирования в антителе можно легко осуществлять путем такого изменения аминокислотной последовательности, которое позволяет создавать или удалять один или несколько сайтов гликозилирования.
Если антитело содержит Fc-область, то можно изменять присоединенный к ней углевод. Нативные антитела, которые продуцируются клетками млекопитающих, как правило, содержат разветвленный биантенный олигосахарид, который, как правило, присоединен с помощью N-связи к Asn297 СН2-домена Fc-области (см., например, Wright и др., TIBTECH 15, 1997, сс. 26-32). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стебле» биантенной олигосахаридной структуры. В некоторых вариантах осуществления изобретения модификации олигосахарида в антителе, предлагаемом в изобретении, можно осуществлять для создания вариантов антитела с определенными улучшенными свойствами. Одним из вариантов осуществления изобретения являются варианты антитела, имеющие углеводную структуру, в которой снижено содержание фукозы, присоединенной (прямо или косвенно) к Fc-области. Например, содержание фукозы в указанной Fc-области может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Содержание фукозы определяют путем расчета среднего содержания фукозы в сахарной цепи на Asn297 относительно суммы всех гликоструктур, присоединенных к Asn297 (например, комплексных, гибридных структур и структур с высоким содержанием маннозы), которое измеряют с помощью MALDI-TOF-масс-спектрометрии согласно методу, описанному, например, в WO 2008/077546. Asn297 обозначает остаток аспарагина, локализованный примерно в положении 297 в Fc-области (EU-нумерация остатков Fc-области); однако вследствие минорных вариаций последовательности антитела Asn297 может располагаться также в положении, находящемся на расстоянии примерно ±3 аминокислоты в прямом или обратном направлении от положения 297, т.е. между положениями 294 и 300. Указанные фукозилированные варианты могут обладать улучшенной ADCC-функцией (см., например, публикации патентов США №№ US 2003/0157108 (Presta L.); US 2004/0093621 (фирма Kyowa Hakko Kogyo Co., Ltd). Примерами публикаций, касающихся «дефукозилированных» вариантов антител или вариантов антител «с дефицитом фукозы» являются: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki и др., J. Mol. Biol. 336, 2004, cc. 1239-1249; Yamane-Ohnuki и др., Biotech. Bioeng. 87, 2004, cc. 614-622). Примерами клеточных линий, которые обладают способностью продуцировать дефукозилированные антитела, являются Lec13 СНО-клетки с дефицитом белкового фукозилирования (Ripka и др., Arch. Biochem. Biophys. 249, 1986, сс.533-545; заявка па патент США 2003/0157108 А1 (на имя Presta L.) и WO 2004/056312 А1 (на имя Adams и др., см., прежде всего, пример 11), и клеточные линии с «выключенным» геном, например, СНО клетки с «выключенным» геном альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki и др., Biotech. Bioeng. 87, 2004, сс. 614-622); Kanda и др., Biotechnol. Bioeng., 94(4), 2006, сс. 680-688; и WO 2003/085107).
Кроме того, описаны варианты антител с бисекционными олигосахаридами, например, в которых биантенный олигосахарид, присоединенный к Fc-области антитела, бисекционнируется с помощью GlcNAc. Указанные варианты антител могут отличаться пониженным фукозилированием и/или повышенной ADCC-функцией. Примеры указанных вариантов антител описаны, например, в WO 2003/011878 (на имя Jean-Mairet и др.); US №6602684 (на имя Umana и др.) и US 2005/0123546 (на имя Umana и др.). Предложены также варианты антител по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Указанные варианты антител могут обладать повышенной CDC-функцией. Указанные варианты антител описаны, например, в WO 1997/30087 (на имя Patel и др.); WO 1998/58964 (на имя Raju S.); и WO 1999/22764 (на имя Raju S.).
Может оказаться предпочтительным модифицировать антитело для снижения степени гликозилирования. В некоторых вариантах осуществления изобретения антитело может быть агликозилированным или дегликозилированным. Антитело может иметь замену в N297, например, N297D/A.
Варианты Fc-области
Согласно конкретным вариантам осуществления изобретения можно интродуцировать в Fc-область антитела, представленного в настоящем описании, одну или несколько аминокислотных модификаций, создавая тем самым вариант Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), включающую аминокислотную модификацию (например, замену) в одном или нескольких аминокислотных положениях.
Некоторые варианты осуществления изобретения относятся к варианту антитела, который обладает пониженный эффекторной функцией например, пониженной или элиминированной CDC, ADCC и/или FcγR-связывающей активностью. В некоторых объектах изобретения предложен вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его перспективным кандидатом для путей применения, для которых является важным время полужизни антитела in vivo, однако некоторые эффекторные функции (такие как комплементзависимая цитотоксичность (CDC) и антитело-обусловленная клеточнозависимая цитотоксичность (ADCC)) не являются необходимыми или являются вредными.
Можно осуществлять анализы цитотоксичности in vitro и/или in vivo для подтверждения снижения/истощения CDC- и/или ADCC-активности. Например, можно осуществлять анализы связывания Fc-рецептора (FcR) для подтверждения того, что у антитела отсутствует способность к связыванию с FcγR (и поэтому, вероятно, отсутствует ADCC-активность). Первичные клетки, опосредующие ADCC, т.е. NK-клетки, экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Данные об экспрессии FcR на гематопоэтических клетках обобщены в таблице 3 на с. 464 у Ravetch и Kinet, Annu. Rev. Immunol. 9, 1991, cc. 457-492. Примерами анализов in vitro ADCC-активности представляющей интерес молекулы являются (но, не ограничиваясь только ими) анализы, описанные в US №5500362 (см., например, Hellstrom I. и др., Proc. Nat'l Acad. Sci. USA 83, 1986, cc. 7059-7063 и Hellstrom I. и др., Proc. Nat'l Acad. Sci. USA 82, 1985, cc. 1499-1502); US №№5821337 (см. Bruggemann M. и др., J. Exp.Med. 166, 1987, cc. 1351-1361). В альтернативном варианте можно применять нерадиоактивные методы анализа (см., например, нерадиоактивный анализ цитотоксичности АСТ1™ на основе проточной цитометрии (фирма CellTechnology, Inc. Маунтин-Вью, шт. Калифорния); и нерадиоактивный анализ цитотоксичности CytoTox 96® (фирма Promega, Мэдисон, шт. Висконсин). Пригодными для таких анализов эффекторными клетками являются мононуклеарные клетки периферической крови (РВМС) и естественные клетки-киллеры (NK). В альтернативном или дополнительном варианте ADCC-активность представляющей интерес молекулы можно оценивать in vivo, например, на животной модели, например, как описано у Clynes и др., Proc.Nat'l. Acad. Sci. USA 95, 1998, cc. 652-656. Можно осуществлять также анализы связывания Clq для подтверждения того, что антитело не может связываться с Clq и поэтому не обладает CDC-активностью (см., например, описание ELISA для оценки связывания Clq и СЗс в WO 2006/029879 и WO 2005/100402). Для оценки активации комплемента можно осуществлять анализ CDC (см., например, Gazzano-Santoro и др., J. Immunol. Methods 202, 1996, сс.163-171; Cragg M.S.и M.J. Glennie, Blood 101, 2003, cc. 1045-1052; и Cragg, Blood 103, 2004, cc. 2738-2743). Определение FcRn-связывания и клиренса/времени полужизни in vivo можно осуществлять также с использованием методов, известных в данной области (см., например, Petkova S.B. и др., Int'l. Immunol. 18(12), 2006, сс.1759-1769); WO 2013/120929 Al).
Антитела с пониженной эффекторной функцией включают антитела с заменой одного или нескольких следующих остатков Fc-области, таких как 238, 265, 269, 270, 297, 327 и 329 (см., например, US №6737056), например, P329G. К указанным мутантам Fc относятся мутанты Fc с заменами в двух или большем количестве из следующих аминокислотных положений: 265, 269, 270, 297 и 327, включая так называемый мутант «DANA» Fc-области, несущий замену остатков 265 и 297 на аланин (US №7332581).
В некоторых объектах изобретения вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые уменьшают FcγR-связывание, например, замены в положениях 234 и 235 Fc-области (EU-нумерация остатков). В одном из объектов изобретения замены представляют собой L234A и L235A (LALA). В некоторых объектах изобретения вариант антитела дополнительно содержит D265A и/или P329G в Fc-области, имеющей происхождение из Fc-области человеческого IgGl. В одном из объектов изобретения замены представляют собой L234A, L235A и P329G (LALA-PG) в Fc-области, имеющей происхождение из Fc-области человеческого IgG1 (см., например, WO 2012/130831). В другом объекте изобретения замены представляют собой L234A, L235A и D265A (LALA-DA) в Fc-области, имеющей происхождение из Fc-области человеческого IgG1.
В других вариантах осуществления изобретения может оказаться возможным применять подтип IgG с пониженной эффекторной функцией, такой как IgG4 или IgG2.
Описаны некоторые варианты антител с повышенной или пониженной способностью связываться с FcR (см., например, US №6737056; WO 2004/056312 и Shields и др., J. Biol. Chem. 276, 2001, сс. 6591-6604).
В некоторых вариантах осуществления изобретения в Fc-области осуществляют изменения, которые приводят к измененной (т.е. либо повышенной, либо пониженной, предпочтительно пониженной) C1q-связывающей активности и/или комплементзависимой цитотоксичности (CDC), например, как описано в US №6194551, WO 99/51642 и у Idusogie и др., J. Immunol. 164, 2000, сс. 4178-4184.
В некоторых вариантах осуществления изобретения можно снижать связывание FcRn, например, для достижения более короткого времени полужизни. В других вариантах осуществления изобретения связывание FcRn может быть нормальным. Например, в некоторых вариантах осуществления изобретения нормальное связывание FcRn можно применять в способах, включающих очищающий агент.
В некоторых объектах изобретения вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые снижают FcRn-связывание, например, замены в положениях 253 и/или 310, и/или 435 Fc-области (EU-нумерация остатков). В некоторых объектах изобретения вариант антитела содержит Fc-область с аминокислотными заменами в положениях 253, 310 и 435. В одном из объектов изобретения замены представляют собой 1253А, Н310А и Н435А в Fc-области, имеющей происхождение из Fc-области человеческого IgG1 (см., например, Grevys А., и др., J. Immunol. 194, 2015, сс. 5497-5508).
В некоторых объектах изобретения вариант антитела содержит Fc-область с одной или несколькими аминокислотными заменами, которые понижают FcRn-связывание, например, замены в положениях 310 и/или 433, и/или 436 Fc-области (EU-нумерация остатков). В некоторых объектах изобретения вариант антитела содержит Fc-область с аминокислотными заменами в положениях 310, 433 и 436. В одном из объектов изобретения замены представляют собой Н310А, Н433А и Y436A в Fc-области, имеющей происхождение из Fc-области человеческого IgG1 (см., например, WO 2014/177460 А1). Например, в некоторых вариантах осуществления изобретения можно применять нормальное связывание FcRn.
Дополнительные примеры, касающиеся других вариантов Fc-области, описаны также у Duncan и Winter, Nature 322, 1988, сс. 738-740; в US №5648260; US №5624821 и в WO 1994/29351.
С-конец тяжелой цепи антитела, указанного в настоящем описании, может представлять собой полный С-конец, заканчивающийся аминокислотными остатками PGK. С-конец тяжелой цепи может представлять собой укороченный С-конец, из которого удалены один или два С-концевых аминокислотных остатка. В одном из предпочтительных объектов изобретения С-конец тяжелой цепи представляет собой укороченный С-конец, заканчивающийся PG.
В одном из варианте всех объектов изобретения, указанных в настоящем описании, антитело, содержащее тяжелую цепь, которая включает С-концевой СН3-домен, указанный в настоящем описании, содержит С-концевой остаток глицина (G446, нумерация аминокислотных положений согласно EU-индексу). Этот вариант все еще строго подпадает под понятие «полноразмерное антитело» или «полноразмерная тяжелая цепь», которое применяют в контексте настоящего описания.
Производные антител
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, можно дополнительно модифицировать таким образом, чтобы оно содержало известные в данной области и легкодоступные дополнительные небелковые фрагменты. Фрагменты, пригодные для дериватизации антитела, включают (но, не ограничиваясь только ими) водорастворимые полимеры. Примерами водорастворимых полимеров являются (но, не ограничиваясь только ими) полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (либо гомополимеры, либо статистические сополимеры) и декстран- или поли(н-винилпиролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры пропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликоль пропиональдегид может иметь преимущество при производстве благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, присоединенных к антителу, может варьироваться и, если присоединено более одного полимера, то они могут представлять собой одинаковые или различные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определять с учетом таких особенностей (но, не ограничиваясь только ими), как конкретные свойства или функции антитела, подлежащие усовершенствованию, предполагается ли применение производного антитела в терапии в определенных условиях, и т.д.
Другим вариантом осуществления изобретения являются конъюгаты антитела и небелкового фрагмента, которые можно избирательно нагревать, воздействуя на него излучением. В одном из вариантов осуществления изобретения небелковый фрагмент представляет собой углеродную нанотрубку (Kam и др., Proc. Natl. Acad. Sci. USA 102, 2005, cc. 11600-11605). Излучение может иметь любую длину волны и включает (но, не ограничиваясь только ими) длины волн, которые не повреждают обычные клетки, но которые нагревают небелковый фрагмент до температуры, при которой уничтожаются клетки, ближайшие к конъюгату: антитело-небелковый фрагмент.
Методы рекомбинации и композиции
Антитела можно получать, используя методы рекомбинации и композиции, например, описанные в US №4816567. Одним из вариантов осуществления изобретения является выделенная нуклеиновая кислота, кодирующая антитело. Указанная нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкие и/или тяжелые цепи антитела). Следующим вариантом осуществления изобретения является(ются) один или несколько векторов (например, экспрессионных векторов), который(е) содержит(ат) указанную нуклеиновую кислоту. Еще одним вариантом осуществления изобретения является клетка-хозяин, которая содержит указанную нуклеиновую кислоту. В одном из указанных вариантов осуществления изобретения клетка-хозяин содержит (например, в результате трансформации указанными векторами): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела.
В случае мультиспецифических антител предложены нуклеиновые кислоты, кодирующие каждый из компонентов тяжелых и легких цепей антитела конкретного формата. Предложен также вектор или набор векторов, содержащий(их) нуклеиновые кислоты.
В одном из вариантов осуществления изобретения клетка-хозяин представляет собой эукариотическую клетку, например, клетку яичника китайского хомячка (СНО) или лимфоидную клетку (например, Y0-, NS0-, Sp20-клетку). Одним из вариантов осуществления изобретения является способ получения антитела, предлагаемого в изобретении, включающий культивирование клетки-хозяина, содержащей нуклеиновую кислоту, которая кодирует антитело, описанное выше, в условиях, пригодных для экспрессии антитела, и необязательно выделение антитела из клетки-хозяина (или среды для культивирования клетки-хозяина).
Для рекомбинантного получения антитела нуклеиновую кислоту, кодирующую антитело, например, описанную выше, выделяют и встраивают в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Указанную нуклеиновую кислоту легко можно выделять и секвенировать с помощью общепринятых процедур (например, путем использования олигонуклеотидных зондов, которые обладают способностью специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела).
Приемлемые клетки-хозяева для клонирования или экспрессии кодирующих антитело векторов включают прокариотические или эукариотические клетки, представленные в настоящем описании. Например, антитела можно получать в бактериях, в частности, когда не требуется гликозилирование и связанная с Fc эффекторная функция. Сведения об экспрессии фрагментов антитела и полипептидов в бактериях см. например, в US №№5648237, 5789199 и 5840523 (см. также у Charlton в: Methods in Molecular Biology, под ред. Lo В.K.С, изд-во Humana Press, Totowa, NJ, т. 248, 2003, cc. 245-254 описание экспрессии фрагментов антител в Е. coli.) После экспрессии антитело можно выделять из пасты бактериальных клеток в виде растворимой фракции и можно дополнительно очищать.
Помимо прокариотических организмов в качестве хозяев, пригодных для клонирования или экспрессии векторов, которые кодируют антитела, можно использовать эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что позволяет получать антитело с частично или полностью человеческой схемой гликозилирования (см. Gerngross, Nat. Biotech. 22, 2004, cc. 1409-1414 и Li и др., Nat. Biotech. 24, 2006, cc. 210-215).
Клетки-хозяева, которые можно использовать для экспрессии гликозилированного антитела, получают также из многоклеточных организмов (беспозвоночных и позвоночных животных). Примерами клеток беспозвоночных являются клетки насекомых, а также можно применять клетки растений. Были выявлены многочисленные штаммы бакуловирусов и соответствующие пригодные для них в качестве хозяев клетки насекомых, прежде всего для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев можно применять также культуры растительных клеток (см., например, US №№5959177, 6040498, 6420548, 7125978 и 6417429 (описание технологии PLANTIBODIES™ для получения антител в трансгенных растениях).
В качестве хозяев можно применять также клетки позвоночных. Например, можно использовать клеточные линии млекопитающих, которые адаптированы к росту в суспензии. Другими примерами приемлемых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная с помощью ОВ40 (COS-7); линия клеток почки эмбриона человека (HEK 293-клетки или 293-клетки, описанные, например, у Graham и др., J. Gen. Virol., 36, 1977, с. 59); клетки почки детеныша хомяка (ВНК); клетки Сертоли мыши (ТМ4-клетки, описанные, например, у Mather, Biol. Reprod., 23, 1980, сс. 243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76,); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени бычьей крысы (BRL 3А); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, описанные, например, у Mather и др., Annals N.Y. Acad. Sci., 383, 1982, сс. 44-68); клетки MRC 5 и клетки FS4. Другими ценными линиями клеток-хозяев млекопитающих являются клетки яичника китайского хомячка (СНО), включая DHFR--СНО-клетки (Urlaub и др., Proc. Natl. Acad. Sci. USA, 77, 1980, с. 4216); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор конкретных линий клеток-хозяев млекопитающих, которые можно применять для производства антител, см., например, у Yazaki и Wu в: Methods in Molecular Biology под ред. В.К.С.Lo, изд-во Humana Press, Totowa, NJ, т. 248, 2003, сс.255-268.
Анализы
Антитела, представленные в настоящем описании, можно идентифицировать, осуществлять их скрининг или характеризовать их физические/химические свойства и/или виды биологической активности с помощью различных анализов, известных в данной области.
Анализы связывания и другие анализы
Согласно одному из объектов изобретения антитело, предлагаемое в изобретении, тестируют в отношении антигенсвязывающей активности с использованием, например, известных методов, таких как ELISA, Вестерн-блоттинг и т.д.
Согласно другому объекту изобретения можно применять анализы в условиях конкуренции для идентификации антитела, которое конкурирует, например, с PRIT-0213 или PRIT-0214, за связывание с Pb-DOTAM или СЕА. В некоторых вариантах осуществления изобретения указанное конкурирующее антитело связывается с тем же самым эпитопом (например, линейным или конформационным эпитопом), с которым связывается антитело PRIT-0213 или PRIT-0214. Подробное описание приведенных в качестве примера методов картирования эпитопа, с которым связывается антитело, представлено у Morris, «Epitope Mapping Protocols)), в: Methods in Molecular Biology, т. 66, изд-во Humana Press, Totowa, NJ, 1996.
В приведенном в качестве примера анализе в условиях конкуренции иммобилизованный антиген инкубируют в растворе, содержащем первое меченое антитело, которое связывается с антигеном (например, PRIT-0213 и PRIT-0214), и второе немеченое антитело, подлежащее тестированию в отношении способности конкурировать с первым антителом за связывание с антигеном. Второе антитело может присутствовать в супернатанте гибридомы. В качестве контроля иммобилизованный антиген инкубируют в растворе, содержащем первое меченое антитело, но не содержащем второе немеченое антитело. После инкубации в условиях, приемлемых для связывания первого антитела с антигеном, избыток несвязанного антитела удаляют и измеряют количество метки, ассоциированной с иммобилизованным антигеном. Если количество метки, ассоциированной с иммобилизованным антигеном, существенно снижается в тестируемом образце по сравнению с контрольным образцом, то это свидетельствует о том, что второе антитело конкурирует с первым антителом за связывание с антигеном (см., например, Harlow и Lane, Antibodies: A Laboratory Manual, глава 14, изд-во Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1988).
Аффинность антител
В некоторых вариантах осуществления изобретения антитело, представленное в настоящем описании, имеет величину константы диссоциации (Kd), составляющую 1 нМ или менее, 550 пМ или менее, 200 пМ или менее, 100 пМ или менее, 50 пМ или менее, 20 пМ или менее, 10 пМ или менее, 5 пМ или менее или 1 пМ или менее, что продемонстрировано в настоящем описании.
В одном из вариантов осуществления величину Kd измеряют с помощью анализа связывания несущего радиоактивную метку антигена (РИА). В одном из вариантов осуществления изобретения РИА осуществляют с использованием Fab-версии представляющего интерес антитела и его антигена. Например, измеряют в растворе аффинность связывания Fab с антигеном путем уравновешивания Fab минимальной концентрацией 125I-меченного антигена в присутствии полученных титрованием разведений немеченого антигена, осуществляя затем «захват» связанного антигена на сенсибилизированном антителом к Fab планшете (см., например, Chen и др., J. Mol Biol 293, 1999, сс. 865-881). Для обеспечения условий, необходимых для проведения анализа, многолуночные планшеты MICROTITER® (фирма Thermo Scientific) сенсибилизируют в течение ночи захватывающим антителом к Fab (фирма Cappel Labs) в концентрации 5 мкг/мл в 50 мМ карбонате натрия (рН 9,6) и затем блокируют с помощью 2% (мас./об.) бычьего сывороточного альбумина в ЗФР в течение 2-5 ч при комнатной температуре (примерно 23°С). В не адсорбирующем планшете (фирма Nunc, №269620) смешивают взятый в концентрации 100 или 26 пМ меченный с помощью 125I антиген с серийными разведениями представляющего интерес Fab (например, пригодного для оценки антитела к VEGF, Fab-12, см. Presta и др., Cancer Res. 57, 1997, сс.4593-4599). Затем представляющий интерес Fab инкубируют в течение ночи; однако инкубацию можно осуществлять в течение более продолжительного периода времени (например, в течение 65 ч) для того, чтобы гарантировать достижение равновесия. После этого смеси переносят в подготовленный для захвата планшет и инкубируют при комнатной температуре (например, в течение 1 ч). Затем раствор удаляют и планшет промывают восемь раз 0,1% полисорбатом 20 (TWEEN-20®) в ЗФР. После просушки планшетов в них добавляют по 150 мкл/лунку сцинтилляционной жидкости ((MICROSCINT-20™; фирма Packard) и осуществляют считывание планшетов с помощью гамма-счетчика TOPCOUNT™ (фирма Packard) в течение 10 мин. Для осуществления анализа связывания в условиях конкуренции выбирают концентрации каждого Fab, обеспечивающие уровень связывания, составляющий менее или равный 20% от максимального.
В другом варианте осуществления изобретения Kd измеряют методом поверхностного плазмонного резонанса BIACORE®. Например, анализ осуществляют с помощью устройства BIACORE®-2000 или BIACORE®-3000 (фирма GE Healthcare Inc., Пискатавей, шт. Нью-Джерси) при 25°С с использованием антигена, иммобилизованного на СМ5-чипах с уровнем иммобилизации, соответствующим ~10 единицам ответа (RU). В одном из вариантов осуществления изобретения биосенсорные чипы из карбоксиметилированного декстрана (СМ5, фирма GE Healthcare Inc.) активируют с помощью гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиимида (EDC) и N-гидроксисукцинимида (NHS) согласно инструкциям поставщика. Антиген разводят10 ацетатом натрия, рН 4,8 до концентрации 5 мкг/мл (~ 0,2 мкМ) перед инъекцией со скоростью потока 5 мкл/мин для достижения уровня иммобилизации, соответствующего примерно 10 единицам ответа (RU) сшитого белка. После инъекции антигена инъецируют 1 М этаноламин для блокады непрореагировавших групп. Для кинетических измерений инъецируют двукратные серийные разведения Fab (от 0,78 до 500 нМ) в ЗФР, дополненном 0,05% полисорбата 20 (TWEEN-20™) в качестве поверхностно-активного вещества (ЗФРТ), при 25°С со скоростью потока примерно 25 мкл/мин. Скорость реакции ассоциации (kon) и реакции диссоциации (koff) рассчитывают с использованием простой модели связывания Ленгмюра 1:1 (программное обеспечение Evaluation версия 3.2, BIACORE®) путем одновременной аппроксимации сенсограмм ассоциации и диссоциации. Константу равновесия реакции диссоциации (Kd) рассчитывают как соотношение koff/kon (см., например, Chen и др., J Mol Biol 293, 1999, cc. 865-881). Если скорость реакции ассоциации превышает 106М-1 с-1 при оценке с помощью описанного выше метода поверхностного плазмонного резонанса, то скорость реакции ассоциации можно определять с помощью метода гашения флуоресценции, который позволяет измерять повышение или снижение интенсивности излучения флуоресценции (длина волны возбуждения 295 нм; длина волны испускания 340 нм, полоса пропускания 16 нм) при 25°С применяемого в концентрации 20 нМ антитела к антигену (Fab-формат) в ЗФР, рН 7,2, в присутствии возрастающих концентраций антигена, осуществляя измерения с помощью спектрометра, такого как спектрофотометр, снабженный блоком для остановки потока (фирма Aviv Instruments), или спектрофотометр SLM-AMINCO™ серии 8000 (фирма ThermoSpectronic), при перемешивании содержимого кюветы.
В другом варианте осуществления изобретения Kd измеряют с помощью SET-анализа (титрование при равновесии в растворе). При осуществлении указанного анализа тестируемые антитела, как правило, вносят в постоянной концентрации и смешивают с серийными разведениями тестируемого антигена. После инкубации для создания равновесия часть свободных антител «захватывают» на сенсибилизированной антигеном поверхности и оценивают с помощью меченного антивидового антитела, как правило, с использованием электрохемилюминисценции (например, согласно методу, описанному у Haenel и др., Analytical Biochemistry 339, 2005, сс. 182-184).
Например, в одном варианте осуществления изобретения 384-луночные стрептавидиновые планшеты (фирма Nunc, Microcoat №11974998001) инкубируют в течение ночи при 4°С с использованием 25 мкл/лунку смеси антиген-биотин-изомер в ЗФР-буфере в концентрации 20 нг/мл. Для уравновешивания образцов антитела свободным антигеном: 0,01-1 нМ антитело титруют с использованием релевантного антигена в серийных разведениях 1:3, 1:2 или 1:1,7, начиная с концентрации антигена 2500 нМ, 500 нМ или 100 нМ. Образцы инкубируют при 4°С в течение ночи в запечатанных полипропиленовых микропланшетах для хранения REMP (фирма Brooks). После инкубации в течение ночи стрептавидиновые планшеты промывают трижды (3х), используя 90 мкл ЗФРТ на лунку. По 15 мкл каждого образца из планшета для уравновешивания переносят в планшет для анализа и инкубируют в течение 15 мин при КТ, с последующими тремя стадиями промывки (по 90 мкл) ЗФРТ-буфером. Детекцию осуществляют, добавляя 25 мкл конъюгата козье антитело к человеческому IgG-POD (фирма Jackson, 109-036-088, 1:4000 в среде OSEP), с последующими шестью стадиями промывки (по 90 мкл) ЗФРТ-буфером. Добавляют в каждую лунку 25 мкл ТМВ-субстрата (фирма Roche Diagnostics GmbH, каталожный №11835033001). Измерения осуществляют при длине волны 370/492 нм с использованием ридера Safire2 (фирма Tecan).
В другом варианте осуществления изобретения Kd измеряют с помощью KinExA-анализа (кинетический анализ исключения). Согласно этому анализу антиген, как правило, титруют в присутствии постоянной концентрации связывающих сайтов антитела, образцам дают уравновеситься и затем быстро пропускают через проточную ячейку, на которой свободные связывающие сайты антитела «захватываются» на покрытых антигеном гранулах, в то время как насыщенный антигеном комплекс антитела отмывается. Гранулы с «захваченным» антителом затем оценивают с помощью меченого антивидового антитела, например, флуоресцентно меченного антитела (Bee и др., PloS One, 7(4), 2012, е36261). Например, в одном из вариантов осуществления изобретения эксперименты с использованием KinExA осуществляют при комнатной температуре (КГ), применяя ЗФР, рН 7,4 в качестве подвижного буфера. Образцы приготавливают в подвижном буфере, дополненном 1 мг/мл БСА («буфер для образца»). Используют скорость потока 0,25 мл/мин. Постоянное количество антитело с концентраций сайтов связывания 5 пМ титруют с использованием двукратных серийных разведений антигена, начиная с концентрации 100 пМ (диапазон концентраций 0,049-100 пМ). Один образец антитела без антигена служит в качестве 100% сигнала (т.е. без ингибирования). Комплексы антиген-антитело инкубируют при КТ в течение по меньшей мере 24 ч, обеспечивая достижение равновесия. Уравновешенные смеси затем пропускают через колонку, содержащую сшитые с антигеном гранулы, в KinExA-системе в объеме 5 мл, что позволяет несвязанному антителу «захватываться» гранулами без нарушения состояния равновесия раствора. Для детекции «захваченного» антитела используют 250 нг/мл конъюгированного с Dylight 650© специфического для человеческого Fc-фрагмента вторичного антитела в буфере для образца. Во всех экспериментах по уравновешиванию каждый образец оценивают с дублированием. Величины KD определяют с помощью нелинейного регрессионного анализа данных с использованием модели односайтового гомогенного связывания, входящей в пакет программ KinExA (версия 4.0.11), используя метод «стандартного анализа».
Терапевтические способы и композиции
Как указано выше, мультиспецифические антитела, предлагаемые в настоящем изобретении, можно применять для любого лечения, для которого требуется доставка радионуклида к мишени. Таким образом, в настоящем изобретении предложено таргетное антитело, такое как мультиспецифическое или биспецифическое антитело, указанное в настоящем описании, для применения в способе лечении. Более конкретно, предложено таргетное антитело (например, мультиспецифическое или биспецифическое антитело), указанное в настоящем описании, для применения в способе претаргетной радиоиммунотерапии. В указанных вариантах осуществления изобретения хелатный Pb предпочтительно представляет собой 212Pb.
Как отмечено выше, лечение может быть направлено на любое состояние, которое можно лечить с использованием цитотоксической активности, направленной на пораженные заболеванием клетки пациента. Лечение предпочтительно представляет собой лечение опухоли или рака. Однако применимость изобретения не ограничена опухолями и раками. Например, лечение может также представлять собой лечение вирусной инфекции или инфекции, вызываемой другим патогенным организмом, например, прокариотическим организмом. Необязательно таргетинг можно применять также касательно Т-клеток для лечения обусловленного Т-клетками аутоиммунного заболевания или Т-клеточных раков крови. Таким образом, состояние, подлежащее лечению, может включать вирусные инфекции, такие как ВИЧ, бешенство, EBV и ассоциированная с вирусом герпеса саркома Капоши, и аутоиммунные заболевания, такие как рассеянный склероз и реакция «трансплантат-против-хозяина».
В контексте настоящего описания понятие «рак» относится как к солидным опухолям, так и к гематологическим видам рака, таким как лимфомы, лимфолейкозы, рак легкого, немелкоклеточный рак легкого (NSCL), альвеолярно-клеточный рак легкого, рак кости, рак поджелудочной железы, рак кожи, рак головы или шеи, кожная или внутриглазная меланома, рак матки, рак яичника, ректальный рак, рак анальной области, рак желудка, гастральный рак, рак ободочной кишки, рак молочной железы, карцинома фаллопиевых труб, карцинома эндометрия, карцинома шейки матки, карцинома влагалища, карцинома вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркома мягких тканей, рак мочеиспускательного канала, рак пениса, рак предстательной железы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточная карцинома, карцинома почечной лоханки, мезотелиома, печеночно-клеточный рак, билиарный рак, неоплазмы центральной нервной системы (ЦНС), опухоли позвоночника, глиома ствола головного мозга, мультиформная глиобластома, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, плоскоклеточные карциномы, аденома гипофиза и саркома Юинга, включая рефрактерные варианты любого из указанных выше видов рака или комбинации одного или нескольких из указанных выше видов рака.
Способ направленного переноса радиоизотопа в ткань или орган для терапии может включать:
I) введение индивидууму мультиспецифического или биспецифического антитела, указанного в настоящем описании, где антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень; и
II) последующее введение индивидууму радионуклида Pb, хелатированного DOTAM или его функциональным вариантом, где радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, связывается с антителом, локализованном на клеточной поверхности.
Необязательно между стадиями (I) и (II) вводят очищающий/блокирующий агент. Очищающий/блокирующий агент может связываться с антигенсвязывающим сайтом, специфическим в отношении Pb-DOTAM, и блокировать последующее связывание хелатного радионуклида. Очищающий агент может содержать DOTAM или его функциональный вариант, образующий хелат иона металла и конъюгированный с очищающим фрагментом.
Примеры приемлемых очищающих фрагментов могут включать фрагменты, которые увеличивают размер и/или гидродинамический радиус молекулы, препятствуют способности молекулы достигать опухоли, без воздействия на способность молекулы связываться с антителом в кровотоке. Примеры фрагментов включают гидрофильные полимеры. Фрагмент может представлять собой полимер или сополимер, например, декстран, декстрин, ПЭГ, полисиаловые кислоты (PSA), гиалуроновую кислоту, гидроксиэтилкрахмал (HES) или поли(2-этил-2-оксазолин) (PEOZ). В других вариантах осуществления изобретения фрагмент может представлять собой неструктурный пептид или белок, такой как XTEN-полипептиды (неструктурные гидрофильные белковые полимеры), гомополимер аминокислот (НАР), полимер пролина-аланина-серина (PAS), эластинподобный пептид (ELP) или желатиноподобный белок (GLK). Приемлемая молекулярная масса полимеров может находиться в диапазоне, например, по меньшей мере 50 кДа, например, между 50 и 2000 кДа. Например, молекулярная масса может составлять 200-800 кДа, необязательно может быть выше 300, 350, 400 или 450 кДа и необязательно может быть ниже 700, 650, 600 или 550 кДа, необязательно может составлять примерно 500 кДа.
В некоторых вариантах осуществления изобретения очищающий агент может представлять собой DOTAM или его функциональный вариант (образующий хелат с ионом металла), конъюгированный с декстраном или его производным, например, дополнительно описанным ниже.
В некоторых вариантах осуществления изобретения массовое соотношение антитела и очищающего агента может находиться в диапазоне от 1:1, 2:1, 3:1 или 4:1 до 20:1, 15:1, 10:1, 8:1, 6:1 или 5:1, например, в диапазоне от 1:1 до 20:1, от 1:1 до 10:1, от 2:1 до 8:1 или от 2:1 до 6:1.
В некоторых вариантах осуществления изобретения очищающий агент можно вводить через несколько часов или дней после обработки мультиспецифическим антителом. В некоторых вариантах осуществления изобретения может оказаться предпочтительным вводить очищающий агент по меньшей мере через 2, 4, 6, 8, 10, 12, 16, 18, 22 или 24 ч после мультиспецифического антитела или по меньшей мере через 1, 2, 3, 4, 5, 6 или 7 дней. В некоторых вариантах осуществления изобретения может оказаться предпочтительным вводить очищающий агент не более чем через 14 дней после антитела, например, не более чем через 10, 9, 8, 7, 6, 5, 4, 3 или 2 дня.
Необязательно очищающий агент вводят в период времени между 4 и 10 днями, 4 и 7 днями, 2 и 7 днями или 2-4 днями после мультиспецифического антитела.
В некоторых вариантах осуществления изобретения радионуклид Pb вводят через несколько минут, часов или дней после очищающего агента. В некоторых вариантах осуществления изобретения может оказаться предпочтительным вводить радионуклид Pb по меньшей мере через 30 мин после очищающего агента и необязательно в пределах 48 ч, 24 ч, 8 ч или 4 ч после введения очищающего агента. В некоторых вариантах осуществления изобретения радионуклид Pb можно вводить через 1 день после введения очищающего агента.
В некоторых вариантах осуществления изобретения антитела, представленные в настоящем описании, можно применять в качестве компонента комбинированной терапии. Например, их можно вводить в комбинации с одним или несколькими химиотерапевтическими агентами: химиотерапевтический агент и антитело можно вводить одновременно или последовательно в любом порядке.
В некоторых вариантах осуществления изобретения антитела, представленные в настоящем описании, можно в дополнительном или альтернативном варианте вводить в комбинации с радиосенсибилизаторами. Радиосенсибилизатор и антитело можно вводить одновременно или последовательно в любом порядке.
Фармацевтические композиции
Фармацевтические композиции антитела к Pb-DOTAM, указанного в настоящем описании, например, мультиспецифического или биспецифического антитела, получают путем смешения указанного антитела, имеющего необходимую степень чистоты, с одним или несколькими необязательными фармацевтически приемлемыми носителями (Remington's Pharmaceutical Sciences, 16-е изд. под ред. Osol А., 1980) в форме лиофилизированных композиций или водных растворов. Фармацевтически приемлемые носители, как правило, являются нетоксичными для реципиентов в используемых дозах и концентрациях, они включают (но, не ограничиваясь только ими): буферы, такие как фосфатный, цитратный буферы, а также буферы на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и мета-крезол); полипептиды с низкой молекулярной массой (содержащие менее приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поли(винилпирролидон); аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТК; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; содержащие металл комплексы (например, комплексы Zn-белок); и/или неионогенные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). В контексте настоящего описания примеры фармацевтически приемлемых носителей включают также диспергирующие агенты интерстициальных лекарственных средств, такие как растворимые активные в нейтральной среде гликопротеины гиалуронидаз (sHASEGP), например, человеческие растворимые гликопротеины гиалуронидазы РН-20, такие как rHuPH20 (HYLENEX®, фирма Baxter International, Inc.). Некоторые примеры sHASEGP и методы их применения, в том числе rHuPH20, описаны в публикациях патентов США №№2005/0260186 и 2006/0104968. Согласно одному из объектов изобретения sHASEGP объединяют с одним или несколькими дополнительными гликозаминогликаназами, такими как хондроитиназы.
Примеры лиофилизированных композиций антител описаны в US №6267958. Водные композиции антител включают композиции, описанные в US №6171586 и WO 2006/044908, последние композиции включают гистидин-ацетатный буфер.
Композиция, указанная в настоящем описании, может содержать также более одного действующего вещества, если это необходимо для конкретного подлежащего лечению показания, предпочтительно вещества с дополнительными видами активности, которые не оказывают отрицательного воздействия друг на друга. Например, может требоваться применение дополнительных химиотерапевтических агентов и/или радиосенсибилизаторов, указанных выше. Такие действующие вещества должны присутствовать в комбинации в количествах, эффективных для достижения поставленной цели.
Действующие вещества можно включать также в микрокапсулы, например, полученные с помощью методов коацервации или межфазной полимеризации, например в гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли(метилметакрилатные) микрокапсулы соответственно, в коллоидные системы введения лекарственного средства (например в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы описаны в Remington's Pharmaceutical Sciences, 16-е изд., под ред. Osol А., 1980.
Можно приготавливать препараты с замедленным высвобождением. Приемлемыми примерами препаратов с замедленным высвобождением являются полупроницаемые матрицы из твердых гидрофобных полимеров, включающие антитело или конъюгат, такие матрицы представляют собой изделия определенной формы, например, пленки или микрокапсулы. Композиции, предназначенные для применения in vivo, как правило, должны быть стерильными. Стерильность можно легко обеспечивать, например, путем фильтрации через стерильные фильтрующие мембраны.
Способы и композиции для диагностирования и детекции
В настоящем изобретении предложено также таргетное антитело, например, мультиспецифическое антитело, указанное в настоящем описании, для применения в способе диагностирования индивидуума. Способ диагностирования может представлять собой способ претаргетной радиоиммуновизуализации, например, для диагностирования индивидуум, у которого предполагается наличие пролиферативного нарушения или инфекционного заболевания. В таких вариантах осуществления изобретения хелатный Pb предпочтительно представляет собой 203Pb.
Способ направленного переноса радиоизотопа в ткань или орган для визуализации может включать:
I) введение индивидууму мультиспецифического или биспецифического антитела, указанного в настоящем описании, где антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень; и
II) последующее введение индивидууму радионуклида Pb, хелатированного DOTAM или его функциональным вариантом, где радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, связывается с антителом, локализованном на клеточной поверхности.
В другом варианте осуществления изобретения мультиспецифическое или биспецифическое антитело, указанное в настоящем описании, может быть связано с хелатным радионуклидом Pb в момент введения.
Необязательно способ может включать также:
III) визуализацию ткани или органа, в которой радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, локализован или ожидается, что будет локализован.
В другом варианте осуществления изобретения способ, предлагаемый в изобретении, может включать визуализацию ткани или органа индивидуума, в котором индивидуум предварительно обработан:
I) мультиспецифическим или биспецифическим антителом, указанным в настоящем описании, где антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень; и
II) радионуклидом Pb, хелатированным DOTAM или его функциональным вариантом, где радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, связывается с антителом, локализованном у индивидуума на клеточной поверхности.
Необязательно между стадиями (I) и (II) вводят очищающий/блокирующий агент. Очищающий агент, режим введения очищающего агента и массовое соотношение антитела и очищающего агента могут соответствовать описанным выше.
Антиген-мишень может представлять собой любой антиген-мишень, указанный в настоящем описании. В некоторых вариантах осуществления изобретения антиген-мишень может представлять собой опухолеспецифический антиген, указанный выше, и визуализация может представлять собой способ визуализации опухоли или опухолей. Индивидуум может иметь опухоль или у него может предполагаться наличие опухоли.
Например, способ может представлять собой способ визуализации опухолей у индивидуума, который имеет или предполагается, что он имеет рак легкого, немелкоклеточный рак легкого (NSCL), альвеолярно-клеточный рак легкого, рак кости, рак поджелудочной железы, рак кожи, рак головы или шеи, кожную или внутриглазную меланому, рак матки, рак яичника, ректальный рак, рак анальной области, рак желудка, гастральный рак, рак ободочной кишки, рак молочной железы, карциному фаллопиевых труб, карциному эндометрия, карциному шейки матки, карциному влагалища, карциному вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркому мягких тканей, рак мочеиспускательного канала, рак пениса, рак предстательной железы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточную карциному, карциному почечной лоханки, мезотелиому, печеночно-клеточный рак, билиарный рак, неоплазмы центральной нервной системы (ЦНС), опухоли позвоночника, глиому ствола головного мозга, мультиформную глиобластому, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, плоскоклеточные карциномы, аденому гипофиза и саркому Юинга, включая рефрактерные варианты любого из указанных выше видов рака или комбинации одного или нескольких из указанных выше видов рака.
Очищающие агенты
Следующим объектом изобретения является разработанный при создании настоящего изобретения новый очищающий агент. Указанный очищающий агент можно применять в способах диагностирования, визуализации или лечения, которые указаны в настоящем описании.
Один из объектов настоящего изобретения относится к очищающему агенту на основе декстрана, который содержит декстран или его производное, конъюгированный/конъюгированное с M-DOTAM или его функциональным вариантом.
В некоторых вариантах осуществления изобретения очищающий агент может представлять собой соединение следующей формулы:
декстран-(линкер-(М-ВОТАМ))х, в которой декстран обозначает декстран или его производное; линкер обозначает связывающий фрагмент; M-DOTAM обозначает DOTAM или его функциональный вариант, включающий ион металла; и х>1.
В некоторых вариантах осуществления изобретения связывающий фрагмент может представлять собой или может содержать одну или несколько двухвалентных функциональных групп, выбранных из мочевинной группы (-NH-C(O)-NH-), замещенной мочевинной группы (-NRX-C(O)-NRX-, в которой одна или обе RX - группы не обозначают Н), тиомочевинной группы (-NH-C(S)-NH-), замещенной тиомочевинной группы (-NRX-C(S)-NRX-, в которой одна или обе RX - группы не обозначают Н), амидной группы (-C(O)-NH-), замещенной амидной группы (-C(O)-NRX-, в которой RXнe обозначает Н), тиоамидной группы (-C(S)-NH-), замещенной амидной группы (-C(S)-NRX-, в которой RX не обозначает Н), триазольной группы или замещенного триазола. В указанных вариантах осуществления изобретения связывающий фрагмент может необязательно содержать одну или несколько дополнительных двухвалентных функциональных групп, таких как алкиленовая группа, ариленовая группа, а гетероариленовая группа, аралкиленовая группа и гетероаралкиленовая группа. Значения заместителя RX не ограничены конкретным значением. В конкретных вариантах осуществления изобретения RX, если присутствует, выбирают из группы, состоящей из C1-С6алкила, С5-С12арила, С5-С12гетероарила и галогрупп.
В конкретных вариантах осуществления изобретения связывающий фрагмент может представлять собой или может содержать одну или несколько двухвалентных функциональных групп, выбранных из мочевинной группы, тиомочевинной группы, амидной группы, тиоамидной группы или триазольной группы.
В предпочтительном варианте осуществления изобретения связывающий фрагмент содержит двухвалентную тиомочевинную функциональную группу или двухвалентную тиоамидную функциональную группу.
В некоторых вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиомочевинную функциональную группу и необязательно замещенную ариленовую группу. В некоторых вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиомочевинную функциональную группу, необязательно замещенную ариленовую группу и необязательно замещенную алкиленовую группу. В конкретных вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиомочевинную функциональную группу, ковалентно связанную через один из ее атомов азота с необязательно замещенной ариленовой группой. В других вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиомочевинную функциональную группу, ковалентно связанную через один из ее атомов азота с необязательно замещенной ариленовой группой, и необязательно замещенная ариленовая группа ковалентно связана с необязательно замещенной алкиленовой группой. В предпочтительных вариантах осуществления изобретения ариленовая группа является незамещенной. В конкретных вариантах осуществления изобретения ариленовая группа представляет собой фениленовую группу. В предпочтительных вариантах осуществления изобретения алкиленовая группа является незамещенной. В конкретных вариантах осуществления изобретения алкиленовая групп представляет собой C1-С6алкиленовую группу. В особенно предпочтительных вариантах осуществления изобретения алкиленовую группу выбирают из метилена и этилена. Если они присутствуют в связывающем фрагменте, то ариленовая группа и алкиленовая группа могут быть незамещенными. В конкретных вариантах осуществления изобретения связывающий фрагмент состоит из двухвалентной тиомочевинной функциональной группы, ковалентно связанной через один из ее атомов азота с ариленовой группой, и ариленовая группа ковалентно связана с алкиленовой группой.
В некоторых вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиоамидную функциональную группу и необязательно замещенную ариленовую группу. В некоторых вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиоамидную функциональную группу, необязательно замещенную ариленовую группу и необязательно замещенную алкиленовую группу. В конкретных вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиоамидую функциональную группу, ковалентно связанную через один из ее атомов азота с необязательно замещенной ариленовой группой. В других вариантах осуществления изобретения связывающий фрагмент содержит двухвалентную тиоамидную функциональную группу, ковалентно связанную через один из ее атомов азота с необязательно замещенной ариленовой группой, и необязательно замещенную ариленовую группу, ковалентно связанную с необязательно замещенной алкиленовой группой. В предпочтительных вариантах осуществления изобретения ариленовая группа является незамещенной. В конкретных вариантах осуществления изобретения ариленовая группа представляет собой фениленовую группу. В предпочтительных вариантах осуществления изобретения алкиленовая группа является незамещенной. В конкретных вариантах осуществления изобретения алкиленовая группа представляет собой C1-С6алкиленовую группу. В особенно предпочтительных вариантах осуществления изобретения алкиленовую группу выбирают из метилена и этилена. Если они присутствуют в связывающем фрагменте, то ариленовая группа и алкиленовая группа могут быть незамещенными. В конкретных вариантах осуществления изобретения связывающий фрагмент состоит из двухвалентной тиоамидной функциональной группы, ковалентно связанной через один из ее атомов азота с ариленовой группой, и ариленовая группа ковалентно связана с алкиленовой группой.
В некоторых вариантах осуществления изобретения связывающий фрагмент может представлять собой или может содержать группу следующей формулы:
в которой у обозначает 1-6 (предпочтительно 1 или 2), * обозначает точку присоединения декстрана или его производного, а ** обозначает точку присоединения кольцевого атома DOTAM или его функционального варианта.
В некоторых вариантах осуществления изобретения связывающий фрагмент может быть образован в результате конъюгации амина (предпочтительно первичного амина) и изоцианата или изотиоцианата. Указанная конъюгация приводит к образованию двухвалентной мочевинной функциональной группы и тиомочевинной функциональной группы соответственно. В указанных вариантах осуществления изобретения, когда изоцианат является одним из реактантов, то связывающий фрагмент может при необходимости содержать двухвалентную мочевинную функциональную группу или двухвалентную амидную функциональную группу. В указанных вариантах осуществления изобретения, когда изотиоцианат является одним из реактантов, то связывающий фрагмент может при необходимости содержать двухвалентную тиомочевинную функциональную группу или двухвалентную тиоамидную функциональную группу.
Предпочтительно х обозначает число, превышающее 1, в результате на каждую молекулу декстрана приходится в среднем более одной молекулы М-DOTAM или его функционального варианта. Например, х может обозначать 2 или более, 5 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, 35 или более, 40 или более или предпочтительно 50 или более. При создании настоящего изобретения было установлено, что улучшенное очищение может достигаться при применении декстрана, меченного несколькими М-DOTAM- группами.
DOTAM или его функциональный вариант может включать только один линкер, для предупреждения перекрестного связывания декстрана.
Производные декстрана, которые могут найти применение в очищающем агенте, включают аминодекстраны, в которых декстран замещен одним или несколькими аминами. Особенно предпочтительно применять аминодекстраны, в которых одна или несколько гидроксильных групп декстрана замещены аминозамещенной карбоксиметиламидной группой. Такие соединения можно получать путем модификации декстрана карбоксиметильной группой (например, путем взаимодействия с хлоруксусной кислотой) и последующего дополнительного взаимодействия с необязательно замещенным диамином (предпочтительно алкилдиамином, таким как а,ю-алкилендиамин, например этилендиамин).
Аминогруппа является точкой присоединения линкера. По меньшей мере 30% доступных аминогрупп могут быть замещены DOT AM или его функциональным вариантом, предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%.
Предпочтительно «декстран» в приведенной выше формуле обозначает аминодекстран, соответствующий декстрану, замещенному одной или несколькими карбоксиметильными группами, которые сами замещены этилендиамином.
Декстран может быть замещен одной или несколькими группами формулы -CH2C(=O)NH(CH2)fNHRF, в которой RF обозначает водород или линкер для DOTAM, и f обозначает 1-6, наиболее предпочтительно 2. Например, группы могут иметь следующую формулу:
в которой волнистой линией обозначена точка присоединения к кислороду на декстране.
Аминодекстран может иметь каркас из преимущественно α(1,6)-связанных гликопиранозильных повторяющихся звеньев, необязательно с разветвлениями других гликопиранозильных звеньев, связанных, например, через α(1,2), α(1,3) или α(1,4) гликозидные связи. По меньшей мере некоторые из гидроксильных групп замещены аминозамещенной карбоксиметиламидной группой, указанной выше (в частности, группой формулы -CH2C(=O)NHCH2CH2NHRF). Иными словами, аминодекстран может содержать звенья следующей формулы:
в которой RG каждый обозначает Н, аминозамещенную карбоксиметиламидную группу (такую как -CH2C(=O)NHCH2CH2NHRF) или связь для дополнительного гликопиранозильного звена, преимущественно через α(1,6)-связь, и в которой пунктирной линией обозначена связь с соседним звеном.
Очищающий агент может включать одно или несколько звеньев представленной ниже формулы:
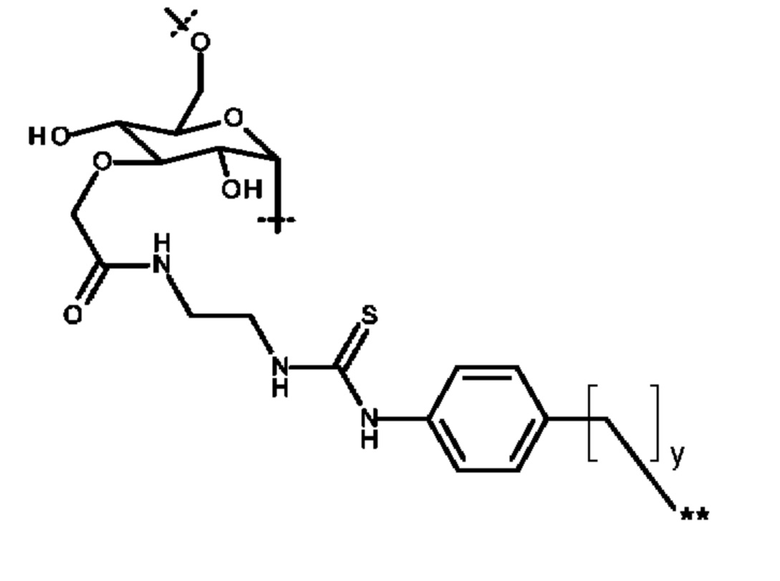
в которой ** обозначает точку присоединения к DOTAM или его функциональному варианту, а у имеет указанные выше значения.
Производные декстрана могут включать декстран или аминодекстран, модифицированный одной или несколькими группами, выбранными из аминокислоты или сахарида, отличного от глюкозы. Например, декстран может быть модифицирован (например, кэпирован) одним или несколькими остатками глутаминовой кислоты или полиглутаминовой кислоты, включая Glu, (Glu)2, (Glu)3 или (Glu)4. В дополнительном или альтернативном варианте декстран может быть модифицирован (например, кэпирован) сахаридом, отличным от глюкозы, таким как N-ацетилгалактозамин (GalNAc), или полисахаридом, образованным из таких сахаридов, таким как три-GalNAc.
В некоторых вариантах осуществления изобретения молекулярная масса компонента, представляющего собой декстран, может составлять по меньшей мере 50 кДа, например, от 50 до 2000 кДа. Например, молекулярная масса может составлять 200-800 кДа, необязательно больше чем 300, 350, 400 или 450 кДа и необязательно меньше чем 700, 650, 600 или 550 кДа, необязательно примерно 500 кДа.
Может оказаться предпочтительным, если количество аминогрупп, выраженное в виде процента от количества глюкозных звеньев декстрана или производного декстрана («насыщение» глюкозных звеньев аминогруппами), может составлять по меньшей мере 0,5%, по меньшей мере 1%, по меньшей мере 2%, по меньшей мере 5%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100%. В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если насыщение декстрана аминогруппами составляет по меньшей мере или примерно 1% или 10%, например, 1%-10%
Может оказаться предпочтительным, если количество групп DOTAM, выраженное в виде процента от количества аминогрупп производного декстрана («насыщение» аминогрупп декстранового компонента с помощью DOTAM) может составлять по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или 100%. В некоторых вариантах осуществления изобретения может оказаться предпочтительным, если насыщение доступных аминогрупп производного декстрана с помощью DOTAM составляет по меньшей мере или примерно 40% или 50%, например, 40%-60%.
Одной из возможных трудностей, связанных с применением очищающих агентов, является возможность того, что они могут проникать в опухоли и связываться с опухолеассоциированными антитела, оказывая отрицательное воздействие на последующее связывание радиолигандов.
При создании настоящего изобретения было обнаружено также, что удовлетворительный клиренс крови может достигаться в сочетании с низким уровнем проникновения очищающего агента в опухоли, при применении очищающего агента на основе декстрана, который имеет I) высокую среднюю молекулярную массу и II) был подвержен отсечению по молекулярной массе, в результате которой были удалены фрагменты, имеющие размер ниже требуемого. Отсечение по молекулярной массе можно применять к декстрану или производному декстрана до стадии конъюгации; и/или применять к очищающему агенту после конъюгации; и/или применять к очищающему агенту после образования комплекса с металлом.
Таким образом, очищающий агент, который можно применять согласно настоящему изобретению, может представлять собой очищающий агент на основе декстрана, содержащий декстран или его производное (например, указанное выше, предпочтительно аминодекстран), конъюгированный/конъюгированное с хелатом металла, в котором I) средняя молекулярная масса декстрана или его производного составляет предпочтительно 200-800 кДа, необязательно превышает 300, 350, 400 или 450 кДа и необязательно является более низкой чем 700, 650, 600 или 550 кДа, необязательно составляет примерно 500 кДа, и II) декстран, производные декстрана или очищающие агенты, имеющие молекулярную массу ниже отсекаемой, удалены, при этом отсечение проводится по молекулярной массе 50 кДа или более, 100 кДа или более, или 200 кДа или более, необязательно по молекулярной массе, которая находится в диапазоне 50-250 кДа или 50-200 кДа, необязательно 100-200 кДа, необязательно около 100 кДа или 150 кДа, или 200 кДа. (Во избежание сомнений следует отметить, что, если отсечение указывается на уровне 50 кДа или выше, то это означает, что отсечение может соответствовать 50 кДа или любой величине, превышающей 50 кДа, но при этом должны быть удалены декстран, производные декстрана или очищающие агенты с молекулярной массой, ниже отсекаемой).
Количество видов с молекулярной массой, ниже отсекаемой, может составлять, например, 5 мас. % или менее, 4 мас. % или менее, 3 мас. % или менее, 2 мас. % или менее, 1 мас. % или менее, 0,5 мас. % или менее, 0,4 мас. % или менее, 0,3 мас. % или менее, 0,2 мас. % или менее, 0,1 мас. % или менее, или 0,01 мас. % или менее, в пересчете на массовый процент очищающего агента. Предпочтительно очищающий агент практически не содержит виды, имеющие молекулярную массу ниже отсекаемой.
Для обеспечения отсечения по молекулярной массе можно применять фильтрацию, например, диафильтрацию, ультрафильтрацию, фильтрацию в тангенциальном потоке или фильтрацию с поперечным потоком. Предпочтительно осуществляют по меньшей мере две стадии фильтрации, необязательно по меньшей мере три. Под «средней молекулярной массой» подразумевается средневзвешенная молекулярная масса, определенная с помощью SEC-MALS-анализа.
Является приемлемым, если включенные в DOTAM или его функциональный вариант металлы присутствуют в виде ионов металлов и если степень окисления варьируется в зависимости от конкретного элемента. Так, специалисту в данной области должно быть очевидно, что, например, под понятие свинец, Pb или 206Pb подпадают ионные формы элемента, в частности, Pb(II).
Присутствующий в очищающем агенте металл может представлять собой стабильный (нерадиоактивный) изотоп свинца или стабильный или практически стабильный изотоп иона другого металла, при условии, что комплекс ион металла-DOTAM распознается с высокой аффинностью антителом. Например, к другим приемлемым металлам могут относиться Zn (Zn2+), Са (Са2+) или 209Bi (Bi2+), последний из которых является радиоактивным, но рассматривается как практически стабильный из-за очень длинного периода полураспада.
Следующим объектом настоящего изобретения является способ получения очищающего агента, включающий конъюгацию декстрана или производного декстрана с DOTAM или его функциональным вариантом или производным, где способ включает получение хелата DOTAM с Pb или другим ионом металла, описанного выше [например, Pb(II)] до и/или после конъюгации DOTAM или его функционального варианта с декстраном.
Следующим объектом настоящего изобретения является способ получения очищающего агента, включающий:
создание конъюгата путем конъюгации DOTAM или его функционального варианта или производного с декстраном или производным декстрана;
в котором перед конъюгацией декстран или производное декстрана подвергают стадии фильтрации для удаления видов с молекулярной массой ниже порога отсечения, например, составляющей 50 кДа или выше, 100 кДа или выше или 200 кДа или выше, необязательно находящейся в диапазоне 50-250 кДа или 50-200 кДа, необязательно 100-200 кДа, например, видов с молекулярной массой ниже 100, 150 или 200 кДа, или где
способ дополнительно включает подвергание конъюгата стадии фильтрации для удаления видов с молекулярной массой ниже порога отсечения, например, составляющей 50 кДа или выше, 100 кДа или выше или 200 кДа или выше, необязательно находящейся в диапазоне 50-250 кДа или 50-200 кДа, необязательно 100-200 кДа, например, видов с молекулярной массой ниже 100, 150 или 200 кДа.
Как указано выше, при создании настоящего изобретения установлено, что целесообразно применять отсечение по молекулярной массе для удаления фрагментов, имеющих размер ниже требуемого. Метод фильтрации может представлять собой, например, диафильтрацию. Специалисту в данной области должно быть очевидно, что понятие «удаление» во фразе «удаление видов с молекулярной массой ниже порога отсечения» является синонимом с «уменьшением количества» и что может сохраняться некоторое количество видов с низкой молекулярной массой в зависимости от конкретного применяемого метода фильтрации. Количество видов, имеющих молекулярную массу, ниже отсекаемой, может составлять, например, 5 мас. % или менее, 4 мас. % или менее, 3 мас. % или менее, 2 мас. % или менее, 1 мас. % или менее, 0,5 мас. % или менее, 0,4 мас. % или менее, 0,3 мас. % или менее, 0,2 мас. % или менее, 0,1 мас. % или менее или 0,01 мас. % или менее в пересчете на массовый процент очищающего агента. Предпочтительно очищающий агент практически не содержит виды, имеющие после фильтрации молекулярную массу ниже порога отсечения.
Функциональный вариант или производное DOTAM может представлять собой указанный/указанное выше, в котором по меньшей мере одна из R1-групп служит в качестве линкерного фрагмента. Например, пригодную группу (линкер-(M-DOTAM)) можно получать путем взаимодействия соединения указанной ниже формулы с указанным выше аминодекстраном:
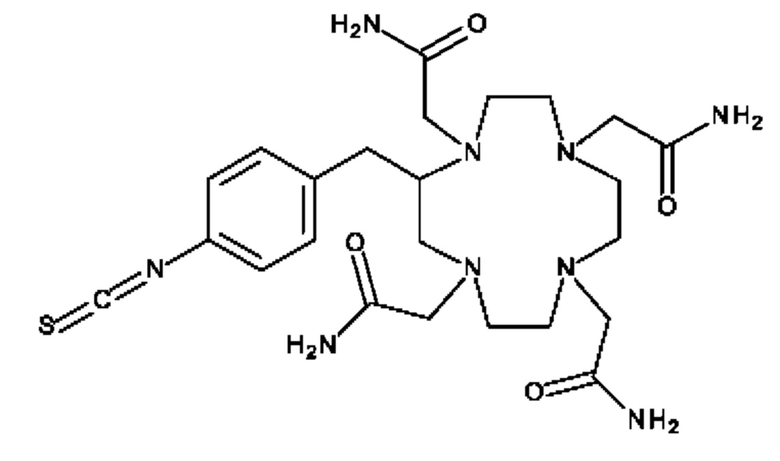
Синтез указанного соединения описан у Chappell и др., Nuclear Medicine and Biology, т. 27, 2000, cc. 93-100, и производные DOTAM поступают в продажу от фирмы Macrocyclics, Inc. (Плано, шт. Техас).
DOTAM или его функциональный вариант можно добавлять в избытке, в результате каждое производное декстрана имеет в среднем больше одной молекулы DOTAM. Среднее количество молекул DOTAM или его функциональных вариантов на каждую молекулу декстрана может быть больше 1, например, 2 или более, 3 или более, 4 или более, 5 или более, 10 или более, 15 или более, 20 или более, 25 или более, 30 или более, 35 или более, 40 или более, 50 или более, 100 или более или предпочтительно 40 или более. При создании настоящего изобретения было установлено, что улучшенного очищения можно достигать путем использования декстрана, конъюгированного с большим количеством групп M-DOTAM.
Предпочтительно декстран имеет среднюю молекулярную массу 200-800 кДа, необязательно больше чем 300, 350, 400 или 450 кДа и необязательно меньше чем 700, 650, 600 или 550 кДа, необязательно примерно 500 кДа.
Способ получения очищающего агента может включать также стадию хелатирования, на которой получают хелат DOTAM или его функционального варианта с ионом металла. Ион металла может представлять собой нерадиоактивный изотоп, например, нерадиоактивный изотоп Pb, Са, Zn или практически стабильный изотоп, такой как 209Bi.
Стадию хелатирования осуществляют перед конъюгацией DOTAM или его функционального варианта с декстраном и/или после конъюгации DOTAM или его функционального варианта с декстраном, но необязательно перед стадией фильтрации. Хелатирование иона металла с помощью DOTAM или его функционального варианта может являться необходимым для гарантии правильного связывания биспецифического антитела с очищающим агентом, например, для гарантии того, что DOTAM или его функциональный вариант принимает правильную конформацию для контакта с антителом.
Когда способ содержит стадию хелатирования, то способ предпочтительно включает также последующую стадию удаления несвязанного металла. Для этой цели можно добавлять дополнительный хелатирующий агент, который можно впоследствии отделять от связанного с декстраном DOTAM или его функционального варианта в процессе стадии фильтрации. Дополнительный хелатирующий агент преимущественно отличает от DOTAM или его функционального варианта. Предпочтительно дополнительный хелатирующий агент имеет более низкую молекулярную массу, чем конъюгат декстран-хелатирующий агент, для облегчения разделения по размеру. Например, дополнительный хелатирующий агент может представлять собой полиаминокарбоновую кислоту, такую как этилендиаминотетрауксусная кислота (ЭДТК) или ее соль.
Предпочтительно способ получения очищающего агента включает:
I) создание конъюгата путем конъюгации DOTAM или его функционального варианта или производного с декстраном или производным декстрана;
II) необязательно удаление низкомолекулярных видов из продукта, полученного на стадии (I);
III) получение хелата конъюгата с ионом металла, например, ионом Pb, Bi, Zn или Са;
IV) добавление дополнительного хелатирующего агента для хелатирования несвязанного иона металла; и
V) осуществление стадии фильтрации для удаления видов с молекулярной массой ниже порога отсечения.
Хелатированный РОТАМ радионуклид Pb
Радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, можно применять в любом из способов диагностирования, визуализации или лечения, указанном в настоящем описании. Является целесообразным, если при применении в указанных способах радионуклид Pb, халатированный DOTAM или его функциональным вариантом, содержится в композиции. В одном из конкретных вариантов осуществления изобретения композиция содержит радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, и DOTAM или его функциональный вариант, не образующий хелат с радионуклидом Pb. Таким образом, другой объект настоящего изобретения относится к указанной композиции и/или к указанной композиции, предназначенной для применения в любом из способов визуализации или лечения, указанных в настоящем описании. Радионуклид Pb, хелатированный DOTAM, касательно применения в указанных способах, может находиться в форме композиции, указанной в настоящем описании.
DOTAM или его функциональный вариант, не образующий хелат с радионуклидом Pb, может представлять собой нехелатный DOTAM или его функциональный вариант. При применении in vivo нехалатный DOTAM или его функциональный вариант могут образовывать комплексы с ионами металлов из окружающей среды, например, с ионами кальция. Указанные ионы кальция, хелатированные DOTAM или его функциональным вариантом, являются фармакологически неактивными и потенциально могут блокировать фармакологически активный радионуклид Pb, хелатированный DOTAM или его вариантом, в мишенях в опухолях и поэтому снижать эффективность лечения и/или требуемый согласно стандарту для визуализации и диагностирования уровень поглощения.
При создании настоящего изобретения было установлено, что путем гашения нехелатного DOTAM или его функционального варианта в определенных условиях можно повышать контроль формирования in vivo хелатного радионуклида Pb и/или можно элиминировать или снижать потенциальную конкуренцию между фармацевтически активным хелатом и фармацевтически неактивным хелатом. Таким образом, в некоторых вариантах осуществления изобретения DOTAM или его функциональный вариант, который не образует хелат с радионуклидом Pb, представляет собой DOTAM или его функциональный вариант, образующий хелат с нерадиоактивным ионом металла.
В некоторых вариантах осуществления изобретения хелатный радионуклид Pb представляет собой 212Pb. В некоторых вариантах осуществления изобретения хелатный радионуклид Pb представляет собой 203Pb.
Считается приемлемым, чтобы включенный в DOTAM или его функциональный вариант радионуклид Pb присутствовал в виде ионов металла и чтобы степень окисления варьировалась в зависимости от конкретного элемента. Так, специалисту в данной области должно быть очевидно, что, например, подразумевается, что под понятие свинец, Pb или 206Pb подпадают ионные формы элемента, в частности, Pb(II).
Нерадиоактивный металл, присутствующий в композиции, может представлять собой стабильный (нерадиоактивный) изотоп свинца, или стабильный или практически стабильный изотоп другого иона металла. Например, другие приемлемые металлы могут представлять собой Gd (Gd2+), Cu (Cu2+), Zn (Zn2+), Ca (Ca2+) или 209Bi (Bi2+), последний из которых является радиоактивным, но рассматривается как практически стабильный из-за очень длинного периода полураспада. В некоторых вариантах осуществления изобретения металл представляет собой Са или Cu. В некоторых вариантах осуществления изобретения металл представляет собой Са.
Функциональный вариант или производное DOTAM может представлять собой указанный/указанное выше.
Следующим объектом настоящего изобретения является способ получения композиции, которая содержит радионуклид Pb, хелатированный DOTAM или его функциональным вариантом, включающий:
I) получение радионуклидов Pb,
II) получение хелата радионуклида Pb DOTAM или\его функционального варианта,
III) получение хелата нехелатного DOTAM или его функционального варианта с нерадиоактивным ионом металла.
Нехелатный DOTAM или его функциональный вариант, полученный на стадии III), представляет собой DOTAM или его функциональный вариант, не образовавший хелат с радионуклидом Pb на стадии II).
Нерадиоактивный ион металла может представлять собой ион Pb, Са, Zn, Gd или Cu. В некоторых вариантах осуществления изобретения ион металла представляет собой ион Са или Cu. В некоторых вариантах осуществления изобретения ион металла представляет собой ион Са, в частности Са2+.
В некоторых вариантах осуществления изобретения на долю нехелатного DOTAM или его функционального варианта, сохраняющегося после осуществления стадии II), приходится по меньшей мере 90 мол. %, по меньшей мере 95 мол. %, по меньшей мере 99 мол. % от DOTAM или функционального варианта, добавленного к радионуклиду РЬ. В одном из конкретных вариантов осуществления изобретения на долю нехелатного DOTAM или его функционального варианта, сохраняющегося после осуществления стадии II), приходится по меньшей мере 99 мол. %.
В некоторых вариантах осуществления изобретения на долю нехелатного DOTAM или его функционального варианта, сохраняющегося после осуществления стадии III), приходится менее 5 мол. %, менее 2 мол. %, менее 1 мол. %, менее 0,1 мол. %, менее 0,01 мол. % от DOTAM или функционального варианта, добавленного к радионуклиду РЬ. В одном из конкретных вариантов осуществления изобретения на долю нехелатного DOTAM или его функционального варианта, сохраняющегося после осуществления стадии III), приходится менее 1 мол. %, менее 0,1 мол. %, менее 0,01 мол. %.
Указанные на стадии а) радионуклиды Pb можно создавать, помещая радиоактивный материал, который расщепляется до представляющего интерес радионуклида Pb, в генератор, в котором радиоактивный материал связывается с твердым материалом. Например, указанный радиоактивный материал, применяемый для получения 212Pb, может представлять собой радий. Затем представляющий интерес радионуклид экстрагируют из генератора в водном растворе, который может содержать радиологические и химические примеси. Водный раствор, содержащий представляющий интерес радионуклид 224Pb и примеси, очищают с помощью жидкостной хроматографии на колонке. Жидкостная хроматография на колонке может представлять собой экстракционную хроматографию или распределительную хроматографию. Экстракционная или распределительная хроматография основаны на разделении элементов, которые должны распределяться между органической фазой или экстрактантом и водной фазой, при этом экстрактант является связанным с инертной подложкой и образует в сочетании с ней стационарную фазу, в то время как водная фаза представляет собой подвижную фазу.
В экстракционной хроматографии можно применять стационарную фазу, которая включает краун-эфир в качестве экстрактанта и, в частности, дициклогексано-18-краун-6 эфир или дибензо-18-крун-6 эфир, в котором циклогексильные или бензильные группы замещены одной или несколькими C1-С2алкильными группами с прямой или разветвленной цепью, в растворе в органическом растворителе, который не смешивается с водой, как правило, в спирте с длинной углеводородной цепью, иными словами, Сх-цепью и более длинной цепью.
В частности, можно использовать стационарную фазу, которая содержит 4,4'(5')ди-ербутилциклоргексано-18-краун-6 эфир в качестве экстрактанта, предпочтительно разведенный в октан-1-оле. Указанная стационарная фаза обладает преимуществом, заключающемся в том, что избирательно удерживает более 99% 212Pb, присутствующего в водном растворе, который содержит от 1,5 до 2,5 моля/л сильной кислоты, которая, как правило, соответствует типам водных растворов, которые применяют для экстракции 212Pb из генератора, содержащего радий-224. Указанный тип стационарной фазы поставляться, например, во флаконах, но также может быть упакован в виде готовых к применению колонках или картриджах для хроматографии, которые поступают в продажу от фирмы TRISKEM International под торговым названием «Pb-смола».
Альтернативно этому, раствор, содержащий требуемый радионуклид и примеси, можно очищать также с помощью ионообменной хроматографии, например, катионообменной хроматографии.
Метод получения и очистки 212Pb описан в WO 2013/174949.
Последовательности
Ниже в таблице представлены некоторые последовательности, указанные в настоящем описании.



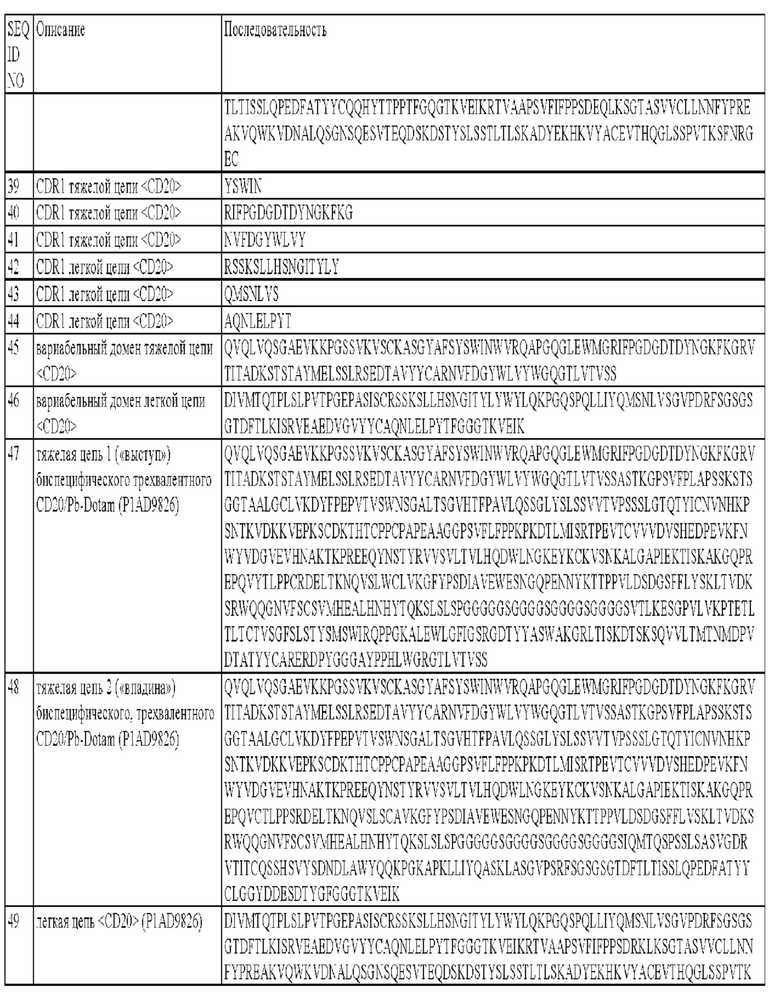

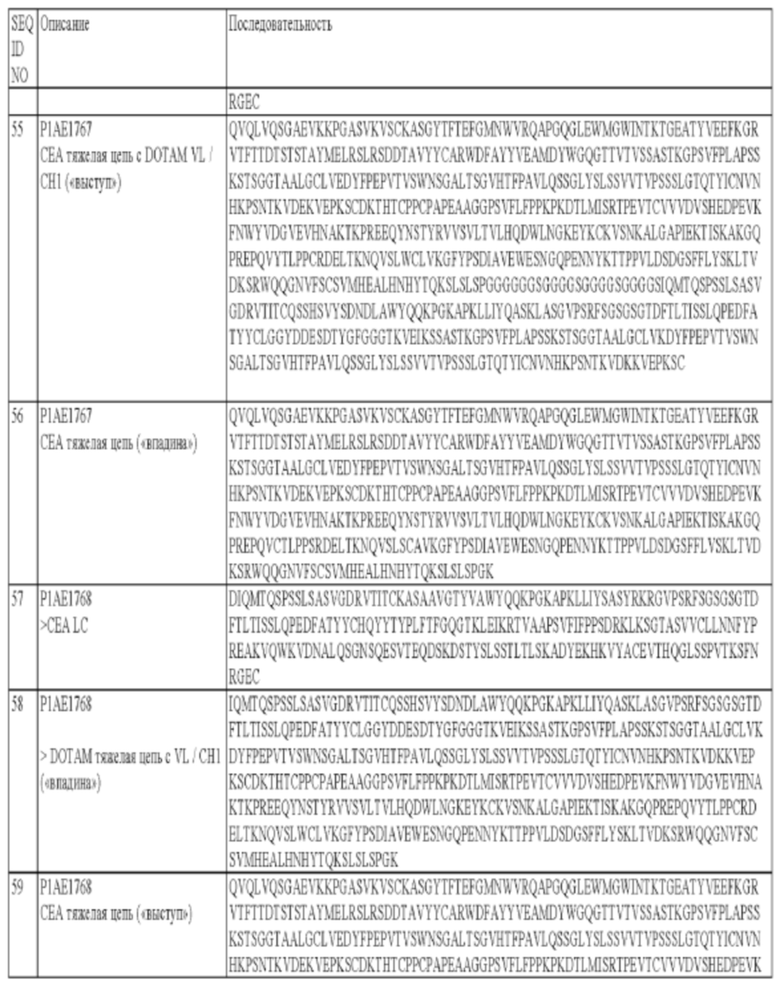
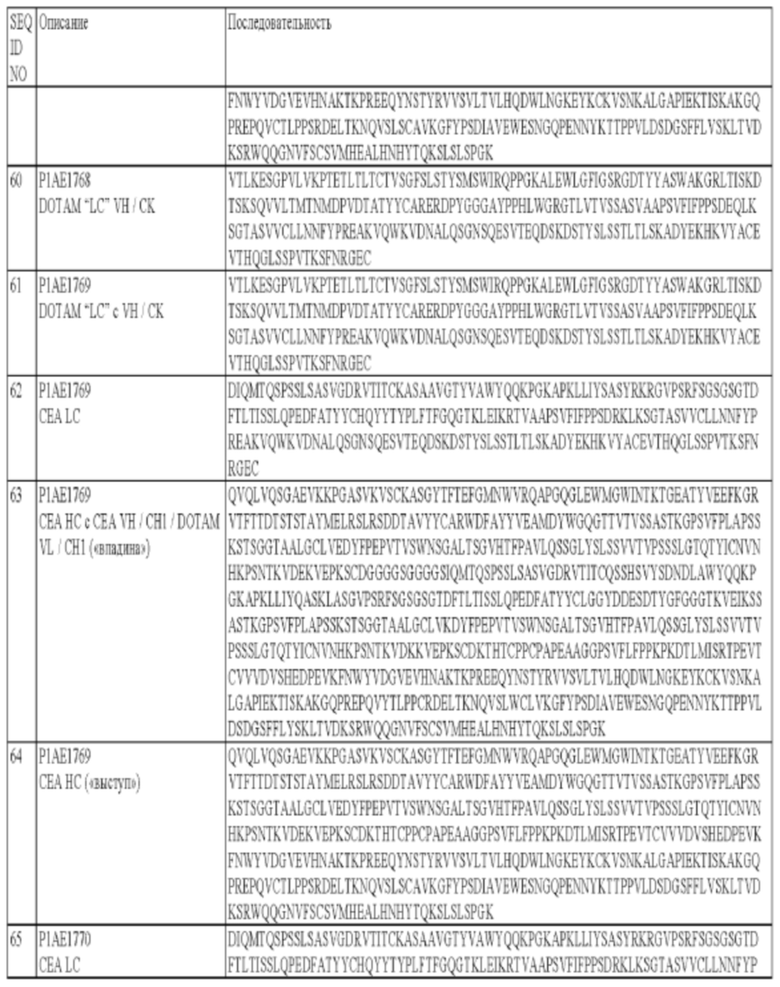
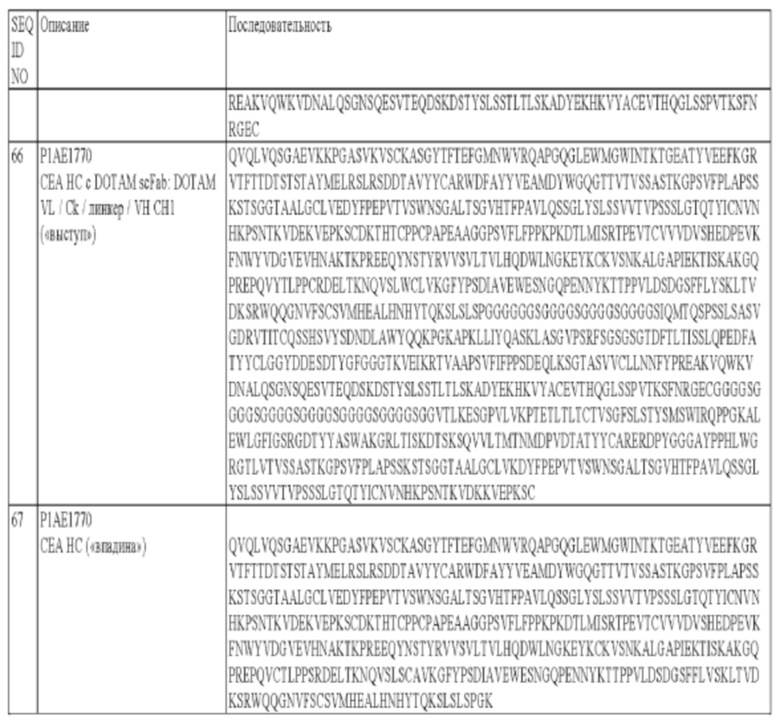
Содержание всех патентов и научной литературы, которые процитированы в настоящем описании, специально полностью включено в настоящее описание в качестве ссылки.
Изобретение дополнительно описано ниже со ссылкой на конкретные примеры. Должно быть очевидно, что можно применять на практике различные другие варианты осуществления изобретения с учетом представленного выше подробного описания.
Примеры
Пример 1: Описание иммунизации
Иммунизация кроликов
Смесь 1:1 двух энантиомерных фракций Pb-DOTAM-алкил-ПЭГ4-KLH (фракция 1 MS2-DOTAM KLH и фракция 2 MS2-DOTAM KLH) применяли для иммунизации новозеландских белых кроликов, несущих локус человеческого иммуноглобулина, которые описаны в WO 2000/46251, WO 2002/12437, WO 2005/007696, WO 2006/047367, US 2007/0033661 и WO 2008/027986. Каждого кролика иммунизировали с использованием 500 мкг иммуногенной смеси, эмульгированной в полном адъюванте Фрейнда, в день 0 путем внутрикожного введения и 500 мкг в каждый из дней 7, 14, 28, 56 путем чередующихся внутримышечных и подкожных введение. Затем кроликов подвергали ежемесячным подкожным иммунизациям с использованием 500 мкг смеси, и получали небольшие образцы крови через 7 дней после иммунизации для определения титров в сыворотке. Более крупные образцы крови (10% от ориентировочного общего объема крови) получали в течение третьего и в течение девятого месяца иммунизации (через 5-7 дней после иммунизации) и выделяли мононуклеарные клетки периферической крови, которые использовали в качестве источника антигенспецифических В-клеток в процессе В-клеточного клонирования (пример 2).
Определение титров в сыворотке (ELISA)
Каждую из двух энантиомерных фракций (Pb-DOTAM (PJRD05.133F1 или PJRD05.133F2) иммобилизовывали на 96-луночном планшете NUNC Maxisorp в концентрации 1 мкг/мл из расчета 100 мкл/лунку в ЗФР, после чего: блокировали планшет с помощью 2% Crotein С в ЗФР, 200 мкл/лунку; вносили с дублированием серийные разведения антисыворотки в 0,5% Crotein С в ЗФР, 100 мкл/лунку; осуществляли детекцию с помощью конъюгированного с HRP ослиного антитела к кроличьему IgG (фирма Jackson Immunoresearch/Dianova 711-036-152; 1/16000) и стрептавидина-HRP, каждый из которых разводили в 0,5% Crotein С в ЗФР, 100 мкл/на лунку. При осуществлении всех стадий планшеты инкубировали в течение 1 ч при 37°С. Между всеми стадиями планшеты промывали 3 раза с помощью 0,05% Твин 20 в ЗФР. Сигнал генерировали путем добавления растворимого субстрата ВМ Blue POD (фирма Roche), 100 мкл/на лунку; и реакцию прекращали путем добавления 1М HCl, 100 мкл/на лунку. Абсорбцию определяли при 450 нм, используя в качестве референс-волны волну длиной 690 нм. Титры определяли в виде разведения антисыворотки, обеспечивающего сигнал, составляющий половину от максимального.
Пример 2: В-клеточное клонирование из крови кроликов
Выделение мононуклеарных клеток периферической крови (РВМС) кроликов
Образцы крови получали из иммунизированных кроликов. Содержащую ЭДТА цельную кровь двукратно разводили с использованием 1×ЗФР (фирма РАА, Пашинг, Австрия) перед центрифугированием в градиенте плотности с использованием лимфоцита млекопитающих (фирма Cedarlane Laboratories, Берлингтон, провинция Онтарио, Канада) согласно спецификациям производителя. РВМС промывали дважды с помощью 1×ЗФР.
Среда EL-4 В5
Использовали среду RPMI 1640 (фирма Pan Biotech, Айденбах, Германия), дополненную 10% FCS (фирма Hyclone, Логан, шт. Юта, США), 2 мМ глутамином, 1% раствором пенициллина/стрептомицина (фирма РАА, Пашинг, Австрия), 2 мМ пируватом натрия, 10 мМ HEPES (фирма PAN Biotech, Айденбах, Германия) и 0,05 мМ b-меркаптоэтанолом (фирма Gibco, Пейсли, Шотландия).
Сенсибилизация планшетов
Стерильные 6-луночные планшеты для культуры клеток сенсибилизировали 2 мкг/мл KLH в карбонатном буфере (0,1 М бикарбонат натрия, 34 мМ динатрийгидрогенкарбоната, рН 9,55) в течение ночи при 4°С. Планшеты перед использованием промывали трижды в стерильном ЗФР. Стерильные покрытые стрептавидином 6-луночные планшеты (фирма Microcoat, Бернрид, Германия) сенсибилизировали смесью 1+1 энантиомеров: изомера А (1 мкг/мл) и изомера Б (1 мкг/мл) биотинилированного ТСМС-Pb-dПЭГ3-биотина в ЗФР в течение 1 ч при комнатной температуре. Перед стадией пэннинга указанные 6-луночные планшеты трижды промывали стерильным ЗФР.
Истощение макрофагов/моноцитов
РВМС высевали на стерильные сенсибилизированные KLH 6-луночные планшеты для истощения макрофагов и моноцитов посредством неспецифической адгезии и для удаления клеток, связывающихся с KLH. Каждую лунку заполняли до максимума 4 мл среды и вплоть до 6×106 РВМС из иммунизированного кролика и давали связываться в течение 1 ч при 37°С и 5% СО2. Клетки, находящиеся в супернатанте (лимфоциты периферической крови (PBL)), применяли на стадии пэннинга антигенов.
Обогащение В-клеток на содержащем Pb ТСМС-энантиомере
В 6-луночные планшеты, сенсибилизированные энантиомерной смесью изомера А и изомера Б ТСМС-Pb-dPEC3-биотина, высевали с плотностью вплоть до 6×10е6 PBL на 4 мл среды и давали связываться с течение 1 ч при 37°С и 5% СО2. Неприкрепившиеся клетки удаляли путем осторожной промывки лунок 1-3 раза с использованием 1×ЗФР. Оставшиеся прикрепленными клетки отделяли путем обработки трипсином в течение 10 мин при 37°С и 5% CO2. Трипсинизацию прекращали, добавляя среду EL-4 В5. Клетки выдерживали на льду до осуществления иммунофлуоресцентного окрашивания.
Иммунофлуоресцентное окрашивание и проточная цитометрия
Анти-IgG-ФИТЦ (фирма AbD Serotec, Дюссельдорф, Германия) применяли для сортировки единичных клеток. Для окрашивания поверхности клетки, полученные на стадии истощения и стадии обогащения, инкубировали с антителом к IgG, конъюгированным с ФИТЦ, в ЗФР и инкубировали в течение 45 мин в темноте при 4°С. После окрашивания РВМС промывали дважды охлажденным на льду ЗФР. В завершение, РВМС ресуспендировали в охлажденном на льду ЗФР и немедленного подвергали FACS-анализу. Добавляли йодид пропидия в концентрации 5 мкг/мл (фирма BD Pharmingen, Сан-Диего, шт. Калифорния, США) перед осуществлением FACS-анализа для разделения мертвых и живых клеток.
Для сортировки единичных клеток применяли устройство Becton Dickinson FACSAria, снабженное компьютером и пакетом программ FACSDiva (фирма BD Biosciences, США).
Культивирование В-клеток
Культивирование кроличьих В-клеток осуществляли согласно методу, описанному у Lightwood и др., J Immunol Methods, 316, 2006, сс. 133-143. В целом, метод состоял в следующем: полученные в результате сортировки единичных клеток кроличьи В-клетки инкубировали в 96-луночных планшетах с использованием 200 мкл/на лунку среды EL-4 В, содержащей клетки Pansorbin® 1:100000) (фирма Calbiochem (Merck), Дармштадт, Германия), 5% супернатанта кроличьих тимоцитов (фирма MicroCoat, Бернрид, Германия) и обработанные гамма-излучением клетки мышиной тимомы EL-4 B5s (5×105 клеток/на лунку), в течение 7 дней при 37°С в инкубаторе. Супернатанты, полученные при культивировании В-клеток, удаляли для скрининга и оставшиеся клетки немедленно собирали и замораживали при -80°С в 100 мкл RLT-буфера (фирма Qiagen, Хильден, Германия).
Пример 3: Экспрессия кроличьего антитела
ПЦР-амплификация V-доменов
Общую РНК получали из лизата В-клеток (ресуспендированного в RLT-буфере (фирма Qiagen, каталожный №79216)), используя набор для РНК NucleoSpin 8/96 (фирма Macherey&Nagel; 740709.4, 740698) согласно протоколу производителя. РНК элюировали с помощью 60 мкл свободной от РНКазы воды. 6 мкл РНК применяли для создания кДНК с использованием реакции с обратной транскриптазой с помощью смеси для синтеза первой цепи (Superscript III First-Strand Synthesis SuperMix) (фирма Invitrogen, 18080-400) и олиго-dT-праймера согласно инструкциям производителя. Все стадии осуществляли с помощью системы Hamilton ML Star. 4 мкл кДНК применяли для амплификации вариабельных областей тяжелой и легкой цепей иммуноглобулина (VH и VL) с помощью AccuPrime Supermix (фирма Invitrogen 12344-040) в конечном объеме 50 мкл, используя праймеры rbHC.up и rbHC.do для тяжелой цепи и rbLC.up и rbLC.do для легкой цепи (таблица 3). Все «прямые» праймеры представляли собой специфические для сигнального пептида праймеры (соответственно VH и VL), а «обратные» праймеры представляли собой специфические для константных областей праймеры (соответственно VH и VL). Условия проведения ПЦР для RbVH+RbVL были следующими: горячий старт при 94°С в течение 5 мин; 35 циклов по 20 с при 94°С, 20 с при 70°С, 45 с при 68°С, и конечное удлинение при 68°С в течение 7 мин.

8 мкл из 50 мкл раствора, в которой осуществляли ПЦР, вносили на 2% гель 48 Е (фирма Invitrogen, G8008-02). Продукты положительных ПЦР-реакций очищали с помощью набора NucleoSpin Extract II (фирма Macherey&Nagel; 740609250) согласно протоколу производителя и элюировали с использованием 5 мкл буфера для элюции. Все стадии очистки осуществляли с помощью системы Hamilton ML Starlet.
Рекомбинантная экспрессия кроличьих моноклональных двухвалентных антител
Для рекомбинантной экспрессии кроличьих моноклональных двухвалентных антител ПЦР-продукты, кодирующие VH или VL, клонировали в виде кДНК в экспрессионных векторах с помощью метода клонирования по выступающим концам (R.S. Haun и др., Biotechniques 13, 1992, сс. 515-518; M.Z. Li и др., Nature Methods 4, 2007, сс. 251-256). Экспрессионные векторы содержали кассету экспрессии, состоящую из 5' CMV-промотора, включая интрон А, и последовательности полиаденилирования 3' BGH. Помимо кассеты экспрессии плазмиды содержали сайт инициации репликации из pUC18 и ген бета-лактамазы, придающий устойчивость к ампициллину для амплификации плазмиды в E.coli. Применяли три варианта основной плазмиды: одну плазмиду, содержащую константную область кроличьего IgG, созданную для включения VH-областей, и две дополнительные плазмиды, содержащие константную область кроличьей или человеческой каппа-LC, для включения VL-областей. Линеризованные экспрессионные плазмиды, кодирующие константную область каппа- или гамма-цепи и VL/VH-вставки, амплифицировали с помощью ПЦР с использованием перекрывающихся праймеров. Очищенные ПЦР-продукты инкубировали с ДНК-полимеразой Т4, которая создавала одноцепочечные выступающие концы. Реакцию прекращали, добавляя dCTP. На следующей стадии плазмиду и вставку объединяли и инкубировали с recA, что индуцировало сайтспецифическую рекомбинацию. Плазмидами, полученными в результате рекомбинации, трансформировали E.coli. На следующий день выросшие колонии собирали и тестировали в отношении правильной рекомбинантной плазмиды после получения плазмиды путем осуществления рестрикционного анализа и ДНК-секвенирования. Для экспрессии антитела выделенными плазмидами, несущими НС и LC, кратковременно совместно трансфектировали клетки FreeStyle HEK293-F (фирма Invitrogen R790-07) в 2 мл (96-луночный планшет), применяя реагент для трансфекции 239-Free (фирма Novagen) согласно процедуре, предложенной поставщиком реагента. Супернатанты собирали через 1 неделю и передавали для очистки.
Пример 4: Селекция кроличьих моноклональных антител
Ниже в таблице вкратце обобщены свойства различных моноклональных двухвалентных кроличьих антител. Антитело PRIT-0128 выбирали в качестве наиболее перспективного кандидата, поскольку оно обладает сопоставимым связыванием с хелатными Pb и Bi, пониженной способностью связываться с другими хелатными металлами и высокой аффинностью (<100 пМ).
SET-анализ (титрование при равновесии в растворе) осуществляли согласно описанному ниже методу.
Приготовление планшета для анализа: 384-луночные стрептавидиновые планшеты (фирма Nunc, Microcoat, №11974998001) инкубировали в течение ночи при 4°С с 25 мкл/на лунку смеси изомеров DOTAM-биотин в ЗФР-буфере в концентрации 20 нг/мл.
Уравновешивание образцов антител к DOTAM с помощью свободных хелатов DOTAM-металл (Pb, Bi, Са, Cu, Zn, Mg, Fe): 0,01-1 нМ антитело титровали с помощью релевантных хелатов DOTAM-металл с использованием стадий разведения 1:3, 1:2 или 1:1,7, начиная с концентрации хелата DOTAM-металл 2500 нМ, 500 нМ или 100 нМ. Образцы инкубировали при 4°С в течение ночи в запечатанных полипропиленовых микропланшетах для хранения REMP (фирма Brooks).
После инкубации в течение ночи стрептавидиновые планшеты промывали 3×90 мл ЗФРТ на лунку. 15 мкл каждого образца из планшета для уравновешивания переносили в планшет для анализа и инкубировали в течение 15 мин при КТ, после чего осуществляли три стадии промывки, используя по 90 мкл ЗФРТ-буфера. Детекцию осуществляли, добавляя 25 мкл козьего антитела к человеческому IgG, конъюгированного с POD (фирма Jackson, 109-036-088, разведение 1:4000 в OSEP), после чего осуществляли три стадии промывки, используя по 90 мкл ЗФРТ-буфера. В каждую лунку добавляли по 25 мкл ТМВ-субстрата (фирма Roche Diagnostics GmbH, каталожный №11835033001). Измерения осуществляли при 370/492 нм с помощью ридера Safire2 (фирма Tecan).
Материалы:
1. Смесь изомеров DOTAM-биотин:
Смесь включала следующие компоненты в концентрации 20 нг/мл:
- Pb-DOTAM-Bn-биотин/ТСМС-Pb-dПЭГ3-биотин, изомер А
- Pb-DOTAM-Bn-биотин/ТСМС-Pb-dПЭГ3-биотин, изомер Б
- Pb-DOTAM-алкил-биотин, изомер А
- Pb-DOTAM-алкил-биотин, изомер Б.
2. ЗФР: DЗФР, фирма PAN, Р04-36500.
3. БСА (BSA): фирма Roche, 10735086001.
4. Твин 20: полисорбат 20 (фирма usb, №20605, 500 мл).
5. ЗФРТ: 10×, фирма Roche, №11666789001/0,1% Твин 20.
6. OSEP: ЗФР (10×, фирма Roche, №11666789001/0,5% БСА (бычий сывороточный альбумин, фракция V, без жирных кислот, фирма Roche, №10735086001)/0,05% Твин 20.
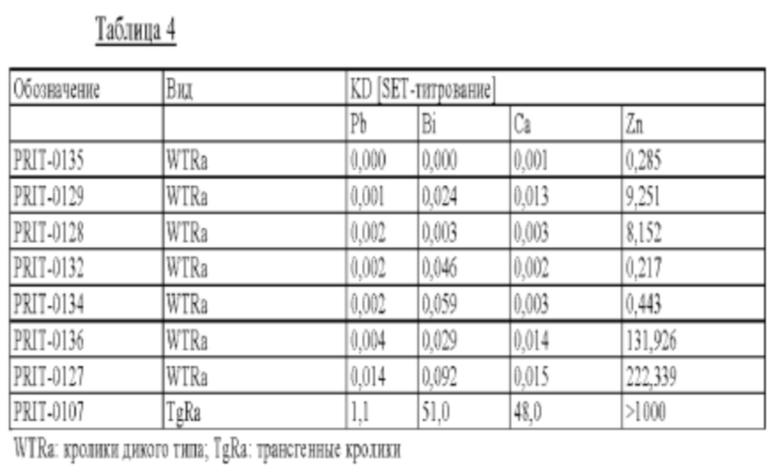
Пример 5: Молекулярная биология
Методы рекомбинантной ДНК
Для манипуляций с ДНК использовали стандартные методы, описанные у Sambrook J. и др., Molecular cloning: A laboratory manual; изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии применяли согласно инструкциям производителей.
Синтез генов и олигонуклеотидов
Требуемые сегменты генов получали путем химического синтеза на фирме Geneart AG (Регенсбург, Германия). Для размножения/амплификации синтезированные фрагменты генов клонировали в плазмидах Е. coli. Последовательности ДНК субклонированных фрагментов генов подтверждали с помощью ДНК-секвенирования. Альтернативно этому, короткие синтетические ДНК-фрагменты собирали путем отжига химически синтезированных олигонуклеотидов или с помощью ПЦР. Соответствующие олигонуклеотиды получали на фирме Metabion GmbH (Планегг-Мартинсрид, Германия).
Определение белка
Концентрацию белка очищенных полипептидов определяли на основе оптической плотности (ОП) при 280 нм, используя коэффициент молярной экстинкции, рассчитанный на основе аминокислотной последовательности полипептида.
Создание плазмид для рекомбинантной экспрессии тяжелых или легких цепей антител
Требуемые белки экспрессировали путем кратковременной трансфекции клеток почки человеческого эмбриона (HEK 293). Для экспрессии требуемого гена/белка (например, тяжелой цепи полноразмерного антитела, легкой цепи полноразмерного антитела или тяжелой цепи полноразмерного антитела, содержащей дополнительный домен (например, вариабельного домена тяжелой или легкой цепи иммуноглобулина на его С-конце) применяли транскрипционную единицу, содержащую следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5'UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность (SS) тяжелой цепи мышиного иммуноглобулина,
- подлежащий экспрессии ген/белок и
- последовательность полиаденилирования бычьего гормона роста (BGH рА).
Добавляли к экспрессионной единице/кассете, включающей требуемый подлежащий экспрессии ген, базовую/стандартную экспрессионную плазмиду для клеток млекопитающих, которая содержала сайт инициации репликации из pUC18, позволяющий осуществлять репликацию этой плазмиды в Е. coli, и ген бета-лактамазы, придающий устойчивость к ампициллину в E.coli.
а) Экспрессионная плазмида для тяжелых цепей антител
Гены, кодирующие тяжелую цепь антитела, включая С-концевые слитые гены, содержащие полную и функциональную тяжелую цепь антитела, с расположенным далее дополнительным V-доменом тяжелой или V-доменом легкой цепи антитела, объединяли путем слияния ДНК-фрагмента, кодирующего соответствующие элементы последовательности (V-домен тяжелой или V-домен легкой цепи), каждый из которых отделен G4Sx4-линкером, с С-концом СН3-домена молекулы человеческого IgG (VH-СН1-шарнир-СН2-СН3-линкер-VH или VH-СН1-шарнир-СН2-СН3-линкер-VL). Молекулы рекомбинантных антител, несущие один VH- и один VL-домен на С-концах двух СН3-доменов соответственно, экспрессировали с использованием технологии knob-into-hole.
Экспрессионные плазмиды для кратковременной экспрессии тяжелой цепи антитела с С-концевым VH- или VL-доменом в клетках HEK293, содержали помимо кассеты экспрессии для фрагмента тяжелой цепи антитела с С-концевым VH- или VL-доменом, сайт инициации репликации из pUC18, позволяющий осуществлять репликацию этой плазмиды в Е. coli, и ген бета-лактамазы, придающий устойчивость к ампициллину в E.coli. Транскрипционная единица слитого гена фрагмента тяжелой цепи антитела с С-концевым VH- или VL-доменом содержала следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5'UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность тяжелой цепи мышиного иммуноглобулина,
- нуклеиновую кислоту, кодирующую тяжелую цепь антитела (VH-CH1-шарнир-СН2-СН3-линкер-VH или VH-СН1-шарнир-СН2-СН3-линкер-VL), и
- последовательность полиаденилирования бычьего гормона роста (BGH рА).
Экспрессионные плазмиды, кодирующие полипептиды/белки всех тяжелых цепей, указанных в таблице 2, конструировали согласно изложенным выше методам.
б) Экспрессионная плазмида для легких цепей антитела
Гены, кодирующие легкие цепи антитела, содержащие полную и функциональную легкую цепь антитела, объединяли путем слияния ДНК-фрагмента, кодирующего соответствующие элементы последовательности.
Экспрессионная плазмида для кратковременной экспрессии легкой цепи содержала помимо фрагмента легкой цепи сайт инициации репликации из pUC18, позволяющий осуществлять репликацию этой плазмиды в Е. coli, и ген бета-лактамазы, придающий устойчивость к ампициллину в E.coli. Транскрипционная единица фрагмента легкой цепи антитела содержала следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5'UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность тяжелой цепи мышиного иммуноглобулина,
- нуклеиновую кислоту, кодирующую легкую цепь антитела (VL-CL),
- последовательность полиаденилирования бычьего гормона роста (BGH рА).
Экспрессионные плазмиды, кодирующие полипептиды/белки всех легких цепей, указанных в таблице 2, конструировали согласно изложенным выше методам.
Схематическое изображение применяемого формата представлено на фиг. 1. Звездочкой обозначены PGLALA-замены: цифрой 1 обозначен связывающий DOTAM агент, а цифрами 2 и 3 обозначены связывающие мишень агенты (в данном случае антитела к СЕА).
Пример 6: Кратковременная экспрессия PRIT-молекул
Кратковременная экспрессия молекул антител
Молекулы антител создавали в кратковременно трансфектированных клетках HEK293 (линия клеток почки человеческого эмбриона, полученная из 293), которую культивировали в среде F17 (фирма Invitrogen Corp.). Для трансфекции применяли трансфектирующий реагент «293-Free» (фирма Novagen). Соответствующие молекулы тяжелых и легких цепей антител, описанные выше, экспрессировали из индивидуальных экспрессионных плазмид. Трансфекции осуществляли согласно инструкциям производителя. Супернатанты клеточных культур, содержащие иммуноглобулин, собирали через три-семь (3-7) дней после трансфекции. Супернатанты хранили при пониженной температуре (например, -80°С) вплоть до очистки.
Общая информация, касающаяся рекомбинантной экспрессии человеческих иммуноглобулинов, например в HEK293-клетках, представлена у: Meissner Р. и др., Biotechnol. Bioeng. 75, 2001, сс. 197-203.
Пример 7: Очистка белков
Собранный супернатант клеточной культуры вносили на колонку (диаметр 1,1 см, длина 5 см), заполненную 5 мл смолы с белком А (смола Mab Select Sure), со скоростью потока 5 мл/мин. После промывки с использованием 20 мл ЗФР-буфера антитело элюировали с использованием 25 мл Na-цитрата, рН 3,0.
Затем значение рН элюата доводили до рН 5,0 с помощью 1 М Трис, рН 9,0 и инкубировали при 4°С в течение ночи.
После центрифугирования в течение 10 мин при 10000×g и фильтрации через фильтр с размером пор 0,2 мкм фильтрат вносили на колонку для гель-фильтрации Супердекс 200 (диаметр 2,6 см, длина 60 см), предварительно уравновешенную буфером, содержащим 20 мМ гистидин, 140 мМ NaCl, рН 6,0, и элюировали таким же буфером.
Полученный после элюции основной пик, содержащий очищенное антитело, собирали и анализировали конечную чистоту.
Пример 8: FACS
MKN-45 клетки открепляли от культурального флакона с помощью трипсина и подсчитывали количество с помощью счетчика клеток Casy. После пеллетирования при 4°С, 300×g клетки ресуспендировали в FACS-буфере (2,5% FCS в ЗФР), доводили количество до 2,0Е+06 клеток/мл, вносили в 96-луночный РР-планшет с V-образным дном (25 мкл/на лунку=5,0Е+04 клеток/на лунку).
FACS-окрашивание с использованием DOTAM-ФИТЦ
Концентрацию первичных СЕА-специфических антител доводили до 40 мкг/мл в FACS-буфере с получением конечной концентрации 10 мкг/мл. RS-CEA-Il2v применяли в качестве референс-антитела. Хелат Pb-DOTAM, меченный ФИТЦ, и первичные антитела применяли в эквимолярных соотношениях. Их смешивали и инкубировали в течение 10 мин при КТ, давая связываться антителам с Pb-DOTAM. Затем 20 мкл полученной смеси добавляли к 25 мкл клеточной суспензии и инкубировали в течение 1 ч при 4°С. Затем клетки промывали дважды FACS-буфером и ресуспендировали, используя 70 мкл/на лунку FACS-буфера, для оценки с помощью устройства FACS Canto (фирма BD, Pharmingen).
FACS-окрашивание с использованием <hu каппа>
Концентрацию первичных СЕА-специфических антител доводили до 20 мкг/мл в FACS-буфере с получением конечной концентрации 10 мкг/мл. RS-CEA-Il2v применяли в качестве референс-антитела. Добавляли 20 мкл к 25 мкл клеточной суспензии и инкубировали в течение 1 ч при 4°С. Затем клетки промывали дважды FACS-буфером. После промывки клетки ресуспендировали в 50 мкл FACS-буфера, содержащего вторичное антитело (<huIgG>-Alexa488, с=10 мкг/мл), и инкубировали в течение 1 ч при 4°С. Затем клетки промывали дважды FACS-буфером и ресуспендировали, используя 70 мкл/на лунку FACS-буфера, для оценки с помощью устройства FACS Canto (фирма BD, Pharmingen).
На фиг. 36 представлен пример, демонстрирующий связывание одного антитела (PRIT-0165) с клетками MKN-4, для обнаружения которого применяли либо вторичное антитело (правая панель, Alexa 488), либо DOTAM-ФИТЦ (левая панель, ФИТЦ-А).
Пример 9: Гуманизация
Для идентификации приемлемого человеческого акцепторного каркасного участка в процессе гуманизации антитела PRIT-0128, связывающего DOTAM, использовали сочетание двух методологий. С одной стороны, применяли классический подход для поиска акцепторного каркасного участка с высокой гомологией последовательности с родительским антителом и последующей трансплантации CDR-участков в указанный акцепторный каркасный участок. Каждое отличие аминокислот в идентифицированных каркасных участках относительно родительского антитела оценивали в отношении воздействия на структурную целостность связывающего агента и всякий раз при необходимости интродуцировали обратные мутации в родительскую последовательность.
С другой стороны, в лаборатории заявителей был разработан инструмент для прогнозирования in silico ориентации VH- и VL-доменов гуманизированных версий друг относительно друга (см. WO 2016/062734). Его осуществляли для виртуальных трансплантаций CDR во все возможные комбинации человеческой зародышевой линии. Результаты сравнивали с ориентацией VH-VL-доменов родительского связывающего агента для выбора комбинаций каркасных участков, близких по геометрии к исходному антителу.
В каждом случае следующие CDR-области родительского антитела трансплантировали в акцепторный каркасный участок (нумерация согласно Кэботу):
VH_CDR1: 31-35
VH_CDR2: 50-65
VH_CDR3: 95-102
VL_CDR1: 24-34
VL_CDR2: 50-56
VL_CDR3: 89-97
Гуманизированные варианты получали в окончательном формате, в котором связывающий DOTAM агент сливали с С-концом Fc таргетирующего опухоль IgG в виде слияния VH/VL Fv (без СН1 и Ck соответственно). Родительский (негуманизированный) связывающий DOTAM агент PRIT-0128, полученный из молекулы, имеющий окончательный формат, обозначали как PRIT-0156.
Включали каркасный участок герцептина из-за его прогнозируемой пригодности для VH/VL и повышенной стабильности каркасного участка. Для всех гуманизированных вариантов VH применяли человеческий J-элемент hJH2. Для всех гуманизированных вариантов VK применяли человеческий J-элемент hJK4.
НС4 представлял собой результат трансплантации PRIT-128 в человеческую зародышевую линию IGHV3-30-02 с одной обратной мутацией A49G (нумерация согласно Кэботу).
Для получения НС5 вариабельной тяжелой цепи CDR трансплантировали в человеческую зародышевую линию hVH_2_26 с A49G в качестве обратной мутации и делецией первой аминокислоты для соответствия исходному N-концу кроличьего антитела.
Трансплантированный в V-область герцептина (полученную из человеческой зародышевой линии hVH3_66), вариант НС7 характеризовался наличием нескольких модификаций в акцепторном каркасном участке: делецией на N-конце Е, A49G, A71R и S93A.
Для получения НС10 CDR PRIT-128 трансплантировали в человеческую зародышевую линию IGHV4_34_01.
При этом N-конец модифицировали, начиная с V2, для соответствия исходному кроличьему антителу, начиная с Q2. Кроме того, G29F и F31L рассматривали в качестве обратной мутации (согласно номенклатуре Кэбота), а также V71R и F78V в каркасном участке 3.
Для легкой цепи LC1 CDR трансплантировали в человеческую зародышевую линию IGKV1_39_01 без какой-либо обратной мутации. В качестве начала выбирали 12 для соответствия исходному кроличьему Ат, начинающемуся с А2.
Вариант легкой цепи LC3 получали путем трансплантации CDR в человеческую зародышевую линию hVK1_5. D1 удаляли путем делеции и обратную мутацию 12A рассматривали в качестве нового N-конца. В качестве дополнительных обратных мутаций рассматривали K42Q и А43Р.
Получали не все возможные комбинации матрицы гуманизации, но выбор определенных комбинаций был основан на таких соображениях как предсказание VH/VL и последующие риски, связанные с данной комбинацией.
Целью гуманизации было получение гуманизированных связывающих агентов, у которых аффинность к DOTAM не снижалась более чем в 10 раз, и которые обладали повышенной стабильностью, если это было возможно. Этой цели удалось достичь для нескольких связывающих агентов, которые обладали сопоставимой или даже более высокой аффинностью к DOTAM а также увеличенной термостабильностью, которую измеряли методом DLS (динамическое рассеяние света) примерно при 10-15°С (см. таблицы 7 и 8, ниже).
Пример 10: Определение kd в условиях равновесия в растворе
Для скрининга большого количества гуманизированных кандидатов в отношении их аффинности к Pb-DOTAM применяли метод титрования при равновесии в растворе (SET).
В таблице 6 приведены детали определения аффинности на основе метода SET отобранных DOTAM-связывающих агентов в отношении Pb-DOTAM. Все антитела, приведенные в таблице 6, представляют собой биспецифические антитела, которые характеризуются двухвалентным связывание с СЕА и одновалентным связыванием с Pb-DOTAM (формат 2:1, см. фиг. 1):
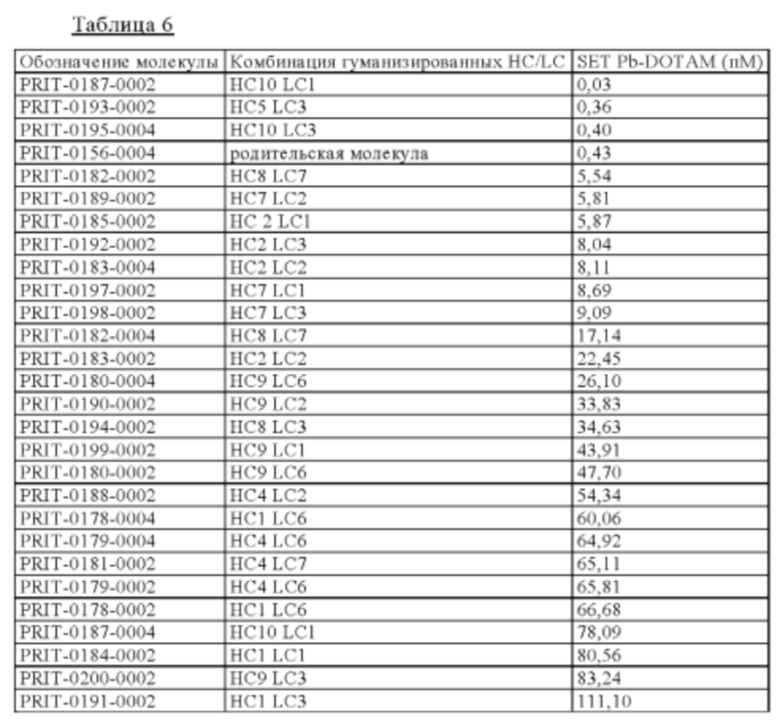
Пример 11: Определение kd на основе метода Kinexa
Для более подробного анализа и определения аффинности ортогональным методом применяли метод Kinexa.
Инструментарий и материалы
Применяли устройство KinExA 3200 фирмы Sapidyne Instruments (Бойсе, шт. Айдахо) с автосэмплером. Полиметилметакрилатные (РММА) гранулы покупали у фирмы Sapidyne, а ЗФР (забуференный фосфатом физиологический раствор), БСА (фракция V, бычий сывороточный альбумин) и антитела к DOTAM получали в лаборатории заявителей (фирма Roche). Конъюгированное с Dylight650® очищенное с помощью аффинной хроматографии козье антитело к человеческому IgG, перекрестно адсорбированное с Fc-фрагментом, покупали у фирмы Bethyl Laboratories (Монтгомери, шт. Техас). Биотинилированные антигены Pb-DOTAM (Pb-DOTAM-алкил-биотин, изомеры А и Б, Pb-DOTAM-Bn-биотин/ТСМС-Pb-dПЭГ3-биотин, изомеры А и Б) и небиотинилированный Pb-DOTAM получали от фирмы AREVA Med (Бетезда, шт. Мэриленд).
Получение покрытых антигеном гранул
На РММА-гранулы наносили покрытие согласно протоколу, указанному в справочнике KinExA для биотинилированных молекул (фирма Sapidyne). В целом, метод состоял в следующем: во-первых, добавляли биотин-БСА (фирма Thermo Scientific) в 1 мл ЗФР (рН 7,4) из расчета по 10 мкг на флакон (200 мг) гранул для нанесения адсорбирующего покрытия. После вращения в течение 2 ч при комнатной температуре супернатант удаляли и гранулы промывали 5 раз 1 мл ЗФР. Во-вторых, к гранулам добавляли 1 мл 100 мкг нейтравидина, биотин-связывающего белка (фирма Thermo Scientific) в ЗФР, содержащем 10 мг/мл БСА, и инкубировали при комнатной температуре еще в течение 2 ч для связывания нейтравидина с гранулами и для создания дополнительных связывающих биотин сайтов для последующего связывания биотинилированных белков. Затем покрытые нейтравидином гранулы промывали 5 раз 1 мл ЗФР. В завершение, на гранулы наносили покрытие с использованием 200 нг/мл смеси биотинилированных изомеров Pb-DOTAM (по 50 нг каждого изомера) в ЗФР и инкубировали еще в течение 2 ч при комнатной температуре. Затем гранулы ресуспендировали в 30 мл ЗФР и сразу же использовали.
Анализы методом KinExA в условиях равновесия
Все эксперименты по методу KinExA осуществляли при комнатной температуре (КТ) с использованием ЗФР, рН 7,4 в качестве подвижного буфера. Образцы приготавливали в подвижном буфере, дополненном 1 мг/мл БСА («буфер для образца»). Скорость потока составляла 0,25 мл/мин. Постоянное количество антитела к DOTAM с концентраций сайтов связывания 5 пМ титровали с использованием двукратных серийных разведений антигена Pb-DOTAM, начиная с концентрации 100 пМ (диапазон концентраций 0,049-100 пМ). Один образец антитела без антигена служил в качестве образца, дающего 100% сигнал (т.е. без ингибирования). Комплексы антиген-антитело инкубировали при КТ в течение по меньшей мере 24 ч, обеспечивая достижение равновесия. Затем уравновешенные смеси пропускали через колонку, содержащую сшитые с Pb-DOTAM гранулы, в KinExA-системе в объеме 5 мл, что позволяло несвязанному антителу «захватываться» гранулами без нарушения состояния равновесия раствора. Для детекции «захваченного» антитела использовали 250 нг/мл конъюгированного с Dylight 650© специфического для человеческого Fc-фрагмента вторичного антитела в буфере для образца. Каждый образец оценивали с дублированием во всех экспериментах по уравновешиванию.
Величины KD определяли с помощью нелинейного регрессионного анализа данных с использованием модели односайтового гомогенного связывания, входящей в пакет программ KinExA (версия 4.0.11), используя метод «стандартного анализа». Программное обеспечение позволяло рассчитывать величины KD и определять 95%-ный доверительный интервал на основе аппроксимации экспериментальных точек теоретической кривой KD. 95%-ный доверительный интервал (Sapidyne TechNote TN207R0) представлен в виде величин KDlow и KDhigh.
Ниже представлены другие примеры величин аффинности, определенных методом Kinexa, для PRIT-213. PRIT-0213 представляет собой ту же самую молекулу, что и PRIT-0186, за исключением другой связывающей СЕА комбинации VH/VL, см. таблицу 8.
PRIT-213
Аффинности CEA-DOTAM BsAb в отношении хелата металл-DOTAM
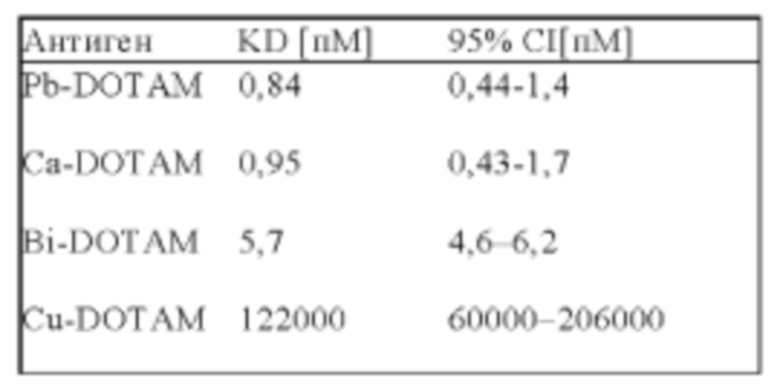
Дополнительные параметры приведены ниже:
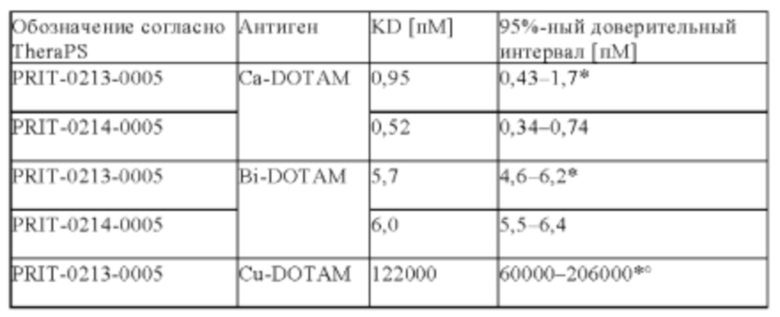
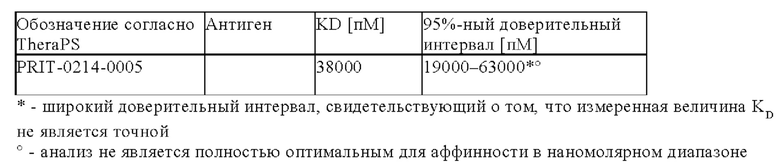
Пример 12: Измерения термостабильности гуманизированных PRIT-молекул
Метод и анализ данных
Различные варианты гуманизированных PRIT-молекул в окончательном формате (в 20 мМ гистидине, 140 мМ NaCl, рН 6,0) разводили в одном и том же буфере до концентрации 1 мг/мл. По 30 мкл каждого образца переносили в 384-луночный планшет фильтровального устройства (наряду с антителом к HER3 в качестве референс-антитела). После центрифугирования при 1000×g в течение 1 мин в лунки наслаивали по 10 мкл парафинового масла. Планшет снова центрифугировали (1000×g в течение 1 мин) и переносили в планшет-ридер для DLS (Dyna Pro PlateReader-II, фирма Wyatt). Температуру повышали, начиная с 25°С, до 79,9°С со скоростью 0,05°С/мин. Рассеянный свет регистрировали использованием программы Dynamics Software (V7.0).
Данные переносили в базу Excel (фирма Microsoft), сортировали в зависимости от образца и температуры и применяли пакет программ надстройки для создания кривых плавления. Температуру, при которой наблюдалось выраженное отклонение от исходного уровня, определяли как «начало агрегации», а точку излома на кривой плавления как «температуру плавления».
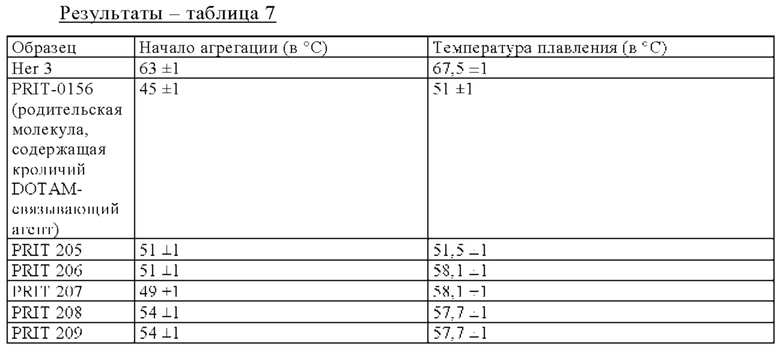

Пример 13: Отбор кандидатов
PRIT-0156 представляет собой антитело в формате 2:1, содержащее кроличий DOTAM-связывающий агент PRIT-0128, объединенный с СЕА-связывающим агентом СН1А1А. PRIT-0178-PRIT-0204 представляют собой гуманизированные варианты одинакового формата с одинаковым СЕА-связывающим агентом. PRIT-0205-PRIT-0221 соответствуют гуманизированным вариантам PRIT-0178-PRIT-0204 в DOTAM-связывающей области, но в них СЕА-связывающий агент заменен на Т84.66.
В приведенной таблице ниже представлено сравнение свойств различных PRIT-молекул. Предпочтительными соединениями являются PRIT-213 и PRIT-214.
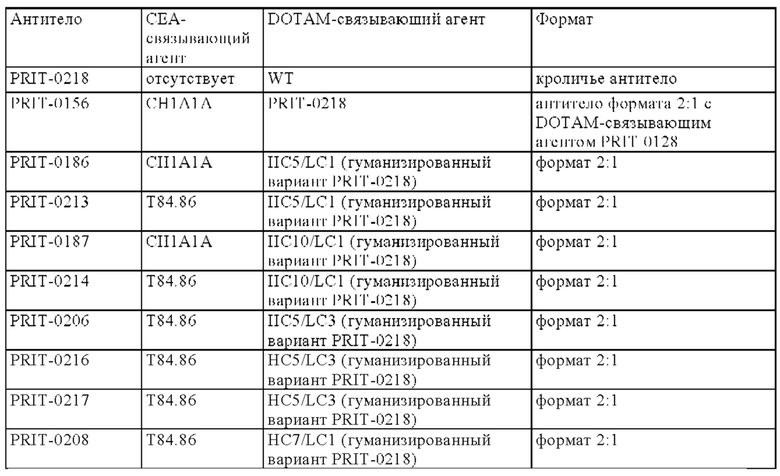
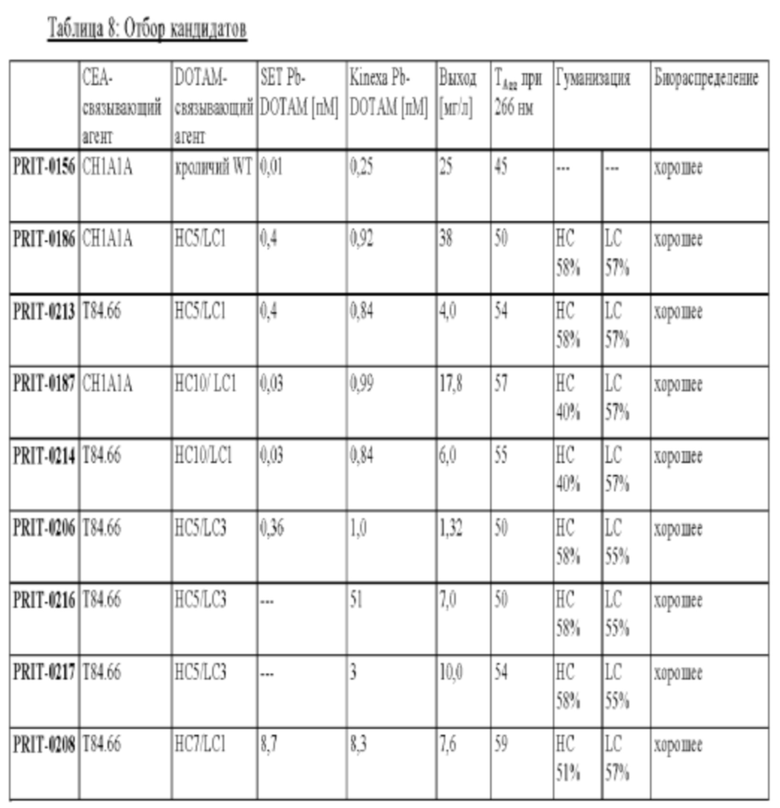
Последовательности
НС5:

LC1:
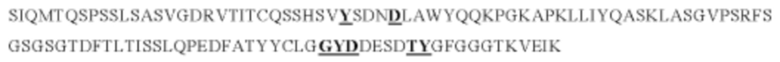
LC3: 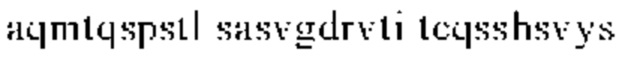
НС7: 
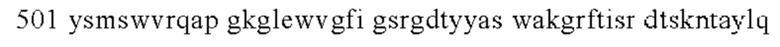
НС10: 
T84.66 VL:
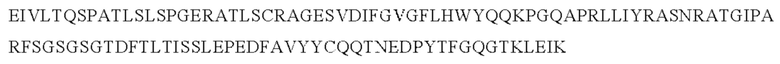
CH1A1A VH:
СН1А1А VL:
Пример 14: Кристаллизация, сбор данных и определение структуры комплекса Fab Р1АА1227 Pb-DOTAM
Для формирования комплекса смешивали Fab, полученный из гуманизированного VH/VL PRIT-0213, который обозначали как Р1АА1227, в концентрации 26 мг/мл с порошкообразным Pb-DOTAM в молярном соотношении 1:4.2. После инкубации в течение 2 ч при 4°С осуществляли начальные опыты по кристаллизации в устройствах для диффузии паров по методу «сидячей» капли при 21°С с использованием экрана JCSG+ (фирма Qiagen, Хильден). Кристаллы формировались в течение 5 дней из 0,2 М (NH4)2SO4, 0,1 М Бис-Трис, рН 5,5, 25 мас./об. % ПЭГ3350. Кристаллы собирали непосредственно с планшета для скрининга без применения дополнительной стадии оптимизации.
Сбор данных и определение структуры. Для сбора данных кристаллы мгновенно замораживали при 100 K в растворе для осаждения, содержащем 10% этиленгликоля. Данные о дифракции собирали при длине волны 1,0000  с использованием детектора PILATUS 6М для электронного пучка X10SA источника синхротронного излучения (Швейцарский источник света (Swiss Light Source) (Виллиген, Швейцария). Данные обрабатывали с помощью XDS (Kabsch W., Acta Cryst. D66, 2010, сс. 133-144) и стандартизовали с помощью SADABS (фирма BRUKER). Кристаллы комплекса относились к пространственной группе С2 с осями элементарной ячейки а=135,63
с использованием детектора PILATUS 6М для электронного пучка X10SA источника синхротронного излучения (Швейцарский источник света (Swiss Light Source) (Виллиген, Швейцария). Данные обрабатывали с помощью XDS (Kabsch W., Acta Cryst. D66, 2010, сс. 133-144) и стандартизовали с помощью SADABS (фирма BRUKER). Кристаллы комплекса относились к пространственной группе С2 с осями элементарной ячейки а=135,63  b=56,42
b=56,42  с=64,52
с=64,52  и β=108,36°, картину дифракции получали с разрешением 1,40
и β=108,36°, картину дифракции получали с разрешением 1,40  Структуру определяли методом молекулярного замещения с помощью программы PHASER (McCoy A.J, Grosse-Kunstleve R.W., Adams P.D., Storoni L.C. и Read R.J., J. Appl. Cryst. 40, 2007, cc. 658-674) с использованием координат разработанной в лаборатории заявителей структуры Fab в качестве поисковой модели. При уточнении реальной пространственной структуры использовали различия в электронной плотности для определения положения Pb-DOTAM и изменения аминокислот в соответствии с различиями последовательностей. Структуры уточняли с помощью программ из пакета ССР4 (Collaborative Computational Project, Number 4 Acta Cryst. D50, 1994, cc. 760-763) и BUSTER (Bricogne G., Blanc E., Brandl M., Flensburg C., Keller P., Paciorek W., Roversi P., Sharff A., Smart O.S., Vonrhein C., Womack T.O, 2011, Buster, версия 2.9.5 Cambridge, United Kingdom: Global Phasing Ltd.). Перестроение вручную осуществляли с помощью программы COOT (Emsley P., Lohkamp В., Scott W.G. и Cowtan K., Acta Cryst D66, 2010, cc. 486-501).
Структуру определяли методом молекулярного замещения с помощью программы PHASER (McCoy A.J, Grosse-Kunstleve R.W., Adams P.D., Storoni L.C. и Read R.J., J. Appl. Cryst. 40, 2007, cc. 658-674) с использованием координат разработанной в лаборатории заявителей структуры Fab в качестве поисковой модели. При уточнении реальной пространственной структуры использовали различия в электронной плотности для определения положения Pb-DOTAM и изменения аминокислот в соответствии с различиями последовательностей. Структуры уточняли с помощью программ из пакета ССР4 (Collaborative Computational Project, Number 4 Acta Cryst. D50, 1994, cc. 760-763) и BUSTER (Bricogne G., Blanc E., Brandl M., Flensburg C., Keller P., Paciorek W., Roversi P., Sharff A., Smart O.S., Vonrhein C., Womack T.O, 2011, Buster, версия 2.9.5 Cambridge, United Kingdom: Global Phasing Ltd.). Перестроение вручную осуществляли с помощью программы COOT (Emsley P., Lohkamp В., Scott W.G. и Cowtan K., Acta Cryst D66, 2010, cc. 486-501).
Собранные данные и полученные при уточнении статистические данные обобщены в таблице 9.
Все графические презентации получали с помощью системы PYMOL (The Pymol Molecular Graphics System, версия 1.7.4.  LLC.).
LLC.).


Структура Fab Р1АА1227 в комплексе с Pb-Dotam
Для детальной характеризации взаимодействия Pb-Dotam с Fab Р1АА1227 при создании изобретения определяли кристаллическую структуру комплекса с разрешением 1,40 Анализ структуры выявил, что основной вклад в связывание Fab Р1АА1227 с Pb-DOTAM вносят CDR1 и CDR3 легкой цепи и CDR2 и CDR3 тяжелой цепи.
Анализ структуры выявил, что основной вклад в связывание Fab Р1АА1227 с Pb-DOTAM вносят CDR1 и CDR3 легкой цепи и CDR2 и CDR3 тяжелой цепи.
Анализ связывающихся поверхностей раздела с помощью программы PISA выявил схему взаимодействия Fab Р1АА1227 с Pb-DOTAM, включающую 3 водородные связи, полярные взаимодействия и контакты, обусловленные Ван-дер-Ваальсовыми силами. Pb-DOTAM связывается в кармане, образованном тяжелой и легкой цепями. Этот карман имеет форму коробки, открытой с одной стороны. Боковые стенки и дно кармана участвуют в неполярных взаимодействиях, в то время как на кромке стенок превалируют полярные взаимодействия. Водородные связи боковой цепи формируются между остатками Glu95 и Asp97 CDR3 тяжелой цепи и карбамоильными атомами азота N7 и N8 DOTAM. Дополнительная водородная связь формируется между основным карбонильным атомом Arg96 и атомом N7 DOTAM. Комплекс дополнительно стабилизируется посредством неполярных взаимодействий Phe50 и Tyr58 CDR2 тяжелых цепей, боковые цепи которых ориентированы по типу «ребро-к грани» относительно азациклододеканового кольца. Легкая цепь в основном формирует «дно» кармана с помощью остатков Gly91-Tyr96 CDR3, обеспечивающих неполярные контакты с тетрациклододекановым кольцом. Asp32 обеспечивает водородную связь с карбамоильным атомом азота N6 DOTAM (нумерация согласно Кэботу).
На фиг. 2 представлена структура Р1АА1227 в комплексе с Pb-DOTAM.
На фиг. 3 представлено схематическое изображение сайта взаимодействия.
В приведенной ниже таблице представлены остатки паратопа в тяжелой цепи, выявленные на основе анализа с помощью программы PISA.
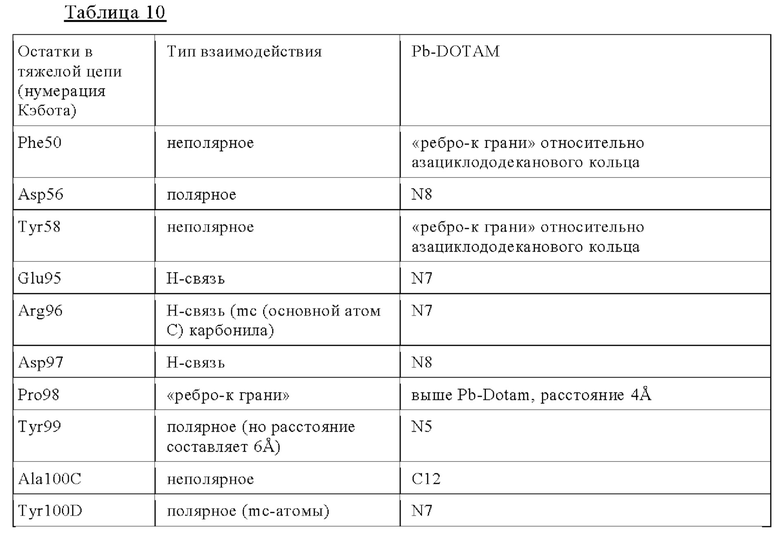
В приведенной ниже таблице представлены остатки паратопа в легкой цепи, выявленные на основе анализа с помощью программы PISA.
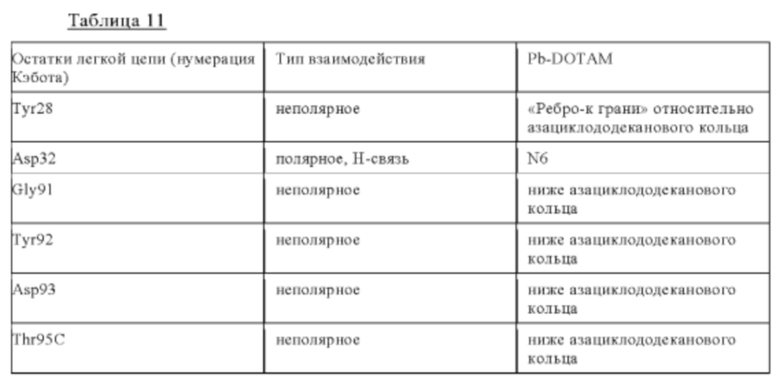

В приведенных ниже последовательностях остатки паратопа подчеркнуты:
> Р1АА1227_НС

> P1AA1227_LC

Пример 15: Биораспределение и эффективность in vivo; материалы и методы
Словарь сокращений, применяемых в приведенных ниже примерах


Материал и методы протоколов
Общие сведения
Все экспериментальные протоколы были рассмотрены и одобрены местными регулирующими органами (
 d'Ethique de
d'Ethique de  Animale du Limousin (CREEAL), Laboratoire
Animale du Limousin (CREEAL), Laboratoire  d'Analyses et de Recherches de la Haute-Vienne). Самок мышей с серьезным комбинированным иммунодефицитом (SCID) (фирма Charles River) содержали в свободных от специфических патогенов условиях с суточными циклами свет/темнота (12 ч/12 ч) в соответствии с руководствами по этике. Никаких манипуляций не осуществляли в течение первой недели после доставки, давая животным акклиматизироваться к новой окружающей среде. Осуществляли ежедневный мониторинг физического состояния и общего состояния здоровья всех мышей.
d'Analyses et de Recherches de la Haute-Vienne). Самок мышей с серьезным комбинированным иммунодефицитом (SCID) (фирма Charles River) содержали в свободных от специфических патогенов условиях с суточными циклами свет/темнота (12 ч/12 ч) в соответствии с руководствами по этике. Никаких манипуляций не осуществляли в течение первой недели после доставки, давая животным акклиматизироваться к новой окружающей среде. Осуществляли ежедневный мониторинг физического состояния и общего состояния здоровья всех мышей.
Объемы опухолей определяли с помощью кронциркуля, осуществляя расчет по следующей формуле: объем=0,5×длина×ширина2. Кровь собирали по окончанию эксперимента из венозного синуса, используя ретро-орбитальное кровопускание, после чего изымали дополнительные ткани для измерений радиоактивности и/или гистологического анализа, в соответствии с протоколом. Неожиданные или аномальные состояния документировали.
Статистический анализ осуществляли с использованием программы GraphPad Prism 6 (фирма GraphPad Software, Inc.) и JMP 8 (фирма SAS Institute Inc.).
Реагенты
Биспецифические антитела получали от фирмы Roche Diagnostics GmbH, Pharma Research Penzberg (Пенцберг, Германия) и хранили при -80°С до дня осуществления инъекции. Затем их подвергали оттаиванию и разводили в стандартном применяемом в качестве наполнителя буфере (20 мМ гистидин/гистидин×HCl, 140 мМ NaCl; рН 6,0).
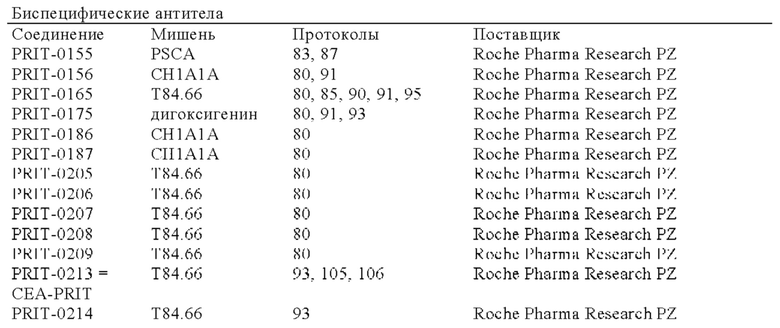
Очищающие реагенты получали от фирмы Macrocyclics (Плано, шт. Техас, США). Их хранили при -80°С до дня осуществления инъекции и подвергали оттаиванию и разводили в ЗФР до требуемой концентрации.
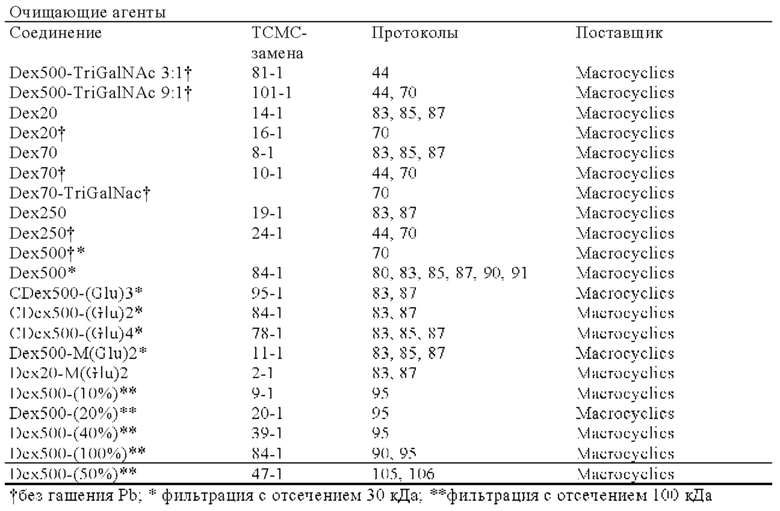
Хелаты DOTAM для радиоактивного мечения получали от фирмы Macrocyclics и выдерживали при 20°С до осуществления радиоактивного мечения. Последующее мечение либо свинцом-203 (203Pb), либо свинцом-212 (212Pb) осуществляли на фирме AREVA Med (Разес, Франция). Мышам инъецировали внутривенно (i.v.) 100 мкл соответствующих растворов Pb-DOTAM, разведенных ЗФР, для получения требуемой дозы РЬ/активной концентрации. 203Pb-DOTAM применяли предварительно связанным с биспецифическими антителами, в то время как 212Pb-DOTAM вводили после PRIT и очищающего агента. Измерения радиоактивности осуществляли с помощью автоматического гамма-счетчика 2470 WIZARD2 (фирма PerkinElmer).

ВхРС3 представляет собой клеточную линию человеческой первичной аденокарциномы поджелудочной железы, которая в естественных условиях экспрессирует СЕА. Клетки ВхРС3 культивировали в среде RPMI-1640 (фирма Gibco, каталожный №42401-018), обогащенной 10% фетальной бычьей сыворотки и 1% GlutaMAX (фирма Gibco, каталожный №35050-061). LS174T представляет собой клеточную линию человеческой колоректальной аденокарциномы, которая в естественных условиях экспрессирует СЕА. Клетки LS174T культивировали в среде DMEM (фирма Gibco, каталожный №42430-082), обогащенной 10% фетальной бычьей сыворотки. MKN45 представляет собой клеточную линию человеческой аденокарциномы желудка, которая в естественных условиях экспрессирует СЕА. Клетки MKN45 культивировали в среде RPMI-1640 (фирма Gibco, каталожный №42401-018), обогащенной 20% фетальной бычьей сыворотки. Ксенотрансплантаты солидных опухолей создавали путем подкожной инъекции в правую боковую область клеток в среде RPMI или DMEM, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №.354230).


Пример 16: Претаргетинг с использованием различных направленных против СЕА биспецифических антител (протокол 80 (а, б, в))
Целью настоящего исследования была оценка накопления РЬ в ксенотрансплантатах аденокарциномы поджелудочной железы у мышей после претаргетинга с использованием различных направленных против СЕА биспецифических антител, для оптимизации режима и отбора наиболее пригодного кандидата для включения в клинические исследования. Эксперименты осуществляли в соответствии с тремя различными протоколами, осуществляя их в следующем порядке I) 80б, II) 80в и III) 80а. Согласно протоколу 80б оценивали поглощение опухолью пяти конструкций полностью гуманизированных биспецифических антител, предварительно связанных с 203Pb-DOTAM. Согласно протоколу 80в оценивали поглощение опухолью конструкций биспецифических антител, предварительно связанных с 203Pb-DOTAM, в три различных момента времени после инъекции для оптимизации периода времени между инъекцией PRIT и инъекцией СА/хелата в режимах претаргетинга. И, наконец, согласно протоколу 80а оценивали поглощение опухолью 212Pb-DOTAM в стандартной схеме претаргетинга, используя пять конструкций полностью гуманизированных биспецифических антител, мишенью которых является либо Т84.66, либо СН1А1А.
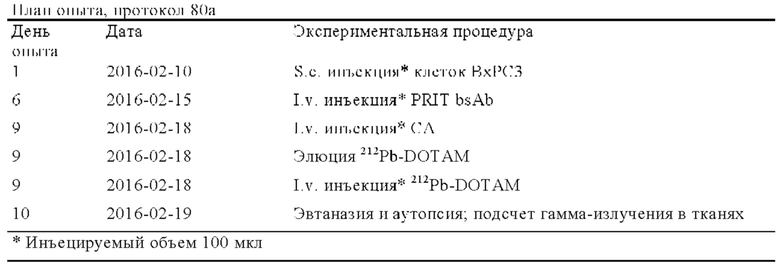


Каждой мыши (возрастом 6-7 недель) инъецировали подкожно (s.c.) клетки ВхРС3 (пассаж 30) в 100 мкл RPMI/Matrigel в правую боковую область. Через пять дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 160-170 мм3. Антитела разводили до конечной концентрации 30 мкг на 100 мкл и затем вводили i.v. либо индивидуально (80а), либо после предварительного связывания с 203Pb-DOTAM (80б, 80в). PRIT-0165 и PRIT-0156 применяли в качестве положительных СЕА-связывающих контролей, таргетирующих Т84.66 и СН1А1А соответственно. PRIT-0175 применяли в качестве не связывающего СЕА контроля.
Согласно протоколу 80а очищающий агент инъецировали внутривенно в концентрации 30 мкг на 100 мкл через три дня после биспецифического антитела, после чего через два часа вводили 212Pb-DOTAM.
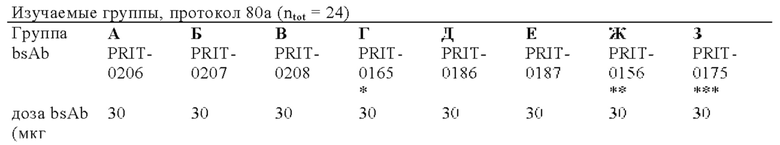


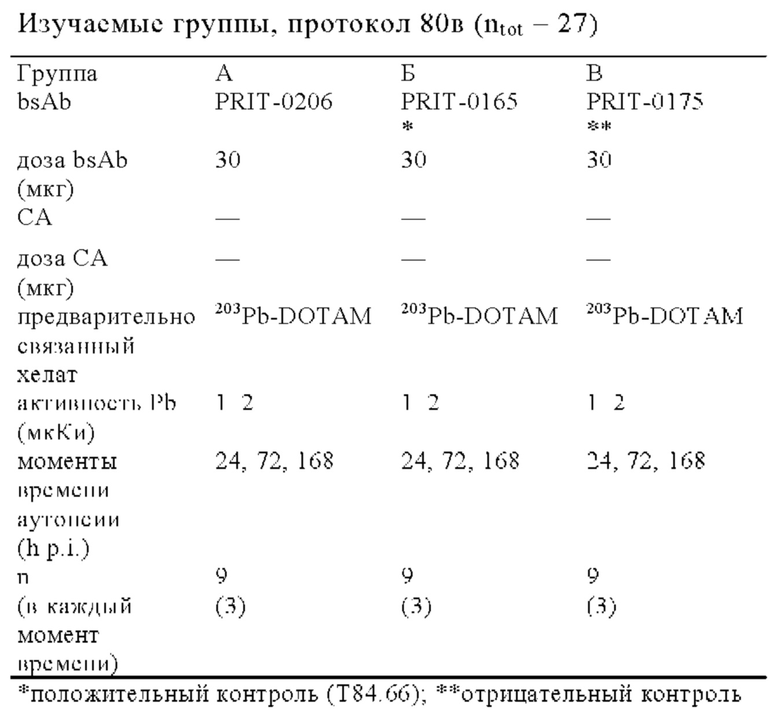
Мышей умерщвляли для изучения биораспределения через 24 ч (80а); 96 ч (80б); или 24, 72 или 168 ч (80в). У всех мышей согласно протоколу 80а изымали: кровь, мочевой пузырь, селезенку, почки, печень, легкие, сердце, мышцы и опухоли. У всех мышей согласно протоколам 80б и 80в изымали: кровь, мочевой пузырь, тонкий кишечник, ободочную кишку, селезенку, поджелудочные железы, почки, печень, легкие, сердце, бедренную кость, мышцы и опухоли. Собранные ткани взвешивали и помещали в пластиковые пробирки для немедленного измерения радиоактивности. Затем рассчитывали процент инъецируемой дозы на грамм ткани (%ID/г), включая коррекции на радиоактивный распад и фоновый уровень.
Результаты, протокол 80а
Все СЕА-связывающие биспецифические антитела обеспечивали специфический таргетинг 212Pb-DOTAM к опухолям, при этом имело место небольшое поглощение или отсутствие поглощения здоровыми тканями через 24 ч после инъекции DOTAM. Для PRIT-0206, PRIT-0207 и PRIT-0208 среднее поглощение опухолью ± SD составляло 8,62±1,05, 7,30±3,84 и 7,75±2,61 %ID/г соответственно, при этом для соответствующего Т84.66-связывающего применяемого в качестве положительного контроля антитела PRIT-0165 поглощение составляло 9,13±1,82 %ID/г. Для CH1A1A-связывающих PRIT-0186 и PRIT-0187 указанные величины в опухолях составляли 17,44±1,39 и 16,50±3,25 %ID/г, а для применяемого для них в качестве положительного контроля PRIT-0156 - 18,98±1,89 %ID/г. Поглощение опухолью не связывающего СЕА антитела PRIT-0175 составляло 0,41±0,42%ID/r.
В целом, биспецифические антитела, таргетирующие СН1А1А, обеспечивали существенно более высокое поглощение опухолями, чем таргетирующие Т84.66 (непарный t-критерий, р<0,0001); таргетинг как СН1А1А, так и Т84.66, обеспечивал существенно более высокое поглощение по сравнению с отрицательным контролем (односторонний дисперсионный анализ, р<0,0001). Результаты представлены на фиг. 4.
Результаты, протокол 80б
Специфический таргетинг опухолей достигался при применении всех предварительно связанных СЕА-связывающих биспецифических антител, хотя величина %ID/г оказалась несколько более низкой в случае полностью гуманизированных версий по сравнению с применяемым в качестве положительного контроля PRIT-0165. Не связывающий СЕА контроль приводил к относительно незначимому накоплению в опухолях.
В целом, рассчитанные величины %ID/г при осуществлении указанной схемы эксперимента, включающей предварительное связывание, достигали уровней, которые примерно в 10 раз превышали уровни, соответствующие PRIT-режимам; при этом выходные данные отражали измеренную активность гамма-излучения и не были обнаружены ошибки вычисления. Важно отметить, что соотношения накопления в опухоли/здоровой ткани оставалось в ожидаемом диапазоне. В частности, соотношение накопления в опухоли/здоровой ткани (±SD, n=3) составляли 6,76±2,95 для PRIT-0205, 7,56±2,27 для PRIT-0206, 9,33±0,91 для PRIT-0207, 10,77±0,84 для PRIT-0208, 11,71±0,84 для PRIT-0209, 10,78±0,88 для PRIT-0165 и 0,85±0,12 для PRIT-0175. Результаты представлены на фиг. 5.
Результаты, протокол 80в
Применение обоих СЕА-связывающих антител приводило к существенному накоплению в опухолях по сравнению с отрицательным контролем. Статистический анализ продемонстрировал отсутствие какого-либо значимого различия между таргетингом опухоли с использованием PRIT-0206 или PRIT-0165 в любые изученные моменты времени, отсутствие какого-либо значимого различия в величинах %ГО/г между 3 и 7 днями при использовании любого из изученных антител (двусторонний дисперсионный анализ, р<0,05). Аналогично результатам, полученным при осуществлении протокола 80б, рассчитанные величины %ID/г в целом были высокими; однако соотношения «опухоль/кровь» оставались в ожидаемом диапазоне. Никаких значимых различий в соотношениях «опухоль/кровь» не обнаружено в дни 1 и 3, но для PRIT-0165 и PRIT-0206 после продолжения опыта в течение 7 дней обнаружено существенное повышение указанного соотношения по сравнению с PRIT-0213 (двусторонний дисперсионный анализ р<0,05). Результаты представлены на 6.
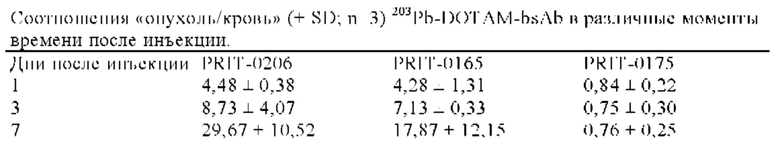
Обобщение результатов и заключение
Все полностью гуманизированные биспецифические антитела, таргетирующие либо Т84.66, либо СН1А1А, обеспечивали существенное накопление радиоактивности в ВхРС3-опухолях, сопоставимое с накоплением соответствующих положительных контролей.
Величина %ID/г 203Pb-DOTAM-bsAb в опухоли не отличалась существенно между днями 3 и 7, но соответствующее соотношение опухоль/кровь оказалось благоприятным в более поздний момент времени из-за снижения радиоактивности в крови.
Пример 17: Биораспределение очищающих агентов (протокол 44)
Целью настоящего опыта было изучение биораспределения отобранных очищающих агентов, обладающих различными свойствами (например, молекулярный каркас, размер и заряд) и, более конкретно, их присутствие и/или накопление в опухолях. Представлялось важным это установить, поскольку очищающие агенты могли потенциально проникать в опухоли, связываться со связанными с опухолью антителами и/или вытеснять их из опухоли, оказывая отрицательное воздействие на последующее связывание радиолигандов.

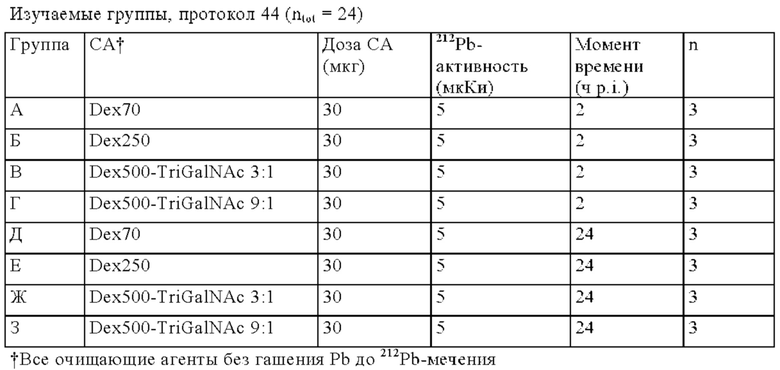
По 30 мкг Dex70, Dex250 и TriGalNac-модифицированных очищающих агентов гасили с помощью 5 мкКи 212Pb и разводили в ЗФР с получением 30 мкг на 100 мкл полного объема для i.v.-инъекции.
Мышей умерщвляли и подвергли аутопсии через 2 или 24 ч после инъекции 212Pb-СА. Кровь, мочевой пузырь, сердце, легкие, печень, селезенку, почки, тонкий кишечник (двенадцатиперстная кишка, тощая кишка, подвздошная кишка), ободочную кишку, поджелудочную железу, желудок, яичники, головной мозг, бедро с костным мозгом и опухоли изымали, взвешивали и измеряли уровень радиоактивности, а затем рассчитывали %ID и %ID/г. Кроме того, получали образцы мочи в момент времени 24 ч.
Результаты, протокол 44
Все очищающие агенты быстро удалялись из кровотока, накапливаясь в основном в печени и ободочной кишке уже через 2 ч после инъекции. Примерно 50% инъецируемого 212Pb обнаружено в печени через 24 ч после инъекции, что продемонстрировано по распределению радиоактивности по органам (%ID). TriGalNAc-модифицированные очищающие агенты накапливались также в определенной степени в селезенке, о чем свидетельствует присутствие молекул TriGalNAc. В целом, небольшая радиоактивность обнаружена в моче через 24 ч. Однако, имеющий самую низкую молекулярную массу очищающий агент (Dex70) экскретировался медленнее, чем ожидалось.
На фиг. 7 продемонстрировано распределение радиоактивности в отобранных тканях через 2 ч после инъекции 212Pb-меченных очищающих агентов в несущих MKN45-опухоли мышах (%ID/г ± SD, n=3).
На фиг. 8 продемонстрировано распределение радиоактивности в отобранных тканях и в моче через 24 ч после инъекции 212Pb-меченных очищающих агентов в несущих MKN45-опухоли мышах (%ID/г ± SD, n=3).
На фиг. 9 продемонстрировано распределение радиоактивности по органам в отобранных тканях и в моче через 24 ч после инъекции 212Pb-меченных очищающих агентов в несущих MKN45-опухоли мышах (%ID ± SD, n=3).
Обобщение результатов и заключение
Применение ни одного из радиоактивно меченных СА не приводило к поглощению опухолью 212Pb, вводимого в дозе 30 мкг на мышь. Модификация Dex500 с помощью TriGalNac не обладала преимуществом по сравнению с не модифицированными очищающими агентами Dex70 и Dex250.
Пример 18: Долговременное биораспределение очищающих агентов (протокол 70)
В настоящем исследовании сравнивали долговременное биораспределение шести различных очищающих агентов. Отслеживание in vivo осуществляли на основе радиоактивного мечения с помощью 203Pb.
План опыта, протокол 70

Изучаемые группы, протокол 70 (ntot=18)

Результаты, протокол 70
На фиг. 10 продемонстрировано распределение радиоактивности в отобранных тканях через 1 неделю после инъекции 203Pb-меченных очищающих агентов в свободных от опухолей мышах (%ID/г ± SD, n=3).
На фиг. 11 продемонстрировано распределение радиоактивности по органам в отобранных тканях через 1 неделю после инъекции 203Pb-меченных очищающих агентов в свободных от опухолей мышах (%ID ± SD, n=3).
Пример 19: Очищающий агент в крови (протоколы 83 и 87)
В настоящем примере представлены результаты двух опытов, из которых первый включал первоначальный скрининг, в котором сравнивали девять различных очищающих агентов на основе декстрана с ЗФР с позиции времени удерживания в крови. Цель состояла в идентификации перспективных кандидатов для последующих экспериментов по претаргетингу, включающему повторяющиеся обработки, разделенные тремя неделями. Гипотеза состояла в том, что очищающие агенты, которые сохраняются в кровотоке в течение пролонгированного времени, связываются с введенными биспецифическими антителами, эффективно блокируя связывание инъецируемого в дальнейшем радиоактивно меченого DOTAM. Очищающие реагенты варьировались по размеру, заряду и загрузке 1,4,7,10-тетракис(карбамоилметил)-1,4,7,10-тетраазациклододеканом (ТСМС).
Во втором опыте оценивали девять очищающих агентов с позиции эффективности в отношении очищения от антител. План эксперимента включал стандартный PRIT-режим на основе однократной инъекции свободным от опухолей мышам, оценку удерживания 212Pb-DOTAM в крови в качестве показателя удерживания bsAb после введения СА.
План опыта, протокол 83
План опыта, протокол 87

Изучаемые группы, протоколы 83 и 87 ntot=30 на протокол)
Изучаемые группы, протоколы 83 и 87 (ntot=30 на протокол)
В момент первой инъекции PRIT возраст мышей составлял 7-9 недель. Все мыши в опыте были свободны от опухолей; поэтому для скрининга можно было использовать любое DOTAM-связывающее биспецифическое антитело. Очищающие агенты вводили внутривенно через 1 день после PRIT-0155, их разводили в ЗФР до конечной концентрации 30 мкг (протокол 83) или 60 мкг (протокол 87) на 100 мкл. Группа П получали ЗФР вместо очищающего агента. Основой соединений в группах А-Г был декстран размером 20, 70, 250 или 500 кДа, с различным ТСМС-замещением. Основой соединений в группах К-О был кэпированный (т.е. с нейтрализацией избытка аминов) декстран-500 (CDex) с различными зарядами (Glu) или декстран-500 и декстран-20 с моноверсиями Glu (M(Glu)). Кэпированный декстран с -(Glu)4, -(Glu)3 и -(Glu)2 соответствовал сильно отрицательному, отрицательному и нейтральному чистому заряду соответственно; моноверсия -M(Glu)2 соответствовала чистому заряду от отрицательного до слегка положительного.
Результаты, протокол 83
Средние уровни радиоактивности (%ID/г) в крови в группах, существенно отличающиеся от радиоактивности в обработанной ЗФР контрольной группе (41,1±1,4%ID/г), свидетельствовали об удерживании очищающего агента в кровотоке через три недели после введения. Три соединения не отличались существенно от контроля: кэпированный Dex500-(Glu)4, Dex500-моно-(Glu)2 и Dex20-моно-(Glu)3; другие отличались в различной степени.
На фиг. 12 представлены данные об уровне радиоактивности в крови через 4 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3). Закрашенный полосками прямоугольник соответствует контролю без СА, с которым сравнивали все реагенты-кандидаты. Звездочками обозначен уровень статистической значимости от более низкого (*) до более высокого (***).
Результаты, протокол 87
Все протестированные очищающие агенты, в целом, хорошо действовали за одним исключением: Dex500-M(Glu)2, для которого величина 8,07 ± 0,61%ID/г сохранялась в крови после 24 ч.
На фиг. 13 продемонстрирован средний уровень радиоактивности в крови через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3). Закрашенный полосками прямоугольник соответствует контролю без СА, с которым сравнивали все реагенты-кандидаты.
Обобщение результатов и заключение
Подвергнутые скринингу реагенты в целом хорошо действовали с точки зрения самоклиренса и обеспечения клиренса антитела из кровотока. Повторяющиеся обработки с трехнедельными интервалами между инъекциями СА доказали возможность их применения без риска связывания 212Pb-DOTAM с биспецифическими антителами. Кроме того, при применении большинства тестируемых соединений клиренс биспецифических антител происходил в пределах 2 ч после инъекции СА.
Пример 20: Проникновение в опухоли очищающих агентов (протокол 85)
Проникновение очищающего агента в опухоль является потенциальной проблемой при осуществлении PRIT-режимов, поскольку проникающие связанные с DOTAM СА-фрагменты могут конкурировать с 212Pb-DOTAM за связывание с опухолевыми клетками, предварительно таргетированными антителом. В данном опыте сравнивали шесть различных очищающих агентов с позиции ингибирования ассоциации 212Pb-DOTAM с опухолями. Кандидатов выбирали на основе результатов основного скрининга СА (протокол 83) для оценки воздействия на ассоциированную с опухолью радиоактивность, обусловленного I) размером декстрана и II) зарядом молекулы.
План опыта, протокол 85

Изучаемые группы, протокол 85 (ntot=18)
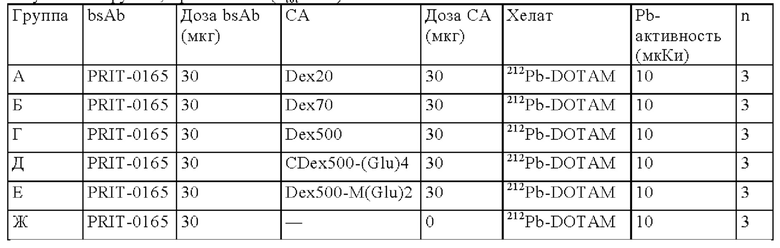
Каждой мыши (возрастом 7 недель) инъецировали s.c. ВхРС3-клетки (пассаж 33) в 100 мкл RPMI/Matrigel в правую боковую область. Через пять дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 200 мм3.
Основой очищающих агентов являлся декстран с молекулярной массой 20, 70 или 500 кДа, с различным ТСМС-замещением. Основой CDex500-(Glu)4 являлся кэпированный (т.е. с нейтрализацией избытка аминов) декстран-500 (CDex) с отрицательным чистым зарядом; Dex500-M(Glu)2 имел моноверсию Glu (M(Glu)) и чистый заряд от нейтрального до слегка положительного. Все соединения вводили внутривенно, 30 мкг на 100 мкл, через 3 дня после PRIT-0165. Группу Ж обрабатывали ЗФР вместо очищающего агента.
Мышей умерщвляли через 24 ч после инъекции 212Pb-DOTAM. Получали образцы крови и опухолей и рассчитывали соответствующие величины %ID/г в зависимости от веса образцов и уровня радиоактивности.
Результаты, протокол 85
Для трех кандидатов характерным являлось очень низкое поглощение или отсутствие поглощения опухолями Dex20 (0,26±0,03%ID/г), Dex70 (4,06±2,06%ID/г) и CDex500-(Glu)4 (2,98±0,73%ID/г), свидетельствующее о значительном проникновении СА в опухоли. В противоположность этому, при применении Dex500-M(Glu)2 обнаружена высокая радиоактивность как в опухолях (69,39 ± 9,70%ID/г), так и в крови (20,68±1,22%ID/г), уровень которого очень близок к уровню, обнаруженному при применении ЗФР в качестве контроля (59,02±15,53 и 27,92±2,38%ID/г в опухолях и крови соответственно), что можно интерпретировать как недостаточность клиренса bsAb. Для Dex500 характерно значительное накопление радиоактивности в опухолях (20,78±3,76%ID/г), что свидетельствовало о низком проникновении в опухоль, в то время как клиренс крови был практически полным (0,17±0,01%ID/г). Тем не менее, поглощение опухолью, достигаемое при применении контроля без СА, свидетельствовало о том, что некоторая часть низкомолекулярных (MW) DOTAM-фрагментов декстрана также присутствовала в партии Dex500.
На фиг. 14 представлены данные об уровне радиоактивности в крови и опухолях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3).
Обобщение результатов и заключение
В указанном эксперименте подтверждена концепция о том, что присутствие имеющих низкую MW видов СА может оказывать существенное влияние на накопление в опухолях 212Pb-DOTAM. Это продемонстрировано также для молекул с широким диапазоном MW, которые присутствовали в любой партии СА, вне зависимости от указанного заданного размера. Из этого следует, что чем больше заданный размер СА, тем меньший риск интродукции фрагментов с низкой MW, которые могут проникать в опухоли. Однако даже для Dex500 определенный уровень проникновения был выявлен, о чем свидетельствовало различие в поглощении опухолью по сравнению с контролем без СА. Поэтому указанный раствор в будущем следует подвергать диафильтрации с использованием более высокого порога отсечения MW для максимально возможного удаления указанных интерферирующих фрагментов. Принято решение о повышении для Dex500 порога отсечения MW с 30 до 100 кДа.
Пример 21: Процесс получения очищающего агента Рех500
Аминодекстран (20,0 г) растворяли в смеси 0,1М Na2CO3 в H2O (400 мл) и 0,1М NaHCO3 в H2O (400 мл). После получения прозрачного бесцветного раствора добавляли при перемешивании пара-SCN-Bn-DOTAM 4HCl (тетрагидрохлорид S-2-(4-изотиоцианатбензил)-1,4,7,10-тетрааза-1,4,7,10-тетра(2-карбамоилметил)циклододекана, 2,03 г). Полученный слегка мутный раствор перемешивали при комнатной температуре в течение 4 ч, после чего реакционную смесь нейтрализовали до достижения рН 6-7 путем добавления 2М HCl. Полученный раствор очищали посредством фильтрации в тангенциальном потоке с отсечением по молекулярной массе 100 кДа (Sartorius Hydrosart, Slice 200 100 кДа 0,02 м2, мембрана на основе стабилизированной целлюлозы, кассета для ультрафильтрации) для удаления примесей с низкой молекулярной массой. Полученный раствор лиофилизировали при пониженном давлении, в результате чего получали 17,9 г требуемого промежуточного продукта.
16,1 г твердого продукта, полученного в результате сушки вымораживанием, растворяли в смеси ОДМ АсОН в H2O (60 мл) и 0,1М NaOAc⋅3H2O (540 мл). К прозрачному бесцветному раствору добавляли Pb(ОАс)2⋅3H2O (744 мг) в виде твердого вещества. Прозрачный бесцветный раствор перемешивали в течение 60 мин, после чего добавляли раствор ксиленолового оранжевого (1% в Н2О, 250 мкл). Пурпурная окраска свидетельствовала о присутствии в растворе свободного Pb(II). Добавляли раствор ЭДТК (0,01М в H2O) до тех пор пока не происходило изменение окраски на желтую (65,2 мл). Полученный раствор очищали посредством фильтрации в тангенциальном потоке с отсечением по молекулярной массе 100 кДа (мембрана Sartorius Hydrosart, Slice 200 100 кДа 0,02 м2 на основе стабилизированной целлюлозы, кассета для ультрафильтрации) для удаления примесей с низкой молекулярной массой. Полученный раствор лиофилизировали при пониженном давлении, в результате чего получали 14,0 г требуемого очищающего агента.
Полученный очищающий агент замещали несколькими Pb-DOTAM-фрагментами, схематически представленными ниже:
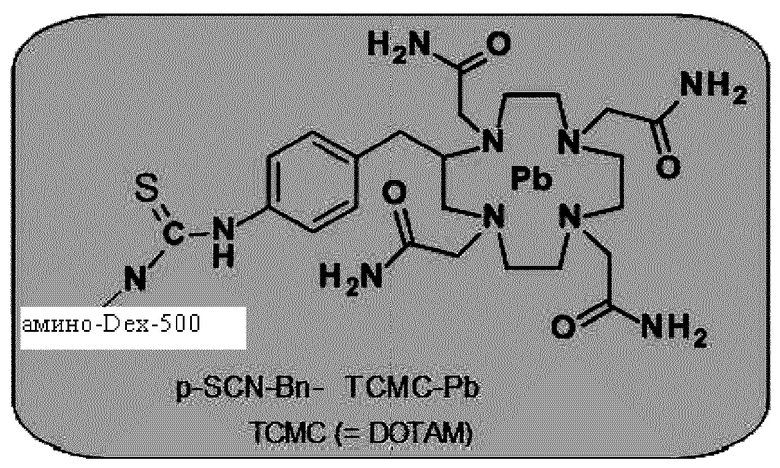
Пример 22: Влияние дозы очищающего агента на накопление в опухоли (протокол 90)
Целью настоящего исследования было изучение влияния дозы очищающего агента на накопление в опухоли на модели подкожной опухоли с учетом гипотезы о том, что большее количество очищающего агента может приводить к более высокой степени проникновения очищающего агента в опухоли, что снижает последующее поглощение меченого хелата в результате блокады DOTAM-связывающего плеча связанных с опухолью биспецифических антител. Однако слишком низкая доза может приводить к менее эффективному накоплению ассоциированной с опухолью радиоактивности из-за более высоких уровней DOTAM-связывающих биспецифических антител в кровотоке.
Кроме того, осуществляли сравнение партий очищающего агента, которые были подвергнуты диафильтрации для удаления компонентов с низкой MW с порогом отсечения либо 30, либо 100 кДа, для снижения проникновения в опухоли связанных с DOTAM фрагментов декстрана.
План опыта, протокол 90

Изучаемые группы, протокол 90 (ntot=24)
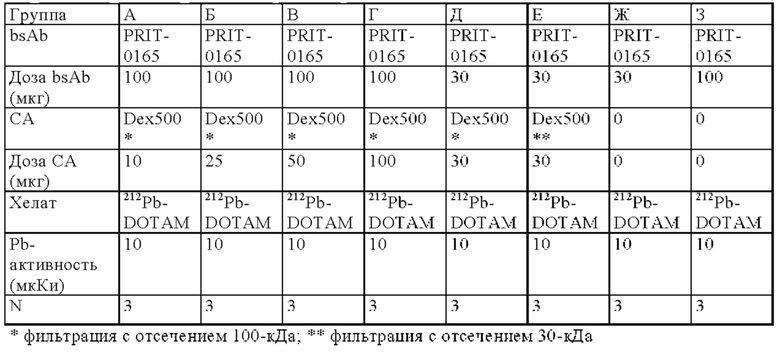
Ксенотрансплантаты солидных опухолей создавали путем подкожной инъекции клеток ВхРС3 (пассаж 30) в среде RPMI, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №354230). Каждой мыши (возрастом 11 недель) инъецировали s.c. в правую боковую область 5×106 клеток в 100 мкл RPMI/Matrigel. Через 5 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 150 мм3.
PRIT-0165 инъецировали i.v. в концентрации либо 30, либо 100 мкг на 100 мкл, затем через четыре дня вводили очищающий агент (группы А-Е) в концентрации 10, 25, 30, 50 или 100 мкг на 100 мкл. 212Pb-DOTAM инъецировали через 2 ч после СА. Группы Ж и З не получали очищающего агента между введениями биспецифического антитела и радиоактивно меченного хелата. Мышей умерщвляли для оценки биораспределения через 24 ч после инъекции 212Pb-DOTAM и изымали кровь, мочевой пузырь, селезенку, почки, печень, легкое, мышцы, хвост и опухоли. Образцы взвешивали и измеряли радиоактивность и затем рассчитывали величины %ID/г для каждого органа, внося коррекции на распад и фоновый уровень.
Результаты, протокол 90
Накопление 212Pb в здоровых тканях оказалось низким во всех группах, которым вводили очищающий агент; в противоположность этому, в группах, которые не получали очищающего агента, в целом, обнаружены значительные уровни радиоактивности.
Данные о биораспределении продемонстрировали влияние различных количеств очищающего агента (порог отсечения 100 кДа). Кривые, полученные для индивидуальных опухолей методом линейной регрессии, характеризовались значительной крутизной наклона, что свидетельствовало о более высоком поглощении опухолью при уменьшении количеств Dex500 (р=0,03, R2=0,31). Воздействие на уровень радиоактивности в крови оказалось очень сильным, величина ID/г составляла 31,5±0,2% при применении ЗФР, а при применении 10 мкг Dex500 величина ID/г составляла 1,6±0,5%.
Две группы обрабатывали PRIT-0165 и Dex500 в одинаковом (1:1) соотношении; либо по 100, либо по 30 мкг каждого агента. Полученные величины %ID/г в опухолях не различались существенно между двумя группами (непарный t-критерий, р=0,726). Важно отметить, что сравнение одинаковых количеств Dex500, который подвергали фильтрации с разными порогами отсечения, выявлено значимое увеличение накопления в опухолях при более высокой отсекаемой MW: 54,9±6,9 вместо 32,5±4,4%ID/г (непарный t-критерий, р=0,009).
На фиг. 15 продемонстрировано распределение 212Pb через 24 ч после инъекции радиоактивно меченого DOTAM (%ID/г ± SD, n=3) при использовании 30 или 100 мкг биспецифического антитела и 10-100 мкг очищающих агентов, подвергнутых фильтрации с порогом отсечения 100- или 30-кДа, или вообще без использования очищающего агента (ЗФР).
На фиг. 16 продемонстрировано влияние возрастающих количеств очищающего агента (0-100 мкг) на активность концентрации 212Pb в крови и опухоли. Опухоли предварительно таргетировали, используя 100 мкг PRIT-0165, с последующим введением через 4 дня Dex500, подвергнутого диафильтрации с порогом отсечения 100-кДа, или ЗФР. 212Pb-DOTAM вводили через 2 ч после СА. Символами обозначена величина %ID/г через 24 ч после инъекции радиоактивного соединения, а линией обозначены результаты, полученные обработкой данных для опухолей методом линейной регрессии.
Обобщение результатов и заключение
В опыте продемонстрировано резкое повышение накопления в опухолях Pb, с сохранением низких уровней радиоактивности в кровотоке. По результатам сделано два основных вывода: 1) диафильтрация с порогом отсечения 100 кДа существенно снижала проникновение в опухоли связанных с DOTAM фрагментов декстрана и 2) соотношение СА и bsAb влияло на результат в большей степени, чем абсолютное количество СА. Оптимальное соотношение «опухоль/кровь» было достигнуто при использовании соотношения антитело : СА 10:1 при введении 100 мкг биспецифического антитела. Кроме того, сделано заключение о том, что все последующие изучения с использованием очищающих агентов на основе Dex500 следует проводить с использованием реагентов, подвергнутых диафильтрации с порогом отсечения 100-кДа.
Пример 23: Влияние на ассоциированную с опухолью радиоактивность (протокол 95)
В данном опыте сравнивали девять различных очищающих агентов на основе декстрана-500 в отношении ингибирования ассоциации 212Pb-DOTAM с опухолями. Более конкретно, в эксперименте оценивали воздействие I) ТСМС-насыщения, II) соотношения очищающий агент/антитело и III) метода получения/очистки на ассоциированную с опухолью радиоактивность. Кроме того, сравнивали поглощение опухолью после инъекции 10 или 30 мкКи 212Pb-DOTAM без претаргетинга или добавления СА, для решения вопроса о том, может ли насыщение опухоли радиоактивностью достигаться уже при низкой дозе.
План опыта, протокол 95

Изучаемые группы, протокол 95 (ntot=27)
Ксенотрансплантаты солидных опухолей создавали путем подкожной инъекции клеток ВхРС3 (пассаж 20) в среде RPMI, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №354230). Каждой мыши (возрастом 8 недель) инъецировали в правую боковую область s.c. 5×106 клеток в 100 мкл RPM1/Matrigel. Через 15 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 210 мм3.
PRIT-0165 (100 мкг на 100 мкл) вводили i.v., с последующим введением через 3 дня очищающих реагентов на основе декстрана-500 (10-170 мкг на 100 мкл) с различной ТСМС-заменой (10-100%). Группы К и Л получали ЗФР вместо очищающего агента. Через 2 ч мышам инъецировали соответствующие растворы 212Pb-DOTAM (10 или 30 мкКи).
Мышей умерщвляли и подвергали аутопсии через 24 ч после инъекции 212Pb-DOTAM. Изымали кровь, мочевой пузырь, селезенку, почки, печень, легкие, мышцы, хвост и опухоли, взвешивали и измеряли уровень радиоактивности и затем рассчитывали величины %ID/г.
Результаты, протокол 95
Данные о биораспределении продемонстрировали выраженную тенденцию зависимости среднего уровня радиоактивности (%ID/г ± SD) в собранных тканях через 24 ч после инъекции 212Pb-DOTAM от вводимого количества, с наличием более высокой концентрации 212Pb в крови и здоровых тканях при уменьшении количества СА. Кроме того, более высокая 212Pb-активность приводила к более высокому накоплению 212Pb в опухоли без соответствующего повышения поглощения здоровыми тканями.
На фиг. 17 показано распределение радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3). Прямоугольники темно-серого и черного цвета соответствуют положительному контролю без СА, с которым сравнивали реагенты-кандидаты.
Односторонний дисперсионный анализ с коррекцией на множественные сравнения продемонстрировал, что с точки зрения соотношения кровь-опухоль только Dex500-(40%) (76,79±33,28, р<0,0001) и Dex500-(100%) (43,26±24,66, р=0.0115) существенно отличались от данных, полученных для отрицательного контроля с 10-мкКи (2,26±0,25). Выявленное различие между очищающим реагентом с наиболее высоким соотношением «опухоль/кровь» (Dex500-(40%)) и ранее применяемым стандартом (Dex500-(100%)), оказалась статистически незначимым (непарный t-критерий, р=0,2336).
На фиг. 18 показано содержание 212Pb в крови и опухолях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3) и соответствующие соотношения «опухоль/кровь». Прямоугольники темно-серого и черного цвета соответствуют положительному контролю без СА, с которым сравнивали реагенты-кандидаты.
Анализ методом линейной регрессии и полиномиальной (кубической) аппроксимации кривых осуществляли для оценки влияния возрастающих количеств очищающего агента (Dex500-(10%)) и ТСМС-загрузки (без СА, Dex500-(10%), Dex500-(20%), Dex500-(40%), Dex500-(100%)) на соотношение «опухоль/кровь». Кривизна наклона линейной кривой оказалась статистически значимой (р<0,0001), свидетельствуя о необходимости в более крупных количествах Dex500-(10%) для увеличения соотношения «опухоль/кровь», в то время как для инъецируемого определенного количества ТСМС в пересчет на 100 мкг Dex500-(10%), максимум обнаружен при соотношении TCMC-к-Dex500 около 60. Важно отметить, что эти результаты не следует учитывать в контексте общих заключений о количествах реагента или ТСМС-насыщения; они пригодны только для применяемых условий.
На фиг. 19 показано соотношение «опухоль/кровь» через 24 ч после инъекции 212Pb-DOTAM в зависимости от количества СА (PJRD08-46) и ТСМС-насыщения (9, 20, 39 или 84 к 1). Пунктирными линиями обозначены результаты аппроксимации соответствующих данных прямой линией по методу линейной регрессии (R2=0,82) и нелинейной кривой (R2=0,74).
В последнем опыте сравнивали применение 212Pb-DOTAM с радиоактивностью 10 и 30 мкКи без инъекции антител или очищающих агентов. Содержание в крови оказалось сходным в двух изучаемых группах, но применение соединения с радиоактивностью 30 мкКи приводило к более выраженному накоплению в опухолях по сравнению с 10 мкКи: 107,74±14,71 против 72,38±10,83%ID/г (± SD, n=3). Различие полученных соотношений «опухоль/кровь» было статистически значимым: 3,20±0,20 против 2,26±0,25 для 30 и 10 мкКи соответственно (непарный t-критерий, р=0,007).
Обобщение результатов и заключение
Таким образом, установлено, что применение 20 мкг Dex500 с 39 ТСМС на молекулу (Dex500-(40%)) приводило к очень высокому накоплению 212Pb в предварительно таргетированных опухолях в сочетании с низким временем удерживания в крови через 24 ч после введения 212Pb-DOTAM. Он представлял собой наиболее эффективный очищающий агент, наряду с 10 мкг ранее известного стандарта СА, т.е. Dex500-(100%), с 84 ТСМС на молекулу. Из-за значительной вариабельности внутри групп и небольшого размера выборки различие для двух реагентов в соотношении «опухоль/кровь» не было статистически значимым, но в среднем свидетельствовало о том, что Dex500-(100%) может являться предпочтительным. Кроме того, было сделано заключение о том, что 30 мкКи 212Pb-DOTAM может приводить к более выраженному накоплению в опухолях по сравнению с 10 мкКи, т.е. при применении более низкой дозы насыщение не достигалось.
Пример 24: Влияние дозы биспецифического антитела на накопление в опухоли (протокол 91)
Целью настоящего исследования было изучение влияния дозы биспецифического антитела на накопление в опухоли на модели подкожной опухоли. Гипотеза состояла о том, что большее количество биспецифического антитела может приводить к более легкому насыщению доступных сайтов связывания на опухолевых клетках, повышая тем самым последующее поглощение меченого хелата. С другой стороны, применение возрастающей дозы может приводить к менее эффективному накоплению ассоциированной с опухолью радиоактивности из-за более высоких уровней в кровотоке биспецифических антител.
План опыта, протокол 91

Изучаемые группы, протокол 91 (ntot=21)
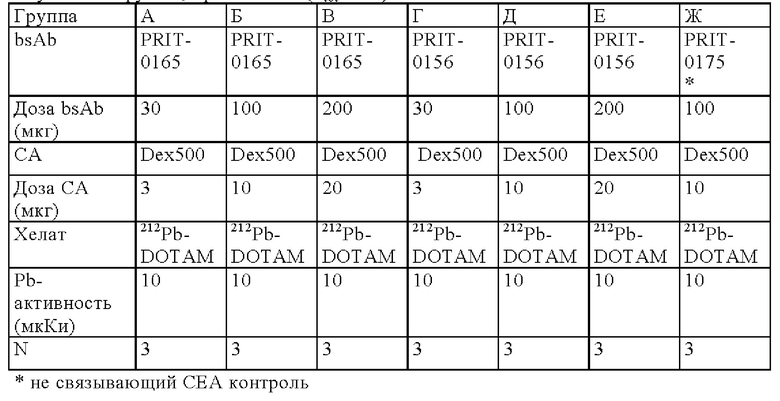
Ксенотрансплантаты солидных опухолей создавали путем подкожной инъекции клеток ВхРС3 (пассаж 34) в среде RPMI, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №. 354230). Каждой мыши (возрастом 7-12 недель) инъецировали в правую боковую область s.c. 5×106 клеток в 100 мкл RPMI/Matrigel. Через 5 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 220 мм3.
Мышам вводили i.v. СЕА-связывающие биспецифические антитела PRIT-0165 или PRIT-0156 в концентрации 30-200 мкг на 100 мкл или 100 мкг не связывающего СЕ А контрольного антитела PRIT-0175. Через четыре дня всем группам инъецировали i.v. очищающий агент в концентрации 3, 10 или 20 мкг на 100 мкл (1/10 от инъецируемой дозы антитела), затем через 2 ч вводили 10 мкКи 212Pb-DOTAM. Мышей умерщвляли через 24 ч и осуществляли аутопсию. Изымали кровь, мочевой пузырь, селезенку, почки, печень, легкие, мышцы, хвост и опухоли. Образцы взвешивали и измеряли уровень радиоактивности и рассчитывали величины %ID/г для каждого органа.
Результаты, протокол 91
Претаргетинг с использованием 100 мкг любого из СЕА-связывающих биспецифических антител приводил к значительному накоплению 212Pb в опухолях по сравнению с применением в количестве 100 мкг не связывающего СЕА контроля (непарный t-критерий, р=0,027 и 0,008 для сравнению с PRIT-0156 и PRIT-0165 соответственно). Никакого существенного различия не обнаружено для поглощения опухолями при повышении дозы антитела от 30 до 200 мкг для любой из СЕА-связывающих конструкций. Однако общие уровни радиоактивности слегка возрастали в большинстве здоровых тканей при применении антител в дозах, превышающих 30 мкг. Важно отметить, что величина %ID/г в крови существенно возрастала для каждого повышения дозы за исключением разницы при применении в дозах 100 и 200 мкг PRIT-0156 (непарный t-критерий, р<0,05).
На фиг. 20 показано распределение 212Pb через 24 ч после инъекции радиоактивно меченого DOTAM (%ID/г ± SD, n=3). Прямоугольниками с белым и серым фоном обозначены варианты, таргетируемые Т84.66 и СН1А1А соответственно; черным прямоугольником обозначены данные для не связывающего СЕА контроля.
Обобщение результатов и заключение
Применение антител во всех трех уровнях доз (30, 100 и 200 мкг) приводило к высокому специфическому поглощению радиоактивности опухолями в случае обеих изученных биспецифических конструкций. Обнаружено отсутствие различия в накоплении в опухолях при возрастании дозы, свидетельствующее о том, что сайты связывания насыщались уже более низкой дозой, или о том, что промежуток времени между введением PRIT и инъекцией очищающего агента (четыре дня) был слишком коротким, для того, чтобы обеспечивать дополнительное накопление в опухоли. Как и ожидалось, уровни радиоактивности в крови повышались с возрастанием дозы, что несколько сужает терапевтическое окно при лечении.
Пример 25: Эффективность CEA-PRIT (протокол 93 (а, б))
В данном опыте оценивали эффективность CEA-PRIT с использованием двух bsAb, являющихся кандидатами для клинических испытаний (PRIT-0213 и PRIT-0214), с использованием параллельно двух подкожных моделей опухолей: ВхРС3 и LS174T. Биспецифические антитела сравнивали одновременно с использованием схем лечения, состоящих из одного и двух циклов.
План опыта, протокол 93а (ВхРС3)
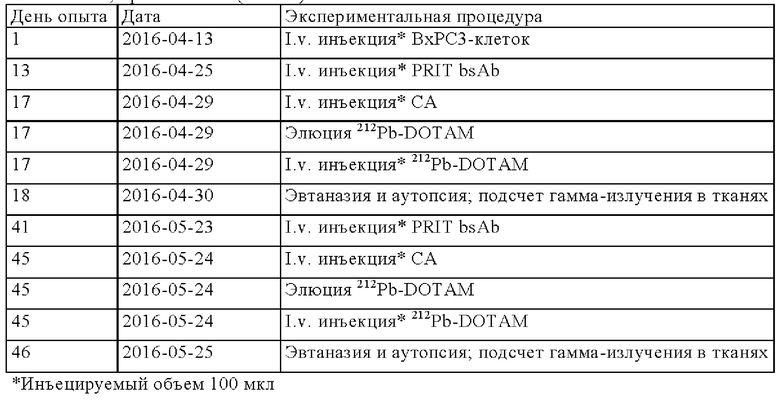
План опыта, протокол 93б (LS174T)
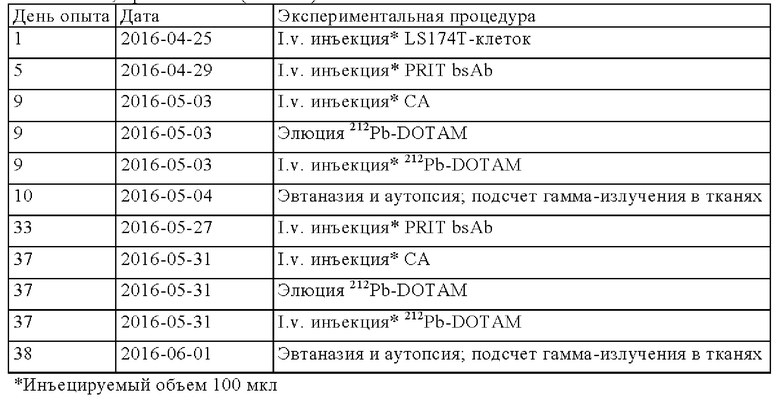
Группы А-Ж включали обработанных мышей, которые предназначались для оценки эффективности, мышей из групп З-М умерщвляли для оценки биораспределения через 24 ч после последней инъекции 212Pb-DOTAM.
Изучаемые группы, протокол 93а (ВхРСЗ; ntot=71)
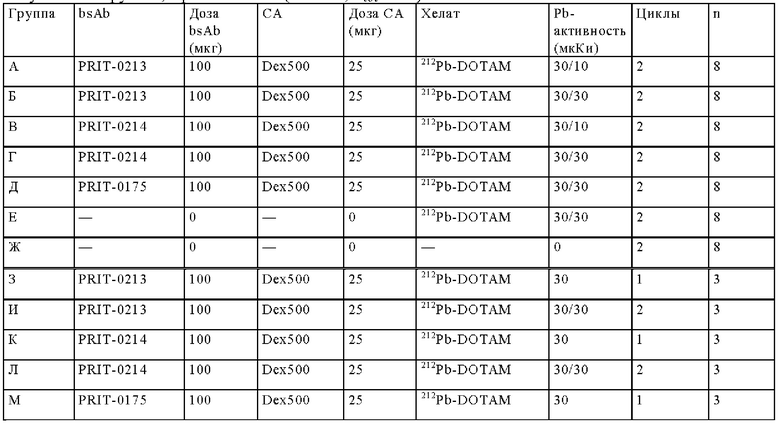
Изучаемые группы (протокол 93б) (LS174T; ntot=71)
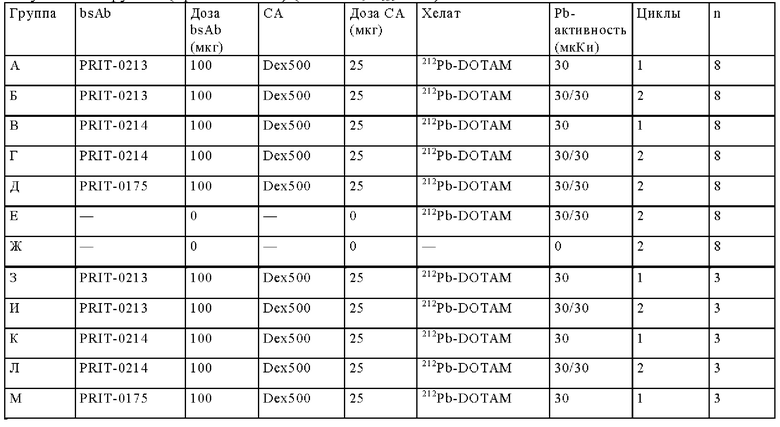
Ксенотрансплантаты солидных опухолей создавали путем подкожной инъекции клеток в среде RPMI (ВхРС3) или DMEM (LS174T), смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №.354230).
Согласно протоколу 93а мышам (возрастом 7 недель) инъецировали s.c. в правую боковую область 5×106 клеток ВхРС3 (пассаж 33) в 100 мкл RPMI/Matrigel. Через 12 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 235 мм3.
Согласно протоколу 93б мышам (возрастом 9 недель) инъецировали в правую боковую область s.c 1×106 клеток LS174T (пассаж 26) в 100 мкл DMEM/Matrigel. Через 3 дня после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 150 мм3.
Каждый цикл лечения начинался с инъекции гуманизированных bsAb (PRIT-0213, PRIT-0214 или PRIT-0175). Через четыре дня вводили очищающий агент (Dex500), а через 2 ч 212Pb-DOTAM. Для минимизации повторного поглощения радиоактивной мочи/экскрементов после цикла лечения 1, клетки заменяли через 4 ч после обработки 212Pb-DOTAM и затем вновь через 24 ч p.i. При осуществлении цикла 2 мышей помещали в клетки с решетчатыми полами через 4 ч после введения 212Pb-DOTAM перед переносом в новые клетки со стандартной подстилкой. Затем все клетки заменяли через 24 ч p.i., как и в случае цикла 1.
Развитие опухолей у мышей отслеживали путем многократных измерений с помощью кронциркуля трижды в неделю. При необходимости осуществляли дополнительное измерение с помощью кронциркуля в промежутках между предусмотренных схемой измерений. Вес тела животных определяли путем многократных измерений согласно аналогичной схеме. Мышей, объем опухолей которых достигал 3000 мм3, немедленно умерщвляли. Другие факторы, учитывающиеся при эвтаназии по этическим соображениям, представляли собой снижение веса тела, статус опухоли (например, изъязвление) и общий вид животного. Всем животным давали влажный корм, начиная с пятого дня после радиоактивной инъекции, если принималось такое решение из-за острой потери веса тела (коллективной или индивидуальной) из-за индуцированной радиацией токсичности, вплоть до существенного восстановления всех особей.
В группах А-Ж в момент эвтаназии изымали следующие органы и ткани: мочевой пузырь, яичник, печень, селезенку, почки, бедра (включая костный мозг), ободочную кишку, тощую кишку, желудок и опухоль. Неожиданные или аномальные состояния фиксировали и фотографировали. Ткани немедленно помещали в 10%-ный нейтральный забуференный формалин (4°С), а затем через 5 дней переносили в ЗФР (4°С). Затем фиксированные в формалине образцы отправляли на фирму Roche Pharma Research and Early Development, Roche Innovation Center Basel, для дополнительной обработки и анализа.
Мышей из групп З, К и М умерщвляли и осуществляли аутопсию через 24 ч после первой и единственной инъекции 212Pb-DOTAM; мышей групп И и Л умерщвляли и осуществляли аутопсию через 24 ч после второй инъекции 212Pb-DOTAM. Изымали следующие органы и ткани: кровь, мочевой пузырь, тонкий кишечник, ободочную кишку, селезенку, поджелудочную железу, почки, печень, легкие, сердце, бедренную кость, мышцы, хвост и опухоль. Собранные образцы взвешивали и измеряли радиоактивность, и затем рассчитывали процент инъецируемой дозы на грамм ткани (%ID/г) с учетом коррекций на распад и фоновый уровень.
Результаты, протокол 93а
Полученные после цикла 1 лечения результаты хорошо соответствовали полученным ранее на этой модели результатам, касающимся поглощения тканями, в которых продемонстрирован высокий уровень накопления в опухолях (25,0±9,7 и 19,5±6,4%ID/г для PRIT-0213 и PRIT-0214 соответственно) и низкое накопление в здоровых тканях. Однако полученные после цикла 2 результаты продемонстрировали другой профиль распределения 212Pb с повышенным уровнем радиоактивности во всех собранных тканях.
На фиг. 21 показано распределение радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SEM, n=3) для циклов лечения 1 и 2 на ВхРС3-модели.
Всем мышам, которым инъецировали 30 мкКи 212Pb-DOTAM обнаружено резкое начальное уменьшение веса тела, которое становилось менее выраженным к 10 дню после первой радиоактивной инъекции. В группах, которым вводили вторую инъекцию соединения с радиоактивностью 30 мкКи, обнаружена более выраженная острая потеря веса, вероятно, из-за медленного клиренса радиоактивности и в результате этого накопления 212Pb в здоровых тканях, что обнаружено при оценке биораспределения в цикле 2. Никакого указанного действия не выявлено в группах, которые во втором цикле получали радиоактивную инъекцию 10 мкКи, после которой потеря веса была умеренной.
На фиг. 22 представлены данные о среднем весе тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на ВхРС3-модели. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана опыта.
На фиг. 23 представлены данные об изменении среднего веса тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на ВхРС3-модели, выраженном в виде процента от начального веса тела. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана опыта.
Во всех группах объем опухолей удваивался к дню 14 19 по сравнению с исходным уровнем. Первую обработку 212Pb-DOTAM осуществляли в день 16, после чего опухоли в группах, обработанных PRIT-0213 и PRIT-0214, продолжали расти в течение примерно 1 недели до наступления регресса объема. Не обнаружен ни один случай полного регресса опухолей, но объемы сохранялись на относительно постоянном низком уровне вплоть до дня 40-47, после чего их объем вновь удваивался по сравнению с исходным объемом. В этот момент времени осуществляли вторую инъекцию 30 мкКи 212Pb-DOTAM (день 44), которая, как обсуждено ранее, приводила к острой потере весе и к необходимости в последующей эвтаназии наиболее пораженных мышей на следующей неделе. У мышей, которых подвергали второму циклу лечения с использованием 10 мкКи вместо 30 мкКи 212Pb-DOTAM (день 55), обнаружена менее выраженная индуцированная радиацией токсичность. Радиоактивная инъекции 10 мкКи приводила к второй фазе регресса опухолей, которая продолжалась примерно от 58 дня до 75 дня. Осуществление PRIT с использованием неспецифического bsAb приводило к существенному, но ограниченному ингибированию роста опухолей по сравнению с обработкой только 212Pb-DOTAM или ЗФР. Сравнение средних значений с использованием метода Даннета продемонстрировало, что все схемы лечения с использованием PRIT-0213 и PRIT-0214 приводили к одинаковым соотношениям «лечение/контроль», во всех случаях обнаружено значимое различие по сравнению с любой контрольной группой с дня 23-26 и далее.
В день 47, в последний день, в который еще были представлены все обрабатываемые группы, ингибирование роста опухолей (TGI) составляло 87,3, 88,8, 84,1, 88,7, 57,5 и 15,8% для групп «PRIT-0213, 30+10 мкКи», «PRIT-0213, 30+30 мкКи», «PRIT-0214, 30+10 мкКи», «PRIT-0214, 30+30 мкКи», «PRIT-0175, 30+30 мкКи и «только DOTAM, 30+30 мкКи» соответственно по сравнению с ЗФР. Последняя мышь из обработанной наполнителем контрольной группы была проанализирована в день 66, в этом момент времени величина TGI составляла 87,8, 84,5 и 87,3% для трех оставшихся групп: «PRIT-0213, 30+10 мкКи», «PRIT-0214, 30+10 мкКи» и «PRIT-0214, 30+30 мкКи» соответственно. Всего шесть мышей выжило к концу эксперимента (день 84): одна из группы «PRIT-0213, 30+10 мкКи», три из группы «PRIT-0214, 30+10 мкКи» и две из группы «PRIT-0214, 30+30 мкКи».
На фиг. 24 представлены данные о росте опухолей, выраженные в виде средних значений со стандартной ошибкой в группах А-Ж, полученные на ВхРС3-модели (n=8). Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана опыта.
На фиг. 25 показаны кривые роста индивидуальных опухолей в группах А-Ж, полученные на ВхРС3-модели. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM.
На фиг. 26 показаны кривые Каплана-Мейера, демонстрирующие выживаемость в группах А-Ж, полученные на ВхРС3-модели (n=8). Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM.
Попарное тестирование осуществляли для выявления групп, в которых имело место существенное различие выживаемости: логранговый критерий (больший вес для более поздних случаев выживания), и критерий Уилкоксона (больший вес для ранних случаев), в обоих случаях с использованием поправки Бонферрони для множественных сравнений. Из-за индуцированной радиацией токсичности в группе «PRIT-0213, 30+30 мкКи» смертность была такой же или хуже, чем в контрольных группах. Соответствующая обработка PRIT-0214 обеспечивала несколько лучшие результаты, увеличивая выживаемость по сравнению с обработкой ЗФР и неспецифическим антителом. В двух группах, обработанных 10 мкКи 212Pb-DOTAM во втором цикле, существенно увеличивалась выживаемость по сравнению со всеми группам, за исключением друг друга и группы «PRIT-0214, 30+30 мкКи».
Парный логранговый критерий (уровень значимости для множественных сравнений = 0,00238)
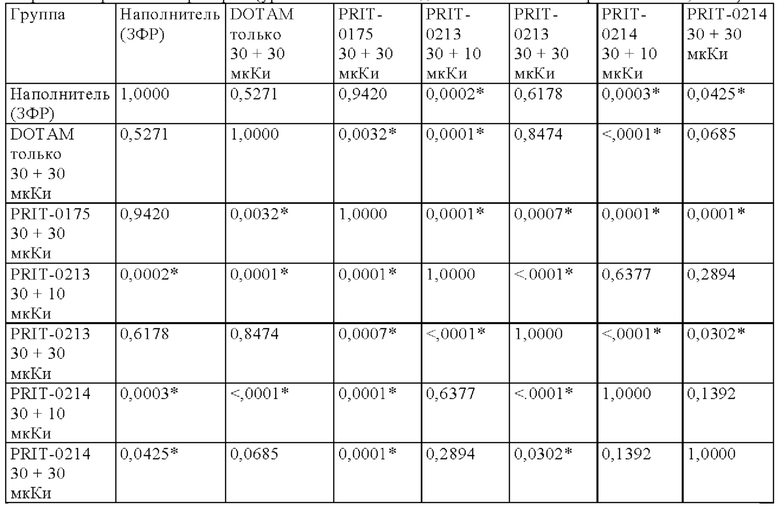
Парный критерий Уилкоксона (уровень значимости для множественных сравнений = 0,00238)
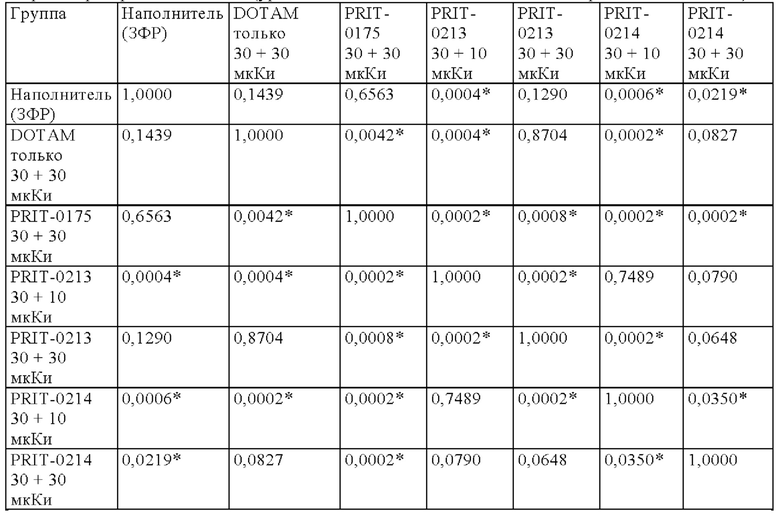
Результаты, протокол 93б
Оба цикла лечения приводили к высокому уровню накопления 212Pb в опухолях и низкому уровню накопления 212Pb в здоровых тканях. Поглощение опухолями после цикла 1 составляло 30,9±2,9 и 21,4±1,9%ID/г для PRIT-0213 и PRIT-0214 соответственно; после цикла 2 соответствующие показатели составляли 33,2±0,7 и 40,1±6,5%ID/г.
На фиг. 27 представлены данные о распределении радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3) в циклах лечения 1 и 2, полученные на LS174T-модели.
У всех мышей, которым инъецировали 212Pb-DOTAM, выявлено умеренное снижение веса тела, сходное после обоих циклов лечения с использованием 30 мкКи. Однако многих животных умерщвляли до окончания опыта из этических соображений из-за плохого статуса опухоли.
На фиг. 28 представлены данные о среднем весе тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на модели LS174T3. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана опыта.
На фиг. 29 представлены данные об изменении среднего веса тела в группах А-Ж (n=8) после обработки CEA-PRIT, полученные на LS174T3-модели, которое выражали в виде процента от начального веса тела. Кривые обрывали после первого случая гибели в каждой группе. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана опыта.
Первую обработку 212Pb-DOTAM осуществляли в день 8. Не обнаружено никакого регресса опухолей по сравнению с исходным уровнем, но в обработанных PRIT-0213 и PRIT-0214 группах средние величины оставались относительно постоянным, увеличение опухолей было очень медленным. Вторую инъекцию 30 мкКи 212Pb-DOTAM осуществляли в день 36; однако из-за быстрого роста опухолей в контрольных группах и ранее описанной проблемы, связанной с плохим статусом опухолей во всех группах, многие мыши уже были умерщвлены к этому моменту времени. У тех животных, которые получали вторую инъекции 30 мкКи 212Pb-DOTAM, восстанавливался или сохранялся контроль опухолей, но не обнаружен полный регресс опухолей. Сравнение средних значений с использованием метода Даннета продемонстрировало, что все схемы лечения с использованием PRIT-0213 и PRIT-0214 приводили к одинаковым соотношениям «лечение/контроль» по сравнению с обработанной ЗФР группой, начиная с дня 14. Кроме того, обработанные PRIT-0175 и DOTAM -контролями группы существенно отличались от обработанного наполнителем контроля, начиная со дня 16 и 22 соответственно.
В день 22, в последний день, в который были еще представлены все обрабатываемые группы, величина TGI составляла 83,1, 88,8, 87,5, 91,0, 53,0 и 64,7% для групп «PRIT-0213, 30 мкКи», «PRIT-0213, 30+30 мкКи», «PRIT-0214, 30 мкКи», «PRIT-0214, 30+30 мкКи», «PRIT-0175, 30+30 мкКи» и «только DOTAM 30+30 мкКи» соответственно по сравнению с ЗФР. Последняя мышь из обработанной наполнителем контрольной группы была проанализирована в день 30, в этом момент времени величина TGI составляла 92,5, 89,5, 90,8, 92,6 и 79,9% для оставшихся групп «PRIT-0213, 30 мкКи», «PRIT-0213, 30+30 мкКи», «PRIT-0214, 30 мкКи», «PRIT-0214, 30+30 мкКи» и «PRIT-0175, 30+30 мкКи» соответственно. Две мыши выжили к концу эксперимента (день 101), обе из группы «PRIT-0214, 30+30 мкКи».
На фиг. 30 представлены данные о росте опухолей, выраженные в виде средних значений со стандартной ошибкой, в группах А-Ж (n=8), полученные на LS 174Т3-модели. Кривые обрывали в каждой группе после первого случая гибели. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM некоторых или всех групп в зависимости от плана опыта.
На фиг. 31 показаны кривые индивидуального роста опухолей в группах А-Ж, полученные на LS 174Т3-модели. Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM.
На фиг. 32 показаны кривые Каплана-Мейера, демонстрирующие выживаемость в группах А-Ж, полученные на LS 174Т3-модели (n=8). Пунктирными вертикальными линиями обозначена обработка 212Pb-DOTAM.
Логранговый критерий и критерий Уилкоксона применяли с использованием поправки Бонферрони для выявления групп, в которых имело место существенное различие выживаемости. Решающее значение имели данные о том, что в группе «PRIT-0214, 30+30 мкКи» существенно повышалась выживаемость по сравнению со всеми группами за исключением «PRIT-0213, 30 мкКи». Однако общая выживаемость в группе «PRIT-0213, 30+30 мкКи» не существенно отличалась от выживаемости в группе «PRIT-0213, 30 мкКи», она лишь слегка отличалась от выживаемости в группе «PRIT-0214, 30+30 мкКи».
Парный логранговый критерий (уровень значимости для множественных сравнений = 0,00238)
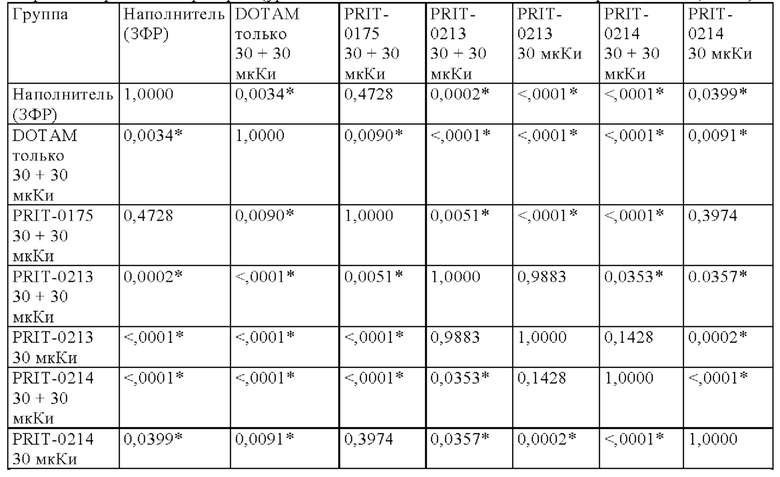
Парный критерий Уилкоксона (уровень значимости для множественных сравнений = 0,00238)
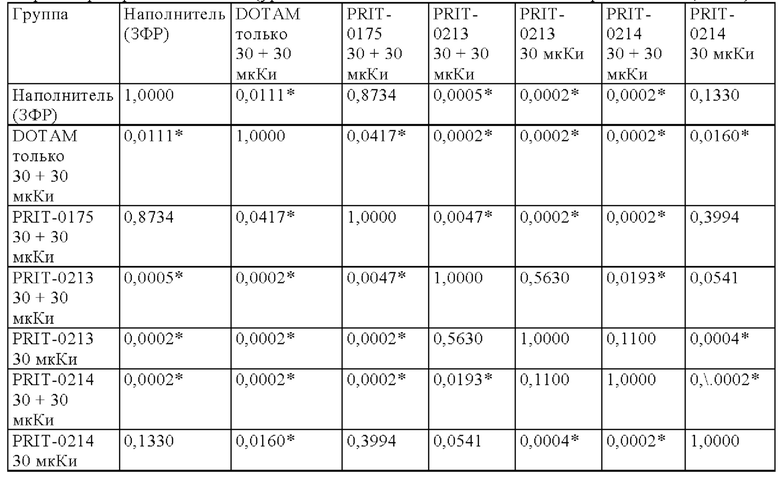
Обобщение результатов и заключение
Осуществление CEA-PRIT с использованием любого из двух биспецифических антител PRIT-0213 и PRIT-0214 в одном или двух циклах лечения приводило к существенному ингибированию роста опухолей и повышению выживаемости на обеих изученных моделях опухолей. Усилия по устранению проблем, связанных с неожиданным биореспределением 212Pb после второго цикла и последующей связанной с радиацией токсичности, выявленном на ВхРС3-модели, позволили сделать заключение о том, что ситуация, вероятно, вызвана не идентифицированной проблемой, связанной с инъекцией. Опыты по изучению эффективности на ВхРС3-модели следует повторить для подтверждения наличия проблемы, не связанной со схемой лечения. Дополнительные эксперименты на LS174T-модели нельзя осуществлять вплоть до решения проблемы с плохим статусом опухолей. Для повышения эффективности второго цикла лечения предполагается укоротить промежуток времени между двумя циклами во избежание повторного роста опухолей. Кроме того, для дополнительного изучения можно рассматривать также включение третьего цикла лечения.
И, наконец, исследования механизмов, лежащих в основе роста опухолей, представляют очень большой интерес для решения вопроса о том, почему, хотя их рост явно ингибируется, не происходит полный регресс опухолей, но вместо этого происходит их повторный рост после некоторого промежутка времени.
Пример 26: Дозы очищающего агента (протокол 105)
В этом исследовании изучали диапазон доз очищающего агента на основе декстрана-500 с 50%-ным ТСМС-насыщением с позиции воздействия на ассоциацию 212Pb-DOTAM с предварительно таргетированными опухолями.
План опыта, протокол 105

Изучаемые группы, протокол 105 (ntot=18)
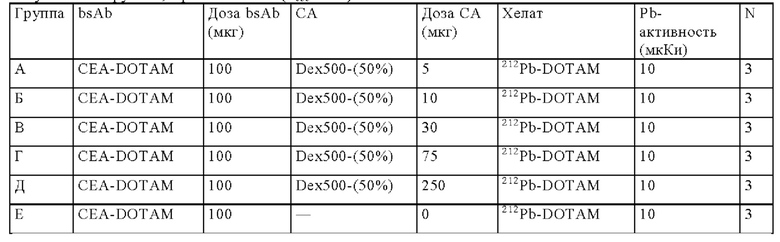
Ксенотрансплантаты солидных опухолей создавали путем подкожной инъекции клеток ВхРС3 (пассаж 26) в среде RPMI, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №.354230). Каждой мыши (возрастом 6 недель) инъецировали s.c. в правую боковую область 5×106 клеток в 100 мкл RPMI/Matrigel. Через 15 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 222 мм3.
CEA-PRIT (100 мкг на 100 мкл) вводили i.v., после чего через три дня вводили Dex500-(50%) (5-250 мкг на 100 мкл). Группа Е получала ЗФР вместо очищающего агента. Через 2 ч мышам инъецировали i.v. 100 мкл 212Pb-DOTAM (10 мкКи).
Мышей умерщвляли и осуществляли аутопсию через 24 ч после инъекции 212Pb-DOTAM. Изымали кровь, мочевой пузырь, селезенку, почки, печень, легкие, мышцы, хвост и опухоли, взвешивали и измеряли уровень радиоактивности и затем рассчитывали величину %ID/г.
Результаты, протокол 105
Накопление в опухолях 212Pb оказалось, в целом, высоким для всех доз СА за исключением 250 мкг. Клиренс крови от CEA-PRIT-биспецифического антитела оказался наиболее эффективным при применении СА в дозах 30, 75 и 250 мкг. Таким образом, наиболее высокие соотношения «опухоль/кровь» достигались при применении 30, 75 и 250 мкг СА: 187,7, 180,2 и 243,5 соответственно. Соответствующее поглощение опухолями 212Pb составляло 91,8±18,9, 70,5±11,1 и 35,2±17,0%ID/г (± SD, n=3).
На фиг. 33 представлены данные о распределении радиоактивности в отобранных тканях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3). Серыми прямоугольниками обозначено накопление в ткани после инъекции в различных количествах СА Dex500-(50%); черный прямоугольник соответствует контролю без СА.
На фиг. 34 представлены данные о содержании 212Pb в крови и опухолях через 24 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3) и соответствующие соотношения «опухоль/кровь». Серыми прямоугольниками обозначено накопление в ткани после инъекции в различных количествах СА Dex500-(50%); черный прямоугольник соответствует контролю без СА.
Обобщение результатов и заключение
На основе данных о накоплении 212Pb в опухолях, о клиренсе крови и соответственно соотношении «опухоль/кровь», установлено, что доза 30 мкг Dex500-(50%) может являться предпочтительной для CEA-PRIT на ВхРС3-модели.
Пример 27: Время нахождении в крови очищающего агента на основе декстрана-500 (протокол 106)
В настоящем исследовании изучали очищающий агент на основе декстрана-500 с 50%-ным ТСМС-насыщением в отношении времени нахождения в крови. Цель состояла в идентификации приемлемой временной рамки для повторяющихся обработок (1-4 недели), гарантирующей, что в кровотоке остается небольшое количество очищающего агента или отсутствует очищающий агент, который может связываться с введенными биспецифическими антителами, эффективно блокируя связывание вводимого путем последующей инъекции радиоактивно меченного DOTAM.
План опыта, протокол 106

План опыта, протокол 106
Изучаемые группы, протокол 106 (ntot=24)
Мышам (возрастом 9 недель) вводили i.v. CEA-PRIT (100 мкг на 100 мкл), затем через один день Dex500-(50%) (25 мкг на 100 мкл). Группам Б, Г, Е и З вводили ЗФР вместо очищающего агента. Через 1, 2, 3 или 4 недели мышам повторно инъецировали i.v. CEA-PRIT (100 мкг на 100 мкл), затем через 1 день 100 мкл 212Pb-DOTAM (10 мкКи).
Мышей умерщвляли через 4 ч после инъекции 212Pb-DOTAM. Получали образцы крови в момент эвтаназии и образцы взвешивали и измеряли уровень радиоактивности. Затем рассчитывали процент инъецированной дозы на грамм крови (%ID/г) с учетом коррекции на распад и фоновый уровень.
Результаты, протокол 106
Не обнаружено статистически значимого различия в среднем уровне радиоактивности между мышами, которым вводили Dex500-(50%) или ЗФР, в любой из изученных моментов времени (1-сторонний дисперсионный анализ, критерий множественных сравнений Сидака, р>0,05).
На фиг. 35 представлены данные об уровне радиоактивности в крови через 4 ч после инъекции 212Pb-DOTAM (%ID/г ± SD, n=3).
Обобщение результатов и заключение
Результаты продемонстрировали, что повторный цикл обработки CEA-PRIT можно начинать уже через 1 неделю после последней инъекции Dex500-(50%) и это не приводит к блокаде связывания вводимого следом 212Pb-DOTAM с СЕА-DOTAM bsAb.
Пример 28: Дополнительные форматы биспецифических антител
Создание плазмид для рекомбинантной экспрессии тяжелых или легких цепей антитела
Требуемые белки экспрессировали путем кратковременной трансфекции клеток почки человеческого эмбриона (HEK 293). Для экспрессии требуемого гена/белка (например, тяжелой цепи полноразмерного антитела, легкой цепи полноразмерного антитела или тяжелой цепи полноразмерного антитела, содержащей дополнительный домен (например, вариабельный домен тяжелой или легкой цепи иммуноглобулина на его С-конце) применяли транскрипционную единицу, содержащую следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5'UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность (SS) тяжелой цепи мышиного иммуноглобулина,
- подлежащий экспрессии ген/белок и
- последовательность полиаденилирования бычьего гормона роста (BGH рА).
В дополнение к экспрессионной единице/кассете, включающей требуемый подлежащий экспрессии ген, базовая/стандартная экспрессионная плазмида для клеток млекопитающих содержала
- сайт инициации репликации из pUC18, позволяющий осуществлять репликацию этой плазмиды в Е. coli, и
- ген бета-лактамазы, придающий устойчивость к ампициллину в E. coli.
Р1АЕ1766
Сборку генов, кодирующих тяжелую цепь антитела, включая С-концевые слитые гены, содержащую полную и функциональную тяжелую цепь антитела с расположенным далее дополнительным VL-CH1-доменом или VH-C-каппа-доменом, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности, каждый из которых был отделен G4Sx4-линкером, с С-концом СН3-домена молекулы человеческого IgG (VH-СН1-шарнир-СН2-СН3-линкер-VL-СН1 или VH-СН1-шарнир-СН2-СН3-линкер-VH-Ck). Рекомбинантные молекулы антител, несущие один VH-CH1-домен и один VH-Ck-домен на С-концах двух СН3-доменов соответственно, экспрессировали с использованием технологии knob-into-hole.
Сборку генов, кодирующих легкую цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Р1АЕ1768
Сборку генов, кодирующих тяжелую цепь антитела, содержащую полную и функциональную тяжелую цепь антитела, таргетирующую либо СЕА, либо DOTAM, осуществляли с использованием технологии knob-into-hole.
Сборку генов, кодирующих легкую цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности. Правильную сборку обеспечивали путем применения технологии CrossMab (обмен CL- и CH1-доменов в одном из плечей).
Р1АЕ1769
Сборку слитого гена осуществляли путем слияния ДНК-фрагмента, кодирующего VH-CH1-домен, с помощью G4Sx4-линкера с N-концом VL-домена молекулы человеческого IgG, содержащей обмен VL-домена на VH домен (в результате чего создавали слитый ген, кодирующий структуру VH-CH1-линкер-VL-CH1-шарнир-CH2-CH3). Рекомбинантные молекулы антитела, несущие один VH-CH1 и не содержащие слияния на N-концах соответственно, экспрессировали с использованием технологии knob-into-hole.
Сборку генов, кодирующих VH-C-каппа-цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Сборку генов, кодирующих легкую цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Правильную сборку обеспечивали путем применения технологии CrossMab.
Р1АЕ1767
Сборку генов, кодирующих тяжелую цепь антитела, включая С-концевые слитые гены, содержащую полную и функциональную тяжелую цепь антитела, с расположенным далее дополнительным V-доменом легкой цепи-СН1-доменом антитела осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности, каждый из которых отделен с помощью G4Sx4-линкера, с С-концом СН3-домена молекулы человеческого IgG (VH-СН1-шарнир-СН2-СН3-линкер-VL-CH1). Рекомбинантные молекулы антитела, несущие один VL-CH1 и не имеющие слияния на С-концах двух СН3-доменов соответственно, экспрессировали с использованием технологии knob-into-hole.
Сборку генов, кодирующих VH-C-каппа-цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Сборку генов, кодирующих легкую цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Правильную сборку обеспечивали путем применения технологии CrossMab.
Р1АЕ1770
Сборку генов, кодирующих тяжелую цепь антитела, включая С-концевые слитые гены, содержащую полную и функциональную тяжелую цепь антитела, с расположенным далее дополнительным V-легкой цепи-С-каппа-цепи-линкер-V-тяжелой цепи-CH1-доменом антитела осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности, каждый из которых отделен с помощью G4Sx4-линкера, с С-концом СН3-домена молекулы человеческого IgG (VH-СН1-шарнир-СН2-СН3-линкер-VL-Ck-линкер-VH-CH1). Рекомбинантные молекулы антитела, несущие один одноцепочечный Fab и не имеющие слияния на С-концах двух СН3-доменов соответственно, экспрессировали с использованием технологии knob-into-hole.
Сборку генов, кодирующих легкую цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Кратковременная экспрессия молекул антитела
Молекулы антител создавали в кратковременно трансфектированных клетках HEK293 (полученных из линии клеток почки человеческого эмбриона 293), которую культивировали в среде F17 (фирма Invitrogen Corp.). Для трансфекции применяли трансфектирующий реагент «293-Free» (фирма Novagen). Соответствующие молекулы тяжелых и легких цепей антител, описанные выше, экспрессировали из индивидуальных экспрессионных плазмид. Трансфекции осуществляли согласно инструкциям производителя. Супернатанты клеточных культур, содержащие иммуноглобулин, собирали через три-семь (3-7) дней после трансфекции. Супернатанты хранили при пониженной температуре (например, при -80°С) вплоть до очистки.
Общая информация, касающаяся рекомбинантной экспрессии человеческих иммуноглобулинов, например в HEK293-клетках, представлена у: Meissner Р. и др., Biotechnol. Bioeng. 75, 2001, сс. 197-203.
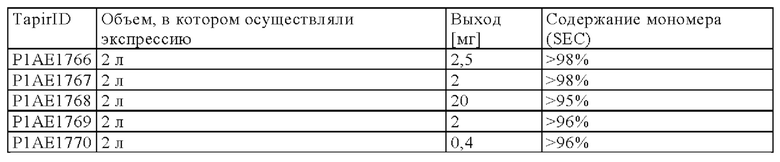
Молекулы очищали с помощью MabSelect Sure (аффинная хроматография) и затем с помощью Супердекс 200 (гель-фильтрация).
Оценка методом Kinexa DOTAM-связывающих свойств различных форматов
Для подробного анализа аффинности применяли метод Kinexa.
Инструментарий и материалы
Применяли устройство KinExA 3200 фирмы Sapidyne Instruments (Бойсе, шт. Айдахо) с автосэмплером. Полиметилметакрилатные (РММА) гранулы покупали у фирмы Sapidyne, а ЗФР (забуференный фосфатом физиологический раствор), БСА (фракция V, бычий сывороточный альбумин) и антитела к DOTAM получали в лаборатории заявителей (фирма Roche). Конъюгированное с Dylight650® очищенное с помощью аффинной хроматографии козье антитело к человеческому IgG, перекрестно адсорбированное с Fc-фрагментом, покупали у фирмы Bethyl Laboratories (Монтгомери, шт. Техас). Биотинилированные антигены Pb-DOTAM (Pb-DOTAM-алкил-биотин, изомеры А и Б, Pb-DOTAM-Bn-биотин/ТСМС-Pb-dПЭГ3-биотин, изомеры А и Б) и небиотинилированный 212Pb-DOTAM получали от фирмы AREVA Med (Бетезда, шт. Мэриленд).
Получение покрытых антигеном гранул
На РММА-гранулы наносили покрытие согласно протоколу, указанному в справочнике KinExA для биотинилированных молекул (фирма Sapidyne). В целом, метод состоял в следующем: во-первых, добавляли биотин-БСА (фирма Thermo Scientific) в 1 мл ЗФР (рН 7,4) из расчета по 10 мкг на флакон (200 мг) гранул для нанесения адсорбирующего покрытия. После вращения в течение 2 ч при комнатной температуре супернатант удаляли и гранулы промывали 5 раз 1 мл ЗФР. Во-вторых, к гранулам добавляли 1 мл 100 мкг нейтравидина, биотин-связывающего белка (фирма Thermo Scientific) в ЗФР, содержащем 10 мг/мл БСА, и инкубировали при комнатной температуре еще в течение 2 ч для связывания нейтравидина с гранулами и для создания дополнительных связывающих биотин сайтов для последующего связывания биотинилированных белков. Затем покрытые нейтравидином гранулы промывали 5 раз 1 мл ЗФР. В завершение, на гранулы наносили покрытие с использованием 200 нг/мл смеси биотинилированных изомеров Pb-DOTAM (по 50 нг каждого изомера) в ЗФР и инкубировали еще в течение 2 ч при комнатной температуре. Затем гранулы ресуспендировали в 30 мл ЗФР и сразу же использовали.
Анализы методом KinExA в условиях равновесия
Все эксперименты по методу KinExA осуществляли при комнатной температуре (КТ) с использованием ЗФР, рН 7,4 в качестве подвижного буфера. Образцы приготавливали в подвижном буфере, дополненном 1 мг/мл БСА («буфер для образца»). Скорость потока составляла 0,25 мл/мин. Постоянное количество антитела к DOTAM с концентраций сайтов связывания 5 пМ титровали с использованием двукратных серийных разведений антигена, начиная с концентрации 100 пМ (диапазон концентраций 0,049-100 пМ). Один образец антитела без антигена служил в качестве образца, дающего 100% сигнал (т.е. без ингибирования). Комплексы антиген-антитело инкубировали при КТ в течение по меньшей мере 24 ч, обеспечивая достижение равновесия. Затем уравновешенные смеси пропускали через колонку, содержащую сшитые с антигеном гранулы, в KinExA-системе в объеме 5 мл, что позволяло несвязанному антителу «захватываться» гранулами без нарушения состояния равновесия раствора. Для детекции «захваченного» антитела использовали 250 нг/мл конъюгированного с Dylight 650© специфического для человеческого Fc-фрагмента вторичного антитела в буфере для образца. Каждый образец оценивали с дублированием во всех экспериментах по уравновешиванию.
Величины KD определяли с помощью нелинейного регрессионного анализа данных с использованием модели односайтового гомогенного связывания, входящей в пакет программ KinExA (версия 4.0.11), используя метод «стандартного анализа». Программное обеспечение позволяло рассчитывать величины KD и определять 95%-ный доверительный интервал на основе аппроксимации экспериментальных точек теоретической кривой KD. 95%-ный доверительный интервал (Sapidyne TechNote TN207R0) представлен в виде величин KDlow и KDhigh.
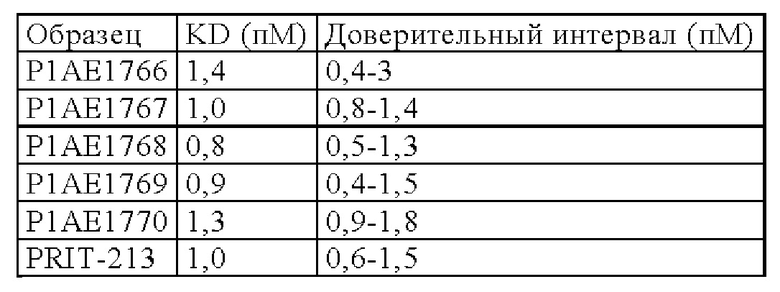
Все конструкции характеризовались сопоставимыми величинами KD, поскольку доверительные интервалы перекрывались.
Последовательности молекул в различных форматах:
Необязательные модификации зарядов выделены жирным шрифтом и подчеркнуты: EQ->RK или KK->ЕЕ
Р1АЕ1766
>СЕА легкая цепь RK
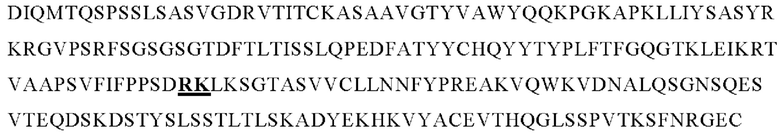
>СЕА тяжелая цепь с DOTAM VL/CH1

>СЕА тяжелая цепь с DOTAM VH/CK

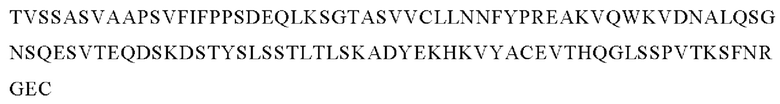
Р1АЕ1767
>DOTAM «LC» с VH/CK

>СЕА легкая цепь RK

>СЕА тяжелая цепь с DOTAM VL/CH1

>СЕА тяжелая цепь

Р1АЕ1768
>СЕА LC

>DOTAM тяжелая цепь с VL/CH1

>СЕА тяжелая цепь
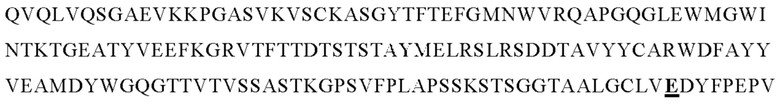

>DOTAM «LC» VH/CK

P1AE1769
DOTAM «LC» с VH/CK

>CEA LC

>СЕА НС с CEA VH/CH1/DOTAM VL/CH1







Пример 29: Биспецифические антитела, связывающиеся с Pb-DOTAM и CD20 или Her2
Создание плазмид для рекомбинантной экспрессии тяжелых или легких цепей антитела
Требуемые белки экспрессировали путем кратковременной трансфекции клеток почки человеческого эмбриона (HEK 293). Для экспрессии требуемого гена/белка (например, тяжелой цепи полноразмерного антитела, легкой цепи полноразмерного антитела или тяжелой цепи полноразмерного антитела, содержащей дополнительный домен (например, вариабельный домен тяжелой или легкой цепи иммуноглобулина на его С-конце) применяли транскрипционную единицу, содержащую следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5'UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность (SS) тяжелой цепи мышиного иммуноглобулина,
- подлежащий экспрессии ген/белок и
- последовательность полиаденилирования бычьего гормона роста (BGH рА).
В дополнение к экспрессионной единице/кассете, включающей требуемый подлежащий экспрессии ген, базовая/стандартная экспрессионная плазмида для клеток млекопитающих содержала
- сайт инициации репликации из pUC18, позволяющий осуществлять репликацию этой плазмиды в Е. coli, и
- ген бета-лактамазы, придающий устойчивость к ампициллину в E.coli.
а) Экспрессионная плазмида для тяжелых цепей антитела
Осуществляли сборку генов, кодирующих тяжелую цепь антитела, включая С-концевые слитые гены, содержащую полную и функциональную тяжелую цепь антитела, с расположенным далее дополнительным V-доменом тяжелой цепи или V-доменом легкой цепи антитела, путем слияния ДНК-фрагмента, кодирующего соответствующие элементы последовательности (V-домен тяжелой или V-домен легкой цепи), каждый отделенный с помощью G4Sx4-линкера, с С-концом СН3-домена молекулы человеческого IgG (VH-СН1-шарнир-СН2-СН3-линкер-VH или VH-СН1-шарнир-СН2-СН3-линкер-VL). Рекомбинантные молекулы антитела, несущие один VH-домен и один VL-домен на С-концах двух СН3-доменов соответственно, экспрессировали с использованием технологии knob-into-hole.
Экспрессионные плазмиды для кратковременной экспрессии тяжелой цепи с С-концевым VH- или VL-доменом антитела в HEK293-клетках содержали помимо кассеты экспрессии для фрагмента тяжелой цепи антитела с С-концевым VH- или VL-доменом сайт инициации репликации из вектора pUC18, обеспечивающий репликацию указанной плазмиды в Е. coli, и ген бета-лактамазы, придающий устойчивость к ампициллину в Е. coli. Транскрипционная единица слитого гена фрагмента тяжелой цепи антитела с С-концевым VH- или VL-доменом содержала следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5'UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность тяжелой цепи мышиного иммуноглобулина,
- нуклеиновую кислоту, кодирующую тяжелую цепь антитела (VH-CH1-шарнир-СН2-СН3-линкер-VH или VH-СН1-шарнир-СН2-СН3-линкер-VL), и
- последовательность полиаденилирования бычьего гормона роста (BGH рА).
Аминокислотная последовательность слитого белка в виде фрагмента тяжелой цепи зрелого антитела с С-концевым VH- или VL-доменом представляет собой



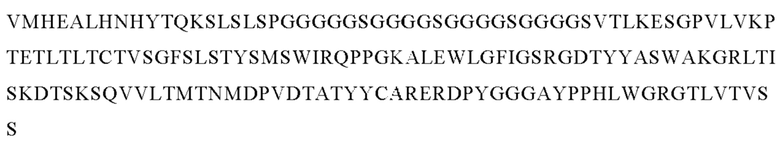
б) Экспрессионная плазмида для легких цепей антитела
Сборку генов, кодирующих легкую цепь антитела, содержащую полную и функциональную легкую цепь антитела, осуществляли путем слияния ДНК-фрагментов, кодирующих соответствующие элементы последовательности.
Экспрессионная плазмида для кратковременной экспрессии легкой цепи антитела содержала помимо фрагмента легкой цепи антитела сайт инициации репликации из вектора pUC18, который обеспечивал репликацию указанной плазмиды в Е. coli, и ген бета-лактамазы, обусловливающий устойчивость к ампициллину в Е. coli. Транскрипционная единица фрагмента легкой цепи антитела содержала следующие функциональные элементы:
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемую область (5' UTR) тяжелой цепи человеческого иммуноглобулина,
- сигнальную последовательность тяжелой цепи мышиного иммуноглобулина,
- нуклеиновую кислоту, кодирующую легкую цепь антитела (VL-CL), и
- последовательность полиаденилирования бычьего гормона роста (BGH рА).

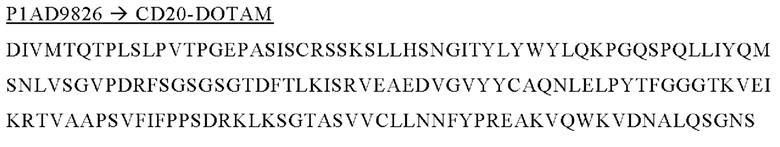
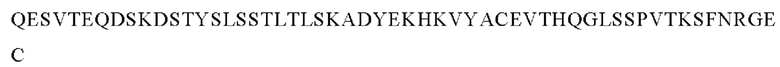
Кратковременная экспрессия молекул антитела
Молекулы антител создавали в кратковременно трансфектированных клетках HEK293 (полученных из линии клеток почки человеческого эмбриона 293), которую культивировали в среде F17 (фирма Invitrogen Corp.). Для трансфекции применяли трансфектирующий реагент «293-Free» (фирма Novagen). Соответствующие молекулы тяжелых и легких цепей антител, описанные выше, экспрессировали из индивидуальных экспрессионных плазмид. Трансфекции осуществляли согласно инструкциям производителя. Супернатанты клеточных культур, содержащие иммуноглобулин, собирали через три-семь (3-7) дней после трансфекции. Супернатанты хранили при пониженной температуре (например, при -80°С) вплоть до очистки.
Общая информация, касающаяся рекомбинантной экспрессии человеческих иммуноглобулинов, например в HEK293-клетках, представлена у: Meissner Р. и др., Biotechnol. Bioeng. 75, 2001, сс. 197-203.
Очистка молекул антител P1AD8926 и P1AD8927
Молекулы PRIT очищали с помощью MabSelect Sure (аффинная хроматография) и затем Супердекс 200 (гель-фильтрация).
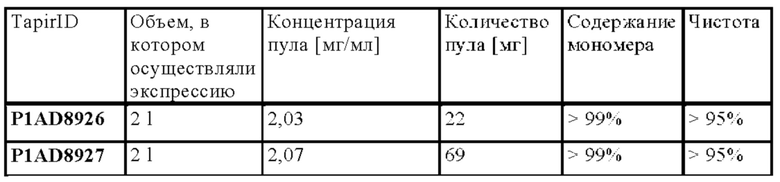
Масс-спектрометрический анализ:
Для подтверждения идентичности молекул PRIT применяли ESI-MC.
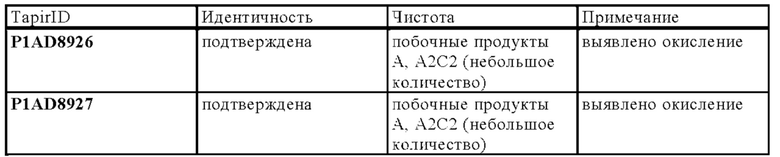
FACS-анализ функциональности P1AD8927
Для оценки функциональности P1AD8927 клетки KPL-4 отделяли от сосуда для культивирования путем обработки аккутазой при 37°С в течение 10 мин. Затем клетки промывали дважды в ЗФР и высевали в 96-луночные планшеты с V-образным дном до достижения конечной плотности 4×106 клеток/лунку.
Антитело предварительно метили Zenon<человеческий IgG>A488, добавляя его к клеткам в концентрациях, указанных на фиг. 38. Затем клетки инкубировали в течение 1 ч на льду, промывали дважды в ЗФР и ресуспендировали в 200 мкл ЗФР/5% FCS для изменений ФИТЦ-флуоресценции с помощью устройства FACS canto.
Результаты представлены на фиг. 38.
Для оценки связывающей способности антитела к DOTAM после связывания с клетками KPL-4 клетки промывали для удаления несвязанного антитела. Затем добавляли Pb-DOTAM-ФИТЦ для детекции антител, связанных с компетентными в отношении связывания с DOTAM клетками (фиг. 39). P1AD8927 характеризовалось зависимым от дозы ФИТЦ-сигналом с учетом коррекции на изотип. Этот эксперимент продемонстрировал, что рассматриваемое антитело обладало функциональностью в отношении связывания с DOTAM.
FACS-анализ функциональности P1AD8926
Для оценки функциональности P1AD8926 клетки Raji промывали дважды в ЗФР и высевали в 96-луночные планшеты с V-образным дном до достижения конечной плотности 4×106 клеток/лунку.
Антитело предварительно метили Zenon<человеческий IgG>A488, добавляя его к клеткам в концентрациях, указанных на фиг. 38. Затем клетки инкубировали в течение 1 ч на льду, промывали дважды в ЗФР и ресуспендировали в 200 мкл ЗФР/5% FCS для изменений ФИТЦ-флуоресценции с помощью устройства FACS canto.
Для оценки связывающей способности антитела к DOTAM после связывания с клетками Raji клетки промывали для удаления несвязанного антитела. Затем добавляли Pb-DOTAM-FITC для детекции антител, связанных с компетентными в отношении связывания с DOTAM клетками (фиг. 41). P1AD8927 характеризовалось зависимым от дозы ФИТЦ-сигналом с учетом коррекции на изотип. Этот эксперимент продемонстрировал, что рассматриваемое антитело обладало функциональностью в отношении связывания с DOTAM.
Пример 30: Материал и методы, применяемые в примерах 31-37
Для опытов, описанных в примерах 31-37, применяли следующий набор материалов и методов.
Трехстадийный PRIT (один цикл)
Стадия 1: Введение BsAb (i.v. или i.p.): BsAb, применяемый в опытах, связывает Pb-DOTAM с высокой аффинностью и таргетирует опухоль, например, посредством СЕА.
Стадия 2: Введение СА (i.v. или i.p.): Для обеспечения эффективного накопления в опухоли 212Pb-DOTAM необходимо нейтрализовать циркулирующий в крови BsAb с использованием СА, который блокирует связывание 212Pb-DOTAM с несвязанным BsAb, без проникновения в опухоль и, следовательно, блокирует предварительно таргетированные сайты. СА вводили сразу как только BsAb накапливался в достаточной степени в опухоли, как правило, через 4-10 дней. СА, представляющий собой Pb-DOTAM-декстран-500, создавали на основе аминодекстрана со средней молекулярной массой 500 кДа, с которым DOTAM конъюгировали через тиомочевинный линкер, как продемонстрировано ниже.
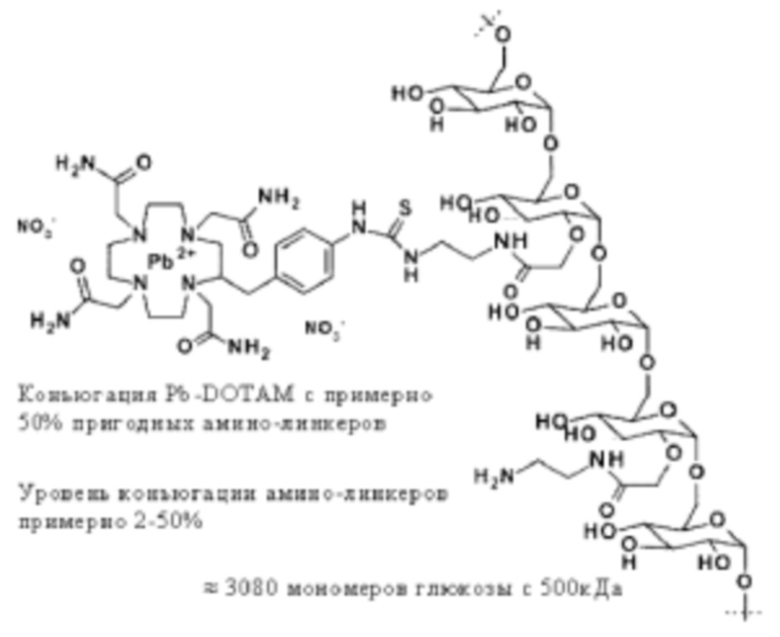
Стадия 3: Введение 212Pb-DOTAM (i.v.): Радиоактивную инъекцию осуществляли на последней стадии, как правило, через 24-48 ч после инъекции СА, давая 212Pb-DOTAM связываться преимущественно с предварительно таргетированными опухолевыми сайтами, эффективно снижая системную экспозиционную дозу радиации.
Общие материалы и методы
Экспериментальные протоколы, которые осуществляли в ARCoLab, были рассмотрены и одобрены местными регулирующими органами (Comité Regional d' Ethique de l'expérimentation Animale du Limousin (CREEAL), Laboratoire départemental d' Analyses et de Recherches de la Haute-Vienne). Мышей с серьезным комбинированным иммунодефицитом (SCID) и CD1-мышей покупали у фирмы Charles River и содержали в свободных от специфических патогенов и условно-патогенных организмов (SOPF) условиях, с суточными циклами свет/темнота (12 ч/12 ч), в соответствии с руководствами по этике. В опыте, проведенном согласно протоколу 121, применяли свободные от специфических патогенов (SPF) условия, а мышей покупали у фирмы Envigo. Никаких манипуляций не осуществляли в течение первых 5 дней после доставки, давая животным акклиматизироваться к новой окружающей среде. Всех мышей контролировали ежедневно клинические симптомы и выявляли нежелательные явления.
Ксенотрансплантаты солидных опухолей создавали путем подкожной (s.c.) инъекции экспрессирующих СЕА опухолевых клеток в среде для культуры клеток, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №354230). Объемы опухолей оценивали вручную с помощью кронциркуля, рассчитывая согласно формуле:
объем = 0,5 × длина × ширина2.
Для минимизации повторного поглощения радиоактивной мочи/экскрементов при осуществлении всех опытов по оценке эффективности мышей помещали в клетки с решетчатыми полами через 4 ч после введения 212Pb-DOTAM перед переносом в новые клетки со стандартной подстилкой. Затем все клетки заменяли через 24 ч после инъекции (p.i.). Указанную процедуру не осуществляли для мышей, подлежащих эвтаназии для изучения биораспределения в течение периода времени вплоть до 24 ч после радиоактивной инъекции.
Вес тела (BW) подопытных животных определяли по меньшей мере 3 раза в неделю, при необходимости осуществлял и дополнительные измерения в зависимости от состояния здоровья. Мышей, у которых потеря BW превышала 25% от их начального BW, или у которых объем опухолей достигал 3000 мм3, немедленно умерщвляли. Другие факторы, которые учитывали для эвтаназии по этическим соображениям, связаны со статусом опухолей (например, некротические области, просачивание крови/жидкости, признаки членовредительства) и общим внешним видом животного (например, состояние шерсти, положение тела, движение). Всем мышам давали влажный корм, начиная с пятого дня после радиоактивной инъекции, если принималось такое решение из-за острой потери BW (коллективной или индивидуальной), вплоть до существенного восстановления всех особей.
Кровь собирали по окончанию эксперимента из венозного синуса посредством ретро-орбитального кровопускания, после чего изымали дополнительные ткани для измерений радиоактивности и/или гистологического анализа, в соответствии с протоколом. Неожиданные или аномальные состояния документировали. Ткани, собранные для фиксации в формалине, немедленно помещали в 10%-ный нейтральный забуференный формалин (4°С), а затем через 5 дней переносили в забуференный фосфатом физиологический раствор (ЗФР, 4°С). Органы и ткани, собранные для оценки биораспределения, взвешивали и измеряли радиоактивность, используя автоматический гамма-счетчик 2470 WIZARD (фирма PerkinElmer), и затем рассчитывали процент инъецируемой дозы на грамм ткани (% ID/г), включая коррекции на распад и фоновый уровень.
Статистический анализ осуществляли с использованием программы GraphPadPrism 6 (фирма GraphPad Software, Inc.) и IMP 8 (фирма SAS Institute Inc.).
Анализ кривых, описывающих ингибирование роста опухолей (TGI), осуществляли на основе средних объемов опухолей с использованием формулы:
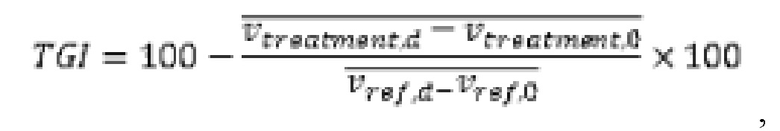
в которой d обозначает день обработки, а 0 обозначает исходное значение. Для обработки референс-группы использовали наполнитель (ЗФР). Регресс опухоли (TR) рассчитывали согласно формуле:

в которой положительные величины свидетельствуют о регрессе опухоли, а величины ниже -1 соответствуют росту, выходящему за пределы удвоенного исходного значения.
Попарное тестирование осуществляли для выявления групп, в которых имело место существенное различие выживаемости: логранговый критерий (больший вес для более поздних случаев выживания), и критерий Уилкоксона (больший вес для ранних случаев), в обоих случаях с использованием поправки Бонферрони для множественных сравнений.
Тестируемые соединения
Соединения, которые использовали в описанных опытах, представлены ниже в таблицах, озаглавленных «Биспецифические антитела», «Очищающие агенты» и «Радиоактивно меченые хелаты».
CEA-DOTAM (PRIT-0213) представляет собой полностью гуманизированное BsAb, мишенью которого является эпитоп Т84. 66 CEA, в то время как DIG-DOTAM (PRIT-0175) представляет собой не связывающее CEA BsAb, которое применяют в качестве отрицательного контроля. Р1АЕ1766, Р1АЕ1767, Р1АЕ1768, Р1АЕ1769 и Р1АЕ1770 представляют собой гуманизированные CEA-DOTAM BsAb, мишенью которых является эпитоп СН1А1А СЕА, описанные выше в примере 28. Конструкции антител хранили при -80°С вплоть до дня инъекции, когда их подвергали оттаиванию и разводили стандартным применяемым в качестве наполнителя буфером (20 мМ гистидин, 140 мМ NaCl; рН 6,0) или 0,9% NaCl до их конечных концентраций для внутривенного (i.v.) или внутрибрюшинного (i.p.) введения соответственно.
СА, представляющие собой Ca-DOTAM-декстран-500 и Pb-DOTAM-декстран-500, хранили при -20°С вплоть до дня инъекции, когда их подвергали оттаиванию и разводили забуференным фосфатом физиологическим раствором (ЗФР) для i.v. - или i.р. -введения.
Хелат DOTAM для радиоактивного мечения получали от фирмы Macrocyclics и выдерживали при -20°С вплоть до осуществления радиоактивного мечения. Pb-DOTAM создавали путем элюции DOTAM из ториевого генератора и последующего гашения с использованием Cu, Са, Zn, Gd или Pb после введения метки. Раствор 212Pb-DOTAM разводили ЗФР или 0,9% NaCl с получением требуемого для i.v.-инъекции уровня активности 212Pb.
Мыши в обрабатываемых наполнителем контрольных группах получали несколько инъекций ЗФР вместо BsAb, СА и 212Pb-DOTAM.
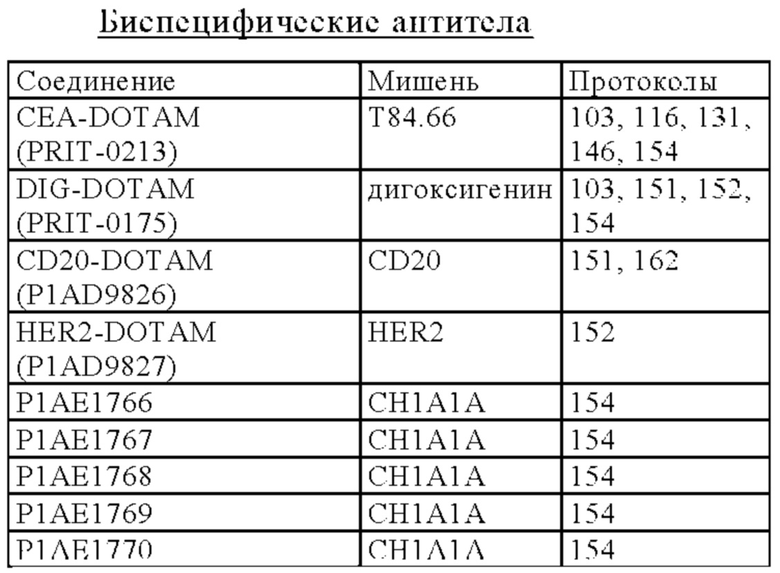

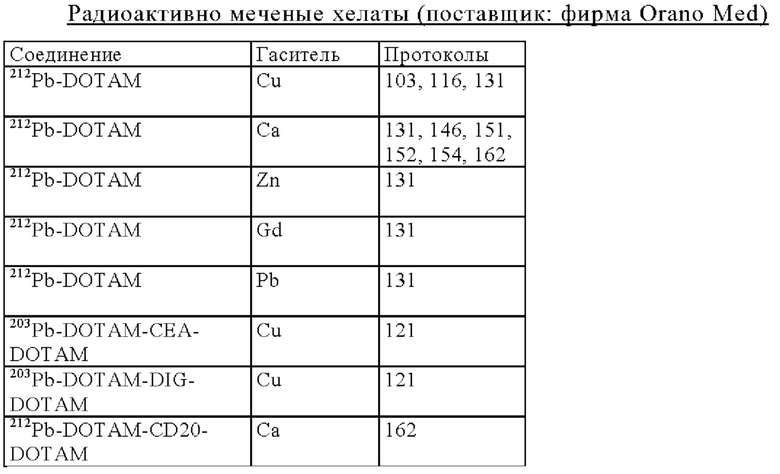
Модели опухолей
Применяемые линии опухолевых клеток и инъецируемое количество для инокуляции мышам описаны ниже в таблице «Линии опухолевых клеток». ВхРС3 представляет собой клеточную линию человеческой первичной аденокарциномы поджелудочной железы, которая в естественных условиях экспрессирует СЕА. Клетки ВхРС3 культивировали в среде RPMI-1640, дополненной GlutaMAX™, HEPES (фирма Gibco каталожный №42401-018), обогащенной 10% фетальной бычьей сыворотки (фирма GE Healthcare Hyclone SH30088.03). HPAF-II (CRL-1997) представляет собой клеточную линию человеческой аденокарциномы поджелудочной железы, которая в естественных условиях экспрессирует СЕА. HPAF-II-клетки культивировали в среде ЕМЕМ (фирма Gibco, 31095-029), обогащенной 10% стандартной фетальной бычьей сыворотки, 1% GlutaMAX 100×, 1% MEM NEAA (минимальная поддерживая среда с заменимыми аминокислотами), 100× (фирма Gibco, 11140-035) и 1% пирувата натрия (100 мМ; фирма Gibco, 11360-070). WSU-DLCL2 представляет собой клеточную линию человеческой В-клеточной лимфомы, которая в естественных условиях экспрессирует CD20. WSU-DLCL2-клетки культивировали в среде RPMI-1640 (фирма Gibco, каталожный №. 72400-021), обогащенной 10% фетальной бычьей сыворотки. NCI-N87 представляет собой клеточную линию рака желудка, которая в естественных условиях экспрессирует HER2. NCI-N87-клетки культивировали в среде RPMI-1640 (фирма Gibco, каталожный №. 72400-021), обогащенной 10% фетальной бычьей сыворотки. Ксенотрансплантаты солидных опухолей создавали в SCID-мышах в день опыта 0 путем подкожной инъекции в правую боковую область клеток в среде RPMI или DMEM, смешанной в соотношении 1:1 с матрицей базальной мембраны Corning® Matrigel® (уменьшенный уровень фактора роста; каталожный №354230).

Экспериментальная часть
Пример 31: Эффективность CEA-PRIT на модели опухоли (протокол 103)
В данном исследовании оценивали эффективность CEA-PRIT с использованием CEA-DOTAM BsAb для лечения созданных путем s.с.-инъекции опухолей ВхРС3 у мышей. Терапевтические средства вводили либо в виде однократной обработки 10 или 30 мкКи 212Pb-DOTAM, либо с использованием трех повторяющихся циклов для каждого из двух уровней радиоактивности. Осуществляли сравнение с PRIT с использованием не связывающего СЕА контрольного антитела (DIG-DOTAM), только CEA-DOTAM BsAb (без радиоактивности) и без обработки (ЗФР). Оценку биораспределения осуществляли после первого и второго циклов для подтверждения таргетинга и клиренса 212Pb-DOTAM, и эффективность лечения оценивали на основе TGI, TR и выживаемости. Осуществляли тщательный мониторинг мышей в процессе всего опыта для оценки переносимости различных схем лечения. План исследования представлен на фиг 42.
План опыта
Ниже в таблицах представлены временной график и план опыта согласно протоколу 103.


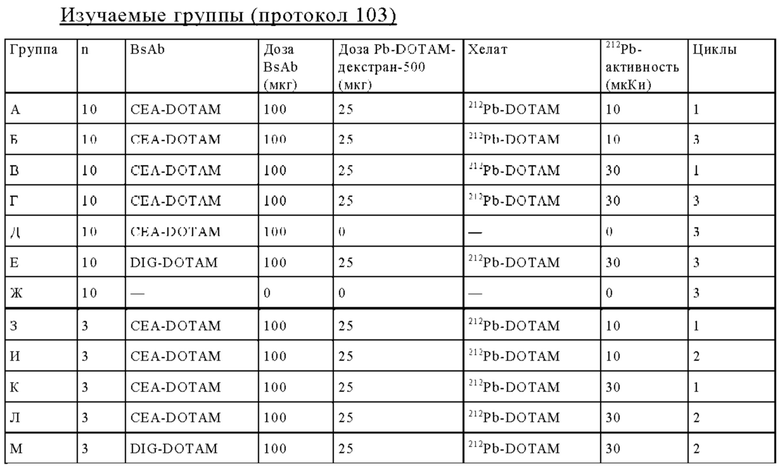
Ксенотрансплантаты солидных опухолей создавали путем s.c. инъекции 5×106 клеток (пассаж 24) в среде RPMI/Matrigel в правую боковую область каждой SCID-мыши в день 0. Через 20 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухолей 290 мм3.
Из-за связанных с логистикой причин начальные обработки осуществляли в течение 2 дней, при этом в первый день осуществляли инъекции BsAb или ЗФР группам В, Г, Д, Е, Ж, К, Л и М, как указано в приведенной выше таблице, а на следующий день осуществляли инъекции BsAb группам А, Б, З и И. Через 4 дня инъецировали СА, а через 2 ч после этого 212-DOTAM или ЗФР. Сначала осуществляли инъекции животным в группах, получающих 30 или 0 мкКи, на следующий день осуществляли инъекции в группах, получающих 10 мкКи. Группы Б, Г, Д, Е, Ж, И, Л и М подвергали нескольким обработкам с 2-недельным интервалом между радиоактивными инъекциями. Все циклы повторных обработок (2-ой и 3-ий) начинали одновременно, т.е. все мыши получали инъекцию BsAb или ЗФР в один и тот же день, с последующей обработкой СА и 212Pb-DOTAM или ЗФР чрез 4 дня. У групп А-Ж в момент эвтаназии изымали следующие органы и ткани: мочевой пузырь, яичники, печень, селезенку, почки, бедро (включая костный мозг), ободочную кишку, тощую кишку, желудок и опухоль.
Мышей в группах Н и J умерщвляли и осуществляли аутопсию через 24 ч после первой для них и единственной инъекции 212Pb-DOTAM; группы И, Л и М умерщвляли и осуществляли аутопсию через 24 ч после второй для них инъекции 212Pb-DOTAM. В момент эвтаназии изымали кровь, мочевой пузырь, тонкий кишечник, ободочную кишку, селезенку, поджелудочную железу, почки, печень, легкие, сердце, бедренную кость, мышцы, хвост и опухоль для измерения радиоактивности.
Результаты
Распределение in vivo 212Pb через 24 ч p.i. продемонстрировало высокий уровень поглощения подкожными ВхРС3-опухолями и низкий уровень накопления в здоровых тканях, что продемонстрировано на фиг. 43. Существенное различие между опухолями, предварительно таргетированными CEA-DOTAM и предварительно таргетированные не связывающим СЕА BsAb DIG-DOT AM, подтвердило высокую специфичность лечения. Односторонний анализ вариантов (односторонний дисперсионный анализ) с критерием множественных сравнений Сидака продемонстрировал отсутствие существенных различий в среднем уровне поглощения опухолью 212Pb ни между радиоактивными инъекциями 1 (50,3% ID/г) и 2 (43,0% ID/г) 10 мкКи, ни между инъекциями 1 (37,6% ГО/г) и 2 (24,1% ID/г) 30 мкКи. Аналогично этому, не обнаружено существенного различия между введением в цикле 1 10 мкКи и в цикле 1 30 мкКи (р>0,05). Однако поглощение опухолями оказалось существенно более низким после 2 циклов введения 30 мкКи по сравнению с 2 циклами, в которых вводили 10 мкКи, также как и при поглощении после 2 циклов 30 мкКи опухолями, предварительно таргетированными DIG-DOTAM (2,9% ID/г), по сравнению с любой из обработок CEA-DOTAM. На панели Б на фиг. 43 продемонстрировано, что величина % ID/г оказалась сопоставимой для опухолей различных размеров в различных обрабатываемых группах.
Кривые среднего роста опухолей и кривые роста индивидуальных опухолей представлены на фиг. 44 и 45 соответственно. Во всех группах объем опухолей удвоился ко дню 20-24. Первую обработку Pb-DOTAM осуществляли в день 24, после чего опухоли в обработанных CEA-DOTAM группах продолжали расти в течение примерно 1 недели до начала их сокращения. Не выявлен полный регресс опухолей, но объемы в группах, которые обрабатывали несколькими соединениями (Б и Г), оставались относительно постоянным вплоть до последней обработки в день 52. Опухоли у мышей, которым однократно инъецировали только либо 10, либо 30 мкКи, начинали уменьшаться в день 44, хотя в группах, которых обрабатывали с использованием более высокой радиоактивности, обнаружена несколько более медленная скорость роста. Осуществление неспецифического PRIT с использованием DIG-DOTAM приводило к статистически значимому, но ограниченному TGI по сравнению с обработкой только CEA-DOTAM или наполнителем. В двух последних контрольных группах развитие опухолей оказалось идентичным.
В день 63, в последний день, в который еще можно было проанализировать все обрабатываемые группы на основе средних, величина TGI составляла 57,2, 89,8, 77,7, 96,6, -6,2 и 67,3% для групп А, Б, В, Г, Д и Е соответственно по сравнению с вариантом без обработки. Последнюю мышь в обработанной наполнителем контрольной группе проанализировали в день 74, в этот момент времени величина TGI составляла 48,5, 83,3, 63,5 и 95,7% для четырех оставшихся групп: А, Б, В и Г соответственно. Опыт заканчивали в день 118 после инъекции клеток путем эвтаназии последней оставшейся мыши (группа Г).
Логранговый критерий и критерий Уилкоксона применяли для выявления групп, в которых имело место существенное различие выживаемости, результаты представлены в двух таблицах ниже. Все на PRIT-режимы основе CEA-DOTAM существенно повышали выживаемость по сравнению с тремя контрольными группами. Общая выживаемость оказалась несколько более высокой после 3 циклов с использованием 10 мкКи по сравнению с одним циклом с использованием 10 мкКи (р=0,0110), но не было обнаружено существенного различия между 3 циклами с использованием 10 мкКи и любым из режимов на основе 30 мкКи при применении претаргетинга CEA-DOTAM. Кривые Каплана-Мейера представлены на фиг. 46.
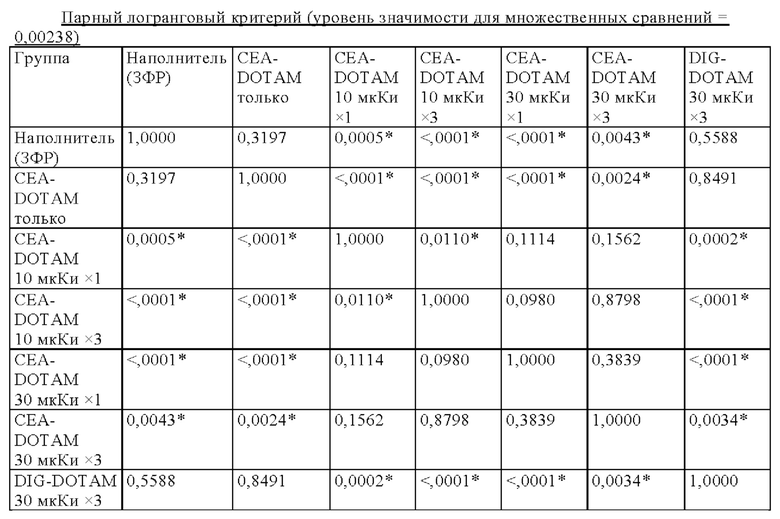
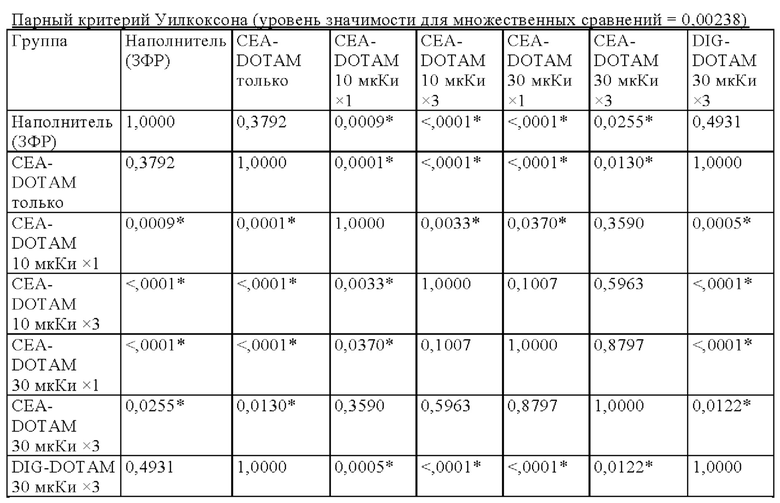
Неблагоприятные явления и токсичность
Неблагоприятные явления, обусловливающие необходимость умерщвлять мышей по этическим соображениям, можно разделить на две группы: 1) статус опухоли и 2) индуцированная радиацией токсичность. Первая относится к некротическим и/или язвенным опухолям, побуждающим мышей «чистить» область опухоли на себе или на соседях по клетке. Мышей, достигших этой стадии, немедленно подвергали эвтаназии для избавления от страданий.
Вторая группа неблагоприятных явлений включает типичные симптомы индуцированной радиацией токсичности, например, потерю BW, диарею и апатию. На фиг. 47 представлено изменение BW в подвергнутых терапии группах. У всех мышей, которым инъецировали 30 мкКи Pb-DOTAM, обнаружено начальное резкое уменьшение BW, которое становилось менее выраженным к 8 дню после первой радиоактивной инъекции. Группы, которым вводили несколько 30 мкКи-инъекций (Г и Е) характеризовались более пролонгированной потерей BW. Потеря BW была более слабой в группах, которые обрабатывали 10 мкКи (А и Б), даже в случае нескольких инъекций. Так, было выявлено выраженное различие между мышами, которых обрабатывали 10 и 30 мкКи 212Pb-DOTAM. Даже после повторных 10 мкКи-обработок у мышей, как правило, не было выявлено основных признаков неблагоприятной токсичности. Мыши, которых подвергали одному циклу обработки 30 мкКи, характеризовались кратковременной потерей веса, но он восстанавливался. Однако второй и третий циклы обработки 30 мкКи приводили к более интенсивной неблагоприятной токсичности, в результате чего многие животные были умерщвлены из-за комбинации потери BW и признаков боли и/или дискомфорта. На основе этих данных можно сделать заключение о том, что 10 мкКи представляет собой безопасный уровень радиоактивности в рассматриваемых условиях, в то время как уровень радиоактивности 30 мкКи близок к максимально переносимому.
Заключение
При создании изобретения было сделано заключение о том, что применение в качестве монотерапевтического режима повторной обработки CEA-PRIT с 10 или 30 мкКи 212Pb-DOTAM обеспечивало существенное повышение выживаемости и TGI, но не приводило ни к полному искоренению опухоли, ни к продолжительному контролю опухоли в рассматриваемых условиях. Полученные результаты позволяют предположить, что для того, чтобы избежать индуцированной радиацией токсичности при поддержании контроля опухоли в течение некоторого времени следует применять 212Pb-DOTAM с уровнем радиоактивности менее 30, но более 10 мкКи.
Пример 32: Выбор пригодного промежутка времени между инъекциями BsAb и СА для CEA-PRIT (протоколы 116 и 121)
Протоколы 116 и 121 разрабатывали для осуществления выбора пригодного промежутка времени между инъекциями BsAb и СА для CEA-PRIT на основе высокого поглощения опухолью и равномерного распределения BsAb в опухоли. Протокол 116 предназначен для оценки распределения BsAb в опухоли в сравнении с экспрессией СЕА при использовании ВхРС3-модели в дни 4, 7 и 14 после i.v. инъекции, для обнаружения которого применяли иммунофлуоресцентное окрашивание. Протокол 121 предназначен для оценки накопления BsAb непосредственно меченного свинцом-203 (203Pb) в опухолях ВхРС3 через 1, 4, 7 и 10 дней. BsAb метили изотопом свинца 203Pb (период полураспада 2,2 дня) для того, чтобы иметь возможность отслеживать процесс в течение более длительного периода времени, чем при использовании 212Pb (период полураспада 10,6 ч).
План опыта
Временные графики и планы протоколов 116 и 121 даны в четырех таблицах, представленных ниже.

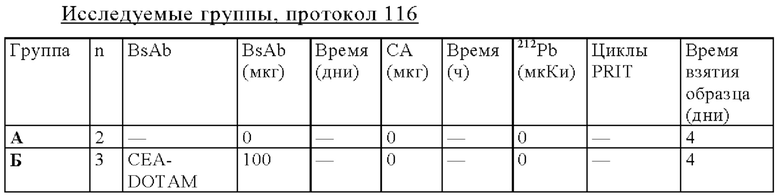
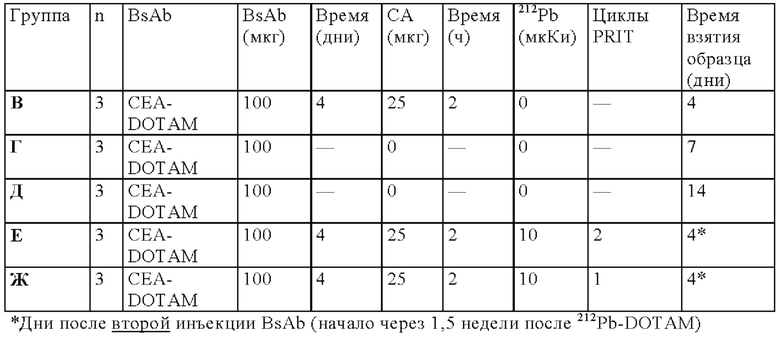

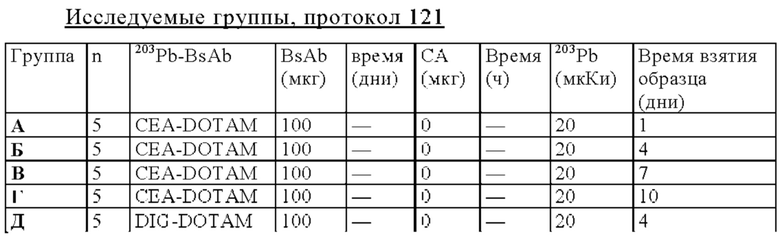
Ксенотрансплантаты солидных опухолей создавали посредством s.c. -инъекции клеток ВхРС3. Через 19 дней после инокуляции мышей согласно протоколу 116 разделяли на экспериментальные группы со средним объемом опухоли 227 мм3. Согласно протоколу 121 средний объем опухоли составлял 203 мм3 через 20 дней.
Иммунофлуоресцентное окрашивание
Согласно протоколу 116 после эвтаназии мышей проводили аутопсию и изымали селезенку, почки, печень, мышцы и опухоль. Собранные ткани разрезали на две части: одну помещали в криоблок, содержащий заливочную среду Tissue-Tek® с оптимальной для приготовления срезов температурой (ОСТ), и помещали на сухой лед для быстрого замораживания, а другую фиксировали в 10% нейтральном забуференном формалине (NBF) в течение 24 ч и затем переносили в ЗФР для хранения до заливки в парафин. Замороженные при ОСТ образцы поддерживали при -80°С до приготовления срезов. Гистологическое окрашивание осуществляли с использованием реагентов, перечисленных в приведенной ниже таблице.
Окрашивание для выявления CEA
С помощью криотома Leica CM 1850 UV получали 12 мкм-срезы замороженных опухолей и предметные стекла хранили при -20°С. Перед окрашиванием предметные стекла подвергали оттаиванию в течение 1 ч при комнатной температуре (КТ) и затем промывали в течение 5 мин ЗФР (1X, рН 7,4), затем в течение 5 мин холодным ацетоном (-20°С) и еще один раз ЗФР в течение 5 мин. Предметные стекла просушивали путем промокания ткани кусочком бумаги и ткань обводили кружочком с помощью гидрофобного карандаша (маркера) (фирма Dako; Agilent). Инкубацию срезов осуществляли с использованием 400 мкл блокирующей сыворотки (козьей) в течение 1 ч при КТ. Блокирующую сыворотку удаляли и к срезам добавляли 200 мкл первичного антитела (кроличий IgG к человеческому СЕА), после чего осуществляли инкубацию в течение ночи при 4°С в темной камере. В день 2 срезы промывали дважды ЗФР в течение 10 мин и добавляли 400 мкл блокирующей сыворотки. После инкубации в течение 1 ч при КТ блокирующую сыворотку удаляли и к срезам добавляли 200 мкл вторичного антитела (меченное Alexa Fluor 488 козье антитело к кроличьему IgG) и осуществляли контрастное окрашивание (краситель фирмы Hoechst), после чего проводили инкубацию в темной камере в течение 2 ч при КТ. Затем предметные стекла промывали дважды ЗФР в течение 10 мин. В завершение, к срезам добавляли флуоресцентную заливочную среду (Dako S3023; фирма Agilent) и предметные стекла накрывали покровным стеклом, после чего сушили на воздухе и хранили в темноте при -20°С. Анализ окрашенных предметных стекол осуществляли с помощью модульного микроскопа Zeiss Axio Scope. A1.
Окрашивание для выявления CEA-DOTAM
Замороженные срезы опухоли хранили и обрабатывали таким же образом, как описано выше для окрашивания СЕА, но применяя ЗФР вместо первичного антитела. Вторичное антитело представляло собой козье антитело к человеческому IgG, меченное Alexa Fluor 555.
Н&Е-окрашивание
Окрашивание гематоксилином и эозином (Н&Е) замороженных срезов опухоли, полученных согласно протоколу 116, осуществляли с использованием автоматической системы для окрашивания предметных стекол Leica Autostainer XL.
Биораспределение
Согласно протоколу 121 мышей умерщвляли для анализа биораспределения и собирали их кровь посредством ретро-орбитального кровопускания перед смертью. Кроме того, после эвтаназии изымали мочевой пузырь, тонкий кишечник, ободочную кишку, селезенку, поджелудочную железу, почки, желудок, печень, легкое, сердце, головной мозг, бедренную кость, мышцы, кожу, хвост и опухоль и измеряли радиоактивность.
Результаты
Иммунофлуоресцентное окрашивание - протокол 116
Необработанный контроль характеризовался высокими уровнями СЕА, равномерно распределенными в собранных опухолях. Как и ожидалось, при окрашивании контроля для выявления CEA-DOTAM BsAb сигнал не был обнаружен.
Опухоли мышей, которым инъецировали CEA-DOTAM BsAb, изымали в дни 4, 7 и 14 p.i. В день 4 BsAb полностью покрывало границы узлов опухолевых клеток, но не проникало полностью в более крупные узлы. Это выявлялось по присутствию более темных окрашенных Hoechst областей, окруженных ярко-красными пограничными слоями клеток. Через 7 дней было видно, что сигнал, обусловленный окрашиванием CEA-DOTAM, становился более равномерно распределенным и было меньше опухолевых клеток, характеризующихся структурой с темными внутренними областями. Еще через одну неделю, в день 14 p.i., распределение флуоресцентного сигнала оставалось сходным с распределением в день 7, но уровень сигнала повсеместно был более низким.
В образцах опухолей из двух групп животных, которых подвергали полному PRIT-циклу, включая облучение с использованием 212Pb-DOTAM, выявлены области некроза, обусловленные индуцированной радиацией гибелью клеток. Полученные данные включали набухание клеток, утрату некоторых клеточных структур (клетки-призраки), определенную степень ядерной атипии и потенциальное увеличение интерстициального фиброза. В опухолях у мышей в группе Е, которым вводили вторую дозу CEA-DOTAM через 2 недели после первой инъекции BsAb, сохранялась высокая и равномерная экспрессия СЕА, а распределение CEA-DOTAM через 4 дня после второй инъекции BsAb было сходным с распределением в не подвергнутых облучению опухолях, т.е. имело место распределение BsAb по всем экспрессирующим СЕА частям опухоли, но проникновение в определенные более крупные структуры, приводящее к возникновению более темных областей на изображениях, было ограниченным. Одновременно собирали соответствующие контрольные образцы в группе Ж, в которой не осуществляли вторую инъекцию BsAb после осуществления начального цикла PRIT. На этих срезах опухоли можно видеть тусклый сигнал, обусловленный окрашиванием CEA-DOTAM, свидетельствующий о том, что определенное количество BsAb все еще оставалось связанным с опухолью спустя 18 дней после инъекции.
Биораспределение - протокол 121
Данные о среднем накоплении 203Pb и клиренсе у несущих опухоль ВхРС3 SCID-мышей представлены на фиг. 48. Клиренс крови был медленным, как это и ожидалось для антитела без применения СА. Не было выявлено неожиданного поглощения меченого BsAb в здоровых органах или тканях. Данные о накоплении в опухоли с течением времени представлены на фиг. 49, начиная со средней величины ID/г 49% через 1 день после инъекции, оно повышалось до 130, 189 и 197% ID/г в дни 4, 7 и 10 соответственно. При применении меченого отрицательного контроля накопление в опухоли не происходило, величина ID/г составляла 3% в день 4 после инъекции.
Статистический анализ осуществляли методом одностороннего дисперсионного анализа с использованием критерия Сидака для множественных сравнений для оценки того, были ли статистически значимыми обнаруженные различия в поглощении опухолью в различные моменты времени. Согласно критерию статистически значимое накопление 203Pb имело место только в период между днями 1 и 7 и между днями 1 и 10; для других моментов времени различия не были статистически значимыми. Анализ, в котором сравнивали данные только для дней 4 и 7 с использованием непарного t-критерия, показал наличие статистически значимого увеличения в день 7 (р=0,0468), в то время как при осуществлении соответствующего анализа, в котором сравнивали данные для дней 7 и 10, значимых различий не было выявлено (р=0,8316).
Заключение
Результаты продемонстрировали наличие общего увеличения поглощения 203Pb-BsAb опухолью в период между днями 4 и 7 после инъекции, но при увеличении интервала времени до 10 дней дальнейшего увеличения не было выявлено. Путем микроскопического исследования установлено, что проникновение BsAb в опухоли повышалось через 7 дней по сравнению с днем 4. В день 14 благоприятного действия не было выявлено, в это время общий флуоресцентный сигнал, по-видимому, снижался без улучшения распределения внутри опухоли. Результаты исследования подтвердили, что целесообразно выбирать интервал времени между введениями BsAb и СА, составляющий 7 дней, а не 4 дня.
Пример 33: Биораспределение (данные о распределении in vivo 212Pb-DOTAM для оценки радиоактивной дозы, поглощенной опухолью и тканями) (протокол 146)
Протокол 146 разрабатывали для получения данных о распределении in vivo 212Pb-DOTAM после PRIT, требующихся для оценки радиоактивной дозы, поглощенной опухолью и здоровыми тканями SCID-мышей, несущих s.c. опухоли ВхРС3. Гашение 212Pb-DOTAM осуществляли с использованием Са (см. пример 34).
Предварительный таргетинг осуществляли путем инъекции CEA-DOTAM BsAb, а затем спустя 7 дней СА. Через 24 ч вводили 212Pb-DOTAM. Группы мышей умерщвляли в несколько моментов времени в период с 5 мин и до 48 ч после радиоактивной инъекции и изымали кровь и органы для измерений радиоактивности. В выбранные моменты времени собирали экскременты с использованием метаболических клеток для оценки скорости экскреции радиоактивного соединения. В завершение рассчитывали поглощенные дозы с помощью принятых методов на основе полученных кривых время-активность.
План опыта
Временной график и план опыта согласно протоколу 146 представлены в приведенных ниже двух таблицах.

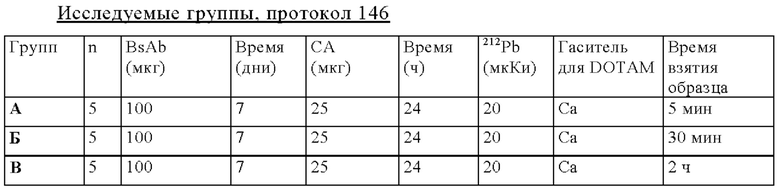
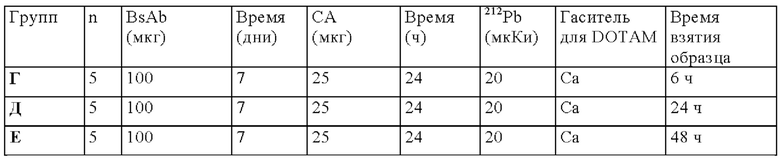
Ксенотрансплантаты солидных опухолей создавали путем s.c. инъекции клеток ВхРС3 SCID-мышам (фирма Envigo) возрастом 8 недель. Через 28 дней после инокуляции мышей разделяли на экспериментальные группы со средним объемом опухоли 310 мм3.
Мышей в группах Д и Е содержали индивидуально (т.е. по одной мыши на клетку) в метаболических клетках с решетчатым полом. Мочу и экскременты собирали в каждой клетке через 2, 6 и 24 ч после радиоактивной инъекции и затем измеряли радиоактивность индивидуальных образцов. Мышей в группе Е переносили в обычную клетку через 24 ч p.i.
Кровь/сыворотку собирали у всех мышей посредством ретро-орбитального кровопускания перед смертью. После эвтаназии дополнительно изымали перечисленные ниже органы и ткани для измерения радиоактивности и расчета % ID/г: мочевой пузырь, матка, тонкий кишечник, ободочная кишка, селезенка, поджелудочная железа, почки, желудок, печень, легкое, сердце, головной мозг, бедренная кость, кожа, мышцы, абдоминальный жир, хвост и опухоль.
Радиационная дозиметрия
Поглощенные дозы рассчитывали по приведенным ниже формулам, которые описаны в техническом проспекте №21 комитета по медицинской внутренней радиационной дозе (MIRD) (Bolch W.E., Eckerman K.F., Sgouros G., и Thomas S.R., J Nucl Med; 50, 2009, cc. 477-484). Формально дозу D(rT), поглощенную тканью- или областью-мишенью rT, рассчитывали в виде интеграла по времени от активности A(rS,t), умноженной на скорость поглощения дозы на единицу активности S(rT ← rS), просуммированного по всем являющимся источниками излучения областям rS. С учетом того, что подавляющая часть энергии, высвобождаемой в результате цепочки распадов 212Pb, обусловлена альфа-излучением и, кроме того, что альфа-излучение обладает намного большей биологической эффективностью (характеризуемой относительной биологической эффективностью, RBE) были сделаны допущения:
1) энергия поглощается локально вследствие короткого диапазона (глубины проникновения) альфа-частиц, это означает, что учитывается только член S(rT ← rT),
2) пренебрегается вкладом, обусловленным бета- и гамма-излучением.
Если Δα представляет собой энергию, высвобождаемую в результате альфа-распадов или более строго сумму средних энергий, высвобождаемых в результате альфа-распадов в цепочке распадов 212Pb, выраженной в единицах J/(мкКи⋅ч), то дозу, поглощенную тканью- или областью-мишенью rT, рассчитывали по формуле:
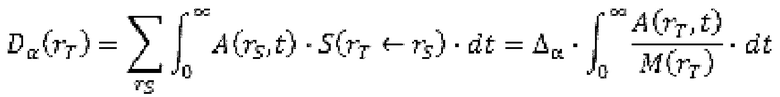
Расчеты основаны на использовании составной (средней) скорректированной на распад радиоактивной концентрации в момент времени t, выраженной в виде [%ID/г]t. На первой стадии ее трансформировали в радиоактивные концентрации, выраженные в μCi на kg веса ткани для ткани-мишени rT:
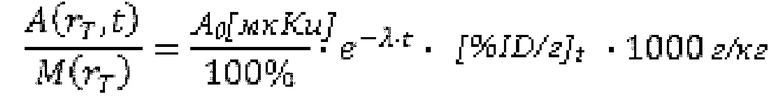
А0[мкКи] обозначает инъецированную активность вещества, а λ обозначает скорость экспоненциального распада 212Pb, 0.0651 ч-1 (что соответствует периоду полураспада 10,6 ч).
На второй стадии радиоактивную концентрацию интегрировали по времени для каждой ткани. Этот расчет осуществляли с использованием «линейно-логарифмического метода трапеций», входящего в программное обеспечение для расчета фармакокинетических характеристик Phoenix WinNonlin 6.4 (фирма Certara USA, Inc., 100 Overlook Center, Suite 101, Princeton, NJ 08540 USA). Применяли тип модели «плазма» с равномерным весом.
На конечной стадии рассчитывали поглощенную дозу путем умножения интегралов по времени от радиоактивной концентрации на высвобождение энергии Δα=170⋅10-6 J/(мкКи⋅ч). Дополнительно рассчитывали с использованием величины RBE (относительная биологическая эффективность) RBE-взвешенные поглощенные дозы, равной 5, как рекомендовано в техническом проспекте MIRD №22 (Sgouros G., Roeske J.C., McDevitt M.R. и др., J Nucl Med; 51, 2010, cc. 311 - 328).
Результаты
Биораспределение
Данные о среднем накоплении и клиренсе 212Pb у несущих опухоли SCID-мышей представлены на фиг. 50. Накопление радиоактивности в опухоли было высоким, величина ID/г составляла 17% уже через 5 мин после инъекции, возрастала до более чем 40% через 6 ч p.i. и оставалась на этом уровне в течение по меньшей мере 48 ч. В здоровых тканях не было выявлено больших различий по сравнению со свободным от опухоли состоянием.
Поглощенные дозы
Средние поглощенные дозы в Гр, рассчитанные согласно протоколу 146, приведены в таблице 24, обе в виде абсолютных значений (RBE=1) и скорректированных на более высокую цитотоксичность альфа-излучателей (RBE=5). Для SCID-мышей средний вес составлял 18,5 г; при активности инъецированного 212Pb, составляющей 20 мкКи, это приводило к стандартизованной величине инъецированной активности, составляющей 1,1 мкКи/г веса тела.
В опухоли поглощенная доза достигала примерно 20 Гр, в то время как в большинстве здоровых тканей за исключением мочевого пузыря поглощенная доза оставалась на уровне существенно ниже 2 Гр. Содержание 212Pb в мочевом пузыре существенно различалось у различных мышей, что свидетельствует о высокой вариабельности для указанной ткани. Скорректированные на RBE дозы приведены для того, чтобы иметь возможность проводить сравнение с радиотерапией с использованием внешнего источника гамма-излучения, это приводило к оценке поглощенной опухолью дозы, составляющей примерно 100 Гр на инъекцию 20 мкКи 212Pb-DOTAM.

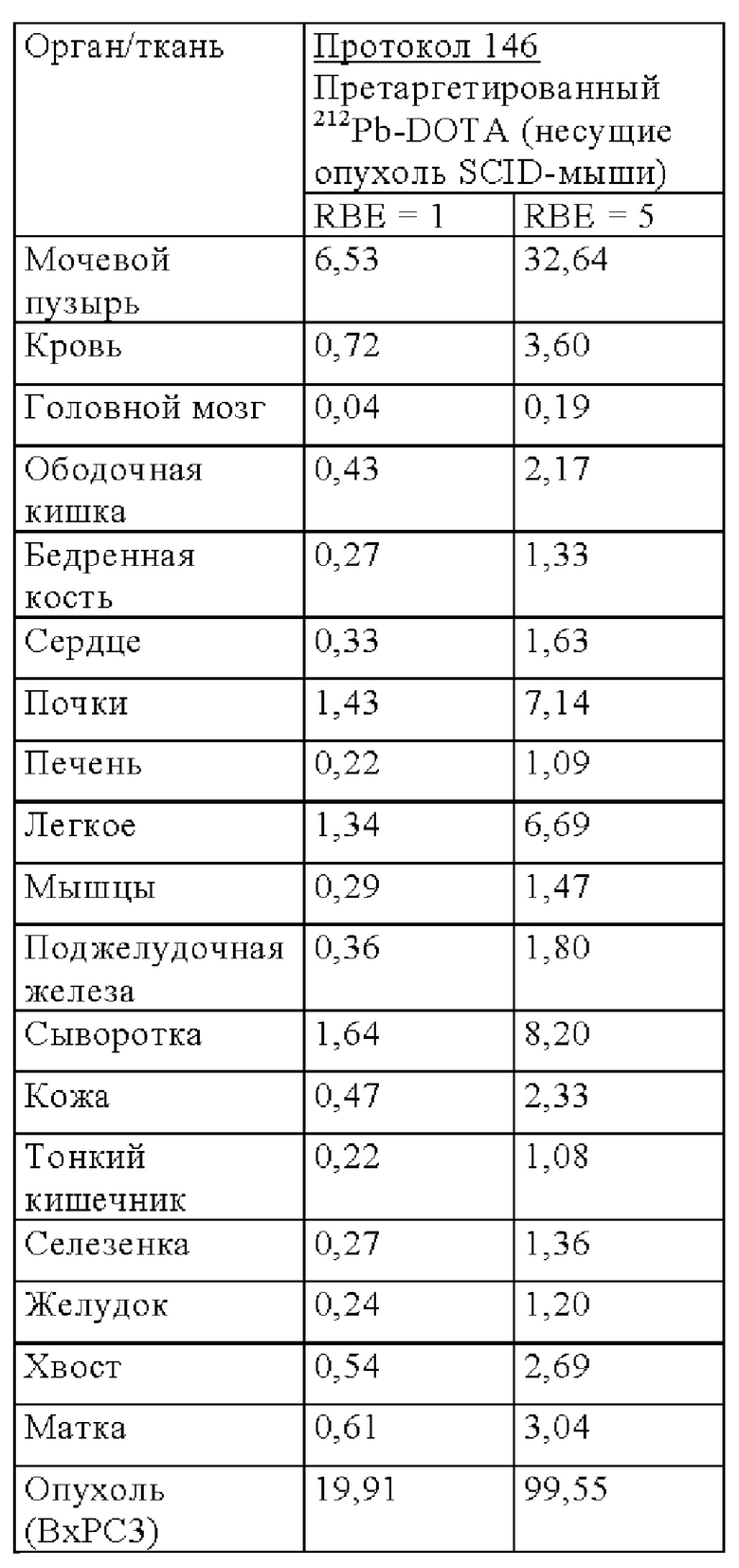
Заключение
В опухоли величина поглощенной дозы достигала примерно 20 Гр, в то время как в большинстве здоровых тканей поглощенная доза оставалась на уровне существенно ниже 2 Гр. В целом, исследование продемонстрировало отсутствие больших рисков ограничивающей дозу токсичности при применении изученного PRIT-режимов.
Пример 34: Гашение нехелатного DOTAM (протокол 131)
Альфа-излучатель 212Pb хелатировали DOTAM после элюции из содержащей торий смолы, получая фармакологически активный 212Pb-DOTAM. После радиоактивного мечения избыток (>99%) свободного нехелатного DOTAM остается в растворе, легко захватывая ионы металла из окружающей среды. После инъекции пациенту он может быстро связываться с находящимся в кровотоке Са2+, формируя фармакологически неактивный Ca-DOTAM. Хотя CEA-DOTAM BsAb создано для того, чтобы связываться преимущественно с Pb-DOTAM, оно может также в сопоставимой степени формировать стабильные комплексы с Ca-DOTAM. Вследствие этого в условиях насыщения может происходить блокирование доступа 212Pb-DOTAM к предварительно таргетированным сайтам опухоли, что может снижать эффективность PRIT-обработки. Поэтому для того, чтобы избежать возможной конкуренции, к процессу элюции 212Pb-DOTAM добавляют стадию гашения, на которой интродуцируют ион металла («X») для контроля формирования «X-DOTAM», характеризующегося существенно более высокой константой диссоциации (Kd). Аффинность DOTAM-связывающего агента в отношении различных хелатов X-DOTAM, по данным измерений в условиях равновесия методом KinExA (кинетический анализ исключения) представлена в приведенной ниже таблице.

Сначала для осуществления гашения был выбран Cu, т.е. для замещения нехелатного DOTAM погашенным неконкурирующим Cu-DOTAM, и в рамках программы CEA-PRIT было проведено большое количество исследований в этих условиях. Однако позднее было обнаружено, что комплекс Cu-DOTAM фактически не обладал стабильностью in vivo, и было предпринято исследование для оценки его стабильности по сравнению с комплексами, содержащими другие гасящие ионы, в человеческой и мышиной сыворотке, плазме и крови. Для этой цели были выбраны, с одной стороны, ионы Pb и Са, обладающие сходной аффинностью, и, с другой стороны, Zn и Cu, обладающие существенно более низкой аффинностью. Исследование продемонстрировало, что Zn-DOTAM обладает существенно более высокой стабильностью по сравнению с Cu-DOTAM в человеческой и мышиной крови, о чем свидетельствовало спонтанное формирование Ca-DOTAM через 35 мин p.i., достигавшее примерно 25% и 30% в мышиной и человеческой крови соответственно после первоначальной инъекции Cu-DOTAM, по сравнению с 0% и 0% после инъекции Zn-DOTAM и 4% и 4% после инъекции Pb-DOTAM.
Протокол 131 был разработан для оценки in vivo гашения DOTAM металлами-кандидатами путем сравнения их воздействия на накопление 212Pb-DOTAM в опухоли и здоровой ткани после предварительного таргетинга СЕА-DOTAM и очищения с помощью Pb-DOTAM-декстран-500. Параллельно с Cu оценивали Zn и Gd, которые образуют обладающий приемлемой стабильностью in vitro комплекс с DOTAM. Кроме того, в качестве контролей применяли Са и стабильный Pb. Схема исследования представлена на фиг. 51.
Временной график и план протокола 131 представлены в приведенных ниже двух таблицах.

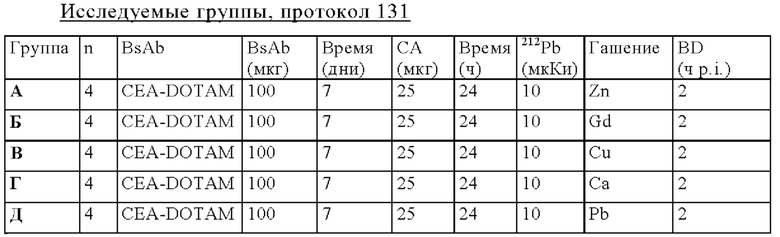
Ксенотрансплантаты солидных опухолей создавали путем s.c.-инъекции клеток ВхРС3 в правую боковую область SCID-мышей возрастом 5-7 недель. Через 18 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухоли 200 мм3. 212Pb-DOTAM инъецировали в день 26 после инокуляции, в этот момент времени средний объем опухоли составлял 350 мм3.
Антитело разводили в 20 мМ His/His-HCl, 140 мМ NaCl, рН 6,0 до конечной концентрации 100 мкг на 100 мкл для i.v.-введения как указано в таблице, озаглавленной «Исследуемые группы, протокол 131», и на фиг. 51. СА подвергали оттаиванию и разводили в ЗФР до конечной концентрации 25 мкг на 100 мкл для i.v.-введения через 7 дней после инъекции BsAb. Еще через 24 ч 212Pb-DOTAM, погашенный с использованием одного из 5 различных металлов, инъецировали i.v. (10 мкКи в 100 мкл 0,9% NaCl).
Мышей умерщвляли для целей оценки биораспределения через 2 ч после инъекции 212Pb-DOTAM и изымали следующие ткани и органы: кровь, мочевой пузырь, селезенку, почки, печень, легкое, мышцы, хвост и опухоль.
Результаты
Данные о среднем накоплении 212Pb и клиренсе во всех собранных тканях через 2 ч после инъекции 212Pb-DOTAM представлены на фиг. 52. Не было выявлено статистически значимых различий в средних величинах % ID/г при использовании любых гасящих металлов по сравнению с применяемым в качестве контроля Pb (двусторонний дисперсионный анализ с критерием Даннета для множественных сравнений). Индивидуальные величины для опухолей и выбранных здоровых тканей представлены на фиг. 53. После гашения с использованием Zn и Gd выявлена большая индивидуальная вариация содержания 212Pb в селезенке и мышцах по сравнению с другими металлами, хотя различие средних величин % ID/г между различными группами обработки не было статистически значимым (односторонний дисперсионный анализ с критерием Тьюки для множественных сравнений).
Заключение
Согласно протоколу 131 оценивали воздействие гашения DOTAM с использованием Zn, Gd или Cu на накопление в опухоли и здоровой ткани предварительно таргетированного 212Pb-DOTAM по сравнению с Са- или Pb-гашением. Не было выявлено различия в поглощении опухолью 212Pb, что свидетельствует об отсутствии существенного блокирования предварительно таргетированных связывающих сайтов с использованием в качестве контролей Са и Pb в рассматриваемых экспериментальных условиях. Была выявлена повышенная вариация содержания 212Pb в селезенке и мышцах после гашения с использованием Zn и Gd, свидетельствующая о том, что комплексы металл-DOTAM, которые конкурируют с 212Pb-DOTAM, могут вносить вклад в нейтрализацию не связанного с СА CEA-DOTAM в нетаргетированных тканях. Гашение с использованием Cu приводило к результатам, сопоставимым с Са-гашением, что дополнительно подтверждает указанную нестабильность in vivo Cu-DOTAM. В конечном итоге для гашения DOTAM был выбран Са по трем основным причинам: более высокий контроль in vivo формирования раствора 212Pb-DOTAM, по сравнению с Cu-гашением; возможность усиления нейтрализации не связанного с СА BsAb, снижающей вариабельность поглощения 212Pb здоровыми тканями; и возможность снижения воздействия гипотетического феномена «обусловленного связывающим сайтом барьера», в результате которого проникновение молекул с высокой аффинностью к антигену ограничивается вследствие того, что они сразу же связываются с легкодоступными мишенями в условиях неполного насыщения.
Пример 35: Оценка CD20-DOTAM и HER2-DOTAM для PRIT (протоколы 151 и 152)
Протоколы 151 и 152 предназначены для оценки связывания 212Pb-DOTAM с подкожными опухолями у мышей предварительно таргетированными полностью гуманизированными BsAb CD20-DOTAM и HER2-DOTAM соответственно. Трехстадийный PRIT осуществляли путем инъекции конструкций антитела, затем через 7 дней Са-DOTAM-декстран-500 в качестве СА и, в завершение, через 24 ч 212Pb-DOTAM. Мышей умерщвляли через 24 ч после радиоактивной инъекции и изымали кровь и органы для измерения радиоактивности.
План опыта
Временные графики и планы протоколов 151 и 152 представлены в приведенных ниже четырех таблицах.


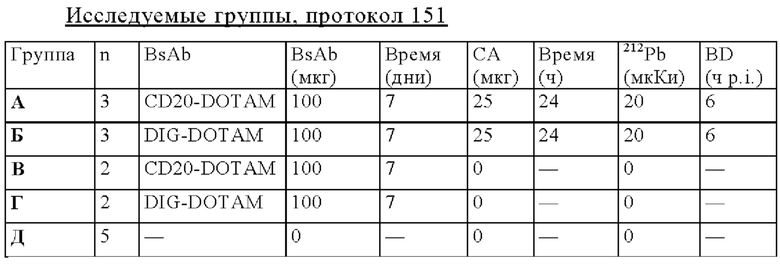

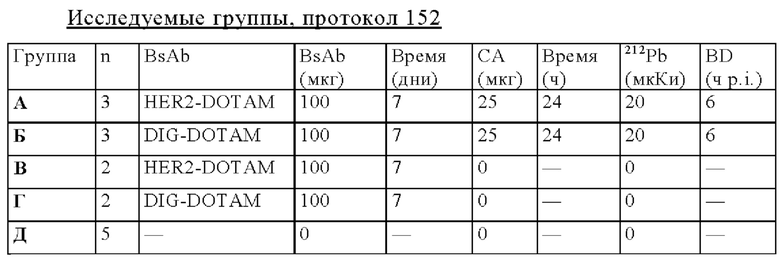
Ксенотрансплантаты солидных опухолей создавали согласно протоколу 151 путем s.с.-инъекции экспрессирующих CD20 клеток человеческой В-клеточной лимфомы WSU-DLCL2 в среде DMEM SCID-мышам возрастом 9 недель. Через 4 дня после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухоли 105 мм3. В день 13 после инокуляции инъецировали 212Pb-DOTAM, в этот момент времени средний объем опухоли составлял 242 мм3.
Согласно протоколу 152 у SCID-мышей возрастом 10 недель создавали ксенотрансплантаты путем s.c. инъекции экспрессирующие HER2 клетки человеческой карциномы желудка NCI-N87 в среде RPMI 1640. Через 14 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухоли 74 мм3. В день 22 после инокуляции инъецировали 212Pb-DOTAM; средний объем опухоли в день 21 составлял 61 мм3.
Предназначенных для исследования биораспределения мышей умерщвляли через 24 ч после радиоактивной инъекции. Перед смертью собирали кровь посредством ретро-орбитального кровопускания под анестезией. Всех мышей после эвтаназии подвергали аутопсии, при этом дополнительно изымали кожу, мочевой пузырь, желудок, тонкий кишечник, ободочную кишку, селезенку, поджелудочную железу, почки, печень, легкое, сердце, бедренную кость, мышцы, хвост и опухоль.
Результаты
Биораспределение - протокол 151
Данные о среднем содержании 212Pb во всех собранных тканях через 24 после инъекции представлены на фиг. 54. При применении в качестве отрицательного контроля BsAb DIG-DOTAM не выявлено накопления радиоактивности в опухолях WSU-DLCL2 (0,4±0,3% ID/г), в то время как при применении CD20-DOTAM BsAb уровень накопления составлял 25,0±3,7% ID/г. Не было выявлено существенного поглощения Pb в здоровых тканях/органах за исключением мочевого пузыря (12,5±4,5% ID/г), почках (3,1±0,3% ID/г) и легком (2,2±0,2% ID/г).
Биораспределение протокол 152
Данные о среднем содержании 212Pb во всех собранных тканях через 24 после инъекции представлены на фиг. 55. При применении в качестве отрицательного контроля BsAb DIG-DOTAM не выявлено накопления радиоактивности в опухолях NCI-N87 (0,8±0,4% ID/г), в то время как при применении HER2-DOTAM BsAb уровень накопления составлял 24,6±1,5% ID/г. Не было выявлено существенного поглощения Pb в здоровых тканях/органах за исключением очень небольших количеств в почках (1,6±0,1% ID/г) и легком (1,4±0,2% ID/г).
Заключение
Результаты исследования согласно протоколам 151 и 152 продемонстрировали наличие специфического таргетинга и in vivo доказали правильность концепции о применении CD20-PRIT и HER2-PRIT на основе разработанного для CEA-PRIT подхода с использованием трехстадийного предварительного таргетинга.
Пример 36: Данные о распределении in vivo и накоплении в опухоли для других форматов (протокол 154)
Протокол 154 разработан для оценки возможности применения 5 новых конструкций CEA-DOTAM BsAb для PRIT. Он разработан для изучения распределения in vivo и накопления в опухоли 212Pb-DOTAM у SCID-мышей, несущих подкожные ксенотрансплантаты HPAF-II. Трехстадийный PRIT-режим осуществляли путем инъекции конструкций CEA-DOTAM, затем через 7 дней Са-DOTAM-декстран-500 в качестве СА и в завершение 212Pb-DOTAM через 24 ч после СА. Мышей умерщвляли через 6 ч после радиоактивной инъекции и изымали кровь и органы для измерений радиоактивности. Осуществляли сравнения с PRIT с использованием в качестве стандарта CEA-DOTAM BsAb и не связывающееся с СЕА BsAb. Схема исследования представлена на фиг. 56.
План опыта
Временной график и план протокола 154 представлены в приведенных ниже двух таблицах.

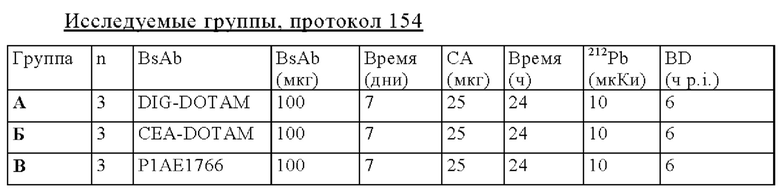
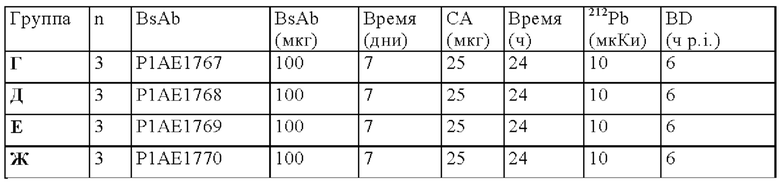
Трансплантаты солидных опухолей создавали путем s.с.-инъекции клеток HPAF-II в правую боковую область мышей возрастом 9 недель. Через 11 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухоли 79 мм3. В день 20 после инокуляции инъецировали 212Pb-DOTAM, в этот момент времени средний объем опухоли составлял 230 мм3.
Все антитела разводили в 20 мМ His/His-HCl, 140 мМ NaCl, рН 6,0 до конечной концентрации 100 мкг на 100 мкл для i.v.-введения согласно таблице, озаглавленной «Исследуемые группы, протокол 154» и фиг. 56. СА подвергали оттаиванию и разводили в ЗФР до конечной концентрации 25 мкг на 100 мкл для i.v.-введения через 7 дней после инъекции BsAb. Еще через 24 ч инъецировали i.v. 212Pb-DOTAM, погашенный Са (10 мкКи в 100 мкл 0,9% NaCl).
Мышей умерщвляли для целей оценки биораспределения через 6 ч после инъекции 212Pb-DOTAM и изымали следующие ткани и органы: кровь, кожу, мочевой пузырь, желудок, тонкий кишечник, ободочную кишку, селезенку, поджелудочную железу, почки, печень, легкое, сердце, бедренную кость, мышцы, головной мозг, хвост и опухоль.
Результаты
Данные о среднем накоплении 212Pb и клиренсе для всех собранных тканей через 6 ч после инъекции представлены на фиг. 57. Данные о содержании в крови и накоплении в опухоли более подробно представлены на фиг. 58. При применении BsAb DIG-DOTAM в качестве отрицательного контроля не было выявлено накопления радиоактивности в опухолях (1,4±0,1% ID/г), в то время как при использовании стандартного CEA-DOTAM BsAb уровень накопления составлял 36,8±3,4% ID/г. Для новых конструкций соответствующие величины составляли 31,8±3,1% ID/г (Р1АЕ1766), 35,0±8,5% ID/г (Р1АЕ1767), 14,2±6,7% ID/г (Р1АЕ1768), 38,8±10,1% ID/г (Р1АЕ1769) и 39,5±6,8% ID/г (Р1АЕ1770). Существенно более низкий уровень накопления в опухоли Р1АЕ1768 можно объяснить одновалентностью указанной конструкции антитела в отношении СЕА.
Не выявлено существенного накопления 212Pb в здоровых тканях/органах за исключением мочевого пузыря, в котором радиоактивность достигала уровней, как правило, обнаруживаемых через 6 ч p.i. для рассматриваемого PRIT-режима.
Заключение
Результаты исследования продемонстрировали in vivo правильность концепции трехстадийного PRIT-режима для всех протестированных конструкций CEA-DOTAM-антитела (Р1АЕ1766, Р1АЕ1767, Р1АЕ1768, Р1АЕ1769 и Р1АЕ1770).
Пример 37: Эффективность CD20-PRIT (протокол 162)
Целью данного исследования было доказательство правильности концепции режима обработки, разработанного для CEA-PRIT, при его применении в отношении другой мишени: кластера дифференцировки 20 (CD20). Терапевтическую эффективность оценивали после осуществления 3 циклов CD20-PRIT на мышах, несущих подкожные опухоли WSU-DLCL2. Осуществляли сравнение с: 1-одностадийным режимом CD20-RIT с использованием BsAb, которые предварительно инкубировали с 212Pb-DOTAM перед осуществлением инъекции; с ритуксимабом, представляющим собой моноклональное антитело типа I против CD20 и являющимся эталоном при лечении заболеваний, характеризующихся экспрессией CD20 (CD20-заболеваний); и с GA101, моноклональным антителом типа II, также направленным против CD20.


Животных обрабатывали согласно схеме эксперимента, проиллюстрированной на фиг. 59. Ксенотрансплантаты солидных опухолей WSU-DLCL2 создавали у SCID-мышей возрастом 8 недель в день исследования 0 путем подкожной инъекции 1,5×105 клеток в правую боковую область. Через 7 дней после инъекции опухолевых клеток мышей разделяли на экспериментальные группы со средним объемом опухоли 144 мм3. В день 15 после инокуляции инъецировали 212Pb-DOTAM; при этом в день 14 средний объем опухоли составлял 306 мм3.
Для оценки эффективности обработок осуществляли наблюдения за мышами в группах А-Ж. При осуществлении PRIT-режима вводимая радиоактивность составляла 20 мкКи на цикл, в то время как соответствующая радиоактивность при осуществлении 1-стадийного RIT-режима составляла 10 мкКи во избежание острой индуцированной радиацией токсичности обусловленной более продолжительным временем удерживания в крови и здоровых тканях при применении указанного режима.
Мышей в группах З, И и К умерщвляли и проводили аутопсию для целей оценки биораспределения через 24 ч после осуществления им первой и единственной инъекции 212Pb-DOTAM или 212Pb-DOTAM-BsAb; животных в группах К и Л умерщвляли и подвергали аутопсии через 24 ч после осуществления им второй и третьей инъекции 212Pb-DOTAM соответственно. У указанных мышей изымали следующие органы и ткани: кровь, мочевой пузырь, селезенку, почки, печень, легкое, мышцы, хвост, кожу и опухоль. Собранные образцы взвешивали и затем измеряли радиоактивность с помощью автоматического гамма-счетчика 2470 WIZARD2 (фирма PerkinElmer), и после этого рассчитывали процент от инъецированной дозы на грамм ткани (%ID/г) с учетом коррекции на распад и фоновый уровень.
Для осуществления PRIT разводили либо CEA-DOTAM BsAb, либо DIG-DOTAM (BsAb, применяемое в качестве отрицательного контроля) в 20 мМ гистидине, 140 мМ NaCl (рН 6,0) до конечной концентрации 100 мкг на 200 мкл для i.р.-введения. Через 7 дней после инъекции вводили i.p. BsAb Ca-DOTAM-декстран-500 в качестве СА (25 мкг на 200 мкл), затем через 24 ч погашенный с использованием Са 212Pb-DOTAM (20 мкКи в 100 мкл).
Мышам, которых обрабатывали согласно 1-стадийному RIT-режиму, вводили предварительно связанный комплекс 212Pb-DOTAM-CD20-DOTAM, полученный путем инкубации 212Pb-DOTAM с CD20-DOTAM BsAb в течение 10 мин при 37°C, после чего инъецировали i.v. BsAb (20 мкКи/20 мкг) в 100 мкл 0,9% NaCl.
Мышей, которых обрабатывали ритуксимабом (MabThera®) или GA101 (обинутузумаб), вводили i.v. соответствующие антитела, разведенные в 20 мММ гистидине, 140 мМ NaCl (рН 6,0) до конечной концентрации 600 мкг в 200 мкл. Инъекции осуществляли в тот же самый день, что и радиоактивные инъекции в группах, которые обрабатывали согласно PRIT- и RIT-режимам.
Предварительные результаты, протокол 162
Исследование еще продолжалось во время написания данного примера, однако уже могут быть представлены предварительные результаты:
Данные о среднем накоплении и клиренсе 212Pb для всех собранных тканей через 24 ч после инъекции для первого цикла обработки представлены на фиг. 60. При применении PRIT в качестве отрицательного контроля поглощения опухолью не выявлено (0,7% ID/г). Поглощение опухолью составляло 22% ID/г как для CD20-PRIT, так и для CD20-RIT.
Среднее развитие опухолей при изученных вариантах обработки представлено на фиг. 61. Опухоли в не обработанной (т.е. обработанной только носителем) группе, обработанной DIG-DOTAM группе и группах, обработанных антителом характеризовались постоянным ростом. В противоположность этому размеры опухолей в CD20-RIT- и CD20-PRIT-группах уменьшались после первого цикла обработки.
Изменение BW во всех подвергнутых терапии группах представлено на фиг. 62. Несколько инъекций 20 мкКи 212Pb-DOTAM хорошо переносились, но инъекции 10 мкКи предварительно связанного комплекса Pb-DOTAM-CD20-DOTAM приводили к существенному резкому уменьшению BW.
Выводы и заключение
Исследование продемонстрировало правильность концепции CD20-PRIT, результаты свидетельствуют об ингибировании роста опухоли, сопоставимом с ингибированием, которое продемонстрировано ранее для CEA-PRIT. CD20-PRIT-режим хорошо переносился, что не имело места в случае обработки согласно 1-стадийному СВ20-RIT-режимом.
Дополнительные варианты осуществления изобретения
Перечисленные ниже пронумерованные положения представляют собой некоторые дополнительные объекты изобретения и варианты осуществления изобретения.
1. Очищающий агент, содержащий декстран или его производное, конъюгированный/конъюгированное с хелатирующим агентом, выбранным из DOTAM и функционального варианта DOTAM, в котором указанный хелатирующий агент образует комплекс с ионом металла.
2. Очищающий агент по п. 1, который представляет собой соединение следующей формулы:
декстран-(линкер-(М-ВОТАМ))х,
в которой
декстран обозначает декстран или его производное;
линкер обозначает связывающий фрагмент;
M-DOTAM обозначает DOTAM или его функциональный вариант, включающий ион металла; и
х≥1.
3. Очищающий агент по п. 1, в котором х обозначает 20 или более, 25 или более, 30 или более, 35 или более, 40 или более или 50 или более.
4. Очищающий агент по п. 2 или п. 3, в котором связывающий фрагмент представляет собой двухвалентную группу следующей формулы:
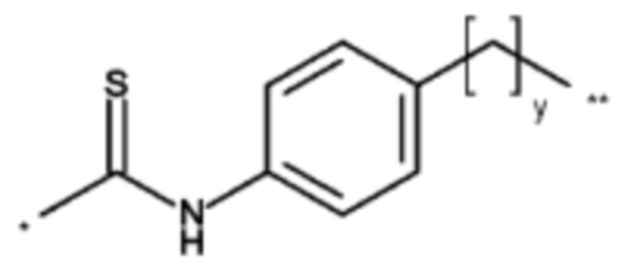
в которой у обозначает 1-6, * обозначает точку присоединения к декстрану и ** обозначает точку присоединения к кольцевому атому в DOT AM или его функциональном варианте.
5. Очищающий агент по одному из предыдущих пунктов, в котором производное декстрана представляет собой аминодекстран, необязательно замещенный одной или несколькими группами, выбранными из аминокислоты и сахарида, отличного от глюкозы.
6. Очищающий агент по одному из предыдущих пунктов, в котором количество групп DOT AM, выраженное в виде процента от количества глюкозных звеньев декстрана или производного декстрана, составляет по меньшей мере 1%, по меньшей мере 1,5%, по меньшей мере 2%, по меньшей мере 2,5%, по меньшей мере 3%, по меньшей мере 5%.
7. Очищающий агент по одному из предыдущих пунктов, в котором средняя молекулярная масса декстрана оставляет 200-800 кДа, необязательно больше чем 300, 350, 400 или 450 кДа и необязательно меньше чем 700, 650, 600 или 550 кДа, необязательно примерно 500 кДа.
8. Очищающий агент по п. 7, в котором компоненты, представляющие собой декстран, или очищающие агенты, имеющие молекулярную массу ниже отсекаемой, удалены, где отсекаемая молекулярная масса составляет 50 кДа или более, 100 кДа или более или 200 кДа или более.
9. Очищающий агент по п. 8, в котором отсекаемая молекулярная масса находится в диапазоне 50-250 кДа или 50-200 кДа, необязательно 100-200 кДа и необязательно составляет примерно 100, 150 или 200 кДа.
10. Очищающий агент по одному из п.п. 7-9, средняя молекулярная масса которого составляет 450-550 кДа, например, примерно 500 кДа.
11. Очищающий агент по одному из предыдущих пунктов, в котором ион металла представляет собой стабильный изотоп или практически стабильный изотоп.
12. Очищающий агент по одному из предыдущих пунктов, в котором ион металла представляет собой ион Pb, Bi или Са.
13. Способ получения очищающего агента, включающий:
конъюгацию декстрана или производного декстрана с DOTAM или его функциональным вариантом или производным, где способ включает получение хелата DOTAM с ионом металла до и/или после конъюгации DOTAM с декстраном.
14. Способ по п. 13, в котором ион металла представляет собой стабильный или практически стабильный ион металла.
15. Способ по п. 13 или п. 14, в котором ион металла представляет собой ион Pb, Bi или Са.
16. Способ по одному из п.п. 13-15, в котором перед конъюгацией декстран или производное декстрана подвергают стадии фильтрации для удаления видов, имеющих молекулярную массу ниже отсекаемой молекулярной массы/порога отсечения, составляющей/составляющего 50 кДа или выше, 100 кДа или выше или 200 кДа или выше, и/или где способ включает подвергание конъюгата стадии фильтрации для удаления видов, имеющих молекулярную массу ниже отсекаемой молекулярной массы/порога отсечения,
составляющей/составляющего 50 кДа или выше, 100 кДа или выше или 200 кДа или выше.
17. Очищающий агент по одному из п.п. 1-12, предназначенный для применения в способе радиоиммунотерапии или радиоиммуновизуализации.
18. Очищающий агент по одному из п.п. 1-12, предназначенный для применения в способе претаргетной радиовизуализации, включающий:
I) введение индивидууму мультиспецифического или биспецифического антитела, которое содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для антигена-мишени, в котором после введения антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень;
II) введение очищающего агента по одному из п.п. 1-11, в котором очищающий агент обладает способностью связываться с антителом в сайте связывания с хелатом Pb-DOTAM и повышает клиренс и/или блокирует антигенсвязывающий сайт антитела, не локализованного на поверхности клетки;
где
способ необязательно дополнительно включает
III) последующее введение комплекса, содержащего DOTAM, образующий хелат с радиоизотопом Pb, в котором указанный комплекс связывается с антителом, локализованным на поверхности клетки; и способ необязательно дополнительно включает
IV) визуализацию ткани или органа, в которой/котором локализован хелатный радионуклид.
19. Очищающий агент по одному из п.п. 1-12, предназначенный для применения в способе претраргетной радиоиммунотерапии, включающий:
I) введение индивидууму мультиспецифического или биспецифического антитела, которое содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для антигена-мишени, в котором после введения антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень;
II) введение очищающего агента по одному из п.п. 1-11, в котором очищающий агент обладает способностью связываться с антителом в сайте связывания с хелатом Pb-DOTAM и повышает клиренс и/или блокирует антигенсвязывающий сайт антитела, не локализованного на поверхности клетки;
где
способ необязательно дополнительно включает
III) последующее введение комплекса, содержащего DOTAM, образующий хелат с радиоизотопом Pb, в котором указанный комплекс связывается с антителом, локализованным на поверхности клетки.
20. Способ претаргетной радиовизуализации, включающий:
I) введение индивидууму мультиспецифического или биспецифического антитела, которое содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для антигена-мишени, в котором после введения антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень;
II) введение очищающего агента по одному из п.п. 1-11, в котором очищающий агент обладает способностью связываться с антителом в сайте связывания с хелатом Pb-DOTAM и повышает клиренс и/или блокирует антигенсвязывающий сайт антитела, не локализованного на поверхности клетки;
где
способ необязательно дополнительно включает
III) последующее введение комплекса, содержащего DOTAM или его функциональный вариант, образующий хелат с радиоизотопом Pb, в котором указанный комплекс связывается с антителом, локализованным на поверхности клетки; и необязательно дополнительно включает
IV) визуализацию ткани или органа, в которой/котором локализован хелатный радионуклид.
21. Способ претаргетной радиоиммунотерапии, включающий:
I) введение индивидууму мультиспецифического или биспецифического антитела, которое содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для антигена-мишени, в котором после введения антитело связывается с антигеном-мишенью и локализуется на поверхности клетки, экспрессирующей антиген-мишень;
II) введение очищающего агента по одному из п.п. 1-11, в котором очищающий агент обладает способностью связываться с антителом в сайте связывания с хелатом Pb-DOTAM и повышает клиренс и/или блокирует антигенсвязывающий сайт антитела, не локализованного на поверхности клетки; необязательно дополнительно включающий
III) последующее введение комплекса, содержащего DOTAM или его функциональный вариант, образующий хелат с радиоизотопом Pb, в котором указанный комплекс связывается с антителом, локализованным на поверхности клетки.
22. Очищающий агент, предназначенный для применения по п. 18 или п. 19, или способ по п. 20 или п. 21, в котором антигенсвязывающий сайт, специфический для халата Pb-DOTAM, содержит по меньшей мере:
а) CDR2 тяжелой цепи, содержащий аминокислотную последовательность FIGSRGDTYYASWAKG (SEQ ID NO: 2) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 2, где указанные замены не включают Phe50, Asp56 и Tyr58 необязательно не включают также Gly52 и/или Arg 54;
б) CDR3 тяжелой цепи, содержащий аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 3, где указанные замены не включают Glu95, Arg96, Asp97, Pro98 и необязательно не включают также Ala100C, Tyr100D и/или Pro100E, и/или необязательно не включают также Tyr99;
в) CDR1 легкой цепи, содержащий аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 4, где указанные замены не включают Tyr28 and Asp32;
г) CDR3 легкой цепи, содержащий аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 6, где указанные замены не включают Gly91, Tyr92, Asp93, Thr95c и Tyr96,
где нумерация соответствует нумерации по Кэботу.
23. Очищающий агент, предназначенный для применения, или способ по п. 22, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит также CDR1 тяжелой цепи и CDR2 легкой цепи.
24. Очищающий агент, предназначенный для применения, или способ по п. 23, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит:
I) CDR1 тяжелой цепи, содержащий аминокислотную последовательность GFSLSTYSMS (SEQ ID NO: 1) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 1, необязательно консервативных замен; и/или
II) CDR2 легкой цепи, содержащий аминокислотную последовательность QASKLAS (SEQ ID NO: 5) или ее вариант, имеющий вплоть до 1, 2 или 3 замен в SEQ ID NO: 5, необязательно консервативных замен.
25. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-24, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит по меньшей мере один, два, три, четыре, пять или шесть CDR, которые выбраны из:
а) CDR1 тяжелой цепи, содержащего аминокислотную последовательность GFSLSTYSMS (SEQ ID NO: 1),
б) CDR2 тяжелой цепи, содержащего аминокислотную последовательность FIGSRGDTYYASWAKWG (SEQ ID NO: 2),
в) CDR3 тяжелой цепи, содержащего аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3),
г) CDR1 легкой цепи, содержащего аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4),
д) CDR2 легкой цепи, содержащего аминокислотную последовательность QASKLAS (SEQ ID NO: 5),
е) CDR3 легкой цепи, содержащего аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6).
26. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-25, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, связывается с тем же эпитопом или с перекрывающимся эпитопом хелата Pb-DOTAM, что и:
I) антитело, которое имеет вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 7, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 8; или
II) антитело, которое имеет вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 9, вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 10.
27. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-26, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, является человеческим, химерным или гуманизированным.
28. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-27, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 7 и SEQ ID NO: 9, или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 7 или SEQ ID NO: 9.
29. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-28, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит вариабельный домен легкой цепи, который содержит аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 8 и SEQ ID NO: 10, или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 8 или 10.
30. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-29, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 7, и вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 8.
31. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-30, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, содержит вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 9, и вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 10.
32. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-30, где связывание антигенсвязывающего сайта, специфического для хелата Pb-DOTAM, с хелатом Pb-DOTAM характеризуется величиной Kd, составляющей 100, 50, 20, 10, 5, 1пМ или менее.
33. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-32, где антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, связывается с хелатом Pb-DOTAM и с хелатом Bi-DOTAM, и где соотношение величин Kd для хелата Bi-DOTAM/хелата Pb-DOTAM находится в диапазоне 0,1-10 или 1-10.
34. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-33, где антиген-мишень представляет собой опухолеспецифический антиген.
35. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-34, где опухолеспецифический антиген выбран из группы, состоящей из СЕА, HER2 и CD20.
36. Очищающий агент, предназначенный для применения, или способ по одному из п.п.18-35, где опухолеспецифический антиген представляет собой карциноэмбриональный антиген (СЕА).
37. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-36, где мультиспецифическое или биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для СЕА, и в котором антигенсвязывающий сайт, специфический для СЕА, содержит тяжелую цепь, которая содержит по меньшей мере один, два или три CDR тяжелой цепи: где:
г) CDR1 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 11,
д) CDR2 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 12,
e) CDR3 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 13;
и/или антигенсвязывающий сайт, специфический для СЕА, содержит легкую цепь, которая содержит по меньшей мере один, два или три CDR легкой цепи: где:
а) CDR1 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 14,
б) CDR2 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 15;
в) CDR3 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 16.
38. Очищающий агент, предназначенный для применения, или способ по одному из п. 37, где антигенсвязывающий сайт для СЕА содержит по меньшей мере один, два, три, четыре, пять или шесть (т.е. все) CDR, выбранные из:
а) CDR1 тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 11;
б) CDR2 тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 12;
в) CDR3 тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 13;
г) CDR1 легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 14;
д) CDR2 легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 15;
е) CDR3 легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 16.
39. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-38, где мультиспецифическое или биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для СЕА, и в котором антигенсвязывающий сайт, специфический для СЕА, содержит
I) вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 17 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 17; и/или
II) вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 18 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 18.
40. Очищающий агент, предназначенный для применения, или способ по п. 39, где антигенсвязывающий сайт, специфический для СЕА, содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 17, и/или вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 18.
41. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-35, где мультиспецифическое или биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для ERBB2, и в котором антигенсвязывающий сайт, специфический для ERBB2, содержит по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29; (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32; и (е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33.
42. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-35 или 41, где мультиспецифическое или биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для ERBB2, и в котором антигенсвязывающий сайт, специфический для ERBB2, содержит
I) вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 34 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 34; и/или
II) вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 35 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 35.
43. Очищающий агент, предназначенный для применения, или способ по 42, где антигенсвязывающий сайт, специфический для ERBB2, содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 34 и/или вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 35.
44. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-35, где мультиспецифическое или биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для CD20, и в котором антигенсвязывающий сайт, специфический для CD20, содержит по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из (a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39; (б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40 (в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41; (г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 42; (д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43; и (е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44.
45. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-35 или 44, где мультиспецифическое или биспецифическое антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для CD20, и в котором антигенсвязывающий сайт, специфический для CD20, содержит
I) вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 45 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 45; и/или
II) вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 46 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 46.
46. Очищающий агент, предназначенный для применения, или способ по 45, где антигенсвязывающий сайт, специфический для CD20, содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 45 и/или вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 46.
47. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 18-46, где мультиспецифическое или биспецифическое антитело содержит Fc-область.
48. Очищающий агент, предназначенный для применения, или способ по. 47, где Fc-область конструируют для снижения эффекторной функции.
49. Очищающий агент, предназначенный для применения, или способ по. 48, где Fc-область конструируют путем замены одного или нескольких остатков 234, 235, 238, 265, 269, 270, 297, 327 и/или 329.
50. Очищающий агент, предназначенный для применения по одному из п.п. 47-49, где мультиспецифическое или биспецифическое антитело содержит полноразмерное антитело, которое содержит первую и вторую тяжелые цепи антитела и первую и вторую легкие цепи антитела, в котором первая тяжелая цепь и первая легкая цепь объединены с образованием антигенсвязывающего сайта для первого антигена и в котором вторая тяжелая цепь и вторая легкая цепь объединены с образованием антигенсвязывающего сайта для второго антигена,
и либо первый, либо второй антиген представляет собой хелат Pb-DOTAM, а другой представляет собой антиген-мишень.
51. Очищающий агент, предназначенный для применения по п. 50, где мультиспецифическое или биспецифическое антитело содержит также дополнительный антигенсвязывающий фрагмент для первого антигена.
52. Очищающий агент, предназначенный для применения по п. 51, где мультиспецифическое или биспецифическое антитело содержит:
полноразмерное антитело, которое содержит первую и вторую тяжелые цепи антитела и первую и вторую легкие цепи антитела, в котором первая тяжелая цепь и первая легкая цепь объединены с образованием Fab, содержащего антигенсвязывающий сайт для первого антигена, и в котором вторая тяжелая цепь и вторая легкая цепь объединены с образованием cross-Fab, содержащего антигенсвязывающий сайт для второго антигена;
и в котором либо первая, либо вторая тяжелая цепь антитела слита через линкер с полипептидом, содержащим СН1- и VH-домены, и указанный первый полипептид объединен со вторым полипептидом, содержащим CL и VL, в результате чего первый и второй полипептиды объединены с образованием Fab, содержащего антигенсвязывающий сайт для первого антигена.
53. Очищающий агент, предназначенный для применения по п. 52, где N-конец тяжелой цепи второго антитела слит через линкер с указанным первым полипептидом.
54. Очищающий агент, предназначенный для применения по одному из п.п. 47-49, где мультиспецифическое или биспецифическое антитело содержит
I) полноразмерное антитело, которое содержит антигенсвязывающий сайт для первого антигена, и
II) по меньшей мере вариабельный домен второй тяжелой цепи и вариабельный домен второй легкой цепи, которые вместе образуют антигенсвязывающий сайт для второго антигена, где либо первый, либо второй антиген представляет собой хелат Pb-DOTAM, а другой представляет собой антиген-мишень.
55. Очищающий агент, предназначенный для применения по п. 54, где мультиспецифическое или биспецифическое антитело содержит полноразмерное антитело, содержащее антигенсвязывающий сайт для первого антигена, в котором N- или С-конец одной из тяжелых цепей сцеплен через полипептидный линкер с первым полипептидом и в котором первый полипептид объединен со вторым полипептидом с образованием Fab или cross-Fab, который содержит связывающий сайт для второго антигена.
56. Очищающий агент, предназначенный для применения по п. 55, где мультиспецифическое или биспецифическое антитело содержит:
I) первый полипептид, состоящий из VH-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VL- и CL-доменов; или
II) первый полипептид, состоящий из VL-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VH- и CL-доменов; или
III) первый полипептид, состоящий из VH-домена и CL-домена, который объединен со вторым полипептидом, состоящим из VL- и СН1-доменов;
в результате чего первый и второй полипептид вместе образуют антигенсвязывающий сайт для второго антигена.
57. Очищающий агент, предназначенный для применения по п. 56, где мультиспецифическое или биспецифическое антитело содержит полноразмерное антитело, которое содержит антигенсвязывающий сайт для первого антигена, в котором С-конец одной из тяжелых цепей сцеплен через полипептидный линкер с первым полипептидом, состоящим из VL-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VH- и CL-доменов.
58. Очищающий агент, предназначенный для применения, или способ по п. 44, где мультиспецифическое или биспецифическое антитело содержит:
а) полноразмерное антитело, которое специфически связывается с первым антигеном и состоит из двух тяжелых цепей антитела и двух легких цепей антитела;
б) полипептид, который состоит из
I) вариабельного домена тяжелой цепи антитела (VH); или
II) вариабельного домена тяжелой цепи антитела (VH) и константного домена антитела (СН1); или
III) вариабельного домена тяжелой цепи антитела (VH) и константного домена легкой цепи антитела (CL),
в котором указанный полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
в) полипептид, который состоит из
I) вариабельного домена легкой цепи антитела (VL); или
II) вариабельного домена легкой цепи антитела (VL) константного домена легкой цепи антитела (CL); или
III) вариабельного домена легкой цепи антитела (VL) и константного домена тяжелой цепи антитела (СН1);
в котором указанный полипептид слит с помощью N-конца VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
и в котором вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для второго антигена,
где либо первый, либо второй антиген представляет собой хелат Pb-DOTAM, а другой представляет собой антиген-мишень.
59. Очищающий агент, предназначенный для применения, или способ по одному из п.п. 50-58, где первый антиген представляет собой антиген-мишень, а второй антиген представляет собой хелат Pb-DOTAM.
60. Очищающий агент, предназначенный для применения, или способ по п. 59, где первый антиген представляет собой СЕА.
61. Очищающий агент, предназначенный для применения, или способ по п. 60, где мультиспецифическое или биспецифическое антитело содержит:
а) полноразмерное антитело, которое специфически связывается с СЕА и состоит из двух тяжелых цепей антитела и двух легких цепей антитела; в котором тяжелые цепи по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичны аминокислотам 1-450 SEQ ID NO: 22 или 23 тяжелой цепи;
и в котором легкие цепи по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичны легкой цепи, имеющей SEQ ID NO 21; и/или
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7; или
II) указанного вариабельного домена тяжелой цепи антитела (VH), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7, и константного домена тяжелой цепи антитела (СН1); или
III) указанного вариабельного домена тяжелой цепи антитела (VH), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 7, и константного домена легкой цепи антитела (CL);
в котором полипептид слит с помощью N-конца VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела; и/или
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену легкой цепи, имеющему SEQ ID NO: 8; или
II) указанного вариабельного домена легкой цепи антитела (VL), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену легкой цепи, имеющему SEQ ID NO: 8, и константного домена легкой цепи антитела (CL); или
III) указанного вариабельного домена легкой цепи антитела (VL), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену легкой цепи, имеющему SEQ ID NO: 8, и константного домена тяжелой цепи антитела (СН1),
в котором указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
в котором вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для хелата Pb-DOTAM.
62. Очищающий агент, предназначенный для применения, или способ по п. 60, где мультиспецифическое или биспецифическое антитело содержит:
а) полноразмерное антитело, которое специфически связывается с СЕА и состоит из двух тяжелых цепей антитела и двух легких цепей антитела;
в котором тяжелые цепи по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичны аминокислотам 1-450 SEQ ID NO: 19 или 20 тяжелой цепи; и в котором легкие цепи по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичны легкой цепи, имеющей SEQ ID NO 21;
б) полипептид, состоящий из
I) вариабельного домена тяжелой цепи антитела (VH), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 9; или
II) указанного вариабельного домена тяжелой цепи антитела (VH), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 9, и константного домена тяжелой цепи антитела (СН1); или
III) указанного вариабельного домена тяжелой цепи антитела (VH), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену тяжелой цепи, имеющему SEQ ID NO: 9, и константного домена легкой цепи антитела (CL);
в котором полипептид слит на N-конце VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела;
в) полипептид, состоящий из
I) вариабельного домена легкой цепи антитела (VL), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену легкой цепи, имеющему SEQ ID NO: 10; или
II) указанного вариабельного домена легкой цепи антитела (VL), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену легкой цепи, имеющему SEQ ID NO: 10, и константного домена легкой цепи антитела (CL); или
III) указанного вариабельного домена легкой цепи антитела (VL), который по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичен вариабельному домену легкой цепи, имеющему SEQ ID NO: 10, и константного домена тяжелой цепи антитела (СН1),
в котором указанный полипептид слит на N-конце VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела;
и в котором вариабельный домен тяжелой цепи антитела пептида, указанного в подпункте (б), и вариабельный домен легкой цепи антитела пептида, указанного в подпункте (в), вместе образуют антигенсвязывающий сайт для хелата Pb-DOTAM.
63. Очищающий агент, предназначенный для применения, или способ по п. 62, где мультиспецифическое или биспецифическое антитело содержит:
I) первую тяжелую цепь, имеющую аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична тяжелой цепи, имеющей SEQ ID NO: 22,
II) вторую тяжелую цепь, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична тяжелой цепи, имеющей SEQ ID NO: 23,
III) две легкие цепи антитела, которые по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичны легкой цепи, имеющей SEQ ID NO: 21.
64. Очищающий агент, предназначенный для применения, или способ по п. 62, где мультиспецифическое или биспецифическое антитело содержит:
I) первую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 22;
II) вторую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 23; и
III) две легкие цепи антитела, которые имеют аминокислотную последовательность SEQ ID NO: 21.
65. Очищающий агент, предназначенный для применения, или способ по п. 62, где мультиспецифическое или биспецифическое антитело содержит:
I) первую тяжелую цепь, имеющую аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична тяжелой цепи, имеющей SEQ ID NO: 19,
II) вторую тяжелую цепь, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентична тяжелой цепи, имеющей SEQ ID NO: 20,
III) две легкие цепи антитела, которые по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% идентичны легкой цепи, имеющей SEQ ID NO: 21.
66. Очищающий агент, предназначенный для применения, или способ по п. 62, где мультиспецифическое или биспецифическое антитело содержит:
I) первую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 19;
II) вторую тяжелую цепь, которая имеет аминокислотную последовательность SEQ ID NO: 20; и
III) две легкие цепи антитела, которые имеют аминокислотную последовательность SEQ ID NO: 21.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф. ХОФФМАНН-ЛЯ РОШ АГ
<120> АНТИТЕЛА К ХЕЛАТНЫМ РАДИОНУКЛИДАМ
<130> 007470503
<150> US 62/658468
<151> 2018-04-16
<160> 82
<170> PatentIn version 3.5
<210> 1
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain CDR1 <Pb-Dotam> PRIT-213
<400> 1
Gly Phe Ser Leu Ser Thr Tyr Ser Met Ser
1 5 10
<210> 2
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain CDR2 <Pb-Dotam> PRIT-213
<400> 2
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 3
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain CDR3 <Pb-Dotam> PRIT-213
<400> 3
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu
1 5 10
<210> 4
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain CDR1 <Pb-Dotam> PRIT-213
<400> 4
Gln Ser Ser His Ser Val Tyr Ser Asp Asn Asp Leu Ala
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain CDR2 <Pb-Dotam> PRIT-213
<400> 5
Gln Ala Ser Lys Leu Ala Ser
1 5
<210> 6
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain CDR3 <Pb-Dotam> PRIT-213
<400> 6
Leu Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly
1 5 10
<210> 7
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain variable domain 1 of <Pb-Dotam> PRIT-213
<400> 7
Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu Thr
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu Thr
65 70 75 80
Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 8
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain variable domain <Pb-Dotam> PRIT-213
<400> 8
Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp
1 5 10 15
Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val Tyr Ser Asp Asn
20 25 30
Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp Glu
85 90 95
Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 9
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain variable domain <Pb-Dotam> PRIT-214
<400> 9
Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Ala Val Tyr Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Val Ser Leu Lys
65 70 75 80
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 10
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain variable domain <Pb-Dotam> PRIT-214
<400> 10
Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp
1 5 10 15
Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val Tyr Ser Asp Asn
20 25 30
Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp Glu
85 90 95
Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 11
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain CDR1 <CEA>
<400> 11
Gly Phe Asn Ile Lys Asp Thr Tyr Met His
1 5 10
<210> 12
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain CDR2 <CEA>
<400> 12
Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe Gln
1 5 10 15
Gly
<210> 13
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain CDR3 <CEA>
<400> 13
Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr
1 5 10
<210> 14
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain CDR1 <CEA>
<400> 14
Arg Ala Gly Glu Ser Val Asp Ile Phe Gly Val Gly Phe Leu His
1 5 10 15
<210> 15
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain CDR2 <CEA>
<400> 15
Arg Ala Ser Asn Arg Ala Thr
1 5
<210> 16
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain CDR3 <CEA>
<400> 16
Gln Gln Thr Asn Glu Asp Pro Tyr Thr
1 5
<210> 17
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain variable domain <CEA> 84.66
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 18
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain variable domain <CEA> 84.66
<400> 18
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Gly Glu Ser Val Asp Ile Phe
20 25 30
Gly Val Gly Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Arg Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Thr Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 19
<211> 591
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain 1 of bispecific, trivalent <CEA/Pb-Dotam> PRIT-214
VH_84.66
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Val Gln Leu Gln Gln Trp Gly Ala Gly Leu
465 470 475 480
Leu Lys Pro Ser Glu Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Phe
485 490 495
Ser Leu Ser Thr Tyr Ser Met Ser Trp Ile Arg Gln Pro Pro Gly Lys
500 505 510
Gly Leu Glu Trp Ile Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr
515 520 525
Ala Ser Trp Ala Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys
530 535 540
Asn Gln Val Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala
545 550 555 560
Val Tyr Tyr Cys Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr
565 570 575
Pro Pro His Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
580 585 590
<210> 20
<211> 581
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain 2 of bispecific, trivalent<CEA/Pb-Dotam> PRIT-214
VL_84.66
<400> 20
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
465 470 475 480
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser His
485 490 495
Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly
500 505 510
Lys Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly
515 520 525
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
530 535 540
Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu
545 550 555 560
Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr
565 570 575
Lys Val Glu Ile Lys
580
<210> 21
<211> 218
<212> PRT
<213> Artificial Sequence
<220>
<223> light chain <CEA> 84.66
<400> 21
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Gly Glu Ser Val Asp Ile Phe
20 25 30
Gly Val Gly Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Arg Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Thr Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 22
<211> 591
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain 1 of bispecific, trivalent <CEA/Pb-Dotam> PRIT-213
VH_84.66 ->knob
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Val Thr Leu Lys Glu Ser Gly Pro Val Leu
465 470 475 480
Val Lys Pro Thr Glu Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe
485 490 495
Ser Leu Ser Thr Tyr Ser Met Ser Trp Ile Arg Gln Pro Pro Gly Lys
500 505 510
Ala Leu Glu Trp Leu Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr
515 520 525
Ala Ser Trp Ala Lys Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys
530 535 540
Ser Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala
545 550 555 560
Thr Tyr Tyr Cys Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr
565 570 575
Pro Pro His Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
580 585 590
<210> 23
<211> 581
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain 2 of bispecific, trivalent <CEA/Pb-Dotam> PRIT-213
VL_84.66 ->hole
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser Leu
465 470 475 480
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser His
485 490 495
Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly
500 505 510
Lys Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly
515 520 525
Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
530 535 540
Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu
545 550 555 560
Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr
565 570 575
Lys Val Glu Ile Lys
580
<210> 24
<211> 581
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain 1 of bispecific, <CEA/Pb-Dotam> Rabbit Dotam _84.66
<400> 24
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Ala Val Leu Thr Gln Thr Pro Ser Pro Val
465 470 475 480
Ser Pro Ala Val Gly Gly Thr Val Thr Ile Ser Cys Gln Ser Ser His
485 490 495
Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Leu Gly
500 505 510
Gln Pro Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly
515 520 525
Val Ser Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu
530 535 540
Thr Ile Ser Gly Val Gln Ser Asp Asp Ala Ala Thr Tyr Tyr Cys Leu
545 550 555 560
Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr
565 570 575
Glu Val Val Val Lys
580
<210> 25
<211> 589
<212> PRT
<213> Artificial Sequence
<220>
<223> heavy chain 2 of bispecific, tetravalent <CEA/Pb-Dotam> Rabbit
Dotam_84.66
<400> 25
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Gln Ser Val Glu Glu Ser Gly Gly Arg Leu
465 470 475 480
Val Thr Pro Gly Thr Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Phe
485 490 495
Ser Leu Ser Thr Tyr Ser Met Ser Trp Val Arg Gln Ala Pro Gly Lys
500 505 510
Gly Leu Glu Trp Ile Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr
515 520 525
Ala Ser Trp Ala Lys Gly Arg Phe Thr Val Ser Arg Thr Ser Thr Thr
530 535 540
Val Asp Leu Lys Ile Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr
545 550 555 560
Phe Cys Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro
565 570 575
His Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
580 585
<210> 26
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 26
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 27
<211> 1379
<212> PRT
<213> Artificial Sequence
<220>
<223> Immunomedics hNM14
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Thr Thr Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile His Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Phe Gly Phe Pro Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr
465 470 475 480
Pro Gly Thr Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu
485 490 495
Ser Thr Tyr Ser Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
500 505 510
Glu Trp Ile Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser
515 520 525
Trp Ala Lys Gly Arg Phe Thr Val Ser Arg Thr Ser Thr Thr Val Asp
530 535 540
Leu Lys Ile Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys
545 550 555 560
Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu
565 570 575
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Glu Val Gln Leu Val
580 585 590
Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser
595 600 605
Cys Ser Ala Ser Gly Phe Asp Phe Thr Thr Tyr Trp Met Ser Trp Val
610 615 620
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile His Pro
625 630 635 640
Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu Lys Asp Arg Phe Thr
645 650 655
Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe Leu Gln Met Asp Ser
660 665 670
Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys Ala Ser Leu Tyr Phe
675 680 685
Gly Phe Pro Trp Phe Ala Tyr Trp Gly Gln Gly Thr Pro Val Thr Val
690 695 700
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
705 710 715 720
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
725 730 735
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
740 745 750
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
755 760 765
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
770 775 780
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
785 790 795 800
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
805 810 815
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
820 825 830
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
835 840 845
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
850 855 860
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
865 870 875 880
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
885 890 895
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
900 905 910
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
915 920 925
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
930 935 940
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly
945 950 955 960
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
965 970 975
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
980 985 990
Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
995 1000 1005
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
1010 1015 1020
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly
1025 1030 1035
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1040 1045 1050
Gly Ser Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Pro Ala
1055 1060 1065
Val Gly Gly Thr Val Thr Ile Ser Cys Gln Ser Ser His Ser Val
1070 1075 1080
Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Leu Gly Gln
1085 1090 1095
Pro Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly
1100 1105 1110
Val Ser Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr
1115 1120 1125
Leu Thr Ile Ser Gly Val Gln Ser Asp Asp Ala Ala Thr Tyr Tyr
1130 1135 1140
Cys Leu Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly
1145 1150 1155
Gly Gly Thr Glu Val Val Val Lys Asp Ile Gln Leu Thr Gln Ser
1160 1165 1170
Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
1175 1180 1185
Cys Lys Ala Ser Gln Asp Val Gly Thr Ser Val Ala Trp Tyr Gln
1190 1195 1200
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Trp Thr Ser
1205 1210 1215
Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser
1220 1225 1230
Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu Asp
1235 1240 1245
Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Leu Tyr Arg Ser Phe
1250 1255 1260
Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
1265 1270 1275
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
1280 1285 1290
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
1295 1300 1305
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
1310 1315 1320
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
1325 1330 1335
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
1340 1345 1350
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
1355 1360 1365
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
1370 1375
<210> 28
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain CDR1 <ERBB2>
<400> 28
Asp Thr Tyr Ile His
1 5
<210> 29
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain CDR2 <ERBB2>
<400> 29
Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 30
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain CDR3 <ERBB2>
<400> 30
Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr
1 5 10
<210> 31
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain CDR1 <ERBB2>
<400> 31
Arg Ala Ser Gln Asp Val Asn Thr Ala Val Ala
1 5 10
<210> 32
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain CDR2 <ERBB2>
<400> 32
Ser Ala Ser Phe Leu Tyr Ser
1 5
<210> 33
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain CDR3 <ERBB2>
<400> 33
Gln Gln His Tyr Thr Thr Pro Pro Thr
1 5
<210> 34
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain variable domain <ERBB2>
<400> 34
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 35
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain variable domain <ERBB2>
<400> 35
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 36
<211> 590
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain 1 (knob) of bispecific, trivalent ERbB2/Pb-Dotam
(P1AD9827)
<400> 36
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val
465 470 475 480
Lys Pro Thr Glu Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser
485 490 495
Leu Ser Thr Tyr Ser Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala
500 505 510
Leu Glu Trp Leu Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala
515 520 525
Ser Trp Ala Lys Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser
530 535 540
Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr
545 550 555 560
Tyr Tyr Cys Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro
565 570 575
Pro His Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
580 585 590
<210> 37
<211> 580
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain 2 (hole) of bispecific, trivalent ERbB2/Pb-Dotam
(P1AD9827)
<400> 37
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
450 455 460
Gly Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
465 470 475 480
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser
485 490 495
Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
500 505 510
Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val
515 520 525
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
530 535 540
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly
545 550 555 560
Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys
565 570 575
Val Glu Ile Lys
580
<210> 38
<211> 214
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain <ErbB2> (P1AD9827)
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 39
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain CDR1 <CD20>
<400> 39
Tyr Ser Trp Ile Asn
1 5
<210> 40
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain CDR2 <CD20>
<400> 40
Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe Lys
1 5 10 15
Gly
<210> 41
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain CDR3 <CD20>
<400> 41
Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr
1 5 10
<210> 42
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain CDR1 <CD20>
<400> 42
Arg Ser Ser Lys Ser Leu Leu His Ser Asn Gly Ile Thr Tyr Leu Tyr
1 5 10 15
<210> 43
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain CDR2 <CD20>
<400> 43
Gln Met Ser Asn Leu Val Ser
1 5
<210> 44
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain CDR3 CD20>
<400> 44
Ala Gln Asn Leu Glu Leu Pro Tyr Thr
1 5
<210> 45
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain variable domain <CD20>
<400> 45
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 46
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain variable domain <CD20>
<400> 46
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Val Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 47
<211> 589
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain 1 (knob) of bispecific, trivalent CD20/Pb-Dotam
(P1AD9826)
<400> 47
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys
465 470 475 480
Pro Thr Glu Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu
485 490 495
Ser Thr Tyr Ser Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu
500 505 510
Glu Trp Leu Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser
515 520 525
Trp Ala Lys Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln
530 535 540
Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr
545 550 555 560
Tyr Cys Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro
565 570 575
His Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
580 585
<210> 48
<211> 579
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy chain 2 (hole) of bispecific, trivalent CD20/Pb-Dotam
(P1AD9826)
<400> 48
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
465 470 475 480
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val
485 490 495
Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala
500 505 510
Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
530 535 540
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly
545 550 555 560
Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val
565 570 575
Glu Ile Lys
<210> 49
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> Light chain <CD20> (P1AD9826)
<400> 49
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Val Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
115 120 125
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 50
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1766 >CEA Light Chain RK
<400> 50
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 51
<211> 687
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1766 CEA Heavy Chain with DOTAM VL / CH1
<400> 51
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser
465 470 475 480
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser
485 490 495
His Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro
500 505 510
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser
515 520 525
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
530 535 540
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
545 550 555 560
Leu Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly
565 570 575
Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
580 585 590
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
595 600 605
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
610 615 620
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
625 630 635 640
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
645 650 655
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
660 665 670
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
675 680 685
<210> 52
<211> 699
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1766 CEA Heavy Chain with DOTAM VH / CK
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Val Thr Leu Lys Glu Ser Gly Pro Val
465 470 475 480
Leu Val Lys Pro Thr Glu Thr Leu Thr Leu Thr Cys Thr Val Ser Gly
485 490 495
Phe Ser Leu Ser Thr Tyr Ser Met Ser Trp Ile Arg Gln Pro Pro Gly
500 505 510
Lys Ala Leu Glu Trp Leu Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr
515 520 525
Tyr Ala Ser Trp Ala Lys Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser
530 535 540
Lys Ser Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr
545 550 555 560
Ala Thr Tyr Tyr Cys Ala Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala
565 570 575
Tyr Pro Pro His Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
580 585 590
Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
595 600 605
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
610 615 620
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
625 630 635 640
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
645 650 655
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
660 665 670
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
675 680 685
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
690 695
<210> 53
<211> 228
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1767 >DOTAM "LC" with VH / CK
<400> 53
Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu Thr
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu Thr
65 70 75 80
Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser
115 120 125
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
130 135 140
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
145 150 155 160
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
165 170 175
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
180 185 190
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
195 200 205
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
210 215 220
Arg Gly Glu Cys
225
<210> 54
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1767 CEA Light Chain RK
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 55
<211> 687
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1767 CEA Heavy Chain with DOTAM VL / CH1 (knob)
<400> 55
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser
465 470 475 480
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser
485 490 495
His Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro
500 505 510
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser
515 520 525
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
530 535 540
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
545 550 555 560
Leu Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly
565 570 575
Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
580 585 590
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
595 600 605
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
610 615 620
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
625 630 635 640
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
645 650 655
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
660 665 670
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
675 680 685
<210> 56
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1767 CEA Heavy Chain (hole)
<400> 56
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 57
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1768 >CEA LC
<400> 57
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 58
<211> 443
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1768> DOTAM Heavy Chain with VL / CH1 (hole)
<400> 58
Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp
1 5 10 15
Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val Tyr Ser Asp Asn
20 25 30
Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp Glu
85 90 95
Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 59
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1768 CEA Heavy chain (knob)
<400> 59
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 60
<211> 228
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1768 DOTAM "LC" VH / CK
<400> 60
Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu Thr
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu Thr
65 70 75 80
Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser
115 120 125
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
130 135 140
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
145 150 155 160
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
165 170 175
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
180 185 190
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
195 200 205
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
210 215 220
Arg Gly Glu Cys
225
<210> 61
<211> 228
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1769 DOTAM "LC" with VH / CK
<400> 61
Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu Thr
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu Thr
65 70 75 80
Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser
115 120 125
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
130 135 140
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
145 150 155 160
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
165 170 175
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
180 185 190
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
195 200 205
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
210 215 220
Arg Gly Glu Cys
225
<210> 62
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1769 CEA LC
<400> 62
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 63
<211> 678
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1769 CEA HC with CEA VH / CH1 / DOTAM VL / CH1 (hole)
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ile Gln Met Thr Gln
225 230 235 240
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
245 250 255
Cys Gln Ser Ser His Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr
260 265 270
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser
275 280 285
Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
290 295 300
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
305 310 315 320
Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly
325 330 335
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr Lys
340 345 350
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
355 360 365
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
370 375 380
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
385 390 395 400
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
405 410 415
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
420 425 430
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
435 440 445
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
450 455 460
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
465 470 475 480
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
485 490 495
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
500 505 510
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
515 520 525
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
530 535 540
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly
545 550 555 560
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
565 570 575
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn
580 585 590
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
595 600 605
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
610 615 620
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
625 630 635 640
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
645 650 655
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
660 665 670
Ser Leu Ser Pro Gly Lys
675
<210> 64
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1769 CEA HC (knob)
<400> 64
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 65
<211> 215
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1770 CEA LC
<400> 65
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 66
<211> 945
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1770 CEA HC with DOTAM scFab: DOTAM VL / Ck / Linker / VH CH1
(knob)
<400> 66
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Ile Gln Met Thr Gln Ser Pro Ser Ser
465 470 475 480
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser
485 490 495
His Ser Val Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro
500 505 510
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser
515 520 525
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
530 535 540
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
545 550 555 560
Leu Gly Gly Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly
565 570 575
Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile
580 585 590
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
595 600 605
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
610 615 620
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
625 630 635 640
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
645 650 655
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
660 665 670
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
675 680 685
Cys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
690 695 700
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
705 710 715 720
Gly Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
725 730 735
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Tyr
740 745 750
Ser Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
755 760 765
Gly Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys
770 775 780
Gly Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
785 790 795 800
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
805 810 815
Arg Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp
820 825 830
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
835 840 845
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
850 855 860
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
865 870 875 880
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
885 890 895
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
900 905 910
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
915 920 925
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
930 935 940
Cys
945
<210> 67
<211> 451
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AE1770 CEA HC (hole)
<400> 67
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 68
<211> 496
<212> PRT
<213> Homo sapiens cartilage protein
<400> 68
Met Arg Val Leu Ser Gly Thr Ser Leu Met Leu Cys Ser Leu Leu Leu
1 5 10 15
Leu Leu Gln Ala Leu Cys Ser Pro Gly Leu Ala Pro Gln Ser Arg Gly
20 25 30
His Leu Cys Arg Thr Arg Pro Thr Asp Leu Val Phe Val Val Asp Ser
35 40 45
Ser Arg Ser Val Arg Pro Val Glu Phe Glu Lys Val Lys Val Phe Leu
50 55 60
Ser Gln Val Ile Glu Ser Leu Asp Val Gly Pro Asn Ala Thr Arg Val
65 70 75 80
Gly Met Val Asn Tyr Ala Ser Thr Val Lys Gln Glu Phe Ser Leu Arg
85 90 95
Ala His Val Ser Lys Ala Ala Leu Leu Gln Ala Val Arg Arg Ile Gln
100 105 110
Pro Leu Ser Thr Gly Thr Met Thr Gly Leu Ala Ile Gln Phe Ala Ile
115 120 125
Thr Lys Ala Phe Gly Asp Ala Glu Gly Gly Arg Ser Arg Ser Pro Asp
130 135 140
Ile Ser Lys Val Val Ile Val Val Thr Asp Gly Arg Pro Gln Asp Ser
145 150 155 160
Val Gln Asp Val Ser Ala Arg Ala Arg Ala Ser Gly Val Glu Leu Phe
165 170 175
Ala Ile Gly Val Gly Ser Val Asp Lys Ala Thr Leu Arg Gln Ile Ala
180 185 190
Ser Glu Pro Gln Asp Glu His Val Asp Tyr Val Glu Ser Tyr Ser Val
195 200 205
Ile Glu Lys Leu Ser Arg Lys Phe Gln Glu Ala Phe Cys Val Val Ser
210 215 220
Asp Leu Cys Ala Thr Gly Asp His Asp Cys Glu Gln Val Cys Ile Ser
225 230 235 240
Ser Pro Gly Ser Tyr Thr Cys Ala Cys His Glu Gly Phe Thr Leu Asn
245 250 255
Ser Asp Gly Lys Thr Cys Asn Val Cys Ser Gly Gly Gly Gly Ser Ser
260 265 270
Ala Thr Asp Leu Val Phe Leu Ile Asp Gly Ser Lys Ser Val Arg Pro
275 280 285
Glu Asn Phe Glu Leu Val Lys Lys Phe Ile Ser Gln Ile Val Asp Thr
290 295 300
Leu Asp Val Ser Asp Lys Leu Ala Gln Val Gly Leu Val Gln Tyr Ser
305 310 315 320
Ser Ser Val Arg Gln Glu Phe Pro Leu Gly Arg Phe His Thr Lys Lys
325 330 335
Asp Ile Lys Ala Ala Val Arg Asn Met Ser Tyr Met Glu Lys Gly Thr
340 345 350
Met Thr Gly Ala Ala Leu Lys Tyr Leu Ile Asp Asn Ser Phe Thr Val
355 360 365
Ser Ser Gly Ala Arg Pro Gly Ala Gln Lys Val Gly Ile Val Phe Thr
370 375 380
Asp Gly Arg Ser Gln Asp Tyr Ile Asn Asp Ala Ala Lys Lys Ala Lys
385 390 395 400
Asp Leu Gly Phe Lys Met Phe Ala Val Gly Val Gly Asn Ala Val Glu
405 410 415
Asp Glu Leu Arg Glu Ile Ala Ser Glu Pro Val Ala Glu His Tyr Phe
420 425 430
Tyr Thr Ala Asp Phe Lys Thr Ile Asn Gln Ile Gly Lys Lys Leu Gln
435 440 445
Lys Lys Ile Cys Val Glu Glu Asp Pro Cys Ala Cys Glu Ser Leu Val
450 455 460
Lys Phe Gln Ala Lys Val Glu Gly Leu Leu Gln Ala Leu Thr Arg Lys
465 470 475 480
Leu Glu Ala Val Ser Lys Arg Leu Ala Ile Leu Glu Asn Thr Val Val
485 490 495
<210> 69
<211> 39
<212> PRT
<213> Artificial Sequence
<220>
<223> trimerization domain sequence
<400> 69
Cys Ala Cys Glu Ser Leu Val Lys Phe Gln Ala Lys Val Glu Gly Leu
1 5 10 15
Leu Gln Ala Leu Thr Arg Lys Leu Glu Ala Val Ser Lys Arg Leu Ala
20 25 30
Ile Leu Glu Asn Thr Val Val
35
<210> 70
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> rbHC.up
<400> 70
aagcttgcca ccatggagac tgggctgcgc tggcttc 37
<210> 71
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> https://protect-eu.mimecast.com/s/kCbWCk5L8Tw8BJC9Dmxl?domain=rbhcf.do
<400> 71
ccattggtga gggtgcccga g 21
<210> 72
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<223> rbLC.up
<400> 72
aagcttgcca ccatggacay gagggccccc actc 34
<210> 73
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> https://protect-eu.mimecast.com/s/05w_ClOgxUM0EYi1ugxe?domain=rblc.do
<400> 73
cagagtrctg ctgaggttgt aggtac 26
<210> 74
<211> 579
<212> PRT
<213> Artificial Sequence
<220>
<223> Immunomedics hNM14
<400> 74
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Thr Thr Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile His Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Phe Gly Phe Pro Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
450 455 460
Gly Gly Gly Ser Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Pro
465 470 475 480
Ala Val Gly Gly Thr Val Thr Ile Ser Cys Gln Ser Ser His Ser Val
485 490 495
Tyr Ser Asp Asn Asp Leu Ala Trp Tyr Gln Gln Lys Leu Gly Gln Pro
500 505 510
Pro Lys Leu Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Ser
515 520 525
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile
530 535 540
Ser Gly Val Gln Ser Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly
545 550 555 560
Tyr Asp Asp Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Glu Val
565 570 575
Val Val Lys
<210> 75
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> LC1
<400> 75
Ser Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val Tyr Ser Asp
20 25 30
Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp
85 90 95
Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 76
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> LC3
<400> 76
Ala Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly Asp
1 5 10 15
Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val Tyr Ser Asp Asn
20 25 30
Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp Glu
85 90 95
Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 77
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> HC7
<400> 77
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
1 5 10 15
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Asp Thr Ser Lys Asn Thr Ala Tyr Leu Gln
65 70 75 80
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 78
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> CH1A1A VH
<400> 78
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 79
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> CH1A1A VL
<400> 79
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 80
<211> 224
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AA1227_HC
<400> 80
Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu Thr
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Tyr Ser
20 25 30
Met Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Gly
35 40 45
Phe Ile Gly Ser Arg Gly Asp Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu Thr
65 70 75 80
Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala Arg
85 90 95
Glu Arg Asp Pro Tyr Gly Gly Gly Ala Tyr Pro Pro His Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 81
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> P1AA1227_LC
<400> 81
Ser Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser His Ser Val Tyr Ser Asp
20 25 30
Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Asp Asp
85 90 95
Glu Ser Asp Thr Tyr Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 82
<211> 213
<212> PRT
<213> Artificial Sequence
<220>
<223> Immunomedics hNM14
<400> 82
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Gly Thr Ser
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Thr Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Leu Tyr Arg Ser
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С РАКОВЫМИ КЛЕТКАМИ И ОБЕСПЕЧИВАЮТ НАПРАВЛЕННЫЙ ПЕРЕНОС РАДИОНУКЛИДОВ К УКАЗАННЫМ КЛЕТКАМ | 2020 |
|
RU2833191C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| МОЛЕКУЛЫ АНТИТЕЛ ПРОТИВ ВИРУСА BK | 2021 |
|
RU2835801C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
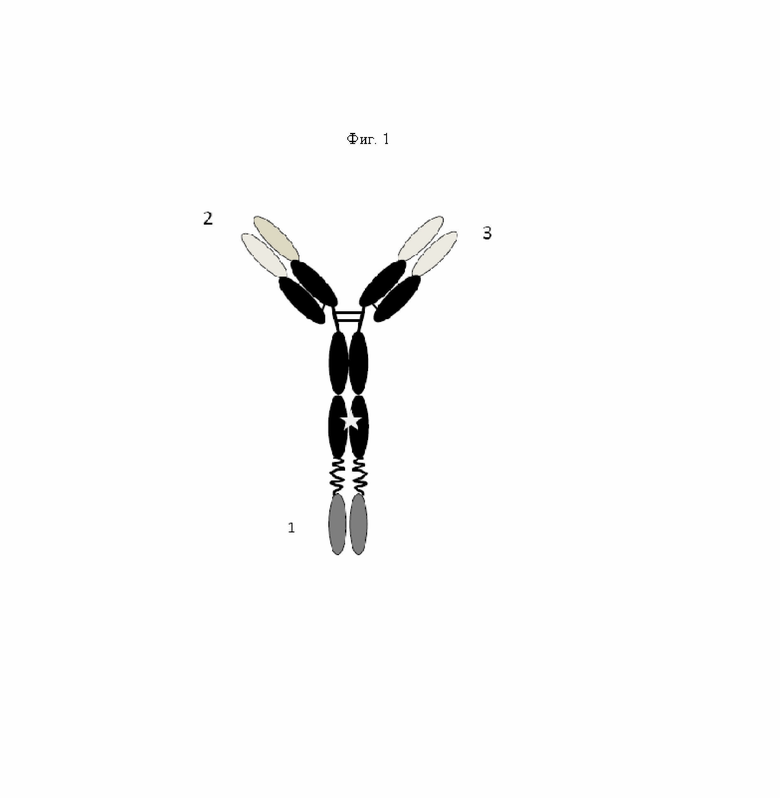

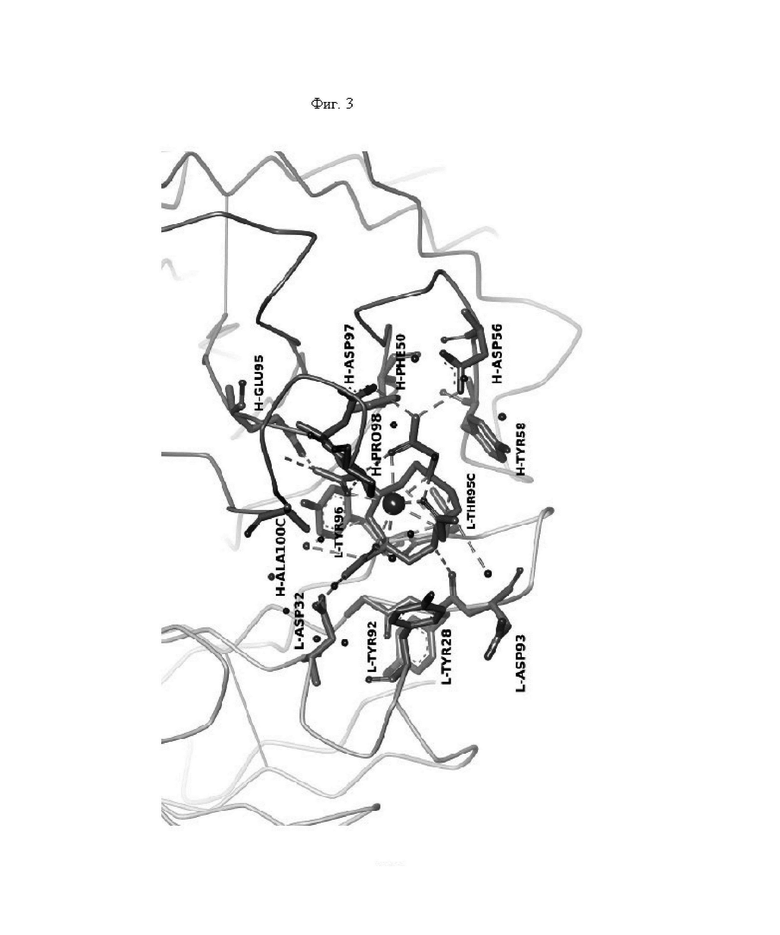
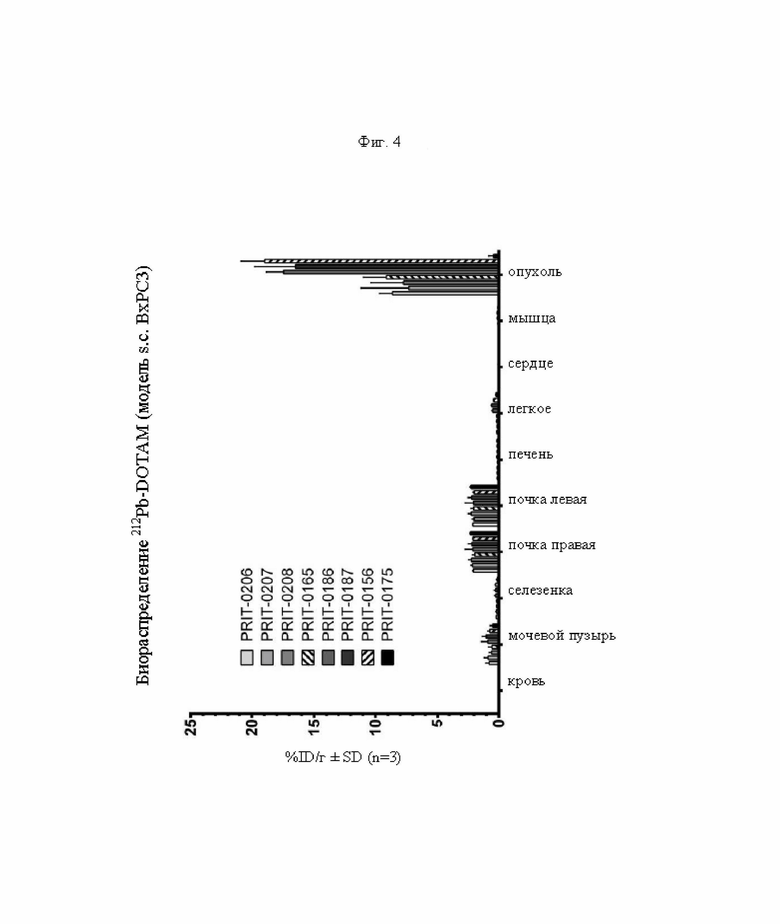
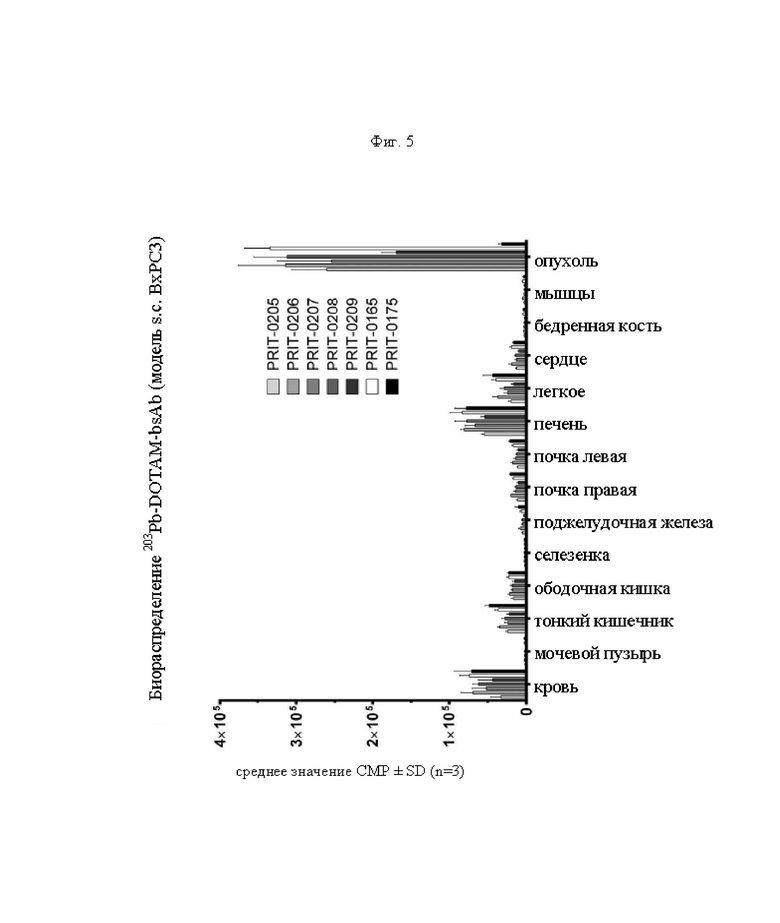
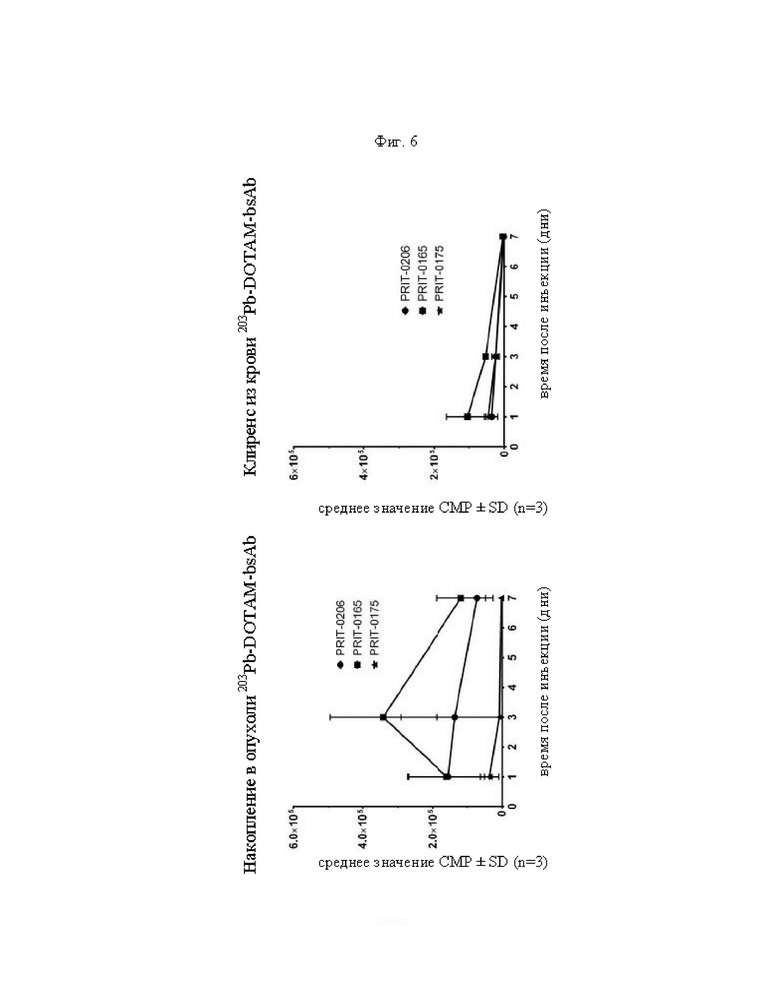
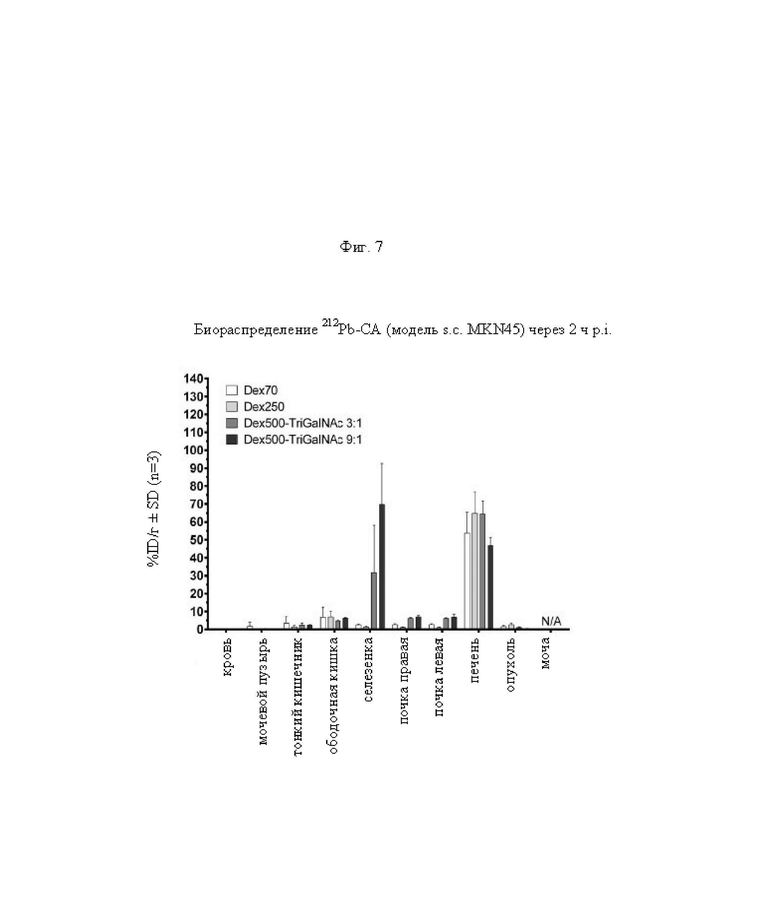
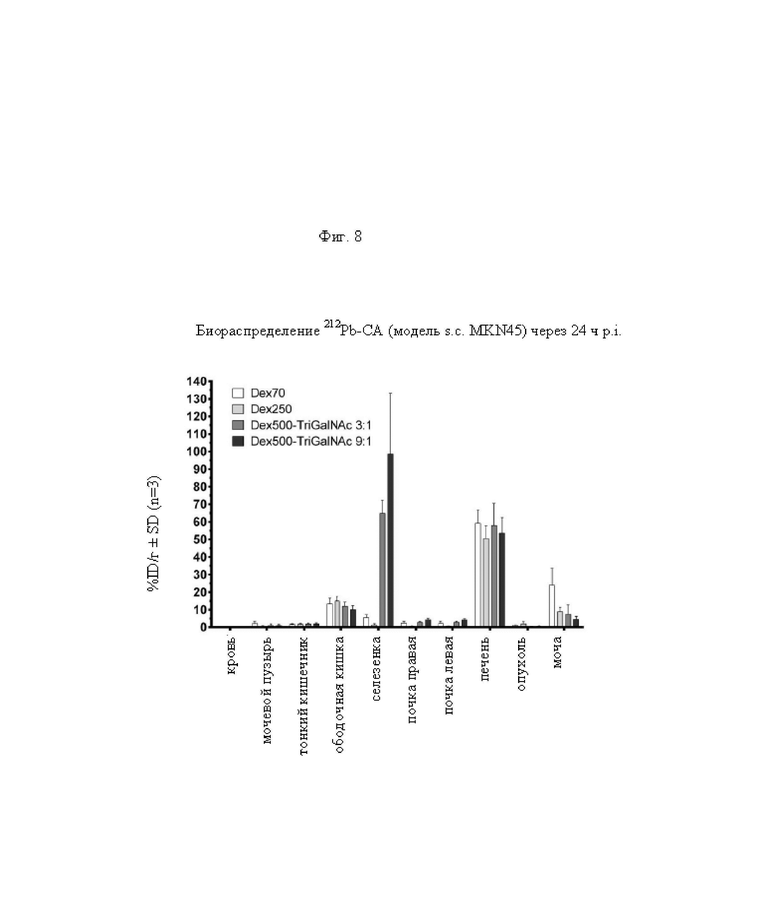
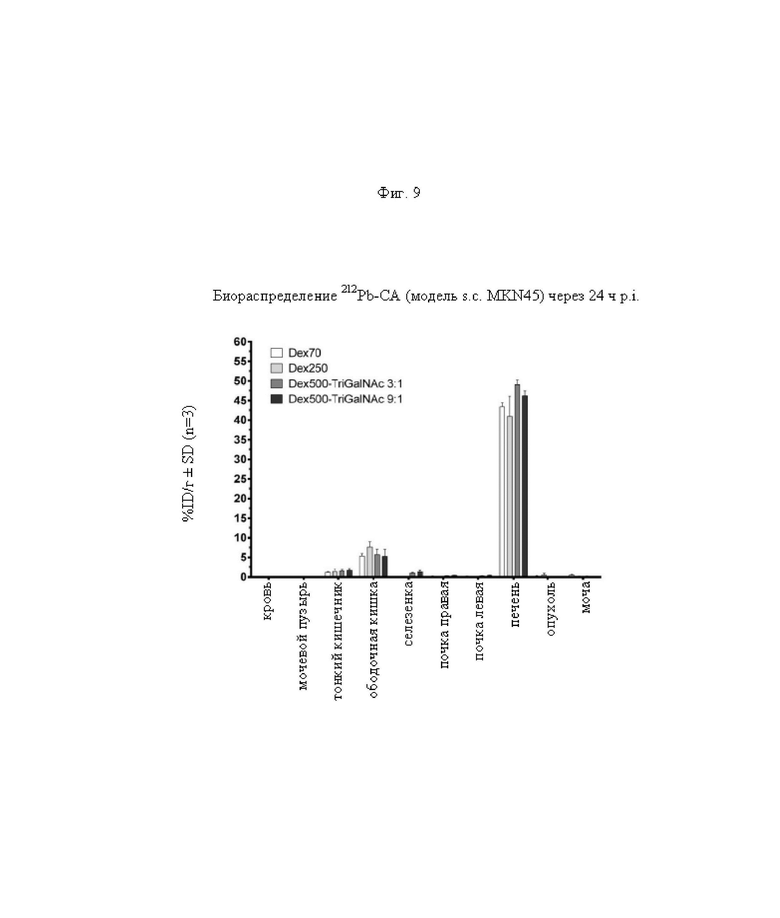
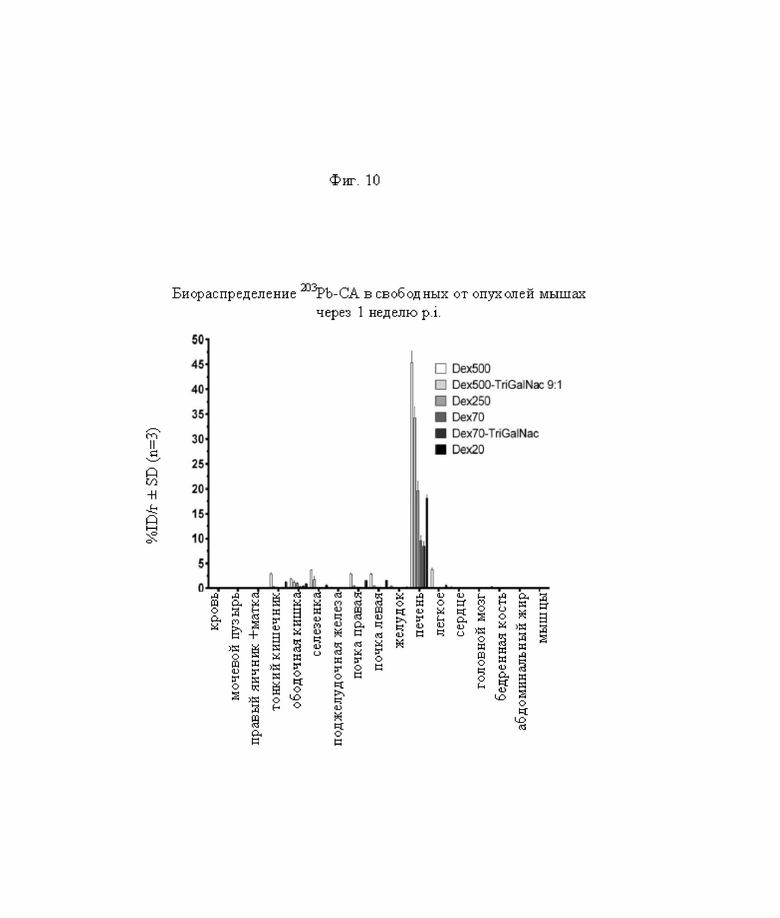
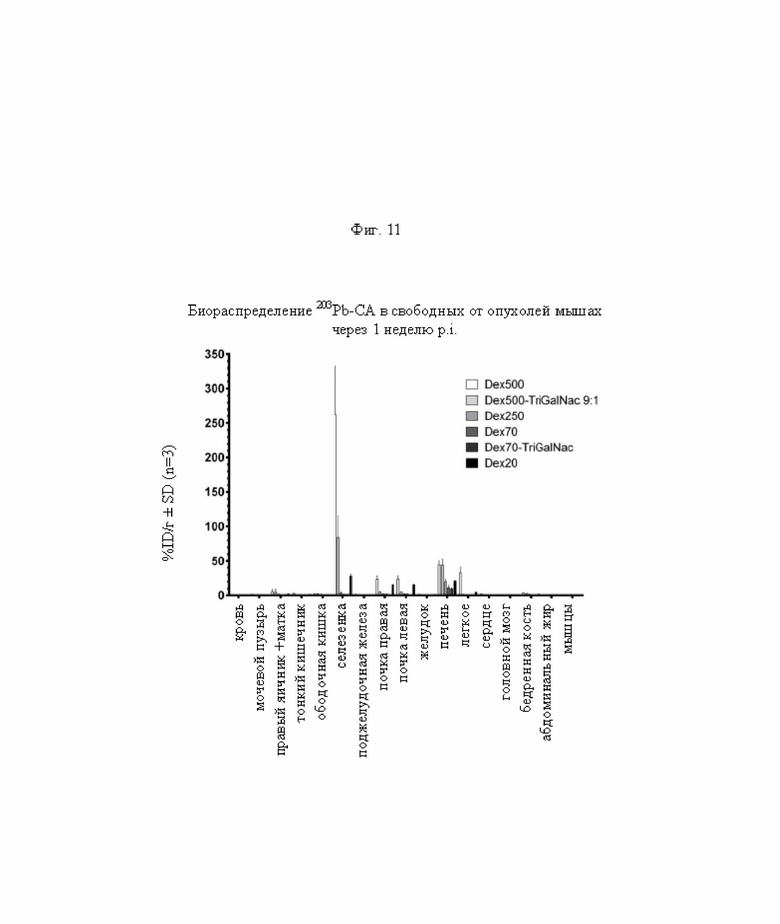
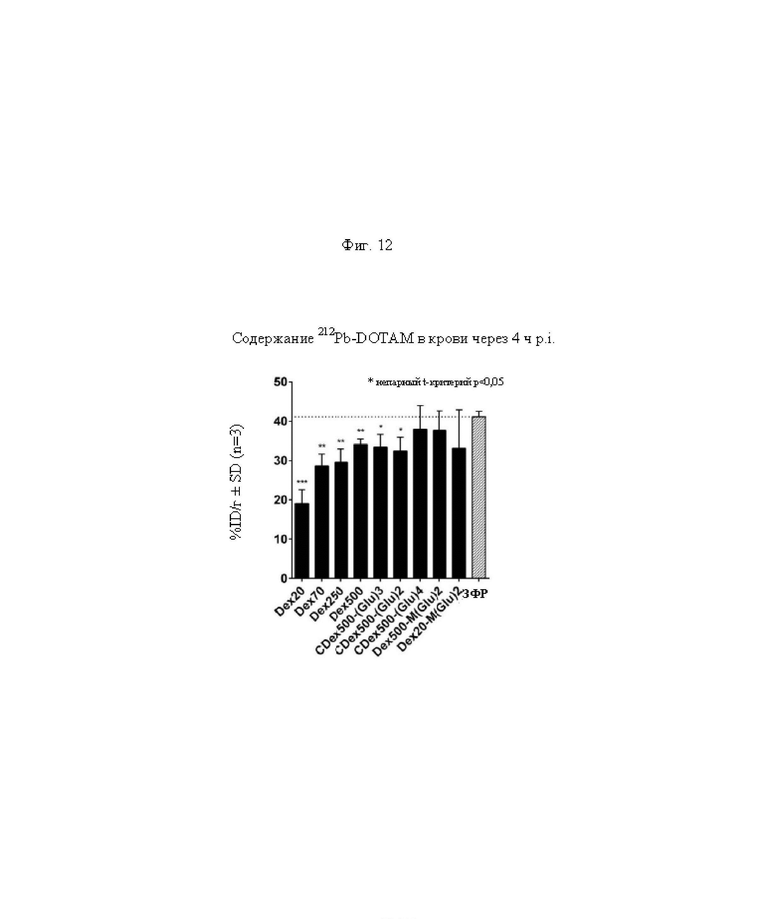
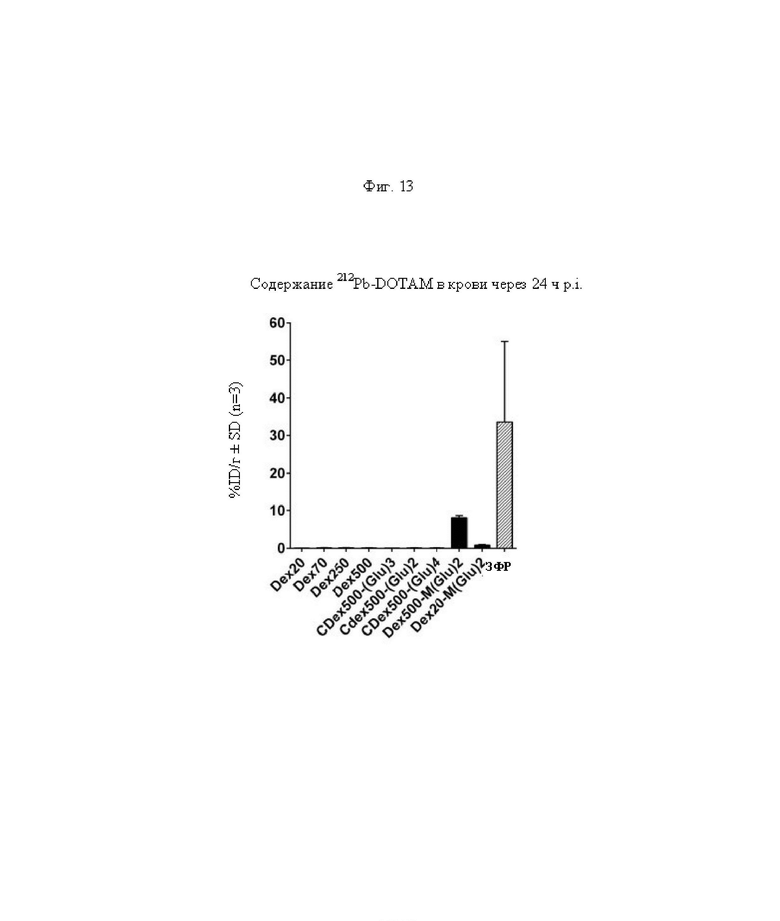
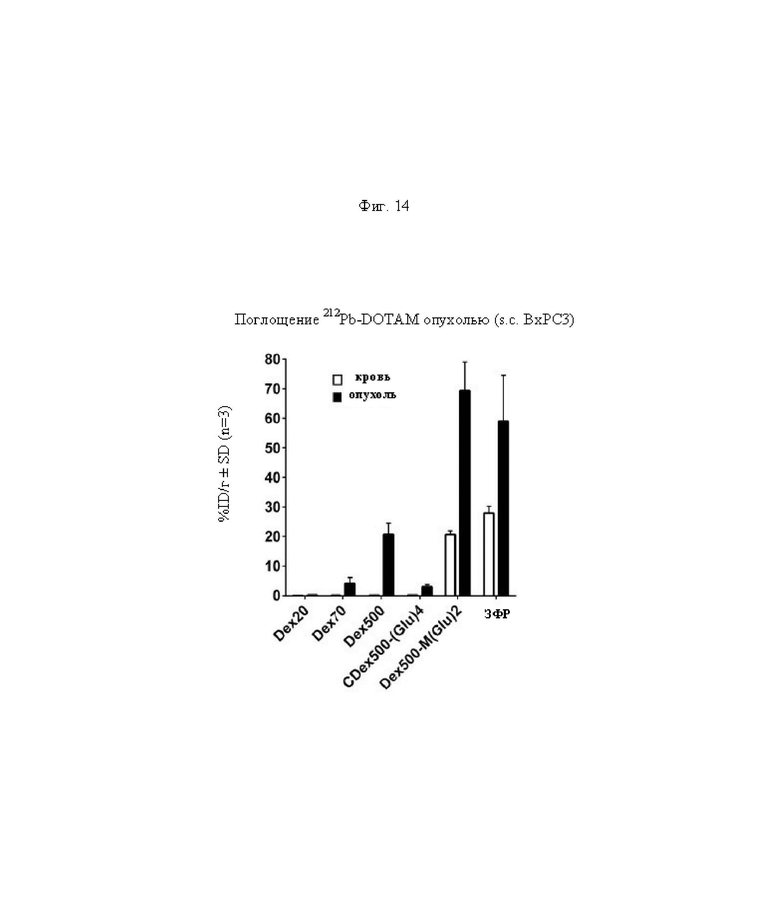
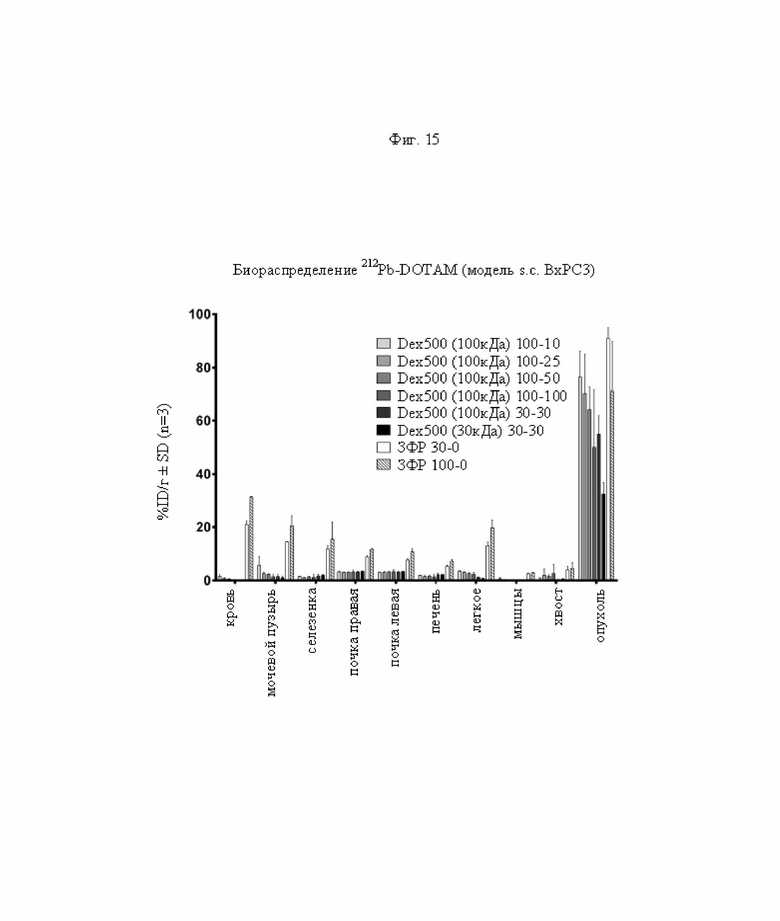
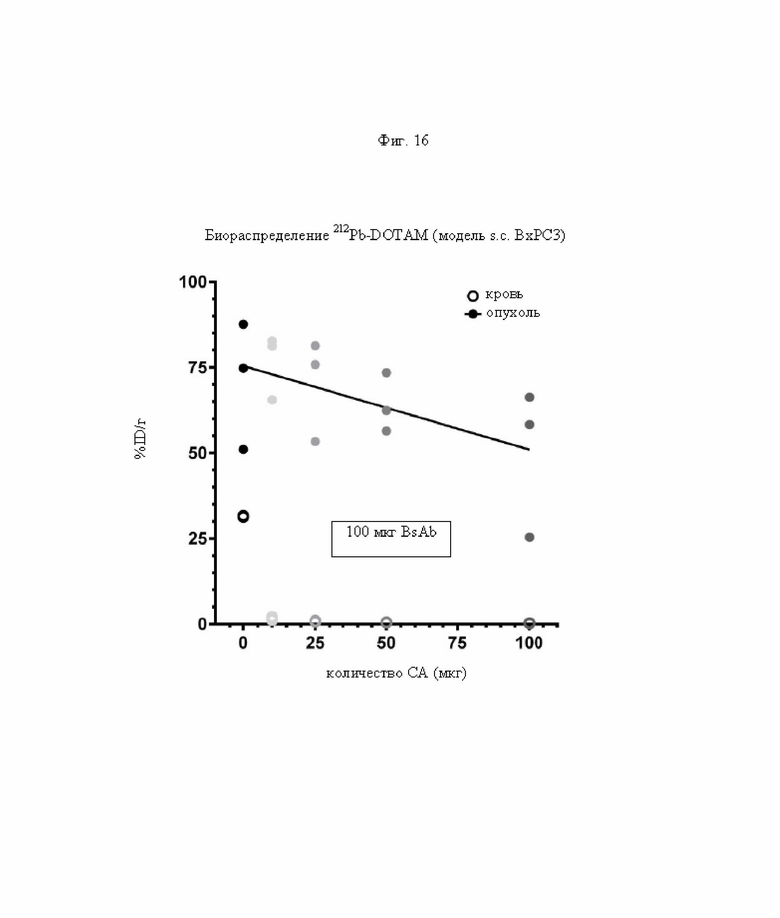
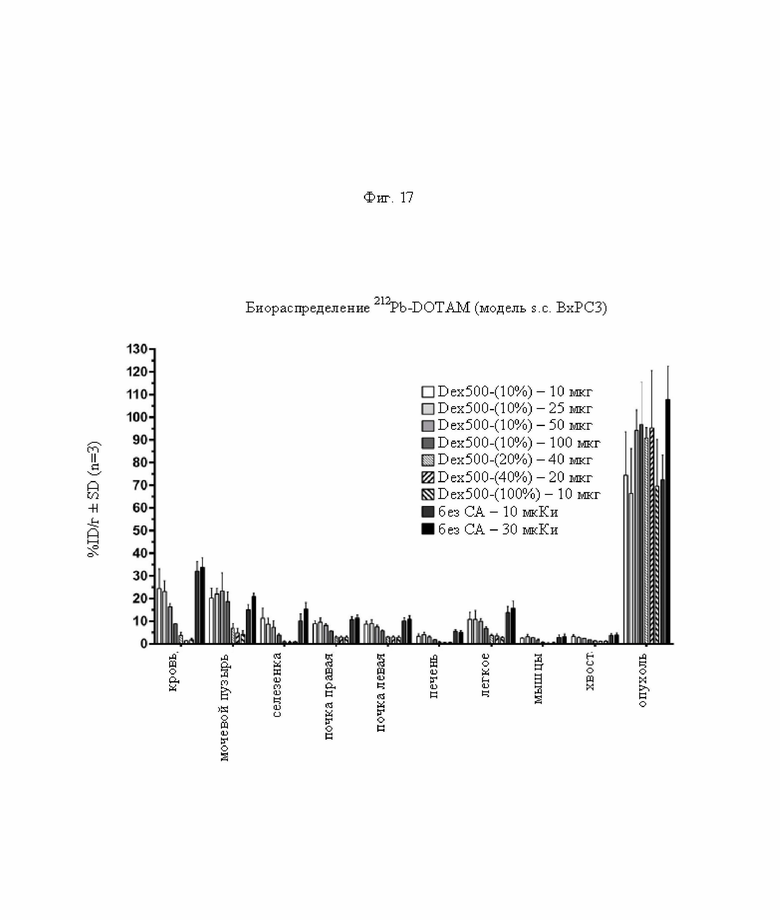
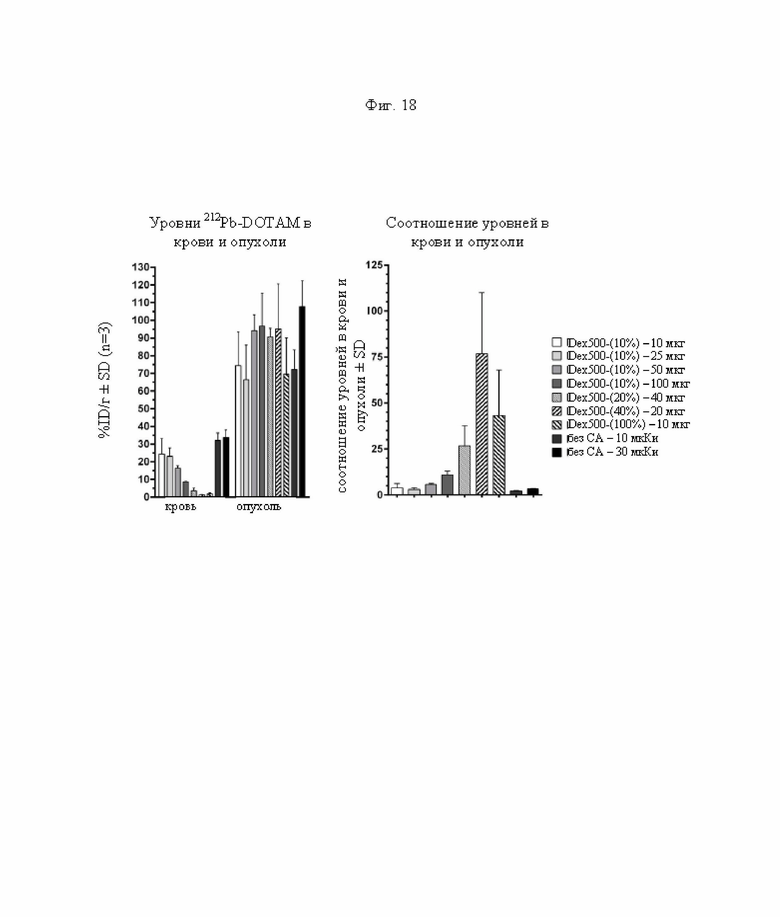
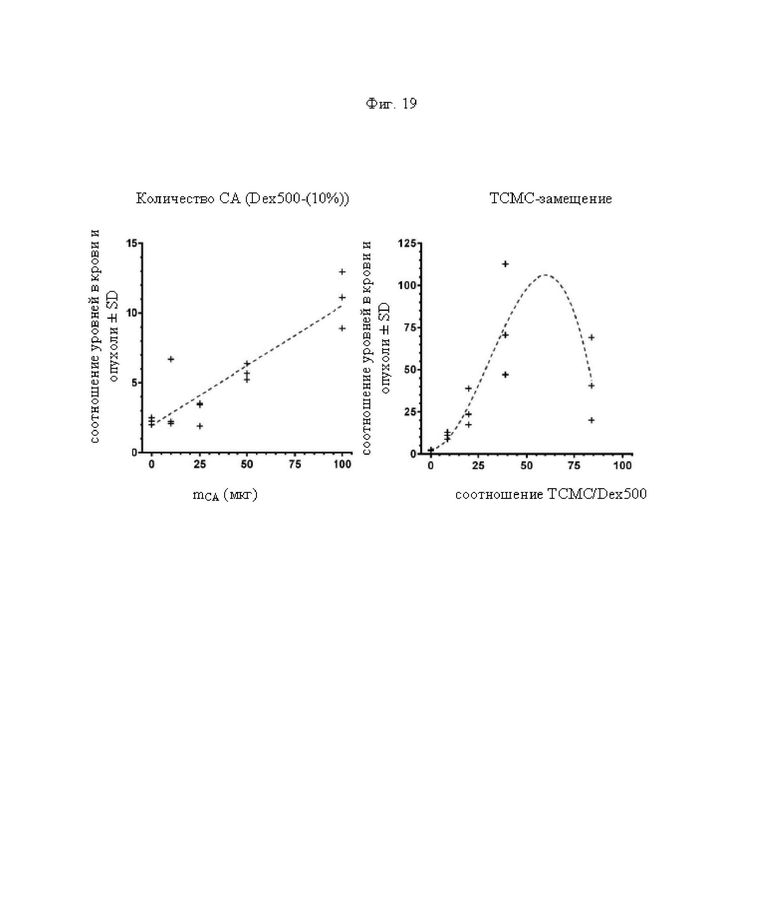
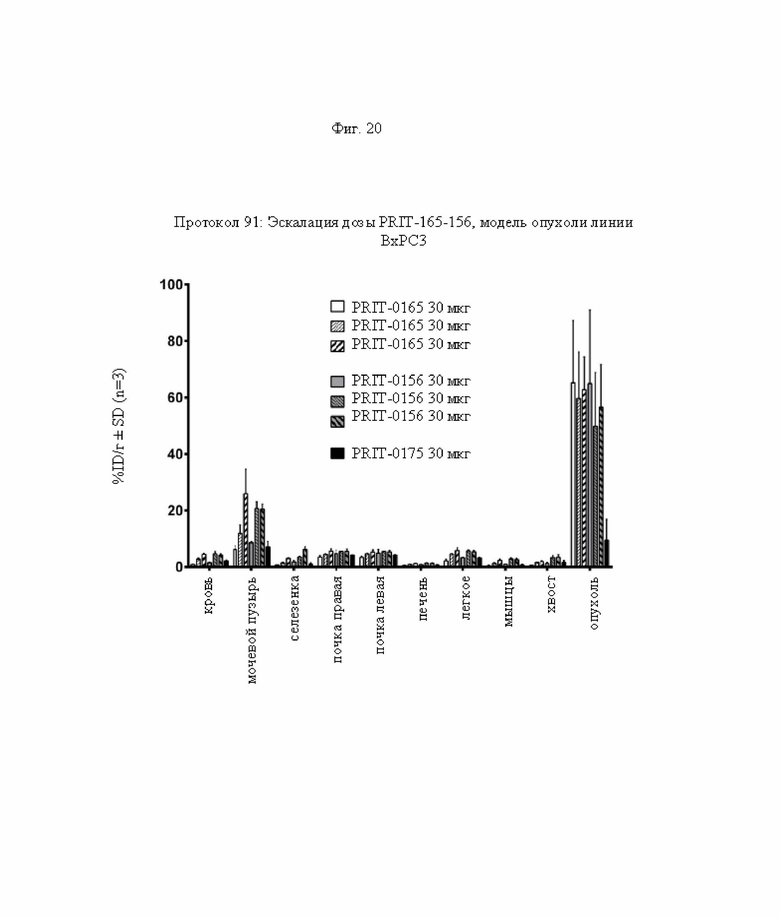
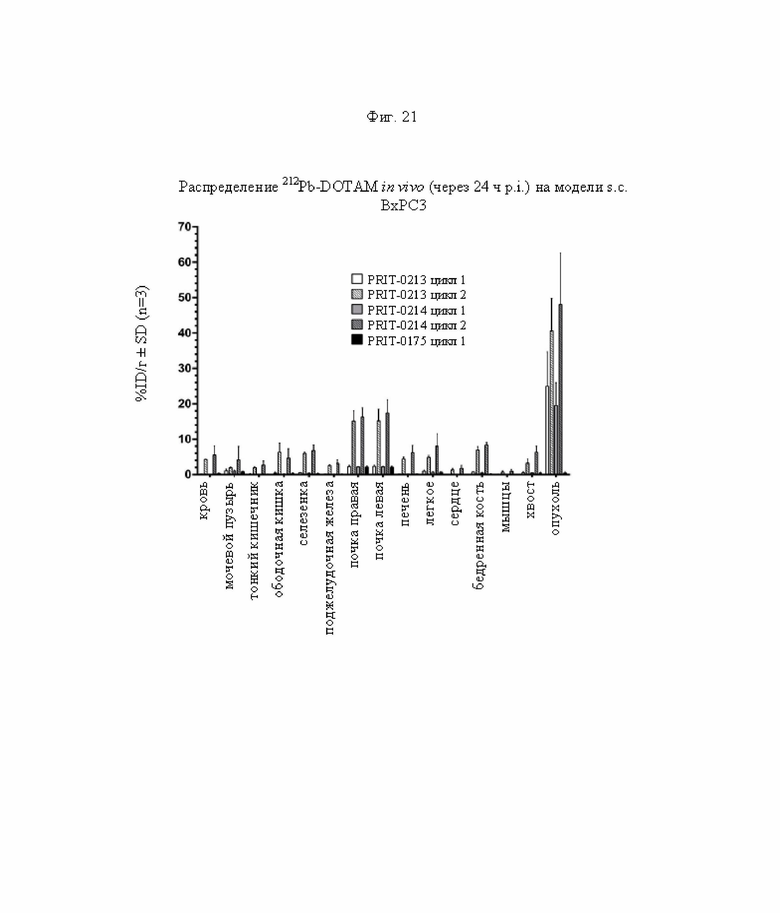
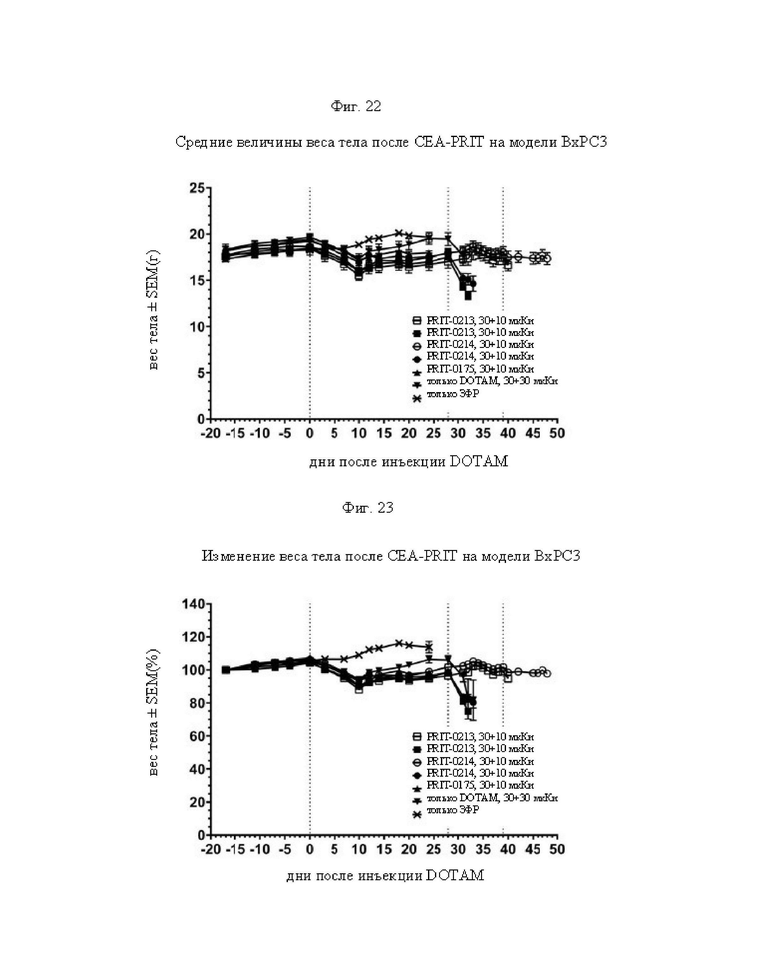
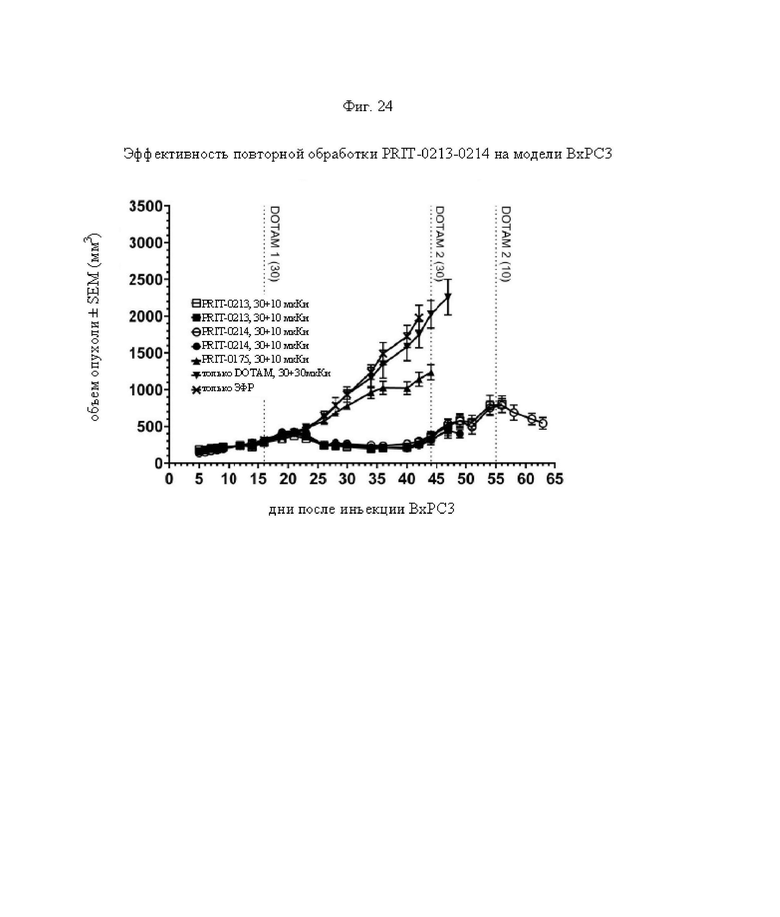
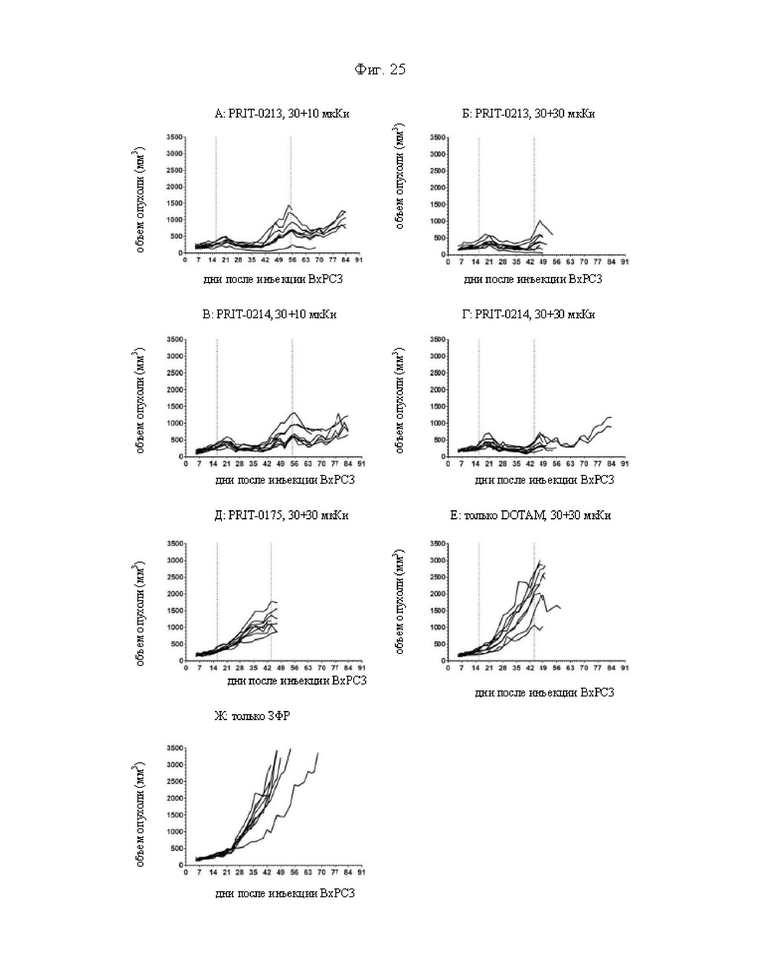
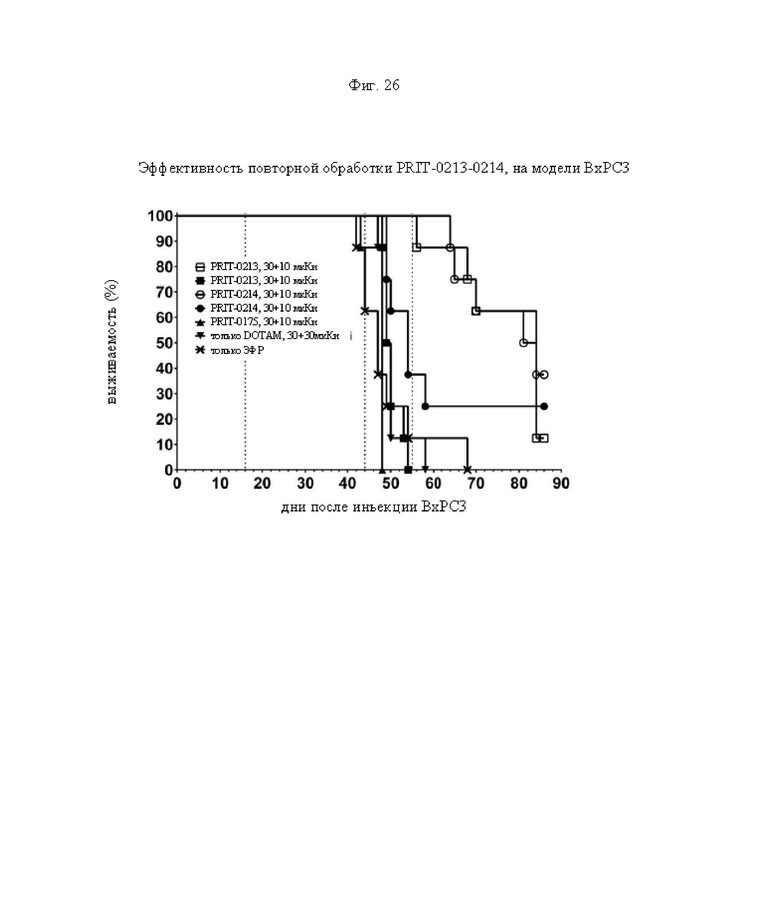
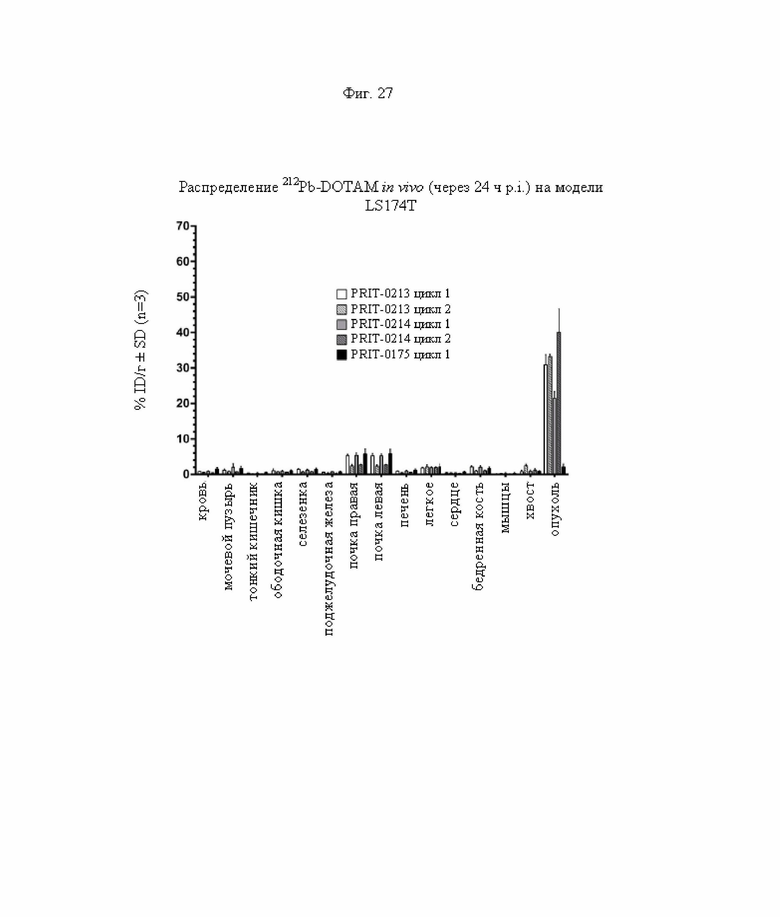
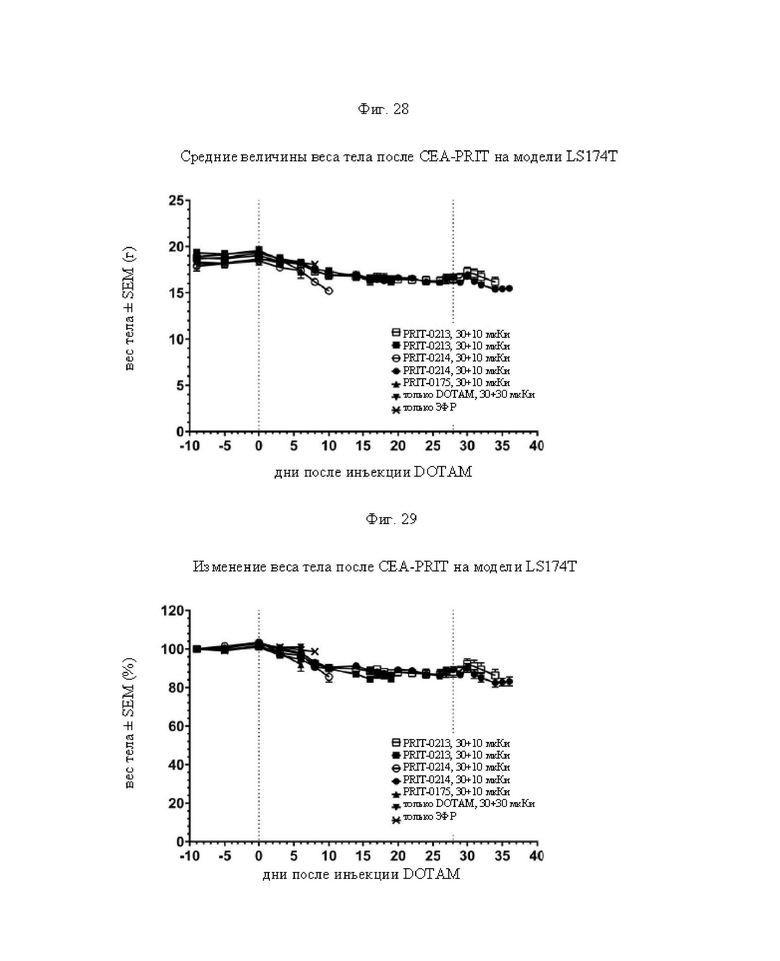
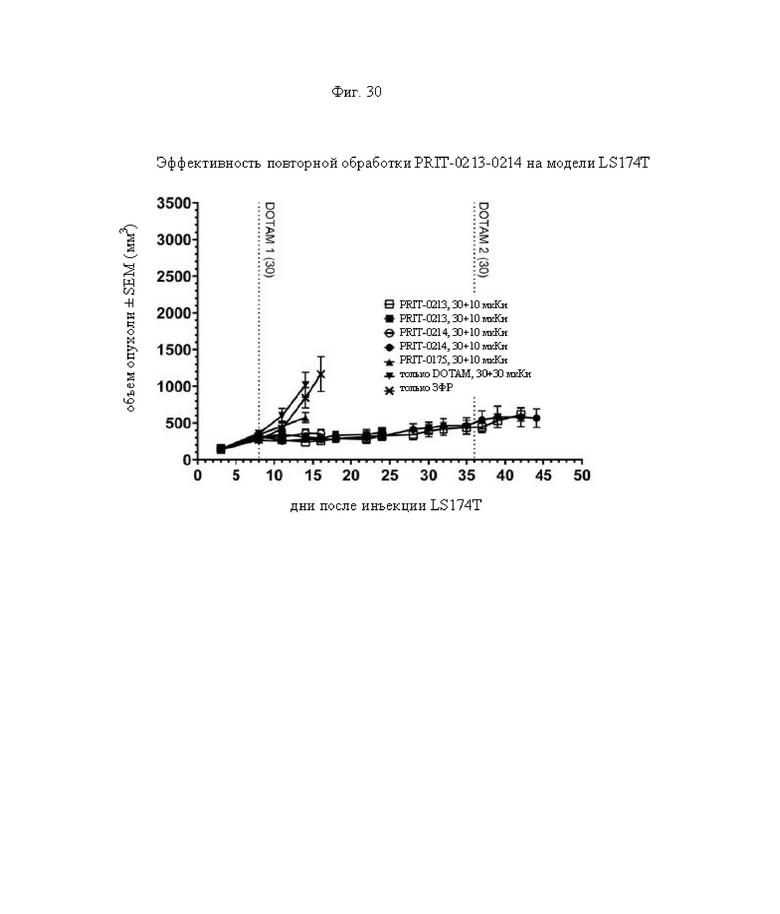
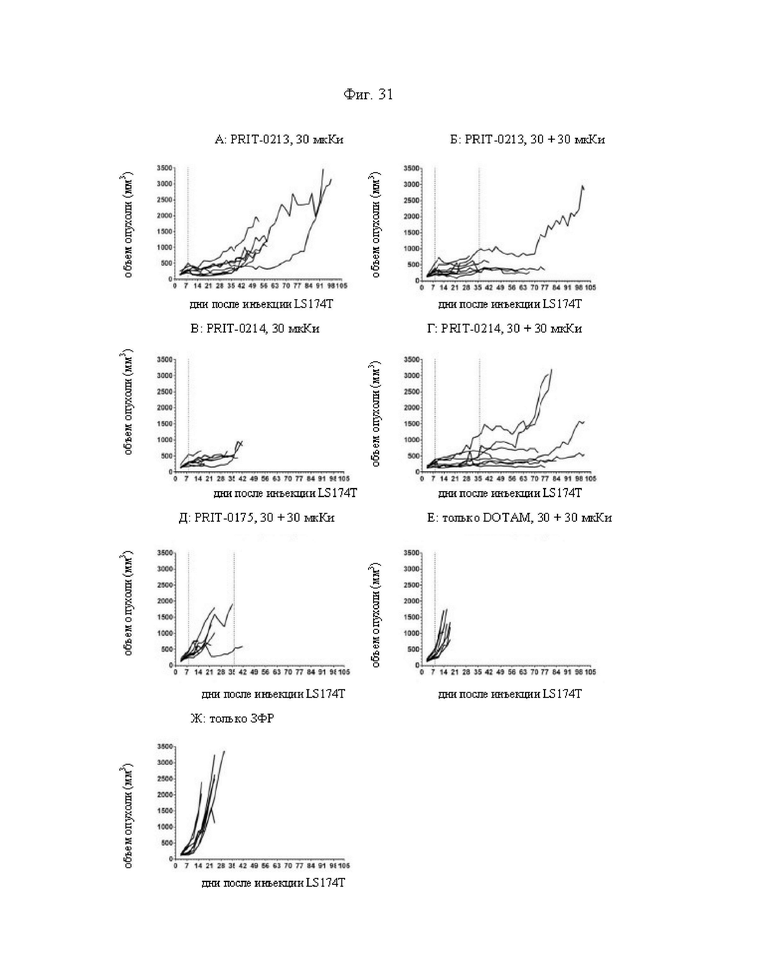
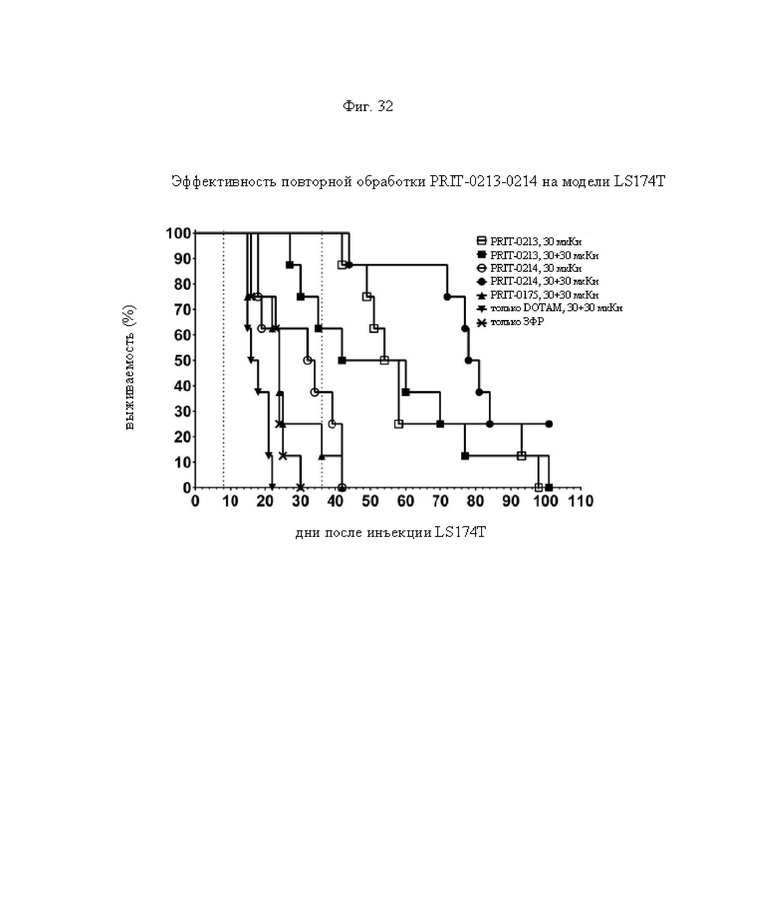
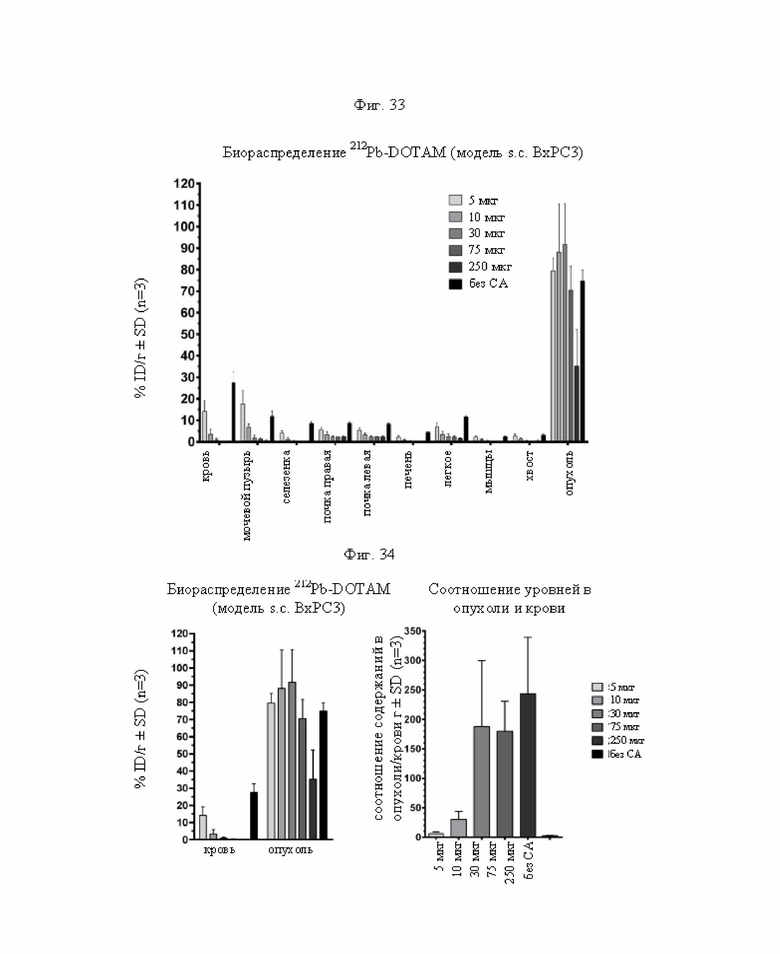
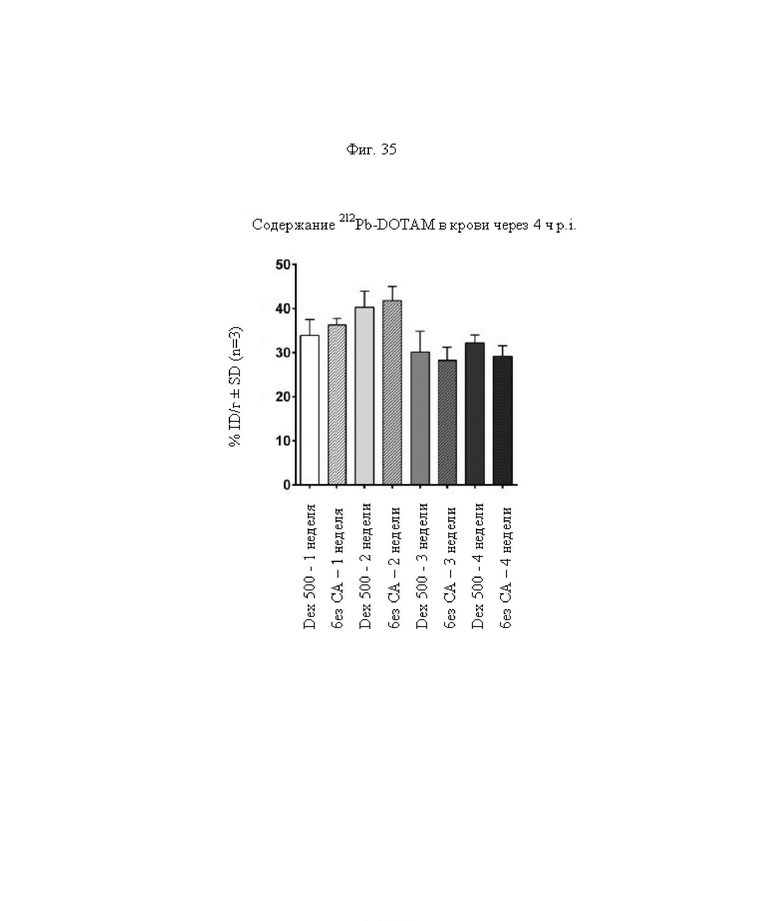
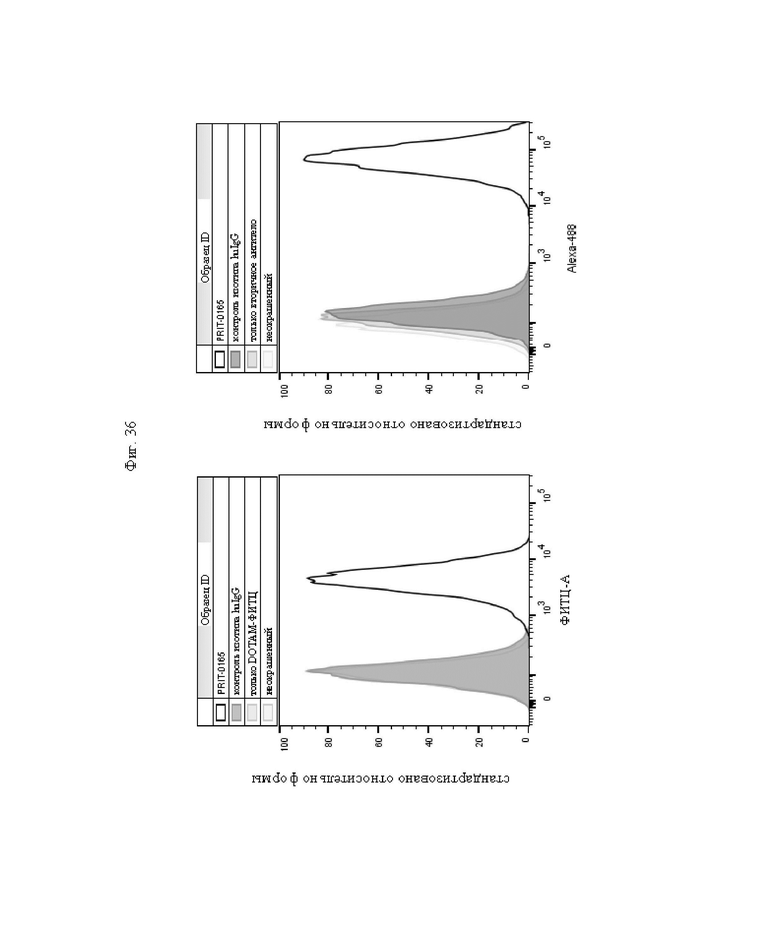
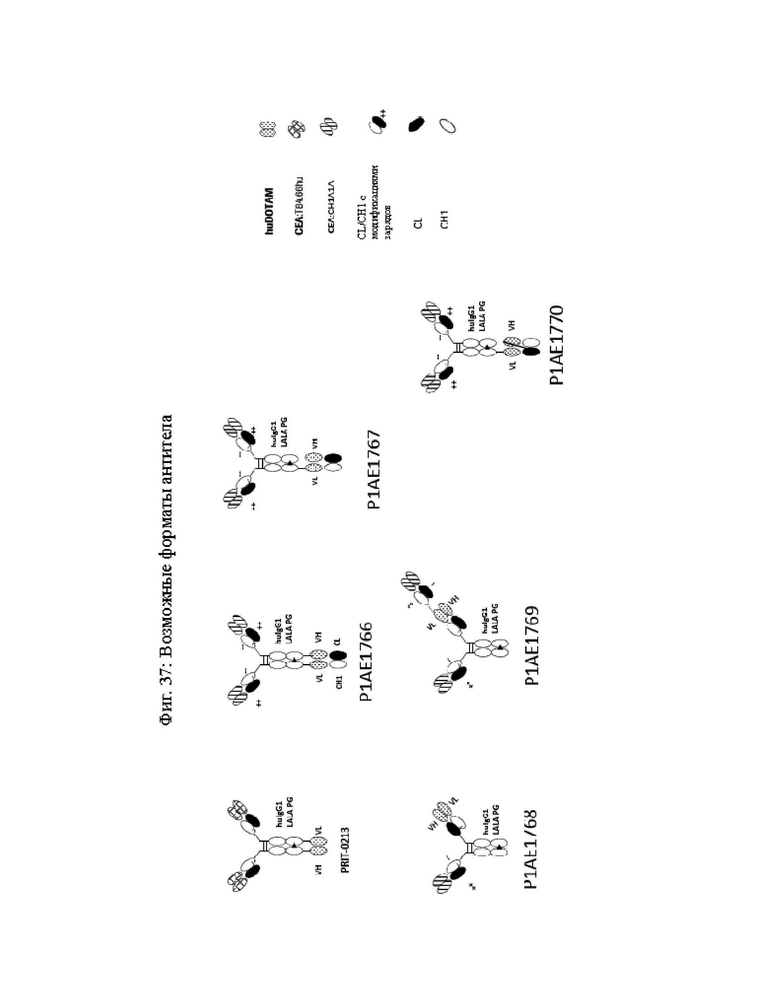
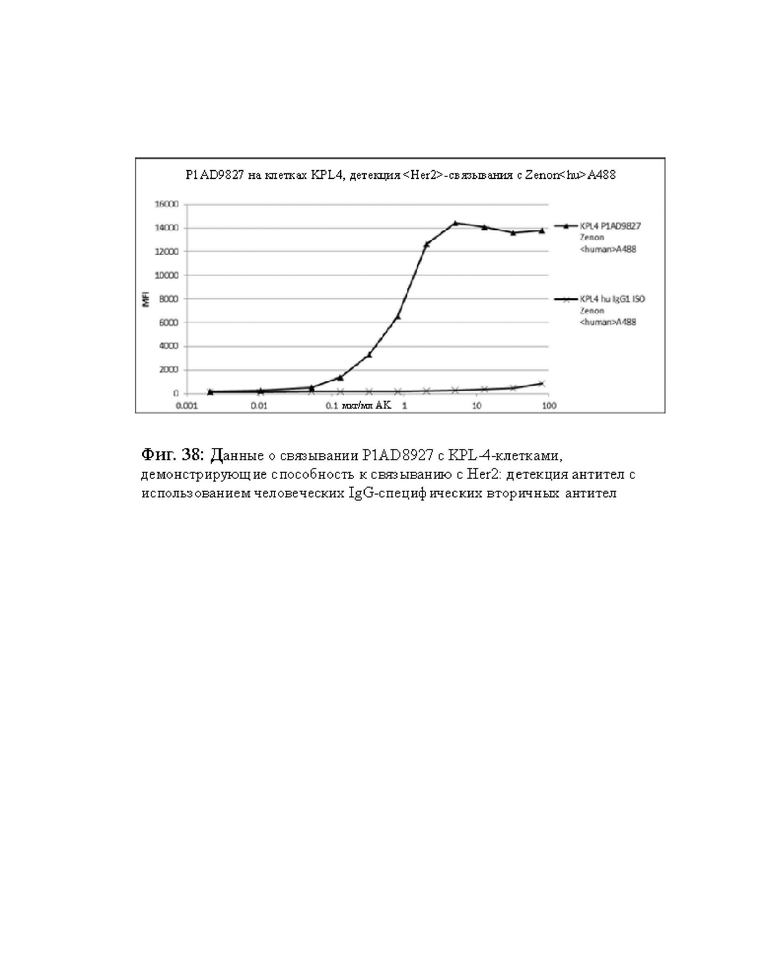


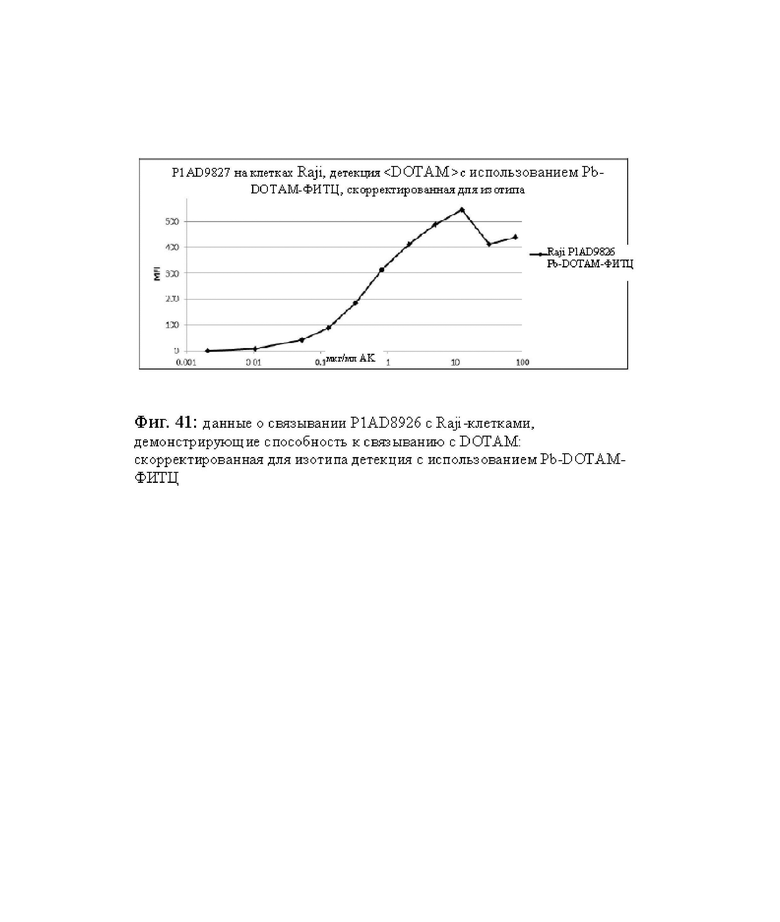
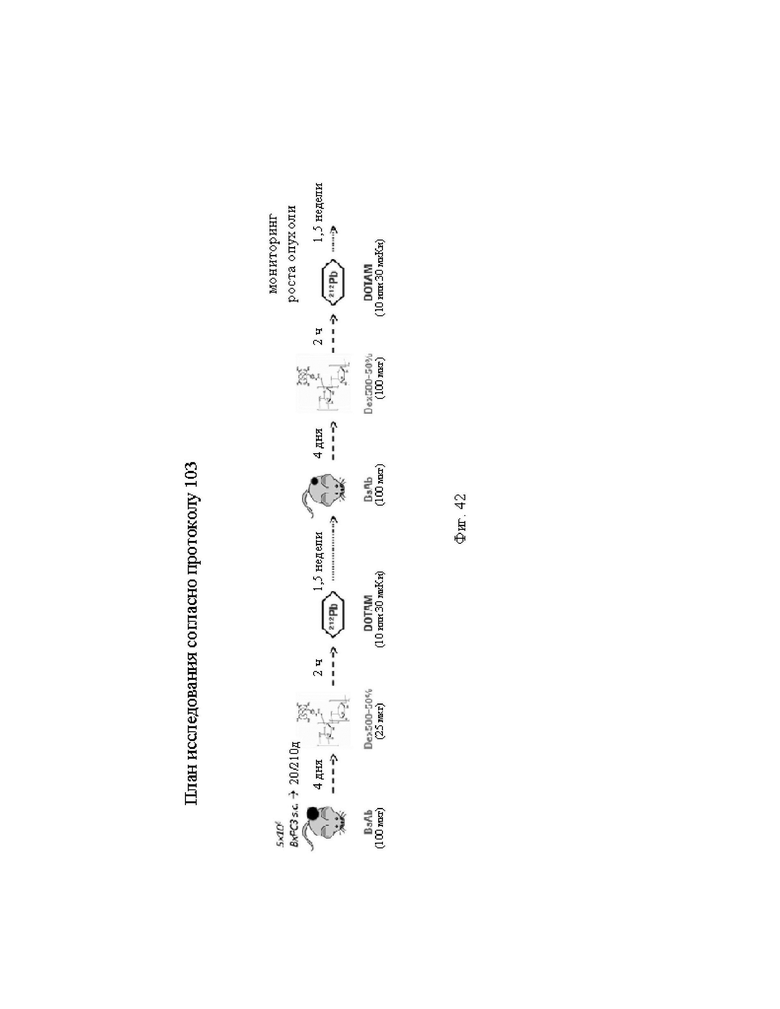
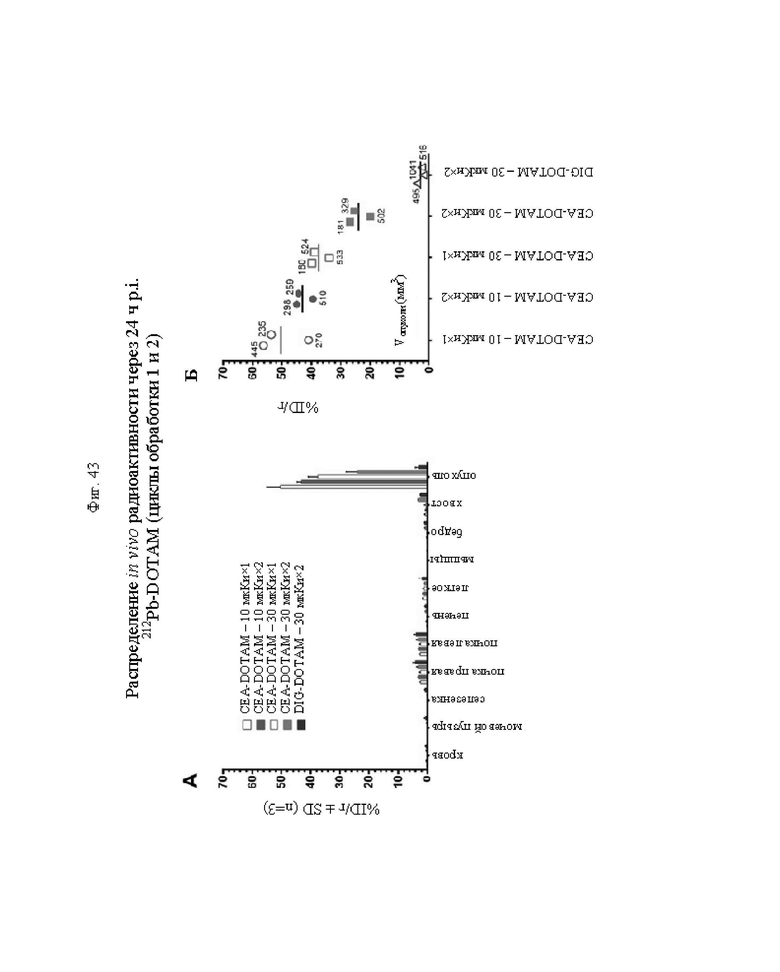
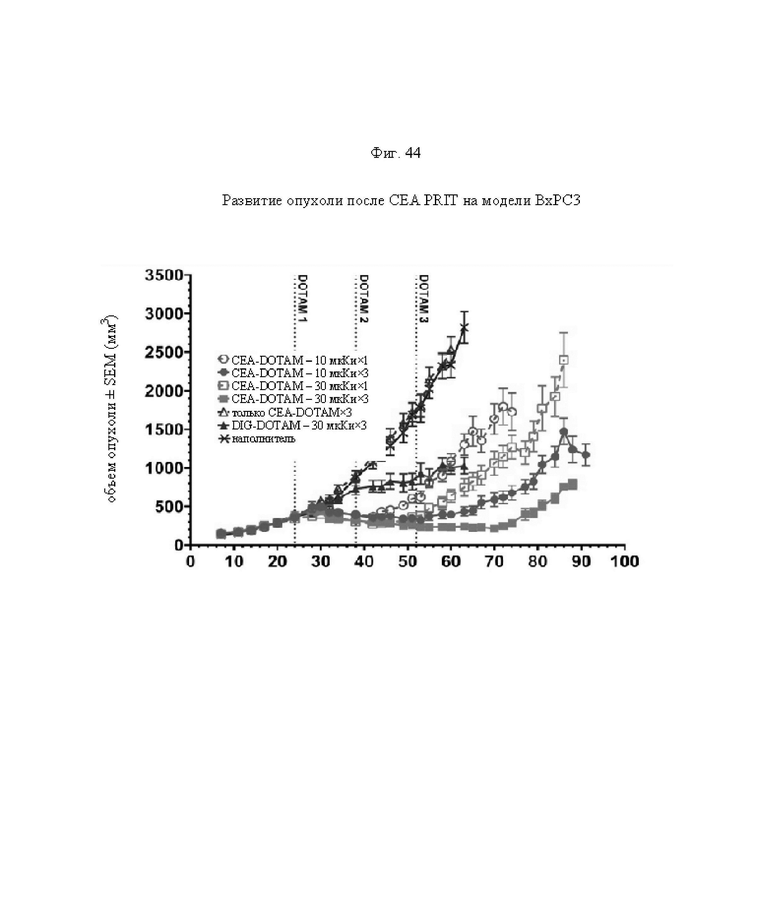

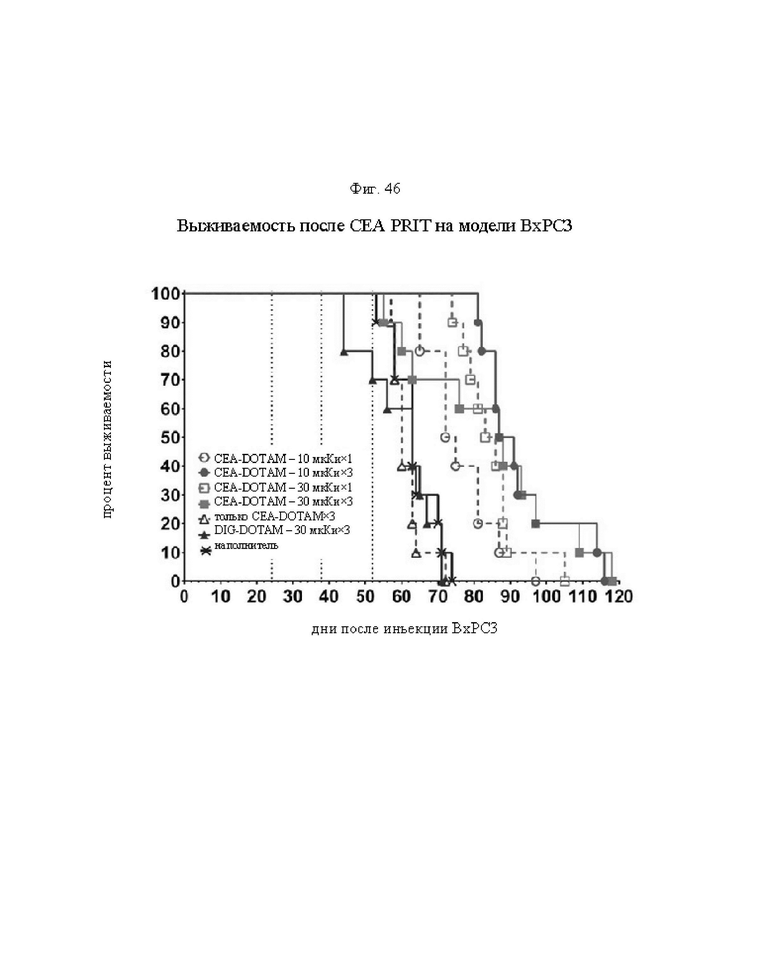
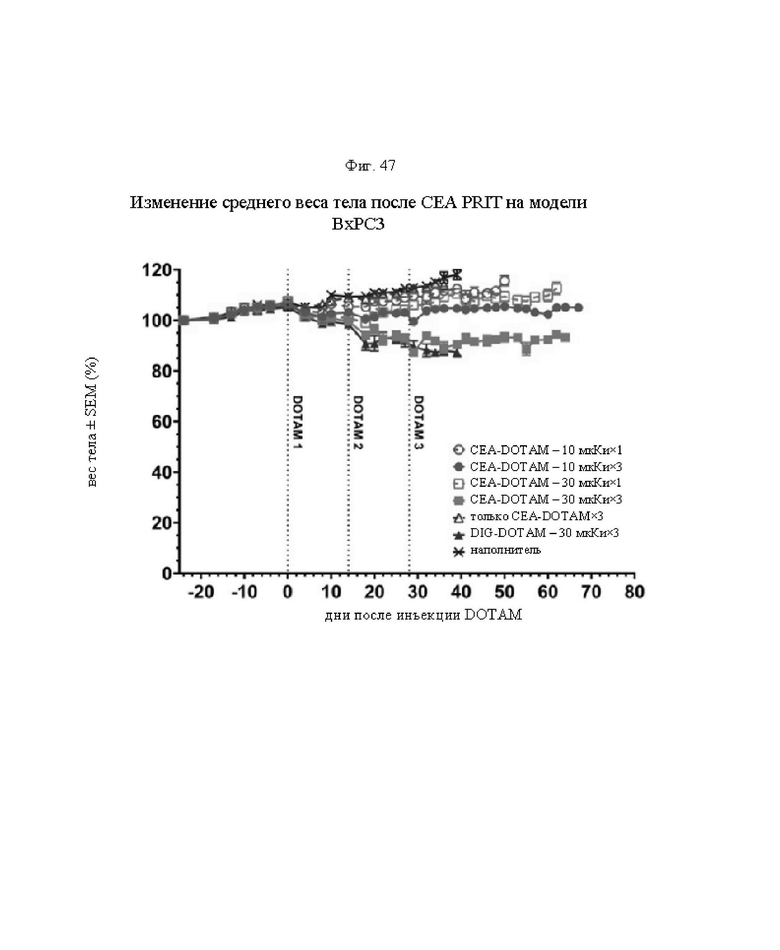
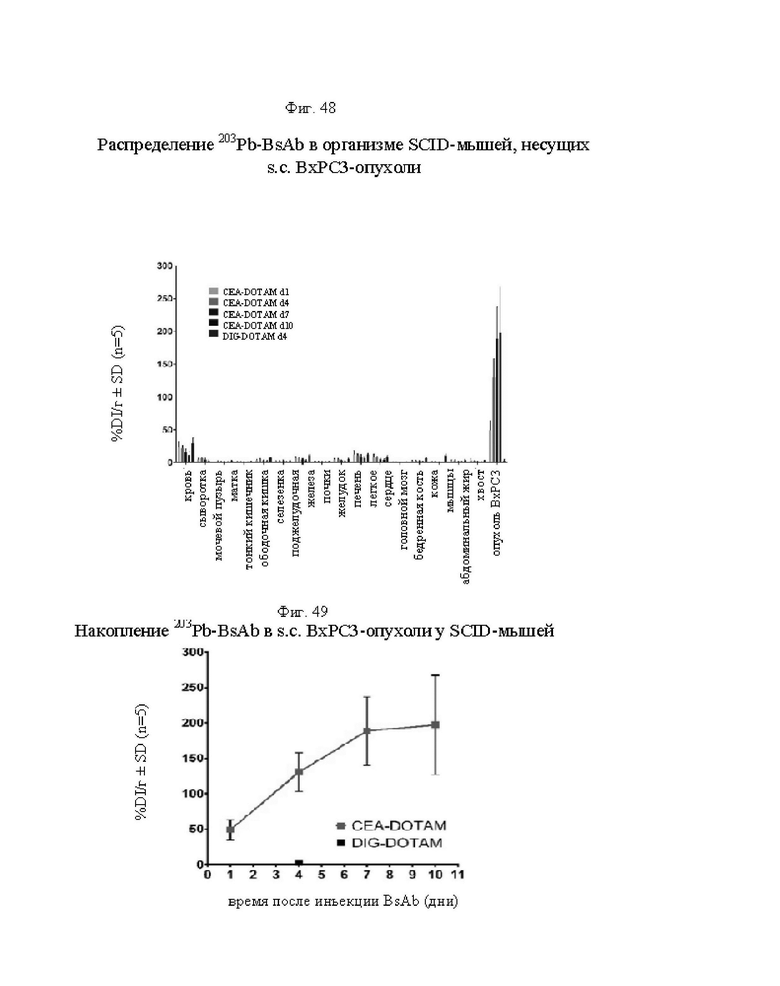
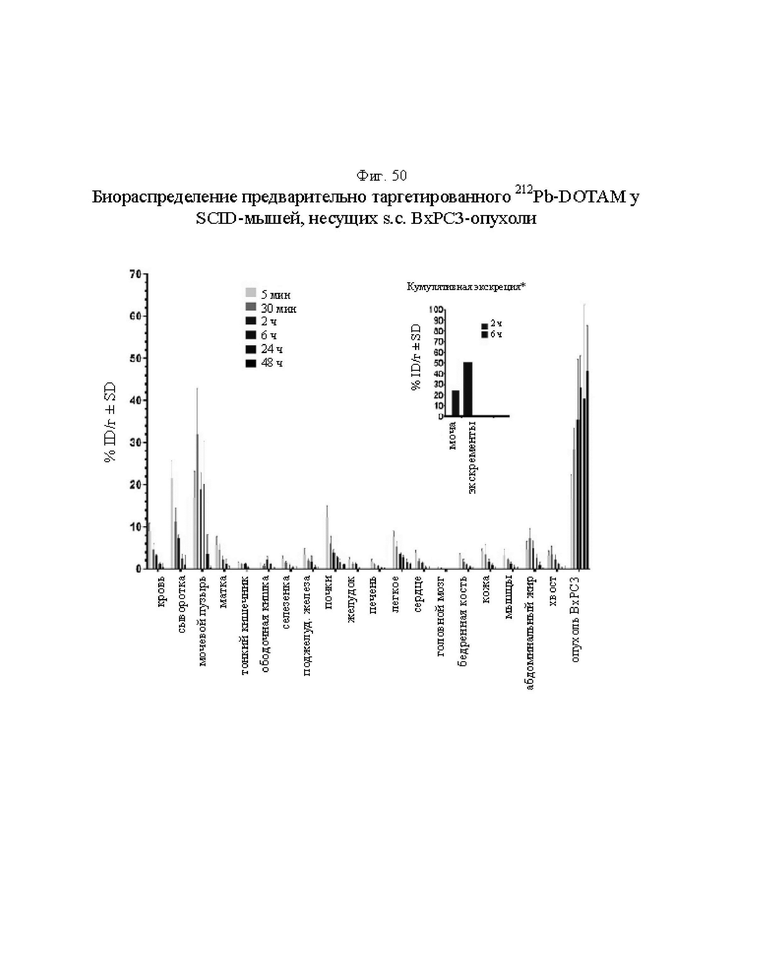
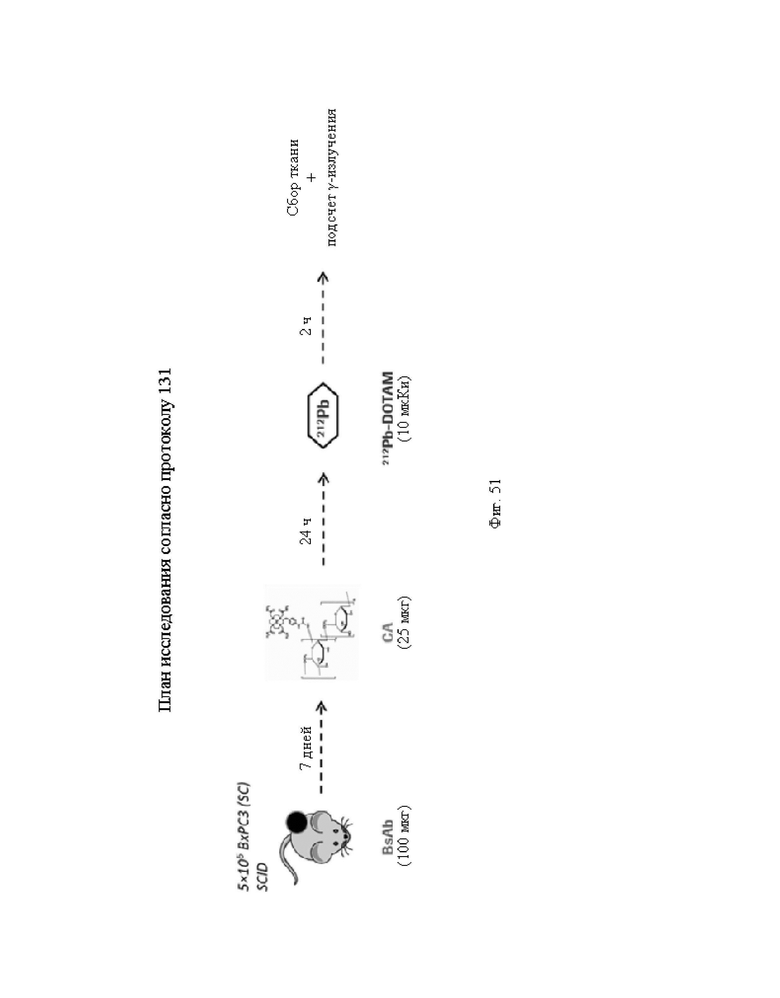
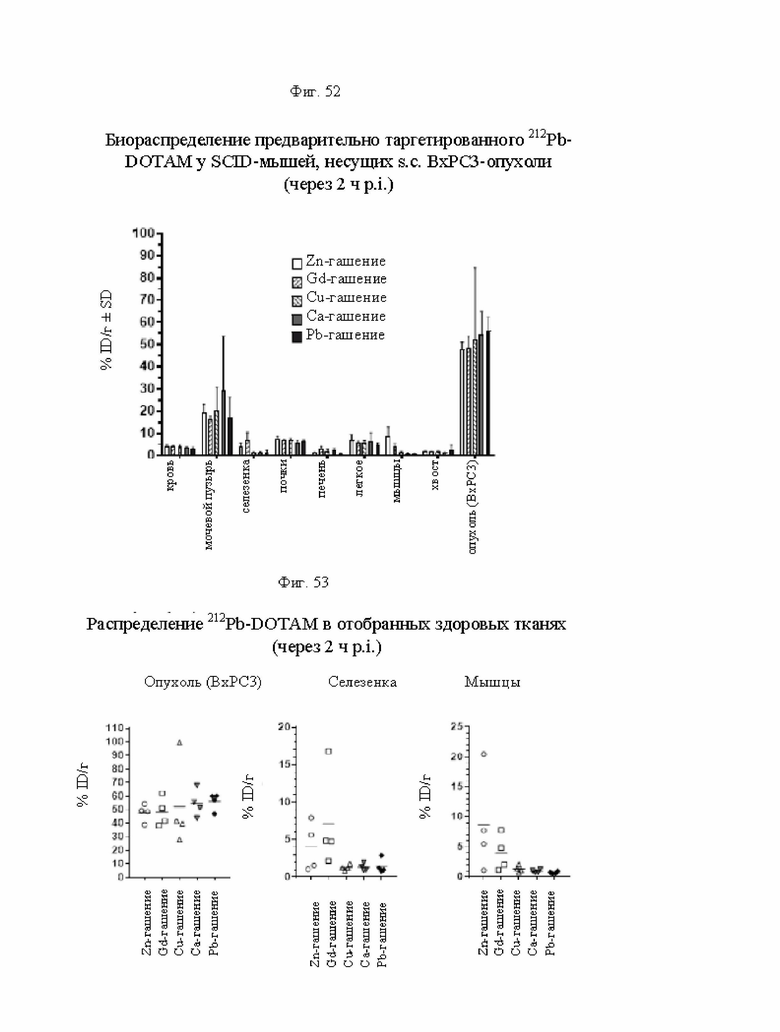
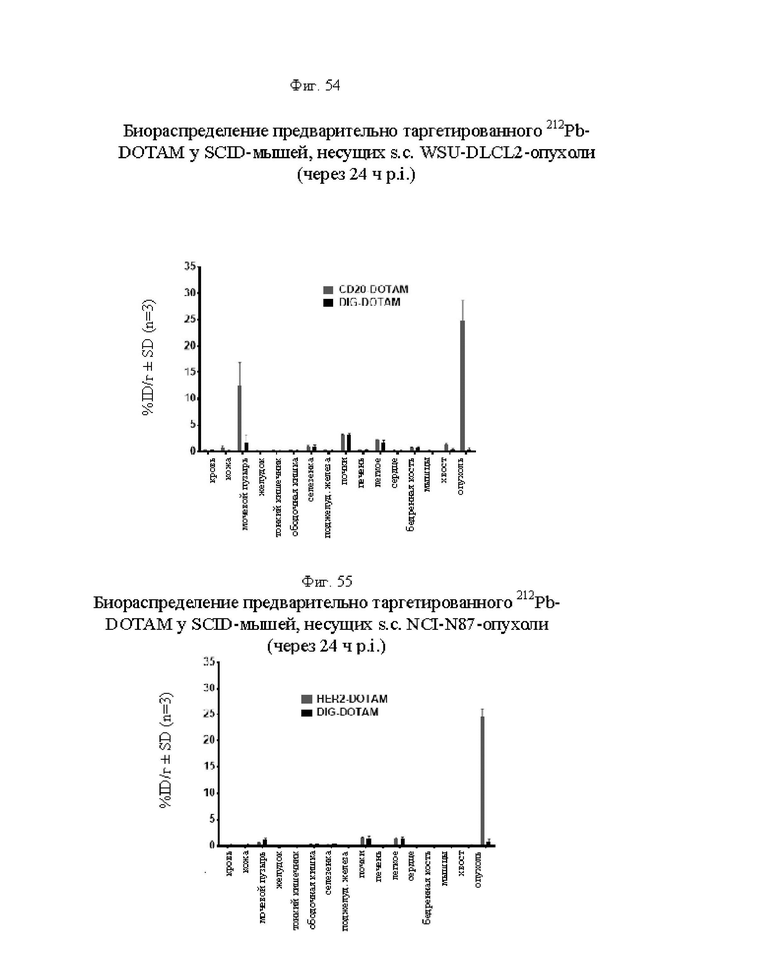
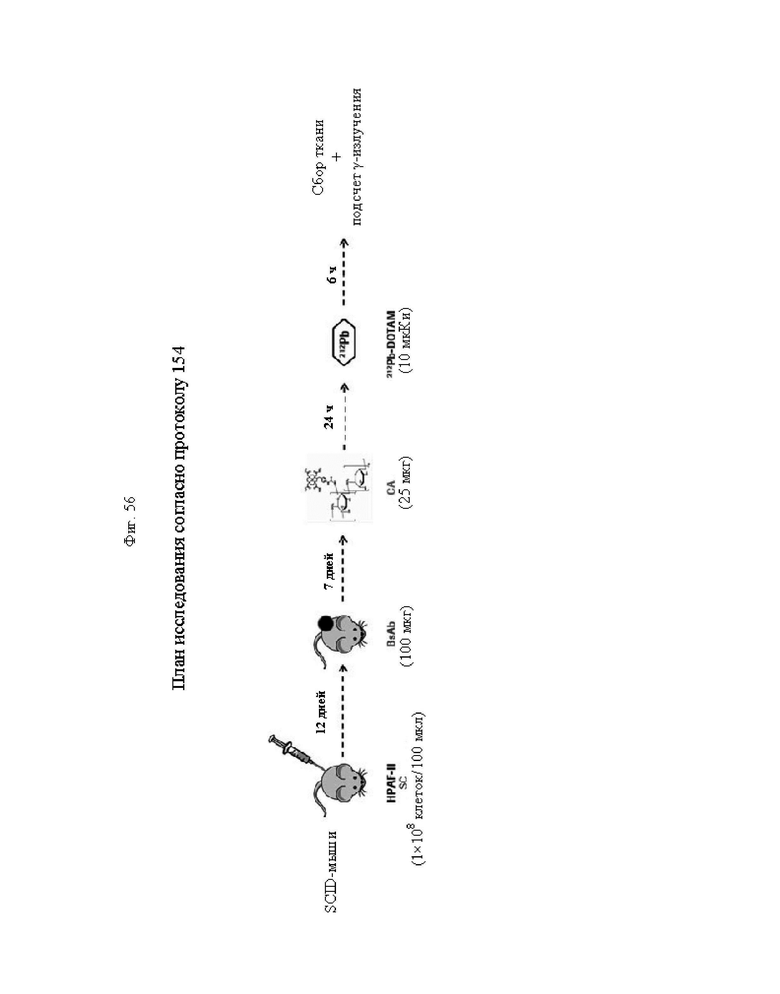
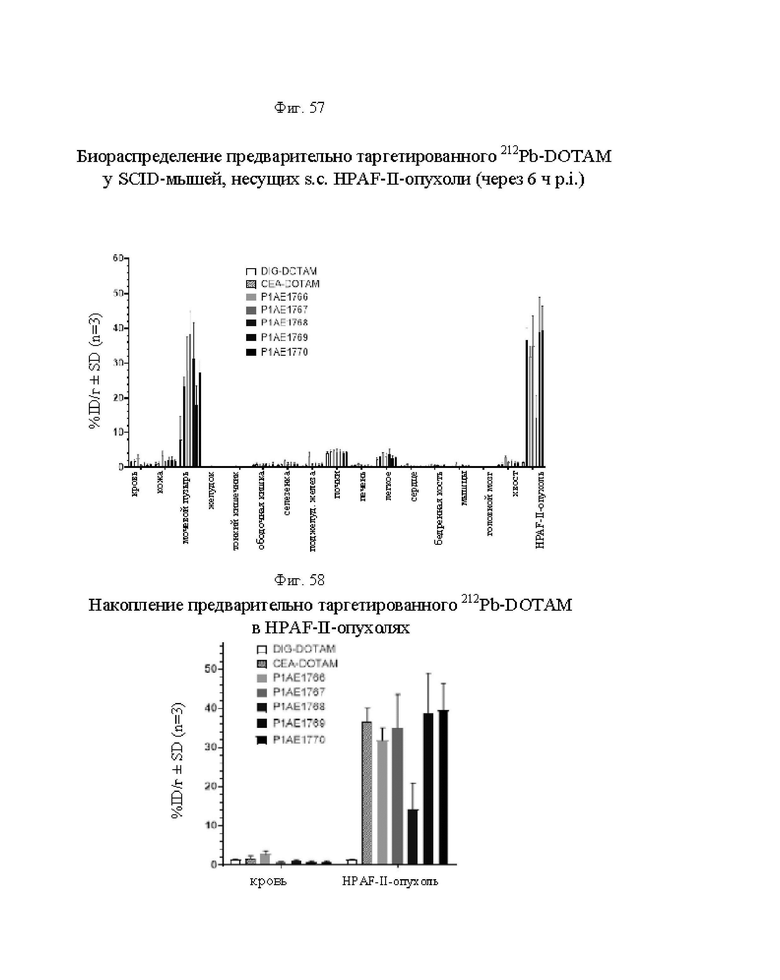
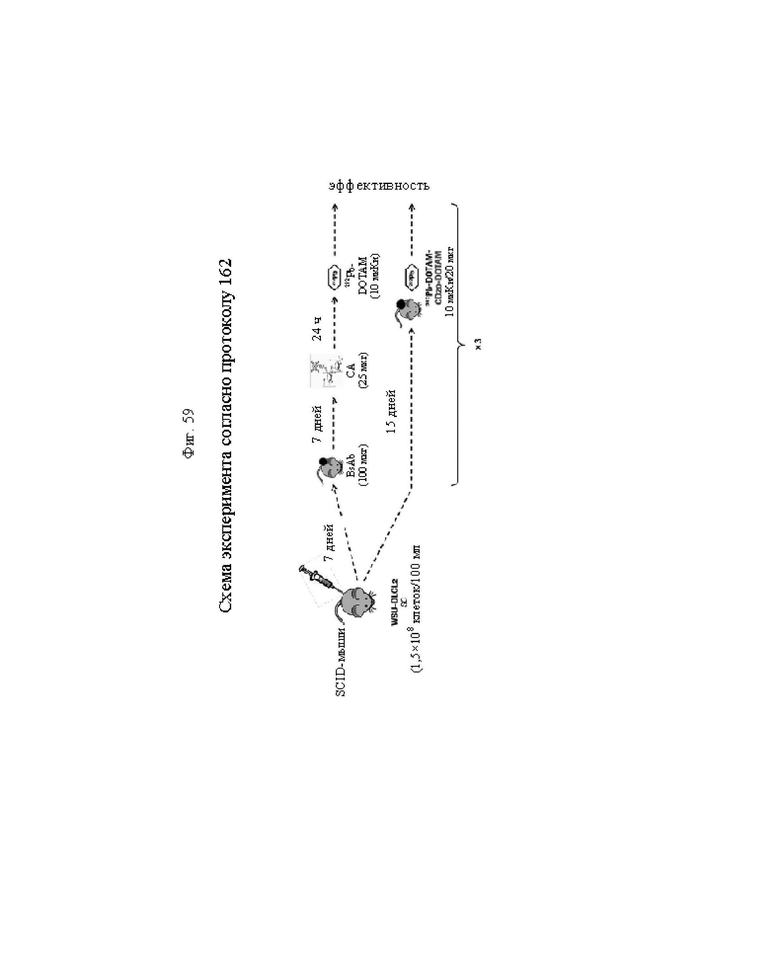
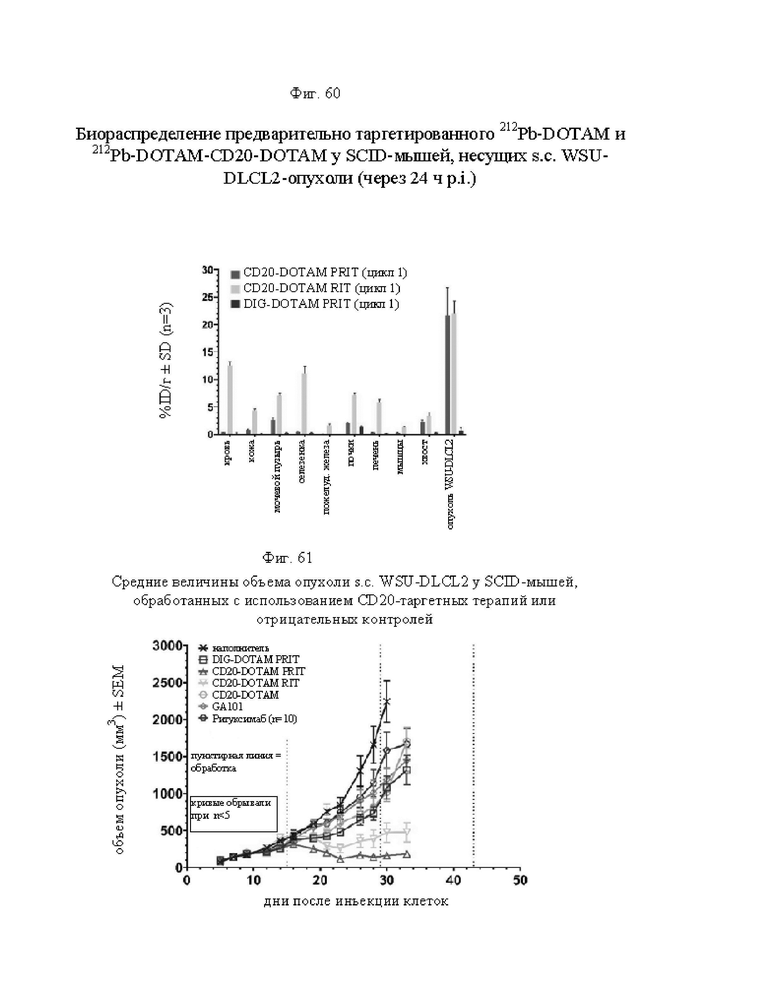
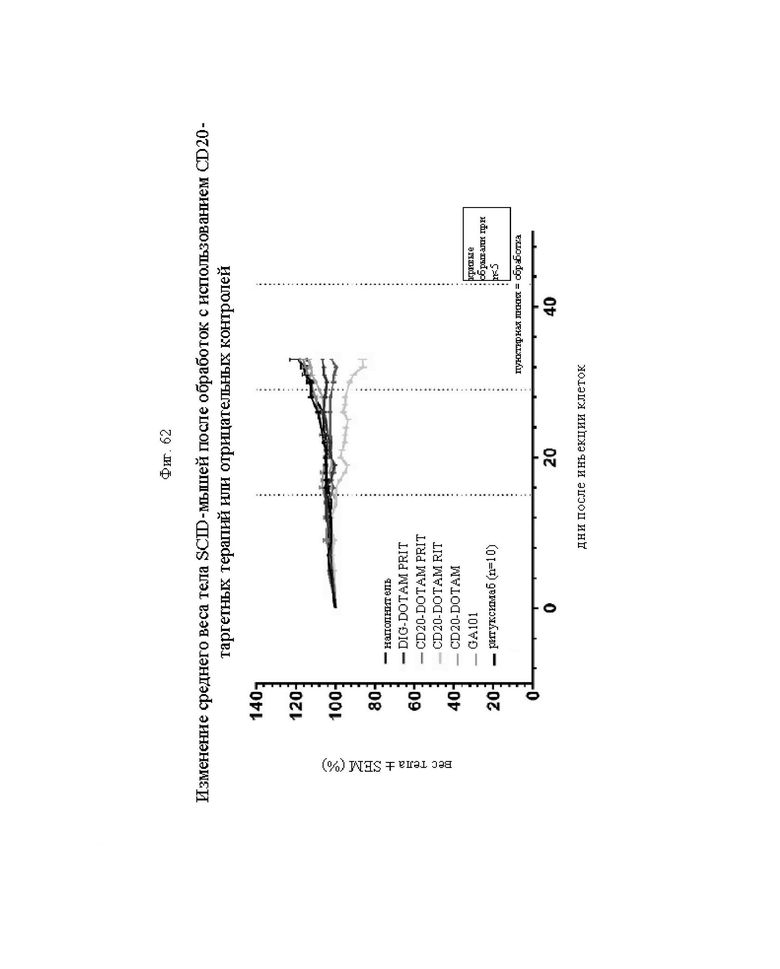
Группа изобретений относится к биотехнологии. Представлены: антитело или антигенсвязывающий фрагмент, специфичные для хелата DOTAM-свинец (Pb), содержащие антигенсвязывающий сайт, биспецифическое антитело, специфичное для хелата DOTAM-свинец (Pb) и опухолеспецифического антигена, выбранного из CEA, HER2 и CD20, и способы их получения. Также раскрыты: выделенные полинуклеотиды, которые кодируют антитела, наборы выделенных полинуклеотидов, экспрессионные вектора, содержащие полинуклеотиды, наборы экспрессионных векторов, клетки-хозяина, содержащие выделенный полинуклеотид или набор выделенных полинуклеотидов, для получения антитела или антигенсвязывающего фрагмента или биспецифического антитела. Изобретение может применяться в таких областях, как радиоиммуновизуализация и радиоиммунотерапия. 14 н. и 20 з.п. ф-лы, 62 ил., 84 табл., 37 пр.
1. Антитело или антигенсвязывающий фрагмент, специфичные для хелата DOTAM-свинец (Pb) и содержащие антигенсвязывающий сайт, который включает:
a) CDR1 тяжелой цепи, содержащий аминокислотную последовательность GFSLSTYSMS (SEQ ID NO: 1);
б) CDR2 тяжелой цепи, содержащий аминокислотную последовательность FIGSRGDTYYASWAKG (SEQ ID NO: 2);
в) CDR3 тяжелой цепи, содержащий аминокислотную последовательность ERDPYGGGAYPPHL (SEQ ID NO: 3);
г) CDR1 легкой цепи, содержащий аминокислотную последовательность QSSHSVYSDNDLA (SEQ ID NO: 4);
д) CDR2 легкой цепи, содержащий аминокислотную последовательность QASKLAS (SEQ ID NO: 5); и
е) CDR3 легкой цепи, содержащий аминокислотную последовательность LGGYDDESDTYG (SEQ ID NO: 6), где нумерация соответствует нумерации по Кэботу, где антитело или антигенсвязывающий фрагмент связывает хелат металла, включающий DOTAM и свинец (Pb).
2. Антитело или антигенсвязывающий фрагмент по п. 1, которое является человеческим, химерным или гуманизированным.
3. Антитело или антигенсвязывающий фрагмент по любому из предыдущих пунктов, в котором
i) антигенсвязывающий сайт содержит вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 7 и SEQ ID NO: 9, или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 7 или SEQ ID NO: 9; и/или
ii) антигенсвязывающий сайт содержит вариабельный домен легкой цепи, который содержит аминокислотную последовательность, выбранную из группы, которая состоит из SEQ ID NO: 8 и SEQ ID NO: 10, или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 8 или 10.
4. Антитело или антигенсвязывающий фрагмент по любому из предыдущих пунктов, в котором
i) антигенсвязывающий сайт содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NО: 7, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 8; или
ii) антигенсвязывающий сайт содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NО: 9, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 10.
5. Антитело или антигенсвязывающий фрагмент по любому из предыдущих пунктов, в котором связывание антигенсвязывающего сайта с хелатом Pb-DOTAM характеризуется величиной Kd, составляющей 100, 50, 20, 10, 5, 1пМ или менее.
6. Антитело или антигенсвязывающий фрагмент по любому из предыдущих пунктов, в котором антигенсвязывающий сайт связывается с хелатом Pb-DOTAM и хелатом Bi-DOTAM, и где соотношение величин Kd для хелата Bi-DOTAM/хелата Pb-DOTAM находится в диапазоне 0,1-10 или 1-10.
7. Антитело или антигенсвязывающий фрагмент по любому из предыдущих пунктов, которое представляет собой цельное антитело или которое представляет собой фрагмент антитела, выбранный из группы, состоящей из молекулы Fv, Fab, Fab', Fab'-SH, F(ab')2, димерного антитела, линейного антитела или одноцепочечного антитела.
8. Биспецифическое антитело, специфичное для хелата DOTAM-свинец (Pb) и опухолеспецифического антигена, выбранного из CEA, HER2 и CD20, включающее антигенсвязывающий сайт антитела или антигенсвязывающий фрагмент по любому из пп. 1-7 и антигенсвязывающий сайт, специфичный для опухолеспецифического антигена, выбранного из CEA, HER2 и CD20.
9. Биспецифическое антитело по п. 8, где антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для CEA, и в котором антигенсвязывающий сайт, специфический для CEA, содержит тяжелую цепь, содержащую по меньшей мере один, два или три CDR тяжелой цепи, где:
а) CDR1 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:11;
б) CDR2 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:12;
в) CDR3 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:13;
и/или антигенсвязывающий сайт, специфический для CEA, содержит легкую цепь, содержащую по меньшей мере один, два или три CDR легкой цепи, где:
г) CDR1 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 14;
д) CDR2 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 15;
е) CDR3 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 16.
10. Биспецифическое антитело по п. 8, где антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для CEA, и в котором антигенсвязывающий сайт, специфический для CEA, содержит
I) вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 17 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 17; и/или
II) вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 18 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 18.
11. Биспецифическое антитело по п. 8, где антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для HER2, и в котором антигенсвязывающий сайт, специфический для HER2, содержит по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из
(a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 28;
(б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 29;
(в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 30;
(г) CDR-L1, который содержит аминокислотную последовательность SEQ ID NO: 31;
(д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 32; и
(е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 33.
12. Биспецифическое антитело по п. 8, где антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для HER2, и в котором антигенсвязывающий сайт, специфический для HER2, содержит
I) вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 34 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 34; и/или
II) вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 35 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 35.
13. Биспецифическое антитело по п. 8, где антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для CD20, и в котором антигенсвязывающий сайт, специфический для CD20, содержит по меньшей мере один, два, три, четыре, пять или шесть CDR, выбранных из
(a) CDR-H1, который содержит аминокислотную последовательность SEQ ID NO: 39;
(б) CDR-H2, который содержит аминокислотную последовательность SEQ ID NO: 40
(в) CDR-H3, который содержит аминокислотную последовательность SEQ ID NO: 41;
(г) CDR-L1 , который содержит аминокислотную последовательность SEQ ID NO: 42;
(д) CDR-L2, который содержит аминокислотную последовательность SEQ ID NO: 43; и
(е) CDR-L3, который содержит аминокислотную последовательность SEQ ID NO: 44.
14. Биспецифическое антитело по п. 8, где антитело содержит по меньшей мере один антигенсвязывающий сайт, специфический для хелата Pb-DOTAM, и по меньшей мере один антигенсвязывающий сайт, специфический для CD20, и в котором антигенсвязывающий сайт, специфический для CD20, содержит
I) вариабельный домен тяжелой цепи, который содержит аминокислотную последовательность SEQ ID NO: 45 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 45; и/или
II) вариабельный домен легкой цепи, который содержит аминокислотную последовательность SEQ ID NO: 46 или ее вариант, содержащий аминокислотную последовательность, которая по меньшей мере на 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% идентична SEQ ID NO: 46.
15. Биспецифическое антитело по любому из пп. 8-14, которое содержит Fc-область.
16. Биспецифическое антитело по п. 15, в котором Fc-область конструируют для снижения эффекторной функции, необязательно путем замены одного или нескольких остатков в положениях 234, 235, 238, 265, 269, 270, 297, 327 и/или 329 Fc-области.
17. Биспецифическое антитело по п. 15 или 16, содержащее
I) полноразмерное антитело, которое содержит антигенсвязывающий сайт для первого антигена, и
II) по меньшей мере вариабельный домен второй тяжелой цепи и вариабельный домен второй легкой цепи, которые вместе образуют антигенсвязывающий сайт для второго антигена, где либо первый, либо второй антиген представляет собой хелат Pb-DOTAM, а другой представляет собой антиген-мишень, где необязательно первый антиген является антиген-мишенью, а второй антиген представляет собой хелат Pb-DOTAM.
18. Биспецифическое антитело по п. 17, содержащее полноразмерное антитело, которое содержит антигенсвязывающий сайт для первого антигена, в котором N-или C-конец одной из тяжелых цепей сцеплен через полипептидный линкер с первым полипептидом, и в котором первый полипептид объединен со вторым полипептидом с образованием Fab или cross-Fab, который содержит связывающий сайт для второго антигена.
19. Биспецифическое антитело по п. 18, которое содержит:
I) первый полипептид, состоящий из VH-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VL- и CL-доменов; или
II) первый полипептид, состоящий из VL-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VH- и CL-доменов; или
III) первый полипептид, состоящий из VH-домена и CL-домена, который объединен со вторым полипептидом, состоящим из VL- и CH1-доменов; в результате чего первый и второй полипептид вместе образуют антигенсвязывающий сайт для второго антигена.
20. Биспецифическое антитело по п. 19, содержащее полноразмерное антитело, которое содержит антигенсвязывающий сайт для первого антигена, в котором C - конец одной из тяжелых цепей сцеплен через полипептидный линкер с первым полипептидом, состоящим из VL-домена и CH1-домена, который объединен со вторым полипептидом, состоящим из VH- и CL-доменов.
21. Выделенный полинуклеотид, который кодирует антитело или антигенсвязывающий фрагмент по любому из пп. 1-7.
22. Набор выделенных полинуклеотидов, кодирующий антитело или антигенсвязывающего фрагмент по любому из пп. 1-7, для получения антитела или антигенсвязывающего фрагмента по любому из пп. 1-7.
23. Экспрессионный вектор, содержащий полинуклеотид по п. 21 или набор выделенных полинуклеотидов по п. 22.
24. Набор экспрессионных векторов, содержащий полинуклеотид по п. 21, или набор полинуклеотидов по п. 22 для получения антитела или антигенсвязывающего фрагмента по любому из пп. 1-7.
25. Клетка-хозяин, содержащая выделенный полинуклеотид по п. 21 или набор выделенных полинуклеотидов по п. 22, необязательно содержащая экспрессионный вектор по п. 23 или набор экспрессионных векторов по п. 24, для получения антитела или антигенсвязывающего фрагмента по любому из пп. 1-7.
26. Клетка-хозяин по п. 25 представляет собой прокариотическую или эукариотическую клетку-хозяина.
27. Способ получения антитела или антигенсвязывающего фрагмента по любому из пп. 1-7, включающий экспрессию антитела или антигенсвязывающего фрагмента в клетке-хозяине по п. 25.
28. Выделенный полинуклеотид, кодирующий биспецифическое антитело по любому из пп. 8-20.
29. Набор выделенных полинуклеотидов, кодирующий биспецифическое антитело по любому из пп. 8-20 для получения биспецифического антитела по любому из пп. 8-20.
30. Экспрессионный вектор, содержащий полинуклеотид по п. 28 или набор полинуклеотидов по п. 29.
31. Набор экспрессионных векторов, содержащий полинуклеотид по п. 28 или набор полинуклеотидов по п. 29, для получения биспецифического антитела по любому из пп. 8-20.
32. Клетка-хозяин, содержащая выделенный полинуклеотид по п. 28 или набор выделенных полинуклеотидов по п. 29, необязательно содержащая вектор экспрессии по п. 30 или набор векторов по п. 31 для получения биспецифического антитела по любому из пп. 8-20.
33. Клетка-хозяин по п. 32, которая является прокариотической или эукариотической клеткой-хозяином.
34. Способ получения биспецифического антитела по любому из пп. 8-20, включающий экспрессию биспецифического антитела из клетки-хозяина по п. 32.
| WO 2016130539 A2, 18.08.2016 | |||
| YAZAKI PAUL J | |||
| et al | |||
| "A series of anti-CEA/anti-DOTA bispecific antibody formats evaluated for pre-targeting: comparison of tumor uptake and blood clearance", PROTEIN ENGINEERING, DESIGN & SELECTION PEDS, vol | |||
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Индукционная катушка | 1920 |
|
SU187A1 |
| CHEAL SARAH et al | |||
| Кулисный парораспределительный механизм | 1920 |
|
SU177A1 |

