ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к мутантной нитрилгидратазе, нуклеиновой кислоте, кодирующей указанную мутантную нитрилгидратазу, вектору экспрессии и транс фор манту, содержащим указанную нуклеиновую кислоту, способу получения указанной мутантной нитрилгидратазы и способу получения амидного соединения.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Нитрилгидратаза представляет собой фермент, обладающий нитрилгидратирующей активностью, превращающий нитрильную группу различных соединений в амидную группу посредством гидратации и используемый на этапе промышленного получения амидных соединений с применением ферментативной реакции.
В последние годы отмечается тенденция к росту спроса на технологии промышленного получения амидных соединений. В свете этих обстоятельств до настоящего времени были проведены различные исследования, направленные на уменьшение производственных расходов на нитрилгидратазу в себестоимости амидных соединений. В частности, применительно к нитрилгидратазе, было несколько сообщений о технологии, относящейся к мутанту, позволяющему повысить показатель активности на единицу массы нитрилгидратазы (опубликованные заявки на патент Японии (Japanese Patent Application Laid-Open, JP-A) №№ H09-275978, 2004-194588 и 2005-160403, международные публикации №№ WO 2004/056990 и 2010/055666, JP-A №№2007-143409 и 2008-253182, и тому подобное).
В данном случае, на типичном этапе получения промышленного амидного соединения с применением ферментативной реакции обычно проводят, в указанном порядке, стадию взаимодействия, включающую проведение реакции синтеза амидного соединения из нитрильного соединения с применением каталитического действия нитрилгидратазы в растворе, рН которого обычно скорректирован до 7-9, и стадию очистки, включающую коррекцию рН раствора до 3,5-6,5 и удаление нитрилгидратазы, присутствующей в растворе, посредством адсорбции с использованием активированного угля при указанном выше рН с получением очищенного амидного соединения (JP-A №2001-270857).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Техническая проблема
Авторы настоящего изобретения провели интенсивные исследования для повышения эффективности получения промышленного амидного соединения с использованием нитрилгидратазы в сравнении с предшествующим уровнем техники. В результате, авторы настоящего изобретения обнаружили, что непрерывное проведение реакции синтеза амидного соединения из нитрильного соединения с применением каталитического действия нитрилгидратазы даже на стадии очистки, проводимой при кислом рН, является эффективным направлением разработки.
Тем не менее, обычно оптимальный рН для нитрилгидратазы дикого типа составляет от 7 до 9, и активность нитрилгидратазы дикого типа значительно снижается в условиях рН от 3,5 до 6,5. Таким образом, при использовании нитрилгидратазы дикого типа ферментативная реакция на стадии очистки не поддерживается на удовлетворительном уровне. Кроме того, есть сообщения о различных мутантах нитрилгидратазы, как описано выше, но их активность значительно снижается в кислых условиях, применяемых на стадии очистки. Таким образом, до настоящего времени не было сообщений о технологии с мутантом нитрилгидратазы, обладающим отличной рН-стабильностью ферментативной активности, катализирующей реакцию синтеза амидного соединения из нитрильного соединения, на стадии очистки в кислых условиях. В условиях относительно сильного закисления, в частности при рН 5,0 или менее, белок более склонен к деактивации, и до настоящего времени не было сообщений о технологии с мутантом нитрилгидратазы, ферментативная активность которого хорошо сохранялась бы даже в условиях такого рН.
Таким образом, согласно изобретению предложена технология с мутантной нитрилгидратазой, имеющей новую точковую мутацию, у которой повышена рН-стабильность ферментативной активности, катализирующей реакцию синтеза амидного соединения из нитрильного соединения. Конкретнее, предложена технология с мутантной нитрилгидратазой, сохраняющей отличную ферментативную активность даже при кислом рН от 3,5 до 6,5. Еще конкретнее, предложена технология с мутантной нитрилгидратазой, сохраняющей отличную ферментативную активность даже при относительно сильном закислении до рН от 3,5 до 5,0.
Решение проблемы
Изобретение включает следующие аспекты.
<1> Мутантная нитрил гидр атаза, имеющая происхождение из Pseudonocardia thermophila и имеющая α-субъединицу и β-субъединицу, содержащая по меньшей мере одну замену аминокислотного остатка, выбранную из группы, состоящей из следующих замен аминокислотных остатков (а)-(е):
(a) замена 40-го аминокислотного остатка от N-конца α-субъединицы на Asn,
(b) замена 43-го аминокислотного остатка от N-конца α-субъединицы на Val,
(c) замена 205-го аминокислотного остатка от N-конца β-субъединицы на Val,
(d) замена 206-го аминокислотного остатка от N-конца β-субъединицы на Gln и
(e) замена 215-го аминокислотного остатка от N-конца β-субъединицы на Asn.
<2> Мутантная нитрилгидратаза, описанная в <1>, содержащая:
две или более замены аминокислотных остатков, выбранные из группы, состоящей из замен аминокислотных остатков (а)-(е).
<3> Мутантная нитрилгидратаза, описанная в <1> или <2>, содержащая: замену аминокислотного остатка (b); и
по меньшей мере одну замену, выбранную из группы, состоящей из замен аминокислотных остатков (а), (с), (d) и (е).
<4> Мутантная нитрилгидратаза, описанная в любом из <1>-<3>, где 36-я аминокислота от N-конца аминокислотной последовательности α-субъединицы представляет собой остаток Trp.
<5> Мутантная нитрилгидратаза, описанная в любом из <1>-<3>, где 36-я аминокислота от N-конца аминокислотной последовательности α-субъединицы представляет собой Met, Ser, Gly или Ala.
<6> Мутантная нитрилгидратаза, описанная в любом из <1>-<5>, где аминокислотная последовательность α-субъединицы удовлетворяет одному или более из следующих условий (1)-(13):
(1) 6-й аминокислотный остаток от N-конца представляет собой Thr или Ala,
(2) 13-й аминокислотный остаток от N-конца представляет собой Leu,
(3) 19-й аминокислотный остаток от N-конца представляет собой Val,
(4) 27-й аминокислотный остаток от N-конца представляет собой Ile,
(5) 48-й аминокислотный остаток от N-конца представляет собой Gln,
(6) 71-й аминокислотный остаток от N-конца представляет собой His,
(7) 92-й аминокислотный остаток от N-конца представляет собой Glu,
(8) 94-й аминокислотный остаток от N-конца представляет собой Ile,
(9) 126-й аминокислотный остаток от N-конца представляет собой Tyr,
(10) 148-й аминокислотный остаток от N-конца представляет собой Asp,
(11) 188-й аминокислотный остаток от N-конца представляет собой Gly,
(12) 197-й аминокислотный остаток от N-конца представляет собой Cys и
(13) 204-й аминокислотный остаток от N-конца представляет собой Arg.
<7> Мутантная нитрилгидратаза, описанная в любом из <1>-<6>, где аминокислотная последовательность β-субъединицы удовлетворяет по меньшей мере одному из следующих условий (15)-(47):
(15) 4-й аминокислотный остаток от N-конца представляет собой Met,
(16) 8-й аминокислотный остаток от N-конца представляет собой Ala,
(17) 10-й аминокислотный остаток от N-конца представляет собой Asp,
(18) 24-й аминокислотный остаток от N-конца представляет собой Ile,
(19) 33-й аминокислотный остаток от N-конца представляет собой Val или Met,
(20) 37-й аминокислотный остаток от N-конца представляет собой Val или Leu,
(21) 40-й аминокислотный остаток от N-конца представляет собой Ile, Val или Leu,
(22) 41-й аминокислотный остаток от N-конца представляет собой Ile,
(23) 46-й аминокислотный остаток от N-конца представляет собой Lys,
(24) 48-й аминокислотный остаток от N-конца представляет собой Val,
(25) 51-й аминокислотный остаток от N-конца представляет собой Val,
(26) 61-й аминокислотный остаток от N-конца представляет собой Val, Gly, Trp, Ser, Leu или Thr,
(27) 79-й аминокислотный остаток от N-конца представляет собой Asn.
(28) 96-й аминокислотный остаток от N-конца представляет собой Arg,
(29) 107-й аминокислотный остаток от N-конца представляет собой Met,
(30) 108-й аминокислотный остаток от N-конца представляет собой Asp или Arg,
(31)110-й аминокислотный остаток от N-конца представляет собой Asn,
(32) 112-й аминокислотный остаток от N-конца представляет собой Val или Не,
(33) 118-й аминокислотный остаток от N-конца представляет собой Val,
(34) 127-й аминокислотный остаток от N-конца представляет собой Ser.
(35) 146-й аминокислотный остаток от N-конца представляет собой Gly.
(36) 150-й аминокислотный остаток от N-конца представляет собой Asn или Ser,
(37) 160-й аминокислотный остаток от N-конца представляет собой Cys, Trp или Met,
(38) 168-й аминокислотный остаток от N-конца представляет собой Glu,
(39) 176-й аминокислотный остаток от N-конца представляет собой Ala. Thr, Met или Cys,
(40) 186-й аминокислотный остаток от N-конца представляет собой.Arg,
(41) 200-й аминокислотный остаток от N-конца представляет собой Glu,
(42) 212-й аминокислотный остаток от N-конца представляет собой Tyr,
(43) 217-й аминокислотный остаток от N-конца представляет собой Val, His, Met, Gly, Ser, Leu или Cys,
(44) 218-й аминокислотный остаток от N-конца представляет собой Met или Ser.
(45) 226-й аминокислотный остаток от N-конца представляет собой Ile,
(46) 230-й аминокислотный остаток от N-конца представляет собой Glu и
(47) 231-й аминокислотный остаток от N-конца представляет собой Val.
<8> Мутантная нитрилгидратаза, описанная в любом из <1>-<7>, содержащая по меньшей мере одну замену аминокислотного остатка, выбранную из группы, состоящей из замен аминокислотных остатков (а)-(е) в любой из следующих нитрил гидр атаз [1]-[49], имеющих происхождение из Pseudonocardia thermophila:
[1] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 2,
[2] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 16, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 33,
[3] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 17, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 33,
[4] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 34,
[5] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 19, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 34,
[6] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную
последовательность SEQ ID NO: 20, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 35,
[7] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 20, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 36,
[8] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 37,
[9] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 38,
[10] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 39,
[11] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 40,
[12] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 41,
[13] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 42,
[14] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 43,
[15] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 22, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 44,
[16] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 23, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 45,
[17] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 24, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 46,
[18] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 25, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 47,
[19] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 48,
[20] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 23, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 49,
[21] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 16, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 50,
[22] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 26, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 51,
[23] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 27, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 52,
[24] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 28, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 53,
[25] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 17, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 54,
[26] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 29, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 55,
[27] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 56,
[28] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 57,
[29] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 58,
[30] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 29, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 59,
[31] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 31, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 60,
[32] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 61,
[33] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 32, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 62,
[34] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 63,
[35] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 64,
[36] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 65,
[37] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 25, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 54,
[38] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 66,
[39] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 67,
[40] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 68,
[41] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 69,
[42] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 70,
[43] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 71,
[44] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 72,
[45] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 73,
[46] нитрилгидратаза, где 36-й аминокислотный остаток от N-конца α-субъединицы любой из нитрилгидратаз [1]-[45] представляет собой остаток Trp,
[47] нитрилгидратаза, имеющая α-субъединицу нитрилгидратазы (А) по любому из [1]-[46] или вариант α-субъединицы, состоящий из аминокислотной последовательности, на 90% или более идентичной α-субъединице, и β-субъединицу нитрилгидратазы (А) или вариант β-субъединицы, состоящий из аминокислотной последовательности, на 90% или более идентичной β-субъединице, где по меньшей мере одна из α-субъединицы или β-субъединицы представляет собой вариант α-субъединицы или вариант β-субъединицы, и
[48] нитрилгидратаза, где общее число присоединенных, замененных, удаленных и/или введенных аминокислотных остатков в α- субъединице нитрилгидратазы (В) по любому из [1]-[46] (за исключением аминокислотных остатков, подлежащих замене (а) и (b)) составляет от 1 до 10, и общее число присоединенных, замененных, удаленных и/или введенных аминокислотных остатков в β-субъединице нитрилгидратазы (В) (за исключением аминокислотных остатков, подлежащих замене (с)-(е)) составляет от 1 до 10.
<9> Нуклеиновая кислота, кодирующая мутантную нитрилгидратазу, описанную в любом из <1>-<8>.
<10> Вектор, содержащий нуклеиновую кислоту, описанную в <9>.
<11> Вектор, описанный в <10>, представляющий собой вектор экспрессии.
<12> Трансформант, содержащий вектор экспрессии, описанный в <11>.
<13> Способ получения мутантной нитрилгидратазы, включающий: культивирование трансформанта, описанного в <12>, в среде; и выделение мутантной нитрилгидратазы, описанной в любом из <1>-<8>, из по меньшей мере одного из культивированного трансформанта и среды.
<14> Мутантная нитрилгидратаза, полученная способом, описанным в <13>.
<15> Способ получения амидного соединения, включающий приведение мутантной нитрилгидратазы, описанной в любом из <1>-<8> и <14>, в контакт с нитрильным соединением.
<16> Способ получения амидного соединения, описанный в <15>, дополнительно включающий удаление примесей из раствора, содержащего амидное соединение, при рН от 3,5 до 6,5.
<17> Способ получения амидного соединения, описанный в <15> или <16>, дополнительно включающий очистку амидного соединения с использованием активированного угля.
Полезные эффекты изобретения
Согласно изобретению может быть предложена технология с мутантной нитрилгидратазой, имеющей новую точковую мутацию, у которой повышена рН-стабильность ферментативной активности, катализирующей реакцию синтеза амидного соединения из нитрильного соединения. Конкретнее, может быть предложена технология с мутантной нитрилгидратазой, сохраняющей отличную ферментативную активность даже при кислом рН от 3,5 до 6,5. Еще конкретнее, может быть предложена технология с мутантной нитрилгидратазой, сохраняющей отличную ферментативную активность даже при относительно сильном закислении до рН от 3,5 до 5,0.
ОПИСАНИЕ ВОПЛОЩЕНИЙ
Мутантная нитрилгидратаза
Согласно изобретению предложена мутантная нитрилгидратаза (далее называемая мутантной нитрилгидратазой А), имеющая происхождение из Pseudonocardia thermophila и имеющая α-субъединицу и β-субъединицу, содержащая по меньшей мере одну замену аминокислотного остатка, выбранную из группы, состоящей из следующих замен аминокислотных остатков (а)-(е):
(a) замена 40-го аминокислотного остатка от N-конца α-субъединицы на Asn,
(b) замена 43-го аминокислотного остатка от N-конца α-субъединицы на Val,
(c) замена 205-го аминокислотного остатка от N-конца β-субъединицы на Val,
(d) замена 206-го аминокислотного остатка от N-конца β-субъединицы на Gln, и
(e) замена 215-го аминокислотного остатка от N-конца β-субъединицы на Asn.
До настоящего времени не было сообщений о мутантах, содержащих по меньшей мере одну или более чем одну из замен аминокислотных остатков (а)-(е), описанных выше. То есть, все замены аминокислотных остатков (а)-(е), которые содержит мутантная нитрилгидратаза, могут быть названы мутациями, сообщений о которых до настоящего времени не было. Кроме того, далее эти замены аминокислотных остатков (а)-(е) названы заменами аминокислотных остатков группы А. Более того, мутантная нитрилгидратаза по изобретению, содержащая мутации, описанные выше, представляет собой фермент, способный демонстрировать ферментативную активность, стабильную в широком диапазоне значений рН, по сравнению с традиционным мутантом нитрилгидратазы, применительно к реакции синтеза амидного соединения из нитрильного соединения. Это подробно объяснено в Примерах.
Попытки получить мутанты нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, уже предпринимались. Тем не менее, эти попытки были направлены главным образом на повышение ферментативной активности в условиях оптимального рН, а ферментативная активность в кислых условиях, в противоположность условиям оптимального рН, при этом не изучалась. Ферментативная активность нитрилгидратазы в кислых условиях, отклоняющихся от оптимальных условий, была впервые открыта авторами настоящего изобретения, и авторы настоящего изобретения обнаружили, что описанные выше новые мутации аминокислотных остатков эффективны.
Далее мутантная нитрилгидратаза по изобретению описана более подробно. Описание нуклеотидной последовательности, представленное в данном изобретении, может быть сосредоточено на последовательности одной цепи, даже если нуклеиновая кислота, содержащая эту нуклеотидную последовательность, образует двойную цепь. Тем не менее, применительно к другим цепям двойной цепи следует использовать последовательности, полученные заменой на соответствующую комплементарную последовательность.
В данном изобретении термин «стадия» включен в этот термин не только тогда, когда предполагаемая цель стадии достигается на независимой стадии, но также тогда, когда стадию нельзя четко отделить от других стадий.
В данном изобретении численный диапазон, указанный с использованием формулировки «от … до», обозначает диапазон, включающий численные значения, описанные до и после формулировки «от … до» как минимальное значение и максимальное значение, соответственно.
В данном изобретении, если не указано иное, в случае присутствия множества веществ, соответствующих каждому компоненту композиции, количество каждого компонента композиции обозначает общее количество множества веществ, присутствующих в композиции.
В данном изобретении понятие нитрилгидратазы, имеющей происхождение из Pseudonocardia thermophila, включает не только нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, которая имеет α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 2, но также модифицированную нитрилгидратазу, которая была модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, в такой степени, что специалист в данной области может рассматривать ее как модифицированную последовательность нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila. Модифицированная нитрил гидр атаза, включенная в понятие нитрилгидратазы, имеющей происхождение из Pseudonocardia thermophila, включает модифицированную нитрилгидратазу, в которой проведены одна или более чем одна модификация относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, выбранные из группы, состоящей из (i) замены одного или более чем одного аминокислотного остатка на другой аминокислотный остаток, (ii) делеции одного или более чем одного аминокислотного остатка, отличных от (а)-(е), (iii) вставки аминокислотных остатков, (iv) присоединения аминокислотного остатка к N-концу и/или С-концу аминокислотной последовательности α-субъединицы и (v) присоединения аминокислотного остатка к N-концу и/или С-концу аминокислотной последовательности β-субъединицы.
В α-субъединице модифицированной нитрилгидратазы число замененных аминокислотных остатков составляет, например, от 1 до 20, от 1 до 15, от 1 до 10, от 1 до 8, от 1 до 7, от 1 до 6, от 1 до 5, от 1 до 4, от 1 до 3, от 1 до 2 или 1. Число замененных аминокислотных остатков может быть равно нулю.
В β-субъединице модифицированной нитрилгидратазы число замененных аминокислотных остатков составляет, например, от 1 до 20, от 1 до 15, от 1 до 10, от 1 до 8, от 1 до 7, от 1 до 6, от 1 до 5, от 1 до 4, от 1 до 3, от 1 до 2 или 1. Число замененных аминокислотных остатков может быть равно нулю.
В α-субъединице модифицированной нитрилгидратазы число удаленных аминокислотных остатков составляет, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число удаленных аминокислотных остатков может быть равно нулю.
В β-субъединице модифицированной нитрилгидратазы число удаленных аминокислотных остатков составляет, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число удаленных аминокислотных остатков может быть равно нулю.
В α-субъединице модифицированной нитрилгидратазы число введенных аминокислотных остатков составляет, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число введенных аминокислотных остатков может быть равно нулю.
В β-субъединице модифицированной нитрилгидратазы число введенных аминокислотных остатков составляет, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число введенных аминокислотных остатков может быть равно нулю.
В α-субъединице модифицированной нитрилгидратазы общее число замененных, удаленных или введенных аминокислотных остатков составляет, например, от 1 до 20, от 1 до 14, от 1 до 8, от 1 до 4, от 1 до 2 или 1. В случае концевого присоединения и тому подобного общее число замененных, удаленных или введенных аминокислотных остатков может быть равно нулю.
В β-субъединице модифицированной нитрилгидратазы общее число замененных, удаленных или введенных аминокислотных остатков составляет, например, от 1 до 20, от 1 до 14, от 1 до 8, от 1 до 4, от 1 до 2 или 1. В случае концевого присоединения и тому подобного общее число замененных, удаленных или введенных аминокислотных остатков может быть равно нулю.
В α-субъединице модифицированной нитрилгидратазы число концевых присоединенных аминокислотных остатков составляет, например, от 1 до 60, от 1 до 40, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3 или 1 на один конец. Число присоединенных аминокислотных остатков может быть равно нулю.
В β-субъединице модифицированной нитрилгидратазы число концевых присоединенных аминокислотных остатков составляет, например, от 1 до 60, от 1 до 40, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3 или 1 на один конец. Число присоединенных аминокислотных остатков может быть равно нулю.
Концевой присоединенный аминокислотный остаток может представлять собой, например, секреторную сигнальную последовательность. Концевой присоединенный аминокислотный остаток может присутствовать только на N-конце, только на С-конце или как на N-конце, так и на С-конце.
Сходство между модифицированной нитрилгидратазой и нитрилгидратазой дикого типа, имеющей происхождение из Pseudonocardia thermophila, может также быть выражено как идентичность последовательности.
Аминокислотная последовательность α-субъединицы модифицированной нитрилгидратазы, например, на 70% или более идентична, на 80% или более идентична, на 85% или более идентична, на 90% или более идентична, на 95% или более идентична, на 96% или более идентична, на 97% или более идентична, на 98% или более идентична или на 99% или более идентична аминокислотной последовательности α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, представленной как SEQ ID NO: 1.
Аминокислотная последовательность β-субъединицы модифицированной нитрилгидратазы, например, на 70% или более идентична, на 80% или более идентична, на 85% или более идентична, на 90% или более идентична, на 95% или более идентична, на 96% или более идентична, на 97% или более идентична, на 98% или более идентична или на 99% или более идентична аминокислотной последовательности β-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, представленной как SEQ ID NO: 2.
Кроме того, выравнивание последовательностей может быть проведено с использованием ClustalW (1.83) и с применением начальных параметров (включая штраф за открытие разрыва 10 и штраф за продолжение разрыва 0,05).
При выравнивании аминокислотной последовательности модифицированной нитрилгидратазы с аминокислотной последовательностью нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, в зависимости от характера модификаций в модифицированной нитрил гидр атазе, возможны случаи, когда расстояние от N-концов субъединиц, определенных, как описано выше, будет различаться, даже если на выравнивании аминокислотные остатки соответствуют друг другу. В таком случае в данном изобретении, применительно к модифицированной нитрил гидр атазе, Х-й аминокислотный остаток от N-конца субъединицы относится к аминокислотному остатку, соответствующему Х-му аминокислотному остатку на выравнивании от N-конца соответствующей субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila. Например, в данном изобретении «40-й аминокислотный остаток от N-конца α-субъединицы», «остаток Asp в 40-м положении от N-конца α-субъединицы» или «остаток Asp, являющийся 40-м аминокислотным остатком от N-конца α-субъединицы» может быть расположен в положении, отличном от 40-го положения от N-конца α-субъединицы в аминокислотной последовательности модифицированной нитрилгидратазы. Например, аминокислотный остаток в 41-м положении от N-конца α-субъединицы модифицированной нитрилгидратазы (не обязательно представляющий собой Asp) может соответствовать, в результате вставки аминокислотного остатка, остатку Asp, являющемуся 40-м аминокислотным остатком от N-конца α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila. В таком случае в описании данного изобретения «40-й аминокислотный остаток от N-конца α-субъединицы», «остаток Asp в 40-м положении от N-конца α-субъединицы» или «остаток Asp, являющийся 40-м аминокислотным остатком от N-конца α-субъединицы» относится к 41-му аминокислотному остатку от N-конца α-субъединицы модифицированной нитрилгидратазы.
В нитрилгидратазе дикого типа, имеющей происхождение из Pseudonocardia thermophila, остаток Met в начале SEQ ID NO: 1 представляет собой 1-й аминокислотный остаток от N-конца аминокислотной последовательности α-субъединицы, 40-й аминокислотный остаток от N-конца аминокислотной последовательности α-субъединицы представляет собой остаток Asp и 43-й аминокислотный остаток от N-конца аминокислотной последовательности α- субъединицы представляет собой остаток Ala. Кроме того, в нитрилгидратазе дикого типа, имеющей происхождение из Pseudonocardia thermophila, остаток Met в начале SEQ ID NO: 2 представляет собой 1-й аминокислотный остаток от N-конца аминокислотной последовательности β-субъединицы, 205-й аминокислотный остаток от N-конца аминокислотной последовательности β-субъединицы представляет собой остаток Gly, 206-й аминокислотный остаток от N-конца аминокислотной последовательности β-субъединицы представляет собой остаток Pro и 215-й аминокислотный остаток от N-конца аминокислотной последовательности β-субъединицы представляет собой остаток Tyr.
Последовательность модифицированной нитрилгидратазы, которая может быть использована в качестве мишени для введения одной или более чем одной из замен аминокислотных остатков (а)-(е), может быть выбрана из последовательностей нитрилгидратазы, зарегистрированных в базе GenBank, предоставляемой Национальным центром биотехнологической информации США (National Center for Biotechnology Information, NCBI), или последовательностей нитрилгидратазы, описанных в известных литературных источниках. Кроме того, последовательность модифицированной нитрилгидратазы может быть выбрана из последовательностей модифицированных нитрилгидратаз, описанных в JP-A №№ Н09-275978, 2004-194588, 2005-160403, WO 2004/056990 или WO 2010/055666, описанных выше. В таком случае, помимо улучшений, описанных в этих патентных документах, может быть получен новый эффект повышения рН-стабильности, благодаря введению одной или более чем одной из замен аминокислотных остатков (а)-(е).
Кроме того, модифицированная нитрилгидратаза, которая может быть использована в качестве мишени для введения одной или более чем одной из замен аминокислотных остатков (а)-(е), может представлять собой нитрилгидратазу, где 36-й аминокислотный остаток от N-конца (аминокислотный остаток, соответствующий остатку Thr на выравнивании в 36-м положении от N-конца аминокислотной последовательности нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila) представляет собой остаток Trp (что далее названо заменой аминокислотного остатка (f)). Наличие замены аминокислотного остатка (f) в дополнение к одной или более чем одной из замен аминокислотных остатков (а)-(е) предпочтительно с точки зрения получения мутантной нитрилгидратазы, обладающей более высокой рН-стабильностью. Иными словами, в присутствие замены аминокислотного остатка (f) эффект замен аминокислотных остатков (а)-(е) обычно более выражен.
Кроме того, например, в WO 2010/055666 раскрыто введение замен аминокислотных остатков (которые далее названы заменами аминокислотных остатков группы В), как описано ниже, в аминокислотную последовательность нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Замены аминокислотных остатков в аминокислотной последовательности α-субъединицы (представленные по положению относительно N-конца α-субъединицы):
замена Leu, являющегося 6-м аминокислотным остатком, на Thr или Ala,
замена Ile, являющегося 13-м аминокислотным остатком, на Leu,
замена Ala, являющегося 19-м аминокислотным остатком, на Val,
замена Met, являющегося 27-м аминокислотным остатком, на Ile,
замена Thr, являющегося 36-м аминокислотным остатком, на Met, Ser, Gly или Ala,
замена Asn, являющегося 48-м аминокислотным остатком, на Gln,
замена Arg, являющегося 71-м аминокислотным остатком, на His,
замена Asp, являющегося 92-м аминокислотным остатком, на Glu,
замена Met, являющегося 94-м аминокислотным остатком, на Ile,
замена Phe, являющегося 126-м аминокислотным остатком, на Tyr,
замена Gly, являющегося 148-м аминокислотным остатком, на Asp,
замена Thr, являющегося 188-м аминокислотным остатком, на Gly,
замена Gly, являющегося 197-м аминокислотным остатком, на Cys и
замена Val, являющегося 204-м аминокислотным остатком, на Arg.
Замены аминокислотных остатков в аминокислотной последовательности β-субъединицы (представленные по положению относительно N-конца β-субъединицы):
замена Val, являющегося 4-м аминокислотным остатком, на Met,
замена Gly, являющегося 8-м аминокислотным остатком, на Ala,
замена Thr, являющегося 10-м аминокислотным остатком, на Asp,
замена Val, являющегося 24-м аминокислотным остатком, на Ile,
замена Ala, являющегося 33-м аминокислотным остатком, на Val или Met,
замена Phe, являющегося 37-м аминокислотным остатком, на Val или Leu,
замена Thr, являющегося 40-м аминокислотным остатком, на Ile, Val или Leu,
замена Phe, являющегося 41-м аминокислотным остатком, на Ile,
замена Met, являющегося 46-м аминокислотным остатком, на Lys,
замена Leu, являющегося 48-м аминокислотным остатком, на Val,
замена Phe, являющегося 51-м аминокислотным остатком, на Val,
замена Ala, являющегося 61-м аминокислотным остатком, на Val, Gly, Trp, Ser, Leu или Thr,
замена His, являющегося 79-м аминокислотным остатком, на Asn,
замена Gin, являющегося 96-м аминокислотным остатком, на Arg,
замена Pro, являющегося 107-м аминокислотным остатком, на Met,
замена Glu, являющегося 108-м аминокислотным остатком, на Asp или Arg,
замена Glu, являющегося 110-м аминокислотным остатком, на Asn,
замена Lys, являющегося 112-м аминокислотным остатком, на Val или Не,
замена Phe, являющегося 118-м аминокислотным остатком, на Val,
замена Leu, являющегося 127-м аминокислотным остатком, на Ser,
замена Arg, являющегося 146-м аминокислотным остатком, на Gly,
замена Ala, являющегося 150-м аминокислотным остатком, на Asn или Ser,
замена Arg, являющегося 160-м аминокислотным остатком, на Cys, Trp или Met,
замена Thr, являющегося 168-м аминокислотным остатком, на Glu,
замена Tyr, являющегося 176-м аминокислотным остатком, на Ala, Thr, Met или Cys,
замена Leu, являющегося 186-м аминокислотным остатком, на Arg,
замена Ala, являющегося 200-м аминокислотным остатком, на Glu,
замена Pro, являющегося 206-м аминокислотным остатком, на Leu,
замена Ser, являющегося 212-м аминокислотным остатком, на Tyr,
замена Asp, являющегося 217-м аминокислотным остатком, на Val, His, Met, Gly, Ser, Leu или Cys,
замена Cys, являющегося 218-м аминокислотным остатком, на Met или Ser,
замена Val, являющегося 226-м аминокислотным остатком, на Ile,
замена Ala, являющегося 230-м аминокислотным остатком, на Glu и
замена Ala, являющегося 231-м аминокислотным остатком, на Val.
Модифицированная нитрилгидратаза, которая может быть использована в качестве мишени для введения одной или более чем одной из замен аминокислотных остатков (а)-(е), может содержать одну или более чем одну замену аминокислотного остатка, выбранные из замен аминокислотных остатков группы В, и, в дополнение к ним, может дополнительно иметь замену аминокислотного остатка (f), по сравнению с аминокислотной последовательностью нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila. Возможно включение двух или более замен аминокислотных остатков в комбинации. Несколько примеров таких комбинаций описаны также в WO 2010/055666. Кроме того, модифицированная нитрилгидратаза должна обладать нитрилгидратазной активностью.
Например, примеры случаев включения трех замен аминокислотных остатков в комбинации могут включать следующее:
комбинация Thr36Ser и Asp92Glu в α-субъединице и Ala33Val в β-субъединице,
комбинация Met94Ile в α-субъединице и Ala61Gly и Ala150Asn в β-субъединице,
комбинация Val4Met, Tyr176Ala и Asp217Val в β-субъединице,
комбинация Ala33Met, His79Asn и Tyr176Thr в β-субъединице и
комбинация Thr40Val, Cys218Met и Val226Ile в β-субъединице, и тому подобное. Кроме того, примеры случаев включения восьми замен аминокислотных остатков в комбинации могут включать, например, следующее:
комбинация Ile 13Leu, Ala19Val, Arg71His и Phe126Tyr в α-субъединице и Phe37Leu, Gln96Arg, Glu108Asp и Ala200Glu в β-субъединице (международная публикация № WO 2010/055666: трансформант №59),
комбинация Leu6Thr, Met27Ile, Thr36Met и Phe126Tyr в α-субъединице и Thr10Asp, Pro107Met, Phe118Val и Ala200Glu в β-субъединице (международная публикация № WO 2010/055666: трансформант №68),
комбинация Leu6Thr, Thr36Met и Phe126Tyr в α-субъединице и Thr10Asp, Phe118Val, Ala200Glu, Pro206Leu и Ala230Glu в β-субъединице (международная публикация № WO 2010/055666: трансформант №92),
комбинация Leu6Thr, Ala19Val и Phe126Tyr в α-субъединице и Leu48Val, His79Asn, Glu108Arg, Ser212Tyr и Ala230Glu в β-субъединице (международная публикация № WO 2010/055666: трансформант №85) и
комбинация Thr36Met, Gly148Asp и Val204Arg в α-субъединице и Phe41Ile, Phe51Val, Glu108Asp, Pro206Leu и Ala230Glu в β-субъединице (международная публикация № WO 2010/055666: трансформант №93) и тому подобное.
Кроме того, аминокислотная последовательность модифицированной нитрилгидратазы, которая может быть использована в качестве мишени для введения одной или более чем одной из замен аминокислотных остатков (а)-(е), может содержать замены аминокислотных остатков, расположенных в положениях, отличных от указанных выше (отличные от замен аминокислотных остатков группы В и замены аминокислотного остатка (f)), по сравнению с аминокислотной последовательностью нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila. В модифицированной нитрилгидратазе, содержащей замену аминокислотного остатка, отличную от замен аминокислотных остатков группы В и замены аминокислотного остатка (f), в дополнение к одной или более чем одной замене аминокислотного остатка группы В и/или замене аминокислотного остатка (f) или без замен аминокислотных остатков группы В и замены аминокислотного остатка (f), введение одной или более чем одной из замен аминокислотных остатков (а)-(е) приводит к эффекту повышения рН-стабильности модифицированной нитрилгидратазы посредством стабилизации ее трехмерной структуры, как описано ниже.
Одна или более чем одна из замен аминокислотных остатков (а)-(е) может быть введена в нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, или модифицированную нитрилгидратазу с получением мутантной нитрилгидратазы по изобретению.
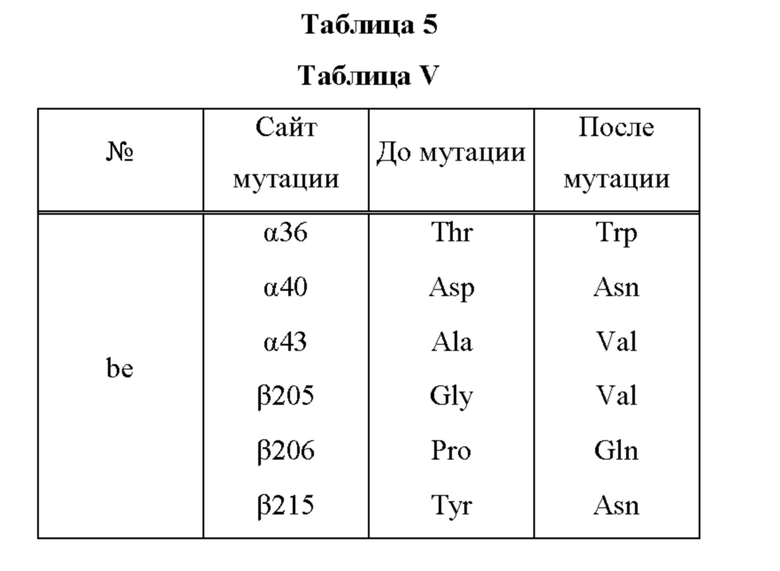
Мутантная нитрилгидратаза может содержать комбинацию двух из замен аминокислотных остатков (а)-(е) и (f), показанную в Таблице I.
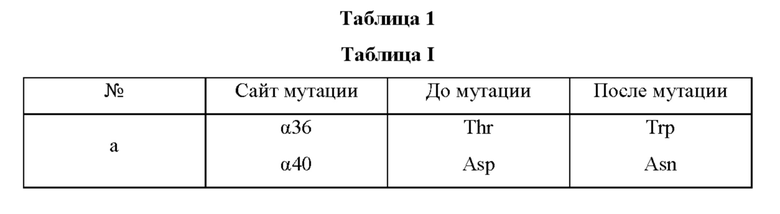

Мутантная нитрилгидратаза может содержать комбинацию трех из замен аминокислотных остатков (а)-(е) и (f), показанную в Таблице II.


Мутантная нитрилгидратаза может содержать комбинацию четырех из замен аминокислотных остатков (а)-(е) и (f), показанную в Таблице III.

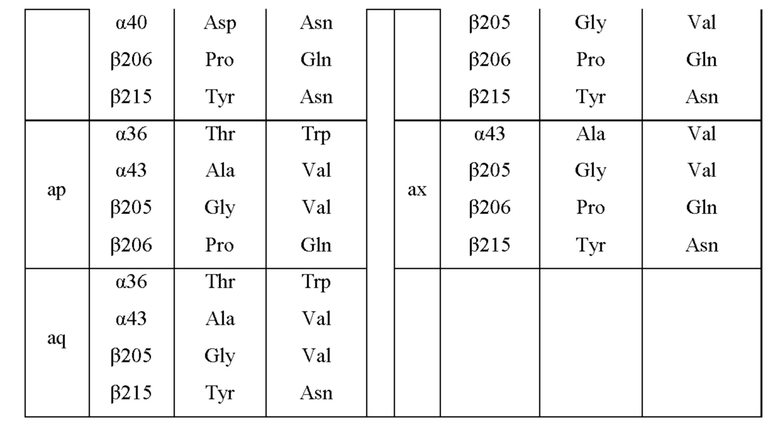
Мутантная нитрилгидратаза может содержать комбинацию пяти из замен аминокислотных остатков (а)-(е) и (f), показанную в Таблице IV.
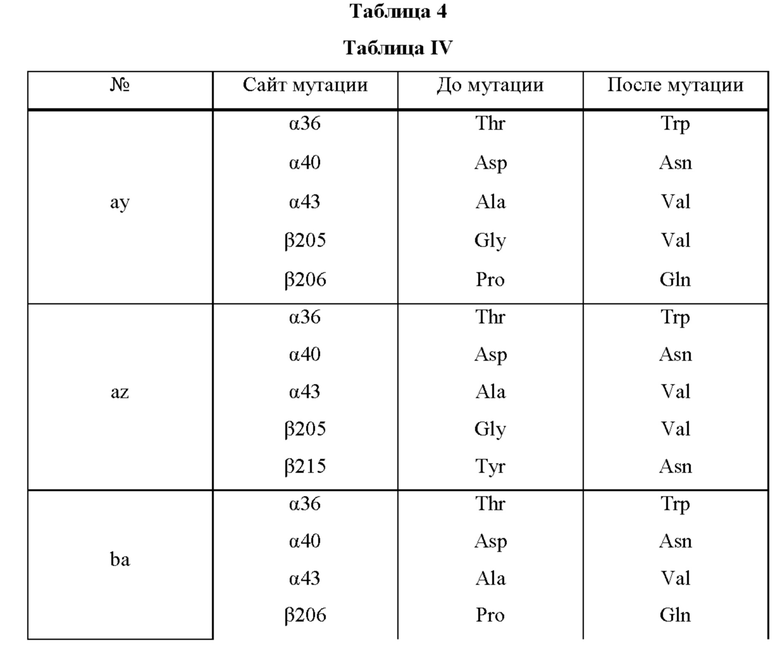
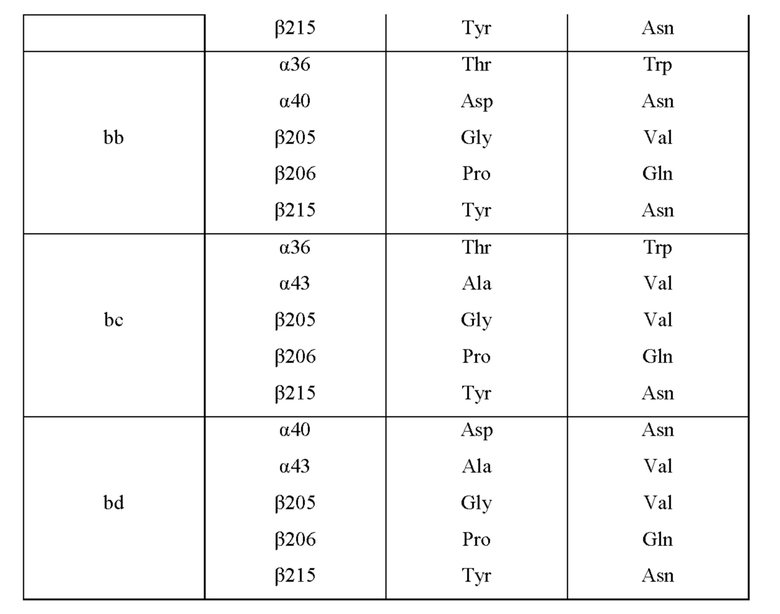
Мутантная нитрилгидратаза может содержать комбинацию всех из замен аминокислотных остатков (а)-(е) и (f) (шесть замен аминокислотных остатков, показанные в Таблице V).
Аминокислотная последовательность модифицированной нитрилгидратазы, которая может быть использована в качестве мишени для введения одной или более чем одной из замен аминокислотных остатков (а)-(е), может представлять собой аминокислотную последовательность модифицированной нитрилгидратазы, полученной в WO 2010/055666, и тому подобное, или последовательность, сходную с ней. Таким образом, мутантная нитрилгидратаза по изобретению может быть получена, например, введением по меньшей мере одной из следующих замен:
замена 40-го аминокислотного остатка от N-конца α-субъединицы на Asn, замена 43-го аминокислотного остатка от N-конца α-субъединицы на Val, замена 205-го аминокислотного остатка от N-конца β-субъединицы на Val, замена 206-го аминокислотного остатка от N-конца β-субъединицы на Gln и замена 215-го аминокислотного остатка от N-конца β-субъединицы на Asn; в любую из следующих нитрилгидратаз (1)-(47) (и названа далее «мутантной нитрилгидратазой В»):
(1) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 2,
(2) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 16, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 33,
(3) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 17, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 33,
(4) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 34,
(5) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 19, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 34,
(6) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 20, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 35,
(7) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 20, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 36,
(8) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 37,
(9) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 38,
(10) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 39,
(11) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 40,
(12) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO:41,
(13) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 42,
(14) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 43,
(15) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 22, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 44,
(16) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 23, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 45,
(17) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 24, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 46,
(18) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 25, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 47,
(19) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 48,
(20) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 23, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 49,
(21) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 16, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 50,
(22) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 26, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 51,
(23) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 27, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 52,
(24) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 28, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 53,
(25) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 17, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 54,
(26) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 29, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 55,
(27) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 56,
(28) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 57,
(29) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 58,
(30) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO:29, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 59,
(31) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 31, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 60,
(32) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 61,
(33) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 32, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 62,
(34) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 63,
(35) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 64,
(36) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 65,
(37) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 25, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 54,
(38) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 66,
(39) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 67,
(40) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 68,
(41) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 69,
(42) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 70,
(43) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 71,
(44) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 72,
(45) нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 73,
(46) нитрилгидратаза, где 36-й аминокислотный остаток от N-конца α-субъединицы любой из нитрилгидратаз (1)-(45) представляет собой остаток Trp,
(47) модифицированная нитрилгидратаза, имеющая вариант α-субъединицы, состоящий из аминокислотных последовательностей α-субъединицы определенной нитрилгидратазы, представляющей собой любую из нитрилгидратаз (1)-(46), или аминокислотных последовательностей, на 70% или более идентичных этим аминокислотным последовательностям, и вариант β-субъединицы, состоящий из аминокислотных последовательностей β-субъединицы определенной нитрилгидратазы или аминокислотных последовательностей, на 70% или более идентичных этим аминокислотным последовательностям, где по меньшей мере одна из аминокислотных последовательностей α-субъединицы и аминокислотных последовательностей β-субъединицы представляет собой аминокислотную последовательность варианта α-субъединицы или варианта β-субъединицы.
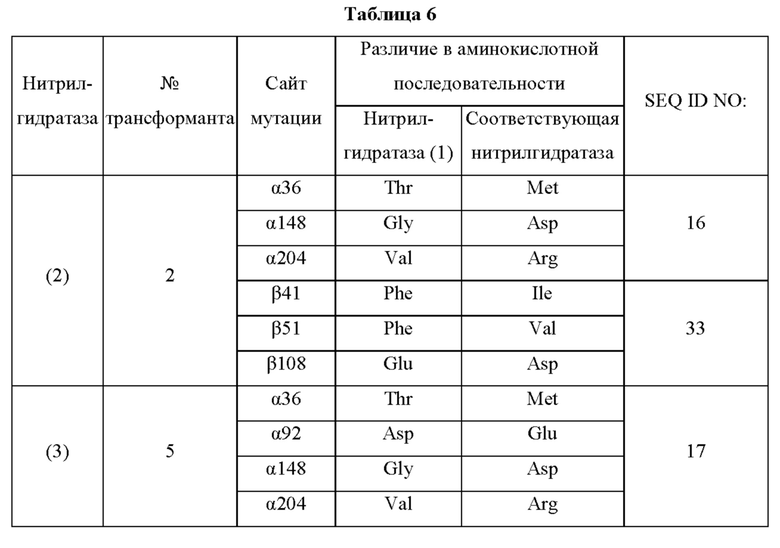
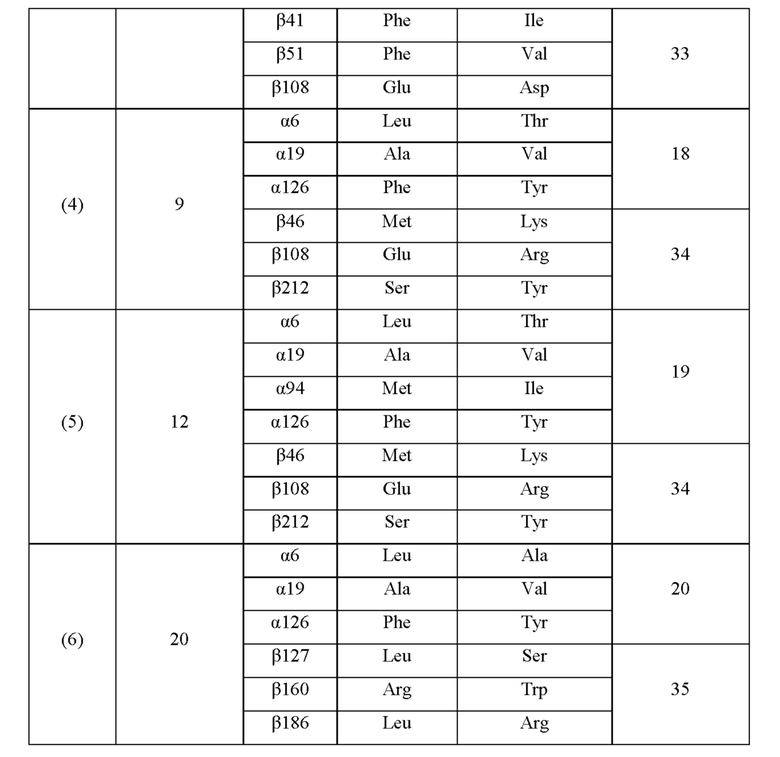
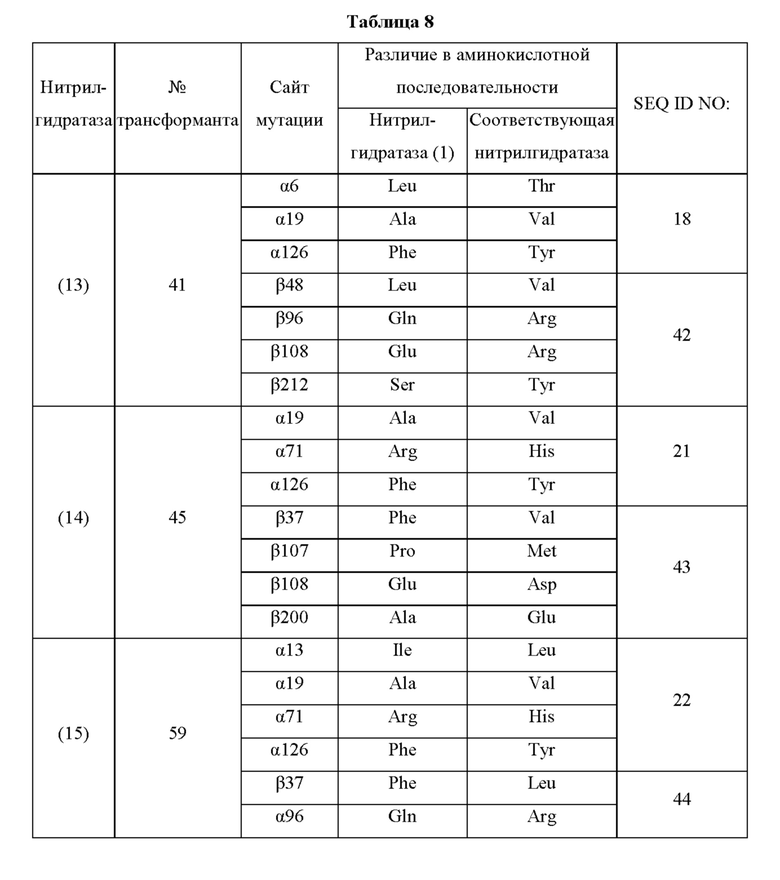
Кроме того, нитрилгидратазы (2)-(45) отличаются от нитрилгидратазы (1), имеющей α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 2, по аминокислотным остаткам, описанным в следующих Таблицах 6-13, и описаны с указанием номера трансформанта в WO 2010/055666. В приведенных ниже Таблицах число в столбце сайта мутации указывает на положение от N-конца аминокислотной последовательности субъединицы. Кроме того, номер трансформанта представляет собой номер, присвоенный в международной публикации WO 2010/055666.



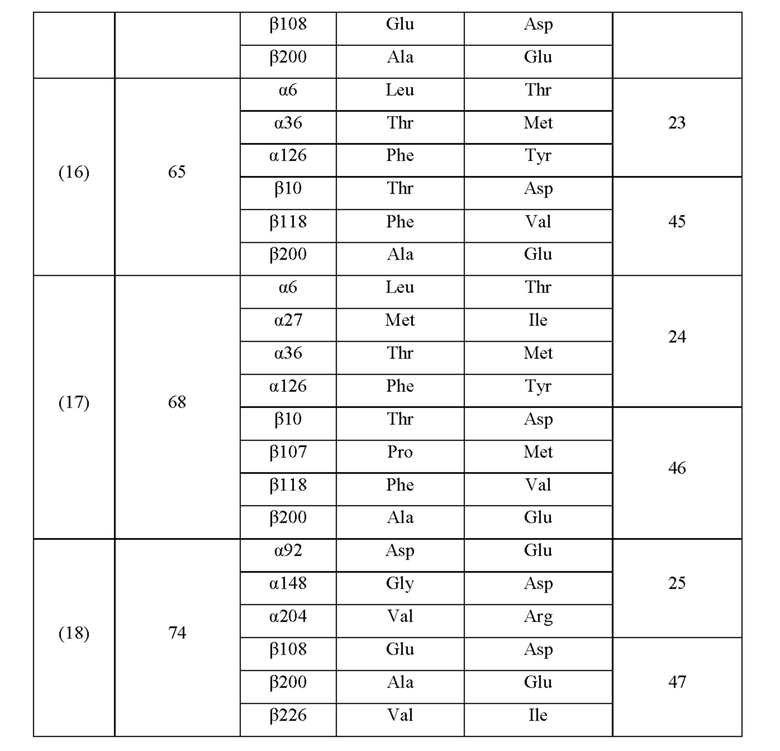

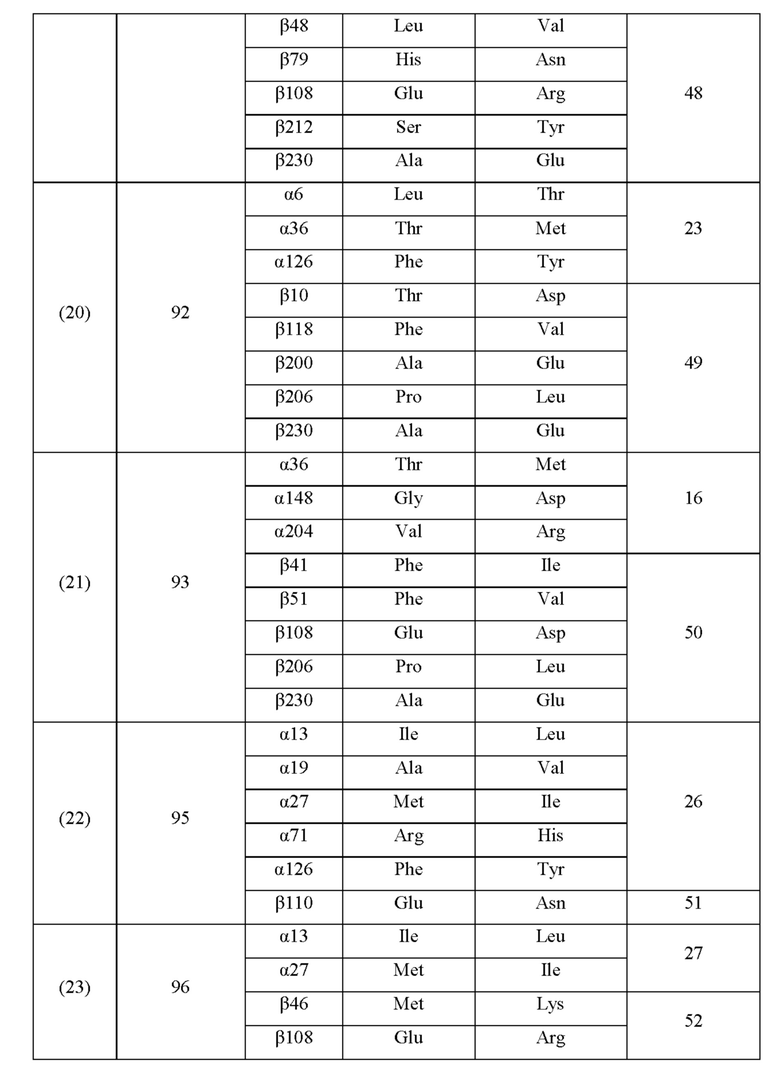

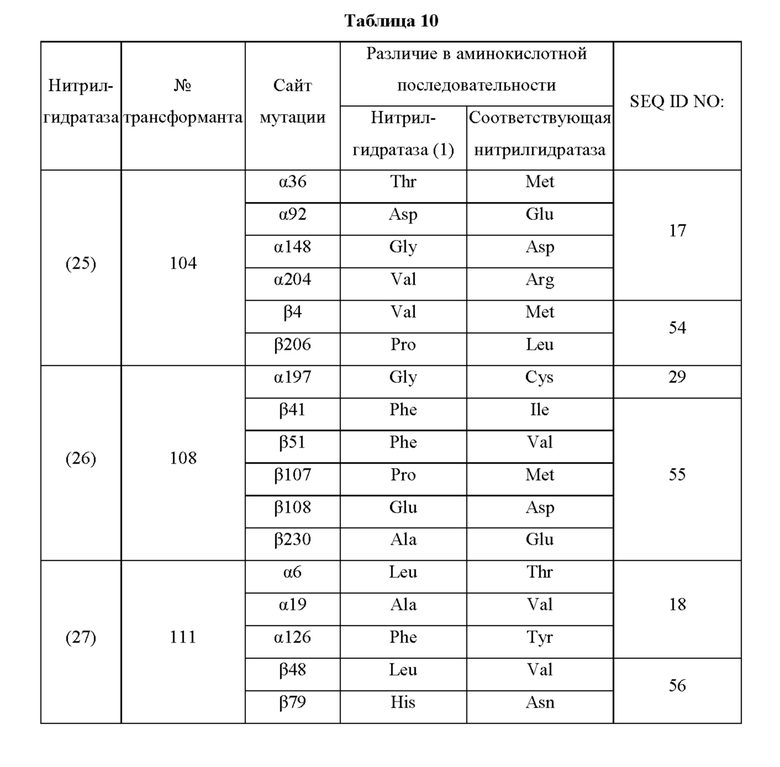
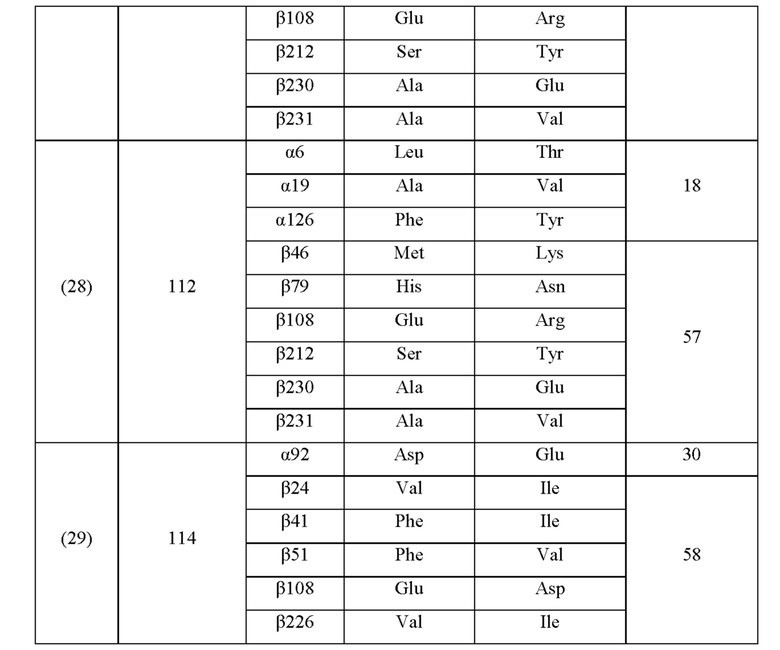
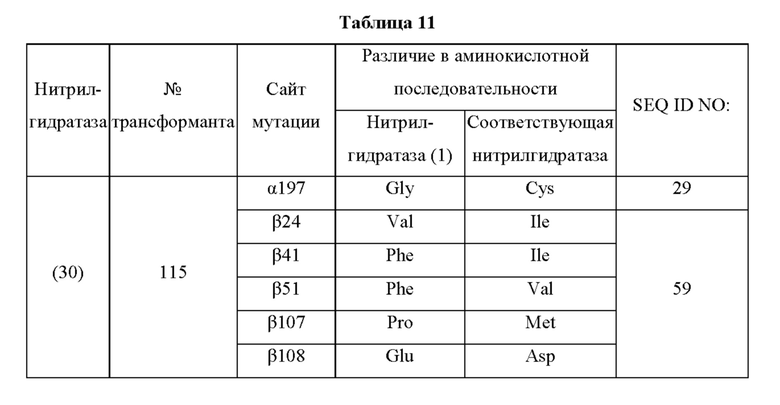
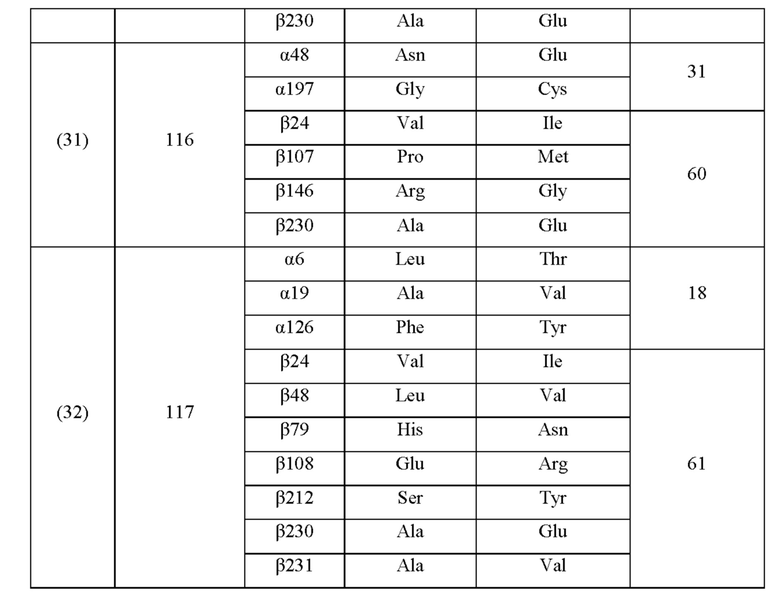
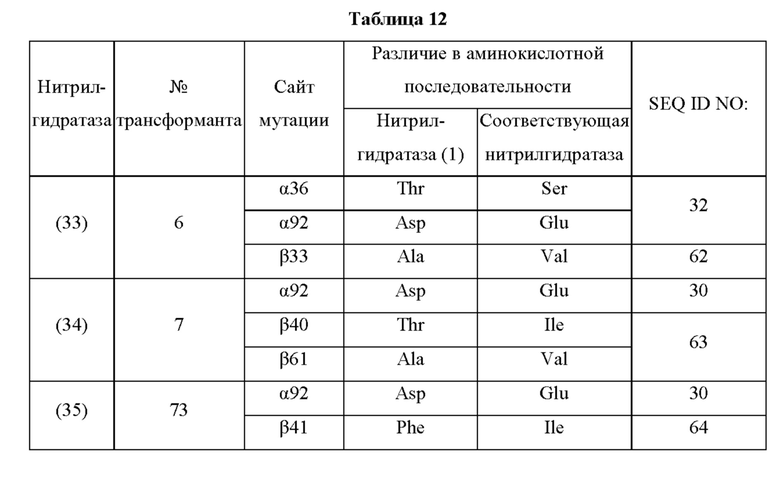
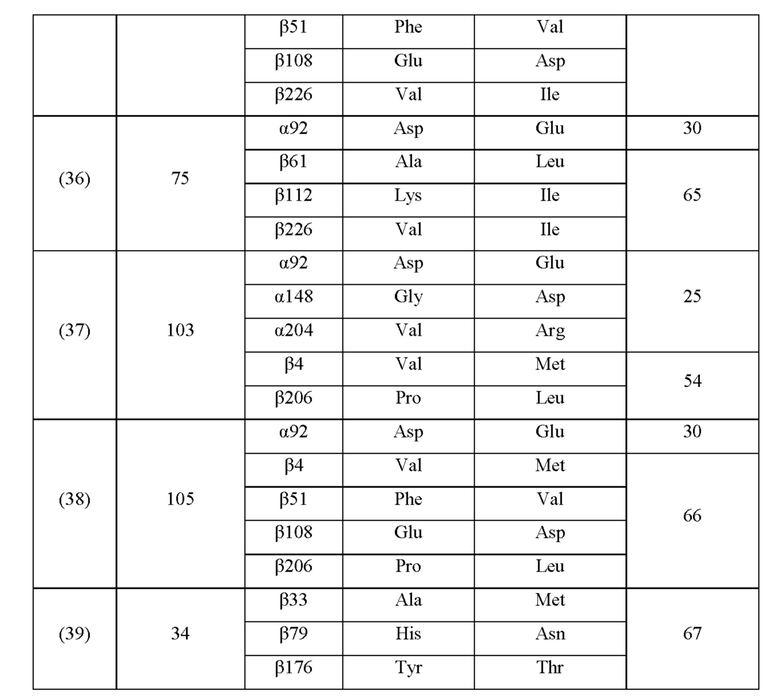
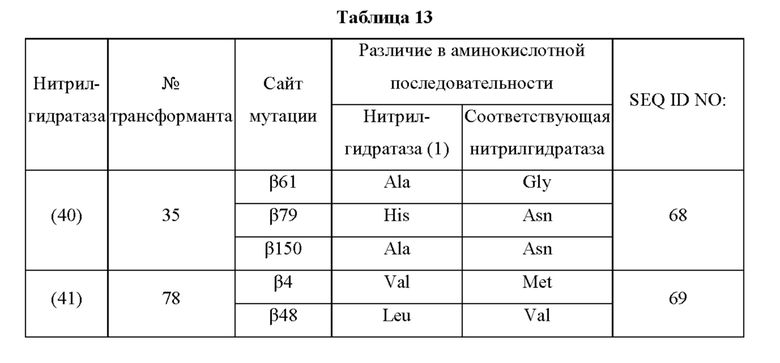
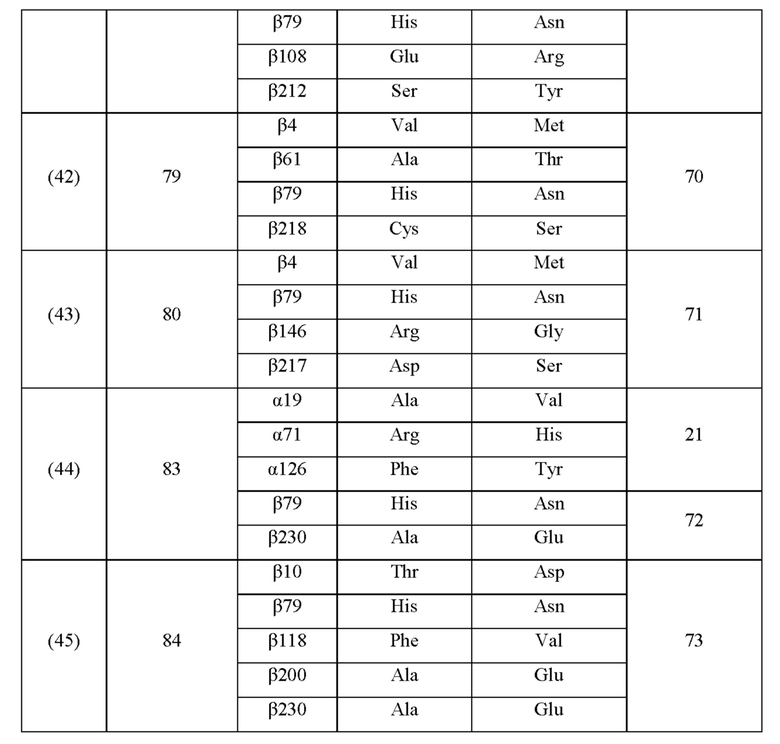
Аминокислотная последовательность модифицированной нитрилгидратазы, которая может быть использована в качестве мишени для введения одной или более чем одной из замен аминокислотных остатков (а)-(е), предпочтительно представляет собой аминокислотную последовательность любой из 44 нитрилгидратаз (2)-(45), описанных выше, и, более предпочтительно, аминокислотную последовательность любой из нитрилгидратаз (3), (11), (18), (19), (25), (27), (28), (29), (32), (33), (34), (35), (36), (37), (38), (39), (40), (41), (42), (43), (44) и (45).
В (47) идентичность аминокислотной последовательности варианта α-субъединицы α-субъединице определенной нитрилгидратазы может составлять 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более. Сходным образом, в (47) идентичность аминокислотной последовательности варианта β-субъединицы β-субъединице определенной нитрилгидратазы может составлять 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более. Кроме того, вариант α-субъединицы относится к α-субъединице, удовлетворяющей идентичности последовательности, определенной выше, и имеющей аминокислотную последовательность, отличающуюся от α-субъединицы определенной нитрилгидратазы. Вариант β-субъединицы относится к β-субъединице, удовлетворяющей идентичности последовательности, определенной выше, и имеющей аминокислотную последовательность, отличающуюся от β-субъединицы определенной нитрилгидратазы.
Описанные выше нитрилгидратазы (2)-(45) представляют собой нитрилгидратазы, активность которых подтверждена в Примерах международной публикации № WO 2010/055666 (трансформанты №№5, 62, 68, 95 и 111), и, кроме того, описанная выше нитрилгидратаза (1) представляет собой нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila. Нитрилгидратазы (2)-(46) могут также быть использованы в качестве мишеней для введения одной или более чем одной из замен аминокислотных остатков (а)-(е). Кроме того, в качестве мишеней для введения одной или более чем одной из замен аминокислотных остатков (а)-(е) могут также быть использованы другие модифицированные нитрилгидратазы, имеющие аминокислотную последовательность, сходную с аминокислотной последовательностью этих нитрилгидратаз. Таким образом, при описании различий аминокислотной последовательности модифицированной нитрилгидратазы и нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila (включая описание степени модификации последовательности и примеров), «нитрилгидратаза дикого типа, имеющая происхождение из Pseudonocardia thermophila)), может быть заменена любой из описанных выше нитрилгидратаз (1)-(46) и может быть использована сама по себе. Кроме того, модифицированная нитрилгидратаза должна обладать нитрилгидратазной активностью.
Например, применительно к заменам аминокислотных остатков и их комбинациям, которые могут быть включены в мутантную нитрилгидратазу В, и 36-му аминокислотному остатку от N-конца α-субъединицы нитрилгидратазы, мутантная нитрилгидратаза В может содержать комбинацию аминокислотных остатков, выбранную из группы, состоящей из комбинаций аминокислотных остатков a-о, указанных в Таблице I выше, комбинаций аминокислотных остатков p-ai, указанных в Таблице II выше, комбинаций аминокислотных остатков aj-ax, описанных в Таблице III выше, комбинаций аминокислотных остатков ay-bd, описанных в Таблице IV выше, и комбинации аминокислотных остатков be, описанной в Таблице V выше. Иными словами, может быть введена комбинация замен аминокислотных остатков относительно аминокислотной последовательности нитрилгидратаз (1)-(46).
Кроме того, например, нитрилгидратаз а (47) может представлять собой модифицированную нитрилгидратазу, в которой проведены одна или более чем одна модификация относительно любой из нитрилгидратаз (1)-(46), выбранные из группы, состоящей из (1) замены одного или более чем одного аминокислотного остатка на другой аминокислотный остаток, (2) делеции одного или более чем одного аминокислотного остатка, отличных от (а)-(е), (3) вставки аминокислотных остатков, (4) присоединения аминокислотного остатка к N-концу и/или С-концу аминокислотной последовательности α-субъединицы и (5) присоединения аминокислотного остатка к N-концу и/или С-концу аминокислотной последовательности β-субъединицы.
В α-субъединице нитрилгидратазы (47) число замененных аминокислотных остатков в случае α-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 20, от 1 до 15, от 1 до 10, от 1 до 8, от 1 до 7, от 1 до 6, от 1 до 5, от 1 до 4, от 1 до 3, от 1 до 2 или 1. Число замененных аминокислотных остатков может быть равно нулю.
В β-субъединице нитрилгидратазы (47) число замененных аминокислотных остатков в случае β-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 20, от 1 до 15, от 1 до 10, от 1 до 8, от 1 до 7, от 1 до 6, от 1 до 5, от 1 до 4, от 1 до 3, от 1 до 2 или 1. Число замененных аминокислотных остатков может быть равно нулю.
В α-субъединице нитрилгидратазы (47) число удаленных аминокислотных остатков в случае α-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число удаленных аминокислотных остатков может быть равно нулю.
В β-субъединице нитрилгидратазы (47) число удаленных аминокислотных остатков в случае β-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число удаленных аминокислотных остатков может быть равно нулю.
В α-субъединице нитрилгидратазы (47) число введенных аминокислотных остатков в случае α-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число введенных аминокислотных остатков может быть равно нулю.
В β-субъединице нитрилгидратазы (47) число введенных аминокислотных остатков в случае β-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 10, от 1 до 7, от 1 до 4, от 1 до 2 или 1. Число введенных аминокислотных остатков может быть равно нулю.
В α-субъединице нитрилгидратазы (47) общее число замененных, удаленных или введенных аминокислотных остатков в случае α-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 20, от 1 до 14, от 1 до 8, от 1 до 4, от 1 до 2 или 1. В случае концевого присоединения и тому подобного общее число замененных, удаленных или введенных аминокислотных остатков может быть равно нулю.
В β-субъединице модифицированной нитрилгидратазы (47) общее число замененных, удаленных или введенных аминокислотных остатков в случае β-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 20, от 1 до 14, от 1 до 8, от 1 до 4, от 1 до 2 или 1. В случае концевого присоединения и тому подобного общее число замененных, удаленных или введенных аминокислотных остатков может быть равно нулю.
В α-субъединице нитрилгидратазы 47 число концевых присоединенных аминокислотных остатков в случае α-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 60, от 1 до 40, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3 или 1 на один конец. Число присоединенных аминокислотных остатков может быть равно нулю.
В β-субъединице нитрилгидратазы (47) число концевых присоединенных аминокислотных остатков в случае β-субъединицы любой из нитрилгидратаз (1)-(46) составляет, в качестве ориентира, например, от 1 до 60, от 1 до 40, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3 или 1 на один конец. Число присоединенных аминокислотных остатков может быть равно нулю.
Нитрилгидратаза (47) может содержать одну или более чем одну замену аминокислотного остатка, выбранные из замен аминокислотных остатков группы В, по сравнению с аминокислотной последовательностью любой из нитрилгидратаз (1)-(46). Возможно включение двух или более замен аминокислотных остатков в комбинации.
В приведенном выше описании вариант α/β-субъединицы нитрилгидратазы (47) не имеет вставок аминокислот или делеций аминокислотных остатков и может иметь только замену или концевое присоединение аминокислотного остатка, по сравнению с аминокислотной последовательностью α/β-субъединицы определенной нитрилгидратазы. Положение аминокислотного остатка в варианте α/β-субъединицы нитрилгидратазы (47) обозначает, как описано выше, положение, соответствующее указанному аминокислотному остатку в α/β-субъединице определенной нитрилгидратазы на выравнивании.
Мутантная нитрилгидратаза по изобретению может быть получена введением по меньшей мере одной из следующих замен аминокислотных остатков относительно определенной нитрилгидратазы, представляющей собой любую из нитрилгидратаз (1)-(46), или относительно модифицированной нитрилгидратазы, имеющей модифицированную α-субъединицу, последовательность которой на 70% или более идентична α-субъединице определенной нитрилгидратазы, и модифицированную β-субъединицу, последовательность которой на 70% или более идентична β-субъединице определенной нитрилгидратазы, но не является самой этой определенной нитрилгидратазой (и названа далее «мутантной нитрилгидратазой С»):
замена аминокислотного остатка, соответствующего 40-му аминокислотному остатку от N-конца α-субъединицы определенной нитрилгидратазы, на Asn,
замена аминокислотного остатка, соответствующего 43-му аминокислотному остатку от N-конца α-субъединицы определенной нитрилгидратазы, на Val,
замена аминокислотного остатка, соответствующего 205-му аминокислотному остатку от N-конца β-субъединицы определенной нитрилгидратазы, на Val,
замена аминокислотного остатка, соответствующего 206-му аминокислотному остатку от N-конца β-субъединицы определенной нитрилгидратазы, на Gln, и
замена аминокислотного остатка, соответствующего 215-му аминокислотному остатку от N-конца β-субъединицы определенной нитрилгидратазы, на Asn.
В приведенном выше описании в случае, когда нитрилгидратаза до введения замен представляет собой модифицированную нитрилгидратазу, «аминокислотный остаток, соответствующий А-му аминокислотному остатку от N-конца α-субъединицы определенной нитрилгидратазы», относится к аминокислотному остатку в аминокислотной последовательности α-субъединицы модифицированной нитрилгидратазы, соответствующему А-му аминокислотному остатку от N-конца аминокислотной последовательности α-субъединицы определенной нитрилгидратазы при выравнивании аминокислотной последовательности α-субъединицы модифицированной нитрилгидратазы с аминокислотной последовательностью α-субъединицы определенной нитрилгидратазы.
Сходным образом, «аминокислотный остаток, соответствующий А-му аминокислотному остатку от N-конца β-субъединицы определенной нитрилгидратазы», относится к аминокислотному остатку в аминокислотной последовательности β-субъединицы модифицированной нитрилгидратазы, соответствующему А-му аминокислотному остатку от N-конца аминокислотной последовательности β-субъединицы определенной нитрилгидратазы при выравнивании аминокислотной последовательности β-субъединицы модифицированной нитрилгидратазы с аминокислотной последовательностью β-субъединицы определенной нитрилгидратазы. Кроме того, в случае, когда нитрилгидратаза до введения замен представляет собой определенную нитрилгидратазу, «аминокислотный остаток, соответствующий А-му аминокислотному остатку от N-конца α-субъединицы определенной нитрилгидратазы», относится к А-му аминокислотному остатку от N-конца α-субъединицы определенной нитрилгидратазы, а «аминокислотный остаток, соответствующий А-му аминокислотному остатку от N-конца β-субъединицы определенной нитрилгидратазы», относится к А-му аминокислотному остатку от N-конца β-субъединицы определенной нитрилгидратазы.
В модифицированной нитрилгидратаз е идентичность последовательности α-субъединицы α-субъединице определенной нитрилгидратазы может составлять 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более. Сходным образом, идентичность
последовательности β-субъединицы β-субъединице определенной нитрилгидратазы может составлять 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более. В частности, в модифицированной нитрилгидратазе идентичность последовательности α-субъединицы α-субъединице определенной нитрилгидратазы предпочтительно составляет 90% или более и, более предпочтительно, 95% или более. Сходным образом, в модифицированной нитрилгидратазе идентичность последовательности β-субъединицы β-субъединице определенной нитрилгидратазы предпочтительно составляет 90% или более и, более предпочтительно, 95% или более. При описании различий аминокислотной последовательности модифицированной нитрилгидратазы и нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila (включая описание степени модификации последовательности и примеров), «нитрилгидратаз а дикого типа, имеющая происхождение из Pseudonocardia thermophila)), может быть заменена определенной нитрилгидратазой и может быть использована сама по себе. В приведенном выше описании аминокислотная последовательность α/β-субъединицы модифицированной нитрилгидратазы не имеет вставок аминокислот или делеций аминокислотных остатков и может иметь только замену или концевое присоединение аминокислотного остатка, по сравнению с аминокислотной последовательностью α/β-субъединицы определенной нитрилгидратазы. Кроме того, модифицированная нитрилгидратаз а должна обладать нитрилгидратазной активностью.
Применительно к заменам аминокислотных остатков и их комбинациям, которые могут быть включены в мутантную нитрил гид ратазу С, и аминокислотному остатку, соответствующему 36-му аминокислотному остатку от N-конца α-субъединицы определенной нитрилгидратазы, мутантная нитрилгидратаз а С может содержать комбинацию аминокислотных остатков, выбранную из группы, состоящей из комбинаций аминокислотных остатков а-о, указанных в Таблице I выше, комбинаций аминокислотных остатков p-ai, указанных в Таблице II выше, комбинаций аминокислотных остатков aj-ax, описанных в Таблице III выше, комбинаций аминокислотных остатков ay-bd, описанных в Таблице IV выше, и комбинации аминокислотных остатков be, описанной в Таблице V выше. Иными словами, может быть введена комбинация замен аминокислотных остатков относительно аминокислотной последовательности определенной нитрилгидратазы или модифицированной нитрилгидратазы.
Кроме того, модифицированная нитрилгидратаза может, например, содержать одну или более чем одну замену аминокислотного остатка, выбранные из замен аминокислотных остатков группы В, по сравнению с аминокислотной последовательностью любой из нитрилгидратаз (1)-(46). Возможно включение двух или более замен аминокислотных остатков в комбинации.
Описанная выше модифицированная нитрилгидратаза может представлять собой нитрил гидр атазу, имеющую полипептид, где к определенной α-субъединице или к ее N-концу и/или С-концу присоединена дополнительная последовательность, и полипептид, где к определенной β-субъединице или к ее N-концу и/или С-концу присоединена дополнительная последовательность. В α-субъединице модифицированной нитрилгидратазы число концевых присоединенных аминокислотных остатков составляет, например, от 1 до 60, от 1 до 40, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3 или 1 на один конец. Число присоединенных аминокислотных остатков может быть равно нулю. В β-субъединице модифицированной нитрилгидратазы число концевых присоединенных аминокислотных остатков составляет, например, от 1 до 60, от 1 до 40, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3 или 1 на один конец. Число присоединенных аминокислотных остатков может быть равно нулю.
В случае присоединения, замены, делеции и/или вставки аминокислотной последовательности (за исключением аминокислотных остатков, подлежащих замене (а) и (b) выше) в α-субъединице модифицированной нитрилгидратазы, общее число замененных, удаленных и/или введенных аминокислотных остатков предпочтительно составляет от 1 до 10 и, более предпочтительно, от 1 до 5. Сходным образом, в случае присоединения, замены, делеции и/или вставки аминокислотной последовательности в α-субъединице модифицированной нитрилгидратазы (за исключением аминокислотных остатков, подлежащих замене (с)-(е) выше) в β-субъединице модифицированной нитрилгидратазы, общее число замененных, удаленных и/или введенных аминокислотных остатков предпочтительно составляет от 1 до 10 и, более предпочтительно, от 1 до 5.
Мутантная нитрилгидратаза по изобретению обычно функционирует посредством объединения двух α-субъединиц, определенных, как описано выше, и двух β-субъединиц, определенных, как описано выше.
В отличие от традиционного мутанта нитрилгидратазы, мутантная нитрилгидратаза по изобретению обладает отличной ферментативной активностью применительно к реакции синтеза амидного соединения из нитрильного соединения, даже в кислых условиях, таких как рН от 3,5 до 6,5 и тому подобное. Повышение стабильности ферментативной активности мутантной нитрилгидратазы по изобретению в кислых условиях особенно заметно в области рН 5,0 или менее. Таким образом, мутантная нитрилгидратаза позволяет в достаточной степени проводить реакцию синтеза амидного соединения, нитрильная связь которого, будучи субстратом, превращается в амидную связь, как на стадии очистки, на которой происходит адсорбция и удаление примесей, таких как белки, присутствующие в растворе, с использованием активированного угля в кислых условиях, таких как рН от 3,5 до 6,5 и тому подобное, с получением посредством этого очищенного амидного соединения, так и на стадии взаимодействия, включающей проведение реакции синтеза амидного соединения из нитрильного соединения, проводимых на этапе получения промышленного амидного соединения с использованием нитрилгидратазы. Таким образом, мутантная нитрилгидратаза, по сравнению с использованием традиционного фермента, позволяет значительно повысить эффективность получения амидного соединения, являющегося желаемым продуктом, на этапе получения промышленного амидного соединения. Кислые условия могут представлять собой рН 5,0 или менее, например, рН от 3,5 до 5,0, для более эффективного удаления ненужных белков.
Кроме того, значение отношения (начальная скорость реакции после обработки кислотой/начальная скорость реакции до обработки кислотой) начальной скорости реакции при использовании мутантной нитрилгидратазы, подвергнутой обработке кислотой (например, при рН 4,0 и 30°С на протяжении 30 часов), к начальной скорости реакции при использовании мутантной нитрилгидратазы, не подвергнутой обработке кислотой, предпочтительно в 1,1 раза или более превышает значение отношения (начальная скорость реакции после обработки кислотой/начальная скорость реакции до обработки кислотой) начальной скорости реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, подвергнутой обработке кислотой, к начальной скорости реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, не подвергнутой обработке кислотой.
Кроме того, рН-стабильность описанной выше мутантной нитрилгидратазы или нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, может быть, например, оценена следующим методом.
Что касается условий взаимодействия, реакцию проводят при температуре 20°С на протяжении 15-60 минут с использованием 50 мМ водного раствора трис-соляной кислоты (рН 8,0), содержащего 2,5% (об./об) акрилонитрила в качестве субстрата. По завершении взаимодействия определяют количество полученного акриламида. Количество акриламида можно анализировать с применением ВЭЖХ. Кроме того, обработка кислотой может быть проведена, например, обработкой при рН приблизительно 4,0 и 30°С на протяжении 30 часов. Конкретно, рН-стабильность может быть оценена способом, описанным в разделе по оценке рН-стабильности в Примерах.
Например, 40 мкл раствора, полученного по завершении культивирования транс фор манта, продуцирующего нитрилгидратазу, суспендируют в 740 мкл 54 мМ водного раствора трис-соляной кислоты (рН 8,0), добавляют 20 мкл акрилонитрила и полученную смесь подвергают взаимодействию на протяжении 15-60 минут с осторожным перемешиванием при 20°С. По завершении взаимодействия проводят анализ реакционного раствора с применением ВЭЖХ для измерения количества (Р) полученного амидного соединения (или оно может быть рассчитано по количеству потребленного нитрильного соединения). Кроме того, 1000 мкл раствора, полученного по завершении культивирования транс форманта, отбирают и центрифугируют для выделения бактериальных клеток, к выделенным бактериальным клеткам добавляют 1000 мкл буфера с 50 мМ лимонной кислотой (рН 4,0) и полученный раствор обрабатывают при 30°С на протяжении 30 часов (обработка кислотой) с перемешиванием. После обработки 780 мкл этой смеси центрифугируют для выделения бактериальных клеток, бактериальные клетки суспендируют в 780 мкл 54 мМ водного раствора трис-соляной кислоты (рН 8,0), добавляют 20 мкл акрилонитрила и полученную смесь подвергают взаимодействию на протяжении 15-60 минут с осторожным перемешиванием при 20°С. По завершении взаимодействия проводят анализ реакционного раствора с применением ВЭЖХ для измерения количества (Q) полученного амидного соединения (или оно может быть рассчитано по количеству потребленного нитрильного соединения). Для каждого условия взаимодействие и анализ проводят по 3 раза или более. Коэффициент (R) получают делением количества (Q) амидного соединения, полученного с использованием нитрилгидратазы после обработки кислотой, к количеству (Р) амидного соединения, полученного с использованием нитрилгидратазы до обработки кислотой. Коэффициент R определяют для нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, и для анализируемой нитрилгидратазы, соответственно, и определяют отношение (анализируемый объект/дикий тип) значения R анализируемой нитрилгидратазы к значению R нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila. Если это отношение превышает 1,0, то рН-стабильность выше, чем у нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Мутантная нитрилгидратаза по изобретению имеет значение R, превышающее соответствующее значение нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, и, предпочтительно, значение R мутантной нитрилгидратазы по изобретению в 1,1 раза или более, более предпочтительно, в 1,2 раза или более, еще более предпочтительно, в 1,3 раза или более, еще более предпочтительно, в 1,4 раза или более превышает значение R нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Кроме того, как описано выше, мутантная нитрилгидратаза представляет собой фермент с отличной рН-стабильностью, то есть мутантная нитрилгидратаза демонстрирует ферментативную активность в широком диапазоне рН, по сравнению с традиционной мутантной нитрилгидратаз ой, применительно к реакции синтеза амидного соединения из нитрильного соединения. В результате рассмотрения авторами изобретения тех факторов, которые повышают рН-стабильность ферментативной активности нитрилгидратазы при заменах аминокислотных остатков (а)-(е), описанных выше, было обнаружено, что все аминокислоты, вовлеченные в замены аминокислотных остатков (а)-(е), расположены вблизи субстратного кармана фермента. Предполагается, что при введении одной или более чем одной из замен аминокислотных остатков (а)-(е), описанных выше, трехмерная (свернутая) структура белка хорошо сохраняется в широком диапазоне рН. Поэтому замены аминокислотных остатков (а)-(е) рассматриваются как группа замен аминокислотных остатков (замены аминокислотных остатков группы А).
Кроме того, в случае мутантной нитрилгидратазы происходит образование димеров, в которых α-субъединица и β-субъединица объединены, образуя основную структурную единицу нитрилгидратазы, а димеры объединены далее с образованием тетрамеров. Цистеиновый остаток, являющийся 111-й аминокислотой от N-конца α-субъединицы, подвергается посттрансляционной модификации с образованием цистеинсульфиновой кислоты (Cys-SOOH), а цистеиновый остаток, являющийся 113-й аминокислотой от N-конца α-субъединицы, подвергается посттрансляционной модификации с образованием цистеинсульфеновой кислоты (Cys-SOH), соответственно, и полипептидная цепь α-субъединицы связывается с атомом кобальта через эти модифицированные аминокислотные остатки, образуя активный центр.
Могут быть использованы плазмида, способная экспрессировать нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, в достаточном количестве, и клеточной линия, трансформированная указанной выше плазмидой, МТ-10822 (регистрационный номер FERM ВР-5785, депонированная в Международном депозитарии патентованных микроорганизмов (International Patent Organism Depositary, NITE-IPOD, 1-1-1 Higashi, Tsukuba, Ibaraki), 7 февраля 1996 г.). Кроме того, модифицированая нитрилгидратаз а может быть получена с применением традиционной генно-манипуляционной технологии. При высокой степени гомологии аминокислотных последовательностей высока вероятность сходства трехмерной структуры белков, независимо от рода и вида штамма, содержащего белок, и, таким образом, одна или более чем одна из замен аминокислотных остатков (а)-(е) оказывают эффект на нитрилгидратазу, имеющую происхождение из Pseudonocardia thermophila, как определено выше, в целом, включая модифицированную нитрилгидратазу. Кроме того, полученная мутантная нитрилгидратаза может обладать более высокой рН-стабильностью, чем до введения одной или более чем одной из замен аминокислотных остатков (а)-(е), где рН-стабильность оценивают по ферментативной активности при рН 4,0 в сравнении с ферментативной активностью при рН 8,0.
Кроме того, мутантная нитрил гидр атаза содержит по меньшей мере одну из замен аминокислотных остатков (а)-(е). Мутантная нитрилгидратаза может содержать только одну из замен аминокислотных остатков (а)-(е) или может содержать две или более замены аминокислотных остатков. Например, примеры комбинаций из двух замен аминокислотных остатков могут включать комбинацию (а) и (b), комбинацию (а) и (с), комбинацию (а) и (d), комбинацию (а) и (е), комбинацию (b) и (с), комбинацию (b) и (d), комбинацию (b) и (е), комбинацию (с) и (d), комбинацию (с) и (е), комбинацию (d) и (е) и комбинацию (d) и (f).
Примеры комбинаций из трех замен аминокислотных остатков могут включать комбинацию (а), (b) и (с), комбинацию (а), (b) и (d), комбинацию (а), (b) и (е), комбинацию (а), (с) и (d), комбинацию (а), (с) и (е), комбинацию (a), (d) и (е), комбинацию (b), (с) и (d), комбинацию (b), (с) и (е), комбинацию (b), (d) и (е) и комбинацию (с), (d) и (е).
Примеры комбинаций из четырех замен аминокислотных остатков могут включать комбинацию (а), (b), (с) и (d), комбинацию (а), (b), (с) и (е), комбинацию (а), (b), (d) и (е), комбинацию (а), (с), (d) и (е) и комбинацию (b), (с), (d) и (е).
Возможна комбинация из пяти замен аминокислотных остатков (а)-(е).
В одном воплощении мутантная нитрилгидратаза содержит одну или более чем одну из замен аминокислотных остатков, выбранных из группы, состоящей из указанной выше замены аминокислотного остатка (b) и замен аминокислотных остатков (а) и (с)-(е). Одна или более чем одна из замен аминокислотных остатков в группе, состоящей из замен аминокислотных остатков (а) и (с)-(е), вместе с указанной выше заменой аминокислотного остатка (b) может представлять собой замену двух аминокислотных остатков, замену трех аминокислотных остатков или замену четырех аминокислотных остатков (то есть все из (а) и (с)-(е)). Кроме того, в этом воплощении мутантная нитрилгидратаза может дополнительно иметь описанную выше замену аминокислотного остатка (f).
Как указано выше, модифицированная нитрилгидратаза, в которую предполагается вводить по меньшей мере одну из замен аминокислотных остатков (а)-(е), может содержать модификацию, такую как аминокислотная замена или тому подобное, относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, или конкретных последовательностей (1)-(5), описанных выше. Таким образом, мутантная нитрилгидратаза может содержать не только замены аминокислотных остатков (а)-(е), но также замену аминокислотного остатка в положении, отличном от замен аминокислотных остатков (а)-(е), по сравнению с нитрилгидратазой дикого типа, имеющей происхождение из Pseudonocardia thermophila, или конкретными последовательностями (1)-(5), описанными выше. Как описано выше, мутантная нитрилгидратаза, содержащая другую замену аминокислотного остатка, может также обладать эффектом, полученным посредством замен аминокислотных остатков группы А, занимающих определенное положение в трехмерной структуре.
В одном воплощении мутантная нитрилгидратаза по изобретению обладает не только повышенной рН-стабильностью, но также повышенной начальной скоростью реакции получения амидного соединения из нитрильного соединения. Иными словами, введение по меньшей мере одной из замен аминокислотных остатков (a)-(e) в нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, или модифицированную нитрилгидратазу приводит к получению нитрилгидратазы, демонстрирующей более высокую начальную скорость реакции, чем до введения замены. Начальную скорость реакции можно, например, определять, суспендируя 40 мкл раствора, полученного по завершении культивирования транс фор манта, продуцирующего нитрилгидратазу, в 740 мкл 54 мМ водного раствора трис-соляной кислоты (рН 8,0), добавляя 20 мкл акрилонитрила, подвергая полученную смесь взаимодействию на протяжении 15-60 минут с осторожным перемешиванием при 20°С и анализируя реакционный раствор с применением ВЭЖХ по завершении взаимодействия. Кроме того, мутантная нитрилгидратаза по изобретению предпочтительно демонстрирует более высокую начальную скорость реакции при рН 8,0, которая, более предпочтительно, в 1,1 раза или более, более предпочтительно, в 1,2 раза или более, более предпочтительно, в 1,3 раза или более и, более предпочтительно, в 1,4 раза или более превышает начальную скорость реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Мутантная нитрилгидратаза может быть получена следующим способом.
Например, может быть получен вектор экспрессии, содержащий ДНК, кодирующую мутантную нитрилгидратазу, произвольная клетка-хозяин может быть трансформирована этим вектором экспрессии с получением трансформанта или трансформированной клеточной линии, и затем трансформант или клеточная линия могут быть культивированы с получением мутантной нитрилгидратазы.
Здесь ген, кодирующий нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, состоит из нуклеотидной последовательности, показанной в SEQ ID NO: 3, и нуклеотидной последовательности, показанной в SEQ ID NO: 4. Кроме того, нуклеотидная последовательность, представленная как SEQ ID NO: 3, соответствует аминокислотной последовательности, состоящей из SEQIDNO:l, а нуклеотидная последовательность, представленная как SEQ ID NO: 4, соответствует аминокислотной последовательности, состоящей из SEQ ID NO: 2.
Более того, ДНК, кодирующая мутантную нитрилгидратазу, имеет по меньшей мере следующие нуклеотидные замены в одном или более чем одном кодоне, соответствующих положениям указанных выше замен аминокислотных остатков (a)-(e) (всего 5 частей), в нуклеотидной последовательности, представленной как SEQ ID NO: 3, или в нуклеотидной последовательности, представленной как SEQ ID NO: 4, а также нуклеотидную замену, соответствующую модифицированной нитрилгидратазе, в других положениях.
Конкретно, например, в случае получения мутанта, где остаток Asp, являющийся 40-м аминокислотным остатком от N-конца α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Asn, используют ДНК, где GAC, представляющий собой нуклеотиды в положениях 118-120 от 5'-конца нуклеотидной последовательности, представленной как SEQ ID NO: 3, заменен на ААС или ААТ.
В случае получения мутанта, где остаток Ala, являющийся 43-м аминокислотным остатком от N-конца α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Val, используют ДНК, где GCC, представляющий собой нуклеотиды в положениях 127-129 от 5'-конца нуклеотидной последовательности, представленной как SEQ ID NO: 3, заменен на GTT, GTC, GTA или GTG.
В случае получения мутанта, где остаток Gly, являющийся 205-м аминокислотным остатком от N-конца β-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Val, используют ДНК, где GGG, представляющий собой нуклеотиды в положениях 613-615 от 5'-конца нуклеотидной последовательности, представленной как SEQ ID NO: 4, заменен на GTT, GTC, GTA или GTG
В случае получения мутанта, где остаток Pro, являющийся 206-м аминокислотным остатком от N-конца β-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Gin, используют ДНК, где GGG, представляющий собой нуклеотиды в положениях 616-618 от 5'-конца нуклеотидной последовательности, представленной как SEQ ID NO: 4, заменен на САА или CAG.
В случае получения мутанта, где остаток Туг, являющийся 215-м аминокислотным остатком от N-конца α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Asn, используют ДНК, где ТАС, представляющий собой нуклеотиды в положениях 643-645 от 5'-конца нуклеотидной последовательности, представленной как SEQ ID NO: 4, заменен на ААС или ААТ.
Кроме того, применительно к замене аминокислотного остатка (f), в случае получения мутанта, где остаток Thr, являющийся 36-м аминокислотным остатком от N-конца α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Trp, используют ДНК, где ACG, представляющий собой нуклеотиды в положениях 106-108 от 5'-конца нуклеотидной последовательности, представленной как SEQ ID NO: 3, заменен на TGG.
Вектор экспрессии содержит ген, кодирующий α-субъединицу мутантной нитрилгидратазы, и ген, кодирующий β-субъединицу мутантной нитрилгидратазы, а также, по необходимости, элементы, позволяющие трансформанту или клеточной линии продуцировать мутантную нитрилгидратазу, такие как регуляторная область, необходимая для экспрессии каждого гена, область, необходимая для автономной репликации, и тому подобное. Клетка-хозяин, в которую вводят вектор экспрессии, может представлять собой любую клетку, например, Escherichia coli.
Регуляторные области, необходимые для экспрессии, могут включать промоторную последовательность (включая операторную последовательность для контроля транскрипции), последовательность связывания рибосом (SD-последовательность), последовательность терминации транскрипции и тому подобное. Конкретные примеры промоторной последовательности могут включать trp-промотор триптофанового оперона, имеющий происхождение из Е. coli, lac-промотор лактозного оперона, PL-промотор и PR-промотор, имеющие происхождение от фага лямбда, и тому подобное. Кроме того, может быть использована искусственно разработанная и модифицированная последовательность, такая как tac-промотор или trc-промотор.
Предпочтительной последовательностью связывания рибосом является TAAGGAGGT, включенная в SEQ ID NO: 74. Что касается порядка расположения последовательностей в регуляторной области вектора экспрессии, предпочтительно, чтобы промоторная последовательность и последовательность связывания рибосом были расположены ближе к 5'-концу относительно гена, кодирующего мутантную нитрилгидратазу, а последовательность терминации транскрипции - ближе к 3'-концу относительно гена, кодирующего мутантную нитрилгидратазу. Кроме того, в зависимости от регуляторной области, возможна экспрессия гена α-субъединицы и гена β-субъединицы мутантной нитрилгидратазы в форме независимых цистронов, соответственно, или в форме полицистрона с общей регуляторной областью.
Примеры вектора, удовлетворяющего описанным выше требованиям, могут включать pBR322, pUC18, pBluescript, pKK223-3 и pSC101, содержащие область, автономно реплицируемую в Е. coli.
В качестве метода конструирования вектора экспрессии по изобретению введением гена, кодирующего мутантную нитрилгидратазу, в вектор вместе с областью, необходимой для экспрессии активности мутантной нитрилгидратазы, метода трансформации желаемой клетки-хозяина вектором экспрессии, метода продукции нитрилгидратазы трансформантом и тому подобного могут быть применены обычные методы или клетки-хозяева, известные в области молекулярной биологии, биотехнологии и генной инженерии, такие как описанные в "Molecular Cloning 3rd Edition" (J. Sambrook, and the like; Cold Spring Harbor Laboratory Press, 2001) и тому подобные.
Трансформанта, полученного трансформацией желаемой клетки-хозяина описанным выше вектором экспрессии, можно культивировать в среде с получением мутантной нитрилгидратазы на основе гена нитрилгидратазы, введенного в вектор экспрессии. Когда клетка-хозяин представляет собой Е. coli, в качестве среды для культивирования трансформанта обычно используют среду LB, среду М9 или тому подобное. Более предпочтительно, среда содержит, в качестве своих компонентов, ионы Fe и ионы Со в количестве 0,1 мкг/мл. Трансформант можно вносить в среду и выращивать при подходящей температуре культивирования (обычно от 20°С до 50°С).
При получении мутантной нитрилгидратазы, обладающей желаемой ферментативной активностью, экспрессией гена, кодирующего мутантную нитрилгидратазу, возможна также экспрессия гена, кодирующего белок, вовлеченный в активацию нитрилгидратазы.
Белок, вовлеченный в активацию нитрилгидратазы, представляет собой белок, экспрессия или отсутствие экспрессии которого непосредственно влияет на активацию нитрилгидратазы, и может включать, в качестве типичного примера, белок (белок, активирующий нитрилгидратазу), вовлеченный в активацию нитрилгидратазы, имеющей происхождение из Pseudonocardia thermophila, описанной в JP-A № H11-253168. Последовательность белка, активирующего нитрилгидратазу, представлена как SEQ ID NO: 75.
Нуклеиновая кислота
Нуклеиновая кислота по изобретению имеет нуклеотидную последовательность, кодирующую аминокислотную последовательность мутантной нитрилгидратазы.
Что касается метода синтеза нуклеотидной последовательности, кодирующей аминокислотную последовательность мутантной нитрилгидратазы, то существуют метод введения точковой мутации в нуклеотидную последовательность, кодирующую соответствующую нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, метод химического синтеза всей нуклеотидной последовательности, включая точковую мутацию, и тому подобные. В качестве метода генерирования мутации в гене с использованием нуклеотидной последовательности, кодирующей нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila, в качестве матрицы могут быть включены, например, метод сайт-специфического мутагенеза (Kramer, W. and frita, Н. J., Methods in Enzymology, vol. 154, P. 350(1987)), метод рекомбинантной ПЦР (PCR Technology, Stockton Press(1989), метод химического синтеза определенной части ДНК, метод обработки гена гидр оке ил амином, метод облучения штамма, содержащего ген, УФ светом, метод обработки гена химическим агентом, таким как нитрозогуанидин, азотистая кислота или тому подобное, метод с использованием имеющегося в продаже набора для мутагенеза и тому подобные. Нуклеиновая кислота по изобретению может представлять собой ДНК или РНК.
Вектор
Конкретных ограничений относительно вектора по изобретению нет, при условии, что он представляет собой вектор, содержащий нуклеиновую кислоту, представленную нуклеотидной последовательностью, кодирующей аминокислотную последовательность мутантной нитрилгидратазы, и он может включать, например, вектор, полученный введением мутантной нитрилгидратазы в известный вектор. Кроме того, вектор может представлять собой фаговый вектор или плазмидный вектор.
Вектор экспрессии
Конкретных ограничений относительно вектора экспрессии по изобретению нет, при условии, что он представляет собой вектор экспрессии, содержащий нуклеиновую кислоту, представленную нуклеотидной последовательностью, кодирующей аминокислотную последовательность мутантной нитрилгидратазы, но плазмидный вектор или фаговый вектор, имеющий показанную ниже структуру, более предпочтителен с точки зрения повышения эффективности трансформации, эффективности трансляции или тому подобного.
Общая структура вектора экспрессии
Вектор экспрессии содержит нуклеотидную последовательность, кодирующую мутантную нитрилгидратазу, и относительно него нет конкретных ограничений, при условии, что вектор экспрессии способен трансформировать клетку-хозяина. По необходимости, вектор экспрессии может содержать, помимо этой нуклеотидной последовательности, нуклеотидную последовательность, составляющую другую область (далее называемую здесь просто «другой областью»). Примеры другой области могут включать регуляторную область, необходимую для того, чтобы трансформант продуцировал мутантную нитрилгидратазу, область, необходимую для автономной репликации, и тому подобное.
Кроме того, с точки зрения облегчения селекции трансформанта, вектор экспрессии может дополнительно содержать нуклеотидную последовательность, кодирующую селекционный ген, который может быть селективным маркером.
Регуляторные области, необходимые для продуцирования мутантной нитрилгидратазы, могут включать промоторную последовательность (включая операторную последовательность для контроля транскрипции), последовательность связывания рибосом (SD-последовательность), последовательность терминации транскрипции и тому подобное.
Вектор экспрессии при использовании дрожжей в качестве клетки-хозяина При использовании дрожжей в качестве клетки-хозяина предпочтительно, чтобы вектор экспрессии содержал промоторную последовательность, помимо нуклеотидной последовательности, кодирующей мутантную нитрилгидратазу. В качестве промоторной последовательности может быть использована любая промоторная последовательность, при условии, что она может обеспечить экспрессию мутантной нитрилгидратазы в трансформанте при использовании дрожжей в качестве клетки-хозяина.
Например, используют такие промоторные последовательности, как промотор алкогольдегидрогеназы (ADH1), промотор фосфоглицераткиназы (PGK1), промотор фактора элонгации пептидной цепи (TEF), промотор глицерол-3-фосфатдегидрогеназы (GPD), промотор галактокиназы (GAL1), промотор металлотионеина (CUP1), промотор ингибирующей кислой фосфатазы (PHO5), промотор глицеральдегид-3-фосфатдегидрогеназы (GAPDH) и тому подобное.
Следует отметить, что источник промоторной последовательности не ограничен дрожжами, используемыми в качестве клетки-хозяина.
Может быть использован экзогенный промотор, такой как промотор цитоме гало вирус a (CMV) или тому подобные. Они могут быть надлежащим образом выбраны, исходя из источника и типа используемого фермента.
Кроме того, вектор экспрессии может содержать секреторный сигнал. В результате, когда трансформант продуцирует мутантную нитрилгидратазу, возможна ее секреция за пределы клетки.
Конкретных ограничений относительно секреторного сигнала нет, при условии, что мутантная нитрилгидратаза может быть секретирована из дрожжей, используемых в качестве клетки-хозяина. С точки зрения эффективности секреции предпочтительно использовать сигнальную последовательность α-фактора, сигнальную последовательность инвертазы, сигнальную последовательность кислой фосфатазы, сигнальную последовательность глюкоамилазы и тому подобное.
Конкретные примеры векторов экспрессии, содержащих промоторную последовательность или секреторный сигнал, как описано выше, могут включать pRS423, pRS424, YEplac195 и тому подобное.
Вектор экспрессии при использовании нитчатых грибов в качестве клетки-хозяина
При использовании нитчатых грибов в качестве клетки-хозяина предпочтительно, чтобы вектор экспрессии содержал промоторную последовательность, помимо нуклеотидной последовательности, кодирующей мутантную нитрилгидратазу. В качестве промоторной последовательности может быть использована любая промоторная последовательность, при условии, что она может обеспечить экспрессию мутантной нитрилгидратазы в трансформанте при использовании нитчатых грибов в качестве клетки-хозяина.
Векторы экспрессии, подходящие для нитчатых грибов, описаны в van den Hondel, С. А. М. J. J. et al. (1991) In: Bennett, J. W. and Lasure, L. L. (eds.) More gene Manipulations in Fungi. Academic Press, pp. 396-428.
Кроме того, могут быть использованы другие широко применяемые векторы экспрессии, такие как pUC18, pBR322, pUC100, pSL1180 (изготавливаемый Pharmacia Inc.), pFB6, Aspergillus pRAX, Trichoderma pTEX и тому подобные.
Вектор экспрессии при использовании прокариот в качестве клетки-хозяина
При использовании прокариот, таких как Е. coli, Bacillus subtilis, Actinomycetes или тому подобные, в качестве клетки-хозяина предпочтительно, чтобы вектор экспрессии содержал промоторную последовательность, помимо нуклеотидной последовательности, кодирующей мутантную нитрилгидратазу. Кроме того, вектор экспрессии может содержать, помимо промоторной последовательности, последовательность связывания рибосом, последовательность терминации транскрипции или тому подобное.
Примеры промоторной последовательности могут включать trp-промотор триптофанового оперона, имеющий происхождение из Е. coli, lac-промотор лактозного оперона, PL-промотор и PR-промотор, имеющие происхождение от фага лямбда, промотор глюконатсинтазы (gnt), имеющий происхождение из Bacillus subtilis, промотор щелочной протеазы (apr), промотор нейтральной протеазы (npr), промотор α-амилазы (amy) и тому подобное.
Кроме того, может также быть использована независимо модифицированная или разработанная промоторная последовательность, такая как tac-промотор.
В качестве последовательности связывания рибосом могут быть включены последовательности, имеющие происхождение из Escherichia coli или Bacillus subtilis, однако конкретных ограничений нет, при условии, что последовательность связывания рибосом представляет собой последовательность, выполняющую свои функции в желаемой клетке-хозяине, такой как Escherichia coli, Bacillus subtilis или тому подобные.
Например, в качестве последовательности связывания рибосом, из последовательностей, комплементарных 3'-концевой области 168-рибосомальной РНК, может быть включена последовательность, полученная ДНК-синтезом консенсусной последовательности протяженностью 4 нуклеотида или более и тому подобные.
Наличие последовательности терминации транскрипции не обязательно, но могут быть использованы последовательности, независимые от β-факторов, такие как терминатор липопротеина, терминатор trp-оперона и тому подобные.
Конкретных ограничений относительно порядка расположения последовательностей этих регуляторных областей в векторе экспрессии нет, но, из соображений эффективности транскрипции, предпочтительно, чтобы были расположены в следующем порядке от 5'-конца: промоторная последовательность; последовательность связывания рибосом; ген, кодирующий целевой белок; и последовательность терминации транскрипции.
Конкретные примеры вектора экспрессии могут включать pBR322, pUC18, Bluescript II SK(+), pKK223-3, pSC101, содержащие область, автономно реплицируемую в Е. coli, и pUB110, pTZ4, рС194, ρ11, ϕ1, ϕ105 и тому подобные, содержащие область, автономно реплицируемую в Bacillus subtilis, которые могут быть использованы в качестве вектора экспрессии.
Кроме того, как пример вектора экспрессии, способного к автономной репликации в двух или более видах хозяев, в качестве вектора экспрессии могут быть использованы pHV14, TRp7, YEp7, pBS7 и тому подобные.
Метод получения трансформанта
Трансформант по изобретению может быть получен известным методом. Например, может быть включен метод, где конструируют вектор экспрессии, содержащий нуклеотидную последовательность, кодирующую мутантную нитрилгидратазу по изобретению, и, по необходимости, другую область, и полученным вектором экспрессии трансформируют желаемую клетку-хозяина, и тому подобные. Конкретно, могут быть применены обычные методы, известные в области молекулярной биологии, биоинженерии и генной инженерии, описанные в Sambrook, J., et. al., "Molecular Cloning A Laboratory Manual, 3rd Edition", Cold Spring Harbor Laboratory Press, (2001), и тому подобные.
Трансформанта по изобретению можно конструировать не только введением вектора экспрессии в клетку-хозяина, но также, по необходимости, введением молчащей мутации таким образом, что кодон, редко используемый в клетке-хозяине, превращается в часто используемый кодон, и тому подобным.
Это может приводить к увеличению количества продуцируемого белка, имеющего происхождение из мутантной нитрилгидратазы, включенной в вектор экспрессии.
Применительно к введению молчащей мутации, в случае, когда ее введение является методом корректировки гена нитрилгидратазы в векторе экспрессии и кодонов сигнальной последовательности внеклеточной секреции гена нитрилгидратазы в соответствии с частотой использования кодонов в клетке-хозяине, конкретных ограничений по методу введения молчащей мутации, точке мутации, типу изменяемого нуклеотида и тому подобному нет.
Способ получения мутантной нитрилгидратазы
Способ получения мутантной нитрилгидратазы по изобретению включает культивирование трансформанта в среде и выделение мутантной нитрилгидратазы из по меньшей мере одного из культивированного трансформанта и среды.
Метод культивирования трансформанта
Условия культивирования трансформанта, полученного трансформацией вектором экспрессии, идентичны условиям культивирования клетки-хозяина до трансформации, и могут быть использованы известные условия.
В качестве среды может быть использована синтетическая среда и натуральная среда при условии, что среда содержит нужное количество источника углерода, источника азота, неорганических веществ и других питательных веществ. В качестве компонентов среды могут быть использованы известные компоненты. Например, могут быть использованы, в подходящей комбинации, органические питательные вещества, такие как мясной экстракт, дрожжевой экстракт, солодовый экстракт, пептон, NZ-амин и картофель, источники углерода, такие как глюкоза, мальтоза, сахароза, крахмал, органические кислоты и тому подобное, источники азота, такие как сульфат аммония, мочевина, хлорид аммония и тому подобное, неорганические питательные вещества, такие как соли фосфорной кислоты, магний, калий, железо и тому подобное, и витамины.
Кроме того, при культивировании трансформанта, трансформированного вектором экспрессии, содержащим селективный маркер, например, в случае, когда селективный маркер придает резистентность к какому-либо лекарственному средству, используют среду, содержащую соответствующее лекарственное средство, в случае, когда селективный маркер придает способность расти в отсутствие какого-либо питательного вещества, используют среду без соответствующего питательного вещества. рН среды может быть выбран так, чтобы входить в диапазон от рН 4 до рН 8.
Культивирование трансформанта можно проводить в жидкой среде, содержащей описанную выше среду, с применением традиционных методов культивирования, таких как культивирование с перемешиванием, культивирование с перемешиванием и аэрацией, постоянная культура, подпитываемая культура и тому подобное.
Условия культивирования могут быть надлежащим образом подобраны с учетом типа трансформанта, культуральной среды и метода культивирования, без каких-либо конкретных ограничений при условии, что они поддерживают рост трансформанта и его способность продуцировать мутантную нитрилгидратазу по изобретению.
Культивирование проводят в аэробных условиях при температуре от 20°С до 45°С и, предпочтительно, от 24°С до 37°С.
Продолжительность периода культивирования до достижения максимального содержания белка, обладающего желаемой активностью мутантной нитрилгидратазы, может составлять от 1 суток до 7 суток.
Стадия выделения мутантной нитрилгидратазы
Стадия выделения мутантной нитрилгидратазы представляет собой стадию выделения мутантной нитрилгидратазы из по меньшей мере одного из культивированного трансформанта и среды после культивирования.
В качестве метода выделения мутантной нитрилгидратазы по изобретению после культивирования трансформированного трансформанта может быть применен метод, широко применяемый в данной области.
В случае секреции мутантной нитрилгидратазы по изобретению за пределы трансформированного трансформанта, раствор неочищенного фермента можно легко получить, проводя центрифугирование, фильтрацию культуры трансформанта или тому подобное. Кроме того, в случае накопления мутантной нитрилгидратазы по изобретению в трансформированном трансформанте, культивированный трансформант выделяют таким методом, как центрифугирование или тому подобное, выделенный трансформант суспендируют в буфере и проводят лизис клеточной мембраны трансформанта с применением известного метода, такого как обработка лизоцимом, замораживание-оттаивание, обработка ультразвуком или тому подобное, получая посредством этого раствор неочищенного фермента.
Раствор неочищенного фермента можно использовать в качестве концентрированного фермента, концентрируя его ультрафильтрацией или тому подобным и добавляя в него консервант или тому подобное. Кроме того, после концентрирования можно получить порошок фермента мутантной нитрилгидратазы, применяя метод распылительной сушки или тому подобное.
В случае необходимости разделения и очистки раствора неочищенного фермента, обладающего активностью выделяемой нитрилгидратазы, могут быть применены, например, высаливание сульфатом аммония или тому подобное, метод преципитации с органическим растворителем с использованием спирта или тому подобное, метод мембранного разделения, такой как диализ, ультрафильтрация или тому подобное, или известный метод хроматографического разделения, такой как ионообменная хроматография, высокоскоростная хроматография с обращенной фазой, аффинная хроматография, гель-фильтрационная хроматография или тому подобное, в подходящей комбинации.
Способ получения амидного соединения по изобретению включает приведение мутантной нитрилгидратазы в контакт с нитрильным соединением. Мутантная нитрилгидратаза катализирует реакцию синтеза амидного соединения из нитрильного соединения. Способ получения амидного соединения описан ниже более подробно.
Сначала культивируют трансформант или клеточную линию, продуцирующие мутантную нитрилгидратазу, и полученный культуральный раствор, клетки или обработанные клеточные продукты приводят в контакт с нитрильным соединением в среде. Таким образом, мутантную нитрилгидратазу приводят в контакт с нитрильным соединением, и мутантная нитрилгидратаза превращает нитрильное соединение в соответствующее амидное соединение. При использовании здесь термин «обработанный клеточный продукт» относится к экстракту или измельченному продукту из трансформанта, продукту выделения из необработанного ферментного препарата, полученному выделением активных нитрилгидратазных фракций экстракта или измельченного продукта, или очищенному ферментному продукту, полученному дальнейшей очисткой или тому подобным, или иммобилизованному продукту, полученному иммобилизацией трансформанта или экстракта, измельченного продукта или продукта выделения из трансформанта с применением подходящего метода.
Температура описанного выше приведения мутантной нитрилгидратазы в контакт с нитрильным соединением предпочтительно входит в диапазон температур, при которых не происходит инактивации мутантной нитрилгидратазы, более предпочтительно, от 0°С до 60°С и, более предпочтительно, от 15°С до 35°С. Например, культуральный раствор, полученный культивированием трансформанта или клеточной линии, продуцирующих мутантную нитрилгидратазу, может быть непосредственно добавлен в водный раствор, содержащий нитрильное соединение, или культуральный раствор может быть центрифугирован для выделения бактериальных клеток, и бактериальные клетки могут быть добавлены в водный раствор, содержащий нитрильное соединение. рН водного раствора во время реакции предпочтительно составляет от 7 до 9 и, более предпочтительно, от 7,5 до 8,5. Нитрильное соединение может присутствовать в водном растворе в концентрации, например, от 0,25% до 20,0% по объему и, более предпочтительно, от 2,0% до 5,0% по объему. Подробности о культивировании трансформанта идентичны описанным в разделе, посвященном способу получения мутантной нитрилгидратазы.
Конкретных ограничений относительно примеров нитрильного соединения нет, при условии, что оно представляет собой соединение, которое может быть субстратом мутантной нитрилгидратазы. Предпочтительно, в качестве типичного примера, нитрильное соединение может включать нитрильное соединение, имеющее от 2 до 4 атомов углерода, такое как ацетонитрил, пропионитрил, акрилонитрил, метакрилонитрил, н-бутиронитрил, изобутиронитрил, кротононитрил, α-гидроксиизобутиронитрил или тому подобное. Поскольку нитрильная группа нитрильного соединения превращается в амидную группу посредством гидратации, акрилонитрил, например, может быть превращен в акриламид.
Затем, добавляя активированный уголь в среду без дезактивации мутантной нитрилгидратазы, проводят очистку полученного амидного соединения от нитрильного соединения с использованием мутантной нитрилгидратазы в качестве катализатора без прекращения реакции синтеза амидного соединения.
Кроме того, реакцию получения амидного соединения из нитрильного соединения в результате приведения его в контакт с мутантной нитрилгидратазой проводят в условиях, соответствующих рН-оптимуму нитрилгидратазы, например, в нейтральных или основных условиях, таких как рН от 7 до 9. По необходимости, рН можно корректировать, например, с использованием основания, такого как аммиак или гидроксид натрия в буфере. В то же время, очистку амидного соединения предпочтительно проводят в кислых условиях, таких как, например, рН от 3,5 до 6,5. Это нужно для эффективного удаления примесей, в частности белков, присутствующих в жидкости, содержащей амидное соединение. рН может быть изменен в кислую сторону с использованием кислоты, такой как уксусная кислота, пропионовая кислота, октановая кислота, валериановая кислота, соляная кислота, акриловая кислота, метакриловая кислота или тому подобное. Таким образом, способ получения амидного соединения предпочтительно включает стадию очистки, на которой происходит удаление примесей из раствора, содержащего полученное амидное соединение, в условиях рН от 3,5 до 6,5. На этой стадии для удаления примесей предпочтительно использовать активированный уголь. То есть, в одном воплощении способ получения амидного соединения по изобретению дополнительно включает очистку амидного соединения с использованием активированного угля. Таким образом, предпочтительно, чтобы ферментативная активность мутантной нитрилгидратазы была выше, чем до введения по меньшей мере одной из замен аминокислотных остатков (а)-(е), предпочтительно в условиях рН от 3,5 до 6,5. рН может составлять 5,0 или менее для более эффективного удаления ненужных белков.
Как описано выше, мутантная нитрилгидратаза по изобретению позволяет достичь высокой эффективности превращения на этапе получения амидного соединения из нитрильного соединения, благодаря повышенной рН-стабильности. Несмотря на эти воплощения, описанные выше, они приведены в качестве примеров, и возможны различные формы изобретения, отличные от описанных выше.
ПРИМЕРЫ
Воплощение изобретения описано более подробно посредством следующих Примеров, но настоящее изобретение никоим образом не ограничено этими Примерами. Кроме того, в следующих Примерах термин «сайт мутации» относится к положению, в котором рассматриваемая аминокислотная последовательность отличается от нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, которая имеет α-субъединицу, имеющую аминокислотную последовательность, представленную как SEQIDNO:l, и β-субъединицу, имеющую аминокислотную последовательность, представленную как SEQ ID NO: 2.
Сравнительный пример 1. Получение нитрилгидратазы (1) дикого типа, имеющей происхождение из Pseudonocardia thermophila
Подготавливали 10 мл жидкой среды LB в пробирке объемом 30 мл и жидкую среду стерилизовали автоклавированием при 121°С на протяжении 20 минут. В жидкую среду добавляли ампициллин до конечной концентрации 100 мкг/мл, после чего вносили одну платиновую петлю клеточной линии МТ-10822 и проводили культивирование при 37°С и 300 об/мин на протяжении приблизительно 20 часов. Затем 1 мл полученного культурального раствора разделяли на фракции в подходящей центрифужной пробирке и бактериальные клетки выделяли центрифугированием (15000 об/мин × 5 минут).
После этого из бактериальных клеток, полученных при выделении, получали плазмиду pPT-DB1 с применением метода щелочной экстракции с SDS. Полученной плазмидой трансформировали компетентные клетки Escherichia coli НВ101 (Toyobo Co., Ltd.) с получением трансформанта (1). Трансформант (1) продуцирует нитрилгидратазу (1), представляющую собой нитрилгидратазу дикого типа, имеющую происхождение из Pseudonocardia thermophila.
Сравнительный пример 2. Получение мутантной нитрилгидратазы (2)
Для получения трансформанта (2), экспрессирующего мутантную нитрилгидратазу, где Asp, являющийся 92-м аминокислотным остатком от N-конца α-субъединицы нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, заменен на Glu, как показано в Таблице 14, проводили сайт-специфический мутагенез с использованием набора «LA PCR in vitro mutagenesis Kit» (называемого далее набором для мутагенеза), изготовленного Takara Shuzo Co., Ltd. Проводили ПЦР с использованием плазмиды экспрессии нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, pPT-DB1 в качестве матрицы.
ПЦР №1 проводили, повторяя 25 циклов тепловой денатурации (98°С) на протяжении 15 секунд, отжига (55°С) на протяжении 30 секунд и реакции элонгации (72°С) на протяжении 120 секунд в системе (состав соответствует условиям, описанным в наборе для мутагенеза) общим объемом 50 мкл, содержащей по 50 пмоль праймера SEQ ID NO: 5 и праймера М13 М4 (последовательность представлена как SEQ ID NO: 12).
ПЦР №2 проводили так же, как ПЦР№1, в системе (состав соответствует условиям, описанным в наборе для мутагенеза) общим объемом 50 мкл, содержащей по 50 пмоль праймера MUT4 (последовательность представлена как SEQIDNO: 14) и праймера М13 RV (последовательность представлена как SEQ ID NO: 13).
По результатам, полученным при проведении анализа продукта амплификации ДНК посредством электрофореза в агарозном геле (концентрация агарозы 1,0 масс. %) с использованием 5 мкл ПЦР-раствора после завершения ПЦР №1 и №2, можно было подтвердить присутствие амплифицированного ДНК-продукта.
Из каждого ПЦР-раствора после завершения реакции удаляли избыток праймера и dNTP с использованием Microcon 100 (Takara Shuzo Co., Ltd.) и затем добавляли ТЕ с получением 50 мкл каждого раствора. Подготавливали в общей сложности 47,5 мкл раствора для отжига (состав основан на условиях, описанных в наборе для мутагенеза), содержащего по 0,5 мкл каждого ТЕ-раствора, и проводили его тепловую денатурацию (98°С) на протяжении 10 минут. Затем смесь охлаждали с постоянной скоростью до 37°С на протяжении 60 минут и непрерывно поддерживали температуру 37°С на протяжении 15 минут, проводя посредством этого отжиг.
В жидкость для отжига добавляли 0,5 мкл TaKaRa LA Taq и проводили тепловую обработку при 72°С на протяжении 3 минут для построения двойных цепей.
Затем добавляли по 50 пмоль праймераМ13 М4 (последовательность представлена как SEQ ID NO: 12) и праймера М13 RV (последовательность представлена как SEQ ID NO: 13) с получением общего объема 50 мкл, после чего проводили ПЦР №3, повторяя 25 циклов тепловой денатурации (98°С) на протяжении 15 секунд, отжига (55°С) на протяжении 30 секунд и реакции элонгации (72°С) на протяжении 120 секунд.
По результатам проведения анализа продукта амплификации ДНК посредством электрофореза в агарозном геле (с использованием агарозы VII типа с низкой температурой плавления, изготовленной Sigma- Aldrich; концентрация агарозы 0,8 масс. %) с использованием 5 мкл ПЦР-раствора после завершения ПЦР №3, можно было подтвердить присутствие амплифицированного ДНК-продукта размером приблизительно 2 т.п.о.
Затем фрагмент ДНК размером приблизительно 2 т.п.о. был вырезан из агарозного геля, вырезанный фрагмент агарозы (приблизительно 1,0 г) тонко измельчали, суспендировали в 1 мл ТЕ-раствора и инкубировали при 55°С на протяжении 1 часа для полного расплавления агарозы.
Этот расплавленный раствор подвергали фенол/хлороформной экстракции и преципитации с этанолом для очистки фрагмента ДНК и, в завершение, полученный фрагмент ДНК растворяли в 10 мкл ТЕ.
Очищенный амплифицированный фрагмент ДНК размером приблизительно 2 т.п.о. расщепляли рестриктазами EcoRI и HindIII, раствор, полученный после обработки рестриктазами, подвергали фенол/хлороформной экстракции и преципитации с этанолом для очистки фрагмента ДНК и, в завершение, полученный фрагмент ДНК растворяли в 10 мкл ТЕ.
Сходным образом, плазмиду экспрессии нитрилгидратазы pPT-DB1 расщепляли рестриктазами EcoRI и HindIII с последующим электрофорезом в агарозном геле (с использованием агарозы VII типа с низкой температурой плавления, изготовленной Sigma-Aldrich; концентрация агарозы 0,7%) и из агарозного геля вырезали фрагмент ДНК размером приблизительно 2,7 т.п.о.
Вырезанные фрагменты агарозы (приблизительно 1,0 г) тонко измельчали, суспендировали в 1 мл ТЕ-раствора и инкубировали при 55°С на протяжении 1 часа для полного расплавления агарозы.
Этот расплавленный раствор подвергали фенол/хлороформной экстракции и преципитации с этанолом для очистки фрагмента ДНК и, в завершение, полученный фрагмент ДНК растворяли в 10 мкл ТЕ.
Полученные таким образом фрагменты ДНК размерами приблизительно 2 т.п.о. и приблизительно 2,7 т.п.о. лигировали с использованием набора для лигирования ДНК (изготовленного Takara Shuzo Co., Ltd.) и трансформировали ими компетентные клетки Е. coli НВ101 (от Toyobo Co., Ltd.) с получением трансформанта (2). Кроме того, с применением метода щелочной экстракции с SDS из бактериальных клеток была получена плазмида, нуклеотидная последовательность ее части, содержащей ген нитрилгидратазы, была определена с использованием ДНК-секвенатора, и было подтверждено, что плазмида pPT-DBl, описанная в Примере 1, имела мутацию, где Asp, являющийся 92-м аминокислотным остатком от N-конца α-субъединицы, был заменен на Glu. Трансформант (2) продуцирует нитрилгидратазу (2), имеющую мутацию, описанную в Таблице 14.
Сравнение рН-стабильности
рН-стабильность при получении амидного соединения с использованием полученных выше трансформанта (2) и трансформанта (1), содержащих pPT-DB1 в качестве своего основного элемента, сравнивали следующим методом. В результате, можно провести оценку относительной рН-стабильности нитрилгидратазы (2) в сравнении с нитрилгидратазой (1).
В пробирке готовили 5 мл жидкой среды LB, содержащей 40 мкг/мл гептагидрата сульфата железа и 10 мкг/мл дигидрата хлорида кобальта и стерилизовали ее автоклавированием при 121°С на протяжении 20 минут.
В жидкую среду добавляли ампициллин до конечной концентрации 100 мкг/мл, после чего вносили одну платиновую петлю каждого трансформанта и проводили культивирование при 37°С и 200 об/мин на протяжении приблизительно 20 часов.
Отбирали 40 мкл раствора, полученного по завершении культивирования, и суспендировали его в 740 мкл 54 мМ водного раствора трис-соляной кислоты (рН 8,0), добавляли 20 мкл акрилонитрила и полученную смесь подвергали взаимодействию на протяжении 15-60 минут с осторожным перемешиванием при 20°С. По завершении взаимодействия реакционный раствор анализировали с применением ВЭЖХ и определяли ферментативную активность по получению амидного соединения из нитрильного соединения. Для каждого трансформанта взаимодействие и анализ проводили три или более раз и вариабельность данных корректировали дозированием и тому подобным.
Сходным образом, 1000 мкл раствора, полученного по завершении культивирования, отбирали и центрифугировали для выделения клеток, к выделенным бактериальным клеткам добавляли 1000 мкл буфера с 50 мМ лимонной кислотой (рН 4,0) и полученный раствор обрабатывали при 30°С на протяжении 30 часов (обработка кислотой) с перемешиванием. После обработки 780 мкл этого раствора центрифугировали для выделения бактериальных клеток, бактериальные клетки суспендировали в 780 мкл 54 мМ водного раствора трис-соляной кислоты (рН 8,0), добавляли 20 мкл акрилонитрила и полученную смесь подвергали взаимодействию на протяжении 15-60 минут с осторожным перемешиванием при 20°С. По завершении взаимодействия реакционный раствор анализировали с применением ВЭЖХ и определяли ферментативную активность по получению амидного соединения из нитрильного соединения. Для каждого трансформанта взаимодействие и анализ проводили три или более раз и вариабельность данных корректировали дозированием и тому подобным.
Условия проведения анализа
Аналитический прибор: JASCO Corporation HPLC
Колонка: YMC Pack ODS-A (150×6,00 мм)
Температура проведения анализа: 40°С
Подвижная фаза: 3%-й ацетонитрил, 10 мМ фосфорная кислота
рН-стабильность оценивали по соотношению активности бактериальных клеток, обработанных буфером с 50 мМ лимонной кислотой (рН 4,0), и ферментативной активности необработанных бактериальных клеток. рН-стабильность трансформанта (2) составляла 0,82 от соответствующего показателя дикого типа (1).
Приведенные выше результаты показывают, что даже если мутация, предположительно, повышает начальную скорость реакции в условиях, оптимальных для нитрилгидратазы (см. международную публикацию № WO 2010/055666), она может не приводить к повышению рН-стабильности.
Пример 1. Получение мутантной нитрилгидратазы (3)
Для получения трансформанта (3), экспрессирующего мутантную нитрилгидратазу, имеющую замену аминокислотного остатка, как показано в Таблице 14, получали плазмиду, и трансформант (3) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 7. Трансформант (3) продуцирует нитрилгидратазу (3), имеющую мутацию, описанную в Таблице 14.
рН-стабильность нитрилгидратазы (3) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 14 ниже.
Пример 2. Получение мутантной нитрилгидратазы (4)
Для получения трансформанта (4), экспрессирующего мутантную нитрилгидратазу, имеющую замену аминокислотного остатка, как показано в Таблице 14, получали плазмиду, и трансформант (4) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 8. Трансформант (4) продуцирует нитрилгидратазу (4), имеющую мутацию, описанную в Таблице 14.
рН-стабильность нитрилгидратазы (4) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 14 ниже.
Пример 3. Получение мутантной нитрилгидратазы (5)
Для получения трансформанта (5), экспрессирующего мутантную нитрилгидратазу, имеющую замену аминокислотного остатка, как показано в Таблице 14, получали плазмиду, и трансформант (5) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 9. Трансформант (5) продуцирует нитрилгидратазу (5), имеющую мутацию, описанную в Таблице 14.
рН-стабильность нитрилгидратазы (5) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 14 ниже.
Пример 4. Получение мутантной нитрилгидратазы (6)
Для получения трансформанта (6), экспрессирующего мутантную нитрилгидратазу, имеющую замену аминокислотного остатка, как показано в Таблице 14, получали плазмиду, и трансформант (6) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 10. Трансформант (6) продуцирует нитрилгидратазу (6), имеющую мутацию, описанную в Таблице 14.
рН-стабильность нитрилгидратазы (6) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 14 ниже.
Пример 5. Получение мутантной нитрилгидратазы (7)
Для получения трансформанта (7), экспрессирующего мутантную нитрилгидратазу, имеющую замену аминокислотного остатка, как показано в Таблице 14, получали плазмиду, и трансформант (7) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 11. Трансформант (7) продуцирует нитрилгидратазу (7), имеющую мутацию, описанную в Таблице 14.
рН-стабильность нитрилгидратазы (7) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 14 ниже.
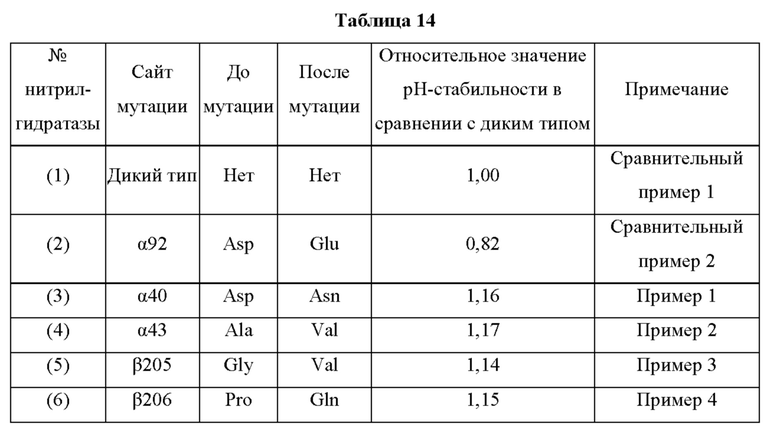

Из результатов, показанных в Таблице 14, видно, что каждая замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила к повышению рН-стабильности нитрилгидратазы сама по себе. В то же время, замена остатка Asp в 92-м положении от N-конца α-субъединицы на Glu, не принадлежащая к заменам аминокислотных остатков группы А, не повышала рН-стабильность по сравнению с нитрилгидратазой дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Пример 6. Получение мутантной нитрилгидратазы (8)
Для получения трансформанта (8), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (8) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 1, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 6. Трансформант (8) продуцирует нитрилгидратазу (8), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (8) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 7. Получение мутантной нитрилгидратазы (9)
Для получения трансформанта (9), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (9) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 2, а вместо праймера SEQIDNO:5 использовали праймер SEQ ID NO: 6. Трансформант (9) продуцирует нитрилгидратазу (9), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (9) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 8. Получение мутантной нитрилгидр атазы (10)
Для получения трансформанта (10), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (10) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 3, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 6. Трансформант (10) продуцирует нитрилгидратазу (10), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (10) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 9. Получение мутантной нитрилгидратазы (11)
Для получения трансформанта (11), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (11) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 4, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 6. Трансформант (11) продуцирует нитрилгидратазу (11), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (11) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 10. Получение мутантной нитрилгидратазы (12)
Для получения трансформанта (12), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (12) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 5, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 6. Трансформант (12) продуцирует нитрилгидратазу (12), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (12) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 11. Получение мутантной нитрилгидратазы (13)
Для получения трансформанта (13), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (13) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 1, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 8. Трансформант (13) продуцирует нитрилгидратазу (13), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (13) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 12. Получение мутантной нитрилгидратазы (14)
Для получения трансформанта (14), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (14) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 1, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 9. Трансформант (14) продуцирует нитрилгидратазу (14), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (14) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 13. Получение мутантной нитрилгидратазы (15)
Для получения трансформанта (15), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (15) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 2, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 9. Трансформант (15) продуцирует нитрилгидратазу (15), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (15) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 14. Получение мутантной нитрилгидратазы (16)
Для получения трансформанта (16), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (16) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 2, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 10. Трансформант (16) продуцирует нитрилгидратазу (16), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (16) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 15. Получение мутантной нитрилгидратазы (17)
Для получения трансформанта (17), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (17) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 2, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 11. Трансформант (17) продуцирует нитрилгидратазу (17), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (17) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 16. Получение мутантной нитрилгидратазы (18)
Для получения трансформанта (18), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (18) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Сравнительному примеру 1, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 15. Трансформант (18) продуцирует нитрилгидратазу (18), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (18) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 17. Получение мутантной нитрилгидратазы (19)
Для получения трансформанта (19), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (19) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 3, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 11. Трансформант (19) продуцирует нитрилгидратазу (19), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (19) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
Пример 18. Получение мутантной нитрилгидратазы (20)
Для получения трансформанта (20), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 15, трансформант (20) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 4, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 11. Трансформант (20) продуцирует нитрилгидратазу (20), имеющую мутации, описанные в Таблице 15.
рН-стабильность нитрилгидратазы (20) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 15 ниже.
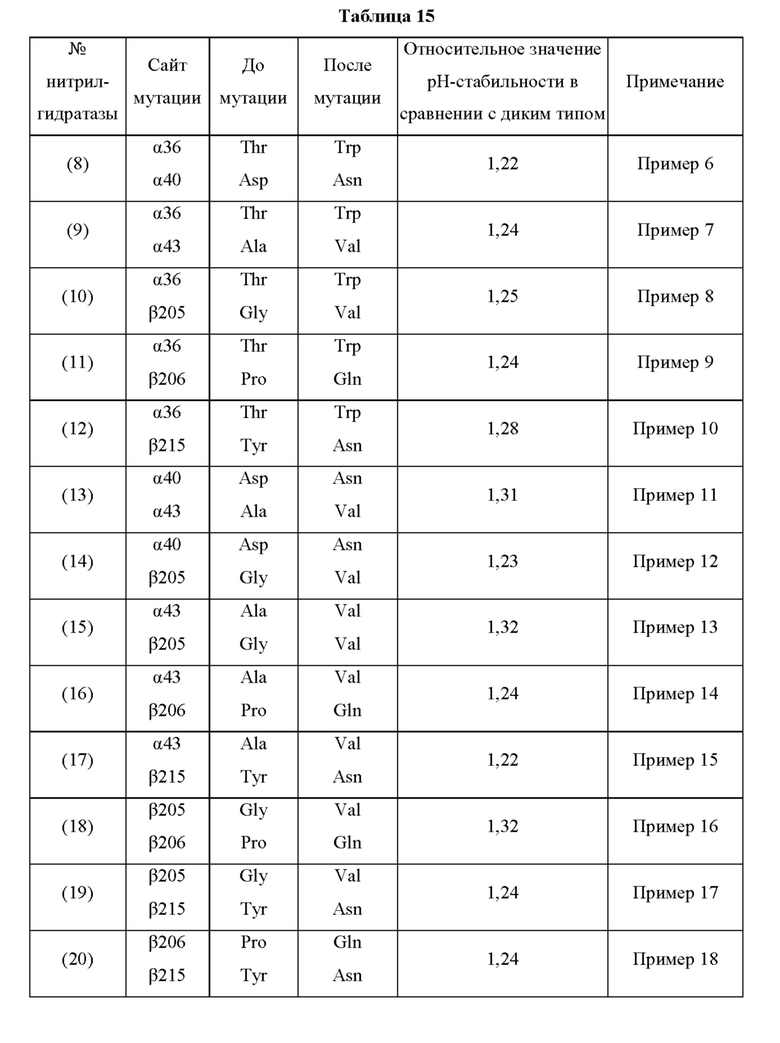
Из результатов, показанных в Таблице 15, видно, что каждая замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила к повышению рН-стабильности нитрилгидратазы даже в случае комбинирования этих замен аминокислотных остатков друг с другом. Кроме того, видно, что в случае комбинирования замен аминокислотных остатков, принадлежащих к заменам аминокислотных остатков группы А, друг с другом или в случае комбинирования этих замен аминокислотных остатков с заменой аминокислотного остатка (f) была тенденция к дальнейшему повышению рН-стабильности.
Пример 19. Получение мутантной нитрилгидратазы (21)
Для получения трансформанта (21), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 16, трансформант (21) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 7, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 9. Трансформант (21) продуцирует нитрилгидратазу (21), имеющую мутации, описанные в Таблице 16.
рН-стабильность нитрилгидратазы (21) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 16 ниже.
Пример 20. Получение мутантной нитрилгидратазы (22)
Для получения трансформанта (22), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 16, трансформант (22) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 11, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 9. Трансформант (22) продуцирует нитрилгидратазу (22), имеющую мутации, описанные в Таблице 16.
рН-стабильность нитрилгидратазы (22) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 16 ниже.
Пример 21. Получение мутантной нитрилгидратазы (23)
Для получения трансформанта (23), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 16, трансформант (23) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 7, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 11. Трансформант (23) продуцирует нитрилгидратазу (23), имеющую мутации, описанные в Таблице 16.
рН-стабильность нитрилгидратазы (23) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 16 ниже.
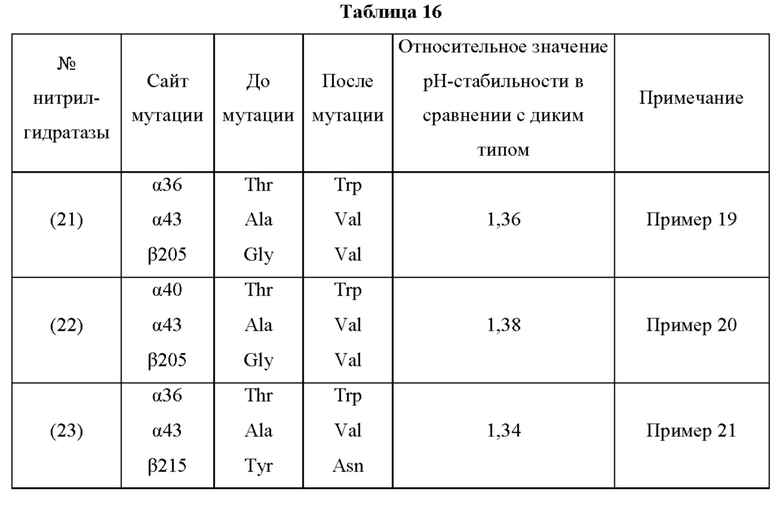
Из результатов, показанных в Таблице 16, видно, что каждая замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила к повышению рН-стабильно ста нитрилгидратазы в случае комбинирования двух или более замен аминокислотных остатков друг с другом и даже в случае их дополнительного комбинирования с заменой аминокислотного остатка (f).
Кроме того, видна тенденция к росту степени стабилизации в зависимости от используемых комбинаций.
Пример 22. Получение мутантной нитрилгидратазы (24)
Для получения трансформанта (24), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 17, трансформант (24) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 7, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 15. Трансформант (24) продуцирует нитрилгидратазу (24), имеющую мутации, описанные в Таблице 17.
рН-стабильность нитрилгидратазы (24) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 17 ниже.
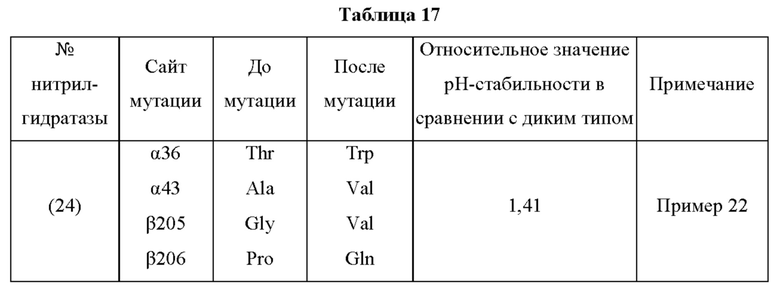
Из результатов, показанных в Таблице 17, видно, что каждая замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила к повышению рН-стабильности нитрилгидратазы даже в случае комбинирования трех или более замен аминокислотных остатков друг с другом. Кроме того, есть тенденция к значительному росту степени этого повышения.
Пример 23. Начальная скорость реакции при использовании мутантной нитрилгидратазы
Начальная скорость реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, и нитрилгидратаз, имеющих замены аминокислотных остатков, показанные в Таблице 18, показана в Таблице 18 как результаты анализа, полученные измерением начальной скорости реакции при оценке получения амидного соединения до обработки кислотой, описанной в разделе «Сравнение рН-стабильности». Значение начальной скорости реакции показано как относительное значение в сравнении с начальной скоростью реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.

Из результатов, показанных в Таблице 18, видно, что каждая замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, также приводила к повышению начальной скорости реакции при использовании соответствующей нитрилгидратазы.
Сравнительный пример 3. Получение мутанта нитрилгидратазы (25)
Трансформант (25) получали таким же образом, как при трансформации, описанной в Сравнительном примере 2, с использованием плазмиды, содержащей мутант нитрилгидратазы с заменами аминокислот в положениях, показанных в Таблице 19 (см. патент Японии №5551081, трансформант №92). Трансформант (25) продуцирует нитрилгидратазу (25), имеющую мутации, описанные в Таблице 19.
рН-стабильность нитрилгидратазы (25) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 19 ниже.
Пример 24. Получение мутанта нитрилгидратазы (26)
Для получения трансформанта (26), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 19, получали плазмиду, и трансформант (26) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Сравнительному примеру 3, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 8. Трансформант (26) продуцирует нитрилгидратазу (26), имеющую мутации, описанные в Таблице 19.
рН-стабильность нитрилгидратазы (26) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 19 ниже.


Из результатов, показанных в Таблице 19, видно, что замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила не только к повышению рН-стабильности нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, но также к повышению рН-стабильности нитрилгидратазы, аминокислотная последовательность которой модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Пример 25. Начальная скорость реакции при использовании мутанта нитрилгидратазы
Начальная скорость реакции при использовании мутантов нитрилгидратазы с заменами аминокислот в положениях, показанных в Таблице 20, показана в Таблице 20 как результаты анализа, полученные измерением начальной скорости реакции при оценке получения амидного соединения до обработки кислотой, описанной в разделе «Сравнение рН-стабильности». Значение начальной скорости реакции показано как относительное значение в сравнении с начальной скоростью реакции при использовании нитрилгидратазы (25).

Из результатов, показанных в Таблице 20, видно, что замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила не только к повышению начальной скорости реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, но также к повышению начальной скорости реакции при использовании нитрилгидратазы, аминокислотная последовательность которой модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Сравнительный пример 4. Получение мутанта нитрилгидратазы (27)
Трансформант (27) получали таким же образом, как при трансформации, описанной в Сравнительном примере 2, с использованием плазмиды, содержащей мутант нитрилгидратазы с заменами аминокислот в положениях, показанных в Таблице 21 (см. патент Японии №5551081, трансформант №114). Трансформант (27) продуцирует нитрилгидратазу (27), имеющую мутации, описанные в Таблице 21.
рН-стабильность нитрилгидратазы (27) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 21 ниже.
Сравнительный пример 26. Получение мутанта нитрилгидратазы (28)
Для получения трансформанта (28), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 21, получали плазмиду, и трансформант (28) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Примеру 4, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 8. Трансформант (28) продуцирует нитрилгидратазу (28), имеющую мутации, описанные в Таблице 21.
рН-стабильность нитрилгидратазы (28) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 21 ниже.


Из результатов, показанных в Таблице 21, видно, что замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила не только к повышению рН-стабильности нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, но также к повышению рН-стабильности нитрилгидратазы, аминокислотная последовательность которой модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Пример 27. Начальная скорость реакции при использовании мутанта нитрилгидратазы
Начальная скорость реакции при использовании мутантов нитрилгидратазы с заменами аминокислот в положениях, показанных в Таблице 21, показана в Таблице 22 как результаты анализа, полученные измерением начальной скорости реакции при оценке получения амидного соединения до обработки кислотой, описанной в разделе «Сравнение рН-стабильности». Значение начальной скорости реакции показано как относительное значение в сравнении с начальной скоростью реакции при использовании нитрилгидратазы (27).
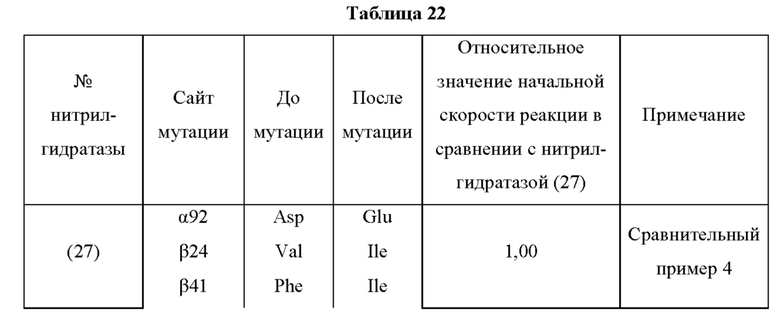

Из результатов, показанных в Таблице 22, видно, что замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила не только к повышению начальной скорости реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, но также к повышению начальной скорости реакции при использовании нитрилгидратазы, аминокислотная последовательность которой модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Сравнительный пример 5. Получение мутанта нитрилгидратазы (29)
Трансформант (29) получали таким же образом, как при трансформации, описанной в Сравнительном примере 2, с использованием плазмиды, содержащей мутант нитрилгидратазы с заменами аминокислот в положениях, показанных в Таблице 23 (см. патент Японии №5551081, трансформант №33). Трансформант (29) продуцирует нитрилгидратазу (29), имеющую мутации, описанные в Таблице 23.
рН-стабильность нитрилгидратазы (29) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 23 ниже.
Пример 28. Получение мутанта нитрилгидратазы (30)
Для получения трансформанта (30), экспрессирующего мутантную нитрилгидратазу, имеющую замены аминокислотных остатков, как показано в Таблице 23, получали плазмиду, и трансформант (30) получали таким же образом, как в Сравнительном примере 2 выше, за исключением того, что в качестве матрицы использовали плазмиду по Сравнительному примеру 5, а вместо праймера SEQ ID NO: 5 использовали праймер SEQ ID NO: 8. Трансформант (30) продуцирует нитрилгидратазу (30), имеющую мутации, описанные в Таблице 23.
рН-стабильность нитрилгидратазы (30) оценивали методом, описанным выше в разделе «Сравнение рН-стабильности». Полученные результаты показаны в Таблице 23 ниже.
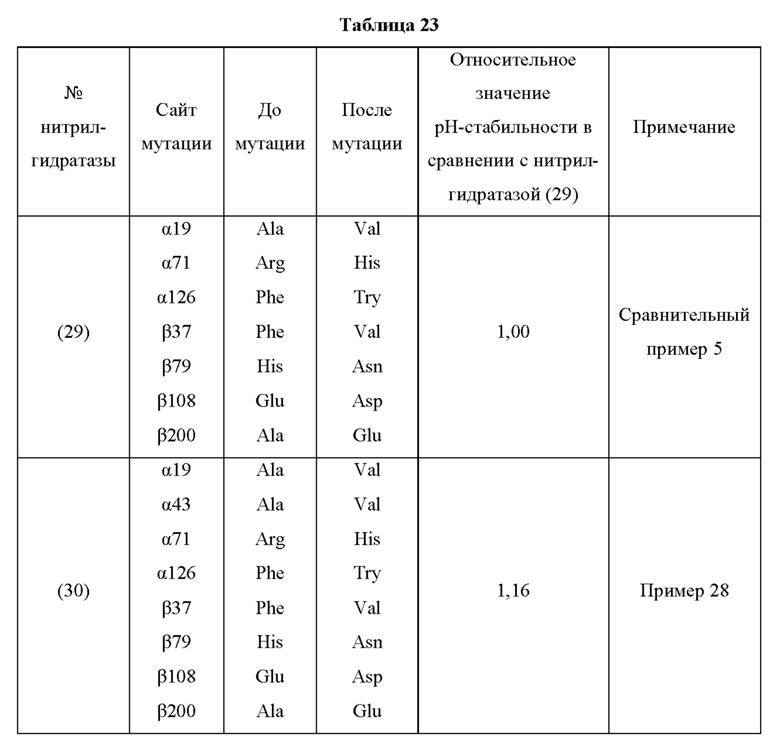
Из результатов, показанных в Таблице 23, видно, что замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила не только к повышению рН-стабильности нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, но также к повышению рН-стабильности нитрилгидратазы, аминокислотная последовательность которой модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Пример 29. Начальная скорость реакции при использовании мутанта нитрилгидратазы
Начальная скорость реакции при использовании мутантов нитрилгидратазы с заменами аминокислот в положениях, показанных в Таблице 23, показана в Таблице 24 как результаты анализа, полученные измерением начальной скорости реакции при оценке получения амидного соединения до обработки кислотой, описанной в разделе «Сравнение рН-стабильности». Значение начальной скорости реакции показано как относительное значение в сравнении с начальной скоростью реакции при использовании нитрилгидратазы (29).


Из результатов, показанных в Таблице 24, видно, что замена аминокислотного остатка, принадлежащая к заменам аминокислотных остатков группы А, приводила не только к повышению начальной скорости реакции при использовании нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila, но также к повышению начальной скорости реакции при использовании нитрилгидратазы, аминокислотная последовательность которой модифицирована относительно нитрилгидратазы дикого типа, имеющей происхождение из Pseudonocardia thermophila.
Описание заявки на патент Японии №2016-256050, поданной 28 декабря 2016 г., полностью включено сюда посредством ссылки.
Все документы, заявки на патенты и технические стандарты, указанные в данном описании, полностью включены сюда посредством ссылки в такой же степени, как в случае, если бы для каждого документа, заявки на патент и технического описания было прямо и по отдельности указано, что они включены посредством ссылки.
Перечень последовательностей
Международная заявка в соответствии с Договором о международной патентной кооперации (International Patent Cooperation Treaty) MT-F03210-JP17047127_24.app.
.
--->
SEQUENCE LISTING
<110> MITSUI CHEMICALS, INC.
<120> Mutant Nitrile Hydratase, Nucleic Acid Encoding the Mutant
Nitrile Hydratase, Expression Vector and Transformant Containing
the Nucleic Acid, Production Method for Muntant Nitrile
Hydratase, and Production Method for Amide Compound
<130> 2017P00694
<150> JP2016-256050
<151> 2016-12-28
<160> 75
<170> PatentIn version 3.5
<210> 1
<211> 205
<212> PRT
<213> Pseudonocardia thermophila
<400> 1
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 2
<211> 233
<212> PRT
<213> Pseudonocardia thermophila
<400> 2
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 3
<211> 618
<212> DNA
<213> Pseudonocardia thermophila
<400> 3
atgaccgaga acatcctgcg caagtcggac gaggagatcc agaaggagat cacggcgcgg 60
gtcaaggccc tggagtcgat gctcatcgaa cagggcatcc tcaccacgtc gatgatcgac 120
cggatggccg agatctacga gaacgaggtc ggcccgcacc tcggcgcgaa ggtcgtcgtg 180
aaggcctgga ccgacccgga gttcaagaag cgtctgctcg ccgacggcac cgaggcctgc 240
aaggagctcg gcatcggcgg cctgcagggc gaggacatga tgtgggtgga gaacaccgac 300
gaggtccacc acgtcgtcgt gtgcacgctc tgctcctgct acccgtggcc ggtgctgggg 360
ctgccgccga actggttcaa ggagccgcag taccgctccc gcgtggtgcg tgagccccgg 420
cagctgctca aggaggagtt cggcttcgag gtcccgccga gcaaggagat caaggtctgg 480
gactccagct ccgagatgcg cttcgtcgtc ctcccgcagc gccccgcggg caccgacggg 540
tggagcgagg aggagctcgc caccctcgtc acccgcgagt cgatgatcgg cgtcgaaccg 600
gcgaaggcgg tcgcgtga 618
<210> 4
<211> 702
<212> DNA
<213> Pseudonocardia thermophila
<400> 4
atgaacggcg tgtacgacgt cggcggcacc gatgggctgg gcccgatcaa ccggcccgcg 60
gacgaaccgg tcttccgcgc cgagtgggag aaggtcgcgt tcgcgatgtt cccggcgacg 120
ttccgggccg gcttcatggg cctggacgag ttccggttcg gcatcgagca gatgaacccg 180
gccgagtacc tcgagtcgcc gtactactgg cactggatcc gcacctacat ccaccacggc 240
gtccgcaccg gcaagatcga tctcgaggag ctggagcgcc gcacgcagta ctaccgggag 300
aaccccgacg ccccgctgcc cgagcacgag cagaagccgg agttgatcga gttcgtcaac 360
caggccgtct acggcgggct gcccgcaagc cgggaggtcg accgaccgcc caagttcaag 420
gagggcgacg tggtgcggtt ctccaccgcg agcccgaagg gccacgcccg gcgcgcgcgg 480
tacgtgcgcg gcaagaccgg gacggtggtc aagcaccacg gcgcgtacat ctacccggac 540
accgccggca acggcctggg cgagtgcccc gagcacctct acaccgtccg cttcacggcc 600
caggagctgt gggggccgga aggggacccg aactccagcg tctactacga ctgctgggag 660
ccctacatcg agctcgtcga cacgaaggcg gccgcggcat ga 702
<210> 5
<211> 24
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 5
cagggcgagg agatgatgtg ggtg 24
<210> 6
<211> 21
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 6
atcctcacct ggtcgatgat c 21
<210> 7
<211> 21
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 7
gtcgatgatc aaccggatgg c 21
<210> 8
<211> 20
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 8
accggatggt cgagatctac 20
<210> 9
<211> 21
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 9
gagctgtggg tgccggaagg g 21
<210> 10
<211> 21
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 10
ctgtgggggc aggaagggga c 21
<210> 11
<211> 21
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 11
ctccagcgtc aactacgact g 21
<210> 12
<211> 17
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 12
gttttcccag tcacgac 17
<210> 13
<211> 17
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 13
caggaaacag ctatgac 17
<210> 14
<211> 20
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 14
ggccagtgcc tagcttacat 20
<210> 15
<211> 20
<212> DNA
<213> Artificial
<220>
<223> Праймер
<400> 15
gctgtgggtg caggaagggg 20
<210> 16
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 16
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Met Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Asp Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Arg Ala
195 200 205
<210> 17
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 17
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Met Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Glu Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Asp Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Arg Ala
195 200 205
<210> 18
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 18
Met Thr Glu Asn Ile Thr Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Val Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 19
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 19
Met Thr Glu Asn Ile Thr Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Val Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Ile Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 20
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 20
Met Thr Glu Asn Ile Ala Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Val Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 21
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 21
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Val Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys His Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 22
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 22
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Leu Gln Lys Glu
1 5 10 15
Ile Thr Val Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys His Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 23
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 23
Met Thr Glu Asn Ile Thr Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Met Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 24
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 24
Met Thr Glu Asn Ile Thr Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Ile Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Met Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 25
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 25
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Glu Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Asp Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Arg Ala
195 200 205
<210> 26
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 26
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Leu Gln Lys Glu
1 5 10 15
Ile Thr Val Arg Val Lys Ala Leu Glu Ser Ile Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys His Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Tyr Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 27
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 27
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Leu Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Ile Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 28
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 28
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Leu Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 29
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 29
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Cys Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 30
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 30
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Glu Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 31
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 31
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Thr Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Glu
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Asp Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Cys Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 32
<211> 205
<212> PRT
<213> Artificial
<220>
<223> Альфа-субъединица модифицированной нитрилгидратазы
<400> 32
Met Thr Glu Asn Ile Leu Arg Lys Ser Asp Glu Glu Ile Gln Lys Glu
1 5 10 15
Ile Thr Ala Arg Val Lys Ala Leu Glu Ser Met Leu Ile Glu Gln Gly
20 25 30
Ile Leu Thr Ser Ser Met Ile Asp Arg Met Ala Glu Ile Tyr Glu Asn
35 40 45
Glu Val Gly Pro His Leu Gly Ala Lys Val Val Val Lys Ala Trp Thr
50 55 60
Asp Pro Glu Phe Lys Lys Arg Leu Leu Ala Asp Gly Thr Glu Ala Cys
65 70 75 80
Lys Glu Leu Gly Ile Gly Gly Leu Gln Gly Glu Glu Met Met Trp Val
85 90 95
Glu Asn Thr Asp Glu Val His His Val Val Val Cys Thr Leu Cys Ser
100 105 110
Cys Tyr Pro Trp Pro Val Leu Gly Leu Pro Pro Asn Trp Phe Lys Glu
115 120 125
Pro Gln Tyr Arg Ser Arg Val Val Arg Glu Pro Arg Gln Leu Leu Lys
130 135 140
Glu Glu Phe Gly Phe Glu Val Pro Pro Ser Lys Glu Ile Lys Val Trp
145 150 155 160
Asp Ser Ser Ser Glu Met Arg Phe Val Val Leu Pro Gln Arg Pro Ala
165 170 175
Gly Thr Asp Gly Trp Ser Glu Glu Glu Leu Ala Thr Leu Val Thr Arg
180 185 190
Glu Ser Met Ile Gly Val Glu Pro Ala Lys Ala Val Ala
195 200 205
<210> 33
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 33
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Ile Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 34
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 34
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Lys Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 35
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 35
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Ser Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Trp
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Arg Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 36
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 36
Met Asn Gly Met Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Ser Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Trp
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Arg Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 37
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 37
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Leu Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 38
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 38
Met Asn Gly Val Tyr Asp Val Ala Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Leu Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 39
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 39
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Val Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 40
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 40
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Val Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 41
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 41
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Val
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 42
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 42
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Val
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Arg
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 43
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 43
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Val Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Met Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 44
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 44
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Leu Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Arg
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 45
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 45
Met Asn Gly Val Tyr Asp Val Gly Gly Asp Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Val Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 46
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 46
Met Asn Gly Val Tyr Asp Val Gly Gly Asp Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Met Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Val Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 47
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 47
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Ile Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 48
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 48
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Val
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 49
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 49
Met Asn Gly Val Tyr Asp Val Gly Gly Asp Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Val Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Leu Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 50
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 50
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Ile Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Leu Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 51
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 51
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Asn Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 52
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 52
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Lys Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Asn Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 53
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 53
Met Asn Gly Val Tyr Asp Val Gly Gly Asp Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Val Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Leu Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Ile Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 54
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 54
Met Asn Gly Met Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Leu Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 55
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 55
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Ile Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Met Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 56
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 56
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Val
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Val Ala Ala
225 230
<210> 57
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 57
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Lys Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Val Ala Ala
225 230
<210> 58
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 58
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Ile Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Ile Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Ile Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 59
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 59
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Ile Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Ile Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Met Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 60
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 60
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Ile Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Met Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Gly Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 61
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 61
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Ile Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Val
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Val Ala Ala
225 230
<210> 62
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 62
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Val Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 63
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 63
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Ile Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Val Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 64
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 64
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Ile Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Ile Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 65
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 65
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Leu Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Ile
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Ile Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 66
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 66
Met Asn Gly Met Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Val Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His His Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Asp His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Leu Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 67
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 67
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Met Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Thr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 68
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 68
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Gly Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Asn Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 69
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 69
Met Asn Gly Met Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Val
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Arg His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Tyr Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 70
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 70
Met Asn Gly Met Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Thr Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Ser Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 71
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 71
Met Asn Gly Met Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Gly Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Ser Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Ala Ala Ala Ala
225 230
<210> 72
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 72
Met Asn Gly Val Tyr Asp Val Gly Gly Thr Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Phe Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Ala Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 73
<211> 233
<212> PRT
<213> Artificial
<220>
<223> Бета-субъединица модифицированной нитрилгидратазы
<400> 73
Met Asn Gly Val Tyr Asp Val Gly Gly Asp Asp Gly Leu Gly Pro Ile
1 5 10 15
Asn Arg Pro Ala Asp Glu Pro Val Phe Arg Ala Glu Trp Glu Lys Val
20 25 30
Ala Phe Ala Met Phe Pro Ala Thr Phe Arg Ala Gly Phe Met Gly Leu
35 40 45
Asp Glu Phe Arg Phe Gly Ile Glu Gln Met Asn Pro Ala Glu Tyr Leu
50 55 60
Glu Ser Pro Tyr Tyr Trp His Trp Ile Arg Thr Tyr Ile His Asn Gly
65 70 75 80
Val Arg Thr Gly Lys Ile Asp Leu Glu Glu Leu Glu Arg Arg Thr Gln
85 90 95
Tyr Tyr Arg Glu Asn Pro Asp Ala Pro Leu Pro Glu His Glu Gln Lys
100 105 110
Pro Glu Leu Ile Glu Val Val Asn Gln Ala Val Tyr Gly Gly Leu Pro
115 120 125
Ala Ser Arg Glu Val Asp Arg Pro Pro Lys Phe Lys Glu Gly Asp Val
130 135 140
Val Arg Phe Ser Thr Ala Ser Pro Lys Gly His Ala Arg Arg Ala Arg
145 150 155 160
Tyr Val Arg Gly Lys Thr Gly Thr Val Val Lys His His Gly Ala Tyr
165 170 175
Ile Tyr Pro Asp Thr Ala Gly Asn Gly Leu Gly Glu Cys Pro Glu His
180 185 190
Leu Tyr Thr Val Arg Phe Thr Glu Gln Glu Leu Trp Gly Pro Glu Gly
195 200 205
Asp Pro Asn Ser Ser Val Tyr Tyr Asp Cys Trp Glu Pro Tyr Ile Glu
210 215 220
Leu Val Asp Thr Lys Glu Ala Ala Ala
225 230
<210> 74
<211> 33
<212> DNA
<213> Artificial
<220>
<223> Олигонуклеотид, действующий как ПЦР-праймер
<400> 74
tacgaattct aaggaggtct cagcatgaac ggc 33
<210> 75
<211> 144
<212> PRT
<213> Pseudonocardia thermophila
<400> 75
Met Ser Ala Glu Ala Lys Val Arg Leu Lys His Cys Pro Thr Ala Glu
1 5 10 15
Asp Arg Ala Ala Ala Asp Ala Leu Leu Ala Gln Leu Pro Gly Gly Asp
20 25 30
Arg Ala Leu Asp Arg Gly Phe Asp Glu Pro Trp Gln Leu Arg Ala Phe
35 40 45
Ala Leu Ala Val Ala Ala Cys Arg Ala Gly Arg Phe Glu Trp Lys Gln
50 55 60
Leu Gln Gln Ala Leu Ile Ser Ser Ile Gly Glu Trp Glu Arg Thr His
65 70 75 80
Asp Leu Asp Asp Pro Ser Trp Ser Tyr Tyr Glu His Phe Val Ala Ala
85 90 95
Leu Glu Ser Val Leu Gly Glu Glu Gly Ile Val Glu Pro Glu Ala Leu
100 105 110
Asp Glu Arg Thr Ala Glu Val Leu Ala Asn Pro Pro Asn Lys Asp His
115 120 125
His Gly Pro His Leu Glu Pro Val Ala Val His Pro Ala Val Arg Ser
130 135 140
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ П-ГИДРОКСИФЕНИЛПИРУВАТДИОКСИГЕНАЗА, НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ЕЕ, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2781830C2 |
| ПОВЫШЕНИЕ РЕАКЦИОННОЙ СПОСОБНОСТИ НИТРИЛГИДРАТАЗЫ С ПОМОЩЬЮ АЛЬДЕГИДА | 2022 |
|
RU2827406C2 |
| ИММУНОКОНЪЮГАТЫ IL2 И МУТАНТНОГО TNF | 2017 |
|
RU2758139C2 |
| МУТАНТНЫЙ БЕЛОК F RSV И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2807742C1 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2819144C2 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| Микроорганизмы для получения орнитина и способ получения орнитина с использованием указанных микроорганизмов | 2016 |
|
RU2721852C1 |
| МУТАНТНЫЙ ПОЛИПЕПТИД ГИДРОКСИФЕНИЛПИРУВАТДИОКСИГЕНАЗА, КОДИРУЮЩИЙ ЕГО ГЕН И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2822892C1 |
| Композиции, относящиеся к мутантному токсину CLOSTRIDIUM DIFFICILE, и способы их применения | 2017 |
|
RU2773880C1 |
Группа изобретений относится к биотехнологии. Предложена мутантная нитрилгидратаза, имеющая происхождение из Pseudonocardia thermophila, имеющая α-субъединицу и β-субъединицу и содержащая по меньшей мере одну замену аминокислотного остатка, выбранную из группы, состоящей из следующих замен аминокислотных остатков (а)-(е): (a) замена аминокислоты, соответствующей 40-му аминокислотному остатку от N-конца SEQ ID NO: 1, на Asn в α-субъединице, (b) замена аминокислоты, соответствующей 43-му аминокислотному остатку от N-конца SEQ ID NO: 1, на Val в α-субъединице, (c) замена аминокислоты, соответствующей 205-му аминокислотному остатку от N-конца SEQ ID NO: 2, на Val в β-субъединице, (d) замена аминокислоты, соответствующей 206-му аминокислотному остатку от N-конца SEQ ID NO: 2, на Gln в β-субъединице и (e) замена аминокислоты, соответствующей 215-му аминокислотному остатку от N-конца SEQ ID NO: 2, на Asn в β-субъединице. Предложены также нуклеиновая кислота, кодирующая мутантную нитрилгидратазу, вектор экспрессии, содержащий нуклеиновую кислоту, трансформант для получения мутантной нитрилгидратазы, содержащий вектор экспрессии. А также способ получения мутантной нитрилгидратазы, включающий культивирование трансформанта в среде и выделение мутантной нитрилгидратазы из по меньшей мере одного из культивированного трансформанта или среды, и способ получения амидного соединения, включающий приведение мутантной нитрилгидратазы в контакт с нитрильным соединением и удаление примесей из раствора, содержащего амидное соединение, при pH от 3,5 до 6,5. Группа изобретений обеспечивает получение мутантной нитрилгидратазы, которая сохраняет отличную ферментативную активность, катализирующую реакцию синтеза амидного соединения из нитрильного соединения даже в кислом диапазоне, при pH от 3,5 до 6,5. 6 н. и 8 з.п. ф-лы, 24 табл., 29 пр.
1. Мутантная нитрилгидратаза, имеющая происхождение из Pseudonocardia thermophila и имеющая α-субъединицу и β-субъединицу, содержащая по меньшей мере одну замену аминокислотного остатка, выбранную из группы, состоящей из следующих замен аминокислотных остатков (а)-(е):
(a) замена аминокислоты, соответствующей 40-му аминокислотному остатку от N-конца SEQ ID NO: 1, на Asn в α-субъединице,
(b) замена аминокислоты, соответствующей 43-му аминокислотному остатку от N-конца SEQ ID NO: 1, на Val в α-субъединице,
(c) замена аминокислоты, соответствующей 205-му аминокислотному остатку от N-конца SEQ ID NO: 2, на Val в β-субъединице,
(d) замена аминокислоты, соответствующей 206-му аминокислотному остатку от N-конца SEQ ID NO: 2, на Gln в β-субъединице, и
(e) замена аминокислоты, соответствующей 215-му аминокислотному остатку от N-конца SEQ ID NO: 2, на Asn в β-субъединице.
2. Мутантная нитрилгидратаза по п. 1, содержащая две или более замены аминокислотных остатков, выбранные из группы, состоящей из замен аминокислотных остатков (а)-(е).
3. Мутантная нитрилгидратаза по п. 1 или 2, содержащая замену аминокислотного остатка (b) и по меньшей мере одну замену, выбранную из группы, состоящей из замен аминокислотных остатков (а), (с), (d) и (е).
4. Мутантная нитрилгидратаза по п. 1 или 2, где аминокислота, соответствующая 36-й аминокислоте от N-конца SEQ ID NO: 1 в аминокислотной последовательности α-субъединицы, представляет собой Trp.
5. Мутантная нитрилгидратаза по п. 1 или 2, где аминокислота, соответствующая 36-й аминокислоте от N-конца SEQ ID NO: 1 в аминокислотной последовательности α-субъединицы, представляет собой Met, Ser, Gly или Ala.
6. Мутантная нитрилгидратаза по п. 1 или 2, где аминокислотная последовательность α-субъединицы удовлетворяет одному или более из следующих условий (1)-(13):
(1) аминокислота, соответствующая 6-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Thr или Ala,
(2) аминокислота, соответствующая 13-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Leu,
(3) аминокислота, соответствующая 19-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Val,
(4) аминокислота, соответствующая 27-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Ile,
(5) аминокислота, соответствующая 48-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Gln,
(6) аминокислота, соответствующая 71-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой His,
(7) аминокислота, соответствующая 92-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Glu,
(8) аминокислота, соответствующая 94-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Ile,
(9) аминокислота, соответствующая 126-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Tyr,
(10) аминокислота, соответствующая 148-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Asp,
(11) аминокислота, соответствующая 188-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Gly,
(12) аминокислота, соответствующая 197-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Cys и
(13) аминокислота, соответствующая 204-му аминокислотному остатку от N-конца SEQ ID NO: 1, представляет собой Arg.
7. Мутантная нитрилгидратаза по п. 1 или 2, где аминокислотная последовательность β-субъединицы удовлетворяет по меньшей мере одному из следующих условий (15)-(47):
(15) аминокислота, соответствующая 4-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Met,
(16) аминокислота, соответствующая 8-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ala,
(17) аминокислота, соответствующая 10-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Asp,
(18) аминокислота, соответствующая 24-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ile,
(19) аминокислота, соответствующая 33-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val или Met,
(20) аминокислота, соответствующая 37-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val или Leu,
(21) аминокислота, соответствующая 40-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ile, Val или Leu,
(22) аминокислота, соответствующая 41-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ile,
(23) аминокислота, соответствующая 46-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Lys,
(24) аминокислота, соответствующая 48-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val,
(25) аминокислота, соответствующая 51-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val,
(26) аминокислота, соответствующая 61-му аминокислотному остатку от N-конца SEQ ID NO: 2 представляет собой Val, Gly, Trp, Ser, Leu или Thr,
(27) аминокислота, соответствующая 79-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Asn,
(28) аминокислота, соответствующая 96-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Arg,
(29) аминокислота, соответствующая 107-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Met,
(30) аминокислота, соответствующая 108-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Asp или Arg,
(31) аминокислота, соответствующая 110-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Asn,
(32) аминокислота, соответствующая 112-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val или Ile,
(33) аминокислота, соответствующая 118-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val,
(34) аминокислота, соответствующая 127-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ser,
(35) аминокислота, соответствующая 146-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Gly,
(36) аминокислота, соответствующая 150-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Asn или Ser,
(37) аминокислота, соответствующая 160-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Cys, Trp или Met,
(38) аминокислота, соответствующая 168-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Glu,
(39) аминокислота, соответствующая 176-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ala, Thr, Met или Cys,
(40) аминокислота, соответствующая 186-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Arg,
(41) аминокислота, соответствующая 200-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Glu,
(42) аминокислота, соответствующая 212 -му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Tyr,
(43) аминокислота, соответствующая 217-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val, His, Met, Gly, Ser, Leu или Cys,
(44) аминокислота, соответствующая 218-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Met или Ser,
(45) аминокислота, соответствующая 226-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Ile,
(46) аминокислота, соответствующая 230-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Glu и
(47) аминокислота, соответствующая 231-му аминокислотному остатку от N-конца SEQ ID NO: 2, представляет собой Val.
8. Мутантная нитрилгидратаза по п. 1 или 2, содержащая по меньшей мере одну замену аминокислотного остатка, выбранную из группы, состоящей из замен аминокислотных остатков (а)-(е) в любой из следующих нитрилгидратаз [1]-[48], имеющих происхождение из Pseudonocardia thermophila:
[1] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 2,
[2] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 16, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 33,
[3] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 17, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 33,
[4] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 34,
[5] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 19, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 34,
[6] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 20, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 35,
[7] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 20, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 36,
[8] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 37,
[9] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 38,
[10] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 39,
[11] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 40,
[12] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 41,
[13] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 42,
[14] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 43,
[15] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 22, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 44,
[16] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 23, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 45,
[17] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 24, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 46,
[18] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 25, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 47,
[19] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 48,
[20] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 23, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 49,
[21] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 16, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 50,
[22] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 26, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 51,
[23] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 27, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 52,
[24] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 28, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 53,
[25] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 17, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 54,
[26] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 29, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 55,
[27] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 56,
[28] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 57,
[29] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 58,
[30] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 29, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 59,
[31] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 31, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 60,
[32] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 18, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 61,
[33] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 32, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 62,
[34] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 63,
[35] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 64,
[36] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 65,
[37] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 25, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 54,
[38] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 30, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 66,
[39] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 67,
[40] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 68,
[41] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 69,
[42] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 70,
[43] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 71,
[44] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 21, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 72,
[45] нитрилгидратаза, имеющая α-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 1, и β-субъединицу, имеющую аминокислотную последовательность SEQ ID NO: 73,
[46] нитрилгидратаза, где 36-й аминокислотный остаток от N-конца α-субъединицы любой из нитрилгидратаз [1]-[45] представляет собой остаток Trp,
[47] нитрилгидратаза, имеющая α-субъединицу нитрилгидратазы (А) по любому из [1]-[46] или вариант α-субъединицы, состоящий из аминокислотной последовательности, на 90% или более идентичной α-субъединице, и β-субъединицу нитрилгидратазы (А) или вариант β-субъединицы, состоящий из аминокислотной последовательности, на 90% или более идентичной β-субъединице, где по меньшей мере одна из α-субъединицы или β-субъединицы представляет собой вариант α-субъединицы или вариант β-субъединицы, и
[48] нитрилгидратаза, где общее число присоединенных, замененных, удаленных и/или введенных аминокислотных остатков или общее число указанных остатков в любой их комбинации в α-субъединице нитрилгидратазы (В) по любому из [1]-[46] (за исключением аминокислотных остатков, подлежащих замене (а) и (b)) составляет от 1 до 10 и общее число присоединенных, замененных, удаленных и/или введенных аминокислотных остатков или общее число указанных остатков в любой их комбинации в β-субъединице нитрилгидратазы (В) (за исключением аминокислотных остатков, подлежащих замене (с)-(е)) составляет от 1 до 10.
9. Нуклеиновая кислота, кодирующая мутантную нитрилгидратазу по п. 1 или 2.
10. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 9.
11. Трансформант для получения мутантной нитрилгидратазы, содержащий вектор экспрессии по п. 10.
12. Способ получения мутантной нитрилгидратазы, включающий:
культивирование трансформанта по п. 11 в среде и
выделение мутантной нитрилгидратазы из по меньшей мере одного из культивированного трансформанта или среды.
13. Способ получения амидного соединения, включающий приведение мутантной нитрилгидратазы по п. 1 или 2 в контакт с нитрильным соединением и удаление примесей из раствора, содержащего амидное соединение, при pH от 3,5 до 6,5.
14. Способ получения амидного соединения по п. 13, дополнительно включающий очистку амидного соединения с использованием активированного угля.
| MIYANACA AKIMASA et al., Mutational and structural analysis of cobalt-containing nitrile hydratase on substrate and metal binding, EUR | |||
| J | |||
| Biochem, 2004, vol.271, p.429-438, fig.1 | |||
| WO 2010055666 A1, 20.05.2010 | |||
| УСТОЙЧИВЫЕ К ЦИАНИДАМ НИТРИЛГИДРАТАЗЫ | 2005 |
|
RU2385876C2 |

