ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к соединениям для применения в генной терапии глазных болезней Более конкретно, настоящее изобретение относится к векторам на основе аденоассоциированного вируса (ААВ) (аденоассоциированным вирусным векторам) для применения для лечения или профилактики возрастной макулярной дегенерации (ВМД, AMD), которые способны доставлять в глаз Фактор I и/или Фактор H или его фрагменты или производные.
УРОВЕНЬ ТЕХНИКИ
Макула - это небольшая область в сетчатке глаза, размером примерно 3-5 миллиметров, находящаяся рядом со зрительным нервом. Она представляет собой чувствительную область сетчатки, которая содержит центральную ямку (фовеа), углубленный участок, обеспечивающий высокую остроту зрения и содержащий высокую концентрацию колбочек, фоторецепторов, отвечающих за цветное зрение.
Возрастная макулярная дегенерация (ВМД) является наиболее распространенной причиной функциональной слепоты в развитых странах у лиц старше 50 лет (Seddon, J M. Epidemiology of age-related macular degeneration. In: Ogden, T E, et al., eds. Ryan S J, ed-in-chief. Retina Vol II. 3rd ed. St. Louis, Mo.: Mosby; 2001:1039-50). ВМД связана с неоваскуляризацией, возникающей в системе хориоидальных сосудов и распространяющейся в субретинальное пространство. Кроме того, ВМД характеризуется прогрессирующей дегенерацией сетчатки, пигментного эпителия сетчатки (ПЭС, RPE) и расположенной ниже хориоидеи (сосудистой ткани, расположенной под ПЭС, между сетчаткой и склерой).
Патогенезу ВМД может способствовать множество факторов, включая оксидативный стресс, воспаление с возможным аутоиммунным компонентом, генетический фон (например, мутации) и экологические или поведенческие факторы, такие как курение и питание.
Этапы характерные для клинического прогрессирования ВМД соответствуют происходящим в макуле изменениям. Отличительной особенностью ранней стадии ВМД являются друзы, представляющие собой скопления под сетчаткой внеклеточного дебриса, видимые на сетчатке при клиническом осмотре глазного дна и на фотографиях глаза в виде желтых пятен. В зависимости от размера выделяют небольшие (<63 мкм), средние (63-124 мкм) и крупные (> 124 мкм) друзы. В зависимости от внешнего вида границы друз при офтальмологическом осмотре их также подразделяют на жесткие или мягкие. В то время как твердые друзы имеют четко определенные границы, границы мягких друз менее четкие и размытые. Одной из основных систем классификации, используемых для этого состояния, является классификация, разработанная в ходе исследования возрастной глазной патологии (AREDS) на основании снимков глазного дна.
Выделяют "сухую" и "влажную" (экссудативную или неоваскулярную) формы ВМД. Сухая форма ВМД является более распространенной чем влажная, однако она может прогрессировать до влажной формы, и в целом ряде случаев эти две формы сосуществуют. Сухая форма ВМД обычно характеризуется прогрессирующим апоптозом клеток в слое ПЭС, вышележащих фоторецепторных клеток, а также часто и нижележащих клеток хориокапиллярного слоя. Сливающиеся области гибели клеток ПЭС, сопровождающейся атрофией вышележащих фоторецепторов, называются географической атрофией. У пациентов с такой формой ВМД происходит медленное и прогрессивное ухудшение центрального зрения.
Влажная форма ВМД характеризуется кровотечением и/или утечкой жидкости из аномальных сосудов, выросших из хориоидальных сосудов (хориокапилляров) под ПЭС и макулой, что может стать причиной внезапной и инвалидизируюшей потери зрения. Согласно имеющимся оценкам, в большинстве случаев потеря зрения у пациентов обусловлена такой хориоидальной неоваскуляризацией (ХНВ) и вызываемыми ею вторичными осложнениями. Подтип неоваскулярной ВМД называется ретинальной ангиоматозной пролиферацией (РАП). В этом случае ангиоматозная пролиферация возникает в сетчатке и распространяется вглубь в субретинальное пространство, в конечном итоге затрагивая в некоторых случаях новые хориоидальные сосуды.
Система комплемента (СК, CS) вовлечена в ранний патогенез ВМД, о чем свидетельствуют компоненты СК в друзах, полученных из глаз пациентов с ВМД. При ВМД было идентифицировано по меньшей мере 129 типов белков, осажденных в друзах, включая различные типы аполипопротеинов (E, B или A-I), несколько видов амилоидных пептидов (P, Aβ или SA-1), TIMP-3, сывороточный альбумин и некоторые белки, связанные с клеточной функцией (например, субъединица β АТФ-синтазы, фагоцитарный рецептор В2 и ретинолдегидрогеназа). Друзы при ВМД также содержат почти все белки комплемента, включая регуляторные белки (CFH, рецептор комплемента 1 (CR1), витронектин и кластерин), продукты активации и деградации СК (C1q, C3, C3a, C3b и C5a) и членов терминального пути СК, содержащего компоненты МАК (MAC) (т.е. 5, 6, 8 (α, β и γ) и 9), в виде отдельных компонентов и в виде комплексов. Скопление друз может активировать СК, вызывать локальное продуцирование воспалительных медиаторов и привлекать лейкоциты, которые, в свою очередь, усиливают очаговое воспаление, присутствующее при ВМД.
Текущие методы лечения ВМД включают фотодинамическую терапию с использованием бензопорфирина (Arch Ophthalmol, 1999; 117:1329-1345) и ряда терапевтических агентов, нацеленных на сигнальный путь, опосредованный через сосудистый эндотелиальный фактор роста (VEGF). Примерами таких терапевтических агентов, нацеленных на VEGF, являются пегаптаниб, препарат на основе аптамера (N Engl J Med. 2004;351:2805-2816), и антитела, такие как ранибизумаб (N Engl J Med. 2006 Oct 5; 355(14):1432-44) и бевацизумаб (BMJ. 2010 Jun 9; 340:c2459). Тем не менее, не все пациенты реагируют на лечение с помощью анти-VEGF антитела, и зрение либо не восстанавливается, либо прогрессирует до диагностируемой слепоты.
Разработан терапевтический способ лечения географической атрофии, который в настоящее время находится на стадии клинического исследования фазы III (исследование MAHALO Genentech/Roche). Лампализумаб является гуманизированным моноклональным антителом, ингибирующим фактор D комплемента, вводимым путем инъекции в стекловидное тело, для остановки скорости прогрессирования географической атрофии. Как показано на Фиг. 1, фактор D является частью цикла обратной связи ("амплификации") C3b компонента. Фактор D присутствует в очень низких концентрациях в сыворотке крови и является важным фактором альтернативного пути. Тем не менее, из-за его небольшого размера (27 кДа), фактор D быстро выводится почками и быстро возобновляется в результате синтеза. Этот метод лечения требует введения ежемесячных инъекций в стекловидное тело.
В настоящее время существует потребность в новых подходах к лечению ВМД.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения предлагают подход для модуляции системы комплемента, который может быть применен, например, для лечения ВМД. Авторы изобретения предоставляют фактор I, доставляемый посредством генной терапии для регуляции цикла обратной связи C3b компонента комплемента путем направленного воздействия на цикл расщепления (Фиг. 1). Восстановление стабильного состояния петли обратной связи альтернативного пути будет вызывать расщепление C3b и iC3b и, таким образом, способствовать удалению основных факторов заболевания в опосредованных комплементом расстройствах, в частности расстройствах, причиной которых является дефект в регуляции альтернативного пути. В качестве альтернативы, можно использовать фактор H.
В одном из аспектов изобретение относится к аденоассоциированному вирусному (AAV) вектору, содержащему нуклеотидную последовательность, кодирующую фактор I или его фрагмент или производное. В другом аспекте изобретение относится к аденоассоциированному вирусному (AAV) вектору, содержащему нуклеотидную последовательность, кодирующую фактор H или его фрагмент или производное. В другом аспекте изобретения предоставляется аденоассоциированный вирусный (AAV) вектор, содержащий нуклеотидную последовательность, кодирующую антитело к фактору D. В другом аспекте изобретение относится к аденоассоциированному вирусному (AAV) вектору, содержащему нуклеотидную последовательность, кодирующую антитело к компоненту комплемента 5 (C5).
В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное, содержит последовательность, выбранную из группы, состоящей из:
(а) нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая имеет по меньшей мере 70% идентичность с SEQ ID NO: 1 или 9;
(b) нуклеотидной последовательности, которая имеет по меньшей мере 70% идентичность с SEQ ID NO: 2 или 8; а также
(c) нуклеотидной последовательности SEQ ID NO: 2 или 8.
Предпочтительно нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное, кодирует белок с природной активностью фактора I (например, белок, представленный SEQ ID NO: 1 или 9). Например, нуклеотидная последовательность, кодирующая фактор I или его фрагмент или ее производное, может кодировать белок, обладающий способностью к процессингу C3b и iC3b, превращая их в неактивные продукты расщепления. Другими словами, фактор I или его фрагмент или производное предпочтительно сохраняет активность, инактивирующую C3b и вызывающую деградацию iC3b.
В предпочтительном варианте осуществления нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное, кодирует белок с активностью, инактивирующей C3b и вызывающей деградацию iC3b.
В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное, включает последовательность, выбранную из группы, состоящей из:
(а) нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая имеет по меньшей мере 70% идентичность с SEQ ID NO: 3;
(b) нуклеотидной последовательности, которая имеет по меньшей мере 70% идентичность с SEQ ID NO: 4; и
(c) нуклеотидной последовательности SEQ ID NO: 4.
Предпочтительно нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное, кодирует белок с природной активностью фактора H (например, белок, представленный SEQ ID NO: 3). Например, нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное, может кодировать белок, обладающий способностью действовать в качестве кофактора для опосредованного фактором I расщепления C3b и увеличивать скорость диссоциации C3-конвертазы и C5-конвертазы.
В другом аспекте изобретения предоставляется клетка, трансфицированная AAV-вектором по изобретению.
В другом аспекте изобретение относится к фармацевтической композиции, содержащей AAV-вектор по изобретению или клетку по изобретению в комбинации с фармацевтически приемлемым носителем, разбавителем или наполнителем. В предпочтительном варианте осуществления фармацевтическая композиция предназначена для интраокулярного введения.
В одном из вариантов осуществления AAV-вектор содержит промотор куриного бета-актина (CBA), например, функционально связанный с нуклеотидной последовательностью, кодирующей фактор I или Н или его фрагмент или производное. В одном из вариантов осуществления AAV-вектор содержит промотор CAG, например, функционально связанный с нуклеотидной последовательностью, кодирующей фактор I или H или его фрагмент или производное. В одном из вариантов осуществления AAV-вектор содержит промотор с нуклеотидной последовательностью SEQ ID NO: 5, например функционально связанный с нуклеотидной последовательностью, кодирующей фактор I или H или его фрагмент или производное.
В одном из вариантов осуществления AAV-вектор содержит энхансерный элемент цитомегаловируса (CMV), например функционально связанный с нуклеотидной последовательностью, кодирующей фактор I или Н или его фрагмент или производное.
В одном из вариантов осуществления AAV-вектор содержит поли-A сигнал бычьего гормона роста, например, функционально связанный с нуклеотидной последовательностью, кодирующей фактор I или H или его фрагмент или производное, предпочтительно поли-A сигнал бычьего гормона роста, имеющий нуклеотидную последовательность SEQ ID NO: 6.
В одном из вариантов осуществления AAV-вектор содержит посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE), например функционально связанный с нуклеотидной последовательностью, кодирующей фактор I или Н или его фрагмент или производное, предпочтительно WPRE, имеющий нуклеотидную последовательность SEQ ID NO: 7.
В предпочтительном варианте осуществления AAV-вектор по изобретению находится в форме вирусной частицы.
В предпочтительном варианте осуществления AAV вирусная частица содержит геном AAV2 и капсидные AAV2 белки. Нуклеотидная последовательность, кодирующая фактор I или Н или его фрагмент или производное, предпочтительно функционально связана с промотором CAG, предпочтительно с промотором с нуклеотидной последовательностью SEQ ID NO: 5.
AAV-вектор, клетку или фармацевтическую композицию по изобретению можно использовать для лечения или профилактики глазных заболеваний.
В одном из варианте осуществления изобретения предоставляется AAV-вектор, клетка или фармацевтическая композиция по изобретению для применения для лечения или профилактики опосредованного комплементом расстройства зрения.
В одном из вариантов осуществления расстройство связано с чрезмерной активностью цикла обратной связи C3b и/или недостаточной активностью цикла расщепления C3b (см. Фиг. 1). В одном из вариантов осуществления расстройство представляет собой возрастную макулярную дегенерацию (ВМД) или диабетическую ретинопатию. В предпочтительном варианте осуществления расстройство представляет собой ВМД, предпочтительно сухую форму ВМД.
В одном из вариантов осуществления применение предназначено для лечения или профилактики расстройства у субъекта:
(а) у которого уровень активности или концентрация фактора I в глазу и/или сыворотке ниже нормального уровня, предпочтительно у которого концентрация или уровень активности в сыворотке эквивалентны 0-30 или 0-20 или 0-10 мкг/мл; и/или
(b) который является гетерозиготным или гомозиготным по SNP, ассоциированным с возрастной макулярной дегенерацией (ВМД), предпочтительно по редкому варианту фактора I.
В одном из вариантов осуществления применение предназначено для лечения или профилактики расстройства у субъекта:
(а) у которого нормальный уровень активности или концентрации фактора I в глазу и/или сыворотке, предпочтительно равный по меньшей мере 30 мкг/мл, например 30-40 мкг/мл в сыворотке; и/или
(b) который несет редкий аллель варианта фактора I.
В другом аспекте изобретение относится к AAV-вектору, клетке или фармацевтической композиции по изобретению для применения для лечения или профилактики возрастной макулярной дегенерации (ВМД). ВМД может быть, например, сухой формой ВМД. В предпочтительном варианте осуществления ВМД представляет собой сухую форму ВМД.
В другом аспекте изобретение относится к AAV-вектору, клетке или фармацевтической композиции по изобретению для применения для лечения или профилактики диабетической ретинопатии.
В одном из вариантов осуществления развитие географической атрофии предотвращают или задерживают. В другом варианте осуществления величина географической атрофии уменьшается.
В одном из вариантов осуществления прогрессирование географической атрофии замедляется. Предпочтительно, увеличение площади географической атрофии снижается по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% за 12 месяцев после введения в подвергшийся лечению глаз субъекта по сравнению с не подвергшимся лечению глазом за тот же период.
В другом аспекте изобретения предоставляется AAV-вектор, клетка или фармацевтическая композиция по изобретению для применения для улучшения или восстановления зрения или остроты зрения, например у субъекта, страдающего расстройством зрения, таким как раскрытое в настоящей заявке расстройство зрения. В другом аспекте изобретения предоставляется AAV-вектор, клетка или фармацевтическая композиция по изобретению для применения для предотвращения потери зрения или потери остроты зрения, например, потери зрения или остроты зрения, связанных с расстройством зрения, таким как раскрытое в настоящей заявке расстройство зрения.
В другом аспекте изобретения предоставляются AAV-вектор, клетка или фармацевтическая композиция по изобретению для применения для улучшения или восстановления скорости чтения у субъекта, например у субъекта, страдающего расстройством зрения, таким как раскрытое в настоящей заявке расстройство зрения. В другом аспекте изобретения предоставляется AAV-вектор, клетка или фармацевтическая композиция по изобретению для применения для предотвращения уменьшения скорости чтения у субъекта, например уменьшения скорости чтения, связанного с расстройством зрения, таким как раскрытое в настоящей заявке расстройство зрения.
В другом аспекте изобретение относится к AAV-вектору, клетке или фармацевтической композиции по изобретению для применения для уменьшения или предотвращения потери фоторецепторов и/или пигментного эпителия сетчатки (ПЭС), например, потери фоторецепторов и/или ПЭС, связанной с расстройством зрения, таким как раскрытое в настоящей заявке расстройство зрения.
В предпочтительном варианте осуществления AAV-вектор, клетку или фармацевтическую композицию для применения в соответствии с изобретением вводят интраокулярно. Авторы изобретения признают, что такое локальное введение терапевтического средства по существу позволяет достичь требуемых уровней фактора комплемента для лечения или профилактики опосредованного комплементом расстройства зрения, например ВМД.
В одном из вариантов осуществления AAV-вектор, клетку или фармацевтическую композицию по изобретению вводят в глаз субъекта путем субретинальной инъекции, инъекции непосредственно в ретину, супрахориоидальное пространство или стекловидное тело.
В особенно предпочтительном варианте осуществления AAV-вектор, клетку или фармацевтическую композицию по изобретению вводят в глаз субъекта путем субретинальной инъекции.
В одном из вариантов осуществления введение AAV-вектора, клетки или фармацевтической композиции по изобретению повышает уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта, в частности в глазу, например, в ПЭС субъекта. В другом варианте осуществления введение AAV-вектора, клетки или фармацевтической композиции по изобретению повышает уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта, в частности в глазу, например в ПЭС, субъекта до уровня, превышающего нормальный уровень в глазу.
В другом аспекте изобретение относится к способу лечения или профилактики опосредованного комплементом расстройства зрения, включающему введение AAV-вектора, клетки или фармацевтической композиции по изобретению нуждающемуся в этом субъекту.
В одном из вариантов осуществления расстройство связано с чрезмерной активностью цикла обратной связи C3b комплемента и/или недостаточной активности цикла расщепления C3b. В одном из вариантов осуществления расстройство представляет собой возрастную макулярную дегенерацию (ВМД) или диабетическую ретинопатию. В предпочтительном варианте осуществления расстройство представляет собой ВМД, предпочтительно сухую форму ВМД.
В другом аспекте изобретение относится к способу лечения или профилактики возрастной макулярной дегенерации (ВМД), включающему введение AAV-вектора, клетки или фармацевтической композиции по изобретению нуждающемуся в этом субъекту. ВМД может представлять собой, например, сухую форму ВМД. В предпочтительном варианте осуществления ВМД представляет собой сухую форму ВМД.
В другом аспекте изобретение относится к способу лечения или профилактики диабетической ретинопатии, включающему введение AAV-вектора, клетки или фармацевтической композиции по изобретению нуждающемуся в этом субъекту.
Субъект, например, может быть диагностирован как имеющий ВМД, или может быть подвержен риску развития ВМД.
В предпочтительном варианте осуществления AAV-вектор, клетку или фармацевтическую композицию вводят интраокулярно.
В одном из вариантов осуществления AAV-вектор, клетку или фармацевтическую композицию вводят в глаз субъекта путем субретинальной инъекции, инъекции непосредственно в ретину, супрахориоидальное или стекловидное тело.
В особенно предпочтительном варианте осуществления AAV-вектор, клетку или фармацевтическую композицию вводят в глаз субъекта путем субретинальной инъекции.
В другом аспекте изобретение относится к применению AAV-вектора, клетки или фармацевтической композиции по изобретению для изготовления лекарственного средства для лечения или профилактики опосредованного комплементом расстройства зрения.
В одном из вариантов осуществления расстройство связано с чрезмерной активностью цикла обратной связи C3b комплемента и/или недостаточной активности цикла расщепления C3b. В одном из вариантов осуществления расстройство представляет собой возрастную макулярную дегенерацию (ВМД) или диабетическую ретинопатию. В предпочтительном варианте осуществления расстройство представляет собой ВМД, предпочтительно сухую форму ВМД.
В другом аспекте изобретение относится к применению AAV-вектора, клетки или фармацевтической композиции по изобретению для изготовления лекарственного средства для лечения или профилактики возрастной макулярной дегенерации (ВМД). В предпочтительном варианте осуществления ВМД представляет собой сухую форму ВМД.
В другом аспекте изобретение относится к применению AAV-вектора, клетки или фармацевтической композиции по изобретению для изготовления лекарственного средства для лечения или профилактики диабетической ретинопатии.
В одном из вариантов осуществления AAV-вектор по изобретению не содержит промотор hAAT.
В одном из вариантов осуществления AAV-вектор по изобретению не содержит энхансер ApoR.
В другом варианте осуществления AAV-вектор по изобретению не содержит два энхансера ApoR.
В одном из вариантов осуществления AAV-вектор по изобретению не содержит геном AAV2 и капсидный белок AAV8, т.е. AAV-вектор по изобретению не является вектором AAV2/8.
В одном из вариантов осуществления AAV-вектор, клетку или фармацевтическую композицию по изобретению не вводят системно. В другом варианте осуществления AAV-вектор, клетку или фармацевтическую композицию по изобретению не вводят внутривенно.
ОПИСАНИЕ РИСУНКОВ
Фиг. 1
Циклы обратной связи (амплификации) и расщепления (даун-регуляции) C3b альтернативного пути комплемента у позвоночных (ʺIʺ=фактор I; ʺHʺ=фактор H; ʺBʺ=фактор B; и ʺDʺ=фактор D).
Фиг. 2
фиг. 2 Агарозный гель, рестрикционное расщепление CFI и CFIco. Полосу 1752 п.о. CFI вырезали и клонировали в pAAV-CBA-WPRE-bGHpA остов.
Фиг. 3
Иммуноблоттинг CFI (Фиг. 3A) и GFP (Фиг. 3B). 3А: CFI проявляется в виде полосы 70 кДа (не уменьшенный) и экспрессируется с одной и той же скоростью после трансфекции ARPE-19 векторами pAAV.CFI или pAAV.CFIco. После трансфекции pAAV-ветора CFI не экспрессировался. В качестве положительного контроля в иммуноблоттинге CFI использовали 10% нормальную человеческую сыворотку (NHS). 3B: Эффективность трансфекции анализировали путем котрансфекции клеток ARPE-19 вектором pCMV.GFP. GFP проявляется в виде полосы 30 кДа, и иммуноблот подтвердил, что клетки были трансфицированы с аналогичной эффективностью.
Фиг. 4
Иммуноблоттинг CFI в супернатанте клеточных линий HEK-293 и ARPE-19, трансдуцированных вирусом. 4A: Супернатант наносили в неденатурирующих условиях, и CFI детектировали с помощью мышиного моноклонального антитела к человеческому CFI (OX21, Thermo Fisher Scientific) и HRP-конъюгированного антитела осла к мышиным IgG (Abcam). CFI и CFIco экспрессировались в обеих клеточных линиях. Трансдукция клеточных линий вектором AAV.GFP служила в качестве отрицательного контроля, тогда как трансдукция 0,5 мкг очищенного от плазмы человеческого CFI (упоминаемого в настоящем описании "CFIpl") (Comptech) служила в качестве положительного контроля. 4B: Супернатант наносили в денатурирующих условиях, и CFI детектировали с помощью козьей антисыворотки к человеческому CFI (Comptech) и пероксидазным кроличьим антителом к козьим IgG (специфичным к цельной молекуле) (Sigma). CFI проявляется в виде полос 80 кДа (про-фермент), 50 кДа (подвергнутая процессингу тяжелая цепь) и 35 кДа (подвергнутая процессингу легкая цепь). AAV.GFP служил в качестве отрицательного контроля, тогда как 0,5 мкг очищенного от плазмы человеческого CFI (Comptech) и 10% нормальная человеческая сыворотка (упоминаемая в настоящем описании "NHS") служили положительным контролем. CFI и CFIco экспрессировались в обеих клеточных линиях.
Фиг. 5
Репрезентативный результат анализа расщепления C3b. Дорожка 1 представляет C3b, инкубированный только с CFH. Дорожка 2 представляет C3b, инкубированный с CFH и CFIpl. C3b расщепленный CFIpl до iC3b и C3dg. Дорожка 3 представляет C3b, инкубированный с CFH и супернатантом клеток HEK-293, трансдуцированных вектором AAV.CFI. Дорожки 4-5 представляют C3b, инкубированный с CFH и иммуноосажденным CFI (обозначенным "IP CFI") из клеток ARPE-19, трансдуцированных либо AAV.CFI (дорожка 4), либо AAV.CFIco (дорожка 5). Дорожки 6-7 представляют C3b, инкубированный с CFH и иммуноосажденным CFI из клеток HEK-293, трансдуцированных либо AAV.CFI (дорожка 6), либо AAV.CFIco (дорожка 7).
Фиг. 6
Иммуноблоттинг CFI, секретируемого клетками ARPE-19, культивированными на фильтрах Transwell. Клетки не были дифференцированы в гексагональные клетки, однако их культивировали в виде конфлюэнтного монослоя клеток, и деление клеток было сведено к минимуму путем добавления 1% сывороточной среды. 6A: Супернатант из обоих компартментов наносили в неденатурирующих условиях, и вестерн-блот-анализ проводили с использованием мышиного моноклонального антитела к CFI (Ap=апикальный компартмент и Bl=базолатеральный компартмент). Показано, что CFI экспрессируется в конфлюэнтном монослое клеток, и что секретируемый белок обнаруживается в обоих компартментах. 6B: Ядра окрашивали красителем Хехст (Hoechst) для подтверждения наличия монослоя клеток (окрашивание проводили после сбора супернатанта).
Фиг. 7
Экспрессия белка CFI в объединенных образцах анализировали методом иммуноблоттинга. CFI экспрессировался на детектируемых уровнях во всех дозах как в случае вектора AAV.CFI, так и в случае вектора AAV.CFIco. β-актин использовали в качестве контроля для нанесения. 7A: 40 мкг белкового лизата наносили в денатурирующих условиях, и CFI детектировали с помощью поликлональной козьей антисыворотки к человеческому CFI. CFI проявляется в виде полос 80 кДа (про-фермент), 50 кДа (подвергнутая процессингу тяжелая цепь) и 35 кДа (подвергнутая процессингу легкая цепь). Эти полосы соответствуют ожидаемому размеру CFI и подтверждают наличие процессинга, т.е. наличие тяжелой и легкой цепи. 7B: Такое же количество белкового лизата также использовали для образцов лизата, полученных из глаз с введенным путем инъекции AAV.CFIco в количестве 109 гк/глаз и из глаз без инъекции. CFI детектировали с помощью мышиного моноклонального антитела к CFI (слева, гель в неденатурирующих условиях) и козьей антисыворотки к CFI (справа, гель в денатурирующих условиях). В геле в неденатурирующих условиях (слева) CFI проявляется в виде полосы при 75 кДа у инъецированных животных, а у животных, в глаза которых не вводили инъекцию, полосы отсутствуют. В геле в денатурирующих условиях (справа) CFI проявляется в виде полос 80 кДа (про-фермент), 50 кДа (подвергнутая процессингу тяжелая цепь) и 35 кДа (подвергнутая процессингу легкая цепь).
Фиг. 8
Анализ экспрессии генов методом количественной ПЦР (кПЦР, qPCR).
Фиг. 9
Локализация hCFI: глаза с введенной ложной инъекцией (A-C), инъекцией AAV.CFI (D-F) и AAV.CFIco (G-H). Срезы сетчатки метили дважды: фибронектином (A, D и G) и hCFI (B, E и H). Ядра окрашивали DAPI, показаны в режиме наложения изображений (C, F и I). Scl: Склера, RPE: пигментный эпителий сетчатки, ОС: внешний сегмент фоторецепторов, IS: внутренний сегмент фоторецепторов, OPL: внешний слой плексиформного слоя, GCL: слой ганглиозных клеток, НФЛ: слой нервных волокон, увеличение: 20X, масштабная метка: 50 мкм.
Фиг. 10
Локализация hCFI: глаза с введенной ложной инъекцией (A-C), инъекцией AAV.CFI (D-F): более высокое увеличение ПЭС. Срезы сетчатки метили дважды: фибронектином (A и D) и hCFI (B и E). Ядра окрашивали DAPI, показаны в режиме наложения изображений (C и F). Scl: Склера, Bru: мембрана Бруха, Cho: Хориокапилляры, RPE: пигментный эпителий сетчатки, IPM: внутренний фоторецепторный матрикс. Увеличение: 189X, масштабная метка: 10 мкм. Окрашивание везикул показано стрелками, микроэлементы ПЭС показаны в нижней части СЭП звездочками.
Фиг. 11
Локализация hCFI: глаза с введенной ложной инъекцией (A-C), инъекцией инъекцией AAV.CFI (D-F): более высокое увеличение фоторецепторных слоев. Срезы сетчатки метили дважды: фибронектином (A и D) и hCFI (B и E). Ядра окрашивали DAPI, показаны в режиме наложения изображений (C и F). IPM: внутренний фоторецепторный матрикс, ОС: внешний сегмент, IS: внутренний сегмент, ONL: внешний ядерный слой. Увеличение: 189X, масштабная метка: 10 мкм.
Фиг. 12
Локализация hCFI: глаза с введенной ложной инъекцией (A-C) и инъекцией AAV.CFI (D-F): более высокое увеличение наружного плексиформного слоя (OPL). Срезы сетчатки метили дважды: фибронектином (A и D) и hCFI (B и E). Ядра окрашивали DAPI, показаны в режиме наложения изображений (C и F). ONL: Внешний ядерный слой, INL: внутренний ядерный слой. Увеличение: 189X, масштабная метка: 10 мкм. Окрашивание горизонтальных клеток показано стрелками.
Фиг. 13
Локализация hCFI: глаза с введенной ложной инъекцией (A-C) и инъекцией AAV.CFI (D-F): более высокое увеличение слоя ганглиозных клеток (GCL). Срезы сетчатки метили дважды: фибронектином (A и D) и hCFI (B и E). Ядра окрашивали DAPI, показаны в режиме наложения изображений (C и F). IPL: Внутренний плексиформный слой, НФЛ: слой нервных волокон. Увеличение: 189X, масштабная метка: 10 мкм.
Фиг. 14
Локализация hCFI: глаза с введенной ложной инъекцией (A-C) и инъекцией AAV.CFI (D-F) и AAV.CFIco (G-H). Цельный препарат пигментного эпителия сетчатки метили дважды: фибронектином (A, D и G) и hCFI (B, E и H). Увеличение: 40X, масштабная метка: 30 мкм.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Система комплемента
Система комплемента является неотъемлемой частью гуморальной иммунной системы и вовлечена в воспалительные процессы тканей, опсонизацию клеток и цитолиз. Она обеспечивает защиту от микроорганизмов и опосредует очистку экзогенного и эндогенного клеточного дебриса из тканей хозяина.
Каскад системы комплемента состоит из четырех путей активации. Все пути в конечном итоге заканчиваются расщеплением фактора С3 и генерацией его активных фрагментов С3а и C3b, что является центральным моментом каскада. C3a представляет собой анафилотоксин, который вызывает ряд хемотаксических и провоспалительных реакций, таких как рекрутирование воспалительных клеток и усиление проницаемости микроциркуляторного русла, тогда как C3b отвечает за опсонизацию чужеродных поверхностей, ковалентно связанных с C3b. Опсонизация активированными фрагментами C3 (C3b и iC3b) выполняет три основные функции: (i) элиминацию клеточного дебриса фагоцитарными клетками (например, макрофагами или клетками микроглии) и стимуляцию адаптивной иммунной системы (B и Т-клетки); (ii) усиление активации комплемента посредством образования поверхностно-связанной C3-конвертазы; и (iii) сборку C5-конвертазы.
Сборка C5-конвертазы ответственна за расщепление C5, что приводит к образованию цитолитического мембранного комплекса атаки (MAC), способного генерировать перфорации в клеточной мембране, тем самым способствуя лизису клеток и устранению ненужных клеток. Благодаря всем этим действиям компонент врожденного иммунитета каскада комплемента поддерживает и активизирует функцию последующих механизмов иммунной системы, которые защищают целостность ткани хозяина. В целом, активация пути системы комплемента приводит к провоспалительному ответу, включая генерацию MAC, который опосредует лизис клеток, высвобождение хемокинов для привлечения воспалительных клеток к месту повреждения и повышение проницаемости капилляров, содействуя экстравазации инфильтрующих лейкоцитов. В физиологических условиях активация комплемента эффективно контролируется скоординированным действием растворимых и связанных с мембраной регуляторных молекул комплемента (CRM). Растворимые регуляторы комплемента, такие как ингибитор C1, ингибитор анафилотоксинов, C4b-связывающий белок (C4BP), фактор H комплемента (CFH), фактор I комплемента (CFI), кластерин и витронектин, ограничивают действие комплемента в тканях человека на нескольких участках каскадной реакции. Кроме того, каждая отдельная клетка защищена от атаки гомологичного комплемента поверхностными белками, такими как рецептор комплемента 1 (CR1, CD35), мембранный кофакторный белок (CD46), и гликозилфосфатидилинозитол-заякоренными белками, такими как фактор ускорения распада (CD55) или молекула CD59. Следует отметить, что клетки и ткани хозяева, которые неадекватно защищены от атаки комплемента, могут быть подвергнуты лизису посторонними клетками.
Изобретение относится к лечению или профилактике опосредованного комплементом расстройства зрения. Например, опосредованное комплементом расстройство может быть расстройством, связанным с дефектом в регуляции альтернативного пути, и, в частности, с чрезмерной активностью цикла обратной связи C3b и/или недостаточной активностью цикла расщепления C3b.
В одном из вариантов осуществления до введения AAV-вектора, клетки или фармацевтической композиция по изобретению у субъекта был низкий (например, ниже нормального уровня) уровень активности фактора I, например низкий уровень активности фактора I в глазу и/или низкий уровень активности фактора I в сыворотке. Недостаточный уровень активности фактора I может быть обусловлен недостаточным уровнем экспрессии нормально функционирующего фактора I или, по меньшей мере, частичной (например, гетерозиготной) экспрессией (при нормальном или субнормальном уровне) нефункционального или недостаточно функционального варианта фактора I. (Такой субъект может нести одну или более копий SNP, ассоциированных с ВМД, например, субъект может быть гомо- или гетерозиготным по одному из редких вариантов фактора I, обсуждаемых ниже.) Таким образом, субъект может иметь низкую концентрацию (например, более низкую, чем обычно) фактора I в глазу и/или сыворотке. В случае человека, нормальный уровень активности фактора I (активности, инактивирующей C3b и вызывающей деградацию iC3b) может быть эквивалентным уровню, обеспечиваемому 30-40 мкг/мл фактора I в сыворотке субъекта. Таким образом, у субъекта с низкой активностью фактора I активность фактора I в сыворотке может соответствовать менее чем 30 мкг/мл и более чем 0 мкг/мл фактора I, например в диапазоне 0-20 или 0-10 мкг/мл (диапазоны концентрации фактора I в сыворотке, в которые могут попадать субъекты с низкой концентрацией фактора I).
Таким образом, субъект, подлежащий лечению по настоящему изобретению, может страдать опосредованным комплементом расстройством зрения, таким как ВМД, более конкретно сухой формой ВМД (например, характеризующейся географической атрофией) или может иметь риск развития такого расстройства. Например, субъект может быть гомозиготным или гетерозиготным в отношении восприимчивости к одному или более SNP, связанных с опосредованным комплементом расстройством.
В одном из вариантов осуществления субъект подвержен риску развития ВМД. Например, субъект может быть гомозиготным или гетерозиготным по генетической предрасположенности из-за наличия одного или более SNP ассоциированных с ВМД, например, нести редкие мутации в факторе I, связанные с прогрессирующим ВМД, которые обычно приводят к сниженным уровням фактора I в сыворотке (Kavanagh et al., Hum Mol Genet. 2015 июль 1, 24 (13):3861-70). В частности, субъект может иметь одну или две копии одного или более из следующих редких вариантов фактора I: rs144082872 (кодирующий P50A); 4:110687847 (кодирующий P64L); rs141853578 (кодирующий G119R); 4:110685721 (кодирующий V152M); 4:110682846 (кодирующий G162D); 4:110682801 (кодирующий N177I); rs146444258 (кодирующий A240G); rs182078921 (кодирующий G287R); rs41278047 (кодирующий K441R); rs121964913 (кодирующий R474).
Изобретение может дополнительно содержать определение, подвержен ли субъект риску развития опосредованного комплементом расстройства (например, ВМД), например, путем определения, является ли субъект гомозиготным или гетерозиготным по генетической предрасположенности, по наличию одного или более SNP, ассоциированных с опосредованным комплементом расстройством (например, путем определения, является ли субъект гомозиготным или гетерозиготным по генетической предрасположенности, по наличию одного или более перечисленных выше редких вариантов фактора I, ассоциированных с ВМД).
Альтернативно, у субъекта может быть нормальный уровень активности или концентрация эндогенного фактора I, например, в глазу и/или сыворотке, и/или может отсутствовать аллель редкого варианта фактора I.
В одном из вариантов осуществления введение AAV-вектора, клетки или фармацевтической композиции по изобретению повышает уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, в глазу субъекта. В другом варианте осуществления введение AAV-вектора, клетки или фармацевтической композиции по изобретению повышает уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, в глазу субъекта до уровня, который превышает нормальный уровень в глазу. Более конкретно, уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, в ПЭС глаза увеличивается.
Понятно, что активность, инактивирующая C3b и вызывающая деградацию iC3b, у субъекта в результате экспрессии фактора I или его фрагмента или производного с помощью AAV-вектора по изобретению может включать активность, инактивирующую C3b и вызывающую деградацию iC3b, эндогенного фактора I субъекта (т.е. фактора I субъекта, который не вырабатывается в результате экспрессии вектора AAV), и активность инактивирующую C3b и вызывающую деградацию iC3b, полученную в результате экспрессии AAV-вектора по изобретению, так что общий уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта будет превышать нормальный уровень.
В одном из вариантов осуществления уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта, например в глазу, увеличивается до уровня, который по меньшей мере на 5%, 10%, 15%, 20% или 25% выше нормального уровня.
В другом варианте осуществления уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта, например в глазу, увеличивается до уровня, который в два раза превышает нормальный уровень или который превышает нормальный уровень на 80%, 60%, 40% или 20%.
Например, уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта, например в глазу, может быть увеличен до уровня, превышающего нормальный уровень на 5-100%, 5-80%, 5-60%, 5-40%, 5-20%, 10-100%, 10-80%, 10-60%, 10-40%, 10-20%, 15-100%, 15-80%, 15-60%, 15-40%, 15-20%, 20-100%, 20-80%, 20-60%, 20-40%, 25-100%, 25-80%, 25-60% или 25-40%.
В одном из вариантов осуществления введение AAV-вектора, клетки или фармацевтической композиции по изобретению не обеспечивает заметного увеличения уровня активности, инактивирующей C3b и вызывающей деградацию iC3b, в глазу субъекта. В другом варианте осуществления введение AAV-вектора, клетки или фармацевтической композиции по изобретению не обеспечивает заметного увеличения уровня активности, инактивирующей C3b и вызывающей деградацию iC3b, в глазу субъекта до уровня, который превышает нормальный уровень в глазу.
В вышеприведенном разделе, за исключением очевидно неприменимых случаев, ссылка на активность фактора I и активность, инактивирующую C3b и вызывающую деградацию iC3b, может быть заменена ссылкой на фактор H и его способность действовать в качестве кофактора для опосредованного фактором I расщепления C3b и для увеличения скорости диссоциация C3-конвертазы и C5-конвертазы, соответственно. В одном из вариантов осуществления до введения AAV-вектора, клетки или фармацевтической композиции по изобретению у субъекта был низкий уровень (например, ниже нормального уровня) активности фактора H, например низкий уровень активности фактора H в глазу и/или низкий уровень активности фактора H в сыворотке. У человека нормальный уровень фактора H может составлять примерно 200-500 мкг/мл в сыворотке субъекта. Таким образом, у субъекта с низкими уровнями фактора H уровни в сыворотке могут быть менее 200 мкг/мл и выше 0 мкг/мл, например 0-100 мкг/мл. Альтернативно, уровень эндогенного фактора H у субъекта, например в глазу и/или сыворотке, может быть нормальным.
Фактор I
Фактор I комплемента (фактор I, CFI), также известный как C3b/C4b инактиватор, представляет собой белок, который у человека кодируется геном CFI.
Фактор I представляет собой сериновую протеазу, которая циркулирует в зимогенподобной форме (Roversi et al., PNAS, 2011; 108 (31):12839-12844) в концентрации ~ 35 мкг/мл (Nilsson et al., Mol Immunol 2011, 48 (14):1611-1620). Белок фактор I представляет собой сильно N-гликозилированный гетеродимер, состоящий из двух полипептидных цепей, связанных одной дисульфидной связью. Тяжелая цепь (50 кДа) содержит N-концевую область; мембраноатакующий комплекс FI (FIMAC); CD5-подобный домен или богатый цистеином домен фагоцитарных рецепторов (SRCR); два домена рецепторов липопротеинов низкой плотности (LDLr); и С-терминальную область неизвестной функции, которая является сайтом изменчивости последовательности, обеспечивающим разнообразие видов (Roversi et al., см. выше). Легкая цепь (38 кДа) содержит домен сериновой протеазы (SP) с консервативными каталитическими остатками (Goldberger et al., J Biol Chem 1987, 262 (21):10065-10071).
Фактор I инактивирует C3b путем его расщепления на iC3b, C3d и C3d,g и C4b аналогичным образом на C4c и C4d. Для нормального функционирования фактора I необходимо наличие кофакторных белков, таких как C4b-связывающий белок (C4BP), фактор комплемента H (CFH), рецептор комплемента 1 (CR1 CD35) и мембранный кофакторный белок (MCP/CD46) (Degn et al., Am J Hum Genet 2011, 88 (6): 689-705).
iC3b не способен связываться с фактором B и, следовательно, не может поддерживать усиление каскада комплемента или активацию через альтернативный путь. Следовательно, после расщепления C3b до iC3b не происходит ни инициация альтернативного пути, ни активация терминального каскада комплемента.
iC3b способен обеспечивать провоспалительное действие посредством связывания с рецептором комплемента 3 (CR3) (CD11b/CD18) на полиморфноядерных лейкоцитах (в основном нейтрофилах), NK-клетках и мононуклеарных фагоцитах, таких как макрофаги.
Фактор I способен осуществлять процессинг iC3b в C3d,g благодаря протеазной активности, требующей наличия кофактора CR1. C3d,g не может связываться с CR3. Поскольку iC3b, взаимодействующий с CR3 рецептором комплемента, является основным механизмом, посредством которого активация комплемента приводит к воспалению, разложение iC3b до C3d,g имеет важное значение для уменьшения воспаления, вызванного комплементом (Lachmann (2009), Adv. Immunol., 104:115-149).
Уникальная способность фактора I как стимулировать расщепление C3b до iC3b, так и ускорять расщепление iC3b, в сочетании с его относительно низкой концентрацией в сыворотке человека, в смысле количества, требуемого для терапевтической эффективности, делают его особенно выгодной мишенью.
В одном из вариантов осуществления полипептид фактора I или его фрагмент или производное способен расщеплять C3b до неактивного продукта разложения. Например, полипептид фактора I или его фрагмент или производное может быть способным расщеплять C3b до iC3b.
В одном из вариантов осуществления полипептид фактора I или его фрагмент или производное способен к процессингу iC3b в неактивный продукт разложения. Например, полипептид фактора I или его фрагмент или производное может быть способен осуществлять процессинг iC3b в C3d,g.
В предпочтительном варианте осуществления полипептид фактора I или его фрагмент или производное способен расщеплять C3b до iC3b и осуществлять процессинг iC3b в C3d,g.
Фрагмент или производное фактора I может сохранять по меньшей мере 50%, 60%, 70%, 80%, 90%, 95% или 100% активность, инактивирующую C3b и вызывающую деградацию iC3b, нативного фактора I. Инактивирующая C3b и вызывающая деградацию iC3b активность фрагмента или производного фактора I и нативного фактора I может быть определена любым подходящим методом, известным специалистам в данной области. Например, измерение протеолитической активности фактора I описано у Hsiung et al. (Biochem. J. (1982), 203, 293-298). Анализ гемолитической и конглютинирующей активностей FI описаны у Lachmann PJ & Hobart MJ (1978) "Complement Technology" in Handbook of Experimental Immunology 3rd edition Ed DM Weir Blackwells Scientific Publications Chapter 5A p17. Более подробное описание, также включающее описание протеолитического анализа, дает Harrison RA (1996) в "Weir's Handbook of Experimental Immunology" 5th Edition Eds; Herzenberg Leonore A'Weir DM, Herzenberg Leonard A & Blackwell C Blackwells Scientific Publications Chapter 75 36-37. Анализ конглютинирующей активности является высокочувствительным и может быть использован для обнаружения как первого (двойного) укорочения, преобразующего фиксированный C3b в iC3b и обеспечивающий способность взаимодействовать с конглютинином; так и для обнаружения конечного укорочения с получением C3dg, начиная с фиксированного iC3b с дальнейшим поиском потери способности взаимодействовать с конглютинином. Гемолитический анализ используется для превращения C3b в iC3b, а протеолитический анализ обнаруживает все укорочения.
В одном из вариантов осуществления фактор I является человеческим фактором I.
Примером человеческого фактора I является белок фактор I человека с номером доступа UniProtKB P05156. Эта приведенная в качестве примера последовательность имеет длину 583 аминокислоты (см. SEQ ID NO:1), из которых аминокислоты 1-18 образуют сигнальную последовательность.
В одном из вариантов осуществления аминокислотная последовательность фактора I представляет собой последовательность, приведенную в SEQ ID NO:1. В одном из вариантов осуществления аминокислотная последовательность фактора I представляет собой последовательность, представленную с 19 по 583 позицию в SEQ ID NO:1.
MKLLHVFLLF LCFHLRFCKV TYTSQEDLVE KKCLAKKYTH LSCDKVFCQP
WQRCIEGTCV CKLPYQCPKN GTAVCATNRR SFPTYCQQKS LECLHPGTKF
LNNGTCTAEG KFSVSLKHGN TDSEGIVEVK LVDQDKTMFI CKSSWSMREA
NVACLDLGFQ QGADTQRRFK LSDLSINSTE CLHVHCRGLE TSLAECTFTK
RRTMGYQDFA DVVCYTQKAD SPMDDFFQCV NGKYISQMKA CDGINDCGDQ
SDELCCKACQ GKGFHCKSGV CIPSQYQCNG EVDCITGEDE VGCAGFASVT
QEETEILTAD MDAERRRIKS LLPKLSCGVK NRMHIRRKRI VGGKRAQLGD
LPWQVAIKDA SGITCGGIYI GGCWILTAAH CLRASKTHRY QIWTTVVDWI
HPDLKRIVIE YVDRIIFHEN YNAGTYQNDI ALIEMKKDGN KKDCELPRSI
PACVPWSPYL FQPNDTCIVS GWGREKDNER VFSLQWGEVK LISNCSKFYG
NRFYEKEMEC AGTYDGSIDA CKGDSGGPLV CMDANNVTYV WGVVSWGENC
GKPEFPGVYT KVANYFDWIS YHVGRPFISQ YNV
(SEQ ID NO: 1)
В одном из вариантов осуществления аминокислотная последовательность фактора I представляет собой последовательность, приведенную в SEQ ID NO:9, которая соответствует номеру доступа NCBI NP_000195. В одном из вариантов осуществления аминокислотная последовательность фактора I представляет собой последовательность, представленную с 19 по 583 позицию в SEQ ID NO:9.
MKLLHVFLLF LCFHLRFCKV TYTSQEDLVE KKCLAKKYTH LSCDKVFCQP WQRCIEGTCV
CKLPYQCPKN GTAVCATNRR SFPTYCQQKS LECLHPGTKF LNNGTCTAEG KFSVSLKHGN
TDSEGIVEVK LVDQDKTMFI CKSSWSMREA NVACLDLGFQ QGADTQRRFK LSDLSINSTE
CLHVHCRGLE TSLAECTFTK RRTMGYQDFA DVVCYTQKAD SPMDDFFQCV NGKYISQMKA
CDGINDCGDQ SDELCCKACQ GKGFHCKSGV CIPSQYQCNG EVDCITGEDE VGCAGFASVA
QEETEILTAD MDAERRRIKS LLPKLSCGVK NRMHIRRKRI VGGKRAQLGD LPWQVAIKDA
SGITCGGIYI GGCWILTAAH CLRASKTHRY QIWTTVVDWI HPDLKRIVIE YVDRIIFHEN
YNAGTYQNDI ALIEMKKDGN KKDCELPRSI PACVPWSPYL FQPNDTCIVS GWGREKDNER
VFSLQWGEVK LISNCSKFYG NRFYEKEMEC AGTYDGSIDA CKGDSGGPLV CMDANNVTYV
WGVVSWGENC GKPEFPGVYT KVANYFDWIS YHVGRPFISQ YNV
(SEQ ID NO: 9)
Примером нуклеотидной последовательности, кодирующей фактор I, является нуклеотидная последовательность, имеющая номер доступа NCBI NM_000204. В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор I, представляет собой нуклеотидную последовательность, имеющую номер доступа NCSCI NM_000204.
В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор I, представляет собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2.
atgaagcttc ttcatgtttt cctgttattt ctgtgcttcc acttaaggtt ttgcaaggtc
acttatacat ctcaagagga tctggtggag aaaaagtgct tagcaaaaaa atatactcac
ctctcctgcg ataaagtctt ctgccagcca tggcagagat gcattgaggg cacctgtgtt
tgtaaactac cgtatcagtg cccaaagaat ggcactgcag tgtgtgcaac taacaggaga
agcttcccaa catactgtca acaaaagagt ttggaatgtc ttcatccagg gacaaagttt
ttaaataacg gaacatgcac agccgaagga aagtttagtg tttccttgaa gcatggaaat
acagattcag agggaatagt tgaagtaaaa cttgtggacc aagataagac aatgttcata
tgcaaaagca gctggagcat gagggaagcc aacgtggcct gccttgacct tgggtttcaa
caaggtgctg atactcaaag aaggtttaag ttgtctgatc tctctataaa ttccactgaa
tgtctacatg tgcattgccg aggattagag accagtttgg ctgaatgtac ttttactaag
agaagaacta tgggttacca ggatttcgct gatgtggttt gttatacaca gaaagcagat
tctccaatgg atgacttctt tcagtgtgtg aatgggaaat acatttctca gatgaaagcc
tgtgatggta tcaatgattg tggagaccaa agtgatgaac tgtgttgtaa agcatgccaa
ggcaaaggct tccattgcaa atcgggtgtt tgcattccaa gccagtatca atgcaatggt
gaggtggact gcattacagg ggaagatgaa gttggctgtg caggctttgc atctgtggct
caagaagaaa cagaaatttt gactgctgac atggatgcag aaagaagacg gataaaatca
ttattaccta aactatcttg tggagttaaa aacagaatgc acattcgaag gaaacgaatt
gtgggaggaa agcgagcaca actgggagac ctcccatggc aggtggcaat taaggatgcc
agtggaatca cctgtggggg aatttatatt ggtggctgtt ggattctgac tgctgcacat
tgtctcagag ccagtaaaac tcatcgttac caaatatgga caacagtagt agactggata
caccccgacc ttaaacgtat agtaattgaa tacgtggata gaattatttt ccatgaaaac
tacaatgcag gcacttacca aaatgacatc gctttgattg aaatgaaaaa agacggaaac
aaaaaagatt gtgagctgcc tcgttccatc cctgcctgtg tcccctggtc tccttaccta
ttccaaccta atgatacatg catcgtttct ggctggggac gagaaaaaga taacgaaaga
gtcttttcac ttcagtgggg tgaagttaaa ctaataagca actgctctaa gttttacgga
aatcgtttct atgaaaaaga aatggaatgt gcaggtacat atgatggttc catcgatgcc
tgtaaagggg actctggagg ccccttagtc tgtatggatg ccaacaatgt gacttatgtc
tggggtgttg tgagttgggg ggaaaactgt ggaaaaccag agttcccagg tgtttacacc
aaagtggcca attattttga ctggattagc taccatgtag gaaggccttt tatttctcag
tacaatgtat aa
(SEQ ID NO: 2)
Нуклеотидные последовательности, используемые в изобретении, могут быть оптимизированы по кодону. Оптимизация кодона ранее была описана в WO 1999/041397 и WO 2001/079518. Разные клетки различаются по использованию конкретных кодонов. Это смещение кодонов соответствует смещению в относительной численности конкретных тРНК в типе клеток. Путем изменения кодонов в последовательности таким образом, чтобы они соответствовали относительной численности соответствующих тРНК, можно усилисть экспрессию. Точно так же можно уменьшить экспрессию путем преднамеренного выбора кодонов, для которых известно, что соответствующие тРНК являются редкими в конкретном типе клеток. Таким образом, имеется дополнительная степень контроля на уровне трансляции.
В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор I, представляет собой нуклеотидную последовательность, приведенную в SEQ ID NO: 8.
ATGAAGCTGCTGCATGTCTTTCTGCTGTTTCTGTGCTTCCATCTGCGGTTCTGTAAAGTGACCTATACTAGCCAGGAGGATCTGGTGGAGAAGAAGTGTCTGGCCAAGAAGTACACACACCTGAGCTGCGACAAGGTGTTCTGTCAGCCTTGGCAGCGGTGCATCGAGGGCACCTGCGTGTGCAAGCTGCCTTACCAGTGCCCAAAGAACGGCACCGCCGTGTGCGCCACAAATCGGAGATCTTTTCCAACATATTGCCAGCAGAAGAGCCTGGAGTGTCTGCACCCCGGCACCAAGTTCCTGAACAATGGCACCTGCACAGCCGAGGGCAAGTTTTCTGTGAGCCTGAAGCACGGCAACACAGATAGCGAGGGCATCGTGGAGGTGAAGCTGGTGGACCAGGATAAGACCATGTTCATCTGTAAGAGCTCCTGGTCCATGAGGGAGGCAAACGTGGCATGCCTGGATCTGGGATTCCAGCAGGGAGCAGACACACAGAGGCGCTTTAAGCTGTCCGACCTGTCTATCAATAGCACCGAGTGCCTGCACGTGCACTGTAGGGGCCTGGAGACATCCCTGGCAGAGTGCACCTTCACAAAGCGGAGAACCATGGGCTACCAGGACTTTGCCGACGTGGTGTGCTATACCCAGAAGGCCGATAGCCCCATGGACGATTTCTTTCAGTGCGTGAACGGCAAGTATATCTCCCAGATGAAGGCCTGCGACGGCATCAATGACTGTGGCGATCAGTCTGACGAGCTGTGCTGTAAGGCCTGTCAGGGCAAGGGCTTCCACTGCAAGAGCGGCGTGTGCATCCCTTCCCAGTACCAGTGCAACGGCGAGGTGGATTGTATCACAGGAGAGGACGAAGTGGGATGCGCAGGATTTGCATCTGTGGCACAGGAGGAGACAGAGATCCTGACAGCCGACATGGATGCCGAGAGGCGCCGGATCAAGTCTCTGCTGCCTAAGCTGAGCTGTGGCGTGAAGAATCGGATGCACATCAGAAGGAAGCGCATCGTGGGAGGCAAGAGGGCACAGCTGGGCGATCTGCCATGGCAGGTGGCCATCAAGGACGCCTCTGGCATCACCTGCGGCGGCATCTACATCGGAGGATGTTGGATCCTGACCGCAGCACACTGCCTGAGAGCAAGCAAGACACACAGGTATCAGATCTGGACCACAGTGGTGGATTGGATCCACCCAGACCTGAAGAGAATCGTGATCGAGTACGTGGATAGGATCATCTTTCACGAGAACTACAATGCCGGCACATATCAGAACGACATCGCCCTGATCGAGATGAAGAAGGATGGCAATAAGAAGGACTGTGAGCTGCCCAGATCCATCCCTGCATGCGTGCCATGGAGCCCCTATCTGTTCCAGCCCAACGATACCTGCATCGTGTCCGGATGGGGAAGGGAGAAGGACAATGAGCGGGTGTTTTCTCTGCAGTGGGGCGAGGTGAAGCTGATCTCCAACTGTTCTAAGTTCTACGGCAATAGGTTTTATGAGAAGGAGATGGAGTGCGCCGGCACCTACGATGGCAGCATCGACGCCTGTAAGGGCGATTCCGGAGGACCACTGGTGTGCATGGACGCAAACAATGTGACATACGTGTGGGGAGTGGTGTCCTGGGGAGAGAACTGCGGCAAGCCAGAGTTCCCCGGCGTATATACCAAGGTGGCCAATTATTTTGATTGGATTTCCTACCACGTCGGCAGGCCCTTTATTTCCCAGTATAATGTCTAA
(SEQ ID NO: 8)
Нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное может содержать, например, нуклеотидную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO: 2 или 8, при этом белок, кодируемый нуклеотидной последовательностью по существу сохраняет функциональную активность белка, представленного SEQ ID NO:1 или 9.
Нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное может содержать, например, нуклеотидную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с последовательностью, представленной с 55 по 1752 позицию в SEQ ID NO: 2 или 8, при этом белок, кодируемый нуклеотидной последовательностью, по существу сохраняет функциональную активность белка, представленного SEQ ID NO: 1 или 9.
Нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное может, например, кодировать аминокислотную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO:1 или 9, при этом аминокислотная последовательность по существу сохраняет функциональную активность белка, представленного SEQ ID NO:1 или 9.
Нуклеотидная последовательность, кодирующая фактор I или его фрагмент или производное, может, например, кодировать аминокислотную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с последовательностью, представленной с 19 по 583 позицию в SEQ ID NO: 1 или 9, при этом аминокислотная последовательность по существу сохраняет функциональную активность белка, представленного SEQ ID NO: 1 или 9.
Преимущество изобретения заключается в том, что фактор I практически невозможно получить в виде очищенного белка. Соответственно, авторы изобретения разработали способ модуляции системы комплемента, например, позволяющий осуществлять лечение возрастной макулярной дегенерации (ВМД) путем введения фактора I в виде AAV-вектора, содержащего нуклеотидную последовательность, кодирующую фактор I. AAV-вектор можно вводить в представляющее интерес место, например глаз, чтобы обеспечить возможность трансляции in situ полипептида фактора I.
Фактор H
Фактор H комплемента (фактор H, CFH) является комплемент-контролирующим белком.
Это большой (155 кДа) растворимый гликопротеин, который присутствует в плазме человека с типичной концентрации 200-300 мкг/мл (Hakobyan et al., 2008; 49 (5):1983-90). Основная функция фактора H - регулировать альтернативный путь системы комплемента.
Фактор H обеспечивает кофакторную активность для опосредованного фактором I расщепления C3b. Фактор H также увеличивает скорость диссоциации комплекса C3bBb (C3-конвертазы) и комплекса (C3b)NBB (C5-конвертаза) и, тем самым, уменьшает активность альтернативного пути комплемента.
Фактор H состоит из модулей 20 комплемент-контролирующих белков (CCP) (также называемых короткими консенсусными повторами или доменами Суши (Sushi)), соединенных друг с другом короткими линкерами (от трех до восьми аминокислотных остатков) и простирающихся от головной до хвостовой части. Каждый из модулей CCP состоит из примерно 60 аминокислот с четырьмя цистеиновыми остатками, соединенными дисульфидами связями по схеме 1-3 2-4, и гидрофобного ядра, образованного вокруг практически инвариантного остатка триптофана. Модули CCP нумеруются от 1 до 20 (от N-конца белка). 1-4 CCP и 19-20 CCP взаимодействуют с C3b, тогда как 7 CCP и 19-20 CCP связываются с GAG и сиаловой кислотой (Schmidt et al., 2008; Journal of Immunology 181 (4): 2610-9).
Было показано, что генная терапия с использованием фактора H может облегчить индуцированную ВМД-подобную патологию у мышей (Cashman et al. (2015) J. Gene Med. 17:229-243). Мышам субретинально вводили совместно: (i) аденовирусный вектор, экспрессирующий C3 компонент комплемента, который, как ранее было показано, связан со многими патологическими признаками ВМД человека; и (ii) аденовирусный вектор, экспрессирующий фактор H. Относительно контрольных животных, получавших GFP вместо фактора H, у мышей, трансформированных фактором H, наблюдалось 91% снижение уровня пролиферации эндотелиальных клеток и 69% ослабление атрофии СЭП. Электроретинография показала улучшенную функцию сетчатки у мышей, получавших фактор H, а иммуноцитохимия родопсина и RPE65 свидетельствовала о сохранении фоторецепторов и СЭП у таких животных.
В одном из вариантов осуществления полипептид фактора H или его фрагмент или производное способен действовать в качестве кофактора для опосредованного фактором I расщепления C3b. В одном из вариантов осуществления полипептид фактора H или его фрагмент или производное способны увеличивать скорость диссоциации C3-конвертазы и C5-конвертазы.
В предпочтительном варианте осуществления полипептид фактора H или его фрагмент или производное способен действовать в качестве кофактора для опосредованного фактором I расщепления C3b и увеличивать скорость диссоциации C3-конвертазы и C5-конвертазы.
В одном из вариантов осуществления фактор H является человеческим фактором H.
В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор I, представляет собой нуклеотидную последовательность с номером доступа UniProtKB P08603. Эта приведенная в качестве примера последовательность имеет длину 1231 аминокислоты (приведена в SEQ ID NO:3), из которых аминокислоты 1-18 образуют сигнальную последовательность.
В одном из вариантов осуществления аминокислотная последовательность фактора H представляет собой последовательность, приведенную в SEQ ID NO: 3. В одном из вариантов осуществления аминокислотная последовательность фактора H представляет собой последовательность, представленную с 19 по 1231 позицию в SEQ ID NO: 3.
MRLLAKIICL MLWAICVAED CNELPPRRNT EILTGSWSDQ TYPEGTQAIY
KCRPGYRSLG NVIMVCRKGE WVALNPLRKC QKRPCGHPGD TPFGTFTLTG
GNVFEYGVKA VYTCNEGYQL LGEINYRECD TDGWTNDIPI CEVVKCLPVT
APENGKIVSS AMEPDREYHF GQAVRFVCNS GYKIEGDEEM HCSDDGFWSK
EKPKCVEISC KSPDVINGSP ISQKIIYKEN ERFQYKCNMG YEYSERGDAV
CTESGWRPLP SCEEKSCDNP YIPNGDYSPL RIKHRTGDEI TYQCRNGFYP
ATRGNTAKCT STGWIPAPRC TLKPCDYPDI KHGGLYHENM RRPYFPVAVG
KYYSYYCDEH FETPSGSYWD HIHCTQDGWS PAVPCLRKCY FPYLENGYNQ
NYGRKFVQGK SIDVACHPGY ALPKAQTTVT CMENGWSPTP RCIRVKTCSK
SSIDIENGFI SESQYTYALK EKAKYQCKLG YVTADGETSG SITCGKDGWS
AQPTCIKSCD IPVFMNARTK NDFTWFKLND TLDYECHDGY ESNTGSTTGS
IVCGYNGWSD LPICYERECE LPKIDVHLVP DRKKDQYKVG EVLKFSCKPG
FTIVGPNSVQ CYHFGLSPDL PICKEQVQSC GPPPELLNGN VKEKTKEEYG
HSEVVEYYCN PRFLMKGPNK IQCVDGEWTT LPVCIVEEST CGDIPELEHG
WAQLSSPPYY YGDSVEFNCS ESFTMIGHRS ITCIHGVWTQ LPQCVAIDKL
KKCKSSNLII LEEHLKNKKE FDHNSNIRYR CRGKEGWIHT VCINGRWDPE
VNCSMAQIQL CPPPPQIPNS HNMTTTLNYR DGEKVSVLCQ ENYLIQEGEE
ITCKDGRWQS IPLCVEKIPC SQPPQIEHGT INSSRSSQES YAHGTKLSYT
CEGGFRISEE NETTCYMGKW SSPPQCEGLP CKSPPEISHG VVAHMSDSYQ
YGEEVTYKCF EGFGIDGPAI AKCLGEKWSH PPSCIKTDCL SLPSFENAIP
MGEKKDVYKA GEQVTYTCAT YYKMDGASNV TCINSRWTGR PTCRDTSCVN
PPTVQNAYIV SRQMSKYPSG ERVRYQCRSP YEMFGDEEVM CLNGNWTEPP
QCKDSTGKCG PPPPIDNGDI TSFPLSVYAP ASSVEYQCQN LYQLEGNKRI
TCRNGQWSEP PKCLHPCVIS REIMENYNIA LRWTAKQKLY SRTGESVEFV
CKRGYRLSSR SHTLRTTCWD GKLEYPTCAK R
(SEQ ID NO: 3)
Примером нуклеотидной последовательности, кодирующей фактор H, является нуклеотидная последовательность с номером доступа NCBI NM_000186. В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор H, представляет собой нуклеотидную последовательность с номером доступа NCSCI NM_000186.
В одном из вариантов осуществления нуклеотидная последовательность, кодирующая фактор H, представляет собой нуклеотидную последовательность, приведенную в SEQ ID NO: 4.
atgagacttc tagcaaagat tatttgcctt atgttatggg ctatttgtgt agcagaagat
tgcaatgaac ttcctccaag aagaaataca gaaattctga caggttcctg gtctgaccaa
acatatccag aaggcaccca ggctatctat aaatgccgcc ctggatatag atctcttgga
aatgtaataa tggtatgcag gaagggagaa tgggttgctc ttaatccatt aaggaaatgt
cagaaaaggc cctgtggaca tcctggagat actccttttg gtacttttac ccttacagga
ggaaatgtgt ttgaatatgg tgtaaaagct gtgtatacat gtaatgaggg gtatcaattg
ctaggtgaga ttaattaccg tgaatgtgac acagatggat ggaccaatga tattcctata
tgtgaagttg tgaagtgttt accagtgaca gcaccagaga atggaaaaat tgtcagtagt
gcaatggaac cagatcggga ataccatttt ggacaagcag tacggtttgt atgtaactca
ggctacaaga ttgaaggaga tgaagaaatg cattgttcag acgatggttt ttggagtaaa
gagaaaccaa agtgtgtgga aatttcatgc aaatccccag atgttataaa tggatctcct
atatctcaga agattattta taaggagaat gaacgatttc aatataaatg taacatgggt
tatgaataca gtgaaagagg agatgctgta tgcactgaat ctggatggcg tccgttgcct
tcatgtgaag aaaaatcatg tgataatcct tatattccaa atggtgacta ctcaccttta
aggattaaac acagaactgg agatgaaatc acgtaccagt gtagaaatgg tttttatcct
gcaacccggg gaaatacagc aaaatgcaca agtactggct ggatacctgc tccgagatgt
accttgaaac cttgtgatta tccagacatt aaacatggag gtctatatca tgagaatatg
cgtagaccat actttccagt agctgtagga aaatattact cctattactg tgatgaacat
tttgagactc cgtcaggaag ttactgggat cacattcatt gcacacaaga tggatggtcg
ccagcagtac catgcctcag aaaatgttat tttccttatt tggaaaatgg atataatcaa
aatcatggaa gaaagtttgt acagggtaaa tctatagacg ttgcctgcca tcctggctac
gctcttccaa aagcgcagac cacagttaca tgtatggaga atggctggtc tcctactccc
agatgcatcc gtgtcaaaac atgttccaaa tcaagtatag atattgagaa tgggtttatt
tctgaatctc agtatacata tgccttaaaa gaaaaagcga aatatcaatg caaactagga
tatgtaacag cagatggtga aacatcagga tcaattacat gtgggaaaga tggatggtca
gctcaaccca cgtgcattaa atcttgtgat atcccagtat ttatgaatgc cagaactaaa
aatgacttca catggtttaa gctgaatgac acattggact atgaatgcca tgatggttat
gaaagcaata ctggaagcac cactggttcc atagtgtgtg gttacaatgg ttggtctgat
ttacccatat gttatgaaag agaatgcgaa cttcctaaaa tagatgtaca cttagttcct
gatcgcaaga aagaccagta taaagttgga gaggtgttga aattctcctg caaaccagga
tttacaatag ttggacctaa ttccgttcag tgctaccact ttggattgtc tcctgacctc
ccaatatgta aagagcaagt acaatcatgt ggtccacctc ctgaactcct caatgggaat
gttaaggaaa aaacgaaaga agaatatgga cacagtgaag tggtggaata ttattgcaat
cctagatttc taatgaaggg acctaataaa attcaatgtg ttgatggaga gtggacaact
ttaccagtgt gtattgtgga ggagagtacc tgtggagata tacctgaact tgaacatggc
tgggcccagc tttcttcccc tccttattac tatggagatt cagtggaatt caattgctca
gaatcattta caatgattgg acacagatca attacgtgta ttcatggagt atggacccaa
cttccccagt gtgtggcaat agataaactt aagaagtgca aatcatcaaa tttaattata
cttgaggaac atttaaaaaa caagaaggaa ttcgatcata attctaacat aaggtacaga
tgtagaggaa aagaaggatg gatacacaca gtctgcataa atggaagatg ggatccagaa
gtgaactgct caatggcaca aatacaatta tgcccacctc cacctcagat tcccaattct
cacaatatga caaccacact gaattatcgg gatggagaaa aagtatctgt tctttgccaa
gaaaattatc taattcagga aggagaagaa attacatgca aagatggaag atggcagtca
ataccactct gtgttgaaaa aattccatgt tcacaaccac ctcagataga acacggaacc
attaattcat ccaggtcttc acaagaaagt tatgcacatg ggactaaatt gagttatact
tgtgagggtg gtttcaggat atctgaagaa aatgaaacaa catgctacat gggaaaatgg
agttctccac ctcagtgtga aggccttcct tgtaaatctc cacctgagat ttctcatggt
gttgtagctc acatgtcaga cagttatcag tatggagaag aagttacgta caaatgtttt
gaaggttttg gaattgatgg gcctgcaatt gcaaaatgct taggagaaaa atggtctcac
cctccatcat gcataaaaac agattgtctc agtttaccta gctttgaaaa tgccataccc
atgggagaga agaaggatgt gtataaggcg ggtgagcaag tgacttacac ttgtgcaaca
tattacaaaa tggatggagc cagtaatgta acatgcatta atagcagatg gacaggaagg
ccaacatgca gagacacctc ctgtgtgaat ccgcccacag tacaaaatgc ttatatagtg
tcgagacaga tgagtaaata tccatctggt gagagagtac gttatcaatg taggagccct
tatgaaatgt ttggggatga agaagtgatg tgtttaaatg gaaactggac ggaaccacct
caatgcaaag attctacagg aaaatgtggg ccccctccac ctattgacaa tggggacatt
acttcattcc cgttgtcagt atatgctcca gcttcatcag ttgagtacca atgccagaac
ttgtatcaac ttgagggtaa caagcgaata acatgtagaa atggacaatg gtcagaacca
ccaaaatgct tacatccgtg tgtaatatcc cgagaaatta tggaaaatta taacatagca
ttaaggtgga cagccaaaca gaagctttat tcgagaacag gtgaatcagt tgaatttgtg
tgtaaacggg gatatcgtct ttcatcacgt tctcacacat tgcgaacaac atgttgggat
gggaaactgg agtatccaac ttgtgcaaaa agatag
(SEQ ID NO: 4)
Нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное может содержать, например, нуклеотидную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO: 4, при этом белок, кодируемый нуклеотидной последовательностью по существу сохраняет функциональную активность белка, приведенного в SEQ ID NO:3.
Нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное может содержать, например, нуклеотидную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с последовательностью, представленной с 55 по 3696 позицию в SEQ ID NO: 4, при этом белок, кодируемый нуклеотидной последовательностью, по существу сохраняет функциональную активность белка, приведенного в SEQ ID NO: 3.
Нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное может, например, кодировать аминокислотную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO:3, при этом аминокислотная последовательность по существу сохраняет функциональную активность белка, приведенного в SEQ ID NO:3.
Нуклеотидная последовательность, кодирующая фактор H или его фрагмент или производное может, например, кодировать аминокислотную последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с последовательностью, представленной с 19 по 1231 позицию в SEQ ID NO:3, при этом аминокислотная последовательность по существу сохраняет функциональную активность белка, приведенного в SEQ ID NO:3.
Фактор D
Фактор D комплемента (фактор D, CFD) участвует в альтернативном пути системы комплемента. Его функцией является расщепление фактора B на факторы Bb и Ba.
Фактор D является членом пептидаз семейства трипсина. Все члены сериновых протеаз семейства химотрипсина имеют очень схожие структуры. Все они, включая фактор D, имеют два антипараллельных домена β-баррелей, где каждый баррель содержит шесть β-нитей с одинаковой типологией во всех ферментах. Основное отличие в структуре остова между фактором D и другими сериновыми протеазами семейства химотрипсина заключается в поверхностных петлях, соединяющих вторичные структурные элементы.
Каскад альтернативного пути активации комплемента инициируется спонтанным гидролизом C3, который в изобилии содержится в плазме крови. Постоянный уровень ("tickover") поддерживается за счет спонтанного расщепления тиоэфирной связи в С3 с образованием С3(Н2О).
Такое изменение формы дает возможность связывать фактор В белка плазмы, что позволяет Фактору D расщеплять фактор B на Ba и Bb. Bb становится частью C3(H2O), образуя C3(H2O)Bb. Этот комплекс также известен как жидкофазная C3-конвертаза. Эта конвертаза, хотя и продуцируется только в небольших количествах, может расщеплять несколько C3-белков на C3a и C3b.
AAV-вектор по настоящему изобретению может содержать нуклеотидную последовательность, которая кодирует антитело к фактору D.
Антитело к фактору D может связываться с фактором D и снижать или подавлять функциональную активность фактора D. Например, антитело к фактору D может уменьшать или предотвращать связывание фактора D с фактором B и/или расщепление фактора B фактором D.
Подходящие антитела к фактору D известны в данной области. Такие антитела включают, без ограничения, лампализумаб.
Компонент комплимента 5
Компонент комплемента 5 (C5) является пятым компонентом комплемента, который играет важную роль в процессах воспаления и гибели клеток. C5 состоит из альфа- и бета-полипептидных цепей, которые связаны дисульфидным мостиком.
C5 расщепляется протеазой C5-конвертазой на фрагменты 5a (C5a) и C5b компонента комплемента. C5b играет важную роль в поздних событиях каскада комплемента, тогда как C5a действует в качестве белка воспалительного процесса. C5 обычно синтезируется гепатоцитами, но его синтез также можно обнаружить в макрофагах, и это может привести к локальному увеличению C5a. C5a обладает хемотаксическими и анафилатоксическими свойствами, он является важным компонентом врожденного иммунитета, но также связан с адаптивным иммунитетом. Усиление продуцирования C5a связано с рядом воспалительных заболеваний.
C5a является анафилатоксином, вызывая повышенную экспрессию молекул адгезии на эндотелии, сокращение гладких мышц и повышенную проницаемость сосудов. C5a-des-Arg (с отсутствующим С-концевым аргинином) является гораздо менее мощным анафилатоксином. Оба C5a и C5a-des-Arg могут вызывать дегрануляцию тучных клеток, высвобождая провоспалительные молекулы гистамина и TNF-α. C5a также является мощным хемоаттрактантом, инициирующим накопление комплемента и фагоцитарных клеток в местах инфекции или рекрутирование антигенпрезентирующих клеток в лимфатические узлы. C5a играет ключевую роль в усилении миграции и адгезии нейтрофилов и моноцитов к стенкам сосудов. Лейкоциты активируются повышением авидности интегрина, усилением липоксигеназного пути и метаболизма арахидоновой кислоты. C5a также модулирует баланс между активирующими и ингибирующими Fc-рецепторами к IgG на лейкоцитах, тем самым усиливая аутоиммунный ответ.
AAV-вектор по настоящему изобретению может содержать нуклеотидную последовательность, которая кодирует анти-С5 антитело.
Анти-С5 антитело может связывать С5 и предотвращать расщепление С5 на C5a и C5b.
Подходящие анти-С5 антитела известны в данной области. Такие антитела включают, без ограничения, экулизумаб.
Антитело
Антитело относится к любой части антитела или его фрагмента, которая сохраняет способность связываться с той же самой антигенной мишенью, что и родительское антитело.
Антитело может быть химерным антителом. Химерные антитела могут быть получены путем трансплантации вариабельных доменов антитела одного вида (например, мыши) в постоянные домены антитела другого вида (например, человека).
Антитело может представлять собой полноразмерное классическое антитело. Например, антитело может представлять собой молекулу IgG, IgM или IgA.
Антитело может быть функциональным фрагментом антитела. Конкретные фрагменты антител включают, без ограничения, (i) Fab-фрагмент, состоящий из доменов VL, VH, CL и CH1, (ii) Fd-фрагмент, состоящий из доменов VH и CH1, (iii) Fv-фрагмент, состоящий из доменов VL и VH одного антитела, (iv) dAb фрагмент, который состоит из одного вариабельного домена, (v) выделенные CDR области, (vi) F(ab')2 фрагменты, бивалентный фрагмент, содержащий два связанных Fab фрагмента (vii) одноцепочечные молекулы Fv (scFv), в которых домен VH и домен VL связаны пептидным линкером, позволяющим двум доменам ассоциировать с образованием антигенсвязывающего сайта, (viii) биспецифические одноцепочечные Fv-димеры и (ix) "диатела" или "триатела", мультивалентные или мультиспецифические фрагменты, сконструированные путем слияния генов. Фрагменты антител могут быть модифицированы. Например, молекулы могут быть стабилизированы путем введения дисульфидных мостиков, связывающих домены VH и VL.
Антитело, описанное в настоящей заявке, может быть мультиспецифическим антителом и, в частности биспецифическим антителом, также иногда называемым "диателом". Это - антитела, которые связываются с двумя (или более) различными антигенами. Антитело может быть минителом. Минитела представляют собой минимизированные антителоподобные белки, содержащие scFv, присоединенный к домену СН3. В некоторых случаях scFv может быть присоединен к Fc-области и может включать часть или весь шарнирный участок.
Антитело может быть доменным антителом (также называемым однодоменным антителом или нанотелом). Это - фрагмент антитела, содержащий один мономерный единичный вариабельный домен антитела. Примеры однодоменных антител включают, без ограничения, фрагменты VHH, впервые обнаруженные у верблюдов, и фрагменты VNAR, впервые обнаруженные у хрящевых рыб. Однодоменные антитела также могут быть получены путем расщепления димерных вариабельных доменов, происходящих из общих молекул IgG, на мономеры.
Антитело может быть синтетическим антителом (также называемым антителом-миметиком). Антитела-миметики включают, без ограничения, аффитела, DARP белки, антикалины, авимеры, версатела и дуокалины.
Возрастная макулярная дегенерация (ВМД)
Этапы характерные для клинического пирования ВМД соответствуют происходящим в макуле изменениям. Отличительной особенностью ранней стадии ВМД являются друзы, представляющие собой скопления под сетчаткой внеклеточного дебриса, видимые на сетчатке при клиническом осмотре глазного дна и на фотографиях глаза в виде желтых пятен. В зависимости от размера выделяют небольшие (<63 мкм), средние (63-124 мкм) и крупные (> 124 мкм) друзы. В зависимости от внешнего вида границы друз при офтальмологическом осмотре их также подразделяют на жесткие или мягкие. В то время как твердые друзы имеют четко определенные границы, границы мягких друз менее четкие и размытые. Одной из основных систем классификации, используемых для этого состояния, является классификация, разработанная в ходе исследования возрастной глазной патологии (AREDS) на основании снимков глазного дна.
Выделяют "сухую" и "влажную" (экссудативную или неоваскулярную) формы ВМД. Сухая форма ВМД является более распространенной чем влажная, однако она может прогрессировать до влажной формы, и в целом ряде случаев эти две формы сосуществуют. Сухая форма ВМД обычно характеризуется прогрессирующим апоптозом клеток в слое ПЭС, вышележащих фоторецепторных клеток, а также часто и нижележащих клеток хориокапиллярного слоя. Сливающиеся области гибели клеток ПЭС, сопровождающейся атрофией вышележащих фоторецепторов, называются географической атрофией. У пациентов с такой формой ВМД происходит медленное и прогрессивное ухудшение центрального зрения.
Влажная форма ВМД характеризуется кровотечением и/или утечкой жидкости из аномальных сосудов, выросших из хориоидальных сосудов (хориокапилляров) под ПЭС и макулой, что может стать причиной внезапной и инвалидизируюшей потери зрения. Согласно имеющимся оценкам, в большинстве случаев потеря зрения у пациентов обусловлена такой хориоидальной неоваскуляризацией (ХНВ) и вызываемыми ею вторичными осложнениями.
Описанное в настоящей заявке лечение или профилактика ВМД может уменьшить или предотвратить проявление описанного выше фенотипа ВМД. Предпочтительно, лечение ВМД позволяет поддерживать или улучшать функцию зрения.
В одном из вариантов осуществления лечение или профилактика ВМД приводит к предотвращению или уменьшению образования географической атрофии. В другом варианте осуществления лечение или профилактика ВМД приводит к замедлению прогрессирования географической атрофии. Например, увеличение площади географической атрофии снижается по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% за 12 месяцев после введения в подвергшийся лечению глаз субъекта по сравнению с не подвергшимся лечению глазом за тот же период. В другом варианте осуществления лечение или профилактика ВМД приводит к лечению географической атрофии, например, к уменьшению величины географической атрофии.
В одном из вариантов осуществления лечение или профилактика ВМД приводит к предотвращению или уменьшению образования друз. В другом варианте осуществления лечение или профилактика ВМД приводит к уменьшению существующих друз, например, к уменьшению размера и/или количества существующих друз.
В одном из вариантов осуществления лечение или профилактика ВМД приводит к предотвращению или уменьшению отложения комплемента. В другом варианте осуществления лечение или профилактика ВМД приводит к уменьшению существующего отложения комплемента.
В одном из вариантов осуществления лечение или профилактика ВМД приводит к улучшению или восстановлению зрения или остроты зрения. В другом варианте осуществления лечение или профилактика ВМД уменьшает скорость потери зрения или остроты зрения.
В одном из вариантов осуществления лечение или предотвращение ВМД приводит к улучшению или восстановлению скорости чтения у субъекта. В другом варианте осуществления лечение или профилактика ВМД уменьшает снижение скорости чтения у субъекта.
В одном из вариантов осуществления лечение или профилактика ВМД приводит к уменьшению или предотвращению потери фоторецепторов и/или пигментного эпителия сетчатки (ПЭС).
Диабетическая ретинопатия
Диабетическая ретинопатия - это состояние, которое характеризуется поражением кровеносных сосудов сетчатки, вызванным высокими уровнями сахара в крови, связанными с диабетом. Если не лечить диабетическую ретинопатию, это может привести к слепоте.
Хотя у субъектов с легкой диабетической ретинопатией может быть хорошее зрение, два типа диабетической ретинопатии, а именно диабетический макулярный отек (DMO) и пролиферативная диабетическая ретинопатия (PDR), могут угрожать зрению субъекта.
Диабетический макулярный отек характеризуется утечкой жидкости из поврежденных кровеносных сосудов в задней части глаза. Просочившаяся жидкость накапливается в макуле, что приводит к отеку и расфокусированному зрению. Это может в конечном итоге привести к плохому центральному зрению и невозможности чтения или вождения. Боковое зрение обычно остается нормальным.
Пролиферативная диабетическая ретинопатия характеризуется закрытием кровеносных сосудов сетчатки, что приводит к росту аномальных хрупких кровеносных сосудов на поверхности сетчатки. Это может привести к потере зрения из-за кровоизлияния в глазу, рубцевания и отслоения сетчатки.
Структура глаза
Представленные в настоящей заявке лекарственные средства могут быть доставлены млекопитающему, предпочтительно в человеческий глаз, с целью лечения или профилактики возрастной макулярной дегенерации (ВМД).
Специалист по лечению заболеваний глаз имеет детальное и полное понимание структуры глаза. Однако ниже описаны структуры, имеющие особое значение для изобретения.
Сетчатка
Сетчатка представляет собой многослойную мембрану, которая выстилает внутреннюю заднюю камеру глаза и воспринимает образ видимого мира, который передается в мозг по зрительному нерву. В направлении изнутри наружу сетчатка глаза содержит слои нейросенсорной сетчатки и пигментный эпителий сетчатки, сосудистую оболочку, расположенную за пигментным эпителием сетчатки.
Нейросенсорная сетчатка и фоторецепторные клетки
Нейросенсорная сетчатка содержит фоторецепторные клетки, которые непосредственно воспринимают свет. Она содержит следующие слои: внутренняя ограничивающая мембрана (ILM); слой нервных волокон; слой ганглиозных клеток; внутренний сетчатый слой; внутренний ядерный слой; наружный сетчатый слой; внешний ядерный слой (ядра фоторецепторов); наружная пограничная мембрана (ELM); и фоторецепторы (внутренний и внешний сегменты палочек и колбочек).
Специалист в данной области имеет детальное понимание фоторецепторных клеток. Упоминая вкратце, фоторецепторные клетки представляют собой специализированные нейроны, расположенные в сетчатке, которые преобразуют свет в биологические сигналы. Фоторецепторные клетки содержат палочки и колбочки, которые по-разному распределены в сетчатке.
Палочки распределены главным образом во внешних частях сетчатки. Они очень чувствительны и обеспечивают зрение при низких уровнях освещенности. В нормальной человеческой сетчатке в среднем содержится примерно 125 миллионов палочек.
Колбочки распределены по всей сетчатке с наибольшей концентрацией в фовеа, углублении в нейросенсорной сетчатке, которая отвечает за центральное зрение, отличающееся высоким разрешением. Колбочки менее чувствительны, чем палочки. Нормальная человеческая сетчатка содержит в среднем примерно 6-7 миллионов колбочек.
Пигментный эпителий сетчатки
Пигментный эпителий сетчатки (ПЭС) представляет собой пигментированный слой клеток, расположенных непосредственно снаружи нейросенсорной сетчатки. ПЭС выполняет ряд функций, включая перенос питательных веществ и других веществ в фоторецепторные клетки, и поглощение рассеянного света для улучшения зрения.
Сосудистая оболочка (хориоидея)
Хориоидея - сосудистый слой, расположенный между ПЭС и внешней склерой глаза. Сосудистая система хориоидеи обеспечивает сетчатку кислородом и питательными веществами.
Аденоассоциированные вирусные (AAV) векторы
В одном из аспектов изобретения предоставляется AAV-вектор, содержащий нуклеотидную последовательность, кодирующую фактор I или его фрагмент или производное и/или фактор H или его фрагмент или производное.
Предпочтительно AAV-вектор находится в форме AAV частицы.
Способы получения и модификации вирусных векторов и вирусных векторных частиц, например, полученных из AAV, хорошо известны в данной области.
AAV-вектор может содержать геном AAV или его фрагмент или производное.
Геном AAV представляет собой полинуклеотидную последовательность, которая кодирует функции, необходимые для получения AAV частицы. Эти функции включают те, которые работают в цикле репликации и упаковки AAV в клетке хозяина, включая инкапсуляцию генома AAV в AAV частицу. Встречающиеся в природе AAV являются дефицитными по репликации и зависят от внешних вспомогательных функций для завершения цикла репликации и упаковки. Соответственно, геном AAV AAV-вектора по изобретению, как правило, является дефицитным по репликации.
Геном AAV может быть одноцепочечным, как положительным, так и отрицательным, или, альтернативно, двухцепочечным. Использование двухцепочечной формы позволяет обходить стадию репликации ДНК в клетке-мишени и, таким образом, ускорять экспрессию трансгена.
Геном AAV может происходить из любого встречающегося в природе серотипа, изолята или клада AAV. Таким образом, геном AAV может представлять собой полный геномом встречающегося в природе AAV. Как известно специалисту в данной области, встречающиеся в природе AAV могут быть классифицированы в соответствии с разными биологическими системами.
Обычно AAV называют по их серотипу. Серотип соответствует вариантному подвиду AAV, который благодаря своему профилю экспрессии поверхностных капсидных антигенов имеет характерную реактивность, которую можно использовать для отличия его от других вариантных подвидов. Как правило, вирус с конкретным серотипом AAV не вступает в перекрестные реакции с нейтрализующими антителами, специфичными к любому другому серотипу AAV.
Серотипы AAV включают AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10 и AAV11, а также рекомбинантные серотипы, такие как Rec2 и Rec3, недавно выделенные из мозга приматов. Любой из этих серотипов AAV может быть использован в изобретении. Таким образом, в одном из вариантов осуществления изобретения предоставляется AAV-векторная частица, которая представляет собой векторную частицу AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, Rec2 или Rec3 AAV.
В одном из вариантов осуществления AAV может относиться к серотипу AAV1, AAV2, AAV5, AAV7, AAV8 или AAV8.
В одном из вариантов осуществления AAV может относиться к серотипу AAV2 или AAV8.
Капсидный белок может представлять собой мутантный капсидный белок, такой как раскрытый в документе WO 2008/124724, включенном в настоящее описание в виде ссылки.
В одном из вариантов осуществления AAV-вектор содержит капсид AAV8 с мутацией Y733F.
Обзор серотипов AAV можно найти в Choi et al. (2005) Curr. Gene Ther. 5: 299-310 и Wu et al. (2006) Molecular Therapy 14: 316-27. Последовательности геномов AAV или элементов геномов AAV, включая ITR последовательности, гены rep или cap, для использования в изобретении, могут быть получены из цельных последовательностей генома AAV со следующими номерами доступа: аденоассоциированный вирус 1 NC_002077, AF063497; аденоассоциированный вирус 2 NC_001401; аденоассоциированный вирус 3 NC_001729; аденоассоциированный вирус 3B NC_001863; аденоассоциированный вирус 4 NC_001829; аденоассоциированный вирус 5 Y18065, AF085716; аденоассоциированный вирус 6 NC_001862; птичий AAV ATCC VR-865 AY186198, AY629583, NC_004828; штамм DA-1 птичьего AAV NC_006263, AY629583; бычий AAV NC_005889, AY388617.
AAV также может упоминаться в терминах кладов или клонов. Это относится к филогенетической взаимосвязи между встречающимися в природе AAV и, как правило, к филогенетической группе AAV, которую можно проследить до общего предка, и включает все его потомство. Кроме того, AAV могут упоминаться в терминах конкретного изолята, т.е. генетического изолята конкретного встречающегося в природе AAV. Термин генетический изолят описывает популяцию AAV, которая подверглась ограниченному генетическому смешению с другими встречающимися в природе AAV, таким образом определяя распознаваемо отличающуюся популяцию на генетическом уровне.
Специалист может выбрать подходящий серотип, клад, клон или изолят AAV для использования в изобретении, исходя из общих сведений о них. Например, была показана эффективная трансдукция капсида AAV5 в фоторецепторы приматов, о чем свидетельствует успешная коррекция наследственного дефекта цветового зрения (Mancuso et al. (2009) Nature 461: 784-7).
Серотип AAV определяет тканевую специфичность инфекции (или тропизм) AAV вируса. Соответственно, предпочтительными серотипами AAV для использования в качестве вводимых пациентам AAV в соответствии с изобретением, являются те, которые имеют естественный тропизм или высокую эффективность инфицирования клеток-мишеней в глазу. В одном из вариантов осуществления серотипы AAV для использования в изобретении представляют собой серотипы, которые трансдуцируют клетки нейросенсорной сетчатки, пигментного эпителия сетчатки и/или сосудистой оболочки.
Как правило, геном AAV полученного естественным образом серотипа, изолята или клада AAV содержит по меньшей мере одну последовательность инвертированного концевого повтора (ITR). ITR последовательность действует в цис-конфигурации, обеспечивая функциональную точку начала репликации, и позволяет встраивать и вырезать вектор из генома клетки. В предпочтительных вариантах осуществления одна или более ITR последовательностей фланкируют нуклеотидные последовательности, кодирующие фактор I и/или фактор H (или их фрагменты или производные). Геном AAV обычно также включает необходимые для упаковки гены, такие как гены rep и/или cap, которые кодируют функции упаковки AAV частицы. Ген rep кодирует один или более белков Rep78, Rep68, Rep52 и Rep40 или их варианты. Ген cap кодирует один или более капсидных белков, таких как VP1, VP2 и VP3 или их варианты. Эти белки составляют капсид AAV частицы. Варианты капсида обсуждаются ниже.
Промотор функционально связан с каждым из генов упаковки. Конкретные примеры таких промоторов включают промоторы p5, p19 и p40 (Laughlin et al., 1979) Proc. Natl. Acad. Sci. USA 76: 5567-5571). Например, промоторы p5 и p19 обычно используют для экспрессии гена rep, тогда как промотор p40 обычно используют для экспрессии гена cap.
Как обсуждалось выше, геном AAV, используемый в AAV-векторе по изобретению, может следовательно представлять собой полный геномом встречающегося в природе AAV. Например, для получения AAV-вектора или векторной частицы in vitro может быть использован вектор, содержащий полный геном AAV. Однако, хотя такой вектор можно в принципе вводить пациентам, на практике это осуществляется редко. Предпочтительно геном AAV дериватизируют для введения пациентам. Такая дериватизация является стандартной в данной области, и в объем изобретения включено использование любого известного производного генома AAV, а также производных, которые могут быть получены методами, известных в данной области. Дериватизация генома AAV и капсида AAV рассмотрена в Coura and Nardi (2007) Virology Journal 4: 99, а в Choi et al. и Wu et al., упомянутых выше.
Производные генома AAV включают любые усеченные или модифицированные формы генома AAV, которые обеспечивают экспрессию трансгена из AAV-вектора по изобретению in vivo. Как правило, геном AAV может быть усеченным в значительной степени так, чтобы он включал минимальную вирусную последовательность, сохраняя при этом указанную выше функцию. Это является предпочтительным из соображения безопасности для снижения риска рекомбинации вектора с вирусом дикого типа, а также для предотвращения инициирования клеточного иммунного ответа из-за наличия вирусных генных белков в клетке-мишени.
Как правило, производное включает по меньшей мере одну последовательность инвертированного концевого повтора (ITR), предпочтительно более одного ITR, например два ITR или более. Один или более ITR могут быть получены из геномов AAV, имеющих разные серотипы, или могут быть химерным или мутантным ITR. Предпочтительный мутантный ITR представляет собой ITR, имеющий делецию trs (сайт концевого разрешения). Эта делеция позволяет продолжать репликацию генома для получения одноцепочечного генома, который содержит как кодирующие, так и комплементарные последовательности, т.е. самокомплементарный геном AAV. Это позволяет обходить репликацию ДНК в клетке-мишени и, таким образом, обеспечивает ускоренную экспрессию трансгена.
Один или несколько ITR предпочтительно фланкируют с обоих концов нуклеотидную последовательность, кодирующую фактор I и/или фактор H (или его фрагмент или производные). Включение одного или более ITR предпочтительнее для образования конкатемера вектора по изобретению в ядре клетки-хозяина, например, после превращения одноцепочечной векторной ДНК в двухцепочечную ДНК в результате действия ДНК полимеразы клетки-хозяина. Образование таких эписомальных конкатемеров защищает векторную конструкцию в течение жизни клетки-хозяина, тем самым обеспечивая длительную экспрессию трансгена in vivo.
В предпочтительных вариантах осуществления элементы ITR являются единственными последовательностями, оставшимися в производном от нативного генома AAV. Таким образом, производное предпочтительно не включает гены rep и/или cap нативного генома и любые другие последовательности нативного генома. Это является предпочтительным по причинам, описанным выше, а также для уменьшения возможности интеграции вектора в геном клетки-хозяина. Кроме того, уменьшение размера генома AAV позволяет повысить гибкость при внедрении в вектор других элементов последовательности (таких как регуляторные элементы) дополнительно к трансгену.
Следовательно, из производного по изобретению могут быть удалены следующие части: одна последовательность инвертированных терминальных повторов (ITR), гены репликации (rep) и капсида (cap). Однако в некоторых вариантах осуществления производные могут дополнительно включать один или более генов rep и/или cap или другие вирусные последовательности генома AAV. Встречающийся в природе AAV способен встраиваться с высокой частотой в определенном месте хромосомы человека 19 с незначительной частотой случайной интеграции, вследствие чего сохранение способности вектора к интеграции допустима в схеме лечения.
Если производное содержит капсидные белки, т.е. VP1, VP2 и/или VP3, оно может быть химерным производным одного или более встречающихся в природе AAV, производным с перетасованными экзонами или производным с модифицированным капсидом. В частности, в объем изобретения также включено предоставление последовательностей капсидных белков из разных серотипов, кладов, клонов или изолятов AAV в пределах одного и того же вектора (т.е. псевдотипированного вектора).
Химерные, перетасованные или с модифицированным капсидом производные обычно выбирают для получения AAV-вектора с одной или более желаемыми функциональными возможностями. Таким образом, эти производные могут проявлять повышенную эффективность доставки гена, сниженную иммуногенность (гуморальную или клеточную), измененный диапазон тропизма и/или улучшенное нацеливание на конкретный тип клеток по сравнению с AAV-вектором, включающим встречающийся в природе геном AAV, такой как геном AAV2. Повышенная эффективность доставки гена может быть достигнута за счет улучшенного связывания рецептора или корецептора на поверхности клетки, улучшенной интернализации, улучшенной миграции внутри клетки и в ядро, улучшенной декапсидации вирусной частицы и улучшенного превращения одноцепочечного генома в двухцепочечную форму. Повышенная эффективность может также относиться к измененному диапазону тропизма или направленного воздействия на конкретную популяцию клеток, благодаря чему доза вектора не разбавляется в результате введения в ткани, если в этом нет необходимости.
Химерные капсидные белки включают белки, полученные в результате рекомбинации между двумя или более кодирующими капсидный белок последовательностями встречающихся в природе серотипов AAV. Их получение возможно, например, с помощью метода спасения генетического маркера, при котором неинфекционные последовательности капсидного белка одного серотипа трансфицируют вместе с последовательностями капсидного белка другого серотипа, с последующим осуществлением направленного отбора для отбора последовательностей капсидных белков, имеющих нужные свойства. Последовательности капсидных белков различных серотипов могут быть изменены путем гомологичной рекомбинации внутри клетки для получения новых химерных капсидных белков.
Химерные капсидные белки также включают белки, которые получают путем конструирования последовательностей капсидных белков для переноса конкретных доменов капсидного белка, поверхностных петель или отдельных аминокислотных остатков между двумя или более капсидными белками, например, между двумя или более капсидными белками разных серотипов.
Перетасованные или химерные капсидные белки также могут быть получены путем перетасовки ДНК или методом ПЦР с внесением ошибок. Гибридные гены капсидных белков AAV могут быть созданы путем случайной фрагментации последовательностей родственных генов AAV, например, генов, которые кодируют капсидные белки из нескольких различных серотипов, с последующей сборкой фрагментов в полимеразной реакции самопраймирования, что также может приводить к кроссоверам в областях гомологии последовательностей. Библиотека гибридных генов AAV, созданная таким образом путем перетасовки генов капсидных белков нескольких серотипов, может быть подвергнута скринингу для идентификации вирусных клонов, имеющих нужные функциональные свойства. Аналогично, можно использовать ПЦР с внесением ошибок для случайного мутирования генов капсидных белков AAV для создания библиотеки различных вариантов, которые затем могут быть подвергнуты отбору по нужному свойству.
Последовательности генов капсидных белков также могут быть генетически модифицированы для введения конкретных делеций, замещений или вставок относительно последовательности нативного дикого типа. В частности, гены капсидных белков могут быть модифицированы путем вставки последовательности неродственного белка или пептида в открытую рамку считывания кодирующей капсидный белок последовательности или на N- и/или С-конце кодирующей капсидный белок последовательности.
Неродственный белок или пептид может преимущественно быть белком, который действует в качестве лиганда для конкретного типа клеток, тем самым обеспечивая улучшенное связывание с клеткой-мишенью или улучшая специфичность нацеливания вектора на конкретную популяцию клеток. Примером может служить использование пептида RGD для блокирования всасывания в пигментный эпителий сетчатки и, таким образом, усиливая трансдукцию окружающих тканей сетчатки (Cronin et al. (2008) ARVO Abstract: D1048). Неродственный белок также может быть белком, который способствует очистке вирусной частицы как части процесса получения, т.е. меткой эпитопа или аффинности. Сайт для вставки обычно выбирают таким образом, чтобы он не интерферировал с другими функциями вирусной частицы, например, интернализацией, миграцией вирусной частицы. Специалист в данной области может определить подходящие сайты для вставки, исходя из общих знаний. Конкретные сайты описаны в Choi et al., ссылки приведены выше.
Изобретение дополнительно включает получение последовательностей генома AAV с другим порядком и конфигурацией относительно нативного генома AAV. В объем изобретения также входит замена одной или более последовательностей AAV или гены с последовательностями из другого вируса или химерные гены, состоящие из последовательностей более чем одного вируса. Такие химерные гены могут состоять из последовательностей, полученных из двух или более родственных вирусных белков вирусов разных видов.
AAV-вектор по изобретению может принимать форму нуклеотидной последовательности, содержащей геном AAV или его производное, и последовательность, кодирующую трансген фактора I и/или фактора H или его производные.
AAV частицы по изобретению включают транскапсидированные формы, в которых геном AAV или производное, имеющее ITR одного серотипа, упакован в капсид другого серотипа. AAV частицы по изобретению также включают мозаичные формы, в которых смесь немодифицированных капсидных белков из двух или более различных серотипов составляет вирусный капсид. AAV частица также включает химически модифицированные формы, несущие лиганды, адсорбированные на поверхности капсида. Например, такие лиганды могут включать антитела для нацеливания на конкретный рецептор на поверхности клетки.
Так, например, AAV частицы по изобретению включают частицы с геномом AAV2 и капсидныыми белками AAV2 (AAV2/2), частицы с геномом AAV2 и капсидными белками AAV5 (AAV2/5), и частицы с геномом AAV2 и капсидными белками AAV8 (AAV2/8), а также частицы с геномом AAV2 и капсидными белками более чем одного серотипа.
AAV-вектор может содержать множество копий (например, 2, 3 и т.д.) раскрытой в настоящей заявке нуклеотидной последовательности.
Промоторы и регуляторные последовательности
AAV-вектор по изобретению может также включать элементы, обеспечивающие экспрессию трансгенов фактора I и/или фактора H (или их фрагментов или производных) in vitro или in vivo. Они могут упоминаться как последовательности контроля экспрессии. Таким образом, AAV-вектор обычно содержит последовательности контроля экспрессии (например, содержащие промоторную последовательность), функционально связанные с нуклеотидной последовательностью, кодирующей трансген.
Может использоваться любой подходящий промотор, выбор которого не представляет никакой сложности для специалиста. Промоторная последовательность может быть конститутивно активной (т.е. функциональной в любой среде клетки-хозяина) или, альтернативно, может быть активной только в конкретной среде клетки-хозяина, что позволяет осуществлять целенаправленную экспрессию трансгена в конкретном типе клеток (например, тканеспецифический промотор). Промотор может иметь индуцируемую экспрессию в ответ на присутствие другого фактора, например фактора, присутствующего в клетке-хозяине. В любом случае, при введении вектора для терапии предпочтительно, чтобы промотор был функциональным в среде клеток-мишеней.
В некоторых вариантах осуществления предпочтительно, чтобы промотор имел специфическую для сетчатки экспрессию, обеспечивая экспрессию трансгена только в популяциях клеток сетчатки. Таким образом, экспрессия с промотора может быть специфичной по отношению к сетчатке, например, ограничиваться только клетками нейросенсорной сетчатки и пигментным эпителием сетчатки.
Предпочтительные промоторы, которые не являются специфичными к клеткам сетчатки, включают промотор куриного бета-актина (CBA), необязательно в сочетании с энхансерным элементом цитомегаловируса (CMV). Примером промотора для использования в изобретении является промотор CAG, например промотор, используемый в кассете экспрессии rAVE (GeneDetect.com). Другой пример промотора для использования в изобретении имеет последовательность:
ATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTG
GAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTG
ACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTG
GCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTCGAGGTGAGCCCCACGTTCTGCTTCACTCTC
CCCATCTCCCCCCCCTCCCCACCCCCAATTTTGTATTTATTTATTTTTTAATTATTTTGTGCAGCGATGG
GGGCGGGGGGGGGGGGGGGGCGCGCGCCAGGCGGGGCGGGGCGGGGCGAGGGGCGGGGCGGGGCGAGGCG
GAGAGGTGCGGCGGCAGCCAATCAGAGCGGCGCGCTCCGAAAGTTTCCTTTTATGGCGAGGCGGCGGCGG
CGGCGGCCCTATAAAAAGCGAAGCGCGCGGCGGGCGGGAGTCGCTGCGCGCTGCCTTCGCCCCGTGCCCC
GCTCCGCCGCCGCCTCGCGCCGCCCGCCCCGGCTCTGACTGACCGCGTTACTCCCACAGGTGAGCGGGCG
GGACGGCCCTTCTCCTCCGGGCTGTAATTAGCGCTTGGTTTAATGACGGCTTGTTTCTTTTCTGTGGCTG
CGTGAAAGCCTTGAGGGGCTCCGGGAGGGCCCTTTGTGCGGGGGGAGCGGCTCGGGGCTGTCCGCGGGGG
GACGGCTGCCTTCGGGGGGGACGGGGCAGGGCGGGGTTCGGCTTCTGGCGTGTGACCGGCGGCTCTAGAG
CCTCTGCTAACCATGTTCATGCCTTCTTCTTTTTCCTACAGCTCCTGGGCAACGTGCTGGTTATTGTGCT
GTCTCATCATTTTGGCAAAGAATT
(SEQ ID NO: 5)
В одном из вариантов осуществления AAV-вектор содержит промотор с нуклеотидной последовательностью SEQ ID NO: 5. В другом варианте осуществления AAV-вектор содержит промотор с нуклеотидной последовательностью, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO: 5, при этом нуклеотидная последовательность по существу сохраняет функциональную активность промотора, приведенного в SEQ ID NO: 5.
Примеры промоторов на основе человеческих последовательностей, которые индуцируют специфическую экспрессию генов, включают родопсинкиназу для палочек и колбочек (Allocca et al. (2007) J. Virol. 81: 11372-80), PR2.1 только для колбочек (Mancuso et al. (2009) Nature 461: 784-7) и/или RPE65 (Bainbridge et al. (2008) N. Engl. J. Med. 358: 2231-9) или VMD2 (Esumi et al. (2004) J. Biol. Chem. 279: 19064-73) для пигментного эпителия сетчатки.
AAV-вектор по изобретению также может содержать одну или более дополнительных регуляторных последовательностей, которые могут действовать до или после транскрипции. Регуляторная последовательность может быть частью нативного трансгенного локуса или может быть гетерологичной регуляторной последовательностью. AAV-вектор по изобретению может содержать части 5'-UTR или 3'-UTR из нативного трансгенного транскрипта.
Регуляторными последовательностями являются любые последовательности, которые облегчают экспрессию трансгена, т.е. действие которых усиливает экспрессию транскрипта, улучшает ядерный экспорт мРНК или повышает ее стабильность. Такие регуляторные последовательности включают, например, энхансерные элементы, пост-транскрипционные регуляторные элементы и сайты полиаденилирования. Предпочтительным сайтом полиаденилирования является поли-A сигнал бычьего гормона роста, который может быть таким, как показано ниже:
TCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCC
TTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGA
GTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAG
CAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTTCTGAGGCGGAAAGAACCAGCTGGGG
(SEQ ID NO: 6)
В одном из вариантов осуществления AAV-вектор содержит сайт полиаденилирования с нуклеотидной последовательностью SEQ ID NO: 6. В другом варианте осуществления AAV-вектор содержит сайт полиаденилирования с нуклеотидной последовательностью, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO: 6, при этом нуклеотидная последовательность по существу сохраняет функциональную активность сайта полиаденилирования, приведенного в SEQ ID NO: 6.
В контексте AAV-вектора по изобретению такие регуляторные последовательности будут цис-действующими. Однако изобретение также включает использование тран-действующих регуляторных последовательностей, расположенных на дополнительных генетических конструкциях.
Предпочтительным посттранскрипционным регуляторным элементом для использования в AAV-векторе по изобретению является посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE) или его вариант. Пример последовательности WPRE показан ниже:
ATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCT
ATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCC
TTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGT
GCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGAC
TTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGG
GCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAATCATCGTCCTTTCCTTGGCTGCTCG
CCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGA
CCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGT
CGGATCTCCCTTTGGGCCGCCTCCCCGC
(SEQ ID NO: 7)
В одном из вариантов осуществления AAV-вектор содержит посттранскрипционный регуляторный элемент с нуклеотидной последовательностью SEQ ID NO: 7. В другом варианте осуществления AAV-вектор содержит посттранскрипционный регуляторный элемент с нуклеотидной последовательностью, которая имеет по меньшей мере 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% или 100% идентичность с SEQ ID NO: 7, при этом нуклеотидная последовательность по существу сохраняет функциональную активность посттранскрипционного регуляторного элемента, приведенного в SEQ ID NO: 7.
Изобретение включает использование любой вариантной последовательности WPRE, которая увеличивает экспрессию трансгена по сравнению с AAV-вектором без WPRE. Предпочтительно вариантные последовательности имеют по меньшей мере 70% идентичность с SEQ ID NO: 7 относительно всей последовательности, более предпочтительно 75, 80, 85, 90 и более предпочтительно по меньшей мере 95%, 96% 97%, 98% или 99% идентичность с SEQ ID NO: 7 относительно всей последовательности.
Другая регуляторная последовательность, которую можно использовать в AAV-векторе по изобретению, представляет собой область связывания с ядерным матриксом (SAR). Специалистам не составит никакого труда выбирать дополнительные регуляторные последовательности.
Способ введения
В предпочтительном варианте осуществления AAV-вектор вводят интраокулярно.
Термин "интраокулярный" относится к внутренней части глаза, поэтому интраокулярное введение относится к введению внутрь глаза субъекта.
В одном из вариантов осуществления AAV-вектор вводят в глаз субъекта путем субретинальной инъекции, инъекции непосредственно в ретину, супрахориоидальное пространство или стекловидное тело. В одном из вариантов осуществления указанное введение выполняют с помощью робота.
Объем вводимой лекарственной композиции может составлять, например, примерно 10-500 мкл, например примерно 50-500, 100-500, 200-500, 300-500, 400-500, 50-250, 100-250, 200-250 или 50-150 мкл. Объем может составлять, например, примерно 10, 50, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мкл. Предпочтительно, объем вводимой лекарственной композиции составляет 100 мкл.
Специалисту в данной области хорошо известны эти способы инъекции, и он способен самостоятельно выполнить субретинальную инъекцию, инъекцию непосредственно в ретину, супрахориоидальное пространство или стекловидное тело.
Предпочтительно, AAV-вектор вводят путем субретинальной инъекции.
В одном из вариантов осуществления AAV-вектор или фармацевтическую композицию, содержащую его, вводят не более одного раза или не более двух раз в течение жизни субъекта.
Субретинальная инъекция
Субретинальные инъекции представляют собой инъекции в субретинальное пространство, т.е. под нейросенсорную сетчатку. Во время субретинальной инъекции вводимый материал направляется внутрь и создает пространство между слоем фоторецепторных клеток и слоем пигментного эпителия сетчатки (ПЭС).
При проведении инъекции посредством ретинотомии с небольшим отверстием может произойти отслоение сетчатки. Отслоенный приподнятый слой сетчатки, который является результатом инъецирования материала, называется "пузырьком".
Отверстие, созданное субретинальной инъекцией, должно быть достаточно малым, чтобы инъецированный раствор после введения не вытекал в больших количествах обратно в стекловидную полость. Такой обратный поток будет особенно проблематичным при введении лекарственного средства, поскольку воздействие лекарственного средства окажется в стороне от целевой зоны. Предпочтительно, инъекция создает самоуплотняемую точку входа в нейросенсорную сетчатку, т.е. после удаления инъекционной иглы отверстие, созданное иглой вновь смыкается настолько, что через него практически не вытекает или вытекает незначительное количество введенного материала.
С целью облегчения процесса на рынке появились специализированные иглы для субретинульной инъекции (например, DECC 41G тефлоновые иглы для субретинальной инъекции, Dutch Ophthalmic Research Center International BV, Zuidland, The Netherlands). Эти иглы предназначены для осуществления субретинальных инъекций.
При отсутствии повреждения сетчатки во время инъекции и при условии использования достаточно малой иглы по существу весь инъецируемый материал остается между отслоенной нейросенсорной сетчаткой и ПЭС в месте локализованного отслоения сетчатки (т.е. без обратного потока в полость стекловидного тела). Действительно, типичное сопротивление пузырька в течение короткого временного интервала указывает на незначительное количество, вытекшее в стекловидное тело. Пузырек может рассасываться в течение более длительного периода времени, пока не абсорбируется введенный материал.
Перед операцией можно выполнить визуализацию глаза, в частности сетчатки, например, с помощью оптической когерентной томографии.
Объем вводимой лекарственной композиции может составлять, например, примерно 10-500 мкл, например примерно 50-500, 100-500, 200-500, 300-500, 400-500, 50-250, 100-250, 200-250 или 50-150 мкл. Объем может составлять, например, примерно 10, 50, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мкл. Предпочтительно, объем вводимой лекарственной композиции составляет 100 мкл. Большие объемы могут увеличить риск отслоения сетчатки, в то время как малые объемы могут быть трудно различимыми.
Двухэтапная субретинальная инъекция
Вектор по изобретению может быть доставлен с повышенной точностью и безопасностью при помощи двухэтапного метода, при котором локализованное отслоение сетчатки создается субретинальной инъекцией первого раствора. Первый раствор не содержит вектор. Затем в субретинальную жидкость пузырька, образованного первой субретинальной инъекцией, вводят вторую субретинальную инъекцию для доставки лекарственного средства, содержащего вектор. Поскольку инъекция, доставляющая лекарственное средство, не используется для отслоения сетчатки, на второй стадии может быть введен определенный объем раствора.
В одном из вариантов осуществления изобретения субретинальная инъекция вектора содержит этапы:
(а) введения раствора субъекту путем субретинальной инъекции в количестве, эффективном для по меньшей мере частичного отделения сетчатки с образованием субретинального пузырька, при этом раствор не содержит вектор; и
(b) введение лекарственной композиции путем субретинальной инъекции в пузырек, образованный на стадии (а), при этом лекарственное средство содержит вектор.
Объем раствора, вводимого на стадии (а) для по меньшей мере частичного отделения сетчатки может составлять, например, примерно 10-1000 мкл, например примерно 50-1000, 100-1000, 250-1000, 500-1000, 10-500, 50-500, 100-500, 250-500 мкл. Объем может составлять, например, примерно 10, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 мкл.
Объем лекарственной композиции, вводимой на стадии (b), может составлять, например, примерно 10-500 мкл, например примерно 50-500, 100-500, 200-500, 300-500, 400-500, 50-250, 100-250, 200-250 или 50-150 мкл. Объем может составлять, например, примерно 10, 50, 100, 150, 200, 250, 300, 350, 400, 450 или 500 мкл. Предпочтительно объем лекарственной композиции, вводимой на стадии (b), составляет 100 мкл. Большие объемы могут увеличить риск отслоения сетчатки, в то время как малые объемы могут быть трудно различимыми.
Раствор, который не включает лекарственное средство (т.е. "раствор" стадии (а)), может быть составлен аналогичным образом и может представлять собой раствор, который содержит лекарственное средство, как описано ниже. Предпочтительным раствором, который не включает лекарственное средство, является сбалансированный солевой раствор (BSS) или аналогичный буферный раствор с рН и осмоляльностью, соответствующими таковым субретинального пространства.
Визуализация сетчатки во время операции
При определенных обстоятельствах, например, при терминальной стадии дегенерации сетчатки идентификация сетчатки затруднена, поскольку она становится тонкой, прозрачной и трудно различимой на фоне разрушенного и сильно пигментированного эпителия, на котором она держится. Использование синего витального красителя (например, Brilliant Peel®, Geuder, MembraneBlue-Dual®, Dorc) может облегчить идентификацию отверстия сетчатки, сделанного для процедуры отслоения сетчатки (например, на этапе (а) в двухэтапном методе субретинальной инъекции по изобретению), так что лекарственное средство можно вводить через то же самое отверстие без риска обратного оттока в стекловидную полость.
Использование синего витального красителя также позволяет идентифицировать любые области сетчатки, где имеется утолщенная внутренняя пограничная мембрана или эпиретинальная мембрана, поскольку инъекция через любую из этих структур препятствует чистому доступу в субретинальное пространство. Кроме того, сокращение любой из этих структур в ближайший послеоперационный период может привести к растяжению входного отверстия сетчатки, что может привести к обратному оттоку лекарственного средства в стекловидную полость.
Инъекция в супрахориоидальное пространство
Вектор по изобретению может быть доставлен в супрахориоидальное пространство с помощью ab externo-подхода, в котором используется микрокатетер (см., например, Peden et al. (2011) PLoS One 6 (2): e17140). В этом методе выполняют лимбальную перитомию конъюнктивы, обнажая голую склеру, а затем склеротомию, обнажая голую сосудистую оболочку. Микрокатетер (например, iTrack 250A компании iScience Interventional, необязательно соединенный с системой освещения, такой как система микроосвещения на основе лазерного диода iLumin (iScience Interventional)), вводят в супрахориоидальное пространство и продвигают вглубь к оптическому диску. После выставления наконечника микрокатетера в нужное положение выполняют инъекцию вектора с образованием пузырька внутри сетчатки и хориоидеи.
Таким образом, в одном из вариантов осуществления вектор доставляется в супрахориоидальное пространство методом, включающим (i) введение микрокатетера в супрахориоидальное пространство; (ii) продвижение микрокатетера в указанном пространстве до тех пор, пока наконечник не окажется рядом с пораженной областью сетчатки; и (iii) инъекцию вектора из наконечника микрокатетера с образованием пузырька.
В одном из вариантов осуществления вышеуказанные процедуры введения выполняются непосредственно роботом.
Фармацевтические композиции и инъекционные растворы
Медикаменты, например AAV-векторы по изобретению, могут быть составлены в виде фармацевтических композиций. Эти композиции могут включать дополнительно к лекарственному средству фармацевтически приемлемый носитель, разбавитель, наполнитель, буфер, стабилизатор или другие материалы, хорошо известные в данной области. Такие материалы должны быть нетоксичными и не должны влиять на эффективность активного ингредиента. Точная природа носителя или другого материала может быть определена квалифицированным специалистом в соответствии со способом введения, например, субретинальной инъекции, инъекции непосредственно в сетчатку, в супрахориоидальное пространство или стекловидное тело.
Фармацевтическая композиция обычно находится в жидком виде. Жидкие фармацевтические композиции обычно включают жидкий носитель, такой как вода, нефть, животные или растительные масла, минеральное масло или синтетическое масло. Могут быть включены физиологический солевой раствор, раствор хлорида магния, декстрозы или другого сахарида, или гликолей, таких как этиленгликоль, пропиленгликоль или полиэтиленгликоль. В некоторых случаях может быть использовано поверхностно-активное вещество, такое как 0,001% плуроновая кислота (PF68).
Для инъекций в месте поражения активный ингредиент может быть в виде водного раствора, который не содержит пирогенных веществ, и имеет подходящий рН, изотоничность и стабильность. Специалист в данной области способен приготовить подходящие растворы, используя, например, изотонические носители, такие как раствор хлорида натрия для инъекций, раствор Рингера для инъекций или Рингер лактат раствор для инъекций. При необходимости могут быть включены консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки.
Для замедленного высвобождения лекарственное средство может быть включено в фармацевтическую композицию, которая находится в лекарственной форме для замедленного высвобождения, например в микрокапсулах, образованных из биосовместимых полимеров или в липосомальных системах доставки, способами, известными в данной области.
Способ лечения
Следует учесть, что все ссылки на лечение, упомянутые в настоящей заявке, включают лечебное, паллиативное и профилактическое лечение; хотя в контексте изобретения ссылки на профилактику чаще связаны с профилактическим лечением. Лечение также может включать остановку прогрессирования заболевания.
Предпочтительным является лечение млекопитающих, в частности людей. Однако в объем изобретения входит как лечение человека, так и ветеринарное лечение.
Варианты, производные, аналоги, гомологи и фрагменты
В дополнение к упомянутым в настоящем описании конкретным белкам и нуклеотидам, изобретение также включает применение вариантов, производных, аналогов, гомологов и их фрагментов.
В контексте изобретения вариант любой заданной последовательности представляет собой последовательность, в которой конкретная последовательность остатков (будь то аминокислотная или нуклеиновая кислота) модифицирована таким образом, что рассматриваемый полипептид или полинуклеотид по существу сохраняет свою функцию. Вариантную последовательность можно получить путем добавления, делеции, замещения, модификации, замены и/или изменения по меньшей мере одного остатка, присутствующего во встречающемся в природе белке.
Используемый в настоящем описании термин "производное" в отношении белков или полипептидов по изобретению включает любое замещение, изменение, модификацию, замену, делецию и/или добавление одного (или более) аминокислотных остатков из или в последовательности, при условии, что полученный в результате белок или полипептид по существу сохраняет по меньшей мере одну из своих эндогенных функций.
Используемый в настоящем описании термин "аналог" в отношении полипептидов или полинуклеотидов включает любой миметик, т.е. химическое соединение, которое обладает по меньшей мере одной из эндогенных функций полипептидов или полинуклеотидов, которые он имитирует.
Как правило, аминокислотные замещения могут включать, например, 1, 2 или 3 до 10 или 20 замещений, при условии, что модифицированная последовательность по существу сохраняет требуемую активность или способность. Аминокислотные замещения могут включать использование не встречающихся в природе аналогов.
Белки, используемые в изобретении, могут также иметь делеции, вставки или замещения аминокислотных остатков, которые образуют молчащие изменения и приводят к функционально эквивалентному белку. Такие аминокислотные замещения могут быть сделаны на основании сходства по полярности, заряду, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков при условии сохранения эндогенной функции. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин; и аминокислоты с незаряженными полярными группами, имеющими сходные значения гидрофильности, включают аспарагин, глутамин, серин, треонин и тирозин.
Консервативные замены могут быть сделаны, например, согласно приведенной ниже таблице. Аминокислоты, находящиеся в одном блоке, в одной ячейке во второй колонке и в одной и той же строке третьей колонки, могут замещать друг друга:
Используемый в настоящем описании термин "гомолог" означает объект, имеющий определенную гомологию с аминокислотной последовательностью дикого типа и нуклеотидной последовательностью дикого типа. Термин "гомология" можно приравнять к "идентичности".
Гомологичная последовательность может включать аминокислотную последовательность, которая может быть по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85% или 90% идентичной, предпочтительно по меньшей мере на 95% или 97% или 99% идентичной последовательности субъекта. Как правило, гомологи содержат такие же активные сайты и т.д., что и аминокислотная последовательность субъекта. Хотя гомологию также можно рассматривать с точки зрения подобия (т.е. аминокислотных остатков, имеющих сходные химические свойства/функции), в контексте изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательности.
Гомологичная последовательность может включать нуклеотидную последовательность, которая может быть по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85% или 90% идентичной, предпочтительно по меньшей мере на 95% или 97% или 99% идентичной последовательности субъекта. Хотя гомологию также можно рассматривать с точки зрения подобия, в контексте изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательности.
Предпочтительно, ссылка на последовательность, которая имеет некий процент идентичности с любой одной из последовательностей SEQ ID NO, подробно описанных в настоящей заявке, относится к последовательности, которая имеет заявленную процент идентичности относительно всей длины этой SEQ ID NO.
Сравнения на определение гомологии можно выполнять визуально или, как принято, с помощью легкодоступных программ сравнения последовательностей. Эти коммерчески доступные компьютерные программы могут рассчитать процент гомологии или идентичность между двумя или более последовательностями.
Процент гомологии может быть рассчитан для непрерывных последовательностей, т.е. одну последовательность выравнивают относительно другой последовательности, и каждую аминокислоту в одной последовательности непосредственно сравнивают с соответствующей аминокислотой в другой последовательности, по одному остатку за раз. Это называется выравниванием "без разрывов (гэпов)". Как правило, такие выравнивания "без разрывов" выполняются только для относительно небольшого количества остатков.
Хотя это очень простой и последовательный метод, в нем не учитывается тот факт, что, например, в паре иным образом идентичных последовательностей одна вставка или делеция в нуклеотидной последовательности может приводить к выпадению из процесса выравнивания последующих кодонов, таким образом потенциально приводя к значительному уменьшению процента гомологии при проведении общего выравнивания. Следовательно, большинство методов сравнения последовательностей предназначены для получения оптимальных выравниваний, в которых учитываются возможные вставки и делеции, без чрезмерных потерь, влияющих на общий показатель гомологии. Это достигается вставкой "разрыва" при выравнивании последовательности для получения максимальной локальной гомологии.
Однако в этих более сложных методах назначаются "штрафы за разрывы" за каждый разрыв, образующийся при выравнивании, и, таким образом, для одинакового количества идентичных аминокислот выравнивание последовательности с минимально возможным количеством разрывов, отражающим более высокое родство между двумя сравниваемыми последовательностями, получит более высокий показатель, чем выравнивание со многими разрывами. Обычно используются стоимость однородных разрывов (ʺaffine gap costsʺ), при которой назначается относительно высокая стоимость за открытие разрыва и меньший по размеру штраф за каждый последующий остаток в разрыве (продление разрыва). Это наиболее часто используемая система оценки разрывов. Высокие штрафы за разрыв, конечно же, будут способствовать получению оптимизированных выравниваний с меньшим количеством разрывов. Большинство программ выравнивания позволяют модифицировать штрафы за разрыв. Однако при использовании такого программного обеспечения для сравнения последовательностей предпочтительно использовать значения по умолчанию. Например, в случае пакета GCG Wisconsin Bestfit штраф за разрыв в аминокислотных последовательностях, назначаемый по умолчанию, составляет -12 для разрыва и -4 за каждое удлинение.
Поэтому при вычислении максимального процента гомологии сначала необходимо создать оптимальное выравнивание с учетом штрафов за разрыв. Подходящей компьютерной программой для проведения такого выравнивания является пакет GCG Wisconsin Bestfit (University of Wisconsin, U.S.A, Devereux et al. (1984) Nucleic Acids Res. 12: 387). Примеры других компьютерных программ, которые могут выполнять сравнение последовательностей, включают, без ограничения, пакет BLAST (см. Ausubel et al. (1999) ibid - Ch. 18), FASTA (Atschul et al. (1990) J. Mol. Biol. 403-410) и набор средств для сравнения GENEWORKS. Как BLAST, так и FASTA доступны для работы как в автономном, так и интерактивном режимах (см. Ausubel et al. (1999) ibid, pages 7-58 to 7-60). Однако для некоторых приложений предпочтительно использовать программу GCG Bestfit. Для сравнения белковых и нуклеотидных последовательностей также доступен новый инструмент, названный BLAST 2 Sequences (см. FEMS Microbiol. Lett. (1999) 174: 247-50, FEMS Microbiol. Lett. (1999) 177: 187-8).
Хотя конечный процент гомологии может быть измерен в терминах идентичности, процесс выравнивания сам по себе обычно не основан на сравнении пар по принципу "все или ничего". Вместо этого обычно используют матрицу оценки взвешенного подобия, которая назначает оценку каждому парному сравнению на основании химического сходства или эволюционного расстояния. Примером такой обычно используемой матрицы является матрица BLOSUM62, которая представляет собой матрицу, используемую по умолчанию в программах BLAST. В программах GCG Wisconsin обычно применяют либо общедоступные значения по умолчанию, либо, при наличии, пользовательскую таблицу сравнения символов (дополнительную информацию см. в руководстве пользователя). Для некоторых приложений предпочтительно использовать общедоступные значения по умолчанию для пакета GCG или, в случае другого программного обеспечения, матрицу по умолчанию, например BLOSUM62.
После получения оптимального выравнивания, вычисленного с помощью компьютерной программы, можно рассчитать процент гомологии, предпочтительно процент идентичности последовательностей. Как правило, это является частью выполняемого программой сравнения последовательностей, и выдается в виде численного результата.
"Фрагменты" фактора I или фактора H полной длины также являются вариантами, и этот термин обычно относится к выбранной области полипептида или полинуклеотида, которая представляет интерес либо с точки зрения функциональности, либо, например, с точки зрения анализа. Таким образом, "фрагмент" относится к аминокислотной последовательности или последовательности нуклеиновой кислоты, которая является частью полноразмерного полипептида или полинуклеотида.
Такие варианты могут быть получены стандартными методами рекомбинантной ДНК, такими как сайт-направленный мутагенез. Если необходимо выполнить вставку, можно создать синтетическую ДНК, кодирующую вставку вместе с 5'- и 3'-фланкирующими областями, соответствующими встречающейся в природе последовательности, расположенными с обеих сторон сайта вставки. Фланкирующие области могут содержать подходящие сайты рестрикции, соответствующие сайтам встречающейся в природе последовательности, с тем, чтобы последовательность могла быть разрезана соответствующим ферментом(ферментами), позволяя осуществить лигирование ДНК в этом разрезе. Затем осуществляется экспрессия ДНК в соответствии с изобретением с получением кодированного белка. Эти методы являются лишь иллюстрацией многочисленных стандартных методов, известных в данной области техники для манипулирования последовательностями ДНК, при этом также можно использовать другие известные методы.
Различные предпочтительные признаки и варианты осуществления настоящего изобретения описаны ниже в виде неограничивающих примеров.
При осуществлении настоящего изобретения использованы, если не указано иное, способы, обычно используемые в области химии, биохимии, молекулярной биологии, микробиологии и иммунологии, которые находятся в пределах компетентности специалиста в данной области техники. Такие методы описаны в литературе. См. например, Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press; Ausubel, F.M. et al. (1995 and periodic supplements) Current Protocols in Molecular Biology, Ch. 9, 13 and 16, John Wiley & Sons; Roe, B., Crabtree, J. and Kahn, A. (1996) DNA Isolation and Sequencing: Essential Techniques, John Wiley & Sons; Polak, J.M. and McGee, J.O'D. (1990) In Situ Hybridization: Principles and Practice, Oxford University Press; Gait, M.J. (1984) Oligonucleotide Synthesis: A Practical Approach, IRL Press; and Lilley, D.M. and Dahlberg, J.E. (1992) Methods in Enzymology: DNA Structures Part A: Synthesis and Physical Analysis of DNA, Academic Press., каждый из которых включен в настоящее описание в виде ссылки.
ПРИМЕРЫ
Пример 1
Клонирование кДНК человеческого CFI и создание экспрессионной кассеты CBA-CFI-WPRE, конструирование pAAV-CBA-CFI-WPRE-bGHpA и упаковка вируса AAV-CFI.
кДНК наиболее распространенного варианта последовательности человеческого CFI загружали из Genbank под номером доступа NM_000204.4. кДНК имеет последовательность SEQ ID NO: 2 и была заказана в виде фрагментов генов GBlocks® компании Integrated DNA Technologies. Также была заказана вторая конструкция CFI, в которой кДНК-последовательность гена CFI оптимизирована по кодону для экспрессии в клетках человека. Оптимизированная по кодону последовательность (ʺCFIcoʺ) имеет последовательность SEQ ID NO: 8 и была заказана в компании GeneWiz (CFIco в плазмиде pUC57).
Эти последовательности кДНК вставляли в плазмиду pAAV в cis-положении, далее упоминаемую как pAM. pAM представляет собой плазмиду, позволяющую создавать большое количество копий, первоначально полученную из pBR322, и которая включает стабилизированные в AAV-2 левые и правые инвертированные концевые повторы, фланкирующие выбранную экспрессионную кассету. В векторах AAV-CFI и AAV-CFIco для стимулирования экспрессии CFI и CFIco использовали модифицированный промотор CBA/CAG (бета-актин цыпленка с энхансером CMV, упоминаемым в настоящем описании как ʺCBAʺ), и на 3'-конце кДНК обеспечивали модифицированную WPRE последовательность и bGH полиA. Эти две плазмиды обозначены как pAAV2.CBA-hCFI-WPRE-bGH, (pAAV-CFI) и pAAV2.CBA-hCFIco-WPRE-bGH, (pAAV-CFIco).
На фиг. 2 показан агарозный гель с рестриктами CFI и CFIco. Полосу 1752 п.о. CFI вырезали и клонировали в остов pAAV-CBA-WPRE-bGHpA.
Сравнение уровней экспрессии CFI вектора pAAV.CFI с вектором pAAV.CFIco, котрансфекция клеточной линиии ARPE-19 векторами pAAV-CFI или pAAV.CFIco и pCMV.GFP и иммуноблоттинг CFI
ARPE-19 (ATCC® CRL-2302™) представляет собой спонтанно возникающую клеточную линию пигментного эпителия сетчатки (ПЭС), полученную в 1986 году Эми Атаки-Кином (Amy Aotaki-Keen) из нормальных глаз 19-летнего мужчины, который умер от травмы головы в автомобильной аварии. Эти клетки образуют устойчивые адгезивные монослои, для которых характерна морфологическая поляризация, наблюдаемая при их нанесении на покрытые ламинином фильтры Transwell-COL в среде с низкой концентрацией сыворотки. Клетки ARPE-19 получали из Американской коллекции типовых культур (АТСС). Клетки выращивали в DMEM/F12 (Thermo Fisher Scientific), дополненной 10% инактивированной нагреванием фетальной бычьей сывороткой (Gibco), 1% 200мМ L-глютамина (Sigma Aldrich) и 1% пенициллина-стрептомицина (Sigma Aldrich, 10000 единиц пенициллина, 10 мг стрептомицина/мл).
Котрансфекцию ARPE-19 выполняли с использованием липофектамина LTX (Life Technologies) в соответствии с протоколом производителя. 0,5 мкг pAAV-CFI или pAAV.CFIco котрансфицировали с 0,5 мкг pCMV.GFP (Plasmid Factory). В качестве отрицательного контроля служила котрансфекция pAAV-CBA-WPRE-bGHpA (pAAV [без трансгенной кассеты]). Через 24 часа после трансфекции клетки промывали PBS (забуференный фосфатом физиологический раствор, pH 7,2, Gibco™) и культивировали в бессывороточной среде для роста в течение 72 часов. Супернатант отбирали, очищали центрифугированием и хранили при -80°C. Клетки ARPE-19 отделяли с помощью TrypLE Express (Gibco) и подсчитывались для обеспечения равного количества клеток в каждой лунке. Осадок замораживали при -20°C в течение 30 минут. Буфер для лизиса 1xRIPA (Merck Millipore), дополненный коктейлем cOmplete™ ингибиторов протеаз без ЭДТА (Roche), добавляли к осадку, и клетки разрушали ультразвуком. Нерастворимый белок центрифугировали, и супернатант (= лизат) хранили при -80°C.
Иммуноблоттинг CFI проводили путем загрузки 30 мкл неразбавленного супернатанта в 4-кратный буфер Laemmli (250 мМ Трис-HCl (pH 6,8), 8% SDS, 40% глицерина и 0,02% бромфенолового синего) и разделением белков в 10% готовом полиакриламидном геле (Bio-Rad). Белки переносили на мембрану PVDF (Bio-Rad) методом полусухого переноса, после чего мембрану блокировали в блокирующем буфере (1xTBS pH 8 [Sigma], 0,05% Твин-20 и 5% сухого обезжиренного молока [Marvel]), CFI детектировали с помощью OX21 (таблица 1) и HRP-конъюгированного антитела к мышиным IgG (таблица 1), разбавленных в блокирующем буфере. Полосы белка визуализировали с помощью субстрата Clarity Western ECL-(Bio-Rad) и анализировали с помощью системы визуализации Odyssey® Fc. Иммуноблоттинг GFP осуществляли путем загрузки 5 мкл неразбавленного супернатанта в 4х буфер Лэммли (Laemmli), дополненный 50 мкл/мл β-меркаптоэтанолом. Иммуноблотинг выполняли, как описано выше. GFP детектировали с помощью антитела к TurboGFP (таблица 1) и HRP-конъюгированного антитела к кроличьим IgG (таблица 1).
Таблица 1. Первичные и вторичные антитела, используемые для иммуноблоттинга или иммунопреципитации
На Фиг. 3 показан иммуноблоттинг CFI (Фиг. 3A) и GFP (Фиг. 3B).
3А: CFI проявляется в виде полосы 70 кДа (не уменьшенный) и экспрессируется с одной и той же скоростью после трансфекции ARPE-19 векторами pAAV.CFI или pAAV.CFIco. После трансфекции pAAV-ветора CFI не экспрессировался. В качестве положительного контроля в иммуноблоттинге CFI использовали 10% нормальную человеческую сыворотку (NHS). 3B: Эффективность трансфекции анализировали путем котрансфекции клеток ARPE-19 вектором pCMV.GFP. GFP проявляется в виде полосы 30 кДа, и иммуноблот подтвердил, что клетки были трансфицированы с аналогичной эффективностью.
Получение AAV.CFI и AAV.CFIco.
Вирус AAV2 получали двойной трансфекцией клеточных линий HEK-293 (ATCC® CRL-1573) или HEK-293T (ATCC® CRL-3216™) (обе адгезивные) векторами pAAV.CFI или pAAV.CFIco и pDG (Plasmid Factory), несущими аденовирусные последовательности-помощники и последовательности упаковки (Rep/Cap). Клетки HEK-293 и HEK-293T выращивали в DMEM (Sigma), дополненной 10% инактивированной нагреванием фетальной бычьей сывороткой (Gibco), 1% 200мМ L-глутамина (Sigma Aldrich) и 1% пенициллина-стрептомицина (Sigma Aldrich, 10000 единиц пенициллина, 10 мг стрептомицина/мл). Клетки собирали через 72 часа и лизировали для очистки вирусных частиц. AAV частицы очищали в градиенте иодиксанола и извлекали из 40% фракции. Вирус концентрировали на центрифужных фильтрах Amicon Ultra-15 (Merck Millipore) и хранили в аликвотах при -80°C. Чистоту вируса оценивали с помощью SDS PAGE, а титр определяли с помощью кПЦР.
Анализ экспрессии AAV.CFI, AAV.CFIco и AAV.GFP in vitro, трансдукция клеточных линий HEK-293 и ARPE-19, иммуноблот супернатанта тканевой культуры для анализа экспрессии CFI
70% конфлюентных линий HEK-293 (ʺ293ʺ) и ARPE-19 трансдуцировали векторами AAV.CFI, AAV.CFIco и AAV.GFP с множественностью инфекции (MOI) 1×104 в нормальной среде для роста, дополненной только 1% инактивированной нагреванием фетальной бычьей сывороткой. Через 7 дней супернатант собирали и очищали центрифугированием. Супернатант хранили в аликвотах при -80°C.
Иммуноблоттинг выполняли, как описано выше, и 30 мкл супернатанта смешивали с 4х буфером Лэммли и загружали в 10% -готовый полиакриламидный гель. CFI детектировали с помощью OX21 (таблица 1) и HRP-конъюгированного антитела к мышиным IgG (таблица 1), разбавленных в блокирующем буфере (когда супернатант загружали в неденатурирующих условиях), или с помощью антисыворотки к человеческому CFI (таблица 1) и конъюгированного с пероксидазой антитела к козьим IgG (цельная молекула) (таблица 1) (когда супернатант загружали в денатурирующих условиях).
На Фиг. 4 показан иммуноблоттинг CFI в супернатанте клеточных линий HEK-293 и ARPE-19, трансдуцированных вирусом. 4A: Супернатант наносили в неденатурирующих условиях, и CFI детектировали с помощью мышиного моноклонального антитела к человеческому CFI (OX21, Thermo Fisher Scientific) и HRP-конъюгированного антитела осла к мышиным IgG (Abcam). CFI и CFIco экспрессировались в обеих клеточных линиях. Трансдукция клеточных линий вектором AAV.GFP служила в качестве отрицательного контроля, тогда как трансдукция 0,5 мкг очищенного от плазмы человеческого CFI (упоминаемого в настоящем описании "CFIpl") (Comptech) служила в качестве положительного контроля. 4B: Супернатант наносили в денатурирующих условиях, и CFI детектировали с помощью козьей антисыворотки к человеческому CFI (Comptech) и пероксидазным кроличьим антителом к козьим IgG (специфичным к цельной молекуле) (Sigma). CFI проявляется в виде полос 80 кДа (про-фермент), 50 кДа (подвергнутая процессингу тяжелая цепь) и 35 кДа (подвергнутая процессингу легкая цепь). AAV.GFP служил в качестве отрицательного контроля, тогда как 0,5 мкг очищенного от плазмы человеческого CFI (Comptech) и 10% нормальная человеческая сыворотка (упоминаемая в настоящем описании "NHS") служили положительным контролем. CFI и CFIco экспрессировались в обеих клеточных линиях.
Качественный анализ экспрессии CFI, анализ расщепления C3b для измерения функциональной активности
Для качественного анализа осуществляли иммунопреципитацию CFI с использованием набора для иммуннопреципитации Pierce (Co-IP) (Thermo Fisher Scientific) в соответствии с протоколом производителя. 30 мкг аффинно-очищенного моноклонального антитела к человеческому CFI (Ox21, Thermo Fisher Scientific) инкубировали в течение 2 часов при комнатной температуре с 25 мкл смолы AminoLink Plus Coupling. Супернатант трансдуцированных клеток HEK-293 или ARPE-19 инкубировали с полученной смолой в течение ночи при 4°C. На следующий день смолу промывали несколько раз, используя буфер IP для лизиса/промывки. Связанный CFI элюировали буфером из набора для элюирования (0,1 М глицина, рН 2,7) и сразу же нейтрализовали 1М Трис, pH 9,5. Иммунопреципитацию CFI оценивали путем измерения поглощения при 280 нм и с помощью анализа SDS PAGE. CFI либо использовали непосредственно в анализе расщепления C3b, либо хранили в аликвотах при -80°C.
В анализе расщепления C3b 1 мг очищенного от плазмы C3b инкубировали в течение 1 часа при 37°C с 0,5 мкг очищенного от плазмы фактора H комплемента (CFH) и либо с очищенным от плазмы фактором I комплемента (CFIpl) (все очищенные от плазмы белки были приобретены у компании Comptech), супернатантом культуры клеток, трансдуцированных вектором AAV.CFI, либо с иммунопреципитированным CFI/CFIco. Для остановки реакции добавляли 4x буфер Лэммли с β-меркаптоэтанолом. Образцы разбавляли и загружали в 10% готовый полиакриламидный гель SDS-PAGE (Bio-Rad). После переноса на мембрану PVDF (Bio-Rad) и блокирования в блокирующем буфере (1xTBS pH 8 [Sigma]/0,05% Твин-20 и 5% сухого обезжиренного молока [Marvel]), детектировали расщепление C3b с помощью биотинилированного клона 9 (Таблица 1) и HRP-конъюгированного экстравидина (таблица 1). Это антитело реагирует с эпитопом на C3g и в денатурирующем SDS PAGE распознает α-цепь C3, α'-цепь C3b, C3α'1-цепь iC3b и C3dg. Поэтому его можно использовать для анализа деградации C3b под действием CFI. CFH служил в качестве кофактора для деградации C3b, и поскольку анализ проводили в условиях низкой ионной силы, он также служил в качестве кофактора для второго расщепления iC3b до C3dg. Используемым буфер представляет собой "буфер для элюции" из набора для коиммуннопреципитации Pierce (Co-IP) (Thermo Fisher Scientific), нейтрализованный 1M Трис, pH 9,5.
На Фиг. 5 показан репрезентативный результат анализа расщепления C3b. Дорожка 1 представляет C3b, инкубированный только с CFH. Дорожка 2 представляет C3b, инкубированный с CFH и CFIpl. C3b расщепленный CFIpl до iC3b и C3dg. Дорожка 3 представляет C3b, инкубированный с CFH и супернатантом клеток HEK-293, трансдуцированных вектором AAV.CFI. Дорожки 4-5 представляют C3b, инкубированный с CFH и иммуноосажденным CFI (обозначенным "IP CFI") из клеток ARPE-19, трансдуцированных либо AAV.CFI (дорожка 4), либо AAV.CFIco (дорожка 5). Дорожки 6-7 представляют C3b, инкубированный с CFH и иммуноосажденным CFI из клеток HEK-293, трансдуцированных либо AAV.CFI (дорожка 6), либо AAV.CFIco (дорожка 7). CFI, секретируемый из трансдуцированных клеточных линий, является функционально активным и расщепляет α-цепь C3b до iC3b. Иммунопреципитация CFI приводит к увеличению общего количества CFI, которое отражается появлением полосы C3dg в дополнение к полосе α1-фрагмента iC3b. Это второе расщепление происходит только в условиях низкой ионной силы и с низкой скоростью, что требует большего количества CFI.
Анализ in vitro экспрессии CFI в трансдуцированных ARPE-19, выращенных на проницаемом фильтре transwell.
Клетки ПЭС представляют собой пигментированные, неделящиеся клетки, для которых характерна морфологическая поляризация при нанесении на покрытые ламинином фильтры Transwell-COL в среде с низкой концентрацией сыворотки. Фильтр Transwell (вставки из полиэфирной мембраны, диаметр пор 0,4 мкм, диаметр мембраны 12 мм) позволяет разделить культуральные лунки на два компартмента, апикальный (верхний) домен, соответствующий стороне монослоя ПЭС, обращенной к сетчатке, и базолатеральный (нижний) домен, соответствующий стороне монослоя ПЭС, обращенного к хориоидальному слою. При выходе из условий низкой сывороточной среды клетки ARPE-19 дифференцируются и становятся гексагональными, слегка пигментированными клетками. Модель transwell можно использовать для оценки экспрессии в покоящихся клетках, что отражает ситуацию in vivo.
Мембраны Transwell покрывали 8 мкг/мл ламинина (Sigma), а клетки ARPE-19 высевали в 10% FBS-среде в виде монослоев на Transwells. Через 48 часов среду заменяли на среду с 1% FBS, и клетки трансдуцировали с MOI 105/клетка (AAV.CFI, AAV.CFIco и AAV.GFP). Через 7 дней супернатант собирали из обоих компартментов и очищали центрифугированием. Из-за разных объемов в верхнем и нижнем компартментах, т.е. 500 мкл среды в верхнем и 1500 мкл в нижнем компартменте, супернатант анализировали, нормализуя до общего объема: 30 мкл супернатанта из нижнего компартмента смешивали с 4х буфером Лэммли, и 10 мкл супернатанта из верхнего компартмента смешивали с 4x буфером Лэммли. Образцы загружали в 10% готовый полиакриламидный гель. CFI детектировали с помощью OX21 (таблица 1) и HRP-конъюгированного антитела к мышиным IgG (таблица 1), разбавленных в блокирующем буфере. Для окрашивания ядер клетки промывали 2x PBS (Gibco) и фиксировали в течение 15 минут 4% параформальдегидом (Sigma) в PBS и пермеабилизировали в течение 45 минут с 1% BSA (Thermo Fisher Scientific)-0,1% Тритон X-100 (Sigma ) в PBS. Хехст (Thermo Fisher Scientific) инкубировали при соотношении 1:5000 в буфере для пермеабилизации в течение 15 минут и оценивали окрашивание ядер.
На фиг. 6 показан иммуноблоттинг CFI, секретируемого из клеток ARPE-19, культивированных на transwell. Клетки не были дифференцированы в гексагональные клетки, однако их культивировали в виде конфлюэнтного монослоя клеток, и деление клеток сбыло сведено к минимуму путем добавления 1% сывороточной среды. 6A: Супернатант из обоих компартментов наносили в неденатурирующих условиях, и вестерн-блот-анализ проводили с использованием мышиного моноклонального антитела к CFI (Ap=апикальный компартмент и Bl=базолатеральный компартмент). Показано, что CFI экспрессируется в конфлюентном монослое клеток, и что секретируемый белок обнаруживается в обоих компартментах. 6B: Ядра окрашивали красителем Хехст (Hoechst) для подтверждения наличия монослоя клеток (окрашивание проводили после сбора супернатанта).
Анализ экспрессии in vivo AAV.CFI и AAV.CFIco, субретинальная инъекция мышам C57/Black 6, анализ экспрессии CFI с помощью иммуноблоттинга, кПЦР и иммуногистохимии
Субретинальные инъекции
Во всех экспериментах использовали 8-10 недельных самок мышей C57BL/6J (Charles River Laboratories). Со всеми животными, использованными в этом исследовании, обращались гуманно в соответствии с правилами министерства внутренних дел Великобритании согласно лицензии проекта 30/3363. Мышей содержали в условиях дневного цикла 12:12 час. день/ночь.
Мышей анестезировали смесью ксилазина (10 мг/кг)/кетамина (80 мг/кг) в стерильном физиологическом растворе; зрачки расширяли фенилэфрина гидрохлоридом (2,5%) и тропикамидом (1%). Проксиметакаин гидрохлорида (0,5%) в виде глазных капель использовали для дополнительной местной анестезии. Перед субретинальной инъекцией выполняли прокол передней камеры, используя стерильные иглы 33G (TSK Laboratory) и карбомерный гель (Viscotears, Novartis Pharmaceuticals Ltd), а для хорошей визуализации дна использовали небольшое круглое покровное стекло. Инъекцию проводили через заднюю поверхность сетчатки, используя 10 мкл шприц NanoFil и 35G скошенную иглу NanoFil (World Precision Instruments Ltd). Анестезию снимали атипамезолом (2 мг/кг) в стерильном физиологическом растворе.
Мышам вводили две разные дозы (107 геномных копий [гк]/глаз и 108 гк/глаз) и две AAV-конструкции (AAV.CFI и AAV.CFIco). Трем мышам вводили третью дозу, 109 гк/глаз AAV.CFIco; этих мышей использовали для калибровки иммуноблотов. Вирус разводили в 0,001% PF68 (Gibco) в PBS (Gibco), и делали ложные инъекции, используя тот же разбавитель. Субретинальные инъекции вводили в 12 глаз на одно состояние.
Подготовка образцов тканей для иммуноблоттинга CFI
Мышей умерщвляли мгновенной дислокацией шейных позвонков через 4 недели после инъекции. Глаза вынимали и готовили следующим образом. Используя препаровальную лупу, делали разрез в роговице. Глаз разделяли на роговицу/радужку (передний сегмент), хрусталик и заднюю стенку глазного бокала. Для цельного образца глазного бокала, заднюю стенку глазного бокала помещали в стерильную пробирку. Некоторые глаза разделяли на сетчатку и ПЭС/хориоидею/склеру, осторожно отделяли слой сетчатки от комплекса ПЭС/хориоидея/склера. Образцы, по три мыши на каждое состояние, объединяли (фиг. 7А) за исключением глаз, в которые вводили 109 гк/глаз AAV.CFIco (фиг. 7B), где образцы от двух мышей объединяли и сравнивали со вторым глазом без инъекции. Все образцы сразу же помещали на сухой лед для предотвращения деградации белка и хранили при -80°C. Для иммуноблоттинга образцы гомогенизировали в буфере для лизиса 1xRIPA (Merck Millipore), дополненном коктейлем ингибитора протеазы cOmplete™ без ЭДТА (Roche), и клетки механически разрушали в ступке с пестиком и обработкой ультразвуком. Нерастворимый белок центрифугировали, и супернатант аликвотировали и хранили при -80°C. Концентрацию белка определяли с помощью набора для анализа белка Pierce BCA (Thermo Fisher Scientific), и 40 мкг белкового лизата загружали в 4х буфер Лэммли (250 мМ Tris-HCl (pH 6,8), 8% SDS, 40% глицерин и 0,02% бромфеноловый синий). Белки разделяли в 10% готовом полиакриламидном геле (Bio-Rad). Белки переносили на мембрану PVDF (Bio-Rad) методом полусухого переноса, после чего мембрану блокировали в блокирующем буфере (1xTBS pH 8 [Sigma], 0,05% Твин-20 и 5% порошковое обезжиренное молоко [Marvel]), CFI детектировали с помощью: a) ослабленных: антисыворотки к человеческому CFI (таблица 1) и HRP-конъюгированного антитела к козьим IgG (таблица 1) или b) неослабленных: OX21 (таблица 1) и HRP-конъюгированного антитела к мышиным IgG (Abcam), разведенных в блокирующем буфере. Для обнаружения β-актина мембрану удаляли с помощью буфера Restore Western Blot Stripping (Thermo Fisher Scientific) и анализировали с использованием анти-β-актина (таблица 1) и HRP-конъюгированного антитела к мышиным IgG (таблица 1), разбавленных в блокирующем буфере. Альтернативно, мышиное анти-β-актин антитело использовали в сочетании с козьей антисывороткой к CFI.
На Фиг. 7 показана экспрессия белка CFI в объединенных образцах, проанализированных методом иммуноблоттинга. CFI экспрессировался на детектируемых уровнях во всех дозах, в обоих случаях, как в случае вектора AAV.CFI, так в случае для вектора AAV.CFIco. β-актин использовали в качестве контроля для нанесения. 7A: 40 мкг белкового лизата наносили в денатурирующих условиях, и CFI детектировали с помощью поликлональной козьей антисыворотки к человеческому CFI. CFI проявляется в виде полос: 80 кДа (про-фермент), 50 кДа (подвергнутая процессингу тяжелая цепь) и 35 кДа (подвергнутая процессингу легкая цепь). Эти полосы соответствуют ожидаемому размеру CFI и подтверждают наличие процессинга, т.е. наличие тяжелой и легкой цепи. 7B: Такое же количество белкового лизата также использовали для образцов лизата, полученных из глаз с введенным AAV путем инъекции в количестве 109 гк/глаз и глаз без инъекции. CFI детектировали с помощью мышиного моноклонального антитела к CFI (слева, гель в неденатурирующих условиях) и козьей антисыворотки к CFI (справа, гель в денатурирующих условиях). В геле в неденатурирующих условиях (слева) CFI проявляется в виде полосы 75 кДа у инъецированных животных, а у животных, в глаза которых не вводили инъекцию, полосы отсутствуют. В геле в денатурирующих условиях (справа) CFI проявляется в виде полос 80 кДа (подвергнутая процессингу тяжелая цепь) и 35 кДа (подвергнутая процессингу легкая цепь).
Подготовка образцов тканей для анализа экспрессии генов
Мышей умерщвляли, как описано выше, и задние стенки глазных бокалов оставляли нетронутыми, или сетчатку и ПЭС отделяли, как описано выше. Образцы тканей сразу помещали в стерильные пробирки, содержащие RNALater (Thermo Fisher Scientific). Образцы хранили при 4°С до дальнейшей обработки. РНК выделяли с помощью набора RNeasy Mini (Qiagen), и ткань гомогенизировали в ступке с пестиком в буфере RLT (с помощью набора). Для гарантии отсутствия геномной ДНК дополнительно проводили комплементарную обработку свободной от РНКазы ДНКазой (Qiagen). 100 нг РНК транскрибировали в обратном направлении в кДНК с использованием набора Superscript III First Strand Synthesis (Thermo Fisher Scientific). После обратной транскрипции реакционный продукт очищали с помощью ПЦР набора для очистки QIAquick PCR Purification (Qiagen). кПЦР выполняли на кДНК из одного глаза на каждое состояние (имитация, 107 гк/глаз CFI, 108 гк/глаз CFI, 107 гк/глаз CFIco и 108 гк/глаз CFIco) с использованием системы обнаружения методом ПЦР в режиме реального времени CFX Connect™ (Bio Rad). В каждую лунку вводили по 1,25 нг кДНК в качестве матрицы для кПЦР реакций с использованием зеленого мастер-микс SYBR (iTaq Universal SYBR Green Supermix, Bio Rad). Для каждого условия выполняли по три повтора; значения Ct получали, используя компьютерную программу. Сравнительный анализ ΔΔCt с помощью мышиного бета-актина в качестве гена хозяина использовали для определения относительной экспрессии CFI для контролей с ложными инъекциями.
На Фиг. 8 показан анализ экспрессии генов методом кПЦР. Как и ожидалось, у ложно-инъекционных мышей отсутствовала экспрессия человеческого CFI или CFIco. Поскольку анализировали только один глаз на каждое состояние, статистический анализ не проводили. CFI экспрессировался из обеих AAV-конструкций, и экспрессия, по-видимому, происходила преимущественно в ПЭС, однако инъекция 108 гк/глаз приводила к увеличению экспрессии мРНК CFI в ткани сетчатки. Предполагается, что в образце глазного бокала количество мРНК CFI на общее количество мРНК глазного бокала будет меньше, чем в ПЭС, потому что во многих областях глаза отсутствует экспрессия CFI; что отражено в результатах.
Подготовка срезов сетчатки для иммуногистохимии
Мышей умерщвляли мгновенной дислокацией шейных позвонков через 4 недели после инъекции. Глаза вынимали и готовили следующим образом. Отверстие делали сразу за зубчатым краем (ora serrata), и глазной бокал помещали в 4% (мас./об.) параформальдегида в 0,12 М фосфатном буфере, рН 7,2 на 1 час при комнатной температуре. Затем удаляли передний сегмент и хрусталик, и готовили срезы глаза или цельный препарат. Глазные бокалы, используемые для срезов, криоконсервировали с увеличением концентраций сахарозы в фосфатном буфере 0,12 М, рН 7,2 (10%, в течение 1 часа при комнатной температуре и 30% в течение ночи при +4°С), заливали OCT (Tissue-tek, Sakura Finetek USA inc, Ca, USA) и замораживали в форме при -80°C. Срезы получали на криостате с толщиной 10 мкм и помещали на предметные стекла (Super-Frost, Thermo Fisher Scientific, Waltham, MA, USA). Предметные стекла сушили на воздухе в течение 2 часов при комнатной температуре и хранили при -80°C. В случае глазных бокалов, используемых для получения цельных препаратов, сетчатку и ПЭС/хориоидея разделяли, осторожно отделяя сетчатку от ПЭС и хранили отдельно в PBS при +4°C.
Иммуноокрашивание
Срезы блокировали и пермеабилизировали путем инкубации при комнатной температуре в течение 60 мин в растворе PBSGT (0,2% (в./об.) желатин, 0,25% (м/м) Тритон X-100 в PBS). Цельные препараты блокировали и пермеабилизировали путем инкубации при комнатной температуре в течение 2 часов в 0,2% (в./об.) желатина, 1,5% (в/в) Тритон X-100 в PBS. Затем срезы и цельные препараты инкубировали с первичными антителами (см. таблицу 1) в разбавленном растворе PBSGT в течение ночи при комнатной температуре. После промывки в растворе PBST (0,1% (об./об.) Тритон X-100 в PBS) срезы и цельные препараты инкубировали со вторичными антителами, связанными с Alexa Fluor594 (Life Technologies, Thermo Fisher Scientific, Waltham, Massachusetts, USA) при разбавлении 1:5000 и окрашивали DAPI в растворе PBSGT в течение 1,5 ч при комнатной температуре. Слайды промывали раствором PBST и затем покрывали средой для заливки (Mowiol, Merck Millipore).
Таблица 2. Первичные антитела, используемые в этом исследовании в иммуногистохимии
На фиг. 9 показана локализация фибронектина и hCFI в срезах сетчатки глаза ложно-инъецированных, инъецированных вектором AAV.CFI и вектором AAV.CFIco. Фибронектин используется в качестве ко-маркера для различения различных слоев сетчатки. В глазах, инъецированных AAV.CFI и AAV.CFIco hCFI локализуется в нескольких компартментах: в небольших количествах в склере (Scl), в больших количествах в ПЭС, наружном (OS) и внутреннем (IS) сегментах фоторецепторов, наружном плексиформном слое (OPL) и предположительно в слое ганглиозных клеток (GCL) и слое нервных волокон (NFL). Небольшой сигнал в GCL+NFL виден в ложно инъецированных глазах, свидетельствуя о том, что антитело не является полностью специфичным к этим слоям.
На фиг. 10-13 показаны при более высоком увеличении различные области, в которых локализуется hCFI, в срезах сетчатки глаз, ложно инъецированных и инъецированных вектором AAV.CFI. Такой же сигнал наблюдается в срезах сетчатки глаз, инъецированных вектором AAV.CFIco (не показан).
На фиг. 10 показаны при более высоком увеличении области СЭП, в срезах сетчатки глаз, ложно инъецированных и инъецированных вектором AAV.CFI. hCFI локализуется во всем слое СЭП и в микроворсинках СЭП. Окрашивание является везикулярным, указывая на локализацию секреторного пути hCFI.
На фиг. 11 показана при более высоком увеличении область фоторецепторов в срезах сетчатки глаз, ложно инъецированных и инъецированных вектором AAV.CFI. hCFI локализуется во внутреннем и внешнем компартментах фоторецепторов, но не присутствует в области ядерно-эндоплазматического ретикулума фоторецепторов.
На фиг. 12 показан при более высоком увеличении внешний плексиформный слой в срезах сетчатки глаз, ложно инъецированных и инъецированных вектором AAV.CFI. hCFI локализуется в клеточных телах, что подтверждено окрашиванием горизонтально расположенных клеток. В приведенных ниже ссылках, показано, как проявляется экспрессия в горизонтально расположенных клетках в иммуногистохимическом анализе в отсутствие маркера в горизонтально расположенных клетках: Poche et al (2009), Development, 136: 2141-51; Cuenca et al (2010), Exp Eye Res., 91: 273-85; Ho et al (2012), PLoS One, 7: e29892.
На фиг. 13 показана при более высоком увеличении область слоя ганглиозных клеток в срезах сетчатки глаз, ложно инъецированных и инъецированных вектором AAV.CFI. Хотя интенсивность мечения hCFI в слое нервных волокон в срезах сетчатки глаз, инъецированных вектором AAV.CFI, по-видимому, выше интенсивности в срезах ложно инъецированных сетчаток глаз, трудно определить, локализован ли hCFI в слое нервных волокон.
На фиг. 14 показана локализация фибронектина и hCFI в ПЭС цельных препаратов глаз, ложно инъецированных или инъецированных векторами AAV.CFI и AAV.CFIco. В глазах, инъецированных как AAV.CFI, так и AAV.CFIco, hCFI локализуется в клетках ПЭС. Окрашивание является прерывистым и, по-видимому, локализуется в везикулах секреторного пути, где осуществляется процессинг hCFI.
Пример 2
Функциональный анализ in vivo способности AAV.CFI или AAV.CFIco уменьшать индуцированные светом повреждения сетчатки
Мышей Albino BALB/c содержали в учреждении для ухода за животными в условиях дневного цикла 12:12 час. день/ночь с неограниченным доступом к пище и воде. Интенсивность внешнего освещения на уровне глаз животных составляла 85±18 люкс. Все эксперименты проводили в соответствии с заявлением ARVO по использованию животных в офтальмологических исследованиях и исследованиях зрения, одобренным Министерством внутренних дел.
Для модели географической атрофии (Bärbel Rohrer; Yao Guo; Kannan Kunchithapautham; Gary S. Gilkeson, Investigative Ophthalmology & Visual Science November 2007, Vol.48, 5282-5289) взрослых мышей (приблизительно 1 год) перемещали в комнату для фототерапии и оставляют на ночь в темноте для адаптации. Животных размещали по две особи на клетку с прозрачными акриловыми стеклами (Plexiglas; Rohm and Haas, Philadelphia, PA) с неограниченным доступом к пище и воде. Индуцируемое светом повреждение вызывали, подвергая животных освещению белым светом интенсивностью до 1000 люкс, обеспечиваемой двумя флуоресцентными лампами 30 Вт (T30T12-CW-RS, General Electric, Piscataway, NJ), подвешенными над клетками на высоте примерно 40 см. Интенсивность света измеряли с помощью измерителя света (Extech Instruments, Waltham, MA), чтобы обеспечить одинаковую яркость всем животным. Это количество света уменьшает количество колбочек до одного ряда в течение 2-3 недель у мышей-альбиносов.
Для изучения роли фактора I комплемента (CFI) при индуцированной светом потери внешних нейронов сетчатки, мыши получали субретинальную инъекцию вектора CFI-AAV.CFI (или AAV.CFIco), контрольного вектора GFP-AAV.GFP или ложную инъекцию. Обычно использовали когорты из 6-8 мышей. После 3-4 недель воздействия AAV, инициировали вызванное светом повреждение, как описано выше. Через 2-3 недели проводили электроретинографию (ERG). Животных анестезировали с помощью ксилазина и кетамина (соответственно 20 и 80 мг/кг), а зрачки расширяли 1 каплей фенилэфрина HCl (2,5%) и атропинсульфата (1%), и помещали на нагретый блок в светонепронецаемой клетке Фарадея, в которой поддерживали температуру на уровне 37°C. Световую стимуляцию обеспечивали с помощью установки ERG, предоставленной Micron IV (Phoenix Labs). Оптический сигнал регулировали механически управляемыми шторами, концентрацией нейтральных частиц, устанавливаемой вручную, и 500 нм полосовым фильтром. Интенсивность света при 10 мс вспышке, предусмотренной в качестве стимула, можно изменять с шагом 0,3 log в пределах от 3,0×105 до 3,0×1011 фотонов/мм2. Скотопические электроретинограммы регистрировали в ответ на однократную стимуляцию увеличивающейся интенсивности света, в виде среднего значения для трех-пяти ответов. Пиковую амплитуду a-волны измеряли от базовой линии до начального падения напряжения, тогда как амплитуду пиковой b-волны измеряли от минимума a-волны до пика положительной b-волны. После ERG получали глаза мышей и обрабатывали для гистологического анализа путем окрашивания гематоксилином и эозином, и определяли толщину наружных ядерных слоев сетчатки.
Предполагается, что снижение активности альтернативного пути комплемента под воздействием CFI-AAV.CFI/AAV.CFIco проявится в неизменившейся функции ERG и приведет к уменьшению потери внешних нейронов сетчатки.
Таким образом, аденоассоциированный вирус является эффективным средством для обеспечения устойчивой экспрессии фактора I комплемента, регулятора альтернативного пути комплемента. Неожиданно было обнаружено, что CFI, доставленный при помощи AAV, экспрессируется, подвергается нормальному процессингу и секретируется в функционально активной форме как в активных, так и в конфлюентных человеческих клетках ПЭС. Эти данные показывают, что человеческий ПЭС содержит про-белковые конвертазы, необходимые для секреции CFI в активной форме. В исследованиях трансляции у животных субретинальная доставка CFI.AAV приводила к легко обнаруживаемой экспрессии подвергнутой процессингу активной формы hCFI, продуцируемой мышиными клетками ПЭС. Иммуноокрашивание также предполагает, что, несмотря на отсутствие внутриклеточной экспрессии hCFI в фоторецепторах мышей, белок присутствовал во внутренних и внешних сегментах фоторецепторов, что свидетельствует о диффузии hCFI, секретируемого ПЭС, который может быть потенциально важным регуляторным элементом пути комплемента в широкой области хориоретины. Учитывая данные, связывающие дисрегуляцию комплемента с возрастной макулярной дегенерацией, этот новый терапевтический подход может сыграть решающую роль в лечении этого состояния ослепления, обеспечивающий продолжительный эффект.
Все упомянутые выше публикации включены в настоящее описание посредством ссылки. Различные модификации и варианты описанных векторов, клеток, композиций, применений и способов по настоящему изобретению будут очевидны специалистам в данной области без отхода от объема и сущности настоящего изобретения. Хотя настоящее изобретение было описано в отношении конкретных предпочтительных вариантов осуществления, следует понимать, что заявленное изобретение никоим образом не ограничивается такими конкретными вариантами осуществления. Действительно, различные модификации описанных режимов осуществления изобретения, которые очевидны специалистам в области биохимии и биотехнологии или смежных областях, также входят в объем приведенных ниже пунктов формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Syncona Management LLP
<120> ГЕННАЯ ТЕРАПИЯ
<130> P108488PCT
<140> PCT/GB2016/053343
<141> 2016-10-27
<150> GB 1519086.1
<151> 2015-10-28
<160> 9
<170> PatentIn version 3.5
<210> 1
<211> 583
<212> Протеин
<213> Homo sapiens
<400> 1
Met Lys Leu Leu His Val Phe Leu Leu Phe Leu Cys Phe His Leu Arg
1 5 10 15
Phe Cys Lys Val Thr Tyr Thr Ser Gln Glu Asp Leu Val Glu Lys Lys
20 25 30
Cys Leu Ala Lys Lys Tyr Thr His Leu Ser Cys Asp Lys Val Phe Cys
35 40 45
Gln Pro Trp Gln Arg Cys Ile Glu Gly Thr Cys Val Cys Lys Leu Pro
50 55 60
Tyr Gln Cys Pro Lys Asn Gly Thr Ala Val Cys Ala Thr Asn Arg Arg
65 70 75 80
Ser Phe Pro Thr Tyr Cys Gln Gln Lys Ser Leu Glu Cys Leu His Pro
85 90 95
Gly Thr Lys Phe Leu Asn Asn Gly Thr Cys Thr Ala Glu Gly Lys Phe
100 105 110
Ser Val Ser Leu Lys His Gly Asn Thr Asp Ser Glu Gly Ile Val Glu
115 120 125
Val Lys Leu Val Asp Gln Asp Lys Thr Met Phe Ile Cys Lys Ser Ser
130 135 140
Trp Ser Met Arg Glu Ala Asn Val Ala Cys Leu Asp Leu Gly Phe Gln
145 150 155 160
Gln Gly Ala Asp Thr Gln Arg Arg Phe Lys Leu Ser Asp Leu Ser Ile
165 170 175
Asn Ser Thr Glu Cys Leu His Val His Cys Arg Gly Leu Glu Thr Ser
180 185 190
Leu Ala Glu Cys Thr Phe Thr Lys Arg Arg Thr Met Gly Tyr Gln Asp
195 200 205
Phe Ala Asp Val Val Cys Tyr Thr Gln Lys Ala Asp Ser Pro Met Asp
210 215 220
Asp Phe Phe Gln Cys Val Asn Gly Lys Tyr Ile Ser Gln Met Lys Ala
225 230 235 240
Cys Asp Gly Ile Asn Asp Cys Gly Asp Gln Ser Asp Glu Leu Cys Cys
245 250 255
Lys Ala Cys Gln Gly Lys Gly Phe His Cys Lys Ser Gly Val Cys Ile
260 265 270
Pro Ser Gln Tyr Gln Cys Asn Gly Glu Val Asp Cys Ile Thr Gly Glu
275 280 285
Asp Glu Val Gly Cys Ala Gly Phe Ala Ser Val Thr Gln Glu Glu Thr
290 295 300
Glu Ile Leu Thr Ala Asp Met Asp Ala Glu Arg Arg Arg Ile Lys Ser
305 310 315 320
Leu Leu Pro Lys Leu Ser Cys Gly Val Lys Asn Arg Met His Ile Arg
325 330 335
Arg Lys Arg Ile Val Gly Gly Lys Arg Ala Gln Leu Gly Asp Leu Pro
340 345 350
Trp Gln Val Ala Ile Lys Asp Ala Ser Gly Ile Thr Cys Gly Gly Ile
355 360 365
Tyr Ile Gly Gly Cys Trp Ile Leu Thr Ala Ala His Cys Leu Arg Ala
370 375 380
Ser Lys Thr His Arg Tyr Gln Ile Trp Thr Thr Val Val Asp Trp Ile
385 390 395 400
His Pro Asp Leu Lys Arg Ile Val Ile Glu Tyr Val Asp Arg Ile Ile
405 410 415
Phe His Glu Asn Tyr Asn Ala Gly Thr Tyr Gln Asn Asp Ile Ala Leu
420 425 430
Ile Glu Met Lys Lys Asp Gly Asn Lys Lys Asp Cys Glu Leu Pro Arg
435 440 445
Ser Ile Pro Ala Cys Val Pro Trp Ser Pro Tyr Leu Phe Gln Pro Asn
450 455 460
Asp Thr Cys Ile Val Ser Gly Trp Gly Arg Glu Lys Asp Asn Glu Arg
465 470 475 480
Val Phe Ser Leu Gln Trp Gly Glu Val Lys Leu Ile Ser Asn Cys Ser
485 490 495
Lys Phe Tyr Gly Asn Arg Phe Tyr Glu Lys Glu Met Glu Cys Ala Gly
500 505 510
Thr Tyr Asp Gly Ser Ile Asp Ala Cys Lys Gly Asp Ser Gly Gly Pro
515 520 525
Leu Val Cys Met Asp Ala Asn Asn Val Thr Tyr Val Trp Gly Val Val
530 535 540
Ser Trp Gly Glu Asn Cys Gly Lys Pro Glu Phe Pro Gly Val Tyr Thr
545 550 555 560
Lys Val Ala Asn Tyr Phe Asp Trp Ile Ser Tyr His Val Gly Arg Pro
565 570 575
Phe Ile Ser Gln Tyr Asn Val
580
<210> 2
<211> 1752
<212> ДНК
<213> Homo sapiens
<400> 2
atgaagcttc ttcatgtttt cctgttattt ctgtgcttcc acttaaggtt ttgcaaggtc 60
acttatacat ctcaagagga tctggtggag aaaaagtgct tagcaaaaaa atatactcac 120
ctctcctgcg ataaagtctt ctgccagcca tggcagagat gcattgaggg cacctgtgtt 180
tgtaaactac cgtatcagtg cccaaagaat ggcactgcag tgtgtgcaac taacaggaga 240
agcttcccaa catactgtca acaaaagagt ttggaatgtc ttcatccagg gacaaagttt 300
ttaaataacg gaacatgcac agccgaagga aagtttagtg tttccttgaa gcatggaaat 360
acagattcag agggaatagt tgaagtaaaa cttgtggacc aagataagac aatgttcata 420
tgcaaaagca gctggagcat gagggaagcc aacgtggcct gccttgacct tgggtttcaa 480
caaggtgctg atactcaaag aaggtttaag ttgtctgatc tctctataaa ttccactgaa 540
tgtctacatg tgcattgccg aggattagag accagtttgg ctgaatgtac ttttactaag 600
agaagaacta tgggttacca ggatttcgct gatgtggttt gttatacaca gaaagcagat 660
tctccaatgg atgacttctt tcagtgtgtg aatgggaaat acatttctca gatgaaagcc 720
tgtgatggta tcaatgattg tggagaccaa agtgatgaac tgtgttgtaa agcatgccaa 780
ggcaaaggct tccattgcaa atcgggtgtt tgcattccaa gccagtatca atgcaatggt 840
gaggtggact gcattacagg ggaagatgaa gttggctgtg caggctttgc atctgtggct 900
caagaagaaa cagaaatttt gactgctgac atggatgcag aaagaagacg gataaaatca 960
ttattaccta aactatcttg tggagttaaa aacagaatgc acattcgaag gaaacgaatt 1020
gtgggaggaa agcgagcaca actgggagac ctcccatggc aggtggcaat taaggatgcc 1080
agtggaatca cctgtggggg aatttatatt ggtggctgtt ggattctgac tgctgcacat 1140
tgtctcagag ccagtaaaac tcatcgttac caaatatgga caacagtagt agactggata 1200
caccccgacc ttaaacgtat agtaattgaa tacgtggata gaattatttt ccatgaaaac 1260
tacaatgcag gcacttacca aaatgacatc gctttgattg aaatgaaaaa agacggaaac 1320
aaaaaagatt gtgagctgcc tcgttccatc cctgcctgtg tcccctggtc tccttaccta 1380
ttccaaccta atgatacatg catcgtttct ggctggggac gagaaaaaga taacgaaaga 1440
gtcttttcac ttcagtgggg tgaagttaaa ctaataagca actgctctaa gttttacgga 1500
aatcgtttct atgaaaaaga aatggaatgt gcaggtacat atgatggttc catcgatgcc 1560
tgtaaagggg actctggagg ccccttagtc tgtatggatg ccaacaatgt gacttatgtc 1620
tggggtgttg tgagttgggg ggaaaactgt ggaaaaccag agttcccagg tgtttacacc 1680
aaagtggcca attattttga ctggattagc taccatgtag gaaggccttt tatttctcag 1740
tacaatgtat aa 1752
<210> 3
<211> 1231
<212> Протеин
<213> Homo sapiens
<400> 3
Met Arg Leu Leu Ala Lys Ile Ile Cys Leu Met Leu Trp Ala Ile Cys
1 5 10 15
Val Ala Glu Asp Cys Asn Glu Leu Pro Pro Arg Arg Asn Thr Glu Ile
20 25 30
Leu Thr Gly Ser Trp Ser Asp Gln Thr Tyr Pro Glu Gly Thr Gln Ala
35 40 45
Ile Tyr Lys Cys Arg Pro Gly Tyr Arg Ser Leu Gly Asn Val Ile Met
50 55 60
Val Cys Arg Lys Gly Glu Trp Val Ala Leu Asn Pro Leu Arg Lys Cys
65 70 75 80
Gln Lys Arg Pro Cys Gly His Pro Gly Asp Thr Pro Phe Gly Thr Phe
85 90 95
Thr Leu Thr Gly Gly Asn Val Phe Glu Tyr Gly Val Lys Ala Val Tyr
100 105 110
Thr Cys Asn Glu Gly Tyr Gln Leu Leu Gly Glu Ile Asn Tyr Arg Glu
115 120 125
Cys Asp Thr Asp Gly Trp Thr Asn Asp Ile Pro Ile Cys Glu Val Val
130 135 140
Lys Cys Leu Pro Val Thr Ala Pro Glu Asn Gly Lys Ile Val Ser Ser
145 150 155 160
Ala Met Glu Pro Asp Arg Glu Tyr His Phe Gly Gln Ala Val Arg Phe
165 170 175
Val Cys Asn Ser Gly Tyr Lys Ile Glu Gly Asp Glu Glu Met His Cys
180 185 190
Ser Asp Asp Gly Phe Trp Ser Lys Glu Lys Pro Lys Cys Val Glu Ile
195 200 205
Ser Cys Lys Ser Pro Asp Val Ile Asn Gly Ser Pro Ile Ser Gln Lys
210 215 220
Ile Ile Tyr Lys Glu Asn Glu Arg Phe Gln Tyr Lys Cys Asn Met Gly
225 230 235 240
Tyr Glu Tyr Ser Glu Arg Gly Asp Ala Val Cys Thr Glu Ser Gly Trp
245 250 255
Arg Pro Leu Pro Ser Cys Glu Glu Lys Ser Cys Asp Asn Pro Tyr Ile
260 265 270
Pro Asn Gly Asp Tyr Ser Pro Leu Arg Ile Lys His Arg Thr Gly Asp
275 280 285
Glu Ile Thr Tyr Gln Cys Arg Asn Gly Phe Tyr Pro Ala Thr Arg Gly
290 295 300
Asn Thr Ala Lys Cys Thr Ser Thr Gly Trp Ile Pro Ala Pro Arg Cys
305 310 315 320
Thr Leu Lys Pro Cys Asp Tyr Pro Asp Ile Lys His Gly Gly Leu Tyr
325 330 335
His Glu Asn Met Arg Arg Pro Tyr Phe Pro Val Ala Val Gly Lys Tyr
340 345 350
Tyr Ser Tyr Tyr Cys Asp Glu His Phe Glu Thr Pro Ser Gly Ser Tyr
355 360 365
Trp Asp His Ile His Cys Thr Gln Asp Gly Trp Ser Pro Ala Val Pro
370 375 380
Cys Leu Arg Lys Cys Tyr Phe Pro Tyr Leu Glu Asn Gly Tyr Asn Gln
385 390 395 400
Asn Tyr Gly Arg Lys Phe Val Gln Gly Lys Ser Ile Asp Val Ala Cys
405 410 415
His Pro Gly Tyr Ala Leu Pro Lys Ala Gln Thr Thr Val Thr Cys Met
420 425 430
Glu Asn Gly Trp Ser Pro Thr Pro Arg Cys Ile Arg Val Lys Thr Cys
435 440 445
Ser Lys Ser Ser Ile Asp Ile Glu Asn Gly Phe Ile Ser Glu Ser Gln
450 455 460
Tyr Thr Tyr Ala Leu Lys Glu Lys Ala Lys Tyr Gln Cys Lys Leu Gly
465 470 475 480
Tyr Val Thr Ala Asp Gly Glu Thr Ser Gly Ser Ile Thr Cys Gly Lys
485 490 495
Asp Gly Trp Ser Ala Gln Pro Thr Cys Ile Lys Ser Cys Asp Ile Pro
500 505 510
Val Phe Met Asn Ala Arg Thr Lys Asn Asp Phe Thr Trp Phe Lys Leu
515 520 525
Asn Asp Thr Leu Asp Tyr Glu Cys His Asp Gly Tyr Glu Ser Asn Thr
530 535 540
Gly Ser Thr Thr Gly Ser Ile Val Cys Gly Tyr Asn Gly Trp Ser Asp
545 550 555 560
Leu Pro Ile Cys Tyr Glu Arg Glu Cys Glu Leu Pro Lys Ile Asp Val
565 570 575
His Leu Val Pro Asp Arg Lys Lys Asp Gln Tyr Lys Val Gly Glu Val
580 585 590
Leu Lys Phe Ser Cys Lys Pro Gly Phe Thr Ile Val Gly Pro Asn Ser
595 600 605
Val Gln Cys Tyr His Phe Gly Leu Ser Pro Asp Leu Pro Ile Cys Lys
610 615 620
Glu Gln Val Gln Ser Cys Gly Pro Pro Pro Glu Leu Leu Asn Gly Asn
625 630 635 640
Val Lys Glu Lys Thr Lys Glu Glu Tyr Gly His Ser Glu Val Val Glu
645 650 655
Tyr Tyr Cys Asn Pro Arg Phe Leu Met Lys Gly Pro Asn Lys Ile Gln
660 665 670
Cys Val Asp Gly Glu Trp Thr Thr Leu Pro Val Cys Ile Val Glu Glu
675 680 685
Ser Thr Cys Gly Asp Ile Pro Glu Leu Glu His Gly Trp Ala Gln Leu
690 695 700
Ser Ser Pro Pro Tyr Tyr Tyr Gly Asp Ser Val Glu Phe Asn Cys Ser
705 710 715 720
Glu Ser Phe Thr Met Ile Gly His Arg Ser Ile Thr Cys Ile His Gly
725 730 735
Val Trp Thr Gln Leu Pro Gln Cys Val Ala Ile Asp Lys Leu Lys Lys
740 745 750
Cys Lys Ser Ser Asn Leu Ile Ile Leu Glu Glu His Leu Lys Asn Lys
755 760 765
Lys Glu Phe Asp His Asn Ser Asn Ile Arg Tyr Arg Cys Arg Gly Lys
770 775 780
Glu Gly Trp Ile His Thr Val Cys Ile Asn Gly Arg Trp Asp Pro Glu
785 790 795 800
Val Asn Cys Ser Met Ala Gln Ile Gln Leu Cys Pro Pro Pro Pro Gln
805 810 815
Ile Pro Asn Ser His Asn Met Thr Thr Thr Leu Asn Tyr Arg Asp Gly
820 825 830
Glu Lys Val Ser Val Leu Cys Gln Glu Asn Tyr Leu Ile Gln Glu Gly
835 840 845
Glu Glu Ile Thr Cys Lys Asp Gly Arg Trp Gln Ser Ile Pro Leu Cys
850 855 860
Val Glu Lys Ile Pro Cys Ser Gln Pro Pro Gln Ile Glu His Gly Thr
865 870 875 880
Ile Asn Ser Ser Arg Ser Ser Gln Glu Ser Tyr Ala His Gly Thr Lys
885 890 895
Leu Ser Tyr Thr Cys Glu Gly Gly Phe Arg Ile Ser Glu Glu Asn Glu
900 905 910
Thr Thr Cys Tyr Met Gly Lys Trp Ser Ser Pro Pro Gln Cys Glu Gly
915 920 925
Leu Pro Cys Lys Ser Pro Pro Glu Ile Ser His Gly Val Val Ala His
930 935 940
Met Ser Asp Ser Tyr Gln Tyr Gly Glu Glu Val Thr Tyr Lys Cys Phe
945 950 955 960
Glu Gly Phe Gly Ile Asp Gly Pro Ala Ile Ala Lys Cys Leu Gly Glu
965 970 975
Lys Trp Ser His Pro Pro Ser Cys Ile Lys Thr Asp Cys Leu Ser Leu
980 985 990
Pro Ser Phe Glu Asn Ala Ile Pro Met Gly Glu Lys Lys Asp Val Tyr
995 1000 1005
Lys Ala Gly Glu Gln Val Thr Tyr Thr Cys Ala Thr Tyr Tyr Lys
1010 1015 1020
Met Asp Gly Ala Ser Asn Val Thr Cys Ile Asn Ser Arg Trp Thr
1025 1030 1035
Gly Arg Pro Thr Cys Arg Asp Thr Ser Cys Val Asn Pro Pro Thr
1040 1045 1050
Val Gln Asn Ala Tyr Ile Val Ser Arg Gln Met Ser Lys Tyr Pro
1055 1060 1065
Ser Gly Glu Arg Val Arg Tyr Gln Cys Arg Ser Pro Tyr Glu Met
1070 1075 1080
Phe Gly Asp Glu Glu Val Met Cys Leu Asn Gly Asn Trp Thr Glu
1085 1090 1095
Pro Pro Gln Cys Lys Asp Ser Thr Gly Lys Cys Gly Pro Pro Pro
1100 1105 1110
Pro Ile Asp Asn Gly Asp Ile Thr Ser Phe Pro Leu Ser Val Tyr
1115 1120 1125
Ala Pro Ala Ser Ser Val Glu Tyr Gln Cys Gln Asn Leu Tyr Gln
1130 1135 1140
Leu Glu Gly Asn Lys Arg Ile Thr Cys Arg Asn Gly Gln Trp Ser
1145 1150 1155
Glu Pro Pro Lys Cys Leu His Pro Cys Val Ile Ser Arg Glu Ile
1160 1165 1170
Met Glu Asn Tyr Asn Ile Ala Leu Arg Trp Thr Ala Lys Gln Lys
1175 1180 1185
Leu Tyr Ser Arg Thr Gly Glu Ser Val Glu Phe Val Cys Lys Arg
1190 1195 1200
Gly Tyr Arg Leu Ser Ser Arg Ser His Thr Leu Arg Thr Thr Cys
1205 1210 1215
Trp Asp Gly Lys Leu Glu Tyr Pro Thr Cys Ala Lys Arg
1220 1225 1230
<210> 4
<211> 3696
<212> ДНК
<213> Homo sapiens
<400> 4
atgagacttc tagcaaagat tatttgcctt atgttatggg ctatttgtgt agcagaagat 60
tgcaatgaac ttcctccaag aagaaataca gaaattctga caggttcctg gtctgaccaa 120
acatatccag aaggcaccca ggctatctat aaatgccgcc ctggatatag atctcttgga 180
aatgtaataa tggtatgcag gaagggagaa tgggttgctc ttaatccatt aaggaaatgt 240
cagaaaaggc cctgtggaca tcctggagat actccttttg gtacttttac ccttacagga 300
ggaaatgtgt ttgaatatgg tgtaaaagct gtgtatacat gtaatgaggg gtatcaattg 360
ctaggtgaga ttaattaccg tgaatgtgac acagatggat ggaccaatga tattcctata 420
tgtgaagttg tgaagtgttt accagtgaca gcaccagaga atggaaaaat tgtcagtagt 480
gcaatggaac cagatcggga ataccatttt ggacaagcag tacggtttgt atgtaactca 540
ggctacaaga ttgaaggaga tgaagaaatg cattgttcag acgatggttt ttggagtaaa 600
gagaaaccaa agtgtgtgga aatttcatgc aaatccccag atgttataaa tggatctcct 660
atatctcaga agattattta taaggagaat gaacgatttc aatataaatg taacatgggt 720
tatgaataca gtgaaagagg agatgctgta tgcactgaat ctggatggcg tccgttgcct 780
tcatgtgaag aaaaatcatg tgataatcct tatattccaa atggtgacta ctcaccttta 840
aggattaaac acagaactgg agatgaaatc acgtaccagt gtagaaatgg tttttatcct 900
gcaacccggg gaaatacagc aaaatgcaca agtactggct ggatacctgc tccgagatgt 960
accttgaaac cttgtgatta tccagacatt aaacatggag gtctatatca tgagaatatg 1020
cgtagaccat actttccagt agctgtagga aaatattact cctattactg tgatgaacat 1080
tttgagactc cgtcaggaag ttactgggat cacattcatt gcacacaaga tggatggtcg 1140
ccagcagtac catgcctcag aaaatgttat tttccttatt tggaaaatgg atataatcaa 1200
aatcatggaa gaaagtttgt acagggtaaa tctatagacg ttgcctgcca tcctggctac 1260
gctcttccaa aagcgcagac cacagttaca tgtatggaga atggctggtc tcctactccc 1320
agatgcatcc gtgtcaaaac atgttccaaa tcaagtatag atattgagaa tgggtttatt 1380
tctgaatctc agtatacata tgccttaaaa gaaaaagcga aatatcaatg caaactagga 1440
tatgtaacag cagatggtga aacatcagga tcaattacat gtgggaaaga tggatggtca 1500
gctcaaccca cgtgcattaa atcttgtgat atcccagtat ttatgaatgc cagaactaaa 1560
aatgacttca catggtttaa gctgaatgac acattggact atgaatgcca tgatggttat 1620
gaaagcaata ctggaagcac cactggttcc atagtgtgtg gttacaatgg ttggtctgat 1680
ttacccatat gttatgaaag agaatgcgaa cttcctaaaa tagatgtaca cttagttcct 1740
gatcgcaaga aagaccagta taaagttgga gaggtgttga aattctcctg caaaccagga 1800
tttacaatag ttggacctaa ttccgttcag tgctaccact ttggattgtc tcctgacctc 1860
ccaatatgta aagagcaagt acaatcatgt ggtccacctc ctgaactcct caatgggaat 1920
gttaaggaaa aaacgaaaga agaatatgga cacagtgaag tggtggaata ttattgcaat 1980
cctagatttc taatgaaggg acctaataaa attcaatgtg ttgatggaga gtggacaact 2040
ttaccagtgt gtattgtgga ggagagtacc tgtggagata tacctgaact tgaacatggc 2100
tgggcccagc tttcttcccc tccttattac tatggagatt cagtggaatt caattgctca 2160
gaatcattta caatgattgg acacagatca attacgtgta ttcatggagt atggacccaa 2220
cttccccagt gtgtggcaat agataaactt aagaagtgca aatcatcaaa tttaattata 2280
cttgaggaac atttaaaaaa caagaaggaa ttcgatcata attctaacat aaggtacaga 2340
tgtagaggaa aagaaggatg gatacacaca gtctgcataa atggaagatg ggatccagaa 2400
gtgaactgct caatggcaca aatacaatta tgcccacctc cacctcagat tcccaattct 2460
cacaatatga caaccacact gaattatcgg gatggagaaa aagtatctgt tctttgccaa 2520
gaaaattatc taattcagga aggagaagaa attacatgca aagatggaag atggcagtca 2580
ataccactct gtgttgaaaa aattccatgt tcacaaccac ctcagataga acacggaacc 2640
attaattcat ccaggtcttc acaagaaagt tatgcacatg ggactaaatt gagttatact 2700
tgtgagggtg gtttcaggat atctgaagaa aatgaaacaa catgctacat gggaaaatgg 2760
agttctccac ctcagtgtga aggccttcct tgtaaatctc cacctgagat ttctcatggt 2820
gttgtagctc acatgtcaga cagttatcag tatggagaag aagttacgta caaatgtttt 2880
gaaggttttg gaattgatgg gcctgcaatt gcaaaatgct taggagaaaa atggtctcac 2940
cctccatcat gcataaaaac agattgtctc agtttaccta gctttgaaaa tgccataccc 3000
atgggagaga agaaggatgt gtataaggcg ggtgagcaag tgacttacac ttgtgcaaca 3060
tattacaaaa tggatggagc cagtaatgta acatgcatta atagcagatg gacaggaagg 3120
ccaacatgca gagacacctc ctgtgtgaat ccgcccacag tacaaaatgc ttatatagtg 3180
tcgagacaga tgagtaaata tccatctggt gagagagtac gttatcaatg taggagccct 3240
tatgaaatgt ttggggatga agaagtgatg tgtttaaatg gaaactggac ggaaccacct 3300
caatgcaaag attctacagg aaaatgtggg ccccctccac ctattgacaa tggggacatt 3360
acttcattcc cgttgtcagt atatgctcca gcttcatcag ttgagtacca atgccagaac 3420
ttgtatcaac ttgagggtaa caagcgaata acatgtagaa atggacaatg gtcagaacca 3480
ccaaaatgct tacatccgtg tgtaatatcc cgagaaatta tggaaaatta taacatagca 3540
ttaaggtgga cagccaaaca gaagctttat tcgagaacag gtgaatcagt tgaatttgtg 3600
tgtaaacggg gatatcgtct ttcatcacgt tctcacacat tgcgaacaac atgttgggat 3660
gggaaactgg agtatccaac ttgtgcaaaa agatag 3696
<210> 5
<211> 934
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Пример промоторной последовательности
<400> 5
attgacgtca ataatgacgt atgttcccat agtaacgcca atagggactt tccattgacg 60
tcaatgggtg gagtatttac ggtaaactgc ccacttggca gtacatcaag tgtatcatat 120
gccaagtacg ccccctattg acgtcaatga cggtaaatgg cccgcctggc attatgccca 180
gtacatgacc ttatgggact ttcctacttg gcagtacatc tacgtattag tcatcgctat 240
taccatggtc gaggtgagcc ccacgttctg cttcactctc cccatctccc ccccctcccc 300
acccccaatt ttgtatttat ttatttttta attattttgt gcagcgatgg gggcgggggg 360
gggggggggg cgcgcgccag gcggggcggg gcggggcgag gggcggggcg gggcgaggcg 420
gagaggtgcg gcggcagcca atcagagcgg cgcgctccga aagtttcctt ttatggcgag 480
gcggcggcgg cggcggccct ataaaaagcg aagcgcgcgg cgggcgggag tcgctgcgcg 540
ctgccttcgc cccgtgcccc gctccgccgc cgcctcgcgc cgcccgcccc ggctctgact 600
gaccgcgtta ctcccacagg tgagcgggcg ggacggccct tctcctccgg gctgtaatta 660
gcgcttggtt taatgacggc ttgtttcttt tctgtggctg cgtgaaagcc ttgaggggct 720
ccgggagggc cctttgtgcg gggggagcgg ctcggggctg tccgcggggg gacggctgcc 780
ttcggggggg acggggcagg gcggggttcg gcttctggcg tgtgaccggc ggctctagag 840
cctctgctaa ccatgttcat gccttcttct ttttcctaca gctcctgggc aacgtgctgg 900
ttattgtgct gtctcatcat tttggcaaag aatt 934
<210> 6
<211> 270
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Поли-A сигнал бычьего гормона роста
<400> 6
tcgctgatca gcctcgactg tgccttctag ttgccagcca tctgttgttt gcccctcccc 60
cgtgccttcc ttgaccctgg aaggtgccac tcccactgtc ctttcctaat aaaatgagga 120
aattgcatcg cattgtctga gtaggtgtca ttctattctg gggggtgggg tggggcagga 180
cagcaagggg gaggattggg aagacaatag caggcatgct ggggatgcgg tgggctctat 240
ggcttctgag gcggaaagaa ccagctgggg 270
<210> 7
<211> 588
<212> ДНК
<213> Вирус гепатита лесных сурков
<400> 7
atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac tatgttgctc 60
cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt gcttcccgta 120
tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat gaggagttgt 180
ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca acccccactg 240
gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc cccctcccta 300
ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg gctcggctgt 360
tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct tggctgctcg 420
cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct tcggccctca 480
atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt ccgcgtcttc 540
gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgc 588
<210> 8
<211> 1752
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая Фактор I
<400> 8
atgaagctgc tgcatgtctt tctgctgttt ctgtgcttcc atctgcggtt ctgtaaagtg 60
acctatacta gccaggagga tctggtggag aagaagtgtc tggccaagaa gtacacacac 120
ctgagctgcg acaaggtgtt ctgtcagcct tggcagcggt gcatcgaggg cacctgcgtg 180
tgcaagctgc cttaccagtg cccaaagaac ggcaccgccg tgtgcgccac aaatcggaga 240
tcttttccaa catattgcca gcagaagagc ctggagtgtc tgcaccccgg caccaagttc 300
ctgaacaatg gcacctgcac agccgagggc aagttttctg tgagcctgaa gcacggcaac 360
acagatagcg agggcatcgt ggaggtgaag ctggtggacc aggataagac catgttcatc 420
tgtaagagct cctggtccat gagggaggca aacgtggcat gcctggatct gggattccag 480
cagggagcag acacacagag gcgctttaag ctgtccgacc tgtctatcaa tagcaccgag 540
tgcctgcacg tgcactgtag gggcctggag acatccctgg cagagtgcac cttcacaaag 600
cggagaacca tgggctacca ggactttgcc gacgtggtgt gctataccca gaaggccgat 660
agccccatgg acgatttctt tcagtgcgtg aacggcaagt atatctccca gatgaaggcc 720
tgcgacggca tcaatgactg tggcgatcag tctgacgagc tgtgctgtaa ggcctgtcag 780
ggcaagggct tccactgcaa gagcggcgtg tgcatccctt cccagtacca gtgcaacggc 840
gaggtggatt gtatcacagg agaggacgaa gtgggatgcg caggatttgc atctgtggca 900
caggaggaga cagagatcct gacagccgac atggatgccg agaggcgccg gatcaagtct 960
ctgctgccta agctgagctg tggcgtgaag aatcggatgc acatcagaag gaagcgcatc 1020
gtgggaggca agagggcaca gctgggcgat ctgccatggc aggtggccat caaggacgcc 1080
tctggcatca cctgcggcgg catctacatc ggaggatgtt ggatcctgac cgcagcacac 1140
tgcctgagag caagcaagac acacaggtat cagatctgga ccacagtggt ggattggatc 1200
cacccagacc tgaagagaat cgtgatcgag tacgtggata ggatcatctt tcacgagaac 1260
tacaatgccg gcacatatca gaacgacatc gccctgatcg agatgaagaa ggatggcaat 1320
aagaaggact gtgagctgcc cagatccatc cctgcatgcg tgccatggag cccctatctg 1380
ttccagccca acgatacctg catcgtgtcc ggatggggaa gggagaagga caatgagcgg 1440
gtgttttctc tgcagtgggg cgaggtgaag ctgatctcca actgttctaa gttctacggc 1500
aataggtttt atgagaagga gatggagtgc gccggcacct acgatggcag catcgacgcc 1560
tgtaagggcg attccggagg accactggtg tgcatggacg caaacaatgt gacatacgtg 1620
tggggagtgg tgtcctgggg agagaactgc ggcaagccag agttccccgg cgtatatacc 1680
aaggtggcca attattttga ttggatttcc taccacgtcg gcaggccctt tatttcccag 1740
tataatgtct aa 1752
<210> 9
<211> 583
<212> Протеин
<213> Homo sapiens
<400> 9
Met Lys Leu Leu His Val Phe Leu Leu Phe Leu Cys Phe His Leu Arg
1 5 10 15
Phe Cys Lys Val Thr Tyr Thr Ser Gln Glu Asp Leu Val Glu Lys Lys
20 25 30
Cys Leu Ala Lys Lys Tyr Thr His Leu Ser Cys Asp Lys Val Phe Cys
35 40 45
Gln Pro Trp Gln Arg Cys Ile Glu Gly Thr Cys Val Cys Lys Leu Pro
50 55 60
Tyr Gln Cys Pro Lys Asn Gly Thr Ala Val Cys Ala Thr Asn Arg Arg
65 70 75 80
Ser Phe Pro Thr Tyr Cys Gln Gln Lys Ser Leu Glu Cys Leu His Pro
85 90 95
Gly Thr Lys Phe Leu Asn Asn Gly Thr Cys Thr Ala Glu Gly Lys Phe
100 105 110
Ser Val Ser Leu Lys His Gly Asn Thr Asp Ser Glu Gly Ile Val Glu
115 120 125
Val Lys Leu Val Asp Gln Asp Lys Thr Met Phe Ile Cys Lys Ser Ser
130 135 140
Trp Ser Met Arg Glu Ala Asn Val Ala Cys Leu Asp Leu Gly Phe Gln
145 150 155 160
Gln Gly Ala Asp Thr Gln Arg Arg Phe Lys Leu Ser Asp Leu Ser Ile
165 170 175
Asn Ser Thr Glu Cys Leu His Val His Cys Arg Gly Leu Glu Thr Ser
180 185 190
Leu Ala Glu Cys Thr Phe Thr Lys Arg Arg Thr Met Gly Tyr Gln Asp
195 200 205
Phe Ala Asp Val Val Cys Tyr Thr Gln Lys Ala Asp Ser Pro Met Asp
210 215 220
Asp Phe Phe Gln Cys Val Asn Gly Lys Tyr Ile Ser Gln Met Lys Ala
225 230 235 240
Cys Asp Gly Ile Asn Asp Cys Gly Asp Gln Ser Asp Glu Leu Cys Cys
245 250 255
Lys Ala Cys Gln Gly Lys Gly Phe His Cys Lys Ser Gly Val Cys Ile
260 265 270
Pro Ser Gln Tyr Gln Cys Asn Gly Glu Val Asp Cys Ile Thr Gly Glu
275 280 285
Asp Glu Val Gly Cys Ala Gly Phe Ala Ser Val Ala Gln Glu Glu Thr
290 295 300
Glu Ile Leu Thr Ala Asp Met Asp Ala Glu Arg Arg Arg Ile Lys Ser
305 310 315 320
Leu Leu Pro Lys Leu Ser Cys Gly Val Lys Asn Arg Met His Ile Arg
325 330 335
Arg Lys Arg Ile Val Gly Gly Lys Arg Ala Gln Leu Gly Asp Leu Pro
340 345 350
Trp Gln Val Ala Ile Lys Asp Ala Ser Gly Ile Thr Cys Gly Gly Ile
355 360 365
Tyr Ile Gly Gly Cys Trp Ile Leu Thr Ala Ala His Cys Leu Arg Ala
370 375 380
Ser Lys Thr His Arg Tyr Gln Ile Trp Thr Thr Val Val Asp Trp Ile
385 390 395 400
His Pro Asp Leu Lys Arg Ile Val Ile Glu Tyr Val Asp Arg Ile Ile
405 410 415
Phe His Glu Asn Tyr Asn Ala Gly Thr Tyr Gln Asn Asp Ile Ala Leu
420 425 430
Ile Glu Met Lys Lys Asp Gly Asn Lys Lys Asp Cys Glu Leu Pro Arg
435 440 445
Ser Ile Pro Ala Cys Val Pro Trp Ser Pro Tyr Leu Phe Gln Pro Asn
450 455 460
Asp Thr Cys Ile Val Ser Gly Trp Gly Arg Glu Lys Asp Asn Glu Arg
465 470 475 480
Val Phe Ser Leu Gln Trp Gly Glu Val Lys Leu Ile Ser Asn Cys Ser
485 490 495
Lys Phe Tyr Gly Asn Arg Phe Tyr Glu Lys Glu Met Glu Cys Ala Gly
500 505 510
Thr Tyr Asp Gly Ser Ile Asp Ala Cys Lys Gly Asp Ser Gly Gly Pro
515 520 525
Leu Val Cys Met Asp Ala Asn Asn Val Thr Tyr Val Trp Gly Val Val
530 535 540
Ser Trp Gly Glu Asn Cys Gly Lys Pro Glu Phe Pro Gly Val Tyr Thr
545 550 555 560
Lys Val Ala Asn Tyr Phe Asp Trp Ile Ser Tyr His Val Gly Arg Pro
565 570 575
Phe Ile Ser Gln Tyr Asn Val
580
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ КОМПЛЕМЕНТ-ОПОСРЕДУЕМЫХ РАССТРОЙСТВ | 2017 |
|
RU2768982C2 |
| СЛИТЫЙ БЕЛОК КОРОТКОЙ ФОРМЫ ФАКТОРА ЖИЗНЕСПОСОБНОСТИ КОЛБОЧЕК, ПОЛУЧЕННОГО ИЗ ПАЛОЧЕК, И ГИДРОФИЛЬНОГО ПЕПТИДА | 2017 |
|
RU2773368C2 |
| ВЕКТОРЫ AAV, НАЦЕЛЕННЫЕ НА ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ | 2015 |
|
RU2727015C2 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ | 2016 |
|
RU2734678C2 |
| ВАРИАНТ AAV, КОМПОЗИЦИИ И СПОСОБЫ, В КОТОРЫХ ОН ИСПОЛЬЗУЕТСЯ, А ТАКЖЕ СПОСОБЫ ЕГО ПРИМЕНЕНИЯ ДЛЯ ПЕРЕНОСА ГЕНОВ В КЛЕТКИ, ОРГАНЫ И ТКАНИ | 2014 |
|
RU2837753C2 |
| КОМПОЗИЦИЯ ДЛЯ ОБЛЕГЧЕНИЯ ИЛИ ЛЕЧЕНИЯ БОЛИ | 2017 |
|
RU2725830C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ НЕЙРОДЕГЕНЕРАТИВНОГО НАРУШЕНИЯ ИЛИ ИНСУЛЬТА | 2018 |
|
RU2757932C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И ВЕКТОРЫ ДЛЯ ГЕНОТЕРАПИИ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА И ДРУГИХ СОСТОЯНИЙ | 2015 |
|
RU2745567C2 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок фактора свёртывания крови IX, и ее применение | 2021 |
|
RU2831751C2 |
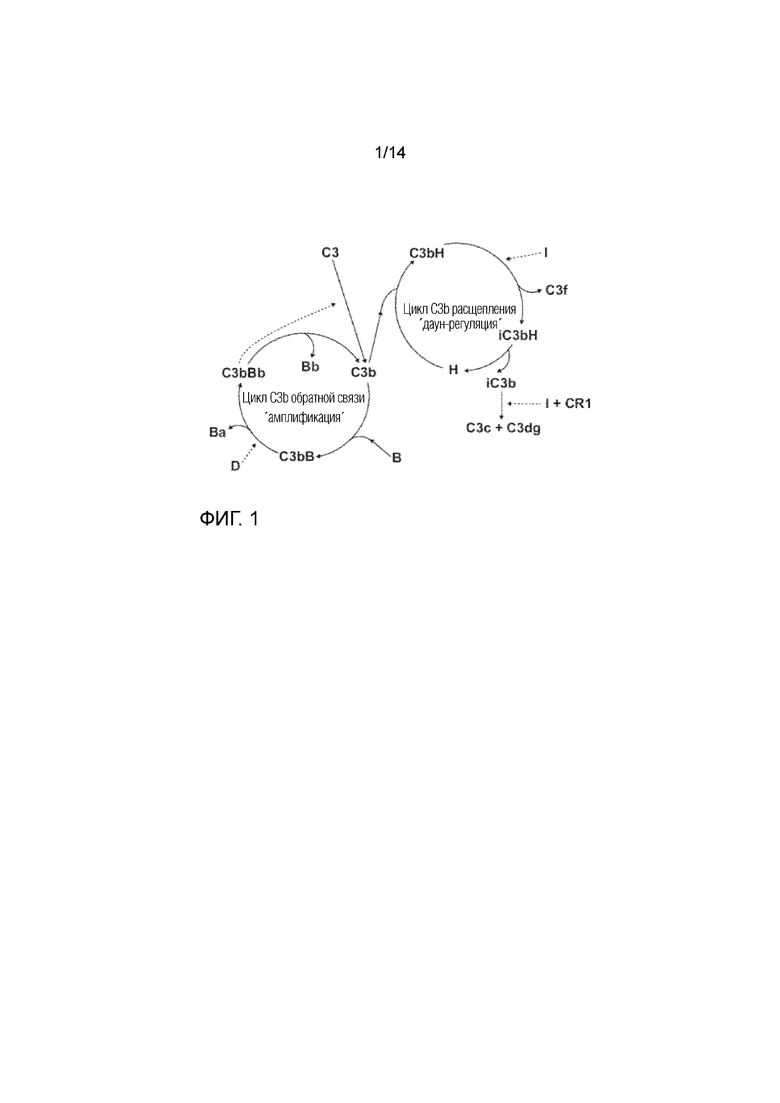
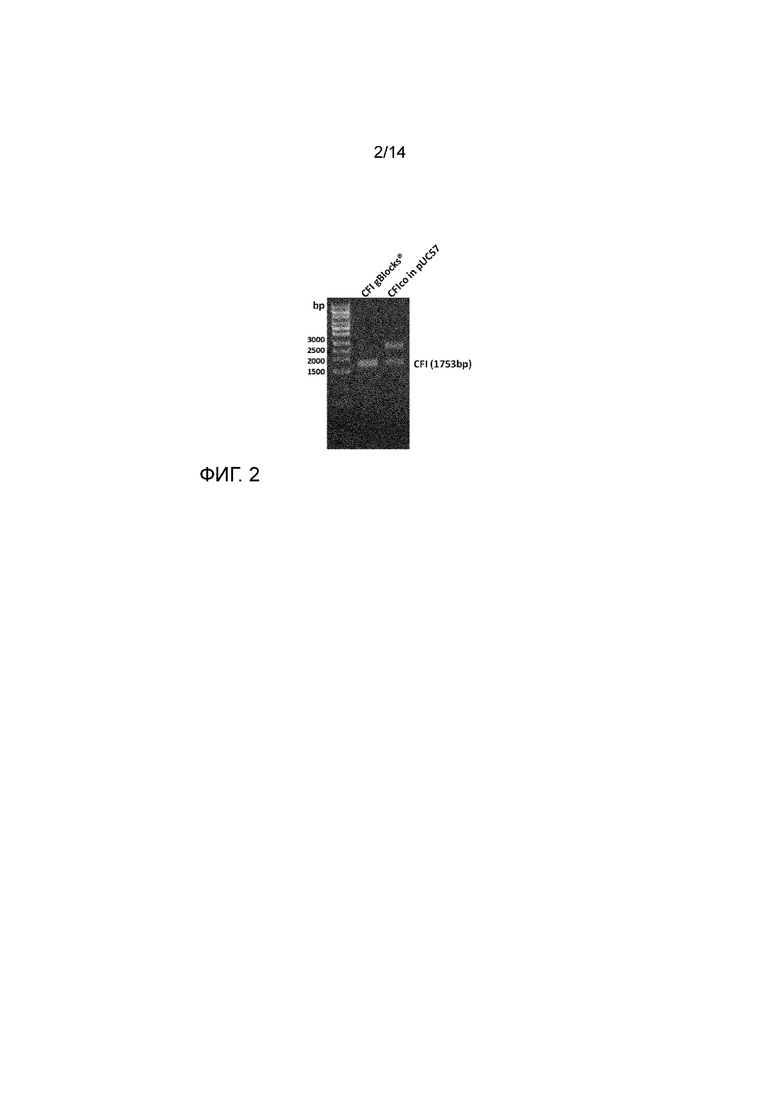
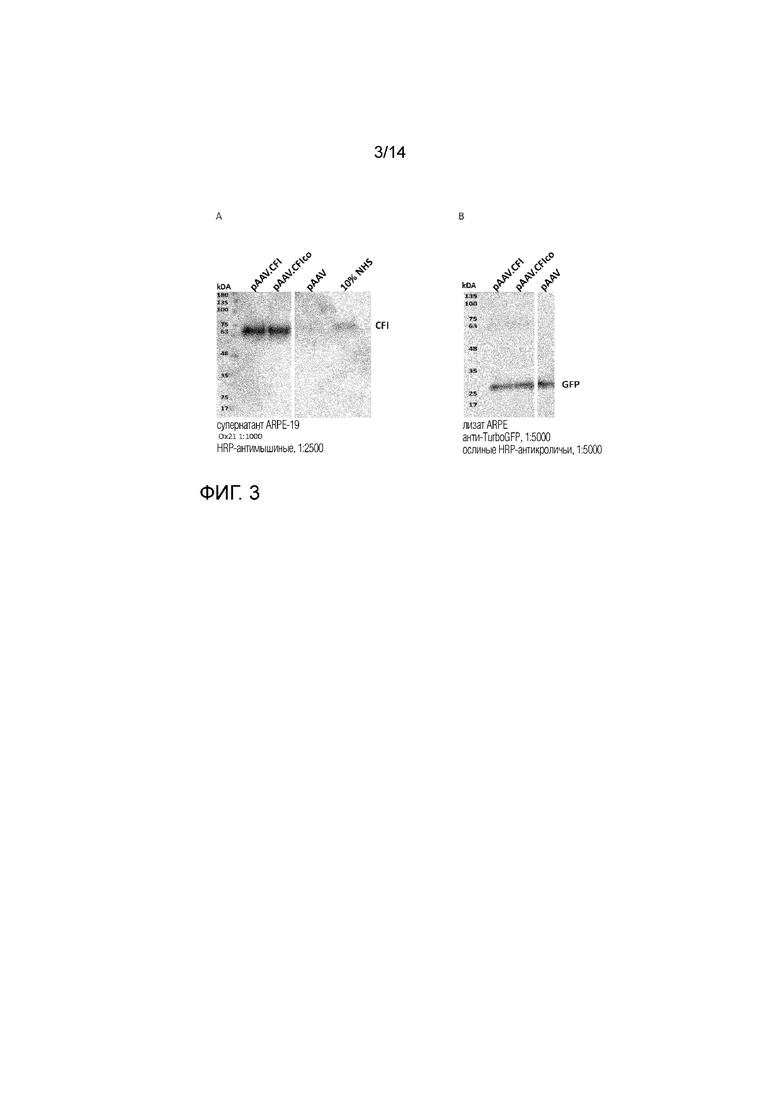


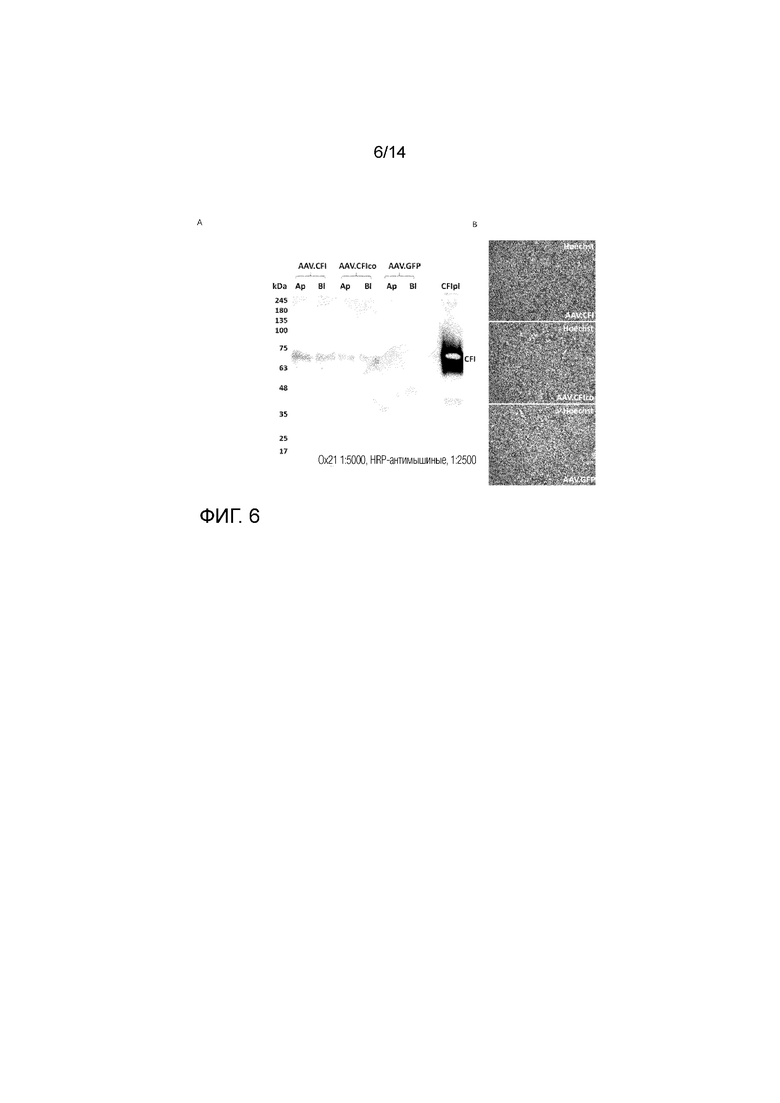
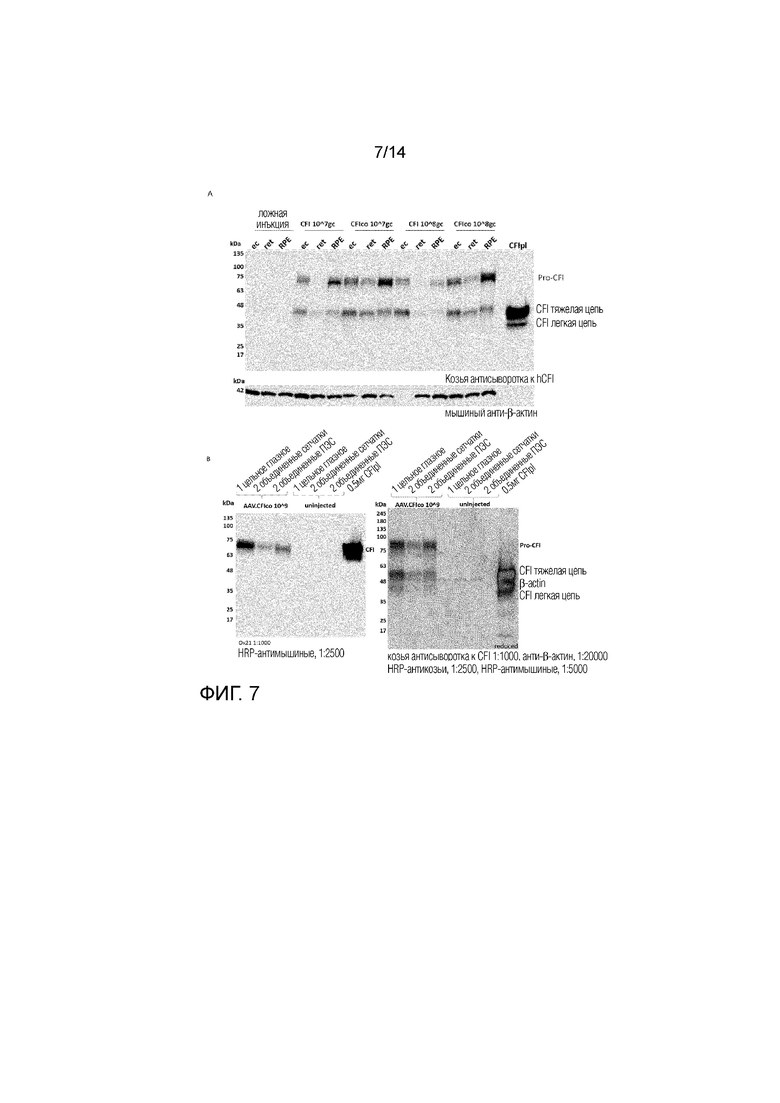
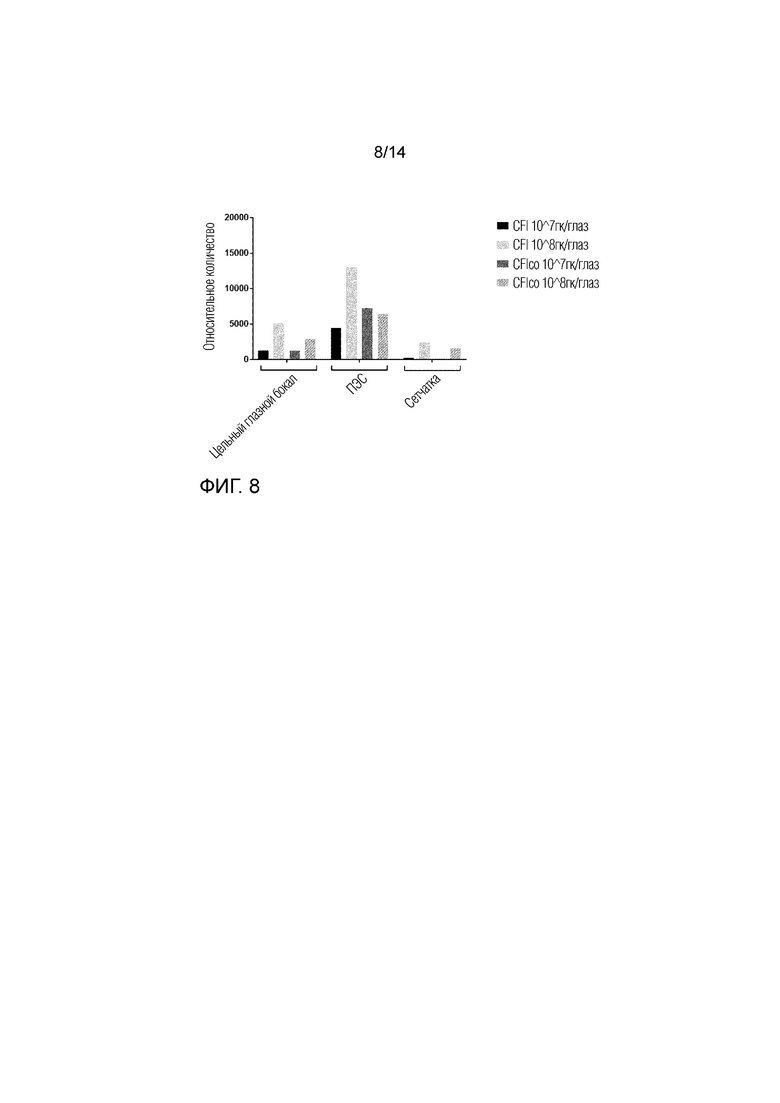
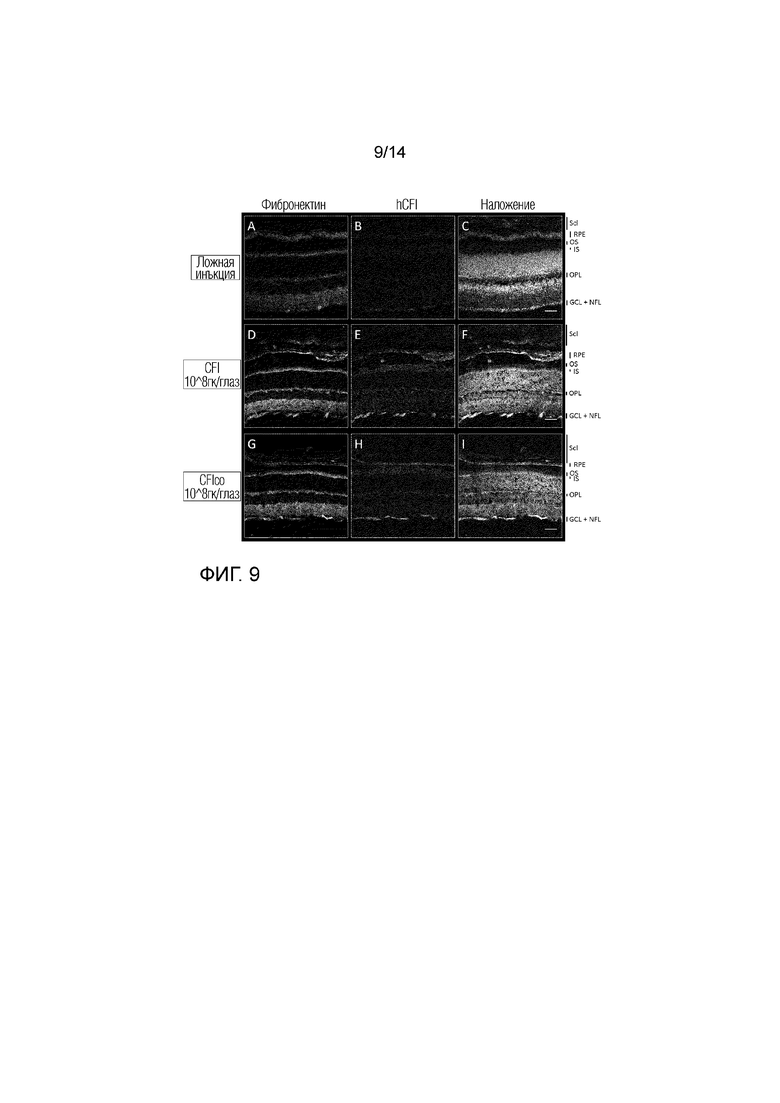
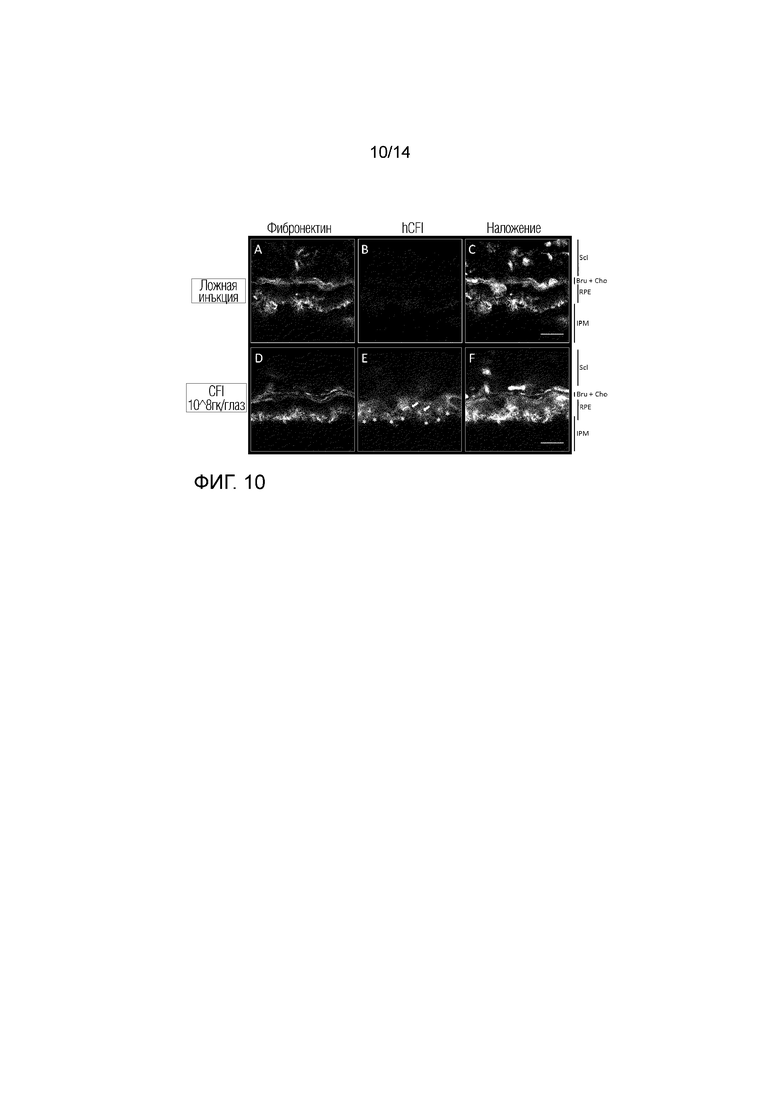
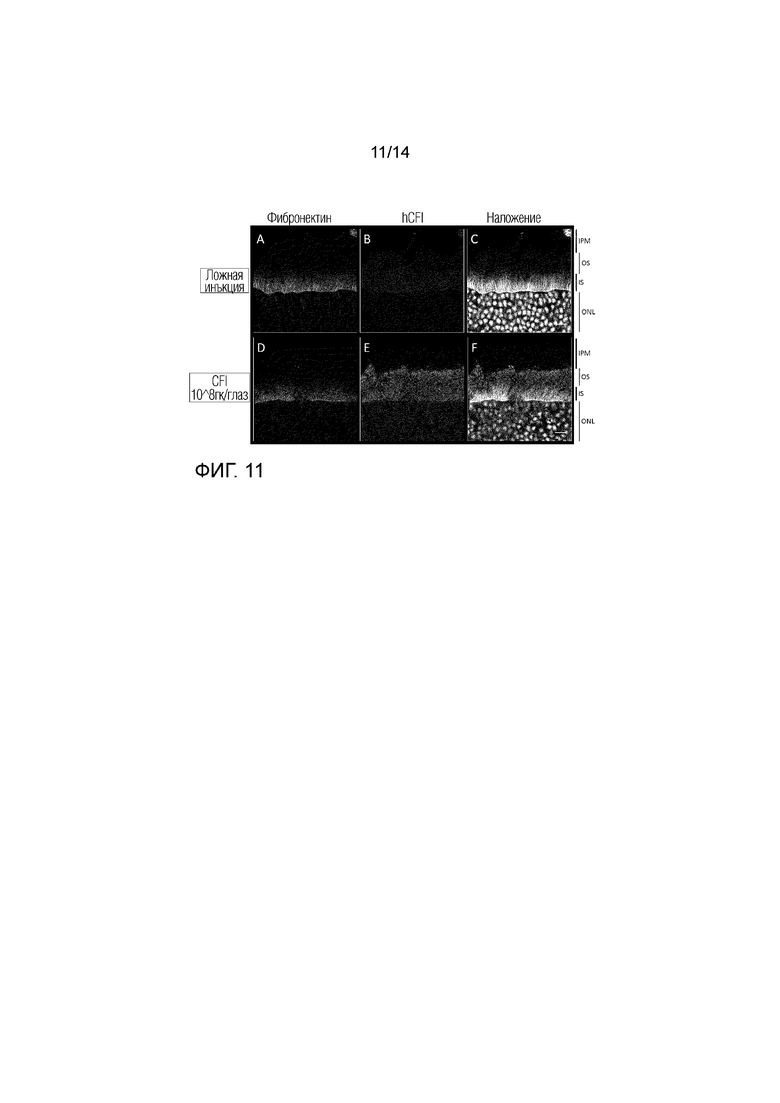

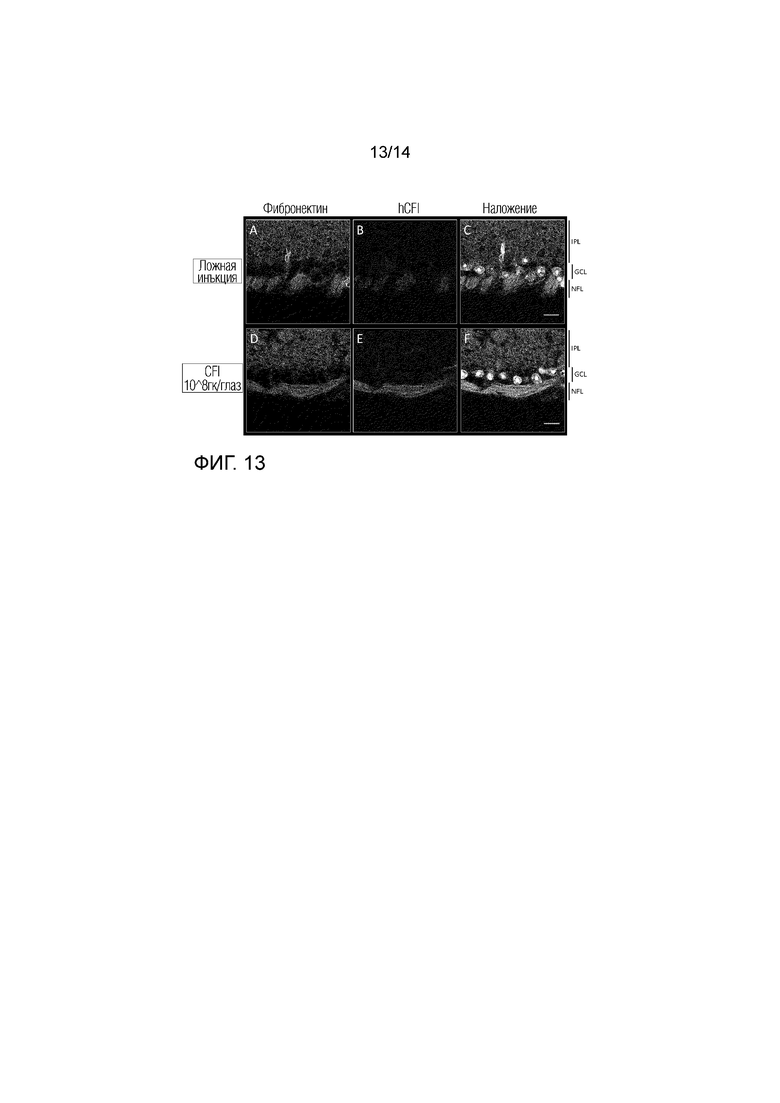

Изобретение относится к биотехнологии. Описан аденоассоциированный вирусный (AAV) вектор для лечения или профилактики опосредованного комплементом расстройства, содержащий нуклеотидную последовательность, кодирующую фактор I или его фрагмент, где фрагмент способен расщеплять C3b до iC3b. Изобретение расширяет арсенал средств для лечения или профилактики опосредованного комплементом расстройства. 5 н. и 22 з.п. ф-лы, 14 ил., 2 табл., 2 пр.
1. Аденоассоциированный вирусный (AAV) вектор для лечения или профилактики опосредованного комплементом расстройства, содержащий нуклеотидную последовательность, кодирующую фактор I или его фрагмент, где фрагмент способен расщеплять C3b до iC3b.
2. AAV-вектор по п.1, в котором нуклеотидная последовательность, кодирующая фактор I или его фрагмент, включает последовательность, выбранную из группы, состоящей из:
(а) нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая имеет по меньшей мере 70% идентичность с SEQ ID NO: 1 или 9;
(b) нуклеотидной последовательности, которая имеет по меньшей мере 70% идентичность с SEQ ID NO: 2 или 8; и
(c) нуклеотидной последовательности SEQ ID NO: 2 или 8.
3. AAV-вектор по п.1 или 2, находящийся в виде вирусной частицы.
4. AAV-вектор по п.3, в котором вирусная частица AAV содержит геном AAV2 и капсидные белки AAV2.
5. AAV-вектор по любому из предыдущих пунктов, в котором нуклеотидная последовательность, кодирующая фактор I или его фрагмент, функционально связана с промотором CAG, предпочтительно с промотором с нуклеотидной последовательностью SEQ ID NO: 5.
6. Клетка для лечения или профилактики опосредованного комплементом расстройства, трансфицированная AAV-вектором по любому из пп.1-5.
7. Фармацевтическая композиция для лечения или профилактики опосредованного комплементом расстройства, содержащая AAV-вектор по любому из пп.1-5 или клетку по п.6 в комбинации с фармацевтически приемлемым носителем, разбавителем или наполнителем.
8. Применение AAV-вектора, клетки или фармацевтической композиции по любому из пп.1-7 для лечения или профилактики опосредованного комплементом расстройства зрения.
9. Применение по п.8, где расстройство представляет собой возрастную макулярную дегенерацию (ВМД) или диабетическую ретинопатию, предпочтительно ВМД.
10. Применение по п.9, где ВМД является сухой формой ВМД.
11. Применение по любому из пп.8-10, где образование географической атрофии предотвращается или замедляется и/или уменьшается величина географической атрофии.
12. Применение по любому из пп.8-10, где прогрессирование географической атрофии замедляется.
13. Применение по п.12, где по меньшей мере на 10% уменьшается увеличение площади географической атрофии за 12 месяцев после введения в подвергшийся лечению глаз субъекта по сравнению с не подвергшимся лечению глазом за тот же период.
14. Применение по любому из пп.8-13, где введение AAV-вектора, клетки или фармацевтической композиции увеличивает уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта или в глазу субъекта, например, в пигментном эпителии сетчатки (ПЭС), необязательно до уровня, который превышает нормальный уровень у субъекта или в его глазу или ПЭС.
15. Применение по любому пп.8-14, где AAV-вектор, клетку или фармацевтическую композицию вводят интраокулярно.
16. Применение по любому из пп.8-15, где AAV-вектор, клетку или фармацевтическую композицию вводят в глаз субъекта путем субретинальной инъекции, инъекции непосредственно в сетчатку, супрахориоидальное пространство или стекловидное тело.
17. Применение по любому из пп.8-16, где AAV-вектор, клетку или фармацевтическую композицию вводят в глаз субъекта путем субретинальной инъекции.
18. Способ лечения или профилактики опосредованного комплементом расстройства зрения, включающий введение нуждающемуся в этом субъекту AAV-вектора, клетки или фармацевтической композиции по любому из пп.1-7.
19. Способ по п.18, в котором расстройство представляет собой возрастную дегенерацию желтого пятна (ВМД) или диабетическую ретинопатию, предпочтительно ВМД.
20. Способ по п.19, в котором ВМД представляет собой сухую форму ВМД.
21. Способ по любому из пп.18-20, в котором образование географической атрофии предотвращается или уменьшается и/или уменьшается величина географической атрофии.
22. Способ по любому из пп.18-20, в котором прогрессирование географической атрофии замедляется.
23. Способ по п.22, в котором по меньшей мере на 10% уменьшается увеличение площади географической атрофии в течение 12 месяцев после введения в подвергшийся лечению глаз субъекта по сравнению с не подвергшимся лечению глазом в течение того же периода.
24. Способ по любому из пп.18-23, в котором введение AAV-вектора, клетки или фармацевтической композиции увеличивает уровень активности, инактивирующей C3b и вызывающей деградацию iC3b, у субъекта или в глазу субъекта, например, в пигментном эпителии сетчатки (ПЭС), необязательно до уровня, который превышает нормальный уровень у субъекта или в его глазу или ПЭС.
25. Способ по любому из пп.18-24, в котором AAV-вектор, клетку или фармацевтическую композицию вводят интраокулярно.
26. Способ по любому из пп.18-25, в котором AAV-вектор, клетку или фармацевтическую композицию вводят в глаз субъекта путем субретинальной инъекции, инъекции непосредственно в сетчатку, супрахориоидальное пространство или стекловидное тело.
27. Способ по любому из пп.18-26, в котором AAV-вектор, клетку или фармацевтическую композицию вводят в глаз субъекта путем субретинальной инъекции.
| WO 2015121501 A1, 20.08.2015 | |||
| WO 2014116880 A1, 31.07.2014 | |||
| GB 904427 D0, 29.04.2009 | |||
| RU 2005138368 A, 27.07.2006. |

