ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
Настоящая заявка испрашивает приоритет по предварительной заявке на патент США №62/293406, поданной 10 февраля 2016 года, которая включена в данный документ посредством ссылки во всей своей полноте.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
В данном документе описаны композиции, наборы, векторы и способы, включающие антитела, направленные на эпитопы, обнаруживаемые в липоарабиноманнане (LAM), липоманнане (LM) и фосфатидил-мио-инозитол-маннозиде 6 (PIM6), для диагностики, предупреждения и лечения инфекций, вызванных Mycobacterium tuberculosis.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка содержит перечень последовательностей, который был подан в формате ASCII с помощью EFS-WEB и настоящим включен посредством ссылки во всей своей полноте. Указанная ASCII-копия, созданная 1 февраля 2017 года, имеет название 096747.00337_ST25.txt, и ее размер составляет 29097 байт.
УРОВЕНЬ ТЕХНИКИ
A. Mycobacterium tuberculosis
Туберкулез (ТВ) остается одним из самых смертоносных инфекционных заболеваний в мире, при этом в настоящее время инфицирована приблизительно 1/3 мирового населения. Согласно Глобальному докладу о туберкулезе ВОЗ за 2014 год: туберкулез, по оценке в 2013 году ТВ развился у 9,0 миллиона людей, а 1,5 миллиона умерли от данного заболевания. Хотя в настоящее время доступны эффективные лекарственные средства для лечения ТВ, они требуют длительного лечения с применением нескольких антибиотиков, и они все больше компрометируются из-за развития штаммов с множественной лекарственной устойчивостью (MDR-TB), которые в настоящее время ответственны за приблизительно 3,5% последних случаев заражения. Эти штаммы намного сложнее лечить, и они характеризуются значительно более низкими уровнями излечения. Также распространяются штаммы ТВ с широкой лекарственной устойчивостью (XDR-TB), которые даже более дорогие и сложные для лечения, чем штаммы MDR-TB, и к настоящему времени о них сообщалось в 100 странах по всему миру. Следовательно необходимы новые подходы для более ранней диагностики и лечения ТВ-инфекций.
В. Липоарабиноманнан (LAM)
Гликолипид липоарабиноманнан (LAM) является главным структурным и антигенным компонентом клеточной стенки представителей комплекса Mycobacterium tuberculosis, и он опосредует целый ряд важных функций, которые ускоряют продуктивную инфекцию и развитие заболевания. LAM также является важной мишенью для иммунодиагностики, служащей для выявления активной инфекции при ТВ, в частности, у пациентов, инфицированных одновременно HIV-1, а также потенциальной вакцинной мишенью. Несмотря на важность LAM в качестве мишени для иммунодиагностики и его важную роль в физиологии и патогенности M.tb при инфицировании, удивительно мало известно о природе гуморального ответа человека на данный антиген. Доступные ранее LAM-специфические моноклональные антитела были получены от мышей, иммунизированных LAM, очищенным либо из Mycobacterium leprae, либо из Mycobacterium tuberculosis, и при этом отсутствовали данные о каких-либо человеческих моноклональных антителах против LAM, которые были индуцированы в ответ либо на иммунизацию, либо на инфекцию, вызванную Mycobacterium tuberculosis.
Липоманнан (LM) представляет собой промежуточный предшественник LAM и содержит фосфатидил-мио-инозитольный домен, модифицированный маннановым доменом, состоящим из α(1→6)-связанного остова Manp, замещенного короткими α(1→2)--маннопиранозильными боковыми цепями, но не содержащего арабинозные боковые цепи.
С. Фосфатидил-мио-инозитол-маннозид 6 (PIM6)
PIM6 является продуктом PIM2, общим предшественником LM и LAM. Коровой частью этих молекул является мио-инозитольная структура, гликозилированная структурной единицей Manp в положениях 2 и 6. В PIM6 структурная единица Manp в положении 6 дополнительно замещена двумя концевыми α-Manp(1→2)-связанными остатками сахара, идентичными маннозному кэпу на ManLAM. Данные молекулы ацилированы с помощью вплоть до 4 жирнокислотных цепей, прикрепленных к группе инозитольной головки и к коровому остатку Man, которые нековалентно заякоривают данные молекулы во внутренней и наружной мембранах клеточной оболочки. Сообщалось, что PIM6 связывается с лектинами С-типа и DC-SIGN, главным рецептором на дендритных клетках, и является сильным агонистом TLR2 и усилителем репликации HIV-1, обладающим сильной противовоспалительной активностью.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В данном документе описаны новые моноклональные антитела (mAb) к LAM и к PIM6/LAM для диагностики и лечения инфекций, вызванных Mycobacterium tuberculosis. Ниже описано выделение и определение характеристик этих новых человеческих антител, специфических в отношении гликолипидов Mycobacterium tuberculosis, включая человеческие mAb, специфические в отношении эпитопов LAM, и человеческое mAb, специфическое в отношении эпитопа, общего для LAM и PIM6.
Соответственно, в данном документе описано человеческое моноклональное антитело к липоарабиноманнану (к LAM) или его антигенсвязывающая часть, которые специфически связываются с эпитопом LAM, включая структуру Ara4, структуру Ara6 или их комбинацию, где антитело к LAM включает CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами. Человеческое моноклональное антитело к LAM или его антигенсвязывающая часть могут включать вариабельный участок тяжелой цепи, включающий аминокислотные последовательности под SEQ ID NO: 21 и SEQ ID NO: 23, и вариабельный участок легкой цепи, включающий аминокислотные последовательности под SEQ ID NO: 24 и SEQ ID NO: 26. Антителом к LAM может быть антитело, представляющее собой scFv-IgG и IgGa или IgM. Примером антитела к LAM является А194.
В данном документе также описано человеческое моноклональное антитело к LAM или его антигенсвязывающая часть, которые специфически связываются с эпитопом LAM, включающим по меньшей мере одно из кэпированной маннозой структуры Ara4 и кэпированной маннозой структуры Ara6. Антитело к LAM может включать CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 7 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 8 или ее антигенными фрагментами, СОЮ вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 9 или SEQ ID NO: 32 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 10 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 11 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 12 или SEQ ID NO: 29 или их антигенными фрагментами. Антитело может включать вариабельный участок тяжелой цепи, включающий аминокислотную последовательность под SEQ ID NO: 43, и вариабельный участок легкой цепи, включающий аминокислотную последовательность под SEQ ID No: 44. Антителом к LAM может быть, например, антитело, представляющее собой IgM или IgA. Примером антитела к LAM является Р3В09.
В данном документе дополнительно описано человеческое моноклональное антитело к LAM или его антигенсвязывающая часть, которые специфически связываются с эпитопом LAM, включающим α-Manp(1→2)-связанную структуру, прикрепленную к невосстанавливающему концу Ara4 или Ara6, где антитело к LAM включает CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 7 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 8 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 9 или ее антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 10 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 11 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 12 или ее антигенными фрагментами. Антителом к LAM (например, Р3В09) может быть, например, антитело, представляющее собой IgM или IgA.
В данном документе еще дополнительно описано человеческое моноклональное антитело к PIM6/LAM или его антигенсвязывающая часть, которые специфически связываются с эпитопом, присутствующим в LAM и PIM6, причем эпитоп содержит по меньшей мере одну полиманнозную структуру. Эпитоп находится в маннановом домене PIM6 и также присутствует в микобактериальном липоманнане (LM). Антитело к PIM6/LAM может включать CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 13 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 14 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 15 или ее антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 16 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 17 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 18 или ее антигенными фрагментами. Например, антитело может включать вариабельный участок тяжелой цепи, включающий аминокислотную последовательность под SEQ ID NO: 47, и вариабельный участок легкой цепи, включающий аминокислотную последовательность под SEQ ID NO: 48. Антителом к PIM6/LAM может быть, например, антитело, представляющее собой IgM, IgA или IgG. Примером антитела к PIM6/LAM является Р95С1.
В данном документе также описан набор для выявления по меньшей мере одного эпитопа LAM. Набор включает (а) по меньшей мере первое антитело к LAM, которое специфически связывается с эпитопом LAM; (b) подложку, с которой связано по меньшей мере первое антитело к LAM; (с) выявляющее антитело, которое специфически связывается с LAM или специфически связывается с по меньшей мере первым антителом к LAM, где выявляющее антитело помечено репортерной молекулой; и (d) буфер. По меньшей мере первое антитело к LAM представляет собой, например, человеческое моноклональное антитело к LAM, как описано в данном документе. Выявляющее антитело может представлять собой, например, второе антитело к LAM, которое специфически связывается с LAM. В некоторых вариантах осуществления по меньшей мере одно из первого антитела к LAM и второго антитела к LAM является антителом, представляющим собой scFv-IgG или IgM, и оно включает CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами. В некоторых вариантах осуществления набора по меньшей мере одно из первого антитела к LAM и второго антитела к LAM включает вариабельный участок тяжелой цепи, включающий аминокислотные последовательности под SEQ ID NO: 21 и SEQ ID NO: 23, и вариабельный участок легкой цепи, включающий аминокислотные последовательности под SEQ ID NO: 24 и SEQ ID NO: 26.
Более того, в данном документе описан способ диагностирования активной туберкулезной инфекции у индивидуума, включающий: (а) получение от индивидуума образца, который включает или предположительно включает LAM; (b) обработку указанного образца для демаскирования эпитопов LAM индивидуума; (с) приведение указанного образца в контакт с по меньшей мере первым антителом, которое специфически связывается с первым эпитопом на указанном LAM; (d) приведение указанного образца в контакт с выявляющим антителом, которое специфически связывается с LAM или специфически связывается с по меньшей мере первым антителом; (е) выявление связывания по меньшей мере первого антитела с указанным первым эпитопом на LAM и (f) диагностирование указанного пациента как имеющего активную туберкулезную инфекцию, при этом связывание по меньшей мере первого антитела с указанным первым эпитопом на LAM является показателем активной туберкулезной инфекции. По меньшей мере первое антитело представляет собой, например, человеческое моноклональное антитело к LAM или человеческое моноклональное антитело к PIM6/LAM, как описано в данном документе. Выявляющее антитело может представлять собой, например, антитело к LAM, которое специфически связывается с LAM. В некоторых вариантах осуществления способа каждое из по меньшей мере первого антитела и выявляющего антитела включает CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами. В некоторых вариантах осуществления способа по меньшей мере одно из первого антитела и выявляющего антитела является антителом, представляющим собой scFv-IgG или IgM, и оно включает участок CDR1 в составе вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами. В некоторых вариантах осуществления индивидуум является человеком.
Также в данном документе описан способ лечения туберкулезной инфекции у индивидуума (например, человека). Способ включает введение указанному индивидууму терапевтически эффективного количества по меньшей мере одного человеческого моноклонального антитела к LAM или человеческого моноклонального антитела к PIM6/LAM, как описано в данном документе. Способ может дополнительно включать введение указанному индивидууму терапевтически эффективного количества по меньшей мере одного антибиотика. Туберкулезная инфекция может представлять собой туберкулезную инфекцию с множественной лекарственной устойчивостью (MDR-TB).
В данном документе дополнительно описаны нуклеотидные последовательности, кодирующие тяжелые цепи и легкие цепи (включая вариабельные участки) антител, описанных в данном документе.
А. Антитела к LAM и антитела к PIM6/LAM
В некоторых вариантах осуществления настоящего изобретения представлены антитело к LAM или его антигенсвязывающая часть. В некоторых вариантах осуществления настоящего изобретения представлены антитело к PIM6/LAM или его антигенсвязывающая часть. Антитело к LAM (или его антигенсвязывающая часть), как описано в данном документе, специфически связываются с эпитопом LAM. Антитело к PIM6/LAM (или его антигенсвязывающая часть), как описано в данном документе, специфически связываются как с эпитопом LAM, так и с эпитопом PIM6. В некоторых вариантах осуществления эпитопы LAM и PIM6 получены из различных видов микобактерий. В дополнительных вариантах осуществления различные виды микобактерий представляют собой вирулентных представителей комплекса Mycobacterium tuberculosis. В еще одних вариантах осуществления видом микобактерий является Mycobacterium tuberculosis. В некоторых вариантах осуществления антитело к LAM или антитело к PIM6/LAM представляют собой моноклональное антитело (mAb). В дополнительных вариантах осуществления антитело к LAM или антитело к PIM6/LAM представляют собой человеческое моноклональное антитело к LAM или человеческое моноклональное антитело к PIM6/LAM соответственно. В других вариантах осуществления антитело к LAM или антитело к PIM6/LAM представляют собой гуманизированное моноклональное антитело к LAM или антитело к PIM6/LAM соответственно. В некоторых вариантах осуществления антитело к LAM связывается со структурами Ara4 и Ara6.
В некоторых вариантах осуществления эпитоп LAM представляет собой некэпированную арабинозную цепь. В некоторых вариантах осуществления эпитоп LAM представляет собой некэпированную или кэпированную единичной маннозой арабинозную цепь с концевой заменой МТХ или без нее.
В некоторых вариантах осуществления эпитоп LAM представляет собой кэпированную маннозой структуру Ara4 и кэпированную маннозой структуру Ara6. В других вариантах осуществления антитело к LAM специфически связывается с α(1→2)-связанной диманнозной структурой, которая может быть присоединена либо к структуре Ara4/Ara6, либо к полиманнозной структуре (ФИГ. 8). В некоторых вариантах осуществления эпитоп PIM6 включает по меньшей мере одну полиманнозную структуру, также присутствующую в микобактериальном липоманнане (LM). В некоторых вариантах осуществления антитело к PIM6/LAM специфически связывается с эпитопом PIM6, который включает по меньшей мере одну полиманнозную структуру в маннановом домене PIM6. В некоторых вариантах осуществления эпитоп LAM включает по меньшей мере одну метилтиоксилозную (МТХ) или метилсульфинилксилофуранозильную (MSX) замену. В некоторых вариантах осуществления эпитоп LAM включает по меньшей мере одну фосфатидил-мио-инозитольную замену (PILAM). В некоторых вариантах осуществления эпитоп LAM представляет собой арабинозную цепь, кэпированную с помощью по меньшей мере одной маннозы, т.е. маннозилированный эпитоп Man-LAM. В дополнительных вариантах осуществления кэпированная арабинозная цепь включает структуры Ara4 и/или Ara6. В некоторых вариантах осуществления эпитоп Man-LAM включает замещенные мономаннозой арабинозные цепи, замещенные диманнозой арабинозные цепи, замещенные триманнозой арабинозные цепи или их комбинации. В некоторых вариантах осуществления эпитоп Man-LAM включает кэпированные диманнозой или триманнозой структуры Ara4 и/или Ara6. В некоторых вариантах осуществления эпитоп Man-LAM представляет собой кэпированную диманнозой Ara6. В некоторых вариантах осуществления антитело к LAM или антитело к PIM6/LAM включают антитело, представляющее собой IgG. В дополнительных вариантах осуществления антитело, представляющее собой IgG, к LAM или антитело к PIM6/LAM включает подкласс IgG1, IgG2 или IgG3. В некоторых вариантах осуществления антитело к LAM или антитело к PIM6/LAM не являются антителом, представляющим собой IgG. В других вариантах осуществления антитело к LAM или антитело к PIM6/LAM включают антитело, представляющее собой IgA. В других вариантах осуществления антитело к LAM или антитело к PIM6/LAM включают антитело, представляющее собой IgM. В некоторых вариантах осуществления антитело к LAM или антитело к PIM6/LAM включают второй изотип, на который было выполнено переключение из изотипа, который был изначально выделен. В некоторых вариантах осуществления антитело к LAM или антитело к PIM6/LAM включают рекомбинантное антитело. В некоторых вариантах осуществления рекомбинантное антитело включает мультивалентное антитело, представляющее собой IgM. В дополнительных вариантах осуществления рекомбинантное антитело включает пентавалентное антитело, представляющее собой IgM. В других вариантах осуществления рекомбинантное антитело включает антитело, представляющее собой ScFv-IgG, в котором одноцепочечный Fv-фрагмент одного антитела присоединен к N-концу тяжелой цепи того же или отличного mAb к LAM. В дополнительных вариантах осуществления рекомбинантное антитело включает мультивалентное антитело, представляющее собой ScFv-IgG. В дополнительных вариантах осуществления рекомбинантное антитело включает гомологичное тетравалентное антитело, представляющее собой ScFv-IgG, в котором scFv-домены получены из вариабельных участков IgG, присутствующего в конструкции. В еще одних вариантах осуществления рекомбинантное антитело включает гетерологичное тетрамерное антитело, представляющее собой scFv-IgG, в котором участки scFv получены из антитела к LAM или антитела к PIM6/LAM, отличающегося от такового, в который включен участок IgG. В некоторых вариантах осуществления scFv-домен включает лидерную последовательность, присоединенную к вариабельному участку тяжелой цепи (VH) второго антитела к LAM или антитела к PIM6/LAM, который присоединен к вариабельному домену легкой цепи (VL) указанного антитела к LAM или антитела к PIM6/LAM. В других вариантах осуществления scFv-домен включает лидерную последовательность, присоединенную к вариабельному участку легкой цепи первого антитела к LAM или антитела к PIM6/LAM, который присоединен к вариабельному участку тяжелой цепи (VH) второго антитела к LAM или антитела к PIM6/LAM. В некоторых вариантах осуществления антитело к LAM представляет собой выделенное антитело к LAM, которое специфически связывается с эпитопом LAM (например, с одной из Ara4 и Ara6 или их комбинаций, с α(1→2)-связанной диманнозной структурой, которая может быть присоединена к одной из структуры Ara4/Ara6, или с полиманнозной структурой). В некоторых вариантах осуществления антитело к LAM не конкурирует с CS-35 и FIND25. В некоторых вариантах осуществления антитело к PIM6/LAM представляет собой выделенное антитело к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в микобактериальном липоманнане (LM).
В некоторых вариантах осуществления антитело к LAM или антитело к PIM6/LAM включает гибкий линкер. В некоторых вариантах осуществления гибкий линкер соединяет соответствующие домены тяжелой и легкой цепи в одноцепочечную молекулу. В некоторых вариантах осуществления гибкий линкер соединяет легкую цепь иммуноглобулина (IgVL) с тяжелой цепью иммуноглобулина (IgVH). В дополнительных вариантах осуществления гибкий линкер соответствует формуле (GGSGG)n (SEQ ID NO: 19), где n представляет собой любое положительное число от 1 до 200 и любые диапазоны между ними, например, от 1 до 5, от 1 до 10, от 1 до 15, от 1 до 25, от 1 до 50, от 5 до 10, от 5 до 25, от 10 до 25, от 10 до 50, от 1 до 100, от 1 до 150 и все промежуточные интервалы.
В некоторых вариантах осуществления антитело к LAM (например, Р30В9, А194-01) имеет по меньшей мере один (например, один, два, три) определяющий комплементарность участок (CDR) (например CDR1, CDR2, CDR3). В некоторых вариантах осуществления CDR1 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 1 или ее антигенных фрагментов. В других вариантах осуществления участок CDR1 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 7 или ее антигенных фрагментов. В других вариантах осуществления участок CDR1 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 13 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR1 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 4 или ее антигенных фрагментов. В других вариантах осуществления участок CDR1 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 10 или ее антигенных фрагментов. В других вариантах осуществления участок CDR1 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 16 или ее антигенных фрагментов.
В некоторых вариантах осуществления CDR2 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 2 или ее антигенных фрагментов. В других вариантах осуществления CDR2 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 8 или ее антигенных фрагментов. В других вариантах осуществления CDR2 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 14 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR2 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 5 или ее антигенных фрагментов. В других вариантах осуществления участок CDR2 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 11 или ее антигенных фрагментов. В других вариантах осуществления участок CDR2 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 17 или ее антигенных фрагментов.
В некоторых вариантах осуществления CDR3 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 3 или ее антигенных фрагментов. В других вариантах осуществления CDR3 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 9 или ее антигенных фрагментов. В других вариантах осуществления CDR3 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 15 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR3 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 6 или ее антигенных фрагментов. В других вариантах осуществления участок CDR3 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 12 или ее антигенных фрагментов. В других вариантах осуществления участок CDR3 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 18 или ее антигенных фрагментов.
В некоторых вариантах осуществления антитело к PIM6/LAM (например, Р95С1) имеет по меньшей мере один (например, один, два, три) CDR (например, CDR1, CDR2, CDR3). В некоторых вариантах осуществления CDR1 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 13 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR1 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 16 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR2 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 14 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR2 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 17 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR3 вариабельного участка легкой цепи состоит фактически из SEQ ID NO: 15 или ее антигенных фрагментов. В некоторых вариантах осуществления CDR3 вариабельного участка тяжелой цепи состоит фактически из SEQ ID NO: 18 или ее антигенных фрагментов.
В. Диагностические наборы и способы
В некоторых вариантах осуществления настоящего изобретения представлены наборы для выявления присутствия LAM и/или PIM6 в биологических жидкостях субъекта-человека. В некоторых вариантах осуществления компоненты данных анализов собраны в виде устройства с горизонтальным потоком (см. World Health Organization 2015, The use of lateral flow lipoarabinomannan assay (LF-LAM) for the diagnosis and screening of active tuberculosis in people living with HIV). В некоторых вариантах осуществления наборы включают первое захватывающее антитело к LAM (например, А194-01, Р30 В9) или к PIM6/LAM (например, Р95С1), второе детекторное (выявляющее) антитело к LAM или к PIM6/LAM, помеченное с помощью репортерной молекулы, подложку, с которой связано первое антитело к LAM или к PIM6/LAM, и буфер. В некоторых вариантах осуществления по меньшей мере одно из первого антитела к LAM или к PIM6/LAM и второго антитела к LAM или к PIM6/LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций, или человеческое моноклональное антитело к PIM6/LAM, которое специфически связывается с маннановым доменом LAM (и липоманнана (LM)). В некоторых вариантах осуществления первое антитело к LAM и второе антитело к LAM связываются с теми же эпитопами LAM, которые присутствуют в виде множественных копий на одной молекуле LAM. В других вариантах осуществления первое антитело к LAM и второе антитело к LAM связываются с различными эпитопами, присутствующими на одной молекуле LAM. Эпитопы LAM и PIM6 могут представлять собой любой из эпитопов LAM и PIM6, описанных в данном документе. В других вариантах осуществления включено третье детекторное (выявляющее) антитело, которое связывается с неконкурирующим сайтом второго антитела. В некоторых вариантах осуществления первое антитело и второе антитело относятся к одному и тому же изотипу. В других вариантах осуществления первое антитело и второе антитело относятся к разным изотипам. В некоторых вариантах осуществления анализа захвата только одно из захватывающего антитела или выявляющего антитела представляет собой антитело к LAM (например, А194-01, Р30В9) или антитело к PIM6/LAM (например, Р95С1), как описано в данном документе.
Антитела, описанные в данном документе, можно применять для дополнительного выявления и применений в диагностике. Например, в одном диагностическом анализе одно или несколько антител, описанных в данном документе (например, А194-01, Р30В9, Р95С1), можно применять для окрашивания тканей, полученных от пациентов, для выявления присутствия LAM в очагах, предположительно содержащих ТВ, или клетках, инфицированных ТВ (например, в гранулемах). Его можно выполнять, например, с помощью одного антитела, описываемого в данном документе (например, А194-01, Р30В9, Р95С1), которое конъюгировано с меткой, которая обеспечивает возможность чувствительного выявления. При таком способе или анализе в инфицированных тканях можно осуществлять выявление PIM6 или родственных молекул с помощью Р95С1. В другом примере Р95С1 можно применять в конкурентном анализе PIM6, в котором происходит конкурирование за захват иммобилизированным Р95С1 между меченой формой PIM6 и растворимым PIM6, присутствующим в биологической жидкости (например, крови или моче) человека с подозрением на заболевание. В отсутствие растворимого PIM6 это будет приводить к захвату сигнала, с которым будет происходить конкуренция в случае присутствия растворимого PIM6 (см. World Health Organization, 2015 Policy Guidance- The use of lateral flow urine lipoarabinomannan assay (LF-LAM) for the diagnosis and screening of active tuberculosis in people living with HIV).
В некоторых вариантах осуществления настоящего изобретения представлены наборы для различения между патогенным представителем комплекса Mycobacterium tuberculosis и непатогенным представителем комплекса Mycobacterium tuberculosis. В некоторых вариантах осуществления антитело к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из структур Ara4 и Ara6 с заменой Man или МТХ-Man или без нее или их комбинациями, или антитело к PIM6/LAM, которое специфически связывает по меньшей мере одну полиманнозную структуру в PIM6 или в маннановом домене LAM. В некоторых вариантах осуществления антитело к LAM специфически связывается с эпитопом Man-LAM, включающим замещенные диманнозой боковые цепи, замещенные триманнозой боковые цепи или их комбинации. В дополнительных вариантах осуществления антитело к LAM специфически связывается с эпитопами Man-LAM, включающими кэпированные диманнозой или триманнозой структуры Ara4 и/или Ara6. В еще одних вариантах осуществления антитело к LAM специфически связывается с кэпированными диманнозой структурами Ara6.
В некоторых вариантах осуществления настоящего изобретения представлены способы диагностирования активной туберкулезной инфекции у индивидуума. В некоторых вариантах осуществления антитело к LAM или к PIM6/LAM можно модифицировать с помощью чувствительной метки и применять для идентификации микобактериального PIM6 или LAM-родственного материала в образце ткани в качестве диагностического критерия ТВ-инфекции и ее локализации. В некоторых вариантах осуществления способ предусматривает захват растворимого LAM и включает стадии: (а) получения от индивидуума образца, который включает LAM; (b) обработки образца для выделения или демаскирования указанного LAM, (с) захвата указанного выделенного или демаскированного LAM с помощью первого антитела к LAM, которое связывается с первым эпитопом на указанном LAM; (d) приведения указанного выделенного или демаскированного LAM в контакт со вторым антителом к LAM, где указанное второе антитело к LAM связывается со вторым эпитопом на указанном LAM; (е) выявления связывания по меньшей мере одного из указанного первого антитела к LAM и указанного второго антитела к LAM с указанным LAM; и (f) диагностирования указанного пациента как имеющего активную туберкулезную инфекцию, где указанное присутствие связывания по меньшей мере одного из указанного первого антитела к LAM и указанного второго антитела к LAM с указанным LAM является показателем активной туберкулезной инфекции. В некоторых вариантах осуществления по меньшей мере одно из первого антитела к LAM и второго антитела к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В некоторых вариантах осуществления по меньшей мере одно из первого и второго антител представляет собой человеческое моноклональное антитело к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене LAM. В дополнительных вариантах осуществления первое антитело и второе антитело относятся к разным изотипам. В некоторых вариантах осуществления по меньшей мере одно из первого антитела и второго антитела представляют собой рекомбинантные антитела. В других вариантах осуществления ни первое антитело, ни второе антитело не представляют собой рекомбинантные антитела. Дополнительно в других вариантах осуществления как первое антитело, так и второе антитело представляют собой рекомбинантные антитела.
В некоторых вариантах осуществления настоящего изобретения представлены способы количественной оценки содержания LAM и/или PIM6, присутствующих в образце. В некоторых вариантах осуществления способ включает стадии: (а) получения образца, который включает LAM и/или PIM6; (b) приведения указанного образца в контакт с антителом к LAM и/или антителом к PIM6; (с) выявления присутствия специфического связывания антитела к LAM с указанным LAM и/или связывания антитела к PIM6/LAM с указанным LAM или указанным PIM6; и (d) количественной оценки содержания LAM или PIM6 в указанном образце. В некоторых вариантах осуществления антитело к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В некоторых вариантах осуществления антитело к PIM6/LAM представляет собой человеческое моноклональное антитело к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6 (например, с по меньшей мере одной полиманнозной структурой в микобактериальном липоманнане (LM)). В некоторых вариантах осуществления количественную оценку указанного содержания LAM и/или PIM6 осуществляют путем сравнения интенсивности сигнала с сигналом у серийно разведенного контрольного образца с известной концентрацией LAM и/или PIM6.
В некоторых вариантах осуществления настоящего изобретения представлены способы диагностирования индивидуума как инфицированного Mycobacterium tuberculosis. В некоторых вариантах осуществления способ включает стадии: (а) получения образца, который включает LAM или PIM6; (b) приведения указанного образца в контакт с антителом к LAM и/или антителом к PIM6, где антитело к LAM специфически связывается с эпитопом LAM, включая Man-LAM, имеющим по меньшей мере одну 5-дезокси-5-метилтиопентофуранозильную (МТХ) замену, а антитело к PIM6/LAM специфически связывается с эпитопом, включающим по меньшей мере одну полиманнозную структуру в маннановом домене LAM, и (с) обнаружения присутствия специфического связывания антитела к LAM с указанным Man-LAM и/или присутствия специфического связывания антитела к PIM6/LAM с указанным PIM6. В некоторых вариантах осуществления антитело к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В некоторых вариантах осуществления антитело к PIM6/LAM представляет собой человеческое моноклональное антитело к PIM6/LAM (например, Р95С1), которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6.
В некоторых вариантах осуществления способ включает стадию амплификации, которая повышает чувствительность способа выявления. Примеры предусматривают получение дополнительных целевых сайтов путем применения набора для амплификации сигнала с помощью тирамида (Perkin-Elmer) или амплификации исходного сигнала путем применения системы амплификации для ELISA (Thermo Fisher).
В некоторых вариантах осуществления настоящего изобретения представлены способы разграничения патогенного представителя комплекса Mycobacterium tuberculosis и непатогенного представителя комплекса Mycobacterium tuberculosis. В некоторых вариантах осуществления способ предусматривает стадии: (а) получения образца, который содержит LAM и/или PIM6; (b) приведения указанного образца в контакт с антителом к LAM, которое специфически связывается с эпитопом Man-LAM, который включает замещенные диманнозой боковые цепи, замещенные триманнозой боковые цепи или их комбинации, или с антителом к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6; и (с) выявления присутствия специфического связывания антитела к LAM с указанным Man-LAM или присутствия специфического связывания антитела к PIM6/LAM с указанной по меньшей мере одной полиманнозной структурой в маннановом домене PIM6, где присутствие указанного специфического связывания является показателем присутствия патогенного представителя комплекса Mycobacterium tuberculosis. В некоторых вариантах осуществления антитело к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В дополнительных вариантах осуществления эпитоп Man-LAM включает кэпированные диманнозой или триманнозой структуры Ara4 и/или Ara6. В еще одних вариантах осуществления эпитоп Man-LAM представляет собой кэпированную диманнозой Ara6. В некоторых вариантах осуществления антитело к PIM6/LAM представляет собой человеческое моноклональное антитело к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6.
С. Терапевтические композиции, способы, вакцины и векторы
В некоторых вариантах осуществления настоящего изобретения представлены способы лечения инфекции, вызванной вирулентным представителем комплекса Mycobacterium tuberculosis, у индивидуума. В некоторых вариантах осуществления способ включает введение терапевтически эффективного количества по меньшей мере одного антитела к LAM или антитела к PIM6/LAM индивидууму, находящемуся под воздействием инфекционного M.tb. В дополнительных вариантах осуществления способ предусматривает введение по меньшей мере одного антибиотика. В некоторых вариантах осуществления ТВ-инфекция является активной. В других вариантах осуществления ТВ-инфекция является латентной. В некоторых вариантах осуществления инфекция вызвана штаммом туберкулеза с множественной лекарственной устойчивостью (MDR). В других вариантах осуществления инфекция вызвана штаммом туберкулеза с широкой лекарственной устойчивостью (XDR).
В некоторых вариантах осуществления настоящего изобретения представлена комбинированная терапия для лечения инфекции, вызванной вирулентным представителем комплекса Mycobacterium tuberculosis, у индивидуума. В некоторых вариантах осуществления способ включает введение терапевтически эффективного количества первого антитела к LAM, которое специфически связывается с первым эпитопом LAM, включающим по меньшей мере одно из незамещенного LAM, мономаннозилированного Man-LAM, MSX-замещенного LAM и их комбинаций, или первого антитела к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6 и LAM; и введение терапевтически эффективного количества второго антитела к LAM, которое специфически связывается со вторым эпитопом LAM, включающим по меньшей мере одно из замещенного диманнозой Man-LAM, замещенного триманнозой Man-LAM и их комбинаций. В некоторых вариантах осуществления первое антитело и второе антитело вводят одновременно (например, в одной композиции или в двух композициях, вводимых в одно и то же время). В других вариантах осуществления первое антитело и второе антитело вводят в разные моменты времени. В некоторых вариантах осуществления по меньшей мере одно из первого антитела к LAM и второго антитела к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В некоторых вариантах осуществления антитело к PIM6/LAM представляет собой человеческое моноклональное антитело к PIM6, которое специфически связывается с по меньшей мере одной полиманнозной структурой в PIM6 и/или в перекрестно реагирующем маннановом домене PIM6 из LAM. В некоторых вариантах осуществления первое антитело к LAM и второе антитело к LAM или антитело к PIM6/LAM относятся к различным изотипам. В некоторых вариантах осуществления по меньшей мере одно из первого антитела к LAM и второго антитела к LAM и антитела к PIM6/LAM представляют собой рекомбинантные антитела. В других вариантах осуществления ни первое антитело к LAM, ни второе антитело к LAM, ни антитело к PIM6/LAM не представляют собой рекомбинантные антитела. Дополнительно в других вариантах осуществления как первое антитело к LAM, так и второе антитело к LAM или антитело к PIM6/LAM представляют собой рекомбинантные антитела. В дополнительных вариантах осуществления способ предусматривает введение по меньшей мере одного антибиотика. В таких вариантах осуществления по меньшей мере один антибиотик можно вводить (например, вводить совместно) одновременно с первым и вторым антителами, или по меньшей мере один антибиотик можно вводить в момент времени, отличный от момента времени введения первого и второго антител. В некоторых вариантах осуществления инфекция является активной. В других вариантах осуществления инфекция является латентной. В некоторых вариантах осуществления инфекция представляет собой туберкулезную инфекцию с множественной лекарственной устойчивостью (MDR). В других вариантах осуществления инфекция представляет собой туберкулезную инфекцию с широкой лекарственной устойчивостью (XDR).
В некоторых вариантах осуществления настоящего изобретения представлены вакцины или фармацевтические композиции для предупреждения инфекции, вызванной вирулентным представителем комплекса Mycobacterium tuberculosis. В некоторых вариантах осуществления настоящего изобретения представлен способ стимуляции иммунного ответа хозяина у пациента, включающий введение указанному пациенту терапевтически эффективного количества антигена LAM и/или антигена PIM6. В некоторых вариантах осуществления данные антигены конъюгированы с иммуногенными белками-носителями и/или вводятся совместно с адъювантом, который эффективно стимулирует иммунный ответ на гликолипидные антигены. В некоторых вариантах осуществления вакцинная или фармацевтическая композиция индуцируют антитело к LAM, которое специфически связывается с эпитопом Man-LAM, и/или антитело к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6. В дополнительных вариантах осуществления эпитоп Man-LAM, присутствующий в вакцинной или фармацевтических композициях, включает кэпированные диманнозой или триманнозой структуры Ara4 и/или Ara6. В еще одних вариантах осуществления эпитоп Man-LAM представляет собой кэпированную диманнозой Ara6. В некоторых вариантах осуществления эпитоп Man-LAM имеет по меньшей мере одну замену МТХ. В некоторых вариантах осуществления антителом к LAM и/или антителом к PIM6/LAM является антитело, представляющее собой IgM. В других вариантах осуществления антитело к LAM и/или антитело к PIM6/LAM представляет собой рекомбинантное антитело.
В некоторых вариантах осуществления настоящего изобретения представлен способ предупреждения инфекции, вызванной вирулентным представителем комплекса Mycobacterium tuberculosis, у индивидуума с помощью пассивного введения защитного антитела. В некоторых вариантах осуществления антитело к LAM представляет собой человеческое моноклональное антитело к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В некоторых вариантах осуществления антитело к PIM6/LAM представляет собой человеческое моноклональное антитело к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6 и LAM. В некоторых вариантах осуществления способ предусматривает введение индивидууму терапевтически эффективного количества антитела к LAM, которое специфически связывается с эпитопом Man-LAM, и/или антитела к PIM6, которое специфически связывается с эпитопом PIM6 (например, эпитопом, общим для PIM6 и LAM). В дополнительных вариантах осуществления подвергаемый нацеливанию эпитоп ManLAM включает кэпированные диманнозой или триманнозой структуры Ara4 и/или Ara6. В еще одних вариантах осуществления эпитоп ManLAM представляет собой кэпированную диманнозой Ara6. В некоторых вариантах осуществления эпитоп ManLAM имеет по меньшей мере одну замену МТХ. В некоторых вариантах осуществления антителом к LAM или антителом к PIM6/LAM является антитело, представляющее собой IgM. В других вариантах осуществления антитело к LAM или антитело к PIM6/LAM представляют собой рекомбинантное антитело.
В некоторых вариантах осуществления настоящего изобретения представлено пассивное введение защитного антитела с помощью рекомбинантных векторов. В одном варианте осуществления рекомбинантные векторы включают первую нуклеиновую кислоту, кодирующую IgVL антитела к LAM, и вторую нуклеиновую кислоту, кодирующую IgVH антитела к LAM, где каждая из нуклеиновых кислот функционально связана с промоторным участком. В некоторых вариантах осуществления по меньшей мере один из IgVL и IgVH получен из человеческого моноклонального антитела к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций. В другом варианте осуществления рекомбинантные векторы включают первую нуклеиновую кислоту, кодирующую IgVL антитела к PIM6/LAM, и вторую нуклеиновую кислоту, кодирующую IgVH антитела к PIM6/LAM, где каждая из нуклеиновых кислот функционально связана с промоторным участком. В некоторых вариантах осуществления рекомбинантные векторы включают дополнительные элементы регуляции транскрипции. В некоторых вариантах осуществления по меньшей мере одна из последовательности первой нуклеиновой кислоты и последовательности второй нуклеиновой кислоты организована в виде оперона. В некоторых вариантах осуществления по меньшей мере одна из последовательности первой нуклеиновой кислоты и последовательности второй нуклеиновой кислоты организована в виде кассеты экспрессии. В некоторых вариантах осуществления последовательность первой нуклеиновой кислоты и последовательность второй нуклеиновой кислоты организованы в виде одной кассеты экспрессии. В некоторых вариантах осуществления первая нуклеиновая кислота и вторая нуклеиновая кислота находятся в одном и том же клонирующем векторе. В других вариантах осуществления первая нуклеиновая кислота и вторая нуклеиновая кислота находятся в разных клонирующих векторах. В некоторых вариантах осуществления экспрессия первой нуклеиновой кислоты и второй нуклеиновой кислоты может быть одновременной. В других вариантах осуществления экспрессия первой нуклеиновой кислоты и второй нуклеиновой кислоты индуцируется отдельно. В некоторых вариантах осуществления экспрессия первой нуклеиновой кислоты может быть отделена временным промежутком от экспрессии второй нуклеиновой кислоты. В некоторых вариантах осуществления рекомбинантный вектор представляет собой плазмиду. В других вариантах осуществления рекомбинантный вектор представляет собой нереплицирующийся вирус. В дополнительных вариантах осуществления рекомбинантный вектор представляет собой аденоассоциированный вирус.
В некоторых вариантах осуществления настоящего изобретения представлен способ лечения инфекции, вызванной вирулентным представителем комплекса Mycobacterium tuberculosis, у индивидуума. В некоторых вариантах осуществления способ включает введение индивидууму первой нуклеиновой кислоты, кодирующей IgVH антитела к LAM, и второй нуклеиновой кислоты, кодирующей IgVL антитела к LAM, где каждая из нуклеиновых кислот функционально связана с промоторным участком. В других вариантах осуществления способ предусматривает введение индивидууму первой нуклеиновой кислоты, кодирующей IgVH антитела к PIM6/LAM, и второй нуклеиновой кислоты, кодирующей IgVL антитела к PIM6/LAM, где каждая из нуклеиновых кислот функционально связана с промоторным участком. В некоторых вариантах осуществления по меньшей мере один из IgVL и IgVH получен из человеческого моноклонального антитела к LAM, которое специфически связывается с одной из Ara4 и Ara6 или их комбинаций, или из человеческого моноклонального антитела к PIM6/LAM, которое специфически связывается с по меньшей мере одной полиманнозной структурой в маннановом домене PIM6. В некоторых вариантах осуществления первая нуклеиновая кислота и вторая нуклеиновая кислота находятся в одном и том же клонирующем векторе. В других вариантах осуществления первая нуклеиновая кислота и вторая нуклеиновая кислота находятся в разных клонирующих векторах. В некоторых вариантах осуществления рекомбинантный вектор представляет собой плазмиду. В других вариантах осуществления рекомбинантный вектор представляет собой нереплицирующийся вирус. В дополнительных вариантах осуществления рекомбинантный вектор представляет собой аденоассоциированный вирус.
Дополнительные варианты осуществления, признаки и преимущества будут совершенно очевидны для специалиста в данной области, исходя из раскрытия, представленного в данном документе. Другие признаки станут более очевидны для лиц, обладающих очевидными познаниями в области техники, относящейся к упаковкам, и из следующего описания и пунктов формулы изобретения. Хотя антитела, композиции, наборы и способы, аналогичные или эквивалентные таковым, описанным в данном документе, можно применять при осуществлении настоящего изобретения на практике или при его тестировании, подходящие антитела, композиции, наборы и способы описаны ниже. Все публикации, патентные заявки и патенты, упомянутые в данном документе, включены посредством ссылки во всей своей полноте. В случае конфликта настоящее описание, включая определения, будет иметь преимущественную силу. Конкретные варианты осуществления, обсуждаемые ниже, служат только для иллюстрации и не предусматриваются как ограничивающие.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГ. 1А - модель формы IgG А194-01 и ее фрагментов, применяемых в анализе конкурентного связывания. Они включают моновалентные структуры scFv и Fab, а также бивалентные димер scFv и природный IgG. 1В - кривые конкурентного связывания, показывающие, что моновалентные формы А194-01 конкурировали с меньшей эффективностью, чем бивалентные формы. 1С - структура формы А194-01 с более высокой валентностью. Она представляет собой гомологичное тетравалентное А194-01 scFv-IgG, которое содержит scFv-домен А194-01, присоединенный к N-концу каждой из нормальных тяжелых цепей.
ФИГ. 2А - связывающая активность форм IgG и IgM Р30 В9, а также IgM, в котором 6-аминокислотная вставка в участке VH была подвергнута делеции, или 9 соматических мутаций в участке VH были возвращены к последовательности ближайшей зародышевой линии, с ManLAM, полученным из Mycobacterium tuberculosis. На реактивность в большей степени влияла 6-аминокислотная вставка, чем 9 соматических аминокислотных мутаций. На ФИГ. 2В и 2С проведено сравнение реактивности форм IgM и IgG Р30В9 и мутантной формы с 6-аминокислотной делецией в тяжелой цепи в отношении ManLAM от Mycobacterium tuberculosis (В) и PILAM от Mycobacterium smegmatis (С). Форма IgM, но не форма IgG, специфически реагирует с ManLAM, полученным из Mycobacterium tuberculosis (2В), но не с PILAM (2С), а реактивность мутантной формы, содержащей Δ6 аминокислот, была в значительной степени снижена в отношении ManLAM и отрицательной в отношении PILAM.
ФИГ. 3 - сравнение реактивности 2 человеческих mAb и 4 мышиных mAb в отношении PILAM на левой панели и в отношении ManLAM, выделенного из штамма H37Rv Mycobacterium tuberculosis, на правой панели. Кривые наносили на график с применением молярных концентраций антител для учета различной молекулярной массы этих реагентов.
ФИГ. 4А - структуры 25 синтетических олигосахаридов, представляющих собой структуры микробных гликанов, родственных мотивам, присутствующим в LAM. Эти структуры присоединяли к белку-носителю из BSA и применяли для проверки эпитопной специфичности. 4В - профили связывания шести LAM-специфических моноклональных антител с панелью из 25 синтетических олигосахаридов. Результаты связывания показаны для трех концентраций, а относительные аффинности антител с этими антигенами показаны с помощью паттерна титрации.
ФИГ. 5 - левая панель - структура IgA1 (A), IgA2 (В) и димерного комплекса из димерного IgA1-J (С). Правая панель- гель SDS-PAGE с очищенными белками Р30В9 IgA1, IgA2 и IgA3, как перед, так и после восстановления с помощью DTT. Позже было установлено, что Р30В9 IgA3 представляет собой артефакт ПЦР с более длинным шарнирным участком.
ФИГ. 6 - кривые связывания различных изотипов Р30В9 с ManLAM, показывающие самую большую активность у формы IgM, а затем у форм IgA, при этом с отсутствием реактивности у формы IgG.
ФИГ. 7 - сравнение эффективности зондов на основе биотинилированного моноклонального антитела при выявлении растворимого ManLAM в анализе захвата с CS-35, в котором указанную концентрацию ManLAM захватывали с помощью CS-35 и выявляли с помощью указанных mAb, меченных биотином.
ФИГ. 8 - кривые связывания Р30В9 с различными кэпированными маннозой структурами Ara4 или с тетра- и пентаманнозными структурами. Преимущественное связывание наблюдали в случае структур 3 (диманноза-Ara4) и 59, которые содержали родственную связь α-Manp(1→2)-Manp.
ФИГ. 9 - титрование моноклональных антител к LAM в отношении различных некэпированных LAM-родственных гликоконъюгатов для определения структурных требований для обеспечения реактивности. 9А - анализ важности связи Ara-α(1→5)-Ara в предпоследнем положении от невосстанавливающего конца последовательности Ara4. 9В - анализ зависимости связи Ara-β(1→2)-Ara в концевом положении последовательности Ara4.
ФИГ. 10 - кривые связывания А194-01 IgG и трех мышиных антител к LAM с различными Ara6-содержащими гликоконъюгатами, показывающие эффект разных кэпирующих мотивов на реактивность антитела.
ФИГ. 11 - исследования конкурентного связывания для измерения способности индивидуальных антител к LAM конкурировать за связывание антитела-зонда с антигеном ManLAM. Антитела были биотинилированными при тестировании в сравнении с антителами от того же вида. Следует отметить неспособность А194-01 конкурировать с биотинилированным Р30 В9.
ФИГ. 12 - конкурирование за связывание моноклональных антител к LAM с LAM, полученным из Mycobacterium tuberculosis (ManLAM), и LAM, полученным из Mycobacterium smegmatis (PILAM). Эффективное конкурирование между FIND25 и Р30В9 за ManLAM согласуется с преобладанием диманноза-замещенной Ara6, в то время как отсутствие конкурирования этих двух mAb с А194 согласуется с их слабой реактивностью с кэпированными диманнозой структурами. Эффективное конкурирование А194 с FIND25 в отношении PILAM согласуется с отсутствием кэпирования диманнозой данной структуры.
ФИГ. 13 - конкурентное связывание биотинилированных моноклональных антител-зондов и избытка немодифицированных антител с природными антигенами LAM и выбранными гликоконъюгатами. 13А - конкурирование за связывание биотинилированных А194-01 IgG, CS-35 и FIND25 с MAnLAM с применением четырех mAb; 13 В - конкурирование за связывание FIND25 с обоими ManLAM и PILAM с применением трех mAb; 13С - конкурирование за связывание Р30 В9 IgM с MAnLAM и двумя синтетическими антигенами на основе гликоконъюгатов с применением четырех mAb.
ФИГ. 14 - сконструированные варианты и/или производные А194-01 реагируют с более широким спектром гликоконъюгатов, включая замещенные ди- и триманнозой формы, которые слабо распознаются изотипом IgG А194-01.
ФИГ. 15 - дифференциальное конкурирование А194-01 IgG и сконструированных вариантов и/или производных А194-01 за связывание ManLAM с FIND25 и Р30В9 IgM. Хотя А194 IgG не конкурирует с Р30В9 или FIND25 за ManLAM, мультимерные формы действительно конкурируют, что согласуется с более широкой эпитопной специфичностью этих форм. Как показано выше, А194 IgG не конкурирует в достаточной степени с FIND25 за PILAM.
ФИГ. 16 - сравнение анализа эффекта кэпирования маннозой на реактивность mAb CS-40, А194-01 и Р30В9. Специфичность связывания антител измеряли с помощью ELISA в отношении специфических гликонъюгатов, содержащих разные маннозные замещения. Титрации антител показаны на 16А, а структуры антигенов на основе манноза-содержащих гликанов показаны на 16В.
ФИГ. 17-17А - гомологичный scFv-IgG. В данном примере как IgG, так и scFv-домены получены из одного и того же антитела. Это приводит к увеличенной валентности (тетравалентный по сравнению с бивалентным), но непосредственно не модифицирует специфичность в отношении мишени. 17В - гетерологичный scFv-IgG. В дополнение к увеличению валентности также вводится более широкая специфичность, что может обеспечивать возможность распознавания отличающихся эпитопов в одной молекуле антигена. 17С - гетерологичный scFv-IgM. В данном составе отличающиеся scFv объединены с конструкцией IgM. Одним примером может быть связывание А194-01 scFv с Р30В9 IgM. В дополнение к увеличению валентности это будет вводить дополнительную эпитопную специфичность, которая может обеспечивать возможность мультивалентного распознавания отличающихся эпитопов, которые не могут распознаваться гомологичным scFv-IgM, и приводить к повышенной аффинности.
ФИГ. 18А-18С - картирование эпитопов, распознаваемых новыми mAb. Эпитопную специфичность Р95С1 сравнивали с таковой двух ранее описанных mAb, А194-01 и Р30В9, и двух новых mAb, Р61Н5 и Р83А8, которые распознают эпитопы, родственные таковым для двух ранее описанных mAb. 18А - реактивность LAM-специфических mAb в отношении молекул-предшественников LAM. Р30В9 и Р61Н5 были специфическими в отношении ManLAM по сравнению с PILAM, в то время как А194-01, Р83А8 и Р95С1 распознавали обе формы LAM. Р95С1 также эффективно связывалось с LM и PIM6. Слабая реактивность остальных mAb в отношении LM и PIM6 обусловлена, по меньшей мере частично, загрязнением этих материалов ManLAM. 18В - реактивность в отношении синтетических гликоконъюгатов, полученных из LAM. 18С - в отличие от ранее известных mAb Р95С1 было единственным антителом, которое не распознавало никакую из полиарабинозных структур, но специфически реагировало с двумя полиманнозными структурами, YB-BSA-05 и YB-BSA-11, которые напоминали структуры, присутствующие в PIM6 и в маннановых доменах в основе LM и LAM.
ФИГ. 19 - эффект переключения изотопа на связывание Р95С1 и Р30 В9 с ManLAM и PI-LAM. В случае Р95С1 все изотопы IgM, IgA и IgG характеризовались сравнимой связывающей активностью с обоими ManLAM и PILAM, в отличие от Р30 В9, которое реагировало только с ManLAM и только в формах IgM и IgA, но не IgG.
ФИГ. 20(А)-20(В) - вестерн-блот анализ перекрестной реактивности Р95С1 с LAM и дополнительными гликолипидами M.tb. 20(A) - очищенные LAM-ассоциированные гликолипиды разделяли на 12% SDS-PAGE геле с последующим окислением и окрашиванием молекул сахара с помощью реактива Шиффа для обнаружения материала, содержащего реакционноспособные гликаны. 20(B) - параллельные блоты исследовали с помощью mAb, А194 IgG1, Р30 В9 IgM и Р95С1 IgM с последующей обработкой вторичными антителами к человеческим IgG и IgM, конъюгированными с щелочной фосфатазой, и обработкой субстратом для проявления цвета bcip/nbt. А194-01 перекрестно реагирует с ManLAM из M.tb и PILAM из M.smeg. Р30 В9 является специфическим в отношении ManLAM из M.tb. Р95С1 распознает оба вида LAM, а также LM и PIM6, выделенные из M.tb. Слабое окрашивание с помощью А194-01 полосок LM и PIM6, которые мигрировали совместно с LAM, по-видимому, обусловлено незначительным загрязнением этих образцов с помощью LAM.
ФИГ. 21 - выравнивания аминокислотных последовательностей с применением последовательностей вариабельных участков тяжелой цепи и легкой цепи А194 и их сравнение с их ближайшими последовательностями зародышевой линии. В верхнем выравнивании первая сверху аминокислотная последовательность (A194-VH) представляет собой последовательность вариабельного участка тяжелой цепи А194 без последовательности CDR3 (SEQ ID NO: 23). Последовательность вариабельного участка тяжелой цепи без CDR3 представляет собой SEQ ID NO: 21. В верхнем выравнивании вторая аминокислотная последовательность (Homsap IGHV3-20*01 зародышевой линии) представляет собой SEQ ID NO: 22. В верхнем выравнивании третья аминокислотная последовательность представляет собой CDR3 вариабельного участка тяжелой цепи А194, и она представляет собой SEQ ID NO: 23. В нижнем выравнивании первая сверху аминокислотная последовательность (А-194-Vk) представляет собой вариабельный участок легкой цепи А194 без последовательности CDR3 (SEQ ID NO: 26). Последовательность вариабельного участка легкой цепи без CDR3 представляет собой SEQ ID NO: 24. В нижнем выравнивании вторая аминокислотная последовательность (Homsap IGKV3-15*01 зародышевой линии) представляет собой SEQ ID NO: 25. В нижнем выравнивании третья последовательность представляет собой CDR3 вариабельного участка легкой цепи А194, и она представляет собой SEQ ID NO: 26.
ФИГ. 22 - Аминокислотные последовательности у последовательностей вариабельных участков тяжелой цепи и легкой цепи P30B9-IgM и их сравнение с их ближайшими последовательностями зародышевой линии. В верхнем выравнивании первая сверху аминокислотная последовательность (P30B9-Vh) представляет собой последовательность вариабельного участка тяжелой цепи P30B9-IgM без последовательности CDR3 (SEQ ID NO: 29). Последовательность вариабельного участка тяжелой цепи без CDR3 представляет собой SEQ ID NO: 27. Вторая аминокислотная последовательность (Homsap IGHV4-34*01 F) представляет собой SEQ ID NO: 28. Третья аминокислотная последовательность представляет собой вариабельный участок тяжелой цепи P30B9-IgM, и она представляет собой SEQ ID NO: 29. В нижнем выравнивании первая сверху аминокислотная последовательность (P30B9-Vk) представляет собой вариабельный участок легкой цепи Р30 В9 без последовательности CDR3 (SEQ ID NO: 32). Последовательность вариабельного участка легкой цепи без CDR3 представляет собой SEQ ID NO: 30. В нижнем выравнивании вторая аминокислотная последовательность (Homsap IGKV1-5*03 зародышевой линии) представляет собой SEQ ID NO: 31. В нижнем выравнивании третья последовательность представляет собой CDR3 вариабельного участка легкой цепи Р30В9, и она представляет собой SEQ ID NO: 32.
ФИГ. 23 - выравнивания аминокислотных последовательностей с применением последовательностей вариабельных участков тяжелой цепи и легкой цепи P95C1-IgM и их сравнение с их ближайшими последовательностями зародышевой линии. В верхнем выравнивании первая сверху аминокислотная последовательность (P95C1-VH) представляет собой последовательность вариабельного участка тяжелой цепи Р95С1 без последовательности CDR3 (SEQ ID NO: 18). Последовательность вариабельного участка тяжелой цепи без CDR3 представляет собой SEQ ID NO: 33. В верхнем выравнивании вторая аминокислотная последовательность (Homsap IGHV4-4*02 зародышевой линии) представляет собой SEQ ID NO: 34. В верхнем выравнивании третья аминокислотная последовательность представляет собой CDR3 вариабельного участка тяжелой цепи P95C1-gM, и она представляет собой SEQ ID NO: 18. В нижнем выравнивании первая сверху аминокислотная последовательность (P95C1-Vk) представляет собой вариабельный участок легкой цепи Р95С1 без последовательности CDR3 (SEQ ID NO: 15). Последовательность вариабельного участка легкой цепи без CDR3 представляет собой SEQ ID NO: 36. В нижнем выравнивании вторая аминокислотная последовательность (Homsap IGKV4-1*01 F зародышевой линии) представляет собой SEQ ID NO: 37. В нижнем выравнивании третья последовательность представляет собой CDR3 вариабельного участка легкой цепи Р95С1, и она представляет собой SEQ ID NO: 15.
ПОДРОБНОЕ ОПИСАНИЕ
А. Определения
Если не указано иное, все технические термины, используемые в данном документе, имеют то же самое значение, которое общеизвестно обычному специалисту в области техники, к которой принадлежит настоящее изобретение.
Антитело к LAM может находиться в одной из множества форм, известных из уровня техники, как раскрыто в данном документе. Помимо всего прочего, антитела определяются антигенами, с которыми они связываются, таким образом, "антитело к LAM" представляет собой любое подобное антитело, которое специфически связывает по меньшей мере один эпитоп липоарабиноманнана (LAM), как описано в данном документе. Из уровня техники понятно, что антитело представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, соединенные межу собой с помощью дисульфидных связей, или его антигенсвязывающую часть. Тяжелую цепь составляют вариабельный участок тяжелой цепи (VH) и константный участок тяжелой цепи (CH1, СН2 и СН3). Легкую цепь составляют вариабельный участок легкой цепи (VL) и константный участок легкой цепи (CL). Вариабельные участки обеих тяжелой и легкой цепей содержат каркасные участки (FWR) и определяющие комплементарность участки (CDR). Четыре участка FWR являются относительно консервативными, в то время как участки CDR (CDR1, CDR2 и CDR3) представляют гипервариабельные участки, и они расположены от NH2-конца до СООН-конца следующим образом: FWR1, CDR1, FWR2, CDR2, FWR3, CDR3, FWR4. Вариабельные участки тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном, в то время как константный участок(участки) в зависимости от изотипа может(могут) опосредовать связывание иммуноглобулина с тканями или факторами хозяина.
Антитело к PIM6/LAM может находиться в одной из множества форм, известных из уровня техники, как раскрыто в данном документе. "Антитело к PIM6/LAM" представляет собой любое такое антитело, которое специфически связывает по меньшей мере один эпитоп, который является общим для фосфатидилинозитол-маннозида 6 (PIM6) и LAM, как описано в данном документе. Человеческое mAb, специфическое в отношении эпитопа, общего для LAM и PIM6, описанных в данном документе, представляет собой Р95С1, которое специфически связывается с по меньшей мере одной полиманнозной структурой в PIM6 и в PIM6-родственном маннановом домене в LM и LAM. Р95С1 связывается с обоими LAM и PIM6, поскольку оно различает одинаковый (общий) эпитоп, и, таким образом, обозначается в данном документе как "антитело к PIM6/LAM" или "моноклональное антитело к PIM6/LAM", "человеческое антитело к PIM6/LAM" или "человеческое моноклональное антитело к PIM6/LAM".
Из уровня техники известно, что можно осуществлять манипуляции с моноклональными и другими антителами и применять методики технологии рекомбинантной ДНК для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела. Такие методики могут включать введение ДНК, кодирующей вариабельный участок иммуноглобулина или CDR антитела, в константные участки или константные участки с каркасными участками другого иммуноглобулина.
Используемый в данном документе термин "антитело" (Ab) используется в самом широком смысле и, в частности, может включать любой иммуноглобулин, природный или полученный частично или полностью синтетическим путем, включая без ограничений моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела и полиреактивные антитела) и фрагменты антител. Таким образом, подразумевается, что термин "антитело", используемый в любом контексте в пределах настоящего описания, включает без ограничений любой специфический связывающий элемент, класс и/или изотип иммуноглобулина (например, IgG1, IgG2a, IgG2b, IgG3, IgG4, IgM, IgA1, IgA2, IgD и IgE) и его биологически релевантный фрагмент или специфический связывающий элемент, включая без ограничений Fab, F(ab')2, scFv (одноцепочечный или родственный объект) и (scFv)2.
Используемый в данном документе термин "фрагменты антитела" может включать фрагменты антитела, которые получены с применением методик, хорошо известных и доступных обычным специалистам в данной области, как рассмотрено в данном документе. Следовательно, термин "антитело" описывает любой полипептид или белок, содержащие часть интактного антитела, такую как антигенсвязывающий или вариабельный участок интактного антитела. Они могут быть получены из природных источников, или они могут быть получены частично или полностью синтетическим путем. Примеры фрагментов антител включают без ограничений Fab-, Fab'-, F(ab')2- и Fv-фрагменты; диатела и линейные антитела. В частности, используемые в данном документе "одноцепочечные Fv" ("sFv" или "scFv") представляют собой фрагменты антител, которые содержат VH- и VL-домены антитела, соединенные в одну полипептидную цепь. Полипептид sFv может дополнительно содержать, например, линкер, такой как гибкий полипептидный линкер между VH- и VL-доменами, который позволяет scFv образовывать требуемую структуру для связывания антигена.
Используемый в данном документе термин "моноклональное антитело" или "mAb" может относится к антителу, полученному из группы в значительной степени однородных антител, т.е. индивидуальные антитела, составляющие группу, являются идентичными за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в минимальных количествах.
Используемые в данном документе термины "варианты", "производные" и/или "варианты и/или производные" могут обозначать антитела, фрагменты антител, рекомбинантные антитела, независимо от того, получены ли они из природных источников или получены частично или полностью синтетическим путем, а также белки, фрагменты белков и полипептиды, в том случае, если вышеупомянутые соединения являются либо структурно подобными, т.е. сохраняют степень идентичности, которая составляет по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% или больше идентичности последовательности с оригинальным немодифицированным антителом, и/или, независимо от идентичности структуры, могут быть функционально подобны исходным немодифицированным антителам к LAM и к PIM6/LAM, то есть они сохраняют способность к специфическому связыванию с по меньшей мере одним эпитопом LAM или с общим эпитопом PIM6/LAM соответственно. Например, такие варианты и/или производные могут включать антитела к LAM или к PIM6/LAM с вариантными Fc-доменами, химерные антитела, слитые белки, биспецифические антитела или другие рекомбинантные антитела. Такие варианты и/или производные антитела могут, но не обязательно, обладать большей специфичностью связывания в отношении одного или нескольких эпитопов LAM или PIM6, и/или могут быть способны связываться с дополнительными эпитопами LAM или PIM6.
Термин "биологический образец" обозначает образец, полученный из организма (например, пациента) или из компонентов (например, клеток) организма. Образец может представлять собой любую биологическую ткань, клетку(клетки) или жидкость. Образец может представлять собой "клинический образец", который представляет собой образец, полученный от субъекта, такого как пациент-человек. Такие образцы включают без ограничений слюну, мокроту, кровь, клетки крови (например, лейкоциты), амниотическую жидкость, плазму, семенную жидкость, костный мозг, а также ткань или образцы тонкоигольной пункционной биопсии, мочу, перитонеальную жидкость и плевральную жидкость или клетки из них. Биологические образцы могут также включать срезы тканей, такие как замороженные срезы, получаемые для гистологических целей. Биологический образец может также обозначаться как "образец от пациента". Биологический образец может также включать в значительной степени очищенный или выделенный белок, мембранный препарат или культуру клеток.
Используемые в данном документе термины "эффективное количество" или "терапевтически эффективное количество" могут обозначать количество соединения или средства, которое способно приводить к требуемому с медицинской точки зрения результату у субъекта, подвергаемого лечению. Способ лечения можно осуществлять in vivo или ex vivo, отдельно или в сочетании с другими лекарственными средствами или терапией. Терапевтически эффективное количество можно вводить в виде одного или нескольких введений, нанесений или дозировок, и при этом не подразумевается ограничение конкретным составом или путем введения.
Используемые в данном документе термины "антигенсвязывающий фрагмент" или "Fab" могут обозначать участок на антителе, который связывается с антигенами. Обычный специалист в данной области будет понимать, что Fab составляют один константный и один вариабельный домен каждой из тяжелой и легкой цепей антитела.
Используемые в данном документе термины "специфическое связывание", "селективное связывание", "селективно связывает" и "специфически связывает" могут обозначать связывание антитела с эпитопом на заданном антигене, а не на других антигенах. Как правило, антитело (i) связывается при равновесной константе диссоциации (KD), составляющей приблизительно менее 10-6 М, как, например, приблизительно менее 10-7 М, 10-8 М, 10-9 М или 10-10 М или даже ниже при определении, например, с помощью технологии поверхностного плазмонного резонанса (SPR) в установке для исследования поверхностного плазмонного резонанса BIACORE® 2000 с применением заданного антигена, например, эпитопа LAM, в качестве аналита и антитела в качестве лиганда, или с помощью анализа Скэтчарда связывания антитела антиген-положительными клетками, и (ii) связывается с заданным антигеном с аффинностью, которая по меньшей мере в два раза превышает аффинность его связывания с неспецифическим антигеном (например, BSA, казеином), отличным от заданного антигена или близкородственного антигена.
Используемые в данном документе термины "консервативные модификации последовательности" или "консервативные замены" могут обозначать аминокислотные модификации, которые значимо не влияют или не изменяют характеристики связывания антитела, содержащего аминокислотную последовательность. Такие консервативные модификации включают аминокислотные замены, добавления и делеции. Модификации можно вводить в антитело по настоящему изобретению с помощью стандартных методик, известных из уровня техники, таких как сайт-направленный мутагенез и опосредованный ПЦР мутагенез. Консервативные аминокислотные замены представляют собой замены, при которых аминокислотный остаток замещается на аминокислотный остаток с подобной боковой цепью. Семейства аминокислотных остатков, имеющих подобные боковые цепи, были определены в уровне техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в пределах участков CDR антитела по настоящему изобретению могут быть замещены другими аминокислотными остатками из того же семейства боковых цепей, и измененное антитело можно тестировать в отношении сохраненной функции с применением функциональных анализов, описанных в данном документе.
Используемый в данном документе термин "идентичность" может обозначать существование общей структуры у двух композиций. В контексте белков термин "идентичность" может обозначать количество (например, выраженное в виде процентной доли) совпадений между двумя или более аминокислотными и/или пептидными последовательностями. В контексте нуклеиновых кислот термин может обозначать количество (например, выраженное в виде процентной доли) совпадений между двумя или более последовательностями нуклеиновой кислоты. Используемый в данном документе процент (%) идентичности между двумя последовательностями эквивалентен проценту идентичности между двумя последовательностями. Процент идентичности между двумя последовательностями представляет собой функцию числа идентичных положений, общих для последовательностей (т.е. % идентичности = число идентичных положений/общее число положений × 100), с учетом числа гэпов и длины каждого гэпа, который нужно вводить для оптимального выравнивания двух последовательностей. Сравнение последовательностей и определение процента идентичности межу двумя последовательностями можно выполнять с применением математического алгоритма. Такая идентичность хорошо описана в уровне техники с помощью инструментов и/или алгоритмов для локального выравнивания, и может включать попарное выравнивание, способы выравнивания множественных последовательностей, способы структурного выравнивания и/или способы филогенетического анализа. Специфические примеры включают следующее. Процент идентичности между двумя аминокислотными последовательностями можно определять с применением алгоритма Э. Мейерса и У. Миллера (Comput. Appl. Biosci., 4:11-17 (1988)), который был встроен в программу ALIGN (версия 2.0), с применением таблицы весов замен остатков РАМ120, штрафа за удлинение гэпа 12 и штрафа за введение гэпа 4. В дополнение, процент идентичности между двумя аминокислотными последовательностями можно определять с применением алгоритма Нидмана и Вунша (J. Mol. Biol. 48:444-453 (1970)), который был встроен в программу GAP в пакете программного обеспечения GCG (доступен на www.gcg.com), с применением либо матрицы Blossum 62, либо матрицы РАМ250, и веса гэпа 16, 14, 12, 10, 8, 6 или 4 и веса длины 1, 2, 3, 4, 5 или 6. Дополнительно или альтернативно белковые последовательности по настоящему изобретению можно также применять в качестве "последовательности запроса" для проведения поиска в открытых базах данных, например, в отношении идентичности родственных последовательностей. Такие поиски можно проводить с применением программы XBLAST (версия 2.0) из Altschul, et al. (1990) J. Mol. Biol. 215:403-10. Белковые поиски BLAST можно проводить с помощью программы XBLAST, балл=50, длина слова=3 для получения аминокислотных последовательностей, гомологичных молекулам антитела по настоящему изобретению. Чтобы получить выравнивания с гэпами для целей сравнения, можно использовать Gapped BLAST, как описано в Altschul et al., (1997) Nucleic Acids Res. 25(17):3389-3402. При использовании программ BLAST и Gapped BLAST можно использовать параметры по умолчанию соответствующих программ (например, XBLAST и NBLAST).
Используемые в данном документе термины "совместное введение", "вводимый совместно" и "в комбинации с" могут обозначать введение по меньшей мере двух средств или терапий субъекту. В некоторых вариантах осуществления совместное введение двух или более средств/терапий является одновременным. В других вариантах осуществления первое средство/терапию вводят перед вторым средством/терапией. Специалисты в данной области поймут, что составы и/или пути введения различных применяемых средств/терапий могут варьировать.
Используемый в данном документе термин "носители" может включать фармацевтически приемлемые носители, наполнители или стабилизаторы, которые являются нетоксичными для клетки или млекопитающего, подвергаемого их воздействию при используемых дозировках и концентрациях. Зачастую физиологически приемлемый носитель представляет собой водный раствор с забуференным рН. Примеры физиологически приемлемых носителей включают без ограничений буферы, такие как фосфатный, цитратный и на основе других органических кислот; антиоксиданты, включая без ограничений аскорбиновую кислоты; низкомолекулярные полипептиды (состоящие из менее чем приблизительно 10 остатков); белки, такие как без ограничений сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как без ограничений поливинилпирролидон; аминокислоты, такие как без ограничений глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая без ограничений глюкозу, маннозу или декстрины; хелатирующие средства, такие как без ограничений EDTA; сахароспирты, такие как без ограничений маннит или сорбит; солеобразующие противоионы, такие как без ограничений натрий; и/или неионные поверхностно-активные вещества, такие как без ограничений TWEEN; полиэтиленгликоль (PEG) и PLURONICS.
Термин "осуществление лечения" или "лечение" заболевание обозначает выполнение протокола, который может включать введение одного или нескольких лекарственных средств пациенту (человеку или другому субъекту) в попытке уменьшить признаки или симптомы заболевания. Уменьшение может происходить до проявления признаков или симптомов заболевания, а также после их проявления. Таким образом, "осуществление лечения" или "лечение" включают "осуществление предупреждения" или "предупреждение" заболевания. Термины "предупреждать" или "осуществление предупреждения" обозначают профилактически и/или превентивные меры, при этом у объекта предупреждают или замедляют целевое патологическое состояние или нарушение. Например, в случае инфекции, вызванной вирулентным штаммом комплекса Mycobacterium tuberculosis, "осуществление предупреждения" или "осуществление предупреждения" могут происходить в ситуации, когда в ходе лечения его проводят заранее, чтобы предупредить или замедлить инфекцию, вызванную вирулентным штаммом комплекса Mycobacterium tuberculosis, как, например, с помощью вакцинации или пассивного введения защитного антитела. Такое "осуществление предупреждения" или "предупреждение" также происходят в случае латентной инфекции, вызванной Mycobacterium tuberculosis, при котором у объекта будет предупреждаться развитие активной инфекции и/или пациент будет избавлен от указанной латентной инфекции. В дополнение, "осуществление лечения" или "лечение" не требуют полного облегчения признаков или симптомов, не требует излечения и, в частности, включают протоколы, которые оказывают только незначительный эффект на пациента.
Термины "пациент", "субъект" и "индивидуум" используются в данном документе взаимозаменяемо и могут относится к биологической системе, в отношении которой применяют лечение. Биологическая система может включать, например, индивидуальную клетку, набор клеток (например, культуру клеток), орган, ткань или многоклеточный организм. "Пациент", "субъект" или "индивидуум" могут относиться к пациенту-человеку, субъекту-человеку 'или индивидууму-человеку или к пациенту, субъекту или индивидууму, отличным от человека.
Используемый в данном документе термин "эпитоп" может относиться к участку антигена, с которым связывается антитело или Т-клетка. "Антиген" обозначает вещество, которое вызывает иммунологическую реакцию или связывается с продуктами такой реакции.
Используемый в данном документе термин "вектор" означает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она была связана. Векторы, способные управлять экспрессией генов, с которыми они функционально связаны, обозначены в данном документе как "векторы экспрессии".
Используемые в данном документе "белок" и "полипептид" используются в качестве синонимов для обозначения любой цепи из аминокислот с пептидными связями, независимо от длины или посттрансляционной модификации, например, гликозилирования или фосфорилирования.
Подразумевается, что термин "меченный" в отношении антитела, нуклеиновой кислоты, пептида, полипептида, клетки или зонда, охватывает прямое мечение антитела, нуклеиновой кислоты, пептида, полипептида, клетки или зонда путем присоединения (т.е. физического связывания) обнаруживаемого вещества с антителом, нуклеиновой кислотой, пептидом, полипептидом, клеткой или зондом.
Используемые в данном документе термины "очищенный" или "выделенный" пептид, полипептид или белок обозначают пептид, полипептид или белок и могут обозначать пептид, полипептид или белок, которые были отделены от других белков, липидов и нуклеиновых кислот, с которыми они ассоциированы в природе. Полипептид/белок могут составлять по меньшей мере 10% (т.е. любую процентную долю от 10% до 100%, например, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 95% и 99%) по сухому весу очищенного препарата. Чистота может быть измерена с помощью любого подходящего стандартного способа, например, с помощью колоночной хроматографии, электрофореза на полиакриламидном геле или ВЭЖХ-анализа. Выделенный полипептид/белок (например, антитела к LAM), описанные в настоящем изобретении, можно получать с помощью методик рекомбинантной ДНК.
В. Mycobacterium tuberculosis
Туберкулез (ТВ) остается одним из самых смертоносных инфекционных заболеваний в мире, при этом в настоящее время инфицирована приблизительно одна треть мирового населения. По оценке у 9,0 миллиона людей развился ТВ в 2013 году, и по оценке 1,5 миллиона людей умерли из-за данного заболевания. Хотя в настоящее время доступны антибиотические средства для лечения, они требуют длительного лечения, и они все больше компрометируются из-за развития штаммов с множественной лекарственной устойчивостью (MDR-TB), которые в настоящее время ответственны за приблизительно 3,5% недавних случаев заражения. Эти штаммы намного сложнее лечить, и они характеризуются значительно более низкими уровнями излечения. Также распространяются штаммы ТВ с широкой лекарственной устойчивостью (XDR-TB), которые даже более дорогие и сложные для лечения, чем штаммы MDR-TB, и к настоящему времени о них сообщалось в 100 странах по всему миру.
Существует издавна установленная парадигма, что иммунитет против ТВ зависит исключительно от клеточных защитных механизмов. Однако, исследования в области ВИЧ указывают на удивительную способность гуморальной иммунной системы человека вырабатывать различные антитела с удивительным размахом и эффективностью нейтрализации, и в настоящем изобретении указана способность гуморальной иммунной системы вырабатывать антитела с высокой аффинностью, которые распознают множественные эпитопы LAM. Это позволяет предположить, что большая часть затруднений в прошлом, связанных с демонстрацией важной роли опосредованной антителами защиты от ТВ, может быть обусловлена ограничениями в качестве и источнике антител, применяемых в прежних исследованиях, и что применение способов по настоящему изобретению для получения более высокоразвитых антител от пациентов-людей с хронической инфекцией может иллюстрировать критическую роль гуморального ответа в иммунитете в отношении ТВ.
Некоторые варианты осуществления настоящего изобретения направлены на способы in vitro культивирования В-клеток памяти из инфицированных людей и молекулярного клонирования вариабельных участков тяжелых (Н) и легких (L) цепей IgG из одиночной клетки. Эти способы можно использовать для получения человеческих моноклональных антител против главного поверхностного антигена LAM. Настоящее изобретение относится к таким антителам и сконструированным производным этих антител, которые обладают уникальной эпитопной специфичностью и свойствами связывания, а также к иммунодиагностическим и иммунотерапевтическим применениям данных антител.
С. Липоарабиноманнан (LAM)
Одной заметной антигенной мишенью антител по настоящему изобретению является поверхностный гликолипид, липоарабиноманнан (LAM), главный структурный компонент клеточной стенки представителей комплекса Mycobacterium tuberculosis. В настоящем изобретении идентифицированы ранее недооцененная гетерогенность в антигенной структуре LAM и гуморальном иммунном ответе против LAM в ответ на инфекцию и иммунизацию. Структура LAM подробно описана в Khoo et al., "Variation in Mannose-capped Terminal Arabinan Motifs of Lipoarabinomannan from Clinical Isolates of Mycobacterium tuberculosis and Mycobacterium avium Complex," Journal of Biological Chemistry Vol 276, No. 6, Feb 9, 2001, включенной в данный документ посредством ссылки во всей своей полноте. Структура LAM является сложной, проявляющей общую трехчленную структуру с четырьмя различимыми структурными доменами; фосфатидилинозитол-липидным якорем (маннозил-фосфатидил-мио-инозитол), α(1→6)-связанным D-маннановым остовом с концевыми α(1→2)-Manp-связанными боковыми цепями, D-арабинановой цепью, содержащей множественные тетра-/гекса-арабинофуранозидные ветви, и различными кэпирующими мотивами. LAM состоит из гетерогенной популяции молекул, которые могут быть разделены на множество изоформ, которые обладают разными биологическими свойствами. Эта гетерогенность обусловлена варьирующими длинами маннановой и арабинозной цепей, различными паттернами ветвления, разным числом таких ветвей и модификацией арабинозных боковых цепей с помощью кэпирования маннозой, добавления МТХ и ацилирования с помощью жирных кислот, сукцинатов и лактатов.
У вирулентных штаммов комплекса Mycobacterium tuberculosis эти молекулы широко кэпированы с помощью моно-, ди- и три-α(1→2)-D-Manp-сахаридных структурных единиц, в то время как у быстрорастущих непатогенных штаммов, подобных М. smegmatis, имеются некэпированные концы или фосфатидил-мио-инозитольные кэпы (PILAM). Согласно проведенной оценке 40-70% невосстанавливающих концов LAM из патогенных штаммов комплекса Mycobacterium tuberculosis кэпированы маннозой, и анализ относительного обилия разных мотивов кэпов у вирулентного клинического штамма МТ103 показал, что диманнозильная структурная единица была главным структурным мотивом (75-80%), в то время как маннозильный и триманнозильный мотивы присутствовали при более низких концентрациях (10-13%). Данное широкое кэпирование может представлять собой уникальный маркер для разграничения вирулентных штаммов комплекса Mycobacterium tuberculosis от невирулентных/непатогенных штаммов, таких как М. smegmatis, и также может обеспечивать возможные антигенные мишени для терапевтического применения антитела к LAM по настоящему изобретению. В дополнение, некоторые из концевых маннозных остатков сахара в ManLAM, обнаруженном в штамме М. tuberculosis, дополнительно модифицированы путем α(1→4)-добавления уникальной структуры, 5-дезокси-5-метил-тио-пентофуранозы (МТХ), которая влияет на иммунную реактивность разных mAb, чувствительных к кэпирующим мотивам, таких как А194-01 и Р30В9; при этом добавление МТХ повышает реактивность в случае А194-01 и снижает реактивность для Р30В9. Данная замена присутствует с низкой частотой, и может представлять собой уникальный маркер для идентификации М. tuberculosis, потенциально даже от других вирулентных представителей комплекса Mycobacterium tuberculosis, как, например, от М. bovis и М. africanum, и также может обеспечивать потенциальную антигенную мишень для терапевтического применения антител к LAM по настоящему изобретению.
Секретируемые формы LAM являются важными мишенями для иммунодиагностических анализов инфекции, вызванной патогенными представителями комплекса М. tuberculosis. В дополнение, значительная совокупность доказательств указывает на то, что LAM представляет собой важный медиатор целого ряда функций, которые содействуют продуктивной инфекции и патогенности. LAM вовлечен в поддержание целостности клеточной стенки и устойчивость к бета-лактамным антибиотикам. Сниженная экспрессия LAM на поверхности бактерий коррелировала с нарушенным попаданием в макрофаги, ингибированием фагосомно-лизосомного слияния, ослаблением макрофагов и повышенной чувствительностью к адаптивному иммунитету, а связывание концевых маннозильных структурных единиц ManLAM с маннозным рецептором на поверхности макрофагов было описано в качестве важнейшей стадии в захвате микобактерий фагоцитирующими клетками. Без ограничения теорией полагают, что ManLAM взаимодействует с лектинами С-типа, такими как молекулы межклеточной адгезии дендритных клеток 3 (ICAM-3)-захватывающего неинтегрина (DC-SIGN), маннозный рецептор макрофага (MMR) и дектин-2 на дендритных клетках. Полагают, что попав внутрь макрофагов, LAM ингибирует фагосомно-лизосомное слияние, которое приводило бы к разрушению бактерий, что, тем самым, обеспечивает возможность персистирования бактерий внутри макрофагов.
LAM также секретируется на поверхности бактерий, и внеклеточный LAM связывается с поверхностными рецепторами дендритной клетки, включая DC-SIGN и дектин-2. Полагают, что эти взаимодействия подавляют функцию дендритных клеток и противодействуют иммунной системе хозяина, способствуя уклонению от распознавания иммунной системой. Поскольку LAM присутствует в относительно больших количествах во время активной инфекции, его можно обнаружить в крови и моче инфицированных пациентов, например, с помощью одного или нескольких антител к LAM по настоящему изобретению. Их можно применять, например, в диагностических наборах и способах, связанных с указанными диагностическими наборами.
D. Антитела к LAM и к PIM6/LAM
Антитела к LAM по настоящему изобретению могут предусматривать выделенные, полученные в результате культивирования клеток или сконструированные варианты и/или производные человеческих моноклональных антител, которые распознают по меньшей мере один эпитоп на липоарабиноманнане (LAM). Антитело к PIM6/LAM (например, Р95С1), как описано в данном документе, специфически связывает по меньшей мере одну полиманнозную структуру в PIM6 и в перекрестно-реагирующем маннановом домене PIM6 из LAM. Антитела к LAM и к PIM6/LAM по настоящему изобретению можно очищать согласно способам, известным из уровня техники. Такие способы могут включать, например, без ограничений аффинную хроматографию, ионообменную хроматографию, хроматографию с иммобилизованными хелатами металлов, тиофильную адсорбцию, физико-химическое фракционирование или другие антиген-специфические аффинные способы, например, способы, включающие лиганды для связывания антител на основе белка A, G и L. Такие очищенные антитела могут обладать или могут не обладать структурными характеристиками, которые отличаются от человеческих моноклональных антител, не подвергнутых очистке. Например, после очистки могут возникать изменения конформационных эпитопов человеческих моноклональных антител. Антитела могут быть связаны с дополнительными молекулами, которые удаляются после очистки. Соответственно, такие очищенные антитела могут характеризоваться или могут не характеризоваться другой функциональной активностью. Антитела к LAM и к PIM6/LAM по настоящему изобретению могут иметь целый ряд структурных модификаций. Например, антитела к LAM и к PIM6/LAM по настоящему изобретению могут быть гликозилированы, ПЭГилированы или иным образом химически модифицированы, чтобы воздействовать на стабильность, функцию, биодоступность, распознавание эпитопа или другую функциональную активность. Антитела к LAM и к PIM6/LAM по настоящему изобретению могут представлять собой сконструированные варианты и/или производные таких антител, описанных ниже, и могут обладать или могут не обладать функциональной или структурной эквивалентностью. Соответственно, такие варианты и/или производные все еще рассматриваются в пределах объема настоящего изобретения при условии, что они получены или сконструированы по меньшей мере частично из выделенного человеческого моноклонального антитела к LAM или к PIM6/LAM, и/или распознают по меньшей мере один эпитоп на LAM.
1. А194-01
В некоторых вариантах осуществления настоящее изобретение направлено на человеческое моноклональное антитело А194-01, включая его варианты и/или производные. А194-01 является специфическим в отношении LAM. А194-01 обладает очень высокой связывающей активностью LAM, например, изотип IgG А194-01 может проявлять 50% максимальную связывающую активность антитела при концентрации, составляющей приблизительно 20 нг/мл, что, таким образом, служит признаком высокой аффинности в отношении LAM. А194-01 было изначально выделено и очищено в виде IgG, однако, А194-01 может существовать в виде целого ряда изотипов, а также сконструированных и рекомбинантных изотипов, включая без ограничений IgG, IgA, IgM, моновалентные одноцепочечные Fv (scFv) фрагменты, Fab-белки, бивалентные scFv-фрагменты, одноцепочечные scFv-фрагменты (мономеры), где индивидуальные вариабельные участки легкой цепи и вариабельные участки тяжелой цепи соединены, например, с помощью гибкого линкера, и димерные scFv-белки, в которых два scFv-мономера соединены друг с другом (ФИГ. 1А). Некоторые конкретные сконструированные варианты и/или производные А194-01 включают без ограничений следующие. Один сконструированный вариант и/или производное А194-01 содержат тетравалентный scFv-IgG, образованный путем соединения А194-01 scFv, нацеленного на антиген, с N-концом А194-01 IgG (ФИГ. 1В, ФИГ. 17), что может повышать аффинность связывания и расширять диапазон распознаваемых эпитопов (примеры этого приведены на ФИГ. 14 и 15). Тетравалентное scFv-IgG может содержать лидерную последовательность-VH-VL-IgG или может содержать лидерную последовательность-VL-VH-IgG. Обычный специалист в данной области будет понимать, что сконструированные варианты и/или производные на основе scFv-IgG могут иметь валентности, больше чем просто тетравалентные. Другой сконструированный вариант и/или производное А194-01 содержат пентавалентный IgM, полученный путем перевода димерного А194-01 IgG на константный домен человеческого IgM, где такой пентавалентный IgM содержит 10 сайтов связывания (ФИГ. 1В). Обычный специалист в данной области будет понимать, что возможны дополнительные комбинации антигенных фрагментов А194-01, и они рассматриваются в пределах объема настоящего изобретения, в частности, те фрагменты антитела, которые имеют определяющие комплементарность участки (CDR), специфические для А194-01.
Изотип IgG А194-01 распознает уникальный и сложный эпитоп, который находится на немодифицированных боковых цепях Ara4 и Ara6 и на боковых цепях, несущих единичную маннозу. Хотя А194-01 не распознает боковые цепи, несущие ди- или триманнозные замены, оно реагирует с такими структурами, если они дополнительно модифицированы с помощью заместителя MSX. Соответственно, изотип IgG А194-01 связывается с PILAM и ManLAM с высокой аффинностью, и также сильно связывается с некэпированными вариантами обеих структур Ara4 и Ara6, и связывается несколько менее сильно с кэпированными единичной маннозой структурами Ara4/Ara6 и замещенными MSX, но слабо или совсем не связывается с дизамещенными и тризамещенными ManLAM (ФИГ. 4). Без ограничения теорией очень сильно отличающийся эффект прикрепления маннозы и MSX к концевой маннозе в мономаннозилированной стуктуре Ara4 может отражать отличие между α(1→2)-связью маннозы и α(1→4)-связью замены MSX. Сконструированные варианты и/или производные А194-01, включая те, которые обладают более высокими валентностями, могут проявлять более широкую эпитопную специфичность, чем А194-01 IgG (ФИГ. 14), и могут дополнительно проявлять усиленную аффинность в отношении LAM (ФИГ. 15). Например, сконструированное А194-01 в виде тетравалентного scFv-IgG и сконструированные изотипы IgA и IgM связываются с обеими структурами Ara4 и Ara6 с более высокой аффинностью, чем изотип IgG А194-01, и, более того, также распознает структуры, кэпированные диманнозой и триманнозой, с которыми изотип IgG связывается в небольшой степени (ФИГ. 14). Поскольку у патогенных видов комплекса Mycobacterium tuberculosis преимущественно наблюдаются структуры, кэпированные диманнозой, эти сконструированные варианты и/или производные А194-01, включая scFv-IgG и изотипы IgM, могут оказаться особенно применимыми для диагностических наборов и способов, а также для терапевтического применения.
Дополнительные сконструированные варианты и/или производные А194-01 включают такие антитела, где Fc-домен IgG1 превращен в IgG3, который является более опсогенным, или путем получения мультимерных вариантов с помощью замены константного домена IgG1 на димерный IgA или пентамерный или гексамерный IgM. Без ограничения теорией это может значительно усиливать авидность антител к LAM путем увеличения гибкости и диапазона бивалентного и мультивалентного связывания, что способствует повышению аффинности (ФИГ. 1). Это имеет потенциально важное клиническое значение, поскольку такие виды лечения будут особенно ценными в случаях воздействия или инфицирования штаммами Myocbacterium tuberculosis с MDR или X-MDR, которые нельзя эффективно лечить с помощью традиционных антибиотиков.

Обычный специалист в данной области будет понимать, что CDR являются очень важными для разнообразия по антигенной специфичности. Обычный специалист в данной области будет также понимать, что CDR3 является наиболее вариабельным из участков CDR, и сам по себе обладает наибольшей важностью, при этом разнообразия в участке CDR3 из вариабельного участка тяжелой цепи достаточно для большей части специфичности антител. Соответственно, в некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 1, 2 и 3 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 1, 2 и 3 соответственно с консервативными модификациями в последовательности. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 95% идентичностью с SEQ ID NO: 1, 2 и 3 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 90% идентичностью с SEQ ID NO: 1, 2 и 3 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 85% идентичностью с SEQ ID NO: 1, 2 и 3 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 80% идентичностью с SEQ ID NO: 1, 2 и 3 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 3. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 3, с консервативными модификациями в последовательности. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 95% идентичностью с SEQ ID NO: 3. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 90% идентичностью с SEQ ID NO: 3. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 85% идентичностью с SEQ ID NO: 3. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 80% идентичностью с SEQ ID NO: 3.
В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 4, 5 и 6 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 4, 5 и 6 соответственно, с консервативными модификациями в последовательности. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 95% идентичностью с SEQ ID NO: 4, 5 и 6 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 90% идентичностью с SEQ ID NO: 4, 5 и 6 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующий вплоть до 85% идентичностью с SEQ ID NO: 4, 5 и 6 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 80% идентичностью с SEQ ID NO: 4, 5 и 6 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 6. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 6, с консервативными модификациями в последовательности. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 95% идентичностью с SEQ ID NO: 6. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 90% идентичностью с SEQ ID NO: 6. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 85% идентичностью с SEQ ID NO: 6. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 80% идентичностью с SEQ ID NO: 6.
В экспериментах, описанных в данном документе, антитело А194-01 экспрессировали путем трансфекции векторами с Н- и L-цепью клеток Expi293 и культивирования их в бессывороточной среде, стандартной для Expi293, в течение нескольких дней. Секретируемое антитело очищали из культурального супернатанта с помощью аффинной хроматографии на колонках путем конъюгации с лигандами для белка А или белка G. Связанные антитела освобождали от лигандов путем обработки с помощью буфера с низким рН (0,2 М глицин-HCl, рН 2,5) и нейтрализацией с помощью 1/50 объема 2 М трис-буфера (рН 8,8). Замену буфера на PBS проводили с помощью диализа или с помощью нескольких раундов концентрирования на центрифужных фильтрах (центрифужные фильтры Amicon Ultra, предел отсечения по молекулярной массе 30000).
Аминокислотные (аа) последовательности и последовательности нуклеиновой кислоты (nt) для последовательностей тяжелой и легкой цепи А194 являются следующими:
nt-последовательность тяжелой цепи А194:

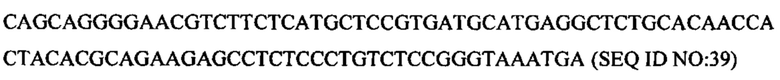
аа-последовательность тяжелой цепи А194:

nt-последовательность легкой цепи А194 (каппа-цепи):

аа-последовательность легкой цепи А194 (каппа-цепи):
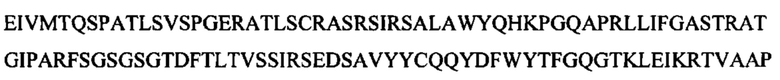

2. Р30В9
В некоторых вариантах осуществления настоящее изобретение направлено на рекомбинантное человеческое моноклональное антитело Р30В9, включая его варианты и/или производные. Р30В9 является специфическим в отношении LAM. Р30В9 было изначально выделено и очищено в виде IgM, однако Р30В9 может существовать в виде целого ряда изотипов, а также сконструированных и рекомбинантных изотипов, включая без ограничений IgM, IgG, IgA, а также их антигенных фрагментов, включая без ограничений моновалентные одно цепочечные Fv- (scFv) фрагменты, Fab-белки, бивалентные scFv-фрагменты, одноцепочечные scFv-фрагменты (мономеры), где индивидуальные вариабельные участки легкой цепи и вариабельные участки тяжелой цепи соединены, например, с помощью гибкого линкера, и димерные scFv-белки, в которых два scFv-мономера соединены друг с другом.
Изотип IgM Р30В9 наиболее эффективно связывается с эпитопами LAM на основе замещенных диманнозой Ara4 и Ara6 со структурами Manp-α(1→2)-Manp-(1→5)-Araf (ФИГ. 4, 16 и 18) хотя другие замещенные Manp-α структуры (например, структура 2, 4 и 59 на ФИГ. 8) также могут распознаваться с более низкими аффинностями. Предпочтительное распознавание Р30В9 кэпированных диманнозой LAM имеет потенциально важное клиническое значение, поскольку сообщалось, что диманнозные кэпы являются доминантной модификацией LAM на вирулентных штаммах комплекса Mycobacterium tuberculosis. Без ограничения теорией полагают, что концевые маннозильные структурные единицы опосредуют связывание LAM вирулентных штаммов комплекса Mycobacterium tuberculosis с человеческими макрофагами и другими иммунными клетками, что приводит к расстройству иммунной функции и развитию стабильной инфекции. Без ограничения теорией полагают, что связывание маннозных кэпов с маннозным рецептором ограничивает фагосомно-лизосомное (P-L) слияние и облегчает выживание бактерии в инфицированных макрофагах. На специфичность Р30В9 в отношении кэпированных диманнозой LAM указывает специфичность данного mAb в отношении гликоконъюгатов, несущих данную структуру, а также тот факт, что изотип IgM Р30В9 специфически связывается с LAM, полученным от любой Mycobacterium tuberculosis, но не с LAM от Mycobacterium smegmatis или Mycobacterium leprae, которые не содержат эпитопы LAM, кэпированные диманнозой. В этом его отличие от изотипа IgG А194-01, которое связывается с PILAM, некэпированными остатками Ara4/Ara6 и эпитопами LAM, кэпированными мономаннозой, все из которых являются обычными для Mycobacterium smegmatis и Mycobacterium leprae. Аналогично изотипу IgM Р30В9 изотип IgM А194-01 способен связываться с эпитопами LAM, кэпированными диманнозой и триманнозой (ФИГ. 14), преимущественно вследствие повышенной авидности связывания.
Следовательно, изотипы IgM Р30В9 могут служить в качестве важного иммунодиагностического реагента для выявления инфекции, вызванной вирулентными представителями комплекса Mycobacterium tuberculosis, и различения указанных вирулентных представителей от других непатогенных видов микобактерий, поскольку оно является специфическим в отношении кэпированного диманнозой LAM. Более того, изотип IgM антитела Р30В9, а также сконструированные варианты и/или производные А194-01 могут обладать иммунотерапевтической активностью, которая ограничивает инфекцию и патогенез вирулентных представителей комплекса Mycobacterium tuberculosis, и могут подходить для применения в терапии, либо в комбинации с традиционными антибиотиками, дополнительными антителами, либо отдельно, или могут применяться в качестве средства для пассивной иммунотерапии. Изотип IgM Р30В9 специфически связывается с ManLAM, полученным от Mycobacterium tuberculosis, с высокой аффинностью (ФИГ. 2А, В), но не с PILAM, полученным от Mycobacterium smegmatis (ФИГ. 2С).
Сконструированные варианты и/или производные Р30В9 могут включать, например, Р30В9, экспрессируемый в виде изотипа IgA, включая димерные IgA1 и IgA2. Без ограничения теорией полагают, что для функционирования Р30В9 требуется поливалентность, поскольку данное антитело было выделено в виде IgM и не является активным, если оно экспрессируется в виде IgG. В настоящем изобретении показано, что Р30В9 является активным в виде сконструированных изотипов IgA, включая димерные IgA1 и IgA2. Это тестировали путем перемещения VH-домена Р30В9 в векторы с IgA1 и IgA2. IgA1 отличается от IgA2, главным образом, присутствием 16-аминокислотной вставки, составленной из повтора 8 аминокислот, богатых пролином, серином и треонином, и модифицированной 3-6 О-связанными олигосахаридами [ФИГ. 5]. Связывающую активность сконструированных форм IgA Р30В9 в отношении ManLAM сравнивали с таковой у форм IgG и IgM. Форма IgM характеризовалась самой высокой активностью, в то время как обе формы IgA также были способны связываться с ManLAM, при этом форма IgA2 демонстрировала более слабую активность, чем форма IgA1, а форма IgG была неактивной в ELISA в отношении ManLAM (ФИГ. 6).

Обычный специалист в данной области будет понимать, что CDR являются очень важными для разнообразия по антигенной специфичности. Обычный специалист в данной области будет также понимать, что CDR3 является наиболее вариабельным из участков CDR, и сам по себе обладает наибольшей важностью, при этом разнообразия в участке CDR3 из вариабельного участка тяжелой цепи достаточно для большей части специфичности антител. Соответственно, в некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 7, 8 и 9 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 7, 8 и 9 соответственно с консервативными модификациями в последовательности. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 95% идентичностью с SEQ ID NO: 7, 8 и 9 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 90% идентичностью с SEQ ID NO: 7, 8 и 9 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 85% идентичностью с SEQ ID NO: 7, 8 и 9 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 80% идентичностью с SEQ ID NO: 7, 8 и 9 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 9. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 9, с консервативными модификациями в последовательности. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 95% идентичностью с SEQ ID NO: 9. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 90% идентичностью с SEQ ID NO: 9. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 85% идентичностью с SEQ ID NO: 9. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 80% идентичностью с SEQ ID NO: 9.
В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 10, 11 и 12 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 10, 11 и 12 соответственно, с консервативными модификациями в последовательности. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 95% идентичностью с SEQ ID NO: 10, 11 и 12 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 90% идентичностью с SEQ ID NO: 10, 11 и 12 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующий вплоть до 85% идентичностью с SEQ ID NO: 10, 11 и 12 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 80% идентичностью с SEQ ID NO: 10, 11 и 12 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 12. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 12, с консервативными модификациями в последовательности. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 95% идентичностью с SEQ ID NO: 12. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 90% идентичностью с SEQ ID NO: 12. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 85% идентичностью с SEQ ID NO: 12. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 80% идентичностью с SEQ ID NO: 12.
В экспериментах, описанных в данном документе, антитело Р30В9 экспрессировали путем трансфекции векторами с Н- и L-цепью клеток Expi293 и культивирования их в бессывороточной среде, стандартной для Expi293, в течение нескольких дней. Секретируемое антитело очищали из культурального супернатанта с помощью аффинной хроматографии на колонках путем конъюгации с лигандами для белка L. Связанное антитело освобождали от лигандов путем обработки с помощью буфера с низким рН (0,2 М глицин-HCl, рН 2,5) и нейтрализацией с помощью 1/50 объема 2 М трис-буфера (рН 8,8). Замену буфера на PBS проводили с помощью диализа или с помощью нескольких раундов концентрирования на центрифужных фильтрах (центрифужные фильтры Amicon Ultra, предел отсечения по молекулярной массе 30000).
Аминокислотные последовательности для тяжелой цепи и легкой цепи Р30В9 и их сравнение с их ближайшей последовательностью зародышевой линии показано на ФИГ. 22. Аминокислотные и нуклеотидные последовательности Р30В9, включая участок CDR3, воспроизведены ниже:
Вариабельный участок тяжелой цепи Р30В9:

Вариабельный участок легкой цепи Р30В9: 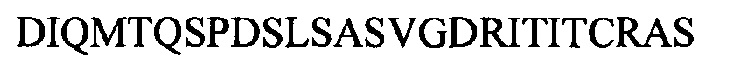

Последовательность ДНК тяжелой цепи Р30В9:

Легкая цепь Р30В9:

3. Р95С1
В некоторых вариантах осуществления настоящее изобретение направлено на рекомбинантное человеческое моноклональное антитело Р95С1, включая его варианты и/или производные. Р95С1 является специфическим в отношении эпитопа, общего для LAM, LM и PIM6. Хотя Р95С1 был изначально выделен и очищен в виде IgM, Р95С1 также является активным, когда оно экспрессируется в виде других изотипов, включая без ограничений формы IgG и IgA.
Обычный специалист в данной области будет понимать, что CDR являются очень важными для разнообразия по антигенной специфичности. Обычный специалист в данной области будет также понимать, что CDR3 является наиболее вариабельным из участков CDR и сам по себе обладает наибольшей важностью, при этом разнообразия в участке CDR3 из вариабельного участка тяжелой цепи достаточно для большей части специфичности антител. Соответственно, в некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 13, 14 и 15 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 13, 14 и 15 соответственно, с консервативными модификациями в последовательности. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 95% идентичностью с SEQ ID NO: 13, 14 и 15 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 90% идентичностью с SEQ ID NO: 13, 14 и 15 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 85% идентичностью с SEQ ID NO: 13, 14 и 15 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка легкой цепи, характеризующиеся вплоть до 80% идентичностью с SEQ ID NO: 13, 14 и 15 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 15. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, как изложено под SEQ ID NO: 15, с консервативными модификациями в последовательности. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 95% идентичностью с SEQ ID NO: 15. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 90% идентичностью с SEQ ID NO: 15. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 85% идентичностью с SEQ ID NO: 15. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка легкой цепи, характеризующийся вплоть до 80% идентичностью с SEQ ID NO: 15.
В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 16, 17 и 18 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 16, 17 и 18 соответственно, с консервативными модификациями в последовательности. В некоторых вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 95% идентичностью с SEQ ID NO: 16, 17 и 18 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 90% идентичностью с SEQ ID NO: 16, 17 и 18 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 85% идентичностью с SEQ ID NO: 16, 17 и 18 соответственно. В других вариантах осуществления антитела к LAM имеют участки CDR1, CDR2 и CDR3 из вариабельного участка тяжелой цепи, характеризующиеся вплоть до 80% идентичностью с SEQ ID NO: 16, 17 и 18 соответственно. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 18. В некоторых вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, как изложено под SEQ ID NO: 18 с консервативными модификациями в последовательности. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 95% идентичностью с SEQ ID NO: 18. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 90% идентичностью с SEQ ID NO: 18. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 85% идентичностью с SEQ ID NO: 18. В других вариантах осуществления антитела к LAM имеют участок CDR3 из вариабельного участка тяжелой цепи, характеризующийся вплоть до 80% идентичностью с SEQ ID NO: 18.

Антитело Р95С1 IgM/κ первоначально было идентифицировано в супернатантах BCL6/Bcl-xL-трансдуцированных В-клеток памяти и клонировано из этих клеток в векторы экспрессии IgM/κ с применением стандартного протокола для RT-PCR. Антитело экспрессировали путем трансфекции векторами с Н- и L-цепью клеток Expi293 и культивирования их в бессывороточной среде, стандартной для Expi293, в течение нескольких дней. Секретируемое антитело очищали из культурального супернатанта с помощью аффинной хроматографии на колонках путем конъюгации с лигандами для белка L. Связанное антитело освобождали от лигандов путем обработки с помощью буфера с низким рН (0,2 М глицин-HCl, рН 2,5) и нейтрализацией с помощью 1/50 объема 2 М трис-буфера (рН 8,8). Замену буфера на PBS проводили с помощью диализа или с помощью нескольких раундов концентрирования на центрифужных фильтрах (центрифужные фильтры Amicon Ultra, предел отсечения по молекулярной массе 30000).
Аминокислотные последовательности для тяжелой цепи и легкой цепи Р95С1 и их сравнение с их ближайшей последовательностью зародышевой линии показано на ФИГ. 23. Аминокислотные и нуклеотидные последовательности Р95С1, включая участок CDR3, воспроизведены ниже:
Вариабельный участок тяжелой цепи Р95С1: 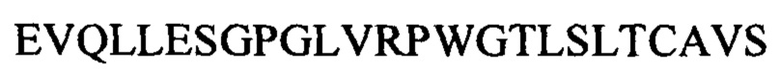

Вариабельный участок легкой цепи Р95С1: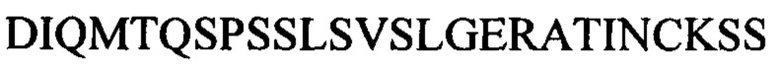
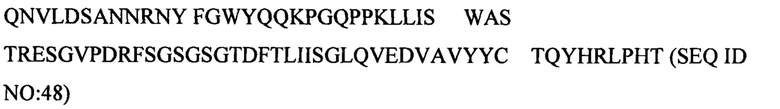
Тяжелая цепь Р95С1:

Легкая цепь Р95С1:

Е. Дополнительные варианты и/или производные
Обычный специалист в данной области будет понимать, что с помощью участков CDR из А194-01, Р30В9 и Р95С1 можно разработать огромное количество сконструированных вариантов и/или производных антител к LAM, раскрытых в данном документе. Например, антитела к LAM по настоящему изобретению могут быть сконструированы в виде химерных антител, гуманизированных антител и химерных/гуманизированных антител, которые проявляют аффинность к одному или нескольким эпитопам LAM. Антитела могут быть сконструированы в виде биспецифических антител или могут быть сконструированы таким образом, что конструкция на основе одного антитела связывается с множественными эпитопами LAM.
Как описано в данном документе, антитела к LAM по настоящему изобретению можно конструировать в виде гомологичных конструкций scFv-IgG или в виде гетерологичных конструкций scFv-IgG. Гомологичные конструкции scFv-IgG из А194-01 подробно описаны в настоящей заявке (ФИГ. 17А). Одним неограничивающим примером гетерологичной конструкции scFv-IgG будет такая, в которой VH- и VL-цепи Р30В9 соединены с А194-01 IgG с помощью линкера (ФИГ. 17В), хотя можно использовать другие VH/VL-цепи, например, из других антител к LAM, таких как мышиные антитела к LAM. Это может обеспечивать возможность распознавания отдельных эпитопов в одной молекуле антигена и может усиливать мультивалентное связывание и приводить к повышенной аффинности. Альтернативно с помощью гетерологичных конструкций scFv-IgG можно получать биспецифические антитела, если дополнительные VH/VL-цепи нацеливаются на антиген, отличный от LAM.
Антитела к LAM по настоящему изобретению можно также конструировать для создания конструкций scFv-IgM, включающих как гомологичные, так и гетерологичные конструкции scFv-IgM. Неограничивающим примером гомологичного scFv-IgM будет конструкция, в которой VH/VL-цепи Р30В9 соединены с Р30В9 IgM. В данной конструкции все сайты связывания будут обладать одинаковой эпитопной специфичностью. Неограничивающим примером гетерологичной конструкции scFv-IgM является конструкция, в которой А194-01 scFv соединен с Р30В9 IgM, в противоположность константному домену IgG [неограничивающий пример [ФИГ. 17С]]. Такая сконструированная вариантная и/или производная конструкция будет сохранять IgM-зависимое распознавание диманнозных эпитопов от исходных mAb Р30В9 и добавлять дополнительную специфичность связывания А194-01 scFv. Это может обеспечивать возможность распознавания уникальных наборов эпитопов и приводить к усиленным аффинностям, что могло бы быть ценным для улучшенных анализов выявления антигена "у постели больного". F. Диагностические наборы и способы
Один вариант осуществления настоящего изобретения относится к диагностическим наборам и способам выявления и/или количественной оценки LAM и/или PIM6 в образце. Как описано в данном документе, антитела к LAM А194-01 и Р30В9, а также антитело к PIM6/LAM, Р95С1, включая его сконструированные варианты и/или производные, может быть эффективным в выявлении и/или количественной оценке содержания LAM и/или PIM6, присутствующих в образце. LAM или PIM6 могут быть получены из любого источника, как, например, из Mycobacterium tuberculosis или Mycobacterium smegmatis, или из образца сыворотки крови или мочи от пациента, например, пациента, инфицированного вирулентным штаммом комплекса Mycobacterium tuberculosis. Например, LAM может представлять собой PILAM, ManLAM или некэпированный/немодифицированный AraLAM из других штаммов микобактерий, таких как М. leprae. Данные штаммы отличаются по природе и степени встречающегося кэпирования, и разные комбинации антител, следовательно, будут характеризоваться разной специфичностью в отношении разных форм, что обеспечивает возможность осуществления некоторого уровня разграничения или типирования. В частности, изотип IgM и сконструированный изотип IgA1 Р30В9, а также сконструированные изотипы IgM и scFv-IgG А194-01 будут хорошо подходить для выявления и/или количественной оценки замещенного диманнозой ManLAM в образце от пациента с ТВ, который в ряде случаев может составлять 80% указанного LAM, и различные изотипы Р30В9 будут особенно эффективными при выявлении и/или количественной оценке LAM, несущего замещенные диманнозой остатки Ara6, которые, как описано в данном документе, особенно широко распространены на LAM, полученном из Mycobacterium tuberculosis. Поскольку эпитоп, распознаваемый Р95С1, является высоко консервативным у всех молекул LAM, данное антитело, при сочетании со вторым антителом с надлежащей специфичностью, будет хорошо подходить для выявления и/или количественной оценки различных типов LAM в образце. Изотип IgG А194-01 очень эффективно связывается с различными формами LAM, в частности, незамещенным LAM, мономаннозилированным LAM и PILAM, и, вследствие этого, будет эффективным в выявлении и/или количественной оценке LAM, полученного из различных штаммов микобактерий. Сконструированные изотипы IgM и scFv-IgG также будут довольно эффективными в выявлении и/или количественной оценке содержания незамещенного LAM, мономаннозилированного LAM и PILAM и дополнительно могут связываться с LAM, замещенным ди- и триманнозой. Это снабжает сконструированные варианты и/или производные А194-01 лучшим эпитопным распознаванием, чем у изотипа IgG А194-01 или изотипа IgM Р30В9, но за счет специфичности в отношении только тех эпитопов LAM, которые являются специфическими для вирулентных штаммов Mycobacterium tuberculosis. В некоторых вариантах осуществления количественную оценку указанной специфичности в отношении LAM и/или PIM6 осуществляют путем сравнения интенсивности сигнала серийно разведенного контрольного образца с известной концентрацией LAM и/или PIM6 в различных анализах прямого связывания или анализах захвата антигена.
Поскольку изотип IgG А194-01, изотипы IgM/IgA Р30В9 и различные изотипы Р95С1 связываются с разными эпитопами LAM, которые по-разному экспрессируются в разных штаммах Mycobacterium tuberculosis, эти конкретные изотипы можно было применять для разграничения происхождения источника LAM; при этом замещенный диманнозой LAM, в частности, замещенные диманнозой остатки Ara6, составляют большинство остатков LAM у вирулентных штаммов Mycobacterium tuberculosis, тогда как остатки незамещенного LAM/PILAM составляют большинство остатков LAM у быстрорастущих невирулентных штаммов, таких как Mycobacterium smegmatis. Например, образцы, содержащие LAM, который связывается только с А194-01 IgG, а не Р30В9 IgM, с большой вероятностью, не получены от вирулентного штамма Mycobacterium tuberculosis, тогда как образцы, которые связываются с обоими P30B0 IgM и А194-01 IgG, с большой вероятностью, действительно получены от вирулентного штамма Mycobacterium tuberculosis или вида микобактерий, в который введен аналогичный кэпирующий мотив.
Поскольку изотипы IgM/IgA Р30В9 являются специфическими в отношении замещенного диманнозой ManLAM, который, как подробно описано в данном документе, является доминантной формой у вирулентных штаммов Mycobacterium tuberculosis, указанные изотипы Р30В9 являются идеальными кандидатами для диагностических наборов и способов для применения в диагностировании пациентов как инфицированных вирулентным штаммом комплекса Mycobacterium tuberculosis. Более того, сконструированные варианты и/или производные IgM и scFv-IgG А194-01 могут подходить для такого применения, поскольку они также распознают замещенные диманнозой и триманнозой эпитопы ManLAM. Такой пациент мог иметь протекающую или активную инфекцию, или инфекция могла быть латентной. Штамм мог характеризоваться множественной лекарственной устойчивостью (MDR) или мог характеризоваться широкой лекарственной устойчивостью (XDR). В частности, касательно пациентов, имеющих латентные инфекции, изменение концентрации LAM в сыворотке крови или моче может иметь особое значение, поскольку повышение концентраций может являться показателем перехода к активной инфекции. Альтернативно снижение концентрации у индивидуума, который имеет активную инфекцию, может служить показателем того, что лечение является эффективным и должно быть продолжено, или повышение концентрации во время лечения может указывать на то, что текущее лечение не является эффективным и должно быть отменено, изменено и/или модифицировано.
Способы диагностирования инфекции могут включать приведение биологического образца, например, крови, плазмы, мочи, мокроты или другой биологической жидкости, от указанного пациента в контакт с по меньшей мере одним антителом к LAM и/или с по меньшей мере одним антителом к PIM6/LAM по настоящему изобретению, в частности, теми антителами к LAM, которые распознают замещенный диманнозой ManLAM, и теми антителами к PIM6/LAM, которые распознают по меньшей мере одну полиманнозную структуру в маннановом домене PIM6. Они включают, например, изотипы IgM и IgA Р30В9, сконструированные изотипы IgA, IgM и scFv-IgG A194-01 и различные изотипы (IgG, IgM, IgA) P95C1.
Антитела, применяемые в качестве выявляющего реагента, могут быть связаны с репортерными молекулами, такими как молекулы, известные из уровня техники. Антитела могут составлять часть набора, например, быть связанными с субстратом или составлять часть сэндвич-анализа. Наборы могут включать первое захватывающее антитело к LAM или к PIM6/LAM, второе детекторное (выявляющее) антитело к LAM или к PIM6/LAM, которое связано с репортерной молекулой, и подложку, с которой связано захватывающее антитело к LAM или к PIM6/LAM. Первое и второе антитела к LAM могут связываться с одними и теми же эпитопами LAM, которые присутствуют в виде множественных копий на одной молекуле LAM, или предпочтительно они могут связываться с разными эпитопами, присутствующими на одной молекуле LAM. Эпитопы LAM и PIM6 могут представлять собой любые из эпитопов, описанных в данном документе. Наборы могут включать третье захватывающее или детекторное (выявляющее) антитело, которое связывается с неконкурирующим сайтом первого и второго антитела. Оно может увеличивать число захваченных молекул и число связанных детекторных молекул, а также силу соответствующего сигнала.
Наборы могут включать инструкции для применения и могут дополнительно содержать различные реагенты, растворители, разбавители и/или фармацевтически приемлемые консерванты. Чувствительность разных меченных биотином моноклональных антител к LAM исследовали в таком анализе [ФИГ. 7]. В данном анализе мышиное антитело к LAM CS-35 применяли для захвата ManLAM из раствора. Данное антитело было выбрано из-за его широкой специфичности. CS-35 (250 нг/лунка) применяли для захвата ManLAM из растворов, содержащих отличающиеся концентрации, и затем разные биотинилированные моноклональные антитела применяли для исследования присутствия ManLAM в лунке для захвата. При применении порогового значения, составляющего 3 х SD фона, наиболее чувствительным зондом было А194-01 IgM, которое приводило к сильному сигналу (1,8 OD) в случае самого высокого разбавления ManLAM (0,016 нг/лунка). Оно превосходило два мышиных антитела FIND, которые ранее считались наилучшими доступными зондами для данного типа анализа.
G. Терапевтические композиции, способы, вакцины и векторы
Один вариант осуществления настоящего изобретения направлен на фармацевтические композиции, содержащие по меньшей мере одно антитело к LAM или антитело к PIM6/LAM по настоящему изобретению, а также способы их применения в осуществлении лечения нуждающегося в этом пациента. Пациент может иметь латентную или активную инфекцию, вызванную вирулентным штаммом Mycobacterium tuberculosis, и, что особенно важно, штамм может характеризоваться множественной лекарственной устойчивостью (MDR) или широкой лекарственной устойчивостью (XDR) в отношении традиционных средств терапии/антибиотиков. Антитела к LAM и к PIM6/LAM, используемые в данных композициях и способах, могут представлять собой любое антитело к LAM или антитело к PIM6/LAM по настоящему изобретению, но, что особенно важно, могут представлять собой такие антитела к LAM, которые распознают кэпированный диманнозой ManLAM, в частности, кэпированные диманнозой остатки Ara6, например, Р30В9 IgM или изотип IgA1/IgA2 и пентавалентное А194-01 IgM или пентавалентный изотип scFv-IgG и различные изотипы Р95С1.
Фармацевтически приемлемая композиция на основе антитела к LAM и/или антитела к PIM6/LAM, подходящая для введения пациенту, будет содержать эффективное количество антитела или антител к LAM или к PIM6/LAM в составе, оба из которых сохраняют биологическую активность, при этом также содействовать максимальной стабильности во время хранения в пределах приемлемого диапазона температур. Фармацевтические композиции могут также включать, в зависимости от требуемого состава, фармацевтически приемлемые разбавители, фармацевтически приемлемые носители и/или фармацевтически приемлемые наполнители, или любую такую среду-носитель, обычно применяемую для составления фармацевтических композиций для введения животному или человеку. Разбавитель выбирают таким образом, чтобы он не воздействовал на биологическую активность комбинации. Примерами таких разбавителей являются дистиллированная вода, физиологический фосфатно-солевой буферный раствор, растворы Рингера, раствор декстрозы и раствор Хенкса. Количество наполнителя, который применим в фармацевтической композиции или составе по настоящему изобретению, представляет собой количество, которое служит для однородного распределения антитела по всей композиции, вследствие чего оно может быть однородно распределено, когда его следует доставлять нуждающемуся в этом субъекту. Он может служить для разбавления антитела до концентрации, которая обеспечивает требуемые благоприятные паллиативный или лечебный результаты, при этом в то же самое время сводит к минимуму любые негативные побочные эффекты, которые могли бы возникнуть из-за слишком высокой концентрации. Он также может обладать консервирующим эффектом. Таким образом, в случае антитела с высокой физиологической активностью будет использоваться больше наполнителя. С другой стороны, в случае любого(любых) активного(активных) ингредиента(ингредиентов), который(которые) проявляет(проявляют) более низкую физиологическую активность, будет использоваться меньшее количество наполнителя.
Фармацевтически приемлемая композиция на основе антитела к LAM и/или антитела к PIM6/LAM может находиться в жидкой форме или твердой форме. Твердый состав обычно является лиофилизированным и переводится в раствор перед введением в качестве или однократного, или многократного введения доз. Составы не следует подвергать воздействию экстремальных температур или рН во избежание термической денатурации. Таким образом, необходимо составлять композицию на основе антитела по настоящему изобретению в пределах биологически релевантного диапазона рН. Раствор, забуференный для поддержания надлежащего диапазона рН во время хранения, требуется, особенно в случае жидких составов, хранящихся в течение более длительных периодов времени между составлением и введением. До настоящего времени как для жидких, так и для твердых составов требовалось хранение при более низких температурах (обычно 2- 8°С), чтобы они сохраняли стабильность в течение более длительных периодов. Составленные композиции на основе антитела, особенно жидкие составы, могут содержать бактериостатик для предупреждения или сведения к минимуму протеолиза во время хранения, включая без ограничений эффективные концентрации (обычно <1% вес/объем) бензилового спирта, фенола, м-крезола, хлорбутанола, метилпарабена и/или пропилпарабена. Бактериостатик может быть противопоказан для некоторых пациентов. Следовательно, лиофилизированный состав может быть восстановлен в раствор, либо содержащий, либо не содержащий такой компонент. В забуференный жидкий или твердый состав на основе антитела можно добавлять дополнительные компоненты, включая без ограничений сахара в качестве криопротектора (включая, но не обязательно ограничиваясь, полигидроксиуглеводы, такие как сорбит, маннит, глицерин и дульцит, и/или дисахариды, такие как сахароза, лактоза, мальтоза или трегалоза) и, в некоторых случаях, соответствующую соль (включая без ограничений NaCl, KCl или LiCl). Такие составы на основе антитела, особенно жидкие составы, подлежащие длительному хранению, будут зависеть от применимого диапазона общей осмолярности, чтобы содействовать длительной стабильности при температуре 2-8°С или выше, и при этом также делать состав применимым для парентеральной инъекции. Эффективный диапазон общей осмолярности (общее число молекул в растворе) составляет от приблизительно 200 мОсм/л до приблизительно 800 мОсм/л. Будет очевидно, что количество криопротектора, такого как сахароза или сорбит, будет зависеть от количества соли в составе, чтобы общая осмолярность раствора оставалась в пределах подходящего диапазона. Следовательно, состав, не содержащий соли, может содержать от приблизительно 5% до приблизительно 25% сахарозы, с предпочтительным диапазоном сахарозы от приблизительно 7% до приблизительно 15%, с особенно предпочтительной концентрацией сахарозы в составе, не содержащем солей, составляющей от 10% до 12%. Альтернативно состав на основе сорбита, не содержащий соли, может содержать сорбит в пределах диапазона от приблизительно 3% до приблизительно 12%, с предпочтительным диапазоном от приблизительно 4% до 7%, и особенно предпочтительный диапазон составляет от приблизительно 5% до приблизительно 6% сорбита в составе, не содержащем соли. Для составов, не содержащих соли, конечно, будут требоваться повышенные диапазоны соответствующего криопротектора, чтобы поддерживать эффективные уровни осмолярности. Такой состав также может содержать бивалентный катион (включая, но не обязательно ограничиваясь, MgCl2, CaCl2 и MnCl2) и не-32-ионное поверхностно-активное вещество (включая, но не обязательно ограничиваясь, полисорбат-80 (Tween 80®), полисорбат-60 (Tween 60®), полисорбат-40 (Tween 40®) и полисорбат-20 (Tween 20®), простые полиоксиэтиленалкиловые эфиры, включая без ограничений Brij 58®, Brij 35®, а также другие, такие как Triton Х-100®, Triton X 114®, NP40®, Span 85 и неионные поверхностно-активные вещества серии Pluronic (например, Pluronic 121)). Любая комбинация таких компонентов, включая возможное включение бактериостатика, может применяться для заполнения антитело-содержащих составов по настоящему изобретению. Композиция на основе антитела по настоящему изобретению также может представлять собой "химическое производное", которое описывает антитело, которое содержит дополнительные химические фрагменты, которые в норме не являются частью молекулы иммуноглобулина (например, пэгилирование). Такие фрагменты могут усиливать растворимость, период полувыведения, всасывание и т.д. основной молекулы. Альтернативно фрагменты могут ослаблять нежелательные побочные эффекты основной молекулы или снижать токсичность основной молекулы.
Специфические варианты осуществления включают микросферы из PLGA, обсуждаемые в данном документе и дополнительно известные из уровня техники, а также неразлагаемые среды-носители на основе полимера, содержащие сополимер полиэтилена и поливинилацетата (PEVAc). Дополнительно обзор о доставке с контролированным высвобождением и локализованной доставке терапевтических продуктов на основе антитела приведен в Grainger, et al., 2004, Expert Opin. Biol. Ther. 4(7): 1029-1044), включенном настоящим посредством ссылки во всей своей полноте. Подходящие микрокапсулы, в которые можно инкапсулировать антитело, также могут включать гидроксиметилцеллюлозу или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, полученные с помощью методик коацервации или с помощью межфазной полимеризации. См. публикацию согласно РСТ WO 99/24061 под заголовком "Method for Producing IGF-1 Sustained-Release Formulations", где белок инкапсулирован в микросферы из PLGA, причем данный литературный источник настоящим включен в данный документ посредством ссылки во всей своей полноте. В дополнение, также можно применять микроэмульсии или системы для доставки коллоидных лекарственных средств, такие как липосомы и альбуминовые микросферы. В других предпочтительных композициях с замедленным высвобождением используется биоадгезив, чтобы сохранить антитело в месте введения. Как отмечено выше, состав с замедленным высвобождением может содержать биоразлагаемый полимер, в котором расположено антитело, что может обеспечивать отсутствие немедленного высвобождения. Неинъецируемые устройства могут описываться в данном документе как "имплантат", "фармацевтический имплантат-депо", "имплантат- депо", "неинъецируемое депо" или каким-либо таким аналогичным термином. Обычные имплантаты-депо могут включать без ограничений твердые биоразлагаемые и небиоразлагаемые полимерные устройства (такие как удлиненное полимерное устройство или устройство в форме коаксиального стержня), а также многочисленные системы-насосы, также известные из уровня техники. Инъецируемые устройства разделяются на болюсные инъекции (высвобождение и рассеивание лекарственного средства после инъекции) и депонирующую инъекцию или инъекцию-депо, которые обеспечивают резервуар для хранения в месте инъекции, обеспечивая возможность замедленного высвобождения биологического средства с течением времени. Имплантат-депо может быть хирургическим путем привязан к точке доставки, чтобы обеспечить соответствующий резервуар для пролонгированного высвобождения антитела с течением времени. Такое устройство будет способно переносить состав на основе лекарственного средства в таких количествах, которые требуются в терапевтических или профилактических целях для лечения на протяжении заранее выбранного периода. Имплантат-депо также может обеспечивать защиту состава от разрушения под действием процессов, происходящих в организме (таких как действие протеаз), на весь период проведения лечения. Как известно из уровня техники, термин "замедленное высвобождение" обозначает постепенное (непрерывное или прерывистое) высвобождение такого антигена из матрицы на основе блок-полимера на протяжении продолжительного периода времени. Независимо от специфического устройства замедленное высвобождение композиции на основе антитела к LAM и/или антитела к PIM6/LAM будет приводить к локальным биологически эффективным концентрациям антитела. Замедленное высвобождение биологического средства(средств) будет осуществляться в течение периода, составляющего один день, несколько дней, неделю или более; но, наиболее приемлемо, в течение месяца или более, или вплоть до приблизительно шести месяцев, в зависимости от состава. Природные или синтетические полимеры, известные из уровня техники, будут применимы в качестве имплантата-депо, что обусловлено их характеристиками, такими как универсальная кинетика разрушения, безопасность и биосовместимость. Эти сополимеры можно подвергать манипуляциям, чтобы модифицировать фармакокинетику активного ингредиента, защитить средство от разрушающего действия ферментов, а также разрушения с течением времени в месте прикрепления или инъекции. Специалист будет понимать, что в уровне техники имеется предостаточно идей для осуществления манипуляций со свойствами этих сополимеров, включая соответствующий процесс получения, применяемые катализаторы и конечную молекулярную массу имплантата-депо или инъекции-депо с замедленным высвобождением. Природные полимеры включают без ограничений белки (например, коллаген, альбумин или желатин); полисахариды (целлюлозу, крахмал, альгинаты, хитин, хитозан, циклодекстрин, декстран, гиалуроновую кислоту) и липиды. Биоразлагаемые синтетические полимеры могут включать без ограничений различные сложные полиэфиры, сополимеры L-глутаминовой кислоты и гамма-этил-L-глутамата (Sidman et al., 1983, Biopolymers 22:547-556), полилактиды ([PLA]; патент США №3773919 и ЕР 058481), полилактат-полигликолат (PLGA), такой как сополимер полилактида и гликолида (см., например, патенты США №№4767628 и 5654008), полигликолид (PG), полиэтиленгликоль (PEG), конъюгаты поли(α-гидроксикислот), сложные полиортоэфиры, полиаспирины, полифосфагены, винилпирролидон, поливиниловый спирт (PVA), PVA-g-PLGA, сополимер PEGT-PBT (полиактивный), метакрилаты, поли(N-изопропилакриламид), РЕО-РРО-РЕО (плюроники), сополимеры PEO-PPO-PAA, PLGA-PEO-PLGA, сложные полиортоэфиры (РОЕ) или любые их комбинации, как описано выше (см., например, патент США №6991654 и заявку на патент США №20050187631, каждый из которых включен в данный документе посредством ссылки во всей своей полноте), гидрогели (см., например, Langer et al., 1981, J. Biomed. Mater. Res. 15:167-277; Langer, 1982, Chem. Tech. 12:98-105), неразлагаемый этилен-винилацетат (например, этилен-винилацетатные диски и сополимер полиэтилена и поливинилацетата)), разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как Lupron Depot™, поли-D-(-)-3-гидроксимасляная кислота (ЕР 133988), гели из гиалуроновой кислоты (см., например, патент США №4636524), суспензии альгиновой кислоты, сложные полиортоэфиры (РОЕ) и т.п. Полилактид (PLA) и его сополимеры с гликолидом (PLGA) были хорошо известны из уровня техники с тех пор, как в коммерческое использование был введен Lupron Depot™, одобренный в 1989 году в качестве первого парентерального состава с замедленным высвобождением с использованием полимеров PLA. Дополнительные примеры продуктов, в которые PLA и PLGA используются в качестве наполнителей для обеспечения замедленного высвобождения активного ингредиента, включают Amidox (PLA; периодонтит), Nutropin Depot (PLGA; с hGH) и Trelstar Depot (PLGA; рак предстательной железы). Другие синтетические полимеры включали без ограничений поли(с-капролактон), поли-3-гидроксибутират, поли(β-яблочную кислоту) и поли(диоксанон)]; полиангидриды, полиуретан (см. WO 2005/013936), полиамиды, циклодекстраны, сложные полиортоэфиры, n-виниловый спирт, полиэтиленоксид/полиэтилентерефталат, полифосфат, полифосфонат, сложный полиортоэфир, полицианакрилат, полиэтиленгликоль, полидигидропиран и полиациталь. Небиоразлагаемые устройства включают без ограничений различные производные целлюлозы (карбоксиметилцеллюлозу, ацетат целлюлозы, ацетат- пропионат целлюлозы, этилцеллюлозу, гидроксипропилметил целлюлозу), имплантаты на основе кремния (полидиметилсилоксан), акриловые полимеры (полиметакрилат, полиметилметакрилат, полигидрокси(этилметилакрилат), а также сополимер полиэтилена и винилацетата, полоксамер, поливинилпирролидон, полоксамин, полипропилен, полиамид, полиацеталь, сложный полиэфир, полиэтилен-хлортрифторэтилен, политетрафторэтилен (PTFE или "Teflon™"), бутадиенстирольный каучук, полиэтилен, полипропилен, полифениленоксид- полистирол, поли-а-хлор-р-ксилол, полиметилпентен, полисульфон и другие родственные биостабильные полимеры. Носители, подходящие для составов-депо с замедленным высвобождением включают без ограничений микросферы, пленки, капсулы, частицы, гели, покрытия, матрицы, облатки, пили или другие фармацевтические композиции для доставки. Примеры таких составов с замедленным высвобождением описаны выше. См. также патенты США №№6953593; 6946146; 6656508; 6541033 и 6451346, содержание каждого из которых включено в данный документ посредством ссылки. Лекарственная форма должна быть способна переносить состав на основе лекарственного средства в таких количествах и концентрации, которые требуются в терапевтических целях для лечения на протяжении заранее выбранного периода, и должна обеспечивать достаточную защиту состава от разрушения под действием процессов, происходящих в организме, на весь период проведения лечения. Например, лекарственная форма может быть окружена внешним слоем, изготовленным из материала, который имеет свойства обеспечивать защиту от разрушения под действием метаболических процессов и риска, например, утечки, разламывания, разрыва или деформации. Это может предупреждать выброс содержимого лекарственной формы неконтролируемым образом в условиях напряжений, которым она будет поддаваться во время применения, например, вследствие физических сил, приложенных к устройству высвобождения лекарственного средства в результате нормального движения в суставе и других движений субъекта, или, например, в устройствах для конвективной доставки лекарственного средства физические силы, ассоциированные с давлением, генерируются внутри резервуара. Резервуар для лекарственного средства или другие средства для удержания или содержания лекарственного средства также должны быть из такого материала, чтобы избежать неожиданных реакций с составом на основе активного средства, и предпочтительно биосовместимыми (например, если лекарственная форма имплантируется, то есть в значительной степени инертными с точки зрения организма субъекта или жидкостей организма). В целом, соответствующее биологическое(биологические) средство(средства) вводят индивидууму на срок от по меньшей мере 12 часов до по меньшей мере недели, и, наиболее приемлемо, с помощью имплантата, разработанного для доставки лекарственного средства, на срок по меньшей мере 10, 20, 30, 100 дней, или по меньшей мере 4 месяца, или по меньшей мере 6 месяцев или более, в случае необходимости. Антитело к LAM и/или антитело к PIM6/LAM может доставляться при таких относительно низких объемных скоростях, например, составляющих от приблизительно 0,001 мл/сутки до 1 мл/сутки, чтобы свести к минимуму нарушения в ткани или травму вблизи места, в которое высвобождается состав. Состав может высвобождаться со скоростью, в зависимости от специфического(специфических) биологического(биологических) средства(средств), при низкой дозе, например, от приблизительно 0,01 мкг/ч или 0,1 мкг/ч, 0,25 мкг/ч, 1 мкг/ч, обычно вплоть до приблизительно 200 мкг/ч, или состав доставляется с низкой объемной скоростью, например, объемная скорость составляет от приблизительно 0,001 мл/сутки до приблизительно 1 мл/сутки, например, от 0,01 микрограмма в сутки вплоть до приблизительно 20 миллиграммов в сутки. Дозировка зависит от целого ряда факторов, таких как активность, биодоступность и токсичность применяемого активного ингредиента (например, IgG-антитела) и потребностей субъекта.
Для in vivo лечения пациента-человека и пациентов, отличных от человека, пациенту вводят или предоставляют фармацевтический состав, включающий по меньшей мере одно антитело к LAM и/или по меньшей мере одно антитело к PIM6/LAM по настоящему изобретению. При применении для in vivo терапии антитела к LAM или к PIM6/LAM по настоящему изобретению вводят пациенту в терапевтически эффективных количествах (т.е. количествах, которые устраняют или снижают общую бактериальную нагрузку). Антитела вводят пациенту-человеку в соответствии с известными способами, такими как внутривенное введение, например, в виде болюса или с помощью непрерывной инфузии на протяжении определенного периода времени, с помощью внутримышечного, внутрибрюшинного, интрацереброспинального, подкожного, интраартикулярного, интрасиновиального, интратекального, перорального, местного или ингаляционного путей. Антитела можно вводить парентерально, если возможно, в сайт с целевыми клетками, или внутривенно. В некоторых вариантах осуществления, антитело вводят с помощью внутривенного или подкожного введения. Терапевтические композиции по настоящему изобретению можно вводить пациенту или субъекту системно, парентерально или локально. Вышеуказанные параметры для оценки успешности лечения и улучшения при заболевании легко измеряются с помощью обычных процедур, хорошо известных лечащему врачу.
Для парентерального введения антитела к-LAM и к PIM6/LAM можно составлять в виде стандартной лекарственной инъецируемой формы (раствор, суспензия, эмульсия) в сочетании с фармацевтически приемлемой средой-носителем для парентерального введения. Примеры таких сред-носителей включают без ограничений воду, солевой раствор, раствор Рингера, раствор декстрозы и 5% человеческий сывороточный альбумин. Неводные среды-носители включают без ограничений нелетучие масла и этилолеат. В качестве носителей можно применять липосомы. Среда-носитель может содержать незначительные количества добавок, таких как вещества, которые улучшают изотоничность и химическую стабильность, такие как, например, буферы и консерванты.
Антитела к LAM и к PIM6/LAM по настоящему изобретению можно вводить хозяину с помощью любого способа, стратегии и/или комбинации, доступных из уровня техники, в количествах, достаточных для обеспечения терапевтического лечения инфекции, вызванной вирулентным штаммом комплекса Mycobacterium tuberculsosis. Эти композиции могут предоставляться индивидууму с помощью разнообразных путей, известных из уровня техники, особенно парентеральных путей, включающих без ограничений парентеральные пути, такие как внутривенное (IV), внутримышечное (IM) или подкожное (SC) введение, при этом IV введение представляет собой норму для введения терапевтических антител в данной области техники. Эти композиции можно вводить в виде отдельных или множественных доз (т.е. введение антитела с перерывами по времени при поддержании стерильного состояния состава на протяжении схемы лечения).
Доза и режим дозирования зависят от множества факторов, которые легко может определить лечащий врач, таких как природа инфекции, например, ее терапевтический индекс, пациент и анамнез пациента. В целом, пациенту вводят терапевтически эффективное количество антитела. В некоторых вариантах осуществления количество вводимого антитела находится в диапазоне от приблизительно 0,01 мг/кг до приблизительно 1000 мг/кг массы тела пациента и в любом диапазоне между ними. В зависимости от типа и тяжести инфекции от приблизительно 0,1 мг/кг до приблизительно 50 мг/кг массы тела (например, приблизительно 0,1-15 мг/кг/доза) антитела является первоначальной кандидатной дозировкой для введения пациенту, вне зависимости от того, например, вводится ли она с помощью одного или нескольких отдельных введений или с помощью непрерывной инфузии. Прогресс данной терапии легко отслеживать с помощью традиционных способов и анализов и исходя из критериев, известных лечащему врачу или другим специалистам в данной области. Вышеуказанные параметры для оценки успешности лечения и улучшения при заболевании легко измеряются с помощью обычных процедур, хорошо известных лечащему врачу.
Эти антитела также можно вводить с помощью генетических векторов, которые экспрессируют парные тяжелые и легкие цепи данного антитела. Это может подразумевать плазмиду, которая эффективно экспрессирует эти гены, или вирусный вектор, такой как аденовирусный вектор или вектор на основе аденоассоциированного вируса (AAV). Эти векторы можно доставлять путем инъекции в мышечную ткань, и в зависимости от дозы они могут приводить к секреции относительно больших количеств секретируемого антитела в кровотоке в течение относительно продолжительного периода времени.
Другие терапевтические схемы можно комбинировать с введением антител к LAM и/или к PIM6/LAM по настоящему изобретению, например, с другим антителом к LAM, включая без ограничений такие антитела к LAM, известные из уровня техники, например, мышиными антителами к LAM или их гуманизированными вариантами, или с фармацевтическим соединением, таким как без ограничений антибиотики. Антибиотики, которые подходят для совместного введения с антителами к LAM и/или к PIM6/LAM по настоящему изобретению, включают без ограничений изониазид, рифампин, рифапентин, этамбутол, пиразинамид, бедаквилин, капреомицин, циклосерин, дексаметазон, канамицин и тинокордин. Комбинированное введение включает совместное введение с применением отдельных составов или единого фармацевтического состава и последовательное введение в любом порядке, где предпочтительно имеется период времени, в течение которого оба (или все) активные средства одновременно проявляют свою биологическую активность. Такая комбинированная терапия может приводить к синергическому терапевтическому эффекту. Вышеуказанные параметры для оценки успешности лечения и улучшения при заболевании легко измеряются с помощью обычных процедур, хорошо известных лечащему врачу.
Согласно другому варианту осуществления в настоящем изобретении представлена пассивная вакцинная или фармацевтическая композиции, включающие по меньшей мере одно антитело к LAM и/или к PIM6/LAM по настоящему изобретению и фармацевтически приемлемый носитель. Согласно одному варианту осуществления вакцинная или фармацевтическая композиции представляют собой композицию, включающую по меньшей мере одно антитело, описанное в данном документе, и фармацевтически приемлемый носитель. Вакцина может включать несколько антител с характеристиками, описанными в данном документе, в любой комбинации и может дополнительно включать другие антитела к LAM, включая антитела по настоящему изобретению и антитела, известные из уровня техники, например, мышиные антитела к LAM или их гуманизированные варианты. Пассивная вакцина может включать один или несколько фармацевтически приемлемых консервантов, носителей и/или наполнителей, которые известны из уровня техники.
Согласно другому варианту осуществления настоящее изобретение охватывает активную вакцинную или фармацевтическую композиции, предусматривающие введение пациенту по меньшей мере одного антигенного эпитопа LAM или PIM6. Конкретный эпитоп, который подлежит использованию, можно определять путем тестирования терапевтической активности антител, описанных в данном патенте, на подходящей животной модели ТВ-инфекции и/или патогенеза. Этим модельным видом может служить мышь, или морская свинка, или кролик, или примат. Например, если А194-01 является наиболее защитным, то будут применять вакцину, несущую форму эпитопа для А194-01, тогда как если Р30В9 является наиболее защитным, замещенные диманнозой остатки Ara6 могут быть более эффективными в выработке соответствующего гуморального ответа. Активная вакцина может включать один или несколько адъювантов, которые известны из уровня техники, например алюмокалиевые квасцы, гидроксид алюминия, фосфат алюминия, парафиновое масло и цитокины, например IL-1, IL-2, IL-12. Активная вакцина может содержать один или несколько фармацевтически приемлемых консервантов, носителей и/или наполнителей, которые известны из уровня техники.
В некоторых вариантах осуществления настоящее изобретение направлено на рекомбинантный вектор, например, плазмиду, содержащую нуклеиновую кислоту, кодирующую тяжелую цепь иммуноглобулина (Ig VH) антитела к LAM или антитела к PIM6/LAM, и вторую нуклеиновую кислоту, кодирующую легкую цепь иммуноглобулина (Ig VL). В других вариантах осуществления первая нуклеиновая кислота и вторая нуклеиновая кислота находятся в двух разных рекомбинантных векторах. Согласно другому варианту осуществления настоящее изобретение охватывает способ лечения туберкулезной инфекции у индивидуума, включающий введение указанному индивидууму первой нуклеиновой кислоты, кодирующей тяжелую цепь иммуноглобулина (Ig VH) антитела к LAM или к PIM6/LAM, и второй нуклеиновой кислоты, кодирующей легкую цепь иммуноглобулина (Ig VL) антитела к LAM или к PIM6/LAM, где каждая из нуклеиновых кислот функционально связана с промоторным участком. Первая нуклеиновая кислота и вторая нуклеиновая кислота могут находиться в одном и том же рекомбинантным векторе или в двух разных рекомбинантных векторах. Рекомбинантным вектором могут быть нереплицирующиеся вирусные векторы, например, аденоассоциированные вирусы (AAV), или могут быть плазмиды. В определенных вариантах осуществления настоящее изобретение направлено на клетку, трансформированную с помощью одного или нескольких векторов, раскрытых в данном документе.
Описанные выше антитела и композиции на основе антител, вакцинные композиции и векторы можно вводить для профилактического и терапевтического лечения инфекции, вызванной вирулентными штаммами комплекса Mycobacterium tuberculosis.
Н. Эквиваленты
Если значение представлено в виде диапазонов, понимается, что каждое промежуточное значение до десятых единицы нижнего пределах, если из контекста явно не следует иного, между верхним и нижним пределом такого диапазона и любое другое такое заявленное или промежуточное значение в таком заявленном диапазоне охватывается настоящим изобретением. Верхний и нижний пределы этих меньших диапазонов, которые могут быть независимо включены в меньшие диапазоны, также охватываются настоящим изобретением, кроме случаев любого специфически исключенного предела в заявленном диапазоне. Если заявленный диапазон включает один или оба передела, диапазоны, исключающие любой из двух таких включенных пределов, также включены в настоящее изобретение.
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же самое значение, которое общеизвестно обычному специалисту в области техники, к которой относится настоящее изобретение. Хотя любые способы и материалы, аналогичные или эквивалентные таковым, описанным в данном документе, также можно применять при осуществлении настоящего изобретения на практике или при его тестировании, предпочтительные способы и материалы описаны в данном документе. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки во всей их полноте.
Используемые в данном документе и прилагаемой формуле изобретения формы единственного числа включают ссылки на формы множественного числа, если из контекста явно не следует обратное.
Термин "приблизительно" обозначает диапазон значений, которые не будет считаться обычным специалистом в данной области как в значительной степени отличающийся от исходных значений. Например, термин "приблизительно" может обозначать значение, которое находится в пределах 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,5%, 0,1%, 0,05% или 0,01% от заявленного значения, а также значения, промежуточные для таких заявленных значений.
Публикации, раскрытые в данном документе, представлены исключительно в виде их раскрытия до даты подачи настоящего изобретения. Ничто в данном документе не следует истолковывать как признание того, что настоящее изобретение не имеет права датировать такую публикацию задним числом с учетом предшествующего изобретения. Кроме того, представленные даты публикации могут отличаться от действительных дат публикации, которые могут нуждаться в независимом подтверждении.
Каждая из заявок и патентов, цитируемых в данном тексте, а также каждый документ или ссылка, патентная или непатентная литература, цитируемые в каждой из заявок и патентов (в том числе во время рассмотрения дела каждого выданного патента; "документы, цитируемые в заявке"), и каждая из заявок согласно РСТ и зарубежных заявок или патентов, соответствующих и/или испрашивающих приоритет на основании любой из этих заявок и патентов, и каждый из документов, цитируемых или упоминаемый в каждом из документов, цитируемых в заявке, настоящим явно включены в данный документ посредством ссылки во всей своей полноте. В более широком смысле, документы или ссылки, цитируемые в данном тексте, либо в списке литературы перед формулой изобретения, либо в самом тексте, и каждый из этих документов или ссылок ("ссылки, цитируемые в данном документе"), а также каждый документ или ссылка, цитируемые в каждой из ссылок, цитируемых в данном документе (включая любые спецификации производителя, инструкции и т.д.), настоящим явно включены в данный документ посредством ссылки.
Следующие неограничивающие примеры служат для дополнительной иллюстрации настоящего изобретения.
ПРИМЕРЫ
ПРИМЕР 1. Способы, описанные в данном документе, применяли для культивирования В-клеток памяти in vitro и для молекулярного клонирования генов вариабельного участка иммуноглобулина с целью выделения нескольких новых человеческих моноклональных антител (rnAb), специфических в отношении LAM. Специалисту в данной области техники будет понятно, что данные способы, описанные в данном документе, можно корректировать для селективной идентификации редких антител с очень высокой аффинностью, которые могут присутствовать на всего лишь 1 из 100000 В-клеток памяти, циркулирующих в крови пациента.
Моноклональные антитела
Мышиные моноклональные антитела: линии клеток гибридомы, продуцирующие LAM-специфические мышиные моноклональные антитела CS-35 и CS-40, полученные из лаборатории доктора Delphi Chatterjee, повторно клонировали до однородности, и антитела очищали посредством хроматографии на основе связывания с белком А.
Антитела 906.41, 906.7, 908.1 и 922.5 были предоставлены доктором John Spencer, и FIND25 и FIND 170 были предоставлены Tobias Broger из FIND.
Антигены
Полученный из Mycobacterium tuberculosis липоарабиноманнан (LAM) H37R.V (NR-14848) и полученный из Mycobacterium smegmatis LAM (NR-14860) получали из Университета штата Колорадо посредством ресурсов BEI. Полученные из LAM гликоконъюгаты синтезировали в Lowary lab.
Анализы посредством ELISA
Man-LAM (H37Rv) и PI-LAM (полученный из Mycobacterium smegmatic) разбавляли в буфере СВС (7,5 мМ карбоната натрия, 17,4 мМ бикарбоната натрия, рН 9,0) и высевали при концентрации 100 нг/лунка в 96-луночные планшеты для ELISA. После инкубирования планшетов в течение ночи при 4°С лунки промывали посредством PBS, pH 7,4, содержащего 0,05% Tween-20 (PBST), затем блокировали посредством 1% BSA (Sigma) в буфере PBS. Промытые посредством PBST планшеты инкубировали в течение 1 часа при 37°C с плазмой крови, полученной от индивидуумов, инфицированных Mycobacterium tuberculosis, и контрольной плазмой крови, разбавленной в среде RPMI, содержащей 2% FBS. Промытые посредством PBST планшеты затем инкубировали в течение 1 часа с разведением 1:1000 конъюгиро ванных с щелочной фосфатазой IgG (γ-специфического) (Millipore), или IgM (μ-специфического) (Millipore), или IgA (α-специфического) козы к антителу человека. После промывания посредством PBST цвет проявляли посредством 50 мкл буфера DEA. OD измеряли при 405 нм посредством спектрофотометра. Титры определяли в качестве кратного разбавления, посредством которого получали OD после вычитания фонового OD, охватывающего покрытые BSA планшеты, и определяли посредством интерполирования с помощью экспоненциальной функции.
Титрование плазмы крови
Два очищенных антигена получали из депозитария BEI, и высевали на ночь при 4°С в 96-луночные планшеты для ELISA при концентрации 2 мкг/мл, и планшеты затем блокировали посредством 1% BSA в 1X PBS. LAM-специфические титры плазмы крови тестировали посредством инкубирования серийно разведенных образцов при 37°С в течение 1 часа, после чего планшеты промывали три раза посредством PBS +0,1% Tween20. Связанное антитело выявляли посредством смеси конъюгированных с щелочной фосфатазой каппа-цепи козы к антителу человека и лямбда-цепи козы к антителу человека при разведении 1:1000 в 1% BSA в PBS и сигналы обнаруживали посредством добавления субстрата, представляющего собой щелочной фосфат, в буфере DEA. Реактивность измеряли в виде OD405 в течение 30 мин.
Субъекты-люди
Пациентов с активной инфекцией, вызванной Mycobacterium tuberculosis, включали в исследование в лечебной практике Lattimore во Всемирном институте туберкулеза. Активную инфекцию определяли посредством подтвержденного культурой туберкулезного заболевания или диагноза клинического туберкулеза. Данная группа включала пациентов с недавним диагнозом туберкулеза, пациентов, которые находились на втором месяце терапии. Неинфицированные пациенты являлись HIV-сероотрицательными, имели отрицательную туберкулиновую кожную пробу, являлись здоровыми добровольцами без вакцинации Bacillus  (BCG) в анамнезе и имели отрицательный ответ в отношении анализа высвобождения интерферона-гамма (IGRA) (Quantiferon Gold In-Tube, Cellestis Inc, Валенсия, Калифорния). Информированное письменное согласие получали от участников, и исследование одобряли в Институтском наблюдательном совете Университета Ратджерса.
(BCG) в анамнезе и имели отрицательный ответ в отношении анализа высвобождения интерферона-гамма (IGRA) (Quantiferon Gold In-Tube, Cellestis Inc, Валенсия, Калифорния). Информированное письменное согласие получали от участников, и исследование одобряли в Институтском наблюдательном совете Университета Ратджерса.

1. Культивирование и выделение A194-01 (изотип IgG)
Человеческое моноклональное антитело к LAM А194-01, изотипа IgG, выделяли из культивированных В-клеток памяти, полученных от инфицированного ТВ пациента, ТВ-194. Особо важным компонентом системы культуры in vitro является наличие подходящих питающих клеток, которые могут обеспечивать стимуляцию посредством CD40L, лиганда для CD40, члена суперсемейства рецептора TNF, который экспрессируется на поверхности В-клеток и играет важную роль в опосредовании зависимого от Т-клеток переключения класса иммуноглобулинов и развитии В-клетки. В-клетки памяти высевали на питающий слой экспрессирующих CD40L клеток, бедных на MS40L. Данные клетки экспрессируют низкий уровень CD40L, и ранее было показано, что они эффективно поддерживают репликацию В-клеток памяти и их созревание в плазматические клетки (Luo, X., et al., Blood, 2009. 113(7). Данные клетки образовывали посредством инфицирования мышиных стромальных клеток MS5, которые обеспечивают фактор роста В-клеток IL-7, с помощью FUW-CD40L, вируса, который трансдуцирует CD40L человека, первоначально полученный от Origene (Роквилл, Мэриленд). В-клетки памяти выделяли с помощью набора для выделения человеческих В-клеток памяти MACS от Miltenyi (№ по кат. 130-093-546). Клетки, отличные от В-клеток, отделяли от РВМС посредством негативного отбора с помощью магнитных гранул, содержащих антитела к маркеру клеточной поверхности CD2, CD3, CD14, CD16, CD36, CD43, CD56, CD66b и гликофорину А. Для дополнительного удаления интактных В-клеток, осуществляли позитивный отбор субпопуляций В-клеток памяти с помощью магнитных гранул, связанных с антителом к маркеру клеточной поверхности CD27, маркеру В-клетки памяти, который также экспрессируется на низких уровнях на плазматических клетках, но не на интактных В-клетках. В присутствии экспрессирующих CD40L питающих клеток, данные условия поддерживают репликацию В-клеток памяти и их дифференциацию в лимфобласты, секретирующие относительно высокие титры Ig в супернатантах культуры.
Культуры повторно подпитывали с недельными интервалами посредством замены половины супернатанта культуры свежими средами. Через 2-3 недели было достаточно В-клеток для получения ~1-5 мкг/мл секретированного антитела. Предполагая наличие 100-1000 отличающихся клонов в каждой лунке, это соответствовало средней концентрации 1-10 нг/мл Ig на клон В-клетки. Данная концентрация является достаточно низкой, и, следовательно, данный способ смещался в сторону антитела с относительно высокими значениями аффинности в отношении целевых антигенов. Примерно 80000 В-клеток памяти очищали из крови данного пациента и культивировали в 96 лунках 96-луночного культурального планшета при исходной плотности ~800 клеток/лунка.
Осуществляли скрининг супернатантов культуры посредством ELISA в отношении наличия антитела к полученному из Mycobacterium tuberculosis LAM. LAM покрывали в концентрации 2 мкг/мл посредством 50 мкл бикарбонатного покрывающего буфера на лунку 96-луночного планшета для ELISA и инкубировали при 4°С в течение ночи. Планшет промывали посредством PBST (0,1% Tween 20 в 1X PBS) 4 раза и блокировали посредством 200 мкл 2% обезжиренного молока в 1X PBS в течение 1 часа при 37°С. 100 мкл супернатанта культуры добавляли в соответствующие лунки планшета для ELISA, содержащие LAM, и инкубировали в течение 1 часа при 37°С. После дополнительных стадий промывания добавляли АР-конъюгированные антитела мыши к Fab человека для выявления связанного антитела человека. После инкубирования в течение получаса при 37°С 100 мкл АР-субстрата в буфере DEA добавляли в лунки для ELISA и колориметрически определяли реактивность посредством измерения поглощения при 405 нм.
Положительный сигнал (OD ~1 через 1 ч.) выявляли только в 1 лунке из 96 лунок, что указывает на низкую распространенность данных клеток в данном образце. Клетки из положительных лунок повторно культивировали при плотности 5-10 клеток/лунка в 10 лунках 96-луночных планшетов и повторно подвергали скринингу в отношении активности к LAM. Эти приводило в результате к ~6 положительным лункам (OD ~1 через 1 ч.), вновь соответствующим низкой частоте LAM-реактивных клеток, и что предполагает, что первоначальная положительная лунка содержала только один LAM-положительный клон В-клетки. Клетки из нескольких положительных субклонов лизировали и применяли для выделения вариабельных участков Н- и L-цепей, которые затем клонировали в векторы, экспрессирующие Н- и L-цепь. Всего 10 различных последовательностей VH и 9 последовательностей VL выделяли из данных лунок, и их затем тестировали в отношении активности посредством трансфицирования отдельных комбинаций в 293 клетки. Из 90 тестированных комбинаций только одна комбинация тяжелой цепи (p9045-IgG1-VH) и легкой цепи (p9044-Vk) обеспечивала положительный сигнал в отношении LAM. Антитела экспрессировали посредством котрансфекции соответствующих плазмид, содержащих тяжелую и легкую цепи, в клетки Expi-292, как описано производителем, и выращивали в бессывороточных средах. Антитела очищали посредством аффинной хроматографии либо на гранулах с белком А (для IgG), либо на гранулах с белком L (для IgG) и элюировали буфером с низким значением рН. Очищенные антитела концентрировали и характеризовали посредством SS-PAGE в отношении размера и чистоты.
2. Выделение и культивирование изотипа IgM Р30В9
Человеческое моноклональное антитело к LAM Р30В9, изотипа IgM, выделяли из культивированных В-клеток памяти, полученных от инфицированного ТВ пациента, ТВ-314. РВМС выделяли из крови пациента ТВ-314 посредством центрифугирования в градиенте фиколла, и ~30000 В-клеток памяти очищали, как описано выше, с помощью набора для выделения человеческих В-клеток памяти MACS от Miltenyi. Очищенные В-клетки памяти культивировали в течение 14 дней посредством высевания 400 клеток/лунка на монослои клеток MS40-L, выращенных в 96-луночных планшетах, в присутствии IL-21 (100 нг/мл), IL-10 (100 нг/мл), IL-2 (10 нг/мл), IL-4 (2 нг/мл) и CpG (1 мкМ), и клеточные супернатанты подергали скринингу посредством ELISA в отношении связывания с H37Rv ManLAM. ManLAM покрывали в концентрации 2 мкг/мл посредством 50 мкл бикарбонатного покрывающего буфера на лунку 96-луночного планшета для ELISA и инкубировали при 4°С в течение ночи. Планшет промывали посредством PBST (0,1% Tween 20 в 1X PBS) 4 раза и блокировали посредством 100 мкл 1% BSA в 1 X PBS в течение 1 часа при 37°С. 0 мкл супернатанта культуры или разбавленного антитела добавляли в соответствующие лунки планшета для ELISA, содержащие LAM, и инкубировали в течение 1 часа при 37°С. осле дополнительных стадий промывания добавляли АР-конъюгированные антитела козы к IgG (H+L) человека для выявления связанного антитела человека. После инкубирования в течение получаса при 37°С 50 мкл АР-субстрата в буфере DEA добавляли в лунки для ELISA и определяли реактивность посредством измерения желтого цвета при 405 нм. Только 1 из 78 лунок давала положительный сигнал при мечении реагентом, представляющим собой вторичное антитело козы к каппа-цепи IgG, IgA, IgM человека. После размножения данную лунку трансдуцировали в отношении BCL6 и Bcl-xL, связанных саморасщепляющейся пептидной последовательностью свиного тесховируса-1 (Р2А), после чего следует репортерный ген GFP под управлением IRES. Два этих гена стабилизируют В-клетки памяти для длительной репликации и обеспечивают культивирование клеток даже после отбора антиген-положительных клеток посредством вовлечения BCR. Ретровирусные векторы псевдотипировали посредством гликопротеина оболочки вируса лейкоза гиббонов (GaLV) с удаленным пептидом R из С-концевого ТМ-домена. Успешная трансдукция первичных В- клеток привела к экспрессии BCL-6, Bcl-xL и маркерного белка GFP. Титры вируса определяли посредством подсчета положительных в отношении GFP клеток 293Т под флюоресцентным микроскопом. Активированные В-клетки трансдуцировали ретровирусным вектором в присутствии полибрена/ретронектина.
После дополнительного размножения трансдуцированные клетки субкультивировали при предельных разведениях в присутствии IL-21 (100 нг/мл) и IL-2 (10 нг/мл). Лунку В9 на планшете 30 (Р30В9) выбирали, исходя из ее высокой LAM-связывающей активности и обнаружения посредством микроскопа присутствия одного клона. Супернатант Р30В9 связывался исключительно с лунками, покрытыми H37Rv-LAM, а не с лунками, покрытыми LAM, полученным из Mycobacterium smegmatic, или альфа-кристаллином. Клетки из данных лунок лизировали и выделяли РНК с применением мини-набора RNeasy (Qiagen) с последующим синтезом кДНК с олиго-(dT) с применением системы синтеза кДНК Superscript III (Invitrogen). Вариабельные участки тяжелой и легкой цепей антитела амплифицировали с применением праймеров Smith-Tiller и клонировали в векторы, экспрессирующие тяжелую и легкую цепи человека. Вариабельный участок тяжелой цепи изначально клонировали в стандартный вектор IgG. Однако при объединении с последовательностью легкой цепи, клонированной в вектор, экспрессирующий каппа-цепь человека, LAM-связывающую активность не выявляли. На этом этапе ManLAM-реактивные антитела, продуцированные в первоначальной стабильно трансдуцированной поликлональной лунке, повторно метили изотип-специфическими реагентами и обнаружили, что они представляют собой исключительно IgM. Последовательность VH Р30В9 последовательно клонировали в вектор, экспрессирующий константный участок Н-цепи IgM, и получали хорошую связывающую активность при котрансфекции с соответствующей каппа-цепью.
3. Определение характеристик эпитопной специфичности А194-01 IgG и Р30В9 IgM, а также антител мыши к LAM, направленных против LAM
А. Для определения эпитопов, которые распознаются А194-01 IgG и Р30В9 IgM, значения связывающей активности указанных антител сравнивали со значениями связывающей активности ряда LAM-специфических моноклональных антител мыши (CS-35, CS-40, FIND25, FIND170 и 900 серий моноклональных антител, представленных 908.1) по сравнению с сериями 25 гликоконъюгатов, в которых синтетические гликаны, представляющие различные структуры, присутствующие в LAM, были конъюгированы с бычьим сывороточным альбумином (фиг. 4А). Они варьировали в размерах от 4 до 26 углеводных колец и представляли собой ряд структурных мотивов, которые, как известно, присутствуют в различных микобактериальных LAM, в том числе ряд полиарабинозных структур, как некэпированных, так и кэпированных фосфоинозитолом, альфа-(1→2)-связанные моно-, ди- и триманнопиранозилманнозные структуры и 5-деокси-5-метилтиопентофуранозильные (МТХ) мотивы и различные кэпированные структуры Ara4 и Ara6.
Шесть отличающихся паттернов реактивности получали посредством данной антигенной панели для данных моноклональных антител (фиг. 4В). Значения относительной аффинности моноклональных антител в отношении данных антигенов указаны профилем титрования; реакции с высокой аффинностью сохраняют высокую реактивность при промежуточном разведении, тогда как низкая аффинность указана быстрым снижением реактивности. Самый широкий паттерн наблюдали для mAb мыши CS-35, которое вступало в реакцию с небольшой аффинностью с LAM, полученным из Mycobacterium tuberculosis, и LAM, полученным из Mycobacterium smegmatis, и распознавало как кэпированные, так и некэпированные структуры, содержащие основные мотивы Ara4 и Ara6, что соответствует известной специфичности данных mAb в отношении мотива β-D-Araƒ-(1→2)-α-D-Araƒ-(1→5)-α-D-Araƒ-(1→5)-α-D-Araf.
Человеческое моноклональное антитело к LAM А194-01 IgG также распознавало большую фракцию данных структур и во многих случаях обладало самой большой аффинностью. А109-01 IgG сильно связывался со всеми некэпированными структурами Ara4 и Ara6 и с кэпированной фосфоинозитолом структурой Ara4, и менее сильно с подгруппой кэпированных маннозой структур. А109-01 IgG хорошо связывался с кэпированными мономаннозой структурами, но очень слабо с ди- и триманнозными структурами, хотя реактивность в отношении последних структур увеличивалась при наличии замены МТХ. Четыре из 900 серий моноклональных антител мыши (представленных 908.1) вступали в реакцию с относительно слабой аффинностью со всеми некэпированными структурами Ara4 и Ara6, но не с какой-либо из кэпированных структур. Два моноклональных антитела мыши от FIND (FIND25, также называемый KI25), сильно связываются со всеми структурами Ara6, независимо от наличия или отсутствия кэпирования, но не распознают какие-либо структуры Ara4. CS-40, которое, как известно, специфично вступает в реакцию с ManLAM, слабо вступает в реакцию с LAM, полученным из Mycobacterium tuberculosis, и преимущественно связывалось с кэпированными мономаннозой структурами Ara4 и Ara6.
Человеческое моноклональное антитело к LAM Р30В9 IgM сильно и с высокой специфичностью реагировало с ManLAM, полученным из Mycobacterium tuberculosis, и с кэпированными диманнозой структурами Ara4 и Ara6, и со значительно более слабой активностью с другими содержащими маннозу структурами. Визуализация данной остаточной активности зависит от условий анализа и проявляется в одних форматах анализа (например, фигуры 4b, 8), но не в других (например, фигуры 16, 18). Без ограничения теорией относительная специфичность Р30В9 IgM в отношении кэпированных диманнозой структур является потенциально клинически значимой, поскольку известно, что концевые маннозильные единицы опосредуют связывание липоарабиноманнана из вирулентных штаммов комплекса Mycobacterium tuberculosis с макрофагами человека, и, кроме того, известно, что диманнозные кэпы являются преобладающей модификацией LAM, полученного из Mycobacterium tuberculosis.
Схожие результаты получали, если эпитопную специфичность А104-01 IgG и Р30В9 IgM дополнительно картировали в микроматричном анализе в отношении большей панели углеводных антигенов. Данная панель включала несколько дополнительных полиманнозных структур, которые распознавались Р30В9 IgM, но не любым из других тестированных антител (фиг. 8). Это соответствовало предпочтению Р30В9 IgM в отношении кэпированных диманнозой структур Ara4 и Ara6, в частности, но не обязательно, таковых, содержащих Man-α(1→2)-Man-α(1→5), связанный с концевой арабинозой. Р30В9 IgM также сильно реагировал с пентаманнозной структурой (59. AS-3-71), которая содержала Man-α(1→2)-Man-α(1→6), но лишь слабо со схожей структурой, содержащей Man-α(1→3)-Man-α(1→6) (50. YB-BSA-18). Несмотря на то, что он предпочитает связь α(1→2), Р30В9 IgM не вступает в реакцию с AS-2-91, тетраманнозной структурой, которая содержала связь Man-α(1→2) наряду с дополнительной маннозой, связанной α(1→6) со второй маннозой. Без ограничения теорией это предполагает, что специфичность изотипа IgM Р30В9 может требовать того, чтобы оба остатка сахара диманнозного мотива не содержали каких-либо дополнительных замен.
В. Более точное титрование для картирования тонкой специфичности данных моноклональных антител в отношении полученных из LAM гликанов показало особо важную роль концевого дисахарида β-D-Araƒ-(1→2)-α-D-Araƒ-(1→5) в распознавании антителом структур Ara4. Структура Ara4 состоит из тетрасахарида β-D-Araf-(1→2)-α-D-Araf-(1→5)-α-D-Araf-(1→5)-α-D-Araf, тогда как структура Ara6 содержит дополнительную дисахаридную ветвь β-D-Araf-(1→2)-α-D-Araf-(1→3) во втором сахаре. Три моноклональных антитела связываются как со структурой Ara4, так и со структурой Ara6, независимо от кэпирования маннозой. Все три моноклональных антитела связываются со структурой Ara4 (YB-8-099) и YB-BSA- 03, соответствующей структуре Ara4 с четырьмя дополнительными остатками сахара α-D-Araƒ-(1→5) на восстанавливающем конце (фиг. 9А). Однако ни одно из моноклональных антител не связывалось с родственным октасахаридом (MJ-LZ-2), который содержал концевой дисахарид β-D-Araf-(1→2)-α-D-Araf-(1→3), соответствующий нижней ветви структуры Ara6. Это указывало на то, что верхняя ветвь структуры Ara6, содержащая связь β-D-Araf-(1→2)-α-D-Araf-(1→5), распознавалась данными моноклональными антителами, а не нижняя ветвь, которая содержала дисахарид β-D-Araf-(1→2)-α-D-Araf-(1→3).
Роль концевой связи β-D-Araf-(1→2) в распознавании антитела исследовали посредством изучения реактивности данных моноклональных антител и Ara6-зависимого антитела FIND25 в отношении трех родственных структур поли-α-D-Araf-(1→5), которые содержали усеченные формы концевого дисахарида (фиг. 9В). Все три структуры также содержали внутреннюю ветвь α-D-Araf-(1→3). YB-BSA-07 завершалась линейной структурой α-D-Araf-(1→5), и была полностью нереактивной со всеми антителами к LAM. YB-BSA-09 содержала дополнительные сахара α-D-Araf, прикрепленные посредством ветви (1→3) на предпоследних остатках сахара двух более длинных ветвей, напоминающих структуру ветви Ara6. Данная структура распознавалась лишь слабо более высокими тестируемыми концентрациями изотипа IgG А194-01 и CS-35. YB-BSA-10 содержала концевые остатки сахара β-D-Araƒ-(1→2) на каждой из ветвей, образуя две полные структуры Ara6 на невосстанавливающих концах полисахарида. Данная структура распознавалась всеми моноклональными антителами с относительными значениями силы связывания, соответствующими значениям их аффинности в отношении природного антигена LAM. Данные анализы показали, что концевой дисахарид β-D-Araf-(1→2)-α-D-Araf-(1→5) являлся особо важным компонентом всех из доступных арабиноза-реактивных LAM-специфических моноклональных антител.
С. Принципиальным различием между патогенными штаммами комплекса Mycobacterium tuberculosis, такими как Mycobacterium tuberculosis и Mycobacterium bovis, и непатогенными быстро растущими штаммами, такими как Mycobacterium smegmatis, является наличие кэпированных маннозой концов на патогенных штаммах. Соответственно, моноклональные антитела, которые являются специфическими в отношении отличающихся маннозилированных структур, могут являться пригодными для структурных исследований и для определения функциональных вкладов данных модификаций. Активности двух моноклональных антител, характеризованных в данном исследовании, CS-35 и FIND25/170, совершенно не зависели от наличия или отсутствия маннозных кэпов. Связывание 900 серий моноклональных антител, с другой стороны, полностью аннулировалось посредством маннозилирования любого типа (фиг. 4).
CS-40, с другой стороны, связывалось лишь слабо с немодифицированным гликаном Ara4 (YB-8-099), но сильно со структурами Ara4 (YB-8-101) и Ara6 (YB-8-149), которые содержали единичные маннозные кэпы. В данном эксперименте применяли модифицированный CS-40, в котором домен тяжелой цепи мыши был замещен константной последовательностью IgG1 человека, поскольку это привело в результате к более чувствительному выявлению связывания по сравнению с природным антителом мыши, применяемым на фиг. 4. Слабая реактивность CS-40 с некэпированной арабинофуранозной структурой была отражена в его слабой реактивности с LAM из М. smegmatis по сравнению с LAM из М. tb. Присоединение α(1→4)-связанного сахара MSX к концевой маннозе (т.е. YB-8-141 и YB-8-149) не оказывало влияния на аффинность связывания, тогда как присоединение второго α(1→2)-связанного маннозного сахара (YB-8-111 и YB-8-125) для получения диманнозного кэпа полностью аннулировало реактивность CS-40 (фиг. 16).
А194-01 обладал более сложным паттерном реактивности. А194-01 связываются сильно с некэпированными арабинофуранозильными боковыми цепями и с кэпированными мономаннозой структурами Ara4 (YB-8-101) и Ara6 (YB-8-123), но с данным mAb вступал в реакцию лишь слабо с кэпированной диманнозой Ara4 (YB-8-111), и даже более слабо с кэпированной триманнозой Ara4 (YB-8-113), и практически не вступал в реакцию с кэпированной диманнозой Ara6 (YB-8-125). Как было видно для CS-40, замена МТХ на мономаннозные структуры (YB-8-141, YB-8-149) не подавляла связывание А104-01, и что представляет особый интерес, добавление MSX значительно улучшало распознавание кэпированных диманнозой и триманнозой структур Ara4 (YB-8-133, YB-8-143). В соответствии с высокой селективностью Р30В9 в отношении ManLAM mAb специфично связывались с кэпированными диманнозой структурами Ara4 (YB-111) и Ara6 (YB-8-125). В отличие от благотворных или благоприятных эффектов замены MSX в отношении связывания CS-40 и А194-01, данная замена приводила в результате к полной потере реактивности Р30В9, как и добавление маннозы с образованием кэпированных триманнозой структур. Данные результаты предполагают, что различные моноклональные антитела распознавали различные участки и структурные аспекты структуры LAM, при этом некоторые связываются только с арабинофуранозными боковыми цепями, а другие связываются с различными уровнями специфичности с кэпирующими мотивами.
Относительные значения специфичности связывания и аффинностей Ara6-реактивных антител сравнивали для иллюстративных гликоконъюгатов (фиг. 11). Общие паттерны соответствовали таковым, полученным для природных антигенов, PILAM и ManLAM (фиг. 3) и в предварительном титровании в отношении гликоконъюгатов (фиг. 4). А194-01 IgG человека обладал более высокой относительной аффинностью в отношении всех из некэпированных структур и в отношении MSX-замещенной Ara6-мономаннозной структуры (YB-8-149), вступал в реакцию с равной аффинностью со структурой Ara6 с единичными маннозными кэпами, но не распознавал структуры с диманнозными или триманнозными кэпами. FIND25 связывалось со схожей или немного более высокой аффинностью, чем CS-35, со всеми структурами, которые несут стандартные структуры Ara6, как в кэпированной, так и в некэпированной формах, но не связывалось с двумя структурами (YB-BSA-06 и YB-BSA-08), в которых одна из ветвей была вытянута на невосстанавливающем конце от точки ветвления. 908.1 связывалось с более слабой аффинностью со всеми из некэпированных структур, в том числе последними двумя, но не распознавало какие-либо из кэпированных маннозой структур 4.
Исследования конкурирования, включающие моноклональные антитела к LAM, обзор
Способность отдельных антител конкурировать за связывание биотинилированных mAb-зондов с LAM определяли титрованием с помощью ELISA. Типичные кривые конкурирования показаны на фиг. 16 для четырех антител к LAM, А194-01, CS-35, FIND25 и Р30В9. Как и ожидалось, все биотинилированные антитела конкурировали с их избыточными количествами их немеченных вариантов. Антитело к LAM мыши 908.6 конкурировало слабо или вообще не конкурировало с другими антителами. Это было связано в некоторой степени со слабой аффинностью данного антитела, но также отражает ограничение связывания 908.6 с некэпированными структурами и предполагает, что кэпированные структуры были доминирующими мишенями в ManLAM, распознаваемом CS-35 и FIND25.
В соответствии с его широкой реактивностью, CS-35 в полной мере конкурировало за связывание всех из антител-зондов, хотя его конкурирование с биотинилированным А194-01 было менее активным, чем с А194-01 как таковым, что соответствовало более низкой аффинности CS-35 в отношении LAM. В то время как CS-35 в полной мере конкурировало с биотинилированным FIND25, FIND25 конкурировало только частично с меченным CS-35 (~74% максимального конкурирования) и даже менее эффективно с А194-01 (~50%). Без ограничения теорией данный результат предположительно отражает наличие структур Ara4, которые распознаются А194-01 и CS-35, но не FIND25, которое связывается исключительно с мотивом Ara6. Тот факт, что FIND25 конкурировало в большинстве случаев связывания с CS-35, предположил, что структуры Ara6 более распространены, чем структуры Ara4. Несмотря на его высокую аффинность, А194-01 конкурировало только с собой, но не с CS-35 либо с FIND25, что дополнительно предполагает, что мишени в LAM, распознаваемые последними двумя антителами, преимущественно состояли из структур (например кэпированных диманнозой и триманнозой структур), которые не распознаются А194-01. В отличие от данного результата А194-01 в полной мере и эффективно конкурировало за связывание FIND25 с неманнозилированной PILAM, что соответствует роли эффективного маннозного кэпирования структур Ara6 в отсутствии конкурирования в ManLAM.
Исследования конкурирования с применением антитела Р30В9 дополнительно подтвердили вывод о том, что подавляющее большинство структур Ara6 в ManLAM были кэпированы диманнозой и что основная масса диманнозных кэпов находилась на структурах Ara6. Р30В9 конкурировало с FIND25 на ~70% и с CS-35 на -80% за связывание с ManLAM, что подтверждает, что большинство структур, распознаваемых данными mAb мыши, также распознавались Р30В9. Связывание Р30В9 с ManLAM эффективно конкурировало само по себе, и как с CS-35, так и с FIND25. Уровень конкурирования Р30В9 с FIND25 был близким к 100%, указывая на то, что, по сути, все диманноза-зависимые сайты связывания Р30В9 были расположены на сайтах Ara6, и несколько на структурах Ara4. Как и ожидалось, А194-01 очень слабо конкурировало за связывание Р30В9 с ManLAM, и 908.7 вообще не конкурировало, что соответствует слабому распознаванию кэпированных диманнозой структур данными антителами. Неспособность последних антител эффективно конкурировать за связывание Р30В9 подтвердило, что данный эффект требует связывания конкурирующего mAb с той же ветвью, что и mAb-зонд, и что связывание с гетерологичными эпитопами, расположенными на смежной ветви той же молекулы, не приводило к эффективному конкурированию. В. Относительные значения аффинности А194-01 IgG и Р30В9 IgM,
полученные посредством конкурентного анализа
Картирование реактивности отдельных моноклональных антител к LAM, в том числе изотипа IgG А194-01 и изотипа IgM Р30В9, в отношении специфических гликановых структур обеспечило получение характеристик распределения указанных специфических гликановых структур в LAM посредством конкурентных исследований антитела (фиг. 10). Посредством данных конкурентных анализов предположили, что для того, чтобы одно антитело конкурировало за связывание второго (биотинилированного, в тех случаях, когда они получены от одного и того же вида) антитела, два эпитопа должны находиться в непосредственной близости друг от друга в нативной молекуле, возможно, но не обязательно, на той же или соседней арабинановой ветви. Данная модель поддерживалась асимметрическими паттернами конкурирования, где, например, биотинилированный IgG А194-01, который связывается с некэпированными, мономаннозилированными и замещенными MSX структурами, как Ara4, так и Ara6, эффективно конкурировал сам по себе и со сконструированными вариантами и/или производными А194-01, но лишь частично с моноклональным антителом FIND25 мыши, которое связывается только со структурами Ara6. С другой стороны, моноклональное антитело CS-35 мыши, которое связывается со всеми структурами Ara4 и Ara6, обеспечивает более полное конкурирование, хотя менее эффективное, предположительно вследствие его относительно низкой аффинности.
Результаты данных анализов выявили некоторые поразительные и неожиданные свойства. Например, изотип IgM Р30В9, который связывается со всеми кэпированными диманнозой структурами ManLAM, сильно и в полной мере конкурировал сам с собой, с CS-35 и FIND170. Без ограничения теорией эффективное конкурирование Р30В9 с моноклональным антителом мыши к LAM FIND25 предполагает, что кэпированные диманнозой структуры в нативном LAM в основном расположены на структурах Ara6, распознаваемых антителами FIND, и существенно не экспрессируются на структурах Ara4. Это повышает важность наличия возможности целенаправленно воздействовать и специфически связываться с кэпированными диманнозой остатками Ara6, поскольку полагают, что кэпирование диманнозой является преобладающей формой LAM, обнаруженной в штаммах комплекса Mycobacterium tuberculosis. Высокоэффективное конкурирование биотинилированного FIND25 со сконструированным вариантом изотипа IgM А194-01 является еще одним свидетельством увеличения распознавания маннозилированных структур изотипом IgM А194-01.
С. Исследования конкурирования А194-01 IgG, и Р30В9 IgM, и антител мыши к LAM за связывание с ManLAM и PILAM
Анализы конкурентного связывания между различными моноклональными антителами к LAM применяли для анализа распределения различных структурных форм в LAM. Изотип IgG A194-01 распознавал немодифицированные боковые цепи, как Ara4, так и Ara6, или цепи, которые содержали единичный маннозный кэп, но не связывался с боковыми цепями с либо диманнозными, либо триманнозными кэпирующими мотивами. Два антитела мыши FIND вступали в реакцию со всеми формами Ara6, но не с какими-либо структурами Ara4. Р30В9 IgM являлся относительно специфическим в отношении структур Ara4 и Ara6, которые содержали диманнозные кэпы.
В соответствии с расширением реактивности в отношении конструкций А194-01 с повышенными валентностями, данные конструкции проявляли повышенную эффективность в отношении конкурентной активности антител. При тестировании способности конкурирования в отношении связывания биотинилированного А194-01 IgG и ManLAM декамерные варианты А194-01 IgM и тетрамерные варианты scFv-IgG конкурировали более эффективно, чем изотип А194-01 IgG как таковой (фиг. 11), что означает увеличение потенциальной терапевтической и диагностической применимости, тогда как мономерные Fab- и scFv-формы конкурировали менее эффективно (фиг. 1). Димерный сконструированный вариант scFv и/или производное А194-01 конкурировали в равной степени, как и изотип А194-01 IgG.
При сравнении эпитопной специфичности сконструированных вариантов и/или производных А194-01 со специфичностью эпитопа А194-01 IgG наблюдали, что они обладали более широкой реактивностью (фиг. 14). В то время как изотип IgG существенно не связывался с замещенными диманнозой (YB-8-123, YB-8-125) и триманнозой (YB-BSA-113, YB-BSA-13) структурами, IgM распознавал данные структуры, и форма scFv-IgG также характеризовалась повышенной активностью в отношении некоторых из этих структур. Поскольку кэпирование диманнозой и, в частности, кэпированная диманнозой Ara6, является преобладающим мотивом LAM в вирулентном Mycobacterium tuberculosis, это предполагает потенциально повышенную применимость данных сконструированных форм А194-01 в терапевтических и диагностических применениях.
Немеченный А194-01 IgG конкурировал в с биотинилированным А194-01 IgG за связывание с LAM, полученным либо из Mycobacterium tuberculosis (ManLAM) (фиг. 12А), либо из Mycobacterium smegmatis (PILAM) (фиг. 12D), в то время как моноклональные антитела FIND 170 и Р30В9 не были способны конкурировать с А194-01 за связывание с любым антигеном (фиг. 12А, D). Это соответствовало преобладающему распознаванию структур Ara4, которые распознавались изотипом А194-01 IgG, но не FIND170, которое является специфическим в отношении Ara6, либо Р30В9 IgM, который зависит от диманнозных кэпирующих остатков. Подобным образом, А194-01 не конкурировало за связывание либо биотинилированного FIND25 (фиг. 12В), либо Р30В9 IgM (фиг. 12С) с ManLAM, что соответствует различным эпитопным специфичностям для этих антител. В отличие от неспособности А194-01 IgG конкурировать с FIND25 за связывание с ManLAM, А194-01 IgG сильно конкурировал, на ~90%, с FIND25 за связывание с PILAM (фиг. 12Е), что соответствует известному отсутствию кэпирования маннозы в PILAM и с высокой аффинности А194-01 в отношении некэпированных структур Ara4 и Ara6.
В отличие от неэффективного конкурирования А194-01 IgG, Р30В9 IgM конкурировал за связывание с FIND25 на ~80%, и FIND 170 конкурировало практически в полной мере за связывание с Р30В9 (фиг. 12В, С). Этот факт неопровержимо свидетельствует о том, что подавляющее большинство структур Ara6, распознаваемых антителами мыши FIND, содержало диманнозные кэпы, и, таким образом, также распознавалось Р30В9 IgM, и что большинство кэпированных диманнозой структур, распознаваемых Р30В9, присутствовали на структурах Ara6. Это предполагает, что кэпированный диманнозой Ara6 является преобладающим иммунологическим мотивом в мотиве LAM из вирулентного Mycobacterium tuberculosis.
D. Дополнительные исследования конкурирования
Дополнительные исследования конкурирования проводили для того, чтобы подчеркнуть тот факт, что LAM является сложным антигеном неопределенной гетерогенности. Определение различных эпитопных специфичностей LAM-реактивных моноклональных антител обеспечило возможность применения анализов конкурентного связывания для изучения распределения различных эпитопов в нативных LAM. Способность различных антител конкурировать за связывание биотинилированных моноклональных антител-зондов с LAM и синтетическими гликоконъюгатами определяли титрованием с помощью ELISA. Типичные кривые конкурирования для трех моноклональных антител к LAM, А194-01 IgG, CS-35 и FIND25, показаны на фиг. 13А. Биотинилированные моноклональные антитела-зонды конкурировали со своими немеченными вариантами при присутствии в большом избытке. Моноклональное антитело мыши 908.6 конкурировало слабо или вообще не конкурировало с другими антителами.
Это было связано в некоторой степени со слабой аффинностью данного антитела, но также отражало ограничение связывания 908.6 с некэпированными структурами и дополнительно предполагало, что копированные маннозой структуры были доминирующими мишенями в ManLAM, распознаваемом CS-35 и FIND25. В соответствии с его широкой реактивностью CS-35 конкурировало за связывание с биотинилированным А194-01 IgG, хотя и менее эффективно, чем А194-01 IgG так таковой, что соответствует более высокой аффинности А104-01 IgG к LAM. CS-35 также в полной мере конкурировало с биотинилированным FIND25, тогда как FIND25 конкурировало только частично с меченным CS-35 (максимальное конкурирование ~74%), и даже менее эффективно с А194-01 IgG (~50%). Данный результат предположительно отражает наличие структур Ara4, которые распознаются А194-01 IgG и CS-35, но не FIND25, которое связывается исключительно со структурами, содержащими остов Ara6. Несмотря на его в целом высокую аффинность в отношении LAM, А194-01 IgG конкурировал только в отношении себя, но не в отношении либо CS-35, либо FIND25. Это предполагало, что в сайтах в LAM, распознаваемых моноклональными антителами мыши, преобладали кэпированные диманнозой и триманнозой структуры, которые не распознавались А194-01 IgG.
Дополнительные исследования конкурирования с применением Р30В9 IgM, который специфически связывается с кэпированным диманнозой ManLAM, дополнительно подтвердили клинически значимый вывод о том, что подавляющее большинство структур Ara6 в ManLAM являлись копированными диманнозой и что основная масса диманнозных кэпов находилась на структурах Ara6 в ManLAM, полученном из Mycobacterium tuberculosis. Р30В9 IgM конкурировал с FIND25 за связывание с ManLAM на ~80% (фиг. 13В), что соответствует большинству структур Ara6, распознаваемых FIND25, которые также при этом распознаются Р30В9 IgM. Р30В9 IgM не конкурировал с FIND25 за связывание с PILAM, что соответствует отсутствию диманнозного эпитопа для Р30В9 в PILAM вследствие отсутствия маннозилирования в PILAM. Более того, А194-01 IgG не конкурировал с FIND25 за связывание с ManLAM, что опять-таки соответствует подавляющему большинству структур Ara6, несущих диманнозные кэпы, которые не распознаются А194-01 IgG. Подтверждая роль маннозилирования в данном эффекте, А194-01 IgG очень эффективно конкурировал с FIND25 за связывание с PILAM, что соответствует высокой аффинности А194-01 в отношении PILAM и отсутствию маннозного кэпирования в данном антигене.
Данные по конкурированию за связывание биотинилированного Р30В9 IgM дополнительно подтвердили этот вывод. Биотинилированный Р30В9 IgM наиболее эффективно конкурировал за связывание сам по себе и с равной эффективностью с CS-35 и FIND25, но лишь слабо и несовершенно с А194-01 IgG (фиг. 13С). Уровень конкурирования с FIND25 был близким к 100%, указывая на то, что, по сути, все диманноза-зависимые сайты связывания Р30В9 IgM были расположены на структурах Ara6. В соответствии с данным определением, CS-35 также конкурировало с Р30В9 за связывание с кэпированной диманнозой Ara4 (YB-8-111) и кэпированной диманнозой Ara6 (YB-8-125), тогда как FIND 170 конкурировало только с последним антигеном, и А194-01 IgG не конкурировал с ними. Общее сходство между кривыми конкурирования для ManLAM и гомогенных гликоконъюгатов YB-8-125 указывало на то, что результаты конкурирования коррелировали с наличием или отсутствием релевантных эпитопов на одной углеводной боковой цепи и что опосредованные стерические эффекты вследствие связывания антитела с более удаленными гетерологичными сайтами играли небольшую роль в конкурировании или совсем не играли никакой роли.
Одним поразительным и неожиданным результатом являлось полное отсутствие конкурирования между А194-01 IgG и CS-35, а также FIND25. Способность CS-35 конкурировать с А194-01 IgG за связывание с ManLAM являлась ожидаемой вследствие распознавания всех мишеней А194-01 IgG посредством CS-35, и менее эффективное конкурирование с CS-35, чем с А194-01 IgG самим по себе, соответствовало относительным значениям аффинности данных антител к ManLAM (фиг. 3). Подобным образом, неполное конкурирование за связывание А194-01 IgG с FIND25 можно объяснить присутствием мишеней Ara4, распознаваемых первым, но не последним антителом.
Эффективное и полное конкурирование за связывание биотинилированного Р30В9 IgM как с CS-35, так и с FIND170 свидетельствует о том, что диманнозные кэпы, распознаваемые Р30В9 IgM, присутствовали почти исключительно на структурах Ara6, распознаваемых посредством FIND mAb, которое имеет клиническое и диагностическое значение. Это поддерживалось относительно эффективной конкуренцией связывания FIND25 посредством Р30В9 IgM, который блокировал ~80% связывающей активности FIND25 в отношении ManLAM, но не имел каких-либо эффектов в отношении PILAM. Это дает веские основания предполагать, что ~80% сайтов Ara6 в ManLAM, распознаваемых FIND25, содержали диманнозные кэпы и что, по сути, все из диманнозных кэпов присутствуют на структурах Ara6, а не на Ara4. Эти результаты, вместе с неспособностью А194-01 IgG конкурировать с CS-35 или FIND25, демонстрируют, что замещенная диманнозой Ara6 являлась преобладающей иммуногенной структурой на ManLAM, полученном из Mycobacterium tuberculosis, и, таким образом, представляет собой крайне важную антигенную мишень.
Исследования ответов LAM-специфических антител в плазме крови пациентов указывают, что в ответе преобладают изотипы IgG2, направленные на линейные структуры Ara4/Ara6, независимо от картирования. Эффективное конкурирование Р30В9 IgM с моноклональными антителами IgG к LAM, специфическими в отношении арабинофураноза-зависимых эпитопов, такими как CS-35 и FIND25, предполагает, что преобладающие ответы IgG2 в отношении таких эпитопов в плазме крови пациента будут также конкурировать в отношении каких-либо диманноза-зависимых ManLAM-специфических антител, которые могут продуцироваться в более низких титрах. Таким образом, даже если последний класс антител может иметь более эффективную антибактериальную активность, вполне вероятно, что эти эффекты могут быть ограничены конкурированием за связывание с преобладающими, нефункциональными антителами IgG2, направленными в отношении арабиноза-зависимых эпитопов, которые присутствуют в сыворотке крови пациента. Без ограничения теорией преобладающий гуморальный ответ может фактически защитить бактерии от потенциальных эффектов редких антител, таких как поливалентные антитела, такие как Р30В9 IgM или сконструированные варианты и/или производные А194-01, такие как пятивалентный изотип IgM или тетравалентный scFv-IgG, которые могут обеспечить иммунный контроль в отношении инфекции и патогенности.
5. Эффекты валентности в отношении связывания А194-01
Открытие факта реактивности зависимости диманноза-реактивного Р30В9 от его изотипа IgM предполагает, что поливалентность может способствовать аффинности антител в отношении LAM. Без ограничения теорией это указывало на то, что одна молекула LAM обладала несколькими сайтами или эпитопами связывания с антителом, что соответствует известной разветвленной структуре и сложности LAM, и что антитела с более высокими валентностями являлись способными связываться с большим количеством сайтом, чем двухвалентные антитела, что приводит в результате к более высоким значениям аффинности. Эффект валентности антитела в отношении эффективности связывания с LAM изучали для различных сконструированных вариантов, и/или производных форм, и/или изотипов человеческого моноклонального антитела А194-01 в анализе конкурентного связывания с применением биотинилированного А194-01 IgG в качестве мишени. Формы антитела включали моновалентный одноцепочечный scFv, в котором участки VH и VL соединены с помощью гибкого пептидного линкера, моновалентный Fab-белок, димерный scFv-белок, в котором два scFv-домена соединены гибким линкером, природный димерный IgG (фиг. 1А) и две формы с высокой валентностью, тетравалентный А194-01 scFv-IgG и пятивалентный (декавалентный для сайтов связывания) изотип IgM (фиг. 1В). Превращение интактного двухвалентного IgG в моновалентный Fab приводило в результате к очень большой потере связывающей активности, с >100-кратным увеличением концентрации, необходимой для конкуренции за 50% связывающую активность биотинилированного А194-01 IgG по сравнению с IgG в отношении самого себя (фиг. 1С). Одна цепь также конкурировала неэффективно, с 33-кратной уменьшением активности. С другой стороны, димер scFv конкурировал со схожей эффективностью, что и изотип IgG А194-01. Без ограничения теорией это предполагало, что эффективное связывание двухвалентных форм А194-01 с LAM требовало присоединения обоих сайтов связывания к смежным мишеням в одной молекуле антигена. Формы с высокой валентностью конкурировали более эффективно, на молярной основе. Это может объясняться исключительно наличием дополнительных сайтов связывания, но может также отражать увеличение аффинности форм с высокой валентностью.
Специфичности как тетрамерного варианта scFv-IgG и/или производного А194-01, так и декамерного изотипа IgM А194-01 сравнивали со специфичностью изотипа IgG A194-01 в отношении панели синтетического гликоконъюгата, описанной выше (фиг. 14). Это показало, что как сконструированный вариант scFv-IgG, так и сконструированный изотип IgM характеризовались более широкими реактивностями в отношении некоторых гликанов, которые изотип IgG распознавал лишь слабо или вообще не распознавал. Повышенное связывание наблюдали для кэпированных диманнозой структур (YB-8-111, YB-8-125, YB-8-133) и для некоторых из кэпированных триманнозой структур (YB-8-113, YB-8-143, YB-BSA- 13) со сконструированным изотипом IgM A194-01, обладающим самой широкой панелью реактивности. Это имеет значительный диагностический и терапевтический потенциал, учитывая значимость кэпирования диманнозой. Интересно отметить, что увеличенная реактивность также наблюдалась для изотипа А194-01 IgM с арабинозной структурой, в которой отсутствовала концевая связь β-D-Araƒ-(1→2) (YB-BSA-09), которая не распознавалась какими-либо из тестируемых моноклональных антител к LAM. Без ограничения теорией это указывало на то, что повышенная валентность приводила к усилению авидности изотипа IgM A194-01 в отношении его основного арабиноза-содержащего эпитопа до такой степени, что подавляющие эффекты терминальной замены были преодолены.
Профили реактивности А194-01 IgG и сконструированных форм IgM и scFv-IgG А194-01 дополнительно анализировали посредством реципрокных конкурентных экспериментов по связыванию в отношении либо ManLAM, либо PILAM. В то время как все три формы перекрестно конкурировали за связывание биотинилированных антител с PILAM и за связывание А194-01 IgG с ManLAM, только формы с высокой валентностью scFv-IgG и IgM А194-01 эффективно конкурировали за связывание модифицированных антител с ManLAM. Эти различия в значениях конкурентной активности соответствовали более широкому профилю связывания сконструированных форм и позволяли предложить, что они связываются с сайтами на LAM, которые не распознавались формой IgG.
Этот вывод дополнительно подтвержадается реципрокными конкурентными исследованиями, в которых различные изотипы А194-01 конкурировали за связывание биотинилированного FIND25 IgG и Р30В9 IgM с ManLAM. В то время как A194-01 IgG не конкурировал с Р30В9 IgM и FIND25 за связывание с ManLAM, и форма scFv-IgG, и форма IgM A194-01 конкурировали практически в полной мере за связывание данных двух моноклональных антител (фиг. 15А, В). Как ожидалось, все три формы А194-01 конкурировали за связывание FIND25 с PILAM (фиг. 15С), что соответствовало отсутствию маннозилирования в PILAM. Данная усиленная активность позволила предположить, что данные модификации повышают потенциальную применимость данного антитела для иммунодиагностических применений и, кроме того, могут также повысить эффективность данных реагентов в иммунотерапевтических целях.
ПРИМЕР 2. Выделение и получение характеристик новых человеческих моноклональных антител, специфических в отношении гликолипидов M.tb. Выделение первого mAb, специфического в отношении эпитопа, общего для LAM и PIM6.
Тяжелую и легкую цепи антитела Р95С1 выделяли из одного клона В-клетки, посредством осуществления скрининга в отношении реактивности с ManLAM, от пациента с латентной туберкулезной инфекцией (LTBI). Р95С1 представляет собой антитело изотипа IgM, которое связывается как с ManLAM, так и PILAM. Это было показано посредством исследований связывания с гликоконъюгатом, которые указывают на то, что Р95С1 не связывается с какими-либо молекулами, которые экспрессируют различные арабинозные боковые цепи, либо некэпированные, либо кэпированные различными маннозными структурами. Единственные распознаваемые структуры представляли собой две полиманнозные структуры, которые содержали структурные мотивы, присутствующие в маннановой основе, которые сохранялись между PIM6 и различными LAM (фиг. 18(A), 18(B), 18(C)). Данную перекрестную реактивность LAM-PIM6 подтверждали посредством вестерн-блот-анализа, показывающего, что в то время как А194-01 и Р30В9 связывались только с LAM, Р95С1 также вступал в реакцию с молекулами-предшественниками гликолипидов LAM, LM и PIM6 (фиг. 20(A), 20(B)).
Хотя Р95С1, как и Р30В9, в естественных условиях экспрессировалось в качестве IgM, в отличие от Р30В9 оно сохраняло реактивность при превращении в изотип IgA или изотип IgG (фиг. 19). Это может быть отражением природы эпитопа или его расположения в маннановом участке молекулы LAM или может быть связано с большим числом мутаций в вариабельных участках Р95С1, соответствующих последовательности более зрелого антитела. Вариабельные участки тяжелой и легкой цепей Р95С1 содержат 19 и 13 аминокислотных точечных мутаций соответственно последовательности их ближайшего антитела эмбрионального типа.
Недавно было показано, что антитела, полученные от индивидуумов с латентным заболеванием, функционально превосходят антитела от индивидуумов с активным туберкулезом в содействии фаголизосомального слияния, активации инфламмасомы и уничтожения макрофагами интернализованных микобактерий (Lu et al. 2016). Таким образом, интерес представляет Р95С1, выделенное из пациентов с LTBI, и оно имеет больше мутаций в своей вариабельной области, чем Р30В9 и два других LAM-специфических mAb, которые были выделены из того же пациента с LTBI, которые характеризуются отличающимися эпитопными специфичностями в отношении ManLAM.
О природе гуморального иммунного ответа человека в отношении инфекции, вызванной М. tb, мало что известно. Хотя широко известно, что поверхностные гликолипиды М. tb способствуют подавлению активности макрофагов и дендритных клеток, существует противоречивая информация о том, является ли кэпированный маннозой липоарабиноманнан (ManLAM) или фосфатидилинозитолманнозид 6 (PIM6) основным иммуноингибирующим поверхностным компонентом М. tb. Этот вопрос дополнительно усложняется часто встречающимся загрязнением очищенных препаратов PIM6 посредством ManLAM и наоборот. Одним из способов решения этого вопроса является тестирование способности антител, специфических в отношении данных двух модуляторов, подавлять данные подавляющие активности. Однако это не было возможным из-за отсутствия хорошо охарактеризованных антител, которые являются специфическими в отношении данных двух антигенов. На сегодняшний день не было обнаружено антител, которые распознают PIM6, и настоящее изобретение описывает первое mAb с высокой аффинностью, которое распознает PIM6. PIM (PIM2 и PIM4) являются предшественниками ManLAM, и существует некоторая структурная взаимосвязь между маннанным доменом ManLAM и полиманнозной структурой PIM6.
Антитела могут выполнять свои функции двумя способами: 1) путем прямого блокирования внедрения в клетки-хозяева и нейтрализации бактериальных продуктов; 2) опосредованно через Fc-опосредованный комплемент и механизмы активации клеток через Fc-рецепторы. На антитело-опосредованную эффекторную функцию сильно влияет изотип антитела. Недавнее исследование показало изотип- зависимые подавляющие ответы антитела человека в отношении М. tb и продемонстрировали, что антитела IgA, но не IgG, специфические в отношении различных поверхностных антигенов М. Tb, могут блокировать захват М. tb эпителиальными клетками легкого, независимо от экспрессии рецепторов IgA Fc. Для тестирования эффекта изотипов Р95С1 в отношении связывания LAM константный участок тяжелой цепи P95C1-IgM замещали CH-IgA и CH-IgG для получения P95C1-IgA и P95C1-IgG. Аффинность связывания изотипов Р95С1 (IgM, IgA, IgG) с ManLAM и PILAM являлась сопоставимой (фиг. 19).
ДРУГИЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ
Любые улучшения можно производить в части или во всех антителах, композициях, наборах и способах. Все ссылочные материалы, в том числе публикации, заявки на патент и патенты, приведенные в данном документе, включены в данный документ посредством ссылки. Применение всех возможных примеров или выражений, означающих пример (например, "такой как"), предусмотренных в данном документе, предназначено для освещения настоящего изобретения и не представляет собой ограничение объема настоящего изобретения, если не указано иное. Любое заявление в данном документе в отношении природы или преимуществ настоящего изобретения или предпочтительных вариантов осуществления не предназначено для ограничения, и прилагаемая формула изобретения не должна считаться ограниченной такими заявлениями. В более общем плане, ни одна формулировка в настоящем описании не должна истолковываться как указание какого-либо незаявленного элемента как существенного для применения на практике настоящего изобретения. Настоящее изобретение включает все модификации и эквиваленты объекта изобретения, изложенного в формуле изобретения, приложенной в данном документе, что разрешено действующим законодательством. Более того, любая комбинация вышеописанных элементов во всех их возможных вариациях охватывается настоящим изобретением, если иное не указано в данном документе или иначе явно противопоказано контекстом.
--->
Перечень последовательностей
<110> Rutgers, The State University of New Jersey
Пинтер, Абрахам Патель,
Деендейл Чоудхари, Алок
<120> НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К LAM И PIM ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ ИНФЕКЦИЙ, ВЫЗВАННЫХ MYCOBACTERIUM TUBERCULOSIS
<130> 096747.00337
<150> 62/293,406
<151> 2016-02-10
<160> 50
<170> PatentIn версия 3.5
<210> 1
<211> 6
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 1
Arg Ser Ile Arg Ser Ala
1 5
<210> 2
<211> 3
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 2
Gly Ala Ser
1
<210> 3
<211> 9
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 3
Gln Gln Tyr Asp Phe Trp Tyr Thr Phe
1 5
<210> 4
<211> 8
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 4
Gly Phe Asn Phe Glu Asp Phe Gly
1 5
<210> 5
<211> 8
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 5
Ile Ser Trp Asn Gly Ala Asn Ile
1 5
<210> 6
<211> 13
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 6
Ile Asp Trp Tyr Arg Asp Asp Tyr Tyr Lys Met Asp Val
1 5 10
<210> 7
<211> 6
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 7
Gln Ser Ile Asn Ser Asn
1 5
<210> 8
<211> 3
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 8
Lys Ala Ser
1
<210> 9
<211> 9
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 9
Gln Gln Tyr Lys Ala Phe Lys Thr Phe
1 5
<210> 10
<211> 8
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 10
Gly Gly Ser Phe Ser Gly Tyr Tyr
1 5
<210> 11
<211> 13
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 11
Phe Asp Leu Gly Gly Ser Ile Thr His Ser Arg Gly Thr
1 5 10
<210> 12
<211> 13
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 12
Arg Gly Leu Ala Met Gly Gly Thr Lys Glu Phe Asp Ser
1 5 10
<210> 13
<211> 12
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 13
Gln Asn Val Leu Asp Ser Ala Asn Asn Arg Asn Tyr
1 5 10
<210> 14
<211> 3
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 14
Trp Ala Ser
1
<210> 15
<211> 9
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 15
Thr Gln Tyr His Arg Leu Pro His Thr
1 5
<210> 16
<211> 9
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 16
Gly Gly Ser Ile Asn Thr Asn Asn Trp
1 5
<210> 17
<211> 7
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 17
Ile His Arg His Gly Asp Thr
1 5
<210> 18
<211> 15
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 18
Cys Pro Leu Gly Tyr Cys Ser Gly Asp Asp Cys His Arg Val Ala
1 5 10 15
<210> 19
<211> 5
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 19
Gly Gly Ser Gly Gly
1 5
<210> 20
<400> 20
000
<210> 21
<211> 96
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 21
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Glu Asp Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Trp Asn Gly Ala Asn Ile Gly Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
<210> 22
<211> 96
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Asn Trp Asn Gly Gly Ser Thr Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr His Cys
85 90 95
<210> 23
<211> 14
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 23
Ala Ile Asp Trp Tyr Arg Asp Asp Tyr Tyr Lys Met Asp Val
1 5 10
<210> 24
<211> 88
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 24
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Ile Arg Ser Ala
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Phe Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Val Ser Ser Ile Arg Ser
65 70 75 80
Glu Asp Ser Ala Val Tyr Tyr Cys
85
<210> 25
<211> 88
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 25
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys
85
<210> 26
<211> 8
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 26
Gln Gln Tyr Asp Phe Trp Tyr Thr
1 5
<210> 27
<211> 101
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 27
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Thr Gly Leu Glu Trp Leu
35 40 45
Gly Glu Phe Asp Leu Gly Gly Ser Ile Thr His Ser Arg Gly Thr Asn
50 55 60
Tyr Asn Pro Ser Leu Lys Ser Arg Val Thr Ile Ser Gly Asp Thr Ser
65 70 75 80
Lys Asn Gln Phe Ser Leu Lys Leu Thr Ser Val Thr Ala Ala Asp Thr
85 90 95
Ala Val Tyr Tyr Cys
100
<210> 28
<211> 95
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 28
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
<210> 29
<211> 14
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 29
Ala Arg Gly Leu Ala Met Gly Gly Thr Lys Glu Phe Asp Ser
1 5 10
<210> 30
<211> 88
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asp Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys
85
<210> 31
<211> 88
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys
85
<210> 32
<211> 8
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 32
Gln Gln Tyr Lys Ala Phe Lys Thr
1 5
<210> 33
<211> 96
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 33
Glu Val Gln Leu Leu Glu Ser Gly Pro Gly Leu Val Arg Pro Trp Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Asn Thr Asn
20 25 30
Asn Trp Trp Ser Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Glu Ile His Arg His Gly Asp Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Arg Arg Val Ser Ile Ser Met Asp Glu Ser Met Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ile Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
<210> 34
<211> 96
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 34
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Asn Trp Trp Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Glu Ile Tyr His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Lys Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
<210> 35
<400> 35
000
<210> 36
<211> 94
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 36
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Asn Val Leu Asp Ser
20 25 30
Ala Asn Asn Arg Asn Tyr Phe Gly Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ile
65 70 75 80
Ile Ser Gly Leu Gln Val Glu Asp Val Ala Val Tyr Tyr Cys
85 90
<210> 37
<211> 94
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 37
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys
85 90
<210> 38
<400> 38
000
<210> 39
<211> 1356
<212> ДНК
<213> homo sapiens
<400> 39
caagtgcagc tgttggagtc tgggggaggt gtggtacggc cgggggggtc cctgagactc 60
tcctgtgcag cctctggatt caactttgaa gattttggca tgagctgggt ccgccaagct 120
ccagggaagg ggctggagtg ggtctctagt attagttgga atggtgctaa tataggctat 180
gtagactctg tgaagggccg attcaccatc tccagagaca acgccaagaa ctccctatat 240
ctgcaaatga acagtctgag agccgaggac acggccttat attactgtgc gatagactgg 300
tacagagacg actactacaa gatggacgtc tggggcaaag ggaccacggt caccgtctcc 360
tcagcctcga ccaagggccc atcggtcttc ccgctagcgc cctcctccaa gagcacctct 420
gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc tgtgacggtc 480
tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc 540
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 600
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag 660
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 720
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 780
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 840
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 900
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 960
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 1020
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 1080
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 1140
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 1200
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1260
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1320
acgcagaaga gcctctccct gtctccgggt aaatga 1356
<210> 40
<211> 451
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 40
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Glu Asp Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Trp Asn Gly Ala Asn Ile Gly Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ile Asp Trp Tyr Arg Asp Asp Tyr Tyr Lys Met Asp Val Trp Gly
100 105 110
Lys Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 41
<211> 642
<212> ДНК
<213> HOMO SAPIENS
<400> 41
gaaatagtga tgacgcagtc tccagccacc ctgtctgtct ctccagggga aagagccacc 60
ctctcctgca gggccagtcg gagtattcgc agcgccttag cctggtacca gcacaaacct 120
ggccaggctc ccaggctcct catctttggt gcatccacca gggccactgg tatcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccgtcagcag catacggtct 240
gaggattctg cagtttatta ctgtcagcag tatgatttct ggtacacttt tggccagggg 300
accaagctgg agatcaaacg aactgtggct gcaccatctg tcttcatctt cccgccatct 360
gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc 420
agagaggcca aagtacagtg gaaggtcgac aacgccctcc aatcgggtaa ctcccaggag 480
agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg 540
agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg 600
agctcgcccg tcacaaagag cttcaacagg ggagagtgtt ag 642
<210> 42
<211> 213
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 42
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Ile Arg Ser Ala
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Phe Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Val Ser Ser Ile Arg Ser
65 70 75 80
Glu Asp Ser Ala Val Tyr Tyr Cys Gln Gln Tyr Asp Phe Trp Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 43
<211> 115
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 43
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Thr Gly Leu Glu Trp Leu
35 40 45
Gly Glu Phe Asp Leu Gly Gly Ser Ile Thr His Ser Arg Gly Thr Asn
50 55 60
Tyr Asn Pro Ser Leu Lys Ser Arg Val Thr Ile Ser Gly Asp Thr Ser
65 70 75 80
Lys Asn Gln Phe Ser Leu Lys Leu Thr Ser Val Thr Ala Ala Asp Thr
85 90 95
Ala Val Tyr Tyr Cys Ala Arg Gly Leu Ala Met Gly Gly Thr Lys Glu
100 105 110
Phe Asp Ser
115
<210> 44
<211> 96
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 44
Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asp Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Lys Ala Phe Lys Thr
85 90 95
<210> 45
<211> 379
<212> ДНК
<213> HOMO SAPIENS
<400> 45
caggtgcagc tacagcagtg gggcgcagga ctgttgaagc cttcggagac cctgtccctc 60
acctgcgctg tctatggtgg gtccttcagt ggttactact ggagctggat ccgccagtcc 120
ccagagacgg ggctggagtg gcttggcgaa ttcgatcttg gtggaagcat cactcatagt 180
agaggcacca actacaaccc gtcgctcaag agtcgagtca ccatctcagg agacacgtcc 240
aagaaccagt tctccctgaa actgacctct gtgaccgccg cggacacggc tgtctattac 300
tgtgcgagag gtttagcaat gggtggaact aaggagtttg actcctgggg ccagggaacc 360
ctggtcaccg tctcctcag 379
<210> 46
<211> 319
<212> ДНК
<213> HOMO SAPIENS
<400> 46
gacatccaga tgacccagtc tccagactcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggccagtca gagtattaat agtaatttgg cctggtatca gcagaaaccg 120
gggaaagccc ctaagctcct gatctataag gcgtctgatt tagaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca ccatcagcag cctgcagcct 240
gatgattttg caacttatta ttgccaacag tataaagcat tcaagacgtt cggccacggg 300
accaaggtgg aaatcaaac 319
<210> 47
<211> 111
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Pro Gly Leu Val Arg Pro Trp Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Asn Thr Asn
20 25 30
Asn Trp Trp Ser Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Glu Ile His Arg His Gly Asp Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Arg Arg Val Ser Ile Ser Met Asp Glu Ser Met Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ile Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Cys Pro Leu Gly Tyr Cys Ser Gly Asp Asp Cys His Arg Val Ala
100 105 110
<210> 48
<211> 103
<212> БЕЛОК
<213> HOMO SAPIENS
<400> 48
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Asn Val Leu Asp Ser
20 25 30
Ala Asn Asn Arg Asn Tyr Phe Gly Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Ser Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ile
65 70 75 80
Ile Ser Gly Leu Gln Val Glu Asp Val Ala Val Tyr Tyr Cys Thr Gln
85 90 95
Tyr His Arg Leu Pro His Thr
100
<210> 49
<211> 367
<212> ДНК
<213> HOMO SAPIENS
<400> 49
gaggtgcagc tcttggagtc gggcccagga ctggtgaggc cttgggggac tctgtccctc 60
acctgcgctg tctctggtgg ctccatcaat actaataact ggtggagttg ggtccgccag 120
tccccgggga aggggctgga gtggattgga gaaatccatc gtcatgggga caccaactac 180
aacccgtcac tcaagaggcg agtctccata tcgatggacg agtccatgaa ccagttctcc 240
ctgaggctta tctctgtgac cgccgcggac acggccgtgt attactgttg tcccctagga 300
tattgtagtg gtgatgactg tcaccgagtt gcctggggcc ggggaatcct ggtcaccgtc 360
tcttcag 367
<210> 50
<211> 340
<212> ДНК
<213> HOMO SAPIENS
<400> 50
gacatccaga tgacccagtc tccatcctcc ctgtctgtgt ctctgggcga gagggccacc 60
atcaactgca agtccagcca gaatgtttta gacagcgcca acaataggaa ctacttcggt 120
tggtaccagc agaaaccagg gcagcctcct aagctgctca tttcctgggc atctacacgg 180
gaatccgggg tccctgaccg attcagtggc agcggctctg ggacagactt cactctcatc 240
atcagcggcc tgcaggttga agatgtggca gtttattact gtacacagta tcatagactt 300
cctcacacct tcggccaagg gacacgactg gaaattaaac 340
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
| Композиции и способы лечения и предупреждения инфекций, вызванных Staphylococcus aureus | 2015 |
|
RU2764981C1 |
| АНТИТЕЛА, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ В-КЛЕТОК, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2766094C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К TIM-3 ЧЕЛОВЕКА | 2019 |
|
RU2777336C1 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К ИНТЕРЛЕЙКИНУ-31 ДЛЯ ВЕТЕРИНАРНОГО ПРИМЕНЕНИЯ | 2019 |
|
RU2795411C2 |
| АНТИТЕЛА АНТИ-TNFRSF25 | 2017 |
|
RU2746314C2 |
| МЕТОДЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СЕРДЕЧНЫХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2830232C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С CD19 | 2019 |
|
RU2824170C2 |
| АНТИТЕЛА ПРОТИВ АЛЬФА-СУБЪЕДИНИЦЫ ИЛ-7R И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2811912C2 |
| Моноклональные антитела к альфа-синуклеину для предотвращения агрегации тау-белка | 2017 |
|
RU2760334C2 |







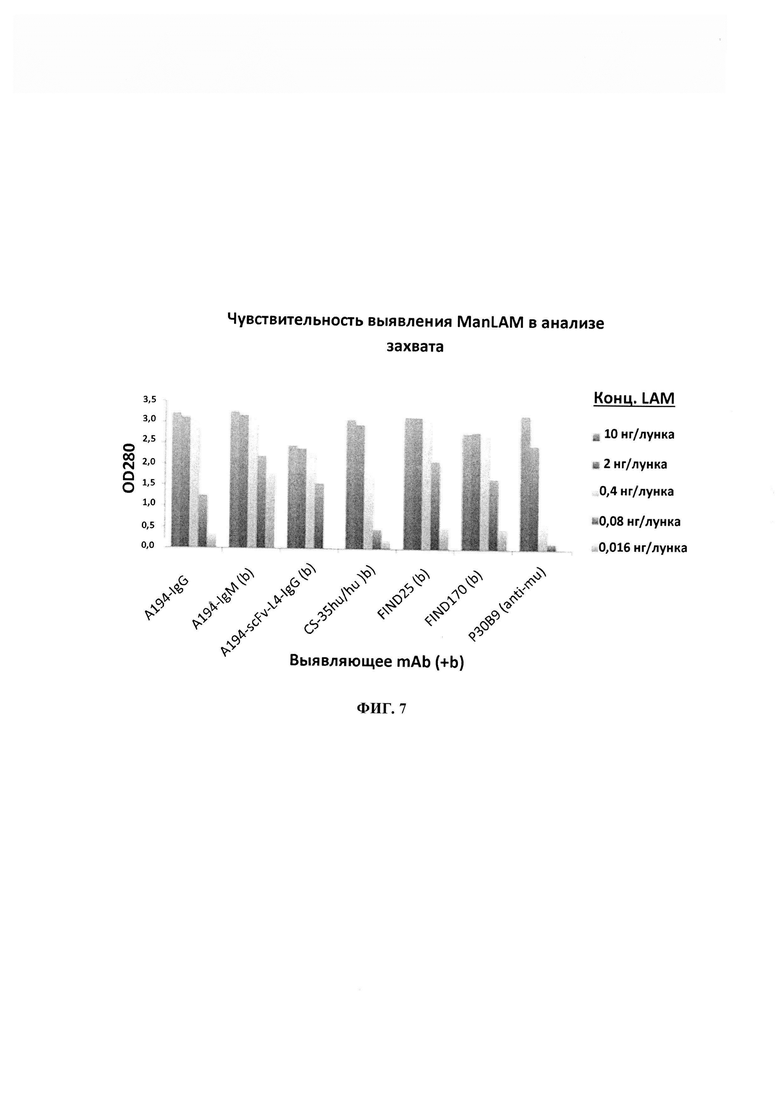
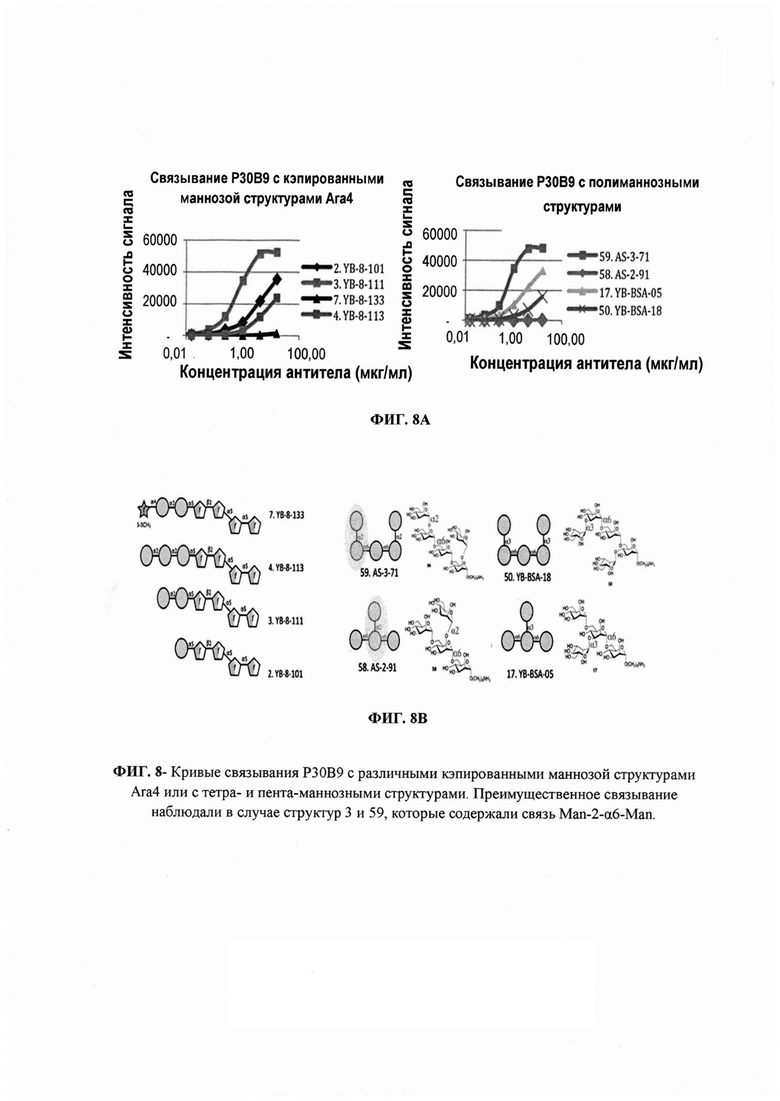
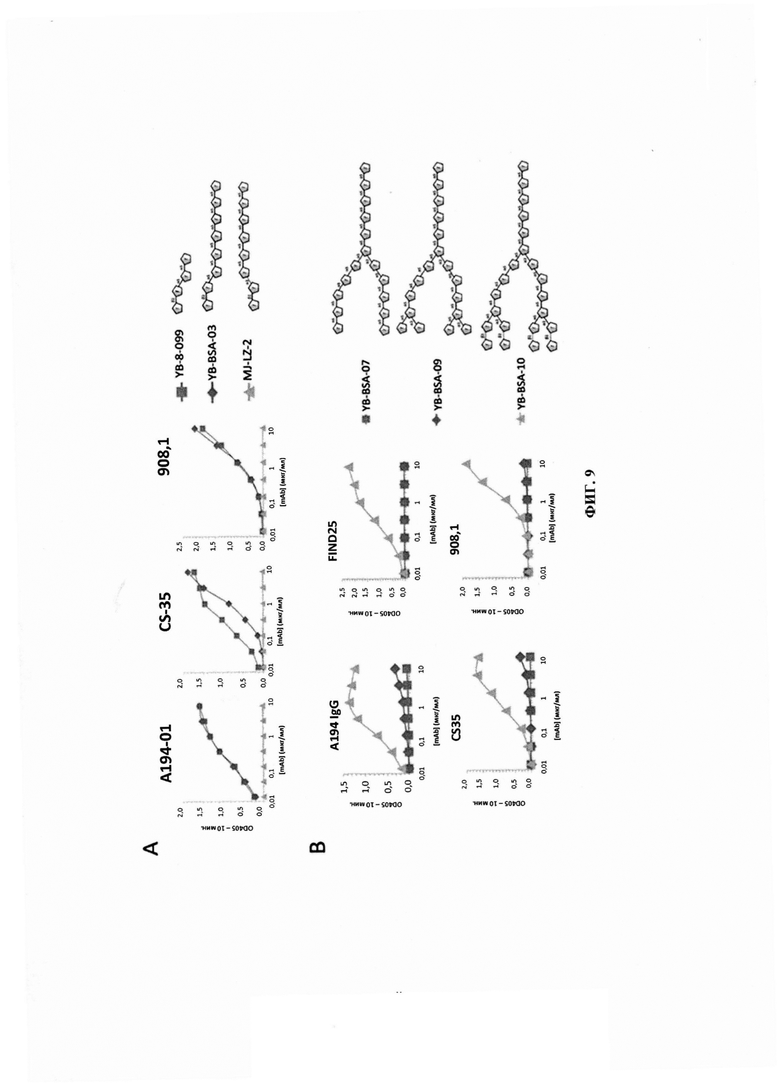

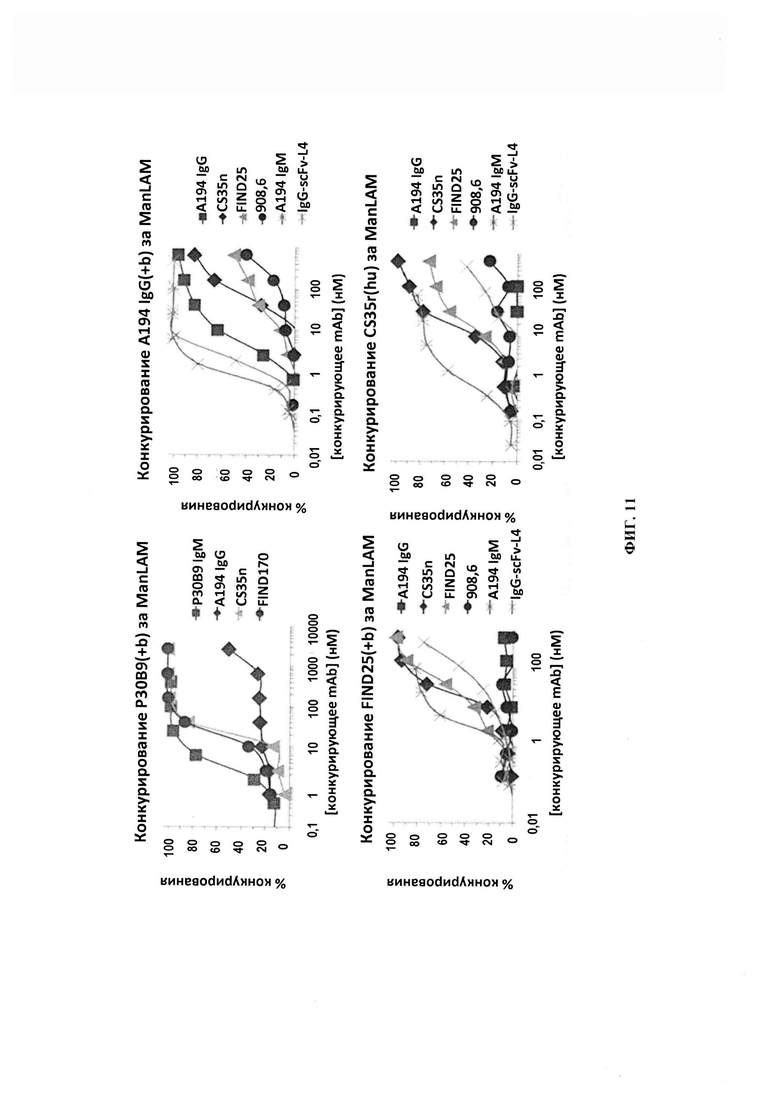
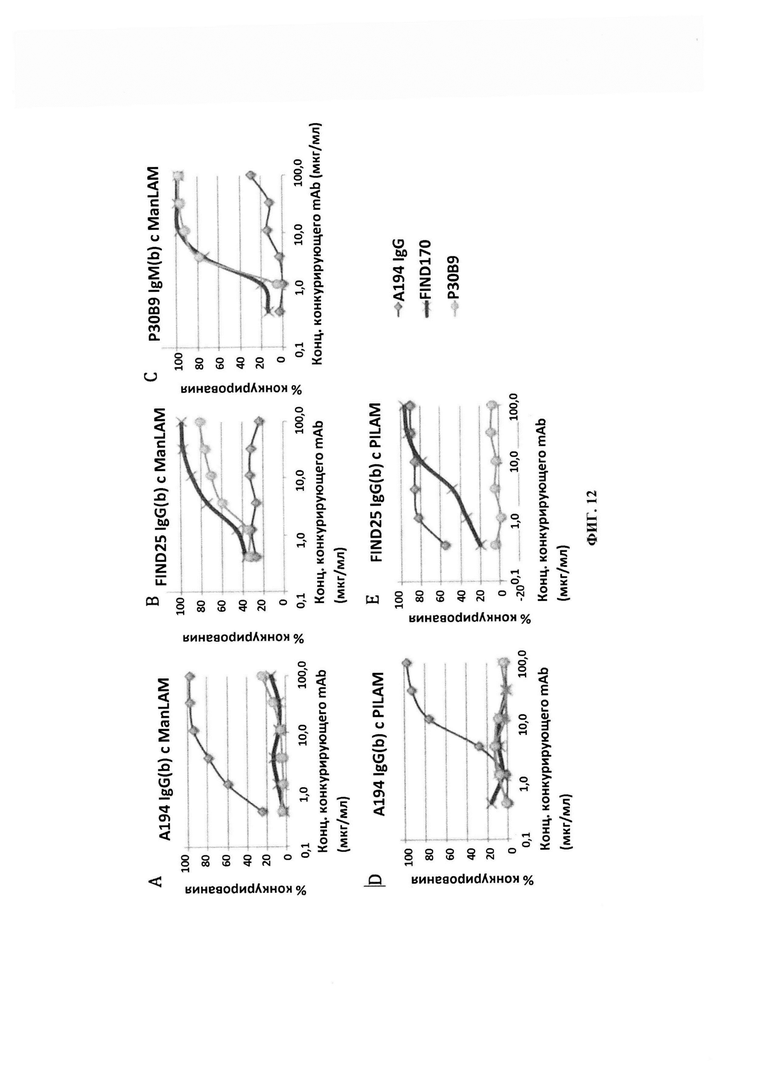

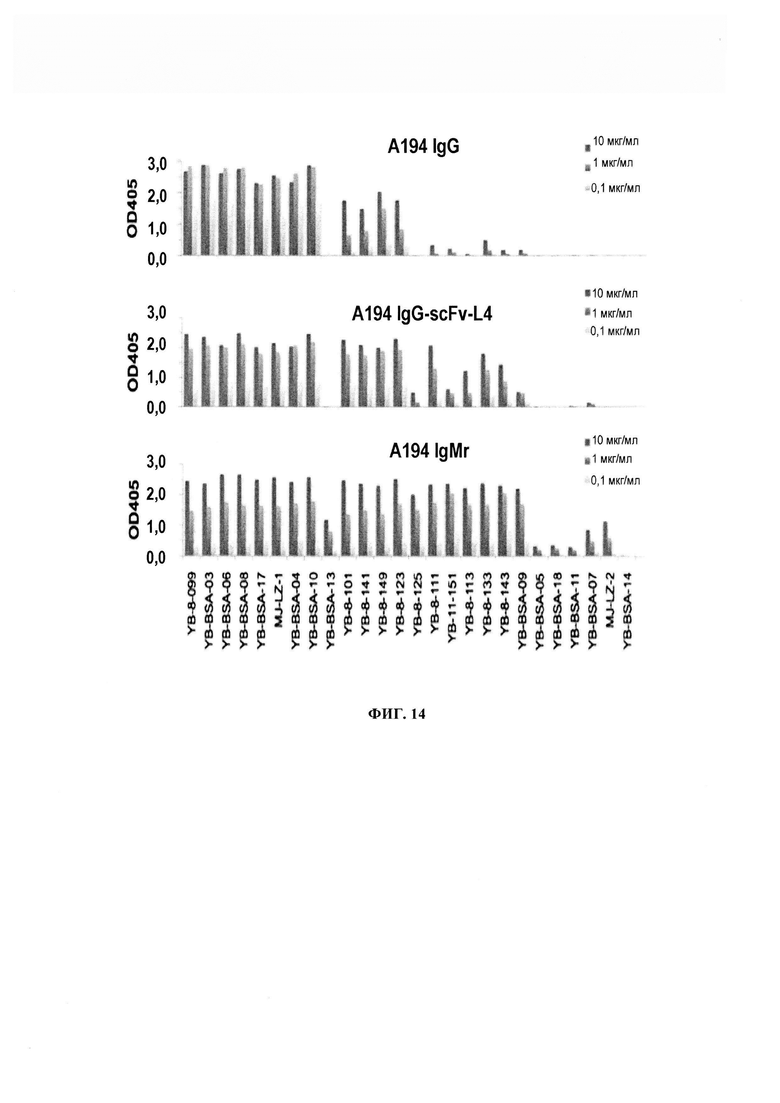
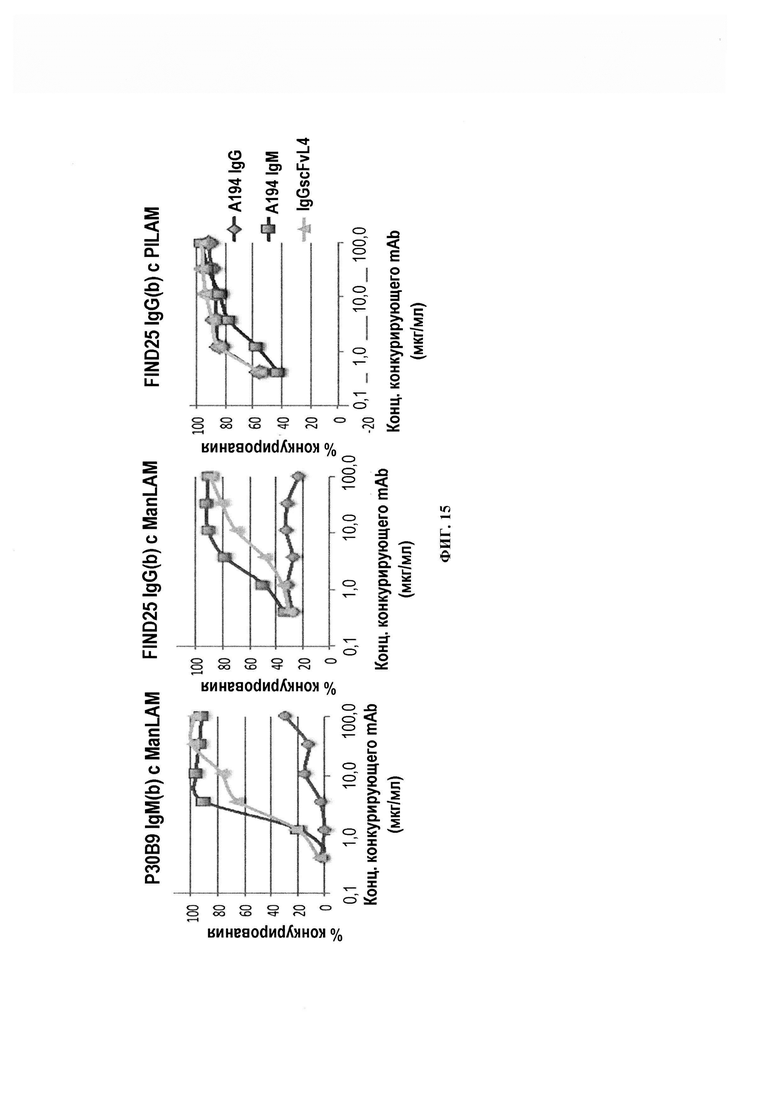




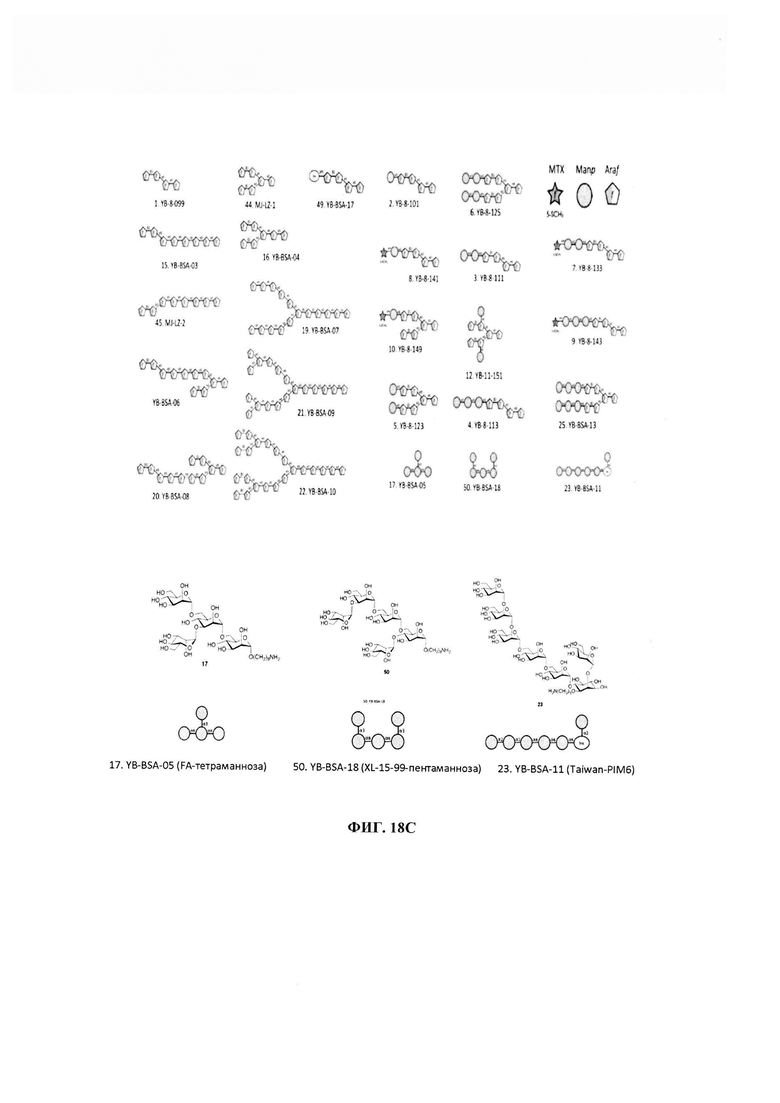
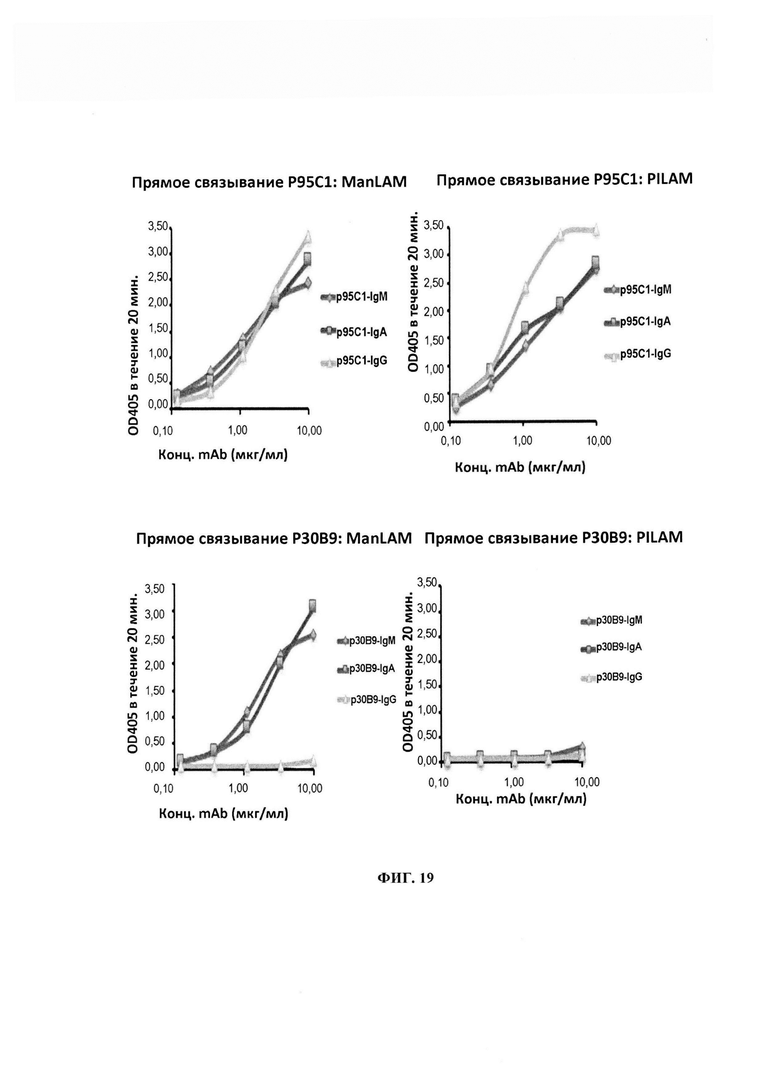

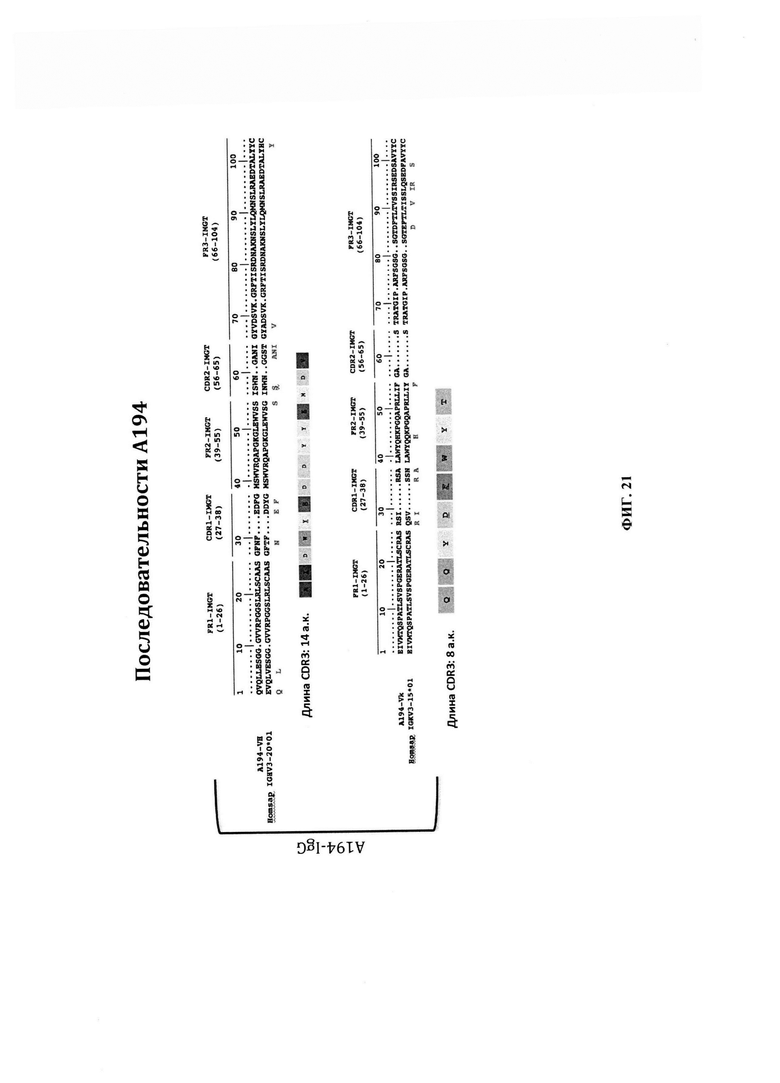


Группа изобретений относится к моноклональным антителам, направленным на эпитопы, обнаруживаемые в пределах липоарабиноманнана (LAM), для диагностики и лечения инфекций, вызванных Mycobacterium tuberculosis. Раскрыто человеческое моноклональное антитело к липоарабиноманнану (к LAM) или его антигенсвязывающая часть, которые специфически связываются с эпитопом LAM, содержащим структуру Ara4, структуру Ara6 или их комбинацию, где антитело к LAM содержит CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами. Также раскрыты набор для выявления по меньшей мере одного эпитопа LAM, способ диагностирования активной туберкулезной инфекции у индивидуума и способ лечения туберкулезной инфекции у индивидуума. Группа изобретений обеспечивает эффективное лечение и диагностику туберкулезной инфекции. 4 н. и 11 з.п. ф-лы, 32 ил., 4 табл., 2 пр.
1. Человеческое моноклональное антитело к липоарабиноманнану (к LAM) или его антигенсвязывающая часть, которые специфически связываются с эпитопом LAM, содержащим структуру Ara4, структуру Ara6 или их комбинацию, для диагностики или лечения инфекций, вызванных Mycobacterium tuberculosis, где антитело к LAM содержит CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами.
2. Человеческое моноклональное антитело к LAM или его антигенсвязывающая часть по п. 1, причем указанное антитело содержит вариабельный участок тяжелой цепи, содержащий аминокислотные последовательности под SEQ ID NO: 21 и SEQ ID NO: 23, и вариабельный участок легкой цепи, содержащий аминокислотные последовательности под SEQ ID NO: 24 и SEQ ID NO: 26.
3. Человеческое моноклональное антитело к LAM или его антигенсвязывающая часть по п. 1, где антителом к LAM является антитело, представляющее собой scFv-IgG, IgА или IgM.
4. Набор для выявления по меньшей мере одного эпитопа LAM, содержащий:
(a) первое антитело к LAM, которое специфически связывается с эпитопом, присутствующим на молекуле LAM;
(b) подложку, с которой связано указанное первое антитело к LAM;
(c) выявляющее антитело, которое специфически связывается с сайтом связывания в молекуле LAM, отличающимся от сайта связывания, связанного указанным первым антителом к LAM, где выявляющее антитело помечено репортерной молекулой; и
(d) буфер,
где либо указанное первое антитело к LAM, либо указанное выявляющее антитело представляет собой человеческое моноклональное антитело к LAM по п. 1.
5. Набор по п. 4, где выявляющее антитело представляет собой второе антитело к LAM, которое специфически связывается с LAM.
6. Набор по п. 4, где по меньшей мере одно из первого антитела к LAM и выявляющего антитела является антителом, представляющим собой scFv-IgG или IgM, и содержит CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO:23 или их антигенными фрагментами.
7. Набор по п. 6, где по меньшей мере одно из первого антитела к LAM и выявляющего антитела содержит вариабельный участок тяжелой цепи, содержащий аминокислотные последовательности под SEQ ID NO: 21 и SEQ ID NO: 23, и вариабельный участок легкой цепи, содержащий аминокислотные последовательности под SEQ ID NO: 24 и SEQ ID NO: 26.
8. Способ диагностирования активной туберкулезной инфекции у индивидуума, предусматривающий:
(a) получение от индивидуума образца, который содержит или предположительно содержит LAM;
(b) приведение указанного образца в контакт с первым антителом, которое специфически связывается с эпитопом, присутствующим на молекуле LAM;
(с) приведение указанного образца в контакт с выявляющим антителом, которое специфически связывается с сайтом связывания на молекуле LAM, отличающимся от сайта связывания, связанного указанным первым антителом;
(d) выявление связывания выявляющего антитела с указанным отличающимся сайтом связывания на молекуле LAM; и
(e) диагностирование указанного пациента как имеющего активную туберкулезную инфекцию,
где захват LAM указанным первым антителом является показателем активной туберкулезной инфекции, и
где по меньшей мере одно из указанных антител представляет собой человеческое моноклональное антитело к LAM по п. 1.
9. Способ по п. 8, где выявляющее антитело представляет собой антитело к LAM по п. 1.
10. Способ по п. 8, где выявляющее антитело представляет собой антитело к LAM, которое специфически связывается с LAM, и где первое антитело или выявляющее антитело содержит CDR1 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами.
11. Способ по п. 8, где выявляющее антитело представляет собой антитело к LAM, которое специфически связывается с LAM, и где первое антитело или выявляющее антитело является антителом, представляющим собой scFv-IgG или IgM, и содержит участок CDR1 в составе вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 1 или ее антигенными фрагментами, CDR2 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 2 или ее антигенными фрагментами, CDR3 вариабельного участка легкой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 3 или SEQ ID NO: 26 или их антигенными фрагментами, CDR1 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 4 или ее антигенными фрагментами, CDR2 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 5 или ее антигенными фрагментами, и CDR3 вариабельного участка тяжелой цепи, характеризующийся по меньшей мере 80% идентичностью с SEQ ID NO: 6 или SEQ ID NO: 23 или их антигенными фрагментами.
12. Способ по п. 8, где индивидуум является человеком.
13. Способ лечения туберкулезной инфекции у индивидуума, предусматривающий введение указанному индивидууму терапевтически эффективного количества человеческого моноклонального антитела к LAM по п. 1.
14. Способ по п. 13, дополнительно предусматривающий введение указанному индивидууму терапевтически эффективного количества по меньшей мере одного антибиотика.
15. Способ по п. 13, где туберкулезная инфекция представляет собой туберкулезную инфекцию с множественной лекарственной устойчивостью (MDR-TB).
| УСТРОЙСТВО ДЛЯ ИЗГОТОВЛЕНИЯ ОБЕЧАЙКИ ЛЕТАТЕЛЬНОГО АППАРАТА ИЗ ТРУБНОЙ ЗАГОТОВКИ ТИТАНОВОГО СПЛАВА В СОСТОЯНИИ СВЕРХПЛАСТИЧНОСТИ | 2023 |
|
RU2821415C1 |
| US 20130309237 A1, 21.11.2013 | |||
| US 20010007660 A1, 12.07.2001 | |||
| Chan E.C | |||
| et al | |||
| The diagnostic targeting of a carbohydrate virulence factor from M.Tuberculosis // Scientific Reports, 2015, V.5, p.10281 | |||
| Термоэлектрическая батарея | 1928 |
|
SU13228A1 |

