Область техники
Настоящая заявка заявляет приоритет по корейской заявке на патент KR 10-2017-0038775, поданной 27 марта 2017 года, и корейской заявке на патент KR 10-2017-0118890 и KR 10-2017-0118917, поданной 15 сентября 2017 года, соответственно, полные описания которых включены в данный документ во всей своей полноте посредством ссылки.
Настоящее изобретение относится к антителу или его фрагменту, специфически связывающемуся с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы, и его применению и, более конкретно, к антителу или его фрагменту, имеющему конкретные последовательности определяющей комплементарность области (CDR - complementary determining region), определенные в настоящем описании изобретения, и специфически связывающемуся с эпитопом, содержащим последовательность SEQ ID NO: 97 в N-конце лизил-тРНК-синтетазы (KRS), к применению данного антитела или его фрагмента для ингибирования метастаза рака, к применению данного антитела или его фрагмента для диагностики рака и к фармацевтической композиции для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Предшествующий уровень техники
Недавние исследования установили, что лизил-тРНК-синтетаза человека (KRS), обычно присутствующая в цитозоле, перемещается к плазматической мембране (клеточной мембране), взаимодействуя с рецептором ламинина, массой 67 кДа (67LR), находящимся на плазматической мембране, стимулируя, таким образом, миграцию опухолевых (или раковых) клеток с влиянием на метастаз рака (Dae Gyu Kim et al., Chemical inhibition of prometastatic lysyl-tRNA synthetaselaminin receptor interaction, Nat Chem Biol. 2014 Jan; 10(1): 2934, Dae Gye Kim et. al. Interaction of two translational components, lysyl-tRNA synthetase and p40/37LRP, in plasma membrane promotes laminin-dependent cell migration, FASEB J. (2012)26, 4142-4159). KRS человека (учетный номер в Genbank NP_005539.1 и т.д.) содержит N-концевой удлиняющий сегмент (1-72), антикодон-связывающий домен (73-209) и каталитический домен (220-597). KRS человека представляет собой фермент, являющийся существенным для синтеза белка, и обычно располагается в пределах мульти-тРНК-синтетазного комплекса (MSC - multi-tRNA synthetase complex) в цитозоле. Однако, после введения сигнала - ламинина р38 MAPK (mitogen-activated protein kinase - митоген-активируемая протеинкиназа) форсфорилирует KRS в положениях остатков Т52, и KRS перемещается к клеточной мембране, где KRS защищает 67LR от убиквитин-опосредованной деградации. Также сообщалось, что KRS, перемещенная к клеточной мембране, ускоряет метастаз рака в результате стабилизации и взаимодействия с 67LR, ассоциированным с метастазом рака.
В то же время, иммунные клетки участвуют в механизме первичной защиты в организме, но об избыточной активации иммунных клеток недавно сообщалось как об одном из главных патогенезов. Повышенная подвижность иммунных клеток обычно наблюдается при активации иммунных клеток воспаления, и, в частности, такая миграция и инвазия иммунных клеток, как сообщается, всесторонне вовлечены в патологию заболевания в следующих заболеваниях.
Например, сердечно-сосудистое заболевание, чьи очаговые поражения встречаются в сердце и главных артериях, включает атеросклероз и коронарную болезнь сердца (Ross R et al., New Engl J Med, 1999:340(2): 115-26, Poli G et al., Redox Biol 2013; 1(1): 125-30, Libby P et al., Circulation 2002;5; 105(9): 1135-43). Атеросклероз представляет собой воспалительное заболевание, провоцируемое холестерином, и вызвано атеромой, образованной холестерином, отложенным на внутренней мембране артерии, и иммунными клетками, мигрирующими из крови во внутреннюю часть артерии. То есть, атерома образуется в результате миграции иммунных клеток, таких как моноциты, к сайту, где окисленный холестерин вызывает воспаление. Образование атеромы делает внутреннюю поверхность кровеносных сосудов шероховатой и утолщает стенку кровеносных сосудов, и, таким образом, внутренний диаметр кровеносных сосудов становится суженным, приводя к нарушениям кровообращения. Разрыв фиброзных мембран, окружающих атерому, вызывает образование тромбов в кровеносных сосудах и кровотечение в атероме, и, таким образом, внутренний диаметр кровеносных сосудов быстро сужается или кровеносные сосуды блокируются. Это происходит главным образом в кровеносных сосудах, поставляющих кровь к сердцу, кровеносных сосудах, поставляющих кровь в мозг, кровеносных сосудах, поставляющих кровь к почкам, и периферических кровеносных сосудах, вызывая, вследствие этого, ишемическую болезнь сердца, ишемическую цереброваскулярную болезнь (инсульт), почечную недостаточность и ишемическое заболевание артерий нижних конечностей. Ранее было известно, что СС хемокиновый лиганд 2 (CCL2, МСР-1), который вызывает воспалительную реакцию в результате индукции миграции моноцитов, играет важную роль в возникновении и развитии таких сердечно-сосудистых заболеваний, и, вследствие этого, были предложены новые средства лечения таких сердечно-сосудистых заболеваний посредством ингибирования действия CCL2 и возникающей в результате миграции моноцитов (Gu L et al., Mol Cell, 1998; 2(2):275-81, Aiello RJ et al., Arterioscler Thromb Vase Biol 1999; 19(6):1518-25, Gosling J1 et al., Clin Invest 1999; 103(6):773-8, Harrington JR et al., Stem Cells 2000; 18(1):65-6, Ikeda U et al., Clin Cardiol 2002; 25(4): 143-7).
Легочная артериальная гипертензия (ЛАГ) относится к Группе 1 в клинической классификационной системе (ESC Guidelines, European Heart Journal 2015) Всемирной организации здравоохранения (ВОЗ) и является редким заболеванием, клинически характеризующимся затруднительным дыханием, увеличением среднего легочного артериального давления (mPAP - mean pulmonary artery pressure, mPAP выше 25 мм рт. ст.) и дисфункцией правого желудочка. Несколько заведомо существующих факторов, таких как наследственность, инфекция и родственные заболевания, задействованы в такой легочной артериальной гипертензии, но иммунный ответ, возникающий в результате повреждения эндотелнальных клеток, как известно, действует в качестве ключевого патологического фактора (Huertas et al., Circulation, 129:1332-1340, 2014). Что касается такого явления, известно, что целый ряд процессов в соответствии с инвазией и дисфункцией иммунных клеток сильно связан с патологическими явлениями, и особенно взаимодействие между иммунными клетками и сосудистыми эндотелиальными клетками, как известно, является важным в ЛАГ. Также сообщалось, что инвазия моноцитов и макрофагов ускоряет развитие синдрома Альпорта.
В заболеваниях, связанных с фиброзом, продолжительные (хронические) воспалительные ответы активируют ранозаживляющий механизм, приводя к фиброзу. После повреждения тканей иммунные клетки воспаления, такие как моноциты/макрофаги, нейтрофилы, эозинофилы и тучные клетки, быстро внедряются в поврежденное место, будучи активированными, и выделяют разные цитокины, которые, в свою очередь, активируют окружающие фибробласты, эпителиальные клетки или гладкомышечные клетки в клетки типа миобластов, и данные клетки типа миобластов образуют и секретируют белки внеклеточного матрикса в больших количествах, вызывая, в конечном итоге, аккумуляцию белков внеклеточного матрикса в больших количествах и приводя к образованию рубцов и фиброзу или гипертрофии тканей (Gurtner GC et al., Trends Cell Biol. 15: 599-607, 2005). Данная патология представляет собой одну из фундаментальных причин следующих явлений: образование рубцов на тканях кожи, вызванное ранами на коже, обусловленными порезами, ожогами, пролежнями и тому подобное; или склерозирующий фиброз печени, почки, сосудистых и легочных тканей. Также показано, что фиброз является основной патологической характеристикой в хронических аутоиммунных заболеваниях, таких как склеродерма, ревматоидный артрит, болезнь Крона, язвенный колит, миелофиброз и системная красная волчанка. Также известно, что активация иммунных клеток воспаления способствует патологии при атопических заболеваниях, астме, ХОБЛ (Хроническая обструктивная болезнь легких), псориазе, келоиде, пролиферативной ретинопатии и т.д.
В особенности, в ранозаживляющем механизме фибробласты, активированные в клетки типа миобластов, называют миофибробластами. Поскольку миофибробласты находятся в центре всех патологических заболеваний, ассоциированных с фиброзом, устранение молекулярных или иммунологических механизмов, вызывающих активность миофибробластов, является ключевым элементом лечения заболевания. Широко известно, что многие типы врожденного иммунитета или приобретенного иммунитета важны в активации и дифференциации фибробластов, и, вследствие этого, устранение воспалительного ответа в месте раны является ключевым фактором в предотвращении ремоделирования тканей в фиброз и поддержании нормальной морфологии тканей. Однако, так как на практике воспалительный ответ не легко устранить, понимание механизмов врожденного и приобретенного иммунитета для нахождения ключевых медиаторов является важным в замедлении фиброза.
В некоторых случаях моноциты, макрофаги и т.п. способствуют заживлению ран, но выделяют активные формы кислорода, азота и т.п., и таким образом оказывают пагубное действие на окружающие клетки. Таким образом, моноциты и макрофаги, если их быстро не удалить, вызывают большее повреждение тканей, приводя к фиброзу. Таким образом, ограничение моноцитов и макрофагов, которые первыми отвечают на ранних стадиях заболевания, считается терапевтической стратегией для разных хронических заболеваний, связанных с воспалением и фиброзом.
Известно, что когда механизм заживления ран запускает ответ в виде фиброза, фактор роста тромбоцитов (PDGF - platelet-derived growth factor), ассоциированный с гемагглютинацией, рекрутирует другие иммунные клетки воспаления к месту раны, и TGF-β1 (transforming growth factor - трансформирующий фактор роста) ускоряет синтез внеклеточного матрикса из местных фибробластов. Однако сообщалось, что факторы, участвующие в гемагглютинации, вызывают фиброз даже тогда, когда факторов недостаточно.
Между тем, тот факт, что Myc-KRS41-597 (ΔN) с делецией 40 концевых остатков в N-концевом удлиняющем сегменте (N-ext) не локализована на плазматической мембране, указывает на то, что область N-ext KRS представляет собой существенную область в перемещении KRS к клеточной мембране. В отношении метастаза рака, в частности, известно, что область N-ext KRS участвует в связывании KRS с 67LR при их взаимодействии. Для использования данного факта в терапевтических или диагностических целях, необходимо специфически нацеливаться на конкретный сайт (особенно, N-ext KRS) в белке KRS в соответствии с характеристиками нескольких доменов, образующих белок KRS.
Однако, несмотря на важность аминоацил-тРНК-синтетазы (ARS - aminoacyl-tRNA synthetases), включая KRS, в качестве биомаркеров, ARS похожи с точки зрения структуры белка, и, таким образом, антитела, полученные посредством иммунизации животных белком ARS, демонстрируют перекрестную реактивность, например, связывания с другими ARS, и во многих случаях даже не получают высокочувствительных антител.
В заболеваниях, вызванных избыточной активацией иммунных клеток, как упомянуто выше, были традиционно предложены целевые факторы для предупреждения транслокации (и инвазии) иммунных клеток, и были сделаны попытки разработки терапевтических способов лечения заболеваний, регулирующих целевые факторы, но сообщается об их соответствующих ограничениях. Таким образом, для эффективного лечения заболевания установление ключевого медиатора и того, какая стратегия будет контролировать ключевой медиатор в смягчении негативных воздействий иммунных клеток, все еще является острой проблемой.
Подробное описание изобретения
Техническая проблема
При исследовании для конструирования антитела, специфически связывающегося с экспонированной во внеклеточную среду N-концевой областью KRS, авторы настоящего изобретения удостоверились в том, что антитела, имеющие конкретные последовательности определяющей комплементарность области (CDR), определенные в настоящем описании изобретения, показали очень высокую специфичность и аффинность связывания с N-концевой областью KRS, а также ингибировали метастаз рака in vivo. Кроме того, авторы настоящего изобретения удостоверились в том, что повышение уровня KRS в клеточной мембране иммунных клеток (моноцитов/макрофагов) является важным патологическим явлением в заболеваниях, связанных с миграцией иммунных клеток и инвазией, и, таким образом, KRS имеет определенную корреляцию с ламинином (особенно, подтип ламинина α4β2γ1), и удостоверились в том, что антитела, связывающиеся с N-концом KRS, предложенные в настоящем изобретении, снижали уровень KRS, повышенный на клеточной мембране иммунных клеток, и фактически ингибировали миграцию и инфильтрацию иммунных клеток и, таким образом, оказывали эффект лечения родственных заболеваний, и, вследствие этого, авторы настоящего изобретения завершили настоящее изобретение.
Таким образом, аспект настоящего изобретения заключается в обеспечении антитела или его фрагмента, специфически связывающегося с эпитопом, содержащим последовательность SEQ ID NO: 97 в N-конце лизил-тРНК-синтетазы (KRS).
Другой аспект настоящего изобретения заключается в обеспечении полинуклеотида, кодирующего антитело или его фрагмент по настоящему изобретению, рекомбинантного экспрессирующего вектора, содержащего данный полинуклеотид, и клетки, трансформированной данным рекомбинантным вектором.
Еще один аспект настоящего изобретения заключается в обеспечении способа получения антитела или его фрагмента, специфически связывающегося с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы (KRS), включающего: (а) трансформацию клеток-хозяев рекомбинантным экспрессирующим вектором; (б) инкубирование трансформированных клеток-хозяев с получением антитела или его фрагмента; и (в) сбор антитела или его фрагмента, полученного в клетках-хозяевах.
Еще один аспект настоящего изобретения заключается в обеспечении фармацевтической композиции, содержащей антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для ингибирования метастаза рака.
Еще один аспект настоящего изобретения заключается в обеспечении фармацевтической композиции, состоящей из антитела или его фрагмента по настоящему изобретению, для ингибирования метастаза рака.
Еще один аспект настоящего изобретения заключается в обеспечении фармацевтической композиции, по существу состоящей из антитела или его фрагмента по настоящему изобретению, для ингибирования метастаза рака.
Еще один аспект настоящего изобретения заключается в обеспечении композиции, содержащей антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для диагностики рака.
Еще один аспект настоящего изобретения заключается в обеспечении композиции, состоящей из антитела или его фрагмента по настоящему изобретению, для диагностики рака.
Еще один аспект настоящего изобретения заключается в обеспечении композиции, по существу состоящей из антитела или его фрагмента по настоящему изобретению, для диагностики рака.
Еще один аспект настоящего изобретения заключается в обеспечении фармацевтической композиции, содержащей антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Еще один аспект настоящего изобретения заключается в обеспечении фармацевтической композиции, состоящей из антитела или его фрагмента по настоящему изобретению, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Еще один аспект настоящего изобретения заключается в обеспечении фармацевтической композиции, по существу состоящей из антитела или его фрагмента по настоящему изобретению, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Еще один аспект настоящего изобретения заключается в обеспечении применения антитела или его фрагмента по настоящему изобретению для получения агента для ингибирования метастаза рака.
Еще один аспект настоящего изобретения заключается в обеспечении способа ингибирования метастаза рака у нуждающегося в этом субъекта, причем способ включает введение антитела или его фрагмента по настоящему изобретению субъекту в количестве, эффективном для ингибирования метастаза рака.
Еще один аспект настоящего изобретения заключается в обеспечении применения антитела или его фрагмента по настоящему изобретению для получения агента для диагностики рака.
Еще один аспект настоящего изобретения заключается в обеспечении способа диагностирования рака у нуждающегося в этом субъекта, причем способ включает введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для диагностирования рака.
Еще один аспект настоящего изобретения заключается в обеспечении применения антитела или его фрагмента по настоящему изобретению для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
Еще один аспект настоящего изобретения заключается в обеспечении способа лечения заболевания, связанного с миграцией иммунных клеток, у нуждающегося в этом субъекта, причем способ включает введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для лечения заболевания, связанного с миграцией иммунных клеток.
Техническое решение
Согласно аспекту настоящего изобретения обеспечено антитело или его фрагмент, специфически связывающееся с эпитопом, содержащим последовательность SEQ ID NO: 97 в N-конце лизил-тРНК-синтетазы (KRS).
Согласно другому аспекту настоящего изобретения обеспечен полинуклеотид, кодирующий кодирующего антитело или его фрагмент по настоящему изобретению, рекомбинантный экспрессирующий вектор, содержащий данный полинуклеотид, и клетка, трансформированная данным рекомбинантным вектором.
Согласно еще одному аспекту настоящего изобретения обеспечен способ получения антитела или его фрагмента, специфически связывающийся с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы (KRS), причем способ включает: (а) трансформацию клеток-хозяев рекомбинантным экспрессирующим вектором; (б) инкубирование трансформированных клеток-хозяев с получением антитела или его фрагмента; и (в) сбор антитела или его фрагмента, полученного в клетках-хозяевах.
Согласно еще одному аспекту настоящего изобретения обеспечена фармацевтическая композиция, содержащая антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для ингибирования метастаза рака.
Согласно еще одному аспекту настоящего изобретения обеспечена фармацевтическая композиция, состоящая из антитела или его фрагмента по настоящему изобретению, для ингибирования метастаза рака.
Согласно еще одному аспекту настоящего изобретения обеспечена фармацевтическая композиция, по существу состоящая из антитела или его фрагмента по настоящему изобретению, для ингибирования метастаза рака.
Согласно еще одному аспекту настоящего изобретения обеспечена композиция, содержащая антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для диагностики рака.
Согласно еще одному аспекту настоящего изобретения обеспечена композиция, состоящая из антитела или его фрагмента по настоящему изобретению, для диагностики рака.
Согласно еще одному аспекту настоящего изобретения обеспечена композиция, по существу состоящая из антитела или его фрагмента по настоящему изобретению в качестве активного вещества, для диагностики рака.
Согласно еще одному аспекту настоящего изобретения обеспечена фармацевтическая композиция, содержащая антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Согласно еще одному аспекту настоящего изобретения обеспечена фармацевтическая композиция, состоящая из антитела или его фрагмента по настоящему изобретению, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Согласно еще одному аспекту настоящего изобретения обеспечена фармацевтическая композиция, по существу состоящая из антитела или его фрагмента по настоящему изобретению, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Согласно еще одному аспекту настоящего изобретения обеспечено применение антитела или его фрагмента по настоящему изобретению для получения агента для ингибирования метастаза рака.
Согласно еще одному аспекту настоящего изобретения обеспечен способ ингибирования метастаза рака у нуждающегося в этом субъекта, причем способ включает введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для ингибирования метастаза рака.
Согласно еще одному аспекту настоящего изобретения обеспечено применение антитела или его фрагмента по настоящему изобретению для получения агента для диагностики рака.
Согласно еще одному аспекту настоящего изобретения обеспечен способ диагностирования рака у нуждающегося в этом субъекта, причем способ включает введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для диагностирования рака.
Согласно еще одному аспекту настоящего изобретения обеспечено применение антитела или его фрагмента по настоящему изобретению для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
Согласно еще одному аспекту настоящего изобретения обеспечен способ лечения заболевания, связанного с миграцией иммунных клеток, у нуждающегося в этом субъекта, причем способ включает введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для лечения заболевания, связанного с миграцией иммунных клеток.
Далее в данном документе настоящее изобретение будет подробно описано.
В том виде, в котором он используется в данном документе, термин «внеклеточно экспонированная N-концевая область лизил-тРНК-синтетазы (KRS)» относится к конкретной последовательности, экспонированной во внеклеточную среду или на поверхности клеточной мембраны, когда KRS, продуцируемая в клетках, перемещается к клеточной мембране (или плазматической мембране), и может обычно относится к частичной или полноразмерной последовательности области из 1-72 аминокислот в N-конце KRS. Кроме того, имеет место сходство последовательностей среди видов в N-концевой области KRS, и главным образом N-концевая область KRS может содержать аминокислотную последовательность, определенную SEQ ID NO: 97. Предпочтительно, N-концевая область KRS содержит последовательность, определенную SEQ ID NO: 75 для человека, последовательность, определенную SEQ ID NO: 113 для мышей, и последовательность, определенную SEQ ID NO: 114 для крыс.
В том виде, в котором он используется в данном документе, термин «KRS» относится к полноразмерному полипептиду, известному как лизил-тРНК-синтетаза, или любой последовательности-фрагменту KRS, содержащему N-концевую область. Как описано выше, антитела или их фрагменты согласно настоящему изобретению специфически выявляют экспонированную во внеклеточную среду N-концевую область KRS и таким образом также могут выявлять приведенный выше полноразмерный полипептид KRS или любую последовательность-фрагмент KRS, содержащий N-концевую область. Конкретная последовательность KRS особым образом не ограничивается, в том случае, если последовательность содержит полипептид, определенный SEQ ID NO: 75, и известна в данной области как лизил-тРНК-синтетаза. Например, KRS по настоящему изобретению включает: последовательность, происходящую из человека (Homo sapiens) и известную как учетный номер в NCBI (база генетических данных) NP_005539.1 или тому подобное; последовательность, происходящую из мыши (Mus musculus) и известную как учетный номер в NCBI (база генетических данных) NP_444322.1 или тому подобное; и последовательность, происходящую из крысы (Rattus norvegicus) и известную как учетный номер в NCBI (база генетических данных) ХР_006255692.1 или тому подобное, и, помимо этого, можно сделать ссылку на следующую информацию о последовательностях, но она не ограничивается следующим: ХР_005004655.1 (морская свинка: Cavia porcellus), ХР_021503253.1 (песчанка, Meriones unguiculatus), ХР_002711778.1 (кролик, Oryctolagus cuniculus), ХР_536777.2 (собака, Canis lupus familiaris), ХР_003126904.2 (свинья, Sus scrofa), ХР_011755768.1 (обезьяна, Масаса nemestrina), ХР_008984479.1 (мартышка, Callithrix jacchus), ХР_019834275.1 (корова, Bos indicus) и ХР_511115.2 (шимпанзе, Pan troglodytes). Наиболее предпочтительно, KRS может представлять собой полипептид, состоящий из аминокислотной последовательности, определенной SEQ ID NO: 76 (учетный номер в Genbank NP_005539.1).
В настоящем изобретении антитело также называется иммуноглобулином (Ig) и представляет собой общий обозначение белков, которые вовлечены в биологический иммунитет посредством селективного действия на антигены. Целое антитело, найденное в природе, обычно состоит из двух пар легкой цепи (LC - light chain) и тяжелой цепи (НС - heavy chain), каждая из которых представляет собой полипептид, состоящий из нескольких доменов, и имеет две пары HC/LC в качестве основной единицы. Существует пять типов тяжелых цепей, составляющих антитела млекопитающих, которые обозначены греческими буквами: α, δ, ε, γ и μ, и разные типы тяжелых цепей составляют разные типы антител: IgA, IgD, IgE, IgG и IgM, соответственно. Существует два типа легких цепей, составляющих антитела млекопитающих, которые обозначаются λ и κ.
Тяжелые и легкие цепи антител структурно подразделяются на вариабельную область и константную область в соответствии с вариабельностью аминокислотной последовательности. Константная область тяжелой цепи состоит из трех или четырех константных областей тяжелой цепи, таких как CH1, СН2 и СН3 (антитела IgA, IgD и IgG) и СН4 (антитела IgE и IgM), в зависимости от типа антитела, и легкая цепь имеет одну константную область CL. Каждая из вариабельных областей тяжелой и легкой цепей состоит из одного домена вариабельной области тяжелой цепи (VH) или вариабельной области легкой цепи (VL). Легкая цепь и тяжелая цепь связаны друг с другом одной ковалентной дисульфидной связью, при этом их вариабельные области и константные области расположены параллельно, и две молекулы тяжелой цепи, которые связаны с легкими цепями, связаны друг с другом двумя ковалентными дисульфидными связями, образуя, таким образом, целое антитело. Целое антитело состоит из двух пар тяжелой и легкой цепей (HC-LC), и таким образом одна молекула целого антитела имеет двухвалентную моноспецифичность, где одна молекула целого антитела связывается с двумя одними и теми же антигенами посредством двух вариабельных областей.
Каждая из вариабельных областей антитела, которые содержат антигенсвязывающие сайты, подразделяется на каркасные области (FR - framework region) с низкой вариабельностью последовательностей и области, определяющие комплементарность (CDR), которые представляют собой определяющие комплементарность области с высокой вариабельностью последовательностей. В VH и VL три CDR и четыре FR расположены в следующем порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4 в направлении от N-конца к С-концу. CDR, которые имеют наивысшую вариабельность последовательностей в вариабельных областях антитела, представляют собой сайты, которые прямо связываются с антигеном, и являются очень важными в специфичности антитела в отношении антигена.
Настоящее изобретение обеспечивает антитело или его фрагмент, специфически связывающиеся с эпитопом, содержащим последовательность SEQ ID NO: 97 в N-конце лизил-тРНК-синтетазы (KRS).
В том виде, в котором он используется в данном документе, термин «эпитоп» относится к белковой детерминанте, способной к специфическому связыванию с антителом. Эпитоп обычно состоит из поверхностных групп молекул, таких как аминокислоты или боковые цепи Сахаров, и обычно имеют специфические характеристики трехмерной структуры, а также специфические характеристики заряда. Конформационные и неконформационные эпитопы отличаются друг от друга тем, что в присутствии денатурирующих растворителей теряется связывание с конформационными эпитопами, но не с неконформационными эпитопами. Эпитоп может содержать аминокислотные остатки, непосредственно участвующие в связывании (также называемые иммуногенным компонентом эпитопа), и другие аминокислотные остатки, не прямо участвующие в связывании, например, аминокислотные остатки, эффективно блокируемые специфическим антигенсвязывающим пептидом (другими словами, аминокислотный остаток, находящийся в пределах области узнавания специфического антигенсвязывающего пептида).
Предпочтительно, эпитоп представляет собой сайт, с которым связывается моноклональное антитело N3 по настоящему изобретению, происходящий из N-концевой последовательности KRS, и его конкретная последовательность особым образом не ограничивается в случае, если последовательность представляет собой непрерывную область, содержащую аминокислоты (klsknelkrrlka), определенные SEQ ID NO: 97, и обычно может состоять из 13-52 аминокислотной последовательности, более предпочтительно 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41 или 42 аминокислотной последовательности, содержащей аминокислотную последовательность SEQ ID NO: 97.
Предпочтительно, эпитоп по настоящему изобретению может включать аминокислотные последовательности, определенные SEQ ID NO: 75, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100 и SEQ ID NO: 101, которые происходят из N-конца KRS человека; аминокислотные последовательности, определенные SEQ ID NO: 113, SEQ ID NO: 103, SEQ ID NO: 104, SEQ ID NO: 105 и SEQ ID NO: 106, которые происходят из N-конца KRS мыши; и аминокислотные последовательности, определенные SEQ ID NO: 114, SEQ ID NO: 108, SEQ ID NO: 109, SEQ ГО NO: 110 и SEQ ГО NO: 111, которые происходят из N-конца KRS крысы. Эпитоп может более предпочтительно представлять собой аминокислотную последовательность в положениях 15-29 в N-концевой области KRS человека, определенную SEQ ID NO: 75 (SEQ ID NO: 75, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100 и SEQ ID NO: 101), и наиболее предпочтительно аминокислотную последовательность в положениях 15-42 в N-концевой области KRS человека, определенную SEQ ID NO: 75 (SEQ ID NO: 101).
«Антитело или его фрагмент, специфически связывающиеся с экспонированной во внеклеточную среду N-концевой областью KRS», предложенные в настоящем изобретении, содержат:
вариабельную область тяжелой цепи (VH), содержащую: определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 13, SEQ ID NO: 25 и SEQ ID NO: 37; определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 3, SEQ ID NO: 15, SEQ ID NO: 27 и SEQ ID NO: 39; и определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 5, SEQ ID NO: 17, SEQ ID NO: 29, и SEQ ID NO: 41; и
вариабельную область легкой цепи (VL), содержащую: определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 19, SEQ ID NO: 31 и SEQ ID NO: 43; определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 9, SEQ ID NO: 21, SEQ ID NO: 33 и SEQ ID NO: 45; и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 11, SEQ ID NO: 23, SEQ ID NO: 35 и SEQ ID NO: 47.
Антитела, состоящие из последовательностей CDR, имеют превосходную способность специфически связываться с экспонированной во внеклеточную среду N-концевой областью KRS. Данный признак хорошо описан в примерах настоящего описания изобретения. В примере настоящего изобретения для конструирования scFv фрагментов (single-chain variable fragment - одноцепочечный вариабельный фрагмент), специфически связывающихся с экспонированной во внеклеточную среду N-концевой областью KRS, проводили в общей сложности пять экспериментальных стадий, начиная с осуществления первичного скрининга посредством скрининга фаговой библиотеки scFv, кончая непрямым ELISA (enzyme-linked immunosorbent assay - твердофазный иммуноферментный анализ) (вторичный скрининг), вестерн-блоттингом (третичный скрининг), иммунопреципитацией (четвертичный скрининг), иммунофлуоресцентным окрашиванием (пятеричный скрининг), для отбора scFv фрагментов, демонстрирующих высокую специфичность связывания и аффинность связывания с точки зрения связывания с N-концом KRS. В общей сложности отбирали 1920 клонов scFv при первичном скрининге посредством осуществления скрининга фаговой библиотеки scFv, а за пять стадий скрининга в конечном итоге отбирали четыре типа фрагментов, scFv N3, scFv N5, scFv N7 и scFv N9, которые обладают наивысшей специфичностью. Кроме того, scFv фрагменты превращали в антитела IgG, конструируя, таким образом, антитела IgGN3, IgG N5, IgG N7 и IgG N9, и данные антитела, как также было проверено, демонстрировали высокую специфичность связывания с точки зрения связывания с N-концом KRS.
Антитела или их фрагменты, специфически связывающиеся с экспонированной во внеклеточную среду N-концевой областью KRS согласно настоящему изобретению, представляют собой антитела, имеющие следующие конформации CDR вариабельных областей тяжелой и легкой цепей, где нижеуказанные (i), (ii), (iii) и (iv) показывают комбинации CDR антител N3, N5, N7 и N9 в соответствующих примерах:
(1) вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ГО NO: 1, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 3, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 11;
(2) вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 13, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 17, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 19, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 23;
(3) вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 27, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 29, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 31, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 33, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 35; и
(4) вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 37, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 39, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 41, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 43, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 45, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 47.
Наиболее предпочтительно, антитела или их фрагменты согласно настоящему изобретению характеризуются содержанием следующих вариабельных областей тяжелой цепи и легкой цепи: в антителах или их фрагментах вариабельная область тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 49 (VH N3), SEQ ID NO: 53 (VH N5), SEQ ID NO: 57 (VH N7) и SEQ ID NO: 61 (VH N9), и вариабельная область легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51 (VLN3), SEQ ID NO: 55 (VL N5), SEQ ID NO: 59 (VL N7) и SEQ ID NO: 63 (VL N9).
Антитело, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), может представлять собой антитело, содержащее тяжелую цепь, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 77, SEQ ID NO: 81, SEQ ID NO: 85 и SEQ ID NO: 89, и легкую цепь, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 79, SEQ ID NO: 83, SEQ ID NO: 87 и SEQ ID NO: 91.
Наиболее предпочтительно, антитела могут представлять собой антитела, содержащие: тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 77, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 79; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 81, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 83; тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 85, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 87; и тяжелую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 89, и легкую цепь, содержащую аминокислотную последовательность, определенную SEQ ID NO: 91.
«Антитело, специфически связывающееся с экспонированной во внеклеточную среду N-концевой областью KRS» согласно настоящему изобретению не ограничивается его типом, в том случае, если антитело имеет указанные выше комбинации CDR или комбинации VH и VL. В качестве конкретного примера, антитело может быть выбрано из группы, состоящей из антител IgG, IgA, IgM, IgE и IgD, и может предпочтительно представлять собой антитело IgG.
Антитела по настоящему изобретению могут представлять собой моноклональные антитела или поликлональные антитела в том случае, если антитела имеют указанные выше комбинации CDR или комбинации VH и VL, которые специфически связываются с N-концевой областью KRS, но предпочтительно представляют собой моноклональные антитела, которые являются группой антител, каждое из которых имеет по существу идентичные аминокислотные последовательности в тяжелой и легкой цепях.
Антитело по настоящему изобретению может происходить из любых животных, включая млекопитающих, включающих человека, и птиц, и может предпочтительно происходить из человека. Однако антитело по настоящему изобретению может представлять собой химерное антитело, включающее часть антитела, происходящую из человека, и часть антитела, происходящую из другого вида животного. То есть, настоящее изобретение включает все химерные антитела, гуманизированные антитела и человеческие антитела, и может предпочтительно представлять собой человеческие антитела.
Кроме того, фрагмент антитела по настоящему изобретению относится к фрагменту антитела, который сохраняет антиген-специфическую способность связывания целого антитела. Предпочтительно, фрагмент сохраняет по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% аффинности связывания с N-концом KRS исходного антитела. Более конкретно, фрагмент может находиться в виде Fab, F(ab)2, Fab', F(ab')2, Fv, диатела, scFv или т.п.
Fab (антигенсвязывающий фрагмент) представляет собой антигенсвязывающий фрагмент антитела и состоит из тяжелой цепи и легкой цепи, причем каждая состоит из одного вариабельного домена и одного константного домена. F(ab')2 представляет собой фрагмент, полученный в результате гидролиза антитела под действием пепсина, и F(ab')2 имеет вид, в котором две молекулы Fab связаны дисульфидными связами в шарнирной области тяжелой цепи. F(ab') представляет собой мономерный фрагмент антитела, в котором шарнир тяжелой цепи добавлен к Fab, отделенному от фрагмента F(ab')2 в результате восстановления его дисульфидных связей. Fv (вариабельный фрагмент) представляет собой фрагмент антитела, состоящий только из соответствующих вариабельных областей тяжелой и легкой цепей. scFv (одноцепочечный вариабельный фрагмент) представляет собой рекомбинантный фрагмент антитела, в котором вариабельная область тяжелой цепи (VH) и вариабельная область легкой цепи (VL) связаны друг с другом через гибкий пептидный линкер. Диатело относится к фрагменту, в котором VH и VL scFv связаны очень коротким линкером, и таким образом не могут быть связаны друг с другом, и связываются с VL и VH другого scFv в таком же виде, соответственно, с образованием димера.
В целях настоящего изобретения фрагмент антитела не ограничивается его структурой или конформацией, в том случае, если фрагмент антитела сохраняет специфичность связывания с N-концевой областью KRS, но может предпочтительно представлять собой scFv. scFv согласно настоящему изобретению имеет конформацию CDR или конформацию VH и VL, специфическую к N-концевой области KRS, и его последовательность особым образом не ограничивается, при условии, что С-конец VH и N-конец VL связаны посредством линкера. Линкер особым образом не ограничивается его типом, в том случае, если он известен как линкер, применяемый для scFv в данной области, но может представлять собой пептид, содержащий аминокислотную последовательность, определенную SEQ ID NO: 65. В частности, scFv по настоящему изобретению может содержать аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 67 (scFv N3), SEQ ID NO: 69 (scFv N5), SEQ ID NO: 71 (scFv N7) и SEQ ID NO: 73 (scFv N9).
Антитело или его фрагмент по настоящему изобретению может содержать консервативную аминокислотную замену (также называемое консервативным вариантом антитела), которая по существу не меняет его биологической активности.
Кроме того, указанное выше антитело или его фрагмент по настоящему изобретению можно конъюгировать с ферментом, флуоресцентным веществом, радиоактивным веществом и белком, но, не ограничиваясь ими. Вместе с тем, способы конъюгирования указанных выше веществ с антителом хорошо известны в данной области.
Настоящее изобретение обеспечивает полинуклеотид, кодирующий указанное выше антитело или его фрагмент согласно настоящему изобретению.
В настоящем описании изобретения полинуклеотид может быть описан как олигонуклеотид или нуклеиновая кислота, и включает: аналоги ДНК или РНК (например, mпептидо-нуклеиновые кислоты и не встречающиеся в природе нуклеотидные аналоги), полученные с использованием молекул ДНК (например, кДНК (комплементарная ДНК) или геномная ДНК), молекул РНК (например, мРНК (матричная РНК)) или нуклеотидных аналогов; и их гибридов. Полипептид может быть одноцепочечным и двуцепочечным.
Полинуклеотид относится к нуклеотидной последовательности, кодирующей антитело, состоящее из тяжелой и легкой цепей, причем каждая имеет конформацию CDR или конформацию VH и VL, специфическую к N-концевой области KRS. Полинуклеотид по настоящему изобретению особым образом не ограничивается его последовательностью, при условии, что последовательность кодирует антитело или его фрагмент по настоящему изобретению. Полинуклеотиды, кодирующие указанные выше последовательности CDR в описанных выше антителах согласно настоящему изобретению, особым образом не ограничиваются их последовательностями, но могут предпочтительно содержать нуклеотидную последовательность, определенную SEQ ID NO: 2 (CDR1 тяжелой цепи), SEQ ID NO: 4 (CDR2 тяжелой цепи), SEQ ID NO: 6 (CDR3 тяжелой цепи), SEQ ID NO: 8 (CDR1 легкой цепи), SEQ ID NO: 10 (CDR2 легкой цепи), SEQ ID NO: 12 (CDR3 легкой цепи), SEQ ID NO: 14 (CDR1 тяжелой цепи), SEQ ID NO: 16 (CDR2 тяжелой цепи), SEQ ID NO: 18 (CDR3 тяжелой цепи), SEQ ID NO: 20 (CDR1 легкой цепи), SEQ ID NO: 22 (CDR2 легкой цепи), SEQ ID NO: 24 (CDR3 легкой цепи), SEQ ID NO: 26 (CDR1 тяжелой цепи), SEQ ID NO: 28 (CDR2 тяжелой цепи), SEQ ID NO: 30 (CDR3 тяжелой цепи), SEQ ID NO: 32 (CDR1 легкой цепи), SEQ ID NO: 34 (CDR2 легкой цепи), SEQ ID NO: 36 (CDR3 легкой цепи), SEQ ID NO: 38 (CDR1 тяжелой цепи), SEQ ID NO: 40 (CDR2 тяжелой цепи), SEQ ID NO: 42 (CDR3 тяжелой цепи), SEQ ID NO: 44 (CDR1 легкой цепи), SEQ ID NO: 46 (CDR2 легкой цепи) или SEQ ID NO: 48 (CDR3 легкой цепи).
Кроме того, полинуклеотиды, кодирующие вышеприведенные VH hVL в антителе по настоящему изобретению, особым образом не ограничиваются их последовательностями, но могут предпочтительно содержать нуклеотидную последовательность, определенную SEQ ID NO: 50 (VH), SEQ ID NO: 52 (VL), SEQ ID NO: 54 (VH), SEQ ID NO: 56 (VL), SEQ ID NO: 58 (VH), SEQ ID NO: 60 (VL), SEQ ID NO: 62 (VH) или SEQ ID NO: 64 (VL).
Кроме того, полинуклеотид, кодирующий фрагмент антитела, может предпочтительно содержать нуклеотидную последовательность любой последовательности, выбранной из группы, состоящей из SEQ ID NO: 68, SEQ ID NO: 70, SEQ ID NO: 72 и SEQ ID NO: 74, которые кодируют scFv фрагменты согласно изобретению.
Полинуклеотиды, кодирующие антитело или его фрагмент по настоящему изобретению, могут быть получены способом, известным в данной области. Например, на основе последовательностей ДНК, кодирующих часть или полноразмерные тяжелые и легкие цепи антитела, или соответствующих аминокислотных последовательностей, полинуклеотиды могут быть синтезированы способами синтеза олигонуклеотидов, которые известны в данной области, например методом полимеразной цепной реакции (ПЦР).
Настоящее изобретение обеспечивает рекомбинантный экспрессирующий вектор, содержащий полинуклеотид, кодирующий антитело или его фрагмент согласно настоящему изобретению.
В том виде, в котором он используется в данном документе, термин «рекомбинантный», используемый взаимозаменяемо с термином «генетическая манипуляция», относится к конструкции гена в виде, который не существует в природе, посредством использования экспериментальных методик молекулярного клонирования, таких как генная трансформация, расщепление или связывание.
В том виде, в котором он используется в данном документе, термин «экспрессия» относится к получению белков или нуклеиновых кислот в клетках.
В том виде, в котором он используется в данном документе, термин «рекомбинантный экспрессирующий вектор» представляет собой вектор, который может экспрессировать целевой белок или нуклеиновую последовательность (РНК) в подходящей клетке-хозяине, и относится к генетической конструкции, содержащей существенные элементы, осуществляющие контроль, которые функционально связаны с возможностью экспрессировать вставку полинуклеотида (гена). Термин «функционально связанный» относится к функциональной связи последовательности, осуществляющей контроль экспрессии нуклеиновой кислоты, и последовательности нуклеиновой кислоты, кодирующей целевой белок или РНК, с возможностью выполнения общих функций, который означает связь между ними с возможностью гена экспрессироваться с помощью данной последовательности, осуществляющей контроль экспрессии. Последовательность, осуществляющая контроль экспрессии, относится к последовательности ДНК, которая контролирует экспрессию функционально связанной полинуклеотидной последовательности в конкретной клетке-хозяине. Такая последовательность, осуществляющая контроль экспрессии, включает промотор для транскрипции, любую последовательность оператора для осуществления контроля транскрипции, последовательность для кодирования соответствующего сайта связывания рибосомы мРНК, последовательность для осуществления контроля терминации транскрипции и трансляции, старт-кодон, стоп-код он, сигнал полиаденилирования А, энхансер и т.п.
Рекомбинантный экспрессирующий вектор по настоящему изобретению особым образом не ограничивается его типом, в том случае, если вектор обычно используется в области клонирования, и примеры рекомбинантного экспрессирующего вектора включают плазмидный вектор, космидный вектор, бактериофаговый вектор и вирусный вектор, но не ограничиваются ими. Примеры плазмиды могут включать плазмиды, происходящие из Escherichia coli (pBR322, pBR325, pUC118, pUC119 и pET-22b(+)), плазмиды, происходящие из Bacillus subtilis (pUB110 и pTP5), и плазмиды, происходящие из дрожжей (YEp13, YEp24 и YCp50), и примеры вируса могут включать: вирусы животных, такие как ретровирус, аденовирус или вирус осповакцины; и вирусы насекомых, такие как бакуловирус.
Под рекомбинантным экспрессирующим вектором по настоящему изобретению подразумевается генетическая конструкция, которая функционально связана таким образом, чтобы мочь экспрессировать в подходящей клетке-хозяине полинуклеотид, кодирующий антитело или его фрагмент, состоящее из тяжелой и легкой цепей, имеющих приведенные выше конформации CDR или VH и VL, способные специфически связываться с N-концевой областью KRS.
Полинуклеотиды, кодирующие тяжелую и легкую цепи антитела согласно настоящему изобретению, могут содержаться в отдельных рекомбинантных экспрессирующих векторах, соответственно, или могут содержаться в одном рекомбинантном экспрессирующем векторе.
Настоящее изобретение обеспечивает клетки, трансформированные описанным выше рекомбинантным экспрессирующим вектором
Клетки по настоящему изобретению особым образом не ограничиваются их типом при условии, что клетки можно использовать для экспрессии полинуклеотида, кодирующего антитело или его фрагмент, содержащегося в рекомбинантном экспрессирующем векторе по настоящему изобретению. Клетки (клетки-хозяева), трансформированные рекомбинантным экспрессирующим вектором согласно настоящему изобретению, могут представлять собой прокариотические клетки (например, Е. coli), эукариотические клетки (например, дрожжи или другие грибы), растительные клетки (например, растительные клетки табака или томата), клетки животных (например, клетки человека, клетки обезьяны, клетки хомяка, клетки крысы, клетки мыши или клетки насекомого) или полученные из них гибридомы. Предпочтительно, клетки могут происходить из млекопитающих, включая человека.
Иллюстративные прокариоты, подходящие для настоящей цели, включают грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, как например, Escherichia, например, Е. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescens, и Shigella, а также Bacilli, например, В. subtilis и В. licheniformis, Pseudomonas, например, P. Aeruginosa и Streptomyces. Клетки по настоящему изобретению особым образом не ограничиваются, при условии, что данные клетки могут экспрессировать вектор по настоящему изобретению, но могут предпочтительно представлять собой Е. coli.
Saccharomyces cerevisiae наиболее часто используется в качестве эукариота для клеток по настоящему изобретению. Однако можно использовать целый ряд других родов, видов и штаммов, но они не ограничиваются, например, Schizosaccharomyces pombe; хозяевами Kluyveromyces, как например, K lactis, K. fragilis (ATCC 12, 424), K. bulgaricus (АТСС 16,045), K. wickeramii (АТСС 24,178), K. waltii (АТСС 56,500), K. drosophilarum (АТСС 36,906), K. Thermotolerans и K. marxianus; yarrowia (ЕР 402,226); Pichia pastoris (ЕР 183,070); Candida; Trichoderma reesia (ЕР 244,234); Neurospora crassa; Schwanniomyces, как например, Schwanniomyces occidentalis; и нитчатыми грибами, например, хозяева Neurospora, Penicillium, Tolypocladium Aspergillus, как например, A. nidulans и A. niger.
Термин «трансформация» относится к модификации генотипа клетки-хозяина благодаря введению чужого полинуклеотида в клетку-хозяина и относится к введению чужого полинуклеотида в клетку-хозяина, вне зависимости от способа, используемого для трансформации. Чужой полинуклеотид, вводимый в клетку-хозяина, встраивается в и поддерживается в геноме клетки-хозяина или поддерживается без встраивания в него, и настоящее изобретение включает оба варианта.
Рекомбинантный экспрессирующий вектор, способный экспрессировать антитело или его фрагмент, специфически связывающиеся с N-концевой областью KRS согласно настоящему изобретению, можно вводить в клетки для получения антитела или его фрагмента, способом, известным в данной области, например, но, не ограничиваясь временной трансфекцией, микроинъекцией, трансдукцией, слиянием клеток, осаждением с помощью фосфата кальция, трансфекцией, опосредованной липосомами, трансфекцией, опосредованной декстраном DEAE (diethylaminoethyl - диэтиламиноэтилдекстран), трансфекцией, опосредованной полибреном, электропорацией, генной пушкой и известными способами введения нуклеиновых кислот в клетки, и, в таком случает, может трансформировать данные клетки.
Настоящее изобретение обеспечивает способ получения антитела или его фрагмента, специфически связывающийся с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы (KRS), включающий:
(а) трансформация клеток-хозяев рекомбинантным экспрессирующим вектором;
(б) инкубирование трансформированных клеток-хозяев с получением антитела или его фрагмента; и
(в) сбор антитела или его фрагмента, полученного в клетках-хозяевах.
На стадии (а), для получения антитела или его фрагмента согласно настоящему изобретению клетки-хозяева трансформируют рекомбинантным экспрессирующим вектором, где полинуклеотид, кодирующий антитело или его фрагмент, функционально связан.
Специалист в данной области может проводить настоящую стадию посредством выбора подходящего способа трансформации в зависимости от выбранных клеток-хозяев и рекомбинантного экспрессирующего вектора, как описано выше. Рекомбинантными экспрессирующими векторами, содержащими нуклеотидные последовательности тяжелой и легкой цепей, можно осуществлять совместную трансформацию в одной и той же клетке-хозяине для обеспечения возможности экспрессии тяжелой и легкой цепей в одной клетке, или рекомбинантными экспрессирующими векторами, содержащими нуклеотидные последовательности тяжелой и легкой цепей, можно трансформировать отдельные клетки-хозяева для обеспечения возможности экспрессии тяжелой и легкой цепей по отдельности.
На стадии (б) трансформированные клетки-хозяева инкубируют с получением полипептидов тяжелой и легкой цепей антитела или его фрагмента антитела согласно настоящему изобретению от рекомбинантного экспрессирующего вектора, введенного в клетки-хозяева.
Составы сред, условия инкубации и время инкубации для инкубирования клеток-хозяев могут быть соответствующим образом выбраны в соответствии со способом, обычно используемым в данной области. Молекулы антитела, продуцируемые в клетке-хозяине, могут аккумулироваться в цитоплазме клетки, могут секретироваться наружу из клетки или в культуральную среду с помощью подходящей сигнальной последовательности, или на них можно нацеливаться с использованием периплазмы или т.п. Также предпочтительно, чтобы антитело согласно настоящему изобретению имело функциональную конформацию в результате повторного сворачивания белка с использованием способа, известного в данной области, таким образом, чтобы сохранялась специфичность связывания с N-концевой областью KRS. В том, что касается получения антитела типа IgG, тяжелая и легкая цепи могут экспрессироваться в отдельных клетках и затем приводиться в контакт друг с другом на отдельной стадии с образованием целого антитела, или тяжелая и легкая цепи могут экспрессироваться водной и той же клетке с образованием целого антитела внутри клетки.
На стадии (в) получают антитело или его фрагмент, продуцируемые в клетках-хозяевах.
Специалист в данной области может должным образом выбирать и контролировать способ сбора, учитывая характеристики полипептидов антитела или его фрагмента, продуцируемых в клетках-хозяевах, характеристики клеток-хозяев, способа экспрессии или наличие нацеливания или отсутствие нацеливания на полипептид. Например, антитело или его фрагмент, секретируемые в культуральную среду, можно собирать посредством получения культуральной среды, в которой культивируют клетки-хозяева, удаляя примеси посредством центрифугирования и т.п. Для выделения, при необходимости, антитела, находящегося в конкретных органеллах или цитоплазме в клетках, из клеток и сбора антитела клетки можно лизировать в той степени, которая не влияет на функциональную структуру антитела или его фрагмента. Полученное антитело можно дополнительно подвергать процессу дополнительного удаления примесей и осуществления концентрирования, посредством хроматографии, фильтрования с использованием фильтра, диализа или т.п.
Полипептид в способе изготовления (получения) по настоящему изобретению может представлять собой собственно антитело или его фрагмент по настоящему изобретению и полипептид, с которым дополнительно связана другая аминокислотная последовательность, отличная от антитела или его фрагмента по настоящему изобретению. В данном случае аминокислотная последовательность может быть освобождена от антитела или его фрагмента по настоящему изобретению посредством использования способа, хорошо известного специалисту в данной области.
Антитело или его фрагмент по настоящему изобретению специфически связывается с N-концевой областью KRS и таким образом является полезным в диагностическом анализе для выявления и количественной оценки белков KRS, например, в определенных клетках, тканях или сыворотке. В особенности, экспонированную во внеклеточную среду N-концевую область KRS можно специфически выявлять без лизиса клеток. Таким образом, настоящее изобретение обеспечивает способ специфического выявления экспонированной во внеклеточную среду N-концевой области лизил-тРНК-синтетазы (KRS), включающий: приведение антитела или его фрагмента в контакт с образцом; и выявление антитела или его фрагмента.
Способ выявления по настоящему изобретению может включать стадию получения образца, который подлежит оцениванию на наличие или отсутствие KRS (или экспонированного во внеклеточную среду N-концевого пептида KRS) и в отношении ее концентрации посредством использования антитела или его фрагмента согласно настоящему изобретению (стадия (1)), перед приведением антитела или его фрагмента согласно настоящему изобретению в контакт с образцом.
Специалист в данной области может подходящим образом выбрать известный способ выявления белка с использованием антитела и получить образец, подходящий для выбранного способа. Кроме того, образец может представлять собой клетки или ткани, полученные с помощью биопсии, крови, цельной крови, сыворотки, плазмы, слюны, спинномозговой жидкости или т.п., который отбирают у субъекта, подлежащего обследованию в отношении наличия или отсутствия рака (особенно рака молочной железы или рака легкого) или метастаза рака. Примеры способа выявления белка с использованием антитела включают вестерн-блоттинг, иммуноблоттинг, дот-блоттинг, иммуногистохимию, твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ, метод конкурентного связывания, иммунопреципитацию и т.п., но не ограничиваются ими. Например, в случае вестерн-блоттинга получение можно осуществлять посредством добавления буфера, подходящего для электрофореза, к образцу или клеточному лизату с последующим кипячением, и в случае иммуногистохимии обработку можно проводить посредством иммобилизации и блокирования клеток или тканевых срезов с последующим блокированием.
Далее, проводят стадию приведения антитела или его фрагмента согласно настоящему изобретению в контакт с образцом, полученным на описанной выше стадии (стадия (2)).
Антитело согласно настоящему изобретению представляет собой антитело или его фрагмент, которые имеют описанные выше конформации CDR или VH и VL и специфически связываются с N-концевой областью KRS, и конкретные типы и их организация последовательностей представляют собой такие, как описано выше.
Антитело или его фрагмент можно метить обычной выявляемой группировкой для их «выявления». Например, антитело или его фрагмент можно метить радиоизотопом или флуоресцентной меткой посредством использования методики, описанной в литературе [Current Protocols in Immunology, Volumes 1 and 2, 1991, Coligen et al., Ed. Wiley-Interscience, New York, N. Y., Pubs]. Кроме того, полезны разные фермент-субстратные метки, и примеры ферментной метки включают следующие метки: люцифераза, как например люцифераза дрозофилы и бактериальная люцифераза (патент США US 4737456), люциферин, 2,3-дигидрофталазин-дион, малатдегидрогеназа, уреаза, пероксидаза, как например, пероксидаза хрена (HRPO - horseradish peroxidase), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизозим, сахарид-оксидаза (например, глюкооксидаза, галактозоксидаза и глюкозо-6-фосфат дегидрогеназа), гетероциклилоксидаза (например, уриказа и ксантиноксидаза), лактопероксидаза, микропероксидаза и т.п. Методики конъюгирования ферментов с антителами описаны, например, в литературе [O'Sullivan et al., 1981, Methods for the Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (J. Langone & H. Van Vunakis, eds.), Academic press, N. Y., 73: 147-166]. Метки могут быть прямо или непрямо конъюгированы с антителами с использованием разных известных методик. Например, антитело можно конъюгировать с биотином и любыми метками, принадлежащими к трем классам широко распространенных категорий, приведенных выше, можно конъюгировать с авидином или наоборот. Биотин может селективно связываться с авидином, и, вследствие этого, данная метка может быть конъюгирована с антителом таким непрямым образом. В качестве альтернативы, для достижения непрямой конъюгации метки с антителом антитело может быть конъюгировано с небольшим гаптеном (например, диоксином), и один из разных типов меток, приведенных выше, может быть конъюгирован с антителом против гаптена (например, антителом против диоксина). Таким образом, можно достигать непрямой конъюгации метки с антителом.
В том виде, в котором он используется в данном документе, термин «приведение в контакт» используют в его общем смысле, и он относится к смешиванию, связыванию или соприкосновению двух или более веществ. Приведение в контакт можно осуществлять in vitro или в другом контейнере, или можно осуществлять на месте, in vivo, у субъекта, в ткани или в клетке.
Далее, проводят стадию выявления антитела или его фрагмента согласно настоящему изобретению из образца после осуществления стадии (2) (стадия(3)).
«Выявление» проводят на комплексе антитела или его фрагмента согласно настоящему изобретению и антигена, причем данный комплекс образуется в образце, и относится к выявлению наличия или отсутствия N-концевого пептида KRS (или белка, включающего пептид, например, KRS) или измерению (включая качественное измерение, количественное измерение или и то и другое) уровня данного пептида. Таким образом, способ выявления по настоящему изобретению может дополнительно включать стадию удаления избыточных антител или их фрагментов, которые не образовали комплекса с N-концевой областью KRS, после осуществления стадии (2) перед стадией (3), описанной позже.
Когда антитело или его фрагмент, используемое на стадии (2), описанной выше, содержит выявляемую группировку, такую как флуоресценция, радиоактивный изотоп или фермент, которая прямо метит антитело или его фрагмент, выявление можно осуществлять способом выявления для соответствующей группировки, известным в данной области. Например, радиоактивность можно измерять, например, посредством сцинтилляционного счета, а флуоресценцию можно количественно оценивать, используя флуориметр.
Когда антитело или его фрагмент, как таковые, используемые на стадии (2), описанной выше, не содержат указанной выше выявляемой группировки, можно проводить непрямое выявление, используя вторичное антитело, меченное флуоресценцией, радиоактивностью, ферментом или т.п. Вторичное антитело связывается с антителом или его фрагментом (первичное антитело) согласно настоящему изобретению.
Недавние исследования установили, что лизил-тРНК-синтетаза (KRS) человека, присутствующая в цитозоле, перемещается к плазматической мембране (клеточной мембране), взаимодействуя с 67-кДа рецептором ламинина (67LR), находящимся на плазматической мембране, стимулируя, таким образом, миграцию опухолевых (или раковых) клеток, влияя на метастаз рака (Dae Gyu Kim et al., Chemical inhibition of prometastatic lysyl-tRNA synthetaselaminin receptor interaction, Nat Chem Biol. 2014 Jan; 10(1): 2934.). В этом случае, N-концевая удлиняющая область KRS (N-ext), как известно, важна для перемещения KRS к мембране клетки. В том, что касается метастаза рака, более конкретно, N-ext область KRS, как известно, участвует в связывании KRS и 67LR при их взаимодействии.
Антитела или их фрагменты согласно настоящему изобретению являются превосходными в способности специфического связывания с N-ext областью KRS. Фактически, антитела и их фрагменты по настоящему изобретению связываются с N-ext областью KRS, и таким образом ингибирует связывание (взаимодействие) с рецептором ламинина, демонстрируя, таким образом, превосходную способность ингибирования метастаза рака. Это хорошо описано в примерах изобретения. В примере в настоящем описании изобретения удостоверились в том, что в результате введения антитела согласно настоящему изобретению в in vivo модели метастаза рака с индуцированным раком, антитело по настоящему изобретению демонстрировало превосходную ингибирующую способность в отношении метастаза рака дозозависимым образом. В особенности, ингибирующая способность антитела по настоящему изобретению в отношении метастаза рака была абсолютно превосходной даже в сравнении с соединением YH 16899, которое, как известно, ингибирует метастаз рака в результате ингибирования взаимодействия рецептора ламинина (67LR) с KRS.
Таким образом, согласно настоящему изобретению предложена фармацевтическая композиция для ингибирования метастаза рака и композиция для диагностики рака, причем каждая из композиций содержит указанное выше антитело или его фрагмент по настоящему изобретению в качестве активного вещества для ингибирования метастаза рака.
Кроме того, согласно настоящему изобретению предложена фармацевтическая композиция для ингибирования метастаза рака и композиция для диагностики рака, причем каждая из данных композиций состоит из указанного выше антитела или его фрагмента по настоящеиу изобертению.
Кроме того, настоящее изобретение обеспечивает фармациевтическую композицию для ингибирования метастаза рака и композиция для диагностики рака, причем каждая композиция по существу состоит из указанного выше антитела или его фрагмента по настоящему изобретению.
Рак особым образом не ограничивается его типом в том случае, если рак известен как злокачественная опухоль в данной области, и его пример может быть выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи и интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, Лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, карциномы щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза. Предпочтительно, рак может представлять собой рак молочной железы или рак легкого.
Настоящее изобретение обеспечивает фармацевтическую композицию, содержащую антитело или его фрагмент по настоящему изобретению в качестве активного вещества, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Кроме того, настоящее изобретение обеспечивает фармацевтическую композицию, состоящей из антитела или его фрагмента по настоящему изобретению, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Кроме того, настоящее изобретение обеспечивает фармацевтическая композиция, по существу состоящей из антитела или его фрагмента по настоящему изобретению, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
В том виде, в котором он используется в данном документе, термин «иммунные клетки» предпочтительно относится к моноцитам или макрофагам.
В том виде, в котором он используется в данном документе, термин «заболевание, связанное с миграцией иммунных клеток» особым образом не ограничивается конкретным его типом в том случае, если в данной области известно, что избыточная миграция (и инвазия) иммунных клеток представляет собой главный патогенез заболевания, и его примеры могут быть выбраны из группы, состоящей из сердечнососудистого заболевания, фиброзного заболевания, хронического воспалительного заболевания и синдрома Альпорта.
Сердечно-сосудистое заболевание особым образом не ограничивается следующими конкретными типами сердечно-сосудистых заболеваний и может быть выбрано из группы, состоящей из легочной артериальной гипертензии, атеросклероза, стенокардии, инфаркта миокарда, ишемической цереброваскулярной болезни, артериосклероза и мезентериального склероза.
Фиброзное заболевание особым образом не ограничивается следующими конкретными типами фиброзных заболеваний и может быть выбрано из группы, состоящей из склеродермы, ревматоидного артрита, болезни Крона, язвенного колита, миелофиброза, фиброза легких, фиброза печени, цирроза печени, фиброза почки, миофиброза, фиброза миокарда, системной красной волчанки, наследственного фиброза, инфекционного фиброза (в особенности фиброза, вызываемого длительно инфекцией), фиброза, вызываемого раздражителем (фиброза, вызываемого повторяющимся воздействием раздражителей, таких как табак и токсичные вещества), фиброза, вызываемого хроническим аутоиммунным заболеванием, фиброза, вызываемого антигенной несовместимостью во время трансплантации органов, фиброза, вызываемого гиперлипидемией, фиброза, вызываемого ожирением, диабетического фиброза, фиброза, вызываемого гипертензией, и окклюзии, вызываемой фиброзом при имплантации стента.
Хроническое воспалительное заболевание может быть выбрано из группы, состоящей из астмы, атопического дерматита, экземы, псориаза, остеоартрита, подагры, псориатического артрита, цирроза, неалкогольного стеатогепатита, хронического обструктивного заболевания легких, ринита, диабетической ретинопатии, диабетической почечной недостаточности, диабетической нейропатии и рассеянного склероза.
Фармацевтическая композиция согласно настоящему изобретению может содержать только антитело или его фрагмент по настоящему изобретению или может дополнительно содержать по меньшей мере один фармацевтически приемлемый носитель. В том виде, в котором он используется в данном документе, термин «фармацевтически приемлемый» относится к нетоксичному составу, который является физиологически приемлемым, не ингибирует действие активного вещества при введении человеку и обычно не вызывает аллергической реакции или похожих реакций, как например, проблемы с желудочно-кишечным трактом или головокружение.
В фармацевтической композиции согласно настоящему изобретению антитело или его фрагмент можно вводить в нескольких пероральных и парентеральных лекарственных формах во время клинического введения. Антитело или его фрагмент при приготовлении можно получать с использованием разбавителя или эксципиента, такого как наполнитель, сухой разбавитель, связующее вещество, увлажнитель, дезинтегратор или поверхностно-активное вещество, которое обычно используют. Твердые композиции для перорального введения включают таблетку, пилюлю, порошок, гранулы, капсулу, пастилку и т.п. Данные твердые композиции могут быть получены посредством смешивания антитела или его фрагмента по настоящему изобретению с по меньшей мере одним эксципиентом, например, крахмалом, карбонатом кальция, сахарозой или лактозой или желатином. Кроме того, помимо простых эксципиентов могут быть использованы смазывающие вещества, такие как стеарат магния и тальк. Жидкие композиции для перорального введения включают суспензию, раствор для внутреннего применения, эмульсию, сироп и тому подобное. Помимо простых разбавителей, которые часто используются, таких как вода и жидкий парафин, в жидких композициях могут содержаться несколько эксципиентов, например, увлажнитель, подсластитель, отдушка, консервант и т.п.
Иллюстративные композиции для парентерального введения включают стерильный водный раствор, неводный растворитель, суспензионный растворитель, эмульсию, лиофилизирующий агент и суппозиторий. Композиция для лечения по настоящему изобретению может быть получена в виде лиофилизата или водного раствора для смешивания и хранения любого физиологически приемлемого носителя, эксципиента или стабилизатора (Remington: The Science and Practice of Pharmacy, 19th Edition, Alfonso, R., ed, Mack Publishing Co. (Easton, PA: 1995)) и антитела с предпочтительной чистотой. Приемлемый носитель, эксципиент или стабилизатор является нетоксичным для пользователя при используемой дозе и концентрации, и их примеры включают: буферы, например, фосфорную кислоту, лимонную кислоту и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярные полипептиды (меньше чем примерно 10 остатков); белки, например, сывороточный альбумин, желатин или иммуноглобулин; гидрофильные полимеры, например, поливинилпирролидон; аминокислоты, например, глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрин; хелатирующие агенты, например, EDT (1,2-ethane dithiol - 1,2-этандитиол); сахароспирты, например, маннит, например, маннит или сорбит; солеобразующие противоионы, например, натрий; и (или) неионные поверхностно-активные вещества, например, твин (Tween), полоксамеры или полиэтиленгликоль (ПЭГ).
Антитело по настоящему изобретению можно вводить в фармацевтически эффективном количестве субъекту, борющемуся с раком или заболеванием, связанным с миграцией иммунных клеток. В том виде, в котором он используется в данном документе, термин «фармацевтически эффективное количество» относится к количеству, показывающему более сильную реакцию, по сравнению с отрицательным контролем, и предпочтительно относится к количеству, достаточному для лечения рака, количеству, достаточному для ингибирования метастаза рака, и количеству, достаточному для лечения заболевания, связанного с миграцией иммунных клеток. Общее эффективное количество антитела или его фрагмента по настоящему изобретению можно вводить пациенту в виде однократной дозы или можно вводить с помощью протокола фракционированного лечения, в котором многократные дозы вводят на протяжении длительного периода времени. Доза антитела или его фрагмента по настоящему изобретению для организма человека может обычно составлять 0,01-100 мг/кг/неделя, предпочтительно 0,1-20 мг/кг/неделя и более предпочтительно 5-10 мг/кг/неделя. Однако, в том, что касается дозы антитела или его фрагмента по настоящему изобретению, его эффективную дозу в отношении пациента определяют при рассмотрении разных факторов, например, пути введения фармацевтической композиции, количества раз, за которое осуществляется лечение, возраста пациента, массы тела, состояния здоровья и пола, тяжести заболевания, рациона и скорости экскреции, и, таким образом, учитывая данный факт, специалист в данной области мог бы определять подходящее эффективное количество антитела или его фрагмента по настоящему изобретению в соответствии с конкретным использованием в качестве ингибитора метастаза раза. Фармацевтическая композиция согласно настоящему изобретению особым образом не ограничивается лекарственной формой, путем введения и способом ее введения с том случае, если композиция демонстрирует эффекты по настоящему изобретению.
Путь введения композиции по настоящему изобретению может представлять собой известный способ введения антитела, например, инъекцию или инфузию посредством внутривенного, внутрибрюшинного, внутричерепного, подкожного, внутримышечного, внутриглазного, внутриартериального, спинномозгового или внутриочагового пути, или инъекцию или инфузию посредством системы с замедленным высвобождением, описанной ниже. Например, антитело по настоящему изобретению можно вводить в организм в целом или местно.
Фармацевтическую композицию по настоящему изобретению можно использовать отдельно или в сочетании с хирургическим вмешательством, гормональной терапией, химиотерапией и способами с использованием регулятора биологической реакции, для предупреждения или лечения рака.
Фармацевтическую композицию по настоящему изобретению также можно использовать отдельно или в сочетании с хирургическим вмешательством, гормональной терапией, химиотерапией и способами с использованием регулятора биологической реакции, для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток.
Диагностику и прогнозирование рака (или метастаза рака) согласно настоящему изобретению можно оценивать посредством выявления белков KRS (особенно, экспонированной во внеклеточную среду N-концевой области KRS) в биологическом образце, и диагностику и прогнозирование заболевания, связанного с миграцией иммунных клеток согласно настоящему изобретению, можно оценивать посредством выявления белков KRS (особенно, экспонированной во внеклеточную среду N-концевой области KRS) в биологическом образце.
В том виде, в котором он используется в данном документе, термин «диагностика» относится к идентификации наличия или характерных признаков патологического состояния. В настоящем изобретении диагностика состоит в том, чтобы идентифицировать возникновение или вероятность (риск) возникновения рака и/или метастаза рака или заболевания, связанного с миграцией иммунных клеток.
Термин «выявление» представляет собой такой, как описано выше, и биологический образец включает кровь и другие жидкие образцы, имеющие биологическое происхождение, биопсийный материал, твердые образцы ткани, такие как культура ткани, или клетки, полученные из нее. Более конкретно, примеры биологического образца могут включать ткани, экстракты, клеточные лизаты, цельную кровь, плазму, сыворотку, слюну, внутриглазную жидкость, спинномозговую жидкость, пот, мочу, молоко, асцитическую жидкость, синовиальную жидкость, перитонеальную жидкость и тому подобное, но не ограничиваются ими. Образец может быть получен из животных, предпочтительно млекопитающих, и наиболее предпочтительно человека. Образец можно предварительно обрабатывать перед применением для выявления. Примеры предварительной обработки могут включать фильтрацию, перегонку, экстракцию, концентрирование, дезактивацию мешающего вещества, добавление реагента и т.п. Кроме того, для выявления можно использовать нуклеиновые кислоты и белки, выделенные из образца.
Антитело или его фрагмент согласно настоящему изобретению могут быть предложены в виде набора для диагностики. Набор особым образом не ограничивается его типом в том случае, если набор известен в данной области как набор для анализа, в котором предложен пептид, имеющий антитело или конкретный связывающий домен в качестве компонента, и его примеры включают набор для вестерн-блоттинга, ELISA, радиоиммунного анализа, радиоиммунодиффузии, радиоиммунодиффузии по Оухтерлони, ракетного иммуноэлектрофореза, иммуногистохимии, анализа на основе иммунопреципитации, анализа на основе фиксации комплемента, FACS (Fluorescence-Activated Cell Sorting - сортировка клеток с активированной флуоресценцией), белкового чипа или т.п.
Антитело или его фрагмент по настоящему изобретению можно использовать в наборе, то есть упакованной комбинации реагентов в заранее определенных количествах с инструкциями по проведению диагностирующего анализа. Когда антитело мечено ферментом, набор может включать субстраты и кофакторы, требующиеся ферменту (например, предшественник субстрата, который предоставляет хромофор или флуорофор). Кроме того, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или лизирующий буфер) и т.п. Относительные количества разных реагентов могут сильно различаться для обеспечения концентраций в растворе реагентов, которые по существу оптимизируют чувствительность анализа. Реагенты могут быть предложены в виде сухих порошков, обычно лиофилизированных, включая эксципиенты, которые при растворении будут обеспечивать получение раствора реагента, имеющего соответствующую концентрацию.
Настоящее изобретение обеспечивает применение антитела или его фрагмента по настоящему изобретению для получения агента для ингибирования метастаза рака.
Настояще изобретение обеспечивает способ ингибирования метастаза рака у нуждающегося в этом субъекта, включающий введение субъекту антитела или его фрагмента по настоящему изобретению количества, эффективного для ингибирования метастаза рака.
Настоящее изобретение обеспечивает применение антитела или его фрагмента по настоящему изобретению для получения агента для диагностики рака.
Настоящее изобретение обеспечивает способ диагностирования рака у нуждающегося в этому субъекта, включающий введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для диагностирования рака.
Настоящее изобретение обеспечивает применение антитела или его фрагмента по настоящему изобретению для получения агента для лечения заболевания, связанного с миграцией иммунных клеток.
Настоящее изобретение обеспечивает способ лечения заболевания, связанного с миграцией иммунных клеток, у нуждающегося в этом субъекта, включающий введение субъекту антитела или его фрагмента по настоящему изобретению в количестве, эффективном для лечения заболевания, связанного с миграцией иммунных клеток.
В том виде, в котором он используется в данном документе, термин «эффективное количество» относится к количеству, демонстрирующему эффект облегчения, лечения, предупреждения, выявления или диагностики рака или эффект ингибирования или уменьшения метастаза рака, и относится к количеству, демонстрирующему эффект облегчения, лечения, предупреждения, выявления или диагностики заболевания, связанного с миграцией иммунных клеток. Термин «субъект» может представлять собой животное, предпочтительно млекопитающее, в особенности животное, включая человека, и может представлять собой клетки, ткань, орган или т.п., происходящие из животного. Субъект может представлять собой пациента, нуждающегося в данном эффекте.
В том виде, в котором он используется в данном документе, термин «лечение» широко относится к облегчению рака, заболевания, связанного с раком, или заболевания, связанного с миграцией иммунных клеток, и может включать излечение или по существу предупреждение такого заболевания или облечение состояния при заболевании, и может включать облегчение, излечение или предупреждение одного или большинства симптомов, являющихся последствием рака или заболевания, связанного с раком, но не ограничивается ими.
В том виде, в котором он используется в данном документе, термин «включающий» используется синонимично с термином «содержащий» или «характеризующийся», и не исключает дополнительных компонентов или стадий, не упомянутых в композиции или способе. Термин «состоящий из» означает исключение дополнительных элементов, стадий или компонентов, не описанных иным образом. Термин «по существу состоящий из» означает включение упомянутых элементов или стадий, а также любого элемента или стадии, которые существенно не влияют на основные характеристики упомянутых элементов или стадий в объеме композиций или способов.
Преимущественные эффекты
Антитела или их фрагменты согласно настоящему изобретению имеют конкретные определяющие комплементарность области (CDR), определенные в настоящем описании изобретения, и абсолютно превосходную способность к специфическому связыванию с экспонированной во внеклеточную среду N-концевой областью KRS. Кроме того, антитела или их фрагменты согласно настоящему изобретению специфически нацелены на N-концевую область KRS in vivo и, таким образом, ингибируют взаимодействие рецептора ламинина и N-концевой области KRS, таким образом, оказывая превосходное влияние на ингибирование метастаза рака, и могут контролировать миграцию иммунных клеток, оказывая, таким образом, очень значительное влияние на предупреждение, облегчение и лечение заболевания, связанного с миграцией иммунных клеток.
Краткое описание графических материалов
На Фиг. 1 показаны результаты селекции посредством вестерн-блоттинга (WB - western blotting), фаговых клонов scFv, связывающихся с полноразмерной последовательностью KRS или N-концевыми фрагментами KRS.
На Фиг. 2 показаны результаты селекции посредством иммунопреципитации (IP - immunoprecipitation), фаговых клонов scFv, связывающихся с полноразмерной последовательностью KRS или N-концевыми фрагментами KRS.
На Фиг. 3а показаны результаты, где клетки, имеющие N-концевую область KRS, экспонированную на клеточной мембране, конструировали посредством экспрессии тус-KRS T52D (активный мутант), и было исследовано связываются ли клоны scFv N3, N5, N7 и N9 с экспонированной областью (KRS: подразумевает myc-KRS T52D).
На Фиг. 3б показано, что в экспрессирующей группе неактивного мутанта (myc-KRS T52D) или WT-KRS (необработанный ламинином) не наблюдали выявляемых сигналов, несмотря на обработку клонами scFv N3, N5, N7 и N9, поскольку N-концевая область KRS не была экспонирована на клеточной мембране.
На Фиг. 3в показано, что когда клетки, трансформированные myc-меченной WT-KRS, T52D и Т52А, обрабатывали ламинином и проводили scFv окрашивание, окрашивание наблюдали в похожих сайтах при мембранной локализации WT-KRS с помощью ламинина и у мутанта T52D, и трансформированная myc была также окрашена в том же сайте, но окрашивания не наблюдали у Т52А (scFv: зеленый, myc: красный).
На Фиг. 4 показаны результаты вестерн-блоттинга, подтверждающие то, связываются ли специфически клоны scFv N3, N5, N7 и N9 с N-концом KRS, посредством использования полноразмерной KRS (обозначаемой F, SEQ ID NO: 76), фрагмента KRS с делецией аминокислот в положениях 1-71 в N-конце (обозначаемого 1) и фрагмента KRS, состоящего из аминокислотных остатков 1-200 в N-конце (обозначаемого 2).
На Фиг. 5а показаны результаты вестерн-блоттинга, подтверждающие способность IgG N3 и IgG N5 в качестве типичных представителей среди антител по настоящему изобретению к связыванию с KRS.
На Фиг. 5б показаны результаты иммунопреципитации, подтверждающие способность IgG N3 и IgG N5 в качестве типичных представителей среди антител по настоящему изобретению к связыванию с KRS.
На Фиг. 6а показаны результаты SPR (Surface Plasmon Resonance - поверхностный плазмонный резонанс), количественно подтверждающие способность IgG N3 к связыванию с N-концом KRS.
На Фиг. 6б показаны результаты SPR, количественно подтверждающие способность IgG N5 к связыванию с N-концом KRS.
На Фиг. 7а показаны результаты, где клетки, трансформированные для экспрессии WT-KRS, обрабатывали ламинином для экспонирования во внеклеточную среду N-концевой области KRS, и затем обрабатывали антителом по настоящему изобретению, IgG N3 для изучения связывания IgG N3 с экспонированной во внеклеточную среду областью посредством иммунофлуоресцентного окрашивания.
На Фиг. 7б показаны результаты, в которых клетки, имеющие экспонированную во внеклеточную среду N-концевую область KRS, конструировали посредством экспрессии T52D-KRS (активный мутант, myc-меченная KRS), и затем обрабатывали антителом по настоящему изобретению, IgGN3, для изучения связывания IgGN3 с экспонированной во внеклеточную среду областью посредством иммунофлуоресцентного окрашивания. Также с помощью эксперимента, наделяя клеточную мембрану проницаемостью, подтверждали, что антитело по настоящему изобретению перемещается в клетки и может связываться с белком KRS, находящимся внутри клеток.
На Фиг. 7в показаны результаты, где клетки, трансформированные для экспрессии WT-KRS, обрабатывали ламинином для экспонирования во внеклеточную среду N-концевой области KRS и затем обрабатывали антителом по настоящему изобретению, IgG N5, для изучения связывания IgG N5 с экспонированной во внеклеточною среду областью посредством иммунофлуоресцентного окрашивания (IgG N5: зеленый).
На Фиг. 8 показаны результаты МТТ-теста (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide - 3-(4,5-диметилтиазол-2-ил)-2,5-дифенил-тетразолия бромид), подтверждающие то, что антитело по настоящему изобретению IgG N3 не обладало цитотоксичностью.
На Фиг. 9а показаны результаты, подтверждающие, что миграция клеток подавлялась обработкой антителом по настоящему изобретению IgG N3.
На Фиг. 9б показаны результаты, подтверждающие, что антитело по настоящему изобретению IgG N3 значимо подавляло миграцию клеток дозозависимым образом.
На Фиг. 10 схематично показана экспериментальная схема конструирования моделей мышиного метастаза рака, введения терапевтического вещества (антитело или YH16899) и наблюдения легочных метастазов в мышиных моделях, в эксперименте с использованием in vivo моделей метастаза рака.
На Фиг. 11 показаны образцы легкого мыши, способные подтвердить то, что раковый метастаз в легких значимо подавлялся введением антитела по настоящему изобретению, IgG N3, в in vivo моделях метастаза рака. Степени прогрессирования и тяжести метастаза рака можно было оценивать по числу и состоянию узелков, образованных в образцах легкого.
На Фиг. 12 показаны результаты, подтверждающие, что образованием узелков в легком значимо подавлялось введением антитела по настоящему изобретению, IgG N3, по сравнению с контролем, в in-vivo моделях метастаза рака (то есть, метастаз рака в легких значимо подавлялся).
На Фиг. 13 сравнительно показаны ткани контрольного легкого в и группе обработки IgG N3 и подтверждено, что значимо большое количество рецепторов ламинина экспрессировалось в местах узелков метастазирования в контрольном легком, по сравнению с группой, обрабатываемой антителом по настоящему изобретению.
На Фиг. 14 показаны образцы контрольного легкого, группы обработки YH16899, группы обработки IgG N3 и показаны результаты, демонстрирующие, что образование узелков в легком значимо подавлялось в группе обработки YH16899 и группе обработки IgGN3, по сравнению с контрольным легким.
На Фиг. 15 показана эффективность ингибирования метастаза легкого (эффективность ингибирования образования узелков в легком) веществ, ингибирующих метастаз рака, в соответствии с концентрацией обработки в группе обработки YH16899 и группе обработки IgG N3.
На Фиг. 16 показаны результаты SPR, количественно подтверждающие способность связывания IgG N3 с фрагментами человеческого (h) N-концевого пептида KRS (F1, F2, F3, F4 и F5) (серые столбцы ниже последовательности показывают способность связывания антитела N3 с соответствующими областями (F1 - F5), и чем темнее столбец, тем сильнее способность связывания).
На Фиг. 17 показаны результаты SPR, количественно подтверждающие способность связывания IgG N3 с фрагментами N-концевого пептида KRS (F1, F3 и F5) человека (h), мыши (m) и крысы (r) (серые столбцы ниже последовательности показывают способность связывания антитела N3 с соответствующими областями (F1 - F5), и чем темней столбец, тем сильнее способность связывания).
На Фиг. 18а показаны результаты сравнения воздействий коллагена, фибронектина и ламинина на миграцию иммунных клеток (моноцит/макрофаг) при анализе миграции в системе трансвелл (transwell), и обеспечены изображения мигрирующих клеток, полученные на микроскопе.
На Фиг. 18б представляет собой график, показывающий число клеток, измеренное (количественно оцененное) на изображениях Фиг. 18а, полученных на микроскопе.
На Фиг. 19а показаны результаты сравнения воздействий разных подтипов ламинина (LN111, LN211, LN221, LN411, LN421, LN511 и LN521) на миграцию иммунных клеток (моноцит/макрофаг) при анализе миграции в системе трансвелл, и предложены изображения мигрирующих клеток, полученные на микроскопе.
На Фиг. 19б показан график, показывающий число клеток, измеренное (количественно оцененное) на изображениях Фиг. 19а, полученных на микроскопе.
На Фиг. 19в показаны результаты вестерн-блоттинга, подтверждающие, что уровень KRS увеличивался на мембране моноцитов/макрофагов в результате обработки LN421.
На Фиг. 20а показаны результаты сравнения ингибирующих эффектов антитела по настоящему изобретению, IgG N3, оказываемых на LN421-специчную миграцию моноцитов/макрофагов при анализе миграции в системе трансвелл, и предложены полученные на микроскопе изображения мигрирующих клеток.
Фиг. 20б представляет собой график, показывающий число клеток, измеренное (количественно определенное) на изображениях Фиг. 20а, полученных на микроскопе.
На Фиг. 20в показаны результаты вестерн-блоттинга, подтверждающие, что уровень KRS, повышенный в результате обработки LN421 в мембране моноцита/макрофага, снижался в результате обработки антителом по настоящему изобретению IgG N3.
На Фиг. 21а показано изменение в конечно-систолическом давлении в правом желудочке (RVESP - right ventricular end-systolic pressure) в результате введения антитела по настоящему изобретению, IgG N3, в моделях легочной артериальной гипертензии (ЛАГ) (имитирующее IgG: отрицательный контроль, Ab 1 мг/кг массы тела: антитело N3 1 мг/кг массы тела, Ab 10 мг/кг массы тела: антитело N3 10 мг/кг массы тела, силденафил: положительный контроль).
На Фиг. 21б показано изменение в конечно-систолическом давлении в левом желудочке (LVESP - change in left ventricular end-systolic pressure) в результате введения антитела по настоящему изобретению, IgG N3, в моделях легочной артериальной гипертензии (ЛАГ) (имитирующее IgG: отрицательный контроль, Ab мг/кг массы тела: IgG N3 1 мг/кг массы тела, Ab 10 мг/кг массы тела: IgG N3 10 мг/кг массы тела, силденафил: положительный контроль).
На Фиг. 22 показаны результаты ИГХ (иммуногистохимического) окрашивания, подтверждающие, что введение антитела по настоящему изобретению, IgGN3, уменьшало миграцию иммунных клеток и инвазию в моделях легочной артериальной гипертензии (ЛАГ).
Способ осуществления изобретения
Далее в данном документе настоящее изобретение будет описано подробно. Однако, следующие примеры предназначены лишь для иллюстрации настоящего изобретения и не предназначены для ограничения объема настоящего изобретения.
Пример 1
Осуществление скрининга библиотеки scFv 1-1.
Осуществление скрининга фагов scFv - первичный скрининг
Для отбора антител scFv, специфически связывающихся только с N-концевой областью KRS (SEQ ID NO: 75), экспонированной во внеклеточную среду при перемещении KRS к клеточной мембране с помощью сигнала ламинина, в полноразмерной последовательности KRS (SEQ ID NO: 76) пэннинг с использованием фагового дисплея осуществляют с использованием фаговой библиотеки scFv, полученной из НА-меченных В-клеток человека. Библиотека фагового дисплея scFv (размер библиотеки: приблизительно 7,6×109, библиотека, полученная проф. Hyunbo Shim), используемая в настоящем эксперименте, раскрыта в корейском патенте KR 10-0961392. Как показано ниже в Таблице 1, полноразмерные последовательности KRS и фрагменты KRS с конкретными разными областями N-конца использовали в качестве белков-антигенов для пэннинга с использованием фагового дисплея.
К иммунологической пробирке, содержащей 1 мл раствора 1X PBS (phosphate buffered saline - фосфатно-солевой буферный раствор), добавляют 1-10 мкг белков-антигенов и инкубируют при 37°С в течение 1 ч при 200 об/мин, покрывая, таким образом, внутреннюю поверхность пробирки антигенами. Раствор антигенов высушивают, и антигены, не участвующие в покрытие, удаляют посредством однократной промывки водопроводной водой. Для предотвращения неспецифического связывания белков-антигенов и фагов, иммунологическую пробирку и библиотеку scFv инкубируют по отдельности с 1X PBST (Phosphate Buffered Saline with Tween - фосфатно-солевой буферный раствор с твином) (0,05% PBS, содержащий твин 20), содержащим 3% обезжиренное молоко, при комнатной температуре в течение 1 ч. После удаления обезжиренного молока из иммунологической пробирки добавляют библиотеку scFv и инкубируют при 150 об/мин в течение 1 ч при 37°С, таким образом, связывая фаги scFv с антигенами. После инкубации фагов scFv в пробирке, несвязанные фаги scFv удаляют посредством двукратной или пятикратной промывки 1X PBST.
Фаги ScFv, специфически связывающиеся с соответствующими антигенами KRS, выделяют на протяжении 10 мин посредством добавления 1 мл триэтиламина (100 ммоль/л) при комнатной температуре и нейтрализуют Tris (tris(hydroxymethyl)aminomethane - трис(гидроксиметил)аминометан) (1 моль/л, рН 7,4). Отфильтрованные фаги scFv добавляют к Е. coli ER2537, культивированной до OD (optical density - оптическая плотность) меньше 1 с последующим инфицированием с инкубацией при 120 об/мин в течение 1 ч и 30 мин при 37°С. Е. coli, инфицируемую фагами, центрифугируют с частичным удалением культурального супернатанта с последующим повторным диспергированием и затем распределяют по чашке, диаметром 15 см, с агарозой, содержащей ампициллин и глюкозу (2%). На следующий день 5 мл среды SB применяли для сбора всех клеток, выросших на чашке, и добавляют глицерин (50%) до половины общего объема с последующим перемешиванием, и затем смесь делят на 1 мл-порции и хранят при минус 80°С (стоковый раствор scFv для пэннинга). Затем, 20 мкл полученного стокового раствора засевают в 20 мл раствора SB с последующей инкубацией и затем осуществляют конструирование в фаговую библиотеку scFv (1 мл) для следующей стадии пэннинга с помощью фагов посредством использования хелперных фагов. Указанную выше процедуру выделения фагов, экспрессирующих scFv, специфические к антигенам, повторяют два или три раза.
1-2. Осуществление скрининга специфически связывающихся антител scFv посредством ELISA - вторичный скрининг
Посредством непрямого ELISA исследуют то, связываются ли фаги с экспрессией scFv, отобранные в Приме 1-1, с приведенной выше полноразмерной KRS или N-концевыми фрагментами.
Продукт scFv, полученный в результате трех циклов пэннинга, разводят и наносят на чашку с агарозой, диаметром 10 см. На следующий день проводят отбор соответствующих колоний и инкубируют в 96-луночном планшете, содержащем 200 мкл среды SB. После того, как колонии проверяют на общий рост, добавляют IPTG (isopropylthiogalactoside - изопропилтиогалактозид) (1 ммоль/л) с последующей инкубацией при 30°С в течение 16 ч, индуцируя, таким образом, получение scFv. На следующий день 96-луночный планшет центрифугируют для выделения исключительно клеток, и затем данные клетки лизируют посредством раствора TES с последующим повторным центрифугированием, отделяя, таким образом, только супернатант. Полученный супернатант подвергают непрямому ELISA для отбора scFv, специфически связывающихся с антигенами. Планшеты покрывают антигенами -полноразмерными KRS или антигенами - фрагментами N-концевой области, соответственно, инкубируют с культуральным супернатантом, содержащим scFv, и затем инкубируют с антителом против HA-HRP (Roche Applied Science) в качестве вторичного антитела. Окрашивание осуществляют с использованием тетраметилбензимидина (ТМВ - tetramethyl benzimidine, Thermo scientific) и затем останавливают, используя H2SO4 (1 моль/л), и показание по поглощению снимают при 450 нм, используя ELISA-ридер. ELISA проводят на контроле (холостая проба) и описанных выше антигенах (Ag), одновременно отбирая только колоний с положительными значениями. Из общего числа 1920 колоний, подвергавшихся ELISA, отбирают 93 колонии в качестве позитивных (см. Таблицу 1).
Таблица 1
1-3. Секвенирование
Последовательности анализируют для того, чтобы исключить одни и те же Ab с перекрывающимися последовательностями CDR из 93 колоний, отобранных посредством ELISA-скрининга в Примере 1-2. Более конкретно, секвенирование проводят следующим способом. После культивирования Е. coli, сохраняющей клоны scFv, фагмиды получали посредством miniprep. Фагмиды секвенируют с использованием праймера Omp (Hye young Yang, et. al., 2009, Mol. Cells 27, 225-235). Полученную таким образом последовательность используют для проверки последовательностей областей CDR фагмид посредством программы Bioedit. Из них исключают клоны с перекрывающимися последовательностями CDR для проверки клонов scFv соответствующих независимых последовательностей CDR.
Таблица 2
В результате скрининга антител с перекрывающимися последовательностями CDR посредством секвенирования, получают 38 клонов scFv с разными последовательностями CDR, как показано в Таблице 2.
1-4. Осуществление скрининга специфически связывающихся антител scFv посредством вестерн-блоттинга - третичный скрининг
Посредством вестерн-блоттинга исследовали то, связываются ли специфически 38 клонов scFv, выделенных в Примере 1-3, с KRS.
scFv позитивные клоны из одной колонии инкубируют в 5 мл канамицин-содержащей среды SB (бактотриптон 30 г, дрожжевой экстракт 20 г, MOPS буфер 10 г/л) для запуска культуры для посева, и после инкубации в течение ночи культуру переносят в 500 мл канамицин-содержащей среды SB. Когда значение OD при 600 нм достигало примерно 0,5, добавляют IPTG до достижения 1 ммоль/л с последующей инкубацией при 30°С в течение ночи с экспрессией, вследствие этого, белков scFv в периплазме Е. coli. На следующий день Е. coli, полученную посредством центрифугирования, суспендируют в буфере 1X TES (50 ммоль/л Tris, 1 ммоль/л ЭДТА (этилендиаминтетрауксусная кислота), 20% сахароза, рН 8,0) и затем добавляют 0,2Х TES в 1,5 раза с последующим перемешиванием, и затем супернатант отбирают посредством центрифугирования, экстрагируя, таким образом, периплазму.
Наконец, к антителам scFv добавляют 5 ммоль/л MgSO4, выделенным из периплазмы, и полученный в результате материал смешивают с шариками Ni-NTA (никель-нитрилоацетат), ранее уравновешенными PBS, с последующим перемешиванием в течение 1 ч в холодильнике для связывания антитела с шариками Ni-NTA. Далее, для достаточной отмывки несвязанных белков PBS осуществляют аффинную хроматографию. После дополнительной достаточной промывки буфером, содержащим 5 ммоль/л имидазол, связанные антитела scFv элюируют, используя 200 ммоль/л имидазоловый буфер. Элюированные антитела диализируют, и их чистоту проверяют посредством электрофореза. Количественную оценку белков проводят посредством анализа на основе BCA (bicinhoninic acid - бицинхониновая кислота), и количество очищенных антител регистрируют, и затем их определенное количество разделяют и затем храняти в замороженном виде.
Как описано выше, антитела scFv, выделенные из периплазмы, используют для исследования с использованием вестерн-блоттинга того, связываются ли антитела scFv с полноразмерной KRS или соответствующими фрагментами N-концевой области KRS. Затем 30 мкг клеточного лизата НСТ116 подвергают электрофоретическому разделению посредством SDS PAGE (ДНС-ПААГ) (sodium dodecyl sulphate polyacrylamide gel electrophoresis - электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия), переносят на мембрану PVDF (polyvinylidene fluoride - поливинилиденфторид) и затем блокируют 3% обезжиренным молоком. Далее, выделенные антитела scFv добавляют в количестве 1,0 мкг/мкл с последующей инкубацией в течение 1 ч. Проводят отмывку от несвязанных антител scFv, и для выявления scFv, связывающиеся с антигенами, инкубируют с вторичными антителами против НА, связанными с пероксидазой хрена (HRP), и проводят сенсибилизацию пленки с использованием реагента ECL в качестве субстрата в темной комнате. Сенсибилизированные полосы сравнивают со стандартными молекулярными маркерами для идентификации полос, соответствующих размерам полноразмерной KRS и соответствующих фрагментов.
Посредством вестерн-блоттинга клоны scFv с чрезвычайно слабыми полосами (бледными полосами) и неспецифическими полосами (двойными полосами) исключают. Таким образом, отбирают 13 клонов scFv, и данные результаты показаны на Фиг. 1.
1-5. Осуществление скрининга специфически связывающихся антител scFv посредством иммунопреципитации - четвертичный скрининг
Иммунопреципитацию проводят для исследования того, связываются ли на самом деле клоны scFv, отобранные в Примере 1-4, с нативной KRS. Очищенные клоны scFv и клеточный лизат НСТ116 подвергают связыванию Ag-Ab, и иммунопреципитацию проводят, используя НА-метку scFv.
Более конкретно, клетки НСТ116 лизируют в буфере 20 ммоль/л Tris-HCl (рН 7,4, лизирующий буфер), содержащем 150 ммоль/л NaCl, 0,5% Triton Х-100, 0,1% SDS и ингибитор протеазы. Каждый scFv (5 мкг) добавляют к 500 мкг клеточного лизата НСТ116 и затем инкубируют при 4°С в течение ночи. Затем, добавляют 30 мкл агарозных шариков с антителом против НА с последующей инкубацией при 4°С в течение 4 ч. Супернатант удаляют посредством центрифугирования. Полученный таким образом преципитат растворяют в SDS-буфере для образца и кипятят в течение 7 мин. Стадию растворения и кипячения повторяют два раза.
Каждый из иммунопреципитированных образцов, полученных посредством описанного выше способа, подвергают электрофоретическому разделению с помощью SDS PAGE, переносят на мембрану PVDF, и затем блокируют 3% обезжиренным молоком. Далее, добавляют поликлональные антитела против KRS (кроличьи, Neomics, Co. Ltd. #NMS-01-0005) с последующей инкубацией в течение 1 ч. После отмывки несвязанных антител добавляют вторичные антитела против кроличьих антител (ThermoFisher Scientific, #31460) с последующей инкубацией. После инкубации с вторичными антителами проводят сенсибилизацию пленки с использованием реагента ECL в качестве субстрата в темной комнате. Сенсибилизированные шарики сравнивают со стандартными молекулярными маркерами для идентификации полос, соответствующих размерам полноразмерной KRS и соответствующих фрагментов.
Таким образом, отбирают 12 клонов scFv, и данные результаты показаны на Фиг. 2.
1-6. Осуществление скрининга специфически связывающихся антител scFv посредством иммунофлуоресценции - пятеричный скрининг
Иммунофлуоресценцию проводят для исследования того, связываются ли фактически клоны scFv, отобранные в Примере 1-5, с N-концевой областью KRS, экспонированной на клеточной мембране. Для воспроизведения явления, в котором KRS экспонирована на мембране, использкют мутант KRS-T52D (активный мутант). В особенности, клетки А549 высевают на стеклянное покровное стекло (1×105 клеток, на основе 12-луночного планшета), и спустя 24 ч, myc-KRS WT/myc-KRS T52D (активный мутант)/myc-KRS Т52А (неактивный мутант) сверхэкспрессировались, соответственно. После инкубации в течение 24 ч, клетки инкубируют с использованием сред, не содержащих сыворотку (среды RPMI 1640) при 37°С в течение 1 ч. Далее, клетки обрабатывают 10 мкг/мл ламинина (L2020; Sigma) и затем инкубируют при 37°С в течение 1 ч. Каждый из векторов myc-KRS T52D (активный мутант) и myc-KRS Т52А (неактивный мутант) конструируют с использованием вектора pcDNA3-myc-KRS WT в качестве остова посредством сайт-направленного мутагенеза (набор для сайт-направленного мутагенеза QuikChange II, Agilent, #200523).
Полученные образцы промывают PBS (4°С), фиксировали посредством обработки 4% параформальдегидом в течение 10 мин и затем промывают. Образцы блокируют блокирующим буфером CAS в течение 10 мин и обрабатывают антителами scFv и Мус в течение 2 ч. Далее, осуществляют отмывку несвязанных антител и инкубируют с вторичными антителами в течение 1 ч в темной комнате. Окрашивание DAPI (4',6-Diamidino -2-phenylindole dihydrochloride - 4', 6-диамидин-2-фенилиндол дигидрохлорид) проводят в течение 10 мин перед заключением.
Как показано на Фиг. 3, результаты экспериментов показали, что было отобрано всего четыре клона scFv (N3, N5, N7 и N9), не связывающихся с неактивным мутантом (Т52А) и WT-KRS (не обработанный ламинином), но связывающихся только с клетками активного мутанта. При проведении обработки ламинином для индукции мембранной локализации WT-KRS и проведении окрашивания scFv, в WT-KRS и мутанте T52D. наблюдают похожие области окрашивания.
Было подтверждено, что данные клоны специфически связываются с клеточной поверхностью (верхняя часть).
1-7. Проверка способности специфического связывания с N-концом KRS
Для исследования того, связываются ли по существу клоны scFv (N3, N5, N7 и N9), отобранные в конечном итоге в Примере 1-6, с N-концом KRS, для проведения вестерн-блоттинга используют полноразмерную KRS (обозначаемую F, SEQ ID NO: 76), фрагмент KRS с делецией в аминокислотах в положениях 1-71 в N-конце (обозначаемый SEQ ID NO: 1) и фрагмент KRS, состоящий из аминокислотных остатков 1-200 в N-конце (обозначаемый SEQ ID NO: 2).
Клетки А549 трансформируют способом, как описано в Примере 1-6, с использованием полинуклеотидов, кодирующих полноразмерную KRS и фрагменты KRS. Далее, клетки лизируют, и проводят вестерн-блоттинг тем же способом, как описано в Примере 1-4.
Как показано на Фиг. 4, результаты подтвердили, что все отобранные клоны scFv (N3, N5, N7 и N9) демонстрируют полосы в единственном фрагменте с аминокислотами в положениях 1-200 в N-конце, одновременно не демонстрируя полос во фрагменте 1 без аминокислот в положениях 1-72 в N-конце. Таким образом, было установлено, что все клоны scFv N3, N5, N7 и N9 специфически связываются в N-концом KRS.
1-8. Секвенирование клонов scFv, специфически связывающихся с N-концом KRS
Клоны scFv (N3, N5, N7 и N9), в конечном итоге отобранные в Примере 1-6, анализируют в отношении их конформации CDR и последовательностей VH и VL. Секвенирование проводят тем же способом, как и описано в Примере 1-3.
По итогам секвенирования scFv N3 состоит из аминокислотной последовательности, определенной SEQ ID NO: 67, которая содержит линкерную последовательность SEQ ID NO: 65 в ее середине. Кроме того, VH N3 состоит из аминокислотной последовательности, определенной SEQ ID NO: 49, и VL N3 состоит из аминокислотной последовательности, определенной SEQ ID NO: 51. По итогам секвенирования соответствующих CDR, содержащихся в VH и VL N3, VH N3 содержит CDR1 тяжелой цепи, определенную SEQ ID NO: 1, CDR2 тяжелой цепи, определенную SEQ ID NO: 3, и CDR3 тяжелой цепи, определенную SEQ ID NO: 5, и VL N3 содержит CDR1 легкой цепи, определенную SEQ ID NO: 7, CDR2 легкой цепи, определенную SEQ ID NO: 9, и CDR3 легкой цепи, определенную SEQ ID NO: 11.
scFv N5 состоит из аминокислотной последовательности, определенной SEQ ID NO: 69, которая содержит линкерную последовательность SEQ ID NO: 65 в ее середине. Кроме того, VH N5 состоит из аминокислотной последовательности, определенной SEQ ID NO: 53, и VL N5 состоит из аминокислотной последовательности, определенной SEQ ID NO: 55. VH N5 содержит CDR1 тяжелой цепи, определенную SEQ ID NO: 13, CDR2 тяжелой цепи, определенную SEQ ID NO: 15, и CDR3 тяжелой цепи, определенную SEQ ID NO: 17, и VL N5 содержат CDR1 легкой цепи, определенную SEQ ID NO: 19, CDR2 легкой цепи, определенную SEQ ID NO: 21, и CDR3 легкой цепи, определенную SEQ ID NO: 23.
scFv N7 состоит из аминокислотной последовательности, определенной SEQ ID NO: 71, которая содержит линкерную последовательность SEQ ID NO: 65 в ее середине. Кроме того, VH N7 состоит из аминокислотной последовательности, определенной SEQ ID NO: 57, и VL N7 состоит из аминокислотной последовательности, определенной SEQ ID NO: 59. VH N7 содержит CDR1 тяжелой цепи, определенную SEQ ID NO: 25, CDR2 тяжелой цепи, определенную SEQ ID NO: 27, и CDR3 тяжелой цепи, определенную SEQ ID NO: 29, и VL N7 содержит CDR1 легкой цепи, определенную SEQ ID NO: 31, CDR2 легкой цепи, определенную SEQ ID NO: 33, и CDR3 легкой цепи, определенную SEQ ID NO: 35.
scFv N9 состоит из аминокислотной последовательности, определенной SEQ ID NO: 73, которая содержит линкерную последовательность SEQ ID NO: 65 в ее середине. Кроме того, VH N9 состоит из аминокислотной последовательности, определенной SEQ ID NO: 61, и VL N9 состоит из аминокислотной последовательности, определенной SEQ ID NO: 63. VH N9 состоит из CDR1 тяжелой цепи, определенной SEQ ID NO: 37, CDR2 тяжелой цепи, определенной SEQ ID NO: 39, и CDR3 тяжелой цепи, определенной SEQ ID NO: 41, и VL N9 содержит CDR1 легкой цепи, определенную SEQ ID NO: 43, CDR2 легкой цепи, определенную SEQ ID NO: 45, и CDR3 легкой цепи, определенную SEQ ID NO: 47.
Пример 2
Превращение антител scFv в антитела IgG и оценка их способности к специфическому связыванию
2-1 Превращение антител scFv в антитела IgG
Сначала, полипептиды, кодирующие cFv, амплифицируют посредством ПЦР с фаговых геномов N3, N5, N7 и N9. Нуклеотидные последовательности праймеров, используемых для амплификации гена области VH антител scFv: Прямой праймер (AGA GAG TGT АСА СТС CCA GGC GGC CGA GGT GCA G, SEQ ID NO: 93), Обратный праймер (CGC CGC TGG GCC СТТ GGT GGA GGC TGA GCT САС GGT GAC CAG, SEQ ID NO: 94). Нуклеотидные последовательности праймеров, используемых для амплификации гена области VL антител scFv: Прямой праймер (AAG CGG CCG ССА CCA TGG GAT GGA GCT GTA ТСА ТСС ТСТ ТСТ TGG TAG САА CAG СТА CAG GTG ТАС ACT ССС AGT CTG TGC TGA СТС AG, SEQ ID NO: 95), Обратный праймер (CGC CGC CGT ACG TAG GAC CGT CAG СТТ GGT, SEQ ID NO: 96)
ПЦР проводят с каждой фаговой ДНК (50 нг) в качестве матрицы, используя праймеры (10 пмоль каждого) в следующих условиях: 95°С/3 мин; 95°С/30 с, 60°С/30 с, 72°С/30 с, 30 циклов; и 72°С/5 мин, амплифицируя, таким образом, ген VH или VL scFv N3, N5, N7 или N9. ПЦР-продукт вставляют в вектор pcDNA3.4, используемый в получении IgG с использованием рестриктаз. Белки тяжелой и легкой цепей IgG кодировались в отдельных плазмидах.
Сконструированные векторы, содержащие ДНК, кодирующие тяжелую и легкую цепи каждого из IgG (далее в данном документе, называемого IgG N3, IgG N5, IgG N7 и IgG N9, соответственно), содержащих вариабельные области scFv, совместно трансформируют вольным образом в клетках 293F для экспрессии тяжелой и легкой цепей совместно в клетках. Трансформированные клетки 293F инкубируют в условиях 37°С и 8% СО2 в течение 7 суток и получают супернатант. Супернатант фильтруют через фильтр с мембраной из ацетата целлюлозы (размер пор 0,22 мкм, Corning) и очищают с использованием колонки с белком A CaptivA™ PriMAB (Repligen, США). Концентрации полученных антител измеряют, используя набор ВСА (Pierce, 23225), и анализируют белки-антитела IgG, полученные в восстановительных и невосстановительных условиях.
2-2. Проверка способности превращенных IgG к связыванию с KRS - Вестерн-блоттинг и иммунопреципитация
KRS-связывающую способность IgG, сконструированных в Примере 2-1, исследуют посредством вестерн-блоттинга (WB) и иммунопреципитации. Вестерн-блоттинг проводят таким же образом, как описано в Примере 1-4, и иммунопреципитацию проводили таким же образом, как описано в Примере 1-5.
Результаты установили, что IgG, сконструированные в настоящем изобретении, связываются с KRS, и на Фиг. 5 показаны данные результаты с использованием IgG N3 и IgG N5 в качестве типичных представителей.
2-3. Проверка способности превращенных IgG специфически связываться с N-концом KRS - SPR
Количественная связывающая способность очищенных белков-антител (IgG N3 и IgG N5) в отношении антигена (KRS 1-207 аа) измеряют, используя биосенсор Biacore 2000 SPR (поверхностный плазмонный резонанс) (GE healthcare, США). После иммобилизации KRS на сенсорном чипе (СМ5, GE healthcare, США), белкам-антителам (6,25-100 нМ), которые подвергают серийному разведению буферным раствором HES (10 ммоль/л HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid - 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота), рН 7,4, 150 ммоль/л NaCl, 3 ммоль/л ЭДТА, 0,005% поверхностно-активное вещество Р20), дают течь со скоростью 30 мкл/мин в течение 3 мин, и 1 моль/л NaCl/20 ммоль/л NaOH дают течь со скоростью 30 мкл/мин в течение 3 мин, вызывая, таким образом, диссоциацию белков, связанных с антигеном. Имитирующий IgG используют в качестве контроля. Конкретные условия эксперимента выглядят следующим образом:
Иммобилизованный антиген: KRS
Иммобилизованный уровень: 185 RU (response unit - единица ответа)
Антитело: IgG N3 и IgG N5
Подвижный буфер: буфер HBS-N
Регенерация: 2 М NaCl, 20 ммоль/л NaOH (поток 30 мкл/мин 1 мин)
Таблица 3
На Таблице 3 показаны кинетические константы скорости и константы равновесия диссоциации, измеренные для IgG N3 и IgG N5 с использованием Biacore 2000 SPR. Аффинность получают на основе кинетических констант скорости (ka и kd) и констант равновесия диссоциации (KD), используя ознакомительную версию программного обеспечения BIA 3.2. На Фиг. 6 показаны результаты SPR в виде графика для IgG N3 и IgG N5, соответственно. На основе Фиг. 6 и Таблицы 3 было подтверждено, что IgG N3 и IgG N5 по настоящему изобретению обладают способностью высокоспецифического связывания с N-концевой областью KRS. В имитирующем IgG в качестве контроля не наблюдали сигналов связывания.
2-4. Проверка способности превращенного IgG к специфическому связыванию с N-концом KRS IgG - иммунофлуоресцентное окрашивание
Для исследования того, связываются ли в действительности IgG, сконструированные в настоящем изобретении, с областью KRS, экспонированной на клеточной мембране, осуществляют иммунофлуоресценцию. Иммунофлуоресцентное окрашивание проводят таким же способом, как описано в Примере 1-6, используя IgG N3 и IgG N5 в качестве типичных представителей.
Как показано на Фиг. 7А, результаты подтвердили, что IgG N3 эффективно специфически связывается с экспонированной во внеклеточную среду N-концевой областью KRS, когда локализацию WT-KRS на мембране индуцируют посредством обработки ламинином. Как показано на Фиг. 7Б, эксперимент с использованием T52D-KRS (активный мутант, myc меченная KRS) также демонстрировал, что антитело по настоящему изобретению эффективно связывается с экспонированной во внеклеточную среду N-концевой областью KRS, и когда экспериментальные клетки становились перемеабилизированными, белки KRS, находящиеся в цитозоле, выявляют при высокой чувствительности. Было также подтверждено, как показано на Фиг. 7В, что IgG N5 благоприятно специфически связывался с экспонированной во внеклеточную среду N-концевой областью KRS, когда локализацию WT-KRS на мембране индуцируют обработкой ламинином.
Таким образом, было установлено, что антитела, предложенные в настоящем изобретении, имеют высокую специфичность связывания с экспонированной во внеклеточную среду N-концевой областью KRS.
Пример 3
Проверка ингибирующего действия на метастаз рака 3-1.
Оценка цитотоксичности
МТТ-тест проводят на IgG N3 в качестве типичного представителя. Клетки А549 высевают в 96-луночные планшеты (5000 клеток/лунка) и инкубируют. Клетки промывают средами, не содержащими сыворотку, и затем обрабатывают человеческим имитирующим IgG и IgG N3 в концентрации 0, 50, 100, 500 нмоль/л (в средах, не содержащих сыворотку). После 24-ч инкубации раствор МТТ добавляют в количестве 50 мкг/лунка с последующей инкубацией в течение 4 ч. После удаления раствора МТТ лунки обрабатывают 100 мкл ДМСО (диметилсульфоксид), и затем поглощение измеряют при 570 нм.
Как показано на Фиг. 8, результаты эксперимента подтвердили, что антитело по настоящему изобретению не демонстрирует цитотоксичности.
3-2. Анализ миграции клеток
Миграцию клеток оценивают, используя 24-луночную камеру Трансвелл с поликарбонатной мембраной (размер пор 8,0 мкм, Costar), как раскрыто в предшествующем уровне техники (Park, S. G. et al., Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. USA 102, 6356-6361 (2005)). В камере Трансвелл нижнюю стенку покрывают 10 мкг ламинина (в желатине) и сушат УФ (ультрафиолетовое излучение). Далее, клетки А549 суспендируют в средах RPEVI, не содержащих сыворотку, и затем помещают в концентрации 1×105 клеток на лунку в верхней камере. Камеру обрабатывают IgG N3 или человеческим имитирующим IgG (контроль) в концентрации 100 нмоль/л или 500 нмоль/л с последующей инкубацией в течение 24 ч. Далее, камеру дважды промывают PBS и обрабатывают 70% МеОН (в PBS) в течение 30 мин. Камеру еще раз дважды промывают PBS и обрабатывают раствором гематоксилина в течение 30 мин. Камеру три раза промывают DW (ДВ) (deionized water - деионизированная вода), и мембрану в камере вырезают и устанавливают на предметное стекло.
Как показано на Фиг. 9А, результаты эксперимента показали, что IgG N3 значительно ингибировал миграцию клеток А549. Результаты эксперимента также показали, что данное ингибирующее действие на миграцию клеток являлось дозозависимым (см. Фиг. 9Б).
3-3. Оценка ингибирующего действия на метастаз рака в in-vivo моделях метастаза рака
Поскольку KRS может ускорять миграцию клеток посредством 67LR, ассоциированного с метастазом рака, животные модели опухоли (рака) конструируют, используя клетки рака молочной железы мыши 4Т-1 (корейский банк клеточных линий), которые легко давали метастазы в легкое. Ортотопические животные модели рака молочной железы конструируют посредством инъецирования 4×104 4Т1 клеток в жировое тело шести мышей BALB/cAnCr 7-недельного возраста (Doo Yeol Biotech).
Опухоль инъецируют в жировое тело молочной железы, и, спустя 10 суток (Сутки 10), раковые ткани подвергают резекции из жирового тела. Спустя одни сутки (Сутки 11), IgG N3 (10 мг/кг) вводят два раза в неделю в течение двух недель с интервалами в 3 суток (всего четыре раза, Сутки 11, 14, 18 и 21) через хвостовую вену, внутривенная инъекция, и также вводят такую же дозу контрольного имитирующего IgG (Thermo #31154). Спустя одну неделю после завершения полной схемы дозирования антитела, то есть, 28 дней после инъекции опухоли, мышей умерщвляют для отбора ткани легкого. При вскрытии ткани легкого легкое наполняют посредством инъекции физиологического раствора в бронх с помощью шприца, собирают и затем хранят в растворе Буэна (Sigma #НТ10132) в течение 24 ч. Далее, осуществляют подсчет метастатических узелков в каждой доле легкого посредством микроскопа.
Как показано на Фиг. 11 и 12, результаты экспериментов подтвердят, что многие узелки были образованы в легком вследствие метастаза рака в контроле, и образование таких узелков значительно подавлялось в группе обработки IgG N3. На Фиг. 13 сравнительно показаны ткани легкого в контроле и группе обработки IgG N3 и подтверждено, что значительное большое количество рецепторов ламинина экспрессировалось в местах метастатических узелков в контроле, по сравнению с группой, обработанной антителом по настоящему изобретению.
3-4. Сравнение эффекта с соединением против метастаза рака (YH16899) в in-vivo моделях метастаза рака
Из приведенной выше литературы «Dae Gyu Kim et al., (2014)» известно, что соединение YH16899 оказывает действие на ингибирование метастаза рака посредством подавления взаимодействия 67LR и KRS. В таком случае, ингибирующее действие на метастаз рака сравнивают у YH16899 и IgG N3, антитела по настоящему изобретению. Конструирование in-vivo моделей опухоли и наблюдение за состоянием метастаза в легком осуществляют тем же способом, как и в Примере 3-3. YH16899 перорально вводят в количестве 100 мг/кг массы тела каждые сутки. IgG N3 внутривенно инъецируют в разных концентрациях (1 мг/кг массы тела, 10 мг/кг массы тела) через мышиные хвосты.
Как показано на Фиг. 14 и 15, результаты эксперимента подтвердили, что количество узелков в легком значимо уменьшалось дозозависимым образом в группе обработки IgG N3, и обработка всего лишь 1 мг/кг массы IgG N3 значимо ингибировала метастаз рака, в сравнении с группами обработки YH16899 (100 мг/кг массы тела).
Пример 4
Проверка сайтов связывания антител против KRS
4-1. Сайты связывания моноклональных антител против KRS с человеческой KRS
Для исследования сайтов связывания IgG N3 среди антител против KRS, сконструированных выше, с человеческой KRS осуществляют поверхностный плазмонный резонанс (SPR), как указано ниже.
Сначала, антитело IgG N3 иммобилизуют на Biacore Т200 (GE Healthcare), оборудованном сенсорным чипом СМ5, серия S (GE Healthcare), с использованием набора для связывания по аминогруппе (GE Healthcare). Затем, пептидам, показанным ниже в Таблице 4, растворенным в растворе PBS в соответствующих концентрациях, дают протечь в течение 60 с. Затем, PBS дают протечь в течение 5 мин. Затем, связывающую способность анализируют посредством Biacore Т200, ознакомительная версия программного обеспечения v2.0 (GE Healthcare).
Таблица 4
Информация о пептидах
Как показано на Фиг. 16, результаты показали, что антитело IgG N3 связывалось с эпитопами F1, F2, F3 и F4, но не с эпитопом F5. Кроме того, способность связывания с эпитопом F4 бала самой сильной, и способность связывания с F3, F2 и F1 усиливалась в указанном порядке.
Данные результаты смогли подтвердить, что основной сайт связывания антитела IgG N3 соответствует аминокислотным остаткам в положениях 15-29 в N-концевой области KRS.
4-2. Межвидовая перекрестная активность моноклонального антитела против KRS
Приведенный выше пример подтвердил сайт связывания антитела IgG N3 против KRS с человеческой KRS, и для исследования того, демонстрирует ли IgG N3 перекрестную активность с другими видами, мышь (m) и крыса (r), осуществляют поверхностный плазмонный резонанс (SPR), как указано ниже.
Таким же образом, как и в экспериментальном способе, описанном выше в Примере 4-1, антитело IgG N3 было иммобилизовано на чипе с использованием набора для связывания с аминогруппой (GE Healthcare). Затем, пептидам, показанным в приведенной выше Таблице 4, растворенным в растворе PBS в соответствующих концентрациях, дают протечь в течение 60 с, и PBS дают протечь в течение 5 мин. Затем, связывающую способность анализируют посредством Biacore Т200, пробный выпуск программного обеспечения v2.0 (GE Healthcare).
Как показано на Фиг. 17, результаты показали, что антитело IgG N3 связывалось с эпитопами F1, F2, F3 и F4 человека (h), мыши (m) и крысы (r), но не с эпитопом F5. Более сильная способность связывания с эпитопом F3, в сравнении с F1, совпадала у человека, мыши и крысы (данные по F2 и F4 не показаны).
Данные результаты смогли подтвердить, что антитело IgG N3 способно к межвидовой перекрестной активности.
Пример 5
Проверка роли сигнала ламинина в миграции и инвазии иммунных клеток Исследуют какое из нескольких веществ внеклеточного матрикса, образующих кровеносные сосуды, ускорял миграцию и инвазию моноцитов/макрофагов. Конкретные экспериментальные способы для анализа миграции в системе Трансвелл с использованием коллагена, фибронектина и ламинина в качестве веществ внеклеточного матрикса, представляли собой такие, как указано ниже. Трансвелл (Corning, #3421-5 мм) покрывают желатином (0,5 мг/мл), и затем клетки RAW 264.7 (1×105 клеток/лунка) засевают в верхнюю камеру. Каждую бессывороточную DMEM (Dulbecco modified Eagle's medium -среда Игла, модифицированная по Дульбекко) (500 мкл), содержащую ламинин (смесь ламининов, Biolamina), фибронектин или коллаген (10 мкг/мл), помещают в нижнюю камеру. Спустя 24 ч, немигрирующие клетки, присутствующие на верхней чести мембраны, удаляют ватными тампонами. Клетки в нижней камере фиксируют посредством обработки 70% метанолом в течение 30 мин и затем окрашивают 50% гематоксилином в течение 30 мин. После окрашивания мембрану берут и устанавливают на предметное стекло, и затем наблюдают за мигрирующими клетками, находящимися на нижней поверхности мембраны, и их количественно оценивают посредством микроскопа высокого разрешения.
Как показано на Фиг. 18а и 18б, результаты эксперимента подтвердили, что из нескольких веществ внеклеточного матрикса, ламинин ускорял миграцию моноцитов/макрофагов наиболее сильным образом.
Пример 6
Воздействие на миграцию и инвазию иммунных клеток, оказываемое подтипами ламинина
Оценивают воздействие подтипов ламинина, оказываемое на миграцию и инвазию иммунных клеток. Анализ миграции в системе Transwell проводят тем же способом, как и в Примере 5, с использованием LN111, LN211, LN221, LN411, LN421, LN511 и LN521 (10 мкг/мл) в качестве разных белков-подтипов ламинина (приобретенных у Biolamina). Специфические последовательности подтипов ламинина могут быть соотнесены с цепочкой α4 SEQ ID NO: 115, цепочкой α2 SEQ ID NO: 121, цепочкой α5 SEQ ID NO: 122, цепочкой β2 SEQ ID NO: 117, цепочкой β1 SEQ ID NO: 123 и цепочкой γ1 SEQ ID NO: 119, в соответствии с цепочками, составляющими соответствующие подтипы ламинина.
Клетки RAW 264.7 (2×106 клеток) инкубируют в течение 18 ч, обрабатывают 1 мкг/мл каждого подтипа ламинина в DMEM, не содержащей сыворотку, и затем собирают в моменты времени 0 ч, 12 ч и 24 ч. Белок клеток RAW 264.7 разделяли на цитозольную и мембранную фракции посредством использования набора для экстракции внутриклеточного протеома ProteoExtract (Calbiotech, cat# 539790). Полученный белок подвергают электрофоретическому разделению, переносят на мембрану PVDF (Milipore) и блокируют 3% обезжиренным молоком. Далее, добавляют поликлональное антитело против KRS (кроличье, Neomics, Co. Ltd. #NMS-01-0005) с последующей инкубацией в течение 1 ч. После отмывки несвязанных антител добавляют вторичные антитела против кроличьих антител (ThermoFisher Scientific, #31460) с последующей инкубацией. После инкубации с вторичными антителами проводят сенсибилизацию пленки с использованием реагента ECL в качестве субстрата в темной комнате. Сенсибилизированные полосы сравнивают со стандартными молекулярными маркерами для идентификации полос, соответствующих размерам KRS. Антитела против Na+/K+ АТФазы (Abeam) и тубулина (Sigma) используют для идентификации маркера плазматической мембраны и цитозоля, соответственно.
Как показано на Фиг. 19а и 196, результаты экспериментов подтвердили, что моноциты/макрофаги мигрируют в результате специфической реакции на подтип α4β2γ1 (LN421) среди всех тестируемых подтипов ламинина. То есть, подтвердили, что моноциты/макрофаги осуществляли миграцию и инвазию, специфически реагируя на LN421. Как показано на Фиг. 19в, подтвердили, что обработка моноцитов/макрофагов LN421 увеличивала количество KRS, выявляемой в области клеточной мембраны, но частично уменьшало количество KRS, выявляемой в области цитозоля. Данные результаты указывают на то, что KRS, которая обычно находится в области цитозоля после экспрессии внутри моноцитов/макрофагов, перемещается в область клеточной мембраны в результате обработки LN421, и что повышение уровня KRS в области мембраны иммунной клетки соответствует важному патологическому явлению в заболеваниях, связанных с миграцией и инвазией иммунных клеток.
Пример 7
Конструирование антитела для снижения уровня KRS клеточной мембраны и проверка контрольного воздействия на миграцию/инвазию иммунных клеток
Воздействие на миграцию и инвазию иммунных клеток исследуют с использованием антитела IgG N3 в качестве типичного представителя среди антител, сконструированных в вышеприведенном Примере 1. Конкретные экспериментальные способы выглядели следующим образом. Трансвелл (Corning, #3421-5 мм) покрывают желатином (0,5 мг/мл), и затем клетки RAW 264.7 (1×105 клеток/лунка) засевают в верхнюю камеру. Бессывороточную DMEM (500 мкл), содержащую ламинин 421 (1 мкг/мл), помещают в нижнюю камеру. Верхнюю камеру обрабатывали каждым антителом в концентрации 100 нМ. Спустя 24 ч, осуществляют иммобилизацию 70% метанолом в течение 30 мин, и затем проводят окрашивание 50% гематоксилином в течение 30 мин. Немигрирующие клетки, находящиеся на верхней части мембраны, удаляют ватными тампонами, и затем мембрану берут и устанавлива на предметном стекле. За мигрирующими клетками, находящимися на нижней поверхности мембраны, наблюдают посредством микроскопа высокого разрешения (Фиг. 20а), и осуществляюти подсчет клеток на полученных изображениях и наносят на график (Фиг. 20б).
Клетки RAW 264.7 обрабатывают ламинином 421 (1 мкг/мл) и антителом (100 нМ), инкубируют в течение 24 ч и собирают. Далее, собранный материал разделяют на мембранную и цитозольную фракции посредством использования набора для экстракции внутриклеточного протеома ProteoExtract (Calbiochem), отбирают образцы и затем подвергают вестерн-блоттингу в отношении KRS. Конкретный способ представлял собой такой, как описано в Примере 6.
Как показано на Фиг. 20а и 20б, результаты эксперимента подтвердили, что антитело по настоящему изобретению эффективно ингибировало специфически LN421-зависимую миграцию моноцитов/макрофагов. Как показано на Фиг. 20в, было подтверждено, что обработка LN421 повышала уровень KRS в клеточный мембране моноцитов/макрофагов, и обработка антителом IgG N3 эффективно снижала уровень KRS в клеточной мембране. Кроме того, было подтверждено, что уровень KRS в цитозоле снижался в группе обработки антителом IgG N3.
Данные результаты подтвердили, что антитело по настоящему изобретению является перспективным в качестве нового терапевтического средства против заболеваний, задействованных в миграции иммунных клеток, таких как моноциты/микрофаги.
Пример 8
Проверка воздействия на заболевание, связанное с миграцией иммунных клеток, в in vivo моделях
Как в примерах, описанных выше, следующий эксперимент проводят с использованием антитела IgG N3 в качестве типичного представителя среди антител по настоящему изобретению.
Способы
1. Конструирование модели легочной артериальной гипертензии (ЛАГ) и введение тестируемого вещества
Для индукции ЛАГ у крыс SD (Sprague Dawley) возраста семь недель (Orient Bio) подкожно инъецируют 60 мг/кг массы тела монокроталина (МСТ - monocrotaline). Далее, крыс делят на четыре группы (пять животных на группу), и им вводят 1 мг/кг массы тела имитирующего человеческого IgG (Thermo Fisher Scientific, отрицательный контроль), 1 мг/кг массы тела IgG N3, 10 мг/кг массы тела IgG N3 10 и 25 мг/кг массы тела силденафила (положительный контроль) в течение трех недель. Все антитела в.в. (внутривенно) инъецируют два раза в неделю, и каждый день перорально вводят силденафил.
2. Измерение кровотока и кровяного давления
Спустя три недели, крыс анестезируюьт изофлураном и им измеряют кровоток и кровяное давление, используя пневматическую систему измерения высокой точности (система измерения кардиоваскулярного давления и объема MPVS, название модели: MPVS Ultra, производитель: Millar Instruments). Конечно-систолическое давление в правом желудочке (RVESP - right ventricular end-systolic pressure), конечно-диастолическое давление в правом желудочке, конечно-систолическое давление в левом желудочке, конечно-диастолическое давление в левом желудочке измеряют с использованием эксклюзивного катетера (катетер для измерения давления у крысы Mikro-Tip, производитель: Millar Instruments). Сердечный выброс измеряют с использованием периваскулярного зонда кровотока (зонды потока Transonic, производитель: Millar Instruments), и данный экспериментальный способ осуществляют таким же способом, как раскрыто в следующей литературе: Pacher Р, Nagayama Т, Mukhopadhyay Р, Batkai S, Kass DA. Measurement of cardiac function using pressure-volume conductance catheter technique in mice and rats. Nat Protoc 2008; 3(9): 1422-34.
3. Иммуногистохимия (ИГХ)
Отобранные легкие фиксируют в параформальдегиде (ПФА) в соответствии с обычным способом и затем осуществляют инфильтрацию парафина и заливают в парафин посредством промывки, дегидратации и очистки. Делают микросрезы парафиновых блоков с тканью легкого крысы до толщины 6 мкм и готовят предметные стекла. Далее, проводят окрашивание, как указано ниже. Сначала образец три раза обрабатывают ксилолом в течение 5 мин, обрабатывают 100% этанолом, 95% этанолом, 90% этанолом и 70% этанолом, и DW в указанном порядке в течение 2 мин и промывают PBS в течение 5 мин. После обработки 0,3% Н2О2 образец дважды промывают PBS в течение 5 мин. Образец погружают в цитратный буфер 0,01 моль/л, нагревают и промывают PBS-T (0,03% твин 20). Далее, образец блокируют (2% БСА (Бычий сывороточный альбумин) и 2% козья сыворотка в PBS) при комнатной температуре в течение 30 мин. Образец окрашивают антителом против CD68 (1:200, клон ED1, Abeam) при 4°С в течение ночи. Образец три раза промывают PBS-T в течение 5 мин и затем обрабатывают набором антител против мышиных антител, меченных полимером-HRP, EnVision (DAKO) при 4°С в течение 1 ч. Образец три раза промывают PBS-T и затем развивают окраску посредством обработки субстратным буфером DAB и хромогеном 20 DAB. Окрашенную ткань обрабатывают гематоксилином Майера (Sigma) в течение 1 мин и затем дважды обрабатывают 70% этанолом, 90% этанолом, 95% этанолом и 100% этанолом в указанном порядке в течение 2 мин каждым. Наконец, ткань три раза обрабатывают ксилолом в течение 5 мин и затем проводят наблюдение в оптический микроскоп.
Результаты
8-1. Проверка изменений кровяного давления и сердечного выброса Модели ЛАГ, которая представляет собой заболевание, имеющее тесную связь между инвазией иммунных клеток и патологическими явлениями, обрабатывали антителом IgG N3 (1 мг/кг массы тела или 10 мг/кг массы тела) в течение 3 недель (в.в., дважды в неделю), и затем в них измеряют конечно-систолическое давление в правом желудочке (RVESP), конечно-диастолическое давление в правом желудочке (RVEDP), конечно-систолическое давление в левом желудочке (LVESP), конечно-диастолическое давление в левом желудочке (LVEDP) и сердечный выброс (СО - cardiac output). Данные Результаты показаны в Таблице 5.
Таблица 5
(n равно 4)
+ Ab N3 1 мг/кг массы тела
(n равно 5)
+ Ab N3 10 мг/кг массы тела
(n равно 5)
(n равно 5)
(мм рт. ст.)
(мм рт. ст.)
(мм рт. ст.)
(мм рт. ст.)
(отсутствует измерение СО для одного животного группы МСТ + имитирующий IgG, умершего от анестезии, и одного животного группы обработки силденафилом, умершего во время хирургической операции)
Легочная артериальная гипертензия вызывает повышение конечного давления в правом желудочке вследствие сужения легочной артерии, приводя к правожелудочковой недостаточности. Кроме того, его компенсаторный механизм нарушается вследствие длительной гипертензии, приводя к гипертрофии правого желудочка с последующей дилатацией правого желудочка. Это приводит к сдавливанию левого желудочка вследствие движения межжелудочковой перегородки, приводя к уменьшению конечно-диастолического объема и сердечного выброса левого желудочка (WooSeok Lee, et al., Clinical Characteristics and Prognostic Factors of Patients with Severe Pulmonary Hypertension, Korean Circulation J 2007; 37:265-270). В конечном итоге, легочная артериальная гипертензия главным образом связана с правым желудочком, но также вовлечена в функции левого желудочка.
Пациенты с ЛАГ показывали повышение RVESP, которое также наблюдают в животных моделях ЛАГ настоящего эксперимента. В связи с этим, как показано на Фиг. 21а, антитело N3 значительно снижало RVESP при обеих его концентрациях и эффективно снижало RVESP, особенно в сравнении с силденафилом, лекарственным средством, представляющимся положительным контролем.
Кроме того, не наблюдают снижения в конечно-систолическом давлении в левом желудочке (LVESP) из-за введения антитела IgG N3, а скорее, как показано на Фиг. 21б, LVESP значительно повышалось в группе, которой вводят антитело по настоящему изобретению. Таким образом, антитело по настоящему изобретению сопоставляют с силденафилом, используемым в качестве существующего терапевтического средства против легочной артериальной гипертензии, где силденафил вызывает дилатацию легочной артерии и дилатацию системных артерий, что, вследствие этого, приводит к риску снижения системного кровяного давления.
То есть, было подтверждено, что антитело по настоящему изобретению демонстрирует тенденцию к оказанию слабого действия на системное артериальное давление, по сравнению с силденафилом, и считается, что данное действие является благоприятной характеристикой терапевтического средства, учитывая, что введение силденафила может иметь риск развития гипотензии в клинических местах. Кроме того, тяжелая легочная артериальная гипертензия вызывает систолическую правожелудочковую недостаточность, которая может сопровождаться низким сердечным выбросом и системной гипотензией.
В свою очередь, ожидается, что лечение для облегчения легочной артериальной гипертензии антителом по настоящему изобретению увеличивает сердечный выброс и системное кровяное давление, нормализуя, таким образом, кровяное давление.
В общем, было подтверждено, что введение антитела по настоящему изобретению снижает риск побочных эффектов существующих терапевтических лекарственных средств и демонстрирует облегчение симптомов ЛАГ и эффекты от лечения.
8-2. Эхокардиография
Данные по D-образному левому желудочку, указывающие на перегрузку давлением в правом желудочке, наблюдают у трех животных в группе введения только МСТ (то есть, модели ЛАГ без введения тестируемого вещества) и трех животных в группе введения МСТ и силденафила, но не наблюдают в группах введения терапевтического антитела.
Кроме того, как показано ниже в Таблице 6, массы тела соответствующих групп увеличивались до похожей степени, без значителного различия. А именно, данных, указывающих на патологические признаки, включая патологическое снижение массы, вызываемое введением терапевтического антитела, не наблюдают.
Таблица 6
(n равно 4)
(n равно 5)
(n равно 5)
(n равно 5)
8-3. Проверка степени миграции моноцитов/макрофагов и инфильтрации
ИГХ-окрашивание проводят в отношении CD68, который является маркером моноцитов/макрофагов, используя ткани легкого каждой группы обработки. Как показано на Фиг. 22, результаты эксперимента подтвердили, что группы с обработкой антителами IgG N3 по настоящему изобретению явным образом уменьшают инфильтрацию моноцитов/макрофагов в ткани легкого, и такой эффект значимо превосходил эффект силденафила.
Промышленная применимость
Как описано выше, антитела или их фрагменты согласно настоящему изобретению имеют конкретные последовательности CDR (определяющая комплементарность область), определенные в настоящем описании изобретения, и абсолютно превосходную способность к специфическому связыванию с экспонированной во внеклеточную среду N-концевой областью KRS, и, таким образом, могут быть использованы в диагностике заболевания (например, рака), которое, как известно, сопровождается характерным поведением KRS. Кроме того, антитела или их фрагменты согласно настоящему изобретению также специфически нацелены на N-концевую область KRS in vivo, и, таким образом, ингибируют взаимодействие рецептора ламинина и N-концевой области, оказывая превосходное ингибирующее действие на метастаз рака, и, таким образом, могут быть использованы в качестве терапевтического средства. Кроме того, антитела или их фрагменты согласно настоящему изобретению могут контролировать миграцию иммунных клеток и таким образом могут быть чрезвычайно преимущественно использованы в предупреждении, облегчении и лечении заболеваний, связанных с миграцией иммунных клеток, и, таким образом, имеют высокую промышленную применимость.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> МЕДИСИНАЛ БАЙОКОНВЕРДЖЕНС РЕСЕРЧ СЕНТР
<120> АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ C N-КОНЦЕВОЙ
ОБЛАСТЬЮ ЛИЗИЛ-тРНК-СИНТЕТАЗЫ, ЭКСПОНИРОВАННОЙ НА
КЛЕТОЧНОЙ МЕМБРАНЕ
<130> OP19-0123/PCT/RU
<150> KR 10-2017-0038775
<151> 2017-03-27
<150> KR 10-2017-0118890
<151> 2017-09-15
<150> KR 10-2017-0118917
<151> 2017-09-15
<150> PCT/KR 2018/003594
<151> 2018-03-27
<160> 123
<170> KoPatentIn 3.0
<210> 1
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VH CDR1
<400> 1
Ser Tyr Asp Met Ser
1 5
<210> 2
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH CDR1
<400> 2
agttatgata tgagc 15
<210> 3
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VH CDR2
<400> 3
Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 4
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH CDR2
<400> 4
gcgatctctt atgataatgg taatacatat tacgctgatt ctgtaaaagg t 51
<210> 5
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VH CDR3
<400> 5
Met Ala Leu Asp Phe Asp Tyr
1 5
<210> 6
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH CDR3
<400> 6
atggcgcttg atttcgacta c 21
<210> 7
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VL CDR1
<400> 7
Ser Ser Ser Asn Ile Gly Ser Asn Tyr Val Thr
1 5 10
<210> 8
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL CDR1
<400> 8
tcttcatcta atattggcag taattatgtc acc 33
<210> 9
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VL CDR2
<400> 9
Asp Asn Ser Asn Arg Pro Ser
1 5
<210> 10
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL CDR2
<400> 10
gataatagta atcggccaag c 21
<210> 11
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VL CDR3
<400> 11
Ala Ser Trp Asp Asp Ser Leu Ser Ala
1 5
<210> 12
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL CDR3
<400> 12
gcttcttggg atgatagcct gagtgct 27
<210> 13
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VH CDR1
<400> 13
Asp Tyr Ala Met Ser
1 5
<210> 14
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VH CDR1
<400> 14
gattatgcta tgagc 15
<210> 15
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VH CDR2
<400> 15
Trp Ile Tyr Ser Gly Ser Gly Asn Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 16
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VH CDR2
<400> 16
tggatctatt ctggtagtgg taataaatat tacgctgatt ctgtaaaagg t 51
<210> 17
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VH CDR3
<400> 17
Met Gly Leu Asp Phe Asp Tyr
1 5
<210> 18
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VH CDR3
<400> 18
atgggtttgg atttcgacta c 21
<210> 19
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VL CDR1
<400> 19
Ser Ser Ser Asn Ile Gly Asn Asn Tyr Val Ser
1 5 10
<210> 20
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VL CDR1
<400> 20
tcttcatcta atattggcaa taattatgtc tcc 33
<210> 21
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VL CDR2
<400> 21
Asp Asp Asn Gln Arg Pro Ser
1 5
<210> 22
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VL CDR2
<400> 22
gatgataatc agcggccaag c 21
<210> 23
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VL CDR3
<400> 23
Gly Thr Trp Asp Asp Ser Leu Ser Ala
1 5
<210> 24
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VL CDR3
<400> 24
ggtacttggg atgatagcct gagtgct 27
<210> 25
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VH CDR1
<400> 25
Asp Tyr Tyr Met Ser
1 5
<210> 26
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VH CDR1
<400> 26
gattattata tgagc 15
<210> 27
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VH CDR2
<400> 27
Gly Ile Tyr Ser Gly Thr Gly Ser Ile Tyr Tyr Ala Asp Ser Val Glu
1 5 10 15
Gly
<210> 28
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VH CDR2
<400> 28
gggatctatt ctggtactgg tagtatatat tacgctgatt ctgtagaagg t 51
<210> 29
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VH CDR3
<400> 29
Pro Pro Tyr His Phe Asp Tyr
1 5
<210> 30
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VH CDR3
<400> 30
cctccgtatc atttcgacta c 21
<210> 31
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VL CDR1
<400> 31
Ser Ser Ser Asn Ile Gly Ser Asn Tyr Val Thr
1 5 10
<210> 32
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VL CDR1
<400> 32
tcttcatcta atattggcag taattatgtc acc 33
<210> 33
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VL CDR2
<400> 33
Ala Asp Ser Asn Arg Pro Ser
1 5
<210> 34
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VL CDR2
<400> 34
gctgatagta atcggccaag c 21
<210> 35
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VL CDR3
<400> 35
Gly Ala Trp Asp Tyr Ser Leu Ser Gly
1 5
<210> 36
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VL CDR3
<400> 36
ggtgcttggg attatagcct gagtggt 27
<210> 37
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VH CDR1
<400> 37
Asn Tyr Asp Met Ser
1 5
<210> 38
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VH CDR1
<400> 38
aattatgata tgagc 15
<210> 39
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VH CDR2
<400> 39
Trp Ile Tyr Ser Gly Asp Ser Ser Lys Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 40
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VH CDR2
<400> 40
tggatctatt ctggtgatag tagtaaatat tacgctgatt ctgtaaaagg t 51
<210> 41
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VH CDR3
<400> 41
Glu Thr Arg Thr Phe Asp Tyr
1 5
<210> 42
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VH CDR3
<400> 42
gagacgcgga cgttcgacta c 21
<210> 43
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VL CDR1
<400> 43
Ser Ser Phe Asn Ile Gly Ser Asn Ala Val Tyr
1 5 10
<210> 44
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VL CDR1
<400> 44
tcttcattta atattggcag taatgctgtc tac 33
<210> 45
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VL CDR2
<400> 45
Tyr Asn Ser Gln Arg Pro Ser
1 5
<210> 46
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VL CDR2
<400> 46
tataatagtc agcggccaag c 21
<210> 47
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VL CDR3
<400> 47
Gly Ser Trp Asp Ala Ser Leu Ser Gly
1 5
<210> 48
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VL CDR3
<400> 48
ggctcttggg atgctagcct gagtggt 27
<210> 49
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VH
<400> 49
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 50
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VH
<400> 50
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 51
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 VL
<400> 51
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 52
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 VL
<400> 52
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtgct tcttgggatg atagcctgag tgcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 53
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VH
<400> 53
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Trp Ile Tyr Ser Gly Ser Gly Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Gly Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 54
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VH
<400> 54
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc gattatgcta tgagctgggt ccgccaggct 120
ccagggaaag ggctggagtg ggtctcatgg atctattctg gtagtggtaa taaatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatgggt 300
ttggatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 55
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 VL
<400> 55
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 56
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 VL
<400> 56
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc aataattatg tctcctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gatgataatc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtggt acttgggatg atagcctgag tgcttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 57
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VH
<400> 57
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Tyr Ser Gly Thr Gly Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Pro Pro Tyr His Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 58
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VH
<400> 58
gaggtgcagc tgttggagtc ggggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc gattattata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaggg atctattctg gtactggtag tatatattac 180
gctgattctg tagaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagacctccg 300
tatcatttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 59
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 VL
<400> 59
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ala Asp Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Trp Asp Tyr Ser Leu
85 90 95
Ser Gly Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 60
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 VL
<400> 60
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gctgatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggacg aggctgatta ttactgtggt gcttgggatt atagcctgag tggttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 61
<211> 116
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VH
<400> 61
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Trp Ile Tyr Ser Gly Asp Ser Ser Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Glu Thr Arg Thr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 62
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VH
<400> 62
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc aattatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatgg atctattctg gtgatagtag taaatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagagagacg 300
cggacgttcg actactgggg ccagggtaca ctggtcaccg tgagctca 348
<210> 63
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 VL
<400> 63
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Phe Asn Ile Gly Ser Asn
20 25 30
Ala Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asn Ser Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ala Ser Leu
85 90 95
Ser Gly Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 64
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 VL
<400> 64
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatt taatattggc agtaatgctg tctactggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat tataatagtc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtggc tcttgggatg ctagcctgag tggttatgtc 300
ttcggcggag gcaccaagct gacggtccta 330
<210> 65
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер для scFv
<400> 65
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 66
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> линкер для scFv
<400> 66
ggtggaggcg gttcaggcgg aggtggatcc ggcggtggcg gatcg 45
<210> 67
<211> 241
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N3 scFv (VH+линкер+VL)
<400> 67
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Trp Asp
210 215 220
Asp Ser Leu Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 68
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N3 scFv (VH+линкер+VL)
<400> 68
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgataata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cggtccgagg atgaggctga ttattactgt 660
gcttcttggg atgatagcct gagtgcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 69
<211> 241
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N5 scFv (VH+линкер+VL)
<400> 69
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Trp Ile Tyr Ser Gly Ser Gly Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Gly Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Asn Asn Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Asp Asp Asn Gln Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp
210 215 220
Asp Ser Leu Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 70
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N5 scFv (VH+линкер+VL)
<400> 70
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc gattatgcta tgagctgggt ccgccaggct 120
ccagggaaag ggctggagtg ggtctcatgg atctattctg gtagtggtaa taaatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatgggt 300
ttggatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcaataatt atgtctcctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgatgata atcagcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cggtccgagg atgaggctga ttattactgt 660
ggtacttggg atgatagcct gagtgcttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 71
<211> 241
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N7 scFv (VH+линкер+VL)
<400> 71
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Tyr Ser Gly Thr Gly Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Pro Pro Tyr His Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile
145 150 155 160
Gly Ser Asn Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Ala Asp Ser Asn Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Trp Asp
210 215 220
Tyr Ser Leu Ser Gly Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 72
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N7 scFv (VH+линкер+VL)
<400> 72
gaggtgcagc tgttggagtc ggggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc gattattata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaggg atctattctg gtactggtag tatatattac 180
gctgattctg tagaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagacctccg 300
tatcatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atctaatatt 480
ggcagtaatt atgtcacctg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tatgctgata gtaatcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cggtccgagg acgaggctga ttattactgt 660
ggtgcttggg attatagcct gagtggttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 73
<211> 241
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N9 scFv (VH+линкер+VL)
<400> 73
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Trp Ile Tyr Ser Gly Asp Ser Ser Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Glu Thr Arg Thr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr
130 135 140
Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Phe Asn Ile
145 150 155 160
Gly Ser Asn Ala Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro
165 170 175
Lys Leu Leu Ile Tyr Tyr Asn Ser Gln Arg Pro Ser Gly Val Pro Asp
180 185 190
Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser
195 200 205
Gly Leu Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp
210 215 220
Ala Ser Leu Ser Gly Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val
225 230 235 240
Leu
<210> 74
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N9 scFv (VH+линкер+VL)
<400> 74
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc aattatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatgg atctattctg gtgatagtag taaatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagagagacg 300
cggacgttcg actactgggg ccagggtaca ctggtcaccg tgagctcagg tggaggcggt 360
tcaggcggag gtggatccgg cggtggcgga tcgcagtctg tgctgactca gccaccctca 420
gcgtctggga cccccgggca gagggtcacc atctcttgta ctggctcttc atttaatatt 480
ggcagtaatg ctgtctactg gtaccagcag ctcccaggaa cggcccccaa actcctcatc 540
tattataata gtcagcggcc aagcggggtc cctgaccgat tctctggctc caagtctggc 600
acctcagcct ccctggccat cagtgggctc cggtccgagg atgaggctga ttattactgt 660
ggctcttggg atgctagcct gagtggttat gtcttcggcg gaggcaccaa gctgacggtc 720
cta 723
<210> 75
<211> 52
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N-концевая область Лизил-тРНК-синтетазы (KRS),
экспонированной на мемебране
<400> 75
Met Ala Ala Val Gln Ala Ala Glu Val Lys Val Asp Gly Ser Glu Pro
1 5 10 15
Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys
20 25 30
Val Ala Glu Lys Glu Ala Lys Gln Lys Glu Leu Ser Glu Lys Gln Leu
35 40 45
Ser Gln Ala Thr
50
<210> 76
<211> 597
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Полноразмерная лизил-тРНК-синтетаза (KRS)
<400> 76
Met Ala Ala Val Gln Ala Ala Glu Val Lys Val Asp Gly Ser Glu Pro
1 5 10 15
Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys
20 25 30
Val Ala Glu Lys Glu Ala Lys Gln Lys Glu Leu Ser Glu Lys Gln Leu
35 40 45
Ser Gln Ala Thr Ala Ala Ala Thr Asn His Thr Thr Asp Asn Gly Val
50 55 60
Gly Pro Glu Glu Glu Ser Val Asp Pro Asn Gln Tyr Tyr Lys Ile Arg
65 70 75 80
Ser Gln Ala Ile His Gln Leu Lys Val Asn Gly Glu Asp Pro Tyr Pro
85 90 95
His Lys Phe His Val Asp Ile Ser Leu Thr Asp Phe Ile Gln Lys Tyr
100 105 110
Ser His Leu Gln Pro Gly Asp His Leu Thr Asp Ile Thr Leu Lys Val
115 120 125
Ala Gly Arg Ile His Ala Lys Arg Ala Ser Gly Gly Lys Leu Ile Phe
130 135 140
Tyr Asp Leu Arg Gly Glu Gly Val Lys Leu Gln Val Met Ala Asn Ser
145 150 155 160
Arg Asn Tyr Lys Ser Glu Glu Glu Phe Ile His Ile Asn Asn Lys Leu
165 170 175
Arg Arg Gly Asp Ile Ile Gly Val Gln Gly Asn Pro Gly Lys Thr Lys
180 185 190
Lys Gly Glu Leu Ser Ile Ile Pro Tyr Glu Ile Thr Leu Leu Ser Pro
195 200 205
Cys Leu His Met Leu Pro His Leu His Phe Gly Leu Lys Asp Lys Glu
210 215 220
Thr Arg Tyr Arg Gln Arg Tyr Leu Asp Leu Ile Leu Asn Asp Phe Val
225 230 235 240
Arg Gln Lys Phe Ile Ile Arg Ser Lys Ile Ile Thr Tyr Ile Arg Ser
245 250 255
Phe Leu Asp Glu Leu Gly Phe Leu Glu Ile Glu Thr Pro Met Met Asn
260 265 270
Ile Ile Pro Gly Gly Ala Val Ala Lys Pro Phe Ile Thr Tyr His Asn
275 280 285
Glu Leu Asp Met Asn Leu Tyr Met Arg Ile Ala Pro Glu Leu Tyr His
290 295 300
Lys Met Leu Val Val Gly Gly Ile Asp Arg Val Tyr Glu Ile Gly Arg
305 310 315 320
Gln Phe Arg Asn Glu Gly Ile Asp Leu Thr His Asn Pro Glu Phe Thr
325 330 335
Thr Cys Glu Phe Tyr Met Ala Tyr Ala Asp Tyr His Asp Leu Met Glu
340 345 350
Ile Thr Glu Lys Met Val Ser Gly Met Val Lys His Ile Thr Gly Ser
355 360 365
Tyr Lys Val Thr Tyr His Pro Asp Gly Pro Glu Gly Gln Ala Tyr Asp
370 375 380
Val Asp Phe Thr Pro Pro Phe Arg Arg Ile Asn Met Val Glu Glu Leu
385 390 395 400
Glu Lys Ala Leu Gly Met Lys Leu Pro Glu Thr Asn Leu Phe Glu Thr
405 410 415
Glu Glu Thr Arg Lys Ile Leu Asp Asp Ile Cys Val Ala Lys Ala Val
420 425 430
Glu Cys Pro Pro Pro Arg Thr Thr Ala Arg Leu Leu Asp Lys Leu Val
435 440 445
Gly Glu Phe Leu Glu Val Thr Cys Ile Asn Pro Thr Phe Ile Cys Asp
450 455 460
His Pro Gln Ile Met Ser Pro Leu Ala Lys Trp His Arg Ser Lys Glu
465 470 475 480
Gly Leu Thr Glu Arg Phe Glu Leu Phe Val Met Lys Lys Glu Ile Cys
485 490 495
Asn Ala Tyr Thr Glu Leu Asn Asp Pro Met Arg Gln Arg Gln Leu Phe
500 505 510
Glu Glu Gln Ala Lys Ala Lys Ala Ala Gly Asp Asp Glu Ala Met Phe
515 520 525
Ile Asp Glu Asn Phe Cys Thr Ala Leu Glu Tyr Gly Leu Pro Pro Thr
530 535 540
Ala Gly Trp Gly Met Gly Ile Asp Arg Val Ala Met Phe Leu Thr Asp
545 550 555 560
Ser Asn Asn Ile Lys Glu Val Leu Leu Phe Pro Ala Met Lys Pro Glu
565 570 575
Asp Lys Lys Glu Asn Val Ala Thr Thr Asp Thr Leu Glu Ser Thr Thr
580 585 590
Val Gly Thr Ser Val
595
<210> 77
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N3 IgG
<400> 77
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Tyr Asp Asn Gly Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Met Ala Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 78
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N3 IgG
<400> 78
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agttatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagcg atctcttatg ataatggtaa tacatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagaatggcg 300
cttgatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 79
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N3 IgG
<400> 79
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Thr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 80
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N3 IgG
<400> 80
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gataatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtgct tcttgggatg atagcctgag tgcttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 81
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N5 IgG
<400> 81
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Trp Ile Tyr Ser Gly Ser Gly Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Gly Leu Asp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 82
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N5 IgG
<400> 82
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc gattatgcta tgagctgggt ccgccaggct 120
ccagggaaag ggctggagtg ggtctcatgg atctattctg gtagtggtaa taaatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagaatgggt 300
ttggatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 83
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N5 IgG
<400> 83
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 84
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N5 IgG
<400> 84
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc aataattatg tctcctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gatgataatc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtggt acttgggatg atagcctgag tgcttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 85
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N7 IgG
<400> 85
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Tyr Ser Gly Thr Gly Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Pro Pro Tyr His Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 86
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N7 IgG
<400> 86
gaggtgcagc tgttggagtc ggggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc gattattata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaggg atctattctg gtactggtag tatatattac 180
gctgattctg tagaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagacctccg 300
tatcatttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 87
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N7 IgG
<400> 87
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 88
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N7 IgG
<400> 88
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatc taatattggc agtaattatg tcacctggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat gctgatagta atcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggacg aggctgatta ttactgtggt gcttgggatt atagcctgag tggttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 89
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N9 IgG
<400> 89
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Trp Ile Tyr Ser Gly Asp Ser Ser Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Ser
85 90 95
Ala Arg Glu Thr Arg Thr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 90
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь N9 IgG
<400> 90
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc aattatgata tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatgg atctattctg gtgatagtag taaatattac 180
gctgattctg taaaaggtcg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactctgc gagagagacg 300
cggacgttcg actactgggg ccagggtaca ctggtcaccg tgagctcagc ctccaccaag 360
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc 420
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc 480
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc 540
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 600
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac 660
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc 720
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 780
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 840
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 900
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 960
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 1020
cagccccgag aaccacaggt gtataccctg cccccatccc gggatgagct gaccaagaac 1080
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1140
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac 1200
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 1260
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 1320
tccctgtccc cgggtaaa 1338
<210> 91
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N9 IgG
<400> 91
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Phe Asn Ile Gly Ser Asn
20 25 30
Ala Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asn Ser Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ala Ser Leu
85 90 95
Ser Gly Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 92
<211> 651
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь N9 IgG
<400> 92
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgtactg gctcttcatt taatattggc agtaatgctg tctactggta ccagcagctc 120
ccaggaacgg cccccaaact cctcatctat tataatagtc agcggccaag cggggtccct 180
gaccgattct ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccgg 240
tccgaggatg aggctgatta ttactgtggc tcttgggatg ctagcctgag tggttatgtc 300
ttcggcggag gcaccaagct gacggtccta cgtacggtgg ctgcaccatc tgtcttcatc 360
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 420
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 480
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 540
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 600
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg t 651
<210> 93
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер scFv VH
<400> 93
agagagtgta cactcccagg cggccgaggt gcag 34
<210> 94
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер scFv VH
<400> 94
cgccgctggg cccttggtgg aggctgagct cacggtgacc ag 42
<210> 95
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер scFv VL
<400> 95
aagcggccgc caccatggga tggagctgta tcatcctctt cttggtagca acagctacag 60
gtgtacactc ccagtctgtg ctgactcag 89
<210> 96
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер scFv VL
<400> 96
cgccgccgta cgtaggaccg tcagcttggt 30
<210> 97
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Основной сайт связывания N3 Ab
<400> 97
Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala
1 5 10
<210> 98
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F1 N3
<400> 98
Met Ala Ala Val Gln Ala Ala Glu Val Lys Val Asp Gly Ser Glu Pro
1 5 10 15
Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala
20 25
<210> 99
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F2 N3
<400> 99
Gln Ala Ala Glu Val Lys Val Asp Gly Ser Glu Pro Lys Leu Ser Lys
1 5 10 15
Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys Val Ala
20 25 30
<210> 100
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F3 N3
<400> 100
Lys Val Asp Gly Ser Glu Pro Lys Leu Ser Lys Asn Glu Leu Lys Arg
1 5 10 15
Arg Leu Lys Ala Glu Lys Lys Val Ala Glu Lys Glu Ala
20 25
<210> 101
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F4 N3
<400> 101
Glu Pro Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu
1 5 10 15
Lys Lys Val Ala Glu Lys Glu Ala Lys Gln Lys Glu
20 25
<210> 102
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F5 N3
<400> 102
Lys Arg Arg Leu Lys Ala Glu Lys Lys Val Ala Glu Lys Glu Ala Lys
1 5 10 15
Gln Lys Glu Leu Ser Glu Lys Gln Leu Ser
20 25
<210> 103
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F1 мыши N3 (mF1)
<400> 103
Met Ala Thr Leu Gln Glu Ser Glu Val Lys Val Asp Gly Glu Gln Lys
1 5 10 15
Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala
20 25
<210> 104
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F2 мыши N3 (mF2)
<400> 104
Gln Glu Ser Glu Val Lys Val Asp Gly Glu Gln Lys Leu Ser Lys Asn
1 5 10 15
Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys Leu Ala
20 25
<210> 105
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F3 мыши N3 (mF3)
<400> 105
Lys Val Asp Gly Glu Gln Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg
1 5 10 15
Leu Lys Ala Glu Lys Lys Leu Ala Glu Lys Glu Ala
20 25
<210> 106
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F4 мыши N3 (mF4)
<400> 106
Gln Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys
1 5 10 15
Lys Leu Ala Glu Lys Glu Ala Lys Gln Lys Glu
20 25
<210> 107
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F5 мыши N3 (mF5)
<400> 107
Arg Arg Leu Lys Ala Glu Lys Lys Leu Ala Glu Lys Glu Ala Lys Gln
1 5 10 15
Lys Glu Leu Ser Glu Lys Gln Leu Asn
20 25
<210> 108
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F1 крысы N3 (rF1)
<400> 108
Met Ala Thr Leu Arg Glu Gly Glu Val Lys Leu Asp Gly Glu Pro Lys
1 5 10 15
Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala
20 25
<210> 109
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F2 крысы N3 (rF2)
<400> 109
Arg Glu Gly Glu Val Lys Leu Asp Gly Glu Pro Lys Leu Ser Lys Asn
1 5 10 15
Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys Leu Ala
20 25
<210> 110
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F3 крысы N3 (rF3)
<400> 110
Lys Leu Asp Gly Glu Pro Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg
1 5 10 15
Leu Lys Ala Glu Lys Lys Leu Ala Glu Lys Glu Ala
20 25
<210> 111
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F4 крысы N3 (rF4)
<400> 111
Pro Lys Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys
1 5 10 15
Lys Leu Ala Glu Lys Glu Ala Lys Gln Lys Glu
20 25
<210> 112
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Эпитоп F5 крысы N3 (rF5)
<400> 112
Arg Arg Leu Lys Ala Glu Lys Lys Leu Ala Glu Lys Glu Ala Lys Gln
1 5 10 15
Lys Glu Leu Ser Glu Lys Gln Leu Asn
20 25
<210> 113
<211> 48
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N-концевая лизил-т-РНК-синтетеза (мышь)
<400> 113
Met Ala Thr Leu Gln Glu Ser Glu Val Lys Val Asp Gly Glu Gln Lys
1 5 10 15
Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys Leu
20 25 30
Ala Glu Lys Glu Ala Lys Gln Lys Glu Leu Ser Glu Lys Gln Leu Asn
35 40 45
<210> 114
<211> 48
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> N-концевая лизил-тРНК-синтетаза (крыса)
<400> 114
Met Ala Thr Leu Arg Glu Gly Glu Val Lys Leu Asp Gly Glu Pro Lys
1 5 10 15
Leu Ser Lys Asn Glu Leu Lys Arg Arg Leu Lys Ala Glu Lys Lys Leu
20 25 30
Ala Glu Lys Glu Ala Lys Gln Lys Glu Leu Ser Glu Lys Gln Leu Asn
35 40 45
<210> 115
<211> 1823
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность подтипа Ламинина
альфа-4 (Человек разумный)
<400> 115
Met Ala Leu Ser Ser Ala Trp Arg Ser Val Leu Pro Leu Trp Leu Leu
1 5 10 15
Trp Ser Ala Ala Cys Ser Arg Ala Ala Ser Gly Asp Asp Asn Ala Phe
20 25 30
Pro Phe Asp Ile Glu Gly Ser Ser Ala Val Gly Arg Gln Asp Pro Pro
35 40 45
Glu Thr Ser Glu Pro Arg Val Ala Leu Gly Arg Leu Pro Pro Ala Ala
50 55 60
Glu Lys Cys Asn Ala Gly Phe Phe His Thr Leu Ser Gly Glu Cys Val
65 70 75 80
Pro Cys Asp Cys Asn Gly Asn Ser Asn Glu Cys Leu Asp Gly Ser Gly
85 90 95
Tyr Cys Val His Cys Gln Arg Asn Thr Thr Gly Glu His Cys Glu Lys
100 105 110
Cys Leu Asp Gly Tyr Ile Gly Asp Ser Ile Arg Gly Ala Pro Gln Phe
115 120 125
Cys Gln Pro Cys Pro Cys Pro Leu Pro His Leu Ala Asn Phe Ala Glu
130 135 140
Ser Cys Tyr Arg Lys Asn Gly Ala Val Arg Cys Ile Cys Asn Glu Asn
145 150 155 160
Tyr Ala Gly Pro Asn Cys Glu Arg Cys Ala Pro Gly Tyr Tyr Gly Asn
165 170 175
Pro Leu Leu Ile Gly Ser Thr Cys Lys Lys Cys Asp Cys Ser Gly Asn
180 185 190
Ser Asp Pro Asn Leu Ile Phe Glu Asp Cys Asp Glu Val Thr Gly Gln
195 200 205
Cys Arg Asn Cys Leu Arg Asn Thr Thr Gly Phe Lys Cys Glu Arg Cys
210 215 220
Ala Pro Gly Tyr Tyr Gly Asp Ala Arg Ile Ala Lys Asn Cys Ala Val
225 230 235 240
Cys Asn Cys Gly Gly Gly Pro Cys Asp Ser Val Thr Gly Glu Cys Leu
245 250 255
Glu Glu Gly Phe Glu Pro Pro Thr Gly Met Asp Cys Pro Thr Ile Ser
260 265 270
Cys Asp Lys Cys Val Trp Asp Leu Thr Asp Ala Leu Arg Leu Ala Ala
275 280 285
Leu Ser Ile Glu Glu Gly Lys Ser Gly Val Leu Ser Val Ser Ser Gly
290 295 300
Ala Ala Ala His Arg His Val Asn Glu Ile Asn Ala Thr Ile Tyr Leu
305 310 315 320
Leu Lys Thr Lys Leu Ser Glu Arg Glu Asn Gln Tyr Ala Leu Arg Lys
325 330 335
Ile Gln Ile Asn Asn Ala Glu Asn Thr Met Lys Ser Leu Leu Ser Asp
340 345 350
Val Glu Glu Leu Val Glu Lys Glu Asn Gln Ala Ser Arg Lys Gly Gln
355 360 365
Leu Val Gln Lys Glu Ser Met Asp Thr Ile Asn His Ala Ser Gln Leu
370 375 380
Val Glu Gln Ala His Asp Met Arg Asp Lys Ile Gln Glu Ile Asn Asn
385 390 395 400
Lys Met Leu Tyr Tyr Gly Glu Glu His Glu Leu Ser Pro Lys Glu Ile
405 410 415
Ser Glu Lys Leu Val Leu Ala Gln Lys Met Leu Glu Glu Ile Arg Ser
420 425 430
Arg Gln Pro Phe Phe Thr Gln Arg Glu Leu Val Asp Glu Glu Ala Asp
435 440 445
Glu Ala Tyr Glu Leu Leu Ser Gln Ala Glu Ser Trp Gln Arg Leu His
450 455 460
Asn Glu Thr Arg Thr Leu Phe Pro Val Val Leu Glu Gln Leu Asp Asp
465 470 475 480
Tyr Asn Ala Lys Leu Ser Asp Leu Gln Glu Ala Leu Asp Gln Ala Leu
485 490 495
Asn Tyr Val Arg Asp Ala Glu Asp Met Asn Arg Ala Thr Ala Ala Arg
500 505 510
Gln Arg Asp His Glu Lys Gln Gln Glu Arg Val Arg Glu Gln Met Glu
515 520 525
Val Val Asn Met Ser Leu Ser Thr Ser Ala Asp Ser Leu Thr Thr Pro
530 535 540
Arg Leu Thr Leu Ser Glu Leu Asp Asp Ile Ile Lys Asn Ala Ser Gly
545 550 555 560
Ile Tyr Ala Glu Ile Asp Gly Ala Lys Ser Glu Leu Gln Val Lys Leu
565 570 575
Ser Asn Leu Ser Asn Leu Ser His Asp Leu Val Gln Glu Ala Ile Asp
580 585 590
His Ala Gln Asp Leu Gln Gln Glu Ala Asn Glu Leu Ser Arg Lys Leu
595 600 605
His Ser Ser Asp Met Asn Gly Leu Val Gln Lys Ala Leu Asp Ala Ser
610 615 620
Asn Val Tyr Glu Asn Ile Val Asn Tyr Val Ser Glu Ala Asn Glu Thr
625 630 635 640
Ala Glu Phe Ala Leu Asn Thr Thr Asp Arg Ile Tyr Asp Ala Val Ser
645 650 655
Gly Ile Asp Thr Gln Ile Ile Tyr His Lys Asp Glu Ser Glu Asn Leu
660 665 670
Leu Asn Gln Ala Arg Glu Leu Gln Ala Lys Ala Glu Ser Ser Ser Asp
675 680 685
Glu Ala Val Ala Asp Thr Ser Arg Arg Val Gly Gly Ala Leu Ala Arg
690 695 700
Lys Ser Ala Leu Lys Thr Arg Leu Ser Asp Ala Val Lys Gln Leu Gln
705 710 715 720
Ala Ala Glu Arg Gly Asp Ala Gln Gln Arg Leu Gly Gln Ser Arg Leu
725 730 735
Ile Thr Glu Glu Ala Asn Arg Thr Thr Met Glu Val Gln Gln Ala Thr
740 745 750
Ala Pro Met Ala Asn Asn Leu Thr Asn Trp Ser Gln Asn Leu Gln His
755 760 765
Phe Asp Ser Ser Ala Tyr Asn Thr Ala Val Asn Ser Ala Arg Asp Ala
770 775 780
Val Arg Asn Leu Thr Glu Val Val Pro Gln Leu Leu Asp Gln Leu Arg
785 790 795 800
Thr Val Glu Gln Lys Arg Pro Ala Ser Asn Val Ser Ala Ser Ile Gln
805 810 815
Arg Ile Arg Glu Leu Ile Ala Gln Thr Arg Ser Val Ala Ser Lys Ile
820 825 830
Gln Val Ser Met Met Phe Asp Gly Gln Ser Ala Val Glu Val His Ser
835 840 845
Arg Thr Ser Met Asp Asp Leu Lys Ala Phe Thr Ser Leu Ser Leu Tyr
850 855 860
Met Lys Pro Pro Val Lys Arg Pro Glu Leu Thr Glu Thr Ala Asp Gln
865 870 875 880
Phe Ile Leu Tyr Leu Gly Ser Lys Asn Ala Lys Lys Glu Tyr Met Gly
885 890 895
Leu Ala Ile Lys Asn Asp Asn Leu Val Tyr Val Tyr Asn Leu Gly Thr
900 905 910
Lys Asp Val Glu Ile Pro Leu Asp Ser Lys Pro Val Ser Ser Trp Pro
915 920 925
Ala Tyr Phe Ser Ile Val Lys Ile Glu Arg Val Gly Lys His Gly Lys
930 935 940
Val Phe Leu Thr Val Pro Ser Leu Ser Ser Thr Ala Glu Glu Lys Phe
945 950 955 960
Ile Lys Lys Gly Glu Phe Ser Gly Asp Asp Ser Leu Leu Asp Leu Asp
965 970 975
Pro Glu Asp Thr Val Phe Tyr Val Gly Gly Val Pro Ser Asn Phe Lys
980 985 990
Leu Pro Thr Ser Leu Asn Leu Pro Gly Phe Val Gly Cys Leu Glu Leu
995 1000 1005
Ala Thr Leu Asn Asn Asp Val Ile Ser Leu Tyr Asn Phe Lys His Ile
1010 1015 1020
Tyr Asn Met Asp Pro Ser Thr Ser Val Pro Cys Ala Arg Asp Lys Leu
1025 1030 1035 1040
Ala Phe Thr Gln Ser Arg Ala Ala Ser Tyr Phe Phe Asp Gly Ser Gly
1045 1050 1055
Tyr Ala Val Val Arg Asp Ile Thr Arg Arg Gly Lys Phe Gly Gln Val
1060 1065 1070
Thr Arg Phe Asp Ile Glu Val Arg Thr Pro Ala Asp Asn Gly Leu Ile
1075 1080 1085
Leu Leu Met Val Asn Gly Ser Met Phe Phe Arg Leu Glu Met Arg Asn
1090 1095 1100
Gly Tyr Leu His Val Phe Tyr Asp Phe Gly Phe Ser Gly Gly Pro Val
1105 1110 1115 1120
His Leu Glu Asp Thr Leu Lys Lys Ala Gln Ile Asn Asp Ala Lys Tyr
1125 1130 1135
His Glu Ile Ser Ile Ile Tyr His Asn Asp Lys Lys Met Ile Leu Val
1140 1145 1150
Val Asp Arg Arg His Val Lys Ser Met Asp Asn Glu Lys Met Lys Ile
1155 1160 1165
Pro Phe Thr Asp Ile Tyr Ile Gly Gly Ala Pro Pro Glu Ile Leu Gln
1170 1175 1180
Ser Arg Ala Leu Arg Ala His Leu Pro Leu Asp Ile Asn Phe Arg Gly
1185 1190 1195 1200
Cys Met Lys Gly Phe Gln Phe Gln Lys Lys Asp Phe Asn Leu Leu Glu
1205 1210 1215
Gln Thr Glu Thr Leu Gly Val Gly Tyr Gly Cys Pro Glu Asp Ser Leu
1220 1225 1230
Ile Ser Arg Arg Ala Tyr Phe Asn Gly Gln Ser Phe Ile Ala Ser Ile
1235 1240 1245
Gln Lys Ile Ser Phe Phe Asp Gly Phe Glu Gly Gly Phe Asn Phe Arg
1250 1255 1260
Thr Leu Gln Pro Asn Gly Leu Leu Phe Tyr Tyr Ala Ser Gly Ser Asp
1265 1270 1275 1280
Val Phe Ser Ile Ser Leu Asp Asn Gly Thr Val Ile Met Asp Val Lys
1285 1290 1295
Gly Ile Lys Val Gln Ser Val Asp Lys Gln Tyr Asn Asp Gly Leu Ser
1300 1305 1310
His Phe Val Ile Ser Ser Val Ser Pro Thr Arg Tyr Glu Leu Ile Val
1315 1320 1325
Asp Lys Ser Arg Val Gly Ser Lys Asn Pro Thr Lys Gly Lys Ile Glu
1330 1335 1340
Gln Thr Gln Ala Ser Glu Lys Lys Phe Tyr Phe Gly Gly Ser Pro Ile
1345 1350 1355 1360
Ser Ala Gln Tyr Ala Asn Phe Thr Gly Cys Ile Ser Asn Ala Tyr Phe
1365 1370 1375
Thr Arg Val Asp Arg Asp Val Glu Val Glu Asp Phe Gln Arg Tyr Thr
1380 1385 1390
Glu Lys Val His Thr Ser Leu Tyr Glu Cys Pro Ile Glu Ser Ser Pro
1395 1400 1405
Leu Phe Leu Leu His Lys Lys Gly Lys Asn Leu Ser Lys Pro Lys Ala
1410 1415 1420
Ser Gln Asn Lys Lys Gly Gly Lys Ser Lys Asp Ala Pro Ser Trp Asp
1425 1430 1435 1440
Pro Val Ala Leu Lys Leu Pro Glu Arg Asn Thr Pro Arg Asn Ser His
1445 1450 1455
Cys His Leu Ser Asn Ser Pro Arg Ala Ile Glu His Ala Tyr Gln Tyr
1460 1465 1470
Gly Gly Thr Ala Asn Ser Arg Gln Glu Phe Glu His Leu Lys Gly Asp
1475 1480 1485
Phe Gly Ala Lys Ser Gln Phe Ser Ile Arg Leu Arg Thr Arg Ser Ser
1490 1495 1500
His Gly Met Ile Phe Tyr Val Ser Asp Gln Glu Glu Asn Asp Phe Met
1505 1510 1515 1520
Thr Leu Phe Leu Ala His Gly Arg Leu Val Tyr Met Phe Asn Val Gly
1525 1530 1535
His Lys Lys Leu Lys Ile Arg Ser Gln Glu Lys Tyr Asn Asp Gly Leu
1540 1545 1550
Trp His Asp Val Ile Phe Ile Arg Glu Arg Ser Ser Gly Arg Leu Val
1555 1560 1565
Ile Asp Gly Leu Arg Val Leu Glu Glu Ser Leu Pro Pro Thr Glu Ala
1570 1575 1580
Thr Trp Lys Ile Lys Gly Pro Ile Tyr Leu Gly Gly Val Ala Pro Gly
1585 1590 1595 1600
Lys Ala Val Lys Asn Val Gln Ile Asn Ser Ile Tyr Ser Phe Ser Gly
1605 1610 1615
Cys Leu Ser Asn Leu Gln Leu Asn Gly Ala Ser Ile Thr Ser Ala Ser
1620 1625 1630
Gln Thr Phe Ser Val Thr Pro Cys Phe Glu Gly Pro Met Glu Thr Gly
1635 1640 1645
Thr Tyr Phe Ser Thr Glu Gly Gly Tyr Val Val Leu Asp Glu Ser Phe
1650 1655 1660
Asn Ile Gly Leu Lys Phe Glu Ile Ala Phe Glu Val Arg Pro Arg Ser
1665 1670 1675 1680
Ser Ser Gly Thr Leu Val His Gly His Ser Val Asn Gly Glu Tyr Leu
1685 1690 1695
Asn Val His Met Lys Asn Gly Gln Val Ile Val Lys Val Asn Asn Gly
1700 1705 1710
Ile Arg Asp Phe Ser Thr Ser Val Thr Pro Lys Gln Ser Leu Cys Asp
1715 1720 1725
Gly Arg Trp His Arg Ile Thr Val Ile Arg Asp Ser Asn Val Val Gln
1730 1735 1740
Leu Asp Val Asp Ser Glu Val Asn His Val Val Gly Pro Leu Asn Pro
1745 1750 1755 1760
Lys Pro Ile Asp His Arg Glu Pro Val Phe Val Gly Gly Val Pro Glu
1765 1770 1775
Ser Leu Leu Thr Pro Arg Leu Ala Pro Ser Lys Pro Phe Thr Gly Cys
1780 1785 1790
Ile Arg His Phe Val Ile Asp Gly His Pro Val Ser Phe Ser Lys Ala
1795 1800 1805
Ala Leu Val Ser Gly Ala Val Ser Ile Asn Ser Cys Pro Ala Ala
1810 1815 1820
<210> 116
<211> 5472
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК последовательность подтипа Ламинина альфа-4
(Человека разумный)
<400> 116
atggctttga gctcagcctg gcgctcggtt ctgcctctgt ggctcctctg gagcgctgcc 60
tgctcccgcg ccgcgtccgg ggacgacaac gcttttcctt ttgacattga agggagctca 120
gcggttggca ggcaagaccc gcctgagacg agcgaacccc gcgtggctct gggacgcctg 180
ccgcctgcgg ccgagaaatg caatgctgga ttctttcaca ccctgtcggg agaatgtgtg 240
ccctgcgact gtaatggcaa ttccaacgag tgtttggacg gctcaggata ctgtgtgcac 300
tgccagcgga acacaacagg agagcactgt gaaaagtgtc tggatggtta tatcggagat 360
tccatcaggg gagcacccca attctgccag ccgtgcccct gtcccctgcc ccacttggcc 420
aattttgcag aatcctgcta taggaaaaat ggagctgttc ggtgcatttg taacgaaaat 480
tatgctggac ctaactgtga aagatgtgct cccggttact atggaaaccc cttactcatt 540
ggaagcacct gtaagaaatg tgactgcagt ggaaattcag atcccaacct gatctttgaa 600
gattgtgatg aagtcactgg ccagtgtagg aattgcttac gcaacaccac cggattcaag 660
tgtgaacgtt gcgctcctgg ctactatggg gacgccagga tagccaagaa ctgtgcagtg 720
tgcaactgcg ggggaggccc atgtgacagt gtaaccggag aatgcttgga agaaggtttt 780
gaacccccta caggcatgga ctgcccaacc ataagctgtg ataagtgcgt ctgggacctg 840
actgatgacc tgcggttagc agcgctctcc atcgaggaag gcaaatccgg ggtgctgagc 900
gtatcctctg gggccgccgc tcataggcac gtgaatgaaa tcaacgccac catctacctc 960
ctcaaaacaa aattgtcaga aagagaaaac caatacgccc taagaaagat acaaatcaac 1020
aatgctgaga acacgatgaa aagccttctg tctgacgtag aggaattagt tgaaaaggaa 1080
aatcaagcct ccagaaaagg acaacttgtt cagaaggaaa gcatggacac cattaaccac 1140
gcaagtcagc tggtagagca agcccatgat atgagggata aaatccaaga gatcaacaac 1200
aagatgctct attatgggga agagcatgaa cttagcccca aggaaatctc tgagaagctg 1260
gtgttggccc agaagatgct tgaagagatt agaagccgtc aaccattttt cacccaacgg 1320
gagctcgtgg atgaggaggc agatgaggct tacgaactac tgagccaggc tgagagctgg 1380
cagcggctgc acaatgagac ccgcactctg tttcctgtcg tcctggagca gctggatgac 1440
tacaatgcta agttgtcaga tctccaggaa gcacttgacc aggcccttaa ctatgtcagg 1500
gatgccgaag acatgaacag ggccacagca gccaggcagc gggaccatga gaaacaacag 1560
gaaagagtga gggaacaaat ggaagtggtg aacatgtctc tgagcacatc tgcggactct 1620
ctgacaacac ctcgtctaac tctttcagaa cttgatgata taataaagaa tgcgtcaggg 1680
atttatgcag aaatagatgg agccaaaagt gaactacaag taaaactatc taacctaagt 1740
aacctcagcc atgatttagt ccaagaagct attgaccatg cacaggacct tcaacaagaa 1800
gctaatgaat tgagcaggaa gttgcacagt tcagatatga acgggctggt acagaaggct 1860
ttggatgcat caaatgtcta tgaaaatatt gttaattatg ttagtgaagc caatgaaaca 1920
gcagaatttg ctttgaacac cactgaccga atttatgatg cggtgagtgg gattgatact 1980
caaatcattt accataaaga tgaaagtgag aacctcctca atcaagccag agaactgcaa 2040
gcaaaggcag agtctagcag tgatgaagca gtggctgaca ctagcaggcg tgtgggtgga 2100
gccctagcaa ggaaaagtgc ccttaaaacc agactcagtg atgccgttaa gcaactacaa 2160
gcagcagaga gaggggatgc ccagcagcgc ctggggcagt ctagactgat caccgaggaa 2220
gccaacagga cgacgatgga ggtgcagcag gccactgccc ccatggccaa caatctaacc 2280
aactggtcac agaatcttca acattttgac tcttctgctt acaacactgc agtgaactct 2340
gctagggatg cagtaagaaa tctgaccgag gttgtccctc agctcctgga tcagcttcgt 2400
acggttgagc agaagcgacc tgcaagcaac gtttctgcca gcatccagag gatccgagag 2460
ctcattgctc agaccagaag tgttgccagc aagatccaag tctccatgat gtttgatggc 2520
cagtcagctg tggaagtgca ctcgagaacc agtatggatg acttaaaggc cttcacgtct 2580
ctgagcctgt acatgaaacc ccctgtgaag cggccggaac tgaccgagac tgcagatcag 2640
tttatcctgt acctcggaag caaaaacgcc aaaaaagagt atatgggtct tgcaatcaaa 2700
aatgataatc tggtatacgt ctataatttg ggaactaaag atgtggagat tcccctggac 2760
tccaagcccg tcagttcctg gcctgcttac ttcagcattg tcaagattga aagggtggga 2820
aaacatggaa aggtgttttt aacagtcccg agtctaagta gcacagcaga ggaaaagttc 2880
attaaaaagg gggaattttc gggagatgac tctctgctgg acctggaccc tgaggacaca 2940
gtgttttatg ttggtggagt gccttccaac ttcaagctcc ctaccagctt aaacctgcct 3000
ggctttgttg gctgcctgga actggccact ttgaataatg atgtgatcag cttgtacaac 3060
tttaagcaca tctataatat ggacccctcc acatcagtgc catgtgcccg agataagctg 3120
gccttcactc agagtcgggc tgccagttac ttcttcgatg gctccggtta tgccgtggtg 3180
agagacatca caaggagagg gaaatttggt caggtgactc gctttgacat agaagttcga 3240
acaccagctg acaacggcct tattctcctg atggtcaatg gaagtatgtt tttcagactg 3300
gaaatgcgca atggttacct acatgtgttc tatgattttg gattcagcgg tggccctgtg 3360
catcttgaag atacgttaaa gaaagctcaa attaatgatg caaaatacca tgagatctca 3420
atcatttacc acaatgataa gaaaatgatc ttggtagttg acagaaggca tgtcaagagc 3480
atggataatg aaaagatgaa aatacctttt acagatatat acattggagg agctcctcca 3540
gaaatcttac aatccagggc cctcagagca caccttcccc tagatatcaa cttcagagga 3600
tgcatgaagg gcttccagtt ccaaaagaag gacttcaatt tactggagca gacagaaacc 3660
ctgggagttg gttatggatg cccagaagac tcacttatat ctcgcagagc atatttcaat 3720
ggacagagct tcattgcttc aattcagaaa atatctttct ttgatggctt tgaaggaggt 3780
tttaatttcc gaacattaca accaaatggg ttactattct attatgcttc agggtcagac 3840
gtgttctcca tctcactgga taatggtact gtcatcatgg atgtaaaggg aatcaaagtt 3900
cagtcagtag ataagcagta caatgatggg ctgtcccact tcgtcattag ctctgtctca 3960
cccacaagat atgaactgat agtagataaa agcagagttg ggagtaagaa tcctaccaaa 4020
gggaaaatag aacagacaca agcaagtgaa aagaagtttt acttcggtgg ctcaccaatc 4080
agtgctcagt atgctaattt cactggctgc ataagtaatg cctactttac cagggtggat 4140
agagatgtgg aggttgaaga tttccaacgg tatactgaaa aggtccacac ttctctttat 4200
gagtgtccca ttgagtcttc accattgttt ctcctccata aaaaaggaaa aaatttatcc 4260
aagcctaaag caagtcagaa taaaaaggga gggaaaagta aagatgcacc ttcatgggat 4320
cctgttgctc tgaaactccc agagcggaat actccaagaa actctcattg ccacctttcc 4380
aacagcccta gagcaataga gcacgcctat caatatggag gaacagccaa cagccgccaa 4440
gagtttgaac acttaaaagg agattttggt gccaaatctc agttttccat tcgtctgaga 4500
actcgttcct cccatggcat gatcttctat gtctcagatc aagaagagaa tgacttcatg 4560
actctatttt tggcccatgg ccgcttggtt tacatgttta atgttggtca caaaaaactg 4620
aagattagaa gccaggagaa atacaatgat ggcctgtggc atgatgtgat atttattcga 4680
gaaaggagca gtggccgact ggtaattgat ggtctccgag tcctagaaga aagtcttcct 4740
cctactgaag ctacctggaa aatcaagggt cccatttatt tgggaggtgt ggctcctgga 4800
aaggctgtga aaaatgttca gattaactcc atctacagtt ttagtggctg tctcagcaat 4860
ctccagctca atggggcctc catcacctct gcttctcaga cattcagtgt gaccccttgc 4920
tttgaaggcc ccatggaaac aggaacttac ttttcaacag aaggaggata cgtggttcta 4980
gatgaatctt tcaatattgg attgaagttt gaaattgcat ttgaagtccg tcccagaagc 5040
agttccggaa ccctggtcca cggccacagt gtcaatgggg agtacctaaa tgttcacatg 5100
aaaaatggac aggtcatagt gaaagtcaat aatggcatca gagatttttc cacctcagtt 5160
acacccaagc agagtctctg tgatggcaga tggcacagaa ttacagttat tagagattct 5220
aatgtggttc agttggatgt ggactctgaa gtgaaccatg tggttggacc cctgaatcca 5280
aaaccaattg atcacaggga gcctgtgttt gttggaggtg ttccagaatc tctactgaca 5340
ccacgcttgg cccccagcaa acccttcaca ggctgcatac gccactttgt gattgatgga 5400
cacccagtga gcttcagtaa agcagccctg gtcagcggcg ccgtaagcat caactcctgt 5460
ccagcagcct ga 5472
<210> 117
<211> 1798
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность подтипа Ламинин
бета-2 (Человек разумный)
<400> 117
Met Glu Leu Thr Ser Arg Glu Arg Gly Arg Gly Gln Pro Leu Pro Trp
1 5 10 15
Glu Leu Arg Leu Gly Leu Leu Leu Ser Val Leu Ala Ala Thr Leu Ala
20 25 30
Gln Ala Pro Ala Pro Asp Val Pro Gly Cys Ser Arg Gly Ser Cys Tyr
35 40 45
Pro Ala Thr Gly Asp Leu Leu Val Gly Arg Ala Asp Arg Leu Thr Ala
50 55 60
Ser Ser Thr Cys Gly Leu Asn Gly Pro Gln Pro Tyr Cys Ile Val Ser
65 70 75 80
His Leu Gln Asp Glu Lys Lys Cys Phe Leu Cys Asp Ser Arg Arg Pro
85 90 95
Phe Ser Ala Arg Asp Asn Pro His Ser His Arg Ile Gln Asn Val Val
100 105 110
Thr Ser Phe Ala Pro Gln Arg Arg Ala Ala Trp Trp Gln Ser Glu Asn
115 120 125
Gly Ile Pro Ala Val Thr Ile Gln Leu Asp Leu Glu Ala Glu Phe His
130 135 140
Phe Thr His Leu Ile Met Thr Phe Lys Thr Phe Arg Pro Ala Ala Met
145 150 155 160
Leu Val Glu Arg Ser Ala Asp Phe Gly Arg Thr Trp His Val Tyr Arg
165 170 175
Tyr Phe Ser Tyr Asp Cys Gly Ala Asp Phe Pro Gly Val Pro Leu Ala
180 185 190
Pro Pro Arg His Trp Asp Asp Val Val Cys Glu Ser Arg Tyr Ser Glu
195 200 205
Ile Glu Pro Ser Thr Glu Gly Glu Val Ile Tyr Arg Val Leu Asp Pro
210 215 220
Ala Ile Pro Ile Pro Asp Pro Tyr Ser Ser Arg Ile Gln Asn Leu Leu
225 230 235 240
Lys Ile Thr Asn Leu Arg Val Asn Leu Thr Arg Leu His Thr Leu Gly
245 250 255
Asp Asn Leu Leu Asp Pro Arg Arg Glu Ile Arg Glu Lys Tyr Tyr Tyr
260 265 270
Ala Leu Tyr Glu Leu Val Val Arg Gly Asn Cys Phe Cys Tyr Gly His
275 280 285
Ala Ser Glu Cys Ala Pro Ala Pro Gly Ala Pro Ala His Ala Glu Gly
290 295 300
Met Val His Gly Ala Cys Ile Cys Lys His Asn Thr Arg Gly Leu Asn
305 310 315 320
Cys Glu Gln Cys Gln Asp Phe Tyr Arg Asp Leu Pro Trp Arg Pro Ala
325 330 335
Glu Asp Gly His Ser His Ala Cys Arg Lys Cys Glu Cys His Gly His
340 345 350
Thr His Ser Cys His Phe Asp Met Ala Val Tyr Leu Ala Ser Gly Asn
355 360 365
Val Ser Gly Gly Val Cys Asp Gly Cys Gln His Asn Thr Ala Gly Arg
370 375 380
His Cys Glu Leu Cys Arg Pro Phe Phe Tyr Arg Asp Pro Thr Lys Asp
385 390 395 400
Leu Arg Asp Pro Ala Val Cys Arg Ser Cys Asp Cys Asp Pro Met Gly
405 410 415
Ser Gln Asp Gly Gly Arg Cys Asp Ser His Asp Asp Pro Ala Leu Gly
420 425 430
Leu Val Ser Gly Gln Cys Arg Cys Lys Glu His Val Val Gly Thr Arg
435 440 445
Cys Gln Gln Cys Arg Asp Gly Phe Phe Gly Leu Ser Ile Ser Asp Arg
450 455 460
Leu Gly Cys Arg Arg Cys Gln Cys Asn Ala Arg Gly Thr Val Pro Gly
465 470 475 480
Ser Thr Pro Cys Asp Pro Asn Ser Gly Ser Cys Tyr Cys Lys Arg Leu
485 490 495
Val Thr Gly Arg Gly Cys Asp Arg Cys Leu Pro Gly His Trp Gly Leu
500 505 510
Ser His Asp Leu Leu Gly Cys Arg Pro Cys Asp Cys Asp Val Gly Gly
515 520 525
Ala Leu Asp Pro Gln Cys Asp Glu Gly Thr Gly Gln Cys His Cys Arg
530 535 540
Gln His Met Val Gly Arg Arg Cys Glu Gln Val Gln Pro Gly Tyr Phe
545 550 555 560
Arg Pro Phe Leu Asp His Leu Ile Trp Glu Ala Glu Asp Thr Arg Gly
565 570 575
Gln Val Leu Asp Val Val Glu Arg Leu Val Thr Pro Gly Glu Thr Pro
580 585 590
Ser Trp Thr Gly Ser Gly Phe Val Arg Leu Gln Glu Gly Gln Thr Leu
595 600 605
Glu Phe Leu Val Ala Ser Val Pro Lys Ala Met Asp Tyr Asp Leu Leu
610 615 620
Leu Arg Leu Glu Pro Gln Val Pro Glu Gln Trp Ala Glu Leu Glu Leu
625 630 635 640
Ile Val Gln Arg Pro Gly Pro Val Pro Ala His Ser Leu Cys Gly His
645 650 655
Leu Val Pro Lys Asp Asp Arg Ile Gln Gly Thr Leu Gln Pro His Ala
660 665 670
Arg Tyr Leu Ile Phe Pro Asn Pro Val Cys Leu Glu Pro Gly Ile Ser
675 680 685
Tyr Lys Leu His Leu Lys Leu Val Arg Thr Gly Gly Ser Ala Gln Pro
690 695 700
Glu Thr Pro Tyr Ser Gly Pro Gly Leu Leu Ile Asp Ser Leu Val Leu
705 710 715 720
Leu Pro Arg Val Leu Val Leu Glu Met Phe Ser Gly Gly Asp Ala Ala
725 730 735
Ala Leu Glu Arg Gln Ala Thr Phe Glu Arg Tyr Gln Cys His Glu Glu
740 745 750
Gly Leu Val Pro Ser Lys Thr Ser Pro Ser Glu Ala Cys Ala Pro Leu
755 760 765
Leu Ile Ser Leu Ser Thr Leu Ile Tyr Asn Gly Ala Leu Pro Cys Gln
770 775 780
Cys Asn Pro Gln Gly Ser Leu Ser Ser Glu Cys Asn Pro His Gly Gly
785 790 795 800
Gln Cys Leu Cys Lys Pro Gly Val Val Gly Arg Arg Cys Asp Leu Cys
805 810 815
Ala Pro Gly Tyr Tyr Gly Phe Gly Pro Thr Gly Cys Gln Ala Cys Gln
820 825 830
Cys Ser His Glu Gly Ala Leu Ser Ser Leu Cys Glu Lys Thr Ser Gly
835 840 845
Gln Cys Leu Cys Arg Thr Gly Ala Phe Gly Leu Arg Cys Asp Arg Cys
850 855 860
Gln Arg Gly Gln Trp Gly Phe Pro Ser Cys Arg Pro Cys Val Cys Asn
865 870 875 880
Gly His Ala Asp Glu Cys Asn Thr His Thr Gly Ala Cys Leu Gly Cys
885 890 895
Arg Asp His Thr Gly Gly Glu His Cys Glu Arg Cys Ile Ala Gly Phe
900 905 910
His Gly Asp Pro Arg Leu Pro Tyr Gly Gly Gln Cys Arg Pro Cys Pro
915 920 925
Cys Pro Glu Gly Pro Gly Ser Gln Arg His Phe Ala Thr Ser Cys His
930 935 940
Gln Asp Glu Tyr Ser Gln Gln Ile Val Cys His Cys Arg Ala Gly Tyr
945 950 955 960
Thr Gly Leu Arg Cys Glu Ala Cys Ala Pro Gly His Phe Gly Asp Pro
965 970 975
Ser Arg Pro Gly Gly Arg Cys Gln Leu Cys Glu Cys Ser Gly Asn Ile
980 985 990
Asp Pro Met Asp Pro Asp Ala Cys Asp Pro His Thr Gly Gln Cys Leu
995 1000 1005
Arg Cys Leu His His Thr Glu Gly Pro His Cys Ala His Cys Lys Pro
1010 1015 1020
Gly Phe His Gly Gln Ala Ala Arg Gln Ser Cys His Arg Cys Thr Cys
1025 1030 1035 1040
Asn Leu Leu Gly Thr Asn Pro Gln Gln Cys Pro Ser Pro Asp Gln Cys
1045 1050 1055
His Cys Asp Pro Ser Ser Gly Gln Cys Pro Cys Leu Pro Asn Val Gln
1060 1065 1070
Gly Pro Ser Cys Asp Arg Cys Ala Pro Asn Phe Trp Asn Leu Thr Ser
1075 1080 1085
Gly His Gly Cys Gln Pro Cys Ala Cys His Pro Ser Arg Ala Arg Gly
1090 1095 1100
Pro Thr Cys Asn Glu Phe Thr Gly Gln Cys His Cys Arg Ala Gly Phe
1105 1110 1115 1120
Gly Gly Arg Thr Cys Ser Glu Cys Gln Glu Leu His Trp Gly Asp Pro
1125 1130 1135
Gly Leu Gln Cys His Ala Cys Asp Cys Asp Ser Arg Gly Ile Asp Thr
1140 1145 1150
Pro Gln Cys His Arg Phe Thr Gly His Cys Ser Cys Arg Pro Gly Val
1155 1160 1165
Ser Gly Val Arg Cys Asp Gln Cys Ala Arg Gly Phe Ser Gly Ile Phe
1170 1175 1180
Pro Ala Cys His Pro Cys His Ala Cys Phe Gly Asp Trp Asp Arg Val
1185 1190 1195 1200
Val Gln Asp Leu Ala Ala Arg Thr Gln Arg Leu Glu Gln Arg Ala Gln
1205 1210 1215
Glu Leu Gln Gln Thr Gly Val Leu Gly Ala Phe Glu Ser Ser Phe Trp
1220 1225 1230
His Met Gln Glu Lys Leu Gly Ile Val Gln Gly Ile Val Gly Ala Arg
1235 1240 1245
Asn Thr Ser Ala Ala Ser Thr Ala Gln Leu Val Glu Ala Thr Glu Glu
1250 1255 1260
Leu Arg Arg Glu Ile Gly Glu Ala Thr Glu His Leu Thr Gln Leu Glu
1265 1270 1275 1280
Ala Asp Leu Thr Asp Val Gln Asp Glu Asn Phe Asn Ala Asn His Ala
1285 1290 1295
Leu Ser Gly Leu Glu Arg Asp Arg Leu Ala Leu Asn Leu Thr Leu Arg
1300 1305 1310
Gln Leu Asp Gln His Leu Asp Leu Leu Lys His Ser Asn Phe Leu Gly
1315 1320 1325
Ala Tyr Asp Ser Ile Arg His Ala His Ser Gln Ser Ala Glu Ala Glu
1330 1335 1340
Arg Arg Ala Asn Thr Ser Ala Leu Ala Val Pro Ser Pro Val Ser Asn
1345 1350 1355 1360
Ser Ala Ser Ala Arg His Arg Thr Glu Ala Leu Met Asp Ala Gln Lys
1365 1370 1375
Glu Asp Phe Asn Ser Lys His Met Ala Asn Gln Arg Ala Leu Gly Lys
1380 1385 1390
Leu Ser Ala His Thr His Thr Leu Ser Leu Thr Asp Ile Asn Glu Leu
1395 1400 1405
Val Cys Gly Ala Pro Gly Asp Ala Pro Cys Ala Thr Ser Pro Cys Gly
1410 1415 1420
Gly Ala Gly Cys Arg Asp Glu Asp Gly Gln Pro Arg Cys Gly Gly Leu
1425 1430 1435 1440
Ser Cys Asn Gly Ala Ala Ala Thr Ala Asp Leu Ala Leu Gly Arg Ala
1445 1450 1455
Arg His Thr Gln Ala Glu Leu Gln Arg Ala Leu Ala Glu Gly Gly Ser
1460 1465 1470
Ile Leu Ser Arg Val Ala Glu Thr Arg Arg Gln Ala Ser Glu Ala Gln
1475 1480 1485
Gln Arg Ala Gln Ala Ala Leu Asp Lys Ala Asn Ala Ser Arg Gly Gln
1490 1495 1500
Val Glu Gln Ala Asn Gln Glu Leu Gln Glu Leu Ile Gln Ser Val Lys
1505 1510 1515 1520
Asp Phe Leu Asn Gln Glu Gly Ala Asp Pro Asp Ser Ile Glu Met Val
1525 1530 1535
Ala Thr Arg Val Leu Glu Leu Ser Ile Pro Ala Ser Ala Glu Gln Ile
1540 1545 1550
Gln His Leu Ala Gly Ala Ile Ala Glu Arg Val Arg Ser Leu Ala Asp
1555 1560 1565
Val Asp Ala Ile Leu Ala Arg Thr Val Gly Asp Val Arg Arg Ala Glu
1570 1575 1580
Gln Leu Leu Gln Asp Ala Arg Arg Ala Arg Ser Trp Ala Glu Asp Glu
1585 1590 1595 1600
Lys Gln Lys Ala Glu Thr Val Gln Ala Ala Leu Glu Glu Ala Gln Arg
1605 1610 1615
Ala Gln Gly Ile Ala Gln Gly Ala Ile Arg Gly Ala Val Ala Asp Thr
1620 1625 1630
Arg Asp Thr Glu Gln Thr Leu Tyr Gln Val Gln Glu Arg Met Ala Gly
1635 1640 1645
Ala Glu Arg Ala Leu Ser Ser Ala Gly Glu Arg Ala Arg Gln Leu Asp
1650 1655 1660
Ala Leu Leu Glu Ala Leu Lys Leu Lys Arg Ala Gly Asn Ser Leu Ala
1665 1670 1675 1680
Ala Ser Thr Ala Glu Glu Thr Ala Gly Ser Ala Gln Gly Arg Ala Gln
1685 1690 1695
Glu Ala Glu Gln Leu Leu Arg Gly Pro Leu Gly Asp Gln Tyr Gln Thr
1700 1705 1710
Val Lys Ala Leu Ala Glu Arg Lys Ala Gln Gly Val Leu Ala Ala Gln
1715 1720 1725
Ala Arg Ala Glu Gln Leu Arg Asp Glu Ala Arg Asp Leu Leu Gln Ala
1730 1735 1740
Ala Gln Asp Lys Leu Gln Arg Leu Gln Glu Leu Glu Gly Thr Tyr Glu
1745 1750 1755 1760
Glu Asn Glu Arg Ala Leu Glu Ser Lys Ala Ala Gln Leu Asp Gly Leu
1765 1770 1775
Glu Ala Arg Met Arg Ser Val Leu Gln Ala Ile Asn Leu Gln Val Gln
1780 1785 1790
Ile Tyr Asn Thr Cys Gln
1795
<210> 118
<211> 5397
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК последовательность подтипа Ламинина бета-2
(Человек разумный)
<400> 118
atggagctga cctcaaggga aagagggagg ggacagcctc tgccctggga acttcgactg 60
ggcctactgc taagcgtgct ggctgccaca ctggcacagg cccctgcccc ggatgtgcct 120
ggctgttcca ggggaagctg ctaccccgcc acgggcgacc tgctggtggg ccgagctgac 180
agactgactg cctcatccac ttgtggcctg aatggccccc agccctactg catcgtcagt 240
cacctgcagg acgaaaagaa gtgcttcctt tgtgactccc ggcgcccctt ctctgctaga 300
gacaacccac acagccatcg catccagaat gtagtcacca gctttgcacc acagcggcgg 360
gcagcctggt ggcagtcaga gaatggtatc cctgcggtca ccatccagct ggacctggag 420
gctgagtttc atttcacaca cctcattatg accttcaaga catttcgccc tgctgccatg 480
ctggtggaac gctcagcaga ctttggccgc acctggcatg tgtaccgata tttctcctat 540
gactgtgggg ctgacttccc aggagtccca ctagcacccc cacggcactg ggatgatgta 600
gtctgtgagt cccgctactc agagattgag ccatccactg aaggcgaggt catctatcgt 660
gtgctggacc ctgccatccc tatcccagac ccctacagct cacggattca gaacctgttg 720
aagatcacca acctacgggt gaacctgact cgtctacaca cgttgggaga caacctactc 780
gacccacgga gggagatccg agagaagtac tactatgccc tctatgagct ggttgtacgt 840
ggcaactgct tctgctacgg acacgcctca gagtgtgcac ccgccccagg ggcaccagcc 900
catgctgagg gcatggtgca cggagcttgc atctgcaaac acaacacacg tggcctcaac 960
tgcgagcagt gtcaggattt ctatcgtgac ctgccctggc gtccggctga ggacggccat 1020
agtcatgcct gtaggaagtg tgagtgccat gggcacaccc acagctgcca cttcgacatg 1080
gccgtatacc tggcatctgg caatgtgagt ggaggtgtgt gtgatggatg tcagcataac 1140
acagctgggc gccactgtga gctctgtcgg cccttcttct accgtgaccc aaccaaggac 1200
ctgcgggatc cggctgtgtg ccgctcctgt gattgtgacc ccatgggttc tcaagacggt 1260
ggtcgctgtg attcccatga tgaccctgca ctgggactgg tctccggcca gtgtcgctgc 1320
aaagaacatg tggtgggcac tcgctgccag caatgccgtg atggcttctt tgggctcagc 1380
atcagtgacc gtctgggctg ccggcgatgt caatgtaatg cacggggcac agtgcctggg 1440
agcactcctt gtgaccccaa cagtggatcc tgttactgca aacgtctagt gactggacgt 1500
ggatgtgacc gctgcctgcc tggccactgg ggcctgagcc acgacctgct cggctgccgc 1560
ccctgtgact gcgacgtggg tggtgctttg gatccccagt gtgatgaggg cacaggtcaa 1620
tgccactgcc gccagcacat ggttgggcga cgctgtgagc aggtgcaacc tggctacttc 1680
cggcccttcc tggaccacct aatttgggag gctgaggaca cccgagggca ggtgctcgat 1740
gtggtggagc gcctggtgac ccccggggaa actccatcct ggactggctc aggcttcgtg 1800
cggctacagg aaggtcagac cctggagttc ctggtggcct ctgtgccgaa ggctatggac 1860
tatgacctgc tgctgcgctt agagccccag gtccctgagc aatgggcaga gttggaactg 1920
attgtgcagc gtccagggcc tgtgcctgcc cacagcctgt gtgggcattt ggtgcccaag 1980
gatgatcgca tccaagggac tctgcaacca catgccaggt acttgatatt tcctaatcct 2040
gtctgccttg agcctggtat ctcctacaag ctgcatctga agctggtacg gacaggggga 2100
agtgcccagc ctgagactcc ctactctgga cctggcctgc tcattgactc gctggtgctg 2160
ctgccccgtg tcctggtgct agagatgttt agtgggggtg atgctgctgc cctggagcgc 2220
caggccacct ttgaacgcta ccaatgccat gaggagggtc tggtgcccag caagacttct 2280
ccctctgagg cctgcgcacc cctcctcatc agcctgtcca ccctcatcta caatggtgcc 2340
ctgccatgtc agtgcaaccc tcaaggttca ctgagttctg agtgcaaccc tcatggtggt 2400
cagtgcctgt gcaagcctgg agtggttggg cgccgctgtg acctctgtgc ccctggctac 2460
tatggctttg gccccacagg ctgtcaagcc tgccagtgca gccacgaggg ggcactcagc 2520
agtctctgtg aaaagaccag tgggcaatgt ctctgtcgaa ctggtgcctt tgggcttcgc 2580
tgtgaccgct gccagcgtgg ccagtgggga ttccctagct gccggccatg tgtctgcaat 2640
gggcatgcag atgagtgcaa cacccacaca ggcgcttgcc tgggctgccg tgatcacaca 2700
gggggtgagc actgtgaaag gtgcattgct ggtttccacg gggacccacg gctgccatat 2760
gggggccagt gccggccctg tccctgtcct gaaggccctg ggagccaacg gcactttgct 2820
acttcttgcc accaggatga atattcccag cagattgtgt gccactgccg ggcaggctat 2880
acggggctgc gatgtgaagc ttgtgcccct gggcactttg gggacccatc aaggccaggt 2940
ggccggtgcc aactgtgtga gtgcagtggg aacattgacc caatggatcc tgatgcctgt 3000
gacccccaca cggggcaatg cctgcgctgt ttacaccaca cagagggtcc acactgtgcc 3060
cactgcaagc ctggcttcca tgggcaggct gcccgacaga gctgtcaccg ctgcacatgc 3120
aacctgctgg gcacaaatcc gcagcagtgc ccatctcctg accagtgcca ctgtgatcca 3180
agcagtgggc agtgcccatg cctccccaat gtccagggcc ctagctgtga ccgctgtgcc 3240
cccaacttct ggaacctcac cagtggccat ggttgccagc cttgtgcctg ccacccaagc 3300
cgggccagag gccccacctg caacgagttc acagggcagt gccactgccg tgccggcttt 3360
ggagggcgga cttgttctga gtgccaagag ctccactggg gagaccctgg gttgcagtgc 3420
catgcctgtg attgtgactc tcgtggaata gatacacctc agtgtcaccg cttcacaggt 3480
cactgcagct gccgcccagg ggtgtctggt gtgcgctgtg accagtgtgc ccgtggcttc 3540
tcaggaatct ttcctgcctg ccatccctgc catgcatgct tcggggattg ggaccgagtg 3600
gtgcaggact tggcagcccg tacacagcgc ctagagcagc gggcgcagga gttgcaacag 3660
acgggtgtgc tgggtgcctt tgagagcagc ttctggcaca tgcaggagaa gctgggcatt 3720
gtgcagggca tcgtaggtgc ccgcaacacc tcagccgcct ccactgcaca gcttgtggag 3780
gccacagagg agctgcggcg tgaaattggg gaggccactg agcacctgac tcagctcgag 3840
gcagacctga cagatgtgca agatgagaac ttcaatgcca accatgcact aagtggtctg 3900
gagcgagata ggcttgcact taatctcaca ctgcggcagc tcgaccagca tcttgacttg 3960
ctcaaacatt caaacttcct gggtgcctat gacagcatcc ggcatgccca tagccagtct 4020
gcagaggcag aacgtcgtgc caatacctca gccctggcag tacctagccc tgtgagcaac 4080
tcggcaagtg ctcggcatcg gacagaggca ctgatggatg ctcagaagga ggacttcaac 4140
agcaaacaca tggccaacca gcgggcactt ggcaagctct ctgcccatac ccacaccctg 4200
agcctgacag acataaatga gctggtgtgt ggggcaccag gggatgcacc ctgtgctaca 4260
agcccttgtg ggggtgccgg ctgtcgagat gaggatgggc agccgcgctg tgggggcctc 4320
agctgcaatg gggcagcggc tacagcagac ctagcactgg gccgggcccg gcacacacag 4380
gcagagctgc agcgggcact ggcagaaggt ggtagcatcc tcagcagagt ggctgagact 4440
cgtcggcagg caagcgaggc acagcagcgg gcccaggcag ccctggacaa ggctaatgct 4500
tccaggggac aggtggaaca ggccaaccag gaacttcaag aacttatcca gagtgtgaag 4560
gacttcctca accaggaggg ggctgatcct gatagcattg aaatggtggc cacacgggtg 4620
ctagagctct ccatcccagc ttcagctgag cagatccagc acctggcggg tgcgattgca 4680
gagcgagtcc ggagcctggc agatgtggat gcgatcctgg cacgtactgt aggagatgtg 4740
cgtcgtgccg agcagctact gcaggatgca cggcgggcaa ggagctgggc tgaggatgag 4800
aaacagaagg cagagacagt acaggcagca ctggaggagg cccagcgggc acagggtatt 4860
gcccagggtg ccatccgggg ggcagtggct gacacacggg acacagagca gaccctgtac 4920
caggtacagg agaggatggc aggtgcagag cgggcactga gctctgcagg tgaaagggct 4980
cggcagttgg atgctctcct ggaggctctg aaattgaaac gggcaggaaa tagtctggca 5040
gcctctacag cagaagaaac ggcaggcagt gcccagggtc gtgcccagga ggctgagcag 5100
ctgctacgcg gtcctctggg tgatcagtac cagacggtga aggccctagc tgagcgcaag 5160
gcccaaggtg tgctggctgc acaggcaagg gcagaacaac tgcgggatga ggctcgggac 5220
ctgttgcaag ccgctcagga caagctgcag cggctacagg aattggaagg cacctatgag 5280
gaaaatgagc gggcactgga gagtaaggca gcccagttgg acgggttgga ggccaggatg 5340
cgcagcgtgc ttcaagccat caacttgcag gtgcagatct acaacacctg ccagtga 5397
<210> 119
<211> 1609
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность подтипа Ламинина
гамма-1 (Человек разумный)
<400> 119
Met Arg Gly Ser His Arg Ala Ala Pro Ala Leu Arg Pro Arg Gly Arg
1 5 10 15
Leu Trp Pro Val Leu Ala Val Leu Ala Ala Ala Ala Ala Ala Gly Cys
20 25 30
Ala Gln Ala Ala Met Asp Glu Cys Thr Asp Glu Gly Gly Arg Pro Gln
35 40 45
Arg Cys Met Pro Glu Phe Val Asn Ala Ala Phe Asn Val Thr Val Val
50 55 60
Ala Thr Asn Thr Cys Gly Thr Pro Pro Glu Glu Tyr Cys Val Gln Thr
65 70 75 80
Gly Val Thr Gly Val Thr Lys Ser Cys His Leu Cys Asp Ala Gly Gln
85 90 95
Pro His Leu Gln His Gly Ala Ala Phe Leu Thr Asp Tyr Asn Asn Gln
100 105 110
Ala Asp Thr Thr Trp Trp Gln Ser Gln Thr Met Leu Ala Gly Val Gln
115 120 125
Tyr Pro Ser Ser Ile Asn Leu Thr Leu His Leu Gly Lys Ala Phe Asp
130 135 140
Ile Thr Tyr Val Arg Leu Lys Phe His Thr Ser Arg Pro Glu Ser Phe
145 150 155 160
Ala Ile Tyr Lys Arg Thr Arg Glu Asp Gly Pro Trp Ile Pro Tyr Gln
165 170 175
Tyr Tyr Ser Gly Ser Cys Glu Asn Thr Tyr Ser Lys Ala Asn Arg Gly
180 185 190
Phe Ile Arg Thr Gly Gly Asp Glu Gln Gln Ala Leu Cys Thr Asp Glu
195 200 205
Phe Ser Asp Ile Ser Pro Leu Thr Gly Gly Asn Val Ala Phe Ser Thr
210 215 220
Leu Glu Gly Arg Pro Ser Ala Tyr Asn Phe Asp Asn Ser Pro Val Leu
225 230 235 240
Gln Glu Trp Val Thr Ala Thr Asp Ile Arg Val Thr Leu Asn Arg Leu
245 250 255
Asn Thr Phe Gly Asp Glu Val Phe Asn Asp Pro Lys Val Leu Lys Ser
260 265 270
Tyr Tyr Tyr Ala Ile Ser Asp Phe Ala Val Gly Gly Arg Cys Lys Cys
275 280 285
Asn Gly His Ala Ser Glu Cys Met Lys Asn Glu Phe Asp Lys Leu Val
290 295 300
Cys Asn Cys Lys His Asn Thr Tyr Gly Val Asp Cys Glu Lys Cys Leu
305 310 315 320
Pro Phe Phe Asn Asp Arg Pro Trp Arg Arg Ala Thr Ala Glu Ser Ala
325 330 335
Ser Glu Cys Leu Pro Cys Asp Cys Asn Gly Arg Ser Gln Glu Cys Tyr
340 345 350
Phe Asp Pro Glu Leu Tyr Arg Ser Thr Gly His Gly Gly His Cys Thr
355 360 365
Asn Cys Gln Asp Asn Thr Asp Gly Ala His Cys Glu Arg Cys Arg Glu
370 375 380
Asn Phe Phe Arg Leu Gly Asn Asn Glu Ala Cys Ser Ser Cys His Cys
385 390 395 400
Ser Pro Val Gly Ser Leu Ser Thr Gln Cys Asp Ser Tyr Gly Arg Cys
405 410 415
Ser Cys Lys Pro Gly Val Met Gly Asp Lys Cys Asp Arg Cys Gln Pro
420 425 430
Gly Phe His Ser Leu Thr Glu Ala Gly Cys Arg Pro Cys Ser Cys Asp
435 440 445
Pro Ser Gly Ser Ile Asp Glu Cys Asn Ile Glu Thr Gly Arg Cys Val
450 455 460
Cys Lys Asp Asn Val Glu Gly Phe Asn Cys Glu Arg Cys Lys Pro Gly
465 470 475 480
Phe Phe Asn Leu Glu Ser Ser Asn Pro Arg Gly Cys Thr Pro Cys Phe
485 490 495
Cys Phe Gly His Ser Ser Val Cys Thr Asn Ala Val Gly Tyr Ser Val
500 505 510
Tyr Ser Ile Ser Ser Thr Phe Gln Ile Asp Glu Asp Gly Trp Arg Ala
515 520 525
Glu Gln Arg Asp Gly Ser Glu Ala Ser Leu Glu Trp Ser Ser Glu Arg
530 535 540
Gln Asp Ile Ala Val Ile Ser Asp Ser Tyr Phe Pro Arg Tyr Phe Ile
545 550 555 560
Ala Pro Ala Lys Phe Leu Gly Lys Gln Val Leu Ser Tyr Gly Gln Asn
565 570 575
Leu Ser Phe Ser Phe Arg Val Asp Arg Arg Asp Thr Arg Leu Ser Ala
580 585 590
Glu Asp Leu Val Leu Glu Gly Ala Gly Leu Arg Val Ser Val Pro Leu
595 600 605
Ile Ala Gln Gly Asn Ser Tyr Pro Ser Glu Thr Thr Val Lys Tyr Val
610 615 620
Phe Arg Leu His Glu Ala Thr Asp Tyr Pro Trp Arg Pro Ala Leu Thr
625 630 635 640
Pro Phe Glu Phe Gln Lys Leu Leu Asn Asn Leu Thr Ser Ile Lys Ile
645 650 655
Arg Gly Thr Tyr Ser Glu Arg Ser Ala Gly Tyr Leu Asp Asp Val Thr
660 665 670
Leu Ala Ser Ala Arg Pro Gly Pro Gly Val Pro Ala Thr Trp Val Glu
675 680 685
Ser Cys Thr Cys Pro Val Gly Tyr Gly Gly Gln Phe Cys Glu Met Cys
690 695 700
Leu Ser Gly Tyr Arg Arg Glu Thr Pro Asn Leu Gly Pro Tyr Ser Pro
705 710 715 720
Cys Val Leu Cys Ala Cys Asn Gly His Ser Glu Thr Cys Asp Pro Glu
725 730 735
Thr Gly Val Cys Asn Cys Arg Asp Asn Thr Ala Gly Pro His Cys Glu
740 745 750
Lys Cys Ser Asp Gly Tyr Tyr Gly Asp Ser Thr Ala Gly Thr Ser Ser
755 760 765
Asp Cys Gln Pro Cys Pro Cys Pro Gly Gly Ser Ser Cys Ala Val Val
770 775 780
Pro Lys Thr Lys Glu Val Val Cys Thr Asn Cys Pro Thr Gly Thr Thr
785 790 795 800
Gly Lys Arg Cys Glu Leu Cys Asp Asp Gly Tyr Phe Gly Asp Pro Leu
805 810 815
Gly Arg Asn Gly Pro Val Arg Leu Cys Arg Leu Cys Gln Cys Ser Asp
820 825 830
Asn Ile Asp Pro Asn Ala Val Gly Asn Cys Asn Arg Leu Thr Gly Glu
835 840 845
Cys Leu Lys Cys Ile Tyr Asn Thr Ala Gly Phe Tyr Cys Asp Arg Cys
850 855 860
Lys Asp Gly Phe Phe Gly Asn Pro Leu Ala Pro Asn Pro Ala Asp Lys
865 870 875 880
Cys Lys Ala Cys Asn Cys Asn Leu Tyr Gly Thr Met Lys Gln Gln Ser
885 890 895
Ser Cys Asn Pro Val Thr Gly Gln Cys Glu Cys Leu Pro His Val Thr
900 905 910
Gly Gln Asp Cys Gly Ala Cys Asp Pro Gly Phe Tyr Asn Leu Gln Ser
915 920 925
Gly Gln Gly Cys Glu Arg Cys Asp Cys His Ala Leu Gly Ser Thr Asn
930 935 940
Gly Gln Cys Asp Ile Arg Thr Gly Gln Cys Glu Cys Gln Pro Gly Ile
945 950 955 960
Thr Gly Gln His Cys Glu Arg Cys Glu Val Asn His Phe Gly Phe Gly
965 970 975
Pro Glu Gly Cys Lys Pro Cys Asp Cys His Pro Glu Gly Ser Leu Ser
980 985 990
Leu Gln Cys Lys Asp Asp Gly Arg Cys Glu Cys Arg Glu Gly Phe Val
995 1000 1005
Gly Asn Arg Cys Asp Gln Cys Glu Glu Asn Tyr Phe Tyr Asn Arg Ser
1010 1015 1020
Trp Pro Gly Cys Gln Glu Cys Pro Ala Cys Tyr Arg Leu Val Lys Asp
1025 1030 1035 1040
Lys Val Ala Asp His Arg Val Lys Leu Gln Glu Leu Glu Ser Leu Ile
1045 1050 1055
Ala Asn Leu Gly Thr Gly Asp Glu Met Val Thr Asp Gln Ala Phe Glu
1060 1065 1070
Asp Arg Leu Lys Glu Ala Glu Arg Glu Val Met Asp Leu Leu Arg Glu
1075 1080 1085
Ala Gln Asp Val Lys Asp Val Asp Gln Asn Leu Met Asp Arg Leu Gln
1090 1095 1100
Arg Val Asn Asn Thr Leu Ser Ser Gln Ile Ser Arg Leu Gln Asn Ile
1105 1110 1115 1120
Arg Asn Thr Ile Glu Glu Thr Gly Asn Leu Ala Glu Gln Ala Arg Ala
1125 1130 1135
His Val Glu Asn Thr Glu Arg Leu Ile Glu Ile Ala Ser Arg Glu Leu
1140 1145 1150
Glu Lys Ala Lys Val Ala Ala Ala Asn Val Ser Val Thr Gln Pro Glu
1155 1160 1165
Ser Thr Gly Asp Pro Asn Asn Met Thr Leu Leu Ala Glu Glu Ala Arg
1170 1175 1180
Lys Leu Ala Glu Arg His Lys Gln Glu Ala Asp Asp Ile Val Arg Val
1185 1190 1195 1200
Ala Lys Thr Ala Asn Asp Thr Ser Thr Glu Ala Tyr Asn Leu Leu Leu
1205 1210 1215
Arg Thr Leu Ala Gly Glu Asn Gln Thr Ala Phe Glu Ile Glu Glu Leu
1220 1225 1230
Asn Arg Lys Tyr Glu Gln Ala Lys Asn Ile Ser Gln Asp Leu Glu Lys
1235 1240 1245
Gln Ala Ala Arg Val His Glu Glu Ala Lys Arg Ala Gly Asp Lys Ala
1250 1255 1260
Val Glu Ile Tyr Ala Ser Val Ala Gln Leu Ser Pro Leu Asp Ser Glu
1265 1270 1275 1280
Thr Leu Glu Asn Glu Ala Asn Asn Ile Lys Met Glu Ala Glu Asn Leu
1285 1290 1295
Glu Gln Leu Ile Asp Gln Lys Leu Lys Asp Tyr Glu Asp Leu Arg Glu
1300 1305 1310
Asp Met Arg Gly Lys Glu Leu Glu Val Lys Asn Leu Leu Glu Lys Gly
1315 1320 1325
Lys Thr Glu Gln Gln Thr Ala Asp Gln Leu Leu Ala Arg Ala Asp Ala
1330 1335 1340
Ala Lys Ala Leu Ala Glu Glu Ala Ala Lys Lys Gly Arg Asp Thr Leu
1345 1350 1355 1360
Gln Glu Ala Asn Asp Ile Leu Asn Asn Leu Lys Asp Phe Asp Arg Arg
1365 1370 1375
Val Asn Asp Asn Lys Thr Ala Ala Glu Glu Ala Leu Arg Lys Ile Pro
1380 1385 1390
Ala Ile Asn Gln Thr Ile Thr Glu Ala Asn Glu Lys Thr Arg Glu Ala
1395 1400 1405
Gln Gln Ala Leu Gly Ser Ala Ala Ala Asp Ala Thr Glu Ala Lys Asn
1410 1415 1420
Lys Ala His Glu Ala Glu Arg Ile Ala Ser Ala Val Gln Lys Asn Ala
1425 1430 1435 1440
Thr Ser Thr Lys Ala Glu Ala Glu Arg Thr Phe Ala Glu Val Thr Asp
1445 1450 1455
Leu Asp Asn Glu Val Asn Asn Met Leu Lys Gln Leu Gln Glu Ala Glu
1460 1465 1470
Lys Glu Leu Lys Arg Lys Gln Asp Asp Ala Asp Gln Asp Met Met Met
1475 1480 1485
Ala Gly Met Ala Ser Gln Ala Ala Gln Glu Ala Glu Ile Asn Ala Arg
1490 1495 1500
Lys Ala Lys Asn Ser Val Thr Ser Leu Leu Ser Ile Ile Asn Asp Leu
1505 1510 1515 1520
Leu Glu Gln Leu Gly Gln Leu Asp Thr Val Asp Leu Asn Lys Leu Asn
1525 1530 1535
Glu Ile Glu Gly Thr Leu Asn Lys Ala Lys Asp Glu Met Lys Val Ser
1540 1545 1550
Asp Leu Asp Arg Lys Val Ser Asp Leu Glu Asn Glu Ala Lys Lys Gln
1555 1560 1565
Glu Ala Ala Ile Met Asp Tyr Asn Arg Asp Ile Glu Glu Ile Met Lys
1570 1575 1580
Asp Ile Arg Asn Leu Glu Asp Ile Arg Lys Thr Leu Pro Ser Gly Cys
1585 1590 1595 1600
Phe Asn Thr Pro Ser Ile Glu Lys Pro
1605
<210> 120
<211> 4830
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК последовательность подтипа Ламинина гамма-1
(Человек разумный)
<400> 120
atgagaggga gccatcgggc cgcgccggcc ctgcggcccc gggggcggct ctggcccgtg 60
ctggccgtgc tggcggcggc cgccgcggcg ggctgtgccc aggcagccat ggacgagtgc 120
acggacgagg gcgggcggcc gcagcgctgc atgcccgagt tcgtcaacgc cgccttcaac 180
gtgactgtgg tggccaccaa cacgtgtggg actccgcccg aggaatactg tgtgcagacc 240
ggggtgaccg gggtcaccaa gtcctgtcac ctgtgcgacg ccgggcagcc ccacctgcag 300
cacggggcag ccttcctgac cgactacaac aaccaggccg acaccacctg gtggcaaagc 360
cagaccatgc tggccggggt gcagtacccc agctccatca acctcacgct gcacctggga 420
aaagcttttg acatcaccta tgtgcgtctc aagttccaca ccagccgccc ggagagcttt 480
gccatttaca agcgcacacg ggaagacggg ccctggattc cttaccagta ctacagtggt 540
tcctgtgaga acacctactc caaggcaaac cgcggcttca tcaggacagg aggggacgag 600
cagcaggcct tgtgtactga tgaattcagt gacatttctc ccctcactgg gggcaacgtg 660
gccttttcta ccctggaagg aaggcccagc gcctataact ttgacaatag ccctgtgctg 720
caggaatggg taactgccac tgacatcaga gtaactctta atcgcctgaa cacttttgga 780
gatgaagtgt ttaacgatcc caaagttctc aagtcctatt attatgccat ctctgatttt 840
gctgtaggtg gcagatgtaa atgtaatgga cacgcaagcg agtgtatgaa gaacgaattt 900
gataagctgg tgtgtaattg caaacataac acatatggag tagactgtga aaagtgtctt 960
cctttcttca atgaccggcc gtggaggagg gcaactgcgg aaagtgccag tgaatgcctg 1020
ccctgtgatt gcaatggtcg atcccaggaa tgctacttcg accctgaact ctatcgttcc 1080
actggccatg ggggccactg taccaactgc caggataaca cagatggcgc ccactgtgag 1140
aggtgccgag agaacttctt ccgccttggc aacaatgaag cctgctcttc atgccactgt 1200
agtcctgtgg gctctctaag cacacagtgt gatagttacg gcagatgcag ctgtaagcca 1260
ggagtgatgg gggacaaatg tgaccgttgc cagcctggat tccattctct cactgaagca 1320
ggatgcaggc catgctcttg tgatccctct ggcagcatag atgaatgtaa tattgaaaca 1380
ggaagatgtg tttgcaaaga caatgtcgaa ggcttcaatt gtgaaagatg caaacctgga 1440
ttttttaatc tggaatcatc taatcctcgg ggttgcacac cctgcttctg ctttgggcat 1500
tcttctgtct gtacaaacgc tgttggctac agtgtttatt ctatctcctc tacctttcag 1560
attgatgagg atgggtggcg tgcggaacag agagatggct ctgaagcatc tctcgagtgg 1620
tcctctgaga ggcaagatat cgccgtgatc tcagacagct actttcctcg gtacttcatt 1680
gctcctgcaa agttcttggg caagcaggtg ttgagttatg gtcagaacct ctccttctcc 1740
tttcgagtgg acaggcgaga tactcgcctc tctgcagaag accttgtgct tgagggagct 1800
ggcttaagag tatctgtacc cttgatcgct cagggcaatt cctatccaag tgagaccact 1860
gtgaagtatg tcttcaggct ccatgaagca acagattacc cttggaggcc tgctcttacc 1920
ccttttgaat ttcagaagct cctaaacaac ttgacctcta tcaagatacg tgggacatac 1980
agtgagagaa gtgctggata tttggatgat gtcaccctgg caagtgctcg tcctgggcct 2040
ggagtccctg caacttgggt ggagtcctgc acctgtcctg tgggatatgg agggcagttt 2100
tgtgagatgt gcctctcagg ttacagaaga gaaactccta atcttggacc atacagtcca 2160
tgtgtgcttt gcgcctgcaa tggacacagc gagacctgtg atcctgagac aggtgtttgt 2220
aactgcagag acaatacggc tggcccgcac tgtgagaagt gcagtgatgg gtactatgga 2280
gattcaactg caggcacctc ctccgattgc caaccctgtc cgtgtcctgg aggttcaagt 2340
tgtgctgttg ttcccaagac aaaggaggtg gtgtgcacca actgtcctac tggcaccact 2400
ggtaagagat gtgagctctg tgatgatggc tactttggag accccctggg tagaaacggc 2460
cctgtgagac tttgccgcct gtgccagtgc agtgacaaca tcgatcccaa tgcagttgga 2520
aattgcaatc gcttgacggg agaatgcctg aagtgcatct ataacactgc tggcttctat 2580
tgtgaccggt gcaaagacgg attttttgga aatcccctgg ctcccaatcc agcagacaaa 2640
tgcaaagcct gcaattgcaa tctgtatggg accatgaagc agcagagcag ctgtaacccc 2700
gtgacggggc agtgtgaatg tttgcctcac gtgactggcc aggactgtgg tgcttgtgac 2760
cctggattct acaatctgca gagtgggcaa ggctgtgaga ggtgtgactg ccatgccttg 2820
ggctccacca atgggcagtg tgacatccgc accggccagt gtgagtgcca gcccggcatc 2880
actggtcagc actgtgagcg ctgtgaggtc aaccactttg ggtttggacc tgaaggctgc 2940
aaaccctgtg actgtcatcc tgagggatct ctttcacttc agtgcaaaga tgatggtcgc 3000
tgtgaatgca gagaaggctt tgtgggaaat cgctgtgacc agtgtgaaga aaactatttc 3060
tacaatcggt cttggcctgg ctgccaggaa tgtccagctt gttaccggct ggtaaaggat 3120
aaggttgctg atcatagagt gaagctccag gaattagaga gtctcatagc aaaccttgga 3180
actggggatg agatggtgac agatcaagcc ttcgaggata gactaaagga agcagagagg 3240
gaagttatgg acctccttcg tgaggcccag gatgtcaaag atgttgacca gaatttgatg 3300
gatcgcctac agagagtgaa taacactctg tccagccaaa ttagccgttt acagaatatc 3360
cggaatacca ttgaagagac tggaaacttg gctgaacaag cgcgtgccca tgtagagaac 3420
acagagcggt tgattgaaat cgcatccaga gaacttgaga aagcaaaagt cgctgctgcc 3480
aatgtgtcag tcactcagcc agaatctaca ggggacccaa acaacatgac tcttttggca 3540
gaagaggctc gaaagcttgc tgaacgtcat aaacaggaag ctgatgacat tgttcgagtg 3600
gcaaagacag ccaatgatac gtcaactgag gcatacaacc tgcttctgag gacactggca 3660
ggagaaaatc aaacagcatt tgagattgaa gagcttaata ggaagtatga acaagcgaag 3720
aacatctcac aggatctgga aaaacaagct gcccgagtac atgaggaggc caaaagggcc 3780
ggtgacaaag ctgtggagat ctatgccagc gtggctcagc tgagcccttt ggactctgag 3840
acactggaga atgaagcaaa taacataaag atggaagctg agaatctgga acaactgatt 3900
gaccagaaat taaaagatta tgaggacctc agagaagata tgagagggaa ggaacttgaa 3960
gtcaagaacc ttctggagaa aggcaagact gaacagcaga ccgcagacca actcctagcc 4020
cgagctgatg ctgccaaggc cctcgctgaa gaagctgcaa agaagggacg ggatacctta 4080
caagaagcta atgacattct caacaacctg aaagattttg ataggcgtgt gaacgataac 4140
aagacggccg cagaggaggc actaaggaag attcctgcca tcaaccagac catcactgaa 4200
gccaatgaaa agaccagaga agcccagcag gccctgggca gtgctgcggc ggatgccaca 4260
gaggccaaga acaaggccca tgaggcggag aggatcgcga gcgctgtcca aaagaatgcc 4320
accagcacca aggcagaagc tgaaagaact tttgcagaag ttacagatct ggataatgag 4380
gtgaacaata tgttgaagca actgcaggaa gcagaaaaag agctaaagag aaaacaagat 4440
gacgctgacc aggacatgat gatggcaggg atggcttcac aggctgctca agaagccgag 4500
atcaatgcca gaaaagccaa aaactctgtt actagcctcc tcagcattat taatgacctc 4560
ttggagcagc tggggcagct ggatacagtg gacctgaata agctaaacga gattgaaggc 4620
accctaaaca aagccaaaga tgaaatgaag gtcagcgatc ttgataggaa agtgtctgac 4680
ctggagaatg aagccaagaa gcaggaggct gccatcatgg actataaccg agatatcgag 4740
gagatcatga aggacattcg caatctggag gacatcagga agaccttacc atctggctgc 4800
ttcaacaccc cgtccattga aaagccctag 4830
<210> 121
<211> 3122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность подтипа Ламинина
альфа-2 (Человек разумный)
<400> 121
Met Pro Gly Ala Ala Gly Val Leu Leu Leu Leu Leu Leu Ser Gly Gly
1 5 10 15
Leu Gly Gly Val Gln Ala Gln Arg Pro Gln Gln Gln Arg Gln Ser Gln
20 25 30
Ala His Gln Gln Arg Gly Leu Phe Pro Ala Val Leu Asn Leu Ala Ser
35 40 45
Asn Ala Leu Ile Thr Thr Asn Ala Thr Cys Gly Glu Lys Gly Pro Glu
50 55 60
Met Tyr Cys Lys Leu Val Glu His Val Pro Gly Gln Pro Val Arg Asn
65 70 75 80
Pro Gln Cys Arg Ile Cys Asn Gln Asn Ser Ser Asn Pro Asn Gln Arg
85 90 95
His Pro Ile Thr Asn Ala Ile Asp Gly Lys Asn Thr Trp Trp Gln Ser
100 105 110
Pro Ser Ile Lys Asn Gly Ile Glu Tyr His Tyr Val Thr Ile Thr Leu
115 120 125
Asp Leu Gln Gln Val Phe Gln Ile Ala Tyr Val Ile Val Lys Ala Ala
130 135 140
Asn Ser Pro Arg Pro Gly Asn Trp Ile Leu Glu Arg Ser Leu Asp Asp
145 150 155 160
Val Glu Tyr Lys Pro Trp Gln Tyr His Ala Val Thr Asp Thr Glu Cys
165 170 175
Leu Thr Leu Tyr Asn Ile Tyr Pro Arg Thr Gly Pro Pro Ser Tyr Ala
180 185 190
Lys Asp Asp Glu Val Ile Cys Thr Ser Phe Tyr Ser Lys Ile His Pro
195 200 205
Leu Glu Asn Gly Glu Ile His Ile Ser Leu Ile Asn Gly Arg Pro Ser
210 215 220
Ala Asp Asp Pro Ser Pro Glu Leu Leu Glu Phe Thr Ser Ala Arg Tyr
225 230 235 240
Ile Arg Leu Arg Phe Gln Arg Ile Arg Thr Leu Asn Ala Asp Leu Met
245 250 255
Met Phe Ala His Lys Asp Pro Arg Glu Ile Asp Pro Ile Val Thr Arg
260 265 270
Arg Tyr Tyr Tyr Ser Val Lys Asp Ile Ser Val Gly Gly Met Cys Ile
275 280 285
Cys Tyr Gly His Ala Arg Ala Cys Pro Leu Asp Pro Ala Thr Asn Lys
290 295 300
Ser Arg Cys Glu Cys Glu His Asn Thr Cys Gly Asp Ser Cys Asp Gln
305 310 315 320
Cys Cys Pro Gly Phe His Gln Lys Pro Trp Arg Ala Gly Thr Phe Leu
325 330 335
Thr Lys Thr Glu Cys Glu Ala Cys Asn Cys His Gly Lys Ala Glu Glu
340 345 350
Cys Tyr Tyr Asp Glu Asn Val Ala Arg Arg Asn Leu Ser Leu Asn Ile
355 360 365
Arg Gly Lys Tyr Ile Gly Gly Gly Val Cys Ile Asn Cys Thr Gln Asn
370 375 380
Thr Ala Gly Ile Asn Cys Glu Thr Cys Thr Asp Gly Phe Phe Arg Pro
385 390 395 400
Lys Gly Val Ser Pro Asn Tyr Pro Arg Pro Cys Gln Pro Cys His Cys
405 410 415
Asp Pro Ile Gly Ser Leu Asn Glu Val Cys Val Lys Asp Glu Lys His
420 425 430
Ala Arg Arg Gly Leu Ala Pro Gly Ser Cys His Cys Lys Thr Gly Phe
435 440 445
Gly Gly Val Ser Cys Asp Arg Cys Ala Arg Gly Tyr Thr Gly Tyr Pro
450 455 460
Asp Cys Lys Ala Cys Asn Cys Ser Gly Leu Gly Ser Lys Asn Glu Asp
465 470 475 480
Pro Cys Phe Gly Pro Cys Ile Cys Lys Glu Asn Val Glu Gly Gly Asp
485 490 495
Cys Ser Arg Cys Lys Ser Gly Phe Phe Asn Leu Gln Glu Asp Asn Trp
500 505 510
Lys Gly Cys Asp Glu Cys Phe Cys Ser Gly Val Ser Asn Arg Cys Gln
515 520 525
Ser Ser Tyr Trp Thr Tyr Gly Lys Ile Gln Asp Met Ser Gly Trp Tyr
530 535 540
Leu Thr Asp Leu Pro Gly Arg Ile Arg Val Ala Pro Gln Gln Asp Asp
545 550 555 560
Leu Asp Ser Pro Gln Gln Ile Ser Ile Ser Asn Ala Glu Ala Arg Gln
565 570 575
Ala Leu Pro His Ser Tyr Tyr Trp Ser Ala Pro Ala Pro Tyr Leu Gly
580 585 590
Asn Lys Leu Pro Ala Val Gly Gly Gln Leu Thr Phe Thr Ile Ser Tyr
595 600 605
Asp Leu Glu Glu Glu Glu Glu Asp Thr Glu Arg Val Leu Gln Leu Met
610 615 620
Ile Ile Leu Glu Gly Asn Asp Leu Ser Ile Ser Thr Ala Gln Asp Glu
625 630 635 640
Val Tyr Leu His Pro Ser Glu Glu His Thr Asn Val Leu Leu Leu Lys
645 650 655
Glu Glu Ser Phe Thr Ile His Gly Thr His Phe Pro Val Arg Arg Lys
660 665 670
Glu Phe Met Thr Val Leu Ala Asn Leu Lys Arg Val Leu Leu Gln Ile
675 680 685
Thr Tyr Ser Phe Gly Met Asp Ala Ile Phe Arg Leu Ser Ser Val Asn
690 695 700
Leu Glu Ser Ala Val Ser Tyr Pro Thr Asp Gly Ser Ile Ala Ala Ala
705 710 715 720
Val Glu Val Cys Gln Cys Pro Pro Gly Tyr Thr Gly Ser Ser Cys Glu
725 730 735
Ser Cys Trp Pro Arg His Arg Arg Val Asn Gly Thr Ile Phe Gly Gly
740 745 750
Ile Cys Glu Pro Cys Gln Cys Phe Gly His Ala Glu Ser Cys Asp Asp
755 760 765
Val Thr Gly Glu Cys Leu Asn Cys Lys Asp His Thr Gly Gly Pro Tyr
770 775 780
Cys Asp Lys Cys Leu Pro Gly Phe Tyr Gly Glu Pro Thr Lys Gly Thr
785 790 795 800
Ser Glu Asp Cys Gln Pro Cys Ala Cys Pro Leu Asn Ile Pro Ser Asn
805 810 815
Asn Phe Ser Pro Thr Cys His Leu Asp Arg Ser Leu Gly Leu Ile Cys
820 825 830
Asp Gly Cys Pro Val Gly Tyr Thr Gly Pro Arg Cys Glu Arg Cys Ala
835 840 845
Glu Gly Tyr Phe Gly Gln Pro Ser Val Pro Gly Gly Ser Cys Gln Pro
850 855 860
Cys Gln Cys Asn Asp Asn Leu Asp Phe Ser Ile Pro Gly Ser Cys Asp
865 870 875 880
Ser Leu Ser Gly Ser Cys Leu Ile Cys Lys Pro Gly Thr Thr Gly Arg
885 890 895
Tyr Cys Glu Leu Cys Ala Asp Gly Tyr Phe Gly Asp Ala Val Asp Ala
900 905 910
Lys Asn Cys Gln Pro Cys Arg Cys Asn Ala Gly Gly Ser Phe Ser Glu
915 920 925
Val Cys His Ser Gln Thr Gly Gln Cys Glu Cys Arg Ala Asn Val Gln
930 935 940
Gly Gln Arg Cys Asp Lys Cys Lys Ala Gly Thr Phe Gly Leu Gln Ser
945 950 955 960
Ala Arg Gly Cys Val Pro Cys Asn Cys Asn Ser Phe Gly Ser Lys Ser
965 970 975
Phe Asp Cys Glu Glu Ser Gly Gln Cys Trp Cys Gln Pro Gly Val Thr
980 985 990
Gly Lys Lys Cys Asp Arg Cys Ala His Gly Tyr Phe Asn Phe Gln Glu
995 1000 1005
Gly Gly Cys Thr Ala Cys Glu Cys Ser His Leu Gly Asn Asn Cys Asp
1010 1015 1020
Pro Lys Thr Gly Arg Cys Ile Cys Pro Pro Asn Thr Ile Gly Glu Lys
1025 1030 1035 1040
Cys Ser Lys Cys Ala Pro Asn Thr Trp Gly His Ser Ile Thr Thr Gly
1045 1050 1055
Cys Lys Ala Cys Asn Cys Ser Thr Val Gly Ser Leu Asp Phe Gln Cys
1060 1065 1070
Asn Val Asn Thr Gly Gln Cys Asn Cys His Pro Lys Phe Ser Gly Ala
1075 1080 1085
Lys Cys Thr Glu Cys Ser Arg Gly His Trp Asn Tyr Pro Arg Cys Asn
1090 1095 1100
Leu Cys Asp Cys Phe Leu Pro Gly Thr Asp Ala Thr Thr Cys Asp Ser
1105 1110 1115 1120
Glu Thr Lys Lys Cys Ser Cys Ser Asp Gln Thr Gly Gln Cys Thr Cys
1125 1130 1135
Lys Val Asn Val Glu Gly Ile His Cys Asp Arg Cys Arg Pro Gly Lys
1140 1145 1150
Phe Gly Leu Asp Ala Lys Asn Pro Leu Gly Cys Ser Ser Cys Tyr Cys
1155 1160 1165
Phe Gly Thr Thr Thr Gln Cys Ser Glu Ala Lys Gly Leu Ile Arg Thr
1170 1175 1180
Trp Val Thr Leu Lys Ala Glu Gln Thr Ile Leu Pro Leu Val Asp Glu
1185 1190 1195 1200
Ala Leu Gln His Thr Thr Thr Lys Gly Ile Val Phe Gln His Pro Glu
1205 1210 1215
Ile Val Ala His Met Asp Leu Met Arg Glu Asp Leu His Leu Glu Pro
1220 1225 1230
Phe Tyr Trp Lys Leu Pro Glu Gln Phe Glu Gly Lys Lys Leu Met Ala
1235 1240 1245
Tyr Gly Gly Lys Leu Lys Tyr Ala Ile Tyr Phe Glu Ala Arg Glu Glu
1250 1255 1260
Thr Gly Phe Ser Thr Tyr Asn Pro Gln Val Ile Ile Arg Gly Gly Thr
1265 1270 1275 1280
Pro Thr His Ala Arg Ile Ile Val Arg His Met Ala Ala Pro Leu Ile
1285 1290 1295
Gly Gln Leu Thr Arg His Glu Ile Glu Met Thr Glu Lys Glu Trp Lys
1300 1305 1310
Tyr Tyr Gly Asp Asp Pro Arg Val His Arg Thr Val Thr Arg Glu Asp
1315 1320 1325
Phe Leu Asp Ile Leu Tyr Asp Ile His Tyr Ile Leu Ile Lys Ala Thr
1330 1335 1340
Tyr Gly Asn Phe Met Arg Gln Ser Arg Ile Ser Glu Ile Ser Met Glu
1345 1350 1355 1360
Val Ala Glu Gln Gly Arg Gly Thr Thr Met Thr Pro Pro Ala Asp Leu
1365 1370 1375
Ile Glu Lys Cys Asp Cys Pro Leu Gly Tyr Ser Gly Leu Ser Cys Glu
1380 1385 1390
Ala Cys Leu Pro Gly Phe Tyr Arg Leu Arg Ser Gln Pro Gly Gly Arg
1395 1400 1405
Thr Pro Gly Pro Thr Leu Gly Thr Cys Val Pro Cys Gln Cys Asn Gly
1410 1415 1420
His Ser Ser Leu Cys Asp Pro Glu Thr Ser Ile Cys Gln Asn Cys Gln
1425 1430 1435 1440
His His Thr Ala Gly Asp Phe Cys Glu Arg Cys Ala Leu Gly Tyr Tyr
1445 1450 1455
Gly Ile Val Lys Gly Leu Pro Asn Asp Cys Gln Gln Cys Ala Cys Pro
1460 1465 1470
Leu Ile Ser Ser Ser Asn Asn Phe Ser Pro Ser Cys Val Ala Glu Gly
1475 1480 1485
Leu Asp Asp Tyr Arg Cys Thr Ala Cys Pro Arg Gly Tyr Glu Gly Gln
1490 1495 1500
Tyr Cys Glu Arg Cys Ala Pro Gly Tyr Thr Gly Ser Pro Gly Asn Pro
1505 1510 1515 1520
Gly Gly Ser Cys Gln Glu Cys Glu Cys Asp Pro Tyr Gly Ser Leu Pro
1525 1530 1535
Val Pro Cys Asp Pro Val Thr Gly Phe Cys Thr Cys Arg Pro Gly Ala
1540 1545 1550
Thr Gly Arg Lys Cys Asp Gly Cys Lys His Trp His Ala Arg Glu Gly
1555 1560 1565
Trp Glu Cys Val Phe Cys Gly Asp Glu Cys Thr Gly Leu Leu Leu Gly
1570 1575 1580
Asp Leu Ala Arg Leu Glu Gln Met Val Met Ser Ile Asn Leu Thr Gly
1585 1590 1595 1600
Pro Leu Pro Ala Pro Tyr Lys Met Leu Tyr Gly Leu Glu Asn Met Thr
1605 1610 1615
Gln Glu Leu Lys His Leu Leu Ser Pro Gln Arg Ala Pro Glu Arg Leu
1620 1625 1630
Ile Gln Leu Ala Glu Gly Asn Leu Asn Thr Leu Val Thr Glu Met Asn
1635 1640 1645
Glu Leu Leu Thr Arg Ala Thr Lys Val Thr Ala Asp Gly Glu Gln Thr
1650 1655 1660
Gly Gln Asp Ala Glu Arg Thr Asn Thr Arg Ala Lys Ser Leu Gly Glu
1665 1670 1675 1680
Phe Ile Lys Glu Leu Ala Arg Asp Ala Glu Ala Val Asn Glu Lys Ala
1685 1690 1695
Ile Lys Leu Asn Glu Thr Leu Gly Thr Arg Asp Glu Ala Phe Glu Arg
1700 1705 1710
Asn Leu Glu Gly Leu Gln Lys Glu Ile Asp Gln Met Ile Lys Glu Leu
1715 1720 1725
Arg Arg Lys Asn Leu Glu Thr Gln Lys Glu Ile Ala Glu Asp Glu Leu
1730 1735 1740
Val Ala Ala Glu Ala Leu Leu Lys Lys Val Lys Lys Leu Phe Gly Glu
1745 1750 1755 1760
Ser Arg Gly Glu Asn Glu Glu Met Glu Lys Asp Leu Arg Glu Lys Leu
1765 1770 1775
Ala Asp Tyr Lys Asn Lys Val Asp Asp Ala Trp Asp Leu Leu Arg Glu
1780 1785 1790
Ala Thr Asp Lys Ile Arg Glu Ala Asn Arg Leu Phe Ala Val Asn Gln
1795 1800 1805
Lys Asn Met Thr Ala Leu Glu Lys Lys Lys Glu Ala Val Glu Ser Gly
1810 1815 1820
Lys Arg Gln Ile Glu Asn Thr Leu Lys Glu Gly Asn Asp Ile Leu Asp
1825 1830 1835 1840
Glu Ala Asn Arg Leu Ala Asp Glu Ile Asn Ser Ile Ile Asp Tyr Val
1845 1850 1855
Glu Asp Ile Gln Thr Lys Leu Pro Pro Met Ser Glu Glu Leu Asn Asp
1860 1865 1870
Lys Ile Asp Asp Leu Ser Gln Glu Ile Lys Asp Arg Lys Leu Ala Glu
1875 1880 1885
Lys Val Ser Gln Ala Glu Ser His Ala Ala Gln Leu Asn Asp Ser Ser
1890 1895 1900
Ala Val Leu Asp Gly Ile Leu Asp Glu Ala Lys Asn Ile Ser Phe Asn
1905 1910 1915 1920
Ala Thr Ala Ala Phe Lys Ala Tyr Ser Asn Ile Lys Asp Tyr Ile Asp
1925 1930 1935
Glu Ala Glu Lys Val Ala Lys Glu Ala Lys Asp Leu Ala His Glu Ala
1940 1945 1950
Thr Lys Leu Ala Thr Gly Pro Arg Gly Leu Leu Lys Glu Asp Ala Lys
1955 1960 1965
Gly Cys Leu Gln Lys Ser Phe Arg Ile Leu Asn Glu Ala Lys Lys Leu
1970 1975 1980
Ala Asn Asp Val Lys Glu Asn Glu Asp His Leu Asn Gly Leu Lys Thr
1985 1990 1995 2000
Arg Ile Glu Asn Ala Asp Ala Arg Asn Gly Asp Leu Leu Arg Thr Leu
2005 2010 2015
Asn Asp Thr Leu Gly Lys Leu Ser Ala Ile Pro Asn Asp Thr Ala Ala
2020 2025 2030
Lys Leu Gln Ala Val Lys Asp Lys Ala Arg Gln Ala Asn Asp Thr Ala
2035 2040 2045
Lys Asp Val Leu Ala Gln Ile Thr Glu Leu His Gln Asn Leu Asp Gly
2050 2055 2060
Leu Lys Lys Asn Tyr Asn Lys Leu Ala Asp Ser Val Ala Lys Thr Asn
2065 2070 2075 2080
Ala Val Val Lys Asp Pro Ser Lys Asn Lys Ile Ile Ala Asp Ala Asp
2085 2090 2095
Ala Thr Val Lys Asn Leu Glu Gln Glu Ala Asp Arg Leu Ile Asp Lys
2100 2105 2110
Leu Lys Pro Ile Lys Glu Leu Glu Asp Asn Leu Lys Lys Asn Ile Ser
2115 2120 2125
Glu Ile Lys Glu Leu Ile Asn Gln Ala Arg Lys Gln Ala Asn Ser Ile
2130 2135 2140
Lys Val Ser Val Ser Ser Gly Gly Asp Cys Ile Arg Thr Tyr Lys Pro
2145 2150 2155 2160
Glu Ile Lys Lys Gly Ser Tyr Asn Asn Ile Val Val Asn Val Lys Thr
2165 2170 2175
Ala Val Ala Asp Asn Leu Leu Phe Tyr Leu Gly Ser Ala Lys Phe Ile
2180 2185 2190
Asp Phe Leu Ala Ile Glu Met Arg Lys Gly Lys Val Ser Phe Leu Trp
2195 2200 2205
Asp Val Gly Ser Gly Val Gly Arg Val Glu Tyr Pro Asp Leu Thr Ile
2210 2215 2220
Asp Asp Ser Tyr Trp Tyr Arg Ile Val Ala Ser Arg Thr Gly Arg Asn
2225 2230 2235 2240
Gly Thr Ile Ser Val Arg Ala Leu Asp Gly Pro Lys Ala Ser Ile Val
2245 2250 2255
Pro Ser Thr His His Ser Thr Ser Pro Pro Gly Tyr Thr Ile Leu Asp
2260 2265 2270
Val Asp Ala Asn Ala Met Leu Phe Val Gly Gly Leu Thr Gly Lys Leu
2275 2280 2285
Lys Lys Ala Asp Ala Val Arg Val Ile Thr Phe Thr Gly Cys Met Gly
2290 2295 2300
Glu Thr Tyr Phe Asp Asn Lys Pro Ile Gly Leu Trp Asn Phe Arg Glu
2305 2310 2315 2320
Lys Glu Gly Asp Cys Lys Gly Cys Thr Val Ser Pro Gln Val Glu Asp
2325 2330 2335
Ser Glu Gly Thr Ile Gln Phe Asp Gly Glu Gly Tyr Ala Leu Val Ser
2340 2345 2350
Arg Pro Ile Arg Trp Tyr Pro Asn Ile Ser Thr Val Met Phe Lys Phe
2355 2360 2365
Arg Thr Phe Ser Ser Ser Ala Leu Leu Met Tyr Leu Ala Thr Arg Asp
2370 2375 2380
Leu Arg Asp Phe Met Ser Val Glu Leu Thr Asp Gly His Ile Lys Val
2385 2390 2395 2400
Ser Tyr Asp Leu Gly Ser Gly Met Ala Ser Val Val Ser Asn Gln Asn
2405 2410 2415
His Asn Asp Gly Lys Trp Lys Ser Phe Thr Leu Ser Arg Ile Gln Lys
2420 2425 2430
Gln Ala Asn Ile Ser Ile Val Asp Ile Asp Thr Asn Gln Glu Glu Asn
2435 2440 2445
Ile Ala Thr Ser Ser Ser Gly Asn Asn Phe Gly Leu Asp Leu Lys Ala
2450 2455 2460
Asp Asp Lys Ile Tyr Phe Gly Gly Leu Pro Thr Leu Arg Asn Leu Ser
2465 2470 2475 2480
Met Lys Ala Arg Pro Glu Val Asn Leu Lys Lys Tyr Ser Gly Cys Leu
2485 2490 2495
Lys Asp Ile Glu Ile Ser Arg Thr Pro Tyr Asn Ile Leu Ser Ser Pro
2500 2505 2510
Asp Tyr Val Gly Val Thr Lys Gly Cys Ser Leu Glu Asn Val Tyr Thr
2515 2520 2525
Val Ser Phe Pro Lys Pro Gly Phe Val Glu Leu Ser Pro Val Pro Ile
2530 2535 2540
Asp Val Gly Thr Glu Ile Asn Leu Ser Phe Ser Thr Lys Asn Glu Ser
2545 2550 2555 2560
Gly Ile Ile Leu Leu Gly Ser Gly Gly Thr Pro Ala Pro Pro Arg Arg
2565 2570 2575
Lys Arg Arg Gln Thr Gly Gln Ala Tyr Tyr Ala Ile Leu Leu Asn Arg
2580 2585 2590
Gly Arg Leu Glu Val His Leu Ser Thr Gly Ala Arg Thr Met Arg Lys
2595 2600 2605
Ile Val Ile Arg Pro Glu Pro Asn Leu Phe His Asp Gly Arg Glu His
2610 2615 2620
Ser Val His Val Glu Arg Thr Arg Gly Ile Phe Thr Val Gln Val Asp
2625 2630 2635 2640
Glu Asn Arg Arg Tyr Met Gln Asn Leu Thr Val Glu Gln Pro Ile Glu
2645 2650 2655
Val Lys Lys Leu Phe Val Gly Gly Ala Pro Pro Glu Phe Gln Pro Ser
2660 2665 2670
Pro Leu Arg Asn Ile Pro Pro Phe Glu Gly Cys Ile Trp Asn Leu Val
2675 2680 2685
Ile Asn Ser Val Pro Met Asp Phe Ala Arg Pro Val Ser Phe Lys Asn
2690 2695 2700
Ala Asp Ile Gly Arg Cys Ala His Gln Lys Leu Arg Glu Asp Glu Asp
2705 2710 2715 2720
Gly Ala Ala Pro Ala Glu Ile Val Ile Gln Pro Glu Pro Val Pro Thr
2725 2730 2735
Pro Ala Phe Pro Thr Pro Thr Pro Val Leu Thr His Gly Pro Cys Ala
2740 2745 2750
Ala Glu Ser Glu Pro Ala Leu Leu Ile Gly Ser Lys Gln Phe Gly Leu
2755 2760 2765
Ser Arg Asn Ser His Ile Ala Ile Ala Phe Asp Asp Thr Lys Val Lys
2770 2775 2780
Asn Arg Leu Thr Ile Glu Leu Glu Val Arg Thr Glu Ala Glu Ser Gly
2785 2790 2795 2800
Leu Leu Phe Tyr Met Ala Arg Ile Asn His Ala Asp Phe Ala Thr Val
2805 2810 2815
Gln Leu Arg Asn Gly Leu Pro Tyr Phe Ser Tyr Asp Leu Gly Ser Gly
2820 2825 2830
Asp Thr His Thr Met Ile Pro Thr Lys Ile Asn Asp Gly Gln Trp His
2835 2840 2845
Lys Ile Lys Ile Met Arg Ser Lys Gln Glu Gly Ile Leu Tyr Val Asp
2850 2855 2860
Gly Ala Ser Asn Arg Thr Ile Ser Pro Lys Lys Ala Asp Ile Leu Asp
2865 2870 2875 2880
Val Val Gly Met Leu Tyr Val Gly Gly Leu Pro Ile Asn Tyr Thr Thr
2885 2890 2895
Arg Arg Ile Gly Pro Val Thr Tyr Ser Ile Asp Gly Cys Val Arg Asn
2900 2905 2910
Leu His Met Ala Glu Ala Pro Ala Asp Leu Glu Gln Pro Thr Ser Ser
2915 2920 2925
Phe His Val Gly Thr Cys Phe Ala Asn Ala Gln Arg Gly Thr Tyr Phe
2930 2935 2940
Asp Gly Thr Gly Phe Ala Lys Ala Val Gly Gly Phe Lys Val Gly Leu
2945 2950 2955 2960
Asp Leu Leu Val Glu Phe Glu Phe Arg Thr Thr Thr Thr Thr Gly Val
2965 2970 2975
Leu Leu Gly Ile Ser Ser Gln Lys Met Asp Gly Met Gly Ile Glu Met
2980 2985 2990
Ile Asp Glu Lys Leu Met Phe His Val Asp Asn Gly Ala Gly Arg Phe
2995 3000 3005
Thr Ala Val Tyr Asp Ala Gly Val Pro Gly His Leu Cys Asp Gly Gln
3010 3015 3020
Trp His Lys Val Thr Ala Asn Lys Ile Lys His Arg Ile Glu Leu Thr
3025 3030 3035 3040
Val Asp Gly Asn Gln Val Glu Ala Gln Ser Pro Asn Pro Ala Ser Thr
3045 3050 3055
Ser Ala Asp Thr Asn Asp Pro Val Phe Val Gly Gly Phe Pro Asp Asp
3060 3065 3070
Leu Lys Gln Phe Gly Leu Thr Thr Ser Ile Pro Phe Arg Gly Cys Ile
3075 3080 3085
Arg Ser Leu Lys Leu Thr Lys Gly Thr Gly Lys Pro Leu Glu Val Asn
3090 3095 3100
Phe Ala Lys Ala Leu Glu Leu Arg Gly Val Gln Pro Val Ser Cys Pro
3105 3110 3115 3120
Ala Asn
<210> 122
<211> 3695
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность подтипа Ламинина
альфа-5 (Человек разумный)
<400> 122
Met Ala Lys Arg Leu Cys Ala Gly Ser Ala Leu Cys Val Arg Gly Pro
1 5 10 15
Arg Gly Pro Ala Pro Leu Leu Leu Val Gly Leu Ala Leu Leu Gly Ala
20 25 30
Ala Arg Ala Arg Glu Glu Ala Gly Gly Gly Phe Ser Leu His Pro Pro
35 40 45
Tyr Phe Asn Leu Ala Glu Gly Ala Arg Ile Ala Ala Ser Ala Thr Cys
50 55 60
Gly Glu Glu Ala Pro Ala Arg Gly Ser Pro Arg Pro Thr Glu Asp Leu
65 70 75 80
Tyr Cys Lys Leu Val Gly Gly Pro Val Ala Gly Gly Asp Pro Asn Gln
85 90 95
Thr Ile Arg Gly Gln Tyr Cys Asp Ile Cys Thr Ala Ala Asn Ser Asn
100 105 110
Lys Ala His Pro Ala Ser Asn Ala Ile Asp Gly Thr Glu Arg Trp Trp
115 120 125
Gln Ser Pro Pro Leu Ser Arg Gly Leu Glu Tyr Asn Glu Val Asn Val
130 135 140
Thr Leu Asp Leu Gly Gln Val Phe His Val Ala Tyr Val Leu Ile Lys
145 150 155 160
Phe Ala Asn Ser Pro Arg Pro Asp Leu Trp Val Leu Glu Arg Ser Met
165 170 175
Asp Phe Gly Arg Thr Tyr Gln Pro Trp Gln Phe Phe Ala Ser Ser Lys
180 185 190
Arg Asp Cys Leu Glu Arg Phe Gly Pro Gln Thr Leu Glu Arg Ile Thr
195 200 205
Arg Asp Asp Ala Ala Ile Cys Thr Thr Glu Tyr Ser Arg Ile Val Pro
210 215 220
Leu Glu Asn Gly Glu Ile Val Val Ser Leu Val Asn Gly Arg Pro Gly
225 230 235 240
Ala Met Asn Phe Ser Tyr Ser Pro Leu Leu Arg Glu Phe Thr Lys Ala
245 250 255
Thr Asn Val Arg Leu Arg Phe Leu Arg Thr Asn Thr Leu Leu Gly His
260 265 270
Leu Met Gly Lys Ala Leu Arg Asp Pro Thr Val Thr Arg Arg Tyr Tyr
275 280 285
Tyr Ser Ile Lys Asp Ile Ser Ile Gly Gly Arg Cys Val Cys His Gly
290 295 300
His Ala Asp Ala Cys Asp Ala Lys Asp Pro Thr Asp Pro Phe Arg Leu
305 310 315 320
Gln Cys Thr Cys Gln His Asn Thr Cys Gly Gly Thr Cys Asp Arg Cys
325 330 335
Cys Pro Gly Phe Asn Gln Gln Pro Trp Lys Pro Ala Thr Ala Asn Ser
340 345 350
Ala Asn Glu Cys Gln Ser Cys Asn Cys Tyr Gly His Ala Thr Asp Cys
355 360 365
Tyr Tyr Asp Pro Glu Val Asp Arg Arg Arg Ala Ser Gln Ser Leu Asp
370 375 380
Gly Thr Tyr Gln Gly Gly Gly Val Cys Ile Asp Cys Gln His His Thr
385 390 395 400
Thr Gly Val Asn Cys Glu Arg Cys Leu Pro Gly Phe Tyr Arg Ser Pro
405 410 415
Asn His Pro Leu Asp Ser Pro His Val Cys Arg Arg Cys Asn Cys Glu
420 425 430
Ser Asp Phe Thr Asp Gly Thr Cys Glu Asp Leu Thr Gly Arg Cys Tyr
435 440 445
Cys Arg Pro Asn Phe Ser Gly Glu Arg Cys Asp Val Cys Ala Glu Gly
450 455 460
Phe Thr Gly Phe Pro Ser Cys Tyr Pro Thr Pro Ser Ser Ser Asn Asp
465 470 475 480
Thr Arg Glu Gln Val Leu Pro Ala Gly Gln Ile Val Asn Cys Asp Cys
485 490 495
Ser Ala Ala Gly Thr Gln Gly Asn Ala Cys Arg Lys Asp Pro Arg Val
500 505 510
Gly Arg Cys Leu Cys Lys Pro Asn Phe Gln Gly Thr His Cys Glu Leu
515 520 525
Cys Ala Pro Gly Phe Tyr Gly Pro Gly Cys Gln Pro Cys Gln Cys Ser
530 535 540
Ser Pro Gly Val Ala Asp Asp Arg Cys Asp Pro Asp Thr Gly Gln Cys
545 550 555 560
Arg Cys Arg Val Gly Phe Glu Gly Ala Thr Cys Asp Arg Cys Ala Pro
565 570 575
Gly Tyr Phe His Phe Pro Leu Cys Gln Leu Cys Gly Cys Ser Pro Ala
580 585 590
Gly Thr Leu Pro Glu Gly Cys Asp Glu Ala Gly Arg Cys Leu Cys Gln
595 600 605
Pro Glu Phe Ala Gly Pro His Cys Asp Arg Cys Arg Pro Gly Tyr His
610 615 620
Gly Phe Pro Asn Cys Gln Ala Cys Thr Cys Asp Pro Arg Gly Ala Leu
625 630 635 640
Asp Gln Leu Cys Gly Ala Gly Gly Leu Cys Arg Cys Arg Pro Gly Tyr
645 650 655
Thr Gly Thr Ala Cys Gln Glu Cys Ser Pro Gly Phe His Gly Phe Pro
660 665 670
Ser Cys Val Pro Cys His Cys Ser Ala Glu Gly Ser Leu His Ala Ala
675 680 685
Cys Asp Pro Arg Ser Gly Gln Cys Ser Cys Arg Pro Arg Val Thr Gly
690 695 700
Leu Arg Cys Asp Thr Cys Val Pro Gly Ala Tyr Asn Phe Pro Tyr Cys
705 710 715 720
Glu Ala Gly Ser Cys His Pro Ala Gly Leu Ala Pro Val Asp Pro Ala
725 730 735
Leu Pro Glu Ala Gln Val Pro Cys Met Cys Arg Ala His Val Glu Gly
740 745 750
Pro Ser Cys Asp Arg Cys Lys Pro Gly Phe Trp Gly Leu Ser Pro Ser
755 760 765
Asn Pro Glu Gly Cys Thr Arg Cys Ser Cys Asp Leu Arg Gly Thr Leu
770 775 780
Gly Gly Val Ala Glu Cys Gln Pro Gly Thr Gly Gln Cys Phe Cys Lys
785 790 795 800
Pro His Val Cys Gly Gln Ala Cys Ala Ser Cys Lys Asp Gly Phe Phe
805 810 815
Gly Leu Asp Gln Ala Asp Tyr Phe Gly Cys Arg Ser Cys Arg Cys Asp
820 825 830
Ile Gly Gly Ala Leu Gly Gln Ser Cys Glu Pro Arg Thr Gly Val Cys
835 840 845
Arg Cys Arg Pro Asn Thr Gln Gly Pro Thr Cys Ser Glu Pro Ala Arg
850 855 860
Asp His Tyr Leu Pro Asp Leu His His Leu Arg Leu Glu Leu Glu Glu
865 870 875 880
Ala Ala Thr Pro Glu Gly His Ala Val Arg Phe Gly Phe Asn Pro Leu
885 890 895
Glu Phe Glu Asn Phe Ser Trp Arg Gly Tyr Ala Gln Met Ala Pro Val
900 905 910
Gln Pro Arg Ile Val Ala Arg Leu Asn Leu Thr Ser Pro Asp Leu Phe
915 920 925
Trp Leu Val Phe Arg Tyr Val Asn Arg Gly Ala Met Ser Val Ser Gly
930 935 940
Arg Val Ser Val Arg Glu Glu Gly Arg Ser Ala Thr Cys Ala Asn Cys
945 950 955 960
Thr Ala Gln Ser Gln Pro Val Ala Phe Pro Pro Ser Thr Glu Pro Ala
965 970 975
Phe Ile Thr Val Pro Gln Arg Gly Phe Gly Glu Pro Phe Val Leu Asn
980 985 990
Pro Gly Thr Trp Ala Leu Arg Val Glu Ala Glu Gly Val Leu Leu Asp
995 1000 1005
Tyr Val Val Leu Leu Pro Ser Ala Tyr Tyr Glu Ala Ala Leu Leu Gln
1010 1015 1020
Leu Arg Val Thr Glu Ala Cys Thr Tyr Arg Pro Ser Ala Gln Gln Ser
1025 1030 1035 1040
Gly Asp Asn Cys Leu Leu Tyr Thr His Leu Pro Leu Asp Gly Phe Pro
1045 1050 1055
Ser Ala Ala Gly Leu Glu Ala Leu Cys Arg Gln Asp Asn Ser Leu Pro
1060 1065 1070
Arg Pro Cys Pro Thr Glu Gln Leu Ser Pro Ser His Pro Pro Leu Ile
1075 1080 1085
Thr Cys Thr Gly Ser Asp Val Asp Val Gln Leu Gln Val Ala Val Pro
1090 1095 1100
Gln Pro Gly Arg Tyr Ala Leu Val Val Glu Tyr Ala Asn Glu Asp Ala
1105 1110 1115 1120
Arg Gln Glu Val Gly Val Ala Val His Thr Pro Gln Arg Ala Pro Gln
1125 1130 1135
Gln Gly Leu Leu Ser Leu His Pro Cys Leu Tyr Ser Thr Leu Cys Arg
1140 1145 1150
Gly Thr Ala Arg Asp Thr Gln Asp His Leu Ala Val Phe His Leu Asp
1155 1160 1165
Ser Glu Ala Ser Val Arg Leu Thr Ala Glu Gln Ala Arg Phe Phe Leu
1170 1175 1180
His Gly Val Thr Leu Val Pro Ile Glu Glu Phe Ser Pro Glu Phe Val
1185 1190 1195 1200
Glu Pro Arg Val Ser Cys Ile Ser Ser His Gly Ala Phe Gly Pro Asn
1205 1210 1215
Ser Ala Ala Cys Leu Pro Ser Arg Phe Pro Lys Pro Pro Gln Pro Ile
1220 1225 1230
Ile Leu Arg Asp Cys Gln Val Ile Pro Leu Pro Pro Gly Leu Pro Leu
1235 1240 1245
Thr His Ala Gln Asp Leu Thr Pro Ala Met Ser Pro Ala Gly Pro Arg
1250 1255 1260
Pro Arg Pro Pro Thr Ala Val Asp Pro Asp Ala Glu Pro Thr Leu Leu
1265 1270 1275 1280
Arg Glu Pro Gln Ala Thr Val Val Phe Thr Thr His Val Pro Thr Leu
1285 1290 1295
Gly Arg Tyr Ala Phe Leu Leu His Gly Tyr Gln Pro Ala His Pro Thr
1300 1305 1310
Phe Pro Val Glu Val Leu Ile Asn Ala Gly Arg Val Trp Gln Gly His
1315 1320 1325
Ala Asn Ala Ser Phe Cys Pro His Gly Tyr Gly Cys Arg Thr Leu Val
1330 1335 1340
Val Cys Glu Gly Gln Ala Leu Leu Asp Val Thr His Ser Glu Leu Thr
1345 1350 1355 1360
Val Thr Val Arg Val Pro Lys Gly Arg Trp Leu Trp Leu Asp Tyr Val
1365 1370 1375
Leu Val Val Pro Glu Asn Val Tyr Ser Phe Gly Tyr Leu Arg Glu Glu
1380 1385 1390
Pro Leu Asp Lys Ser Tyr Asp Phe Ile Ser His Cys Ala Ala Gln Gly
1395 1400 1405
Tyr His Ile Ser Pro Ser Ser Ser Ser Leu Phe Cys Arg Asn Ala Ala
1410 1415 1420
Ala Ser Leu Ser Leu Phe Tyr Asn Asn Gly Ala Arg Pro Cys Gly Cys
1425 1430 1435 1440
His Glu Val Gly Ala Thr Gly Pro Thr Cys Glu Pro Phe Gly Gly Gln
1445 1450 1455
Cys Pro Cys His Ala His Val Ile Gly Arg Asp Cys Ser Arg Cys Ala
1460 1465 1470
Thr Gly Tyr Trp Gly Phe Pro Asn Cys Arg Pro Cys Asp Cys Gly Ala
1475 1480 1485
Arg Leu Cys Asp Glu Leu Thr Gly Gln Cys Ile Cys Pro Pro Arg Thr
1490 1495 1500
Ile Pro Pro Asp Cys Leu Leu Cys Gln Pro Gln Thr Phe Gly Cys His
1505 1510 1515 1520
Pro Leu Val Gly Cys Glu Glu Cys Asn Cys Ser Gly Pro Gly Ile Gln
1525 1530 1535
Glu Leu Thr Asp Pro Thr Cys Asp Thr Asp Ser Gly Gln Cys Lys Cys
1540 1545 1550
Arg Pro Asn Val Thr Gly Arg Arg Cys Asp Thr Cys Ser Pro Gly Phe
1555 1560 1565
His Gly Tyr Pro Arg Cys Arg Pro Cys Asp Cys His Glu Ala Gly Thr
1570 1575 1580
Ala Pro Gly Val Cys Asp Pro Leu Thr Gly Gln Cys Tyr Cys Lys Glu
1585 1590 1595 1600
Asn Val Gln Gly Pro Lys Cys Asp Gln Cys Ser Leu Gly Thr Phe Ser
1605 1610 1615
Leu Asp Ala Ala Asn Pro Lys Gly Cys Thr Arg Cys Phe Cys Phe Gly
1620 1625 1630
Ala Thr Glu Arg Cys Arg Ser Ser Ser Tyr Thr Arg Gln Glu Phe Val
1635 1640 1645
Asp Met Glu Gly Trp Val Leu Leu Ser Thr Asp Arg Gln Val Val Pro
1650 1655 1660
His Glu Arg Gln Pro Gly Thr Glu Met Leu Arg Ala Asp Leu Arg His
1665 1670 1675 1680
Val Pro Glu Ala Val Pro Glu Ala Phe Pro Glu Leu Tyr Trp Gln Ala
1685 1690 1695
Pro Pro Ser Tyr Leu Gly Asp Arg Val Ser Ser Tyr Gly Gly Thr Leu
1700 1705 1710
Arg Tyr Glu Leu His Ser Glu Thr Gln Arg Gly Asp Val Phe Val Pro
1715 1720 1725
Met Glu Ser Arg Pro Asp Val Val Leu Gln Gly Asn Gln Met Ser Ile
1730 1735 1740
Thr Phe Leu Glu Pro Ala Tyr Pro Thr Pro Gly His Val His Arg Gly
1745 1750 1755 1760
Gln Leu Gln Leu Val Glu Gly Asn Phe Arg His Thr Glu Thr Arg Asn
1765 1770 1775
Thr Val Ser Arg Glu Glu Leu Met Met Val Leu Ala Ser Leu Glu Gln
1780 1785 1790
Leu Gln Ile Arg Ala Leu Phe Ser Gln Ile Ser Ser Ala Val Phe Leu
1795 1800 1805
Arg Arg Val Ala Leu Glu Val Ala Ser Pro Ala Gly Gln Gly Ala Leu
1810 1815 1820
Ala Ser Asn Val Glu Leu Cys Leu Cys Pro Ala Ser Tyr Arg Gly Asp
1825 1830 1835 1840
Ser Cys Gln Glu Cys Ala Pro Gly Phe Tyr Arg Asp Val Lys Gly Leu
1845 1850 1855
Phe Leu Gly Arg Cys Val Pro Cys Gln Cys His Gly His Ser Asp Arg
1860 1865 1870
Cys Leu Pro Gly Ser Gly Val Cys Val Asp Cys Gln His Asn Thr Glu
1875 1880 1885
Gly Ala His Cys Glu Arg Cys Gln Ala Gly Phe Val Ser Ser Arg Asp
1890 1895 1900
Asp Pro Ser Ala Pro Cys Val Ser Cys Pro Cys Pro Leu Ser Val Pro
1905 1910 1915 1920
Ser Asn Asn Phe Ala Glu Gly Cys Val Leu Arg Gly Gly Arg Thr Gln
1925 1930 1935
Cys Leu Cys Lys Pro Gly Tyr Ala Gly Ala Ser Cys Glu Arg Cys Ala
1940 1945 1950
Pro Gly Phe Phe Gly Asn Pro Leu Val Leu Gly Ser Ser Cys Gln Pro
1955 1960 1965
Cys Asp Cys Ser Gly Asn Gly Asp Pro Asn Leu Leu Phe Ser Asp Cys
1970 1975 1980
Asp Pro Leu Thr Gly Ala Cys Arg Gly Cys Leu Arg His Thr Thr Gly
1985 1990 1995 2000
Pro Arg Cys Glu Ile Cys Ala Pro Gly Phe Tyr Gly Asn Ala Leu Leu
2005 2010 2015
Pro Gly Asn Cys Thr Arg Cys Asp Cys Thr Pro Cys Gly Thr Glu Ala
2020 2025 2030
Cys Asp Pro His Ser Gly His Cys Leu Cys Lys Ala Gly Val Thr Gly
2035 2040 2045
Arg Arg Cys Asp Arg Cys Gln Glu Gly His Phe Gly Phe Asp Gly Cys
2050 2055 2060
Gly Gly Cys Arg Pro Cys Ala Cys Gly Pro Ala Ala Glu Gly Ser Glu
2065 2070 2075 2080
Cys His Pro Gln Ser Gly Gln Cys His Cys Arg Pro Gly Thr Met Gly
2085 2090 2095
Pro Gln Cys Arg Glu Cys Ala Pro Gly Tyr Trp Gly Leu Pro Glu Gln
2100 2105 2110
Gly Cys Arg Arg Cys Gln Cys Pro Gly Gly Arg Cys Asp Pro His Thr
2115 2120 2125
Gly Arg Cys Asn Cys Pro Pro Gly Leu Ser Gly Glu Arg Cys Asp Thr
2130 2135 2140
Cys Ser Gln Gln His Gln Val Pro Val Pro Gly Gly Pro Val Gly His
2145 2150 2155 2160
Ser Ile His Cys Glu Val Cys Asp His Cys Val Val Leu Leu Leu Asp
2165 2170 2175
Asp Leu Glu Arg Ala Gly Ala Leu Leu Pro Ala Ile His Glu Gln Leu
2180 2185 2190
Arg Gly Ile Asn Ala Ser Ser Met Ala Trp Ala Arg Leu His Arg Leu
2195 2200 2205
Asn Ala Ser Ile Ala Asp Leu Gln Ser Gln Leu Arg Ser Pro Leu Gly
2210 2215 2220
Pro Arg His Glu Thr Ala Gln Gln Leu Glu Val Leu Glu Gln Gln Ser
2225 2230 2235 2240
Thr Ser Leu Gly Gln Asp Ala Arg Arg Leu Gly Gly Gln Ala Val Gly
2245 2250 2255
Thr Arg Asp Gln Ala Ser Gln Leu Leu Ala Gly Thr Glu Ala Thr Leu
2260 2265 2270
Gly His Ala Lys Thr Leu Leu Ala Ala Ile Arg Ala Val Asp Arg Thr
2275 2280 2285
Leu Ser Glu Leu Met Ser Gln Thr Gly His Leu Gly Leu Ala Asn Ala
2290 2295 2300
Ser Ala Pro Ser Gly Glu Gln Leu Leu Arg Thr Leu Ala Glu Val Glu
2305 2310 2315 2320
Arg Leu Leu Trp Glu Met Arg Ala Arg Asp Leu Gly Ala Pro Gln Ala
2325 2330 2335
Ala Ala Glu Ala Glu Leu Ala Ala Ala Gln Arg Leu Leu Ala Arg Val
2340 2345 2350
Gln Glu Gln Leu Ser Ser Leu Trp Glu Glu Asn Gln Ala Leu Ala Thr
2355 2360 2365
Gln Thr Arg Asp Arg Leu Ala Gln His Glu Ala Gly Leu Met Asp Leu
2370 2375 2380
Arg Glu Ala Leu Asn Arg Ala Val Asp Ala Thr Arg Glu Ala Gln Glu
2385 2390 2395 2400
Leu Asn Ser Arg Asn Gln Glu Arg Leu Glu Glu Ala Leu Gln Arg Lys
2405 2410 2415
Gln Glu Leu Ser Arg Asp Asn Ala Thr Leu Gln Ala Thr Leu His Ala
2420 2425 2430
Ala Arg Asp Thr Leu Ala Ser Val Phe Arg Leu Leu His Ser Leu Asp
2435 2440 2445
Gln Ala Lys Glu Glu Leu Glu Arg Leu Ala Ala Ser Leu Asp Gly Ala
2450 2455 2460
Arg Thr Pro Leu Leu Gln Arg Met Gln Thr Phe Ser Pro Ala Gly Ser
2465 2470 2475 2480
Lys Leu Arg Leu Val Glu Ala Ala Glu Ala His Ala Gln Gln Leu Gly
2485 2490 2495
Gln Leu Ala Leu Asn Leu Ser Ser Ile Ile Leu Asp Val Asn Gln Asp
2500 2505 2510
Arg Leu Thr Gln Arg Ala Ile Glu Ala Ser Asn Ala Tyr Ser Arg Ile
2515 2520 2525
Leu Gln Ala Val Gln Ala Ala Glu Asp Ala Ala Gly Gln Ala Leu Gln
2530 2535 2540
Gln Ala Asp His Thr Trp Ala Thr Val Val Arg Gln Gly Leu Val Asp
2545 2550 2555 2560
Arg Ala Gln Gln Leu Leu Ala Asn Ser Thr Ala Leu Glu Glu Ala Met
2565 2570 2575
Leu Gln Glu Gln Gln Arg Leu Gly Leu Val Trp Ala Ala Leu Gln Gly
2580 2585 2590
Ala Arg Thr Gln Leu Arg Asp Val Arg Ala Lys Lys Asp Gln Leu Glu
2595 2600 2605
Ala His Ile Gln Ala Ala Gln Ala Met Leu Ala Met Asp Thr Asp Glu
2610 2615 2620
Thr Ser Lys Lys Ile Ala His Ala Lys Ala Val Ala Ala Glu Ala Gln
2625 2630 2635 2640
Asp Thr Ala Thr Arg Val Gln Ser Gln Leu Gln Ala Met Gln Glu Asn
2645 2650 2655
Val Glu Arg Trp Gln Gly Gln Tyr Glu Gly Leu Arg Gly Gln Asp Leu
2660 2665 2670
Gly Gln Ala Val Leu Asp Ala Gly His Ser Val Ser Thr Leu Glu Lys
2675 2680 2685
Thr Leu Pro Gln Leu Leu Ala Lys Leu Ser Ile Leu Glu Asn Arg Gly
2690 2695 2700
Val His Asn Ala Ser Leu Ala Leu Ser Ala Ser Ile Gly Arg Val Arg
2705 2710 2715 2720
Glu Leu Ile Ala Gln Ala Arg Gly Ala Ala Ser Lys Val Lys Val Pro
2725 2730 2735
Met Lys Phe Asn Gly Arg Ser Gly Val Gln Leu Arg Thr Pro Arg Asp
2740 2745 2750
Leu Ala Asp Leu Ala Ala Tyr Thr Ala Leu Lys Phe Tyr Leu Gln Gly
2755 2760 2765
Pro Glu Pro Glu Pro Gly Gln Gly Thr Glu Asp Arg Phe Val Met Tyr
2770 2775 2780
Met Gly Ser Arg Gln Ala Thr Gly Asp Tyr Met Gly Val Ser Leu Arg
2785 2790 2795 2800
Asp Lys Lys Val His Trp Val Tyr Gln Leu Gly Glu Ala Gly Pro Ala
2805 2810 2815
Val Leu Ser Ile Asp Glu Asp Ile Gly Glu Gln Phe Ala Ala Val Ser
2820 2825 2830
Leu Asp Arg Thr Leu Gln Phe Gly His Met Ser Val Thr Val Glu Arg
2835 2840 2845
Gln Met Ile Gln Glu Thr Lys Gly Asp Thr Val Ala Pro Gly Ala Glu
2850 2855 2860
Gly Leu Leu Asn Leu Arg Pro Asp Asp Phe Val Phe Tyr Val Gly Gly
2865 2870 2875 2880
Tyr Pro Ser Thr Phe Thr Pro Pro Pro Leu Leu Arg Phe Pro Gly Tyr
2885 2890 2895
Arg Gly Cys Ile Glu Met Asp Thr Leu Asn Glu Glu Val Val Ser Leu
2900 2905 2910
Tyr Asn Phe Glu Arg Thr Phe Gln Leu Asp Thr Ala Val Asp Arg Pro
2915 2920 2925
Cys Ala Arg Ser Lys Ser Thr Gly Asp Pro Trp Leu Thr Asp Gly Ser
2930 2935 2940
Tyr Leu Asp Gly Thr Gly Phe Ala Arg Ile Ser Phe Asp Ser Gln Ile
2945 2950 2955 2960
Ser Thr Thr Lys Arg Phe Glu Gln Glu Leu Arg Leu Val Ser Tyr Ser
2965 2970 2975
Gly Val Leu Phe Phe Leu Lys Gln Gln Ser Gln Phe Leu Cys Leu Ala
2980 2985 2990
Val Gln Glu Gly Ser Leu Val Leu Leu Tyr Asp Phe Gly Ala Gly Leu
2995 3000 3005
Lys Lys Ala Val Pro Leu Gln Pro Pro Pro Pro Leu Thr Ser Ala Ser
3010 3015 3020
Lys Ala Ile Gln Val Phe Leu Leu Gly Gly Ser Arg Lys Arg Val Leu
3025 3030 3035 3040
Val Arg Val Glu Arg Ala Thr Val Tyr Ser Val Glu Gln Asp Asn Asp
3045 3050 3055
Leu Glu Leu Ala Asp Ala Tyr Tyr Leu Gly Gly Val Pro Pro Asp Gln
3060 3065 3070
Leu Pro Pro Ser Leu Arg Arg Leu Phe Pro Thr Gly Gly Ser Val Arg
3075 3080 3085
Gly Cys Val Lys Gly Ile Lys Ala Leu Gly Lys Tyr Val Asp Leu Lys
3090 3095 3100
Arg Leu Asn Thr Thr Gly Val Ser Ala Gly Cys Thr Ala Asp Leu Leu
3105 3110 3115 3120
Val Gly Arg Ala Met Thr Phe His Gly His Gly Phe Leu Arg Leu Ala
3125 3130 3135
Leu Ser Asn Val Ala Pro Leu Thr Gly Asn Val Tyr Ser Gly Phe Gly
3140 3145 3150
Phe His Ser Ala Gln Asp Ser Ala Leu Leu Tyr Tyr Arg Ala Ser Pro
3155 3160 3165
Asp Gly Leu Cys Gln Val Ser Leu Gln Gln Gly Arg Val Ser Leu Gln
3170 3175 3180
Leu Leu Arg Thr Glu Val Lys Thr Gln Ala Gly Phe Ala Asp Gly Ala
3185 3190 3195 3200
Pro His Tyr Val Ala Phe Tyr Ser Asn Ala Thr Gly Val Trp Leu Tyr
3205 3210 3215
Val Asp Asp Gln Leu Gln Gln Met Lys Pro His Arg Gly Pro Pro Pro
3220 3225 3230
Glu Leu Gln Pro Gln Pro Glu Gly Pro Pro Arg Leu Leu Leu Gly Gly
3235 3240 3245
Leu Pro Glu Ser Gly Thr Ile Tyr Asn Phe Ser Gly Cys Ile Ser Asn
3250 3255 3260
Val Phe Val Gln Arg Leu Leu Gly Pro Gln Arg Val Phe Asp Leu Gln
3265 3270 3275 3280
Gln Asn Leu Gly Ser Val Asn Val Ser Thr Gly Cys Ala Pro Ala Leu
3285 3290 3295
Gln Ala Gln Thr Pro Gly Leu Gly Pro Arg Gly Leu Gln Ala Thr Ala
3300 3305 3310
Arg Lys Ala Ser Arg Arg Ser Arg Gln Pro Ala Arg His Pro Ala Cys
3315 3320 3325
Met Leu Pro Pro His Leu Arg Thr Thr Arg Asp Ser Tyr Gln Phe Gly
3330 3335 3340
Gly Ser Leu Ser Ser His Leu Glu Phe Val Gly Ile Leu Ala Arg His
3345 3350 3355 3360
Arg Asn Trp Pro Ser Leu Ser Met His Val Leu Pro Arg Ser Ser Arg
3365 3370 3375
Gly Leu Leu Leu Phe Thr Ala Arg Leu Arg Pro Gly Ser Pro Ser Leu
3380 3385 3390
Ala Leu Phe Leu Ser Asn Gly His Phe Val Ala Gln Met Glu Gly Leu
3395 3400 3405
Gly Thr Arg Leu Arg Ala Gln Ser Arg Gln Arg Ser Arg Pro Gly Arg
3410 3415 3420
Trp His Lys Val Ser Val Arg Trp Glu Lys Asn Arg Ile Leu Leu Val
3425 3430 3435 3440
Thr Asp Gly Ala Arg Ala Trp Ser Gln Glu Gly Pro His Arg Gln His
3445 3450 3455
Gln Gly Ala Glu His Pro Gln Pro His Thr Leu Phe Val Gly Gly Leu
3460 3465 3470
Pro Ala Ser Ser His Ser Ser Lys Leu Pro Val Thr Val Gly Phe Ser
3475 3480 3485
Gly Cys Val Lys Arg Leu Arg Leu His Gly Arg Pro Leu Gly Ala Pro
3490 3495 3500
Thr Arg Met Ala Gly Val Thr Pro Cys Ile Leu Gly Pro Leu Glu Ala
3505 3510 3515 3520
Gly Leu Phe Phe Pro Gly Ser Gly Gly Val Ile Thr Leu Asp Leu Pro
3525 3530 3535
Gly Ala Thr Leu Pro Asp Val Gly Leu Glu Leu Glu Val Arg Pro Leu
3540 3545 3550
Ala Val Thr Gly Leu Ile Phe His Leu Gly Gln Ala Arg Thr Pro Pro
3555 3560 3565
Tyr Leu Gln Leu Gln Val Thr Glu Lys Gln Val Leu Leu Arg Ala Asp
3570 3575 3580
Asp Gly Ala Gly Glu Phe Ser Thr Ser Val Thr Arg Pro Ser Val Leu
3585 3590 3595 3600
Cys Asp Gly Gln Trp His Arg Leu Ala Val Met Lys Ser Gly Asn Val
3605 3610 3615
Leu Arg Leu Glu Val Asp Ala Gln Ser Asn His Thr Val Gly Pro Leu
3620 3625 3630
Leu Ala Ala Ala Ala Gly Ala Pro Ala Pro Leu Tyr Leu Gly Gly Leu
3635 3640 3645
Pro Glu Pro Met Ala Val Gln Pro Trp Pro Pro Ala Tyr Cys Gly Cys
3650 3655 3660
Met Arg Arg Leu Ala Val Asn Arg Ser Pro Val Ala Met Thr Arg Ser
3665 3670 3675 3680
Val Glu Val His Gly Ala Val Gly Ala Ser Gly Cys Pro Ala Ala
3685 3690 3695
<210> 123
<211> 1786
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность подтипа Ламинина
бета-1 (Человек разумный)
<400> 123
Met Gly Leu Leu Gln Leu Leu Ala Phe Ser Phe Leu Ala Leu Cys Arg
1 5 10 15
Ala Arg Val Arg Ala Gln Glu Pro Glu Phe Ser Tyr Gly Cys Ala Glu
20 25 30
Gly Ser Cys Tyr Pro Ala Thr Gly Asp Leu Leu Ile Gly Arg Ala Gln
35 40 45
Lys Leu Ser Val Thr Ser Thr Cys Gly Leu His Lys Pro Glu Pro Tyr
50 55 60
Cys Ile Val Ser His Leu Gln Glu Asp Lys Lys Cys Phe Ile Cys Asn
65 70 75 80
Ser Gln Asp Pro Tyr His Glu Thr Leu Asn Pro Asp Ser His Leu Ile
85 90 95
Glu Asn Val Val Thr Thr Phe Ala Pro Asn Arg Leu Lys Ile Trp Trp
100 105 110
Gln Ser Glu Asn Gly Val Glu Asn Val Thr Ile Gln Leu Asp Leu Glu
115 120 125
Ala Glu Phe His Phe Thr His Leu Ile Met Thr Phe Lys Thr Phe Arg
130 135 140
Pro Ala Ala Met Leu Ile Glu Arg Ser Ser Asp Phe Gly Lys Thr Trp
145 150 155 160
Gly Val Tyr Arg Tyr Phe Ala Tyr Asp Cys Glu Ala Ser Phe Pro Gly
165 170 175
Ile Ser Thr Gly Pro Met Lys Lys Val Asp Asp Ile Ile Cys Asp Ser
180 185 190
Arg Tyr Ser Asp Ile Glu Pro Ser Thr Glu Gly Glu Val Ile Phe Arg
195 200 205
Ala Leu Asp Pro Ala Phe Lys Ile Glu Asp Pro Tyr Ser Pro Arg Ile
210 215 220
Gln Asn Leu Leu Lys Ile Thr Asn Leu Arg Ile Lys Phe Val Lys Leu
225 230 235 240
His Thr Leu Gly Asp Asn Leu Leu Asp Ser Arg Met Glu Ile Arg Glu
245 250 255
Lys Tyr Tyr Tyr Ala Val Tyr Asp Met Val Val Arg Gly Asn Cys Phe
260 265 270
Cys Tyr Gly His Ala Ser Glu Cys Ala Pro Val Asp Gly Phe Asn Glu
275 280 285
Glu Val Glu Gly Met Val His Gly His Cys Met Cys Arg His Asn Thr
290 295 300
Lys Gly Leu Asn Cys Glu Leu Cys Met Asp Phe Tyr His Asp Leu Pro
305 310 315 320
Trp Arg Pro Ala Glu Gly Arg Asn Ser Asn Ala Cys Lys Lys Cys Asn
325 330 335
Cys Asn Glu His Ser Ile Ser Cys His Phe Asp Met Ala Val Tyr Leu
340 345 350
Ala Thr Gly Asn Val Ser Gly Gly Val Cys Asp Asp Cys Gln His Asn
355 360 365
Thr Met Gly Arg Asn Cys Glu Gln Cys Lys Pro Phe Tyr Tyr Gln His
370 375 380
Pro Glu Arg Asp Ile Arg Asp Pro Asn Phe Cys Glu Arg Cys Thr Cys
385 390 395 400
Asp Pro Ala Gly Ser Gln Asn Glu Gly Ile Cys Asp Ser Tyr Thr Asp
405 410 415
Phe Ser Thr Gly Leu Ile Ala Gly Gln Cys Arg Cys Lys Leu Asn Val
420 425 430
Glu Gly Glu His Cys Asp Val Cys Lys Glu Gly Phe Tyr Asp Leu Ser
435 440 445
Ser Glu Asp Pro Phe Gly Cys Lys Ser Cys Ala Cys Asn Pro Leu Gly
450 455 460
Thr Ile Pro Gly Gly Asn Pro Cys Asp Ser Glu Thr Gly His Cys Tyr
465 470 475 480
Cys Lys Arg Leu Val Thr Gly Gln His Cys Asp Gln Cys Leu Pro Glu
485 490 495
His Trp Gly Leu Ser Asn Asp Leu Asp Gly Cys Arg Pro Cys Asp Cys
500 505 510
Asp Leu Gly Gly Ala Leu Asn Asn Ser Cys Phe Ala Glu Ser Gly Gln
515 520 525
Cys Ser Cys Arg Pro His Met Ile Gly Arg Gln Cys Asn Glu Val Glu
530 535 540
Pro Gly Tyr Tyr Phe Ala Thr Leu Asp His Tyr Leu Tyr Glu Ala Glu
545 550 555 560
Glu Ala Asn Leu Gly Pro Gly Val Ser Ile Val Glu Arg Gln Tyr Ile
565 570 575
Gln Asp Arg Ile Pro Ser Trp Thr Gly Ala Gly Phe Val Arg Val Pro
580 585 590
Glu Gly Ala Tyr Leu Glu Phe Phe Ile Asp Asn Ile Pro Tyr Ser Met
595 600 605
Glu Tyr Asp Ile Leu Ile Arg Tyr Glu Pro Gln Leu Pro Asp His Trp
610 615 620
Glu Lys Ala Val Ile Thr Val Gln Arg Pro Gly Arg Ile Pro Thr Ser
625 630 635 640
Ser Arg Cys Gly Asn Thr Ile Pro Asp Asp Asp Asn Gln Val Val Ser
645 650 655
Leu Ser Pro Gly Ser Arg Tyr Val Val Leu Pro Arg Pro Val Cys Phe
660 665 670
Glu Lys Gly Thr Asn Tyr Thr Val Arg Leu Glu Leu Pro Gln Tyr Thr
675 680 685
Ser Ser Asp Ser Asp Val Glu Ser Pro Tyr Thr Leu Ile Asp Ser Leu
690 695 700
Val Leu Met Pro Tyr Cys Lys Ser Leu Asp Ile Phe Thr Val Gly Gly
705 710 715 720
Ser Gly Asp Gly Val Val Thr Asn Ser Ala Trp Glu Thr Phe Gln Arg
725 730 735
Tyr Arg Cys Leu Glu Asn Ser Arg Ser Val Val Lys Thr Pro Met Thr
740 745 750
Asp Val Cys Arg Asn Ile Ile Phe Ser Ile Ser Ala Leu Leu His Gln
755 760 765
Thr Gly Leu Ala Cys Glu Cys Asp Pro Gln Gly Ser Leu Ser Ser Val
770 775 780
Cys Asp Pro Asn Gly Gly Gln Cys Gln Cys Arg Pro Asn Val Val Gly
785 790 795 800
Arg Thr Cys Asn Arg Cys Ala Pro Gly Thr Phe Gly Phe Gly Pro Ser
805 810 815
Gly Cys Lys Pro Cys Glu Cys His Leu Gln Gly Ser Val Asn Ala Phe
820 825 830
Cys Asn Pro Val Thr Gly Gln Cys His Cys Phe Gln Gly Val Tyr Ala
835 840 845
Arg Gln Cys Asp Arg Cys Leu Pro Gly His Trp Gly Phe Pro Ser Cys
850 855 860
Gln Pro Cys Gln Cys Asn Gly His Ala Asp Asp Cys Asp Pro Val Thr
865 870 875 880
Gly Glu Cys Leu Asn Cys Gln Asp Tyr Thr Met Gly His Asn Cys Glu
885 890 895
Arg Cys Leu Ala Gly Tyr Tyr Gly Asp Pro Ile Ile Gly Ser Gly Asp
900 905 910
His Cys Arg Pro Cys Pro Cys Pro Asp Gly Pro Asp Ser Gly Arg Gln
915 920 925
Phe Ala Arg Ser Cys Tyr Gln Asp Pro Val Thr Leu Gln Leu Ala Cys
930 935 940
Val Cys Asp Pro Gly Tyr Ile Gly Ser Arg Cys Asp Asp Cys Ala Ser
945 950 955 960
Gly Tyr Phe Gly Asn Pro Ser Glu Val Gly Gly Ser Cys Gln Pro Cys
965 970 975
Gln Cys His Asn Asn Ile Asp Thr Thr Asp Pro Glu Ala Cys Asp Lys
980 985 990
Glu Thr Gly Arg Cys Leu Lys Cys Leu Tyr His Thr Glu Gly Glu His
995 1000 1005
Cys Gln Phe Cys Arg Phe Gly Tyr Tyr Gly Asp Ala Leu Gln Gln Asp
1010 1015 1020
Cys Arg Lys Cys Val Cys Asn Tyr Leu Gly Thr Val Gln Glu His Cys
1025 1030 1035 1040
Asn Gly Ser Asp Cys Gln Cys Asp Lys Ala Thr Gly Gln Cys Leu Cys
1045 1050 1055
Leu Pro Asn Val Ile Gly Gln Asn Cys Asp Arg Cys Ala Pro Asn Thr
1060 1065 1070
Trp Gln Leu Ala Ser Gly Thr Gly Cys Asp Pro Cys Asn Cys Asn Ala
1075 1080 1085
Ala His Ser Phe Gly Pro Ser Cys Asn Glu Phe Thr Gly Gln Cys Gln
1090 1095 1100
Cys Met Pro Gly Phe Gly Gly Arg Thr Cys Ser Glu Cys Gln Glu Leu
1105 1110 1115 1120
Phe Trp Gly Asp Pro Asp Val Glu Cys Arg Ala Cys Asp Cys Asp Pro
1125 1130 1135
Arg Gly Ile Glu Thr Pro Gln Cys Asp Gln Ser Thr Gly Gln Cys Val
1140 1145 1150
Cys Val Glu Gly Val Glu Gly Pro Arg Cys Asp Lys Cys Thr Arg Gly
1155 1160 1165
Tyr Ser Gly Val Phe Pro Asp Cys Thr Pro Cys His Gln Cys Phe Ala
1170 1175 1180
Leu Trp Asp Val Ile Ile Ala Glu Leu Thr Asn Arg Thr His Arg Phe
1185 1190 1195 1200
Leu Glu Lys Ala Lys Ala Leu Lys Ile Ser Gly Val Ile Gly Pro Tyr
1205 1210 1215
Arg Glu Thr Val Asp Ser Val Glu Arg Lys Val Ser Glu Ile Lys Asp
1220 1225 1230
Ile Leu Ala Gln Ser Pro Ala Ala Glu Pro Leu Lys Asn Ile Gly Asn
1235 1240 1245
Leu Phe Glu Glu Ala Glu Lys Leu Ile Lys Asp Val Thr Glu Met Met
1250 1255 1260
Ala Gln Val Glu Val Lys Leu Ser Asp Thr Thr Ser Gln Ser Asn Ser
1265 1270 1275 1280
Thr Ala Lys Glu Leu Asp Ser Leu Gln Thr Glu Ala Glu Ser Leu Asp
1285 1290 1295
Asn Thr Val Lys Glu Leu Ala Glu Gln Leu Glu Phe Ile Lys Asn Ser
1300 1305 1310
Asp Ile Arg Gly Ala Leu Asp Ser Ile Thr Lys Tyr Phe Gln Met Ser
1315 1320 1325
Leu Glu Ala Glu Glu Arg Val Asn Ala Ser Thr Thr Glu Pro Asn Ser
1330 1335 1340
Thr Val Glu Gln Ser Ala Leu Met Arg Asp Arg Val Glu Asp Val Met
1345 1350 1355 1360
Met Glu Arg Glu Ser Gln Phe Lys Glu Lys Gln Glu Glu Gln Ala Arg
1365 1370 1375
Leu Leu Asp Glu Leu Ala Gly Lys Leu Gln Ser Leu Asp Leu Ser Ala
1380 1385 1390
Ala Ala Glu Met Thr Cys Gly Thr Pro Pro Gly Ala Ser Cys Ser Glu
1395 1400 1405
Thr Glu Cys Gly Gly Pro Asn Cys Arg Thr Asp Glu Gly Glu Arg Lys
1410 1415 1420
Cys Gly Gly Pro Gly Cys Gly Gly Leu Val Thr Val Ala His Asn Ala
1425 1430 1435 1440
Trp Gln Lys Ala Met Asp Leu Asp Gln Asp Val Leu Ser Ala Leu Ala
1445 1450 1455
Glu Val Glu Gln Leu Ser Lys Met Val Ser Glu Ala Lys Leu Arg Ala
1460 1465 1470
Asp Glu Ala Lys Gln Ser Ala Glu Asp Ile Leu Leu Lys Thr Asn Ala
1475 1480 1485
Thr Lys Glu Lys Met Asp Lys Ser Asn Glu Glu Leu Arg Asn Leu Ile
1490 1495 1500
Lys Gln Ile Arg Asn Phe Leu Thr Gln Asp Ser Ala Asp Leu Asp Ser
1505 1510 1515 1520
Ile Glu Ala Val Ala Asn Glu Val Leu Lys Met Glu Met Pro Ser Thr
1525 1530 1535
Pro Gln Gln Leu Gln Asn Leu Thr Glu Asp Ile Arg Glu Arg Val Glu
1540 1545 1550
Ser Leu Ser Gln Val Glu Val Ile Leu Gln His Ser Ala Ala Asp Ile
1555 1560 1565
Ala Arg Ala Glu Met Leu Leu Glu Glu Ala Lys Arg Ala Ser Lys Ser
1570 1575 1580
Ala Thr Asp Val Lys Val Thr Ala Asp Met Val Lys Glu Ala Leu Glu
1585 1590 1595 1600
Glu Ala Glu Lys Ala Gln Val Ala Ala Glu Lys Ala Ile Lys Gln Ala
1605 1610 1615
Asp Glu Asp Ile Gln Gly Thr Gln Asn Leu Leu Thr Ser Ile Glu Ser
1620 1625 1630
Glu Thr Ala Ala Ser Glu Glu Thr Leu Phe Asn Ala Ser Gln Arg Ile
1635 1640 1645
Ser Glu Leu Glu Arg Asn Val Glu Glu Leu Lys Arg Lys Ala Ala Gln
1650 1655 1660
Asn Ser Gly Glu Ala Glu Tyr Ile Glu Lys Val Val Tyr Thr Val Lys
1665 1670 1675 1680
Gln Ser Ala Glu Asp Val Lys Lys Thr Leu Asp Gly Glu Leu Asp Glu
1685 1690 1695
Lys Tyr Lys Lys Val Glu Asn Leu Ile Ala Lys Lys Thr Glu Glu Ser
1700 1705 1710
Ala Asp Ala Arg Arg Lys Ala Glu Met Leu Gln Asn Glu Ala Lys Thr
1715 1720 1725
Leu Leu Ala Gln Ala Asn Ser Lys Leu Gln Leu Leu Lys Asp Leu Glu
1730 1735 1740
Arg Lys Tyr Glu Asp Asn Gln Arg Tyr Leu Glu Asp Lys Ala Gln Glu
1745 1750 1755 1760
Leu Ala Arg Leu Glu Gly Glu Val Arg Ser Leu Leu Lys Asp Ile Ser
1765 1770 1775
Gln Lys Val Ala Val Tyr Ser Thr Cys Leu
1780 1785
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ДЛЯ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С N-КОНЦЕВЫМ ДОМЕНОМ ЛИЗИЛ-ТРНК-СИНТЕТАЗЫ, ЭКСПОНИРОВАННЫМ НА ВНЕКЛЕТОЧНОЙ МЕМБРАНЕ | 2019 |
|
RU2781304C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО, СПЕЦИФИЧНО СВЯЗЫВАЮЩЕЕСЯ С N-КОНЦОМ ЛИЗИЛ-тРНК-СИНТЕТАЗЫ, В КАЧЕСТВЕ ЭФФЕКТИВНОГО ИНГРЕДИЕНТА ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ, ОБУСЛОВЛЕННОГО МИГРАЦИЕЙ КЛЕТОК | 2018 |
|
RU2749591C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| АНТИТЕЛО К B7-H4, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2792748C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| АНТИТЕЛО ПРОТИВ КЛАУДИНА 18.2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2822550C2 |
| НОВОЕ АНТИ-С-МЕТ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2751720C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |

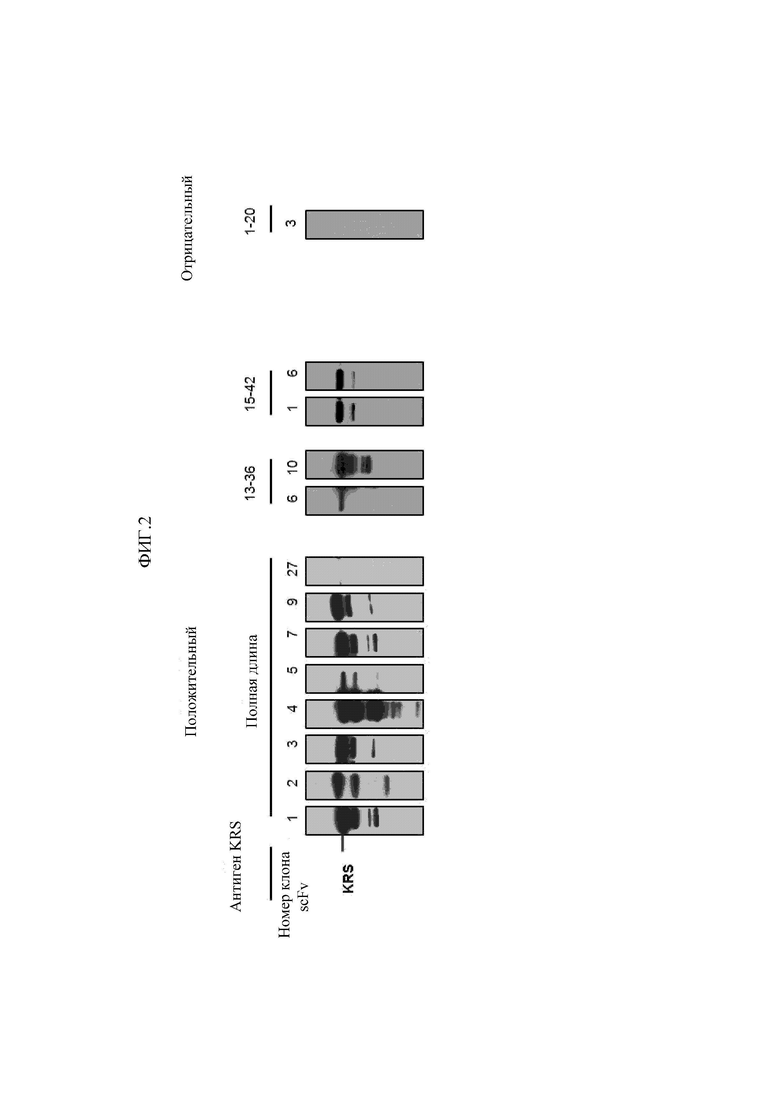
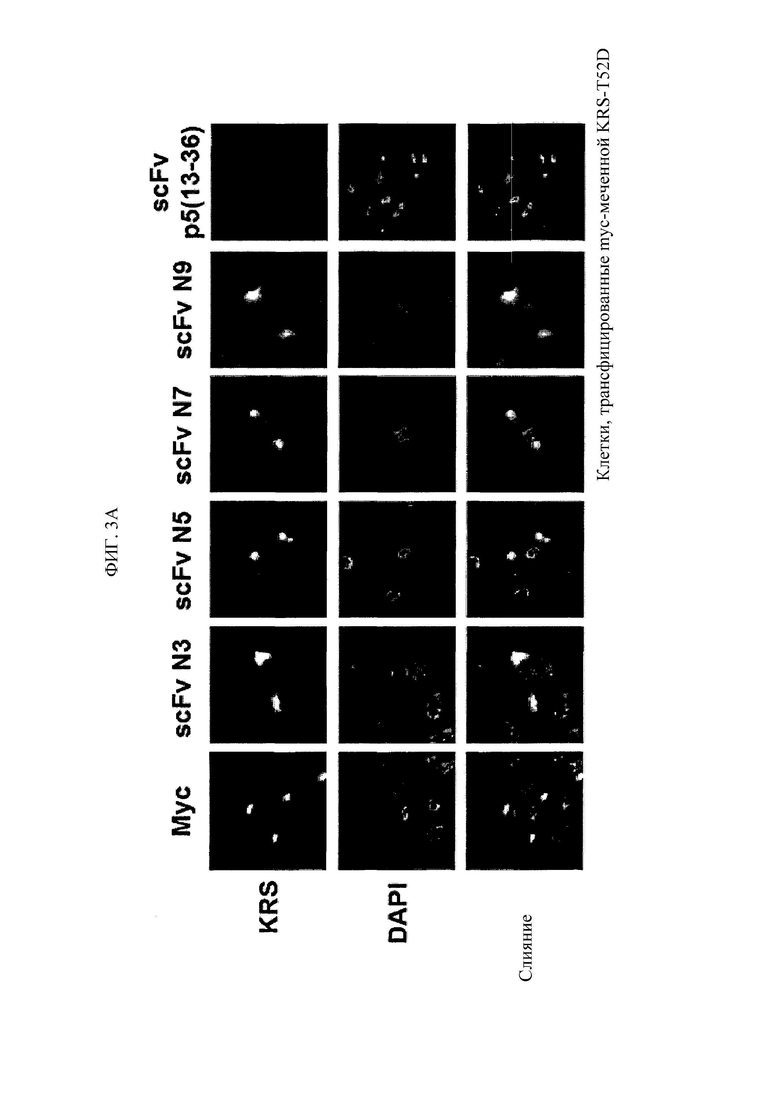

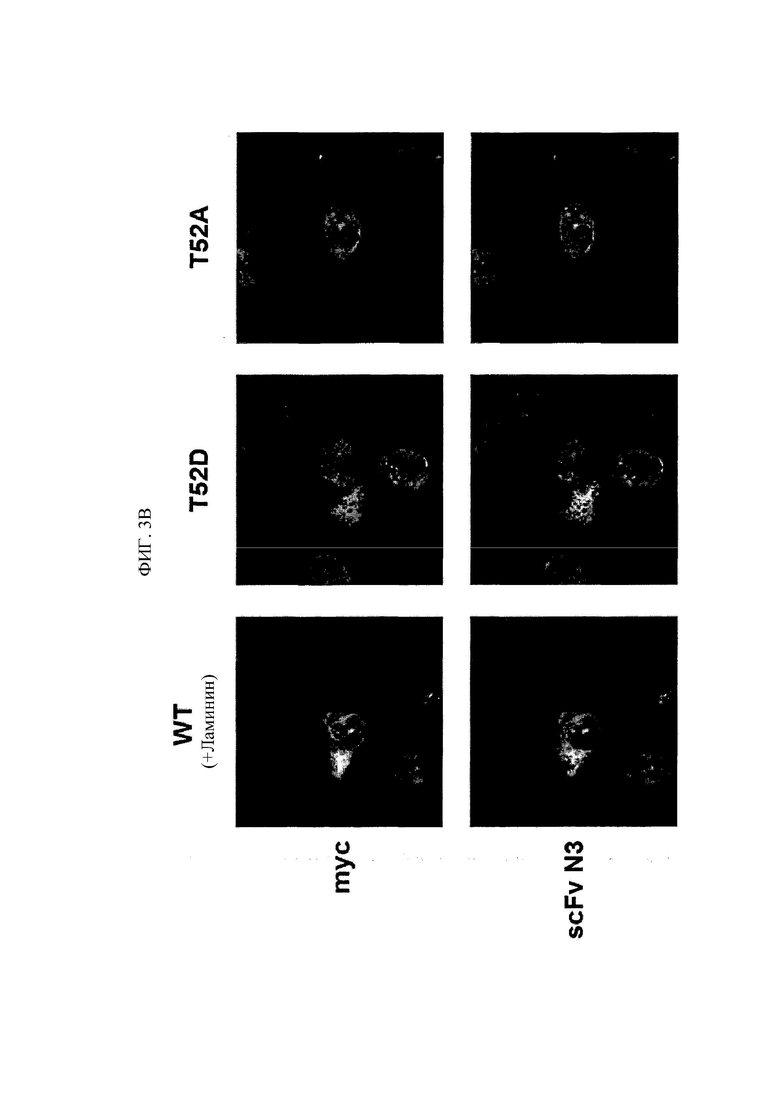

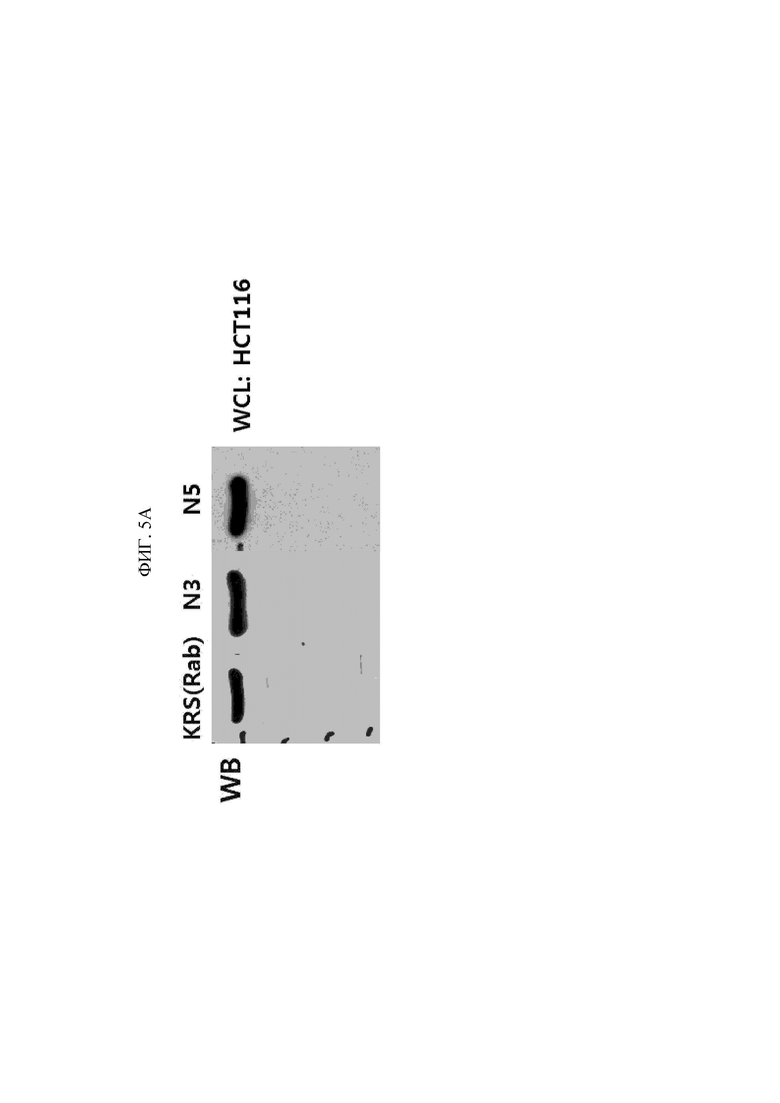

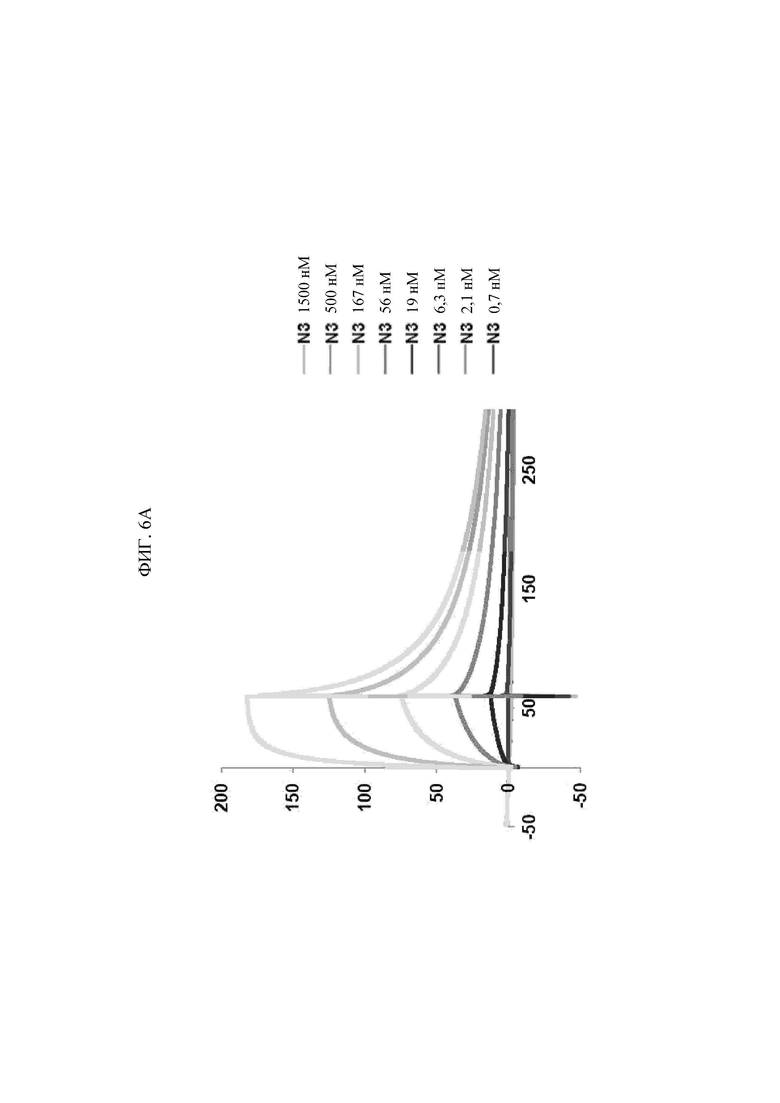
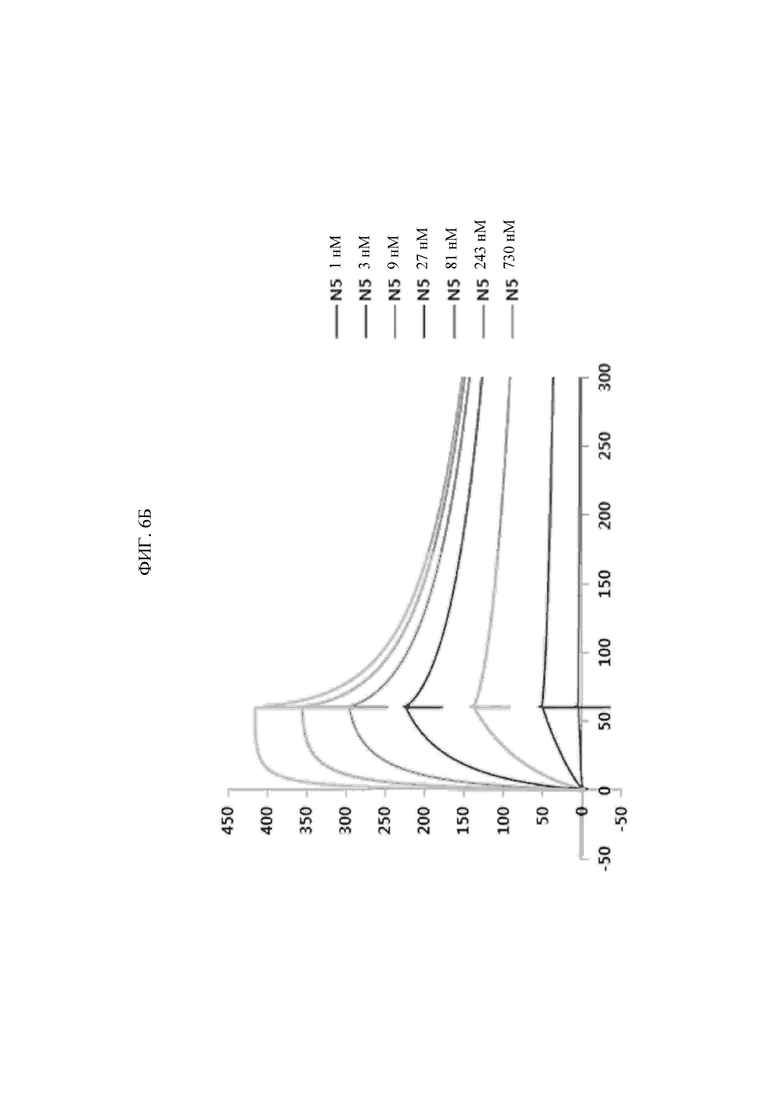
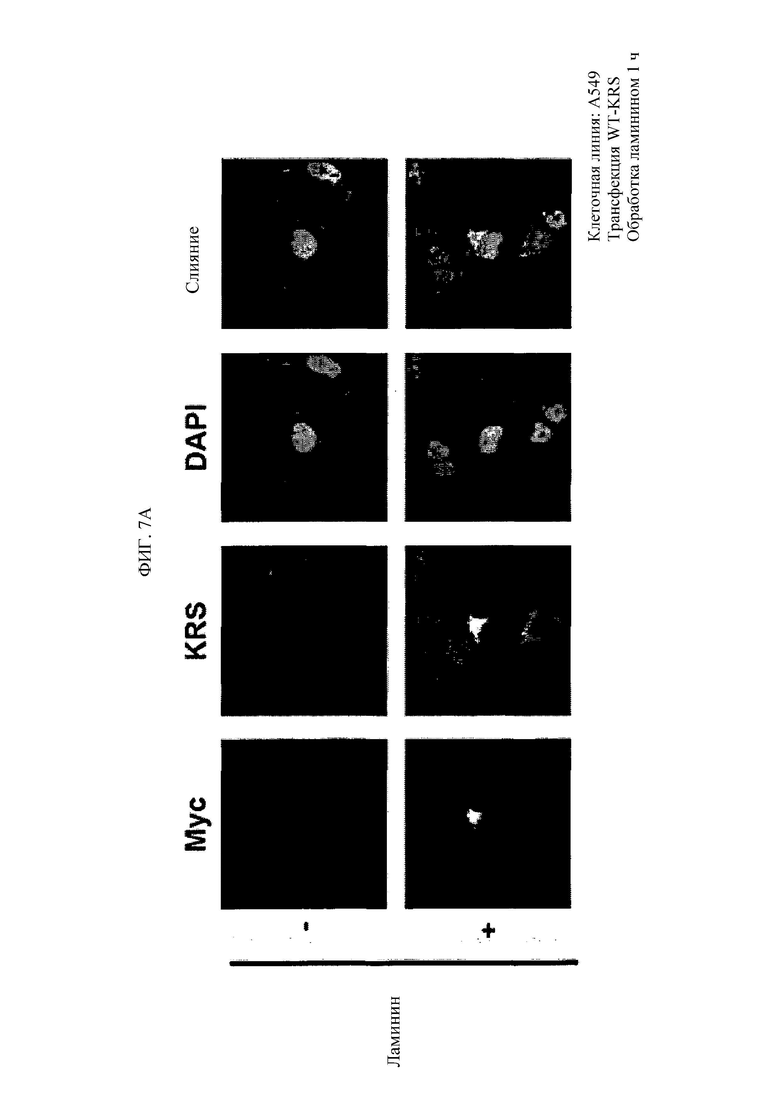
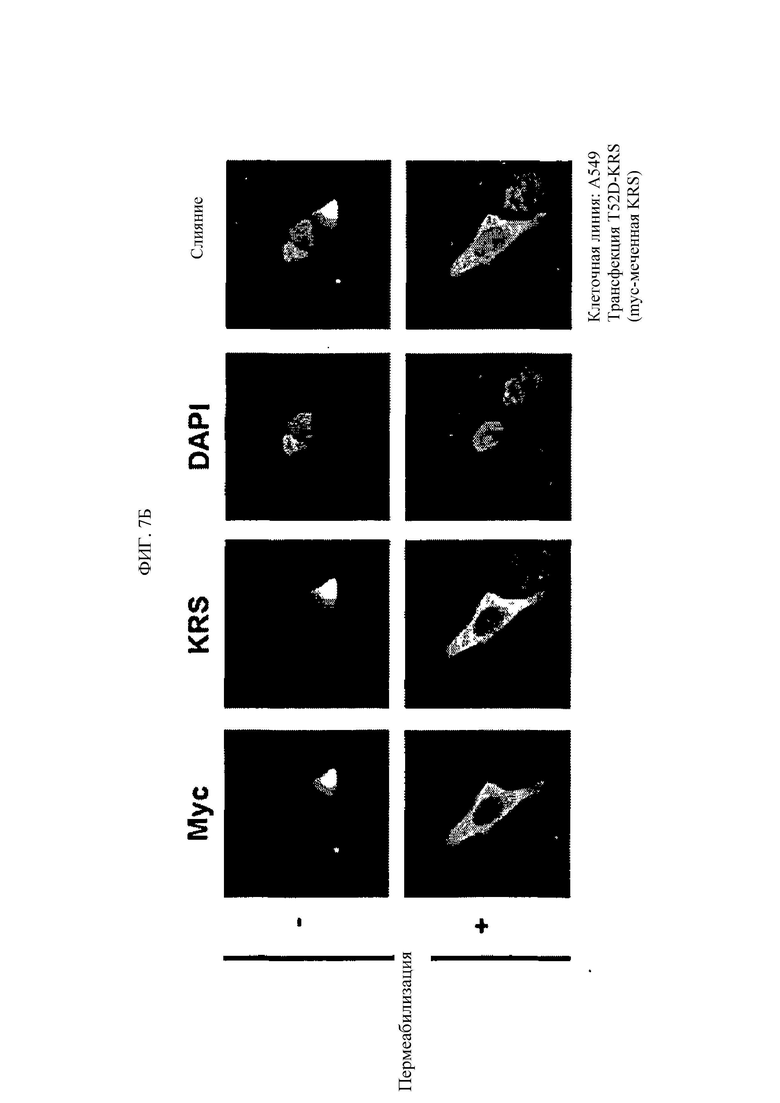
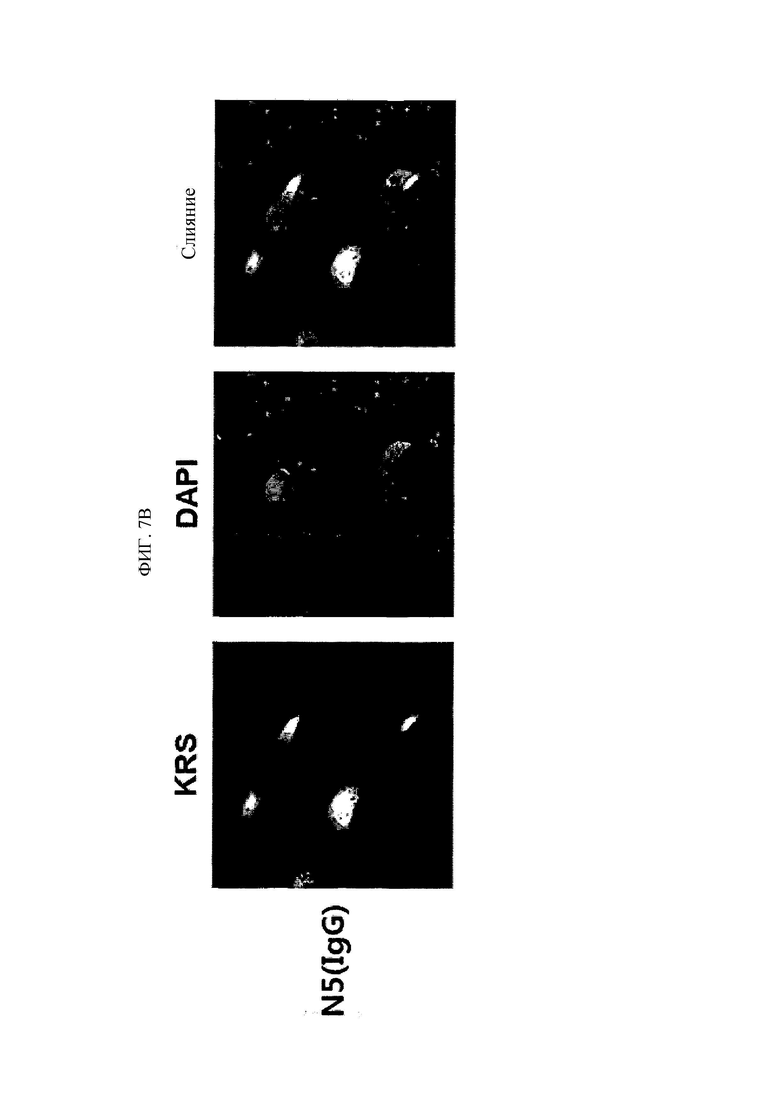
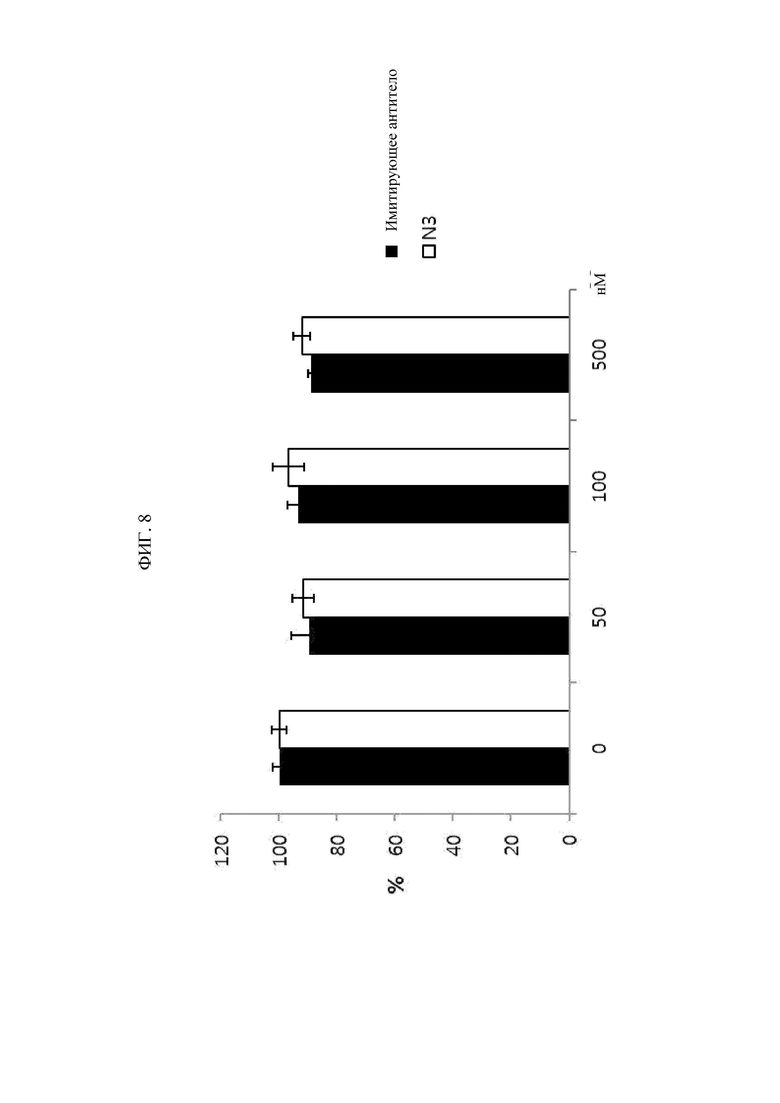
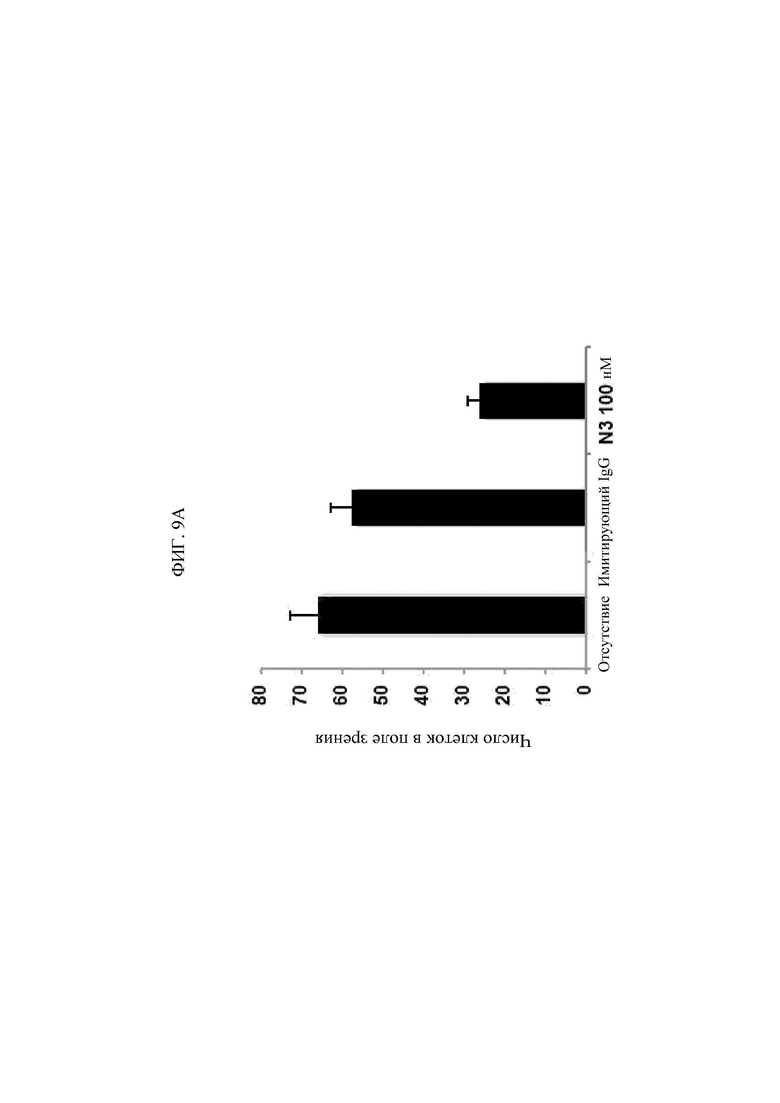
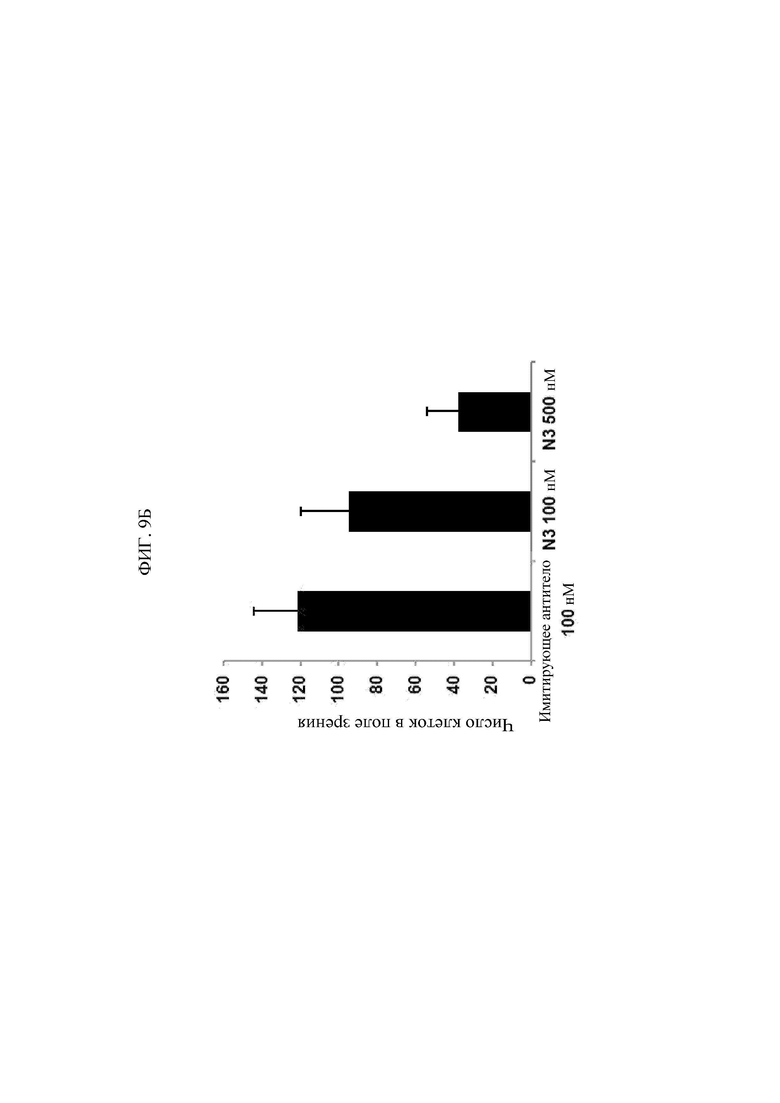
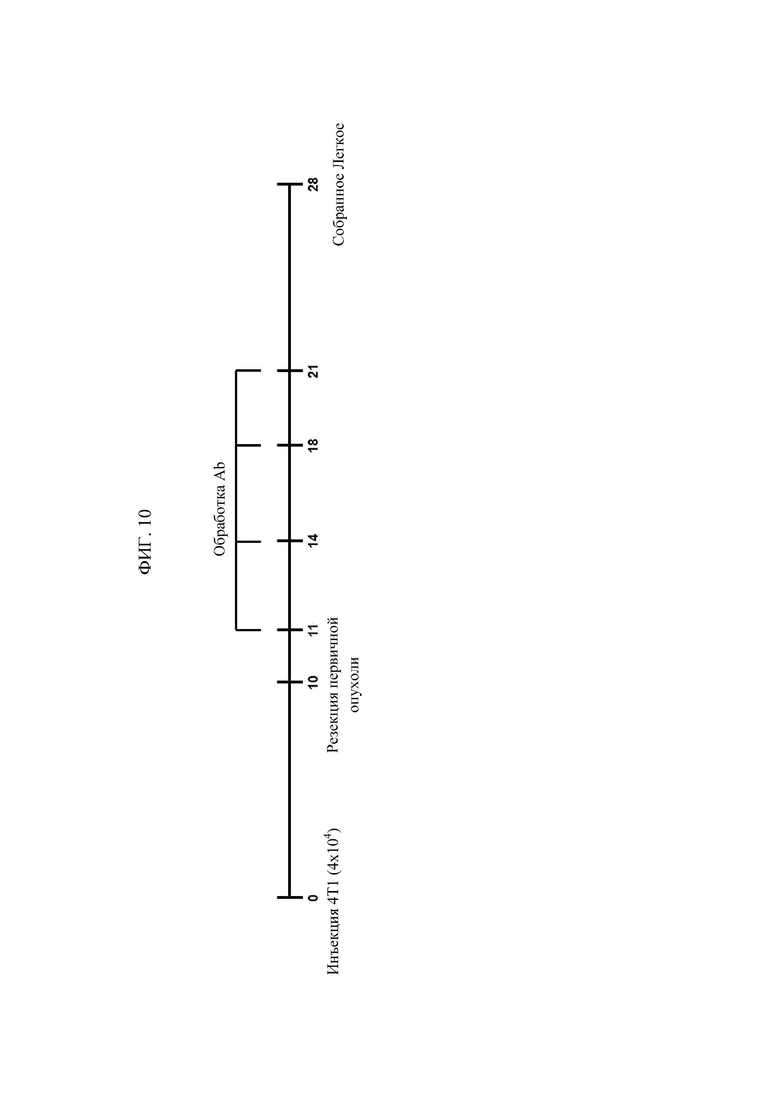
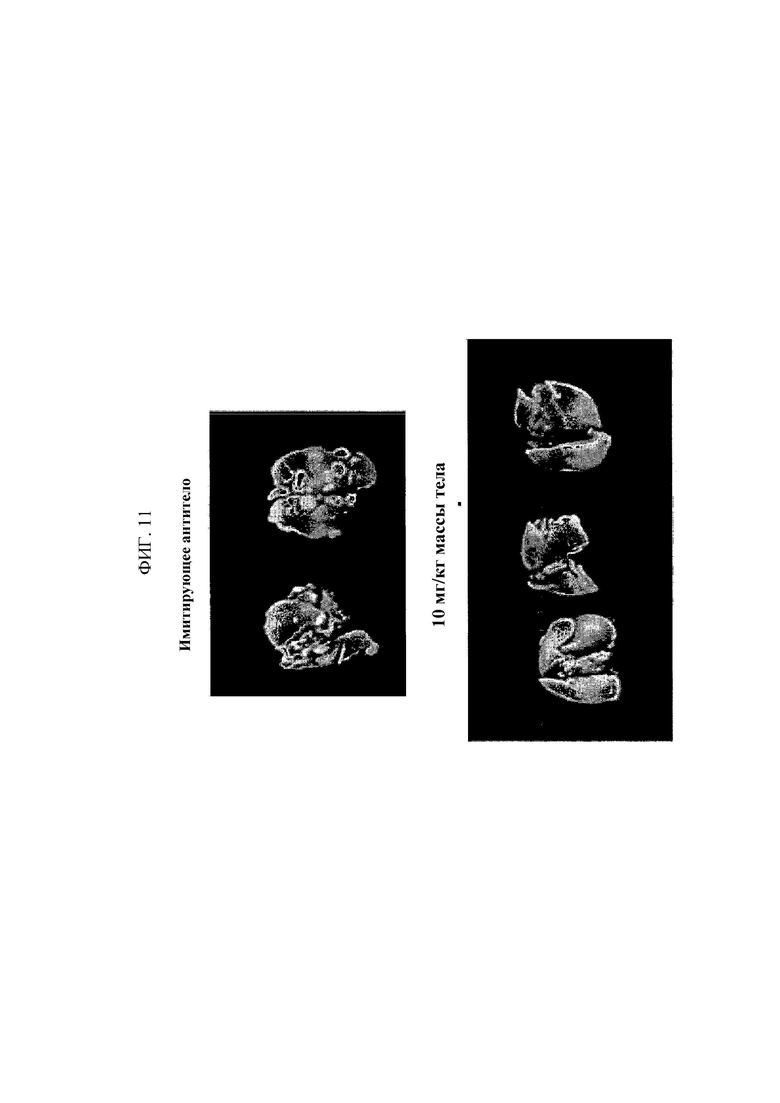
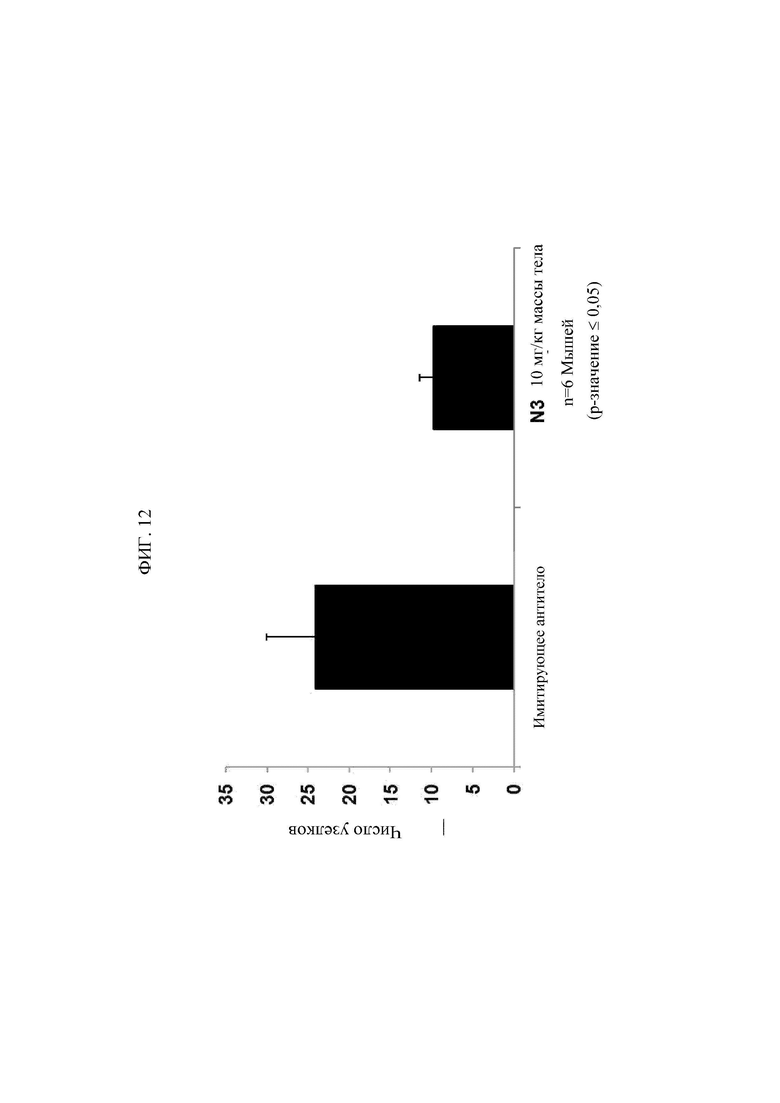
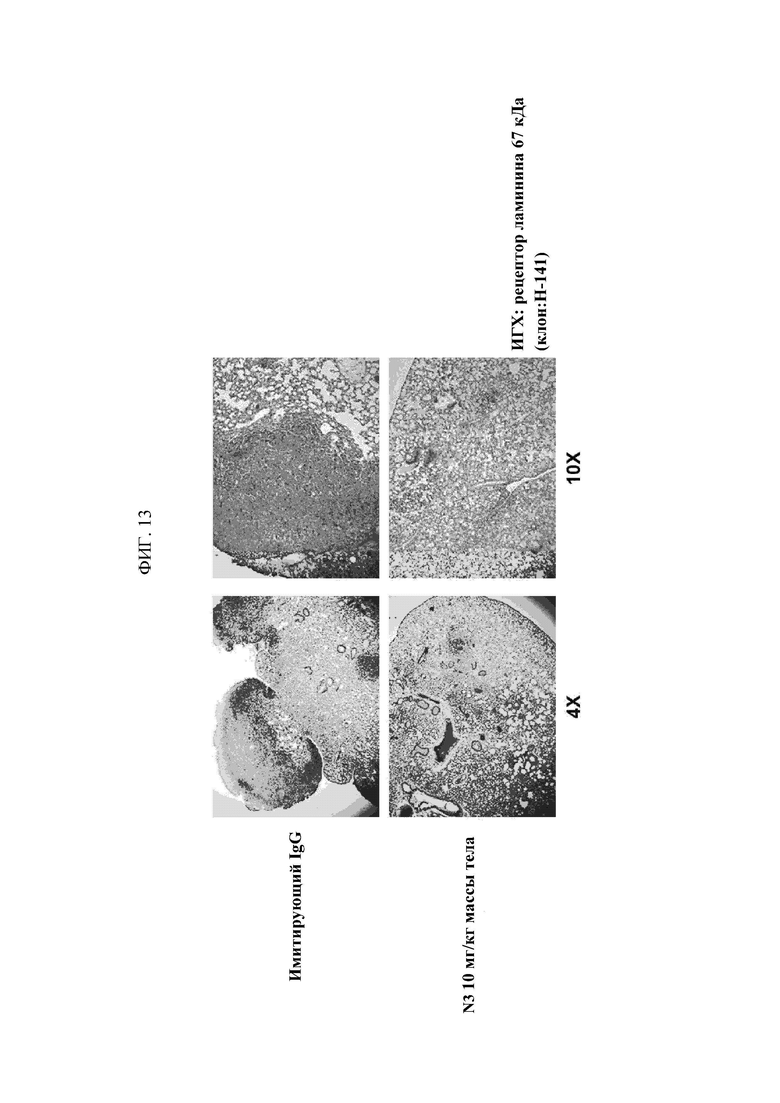

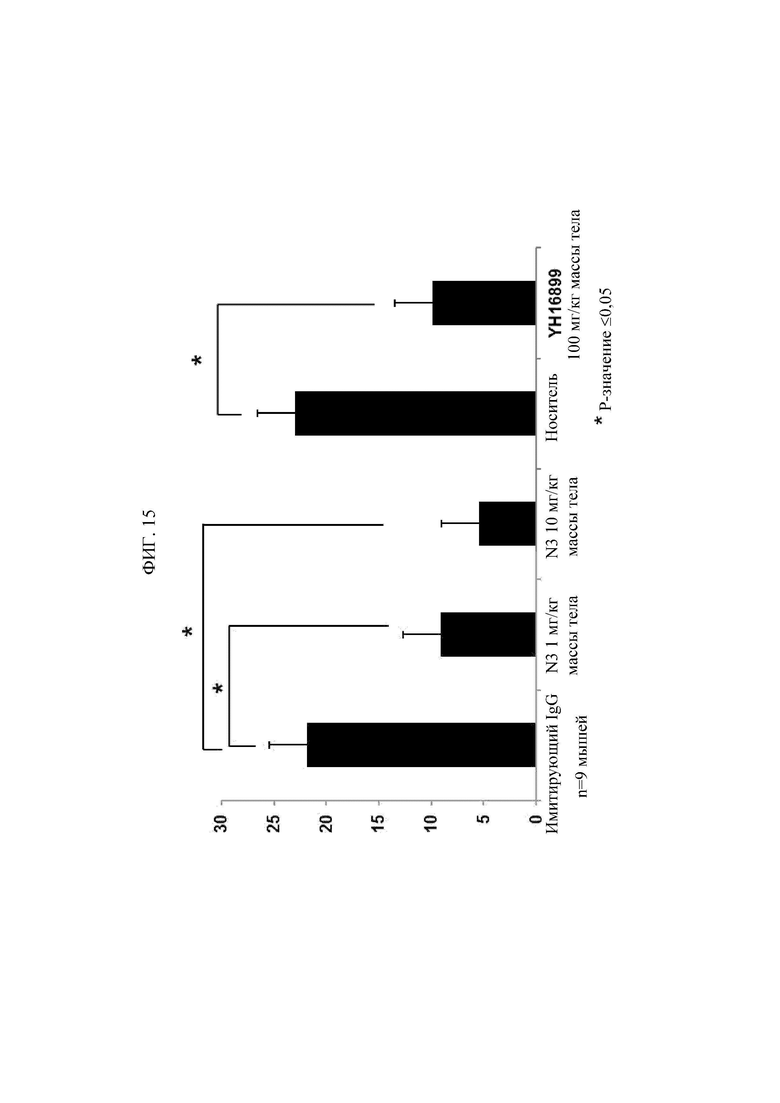
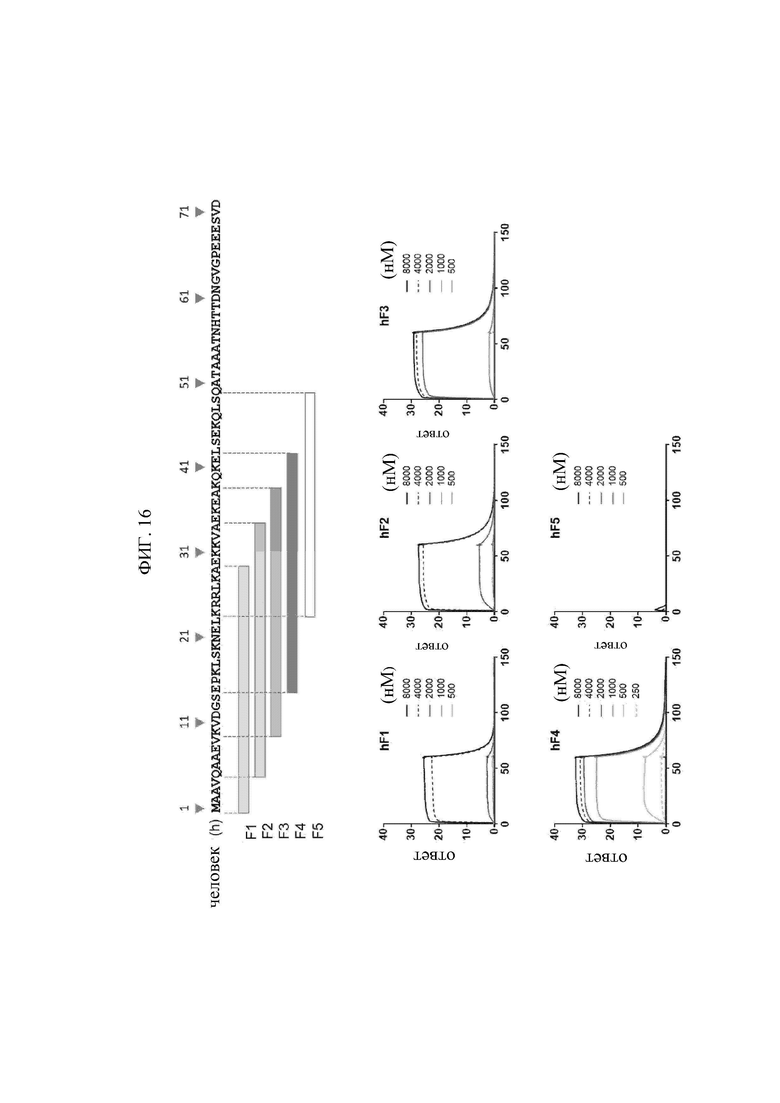
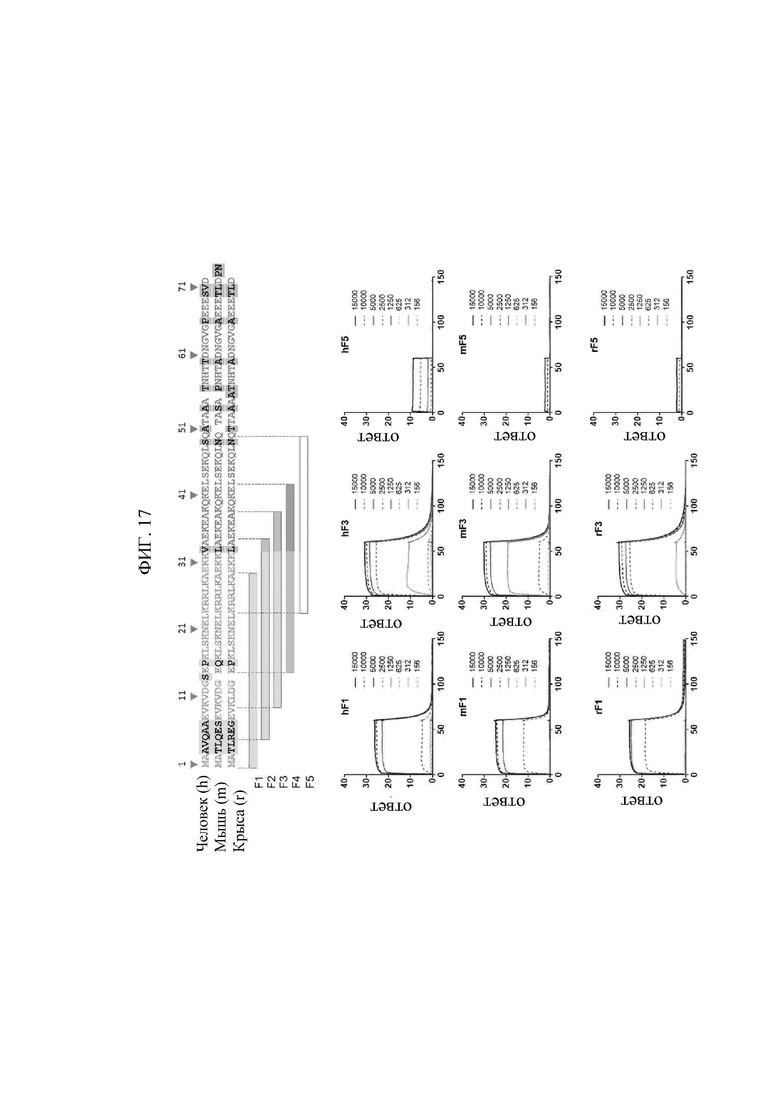

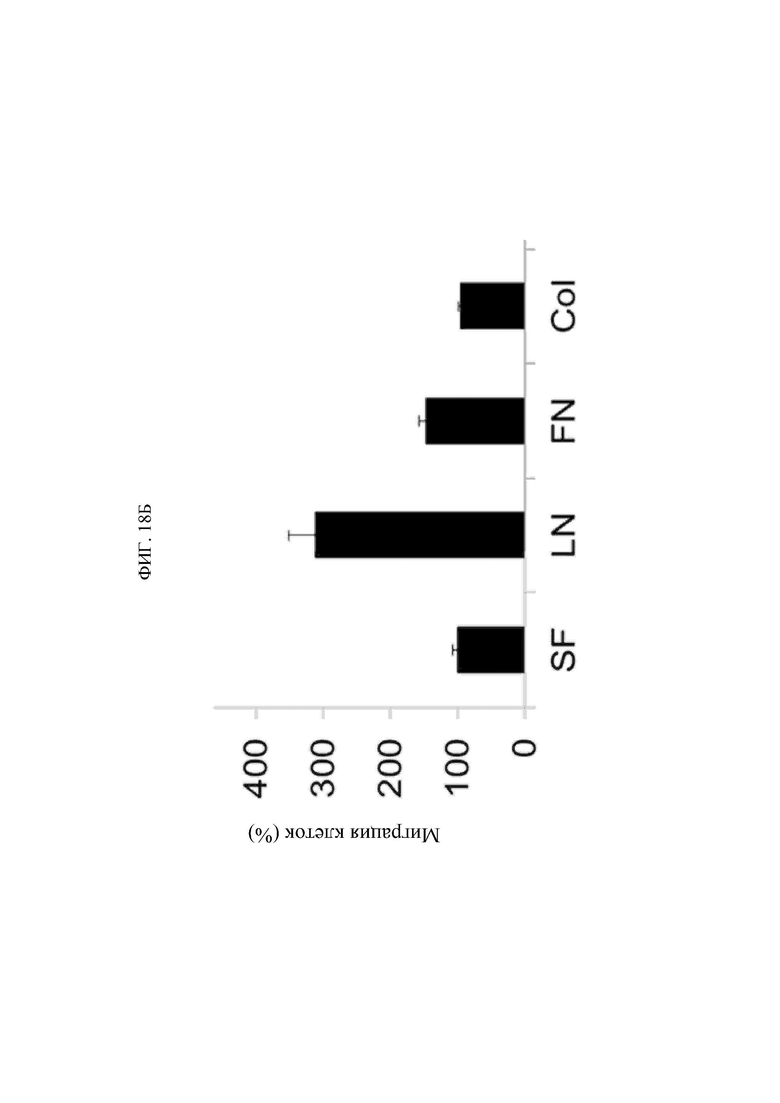
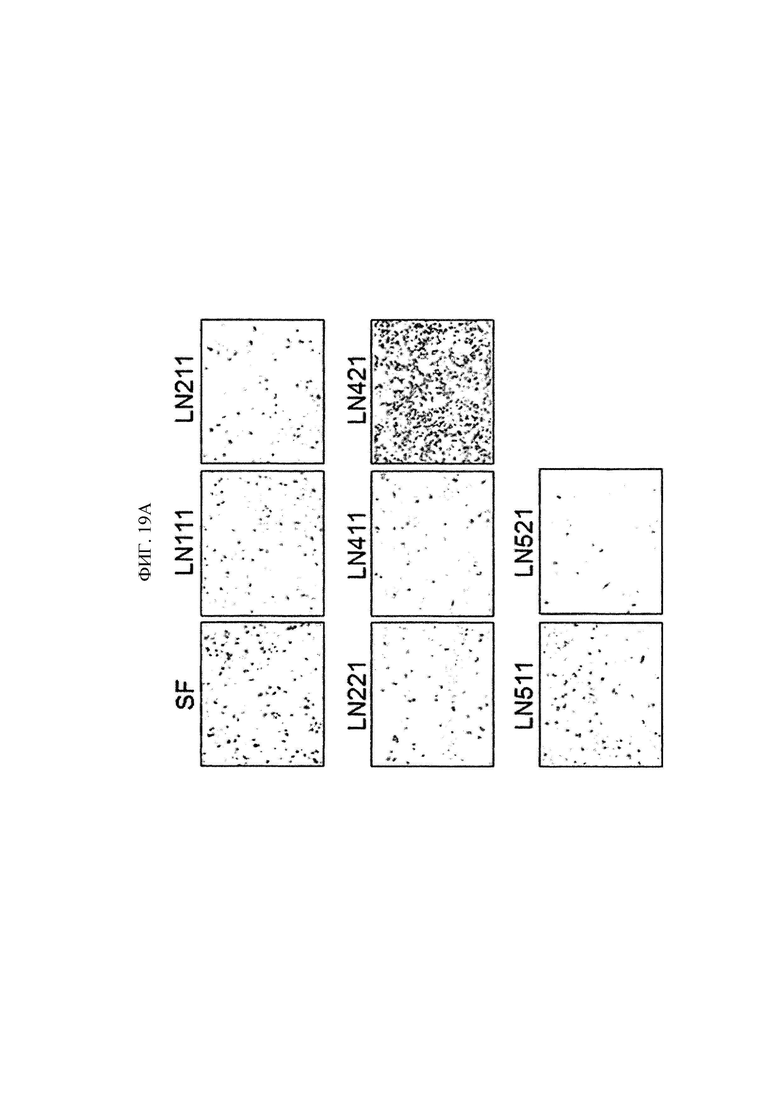
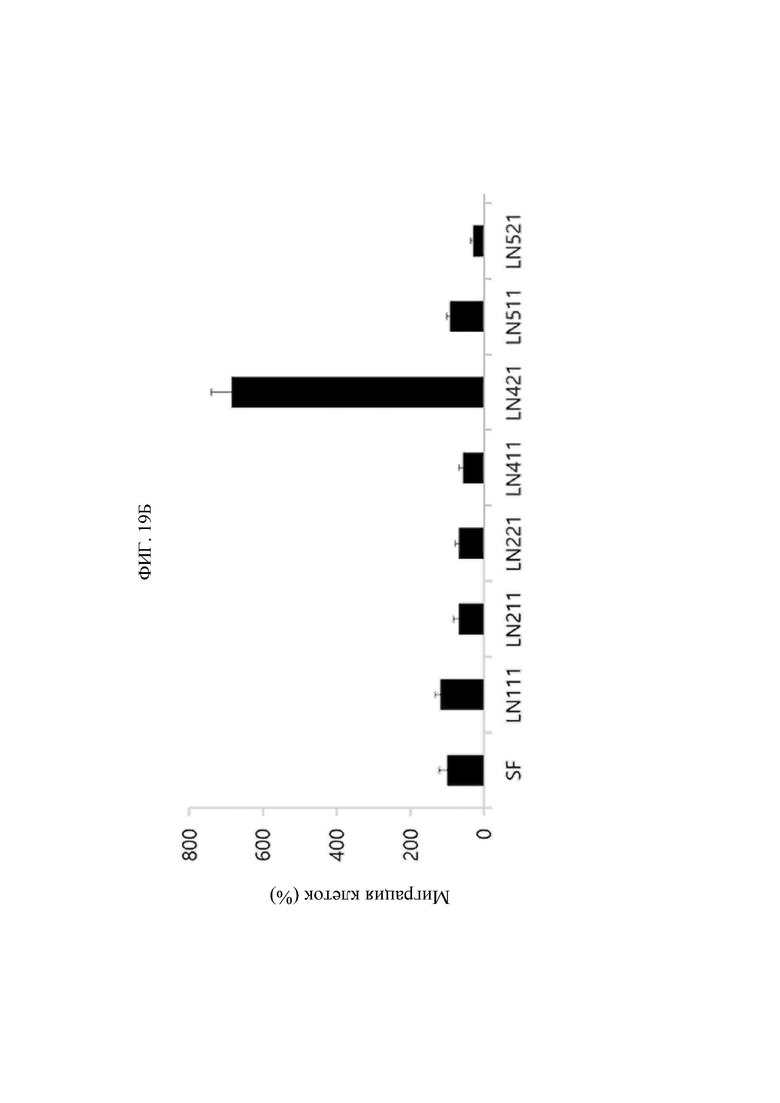
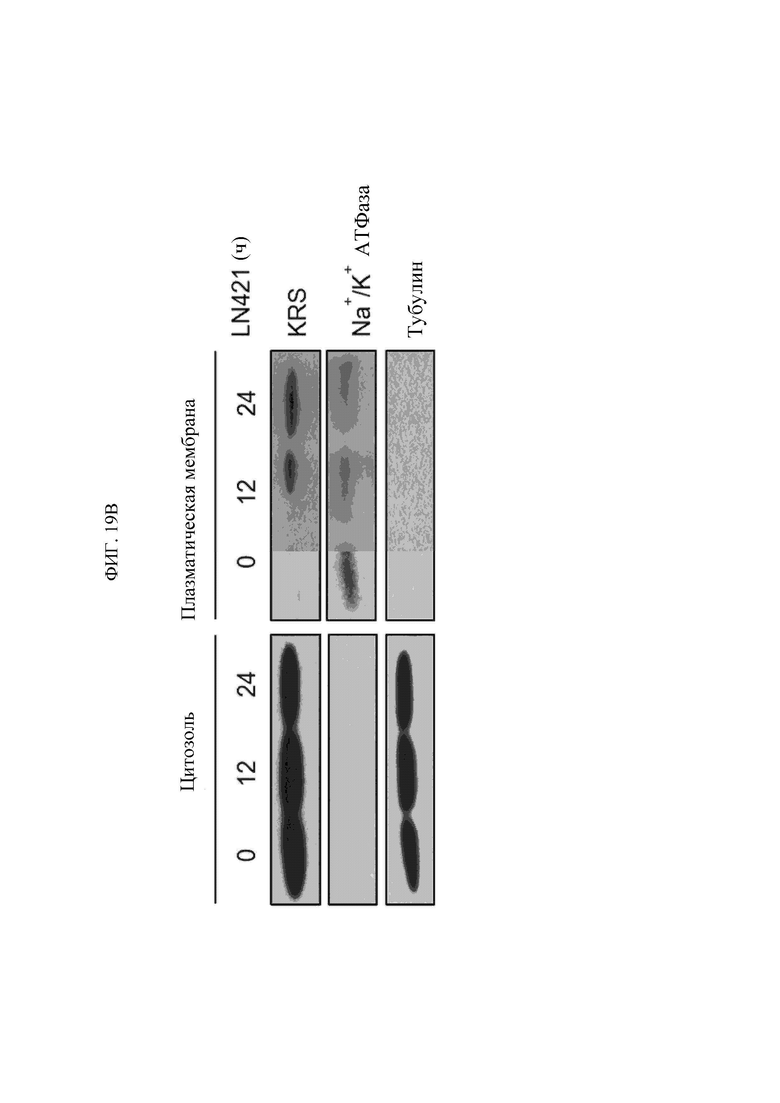

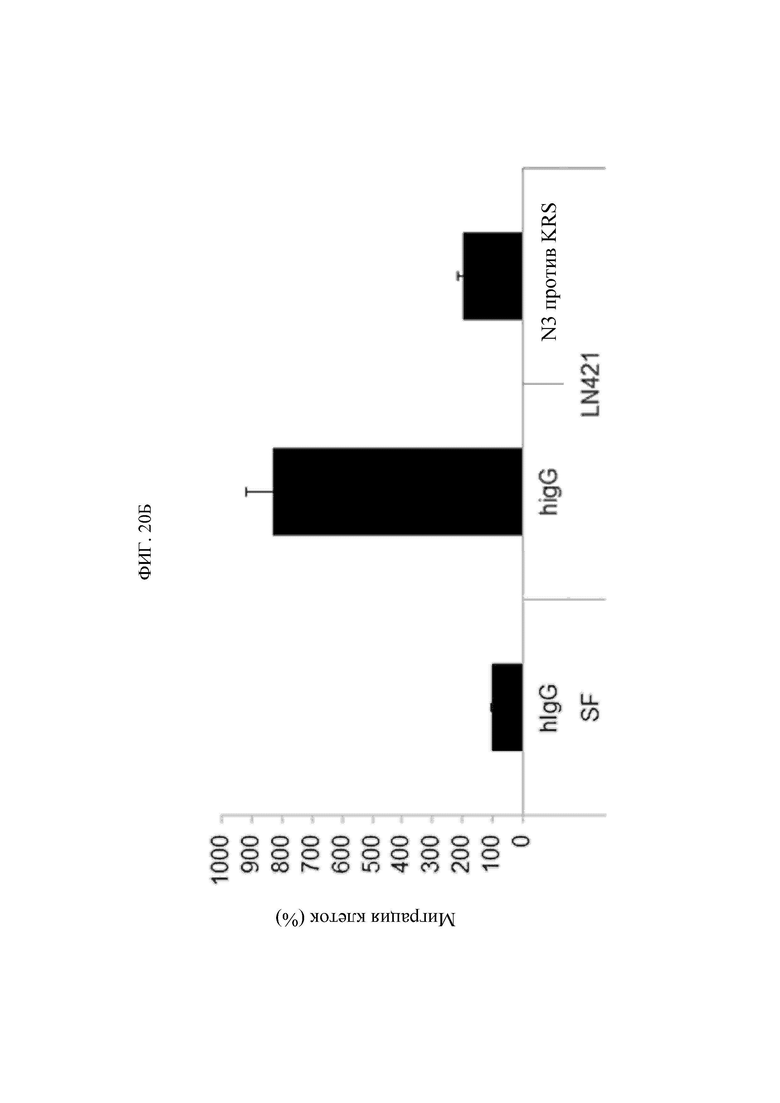
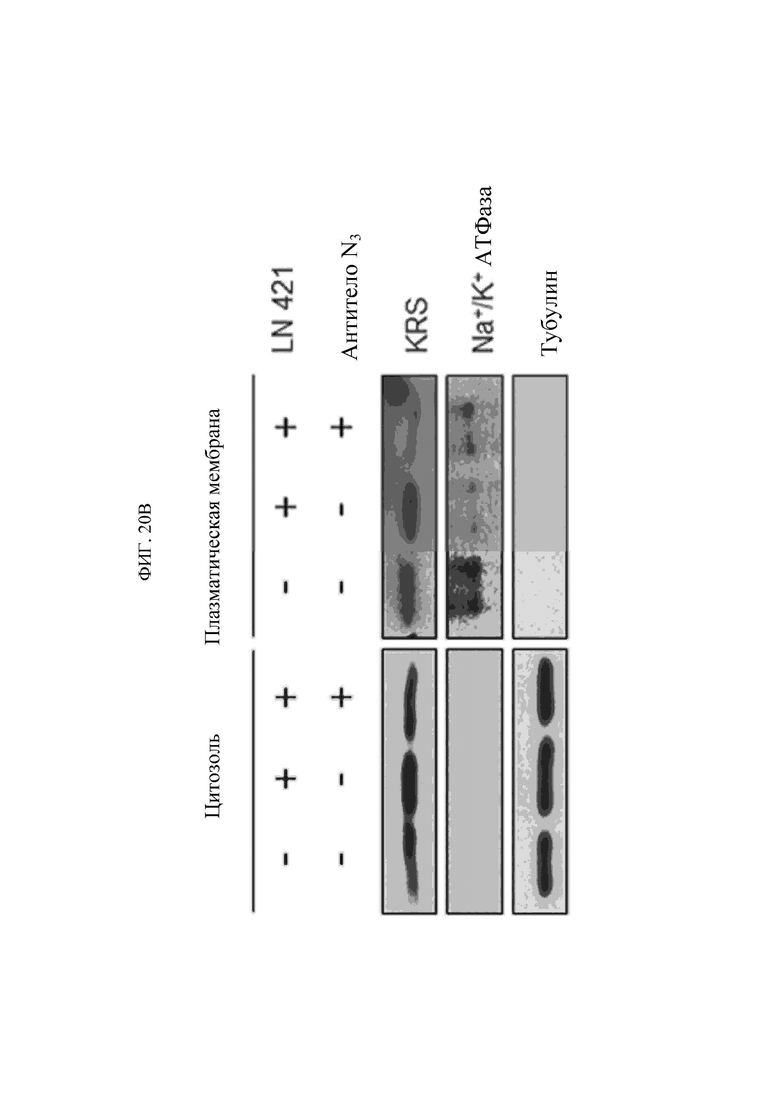
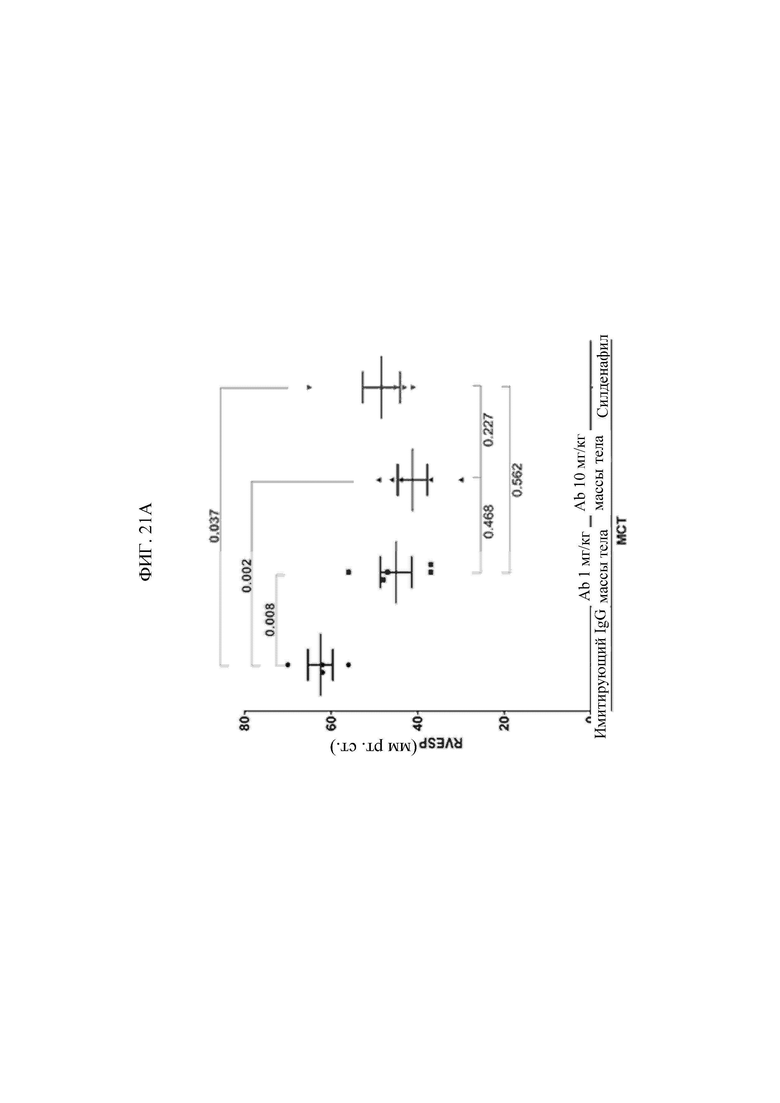
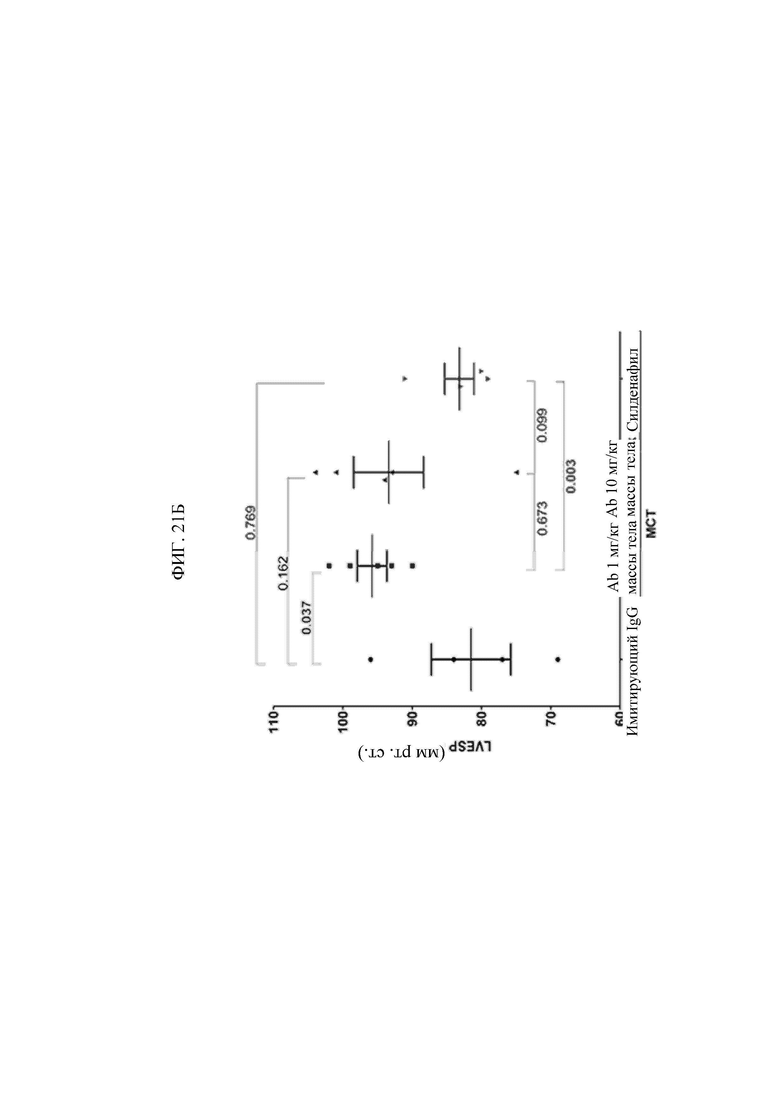

Группа изобретений относится к области медицины, а именно к антителу или его фрагменту, которое специфически связывается с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы (KRS) и содержит: вариабельную область тяжелой цепи (VH), содержащую: определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 13, SEQ ID NO: 25 и SEQ ID NO: 37; определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 3, SEQ ID NO: 15, SEQ ID NO: 27 и SEQ ID NO: 39; и определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 5, SEQ ID NO: 17, SEQ ID NO: 29 и SEQ ID NO: 41; и вариабельную область легкой цепи (VL), содержащую: определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 19, SEQ ID NO: 31 и SEQ ID NO: 43; определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 9, SEQ ID NO: 21, SEQ ID NO: 33 и SEQ ID NO: 45; и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 11, SEQ ID NO: 23, SEQ ID NO: 35 и SEQ ID NO: 47, относится к полинуклеотиду, кодирующему антитело или его фрагмент, относится к рекомбинантному экспрессирующему вектору, содержащему полинуклеотид, также относится к клетке, трансформированной рекомбинантным вектором, относится к способу получения антитела или его фрагмента, относится к способу специфического выявления экспонированной во внеклеточную среду N-концевой области лизил-тРНК-синтетазы (KRS), относится к фармацевтической композиции для ингибирования метастаза рака и к композиции для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток, также относится к способу лечения заболевания, связанного с миграцией иммунных клеток. Группа изобретений обеспечивает получение антитела или его фрагмента, специфически связывающегося с эпитопом, содержащим последовательность SEQ ID NO: 97 в N-конце лизил-тРНК-синтетазы (KRS), где благодаря связыванию с указанным эпитопом антитела снижали уровень KRS, повышенный на клеточной мембране иммунных клеток, и фактически ингибировали миграцию и инфильтрацию иммунных клеток, оказывая эффект лечения связанных с таковой миграцией заболеваний. 15 н. и 5 з.п. ф-лы, 6 табл., 35 ил., 8 пр.
1. Антитело или его фрагмент, которое специфически связывается с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы (KRS) и содержит:
вариабельную область тяжелой цепи (VH), содержащую: определяющую комплементарность область 1 (CDR1) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 13, SEQ ID NO: 25 и SEQ ID NO: 37; определяющую комплементарность область 2 (CDR2) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 3, SEQ ID NO: 15, SEQ ID NO: 27 и SEQ ID NO: 39; и определяющую комплементарность область 3 (CDR3) тяжелой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 5, SEQ ID NO: 17, SEQ ID NO: 29, и SEQ ID NO: 41; и
вариабельную область легкой цепи (VL), содержащую: определяющую комплементарность область 1 (CDR1) легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 19, SEQ ID NO: 31 и SEQ ID NO: 43; определяющую комплементарность область 2 (CDR2) легкой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 9, SEQ ID NO: 21, SEQ ID NO: 33 и SEQ ID NO: 45; и определяющую комплементарность область 3 (CDR3) легкой цепи, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 11, SEQ ID NO: 23, SEQ ID NO: 35 и SEQ ID NO: 47.
2. Антитело или его фрагмент по п. 1, где антитело или его фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, которые выбраны из группы, состоящей из следующих вариабельных областей:
вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 1, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 3, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 5, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 7, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 9, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 11;
вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 13, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 15, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 17, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 19, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 21, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 23;
вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 25, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 27, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 29, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 31, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 33, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 35; и
вариабельная область тяжелой цепи, содержащая определяющую комплементарность область 1 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 37, определяющую комплементарность область 2 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 39, и определяющую комплементарность область 3 тяжелой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 41, и вариабельная область легкой цепи, содержащая определяющую комплементарность область 1 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 43, определяющую комплементарность область 2 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 45, и определяющую комплементарность область 3 легкой цепи, содержащую аминокислотную последовательность, определенную SEQ ID NO: 47.
3. Антитело или его фрагмент по п. 1, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 49, SEQ ID NO: 53, SEQ ID NO: 57 и SEQ ID NO: 61, и вариабельная область легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 51, SEQ ID NO: 55, SEQ ID NO: 59 и SEQ ID NO: 63.
4. Антитело или его фрагмент по п. 1, где антитело выбрано из группы, состоящей из IgG, IgA, IgM, IgE и IgD, и фрагмент выбран из группы, состоящей из диатела, Fab, Fab', F(ab)2, F(ab')2, Fv и scFv.
5. Антитело или его фрагмент по п. 4, где scFv содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 67, SEQ ID NO: 69, SEQ ID NO: 71 и SEQ ID NO: 73.
6. Полинуклеотид, кодирующий антитело или его фрагмент по любому из пп. 1-5.
7. Рекомбинантный экспрессирующий вектор, содержащий полинуклеотид по п. 6.
8. Клетка, трансформированная рекомбинантным вектором по п. 7.
9. Способ получения антитела или его фрагмента, специфически связывающегося с экспонированной во внеклеточную среду N-концевой областью лизил-тРНК-синтетазы (KRS), включающий:
(a) трансформирование клеток-хозяев рекомбинантным экспрессирующим вектором по п. 7;
(б) инкубирование трансформированных клеток-хозяев с получением антитела или его фрагмента; и
(в) сбор антитела или его фрагмента, полученного в клетках-хозяевах.
10. Способ специфического выявления экспонированной во внеклеточную среду N-концевой области лизил-тРНК-синтетазы (KRS), включающий:
приведение антитела или его фрагмента по п. 1 в контакт с образцом; и
выявление антитела или его фрагмента.
11. Фармацевтическая композиция для ингибирования метастаза рака, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного вещества, где рак выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи или интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза.
12. Композиция по п. 11, где рак представляет собой рак молочной железы или рак легкого.
13. Композиция для диагностики рака, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного вещества, где рак выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи или интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза.
14. Фармацевтическая композиция для предупреждения или лечения заболевания, связанного с миграцией иммунных клеток, содержащая эффективное количество антитела или его фрагмента по п. 1 в качестве активного вещества,
где заболевание, связанное с миграцией иммунных клеток, выбрано из группы, состоящей из сердечно-сосудистого заболевания, фиброзного заболевания, хронического воспалительного заболевания и синдрома Альпорта, где сердечно-сосудистое заболевание выбрано из группы, состоящей из легочной артериальной гипертензии, атеросклероза, стенокардии, инфаркта миокарда, ишемической цереброваскулярной болезни, артериосклероза и мезентериального склероза,
где фиброзное заболевание выбрано из группы, состоящей из склеродермии, ревматоидного артрита, болезни Крона, язвенного колита, миелофиброза, фиброза легкого, фиброза печени, цирроза печени, фиброза почки, миофиброза, фиброза миокарда, системной красной волчанки, наследственного фиброза, инфекционного фиброза, фиброза, вызываемого воздействием раздражителей, фиброза, вызываемого хроническим аутоиммунным заболеванием, фиброза, вызываемого антигенной несовместимостью во время трансплантации органов, фиброза, вызываемого гиперлипидемией, фиброза, вызываемого ожирением, диабетического фиброза, фиброза, вызываемого гипертензией, и окклюзии, вызываемой фиброзом во время имплантации стента,
где хроническое воспалительное заболевание выбрано из группы, состоящей из астмы, атопического дерматита, экземы, псориаза, остеоартрита, подагры, псориатического артрита, цирроза, неалкогольного стеатогепатита, хронического обструктивного заболевания легких, ринита, диабетической ретинопатии, диабетической почечной недостаточности, диабетической нейропатии и рассеянного склероза.
15. Применение эффективного количества антитела или его фрагмента по п. 1 для получения агента для ингибирования метастаза рака, где рак выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи или интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза.
16. Способ ингибирования метастаза рака у нуждающегося в этом субъекта, включающий введение эффективного количества антитела или его фрагмента по п. 1 указанному субъекту для ингибирования метастаза рака, где рак выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи или интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза.
17. Применение эффективного количества антитела или его фрагмента по п. 1 для получения агента для диагностики рака, где рак выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи или интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза.
18. Способ диагностирования рака у нуждающегося в этом субъекта, включающий введение указанному субъекту эффективного количества антитела или его фрагмента по п. 1 для диагностирования рака, где рак выбран из группы, состоящей из рака молочной железы, рака толстой кишки, рака легкого, мелкоклеточного рака легкого, рака желудка, рака печени, рака крови, рака кости, рака поджелудочной железы, рака кожи, рака головы или шеи, меланомы кожи или интраокулярной меланомы, рака матки, рака яичника, ректального рака, рака анального канала, рака ободочной кишки, рака молочной железы, карциномы фаллопиевой трубы, карциномы эндометрия, рака шейки матки, рака влагалища, карциномы вульвы, лимфомы Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, карциномы паращитовидной железы, рака надпочечника, саркомы мягких тканей, рака матки, рака пениса, рака предстательной железы, хронического или острого лейкоза, лимфоцитарной лимфомы, рака мочевого пузыря, рака почки или уретры, почечно-клеточной карциномы, карциномы почечной лоханки, опухоли ЦНС (центральная нервная система), первичной лимфомы ЦНС, опухоли спинного мозга, глиомы ствола головного мозга и аденомы гипофиза.
19. Применение эффективного количества антитела или его фрагмента по п. 1 для получения агента для лечения заболевания, связанного с миграцией иммунных клеток,
где заболевание, связанное с миграцией иммунных клеток, выбрано из группы, состоящей из сердечно-сосудистого заболевания, фиброзного заболевания, хронического воспалительного заболевания и синдрома Альпорта,
где сердечно-сосудистое заболевание выбрано из группы, состоящей из легочной артериальной гипертензии, атеросклероза, стенокардии, инфаркта миокарда, ишемической цереброваскулярной болезни, артериосклероза и мезентериального склероза,
где фиброзное заболевание выбрано из группы, состоящей из склеродермии, ревматоидного артрита, болезни Крона, язвенного колита, миелофиброза, фиброза легкого, фиброза печени, цирроза печени, фиброза почки, миофиброза, фиброза миокарда, системной красной волчанки, наследственного фиброза, инфекционного фиброза, фиброза, вызываемого воздействием раздражителей, фиброза, вызываемого хроническим аутоиммунным заболеванием, фиброза, вызываемого антигенной несовместимостью во время трансплантации органов, фиброза, вызываемого гиперлипидемией, фиброза, вызываемого ожирением, диабетического фиброза, фиброза, вызываемого гипертензией, и окклюзии, вызываемой фиброзом во время имплантации стента,
где хроническое воспалительное заболевание выбрано из группы, состоящей из астмы, атопического дерматита, экземы, псориаза, остеоартрита, подагры, псориатического артрита, цирроза, неалкогольного стеатогепатита, хронического обструктивного заболевания легких, ринита, диабетической ретинопатии, диабетической почечной недостаточности, диабетической нейропатии и рассеянного склероза.
20. Способ лечения заболевания, связанного с миграцией иммунных клеток, у нуждающегося в этом субъекта, включающий введение указанному субъекту эффективного количества антитела или его фрагмента по п. 1 для лечения заболевания, связанного с миграцией иммунных клеток,
где заболевание, связанное с миграцией иммунных клеток, выбрано из группы, состоящей из сердечно-сосудистого заболевания, фиброзного заболевания, хронического воспалительного заболевания и синдрома Альпорта,
где сердечно-сосудистое заболевание выбрано из группы, состоящей из легочной артериальной гипертензии, атеросклероза, стенокардии, инфаркта миокарда, ишемической цереброваскулярной болезни, артериосклероза и мезентериального склероза,
где фиброзное заболевание выбрано из группы, состоящей из склеродермии, ревматоидного артрита, болезни Крона, язвенного колита, миелофиброза, фиброза легкого, фиброза печени, цирроза печени, фиброза почки, миофиброза, фиброза миокарда, системной красной волчанки, наследственного фиброза, инфекционного фиброза, фиброза, вызываемого воздействием раздражителей, фиброза, вызываемого хроническим аутоиммунным заболеванием, фиброза, вызываемого антигенной несовместимостью во время трансплантации органов, фиброза, вызываемого гиперлипидемией, фиброза, вызываемого ожирением, диабетического фиброза, фиброза, вызываемого гипертензией, и окклюзии, вызываемой фиброзом во время имплантации стента,
где хроническое воспалительное заболевание выбрано из группы, состоящей из астмы, атопического дерматита, экземы, псориаза, остеоартрита, подагры, псориатического артрита, цирроза, неалкогольного стеатогепатита, хронического обструктивного заболевания легких, ринита, диабетической ретинопатии, диабетической почечной недостаточности, диабетической нейропатии и рассеянного склероза.
| WO 2011153277 A2, 08.12.2011 | |||
| Якубке Х.-Д и др | |||
| Аминокислоты, пептиды, белки, М: Мир, 1985, с.92-94. |

