Область изобретения
Настоящее изобретение относится к варианту аспартокиназы, микроорганизму, содержащему вариант, и способу получения L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты с использованием микроорганизма.
Предшествующий уровень техники
Микроорганизм рода Corynebacterium, в частности Corynebacterium glutamicum, представляет собой грамположительный микроорганизм, который широко используется в получении L-аминокислот и других полезных веществ. Были проведены различные исследования для разработки микроорганизмов с высокоэффективной продуктивностью и технологий процессов ферментации для получения L-аминокислот и других полезных веществ. Например, в основном были использованы способы более точного приближения к веществам-мишеням путем повышения экспрессии генов, кодирующих ферменты, вовлеченные в биосинтез L-лизина, или путем нокаута генов, не являющихся необходимыми для биосинтеза (патент Кореи №10-0838038).
Между тем, среди L-аминокислот, L-лизин, L-треонин, L-метионин, L-изолейцин и L-глицин представляют собой аминокислоты, имеющие происхождение из аспартата (Asp), и эти аминокислоты обычно используют аспартилфосфат (Арр), полученный из Asp посредством аспартокиназы (LysC, Е.С. 2.7.2.4) (Фиг. 1). Поэтому для получения аминокислот посредством способа микробной ферментации, необходимо поддерживать активность ферментов, используемых в биосинтетическом пути, на конкретном уровне или выше и, таким образом, для этого было выполнено интенсивное исследование.
В частности известно, что активность LysC, которая действует в качестве первого фермента в пути биосинтеза аминокислот, имеющих происхождение из аспартата, регулируется ингибированием L-лизина и L-треонина по типу обратной связи (J Mol Biol. 2007 Apr 27; 368(2): 521-36. Epub 2007 Feb 20). В связи с этим, несмотря на то, что были поданы заявки, имеющие отношение к ингибированию по типу обратной связи (патент США № US 8062869 В и патент Японии № JP 3473042 В), по-прежнему требуется продолжение исследований для повышения продуктивности продуктов, имеющих происхождение из аспартата.
При таких обстоятельствах, авторы изобретения завершили настоящее изобретение путем подтверждения того, что продуктивность L-аминокислот, имеющих происхождение из аспартата, или производных этих аминокислот улучшается при применении нового варианта аспартокиназы.
Техническая задача
Задача настоящего изобретения заключается в том, чтобы предложить вариант аспартокиназы, содержащий одну или более чем одну аминокислотную замену в аминокислотной последовательности SEQ ID NO: 1, где аминокислотная замена включает замену аминокислоты в положении 377 в аминокислотной последовательности SEQ ID NO: 1 L-лизином или L-метионином.
Задача настоящего изобретения также заключается в том, чтобы предложить полинуклеотид, кодирующий вариант.
Задача настоящего изобретения также заключается в том, чтобы предложить микроорганизм рода Corynebacterium, продуцирующий L-аминокислоту, имеющую происхождение из аспартата, или производное этой аминокислоты, которое содержит вариант аспартокиназы или имеет усиленную его активность.
Задача настоящего изобретения также заключается в том, чтобы предложить способ получения L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты, включающий: культивирование микроорганизма в среде и выделение L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты из культивированного микроорганизма или культуральной среды.
Задача настоящего изобретения также заключается в том, чтобы предложить способ получения метионина, включающий: культивирование микроорганизма в среде; получение ацетилгомосерина или сукцинилгомосерина из культивированного микроорганизма или культуральной среды; и превращение ацетилгомосерина или сукцинилгомосерина в метионин.
Техническое решение
Далее настоящее изобретение будет описано подробно.
Между тем, каждое из пояснений и типичных воплощений, раскрытых здесь, может быть применено к другим пояснениям и типичным воплощениям. То есть, все комбинации различных факторов, раскрытых здесь, принадлежат объему настоящего изобретения. Кроме того, объем настоящего изобретения не следует ограничивать конкретным описанием изобретения, предложенным ниже.
Дополнительно, специалист в данной области техники сможет распознать или подтвердить на основе обычного экспериментирования многие эквиваленты для конкретных воплощений настоящего изобретения, описанных в данной заявке, и такие эквиваленты предназначены для включения в настоящее изобретение.
Для выполнения задач, приведенных выше, в аспекте настоящего изобретения предложен вариант аспартокиназы, содержащий одну или более чем одну аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1, где аминокислотная замена включает замену аминокислоты в положении 377 другой аминокислотой. Более конкретно, задача настоящего изобретения заключается в том, чтобы предложить вариант аспартокиназы, где аминокислота в положении 377 в аминокислотной последовательности SEQ ID NO: 1 замещается L-лизином или L-метионином.
Как его используют здесь, термин «аспартокиназа» относится к ферменту, который катализирует фосфорилирование аминокислоты, аспартата и действует на первой стадии биосинтеза трех незаменимых аминокислот, L-метионина, L-лизина и L-треонина, которые известны как семейство аспартата.
В настоящем изобретении термин "аспартокиназа" можно использовать взаимозаменяемо с «LysC» или «белком LysC». Белок LysC может быть получен из GenBank Национального Центра Биотехнологической Информации (NCBI), известной базы данных. Белок LysC может представлять собой LysC, полученный из рода Corynebacterium, и, более конкретно, может представлять собой полипептид, имеющий аминокислотную последовательность SEQ ID NO: 1, который имеет происхождение из Corynebacterium glutamicum, но не ограничиваясь этим. Кроме того, LysC в настоящем изобретении может представлять собой полипептид, состоящий из аминокислотной последовательности SEQ ID NO: 1 или аминокислотной последовательности, имеющей гомологию или идентичность с ней 80%, 85%, 90%, 95% или 97%, или выше. Дополнительно, очевидно, что полипептид, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой части последовательности, также включается в объем настоящего изобретения до тех пор, пока аминокислотная последовательность имеет такую же гомологию или идентичность и проявляет эффект, соответствующий таковому у этого полипептида.
В настоящем изобретении в способе получения аспартокиназы (LysC) можно использовать различные способы, хорошо известные в данной области техники. Примеры таких способов включают техники синтеза генов, включая оптимизацию кодонов так, чтобы получать ферменты с высокой эффективностью у микроорганизма рода Corynebacterium, который обычно используют для экспрессии ферментов, и способы скрининга источников полезных ферментов с использованием биоинформационных способов на основе метагенома микроорганизмов, но способы не ограничиваются этим.
Как его используют здесь, термин «вариант» представляет собой полипептид, который по меньшей мере одной аминокислотой в консервативных заменах и/или модификациях отличается от приведенного полипептида таким образом, что функции или свойства полипептида сохраняются. Вариант полипептидов отличается от идентифицированной последовательности заменой, делецией или вставкой нескольких аминокислот. Обычно такие варианты можно идентифицировать посредством модификации одной из приведенных выше полипептидных последовательностей и оценки свойств модифицированного полипептида. Другими словами, способность варианта можно повышать, оставлять прежней или ослаблять по сравнению с нативным белком. Такие варианты обычно можно идентифицировать посредством модификации одной из приведенных выше полипептидных последовательностей и оценки реакционной способности модифицированного полипептида. Кроме того, часть вариантов может включать такие, у которых была удалена одна или более частей, таких как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, у которых была удалена небольшая часть от N-и/или С-конца зрелого белка.
Как его используют здесь, «консервативная замена» означает, что аминокислота замещена другой аминокислотой, которая имеет подобную структуру и/или химические свойства. Например, варианты могут иметь по меньшей мере одну консервативную замену при сохранении еще одной биологической активности. Такие аминокислотные замены обычно могут быть сделаны на основе подобия полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; отрицательно заряженные (кислые) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; ароматические аминокислоты включают фенилаланин, триптофан и трирозин; и гидрофобные аминокислоты включают аланин, валин, изолейцин, лейцин, метионин, фенилаланин, тирозин и триптофан. Обычно консервативные замены практически не влияют на активность полученного полипептида.
Варианты также могут содержать другие модификации, включая делецию или вставку аминокислот, которые имеют минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид можно конъюгировать с сигнальной (или лидерной) последовательностью в N-терминальном конце белка, которая посттрансляционно или посттрансляционно направляет перенос белка. Полипептид также можно конъюгировать с линкерной или другой последовательностью для облегчения синтеза, очистки или идентификации полипептида.
Например, термин «вариант аспартокиназы», заявленный здесь, более конкретно может относиться к модифицированному полипептиду LysC, имеющему аминокислотную последовательность SEQ ID NO: 1, который получен из Corynebacterium sp., и может содержать вариант, содержащий одну или более аминокислотных замен в аминокислотной последовательности SEQ ID NO: 1. Более конкретно вариант может включать замену аминокислотного остатка в положении 377 другой аминокислотой в аминокислотной последовательности SEQ ID NO: 1. «Другую аминокислоту» не ограничивают до тех пор, пока она представляет собой аминокислоту, но не L-лейцин, которая представлет собой аминокислоту в положении 377. Более конкретно, например, аминокислотный остаток в положении 377 может быть замещен лизином или метионином. L-Лизин представляет собой пример основной аминокислоты, и основная аминокислота может представлять собой одну из L-лизина, L-аргинина и L-гистидина. L-Метионин представляет собой пример неполярной аминокислоты, и неполярная аминокислота может представлять собой одну из L-метионина, L-фенилаланина, L-аланина, L-цистеина, L-глицина, L-изолейцина, L-лейцина, L-пролина, L-триптофана и L-валина. Однако не ограничиваются этим.
Как его используют здесь, термин «вариант аспартокиназы» можно использовать взаимозаменяемо с «модифицированной аспартокиназой» или «модифицированной LysC».
Такой вариант аспартокиназы характеризуется наличием повышенной активности аспартокиназы по сравнению с полипептидом, имеющим активность аспартокиназы SEQ ID NO: 1.
Кроме того, даже если он описан здесь как «белок, имеющий аминокислотную последовательность конкретного номера последовательности», очевидно, что полипептид, имеющий аминокислотную последовательность с делецией, модификацией, заменой или вставкой части последовательности, также включен в объем настоящего изобретения до тех пор, пока белок имеет эффект, идентичный или эквивалентный эффекту пептида, состоящего из аминокислотной последовательности соотвествующего номера последовательности. Более конкретно, такой белок не исключает вставку последовательностей, которые не изменяют функцию, в белок в начало или конец аминокислотной последовательности соответствующего номера последовательности, природную мутацию, консервативную замену или ее синонимическую мутацию. Кроме того, очевидно, что белок, имеющий вставку такой последовательности или мутацию, также включается в объем настоящего изобретения.
Как его используют здесь, термин «консервативная замена» относится к замене конкретного аминокислотного остатка на аминокислотный остаток, имеющий подобную боковую цепь. Семейства аминокислотных остатков, имеющих подобные боковые цепи, хорошо известны в данном уровне техники. Примеры таких семейств могут включать аминокислоты, имеющие основную боковую цепь (например лизин, аргинин, гистидин), аминокислоты, имеющие кислую боковую цепь (например аспарагиновая кислота, глутаминовая кислота), аминокислоты, имеющие незаряженную полярную боковую цепь (например аспарагин, глутамин, серин, треонин, тирозин, цистеин), аминокислоты, имеющие неполярную боковую цепь (например глицин, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), аминокислоты, имеющие разветвленную боковую цепь в положении β (например треонин, валин, изолейцин), аминокислоты, имеющие ароматическую боковую цепь (например тирозин, фенилаланин, триптофан, гистидин), аминокислоты, имеющие боковую цепь (например серин, треонин, тирозин), содержащую гидроксильную группу (например спиртовую, фенольную), и аминокислоты, имеющие серосодержащую боковую цепь (например цистеин, метионин). Предпочтительно, консервативная замена аминокислот может представлять собой замену между аспарагиновой кислотой и глутаминовой кислотой, замену между аргинином, лизином и гистидином, замену между триптофаном и фенилаланином, замену между фенилаланином и валином, замену между лейцином, изолейцином и аланином и замену между глицином и аланином.
Как его используют здесь, термин «гомология» или «идентичность» относится к степени релевантности между двумя данными аминокислотными последовательностями или нуклеотидными последовательностями и может быть выражен в процентах.
Термины «гомология» и «идентичность» часто используют взаимозаменяемо друг с другом.
Гомологию последовательностей или идентичность консервативных полинуклеотидных или полипептидных последовательностей можно определять посредством стандартных алгоритмов выравнивания и можно использовать со штрафом за разрыв в последовательности ("gap penalty"), установленным используемой программой. По существу, обычно ожидается, что гомологичные или идентичные полинуклеотиды или полипептиды гибридизуются по меньшей мере примерно на 50%, примерно на 60%, примерно на 70%, примерно на 80% или примерно на 90% всей длины полинуклеотидов-мишеней или полипептидов в умеренных или строгих условиях. Полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов, также рассматриваются в отношении гибридизации.
Имеют ли две любые полинуклеотидные или полипептидные последовательности гомологию, сходство или идентичность друг с другом, можно определить с использованием известного компьютерного алгоритма, такого как программа "FASTA" (например Pearson et al., (1988) Proc. Natl. Acad. Sci. USA 85: 2444), используя параметры штрафов. Альтернативно, это может быть определено с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), который выполняют в программе Нидлмана пакета программ EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (предпочтительно, версия 5.0.0 или версии после этого) (GCG program package (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL., J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, and [CARILLO ETA/.](1988) SIAM J Applied Math 48: 1073). Например, гомология или идентичность могут быть определены с использованием BLAST или ClustalW Национального Центра Биотехнологической Информации (NCBI).
Гомология, сходство или идентичность полинуклеотидных или полипептидных последовательностей может быть определена путем сравнения информации о последовательностях, используя, например, компьютерную программу GAP (например Smith and Waterman, Adv. Appl. Math (1981) 2:482), которая была опубликована. Таким образом, программа GAP определяет гомологию или идентичность в виде значения, полученного путем деления количества сходным образом выровненных символов (то есть нуклеотидов или аминокислот) на общее количество символов в более короткой из двух последовательностей. Параметры штрафов для программы GAP могут включать (1) унарную матрицу сравнения (содержащую значение 1 при идентичности и 0 при отсутствии идентичности) и взвешенную матрицу сравнения Gribskov et at. (1986), Nucl. Acids Res. 14:6745, как раскрыто в Schwartz и Dayhoff, eds., Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, pp. 353-358, 1979; (2) штраф 3,0 за каждый разрыв и дополнительньй штраф 0,10 за каждый символ в каждом разрыве (или штраф за открытие разрыва 10 и штраф за продление разрыва 0,5); и (3) отсутствие штрафа для концевых разрывов. Таким образом, как его используют здесь, термин "гомология" или "идентичность" относится к релевантности между последовательностями.
В другом аспекте настоящего изобретения предложен полинуклеотид, кодирующий вариант аспартокиназы.
Вариант аспартокиназы такой, как описано выше.
Как его используют здесь, термин "полинуклеотид" относится к нуклеотидному полимеру, состоящему из мономеров-нуклеотидов, ковалентно связанных в цепь, и примеры их представляют собой нити ДНК или РНК, имеющие предопределенную или большую длину. Более конкретно, полинуклеотид относится к фрагменту полинуклеотида, кодирующему модифицированный полипептид.
Полинуклеотид, кодирующий вариант аспартокиназы, может представлять собой полинуклеотид, кодирующий полипептид, содержащий одну или более чем одну аминокислотную замену в аминокислотной последовательности SEQ ID NO: 1, в котором аминокислотная замена включает замену аминокислотного остатка в положении 377 другой аминокислотой. Более конкретно, в качестве репрезентативного примера, полинуклеотид, кодирующий вариант аспартокиназы, может представлять собой полинуклеотид, кодирующий полипептид, в котором аминокислотный остаток в положении 377 замещен другой аминокислотой, L-лизином или L-метионином, в аминокислотной последовательности SEQ ID NO: 1. Еще более конкретно, полинуклеотид может представлять собой полинуклеотид, кодирующий белок LysC, имеющий аминокислотную последовательность, состоящую из SEQ ID NO: 3 или SEQ ID NO: 5. Например, полинуклеотид может представлять собой полинуклеотид, состоящий из нуклеотидной последовательности SEQ ID NO: 4 или SEQ ID NO: 6. В кодирующем участке могут быть сделаны различные модификации, при условии, что они не изменяют аминокислотную последовательность белка, экспрессирующегося из кодирующего участка, из-за вырожденности кодона или принимая во внимание кодоны, предоставленные организмом, в котором белок должен экспрессироваться.
Следовательно, очевидно, что из-за вырожденности кодона полипептид, состоящий из аминокислотной последовательности SEQ ID NO: 3 или SEQ ID NO: 5, или полинуклеотид, способный транслироваться в полинуклеотиде, имеющем с ним гомологию или идентичность, также может быть включен в объем настоящего изобретения. С другой стороны, зонд, который может быть получен из известной последовательности гена, например, последовательности, кодирующей белок, имеющий активность фермента, состоящего из аминокислотной последовательности SEQ ID NO: 1, путем гибридизации в жестких условиях с комплементарной последовательностью всей или части полинуклеотидной последовательности, может быть включен без ограничения.
"Жесткие условия" относятся к условиям, которые делают возможной конкретную гибридизацию между полинуклеотидами. Такие условия более конкретно раскрыты в литературе (например J. Sambrook et al.). Например, жесткие условия могут включать условия, при которых гены, имеющие высокую гомологию или идентичность (например 80% или более, 85% или более, конкретно 90% или более, еще более конкретно 95% или более, еще более конкретно 97% или более, и еще более конкретно 99% или более) могут гибридизироваться между собой, тогда как гены, имеющие более низкую гомологию или идентичность, не могут гибридизоваться друг с другом; или условия для обычной саузерн гибридизации (то есть условия для промывания один и, более точно, два или три раза при концентрации солей и температуре, соответствующих 60°С, 1×SSC (раствор цитрата натрия и хлорида натрия) и 0,1% SDS (додецилсульфат натрия); более конкретно при 60°С, 0,1×SSC и 0,1% SDS; и еще более конкретно при 68°С, 0,1×SSC и 0,1% SDS).
Гибридизация требует, чтобы две нуклеиновых кислоты имели комплементарные последовательности, хотя возможные ошибки спаривания между основаниями, зависящие от строгости гибридизации. Термин "комплементарный" используют для описания отношений между нуклеотидными основаниями, которые способны гибридизоваться друг с другом. Например, что касается ДНК, аденозин является комплементарным тимину и цитозин является комплементарным гуанину. Следовательно, настоящее изобретение также может включать по существу подобные последовательности нуклеиновых кислот, а также выделенные фрагменты нуклеиновых кислот, комплементарные всей последовательности.
Более конкретно, полинуклеотид, имеющий гомологию или идентичность, может быть обнаружен с использованием условий гибридизации, включающих стадию гибридизации при значении Тпл 55°С и использование описанных выше условий. Кроме того, значение Тпл может представлять собой 60°С, 63°С или 65°С, но не ограничиваться этим. Специалист в данной области техники может надлежащим образом регулировать значение Тпл в зависимости от свой цели.
Целесообразная жесткость гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов и изменяемые величины хорошо известны в данной области техники (Sambrook et al., выше, 9.50-9.51, 11.7-11.8).
В еще одном аспекте настоящего изобретения предложена клетка-хозяин, содержащая вариант аспартокиназы и микроорганизм, трансформированный с помощью вектора, содержащего полинуклеотид, кодирующий вариант аспартокиназы. Более конкретно, в настоящем изобретении предложен микроорганизм рода Corynebacterium, продуцирующий L-аминокислоту имеющую происхождение из аспартата, или производное этой аминокислоты, которое содержит вариант аспартокиназы.
Микроорганизм, содержащий вариант аспартокиназы по настоящему изобретению, характеризуется тем, что активность аспартокиназы повышена по сравнению с микроорганизмом дикого типа или немодифицированным микроорганизмом.
Как его используют здесь, термин «повышение активности» означает, что активность белка введена, или что активность повышена по сравнению с эндогенной активностью или активность микроорганизма до модификации. «Ввод» активности означает, что активность конкретного белка, которого микроорганизм изначально не имел, экспрессируется естественным путем или искусственно. «Эндогенная активность» относится к активности конкретного белка, который изначально присутствует в родительском штамме до трансформации, когда микробные признаки изменяются вследствие генетической вариации посредством природных или искусственных факторов.
Как его используют здесь, термин «вектор» относится к конструкции ДНК, содержащей нуклеотидную последовательность полинуклеотида, кодирующего белок-мишень, функционально связанную с соответствующей регуляторной последовательностью для экспрессии белка-мишени в соответствующем хозяине. Регуляторная последовательность может включать промотор, который может инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность для кодирования подходящего мРНК-сайта связывания рибосомы, и последовательность для регулирования терминации транскрипции и трансляции. Вектор может быть трансфицирован в подходящего хозяина и затем может реплицироваться или функционировать независимо от генома хозяина и может быть интегрирован в геном самостоятельно.
Вектор, используемый в настоящем изобретении, конкретно не ограничен, при условии, что он может экспрессироваться в хозяине, и можно использовать любой вектор, который известен в данной области техники. Примеры обычно используемых векторов представляют собой плазмиду, космиду, вирус и бактериофаг в естественном состоянии или рекомбинантном состоянии. Например, pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A можно использовать в качестве фагового вектора или космидного вектора; и систему pBR, систему pUC, систему pBluescriptII, систему pGEM, систему pTZ, систему pCL и систему рЕТ можно использовать в качестве плазмидного вектора. Более конкретно, можно использовать векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 и pCC1BAC, но не ограничиваясь этим.
Вектор, который можно использовать в настоящем изобретении, не ограничивается конкретно, и может быть использован известный вектор экспрессии. Кроме того, полинуклеотид, кодирующий белок-мишень, может быть вставлен в хромосому посредством вектора для вставки во внутриклеточную хромосому. Вставка полинуклеотида в хромосому может быть выполнена посредством любого способа, известного в данной области техники, такого как гомологичная рекомбинация, без конкретного ограничения этим. Вектор может дополнительно включать селективный маркер для индикации вставки вектора в хромосому хозяина. Селективный маркер используют для отбора клеток, трансформированных с помощью вектора, то есть для подтверждения того, вставлена ли молекула-мишень нуклеиновой кислоты. Таким образом, селективный маркер может наделить клетку способностью проявлять резистентность к лекарственному средству, резистентность к цитотоксическому агенту, аутотрофию или экспрессию селектируемого фенотипа, такую как экспрессия поверхностного белка. Трансформированные клетки могут быть выбраны в присутствии селективного агента, поскольку только клетки, которые экспрессируют селективный маркер, остаются живыми или показывают другой фенотип.
Как его используют здесь, термин "трансформация" относится к вводу вектора, включающего полинуклеотид, кодирующий белок-мишень, в клетку-хозяина таким образом, чтобы белок, кодированный полинуклеотидом, экспрессировался в клетке-хозяине. До тех пор, пока трансформированный полинуклеотид может экспрессироваться в клетке-хозяине, он может либо интегрироваться в хромосому или помещаться в хромосому клетки-хозяина, либо существовать внехромосомно. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие белок-мишень. Полинуклеотид можно вводить в любой форме до тех пор, пока он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все элементы, необходимые для его автономной экспрессии. Экспрессионная кассета может включать промотор, функционально связанный с полинуклеотидом, терминатор транскрипции, сайты связывания рибосомы или терминатор трансляции. Экспрессионная кассета может быть представлена в форме самореплицируемого вектора экспрессии. Кроме того, полинуклеотид как таковой может быть введен в клетку-хозяина и функционально связывай с последовательностями, необходимыми для экспрессии в клетке-хозяине, но не ограничиваясь этим. Способ трансформации включает любой способ ввода нуклеиновой кислоты в клетку и может быть выполнен путем выбора подходящего стандартного способа, известного в данном уровне техники, в зависимости от клетки-хозяина. Примеры способа включают электропорацию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), микроинъекцию, способ с полиэтиленгликолем (PEG), способ с ДЭАЭ-декстраном, способ с катионными липосомами, способ с использованием ацетат лития-ДМСО и так далее, но не ограничиваются этим.
Дополнительно, термин «функциональное связывание» означает, что полинуклеотидная последовательность фунционально связана с последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего белок-мишень по настоящему изобретению. Функциональное связывание может быть получено с использованием способа рекомбинации генов, известного в данном уровне техники, и сайт-специфическое расщепление и связывание ДНК могут быть получены, используя известные лиазу и лигазу, но не ограничивается этим.
Как его используют здесь, термин «микроорганизм, продуцирующий L-аминокислоту, имеющую происхождение из аспартата, или производное этой аминокислоты» относится к микроорганизму, естественно продуцирующему L-аминокислоту, имеющую происхождение из аспартата, или производное этой аминокислоты, или относится к микроорганизму, у которого продуктивность L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты обеспечена родительским штаммом, не имеющим способности продуцировать L-аминокислоту, имеющую происхождение из аспартата, или производное этой аминокислоты.
Как его используют здесь, термин «L-аминокислота, имеющая происхождение из аспартата, или производное этой аминокислоты» относится к веществу, способному биосинтезироваться, используя аспарагиновую кислоту (аспартат) в качестве предшественника, и может использоваться взаимозаменяемо с термином «продукт, имеющий происхождение из аспартата». «Производное аминокислоты» относится к веществу, которое может быть получено из L-аминокислоты, и веществу, включающему предшественник L-аминокислоты. Кроме того, производное аминокислоты не ограничивается до тех пор, пока оно представляет собой вещество, которое может быть получено путем биосинтеза с использованием аспартата в качестве предшественника. Более конкретно, вещества, синтезированные обычно с использованием ацетилфосфата, включены без ограничения. Например, производное аминокислоты может представлять собой L-лизин, L-треонин, L-метионин, L-глицин, гомосерин, O-ацетилгомосерин, O-сукцинилгомосерин, O-фосфогомосерин, L-изолейцин и кадаверин, но не ограничиваясь этим.
Кадаверин можно прямо биосинтезировать, используя аспартат в качестве предшественника, или можно получать из лизина посредством лизиндекарбоксилазы. L-метионин можно прямо биосинтезировать, используя аспартат в качестве предшественника, или можно получать путем превращения из O-ацетилгомосерина или O-сукцинилгомосерина.
Как его используют здесь, термин «аспартат (аспарагиновая кислота)» сокращается как Asp или D, и относится к аспарагиновой кислоте, которая представляет собой α-аминокислоту используемую в биосинтезе белков. Подобно всем другим аминокислотам, аспартат включает аминогруппу и карбоксильную группу. Обычно, аспартат получают в виде аспартилфосфата посредством аспартокиназы (LysC), и затем он превращается в клетке в L-лизин, L-метионин, L-гомосерин, L-треонин, L-изолейцин и так далее.
Для повышения биосинтеза продукта, имеющего происхождение из аспартата, можно использовать вариант аспартокиназы по настоящему изобретению. Например, для повышения биосинтеза L-лизина, L-треонина, L-метионина, L-глицина, гомосерина, O-ацетилгомосерина, O-сукцинилгомосерина, O-фосфогомосерина, L-изолейцина и кадаверина может быть введен вариант аспартокиназы по настоящему изобретению или может быть повышена его активность. Кроме того, продуктивность продукта, имеющего происхождение из аспартата, в дальнейшем может быть повышена в результате ввода или повышения активности конкретного белка или путем инактивации активности конкретного белка.
Способ ввода, повышения и инактивации активности конкретного белка может быть выполнен с использованием способа, известного в данном уровне техники, в зависимости от характеристик микроорганизмов.
Микроорганизм, продуцирующий продукт, имеющий происхождение из аспартата, L-аминокислоту или производное этой аминокислоты, может представлять собой любой микроорганизм, способный продуцировать «аминокислоту», имеющую происхождение из аспартата, или производное этой аминокислоты путем включения варианта аспартата по настоящему изобретению. Конкретный пример этого может включать Escherichia sp., Serratia sp., Erwinia sp., Enterobacteria sp., Salmonella sp., Streptomyces sp., Pseudomonas sp., Brevibacterium sp. или Corynebacterium sp., и еще более конкретно микроорганизм может представлять собой Corynebacterium glutamicum, но не ограничиваясь этим.
В еще одном аспекте настоящего изобретения предложен способ получения L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты, включающий: культивирование описанного выше микроорганизма; и выделение L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты из культивированного микроорганизма или культуральной среды.
Способ получения L-аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты легко может быть определен специалистом в данной области техники в оптимизированных условиях культивирования и условиях активности фермента, известных в данном уровне техники. Более точно, хотя конкретно не ограничивается, стадия культивирования микроорганизма может быть выполнена путем периодического культивирования, непрерывного культивирования и периодического культивирования с подпиткой, известных в данном уровне техники. Здесь условия культивирования не ограничиваются конкретно, но оптимальный рН (например рН от 5 до 9, более конкретно рН от 6 до 8 и еще более конкретно рН 6,8) можно поддерживать путем использования основного химического вещества (например гидроксида натрия, гидроксида калия или аммиака) или кислого химического вещества (например фосфорной кислоты или серной кислоты). Кроме того, аэробные условия можно поддерживать путем добавления кислорода или кислородсодержащей газовой смеси в культуру клеток. Температура культивирования может поддерживаться при от 20°С до 45°С и более конкретно при от 25°С до 40°С, и культивирование может выполняться в течение примерно от 10 часов до 160 часов, но не ограничивается этим. Аминокислота, имеющая происхождение из аспартата, или производное этой аминокислоты, полученные путем культивирования, могут выделяться в среду или могут оставаться в клетках.
Дополнительно, среда, подлежащая использованию в культивировании, может включать сахар и углевод (например глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассу, крахмал и целлюлозу), масло и жир (например соевое масло, масло семян подсолнечника, арахисовое масло и кокосовое масло), жирную кислоту (например пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирт (например глицерин и этиловый спирт) и органическую кислоту (например уксусную кислоту) по отдельности или в комбинации в качестве источника углерода; азотсодержащее органическое соединение (например пептон, дрожжевой экстракт, мясной сок, солодовый экстракт, кукурузный раствор, порошок соевого шрота и мочевину) или неорганическое соединение (например сульфат аммония, хлорид аммония, фосфат аммония, аммония карбонат и нитрат аммония) по отдельности или в комбинации в качестве источника азота; и первичный кислый фосфат калия, гидрофосфат калия или натрий-содержащую соль, соответствующую этим по отдельности или в комбинации в качестве источника фосфора; но не ограничиваются этим. Кроме того, среда может содержать необходимые для обеспечивания роста вещества, такие как соли других металлов (например сульфат магния или сульфат железа), аминокислоты и витамины.
Способ выделения аминокислоты, имеющей происхождение из аспартата, или производного этой аминокислоты, полученного в способе культивирования по настоящему изобретению, можно выполнять для получения целевой аминокислоты из культуральной среды путем использования подходящего способа, известного в данной области техники, в зависимости от способа культивирования. Например, можно использовать центрифугирование, фильтрацию, анионообменную хроматографию, кристаллизацию и ВЭЖХ, и целевая аминокислота, имеющая происхождение из аспартата, или производное этой аминокислоты может быть выделено из среды или микроорганизма посредством подходящего способа, известного в данном уровне техники.
Дополнительно, способ выделения может дополнительно включать стадию очистки и может быть выполнен с использованием подходящего способа, известного в данном уровне техники.
В еще одном аспекте настоящего изобретения предложен способ получения L-метионина, включающий: культивирование микроорганизма по настоящему изобретению; продуцирование O-ацетилгомосерина или O-сукцинилгомосерина из культивированного микроорганизма или культуральной среды; и превращение O-ацетилгомосерина или O-сукцинилгомосерина в L-метионин.
Более конкретно, «стадия превращения в L-метионин» может включать стадию взаимодействия O-ацетилгомосерина или O-сукцинилгомосерина с O-ацетилгомосерин-сульфгидрилазой или O-сукцинилгомосерин-сульфгидрилазой в присутствии сульфида. «O-ацетилгомосерин или O-сукцинилгомосерин» может представлять собой ферментативый бульон, содержащий O-ацетилгомосерин или O-сукцинилгомосерин, продуцированный микроорганизмом по настоящему изобретению, или может быть представлен в очищенной форме. Кроме того, «сульфид» может представлять собой, например, метилмеркаптан, и метилмеркаптан может относиться к форме метилмеркаптида натрия (CH3S-Na), а также метилмеркаптана (CH3SH) в газообразном или сжиженном состоянии и метилмеркаптана, содержащего диметилсульфид (DMS) в форме, описанной в публикации международной заявки на патент № WO 2010/098629; и метил меркаптан также может относиться к производному метилмеркаптана, содержащему форму, способную обеспечивать присутствие атомов серы. Кроме того, «O-ацетилгомосерин-сульфгидрилаза или (O-сукцинилгомосерин-сульфгидрилаза» может представлять собой ферментативый бульон микроорганизма, продуцирующего то же самое, или может представлять собой очищенную форму.
Способ получения L-метионина может быть без труда определен специалистом в данной области техники в оптимизированных условиях культивирования и условиях активности фермента, известных в данном уровне техники. Конкретный способ культивирования и среда такие, как описано выше.
Полезные эффекты изобретения
С помощью микроорганизма, содержащего вариант аспартокиназы по настоящему изобретению, можно получать L-аминокислоту, имеющую происхождение из аспартата, или продукт, имеющий происхождение из аспартата, с высоким выходом без подавления роста клеток-хозяев по сравнению с микроорганизмом, включающим LysC дикого типа.
Краткое описание графических материалов
На Фиг. 1 показан путь биосинтеза аминокислоты, имеющей происхождение из аспартата, и производного этой аминокислоты у Corynebacterium glutamicum.
Подробное описание изобретения
Далее настоящее изобретение будет описно подробно с сопутствующими типичными воплощениями. Однако типичные воплощения, раскрытые здесь, предназначены только для иллюстративных целей и их не следует рассматривать как ограничивающие объем настоящего изобретения.
Пример 1: Получение штамма с введенным вариантом lysC
Вариацию, активность которой может быть усилена по сравнению с диким типом, отбирали посредством структурного моделирования аспартокиназы и штамм с введенным вариантом получали, как описано ниже.
В гене lysC (SEQ ID NO: 2), кодирующем аспартокиназу (SEQ ID NO: 1), полученную из Corynebacterium glutamicum АТСС13032 (далее обозначаемого как WT), аминокислоту в положении 377 отбирали в качестве сайта вариации, и L-лизин, который представляет собой основную аминокислоту, и L-метионин, который представляет собой неполярную аминокислоту, отбирали в качестве репрезентативных примеров других аминокислот для замены.
Для получения вектора, вводимого с вариацией, сосредотачиваясь на сайте вариации, разрабатывали пару праймеров (SEQ ID NO: 7 и 8 или SEQ ID NO: 7 и 10) для амплификации участка выше по ходу транскрипции и пару праймеров (SEQ ID NO: 9 и 12 или SEQ ID NO: 11 и 12) для амплификации участка ниже по ходу транскрипции (Таблица 1). В праймеры SEQ ID NO: 7 и 12 вставляли сайт фермент рестрикции XbaI (подчеркнуто) на каждом конце. Затем в пару праймеров SEQ ID NO: 8 и 9 или пару праймеров SEQ ID NO: 10 и 11 помещали вариацию замены нуклеотида (подчеркнут) в сайте, предназначенном для кроссинговера друг с другом.

ПЦР выполняли с помощью праймеров SEQ ID NO: 7 и 8, SEQ ID NO: 7 и 10, SEQ ID NO: 9 и 12, или SEQ ID NO: 11 и 12 с использованием хромосомы WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 30 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате, сосредотачиваясь на варианте гена lysC, получали фрагмент ДНК (509 п.н.) участка выше по ходу транскрипции и фрагмент ДНК (520 п.н.) участка ниже по ходу транскрипции, соответственно.
ПЦР выполняли с праймерами SEQ ID NO: 7 и 12 с использованием двух амплифицированных фрагментов ДНК в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате амплифицировали фрагмент ДНК (1011 п.н.), включающий вариацию (SEQ ID NO: 4) гена lysC, кодирующего вариант аспартокиназы (SEQ ID NO: 3), в котором лейцин в положении 377 замещен лизином. Кроме того, для того, чтобы подтвердить важность аминокислоты в положении 377, получали фрагмент ДНК (1011 п.н.), включающий вариацию (SEQ ID NO: 6) гена lysC, кодирующего вариант аспартокиназы (SEQ ID NO: 5), в котором лейцин в положении 377 замещен метионином.
Вектор pDZ (патент Кореи №0924065), который не может реплицироваться у Corynebacterium glutamicum, и фрагмент ДНК (1011 п.н.) обрабатывали ферментом рестрикции, XbaI, и соединяли их с использованием ДНК-лигазы и затем клонировали для получения плазмид. Плазмиды были названы pDZ-lysC(L377K) и pDZ-lysC(L377M).
Каждый из векторов pDZ-lysC(L377K) и pDZ-lysC(L377M) трансформировали в WT с использованием способа электрического импульса (Appl. Microbiol. Biotechnol. (1999) 52:541-545), и затем получали штаммы-трансформанты в среде LB, содержащей канамицин (25 мг/л). WT::lysC(L377K) и WT::lysC(L377M), получали штаммы, у которых вариацию нуклеотида вводят в ген lysC посредством фрагментов ДНК, вставленных в хромосому с помощью процесса вторичной рекомбинации (кроссинговера), и назвали их Corynebacterium glutamicum СА01-2307 и CA01-2308, соответственно. СА01-2307 и CA01-2308 депонировали в Корейском Центре Культур Микроорганизмов, который представляет собой международный орган депонирования в соответствии с Будапештским договором, 29 марта 2017, с присвоением им номеров доступа KCCM12000P и KCCM12001P.
Пример 2: Подтверждение способности штамма с введенным вариантом lysC продуцировать полученную из аспартата аминокислоту
С целью сравнения способностей штаммов CA01-2307 и СА01-2308, полученных в Примере 1, и штамма WT в продуцировании главных полученных из аспартата аминокислот штаммы культивировали с использованием следующего способа и анализировали компоненты в культуральной среде.
Каждый штамм инокулировали в колбы с угловыми перегородками (250 мл), содержащие посевную среду (25 мл), и культивировали при 37°С в течение 20 часов со встряхиванием при 200 об/мин. Раствор с посевной культурой(1 мл) инокулировали в колбу с угловыми перегородками (250 мл), содержащую продукционную среду (24 мл), и затем культивировали при 37°С в течение 24 часов со встряхиванием при 200 об/мин. Концентрации L-лизина и L-треонина, которые представляют собой репрезентативные аминокислоты, имеющие происхождение из L-аспартата и аспартата, анализировали посредством ВЭЖХ и проанализированные концентрации показаны в Таблице 2.
Посевная среда (рН 7,0)
Глюкоза (20 г), пептон (10 г), дрожжевой экстракт (5 г), мочевина (1,5 г), KH2PO4 (4 г), K2HPO4 (8 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамин HCl (1000 мкг), пантотенат кальция (2000 мкг), никотинамид (2000 мкг) (на 1 л дистиллированной воды)
Продукционная среда (рН 7,0)
Глюкоза (100 г), (NH4)2SO4 (40 г), соевый белок (2,5 г), сухие вещества кукурузного экстракта (5 г), мочевина (3 г), KH2PO4 (1 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамина хлорид (1000 мкг), пантотенат кальция (2000 мкг), никотинамид (3000 мкг), СаСО3 (30 г) (на 1 л дистиллированной воды)
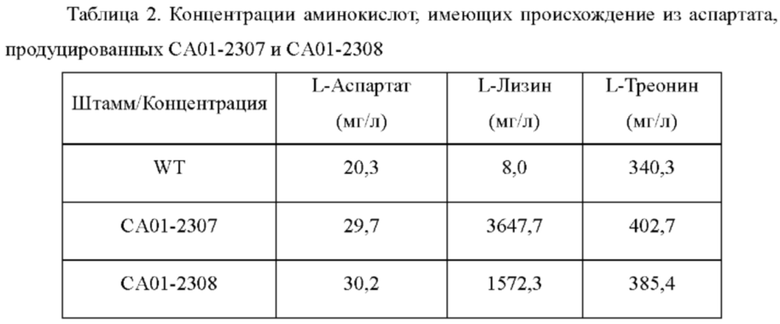
В результате анализа концентраций аминокислот, имеющих происхождение из аспартата, было подтверждено, что когда вводили вариацию lysC, концентрация L-лизина резко повышалась и концентрации L-аспартата и L-треонина также повышались по сравнению с WT. На основании приведенных выше результатов штаммы СА01-2307, СА01-2308 и WT культивировали таким же образом, как описано выше, для подробного сравнения их способностей продуцировать лизин. Затем концентрацию L-лизина в культуральной среде анализировали таким же образом, как описано выше (Таблица 3).
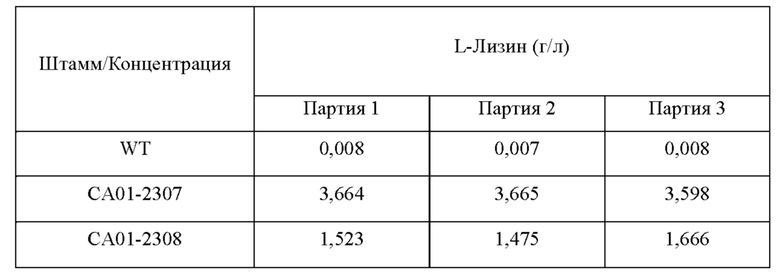
В результате анализа концентрации L-лизина было подтверждено, что продуктивность по L-лизину, продуцированному СА01-2307, включающему вариацию lysC(L377K), значительно повышалась, как и в предыдущем оценивании, по сравнению с таковым, продуцированным штаммом WT. Кроме того, было подтверждено, что продуктивность по L-лизину, продуцированному САО 1-2308, включающим вариацию lysC(L377M), также повышалась по сравнению с таковым, продуцированным штаммом WT. На основании приведенных выше результатов было подтверждено, что продуктивность по L-лизину у аминокислот, имеющих происхождение из аспартата, значительно повышалась благодаря варианту аспартокиназы (SEQ ID NO: 3), в котором лейцин в положении 377, который выбран в настоящем изобретении, замещен лизином и благодаря варианту аспартокиназы (SEQ ID NO: 5), в котором лейцин в положении 377 замещен метионином.
Пример 3: Получение штамма с повышенным продуцированием L-треонина и подтверждение продуктивности по L-треонину
Для четкого подтверждения изменения продуктивности по L-треонину вследствие ввода вариации lysC(L377K), которая имеет более высокую продуктивность по лизину, подтвержденную в Примере 2, вариацию вводили в ген, кодирующий гомосерин-дегидрогеназу, продуцирующую гомосерин, который представляет собой общее промежуточное соединение в биосинтетическом пути производных L-треонина, L-изолейцина, L-метионина и гомосерина. Более конкретно, штамм получали посредством ввода вариации hom(G378E), известной в данной области техники (Appl. Microbiol. Biotechnol. 45, 612-620 (1996)), в штамм СА01-2307, который был получен в Примере 1. Кроме того, штамм, в котором вариацию hom(С378Е) вводят в WT, также получали в качестве контрольной группы. Для получения вектора, вводящего hom(G378E), ПЦР выполняли с SEQ ID NO: 13 и 14 и SEQ ID NO: 15 и 16 с использованием геномной ДНК WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 30 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут.
В результате, сосредотачиваясь на варианте гена hom, получали фрагмент ДНК (220 п.н.) участка выше по ходу транскрипции и фрагмент ДНК (220 п.н.) участка ниже по ходу транскрипции. ПЦР выполняли с SEQ ID NO: 13 и 16 с использованием этих двух продуктов ПЦР в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 30 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате амплифицировали фрагмент ДНК (440 п.н.), включающий вариацию гена hom.

Вектор pDZ, который ранее использовали в Примере 1, и фрагмент ДНК (440 п.н.) обрабатывали ферментом рестрикции XbaI и соединяли их, используя ДНК-лигазу, и затем клонировали для получения плазмиды. Плазмида была названа pDZ-hom(G378E).
Вектор pDZ-hom(G378E) вводили в штаммы WT и САО 1-2307, используя способ электрического импульса (Appl. Microbiol. Biotechnol. (1999, 52:541-545)), и затем получали штаммы-трансформанты в селективной среде, содержащей канамицин (25 мг/л). Получали штаммы WT::hom(G378E) и САО 1-2307::hom(G378E), у которых вариация нуклеотида введена в ген hom посредством фрагмента ДНК, вставленного в хромосому с помощью процесса вторичной рекомбинации (кроссинговера). Для сравнения способностей WT::hom(G378E) и СА01-2307::hom(G378E) к продуцированию треонина, штаммы культивировали таким же образом, как в Примере 2, и анализировали концентрацию треонина в культуральной среде.
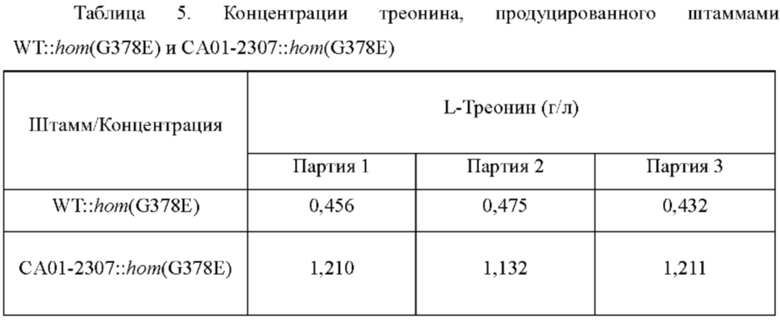
В результате анализа концентрации L-треонина было подтверждено, что продуктивность по L-треонину резко повышалась у штамма, включающего вариацию lysC (Таблица 5).
Пример 4: Получение штамма с повышенным продуцированием L-изолейцина и сравнение продуктивности по L-изолейцину
С целью подтверждения влияния введения вариации lysC(L377K) на продуктивность по L-изолейцину получали вектор для усиления экспрессии вариации ilvA(V323A) гена (Appl. Enviro. Microbiol., Dec. 1996, p. 4345-4351), кодирующего L-треонин-дегидратазу, который известен в данном уровне техники.
Для получения вектора, вводимого с вариантом гена ilvA, сосредотачиваясь на сайте вариации, разрабатывали пару праймеров (SEQ ID NO: 17 и 18) для амплификации участка выше по ходу транскрипции и пару праймеров (SEQ ID NO: 19 и 20) для амплификации участка ниже по ходу транскрипции. В праймерах SEQ ID NO: и 17 и 20 сайт фермента рестрикции BamHI (подчеркнут) вставляли в каждый конец. Затем в праймеры SEQ ID NO: 18 и 19 помещали вариацию нуклеотидной замены (подчеркнуто) в сайтах, предназначенных для кроссинговера друг с другом.


ПЦР выполняли с праймерами SEQ ID NO: 17 и 18 и SEQ ID NO: 19 и 20, используя хромосому WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 30 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате, сосредотачиваясь на варианте гена ilvA, получали фрагмент ДНК (627 п.н.) участка выше по ходу транскрипции и фрагмент ДНК (608 п.н.) участка ниже по ходу транскрипции.
ПЦР выполняли с праймерами SEQ ID NO: 17 и 20, используя два амплифицированных фрагмента ДНК в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате фрагмент ДНК (1217 п.н.), включающий вариацию гена ilvA, кодирующего вариант IlvA, в котором валин в положении 323 замещен аланином, был амплифицирован.
Вектор pECCG117 (патент Кореи №10-0057684) и ДНК фрагмент (1011 п.н.) обрабатывали ферментом рестрикции BamHI и соединяли их, используя ДНК-лигазу, и затем клонировали для получения плазмиды. Плазмида была названа pECCG117-/ilvA(V323A).
Вектор pECCG117-ilvA (V323A) вводили в штаммы WT::hom(G378E) и СА01-2307::hom(G378E), которые были получены в Примере 3 с использованием способа электрического импульса, и затем размазывали по селективной среде, содержащей канамицин (25 мг/л), для получения штаммов-трансформантов.
Штаммы культивировали таким же образом, как в способе культивирования в колбе, показанном в Примере 2, и анализировали концентрацию L-изолейцина в культуральной среде (Таблица 7).
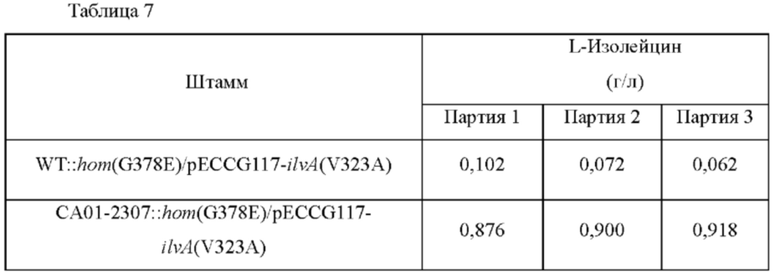
В результате анализа концентрации L-изолейцина было подтверждено, что продуктивность по L-изолейцину значительно повышалась у штамма, включающего вариацию lysC.
Пример 5: Получение штамма с усилением О-ацетилгомосерина и сравнение продуктивности по О-ацетилгомосерину
Для определения влияния ввода вариации lysC(L377K) на продуцирование О-ацетилгомосерина ген metB, который кодирует цистатионин гамма-синтазу, вовлеченную в путь деградации О-ацетилгомосерина, и ген metY, который кодирует О-ацетилгомосерин (тиол)-лиазу, делетировали и затем ген metX, кодирующий гомосерин-O-ацетилтрансферазу, который представляет собой фермент биосинтеза О-ацетилгомосерин, сверхэкспрессировался и посредством этого получали штамм, продуцирующий О-ацетилгомосерин. Сначала для делетирования гена metB, основываясь на информации о нуклеотидной последовательности гена metB, полученного из WT, разрабатывали пару праймеров (SEQ ID NO: 21 и 22) для амплификации участка выше по ходу транскрипции гена metB и пару праймеров (SEQ ID NO: 23 и 24) для амплификации 3' ниже по ходу транскрипции.


ПЦР выполняли с праймерами SEQ ID NO: 21 и 22, SEQ ID NO: 23 и 24, используя хромосому WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд, полимеризация при 72°С в течение 90 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате получали фрагмент ДНК (450 п.н.) участка выше по ходу транскрипции гена metB и фрагмент ДНК (467 п.н.) участка ниже по ходу транскрипции.
ПЦР выполняли с праймерами SEQ ID NO: 21 и 24, используя два амплифицированных фрагмента ДНК в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 3 минут. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате центральный участок гена metB был делетирован и, таким образом, был амплифицирован фрагмент ДНК (917 п.н.), включающий только концы выше и ниже по ходу транскрипции.
Вектор pDZ и фрагмент ДНК (917 п.н.) обрабатывали ферментом рестрикции XbaI и соединяли их с использованием ДНК-лигазы и затем клонировали с получением плазмиды. Плазмида была названа pDZ-ΔmetB.
Вектор pDZ-ΔmetB вводили в штаммы WT::hom(G378E) и СА01-2307::hom(G378E), которые получали в Примере 3, используя способ электрического импульса, и затем получали штаммы-трансформанты на селективной среде, содержащей канамицин (25 мг/л). Получали штаммы WT::hom(G378E)ΔmetB и СА01-2307::hom(G378E)ΔmetB, у которых ген metB делетирован посредством фрагментов ДНК, вставленных в хромосому с помощью процесса вторичной рекомбинации (кроссинговера).
Для делетирования гена metY, вовлеченного в другой путь деградации О-ацетилгомосерина, основываясь на информации о нуклеотидной последовательности гена metY, полученного из WT, разрабатывали пару праймеров (SEQ ID NO: 25 и 26) для амплификации участка выше по ходу траскрипции гена metY и пару праймеров (SEQ ID NO: 27 и 28) для амплификации участка ниже по ходу траскрипции.

ПЦР выполняли с SEQ ID NO: 25 и 26, SEQ ID NO: 27 и 28, используя хромосому WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд, полимеризация при 72°С в течение 90 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате получали фрагмент ДНК (512 п.н.) участка выше по ходу транскрипции гена metY и фрагмент ДНК (520 п.н.) участка ниже по ходу транскрипции.
ПЦР выполняли с праймерами SEQ ID NO: 25 и 28, используя два амплифицированных фрагмента ДНК в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 3 минут. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате центральный участок гена metY был делетирован и таким образом был амплифицирован фрагмент ДНК (1032 п.н.), включающий только выше и ниже по ходу транскрипции.
Вектор pDZ и ДНК фрагмент (1032 п.н.) обрабатывали ферментом рестрикции XbaI и соединяли их, используя ДНК-лигазу, и затем клонировали для получения плазмиды. Плазмида была названа pDZ-ΔmetY.
Вектор pDZ-ΔmetY вводили в каждый из штаммов WT::hom(G378E) ΔmetB и СА01-2307::hom(G378E)ΔmetB, которые были получены выше, используя способ электрического импульса, и затем получали штаммы-трансформанты в селективной среде, содержащей канамицин (25 мг/л). Получали WT::hom(G378E)ΔmetBΔmetY и СА01-2307::hom(G378E)ΔmetBΔmetY, штаммы, у которых ген metY был делетирован посредством фрагментов ДНК, вставленных в хромосому с помощью процесса вторичной рекомбинации (кроссинговера).
Для максимального увеличения продуцирования О-ацетилгомосерина получали вектор, усиливающий экспрессию гена metX. Для амплификации гена, кодирующего гомосерин-O-ацетилтрансферазу (MetX), на основе описанных последовательностей, полученных из WT, разрабатывали вектор посредством вставки участка фермента рестрикции BamHI в оба конца праймеров (SEQ ID NO: 29 и 30) для амплификации от промоторного участка (примерно 300 п.н. выше кодона инициации) до терминаторного участка (примерно 100 п.н. ниже кодона терминации).

ПЦР выполняли с праймерами SEQ ID NO: 25 и 26, используя хромосому WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 90 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате получали фрагмент ДНК (1546 п.н.), включающий ген metX.
Вектор pECCG117 (патент Кореи №10-0057684) и фрагмент ДНК metX обрабатывали ферментом рестрикции BamHI и соединяли их, используя ДНК-лигазу, и затем клонировали для получения плазмиды. Плазмида была названа pECCG117-metX.
Вектор pECCG117-metX вводили в штаммы WT::hom(G378E)ΔmetBΔmetY и СА01-2307::hom(G378E)ΔmetBΔmetY, которые были получены выше, используя способ электрического импульса, и затем размазывали на селективной среде, содержащей канамицин (25 мг/л), для получения штаммов-трансформантов.
Для сравнения способностей штаммов, полученных выше, к продуцированию О-ацетилгомосерина (далее обозначается О-АН), штаммы культивировали, используя способ ниже, и анализировали концентрацию О-ацетилгомосерина в культуральной среде.
Одну платиновую петлю штаммов инокулировали в колбы с угловыми перегородками (250 мл), содержащие О-АН продукционная среда (25 мл), и культивированного при 37°С в течение 20 часов со встряхиванием при 200 об/мин. Концентрацию О-ацетилгомосерина анализировали посредством ВЭЖХ, и проанализированные концентрации показаны в Таблице 11.
Продукционная среда О-АН (рН 7,0)
Глюкоза (100 г), (NH4)2SO4 (40 г), соевый белок (2,5 г), сухие вещества кукурузного экстракта (5 г), мочевина (3 г), KH2PO4 (1 г), MgSO4⋅7H2O (0,5 г), биотин (100 мкг), тиамина хлорид (1000 мкг), пантотенат кальция (2000 мкг), никотинамид (3000 мкг), СаСО3 (30 г), L-метионин (0,3 г) (в 1 л дистиллированной воды).
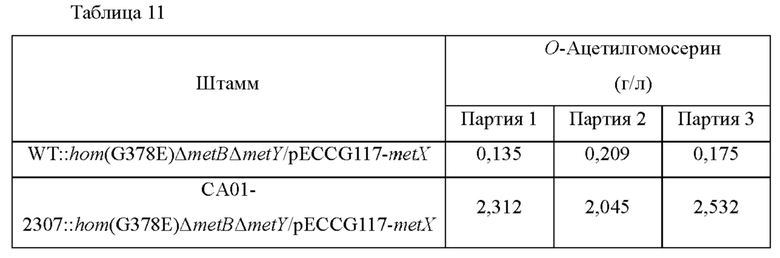
В результате анализа концентрации О-ацетилгомосерина, как показано в таблице, приведенной выше, было подтверждено, что концентрация О-ацетилгомосерина, продуцированного посредством вариации lysC, была повышена.
Пример 6: Получение штамма с усилением О-ацетилгомосерина и сравнение продуктивности по О-сукцинилгомосерину
Для определения влияния ввода вариации lysC(L377K) на продуцирование О-сукцинилгомосерина был получен штамм, продуцирующий О-сукцинилгомосерин. Так как Corynebacterium glutamicum дикого типа по природе не продуцирует О-сукцинилгомосерин, штамм, который был получен в Примере 5, модифицировали для получения активности О-сукцинилтрансферазы (MetX) путем замены аминокислоты в субстрат-связывающем участке О-ацетилтрансферазы с целью продуцирования О-сукцинилгомосерина. Таким образом, на основе последовательностей гена metX, полученного из WT, разрабатывали пару праймеров (SEQ ID NO: 31 и 32) для получения вектора для ввода вариации metX.

ПЦР выполняли с праймерами SEQ ID NO: 27 и 28, используя хромосому WT в качестве матрицы. После денатурации при 95°С в течение 5 минут выполняли в общей сложности 30 циклов ПЦР в следующих условиях: 95°С денатурация в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 90 секунд. После этого выполняли реакцию полимеризации при 72°С в течение 7 минут. В результате получали фрагмент ДНК (1146 п.н.) кодирующего участка гена metX.
Вектор pDZ и фрагмент ДНК (1146 п.н.) обрабатывали ферментом рестрикции XbaI и соединяли их, используя ДНК-лигазу, и затем клонировали для получения плазмиды. Плазмида была названа pDZ-metX. На основе вектора pDZ-metX для создания вектора с вариациями, в котором аминокислота в положении 176 замещена аспарагином и аминокислота в положении 313 замещена аргинином, разрабатывали пару праймеров (SEQ ID NO: 33 и 34), которые вызывают вариацию аминокислоты в положении 176, и пару праймеров (SEQ ID NO: и 35 и 36), которые вызывают вариацию аминокислоты в положении 313.
Вариант гена metX получали с использованием набора для сайт направленного мутагенеза (Stratagene, USA) вместе с каждым из праймеров, приведенных выше. Трансформант с плазмидой на основе существующей плазмиды дикого типа pDZ-metX был назван pDZ-metX (Q176N, L313R). Вектор pDZ-metX(Q176N, L313R), который был получен выше, трансформировали в штаммы WT::hom(G378E)ΔmetBΔmetY и СА01-2307::hom(G378E)ΔmetBΔmetY, полученные в Примере 5, с использованием способа электрического импульса и затем штаммы-трансформанты получали в селективной среде, содержащей канамицин (25 мг/л). Получали штаммы WT::hom(G378E)metX(Q176N, L313R)ΔmetBΔmetY и СА01-2307::hom(G378E)metX(Q176N, L313R)ΔmetBΔmetY, у которых ген metX замещен на metX(Q176N, L313R) посредством фрагмента ДНК, вставленного в хромосому с помощью процесса вторичной рекомбинации (кроссинговера).
Для максимального увеличения продуцирования О-сукцинилгомосерина получали вектор для усиления экспрессии гена-варианта metX(Q176N, L313R). На основе pECCG117-metY, полученного в Примере 5, получали вектор, сверхэкспрессирующий вариант metX, с использованием праймеров SEQ ID NO: 33-36, в котором вариант аминокислоты в положении 176, который имеет активность О-сукцинилтрансферазы, замещен аспарагином и аминокислота в положении 313 замещен аргинином. Более конкретно, вектор получали с использованием набора для сайтнаправленного мутагенеза (Stratagene, USA) вместе с каждым из праймеров и полученный вектор был назван pECCG117-metX(Q176N, L313R).
pECCG117-metX(Q176N, L313R) и пустой вектор pECCG117 вводили в WT::hom(G378E)metX(Q176N, L313R)ΔmetBΔmetY и СА01-2307::hom(G378E)metX(Q176N, L313R)ΔmetBΔmetY, которые представляют собой полученные выше штаммы, продуцирующие О-сукцинилгомосерин, с использованием способа электрического импульса, и размазанные на селективной среде, содержащей канамицин (25 мг/л) для получения штамма-трансформанта.
Для сравнения способности штамма к продуцированию О-сукцинилгомосерина, штамм культивировали с использованием следующих способов и анализировали концентрацию О-сукцинилгомосерина в культуральной среде.
Одну платиновую петлю штамма инокулировали в колбу с угловыми перегородками (250 мл), содержащую 25 мл продукционной среды О-АН такого же состава, используемой в Примере 5, и культивировали при 37°С в течение 20 часов со встряхиванием при 200 об/мин. Концентрацию О-сукцинилгомосерина анализировали посредством ВЭЖХ; и проанализированные концентрации показаны в Таблице 14.
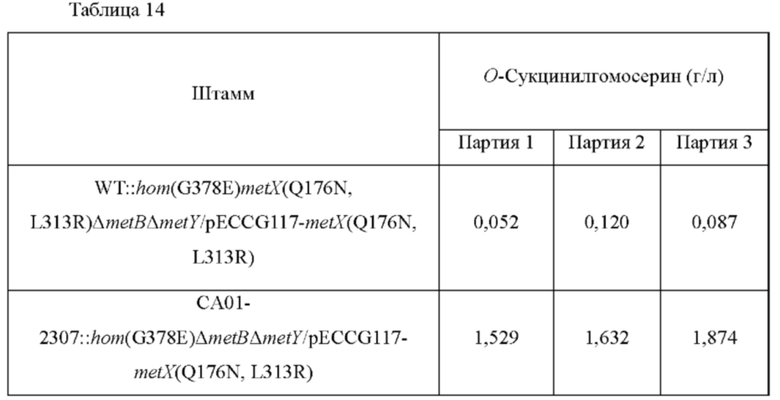
В результате анализа концентрации O-сукцинилгомосерина, как показано в приведенной выше таблице, было подтверждено, что продуцирование О-сукцинилгомосерина, полученное посредством вариации lysC, было повышенным.
Такие результаты указывают на то, что вариант по настоящему изобретению может повышать продуцирование аминокислоты, имеющей происхождение из аспартата, и/или ее производного.
Исходя из изложенного выше, специалист в области техники, к которой принадлежит данное изобретение, понимает, что настоящее изобретение может быть воплощено в других конкретных формах без отступления от технической идеи или существенных признаков настоящего изобретения. В связи с этим, типичные воплощения, раскрытые здесь, предназначаются только для иллюстративных целей и не должны считаться ограничивающими объем настоящего изобретения. Напротив, настоящее изобретение предназначено охватывать не только типичные воплощения, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящего изобретения, который определен прилагаемой формулой.
--->
<110> CJ CheilJedang Corporation
<120> Novel aspartokinase variant and method of producing L-amino acid using the same
<130> OPA18147
<150> 10-2017-0083437
<151> 2017-06-30
<160> 36
<170> KoPatentIn 3.0
<210> 1
<211> 421
<212> PRT
<213> Corynebacterium glutamicum
<400> 1
Val Ala Leu Val Val Gln Lys Tyr Gly Gly Ser Ser Leu Glu Ser Ala
1 5 10 15
Glu Arg Ile Arg Asn Val Ala Glu Arg Ile Val Ala Thr Lys Lys Ala
20 25 30
Gly Asn Asp Val Val Val Val Cys Ser Ala Met Gly Asp Thr Thr Asp
35 40 45
Glu Leu Leu Glu Leu Ala Ala Ala Val Asn Pro Val Pro Pro Ala Arg
50 55 60
Glu Met Asp Met Leu Leu Thr Ala Gly Glu Arg Ile Ser Asn Ala Leu
65 70 75 80
Val Ala Met Ala Ile Glu Ser Leu Gly Ala Glu Ala Gln Ser Phe Thr
85 90 95
Gly Ser Gln Ala Gly Val Leu Thr Thr Glu Arg His Gly Asn Ala Arg
100 105 110
Ile Val Asp Val Thr Pro Gly Arg Val Arg Glu Ala Leu Asp Glu Gly
115 120 125
Lys Ile Cys Ile Val Ala Gly Phe Gln Gly Val Asn Lys Glu Thr Arg
130 135 140
Asp Val Thr Thr Leu Gly Arg Gly Gly Ser Asp Thr Thr Ala Val Ala
145 150 155 160
Leu Ala Ala Ala Leu Asn Ala Asp Val Cys Glu Ile Tyr Ser Asp Val
165 170 175
Asp Gly Val Tyr Thr Ala Asp Pro Arg Ile Val Pro Asn Ala Gln Lys
180 185 190
Leu Glu Lys Leu Ser Phe Glu Glu Met Leu Glu Leu Ala Ala Val Gly
195 200 205
Ser Lys Ile Leu Val Leu Arg Ser Val Glu Tyr Ala Arg Ala Phe Asn
210 215 220
Val Pro Leu Arg Val Arg Ser Ser Tyr Ser Asn Asp Pro Gly Thr Leu
225 230 235 240
Ile Ala Gly Ser Met Glu Asp Ile Pro Val Glu Glu Ala Val Leu Thr
245 250 255
Gly Val Ala Thr Asp Lys Ser Glu Ala Lys Val Thr Val Leu Gly Ile
260 265 270
Ser Asp Lys Pro Gly Glu Ala Ala Lys Val Phe Arg Ala Leu Ala Asp
275 280 285
Ala Glu Ile Asn Ile Asp Met Val Leu Gln Asn Val Ser Ser Val Glu
290 295 300
Asp Gly Thr Thr Asp Ile Thr Phe Thr Cys Pro Arg Ser Asp Gly Arg
305 310 315 320
Arg Ala Met Glu Ile Leu Lys Lys Leu Gln Val Gln Gly Asn Trp Thr
325 330 335
Asn Val Leu Tyr Asp Asp Gln Val Gly Lys Val Ser Leu Val Gly Ala
340 345 350
Gly Met Lys Ser His Pro Gly Val Thr Ala Glu Phe Met Glu Ala Leu
355 360 365
Arg Asp Val Asn Val Asn Ile Glu Leu Ile Ser Thr Ser Glu Ile Arg
370 375 380
Ile Ser Val Leu Ile Arg Glu Asp Asp Leu Asp Ala Ala Ala Arg Ala
385 390 395 400
Leu His Glu Gln Phe Gln Leu Gly Gly Glu Asp Glu Ala Val Val Tyr
405 410 415
Ala Gly Thr Gly Arg
420
<210> 2
<211> 1266
<212> DNA
<213> Corynebacterium glutamicum
<400> 2
gtggccctgg tcgtacagaa atatggcggt tcctcgcttg agagtgcgga acgcattaga 60
aacgtcgctg aacggatcgt tgccaccaag aaggctggaa atgatgtcgt ggttgtctgc 120
tccgcaatgg gagacaccac ggatgaactt ctagaacttg cagcggcagt gaatcccgtt 180
ccgccagctc gtgaaatgga tatgctcctg actgctggtg agcgtatttc taacgctctc 240
gtcgccatgg ctattgagtc ccttggcgca gaagcccaat ctttcacggg ctctcaggct 300
ggtgtgctca ccaccgagcg ccacggaaac gcacgcattg ttgatgtcac tccaggtcgt 360
gtgcgtgaag cactcgatga gggcaagatc tgcattgttg ctggtttcca gggtgttaat 420
aaagaaaccc gcgatgtcac cacgttgggt cgtggtggtt ctgacaccac tgcagttgcg 480
ttggcagctg ctttgaacgc tgatgtgtgt gagatttact cggacgttga cggtgtgtat 540
accgctgacc cgcgcatcgt tcctaatgca cagaagctgg aaaagctcag cttcgaagaa 600
atgctggaac ttgctgctgt tggctccaag attttggtgc tgcgcagtgt tgaatacgct 660
cgtgcattca atgtgccact tcgcgtacgc tcgtcttata gtaatgatcc cggcactttg 720
attgccggct ctatggagga tattcctgtg gaagaagcag tccttaccgg tgtcgcaacc 780
gacaagtccg aagccaaagt aaccgttctg ggtatttccg ataagccagg cgaggctgcg 840
aaggttttcc gtgcgttggc tgatgcagaa atcaacattg acatggttct gcagaacgtc 900
tcttctgtag aagacggcac caccgacatc accttcacct gccctcgttc cgacggccgc 960
cgcgcgatgg agatcttgaa gaagcttcag gttcagggca actggaccaa tgtgctttac 1020
gacgaccagg tcggcaaagt ctccctcgtg ggtgctggca tgaagtctca cccaggtgtt 1080
accgcagagt tcatggaagc tctgcgcgat gtcaacgtga acatcgaatt gatttccacc 1140
tctgagattc gtatttccgt gctgatccgt gaagatgatc tggatgctgc tgcacgtgca 1200
ttgcatgagc agttccagct gggcggcgaa gacgaagccg tcgtttatgc aggcaccgga 1260
cgctaa 1266
<210> 3
<211> 421
<212> PRT
<213> Corynebacterium glutamicum
<400> 3
Val Ala Leu Val Val Gln Lys Tyr Gly Gly Ser Ser Leu Glu Ser Ala
1 5 10 15
Glu Arg Ile Arg Asn Val Ala Glu Arg Ile Val Ala Thr Lys Lys Ala
20 25 30
Gly Asn Asp Val Val Val Val Cys Ser Ala Met Gly Asp Thr Thr Asp
35 40 45
Glu Leu Leu Glu Leu Ala Ala Ala Val Asn Pro Val Pro Pro Ala Arg
50 55 60
Glu Met Asp Met Leu Leu Thr Ala Gly Glu Arg Ile Ser Asn Ala Leu
65 70 75 80
Val Ala Met Ala Ile Glu Ser Leu Gly Ala Glu Ala Gln Ser Phe Thr
85 90 95
Gly Ser Gln Ala Gly Val Leu Thr Thr Glu Arg His Gly Asn Ala Arg
100 105 110
Ile Val Asp Val Thr Pro Gly Arg Val Arg Glu Ala Leu Asp Glu Gly
115 120 125
Lys Ile Cys Ile Val Ala Gly Phe Gln Gly Val Asn Lys Glu Thr Arg
130 135 140
Asp Val Thr Thr Leu Gly Arg Gly Gly Ser Asp Thr Thr Ala Val Ala
145 150 155 160
Leu Ala Ala Ala Leu Asn Ala Asp Val Cys Glu Ile Tyr Ser Asp Val
165 170 175
Asp Gly Val Tyr Thr Ala Asp Pro Arg Ile Val Pro Asn Ala Gln Lys
180 185 190
Leu Glu Lys Leu Ser Phe Glu Glu Met Leu Glu Leu Ala Ala Val Gly
195 200 205
Ser Lys Ile Leu Val Leu Arg Ser Val Glu Tyr Ala Arg Ala Phe Asn
210 215 220
Val Pro Leu Arg Val Arg Ser Ser Tyr Ser Asn Asp Pro Gly Thr Leu
225 230 235 240
Ile Ala Gly Ser Met Glu Asp Ile Pro Val Glu Glu Ala Val Leu Thr
245 250 255
Gly Val Ala Thr Asp Lys Ser Glu Ala Lys Val Thr Val Leu Gly Ile
260 265 270
Ser Asp Lys Pro Gly Glu Ala Ala Lys Val Phe Arg Ala Leu Ala Asp
275 280 285
Ala Glu Ile Asn Ile Asp Met Val Leu Gln Asn Val Ser Ser Val Glu
290 295 300
Asp Gly Thr Thr Asp Ile Thr Phe Thr Cys Pro Arg Ser Asp Gly Arg
305 310 315 320
Arg Ala Met Glu Ile Leu Lys Lys Leu Gln Val Gln Gly Asn Trp Thr
325 330 335
Asn Val Leu Tyr Asp Asp Gln Val Gly Lys Val Ser Leu Val Gly Ala
340 345 350
Gly Met Lys Ser His Pro Gly Val Thr Ala Glu Phe Met Glu Ala Leu
355 360 365
Arg Asp Val Asn Val Asn Ile Glu Lys Ile Ser Thr Ser Glu Ile Arg
370 375 380
Ile Ser Val Leu Ile Arg Glu Asp Asp Leu Asp Ala Ala Ala Arg Ala
385 390 395 400
Leu His Glu Gln Phe Gln Leu Gly Gly Glu Asp Glu Ala Val Val Tyr
405 410 415
Ala Gly Thr Gly Arg
420
<210> 4
<211> 1266
<212> DNA
<213> Corynebacterium glutamicum
<400> 4
gtggccctgg tcgtacagaa atatggcggt tcctcgcttg agagtgcgga acgcattaga 60
aacgtcgctg aacggatcgt tgccaccaag aaggctggaa atgatgtcgt ggttgtctgc 120
tccgcaatgg gagacaccac ggatgaactt ctagaacttg cagcggcagt gaatcccgtt 180
ccgccagctc gtgaaatgga tatgctcctg actgctggtg agcgtatttc taacgctctc 240
gtcgccatgg ctattgagtc ccttggcgca gaagcccaat ctttcacggg ctctcaggct 300
ggtgtgctca ccaccgagcg ccacggaaac gcacgcattg ttgatgtcac tccaggtcgt 360
gtgcgtgaag cactcgatga gggcaagatc tgcattgttg ctggtttcca gggtgttaat 420
aaagaaaccc gcgatgtcac cacgttgggt cgtggtggtt ctgacaccac tgcagttgcg 480
ttggcagctg ctttgaacgc tgatgtgtgt gagatttact cggacgttga cggtgtgtat 540
accgctgacc cgcgcatcgt tcctaatgca cagaagctgg aaaagctcag cttcgaagaa 600
atgctggaac ttgctgctgt tggctccaag attttggtgc tgcgcagtgt tgaatacgct 660
cgtgcattca atgtgccact tcgcgtacgc tcgtcttata gtaatgatcc cggcactttg 720
attgccggct ctatggagga tattcctgtg gaagaagcag tccttaccgg tgtcgcaacc 780
gacaagtccg aagccaaagt aaccgttctg ggtatttccg ataagccagg cgaggctgcg 840
aaggttttcc gtgcgttggc tgatgcagaa atcaacattg acatggttct gcagaacgtc 900
tcttctgtag aagacggcac caccgacatc accttcacct gccctcgttc cgacggccgc 960
cgcgcgatgg agatcttgaa gaagcttcag gttcagggca actggaccaa tgtgctttac 1020
gacgaccagg tcggcaaagt ctccctcgtg ggtgctggca tgaagtctca cccaggtgtt 1080
accgcagagt tcatggaagc tctgcgcgat gtcaacgtga acatcgaaaa gatttccacc 1140
tctgagattc gtatttccgt gctgatccgt gaagatgatc tggatgctgc tgcacgtgca 1200
ttgcatgagc agttccagct gggcggcgaa gacgaagccg tcgtttatgc aggcaccgga 1260
cgctaa 1266
<210> 5
<211> 421
<212> PRT
<213> Corynebacterium glutamicum
<400> 5
Val Ala Leu Val Val Gln Lys Tyr Gly Gly Ser Ser Leu Glu Ser Ala
1 5 10 15
Glu Arg Ile Arg Asn Val Ala Glu Arg Ile Val Ala Thr Lys Lys Ala
20 25 30
Gly Asn Asp Val Val Val Val Cys Ser Ala Met Gly Asp Thr Thr Asp
35 40 45
Glu Leu Leu Glu Leu Ala Ala Ala Val Asn Pro Val Pro Pro Ala Arg
50 55 60
Glu Met Asp Met Leu Leu Thr Ala Gly Glu Arg Ile Ser Asn Ala Leu
65 70 75 80
Val Ala Met Ala Ile Glu Ser Leu Gly Ala Glu Ala Gln Ser Phe Thr
85 90 95
Gly Ser Gln Ala Gly Val Leu Thr Thr Glu Arg His Gly Asn Ala Arg
100 105 110
Ile Val Asp Val Thr Pro Gly Arg Val Arg Glu Ala Leu Asp Glu Gly
115 120 125
Lys Ile Cys Ile Val Ala Gly Phe Gln Gly Val Asn Lys Glu Thr Arg
130 135 140
Asp Val Thr Thr Leu Gly Arg Gly Gly Ser Asp Thr Thr Ala Val Ala
145 150 155 160
Leu Ala Ala Ala Leu Asn Ala Asp Val Cys Glu Ile Tyr Ser Asp Val
165 170 175
Asp Gly Val Tyr Thr Ala Asp Pro Arg Ile Val Pro Asn Ala Gln Lys
180 185 190
Leu Glu Lys Leu Ser Phe Glu Glu Met Leu Glu Leu Ala Ala Val Gly
195 200 205
Ser Lys Ile Leu Val Leu Arg Ser Val Glu Tyr Ala Arg Ala Phe Asn
210 215 220
Val Pro Leu Arg Val Arg Ser Ser Tyr Ser Asn Asp Pro Gly Thr Leu
225 230 235 240
Ile Ala Gly Ser Met Glu Asp Ile Pro Val Glu Glu Ala Val Leu Thr
245 250 255
Gly Val Ala Thr Asp Lys Ser Glu Ala Lys Val Thr Val Leu Gly Ile
260 265 270
Ser Asp Lys Pro Gly Glu Ala Ala Lys Val Phe Arg Ala Leu Ala Asp
275 280 285
Ala Glu Ile Asn Ile Asp Met Val Leu Gln Asn Val Ser Ser Val Glu
290 295 300
Asp Gly Thr Thr Asp Ile Thr Phe Thr Cys Pro Arg Ser Asp Gly Arg
305 310 315 320
Arg Ala Met Glu Ile Leu Lys Lys Leu Gln Val Gln Gly Asn Trp Thr
325 330 335
Asn Val Leu Tyr Asp Asp Gln Val Gly Lys Val Ser Leu Val Gly Ala
340 345 350
Gly Met Lys Ser His Pro Gly Val Thr Ala Glu Phe Met Glu Ala Leu
355 360 365
Arg Asp Val Asn Val Asn Ile Glu Met Ile Ser Thr Ser Glu Ile Arg
370 375 380
Ile Ser Val Leu Ile Arg Glu Asp Asp Leu Asp Ala Ala Ala Arg Ala
385 390 395 400
Leu His Glu Gln Phe Gln Leu Gly Gly Glu Asp Glu Ala Val Val Tyr
405 410 415
Ala Gly Thr Gly Arg
420
<210> 6
<211> 1266
<212> DNA
<213> Corynebacterium glutamicum
<400> 6
gtggccctgg tcgtacagaa atatggcggt tcctcgcttg agagtgcgga acgcattaga 60
aacgtcgctg aacggatcgt tgccaccaag aaggctggaa atgatgtcgt ggttgtctgc 120
tccgcaatgg gagacaccac ggatgaactt ctagaacttg cagcggcagt gaatcccgtt 180
ccgccagctc gtgaaatgga tatgctcctg actgctggtg agcgtatttc taacgctctc 240
gtcgccatgg ctattgagtc ccttggcgca gaagcccaat ctttcacggg ctctcaggct 300
ggtgtgctca ccaccgagcg ccacggaaac gcacgcattg ttgatgtcac tccaggtcgt 360
gtgcgtgaag cactcgatga gggcaagatc tgcattgttg ctggtttcca gggtgttaat 420
aaagaaaccc gcgatgtcac cacgttgggt cgtggtggtt ctgacaccac tgcagttgcg 480
ttggcagctg ctttgaacgc tgatgtgtgt gagatttact cggacgttga cggtgtgtat 540
accgctgacc cgcgcatcgt tcctaatgca cagaagctgg aaaagctcag cttcgaagaa 600
atgctggaac ttgctgctgt tggctccaag attttggtgc tgcgcagtgt tgaatacgct 660
cgtgcattca atgtgccact tcgcgtacgc tcgtcttata gtaatgatcc cggcactttg 720
attgccggct ctatggagga tattcctgtg gaagaagcag tccttaccgg tgtcgcaacc 780
gacaagtccg aagccaaagt aaccgttctg ggtatttccg ataagccagg cgaggctgcg 840
aaggttttcc gtgcgttggc tgatgcagaa atcaacattg acatggttct gcagaacgtc 900
tcttctgtag aagacggcac caccgacatc accttcacct gccctcgttc cgacggccgc 960
cgcgcgatgg agatcttgaa gaagcttcag gttcagggca actggaccaa tgtgctttac 1020
gacgaccagg tcggcaaagt ctccctcgtg ggtgctggca tgaagtctca cccaggtgtt 1080
accgcagagt tcatggaagc tctgcgcgat gtcaacgtga acatcgaaat gatttccacc 1140
tctgagattc gtatttccgt gctgatccgt gaagatgatc tggatgctgc tgcacgtgca 1200
ttgcatgagc agttccagct gggcggcgaa gacgaagccg tcgtttatgc aggcaccgga 1260
cgctaa 1266
<210> 7
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 1
<400> 7
tcctctagag ctgcgcagtg ttgaatacg 29
<210> 8
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 2
<400> 8
tggaaatctt ttcgatgttc acgttgacat 30
<210> 9
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 3
<400> 9
acatcgaaaa gatttccacc tctgagattc 30
<210> 10
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 4
<400> 10
tggaaatcat ttcgatgttc acgttgacat 30
<210> 11
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 5
<400> 11
acatcgaaat gatttccacc tctgagattc 30
<210> 12
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 6
<400> 12
gactctagag ttcacctcag agacgatta 29
<210> 13
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 7
<400> 13
tcctctagac tggtcgcctg atgttctac 29
<210> 14
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 8
<400> 14
gccaaaacct ccacgcgatc 20
<210> 15
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 9
<400> 15
atcgcgtgga ggttttggct 20
<210> 16
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 10
<400> 16
gactctagat tagtcccttt cgaggcgga 29
<210> 17
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 11
<400> 17
acggatccca gactccaaag caaaagcg 28
<210> 18
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 12
<400> 18
acaccacggc agaaccaggt gcaaaggaca 30
<210> 19
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 13
<400> 19
ctggttctgc cgtggtgtgc atcatctctg 30
<210> 20
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 14
<400> 20
acggatccaa ccaaacttgc tcacactc 28
<210> 21
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 15
<400> 21
tctagatgcg ctgattatct cacc 24
<210> 22
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 16
<400> 22
actggtgggt catggttgca tatgagatca actcctgtaa 40
<210> 23
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 17
<400> 23
ttacaggagt tgatctcata tgcaaccatg acccaccagt 40
<210> 24
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 18
<400> 24
tctagacctt gaagttcttg actg 24
<210> 25
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 19
<400> 25
tctagaagta gcgttgctgt acac 24
<210> 26
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 20
<400> 26
atcaatggtc tcgatgccca tatggcattt ggaggtcctt aag 43
<210> 27
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 21
<400> 27
cttaaggacc tccaaatgcc atatgggcat cgagaccatt gat 43
<210> 28
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 22
<400> 28
tctagatgga accgttgcaa ccac 24
<210> 29
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 23
<400> 29
ggatcccctc gttgttcacc cagcaacc 28
<210> 30
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 24
<400> 30
ggatcccaaa gtcacaacta cttatgttag 30
<210> 31
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 25
<400> 31
tctagaatgc ccaccctcgc gccttc 26
<210> 32
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 26
<400> 32
tctagattag atgtagaact cgatg 25
<210> 33
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 27
<400> 33
acgcgccagc gcctggaaca tcggcattca atccg 35
<210> 34
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 28
<400> 34
cggattgaat gccgatgttc caggcgctgg cgcgt 35
<210> 35
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 29
<400> 35
gtagataccg atattcggta cccctaccac cag 33
<210> 36
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> праймер 30
<400> 36
ctggtggtag gggtaccgaa tatcggtatc tac 33
<---
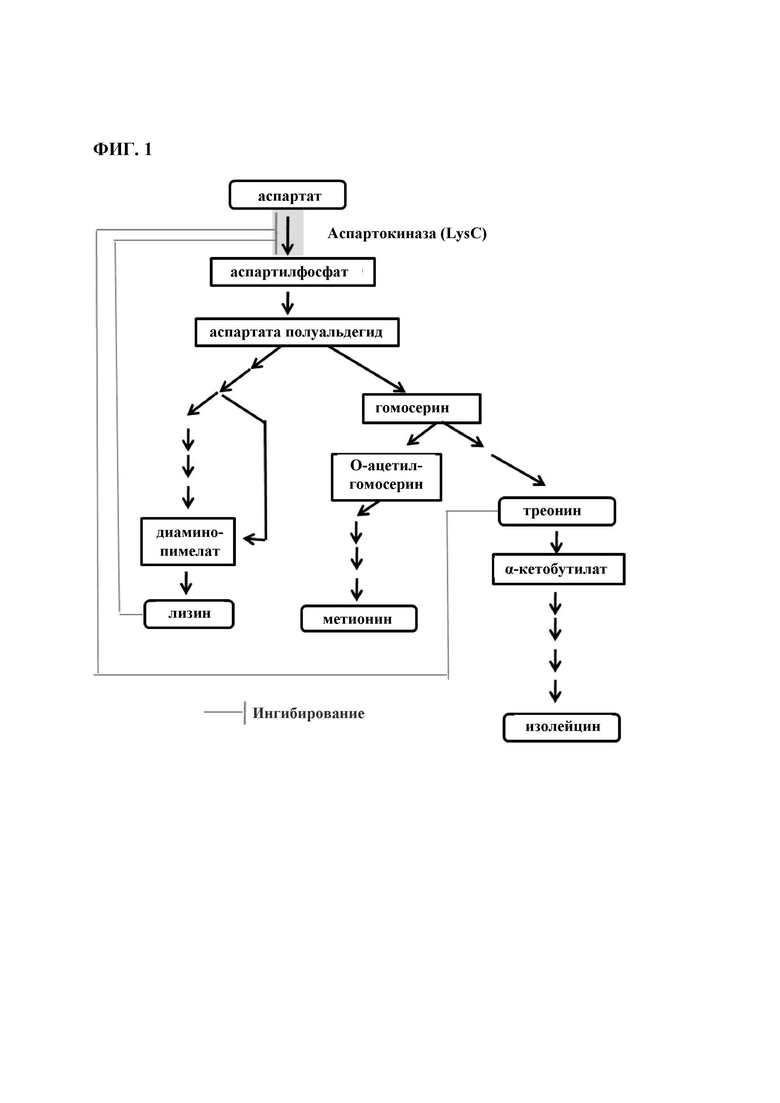
Изобретение относится к биотехнологии и представляет собой аспартакиназу, где аминокислота в положении 377 в аминокислотной последовательности SEQ ID NO: 1 заменена L-лизином или L-метионином. Изобретение относится также к микроорганизму рода Corynebacterium, продуцирующему L-аминокислоту, имеющую происхождение из аспартата, или производное этой аминокислоты, содержащему заявленную аспартокиназу. Изобретение позволяет получать L-аминокислоты, имеющие происхождение из аспартата, с высокой степенью эффективности. 6 н. и 7 з.п. ф-лы, 1 ил., 14 табл., 6 пр.
1. Аспартокиназа, где аминокислота в положении 377 в аминокислотной последовательности SEQ ID NO: 1 заменена L-лизином или L-метионином.
2. Полинуклеотид, кодирующий аспартокиназу по п. 1.
3. Микроорганизм рода Corynebacterium, продуцирующий L-аминокислоту, имеющую происхождение из аспартата, или производное этой аминокислоты, содержащий аспартокиназу по п. 1.
4. Микроорганизм по п. 3, где L-аминокислота, имеющая происхождение из аспартата, или производное этой аминокислоты выбраны из группы, состоящей из лизина, треонина, метионина, гомосерина, O-ацетилгомосерина, O-сукцинилгомосерина, изолейцина и кадаверина.
5. Микроорганизм по п. 3, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
6. Способ получения L-аминокислоты, имеющей происхождение из аспартата, включающий:
культивирование микроорганизма по п. 3 в среде; и выделение L-аминокислоты, имеющей происхождение из аспартата, из культивированного микроорганизма или культуральной среды.
7. Способ по п. 6, где L-аминокислота, имеющая происхождение из аспартата, выбрана из группы, состоящей из лизина, треонина, метионина, гомосерина и изолейцина.
8. Способ по п. 6, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
9. Способ получения производного L-аминокислоты, имеющей происхождение из аспартата, включающий:
культивирование микроорганизма по п. 3 в среде; и выделение указанного производного из культивированного микроорганизма или культуральной среды, где указанное производное представляет собой любое вещество, полученное путем биосинтеза с использованием аспартата в качестве предшественника, включая вещества, полученные из L-аминокислоты, и предшественники L-аминокислоты.
10. Способ по п. 9, где указанное производное выбрано из группы, состоящей из O-ацетилгомосерина, O-сукцинилгомосерина и кадаверина.
11. Способ по п. 9, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
12. Способ получения L-метионина, включающий:
культивирование микроорганизма по п. 3 в среде;
получение O-ацетилгомосерина или O-сукцинилгомосерина из культивированного микроорганизма или культуральной среды; и
превращение O-ацетилгомосерина или O-сукцинилгомосерина в L-метионин.
13. Способ по п. 12, где микроорганизм рода Corynebacterium представляет собой Corynebacterium glutamicum.
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 2000 |
|
RU2224793C1 |
| US 6221636 B1, 24.04.2001 | |||
| EP 699759 B1, 26.03.2003 | |||
| DE 69332805 D1, 30.04.2003 | |||
| DE 69330518 D1, 06.09.2001. | |||

