ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка претендует на приоритет предварительной патентной заявки США №62/194,355, поданной 20 июля 2015 года, содержание которой включено в данный документ путем ссылки во всей полноте.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0002] Данное изобретение касается адъювантов для вакцины.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[0003] В области вакцинологии антигены вводятся хозяину, таким образом, чтобы стимулировать иммунный ответ на антиген, и, таким образом на потенциальный патоген. Индуцирование иммунного ответа зависит от многих факторов, среди которых, как считают, являются химический состав, характеристики и конфигурация антигена, состояние здоровья и состояние иммунной системы хозяина, способ доставки и введения антигена.
[0004] Иммунный ответ имеет много аспектов, некоторые из который продемонстрированы клетками иммунной системы, (например, дендритными клетками, В-лимфоцитами, Т-лимфоцитами, макрофагами, и плазматическими клетками). Клетки иммунной системы принимают участие в иммунном ответе за счет взаимодействия с антигенами или другими клетками иммунной системы, высвобождения цитокинов и реактивности к данным цитокинам. Адаптивный (приобретенный) иммунный ответ условно (но без достаточных оснований) делится на две основные категории - гуморальный и клеточно-опосредованный. Гуморальный компонент иммунного ответа включает продуцирование антител специфических к антигену. Клеточно-опосредованный компонент включает генерирование гиперчувствительности замедленного типа и цитотоксических эффекторных Т-клеток, специфических к антигену.
[0005] Адъюванты представляют собой вещества, используемые для потенциирования иммунного ответа, когда их применяют в сочетании с антигеном. Применение адъюванта в протоколе вакцинации может, например, вызвать иммунный ответ, который является более быстрым или значительно большим, чем тот, который может быть вызван антигеном самостоятельно. Кроме того, адъюванты могут использоваться для направления иммунного ответа на специфические иммунологические пути и служат средствами доставки антигена.
[0006] Липосомы и липосомальные композиции являются примерами адъювантов. Как правило, липосомы могут быть нагружены антигеном(ами) и/или другими иммуномодулирующими соединениями, или липосомы сами по себе могут служить в качестве самостоятельных адъювантов. Антигены и/или другие иммуностимулирующие соединения могут быть инкапсулированными внутри липосомы, и/или они могут быть присоединенными к липосоме или включенными в липидный бислой.
[0007] Факторы, влияющие на приемлемость предоставленной липосомы в качестве средства доставки в представлении предоставленной системы, остаются непонятными. Таким образом, все еще существует потребность в средствах доставки, которые обеспечивают улучшенную эффективность. Такая улучшенная доставка особенно касается введения молекул, стимулирующих и/или вызывающих иммунный ответ, например, антигенов и иммуномодуляторов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0008] Настоящее изобретение касается адъювантов для усиления эффективности вакцины.
[0009] В некоторых аспектах изобретение предусматривает липосому, содержащую внешнюю липидную двухслойную мембрану и внутреннюю камеру, где внешняя липидная двухслойная мембрана содержит: четвертичное аммониевое соединение; стерин; фосфолипид; и гликолипид формулы I:
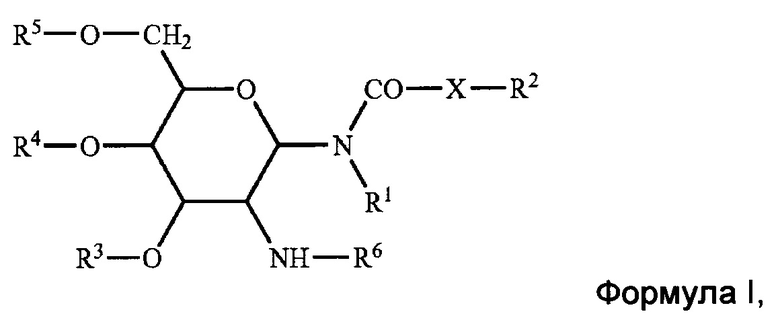
в которой R1 и R2 независимо представляют собой водород, или насыщенный алкильный радикал, имеющий вплоть до 20 атомов углерода; X представляет собой -СН2-, -О- или -NH-; R2 представляет собой водород, или насыщенный или ненасыщенный алкильный радикал, имеющий вплоть до 20 атомов углерода; R3, R4, и R5 независимо представляют собой водород, -SO42-, -PO42-, -СОС1-10 алкил; R6 представляет собой L-аланил, L-альфа-аминобутил, L-аргинил, L-аспаргинил, L-аспартил, L-цистеинил, L-глутамил, L-глицил, L-гистидил, L-гидроксипропил, L-изолейцил, L-лейцил, L-лизил, L-метионил, L-орнитинил, L-фенилаланил, L-пролил, L-серил, L-треонил, L-тирозил, L-триптофанил, и L-валил или их D-изомеры.
[0010] В некоторых вариантах осуществления, четвертичное аммониевое соединение представляет собой DDA, стерин представляет собой холестерин, фосфолипид представляет собой лецитин, и гликолипид представляет собой N-(2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-октадецилдодеканоиламид или его ацетат.
[0011] В некоторых вариантах осуществления, липосома фактически не содержит сапонинов.
[0012] В некоторых вариантах осуществления, липосома дополнительно включает иммуностимулирующий олигонуклеотид, выбранный из группы, состоящей из иммуностимулирующего рибонуклеотида, CpG олигодезоксирибонуклеотида, и их комбинации. В некоторых вариантах осуществления, липосома не содержит CpG олигодезоксирибонуклеотид.
[0013] В некоторых вариантах осуществления, иммуностимулирующий олигонуклеотид вводят во внутреннюю камеру липосомы. В других вариантах осуществления, иммуностимулирующий олигонуклеотид ассоциируется с внешней поверхностью липосомы.
[0014] В некоторых вариантах осуществления, указанный иммуностимулирующий олигонуклеотид включает какую-либо одну из SEQ ID NO 1-14.
[0015] В некоторых аспектах, изобретение предполагает адъювантную композицию, содержащую липосомы, как описано в данном документе.
[0016] В некоторых вариантах осуществления, адъювантная композиция фактически не содержит сапонинов. В некоторых вариантах осуществления, адъювантная композиция фактически не содержит CpG олигодезоксирибонуклеотид.
[0017] В некоторых аспектах, изобретение предполагает вакцинную композицию, содержащую эффективное количество антигенного компонента и адъювантную композицию, как описано в данном документе.
[0018] В некоторых вариантах осуществления, вакцинная композиция фактически не содержит сапонинов.
[0019] В некоторых вариантах осуществления, вакцинная композиция фактически не содержит CpG. В некоторых вариантах осуществления, антигенный компонент фактически свободной от CpG вакцинной композиции содержит вирус с (-)оцРНК.
[0020] В некоторых вариантах осуществления, вирус с (-)оцРНК представляет собой вирус гриппа. В некоторых вариантах осуществления, вирус гриппа представляет собой вирус свиного гриппа.
[0021] В некоторых вариантах осуществления, антигенный компонент вводят во внутреннюю камеру липосомы.
[0022] Антигенный компонент, в выбранных вариантах осуществления, приемлемых для крупного рогатого скота, может включать инактивированные вирусы BVDV-1 и/или BVDV-2 (и BHV-1). В других вариантах осуществления, особенно приемлемых для сельскохозяйственных птиц, антигенный компонент включает профилин.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0023] Определения:
[0024] Термины ‘около’ или ‘приблизительно’, когда они используются в связи с измеряемой числовой переменной, относятся к указанному значению переменной и ко всем значениям переменной, находящимся в экспериментальной погрешности указанного значения (например, в пределах 95% доверительного интервала для среднего значения) или в пределах 10 процентов от указанного значения, в зависимости оттого, которое является большим, за исключением случаев, когда ‘около’ используется по отношению к временным интервалам в неделях, когда "около 3 недель", составляет от 17 до 25 дней, и когда от приблизительно 2 до приблизительно 4 недель составляет от 10 до 40 дней.
[0025] Термин ‘сопутствующая повышенная температура’ касается повышения температуры у вакцинированного животного в пределах одного дня вакцинации. В случае крупного рогатого скота, термин касается ректальной температуры свыше 103,5°F.
[0026] Термин ‘антиген’ в сочетании с данным видом касается патогенов, вызывающих инфекционное заболевание у указанного вида, или компонентов данных патогенов. Таким образом, например, термин ‘антигены крупного рогатого скота’ касается патогенов, способных вызвать инфекционные заболевания у крупного рогатого скота, или компонентов данных патогенов.
[0027] Термин ‘фактически состоит из’ и подобный, применяемый к липосомам и адъювантным композициям по представленному изобретению, касается композиций, не содержащих дополнительных адъювантных или иммуномодулирующих агентов в количествах, при которых указанный агент проявляет адъювантные или иммуномодулирующие эффекты, которые могут быть измерены.
[0028] Термины ‘фактически не содержит сапонин’, ‘фактически является свободным от сапонина’ и подобные им, касаются композиции, не содержащей сапонин в количествах, при которых сапонин проявляет адъювантные или иммуномодулирующие эффекты, которые могут быть измерены. В некоторых вариантах осуществления, фактически свободные от сапонина композиции содержат сапонин в количестве недостаточном для того, чтобы вызвать системный иммунный ответ, такой как повышенная температура. В некоторых вариантах осуществления, фактически свободные от сапонина композиции не содержат никакого сапонина или содержат сапонин в пределах детектирования или ниже.
[0029] Аналогичным образом, термины ‘фактически не содержит CpG дезоксирибонуклеотид’, ‘фактически является свободным от CpG дезоксирибонуклеотида’ и подобные им касаются композиции, не содержащей CpG дезоксирибонуклеотид в количествах, при которых CpG дезоксирибонуклеотид проявляет адъювантные или иммуномодулирующие эффекты, которые могут быть измерены. В некоторых вариантах осуществления, фактически свободные от CpG дезоксирибонуклеотида композиции не содержат никакого CpG дезоксирибонуклеотида или содержат CpG дезоксирибонуклеотид в пределах детектирования или ниже. Термины ‘фактически не содержит CpG дезоксирибонуклеотид’, ‘фактически является свободным от CpG дезоксирибонуклеотида’ и подобные им исключают вакцины, в которых CpG дезоксирибонуклеотид естественно присутствует в антигене.
[0030] Термин ‘иммуностимулирующая молекула’ касается молекулы, усиливающей иммунный ответ.
[0031] Термин ‘липосома’ касается микроскопической сферической частицы, образованной липидным бислоем, окружающим водную камеру.
[0032] Термин ‘парентеральное введение’ касается введения вещества, такого как вакцина, в организм субъекта через или путем, не включающим желудочно-кишечный тракт. Парентеральное введение включает подкожное, внутримышечное, транскутанное, интрадермальное, внутрибрюшное, внутриглазное и внутривенное введение.
[0033] Термины ‘терапевтически эффективное количество’ и ‘эффективное количество’ касаются количества антигена или вакцины, которые будут индуцировать иммунный ответ у субъекта, получающего антиген или вакцину, адекватную для, предотвращения или уменьшения признаков или симптомов заболевания, включая неблагоприятные последствия для здоровья или их осложнения, вызванные инфицированием патогеном, таким как вирус или бактерия. Индуцированным может быть гуморальный иммунитет или клеточно-опосредованный иммунитет или как гуморальный, так и клеточно-опосредованный иммунитет. Иммуногенность и эффективность вакцины у животного могут быть оценены, например, опосредованно путем измерения титров антител, анализов пролиферации лимфоцитов, анализов ELISPOT IFN гамма, анализов цитотоксических Т-клеток или непосредственно путем наблюдения за признаками и симптомами после обнаружения штаммов немутантного типа. Защитный иммунитет, предоставляемый вакциной, может быть оценен путем измерения, например, уменьшения клинических признаков, таких как смертность, заболеваемость, повышенная температура, вирусемия, влияние на клиническую патологию, общее физическое состояние, общее состояние здоровья и активность субъекта. Количество вакцины, являющейся терапевтически эффективной, может варьировать в зависимости от конкретного используемого адъюванта, конкретного используемого антигена или состояния субъекта, и может быть определено квалифицированным специалистом в данной области.
[0034] Изобретение предполагает, в частности, липосомы, содержащие внутреннюю камеру и внешнюю мембрану. Липосомы могут иметь средний размер частиц от 50 до 500 нм. В некоторых не ограничивающих вариантах осуществления, средний размер частиц липосом составляет 100-500 нм, или 150-450 нм, или 150-250 нм, или 300-400 нм, или 250-300 нм. В некоторых вариантах осуществления, мембрана содержит четвертичное аминное соединение, фосфолипид, стерин, и гликолипид. В некоторых вариантах осуществления, липосома фактически не содержит сапонин.
[0035] В некоторых вариантах осуществления, внешняя мембрана фактически состоит из или состоит из четвертичного аминного соединения, фосфолипида, стерина, и гликолипида. В других вариантах осуществления, внешняя камера липосомы не содержит никаких иммуностимулирующих олигонуклеотидов и/или других иммуномодулирующих соединений. Таким образом, в таких вариантах осуществления, липосома фактически состоит из, или состоит из внутренней камеры, фактически состоящей из или состоящей из иммунологически инертного водного носителя, где указанная внутренняя камера окружена внешней мембраной, фактически состоящей из, или состоящей из четвертичного аминного соединения, фосфолипида, стерина, и гликолипида.
[0036] Четвертичные аминные соединения представляют собой соединения на основе аммония с четырьмя углеводородными группами. На практике, углеводородные группы, как правило, ограничиваются алкильной или арильной группами. В наборе вариантов осуществления, четвертичные аминные соединения состоят из четырех алкильных цепей, две из которых представляют собой С10-С20 алкилы, и две, оставшиеся, представляют собой С1-С4 алкилы. В некоторых вариантах осуществления, четвертичный амин представляет собой диметилдиоктадециламмония (DDA) бромид, хлорид или другой фармацевтически приемлемый противоион.
[0037] Стерины имеют общее химическое ядро, представляющее собой стероидную(ые) кольцевую(ые) структуру[ы], содержащее гидроксильную (ОН) группу, как правило, присоединенную к углероду-3. Углеводородная цепь жирнокислотного заместителя варьирует в длину, как правило, от 16 до 20 атомов углерода, и может быть насыщенной или ненасыщенной. Стерины, как правило, содержат одну или несколько двойных связей в кольцевой структуре и также различные заместители, присоединенные к кольцам. Стерины и их жирнокислотные сложные эфиры фактически нерастворимы в воде. Принимая во внимание данные химические сходства, существует, таким образом, вероятность того, что стерины, имеющие данное химическое ядро, будут иметь подобные свойства, при использовании в вакцинных композициях по представленному изобретению. Стерины хорошо известны в данной области из уровня техники и могут быть коммерчески приобретенными. Например холестерин раскрыт в Merck Index, 12th Ed., P. 369. Приемлемые стерины включают, но не ограничиваются этим, β-ситостерин, стигмастерин, эргостерин, эргокальциферол, и холестерин.
[0038] Приемлемые гликолипиды, как правило, представляют собой те, что активируют ответ Th2. Гликолипиды включают, но не ограничиваются этим, те, что охватываются формулой I, и которые, как правило, описаны в патентной публикации США 20070196384 (Ramasamy et al.).
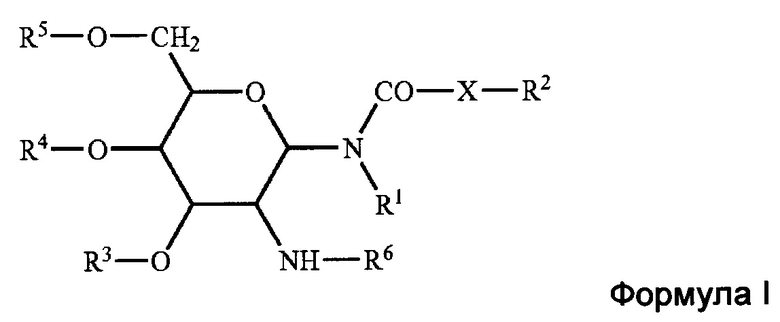
[0039] В структурной формуле I, R1 и R2 независимо представляют собой водород, или насыщенный алкильный радикал, содержащий вплоть до 20 атомов углерода; X представляет собой -СН2-, -О- или -NH-; R2 представляет собой водород, или насыщенный или ненасыщенный алкильный радикал, содержащий вплоть до 20 атомов углерода; R3, R4, и R5 независимо представляют собой водород, -SO42-, -PO42-, -СОС1-10 алкил; R6 представляет собой L-аланил, L-альфа-аминобутил, L-аргинил, L-аспаргинил, L-аспартил, L-цистеинил, L-глутамил, L-глицил, L-гистидил, L-гидроксипропил, L-изолейцил, L-лейцил, L-лизил, L-метионил, L-орнитинил, L-фенилаланил, L-пролил, L-серил, L-треонил, L-тирозил, L-триптофанил, и L-валил или их D-изомеры.
[0040] Примеры гликолипида включают, но не ограничиваются этим, N-(2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-октадецилдодеканоиламид (BayR®1005, или R1005) или его соль (например, ацетатную).
[0041] Лецитин может быть получен в виде смеси фосфатидов и триглицеридов путем промывания водой неочищенных растительных масел, отделения и сушки полученных в результате гидратированных смол. Очищенный продукт может быть получен путем фракционирования смеси нерастворимых в ацетоне фосфолипидов и гликолипидов, оставшихся после удаления триглицеридов, и растительного масла за счет промывания ацетоном. Альтернативно, лецитин может быть получен из различных коммерческих источников.
[0042] Другие приемлемые фосфолипиды включают фосфатидилхолин, фосфатидилглицерин, фосфатидилинозитол, фосфатидилсерин, ацилфосфатидилэтаноламин, дифосфатидилглицерин, лизофосфатидилэтаноламин, лизофосфатидилхолин, фосфатидиновую кислоту, кардиолипин, и фосфатидилэтаноламин. Фосфолипиды могут быть выделены из природных источников или синтезированы общепринятыми способами.
[0043] Липосомы, как описано в данном документе, позволяют использовать гибкие соотношения элементов внешней мембраны. В некоторых вариантах осуществления, массовые соотношения четвертичного аммониевого соединения: стерина фосфолипида : гликолипида составляют 1:0,75-1,25:1,5-2,5:1,5-2,5, соответственно. В некоторых вариантах осуществления, массовые соотношения четвертичного аммониевого соединения: стерина : фосфолипида : гликолипида составляют 1:1:2:2.
[0044] В других вариантах осуществления, общая масса четвертичного аммониевого соединения и стерина составляет приблизительно половину (например, 40%, 45%, 50%, 55%, 60%) от общее массы гликолипида и фосфолипида, при условии, что четвертичное аммониевое соединение составляет, по меньшей мере, приблизительно 5% масс./масс. от общее массы данных четырех соединений (четвертичного аммониевого соединения, стерина, фосфолипида и гликолипида), и гликолипид составляет, по меньшей мере, приблизительно 20% масс./масс. от общей массы данных четырех соединений.
[0045] В некоторых вариантах осуществления, общая масса четвертичного аммониевого соединения и стерина составляет приблизительно 10-40% (например, приблизительно 10%, приблизительно 15%, приблизительно 20%, приблизительно 25%, приблизительно 30%, приблизительно 33,3%, приблизительно 35%, приблизительно 40%) от общее массы гликолипида и фосфолипида, при условии, что четвертичное аммониевое соединение составляет, по меньшей мере, приблизительно 5% масс./масс. от общее массы данных четырех соединений (четвертичного аммониевого соединения, стерина, фосфолипида и гликолипида), и гликолипид составляет, по меньшей мере, приблизительно 20% масс./масс. от общее массы данных четырех соединений.
[0046] В некоторых вариантах осуществления, иммунологически эффективное количество липосом по представленному изобретению может быть введено в качестве адъюванта. В некоторых вариантах осуществления, изобретение предполагает вакцинную комбинацию, содержащую иммунологически эффективное количество адъювантной композиции и антигенный компонент, как описано ниже.
[0047] Масса вида пациента, в конечном итоге, определяет дозу адъювантной композиции по представленному изобретению.
[0048] В некоторых вариантах осуществления, приемлемая для крупного рогатого скота, лошадей и взрослых свиней, одна доза содержит эквивалент 1000-3000 мкг внешнего мембранного компонента (то есть, общая масса четвертичного аммониевого соединения, стерина, фосфолипида и гликолипида), или эквивалент 1000-2000 мкг, или эквивалент 1000-1500 мкг, или эквивалент 1300-1800 мкг, или эквивалент 1500-2000 мкг.
[0049] Масса липосомальной композиции не может быть равна массе мембранного компонента вследствие присутствия внутренней камеры, которая может содержать иммуностимулирующий олигонуклеотид, антигенный компонент, другие иммуномодуляторы, и тому подобное. Применение эквивалентов к липосомальному мембранному компоненту позволяет проводить равномерное дозирование. Схема дозирования, приведенная в данном отношении, гарантирует, что животное крупного рогатого скота получает, по меньшей мере, 200 мкг гликолипида и приблизительно 50 мкг четвертичного аммониевого соединения.
[0050] В некоторых вариантах осуществления приемлемых для овец и коз, одна доза содержит эквивалент 300-1000 мкг внешнего мембранного компонента, например, эквивалент 300-500 мкг, или эквивалент 400-500 мкг, или эквивалент 400-1000 мкг, или эквивалент 500-1000 мкг, или эквивалент 600-1000 мкг, или эквивалент 600-800 мкг.
[0051] В некоторых вариантах осуществления приемлемых для поросят, собак и кошек, одна доза содержит эквивалент 100-400 мкг внешнего мембранного компонента, или эквивалент 100-200 мкг, или эквивалент 100-150 мкг, или эквивалент 130-180 мкг, или эквивалент 150-200 мкг.
[0052] В некоторых вариантах осуществления приемлемых для птиц, одна доза содержит эквивалент 50-200 мкг внешнего мембранного компонента, или эквивалент 50-100 мкг, или эквивалент 50-75 мкг, или эквивалент 65-90 мкг, или эквивалент 75-100 мкг, или эквивалент 75-150 мкг.
[0053] Внутреннее соединение липосомы может содержать антигены или другие иммуномодулирующие молекулы. В некоторых вариантах осуществления, такие иммуномодулирующие молекулы, приемлемые для внутренней камеры, включают, но не ограничиваются этим, экстракты антигенов, субъединицы, синтетические вещества, целые клетки или вирусы.
[0054] В некоторых вариантах осуществления, активные фармацевтические вещества могут быть запакованны вовнутрь липосомы.
[0055] Иммуномодуляторы, которые могут быть упакованы, также включают, но не ограничиваются этим, гмлТ, MPLA, альфа-Gal-Cer, токсин холеры, LPS, липотейхоевые кислоты, поли I:C, флагелин, зимозан, хитин и модифицированные хитиновые формы, бета-глюканы, авридин, инулин и модифицированные инулиновые формы, этилен оксиянтарные ангидриды, плюрониловые вещества, такие как L121 и L141, агонист CD40, агонист TLR5, а также какой-либо агонист TLR, GM-CSF.
[0056] В некоторых вариантах осуществления, липосомы могут нести различные молекулы, которые могут использоваться в качестве маркеров, включая, но не ограничиваются этим, OspA, OspC, пертактин и другие.
[0057] В некоторых вариантах осуществления, адъювантная композиция по представленному изобретению дополнительно содержит иммуностимулирующие олигонуклеотиды, такие как, например, CpG олигодезоксирибонуклеотиды или иммуностимулирующие олигорибонуклеотиды (ORNs), или их химеры. Приемлемые неограничивающие примеры CpG олигодезоксирибонуклеотидов проиллюстрированы в SEQ ID NO 1-10, приемлемые неограничивающие примеры ORN представлены в SEQ ID NO 11-13, и приемлемый неограничивающий пример химерного иммуностимулирующего олигонуклеотида представлен в SEQ ID NO: 14.
[0058] Данные иммуностимулирующие олигонуклеотиды, в некоторых вариантах осуществления, присутствуют во внутренней камере липосомы.
[0059] В некоторых вариантах осуществления, иммуномодулирующие олигонуклеотиды ассоциированы с наружной поверхностью липосомы. Ассоциация может происходить за счет водородных связей, электростатических связей, липофильных связей, Ван-дер-Ваальских сил, и подобного.
[0060] В некоторых вариантах осуществления, отрицательно заряженный иммуностимулирующий олигонуклеотид ассоциирован с наружной поверхностью липосомы за счет взаимодействия с положительно заряженным четвертичным атомом азота в четвертичном аммониевом соединении.
[0061] CpG олигодезоксирибонуклеотиды (также называют CpG дезоксирибонуклеотиды или CpG ODN) представляют собой недавно описанный класс фармакотерапевтических агентов, характеризующихся присутствием неметилированного CG динуклеотида в контекстах специфической нуклеотидной последовательности (CpG мотив). (Hansel ТТ, Barnes PJ (eds): New Drugs for Asthma, Allergy та COPD. Prog Respir Res. Basel, Karger, 2001, vol 31, pp 229-232, включенный в данный документ в виде ссылки). Данные мотивы CpG не наблюдаются в эукариотической ДНК, в которой CG динуклеотиды подавляются и, если присутствуют, как правило, метилируются, но присутствуют в бактериальной ДНК, которой они обеспечивают иммуностимулирующие свойства.
[0062] В некоторых вариантах осуществления, адъюванты по представленному изобретению используют так называемый иммуностимулирующий олигонуклеотид, более предпочтительно, модифицированные иммуностимулирующие олигонуклеотиды Р-класса, еще более предпочтительно, Е-модифицированные олигонуклеотиды Р-класса. Иммуностимулирующие олигонуклеотиды Р - класса представляют собой олигодезоксирибонуклеотид CpG, характеризующийся присутствием палиндромов, как правило, длиной 6-20 нуклеотидов. Олигонуклеотиды Р-класса обладают способностью спонтанно самособираться в конкатемеры, или in vitro, и/или in vivo. Данные олигонуклеотиды, в точном значении, являются одноцепочечными, но присутствие палиндромов позволяет образование конкатемеров или, возможно, стволово-петлевых структур. Общая длина иммуностимулирующих олигонуклеотидов Р-класса составляет от 19 до 100 нуклеотидов, например, 19-30 нуклеотидов, 30-40 нуклеотидов, 40-50 нуклеотидов, 50-60 нуклеотидов, 60-70 нуклеотидов, 70-80 нуклеотидов, 80-90 нуклеотидов, 90-100 нуклеотидов.
[0063] В одном аспекте изобретения иммуностимулирующий олигонуклеотид содержит 5' TLR активационный домен и, по меньшей мере, два палиндромических участка, причем один палиндромический участок представляет собой 5' палиндромический участок из, по меньшей мере, 6 нуклеотидов в длину, и соединенный с 3' палиндромическим участком из, по меньшей мере, 8 нуклеотидов в длину, или непосредственно, или через спейсер.
[0064] Иммуностимулирующие олигонуклеотиды Р-класса могут быть модифицированными согласно методикам, известным в данной области. Например, J-модификация относится к йод-модифицированным нуклеотидам. Е-модификация относятся к этил-модифицированному(ым) нуклеотиду(ам). Таким образом, Е-модифицированные иммуностимулирующие олигонуклеотиды Р-класса представляют собой иммуностимулирующие олигонуклеотиды Р-класса, в которых, по меньшей мере, один нуклеотид (предпочтительно 5' нуклеотид) является этилированным. Дополнительные модификации включают присоединение 6-нитробензимидазола, О-метилирование, модификацию с пронил-dU, инозиновую модификацию, присоединение 2-бромвинила (предпочтительно, куридину).
[0065] Иммуностимулирующие олигонуклеотиды Р-класса также могут содержать модифицированную межнуклеотидную связь, включающую, но не ограничиваются этим, фосфодисложноэфирные связи и фосфоротиоатные связи. Олигонуклеотиды по представленному изобретению могут быть синтезированы или получены из коммерческих источников.
[0066] Олигонуклеотиды Р-класса и модифицированные олигонуклеотиды Р-класса, кроме того, раскрыты в опубликованной заявке РСТ № WO 2008/068638, опубликованной 12 июня 2008 года. Приемлемые неограничивающие примеры модифицированных иммуностимулирующих олигонуклеотидов Р-класса представлены ниже (в SEQ ID NO 1-10, "*" касается фосфоротиоатной связи и "_" касается фосфодисложноэфирной связи). В SEQ ID NO 11-14, все связи представляют собой или фосфодисложноэфирные или фосфоротиоатные связи.

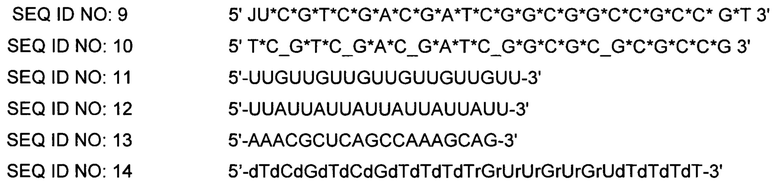
[0067] Доза иммуностимулирующего олигонуклеотида для применения в адъювантных композициях в конечном итоге зависит от предназначенного вида.
[0068] Например, в некоторых вариантах осуществления приемлемых для крупного рогатого скота, овец или взрослых свиней, одна доза адъювантной композиции по представленному изобретению будет содержать от приблизительно 50 до 400 мкг (например, 50-300, или 100-250 мкг, или от приблизительно 50 до приблизительно 100 мкг для взрослых свиней и от приблизительно 100 до приблизительно 250 мкг для крупного рогатого скота) иммуностимулирующего олигонуклеотида.
[0069] В некоторых вариантах осуществления, приемлемых для животных-компаньонов или поросят, одна доза адъювантной композиции по представленному изобретению будет содержать от приблизительно 5 до 100 мкг (например, 10-80 мкг, или 20-50 мкг) иммуностимулирующего олигонуклеотида.
[0070] В некоторых вариантах осуществления приемлемых для домашней птицы, одна доза адъювантной композиции по представленному изобретению будет содержать от приблизительно 0,1 до приблизительно 5 мкг (например, 0,5-3 мкг, или 0,9-1,1 мкг) иммуностимулирующего олигонуклеотида.
[0071] Способы получения липосом хорошо известны в данной области. Коротко говоря, компоненты липосомы растворяют и смешивают в органическом растворителе, например, метиленхлориде, и затем растворитель удаляют путем высушивания для получения пленки. Пленку позже регидратируют, используя водную среду (например, воду или буфер), необязательно содержащую соединения, которые должны быть включены во внутреннюю камеру липосом. В различных вариантах осуществления, соединения могут включать иммуностимулирующие олигонуклеотиды, другие иммуномодуляторы, и/или антигенный компонент.
[0072] Стадия регидратации сопровождается обработкой ультразвуком и/или экструзией для уменьшения размера пузырьков, образующихся при стадии регидратации.
[0073] Существуют две основные методики обработки ультразвуком: зондовая/ наконечниковая обработка ультразвуком и обработка ультразвуком в ванне. Зондовая/наконечниковая обработка ультразвуком имеет высокую энергию потребления, которая приводит к значительной генерации тепла, поэтому необходимо применение ледяной бани для поддержания температуры липосомальной дисперсии для предотвращения разложения липина. Альтернативно, энергия ультразвука может быть опосредованно разделена на липосомальную суспензию, используя ультразвуковую ванну, где температуру легче контролировать, но потери энергии сравнительно высоки. Обработка ультразвуком, как правило, дает небольшие пузырьки (~10 нм), которые спонтанно сливаются в течение времени, ослабляя стресс высокой кривизны мембраны.
[0074] Способ экструдирования включает пропускание липосомальной суспензии через мембрану с определенным размером пор. Данный способ является предпочтительным, поскольку определенный размер пор способствует однородности размера частиц в пределах липосомальной популяции, хотя экструдирования ниже температуры липидного перехода может быть сложным вследствие жесткости мембраны. Липосомальные суспензии часто подвергают экструзии несколько раз, для достижения низкой полидисперсности в конечном продукте.
[0075] Предпочтительным может быть получить стабильный при хранении препарат из липосом. В некоторых вариантах осуществления, такой стабильный при хранении препарат из липосом создают путем высушивания при замораживании. Коротко говоря, сухую пленку, описанную выше, регидратируют в водном буфере, содержащем криопротектор и лиопротектор, такой как сахароза, трегалоза, или их комбинации. В других вариантах осуществления, криопротектор и лиопротектор добавляют после стадии регидратации. Регидратированный препарат затем лиофилизируют, используя способы хорошо известные в данной области. Полученный в результате лиофилизированный препарат является стабильным при хранении. В нужное время, он может быть регидратирован приемлемым буфером.
[0076] Дополнительные иммуномодуляторы, включая, но не ограничиваются этим, иммуностимулирующие олигонуклеотиды и антиген(ы) могут добавляться или перед высушиванием при замораживании, или во время конечного приготовления.
[0077] В некоторых вариантах осуществления, антигены смешиваются с липосомальной композицией после того, как липосомы по представленному изобретению восстановлены. В других вариантах осуществления, липосомы, дополнительные иммуномодуляторы и антигенный компонент получают и сушат вместе.
[0078] В некоторых вариантах осуществления, дополнительные иммуностимулирующие соединения присутствуют в композициях по представленному изобретению. Такие дополнительные иммуностимулирующие соединения могут присутствовать во внутренней камере липосом, и/или ассоциироваться с внешней поверхностью липосом, и/или независимо от липосом, в адъювантных композициях по представленному изобретению.
[0079] Приемлемые неограничивающие примеры таких дополнительных иммуностимулирующих соединений включают, но не ограничиваются этим, несколько классов адъювантов, таких как минеральные соли, например, алюмокалиевые квасцы, гидроксид алюминия, фосфат алюминия и фосфат кальция; поверхностно-активные агенты и микрочастицы, например, неионные блок-полимерные поверхностно-активные вещества, виросомы, сапонины (например, Quil A, QS-21 и GPI-0100), протеосомы, иммунные стимулирующие комплексы, со-хелаты, пиридин, витамин А, витамин Е; бактериальные продукты, такие как RIBI адъювантная система (Ribi Inc.), каркас клеточной стенки Mycobacterium phlei (Detox®), мурамил дипептиды (MDP) и трипептиды (МТР), монофосфориллипид A (MPLA), бацила Кальметта - Герена (БЦЖ), тепловые лабильные энтеротоксины Е. coli, холерный токсин, трегалозы димиколат, цитокины и гормоны, например, интерлейкины (IL-1, IL-2, IL-6, IL-12, IL-15, IL-18), гранулоцитарно-макрофагальный колониестимулирующий фактор, дегидроэпиандростерон, 1,25-дигидрокси витамин D3; полианионы, например, декстран; полиакрилы (например, полиметилметакрилат, КАРБОПОЛ®934Р); носители, например, столбнячный анатоксин, дифтерийный анатоксин, субединица В холерного токсина, мутантный тепловой лабильный энтеротоксин энтеротоксигенной Е. coli (гмлТ), протеины теплового шока; эмульсии масло-в-воде, например, AMPHIGEN® (Hydronics, USA); поликатионные носители (например, DEAE декстран или QAE декстран), и эмульсии вода-в-масле, такие как, например, и неполные адъюванты Фрейнда.
[0080] Другие приемлемые иммуномодуляторы включают Альфа-Gal-Cer. LPS, липотейхоевые кислоты, поли I:C, флагелин, зимозан, хитин и модифицированные хитиновые формы, бета-глюканы, авридин, инулин и модифицированные инулиновые формы, этиленоксиянтарные ангидриды, плюрониловые вещества, такие как L121 и L141, агонист CD40, агонист TLR5, а также какой-либо агонист TLR агонист.
Антигены и заболевания
[0081] В некоторых вариантах осуществления, липосомальная адъювантная композиция по представленному изобретению может быть скомбинирована с антигенным компонентом, таким образом образуя вакцинную композицию по представленному изобретению. Антигенный компонент вакцин по представленному изобретению может присутствовать во внутренней камере липосом, и/или быть ассоциированным с внешней поверхностью липосом, и/или независимо от липосом.
[0082] В некоторых вариантах осуществления, вакцинная композиция фактически не содержит сапонин. В дополнительных вариантах осуществления, вакцинная композиция фактически не содержит дезоксирибонуклеотид CpG.
[0083] Предпочтительными являются варианты осуществления, в которых вакцинная композиция фактически не содержит дезоксирибонуклеотид CpG, если антиген в вакцине содержит цельный вирус с оцРНК (вирус с или (+)оцРНК, или (-)оцРНК), последовательности которых являются иммуностимулирующими, несмотря на направленный TLR7/8. Такие стимулирующие последовательности TLR 7/8 включают полии или GU-обогащенные оцРНК последовательности. Heil F, Hemmi Н. et al., 2004. Science 303(5663):1526-9. Diebold SS., Kaisho T. et al., 2004. Science 303(5663): 1529-31.
[0084] Вирусы, содержащие такие последовательности включают, но не ограничиваются этим, различные вирусы гриппа (например, вирус гриппа крупного рогатого скота, вирус гриппа собак, вирус гриппа коней, вирус свиного гриппа и подобные). В конкретных предпочтительных вариантах осуществления, антигенный компонент вакцины фактически свободный от CpG ODN содержит вирус гриппа.
[0085] Антигенный компонент может включать, в различных иллюстративных вариантах осуществления, антигены крупного рогатого скота, козьи антигены, свиные антигены, антигены домашней птицы, антигены коней, собачьи антигены, антигены коней и кошачьи антигены.
[0086] Антигены могут представлять собой какое-либо из широкого диапазона веществ, способных к выработке желаемого иммунного ответа у субъекта, включая, но не ограничиваются этим, один или несколько вирусов (инактивированных, ослабленных, и модифицированных живых), бактерий, паразитов, нуклеотидов (включая, но не ограничиваются этим, антигены на основе нуклеиновой кислоты, например, ДНК вакцины или мРНК вакцины), полинуклеотидов, пептидов, полипептидов, рекомбинантных протеинов, синтетических пептидов, протеиновых экстрактов, клеток (включая, опухолевые клетки), тканей, полисахаридов, углеводов, жирных кислот, липотейхоевой кислоты, пептидогликанов, липидов, или гликолипидов, индивидуально или в какой-либо их комбинации.
[0087] Антигены, использующиеся с адъювантами по изобретению также включают иммуногенные фрагменты нуклеотидов, полинуклеотидов, пептидов, полипептидов, которые могут быть выделены из организмов, указанных в данном документе, или химически или биологически изготовлены.
[0088] Живые, модифицированные живые и ослабленные вирусные штаммы, не вызывающие заболевания у субъекта были выделены в невирулентной форме или были ослаблены, используя способы, хорошо известные в данной области, включая серийный пассаж в приемлемой клеточной линии или влияние ультрафиолетового света или химического мутагена. Инактивированные или убитые вирусные штаммы представляют собой те, которые были инактивированы, используя способы, известные квалифицированному специалисту в данной области, включая обработку формалином, бета-пропиолактоном (BPL), пероксидами, бинарным этиленимином (BEI), стерилизующим излучением, теплом или другими такими способами.
[0089] Два или более антигенов могут быть совмещенными для получения поливалентой композиции, которая может защитить субъект против широкого диапазона заболеваний, вызывающихся патогенами. В настоящее время, коммерческие производители вакцин, а также конечные потребители отдают предпочтение поливалентным вакцинным продуктам. Несмотря на то, что обычные адъюванты часто ограничиваются различными антигенами, с которыми они могут эффективно использоваться (или моновалентно, или поливалентно), адъюванты, описанные в данном документе, могут эффективно использоваться с широким диапазоном антигенов, как моновалентно, так и поливалентно. Таким образом, антигены, описанные в данном документе, могут сочетаться в единую композицию, содержащую адъюванты, описанные в данном документе.
[0090] Примеры бактерий, которые могут использоваться, в качестве антигенов с адъювантными композициями, описанными в данном документе, включают, но не ограничиваются этим, Acinetobacter calcoaceticus, Acetobacter paseruianus, Actinobacillus pleuropneumoniae, Aeromonas hydrophila, Alicyclobacillus acidocaldarius, Arhaeglobus fulgidus, Bacillus pumilus, Bacillus stearothermophillus, Bacillus subtilis, Bacillus thermocatenulatus, Bordetella bronchiseptica, Burkholderia cepacia, Burkholderia glumae, Campylobacter coli, Campylobacter fetus, Campylobacter jejuni, Campylobacter hyointestinalis, Chlamydia psittaci, Chlamydia trachomatis, Chlamydophila spp., Chromobacterium viscosum, Erysipelothrix rhusiopathieae, Listeria monocytogenes, Ehrlichia canis, Escherichia coli, Haemophilus influenzae, Haemophilus somnus, Helicobacter suis, Lawsonia intracellularis, Legionella pneumophilia, Moraxellsa sp., Mycobactrium bovis, Mycoplasma hyopneumoniae, Mycoplasma mycoides subsp. mycoides LC, Clostridium perfringens, Odoribacter denticanis, Pasteurella (Mannheimia) haemolytica, Pasteurella multocida, Photorhabdus luminescens, Porphyromonas gulae, Porphyromonas gingivalis, Porphyromonas salivosa, Propionibacterium acnes, Proteus vulgaris, Pseudomonas wisconsinensis, Pseudomonas aeruginosa, Pseudomonas fluorescens C9, Pseudomonas fluorescens SIKW1, Pseudomonas fragi, Pseudomonas luteola, Pseudomonas oleovorans, Pseudomonas sp B11-1, Alcaliges eutrophus, Psychrobacter immobilis, Rickettsia prowazekii, Rickettsia rickettsii, Salmonella enterica все серологические варианты, включая, например: Salmonella enterica Typhimurium, Salmonella enterica Bongori, Salmonella enterica Dublin, Salmonella enterica Choleraesuis, и Salmonella enterica Newport, Serratia marcescens, Spirluina platensis, Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus hyicus, Streptomyces albus, Streptomyces cinnamoneus, Streptococcus uberis, Streptococcus suis, Streptomyces exfoliates, Streptomyces scabies, Sulfolobus acidocaldarius, Syechocystis sp., Vibrio cholerae, Borrelia burgdorferi, Treponema denticola, Treponema minutum, Treponema phagedenis, Treponema refringens, Treponema vincentii, Treponema palladium, Trueperella pyogenes и Leptospira species, такие как известные патогены Leptospira canicola, Leptospira grippotyposa, Leptospira hardjo, Leptospira borgpetersenii hardjo-bovis, Leptospira borgpetersenii hardjo-prajitno, Leptospira interrogans, Leptospira icterohaemorrhagiae, Leptospira pomona, и Leptospira bratislava, и их комбинации.
[0091] Как инактивированные вирусы, так и ослабленные живые вирусы могут использоваться в адъювантных композициях. Некоторые примеры вирусов, которые могут использоваться в качестве антигенов, включают, но не ограничиваются этим, птичьи герпесвирусы, герпесвирусы крупного рогатого скота, собачьи герпесвирусы, герпесвирусы коней, вирус кошачьего вирусного ринотрахеита, вирус болезни Марека, овечьи герпесвирусы, свиные герпесвирусы, вирус эпизоотической диареи свиней (PEDv), вирус псевдобешенства, птичьи парамиксовирусы, респираторно-синцитиальный вирус крупного рогатого скота, вирус чумки собак, собачий паравирус гриппа, собачий аденовирус, собачий парвовирус, вирус парагриппа 3 крупного рогатого скота, овечий вирус парагриппа 3, вирус чумы, вирус пограничной болезни овец, вирус вирусной диареи - болезни слизистых крупного рогатого скота (BVDV), BVDV типа I, BVDV типа II, вирус классической чумы свиней, птичий вирус лейкоза, вирус иммунодефицита крупного рогатого скота, вирус лейкоза крупного рогатого скота, туберкулез крупного рогатого скота, вирус инфекционной анемии лошадей, вирус иммунодефицита кошек, вирус лейкоза кошачьих (FeLV), вирус ньюкаслской болезни, овечий вирус прогрессирующей пневмонии, овечий вирус легочной аденокарциномы, собачий коронавирус (CCV), пантропный CCV, собачий респираторный коронавирус, коронавирус крупного рогатого скота, кошачий калицивирус, кошачий кишечный коронавирус, кошачий вирус инфекционного перитонита, свиной вирус эпидемической диареи, свиной вирус гемагглютинирующего энцефаломиелита, свиной парвовирус, свиной цирковирус (PCV) типа I, PCV типа II, свиной вирус репродуктивный и респираторный синдром (PRRS), вирус трансмиссивного гастроэнтерита, корона вирус индюков, вирус эфемерной лихорадки крупного рогатого скота, вирус бешенства, ротовирус, вирус везикулярного стоматита, лентивирус, птичий грипп, риновирусы, вирус гриппа лошадей, вирус свиного гриппа, вирус собачьего гриппа, вирус кошачьего гриппа, вирус гриппа человека, вирус восточного энцефалита лошадей (ЕЕЕ), вирус венесуэльского энцефалита лошадей, вирус западного Нила, вирус западного энцефалита лошадей, вирус иммунодефицита человека, вирус папилломы человека, вирус ветряной оспы, вирус гепатита В, риновирус, вирус кори и их комбинации.
[0092] Примеры пептидных антигенов включают Bordetella bronchiseptica р68, GnRH, пептиды IgE, Fel d1, и раковые антигены, и их комбинации. Примеры других антигенов включают нуклеотиды, углеводы, липиды, гликолипиды, пептиды, жирные кислоты, липотейхоевую и тейхоевую кислоту, и пептидогликаны, и их комбинации.
[0093] Примеры паразитов, которые могут использоваться в качестве антигенов с адъювантными композициями, описанными в данном документе, включают, но не ограничиваются этим, Anaplasma, Fasciola hepatica (печеночная двуустка), Coccidia, Eimeria spp., Neospora caninum, Toxoplasma gondii, Giardia, Dirofilaria (сердечный гельминт), Ancylostoma (анкилостоматиды), Cooperia, Haemonchus contortus (гемонхус), Ostertagia ostertagi (желудочный гельминт), Dictyocaulus viviparous (легочные гельминты), Trypanosoma spp., Leishmania spp., Trichomonas spp., Cryptosporidium parvum, Babesia, Schistosoma, Taenia, Strongyloides, Ascaris, Trichinella, Sarcocystis, Hammondia, и Isopsora, и их комбинации. Кроме того, рассматриваются внешние паразиты, включая, но не ограничиваются этим, клещи, включая виды Ixodes, Rhipicephalus, Dermacentor, Amblyomma, Boophilus, Hyalomma, и Haemaphysalis, и их комбинации.
[0094] Количество антигена, используемого для индуцирования иммунного ответа будут варьироваться в значительной степени в зависимости от используемого антигена, субъекта, и уровня желаемого ответа, и может быть определено квалифицированным специалистом в данной области. Для вакцин, содержащих модифицированные живые вирусы или ослабленные вирусы, терапевтически эффективное количество антигена, как правило, находится в диапазоне от приблизительно 102 инфекционной дозы в тканевой культуре (TCID)50 до приблизительно 1010 TCID50, включительно. Для многих таких вирусов, терапевтически эффективная доза, как правило, находится в диапазоне от приблизительно 102 TCID50 до приблизительно 108 TCID50, включительно. В некоторых вариантах осуществления, диапазоны терапевтически эффективных доз составляют от приблизительно 103 TCID50 до приблизительно 106 TCID50, включительно. В некоторых других вариантах осуществления, диапазоны терапевтически эффективных доз составляют от приблизительно 104 TCID50 до приблизительно 105 TCID50, включительно.
[0095] Для вакцин, содержащих инактивированные вирусы, терапевтически эффективное количество антигена, как правило, составляет по меньшей мере приблизительно 100 относительных единиц на дозу, и часто в диапазоне от приблизительно 1000 до приблизительно 4500 относительных единиц на дозу, включительно. В других вариантах осуществления, терапевтически эффективное количество антигена находится в диапазоне от приблизительно 250 до приблизительно 4000 относительных единиц на дозу, включительно, от приблизительно 500 до приблизительно 3000 относительных единиц на дозу, включительно, от приблизительно 750 до приблизительно 2000 относительных единиц на дозу, включительно, или от приблизительно 1000 до приблизительно 1500 относительных единиц на дозу, включительно.
[0096] Терапевтически эффективное количество антигена в вакцинах, содержащих инактивированные вирусы, также могут измеряться по отношению к относительной активности (RP) на мл. Терапевтически эффективное количество часто находится в диапазоне от приблизительно 0,1 до приблизительно 50 RP на мл, включительно. В других вариантах осуществления, терапевтически эффективное количество антигена находится в диапазоне от приблизительно 0,5 до приблизительно 30 RP на мл, включительно, от приблизительно 1 до приблизительно 25 RP на мл, включительно, от приблизительно 2 до приблизительно 20 RP на мл, включительно, от приблизительно 3 до приблизительно 15 RP на мл, включительно, или от приблизительно 5 до приблизительно 10 RP на мл, включительно.
[0097] Количество клеток для бактериального антигена, вводимого в вакцину, находится в диапазоне от приблизительно 1×106 до приблизительно 5×1010 колониеобразующих единиц (КОЕ) на дозу, включительно. В других вариантах осуществления, количество клеток находится в диапазоне от приблизительно 1×107 до 5×1010 КОЕ/доза, включительно, или от приблизительно 1×108 до 5×1010 КОЕ/доза, включительно. В еще других вариантах осуществления, количество клеток находится в диапазоне от приблизительно 1×102 до 5×1010 КОЕ/доза, включительно, или от приблизительно 1×104 до 5×109 КОЕ/доза, включительно, или от приблизительно 1×105 до 5×109 КОЕ/доза, включительно, или от приблизительно 1×106 до 5×109 КОЕ/доза, включительно, или от приблизительно 1×106 до 5×108 КОЕ/доза, включительно, или от приблизительно 1×107 до 5×109 КОЕ/доза, включительно.
[0098] Количество клеток для паразитарного антигена, вводимого в вакцину, находится в диапазоне от приблизительно 1×102 до приблизительно 1×1010 на дозу, включительно. В других вариантах осуществления, количество клеток находится в диапазоне от приблизительно 1×103 до приблизительно 1×109 на дозу, включительно, или от приблизительно 1×104 до приблизительно 1×108 на дозу, включительно, или от приблизительно 1×105 до приблизительно 1×107 на дозу, включительно, или от приблизительно 1×106 до приблизительно 1×108 на дозу, включительно.
Эксципиенты
[0099] Адъювантная композиция и/или вакцинные композиции могут включать фармацевтически приемлемый носитель. Как используется в данном документе, "фармацевтически-приемлемый носитель" включает какой-либо и все растворители, дисперсионные среды, покрытия, стабилизирующие агенты, разбавители, консерванты, антибактериальные и противогрибковые агенты, изотонические агенты, агенты, замедляющие адсорбцию, и подобные. Носитель(и) должен/должны быть "приемлемым(и)" в смысле совместимости с другими компонентами композиций и не вредными относительно субъекта. Как правило, носители должны быть стерильными но не содержать пирогенов, и их выбирают в зависимости от способа введения, который будет использоваться. Квалифицированным специалистам в данной области хорошо известно, что для предпочтительных композиций фармацевтически приемлемый носитель, содержащийся в композициях представляет собой те фармацевтические носители, которые утверждены в соответствии с нормативными актами, опубликованными Министерством сельского хозяйства Соединенных Штатов (США) или Администрацией США по пищевым продуктам и лекарственным веществам, или аналогичным государственным агентством в других странах кроме США. Таким образом, фармацевтически приемлемый носитель для коммерческого производства композиций представляет собой носитель, который уже утвержден, или будет утвержден, соответствующим государственным органом в США или в иностранном государстве.
Введение композиций
[00100] Размеры доз композиций, как правило, находятся в диапазоне от приблизительно 1 мл до приблизительно 5 мл, включительно, в зависимости от субъекта и антигена. Например, для собачьих или кошачьих, как правило, используемой является доза приблизительно 1 мл, тогда как у крупного рогатого скота, как правило, используемой является доза приблизительно 2-5 мл. Однако, данные адъюванты также могут быть сформулированными в микродозах, при этом использоваться могут дозы приблизительно 100 мкл.
[00101] Способы введения для адъювантных композиций включают парентеральное, пероральное, ороназальное, интраназальное, интратрахеальное, подкожное, внутримышечное, транскутанное, интрадермальное, внутрибрюшное, внутриглазное, внутривенное введение и введение в яйцо. Какое-либо приемлемое устройство могут использовать для введения композиций, включая шприцы, капельницы, безигольные инъекционные устройства, пластыри, и подобные. Способ и устройство, которые выбирают для применения будут зависеть от композиции адъюванта, антигена и субъекта, и они хорошо известны квалифицированному специалисту.
Применение композиций
[00102] Адъювантные композиции, описанные в данном документе, являются легкими относительно изготовления и стабильными в течение, по меньшей мере, 18 месяцев при 4°C. Препараты могут быть стабильными, например, в течение приблизительно 18 месяцев, или от приблизительно 18 до приблизительно 24 месяцев при 4°C. В другом варианте осуществления препараты являются стабильными в течение, по меньшей мере, приблизительно 24 месяцев при 4°C. Ускоренные процедуры исследования также показывают, что препараты, описанные в данном документе, являются стабильными в течение, по меньшей мере, двух недель при 37°C, что соответствует приблизительно 24 месяцам при 4°C.
[00103] Адъювантные композиции, описанные в данном документе, могут безопасно и эффективно быть введены широкому кругу субъектов. Неожиданно было установлено, что адъювантные композиции, описанные в данном документе, демонстрируют улучшение безопасности по сравнению с другими адъювантными композициями.
[00104] Следующие примеры представлены в качестве иллюстративных вариантов осуществления, но они не должны рассматриваться как ограничивающие объем изобретения. Много изменений, вариантов, модификаций и других использований и применений данного изобретения будут очевидны для квалифицированного специалиста в данной области.
ПРИМЕРЫ
Пример 1 - Характеристика липосом
[00105] DCRL липосомы (содержащие DDA, холестерин, Bay®R1005 и лецитин) доводили до конечной концентрации 50, 50, 100 и 100 мкг/мл DDA, холестерина, R1005 и соевого лецитина, соответственно.
[00106] Соединения добавляли в 250 мл круглодонную колбу и заполняли до конечного объема 5 мл хлороформа безводного класса (Sigma-Aldrich, Poole, Dorset, UK), используя 1 мл растворителя пипеточным дозатором Safe™ (Sigma-Aldrich). Растворитель удаляли, используя роторный испаритель ( , Flawil, Switzerland) при 55°C в вакууме (KNF Neuberger, Witney, Oxfordshire, UK) в течение 1 часа до образования сухой белой липидной пленки. Липосомы регидратировали или двойной дистиллированной водой (ddH2O) из собственного модифицированного очистителя воды Option 3, включая обратный осмос для сверхвысокой чистоты (ELGA LabWater, Wycombe, Buckinghameshire, UK), или фосфатным солевым буфером (PBS; Sigma-Aldrich), восстановленным из таблеток с ddH2O.
, Flawil, Switzerland) при 55°C в вакууме (KNF Neuberger, Witney, Oxfordshire, UK) в течение 1 часа до образования сухой белой липидной пленки. Липосомы регидратировали или двойной дистиллированной водой (ddH2O) из собственного модифицированного очистителя воды Option 3, включая обратный осмос для сверхвысокой чистоты (ELGA LabWater, Wycombe, Buckinghameshire, UK), или фосфатным солевым буфером (PBS; Sigma-Aldrich), восстановленным из таблеток с ddH2O.
[00107] После регидратации, липосомы обрабатывали насадкой пипетки с шириной отверстия 1 мл (VWR, Lutterworth, Leicesterhire, UK) для уменьшения фрагментации. Затем, липосомальную суспензию обрабатывали ультразвуком в ванне (Sarose Scientific Instruments, Perivale, Middlesex, UK) в течение 1 часа и/или экструдировали вплоть до 3-х раз через 100 нм поликарбонатную нуклеопоровую трековую мембрану Whatman® (Sigma-Aldrich) с использованием мини-экструдерной кюветы, оснащенной двумя 1 мл шприцами. Поликарбонатная мембрана со стороны контактировала с 10 мм подложкой фильтра.
[00108] Обработанные ультразвуком DCRL липосомы (средний диаметр - 1,14 мкм) были значительно больше (в 5,44-раз), чем экструдированные липосомы. Обработка ультразвуком с последующей экструзией в значительной степени не влияла на размер пузырьков по сравнению с экструдированными образцами (209,6 нм). Полидисперсность экструдированных липосом была в 2,10 раз ниже, чем обработанных ультразвуком липосом. Комбинация обработки ультразвуком и экструзии в значительной степени повышала DCRL полидисперсность в 1,42 раза по сравнению с экструдированными липосомами.
[00109] Для оценки коллоидной стабильности липосом в водном растворе, DCRL липосомы, гидратированные в ddH2O, оценивали в течение 4 недель. Следует отметить, что приблизительный термодинамический эквивалент 2 годов при 4°C представляет собой хранение в течение 4 недель при 37°C, таким образом, мотивируя оценку стабильности продукта как при 4, так и при 37°C. Оценивали как пустые ("-ODN") DCRL липосомы, так и DCRL липосомы, нагруженные иллюстративным иммуностимулирующим олигонуклеотидом CpG ODN ("+ODN"), для определения влияния анионных нуклеиновых кислот на катионную липосомальную коллоидную стабильность. В данном документе, ODN нагружали за счет регидратирования липидной пленки раствором ddH2O и ODN. Размер и полидисперсность DCRL липосом, гидратированных в ddH2O (177 нм, 0,24) были статистически подобны тем, которые гидратированы в PBS (210 нм, 0,17).
[00110] При 4°C, +/-ODN DCRL липосомы претерпели незначительное увеличение в диаметре в 1,10- и 1,01-раз через 28 дней, соответственно. Липосомы, содержащие ODN (+ODN липосомы) (417 нм) были в значительной степени больше, чем липосомы, не содержащие ODN (-ODN липосомы) (177 нм). Аналогичным образом, в незначительной степени уменьшалась полидисперсность в 1,08- и 1,03-раза для +/-ODN липосом через 28 дней, соответственно, хотя полидисперсность на 28 день в значительной степени была более высокой в +ODN липосомах (в 1,23 раза) по сравнению с -ODN противоположностями.
[00111] Противоположно этому, агрегационное поведение липосом наблюдалось через 28 дней при хранении при 37°C. +/-ODN липосомы претерпели значительное увеличение в диаметре в 3,24 и 2,00 раза через 28 дней, соответственно, но стабилизировались после 21-ого дня. Хотя +ODN липосомы были большими (в 1,46 раз на 28-й день), чем -ODN липосомы вследствие ODN-опосредованной компенсации заряда и потери коллоидной стабильности, тенденции стабильности в течение времени были качественно подобными. Однако дестабилизация +ODN липосом возникла после 7-го дня, тога как у -ODN липосом, стабильность сохранялась до после 14-го дня. Кроме того, полидисперсность +/-ODN липосом была статистически подобной через 28 дней, претерпев значительное уменьшение в 1,92 и 1,50 раза, соответственно, относительно полидисперсности по сравнению с пузырьками на 0-й день.
[00112] Данные результаты свидетельствуют о необходимости разработки технологий для сохранения липосомы и также свидетельствуют о том, что более мощные способы обработки ультразвуком и/или экструзией, в итоге приводящие сначала к меньшему размеру липосомы, могут компенсировать начальную агрегацию липосом.
[00113] Сравнивали размеры липосом, полученных с использованием микрофлюидизации (Microtiuidics Corp., Model 110EH) и обработки ультразвуком. Когда стадию обработки ультразвуком заменяли на микрофлюидизацию, формулировка давала липосомы, имеющие средний диаметр приблизительно 59 нм.
[00114] Данные результаты показывают, что липосомы, полученные с использованием микрофлюидизации, могут минимизировать агрегацию липосом, которые могут потенциально быть вызваны за счет добавления ODN или ORN.
Пример 2 - Стабильность лиофилизированных липосом
[00115] Лиофилизацию проводили с использованием сублимационной сушилки Dura-Stop (SP Scientific, Ipswich, Suffolk, UK). Образцами наполняли 10 мл цилиндрические типа I стеклянные флаконы для высушивания при замораживании (Schott, Stafford, Staffordshire, UK) перед тем, как проводить лиофилизацию. Параметры лиофилизационного цикла были следующими. Образцы замораживали в течение 30 минут при 5°C, 30 минут при -5°C и 60 минут при -40°C, все со скоростью линейного изменения 1,00°C/мин. Первоначальную сушку проводили в течение 59 минут при -37°C, 59 минут при -28°C, 59 минут при -23°C и 552 минуты при -21°C, все со скоростью линейного изменения 0,50°C/мин. Вторичную сушку проводили при 20°C в течение 360 минут со скоростью линейного изменения 0,10°C/мин. Всю сушку проводили при давлении камеры 57 мТорр. Альтернативный цикл первоначальной сушки проводили при 57 мТорр в течение 59 минут при -38°C, 59 минут при -38°C, 59 минут при -37°C и 552 минут при -35°C, все со скоростью линейного изменения 0,50°C/мин. Лиофилизированные образцы герметично закрывали 14 мм фармацевтического класса бутилкаучуковыми пробками для высушивания при замораживании (Fisher Scientific, Loughborough, Leicestershire, UK) и Parafilm M® (Sigma-Aldrich).
[00116] Лиопротекторы D-манит, D-(+)-глюкозу, D-(-)-фруктозу, сахарозу, D-(+)-трегалозы дигидрат (Sigma-Aldrich) или D(+)-маннозу (Acros Organics) растворяли в ddH2O или PBS и добавляли в липосомную суспензию до конечной концентрации 2-4% масс/об. перед лиофилизацией, как указано.
[00117] При отсутствии сахаров, ddH2O- и PBS-регидратированные липосомы составляли -210 нм перед лиофилизацией. Без лиопротекторов, лиофилизация повышала размер липосомы в 3,55- и 6,05-раз в ddH2O- и PBS-регидратированных образцах, соответственно. DCRL липосомы, регидратированные в ddH2O или PBS, не претерпевали значительные изменения размера после добавления 3% сахарозы, 4% манита или их комбинации, что свидетельствует о том, что агрегация, наблюдающаяся после лиофилизации, не была вызвана непосредственно за счет добавления лиопротектора(ов).
[00118] Для липосом, регидратированных в PBS, размер липосом в значительной степени увеличивался (в 2,00 раза) только в образцах без лиопротектора после цикла замораживания. Однако, после полного цикла лиофилизации, размер липосомы в значительной степени увеличивался в 6,05-, 3,03-, 5,15- и 4,13-раз для образцов без лиопротектора, с 3% сахарозы, с 4% манита и с комбинацией, соответственно. Аналогичным образом, полидисперсность в значительной степени повышалась в 2,61-, 2,61-, 2,85- и 2,79-раз для DCRL без лиопротектора, с 3% сахарозы, с 4% манита и с комбинацией, соответственно, по сравнению с предварительно лиофилизированными контролями.
[00119] Для липосом, регидратированных в ddH2O, размер пузырька после цикла замораживания увеличивался в значительной степени в течение предыдущей лиофилизации контролей в 17,90- и 11,45 раз для липосом без лиопротекторов и с 4% манита, соответственно, тогда как липосомы с 3% сахарозы или комбинацией с 3% сахарозы и 4% манита претерпели незначительные изменения. После полного цикла лиофилизации, размеры липосом умеренно изменились в 3,55-, 0,88-, 3,48- и 1,47 - раз для образцов без лиопротектора, с 3% сахарозы, с 4% манита и с комбинацией, соответственно.
[00120] Данные результаты показывают, что липосомы, регидратированные в ddH2O, имеют лучшую коллоидную стабильность по сравнению с теми, которые регидратированы в PBS, как проиллюстрировано неизмененным размером пузырька после лиофилизации и что 3% сахарозы могут быть более идеальным лиопротектором для обеспечения хорошей коллоидной стабильности.
[00121] Относительно оптимизации схемы лиофилизации для ddH2O-регидратированных DCRL липосом, оценивали влияние 6 известных сахарных лиопротекторов (индивидуально и в комбинации) в концентрации 2% мас./об. на размер пузырька и полидисперсность.
[00122] Сахарозу, глюкозу, манит, трегалозу, фруктозу и маннозу исследовали в качестве лиопротекторов. Только 2% манита в качестве лиопротектора в значительной степени повышала размер пузырька (в 39,66 раза) и полидисперсность (в 2,95 раза). Однако, несмотря на то, что лиофилизация со всеми другими сахарами не привела в результате к значительному изменению размера, продукты лиофилизационно защищенные с использованием глюкозы, фруктозы и/или маннозы опадали в водонепроницаемые, плотные слои сахара и липосом, которые трудно было регидратировать. Согласно литературе, липосомы, лиофилизационно защищенные с использованием сахарозы или трегалозы, предотвращали изменение размеров частиц и в результате приводили к таблетке, которая могла бы быть восстановленной в липосомальную дисперсию с хорошей коллоидной стабильностью.
[00123] DCRL также был лиофилизированным с комбинациями сахаров. Данное исследование показало, что только 2% сахарозы, 2% трегалозы и 2% сахарозы/2% трегалозы поддерживают хорошую коллоидную стабильность после лиофилизации и достигали фармацевтически элегантной структуры таблетки. Однако, комбинация с 2% сахарозы/2% трегалозы, как оказалось, не была предпочтительной с точки зрения стабильности размера липосом и полидисперсности по сравнению с лиофилизационной защитой с 2% сахарозы или 2% трегалозы самостоятельно.
Пример 3 - Вакцина IBR/BVDV
[00124] Как сообщалось, адъювант Quil-A вызывает системный иммунный ответ, часто сопровождающийся временной повышенной температурой. Такая повышенная температура, как считается, связана с уменьшением молочной продуктивности у лактирующих коров. Целью данного исследования было оценить влияние липосом, не содержащих сапонин на иммунный ответ, вызванный липосомами и возможные побочные эффекты, вызванные адъювантами.
[00125] Для данного исследования использовались восьмимесячного возраста голштинские телята-бычки. Потенциальные исследуемые животные были серологически проверенными, и те, у которых титры нейтрализации сыворотки (SN) <1:2 для BVDV-1 и BVDV-2 были приемлемыми, засчитывались в исследование. Кроме того, животные не были персистентно инфицированными (PI) BVDV, как определялось, применяя биопсию с перфоратором для взятия проб тканей ушной раковины и иммуногистохимии. Животные находились в среде, в которой контролировались температура и влажность и получали коммерческое действие и воду городской системы по желанию.
[00126] Акклиматизация началась приблизительно за неделю до 0-го дня исследования. Телята получали DRAXXIN® и DECTOMAX® перед загрузкой.
[00127] Исследуемые животные, теряющие свое значение, становились поврежденными или умирали из-за обстоятельств, не связанных с исследованиями, были исключены из исследования и соответственно анализа данных. Животные, потерявшие свое значение, подвергались эвтаназии.
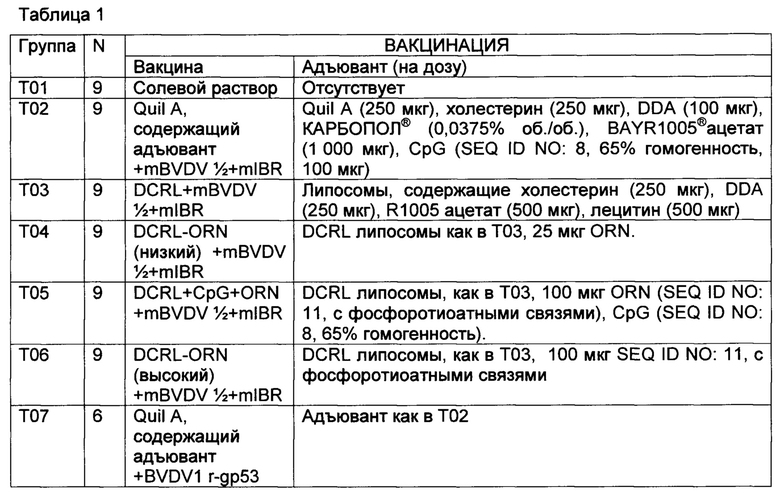
[00128] Животных вакцинировали на 0-й день и на 28-й день путем подкожного введения 2 мл вакцины, как подытоживается в таблице 1.
[00129] На 49-й день животные были контрольно заражены нецитопатическим вирусом вирусной диареи - заболевание слизистых крупного рогатого скота типа 2 (штамм 24515). Каждое животное получало приблизительно 4,88 log10TCID50 на дозу в 5 мл, которую вводили интраназально (2,5 мл на ноздрю), используя аэрозольный ингалятор с сжатым газом.
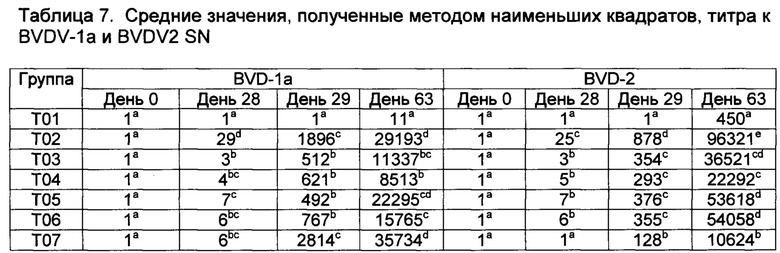
[00130] В группах Т02-Т06, антигенный компонент содержал 4500 Ус.ед./вирус предварительно инактивированного модифицированного BVDV 1/2 и модифицированную IBR (8,0 log10TCID50).
[00131] Животных контрольно заражали на 48-й день, применяя 5 мл BVDV-2 штамма 24515 (4,88 Log10TCID50 на дозу) интраназально (2,5 мл на ноздрю), используя аэрозольный ингалятор с сжатым газом.
[00132] Средние значения ректальной температуры, полученные способом наименьших квадратов, и диапазоны после первой и второй вакцинации показаны в таблице 2.
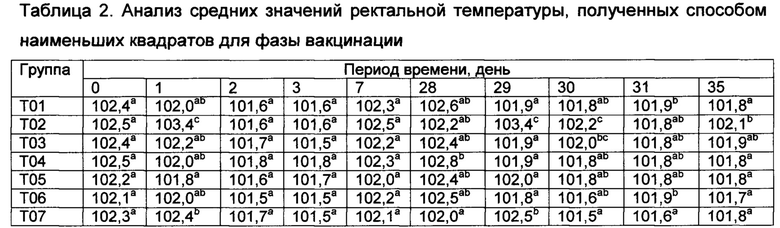
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00133] Ректальные температуры для некоторых субъектов из Т02 и Т07 показали временное повышение на следующий день как после первой, так и второй вакцинации. Повышение ректальных температур у животных в группе Т02 было в значительной степени более высоким (Р≤0,10), чем в других группах (Т01, Т013, Т04, Т05, Т06, и Т07) в течение первого дня после первой вакцинации.
[00134] После первой вакцинации, четыре из 9 животных в группе Т02 имели температуры более высокие, чем 103,5°F (три имели температуры, превышавшие 104,0°F). В отличие от этого, ни одно из животных в группе Т05 не имело температуру, превышающую 102,5°F в течение первого дня после первой вакцинации (данные не показаны).
[00135] После второй вакцинации повышение ректальных температур в группах Т02 и Т07 было в значительной степени (Р≤0,10) более высоким, чем у контрольных и вакцинированных (T03, Т04, Т05, и Т06), однако, повышенные ректальные температуры были кратковременными (Таблица 2). Индивидуальные ответы в группе Т02 находились в диапазоне от 101,1 до 104,5°F. Пять из девяти животных в группе Т02 имели температуры, превышающие 103,5°F (и другие животные имели температуру 103,5°F) после второй вакцинации. Только три из девяти животных в группе Т02 имели температуру ниже 103,5°F в течение первого дня после второй вакцинации. В отличие от этого, ни одна из композиций DCRL не вызывала повышение ректальных температур, превышающую 103,7°F. Индивидуальные ответы в группе Т05 находились в диапазоне от 101,4 до 102,6°F (данные не показаны).
[00136] Фебрильная реакция наблюдалась (>104,5°F) у 8 из 8 животных в группе Т01 после контрольного заражения; это отвечает критериям результатов для успешного контрольного заражения. Ректальные температуры во всех вакцинированных группах были в значительной степени более низкими (Р≤0,10) на 53-й день исследования по сравнению с контролями (через 4 дня после контрольного заражения), и появились некоторые отличия в последующие дни (данные не показаны). В контрольной группе, в ответ на контрольное заражение, наблюдалось типичное двухфазное повышение ректальной температуры.
[00137] Присутствие клинического заболевания BVDV (животное должно было иметь клиническую оценку ≥2) после контрольного заражения определялось соответственно следующей системе оценки:
0 - никаких клинических признаков
1 - клинические признаки в целом не являются специфическими для острой инфекции BVD. Клинические признаки могут включать выделения из носа, ненормальное дыхание и легкую вялость.
2 - клинические признаки в целом являются умеренными по степени и специфическими для острой инфекции BVD. Клинические признаки могут включать выделения из носа, ненормальное дыхание, вялость, похудание, выделения из глаз, повышенное слюноотделение, диарею, дегидратацию, хромоту и/или нежелание двигаться.
3 - клинические признаки в целом являются тяжелыми по степени и характерными для острой инфекции BVD. Клинические признаки могут включать выделения из носа, ненормальное дыхание, вялость, похудание, выделения из глаз, повышенное слюноотделение, диарею, повышенное травмирование, дегидратации, лежачее положение, хромоту и/или нежелание двигаться.
[00138] Не было никаких существенных различий между контрольной группой и вакцинированными группами.
[00139] Лейкопения
[00140] Результат исследования соответствовал критериям достоверного исследования, поскольку 100% из Т01 (контроли) имели лейкопению при применении ≥40% уменьшения и 75% контролей имели <4000 клетки/мкл. Не было существенных различий между количеством животных, у которых развивалась лейкопения ≥40% уменьшение количества белых кровяных клеток по сравнению с фоном после контрольного заражения в Т01 по сравнению с вакцинированными группами лечения Т02, Т03, Т04, Т05, Т06, и Т07 (Р≤0,10). Однако, клиническая лейкопения (<4000 клетки/мкл), которая является более релевантным определением, была обнаружена у 6 из 8 (75%), у 2 из 9 (22,2%), у 1 из 8 (12,5%), у 3 из 9 (33,3%), у 4 из 9 (44,4%), у 4 из 9 (44,4%), и у 0 из 6 в Т01, Т02, Т03, Т04, Т05, Т06, и Т07, соответственно (Таблица 3). Продолжительность лейкопении была значительно длиннее (Р≤0,10) в группе Т01 при применении ≥40% уменьшения по сравнению со всеми вакцинированными и значительно длиннее (Р≤0,10) при применении <4000 мкл по сравнению с вакцинированными группами Т02, Т03, Т05, Т06, и Т07 (Таблица 4).


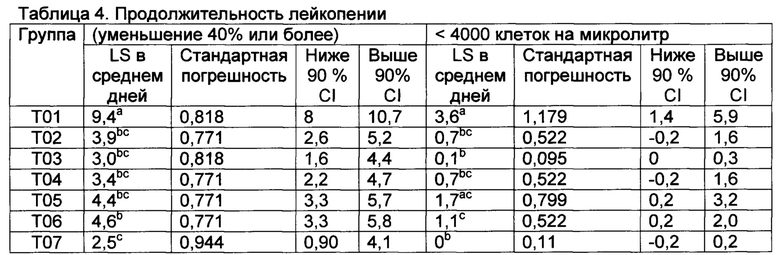
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00141] Вирусемия
[00142] Все экспериментальные вакцины защищали против вирусемии. В некоторых группах существовала полная защита (Т02 и Т05 - обе, содержат CpG) и в других - частичная (T03, Т04, Т06, и Т07). Количество зараженных вирусом животных в Т01 была в значительной степени более высокой, чем количество в Т02, Т03, Т04, Т05, Т06 и Т07 (Р≤0,10) (Таблица 5). В группах Т02 и Т05 никакой вирусемии не наблюдалось. Однако существовала разница между количеством зараженных вирусом животных в группах Т03, Т04, и Т06, которые не содержали никакого ORN, низкую дозу ORN и высокую дозу ORN по сравнению с контролями.
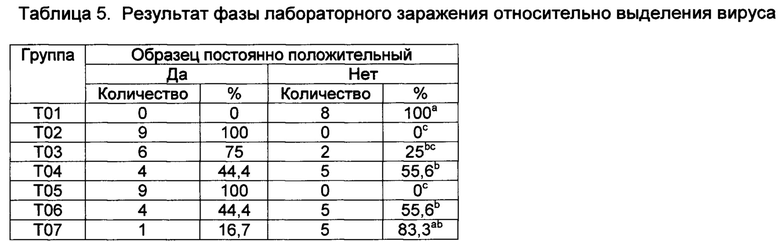
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00143] Продолжительность вирусемии для группы Т01 была в значительной степени большей чем во всех вакцинированных группах (от Т02 по Т07) (Р≤0,1) (Таблица 6). Телята в Т07 получали вакцину, содержащую Quil-A, который содержит адъювант + рекомбинантный gp53 антиген вируса BVDV-1, и были также частично защищенными и имели в значительной степени более короткую продолжительность вирусемии по сравнению с Т01. Для данных телят контрольное заражение происходило другим биотипом вируса BVDV (тип 2), отражающее то, что существовала частичная перекрестная защита между антигеном BVDV-1 gp53 и штаммом контрольного заражения.
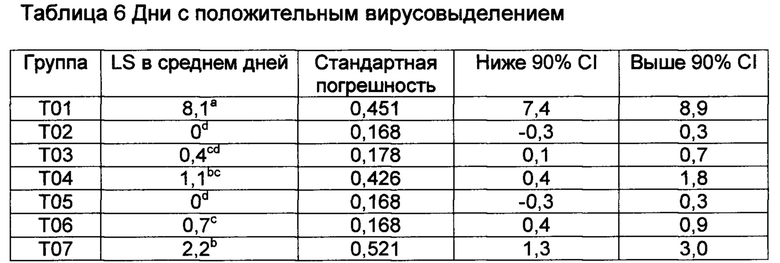
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00144] SN Титры
[00145] На 49-й день, перед контрольным заражением все группы животных с Т02 по Т07 имели SN титр к BVDV-1, BVDV-2, и только группы с Т02 по Т06 имели антитела к антигену IBR ≥ 1:8. Все вакцины, как было признано, имели адекватную эффективность, поскольку они соответствовали требованиям ≥1:8. После первой и второй вакцинации средние значения, полученные методом наименьших квадратов, SN титров к BVDV-1 и 2 (на 28-й и 49-й день) представлены в таблице 7. Все вакцинированные животные имели в значительной степени более высокие SN титры, чем контрольная группа, и существовали значительные различия между SN титрами вакцинированных групп. Липосомальные вакцины (от Т03 до Т06) индуцировали в значительной степени более низкие SN титры к BVDV-1 и 2, чем вакцины, в которые была добавлена в качестве адъюванта композиция, содержащая Quil А (Т02). Добавление ORNS или CpG к данным препаратам не улучшало данные ответа. Вакцина gp53, в которую была добавлена в качестве адъюванта композиция, содержащая Quil А, также индуцировала высокие SN титры на 49-ый день к антигену BVDV-1, но более низкий к антигену BVDV-2. SN титры к BVDV-1 группы Т07 в значительной степени не отличались от титров Т02, но были титры к BVDV-2. После контрольного заражения BVDV-2, SN титры всех вакцинированных групп повышались, тогда как у телят из группы Т01 развивались первичные ответы на антитела (на 63-ий день).
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
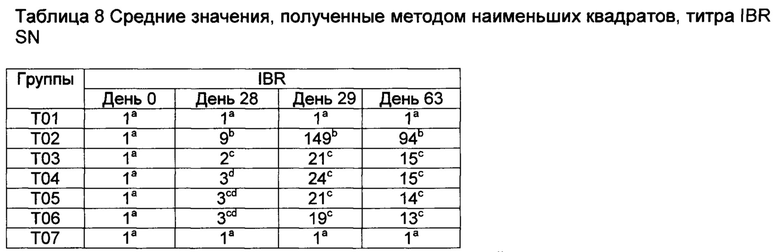
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00146] Реакции в месте инъекции
[00147] Средние значения реакции в месте инъекции, полученные методом наименьших квадратов, (ISR) как для левой, так и для правой стороны шеи, показаны в таблицах 9 и 10. В 1-ый день, небольшие реакции наблюдались во всех шести группах вакцинации, но опускались. Наибольшие реакции на местах наблюдались у Т02 и Т07 на 29-ый, 41,8-й и 16,3-й день, соответственно. Существовали значительные различия между реакциями в месте инъекции в определенные дни. Все вакцины были безопасными, поскольку реакции в месте инъекции быстро рассасываются.
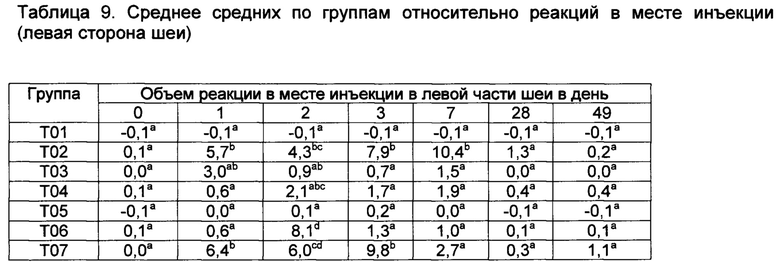
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
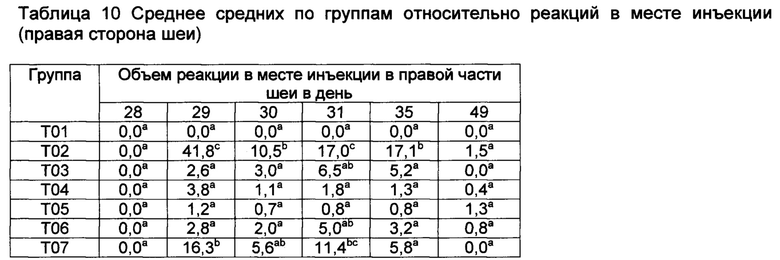
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00148] Клеточно-опосредованный иммунитет
[00149] Анализ ELISPOT BVDV IFN гамма имел высокий фон в группе Т01 перед контрольным заражением и, поэтому, считается ненадежным для демонстрации индуцирования ответов CMI на данные вакцины к данному антигену (данные не показаны). Однако анализы на антигены IBR были отражением индуцирования CMI соответствующими вакцинами (смотрите таблицы 11-13, ответы на убитый антиген IBR, вторичные иммунные ответы на пептиды gB и gD IBR в группах с Т02 по Т06). В целом наиболее высокие ответы в группе вакцинации Т02 и группе Т05, которые имели CpG в составе. После контрольного заражения ответы не улучшались, поскольку контрольное заражение представляло собой контрольное заражение вирусом BVDV. Возможно, подобные ответы CMI были индуцированы вакцинами относительно антигена BVDV.
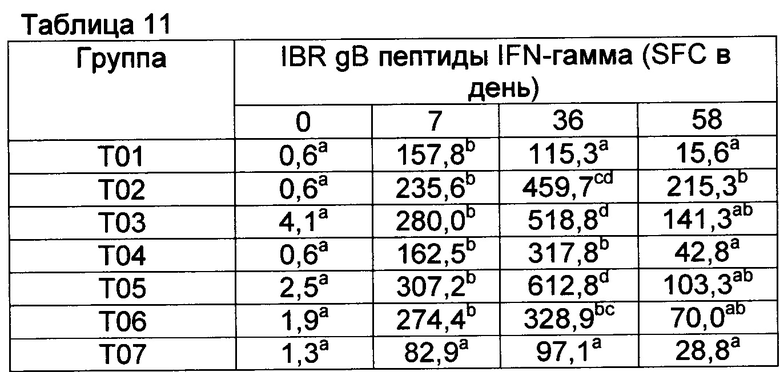
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)

Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
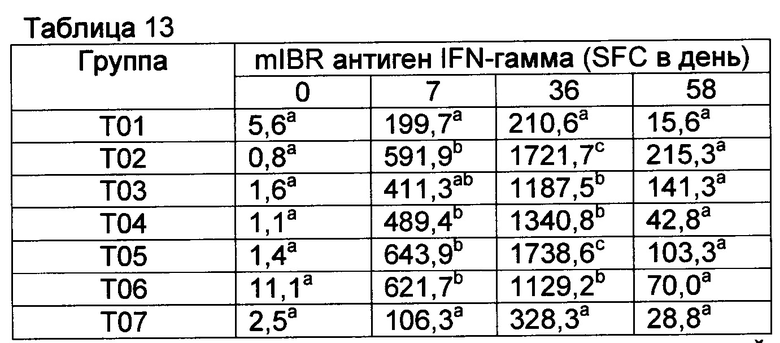
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00150] Эффект вакцины наблюдался на развитие и продолжительность вирусемии. Все экспериментальные вакцины защищали против вирусемии. Полная защита наблюдалась в некоторых группах (Т02 и Т05, содержащие CpG), тогда как в других группах (T03, Т04, Т06 и Т07) наблюдалась частичная защита. Профили ректальной температуры свидетельствуют о транзиторном повышении в первый день после как первой, так и второй вакцинации для групп обработки Т02 и Т07 по сравнению с контрольной Т01. Липосомальные вакцины индуцировали более низкое повышение ректальной температуры по сравнению с Т02 и Т07, которые имели Quil-A, содержащий адъювант. Данный Quil А, содержащий адъювант, как правило, индуцирует однодневное повышение температуры непосредственно после вакцинации. Последнее является особенно важным для молочных коров, так как повышенная температура, сопровождающая вакцинацию, часто в результате приводит к неприемлемому падению количества молока. Липосомальные композиции индуцировали стабильное антитело, а также Т-клеточные ответы (даже липосомы без дополнительных иммуномодуляторов). Липосомальные вакцины индуцировали в значительной степени более низкие SN титры к BVDV-1 и 2 и антиген IBR по сравнению с вакцинами, содержащими Quil А, хотя достигнутые уровни рассматривались в защитном диапазоне для данных соответствующих заболеваний (титры SN BVDV-1 - 1: 256-512; BVDV-2±256 или больше; титр SN IBR - 1:32).
[00151] В конечном итоге, липосомальные композиции обеспечивают безопасную и новую возможность для доставки убитых антигенов для заболеваний, нуждающихся не только в антителах, но и CMI, а также для защиты. Композиции сравнивали с вакцинами, содержащими Quil-A, относительно эффективности. Влияние CpG на эффективность было воспроизводимым, однако, ORN не было эффективным в повышении иммунитета против контрольного заражения BVDV.
Пример 4 - Вирус свиного гриппа
[00152] Целью данного исследования было оценить иммунный ответ и эффективность нескольких SIV-вакцин (Убитый вирус, pH1N1 & H3N2), содержащих новые адъювантные композиции и иммуномодуляторы. Эффективность определяли как по иммунологическим параметрам (гуморальный и клеточный), так и по конечным точкам эффективности, включая клинические признаки, вирусемию, бессимптомное вирусовыделение и поражение легких.
[00153]
[00154] Животные.
[00155] Для данного исследования использовали трехнедельного возраста (21 +/- 3 дней) свиней обоих полов. Животные не имели в анамнезе влияния PRRSV, Mycoplasma hyopneumoniae. Животные или их производительности не имели в анамнезе вакцинации против или влияния какого-либо серотипа SIV. Животные прибывали на место за от четырех до семи дней перед вакцинацией и получали стандартный рацион с водой по желанию.
[00156] Свиньи были вакцинированы композициями из таблицы 14 на 0-ой день (левая сторона шеи) и 21-ый (правая сторона шеи) и проведено контрольное заражение вирусом H3N2 IN/12 на 35-ый день.

*4% субраствор на основе сквалана, разбавленный в 10 раз для конечного продукта (0,4% масло)
[00157] Сыворотку крови собирали для анализа ингибирования гемагглютинации (HAI) на 35-ый и 40-ый день, и цельную кровь собирали для анализа ELISPOT (IFNγ) на 28-й, 35-й и 40-й день. Свиней подвергали эвтаназии на 40-й день (через 5 дней после контрольного заражения), и оценивали процент закрепления для каждой части (левой краниальной, левой средней, левой каудальной, правой краниальной, правой средней, правой каудальной и придаточной) и записывали как процент от 0 до 100%.
[00158] Ни одна из вакцинаций в итоге не приводила к системным побочным эффектам или неприемлемым реакциям в месте инъекции (данные не показаны).
[00159] HAI титры показаны в таблице 15.
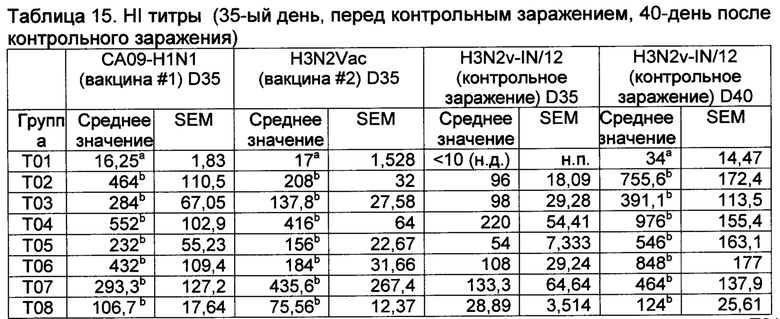
а Значения с различными верхними индексами в значительной степени отличаются от Т01 (р≤0,10) Неспаренные Т исследования с коррекцией Уэлша (GraphPad Prism, Версия 6.04). н.д., не детектируемый; н.п., не применяемый.
[00160] Оценки поражения легких представлены в таблице 16.
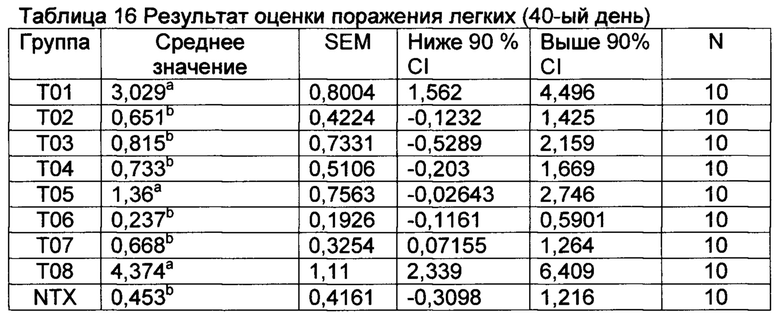
а Значения с различными верхними индексами в значительной степени отличаются от Т01 (р≤0,10) Неспаренные Т исследования с коррекцией Уэлша (GraphPad Prism, Версия 6.04). NTX представляет собой группу невакцинированных, контрольно незараженных свиней.
[00161] Группа Т04 имела самый высокий титр HAI к одному из вакцинных штаммов (H1N1), тогда как Т07 имела самый высокий титр к другому штамму (Н3Н2), несмотря на то, что титры в группах Т02 - Т08 были в значительной степени более высокими, чем в контрольной группе Т01.
[00162] Все вакцины (за исключением Т08) имели тенденцию уменьшать LLS через 5 дней после контрольного заражения. Группа Т06 (ТХО) имела тенденцию демонстрировать наиболее низкие показатели поражения легких.
[00163] Клетки, секретирующие пептид-специфический IFN-гамма к протеинам NP гриппа, и клетки, секретирующие IFN-гамма специфический к цельному вирусу гриппа (обе определены с использованием ELISPOT), представлены в таблицах 19 и 20 соответственно. Пептиды в пулах являются 16-мерными с 12-аминокислотным перекрытием. Четыре пула содержат последовательность NP пептида с N-конца (пул 1) до С-конца (пул 4). Последовательности пептидов представлены в таблицах 17 и 18.
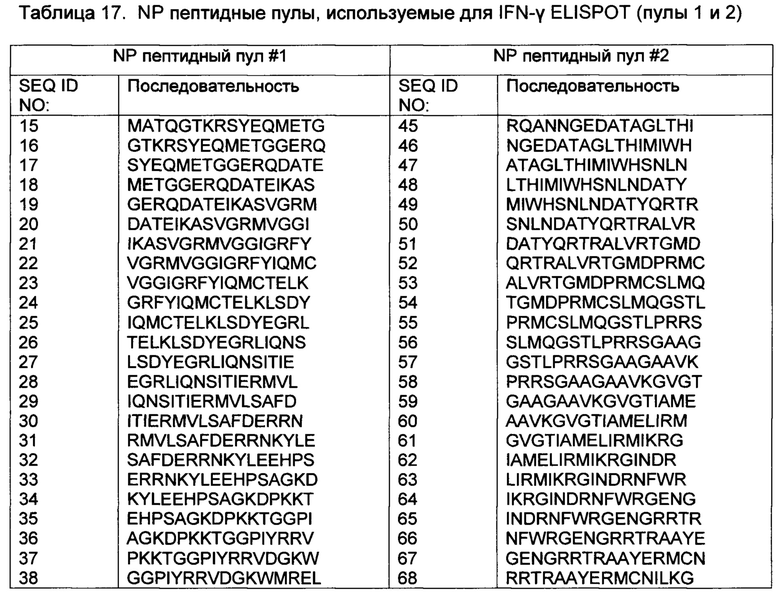
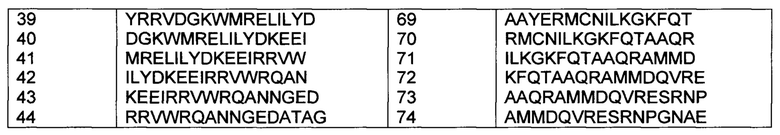
[00164]

а Значения с различными верхними индексами в значительной степени отличаются от Т01 (р≤0,10); Неспаренные Т исследования с коррекцией Уэлша (GraphPad Prism, Версия 6.04).
[00165] NP протеин является, как правило, высоко консервативным среди SIV штаммов, и, поэтому, эффективный ответ CMI, скорее всего, приводит к лучшей эффективности перекрестной защиты.
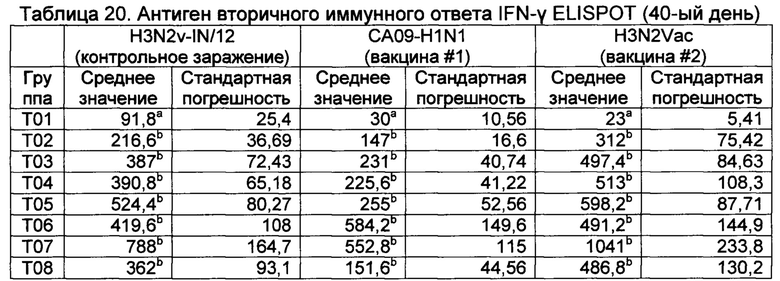
а Значения с различными верхними индексами в значительной степени отличаются от Т01 (р≤0,10); Неспаренные Т исследования с коррекцией Уэлша (GraphPad Prism, Версия 6,04).
[00166] Группа Т07 (DCRL-липосомы) обеспечивала наиболее высокие ответы IFN гамма к трем из четырех пептидных пулов NP протеина и к двум из трех целых вирусов, которые исследовали. Единственной группой, в которой наблюдался ответ IFN-γ после вакцинации (предварительное контрольное заражение), была Т06 (ТХО) (данные не показаны).
[00167] Добавление агониста CD40 и CpG однотипно имели тенденцию снижать титры, а также увеличивать LLS, особенно в контексте липосомальной композиции.
[00168] Взятые в целом, данные результаты свидетельствуют о том, что вакцина, в которую добавлены в качестве адъюванта DCRL липосомы, столь же эффективна в качестве препарат Т02 (экспериментальная композиция, аналогичная коммерческой вакцине) относительно снижения показателей поражения легких. В то же время, DCRL липосомы являются намного более эффективными в активировании ответа CMI, чем другие вакцины, которые исследуются, таким образом, возможно, обеспечивают более широкий перекрестный защитный потенциал и повышают продолжительность иммунитета, чем коммерческий продукт.
Пример 5 Развитие иммунитета к Eimeria maxima после вакцинации профилином
[00169] Цель исследования заключалась в оценке влияния различных адъювантов (собственные разработки Zoetis) на развитие иммунитета к Eimeria maxima после вакцинации профилином.
[00170] Цыплята, только что вылупившиеся из яйца были приобретены из инкубатора Лонгенкера (Longenecker’s hatchery) Elizabethtown, РА. Цыплят обеспечивали кормом и водой по желанию. Птиц держали в брудерных гнездах в оснащении, которое не содержало Eimeria, и переносили в большие висячие клетки в отдельном месте, где они были инфицированы и содержались до конца экспериментального периода.
[00171] Очищенный профилин (50 мкг/доза) смешивали с различными адъювантами, как представлено в таблице 21.
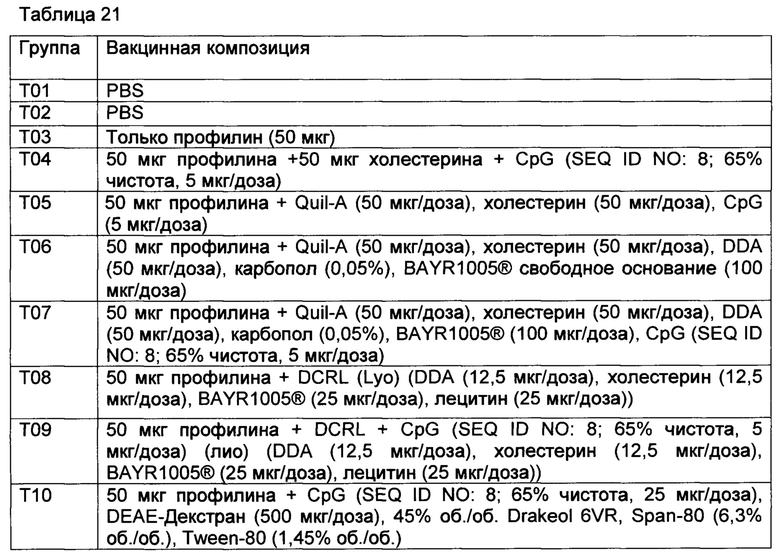
[00172] Птицы были иммунизированы двумя подкожными инъекциями (0,5 мл на дозу) профилина плюс адъювант в 1-й день и затем на 7-й день жизни. Через семь дней после второй иммунизации (14 дневного возраста), птицы (за исключением группы Т01) контролируемо заражали 1×104 спорулированного ооциста Eimeria maxima.
[00173] Увеличение массы тела определяли на 6-й день и на 15-й день после контрольного заражения. Кроме того, определялись ооцисты, которые выделялись в фекалии с 6-9-го дня после контрольного заражения. Сывороточные антитела против профилина оценивали с помощью ELISA.
[00174] Контрольные группы: инъекции PBS и отсутствие контрольного заражения (невакцинированный без контрольного заражения контроль), инъекции PBS и контрольное заражение на 14-й день (невакцинированный с контрольным заражением контроль), профилин/без адъюванта и контрольное заражения на 14-й день (антигенный контроль).
[00175] Для оценки результатов поражения, шесть птиц/группа были эвтанизированы через 6 дней после контрольного заражения. Приблизительно 20 см кишечных сегментов, простирающихся на 10 см спереди и сзади дивертикула, были получены и разрезаны продольно. Содержимое кишечника аккуратно удалялось. Оценка в диапазоне от 0 до 4 предоставлялась в зависимости от тяжести поражения.
[00176] Для оценки продуцирования ооцистов в фекалиях, фекалии из каждой группы (8 птиц/группа, 4 клетки из 2 птиц/клетка) собирали отдельно через от 6 до 9 дней после контрольного заражения. Начиная с 6 дня после контрольного заражения, были установлены сборные клетки, и смотрителям животных было поручено не очищать фекалии. Фекальные испражнения собирали из каждой клетки сбора ооцистов, в которых держали по 2 птицы на клетку. Поддоны для сбора помещали под каждую клетку на 3 дня, начиная с 6 дня после инъекции, и фекальный материал собирали в большие пластиковые баночки. Фекальные испражнения в каждой баночке измельчали в блендере с водой, и два по 35 мл случайных образцов были взяты из каждого образца. Для подсчета ооцистов кокцидии, сначала были сделаны различные разбавления для определения оптимального разбавления для подсчета ооцистов для каждого образца. Ооцисты подсчитывали микроскопически. Общее количество выделения ооцистов на цыпленка рассчитывали, используя формулу: общее количество ооцистов/птица = (количество ооцистов × коэффициент разбавления × объем образца фекалий/объем камеры для подсчета)/ количество птиц на клетку.
[00177] Образцы крови (4 птицы/группа) собирали на 9-й день после контрольного заражения Eimeria для измерения ответа на антитела. Образцам крови давали свертываться при 4°C в течение 4 часов, и сыворотки отделялись. Образцы сыворотки исследовали на антитела против Eimeria, используя ELISA. Коротко говоря, микротитровальные планшеты покрывались в течение ночи 200 нг/лунка рекомбинантного кокцидиального антигена, промывали PBS-0,05% Tween, и блокировали PBS 1% BSA. Добавляли сывороточные разбавления, инкубировали при постоянном осторожном встряхивании, промывали, и связанные Ab детектировали с использованием конъюгированного с пероксидазой кроличьего анти-куриного IgG (Sigma) и специфического к пероксидазе субстрата. Оптическую плотность (OD) измеряли при 450 нм, используя микропланшетный ридер (Bio-Rad, Richmond, СА).
[00178] Все значения выражаются в виде среднего значения ± SEM. Различие между средними значениями считалась значительной при р<0,05.
[00179] Отрицательный контроль и контроль с контрольным заражением в значительной степени отличались через 6 дней, но не через 15 дней после инфицирования. Через 6 дней после контрольного заражения потеря массы в контроле с контрольным заражением по сравнению с отрицательным контролем составляла приблизительно 15%. Ни одна обработка не отличалась ни от адъювантного контроля или от контроля с контрольным заражением, но обработки Т05, Т06 и Т08 также не отличались от отрицательного контроля.
[00180] Чуть меньше 60% птиц имели оценку поражения 2 или более высокую. Ни одна композиция в значительной мере не отличалась от контроля контрольного заражения. Группа Т09 была самым неудачным исполнителем. Т04 была лучшим исполнителем.
[00181] Как правило, говоря об уменьшении <1 log на выходе не считается биологически релевантным, но все еще может свидетельствовать об активном иммунитете. Только композиции, которые давали в значительной степени меньше, чем контроль с контрольным заражением, были Т05 и Т09.
[00182] Ответ на антитело, как правило, не считается имеющим отношение к иммунитету относительно кокцидиоза, поэтому не может коррелировать с другими критериями. В указанных выше экспериментах в группе Т07 вызывался ответ на антитело, который был более высоким, чем ответ, вызванный либо отрицательным (у невакцинированных, неинфицированных птиц), положительным контролем (у невакцинированных, инфицированных птиц), или птиц, вакцинированных только антигеном. В группах Т05, Т06 и Т08-Т10 вызывались более высокие ответы на антитело, чем у отрицательного контроля (у невакцинированных, неинфицированных птиц).
[00183] Результаты просуммированы в таблице 22. Повышение массы тела, уровень антител в сыворотке (выраженный как % неинфицированного контроля) и показатели поражения, продуцирования ооцистов (выраженный как % инфицированного контроля) после иммунизации птиц двумя подкожными инъекциями профилина плюс различные адъюванты в 1-й день и 7-й день после рождения и последующее контрольное заражение 1×104 ооцистами Eimeria maxima в возрасте 14 дней.
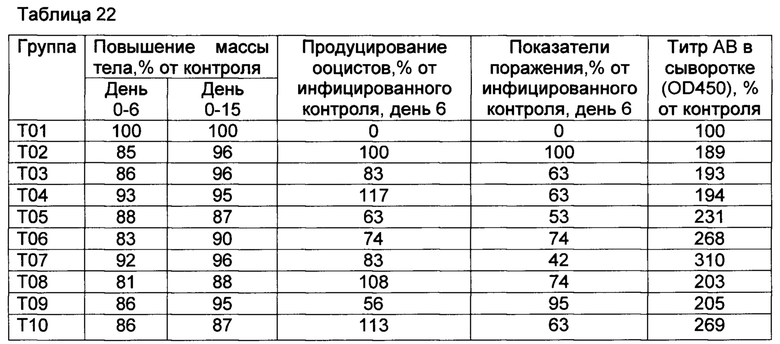
Пример 6 - Вакцина BVDV/IBR, в которую в качестве адъюванта добавлено DCRL + CpG
[00184] В данном примере оценивали адъювантную эффективность DCRL липосом с CpG (без ORN). Кроме того, сравнивались различные способы нагрузки липосом.
[00185] Экспериментальная модель была следующей:
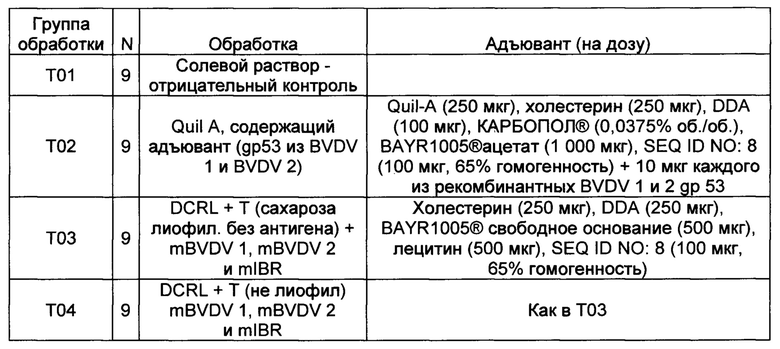
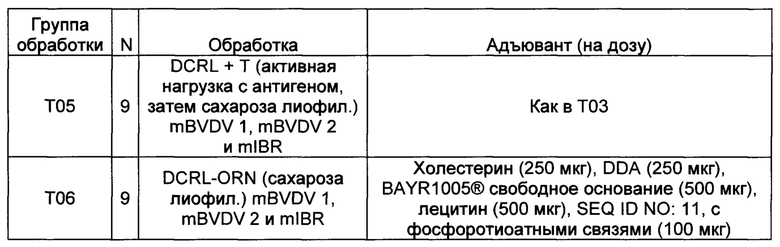
[00186] Вакцины в группах Т03-Т06 также содержали mBVD1 (4 500 усл.ед./доза) + BVDV1 (4500 усл. од.), BVDV 2 (4500 усл. ед.), mIBR 108log10 TCID50 (предварительно инактивированная доза).
[00187] Здоровые голштинские телята (в возрасте 6-7 месяцев, серонегативные относительно BVDV1, BVDV2 и IBR) были включены в данное исследование (n=9 на группу обработки). Телятам вводили 2 мл назначенной вакцины подкожно на 0-й и 28-й день. Вирулентное BVDV2 контрольное заражение (4 мл (5,14 Log10TCID50 на дозу) интраназально) проводили на 49-ый день. Проводили клинические наблюдения, и измеряли места инъекций и ректальные температуры от начала до конца каждой вакцинации. Образцы крови собирали для подсчета белых кровяных клеток, вирусемии и серологии через 63 дня.
[00188] Все телята в группах Т02, Т03, Т04 и Т06 достигали титров 1: 8 против BVDV-1a и BVDV2 перед контрольным заражением, по сравнению с 22,1 и 11,1% телят в Т05. Считается, что благодаря активной нагрузке, используемой в Т05, большинство антигенов была удалена из вакцинного препарата. Таким образом, животные в группе Т05 получали меньше антигена, чем животные в группах Т03, Т04 и Т06. Поэтому интерпретация наблюдений за группой Т05 должна истолковываться осторожно.
[00189] Несмотря на вышесказанное, титры BVDV-1a и BVDV2 для Т05 были в значительной степени более низкими, чем для других вакцинированных групп на 28-й, 49-й и 63-й день. Некоторые телята в Т02 или Т05 не достигали титров 1: 8 против BHV перед контрольным заражением, и данные группы также имели в значительной степени более низкие титры по сравнению с другими вакцинированными группами на 28-й, 49-й и 63-й день. Средние значения, полученные методом наименьших квадратов, для gp53-1 SFC были в значительной степени более высокими для Т02 по сравнению со всеми другими группами обработки на 0-й, 35-й и 56-й день, и средние значения, полученные методом наименьших квадратов, для gp53-2 SFC были в значительной степени более высокими для Т02 на 35-й и 56-й день.
[00190] Вирусемия наблюдалась у всех телят в Т01 и Т05. Ни у кого из телят в группе Т02 не развилась вирусемия после контрольного заражения. В группах Т03, Т04 и Т06 у 22,2, 33,3 и 33,3% телят развилась вирусемия; данные группы в значительной степени не отличались от Т02. Продолжительность вирусемии была в значительной степени более длинной в Т01 и Т05 по сравнению со всеми другими группами обработки. При определении лейкопении по 40% снижению уровня WBC от исходного, 22,2% телят в группе Т02 были лейкопеническими после контрольного заражения, тогда как лейкопеническими были все телята во всех группах. Телята в группах Т01 и Т05 имели в значительной степени более длительную продолжительность лейкопении по сравнению со всеми другими группами (8,6 дней, 0 дней, 0,2 дня, 0,7 дней, 6,7 дней, и 0,8 дней для групп Т01 - Т06, соответственно). Только легкие признаки клинического заболевания наблюдались после контрольного заражения в данном исследовании. В контрольной группе Т01, 22,2% телят имели клинические оценки ≥2 после контрольного заражения, тогда как 11,1% телят в группе Т05 имели клинические оценки ≥2 после контрольного заражения. Ни один из телят в какой-либо другой группе обработки не имел клинические оценки ≥2 после контрольного заражения.
[00191] Временные повышенные температуры наблюдались в Т02 в 1-й и 29-й день. Температуры всех других телят были нормальными в течение всего исследования.
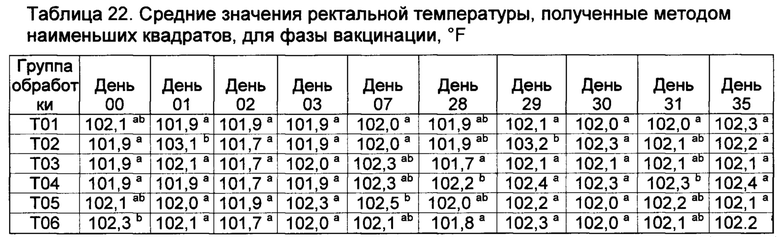
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
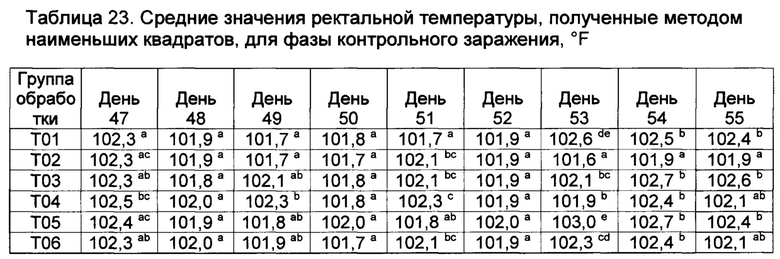
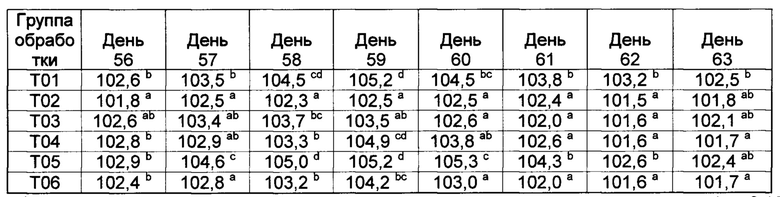
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00192] Анализ отдельных животных показал, что в группе Т02 (в которой добавлены в качестве адъюванта Quil А, содержащий адъювант), четыре животных имели температуру свыше 103,5 F после первой инъекции и два животных имели температуру свыше 103,5 F после второй инъекции. Кроме того, одно животное имело температуру 103,2 F в 1-й день (через день после первой вакцинации) и пять имели температуры в диапазоне 103,0-103,4 F на 29-й день (через день после второй вакцинации). В отличие от этого, ни одно из животных, вакцинированных с адъювантами, не содержащими Quil А, не имело температуру свыше 103,5 F или после первой или после второй вакцинации. Двое имели температуры в диапазоне 103,0-103,4 F.
[00193] В данном исследовании не наблюдалось никаких реакций в месте инъекции >200 см3. Группа Т02 (в которой добавлены в качестве адъюванта Quil А, содержащий адъювант) имела в значительной степени большие реакции в месте инъекции во 2-й, 7-й и 29-й день по сравнению со всеми другими группами обработки. Измеряемые реакции в месте инъекции еще присутствовали на 49-й день в Т02 (вторая инъекция) и Т06 (первая инъекция).
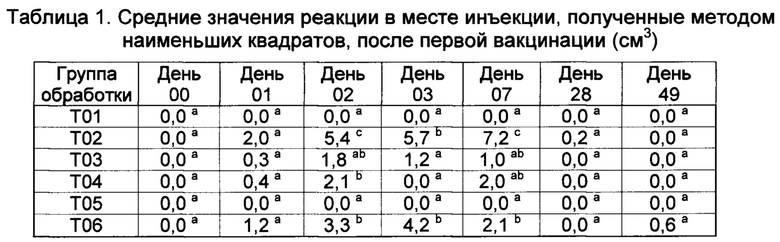
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
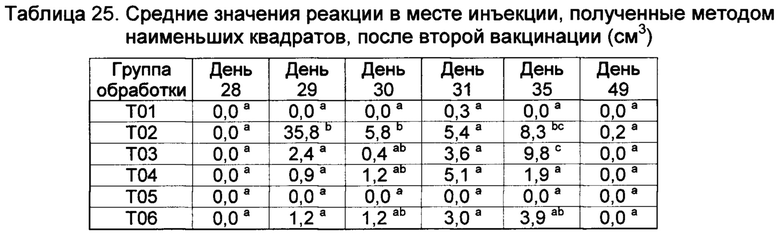
Значения с различными верхними индексами в значительной степени отличаются (р≤0,10)
[00194] В целом, вакцинные кандидаты Т02, Т03, Т04 и Т06 соответствовали критериям результата для обеспечения титров BVDV2>1:8 у всех телят. Кандидат на вакцину Т02 также обеспечил 100% защиту против вирусемии и полную защиту против лейкопении (как измерено по 40% снижению уровня WBC). Телята в данной группе подверглись временному повышению температуры и реакции в месте инъекции.
[00195] Все публикации, приводимые в описании, как патентные публикации, так и не патентные публикации, свидетельствующие об уровне квалифицированности специалистов в области, к которой данное изобретение относится. Все данные публикации включены в настоящий документ в полном объеме в виде ссылки в той же степени, как в случае, когда каждая отдельная публикация была бы конкретно и индивидуально указана как включенная в виде ссылки.
[00196] Несмотря на то, что изобретение в настоящем документе описывается со ссылкой на конкретные варианты осуществления, следует понимать, что данные варианты осуществления являются только иллюстрацией принципов и применений представленного изобретения. Поэтому следует понимать, что многочисленные модификации могут быть сделаны в иллюстративных вариантах осуществления, и что другие варианты могут быть разработаны, не отходя от замысла и объема настоящего изобретения, как определено в следующей формуле изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Zoetis Services LLC
Dominowski, Paul J
Mwangi, Duncan
Rai, Sharath K
Foss, Dennis L
Godbee, Traci K
Sly, Laurel M
Mahan, Suman
Vora, Shaunak
<120> ЛИПОСОМАЛЬНЫЕ АДЪЮВАНТНЫЕ КОМПОЗИЦИИ
<130> ZP000111
<150> US 62/194,355
<151> 2015-07-20
<160> 136
<170> PatentIn version 3.5
<210> 1
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG олигонуклеотид
<400> 1
tcgtcgacga tcggcgcgcg ccg 23
<210> 2
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG олигонуклеотид
<400> 2
tcgacgtcga tcggcgcgcg ccg 23
<210> 3
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG олигонуклеотид
<400> 3
tcgacgtcga tcggcgcgcg ccgt 24
<210> 4
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG олигонуклеотид
<220>
<221> другой_признак
<222> (1)..(1)
<223> 5'-Йод-2'-дезоксиуридин
<400> 4
ncgacgtcga tcggcgcgcg ccg 23
<210> 5
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG олигонуклеотид
<220>
<221> другой_признак
<222> (1)..(1)
<223> 5'-Йод-2'-дезоксиуридин
<400> 5
ncgacgtcga tcggcgcgcg ccgt 24
<210> 6
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG Олигонуклеотид
<220>
<221> другой_признак
<222> (1)..(1)
<223> 5'-Йод-2'-дезоксиуридин
<400> 6
ncgacgtcga tcggcgcgcg ccgt 24
<210> 7
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG олигонуклеотид
<220>
<221> другой_признак
<222> (1)..(1)
<223> 5'-Этил-2'-дезоксиуридин
<400> 7
ncgacgtcga tcggcgcgcg ccg 23
<210> 8
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG Олигонуклеотид
<220>
<221> другой_признак
<222> (1)..(1)
<223> 5'-Йод-2'-дезоксиуридин
<400> 8
ncgtcgacga tcggcggccg ccgt 24
<210> 9
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG Олигонуклеотид
<220>
<221> другой_признак
<222> (1)..(1)
<223> 5'-Йод-2'-дезоксиуридин
<400> 9
ncgtcgacga tcggcggccg ccgt 24
<210> 10
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CpG Олигонуклеотид
<400> 10
tcgtcgacga tcggcgcgcg ccg 23
<210> 11
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> иммуностимулирующий олигорибонуклеотид
<400> 11
uuguuguugu uguuguuguu 20
<210> 12
<211> 20
<212> РНК
<213> Искусственная последовательность
<220>
<223> Иммуностимулирующий олигорибонуклеотид
<400> 12
uuauuauuau uauuauuauu 20
<210> 13
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> Иммуностимулирующий олигорибонуклеотид
<400> 13
aaacgcucag ccaaagcag 19
<210> 14
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК/РНК гибрид
<220>
<221> другой_признак
<222> (11)..(17)
<223> рибонуклеотиди
<400> 14
tcgtcgtttt guuguguttt t 21
<210> 15
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 15
Met Ala Thr Gln Gly Thr Lys Arg Ser Tyr Glu Gln Met Glu Thr Gly
1 5 10 15
<210> 16
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 16
Gly Thr Lys Arg Ser Tyr Glu Gln Met Glu Thr Gly Gly Glu Arg Gln
1 5 10 15
<210> 17
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 17
Ser Tyr Glu Gln Met Glu Thr Gly Gly Glu Arg Gln Asp Ala Thr Glu
1 5 10 15
<210> 18
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 18
Met Glu Thr Gly Gly Glu Arg Gln Asp Ala Thr Glu Ile Lys Ala Ser
1 5 10 15
<210> 19
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 19
Gly Glu Arg Gln Asp Ala Thr Glu Ile Lys Ala Ser Val Gly Arg Met
1 5 10 15
<210> 20
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 20
Asp Ala Thr Glu Ile Lys Ala Ser Val Gly Arg Met Val Gly Gly Ile
1 5 10 15
<210> 21
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 21
Ile Lys Ala Ser Val Gly Arg Met Val Gly Gly Ile Gly Arg Phe Tyr
1 5 10 15
<210> 22
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 22
Val Gly Arg Met Val Gly Gly Ile Gly Arg Phe Tyr Ile Gln Met Cys
1 5 10 15
<210> 23
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 23
Val Gly Gly Ile Gly Arg Phe Tyr Ile Gln Met Cys Thr Glu Leu Lys
1 5 10 15
<210> 24
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 24
Gly Arg Phe Tyr Ile Gln Met Cys Thr Glu Leu Lys Leu Ser Asp Tyr
1 5 10 15
<210> 25
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 25
Ile Gln Met Cys Thr Glu Leu Lys Leu Ser Asp Tyr Glu Gly Arg Leu
1 5 10 15
<210> 26
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 26
Thr Glu Leu Lys Leu Ser Asp Tyr Glu Gly Arg Leu Ile Gln Asn Ser
1 5 10 15
<210> 27
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 27
Leu Ser Asp Tyr Glu Gly Arg Leu Ile Gln Asn Ser Ile Thr Ile Glu
1 5 10 15
<210> 28
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 28
Glu Gly Arg Leu Ile Gln Asn Ser Ile Thr Ile Glu Arg Met Val Leu
1 5 10 15
<210> 29
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 29
Ile Gln Asn Ser Ile Thr Ile Glu Arg Met Val Leu Ser Ala Phe Asp
1 5 10 15
<210> 30
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 30
Ile Thr Ile Glu Arg Met Val Leu Ser Ala Phe Asp Glu Arg Arg Asn
1 5 10 15
<210> 31
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 31
Arg Met Val Leu Ser Ala Phe Asp Glu Arg Arg Asn Lys Tyr Leu Glu
1 5 10 15
<210> 32
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 32
Ser Ala Phe Asp Glu Arg Arg Asn Lys Tyr Leu Glu Glu His Pro Ser
1 5 10 15
<210> 33
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 33
Glu Arg Arg Asn Lys Tyr Leu Glu Glu His Pro Ser Ala Gly Lys Asp
1 5 10 15
<210> 34
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 34
Lys Tyr Leu Glu Glu His Pro Ser Ala Gly Lys Asp Pro Lys Lys Thr
1 5 10 15
<210> 35
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 35
Glu His Pro Ser Ala Gly Lys Asp Pro Lys Lys Thr Gly Gly Pro Ile
1 5 10 15
<210> 36
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 36
Ala Gly Lys Asp Pro Lys Lys Thr Gly Gly Pro Ile Tyr Arg Arg Val
1 5 10 15
<210> 37
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 37
Pro Lys Lys Thr Gly Gly Pro Ile Tyr Arg Arg Val Asp Gly Lys Trp
1 5 10 15
<210> 38
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 38
Gly Gly Pro Ile Tyr Arg Arg Val Asp Gly Lys Trp Met Arg Glu Leu
1 5 10 15
<210> 39
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 39
Tyr Arg Arg Val Asp Gly Lys Trp Met Arg Glu Leu Ile Leu Tyr Asp
1 5 10 15
<210> 40
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 40
Asp Gly Lys Trp Met Arg Glu Leu Ile Leu Tyr Asp Lys Glu Glu Ile
1 5 10 15
<210> 41
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 41
Met Arg Glu Leu Ile Leu Tyr Asp Lys Glu Glu Ile Arg Arg Val Trp
1 5 10 15
<210> 42
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 42
Ile Leu Tyr Asp Lys Glu Glu Ile Arg Arg Val Trp Arg Gln Ala Asn
1 5 10 15
<210> 43
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 43
Lys Glu Glu Ile Arg Arg Val Trp Arg Gln Ala Asn Asn Gly Glu Asp
1 5 10 15
<210> 44
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 44
Arg Arg Val Trp Arg Gln Ala Asn Asn Gly Glu Asp Ala Thr Ala Gly
1 5 10 15
<210> 45
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 45
Arg Gln Ala Asn Asn Gly Glu Asp Ala Thr Ala Gly Leu Thr His Ile
1 5 10 15
<210> 46
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 46
Asn Gly Glu Asp Ala Thr Ala Gly Leu Thr His Ile Met Ile Trp His
1 5 10 15
<210> 47
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 47
Ala Thr Ala Gly Leu Thr His Ile Met Ile Trp His Ser Asn Leu Asn
1 5 10 15
<210> 48
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 48
Leu Thr His Ile Met Ile Trp His Ser Asn Leu Asn Asp Ala Thr Tyr
1 5 10 15
<210> 49
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 49
Met Ile Trp His Ser Asn Leu Asn Asp Ala Thr Tyr Gln Arg Thr Arg
1 5 10 15
<210> 50
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 50
Ser Asn Leu Asn Asp Ala Thr Tyr Gln Arg Thr Arg Ala Leu Val Arg
1 5 10 15
<210> 51
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 51
Asp Ala Thr Tyr Gln Arg Thr Arg Ala Leu Val Arg Thr Gly Met Asp
1 5 10 15
<210> 52
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 52
Gln Arg Thr Arg Ala Leu Val Arg Thr Gly Met Asp Pro Arg Met Cys
1 5 10 15
<210> 53
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 53
Ala Leu Val Arg Thr Gly Met Asp Pro Arg Met Cys Ser Leu Met Gln
1 5 10 15
<210> 54
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 54
Thr Gly Met Asp Pro Arg Met Cys Ser Leu Met Gln Gly Ser Thr Leu
1 5 10 15
<210> 55
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 55
Pro Arg Met Cys Ser Leu Met Gln Gly Ser Thr Leu Pro Arg Arg Ser
1 5 10 15
<210> 56
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 56
Ser Leu Met Gln Gly Ser Thr Leu Pro Arg Arg Ser Gly Ala Ala Gly
1 5 10 15
<210> 57
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 57
Gly Ser Thr Leu Pro Arg Arg Ser Gly Ala Ala Gly Ala Ala Val Lys
1 5 10 15
<210> 58
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 58
Pro Arg Arg Ser Gly Ala Ala Gly Ala Ala Val Lys Gly Val Gly Thr
1 5 10 15
<210> 59
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 59
Gly Ala Ala Gly Ala Ala Val Lys Gly Val Gly Thr Ile Ala Met Glu
1 5 10 15
<210> 60
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 60
Ala Ala Val Lys Gly Val Gly Thr Ile Ala Met Glu Leu Ile Arg Met
1 5 10 15
<210> 61
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 61
Gly Val Gly Thr Ile Ala Met Glu Leu Ile Arg Met Ile Lys Arg Gly
1 5 10 15
<210> 62
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 62
Ile Ala Met Glu Leu Ile Arg Met Ile Lys Arg Gly Ile Asn Asp Arg
1 5 10 15
<210> 63
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 63
Leu Ile Arg Met Ile Lys Arg Gly Ile Asn Asp Arg Asn Phe Trp Arg
1 5 10 15
<210> 64
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 64
Ile Lys Arg Gly Ile Asn Asp Arg Asn Phe Trp Arg Gly Glu Asn Gly
1 5 10 15
<210> 65
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 65
Ile Asn Asp Arg Asn Phe Trp Arg Gly Glu Asn Gly Arg Arg Thr Arg
1 5 10 15
<210> 66
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 66
Asn Phe Trp Arg Gly Glu Asn Gly Arg Arg Thr Arg Ala Ala Tyr Glu
1 5 10 15
<210> 67
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 67
Gly Glu Asn Gly Arg Arg Thr Arg Ala Ala Tyr Glu Arg Met Cys Asn
1 5 10 15
<210> 68
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 68
Arg Arg Thr Arg Ala Ala Tyr Glu Arg Met Cys Asn Ile Leu Lys Gly
1 5 10 15
<210> 69
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 69
Ala Ala Tyr Glu Arg Met Cys Asn Ile Leu Lys Gly Lys Phe Gln Thr
1 5 10 15
<210> 70
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 70
Arg Met Cys Asn Ile Leu Lys Gly Lys Phe Gln Thr Ala Ala Gln Arg
1 5 10 15
<210> 71
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 71
Ile Leu Lys Gly Lys Phe Gln Thr Ala Ala Gln Arg Ala Met Met Asp
1 5 10 15
<210> 72
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 72
Lys Phe Gln Thr Ala Ala Gln Arg Ala Met Met Asp Gln Val Arg Glu
1 5 10 15
<210> 73
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 73
Ala Ala Gln Arg Ala Met Met Asp Gln Val Arg Glu Ser Arg Asn Pro
1 5 10 15
<210> 74
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 74
Ala Met Met Asp Gln Val Arg Glu Ser Arg Asn Pro Gly Asn Ala Glu
1 5 10 15
<210> 75
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 75
Gln Val Arg Glu Ser Arg Asn Pro Gly Asn Ala Glu Ile Glu Asp Leu
1 5 10 15
<210> 76
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 76
Ser Arg Asn Pro Gly Asn Ala Glu Ile Glu Asp Leu Ile Phe Leu Ala
1 5 10 15
<210> 77
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 77
Gly Asn Ala Glu Ile Glu Asp Leu Ile Phe Leu Ala Arg Ser Ala Leu
1 5 10 15
<210> 78
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 78
Ile Glu Asp Leu Ile Phe Leu Ala Arg Ser Ala Leu Val Leu Arg Gly
1 5 10 15
<210> 79
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 79
Ile Phe Leu Ala Arg Ser Ala Leu Val Leu Arg Gly Ser Val Ala His
1 5 10 15
<210> 80
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 80
Arg Ser Ala Leu Val Leu Arg Gly Ser Val Ala His Lys Ser Cys Leu
1 5 10 15
<210> 81
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 81
Val Leu Arg Gly Ser Val Ala His Lys Ser Cys Leu Pro Ala Cys Val
1 5 10 15
<210> 82
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 82
Ser Val Ala His Lys Ser Cys Leu Pro Ala Cys Val Tyr Gly Leu Ala
1 5 10 15
<210> 83
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 83
Lys Ser Cys Leu Pro Ala Cys Val Tyr Gly Leu Ala Val Ala Ser Gly
1 5 10 15
<210> 84
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 84
Pro Ala Cys Val Tyr Gly Leu Ala Val Ala Ser Gly His Asp Phe Glu
1 5 10 15
<210> 85
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 85
Tyr Gly Leu Ala Val Ala Ser Gly His Asp Phe Glu Arg Glu Gly Tyr
1 5 10 15
<210> 86
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 86
Val Ala Ser Gly His Asp Phe Glu Arg Glu Gly Tyr Ser Leu Val Gly
1 5 10 15
<210> 87
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 87
His Asp Phe Glu Arg Glu Gly Tyr Ser Leu Val Gly Ile Asp Pro Phe
1 5 10 15
<210> 88
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 88
Arg Glu Gly Tyr Ser Leu Val Gly Ile Asp Pro Phe Lys Leu Leu Gln
1 5 10 15
<210> 89
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 89
Ser Leu Val Gly Ile Asp Pro Phe Lys Leu Leu Gln Asn Ser Gln Val
1 5 10 15
<210> 90
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 90
Ile Asp Pro Phe Lys Leu Leu Gln Asn Ser Gln Val Phe Ser Leu Ile
1 5 10 15
<210> 91
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 91
Lys Leu Leu Gln Asn Ser Gln Val Phe Ser Leu Ile Arg Pro Asn Glu
1 5 10 15
<210> 92
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 92
Asn Ser Gln Val Phe Ser Leu Ile Arg Pro Asn Glu Asn Pro Ala His
1 5 10 15
<210> 93
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 93
Phe Ser Leu Ile Arg Pro Asn Glu Asn Pro Ala His Lys Ser Gln Leu
1 5 10 15
<210> 94
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 94
Arg Pro Asn Glu Asn Pro Ala His Lys Ser Gln Leu Val Trp Met Ala
1 5 10 15
<210> 95
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 95
Asn Pro Ala His Lys Ser Gln Leu Val Trp Met Ala Cys His Ser Ala
1 5 10 15
<210> 96
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 96
Lys Ser Gln Leu Val Trp Met Ala Cys His Ser Ala Ala Phe Glu Asp
1 5 10 15
<210> 97
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 97
Val Trp Met Ala Cys His Ser Ala Ala Phe Glu Asp Leu Arg Val Ser
1 5 10 15
<210> 98
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 98
Cys His Ser Ala Ala Phe Glu Asp Leu Arg Val Ser Ser Phe Ile Arg
1 5 10 15
<210> 99
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 99
Ala Phe Glu Asp Leu Arg Val Ser Ser Phe Ile Arg Gly Lys Lys Val
1 5 10 15
<210> 100
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 100
Leu Arg Val Ser Ser Phe Ile Arg Gly Lys Lys Val Ile Pro Arg Gly
1 5 10 15
<210> 101
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 101
Ser Phe Ile Arg Gly Lys Lys Val Ile Pro Arg Gly Lys Leu Ser Thr
1 5 10 15
<210> 102
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 102
Gly Lys Lys Val Ile Pro Arg Gly Lys Leu Ser Thr Arg Gly Val Gln
1 5 10 15
<210> 103
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 103
Ile Pro Arg Gly Lys Leu Ser Thr Arg Gly Val Gln Ile Ala Ser Asn
1 5 10 15
<210> 104
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 104
Lys Leu Ser Thr Arg Gly Val Gln Ile Ala Ser Asn Glu Asn Val Glu
1 5 10 15
<210> 105
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 105
Arg Gly Val Gln Ile Ala Ser Asn Glu Asn Val Glu Ala Met Asp Ser
1 5 10 15
<210> 106
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 106
Ile Ala Ser Asn Glu Asn Val Glu Ala Met Asp Ser Asn Thr Leu Glu
1 5 10 15
<210> 107
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 107
Glu Asn Val Glu Ala Met Asp Ser Asn Thr Leu Glu Leu Arg Ser Arg
1 5 10 15
<210> 108
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 108
Ala Met Asp Ser Asn Thr Leu Glu Leu Arg Ser Arg Tyr Trp Ala Ile
1 5 10 15
<210> 109
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 109
Asn Thr Leu Glu Leu Arg Ser Arg Tyr Trp Ala Ile Arg Thr Arg Ser
1 5 10 15
<210> 110
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 110
Leu Arg Ser Arg Tyr Trp Ala Ile Arg Thr Arg Ser Gly Gly Asn Thr
1 5 10 15
<210> 111
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 111
Tyr Trp Ala Ile Arg Thr Arg Ser Gly Gly Asn Thr Asn Gln Gln Arg
1 5 10 15
<210> 112
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 112
Arg Thr Arg Ser Gly Gly Asn Thr Asn Gln Gln Arg Ala Ser Ala Gly
1 5 10 15
<210> 113
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 113
Gly Gly Asn Thr Asn Gln Gln Arg Ala Ser Ala Gly Gln Ile Ser Val
1 5 10 15
<210> 114
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 114
Asn Gln Gln Arg Ala Ser Ala Gly Gln Ile Ser Val Gln Pro Thr Phe
1 5 10 15
<210> 115
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 115
Ala Ser Ala Gly Gln Ile Ser Val Gln Pro Thr Phe Ser Val Gln Arg
1 5 10 15
<210> 116
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 116
Gln Ile Ser Val Gln Pro Thr Phe Ser Val Gln Arg Asn Leu Pro Phe
1 5 10 15
<210> 117
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 117
Gln Pro Thr Phe Ser Val Gln Arg Asn Leu Pro Phe Glu Arg Ala Thr
1 5 10 15
<210> 118
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 118
Ser Val Gln Arg Asn Leu Pro Phe Glu Arg Ala Thr Ile Met Ala Ala
1 5 10 15
<210> 119
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппау
<400> 119
Asn Leu Pro Phe Glu Arg Ala Thr Ile Met Ala Ala Phe Ser Gly Asn
1 5 10 15
<210> 120
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 120
Glu Arg Ala Thr Ile Met Ala Ala Phe Ser Gly Asn Asn Glu Gly Arg
1 5 10 15
<210> 121
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 121
Ile Met Ala Ala Phe Ser Gly Asn Asn Glu Gly Arg Thr Ser Asp Met
1 5 10 15
<210> 122
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 122
Phe Ser Gly Asn Asn Glu Gly Arg Thr Ser Asp Met Arg Thr Glu Val
1 5 10 15
<210> 123
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 123
Asn Glu Gly Arg Thr Ser Asp Met Arg Thr Glu Val Ile Arg Met Met
1 5 10 15
<210> 124
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 124
Thr Ser Asp Met Arg Thr Glu Val Ile Arg Met Met Glu Ser Ala Lys
1 5 10 15
<210> 125
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 125
Arg Thr Glu Val Ile Arg Met Met Glu Ser Ala Lys Pro Glu Asp Leu
1 5 10 15
<210> 126
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 126
Ile Arg Met Met Glu Ser Ala Lys Pro Glu Asp Leu Ser Phe Gln Gly
1 5 10 15
<210> 127
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 127
Glu Ser Ala Lys Pro Glu Asp Leu Ser Phe Gln Gly Arg Gly Val Phe
1 5 10 15
<210> 128
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 128
Pro Glu Asp Leu Ser Phe Gln Gly Arg Gly Val Phe Glu Leu Ser Asp
1 5 10 15
<210> 129
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 129
Ser Phe Gln Gly Arg Gly Val Phe Glu Leu Ser Asp Glu Lys Ala Thr
1 5 10 15
<210> 130
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 130
Arg Gly Val Phe Glu Leu Ser Asp Glu Lys Ala Thr Ser Pro Ile Val
1 5 10 15
<210> 131
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 131
Glu Leu Ser Asp Glu Lys Ala Thr Ser Pro Ile Val Pro Ser Phe Asp
1 5 10 15
<210> 132
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 132
Glu Lys Ala Thr Ser Pro Ile Val Pro Ser Phe Asp Met Ser Asn Glu
1 5 10 15
<210> 133
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 133
Ser Pro Ile Val Pro Ser Phe Asp Met Ser Asn Glu Gly Ser Tyr Phe
1 5 10 15
<210> 134
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 134
Pro Ser Phe Asp Met Ser Asn Glu Gly Ser Tyr Phe Phe Gly Asp Asn
1 5 10 15
<210> 135
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 135
Met Ser Asn Glu Gly Ser Tyr Phe Phe Gly Asp Asn Ala Glu Glu Tyr
1 5 10 15
<210> 136
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Фрагмент протеина NP вируса свиного гриппа
<400> 136
Gly Ser Tyr Phe Phe Gly Asp Asn Ala Glu Glu Tyr Asp Ser
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИСКУССТВЕННЫЕ НЕИЗБИРАТЕЛЬНЫЕ ЭПИТОПЫ КЛЕТОК Т-ХЕЛПЕРОВ КАК ИММУННЫЕ СТИМУЛЯТОРЫ ДЛЯ СИНТЕТИЧЕСКИХ ПЕПТИДНЫХ ИММУНОГЕНОВ | 2019 |
|
RU2839585C2 |
| Выделенный рекомбинантный вирус на основе вируса гриппа для индукции специфического иммунитета к вирусу гриппа и/или профилактики заболеваний, вызванных вирусом гриппа | 2021 |
|
RU2813150C2 |
| АНТИТЕЛА С ПРИВИТЫМ НАТРИЙУРЕТИЧЕСКИМ ПЕПТИДОМ С-ТИПА | 2019 |
|
RU2828786C2 |
| МУЛЬТИВАЛЕНТНЫЕ И МУЛЬТИСПЕЦИФИЧЕСКИЕ 41BB-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2017 |
|
RU2789648C2 |
| МУЛЬТИВАЛЕНТНЫЕ И МУЛЬТИСПЕЦИФИЧЕСКИЕ ГИБРИДНЫЕ БЕЛКИ, СВЯЗЫВАЮЩИЕСЯ С DR5 | 2016 |
|
RU2748620C2 |
| АНТИТЕЛА С ПРИВИТЫМ МОЗГОВЫМ НАТРИЙУРЕТИЧЕСКИМ ПЕПТИДОМ | 2019 |
|
RU2839970C2 |
| АНТИТЕЛА С ПРИВИТЫМ ПРЕДСЕРДНЫМ НАТРИЙУРЕТИЧЕСКИМ ПЕПТИДОМ | 2019 |
|
RU2822433C2 |
| ИММУНОМОДУЛИРУЮЩИЕ СЛИТЫЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2755227C2 |
| АНТИТЕЛА, ПОДХОДЯЩИЕ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ ГРИППА, И ИХ КОМПОЗИЦИИ, КОМБИНАЦИИ И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2720282C1 |
| СЛИТЫЕ БЕЛКИ С АЛЬБУМИН-СВЯЗЫВАЮЩИМИ ДОМЕНАМИ | 2018 |
|
RU2786444C2 |
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая липосому для усиления эффективности вакцины, по сути свободную от сапонина, адъювантную композицию для усиления эффективности вакцины, содержащую вышеуказанную липосому, вакцинную композицию для потенциирования иммунного ответа, содержащую вышеуказанную липосому, способ индуцирования иммунного ответа против BVDV у крупного рогатого скота, способ предупреждения выделения ооциста Eimeria у животных - домашней птицы, инфицированной Eimeria, способ получения вышеуказанной липосомы. В одном из вариантов реализации, липосома является по сути свободной от сапонина, причем сапонин или отсутствует, или присутствует в количестве меньшем, чем определяемое, и содержит внешнюю липидную двухслойную мембрану и внутреннюю камеру. Изобретение расширяет арсенал средств для усиления индуцирования имунного ответа. 6 н. и 17 з.п. ф-лы, 25 табл., 6 пр.
1. Липосома для усиления эффективности вакцины, по сути свободная от сапонина, причем сапонин или отсутствует, или присутствует в количестве меньшем, чем определяемое, содержащая внешнюю липидную двухслойную мембрану и внутреннюю камеру, где внешняя мембрана содержит:
a) четвертичное аммониевое соединение, состоящее из четырех алкильных цепей, две из которых представляют собой С10-С20алкилы, и оставшиеся две представляют собой С1-С4алкилы;
b) стерин, выбранный из группы, состоящей из β-ситостерина, стигмастерина, эргостерина, эргокальциферола и холестерина;
c) фосфолипид; и
d) гликолипид формулы I:
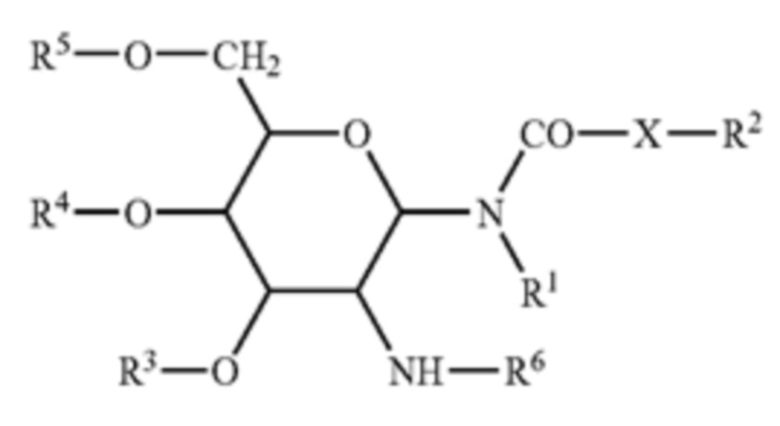
Формула I
в котором, R1 и R2 независимо представляют собой водород, или насыщенный алкильный радикал, имеющий вплоть до 20 атомов углерода; X представляет собой -CH2-, -O- или -NH-; R3, R4 и R5 независимо представляют собой водород, -SO42-, -PO42-, -COC1-10 алкил; R6 представляет собой L-аланил, L-альфа-аминобутил, L-аргинил, L-аспаргинил, L-аспартил, L-цистеинил, L-глутамил, L-глицил, L-гистидил, L-гидроксипропил, L-изолейцил, L-лейцил, L-лизил, L-метионил, L-орнитинил, L-фенилаланил, L-пролил, L-серил, L-треонил, L-тирозил, L-триптофанил, и L-валил или их D-изомеры.
2. Липосома по п. 1, где четвертичное аммониевое соединение представляет собой DDA, стерин представляет собой холестерин, и гликолипид представляет собой N-(2-дезокси-2-L-лейциламино-β-D-глюкопиранозил)-N-октадецилдодеканоиламид или его соль.
3. Липосома по п. 1, дополнительно содержащая, иммуностимулирующий олигонуклеотид, выбранный из группы, состоящей из иммуностимулирующего рибонуклеотида, CpG олигодезоксирибонуклеотида, и их комбинации.
4. Липосома по п. 3, в которой указанный иммуностимулирующий олигонуклеотид содержит какую-либо одну из SEQ ID NO 1-14.
5. Липосома, по сути свободная от сапонина по п. 1, в которой массовые соотношения - четвертичное аммониевое соединение : стерин : фосфолипид : гликолипид составляют 1 : 0,75-1,25 : 1,5-2,5 : 1,5-2,5 соответственно.
6. Липосома, по сути свободная от сапонина по п. 5, в которой массовое соотношение - четвертичное аммониевое соединение : стерин : фосфолипид : гликолипид составляет 1 : 1 : 2 : 2.
7. Липосома, по сути свободная от сапонина по п. 1, в которой
а) общая масса четвертичного аммониевого соединения и стерина составляет около 10-60% от общей массы гликолипида и фосфолипида;
b) четвертичное аммониевое соединение составляет, по меньшей мере, около 5% мас./мас. от общей массы четвертичного аммониевого соединения, стерина, фосфолипида и гликолипида; и
с) гликолипид составляет, по меньшей мере, около 20% мас./мас. от общей массы четвертичного аммониевого соединения, стерина, фосфолипида и гликолипида.
8. Адъювантная композиция для усиления эффективности вакцины, содержащая эффективное количество липосомы по п. 1.
9. Вакцинная композиция для потенциирования иммунного ответа, содержащая эффективное количество антигенного компонента и иммунологически эффективное количество адъювантной композиции по п. 8.
10. Вакцинная композиция по п. 9, где указанная вакцинная композиция фактически не содержит сапонин.
11. Вакцинная композиция по п. 9, в которой антигенный компонент выбран из группы, состоящей из антигенов крупного рогатого скота, козьих антигенов, свиных антигенов, антигенов птицы, антигенов коней, собачьих антигенов и кошачьих антигенов.
12. Вакцинная композиция по п. 9, в которой иммуностимулирующий олигонуклеотид содержит CpG олигодезоксирибонуклеотид.
13. Вакцинная композиция по п. 9, в которой антигенный компонент содержит IBR (вирус инфекционного ринотрахеита крупного рогатого скота), BVDV-1 (вирус типа 1 вирусной диареи крупного рогатого скота) и BVDV-2 (вирус типа 2 вирусной диареи крупного рогатого скота), и где вакцинная композиция фактически не содержит сапонин.
14. Вакцинная композиция по п. 11, в которой антиген содержит антиген птицы.
15. Вакцинная композиция по п. 14, в которой антиген представляет собой профилин.
16. Вакцинная композиция по п. 11, в которой антигенный компонент содержит вирус с оцРНК, и где вакцинная композиция фактически не содержит CpG олигодезоксирибонуклеотид.
17. Вакцинная композиция по п. 16, в которой вирус с оцРНК представляет собой вирус гриппа.
18. Вакцинная композиция по п. 17, в которой вирус гриппа представляет собой инактивированный вирус свиного гриппа (SIV).
19. Способ индуцирования иммунного ответа против BVDV у крупного рогатого скота, который включает введение указанному крупному рогатому скоту вакцинной композиции по п. 13.
20. Способ по п. 19, в котором указанный иммунный ответ индуцируется без сопутствующей повышенной температуры.
21. Способ предупреждения выделения ооциста Eimeria у животных - домашней птицы, инфицированной Eimeria, который включает введение указанному животному вакцинной композиции по п. 15 перед указанным инфицированием.
22. Способ получения липосомы по п. 1, где способ включает:
a) растворение в органическом растворителе четвертичного аммониевого соединения, стерина, фосфолипида и гликолипида формулы I:
Формула I
в котором, R1 и R2 независимо представляют собой водород, или насыщенный алкильный радикал, имеющий вплоть до 20 атомов углерода; X представляет собой -CH2-, -O- или -NH-; R3, R4, и R5 независимо представляют собой водород, -SO42-, -PO42-, -COC1-10 алкил; R6 представляет собой L-аланил, L-альфа-аминобутил, L-аргинил, L-аспаргинил, L-аспартил, L-цистеинил, L-глутамил, L-глицил, L-гистидил, L-гидроксипропил, L-изолейцил, L-лейцил, L-лизил, L-метионил, L-орнитинил, L-фенилаланил, L-пролил, L-серил, L-треонил, L-тирозил, L-триптофанил, и L-валил или их D-изомеры;
b) удаление органического растворителя и образование пленки;
c) регидратирование пленки в водном растворителе, таким образом, образуя регидратированную композицию;
d) микрофлюидизирование регидратированной композиции.
23. Способ по п. 22, в котором водный растворитель содержит иммуностимулирующий олигонуклеотид, выбранный из группы, состоящей из иммуностимулирующего рибонуклеотида, CpG олигодезоксири-бонуклеотида, и их комбинации.
| Паровозный поршень, останавливающийся при езде без пара | 1924 |
|
SU839A1 |
| US 5741517 A1, 21.04.1998 | |||
| Аналоговый преобразователь информации | 1977 |
|
SU750379A1 |
| УСОВЕРШЕНСТВОВАННЫЕ ЛИПОСОМЫ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2482837C2 |
| НОВЫЕ ГЛИКОЛИПИДНЫЕ АДЪЮВАНТНЫЕ КОМПОЗИЦИИ | 2007 |
|
RU2392965C2 |

