ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области рекомбинантных моноклональных антител и их применения в клинических и научных методиках, включая диагностические методики. Согласно настоящему изобретению, также предложены выделенные антитела против IL-31 в форме ветеринарных композиций, полезных для лечения расстройства, опосредованного IL-31, у млекопитающего, такого как кошка, собака или лошадь.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Атопический дерматит определен рабочей группой Американского колледжа ветеринарной дерматологии (American College of Veterinary Dermatology) как «воспалительное аллергическое заболевание кожи, сопровождающееся зудом, обусловленное генетической предрасположенностью и имеющее характерные клинические признаки» (Olivry, et at. Veterinary Immunology and Immunopathology 2001; 81: 143-146). Согласно этой рабочей группе, у собак данное заболевание связано с аллергенспецифическими IgE (Olivry, et al. 2001 supra; Marsella & Olivry Clinics in Dermatology 2003; 21: 122-133). Симптомами, которые наиболее заметны для владельцев животных и вызывают у них наибольшее беспокойство, являются сильный зуд, сопровождающийся вторичным облысением и эритемой.
Факторы, которые могут быть вовлечены в развитие аллергического дерматита, многочисленны и малопонятны. Атопический дерматит могут провоцировать компоненты корма (Picco, et al. Vet Dermatol. 2008; 19: 150-155), а также аллергены внешней среды, такие как блохи, пылевые клещи, амброзия, растительные экстракты и так далее. Также важна роль наследственных факторов. Несмотря на отсутствие подтвержденной предрасположенности у каких-либо пород, полагают, что наследственность некоторым образом усиливает предрасположенность к развитию атопического дерматита (Sousa & Marsella Veterinary Immunology and Immunopathology 2001; 81: 153-157; Schwartzman, etal. Clin. Exp. Immunol. 1971; 9: 549-569).
Согласно оценкам, атопический дерматит есть у 10% всей популяции собак (Marsella & Olivry 2003 supra; Scott, et al. Canadian Veterinary Journal 2002; 43: 601-603; Hillier Veterinary Immunology and Immunopathology 2001; 81: 147-151). Во всем мире приблизительно 4,5 миллиона собак поражены этим хроническим и пожизненным заболеванием. По-видимому, заболеваемость атопическим дерматитом растет. Были подозрения на предрасположенность, обусловленную породой и полом, но она может значительно варьировать в зависимости от географической области (Hillier, 2001 supra; Picco, et al. 2008 supra).
У кошек аллергический дерматит представляет собой воспалительное состояние кожи, сопровождающееся зудом, которое, как полагают, обусловлено аномальным ответом иммунной системы на вещества, не вызывающие реакции у здоровых кошек. Наиболее распространенным признаком аллергического дерматита у кошек является хронический рецидивирующий зуд. Частые клинические проявления аллергического дерматита у кошек включают спонтанное облысение, милиарный дерматит, эозинофильные гранулематозные комплексы (включая бляшки, гранулемы и индолентные язвы) и фокальный зуд в области головы и шеи, характеризующийся экскориациями, эрозиями и/или язвами. Имеющиеся данные не демонстрируют предрасположенности, обусловленной породой и полом, и, по-видимому, к развитию заболевания более склонны молодые кошки (Hobi et al. Vet Dermatol 2011 22: 406-413; Ravens et al. Vet Dermatol 2014; 25: 95-102; Buckely In Practice 2017; 39: 242-254).
В настоящее время варианты лечения кошек, у которых диагностирован аллергический дерматит, зависят от выраженности клинических признаков заболевания, его продолжительности и предпочтений владельца животного и включают аллергенспецифическую иммунотерапию и противозудные лекарственные средства, такие как глюкокортикоиды и циклоспорины (Buckley, supra). Иммунотерапевтическое лечение эффективно у некоторых пациентов, но требует частых инъекций, а до клинического улучшения может проходить 6-9 месяцев (Buckley, supra). Иммуносупрессивные лекарственные средства, такие как глюкокортикоиды и циклоспорины, обычно эффективны, тем не менее, их длительное применение часто приводит к нежелательным эффектам.
У лошадей атопический дерматит рассматривается как возможная причина зуда. Все больше данных указывают на роль аллергенов внешней среды при атопическом дерматите у лошадей. Заболевание может быть сезонным или несезонным, в зависимости от вовлеченного аллергена (аллергенов). Данных о предрасположенности, обусловленной возрастом, породой и полом, немного. В предварительном исследовании, проведенном в Школе ветеринарной медицины Калифорнийского университета в Дейвисе (School of Veterinary Medicine, University of California, Davis (SVM-UCD)), медиана возраста начала заболевания составляла 6,5 лет, наиболее распространенной породой были чистокровные верховые лошади, составлявшие 25% изученных лошадей, и у коней (обычно меринов) заболевание возникало почти в два раза чаще, чем у кобыл; тем не менее, эти данные были получены с использованием всего 24 лошадей и пока не сопоставлены с общей популяцией больных животных. Наиболее частым клиническим признаком атопического дерматита у лошадей является зуд, локализованный обычно в области морды, дистальных отделах конечностей или на туловище. Возможны облысение, эритема, крапивница и папулы. Крапивница может быть весьма выраженной, но не сопровождаться зудом. У лошадей возможна семейная предрасположенность к уртикарному атопическому дерматиту. Лошади могут страдать от вторичной пиодермии, типичными проявлениями которой являются избыточное шелушение, небольшие эпидермальные «венчики» или папулы, покрытые корками («милиарный дерматит»). Диагностика атопического дерматита основана на клинических признаках и исключении других диагнозов, в особенности гиперчувствительности к насекомым (Culicoides) (White Clin Tech Equine Pract 2005; 4: 311-313; Fadok Vet Clin Equine 2013; 29 541-550). В настоящее время лечение атопического дерматита у лошадей проводят как симптоматически, подавляя воспаление и зуд, спровоцированные аллергическим ответом, так и воздействуя на конкретную причину (то есть определяя задействованные аллергены и изготавливая аллергенспецифическую вакцину). Симптоматический подход обычно необходим для краткосрочной терапии в целях обеспечения комфорта пациента и минимизации самотравмирования. Этот подход основан на применении комбинации местной и системной терапии, включая антигистаминные средства, незаменимые жирные кислоты, пентоксифиллин и глюкокортикоиды. Основной подход к контролю аллергии на элементы внешней среды включает определение аллергенов, запускающих реакцию гиперчувствительности. Большинство дерматологов признают, что аллергенспецифическая иммунотерапия может быть полезной для лошадей, страдающих атопией. Тем не менее, у большинства лошадей улучшение наступает, как правило, лишь после первых 6 месяцев иммунотерапии (Marsella Vet Clin Equine 2013; 29: 551-557). Кроме того, длительное применение иммуносупрессивных лекарственных средств у лошадей может приводить к нежелательным эффектам.
Показано, что интерлейкин-31 (IL-31), цитокин, продуцируемый Т-хелперными клетками 2-го типа, приводит к зуду у людей, мышей и собак (Bieber N Engl J Med 2008; 358: 1483-1494; Dillon et al. Nat Immunol 2004; 5:752-60; патент США №8,790,651, выданный Bammert et at; Gonzalez et al. Vet Dermatl. 2013; 24(1): 48-53). IL-31 связывается с корецептором, состоящим из рецептора A IL-31 (IL-31RA) и рецептора онкостатина М (OSMR) (Dillon et al. 2004 supra и Bilsborough et al. J Allergy Clin Immunol. 2006 117(2):418-25). Активация рецепторов приводит к фосфорилированию STAT через рецептор (рецепторы) JAK. Было показано, что указанный корецептор экспрессирован в макрофагах, кератиноцитах и ганглиях задних корешков.
Недавно было обнаружено, что IL-31 вовлечен в дерматит, кожный зуд, аллергию и гиперчувствительность дыхательных путей. Показано, что Cytopoint®, собачье моноклональное антитело против IL-31, изготавливаемое Zoetis Inc. (Парсиппани, штат Нью-Джерси), уменьшает зуд и кожные очаги у собак с атопическим дерматитом (Gonzalez et al. 2013 supra, Michels etal. Vet Dermatol. 2016; Dec; 27(6): 478-e129). Было бы желательным обеспечить другие антитела против IL-31 для предотвращения и лечения расстройств, опосредованных IL-31, у млекопитающих в ветеринарии. С учетом неудовлетворенной на данный момент потребности в безопасных и эффективных альтернативных вариантов лечения атопических и аллергических дерматозов у кошек и лошадей, было бы особенно желательным обеспечить кошачьи и лошадиные антитела против IL-31 для уменьшения зуда и кожных очагов у кошек и лошадей с атопическим дерматитом.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном воплощении согласно настоящему изобретению предложены моноклональное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с областью белка IL-31 млекопитающего, вовлеченной во взаимодействие белка IL-31 с его корецептором, где на связывание указанного антитела с указанной областью влияют мутации в области связывания эпитопа 15Н05, выбранной из по меньшей мере одного из следующего: (а) области приблизительно между аминокислотными остатками 124 и 135 последовательности кошачьего IL-31, представленной в SEQ ID NO:157 (Feline_IL31_wildtype); (b) области приблизительно между аминокислотными остатками 124 и 135 последовательности собачьего IL-31, представленной в SEQ ID NO: 155 (Canine_IL31); и (с) области приблизительно между аминокислотными остатками 118 и 129 последовательности лошадиного IL-31, представленной в SEQ ID NO: 165 (Equine_IL31).
В одном воплощении указанные выше мутации в области связывания эпитопа 15Н05 выбраны из по меньшей мере одного из следующего: (а) мутанта, где положения 126 и 128 SEQ ID NO: 157 заменены на аланин; (b) мутанта, где положения 126 и 128 SEQ ID NO: 155 заменены на аланин; и (с) мутанта, где положения 120 и 122 SEQ ID NO: 165 заменены на аланин.
В одном воплощении моноклональное антитело по настоящему изобретению связывается с областью эпитопа 15Н05. То есть, в одном воплощении согласно настоящему изобретению предложены моноклональное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с областью белка IL-31 млекопитающего, вовлеченной во взаимодействие белка IL-31 с его корецептором, где область связывания представляет собой область связывания эпитопа 15Н05, выбранную из по меньшей мере одного из следующего: (а) области приблизительно между аминокислотными остатками 124 и 135 последовательности кошачьего IL-31, представленной в SEQ ID NO: 157 (Feline_IL31_wildtype); (b) области приблизительно между аминокислотными остатками 124 и 135 последовательности собачьего IL-31, представленной в SEQ ID NO: 155 (Canine_IL31); и (с) области приблизительно между аминокислотными остатками 118 и 129 последовательности лошадиного IL-31, представленной в SEQ ID NO: 165 (Equine_IL31).
В одном воплощении IL-31 млекопитающего, с которым специфически связываются антитело или его антигенсвязывающий фрагмент, представляет собой кошачий IL-31, где антитело связывается с областью приблизительно между аминокислотными остатками 125 и 134 последовательности кошачьего IL-31, представленной в SEQ ID NO: 157 (Feline_IL31_wildtype). В некоторых воплощениях антитело, связывающееся с данной областью кошачьего IL-31, содержит цепь VL, содержащую изменения в каркасной области 2 (FW2), выбранные из следующего: аспарагина вместо лизина в положении 42, изолейцина вместо валина в положении 43, валина вместо лейцина в положении 46, аспарагина вместо лизина в положении 49 и их комбинаций, где положения соответствуют нумерации SEQ ID NO: 127 (FEL_15Н05_VL1).
В одном воплощении IL-31 млекопитающего, с которым специфически связываются антитело или его антигенсвязывающий фрагмент, представляет собой собачий IL-31, где антитело связывается с областью приблизительно между аминокислотными остатками 125 и 134 последовательности собачьего IL-31, представленной в SEQ ID NO: 155 (Canine_IL31).
В другом воплощении IL-31 млекопитающего, с которым специфически связываются антитело или его антигенсвязывающий фрагмент, представляет собой лошадиный IL-31, где антитело связывается с областью приблизительно между аминокислотными остатками 117 и 128 последовательности лошадиного IL-31, представленной в SEQ ID NO: 165 (Equine_IL31).
В одном воплощении моноклональное антитело или его антигенсвязывающий фрагмент содержат следующие комбинации последовательностей гипервариабельных участков (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) (VH-CDR1) SYTIH (SEQ ID NO: 1), VH-CDR2 NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 WGFKYDGEWSFDV (SEQ ID NO: 3), CDR1 вариабельной области легкой цепи (VL) (VL-CDR1) RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 KASNLHI (SEQ ID NO: 5) и VL-CDR3 LQSQTYPLT (SEQ ID NO: 6); или
2) вариант (1), отличающийся от исходного антитела 15Н05 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В одном воплощении вариант антитела 15Н05/1505 содержит замену в одном или более чем одном из следующих положений в CDR: остаток 4 (I) SEQ ID NO: 1; остатки 1-3 (NIN), 5-7 (TSG), 9-11 (TEN) и 13 (Q) SEQ ID NO: 2; остатки 4 (K), 6(D) и 13 (V) SEQ ID NO: 3 в CDR1, 2 и 3 тяжелой цепи, соответственно, и остатки 3-7 (SQGIS) SEQ ID NO:4, остатки 3 (S) и 5 (L) SEQ ID NO: 5 и остатки 4(Q), 5 (Т) и 9 (Т) SEQ ID NO: 6 в CDRL1, 2 и 3, соответственно. В одном воплощении одна или более чем одна из этих замен представляют собой консервативные аминокислотные замены.
В одном воплощении моноклональное антитело 15Н05, указанное выше, содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую FEL_15Н05_VL1_FW2:
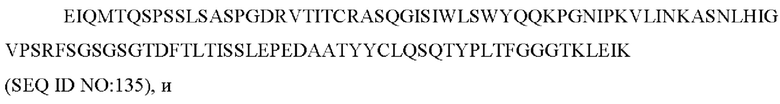
b) вариабельную область тяжелой цепи, содержащую FEL_15Н05_VH1:
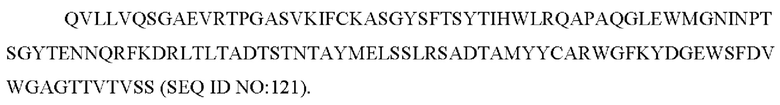
Согласно настоящему изобретению также предложены моноклональное антитело или его антигенсвязывающий фрагмент, содержащие следующие комбинации последовательностей гипервариабельных участков (CDR):
1) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) (VH-CDR1) SYGMS (SEQ ID NO: 13), VH-CDR2 HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 VYTTLAAFWTDNFDY (SEQ ID NO: 15), CDR1 вариабельной области легкой цепи (VL) (VL-CDR1) SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 SDGNRPS (SEQ ID NO: 17) и VL-CDR3 QSFDTTLDAYV (SEQ ID NO: 18);
2) антитело ZIL8: VH-CDR1 DYAMS (SEQ ID NO: 19), VH-CDR2 GIDSVGSGTSYADAVKG (SEQ ID NO: 20), VH-CDR3 GFPGSFEH (SEQ ID NO: 21), VL-CDR1 TGSSSNIGSGYVG (SEQ ID NO:22), VL-CDR2 YNSDRPS (SEQ ID NO: 23), VL-CDR3 SVYDRTFNAV (SEQ ID NO: 24);
3) антитело ZIL9: VH-CDR1 SYDMT (SEQ ID NO: 25), VH-CDR2 DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 RDTERPS (SEQ ID NO: 29), VL-CDR3 ESAVDTGTLV (SEQ ID NO: 30);
4) антитело ZIL11: VH-CDR1 TYVMN (SEQ ID NO: 31), VH-CDR2 SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 SMVGPFDY (SEQ ID NO: 33), VL-CDR1 SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 KDTERPS (SEQ ID NO: 35), VL-CDR3 ESAVSSDTIV (SEQ ID NO: 36);
5) антитело ZIL69: VH-CDR1 SYAMK (SEQ ID NO: 37), VH-CDR2 TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 KDTERPS (SEQ ID NO: 41), VL-CDR3 ESAVSSETNV (SEQ ID NO: 42);
6) антитело ZIL94: VH-CDR1 TYFMS (SEQ ID NO: 43), VH-CDR2 LISSDGSGTYYADAVKG (SEQ ID NO: 44), VH-CDR3 FWRAFND (SEQ ID NO: 45), VL-CDR1 GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 DTGSRPS (SEQ ID NO: 47), VL-CDR3 SLYTDSDILV (SEQ ID NO: 48);
7) антитело ZIL154: VH-CDR1 DRGMS (SEQ ID NO: 49), VH-CDR2 YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 WDGSSFDY (SEQ ID NO: 51), VL-CDR1 KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 KVSNRDP (SEQ ID NO: 53), VL-CDR3 MQAIHFPLT (SEQ ID NO: 54);
8) антитело ZIL159: VH-CDR1 SYVMT (SEQ ID NO: 55), VH-CDR2 GINSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 KDTERPS (SEQ ID NO: 59), VL-CDR3 KSAVSIDVGV (SEQ ID NO: 60);
9) антитело ZIL171: VH-CDR1 TYVMN (SEQ ID NO: 61), VH-CDR2 SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 SMVGPFDY (SEQ ID NO: 63), VL-CDR1 SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 KDTERPS (SEQ ID NO: 65), VL-CDR3 ESAVSSDTIV (SEQ ID NO: 66);
10) антитело 04H07: VH-CDR1 SYWMN (SEQ ID NO: 200), VH-CDR2 MIDPSDSEIHYNQVFKD (SEQ ID NO: 201), VH-CDR3 QDIVTTVDY (SEQ ID NO: 202), VL-CDR1 KSSQSLLYSINQKNHLA (SEQ ID NO: 203), VL-CDR2 WASTRES (SEQ ID NO: 204), VL-CDR3 QQGYTYPFT (SEQ ID NO: 205);
11) антитело 06A09: VH-CDR1 SYWMN (SEQ ID NO: 206), VH-CDR2 MIDPSDSETHYNQIFRD (SEQ ID NO: 207), VH-CDR3 QDIVTTVDY (SEQ ID NO: 208), VL-CDR1 KSSQSLLYSINQKNFLA (SEQ ID NO: 209), VL-CDR2 WASTRES (SEQ ID NO: 210), VL-CDR3 QQHYGYPFT (SEQ ID NO: 211); или
12) вариант (1)-(11), отличающийся от соответствующего исходного антитела ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В некоторых воплощениях настоящего изобретения:
1) антитело ZIL1 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL1_VL:
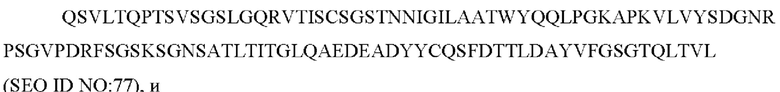
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL1_VH:
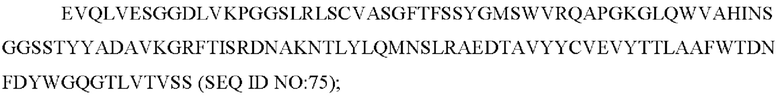
2) антитело ZIL8 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL8_VL:
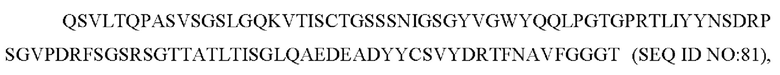
и
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL8_VH:
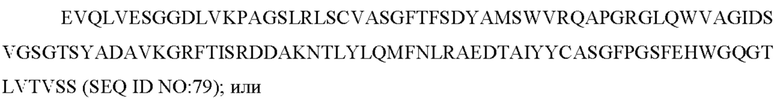
содержит по меньшей мере одно из следующего:
c) вариабельную область легкой цепи, содержащую ZTS_5864_VL:
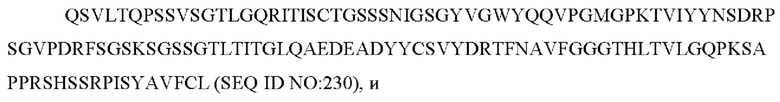
d) вариабельную область тяжелой цепи, содержащую ZTS_5864_VH:
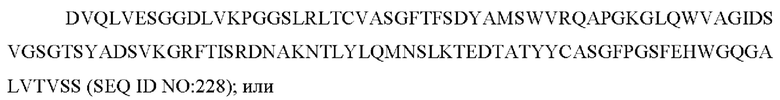
содержит по меньшей мере одно из следующего:
e) вариабельную область легкой цепи, содержащую ZTS 5865 VL:
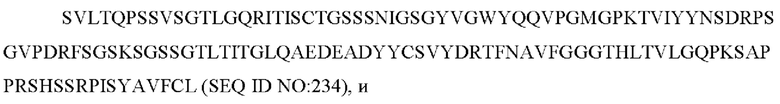
f) вариабельную область тяжелой цепи, содержащую ZTS 5865 VH:
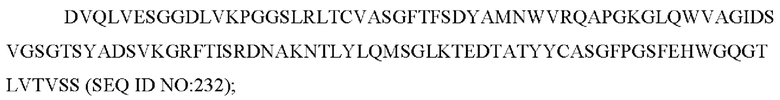
3) антитело ZIL9 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL9_VL:
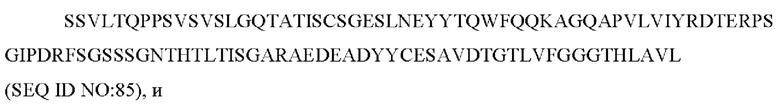
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL9_VH:
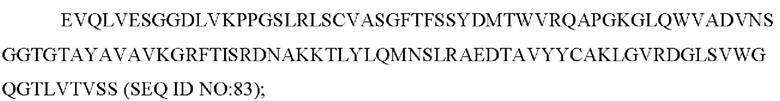
4) антитело ZIL11 содержит по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащую CAN-ZIL11_VL:

b) вариабельную область тяжелой цепи, содержащую CAN-ZIL11_VH:

5) антитело ZIL69 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL69_VL:

b) вариабельную область тяжелой цепи, содержащую CAN-ZIL69_VH:
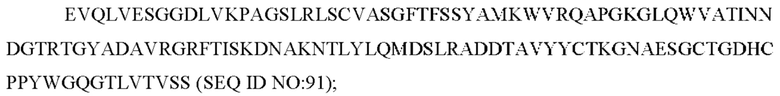
6) антитело ZIL94 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL94_VL:
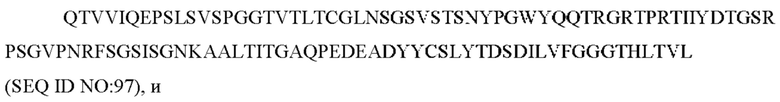
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL94_VH:
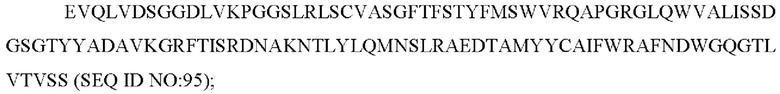
7) антитело ZIL154 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL154_VL:
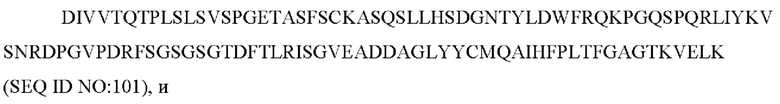
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL154_VH:
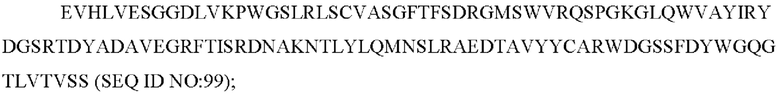
8) антитело ZIL159 содержит по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащую CAN-ZIL159_VL:
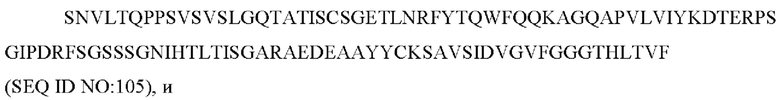
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL159_VH:
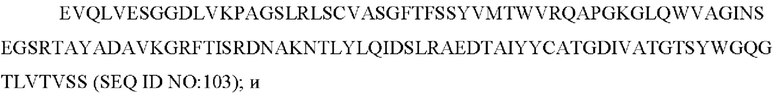
9) антитело ZIL171 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL171_VL:
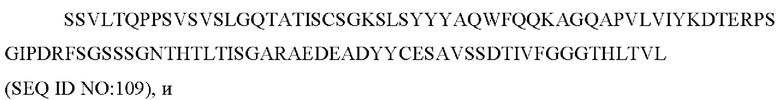
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL171VH:

10) антитело 04Н07 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую Mu_04H07_VL:
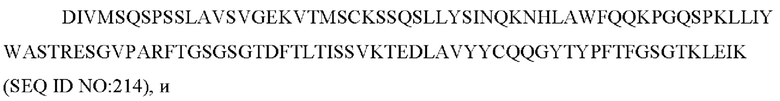
b) вариабельную область тяжелой цепи, содержащую Mu_04H07_VH:

11) антитело 06А09 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую Mu_06А09_VL:

b) вариабельную область тяжелой цепи, содержащую Mu_06А09_VH:
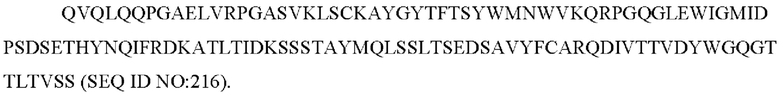
В одном воплощении моноклональное антитело или его антигенсвязывающий фрагмент по настоящему изобретению ослабляет, ингибирует или нейтрализует состояние, сопровождающееся зудом, или аллергическое состояние, опосредованные IL-31, у млекопитающего. В одном воплощении такое млекопитающее выбрано из собаки, кошки или лошади.
В некоторых воплощениях моноклональное антитело представляет собой химерное антитело. В других воплощениях антитело представляет собой канинизированное, фелинизированное, эквинизированное, полностью собачье, полностью кошачье или полностью лошадиное антитело.
Согласно настоящему изобретению также предложена ветеринарная композиция, содержащая терапевтически эффективное количество по меньшей мере одного антитела или его антигенсвязывающей части, описанных выше.
Также предложен способ лечения расстройства, опосредованного IL-31, у субъекта, включающий введение субъекту терапевтически эффективного количества по меньшей мере одного антитела или его антигенсвязывающей части, описанных выше.
В одном воплощении расстройство, опосредованное IL-31, представляет собой состояние, сопровождающееся зудом, или аллергическое состояние. В некоторых воплощениях состояние, сопровождающееся зудом, или аллергическое состояние представляют собой состояние, сопровождающееся зудом, выбранное из атопического дерматита, экземы, псориаза, склеродермии и зуда. В других воплощениях состояние, сопровождающееся зудом, или аллергическое состояние представляют собой аллергическое состояние, выбранное из группы, состоящей из аллергического дерматита, летней экземы, крапивницы, запала, воспалительного заболевания дыхательных путей, рецидивирующей обструкции дыхательных путей, гиперреактивности дыхательных путей, хронической обструктивной болезни легких и воспалительных процессов, являющихся результатами аутоиммунитета.
В других воплощениях расстройство, опосредованное IL-31, представляет собой прогрессирование опухоли. В некоторых воплощениях расстройство, опосредованное IL-31, представляет собой эозинофильное заболевание или мастоцитомы.
Кроме того, предложен способ ингибирования активности IL-31 у млекопитающего, включающий введение млекопитающему антитела или его антигенсвязывающей части, как описано выше.
Также предложены антитело или его антигенсвязывающий фрагмент, описанные выше, для применения в лечении млекопитающего с расстройством, опосредованным IL-31.
Кроме того, предложено применение антитела или его антигенсвязывающего фрагмента, описанных выше, для лечения млекопитающего с расстройством, опосредованным IL-31.
Также предложен способ выявления IL-31, включающий: инкубирование образца, содержащего IL-31, в присутствии антитела или его антигенсвязывающей части, описанных выше; и выявление антитела, связанного с IL-31, в образце. В одном воплощении способ дополнительно включает количественное определение IL-31 в образце.
Согласно настоящему изобретению также предложена клетка-хозяин, продуцирующая моноклональное антитело или его антигенсвязывающую часть, содержащие по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR):
1) антитело 15Н05: CDR1 вариабельной области тяжелой цепи (VH) (VH-CDR1) SYTIH (SEQ ID NO: 1), VH-CDR2 NINPTSGYTENNQRFKD (SEQ ID NO: 2), VH-CDR3 WGFKYDGEWSFDV (SEQ ID NO: 3), CDR1 вариабельной области легкой цепи (VL) (VL-CDR1) RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 KASNLHI (SEQ ID NO: 5) и VL-CDR3 LQSQTYPLT (SEQ ID NO: 6);
2) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) (VH-CDR1) SYGMS (SEQ ID NO: 13), VH-CDR2 HINSGGSSTYYADAVKG (SEQ ID NO: 14), VH-CDR3 VYTTLAAFWTDNFDY (SEQ ID NO: 15), CDR1 вариабельной области легкой цепи (VL) (VL-CDR1) SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 SDGNRPS (SEQ ID NO: 17) и VL-CDR3 QSFDTTLDAYV (SEQ ID NO: 18);
3) антитело ZIL8: VH-CDR1 DYAMS (SEQ ID NO: 19), VH-CDR2 GIDSVGSGTSYADAVKG (SEQ ID NO: 20), VH-CDR3 GFPGSFEH (SEQ ID NO: 21), VL-CDR1 TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 YNSDRPS (SEQ ID NO: 23), VL-CDR3 SVYDRTFNAV (SEQ ID NO: 24);
4) антитело ZIL9: VH-CDR1 SYDMT (SEQ ID NO: 25), VH-CDR2 DVNSGGTGTAYAVAVKG (SEQ ID NO: 26), VH-CDR3 LGVRDGLSV (SEQ ID NO: 27), VL-CDR1 SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 RDTERPS (SEQ ID NO: 29), VL-CDR3 ESAVDTGTLV (SEQ ID NO: 30);
5) антитело ZIL11: VH-CDR1 TYVMN (SEQ ID NO: 31), VH-CDR2 SINGGGSSPTYADAVRG (SEQ ID NO: 32), VH-CDR3 SMVGPFDY (SEQ ID NO: 33), VL-CDR1 SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 KDTERPS (SEQ ID NO: 35), VL-CDR3 ESAVSSDTIV (SEQ ID NO: 36);
6) антитело ZIL69: VH-CDR1 SYAMK (SEQ ID NO: 37), VH-CDR2 TINNDGTRTGYADAVRG (SEQ ID NO: 38), VH-CDR3 GNAESGCTGDHCPPY (SEQ ID NO: 39), VL-CDR1 SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 KDTERPS (SEQ ID NO: 41), VL-CDR3 ESAVSSETNV (SEQ ID NO: 42);
7) антитело ZIL94: VH-CDR1 TYFMS (SEQ ID NO: 43), VH-CDR2 LISSDGSGTYYADAVKG (SEQ ID NO: 44), VH-CDR3 FWRAFND (SEQ ID NO: 45), VL-CDR1 GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 DTGSRPS (SEQ ID NO: 47), VL-CDR3 SLYTDSDILV (SEQ ID NO: 48);
8) антитело ZIL154: VH-CDR1 DRGMS (SEQ ID NO: 49), VH-CDR2 YIRYDGSRTDYADAVEG (SEQ ID NO: 50), VH-CDR3 WDGSSFDY (SEQ ID NO: 51), VL-CDR1 KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 KVSNRDP (SEQ ID NO: 53), VL-CDR3 MQAIHFPLT (SEQ ID NO: 54);
9) антитело ZIL159: VH-CDR1 SYVMT (SEQ ID NO: 55), VH-CDR2 GINSEGSRTAYADAVKG (SEQ ID NO: 56), VH-CDR3 GDIVATGTSY (SEQ ID NO: 57), VL-CDR1 SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 KDTERPS (SEQ ID NO: 59), VL-CDR3 KSAVSIDVGV (SEQ ID NO: 60);
10) антитело ZIL171: VH-CDR1 TYVMN (SEQ ID NO: 61), VH-CDR2 SINGGGSSPTYADAVRG (SEQ ID NO: 62), VH-CDR3 SMVGPFDY (SEQ ID NO: 63), VL-CDR1 SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 KDTERPS (SEQ ID NO: 65), VL-CDR3 ESAVSSDTIV (SEQ ID NO: 66);
11) антитело 04H07: VH-CDR1 SYWMN (SEQ ID NO: 200), VH-CDR2 MIDPSDSEIHYNQVFKD (SEQ ID NO: 201), VH-CDR3 QDIVTTVDY (SEQ ID NO: 202), VL-CDR1 KSSQSLLYSINQKNHLA (SEQ ID NO: 203), VL-CDR2 WASTRES (SEQ ID NO: 204), VL-CDR3 QQGYTYPFT (SEQ ID NO: 205);
12) антитело 06A09: VH-CDR1 SYWMN (SEQ ID NO: 206), VH-CDR2 MIDPSDSETHYNQIFRD (SEQ ID NO: 207), VH-CDR3 QDIVTTVDY (SEQ ID NO: 208), VL-CDR1 KSSQSLLYSINQKNFLA (SEQ ID NO: 209), VL-CDR2 WASTRES (SEQ ID NO: 210), VL-CDR3 QQHYGYPFT (SEQ ID NO: 211); или
13) вариант (1)-(12), отличающийся от соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
Также предложен способ получения антитела, включающий культивирование клетки-хозяина, описанной выше, в условиях, приводящих к продукции антитела, и выделение антитела из клетки-хозяина или культуральной среды клетки-хозяина.
Далее описаны выделенные нуклеиновые кислоты по настоящему изобретению. Такие нуклеиновые кислоты могут содержать нуклеиновокислотную последовательность, кодирующую описанные выше последовательности CDR вариабельной области тяжелой или вариабельной области легкой цепи. Альтернативно, выделенная нуклеиновая кислота по настоящему изобретению может содержать нуклеиновокислотную последовательность, кодирующую CDR вариабельной области как тяжелой, так и легкой цепи.
В одном воплощении согласно изобретению предложена выделенная нуклеиновая кислота, содержащая нуклеиновокислотную последовательность, кодирующую по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области тяжелой цепи:
1) 15Н05: CDR1 вариабельной области тяжелой цепи (VH) (VH-CDR1) SYTIH (SEQ ID NO: 1), VH-CDR2 NINPTSGYTENNQRFKD (SEQ ID NO: 2) и VH-CDR3 WGFKYDGEWSFDV (SEQ ID NO: 3);
2) ZIL1: VH-CDR1 SYGMS (SEQ ID NO: 13), VH-CDR2 HINSGGSSTYYADAVKG (SEQ ID NO: 14) и VH-CDR3 VYTTLAAFWTDNFDY (SEQ ID NO: 15);
3) ZIL8: VH-CDR1 DYAMS (SEQ ID NO: 19), VH-CDR2 GIDSVGSGTSYADAVKG (SEQ ID NO: 20) и VH-CDR3 GFPGSFEH (SEQ ID NO: 21);
4) ZIL9: VH-CDR1 SYDMT (SEQ ID NO: 25), VH-CDR2 DVNSGGTGTAYAVAVKG (SEQ ID NO: 26) и VH-CDR3 LGVRDGLSV (SEQ ID NO: 27);
5) ZIL11: VH-CDR1 TYVMN (SEQ ID NO: 31), VH-CDR2 SINGGGSSPTYADAVRG (SEQ ID NO: 32) и VH-CDR3 SMVGPFDY (SEQ ID NO: 33);
6) ZIL69: VH-CDR1 SYAMK (SEQ ID NO: 37), VH-CDR2 TINNDGTRTGYADAVRG (SEQ ID NO: 38) и VH-CDR3 GNAESGCTGDHCPPY (SEQ ID NO: 39);
7) ZIL94: VH-CDR1 TYFMS (SEQ ID NO: 43), VH-CDR2 LISSDGSGTYYADAVKG (SEQ ID NO: 44) и VH-CDR3 FWRAFND (SEQ ID NO: 45);
8) ZIL154: VH-CDR1 DRGMS (SEQ ID NO: 49), VH-CDR2 YIRYDGSRTDYADAVEG (SEQ ID NO: 50) и VH-CDR3 WDGSSFDY (SEQ ID NO: 51);
9) ZIL159: VH-CDR1 SYVMT (SEQ ID NO: 55), VH-CDR2 GINSEGSRTAYADAVKG (SEQ ID NO: 56) и VH-CDR3 GDIVATGTSY (SEQ ID NO: 57);
10) ZIL171: VH-CDR1 TYVMN (SEQ ID NO: 61), VH-CDR2 SINGGGSSPTYADAVRG (SEQ ID NO: 62) и VH-CDR3 SMVGPFDY (SEQ ID NO: 63);
11) 04H07: VH-CDR1 SYWMN (SEQ ID NO: 200), VH-CDR2 MIDPSDSEIHYNQVFKD (SEQ ID NO: 201) и VH-CDR3 QDIVTTVDY (SEQ ID NO: 202);
12) 06A09: VH-CDR1 SYWMN (SEQ ID NO: 206), VH-CDR2 MIDPSDSETHYNQIFRD (SEQ ID NO: 207) и VH-CDR3 QDIVTTVDY (SEQ ID NO: 208); или
13) вариант (1)-(12), отличающийся от CDR соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH.
В одном воплощении выделенная нуклеиновая кислота, описанная выше, может дополнительно содержать нуклеиновокислотную последовательность, кодирующую по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области легкой цепи:
1) 15Н05: CDR1 вариабельной области легкой цепи (VL)(VL-CDR1) RASQGISIWLS (SEQ ID NO: 4), VL-CDR2 KASNLHI (SEQ ID NO: 5) и VL-CDR3 LQSQTYPLT (SEQ ID NO: 6);
2) ZIL1: VL-CDR1 SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 SDGNRPS (SEQ ID NO: 17) и VL-CDR3 QSFDTTLDAYV (SEQ ID NO: 18);
3) ZIL8: VL-CDR1 TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 YNSDRPS (SEQ ID NO: 23) и VL-CDR3 SVYDRTFNAV (SEQ ID NO: 24);
4) ZIL9: VL-CDR1 SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 RDTERPS (SEQ ID NO: 29) и VL-CDR3 ESAVDTGTLV (SEQ ID NO: 30);
5) ZIL11: VL-CDR1 SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 KDTERPS (SEQ ID NO: 35) и VL-CDR3 ESAVSSDTIV (SEQ ID NO: 36);
6) ZIL69: VL-CDR1 SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 KDTERPS (SEQ ID NO: 41) и VL-CDR3 ESAVSSETNV (SEQ ID NO: 42);
7) ZIL94: VL-CDR1 GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 DTGSRPS (SEQ ID NO: 47) и VL-CDR3 SLYTDSDILV (SEQ ID NO: 48);
8) ZIL154: VL-CDR1 KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 KVSNRDP (SEQ ID NO: 53) и VL-CDR3 MQAIHFPLT (SEQ ID NO: 54);
9) ZIL159: VL-CDR1 SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 KDTERPS (SEQ ID NO: 59) и VL-CDR3 KSAVSIDVGV (SEQ ID NO: 60);
10) ZIL171: VL-CDR1 SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 KDTERPS (SEQ ID NO: 65) и VL-CDR3 ESAVSSDTIV (SEQ ID NO: 66);
11) 04H07: VL-CDR1 KSSQSLLYSINQKNHLA (SEQ ID NO: 203), VL-CDR2 WASTRES (SEQ ID NO: 204), VL-CDR3 QQGYTYPFT (SEQ ID NO: 205);
12) 06A09: VL-CDR1 KSSQSLLYSINQKNFLA (SEQ ID NO: 209), VL-CDR2 WASTRES (SEQ ID NO: 210), VL-CDR3 QQHYGYPFT (SEQ ID NO: 211); или
13) вариант (1)-(12), отличающийся от CDR соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В одном воплощении согласно изобретению предложена выделенная нуклеиновая кислота, содержащая нуклеиновокислотную последовательность, кодирующую по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области легкой цепи:
1) 15Н05: CDR1 вариабельной области легкой цепи (VL)(VL-CDR1) RASQGISIWLS (SEQIDNO:4), VL-CDR2 KASNLHI (SEQ ID NO: 5) и VL-CDR3 LQSQTYPLT (SEQ ID NO: 6);
2) ZIL1: VL-CDR1 SGSTNNIGILAAT (SEQ ID NO: 16), VL-CDR2 SDGNRPS (SEQ ID NO: 17) и VL-CDR3 QSFDTTLDAYV (SEQ ID NO: 18);
3) ZIL8: VL-CDR1 TGSSSNIGSGYVG (SEQ ID NO: 22), VL-CDR2 YNSDRPS (SEQ ID NO: 23) и VL-CDR3 SVYDRTFNAV (SEQ ID NO: 24);
4) ZIL9: VL-CDR1 SGESLNEYYTQ (SEQ ID NO: 28), VL-CDR2 RDTERPS (SEQ ID NO: 29) и VL-CDR3 ESAVDTGTLV (SEQ ID NO: 30);
5) ZIL11: VL-CDR1 SGESLSNYYAQ (SEQ ID NO: 34), VL-CDR2 KDTERPS (SEQ ID NO: 35) и VL-CDR3 ESAVSSDTIV (SEQ ID NO: 36);
6) ZIL69: VL-CDR1 SGESLNKYYAQ (SEQ ID NO: 40), VL-CDR2 KDTERPS (SEQ ID NO: 41) и VL-CDR3 ESAVSSETNV (SEQ ID NO: 42);
7) ZIL94: VL-CDR1 GLNSGSVSTSNYPG (SEQ ID NO: 46), VL-CDR2 DTGSRPS (SEQ ID NO: 47) и VL-CDR3 SLYTDSDILV (SEQ ID NO: 48);
8) ZIL154: VL-CDR1 KASQSLLHSDGNTYLD (SEQ ID NO: 52), VL-CDR2 KVSNRDP (SEQ ID NO: 53) и VL-CDR3 MQAIHFPLT (SEQ ID NO: 54);
9) ZIL159: VL-CDR1 SGETLNRFYTQ (SEQ ID NO: 58), VL-CDR2 KDTERPS (SEQ ID NO: 59) и VL-CDR3 KSAVSIDVGV (SEQ ID NO: 60);
10) ZIL171: VL-CDR1 SGKSLSYYYAQ (SEQ ID NO: 64), VL-CDR2 KDTERPS (SEQ ID NO: 65) и VL-CDR3 ESAVSSDTIV (SEQ ID NO:66);
11) 04H07: VL-CDR1 KSSQSLLYSINQKNHLA (SEQ ID NO: 203), VL-CDR2 WASTRES (SEQ ID NO: 204) и VL-CDR3 QQGYTYPFT (SEQ ID NO: 205);
12) 06A09: VL-CDR1 KSSQSLLYSINQKNFLA (SEQ ID NO: 209), VL-CDR2 WASTRES (SEQ ID NO: 210) и VL-CDR3 QQHYGYPFT (SEQ ID NO: 211); или
13) вариант (1)-(12), отличающийся от CDR соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VL.
Кроме того, согласно настоящему изобретению предложен вектор, содержащий по меньшей мере одну из нуклеиновых кислот, описанных выше.
Согласно настоящему изобретению, также предложен способ улучшения однородности и/или характеристик кошачьего антитела, включающий: экспрессию нуклеотидной последовательности, кодирующей легкую цепь каппа кошачьего IgG, и нуклеотидной последовательности, кодирующей тяжелую цепь кошачьего IgG, в клетке-хозяине с получением кошачьего антитела, где нуклеотидная последовательность, кодирующая легкую цепь каппа кошачьего IgG, содержит нуклеотидную последовательность константной области легкой цепи каппа, в которой последовательность, кодирующая С-концевую последовательность QRE, в иных случаях присутствующую в константной области легкой цепи каппа кошачьего IgG дикого типа, модифицирована и/или удалена. Такие модификации могут включать модификации нуклеотидной последовательности, приводящие, например, к делециям, заменам или добавлениям одной или более чем одной аминокислоты на С-конце.
В одном воплощении способ улучшения однородности и/или характеристик кошачьего антитела включает:
a) предоставление нуклеотидной последовательности, кодирующей константную область легкой цепи каппа кошачьего антитела IgG дикого типа, содержащую С-концевую аминокислотную последовательность QRE;
b) удаление и/или модификацию последовательности, кодирующей С-концевую QRE, в нуклеотидной последовательности, предоставленной в (а), с получением измененной нуклеотидной последовательности константной области легкой цепи каппа;
c) объединение измененной нуклеотидной последовательности константной области легкой цепи каппа, полученной в (b), с нуклеотидной последовательностью, кодирующей вариабельную область легкой цепи каппа кошачьего IgG, с получением нуклеотидной последовательности, кодирующей полную легкую цепь каппа кошачьего IgG; и
d) экспрессию нуклеотидной последовательности, кодирующей полную легкую цепь каппа кошачьего IgG, полученной в (с), и нуклеотидной последовательности, кодирующей тяжелую цепь кошачьего IgG, в клетке-хозяине с получением кошачьего антитела, в котором С-концевая последовательность QRE, в иных случаях присутствующая в константной области легкой цепи каппа кошачьего IgG дикого типа, модифицирована и/или удалена.
В одном воплощении улучшение однородности и/или характеристик кошачьего антитела включает снижение уровней свободной легкой цепи каппа IgG и, посредством этого, повышение процентного содержания мономера интактного кошачьего антитела IgG.
В одном воплощении нуклеотидная последовательность, кодирующая легкую цепь каппа кошачьего IgG, и нуклеотидная последовательность, кодирующая тяжелую цепь кошачьего IgG, присутствуют в одном и том же векторе, используемом для трансформации клетки-хозяина. В другом воплощении нуклеотидная последовательность, кодирующая легкую цепь каппа кошачьего IgG, и нуклеотидная последовательность, кодирующая тяжелую цепь кошачьего IgG, присутствуют в отдельных векторах, используемых для трансформации клетки-хозяина.
В одном воплощении способа улучшения однородности и/или характеристик кошачьего антитела, кошачье антитело специфически связывается с мишенью, вовлеченной в расстройство, опосредованное цитокином и/или фактором роста. В одном конкретном воплощении кошачье антитело специфически связывается с кошачьим IL-31 или кошачьим NGF.
В одном воплощении кошачье антитело содержит константную область легкой цепи каппа, имеющую последовательность:
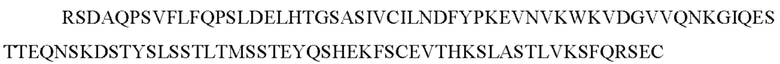 (SEQ ID NO: 186) или ее вариант. Такие варианты могут включать, например, добавление или модификацию одного или более чем одного аминокислотного остатка на С-конце SEQ ID NO: 186.
(SEQ ID NO: 186) или ее вариант. Такие варианты могут включать, например, добавление или модификацию одного или более чем одного аминокислотного остатка на С-конце SEQ ID NO: 186.
Кроме того, согласно настоящему изобретению предложен способ улучшения однородности и/или характеристик собачьего антитела, включающий: экспрессию нуклеотидной последовательности, кодирующей легкую цепь каппа собачьего IgG, и нуклеотидной последовательности, кодирующей тяжелую цепь собачьего IgG, в клетке-хозяине с получением собачьего антитела, где нуклеотидная последовательность, кодирующая легкую цепь каппа собачьего IgG, содержит нуклеотидную последовательность константной области легкой цепи каппа, в которой последовательность, кодирующая С-концевую последовательность QRVD, в иных случаях присутствующую в константной области легкой цепи каппа собачьего IgG дикого типа (Canine LC Kappa wt, SEQ ID NO: 194), модифицирована и/или удалена. Такие модификации могут включать модификации нуклеотидной последовательности, приводящие, например, к делециям, заменам или добавлениям одной или более чем одной аминокислоты на С-конце.
В одном воплощении способ улучшения однородности и/или характеристик собачьего антитела включает:
a) предоставление нуклеотидной последовательности, кодирующей константную область легкой цепи каппа собачьего антитела IgG дикого типа, содержащую С-концевую аминокислотную последовательность QRVD;
b) удаление и/или модификацию последовательности, кодирующей С-концевую QRVD, в нуклеотидной последовательности, предоставленной в (а), с получением измененной нуклеотидной последовательности константной области легкой цепи каппа;
c) объединение измененной нуклеотидной последовательности константной области легкой цепи каппа, полученной в (b), с нуклеотидной последовательностью, кодирующей вариабельную область легкой цепи каппа собачьего IgG, с получением нуклеотидной последовательности, кодирующей полную легкую цепь каппа собачьего IgG; и
d) экспрессию нуклеотидной последовательности, кодирующей полную легкую цепь каппа собачьего IgG, полученной в (с), и нуклеотидной последовательности, кодирующей тяжелую цепь собачьего IgG, в клетке-хозяине с получением собачьего антитела, в котором С-концевая последовательность QRVD, в иных случаях присутствующая в константной области легкой цепи каппа собачьего IgG дикого типа, модифицирована и/или удалена.
В одном воплощении улучшение однородности и/или характеристик собачьего антитела включает снижение уровней свободной легкой цепи каппа IgG и, посредством этого, повышение процентного содержания мономера интактного собачьего антитела IgG.
В одном воплощении нуклеотидная последовательность, кодирующая легкую цепь каппа собачьего IgG, и нуклеотидная последовательность, кодирующая тяжелую цепь собачьего IgG, присутствуют в одном и том же векторе, используемом для трансформации клетки-хозяина. В другом воплощении нуклеотидная последовательность, кодирующая легкую цепь каппа собачьего IgG, и нуклеотидная последовательность, кодирующая тяжелую цепь собачьего IgG, присутствуют в раздельных векторах, используемых для трансформации клетки-хозяина.
В одном воплощении способа улучшения однородности и/или характеристик собачьего антитела собачье антитело специфически связывается с мишенью, вовлеченной в расстройство, опосредованное цитокином и/или фактором роста. В одном конкретном воплощении собачье антитело специфически связывается с собачьим IL-31.
В одном воплощении способа улучшения однородности и/или характеристик собачьего антитела собачье антитело содержит константную область легкой цепи каппа, имеющую последовательность:
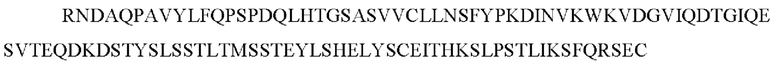 (SEQ ID NO: 179) или ее вариант. Такие варианты могут включать, например, добавление или модификацию одного или более чем одного аминокислотного остатка на С-конце SEQ ID NO: 179.
(SEQ ID NO: 179) или ее вариант. Такие варианты могут включать, например, добавление или модификацию одного или более чем одного аминокислотного остатка на С-конце SEQ ID NO: 179.
Измененные константные области легкой цепи каппа, описанные здесь, могут быть использованы в сочетании с любым числом кошачьих и собачьих антител, включая, например, без ограничения, любые из собачьих или кошачьих антител, описанных в описании и формуле данного изобретения. Предполагается, что собачьи и кошачьи антитела, имеющие мишени, отличные от IL-31, можно также подходящим образом комбинировать с измененными константными областями легкой цепи каппа, раскрытыми здесь. Настоящее изобретение включает любое кошачье или собачье антитело, содержащее такие измененные константные области легкой цепи каппа, раскрытые здесь, поскольку, на основании описания и формулы настоящего изобретения, логично предположить, что такие антитела имеют улучшенную однородность и/или характеристики.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 представлено выравнивание, показывающее консервативность аминокислотной последовательности IL-31 различных видов. В частности, показано сравнение SEQ ID NO: 155 (собачий IL-31), SEQ ID NO: 157 (кошачий IL-31), SEQ ID NO: 165 (лошадиный IL-31) и SEQ ID NO: 181 (человеческий IL-31). Также указан процент идентичности аминокислотной последовательности собачьего, кошачьего, лошадиного и человеческого IL-31.
На Фиг. 2 приведены подробные данные по аффинности, с которой каждое из антител-кандидатов с CDR мышиного происхождения связываются с кошачьим и собачьим IL-31, полученные с применением поверхностного плазмонного резонанса (SPR) и системы Biacore (Biacore Life Sciences (GE Healthcare), Уппсала, Швеция).
На Фиг. 3 представлена таблица, в которой показана активность(IC50 (мкг/мл)) антител-кандидатов с CDR мышиного происхождения, измеренная в анализах с собачьими и кошачьими клетками. В частности, была проведена оценка способности антител-кандидатов ингибировать фосфорилирование STAT, опосредованное IL-31, в собачьих (DH-82) или кошачьих (FCWF4) макрофагоподобных клетках.
На Фиг. 4 показаны результаты, полученные применительно к связыванию моноклональных антител-кандидатов с CDR собачьего происхождения с различными белками с применением как метода непрямого ELISA, так и метода Biacore. В случае непрямого ELISA оценивали связывание (OD при ELISA) с кошачьим IL-31 дикого типа и мутантом кошачьего IL-31 15Н05, имевшим мутации в области эпитопа моноклонального антитела 15Н05. Для подтверждения связывания проводили анализ Biacore, используя поверхности с белками собачьего, кошачьего, лошадиного, человеческого IL-31, мутанта кошачьего IL-31 15Н05 и мутанта кошачьего IL-31 11Е12 при единственной анализируемой концентрации антитела. Мутант кошачьего IL-31 11Е 12 имел мутации в области эпитопа моноклонального антитела 11Е12.
Фиг. 5. На Фиг. 5А показано выравнивание с последовательностью VL мышиного антитела 11Е12 (SEQ ID NO: 73) при сравнении последовательностей раскрытого ранее канинизированного 11Е12, обозначенных как Can_11Е12_VL_cUn_1 (SEQ ID NO: 182) и CAN_11E12_VL_cUn_FW2 (SEQ ID NO: 184), с фелинизированными вариантами, обозначенными как FEL_11Е12_VL1 (SEQ ID NO: 113) и FEL_11E12_VL1_FW2 (SEQ ID NO: 117). Под выравниванием на Фиг. 5A отмечены точки, показывающие положения важных изменений в Fel_11E12_VL1, которые были необходимы для восстановления аффинности этого антитела в отношении белка IL-31. На Фиг. 5В показано выравнивание последовательности VL мышиного антитела 15Н05, обозначенной здесь как MU_15H05_VL (SEQ ID NO: 69), с последовательностями VL фелинизированного 15Н05, обозначенными здесь как FE1_15H05_VL1 (SEQ ID NO: 127) и FEl_15Н05_VL FW2 (SEQ ID NO: 135). Точки под выравниванием на Фиг. 5В указывают на необходимые изменения в VL фелинизированного 15Н05 (Fel_15Н05_VL1), которые были нужны не только для восстановления, но и для улучшения его аффинности в отношении собачьего и кошачьего IL-31 по сравнению с мышиными и химерными формами этого антитела.
Фиг. 6. На Фиг. 6А показано выравнивание кошачьего IL-31 дикого типа (SEQ ID NO: 157) с мутантами 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161), где выделены положения аланиновых замен. На Фиг. 6В показана гомологичная модель кошачьего IL-31, где выделены положения двух аминокислот, вовлеченных в связывание с антителами 11Е12 (сайт 1) и 15Н05 (сайт 2). На Фиг. 6С представлен график, на котором показаны результаты, полученные применительно к связыванию моноклональных антител 11Е12 и 15Н05 с кошачьим IL-31 дикого типа и мутантными белками IL-31 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161) при использовании дикого типа и этих мутантов в качестве сорбируемых антигенов.
На Фиг. 7 представлены графики, на которых показаны конкурентные анализы связывания mAb 15Н05 и 11Е12 с применением Biacore. На Фиг. 7А показаны данные по конкурентному связыванию мышиных антител 15Н05 и 11Е12 с собачьим IL-31. На Фиг. 7В показаны данные по конкурентному связыванию антител 15Н05 и 11Е12 с поверхностью с кошачьим IL-31.
На Фиг. 8 представлен график, на котором показаны результаты, полученные применительно к связыванию отдельных рецепторных субъединиц OSMR и IL-31Ra с кошачьим IL-31 дикого типа и мутантными белками IL-31 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161) при использовании дикого типа и этих мутантов в качестве сорбируемых антигенов.
На Фиг. 9 представлен график, на котором показана предварительная эффективность химерного мышиного-кошачьего 11Е12, химерного мышиного-кошачьего 15Н05 и фелинизированного 11Е12 (Feline 11Е12 1.1) в модели зуда, индуцированного IL-31, у кошек.
На Фиг. 10 представлены графики, на которых показана оценка эффективности фелинизированного антитела против IL-31 15Н05, названного ZTS-361, in vivo в модели провокации зуда у кошек. На Фиг. 10А показаны исходные поведенческие признаки зуда до провокации в группах плацебо-наполнителя Т01 и антитела ZTS-361 Т02 от -7 суток до 28 суток, где нулевыми сутками являются сутки введения антитела в группе Т02. На Фиг. 10В показана эффективность антитела ZTS-361, демонстрирующего существенное уменьшение зуда, наблюдаемого на 7 сутки (р менее 0,0001), 21 сутки (р менее 0,0027) и 28 сутки (р менее 0,0238) после провокации с использованием IL-31 по сравнению с контрольным плацебо-наполнителем.
Фиг. 11. На Фиг. 11А представлен график, на котором показаны уровни IL-31 в плазме у собак с атопическим и аллергическим дерматитом, принадлежащих клиентам, в сравнении с нормальными лабораторными животными. На Фиг. 11В представлен график, на котором показаны результаты недавнего исследования по определению уровней IL-31 в сыворотке у кошек с предположительным диагнозом аллергического дерматита (АД) из нескольких разных географических областей США. На Фиг. 11С представлен график, на котором показан фармакокинетический профиль собачьего IL-31 у собак после его подкожного введения в дозе 1,75 мкг/кг.
На Фиг. 12 показан невосстанавливающий 4-12% SDS-PAGE при сравнении дорожки 1, на которой представлено ZTS-361, тяжелая цепь которого представляет собой (SEQ ID NO:121; FEL_15H05_VH1), объединенную с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_HC_AlleleA_1). Легкая цепь ZTS-361 представляет собой (SEQ ID NO:135; FEL-15H05-VL1 FW2), объединенную с константной областью легкой цепи IgG (SEQ ID NO: 175; Feline LC Kappa G minus). На дорожке 2 представлено мышиное 15H05, тяжелая цепь которого представляет собой (SEQ ID NO: 67; MU_15H05_VH), объединенную с константной областью тяжелой цепи мышиного IgG (SEQ ID NO: 188; Mouse_НС_IgG1). Легкая цепь мышиного 15Н05 представляет собой (SEQ ID NO: 69; MU_15Н05_VL), объединенную с константной областью легкой цепи мышиного IgG (SEQ ID NO: 190; Mouse_LC_Kappa). Интактное (Intact) относится к IgG с двумя тяжелыми цепями и двумя легкими цепями, удерживаемыми вместе межцепочечными дисульфидными связями, и предполагаемой молекулярной массой приблизительно 150 кДа. HHL относится к «Heavy Heavy Light (тяжелая тяжелая легкая)» и представляет собой IgG без одной легкой цепи с предполагаемой молекулярной массой приблизительно 125 кДа. НН относится к «Heavy Heavy (тяжелая тяжелая)» и представляет собой IgG без обеих легких цепей с предполагаемой молекулярной массой приблизительно 100 кДа. HL относится к «Heavy Light (тяжелая легкая)» и представляет собой IgG с одной тяжелой цепью, одной легкой цепью и предполагаемой молекулярной массой приблизительно 75 кДа. L относится к «Light (легкая)» и представляет собой IgG с одной легкой цепью и предполагаемой молекулярной массой приблизительно 25 кДа, также называемый здесь свободной легкой цепью.
Фиг. 13. На Фиг. 13А показаны результаты невосстанавливающего капиллярного гель-электрофореза (NR-CGE) при сравнении IgG из стабильных клеточных линий, экспрессирующих ZTS-361 или мышиное 15Н05 (каждое из которых описано выше). Процент мономера и второстепенных разновидностей рассчитывали по экспериментальным показаниям, полученным с использованием NR-CGE и показанным в виде электрофореграмм на Фиг. 13В. Площадь с поправкой на время (time corrected area, ТСА) определяют как отношение площади отдельного пика по показаниям прибора ко времени выхода этого пика. Общую ТСА определяют как сумму ТСА всех пиков, превышающих или равных 0,3%. Процент мономера интактного IgG (% Monomer) и отдельных фрагментов (%HHL и %L) рассчитывают по их индивидуальным ТСА как процент от общей ТСА. Процент фрагментов (% Fragments) представляет собой сумму площадей всех пиков, молекулярная масса которых меньше, чем у интактного IgG.
Фиг. 14. На Фиг. 14А показан невосстанавливающий 4-12% SDS-PAGE при сравнении IgG из отдельных стабильных СНО-клонов ZTS-361 (описанных выше и в Разделе 1.9 раздела «Примеры»). На дорожках 1 и 8 представлен стандартный образец IgG для сравнения. Процент мономера рассчитывают денситометрическим анализом каждой полосы, перемещающейся в соответствии с предполагаемой молекулярной массой, составляющей приблизительно 150 кДа, с использованием программного обеспечения BioRad VersaDoc. Процент фрагментов (% Fragments) представляет собой сумму отдельных полос с меньшей молекулярной массой. На Фиг. 14В показаны результаты невосстанавливающего капиллярного гель-электрофореза (NR-CGE) при сравнении IgG из отдельных стабильных СНО-клонов ZTS-361. Площадь с поправкой на время (time corrected area, ТСА) определяют как отношение площади отдельного пика по показаниям прибора ко времени выхода этого пика. Общую ТСА определяют как сумму ТСА всех пиков, превышающих или равных 0,3%. Процент мономера интактного IgG (% Monomer) и процент фрагментов (% Fragments) рассчитывают по их индивидуальным ТСА как процент от общей ТСА. Процент фрагментов (% Fragments) представляет собой сумму площадей всех пиков, молекулярная масса которых меньше, чем у интактного IgG.
На Фиг. 15 показаны С-концевые аминокислоты белка константной области легкой цепи каппа Ig указанного вида. В Canine LC kappa wt показанные С-концевые аминокислотные остатки представляют собой положения 103-109 SEQ ID NO: 194, а показанные нуклеотидные остатки представляют собой положения 307-330 SEQ ID NO: 195; в Feline LC kappa G minus (G-) показанные С-концевые аминокислотные остатки представляют собой положения 105-110 SEQ ID NO: 175, а показанные нуклеотидные остатки представляют собой положения 313-330 SEQ ID NO: 176; в Pig LC kappa показанные С-концевые аминокислотные остатки представляют собой положения 104-108 SEQ ID NO: 196, а показанные нуклеотидные остатки представляют собой положения 310-327 SEQ ID NO: 197; в Mink LC kappa показанные С-концевые аминокислотные остатки представляют собой положения 105-108 SEQ ID NO: 198, а показанные нуклеотидные остатки представляют собой положения 313-327 SEQ ID NO: 199; в Human LC kappa показанные С-концевые аминокислотные остатки представляют собой положения 102-106 SEQ ID NO: 192, а показанные нуклеотидные остатки представляют собой положения 304-321 SEQ ID NO: 193; в Mouse LC kappa показанные С-концевые аминокислотные остатки представляют собой положения 102-106 SEQ ID NO: 190, а показанные нуклеотидные остатки представляют собой положения 304-321 SEQ ID NO: 191. Серым прямоугольником показано положение С-концевого цистеина, образующего межцепочечную дисульфидную связь с константной областью тяжелой цепи IgG, необходимую для интактного антитела. Серым треугольником выделено увеличение процента использования легкой цепи каппа в IgG указанных видов, то есть собаки (Canine), кошки (Feline) и свиньи (Pig) (Aran et al. 1996 Zentralbl Veterinarmed. Nov; 43(9):573-6), норки (Mink) (Bovkun et al. 1993 Eur J Immunol. Aug; 23(8): 1929-34), мыши (Mouse) (Woloschak et al. 1987 Mol Immunol. Jul; 24(7):751-7) и человека (Human) (Barandun et al. 1976 Blood. Jan; 47(1):79-89). Нуклеотидами, показанными светло-серым цветом, выделены кодоны, в которых замена одного нуклеотида приводит к стоп-кодону.
Фиг. 16. На Фиг. 16А представлено пиктографическое изображение кошачьего IgG, на котором выделены относительные положения предполагаемых межцепочечных и внутрицепочечных дисульфидных связей. Показанный аминокислотный остаток CYS15 представляет собой положение 15 Feline НС Allele A wt (SEQ ID NO: 171) и Feline НС AlleleA 1 (SEQ ID NO: 173), а показанные нуклеотидные остатки представляют собой положения 43-45 Feline НС AlleleA wt (SEQ ID NO: 172) и 43-45 Feline НС AlleleA 1 (SEQ ID NO: 174), соответственно. Показанный аминокислотный остаток CYS107 представляет собой положение 107 Feline LC Kappa G minus (SEQ ID NO: 175), a показанные нуклеотидные остатки представляют собой положения 319-321 Feline LC Kappa G minus (SEQ ID NO: 176). На Фиг. 16В представлена гомологичная модель ZTS-361, где выделены положения CYS15 и CYS107, описанных выше. На Фиг. 16С представлено увеличенное изображение области, обведенной кругом на Фиг. 16В, с повторным выделением положений двух цистеинов, обеспечивающих межцепочечное спаривание кошачьих тяжелых и легких цепей. Показанные оболочки с сетчатой поверхностью представляют собой расчетный электростатический вклад остатков константной области легкой цепи каппа QRE, следующих непосредственно после CYS107, показанных в Feline LC kappa G- на Фиг. 15.
Фиг. 17. На Фиг. 17А показаны идентификационные номера последовательностей, соответствующие тяжелым и легким цепям, использованным для создания стабильных клеточных линий СНО, продуцирующих антитела ZTS-361 и ZTS-1505 (описанных выше и/или в Разделе 1.9 раздела «Примеры»). Выделена Feline LC Kappa G minus QRE minus (SEQ ID NO: 186), которой соответствует нуклеотидная последовательность Feline LC Kappa G minus QRE minus (SEQ ID NO: 187). На Фиг. 17В показаны результаты невосстанавливающего капиллярного гель-электрофореза (NR-CGE) при сравнении IgG из отдельных стабильных СНО-клонов ZTS-1505. Площадь с поправкой на время (time corrected area, ТСА) определяют как отношение площади отдельного пика по показаниям прибора ко времени выхода этого пика. Общую ТСА определяют как сумму ТСА всех пиков, превышающих или равных 0,3%. Процент мономера интактного IgG (% Monomer) и процент фрагментов (% Fragments) рассчитывают по их индивидуальным ТСА как процент от общей ТСА. Процент фрагментов (% Fragments) представляет собой сумму площадей всех пиков, молекулярная масса которых меньше, чем у интактного IgG. На Фиг. 17С показано сравнение одного стабильного СНО-клона, продуцирующего антитело ZTS-361, с одним стабильным СНО-клоном, продуцирующим ZTS-1505. Сравнение двух стабильных клонов проводили при 8 независимых вариантах условий культивирования, обозначенных как A-G. Указан процент вариабельности каждой культуры через 14 суток культивирования. Титр указывает на количество антитела, продуцируемого через 14 суток культивирования каждым стабильным клоном при соответствующих условиях культивирования. Указан процент мономера, рассчитанный по результатам NR-CGE для каждого клона, выращенного с применением различных условий культивирования.
Фиг. 18. На Фиг. 18А показан процент идентичности при сравнении вариабельных областей антител против кошачьего NGF и против IL-31, рассчитанный с использованием программного обеспечения ClustalW. На Фиг. 18В и 18С показано выравнивание вариабельных тяжелых и легких цепей, соответственно, антител против кошачьего IL-31 и NGF, где CDR выделены рамками.
На Фиг. 19 показаны результаты NR-CGE при сравнении антител против кошачьего IL-31 и против кошачьего NGF с и без модификации С-конца константной цепи каппа.
На Фиг. 20 показано выравнивание последовательностей кошачьего и лошадиного IL-31 с использованием ClustalW.
На Фиг. 21 показано выравнивание вариабельных областей тяжелых (Фиг. 21А) и легких (Фиг. 21В) цепей антител 04Н07 и 06А09 в сравнении с мышиным антителом 15Н05 с использованием ClustalW. Для сравнения расположение каждого из шести CDR выделено рамками.
На Фиг. 22 представлена сенсограмма Biacore, на которой показан средний профиль антитела против IL-31 ZTS-1505 плюс/минус 3 стандартных отклонения, использованный для определения порогового ответа для скрининга мутантов с аланиновыми заменами в CDR.
На Фиг. 23 показаны результаты мутагенеза с использованием аланиновых замен в CDR тяжелой цепи антитела ZTS-1505 по связыванию с IL-31 и ингибированию pSTAT-сигналов, опосредованных IL-31, в сравнении с антителом дикого типа.
На Фиг. 24 показаны результаты мутагенеза с использованием аланиновых замен в CDR легкой цепи антитела ZTS-1505 по связыванию с IL-31 и ингибированию pSTAT-сигналов, опосредованных IL-31, в сравнении с антителом дикого типа.
Фиг. 25. На Фиг. 25А и 25В показана аффинность связывания двух фелинизированных антител, обозначенных здесь как ZTS-5864 и ZTS-5865, и их активность в отношении клеток, соответственно.
На Фиг. 26 представлен график, на котором показаны результаты оценки эффективности фелинизированного антитела против IL-31 ZTS-5864 in vivo в модели провокации зуда у кошек.
Антитела, описанные на Фиг. 12, 13 и 14, были получены при условиях культивирования, эквивалентных условиям культивирования «А» на Фиг. 17С.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь MU_15Н05_VH_CDR1;
SEQ ID NO: 2 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь MU_15Н05_VH_CDR2;
SEQ ID NO: 3 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь MU_15H05_VH_CDR3;
SEQ ID NO: 4 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь MU_15Н05_VL_CDR1;
SEQ ID NO: 5 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь MU_15H05_VL_CDR2;
SEQ ID NO: 6 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь MU_15Н05_VL_CDR3;
SEQ ID NO: 7 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь 11E12-VH-CDR1;
SEQ ID NO: 8 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь 11E12-VH-CDR2;
SEQ ID NO: 9 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь 11E12-VH-CDR3;
SEQ ID NO: 10 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь 11E12-VL-CDR1;
SEQ ID NO: 11 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь 11E12-VL-CDR2;
SEQ ID NO: 12 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь 11E12-VL-CDR3;
SEQ ID NO: 13 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL1_VH_CDR1;
SEQ ID NO: 14 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL1_VH_CDR2;
SEQ ID NO: 15 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL1_VH_CDR3;
SEQ ID NO: 16 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL1_VL_CDR1;
SEQ ID NO: 17 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL1_VL_CDR2;
SEQ ID NO: 18 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL1_VL_CDR3;
SEQ ID NO: 19 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL8_VH_CDR1;
SEQ ID NO: 20 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL8_VH_CDR2;
SEQ ID NO: 21 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL8_VH_CDR3;
SEQ ID NO: 22 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL8_VL_CDR1;
SEQ ID NO: 23 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL8_VL_CDR2;
SEQ ID NO: 24 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL8_VL_CDR3;
SEQ ID NO: 25 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL9_VH_CDR1;
SEQ ID NO: 26 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL9_VH_CDR2;
SEQ ID NO: 27 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL9_VH_CDR3;
SEQ ID NO: 28 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL9_VL_CDR1;
SEQ ID NO: 29 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL9_VL_CDR2;
SEQ ID NO: 30 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL9_VL_CDR3;
SEQ ID NO: 31 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL11_VH_CDR1;
SEQ ID NO: 32 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL11_VH_CDR2;
SEQ ID NO: 33 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL11_VH_CDR3;
SEQ ID NO: 34 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL11_VL_CDR1;
SEQ ID NO: 35 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL11_VL_CDR2;
SEQ ID NO: 36 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL11_VL_CDR3;
SEQ ID NO: 37 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL69_VH_CDR1;
SEQ ID NO: 38 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL69_VH_CDR2;
SEQ ID NO: 39 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL69_VH_CDR3;
SEQ ID NO: 40 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL69_VL_CDR1;
SEQ ID NO: 41 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL69_VL_CDR2;
SEQ ID NO: 42 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL69_VL_CDR3;
SEQ ID NO: 43 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL94_VH_CDR1;
SEQ ID NO: 44 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL94_VH_CDR2;
SEQ ID NO: 45 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL94_VH_CDR3;
SEQ ID NO: 46 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL94_VL_CDR1;
SEQ ID NO: 47 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL94_VL_CDR2;
SEQ ID NO: 48 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL94_VL_CDR3;
SEQ ID NO: 49 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL154_VH_CDR1;
SEQ ID NO: 50 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL154_VH_CDR2;
SEQ ID NO: 51 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL154_VH_CDR3;
SEQ ID NO: 52 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL154_VL_CDR1;
SEQ ID NO: 53 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL154_VL_CDR2;
SEQ ID NO: 54 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL154_VL_CDR3;
SEQ ID NO: 55 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL159_VH_CDR1;
SEQ ID NO: 56 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL159_VH_CDR2;
SEQ ID NO: 57 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL159_VH_CDR3;
SEQ ID NO: 58 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL159_VL_CDR1;
SEQ ID NO: 59 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL159_VL_CDR2;
SEQ ID NO: 60 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL159_VL_CDR3;
SEQ ID NO: 61 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL171_VH_CDR 1;
SEQ ID NO: 62 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL171_VH_CDR2;
SEQ ID NO: 63 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь CAN_ZIL171_VH_CDR3;
SEQ ID NO: 64 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь CAN_ZIL171_VL_CDR 1;
SEQ ID NO: 65 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь CAN_ZIL171_VL_CDR2;
SEQ ID NO: 66 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь CAN_ZIL171_VL_CDR3;
SEQ ID NO: 67 представляет собой вариабельную область тяжелой цепи, называемую здесь MU_15H05_VH;
SEQ ID NO: 68 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь MU_15H05_VH;
SEQ ID NO: 69 представляет собой вариабельную область легкой цепи, называемую здесь MU_15Н05_VL;
SEQ ID NO: 70 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь MU_15H05_VL;
SEQ ID NO: 71 представляет собой вариабельную область тяжелой цепи, называемую здесь MU-11E12-VH;
SEQ ID NO: 72 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь MU-11E12-VH;
SEQ ID NO: 73 представляет собой вариабельную область легкой цепи, называемую здесь MU-11E12-VL;
SEQ ID NO: 74 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь MU-11E12-VL;
SEQ ID NO: 75 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL1_VH;
SEQ ID NO: 76 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL1_VH;
SEQ ID NO: 77 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL1_VL;
SEQ ID NO: 78 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL1_VL;
SEQ ID NO:79 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL8_VH;
SEQ ID NO: 80 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL8_VH;
SEQ ID NO: 81 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL8_VL;
SEQ ID NO: 82 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL8_VL;
SEQ ID NO: 83 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL9_VH;
SEQ ID NO: 84 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL9_VH;
SEQ ID NO: 85 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL9_VL;
SEQ ID NO: 86 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL9_VL;
SEQ ID NO: 87 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL11_VH;
SEQ ID NO: 88 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL11_VH;
SEQ ID NO: 89 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL11_VL;
SEQ ID NO: 90 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL11_VL;
SEQ ID NO: 91 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL69_VH;
SEQ ID NO: 92 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL69_VH;
SEQ ID NO: 93 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL69_VL;
SEQ ID NO: 94 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL69_VL;
SEQ ID NO: 95 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL94_VH;
SEQ ID NO: 96 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL94_VH;
SEQ ID NO: 97 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL94_VL;
SEQ ID NO: 98 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL94_VL;
SEQ ID NO: 99 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL154_H;
SEQ ID NO: 100 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL154_VH;
SEQ ID NO: 101 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL154_VL;
SEQ ID NO: 102 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL154_VL;
SEQ ID NO: 103 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL159_VH;
SEQ ID NO: 104 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL159_VH;
SEQ ID NO: 105 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL159_VL;
SEQ ID NO: 106 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL159_VL;
SEQ ID NO: 107 представляет собой вариабельную область тяжелой цепи, называемую здесь CAN-ZIL171_VH;
SEQ ID NO: 108 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь CAN-ZIL171_VH;
SEQ ID NO: 109 представляет собой вариабельную область легкой цепи, называемую здесь CAN-ZIL171_VL;
SEQ ID NO: 110 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь CAN-ZIL171_VL;
SEQ ID NO: 111 представляет собой вариабельную область тяжелой цепи, называемую здесь FEL_11E12_VH1;
SEQ ID NO: 112 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь FEL_11Е12_VH1;
SEQ ID NO: 113 представляет собой вариабельную область легкой цепи, называемую здесь FEL_11Е12_VL1;
SEQ ID NO: 114 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_11E12_VL1;
SEQ ID NO: 115 представляет собой вариабельную область легкой цепи, называемую здесь FEL_11Е12_VL2;
SEQ ID NO: 116 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_11E12_VL2;
SEQ ID NO: 117 представляет собой вариабельную область легкой цепи, называемую здесь FEL_11Е12_VL1_FW2;
SEQ ID NO: 118 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_11Е12_VL1_FW2;
SEQ ID NO: 119 представляет собой вариабельную область легкой цепи, называемую здесь FEL_11Е12_VL1_K46Q;
SEQ ID NO: 120 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_11E12_VL1_K46Q;
SEQ ID NO: 121 представляет собой вариабельную область тяжелой цепи, называемую здесь FEL_15Н05_VH1;
SEQ ID NO: 122 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь FEL_15H05_VH1;
SEQ ID NO: 123 представляет собой вариабельную область тяжелой цепи, называемую здесь FEL_15Н05_VH2;
SEQ ID NO: 124 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь FEL_15Н05_VH2;
SEQ ID NO: 125 представляет собой вариабельную область тяжелой цепи, называемую здесь FEL_15H05_VH3;
SEQ ID NO: 126 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь FEL_15Н05_VH3;
SEQ ID NO: 127 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1;
SEQ ID NO: 128 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1;
SEQ ID NO: 129 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL2;
SEQ ID NO: 130 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15H05_VL2;
SEQ ID NO: 131 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL3;
SEQ ID NO: 132 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15H05_VL3;
SEQ ID NO: 133 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW1;
SEQ ID NO: 134 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW1;
SEQ ID NO: 135 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2;
SEQ ID NO: 136 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2;
SEQ ID NO: 137 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW3;
SEQ ID NO: 138 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW3;
SEQ ID NO: 139 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW1_FW2;
SEQ ID NO: 140 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW1_FW2;
SEQ ID NO: 141 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW1_FW3;
SEQ ID NO: 142 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW1_FW3;
SEQ ID NO: 143 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2_FW3;
SEQ ID NO: 144 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2_FW3;
SEQ ID NO: 145 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2_K42N;
SEQ ID NO: 146 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2_K42N;
SEQ ID NO: 147 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2_V43I;
SEQ ID NO: 148 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2_V43I;
SEQ ID NO: 149 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2_L46V;
SEQ ID NO: 150 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2_L46V;
SEQ ID NO: 151 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2_Y49N;
SEQ ID NO: 152 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2_Y49N;
SEQ ID NO: 153 представляет собой вариабельную область легкой цепи, называемую здесь FEL_15Н05_VL1_FW2_K42N_V43I;
SEQ ID NO: 154 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь FEL_15H05_VL1_FW2_K42N_V43I;
SEQ ID NO: 155 представляет собой аминокислотную последовательность белка собачьего IL-31, называемую здесь Canine_IL31;
SEQ ID NO: 156 представляет собой нуклеотидную последовательность, кодирующую белок собачьего IL-31, называемую здесь Canine_IL31;
SEQ ID NO: 157 представляет собой аминокислотную последовательность, называемую здесь Feline_IL31_wildtype, которая соответствует белку кошачьего IL-31 дикого типа с С-концевой His-меткой;
SEQ ID NO: 158 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, называемую здесь Feline_IL31_wildtype;
SEQ ID NO: 159 представляет собой аминокислотную последовательность, называемую здесь Feline_IL_31_E_coli, которая соответствует белку кошачьего IL-31 с N-концевой His-меткой;
SEQ ID NO: 160 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, называемую здесь Feline_IL_31_E_coli;
SEQ ID NO: 161 представляет собой аминокислотную последовательность, называемую здесь Feline_IL31_11Е12_mutant, которая соответствует мутантному белку кошачьего IL-31 НЕ 12 с С-концевой His-меткой;
SEQ ID NO: 162 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, называемую здесь Feline_IL31_11E12_mutant;
SEQ ID NO: 163 представляет собой аминокислотную последовательность, называемую здесь Feline_IL31_15H05_mutant, которая соответствует мутантному белку кошачьего IL-31 15Н05 с С-концевой His-меткой;
SEQ ID NO: 164 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, называемую здесь Feline_IL31_15H05_mutant;
SEQ ID NO: 165 представляет собой аминокислотную последовательность белка лошадиного IL-31, называемую здесь Equine_IL31;
SEQ ID NO: 166 представляет собой нуклеотидную последовательность, кодирующую белок лошадиного IL-31, называемую здесь Equine_IL31;
SEQ ID NO: 167 представляет собой аминокислотную последовательность, называемую здесь Feline_OSMR_hIgG1_Fc, которая соответствует внеклеточному домену кошачьего OSMR, слитому с Fc человеческого IgG1;
SEQ ID NO: 168 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, называемую здесь Feline_OSMR_hIgG1_Fc;
SEQ ID NO: 169 представляет собой аминокислотную последовательность, называемую здесь Feline_IL31Ra_HIgG1_Fc_X1_Fn3, которая соответствует кошачьему IL-31Ra, слитому с Fc человеческого IgG1;
SEQ ID NO: 170 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, называемую здесь Feline_IL31Ra_HIgG1_Fc_X1_Fn3;
SEQ ID NO: 171 представляет собой кошачью тяжелую цепь, называемую здесь Feline_HC_AlleleA_wt;
SEQ ID NO: 172 представляет собой нуклеотидную последовательность, кодирующую кошачью тяжелую цепь, называемую здесь Feline_НС_AlleleA_wt;
SEQ ID NO: 173 представляет собой кошачью тяжелую цепь, называемую здесь Feline_НС_AlleleA_1, которая была модифицирована для замены М, L и G в положениях 120, 121 и 123, соответственно, последовательности дикого типа SEQ ID NO: 171 на аланин (А) с целью устранения эффекторной функции антитела;
SEQ ID NO: 174 представляет собой нуклеотидную последовательность, кодирующую кошачью тяжелую цепь, называемую здесь Feline_HC_AlleleA_1;
SEQ ID NO: 175 представляет собой кошачью легкую цепь каппа, называемую здесь Feline_LC_Kappa_G_minus, которая была модифицирована с устранением гликозилирования (G-) в положении 103 таким образом, что N, обычно присутствующий в этом положении кошачьей легкой цепи каппа дикого типа, был изменен на Q;
SEQ ID NO: 176 представляет собой нуклеотидную последовательность, кодирующую аминокислотную последовательность кошачьей легкой цепи каппа, называемую здесь Feline_LC_Kappa_G_minus;
SEQ ID NO: 177 представляет собой собачью тяжелую цепь, называемую здесь Canine_НС_65_1;
SEQ ID NO: 178 представляет собой нуклеотидную последовательность, кодирующую собачью тяжелую цепь, называемую здесь Canine_HC_65_l;
SEQ ID NO: 179 представляет собой собачью легкую цепь каппа, называемую здесь Canine_LC_Kappa;
SEQ ID NO: 180 представляет собой нуклеотидную последовательность, кодирующую собачью легкую цепь каппа, называемую здесь Canine_LC_Kappa;
SEQ ID NO: 181 представляет собой аминокислотную последовательность человеческого IL-31.
SEQ ID NO: 182 представляет собой последовательность вариабельной области легкой цепи mAb, называемую здесь Can_11Е12_VL_cUn_1;
SEQ ID NO: 183 представляет собой нуклеотидную последовательность, кодирующую последовательность вариабельной области легкой цепи mAb, называемую здесь Can_11E12_VL_cUn_l;
SEQ ID NO: 184 представляет собой последовательность вариабельной области легкой цепи mAb, называемую здесь Can_11Е12_VL_cUn_FW2;
SEQ ID NO: 185 представляет собой нуклеотидную последовательность, кодирующую последовательность вариабельной области легкой цепи mAb, называемую здесь Can_11Е12_VL_cUn_FW2;
SEQ ID NO: 186 представляет собой кошачью легкую цепь каппа, называемую здесь Feline_LC_Kappa_G_minus_QRE_minus, которая была модифицирована с (1) устранением гликозилирования (G-) в положении 103 таким образом, что N, обычно присутствующий в этом положении кошачьей легкой цепи каппа дикого типа, был изменен на Q, и (2) удалением С-концевой QRE по сравнению с диким типом;
SEQ ID NO: 187 представляет собой нуклеотидную последовательность, кодирующую кошачью легкую цепь каппа, называемую здесь Feline_LC_Kappa_G_minus_QRE_minus;
SEQ ID NO: 188 представляет собой мышиную тяжелую цепь, обозначаемую здесь как Mouse_HC_IgG1;
SEQ ID NO: 189 представляет собой нуклеотидную последовательность, кодирующую мышиную тяжелую цепь, обозначаемую здесь как Mouse_НС_IgG1;
SEQ ID NO: 190 представляет собой мышиную легкую цепь каппа, обозначаемую здесь как Mouse_LC_Kappa;
SEQ ID NO: 191 представляет собой нуклеотидную последовательность, кодирующую мышиную легкую цепь каппа, обозначаемую здесь как Mouse_LC_Kappa;
SEQ ID NO: 192 представляет собой человеческую легкую цепь каппа, обозначаемую здесь как Human_LC_Kappa;
SEQ ID NO: 193 представляет собой нуклеотидную последовательность, кодирующую человеческую легкую цепь каппа, обозначаемую здесь как Human_LC_Kappa;
SEQ ID NO: 194 представляет собой собачью легкую цепь каппа дикого типа, обозначаемую здесь как Canine_LC_Kappa_wt;
SEQ ID NO: 195 представляет собой нуклеотидную последовательность, кодирующую собачью легкую цепь каппа дикого типа, обозначаемую здесь как Canine_LC_Kappa_wt;
SEQ ID NO: 196 представляет собой свиную легкую цепь каппа, обозначаемую здесь как Pig_LC_Kappa;
SEQ ID NO: 197 представляет собой нуклеотидную последовательность, кодирующую свиную легкую цепь каппа, обозначаемую здесь как Pig_LC_Kappa;
SEQ ID NO: 198 представляет собой легкую цепь каппа норки, обозначаемую здесь как Mink_LC_Kappa;
SEQ ID NO: 199 представляет собой нуклеотидную последовательность, кодирующую легкую цепь каппа норки, обозначаемую здесь как Mink_LC_Kappa;
SEQ ID NO: 200 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь Mu_04Н07_VH_CDR1;
SEQ ID NO: 201 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь Mu_04H07_VH_CDR2;
SEQ ID NO: 202 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь Mu_04Н07_VH_CDR3;
SEQ ID NO: 203 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь Mu_04H07_VL_CDR1;
SEQ ID NO: 204 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь Mu_04Н07_VL_CDR2;
SEQ ID NO: 205 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь Mu_04Н07_VL_CDR3;
SEQ ID NO: 206 представляет собой CDR1 вариабельной области тяжелой цепи, называемый здесь Mu_06A09_VH_CDR1;
SEQ ID NO: 207 представляет собой CDR2 вариабельной области тяжелой цепи, называемый здесь Mu_06А0_ VH_CDR2;
SEQ ID NO: 208 представляет собой CDR3 вариабельной области тяжелой цепи, называемый здесь Mu_06A09_VH_CDR3;
SEQ ID NO: 209 представляет собой CDR1 вариабельной области легкой цепи, называемый здесь Mu_06А09_VL_CD1;
SEQ ID NO: 210 представляет собой CDR2 вариабельной области легкой цепи, называемый здесь Mu_06А09_VL_CD2;
SEQ ID NO: 211 представляет собой CDR3 вариабельной области легкой цепи, называемый здесь Mu_06A09_VL_CDR3;
SEQIDNO:212 представляет собой вариабельную область тяжелой цепи, называемую здесь Mu_04Н07_VH;
SEQ ID NO: 213 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь Mu_04H07_VH;
SEQ ID NO:214 представляет собой вариабельную область легкой цепи, называемую здесь Mu 04Н07 VL;
SEQ ID NO: 215 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь Mu_04H07_VL;
SEQ ID NO:216 представляет собой вариабельную область тяжелой цепи, называемую здесь Mu_06A09_VH;
SEQ ID NO:217 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь Mu_06А09_VH;
SEQ ID NO: 218 представляет собой вариабельную область легкой цепи, называемую здесь Mu_06A09_VL;
SEQ ID NO:219 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь Mu_06А09_VL;
SEQ ID NO: 220 представляет собой вариабельную область тяжелой цепи, называемую здесь ZTS_768_VH;
SEQ ID NO: 221 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь ZTS_768_VH;
SEQ ID NO: 222 представляет собой вариабельную область легкой цепи, называемую здесь ZTS_768_VL;
SEQ ID NO: 223 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь ZTS_768_VL;
SEQ ID NO: 224 представляет собой вариабельную область тяжелой цепи, называемую здесь ZTS_943_VH;
SEQ ID NO: 225 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь ZTS_943_VH;
SEQ ID NO: 226 представляет собой вариабельную область легкой цепи, называемую здесь ZTS_943_VL;
SEQ ID NO: 227 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь ZTS_943_VL;
SEQ ID NO: 228 представляет собой вариабельную область тяжелой цепи, называемую здесь ZTS_5864_VH;
SEQ ID NO: 229 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь ZTS_5864_VH;
SEQ ID NO: 230 представляет собой вариабельную область легкой цепи, называемую здесь ZTS_5864_VL;
SEQ ID NO: 231 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь ZTS_5864_VL;
SEQ ID NO: 232 представляет собой вариабельную область тяжелой цепи, называемую здесь ZTS_5865_VH;
SEQ ID NO: 233 представляет собой нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, называемую здесь ZTS_5865_VH;
SEQ ID NO: 234 представляет собой вариабельную область легкой цепи, называемую здесь ZTS 5865 VL;
SEQ ID NO: 235 представляет собой нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, называемую здесь ZTS_5865_VL;
SEQ ID NO: 236 представляет собой аминокислотную последовательность кошачьей легкой цепи каппа, называемую здесь Feline LC Lambda;
SEQ ID NO: 237 представляет собой нуклеотидную последовательность, кодирующую кошачью легкую цепь каппа, называемую здесь Feline_LC_Lambda.
ОПРЕДЕЛЕНИЯ
Перед подробным описанием настоящего изобретения будут определены некоторые термины, используемые в контексте настоящего изобретения. По необходимости, в дополнение к этим терминам, в других разделах заявки определены другие термины. Если здесь прямо не определено иное, термины, относящиеся к рассматриваемой области, использованы в данном описании в значениях, принятых в данной области.
Если контекстом ясно не продиктовано иное, при использовании в данном описании и формуле изобретения формы единственного числа включают множественное число. Например, ссылка на «антитело» включает множество таких антител.
При использовании здесь термин «содержащий/включающий» предназначен для обозначения того, что композиции и способы включают указанные элементы, не исключая других.
При использовании здесь «эпитоп» относится к антигенной детерминанте, распознаваемой CDR антитела. Иными словами, эпитоп относится к той части любой молекулы, которая способна быть распознанной и связанной антителом. Если не указано иное, при использовании здесь термин «эпитоп» относится к области IL-31, с которой взаимодействует агент против IL-31.
«Антиген» представляет собой молекулу или часть молекулы, способные быть связанными антителом, которые дополнительно способны быть распознанными и связанными антителом (область связывания соответствующего антитела может быть названа паратопом). Обычно эпитопы состоят из химически активных поверхностных групп молекул, например, аминокислот или сахарных боковых цепей, и имеют специфические трехмерные структурные характеристики, а также специфические зарядные характеристики. Эпитопы представляют собой антигенные детерминанты на белке, распознаваемые иммунной системой. Компонентами иммунной системы, распознающими эпитопы, являются антитела, Т-клетки и В-клетки. Представление Т-клеточных эпитопов происходит на поверхности антиген-представляющих клеток (АПК), а их длина обычно составляет 8-11 (МНС класса I) или 15 и более (МНС класса II) аминокислот. Распознавание представленного комплекса МНС-пептид Т-клетками критически важно для их активации. Эти механизмы позволяют надлежащим образом распознавать свои и «чужие» белки, такие как бактерии и вирусы. Вклад во взаимодействие со связывающей бороздой на АПК и последующее распознавание Т-клеточным рецептором вносят независимые аминокислотные остатки, которые могут не быть смежными (Janeway, Travers, Walport, Immunobiology: The Immune System in Health and Disease. 5th edition New York: Garland Science; 2001). Эпитопы, распознаваемые растворимыми антителами и В-клеточными рецепторами, связанными с клеточной поверхностью, значительно варьируют по длине и степени непрерывности (Sivalingam and Shepherd, Immunol. 2012; 51(3-4): 304-309). Кроме того, даже у линейных эпитопов или эпитопов, расположенных на непрерывном участке белковой последовательности, часто будут несмежные аминокислоты, являющиеся ключевыми точками контакта с паратопами антитела или В-клеточным рецептором. Эпитопы, распознаваемые антителами и В-клетками, могут представлять собой конформационные эпитопы с аминокислотами, составляющими общую область контакта на белке в трехмерном пространстве, и зависят от свойств третичной и четвертичной структуры белка. Эти остатки часто расположены в пространственно разделенных областях первичной аминокислотной последовательности.
При использовании здесь «мимотоп» представляет собой линейный или ограниченный пептид, имитирующий эпитоп антигена. Мимотоп может иметь первичную аминокислотную последовательность, способную индуцировать эффекторный Т-клеточный ответ, и/или трехмерную структуру, необходимую для связывания с В-клетками, приводящего к созреванию приобретенного иммунологического ответа у животного. Антитело к заданному эпитопу антигена будет распознавать мимотоп, имитирующий этот эпитоп.
Термин «специфически» в контексте связывания антитела относится к высокой авидности и/или высокой аффинности связывания антитела со специфическим антигеном, то есть полипептидом, или эпитопом. Во многих воплощениях специфический антиген представляет собой антиген (или фрагмент или субфракцию антигена), используемый для иммунизации животного-хозяина, от которого выделяют клетки, продуцирующие антитело. Специфическое связывание антитела с антигеном сильнее связывания того же антитела с другими антигенами. Антитела, специфически связывающиеся с полипептидом, могут быть способны связываться с другими полипептидами на слабом, но поддающемся выявлению уровне (например, 10% или менее от показанного связывания с интересующим полипептидом). Такое слабое связывание или фоновое связывание легко отличить от специфического связывания антитела с соответствующим полипептидом, например, используя подходящие контроли. Обычно аффинность связывания специфических антител с антигеном соответствует KD 10-7 М или менее, например, 10-8 М или менее (например, 10-9 М или менее, 10-10 или менее, 10-11 или менее, 10-12 или менее, или 10-13 или менее, и так далее).
При использовании здесь термин «антитело» относится к интактному иммуноглобулину, имеющему две легкие и две тяжелые цепи. Таким образом, отдельные выделенные антитело или фрагмент могут представлять собой поликлональное антитело, моноклональное антитело, синтетическое антитело, рекомбинантное антитело, химерное антитело, гетерохимерное антитело, канинизированное антитело, фелинизированное антитело, полностью собачье антитело, полностью кошачье антитело или полностью лошадиное антитело. Термин «антитело» предпочтительно относится к моноклональным антителам, их фрагментам и их эквивалентам по иммунологическому связыванию, которые могут связываться с белком IL-31 и его фрагментами. Термин антитело использован для обозначения как однородных молекул, так и смеси, такой как продукт сыворотки, содержащей множество различных молекул.
«Нативные антитела» и «нативные иммуноглобулины» обычно представляют собой гетеротетрамерные гликопротеины массой приблизительно 150000 Да, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как число дисульфидных связей между тяжелыми цепями иммуноглобулинов разных изотипов варьирует. В каждой тяжелой и легкой цепи также есть регулярно повторяющиеся внутрицепочечные дисульфидные связи. Каждая тяжелая цепь имеет на одном конце вариабельный домен (VH), за которым следуют несколько константных доменов. Каждая легкая цепь имеет вариабельный домен на одном конце (VL) и константный домен на ее другом конце; константный домен легкой цепи выровнен с первым константным доменом тяжелой цепи, а вариабельный домен легкой цепи выровнен с вариабельным доменом тяжелой цепи. Полагают, что определенные аминокислотные остатки формируют поверхности вариабельных доменов легкой и тяжелой цепи, обращенные друг к другу.
Термин «фрагмент антитела» относится к структуре, меньшей, чем интактное антитело, включая, без ограничения, выделенную отдельную цепь антитела, Fv-конструкцию, Fab-конструкцию, Fc-конструкцию, последовательность вариабельной области легкой цепи или гипервариабельного участка (CDR) и так далее.
Термин «вариабельная» область включает каркасные области и CDR (также известные как гипервариабельные участки) и относится к тому факту, что определенные положения вариабельных доменов разных антител значительно различаются по последовательности и задействованы в специфическим связывании каждого конкретного антитела с его конкретным антигеном. Тем не менее, распределение вариабельности в вариабельных доменах антител неравномерно. В вариабельных доменах как легкой цепи, так и тяжелой цепи она сосредоточена в трех сегментах, называемых гипервариабельными участками. Более консервативные части вариабельных доменов называют каркасными областями (FR). Каждый вариабельный домен нативной тяжелой и легкой цепи содержит несколько FR, большей частью имеющих β-складчатую конфигурацию и соединенных тремя гипервариабельными участками, которые образуют петли, соединяющие эти β-складчатые структуры и, в некоторых случаях, входящие в их состав. В каждой цепи FR удерживают гипервариабельные участки в непосредственной близости друг к другу, и они, вместе с гипервариабельными участками другой цепи, участвуют в образовании антигенсвязывающего сайта антитела (см. Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), pages 647-669). Константные домены не принимают непосредственного участия в связывании антитела с антигеном, но выполняют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности.
При использовании здесь термин «гипервариабельный участок» относится к аминокислотным остаткам антитела, обеспечивающим связывание с антигеном. Гипервариабельный участок содержит аминокислотные остатки «гипервариабельного участка» или «CDR» (Kabat et al. (1991) выше) и/или остатки «гипервариабельной петли» (Chothia and Lesk J. Mol. Biol. 196:901-917 (1987)). Остатки вариабельных доменов, не являющиеся остатками гипервариабельных участков, как определено здесь, представляют собой остатки «каркасных областей» или «FR».
Расщепление антител папаином приводит к получению двух идентичных антигенсвязывающих фрагментов, называемых «Fab-фрагментами», у каждого из которых есть один антигенсвязывающий сайт, и остаточного «Fc-фрагмента», название которого отражает его способность к быстрой кристаллизации. При обработке пепсином образуется F(ab')2-фрагмент, имеющий два антигенсвязывающих сайта и способность к перекрестному связыванию антигена.
«Fv» представляет собой минимальный фрагмент антитела, содержащий полный антигенраспознающий и антигенсвязывающий сайт. Эта область состоит из димера одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, связанных тесными нековалентными взаимодействиями. Именно в этой конфигурации три гипервариабельных участка каждого вариабельного домена взаимодействуют, определяя антигенсвязывающий сайт на поверхности димера VH-VL. Вместе эти шесть гипервариабельных участков обеспечивают специфичность связывания антитела с антигеном. Тем не менее, даже отдельный вариабельный домен (или половина Fv, содержащая только три гипервариабельных участка, специфичных в отношении антигена) обладает способностью к распознаванию и связыванию антигена, однако с меньшей аффинностью, чем целый связывающий сайт.
Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (СН1) тяжелой цепи. Fab'-фрагменты отличаются от Fab-фрагментов добавлением нескольких остатков на С-конце домена СН1 тяжелой цепи, включая один или более чем один цистеин шарнирной области антитела. Fab'-SH обозначает здесь Fab', в котором цистеиновый остаток (остатки) константного домена имеют свободную тиоловую группу. F(ab')2-фрагменты антител были исходно получены как пары Fab'-фрагментов с цистеинами шарнирной области между ними. Также известны другие химические сочетания фрагментов антител.
«Легкие цепи» антител (иммуноглобулинов) любого вида позвоночных могут быть отнесены к одному из двух четко различающихся типов, называемых каппа (κ) и лямбда (λ), исходя из аминокислотных последовательностей их константных доменов.
В зависимости от аминокислотных последовательностей константных доменов их тяжелых цепей, иммуноглобулины могут быть отнесены к различным классам. В настоящее время существует пять основных классов иммуноглобулинов, IgA, IgD, IgE, IgG, и IgM, и некоторые из них могут быть разделены далее на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA и IgA2 (как их принято обозначать у мышей и человека). Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, называют альфа, дельта, эпсилон, гамма и мю, соответственно. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны у многих видов. Распространенность отдельных изотипов и функциональная активность, связанная с этими константными доменами, видоспецифичны и нуждаются в экспериментальном определении.
«Моноклональное антитело», как определено здесь, представляет собой антитело, продуцируемое одним клоном клеток (конкретно, одним клоном таких клеток, как гибридомные клетки), и поэтому является одним чистым однородным типом антитела. Все моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к одному клону клеток, одной клетке и потомству этой клетки.
«Полностью собачье антитело», как определено здесь, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно клеточной линией СНО), и поэтому является одним чистым однородным типом антитела. Идентифицированные антитела из отдельных В-клеток иммунизированных млекопитающих, таких как собаки, получают в форме рекомбинантных белков IgG после определения последовательностей их вариабельных доменов. Перенос этих вариабельных доменов на собачьи константные домены (константную область тяжелой цепи и константную область легкой цепи каппа или лямбда) приводит к получению рекомбинантных полностью собачьих антител. Все полностью собачьи моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к одному клону клеток, одной клетке и потомству этой клетки.
«Полностью кошачье антитело», как определено здесь, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно клеточной линией СНО), и поэтому является одним чистым однородным типом антитела. Идентифицированные антитела из отдельных В-клеток иммунизированных млекопитающих, таких как собаки, получают в форме рекомбинантных белков IgG после определения последовательностей их вариабельных доменов. Перенос этих вариабельных доменов на кошачьи константные домены (константную тяжелую цепь и константную область легкой цепи каппа или лямбда) приводит к получению рекомбинантных полностью кошачьих антител. Все полностью кошачьи моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к одному клону клеток, одной клетке и потомству этой клетки.
«Полностью лошадиное антитело», как определено здесь, представляет собой моноклональное антитело, продуцируемое клоном клеток (обычно клеточной линией СНО), и поэтому является одним чистым однородным типом антитела. Идентифицированные антитела из отдельных В-клеток иммунизированных млекопитающих, таких как собаки, получают в форме рекомбинантных белков IgG после определения последовательностей их вариабельных доменов. Перенос этих вариабельных доменов на лошадиные константные домены (константную область тяжелой цепи и константную область легкой цепи каппа или лямбда) приводит к получению рекомбинантных полностью лошадиных антител. Все полностью лошадиные моноклональные антитела, полученные из одного и того же клона, идентичны и имеют одинаковую антигенную специфичность. Термин «моноклональный» относится к одному клону клеток, одной клетке и потомству этой клетки.
Здесь моноклональные антитела включают, в частности, «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител, имеющих происхождение от определенного вида, в то время как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям антител, имеющих происхождение от другого вида, а также фрагменты таких антител, при условии, что они демонстрируют желаемую биологическую активность. Обычно химерные антитела представляют собой антитела, гены легких и тяжелых цепей которых были сконструированы, обычно методами генной инженерии, из генов вариабельных и константных областей антител, принадлежащих к разным видам. Например, вариабельные сегменты генов мышиного моноклонального антитела могут быть объединены с собачьими константными сегментами. В одном воплощении химерного мышиного-собачьего IgG антигенсвязывающий сайт имеет происхождение от мыши, в то время как Fc-часть является собачьей.
«Канинизированные» формы антител, не являющихся собачьими (например, мышиных), представляют собой антитела, полученные методами генной инженерии, содержащие минимальную последовательность, имеющую происхождение от иммуноглобулина, не являющегося собачьим. Канинизированные антитела представляют собой последовательности собачьих иммуноглобулинов (антитело-реципиент), в которых остатки гипервариабельных участков реципиента заменены остатками гипервариабельных участков вида, не являющегося собакой (антитело-донор), такого как мышь, имеющих желаемую специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) последовательностей собачьих иммуноглобулинов заменены соответствующими остатками вида, не являющегося собакой. Кроме того, канинизированные антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре. Эти модификации проводят, чтобы дополнительно улучшить свойства антитела. В целом, канинизированное антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные участки соответствуют гипервариабельным участкам последовательности иммуноглобулина, не являющегося собачьим, а все или практически все FR представляют собой FR последовательности собачьего иммуноглобулина. Возможно, канинизированное антитело будет также содержать полную константную область (Fc) иммуноглобулина, обычно константную область (Fc) последовательности собачьего иммуноглобулина, или по меньшей мере ее часть. В одном воплощении видовой адаптации или канинизации мышиного IgG мышиные CDR переносят на собачьи каркасные области.
«Фелинизированные» формы антител, не являющихся кошачьими (например, мышиных), представляют собой антитела, полученные методами генной инженерии, содержащие минимальную последовательность, имеющую происхождение от иммуноглобулина, не являющегося кошачьим. Фелинизированные антитела представляют собой последовательности кошачьих иммуноглобулинов (антитело-реципиент), в которых остатки гипервариабельных участков реципиента заменены остатками гипервариабельных участков вида, не являющегося кошкой (антитело-донор), такого как мышь, имеющих желаемую специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) последовательностей кошачьих иммуноглобулинов заменены соответствующими остатками вида, не являющегося кошкой. Кроме того, фелинизированные антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре. Эти модификации проводят, чтобы дополнительно улучшить свойства антитела. В целом, фелинизированное антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные участки соответствуют гипервариабельным участкам последовательности иммуноглобулина, не являющегося кошачьим, а все или практически все FR представляют собой FR последовательности кошачьего иммуноглобулина. Возможно, фелинизированное антитело будет также содержать полную константную область (Fc) иммуноглобулина, обычно константную область (Fc) последовательности кошачьего иммуноглобулина, или по меньшей мере ее часть.
«Эквинизированные» формы антител, не являющихся лошадиными (например, мышиных), представляют собой антитела, полученные методами генной инженерии, содержащие минимальную последовательность, имеющую происхождение от иммуноглобулина, не являющегося лошадиным. Эквинизированные антитела представляют собой последовательности лошадиных иммуноглобулинов (антитело-реципиент), в которых остатки гипервариабельных участков реципиента заменены остатками гипервариабельных участков вида, не являющегося лошадью (антитело-донор), такого как мышь, имеющих желаемую специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) последовательностей лошадиных иммуноглобулинов заменены соответствующими остатками вида, не являющегося лошадью. Кроме того, эквинизированные антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре. Эти модификации проводят, чтобы дополнительно улучшить свойства антитела. В целом, эквинизированное антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные участки соответствуют гипервариабельным участкам последовательности иммуноглобулина, не являющегося лошадиным, а все или практически все FR представляют собой FR последовательности лошадиного иммуноглобулина. Возможно, эквинизированное антитело будет также содержать полную константную область (Fc) иммуноглобулина, обычно константную область (Fc) последовательности лошадиного иммуноглобулина, или по меньшей мере ее часть.
«Полностью собачьи» антитела представляют собой антитела, полученные методами генной инженерии, не содержащие последовательностей, имеющих происхождение от иммуноглобулинов, не являющихся собачьими. Полностью собачьи антитела представляют собой последовательности собачьих иммуноглобулинов (антитело-реципиент), в которых остатки гипервариабельных участков имеют происхождение от встречающегося в природе собачьего антитела (антитело-донор), имеющего желаемую специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) последовательностей собачьих иммуноглобулинов заменены соответствующими остатками вида, не являющегося собакой. Кроме того, полностью собачьи антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре, например, включая, без ограничения, изменения в CDR для модификации аффинности. Эти модификации проводят, чтобы дополнительно улучшить свойства антитела. В целом, полностью собачье антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные участки соответствуют гипервариабельным участкам последовательности собачьего иммуноглобулина и все или практически все FR представляют собой FR последовательности собачьего иммуноглобулина. Возможно, полностью собачье антитело будет также содержать полную константную область (Fc) иммуноглобулина, обычно константную область (Fc) последовательности собачьего иммуноглобулина, или по меньшей мере ее часть.
«Полностью кошачьи» антитела представляют собой антитела, полученные методами генной инженерии, не содержащие последовательностей, имеющих происхождение от иммуноглобулинов, не являющихся кошачьими. Полностью кошачьи антитела представляют собой последовательности кошачьих иммуноглобулинов (антитело-реципиент), в которых остатки гипервариабельных участков имеют происхождение от встречающегося в природе кошачьего антитела (антитело-донор), имеющего желаемую специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) последовательностей кошачьих иммуноглобулинов заменены соответствующими остатками вида, не являющегося кошкой. Кроме того, полностью кошачьи антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре, например, включая, без ограничения, изменения в CDR для модификации аффинности. Эти модификации проводят, чтобы дополнительно улучшить свойства антитела. В целом, полностью кошачье антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные участки соответствуют гипервариабельным участкам последовательности кошачьего иммуноглобулина и все или практически все FR представляют собой FR последовательности кошачьего иммуноглобулина. Возможно, полностью кошачье антитело будет также содержать полную константную область (Fc) иммуноглобулина, обычно константную область (Fc) последовательности кошачьего иммуноглобулина, или по меньшей мере ее часть.
«Полностью лошадиные» антитела представляют собой антитела, полученные методами генной инженерии, не содержащие последовательностей, имеющих происхождение от иммуноглобулинов, не являющихся лошадиными. Полностью лошадиные антитела представляют собой последовательности лошадиных иммуноглобулинов (антитело-реципиент), в которых остатки гипервариабельных участков имеют происхождение от встречающегося в природе лошадиного антитела (антитело-донор), имеющего желаемую специфичность, аффинность и активность. В некоторых случаях, остатки каркасных областей (FR) последовательностей лошадиных иммуноглобулинов заменены соответствующими остатками вида, не являющегося лошадью. Кроме того, полностью лошадиные антитела могут содержать остатки, которых нет в антителе-реципиенте или в антителе-доноре, например, включая, без ограничения, изменения в CDR для модификации аффинности. Эти модификации проводят, чтобы дополнительно улучшить свойства антитела. В целом, полностью лошадиное антитело будет содержать практически все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или практически все гипервариабельные участки соответствуют гипервариабельным участкам последовательности лошадиного иммуноглобулина и все или практически все FR представляют собой FR последовательности лошадиного иммуноглобулина. Возможно, полностью лошадиное антитело будет также содержать полную константную область (Fc) иммуноглобулина, обычно константную область (Fc) последовательности лошадиного иммуноглобулина, или по меньшей мере ее часть.
Термин «гетерохимерный», как определено здесь, относится к антителу, в котором одна из цепей антитела (тяжелая или легкая) канинизирована, фелинизирована или эквинизирована, в то время как другая является химерной. В одном воплощении фелинизированная вариабельная область тяжелой цепи (где все CDR мышиные, а все FR кошачьи) спарена с химерной вариабельной областью легкой цепью (где все CDR мышиные и все FR мышиные). В данном воплощении вариабельная область тяжелой цепи и вариабельная область легкой цепи слиты с кошачьей константной областью.
«Вариант» антитела против IL-31 относится здесь к молекуле, отличающейся по аминокислотной последовательности от аминокислотной последовательности «исходного» антитела против IL-31 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка в последовательности исходного антитела и сохраняющей по меньшей мере одну желаемую активность исходного антитела против IL-31. Желаемые активности могут включать способность к специфическому связыванию с антигеном, способность к уменьшению, ингибированию или нейтрализации активности IL-31 у животного и способность к ингибированию pSTAT-сигналов, опосредованных IL-31, в клеточном анализе. В одном воплощении вариант содержит одну или более чем одну аминокислотную замену в одной или более чем одной гипервариабельной и/или каркасной области исходного антитела. Например, вариант может содержать по меньшей мере одну, например, от приблизительно одной до приблизительно десяти и предпочтительно от приблизительно двух до приблизительно пяти замен в одной или более чем одной гипервариабельной и/или каркасной области исходного антитела. Обычно вариант будет иметь аминокислотную последовательность, по меньшей мере на 50% идентичную, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95% идентичную аминокислотным последовательностям вариабельных доменов тяжелой или легкой цепи исходного антитела. Идентичность или гомология применительно к данной последовательности определены здесь как процент аминокислотных остатков в последовательности-кандидате, идентичных остаткам исходного антитела, после выравнивания последовательностей и, по необходимости, введения разрывов для достижения максимального процента идентичности последовательностей. N-концевые, С-концевые или внутренние удлинения, делеции или вставки в последовательности антитела не следует рассматривать как влияющие на идентичность или гомологию последовательностей. Вариант сохраняет способность к связыванию с IL-31 и предпочтительно имеет желаемые активности, превосходящие соответствующие активности исходного антитела. Например, вариант может иметь более высокую аффинность связывания, усиленную способность к уменьшению, ингибированию или нейтрализации активности IL-31 у животного и/или усиленную способность к ингибированию pSTAT-сигналов, опосредованных IL-31, в клеточном анализе.
«Вариант» нуклеиновой кислоты относится здесь к молекуле, последовательность которой отличается от «исходной» нуклеиновой кислоты. Расхождение полинуклеотидных последовательностей может быть результатом мутационных изменений, таких как делеции, замены или добавления одного или более чем одного нуклеотида. Каждое из этих изменений может присутствовать в заданной последовательности само по себе или в комбинации, один или более чем один раз.
Здесь «исходное» антитело представляет собой антитело, кодируемое аминокислотной последовательностью, используемой для получения варианта. В одном воплощении исходное антитело имеет собачью каркасную область и, в случае ее (их) присутствия, константную область (области) собачьего антитела. Например, исходное антитело может представлять собой канинизированное или собачье антитело. В качестве другого примера, исходное антитело может представлять собой фелинизированное или кошачье антитело. В качестве еще одного примера, исходное антитело может представлять собой эквинизированное или лошадиное антитело. В качестве еще одного примера, исходное антитело представляет собой мышиное моноклональное антитело.
Термины «антигенсвязывающая область», «антигенсвязывающий фрагмент» и тому подобные, используемые в данном описании и формуле изобретения, относятся к той части молекулы антитела, которая содержит аминокислотные остатки, взаимодействующие с антигеном и придающие антителу его специфичность и аффинность в отношении антигена. Область связывания антитела содержит «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков. Антигенсвязывающий фрагмент антитела по настоящему изобретению может альтернативно быть назван здесь, например, пептидом или полипептидом, специфичным в отношении IL-31, или пептидом или полипептидом против IL-31.
Термин «выделенный» означает, что вещество (например, антитело или нуклеиновая кислота) отделено и/или выделено из компонента его естественного окружения. Примесные компоненты его естественного окружения являются веществами, которые будут препятствовать диагностическому или терапевтическому применению вещества, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. Применительно к нуклеиновой кислоте выделенная нуклеиновая кислота может включать нуклеиновую кислоту, отделенную от 5'-3'-последовательностей, с которыми она обычно связана в хромосоме. В предпочтительных воплощениях вещество будет очищено до более чем 95% по массе вещества и наиболее предпочтительно до более чем 99% по массе. Выделенное вещество включает вещество in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент естественного окружения вещества будет отсутствовать. Однако обычно выделенное вещество будет получено посредством по меньшей мере одной стадии очистки.
При использовании здесь слово «метка» относится к выявляемым соединению или композиции, конъюгированным прямым или непрямым образом с антителом или нуклеиновой кислотой. Сама метка может быть выявляемой сама по себе (например, радио изотопные метки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать выявляемое химическое изменение соединения-субстрата или композиции-субстрата.
Термины «нуклеиновая кислота», «полинуклеотид», «молекула нуклеиновой кислоты» и тому подобные могут быть использованы здесь взаимозаменяемо для обозначения серии нуклеотидных оснований (называемых также «нуклеотидами») в ДНК и РНК. Нуклеиновая кислота может содержать дезоксирибонуклеотиды, рибонуклеотиды и/или их аналоги. Термин «нуклеиновая кислота» включает, например, одноцепочечные и двуцепочечные молекулы. Нуклеиновая кислота может представлять собой, например, ген или фрагмент гена, экзоны, интроны, молекулу ДНК (например, кДНК), молекулу РНК (например, мРНК), рекомбинантные нуклеиновые кислоты, плазмиды и другие векторы, праймеры и зонды. Включены как 5'-3'-полинуклеотиды (смысловые), так и 3'-5'-полинуклеотиды (антисмысловые).
«Субъект» или «пациент» относится к животному, нуждающемуся в лечении, которое может быть проведено молекулами по изобретению. Животные, которых можно лечить согласно изобретению, включают позвоночных, при этом особенно предпочтительными примерами являются млекопитающие, такие как собаки, кошки и лошади.
«Терапевтически эффективное количество» (или «эффективное количество») относится к количеству активного ингредиента, например, агента по изобретению, достаточному для получения полезных или желаемых результатов при введении субъекту или пациенту. Эффективное количество может быть введено с применением одного или более чем одного введения, нанесения или дозы. Терапевтически эффективное количество композиции по изобретению может быть легко определено специалистом в данной области. В контексте данного изобретения «терапевтически эффективное количество» представляет собой количество, приводящее к объективно измеряемому изменению одного или более чем одного параметра, связанного с лечением состояния, сопровождающегося зудом, или аллергического состояния, включая клиническое улучшение симптомов. Разумеется, терапевтически эффективное количество будет варьировать в зависимости от конкретного субъекта и состояния, в отношении которого проводят лечение, массы тела и возраста субъекта, тяжести болезненного состояния, конкретного выбранного соединения, применяемой схемы введения, времени введения, способа введения и тому подобного, и все это может быть легко определено специалистом в данной области.
При использовании здесь термин «терапевтический» охватывает весь спектр вариантов лечения заболевания или расстройства. «Терапевтический» агент по изобретению может действовать профилактическим или превентивным образом, в том числе включающим методики, разработанные для воздействия на животных, которые могут быть идентифицированы как подверженные риску (фармакогенетика), или облегчающим или излечивающим образом или может действовать, уменьшая скорость или степень прогрессирования по меньшей мере одного симптома заболевания или расстройства, по поводу которых проводят лечение.
«Лечение», «лечить» и тому подобное относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. Животные, нуждающиеся в лечении, включают как тех, у которых уже есть расстройство, так и тех, у которых следует предотвратить расстройство. Термин «лечение» заболевания или расстройства включает предотвращение или защиту от заболевания или расстройства (то есть обеспечение отсутствия развития клинических симптомов), подавление заболевания или расстройства (то есть остановку или сдерживание развития клинических симптомов) и/или ослабление заболевания или расстройства (то есть обеспечение обратного развития клинических симптомов). Следует понимать, что разграничение «предотвращения» и «сдерживания» заболевания или расстройства не всегда возможно, поскольку последние индуцирующие событие или события могут быть неизвестными или латентными. Соответственно, термин профилактика следует понимать как тип «лечения», охватывающий как «предотвращение», так и «сдерживание». Таким образом, термин «лечение» включает «профилактику».
Термин «аллергическое состояние» определен здесь как расстройство или заболевание, вызванное взаимодействием иммунной системы с веществом, являющимся для организма чужеродным. Это чужеродное вещество называют «аллергеном». Распространенные аллергены включают аэроаллергены, такие как пыльца, пыль, плесень, белки пылевых клещей, слюну, попадающую в организм при укусах насекомых, и так далее. Примеры аллергических состояний включают, без ограничения, следующее: аллергический дерматит, летнюю экзему, крапивницу, запал лошадей, воспалительное заболевание дыхательных путей, рецидивирующую обструкцию дыхательных путей, гиперреактивность дыхательных путей, хроническую обструктивную болезнь легких и воспалительные процессы, являющиеся результатами аутоиммунитета, такие как синдром раздраженного кишечника (СРК).
Термин «состояние, сопровождающееся зудом», определен здесь как заболевание или расстройство, характеризующиеся интенсивным ощущением зуда, побуждающим тереть или расчесывать кожу, чтобы получить облегчение. Примеры состояний, сопровождающихся зудом, включают, без ограничения, следующее: атопический дерматит, аллергический дерматит, экзему, псориаз, склеродермию и зуд.
При использовании здесь термины «клетка», «клеточная линия» и «клеточная культура» могут быть использованы взаимозаменяемо. Все эти термины также включают их потомство, представляющее собой все и любые последующие поколения. Следует понимать, что все потомство может не быть идентичным из-за преднамеренных или непреднамеренных мутаций. В контексте экспрессии гетерологичной нуклеиновокислотной последовательности «клетка-хозяин» относится к прокариотической или эукариотической клетке (например, бактериальным клеткам, дрожжевым клеткам, клеткам млекопитающих и клеткам насекомых), независимо от их нахождения in vitro или in vivo. Например, клетки-хозяева могут находиться в трансгенном животном. Клетка-хозяин может быть использована в качестве реципиента векторов и может включать любой организм, поддающийся трансформации, способный реплицировать вектор и/или экспрессировать гетерологичную нуклеиновую кислоту, кодируемую вектором.
Термин «композиция» предназначен для обозначения комбинации активного агента и другого соединения или другой композиции, которые могут быть инертными (например, метка) или активными, такими как адъювант.
Как определено здесь, фармацевтически приемлемые носители, подходящие для использования в изобретении, хорошо известны специалистам в данной области. Такие носители включают, без ограничения, воду, физиологический раствор, забуференный физиологический раствор, фосфатный буфер, спиртовые/водные растворы, эмульсии или суспензии. Согласно общепринятым методикам, могут быть добавлены другие часто используемые разбавители, адъюванты и эксципиенты. Такие носители могут включать этанол, полиолы и их подходящие смеси, растительные масла и органические сложные эфиры для инъекций. Могут также быть использованы буферы и агенты для коррекции рН. Буферы включают, без ограничения, соли, полученные из органической кислоты или основания. Типичные буферы включают, без ограничения, соли органических кислот, такие как соли лимонной кислоты, например, цитраты, аскорбиновой кислоты, глюконовой кислоты, гистидина-HCl, угольной кислоты, винной кислоты, янтарной кислоты, уксусной кислоты или фталевой кислоты, трис, гидрохлорид трометамина или фосфатные буферы. Носители для парентерального введения могут включать раствор хлорида натрия, декстрозу Рингера, декстрозу, трегалозу, сахарозу и хлорид натрия, лактат Рингера или нелетучие масла. Носители для внутривенного введения могут включать восполнители жидкости и питательных веществ, такие как восполнители на основе декстрозы Рингера и тому подобное. В фармацевтических носителях могут также присутствовать консерванты и другие добавки, такие как, например, антимикробные агенты, антиоксиданты, хелатирующие агенты (например, ЭДТА), инертные газы и тому подобное. Настоящее изобретение не ограничено выбором носителя. Изготовление этих фармацевтически приемлемых композиций из описанных выше компонентов, имеющих подходящие рН, изотоничность, стабильность и другие общепринятые характеристики, входит в компетенцию специалиста в данной области. См., например, такие публикации, как Remington: The Science and Practice of Pharmacy, 20th ed, Lippincott Williams & Wilkins, publ., 2000, и The Handbook of Pharmaceutical Excipients, 4.sup.th edit., eds. R. C. Rowe et al., APhA Publications, 2003.
Термин «консервативная аминокислотная замена» указывает на любую аминокислотную замену заданного аминокислотного остатка, где заменяющий остаток химически настолько сходен с заданным остатком, что замена не приводит к существенному снижению функции (например, ферментативной активности) полипептида. Консервативные аминокислотные замены хорошо известны в данной области, и их примеры описаны, например, в патентах США №№6,790,639, 6,774,107, 6,194,167 или 5,350,576. В предпочтительном воплощении консервативная аминокислотная замена будет представлять собой любую замену, происходящую в пределах одной из следующих шести групп:
• 1. малые алифатические практически неполярные остатки: Ala, Gly, Pro, Ser и Thr;
• 2. большие алифатические неполярные остатки: Ile, Leu и Val; Met;
• 3. полярные отрицательно заряженные остатки и их амиды: Asp и Glu;
• 4. амиды полярных отрицательно заряженных остатков: Asn и Gln; His;
• 5. полярные положительно заряженные остатки: Arg и Lys; His; и
• 6. большие ароматические остатки: Trp и Tyr; Phe.
В предпочтительном воплощении консервативная аминокислотная замена будет представлять собой любую из следующих замен, перечисленных как пары «нативный остаток (консервативная замена)»: Ala (Ser); Arg (Lys); Asn (Gln; His); Asp (Glu); Gln (Asn); Glu (Asp); Gly (Pro); His (Asn; Gln); Ile (Leu; Val); Leu (Ile; Val); Lys (Arg; Gln; Glu); Met (Leu; Ile); Phe (Met; Leu; Tyr); Ser (Thr); Thr (Ser); Trp (Tyr); Tyr (Trp; Phe); и Val (Ile; Leu).
Точно так же, как полипептид может содержать консервативную аминокислотную замену (замены), соответствующий ему полинуклеотид может содержать консервативную замену (замены) кодона. Замену кодона считают консервативной, если при экспрессии она приводит к консервативной аминокислотной замене, как описано выше. В полинуклеотидах по настоящему изобретению может также быть полезна вырожденная замена кодона, которая не приводит к аминокислотной замене. Таким образом, например, возможна мутация полинуклеотида, кодирующего выбранный полипептид, полезный в воплощении настоящего изобретения, посредством вырожденной замены кодона для приближения частоты использования кодонов к той, которую демонстрирует экспрессионная клетка-хозяин, подлежащая трансформации указанным полинуклеотидом, или для иного улучшения его экспрессии.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следует понимать, что данное изобретение не ограничено конкретными методами, протоколами, реагентами и так далее, описанными здесь, и может, как таковое, варьировать. Термины, использованные здесь, предназначены лишь для описания конкретных воплощений и не ограничивают объем настоящего изобретения, определенный исключительно формулой изобретения.
Если не определено иное, научные и технические термины, использованные в связи с антителами, описанными здесь, имеют то значение, в котором их обычно понимают специалисты в данной области. Кроме того, если контекст не требует иного, формы единственного числа включают множественное число, и формы множественного числа включают единственное число. В целом, номенклатура, использованная в связи с культивированием клеток и тканей, молекулярной биологией, а также химией и гибридизацией белков и олиго- или полинуклеотидов, и соответствующие методики, описанные здесь, хорошо известны и широко применяются в данной области.
Для рекомбинантных ДНК, синтеза олигонуклеотидов, культивирования тканей и трансфекции применяют стандартные методики (например, электропорацию, липофекцию). Ферментативные реакции и методики очистки осуществляют в соответствии с описаниями изготовителя или так, как их обычно осуществляют в данной области или как описано здесь. Описанные выше методики обычно осуществляют, следуя общепринятым методам, хорошо известным в данной области и описанным в различных общих и более специализированных источниках, цитируемых и обсуждаемых в настоящем описании. См., например, Sambrook et al. MOLECULAR CLONING: LAB. MANUAL (3rd ed., Cold Spring Harbor Lab. Press, Cold Spring Harbor, N.Y., 2001) и Ausubel et al. Current Protocols in Molecular Biology (New York: Greene Publishing Association/Wiley Interscience), 1993. Номенклатура, использованная в связи с аналитической химией, синтетической органической химией и медицинской и фармацевтической химией, и соответствующие лабораторные методики, описанные здесь, хорошо известны и широко применяются в данной области. Для химического синтеза, химического анализа, изготовления фармацевтических препаратов, композиций, их доставки и лечения пациентов применяют стандартные методики.
За исключением рабочих примеров или иных случаев, где это указано, все числа, выражающие количество ингредиентов или условия проведения реакций, используемые здесь, следует всегда понимать как модифицированные термином «приблизительно».
Все указанные патенты и иные публикации прямо включены сюда посредством ссылки с целью описания и раскрытия, например, методов, описанных в таких публикациях, которые могут быть применены в связи с настоящим изобретением. Эти публикации приведены исключительно в силу их раскрытия до даты подачи настоящей заявки.
Согласно настоящему изобретению предложены рекомбинантные моноклональные антитела и пептиды и их применение в клинических и научных методиках, включая диагностические методики.
Появление молекулярно-биологических методов и рекомбинантных технологий привело к возможности рекомбинантного получения антител и антителоподобных молекул и, таким образом, создания генных последовательностей, кодирующих определенные аминокислотные последовательности, присутствующие в полипептидной структуре антител. Такие антитела могут быть получены клонированием генных последовательностей, кодирующих полипептидные цепи указанных антител, или прямым синтезом указанных полипептидных цепей и сборкой синтезированных цепей с образованием активных тетрамерных (H2L2) структур, обладающих аффинностью в отношении определенных эпитопов и антигенных детерминант. Это позволило быстро получать антитела, имеющие последовательности, характерные для нейтрализующих антител от различных видов и источников.
Независимо от источника антител, или особенностей их рекомбинантного конструирования, или особенностей их синтеза, in vitro или in vivo, использования трансгенных животных, больших клеточных культур лабораторного или коммерческого размера, использования трансгенных растений или применения прямого химического синтеза без использования живых организмов на любой стадии процесса, все антитела имеют сходную общую 3-мерную структуру. Эту структуру часто обозначают как H2L2, и это указывает на тот факт, что антитела обычно содержат две легкие (L) аминокислотные цепи и 2 тяжелые (Н) аминокислотные цепи. Обе цепи имеют области, способные взаимодействовать со структурно комплементарной антигенной мишенью. Области, взаимодействующие с мишенью, называют «вариабельными» или «V»-областями, и они характеризуются различиями в аминокислотной последовательности антител с разной антигенной специфичностью. Вариабельные области Н- или L-цепей содержат аминокислотные последовательности, способные специфически связываться с антигенными мишенями.
Как указано выше, термины «антигенсвязывающая область», «антигенсвязывающий фрагмент» и тому подобные, используемые в данном описании и формуле изобретения, относятся к той части молекулы антитела, которая содержит аминокислотные остатки, взаимодействующие с антигеном и придающие антителу его специфичность и аффинность в отношении антигена. Область связывания антитела содержит «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков. Антигенсвязывающий фрагмент антитела по настоящему изобретению может быть названа здесь, например, пептидом или полипептидом, специфичным в отношении IL-31, или пептидом или полипептидом против IL-31.
В пределах вариабельных областей Н- или L-цепей, обеспечивающих антигенсвязывающие области, есть последовательности меньших размеров, называемые «гипервариабельными» ввиду их крайней вариабельности у антител разной специфичности. Такие гипервариабельные области также называют «гипервариабельными участками» или «CDR»-участками. Эти CDR-участки обеспечивают основную специфичность антитела в отношении структуры определенной антигенной детерминанты.
CDR представляют собой несмежные участки из аминокислот в пределах вариабельных областей, но обнаружено, что, независимо от видовой принадлежности, эти критически важные аминокислотные последовательности расположены в вариабельных областях тяжелой и легкой цепи сходным образом. Вариабельные области тяжелых и легких цепей всех антител имеют три CDR-участка, несмежных друг с другом.
У всех видов млекопитающих пептиды антител содержат константные (то есть высоко консервативные) и вариабельные области, и, в пределах вариабельных областей, есть CDR и так называемые «каркасные области», образованные аминокислотными последовательностями, расположенными в пределах вариабельной области тяжелой или легкой цепи, но за пределами CDR.
Что касается антигенной детерминанты, распознаваемой CDR-участками антитела, ее называют также «эпитопом». Иными словами, эпитоп относится к той части любой молекулы, которая способна быть распознанной и связанной антителом (область связывания соответствующего антитела может быть названа паратопом).
«Антиген» представляет собой молекулу или часть молекулы, способные быть связанными антителом, которые дополнительно способны индуцировать у животного выработку антитела, способного к связыванию с эпитопом данного антигена. Антиген может иметь один или более чем один эпитоп. Предполагается, что специфические взаимодействие, упомянутое выше, указывает на то, что антиген будет высокоселективным образом взаимодействовать с соответствующим ему антителом, а не с множеством других антител, которые могут быть индуцированы другими антигенами.
Предполагается, что антитела по настоящему изобретению включают как интактные молекулы иммуноглобулинов, так и их части, фрагменты, пептиды и производные, такие как, например, Fab, Fab', F(ab')2, Fv, Fse, CDR-участки, паратопы или любая часть (например, полипептид) или пептидная последовательность антитела, способная связываться с антигеном или эпитопом. Антитело считают «способным связываться» или «способным к связыванию» с молекулой, если оно способно специфически взаимодействовать с молекулой и, посредством этого, обеспечивать связывание молекулы с антителом.
Антитела по настоящему изобретению также включают химерные антитела, гетерохимерные антитела, канинизированные антитела, фелинизированные антитела или эквинизированные антитела, а также их фрагменты, части, пептиды или производные, полученные любой известной методикой, такой как, без ограничения, ферментативное расщепление, пептидный синтез или рекомбинантные методики. Такие антитела по настоящему изобретению способны специфически связываться по меньшей мере с одним из собачьего IL-31 или кошачьего IL-31. Фрагменты или части антител могут быть лишены Fc-фрагмента интактного антитела и подвержены более быстрому клиренсу из циркуляции и могут иметь меньшее неспецифическое связывание с тканями по сравнению с интактным антителом. Примерами фрагментов антител могут быть фрагменты, полученные из интактных антител с применением методов, хорошо известных в данной области, например, протеолитическим расщеплением такими ферментами, как папаин (с получением Fab-фрагментов) или пепсин (с получением Р(ab')2-фрагментов). См., например, Wahl et al., 24 J. Nucl. Med. 316-25 (1983). Части антител могут быть получены любым из указанных выше методов или могут быть получены экспрессией части рекомбинантной молекулы. Например, CDR-участок (CDR-участки) рекомбинантного антитела может быть выделен и субклонирован в подходящий вектор экспрессии. См., например, патент США №6,680,053.
Термины «антигенсвязывающая область», «антигенсвязывающий фрагмент» и тому подобные, используемые в данном описании и формуле изобретения, относятся к той части молекулы антитела, которая содержит аминокислотные остатки, взаимодействующие с антигеном и придающие антителу его специфичность и аффинность в отношении антигена. Область связывания антитела содержит «каркасные» аминокислотные остатки, необходимые для поддержания правильной конформации антигенсвязывающих остатков. Антигенсвязывающий фрагмент антитела по настоящему изобретению может альтернативно быть назван здесь, например, пептидом или полипептидом, специфичным в отношении IL-31, или пептидом или полипептидом против IL-31.
Нуклеотидные и аминокислотные последовательности клонов 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 и 06А09
В некоторых воплощениях согласно настоящему изобретению предложены новые моноклональные антитела, специфически связывающиеся по меньшей мере с одним из собачьего IL-31, кошачьего IL-31 или лошадиного IL-31. В одном воплощении моноклональное антитело по изобретению связывается с собачьим IL-31, кошачьим IL-31 или лошадиным IL-31 и предотвращает их связывание со своими корецепторными комплексами, содержащими рецептор A IL-31 (IL-31Ra) и рецептор, специфичный в отношении онкостатина М (OsmR или IL-31Rb), и активацию ими этих комплексов. Моноклональные антитела по настоящему изобретению обозначены здесь как «15Н05», «ZIL1», «ZIL8», «ZIL9», «ZIL11», «ZIL69», «ZIL94», «ZIL154», «ZIL159», «ZIL171», «04Н07» и «06А09», что указывает на номера, присвоенные их клонам. Здесь «15Н05», «ZIL1», «ZIL8», «ZIL9», «ZIL11», «ZIL69», «ZIL94», «ZIL154», «ZIL159», «ZIL171», «04Н07» и «06А09» также относятся к части моноклонального антитела, паратопу или CDR, специфически связывающимся с эпитопами IL-31, обозначенными как 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 и 06А09 ввиду их способности к связыванию с антителами 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 и 06А09, соответственно. Эти же названия могут быть использованы для обозначения ряда рекомбинантных, химерных, гетерохимерных, канинизированных, фелинизированных, эквинизированных, полностью собачьих, полностью кошачьих и/или полностью лошадиных форм 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 и 06А09, описанных здесь. В некоторых воплощениях 15Н05 может быть альтернативно названо здесь 1505, по меньшей мере ввиду того, что они имеют одинаковые CDR.
В одном воплощении согласно настоящему изобретению предложены моноклональное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с областью белка IL-31 млекопитающего, вовлеченной во взаимодействие белка IL-31 с его корецептором, где на связывание указанного антитела с указанной областью влияют мутации в области связывания эпитопа 15Н05, выбранной из по меньшей мере одного из следующего: (а) области приблизительно между аминокислотными остатками 124 и 135 последовательности кошачьего IL-31, представленной в SEQ ID NO: 157 (Feline_IL31_wildtype); (b) области приблизительно между аминокислотными остатками 124 и 135 последовательности собачьего IL-31, представленной в SEQ ID NO: 155 (Canine_IL31); и (с) области приблизительно между аминокислотными остатками 118 и 129 последовательности лошадиного IL-31, представленной в SEQ ID NO: 165 (Equine_IL31). В одном воплощении мутации в области связывания эпитопа 15Н05 выбраны из по меньшей мере одного из следующего: (а) мутанта, где положения 126 и 128 SEQ ID NO: 157 заменены на аланин; (b) мутанта, где положения 126 и 128 SEQ ID NO: 155 заменены на аланин; и (с) мутанта, где положения 120 и 122 SEQ ID NO: 165 заменены на аланин.
В одном конкретном воплощении антитело по настоящему изобретению связывается с областью связывания эпитопа 15Н05, описанной выше. То есть, в одном воплощении согласно настоящему изобретению предложены моноклональное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с областью белка IL-31 млекопитающего, вовлеченной во взаимодействие белка IL-31 с его корецептором, где область связывания выбрана из по меньшей мере одного из следующего: (а) области приблизительно между аминокислотными остатками 124 и 135 последовательности кошачьего IL-31, представленной в SEQ ID NO: 157 (Feline_IL31_wildtype); (b) области приблизительно между аминокислотными остатками 124 и 135 последовательности собачьего IL-31, представленной в SEQ ID NO: 155 (Canine_IL31); и (с) области приблизительно между аминокислотными остатками 118 и 129 последовательности лошадиного IL-31, представленной в SEQ ID NO: 165 (Equine_IL31).
В одном воплощении IL-31 млекопитающего, с которым специфически связываются антитело или его антигенсвязывающий фрагмент, представляет собой кошачий IL-31, где антитело связывается с областью приблизительно между аминокислотными остатками 125 и 134 последовательности кошачьего IL-31, представленной в SEQ ID NO: 157 (Feline_IL3_wildtype). В некоторых воплощениях антитело, связывающееся с кошачьим IL-31, содержит цепь VL, содержащую изменения в каркасной области 2 (FW2), выбранные из следующего: аспарагина вместо лизина в положении 42, изолейцина вместо валина в положении 43, валина вместо лейцина в положении 46, аспарагина вместо лизина в положении 49 и их комбинаций, где положения соответствуют нумерации SEQ ID NO: 127 (FEL_15H05V_L1).
В одном воплощении моноклональное антитело или его антигенсвязывающий фрагмент содержат следующие комбинации последовательностей гипервариабельных участков (CDR):
1) антитело 15Н05: 


2) вариант (1), отличающийся от исходного антитела 15Н05 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В одном воплощении антитело 15Н05, описанное выше, содержит по меньшей мере одну из следующих вариабельных областей тяжелых и/или легких цепей:
a) вариабельную область легкой цепи, содержащую FEL_15H05_VL1_FW2: 
b) вариабельную область тяжелой цепи, содержащую FEL_15Н05_VH1: 
В другом воплощении моноклональное антитело или его антигенсвязывающий фрагмент содержат следующие комбинации последовательностей гипервариабельных участков (CDR):
1) антитело ZIL1: CDR1 вариабельной области тяжелой цепи (VH) (VH-CDR1) 
2) антитело ZIL8: 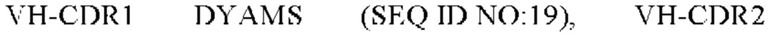

3) антитело ZIL9: 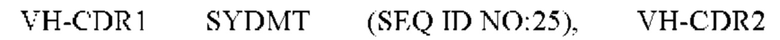

4) антитело ZIL11: 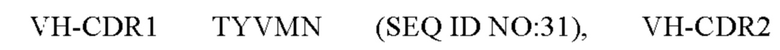

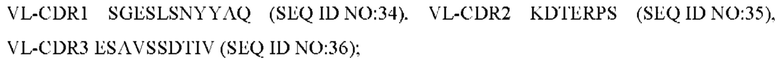
5) антитело ZIL69: 

6) антитело ZIL94: 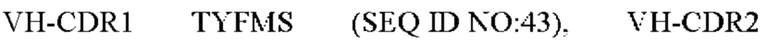
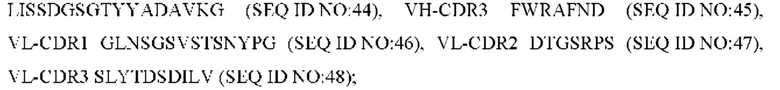
7) антитело ZIL15 4: 

8) антитело ZIL159: 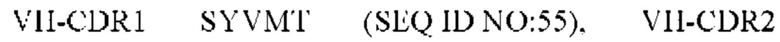

9) антитело ZIL171: 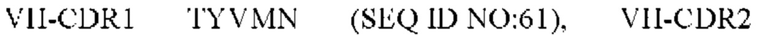

10) антитело 04Н07: 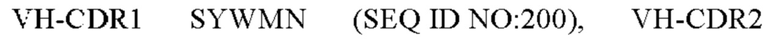

11) антитело 06А09: 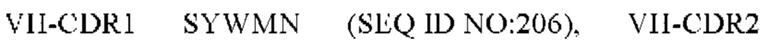

12) вариант (1)-(11), отличающийся от соответствующего исходного антитела ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В некоторых воплощениях настоящего изобретения:
1) антитело ZIL1 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL_1VL:
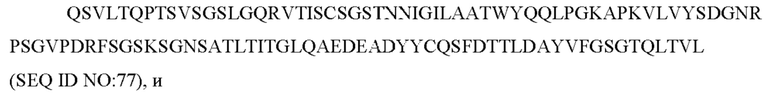
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL1_VH:
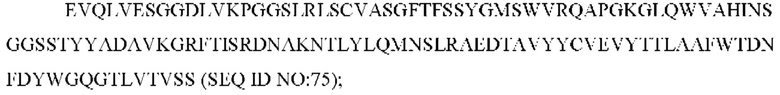
2) антитело ZIL8 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL8_VL:
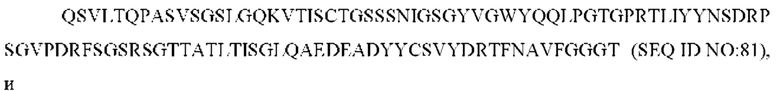
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL8-VH:
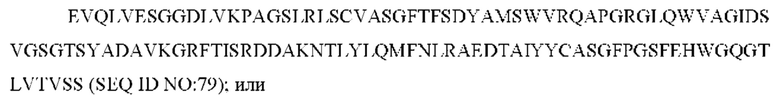
антитело ZIL8 содержит по меньшей мере одно из следующего:
c) вариабельную область легкой цепи, содержащую ZTS_5864_VL:

d) вариабельную область тяжелой цепи, содержащую ZTS 5864 VH:
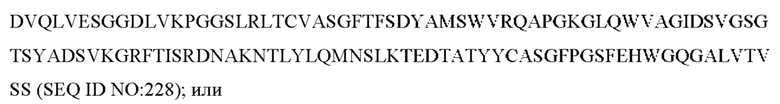
антитело ZIL8 содержит по меньшей мере одно из следующего:
e) вариабельную область легкой цепи, содержащую ZTS_5865_VL:
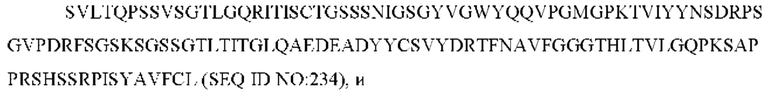
f) вариабельную область тяжелой цепи, содержащую ZTS_5865_VH:
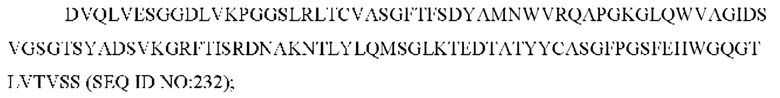
3) антитело ZIL9 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL9_VL:

b) вариабельную область тяжелой цепи, содержащую CAN-ZIL9_VH:
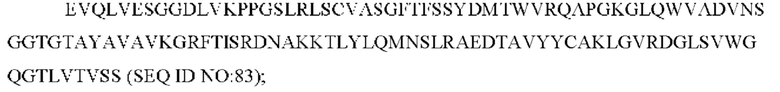
4) антитело ZIL11 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL11_VL:
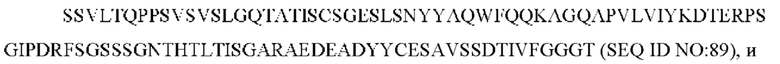
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL11_VH:

5) антитело ZIL69 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL69_VL:

b) вариабельную область тяжелой цепи, содержащую CAN-ZIL69_VH:
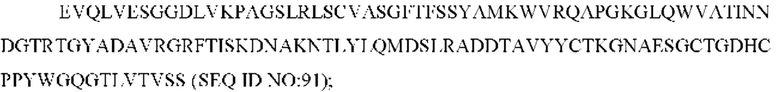
6) антитело ZIL94 содержит по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащую CAN-ZIL94_VL:
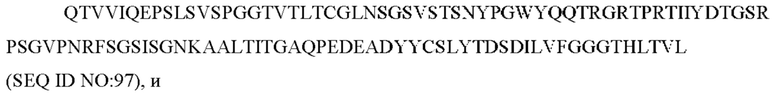
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL94_VH:
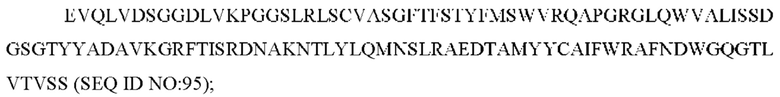
7) антитело ZIL154 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL154_VL:

b) вариабельную область тяжелой цепи, содержащую CAN-ZIL154_VH:
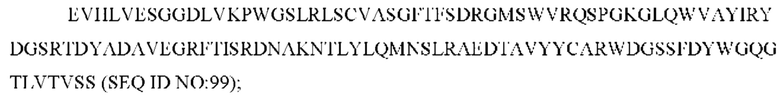
8) антитело ZIL159 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL159_VL:
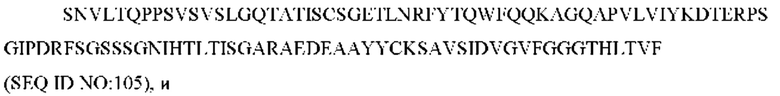
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL159_VH:
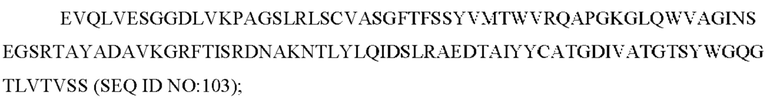
9) антитело ZIL171 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую CAN-ZIL171VL:
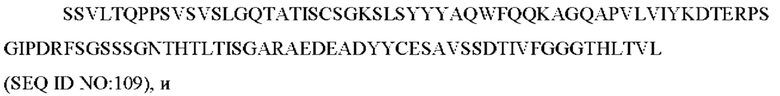
b) вариабельную область тяжелой цепи, содержащую CAN-ZIL_171_VH:
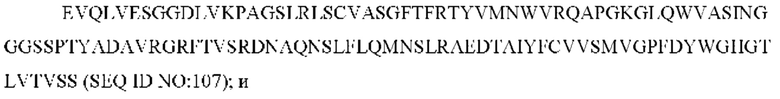
10) антитело 04Н07 содержит по меньшей мере одно из следующего:
а) вариабельную область легкой цепи, содержащую Mu_04H07_VL:

b) вариабельную область тяжелой цепи, содержащую Mu_04H07_VH:
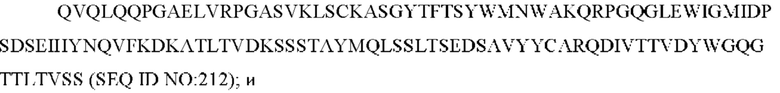
11) антитело 06А09 содержит по меньшей мере одно из следующего:
a) вариабельную область легкой цепи, содержащую Mu_06А09_VL:
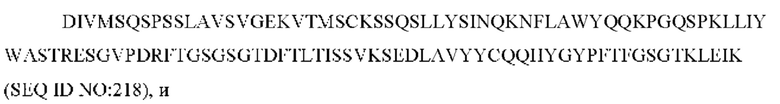
b) вариабельную область тяжелой цепи, содержащую Mu_06A09_VH:
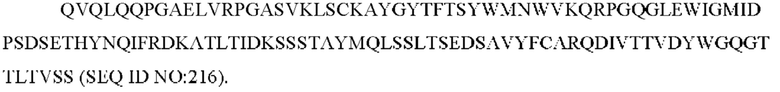
В других воплощениях согласно изобретению предложена клетка-хозяин, продуцирующая антитело, описанное выше.
Объем настоящего изобретения также включает нуклеотидные последовательности, кодирующие вариабельные области легких и тяжелых цепей антитела против IL-31 по настоящему изобретению. Объем изобретения также включает любую нуклеотидную последовательность, кодирующую аминокислотную последовательность 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07, 06А09 или их полипептиды или пептиды, специфичные в отношении IL-31.
В некоторых воплощениях согласно изобретению предложена выделенная нуклеиновая кислота, содержащая нуклеиновокислотную последовательность, кодирующую по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области тяжелой цепи:
1) 15Н05: CDR1 вариабельной области тяжелой цепи 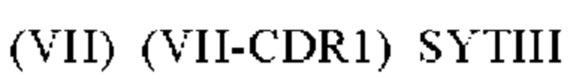

2) 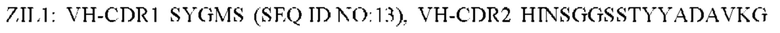
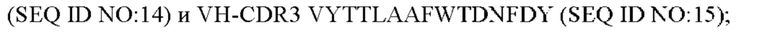
3) ZIL8: 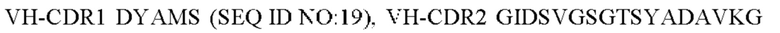
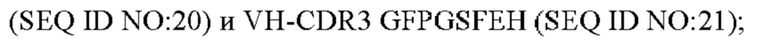
4) ZIL9: 
5) ZIL11: 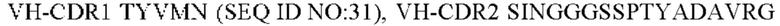
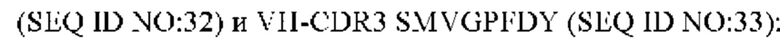
6) ZIL69: 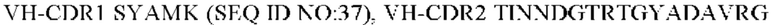

7) ZIL94: 
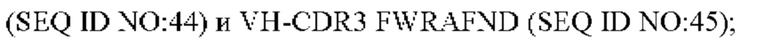
8) ZIL154: 

9) ZIL159: 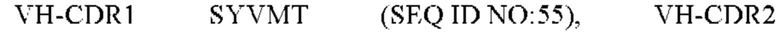

10) ZIL171: 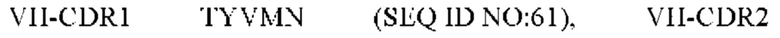
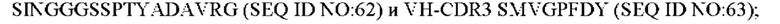
11) 04H07: 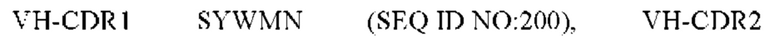
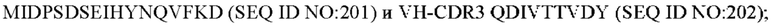
12) 06A09: 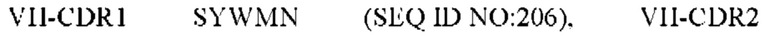
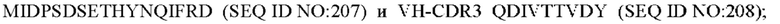 или
или
13) вариант (1)-(12), отличающийся от CDR соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3_VH.
В одном воплощении выделенная нуклеиновая кислота, описанная выше, может дополнительно содержать нуклеиновокислотную последовательность, кодирующую по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области легкой цепи:
1) 15Н05: 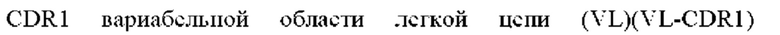

2) ZIL1: 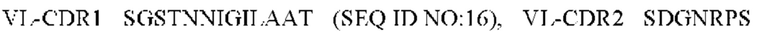
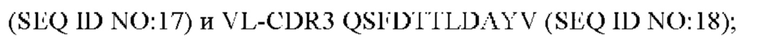
3) ZIL8: 
4) ZIL9: 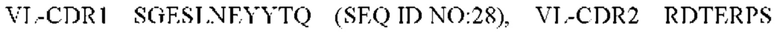
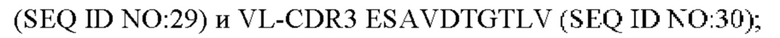
5) ZIL11: 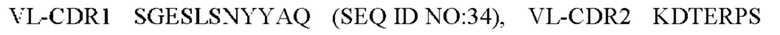
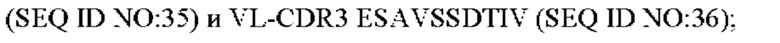
6) ZIL69: 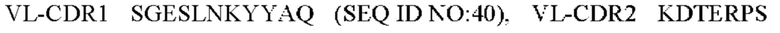
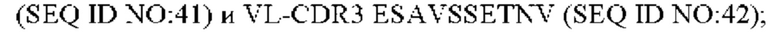
7) ZIL94: 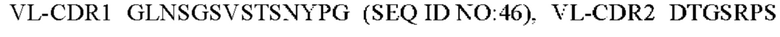
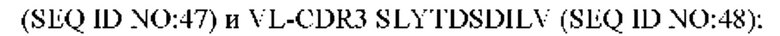
8) ZIL154: 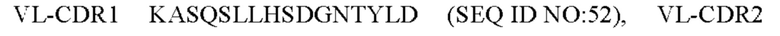
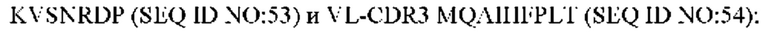
9) ZIL159: 
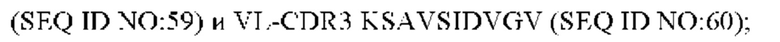
10) ZIL171: 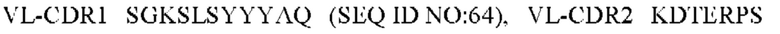
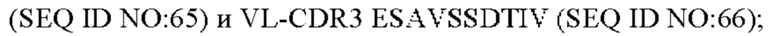
11) 04H07: 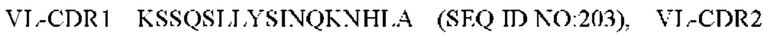
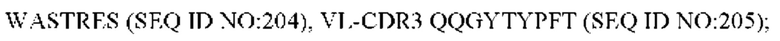
12) 06A09: 
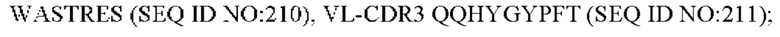 или
или
13) вариант (1)-(12), отличающийся от CDR соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VH или VL.
В одном воплощении согласно изобретению предложена выделенная нуклеиновая кислота, содержащая нуклеиновокислотную последовательность, кодирующую по меньшей мере одну из следующих комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области легкой цепи:
1) 15Н05: 

2) ZIL1: 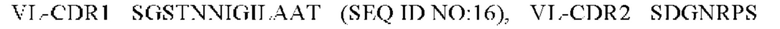
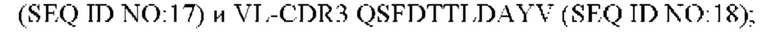
3) ZIL8: 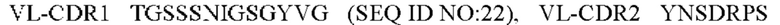
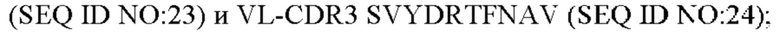
4) ZIL9: 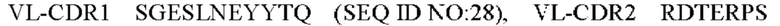
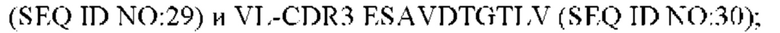
5) ZIL11: 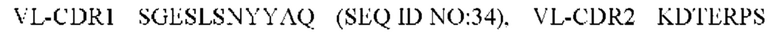
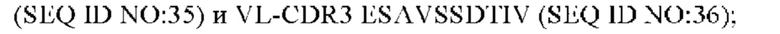
6) ZIL69: 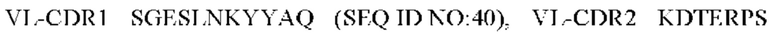

7) ZIL94: 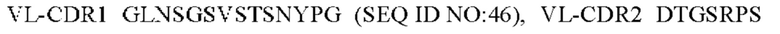
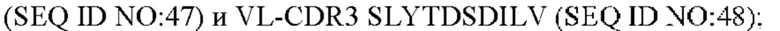
8) ZIL154: 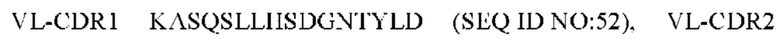
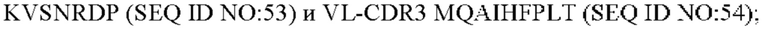
9) ZIL159: 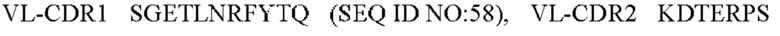
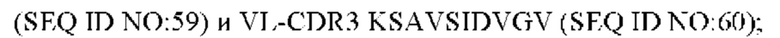
10) ZIL171: 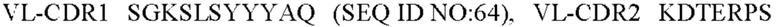
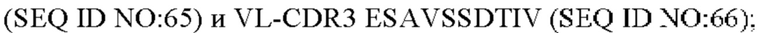
11) 04H07: 

12) 06A09: 
 или
или
13) вариант (1)-(12), отличающийся от CDR соответствующего исходного антитела 15Н05, ZIL1, ZIL8, ZIL9, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159, ZIL171, 04Н07 или 06А09 добавлением, делецией и/или заменой одного или более чем одного аминокислотного остатка по меньшей мере в одном из CDR1, CDR2 или CDR3 VL.
Кроме того, согласно настоящему изобретению предложен вектор, содержащий по меньшей мере одну из нуклеиновых кислот, описанных выше.
Как будет описано более подробно ниже, нуклеиновокислотная последовательность, кодирующая по меньшей мере одну из описанных выше комбинаций последовательностей гипервариабельных участков (CDR) вариабельной области тяжелой цепи, может присутствовать в одном и том же векторе с нуклеиновокислотной последовательностью, кодирующей по меньшей мере одну из описанных выше комбинаций последовательностей CDR вариабельной области легкой цепи. Альтернативно, нуклеиновокислотная последовательность, кодирующая по меньшей мере одну из описанных выше комбинаций последовательностей CDR вариабельной области легкой цепи, и нуклеиновокислотная последовательность, кодирующая по меньшей мере одну из описанных выше комбинаций последовательностей CDR вариабельной области тяжелой цепи, могут присутствовать в разных векторах.
Ввиду вырожденности генетического кода для кодирования конкретной аминокислоты может быть использован более чем один кодон. С использованием генетического кода могут быть определены одна или несколько разных нуклеотидных последовательностей, каждая из которых будет способна кодировать указанную аминокислоту. Вероятность того, что конкретный олигонуклеотид действительно будет составлять фактическую кодирующую последовательность XXX, может быть оценена с учетом закономерностей аномального спаривания оснований и частоты фактического использования конкретного кодона (для кодирования конкретной аминокислоты) в эукариотических или прокариотических клетках, экспрессирующих антитело против IL-31 или его часть, специфичную в отношении IL-31. Такие «правила использования кодонов» раскрыты Lathe, et al., 183 J. Molec. Biol. 1-12 (1985). С применением «правил использования кодонов» по Lathe может быть определена одна нуклеотидная последовательность или группа нуклеотидных последовательностей, содержащих теоретическую «наиболее вероятную» нуклеотидную последовательность, способную кодировать последовательности против IL-31. Кроме того, подразумевается, что области, кодирующие антитело, для использования в настоящем изобретении могут также быть обеспечены изменением существующих генов антител с применением стандартных молекулярно-биологических методик, приводящих к получению вариантов (агонистов) антител и пептидов, описанных здесь. Такие варианты включают, без ограничения, делеции, добавления и замены в аминокислотной последовательности антител против IL-31 или полипептидов или пептидов, специфичных в отношении IL-31, включая CDR-участки антитела. Например, могут быть заменены остатки, не являющиеся критически важными для связывания с антигеном, расположенные в CDR-участках или других областях антитела. Примеры типов экспериментов, применяемых для оценки того, являются ли конкретные остатки критически важными для связывания с антигеном, описаны в Разделе 1.21 раздела «Примеры» ниже. В одном воплощении одна или более чем одна замена представляют собой консервативные аминокислотные замены, описанные здесь более подробно. Тем не менее, варианты антител по настоящему изобретению, включая варианты CDR, не ограничены консервативными аминокислотными заменами.
Например, один класс замен представляет собой консервативные аминокислотные замены. Такие замены представляют собой замены заданной аминокислоты в пептиде антитела против IL-31 другой аминокислотой со сходными характеристиками. Консервативными обычно считают замены одной аминокислоты (остатка) на другую аминокислоту (остаток) в пределах следующих групп: алифатические аминокислоты Ala, Val, Leu и Ile; гидроксильные остатки Ser и Thr; кислые остатки Asp и Glu; амидные остатки Asn и Gln; основные остатки Lys и Arg, ароматические остатки Phe, Tyr; и тому подобное. Информация относительно того, какие аминокислотные изменения, вероятно, не приведут к изменениям фенотипа, представлена в Bowie et al., 247 Science 1306-10 (1990).
Вариант или агонист антител против IL-31 или полипептидов или пептидов, специфичных в отношении IL-31, может быть полностью функциональным или может быть лишен одной или более чем одной функциональной активности. Полностью функциональные варианты обычно содержат только консервативные изменения или изменения остатков или областей, не являющихся критически важными. Функциональные варианты могут также содержать замены сходных аминокислот, не приводящие к функциональным изменениям или приводящие к незначительным функциональным изменениям. Альтернативно, такие замены могут в некоторой степени влиять на функции положительным или отрицательным образом. Нефункциональные варианты обычно содержат одну или более чем одну неконсервативную аминокислотную замену, делецию, вставку, инверсию или усечение или замену, вставку, инверсию или делецию критически важного остатка или критически важной области.
Аминокислоты, важные для выполнения функции, могут быть определены методами, известными в данной области, такими как сайт-направленный мутагенез или мутагенез с применением аланинового сканирования. Cunningham et al., 244 Science 1081-85 (1989). Последняя методика включает введение одиночных аланиновых мутаций по каждому остатку молекулы. Полученные мутантные молекулы затем анализируют на предмет их биологической активности, такой как связывание эпитопов или ADCC-активность in vitro. Сайты, критически важные для связывания лиганда с рецептором, могут также быть определены структурным анализом, таким как кристаллография, ядерный магнитный резонанс или фотоаффинное мечение. Smith et al., 224 J. Mol. Biol. 899-904 (1992); de Vos etal, 255 Science 306-12 (1992).
Более того, полипептиды часто содержат аминокислоты, отличные от двадцати «встречающихся в природе» аминокислот. Кроме того, многие аминокислоты, включая концевые аминокислоты, могут быть модифицированы естественными процессами, такими как процессинг и другие посттрансляционные модификации, или методиками химической модификации, хорошо известными в данной области. Известные модификации включают, без ограничения, ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение гемовой группировки, ковалентное присоединение нуклеотида или нуклеотидного производного, ковалентное присоединение липида или липидного производного, ковалентное присоединение фосфатидилинозита, поперечное связывание, циклизацию, образование дисульфидных связей, деметилирование, образование поперечных ковалентных связей, образование цистина, образование пироглутамата, формилирование, гамма-карбоксилирование, гликозилирование, йодирование, метилирование, миристоилирование, окисление, протеолитический процессинг, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование, присоединение аминокислот к белкам, опосредованное транспортными РНК, такое как аргинилирование, и убиквтинирование.
Такие модификации хорошо известны специалистам в данной области и подробно описаны в научной литературе. Некоторые особенно распространенные модификации, такие как, например, гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и АДФ-рибозилирование, описаны в большинстве основополагающих публикаций, таких как Proteins-Structure and Molecular Properties (2nd ed., Т.E. Creighton, W.H. Freeman & Co., NY, 1993). По этой теме есть много подробных обзоров, такие как Wold, Posttranslational Covalent Modification of proteins, 1-12 (Johnson, ed., Academic Press, NY, 1983), Seifter et al. 182 Meth. Enzymol. 626-46 (1990) и Rattan et al. 663 Ann. NY Acad. Sci. 48-62 (1992).
Соответственно, антитела, полипептиды и пептиды, специфичные в отношении IL-31, по настоящему изобретению также охватывают производные или аналоги, в которых заменяющий аминокислотный остаток отличен от остатков, кодируемых генетическим кодом.
Сходным образом, добавления и замены в аминокислотной последовательности, а также варианты и модификации, описанные выше, могут быть в равной степени применимы к аминокислотной последовательности антигена и/или эпитопа IL-31 или их пептидам и, таким образом, включены в настоящее изобретение. Как указано выше, гены, кодирующие моноклональное антитело по настоящему изобретению, особенно эффективны в распознавании IL-31.
Производные антител
Объем данного изобретения включает производные антител. «Производное» антитела содержит дополнительные химические группировки, не являющиеся обычно частью белка. Объем данного изобретения включает ковалентные модификации белков. Такие модификации могут быть введены в молекулу посредством взаимодействия целевых аминокислотных остатков антитела с органическим дериватизирующим агентом, способным взаимодействовать с выбранными боковыми цепями или концевыми остатками. Например, дериватизация бифункциональными агентами, хорошо известная в данной области, полезна для перекрестного связывания антитела или фрагмента с матрицей водонерастворимой подложки или с другими макромолекулярными носителями.
Производные также включают: радиоактивно меченые моноклональные антитела, меченые, например, радиоактивными йодом (125I, 131I), углеродом (14С), серой (35S), индием (111In, тритием (3Н) или тому подобным; конъюгаты моноклональных антител с биотином или авидином, с ферментами, такими как пероксидаза хрена, щелочная фосфатаза, бета-D-галактозидаза, глюкозооксидаза, глюкоамилаза, ангидраза карбоновых кислот, ацетилхолинэстераза, лизоцим, малатдегидрогеназа или глюкозо-6-фосфатдегидрогеназа; а также конъюгаты моноклональных антител с биолюминесцентными агентами (такими как люцифераза), хемилюминесцентными агентами (такими как сложные эфиры акридина) или флуоресцентными агентами (такими как фикобилипротеины).
Другое бифункциональное производное антитела по настоящему изобретению представляет собой биспецифические антитело, полученное объединением частей двух раздельных антител, распознающих две разные антигенные группы. Это может быть достигнуто поперечным связыванием или рекомбинантными методиками. Кроме того, в антитело или его часть могут быть добавлены группировки, увеличивающие период полувыведения in vivo (например, увеличивая время клиренса из кровотока). Такие методики включают, например, добавление ПЭГ-группировок (также называемое пегилированием) и хорошо известны в данной области. См. публикацию заявки на патент США №20030031671.
Рекомбинантная экспрессия антител
В некоторых воплощениях нуклеиновые кислоты, кодирующие соответствующее моноклональное антитело, вводят непосредственно в клетку-хозяина, и клетку инкубируют в условиях, достаточных для индукции экспрессии кодируемого антитела. В типичных случаях после введения соответствующих нуклеиновых кислот в клетку ее инкубируют, обычно при 37°С, иногда проводя селекцию, на протяжении периода продолжительностью приблизительно 1-24 часа для обеспечения возможности экспрессии антитела. В одном воплощении происходит секреция антитела в супернатант среды, в которой выращивают клетку.
Традиционно моноклональные антитела получают в форме нативных молекул в мышиных гибридомных линиях. В дополнение к этой технологии, согласно настоящему изобретению предложена рекомбинантная экспрессия ДНК моноклональных антител. Это позволяет получать канинизированные, фелинизированные, эквинизированные, полностью собачьи, полностью кошачьи и полностью лошадиные антитела, а также ряд производных антител и слитых белков у выбранного вида-хозяина.
Возможна рекомбинация нуклеиновокислотной последовательности, кодирующей по меньшей мере одно антитело, часть или полипептид против IL-31 по настоящему изобретению с векторной ДНК в соответствии с общепринятыми методиками, включая тупые концы или ступенчатые концы для лигирования, расщепление рестриктазами с получением подходящих концов, соответствующее заполнение липких концов, обработку щелочной фосфатазой во избежание нежелательного соединения и лигирование подходящими лигазами. Методики для проведения таких манипуляций раскрыты, например, в Maniatis et al., MOLECULAR CLONING, LAB. MANUAL (Cold Spring Harbor Lab. Press, NY, 1982 and 1989), и Ausubel et al. 1993 supra и могут быть применены для конструирования нуклеиновокислотных последовательностей, кодирующих молекулу моноклонального антитела или его антигенсвязывающую область.
Молекулу нуклеиновой кислоты, такую как ДНК, считают «способной экспрессировать» полипептид, если она содержит нуклеотидные последовательности, содержащие информацию, необходимую для регуляции транскрипции и трансляции, и такие последовательности «функционально связаны» с нуклеотидными последовательностями, кодирующими полипептид. Функциональная связь представляет собой связь, при которой регуляторные последовательности ДНК и последовательность ДНК, которую следует экспрессировать, соединены таким образом, который позволяет экспрессировать ген в виде пептидов или частей антител против IL-31 в количествах, поддающихся выделению. Конкретная природа регуляторных областей, необходимых для экспрессии генов, может варьировать от организма к организму, что хорошо известно в аналогичных областях. См., например, Sambrook et al., 2001 supra; Ausubel et al., 1993 supra.
Соответственно, настоящее изобретение охватывает экспрессию антитела против IL-31 или пептида или полипептида, специфичных в отношении IL-31, в прокариотических или эукариотических клетках. Подходящие хозяева включают бактериальных или эукариотических хозяев, включая клетки бактерий, дрожжей, насекомых, грибов, птиц и млекопитающих in vivo или in situ или клетки-хозяева, имеющие происхождение от млекопитающих, насекомых, птиц или дрожжей. Клетка или ткань млекопитающего могут иметь происхождение от человека, примата, хомяка, кролика, грызуна, коровы, свиньи, овцы, лошади, козы, собаки или кошки, но может быть использована клетка любого другого млекопитающего.
В одном воплощении вводимая нуклеотидная последовательность будет включена в плазмидный или вирусный вектор, способный к автономной репликации в реципиенте-хозяине. С этой целью может быть использован любой из широкого спектра векторов. См., например, Ausubel et al., 1993 supra. Факторы, важные для выбора конкретного плазмидного или вирусного вектора, включают: простоту распознавания и отбора клеток-реципиентов, содержащих вектор, от тех клеток-реципиентов, которые не содержат вектор; желаемое число копий вектора у конкретного хозяина; и желательность возможности перехода вектора между клетками-хозяевами разных видов.
Примеры прокариотических векторов, известных в данной области, включают плазмиды, такие как плазмиды, способные к репликации в Е. coli (такие как, например, pBR322, ColE1, pSC101, pACYC 184,.pi.VX). Такие плазмиды раскрыты, например, в Maniatis et al., 1989 supra; Ausubel et al., 1993 supra. Плазмиды Bacillus включают pC194, pC221, pT127 и так далее. Такие плазмиды раскрыты в Gryczan, in THE MOLEC. BIO. OF THE BACILLI 307-329 (Academic Press, NY, 1982). Подходящие плазмиды Streptomyces включают pIJ101 (Kendall et al., 169 J. Bacteriol. 4177-83 (1987)) и бактериофаги Streptomyces, такие как.phi.C31 (Chater et al., in SIXTH INT'L SYMPOSIUM ON ACTINOMYCETALES BIO. 45-54 (Akademiai Kaido, Budapest, Hungary 1986)). Обзор плазмид Pseudomonas представлен в John et at, 8 Rev. Infect. Dis. 693-704 (1986), Izaki, 33 Jpn. J. Bacteriol. 729-42 (1978), и Ausubel et al., 1993 supra.
Альтернативно, элементы экспрессии генов, полезные для экспрессии кДНК, кодирующей антитела или пептиды против IL-31, включают, без ограничения: (а) вирусные промоторы транскрипции и их энхансерные элементы, такие как ранний промотор SV40 (Okayama et al., 3 Mol. Cell. Biol. 280 (1983)), LTR вируса саркомы Рауса (Gorman et al., 79 Proc. Natl. Acad. Sci., USA 6777 (1982)) и LTR вируса мышиного лейкоза Молони (Grosschedl et al., 41 Cell 885 (1985)); (b) области сплайсинга и сайты полиаденилирования, такие как области и сайты, имеющие происхождение от поздней области SV40 (Okayarea et al., МСВ, 3: 280 (1983); и (с) сайты полиаденилирования, такие как у SV40 (Okayama et al., 1983, supra).
кДНК-гены иммуноглобулинов могут быть экспрессированы, как описано Weidle et al., 51(1) Gene 21-29 (1987), с использованием, в качестве экспрессионных элементов, раннего промотора SV40 и его энхансера, энхансеров промотора Н-цепи мышиных иммуноглобулинов, поздней области сплайсинга мРНК SV40, вставочной последовательности кроличьего S-глобина, сайтов полиаденилирования иммуноглобулинов и кроличьего S-глобина и элементов полиаденилирования SV40.
Для генов иммуноглобулинов, представленных частично кДНК, частично геномной ДНК (Whittle et al., 1 Protein Engin. 499-505 (1987)), транскрипционный промотор может представлять собой промотор цитомегаловируса человека, энхансеры промотора могут представлять собой энхансеры цитомегаловируса и мышиных/человеческих иммуноглобулинов, а области сплайсинга мРНК и полиаденилирования могут представлять собой нативные хромосомные последовательности иммуноглобулинов.
В одном воплощении для экспрессии кДНК-генов в клетках грызуна транскрипционный промотор представляет собой вирусную LTR-последовательность, энхансеры транскрипционного промотора представляют собой энхансер тяжелой цепи мышиного иммуноглобулина и/или вирусный LTR-энхансер, область сплайсинга содержит интрон размером более 31 п.о., а области полиаденилирования и терминации транскрипции имеют происхождение от нативной хромосомной последовательности, соответствующей синтезируемой цепи иммуноглобулина. В других воплощениях с указанными выше экспрессионными элементами комбинируют последовательности кДНК, кодирующие другие белки, для обеспечения экспрессии белков в клетках млекопитающих.
Каждый слитый ген может быть собран в векторе экспрессии или введен в вектор экспрессии. Затем клетки-реципиенты, способные экспрессировать ген химерной цепи иммуноглобулина, трансфицируют одним геном, кодирующим пептид против IL-31, или химерную Н-цепь, или химерную L-цепь, или котрансфицируют генами химерной Н-цепи и химерной L-цепи. Трансфицированные клетки-реципиенты культивируют в условиях, позволяющих экспрессировать введенные гены, и экспрессированные цепи иммуноглобулинов, интактные антитела или фрагменты выделяют из культуры.
В одном воплощении слитые гены, кодирующие пептид против IL-31, или химерные Н- и L-цепи, или их части, собирают в раздельных векторах экспрессии, которые затем используют для котрансфекции клетки-реципиента. Альтернативно, слитые гены, кодирующие химерные Н- и L-цепи, могут быть собраны на одном и том же векторе экспрессии.
Линия клеток-реципиентов для трансфекции векторами экспрессии и получения химерного антитела может представлять собой клетки миеломы. Клетки миеломы могут обеспечивать синтез, сборку и секрецию иммуноглобулинов, кодируемых генами иммуноглобулинов, использованными при трансфекции, и обладают механизмом гликозилирования иммуноглобулинов. Клетки миеломы можно выращивать в культуре или в брюшинной полости мыши, в случае чего секретированый иммуноглобулин может быть получен из асцитической жидкости. Другие подходящие клетки-реципиенты включают лимфоидные клетки, такие как В-лимфоциты, имеющие происхождение от человека или вида, не являющегося человеком, гибридомные клетки, имеющие происхождение от человека или вида, не являющегося человеком, или межвидовые гетерогибридомные клетки.
Вектор экспрессии, содержащий конструкцию химерного, канинизированного, фелинизированного, эквинизированного, полностью собачьего, полностью кошачьего или полностью лошадиного антитела против IL-31 или полипептид или пептид, специфичные в отношении IL-31 (например, антигенсвязывающую часть антитела, описанного здесь), по настоящему изобретению, может быть введен в соответствующую клетку-хозяина любым из множества подходящих способов, включая такие биохимические способы, как трансформация, трансфекция, конъюгация, слияние протопластов, преципитация с фосфатом кальция и применение поликатионов, таких как диэтиламиноэтилдекстран (DEAE-декстран), и такие механические способы, как электропорация, прямая микроинъекция и бомбардировка микрочастицами. Johnston et al., 240 Science 1538-1541 (1988).
Дрожжи могут обеспечить существенные преимущества над бактериями при получении Н- и L-цепей иммуноглобулинов. В дрожжах происходят посттрансляционные модификации пептидов, включая гликозилирование. В настоящее время существует ряд методик рекомбинантных ДНК, в которых используются последовательности сильных промоторов и плазмиды с большим числом копий, которые могут быть применены для получения желаемых белков в дрожжах. Дрожжи распознают лидерные последовательности продуктов клонированных генов млекопитающих и секретируют пептиды, содержащие лидерные последовательности (то есть препептиды). Hitzman et al., 11th Int'l Conference on Yeast, Genetics & Molec. Biol. (Montpelier, France, 1982).
Дрожжевые системы экспрессии генов можно регулярно оценивать на предмет уровней продукции, секреции и стабильности пептидов против IL-31, антитела и собранных мышиных и химерных, гетерохимерных, канинизированных, фелинизированных, эквинизированных, полностью собачьих, полностью кошачьих или полностью лошадиных антител, их фрагментов и областей. Можно использовать любую из ряда дрожжевых систем экспрессии генов, включающую промоторные и терминационные элементы активно экспрессируемых генов, кодирующих гликолитические ферменты, продуцируемые в больших количествах при выращивании дрожжей в средах с высоким содержанием глюкозы. Известные гликолитические гены могут также обеспечивать очень эффективные сигналы контроля транскрипции. Например, могут быть использованы промоторные и терминаторные сигналы гена фосфоглицераткиназы (ФГК). Для оценки оптимальной экспрессии плазмид для экспрессии клонированных кДНК иммуноглобулинов у дрожжей могут быть применены несколько методик. См. Vol. II DNA Cloning, 45-66, (Glover, ed.,) IRL Press, Oxford, UK (1985).
В качестве хозяев для получения молекул антител или пептидов, описанных в данном изобретении, могут также быть использованы бактериальные штаммы. Вместе с этими бактериальными хозяевами используют плазмидные векторы, содержащие репликон и контрольные последовательности, имеющие происхождение от вида, совместимого с клеткой-хозяином. Вектор содержит сайт репликации, а также определенные гены, способные обеспечивать фенотипическую селекцию трансформированных клеток. Для оценки экспрессии плазмид для получения мышиных, химерных, гетерохимерных, канинизированных, фелинизированных, эквинизированных, полностью собачьих, полностью кошачьих или полностью лошадиных антител, фрагментов и областей или цепей антител, кодируемых клонированными кДНК иммуноглобулинов, у бактерий могут быть применены несколько методик (см. Glover, 1985 supra; Ausubel, 1993 supra; Sambrook, 2001 supra; Colligan et al., eds. Current Protocols in Immunology, John Wiley & Sons, NY, NY (1994-2001); Colligan et al., eds. Current Protocols in Protein Science, John Wiley & Sons, NY, NY (1997-2001)).
Клетки-хозяева, являющиеся клетками млекопитающих, можно выращивать in vitro или in vivo. Клетки млекопитающих обеспечивают посттрансляционные модификации белковых молекул иммуноглобулинов, включая удаление лидерных пептидов, фолдинг и сборку Н- и L-цепей, гликозилирование молекул антител и секрецию функциональных белков антител.
Клетки млекопитающих, которые могут быть полезны в качестве хозяев для получения белков антител, помимо клеток лимфоидного происхождения, описанных выше, включают клетки, имеющие происхождение от фибробластов, такие как клетки Vero (АТСС CRL 81) или СНО-K1 (АТСС CRL 61).
Для экспрессии клонированных генов Н- и L-цепей пептидов против IL-31 в клетках млекопитающих доступно множество векторных систем (см. Glover, 1985 supra). Для получения полных антител H2L2 могут быть применены различные методики. Возможна одновременная экспрессия Н- и L-цепей в одних и тех же клетках, позволяющая достичь внутриклеточного объединения и связывания Н- и L-цепей с образованием полных тетрамерных антител H2L2 и/или пептидов против IL-31. Одновременная экспрессия возможна при использовании одинаковых или разных плазмид у одного и того же хозяина. Гены Н- и L-цепей и/или пептидов против IL-31 могут быть помещены в одну и ту же плазмиду, которой затем трансфицируют клетки, проводя, таким образом, непосредственный отбор клеток, экспрессирующих обе цепи. Альтернативно, клетки могут сначала быть трансфицированы плазмидой, кодирующей одну цепь, например, L-цепь, с последующей трансфекцией полученной клеточной линии плазмидой с Н-цепью, содержащей второй селектируемый маркер. Клеточные линии, продуцирующие пептиды и/или молекулы H2L2 против IL-31 любым из указанных способов, могут быть трансфицированы плазмидами, кодирующими дополнительные копии пептидов, Н-, L- или Н- и L-цепей в сочетании с дополнительными селектируемыми маркерами, для получения клеточных линий с улучшенными свойствами, такими как повышенная продукция собранных молекул антител H2L2 или повышенная стабильность трансфицированных клеточных линий.
Для длительного получения рекомбинантных антител с высоким выходом может быть применена стабильная экспрессия. Например, могут быть получены клеточные линии, стабильно экспрессирующие молекулу антитела. Вместо использования векторов экспрессии, содержащих вирусные репликаторы, клетки-хозяева могут быть трансформированы с использованием экспрессионных кассет иммуноглобулинов и селектируемого маркера. Клетки, полученные после введения чужеродной ДНК, можно выращивать на протяжении 1-2 суток в обогащенной среде и затем переносить их на селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к селекции и позволяет клеткам стабильно интегрировать плазмиду в хромосому и расти, образуя фокусы, которые, в свою очередь, могут быть клонированы и размножены с получением клеточных линий. Полученные таким образом клеточные линии могут быть особенно полезны при скрининге и оценке соединений/компонентов, непосредственно или косвенно взаимодействующих с молекулой антитела.
После получения антитела по изобретению оно может быть очищено любым методом очистки молекул иммуноглобулинов, известным в данной области, например, хроматографией (например, ионообменной, аффинной, особенно аффинной хроматографией с использованием специфического антигена после белка А, и гель-фильтрацией на колонках), центрифугированием, по дифференциальной растворимости или любой другой стандартной методикой очистки белков. Многие воплощения включают секрецию антител в культуральную среду и их выделение из культуральной среды.
Фармацевтические применения
Антитела против IL-31 или полипептиды или пептиды, специфичные в отношении IL-31, по настоящему изобретению могут быть использованы, например, в лечении состояний, сопровождающихся зудом, и/или аллергических состояний у домашних животных, таких как собаки, кошки и лошади. В одном воплощении такие полипептиды или пептиды содержат антигенсвязывающий фрагмент антител против IL-31, описанных здесь. Конкретнее, согласно изобретению также предложена фармацевтическая композиция, содержащая фармацевтически приемлемый носитель или разбавитель и, в качестве активного ингредиента, антитело, или полипептид, или пептид по изобретению. Антитело может представлять собой химерное, гетерохимерное, канинизированное, фелинизированное, эквинизированное, полностью лошадиное, полностью кошачье или полностью лошадиное антитело по настоящему изобретению. Также предусмотрены интактные иммуноглобулины или их связывающие фрагменты, такие как Fab. Антитело и его фармацевтические композиции по данному изобретению могут быть введены парентерально, например, подкожно, внутримышечно или внутривенно.
Антитела против IL-31, и/или полипептиды, специфичные в отношении IL-31, и/или пептиды, специфичные в отношении IL-31, по настоящему изобретению могут быть введены как отдельные терапевтические агенты или в комбинации с другими терапевтическими агентами. Они могут быть введены сами по себе, но обычно их вводят с фармацевтическим носителем, выбранным, исходя из выбранного пути введения и стандартной фармацевтической практики.
Введение антител, раскрытых здесь, может быть осуществлено любым подходящим способом, включая парентеральную инъекцию (такую как внутрибрюшинная, подкожная или внутримышечная инъекция), пероральное или местное введение антител (обычно в составе фармацевтической композиции) на поверхность дыхательных путей. Местное введение на поверхность дыхательных путей может быть осуществлено посредством интраназального введения (например, с использованием капельницы, тампона или ингалятора). Местное введение антител на поверхность дыхательных путей может также быть осуществлено посредством ингаляционного введения, такого как получение пригодных для вдыхания частиц фармацевтической композиции (включая как твердые, так и жидкие частицы), содержащей антитела, в форме аэрозольной суспензии с последующим вдыханием этих пригодных для вдыхания частиц субъектом. Способы и устройства для введения пригодных для вдыхания частиц фармацевтических композиций хорошо известны, и может быть применена любая общепринятая методика. Пероральное введение может быть осуществлено, например, в форме жидкой или твердой композиции, пригодной для приема внутрь.
В некоторых желаемых воплощениях антитела вводят парентеральной инъекцией. Для парентерального введения антитела, или полипептиды, или пептиды против IL-31 могут быть изготовлены в форме раствора, суспензии, эмульсии или лиофилизированного порошка вместе с фармацевтически приемлемым наполнителем для парентерального введения. Например, наполнитель может представлять собой раствор антитела или его смесь, растворенные в приемлемом носителе, таком как водный носитель; такими наполнителями являются вода, физиологический раствор, раствор Рингера, раствор декстрозы, раствор трегалозы или сахарозы, или 5%-й сывороточный альбумин, 0,4%-й физиологический раствор, 0,3%-й глицин и тому подобное. Также могут быть использованы липосомы и неводные наполнители, такие как нелетучие масла. Эти растворы стерильны и обычно не содержат твердых частиц. Эти композиции можно стерилизовать общепринятыми и хорошо известными методиками стерилизации. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, необходимые для создания условий, близких к физиологическим, такие как агенты для коррекции рН и буферные агенты, агенты для коррекции тоничности и тому подобное, например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и так далее. Концентрация антитела в этих композициях может варьировать в широких пределах, например от менее чем приблизительно 0,5%, обычно от 1% или по меньшей мере приблизительно 1%, до 15% или 20% по массе, и ее будут выбирать, исходя в первую очередь из объемов жидкостей, значений вязкости и так далее, в соответствии с конкретным выбранным способом введения. Наполнитель или лиофилизированный порошок может содержать добавки, которые поддерживают изотоничность (например, хлорид натрия, маннит) и химическую стабильность (например, буферы и консерванты). Композицию стерилизуют широко применяемыми методиками.
Существующие способы изготовления парентерально вводимых композиций будут известны или очевидны специалистам в данной области и описаны более подробно, например, в REMINGTON'S PHARMA. SCI. (15th ed., Mack Pub. Co., Easton, Pa., 1980).
Антитела по данному изобретению могут быть лиофилизированы для хранения и восстановлены в подходящем носителе перед применением. Показано, что эта методика эффективна при использовании обычных иммунноглобулинов. Могут быть применены любые подходящие методики лиофилизации и восстановления. Специалистам в данной области будет ясно, что лиофилизация и восстановление могут привести к той или иной степени потери активности антител и что это может привести к необходимости коррекции применяемых уровней.
Композиции, содержащие настоящие антитела или их смесь, могут быть введены для предупреждения рецидива и/или терапевтического лечения существующего заболевания. Подходящие фармацевтические носители описаны в последнем издании REMINGTON'S PHARMACEUTICAL SCIENCES, стандартном руководстве в данной области.
При терапевтическом применении композиции вводят субъекту, уже страдающему от заболевания, в количестве, достаточном для излечения или по меньшей мере частичной остановки развития или облегчения заболевания и его осложнений. Количество, адекватное для достижения этого, определяют как «терапевтически эффективную дозу» или «терапевтически эффективное количество». Количества, эффективные для такого применения, будут зависеть от тяжести заболевания и общего состояния собственной иммунной системы субъекта, но обычно входят в диапазон от приблизительно 0,1 мг антитела на кг массы тела до приблизительно 15 мг антитела на кг массы тела, предпочтительно от приблизительно 0,3 мг антитела на кг массы тела до приблизительно 12 мг антитела на кг массы тела. В одном воплощении терапевтически эффективное количество будет обеспечивать эффективность в течение по меньшей мере одного месяца при дозе до 12 мг/кг массы тела. Минимальное содержание посторонних веществ и уменьшение вероятности отторжения «чужеродного вещества», достигаемые благодаря присутствию настоящих антител, подобных собачьим, кошачьим и лошадиным антителам, могут обеспечить возможность введения этих антител в существенном избытке.
Разумеется, вводимая доза будет варьировать в зависимости от известных факторов, таких как фармакодинамические характеристики конкретного агента, а также способ и путь его введения, возраст, состояние здоровья и масса тела реципиента, характер и выраженность симптомов, сопутствующее лечение, частота введения и желаемый эффект.
В качестве неограничивающего примера, лечение патологии, опосредованной IL-31, у собак, кошек или лошадей может быть проведено в форме введения антител против IL-31 по настоящему изобретению один раз в две недели или один раз в месяц в диапазоне доз, описанном выше.
Примерами антител для терапевтического применения у собак, кошек или лошадей являются высокоаффиные (они могут также быть высокоавидными) антитела и их фрагменты, области и производные, имеющие высокую активность против IL-31 in vivo, по настоящему изобретению. Фрагменты и области антител могут быть альтернативно названы здесь полипептидами или пептидами по настоящему изобретению, содержащими антигенсвязывающую часть антител против IL-31.
Однократное или многократное введение композиций может быть осуществлено при уровнях и типе дозы, выбранных лечащим ветеринаром. В любом случае, фармацевтические композиции должны обеспечивать количество антитела или антител по данному изобретению, достаточное для эффективного лечения субъекта.
Диагностические применения
Согласно настоящему изобретению также предложены указанные выше антитела, полипептиды и/или пептиды против IL-31 для использования в диагностических методах выявления IL-31 у домашних животных с состоянием, сопровождающимся зудом, и/или аллергическим состоянием, или подозрением на наличие этих состояний.
Антитела, полипептиды и/или пептиды против IL-31 по настоящему изобретению полезны для иммунологических анализов, позволяющих выявлять или определять количество IL-31 или антител против IL-31 в образце. Иммунологический анализ на предмет IL-31 обычно включает инкубацию клинического или биологического образца в присутствии высокоаффинного (или высокоавидного) антитела, полипептида или пептида против IL-31 по настоящему изобретению, имеющего метку, поддающуюся выявлению, и способного селективно связываться с IL-31, и выявление меченого полипептида, пептида или антитела, связанных в образце. Различные методики клинических анализов хорошо известны в данной области. См., например, IMMUNOASSAYS FOR THE 80'S (Voller et al., eds., Univ. Park, 1981). Такие образцы включают образцы тканевых биоптатов, крови, сыворотки и кала или жидкости, полученные у субъектов-животных и анализируемые посредством ELISA, как описано ниже.
В некоторых воплощениях связывание антигена с антителом выявляют без использования твердой подложки. Например, связывание антигена с антителом можно выявлять в жидкостном формате.
В других воплощениях антитело, полипептид или пептид против IL-31 могут, например, быть фиксированы на нитроцеллюлозе или другой твердой подложке, способной иммобилизовать клетки, клеточные частицы или растворимые белки. Подложку можно затем промыть подходящими буферами с последующей обработкой полипептидом, пептидом или антителом, специфичным в отношении IL-31, имеющими метку, поддающуюся выявлению. После этого твердую подложку можно промыть буфером во второй раз для удаления несвязанных полипептида, пептида или антитела. Затем, на известных стадиях метода, можно определить количество связанной метки на твердой подложке.
«Твердая подложка» или «носитель» относится к любой подложке, способной связывать полипептид, пептид, антиген или антитело. Хорошо известные подложки или носители включают стекло, полистирол, полипропилен, полиэтилен, поливинилиденфторид (PVDF), декстран, нейлон, амилозы, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Для задач настоящего изобретения носитель должен быть в некоторой степени растворимым или нерастворимым. Вещество подложки может иметь практически любую возможную структурную конфигурацию при условии, что связанная с ним молекула сохраняет способность к связыванию с IL-31 или антителом против IL-31. Таким образом, конфигурация подложки может быть сферической, как у гранулы, или цилиндрической, как у внутренней поверхности пробирки или внешней поверхности стержня. Альтернативно, поверхность может быть плоской, такой как лист, чашка для культивирования, тест-полоска и так далее. Например, подложки могут содержать полистироловые гранулы. Специалистам в данной области будут известны многие другие подходящие носители для связывания антитела, полипептида, пептида или антигена, или они смогут определить такие носители посредством рутинных экспериментов.
На хорошо известных стадиях метода может быть определена связывающая активность заданной партии полипептида, пептида и/или антитела против IL-31. Специалисты в данной области могут определить оптимальные рабочие условия для проведения анализа посредством рутинных экспериментов.
Мечение полипептида, пептида и/или антитела, специфичных в отношении IL-31, меткой, поддающейся выявлению, может быть осуществлено посредством их связывания с ферментом для использования в ферментном иммунологическом анализе (EIA) или твердофазном иммуноферментном анализе (ELISA). Связанный фермент взаимодействует с доступным субстратом, приводя к образованию химической группировки, которая может быть выявлена, например, спектрофотометрическими, флуориметрическими или визуальными методами. Ферменты, которые могут быть использованы для мечения антител, специфичных в отношении IL-31, по настоящему изобретению меткой, поддающейся выявлению, включают, без ограничения, малатдегидрогеназу, стафилококковую нуклеазу, дельта-5-стероидизомеразу, дрожжевую алкогольдегидрогеназу, альфа-глицерофосфатдегидрогеназу, триозофосфатизомеразу, пероксидазу хрена, щелочную фосфатазу, аспарагиназу, глюкозооксидазу, бета-галактозидазу, рибонуклеазу, уреазу, каталазу, глюкозо-6-фосфатдегидрогеназу, глюкоамилазу и ацетилхолинэстеразу.
Радиоактивное мечение антител, специфичных в отношении IL-31, позволяет выявлять IL-31 с применением радиоиммунного анализа (RIA). См. Work et al., LAB. TECHNIQUES & BIOCHEM. IN MOLEC. Bio. (No. Holland Pub. Co., NY, 1978). Радиоактивный изотоп может быть выявлен такими методами, как применение гамма-счетчика или сцинтилляционного счетчика, или авторадиографией. Изотопы, особенно полезные для задач настоящего изобретения, включают 3H, 125I, 131I, 35S, 14C и 125I.
Также возможно мечение антител, специфичных в отношении IL-31, флуоресцентным соединением. При воздействии света с соответствующей длиной волны на флуоресцентно меченое антитело его присутствие можно выявить по флуоресценции. Соединениями, чаще всего используемыми для флуоресцентного мечения, являются изотиоцианат флуоресцеина, родамин, фикоэритрин, фикоцианин, аллофикоцианин, офталевый альдегид и флуорескамин.
Мечение антител, специфичных в отношении IL-31, меткой, поддающейся выявлению, может также быть осуществлено с использованием флуоресцентных металлов, таких как 125Eu или другие металлы группы лантаноидов. Эти металлы могут быть присоединены к антителу, специфическому в отношении IL-31, с использованием таких металлохелатирующих групп, как диэтилентриаминпентауксусная кислота (ДТПА) или этилендиаминтетрауксусная кислота (ЭДТА).
Кроме того, мечение антител, специфичных в отношении IL-31, меткой, поддающейся выявлению, может быть осуществлено сочетанием с хемилюминесцентным соединением. Присутствие хемилюминесцентно меченого антитела затем определяют, выявляя люминесценцию, возникающую при химическом взаимодействии. Примерами соединений, полезных для хемилюминесцентного мечения, являются люминол, изолюминол, тероматический (theromatic) сложный эфир акридиния, имидазол, соль акридиния и сложный эфир щавелевой кислоты.
Сходным образом, для мечения антитела, его части, фрагмента, полипептида или производного, специфичных в отношении IL-31, по настоящему изобретению может быть использовано биолюминесцентное соединение. Биолюминесценция представляет собой тип хемилюминесценции в биологических системах, где эффективность хемилюминесцентного взаимодействия повышена каталитическим белком. Присутствие биолюминесцентного белка определяют, выявляя люминесценцию. Важными биолюминесцентными соединениями, используемыми для мечения, являются люциферин, люцифераза и экворин.
Выявление антитела, его части, фрагмента, полипептида или производного, специфичных в отношении IL-31, может быть осуществлено сцинтилляционным счетчиком, например, если метка, поддающаяся выявлению, представляет собой радиоактивный гамма-излучатель, или флуориметром, например, если метка представляет собой флуоресцентное вещество. В случае ферментной метки выявление может быть осуществлено колориметрическими методами с использованием субстрата соответствующего фермента. Выявление может также быть осуществлено визуальным сравнением степени ферментативного взаимодействия субстрата со стандартными образцами, полученными сходным образом.
Для задач настоящего изобретения IL-31, выявляемый указанными выше анализами, может присутствовать в биологическом образце. Может быть использован любой образец, содержащий IL-31. Например, образец представляет собой биологическую жидкость, такую как, например, кровь, сыворотка, лимфа, моча, кал, воспалительный экссудат, спинномозговая жидкость, амниотическая жидкость, экстракт или гомогенат ткани и тому подобное. Изобретение не ограничено анализами с использованием только этих образцов, однако в свете настоящего описания специалист в данной области сможет определить подходящие условия, позволяющие использовать другие образцы.
Выявление in situ может быть осуществлено посредством получения гистологического образца от субъекта-животного и доставки комбинации меченых антител по настоящему изобретению к такому образцу. Антитело (или его часть) может быть доставлено нанесением или наложением меченого антитела (или его части) на биологический образец. Применяя такую методику, можно определять не только присутствие IL-31, но и распределение IL-31 в анализируемой ткани. При применении настоящего изобретения специалисту в данной области будет ясно, что любой из широкого спектра гистологических методов (таких как методики окрашивания) может быть модифицирован для проведения такого выявления in situ.
Антитело, фрагмент или производное по настоящему изобретению могут быть адаптированы для использования в иммунометрическом анализе, также известном как «двухсайтовый» или «сэндвич»-анализ. Типичный иммунометрический анализ включает связывание определенного количества немеченого антитела (или фрагмента антитела) с твердой подложкой, нерастворимой в анализируемой жидкости, добавление определенного количества растворимого антитела, имеющего метку, поддающуюся выявлению, и/или определение количества тройного комплекса, образованного антителом, фиксированным на твердой фазе, антигеном и меченым антителом.
Антитела могут быть использованы для количественного или качественного выявления IL-31 в образце или для выявления клеток, экспрессирующих IL-31. Это может быть осуществлено иммунофлуоресцентными методиками с использованием флуоресцентно меченого антитела (см. ниже) в сочетании с флуоресцентной микроскопией, проточно-цитометрическим или флуориметрическим выявлением. Для диагностических задач антитела могут быть мечеными или немечеными. Немеченые антитела могут быть использованы в комбинации с другими мечеными антителами (вторичными антителами), способными взаимодействовать с антителом, такими как антитела, специфичные в отношении константных областей собачьих или кошачьих иммуноглобулинов. Альтернативно, возможно прямое мечение антител. Может быть использован широкий спектр меток, таких как радионуклиды, флуоресцентные метки, ферменты, субстраты ферментов, кофакторы ферментов, ингибиторы ферментов, лиганды (в частности, гаптены) и так далее. Доступно множество типов иммунологических анализов, таких как анализы, обсужденные ранее, и они хорошо известны специалистам в данной области.
В одном воплощении диагностический метод выявления IL-31 представляет собой латеральный проточный иммуноанализ. Он также известен как иммунохроматографический анализ, Rapid ImmunoMigration (RIM™) или анализ на тест-полосках. В сущности, латеральные проточные иммуноанализы представляют собой иммунологические анализы, адаптированные для проведения вдоль одной оси, чтобы их можно было применять в формате тест-полосок. Разработан ряд вариантов данной технологии, которые применены в коммерческих продуктах, но все они работают по одному и тому же основному принципу. Типичная тест-полоска состоит из следующих компонентов: (1) зона для образца- абсорбирующая зона, на которую наносят анализируемый образец; (2) зона для конъюгата или реагента - эта зона содержит антитела, специфичные в отношении целевого аналита, конъюгированные с окрашенными частицами (обычно частицами коллоидного золота или латексными микросферами); (3) мембрана для проведения реакции - обычно мембрана из гидрофобной нитроцеллюлозы или ацетата целлюлозы, на которой иммобилизованы антитела против целевого аналита, расположенные по линии, пересекающей мембрану, образуя зону захвата или тест-линию (также может присутствовать контрольная зона, содержащая антитела, специфичные к конъюгированным антителам); и (4) тампон или резервуар для отходов - еще одна абсорбирующая зона, предназначенная для перемещения образца по мембране для проведения реакции под действием капиллярных сил и его сбора. Обычно компоненты полоски зафиксированы на инертном материале подложки и могут быть представлены в формате простой тест-полоски или в пластиковом корпусе с отверстием для образца и реакционным окном с зоной захвата и контрольной зоной.
Существуют два основных типа латерального проточного иммуно анализа, применяемые в микробиологических исследованиях: сэндвич-анализы с двумя антителами и конкурентные анализы. В формате сэндвич-анализа с двумя антителами образец перемещается из зоны для образца через зону для конъюгата, где любой целевой аналит, присутствующий в образце, будет связываться с конъюгатом. Затем образец продолжает перемещаться по мембране, пока не достигнет зоны захвата, где комплекс «целевой аналит/ конъюгат» будет связываться с иммобилизованными антителами, образуя видимую линию на мембране. Затем образец перемещается дальше по полоске, пока не достигнет контрольной зоны, где будет происходить связывание избытка конъюгата с образованием второй видимой линии на мембране. Эта контрольная линия указывает на то, что образец прошел по мембране надлежащим образом. Две отчетливые линии на мембране указывают на положительный результат. Единственная линия в контрольной зоне указывает на отрицательный результат. Конкурентные анализы отличаются от формата сэндвич-анализа с двумя антителами тем, что зона для конъюгата содержит антитела, которые уже связаны с целевым аналитом или с его аналогом. Если в образце присутствует целевой аналит, то вследствие этого он не будет связываться с конъюгатом и останется немеченым. После того, как образец переместится по мембране и достигнет зоны захвата, избыток немеченого аналита будет связываться с иммобилизованными антителами и блокировать захват конъюгата, поэтому образования видимой линии не будет. Затем несвязанный конъюгат будет связываться с антителами в контрольной зоне, образуя видимую контрольную линию. Единственная контрольная линия на мембране указывает на положительный результат. Две видимые линии в зоне захвата и контрольной зоне указывают на отрицательный результат. Однако если избытка немеченого целевого аналита нет, в зоне захвата возможно образование слабо выраженной линии, что указывает на неопределенный результат. Существует целый ряд вариантов латеральной проточной технологии. В зависимости от целевого аналита зона захвата на мембране может содержать иммобилизованные антигены или ферменты, а не антитела. Также возможно использование нескольких зон захвата для проведения множественного анализа. Например, были разработаны коммерческие тест-полоски, позволяющие выявлять оба токсина Шига энтерогеморрагической Е. coli, ST1 и ST2, по отдельности в одном и том же образце.
Важно, что антитела по настоящему изобретению могут быть полезны в диагностике зуда и/или аллергии у собак, кошек или лошадей. Конкретнее, антитело по настоящему изобретению позволяет определять сверхэкспрессию IL-31 у домашних животных. Таким образом, антитело по настоящему изобретению может стать важным инструментом для иммуногистохимии. В одном воплощении здесь предложена методика анализа, где мимотоп (пептид) IL-31 используют для захвата антитела по настоящему изобретению, меченного для выявления в анализе. Аффинность этого захваченного антитела в отношении присоединенного мимотопа будет ниже его аффинности в отношении нативного циркулирующего IL-31 у вида-хозяина. В данном воплощении жидкость, полученную от вида-хозяина, инкубируют с комплексом меченого антитела и мимотопа, связанным с твердой поверхностью. IL-31, присутствующий в анализируемой жидкости, имеющей происхождение от вида-хозяина, будет иметь более высокую аффинность в отношении антитела, приводя, таким образом, к высвобождению меченого антитела с твердой поверхности, после чего оно может быть удалено во время стадий промывки. Поэтому уровень IL-31 в анализируемой жидкости можно соотнести с ослаблением сигнала, происходящим на поверхности, связанной с мимотопом. Предполагается, что такой анализ будет полезен при измерении IL-31 в исследовательских или клинических условиях для применения в качестве диагностического анализа.
Антитела по настоящему изобретению могут быть использованы в матрицах с антителами, особенно подходящих для оценки профилей экспрессии генов.
Наборы
Объем настоящего изобретения также включает наборы для практического применения соответствующих способов. Наборы содержат по меньшей мере одно или более чем одно из антител по настоящему изобретению, нуклеиновую кислоту, кодирующую указанное антитело/антитела, или клетку, содержащую указанную нуклеиновую кислоту. В одном из воплощений антитело по настоящему изобретению может быть представлено в контейнере, обычно в лиофилизированной форме. Антитела, которые могут быть конъюгированы с меткой или токсином или быть неконъюгированными, обычно включены в наборы с буферами, такими как трис, фосфатный, карбонатный и так далее, стабилизаторами, биоцидами, инертными белками, например, сывороточным альбумином, или тому подобным. В большинстве случаев эти вещества будут присутствовать в количестве менее 5 масс. %, исходя из количества активного антитела, и обычно присутствуют в общем количестве по меньшей мере приблизительно 0,001 масс. %, снова исходя из концентрации антитела. Во многих случаях будет желательным включить инертный наполнитель или эксципиент для разбавления активных ингредиентов, где эксципиент может присутствовать в количестве приблизительно от 1% до 99 масс. %. всей композиции. Если в анализе используют вторичное антитело, способное связываться с первичным антителом, оно будет обычно представлено в отдельном флаконе. Вторичное антитело обычно конъюгировано с меткой и включено в композицию, аналогичную композициям антител, описанным выше. В большинстве случаев набор будет также содержать инструкции по применению.
В одном из воплощений набор по настоящему изобретению представляет собой набор тест-полосок (набор для латерального проточного иммуноанализа), полезный для выявления белка собачьего, кошачьего или лошадиного IL-31 в образце. Такая тест-полоска обычно включает: зону для образца, на которую наносят анализируемый образец; зону для конъюгата или реагента, содержащую антитело, специфические в отношении собачьего, кошачьего или лошадиного IL-31, конъюгированное с окрашенными частицами (обычно частицами коллоидного золота); мембрану для проведения реакции, на которой иммобилизованы антитела против IL-31, расположенные по линии, пересекающей мембрану, образуя зону захвата или тест-линию (также может присутствовать контрольная зона, содержащая антитела, специфичные к конъюгированным антителам); и еще одну абсорбирующую зону, предназначенную для перемещения образца по мембране для проведения реакции под действием капиллярных сил и его сбора. В большинстве случаев набор тест-полосок будет также содержать инструкции по применению.
Способы улучшения однородности и/или характеристик кошачьего или собачьего антитела
Такие способы описаны выше в разделе «Краткое изложение сущности изобретения», а также в Примерах и графических материалах настоящей заявки.
Авторы настоящего изобретения неожиданно обнаружили, что удаление или модификация С-конца константной области легкой цепи каппа тех видов животных, у которых нативные последовательности эмбрионального типа кодируют дополнительные остатки, расположенные дальше концевого цистеина легкой цепи, полезным образом сказывается как на получении однородных рекомбинантных антител этих видов, так и на количестве антитела, продуцируемого стабильной клеточной линией (например, повышая выход). Результаты, описанные здесь, подтверждают, что дополнительные аминокислотные остатки, расположенные дальше концевого цистеина кошачьей (и собачьей) легкой цепи каппа, снижают эффективность спаривания с тяжелой цепью, приводя к неправильному спариванию и неудовлетворительным результатам получения антител.
В одном воплощении согласно настоящему изобретению предложен способ улучшения однородности и/или характеристик кошачьего антитела. Данный способ включает экспрессию нуклеотидной последовательности, кодирующей легкую цепь каппа кошачьего IgG, и нуклеотидной последовательности, кодирующей тяжелую цепь кошачьего IgG, в клетке-хозяине с получением кошачьего антитела, где нуклеотидная последовательность, кодирующая легкую цепь каппа кошачьего IgG, содержит нуклеотидную последовательность константной области легкой цепи каппа, в которой последовательность, кодирующая С-концевую последовательность QRE, в иных случаях присутствующую в константной области легкой цепи каппа кошачьего IgG дикого типа, модифицирована и/или удалена. С учетом С-концевых аминокислотных остатков константного домена легкой цепи каппа кошачьего иммуноглобулина, показанных на Фиг. 15, в одном воплощении согласно настоящему изобретению предложено удаление С-концевой «QRE», расположенной сразу после CYS107 последовательности кошачьей легкой цепи каппа SEQ ID NO: 175. Было обнаружено, что данная модификация улучшает получение мономерного рекомбинантного кошачьего IgG. Тем не менее, настоящее изобретение не ограничено указанной модификацией. Например, даже присоединение трех дополнительных следующих друг за другом аминокислот к С-концу цистеина в положении 107 вместо нативной QRE может быть приемлемым, если эти аминокислоты оказывают минимальное влияние на электростатический заряд.
Кроме того, согласно настоящему изобретению предложен способ улучшения однородности и/или характеристик собачьего антитела. Данный способ включает экспрессию нуклеотидной последовательности, кодирующей легкую цепь каппа собачьего IgG, и нуклеотидной последовательности, кодирующей тяжелую цепь собачьего IgG, в клетке-хозяине с получением собачьего антитела, где нуклеотидная последовательность, кодирующая легкую цепь каппа собачьего IgG, содержит нуклеотидную последовательность константной области легкой цепи каппа, в которой последовательность, кодирующая С-концевую последовательность QRVD, в иных случаях присутствующую в константной области легкой цепи каппа собачьего IgG дикого типа, модифицирована и/или удалена. С учетом С-концевых аминокислотных остатков константного домена легкой цепи каппа собачьего иммуноглобулина, показанных на Фиг. 15, в одном воплощении согласно настоящему изобретению предложено удаление С-концевой «QRVD», расположенной сразу после CYS105 последовательности собачьей легкой цепи каппа SEQ ID NO: 194. Тем не менее, настоящее изобретение не ограничено указанной модификацией. Например, даже присоединение трех дополнительных следующих друг за другом аминокислот к С-концу цистеина в положении 105 вместо нативной QRVD может быть приемлемым, если эти аминокислоты оказывают минимальное влияние на электростатический заряд.
Результаты, представленные здесь, ясно демонстрируют, что описанные выше способы применимы к структурно различным антителам, распознающим совершенно разные мишени, и поэтому вероятно, что эти модификации будут применимы к широкому спектру кошачьих антител, включая, без ограничения, антитела против IL-31 и против NGF, а также к другим антителам млекопитающих, имеющим дополнительные С-концевые аминокислоты в константной области легкой цепи каппа. Без ограничения какой-либо теорией, эта модификация легкой цепи, по-видимому, приводит к более точному спариванию цепей иммуноглобулинов при их индуцированной продукции стабильными клеточными линиями СНО, результатами чего являются увеличение количества мономерного IgG и, возможно, повышение общего выхода антитела. Оба эти результата крайне желательны с точки зрения изготовления лекарственных средств, содержащих антитела, для коммерческого применения.
В одном воплощении любое из антител против IL-31, раскрытых здесь, может содержать делеции и/или модификации константной области легкой цепи каппа, раскрытые здесь. Например, в одном воплощении кошачье антитело по настоящему изобретению содержит константную область легкой цепи каппа, где «QRE», обычно присутствующая на С-конце константной области легкой цепи каппа, удалена и, возможно, заменена тремя или менее чем тремя дополнительными аминокислотами, оказывающими минимальное влияние на электростатический заряд. В одном конкретном воплощении кошачье антитело по настоящему изобретению содержит кошачью легкую цепь каппа, имеющую последовательность RSDAQPSVFLFQPSLDELHTGSASIVCILNDFYPKEVNVKWKVDGVVQNKGIQESTTEQNSKDSTYSLSSTLTMSSTEYQSHEKFSCEVTHKSLASTLVKSFQRSEC (SEQ ID NO: 186) или ее вариант.
Теперь изобретение будет описано далее посредством неограничивающих примеров, приведенных ниже. В разделе «Примеры», приведенном ниже, и в графических материалах любые данные, представленные по антителам, обозначение которых содержит «11Е12», предназначены для сравнения с антителами по настоящему изобретению.
ПРИМЕРЫ
1.1. Получение собачьего интерлейкина-31 (cIL-31) из клеток яичника китайского хомячка (СНО)
Консервативность аминокислотной последовательности белка интерлейкина-31 у гомологичных видов варьирует (Фиг. 1), но полагают, что его структурная архитектура аналогична структурной архитектуре других представителей семейства цитокинов I типа (Boulay et al. 2003, Immunity. Aug; 19(2):159-632003; Dillon et al. 2004 Nat Immunol. Jul; 5(7):752-60). Данная топология восходящих и нисходящих пучков важна для механизма распознавания рецепторов, общего для этих цитокинов (Dillon et al. supra, Comelissen et al. 2012 Eur J Cell Biol. Jun-Jul; 91(6-7):552-66). Вариабельность последовательностей белков IL-31 разных видов не позволяет предсказать, будут ли антитела, выработанные против одного вида, перекрестно взаимодействовать с другими видами, из-за различий характера эпитопов и местного аминокислотного состава. Поэтому в данной работе были рассмотрены многие формы белка IL-31, представляющие множество видов и систем экспрессии. Белок собачьего IL-31 (cIL-31) получали для использования в качестве иммуногена и реагента для анализа аффинности и активности выбранных антител. Рекомбинантный cIL-31 получали в клетках СНО с использованием системы CHROMOS АСЕ (Artificial Chromosome Expression) (Chromos Molecular Systems, Inc., Бернаби, Британская Колумбия) с получением секретированного белка собачьего IL-31, имеющего последовательность (SEQ ID NO: 155; Canine_IL31), которой соответствует нуклеотидная последовательность (SEQ ID NO: 156; Canine_IL31). Получали кондиционированную среду из 400 мл клеточной культуры (клеточная линия СНО) и проводили ее диализ против 10 объемов QA-буфера (20 мМ трис, рН 8,0, 20 мМ NaCl) на протяжении 4,5 часа. После диализа среду фильтровали через 0,2 мкм фильтр и загружали на колонку SOURCE™ Q (GE Healthcare, Уппсала, Швеция), предварительно уравновешенную QA-буфером, со скоростью 1 мл/мин. Белок элюировали с применением многоступенчатого линейного градиента. Большая часть cIL-31 оставалась в проточной (FT) фракции, небольшое количество cIL-31 элюировали в начале градиента. Идентичность белка была подтверждена ранее вестерн-иммуноблоттингом и масс-спектрометрическим (MS) анализом продуктов расщепления трипсином. Белок в FT-фракции концентрировали в 4-5 раз и проводили его диализ в течение ночи против забуференного фосфатом физиологического раствора (PBS) при 4°С. Проверяли стабильность белка после диализа в PBS. Преципитации и протеолиза по прошествии нескольких суток при 4°С не было. Эксперименты с дегликозилированием с использованием N-гликозидазы F привели к получению белка, который образовывал одну полосу массой приблизительно 15 кДа при SDS-PAGE. Концентрацию белка определяли с применением бицинхонинового анализа (ВСА-анализа) с бычьим сывороточным альбумином (BSA) в качестве стандарта (ThermoFisher Scientific, Inc., Рокфорд, штат Иллинойс). Полученный раствор белка разделяли на аликвоты, быстро замораживали (жидкий N2) и хранили при -80°С.
1.2. Транзиторная экспрессия кошачьего интерлейкина-31 (fIL-31) дикого типа и мутантного fIL-31 в клетках СНО
Для облегчения идентификации антител с подходящими эпитопсвязывающими свойствами белки кошачьего IL-31 дикого типа и мутантного кошачьего IL-31 экспрессировали в системе экспрессии с клетками млекопитающего для их получения, очистки и оценки в анализах аффинности и клеточных анализах. Сайт связывания антитела 11Е12 на IL-31 был описан ранее (патент США №8,790,651, выданный Bammert, et al.). Определение характеристик нового сайта связывания на IL-31, распознаваемого антителом 15Н05, описано здесь. Обозначение «дикий тип» соответствует полноразмерному белку кошачьего IL-31 без изменений нативных аминокислотных остатков. Мутантные белки были обозначены по названиям соответствующих им антител (11Е12 и 15Н05), указывающим на мутации аминокислот белка IL-31, которые (при их изменении) влияют на его связывание с соответствующим антителом. Определение подходящих мутаций, необходимых для белка кошачьего IL-31 15Н05, описано ниже в Разделе 1.10. Задача состояла в изменении аминокислот в эпитопе IL-31 и подтверждении снижения связывания с соответствующим антителом. Затем при скрининге можно провести сравнение и определить возможное связывание новых антител-кандидатов с белком дикого типа, но не с мутантом. Затем отобранные новые антитела можно разделить на группы по связыванию с таким же эпитопом, что антитело 11Е12 или 15Н05, или со сходным эпитопом.
Проводили оптимизацию кодонов и синтез экспрессионных конструкций для экспрессии в клетках яичника китайского хомячка (СНО). Синтезированные гены клонировали в pD2529 (ATUM-вектор) для транзиторной экспрессии. Белок кошачьего IL-31 дикого типа представлен в (SEQ ID NO: 157; Feline_IL31_wildtype), которой соответствует нуклеотидная последовательность (SEQ ID NO: 158; Feline_IL31_wildtype). Белок мутантного кошачьего IL-31 11Е12 представлен в (SEQ ID NO: 161; Feline_IL31_11E12_mutant), которой соответствует нуклеотидная последовательность (SEQ ID NO: 162; Feline_IL31_11Е12_mutant). Белок мутантного кошачьего IL-31 15Н05 представлен в (SEQ ID NO: 163; Feline_IL31_15H05_mutant), которой соответствует нуклеотидная последовательность (SEQ ID NO: 164; Feline_IL31_15H05_mutant). Рекомбинантные белки кошачьего IL-31 экспрессировали в клетках ExpiCHO-S™ (ThermoFisher Scientific, Inc., Рокфорд, штат Иллинойс), следуя протоколу изготовителя с максимальным титром для транзиторной экспрессии в СНО. Через двенадцать суток после трансфекции клетки центрифугировали и фильтровали для получения секретированного белка в кондиционированной среде. Для каждой конструкции (дикого типа и мутантов) 120 мл кондиционированной среды (из культуры клеток СНО, профильтрованной через 0,2 мкм фильтр) доводили до 30 мСм/см добавлением NaCl, 5 мМ имидазола и рН 7,4. Каждый образец среды объединяли с 5 мл смолы HisPur Cobalt (ThermoFisher Scientific, Inc., Рокфорд, штат Иллинойс), предварительно уравновешенной с использованием 5 мМ имидазола, 20 мМ фосфата натрия, 300 мМ NaCl, рН 7,4. Каждому образцу и смоле давали перемешиваться при 4°С в течение ночи. Смолы собирали (и отделяли от несвязанной фракции) пропусканием через BioRad Econcolumns (Bio-Rad, Геркулес, штат Калифорния). Смолы промывали, используя 5×5 мл буфера (как описано выше), и затем проводили элюирование с использованием 5×5 мл 500 мМ имидазола в том же буфере. Фракции анализировали посредством SDS-PAGE. Концентрацию измеряли ВСА-анализом белка с применением стандартных методов.
1.3 Получение кошачьего интерлейкина-31 (fIL-31) из E. coli
Рекомбинантный белок кошачьего IL-31 получали в экспрессионном хозяине Е. coli для использования в качестве аналитического реагента и для исследований с провокацией in vivo для индукции зуда у кошек. Ген, представляющий кошачий IL-31, синтезировали для оптимальной экспрессии в Е. coli. Получали экспрессионные конструкции с геном полноразмерного кошачьего IL-31, содержащего N-концевую 6-His-метку для выявления и очистки. Данный белок кошачьего IL-31 представлен в (SEQ ID NO: 159; Feline_IL-31_E_coli), которой соответствует нуклеотидная последовательность (SEQ ID NO: 160; Feline_IL-31_Е_coli). Плазмиды с подтвержденной последовательностью использовали для трансформации Е. coli BL21 DE3 (Invitrogen Corp., Карлсбад, штат Калифорния) и проводили последующую экспрессию белков.
Клеточную пасту (262,3 г) из Е. coli лизировали следующим образом. Клеточную пасту ресуспендировали в 500 мл 50 мМ трис, рН 8, фильтровали через фильтр с нержавеющей сеткой для удаления частиц и затем лизировали двойным пропусканием через микрофлюидизатор при 1300 psi (8,963 МПа). Полученный лизат (объемом приблизительно 1200 мл) разделяли на четыре бутылки и центрифугировали при 12000g на протяжении 20 минут при 10°С. Супернатант удаляли декантированием. Каждый осадок промывали суспендированием в 300 мл 5 мМ ЭДТА, 0,5% Triton Х-100, рН 9.0, с последующим центрифугированием при 12000g на протяжении 50 минут при 10°С. Супернатант удаляли декантированием. Промытые осадки хранили при -20°С до фолдинга и выделения.
Перед выделением один из осадков промывали водой для удаления остаточного детергента и затем центрифугировали при 10000 g на протяжении 20 минут при 4°С. Супернатант снова декантировали. В завершение, промытый осадок солюбилизировали в 60 мл 50 мМ фосфата натрия, 300 мМ NaCl, 6 М гуанидина-HCl, 5 мМ имидазола, рН 7,4. Осадку давали перемешиваться при комнатной температуре на протяжении приблизительно 25 минут перед еще одним центрифугированием при 10000g на протяжении 20 минут при 4°С. В этот раз супернатант декантировали и сохраняли для дальнейшей обработки. Осадок (ресуспендированный в воде до исходного объема) оставляли только для SDS-PAGE. Перед фолдингом проводили IMAC (афинная хроматография с использованием иммобилизованных металлов) неочищенного супернатанта для повышения степени чистоты. В данном случае 15 мл Ni-NTA Superflow (Qiagen Еле, Джермантаун, штат Мэриленд, № по каталогу 30450, предварительно уравновешенный в том же буфере) добавляли к осветленному супернатанту и давали перемешиваться при комнатной температуре в течение приблизительно 90 минут. Несвязанную фракцию декантировали и оставляли для SDS-PAGE. Смолу для IMAC промывали с использованием 5 мМ имидазола, 50 мМ фосфата натрия, 300 мМ NaCl, 6 М гуанидина-HCl, рН 7,4 (идентичного солюбилизационному буферу). Элюирование (сначала 7,5 мл и затем несколько раз по 15 мл с мониторингом элюирования белка анализом Бредфорда) проводили с использованием 200 мМ имидазола, 50 мМ фосфата натрия, 300 мМ NaCl, 6 М гуанидина-HCl, рН 7,4. Элюированные фракции, содержавшие белок (согласно анализу Бредфорда), объединяли (125 мл) для дальнейшей обработки.
Фолдинг белка IL-31 проводили следующим образом. IL-31 восстанавливали добавлением дитиотреитола до конечной концентрации 10 мМ и давали перемешиваться при комнатной температуре на протяжении 2 часов. Затем разведенный образец вносили по каплям в 2500 мл (20-кратный объем) PBS с 1 М NaCl при быстром перемешивании. При этом теоретическая концентрация мочевины должна была составлять приблизительно 0,4 М. Остаток мочевины удаляли медленным диализом против 3 смен PBS (по 4 л каждая) при 4°С в течение ночи. После диализа образец фильтровали через 0,2 мкм фильтр для удаления любого белка, не прошедшего фолдинг, или осажденного белка.
Проводили дальнейшую очистку образца посредством второго раунда IMAC, в этот раз проводя элюирование с линейным градиентом. К каждой партии образца добавляли пятнадцать мл смолы Ni-NTA Superflow и давали им связываться при перемешивании (погружной магнитной мешалкой) в течение ночи при 4°С. Несвязанную фракцию снова декантировали и оставляли. Смолу Ni-NTA Superflow помещали в колонку XK16 (GE Healthcare Lifesciences, Марлборо, штат Массачусетс) и соединяли с фирменной хроматографической системой AKTA (GE Healthcare Lifesciences, Марлборо, штат Массачусетс). Затем колонку промывали с использованием 50 мМ трис, 300 мМ NaCl, рН 8,2, после чего проводили элюирование с использованием 150 мл линейного градиента от 0 до 500 мМ имидазола в промывочном буфере. Фракции анализировали посредством SDS-PAGE. Фракции с достаточной степенью чистоты IL-31 объединяли и проводили еще одну замену буфера посредством диализа против 3 смен PBS (по 2 л каждая) при 4°С в течение ночи. В завершение, после диализа получали образец, прошедший фолдинг и очистку, стерилизовали его фильтрацией, измеряли концентрацию, разделяли на аликвоты, быстро замораживали на бане с сухим льдом/изопропанолом и хранили при -80°С.
1.4. Метод определения аффинности антител против IL-31 в отношении IL-31 с применением поверхностного плазмонного резонанса
Аффинность, с которой mAb-кандидаты связываются с кошачьим и собачьим IL-31, определяли с применением поверхностного плазмонного резонанса (SPR) и системы Biacore (Biacore Life Sciences (GE Healthcare), Уппсала, Швеция). Во избежание различий аффинности, связанных с различиями в подготовке поверхностей, возможных при иммобилизации антител на поверхностях, применяли методику с прямой конъюгацией IL-31 на поверхности. Иммобилизацию проводили аминным сочетанием 5 мкг/мл IL-31 с применением N-гидроксисукцинимидной (NHS)/1-этил-3-(3-диметиламинопропил)карбодиимидной (EDC) химии. Чипы гасили этаноламином и оценивали аффинность, с которой все mAb-кандидаты связывались с иммобилизованным IL-31. Все кривые адаптировали к модели 1:1. Константы аффинности (KD) менее 1×10-11 М (1Е-11 М) не достигают нижнего предела количественного определения прибора. Результаты измерения аффинности описаны здесь.
1.5. Метод определения активности антител против IL-31, оцениваемой по ингибированию pSTAT3-сигналов, индуцированных собачьим и кошачьим IL-31, в собачьих и кошачьих макрофагах
Для идентификации кандидатов, обладающих ингибирующей активностью, антитела оценивали на предмет их способности влиять на фосфорилирование STAT3, опосредованное IL-31, в анализе с собачьими и кошачьими клетками. Фосфорилирование STAT3 определяли в собачьих макрофагоподобных клетках DH-82 (АТСС® CRL-10389™) или кошачьих макрофагоподобных клетках FCWF4 (АТСС CRL-2787). Клетки DH82 и FCWF4 примировали собачьим интерфероном-гамма (R&D Systems, Миннеаполис, штат Миннесота), 10 нг/мл на протяжении 24 часов, или кошачьим интерфероном-гамма (R&D Systems, Миннеаполис, штат Миннесота), 125 нг/мл на протяжении 96 часов, соответственно, для повышения экспрессии рецепторов. Оба типа клеток подвергали сывороточному голоданию на протяжении 2 часов перед обработкой IL-31 и mAb. С применением двух независимых методов проводили оценку всех mAb-кандидатов на предмет их способности ингибировать фосфорилирование STAT3, индуцированное 1 мкг/мл собачьего или 0,2 мкг/мл кошачьего IL-31. Также проводили анализы для демонстрации перекрестной реактивности собачьих и кошачьих цитокинов и перекрестной функциональности способности антител ингибировать сигналы у обоих видов. Для обеспечения образования комплекса перед стимуляцией клеток проводили одночасовую совместную инкубацию mAb и цитокина IL-31. Стимуляцию клеток IL-31 проводили на протяжении пяти минут. Фосфорилирование STAT3 измеряли с применением технологии AlphaLISA SureFire ULTRA™ (Perkin Elmer, Уолтем, штат Массачусетс). В случаях, когда концентрация и степень чистоты антитела были неизвестны, проводили количественный анализ гибридомных супернатантов на предмет их способности ингибировать фосфорилирование STAT3 после 1 часа совместной инкубации с 1 мкг/мл собачьего или 0,2 мкг/мл кошачьего IL-31. Активность отдельных моноклональных антител, определенную по их способности ингибировать фосфорилирование STAT3, опосредованное IL-31, в этих анализах, рассматривали как ключевой критерий отбора для дальнейшего совершенствования отобранных антител. Термин «активность» относится к значению IC50, рассчитанному по результатам этих анализов и представляющему собой концентрацию антитела, при которой сигналы, индуцированные IL-31, снижаются до половины их максимального значения. Повышенная активность, описанная здесь, коррелирует с меньшим значением IC50.
1.6. Идентификация мышиных и собачьих моноклинальных антител, распознающих собачий и кошачий интерлейкин-31 (IL-31)
Для идентификации антител мышей и собак иммунизировали рекомбинантным собачьим IL-31 (SEQ ID NO: 155). Титры антител в сыворотке иммунизированных животных определяли с применением твердофазного иммуноферментного анализа (ELISA). Собачий или кошачий IL-31 (50 нг на лунку) иммобилизовали на полистироловых микропланшетах и использовали в качестве антигена захвата. Сыворотку иммунизированных животных разводили в забуференном фосфатом физиологическом растворе с 0,05% Tween-20 (PBST). Присутствие антител против IL-31 выявляли с использованием подходящего вторичного антитела, меченного HRP. После добавления хромогенного субстрата (SureBlue Reserve ТМВ 1-Component Microwell Peroxidase Substrate, KPL, Inc., Гейтерсберг, штат Мэриленд) и десяти минут инкубации при комнатной температуре (к.т.) реакцию останавливали добавлением 100 мкл 0,1 Н HCl. Поглощение каждой лунки определяли как оптическую плотность (OD) при 450 нм. Отбор антител по их способности связываться с собачьим и кошачьим IL-31 проводили с применением ELISA. В некоторых случаях при отборе определяли дополнительные характеристики антител с применением ELISA с мутантной формой белка кошачьего IL-31 в качестве антигена захвата. Клетки, продуцирующие антитела с желаемыми связывающими и ингибирующими свойствами, отбирали для анализа последовательности РНК-транскриптов, соответствующих вариабельным тяжелым (VH) и вариабельным легким (VL) цепям IgG.
В случае мышиных антител для слияния использовали донорные спленоциты единственной ответившей мыши CF-1 и проводили скрининг гибридомных супернатантов на предмет антител, связывавшихся с белками собачьего или кошачьего IL-31, посредством ELISA. Это привело к идентификации одного мышиного антитела, Mu-15Н05, имевшего субнаномолярную аффинность в отношении IL-31 обоих видов (Фиг. 2А). Далее мышиное антитело против IL-31 15Н05 субклонировали для получения гибридомы, продуцирующей однородное антитело, и для секвенирования вариабельной области тяжелой и вариабельной области легкой цепей. Вариабельные последовательности мышиного антитела против IL-31, определенные для антитела 15Н05, представляют собой следующее: вариабельная область тяжелой цепи 15Н05 (SEQ ID NO: 67; MU-15H05-VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 68; MU-15H05-VH), вариабельная область легкой цепи 15Н05 (SEQ ID NO: 69; MU-15H05-VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 70; MU-15H05-VL).
Помимо мышиного антитела 15Н05, также рассматривалось антитело мышиного происхождения 11Е12, описанное ранее в патенте США №8,790,651, выданном Bammert, et al. Здесь описаны данные, показывающие способность антитела 11Е12 связываться с белками как собачьего, так и кошачьего IL-31 с высокой аффинностью. Способность 11Е12 связываться с кошачьим IL-31 сделало это антитело подходящим кандидатом для фелинизации и возможного терапевтического применения у кошек. Вариабельные последовательности мышиного антитела против IL-31, определенные ранее для антитела 11Е12, представляют собой следующее: вариабельная область тяжелой цепи 11Е12 (SEQ ID NO: 71; MU-11E12-VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 72; MU-11E12-VH), вариабельная область легкой цепи 15Н05 (SEQ ID NO: 73; MU-11E12-VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 74; MU-11E12-VL).
Собаки с повышенными титрами антител против IL-31 после вакцинации были выбраны для анализа В-клеточных популяций, продуцирующих антитела с желаемыми фенотипами. В-клетки для дальнейшего анализа получали из МКПК, костного мозга, селезенки или лимфатических узлов. Одиночные В-клетки разносили по отдельным лункам и анализировали на предмет секретируемых IgG, способных связываться с собачьим IL-31 дикого типа и мутантными формами собачьего IL-31 11Е12 и 15Н05 (AbCellera, Ванкувер, Британская Колумбия), с применением методов, описанных в US 2012/0009671 A1, US 2016/0252495 A1, US 9,188,593, WO 2015/176162 A9 и WO 2016/123692 A1.
Эта методика скрининга основана на известных областях белка IL-31, критически важных для связывания и трансдукции сигналов через его корецепторный комплекс. Отбор этих мутантных белков для скрининга описан в Разделе 1.2 данной заявки. Секвенирование вариабельных доменов тяжелых и легких цепей IgG проводили после реакции ОТ-ПЦР от отдельных В-клеток-кандидатов. Этот скрининг привел к идентификации девяти собачьих антител, отобранных для дальнейшего анализа. Вариабельные последовательности этих собачьих антител против IL-31 представляют собой следующее: вариабельная область тяжелой цепи ZIL1 (SEQ ID NO: 75; CAN-ZIL1_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 76; CAN-ZIL1_VH), вариабельная область легкой цепи ZIL1 (SEQ ID NO: 77; CAN-ZIL1_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 78; CAN-ZIL1_)VL); вариабельная область тяжелой цепи ZIL8 (SEQ ID NO: 79; CAN-ZIL8_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 80; CAN-ZIL8_VH), вариабельная область легкой цепи ZIL8 (SEQ ID NO: 81; CAN-ZIL8_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 82; CAN-ZIL8_VL); вариабельная область тяжелой цепи ZIL9 (SEQ ID NO: 83; CAN-ZIL9_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 84; CAN-ZIL9_VH), вариабельная область легкой цепи ZIL9 (SEQ ID NO: 85; CAN-ZIL9_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 86; CAN-ZIL9_VL); вариабельная область тяжелой цепи ZIL11 (SEQ ID NO: 87; CAN-ZIL11_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 88; CAN-ZIL11_VH), вариабельная область легкой цепи ZIL11 (SEQ ID NO: 89; CAN-ZIL11_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 90; CAN-ZIL11_VL); вариабельная область тяжелой цепи ZIL69 (SEQ ID NO: 91; CAN-ZIL69_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 92; CAN-ZIL69_VH), вариабельная область легкой цепи ZIL69 (SEQ ID NO: 93; CAN-ZIL69_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 94; CAN-ZIL69_VL); вариабельная область тяжелой цепи ZIL94 (SEQ ID NO: 95; CAN-ZIL94_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 96; CAN-ZIL94_VH), вариабельная область легкой цепи ZIL94 (SEQ ID NO: 97; CAN-ZIL94_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 98; CAN-ZIL94_VL); вариабельная область тяжелой цепи ZIL154 (SEQ ID NO: 99; CAN-ZIL154_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 100; CAN-ZIL154_VH), вариабельная область легкой цепи ZIL154 (SEQ ID NO: 101; CAN-ZIL154_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 102; CAN-ZIL154_VL); вариабельная область тяжелой цепи ZIL159 (SEQ ID NO: 103; CAN-ZIL159_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 104; CAN-ZIL159_VH), вариабельная область легкой цепи ZIL159 (SEQ ID NO: 105; CAN-ZIL159_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 106; CAN-ZIL159_VL); вариабельная область тяжелой цепи ZIL171 (SEQ ID NO: 107; CAN-ZIL171_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 108; CAN-ZIL171_VH), вариабельная область легкой цепи ZIL171 (SEQ ID NO: 109; CAN-ZIL171_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 110; CAN-ZIL171_VL).
Указанные выше девять моноклональных антител, которые были отобраны для дальнейшего изучения, могут быть обозначены в других частях данного описания, графических материалах или формуле изобретения как ZIL1, ZIL8, ZIL8, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171.
1.7. Конструирование рекомбинантных химерных и полностью собачьих антител
Вариабельные домены антитела обеспечивают его связывание с антигеном. Ожидают, что перенос полноразмерного вариабельного домена на соответствующую константную область окажет незначительное влияние или не повлияет на способность антитела связываться с иммуногеном IL-31. Для одновременного подтверждения правильности определения последовательности вариабельных областей тяжелых и легких цепей и получения однородного материала были разработаны векторы экспрессии для получения рекомбинантных химерных и полностью собачьих антител в системах экспрессии с клетками млекопитающих. Химерные антитела, описанные здесь, состоят из вариабельной последовательности (как CDR, так и каркасных областей) антитела вида-хозяина, перенесенных на соответствующие константные области тяжелой и легкой цепи молекулы кошачьего или собачьего IgG (например, мышиную вариабельную область с собачьей константной областью называют химерным мышиным-собачьим антителом). Полностью собачьи антитела, описанные здесь, состоят из вариабельной последовательности (как CDR, так и каркасных областей) антитела вида-хозяина (собаки), перенесенных на соответствующие константные области тяжелой и легкой цепи молекулы собачьего IgG. Для последовательностей вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) отобранных антител конструировали синтетические последовательности ДНК. Эти последовательности содержат уникальные рестриктазные сайты, консенсусную последовательность Козак и N-концевую секреторную лидерную последовательность для облегчения экспрессии и секреции рекомбинантного антитела из линии клеток млекопитающего.
Для мышиных-кошачьих химерных антител каждую соответствующую вариабельную область клонировали в плазмиду для экспрессии в клетках млекопитающих, содержащую константную область тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_HC_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_HC_AlleleA_1), или константную область легкой цепи кошачьего IgG (SEQ ID NO: 175; Feline_LC_Kappa_G_minus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 176; Feline_LC_Kappa_G_minus). Для мышиных-собачьих химерных антител или полностью собачьих антител каждую мышиную или собачью вариабельную область клонировали в плазмиду для экспрессии в клетках млекопитающих, содержащую константную область тяжелой цепи собачьего IgG (SEQ ID NO: 177; Canine_HC_65_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 178; Canine_НС_65_1), или константную область легкой цепи собачьего IgG (SEQ ID NO: 179; Canine_LC_Kappa), которой соответствует нуклеотидная последовательность (SEQ ID NO: 180; Canine_LC_Kappa). Плазмидами, кодирующими каждую тяжелую и легкую цепь, под контролем CMV-промотора котрансфицировали клетки HEK 293 с применением стандартных методов. После шести суток экспрессии химерные mAb очищали из 50 мл супернатантов транзиторно трансфицированных клеток HEK293FS, используя смолу с белком А MabSelect Sure (GE Healthcare, Уппсала, Швеция) в соответствии со стандартными методами очистки белков. Элюированные фракции объединяли, концентрировали до приблизительно 500 мкл с использованием центрифуги Nanosep Omega (Pall Corp., Порт-Вашингтон, штат Нью-Йорк) с номинальным отсечением по молекулярной массе 10000, подвергали диализу в течение ночи при 4°С в 1x PBS, рН 7,2, и хранении при 4°С. Аффинность и активность отобранных рекомбинантных антител в клеточных анализах описаны ниже.
На Фиг. 2 подробно показана аффинность антител с CDR мышиного происхождения при применении Biacore. На Фиг. 2а показана аффинность мышиных антител против IL-31 11Е12 и 15Н05 и соответствующие аффинности кошачьих и собачьих химерных форм в отношении поверхностей с кошачьим и собачьим IL-31. Эти данные подтверждают правильность последовательности обоих мышиных антител и указывают на то, что превращение в химерные формы приводит к антителам с эквивалентной или более высокой аффинностью, по сравнению с исходными мышиными антителами, за исключением мышиного-кошачьего химерного антитела 15Н05, аффинность которого в отношении IL-31 обоих видов была в некоторой степени снижена в результате превращения в химерную форму.
Полностью мышиные и химерные формы антител 11Е12 и 15Н05 были также проанализированы на предмет активности в анализах с собачьими и кошачьими клетками, описанных в Разделе 1.5. Результаты этих анализов показаны на Фиг. 3. Мышиные антитела 11Е12 и 15Н05 анализировали на предмет активности против собачьих и кошачьих типов клеток с использованием как собачьего, так и кошачьего IL-31 для стимуляции сигналов. При использовании кошачьего цитокина активность обоих мышиных антител против собачьих и кошачьих клеток была сопоставимой, за исключением 15Н05 против кошачьего IL-31 у кошачьих клеток FCWF4, где показано незначительное повышение IC50. Мышиное 15Н05 было способно блокировать сигналы собачьего IL-31 как в кошачьих, так и в собачьих клетках, при этом его активность в анализе с собачьими клетками была несколько выше. Эти результаты показывают, что соответствующие эпитопы, распознаваемые этими антителами, есть как на собачьем, так и кошачьем IL-31 и что связывание этих антител способно нейтрализовать рецептор-опосредованные клеточные сигналы в соответствующих клеточных линиях обоих видов.
На Фиг. 3 также описана активность отобранных химерных антител в обоих клеточных анализах. Превращение мышиных антител в кошачьи и собачьи химерные антитела оказывало минимальное влияние на активность против кошачьего IL-31 в анализе активности с кошачьими клетками (диапазон IC50 1,15-3,45 мкг/мл). Сходные результаты были отмечены при анализе этих химерных антител против сигналов кошачьего IL-31 в собачьих клетках линии DH82, при этом наблюдали незначительное повышение активности (IC50 0,71 мкг/мл) мышиного-собачьего химерного антитела 15Н05. В целом, было отмечено повышение значений IC50 против собачьего IL-31 как у собачьих, так и у кошачьих типов клеток. В анализе данного формата активность мышиного-кошачьего химерного антитела 15Н05 была несколько ниже активности мышиной-собачьей формы (IC50 28,61 против 12,49 мкг/мл). Аналогично мышиным антителам, превращение в собачьи и кошачьи химерные формы приводило к минимальным изменениям активности.
Антитела, описанные выше, которые были идентифицированы из отдельных В-клеток иммунизированных собак, конструировали в форме рекомбинантных белков IgG после определения последовательностей их вариабельных доменов. Перенос этих вариабельных доменов на Fc собачьей тяжелой цепи (изотип 65 1) приводил к получению рекомбинантных полностью собачьих антител. Представляло интерес идентифицировать дополнительные собачьи антитела, которые связывались бы с кошачьим IL-31 дикого типа и связывание которых с мутантом кошачьего IL-31 15Н05 было бы снижено (то есть таких, которые были бы направлены на эпитоп 15Н05). Такие антитела, полученные из этого альтернативного источника (собака против мыши), обеспечивают паратопы (часть антитела, распознающая белок IL-31, содержащая CDR), распознающие эпитоп 15Н05, повышая, таким образом, разнообразие антител с различными физическими свойствами для селекции.
На Фиг. 4 показаны результаты, полученные применительно к связыванию этих рекомбинантных собачьих антител с различными белками с применением как метода ELISA, так и метода Biacore. При непрямом ELISA оценивали связывание антител с белками кошачьего IL-31 дикого типа и мутантного кошачьего IL-31 15Н05. Все девять собачьих моноклональных антител (ZIL1, ZIL8, ZIL8, ZIL11, ZIL69, ZIL94, ZIL154, ZIL159 и ZIL171) были способны связываться с кошачьим IL-31 дикого типа, и на это связывание влияли мутации в области эпитопа mAb 15Н05, что подтверждает правильное определение фенотипа связывания при начальном скрининге, примененном для их идентификации. В сравнении с этим, антитело 11Е12 связывалось с кошачьим IL-31 дикого типа, и на это связывание не влияли мутации в области эпитопа 15Н05, на что указывают данные, представленные на Фиг. 4. Для подтверждения связывания проводили анализ Biacore, используя поверхности с белками собачьего, кошачьего, лошадиного, человеческого IL-31, мутанта кошачьего IL-31 15Н05 и мутанта кошачьего IL-31 11Е12 при единственной анализируемой концентрации антитела. Аналогично данным, полученным при ELISA, все антитела связывались с кошачьим IL-31 дикого типа. В соответствии с данными, описанными выше в этом разделе, как мышиное антитело 11Е12, так и мышиное антитело 15Н05 связывались с поверхностями с собачьим и кошачьим IL-31. Это свойство двойного связывания было показано еще у трех антител: ZIL69 (частичное связывание с собачьим белком), ZIL94 и ZIL159. Из этой группы из девяти полностью собачьих антител только ZIL1 и ZIL9 обладали перекрестной реактивностью в отношении лошадиного IL-31. Следует отметить, что из всех антител, проанализированных здесь, антитело 15Н05 было единственным связывавшимся с собачьим, кошачьим и лошадиным IL-31, указывая на некоторую степень консервативности эпитопов у этих трех видов. В отличие от этого, ни одно из антител, описанных здесь, не связывалось с человеческим IL-31. Для верификации результатов ELISA, указывающих на дифференциальное связывание антител с кошачьим IL-31 дикого типа и двумя белками с мутациями в эпитопах 15Н05 (мутант 15Н05) или 11Е12 (мутант 11Е12), использовали дополнительные Biacore-поверхности. Как и следовало ожидать, мышиное антитело 11Е12 связывалось с мутантом IL-31 15Н05 и не связывалось с мутантом IL-31 11Е12 из-за мутаций в соответствующем эпитопе. Сходным образом, 15Н05 не связывалось с мутантом 15Н05 и сохраняло связывание с мутантом IL-31 11Е12, дополнительно подтверждая, что эти два антитела связываются с раздельными эпитопами. В подтверждение результатов ELISA, все полностью собачьи антитела были чувствительны к мутациям в 15Н05, за исключением ZIL94, ZIL154 и ZIL171 (частично чувствительные). Различия результатов могут быть связаны с различиями двух методов анализа. Кроме того, было показано, что мутации в 11Е12 приводили к снижению связывания трех антител: ZIL1 (частичное снижение), ZIL8 и ZIL159. Эти результаты показывают, что эпитоп, распознаваемый этими антителами, подвержен влиянию изменений в обеих областях белка IL-31. Вместе взятые, эти результаты способствуют определению характеристик девяти антител, имеющих происхождение от собачьих В-клеток, связывающихся с областью белка кошачьего IL-31, распознаваемой антителом 15Н05.
1.8. Фелинизация мышиных антител 11Е12 и 15Н05 и оптимизация аффинности связывания
Выработка противолекарственных антител (ADA) может приводить к снижению эффективности любых биотерапевтических белков, включая моноклональные антитела. Всесторонний анализ литературы показал, что видовая адаптация моноклональных антител позволяет снизить склонность mAb к иммуногенности, несмотря на то, что можно найти примеры иммуногенных полностью человеческих mAb и неиммуногенных химерных mAb. Для снижения рисков, связанных с выработкой ADA, при применении моноклональных антител против IL-31, предложенных здесь, была применена методика фелинизации. Эта методика фелинизации основана на идентификации кошачьей последовательности антитела эмбрионального типа, наиболее подходящей для переноса CDR. После тщательного анализа всех доступных кошачьих последовательностей эмбрионального типа, как тяжелой цепи, так и легкой цепи, кандидаты эмбрионального типа были отобраны, исходя из их гомологии мышиным mAb, и CDR мышиных mAb-предшественников были использованы для замены нативных кошачьих CDR. Задача состояла в сохранении высокой аффинности и активности в отношении клеток при использовании каркасных областей кошачьих антител для минимизации иммуногенного потенциала in vivo. Фелинизированные mAb экспрессировали и характеризовали на предмет их аффинности в отношении кошачьего IL-31 и их активности в клеточных анализах. В случае, если фелинизированное антитело утрачивало способность к связыванию с IL-31, проводили систематический анализ для определения (1) цепи, приведшей к потере функции, (2) каркасной области, приведшей к потере функции, и (3) аминокислоты (аминокислот), приведшей к потере функции.
Получали синтетические нуклеотидные конструкции, соответствовавшие фелинизированным вариабельным тяжелым и легким цепям mAb 11Е12 и 15Н05. После субклонирования каждой вариабельной цепи в плазмиды, содержавшие соответствующие константные области тяжелой цепи или легкой цепи каппа, клетки HEK 293 котрансфицировали полученными плазмидами для экспрессии антител. Исходные попытки фелинизации антитела 11Е12 были сосредоточены на использовании каркасных областей одной кошачьей VH (SEQ ID NO: 111; FEL_11E12_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 112; FEL_11E12_VH1), при их независимом сочетании с каркасными областями VL (SEQ ID NO: 113; FEL_11E12_VL1), которой соответствует нуклеотидная последовательность (SEQ ID NO:114; FEL_11E12_VL1), и (SEQ ID NO: 115; FEL_11E12_VL2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 116; FEL_11E12_VL2), с получением Feline 11E12 1.1 и Feline 11E12 1.2, соответственно. Эта попытка видовой адаптации привела к снижению аффинности у Feline 11Е12 1.1 в отношении белков как кошачьего, так и собачьего IL-31 и полной потере связывания у mAb Feline 11Е12 1.2 по сравнению с мышиной формой этого антитела (Фиг. 2b). Активность этих антител после видовой адаптации оценивали в анализах с собачьими клетками DH82 и кошачьими клетками FCWF4 с использованием цитокина кошачьего IL-31. Активность фелинизированного 11Е12 1.1 против кошачьего IL-31 в анализе с кошачьими клетками FCWF была снижена приблизительно в два раза по сравнению с мышиной формой этого антитела. Аналогично полной потере аффинности фелинизированного 11Е12 1.2, у этого антитела наблюдали полную потерю активности в отношении клеток (Фиг. 3). Исходя из предшествующего опыта канинизации ортолога mAb 11Е12, сходная методика была применена для восстановления аффинности, сниженной при фелинизации (патент США №8,790,651, выданный Bammert, et al.). Была проведена замена фелинизированной каркасной области 2 (FW2) Feline 11Е12 VL1 на мышиную FW2 (SEQ ID NO: 73; Mu_11E12_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 74; Mu_11E12_VL), с получением Feline 11E12 VL1 FW2. Кроме того, была проведена одиночная замена в положении 46 кошачьей VL (K46Q) с получением (SEQ ID NO: 119; FEL_11Е12_VL1_K46Q), которой соответствует нуклеотидная последовательность (SEQ ID NO: 120; FEL_11E12_VL1_K46Q). Сочетание указанных выше VL с Fel_11E12_VH1 приводило к получению Feline 11Е12 1.1 FW2 и Feline 11Е12 1.1 K46Q, соответственно. Изменение FW2 приводило к восстановлению аффинности Feline 11Е12 1.1 FW2 в отношении белка кошачьего IL-31 с KD, эквивалентной KD мышиной и химерной формы (Фиг. 2А и 2В). Тем не менее, эти изменения отрицательно влияли на аффинность Feline 11Е12 1.1 FW2 в отношении белка собачьего IL-31, указывая на явные различия способности антитела 11Е12 к связыванию с этим эпитопом на кошачьем и собачьем цитокине. Одиночная аминокислотная замена в Feline 11Е12 1.1 K46Q не влияла на аффинность этого антитела. Повышенная аффинность антитела 11Е12 1.1 FW2 в отношении белка кошачьего IL-31 привела к росту его активности против кошачьего цитокина в анализе с собачьими клетками DH82 (Фиг. 3).
Попытки фелинизации мышиного антитела 15Н05 были сосредоточены на комбинациях каркасных областей трех кошачьих VH с каркасными областями трех кошачьих VL с получением в общей сложности 9 фелинизированных mAb. FEL_15H05_VH1 (SEQ ID NO: 121; FEL_15H05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15H05_VH1), комбинировали с (SEQ ID NO: 127; FEL_15Н05_VL1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 128; FEL_15Н05_VL1), (SEQ ID NO: 129; FEL_15H05_VL2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 130; FEL_15H05_VL2), и (SEQ ID NO: 131; FEL_15H05_VL3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 132; FEL_15H05_VL3) с получением Feline 15H05 1.1, Feline 15H05 1.2 и Feline 15H05 1.3, соответственно. FEL_15H05_VH2 (SEQ ID NO: 123; FEL_15H05_VH2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 124; FEL_15H05_VH2), комбинировали с (SEQ ID NO: 127; FEL_15H05_VL1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 128; FEL_15H05_VL1), (SEQ ID NO: 129; FEL_15H05_VL2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 130; FEL_15H05_VL2), и (SEQ ID NO: 131; FEL_15H05_VL3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 132; FEL_15H05_VL3) с получением Feline 15H05 2.1, Feline 15H05 2.2 и Feline 15H05 2.3, соответственно. FEL_15H05_VH3 (SEQ ID NO: 125; FEL_15H05_VH3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 126; FEL_15H05_VH3), комбинировали с (SEQ ID NO: 127; FEL_15H05_VL1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 128; FEL_15H05_VL1), (SEQ ID NO: 129; FEL_15H05_VL2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 130; FEL_15H05_VL2), и (SEQ ID NO: 131; FEL_15H05_VL3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 132; FEL_15H05_VL3) с получением Feline 15H05 3.1, Feline 15H05 3.2 и Feline 15H05 3.3, соответственно. Аналогично антителу 11E12, первая попытка фелинизации антитела 15Н05 привела к снижению аффинности в отношении белка кошачьего IL-31 по сравнению с мышиным 15Н05 и к нейтральному эффекту по сравнению с химерным мышиным-кошачьим 15Н05 (Фиг. 2А и 2С). Аналогично связыванию фелинизированного антитела 11Е12 с собачьим IL-31, определенные комбинации кошачьих каркасных областей VH и VL в 15Н05 оказывали нейтральное или положительное влияние на аффинность в отношении собачьего IL-31 (см. Фиг. 2С: Feline 15Н05 1.1, 2.2 и 3.2).
Для восстановления аффинности фелинизированного антитела 15Н05 VH каждого фелинизированного 15Н05 сочетали с VL мышиного 15Н05 с получением гетерохимерных антител. FEL_15Н05_VH1 (SEQ ID NO: 121; FEL_15Н05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15H05_VH1), комбинировали с MU_15H05_VL (SEQ ID NO: 69; MU_15H05_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 70; MU_15Н05_VL), с получением Feline 15Н05 VH1 mouse VL. FEL_15Н05_VH2 (SEQ ID NO: 123; FEL_15H05_VH2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 124; FEL_15H05_VH2), комбинировали с MU_15H05_VL (SEQ ID NO: 69; MU_15H05_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 70; MU_15H05_VL), с получением Feline 15Н05 VH2 mouse VL. FEL_15H05_VH3 (SEQ ID NO: 125; FEL_15H05_VH3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 126; FEL_15H05_VH3), комбинировали с MU_15H05_VL (SEQ ID NO: 69; MU_15H05_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 70; MU_15H05_VL), с получением Feline 15Н05 VH3 mouse VL. Эти гетерохимерные антитела с фелинизированными VH и мышиными VL анализировали на предмет их аффинности в отношении собачьего и кошачьего IL-31. Сочетание VH1 и VH3 фелинизированного 15Н05 с VL мышиного 15Н05 приводило к восстановлению аффинности в отношении кошачьего IL-31 до уровня, эквивалентного или превышающего аффинность мышиной и химерной формы. Эта тенденция к улучшению аффинности была также отмечена при использовании белка собачьего IL-31 (Фиг. 2А и 2С).
Для дополнительного уточнения положений каркасных областей 15Н05, приводящих к снижению аффинности, один фелинизированный VH 15Н05 (FEL_15Н05_VH1) использовали для сочетания с отдельными заменами в каркасных областях VL мышиного 15Н05. FEL_15Н05_VH1 (SEQ ID NO: 122; FEL_15Н05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 123; FEL_15H05_VH1), комбинировали независимым образом с FEL_15H05_VL1-FW1 (SEQ ID NO: 133; FEL_15H05_VL1_FW1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 134; FEL_15H05_VL1_FW1), FEL_15H05_VL1_FW2 (SEQ ID NO: 135; FEL_15H05_VL1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 136; FEL_15H05_VL1_FW2), и FEL_15H05_VL1_FW3 (SEQ ID NO: 137; FEL_15H05_VL1_FW3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 138; FEL_15H05_VL1_FW3), с получением Feline 15H05 1.1 FW1, Feline 15H05 1.1 FW2 и Feline 15H05 1.1 FW3, соответственно. Замена с введением FW1 мышиного 15Н05 в Feline 15Н05 1.1 снижала аффинность как в отношении кошачьего, так и в отношении собачьего IL-31, тем не менее, при замене с введением мышиной FW2 или FW3 в Feline 15Н05 1.1 была получена отличная аффинность в отношении собачьего и кошачьего IL-31, при этом замена FW2 приводила к большему росту аффинности в отношении обоих видов (Фиг. 2С). Были проведены дополнительные попарные замены каркасных областей для определения степени модуляции аффинности при применении данной методики. FEL_15H05_VH1 (SEQ ID NO: 121; FEL_15Н05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15H05_VH1), комбинировали независимым образом с FEL_15Н05_VL1_FW1_2 (SEQ ID NO: 139; FEL_15Н05_VL1_FW1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 140; FEL_15Н05_VL1_FW1_FW2), FEL_15H05_VL1_FW2_3 (SEQ ID NO: 143; FEL_15H05_VL1_FW2_FW3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 144; FEL_15H05_VL1_FW2_FW3), и FEL_15H05_VL1_FW1_3 (SEQ ID NO: 141; FEL_15H05_VL1_FW1_FW3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 142; FEL_15H05_VL1_FW1_FW3), с получением Feline 15H05 1.1 FW1_2, Feline 15H05 1.1 FW2_3 и Feline 15H05 1.1 FW1_3. Интересно, что замена только мышиной FW1 приводила к снижению аффинности, в то время как комбинации FW1 с FW2 или FW3 приводили к хорошей аффинности как в отношении кошачьего, так и в отношении собачьего IL-31 (Фиг. 2С).
В завершение, была предпринята попытка минимизировать число обратных мутаций в кошачьих каркасных областях, начиная с наиболее перспективных комбинаций фелинизированных последовательностей VH и VL. Для этого FEL_15Н05_VH1 (SEQ ID NO: 121; FEL_15Н05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15H05_VH1) комбинировали независимым образом с FEL_15H05_VL1_FW2_K42N (SEQ ID NO: 145; FEL_15H05_VL1_FW2_K42N), которой соответствует нуклеотидная последовательность (SEQ ID NO: 146; FEL_15H05_VL1_FW2_K42N), FEL_15H05_VL1_FW2_V43I (SEQ ID NO: 147; FEL_15H05_VL1_FW2_V43I), которой соответствует нуклеотидная последовательность (SEQ ID NO: 148; FEL_15H05_VL1_FW2_V43I), FEL_15H05_VL1_FW2_L46V (SEQ ID NO: 149; FEL_15H05_VL1_FW2_L46V), которой соответствует нуклеотидная последовательность (SEQ ID NO: 150; FEL_15H05_VL1_FW2_L46V), FEL_15H05_VL1_FW2_Y49N (SEQ ID NO: 151; FEL_15H05_VL1_FW2_Y49N), которой соответствует нуклеотидная последовательность (SEQ ID NO: 152; FEL_15H05_VL1_FW2_Y49N), и FEL_15H05_VL1_FW2_K42N_V43I (SEQ ID NO: 153; FEL_15H05_VL1_FW2_K42N_V43I), которой соответствует нуклеотидная последовательность (SEQ ID NO: 154; FEL_15H05_VL1_FW2_K42N_V43I) с получением Feline 15H05 1.1 K42N, Feline 15H05 1.1 V43I, Feline 15H05 1.1 L46V, Feline 15H05 1.1 Y49N и Feline 15H05 1.1 K42N_V43I, соответственно. В то время как замена с введением всей мышиной каркасной области FW2 в Felinized 15Н05 VL1 приводила к получению антитела с отличной аффинностью в отношении собачьего и кошачьего IL-31 (Фиг. 2С, Feline 15Н05 1.1 FW2), отдельные обратные мутации аминокислотных остатков FW2 оказывали нейтральный или отрицательный эффект, указывая на необходимость всех 4 замен для поддержания оптимальной третичной структуры, обеспечивающей расположение CDR на эпитопе IL-31. Повышение аффинности фелинизированного 15Н05 1.1 FW2 в отношении кошачьего и собачьего IL-31 привела к выбору этого антитела для дальнейшей работы.
На Фиг. 5А показано выравнивание с последовательностью VL мышиного антитела 11Е12 при сравнении последовательности упомянутого ранее канинизированного 11Е12 с фелинизированными вариантами. Под выравниванием отмечены точки, показывающие положения важных изменений в Fel_11E12_VL1, которые были необходимы для восстановления аффинности этого антитела в отношении белка IL-31. Сходным образом, на Фиг. 5В показаны необходимые изменения в VL фелинизированного 15Н05 (Fel_15Н05_VL1), которые были нужны не только для восстановления, но и для улучшения его аффинности в отношении собачьего и кошачьего IL-31 по сравнению с мышиной и химерной формой этого антитела.
1.9. Получение клеточных линий, экспрессирующих фелинизированные антитела с глутаминсинтетазных (GS) плазмид
Фелинизированное 15Н05 1.1 FW2 (Felinized 15Н05 1.1 FW2) было выбрано в качестве кандидата для получения стабильных клеточных линий, которые будут продуцировать однородное антитело для дальнейшего определения его характеристик. Гены, кодирующие фелинизированные тяжелые и легкие цепи, для получения клеточных линий клонировали в GS-плазмиды рЕЕ 6.4 и рЕЕ 12.4, соответственно (Lonza, Базель, Швейцария). Полученные плазмиды расщепляли по протоколу изготовителя и лигировали друг с другом, получая единую плазмиду для экспрессии в клетках млекопитающих. Для ZTS-927 тяжелая цепь представляет собой (SEQ ID NO: 121; FEL_15H05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15Н05_VH1), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 171; Feline_HC_AlleleA_wt), которой соответствует нуклеотидная последовательность (SEQ ID NO: 172; Feline_HC_AlleleA_wt). Для ZTS-927 легкая цепь представляет собой (SEQ ID NO: 135; FEL-15H05-VL1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 136; FEL-15H05-VL1_FW2), в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 175; Feline_LC_Kappa_Gminus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 176; Feline_LC_Kappa_G_minus). Для ZTS-361 тяжелая цепь представляет собой (SEQ ID NO: 121; FEL_15H05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15H05_VH1), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_НС_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_НС_AlleleA_1). Для ZTS-361 легкая цепь представляет собой (SEQ ID NO: 135; FEL-15H05-VL1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 136; FEL-15H05-VL1_FW2), в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 175; Feline_LC_Kappa_G_minus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 176; Feline_LC_Kappa_G_minus). Для ZTS-1505 тяжелая цепь представляет собой (SEQ ID NO: 121; FEL_15H05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15H05_VH1), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_НС_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_HC_AlleleA_1). Для ZTS-1505 легкая цепь представляет собой (SEQ ID NO: 135; FEL-15H05-VL1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 136; FEL-15H05-VL1_FW2), в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 186; Feline_LC_Kappa_G_minus_QRE_minus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 187; Feline_LC_Kappa_G_minus_QRE_minus). Данные по аффинности и активности ZTS-1505 описаны ниже в Разделе 1.18 данного раздела «Примеры».
Для демонстрации транзиторной продукции антитела каждую плазмиду использовали для трансфекции клеток HEK 293 и проводили экспрессию в культурах разного размера. Белок выделяли из кондиционированной среды HEK с применением хроматографии с белом А в соответствии со стандартными методами очистки белков. Среду загружали на хроматографическую смолу и проводили элюирование со сдвигом рН. Перед использованием элюированного белка его подвергали коррекции рН, диализу и стерилизующей фильтрации. ZTS-361 впоследствии использовали в модели зуда у кошек для оценки эффективности in vivo.
Антитела, полученные с одной GS-плазмиды, ZTS-927 и ZTS-361, анализировали на предмет аффинности и активности. На Фиг. 2D показаны результаты оценки аффинности этих антител с применением Biacore. Аффинность ZTS-927 и ZTS-361 в отношении кошачьего IL-31 в высокой степени соответствует аффинности мышиной и химерной формы их мышиного mAb-предшественника 15Н05. Активность этих двух антител определяли против собачьего и кошачьего IL-31 с применением анализов как с собачьими, так и с кошачьими клетками (Фиг. 3). Аналогично предшествующим наблюдениям, значения IC50 были пропорционально выше при использовании собачьей формы IL-31 с клетками обоих типов. Значения IC50 ZTS-927 и ZTS-361 против кошачьего IL-31 также в высокой степени соответствовали значениям IC50 химерной и мышиной форм антитела, указывая на то, что конечный фелинизированный вариант mAb 15Н05, полученный с одной GS-плазмиды, был подходящим для разработки клеточной линии.
Для получения стабильной клеточной линии, продуцирующей антитела-кандидаты, GS-плазмиду линеаризовали перед трансфекцией, используя рестриктазу PvuI, расщепляющую остов плазмиды в единственном сайте. Клетки GS-CHOK1SV (клон 144Е12) трансфицировали линеаризованной плазмидной ДНК посредством электропорации. После трансфекции клетки высевали в 48-луночные планшеты (48WP) для получения стабильных пулов. При по меньшей мере 50% смыкания монослоя в 48-луночных планшетах 100 мкл супернатанта анализировали на предмет экспрессии IgG с применением ForteBio Octet и биосенсоров с белком A (Pall ForteBio, Фримонт, штат Калифорния). Клоны с наилучшей экспрессией масштабировали в 6-луночных планшетах (6WP), а затем во встряхиваемых колбах (SF) объемом 125 мл. После адаптации клеток к суспензионной культуре в колбах объемом 125 мл по 2 колбы пула каждой клеточной линии сохраняли для длительного хранения. Поскольку промышленные клеточные линии должны быть клональными, 3 пула с наиболее высокой экспрессией были субклонированы серийным разведением в 96-луночных культуральных планшетах. Для подтверждения клональности и во избежание второго раунда серийного разведения 96-луночные планшеты визуализировали с использованием устройства Molecular Devices Clone-Select Imager (CSI) (Molecular Devices LLC, Сан-Хосе, штат Калифорния), позволяющего получить изображения отдельных клеток и их последующего роста. Отбор клонов проводили, исходя из успешных CSI-изображений, роста и продукции антител в 96-луночных планшетах.
Для оценки роста и продуктивности клеточных культур проводили дальнейшую оценку пулов с наибольшей экспрессией в подпитываемой культуре во встряхиваемых колбах объемом 125 мл на протяжении 14 суток. Посев клеток проводили с использованием сред-платформ и подпиток, состоявших из CD СНО от Life Technologies с 4 аминокислотами, собственной подпитки CDF v6.2 и 10%-й глюкозы. После 14 суток в подпитываемой культуре пулы центрифугировали и полученное mAb в CD СНО выделяли фильтрацией супернатанта через 0,20 мкм полиэфирсульфоновую мембрану (PES) перед очисткой.
Типичная очистка состоит из двух литров кондиционированной среды (из культуры клеток СНО, профильтрованной через 0,2 мкм фильтр), загружаемых на колонку MabSelect (GE healthcare, № по каталогу 17-5199-02) объемом 235 мл. Колонку предварительно уравновешивали с использованием PBS. Образец загружали при времени удержания более 2,5 минуты. После загрузки колонку промывали еще раз с использованием PBS и затем 25 мМ ацетатом натрия с рН, близким к нейтральному. Проводили элюирование с использованием 25 мМ уксусной кислоты, рН 3,6, и затем десорбцию с использованием 250 мМ уксусной кислоты, 250 мМ хлорида натрия, рН приблизительно 2,2. Фракции (50 мл) собирали на стадиях элюирования и десорбции. Проводили постоянный мониторинг поглощения УФ при А280. Пиковые фракции объединяли, рН корректировали приблизительно до 5,5 добавлением 20 мМ ацетата натрия и затем проводили диализ против трех смен буфера. Диализат собирали, стерилизовали фильтрацией и хранили при 4°С.
1.10. Идентификация эпитопа IL-31, распознаваемого антителом 15H05
Знание эпитопа IL-31, распознаваемого антителом, критически важно для понимания механизма, посредством которого оно нейтрализует цитокин, не позволяя ему связываться с корецептором IL-31Ra:OSMR. Кроме того, знание эпитопа позволяет (без ограничения) оптимизировать аффинность связывания антитела и разрабатывать пептидные миметики эпитопа (мимотопы), которые могут быть очень полезны в качестве аналитических реагентов для захвата и как субъединичные вакцины для индукции иммунного ответа, направленного на соответствующую мишень. Для идентификации и оптимизации пептида, способного связываться с паратопом mAb 15Н05 применяли многостадийную методику с использованием технологии CLIPS (Chemical Linkage of Peptides onto Scaffolds (химическое связывание пептидов с каркасами)) (Timmerman et al. J Mol Recognit. 2007; 20(5): 283-299) (Pepscan, Лелистад, Нидерланды). Аффинность mAb 15H05 в отношении белков как собачьего, так и кошачьего IL-31 высока (Фиг. 2, MU-15H05), поэтому первичная последовательность IL-31 обоих видов была сочтена важной для указанной выше задачи. Была получена библиотека пептидов на микроматрице, соответствующая белку собачьего IL-31, и она была использована для идентификации пептидов, способных связываться с mAb 15Н05, с применением непрямого ELISA. После идентификации пептидов, первичные аминокислотные последовательности которых соответствовали области связывания mAb 15Н05 на IL-31, проводили сфокусированный анализ полной замены с использованием пептидов, соответствующих сегменту IL-31, и с заменой каждой из 12 аминокислот в данной области связывания mAb 15Н05 на 19 других возможных аминокислот в каждом положении. Этот анализ был необходим для идентификации ключевых аминокислотных остатков IL-31, вовлеченных в связывание с mAb 15Н05, а также продемонстрировал, где замены в собачьей первичной последовательности приводят к усилению связывания с антителом.
Аминокислоты белка собачьего IL-31, распознаваемые антителом 11Е12, были описаны ранее (патент США №8,790,651, выданный Bammert, et al.). В нем описан мутационный анализ белка собачьего IL-31, где показаны положения белка собачьего IL-31, влияющие на связывание с mAb 11Е12 при их замене на аланин. Исходя из анализа полной замены, описанного для mAb 15Н05 выше, и известного ранее эпитопа, связываемого 11Е12, были получены мутантные формы кошачьего IL-31, где два ключевых остатка эпитопов, распознаваемых каждым антителом, были заменены на аланин (мутанты описаны в Разделе 1.2 выше). Мутации каждого эпитопа были названы в соответствии с антителом, распознающим сайт мутации (мутанты 11Е12 и 15Н05 против нативной последовательности белка дикого типа).
На Фиг. 6 А показано выравнивание кошачьего IL-31 дикого типа (SEQ ID NO: 157) с мутантами 15Н05 (SEQ ID NO: 163) и 11Е12 (SEQ ID NO: 161), где выделены положения аланиновых замен. IL-31 принадлежит к семейству цитокинов IL-6 с четырьмя спиральными пучками, обладающими восходящей и нисходящей архитектурой (база данных CATH, Dawson et al. 2017 Nucleic Acids Res. 2017 Jan 4; 45 (Database issue): D289 D295). Его гомологичная модель была получена на основе структуры человеческого IL-6 1Р9М (Boulanger et al. 2003 Science. Jun 27; 300(5628):2101-4) с использованием программного обеспечения МОЕ (Chemical Computing Group, Монреаль, Квебек, Канада). На Фиг. 6В показана гомологичная модель кошачьего IL-31, где выделены положения аминокислот, вовлеченных в связывание с антителами 11Е12 (сайт 1) и 15Н05 (сайт 2). Сайты связывания каждого антитела, по-видимому, расположены в разных участках молекулы белка IL-31.
Для определения влияния на способность mAb 11Е12 и 15Н05 к связыванию с этими мутантными формами кошачьего IL-31 проводили непрямой ELISA с использованием данных мутантов, сорбированных непосредственно на планшет для иммунологического анализа. На Фиг. 6С показаны результаты данного ELISA, демонстрирующие, что mAb 11Е12 и 15Н05 способны связываться с кошачьим IL-31 дикого типа в анализе такого формата. При использовании мутанта 11Е12 в качестве белка для захвата связывание mAb 11Е12 значительно ослаблено, а связывание mAb 15Н05 частично ослаблено. Предшествующий анализ эпитопа 11Е12 на собачьем IL-31 (описанный в патенте США №8,790,651, выданном Bammert, et al.) показал, что при аналиновых мутациях на связывание с mAb влияют 4 аминокислотных остатка, поэтому в данном случае мутаций 2 остатков может быть недостаточно для полного устранения высокоаффинного связывания mAb 11Е12 при применении данного формата ELISA. Незначительное ослабление связывания mAb 15Н05 с мутантом 11Е12, вероятно, обусловлено трансляционными эффектами мутаций от смещения двух передних спиралей, влияющего на сайт связывания 15Н05. Мутации, предназначенные для воздействия на связывание mAb 15Н05 (мутант 15Н05), демонстрируют полную потерю способности mAb 1505 к связыванию с данным мутантом IL-31 при ELISA. В отличие от мутанта 11Е12, изменения в случайной спирали, распознаваемые mAb 15Н05 (мутант 15Н05), не влияют на связывание mAb 11Е12, дополнительно подтверждая, что эти два эпитопа отделены друг от друга (Фиг. 6С).
1.11. Конкурентный анализ связывания mAb 15H05 и 11Е12 с применением Biacore
Для дальнейшего определения характеристик эпитопов IL-31, связываемых mAb 15Н05 и 11Е12, проводили эксперименты с блокировкой с применением Biacore, где получали поверхность, содержащую белок IL-31, с последующим последовательным добавлением антител. На Фиг. 7 показано относительное связывание каждого антитела с IL-31 после захвата 11Е12 или 15Н05. Колонки, отмеченные HBS-EP (аналитический буфер), указывают на максимальный сигнал, полученный при самостоятельном связывании каждого антитела с поверхностью с IL-31 без конкуренции. На Фиг. 7А показаны данные по конкурентному связыванию мышиных антител 15Н05 и 11Е12 с собачьим IL-31. Эти результаты ясно показывают, что антитела 15Н05 и 11Е12 способны связываться с собачьим IL-31 в присутствии друг друга, указывая на распознавание ими разных эпитопов данного белка. Сенсограммы, связанные с Фиг. 7А, показывают очень медленную кинетику диссоциации обоих антител на этой новообразованной поверхности Biacore и, следовательно, невозможность занятия новых сайтов связывания при добавлении того же антитела (данные не показаны).
На Фиг. 7 В показаны данные по конкурентному связыванию антител 15Н05 и 11Е12 с поверхностью с кошачьим IL-31, которые снова указывают на отсутствие перекрывания распознаваемых ими эпитопов. Связывание дополнительного антитела в присутствии того же антитела является результатом повышенной скорости диссоциации из-за худшего качества использованной поверхности. Видны повышенные значения скорости диссоциации, которые можно сравнить со значениями Kd, полученными с использованием новообразованных поверхностей с кошачьим IL-31, показанными на Фиг. 2.
Эти результаты являются дополнительным подтверждением данных, полученных при картировании эпитопов, представленных в Разделе 1.10, и указывают на то, что CDR, присутствующие в антителах MU-15H05 и MU-11E12, распознают раздельные эпитопы. Эпитоп, распознаваемый антителом 15Н05, отличается от эпитопа антитела 11Е12, описанного в патенте США №8,790,651, выданном Bammert, et al., и является новой мишенью на белке IL-31 для нейтрализации активности этого цитокина у многих видов. Эти данные подчеркивают пространственную разделенность сайтов связывания, описанную в гомологичной модели кошачьего IL-31 (Фиг. 6В), и подтверждают гипотезу, согласно которой эта поверхность цитокина критически важна для взаимодействия с рецепторным комплексом IL-31Ra:OSMR.
1.12. Синтез и определение характеристик растворимого корецептора кошачьего IL-31 (IL-31RA и OSMR)
Было показано, что гетерохимерный корецепторный комплекс человеческого IL-31, состоящий из субъединиц IL31Ra и OSMR, необходим для внутриклеточной активации пути JAK-STAT, опосредованной IL-31, и вовлечен в атопические заболевания кожи (Dillon etal. 2004 Nat Immunol. Jul; 5(7):752-60, Dreuw et al. 2004 J Biol Chem. 279:36112-36120; и Diveu et al. 2004 Eur Cytokine Netw. 15:291-302). Позднее человеческая субъединица IL-31 Ra была описана как первичное событие связывания, происходящее при контакте IL-31 с рецепторами клеточной поверхности, и это событие является необходимым условием привлечения OSMR с последующим образованием высокоаффинного рецепторного комплекса (Le Saux etal. 2010 J Biol Chem. Jan 29;285(5):3470-7). Здесь авторы изобретения описывают данные, согласно которым кошачий IL-31 способен к независимому связыванию как с OSMR так и с IL-31Ra. Это новое наблюдение, имеющее большое значение для понимания того, как белок IL-31 взаимодействует с корецептором IL-31Ra:OSMR, и биологической роли IL-31 при его независимом взаимодействии с отдельными субъединицами.
Для понимания того, как IL-31 связывается со своим корецептором, и для описания ингибирующих свойств идентифицированных антител были синтезированы две формы рецептора. Обе отдельные субъединицы рецептора IL-31 IL-31Ra (SEQ ID NO: 169; Feline_IL31Ra_HIgG1_Fc_X1_Fn3), которой соответствует нуклеотидная последовательность (SEQ ID NO: 170; Feline_IL31Ra_HIgG1_Fc_X1 Fn3), и OSMR (SEQ ID NO: 167; Feline_OSMR_hIgG1_Fc) которой соответствует нуклеотидная последовательность (SEQ ID NO: 168; Feline_OSMR_hIgG1_ Fc), конструировали в форме слитых белков с Fc человеческого IgG1. На основе гомологии с человеческими гомологами были идентифицированы цитокинсвязывающий домен, домен фибронектина III и иммуноглобулиноподобный домен. Для оценки отдельных субъединиц рецептора внеклеточные домены OSMR и IL-31Ra (с его предполагаемым N-концевым проксимальным доменом фибронектина III) получали в форме слитых белков с Fc человеческого IgG1, в обоих случаях с использованием их нативных сигнальных пептидов. Все синтетические кассеты клонировали в pcDNA3.1, экспрессировали в системе ExpiCHO и очищали, как описано выше.
Для анализа способности этих форм рецепторов к связыванию с белками IL-31 дикого типа и мутантными белками IL-31 проводили непрямой ELISA, сорбируя по 100 мкл каждого соответствующего белка на планшет Immulon 2НВ (1 мкг/мл) в течение ночи в карбонатном/бикарбонатном буфере (Sigma С3041-100САР) при 4°С. Затем планшеты для ELISA блокировали блокирующим буфером с 5% NFDM в PBST на протяжении 1 часа при комнатной температуре с последующим связыванием с каждой рецепторной конструкцией в нескольких концентрациях при комнатной температуре на протяжении 1 часа. После промывки с использованием PBST присутствие связанного рецептора (слитого с Fc) определяли с использованием мышиного антитела против человеческого IgG1 (Lifetech А10684, разведение 1:500) на протяжении 1 часа при комнатной температуре. Лунки промывали еще раз с использованием PBST и проявляли 3,3',5,5'-тетраметилбензидиновым (ТМВ) субстратом для микролунок KPL Sureblue. На Фиг. 8 показаны результаты этого непрямого ELISA с использованием белка кошачьего IL-31 дикого типа и мутантных форм белка кошачьего IL-31 для захвата. Эти данные демонстрируют способность кошачьего IL-31 дикого типа к независимому связыванию с субъединицами рецептора IL-31Ra и OSMR. Эти наблюдения отличаются от предшествующих сообщений о первичном связывании белка IL-31 с субъединицей IL-31Ra с последующим привлечением OSMR к соответствующему сайту. Поскольку биологическая роль IL-31 еще не определена, очень важно понять динамику его связывания с рецепторами и возможные последствия ослабления его роли при таких заболеваниях, как атопический дерматит. По этой причине эти данные учитывались при определении характеристик антител, связывающихся с эпитопами, способными снизить способность IL-31 к распознаванию IL-31Ra и OSMR.
В Разделе 1.2 авторы изобретения описывают ослабленное связывание антител 11Е 12 и 15Н05 с мутантами, в которых ключевые аминокислоты сайтов связывания заменены на аланин (мутанты 11Е12 и 15Н05, соответственно). Поэтому представляло большой интерес понять влияние этих мутаций на способность к связыванию с отдельными рецепторными субъединицами IL-31Ra и OSMR. На Фиг. 8 показано, что мутации в любом из сайтов связывания 11Е12 или 15Н05 приводят к полной утрате способности к связыванию с IL-31Ra и OSMR, указывая на то, что оба антитела связываются с эпитопами, необходимыми для взаимодействия IL-31 с обеими рецепторными субъединицами. Отсутствие связывания могло бы также быть обусловлено изменениями конформации IL-31 в результате мутаций, однако эти мутанты все еще способны связываться с антителом, указывая на то, что отсутствие связывания вызвано другими причинами. Эти ключевые данные подтверждают способность антител 11Е12 и 15Н05 (и их производных) к распознаванию на IL-31 таких эпитопов, которые нейтрализуют способность этого цитокина к передаче сигналов через его корецептор, а также блокируют связывание этого цитокина с клетками через любой из рассматриваемых рецепторов. Эти данные подтверждают идентификацию антител, способных удалять IL-31 из циркуляции и лишать его способности к связыванию с поверхностными клеточными или растворимыми формами рецепторов.
1.13. Оценка химерных антител in vivo в модели с провокацией зуда у кошек с использованием IL-31
Способность антитела к эффективной нейтрализации его мишени можно оценивать in vitro, изучая связывание с соответствующим эпитопом на белке-мишени с подходящей аффинностью и активность в клеточных анализах, что позволяет проводить экстраполяцию на активность in vivo. Выше описаны шаги, предпринятые для определения характеристик двух серий антител, полученных из мышиных mAb-предшественников 11Е12 и 15Н05. В Разделе 1.7 описано получение мышиных-кошачьих химерных форм mAb 11Е12 и 15Н05, при этом полученная аффинность в отношении собачьего и кошачьего IL-31 сопоставима с исходным мышиным моноклональным антителом (Фиг. 2А). Мышиные-кошачьи химерные формы 11Е12 и 15Н05 также имели сопоставимые значения IC50, демонстрируя ингибирование сигналов pSTAT3, индуцированных кошачьим IL-31, в собачьих и кошачьих макрофагах (Фиг. 3). В процессе фелинизации, описанной в Разделе 1.8, мышиное mAb 11Е 12 было превращено в фелинизированный вариант (Feline 11Е12 1.1) с последующим снижением аффинности в отношении собачьего и кошачьего IL-31 (Фиг. 3) и снижением активности против сигналов кошачьего IL-31 в собачьих и кошачьих клетках (Фиг. 3). Перед оптимизацией фелинизированных антител 11Е12 и 15Н05, описанной в Разделе 1.8, представляло интерес понять способность этих предварительно фелинизированных и химерных форм нейтрализовать активность кошачьего IL-31 по индукции зуда в модели с провокацией зуда у кошек. Представляли интерес фармакодинамический эффект этих различных антител на нейтрализацию зуда и понимание любой его корреляции с аффинностью, активностью в отношении клеток или распознаванием эпитопов, которые могут влиять на эффективность. Анализ диапазона активности в отношении клеток, коррелирующей с эффективностью in vivo в модели с провокацией зуда, может позволить прогнозировать дальнейшую оптимизацию с необходимыми in vitro-анализами Для анализа предварительной эффективности химерного мышиного-кошачьего 11Е12, химерного мышиного-кошачьего 15Н05 и фелинизированного 11Е12 (Feline 11Е12 1.1) была разработана модель зуда, индуцированного IL-31, у кошек. После внутривенного введения 0,5 мкг/мл кошачьего IL-31 (SEQ ID NO: 159; Feline_IL-31_E_coli), которому соответствует нуклеотидная последовательность (SEQ ID NO: 160; Feline_IL-31_E_coli), у кошек будут возникать преходящие поведенческие признаки зуда, включающие (без ограничения) лизание, покусывание, расчесывание и мотание головой или телом. Трение о клетку не рассматривалось как проявление зуда. Наблюдение на предмет зуда проводит квалифицированный исследователь на протяжении 30 минут до введения белка IL-31 и на протяжении 1 часа после него. Для данного исследования контрольную провокацию кошачьим IL-31 проводили не ранее, чем за 1 месяц до введения антитела. На нулевые сутки дозу 0,5 мг/кг антитела объединяли с 0,5 мкг/кг кошачьего IL-31 при комнатной температуре на 60 минут перед инъекцией комплекса, прошедшего предварительное связывание, каждому животному. Для контроля был включен контроль «без mAb». Использованная доза mAb соответствует большому молярному избытку антитела относительно цитокина. Описанный мониторинг проявлений зуда проводили на 0, 7 и 21 сутки. Результаты, представленные на Фиг. 9, показывают существенное улучшение (р менее 0,05) показателей зуда при использовании химерного мышиного-кошачьего mAb 15Н05 на 0, 7 и 21 сутки по сравнению с плацебо-контролем. Несмотря на то, что химерное мышиное-кошачье 11Е12 показало начальную тенденцию к эффективности на нулевые сутки, оно не обеспечивало значимого уменьшения зуда по сравнению с плацебо-наполнителем ни в одной временной точке. Кошачье 11Е12 1.1 (Feline 11Е12 1.1) не уменьшало зуд на нулевые сутки и не показало тенденции к эффективности при сравнении с плацебо-наполнителем, поэтому дальнейшие провокации с использованием IL-31 на 7 и 21 сутки не проводились.
Вместе взятые, эти результаты показывают четкое различие между активностью этих антител и отсутствие эффективности кошачьего 11Е12 1.1 в предотвращении поведенческих признаков зуда, индуцированного IL-31, у кошек. Снижение аффинности и активности кошачьего 11Е12 1.1, вероятно, привело к его неэффективности in vivo. При сравнении результатов по эффективности химерного мышиного-кошачьего 11Е12 и химерного мышиного-кошачьего 15Н05 различия менее выражены. Значения Kd химерных форм обоих mAb сопоставимы с их мышиными предшественниками, при этом аффинность мышиного-кошачьего 11Е12 несколько выше как в отношении кошачьего, так и в отношении собачьего IL-31 (Фиг. 2А). Тем не менее, это повышение аффинности не переходит непосредственно в повышение активности, поскольку у химерного мышиного-кошачьего 15Н05 IC50 против pSTAT3-сигналов, индуцированных кошачьим IL-31, в кошачьих клетках FCWF4 приблизительно в 2 раза выше, чем у химерного мышиного-кошачьего 11Е12 (Фиг. 3). Эти данные показывают, что распознавание кошачьего IL-31 гипервариабельными участками антитела 15Н05 приводит к большей нейтрализации способности данного цитокина к передаче сигналов через его корецептор, что, в свою очередь, делает это антитело более эффективным при блокаде зуда у кошек. Различия IC50, наблюдаемые в этих клеточных анализах, делают их перспективным средством прогнозирования эффективности in vivo и выявления незначительных различий в распознавании эпитопов антителами из одной серии и из разных серий.
1.14. Оценка эффективности фелинизированных антител против IL-31 15Н05 in vivo в модели с провокацией зуда у кошек
Исходя из положительных результатов по эффективности химерного мышиного-кошачьего 15Н05, описанных выше, проводили дальнейшую работу для повышения аффинности и активности фелинизированных 15Н05 (описанных выше в Разделе 1.8). Систематические замены каркасных областей вариабельной области легкой цепи кошачьего антитела 15Н05 1.1 (Feline 15Н05 1.1) привели к идентификации Feline 15Н05 1.1 FW2 с повышенной аффинностью как в отношении кошачьего, так и в отношении собачьего IL-31 по сравнению с мышиным 15Н05 (Фиг. 2). Включение тяжелых и легких цепей Feline 15Н05 1.1 FW2 в одну плазмиду привело, после их продукции в системах экспрессии HEK и СНО, к получению антител ZTS-927 и ZTS-361. Аффинность и активность обоих антител, полученных при экспрессии с одной плазмиды, также описаны на Фиг. 2 и 3, соответственно.
Проводили оценку полностью фелинизированного mAb против кошачьего IL-31 ZTS-361 на предмет его способности нейтрализовать поведенческие проявления зуда в модели зуда, индуцированного IL-31, in vivo. На Фиг. 10А показаны исходные поведенческие признаки зуда до провокации в группах плацебо-наполнителя Т01 и антитела ZTS-361 Т02 от -7 суток до 28 суток, где нулевыми сутками являются сутки введения антитела в группе Т02. Как показано на этом графике поведенческие признаки зуда, оцениваемые в группах Т01 и Т02 до введения IL-31, варьировали незначительно, при этом число проявлений зуда, отмеченных за период наблюдения продолжительностью 30 минут, составляло от 0 до 10. Данное исследование отличалось от предварительной модели с провокацией у кошек, описанной выше в Разделе 1.13, тем, что на нулевые сутки кошкам подкожно вводили 4 мг/кг ZTS-361 без его объединения с кошачьим IL-31 для получения комплекса, прошедшего предварительное связывание. Это позволяет провести более точную оценку эффективности, поскольку к моменту первого введения IL-31 антитело ZTS-361 будет циркулировать на протяжении семи суток и сможет связать и нейтрализовать циркулирующий IL-31 только при достаточной экспозиции антитела.
Для данного исследования поведенческие признаки зуда оценивали на 7, 21, и 28 сутки на протяжении 1 часа после внутривенного введения белка IL-31 в дозе 0,5 мкг/кг.На Фиг. 10 В показана эффективность антитела ZTS-361, демонстрирующего существенное уменьшение зуда, наблюдаемого на 7 сутки (р менее 0,0001), 21 сутки (р менее 0,0027) и 28 сутки (р менее 0,0238) после провокации с использованием IL-31, по сравнению с контрольным плацебо-наполнителем. Данные, полученные в этой провокационной модели, подтверждают предшествующие наблюдения, демонстрирующие эффективность химерного мышиного-кошачьего 15Н05, и подтверждают его активность в отношении клеток и значение эпитопа кошачьего IL-31, распознаваемого CDR 15Н05. Кроме того, эти данные подтверждают способность антитела ZTS-361 нейтрализовать зуд, индуцированный кошачьим IL-31 in vivo, и показывают, что это антитело может быть использовано в качестве терапевтического агента при лечении заболевания, опосредованного IL-31, у кошек, включая атопический дерматит.
Полученные недавно данные по изучению уровней IL-31 в плазме у животных, принадлежащих клиентам, указывают на увеличение количества данного цитокина в циркулирующей крови собак с атопическим и аллергическим дерматитом по сравнению с нормальными лабораторными биглями (Фиг. 11А). Недавно было проведено исследование для определения уровней IL-31 в сыворотке у кошек с предположительным диагнозом аллергического дерматита (АД) из нескольких разных географических областей США. На Фиг. 11 В показаны результаты данного исследования, показывающие, что, аналогично собакам с атопическим и аллергическим дерматитом, у 73 кошек с таким предположительным диагнозом, включенных в исследование, средние уровни циркулирующего IL-31 составляли 8799 фг/мл, по сравнению с 205 фг/мл у 17 контрольных кошек того же возраста. Для понимания уровней собачьего IL-31 в исследовании по разработке с применением предшествующей модели, анализировали фармакокинетический профиль собачьего IL-31 у собак после его подкожного введения в дозе 1,75 мкг/кг. На Фиг. 11С показаны пиковые уровни в плазме в течение первого часа с достижением максимального уровня, составлявшего приблизительно 30 нг/мл, и поддержанием уровня, составлявшего приблизительно 400 пг/мл, через три часа. Исходя из этих данных, логично предположить, что количество кошачьего IL-31 в циркулирующей крови после его внутривенного введения в дозе 0,5 мкг/мл, применяемое в данной модели у кошек, будет значительно превышать его количество, наблюдаемое при естественном заболевании у собак и кошек.
1.15. Аналитические методы, применяемые для усовершенствования ведущих фелинизированных антител против IL-31
При разработке клеточных линий применяют множество аналитических методов для обеспечения возможности стабильного получения терапевтического антитела. Пристальное внимание к аналитическим методам, обеспечивающим (без ограничения) подлинность, чистоту и активность продукта, критически важно для стабильного получения ведущих моноклональных антител и тесной корреляции с его активностью и безопасностью у животных целевого вида. Поскольку антитела представляют собой гомодимеры гетеродимерных единиц, удерживаемых друг рядом с другом межцепочечными дисульфидными связями, любое нарушение их спаривания может привести к неоднородности белкового лекарственного препарата. Двумя аналитическими методами, хорошо приспособленными для мониторинга диссоциации антител, являются невосстанавливающий (NR) электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) и невосстанавливающий (NR) капиллярный гель-электрофорез (CGE).
NR SDS-PAGE является удобным методом качественного определения массы отдельных белковых разновидностей в анализируемом образце. SDS вступает в гидрофобное взаимодействие с белками, равномерно придавая им суммарный отрицательный заряд, что позволяет проводить разделение отдельных компонентов по их массе. После электрофоретического разделения в полиакриламидном геле белки окрашивают красителем, таким как кумасси голубой, что позволяет проводить их выявление. Нелинейность окрашивания препятствует определению абсолютного количества белка в отдельных полосах, но позволяет оценивать его с применением программного обеспечения для денситометрического анализа (VersaDoc, Bio-Rad). Капиллярный гель-электрофорез, широко известный как CGE, также включает обработку белков с использованием SDS, которая приводит к равномерному отрицательному заряду и диссоциации нековалентных белковых комплексов. При наличии электрического поля белки, связанные с SDS, перемещаются к аноду, и их выявляют с применением поглощения ультрафиолетового излучения при фиксированной длине волны 220 нм. Разделение основано на размере компонентов образца в капилляре, заполненном заменяемой матрицей с фильтрующим гелем из SDS-полимера. При невосстанавливающем CGE добавляют алкилирующий агент йодацетамид (IAM) для минимизации перетасовки дисульфидных связей во время подготовки образцов. Интактный IgG отделяют от любых фрагментированных разновидностей, что позволяет проводить количественную оценку степени чистоты. В программном обеспечении для CGE-анализа (примерами которого являются Empower или 32 Karat) применяют площадь с поправкой на время (time corrected area, ТСА), которую определяют как отношение площади отдельного пика ко времени выхода этого пика. Общую ТСА определяют как сумму ТСА всех пиков, превышающих или равных 0,3%. Таким образом, процент мономерного интактного IgG и второстепенных разновидностей могут быть рассчитаны по их индивидуальным ТСА как процент от общей ТСА.
С учетом перспективных данных по эффективности ZTS-361 in vivo, описанных для модели зуда у кошек (Раздел 1.14), было проведено дальнейшее определение характеристик разных партий этого антитела с применением этих описанных аналитических методов. При этом было установлено, что стабильные пулы, экспрессировавшие фелинизированное антитело против IL-31 (ZTS-361), имели повышенные уровни разновидностей с меньшей молекулярной массой (включая свободные легкие цепи), видимые при окрашиванием кумасси при SDS-PAGE, по сравнению с мышиной гибридомой-предшественником 15Н05 (Фиг. 12). Интактное (Intact) или интактный мономер относятся здесь к IgG с двумя тяжелыми цепями и двумя легкими цепями, удерживаемыми вместе межцепочечными дисульфидными связями, и предполагаемой молекулярной массой приблизительно 150 кДа. HHL относится здесь к «Heavy Heavy Light (тяжелая тяжелая легкая)», IgG без одной легкой цепи с предполагаемой молекулярной массой приблизительно 125 кДа. НН относится здесь к «Heavy Heavy (тяжелая тяжелая)», IgG без обеих легких цепей с предполагаемой молекулярной массой приблизительно 100 кДа. HL относится здесь к «Heavy Light (тяжелая легкая)», IgG с одной тяжелой цепью, одной легкой цепью и предполагаемой молекулярной массой приблизительно 75 кДа. L относится здесь к «Light (легкая)», IgG с одной легкой цепью и предполагаемой молекулярной массой приблизительно 25 кДа, также называемому здесь свободной легкой цепью. Количественная оценка того же материала с применением NR-CGE выявила значительное уменьшение доли интактного мономерного IgG (83%) ZTS-361 по сравнению с мышиным предшественником 15Н05 (94,7%) (Фиг. 13а). На Фиг. 13 В показаны электрофореграммы после разделения образцов посредством NR-CGE. Данные этих пиков при разном времени удерживания использовали для количественной оценки процента от общей ТСА для образца 1 (ZTS-361) и образца 2 (мышиное 15Н05). Сумму второстепенных пиков с молекулярной массой, меньшей, чем у основного пика интактного IgG, использовали для расчета процента фрагментов, указанного на Фиг. 13А (% Fragments). Стабильные CHO-пулы, продуцировавшие ZTS-361, приводили к тому, что 17% конечного полученного антитела были представлены в виде фрагментированной формы фелинизированного IgG, по сравнению с 5,3% в случае мышиной гибридомы 15Н05.
Для облегчения понимания этого явления получали отдельные клональные изоляты клеток СНО, имевшие происхождение от клонального пула ZTS-361, для выяснения того, варьировал ли процент мономера интактного IgG у отдельных клонов. На Фиг. 14А показан NR SDS-PAGE очищенного антитела, имеющего происхождение от культур 8 отдельных клонов, стабильно экспрессирующих ZTS-361, с окрашиванием кумасси. Для сравнения на дорожках, отмеченных 1 и 8, представлено контрольное антитело с известным высоким процентом мономера интактного IgG (приблизительно 97%). Справа показан количественный денситометрический анализ интенсивности полос. У разных клонов процент интактного мономера варьировал от 80,2% до 86% при среднем значении приблизительно 82%. Сходным образом, процент фрагментов, определенный у отдельных клонов, составлял от 14,0% до 19,5% при среднем значении 17,5%. На Фиг. 14В показана количественная оценка этих отдельных клонов с применением NR-CGE. При применении этого метода была отмечена меньшая вариабельность со средним процентом интактного мономера у 8 проанализированных клонов 86,3% и процентом разновидностей с меньшей молекулярной массой 13,7%. Несмотря на отмеченную высокую однородность процента мономера интактного IgG для ZTS-361 из отдельных клонов, представляло интерес понять, почему общий уровень мономера интактного IgG был меньше, чем у мышиного варианта этого антитела. Важно отметить, что получение антител, включая, без ограничения, фелинизированные антитела, в системах для транзиторной экспрессии (примерами которых являются клетки HEK и СНО) приводило к получению IgG с высоким процентом мономерной формы (от приблизительно 88% до приблизительно 92%) (данные не показаны). При этом количество антител, получаемое из этих транзиторных культур, значительно меньше количества, получаемого из стабильной линии СНО. Без ограничения какой-либо теорией, появление фрагментированных разновидностей антитела, наблюдаемое в случае фелинизированных антител и антител других видов, может быть обусловлено теми условиями, при которых клетка-хозяин продуцирует исключительно большое количество антитела, и присущими ограничениями условий культивирования и/или молекулярного состава антитела.
1.16. Рассмотрение первичных аминокислотных последовательностей константного домена легкой цепи каппа IgG нескольких видов млекопитающих
Анализ потенциальных ограничений фелинизированного антитела ZTS-361 был начат с рассмотрения первичной последовательности антитела. ZTS-361 состоит из тяжелой цепи, содержащей вариабельную область (SEQ ID NO: 121; FEL_15H05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15Н05_VH1), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_НС_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_НС_AlleleA_1), и легкой цепи, содержащей вариабельную область (SEQ ID NO: 135; FEL-15H05-VL1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO: 136; FEL-15H05-VL1_FW2), в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 175; Feline LC Kappa G minus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 176; Feline_LC_Kappa_G_minus). Функциональные свойства встречающихся в природе константных областей тяжелых цепей кошачьих антител описаны ранее Strietzel et al. (2014 Veterinary Immunology and Immunopathology April 15; 158(3-4): 214-223). Здесь авторы изобретения описывают клонирование и экспрессию фелинизированного антитела ZTS-361 с использованием константной области тяжелой цепи (SEQ ID NO: 171; Feline_HC_AlleleA_l). SEQ ID NO: 171 Feline НС AlleleA 1 соответствует кошачьему IgG1a в Strietzel et al. 2014 supra и, по-видимому, функционально эквивалентна человеческому IgG1. Сравнение этих функциональных свойств и выравнивание этой константной области тяжелой цепи с другими константными областями широкого спектра видов не выявило никаких очевидных областей, которые вызывали бы беспокойство, приводя к недостаточному образованию мономера интактного IgG (данные не показаны).
Сходный анализ константной области цепи каппа, использованной в ZTS-361 (SEQ ID NO: 175; Feline_LC_Kappa_G_minus), позволяет выявить уникальный аспект разнообразия константных областей легких цепей каппа, наблюдаемого по меньшей мере в кошачьих и собачьих последовательностях. Различные виды используют легкую цепь каппа в своем репертуаре иммуноглобулинов с разной частотой. На Фиг. 15 показаны типичные С-концевые аминокислоты и соответствующие нуклеотиды нескольких константных областей легких цепей каппа от указанных видов. Процент использования легкой цепи каппа в IgG указанных видов приведен по следующим источникам: собака (Canine), кошка (Feline) и свинья (Pig) (Aran et al. 1996 Zentralbl Veterinarmed. Nov; 43(9):573-6), норка (Mink) (Bovkun et al. 1993 Eur J Immunol. Aug; 23(8): 1929-34), мышь (Mouse) (Woloschak et al. 1987 Mol Immunol. Jul; 24(7):751-7) и человек (Human) (Barandun et al. 1976 Blood. Jan; 47(l):79-89). Обычно моноклональные антитела получают с использованием мышей. Как показано на Фиг. 15, мыши используют легкую цепь каппа приблизительно в 95% случаев по сравнению с легкой цепью лямбда. В отличие от этого, собаки и кошки используют в своем репертуаре иммуноглобулинов преимущественно легкую цепь лямбда (9% и 8%, соответственно). Для сравнения, два млекопитающих, не являющиеся людьми (свинья и норка), и человек демонстрируют сбалансированное использование легких цепей каппа и лямбда (50%, 46% и приблизительно 50%, соответственно). На приведенном изображении показано и выровнено положение большинства С-концевых цистеинов у разных видов. Этот цистеин критически важен для образования четвертичного комплекса и формирования структуры интактного IgG, поскольку он участвует в образовании межцепочечной ковалентной дисульфидной связи с константной тяжелой цепью. Без ограничения какой-либо теорией, крайне интересно отметить, что по меньшей мере собачьи и кошачьи легкие цепи каппа содержат несколько аминокислот после концевого цистеина. Эти дополнительные аминокислотные остатки имеют как полярность (глутамин), так и заряд (аргинин, аспарагиновая и глутаминовая кислоты). Эти остатки обычно расположены там, где они участвуют во взаимодействиях через водородные связи, включая, без ограничения, взаимодействия с внешней водной средой. Природа и расположение этих дополнительных остатков за концевым цистеином может препятствовать образованию межцепочечной дисульфидной связи, необходимой для образования гетеродимера IgG. У двух млекопитающих (свинья и норка) дополнительных остатков после С-концевого цистеина меньше, и они используют легкие цепи каппа и лямбда в приблизительно эквивалентных соотношениях (50% и 46%, соответственно). С применением транзиторной экспрессии из клеток НЕК была получена серия рекомбинантных форм ZTS-361 с изменениями и делециями аминокислот на С-конце константной области легкой цепи каппа, расположенных сразу после цистеина в аминокислотном положении 107 SEQ ID NO: 175, и процент мономерного IgG в них был проанализирован посредством NR-CGE (данные не показаны). Как указано ранее, получение неспаренной легкой цепи и комплексов IgG с меньшей молекулярной массой более вероятно в условиях сверхпродукции антитела стабильными клональными клетками. Тем не менее, NR-CGE позволяет выявить различия в количестве мономера интактного IgG против примесей с меньшей молекулярной массой из культур, полученных транзиторной трансфекцией клеток (например, HEK и СНО). Это позволяет проводить качественную оценку процента мономерного IgG, полученного из транзиторных культур. Это качественное использование NR-CGE применительно к транзиторным культурам позволило проанализировать множество модификаций и делеций С-конца константной цепи каппа ZTS-361 на предмет процента мономерного IgG (данные не показаны). Без ограничения какой-либо теорией, удаление остатков QRE с С-конца кошачьей константной области цепи каппа, по-видимому, является наиболее оптимальным для получения мономерного рекомбинантного кошачьего IgG. Возможно присоединение других остатков к С-концу. В целом, присоединение одной или двух дополнительных аминокислот после цистеина в положении 107 Feline LC Kappa G minus (SEQ ID NO: 175), по-видимому, является приемлемым, исходя из качественной оценки процента мономера после транзиторной продукции IgG. При применении тех же качественных анализов видно, что даже присоединение трех дополнительных следующих друг за другом аминокислот к С-концу цистеина в положении 107 вместо нативных аминокислотных остатков QRE может быть приемлемым, если эти аминокислоты оказывают минимальное влияние на электростатический заряд. Здесь следует отметить, что число и химические свойства дополнительных аминокислот после цистеина в положении 107 Feline LC Kappa G minus (SEQ ID NO: 175) будут влиять на образование межцепочечной дисульфидной связи между константной областью легкой цепью каппа и соответствующей константной областью тяжелой цепью иммуноглобулина. Можно предположить, что модификации и/или делеций в этой области могут благоприятное влиять на получение однородной интактной формы IgG из стабильной рекомбинантной клеточной линии.
С-концевой аминокислотой человеческих и мышиных легких цепей каппа является концевой цистеин, поэтому в них нет дополнительных остатков, которые могли бы взаимодействовать с внешней средой при образовании дисульфидной связи с константной областью тяжелой цепи. Поэтому здесь предполагается, что эти дополнительные С-концевые остатки, присутствующие по меньшей мере в кошачьих и собачьих константных областях легких цепей каппа, ограничивают возможность образования этой дисульфидной связи, что может обычно быть незаметным в естественных условиях ввиду малого количества этих разновидностей антител, но может иметь большое значение при сверхпродукции этих форм, прошедших видовую адаптацию, в лабораторных условиях. Такие ограничения могут включать отсутствие образования дисульфидной связи между С-концом легкой цепи каппа и константной областью тяжелой цепи, что приводит к присутствию разновидностей HHL, НН, HL и L, описанных ранее, при получении рекомбинантного антитела из стабильной клеточной линии. Эти разновидности с меньшей молекулярной массой нежелательны при получении однородного лекарственного препарата, и их присутствие может быть проблемой с точки зрения качества и безопасности.
К этому наблюдению также относятся кодоны, кодирующие аминокислотные остатки данной области, показанные под буквой каждой аминокислоты на Фиг. 15. У млекопитающих есть три стоп-кодона, являющиеся сигналами терминации трансляции полипептида в рибосоме (ТАА, TGA и TAG). Без ограничения какой-либо конкретной теорией, было отмечено, что большинство кодонов, кодирующих аминокислоты, расположенные после С-концевого цистеина (и сам С-концевой цистеин), отличаются от стоп-кодонов лишь одним нуклеотидом (Фиг. 15, светло-серые буквы в кодонах). Авторы изобретения предполагают, что соматические мутации в этих различных нуклеотидных положениях могли обеспечить подбор оптимальных длины и аминокислотного состава константной области легкой цепи каппа иммуноглобулина для эффективной экспрессии и правильного спаривания цепей антитела IgG каппа. При оценке возможности присутствия соответствующей аминокислоты, которая может быть расположена на С-конце рядом с концевым цистеином, нельзя необходимо учитывать тонкие различия. Здесь следует отметить наблюдение, согласно которому присоединение остатков ЕА к легкой цепи свиньи или Q к легкой цепи норки может не оказывать существенного отрицательного влияния на образование межцепочечной дисульфидной связи и экспрессию интактной молекулы IgG. Эти виды демонстрируют эквивалентное использование легких цепей каппа и лямбда, и эти присоединения являются для них приемлемыми. Применительно, без ограничения, к С-концевым остаткам собачьих и кошачьих легких цепей каппа полагают, что расстояние от С-концевого цистеина и заряженная природа остатков аргинина и аспарагиновой и глутаминовой кислот оказывают существенное влияние на способность кошачьих и собачьих антител к эффективному и правильному образованию ковалентной дисульфидной связи с соответствующей тяжелой цепью.
1.17. Анализ четвертичной структуры обращенных друг к другу поверхностей кошачьей тяжелой цепи и легкой цепи каппа
На Фиг. 16А представлено пиктографическое изображение предполагаемой структуры кошачьего IgG, тяжелые и легкие цепи которого эквивалентны ZTS-361 (подтверждающие аналитические данные не показаны). Для упрощения положения внутрицепочечных и межцепочечных дисульфидных связей с выделением цистеина тяжелой цепи (CYS15) и цистеина легкой цепи (CYS107) показаны только в одной ветви структуры. Показанный аминокислотный остаток CYS15 представляет собой положение 15 Feline НС AlleleA wt (SEQ ID NO: 171) и Feline НС AlleleA 1 (SEQ ID NO: 173), а показанные нуклеотидные остатки представляют собой положения 43-45 Feline НС AlleleA wt (SEQ ID NO: 172) и 43-45 Feline НС AlleleA 1 (SEQ ID NO: 174), соответственно. Показанный аминокислотный остаток CYS107 представляет собой положение 107 Feline LC Kappa G minus (SEQ ID NO: 175) и Feline LC Kappa G minus QRE minus (SEQ ID NO: 186), а показанные нуклеотидные остатки представляют собой положения 319-321 Feline LC Kappa G minus (SEQ ID NO: 176) и 319-321 Feline LC Kappa G minus QRE minus (SEQ ID NO: 187). Как указано ранее, состав кошачьего ZTS-361 функционально эквивалентен человеческому IgG1; тем не менее, характер его паттерна дисульфидных связей более сходен с человеческим IgG2 (данные не показаны). На Фиг. 16 В показана гомологичная модель, соответствующая двум Р(ab')2-ветвям ZTS-361, где приблизительное расположение CYS15 и CYS107 для наглядности выделено с применением программного обеспечения МОЕ (Chemical Computing Group, Монреаль, Квебек, Канада). На Фиг. 16С показан вклад дополнительных электростатических зарядов в местную среду, обусловленное остатками константной легкой цепи каппа QRE, расположенными сразу после CYS107. Заряд этих трех аминокислот показан в виде оболочек с сетчатой поверхностью. В гомологичных моделях области со случайной спиральной структурой показаны менее точно, чем области с упорядоченными элементами вторичной структуры, такими как альфа-спирали или области упорядоченной бета-складчатости. Тем не менее, это не имеет существенного значения для CYS15 тяжелой цепи, поскольку эта область константного домена IgG хорошо определена большим объемом структурных данных и упорядоченным характером этой области с консервативной структурой IgG. Сходным образом, положение CYS107 определено в структурах множества антител, что позволяет оценить его близость к CYS15 тяжелой цепи. В данной модели присоединение остатков после концевого цистеина легкой цепи можно определить, только исходя из геометрии соседних остатков и расчета местных энергетических минимумов. Такое отображение кошачьей легкой цепи каппа ZTS-361 является наиболее оптимальной рабочей моделью для разработки экспериментальной гипотезы. Вместе взятые, эти результаты показывают, что дополнительные аминокислотные остатки, расположенные дальше концевого цистеина кошачьей легкой цепи каппа (и, вероятно, легкой цепи каппа других видов), снижают эффективность спаривания с тяжелой цепью, приводя к высокой вероятности неправильного спаривания и неудовлетворительным результатам получения антител.
1.18. Получение фелинизированного антитела против IL-31 ZTS-1505 с модифицированным С-концом константной области легкой цепи каппа
В связи с возможными ограничениями при стабильном получении однородного препарата антитела ZTS-361 было сочтено необходимым устранить снижение процента получаемого мономера. В том числе, но без ограничения, с этой целью было получено ZTS-1505, тяжелая цепь которого содержит вариабельную область (SEQ ID NO: 121; FEL_15Н05_VH1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 122; FEL_15Н05_VH1), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_НС_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_НС_ AlleleA_1). В случае ZTS-1505 легкая цепь содержит вариабельную область (SEQ ID NO:135; FEL-15H05-VL1_FW2), которой соответствует нуклеотидная последовательность (SEQ ID NO:136; FEL-15H05-VL1_FW2), в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 186; Feline_LC_Kappa_G_minus_QRE_minus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 187; Feline_LC_Kappa_G_minus_QRE_minus). ZTS-1505 идентично ZTS-361, за исключением удаления трех дополнительных остатков QRE с самого С-конца константной области легкой цепи каппа во избежание нежелательных эффектов получения антитела с разновидностями с меньшей молекулярной массой в результате неэффективного спаривания с легкой цепью каппа. Предполагается, что удаление этих остатков полезно для получения мономерного IgG. Тем не менее, настоящее изобретение не обязательно ограничено этой одной модификацией легкой цепи каппа, поскольку были проведены и другие изменения в легкой цепи каппа, которые, согласно предварительным экспериментальным данным, оказывали нейтральный или даже положительный эффект при транзиторной экспрессии (данные не показаны).
На Фиг. 17А описаны вариабельные и константные последовательности, использованные для конструирования антител ZTS-361 и ZTS-1505, с выделением отличий легкой цепи, использованной для ZTS-1505. Получение стабильной клеточной линии СНО, экспрессирующей ZTS-1505, описано в Разделе 1.9 данной заявки. Оценка качества антитела представляла большой интерес, поэтому отдельные клоны стабильно трансфицированных СНО изучали на предмет их способности продуцировать мономерное антитело. На Фиг. 17В показаны результаты количественной оценки этих отдельных клонов с применением NR-CGE. Были проанализированы 23 отдельных стабильных клона СНО, и при этом была отмечена высокая степень однородности процента полученного мономера с его средним значением 90,3%. Это соответствует росту среднего процента полученного мономера на 4% по сравнению с отдельными клональными изолятами ZTS-361 (Фиг. 14 В) и на 7,3% по сравнению со стабильным пулом (Фиг. 13А). На Фиг. 17С показано прямое сравнение отдельных стабильных клеточных линии СНО, экспрессирующих антитела ZTS-361 и ZTS-1505, при различных условиях культивирования. Следует отметить, что антитела, описанные на Фиг. 12, 13 и 14, были получены при условиях культивирования, эквивалентных условиям культивирования «А» на Фиг. 17С, и эти условия описаны в Разделе 1.9 данной заявки. Обе клеточные линии выращивали на протяжении 14 суток и определяли жизнеспособность, титр (выход антитела в г/л) и процент мономера по результатам NR-CGE. При всех условиях обе клеточные линии демонстрировали сходный процент жизнеспособности. В случае ZTS-1505 процент мономера по результатам NR-CGE был на 4-5% выше, чем у ZTS-361, еще раз указывая на то, что константная область легкой цепи каппа ZTS-1505 без дополнительных остатков QRE оказывала положительный эффект на соединение кошачьей константной области тяжелой цепи с легкой цепью каппа. Еще более выраженным было увеличение количества полученного антитела ZTS-1505 по сравнению с ZTS-361. В случае ZTS-1505 стабильные клетки СНО продуцировали в среднем в 3,5 раза больше антитела, чем в случае ZTS-361, имеющего константную область легкой цепи каппа дикого типа.
Для того чтобы убедиться, что модификация константной области легкой цепи каппа не влияла на аффинность и активность ZTS-1505, проводили сравнительный анализ с применением Biacore и ингибирования pSTAT-сигналов, опосредованных IL-31 в собачьих и кошачьих клетках. В Таблице 1 ниже описаны результаты анализа с использованием нескольких видов белка IL-31 в качестве поверхности захвата. Эти результаты показывают почти идентичную аффинность ZTS-1505, имеющего модификацию константной области легкой цепи каппа, и ZTS-361, имеющего С-конец константной области цепи каппа дикого типа, в отношении белков собачьего, кошачьего и лошадиного IL-31. Также здесь описано предполагаемое связывание ZTS-1505 с кошачьими мутантами 15Н05 и 11Е12, описанными в Разделе 1.2 данной заявки. Модификация С-конца константной области цепи каппа не меняет селективности в отношении эпитопа, что продемонстрировано отсутствием связывания ZTS-1505 с кошачьим мутантным белком 15Н05 и сохранением связывания с кошачьим мутантом НЕ 12, аналогично ZTS-361.
140
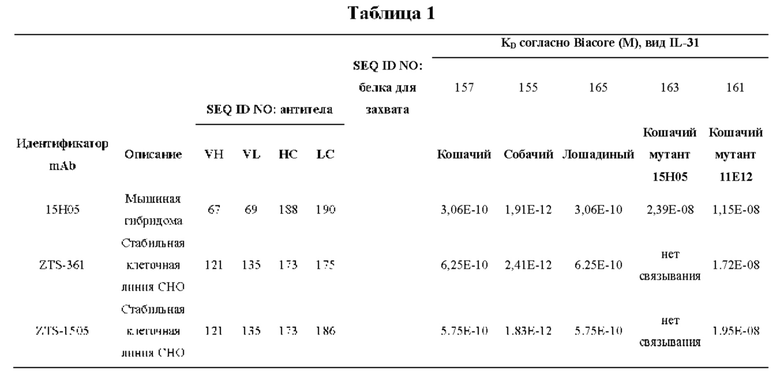
В Таблице 2 ниже показаны данные по активности ZTS-1505 в отношении клеток в сравнении с ZTS-361. Модификация константной области легкой цепи каппа ZTS-1505 не влияла на способность данного антитела к ингибированию клеточных pSTAT-сигналов, индуцированных собачьим и кошачьим IL-31 в собачьих клетках DH82 и кошачьих клетках FCWF-4, соответственно, на что указывают сопоставимые значения IC50.

Вместе взятые, эти результаты показывают, что удаление или модификация С-конца легкой цепи каппа тех видов, у которых нативные последовательности эмбрионального типа кодируют эти дополнительные остатки, полезным образом сказываются как на получении однородных рекомбинантных антител этих видов, так и на количестве антитела, продуцируемого стабильной клеточной линией (например, повышая выход). Кроме того, такие модификации, повышая качество и количество получаемого антитела, не оказывают негативного влияния на аффинность и активность антитела в отношении его белка-мишени IL-31. Здесь следует отметить, что эти факторы могут быть полезны для получения рекомбинантных антител для терапевтического применения у многих видов.
1.19. Подтверждение результатов NR-CGE с использованием mAb против IL-31 1505 и дополнительная демонстрация полезности модифицированного С-конца константной области цепи каппа с использованием антитела против NGF
Представляло интерес определить, применимо ли повышение процента мономерного IgG к получению других антител, чьи белки-мишени отличны от IL-31. Для этого получали дополнительный набор пулов стабильных клеточных линий СНО с фелинизированными антителами, распознающими кошачий фактор роста нервов бета (NGF) (ZTS-768 и ZTS-943). В последовательности антитела ZTS-768 тяжелая цепь представляет собой (SEQ ID NO: 220; ZTS 768 VH), которой соответствует нуклеотидная последовательность (SEQ ID NO:221; ZTS 768 VH), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 171; Feline НС AlleleA wt), которой соответствует нуклеотидная последовательность (SEQ ID NO: 172; Feline НС AlleleA wt). В последовательности антитела ZTS-943 тяжелая цепь представляет собой (SEQ ID NO: 224; ZTS 943 VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 225; ZTS 943 VH), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 171; Feline НС AlleleA wt), которой соответствует нуклеотидная последовательность (SEQ ID NO: 172; FelineHCAlleleAwt). В случае ZTS-768 вариабельная область легкой цепи представляет собой (SEQ ID NO: 222; ZTS 768 VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 223; ZTS 768 VL). В случае ZTS-768, данная вариабельная область легкой цепи представлена в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 175; FelineLCKappaGminus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 176; Feline LC Kappa G minus). В случае ZTS-943 вариабельная область легкой цепи представляет собой (SEQ ID NO: 226; ZTS 943 VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 227; ZTS 943 VL), в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 186; Feline LC Kappa G minus QRE minus), которой соответствует нуклеотидная последовательность (SEQ ID NO: 187; FelineLCKappaGminusQREminus). Для прямого сравнения с антителами против IL-31 также получали стабильные пулы ZTS-361 и ZTS-1505 с применением идентичных условий для культивирования и очистки антител против NGF.
После получения и очистки четырех антител, описанных выше, применяли NR-CGE для определения процента мономерного интактного IgG и второстепенных разновидностей. Сравнение этих антител, полученных и очищенных с применением идентичных методов, позволило оценить полезность включения модифицированного С-конца константной области цепи каппа в структурно различные антитела, распознающие разные белки-мишени. На Фиг. 18А показан процент идентичности при сравнении вариабельных областей антител против кошачьего NGF и против IL-31, рассчитанный с использованием программного обеспечения ClustalW. На Фиг. 18В и 18С показано выравнивание вариабельных областей тяжелых и легких цепей, соответственно, антител против кошачьего IL-31 и NGF, где CDR выделены рамками. Очевидно, что антитела против IL-31 и против NGF отличаются друг от друга, не демонстрируя общей идентичности, особенно в антигенсвязывающих областях, выделенных как CDR.
На Фиг. 19 показаны результаты NR-CGE при сравнении антител против кошачьего IL-31 и против кошачьего NGF с и без модификации С-конца константной области цепи каппа. Аналогично полученным ранее данным, антитело против IL-31, содержащее легкую цепь каппа дикого типа (ZTS-361), имело 80,74% мономерного IgG, при этом наиболее многочисленная второстепенная разновидность, HHL, составляла 8,71%. После удаления трех С-концевых остатков с получением ZTS-1505 снова наблюдали четкое улучшение процента мономера (89,17% мономера при 5,9% HHL). При NR-CGE-анализе очищенного IgG из mAb против кошачьего NGF, содержавшего легкую цепь каппа дикого типа (ZTS-768), наблюдали сходное количество выделенного мономера и второстепенных разновидностей (80,81% мономера при 12,33% HHL) по сравнению с антителом против IL-31 ZTS-361. Следует отметить, что после удаления С-концевых остатков из антитела против кошачьего NGF наблюдали такой же паттерн с получением 88,59% мономера, при этом наиболее многочисленная второстепенная разновидность, HHL, составляла 5,98%.
Эти результаты указывают на наличие структурного различия между белком кошачьего IgG, содержащим аминокислотные остатки дикого типа (QRE) на С-конце легкой цепи каппа, и IgG, из которого эти остатки удалены. Результаты экспериментов, описанные здесь, показывают, что применение этой модификации легкой цепи каппа приводит к получению мономерных кошачьих IgG с меньшим количеством примесных второстепенных разновидностей. В дополнение к этому, результаты, описанные здесь, ясно демонстрируют, что данный способ применим к структурно различным антителам, распознающим совершенно разные мишени, и поэтому вероятно, что эта модификация будет применима к широкому спектру кошачьих антител, а также к другим антителам млекопитающих, имеющим дополнительные С-концевые аминокислоты в константной области легкой цепи каппа. Без ограничения какой-либо теорией, эта модификация легкой цепи, по-видимому, приводит к более точному спариванию цепей иммуноглобулинов при их индуцированной продукции стабильными клеточными линиями СНО, результатами чего являются увеличение количества мономерного IgG и, возможно, повышение общего выхода антитела. Оба эти результата крайне желательны с точки зрения изготовления лекарственных средств, содержащих антитела, для коммерческого применения.
1.20. Идентификация антител против лошадиного IL-31, связывающихся с эквивалентной областью белка лошадиного IL-31, по сравнению со связыванием mAb 15Н05 с кошачьим IL-31
С учетом перспективной in vivo эффективности в модели зуда у кошек при использовании антител из мышиной линии 15Н05, описанных здесь, было желательным идентифицировать новые субстраты-антитела, связывающиеся со сходной областью лошадиного ортолога белка IL-31. С этой целью для идентификации антител, связывающихся с лошадиным IL-31, мышей иммунизировали рекомбинантным лошадиным IL-31 (SEQ ID NO: 165).
Титры антител в сыворотке иммунизированных животных определяли с применением ELISA, как описано ранее. Для слияния использовали донорные спленоциты единственной ответившей мыши и проводили скрининг гибридомных супернатантов на предмет антител, связывавшихся с белком лошадиного IL-31, посредством ELISA. Это привело к идентификации двух мышиных антител, связывавшихся с областью белка лошадиного IL-31, сопоставимой с сайтом связывания антитела 15Н05 на кошачьем IL-31. В Разделе 1.10 данной заявки описано определение характеристик сайта связывания антитела 15Н05 с иллюстрацией приблизительного сайта связывания на гомологичной модели кошачьего IL-31, представленной на Фиг. 6В. На Фиг. 20 показано выравнивание кошачьего IL-31 дикого типа (SEQ ID NO: 157) с лошадиным IL-31 (SEQ ID NO: 165) с применением программного обеспечения ClustalW. Стрелками над выравниванием показаны остатки Р126 и D128, описанные в Разделе 1.10 как входящие в область связывания антитела 15Н05 с кошачьим IL-31 (сайт 2) (Фиг. 6 В). Этими двумя антителами против лошадиного IL-31, имеющими этот общий сайт связывания в белке лошадиного IL-31 являются 04Н07 и 06А09.
Далее 04Н07 и 06А09 против лошадиного IL-31 субклонировали для получения гибридомы, продуцирующей однородное антитело, и для секвенирования их вариабельных областей тяжелых и легких цепей. Последовательности вариабельных областей мышиного антитела против IL-31, определенные для антитела 04Н07, представляют собой следующее: вариабельная область тяжелой цепи 04Н07 (SEQ ID NO: 212; Mu_04Н07_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 213; Mu_04H07_VH), вариабельная область легкой цепи 15Н05 (SEQ ID NO: 214; Mu_04H07_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 215; Mu_04H07_VL). Вариабельная область тяжелой цепи 06А09 (SEQ ID NO: 216; Mu_06A09_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 217; Mu_06A09_VH), вариабельная область легкой цепи 06А09 (SEQ ID NO: 218; Mu_06A09_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 219; Mu_06A09_VL). На Фиг. 21 показано выравнивание вариабельных тяжелых (Фиг. 21А) и легких (Фиг. 21 В) цепей 04Н07 и 06А09 в сравнении с мышиным антителом 15Н05 с использованием Clusta1W. Для сравнения расположение каждого из шести CDR выделено рамками. Антитела против лошадиного IL-31 04Н07 и 06А09 очень похожи друг на друга и, вероятно, происходят от общей клональной линии. Антитело против кошачьего IL-31 15Н05 явно отличается по аминокислотной последовательности CDR и длине CDRH3 и CDRL1 от 04Н07 и 06А09 против лошадиного IL-31. Интересно, что сайт связывания антитела 15Н05 с кошачьим IL-31 сохраняется и в лошадином белке (стрелки на Фиг. 20). Эти результаты являются еще одним примером того, что структурно различные CDR могут распознавать общую эпитопную область на двух ортологах IL-31.
1.21. Аланиновое сканирование CDR антитела против IL-31 ZTS-1505
Область антитела, обеспечивающая распознавание антигена, представляет собой паратоп. Паратоп образован комбинацией аминокислот гипервариабельных участков (CDR) вариабельных областей тяжелой и легкой цепи. Связывание антитела с антигеном часто опосредовано боковыми цепями остатков CDR и боковыми цепями или углеводными группировками антигена. Для содействия определению критически важных боковых цепей, вовлеченных в распознавание антигена антителом, проводили мутагенез с аланиновым сканированием по каждому остатку CDR как тяжелой, так и легкой цепи. Затем проводили раздельный анализ этих мутантов на предмет их способности к связыванию с кошачьим IL-31 с применением Biacore и их оценку на предмет ингибирования сигналов, опосредованных IL-31, в анализе с клетками FCWF-4.
Для определения относительной аффинности мутантных mAb, полученных при аланиновом сканировании, в сравнении с исходным mAb определяли профили связывания с чипами, покрытыми кошачьим IL-31, при 100 нМ с использованием Biacore Т200. Среднее число единиц ответа после четырех повторов, полученное для исходного mAb, плюс/минус 3 стандартных отклонения использовали для расчета параметров, определяющих пороговое число единиц ответа, включая скорости ассоциации и диссоциации при связывании антитела (Фиг. 22). Процент точек данных для каждого мутанта, соответствовавших этому пороговому значению, затем использовали для определения «% показателя сходства» (Фиг. 23). Показатель сходства в результате аланиновых замен по каждому положению CDR тяжелой и легкой цепи антитела ZTS-1505 показан на Фиг. 23 и 24, соответственно.
Для определения относительной активности мутантных антител ZTS-1505 в клеточном анализе каждое антитело оценивали на предмет его способности ингибировать фосфорилирование STAT, опосредованное IL-31, в клетках FCWF-4 при концентрации 15 мкг/мл. Относительное ингибирование фосфорилирования STAT определяли, оценивая % ингибирования фосфорилирования STAT каждым мутантом в сравнении с исходным антителом. Результаты замены каждого положения в CDR тяжелой и легкой цепи ZTS-1505 на аланин показаны как «процент ингибирования в сравнении с исходным антителом» на Фиг. 23 и 24, соответственно. Оценивая эффект отдельных аланиновых замен на аффинность связывания и активность в отношении клеток, можно оценить роль боковых цепей каждого аминокислотного остатка CDR в распознавании антигена. Остатки CDR, которые могут быть заменены на аланин с сохранением активности, вероятно, могут быть подвержены множеству аминокислотных замен и не являются критически важными для распознавания антигена. Исходя из этих данных, по меньшей мере остаток 4(1) SEQ ID NO: 1, остатки 1-3 (NIN), 5-7 (TSG), 9-11 (TEN) и 13 (Q) SEQ ID NO: 2 и остатки 4 (K), 6 (D) и 13 (V) SEQ ID NO: 3 в CDR1, 2 и 3 тяжелой цепи, соответственно, не являются критически важными для связывания с антигеном. Остатки CDR легкой цепи, не являющиеся критически важными для связывания, включают остатки 3-7 (SQGIS) SEQ ID NO: 4, остатки 3 (S) и 5 (L) SEQ ID NO: 5 и остатки 4 (Q), 5 (Т) и 9 (Т) SEQ ID NO: 6 из CDRL1, 2 и 3, соответственно.
1.22. Аффинность связывания двух фелинизированных вариантов антитела ZIL8 и их активность в отношении клеток
Выше, в Разделе 1.6 данного раздела «Примеры», описана идентификация собачьего антитела, распознающего кошачий IL-31, названного ZIL8. Начальный скрининг показал, что данное антитело способно связываться с белком кошачьего IL-31, однако на его связывание влияет мутация 15Н05. Поэтому представляло интерес получить фелинизированную форму данного антитела для применения в качестве терапевтического средства у кошек. Для этого использовали методику фелинизации, описанную ранее в Разделе 1.8 данной заявки. При переносе CDR ZIL8 на подходящие кошачьи каркасные области были получены антитела ZTS-5864 и ZTS-5865. Последовательность тяжелой цепи антитела ZTS-5864 представляет собой (SEQ ID NO: 228; ZTS_5864_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 229; ZTS_5864_VH), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_HC_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_НС_AlleleA_1). В случае ZTS-5864 вариабельная область легкой цепи представляет собой (SEQ ID NO: 230; ZTS_5864_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO:231; ZTS_5864_VL). В случае ZTS-5864, данная вариабельная область легкой цепи представлена в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 236; Feline_LC_ Lambda), которой соответствует нуклеотидная последовательность (SEQ ID NO: 237; Feline_LC_ Lambda). Последовательность тяжелой цепи антитела ZTS-5865 представляет собой (SEQ ID NO: 232; ZTS_5865_VH), которой соответствует нуклеотидная последовательность (SEQ ID NO: 233; ZTS_5865_VH), в комбинации с константной областью тяжелой цепи кошачьего IgG (SEQ ID NO: 173; Feline_HC_AlleleA_1), которой соответствует нуклеотидная последовательность (SEQ ID NO: 174; Feline_НС_AlleleA_1). В случае ZTS-5865 вариабельная область легкой цепи представляет собой (SEQ ID NO: 234; ZTS_5865_VL), которой соответствует нуклеотидная последовательность (SEQ ID NO: 235; ZTS_5865_VL). В случае ZTS-5865, данная вариабельная область легкой цепи представлена в комбинации с константной областью легкой цепи кошачьего IgG (SEQ ID NO: 236; Feline LC Lambda), которой соответствует нуклеотидная последовательность (SEQ ID NO: 237; Feline_LC_Lambda).
На Фиг. 25A и 25В показана аффинность двух фелинизированных антител ZTS-5864 и ZTS-5865 и их активность в отношении клеток. Оба антитела имеют высокую аффинность в отношении кошачьего IL-31, при этом аффинность (Kd (М)) ZTS-5864 приблизительно в 4 раза выше аффинности ZTS-5865. Активность в отношении клеток оценивали с использованием кошачьего IL-31 для стимуляции pSTAT3-сигналов в клетках FCFW4. Значения IC50 рассчитывали для каждого антитела, как описано ранее в данной заявке. Активность ZTS-5864 приблизительно в 3 раза выше при сравнении его значения IC50 c IC50 ZTS-5865. Следует отметить, что оба антитела рассматриваются как активные, при этом их IC50 входят в диапазон IC50 антител линии 15Н05, описанных здесь ранее (Фиг. 3). Значимость этой активности в отношении pSTAT3-сигналов в кошачьих клетках FCFW4 была продемонстрирована ранее положительными результатами по эффективности in vivo при использовании химерных и фелинизированных антител в модели зуда у кошек (Фиг. 9 и Фиг. 10).
1.23. Оценка эффективности фелинизированного антитела против IL-31 ZTS-5864 in vivo в модели с провокацией зуда у кошек
Эффективность ZTS-5864 in vivo оценивали в модели зуда, индуцированного IL-31, у кошек, как описано ранее в Разделе 1.14 данного раздела «Примеры». Результаты этого исследования показаны на Фиг. 26. До введения (7 сутки) показано, что при провокации с использованием IL-31 до введения препаратов у животных из групп Т01 и Т02 развивается одинаковый ответ в виде зуда. Во время исследования, вплоть до 56 суток, в группе Т01 зуд почти не менялся. В отличие от этого, подкожное введение ZTS-5864 (3,0 мг/кг) на нулевые сутки ослабляло зуд после провокации с использованием IL-31 на протяжении всего исследования, вплоть до 56 суток (Фиг. 26, Т02). Сохранение снижения среднего показателя зуда в группе Т02 на 56 сутки указывает на вероятное сохранение эффективности антитела после этой временной точки. Эти результаты подчеркивают надежность критериев, использованных для отбора антител против кошачьего IL-31 в данной заявке. Эти результаты дополнительно обосновывают позиционирование этих антител, прошедших видовую адаптацию, в качестве терапевтических средств для лечения расстройств, опосредованных IL-31, у кошек.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Zoetis Services LLC
Bammert, Gary F.
Dunham, Steven A.
<120> INTERLEUKIN-31 MONOCLONAL ANTIBODIES FOR VETERINARY USE
<130> ZP000226A
<150> 62/643940
<151> 2018-03-16
<160> 237
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Mus musculus
<400> 1
Ser Tyr Thr Ile His
1 5
<210> 2
<211> 17
<212> PRT
<213> Mus musculus
<400> 2
Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe Lys
1 5 10 15
Asp
<210> 3
<211> 13
<212> PRT
<213> Mus musculus
<400> 3
Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val
1 5 10
<210> 4
<211> 11
<212> PRT
<213> Mus musculus
<400> 4
Arg Ala Ser Gln Gly Ile Ser Ile Trp Leu Ser
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Mus musculus
<400> 5
Lys Ala Ser Asn Leu His Ile
1 5
<210> 6
<211> 9
<212> PRT
<213> Mus musculus
<400> 6
Leu Gln Ser Gln Thr Tyr Pro Leu Thr
1 5
<210> 7
<211> 5
<212> PRT
<213> Mus musculus
<400> 7
Tyr Tyr Asp Ile Asn
1 5
<210> 8
<211> 17
<212> PRT
<213> Mus musculus
<400> 8
Trp Ile Phe Pro Gly Asp Gly Gly Thr Lys Tyr Asn Glu Thr Phe Lys
1 5 10 15
Gly
<210> 9
<211> 14
<212> PRT
<213> Mus musculus
<400> 9
Ala Arg Gly Gly Thr Ser Val Ile Arg Asp Ala Met Asp Tyr
1 5 10
<210> 10
<211> 15
<212> PRT
<213> Mus musculus
<400> 10
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Ile Ser Phe Met His
1 5 10 15
<210> 11
<211> 7
<212> PRT
<213> Mus musculus
<400> 11
Arg Ala Ser Asn Leu Glu Ser
1 5
<210> 12
<211> 9
<212> PRT
<213> Mus musculus
<400> 12
Gln Gln Ser Asn Lys Asp Pro Leu Thr
1 5
<210> 13
<211> 5
<212> PRT
<213> Canis familiaris
<400> 13
Ser Tyr Gly Met Ser
1 5
<210> 14
<211> 17
<212> PRT
<213> Canis familiaris
<400> 14
His Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 15
<211> 15
<212> PRT
<213> Canis familiaris
<400> 15
Val Tyr Thr Thr Leu Ala Ala Phe Trp Thr Asp Asn Phe Asp Tyr
1 5 10 15
<210> 16
<211> 13
<212> PRT
<213> Canis familiaris
<400> 16
Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu Ala Ala Thr
1 5 10
<210> 17
<211> 7
<212> PRT
<213> Canis familiaris
<400> 17
Ser Asp Gly Asn Arg Pro Ser
1 5
<210> 18
<211> 11
<212> PRT
<213> Canis familiaris
<400> 18
Gln Ser Phe Asp Thr Thr Leu Asp Ala Tyr Val
1 5 10
<210> 19
<211> 5
<212> PRT
<213> Canis familiaris
<400> 19
Asp Tyr Ala Met Ser
1 5
<210> 20
<211> 17
<212> PRT
<213> Canis familiaris
<400> 20
Gly Ile Asp Ser Val Gly Ser Gly Thr Ser Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 21
<211> 8
<212> PRT
<213> Canis familiaris
<400> 21
Gly Phe Pro Gly Ser Phe Glu His
1 5
<210> 22
<211> 13
<212> PRT
<213> Canis familiaris
<400> 22
Thr Gly Ser Ser Ser Asn Ile Gly Ser Gly Tyr Val Gly
1 5 10
<210> 23
<211> 7
<212> PRT
<213> Canis familiaris
<400> 23
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 24
<211> 10
<212> PRT
<213> Canis familiaris
<400> 24
Ser Val Tyr Asp Arg Thr Phe Asn Ala Val
1 5 10
<210> 25
<211> 5
<212> PRT
<213> Canis familiaris
<400> 25
Ser Tyr Asp Met Thr
1 5
<210> 26
<211> 17
<212> PRT
<213> Canis familiaris
<400> 26
Asp Val Asn Ser Gly Gly Thr Gly Thr Ala Tyr Ala Val Ala Val Lys
1 5 10 15
Gly
<210> 27
<211> 9
<212> PRT
<213> Canis familiaris
<400> 27
Leu Gly Val Arg Asp Gly Leu Ser Val
1 5
<210> 28
<211> 11
<212> PRT
<213> Canis familiaris
<400> 28
Ser Gly Glu Ser Leu Asn Glu Tyr Tyr Thr Gln
1 5 10
<210> 29
<211> 7
<212> PRT
<213> Canis familiaris
<400> 29
Arg Asp Thr Glu Arg Pro Ser
1 5
<210> 30
<211> 10
<212> PRT
<213> Canis familiaris
<400> 30
Glu Ser Ala Val Asp Thr Gly Thr Leu Val
1 5 10
<210> 31
<211> 5
<212> PRT
<213> Canis familiaris
<400> 31
Thr Tyr Val Met Asn
1 5
<210> 32
<211> 17
<212> PRT
<213> Canis familiaris
<400> 32
Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val Arg
1 5 10 15
Gly
<210> 33
<211> 8
<212> PRT
<213> Canis familiaris
<400> 33
Ser Met Val Gly Pro Phe Asp Tyr
1 5
<210> 34
<211> 11
<212> PRT
<213> Canis familiaris
<400> 34
Ser Gly Glu Ser Leu Ser Asn Tyr Tyr Ala Gln
1 5 10
<210> 35
<211> 7
<212> PRT
<213> Canis familiaris
<400> 35
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 36
<211> 10
<212> PRT
<213> Canis familiaris
<400> 36
Glu Ser Ala Val Ser Ser Asp Thr Ile Val
1 5 10
<210> 37
<211> 5
<212> PRT
<213> Canis familiaris
<400> 37
Ser Tyr Ala Met Lys
1 5
<210> 38
<211> 17
<212> PRT
<213> Canis familiaris
<400> 38
Thr Ile Asn Asn Asp Gly Thr Arg Thr Gly Tyr Ala Asp Ala Val Arg
1 5 10 15
Gly
<210> 39
<211> 15
<212> PRT
<213> Canis familiaris
<400> 39
Gly Asn Ala Glu Ser Gly Cys Thr Gly Asp His Cys Pro Pro Tyr
1 5 10 15
<210> 40
<211> 11
<212> PRT
<213> Canis familiaris
<400> 40
Ser Gly Glu Ser Leu Asn Lys Tyr Tyr Ala Gln
1 5 10
<210> 41
<211> 7
<212> PRT
<213> Canis familiaris
<400> 41
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 42
<211> 10
<212> PRT
<213> Canis familiaris
<400> 42
Glu Ser Ala Val Ser Ser Glu Thr Asn Val
1 5 10
<210> 43
<211> 5
<212> PRT
<213> Canis familiaris
<400> 43
Thr Tyr Phe Met Ser
1 5
<210> 44
<211> 17
<212> PRT
<213> Canis familiaris
<400> 44
Leu Ile Ser Ser Asp Gly Ser Gly Thr Tyr Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 45
<211> 7
<212> PRT
<213> Canis familiaris
<400> 45
Phe Trp Arg Ala Phe Asn Asp
1 5
<210> 46
<211> 14
<212> PRT
<213> Canis familiaris
<400> 46
Gly Leu Asn Ser Gly Ser Val Ser Thr Ser Asn Tyr Pro Gly
1 5 10
<210> 47
<211> 7
<212> PRT
<213> Canis familiaris
<400> 47
Asp Thr Gly Ser Arg Pro Ser
1 5
<210> 48
<211> 10
<212> PRT
<213> Canis familiaris
<400> 48
Ser Leu Tyr Thr Asp Ser Asp Ile Leu Val
1 5 10
<210> 49
<211> 5
<212> PRT
<213> Canis familiaris
<400> 49
Asp Arg Gly Met Ser
1 5
<210> 50
<211> 17
<212> PRT
<213> Canis familiaris
<400> 50
Tyr Ile Arg Tyr Asp Gly Ser Arg Thr Asp Tyr Ala Asp Ala Val Glu
1 5 10 15
Gly
<210> 51
<211> 8
<212> PRT
<213> Canis familiaris
<400> 51
Trp Asp Gly Ser Ser Phe Asp Tyr
1 5
<210> 52
<211> 16
<212> PRT
<213> Canis familiaris
<400> 52
Lys Ala Ser Gln Ser Leu Leu His Ser Asp Gly Asn Thr Tyr Leu Asp
1 5 10 15
<210> 53
<211> 7
<212> PRT
<213> Canis familiaris
<400> 53
Lys Val Ser Asn Arg Asp Pro
1 5
<210> 54
<211> 9
<212> PRT
<213> Canis familiaris
<400> 54
Met Gln Ala Ile His Phe Pro Leu Thr
1 5
<210> 55
<211> 5
<212> PRT
<213> Canis familiaris
<400> 55
Ser Tyr Val Met Thr
1 5
<210> 56
<211> 17
<212> PRT
<213> Canis familiaris
<400> 56
Gly Ile Asn Ser Glu Gly Ser Arg Thr Ala Tyr Ala Asp Ala Val Lys
1 5 10 15
Gly
<210> 57
<211> 10
<212> PRT
<213> Canis familiaris
<400> 57
Gly Asp Ile Val Ala Thr Gly Thr Ser Tyr
1 5 10
<210> 58
<211> 11
<212> PRT
<213> Canis familiaris
<400> 58
Ser Gly Glu Thr Leu Asn Arg Phe Tyr Thr Gln
1 5 10
<210> 59
<211> 7
<212> PRT
<213> Canis familiaris
<400> 59
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 60
<211> 10
<212> PRT
<213> Canis familiaris
<400> 60
Lys Ser Ala Val Ser Ile Asp Val Gly Val
1 5 10
<210> 61
<211> 5
<212> PRT
<213> Canis familiaris
<400> 61
Thr Tyr Val Met Asn
1 5
<210> 62
<211> 17
<212> PRT
<213> Canis familiaris
<400> 62
Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val Arg
1 5 10 15
Gly
<210> 63
<211> 8
<212> PRT
<213> Canis familiaris
<400> 63
Ser Met Val Gly Pro Phe Asp Tyr
1 5
<210> 64
<211> 11
<212> PRT
<213> Canis familiaris
<400> 64
Ser Gly Lys Ser Leu Ser Tyr Tyr Tyr Ala Gln
1 5 10
<210> 65
<211> 7
<212> PRT
<213> Canis familiaris
<400> 65
Lys Asp Thr Glu Arg Pro Ser
1 5
<210> 66
<211> 10
<212> PRT
<213> Canis familiaris
<400> 66
Glu Ser Ala Val Ser Ser Asp Thr Ile Val
1 5 10
<210> 67
<211> 122
<212> PRT
<213> Mus musculus
<400> 67
Gln Val Gln Leu Gln Gln Ser Ala Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Lys Thr Thr Leu Thr Val Asp Arg Ser Ser Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu His Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 68
<211> 366
<212> DNA
<213> Mus musculus
<400> 68
caggtccagc tgcagcagtc tgcagctgaa ctggcaagac ctggggcctc agtgaagatg 60
tcctgcaaga cttctggcta cacatttact tcctacacga tacactggat aaaacagagg 120
cctggacagg gtctggaatg gattggaaac attaatccca ccagtggata cactgagaac 180
aatcagaggt tcaaggacaa gaccacattg actgtagaca gatcctccaa cacagcctat 240
ttgcaactgc acagcctgac atctgaggac tctgcggtct atttctgtgc aagatggggc 300
tttaaatatg acggagaatg gtccttcgat gtctggggcg cagggaccac ggtcaccgtc 360
tcctca 366
<210> 69
<211> 107
<212> PRT
<213> Mus musculus
<400> 69
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn
100 105
<210> 70
<211> 321
<212> DNA
<213> Mus musculus
<400> 70
gacatccaaa tgaaccagtc tccatccagt ctgtctgcat ccctcggaga cacaatcacc 60
gtcacttgcc gtgccagtca gggcatcagt atttggttaa gctggtacca gcagaaacca 120
ggaaatattc ctaaagtatt gatcaataag gcttccaact tgcacatagg agtcccacca 180
aggtttagtg gcagtggatc tggaacacat ttcacattaa ctatcaccag cctacagcct 240
gaagacattg ccacttacta ctgtctacag agtcaaactt atcctctcac gttcggaggg 300
gggaccaagc tggaaataaa c 321
<210> 71
<211> 121
<212> PRT
<213> Mus musculus
<400> 71
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Lys Tyr Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Phe Pro Gly Asp Gly Gly Thr Lys Tyr Asn Glu Thr Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Arg Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Thr Ser Val Ile Arg Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 72
<211> 363
<212> DNA
<213> Mus musculus
<400> 72
caggttcagc tgcagcagtc tggagctgaa ctggtaaagc ctggggcttc agtgaagttg 60
tcctgcaagg cttctggcta caccttcaaa tactatgata taaactgggt gaggcagagg 120
cctgaacagg gacttgagtg gattggatgg atttttcctg gagatggtgg tactaagtac 180
aatgagacgt tcaagggcaa ggccacactg actacagaca aatcctccag cacagcctac 240
atgcagctca gcaggctgac atctgaggac tctgctgtct atttctgtgc aagagggggg 300
acttcggtga taagggatgc tatggactac tggggtcaag gaacctcagt caccgtctcc 360
tca 363
<210> 73
<211> 111
<212> PRT
<213> Mus musculus
<400> 73
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asn
65 70 75 80
Pro Val Glu Thr Asp Asp Val Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 74
<211> 333
<212> DNA
<213> Mus musculus
<400> 74
gacattgtgc tgacccaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atctcctgca gagccagcga aagtgttgat aattatggca ttagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatc gtgcatccaa cctagaatct 180
gggatccctg ccaggttcag tggcagtggg tctaggacag acttcaccct caccattaat 240
cctgtggaga ctgatgatgt tgcaacctat tactgtcagc aaagtaataa ggatccgctc 300
acgttcggtg ctgggaccaa gctggagctg aaa 333
<210> 75
<211> 124
<212> PRT
<213> Canis familiaris
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala His Ile Asn Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Glu Val Tyr Thr Thr Leu Ala Ala Phe Trp Thr Asp Asn Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 76
<211> 372
<212> DNA
<213> Canis familiaris
<400> 76
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctggggggtc cctgagactc 60
tcctgtgtgg cttctggatt caccttcagt agttatggca tgagctgggt ccgccaggct 120
ccagggaagg ggctgcagtg ggtcgcacac attaacagtg gtggaagtag cacatactac 180
gcagacgctg tgaagggacg attcaccatc tccagagaca acgccaagaa cacgctctat 240
ctgcagatga acagcctgag agctgaggac acggccgtct attactgtgt ggaggtttac 300
actacgttag ctgcattctg gacagacaat tttgactact ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 77
<211> 110
<212> PRT
<213> Canis familiaris
<400> 77
Gln Ser Val Leu Thr Gln Pro Thr Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Asn Asn Ile Gly Ile Leu
20 25 30
Ala Ala Thr Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Val Leu
35 40 45
Val Tyr Ser Asp Gly Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Asn Ser Ala Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Asp Ala Tyr Val Phe Gly Ser Gly Thr Gln Leu Thr Val Leu
100 105 110
<210> 78
<211> 330
<212> DNA
<213> Canis familiaris
<400> 78
cagtctgtgc tgactcagcc gacctcagtg tcggggtccc ttggccagag ggtcaccatc 60
tcctgctctg gaagcacgaa caacatcggt attcttgctg cgacctggta ccaacaactc 120
ccaggaaagg cccctaaagt cctcgtgtac agtgatggga atcgaccgtc aggggtccct 180
gaccggtttt ccggctccaa gtctggcaac tcagccaccc tgaccatcac tgggcttcag 240
gctgaggacg aggctgatta ttactgccag tcctttgata ccacgcttga tgcttacgtg 300
ttcggctcag gaacccaact gaccgtcctt 330
<210> 79
<211> 117
<212> PRT
<213> Canis familiaris
<400> 79
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Gln Trp Val
35 40 45
Ala Gly Ile Asp Ser Val Gly Ser Gly Thr Ser Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Phe Asn Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Ser Gly Phe Pro Gly Ser Phe Glu His Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 80
<211> 351
<212> DNA
<213> Canis familiaris
<400> 80
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagt gactatgcca tgagctgggt ccgccaggct 120
cctgggaggg gactgcagtg ggtcgcaggt attgacagtg ttggaagtgg cacaagctac 180
gcagacgctg tgaagggccg attcacaatc tccagagacg acgccaagaa cacactgtat 240
ctgcagatgt tcaacctgag agccgaggac acggccatat attactgtgc gagcgggttc 300
cctgggtcct ttgagcactg gggccagggc accctggtca ccgtctcctc a 351
<210> 81
<211> 104
<212> PRT
<213> Canis familiaris
<400> 81
Gln Ser Val Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Leu Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Gly
20 25 30
Tyr Val Gly Trp Tyr Gln Gln Leu Pro Gly Thr Gly Pro Arg Thr Leu
35 40 45
Ile Tyr Tyr Asn Ser Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Arg Ser Gly Thr Thr Ala Thr Leu Thr Ile Ser Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Val Tyr Asp Arg Thr Phe
85 90 95
Asn Ala Val Phe Gly Gly Gly Thr
100
<210> 82
<211> 327
<212> DNA
<213> Canis familiaris
<400> 82
cagtctgtac tgactcagcc ggcctcagtg tctgggtccc tgggccagaa ggtcaccatc 60
tcctgcactg gaagtagttc caacattggt agtggttatg tgggctggta ccagcagctc 120
ccaggaacag gccccagaac cctcatctat tataacagtg accgaccttc gggggtcccc 180
gatcgattct ctggctccag gtcaggcacc acagcaaccc tgaccatctc tggactccag 240
gctgaggacg aggctgatta ttactgctca gtatatgaca ggactttcaa tgctgtgttc 300
ggcggaggca cccacctgac cgtcctc 327
<210> 83
<211> 118
<212> PRT
<213> Canis familiaris
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Pro Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Asp Val Asn Ser Gly Gly Thr Gly Thr Ala Tyr Ala Val Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Leu Gly Val Arg Asp Gly Leu Ser Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 84
<211> 354
<212> DNA
<213> Canis familiaris
<400> 84
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctccagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagc agttatgaca tgacctgggt ccgccaggct 120
cctgggaagg gactgcagtg ggtcgcagat gttaacagtg gtggaactgg cacggcctac 180
gcagtcgctg tgaagggccg attcaccatc tccagagaca acgccaagaa aacactctat 240
ttacagatga acagcctgag agccgaagac acggccgttt attattgtgc gaaactaggt 300
gtgagagatg gtctttctgt ctggggccag ggcaccctgg tcaccgtctc ctcg 354
<210> 85
<211> 107
<212> PRT
<213> Canis familiaris
<400> 85
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Ser Leu Asn Glu Tyr Tyr Thr
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Asp Thr Gly Thr Leu
85 90 95
Val Phe Gly Gly Gly Thr His Leu Ala Val Leu
100 105
<210> 86
<211> 321
<212> DNA
<213> Canis familiaris
<400> 86
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tgggacagac agcaaccatc 60
tcctgctctg gagagagtct gaatgaatat tatacacaat ggttccagca gaaggcaggc 120
caagcccctg tcttggtcat atatagggac actgagcggc cctctgggat ccctgaccga 180
ttctctggct ccagttcagg gaacacacac accctaacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg cgagtcagcg gtcgacactg gaacccttgt ctttggcgga 300
ggcacccacc tggccgtcct c 321
<210> 87
<211> 117
<212> PRT
<213> Canis familiaris
<400> 87
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Val Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Gln Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Val Val Ser Met Val Gly Pro Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 88
<211> 351
<212> DNA
<213> Canis familiaris
<400> 88
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagg acctatgtca tgaactgggt ccgccaggct 120
cctgggaagg ggctgcaatg ggtcgcaagt attaacggtg gtggaagtag cccaacctac 180
gcagacgctg tgaggggccg attcaccgtc tccagggaca acgcccagaa ctcactgttt 240
ctgcagatga acagcctgag agccgaggac acagccgtgt atttttgtgt cgtgtcgatg 300
gttgggccct tcgactactg gggccaaggg accctggtca ccgtgtcctc a 351
<210> 89
<211> 102
<212> PRT
<213> Canis familiaris
<400> 89
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Ser Leu Ser Asn Tyr Tyr Ala
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Ser Ser Asp Thr Ile
85 90 95
Val Phe Gly Gly Gly Thr
100
<210> 90
<211> 321
<212> DNA
<213> Canis familiaris
<400> 90
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tggggcagac agcaaccatc 60
tcctgctctg gagagagtct gagtaactat tatgcacaat ggttccagca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctctggct ccagttcagg gaacacacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg tgagtcagca gtcagttctg atactattgt gttcggcgga 300
ggcacccacc tgaccgtcct c 321
<210> 91
<211> 124
<212> PRT
<213> Canis familiaris
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Lys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Thr Ile Asn Asn Asp Gly Thr Arg Thr Gly Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Lys Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Lys Gly Asn Ala Glu Ser Gly Cys Thr Gly Asp His Cys Pro Pro
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 92
<211> 372
<212> DNA
<213> Canis familiaris
<400> 92
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcggggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagt agttatgcca tgaaatgggt ccgccaggct 120
cctgggaagg ggctgcagtg ggtcgcgact attaacaatg atggaaccag aacaggctac 180
gcagacgctg tgaggggccg attcaccatc tccaaagaca acgccaaaaa cacactgtat 240
ctgcagatgg acagcctgag agccgacgac acggccgtct attactgtac aaagggcaat 300
gccgaatccg gctgtactgg tgatcactgt cctccctact ggggccaggg aaccctggtc 360
accgtctcct ca 372
<210> 93
<211> 107
<212> PRT
<213> Canis familiaris
<400> 93
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Ser Leu Asn Lys Tyr Tyr Ala
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ala Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Ser Ser Glu Thr Asn
85 90 95
Val Phe Gly Ser Gly Thr Gln Leu Thr Val Leu
100 105
<210> 94
<211> 321
<212> DNA
<213> Canis familiaris
<400> 94
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tgggacagac agcaaccatc 60
tcctgctctg gagagagtct gaataaatat tatgcacaat ggttccaaca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctccggct ccagtgcagg caacacacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg cgagtcagca gtcagttctg aaactaacgt gttcggctca 300
ggaacccaac tgaccgtcct t 321
<210> 95
<211> 116
<212> PRT
<213> Canis familiaris
<400> 95
Glu Val Gln Leu Val Asp Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Phe Met Ser Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Gln Trp Val
35 40 45
Ala Leu Ile Ser Ser Asp Gly Ser Gly Thr Tyr Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Ile Phe Trp Arg Ala Phe Asn Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 96
<211> 348
<212> DNA
<213> Canis familiaris
<400> 96
gaggtacaac tggtggactc tgggggagac ctggtgaagc ctggggggtc cctgagactc 60
tcctgtgtgg cctctggatt caccttcagt acctacttca tgtcctgggt ccgccaggct 120
ccagggaggg ggcttcagtg ggtcgcactt attagcagtg atggaagtgg cacatactac 180
gcagacgctg tgaagggccg attcaccatc tccagagaca atgccaagaa cacgctgtat 240
ctgcagatga acagcctgag agccgaggac acggctatgt attactgtgc gatattctgg 300
cgggccttta acgactgggg ccagggcacc ctggtcaccg tctcctca 348
<210> 97
<211> 110
<212> PRT
<213> Canis familiaris
<400> 97
Gln Thr Val Val Ile Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Asn Ser Gly Ser Val Ser Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Tyr Gln Gln Thr Arg Gly Arg Thr Pro Arg Thr
35 40 45
Ile Ile Tyr Asp Thr Gly Ser Arg Pro Ser Gly Val Pro Asn Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Thr Asp Ser
85 90 95
Asp Ile Leu Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 98
<211> 330
<212> DNA
<213> Canis familiaris
<400> 98
cagactgtgg taatccagga gccatcactc tcagtgtctc caggagggac agtcacactc 60
acatgtggcc tcaactctgg gtcagtctcc acaagtaatt accctggctg gtaccagcag 120
acccgaggcc ggactcctcg cacgattatc tacgacacag gcagtcgccc ctctggggtc 180
cctaatcgct tctccggatc catctctgga aacaaagccg ccctcaccat cacaggagcc 240
cagcccgagg atgaggctga ctattactgt tccttatata cggatagtga cattcttgtt 300
ttcggcggag gcacccacct gaccgtcctc 330
<210> 99
<211> 117
<212> PRT
<213> Canis familiaris
<400> 99
Glu Val His Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Trp Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Arg
20 25 30
Gly Met Ser Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Tyr Ile Arg Tyr Asp Gly Ser Arg Thr Asp Tyr Ala Asp Ala Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Gly Ser Ser Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 100
<211> 351
<212> DNA
<213> Canis familiaris
<400> 100
gaggtgcatt tggtggagtc tgggggagac ctggtgaagc cttgggggtc cttgagactg 60
tcctgtgtgg cctctggatt cacctttagt gatcgtggca tgagctgggt ccgtcagtct 120
ccagggaagg ggctgcagtg ggtcgcatat attaggtatg atgggagtag gacagactac 180
gcagacgctg tggagggccg attcaccatc tccagagaca acgccaagaa cacgctctac 240
ctgcagatga acagcctgag agccgaggac acggccgtgt attactgtgc gagatgggac 300
ggtagttctt ttgactattg gggccagggc accctggtca ccgtctcctc a 351
<210> 101
<211> 112
<212> PRT
<213> Canis familiaris
<400> 101
Asp Ile Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Thr Ala Ser Phe Ser Cys Lys Ala Ser Gln Ser Leu Leu His Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Asp Trp Phe Arg Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Arg Leu Ile Tyr Lys Val Ser Asn Arg Asp Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Gly Val Glu Ala Asp Asp Ala Gly Leu Tyr Tyr Cys Met Gln Ala
85 90 95
Ile His Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Val Glu Leu Lys
100 105 110
<210> 102
<211> 336
<212> DNA
<213> Canis familiaris
<400> 102
gatattgtcg tgacacagac cccgctgtcc ctgtccgtca gccctggaga gactgcctcc 60
ttctcctgca aggccagtca gagcctcctg cacagtgatg gaaacacgta tttggattgg 120
ttccgacaga agccaggcca gtctccacag cgtttgatct acaaggtctc caacagagac 180
cctggggtcc cagacaggtt cagtggcagc gggtcaggga cagatttcac cctgagaatc 240
agcggagtgg aggctgacga tgctggactt tattactgca tgcaagcaat acactttcct 300
ctgacgttcg gagcaggaac caaggtggag ctcaaa 336
<210> 103
<211> 119
<212> PRT
<213> Canis familiaris
<400> 103
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Val Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Gly Ile Asn Ser Glu Gly Ser Arg Thr Ala Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Ile Asp Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Thr Gly Asp Ile Val Ala Thr Gly Thr Ser Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 104
<211> 357
<212> DNA
<213> Canis familiaris
<400> 104
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagt agttatgtca tgacctgggt ccgccaggct 120
cctgggaagg gactgcagtg ggtcgcaggc attaatagtg aggggagtag gacagcctac 180
gcagacgctg tgaagggccg attcaccatc tccagagaca acgccaagaa tacactttat 240
ctacaaatag acagcctgag agccgaggac acggccatat attactgtgc gacaggcgat 300
atagtagcga ctggtacttc gtattggggc cagggcaccc tggtcaccgt ctcctca 357
<210> 105
<211> 107
<212> PRT
<213> Canis familiaris
<400> 105
Ser Asn Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Glu Thr Leu Asn Arg Phe Tyr Thr
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Ile His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Ala Tyr Tyr Cys Lys Ser Ala Val Ser Ile Asp Val Gly
85 90 95
Val Phe Gly Gly Gly Thr His Leu Thr Val Phe
100 105
<210> 106
<211> 321
<212> DNA
<213> Canis familiaris
<400> 106
tccaatgtac tgactcagcc tccctcggta tcagtgtctc tgggacagac agcaaccatc 60
tcctgctctg gagagactct gaatagattt tatacacaat ggttccagca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctccggct ccagttcagg gaacatacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg cctattactg caagtcagca gtcagtattg atgttggtgt gttcggcgga 300
ggcacccacc tgaccgtctt c 321
<210> 107
<211> 117
<212> PRT
<213> Canis familiaris
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Ala Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Val Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Ser Ile Asn Gly Gly Gly Ser Ser Pro Thr Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Gln Asn Ser Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Phe Cys
85 90 95
Val Val Ser Met Val Gly Pro Phe Asp Tyr Trp Gly His Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 108
<211> 351
<212> DNA
<213> Canis familiaris
<400> 108
gaggtgcagc tggtggagtc tgggggagac ctggtgaagc ctgcagggtc cctgagactg 60
tcctgtgtgg cctctggatt caccttcagg acctatgtca tgaactgggt ccgccaggct 120
cctgggaagg ggctgcaatg ggtcgcaagt attaacggtg gtggaagtag cccaacctac 180
gcagacgctg tgaggggccg attcaccgtc tccagggaca acgcccagaa ctcactgttt 240
ctgcagatga acagcctgag agccgaggac acagccatat atttttgtgt cgtgtcgatg 300
gttgggccct tcgactactg gggccatggg accctggtca ccgtgtcctc a 351
<210> 109
<211> 107
<212> PRT
<213> Canis familiaris
<400> 109
Ser Ser Val Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Ser Gly Lys Ser Leu Ser Tyr Tyr Tyr Ala
20 25 30
Gln Trp Phe Gln Gln Lys Ala Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Thr Glu Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr His Thr Leu Thr Ile Ser Gly Ala Arg Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Glu Ser Ala Val Ser Ser Asp Thr Ile
85 90 95
Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 110
<211> 321
<212> DNA
<213> Canis familiaris
<400> 110
tccagtgtgc tgactcagcc tccctcggta tcagtgtctc tggggcagac agcaaccatc 60
tcctgctctg gaaagagtct gagttactat tatgcacaat ggttccagca gaaggcaggc 120
caagcccctg tgttggtcat atataaggac actgagcggc cctctgggat ccctgaccga 180
ttctctggct ccagttcagg gaacacacac accctgacca tcagcggggc tcgggccgag 240
gacgaggctg actattactg tgagtcagca gtcagttctg atactattgt gttcggcgga 300
ggcacccacc tgaccgtcct c 321
<210> 111
<211> 121
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Mus musculus и Felis catus
<400> 111
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Ser Phe Thr Tyr Tyr
20 25 30
Asp Ile Asn Trp Leu Arg Gln Ala Pro Glu Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Phe Pro Gly Asp Gly Gly Thr Lys Tyr Asn Glu Thr Phe
50 55 60
Lys Gly Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Thr Ser Val Ile Arg Asp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Ala Leu Val Thr Val Ser Ser
115 120
<210> 112
<211> 363
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, от Mus
musculus и Felis catus
<400> 112
caggtgctgc tggtccagtc aggagcagag gtaaaaaagc ccggggcgag tgtcaagatt 60
ttctgtaagg cctccggata ctcttttacg tattacgata ttaactggct tcgccaggcc 120
cctgagcagg ggctcgaatg gatgggttgg atattccccg gagatggggg aaccaagtac 180
aacgaaacct tcaaggggag gctgaccctg actgcagata ccagcacgaa cacagtgtat 240
atggagttgt cctcactgcg atctgctgat actgccatgt actactgcgc tcgcggcggc 300
acttcagtta tcagggatgc catggactat tgggggcagg gcgcactcgt cactgtctcg 360
agc 363
<210> 113
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 113
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 114
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 114
gaaatccaga tgacacaatc tcccagctcc ctcagcgcat ctcctggcga cagggtaacc 60
atcacctgcc gcgccagcga gtcagtagac aactatggca tatccttcat gcactggtat 120
caacaaaagc ccgggaaagt ccccaaactg ttgatttaca gagcaagcaa tctcgagtca 180
ggagtcccat ctcgcttctc tggttccggt tccggaaccg acttcactct gacaatttct 240
tctctggagc ccgaggatgc cgctacatat tactgtcagc aaagcaataa agatccactg 300
accttcggac agggtaccaa gctggagatc aaa 333
<210> 115
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 115
Glu Val Val Leu Thr Gln Ser Ser Ala Phe Leu Ser Arg Thr Leu Lys
1 5 10 15
Glu Lys Ala Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Asn Gln Ala Pro
35 40 45
Lys Leu Leu Val Lys Arg Ala Ser Asn Leu Glu Ser Gly Ala Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Pro Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 116
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 116
gaggtggtgc tgactcagag tagcgcgttt ctgtctcgga ccctgaaaga gaaagctacc 60
atcacgtgca gggcaagcga gagcgtggac aactatggta tcagcttcat gcattggtat 120
cagcagaaac ctaatcaggc gcctaagctg ctcgtgaaaa gagcctccaa ccttgagagc 180
ggcgcaccat caaggttttc aggaagtggc agcgggacag acttcaccct tacaatctct 240
agtccagagc cggaggacgc agctacctac tattgccagc aatccaataa agacccgttg 300
acattcggcc aaggtacc 318
<210> 117
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 117
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 118
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 118
gagatccaga tgacccagtc tccatcctca ctgagtgcta gcccggggga tcgagtgact 60
ataacatgtc gggccagtga atcagtggac aactatggaa tcagttttat gcactggtat 120
cagcagaagc ccggccagcc accgaagctg ttgatttatc gcgcaagcaa tctggagtca 180
ggagtgccct ctagattttc tgggagcggt tctggcacag atttcacact cacaatatca 240
tccttggaac cggaagacgc agccacatac tattgccagc agagtaacaa ggaccctttg 300
acttttggcc agggtacc 318
<210> 119
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 119
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Val Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Gln Gly Thr
100 105
<210> 120
<211> 318
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 120
gagatccaga tgacccagtc tccatcctca ctgagtgcta gcccggggga tcgagtgact 60
ataacatgtc gggccagtga atcagtggac aactatggaa tcagttttat gcactggtat 120
cagcagaagc ccggccaggt cccgaagctg ttgatttatc gcgcaagcaa tctggagtca 180
ggagtgccct ctagattttc tgggagcggt tctggcacag atttcacact cacaatatca 240
tccttggaac cggaagacgc agccacatac tattgccagc agagtaacaa ggaccctttg 300
acttttggcc agggtacc 318
<210> 121
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Mus musculus и Felis catus
<400> 121
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Arg Thr Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Leu Arg Gln Ala Pro Ala Gln Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 122
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, от Mus
musculus и Felis catus
<400> 122
caagtgcttc tggtgcaaag cggggcggaa gttaggaccc caggagcctc agtaaaaatt 60
ttttgtaagg catccggcta cagtttcacc agctacacta ttcactggct gaggcaggcc 120
ccggcccaag ggctggagtg gatgggaaat atcaatccca cgtctggcta tacagagaat 180
aaccaaaggt ttaaggatag gctgactctg acagctgaca catcaaccaa tacggcatac 240
atggagctct cctctctccg gagtgccgac accgccatgt actactgtgc tcggtggggg 300
tttaaatacg atggcgagtg gagcttcgac gtgtggggcg cgggcacaac cgtgaccgtc 360
tcgagc 366
<210> 123
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Mus musculus и Felis catus
<400> 123
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Arg Thr Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Gly Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Ser Pro Ala Gln Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 124
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, от Mus
musculus и Felis catus
<400> 124
caggtgctgc tcgtgcagag cggagccgaa gtgaggacac ccggtgcgag tgtaaaaatt 60
ttttgcaagg caagcggcta cgggtttaca tcctatacca tccactgggt gaggcagtcc 120
ccagcgcagg gacttgaatg gatgggaaat attaatccaa caagcgggta tactgaaaac 180
aaccaaagat ttaaggacag actgacactc accgcagata catctacaaa tacagcctac 240
atggagttgt cttccctgcg gagtgccgac acggctatgt actactgtgc tcggtggggg 300
tttaagtatg atggcgaatg gtccttcgac gtctggggag ctggaaccac cgtgaccgtc 360
tcgagc 366
<210> 125
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Mus musculus и Felis catus
<400> 125
Gln Val Leu Leu Val Gln Ser Gly Ala Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Phe Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Thr Ile His Trp Leu Arg Gln Ala Pro Glu Gln Gly Leu Glu Trp Met
35 40 45
Gly Asn Ile Asn Pro Thr Ser Gly Tyr Thr Glu Asn Asn Gln Arg Phe
50 55 60
Lys Asp Arg Leu Thr Leu Thr Ala Asp Thr Ser Thr Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Ala Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Phe Lys Tyr Asp Gly Glu Trp Ser Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 126
<211> 366
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, от Mus
musculus и Felis catus
<400> 126
caggtcctct tggttcaaag cggagccgaa gtccgaaaac cgggtgcctc agtgaaaatc 60
ttctgtaagg cctccggcta tagtttcacg agttacacaa tccactggct gcgacaggca 120
ccagagcagg gactggagtg gatgggaaat ataaatccga cgtctgggta cacagaaaac 180
aaccagagat tcaaggatag attgacactg accgcggata ctagtacaaa tacggcttac 240
atggaactgt cctcactccg gtcagccgac accgccatgt attactgtgc tcgctggggg 300
ttcaagtatg atggagagtg gagcttcgac gtatggggag ccggaaccac tgtgaccgtc 360
tcgagc 366
<210> 127
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 127
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 128
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 128
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 129
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 129
Glu Ile Thr Met Thr Gln Ser Pro Gly Ser Leu Ala Gly Ser Pro Gly
1 5 10 15
Gln Gln Val Thr Met Asn Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln His Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Leu Gln Ala
65 70 75 80
Glu Asp Val Ala Ser Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 130
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 130
gaaattacca tgacacaaag ccccggctcc ctggccggct cccccggaca gcaagtgacc 60
atgaattgtc gggccagcca gggaatttct atatggctct cttggtatca gcaaaaaccc 120
ggacagcacc ctaaacttct gatctacaaa gcaagtaact tgcacatcgg cgtccctgat 180
cgattcagtg gctcaggttc cggtacagat tttactctta ccatcagcaa tctgcaggct 240
gaggatgtgg caagctatta ctgtctccaa agtcagactt accctctgac atttgggggc 300
ggtaccaagc tggagatcaa a 321
<210> 131
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 131
Glu Val Val Leu Thr Gln Ser Ser Ala Phe Leu Ser Arg Thr Leu Lys
1 5 10 15
Glu Lys Ala Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Asn Gln Ala Pro Lys Leu Leu Val
35 40 45
Lys Lys Ala Ser Asn Leu His Ile Gly Ala Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Pro Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Gly Asp Gln
100 105
<210> 132
<211> 322
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 132
gaggtagtgc tgactcagtc ctccgccttc ttgtcaagaa ctctcaaaga gaaagcaaca 60
atcacttgtc gggcgtctca agggatatca atttggctga gctggtatca gcagaaacca 120
aatcaagcgc cgaaactgct ggtgaagaag gcctccaatc tccacattgg cgcacccagc 180
aggttttccg gcagtggctc tggcacagat ttcactctga ccatcagctc acccgagccc 240
gaagacgccg ctacatacta ttgcttgcaa tcccagacat accccctgac ttttggggga 300
ggtaccaagc tgggagatca aa 322
<210> 133
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 133
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 134
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 134
gacattcaga tgaatcagtc tcctagctca ctgtcagcca gccttggaga caccattaca 60
gtcacttgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 135
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 135
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 136
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 136
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccg 120
ggcaatatcc caaaggtgct gattaacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 137
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 137
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 138
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 138
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg ggtcccccca 180
aggttcagcg gatctggatc cgggacccac tttactctga ccataacaag cctgcagcct 240
gaagacattg ctacctatta ctgcctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 139
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 139
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 140
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 140
gacattcaga tgaatcagtc tcctagctca ctgtcagcca gccttggaga caccattaca 60
gtcacttgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccg 120
ggcaatatcc caaaggtgct gattaacaag gcctctaacc tgcacattgg cgtgccaagc 180
cgattcagcg ggagcggaag tggcaccgac tttactctga cgatcagttc actggagccc 240
gaggacgctg caacatacta ttgtctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 141
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 141
Asp Ile Gln Met Asn Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Thr Ile Thr Val Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 142
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 142
gacattcaga tgaatcagtc tcctagctca ctgtcagcca gccttggaga caccattaca 60
gtcacttgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccc 120
ggaaaggtcc cgaaactctt gatctacaag gcctctaacc tgcacattgg ggtcccccca 180
aggttcagcg gatctggatc cgggacccac tttactctga ccataacaag cctgcagcct 240
gaagacattg ctacctatta ctgcctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 143
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 143
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Val Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Thr Leu Thr Ile Thr Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 144
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 144
gaaattcaga tgactcaatc accttcatct ttgagtgcat cacccggaga tagagttaca 60
atcacctgca gggcgagtca agggatctcc atatggttgt catggtatca gcagaaaccg 120
ggcaatatcc caaaggtgct gattaacaag gcctctaacc tgcacattgg ggtcccccca 180
aggttcagcg gatctggatc cgggacccac tttactctga ccataacaag cctgcagcct 240
gaagacattg ctacctatta ctgcctgcaa tctcagacct accctctcac ctttggagga 300
ggtaccaagc tggagatcaa a 321
<210> 145
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 145
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Val Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 146
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 146
gaaatccaga tgacacagtc ccccagtagc ctttccgctt caccgggcga tagagtcact 60
attacgtgca gggcctccca gggtatttct atctggctga gctggtatca gcagaagccc 120
ggtaatgtgc caaagctctt gatctacaag gcatctaacc ttcatatcgg agtgccctca 180
agatttagtg ggtcaggcag cggaaccgat ttcacattga ccattagttc tctggaacca 240
gaggacgctg ccacttacta ctgcctgcag tcccaaacat accctttgac ttttgggggg 300
ggtaccaagc tggagatcaa a 321
<210> 147
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 147
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 148
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 148
gagattcaga tgacccagag cccatcaagc ctctccgctt cccccggaga ccgggtgacc 60
atcacatgca gagcttcaca gggaatctca atctggctca gctggtatca gcagaagcca 120
ggcaagattc cgaagttgct tatctataag gccagtaacc tgcatatcgg agttccatca 180
agattcagtg gtagcggaag tgggacagat ttcactctca ccatcagctc cctcgaacca 240
gaggacgctg caacttacta ctgcctgcag tcccagacat atccacttac tttcggcggg 300
ggtaccaagc tggagatcaa a 321
<210> 149
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 149
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Val Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 150
<211> 322
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 150
gagattcaga tgactcagag cccatctagt ctctctgcat ctcccggaga cagagttacg 60
atcacctgca gggctagcca agggatatca atttggctgt cctggtatca gcaaaaacct 120
ggcaaagtgc caaaggtctt gatttacaaa gcatccaatt tgcacatcgg cgtccctagt 180
cgcttttccg ggtctggtag cggcaccgac ttcaccctca ccataagctc actcgagccg 240
gaagatgccg ctacttacta ttgcctgcag tctcagactt accccctgac tttcggcgga 300
ggtaccaagc tggagatcaa ac 322
<210> 151
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 151
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Asn Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 152
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 152
gagatccaga tgacgcagag ccctagcagc ctctctgcat ccccaggaga cagagtaaca 60
attacctgtc gcgccagcca gggaatatct atatggctgt catggtatca acagaaaccg 120
ggaaaggttc caaagctctt gatcaataag gctagcaatc tgcatattgg agtgccctcc 180
cgcttctctg gtagcggaag tggcacagat ttcaccctga ccattagtag tctggagcct 240
gaggatgcgg ccacctacta ctgcctccag tcccaaacct atcccctgac cttcggagga 300
ggtaccaagc tggagatcaa a 321
<210> 153
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 153
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ile Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Asn Ile Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Asn Leu His Ile Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gln Ser Gln Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 154
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 154
gaaattcaga tgactcagag tcctagcagc ctgtccgcaa gcccaggtga ccgagtcacc 60
ataacctgca gggccagtca ggggatctcc atatggctct cttggtatca acagaaaccc 120
ggcaatatcc ctaagctcct gatttataaa gcgtcaaatc tgcatatcgg ggtgccatca 180
agattctctg ggtccggctc aggaaccgac tttaccctga ccatttcttc tctcgaaccc 240
gaggatgccg ccacctatta ttgccttcaa agccagacat acccattgac cttcggcggc 300
ggtaccaagc tggagatcaa a 321
<210> 155
<211> 159
<212> PRT
<213> Canis familiaris
<400> 155
Met Leu Ser His Thr Gly Pro Ser Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Ser Met Glu Thr Leu Leu Ser Ser His Met Ala Pro Thr His Gln Leu
20 25 30
Pro Pro Ser Asp Val Arg Lys Ile Ile Leu Glu Leu Gln Pro Leu Ser
35 40 45
Arg Gly Leu Leu Glu Asp Tyr Gln Lys Lys Glu Thr Gly Val Pro Glu
50 55 60
Ser Asn Arg Thr Leu Leu Leu Cys Leu Thr Ser Asp Ser Gln Pro Pro
65 70 75 80
Arg Leu Asn Ser Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Ile Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln His Glu Pro Glu Thr Glu Ile Ser Val Pro Ala Asp
115 120 125
Thr Phe Glu Cys Lys Ser Phe Ile Leu Thr Ile Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu Ser Val Phe Lys Ser Leu Asn Ser Gly Pro Gln
145 150 155
<210> 156
<211> 477
<212> DNA
<213> Canis familiaris
<400> 156
atgctctccc acacaggacc atccaggttt gccctgttcc tgctctgctc tatggaaacc 60
ttgctgtcct cccatatggc acccacccat cagctaccac caagtgatgt acgaaaaatc 120
atcttggaat tacagccctt gtcgagggga cttttggaag actatcagaa gaaagagaca 180
ggggtgccag aatccaaccg taccttgctg ctgtgtctca cctctgattc ccaaccacca 240
cgcctcaaca gctcagccat cttgccttat ttcagggcaa tcagaccatt atcagataag 300
aacattattg ataaaatcat agaacagctt gacaaactca aatttcaaca tgaaccagaa 360
acagaaattt ctgtgcctgc agatactttt gaatgtaaaa gcttcatctt gacgatttta 420
cagcagttct cggcgtgcct ggaaagtgtg tttaagtcac taaactctgg acctcag 477
<210> 157
<211> 165
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, соответствующая кошачьему IL-31
дикого типа с C-концевой His-меткой
<400> 157
Met Leu Ser His Ala Gly Pro Ala Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Cys Met Glu Thr Leu Leu Pro Ser His Met Ala Pro Ala His Arg Leu
20 25 30
Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu Leu Arg Pro Met Ser
35 40 45
Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu Ile Gly Leu Pro Glu
50 55 60
Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser Asp Ser Gln Leu Pro
65 70 75 80
His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Thr Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val Ser Met Pro Ala Asp
115 120 125
Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn Ser Gly Pro Gln His
145 150 155 160
His His His His His
165
<210> 158
<211> 495
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, соответствующая гену кошачьего
IL-31 дикого типа с кодируемой C-концевой His-меткой
<400> 158
atgctttcac acgctggacc agcccgattc gccctcttcc tcctctgctg tatggagact 60
ctgttgccgt cccacatggc cccggcacat aggctgcagc cgtctgacat ccggaagatc 120
attctcgaac ttcgccccat gtcgaagggg ttgctgcaag actacctgaa gaaggagatc 180
ggcctgcccg aaagcaacca ctcctcgctg ccttgcctgt caagcgattc ccagctgccc 240
cacattaacg gttccgccat cctcccgtac ttccgggcca tcagaccact gtcggacaag 300
aacaccatcg acaagatcat tgaacagctg gacaagctga agtttcagcg cgagcctgaa 360
gccaaagtgt ccatgcccgc cgataacttc gagcggaaga atttcattct cgcggtgctg 420
cagcagttct ccgcgtgcct ggagcacgtc ctgcaatccc tgaacagcgg acctcagcac 480
caccatcacc accat 495
<210> 159
<211> 148
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, соответствующая кошачьему IL-31
N-концевой His-меткой
<400> 159
Met Arg Gly Ser His His His His His His Gly Ser Ser His Met Ala
1 5 10 15
Pro Ala His Arg Leu Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu
20 25 30
Leu Arg Pro Met Ser Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu
35 40 45
Ile Gly Leu Pro Glu Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser
50 55 60
Asp Ser Gln Leu Pro His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe
65 70 75 80
Arg Ala Ile Arg Pro Leu Ser Asp Lys Asn Thr Ile Asp Lys Ile Ile
85 90 95
Glu Gln Leu Asp Lys Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val
100 105 110
Ser Met Pro Ala Asp Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val
115 120 125
Leu Gln Gln Phe Ser Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn
130 135 140
Ser Gly Pro Gln
145
<210> 160
<211> 444
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, соответствующая гену кошачьего
IL-31 с кодируемой N-концевой His-меткой
<400> 160
atgagaggat cccatcacca tcaccaccac ggctcatctc atatggcccc cgcacatcgc 60
ctgcagccga gtgacattcg taaaattatc ttggagctgc gcccgatgtc caagggctta 120
ctgcaggatt atctgaagaa agagatcggg ctgcctgaaa gcaaccatag tagcctgccg 180
tgtttatcgt ctgatagcca gttaccacac atcaatggct ctgcgatttt gccctacttt 240
cgcgccatcc gtccgctgtc cgataaaaat accatcgaca aaattatcga acaactggat 300
aaattgaagt ttcagcgcga gcctgaagcg aaagtttcga tgccagcmga taacttcgaa 360
cgcaaaaact ttattttagc ggtgttgcag cagttttctg cctgtctgga acacgtgctc 420
cagtcactca atagtgggcc acaa 444
<210> 161
<211> 165
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, соответствующая мутантному
белку кошачьего IL-31 11E12 с C-концевой His-меткой
<400> 161
Met Leu Ser His Ala Gly Pro Ala Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Cys Met Glu Thr Leu Leu Pro Ser His Met Ala Pro Ala His Arg Leu
20 25 30
Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu Leu Arg Pro Met Ser
35 40 45
Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu Ile Gly Leu Pro Glu
50 55 60
Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser Asp Ser Gln Leu Pro
65 70 75 80
His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Thr Ile Ala Lys Ile Ala Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val Ser Met Pro Ala Asp
115 120 125
Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn Ser Gly Pro Gln His
145 150 155 160
His His His His His
165
<210> 162
<211> 495
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, соответствующая гену мутантного
кошачьего IL-31 11E12 с кодируемой C-концевой His-меткой
<400> 162
atgctctctc acgccggtcc tgcccggttc gcactgttcc tcctctgttg catggagact 60
ctgcttccct cccacatggc accggcccat agactgcagc cgtccgacat cagaaagatc 120
atccttgaat tgcgccctat gagcaagggg ctgctgcagg attacctgaa aaaggagatc 180
ggcctgccgg aatcgaacca cagctcactg ccatgcctgt cctccgactc gcaactgccc 240
cacatcaatg gatccgccat tctgccgtac ttccgcgcta ttcggcctct ctccgacaag 300
aacaccatcg ccaagattgc cgagcagctg gataagctga agttccagag ggagccagaa 360
gccaaggtgt ccatgcccgc tgacaacttc gagcggaaga actttatcct cgcggtgctg 420
cagcagttct cagcgtgcct cgaacacgtc ttgcaaagcc tgaactcggg accccagcac 480
caccaccatc atcac 495
<210> 163
<211> 165
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность, соответствующая мутантному
белку кошачьего IL-31 15H05 с C-концевой His-меткой
<400> 163
Met Leu Ser His Ala Gly Pro Ala Arg Phe Ala Leu Phe Leu Leu Cys
1 5 10 15
Cys Met Glu Thr Leu Leu Pro Ser His Met Ala Pro Ala His Arg Leu
20 25 30
Gln Pro Ser Asp Ile Arg Lys Ile Ile Leu Glu Leu Arg Pro Met Ser
35 40 45
Lys Gly Leu Leu Gln Asp Tyr Leu Lys Lys Glu Ile Gly Leu Pro Glu
50 55 60
Ser Asn His Ser Ser Leu Pro Cys Leu Ser Ser Asp Ser Gln Leu Pro
65 70 75 80
His Ile Asn Gly Ser Ala Ile Leu Pro Tyr Phe Arg Ala Ile Arg Pro
85 90 95
Leu Ser Asp Lys Asn Thr Ile Asp Lys Ile Ile Glu Gln Leu Asp Lys
100 105 110
Leu Lys Phe Gln Arg Glu Pro Glu Ala Lys Val Ser Met Ala Ala Ala
115 120 125
Asn Phe Glu Arg Lys Asn Phe Ile Leu Ala Val Leu Gln Gln Phe Ser
130 135 140
Ala Cys Leu Glu His Val Leu Gln Ser Leu Asn Ser Gly Pro Gln His
145 150 155 160
His His His His His
165
<210> 164
<211> 495
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, соответствующая гену мутантного
кошачьего IL-31 15H05 с кодируемой C-концевой His-меткой
<400> 164
atgctctctc acgccggtcc tgcccggttc gcactgttcc tcctctgttg catggagact 60
ctgcttccct cccacatggc accggcccat agactgcagc cgtccgacat cagaaagatc 120
atccttgaat tgcgccctat gagcaagggg ctgctgcagg attacctgaa aaaggagatc 180
ggcctgccgg aatcgaacca cagctcactg ccatgcctgt cctccgactc gcaactgccc 240
cacatcaatg gatccgccat tctgccgtac ttccgcgcta ttcggcctct ctccgacaag 300
aacaccatcg acaagattat tgagcagctg gataagctga agttccagag ggagccagaa 360
gccaaggtgt ccatggccgc tgccaacttc gagcggaaga actttatcct cgcggtgctg 420
cagcagttct cagcgtgcct cgaacacgtc ttgcaaagcc tgaactcggg accccagcac 480
caccaccatc atcac 495
<210> 165
<211> 152
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность белка лошадиного IL-31 с
C-концевой His-меткой
<400> 165
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gly Pro Ile Tyr Gln Leu Gln Pro Lys Glu Ile Gln Ala
20 25 30
Ile Ile Val Glu Leu Gln Asn Leu Ser Lys Lys Leu Leu Asp Asp Tyr
35 40 45
Leu Asn Lys Glu Lys Gly Val Gln Lys Phe Asp Ser Asp Leu Pro Ser
50 55 60
Cys Phe Thr Ser Asp Ser Gln Ala Pro Gly Asn Ile Asn Ser Ser Ala
65 70 75 80
Ile Leu Pro Tyr Phe Lys Ala Ile Ser Pro Ser Leu Asn Asn Asp Lys
85 90 95
Ser Leu Tyr Ile Ile Glu Gln Leu Asp Lys Leu Asn Phe Gln Asn Ala
100 105 110
Pro Glu Thr Glu Val Ser Met Pro Thr Asp Asn Phe Glu Arg Lys Arg
115 120 125
Phe Ile Leu Thr Ile Leu Arg Trp Phe Ser Asn Cys Leu Glu His Arg
130 135 140
Ala Gln His His His His His His
145 150
<210> 166
<211> 456
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность гена лошадиного IL-31 с
кодируемой C-концевой His-меткой
<400> 166
atgggctggt cctgcatcat tctgtttctg gtggccacag ccaccggcgt gcactctgga 60
cctatctatc agctgcagcc caaagagatc caggccatca tcgtggaact gcagaacctg 120
agcaagaagc tgctggacga ctacctgaac aaagaaaagg gcgtgcagaa gttcgacagc 180
gacctgccta gctgcttcac cagcgattct caggcccctg gcaacatcaa cagcagcgcc 240
atcctgcctt acttcaaggc catctctccc agcctgaaca acgacaagag cctgtacatc 300
atcgagcagc tggacaagct gaacttccag aacgcccctg aaaccgaggt gtccatgcct 360
accgacaact tcgagcggaa gcggttcatc ctgaccatcc tgcggtggtt cagcaactgc 420
ctggaacaca gagcccagca ccaccaccat caccat 456
<210> 167
<211> 652
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность внеклеточного домена кошачьего
OSMR, слитого с Fc человеческого IgG1
<400> 167
Met Ala Leu Phe Ser Ala Phe Gln Thr Thr Phe Leu Leu Ala Leu Leu
1 5 10 15
Ser Leu Lys Thr Tyr Gln Ser Glu Val Leu Ser Glu Pro Leu Ser Leu
20 25 30
Ala Pro Glu Ser Leu Glu Val Ser Ile Asp Ser Ala Arg Gln Cys Leu
35 40 45
His Leu Lys Trp Ser Val His Asn Leu Ala Tyr His Gln Glu Leu Lys
50 55 60
Met Val Phe Gln Ile Glu Ile Ser Arg Ile Lys Thr Ser Asn Val Ile
65 70 75 80
Trp Val Glu Asn Tyr Ser Thr Thr Val Lys Arg Asn Gln Val Leu Arg
85 90 95
Trp Ser Trp Glu Ser Lys Leu Pro Leu Glu Cys Ala Lys His Ser Val
100 105 110
Arg Met Arg Gly Ala Val Asp Asp Ala Gln Val Pro Glu Leu Arg Phe
115 120 125
Trp Ser Asn Trp Thr Ser Trp Glu Glu Val Asp Val Gln Ser Ser Leu
130 135 140
Gly His Asp Pro Leu Phe Val Phe Pro Lys Asp Lys Leu Val Glu Glu
145 150 155 160
Gly Ser Asn Val Thr Ile Cys Tyr Val Ser Arg Ser His Gln Asn Asn
165 170 175
Ile Ser Cys Tyr Leu Glu Gly Val Arg Met His Gly Glu Gln Leu Asp
180 185 190
Pro Asn Val Cys Val Phe His Leu Lys Asn Val Pro Phe Ile Arg Glu
195 200 205
Thr Gly Thr Asn Ile Tyr Cys Lys Ala Asp Gln Gly Asp Val Ile Lys
210 215 220
Gly Ile Val Leu Phe Val Ser Lys Val Phe Glu Glu Pro Lys Asp Phe
225 230 235 240
Ser Cys Glu Thr Arg Asp Leu Lys Thr Leu Asn Cys Thr Trp Ala Pro
245 250 255
Gly Ser Asp Ala Gly Leu Leu Thr Gln Leu Ser Gln Ser Tyr Thr Leu
260 265 270
Phe Glu Ser Phe Ser Gly Lys Lys Thr Leu Cys Lys His Lys Ser Trp
275 280 285
Cys Asn Trp Gln Val Ser Pro Asp Ser Gln Glu Met Tyr Asn Phe Thr
290 295 300
Leu Thr Ala Glu Asn Tyr Leu Arg Lys Arg Ser Val His Leu Leu Phe
305 310 315 320
Asn Leu Thr His Arg Val His Pro Met Ala Pro Phe Asn Val Phe Val
325 330 335
Lys Asn Val Ser Ala Thr Asn Ala Thr Met Thr Trp Lys Val His Ser
340 345 350
Ile Gly Asn Tyr Ser Thr Leu Leu Cys Gln Ile Glu Leu Asp Gly Glu
355 360 365
Gly Lys Val Ile Gln Lys Gln Asn Val Ser Val Lys Val Asn Gly Lys
370 375 380
His Leu Met Lys Lys Leu Glu Pro Ser Thr Glu Tyr Ala Ala Gln Val
385 390 395 400
Arg Cys Ala Asn Ala Asn His Phe Trp Lys Trp Ser Glu Trp Thr Arg
405 410 415
Arg Asn Phe Thr Thr Ala Glu Ala Ala Asp Lys Thr His Thr Cys Pro
420 425 430
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
435 440 445
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
450 455 460
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
465 470 475 480
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
485 490 495
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
500 505 510
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
515 520 525
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
530 535 540
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
545 550 555 560
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
565 570 575
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
580 585 590
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
595 600 605
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
610 615 620
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
625 630 635 640
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
645 650
<210> 168
<211> 2076
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая внеклеточный домен
кошачьего OSMR, слитый с Fc человеческого IgG1
<400> 168
atggccctgt tcagcgcctt ccagaccacc ttcctgctgg ccctgctgag cctgaaaacc 60
taccagagcg aggtgctgag cgagcccctg tctctggccc ctgagagcct ggaagtgtcc 120
atcgacagcg ccagacagtg cctgcacctg aagtggagcg tgcacaacct ggcctaccac 180
caggaactga agatggtgtt ccagatcgag atcagccgga tcaagaccag caacgtgatc 240
tgggtggaaa actacagcac caccgtgaag cggaaccagg tgctgcggtg gtcctgggag 300
tctaagctgc ctctggaatg cgccaagcac agcgtgcgga tgagaggcgc cgtggatgat 360
gcccaggtgc ccgagctgag attctggtcc aactggacct cctgggaaga ggtggacgtg 420
cagtctagcc tgggccacga ccccctgttc gtgttcccca aggacaagct ggtggaagag 480
ggctccaacg tgaccatctg ctacgtgtcc agaagccacc agaacaacat cagctgctac 540
ctggaaggcg tgcgcatgca cggcgagcag ctggacccta acgtgtgcgt gttccacctg 600
aagaacgtgc ccttcatcag agagacaggc accaacatct actgcaaggc cgaccagggc 660
gacgtgatca agggcatcgt gctgtttgtg tccaaggtgt tcgaggaacc caaggacttc 720
agctgcgaga cacgggatct gaaaaccctg aactgtacct gggcccctgg ctccgatgcc 780
ggactgctga ctcagctgtc ccagagctac accctgttcg agagcttcag cggcaaaaag 840
accctgtgca agcacaagag ctggtgcaac tggcaagtgt cccccgatag ccaggaaatg 900
tacaacttca ccctgaccgc cgagaactac ctgcggaaga gatccgtgca tctgctgttc 960
aacctgaccc acagagtgca ccccatggcc cccttcaacg tgttcgtgaa gaatgtgtcc 1020
gccaccaacg ccaccatgac atggaaggtg cacagcatcg gcaactactc caccctgctg 1080
tgtcagatcg agctggacgg cgagggcaaa gtgatccaga aacagaacgt gtcagtgaaa 1140
gtgaacggca agcacctgat gaagaagctg gaacccagca ccgagtacgc cgcccaggtg 1200
cgctgtgcca acgccaacca cttctggaag tggagtgaat ggacccggcg gaacttcacc 1260
acagccgaag ccgccgctga gaacgaggtg tccacaccta tgcaggccct gaccaccaac 1320
aaggacgacg acaacatcct gttccgggac tccgccaatg ccaccagcct gcctgtgcag 1380
gatagcagct ctgtgctgcc cgccaagccc gagaacatct cctgcgtgtt ctactacgag 1440
gaaaacttca cttgcacctg gtcccccgag aaagaggcca gctacacctg gtacaaagtg 1500
aagagaacct acagctacgg ctacaagagc gacatctgcc ccagcgacaa cagcaccaga 1560
ggcaaccaca ccttctgcag ctttctgccc cccaccatca ccaaccccga caactacacc 1620
atccaggtgg aagcccagaa cgccgacggc atcatcaagt ccgacatcac ccactggtcc 1680
ctggacgcca tcacaaagat cgagcccccc gagatcttct ccgtgaagcc tgtgctgggc 1740
gtgaagagga tggtgcagat caagtggatc cggcccgtgc tggccccagt gtctagcacc 1800
ctgaagtaca ccctgcggtt caagaccgtg aacagcgcct actggatgga agtgaatttc 1860
accaaagagg acatcgaccg ggacgagaca tacaatctga ccggactgca ggccttcaca 1920
gagtacgtgc tggctctgag atgcgccacc aaagaatcca tgttttggag cggctggtcc 1980
caggaaaaga tgggcaccac cgaagagggt aagcctatcc ctaaccctct cctcggtctc 2040
gattctacgc gtaccggtca tcatcaccat caccat 2076
<210> 169
<211> 474
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность кошачьего IL-31-Ra, слитого с Fc
человеческого IgG1
<400> 169
Met Met Trp Pro Gln Val Trp Gly Leu Glu Ile Gln Phe Ser Pro Gln
1 5 10 15
Pro Ala Cys Ile Asp Leu Gly Met Met Trp Ala His Ala Leu Trp Thr
20 25 30
Leu Leu Leu Leu Cys Lys Phe Ser Leu Ala Val Leu Pro Ala Lys Pro
35 40 45
Glu Asn Ile Ser Cys Val Phe Tyr Tyr Glu Glu Asn Phe Thr Cys Thr
50 55 60
Trp Ser Pro Glu Lys Glu Ala Ser Tyr Thr Trp Tyr Lys Val Lys Arg
65 70 75 80
Thr Tyr Ser Tyr Gly Tyr Lys Ser Asp Ile Cys Pro Ser Asp Asn Ser
85 90 95
Thr Arg Gly Asn His Thr Phe Cys Ser Phe Leu Pro Pro Thr Ile Thr
100 105 110
Asn Pro Asp Asn Tyr Thr Ile Gln Val Glu Ala Gln Asn Ala Asp Gly
115 120 125
Ile Ile Lys Ser Asp Ile Thr His Trp Ser Leu Asp Ala Ile Thr Lys
130 135 140
Ile Glu Pro Pro Glu Ile Phe Ser Val Lys Pro Val Leu Gly Val Lys
145 150 155 160
Arg Met Val Gln Ile Lys Trp Ile Arg Pro Val Leu Ala Pro Val Ser
165 170 175
Ser Thr Leu Lys Tyr Thr Leu Arg Phe Lys Thr Val Asn Ser Ala Tyr
180 185 190
Trp Met Glu Val Asn Phe Thr Lys Glu Asp Ile Asp Arg Asp Glu Thr
195 200 205
Tyr Asn Leu Thr Gly Leu Gln Ala Phe Thr Glu Tyr Val Leu Ala Leu
210 215 220
Arg Cys Ala Thr Lys Glu Ser Met Phe Trp Ser Gly Trp Ser Gln Glu
225 230 235 240
Lys Met Gly Thr Thr Glu Glu Asp Lys Thr His Thr Cys Pro Pro Cys
245 250 255
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
260 265 270
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
275 280 285
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
290 295 300
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
305 310 315 320
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
325 330 335
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
340 345 350
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
355 360 365
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
370 375 380
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
385 390 395 400
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
405 410 415
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
420 425 430
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
435 440 445
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
450 455 460
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 170
<211> 1422
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая кошачий IL-31 Ra,
слитый с Fc человеческого IgG1
<400> 170
atgatgtggc cacaagtgtg gggcctggag atccagttca gcccccagcc tgcctgcatc 60
gatctgggca tgatgtgggc tcacgctctg tggaccctgc tgctgctgtg caagttttcc 120
ctggccgtgc tgcccgctaa gcctgagaac atcagctgcg tgttctacta tgaggagaac 180
ttcacctgta catggtcccc cgagaaggag gctagctata cctggtacaa ggtgaagaga 240
acatacagct atggctacaa gtctgatatc tgccccagcg acaactctac ccgcggcaat 300
cacacattct gttcttttct gccccctacc atcacaaacc ctgataatta taccatccag 360
gtggaggccc agaacgctga tggcatcatc aagtctgaca tcacccattg gtccctggac 420
gccatcacaa agatcgagcc acccgagatt ttctccgtga agcccgtgct gggcgtgaag 480
aggatggtgc agatcaagtg gatcaggcct gtgctggctc cagtgtccag caccctgaag 540
tatacactga gattcaagac cgtgaactcc gcttactgga tggaggtgaa cttcaccaag 600
gaggacatcg atagggacga gacctataat ctgacaggcc tgcaggcctt caccgagtac 660
gtgctggccc tgaggtgcgc tacaaaggag tccatgtttt ggtccggctg gagccaggag 720
aagatgggca ccacagagga ggataagacc cacacatgcc ctccatgtcc agctccagag 780
ctgctgggag gaccaagcgt gttcctgttt ccacctaagc ctaaggacac cctgatgatc 840
tctcgcaccc ctgaggtgac atgcgtggtg gtggacgtgt cccacgagga cccagaggtg 900
aagtttaact ggtatgtgga tggcgtggag gtgcataatg ccaagaccaa gcctagagag 960
gagcagtata acagcaccta ccgcgtggtg tctgtgctga cagtgctgca tcaggactgg 1020
ctgaacggca aggagtacaa gtgcaaggtg agcaataagg ccctgcctgc tccaatcgag 1080
aagaccatct ctaaggctaa gggacagcca agggagccac aggtgtatac actgccaccc 1140
agccgggagg agatgaccaa gaaccaggtg tctctgacat gtctggtgaa gggcttctac 1200
ccatctgata tcgctgtgga gtgggagtcc aatggccagc ccgagaacaa ttataagacc 1260
acacctccag tgctggattc tgacggctcc ttctttctgt actccaagct gaccgtggac 1320
aagagcaggt ggcagcaggg caacgtgttt tcttgctccg tgatgcatga ggctctgcac 1380
aatcattaca cacagaagag cctgtctctg tccccaggca ag 1422
<210> 171
<211> 335
<212> PRT
<213> Felis catus
<400> 171
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Thr Thr Ser Gly Ala Thr Val Ala Leu Ala Cys Leu Val Leu Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ala Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Leu Ser Asp Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Arg Lys Thr Asp His Pro Pro Gly Pro Lys Pro Cys Asp Cys
100 105 110
Pro Lys Cys Pro Pro Pro Glu Met Leu Gly Gly Pro Ser Ile Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Ser Ile Ser Arg Thr Pro Glu
130 135 140
Val Thr Cys Leu Val Val Asp Leu Gly Pro Asp Asp Ser Asp Val Gln
145 150 155 160
Ile Thr Trp Phe Val Asp Asn Thr Gln Val Tyr Thr Ala Lys Thr Ser
165 170 175
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Leu His Gln Asp Trp Leu Lys Gly Lys Glu Phe Lys Cys Lys
195 200 205
Val Asn Ser Lys Ser Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Lys Gly Gln Pro His Glu Pro Gln Val Tyr Val Leu Pro Pro Ala
225 230 235 240
Gln Glu Glu Leu Ser Arg Asn Lys Val Ser Val Thr Cys Leu Ile Lys
245 250 255
Ser Phe His Pro Pro Asp Ile Ala Val Glu Trp Glu Ile Thr Gly Gln
260 265 270
Pro Glu Pro Glu Asn Asn Tyr Arg Thr Thr Pro Pro Gln Leu Asp Ser
275 280 285
Asp Gly Thr Tyr Phe Val Tyr Ser Lys Leu Ser Val Asp Arg Ser His
290 295 300
Trp Gln Arg Gly Asn Thr Tyr Thr Cys Ser Val Ser His Glu Ala Leu
305 310 315 320
His Ser His His Thr Gln Lys Ser Leu Thr Gln Ser Pro Gly Lys
325 330 335
<210> 172
<211> 1005
<212> DNA
<213> Felis catus
<400> 172
gcctccacca cggccccatc ggtgttccca ctggccccca gctgcgggac cacatctggc 60
gccaccgtgg ccctggcctg cctggtgtta ggctacttcc ctgagccggt gaccgtgtcc 120
tggaactccg gcgccctgac cagcggtgtg cacaccttcc cggccgtcct gcaggcctcg 180
gggctgtact ctctcagcag catggtgaca gtgccctcca gcaggtggct cagtgacacc 240
ttcacctgca acgtggccca cccgcccagc aacaccaagg tggacaagac cgtgcgcaaa 300
acagaccacc caccgggacc caaaccctgc gactgtccca aatgcccacc ccctgagatg 360
cttggaggac cgtccatctt catcttcccc ccaaaaccca aggacaccct ctcgatttcc 420
cggacgcccg aggtcacatg cttggtggtg gacttgggcc cagatgactc cgatgtccag 480
atcacatggt ttgtggataa cacccaggtg tacacagcca agacgagtcc gcgtgaggag 540
cagttcaaca gcacctaccg tgtggtcagt gtcctcccca tcctacacca ggactggctc 600
aaggggaagg agttcaagtg caaggtcaac agcaaatccc tcccctcccc catcgagagg 660
accatctcca aggccaaagg acagccccac gagccccagg tgtacgtcct gcctccagcc 720
caggaggagc tcagcaggaa caaagtcagt gtgacctgcc tgatcaaatc cttccacccg 780
cctgacattg ccgtcgagtg ggagatcacc ggacagccgg agccagagaa caactaccgg 840
acgaccccgc cccagctgga cagcgacggg acctacttcg tgtacagcaa gctctcggtg 900
gacaggtccc actggcagag gggaaacacc tacacctgct cggtgtcaca cgaagctctg 960
cacagccacc acacacagaa atccctcacc cagtctccgg gtaaa 1005
<210> 173
<211> 335
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность кошачьей тяжелой цепи,
модифицированной для модуляции эффекторной функции антитела
<400> 173
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Thr Thr Ser Gly Ala Thr Val Ala Leu Ala Cys Leu Val Leu Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ala Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Leu Ser Asp Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Arg Lys Thr Asp His Pro Pro Gly Pro Lys Pro Cys Asp Cys
100 105 110
Pro Lys Cys Pro Pro Pro Glu Ala Ala Gly Ala Pro Ser Ile Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Ser Ile Ser Arg Thr Pro Glu
130 135 140
Val Thr Cys Leu Val Val Asp Leu Gly Pro Asp Asp Ser Asp Val Gln
145 150 155 160
Ile Thr Trp Phe Val Asp Asn Thr Gln Val Tyr Thr Ala Lys Thr Ser
165 170 175
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Leu His Gln Asp Trp Leu Lys Gly Lys Glu Phe Lys Cys Lys
195 200 205
Val Asn Ser Lys Ser Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Lys Gly Gln Pro His Glu Pro Gln Val Tyr Val Leu Pro Pro Ala
225 230 235 240
Gln Glu Glu Leu Ser Arg Asn Lys Val Ser Val Thr Cys Leu Ile Lys
245 250 255
Ser Phe His Pro Pro Asp Ile Ala Val Glu Trp Glu Ile Thr Gly Gln
260 265 270
Pro Glu Pro Glu Asn Asn Tyr Arg Thr Thr Pro Pro Gln Leu Asp Ser
275 280 285
Asp Gly Thr Tyr Phe Val Tyr Ser Lys Leu Ser Val Asp Arg Ser His
290 295 300
Trp Gln Arg Gly Asn Thr Tyr Thr Cys Ser Val Ser His Glu Ala Leu
305 310 315 320
His Ser His His Thr Gln Lys Ser Leu Thr Gln Ser Pro Gly Lys
325 330 335
<210> 174
<211> 1005
<212> DNA
<213> Artificial Sequence
<220>
<223> Нкулеотидная последовательность, кодирующая кошачью тяжелую цепь,
модифицированную для модуляции эффекторной функции антитела
<400> 174
gcctccacca cggccccatc ggtgttccca ctggccccca gctgcgggac cacatctggc 60
gccaccgtgg ccctggcctg cctggtgtta ggctacttcc ctgagccggt gaccgtgtcc 120
tggaactccg gcgccctgac cagcggtgtg cacaccttcc cggccgtcct gcaggcctcg 180
gggctgtact ctctcagcag catggtgaca gtgccctcca gcaggtggct cagtgacacc 240
ttcacctgca acgtggccca cccgcccagc aacaccaagg tggacaagac cgtgcgcaaa 300
acagaccacc caccgggacc caaaccctgc gactgtccca aatgcccacc ccctgaggcg 360
gctggagcac cgtccatctt catcttcccc ccaaaaccca aggacaccct ctcgatttcc 420
cggacgcccg aggtcacatg cttggtggtg gacttgggcc cagatgactc cgatgtccag 480
atcacatggt ttgtggataa cacccaggtg tacacagcca agacgagtcc gcgtgaggag 540
cagttcaaca gcacctaccg tgtggtcagt gtcctcccca tcctacacca ggactggctc 600
aaggggaagg agttcaagtg caaggtcaac agcaaatccc tcccctcccc catcgagagg 660
accatctcca aggccaaagg acagccccac gagccccagg tgtacgtcct gcctccagcc 720
caggaggagc tcagcaggaa caaagtcagt gtgacctgcc tgatcaaatc cttccacccg 780
cctgacattg ccgtcgagtg ggagatcacc ggacagccgg agccagagaa caactaccgg 840
acgaccccgc cccagctgga cagcgacggg acctacttcg tgtacagcaa gctctcggtg 900
gacaggtccc actggcagag gggaaacacc tacacctgct cggtgtcaca cgaagctctg 960
cacagccacc acacacagaa atccctcacc cagtctccgg gtaaa 1005
<210> 175
<211> 110
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность кошачьей легкой цепи каппа,
модифицированной устранением гликозилирования (G-) в положении 103
<400> 175
Arg Ser Asp Ala Gln Pro Ser Val Phe Leu Phe Gln Pro Ser Leu Asp
1 5 10 15
Glu Leu His Thr Gly Ser Ala Ser Ile Val Cys Ile Leu Asn Asp Phe
20 25 30
Tyr Pro Lys Glu Val Asn Val Lys Trp Lys Val Asp Gly Val Val Gln
35 40 45
Asn Lys Gly Ile Gln Glu Ser Thr Thr Glu Gln Asn Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Gln
65 70 75 80
Ser His Glu Lys Phe Ser Cys Glu Val Thr His Lys Ser Leu Ala Ser
85 90 95
Thr Leu Val Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Glu
100 105 110
<210> 176
<211> 330
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая кошачью легкую цепь
каппа, модифицированную устранением гликозилирования (G-)
<400> 176
cggagtgatg ctcagccatc tgtctttctc ttccaaccat ctctggacga gttacataca 60
ggaagtgcct ctatcgtgtg catattgaat gacttctacc ccaaagaggt caatgtcaag 120
tggaaagtgg atggcgtagt ccaaaacaaa ggcatccagg agagcaccac agagcagaac 180
agcaaggaca gcacctacag cctcagcagc accctgacga tgtccagtac ggagtaccaa 240
agtcatgaaa agttctcctg cgaggtcact cacaagagcc tggcctccac cctcgtcaag 300
agcttccaga ggagcgagtg tcagagagag 330
<210> 177
<211> 335
<212> PRT
<213> Canis familiaris
<400> 177
Ala Ser Thr Thr Ala Pro Ser Val Phe Pro Leu Ala Pro Ser Cys Gly
1 5 10 15
Ser Thr Ser Gly Ser Thr Val Ala Leu Ala Cys Leu Val Ser Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ser Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ser Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Met Val Thr Val Pro Ser Ser Arg Trp Pro Ser Glu Thr
65 70 75 80
Phe Thr Cys Asn Val Ala His Pro Ala Ser Lys Thr Lys Val Asp Lys
85 90 95
Pro Val Pro Lys Arg Glu Asn Gly Arg Val Pro Arg Pro Pro Asp Cys
100 105 110
Pro Lys Cys Pro Ala Pro Glu Met Leu Gly Gly Pro Ser Val Phe Ile
115 120 125
Phe Pro Pro Lys Pro Lys Asp Thr Leu Leu Ile Ala Arg Thr Pro Glu
130 135 140
Val Thr Cys Val Val Val Asp Leu Asp Pro Glu Asp Pro Glu Val Gln
145 150 155 160
Ile Ser Trp Phe Val Asp Gly Lys Gln Met Gln Thr Ala Lys Thr Gln
165 170 175
Pro Arg Glu Glu Gln Phe Asn Gly Thr Tyr Arg Val Val Ser Val Leu
180 185 190
Pro Ile Gly His Gln Asp Trp Leu Lys Gly Lys Gln Phe Thr Cys Lys
195 200 205
Val Asn Asn Lys Ala Leu Pro Ser Pro Ile Glu Arg Thr Ile Ser Lys
210 215 220
Ala Arg Gly Gln Ala His Gln Pro Ser Val Tyr Val Leu Pro Pro Ser
225 230 235 240
Arg Glu Glu Leu Ser Lys Asn Thr Val Ser Leu Thr Cys Leu Ile Lys
245 250 255
Asp Phe Phe Pro Pro Asp Ile Asp Val Glu Trp Gln Ser Asn Gly Gln
260 265 270
Gln Glu Pro Glu Ser Lys Tyr Arg Thr Thr Pro Pro Gln Leu Asp Glu
275 280 285
Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Ser Val Asp Lys Ser Arg
290 295 300
Trp Gln Arg Gly Asp Thr Phe Ile Cys Ala Val Met His Glu Ala Leu
305 310 315 320
His Asn His Tyr Thr Gln Glu Ser Leu Ser His Ser Pro Gly Lys
325 330 335
<210> 178
<211> 1005
<212> DNA
<213> Canis familiaris
<400> 178
gcctcaacaa ctgctcctag cgtgtttccc ctggccccta gctgcggaag tacctcaggc 60
agcacagtgg ccctggcttg tctggtgtct ggatatttcc ctgagccagt gaccgtgagt 120
tggaacagcg gctctctgac ctccggggtg cacacatttc catctgtgct gcagtctagt 180
ggcctgtact ccctgtcaag catggtgact gtgccttcct ctaggtggcc atcagaaact 240
ttcacctgca acgtggccca tcccgccagc aagaccaaag tggacaagcc cgtgcctaaa 300
agggagaatg gaagggtgcc aagaccacct gattgcccta agtgtccagc tccagaagcg 360
gcgggagcac caagcgtgtt catctttcca cccaagccca aagacacact gctgattgct 420
agaactcccg aggtgacctg cgtggtggtg gacctggatc cagaggaccc cgaagtgcag 480
atctcctggt tcgtggatgg gaagcagatg cagacagcca aaactcagcc tcgggaggaa 540
cagtttaacg gaacctatag agtggtgtct gtgctgccaa ttggacacca ggactggctg 600
aagggcaaac agtttacatg caaggtgaac aacaaggccc tgcctagtcc aatcgagagg 660
actatttcaa aagctagggg acaggctcat cagccttccg tgtatgtgct gcctccatcc 720
cgggaggaac tgtctaagaa cacagtgagt ctgacttgtc tgatcaaaga tttctttccc 780
cctgacattg atgtggagtg gcagagcaat gggcagcagg agccagaatc caagtacaga 840
accacaccac cccagctgga cgaagatggc tcctatttcc tgtacagtaa gctgtcagtg 900
gacaaatcta ggtggcagcg cggggatacc tttatctgcg ccgtgatgca cgaggctctg 960
cacaatcatt acacacaaga aagtctgtca catagccccg gcaag 1005
<210> 179
<211> 106
<212> PRT
<213> Canis familiaris
<400> 179
Arg Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp
1 5 10 15
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe
20 25 30
Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln
35 40 45
Asp Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser
65 70 75 80
His Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr
85 90 95
Leu Ile Lys Ser Phe Gln Arg Ser Glu Cys
100 105
<210> 180
<211> 318
<212> DNA
<213> Canis familiaris
<400> 180
aggaacgacg cccagcctgc tgtgtatctg tttcagccct cccctgatca gctgcacact 60
ggctctgcta gtgtggtgtg tctgctgaac agcttctacc caaaggatat caatgtgaag 120
tggaaagtgg acggcgtgat ccaggatact gggattcagg agtccgtgac cgaacaggac 180
aaagattcaa catatagcct gagctccact ctgaccatgt ctagtaccga gtacctgagc 240
cacgaactgt attcctgcga gatcactcat aagtccctgc cctctaccct gatcaagagc 300
ttccagagat cagagtgt 318
<210> 181
<211> 164
<212> PRT
<213> Homo sapiens
<400> 181
Met Ala Ser His Ser Gly Pro Ser Thr Ser Val Leu Phe Leu Phe Cys
1 5 10 15
Cys Leu Gly Gly Trp Leu Ala Ser His Thr Leu Pro Val Arg Leu Leu
20 25 30
Arg Pro Ser Asp Asp Val Gln Lys Ile Val Glu Glu Leu Gln Ser Leu
35 40 45
Ser Lys Met Leu Leu Lys Asp Val Glu Glu Glu Lys Gly Val Leu Val
50 55 60
Ser Gln Asn Tyr Thr Leu Pro Cys Leu Ser Pro Asp Ala Gln Pro Pro
65 70 75 80
Asn Asn Ile His Ser Pro Ala Ile Arg Ala Tyr Leu Lys Thr Ile Arg
85 90 95
Gln Leu Asp Asn Lys Ser Val Ile Asp Glu Ile Ile Glu His Leu Asp
100 105 110
Lys Leu Ile Phe Gln Asp Ala Pro Glu Thr Asn Ile Ser Val Pro Thr
115 120 125
Asp Thr His Glu Cys Lys Arg Phe Ile Leu Thr Ile Ser Gln Gln Phe
130 135 140
Ser Glu Cys Met Asp Leu Ala Leu Lys Ser Leu Thr Ser Gly Ala Gln
145 150 155 160
Gln Ala Thr Thr
<210> 182
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность канинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Canis
<400> 182
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro
35 40 45
Gln Arg Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser
65 70 75 80
Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 183
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
канинизированной вариабельной области легкой цепи mAb от Mus
musculus и Canis
<400> 183
gacatcgtga tgacccagac ccccctgagc ctgagcgtgt cccctggcga gcctgccagc 60
atcagctgca gagccagcga gagcgtggac aactacggca tcagcttcat gcactggttc 120
cagcagaagc ccggccagag cccccagcgg ctgatctaca gagccagcaa cctggaaagc 180
ggcgtgcccg atcggtttag cggctctggc agcggcaccg acttcaccct gcggatctct 240
cgggtggaag ccgatgacgc cggagtgtac tactgccagc agagcaacaa ggaccccctg 300
acctttggcg ccggtaccaa gctggagatc aag 333
<210> 184
<211> 111
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность канинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Canis
<400> 184
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Ile Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Arg Ala Ser Asn Leu Glu Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile Ser
65 70 75 80
Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Lys Asp Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 185
<211> 333
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
канинизированной вариабельной области легкой цепи mAb от Mus
musculus и Canis
<400> 185
gatatagtga tgacacaaac tcctctcagt ctttccgtat caccgggaga accggcttcc 60
atttcctgtc gggcctcaga gtctgtggac aactacggga tatccttcat gcactggtat 120
cagcagaaac ccggccagcc ccctaaactc cttatttaca gggccagtaa tctggaaagc 180
ggtgtgcccg atcgatttag cggttccggg agcggcacag atttcaccct gcgaatctct 240
agagttgaag cggatgatgc aggagtatat tactgccagc aatccaataa ggatcccctt 300
acattcggcg cgggtaccaa gctggagatc aag 333
<210> 186
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность кошачьей легкой цепи каппа G- с
модифицированным C-концом
<400> 186
Arg Ser Asp Ala Gln Pro Ser Val Phe Leu Phe Gln Pro Ser Leu Asp
1 5 10 15
Glu Leu His Thr Gly Ser Ala Ser Ile Val Cys Ile Leu Asn Asp Phe
20 25 30
Tyr Pro Lys Glu Val Asn Val Lys Trp Lys Val Asp Gly Val Val Gln
35 40 45
Asn Lys Gly Ile Gln Glu Ser Thr Thr Glu Gln Asn Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Gln
65 70 75 80
Ser His Glu Lys Phe Ser Cys Glu Val Thr His Lys Ser Leu Ala Ser
85 90 95
Thr Leu Val Lys Ser Phe Gln Arg Ser Glu Cys
100 105
<210> 187
<211> 324
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая кошачью цепь каппа G-
с модифицированным C-концом
<400> 187
cggagtgatg ctcagccatc tgtctttctc ttccaaccat ctctggacga gttacataca 60
ggaagtgcct ctatcgtgtg catattgaat gacttctacc ccaaagaggt caatgtcaag 120
tggaaagtgg atggcgtagt ccaaaacaaa ggcatccagg agagcaccac agagcagaac 180
agcaaggaca gcacctacag cctcagcagc accctgacga tgtccagtac ggagtaccaa 240
agtcatgaaa agttctcctg cgaggtcact cacaagagcc tggcctccac cctcgtcaag 300
agcttccaga ggagcgagtg ttag 324
<210> 188
<211> 323
<212> PRT
<213> Mus musculus
<400> 188
Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala
1 5 10 15
Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe
20 25 30
Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly
35 40 45
Val His Thr Phe Pro Ala Val Leu Glu Ser Asp Leu Tyr Thr Leu Ser
50 55 60
Ser Ser Val Thr Val Pro Ser Ser Pro Arg Pro Ser Glu Thr Val Thr
65 70 75 80
Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile
85 90 95
Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu
100 105 110
Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr
115 120 125
Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys
130 135 140
Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val
145 150 155 160
His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe
165 170 175
Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly
180 185 190
Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile
195 200 205
Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val
210 215 220
Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser
225 230 235 240
Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu
245 250 255
Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro
260 265 270
Ile Met Asn Thr Asn Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val
275 280 285
Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu
290 295 300
His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser
305 310 315 320
Pro Gly Lys
<210> 189
<211> 971
<212> DNA
<213> Mus musculus
<400> 189
ccaaaacgac acccccatct gtctatccac tggcccctgg atctgctgcc caaactaact 60
ccatggtgac cctgggatgc ctggtcaagg gctatttccc tgagccagtg acagtgacct 120
ggaactctgg atccctgtcc agcggtgtgc acaccttccc agctgtcctg gagtctgacc 180
tctacactct gagcagctca gtgactgtcc cctccagccc tcggcccagc gagaccgtca 240
cctgcaacgt tgcccacccg gccagcagca ccaaggtgga caagaaaatt gtgcccaggg 300
attgtggttg taagccttgc atatgtacag tcccagaagt atcatctgtc ttcatcttcc 360
ccccaaagcc caaggatgtg ctcaccatta ctctgactcc taaggtcacg tgtgttgtgg 420
tagacatcag caaggatgat cccgaggtcc agttcagctg gtttgtagat gatgtggagg 480
tgcacacagc tcagacgcaa ccccgggagg agcagttcaa cagcactttc cgctcagtca 540
gtgaacttcc catcatgcac caggactggc tcaatggcaa ggagttcaaa tgcagggtca 600
acagtgcagc tttccctgcc cccatcgaga aaaccatctc caaaaccaaa ggcagaccga 660
aggctccaca ggtgtacacc attccacctc ccaaggagca gatggccaag gataaagtca 720
gtctgacctg catgataaca gacttcttcc ctgaagacat tactgtggag tggcagtgga 780
atgggcagcc agcggagaac tacaagaaca ctcagcccat catgaacacg aatggctctt 840
acttcgtcta cagcaagctc aatgtgcaga agagcaactg ggaggcagga aatactttca 900
cctgctctgt gttacatgag ggcctgcaca accaccatac tgagaagagc ctctcccact 960
ctcctggtaa a 971
<210> 190
<211> 106
<212> PRT
<213> Mus musculus
<400> 190
Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln
1 5 10 15
Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr
20 25 30
Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln
35 40 45
Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg
65 70 75 80
His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro
85 90 95
Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
100 105
<210> 191
<211> 321
<212> DNA
<213> Mus musculus
<400> 191
gctgatgctg caccaactgt atccatcttc ccaccatcca gtgagcagtt aacatctgga 60
ggtgcctcag tcgtgtgctt cttgaacaac ttctacccca aagacatcaa tgtcaagtgg 120
aagattgatg gcagtgaacg acaaaatggc gtcctgaaca gttggactga tcaggacagc 180
aaagacagca cctacagcat gagcagcacc ctcacgttga ccaaggacga gtatgaacga 240
cataacagct atacctgtga ggccactcac aagacatcaa cttcacccat tgtcaagagc 300
ttcaacagga atgagtgtta g 321
<210> 192
<211> 106
<212> PRT
<213> Homo sapiens
<400> 192
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 193
<211> 321
<212> DNA
<213> Homo sapiens
<400> 193
actgtggctg caccatctgt cttcatcttc ccgccatctg atgagcagtt gaaatctgga 60
actgcctctg ttgtgtgcct gctgaataac ttctatccca gagaggccaa agtacagtgg 120
aaggtggata acgccctcca atcgggtaac tcccaggaga gtgtcacaga gcaggacagc 180
aaggacagca cctacagcct cagcagcacc ctgacgctga gcaaagcaga ctacgagaaa 240
cacaaagtct acgcctgcga agtcacccat cagggcctga gctcgcccgt cacaaagagc 300
ttcaacaggg gagagtgtta g 321
<210> 194
<211> 109
<212> PRT
<213> Canis familiaris
<400> 194
Asn Asp Ala Gln Pro Ala Val Tyr Leu Phe Gln Pro Ser Pro Asp Gln
1 5 10 15
Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Ser Phe Tyr
20 25 30
Pro Lys Asp Ile Asn Val Lys Trp Lys Val Asp Gly Val Ile Gln Asp
35 40 45
Thr Gly Ile Gln Glu Ser Val Thr Glu Gln Asp Lys Asp Ser Thr Tyr
50 55 60
Ser Leu Ser Ser Thr Leu Thr Met Ser Ser Thr Glu Tyr Leu Ser His
65 70 75 80
Glu Leu Tyr Ser Cys Glu Ile Thr His Lys Ser Leu Pro Ser Thr Leu
85 90 95
Ile Lys Ser Phe Gln Arg Ser Glu Cys Gln Arg Val Asp
100 105
<210> 195
<211> 330
<212> DNA
<213> Canis familiaris
<400> 195
aatgatgccc agccagccgt ctatttgttc caaccatctc cagaccagtt acacacagga 60
agtgcctctg ttgtgtgctt gctgaatagc ttctacccca aagacatcaa tgtcaagtgg 120
aaagtggatg gtgtcatcca agacacaggc atccaggaaa gtgtcacaga gcaggacaag 180
gacagtacct acagcctcag cagcaccctg acgatgtcca gtactgagta cctaagtcat 240
gagttgtact cctgtgagat cactcacaag agcctgccct ccaccctcat caagagcttc 300
caaaggagcg agtgtcagag agtggactaa 330
<210> 196
<211> 108
<212> PRT
<213> Sus scrofa
<400> 196
Ala Asp Ala Lys Pro Ser Val Phe Ile Phe Pro Pro Ser Lys Glu Gln
1 5 10 15
Leu Glu Thr Gln Thr Val Ser Val Val Cys Leu Leu Asn Ser Phe Phe
20 25 30
Pro Arg Glu Val Asn Val Lys Trp Lys Val Asp Gly Val Val Gln Ser
35 40 45
Ser Gly Ile Leu Asp Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Ser Leu Pro Thr Ser Gln Tyr Leu Ser
65 70 75 80
His Asn Leu Tyr Ser Cys Glu Val Thr His Lys Thr Leu Ala Ser Pro
85 90 95
Leu Val Lys Ser Phe Ser Arg Asn Glu Cys Glu Ala
100 105
<210> 197
<211> 327
<212> DNA
<213> Sus scrofa
<400> 197
gctgatgcca agccatccgt cttcatcttc ccgccatcga aggagcagtt agagacccaa 60
actgtctctg tggtgtgctt gctcaatagc ttcttcccca gagaagtcaa tgtcaagtgg 120
aaagtggatg gggtggtcca aagcagtggc atcctggata gtgtcacaga gcaggacagc 180
aaggacagca cctacagcct cagcagcacc ctctcgctgc ccacgtcaca gtacctaagt 240
cataatttat attcctgtga ggtcacccac aagaccctgg cctcccctct ggtcaaaagc 300
ttcagcagga acgagtgtga ggcttag 327
<210> 198
<211> 108
<212> PRT
<213> Mustela vison
<400> 198
Arg Asn Asp Ala Gln Pro Ser Val Phe Leu Phe Gln Pro Ser Gln Asp
1 5 10 15
Gln Leu His Thr Gly Ser Ala Ser Val Val Cys Leu Leu Asn Gly Phe
20 25 30
Tyr Pro Lys Glu Val Thr Val Lys Trp Met Val Asp Gly Val Thr Lys
35 40 45
Asn Thr Gly Ile Leu Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Ile Pro Ser Thr Glu Tyr Leu
65 70 75 80
Ser His Glu Thr Tyr Ser Cys Glu Val Thr His Lys Ser Leu Ser Ser
85 90 95
Pro Leu Val Lys Ser Phe Gln Arg Ser Glu Cys Gln
100 105
<210> 199
<211> 327
<212> DNA
<213> Mustela vison
<400> 199
cggaatgatg cccagccatc cgtctttttg ttccaaccat ctcaggacca gttacatacc 60
ggcagtgcct ctgtcgtgtg cttgctgaat ggcttctacc ccaaagaagt cactgtcaaa 120
tggatggttg atggtgttac caaaaacaca ggcatcctag aaagtgtcac agaacaagac 180
agcaaggaca gcacctacag cctcagcagc accctgacga tccccagtac ggagtaccta 240
agtcatgaga cgtactcctg tgaggtcact cacaagagcc tgtcctcccc tcttgtcaag 300
agcttccaaa ggagcgagtg ccaatga 327
<210> 200
<211> 5
<212> PRT
<213> Mus musculus
<400> 200
Ser Tyr Trp Met Asn
1 5
<210> 201
<211> 17
<212> PRT
<213> Mus musculus
<400> 201
Met Ile Asp Pro Ser Asp Ser Glu Ile His Tyr Asn Gln Val Phe Lys
1 5 10 15
Asp
<210> 202
<211> 9
<212> PRT
<213> Mus musculus
<400> 202
Gln Asp Ile Val Thr Thr Val Asp Tyr
1 5
<210> 203
<211> 17
<212> PRT
<213> Mus musculus
<400> 203
Lys Ser Ser Gln Ser Leu Leu Tyr Ser Ile Asn Gln Lys Asn His Leu
1 5 10 15
Ala
<210> 204
<211> 7
<212> PRT
<213> Mus musculus
<400> 204
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 205
<211> 9
<212> PRT
<213> Mus musculus
<400> 205
Gln Gln Gly Tyr Thr Tyr Pro Phe Thr
1 5
<210> 206
<211> 5
<212> PRT
<213> Mus musculus
<400> 206
Ser Tyr Trp Met Asn
1 5
<210> 207
<211> 17
<212> PRT
<213> Mus musculus
<400> 207
Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn Gln Ile Phe Arg
1 5 10 15
Asp
<210> 208
<211> 9
<212> PRT
<213> Mus musculus
<400> 208
Gln Asp Ile Val Thr Thr Val Asp Tyr
1 5
<210> 209
<211> 17
<212> PRT
<213> Mus musculus
<400> 209
Lys Ser Ser Gln Ser Leu Leu Tyr Ser Ile Asn Gln Lys Asn Phe Leu
1 5 10 15
Ala
<210> 210
<211> 7
<212> PRT
<213> Mus musculus
<400> 210
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 211
<211> 9
<212> PRT
<213> Mus musculus
<400> 211
Gln Gln His Tyr Gly Tyr Pro Phe Thr
1 5
<210> 212
<211> 118
<212> PRT
<213> Mus musculus
<400> 212
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met Asn Trp Ala Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asp Pro Ser Asp Ser Glu Ile His Tyr Asn Gln Val Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Ile Val Thr Thr Val Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 213
<211> 354
<212> DNA
<213> Mus musculus
<400> 213
caggtccaac tgcagcagcc tggggctgag ctggtgaggc ctggggcttc agtgaagctg 60
tcctgcaagg cttctggcta caccttcacc agctactgga tgaactgggc gaagcagagg 120
cctggacaag gccttgaatg gattggtatg attgatcctt cagacagtga aattcactac 180
aatcaagtgt tcaaggacaa ggccacattg actgtagaca aatcctccag cacagcctac 240
atgcagctca gcagcctgac atctgaggac tctgcggtct attactgtgc aagacaagat 300
atagtgacta cagttgacta ctggggccaa ggcaccactc tcacagtctc ctca 354
<210> 214
<211> 113
<212> PRT
<213> Mus musculus
<400> 214
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ile Asn Gln Lys Asn His Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ala Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Thr Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Gly Tyr Thr Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 215
<211> 339
<212> DNA
<213> Mus musculus
<400> 215
gacattgtga tgtcacagtc tccatcctcc ctagctgtgt cagttggaga gaaggttaca 60
atgagctgca agtccagtca gagcctttta tatagtatca atcaaaagaa ccacttggcc 120
tggttccagc agaaaccagg gcagtctcct aaactgctga tttactgggc atccactagg 180
gaatctgggg tccctgctcg cttcacaggc agtggatctg gaacagattt cactctcacc 240
atcagcagtg tgaagactga agacctggca gtttattact gtcagcaagg ttatacctac 300
ccattcacgt tcggctcggg gacaaagttg gaaataaaa 339
<210> 216
<211> 118
<212> PRT
<213> Mus musculus
<400> 216
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Tyr Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asp Pro Ser Asp Ser Glu Thr His Tyr Asn Gln Ile Phe
50 55 60
Arg Asp Lys Ala Thr Leu Thr Ile Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gln Asp Ile Val Thr Thr Val Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 217
<211> 354
<212> DNA
<213> Mus musculus
<400> 217
caggtccaat tgcagcagcc tggggctgag ctggtgaggc ctggggcttc agtgaagctg 60
tcctgcaagg cctatggcta caccttcacc agttactgga tgaactgggt gaaacagagg 120
cctggacaag gccttgaatg gattggtatg attgatcctt cagacagtga aactcactac 180
aatcaaatat tcagggacaa ggccacattg actatagaca aatcctccag cacagcctac 240
atgcagctca gcagcctgac atctgaggac tctgcggtct atttctgtgc aagacaagat 300
atagtgacta cagttgacta ctggggccag ggcaccactc tcacagtctc ctca 354
<210> 218
<211> 113
<212> PRT
<213> Mus musculus
<400> 218
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ile Asn Gln Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ser Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
His Tyr Gly Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 219
<211> 339
<212> DNA
<213> Mus musculus
<400> 219
gacattgtga tgtcacagtc tccatcctcc ctagctgtgt cagttggaga gaaggttact 60
atgagctgca agtccagtca gagcctttta tatagtatca atcaaaagaa cttcttggcc 120
tggtaccagc agaaaccagg gcagtctcct aaactgctga tttactgggc atccactagg 180
gaatctgggg tccctgatcg cttcacaggc agtggatctg ggacagattt cactctcacc 240
atcagcagtg tgaagtctga agacctggca gtttattact gtcagcaaca ttatggctat 300
ccattcacgt tcggctcggg gacaaagttg gaaataaaa 339
<210> 220
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Mus musculus и Felis catus
<400> 220
Asp Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Thr Cys Val Ala Ser Gly Phe Thr Tyr Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Arg Ile Asp Pro Tyr Gly Gly Gly Thr Lys His Asn Glu Lys Phe
50 55 60
Lys Arg Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Val Arg Ser Gly Tyr Asp Tyr Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 221
<211> 354
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, от Mus
musculus и Felis catus
<400> 221
gacgtgcaac tggtggagag tggaggggac cttgtaaagc caggcgggtc cctgcgcctg 60
acctgtgtag ccagcggctt cacttactcc aactattgga tgcactgggt cagacaggcc 120
cccggaaaag ggcttcagtg ggtggcaagg atcgatccct acggaggagg aacgaagcat 180
aacgagaagt tcaagcggag gtttactatc agtagagaca acgcgaaaaa tacactgtac 240
ctgcagatga atagtcttaa gacagaggat accgcgacct actattgcgt cagatccggc 300
tatgactatt actttgacgt ttggggacag ggcacactgg tcaccgtctc gagc 354
<210> 222
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 222
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Phe
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Ala Asn Thr Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln His His Phe Gly Thr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 223
<211> 321
<212> PRT
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 223
Gly Ala Ala Ala Thr Thr Cys Ala Ala Ala Thr Gly Ala Cys Cys Cys
1 5 10 15
Ala Gly Thr Cys Ala Cys Cys Cys Ala Gly Thr Thr Cys Cys Thr Thr
20 25 30
Gly Thr Cys Cys Gly Cys Cys Ala Gly Thr Cys Cys Cys Gly Gly Ala
35 40 45
Gly Ala Thr Cys Gly Cys Gly Thr Cys Ala Cys Cys Ala Thr Ala Ala
50 55 60
Cys Cys Thr Gly Cys Cys Gly Gly Gly Cys Cys Ala Gly Thr Gly Ala
65 70 75 80
Ala Ala Ala Thr Ala Thr Thr Thr Ala Thr Thr Cys Ala Thr Thr Cys
85 90 95
Cys Thr Cys Gly Cys Gly Thr Gly Gly Thr Ala Thr Cys Ala Gly Cys
100 105 110
Ala Gly Ala Ala Gly Cys Cys Ala Gly Gly Cys Ala Ala Gly Gly Thr
115 120 125
Gly Cys Cys Gly Ala Ala Gly Cys Thr Gly Cys Thr Gly Ala Thr Ala
130 135 140
Thr Ala Cys Ala Ala Cys Gly Cys Thr Ala Ala Cys Ala Cys Gly Thr
145 150 155 160
Thr Gly Gly Cys Cys Gly Ala Gly Gly Gly Gly Gly Thr Gly Cys Cys
165 170 175
Cys Ala Gly Thr Ala Gly Ala Thr Thr Cys Ala Gly Cys Gly Gly Gly
180 185 190
Thr Cys Cys Gly Gly Cys Ala Gly Thr Gly Gly Ala Ala Cys Cys Gly
195 200 205
Ala Thr Thr Thr Cys Ala Cys Cys Cys Thr Gly Ala Cys Ala Ala Thr
210 215 220
Thr Thr Cys Ala Ala Gly Cys Cys Thr Gly Gly Ala Ala Cys Cys Ala
225 230 235 240
Gly Ala Ala Gly Ala Cys Gly Cys Cys Gly Cys Cys Ala Cys Ala Thr
245 250 255
Ala Cys Thr Ala Cys Thr Gly Cys Cys Ala Gly Cys Ala Thr Cys Ala
260 265 270
Cys Thr Thr Thr Gly Gly Gly Ala Cys Cys Cys Cys Thr Thr Thr Cys
275 280 285
Ala Cys Thr Thr Thr Cys Gly Gly Ala Thr Cys Cys Gly Gly Thr Ala
290 295 300
Cys Cys Ala Ala Gly Cys Thr Gly Gly Ala Gly Ala Thr Cys Ala Ala
305 310 315 320
Ala
<210> 224
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Mus musculus и Felis catus
<400> 224
Asp Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Thr Cys Val Ala Ser Gly Phe Thr Tyr Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Arg Ile Asp Pro Tyr Gly Gly Gly Thr Lys His Asn Glu Lys Phe
50 55 60
Lys Arg Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Val Arg Ser Gly Tyr Asp Tyr Tyr Phe Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 225
<211> 354
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, от Mus
musculus и Felis catus
<400> 225
gacgtgcaac tggtggagag tggaggggac cttgtaaagc caggcgggtc cctgcgcctg 60
acctgtgtag ccagcggctt cacttactcc aactattgga tgcactgggt cagacaggcc 120
cccggaaaag ggcttcagtg ggtggcaagg atcgatccct acggaggagg aacgaagcat 180
aacgagaagt tcaagcggag gtttactatc agtagagaca acgcgaaaaa tacactgtac 240
ctgcagatga atagtcttaa gacagaggat accgcgacct actattgcgt cagatccggc 300
tatgactatt actttgacgt ttggggacag ggcacactgg tcaccgtctc gagc 354
<210> 226
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Mus musculus и Felis catus
<400> 226
Glu Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Pro Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Phe
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Ala Asn Thr Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln His His Phe Gly Thr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 227
<211> 321
<212> PRT
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, от Mus
musculus и Felis catus
<400> 227
Gly Ala Ala Ala Thr Thr Cys Ala Ala Ala Thr Gly Ala Cys Cys Cys
1 5 10 15
Ala Gly Thr Cys Ala Cys Cys Cys Ala Gly Thr Thr Cys Cys Thr Thr
20 25 30
Gly Thr Cys Cys Gly Cys Cys Ala Gly Thr Cys Cys Cys Gly Gly Ala
35 40 45
Gly Ala Thr Cys Gly Cys Gly Thr Cys Ala Cys Cys Ala Thr Ala Ala
50 55 60
Cys Cys Thr Gly Cys Cys Gly Gly Gly Cys Cys Ala Gly Thr Gly Ala
65 70 75 80
Ala Ala Ala Thr Ala Thr Thr Thr Ala Thr Thr Cys Ala Thr Thr Cys
85 90 95
Cys Thr Cys Gly Cys Gly Thr Gly Gly Thr Ala Thr Cys Ala Gly Cys
100 105 110
Ala Gly Ala Ala Gly Cys Cys Ala Gly Gly Cys Ala Ala Gly Gly Thr
115 120 125
Gly Cys Cys Gly Ala Ala Gly Cys Thr Gly Cys Thr Gly Ala Thr Ala
130 135 140
Thr Ala Cys Ala Ala Cys Gly Cys Thr Ala Ala Cys Ala Cys Gly Thr
145 150 155 160
Thr Gly Gly Cys Cys Gly Ala Gly Gly Gly Gly Gly Thr Gly Cys Cys
165 170 175
Cys Ala Gly Thr Ala Gly Ala Thr Thr Cys Ala Gly Cys Gly Gly Gly
180 185 190
Thr Cys Cys Gly Gly Cys Ala Gly Thr Gly Gly Ala Ala Cys Cys Gly
195 200 205
Ala Thr Thr Thr Cys Ala Cys Cys Cys Thr Gly Ala Cys Ala Ala Thr
210 215 220
Thr Thr Cys Ala Ala Gly Cys Cys Thr Gly Gly Ala Ala Cys Cys Ala
225 230 235 240
Gly Ala Ala Gly Ala Cys Gly Cys Cys Gly Cys Cys Ala Cys Ala Thr
245 250 255
Ala Cys Thr Ala Cys Thr Gly Cys Cys Ala Gly Cys Ala Thr Cys Ala
260 265 270
Cys Thr Thr Thr Gly Gly Gly Ala Cys Cys Cys Cys Thr Thr Thr Cys
275 280 285
Ala Cys Thr Thr Thr Cys Gly Gly Ala Thr Cys Cys Gly Gly Thr Ala
290 295 300
Cys Cys Ala Ala Gly Cys Thr Gly Gly Ala Gly Ala Thr Cys Ala Ala
305 310 315 320
Ala
<210> 228
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Canis familiaris и Felis catus
<400> 228
Asp Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Thr Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Gly Ile Asp Ser Val Gly Ser Gly Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Ser Gly Phe Pro Gly Ser Phe Glu His Trp Gly Gln Gly Ala Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 229
<211> 351
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, Canis
familiaris и Felis catus
<400> 229
gatgtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
acctgcgtgg cgagcggctt tacctttagc gattatgcga tgagctgggt gcgccaggcg 120
ccgggcaaag gcctgcagtg ggtggcgggc attgatagcg tgggcagcgg caccagctat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgtat 240
ctgcagatga acagcctgaa aaccgaagat accgcgacct attattgcgc gagcggcttt 300
ccgggcagct ttgaacattg gggccagggc gcgctggtga ccgtgagcag c 351
<210> 230
<211> 132
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Canis familiaris и Felis catus
<400> 230
Gln Ser Val Leu Thr Gln Pro Ser Ser Val Ser Gly Thr Leu Gly Gln
1 5 10 15
Arg Ile Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Gly
20 25 30
Tyr Val Gly Trp Tyr Gln Gln Val Pro Gly Met Gly Pro Lys Thr Val
35 40 45
Ile Tyr Tyr Asn Ser Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ser Ser Gly Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Val Tyr Asp Arg Thr Phe
85 90 95
Asn Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ser Ala Pro Pro Arg Ser His Ser Ser Arg Pro Ile Ser Tyr Ala
115 120 125
Val Phe Cys Leu
130
<210> 231
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, Canis
familiaris и Felis catus
<400> 231
cagagcgtgc tgacccagcc gagcagcgtg agcggcaccc tgggccagcg cattaccatt 60
agctgcaccg gcagcagcag caacattggc agcggctatg tgggctggta tcagcaggtg 120
ccgggcatgg gcccgaaaac cgtgatttat tataacagcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcagc agcggcaccc tgaccattac cggcctgcag 240
gcggaagatg aagcggatta ttattgcagc gtgtatgatc gcacctttaa cgcggtgttt 300
ggcggcggca cccatctgac cgtgctg 327
<210> 232
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области тяжелой
цепи mAb, от Canis familiaris и Felis catus
<400> 232
Asp Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Thr Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Gln Trp Val
35 40 45
Ala Gly Ile Asp Ser Val Gly Ser Gly Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Gly Leu Lys Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Ser Gly Phe Pro Gly Ser Phe Glu His Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 233
<211> 351
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области тяжелой цепи mAb, Canis
familiaris и Felis catus
<400> 233
gatgtgcagc tggtggaaag cggcggcgat ctggtgaaac cgggcggcag cctgcgcctg 60
acctgcgtgg cgagcggctt tacctttagc gattatgcga tgaactgggt gcgccaggcg 120
ccgggcaaag gcctgcagtg ggtggcgggc attgatagcg tgggcagcgg caccagctat 180
gcggatagcg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgtat 240
ctgcagatga gcggcctgaa aaccgaagat accgcgacct attattgcgc gagcggcttt 300
ccgggcagct ttgaacattg gggccagggc accctggtga ccgtgagcag c 351
<210> 234
<211> 132
<212> PRT
<213> Artificial Sequence
<220>
<223> Последовательность фелинизированной вариабельной области легкой
цепи mAb, от Canis familiaris и Felis catus
<400> 234
Gln Ser Val Leu Thr Gln Pro Ser Ser Val Ser Gly Thr Leu Gly Gln
1 5 10 15
Arg Ile Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Gly
20 25 30
Tyr Val Gly Trp Tyr Gln Gln Val Pro Gly Met Gly Pro Lys Thr Val
35 40 45
Ile Tyr Tyr Asn Ser Asp Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ser Ser Gly Thr Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Val Tyr Asp Arg Thr Phe
85 90 95
Asn Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ser Ala Pro Pro Arg Ser His Ser Ser Arg Pro Ile Ser Tyr Ala
115 120 125
Val Phe Cys Leu
130
<210> 235
<211> 327
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность, кодирующая последовательность
фелинизированной вариабельной области легкой цепи mAb, Canis
familiaris и Felis catus
<400> 235
cagagcgtgc tgacccagcc gagcagcgtg agcggcaccc tgggccagcg cattaccatt 60
agctgcaccg gcagcagcag caacattggc agcggctatg tgggctggta tcagcaggtg 120
ccgggcatgg gcccgaaaac cgtgatttat tataacagcg atcgcccgag cggcgtgccg 180
gatcgcttta gcggcagcaa aagcggcagc agcggcaccc tgaccattac cggcctgcag 240
gcggaagatg aagcggatta ttattgcagc gtgtatgatc gcacctttaa cgcggtgttt 300
ggcggcggca cccatctgac cgtgctg 327
<210> 236
<211> 106
<212> PRT
<213> Felis catus
<400> 236
Gly Gln Pro Lys Ser Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Asn
1 5 10 15
Glu Glu Leu Ser Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Ser Gly Leu Thr Val Ala Trp Lys Ala Asp Gly Thr Pro
35 40 45
Ile Thr Gln Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Ser Pro Asn Glu Trp Lys
65 70 75 80
Ser Arg Ser Arg Phe Thr Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Asn Val Val Pro Ala Glu Cys Ser
100 105
<210> 237
<211> 318
<212> DNA
<213> Felis catus
<400> 237
ggccagccca agagcgctcc ctccgtgacc ctgttccccc caagcaacga ggaactgagc 60
gccaacaagg ccaccctggt gtgcctgatc agcgacttct accccagcgg cctgaccgtg 120
gcctggaagg ccgatggcac ccctatcacc cagggcgtgg aaaccaccaa gcccagcaag 180
cagagcaaca acaaatacgc cgccagcagc tacctgagcc tgagccccaa cgagtggaag 240
tcccggtccc ggttcacatg ccaggtgaca cacgagggca gcaccgtgga aaagaacgtg 300
gtgcccgccg agtgcagc 318
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| ПЕПТИДНЫЕ ВАКЦИНЫ ПРОТИВ ИНТЕРЛЕЙКИНА-31 | 2019 |
|
RU2786441C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2018 |
|
RU2795485C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИТЕЛА К ПАРВОВИРУСУ ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2830441C2 |
| АНТИТЕЛО ДЛЯ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА 4 | 2017 |
|
RU2774446C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2838943C1 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |






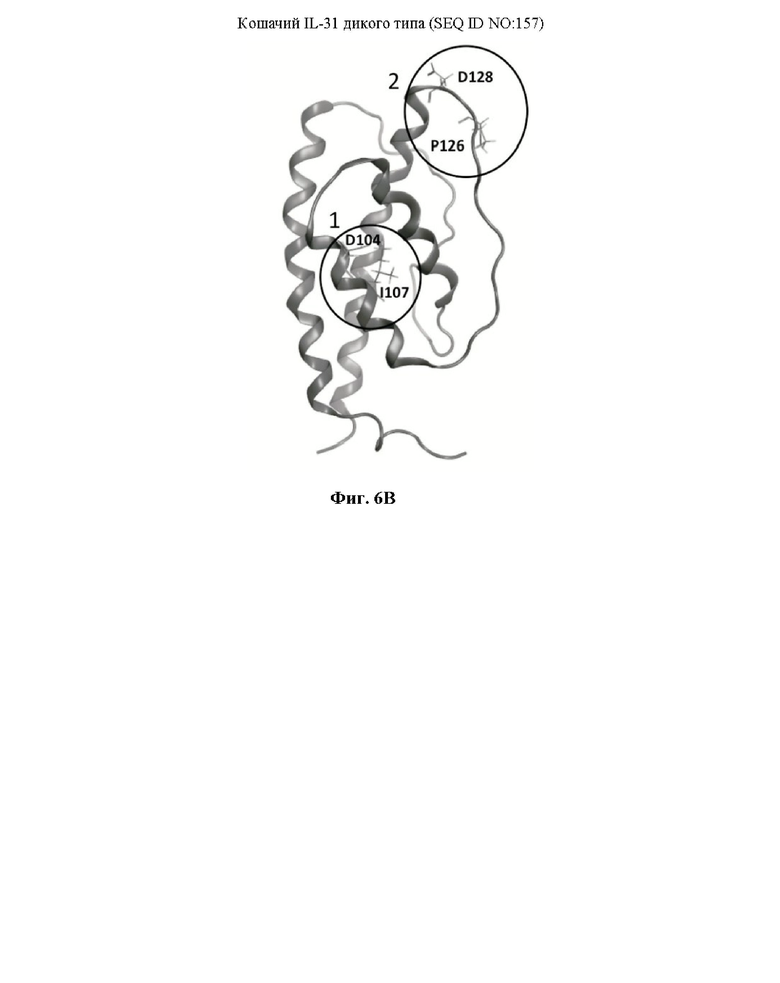
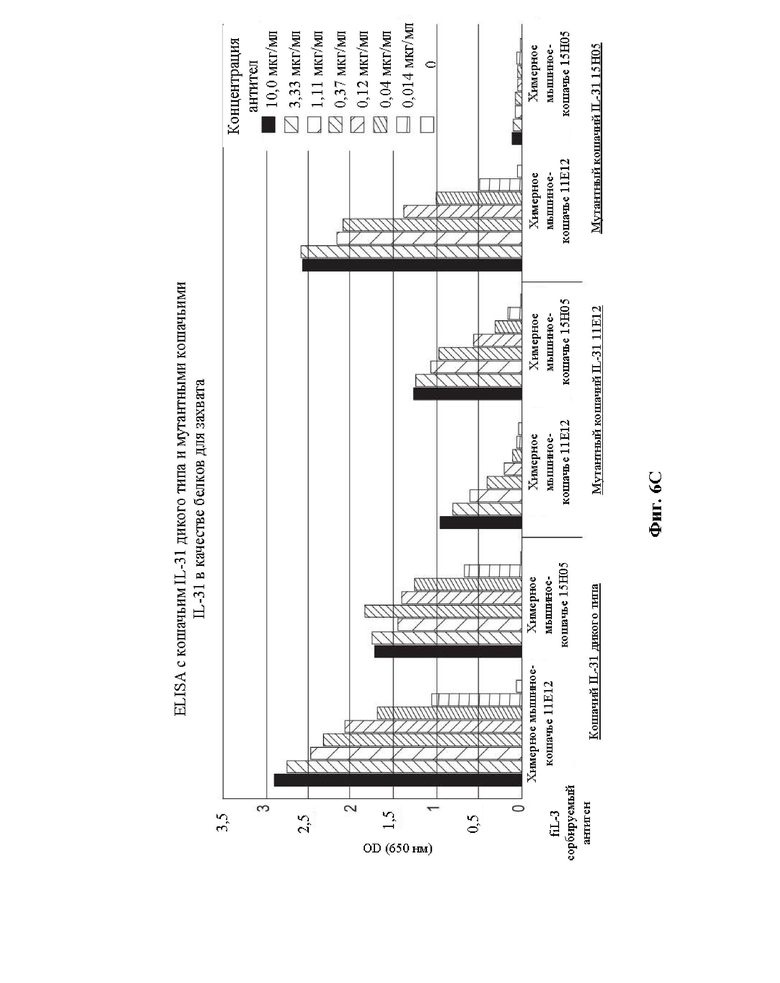
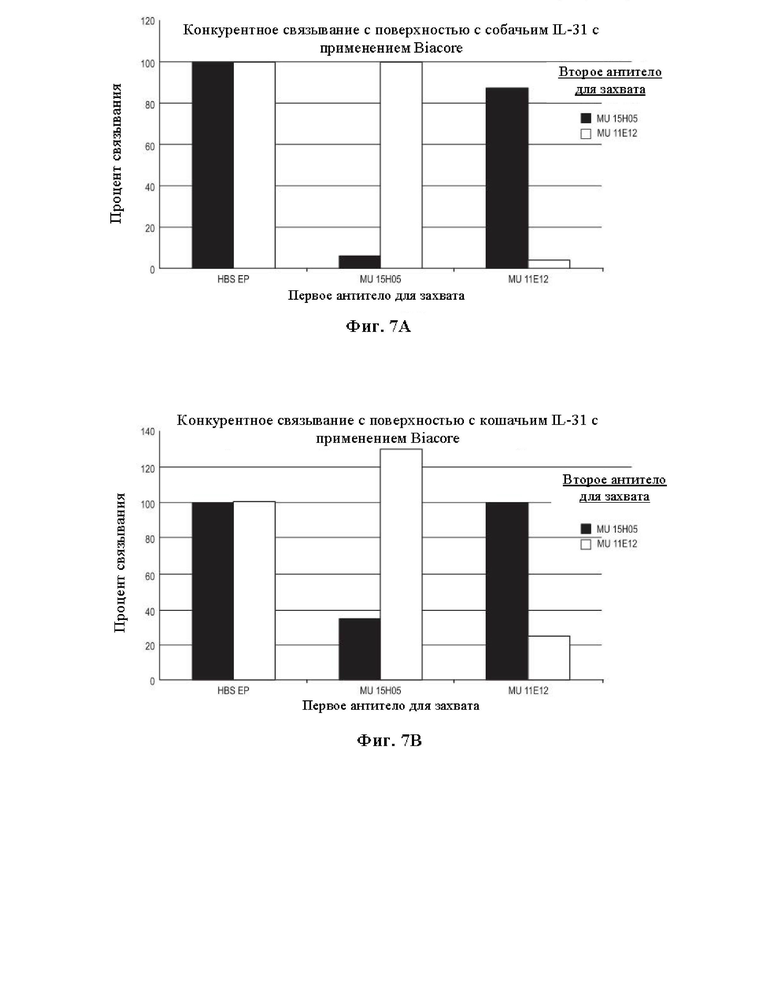
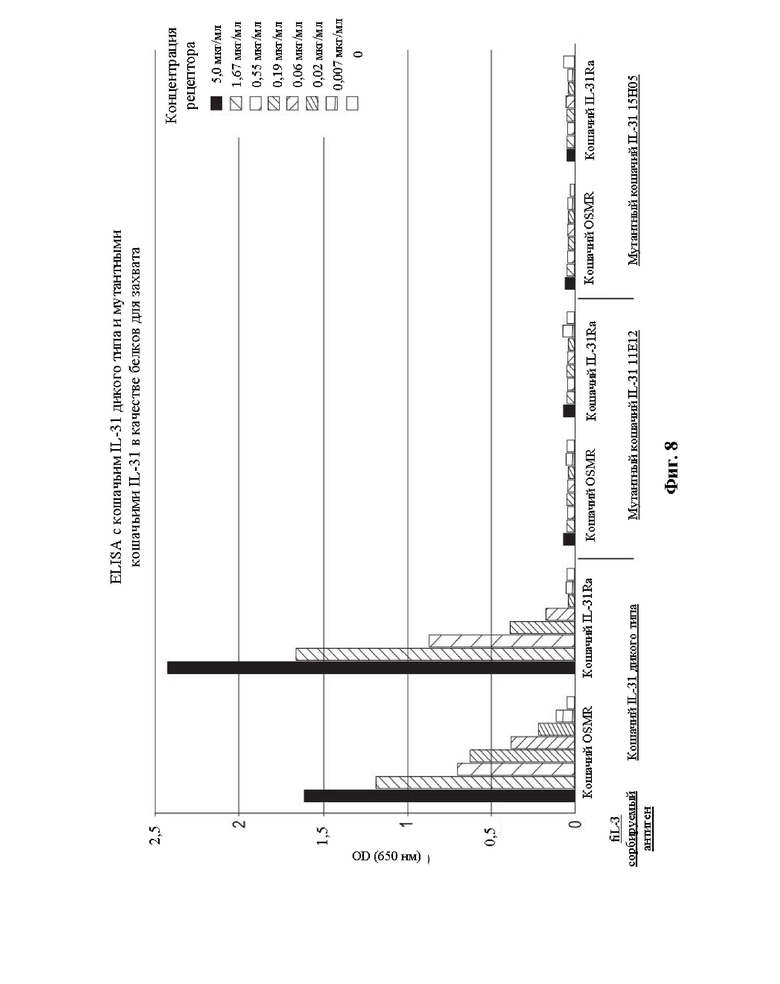
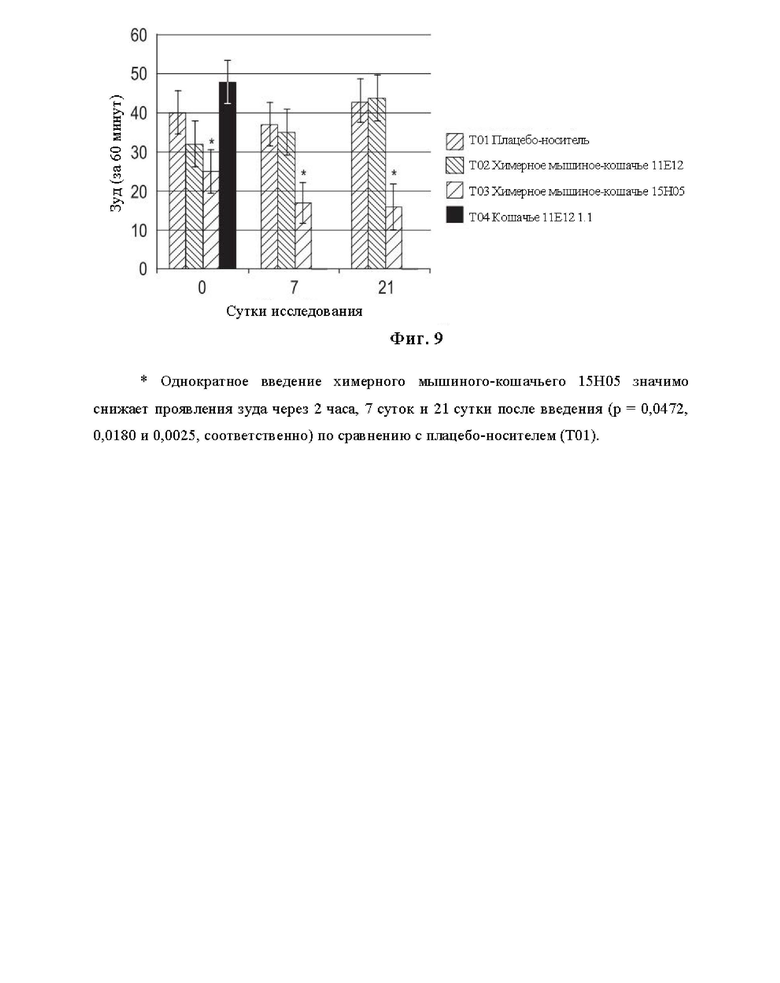
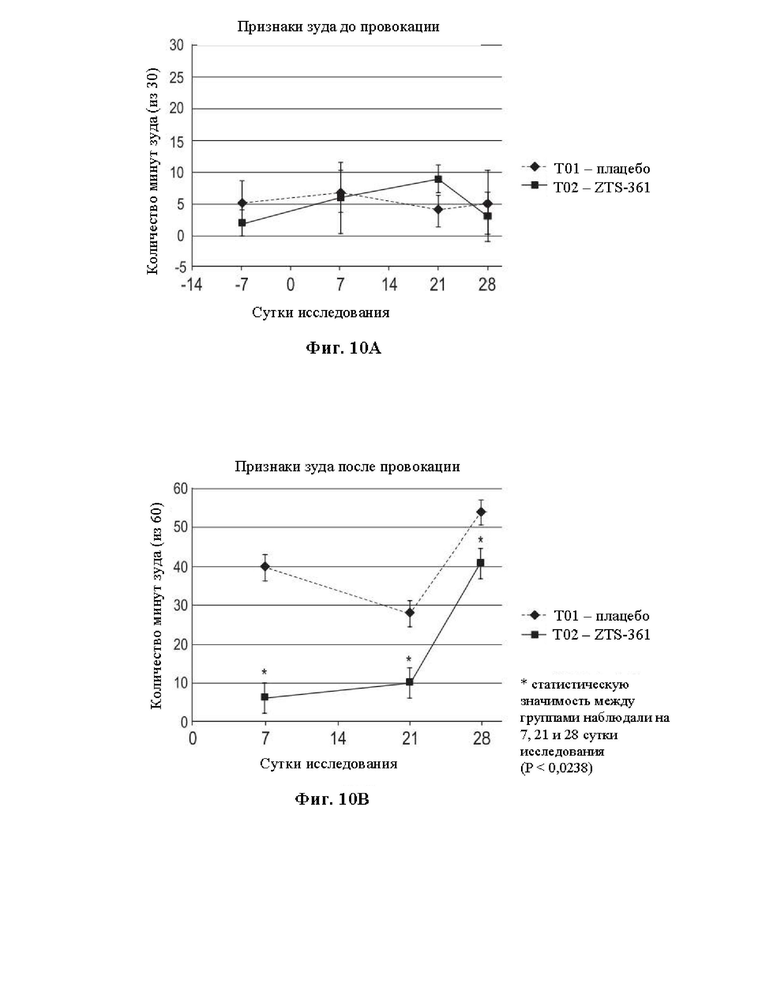

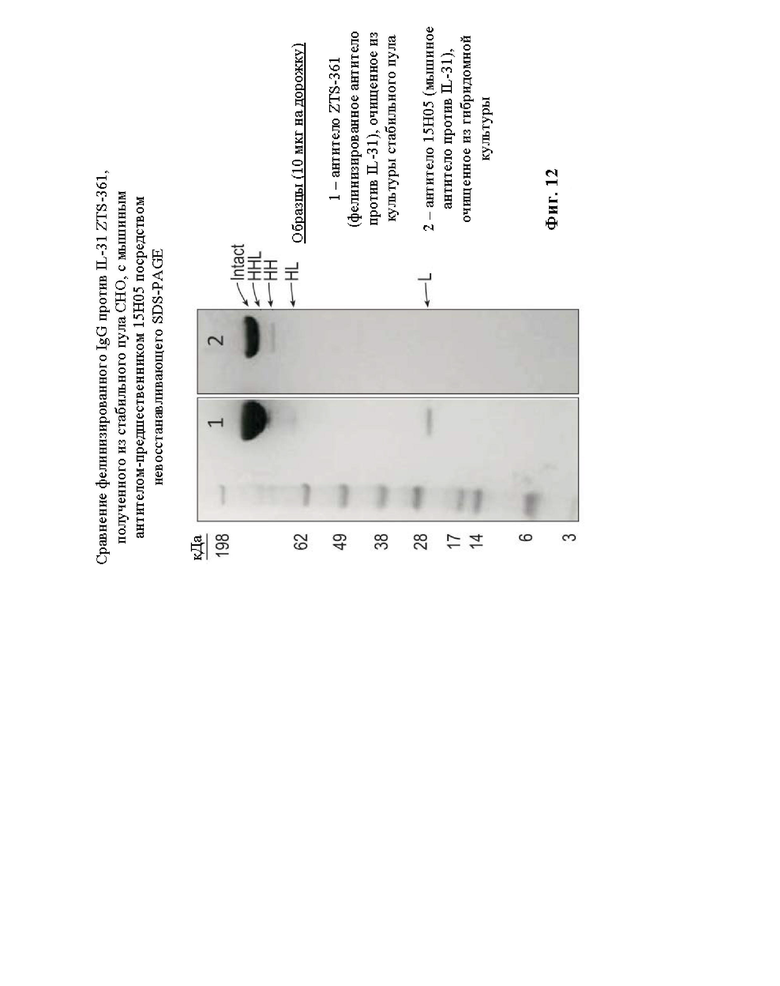
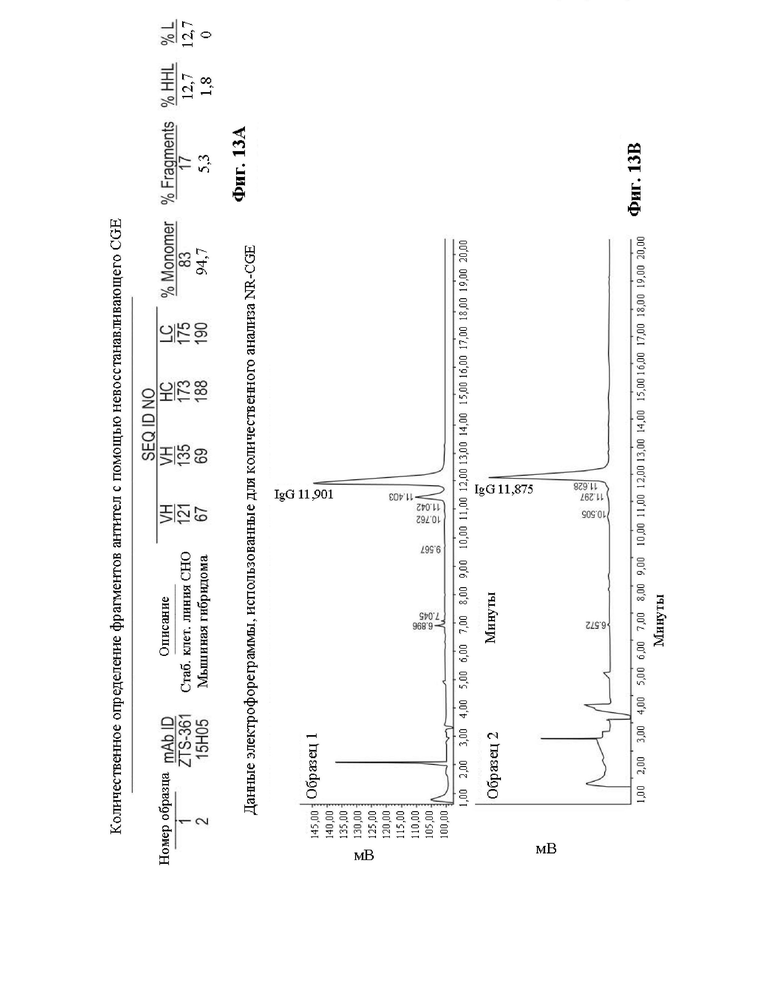


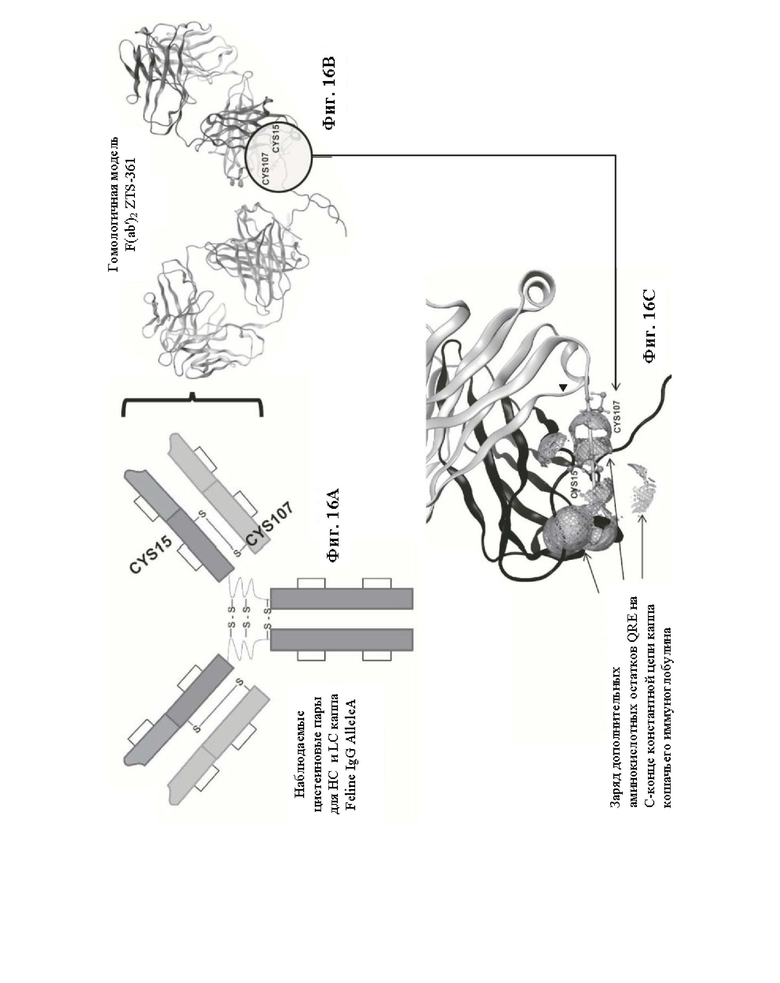

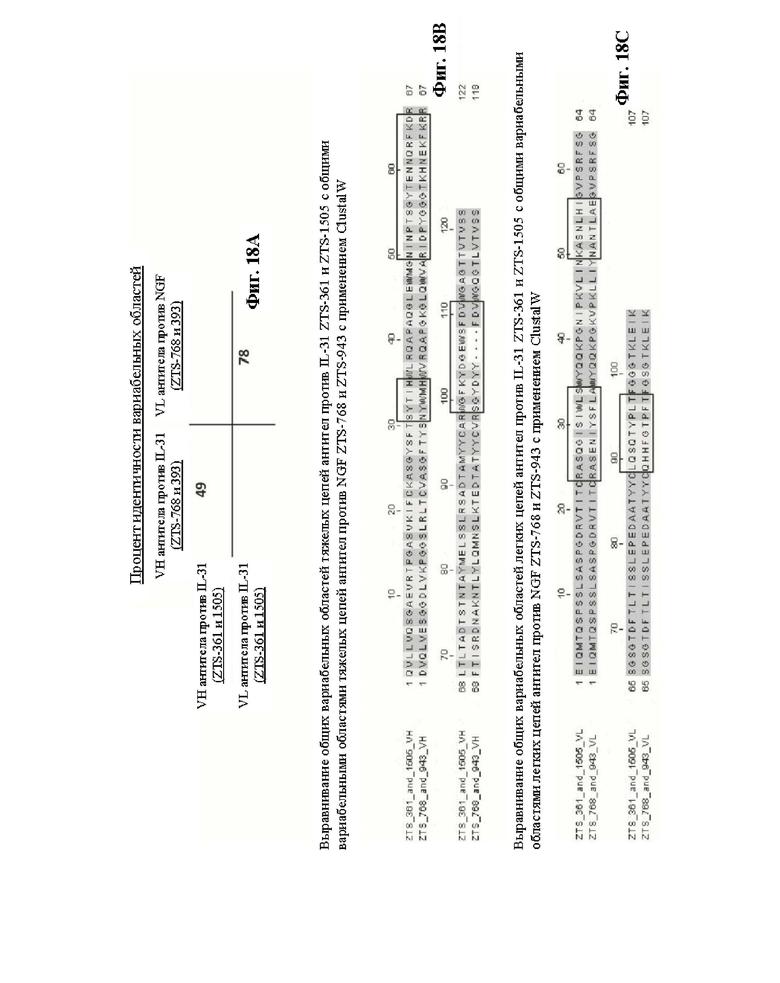




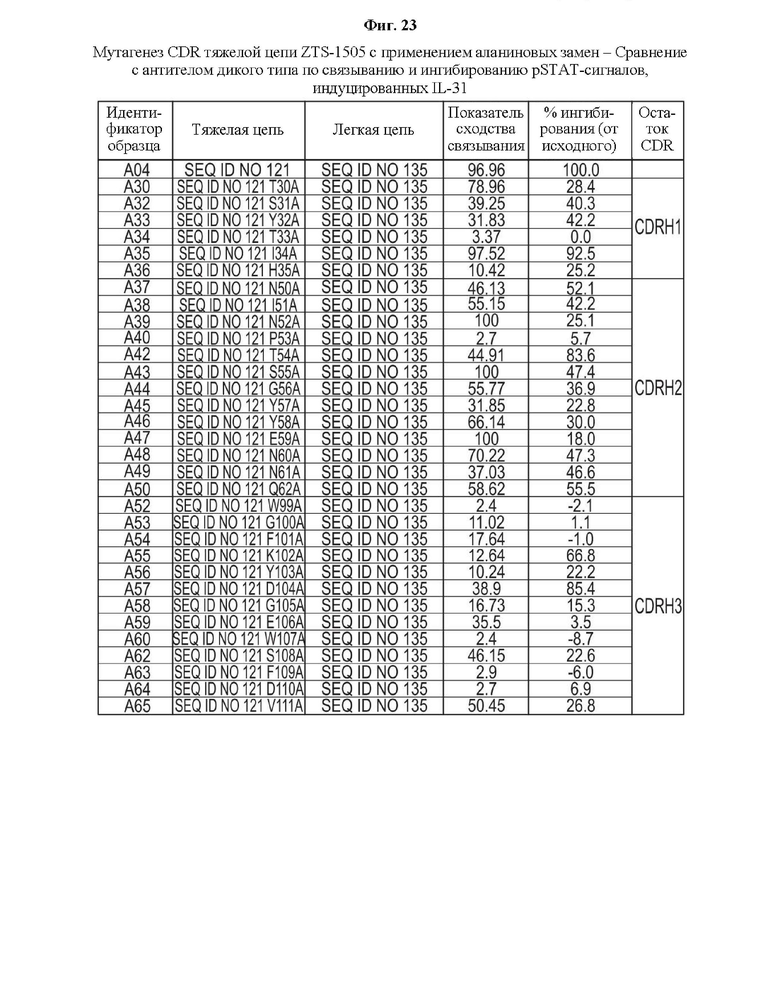
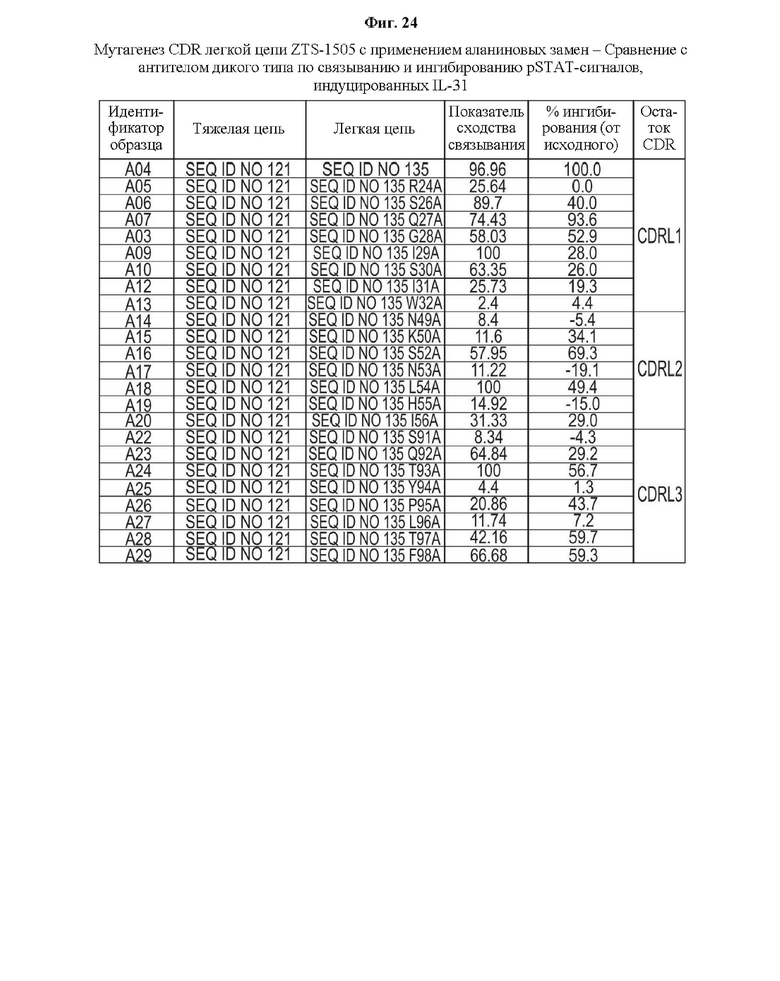
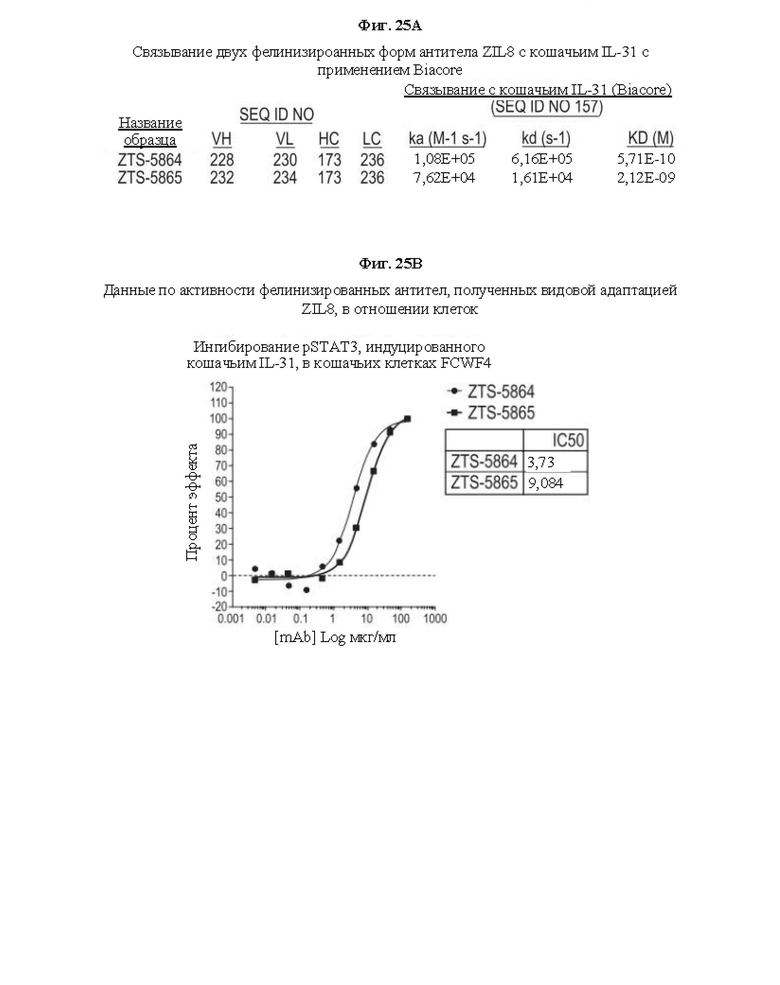
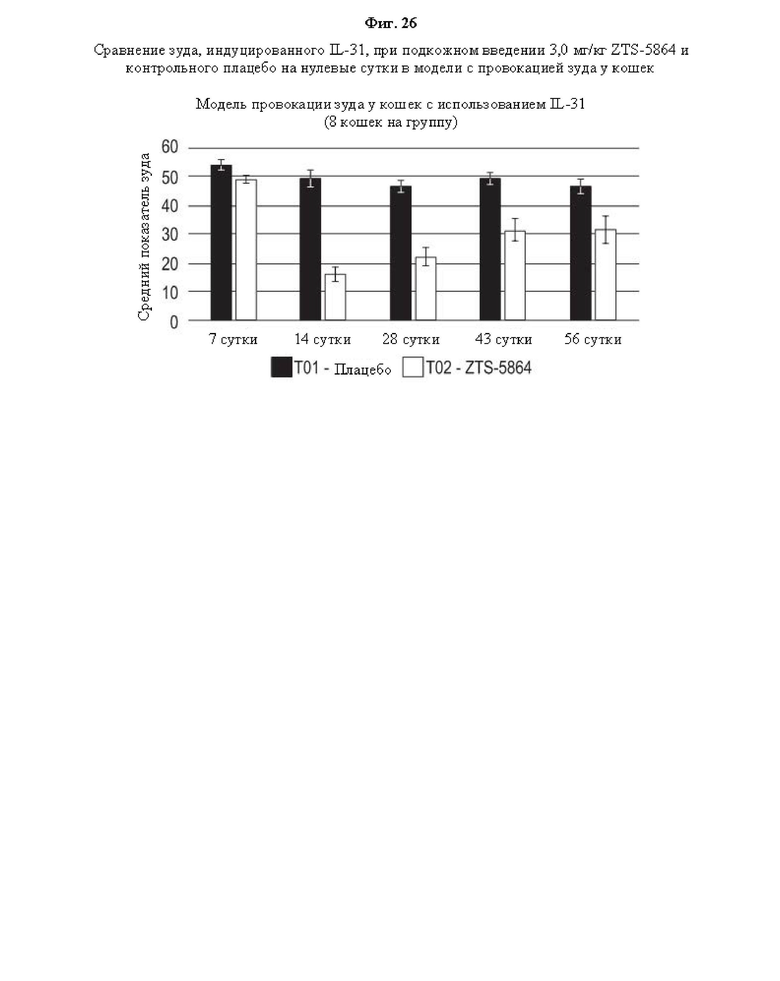
Настоящее изобретение относится к биотехнологии и иммунологии, в частности к моноклональным антителам против IL-31. Также раскрыты композиции, содержащие такие антитела, а также нуклеиновые кислоты, векторы и клетки-хозяева для получения таких антител. Настоящее изобретение обеспечивает новые антитела против IL-31 для предотвращения и лечения расстройств, опосредованных IL-31, у млекопитающих в ветеринарии, в частности для безопасных и эффективных альтернативных вариантов лечения атопических и аллергических дерматозов у кошек и лошадей. 14 н. и 15 з.п. ф-лы, 26 ил., 2 табл., 20 пр.
1. Антитело против IL-31, содержащее:
VH-CDR1 (гипервариабельный участок 1 вариабельной области тяжелой цепи), содержащий аминокислотную последовательность, представленную в SEQ ID NO: 1, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 3, VL-CDR1 (гипервариабельный участок 1 вариабельной области легкой цепи), содержащий аминокислотную последовательность, представленную в SEQ ID NO: 4, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 5, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 6.
2. Антитело по п. 1, которое связывается с кошачьим IL-31 и содержит VL цепь (вариабельная область легкой цепи), содержащую изменения в каркасной области 2 (FW2), выбранные из группы, состоящей из аспарагина вместо лизина в положении 42, изолейцина вместо валина в положении 43, валина вместо лейцина в положении 46, аспарагина вместо лизина в положении 49 и их комбинаций.
3. Антитело против IL-31, содержащее:
1) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 13, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 14, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 15, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 16, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 17, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 18;
2) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 19, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 20, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO:21, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 22, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 23, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO :24;
3) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 25, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 26, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 27, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 28, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 29, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 30;
4) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 31, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 32, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 33, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 34, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 35, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 36;
5) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 37, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 38, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 39, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 40, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 41, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 42;
6) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 43, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 44, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 45, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 46, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 47, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 48;
7) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 49, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 50, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 51, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 52, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 53, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 54;
8) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 55, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 56, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 57, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 58, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 59, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 60; или
9) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 61, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 62, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 63, VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 64, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 65, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 66.
4. Антитело по любому из пп. 1-3, которое ослабляет, ингибирует или нейтрализует состояние, сопровождающееся зудом, или аллергическое состояние, опосредованные IL-31, у млекопитающего.
5. Антитело по п. 4, где млекопитающее выбрано из группы, состоящей из собаки, кошки и лошади.
6. Антитело по любому из пп. 1-5, представляющее собой химерное антитело.
7. Антитело по любому из пп. 1-6, представляющее собой канинизированное, фелинизированное, эквинизированное, полностью собачье, полностью кошачье или полностью лошадиное антитело.
8. Антитело по п. 1, содержащее по меньшей мере одно из группы, состоящей из:
VL цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 135, и VH цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 121.
9. Антитело по п. 3, содержащее:
1) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 77, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 75;
2) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 81, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 79;
3) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 85, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 83;
4) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 89, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 87;
5) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 93, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 91;
6) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 97, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 95;
7) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 101, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 99;
8) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 105, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 103;
9) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 109, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 107;
10) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 230, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 228; или
11) VL цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 234, и VH цепь, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 232.
10. Применение антитела по любому из пп. 1, 3, 8 и 9 для лечения расстройства, опосредованного IL-31, у субъекта.
11. Применение по п. 10, где расстройство, опосредованное IL-31, представляет собой состояние, сопровождающееся зудом, или аллергическое состояние.
12. Применение по п. 11, где состояние, сопровождающееся зудом, или аллергическое состояние представляют собой состояние, сопровождающееся зудом, выбранное из группы, состоящей из атопического дерматита, экземы, псориаза, склеродермии и зуда.
13. Применение по п. 12, где состояние, сопровождающееся зудом, или аллергическое состояние представляют собой аллергическое состояние, выбранное из группы, состоящей из аллергического дерматита, летней экземы, крапивницы, запала лошадей, воспалительного заболевания дыхательных путей, рецидивирующей обструкции дыхательных путей, гиперреактивности дыхательных путей, хронической обструктивной болезни легких и воспалительных процессов, являющихся результатами аутоиммунитета.
14. Применение по п. 10, где расстройство, опосредованное IL-31, представляет собой прогрессирование опухоли.
15. Применение по п. 14, где расстройство, опосредованное IL-31, представляет собой эозинофильное заболевание или мастоцитомы.
16. Применение антитела по любому из пп. 1, 3, 8 и 9 для ингибирования активности IL-31 у млекопитающего.
17. Ветеринарная композиция для лечения расстройства, опосредованного IL-31, у млекопитающего, выбранного из группы, состоящей из собаки, кошки и лошади, содержащая терапевтически эффективное количество антитела по любому из пп. 1, 3, 8 и 9 и приемлемый носитель, разбавитель или эксципиент.
18. Антитело по любому из пп. 1, 3, 8 и 9 для применения в лечении млекопитающего с расстройством, опосредованным IL-31.
19. Применение антитела по любому из пп. 1, 3, 8 и 9 в изготовлении лекарственного средства для лечения млекопитающего с расстройством, опосредованным IL-31.
20. Способ выявления IL-31, включающий:
(a) инкубирование образца, содержащего IL-31, в присутствии антитела по любому из пп. 1, 3, 8 и 9; и
(b) выявление антитела, связанного с IL-31, в образце.
21. Способ по п. 20, дополнительно включающий количественное определение IL-31 в образце.
22. Выделенная нуклеиновая кислота для получения моноклонального антитела по любому из пп. 1-9, содержащая нуклеиновокислотную последовательность, кодирующую:
(1) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 13, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 14, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 15;
(2) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 19, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 20, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 21;
(3) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 25, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 26, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 27;
(4) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 31, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 32, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 33;
(5) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 37, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 38, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 39;
(6) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 43, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 44, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 45;
(7) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 49, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 50, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 51;
(8) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 55, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 56, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 57;
(9) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 61, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 62, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 63; или
(10) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 1, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2, и VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 3.
23. Выделенная нуклеиновая кислота для получения моноклонального антитела по любому из пп. 1-9, содержащая нуклеиновокислотную последовательность, кодирующую:
(1) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 13, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 14, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 15; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 16, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 17, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 18;
(2) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 19, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 20, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 21; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 22, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 23, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 24;
(3) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 25, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 26, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 27; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 28, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 29, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 30;
(4) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 31, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 32, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 33; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 34, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 35, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 36;
(5) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 37, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 38, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 39; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 40, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 41, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 42;
(6) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 43, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 44, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 45; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 46, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 47, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 48;
(7) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 49, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 50, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 51; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 52, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 53, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 54;
(8) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 55, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 56, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 57; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 58, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 59, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 60;
(9) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 61, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 62, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 63; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 64, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 65, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 66; или
(10) VH-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 1, VH-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 2, VH-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 3; VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 4, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 5, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 6.
24. Выделенная нуклеиновая кислота для получения моноклонального антитела по любому из пп. 1-9, содержащая нуклеиновокислотную последовательность, кодирующую:
(1) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 16, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 17, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 18;
(2) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 22, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 23, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 24;
(3) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 28, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 29, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 30;
(4) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 34, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 35, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 36;
(5) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 40, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 41, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 42;
(6) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 46, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 47, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 48;
(7) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 52, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 53, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 54;
(8) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 58, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 59, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 60;
(9) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 64, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 65, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 66; или
(10) VL-CDR1, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 4, VL-CDR2, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 5, и VL-CDR3, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 6.
25. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 22.
26. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 23.
27. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 24.
28. Клетка-хозяин, продуцирующая моноклональное антитело по любому из пп. 1, 3, 8 и 9, содержащая вектор по п. 25 и вектор по п. 27 или вектор по п. 26.
29. Способ получения антитела по любому из пп. 1, 3, 8 и 9, включающий культивирование клетки-хозяина по п. 28 в условиях, приводящих к продуцированию антитела, и выделение антитела из клетки-хозяина или культуральной среды клетки-хозяина.
| US 2013022616 A1, 24.01.2013 | |||
| MARIUZZA R.A | |||
| The structural basis of antigen-antibody recognition, Ann | |||
| Rev | |||
| Biophys | |||
| Biophys | |||
| Chem., 1987, Vol | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| КАМАЕВ А.В | |||
| и др | |||
| Зуд при атопическом дерматите у детей: известные механизмы и возможности его долгосрочного контроля, РМЖ, No.3, 2015, 142-146. | |||

