Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, специфично связывающемуся с рецептором фактора роста гепатоцитов человека (c-Met), и к содержащей их композиции для предотвращения или лечения рака.
Уровень техники
Рецепторные тирозинкиназы (RTK) действуют в качестве жизненно важного модулятора роста, дифференцировки, неоваскуляризации, восстановления тканей и т.д. Помимо таких общих физиологических процессов, аномальная экспрессия определенных RTK связана с развитием и прогрессированием многих видов рака. Таким образом, такие RTK считаются перспективными мишенями для лекарственных средств при лечении рака.
Рецептор фактора роста гепатоцитов (HGFR; c-Met), который представляет собой разновидность RTK, является рецептором на поверхности клеток, обладающим сродством к фактору роста гепатоцитов, известному как рассеивающий фактор (HGF/SF) (Laird AD et al., Expert. Opin. Investig. Drugs 12: 51-64 (2003)). Известно, что аномальная активация c-Met белком HGF, которая является одним из репрезентативных онкогенных механизмов, связана с пролиферацией опухоли, ингибированием апоптоза, неоваскуляризацией, инвазией, метастазированием и т.п. (Bottaro DP et al., Science 251: 802- 804 (1991), Day RM et al., Oncogene 18: 3399-3406 (1999)). Также сообщается, что аномальная активация и амплификация c-Met мутацией c-Met связана с различными видами рака, такими как рак легких, рак толстой кишки, рак головы и шеи, рак желудка, рак молочной железы и т.д., и она также участвует в повышении агрессивности опухоли и ее неблагоприятном прогнозе (Lefebvre J et al., FASEB J 26: 1387-1399 (2012), Liu X et al., Trends Mol Med 16: 37-45 (2010), Smolen GA et al., Proc Natl Acad Sci USA 103: 2316-2321 (2006), Foveau B et al., Mol Biol Cell 20: 2495-2507 (2009)).
Таким образом, c-Met привлек большое внимание в качестве антигена-мишени для лечения таких различных видов рака, и были предложены различные подходы для ингибирования экспрессии и активности c-Met. В качестве c-Met-специфичных низкомолекулярных ингибиторов тирозинкиназы, которые известны в настоящее время, существуют Тивантиниб (ArQule), INC280 (Novartis), AMG337 (Amgen) и т.д. И Рилотумумаб (Amgen), Фиклатузумаб (AVEP Pharmaceuticals), HuL2G7 (Galaxy Biotech) и т.д. были разработаны в качестве HGF-специфичных моноклональных антител, который является лигандом c-Met. Кроме того, в качестве антагониста моноклонального антитела, которое специфично по отношению к c-Met, существует Онартузумаб разработки фирмы Genentech (WO 2006/015371), находящийся в фазе III клинических испытаний, Эмибетузумаб от фирмы Lilly (WO 2010/059654), находящийся в фазе II клинических испытаний, SAIT-301 (US 2014154251), находящийся в фазе I клинических испытаний, ABT-700 (Wang J et al., BMC Cancer. 16: 105-118 (2016)) и т.д. Онартузумаб представляет собой моновалентное антагонистическое антитело, полученное из двухвалентного моноклонального антитела (5D5), который действует на c-Met в качестве агента (Mark Merchant, et al., Proc Natl Acad Sci US A. 110 (32): E2987-E299 (2013)). Таким образом, были разработаны различные лекарственные средства в отношении c-Met, но c-Met связан с возникновением и прогрессированием различных видов рака, как описано выше, поэтому существует постоянная потребность в разработке нового терапевтического средства, способного лечить рак, путем направленного воздействия на c-Met.
Раскрытие изобретения
Техническая проблема
Авторы настоящего изобретения разработали новое анти-c-Met-антитело, связывающееся с c-Met с высокой аффинностью, и также определили, что такое анти-c-Met-антитело, его химера и гуманизированные и оптимизированные по аффинности антитела заметно ингибируют пролиферацию опухолевых клеток и обладают превосходным противоопухолевым эффектом, завершив, таким образом, создание настоящего изобретения.
Решение проблемы
Одной из целей настоящего изобретения является создание антитела или его антигенсвязывающего фрагмента, которое специфично связывается с рецептором фактора роста гепатоцитов (c-Met).
Другой целью настоящего изобретения является молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент, экспрессионный вектор, содержащий эту молекулу нуклеиновой кислоты, клетка-хозяин, содержащая введенный в нее экспрессионный вектор, способ получения антитела или его антигенсвязывающий фрагмент с использованием этой клетки-хозяина.
Еще одной целью настоящего изобретения является композиция для обнаружения c-Met, содержащая антитело или его антигенсвязывающий фрагмент, набор для обнаружения, содержащий эту композицию, и способ обнаружения антигена c-Met с его использованием.
Еще одной целью настоящего изобретения является создание композиции для предотвращения или лечения рака, содержащей антитело или его антигенсвязывающий фрагмент.
Полезный эффект изобретения
Антитело или его антигенсвязывающий фрагмент настоящего изобретения, которое специфично связывается с рецептором фактора роста гепатоцитов (c-Met), имеет новую последовательность и демонстрирует превосходную ингибирующую активность в отношении пролиферации раковых клеток и удивительно превосходную противораковую активность даже в небольших количествах, что позволяет эффективно предотвращать или лечить такие заболевания, как рак.
Краткое описание чертежей
На Фигуре 1 показаны результаты теста in vitro активности ингибирования пролиферации опухолевых клеток гибридомного антитела c-Met настоящего изобретения.
На Фигуре 2 показана схема вектора для экспрессии отдельного транскриптома для дисплея scFv.
На Фигуре 3 показаны результаты анализа активности ингибирования пролиферации опухолевых клеток оптимизированного по аффинности антитела hu8C4 настоящего изобретения.
На Фигуре 4 показаны результаты анализа активности ингибирования пролиферации опухолевых клеток биспецифичного антитела настоящего изобретения.
На Фигуре 5 показаны результаты анализа активности ингибирования пролиферации опухолевых клеток биспецифичного антитела настоящего изобретения.
На Фигуре 6 показаны результаты сравнения активности ингибирования пролиферации опухолевых клеток между биспецифичным антителом настоящего изобретения и комбинированной терапией в отношении клеточных линий U-87 MG (глиоблатома), NCI-H292 (НМРЛ), NCI-H1648 (НМРЛ) и NCI-H596 (НМРЛ).
На Фигуре 7 показаны результаты сравнения активности ингибирования пролиферации опухолевых клеток между биспецифичным антителом настоящего изобретения и комбинированной терапией в отношении клеточных линий LS174T (рак толстой кишки), BT20 (ТНРМЖ) и KP4 (рак поджелудочной железы).
На Фигуре 8 показаны результаты сравнения активности ингибирования пролиферации опухолевых клеток между биспецифичным антителом настоящего изобретения и комбинированной терапией в отношении клеточных линий HCC827 (НМРЛ) и NCI-H596 (НМРЛ).
На Фигуре 9 показаны результаты измерения связывающей способности анти-c-Met антитела и биспецифичного антитела настоящего изобретения в отношении различных видов антигенов c-Met и EGFR методом ИФА.
На Фигуре 10 показаны результаты измерения эффекта снижения уровня рецептора биспецифичным антителом настоящего изобретения в клеточной линии NCI-H820 (НМРЛ).
На Фигуре 11 показаны результаты измерения ингибирования фосфорилирования c-Met и EGFR анти-c-Met антителом и биспецифичным антителом настоящего изобретения в клеточной линии NCI-H820 (НМРЛ).
На Фигуре 12 показаны результаты измерения противоракового действия биспецифичного антитела настоящего изобретения на модели ксенотрансплантата клеток MG-U-87 (глиобластома).
На Фигуре 13 показаны результаты измерения противоракового действия биспецифичного антитела настоящего изобретения на модели ксенотрансплантата клеток NCI-H820 (НМРЛ).
На Фигуре 14 показаны результаты анализа активности ингибирования пролиферации опухолевых клеток, путем обработки анти-c-Met-антителом настоящего изобретения и анти-HER2-антителом, комбинированной терапией в отношении клеточной линии NCI-H2170 (НМРЛ).
На Фигуре 15 показаны результаты измерения противоракового действия комбинированной терапии с анти-c-Met-антителом настоящего изобретения и анти-HER2-антителом на модели ксенотрансплантата клеток NCI-H2170 (НМРЛ).
На Фигуре 16 показаны результаты измерения противоракового действия биспецифичного антитела настоящего изобретения на модели ксенотрансплантата клеток NCI-H596 (НМРЛ).
На Фигуре 17 показаны результаты измерения противоракового действия биспецифичного антитела настоящего изобретения на модели ксенотрансплантата клеток EBC-1 (НМРЛ).
На Фигуре 18 показаны результаты определения количества c-Met на поверхности клеток, измеренного после обработки клеточной линии HCC827 биспецифичным антителом (hu8C4 x scFv-фрагмент вектибикс) и т.д.
На Фигуре 19 показаны результаты определения количества EGFR на поверхности клеток, измеренного после обработки клеточной линии HCC827 биспецифичным антителом (hu8C4 x scFv-фрагмент вектибикс) и т.д.
На Фигуре 20 показаны результаты определения эпитопа биспецифичного антитела, проанализированного при помощи масс-спектрометрии водородно-дейтериевого обмена (HDX-MS), в третичной структуре.
Лучший вариант осуществления изобретения
Далее настоящее изобретение более подробно будет описано следующим образом. Между тем, каждое описание и вариант осуществления изобретения, раскрытые в настоящей заявке, могут также применяться соответственно к другим описаниям и вариантам осуществления изобретения. Другими словами, все комбинации различных элементов, раскрытых в настоящем изобретении, находятся в пределах объема настоящего изобретения. Кроме того, объем настоящего изобретения не может быть ограничен приведенным ниже подробным описанием.
Для достижения вышеуказанных целей в одном аспекте настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое специфично связывается с рецептором фактора роста гепатоцитов (c-Met).
Антитело или его антигенсвязывающий фрагмент настоящего изобретения, специфично связывающееся с c-Met, связывается с c-Met с высокой аффинностью для ингибирования его экспрессии или активности, таким образом, демонстрируя превосходную активность ингибирования пролиферации опухолевых клеток, так что антитело отдельно или с общепринятыми фармацевтически приемлемыми носителями, другими противораковыми лекарственными средствами, противораковыми адъювантами и т.д. может эффективно применяться в качестве противораковой композиции для предотвращения или лечения рака.
В настоящем изобретении термин «антитело» означает молекулу белка, служащую в качестве рецептора для специфичного распознавания антигена, включающую в себя молекулу иммуноглобулина, обладающую иммунологической реакционной способностью с определенным антигеном. Примеры такой молекулы могут включать в себя моноклональное антитело, поликлональное антитело, полноразмерные антитела и все фрагменты антител. Также термин может включать в себя бивалентную или биспецифичную молекулу (например, биспецифичное антитело), диатело, триатело или тетратело.
В настоящем изобретении термин «моноклональное антитело» относится к молекуле антитела из композиции, состоящей из молекул одного типа, полученной из в основном одной и той же популяции антител, где такое моноклональное антитело обладает специфичностью и аффинностью связывания по отношению к одному определенному эпитопу. В настоящем изобретении термин «полноразмерное антитело» означает структуру с двумя полноразмерными легкими цепями и двумя полноразмерными тяжелыми цепями, где каждая из легких цепей связана с тяжелой цепью дисульфидной связью. Константная область тяжелой цепи имеет типы гамма (γ), мю (μ), альфа (α), дельта (δ) и эпсилон (ε), а также гамма1 (γ1), гамма2 (γ2), гамма3 (γ3) ), гамма4 (γ4), альфа1 (α1) и альфа2 (α2) в качестве подклассов. Константная область легкой цепи имеет типы каппа (κ) и лямбда (λ). IgG содержит IgG1, IgG2, IgG3 и IgG4 в качестве подтипа.
В настоящем изобретении термины «фрагмент», «фрагмент антитела» и «антигенсвязывающий фрагмент» относятся к любым фрагментам антитела настоящего изобретения, обладающим антигенсвязывающей функцией антитела, где такие термины используются взаимозаменяемо друг с другом. Типичные антигенсвязывающие фрагменты включают в себя Fab, Fab', F(ab')2, Fv и тому подобное, но не ограничиваются ими.
Указанный фрагмент Fab имеет структуру с вариабельной областью легкой и тяжелой цепей, константной областью легкой цепи и первой константной областью тяжелой цепи (доменом CH1), а также имеет один антигенсвязывающий сайт. Антигенсвязывающий фрагмент молекулы антитела или фрагмент антитела означает фрагмент, обладающий антигенсвязывающей функцией, и Fab' отличается от Fab тем, что первый имеет шарнирную область, содержащую один или несколько остатков цистеина на С-конце СН1 домена тяжелой цепи. Антитело F(ab')2 создано таким образом, что цистеиновый остаток шарнирной области Fab' образует дисульфидную связь. Fv представляет собой минимальный фрагмент антитела, имеющий только вариабельную область тяжелой цепи и вариабельную область легкой цепи, и рекомбинантная технология для создания фрагментов Fv раскрыта в публикациях Международных патентных заявок РСТ WO 88/10649, WO 88/106630, WO 88/07085, WO 88/07086, WO 88/09344 и тому подобное. Двухцепочечный Fv образуется таким образом, что вариабельная область тяжелой цепи и вариабельная область легкой цепи связаны друг с другом нековалентной связью, тогда как одноцепочечный Fv образуется таким образом, что вариабельная область тяжелой цепи и одноцепочечная вариабельная область обычно связаны друг с другом либо ковалентной связью через пептидный линкер, либо непосредственно связаны на С-конце, образуя таким образом структуру, подобную димеру, как показано в двухцепочечном Fv. Такой фрагмент антитела может быть получен с использованием гидролазы белков (например, Fab может быть получен путем проведения рестрикционного расщепления целого антитела папаином, а фрагмент F(ab')2 может быть получен путем выполнения расщепления его пепсином) или может быть получен с помощью технологии рекомбинации генов, но не ограничивается этим.
В частности, в настоящем изобретении может быть предусмотрено, что антитело, специфически связывающееся с c-Met, представляет собой:
(а) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 1; CDR2 легкой цепи, представленную SEQ ID NO: 2; CDR3 легкой цепи, представленную SEQ ID NO: 3, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 7; CDR2 тяжелой цепи, представленную SEQ ID NO: 8; и CDR3 тяжелой цепи, представленную SEQ ID NO: 9;
(b) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 4; CDR2 легкой цепи, представленную SEQ ID NO: 5; CDR3 легкой цепи, представленную SEQ ID NO: 6, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 10; CDR2 тяжелой цепи, представленную SEQ ID NO: 11; и CDR3 тяжелой цепи, представленную SEQ ID NO: 12; или
(с) эти антитела, оптимизированные по аффинности.
В настоящем изобретении термин «тяжелая цепь» может включать в себя как полноразмерную тяжелую цепь, так и ее фрагмент, содержащий домен VH вариабельной области с аминокислотной последовательностью, имеющей последовательность вариабельной области, достаточную для придания специфичности к антигену, а также три домена константной области CH1, CH2 и CH3. Также в настоящем изобретении термин «легкая цепь» может включать в себя как полноразмерную легкую цепь, так и ее фрагмент, содержащий домен VL вариабельной области с аминокислотной последовательностью, имеющей последовательность вариабельной области, достаточную для придания специфичности к антигену, а также домен константной области CL.
В настоящем изобретении антитело может содержать как мышиное антитело, полученное от мыши, так и его мутант, в котором часть аминокислотной последовательности исходного антитела замещена, добавлена и/или удалена для улучшения аффинности, иммунности, и т.д. антитела. В качестве неограничивающего примера, мутант может содержать химерное антитело, гуманизированное антитело, оптимизированное по аффинности антитело и т.д.. В настоящем изобретении термин «мутант» полностью относится к антителу, в котором часть аминокислотной последовательности CDR исходного антитела мутирована (замещена, добавлена или удалена) при условии наличия такой же CDR, что и у исходного антитела, или нацеливания на тот же эпитоп, что и у исходного антитела. Такой мутант может быть соответствующим образом оптимизирован специалистами в данной области техники для улучшения аффинности, иммунности и тому подобного антитела в пределах объема поддержания способности к связыванию с тем же эпитопом.
Другими словами, антитело или его антигенсвязывающий фрагмент настоящего изобретения могут содержать описанную здесь последовательность анти-c-Met-антитела, а также ее биологические эквиваленты с областью специфичного распознавания c-Met. Например, может быть сделано дополнительное изменение в аминокислотной последовательности антитела, чтобы дополнительно улучшить аффинность связывания и/или другие биологические характеристики антитела. Такое изменение включает в себя, например, удаление, вставку и/или замену остатка аминокислотной последовательности антитела. Такая аминокислотная мутация создается на основе относительного сходства заместителя боковой цепи аминокислоты, например, гидрофобности, гидрофильности, заряда, размера и т.д. Анализируя размер, форму и тип заместителя боковой цепи аминокислоты, можно увидеть, что аргинин, лизин и гистидин являются положительно заряженными остатками; аланин, глицин и серин имеют одинаковый размер; и фенилаланин, триптофан и тирозин имеют сходную форму. Таким образом, исходя из таких соображений, можно видеть, что аргинин, лизин и гистидин; аланин, глицин и серин; и фенилаланин, триптофан и тирозин являются биологически функциональными эквивалентами.
В настоящем изобретении термин «химерное антитело» представляет собой антитело, образованное таким образом, что вариабельная область мышиного антитела рекомбинируется с константной областью человеческого антитела, что приводит к значительно улучшенной иммунной реакции по сравнению с мышиным антителом.
В настоящем изобретении термин «гуманизированное антитело» означает антитело, сформированное таким образом, что белковая последовательность антитела, полученного из других видов животных, отличных от человека, модифицирована так, чтобы быть сходной с последовательностью мутантного антитела, естественным образом полученного от человека. Например, гуманизированное антитело может быть создано путем получения гуманизированной вариабельной области путем рекомбинации CDR, полученной от мыши, с FR, полученным из человеческого антитела, и затем путем рекомбинации его с константной областью предпочтительного человеческого антитела. Однако простая прививка только CDR приводит к низкой аффинности гуманизированного антитела, поэтому некоторые ключевые аминокислотные остатки FR, которые, как считается, могут влиять на трехмерную структуру CDR, могут обеспечить аффинность CDR мышиного антитела, и достичь, таким образом, того же уровня аффинности, что и у исходного мышиного антитела.
В настоящем изобретении термин «оптимизированное по аффинности антитело», которое представляет собой мутант, образованный таким образом, что часть последовательности CDR определенного антитела замещена, добавлена или удалена, означает антитело с улучшенной аффинностью связывания с антигеном при связывании с тем же эпитопом антигена, с которым связывается определенное антитело. В частности, термин «оптимизированное по аффинности антитело» настоящего изобретения относится к мутантному антителу, связывающемуся с тем же эпитопом, что и: (а) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 1; CDR2 легкой цепи, представленную SEQ ID NO: 2; CDR3 легкой цепи, представленную SEQ ID NO: 3, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 7; CDR2 тяжелой цепи, представленную SEQ ID NO: 8; CDR3 тяжелой цепи, представленную SEQ ID NO: 9; или (b) антитело, содержащее вариабельную область легкой цепи, содержащее CDR1 легкой цепи, представленную SEQ ID NO: 4; CDR2 легкой цепи, представленную SEQ ID NO: 5; CDR3 легкой цепи, представленную SEQ ID NO: 6, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 10; CDR2 тяжелой цепи, представленную SEQ ID NO: 11; CDR3 тяжелой цепи, представленную SEQ ID NO: 12. Специалист в данной области техники может получить оптимизированное по аффинности антитело, используя известную технологию, основанную на определенных последовательностях CDR легкой цепи и тяжелой цепи. Например, оптимизированное по аффинности антитело настоящего изобретения может быть получено с помощью фагового дисплея. В настоящем изобретении термин «фаговый дисплей» относится к технологии, которая отображает мутантный полипептид в виде белка слитого с, по меньшей мере, частью белка оболочки, на фаге, например, на поверхности частиц филаментного фага. Полезность фагового дисплея заключается в том, что он нацелен на большую библиотеку рандомизированных мутантов белка, что позволяет быстро и эффективно классифицировать последовательности, связывающиеся с антигеном-мишенью с высокой аффинностью. Отображение библиотеки пептидов и белков на фаге было использовано для скрининга миллионов полипептидов, чтобы увидеть полипептид с характеристикой специфичного связывания.
В одном примере варианта осуществления настоящего изобретения предполагается, что антитело представляет собой антитело, содержащее: (a) вариабельную область легкой цепи, представленную SEQ ID NO: 13, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 15; или (b) вариабельную область легкой цепи, представленную SEQ ID NO: 14, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 16. В качестве неограничивающего примера может быть предусмотрено, что указанное антитело представляет собой антитело, содержащее: (а) вариабельную область легкой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 17, и вариабельную область тяжелой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 19; или (b) вариабельную область легкой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 18, и вариабельную область тяжелой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 20.
В соответствии с одним конкретным вариантом осуществления настоящего изобретения группа клеток гибридомы была получена от мыши, где слитый белок домена Sema c-Met человека с Fc представляет собой антиген, из которой анти-c-Met антитело специфично связывающееся с c-Met было выбрано путем скрининга методом ИФА с использованием слитого белка c-Met/His в качестве антигена. Выбранное антитело и его химерное антитело обладают ингибирующей активностью в отношении пролиферации опухолевых клеток, которая равна или даже выше, чем коммерчески доступные известные антитела LY2875358 и OA-5D5 (Таблица 3 и Фигура 1), и таким образом, его применение является очень эффективным для предотвращения или лечение рака.
В другом примере варианта осуществления настоящего изобретения предполагается, что антитело содержит:
(а) вариабельную область легкой цепи, представленную SEQ ID NO: 21, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 23; (b) вариабельную область легкой цепи, представленную SEQ ID NO: 22, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 24; (c) вариабельную область легкой цепи, представленную SEQ ID NO: 29, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 31; или (d) вариабельную область легкой цепи, представленную SEQ ID NO: 30, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 32. В качестве неограничивающего примера может быть предусмотрено, что антитело представляет собой антитело, содержащее: (a) a вариабельную область легкой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 25, и вариабельную область тяжелой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 27; (b) вариабельную область легкой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 26, и вариабельную область тяжелой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 28; (c) вариабельную область легкой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 33, и вариабельную область тяжелой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 35; или (d) вариабельную область легкой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 34, и вариабельную область тяжелой цепи, кодируемую нуклеотидом, представленным SEQ ID NO: 36. Также может быть предусмотрено, что антитело содержит шарнирную область, представленную одной из последовательностей SEQ ID NO: 37 - SEQ ID NO: 44.
В одном конкретном варианте осуществления настоящего изобретения было получено гуманизированное антитело, содержащее CDR антитела, полученные посредством отбора с помощью фагового дисплея, и было установлено, что такое антитело обладало противораковой активностью, которая была аналогична активности химерного антитела настоящего изобретения (Примеры 2 и 3). Кроме того, в другом конкретном варианте осуществления настоящего изобретения ингибирующую активность антитела в отношении пролиферации опухолевых клеток оценивали в зависимости от последовательности шарнирной области, и было установлено, что пролиферация большинства опухолевых клеток эффективно ингибировалась даже при некоторой разнице в активности в зависимости от разницы последовательности шарнирной области (Таблица 7).
В еще одном неограничивающем примерном варианте осуществления настоящего изобретения может быть предусмотрено, что оптимизированное по аффинности антитело в случае гуманизированного антитела представляет собой антитело, где одна или несколько аминокислот замещены в антителе, содержащем: вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 1; CDR2 легкой цепи, представленную SEQ ID NO: 2; CDR3 легкой цепи, представленную SEQ ID NO: 3, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 7; CDR2 тяжелой цепи, представленную SEQ ID NO: 8; CDR3 тяжелой цепи, представленную SEQ ID NO: 9, и где (i) G в первом положении CDR1 легкой цепи замещен A, E, K, L, N, R, S, V или W; А во втором положении этой CDR замещен С, G, I, Р, S, Т или V; S в третьем положении CDR цепи замещен G, M, N, P, Q, R, S или T; Е в 4-ом положении этой CDR замещен А, D, F, G, Н, К, М, Q, R, S, Т или V; N в 5-ом положении этой CDR замещен A, D, E, G, K, L, P, Q, R, S, T или V; I в 6-ом положении этой CDR замещен A, F, L, M, Q, R, S, T или V; Y в 7-ом положении этой CDR замещен F, H, R или V; или G в 8-ом положении этой CDR замещен D, F, H, M, N, R, S, T или V; (ii) G в первом положении CDR2 легкой цепи замещен D, F, H, K, P, Q, S, V или Y; Т в третьем положении этой CDR замещен Q; или N в 4-ом положении этой CDR замещен G; (iii) Q в первом положении CDR3 легкой цепи замещен Е, G, I, М или N; N во втором положении этой CDR замещен A, D, E, H, L, Q, S или T; V в третьем положении этой CDR замещен I, L, M, N, Q, S или T; L в 4-ом положении этой CDR замещен F, Н, I, М, R, S, V, W или Y; S в 5-ом положении этой CDR замещен C, D, E, F, G, H, K, L, N, Q, R, T, V или Y; S в 6-ом положении этой CDR замещен D, E, F, G, H, I, L, M, N, P, Q, R, T, V или Y; P в 7-ом положении этой CDR замещен A, D, E, G, N, Q, S или V; Y в 8-ом положении этой CDR замещен E, F, L, M или Q; или Т в 9-ом положении этой CDR замещен D, F, G, I, L, N, S, V, W или Y; (iv) D в первом положении CDR1 тяжелой цепи замещен G или Q; Y во втором положении этой CDR замещен Q; или I в 4-м положении этой CDR замещен А или Q; (v) F в третьем положении CDR2 тяжелой цепи замещен D, E, W или Y; G в 5-ом положении этой CDR замещен D, H или Y; S в 6-ом положении этой CDR замещен F, P, W или Y; G в 7-ом положении этой CDR замещен A, F, L, N или T; N в 8-ом положении это CDR замещен F, P, S, T или Y; Т в 9-ом положении этой CDR замещен A, D, E, F, G, H, L, P, S или V; Н в 10-ом положении этой CDR замещен А, D, F, М, R, S, Т, V, W или Y; F в 11-ом положении этой CDR замещен G, H, I, L, M, N, P, Q, V или Y; S в 12-ом положении этой CDR замещен A, D, G, H, I, L, P, T или V; А в 13-ом положении этой CDR замещен D, E, F, G, H, I, K, L, M, P, R, S, T, V или Y; R в 14-ом положении этой CDR замещен A, E, G, H, L, N, P, Q, S, W или Y; F в 15-ом положении этой CDR замещен D, E, G, L, M, P, R, S, V или W; K в 16-ом положении этой CDR замещен A, E, F, G, H, L, R, S, T, V или Y; или G в 17-ом положении этой CDR замещен E, F, H, L, M, N, P, Q, R, S, T, V или W; или (vi) G в первом положении CDR3 тяжелой цепи замещен E, F, H, N, Q, V или W; D во втором положении этой CDR замещен E; Y в третьем положении этой CDR замещен L, Q, T или V; G в 4-ом положении этой CDR замещен W; F в 5-ом положении этой CDR замещен L или Y; L в 6-ом положении этой CDR замещен Q, S или Y; или Y в 7-ом положении этой CDR замещен C, L, M, N или Q. Здесь может быть предусмотрено, что CDR1 легкой цепи содержит от 0 до 5 замен, CDR2 легкой цепи содержит от 0 до 1 замены, CDR3 легкой цепи содержит от 0 до 7 замен, CDR1 тяжелой цепи содержит от 0 до 1 замены, CDR2 тяжелой цепи содержит от 0 до 11 замен, и CDR3 тяжелой цепи содержит от 0 до 6 замен.
В частности, в еще одном примерном варианте осуществления настоящего изобретения может быть предусмотрено, что оптимизированное по аффинности антитело содержит вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную любой из SEQ ID NO: 1 и SEQ ID NO: 229 - SEQ ID NO: 268; CDR2 легкой цепи, представленную любой из SEQ ID NO: 2, SEQ ID NO: 182 - SEQ ID NO: 190, SEQ ID NO: 227 и SEQ ID NO: 228; CDR3 легкой цепи, представленную любой из SEQ ID NO: 3, SEQ ID NO: 142 - SEQ ID NO: 181, SEQ ID NO: 191 - SEQ ID NO: 226 и SEQ ID NO: 269 - SEQ ID NO: 301; и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную любой из SEQ ID NO: 7 и SEQ ID NO: 108 - SEQ ID NO: 112; CDR2 тяжелой цепи, представленную любой из SEQ ID NO: 8, SEQ ID NO: 54 - SEQ ID NO: 63, SEQ ID NO: 72 - SEQ ID NO: 107 и SEQ ID NO: 118 - SEQ ID NO: 141; CDR3 тяжелой цепи, представленную любой из SEQ ID NO: 9, SEQ ID NO: 64 - SEQ ID NO: 71 и SEQ ID NO: 113 - SEQ ID NO: 117, более конкретно, содержащую вариабельную область легкой цепи, представленную любой из SEQ ID NO: 21 и SEQ ID NO: 306 - SEQ ID NO: 311, и вариабельную области тяжелой цепи, представленную любой из SEQ ID NO: 23 и SEQ ID NO: 302 - SEQ ID NO: 305, и еще более конкретно, включающую в себя: (а) вариабельную область легкой цепи, представленную SEQ ID NO: 21, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 302; (b) вариабельную область легкой цепи, представленную SEQ ID NO: 21, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 305; (c) вариабельная область легкой цепи, представленную SEQ ID NO: 310, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 23; (d) вариабельную область легкой цепи, представленную SEQ ID NO: 308, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 305; (e) вариабельную область легкой цепи, представленную SEQ ID NO: 306, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 303; (f) вариабельную область легкой цепи, представленную SEQ ID NO: 307, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304; (g) вариабельную область легкой цепи, представленную SEQ ID NO: 308, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304; (h) вариабельную область легкой цепи, представленную SEQ ID NO: 309, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304; (i) вариабельную область легкой цепи, представленную SEQ ID NO: 311, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304; или (j) вариабельную область легкой цепи, представленную SEQ ID NO: 306, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 302, но не ограничивается этим.
В одном конкретном варианте осуществления настоящего изобретения был использован метод конкурентного отбора для отбора антитела с более высокой аффинностью, чем у гуманизированного антитела, что позволило получить ряд антител, оптимизированных по аффинности (Таблицы 8-10 и 12). Оптимизированное по аффинности антитело обладает эффектом ингибирования пролиферации опухолевых клеток, который в 4,3-28,5 раз выше, чем у гуманизированного антитела (Таблица 11, 13 и Фигура 3).
В настоящем изобретении может быть предусмотрено, что антитело представляет собой антитело или его антигенсвязывающий фрагмент, дополнительно специфично связывающееся с рецептором эпидермального фактора роста (EGFR) в дополнение к специфичному связыванию с c-Met.
Известно, что EGFR, одна из тирозинкиназ ErbB, аномально активируется во многих эпидермальных клеточных опухолях, включающих в себя немелкоклеточный рак легкого, вызывает пролиферацию клеток, инвазию, метастазирование и ангиогенез и увеличивает выживаемость клеток. Гефитиниб (Iressa), элотиниб (Tarceva) и осимертиниб (Tagrisso), которые являются ингибиторами тирозинкиназы EGFR, используются в качестве типичного терапевтического средства для лечения рака легких; и цетуксимаб (Erbitux) и панитумумаб (Vectibix), которые являются антителами, направленными против EGFR, используются в качестве терапевтического средства для лечения рака толстой кишки (Yewale C et al., Biomaterials. 2013 34 (34): 8690-707 (2013), Deric L. Wheeler et al., Nature Reviews Clinical Oncology 7, 493-507 (2010)).
Такие терапевтические агенты, нацеленные на EGFR, вызывают устойчивость за год до и после лечения, при этом амплификация c-Met, мутация и HGF-индуцированная активация известны как ключевой механизм устойчивости (Simona Corso Cancer Discovery 3: 978-992 (2013), Curtis R Chong et al., Nature Medicine, 19, 1389-1400 (2013). Также сообщается, что EGFR и c-Met одновременно экспрессируются в различных опухолевых клетках, причем при ингибировании EGFR активируется c-Met, таким образом, быстро развивается устойчивость к ингибиторам тирозинкиназы EGFR (Engelman, JA, et al., Science, 316: 1039-43 (2007)).
Основываясь на таком механизме, в настоящее время на стадии клинических испытаний проводят однократное лечение одним препаратом, нацеленным на c-Met, и комбинированное лечение с препаратом, нацеленным на EGFR, но их эффективность еще не подтверждена в качестве терапевтического средства, и существует необходимость в разработке терапевтического агента для связанных с c-Met раковых опухолей, известных как основная причина резистентности. Соответственно, авторы настоящего изобретения получили биспецифичное в отношении c-Met/EGFR антитело на основе антитела, описанного выше. Биспецифичное антитело не только эффективно ингибирует пролиферацию опухолевых клеток, которые устойчивы к существующим терапевтическим агентам, нацеленным на EGFR, но также проявляет превосходную ингибирующую пролиферацию активность в отношении опухолевых клеток, что, таким образом, эффективно при лечении заболеваний, таких как виды рака, опосредованные различными механизмами с участием c-Met.
Может быть предусмотрено, что биспецифичное антитело конструируется таким образом, что антитело или его антигенсвязывающий фрагмент, специфически связывающееся с EGFR, связано с одним концом легкой цепи или тяжелой цепи антитела, специфичного в отношении c-Met, например, связано с С-концом тяжелой цепи, но не ограничивается этим.
Может быть предусмотрено, что связывающий фрагмент, специфически связывающийся с EGFR, представляет собой Fab, Fab', F(ab')2 или Fv.
В одном примере варианта осуществления настоящего изобретения может быть предусмотрено, что Fv представляет собой фрагмент scFv, где фрагмент scFv связан линкером, способным связывать фрагмент scFv с одним концом легкой цепи или тяжелой цепи анти-c-Met антитела. В одном примере варианта осуществления настоящего изобретения антитело, специфически связывающееся с EGFR, дополнительно получают путем связывания с линкером, представленным SEQ ID NO: 312.
Может быть предусмотрено, что фрагмент scFv анти-EGFR антитела представляет собой scFv анти-EGFR антитела, способный специфично связываться с EGFR, известный в данной области техники, который, например, представляет собой без ограничений эрбитукс, вектибикс, портразза, ТераЦИМ или тому подобное.
В одном примерном варианте осуществления настоящего изобретения может быть предусмотрено, что scFv анти-EGFR антитела представляет собой scFv-фрагмент эрбитукс или вектибикс, в частности scFv анти-EGFR антитела содержит аминокислотную последовательность, представленную SEQ ID NO: 313 или SEQ ID NO: 314, где scFv-фрагмент вектибикс содержит аминокислотную последовательность, представленную SEQ ID NO: 315, но не ограничивается этим.
В соответствии с одним конкретным вариантом осуществления настоящего изобретения, в результате идентификации ингибирующей пролиферацию опухолевых клеток активности биспецифичного антитела было установлено, что это антитело улучшенной эффективностью активности ингибирования опухоли, по сравнением с оптимизированным антителом hu8C4 (Таблицы 16 и 17, и Фигуры 4, 5, 16 и 17). В частности, было установлено, что антитело настоящего изобретения обладает улучшенным ингибирующим пролиферацию клеток действием даже на клеточные линии NCI-H292 и NCI-H1648, в которых c-Met и EGFR экспрессируются в норме (Таблицы 17 и 19 и Фигура 6). На основании таких результатов можно видеть, что противораковое действие антитела настоящего изобретения не ограничивается только аномалией экспрессии c-Met или наличием или отсутствием мутации c-Met и т.д.
Кроме того, было установлено, что биспецифичное антитело настоящего изобретения обладает улучшенной способностью ингибировать пролиферацию опухолевых клеток, по сравнению с комбинированной терапией двумя антителами (Таблицы 18-21 и Фигуры 6-8). Кроме того, в результате выявления влияния биспецифичного антитела настоящего изобретения на активность антигенов и пути передачи сигнала было установлено, что биспецифичное антитело настоящего изобретения обладает более высокой эффективностью ингибирования передачи сигнала, чем одно антитело. (Фигура 11).
Может быть предусмотрено, что антитело или его антигенсвязывающий фрагмент настоящего изобретения связывается с областью эпитопа, представленной аминокислотной последовательностью, выбранной из группы, представленной SEQ ID NO: 331, SEQ ID NO: 332, SEQ ID NO: 333 и/или SEQ ID NO: 334. Оптимизированное по аффинности антитело, полученное на основе определенного антитела (эталонного антитела), характеризуется высокой гомологией с последовательностями CDR легкой цепи и тяжелой цепи вариабельной области эталонного антитела, и таким образом, связывается с той же областью эпитопа, что и эталонное антитело, так что такое оптимизированное по аффинности антитело может обладать всеми биологическими характеристиками, такими как фармацевтический механизм и фармацевтическая эффективность, обусловленными сайтом связывания, специфичностью и антителом, и демонстрировать улучшенное действие на аффинность связывания, по сравнению с эталонным антителом.
Область эпитопа соответственно означает, например, YVSKPGAQL (SEQ ID NO: 331) в положениях 321-329, IGASLNDDI (SEQ ID NO: 332) в положениях 333-341, PIKYVND (SEQ ID NO: 333) в положениях 366-372 и QVVVSRSGPST (SEQ ID NO: 334) в положениях 464-474 от N-конца эталонного антигена c-Met (SEQ ID NO: 335), где последовательность антигена c-Met, связывающаяся с антителом настоящего изобретения или его антигенсвязывающим фрагментом, содержит частичную мутацию (замену, добавление или удаление), или связывающий антиген существует в форме фрагмента, предшественника или подтипа c-Met, и, таким образом, его сайты или последовательности связывания могут, соответственно, несколько изменяться. Тем не менее, специалист в данной области техники может четко определить положение и последовательность, с которой связывается антитело настоящего изобретения или его антигенсвязывающий фрагмент, основываясь на информации о последовательности эпитопа эталонного антигена c-Met.
В одном конкретном варианте осуществления настоящего изобретения было идентифицировано, что биспецифичное антитело hu8C4 × scFv-фрагмент вектибикс настоящего изобретения связывается с 4 эпитопными областями Y321 - L329 (SEQ ID NO: 331), I333 - I341 (SEQ ID NO: 332), P366 - D372 (SEQ ID NO: 333) и Q464 - S474 (SEQ ID NO: 334) β-цепи домена Sema с-Met человека (Таблица 28).
Термин «антитело или его антигенсвязывающий фрагмент, специфично связывающийся с c-Met» настоящего изобретения означает антитело, связывающееся с c-Met человека с KD 1×10-7 М или менее. Может быть предусмотрено, что антитело или его антигенсвязывающий фрагмент связывается с c-Met человека, например, с KD 5×10-8 М или менее, KD 1×10-8 М или менее, KD 5×10-9 М или менее или KD 1×10-9 М или менее, но не ограничивается этим.
В одном конкретном варианте осуществления настоящего изобретения было прямо идентифицировано, что антитело или его антигенсвязывающие фрагменты настоящего изобретения имели высокую аффинность связывания с антигеном c-Met путем идентификации аффинности связывания hu8C4, hu8C4 AH71 и hu8C4 × scFv-фрагмент вектибикс с ВКД c-Met, и, идентифицированное таким образом значение KD составило 3,173×10-10, 9,993×10-11 и 2,78×10-10, соответственно (Таблица 22). Было установлено, что антитело или его антигенсвязывающий фрагмент настоящего изобретения обладают перекрестной реактивностью по отношению к антигену c-Met макака-крабоеда, которая является обезьяной (Таблица 22), но не связывается с антигенами, полученными от других животных (например, грызунов) (Фигура 9). Также было установлено, что антитело или его антигенсвязывающий фрагмент настоящего изобретения не связываются с другими рецепторами на поверхности клеток, кроме c-Met (Таблица 24). Таким образом, из приведенных выше результатов видно, что антитело или его антигенсвязывающий фрагмент настоящего изобретению проявляли специфичность связывания с антигеном c-Met человека и обезьян.
Используемый здесь термин «константа связывания (Kon)» означает коэффициент связывания определенного взаимодействия антитело-антиген, а термин «константа диссоциации (Koff)» означает коэффициент диссоциации определенного взаимодействия антитело-антиген. Кроме того, в настоящем изобретении термин «аффинность к антигену (KD)» означает отношение Koff:Kon (то есть, Koff/Kon) указанное как молярная концентрация (M). Может быть предусмотрено, что значение KD для антитела измеряют с использованием метода, общеизвестного в данной области техники. Например, в качестве способа измерения значения KD антитела может применяться, без ограничений, анализ поверхностного плазмонного резонанса с использованием системы Biocore™.
В другом аспекте настоящее изобретение относится к способу получения молекулы нуклеиновой кислоты, кодирующей антитело или его антигенсвязывающий фрагмент, экспрессионному вектору, содержащему эту молекулу нуклеиновой кислоты, клетке-хозяину, в которую введен этот экспрессионный вектор, антителу, использующему эту клетку-хозяин, или его антигенсвязывающему фрагменту.
Антитело и его антигенсвязывающий фрагмент являются такими, как описано выше.
Используемый здесь термин «молекула нуклеиновой кислоты» имеет значение, которое полностью включает в себя молекулы ДНК и РНК, где нуклеотид, основная составляющая единица в молекуле нуклеиновой кислоты, содержит не только природный нуклеотид, но также аналог, в котором модифицирована часть сахара или основания (Scheit, Nucleotide Analogs, John Wiley, New York (1980); Uhlman and Peyman, Chemical Reviews, (1990) 90: 543-584). Последовательность молекулы нуклеиновой кислоты для кодирования вариабельных областей тяжелой цепи и легкой цепи настоящего изобретения может быть модифицирована, где модификация включает в себя добавление, удаление или неконсервативную или консервативную замену нуклеотида.
Следует понимать, что молекула нуклеиновой кислоты настоящего изобретения также содержит нуклеотидную последовательность, обладающую существенной идентичностью с вышеупомянутой нуклеотидной последовательностью. В настоящем изобретении в случае выравнивания наиболее подходящим образом вышеупомянутой нуклеотидной последовательности настоящего изобретения с любыми другими последовательностями и анализа выровненных последовательностей с помощью алгоритма, обычно используемого в данной области техники, существенная идентичность означает нуклеотидную последовательность, которая имеет минимальную гомологию 80%, предпочтительно, минимальную гомологию 90%, более предпочтительно, минимальную гомологию 95%.
Используемый здесь термин «вектор» означает средство для экспрессии гена-мишени в клетке-хозяине, и включает в себя без ограничений плазмидный вектор; космидный вектор; и вирусный вектор, такой как бактериофаговый вектор, аденовирусный вектор, ретровирусный вектор и адено-родственный вирус, в частности плазмидный вектор.
В векторе настоящего изобретения может быть предусмотрено, что молекула нуклеиновой кислоты, кодирующая вариабельную область легкой цепи, и молекула нуклеиновой кислоты, кодирующая вариабельную область тяжелой цепи, функционально связаны с промотором.
В настоящем изобретении термин «функционально связанный» означает функциональное связывание между последовательностью, регулирующей экспрессию нуклеиновой кислоты, (например, промотором, сигнальной последовательностью или последовательностью в сайте связывания фактора регуляции транскрипции) и другой последовательностью нуклеиновой кислоты, благодаря чему регуляторная последовательность контролирует транскрипцию и/или декодирование другой последовательности нуклеиновой кислоты.
Рекомбинантная векторная система настоящего изобретения может быть создана различными способами, известными в данной области техники. Например, такие способы подробно описаны в книге Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (2001), содержание которой включено в настоящее описание путем ссылки.
Вектор настоящего изобретения, как правило, может быть сконструирован как вектор для клонирования или вектор для экспрессии. Также вектор настоящего изобретения может быть сконструирован таким образом, что его хозяином является прокариотическая клетка или эукариотическая клетка.
Например, если вектор настоящего изобретения является экспрессионным вектором, а хозяином является прокариотическая клетка, то он обычно содержит мощные промоторы, способные выполнять транскрипцию (например, промотор tac, промотор lac, промотор lacUV5, промотор lpp, промотор pLλ, промотор pRλ, промотор rac5, промотор amp, промотор recA, промотор SP6, промотор trp, промотор T7 и тому подобное), сайт связывания рибосомы для начала декодирования и последовательности терминации транскрипции/декодирования. Если в качестве клетки-хозяина используется E.coli (например, штамм HB101, BL21, DH5α и т.д.), то промоторная и операторная части пути биосинтеза триптофана E.coli (Yanofsky, C., J. Bacteriol., (1984) 158: 1018-1024), и левый промотор фага λ (промотор pLλ, Herskowitz, I. and Hagen, D., Ann. Rev. Genet., (1980) 14: 399-445) могут использоваться в качестве регуляторной части. Если в качестве клетки-хозяина используется Bacillus sp., то промотора гена белка токсина Bacillus thuringiensis (Appl. Environ. Microbiol. (1998) 64: 3932-3938; Mol. Gen. Genet. (1996) 250: 734-741) или любые промоторы экспрессируемые в Bacillus sp. могут быть использован в качестве регуляторной части.
Между тем, рекомбинантный вектор настоящего изобретения может быть получен путем манипулирования с плазмидой (например, pCL, pSC101, pGV1106, pACYC177, ColE1, pKT230, pME290, pBR322, pUC8/9, pUC6, pBD9, pHC79, pIJ61, pVAF1, pLAR, серия pGEX, серия pET, pUC19 и тому подобное), фагом (например, λgt4⋅λB, λ-Харон, λΔz1, M13 и тому подобное) или вирусом (например, SV40 и т.д.), часто используемыми в данной области техники.
Между тем, если вектор настоящего изобретения представляет собой экспрессионный вектор, а хозяином является эукариотическая клетка, то можно использовать промоторы, полученные из генома клеток млекопитающих (например, промотор металлотионеина, промотор β-актина, промотор гемоглобина человека и промотор креатина мышц человека) или промоторы, полученные из вируса млекопитающего (например, поздний промотор аденовируса, промотор вируса коровьей оспы 7.5K, промотор SV40, промотор цитомегаловируса (CMV), промотор tk HSV, промотор вируса опухоли молочной железы мыши (MMTV), промотор LTR ВИЧ, промотор вируса Молони, промотор вируса Эпштейна-Барр (EBV) и промотор вируса саркомы Рауса (RSV), где они обычно имеют последовательность полиаденилирования в качестве последовательности терминации транскрипции. В частности, рекомбинантный вектор настоящего изобретения содержит промотор CMV.
Рекомбинантный вектор настоящего изобретения может быть слит с другими последовательностями для облегчения очистки экспрессируемого ими антитела. В качестве примеров слитых последовательностей можно привести глутатион-S-трансферазу (Pharmacia, США), мальтозосвязывающий белок (NEB, США), FLAG (IBI, США), 6× His (гексагистидин; Quiagen, США) и тому подобное. Также белок, экспрессируемый вектором настоящего изобретения, представляет собой антитело, и таким образом, экспрессированное антитело может быть легко очищено через колонку с белком А и т.д. без дополнительной последовательности для очистки.
Между тем, рекомбинантный вектор настоящего изобретения содержит ген устойчивости к антибиотикам, обычно используемый в данной области техники в качестве селективного маркера, где он может включать в себя, например, гены устойчивости к ампициллину, гентамицину, карбенициллину, хлорамфениколу, стрептомицину, канамицину, гененицину, неомицину и тетрациклину.
Вектор для экспрессии антитела настоящего изобретения может существовать как векторная система, в которой легкая цепь и тяжелая цепь одновременно экспрессируются в одном векторе, так и как система, в которой легкая цепь и тяжелая цепь соответственно экспрессируются в отдельном векторе. В последнем случае два вектора могут быть введены в клетку-хозяин, например, путем котрансформации или направленной трансформации. Котрансформация представляет собой метод отбора клеток, которые экспрессируют как легкие, так и тяжелые цепи, после одновременного введения каждой векторной ДНК, кодирующей легкую и тяжелую цепи в клетку-хозяин. Направленная трансформация представляет собой метод отбора клетки, трансформированной вектором, содержащим легкую (или тяжелую) цепь, и трансформации выбранной клетки снова вектором, содержащим тяжелую (или легкую) цепь, чтобы окончательно отобрать клетку, которая экспрессирует как легкую, так и тяжелую цепи.
Можно использовать любые клетки-хозяева, известные в данной области техники, при условии, что они способны стабильно и непрерывно клонировать и экспрессировать вектор настоящего изобретения, такие клетки-хозяева могут включать в себя без ограничений штаммы Bacillus sp., такие как Escherichia coli, Bacillus subtilis и Bacillus thuringiensis, и прокариотические клетки-хозяева, такие как Streptomyces, Pseudomonas (например, Pseudomonas putida), Proteus mirabilis или Staphylococcus (например, Staphylococcus carnosus).
В качестве подходящих для вектора эукариотических клеток-хозяев могут быть мицеты, такие как виды Aspergillus, дрожжи, такие как Pichia pastoris, Saccharomyces cerevisiae, Schizosaccharomyces и Neurospora crassa, другие низшие эукариотические клетки, клетки высших эукариот, такие как клетки, полученные от насекомых, и клетки полученные из растений или млекопитающих.
В частности, клетки-хозяева могут представлять собой без ограничений клетки COS7 (клетки почки обезьяны), клетки NSO, клетки SP2/0, клетки яичника китайского хомячка (CHO), W138, клетки почки детеныша хомяка (BHK), MDCK, клеточные линии миеломы, клетки HuT78 или клетки293, в частности клетки СНО.
В настоящем изобретении «трансформация» и/или «трансфекция» в клетки-хозяева могут быть выполнены путем выбора подходящей стандартной технологии, совместимой с клетками-хозяевами, известной в данной области техники, включая любые способы введения нуклеиновой кислоты в организмы, клетки, ткани или органы. Эти способы включают в себя без ограничений электропорацию, плазмогамию, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), перемешивание с использованием карбидокремниевого волокна, трансформацию, опосредованную агробактериями, ПЭГ, декстрансульфат, липофектамин, трансформацию, опосредованную сушкой/подавлением, и тому подобное.
В настоящем изобретении способ получения антитела или его антигенсвязывающего фрагмента с использованием клетки-хозяина может, в частности, включать в себя этапы: (а) культивирования клетки-хозяина, трансформированной рекомбинантным вектором настоящего изобретения; и (b) экспрессии анти-c-Met-антитела или его антигенсвязывающего фрагмента в клетке-хозяине.
При получении указанного выше антитела культивирование трансформированной клетки-хозяина можно проводить в подходящей среде и в условиях культивирования, известных в данной области техники. Такой процесс культивирования может быть легко оптимизирован в соответствии с выбранным штаммом специалистами в данной области техники. Такой способ культивирования раскрыт в различных документах (например, James M. Lee, Biochemical Engineering, Prentice-Hall International Editions, 138-176). Клеточную культуру разделяют на суспензионную культуру и прикрепленную культуру в зависимости от типа роста клеток, и периодическую культуру, подпитываемую культуру и непрерывную культуру в зависимости от способа культивирования. Среда, используемая в культуре, должна соответствующим образом удовлетворять требованиям определенного штамма.
При культивировании клеток животных среда содержит различные источники углерода, источники азота и микроэлементы. Примеры используемых источников углерода могут включать в себя углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота, где такие источники углерода могут использоваться отдельно или в комбинации.
Источники азота, которые могут быть использованы в настоящем изобретении, могут включать в себя, например, источники органического азота, такие как пептон, дрожжевой экстракт, мясной сок, солодовый экстракт, кукурузный сироп (CSL) и сою-пшеницу, и источники неорганического азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония, где такие источники азота могут использоваться по отдельности или в комбинации. В качестве источника фосфора среда может содержать дигидрофосфат калия, гидрофосфат дикалия и соответствующую ему натрийсодержащую соль. Также среда может содержать соли металлов, такие как сульфат магния или сульфат железа. Кроме того, среда может содержать аминокислоты, витамины, соответствующие предшественники и тому подобное.
Во время культивирования химические соединения, такие как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, добавляются в продукт культивирования соответствующим образом для регулирования значения рН продукта культивирования. Кроме того, во время культивирования образование пузырьков может быть подавлено с помощью пеногасителя, такого как сложный эфир полигликоля жирной кислоты. Кроме того, кислород или кислородсодержащий газ (например, воздух) вводят в продукт культивирования, чтобы поддерживать аэробное состояние продукта культивирования. Температура продукта культивирования обычно составляет от 20 до 45°C, предпочтительно от 25 до 40°C.
Способ получения может дополнительно включать в себя этап: (c) сбора анти-c-Met антитела или его антигенсвязывающего фрагмента, экспрессированного в клетке-хозяине. Антитело, полученное культивированием трансформированной клетки-хозяина, может применяться в неочищенном состоянии или подвергаться дополнительной очистке для получения очищенного антитела с высокой чистотой с использованием различных общепринятых методов, например, диализа, солевого осаждения, хроматографии и тому подобного. Из этих методов чаще всего используется метод хроматографии, в котором тип и порядок колонки могут быть выбраны из ионообменной хроматографии, эксклюзионной хроматографии, аффинной хроматографии и т.д. в зависимости от характеристик антител, метода культивирования и т.д..
В другом аспекте настоящее изобретение относится к композиции для обнаружения c-Met, содержащей антитело или его антигенсвязывающий фрагмент, набору для обнаружения, содержащему ее, и способу обнаружения c-Met при помощи антитела с использованием этого набора.
Композиция для обнаружения c-Met и набор, содержащий ту же форму комплекса антиген-антитело, действуют таким образом, что антитело, специфично связывающееся с c-Met или его антигенсвязывающий фрагмент, вступает в контакт с образцом, таким образом, эффективно обнаруживая c-Met.
Используемый здесь термин «комплекс антиген-антитело» означает конъюгат между c-Met и антителом для его распознавания для идентификации опухоли или раковой клетки, экспрессирующей c-Met в образце.
Способ количественного определения антигена c-Met с использованием композиции для обнаружения c-Met и с использованием набора, содержащего ее, может быть выполнен путем идентификации образования комплекса антиген-антитело, при этом идентификация образования комплекса антиген-антитело может быть выполняется с помощью иммуноферментного анализа (ИФА), вестерн-блоттинга, иммунофлюоресценции, иммуногистохимического окрашивания, проточной цитометрии, иммуноцитохимии, радиоиммуноанализа (РИА), иммунопреципитационного анализа, иммунодиффузионного анализа, анализа фиксации комплемента, белкового чипа и т.д., но не ограничивается этим. ИФА включает в себя различные методы ИФА, такие как прямой ИФА с использованием меченого антитела для распознавания антигена, прикрепленного к твердой подложке; непрямой ИФА с использованием меченого вторичного антитела для распознавания захватывающего антитела в комплексе антитела для распознавания антигена, прикрепленного к твердой подложке; прямой сэндвич-ИФА с использованием другого меченого антитела для распознавания антигена в комплексе антитела и антигена, прикрепленного к твердой подложке; непрямой сэндвич-ИФА с использованием меченого вторичного антитела для реакции с другим антителом для распознавания антигена в комплексе антитела и антигена, прикрепленного к твердой подложке, и затем распознавания такого антитела и т.д.
В качестве метки для качественного или количественного определения образования комплекса антиген-антитело используют без ограничений фермент, флуоресцентный материал, лиганд, светящийся материал, микрочастицу, окислительно-восстановительную молекулу, радиоизотоп и тому подобное. В качестве ферментов используют без ограничений β-глюкуронидазу, β-D-глюкозидазу, β-D-галактозидазу, уреазу, пероксидазу, щелочную фосфатазу, ацетилхолинэстеразу, глюкозооксидазу, гексокиназу и ГДФазу, РНКазу, глюкозооксидазу и люциферазу, фосфофруктокиназу, фосфоенолпируваткарбоксилазу, аспартатаминотрансферазу, фосфоенолпируватдекарбоксилазу, β-лактамазу и т.д..
В другом аспекте настоящее изобретение относится к композиции для предотвращения или лечения рака, содержащей антитело или его антигенсвязывающий фрагмент настоящего изобретения.
В еще одном аспекте настоящее изобретение относится к способу предотвращения или лечения рака, включающему в себя этап введения композиции, содержащей антитело или его антигенсвязывающий фрагмент настоящего изобретения, индивидууму, которому угрожает развитие рака или который имеет его.
В еще одном аспекте настоящее изобретение относится к применению лечения рака и применению получения противоракового лекарственного средства в отношении композиции, содержащей антитело или его антигенсвязывающий фрагмент настоящего изобретения.
Антитело и его антигенсвязывающий фрагмент являются такими, как описано выше.
Антитело или его антигенсвязывающий фрагмент настоящего изобретения способны связываться только с c-Met или комбинацией c-Met и EGFR с высокой аффинностью для ингибирования роста раковых клеток, так что антитело отдельно или в сочетании с обычными фармацевтически приемлемыми носителями может применяться для лечения, предотвращения и диагностики гиперпролиферативных заболеваний, таких как рак.
В настоящем изобретении термин «предотвращение» означает все действия, которые предотвращают или задерживают возникновение или рецидив заболевания, такого как рак и т.д., путем введения композиции настоящего изобретения, а термин «лечение» означает ингибирование развития таких заболеваний, как рак, уменьшение рака или удаление рака.
Может быть предусмотрено, что рак, заболевание, на которое воздействуют при помощи композиции настоящего изобретения, представляет собой без ограничений, в частности, рак легких, рак желудка, рак толстой кишки, рак прямой кишки, трижды негативный рак молочной железы (TNBC), глиобластому, рак поджелудочной железы, рак головы и шеи, рак молочной железы, рак яичников, рак почки, рак мочевого пузыря, рак предстательной железы, соленому, опухоль слюнной железы или рак щитовидной железы, более конкретно рак легких, рак желудка, рак толстой кишки, рак прямой кишки, трижды негативный рак молочной железы (TNBC), глиобластому, рак поджелудочной железы рак головы и шеи, рак молочной железы и, еще более конкретно, рак легких, рак желудка, рак толстой кишки, рак прямой кишки, трижды негативный рак молочной железы (TNBC), глиобластому, рак поджелудочной железы, рак головы и шеи. В настоящем изобретении может быть предусмотрено, что рак представляет собой рак, который вызван, в частности, избыточной экспрессией, амплификацией, мутацией или активацией c-Met, но не ограничивается ими. Другими словами, композиция, содержащая антитело или его связывающий фрагмент настоящего изобретения, оказывает ингибирующее действие на пролиферацию всех раковых опухолей независимо от аномальной экспрессии или мутации c-Met, так что фармацевтическое применение настоящего изобретения не ограничивается аспектом экспрессии или наличием или отсутствием мутации c-Met.
Композиция может находиться в форме фармацевтической композиции, квази-лекарственной композиции и композиции для здорового питания.
Композиция настоящего изобретения для предотвращения или лечения рака может дополнительно содержать фармацевтически приемлемый носитель. Фармацевтически приемлемый носитель представляет собой носитель, обычно используемый при приготовлении композиции, и включает в себя без ограничений лактозу, декстрозу, сахарозу, сорбит, маннит, крахмал, гуммиарабик, фосфат кальция, альгинат, желатин, силикат кальция, микрокристаллическую целлюлозу, поливинилпирролидон, целлюлозу, воду, сироп, метилцеллюлозу, метилгидроксибензоат, пропилгидроксибензоат, тальк, стеарат магния, минеральное масло и тому подобное. Помимо ингредиентов, композиция настоящего изобретения для предотвращения или лечения рака может дополнительно содержать смазывающее вещество, увлажнитель, подсластитель, ароматизатор, эмульгатор, суспендирующий агент, консервант и т.д. Подходящие фармацевтически приемлемые носители и препараты подробно описаны в Remington's Pharmaceutical Sciences (19th ed., 1995).
Композиция настоящего изобретения может быть введена перорально или парентерально, где парентеральное введение может быть выполнено путем внутривенной инфузии, подкожной инфузии, внутримышечной инъекции, внутрибрюшинной инъекции, эндотелиального введения, местного введения, интраназального введения, внутрилегочного введения, ректального введения и тому подобного. Во время перорального введения белок или пептид переваривается, поэтому пероральная композиция может быть составлена таким образом, что ее активное лекарственное средство было покрыто или защищено от разложения в желудке. Композиция настоящего изобретения может вводиться с помощью заранее определенного устройства, через которое активное вещество может перемещаться в клетку-мишень.
Подходящая дозировка композиции настоящего изобретения для предотвращения или лечения рака варьируется в зависимости от таких факторов, как способ введения, тип введения, возраст пациента, масса тела, пол, патологическое состояние, пища, время введения, путь введения, скорость выведения и чувствительность к ответной реакции, с учетом которых обычный квалифицированный врач может легко определить и назначить эффективную дозу для желаемого лечения или предотвращения. В соответствии с одним примерным вариантом осуществления настоящего изобретения суточная доза фармацевтической композиции настоящего изобретения может составлять 0,001-100 мкг/л или более. В настоящем описании термин «эффективная фармацевтическая доза» означает количество, достаточное для лечения, предотвращения и диагностики таких заболеваний, как рак.
Композиция настоящего изобретения для предотвращения или лечения рака может быть получена в виде препарата с использованием фармацевтически приемлемых носителей и/или вспомогательных веществ способом, который может быть легко осуществлен специалистами в области техники, к которой относится настоящее изобретение, так, что такая композиция может быть приготовлена в форме одной дозы или приготовлена в форме нескольких доз, помещенных в контейнер. При этом лекарственная форма может быть в форме раствора в масле или водной среде, суспензии или эмульсии или в форме экстракта, порошка, суппозитория, порошкообразного лекарственного средства, гранулы, таблетки или капсулы и может дополнительно содержать диспергирующий агент или стабилизатор.
Композиция настоящего изобретения может вводиться в виде отдельного терапевтического агента или вводиться в комбинации с другими терапевтическими агентами и может вводиться последовательно или одновременно с обычными терапевтическими агентами.
Антитело или его антигенсвязывающий фрагмент настоящего изобретения можно использовать для лечения рака таким образом, что его вводят in vivo в форме конъюгата антитело-терапевтический агент (функциональная молекула) и биспецифичное антитело-терапевтический агент (функциональная молекула), которые являются такими как описанные выше. Подходящие и желательные различные условия для нацеливания лекарственного средства на конкретную мишень описаны в документах, например, в Trouet et al., Plenum Press, New York and London, (1982) 19-30.
В соответствии с одним конкретным вариантом осуществления настоящего изобретения, в результате выявления противоопухолевой активности композиции настоящего изобретения для предотвращения или лечения рака на модели ксенотрансплантата мыши, было установлено, что ее эффективность по ингибированию активности опухоли была значительно лучше по сравнению с контрольной группой (Фигуры 12 и 13).
Белок c-Met, на который нацелено антитело или его антигенсвязывающий фрагмент, включенное в композицию настоящего изобретения, представляет собой молекулу, экспрессируемую на поверхности раковых клеток, поэтому его можно использовать для предотвращения, лечения и диагностики связанного с c-Met рака, таким образом, что функциональная молекула дополнительно связывается с антителом настоящего изобретения или вводится в комбинации с ним. Функциональная молекула может содержать химическое вещество, радиоактивный нуклид, иммунотерапевтический агент, цитокин, хемокин, токсин, биотический агент, ингибитор фермента и тому подобное.
Функциональная молекула, способная связываться с антителом или его фрагментом настоящего изобретения, приводя к образованию конъюгата антитело-лекарственное средство (АЛС), может представлять собой без ограничений химическое вещество, цитокин или хемокин. Химическое вещество может представлять собой, без ограничений, например, противоопухолевое лекарственное средство, в частности, ацивицин, акларубицин, акодазол, акроницин, адозелезин, аланозин, алдеслейкин, аллопуринол натрия, альтретамин, аминоглутетимид, амонафид, амплиген, амсакрин, андрогены, ангуидин, афидиколин глицинат, асалей, аспарагиназу, 5-азацитидин, азатиоприн, бациллу Кальмета-Герена (БЦЖ), антифол Бейкера, бета-2-диокситиогуанозин, бисантрен HCl, блеомицин сульфат, бульсуфан, бутионсульфоксимин, BWA773U82, BW502U83/HCl, BW7U85 мезилат, церацемид, карбетимер, карбоплатин, кармустин, хлорамбуцил, хлорхиноксалин-сульфонамид, хлорозотоцин, хромомицин A3, цисплатин, кладрибин, кортикостероид, Corynebacterium parvum, СРТ-11, криснатол, циклоцитидин, циклофосфамид, цитарабин, цитембена, дабис малеат, декарбазин, дактиномицин, даунорубицин HCl, диазауридин, дексразоксан, диангидрогалактион, диазихон, дибромодулцитол, дидемнин В, диэтилдитиокарбамат, дигликоальдегид, дигидро-5-азацитидин, доксорубицин, эхиномицин, дедатрексат, эделфосин, эфлорнитин, раствор Эллиота, элсамитруцин, эпирубицин, эсорубицин, эстрамустинфосфат, эстрогены, этанидазол, этиофос, этопозид, фадразол, фазарабин, фенретинид, филграстим, финастерид, флавон уксусной кислоты, флоксуридин, флударабинфосфат, 5-фторурацил, Флуозол™, флутамид, галлия нитрат, гемцитабин, гозерелинацетат, гепсульфам, гексаметилендиизоцианат бисацетамид, гомохаррингтонин, гидразин сульфат, 4-гидроксиандростенедион, гидрозимочевина, идарубицин HCl, ифосфамид, 4-ипомеанол, ипроплатин, изотретиноин, лейковорин кальция, ацетат лейпролида, левамизол, липосомальный даунорубицин, липосомный инкапсулированный доксорубицин, ломустин, лонидамин, майтанзин, гидрохлорид мехлорэтамин, мелфалан, меногарил, мербарон, 6-меркаптопурин, месна, извлеченный метанолом остаток бациллы Кальметта-Герена, метотрексат, N-метилформамид, мифепристон, митогуазон, митомицин-C, митотан, митоксантрона гидрохлорид, моноцитарно/макрофагальный колониестимулирующий фактор, набилон, нафоксидин, неокарзиностатин, октреотида ацетат, ормаплатин, оксалиплатин, паклитаксел, пала, пентостатин, пиперазиндион, пипоброман, пирарубицин, пиритрексим, пироксантрон гидрохлорид, PIXY-321, пликамицин, порфимер натрия, преднимустин, прокарбазин, прогестины, пиразофурин, разоксан, сарграмостим, семустин, спирогерманиум, спиромустин, стрептонигрин, стрептозоцин, сулофенур, сурамин натрия, тамоксифен, таксотер, тегафур, тенипозид, терефталамидин, тероксирон, тиогуанин, тиотэф, инъекция тимидина, тиазофурин, топотекан, торемифен, третиноин, трифлуоперазин гидрохлорид, трифлуридин, триметрексат, фактор некроза опухоли (ФНО), урацилиприт, винбластина сульфат, винкристина сульфат, виндезин, винорелбин, винзолидин, Йоши 864, зорубицин, цитозин арабинозид, этопозид, мелфалан, таксотер, таксол и их смеси.
Выполнение изобретения
Далее настоящее изобретение будет описано более подробно путем приведения Примеров. Следующие Примеры приведены только с целью более подробной иллюстрации настоящего изобретения. Таким образом, в соответствии с целью настоящего изобретения специалистам в данной области техники будет очевидно, что эти Примеры не истолковываются как ограничивающие объем настоящего изобретения.
Пример 1. Получение гибридомной клетки для продуцирования c-Met-специфичного антитела и идентификация его активности, ингибирующей пролиферацию опухолевых клеток
(1) Получение и отбор линий гибридомных клеток для получения моноклонального антитела к белку c-Met
Слитый белок домена Sema c-Met и Fc человека (собственного производства) вводили мышам внутрибрюшинно в качестве антигена для получения иммунизированной мыши, необходимой для получения линии гибридомных клеток посредством иммунизации животных. Скрининг проводили методом ИФА с использованием человеческого слитого белка c-Met/His в качестве антигена, чтобы выбрать гибридомную клетку, специфично отвечающую на белок c-Met, только из группы гибридомных клеток.
(2) c-Met антитело
Аминокислотные последовательности CDR легкой цепи и тяжелой цепи мышиного антитела, полученного из отобранных гибридомных клеточных линий, показаны в Таблицах 1 и 2, соответственно.
Таблица 1
CDR легкой цепи гибридомы
Таблица 2
CDR тяжелой цепи гибридомы
(3) In vitro активность ингибирования пролиферации опухолевых клеток гибридомного анти-c-Met антитела
Что касается c-Met-специфичного мышиного антитела, полученного из линии гибридомных клеток, а также химерного антитела, полученного путем слияния антитела с константными областями тяжелой цепи и легкой цепи человека, то их активность ингибирования пролиферации опухолевых клеток тестировали в клеточной линии глиобластомы человека U-87 MG и клеточной линии рака желудка человека MKN45.
В частности, клетки U-87 MG (ATCC, #HTB14) разводили в культуральной среде EMEM (ATCC, #30-2003), содержащей 10% (об./об.) ФБС, 100 Ед./500 мл пенициллина и 100 мкг/500 мл стрептомицина (Invitrogen, #15140-122), после чего полученные клетки добавляли по 100 мкл в каждую лунку 96-луночного планшета в концентрации 2,5×103 клеток, и культивировали планшет при температуре 37°C, 95% относительной влажности и 5% (об./об.) СО2 в течение 18-24 часов. Среду для культивирования клеток удаляли из каждой лунки, после чего в каждую лунку добавляли по 100 мкл среды EMEM, содержащей 2% (об./об.) ФБС, и антитело, полученное в 2X от конечной концентрации (100 нМ), последовательно разбавляли в соотношении 1/10, так что полученные клетки добавляли по 100 мкл в каждую лунку в шести концентрациях (т.е. 200 нМ, 20 нМ, 2 нМ, 200 пМ, 20 пМ и 2 пМ) для каждого антитела. Затем планшет культивировали в течение 5 суток при температуре 37°C, относительной влажности 95% и 5% (об./об.) СО2, после чего в последний день полученные клетки фиксировали 10%-ным раствором ТХУ (трихлоруксусной кислоты; Sigma, #T0699). Полученные фиксированные клетки окрашивали в течение 25 минут следующим образом: в каждую лунку добавляли 80 мкл 0,4% раствора SRB (сульфородамина В), после чего полученные клетки 5 раз промывали 1% раствором уксусной кислоты. Затем в каждую лунку высушенного планшета вносили 150 мкл 10 мМ раствора триса для растворения красителя SRB, после чего его оптическую плотность измеряли при длине волны 540 нм с использованием устройства для считывания микропланшетов.
Кроме того, клеточные линии MKN45 (# JCRB0254) разводили в среде RPMI-1640 (Gibco, # A10491), содержащей 10% (об./об.) ФБС, после чего полученные клеточные линии разделяли по 2,5×103 клеток в каждую лунку 96-луночного планшета, и культивировали полученный планшет в течение ночи при температуре 37°C, и 5% CO2. Затем среду каждой лунки планшета заменяли 100 мкл среды RPMI-1640, содержащей 1% (об./об.) ФБС, после чего тестируемое антитело последовательно разбавляли в соотношении 1/10 (т.е. 100 нМ, 10 нМ, 1 нМ, 100 пМ, 10 пМ и 1 пМ) для достижения концентрации 1 пМ от конечной концентрации 100 нМ, полученное в результате антитело добавляли по 100 мкл в каждую лунку. Затем планшет культивировали в течение 5 суток при температуре 37°C, и 5% CO2, после чего из него удаляли среду, в каждую лунку вносили 200 мкл раствора ТХУ для фиксации клеток. Как показано в тесте на клетках MGU87, клетки планшета окрашивали в соответствии с традиционным методом колориметрического анализа SRB, после чего измеряли оптическую плотность каждой лунки при длине волны 540 нм с использованием устройства для считывания микропланшетов. Результаты клеточных линий U87MG и MKN45 показаны в Таблице 3 и на Фигуре 1.
Таблица 3
Результаты in vitro определения активности ингибирования пролиферации опухолевых клеток гибридомного анти-c-Met антитела
Как видно из Таблицы 3 и Фигуры 1 выше, все анти-c-Met антитела 8C4, 5G3 и их химерные антитела настоящего изобретения обладают активностью ингибирования пролиферации опухолевых клеток, которая равна или превышает активность у известных анти-c-Met антител LY2875358 и OA-5D5 (контрольная группа). Таким образом, антитела 8C4, 5G3 и их мутанты, такие как химерные антитела, гуманизированные антитела и оптимизированные по аффинности антитела к антигену настоящего изобретения, могут быь очень эффективно применяться для предотвращения или лечения рака, связанного с c-Met.
Конкретные консенсусные последовательности для вариабельных областей легкой цепи и тяжелой цепи антител 8C4, 5G3 настоящего изобретения показаны в следующей Таблице 4.
Таблица 4
Консенсусные SEQ ID NOs для вариабельных областей легкой цепи и тяжелой цепи антител 8C4, 5G3
Пример 2. Получение гуманизированного антитела из антитела 8С4 и идентификация его активности ингибирования пролиферации опухолевых клеток in vitro
В качестве одного примера, мышиное антитело 8C4 было гуманизировано, и была проведена оценка его активности ингибирования пролиферации опухолевых клеток in vitro, чтобы дополнительно идентифицировать действие антитела, полученного в настоящем изобретении.
Для гуманизированной конструкции тяжелых цепей антител 8C4 ген человеческой зародышевой линии, имеющий высокую гомологию с геном в вариабельной области тяжелой цепи мышиного антитела 8C4, сначала анализировали с помощью программы Ig Blast (http: //www.ncbi.nlm.nih.gov/igblast/). В результате установили, что IGHV3-23 обладает 48% гомологии с антителом 8C4 на аминокислотном уровне, а также установили, что IGHV3-11 обладает 46% гомологии с антителом 8C4 на аминокислотном уровне.
CDR-H1, CDR-H2 и CDR-H3 мышиного антитела 8C4 определяли в соответствии с нумерацией по Кабату, и hu8C4-1 получали таким образом, что CDR часть мышиного антитела 8C4 вводили в каркасную область IGHV3-23. При этом аминокислоты в положениях 48 (V→I), 49 (S→G), 71 (R→A), 73 (N→K), 78 (L→A) и 94 (K→G) были подвергнуты обратной мутации в исходную аминокислотную последовательность мышиного антитела 8C4 для окончательного образования тяжелой цепи hu8C4-1. В случае hu8C4-2, CDR часть мышиного антитела 8C4 вводили в каркасную область IGHV3-11, и аминокислоты в положениях 48 (V→I), 49 (S→G), 71 (R→A), 73 (N→K), 78 (L→A) и 94 (R→G) были подвергнуты обратной мутации в исходную аминокислотную последовательность мышиного антитела 8C4 для окончательного образования тяжелой цепи hu8C4-2.
В случае легкой цепи антитела 8C4 для гуманизированной конструкции ген зародышевой линии человека, имеющий высокую гомологию с геном в вариабельной области легкой цепи мышиного антитела 8C4, анализировали с помощью программы Ig Blast (http://www.ncbi.nlm.nih.gov/igblast/). В результате было установлено, что IGKV1-27 обладает гомологией 65,3% с антителом 8C4 на аминокислотном уровне, и что IGKV1-33 обладает гомологией 64,2% с антителом 8C4 на аминокислотном уровне.
CDR-L1, CDR-L2 и CDR-L3 мышиного антитела 8C4 определяли в соответствии с нумерацией по Кабату, и CRD часть мышиного антитела 8C4 вводили в каркасную область IGKV1-33 и каркасную область IGKV1-27, получая, таким образом, hu8C4-1 и hu8C4-2, соответственно. При этом аминокислоты в положении 69 (T→R), как в hu8C4-1, так и в hu8C4-2 подвергли обратной мутации в исходную аминокислотную последовательность мышиного антитела 8C4.
Гуманизированное антитело 8C4 экспрессировали в клетке 293T с использованием вектора pCLS05 (патент Кореи регистрационный номер 10-1420274). Что касается полученных таким образом гуманизированных антител в форме IgG1, то определение того, обладают ли они активностью ингибирования пролиферации опухолевых клеток в клеточной линии глиобластомы человека U-87 MG, проводили тем же способом, который описан выше в Примере 1.
В результате было установлено, что значения ИК50 для hu8C4-1 и hu8C4-2 составляли 30 нМ и 24,6 нМ соответственно, что указывает на уровень противораковой активности, сходный с этим уровнем для химерного антитела 8C4 (ИК50=32,4 нМ).
Конкретные консенсусные последовательности для вариабельных областей легкой цепи и тяжелой цепи гуманизированных антител hu8C4-1 и hu8C4-2 показаны в Таблице 5.
Таблица 5
Консенсусные SEQ ID NOs для вариабельных областей легкой цепи и тяжелой цепи гуманизированных антител hu8C4-1 и hu8C4-2
Пример 3. Получение гуманизированного антитела из антитела 5G3 и определение его активности ингибирования пролиферации опухолевых клеток in vitro
Затем мышиное антитело 5G3 настоящего изобретения гуманизировали для определения его активности ингибирования пролиферации опухолевых клеток in vitro.
В частности, для конструирования тяжелой цепи hu5G3-1 ген зародышевой линии человека, имеющий самую высокую гомологию с геном в вариабельной области тяжелой цепи мышиного антитела 5G3, сначала анализировали с помощью программы Ig Blast (http://www.ncbi.nlm.nih.gov/igblast/). В результате было установлено, что IGHV1-46 обладает 67,3% гомологии с антителом 5G3 на аминокислотном уровне. CDR-H1, CDR-H2 и CDR-H3 мышиного антитела 5G3 определяли в соответствии с нумерацией по Кабату, и CRD часть мышиного антитела 5G3 вводили в каркасную область IGHV1-46. При этом аминокислоты в положениях 48 (M→I), 69 (M→L), 71 (R→A), 73 (T→K) и 78 (V→A) были подвергнуты обратной мутации в исходную аминокислотную последовательность мышиного антитела 5G3. Таким образом, получали тяжелую цепь hu5G3-1.
Для легкой цепи hu5G3-1 прививку CDR проводили в гене IGKV3-20, обладающим 63,5% гомологии с антителом 5G3 и аминокислоты в положениях 43 (A→S), 60 (D→A) и 71 (F→N) были подвергнуты обратной мутации для получения легкой цепи hu5G3-1.
Кроме того, для конструирования тяжелой цепи hu5G3-2 CDR-H1, CDR-H2 и CDR-H3 мышиного антитела 5G3, определенные в соответствии с нумерацией по Кабату, вводили с использованием подтипа VH3, который, как общеизвестно, является наиболее стабильным. При этом аминокислоты в положениях 67 (F→A), 69 (I→L), 73 (T→K), 90 (Y→F) и 94 (T→R) были подвергнуты обратной мутации в исходную аминокислотную последовательность мышиного антитела 5G3. Таким образом, получали тяжелую цепь hu5G3-2.
Для легкой цепи hu5G3-2 прививку CDR проводили в гене IGVK, который, как было известно, стабильно формирует структуру с подтипом VH3, и обратную мутацию не проводили.
Гуманизированное антитело 5G3 экспрессировали в клетке 293T с использованием вектора pCLS05 (патент Кореи регистрационный номер 10-1420274). Что касается полученных таким образом гуманизированных антител в форме IgG2, то определение того, обладают ли они активностью ингибирования пролиферации опухолевых клеток в клеточной линии рака желудка человека MKN45, проводили тем же способом, который описан выше в Примере 1.
В результате было установлено, что значения ИК50 для hu5G3-1 и hu5G3-2 составляли 0,52 нМ и 0,5 нМ соответственно, что указывает на уровень противораковой активности, аналогичный уровню активности химерного антитела 5G3 (ИК50=0,41 нМ).
Консенсусные последовательности для вариабельных областей легкой цепи и тяжелой цепи гуманизированных антител hu5G3-1 и hu5G3-2 показаны в Таблице 6.
Таблица 6
Консенсусные SEQ ID NOs для вариабельных областей легкой цепи и тяжелой цепи гуманизированных антител hu5G3-1 и hu5G3-2
Пример 4. Получение мутанта шарнирной области и проверка его активности ингибирования пролиферации опухолевых клеток
Затем проводили оценку активности ингибирования пролиферации опухолевых клеток в отношении последовательности шарнирной области константной области тяжелой цепи человеческого IgG1.
Прежде всего, шарнирная область константной области тяжелой цепи человеческого IgG1 имела аминокислотную последовательность «EPKSCDKTHTCPPCP (SEQ ID NO: 37)», которая была заменена для получения мутанта шарнирной области, имеющего аминокислотную последовательность SEQ ID NO: С 38 - SEQ ID NO: 44.
Полученные мутанты соответственно клонировали в вектор, содержащий вариабельную область тяжелой цепи гуманизированных антител hu8C4-1, hu8C4-2, полученных в Примере 2 выше. Активность ингибирования пролиферации опухолевых клеток in vitro в зависимости от последовательности шарнирной области определяли на клетках U-87 MG тем же способом, который описан выше в Примере 1.
Кроме того, действие гуманизированного антитела 8C4 в отношении клеточной линии немелкоклеточного рака легких NCI-H1993 (ATCC, #CRL-5909) анализировали следующим образом. Клеточные линии NCI-H1993 разбавляли в среде RPMI-1640 (Gibco, #A10491), содержащей 10% (об./об.) ФБС, после чего полученные клеточные линии распределяли по 3,0×103 клеток в каждую лунку 96-луночного планшета. Полученный таким образом планшет культивировали в течение ночи при температуре 37°C и 5% СО2. После этого среду каждой лунки планшета заменяли 100 мкл среды RPMI-1640, содержащей 2% (об./об.) ФБС, после чего тестируемое антитело последовательно разбавляли в соотношении 1/10 (т.е. 100 нМ, 10 нМ, 1 нМ, 100 пМ, 10 пМ и 1 пМ) для достижения концентрации 0,001 нМ от конечной концентрации 100 нМ, и полученное таким образом антитело добавляли по 100 мкл в каждую лунку. Затем планшет культивировали в течение 5 суток при температуре 37°C и 5% СО2, после чего из него удаляли среду и в каждую лунку вносили 200 мкл раствора ТХУ (Sigma, #T0699) для фиксации клеток. Кроме того, клетки планшета окрашивали в соответствии с традиционным методом колориметрического анализа SRB, после чего измеряли оптическую плотность каждой лунки при длине волны 540 нм с использованием устройства для считывания микропланшетов.
Результаты hu8C4-1 для U-87 MG и NCI-H1993 (ATCC, #CRL-5909) показаны в Таблице 7.
Таблица 7
Мутантные последовательности в шарнирной области и результаты оценки ингибирования пролиферации опухолевых клеток in vitro
Как видно из Таблицы 7, у антитела hu8C4 существует некоторое различие в активности ингибирования пролиферации опухолевых клеток в зависимости от различий шарнирной последовательности, но было установлено, что такое антитело эффективно ингибирует пролиферацию большинства опухолевых клеток. Соответственно, в дальнейшем гуманизированное антитело IgG1, имеющее репрезентативную шарнирную область SEQ ID NO: 38 в hu8C4-1, было названо hu8C4, и на его основе было получено оптимизированное по аффинности антитело, чтобы оценить его действие.
Пример 5. Получение оптимизированного по аффинности антитела hu8C4 и оценка его активности ингибирования пролиферацию опухолевых клеток in vitro
Чтобы получить оптимизированное по аффинности антитело hu8C4, сначала получали библиотеку scFv фагового дисплея, с использованием фагмидного вектора, отображающего в комбинированной форме scFv и pIII, схематическая структура этот вектора показана на Фигуре 2. Фагмидный вектор содержит фрагмент scFv антитела под контролем IPTG-индуктивного lac-промотора, где используемой линкерной последовательностью является GGGGS GGGGS GGGGS (SEQ NO: 53).
Затем для индуцирования разнообразия в CDR домене тяжелой цепи и легкой цепи hu8C4 использовали индуцирующий мутации олигонуклеотид, имеющий NNK-кодон. Соответственно, была приготовлена библиотека scFv hu8C4 со слиянием His, HA и pIII, после чего из полученной библиотеки антител было выбрано c-Met-специфичное антитело человека.
В частности, был использован метод конкурентного отбора для отбора антител с улучшенной аффинностью. Человеческий c-Met-антиген связывали в соответствии с рекомендациями производителя в Dynabeads® M-280 (Thermo Fisher Scientific, 11205D). Частицы со связавшимся с ними антигеном блокировали в течение 2 часов суперблокирующим трис-буферным солевым раствором (TBS, Pierce). Также рекомбинантный фаг выращивали в течение ночи температуре 37°C, а затем рекомбинантный фаг центрифугировали и фаг из его супернатанта блокировали суперблокирующим TBS, 0,05% Твин 20 в течение 2 часов. Затем частицы промывали ФСБ, содержащим 0,05% Твин 20. К промытым частицам добавляли раствор заблокированного фага, после чего полученные частицы инкубировали в ротаторе в течение 2 часов для связывания фага, полученные в результате частицы промывали ФСБ, содержащим 0,05% Твин 20. Затем в 1 мл ФСБ, содержащего 0,05% Твин 20, добавляли человеческий c-Met-антиген, после чего полученный антиген инкубировали в ротаторе в течение 24 часов (Rouet R et al. (2012) Nat Protoc. 7: 364-373). После этого связавшийся с частицами фаг элюировали 100 мМ триэтаноламином в течение 5 минут, после чего элюент нейтрализовали 0,5 М Трис/Cl (рН 7,2). Элюированную нейтрализующую жидкость фага заражели E.coli TG1.
Отдельный клон, отобранный в ходе эксперимента, выращивали в 96-луночном формате в 200 мкл 2-кратного бульона YT с добавлением карбенициллина и ампициллина, после чего его культуральный супернатант непосредственно использовали для ИФА для отбора отображенного фагом scFv связавшегося с планшетом, покрытым белком-мишенью. Аминокислотные последовательности областей CDR легкой цепи и тяжелой цепи детектируемого антитела показаны в Таблицах 8 и 9, а типичные аминокислотные последовательности вариабельных областей легкой цепи и тяжелой цепи оптимизированного по аффинности антитела показаны в Таблице 10.
Таблица 8
Список последовательностей CDR тяжелой цепи
Таблица 9
Таблица 10
Список последовательностей вариабельных областей легкой цепи и тяжелой цепи оптимизированного по аффинности антитела
Кроме того, оценку активности ингибирования пролиферации in vitro, проводили на клеточной линии MG U-87 с использованием части оптимизированных по аффинности антител, результаты этой оценки показаны в Таблице 11.
Таблица 11
Активность ингибирования пролиферации опухолевых клеток in vitro легкой цепи и тяжелой цепи оптимизированных по аффинности антител hu8C4
Анализ ингибирования пролиферации клеток, ИК50 (нМ)
Как видно из Таблицы 11, было установлено, что ИК50 активности ингибирования пролиферации опухолевых клеток оптимизированного по аффинности антитела hu8C4 в отношении клеток MG U-87 составляла 5,0-18 нМ, и его эффективность увеличилась в 4,3-9,8 раза по сравнению с исходным антителом hu8C4. Приведенные выше результаты представляют собой тест, выполненный для части антител, имеющих аминокислотную последовательность, представленную в Таблицах 8-10, у которых была оптимизирована аффинность исходного антитела hu8C4, и все антитела были отобраны на основе аффинности к антигену в процессе отбора. Таким образом, ожидается, что даже в случае остальных оптимизированных по аффинности антител, а также антител с комбинацией представленных CDR вариабельных областей тяжелой цепи и легкой цепи, может иметь место практически аналогичный эффект.
Для дополнительного эксперимента получали 10 видов оптимизированных по аффинности антител путем объединения вариабельных областей легкой цепи и тяжелой цепи. Конкретная комбинация последовательностей легкой цепи и тяжелой цепи показана в Таблице 12.
Таблица 12
Список комбинированных последовательностей вариабельной области оптимизированного по аффинности антитела
Затем активность ингибирования пролиферации опухолевых клеток оценивали тем же способом, который описан в Примере 1 выше, где результаты этого эксперимента показаны в Таблице 13 и на Фигуре 3.
Таблица 13
Активность ингибирования пролиферации опухолевых клеток оптимизированными по аффинности антителами in vitro
Как видно из приведенной выше Таблицы 13, было установлено, что антитело hu8C4, а также 10 типов ключевых антител с комбинацией вариабельных областей легкой цепи и тяжелой цепи его оптимизированного по аффинности антитела также проявляли активность ингибирования пролиферации опухолевых клеток. В частности, ИК50 для 10 типов антител составляла 1,7-3,3 нМ, и было установлено, что они обладают ингибирующим действием на пролиферацию опухолевых клеток, которое в 9,2-28,5 раза сильнее, чем у исходного антитела hu8C4.
Пример 6. Получение биспецифичного антитела и активность ингибиторования пролиферации опухолевых клеток in vitro
Для получения биспецифичного антитела, специфически связывающегося с c-Met и EGFR, scFv-фрагменты эрбитукс и вектибикс, которые, как известно, специфично связываются с EGFR, соединяли соответственно с C-концом тяжелой цепи анти-c-Met антитела настоящего изобретения с помощью линкера GGGGSGGGGGS. (SEQ ID NO: 312).
Для повышения стабильности scFv 44-й остаток тяжелой цепи и 100-й остаток легкой цепи заменяли цистином (Reiter Y. et al., Biochemistry 33 (18): 5451-5459 (1994)). Последовательности scFv эрбитукс и вектибикс, аминокислотные последовательности тяжелой цепи биспецифичного антитела и комбинация вариабельных областей биспецифичного антитела показаны в следующих Таблицах 14 и 15.
Таблица 14
Список аминокислотных последовательностей анти-EGFR антитела для получения биспецифичного антитела, а также биспецифичного антитела
Таблица 15
Список комбинированных последовательностей вариабельной области биспецифичного антитела
Затем противораковую эффективность биспецифичного антитела, связанного с scFv-фрагментами Эрбитукс и Вектибикс, оценивали in vitro в линии опухолевых клеток U-87 MG тем же способом, который описан в Примере 1.
Также активность ингибирования пролиферации опухолевых клеток оценивали с использованием линий клеток рака легкого NCI-H1993, NCI-H292 и NCI-H820. В частности, клеточная линия NCI-H1993 (ATCC, #CRL-5909) имела сверхэкспрессию гена c-Met, клеточная линия NCI-H292 (ATCC, #CRL-1848) имела нормальную экспрессию EGFR и c-Met, и клеточная линия NCI-H820 (ATCC, #HTB-181) имела мутацию в положении 790 аминокислотной последовательности EGFR, которая заключалась в замене треонина (T) на метионин (M). Оценку активности ингибирования пролиферации опухолевых клеток осуществляли следующим способом. Каждую клеточную линию разводили в среде RPMI-1640 (Gibco, #A10491), содержащей 10% (об./об.) ФБС, после чего полученные клеточные линии распределяли по 2,0×103 клеток в каждую лунку 96-луночного планшета, полученный таким образом планшет культивировали в течение ночи при температуре 37°C, и 5% CO2. Затем среду в каждой лунке планшета заменяли 100 мкл бессывороточной среды, после чего полученный планшет культивировали при температуре 37°C, и 5% CO2 в течение 18 часов. После этого среду заменяли на 100 мкл среды RPMI-1640, содержащей 2% (об./об.) ФБС или 50 нг/мл HGF, после чего тестируемое антитело последовательно разбавляли в соотношении 1/10 (т.е. 100 нМ, 10 нМ, 1 нМ, 100 пМ, 10 пМ и 1 пМ) для достижения концентрации 0,001 нМ от конечной концентрации 100 нМ, полученное в результате антитело добавляли по 100 мкл в каждую лунку. Затем планшет культивировали в течение 5 суток при температуре 37°C, и 5% CO2, после чего из него удаляли среду, в каждую лунку вносили 200 мкл раствора ТХУ для фиксации клеток. Кроме того, клетки планшета окрашивали в соответствии с традиционным методом колориметрического анализа SRB, после чего измеряли оптическую плотность каждой лунки при длине волны 540 нм с использованием устройства для считывания микропланшетов.
Результаты активности ингибирования пролиферацию в каждой из вышеперечисленных клеточных линий представлены в Таблицах 16 и 17 и на Фигуре 4 и 5.
Таблица 16
Активность ингибирования пролиферации опухолевых клеток биспецифичных антител in vitro
Таблица 17
Ингибирование пролиферации клеточной линии рака легкого биспецифичным антителом in vitro
В результате не было обнаружено различий в эффективности между биспецифичными антителами, полученными этим способом из линии опухолевых клеток U-87 MG, и было установлено, что их эффективность по ингибированию активности была примерно в 15 раз выше, чем у ИК50 оптимизированного hu8C4 антитела. Также, в результате оценки активности ингибирования пролиферации опухолевых клеток с использованием клеточных линий рака легких NCI-H1993, NCI-H292 и NCI-H820 было установлено, что между полученными биспецифичными антителами не было различий в эффективности.
Такие результаты свидетельствуют о том, что антитело настоящего изобретения оказывает ингибирующее действие на пролиферацию на все типы рака, независимо от сверхэкспрессии или мутации c-Met и EGFR, и, следовательно, может эффективно применяться при лечении этих типов рака.
Пример 7. Сравнительная оценка in vitro активности ингибирования пролиферации опухолевых клеток биспецифичного антитела по сравнению с комбинированной терапией
Восемь типов рака были использованы для сравнения активности ингибирования пролиферации опухолевых клеток между комбинированной терапией каждого антитела, нацеленного на c-Met и EGFR, соответственно, и биспецифичным антителом настоящего изобретения.
В частности, активность ингибирования пролиферации опухолевых клеток оценивали в клеточной линии рака легкого NCI-H292 (ATCC, #CRL-1848), в клеточной линии HGF-аутокринной глиобластомы U-87 MG (ATCC, #HTB-14), клеточных линиях рака легкого NCI-H1648 (ATCC #CRL-5882) и NCI-H596 (ATCC #HTB-178), HCC827 (ATCC, #CRL2868), клеточной линии рака толстой кишки LS174T (ATCC, #CL-188), клеточной линии трижды негативного рака молочной железы (TNBC) BT20 (ATCC, #HTB-19) и клеточной линии рака поджелудочной железы KP4 (JCRB, #RCB1005). Клеточная линия NCI-H1648 характеризуется нормальной экспрессией EGFR и c-Met, клеточная линия NCI-H596 характеризуется делецией определенной последовательности экзона 14 гена MET, и клеточная линия HCC827 характеризуется делецией определенной последовательности экзона 19 гена EGFR. Кроме того, клеточная линия LS174T имеет мутацию KRAS, а KP4 характеризуется аутокринингом HGF.
Клеточную линию MG U-87 оценивали способом, описанным в Примере 1, а клеточную линию NCI-H292 оценивали способом, описанным в Примере 6. Кроме того, клеточные линии NCI-H1648, NCI-H596 и HCC827 разводили в среде RPMI-1640 (Gibco, #A10491), содержащей 10% (об./об.) ФБС, после чего полученные клеточные линии распределяли по 2,0×103 клеток в каждую лунку 96-луночного планшета. Клеточную линию LS174T разводили в среде DMEM (Gibco, # 11995-065), содержащей 10% (об./об.) FBS, после чего полученные клеточные линии распределяли по 2,0×103 клеток. Клеточную линию BT20 разводили в среде EMEM (ATCC, № 30-2003), содержащей 10% (об./об.) ФБС, после чего полученные клеточные линии распределяли по 3,0×103 клеток. И клеточную линию KP4 разводили в среде RPMI-1640 (Gibco, # A10491), содержащей 10% (об./об.) ФБС, после чего полученные клеточные линии распределяли по 1,5×103 клеток, полученный таким образом планшет культивировали в течение ночи при температуре 37°C, и 5% CO2. Затем среду в каждой лунке планшета заменяли 100 мкл бессывороточной среды, после чего полученный планшет культивировали при температуре 37°C, и 5% CO2 в течение 18 часов. После этого среду заменяли на 100 мкл среды RPMI-1640, содержащей 2% (об./об.) ФБС или 50 нг/мл HGF, после чего тестируемое антитело последовательно разбавляли в соотношении 1/10 (т.е. 100 нМ, 10 нМ, 1 нМ, 100 пМ, 10 пМ и 1 пМ) для достижения концентрации 1 пМ от конечной концентрации 100 нМ, полученное в результате антитело добавляли по 100 мкл в каждую лунку. Затем планшет культивировали в течение 5 суток при температуре 37°C, и 5% CO2, после чего из него удаляли среду, в каждую лунку вносили 200 мкл раствора ТХУ для фиксации клеток. Кроме того, клетки планшета окрашивали в соответствии с традиционным методом колориметрического анализа SRB, после чего измеряли оптическую плотность каждой лунки при длине волны 540 нм с использованием устройства для считывания микропланшетов.
Результаты этого Примера показаны в Таблицах 18-21 и на Фигурах 6-8
Таблица 18
Сравнительная оценка активности ингибирования пролиферации опухолевых клеток in vitro между комбинированной терапией и биспецифичным антителом в клеточных линиях U-87 MG и NCI-H292
Таблица 19
Сравнительная оценка активности ингибирования пролиферации опухолевых клеток in vitro между комбинированной терапией и биспецифичным антителом в клеточных линиях NCI-H1648 и NCI-H596
Таблица 20
Сравнительная оценка активности ингибирования пролиферации опухолевых клеток in vitro между комбинированной терапией и биспецифичным антителом в клеточных линиях LS174T, BT20 и KP4
Таблица 21
Сравнительная оценка активности ингибирования пролиферации опухолевых клеток in vitro между комбинированной терапией и биспецифичным антителом в клеточных линиях HCC827 и NCI-H596
В результате было установлено, что способность биспецифичного антитела настоящего изобретения ингибировать пролиферацию опухолевых клеток оказалась выше, чем у hu8C4, препарата Вектибикс или комбинированной терапии двух антител в каждой из 8 типов опухолевых клеточных линий. Также было установлено, что оно обладает более высокой способностью ингибировать пролиферацию опухолевых клеток в клеточных линиях U-87MG, NCI-H292, BT20 и KP4 по сравнению с антителом EM1-MAb (Janssen), используемым в качестве контрольного биспецифичного антитела.
Кроме того, было установлено, что и hu8C4, и hu8C4 × Вектибикс scFv обладали более высокой способностью ингибировать пролиферацию опухолевых клеток в клеточных линиях U-87MG по сравнению с контрольным антителом, в качестве которого использовали антитела LA480 (Lilly), OA-5D5 (Genentech) и AbF46 (Samsung), которые представляли собой анти-c-Met антитела.
Кроме того, препарат Тарцева, ингибитор тирозинкиназы EGFR в клеточной линии HCC827, показал устойчивость в условиях обработки HGF, но было установлено, что он проявляет повышенную способность ингибировать пролиферацию опухолевых клеток в таких условиях при применении в комбинации Тарцева, hu8C4, hu8C4 × Вектибикс scFv или ингибиторы c-Met.
Кроме того, в результате сравнения различных ингибиторов EGFR и ингибиторов c-Met в клеточной линии NCI-H596 было установлено, что способность hu8C4 × Вектибикс scFv ингибировать пролиферацию опухолевых клеток была выше по сравнению с препаратом, действие которого направлено только на EGFR или только на c-Met.
Пример 8. Измерение способности к связыванию с ВКД (BIAcore)
Затем для измерения способности связывания анти-c-Met антитела настоящего изобретения с внеклеточным доменом (ВКД), связывание анти-c-Met антитела и биспецифичного антитела с ВКД c-Met и ВКД EGFR измеряли для ВКД человека и макака-крабоеда при помощи BIAcore анализа.
В частности, использовали ВКД c-Met человека (ACROBiosystems, MET-H5227), ВКД c-Met макака-крабоеда (SiNo. Biological, 90304-C08H), ВКД EGFR человека (ACROBiosystems, EGR-H5285) и ВКД EGFR макака-крабоеда (SiNo. Biological, 90285-C08B).
Прежде всего, чтобы захватить анти-c-Met антитело и биспецифичное антитело, Fc-специфичное антитело против IgG человека (SouthernBiotech, 2047-01) прикрепляли к сенсорному чипу CM5 на уровне 10000 единиц ответа. Антитела разводили в буфере HBS-EP (0,01 М HEPES, рН 7,4, 0,15 М NaCl, 3 мМ ЭДТА и 0,005% (об./об.) ПАВ P20) в концентрации 1-2 мкг/мл, после чего полученные антитела вводили в чип CM5 с фиксированным на нем Fc-специфичным антителом против IgG человека при скорости потока 30 л/мин в течение 10-120 секунд, а затем производили захват в диапазоне 150-200 единиц ответа. Каждый антиген использовали после разбавления до концентрации 10, 5, 2,5, 1,25, 0,625, 0,3125 и 0,15625 нМ, после чего полученные антигены последовательно инъецировали, начиная с более низкой концентрации. Затем полученные антигены вводили со скоростью потока 30 л/мин в течение 5 минут для осуществления связывания, после чего вводили рабочий буфер в течение 10-15 минут для проведения диссоциации. 15 мкл 10 мМ буфера глицин-HCl (рН 1,5) использовали для восстановления чипа. Скорость связывания и диссоциации для каждого цикла оценивали с использованием модели связывания Ленгмюра 1:1 и программного обеспечения BIAevaluation в версии 4.1. Данные Biacore анализа приведены в Таблицах 22 и 23.
Таблица 22
Измерение аффинности к ВКД c-Met
Таблица 23
Измерение аффинности к ВКД EGFR
Эти данные были использованы для доказательства того, что биспецифичные антитела hu8C4, hu8C4 × Вектибикс scFv настоящего изобретения связываются с ВКД c-Met человека и макака-крабоеда с очень хорошей аффинностью.
Пример 9. Измерение способности связывания анти-c-Met антитела с ВКД c-Met, ВКД EGFR у различных видов животных (ИФА)
Связывание анти- c-Met антитела и биспецифичного антитела с ВКД c-Met и ВКД EGFR у мыши, макака-крабоеда и человека измеряли с помощью ИФА.
В частности, антигены c-Met мыши (SiNo. Biological Inc, 50622-M08H), c-Met макака-крабоеда (SiNo. Biological Inc, 90304-C08H), c-Met человека (R & D Systems, 358-MT), EGFR мыши (SiNo Biological Inc, 51091-M08H), EGFR макака-крабоеда (SiNo. Biological, 90285-C08B) и EGFR человека (Abcam, 155639) наносили на 96-луночный планшет в концентрации 2 мкг/мл, после чего полученный планшет инкубировали при температуре 4°C в течение ночи. После блокирования при комнатной температуре в течение 1 часа биспецифичное антитело hu8C4 × Вектибикс scFv последовательно разводили в соотношении 1/5 от 100 нМ для измерения его способности связывания в 7 секциях концентрации (т.е. 100 нМ, 20 нМ, 4 нМ, 800 пм, 160 пм, 32 пм и 6,4 пм).
После связывания биспецифичного антитела hu8C4 × Вектибикс scFv при комнатной температуре в течение 1 часа антитело, специфичное к фрагменту F(ab')2 человеческого IgG, конъюгированное с пероксидазой хрена (HRP) (Jackson Immunoresearch, 109-035-097), разводили в соотношении 1:2500, после чего полученное антитело инкубировали при комнатной температуре в течение 1 часа. Формирование цвета осуществляли с использованием раствора TMB (Sigma, T4444), и значение цвета измеряли при оптической плотности 450 нм. Результаты ИФА показаны на Фигуре 9.
В результате было установлено, что моноспецифичное антитело hu8C4 и биспецифичное антитело hu8C4 × Вектибикс scFv не связываются с c-Met мыши и EGFR мыши, но связываются с c-Mets и EGFR обезьяны и человека. Также было установлено, в случае антитела против IgG человека, используемого в качестве отрицательного контроля, связывание вообще отсутствовало. Приведенные выше результаты свидетельствуют о том, что анти-c-Met антитело настоящего изобретения специфично только для c-Mets и EGFR человека и обезьяны.
Пример 10. Перекрестная реактивность анти-c-Met антитела с различными рецепторам на поверхности клеток
Специфичность антитела hu8C4, специфически связывающегося с c-Met согласно настоящему изобретению, а также его перекрестную реактивность с другими антигенами рецепторных тирозинкиназ анализировали методом непрямого ИФА, и 5 антигенов FGF R3, VEGFR R2, IGF IR, PDGF R и RON были выбраны из ключевых рецепторных тирозинкиназ для проведения этого анализа.
В этом примере в качестве антигенов использовали человеческий химерный белок c-Met Fc (R & D Systems, 358-MT_CF), человеческий химерный белок FGF R3 (IIIc) Fc (R & D Systems, 766-FR), IGF-I R человека(R & D systems, 391-GR-050), человеческий химерный белок PDGF Rβ Fc (R & D Systems, 385-PR_CF), человеческий химерный белок VEGF R2 человека (R & D Systems, 357-KD_CF) и MS/R/Ron человека (R & D Systems, 1947-MS-050).
Каждый антиген разбавляли в 0,05 М карбонат-бикарбонатном буфере (Sigma, C3041) в концентрации 1 мкг/мл, после чего полученный антиген добавляли в каждую лунку 96-луночного планшета (Corning, #2592), полученный таким образом планшет покрывали при температуре 4°C в течение ночи. Планшет один раз промывали буфером TBS-T, после чего TBS-T, содержащий 4% обезжиренного молока, добавляли по 200 мкл в каждую лунку полученного планшета, чтобы ингибировать неспецифическое связывание, полученный планшет инкубировали при температуре 37°C в течение 1 часа. Затем планшет один раз промывали буфером TBS-T, после чего первичное антитело последовательно разводили в буфере TBS-T, содержащем 2% обезжиренного молока, начиная от максимальной концентрации 30 нМ до 0,002 нМ, полученное антитело добавляли по 100 мкл в каждую лунку, и инкубировали при температуре 37°C в течение 2 часов. После трехкратной промывки буфером TBS-T конъюгат антитела против человеческой каппа области легкой цепи и пероксидазы (Sigma, A7164) разводили в соотношении 1:5000 и использовали в качестве вторичного антитела, после чего полученное антитело добавляли по 100 мкл в каждую лунку и инкубировали при температуре 37°C в течение 1 часа. Затем, после трехкратной промывки буфером TBS-T, в каждую лунку добавляли по 100 мкл раствора ТМB (Sigma, T4444) для проведения реакции проявления цвета, после чего в каждую лунку добавляли по 50 мкл 2Н раствора сульфата аммония, чтобы остановить реакцию. Оптическую плотность измеряли на основе значения, полученного при длине волны 450 нм, используя считывающее устройство для микропланшетов и эталонную длину волны 570 нм. Степень связывания анти-c-Met-антитела с каждым антигеном была пропорциональна величине сигнала оптической плотности, результаты которого приведены в Таблице 24.
Таблица 24
Специфичность связывания анти-c-Met антитела hu8C4 с различными антигенами
Как видно из Таблицы 24, антитело hu8C4 настоящего изобретения преимущественно связывается с c-Met, и было установлено, что оно практически не связывается с другими антигенами FGF R3, VEGFR R2, IGF IR, PDGF R и RON.
Пример 11. Активность интернализации in vitro анти-c-Met антитела и ингибирующая активность уровня c-Met биспецифичного антитела
Было установлено, что анти-c-Met антитело настоящего изобретения обладает активностью интернализации in vitro в опухолевых клетках, а также влияет на снижение уровня рецептора биспецифичным антителом, способным одновременно связываться с c-Met и EGFR.
Прежде всего, интернализация антител происходит за счет физиологической активности нормального рецептора, где при связывании со специфичным лигандом рецептор, обычно экспрессируемый вне клеток, активируется посредством гомо- или гетеродимеризации и вызывает опосредованный рецептором эндоцитоз. Антитело, специфичное к рецептору клетки, обладает способностью вызывать такое явление и интернализуется в клетку, вызывая эндоцитоз, индуцируя, таким образом, разложение рецептора, уменьшая степень его экспрессии и, возможно, ингибируя передачу сигнала посредством определенного рецептора. Количество антител, связанных вне клеток, может быть обнаружено с помощью устройства для сортировки флуоресцентно-активированных клеток (FACS), определяя таким образом количество антител, интернализованных внутри клеток. В случае связывания антител с использованием связанного с ФИТЦ антитела против человеческой каппа области легкой цепи в качестве вторичного антитела для легкой цепи измеряемого антитела, можно количественно измерить количество антител, которые не являются интернализованными, но остаются связанными с рецептором-мишенью вне клеток, таким образом, идентифицируя количество интернализованных антител соответственно. Можно измерить фоновый сигнал с помощью неспецифичного связывания антитела, используемого в тесте, с использованием антитела против IgG человека, неспецифического к антигену, таким образом, измеряя флуоресцентный сигнал посредством фактического специфичного связывания.
В этом примере клеточную линию MKN45 (# JCRB0254), которая представляла собой клеточную линию рака желудка, использовали для идентификации in vitro интернализующей активности анти-c-Met антитела внутри опухолевых клеток. MKN45 экспрессирует рецептор c-Met на высоком уровне за счет амплификации гена MET, так что фосфорилирование рецептора c-Met индуцируется HGF-независимым путем. Проводили следующее испытание, чтобы установить, интернализуется ли рецептор c-Met в клетку анти-c-Met антителом hu8C4, снижая таким образом уровень его экспрессии.
Прежде всего, клеточные линии рака желудка MKN45 распределяли по 5,0×105 клеток в каждую лунку 6-луночного планшета, содержащую среду RPMI-1640 (2 мл), содержащую 10% (об./об.) ФБС, после чего планшет культивировали при температуре 37°C, 95% относительной влажности и 5% СО2 в течение 24 часов. Анализируемое анти-c-Met антитело, а также анти-IgG антитело (контроль) разбавляли до конечной концентрации 100 нМ, после чего полученные антитела инкубировали в течение ночи. Планшет, который использовали в качестве неинтернализованной контрольной группы, обрабатывали анти-c-Met антителом и анти-IgG человека антителом (контрольная группа), после чего полученный планшет инкубировали при температуре 4°C в течение 1 часа. Затем клетки каждой лунки собирали в 1 мл не содержащего ферментов буфера для диссоциации клеток (Gibco, #13151), после чего собранные клетки дважды промывали холодным ФСБ. В качестве вторичного антитела связанное с ФИТЦ антитело против человеческой каппа области легкой цепи (LSBio #LS-C60539) разбавляли в соотношении 1:2000, после чего в него добавляли полученное антитело и инкубировали при температуре 4°C в течение 1 часа. Затем клетки дважды промывали ФСБ, после чего полученные клетки фиксировали 100 мкл реагента BD Cytofix (BD, #554655) и промывали один раз ФСБ, среднее геометрическое значение интенсивности флуоресценции (СИФ) ФИТЦ, степень флуоресцентного окрашивания, измеряли с использованием анализатора паренхиматозных клеток BD FACS Canto II. Количество антител, связанных вне клеток, было получено с помощью следующей формулы, результаты этих вычислений приведены в Таблице 25.
Поверхностно-связанное антитело (%) = [(СИФ[37°C эксп.] - СИФ[IgG контроль]) / (СИФ[4°C контроль] - СИФ[IgG контроль])] × 100
Таблица 25
Измерение интернализации антитела hu8C4 и контрольного антитела OA-5D5 в клеточной линии рака желудка MKN45
Как видно из приведенной выше Таблицы 25, показано, что OA-5D5, анти-c-Met антитело, используемое в качестве контроля, практически не интернализировалось в клетки, в то время как антитело hu8C4 настоящего изобретения интернализировалось примерно на 40% или более в клетки клеточной линии рака желудка MKN45. То есть показано, что антитело hu8C4 значительно снижает уровень экспрессии рецептора c-Met.
Затем был проведен тест для измерения уровня рецептора у клеточной линии рака легкого NCI-H820, чтобы идентифицировать эффект снижения уровня рецептора биспецифичным антителом, способным одновременно связываться с рецептором c-Met и рецептором EGFR. Клеточная линия NCI-H820 представляет собой клеточную линию, подходящую для измерения эффекта снижения уровня рецептора биспецифичным антителом против c-Met и EGFR, поскольку в ней рецептор c-Met экспрессируется на уровне примерно 83000 единиц СССА (связывающая способность специфичного антитела) и рецептор EGFR экспрессируется на уровне примерно 74000 единиц СССА.
Прежде всего, клеточные линии NCI-H820 распределяли по 1,0×105 клеток в каждую лунку 6-луночного планшета со средой RPMI-1640 (2 мл), содержащей 10% (об./об.) ФБС, после чего полученный планшет культивировали в течение ночи при температуре 37°C, относительной влажности 95% и 5% СО2 в течение 24 часов. Затем среду заменяли бессывороточной средой, после чего полученный планшет культивировали в течение ночи при температуре 37°C, относительной влажности 95% и 5% СО2 в течение 24 часов. Затем анти-c-Met антитело, биспецифичное антитело против c-Met и EGFR, анти-EGFR антитело и антитело против IgG человека в качестве контроля, которые должны были быть проанализированы, разводили и обрабатывали в среде, содержащей 2% ФБС до достижения конечной концентрации 10 нМ, после чего полученные антитела культивировали в течение 5 суток. После этого клетки каждой лунки собирали в 1 мл не содержащего ферментов буфера для диссоциации клеток, после чего собранные клетки дважды промывали холодным ФСБ. Затем в каждую лунку добавляли по 10 мкл козьего F(ab`)2 фрагмента против мышиного IgG-CSF (R & D Systems Cat. #F0103B) в качестве вторичного антитела и инкубировали при температуре 4°C в течение 1 часа. Затем клетки дважды промывали ФСБ, после чего полученные клетки фиксировали 100 мкл реагента BD Cytofix (BD, #554655) и промывали один раз ФСБ, среднее геометрическое значение интенсивности флуоресценции (СИФ) ФИТЦ, степень флуоресцентного окрашивания, измеряли с использованием анализатора паренхиматозных клеток BD FACS Canto II.
В результате, при обработке анти-c-Met-антителом hu8C4 уровень рецептора EGFR снижался незначительно, а уровень рецептора c-Met заметно снижался до уровня 2% (Фигура 10). Кроме того, анти-EGFR-антитело Вектибикс снижало уровень рецептора EGFR до уровня примерно 83%, но уровень рецептора c-Met снижался незначительно. Напротив, в случае обработки биспецифичным антителом hu8C4 × Вектибикс настоящего изобретения, одновременно связывающимся с рецепторами c-Met и EGFR, было установлено, что уровень рецептора EGFR снижался до уровня примерно 21%, а уровень рецептора c-Met снижался до уровня примерно 4%, соответственно.
Таким образом, было установлено, что биспецифичное антитело hu8C4 × Вектибикс настоящего изобретения значительно снижает уровень экспрессии рецепторов c-Met и EGFR одновременно.
Пример 12. Идентификация активности ингибирования сигнала через c-Met и EGFR биспецифичного антитела in vitro
Затем проводили эксперимент с использованием клеточной линии NCI-H820, чтобы выявить влияние биспецифичного антитела настоящего изобретения на активность антигена и участников пути передачи сигнала.
Прежде всего, клеточные линии NCI-H820 распределяли по 1,0×105 клеток в каждую лунку 6-луночного планшета, после чего полученный планшет культивировали в течение ночи в течение ночи при температуре 37°C и 5% СО2, затем среду заменяли бессывороточной средой и снова культивировали в течение ночи. Антитело разводили и обрабатывали в бессывороточной среде в концентрации 100 нМ, после чего полученное антитело инкубировали в течение 24 часов, после чего за 15 минут до сбора клеток добавляли HGF (Gibco, PHG0254) и EGF (R & D Systems, 236-EG-200) в концентрации 50 нг/мл и 10 нг/мл, соответственно. Затем клетки разбавляли буфером для разбавления для проведения сбора клеток, после чего концентрацию белка определяли количественно с использованием метода анализа Лоури. 20 мкг белка загружали в каждую лунку геля и проводили разделение в ДСН-ПААГ, после чего проводили блоттинг на нитроцеллюлозной мембране. После блокирования мембраны все первичные антитела разбавляли и проводили реакцию в соотношении 1:1000, после чего связанное с HRP анти-кроличье антитело, используемое в качестве вторичного антитела, разбавляли в соотношении 1:5000 и проводили реакцию. Затем антитела, абсорбированные на мембране, обрабатывали реагентами для усиления хемилюминесценции (ECL), после чего полученные антитела измеряли с использованием прибора LC-3000.
В результате, как видно на Фигуре 11, при обработке биспецифичным антителом hu8C4 × Вектибикс scFv фосфорилирование EGFR, фосфорилирование Erk и фосфорилирование Akt заметно снижалось в большей степени, чем при обработке отдельно антителом hu8C4 или отдельно Вектибикс.
Таким образом, биспецифичное антитело hu8C4 × Вектибикс scFv настоящего изобретения может снижать активность рецептора, такого как EGFR, Erk, Akt и т.д., и молекул, участвующих в передачи сигнала на более низкие уровни в клеточной линии NCI-H820. В результате показано, что антитело настоящего изобретения обладает эффективностью за счет ингибирования пути передачи сигнала.
Пример 13. Идентификация активности ингибирования пролиферации опухолевых клеток, на мышиной модели ксенотрансплантата U-87 MG
Эксперимент проводили репрезентативно с использованием антитела hu8C4 IgG2 × Вектибикс scFv, чтобы идентифицировать активность ингибирования пролиферации опухолевых клеток с помощью биспецифичного антитела настоящего изобретения на HGF-зависимой модели ксенотрансплантата клеток U-87 MG.
Прежде всего, клеточные линии U-87 MG глиобластомы человека культивировали при температуре 37°C и 5% СО2 с использованием среды EMEM (ATCC® 30-2003™), содержащей L-глутамин (300 мг/л), 25 мМ HEPES, 25 мМ NaHCO3, 10% инактивированной нагреванием ФБС и тому подобное. Затем клетки U-87 MG подкожно инокулировали в объемк 200 мкл в бок самца бестимусной мыши 6-8-недельного возраста (Harlan) в количестве 1×107 клеток на мышь. После определения того, что объем опухоли, образовавшейся через 25 дней после инокуляции, достиг 60-130 мм3, животных распределяли по группам, после этого им внутрибрюшинно вводили исследуемый материал один раз в неделю в течение 4 недель (всего 5 раз в дни: 0, 7, 14, 21 и 28). Исследуемый материал вводили в количестве 5 мг/кг, а объем опухоли и массу мыши измеряли два раза в неделю. При анализе данных, сравнение между контрольной группой, получавшей вспомогательное вещество, и группой, которой вводили исследуемый материал, обычно проводили с использованием t-критерия Стьюдента, а в качестве статистического метода использовали программу Origin Pro 8.5. «Максимальный процент ингибирования» означает % ингибирования роста опухоли по сравнению с контрольной группой, получавшей растворитель.
В результате группа, которой вводили 3,5 мг/кг и 6,8 мг/кг hu8C4 IgG2 × Вектибикс scFv, имела максимальное ингибирование 96% для объема опухоли по сравнению с контрольной группой, получавшей с растворитель, а группа, которой вводили 1,5 мг/кг этого антитела, имела максимальное ингибирование 80%, таким образом, уменьшая объем опухоли до значимого уровня с 7-го дня после введения до последнего дня испытания (р<0,01) (Фигура 12). Кроме того, при сравнении с BsAB-01 в качестве группы положительного контроля биспецифичное антитело настоящего изобретения снижало рост опухоли до значимого уровня (р<0,01).
Таким образом, на основе приведенных выше результатов было установлено, что биспецифичное антитело настоящего изобретения значительно снижает рост опухоли, и таким образом, обладает очень хорошей противоопухолевой эффективностью.
Пример 14. Идентификация активности ингибирования пролиферации опухолевых клеток на мышиной модели ксенотрансплантата NCI-H820
Клеточная линия NCI-H820, которая представляет собой клеточную линию, имеющую мутацию в положении 790 аминокислотной последовательности EGFR, которая заключается в замене треонина (T) на метионин (M), а также имеющую амплифицированный ген MET, известна как резистентная клеточная линия AZD9291 (осимертиниб, TAGRISSO®), который является ингибитором тирозинкиназы EGFR третьего поколения (Darren AE Cross et al., Cancer Discov. 4(9): 1046-1061 (2014). Оценку проводили на мышиной модели ксенотрансплантата NCI-H820 путем репрезентативного использования биспецифичного антитела настоящего изобретения hu8C4 × Вектибикс scFv, чтобы определить активность ингибирования пролиферации опухолевых клеток биспецифичного антитела в клеточной линии NCI-H820, имеющей устойчивость к такому ингибитору тирозинкиназы EGFR.
В частности, мышь, используемая в этом примере, представляла собой 6-недельного самца (Jackson Laboratory, STOCK Hgftm1.1 (HGF) Aveo Prkdcscid/J), у которого был удален мышиный ген HGF, и который был трансформирован человеческим геном HGF для экспрессии. Клеточную линию NCI-H820 (ATCC, #HTB-181) помещали в колбу для культивирования клеток вместе со средой RPMI1640, содержащей 10% ФБС, после чего полученную колбу культивировали при температуре 37°C и 5% СО2. Затем полученные клетки промывали ФСБ и 2,5% трипсин-ЭДТА (Gibco, 15090) разбавляли в 10 раз, после чего его добавляли к клеткам, чтобы разделить их. После этого проводили центрифугирование (1000 об/мин, 5 минут), чтобы избавиться от супернатанта и получить клеточную суспензию в новой среде. Затем с помощью микроскопа определяли жизнеспособность клеток, после чего полученные клетки разводили в бессывороточной среде в концентрации 5,0×107 клеток/мл, получая, таким образом, клеточные линии. Полученные клеточные линии вводили мышам подкожно в количестве 0,1 мл на животное. После введения, когда размер опухоли в области с трансплантированными в нее клеточными линиями достигал примерно 100-150 мм3, клеточные линии распределяли так, чтобы размер опухоли в каждой группе можно было равномерно распределить в соответствии с ранжированным размером опухоли. Затем онкогенез идентифицировали дважды в неделю с 7-го дня после начала введения клеток до 28-го дня после дня группировки (день начала введения тестируемого материала) и после завершения введения тестируемого материала, после чего большую ось опухоли и малую ось измеряли штангенциркулем, вычисляя, таким образом, размер опухоли (ab2/2 (a: длина большой оси, b: длина малой оси)). Статистический анализ выполняли при помощи программы Prism 5.03 (GraphPad Software Inc., Сан-Диего, Калифорния, США). Если значение p было меньше 0,05, оно считалось статистически значимым.
В результате во всех группах, которым вводили антитело hu8C4 × Вектибикс scFv с 4-го дня после начала введения исследуемого материала до 28-го дня его введения, было показано, что у него активность ингибирования пролиферации опухоли была значительно выше, чем у контрольной группы, получавшей растворитель (р<0,001), и также было установлено, что коэффициент ингибирования опухоли составляет максимум 100% (Фигура 13). С другой стороны, антитело AZD9291 (Selleckchem), используемое в качестве положительного контроля, не показало значимых отличий от контрольной группы, получавшей растворитель.
Пример 15. Идентификация активности ингибирования пролиферации опухолевых клеток in vitro при комбинированном введении анти-c-Met антитела 5G3 и анти-HER2-антитела
In vitro оценку активности ингибирования пролиферации клеток проводили на клеточной линии NCI-H2170, чтобы оценить активность ингибирования пролиферации опухолевых клеток комбинацией анти-c-Met антитела 5G3 настоящего изобретения и анти-HER2 антитела. Клеточная линия NCI-H2170 (ATCC #CRL-5928) представляет собой линию опухолевых клеток немелкоклеточного рака легкого (НМРЛ), в которой по результатам измерения уровня рецептора EGFR, этот рецептор экспрессировался на уровне примерно 2700 единиц СССА (связывающая способность специфичного антитела, тогда как c-Met экспрессировался на уровне примерно 11000 единиц СССА.
В частности, клетки NCI-H2170 разводили в культуральной среде RPMI-1640, содержащей 10% (об./об.) ФБС, после чего полученные клетки вносили по 100 мкл в планшет при концентрации 3,0×103 клеток на лунку, полученный планшет культивировали при температуре 37°C, 95% относительной влажности и 5% (об./об.) СО2 в течение 18-24 часов. Затем из каждой лунки удаляли среду для культивирования клеток, после чего в каждую лунку добавляли по 100 мкл среды RPMI-1640, содержащей 2% (об./об.) ФБС. После этого антитела, полученные в 2-кратной конечной концентрации (100 нМ), последовательно разводили в соотношении 1/10, и полученные антитела добавляли по 100 мкл в каждую лунку в шести концентрациях (то есть 200 нМ, 20 нМ, 2 нМ, 200 пМ, 20 пМ и 2 пМ) для каждого антитела. Планшет культивировали в течение 5 суток при температуре 37°C, 95% относительной влажности и 5% (об./об.) СО2, после чего в каждую лунку в последний день добавляли 20% раствора WST-8 (CCK-8, Dojindo), чтобы осуществить проявление цвета в течение 1-2 часов, оптическую плотность измеряли при длине волны 450 нм с помощью устройства для считывания микропланшетов.
Результаты активности ингибирования пролиферации клеток показаны в Таблице 26 и на Фигуре 14.
Таблица 26
In vitro активность ингибирования пролиферации опухолевых клеток комбинированной терапией анти-c-Met-антителом и анти-HER2-антителом
Из Таблицы 26 видно, что комбинированная обработка антителом 5G3 и антителом A091 (патент Кореи регистрационный номер 10-1515535) в качестве анти-HER2 антитела обладает большей способностью ингибировать пролиферацию опухолевых клеток, чем обработка линии опухолевых клеток NCI-H2170 только одним из этих антител.
Пример 16. Идентификация активности ингибирования пролиферации опухолевых клеток in vivo комбинированным введением анти c-Met антитела 5G3 и анти-HER2 антитела на мышиной модели ксенотрансплантата линии клеток рака легкого человека NCI-H2170
Эксперимент по противораковой активности проводили на мышиной модели ксенотрансплантата линии клеток рака легкого NCI-H2170, чтобы определить совместную эффективность анти-HER2 антитела и анти-c-Met антитела.
В частности, в этом Примере размер опухоли мыши измеряли тем же способом, который описан в Примере 14, с использованием той же мыши, как описано выше в Примере 13. Результаты оценки противоопухолевой эффективности комбинацией A091 и 5G3 на мышиной модели ксенотрансплантата NCI-H2170 в качестве клеток опухоли легкого показаны на Фигуре 15.
В результате, в случае проведения однократного введения одного A091 или комбинации A091 и 5G3, объем опухоли уменьшался до значимого уровня по сравнению с контрольной группой, получавшей растворитель, с 14-го дня после введения (p<0,05). Кроме того, группа, которой вводили комбинацию A091 и 5G3, показала значимое уменьшение объема опухоли по сравнению с группой, которой вводили только A091 или группой, которой вводили BsAB02 (US 2010/0254988 A1) в качестве контрольного биспецифичного антитела (p<0,01).
Пример 17. Идентификация активности ингибирования пролиферации опухолевых клеток на мышиной модели ксенотрансплантата NCI-H596
Поскольку клеточная линия NCI-H596 представляла собой клеточную линию рака легкого с мутацией в экзоне 14 c-Met, то был проведен эксперимент н на мышиной модели ксенотрансплантата NCI-H596 для выявления противоракового эффекта hu8C4 × Вектибикс scFv.
В этом Примере размер опухоли мыши измеряли с использованием той же мыши и того же метода, которые описаны выше в Примере 14.
Результаты оценки противораковой эффективности после введения hu8C4 × Вектибикс scFv один или два раза в неделю в течение всего 4 недель на модели ксенотрансплантата NCI-H596 в качестве клетки опухоли легкого показаны на Фигуре 16.
В результате измерения размера опухоли уровень размера опухоли в группе, которой вводили hu8C4 × Вектибикс scFv в дозе 10 мг/кг два раза в неделю, показал статистически значимое различие по сравнению с контрольной группой с 11-го дня после начала введения исследуемого материала до конца эксперимента, и уровни размеров опухолей в группе, которой вводили hu8C4 × Вектибикс scFv в дозе 5 мг/кг два раза в неделю, и в группе, которой вводили hu8C4 × Вектибикс scFv 10 мг/кг один раз в неделю, также были значительно ниже по сравнению с контрольной группе с 18-го дня после начала введения исследуемого материала. Кроме того, уровень размера опухоли в группе, которой вводили исследуемый материал, имел тенденцию к изменению в зависимости от дозы исследуемого материала, и размер опухоли в опытной группе был ниже по сравнению с контрольной группой даже после последнего дня введения тестового материала (день 28).
Пример 18. Идентификация активности ингибирования пролиферации опухолевых клеток на мышиной модели ксенотрансплантата EBC-1
Поскольку EBC-1 представлял собой клеточную линию рака легкого с амплификацией гена c-Met, то был проведен эксперимент на мышиной модели ксенотрансплантата EBC-1 с целью выявления противоракового эффекта hu8C4 × Вектибикс scFv.
Мышь, используемая в этом Примере, представляла собой шестинедельную самку бестимусной голой мыши (Harlan). Клеточные линии EBC-1 (JCRB, #JCRB0820) помещали в колбу для культивирования клеток вместе со средой EMEM, содержащей 10% ФБС, после чего полученные клеточные линии культивировали при температуре 37°C и 5% СО2. Клеточные линии готовили таким образом, что полученные клеточные линии разводили в бессывороточной среде в концентрации 5,0×107 клеток/мл, после чего клеточные линии вводили мышам подкожно в количестве 0,1 мл на животное. Когда размер опухоли в области с трансплантированными в нее клеточными линиями достигал примерно 100-150 мм3, антитело hu8C4 × Вектибикс scFv вводили один или два раза в неделю в течение всего 4 недель, после чего размер опухоли мыши измеряли тем же методом, который описан в Примере 14.
Результаты оценки противораковой эффективности hu8C4 × Вектибикс scFv на модели ксенотрансплантата EBC-1 в качестве клетки рака легкого показаны на Фигуре 17.
В результате измерения размера опухоли уровень размера опухоли в группе, которой вводили hu8C4 × Вектибикс scFv в дозе 10 мг/кг два раза в неделю, показал статистически значимое различие по сравнению с контрольной группой с 7-го дня после начала введения исследуемого материала до 56-го дня после начала введения исследуемого материала. Группа, которой вводили hu8C4 × Вектибикс scFv в дозе 5 мг/кг два раза в неделю, и группа, которой вводили это антитело один раз в неделю, показали значимый низкий уровень по сравнению с контрольной группой с 18-го дня после начала введения исследуемого материала. Кроме того, уровень размера опухоли в группе, которой вводили исследуемый материал, имел тенденцию к изменению в зависимости от дозы исследуемого материала, и уровень размера опухоли в группе, которой вводили hu8C4 × Вектибикс scFv в дозе 10 мг/кг два раза в неделю в течение периода наблюдения после последнего дня (день 28) введения тестируемого материала был значимо ниже по сравнению с контрольной группой до 56-го дня после начала введения исследуемого материала. В частности, было обнаружено, что одно животное в группе, которой вводили hu8C4 × Вектибикс scFv в дозе 10 мг/кг два раза в неделю, имело полный ответ на 18-й день после начала введения исследуемого материала.
Пример 19. Эффект снижения c-Met и EGFR на поверхности раковых клеток биспецифичным антителом
Эффект снижения c-Met и EGFR на поверхности опухолевых клеток in vitro биспецифичным антителом (hu8C4 × Вектибикс scFv) настоящего изобретения идентифицировали и сравнивали с действием анти-c-Met антитела (hu8C4) настоящего изобретения, Вектибикса, комбинации анти-c-Met/EGFR антител и других антител.
Рецептор, обычно расположенный на клеточной мембране, интернализуется в клетку при связывании с антителом, таким образом, его количество, расположенное на клеточной мембране, уменьшается. Уменьшение рецептора на такой клеточной мембране вызывает ингибирование активации рецептора и уменьшение его последующего сигнала при связывании с лигандом.
В этом примере клеточную линию аденокарциномы легкого HCC827 использовали для наблюдения снижения c-Met и EGFR на клеточной мембране. HCC827 имеет делеционную мутацию EGFR E746-A750 и сверхэкспрессирует c-Met. HCC827 обрабатывали биспецифичным антителом (hu8C4 × Вектибикс scFv) настоящего изобретения и другими антителами, после чего проводили иммунофлуоресцентное окрашивание антителом, специфичным к c-Met и EGFR, полученную клеточную линию анализировали с использованием сортировки флуоресцентно-активированных клеток, измеряя, таким образом, количество c-Met и EGFR на поверхности клеток. Подробное описание этого способа приводится ниже.
Прежде всего, клетки HCC827 (ATCC® CRL-2868™) распределяли по 3,0×105 клеток в каждую лунку 6-луночного планшета, содержащего среду RPMI-1640 (2 мл), содержащую 10% (об./об.) ФБС, затем планшет культивировали при температуре 37°C, относительной влажности 95% и 5% СО2 в течение 24 часов. Биспецифичное антитело (hu8C4 × Вектибикс scFv) настоящего изобретения, антитело c-Met (hu8C4) настоящего изобретения, Вектибикс, смесь антитела c-Met (hu8C4) настоящего изобретения и Вектибикс, C-EM1 и LA480 разбавляли до конечной концентрации 100 нМ, после чего полученные антитела обрабатывали и инкубировали в течение 18 часов. В планшет, который использовали в качестве неубывающей контрольной группы с c-Met и EGFR вносили антитело к IgG человека и инкубировали в течение 18 часов. Затем клетки каждой лунки собирали в 500 мкл не содержащего ферментов буфера для диссоциации клеток (Gibco, № 13151), после чего клетки отделяли от не содержащего ферментов буфера для диссоциации клеток с помощью центробежного сепаратора для удаления не содержащего ферментов буфера для диссоциации клеток. Для иммунофлуоресцентного окрашивания козье анти-c-Met антитело (R & D Systems, AF276), козье анти-EGFR антитело (R & D Systems, AF231) или неспецифичное козье антитело для измерения количества окрашивания смешивали по 2 мкг соответственно с 200 мкл холодного ФСБ, содержащего 2% (об./об.) ФБС, после чего полученные антитела вносили в каждую лунку, и полученный планшет инкубировали при температуре 4°C в течение 1 часа. Затем полученный планшет дважды промывали холодным ФСБ, содержащим 2% (об./об.) ФБС. Антитело ALEXA488 связывали в качестве вторичного антитела, после чего 1 мкл полученного от осла антитела (Thermo Fisher, A-11055), связывающегося с козьим антителом, разбавляли 200 мкл холодного ФСБ, содержащего 2% (об./об.) ФБС, и использовали полученное антитело. После реакции со вторичным антителом при температуре 4°C в течение 1 часа полученные клетки дважды промывали холодным ФСБ, содержащим 2% (об./об.) ФБС, после чего полученные клетки фиксировали, используя 200 мкл реагента BD Cytofix (BD, #554655). После однократной промывки ФСБ, с помощью сортировщика флуоресцентно-активированных клеток BD FACS Canto II измеряли среднее геометрическое значение интенсивности флуоресценции (СИФ) ALEXA488, степень флуоресцентного окрашивания. Количество c-Met и EGFR, расположенных на клеточной мембране, вычисляли как среднее геометрическое значение интенсивности флуоресценции (СИФ) по следующей формуле. Что касается значений, полученных после трехкратного выполнения теста, их среднее значение и стандартное отклонение показаны в Таблице 27 и на Фигуре 18 и 19.
Поверхностное количество c-Met или EGFR=гео СИФ[опытная группа] - гео СИФ[неспецифичное козье антитело]
Таблица 27
Количество c-Met и EGFR на поверхности клеток, измеренное после обработки клеточной линии HCC827 биспецифичным антителом (hu8C4 × Вектибикс scFv) и т.д.
Как видно из Таблицы 27 выше, все антитела, связывающиеся с c-Met, снижали количество c-Met на поверхности клеток на 40~70%, в то время как антитела, связывающиеся с EGFR, демонстрировали незначительный эффект снижения на 10-15%. Кроме того, с точки зрения эффекта снижения c-Met, hu8C4, комбинация hu8C4+Вектибикс, C-EM1 и C-LA480, снижали уровень c-Met на поверхности клеток примерно на 40% или около того, в то время как антитело hu8C4 × Вектибикс scFv снижало уровень c-Met на поверхности клеток на 70%, демонстрируя тем самым большую эффективность снижения c-Met на поверхности клеток, чем другие антитела и комбинации антител.
Приведенные выше результаты показывают, что биспецифичное антитело (hu8C4 × Вектибикс scFv) настоящего изобретения значительно снижает количество c-Met на поверхности клеток.
Пример 20. Картирование эпитопов
Чтобы выяснить с каким эпитопом c-Met-антигена человека взаимодействует биспецифичное антитело (hu8C4 × Вектибикс scFv) настоящего изобретения, проводили анализ, который был поручен группе поддержки разработки молекулярного моделирования Фонда медицинских инноваций Osong (KBIO, Корея). Анализ проводился методом масс-спектрометрия водородно-дейтериевого обмена (HDX-MS).
Домен Sema c-Met состоит из двух цепей α/β, и проводили идентификацию покрытия для каждой из этих двух цепей. Из-за присутствия ряда дисульфидных связей в образце покрытие пептидов было оптимизировано путем регулирования времени выдержки при охлаждении, концентрации TCEP, концентрации пепсина и т.д. Наконец, эксперимент был проведен в условиях буфера для остановки реакции, содержащего 100 мМ K.Фосфата, 125 мМ ТСЕР, 0,5 М гуанидин-HCl и имеющего рН 2,66.
Антигены и антитела готовили в концентрации 3,3 мг/мл и 65 мг/мл соответственно, и за 3 часа до эксперимента связывали 37 пмоль антигенов cMET и 36 пмоль антител. Реакцию с буфером для мечения дейтерием проводили в течение 0, 0,33, 10, 60 и 240 минут. Мечение останавливали с помощью буфера для остановки реакции в соответствии с каждым временем мечения и проводили встряхивание на вортексе, после чего образцы немедленно замораживали в жидком азоте и хранили при температуре -80°C до анализа. Полученные антигены и антитела загружали в колонку с пепсином и анализировали с помощью масс-спектрометра (МС).
В результате анализа было установлено, что биспецифичное антитело (hu8C4 × Вектибикс scFv) настоящего изобретения связывается с трехмерной формой эпитопов в 4 областях Y321 - L329 (SEQ. No. 331), I333 - I341 (SEQ. No. 332), P366-D372 (SEQ. No. 333) и Q464-S474 (SEQ. No. 334) домена Sema β-цепи человеческого c-Met (Таблица 28). Мечение было выполнено для третичной структуры человеческого c-Met-антигена (PDB No. 4K3J) с использованием программы PyMOL, результаты анализа показаны на Фигуре 20.
Таблица 28
Аминокислотная последовательность эпитопной области
Из приведенных выше результатов можно заключить, что мышиное антитело, гуманизированное антитело, оптимизированное по аффинности антитело или его антигенсвязывающие фрагменты настоящего изобретения, специфично связывающиеся с c-Met, избирательно действуют на c-Met, и они обладают превосходной активностью ингибирования пролиферации раковых клеток, а также превосходной противораковой активностью даже в небольшом количестве, эффективно предотвращая или излечивая рак.
Хотя конкретные части настоящего изобретения были подробно описаны выше, для специалистов в данной области техники очевидно, что такие подробные описания приводятся только для иллюстрации примерных вариантов осуществления изобретения, но не предназначены для ограничения объема настоящего изобретения. Таким образом, следует понимать, что существенный объем настоящего изобретения определяется прилагаемой формулой изобретения и ее эквивалентами.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CHONG KUN DANG PHARMACEUTICAL CORP.
<120> Новое анти-c-met антитело и его применение
<130> P18026-CKD
<150> KR 10-2017-0067106
<151> 2017-05-30
<160> 335
<170> Патентная версия 3.0
<210> 1
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 CDR 1 легкой цепи
<400> 1
Gly Ala Ser Glu Asn Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 2
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 CDR 2 легкой цепи
<400> 2
Gly Ala Thr Asn Leu Ala Asp
1 5
<210> 3
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 CDR 3 легкой цепи
<400> 3
Gln Asn Val Leu Ser Ser Pro Tyr Thr
1 5
<210> 4
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 CDR 1 легкой цепи
<400> 4
Ser Ala Thr Ser Ser Val Arg Tyr Met Tyr
1 5 10
<210> 5
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 CDR 2 легкой цепи
<400> 5
Asp Thr Ser Asn Leu Ala Ser
1 5
<210> 6
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 CDR 3 легкой цепи
<400> 6
Gln Gln Trp Ser Ser Tyr Pro Arg Thr
1 5
<210> 7
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 CDR 1 тяжелой цепи
<400> 7
Asp Tyr Tyr Ile Asn
1 5
<210> 8
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 CDR 2 тяжелой цепи
<400> 8
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 9
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 CDR 3 тяжелой цепи
<400> 9
Gly Asp Tyr Gly Phe Leu Tyr
1 5
<210> 10
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 CDR 1 тяжелой цепи
<400> 10
Asp Tyr Thr Leu His
1 5
<210> 11
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 CDR 2 тяжелой цепи
<400> 11
Tyr Ile Asn Pro Tyr Ser Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 12
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 CDR 3 тяжелой цепи
<400> 12
Gly His Met Asp Tyr
1 5
<210> 13
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 вариабельная область легкой цепи
<400> 13
Asp Ile Leu Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Gly Ala Ser Glu Asn Ile Tyr Gly Ala
20 25 30
Leu Asn Trp Tyr Gln Arg Lys Gln Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Met Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Gln Phe Ser Leu Lys Ile Thr Ser Leu His Pro
65 70 75 80
Asp Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 14
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 вариабельная область легкой цепи
<400> 14
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Thr Ser Ser Val Arg Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Gly Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Asn Ser Leu Thr Ile Ser Arg Leu Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro Arg Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 15
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 вариабельная область тяжелой цепи
<400> 15
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Lys Gln Gly Thr Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Thr Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Arg Gly Thr Leu Val
100 105 110
Thr Val Ser Ala
115
<210> 16
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 вариабельная область тяжелой цепи
<400> 16
Gln Gly Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Thr Leu His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Gly Leu Thr Ser Glu Asp Ser Ala Val Phe Tyr Cys
85 90 95
Ala Arg Gly His Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val
100 105 110
Ser Ser
<210> 17
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 вариабельная область легкой цепи
<400> 17
gatattctga tgacccagtc tccagcttca ctgtctgcat ctgtgggaga aactgtcacc 60
atcacatgtg gagcaagtga gaatatttac ggtgctttaa attggtatca gcgaaaacag 120
ggaaaatctc ctcagctcct gatctatggt gcaaccaact tggcagatgg catgtcatcg 180
aggttcagtg gcagtgggtc tggtagacag ttttctctca agatcactag cctgcatcct 240
gacgatgttg caacgtatta ctgtcaaaat gtgctaagta gtccgtacac gttcggaggg 300
gggaccaagc tggaaatcaa a 321
<210> 18
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 вариабельная область легкой цепи
<400> 18
caaattgttc tcacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccacctc aagtgtacgt tacatgtact ggtaccagca gaagccagga 120
tcctccccca gactcctgat ttatgacaca tccaacctgg cttctggagt ccctggtcgc 180
ttcagcggca gtgggtctgg gacctctaac tctctcacaa tcagccgatt ggaggctgaa 240
gatgctgcca cttattactg ccagcagtgg agtagttacc cacggacgtt cggtggaggc 300
accaagctgg aaatcaaa 318
<210> 19
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> гибридома 8C4 вариабельная область тяжелой цепи
<400> 19
gaggttcagc tgcagcagtc tggagctgag ctggcgaggc ccggggcttc agtgaagctg 60
tcctgcaagg cttctggcta caccttcagt gactactata taaactgggt gaagcagggg 120
actggacagg gccttgagtg gattggagag atttttcctg gaagtggaaa tactcacttc 180
agtgcgaggt tcaagggcaa ggccacactg actgcagaca aatcctccag cacagcctac 240
atgcagctca gcagcctgac atctacggac tctgcagtct atttctgtgc cgggggtgac 300
tacgggtttc tttactgggg ccgagggact ctggtcactg tctctgca 348
<210> 20
<211> 342
<212> ДНК
<213> Искусственная последовательность
<220>
<223> гибридома 5G3 вариабельная область тяжелой цепи
<400> 20
cagggccagc tgcagcagtc tggggctgaa ctggcaagac ctggggcctc agtgaagatg 60
tcctgcaagg cttctggcta cacctttact gactacacgc tgcactgggt aaaacagagg 120
cctggacagg gtctggaatg gattggatac attaatcctt acagtggtta tactaattac 180
aatcagaaat tcaaggacaa ggccacattg actgcagaca aatcctccag cacagcctac 240
atgcaactga gcggcctgac atctgaagac tctgcagtct tttattgtgc aagaggacat 300
atggactact ggggtcaagg aacctcagtc accgtctcct ca 342
<210> 21
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4-1 вариабельная область легкой цепи
<400> 21
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Glu Asn Ile Tyr Gly Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 22
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4-2 вариабельная область легкой цепи
<400> 22
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Glu Asn Ile Tyr Gly Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 23
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4-1 вариабельная область тяжелой цепи
<400> 23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 24
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4-2 вариабельная область тяжелой цепи
<400> 24
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Ile Ser Ala Asp Lys Ala Lys Asn Ser Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 25
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu8C4-1 вариабельная область легкой цепи
<400> 25
gatatccaga tgacccagtc tcccagcagt ctttccgctt ctgtgggtga tagggtgacg 60
ataacttgcg gagcaagtga gaatatttac ggtgctttaa attggtacca gcagaagcct 120
gggaaagctc caaagctgct gatctatggt gcaaccaact tggcagatgg cgtccctagc 180
aggttcagcg gcagtggaag cggcagagac ttcactttca caatctcctc cctgcaaccc 240
gaggacattg caacctacta ttgtcaaaat gtgctaagta gtccgtacac gtttggccag 300
ggaaccaagg ttgaaattaa a 321
<210> 26
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu8C4-2 вариабельная область легкой цепи
<400> 26
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcg gagcaagtga gaatatttac ggtgctttaa attggtatca gcagaaacca 120
gggaaagttc ctaagctcct gatctatggt gcaaccaact tggcagatgg ggtcccatct 180
cggttcagtg gcagtggatc tgggcgagat ttcactctca ccatcagcag cctgcagcct 240
gaagatgttg caacttatta ctgtcaaaat gtgctaagta gtccgtacac gtttggccag 300
ggaaccaagg ttgaaattaa a 321
<210> 27
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu8C4-1 вариабельная область тяжелой цепи
<400> 27
gaggttcagt tagtggaatc cggaggagga ctggtgcagc ctggtggaag tttgaggctg 60
tcatgcgcag ccagtggcta caccttcagt gactactata taaactgggt aagacaggct 120
cccggaaaag ggctggagtg gattggagag atttttcctg gaagtggaaa tactcacttc 180
agtgcgaggt tcaagggccg agccaccctc tcagcagaca aaagcaagaa taccgcctat 240
ctgcagatga atagccttcg cgcagaagat actgccgtgt attactgtgc cgggggtgac 300
tacgggtttc tttactgggg acagggcacc ttggtgacag tctcttct 348
<210> 28
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu8C4-2 вариабельная область тяжелой цепи
<400> 28
caggttcagt tagtggaatc cggaggagga ctggtgaagc ctggtggaag tttgaggctg 60
tcatgcgcag ccagtggcta caccttcagt gactactata taaactggat cagacaggct 120
cccggaaaag ggctggagtg gattggagag atttttcctg gaagtggaaa tactcacttc 180
agtgcgaggt tcaagggccg agccaccatc tcagcagaca aagcgaagaa tagcgcctat 240
ctgcagatga atagccttcg cgcagaagat actgccgtgt attactgtgc cgggggtgac 300
tacgggtttc tttactgggg acagggcacc ttggtgacag tctcttct 348
<210> 29
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu5G3-1 вариабельная область легкой цепи
<400> 29
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Thr Ser Ser Val Arg Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Asn Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro Arg Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 30
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu5G3-2 вариабельная область легкой цепи
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Thr Ser Ser Val Arg Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Tyr Pro Arg Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 31
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu5G3-1 вариабельная область тяжелой цепи
<400> 31
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Thr Leu His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly His Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 32
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu5G3-2 вариабельная область тяжелой цепи
<400> 32
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Thr Leu His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Tyr Ile Asn Pro Tyr Ser Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Phe Tyr Cys
85 90 95
Ala Arg Gly His Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 33
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu5G3-1 вариабельная область легкой цепи
<400> 33
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gtgccacctc aagtgtacgt tacatgtact ggtaccagca gaaacctggc 120
cagtctccca ggctcctcat ctatgacaca tccaacctgg cttctggcat cccagcaagg 180
ttcagtggca gtgggtctgg gacagacaac actctcacca tcagcagact ggagcctgaa 240
gattttgcag tttattactg tcagcagtgg agtagttacc cacggacgtt cggcggaggg 300
accaaggtgg agatcaaa 318
<210> 34
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu5G3-2 вариабельная область легкой цепи
<400> 34
gacatccaga tgactcagag tccctcttct ctgtctgcct cagtgggaga tcgggtcaca 60
atcacatgtt cagcaacaag ctcagtgcga tacatgtatt ggtaccagca gaagccaggc 120
aaagccccaa agctgctgat ctatgacaca tctaatctgg ccagcggcgt cccatctcgc 180
ttctcaggct ccggaagcgg tactgatttt accctgacta tttcttcctt gcagcctgag 240
gacttcgcaa cctattattg ccagcagtgg tctagctacc ctcgcacatt cggccaggga 300
accaaggtcg aaattaaa 318
<210> 35
<211> 342
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu5G3-1 вариабельная область тяжелой цепи
<400> 35
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttcacc gactacacgc tgcactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gataggatac attaatcctt acagtggtta tactaattac 180
aatcagaaat tcaaggacag agtcaccttg accgcagaca aatccacgag cacagcctac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtgc tagaggacat 300
atggactact ggggccaagg aaccctggtc accgtctcct ca 342
<210> 36
<211> 342
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hu5G3-2 вариабельная область тяжелой цепи
<400> 36
gaagtccaac ttgtggagtc aggaggcggg ctcgtgcagc caggcggatc attgcgactt 60
tcttgtgctg cctcagggta caccttcact gattatacct tgcattgggt tcgccaagca 120
cccggtaagg gtctcgaatg ggtaggatac attaatccat acagcggcta caccaactac 180
aaccagaaat tcaaagacag ggctaccctt agtgccgaca agtctaagaa caccgcctac 240
cttcagatga actcccttag agccgaggat actgctgtgt tttattgcgc taggggtcat 300
atggactact ggggacaggg gaccttggtg actgtgtctt cc 342
<210> 37
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Шарнирная область константной области IgG1 CH
<400> 37
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 38
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 38
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 39
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 39
Glu Cys Cys Val Glu Cys Pro Pro Cys Pro
1 5 10
<210> 40
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 40
Glu Arg Lys Cys Cys Cys Pro Pro Cys Pro
1 5 10
<210> 41
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 41
Glu Cys Cys Cys Pro Pro Cys Pro
1 5
<210> 42
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 42
Glu Lys Cys Cys Val Glu Cys Pro Pro Cys Pro
1 5 10
<210> 43
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 43
Glu Arg Lys Cys Cys Val Cys Pro Pro Cys Pro
1 5 10
<210> 44
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 44
Glu Lys Cys Cys Val Cys Pro Pro Cys Pro
1 5 10
<210> 45
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Шарнирная область константной области IgG1 CH
<400> 45
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gccca 45
<210> 46
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 46
gagcgaaaat gttgtgtcga gtgcccaccg tgccca 36
<210> 47
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 47
gagtgttgtg tcgagtgccc accgtgccca 30
<210> 48
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 48
gagcgaaaat gttgttgccc accgtgccca 30
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 49
gagtgttgtt gcccaccgtg ccca 24
<210> 50
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 50
gagaaatgtt gtgtcgagtg cccaccgtgc cca 33
<210> 51
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 51
gagcgaaaat gttgtgtctg cccaccgtgc cca 33
<210> 52
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариант шарнирной области IgG1 CH
<400> 52
gagaaatgtt gtgtctgccc accgtgccca 30
<210> 53
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 53
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 54
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 54
Glu Ile Asp Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 55
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 55
Glu Ile Glu Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 56
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 56
Glu Ile Trp Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 57
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 57
Glu Ile Tyr Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 58
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 58
Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 59
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 59
Glu Ile Phe Pro Gly Tyr Gly Asn Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 60
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 60
Glu Ile Phe Pro Gly Ser Gly Tyr Thr His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 61
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 61
Glu Ile Phe Pro Gly Ser Gly Asn Thr Trp Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 62
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 62
Glu Ile Phe Pro Gly Ser Gly Asn Thr Tyr Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 63
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 63
Glu Ile Phe Pro Gly Trp Gly Asn Thr Tyr Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 64
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 64
Gln Asp Tyr Gly Phe Leu Tyr
1 5
<210> 65
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 65
Glu Asp Tyr Gly Phe Leu Tyr
1 5
<210> 66
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 66
His Asp Tyr Gly Phe Leu Tyr
1 5
<210> 67
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 67
Asn Asp Tyr Gly Phe Leu Tyr
1 5
<210> 68
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 68
Val Glu Leu Gly Phe Leu Tyr
1 5
<210> 69
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 69
Phe Glu Thr Gly Tyr Tyr Leu
1 5
<210> 70
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 70
Gly Glu Tyr Gly Tyr Gln Asn
1 5
<210> 71
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 71
Trp Glu Tyr Gly Leu Ser Met
1 5
<210> 72
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 72
Glu Ile Phe Pro His Phe Thr Ser Asp His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 73
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 73
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Trp Met Gly
1 5 10 15
Thr
<210> 74
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 74
Glu Ile Phe Pro Gly Ser Gly Asn Glu Ser Val Ser Phe Arg Phe Lys
1 5 10 15
Gly
<210> 75
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 75
Glu Ile Phe Pro Gly Ser Gly Asn Ser Ala Val Ile Ser Arg Phe Lys
1 5 10 15
Gly
<210> 76
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 76
Glu Ile Phe Pro Gly Ser Gly Asn His Thr Val Val Arg Arg Phe Lys
1 5 10 15
Gly
<210> 77
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 77
Glu Ile Phe Pro Gly Ser Gly Asn Leu Ser Met His Gly Arg Phe Lys
1 5 10 15
Gly
<210> 78
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 78
Glu Ile Phe Pro Gly Ser Gly Asn His Thr Pro Val Phe Arg Phe Lys
1 5 10 15
Gly
<210> 79
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 79
Glu Ile Phe Pro Gly Ser Gly Asn Pro Phe Leu Thr Ile Arg Phe Lys
1 5 10 15
Gly
<210> 80
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 80
Glu Ile Phe Pro Gly Ser Gly Asn Ser His Val Val Ser Arg Phe Lys
1 5 10 15
Gly
<210> 81
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 81
Glu Ile Phe Pro Gly Ser Gly Asn Leu Ser Gly Ile Arg Ser Phe Lys
1 5 10 15
Gly
<210> 82
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 82
Glu Ile Phe Pro Gly Ser Gly Asn Phe Phe His Gly Lys Arg Phe Lys
1 5 10 15
Gly
<210> 83
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 83
Glu Ile Phe Pro Gly Ser Gly Asn Pro Arg Leu Gly Ala Arg Phe Lys
1 5 10 15
Gly
<210> 84
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 84
Glu Ile Phe Pro Gly Ser Gly Asn Val Ser Gln Val Glu Arg Phe Lys
1 5 10 15
Gly
<210> 85
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 85
Glu Ile Phe Pro Gly Ser Gly Asn Phe His Gly Ala Ser Arg Phe Lys
1 5 10 15
Gly
<210> 86
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 86
Glu Ile Phe Pro Gly Ser Gly Asn Val Val Gly Gly Tyr Arg Phe Lys
1 5 10 15
Gly
<210> 87
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 87
Glu Ile Phe Pro Gly Ser Gly Asn Pro Met Tyr Asp Glu Arg Phe Lys
1 5 10 15
Gly
<210> 88
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 88
Glu Ile Phe Pro Gly Ser Gly Asn Ala Asp Leu Thr Ile Arg Phe Lys
1 5 10 15
Gly
<210> 89
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 89
Glu Ile Phe Pro Gly Ser Gly Asn Ser Thr Asn Leu Tyr Arg Phe Lys
1 5 10 15
Gly
<210> 90
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 90
Glu Ile Phe Pro Gly Ser Gly Asn Leu Asp Ile Pro Pro Arg Phe Lys
1 5 10 15
Gly
<210> 91
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 91
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ser Ala Pro Leu
1 5 10 15
Pro
<210> 92
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 92
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ser Glu Phe Val
1 5 10 15
Ser
<210> 93
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 93
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Met Ser Glu Ser
1 5 10 15
Phe
<210> 94
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 94
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Asp Gly Ser Arg
1 5 10 15
Asn
<210> 95
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 95
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ser Ser Val Ser
1 5 10 15
Arg
<210> 96
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 96
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Arg Ser Val Ser
1 5 10 15
Gly
<210> 97
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 97
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Gly Leu Ser Glu
1 5 10 15
Val
<210> 98
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 98
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser His Tyr Trp Ala
1 5 10 15
Ser
<210> 99
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 99
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Thr Gly Leu Thr
1 5 10 15
Gln
<210> 100
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 100
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Arg His Arg Leu
1 5 10 15
His
<210> 101
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 101
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Val Pro Arg Ser
1 5 10 15
Met
<210> 102
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 102
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Leu Gln Asp Tyr
1 5 10 15
Leu
<210> 103
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 103
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Asp Gly Val Ser
1 5 10 15
Ser
<210> 104
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 104
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Met Gln Gly Ser
1 5 10 15
Glu
<210> 105
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 105
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Gly Asn Val His
1 5 10 15
Trp
<210> 106
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 106
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 107
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 107
Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Leu Arg Met Phe
1 5 10 15
Pro
<210> 108
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR1
<400> 108
Asp Tyr Tyr Ala Asn
1 5
<210> 109
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR1
<400> 109
Gly Tyr Tyr Ile Asn
1 5
<210> 110
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR1
<400> 110
Gln Tyr Tyr Ile Asn
1 5
<210> 111
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR1
<400> 111
Asp Gln Tyr Ile Asn
1 5
<210> 112
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR1
<400> 112
Asp Tyr Tyr Gln Asn
1 5
<210> 113
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 113
Gly Asp Val Gly Phe Leu Tyr
1 5
<210> 114
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 114
Gly Asp Tyr Gly Phe Gln Tyr
1 5
<210> 115
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 115
Gly Asp Tyr Gly Phe Leu Gln
1 5
<210> 116
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 116
Gly Asp Gln Trp Leu Leu Cys
1 5
<210> 117
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR3
<400> 117
Trp Asp Tyr Gly Phe Leu Tyr
1 5
<210> 118
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 118
Glu Ile Phe Pro Asp Ser Ala Pro Ser His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 119
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 119
Glu Ile Phe Pro Tyr Phe Leu Pro Pro His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 120
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 120
Glu Ile Phe Pro Gly Pro Phe Thr Pro His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 121
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 121
Glu Ile Phe Pro Gly Ser Asn Phe Gly His Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 122
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 122
Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 123
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 123
Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe Lys
1 5 10 15
Gly
<210> 124
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 124
Glu Ile Phe Pro Gly Tyr Gly Asn Thr Tyr Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 125
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 125
Glu Ile Phe Pro Gly Tyr Gly Asn Thr His Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 126
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 126
Glu Ile Phe Pro Gly Tyr Gly Asn Ser His Val Val Ser Arg Phe Lys
1 5 10 15
Gly
<210> 127
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 127
Glu Ile Phe Pro Gly Ser Gly Asn Thr Tyr Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 128
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 128
Glu Ile Phe Pro Gly Ser Gly Asn Ser His Val Val Arg Ser Pro Thr
1 5 10 15
Pro
<210> 129
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 129
Glu Ile Phe Pro Gly Ser Gly Asn Ser His Val Val Ser Ser Pro Thr
1 5 10 15
Pro
<210> 130
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 130
Glu Ile Phe Pro Asp Ser Ala Pro Ser Tyr Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 131
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 131
Glu Ile Phe Pro Gly Pro Phe Thr Pro Tyr Phe Ser Ala Arg Phe Lys
1 5 10 15
Gly
<210> 132
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 132
Glu Ile Phe Pro Gly Ser Asn Phe Gly Tyr Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 133
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 133
Glu Ile Phe Pro Asp Ser Ala Pro Ser His Val Val Ser Arg Phe Lys
1 5 10 15
Gly
<210> 134
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 134
Glu Ile Phe Pro Gly Pro Phe Thr Ser His Val Val Ser Arg Phe Lys
1 5 10 15
Gly
<210> 135
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 135
Glu Ile Phe Pro Gly Ser Asn Phe Ser His Val Val Ser Arg Phe Lys
1 5 10 15
Gly
<210> 136
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 136
Glu Ile Phe Pro Asp Ser Ala Pro Ser His Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 137
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 137
Glu Ile Phe Pro Gly Pro Phe Thr Pro His Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 138
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 138
Glu Ile Phe Pro Gly Ser Asn Phe Gly His Phe Ser Arg Ser Pro Thr
1 5 10 15
Pro
<210> 139
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 139
Glu Ile Phe Pro Asp Ser Ala Pro Ser His Val Val Ser Ser Pro Thr
1 5 10 15
Pro
<210> 140
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 140
Glu Ile Phe Pro Gly Pro Phe Thr Ser His Val Val Ser Ser Pro Thr
1 5 10 15
Pro
<210> 141
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CH CDR2
<400> 141
Glu Ile Phe Pro Gly Ser Asn Phe Ser His Val Val Ser Ser Pro Thr
1 5 10 15
Pro
<210> 142
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 142
Gln Asn Val Trp Ser Ser Pro Tyr Thr
1 5
<210> 143
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 143
Gln Asn Val Leu Asn Ser Pro Tyr Thr
1 5
<210> 144
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 144
Gln Asn Val Leu Glu Ser Pro Tyr Thr
1 5
<210> 145
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 145
Gln Asn Val Leu Lys Ser Pro Tyr Thr
1 5
<210> 146
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 146
Gln Asn Val Leu Tyr Ser Pro Tyr Thr
1 5
<210> 147
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 147
Gln Asn Val Leu Ser Arg Pro Tyr Thr
1 5
<210> 148
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 148
Gln Asn Val Leu Ser Ser Pro Glu Thr
1 5
<210> 149
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 149
Gln Asn Val Leu Ser Glu Pro Tyr Thr
1 5
<210> 150
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 150
Gln Asn Val Leu Glu Ser Pro Glu Thr
1 5
<210> 151
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 151
Gln Asn Val Leu Ser Val Pro Glu Thr
1 5
<210> 152
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 152
Gln Asn Val Leu Ser Leu Pro Glu Thr
1 5
<210> 153
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 153
Gln Asn Val Leu Ser Ile Pro Glu Thr
1 5
<210> 154
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 154
Gln Asn Val Leu Ser Met Pro Glu Thr
1 5
<210> 155
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 155
Gln Asn Ile Leu Ser Ser Pro Glu Thr
1 5
<210> 156
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 156
Gln Asn Leu Ile Ser Ser Pro Glu Thr
1 5
<210> 157
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 157
Gln Asn Met Ile Ser Ser Pro Glu Thr
1 5
<210> 158
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 158
Gln Asn Ile Ile Ser Leu Pro Glu Thr
1 5
<210> 159
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 159
Gln Asn Ile Ile Ser Ile Pro Glu Thr
1 5
<210> 160
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 160
Gln Asn Ser Leu Ser Ser Pro Glu Thr
1 5
<210> 161
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 161
Gln Asn Thr Leu Ser Ser Pro Glu Thr
1 5
<210> 162
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 162
Gln Asn Val Ser Ser Ser Pro Glu Thr
1 5
<210> 163
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 163
Gln Asn Val Ile Ser Ser Pro Glu Thr
1 5
<210> 164
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 164
Gln Asn Val Phe Ser Ser Pro Glu Thr
1 5
<210> 165
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 165
Gln Asn Val Tyr Ser Ser Pro Glu Thr
1 5
<210> 166
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 166
Gln Asn Val Arg Ser Ser Pro Glu Thr
1 5
<210> 167
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 167
Gln Asn Leu Val Ser Ser Pro Glu Thr
1 5
<210> 168
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 168
Gln Asn Leu Met Ser Ser Pro Glu Thr
1 5
<210> 169
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 169
Gln Asn Ile Met Ser Ser Pro Glu Thr
1 5
<210> 170
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 170
Gln Asn Val His Ser Ser Pro Glu Thr
1 5
<210> 171
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 171
Gln Asn Val Met Ser Ser Pro Glu Thr
1 5
<210> 172
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 172
Gln Asn Leu Leu Ser Ser Pro Glu Thr
1 5
<210> 173
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 173
Gln Ser Val Leu Phe Ser Pro Phe Ser
1 5
<210> 174
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 174
Gln Gln Val Leu Phe Phe Pro Glu Thr
1 5
<210> 175
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 175
Gln Asn Leu Leu Ser Pro Ser Phe Tyr
1 5
<210> 176
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 176
Gln Ser Val Leu Phe Ser Pro Phe Thr
1 5
<210> 177
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 177
Gln Asn Ile Leu Ser Ser Pro Leu Phe
1 5
<210> 178
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 178
Gln Asn Thr Leu His Tyr Ser Leu Val
1 5
<210> 179
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 179
Gln Gln Val Leu Phe Phe Pro Leu Leu
1 5
<210> 180
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 180
Gln Gln Val Leu Asp Phe Val Phe Tyr
1 5
<210> 181
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 181
Gln Asn Val Val Ser Ser Pro Glu Thr
1 5
<210> 182
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 182
Asp Ala Thr Asn Leu Ala Asp
1 5
<210> 183
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 183
Phe Ala Thr Asn Leu Ala Asp
1 5
<210> 184
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 184
His Ala Thr Asn Leu Ala Asp
1 5
<210> 185
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 185
Lys Ala Thr Asn Leu Ala Asp
1 5
<210> 186
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 186
Pro Ala Thr Asn Leu Ala Asp
1 5
<210> 187
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 187
Gln Ala Thr Asn Leu Ala Asp
1 5
<210> 188
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 188
Ser Ala Thr Asn Leu Ala Asp
1 5
<210> 189
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 189
Val Ala Thr Asn Leu Ala Asp
1 5
<210> 190
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 190
Tyr Ala Thr Asn Leu Ala Asp
1 5
<210> 191
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 191
Ile Thr Val Leu Ser Pro Pro Tyr Thr
1 5
<210> 192
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 192
Gln Asn Asn Leu Val Pro Pro Phe Asn
1 5
<210> 193
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 193
Gln His Val Leu Phe Leu Pro Tyr Val
1 5
<210> 194
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 194
Gln Ala Val Leu Thr Asn Ala Tyr Thr
1 5
<210> 195
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 195
Gln Asn Val Leu Arg Val Gly Tyr Leu
1 5
<210> 196
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 196
Gln Ser Val Leu Arg Val Gly Tyr Leu
1 5
<210> 197
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 197
Gln Asn Ile Ile Ser Ser Pro Tyr Thr
1 5
<210> 198
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 198
Gln Gln Val Leu Cys Glu Ser Phe Leu
1 5
<210> 199
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 199
Gln Asn Val Leu Ser Gln Ser Leu Leu
1 5
<210> 200
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 200
Gln Asn Val Leu Gln Pro Ser Tyr Leu
1 5
<210> 201
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 201
Gln Asn Leu Leu Phe Gln Pro Leu Ser
1 5
<210> 202
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 202
Gln Asn Val Leu Phe Gln Pro Leu Val
1 5
<210> 203
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 203
Gln Asn Gln Leu Asp Pro Ser Leu Phe
1 5
<210> 204
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 204
Met Asp Val Leu Glu Ser Pro Tyr Thr
1 5
<210> 205
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 205
Gln Ala Leu Leu Leu Ser Pro Tyr Thr
1 5
<210> 206
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 206
Gln Gln Leu Leu Glu Ser Pro Tyr Thr
1 5
<210> 207
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 207
Asn Leu Thr Leu Val Ser Pro Tyr Thr
1 5
<210> 208
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 208
Gly Asn Ile Leu Asp Ser Pro Tyr Thr
1 5
<210> 209
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 209
Glu Gln Val Leu Leu Ser Pro Tyr Thr
1 5
<210> 210
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 210
Asn Asn Leu Leu Asp Ser Pro Tyr Thr
1 5
<210> 211
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 211
Glu Glu Val Leu Ser Ser Pro Tyr Thr
1 5
<210> 212
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 212
Gln Asn Ile Leu Phe Val Asp Tyr Thr
1 5
<210> 213
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 213
Gln Asn Val Leu His Leu Asn Tyr Thr
1 5
<210> 214
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 214
Gln Asn Val Leu Gln Thr Pro Tyr Thr
1 5
<210> 215
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 215
Gln Asn Ile Leu His Pro Gly Tyr Thr
1 5
<210> 216
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 216
Gln Asn Val Leu Thr Arg Gly Tyr Thr
1 5
<210> 217
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 217
Glu Asn Ile Leu Tyr Ser Pro Tyr Thr
1 5
<210> 218
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 218
Gln Asn Val Leu Gly Gly Gly Gln Gly
1 5
<210> 219
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 219
Gln Asn Val Leu Glu His Pro Leu Ile
1 5
<210> 220
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 220
Gln Asn Val Leu Asp Asp Pro Phe Asp
1 5
<210> 221
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 221
Gln Asn Val Leu Asp Phe Pro Leu Leu
1 5
<210> 222
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 222
Gln Asn Val Leu Tyr Pro Ser Leu Val
1 5
<210> 223
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 223
Gln Asn Val Leu Phe Asp Gln Gln Ser
1 5
<210> 224
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 224
Gln Asn Val Leu Ser Asn Glu Glu Thr
1 5
<210> 225
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 225
Gln Asn Val Leu Lys His Pro Tyr Thr
1 5
<210> 226
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 226
Gln Asn Val Leu Ser Pro Gly Met Trp
1 5
<210> 227
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 227
Gly Ala Thr Gly Leu Ala Asp
1 5
<210> 228
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR2
<400> 228
Gly Ala Gln Asn Leu Ala Asp
1 5
<210> 229
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 229
Gly Ser Ser Arg Ser Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 230
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 230
Arg Ala Gly Arg Ser Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 231
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 231
Leu Gly Arg Arg Gly Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 232
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 232
Glu Val Gln Val Gly Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 233
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 233
Arg Pro Ser Glu Lys Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 234
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 234
Arg Ala Ser Ala Val Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 235
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 235
Lys Thr Gly Asp Leu Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 236
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 236
Ser Cys Arg Val Pro Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 237
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 237
Val Ala Ser Arg Gly Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 238
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 238
Arg Gly Arg Gln Asn Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 239
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 239
Ala Ala Pro Arg Gly Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 240
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 240
Ser Ala Pro Phe Lys Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 241
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 241
Leu Gly Met Asp Asp Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 242
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 242
Asn Val Arg Arg Gly Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 243
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 243
Asn Thr Ser Gly Arg Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 244
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 244
Leu Val Ser Arg Pro Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 245
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 245
Trp Thr Asn Arg Pro Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 246
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 246
Arg Ile Pro Ser Ala Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 247
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 247
Gly Ala Thr Arg Gly Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 248
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 248
Glu Gly Gly Ser Pro Ile Tyr Gly Ala Leu Asn
1 5 10
<210> 249
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 249
Gly Ala Ser Arg Gly Met Phe Arg Ala Leu Asn
1 5 10
<210> 250
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 250
Gly Ala Ser Gly Leu Val Phe Ser Ala Leu Asn
1 5 10
<210> 251
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 251
Gly Ala Ser Arg Gly Thr His Met Ala Leu Asn
1 5 10
<210> 252
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 252
Gly Ala Ser Ser Arg Phe His Asn Ala Leu Asn
1 5 10
<210> 253
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 253
Gly Ala Ser Arg Thr Ala Phe Thr Ala Leu Asn
1 5 10
<210> 254
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 254
Gly Ala Ser Arg Ser Thr Phe Ser Ala Leu Asn
1 5 10
<210> 255
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 255
Gly Ala Ser Gly Pro Met Phe Asp Ala Leu Asn
1 5 10
<210> 256
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 256
Gly Ala Ser His Asp Leu Tyr Gly Ala Leu Asn
1 5 10
<210> 257
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 257
Gly Ala Ser Gly Thr Leu Phe Gly Ala Leu Asn
1 5 10
<210> 258
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 258
Gly Ala Ser Lys Ala Ala Phe Gly Ala Leu Asn
1 5 10
<210> 259
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 259
Gly Ala Ser Glu Gly Ile Val Gly Ala Leu Asn
1 5 10
<210> 260
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 260
Gly Ala Ser His Glu Ile His Val Ala Leu Asn
1 5 10
<210> 261
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 261
Gly Ala Ser Arg Gly Val Phe Gly Ala Leu Asn
1 5 10
<210> 262
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 262
Gly Ala Ser Gly Arg Val Arg Gly Ala Leu Asn
1 5 10
<210> 263
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 263
Gly Ala Ser Thr Gly Ser Phe Ser Ala Leu Asn
1 5 10
<210> 264
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 264
Gly Ala Ser Gly Asn Ser Phe Asp Ala Leu Asn
1 5 10
<210> 265
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 265
Gly Ala Ser Glu Gln Ser Tyr Phe Ala Leu Asn
1 5 10
<210> 266
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 266
Gly Ala Ser Phe Arg Gln Phe Ser Ala Leu Asn
1 5 10
<210> 267
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 267
Gly Ala Ser Ala Pro Arg His Ser Ala Leu Asn
1 5 10
<210> 268
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR1
<400> 268
Gly Ala Ser Met Pro Leu Phe His Ala Leu Asn
1 5 10
<210> 269
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 269
Gln Asn Ile Leu Ser Ser Pro Tyr Thr
1 5
<210> 270
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 270
Gln Asn Val Leu Ser Met Pro Tyr Thr
1 5
<210> 271
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 271
Gln Asn Val Leu Ser Glu Pro Glu Thr
1 5
<210> 272
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 272
Gln Asn Val Leu Tyr Ser Pro Glu Thr
1 5
<210> 273
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 273
Gln Asn Val Leu Glu Glu Pro Tyr Thr
1 5
<210> 274
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 274
Gln Asn Val Leu Glu Leu Pro Glu Thr
1 5
<210> 275
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 275
Gln Asn Val Leu Glu Met Pro Glu Thr
1 5
<210> 276
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 276
Gln Asn Ile Leu Glu Ser Pro Glu Thr
1 5
<210> 277
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 277
Gln Asn Val Ile Glu Ser Pro Glu Thr
1 5
<210> 278
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 278
Gln Asn Val Met Glu Ser Pro Glu Thr
1 5
<210> 279
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 279
Gln Asn Leu Leu Glu Ser Pro Glu Thr
1 5
<210> 280
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 280
Gln Asn Val Leu Tyr Glu Pro Tyr Thr
1 5
<210> 281
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 281
Gln Asn Ile Leu Ser Glu Pro Glu Thr
1 5
<210> 282
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 282
Gln Asn Val Ile Ser Glu Pro Glu Thr
1 5
<210> 283
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 283
Gln Asn Val Met Ser Glu Pro Glu Thr
1 5
<210> 284
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 284
Gln Asn Leu Leu Ser Glu Pro Glu Thr
1 5
<210> 285
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 285
Gln Ser Val Leu Phe Glu Pro Phe Ser
1 5
<210> 286
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 286
Gln Ser Val Leu Phe Glu Pro Phe Thr
1 5
<210> 287
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 287
Gln Asn Ile Leu Tyr Ser Pro Glu Thr
1 5
<210> 288
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 288
Gln Asn Ile Leu Ser Leu Pro Glu Thr
1 5
<210> 289
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 289
Gln Asn Ile Leu Ser Met Pro Glu Thr
1 5
<210> 290
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 290
Gln Asn Val Leu Tyr Met Pro Glu Thr
1 5
<210> 291
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 291
Gln Asn Val Ile Ser Met Pro Glu Thr
1 5
<210> 292
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 292
Gln Asn Val Met Ser Met Pro Glu Thr
1 5
<210> 293
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 293
Gln Asn Leu Leu Ser Met Pro Glu Thr
1 5
<210> 294
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 294
Gln Asn Ile Ile Ser Ser Pro Glu Thr
1 5
<210> 295
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 295
Gln Asn Val Leu Tyr Leu Pro Glu Thr
1 5
<210> 296
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 296
Gln Asn Val Ile Tyr Ser Pro Glu Thr
1 5
<210> 297
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 297
Gln Asn Val Met Tyr Ser Pro Glu Thr
1 5
<210> 298
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 298
Gln Asn Leu Leu Tyr Ser Pro Glu Thr
1 5
<210> 299
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 299
Gln Asn Val Ile Ser Leu Pro Glu Thr
1 5
<210> 300
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 300
Gln Asn Val Met Ser Leu Pro Glu Thr
1 5
<210> 301
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 CL CDR3
<400> 301
Gln Asn Leu Leu Ser Leu Pro Glu Thr
1 5
<210> 302
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> AH71
<400> 302
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 303
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> AH72
<400> 303
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Arg Ser Pro
50 55 60
Thr Pro Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 304
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> AH73
<400> 304
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 305
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> AH85
<400> 305
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 306
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AL130
<400> 306
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Met Pro Leu Phe His Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 307
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AL135
<400> 307
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Glu Asn Ile Tyr Gly Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Glu Glu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 308
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AL165
<400> 308
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Glu Asn Ile Tyr Gly Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Gly Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Glu Ser Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 309
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AL166
<400> 309
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Glu Asn Ile Tyr Gly Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Gly Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Glu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 310
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AL194
<400> 310
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Met Pro Leu Phe His Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Ser Ile Pro Glu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 311
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AL195
<400> 311
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gly Ala Ser Arg Ser Thr Phe Ser Ala
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Thr Asn Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Asn Val Leu Glu Glu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 312
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер
<400> 312
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 313
<211> 246
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эрбитукс scFv HL
<400> 313
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Cys Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Leu Leu Thr
130 135 140
Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly Glu Arg Val Ser Phe
145 150 155 160
Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn Ile His Trp Tyr Gln
165 170 175
Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile Lys Tyr Ala Ser Glu
180 185 190
Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser Glu Asp Ile Ala Asp
210 215 220
Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr Thr Phe Gly Cys Gly
225 230 235 240
Thr Lys Leu Glu Leu Lys
245
<210> 314
<211> 246
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эрбитукс scFv LH
<400> 314
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
115 120 125
Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln Ser
130 135 140
Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr Gly
145 150 155 160
Val His Trp Val Arg Gln Ser Pro Gly Lys Cys Leu Glu Trp Leu Gly
165 170 175
Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr Ser
180 185 190
Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe Lys
195 200 205
Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala Arg
210 215 220
Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ala
245
<210> 315
<211> 246
<212> Белок
<213> Искусственная последовательность
<220>
<223> Вектибикс scFv
<400> 315
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro Gly Lys Cys Leu Glu
35 40 45
Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr Ser Lys Thr Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr
130 135 140
Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
145 150 155 160
Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn Trp Tyr Gln
165 170 175
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp Ala Ser Asn
180 185 190
Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu Asp Ile Ala Thr
210 215 220
Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu Ala Phe Gly Cys Gly
225 230 235 240
Thr Lys Val Glu Ile Lys
245
<210> 316
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 x Эрбитукс scFv HL
<400> 316
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu
450 455 460
Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe
465 470 475 480
Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys
485 490 495
Cys Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr
500 505 510
Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys
515 520 525
Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala
530 535 540
Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala
545 550 555 560
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
595 600 605
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
610 615 620
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
625 630 635 640
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
660 665 670
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
675 680 685
Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
690 695
<210> 317
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH71 x Эрбитукс scFv HL
<400> 317
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu
450 455 460
Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe
465 470 475 480
Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys
485 490 495
Cys Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr
500 505 510
Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys
515 520 525
Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala
530 535 540
Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala
545 550 555 560
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
595 600 605
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
610 615 620
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
625 630 635 640
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
660 665 670
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
675 680 685
Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
690 695
<210> 318
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH72 x Эрбитукс scFv HL
<400> 318
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Arg Ser Pro
50 55 60
Thr Pro Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu
450 455 460
Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe
465 470 475 480
Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys
485 490 495
Cys Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr
500 505 510
Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys
515 520 525
Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala
530 535 540
Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala
545 550 555 560
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
595 600 605
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
610 615 620
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
625 630 635 640
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
660 665 670
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
675 680 685
Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
690 695
<210> 319
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH73 x Эрбитукс scFv HL
<400> 319
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu
450 455 460
Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe
465 470 475 480
Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys
485 490 495
Cys Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr
500 505 510
Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys
515 520 525
Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala
530 535 540
Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala
545 550 555 560
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
595 600 605
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
610 615 620
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
625 630 635 640
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
660 665 670
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
675 680 685
Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
690 695
<210> 320
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH85 x Эрбитукс scFv HL
<400> 320
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu
450 455 460
Val Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe
465 470 475 480
Ser Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys
485 490 495
Cys Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr
500 505 510
Asn Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys
515 520 525
Ser Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala
530 535 540
Ile Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala
545 550 555 560
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
595 600 605
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
610 615 620
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
625 630 635 640
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
660 665 670
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
675 680 685
Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
690 695
<210> 321
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 x Эрбитукс scFv LH
<400> 321
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu
450 455 460
Ser Val Ser Pro Gly Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln
465 470 475 480
Ser Ile Gly Thr Asn Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser
485 490 495
Pro Arg Leu Leu Ile Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro
500 505 510
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile
515 520 525
Asn Ser Val Glu Ser Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn
530 535 540
Asn Asn Trp Pro Thr Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
545 550 555 560
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
565 570 575
Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val
580 585 590
Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser
595 600 605
Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Cys
610 615 620
Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn
625 630 635 640
Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser
645 650 655
Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile
660 665 670
Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr
675 680 685
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
690 695
<210> 322
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH71 x Эрбитукс scFv LH
<400> 322
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu
450 455 460
Ser Val Ser Pro Gly Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln
465 470 475 480
Ser Ile Gly Thr Asn Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser
485 490 495
Pro Arg Leu Leu Ile Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro
500 505 510
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile
515 520 525
Asn Ser Val Glu Ser Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn
530 535 540
Asn Asn Trp Pro Thr Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
545 550 555 560
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
565 570 575
Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val
580 585 590
Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser
595 600 605
Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Cys
610 615 620
Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn
625 630 635 640
Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser
645 650 655
Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile
660 665 670
Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr
675 680 685
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
690 695
<210> 323
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH72 x Эрбитукс scFv LH
<400> 323
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Arg Ser Pro
50 55 60
Thr Pro Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu
450 455 460
Ser Val Ser Pro Gly Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln
465 470 475 480
Ser Ile Gly Thr Asn Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser
485 490 495
Pro Arg Leu Leu Ile Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro
500 505 510
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile
515 520 525
Asn Ser Val Glu Ser Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn
530 535 540
Asn Asn Trp Pro Thr Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
545 550 555 560
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
565 570 575
Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val
580 585 590
Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser
595 600 605
Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Cys
610 615 620
Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn
625 630 635 640
Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser
645 650 655
Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile
660 665 670
Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr
675 680 685
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
690 695
<210> 324
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH73 x Эрбитукс scFv LH
<400> 324
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu
450 455 460
Ser Val Ser Pro Gly Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln
465 470 475 480
Ser Ile Gly Thr Asn Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser
485 490 495
Pro Arg Leu Leu Ile Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro
500 505 510
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile
515 520 525
Asn Ser Val Glu Ser Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn
530 535 540
Asn Asn Trp Pro Thr Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
545 550 555 560
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
565 570 575
Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val
580 585 590
Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser
595 600 605
Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Cys
610 615 620
Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn
625 630 635 640
Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser
645 650 655
Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile
660 665 670
Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr
675 680 685
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
690 695
<210> 325
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH85 x Эрбитукс scFv LH
<400> 325
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu
450 455 460
Ser Val Ser Pro Gly Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln
465 470 475 480
Ser Ile Gly Thr Asn Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser
485 490 495
Pro Arg Leu Leu Ile Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro
500 505 510
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile
515 520 525
Asn Ser Val Glu Ser Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn
530 535 540
Asn Asn Trp Pro Thr Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys
545 550 555 560
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
565 570 575
Gly Gly Gly Ser Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val
580 585 590
Gln Pro Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser
595 600 605
Leu Thr Asn Tyr Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Cys
610 615 620
Leu Glu Trp Leu Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn
625 630 635 640
Thr Pro Phe Thr Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser
645 650 655
Gln Val Phe Phe Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile
660 665 670
Tyr Tyr Cys Ala Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr
675 680 685
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala
690 695
<210> 326
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 x Вектибикс scFv
<400> 326
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Ser Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu
450 455 460
Val Lys Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly
465 470 475 480
Ser Val Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro
485 490 495
Gly Lys Cys Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr
500 505 510
Asn Tyr Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr
515 520 525
Ser Lys Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp
530 535 540
Thr Ala Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp
545 550 555 560
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
595 600 605
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
610 615 620
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
625 630 635 640
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
660 665 670
Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu
675 680 685
Ala Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
690 695
<210> 327
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH71 x Вектибикс scFv
<400> 327
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Ala Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu
450 455 460
Val Lys Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly
465 470 475 480
Ser Val Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro
485 490 495
Gly Lys Cys Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr
500 505 510
Asn Tyr Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr
515 520 525
Ser Lys Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp
530 535 540
Thr Ala Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp
545 550 555 560
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
595 600 605
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
610 615 620
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
625 630 635 640
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
660 665 670
Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu
675 680 685
Ala Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
690 695
<210> 328
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH72 x Вектибикс scFv
<400> 328
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Thr His Phe Ser Arg Ser Pro
50 55 60
Thr Pro Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu
450 455 460
Val Lys Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly
465 470 475 480
Ser Val Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro
485 490 495
Gly Lys Cys Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr
500 505 510
Asn Tyr Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr
515 520 525
Ser Lys Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp
530 535 540
Thr Ala Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp
545 550 555 560
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
595 600 605
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
610 615 620
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
625 630 635 640
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
660 665 670
Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu
675 680 685
Ala Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
690 695
<210> 329
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH73 x Вектибикс scFv
<400> 329
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu
450 455 460
Val Lys Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly
465 470 475 480
Ser Val Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro
485 490 495
Gly Lys Cys Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr
500 505 510
Asn Tyr Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr
515 520 525
Ser Lys Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp
530 535 540
Thr Ala Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp
545 550 555 560
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
595 600 605
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
610 615 620
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
625 630 635 640
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
660 665 670
Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu
675 680 685
Ala Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
690 695
<210> 330
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> hu8C4 AH85 x Вектибикс scFv
<400> 330
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Ser Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Phe Pro Gly Trp Gly Asn Ser His Val Val Ser Arg Phe
50 55 60
Lys Gly Arg Ala Thr Leu Ser Ala Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gln Asp Tyr Gly Phe Leu Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu
450 455 460
Val Lys Pro Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly
465 470 475 480
Ser Val Ser Ser Gly Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro
485 490 495
Gly Lys Cys Leu Glu Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr
500 505 510
Asn Tyr Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr
515 520 525
Ser Lys Thr Gln Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp
530 535 540
Thr Ala Ile Tyr Tyr Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp
545 550 555 560
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Gly Gly
565 570 575
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
580 585 590
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
595 600 605
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
610 615 620
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
625 630 635 640
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
645 650 655
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
660 665 670
Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu
675 680 685
Ala Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
690 695
<210> 331
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эпитопная область Y321~L329
<400> 331
Tyr Val Ser Lys Pro Gly Ala Gln Leu
1 5
<210> 332
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эпитопная область I333~I341
<400> 332
Ile Gly Ala Ser Leu Asn Asp Asp Ile
1 5
<210> 333
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эпитопная область P366~D372
<400> 333
Pro Ile Lys Tyr Val Asn Asp
1 5
<210> 334
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Эпитопная область Q464~S474
<400> 334
Gln Val Val Val Ser Arg Ser Gly Pro Ser Thr
1 5 10
<210> 335
<211> 1390
<212> Белок
<213> Искусственная последовательность
<220>
<223> эталонная последовательность c-Met
<400> 335
Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu Leu Phe
1 5 10 15
Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala Leu Ala Lys
20 25 30
Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro Asn Phe Thr Ala
35 40 45
Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu His His Ile Phe Leu
50 55 60
Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn Glu Glu Asp Leu Gln Lys
65 70 75 80
Val Ala Glu Tyr Lys Thr Gly Pro Val Leu Glu His Pro Asp Cys Phe
85 90 95
Pro Cys Gln Asp Cys Ser Ser Lys Ala Asn Leu Ser Gly Gly Val Trp
100 105 110
Lys Asp Asn Ile Asn Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp
115 120 125
Gln Leu Ile Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His
130 135 140
Val Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys
145 150 155 160
Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys Val
165 170 175
Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys Asp Arg Phe
180 185 190
Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser Ser Tyr Phe Pro Asp
195 200 205
His Pro Leu His Ser Ile Ser Val Arg Arg Leu Lys Glu Thr Lys Asp
210 215 220
Gly Phe Met Phe Leu Thr Asp Gln Ser Tyr Ile Asp Val Leu Pro Glu
225 230 235 240
Phe Arg Asp Ser Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn
245 250 255
Asn Phe Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln
260 265 270
Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285
His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu Lys Arg
290 295 300
Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile Leu Gln Ala Ala
305 310 315 320
Tyr Val Ser Lys Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly Ala Ser
325 330 335
Leu Asn Asp Asp Ile Leu Phe Gly Val Phe Ala Gln Ser Lys Pro Asp
340 345 350
Ser Ala Glu Pro Met Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys
355 360 365
Tyr Val Asn Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg
370 375 380
Cys Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg
385 390 395 400
Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu Tyr
405 410 415
Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu Phe Met Gly
420 425 430
Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser Thr Phe Ile Lys Gly
435 440 445
Asp Leu Thr Ile Ala Asn Leu Gly Thr Ser Glu Gly Arg Phe Met Gln
450 455 460
Val Val Val Ser Arg Ser Gly Pro Ser Thr Pro His Val Asn Phe Leu
465 470 475 480
Leu Asp Ser His Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu
485 490 495
Asn Gln Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys
500 505 510
Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525
Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys
530 535 540
Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile
545 550 555 560
Cys Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu
565 570 575
Gly Gly Thr Arg Leu Thr Ile Cys Gly Trp Asp Phe Gly Phe Arg Arg
580 585 590
Asn Asn Lys Phe Asp Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu
595 600 605
Ser Cys Thr Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys
610 615 620
Thr Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile
625 630 635 640
Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val Asp
645 650 655
Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met Ala Gly Gly
660 665 670
Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn Ser Gly Asn Ser Arg
675 680 685
His Ile Ser Ile Gly Gly Lys Thr Cys Thr Leu Lys Ser Val Ser Asn
690 695 700
Ser Ile Leu Glu Cys Tyr Thr Pro Ala Gln Thr Ile Ser Thr Glu Phe
705 710 715 720
Ala Val Lys Leu Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe
725 730 735
Ser Tyr Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser
740 745 750
Phe Ile Ser Gly Gly Ser Thr Ile Thr Gly Val Gly Lys Asn Leu Asn
755 760 765
Ser Val Ser Val Pro Arg Met Val Ile Asn Val His Glu Ala Gly Arg
770 775 780
Asn Phe Thr Val Ala Cys Gln His Arg Ser Asn Ser Glu Ile Ile Cys
785 790 795 800
Cys Thr Thr Pro Ser Leu Gln Gln Leu Asn Leu Gln Leu Pro Leu Lys
805 810 815
Thr Lys Ala Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr Phe Asp
820 825 830
Leu Ile Tyr Val His Asn Pro Val Phe Lys Pro Phe Glu Lys Pro Val
835 840 845
Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly Asn Asp
850 855 860
Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val Gly Asn Lys
865 870 875 880
Ser Cys Glu Asn Ile His Leu His Ser Glu Ala Val Leu Cys Thr Val
885 890 895
Pro Asn Asp Leu Leu Lys Leu Asn Ser Glu Leu Asn Ile Glu Trp Lys
900 905 910
Gln Ala Ile Ser Ser Thr Val Leu Gly Lys Val Ile Val Gln Pro Asp
915 920 925
Gln Asn Phe Thr Gly Leu Ile Ala Gly Val Val Ser Ile Ser Thr Ala
930 935 940
Leu Leu Leu Leu Leu Gly Phe Phe Leu Trp Leu Lys Lys Arg Lys Gln
945 950 955 960
Ile Lys Asp Leu Gly Ser Glu Leu Val Arg Tyr Asp Ala Arg Val His
965 970 975
Thr Pro His Leu Asp Arg Leu Val Ser Ala Arg Ser Val Ser Pro Thr
980 985 990
Thr Glu Met Val Ser Asn Glu Ser Val Asp Tyr Arg Ala Thr Phe Pro
995 1000 1005
Glu Asp Gln Phe Pro Asn Ser Ser Gln Asn Gly Ser Cys Arg Gln Val
1010 1015 1020
Gln Tyr Pro Leu Thr Asp Met Ser Pro Ile Leu Thr Ser Gly Asp Ser
1025 1030 1035 1040
Asp Ile Ser Ser Pro Leu Leu Gln Asn Thr Val His Ile Asp Leu Ser
1045 1050 1055
Ala Leu Asn Pro Glu Leu Val Gln Ala Val Gln His Val Val Ile Gly
1060 1065 1070
Pro Ser Ser Leu Ile Val His Phe Asn Glu Val Ile Gly Arg Gly His
1075 1080 1085
Phe Gly Cys Val Tyr His Gly Thr Leu Leu Asp Asn Asp Gly Lys Lys
1090 1095 1100
Ile His Cys Ala Val Lys Ser Leu Asn Arg Ile Thr Asp Ile Gly Glu
1105 1110 1115 1120
Val Ser Gln Phe Leu Thr Glu Gly Ile Ile Met Lys Asp Phe Ser His
1125 1130 1135
Pro Asn Val Leu Ser Leu Leu Gly Ile Cys Leu Arg Ser Glu Gly Ser
1140 1145 1150
Pro Leu Val Val Leu Pro Tyr Met Lys His Gly Asp Leu Arg Asn Phe
1155 1160 1165
Ile Arg Asn Glu Thr His Asn Pro Thr Val Lys Asp Leu Ile Gly Phe
1170 1175 1180
Gly Leu Gln Val Ala Lys Gly Met Lys Tyr Leu Ala Ser Lys Lys Phe
1185 1190 1195 1200
Val His Arg Asp Leu Ala Ala Arg Asn Cys Met Leu Asp Glu Lys Phe
1205 1210 1215
Thr Val Lys Val Ala Asp Phe Gly Leu Ala Arg Asp Met Tyr Asp Lys
1220 1225 1230
Glu Tyr Tyr Ser Val His Asn Lys Thr Gly Ala Lys Leu Pro Val Lys
1235 1240 1245
Trp Met Ala Leu Glu Ser Leu Gln Thr Gln Lys Phe Thr Thr Lys Ser
1250 1255 1260
Asp Val Trp Ser Phe Gly Val Leu Leu Trp Glu Leu Met Thr Arg Gly
1265 1270 1275 1280
Ala Pro Pro Tyr Pro Asp Val Asn Thr Phe Asp Ile Thr Val Tyr Leu
1285 1290 1295
Leu Gln Gly Arg Arg Leu Leu Gln Pro Glu Tyr Cys Pro Asp Pro Leu
1300 1305 1310
Tyr Glu Val Met Leu Lys Cys Trp His Pro Lys Ala Glu Met Arg Pro
1315 1320 1325
Ser Phe Ser Glu Leu Val Ser Arg Ile Ser Ala Ile Phe Ser Thr Phe
1330 1335 1340
Ile Gly Glu His Tyr Val His Val Asn Ala Thr Tyr Val Asn Val Lys
1345 1350 1355 1360
Cys Val Ala Pro Tyr Pro Ser Leu Leu Ser Ser Glu Asp Asn Ala Asp
1365 1370 1375
Asp Glu Val Asp Thr Arg Pro Ala Ser Phe Trp Glu Thr Ser
1380 1385 1390
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ МОДИФИЦИРОВАННЫЙ СЛИТЫЙ БЕЛОК FC-ФРАГМЕНТА ИММУНОГЛОБУЛИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800919C2 |
| АНТИ-CEACAM1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2754876C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| СЛИТЫЙ БЕЛОК GDF15 ДЛИТЕЛЬНОГО ДЕЙСТВИЯ И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2836280C1 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| АНТИТЕЛА ПРОТИВ ЛИГАНДА 1 ПРОГРАММИРУЕМОЙ ГИБЕЛИ И ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2786235C1 |
| ПОЛИПЕПТИД, СПЕЦИФИЧНЫЙ К МУЦИНУ-1, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2805840C1 |
| МОДИФИЦИРОВАННЫЕ МИТОХОНДРИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2811467C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
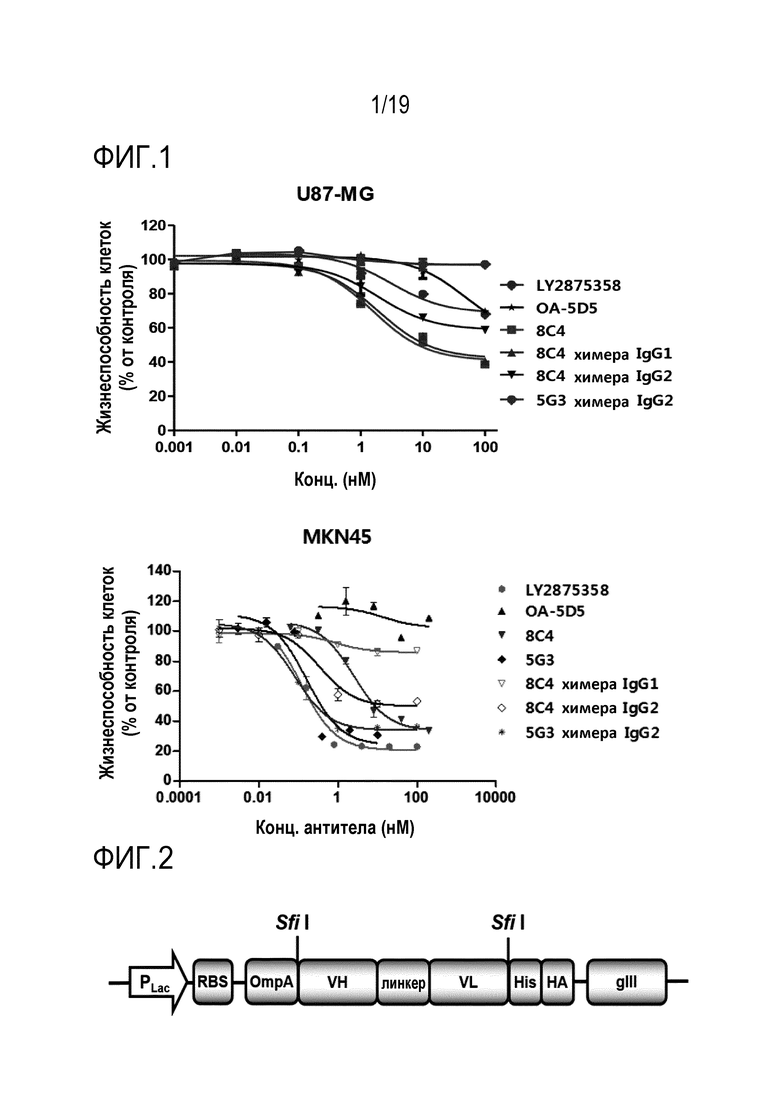
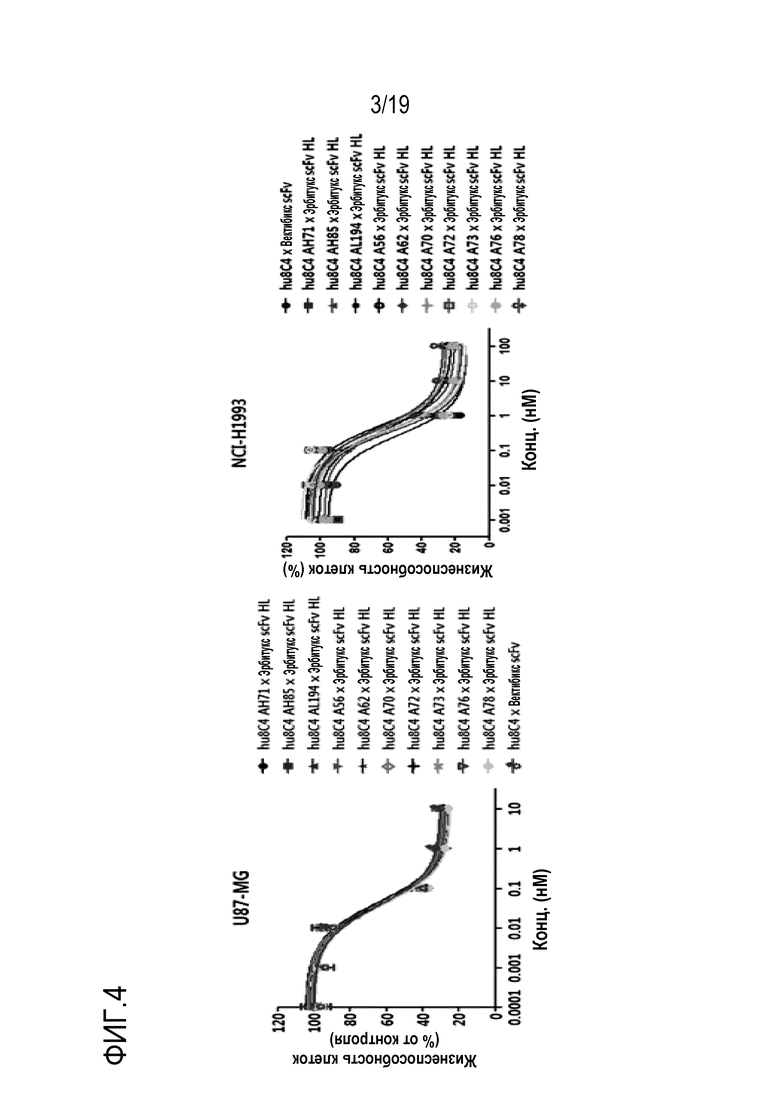
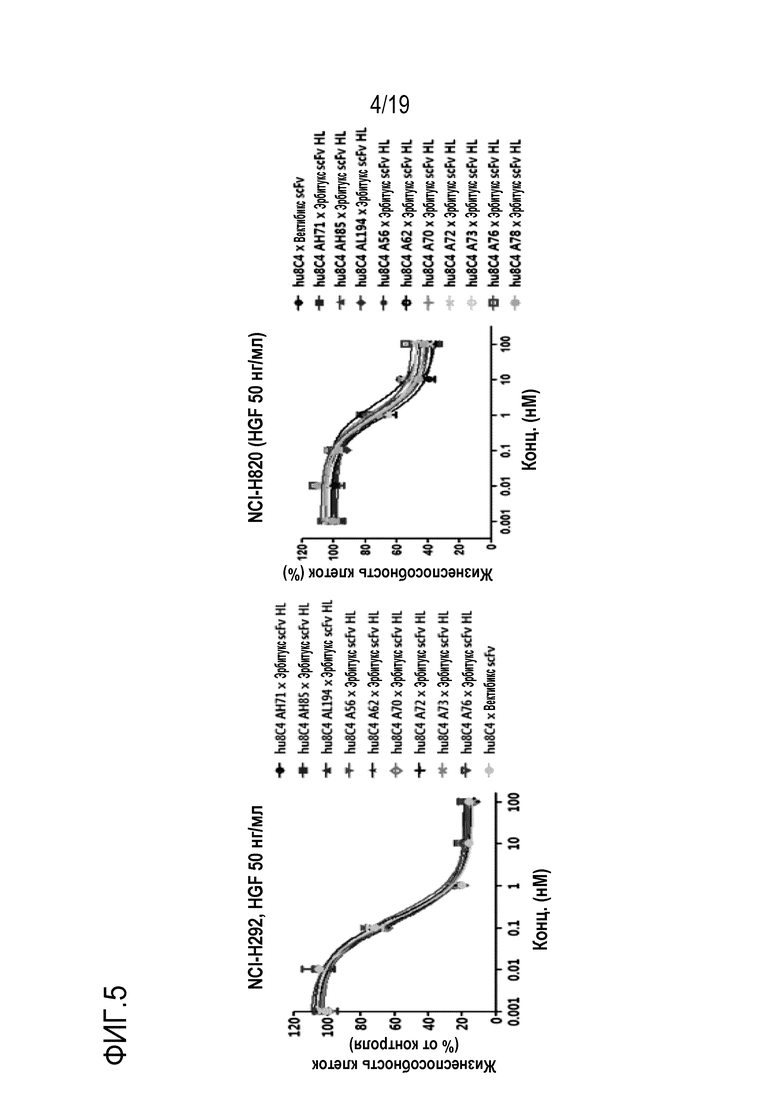
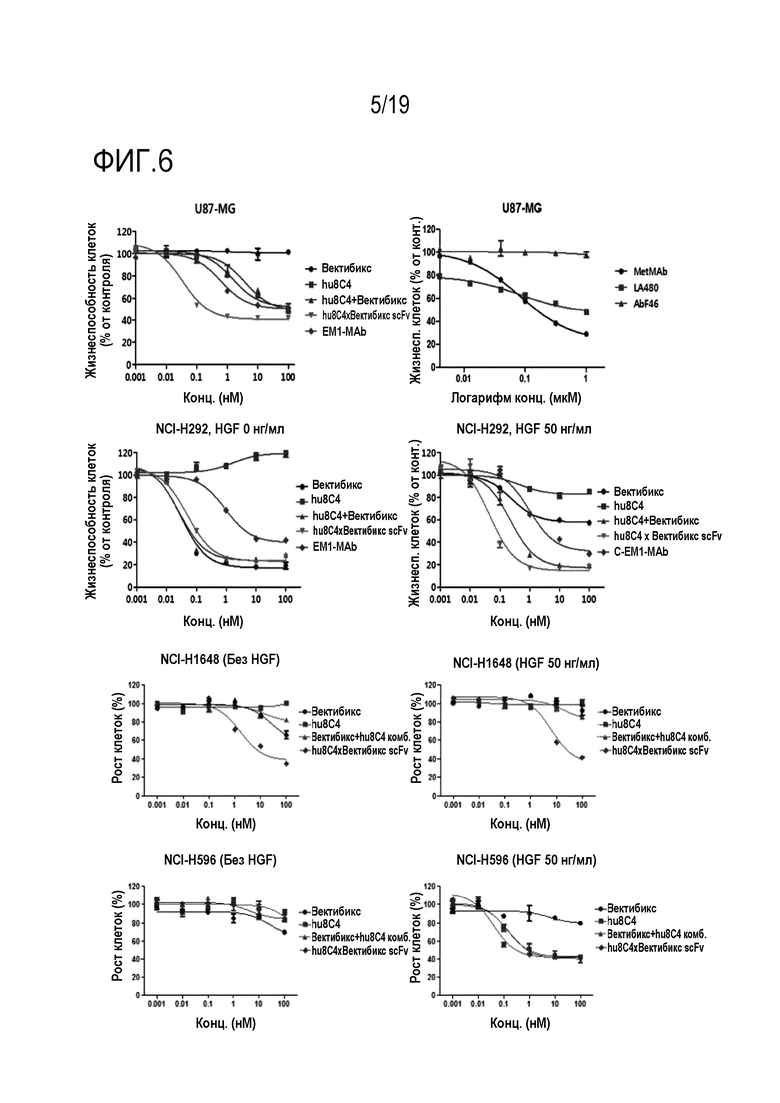
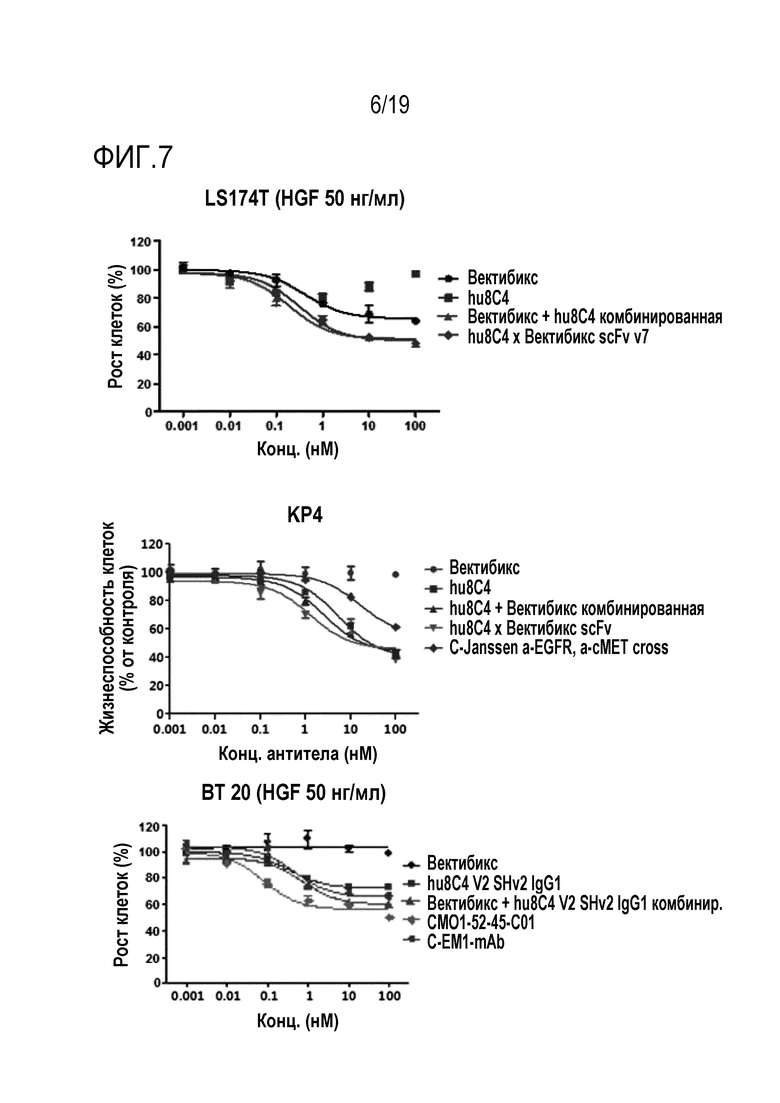

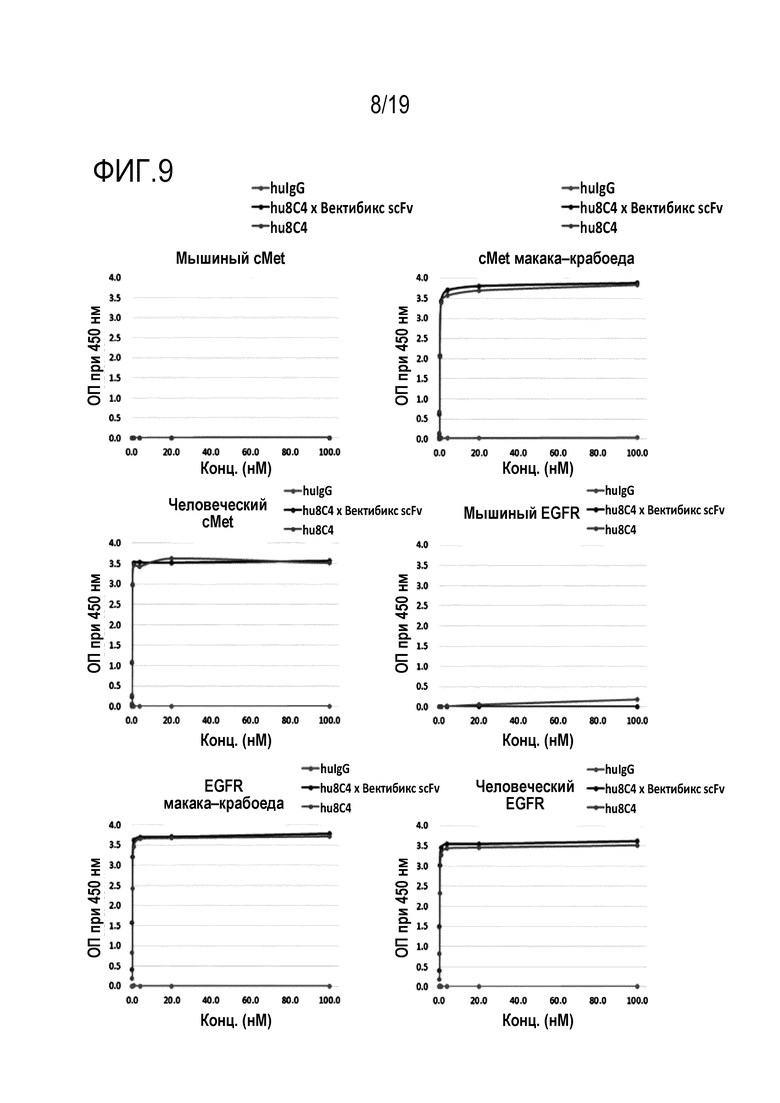
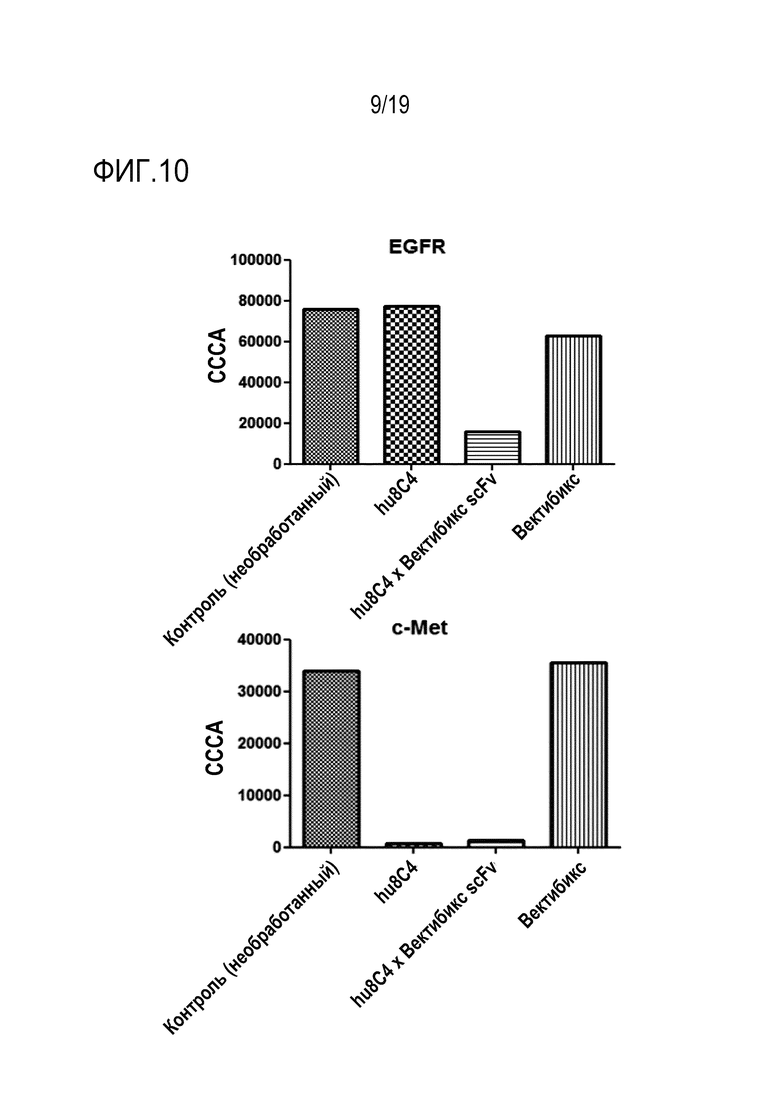
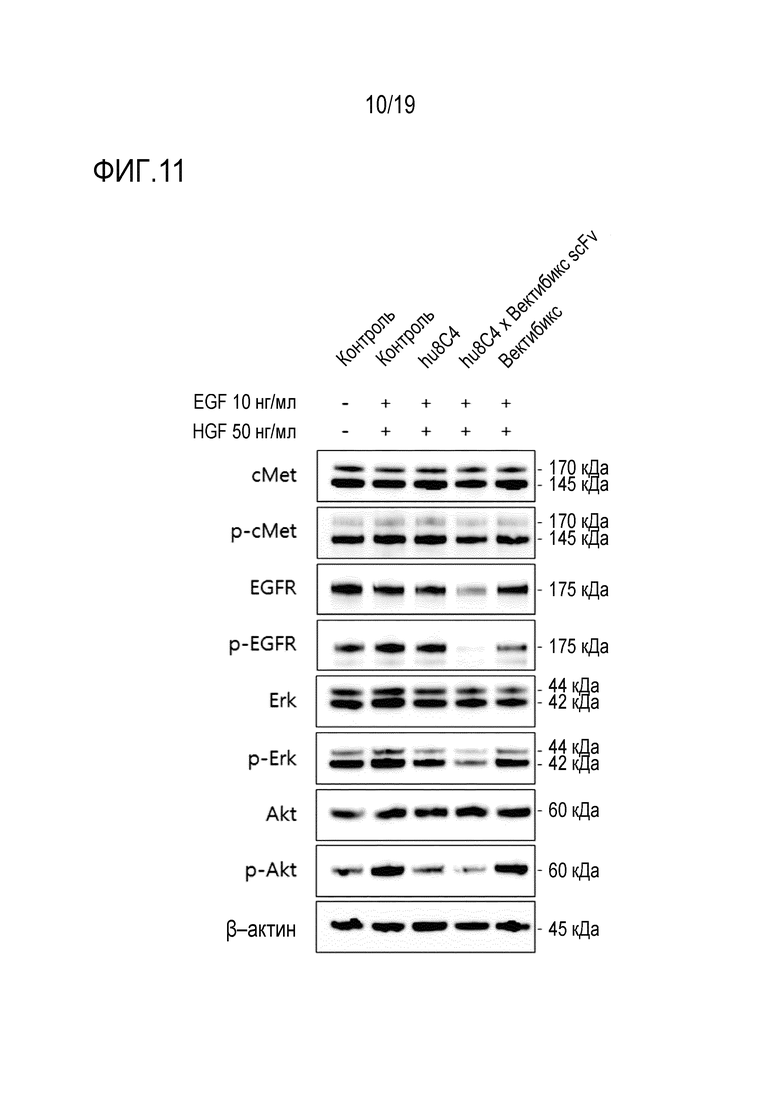
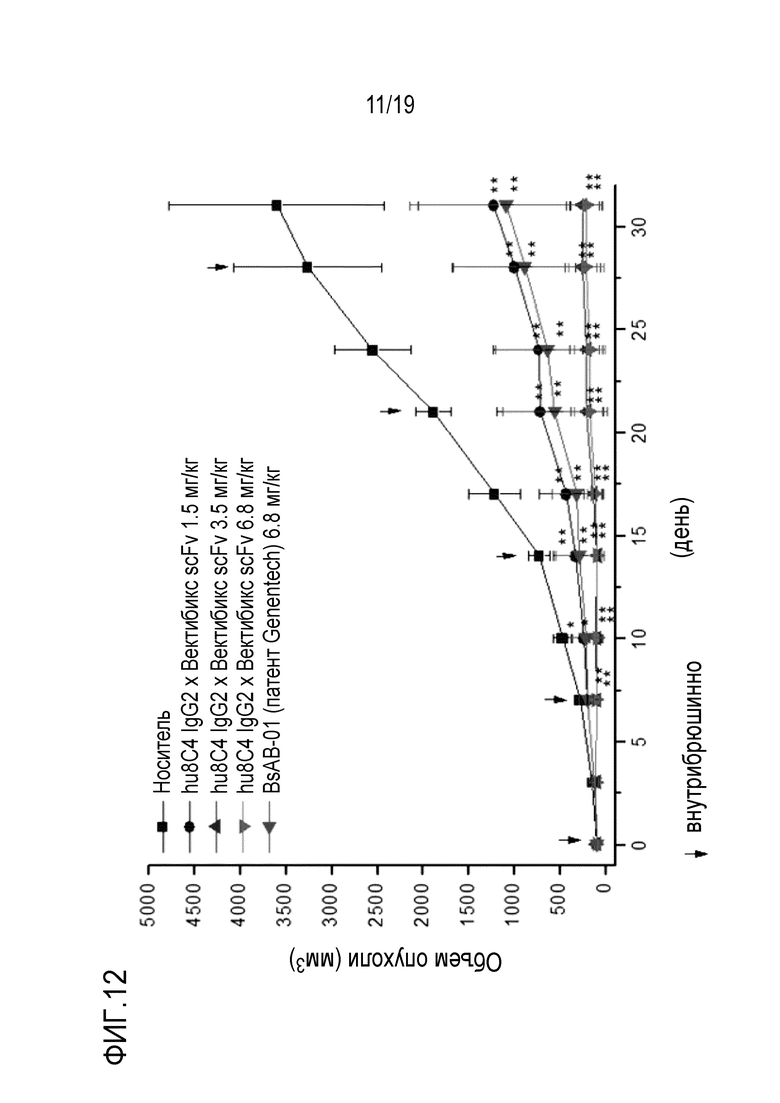
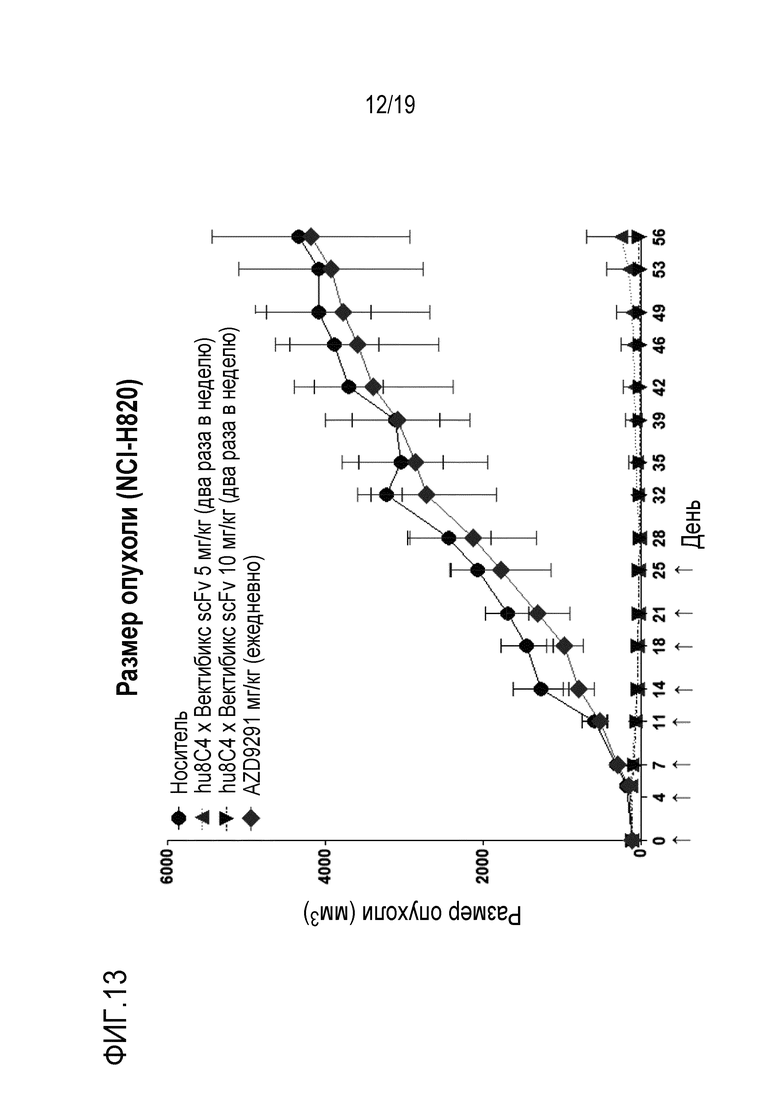
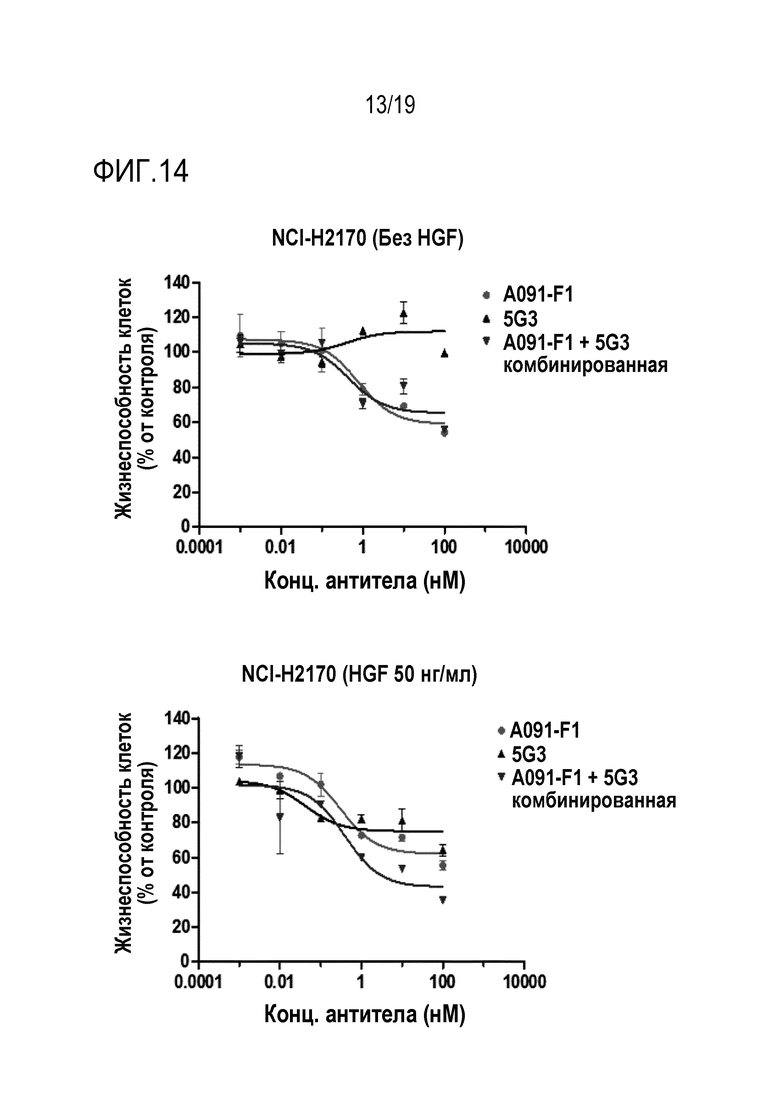
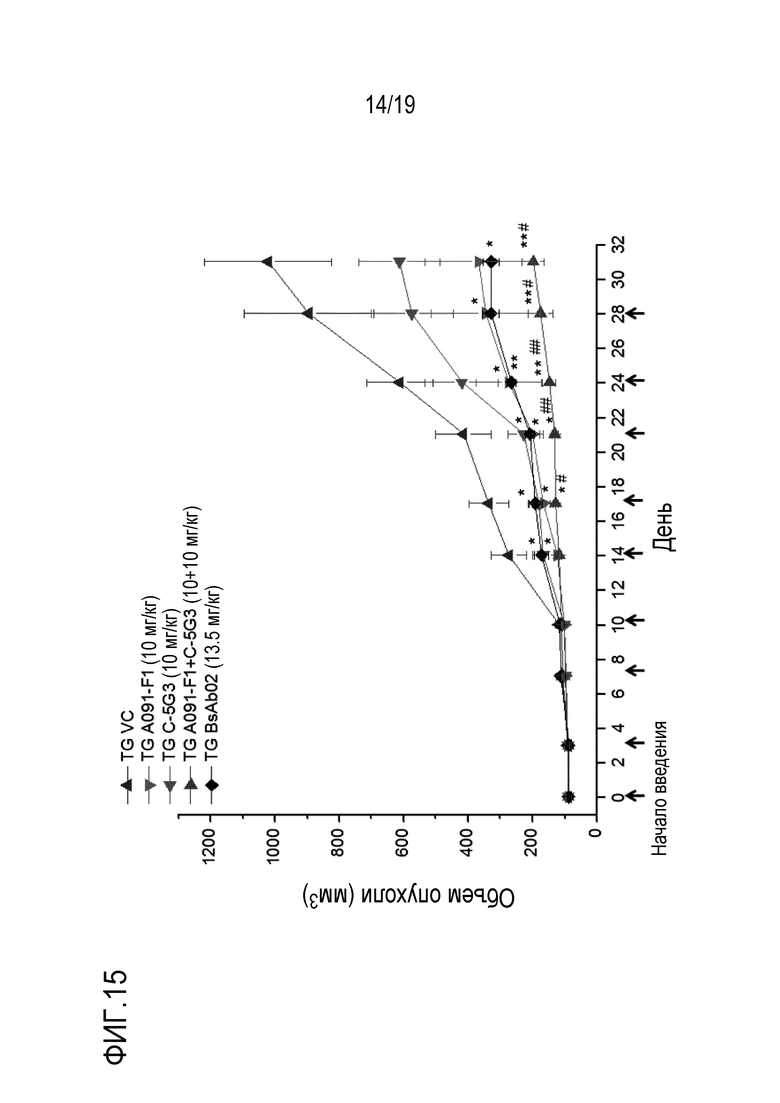
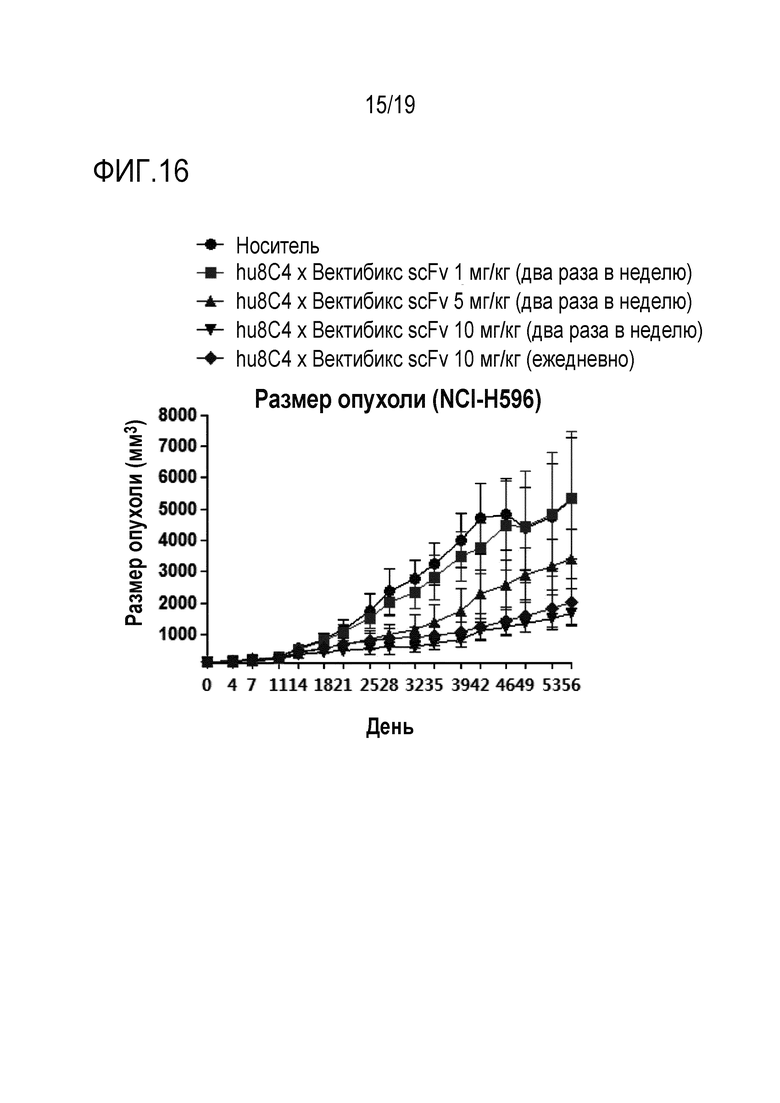
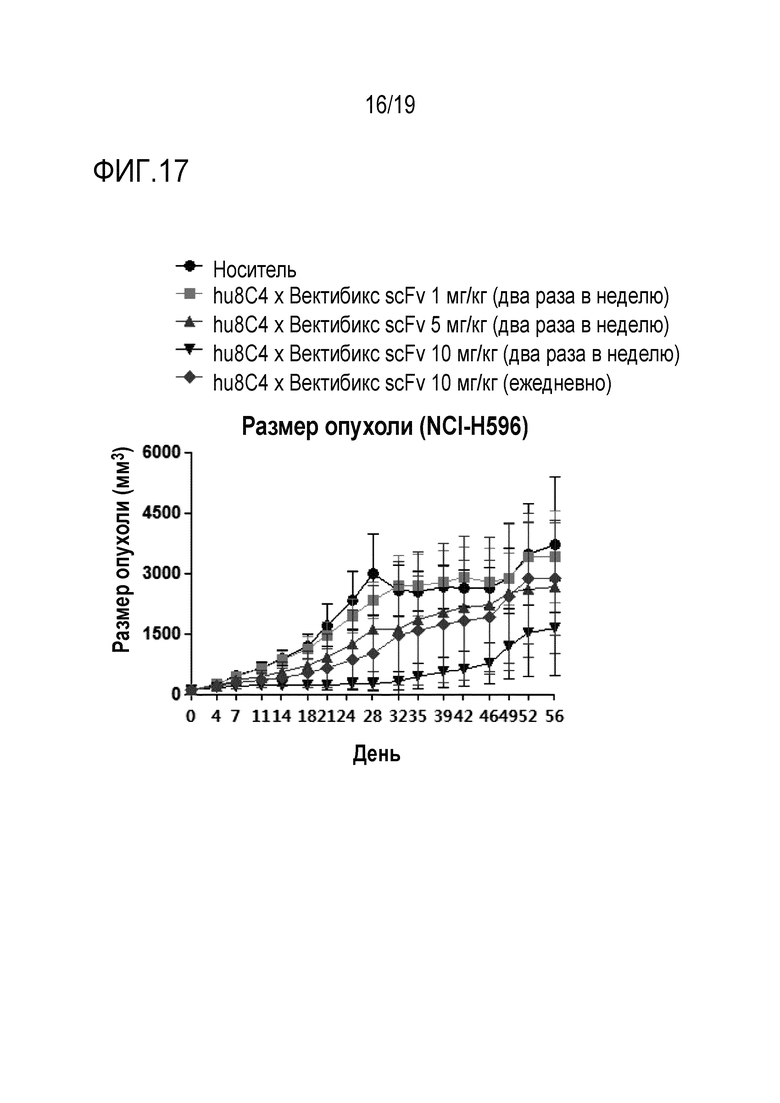
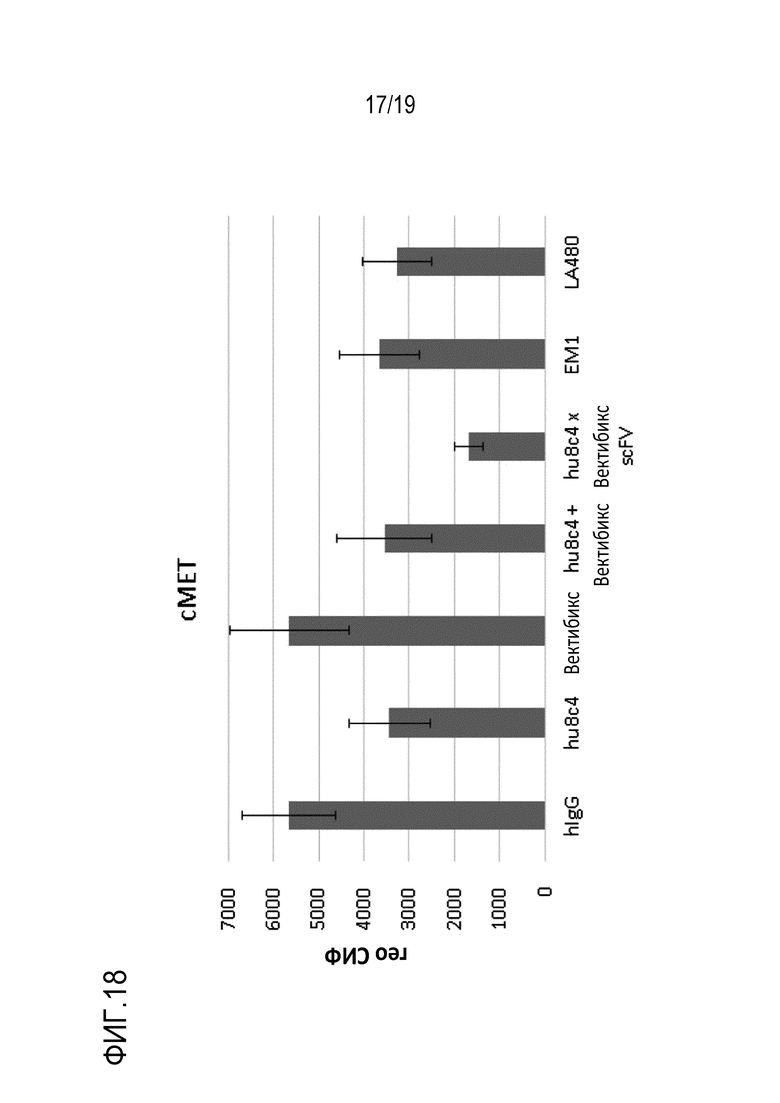
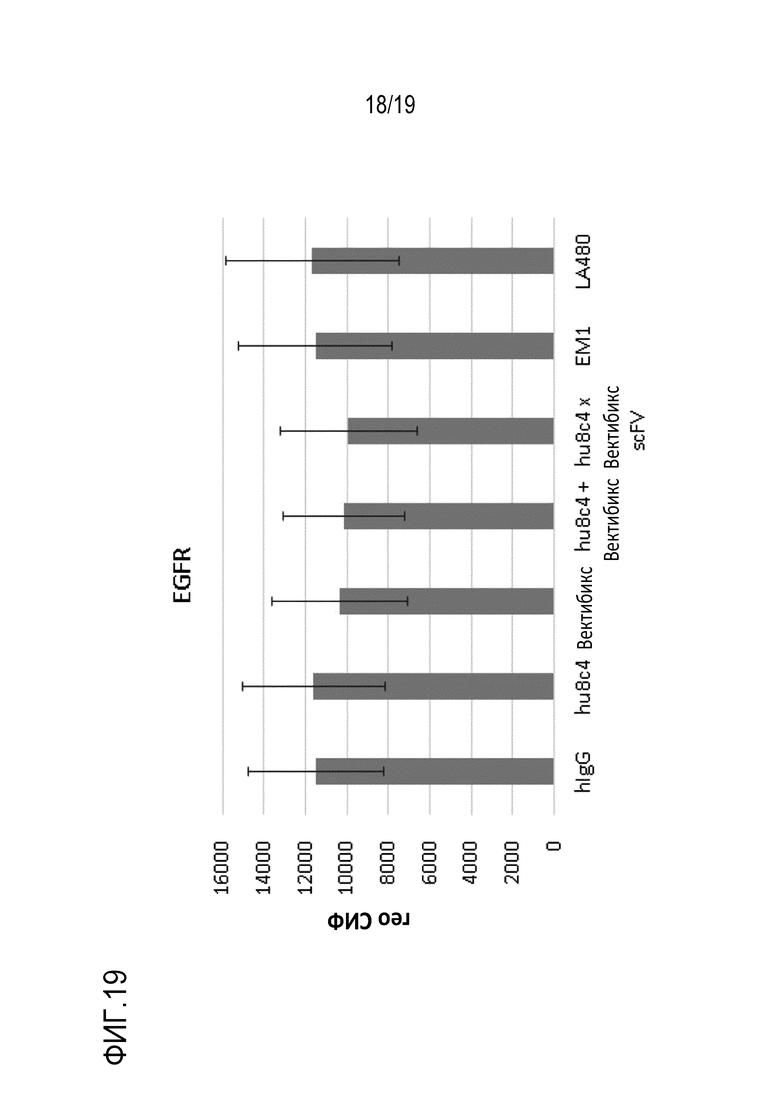
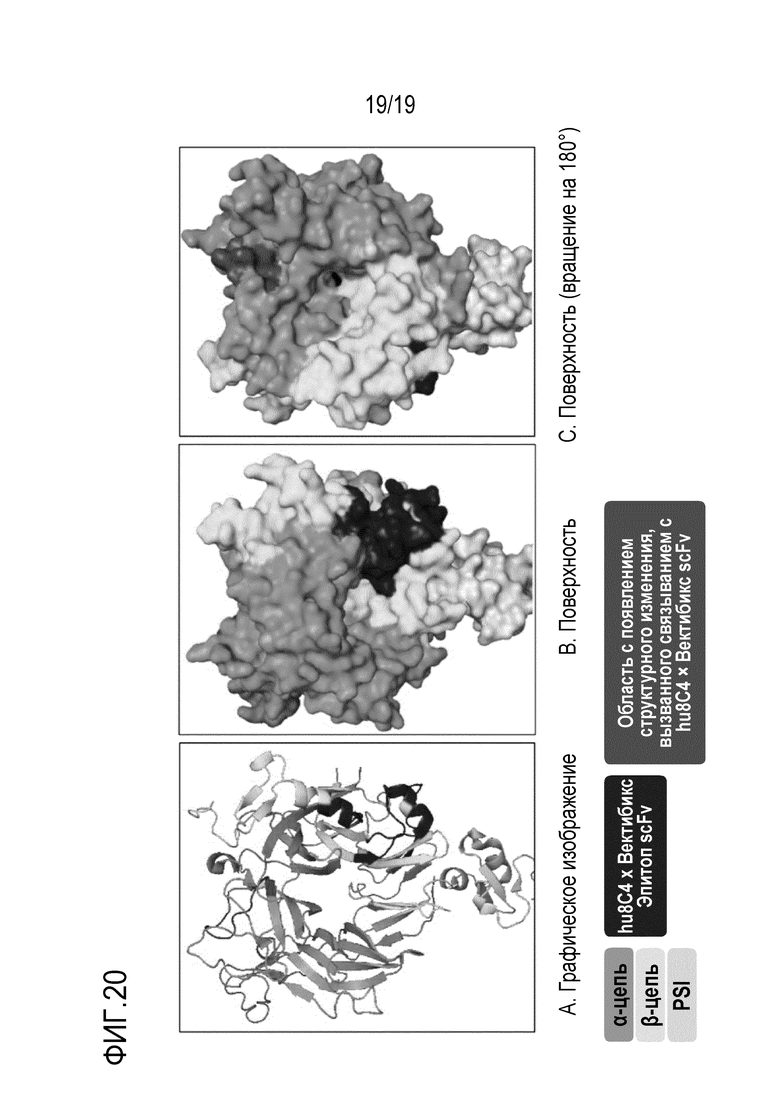
Изобретение относится к области биохимии, в частности к антителу или его антигенсвязывающему фрагменту, которые специфично связываются с рецептором фактора роста гепатоцитов (c-Met). Также раскрыты молекула нуклеиновой кислоты, кодирующая указанное антитело, экспрессионный вектор и клетка-хозяин, содержащие указанную молекулу нуклеиновой кислоты, композиции для обнаружения c-Met и для предотвращения или лечения рака, содержащие указанное антитело. Раскрыты способы обнаружения антигена c-Met с помощью указанного антитела, способ лечения или предотвращения рака с помощью указанного антитела. Изобретение позволяет эффективно лечить рак, ассоциированный с сверхэкспрессией c-Met. 10 н. и 17 з.п. ф-лы, 20 пр., 28 табл., 20 ил.
1. Антитело или его антигенсвязывающий фрагмент, которые специфично связываются с рецептором фактора роста гепатоцитов (c-Met), где антитело представляет собой
(а) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 1; CDR2 легкой цепи, представленную SEQ ID NO: 2; CDR3 легкой цепи, представленную SEQ ID NO: 3, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 7; CDR2 тяжелой цепи, представленную SEQ ID NO: 8; и CDR3 тяжелой цепи, представленную SEQ ID NO: 9;
(b) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 4; CDR2 легкой цепи, представленную SEQ ID NO: 5; CDR3 легкой цепи, представленную SEQ ID NO: 6, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 10; CDR2 тяжелой цепи, представленную SEQ ID NO: 11; CDR3 тяжелой цепи, представленную SEQ ID NO: 12; или
(c) их оптимизированные по аффинности антитела, где оптимизированное по аффинности антитело содержит:
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную любой из SEQ ID NO: 1 и SEQ ID NO: 229 – SEQ ID NO: 268; CDR2 легкой цепи, представленную любой из SEQ ID NO: 2, SEQ ID NO: 182 – SEQ ID NO: 190, SEQ ID NO: 227 и SEQ ID NO: 228; CDR3 легкой цепи, представленную любой из SEQ ID NO: 3, SEQ ID NO: 142 – SEQ ID NO: 181, SEQ ID NO: 191 – SEQ ID NO: 226 и SEQ ID NO: 269 – SEQ ID NO: 301; и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную любой из SEQ ID NO: 7 и SEQ ID NO: 108 – SEQ ID NO: 112; CDR2 тяжелой цепи, представленную любой из SEQ ID NO: 8, SEQ ID NO: 54 – SEQ ID NO: 63, SEQ ID NO: 72 – SEQ ID NO: 107 и SEQ ID NO: 118 – SEQ ID NO: 141; CDR3 тяжелой цепи, представленную любой из SEQ ID NO: 9, SEQ ID NO: 64 – SEQ ID NO: 71 и SEQ ID NO: 113 – SEQ ID NO: 117.
2. Антитело или его антигенсвязывающий фрагмент по п.1, где антитело содержит: (а) вариабельную область легкой цепи, представленную SEQ ID NO: 13, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 15; или (b) вариабельную область легкой цепи, представленную SEQ ID NO: 14, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 16.
3. Антитело или его антигенсвязывающий фрагмент по п.1, где антитело содержит:
(а) вариабельную область легкой цепи, представленную SEQ ID NO: 21, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 23;
(b) вариабельную область легкой цепи, представленную SEQ ID NO: 22, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 24;
(c) вариабельную область легкой цепи, представленную SEQ ID NO: 29, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 31; или
(d) вариабельную область легкой цепи, представленную SEQ ID NO: 30, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 32
4. Антитело или его антигенсвязывающий фрагмент по п.1, где антитело содержит шарнирную область, представленную любой из последовательностей от SEQ ID NO: 37 до SEQ ID NO: 44.
5. Антитело или его антигенсвязывающий фрагмент по п.1, где оптимизированное по аффинности антитело содержит вариабельную область легкой цепи, представленную любой из SEQ ID NO: 21 и SEQ ID NO: 306 – SEQ ID NO: 311, и вариабельную области тяжелой цепи, представленную любой из SEQ ID NO: 23 и SEQ ID NO: 302 – SEQ ID NO: 305.
6. Антитело или его антигенсвязывающий фрагмент по п.5, где оптимизированное по аффинности антитело содержит:
(а) вариабельную область легкой цепи, представленную SEQ ID NO: 21, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 302;
(b) вариабельную область легкой цепи, представленную SEQ ID NO: 21, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 305;
(c) вариабельная область легкой цепи, представленную SEQ ID NO: 310, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 23;
(d) вариабельную область легкой цепи, представленную SEQ ID NO: 308, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 305;
(e) вариабельную область легкой цепи, представленную SEQ ID NO: 306, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 303;
(f) вариабельную область легкой цепи, представленную SEQ ID NO: 307, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304;
(g) вариабельную область легкой цепи, представленную SEQ ID NO: 308, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304;
(h) вариабельную область легкой цепи, представленную SEQ ID NO: 309, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304;
(i) вариабельную область легкой цепи, представленную SEQ ID NO: 311, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 304; или
(j) вариабельную область легкой цепи, представленную SEQ ID NO: 306, и вариабельную область тяжелой цепи, представленную SEQ ID NO: 302.
7. Антитело или его антигенсвязывающий фрагмент, который специфично связывается с рецептором фактора роста гепатоцитов (c–Met), где антитело представляет собой:
(а) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 1; CDR2 легкой цепи, представленную SEQ ID NO: 2; CDR3 легкой цепи, представленную SEQ ID NO: 3, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 7; CDR2 тяжелой цепи, представленную SEQ ID NO: 8; и CDR3 тяжелой цепи, представленную SEQ ID NO: 9;
(b) антитело, содержащее вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную SEQ ID NO: 4; CDR2 легкой цепи, представленную SEQ ID NO: 5; CDR3 легкой цепи, представленную SEQ ID NO: 6, и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную SEQ ID NO: 10; CDR2 тяжелой цепи, представленную SEQ ID NO: 11; CDR3 тяжелой цепи, представленную SEQ ID NO: 12; или
(c) их оптимизированные по аффинности антитела, где оптимизированное по аффинности антитело содержит:
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, представленную любой из SEQ ID NO: 1 и SEQ ID NO: 229 – SEQ ID NO: 268; CDR2 легкой цепи, представленную любой из SEQ ID NO: 2, SEQ ID NO: 182 – SEQ ID NO: 190, SEQ ID NO: 227 и SEQ ID NO: 228; CDR3 легкой цепи, представленную любой из SEQ ID NO: 3, SEQ ID NO: 142 – SEQ ID NO: 181, SEQ ID NO: 191 – SEQ ID NO: 226 и SEQ ID NO: 269 – SEQ ID NO: 301; и вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, представленную любой из SEQ ID NO: 7 и SEQ ID NO: 108 – SEQ ID NO: 112; CDR2 тяжелой цепи, представленную любой из SEQ ID NO: 8, SEQ ID NO: 54 – SEQ ID NO: 63, SEQ ID NO: 72 – SEQ ID NO: 107 и SEQ ID NO: 118 – SEQ ID NO: 141; CDR3 тяжелой цепи, представленную любой из SEQ ID NO: 9, SEQ ID NO: 64 – SEQ ID NO: 71 и SEQ ID NO: 113 – SEQ ID NO: 117,
где антитело дополнительно специфично связывается с рецептором эпидермального фактора роста (EGFR).
8. Антитело или его антигенсвязывающий фрагмент по п.7, где антитело или его антигенсвязывающий фрагмент, специфически связывающееся с EGFR, связано с одним концом легкой цепи или тяжелой цепи антитела, специфичного в отношении c–Met.
9. Антитело или его антигенсвязывающий фрагмент по п.8, где антигенсвязывающий фрагмент, связывающийся с EGFR, представляет собой Fab, Fab', F(ab')2 или Fv.
10. Антитело или его антигенсвязывающий фрагмент по п.9, где Fv представляет собой один или несколько scFv–фрагментов, выбранных из группы, состоящей из эрбитукса, вектибикса, портраззы и ТераЦИМа.
11. Антитело или его антигенсвязывающий фрагмент по п.10, где scFv–фрагмент эрбитукс содержит аминокислотную последовательность, представленную SEQ ID NO: 313 или SEQ ID NO: 314.
12. Антитело или его антигенсвязывающий фрагмент по п.10, где scFv–фрагмент вектибикс содержит аминокислотную последовательность, представленную SEQ ID NO: 315.
13. Антитело или его антигенсвязывающий фрагмент по п. 8, где антитело или антигенсвязывающий фрагмент, связывающиеся с EGFR, содержат аминокислотную последовательность, представленную SEQ ID NO: 313 или SEQ ID NO: 314.
14. Антитело или его антигенсвязывающий фрагмент по п.8, где антитело или его антигенсвязывающий фрагмент связан линкером, представленным SEQ ID NO: 312.
15. Антитело или его антигенсвязывающий фрагмент по п.7, где антитело содержит: вариабельную область легкой цепи, представленную любой из SEQ ID NO: 21 и SEQ ID NO: 306 - SEQ ID NO: 311; и вариабельную область тяжелой цепи, представленную любой из SEQ ID NO: 316 - SEQ ID NO: 330.
16. Антитело или его антигенсвязывающий фрагмент по п.1, где антигенсвязывающий фрагмент представляет собой Fab, Fab', F(ab')2 или Fv.
17. Молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1–16.
18. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п.17.
19. Клетка–хозяин для получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-16, имеющая введенный в нее экспрессионный вектор по п.18.
20. Способ получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-16 с использованием клетки–хозяина по п.19.
21. Композиция для обнаружения c–Met, содержащая эффективную фармацевтическую дозу антитела или его антигенсвязывающего фрагмента по любому из пп. 1–16, которое специфически связывается с с-Met, таким образом обнаруживая с-Met.
22. Способ обнаружения антигена c–Met с использованием антитела или его антигенсвязывающего фрагмента по любому из пп.1–16, которое специфически связывается с с-Met, таким образом обнаруживая с-Met.
23. Композиция для предотвращения или лечения рака, содержащая эффективную фармацевтическую дозу антитела или антигенсвязывающего фрагмента по любому из пп. 1–16, где рак вызван сверхэкспрессией, амплификацией, мутацией или активацией c–Met.
24. Композиция для предотвращения или лечения рака по п.23, где антитело или его антигенсвязывающий фрагмент связывается с c–Met для ингибирования активности рецептора.
25. Композиция для предотвращения или лечения рака по п.24, где антитело или его антигенсвязывающий фрагмент дополнительно связывается с EGFR для ингибирования активности рецептора.
26. Композиция для предотвращения или лечения рака по п.23, где рак выбран из группы, состоящей из рака легкого, рака желудка, рака толстой кишки, рака прямой кишки, трижды негативного рака молочной железы (TNBC), глиобластомы, рака поджелудочной железы, рака головы и шеи, рака молочной железы, рака яичников, рака печени, рака почки, рака мочевого пузыря, рака предстательной железы, рак головного мозга, рак матки, соленомы, рака щитовидной железы, острого миелоидного лейкоза, хронического миелоидного лейкоза, миеломы, множественной миеломы, меланомы, лимфомы и рака коры надпочечников.
27. Способ лечения или предотвращения рака, включающий введение терапевтически эффективного количества композиции, содержащей антитело или его антигенсвязывающий фрагмент по любому из пп. 1–16.
| СПОСОБЫ И КОМПОЗИЦИЯ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2010 |
|
RU2585488C2 |
| US 20150299305 A1, 22.10.2015 | |||
| Способ изготовления искусственной вощины с отстроенными стенками ячеек | 1929 |
|
SU20398A1 |

