ССЫЛКА НА СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Список последовательностей, поданный в электронной форме в форме текстового файла Annex C/ST.25 и имеющий ссылку на файл WOC-019PCT, составляет часть описания.
УРОВЕНЬ ТЕХНИКИ
RdCVF представляет собой подобный тиоредоксину белок, специфически экспрессируемый палочковыми фоторецепторными клетками в сетчатке ( et al. (2004) Nature Genetics 36:755-759 и дополнительная информация). Два различных гена RdCVF обнаружены у человека, и они обозначены RdCVF1 и RdCVF2. Оба гена RdCVF кодируют два продукта посредством альтернативного сплайсинга: полноразмерный белок и белок с C-концевым пост-транскрипционным укорочением, известные как RdCVF-длинный (RdCVFL) и RdCVF-короткий (RdCVFS), соответственно. Подобный нуклеоредоксину ген-1 (Nxnl1) кодирует длинную и короткую форму RdCVF посредством механизма альтернативного сплайсинга. Нокаут Nxnl1 приводит к прогрессивной потере палочек и колбочек у мышей, что позволяет предполагать на генетическом уровне, что этот ген является абсолютно необходимым для выживания фоторецепторных клеток и поддержания надлежащей физиологии и функции сетчатки.
et al. (2004) Nature Genetics 36:755-759 и дополнительная информация). Два различных гена RdCVF обнаружены у человека, и они обозначены RdCVF1 и RdCVF2. Оба гена RdCVF кодируют два продукта посредством альтернативного сплайсинга: полноразмерный белок и белок с C-концевым пост-транскрипционным укорочением, известные как RdCVF-длинный (RdCVFL) и RdCVF-короткий (RdCVFS), соответственно. Подобный нуклеоредоксину ген-1 (Nxnl1) кодирует длинную и короткую форму RdCVF посредством механизма альтернативного сплайсинга. Нокаут Nxnl1 приводит к прогрессивной потере палочек и колбочек у мышей, что позволяет предполагать на генетическом уровне, что этот ген является абсолютно необходимым для выживания фоторецепторных клеток и поддержания надлежащей физиологии и функции сетчатки.
RdCVFS описан как секретируемый трофический фактор для стимуляции выживания колбочек, и RdCVFL - как активный в окислительно-восстановительных реакциях фермент, который взаимодействует с внутриклеточными белками ( et al. (2010) Sci Transl Med. 2(26): 26ps16). Например, tau описан как партнер по связыванию для RdCVF-L, и tau является исключительно внутриклеточным (Fridlich et al. (2009) Molecular & Cellular Proteomics 8(6):1206-18).
Обнаружено, что индивидуумы, страдающие некоторыми дистрофиями сетчатки, имели более низкие уровни RdCVF в глазах, чем индивидуумы без дистрофии сетчатки (Публикация PCT WO 02/081513).
Показано, что различные формы белка RdCVF могут способствовать выживанию колбочковых фоторецепторных клеток in vitro и in vivo. Например, внутриглазные инъекции короткой формы белка RdCVF1 человека (RdCVF1S) не только спасают колбочковые клетки от дегенерации, но также сохраняют их функцию в моделях наследственной дегенерации сетчатки на животных (Yang et al. (2009) Mol Therapy 17:787-795). Однако, демонстрация защитного эффекта in vivo на колбочковые клетки этого белка требовала множества внутриглазных инъекций.
Пигментный ретинит (RP) представляет собой дегенеративное заболевание сетчатки глаза, характеризующееся прогрессирующей дегенерацией палочек с последующей вторичной потерей колбочек. RP является лидирующей причиной наследственной слепоты, поражающей приблизительно 100000 пациентов с 2000 новых случаев в год только в США. RP поражает все этнические группы. Более чем 1,5 миллиона человек поражены RP во всем мире. К сожалению, для пациентов не существует эффективной терапии или одобренной терапии RP. Таким образом, RP остается неудовлетворенной срочной медицинской необходимостью.
Поскольку RP является хроническим дегенеративным заболеванием сетчатки с клиническим течением на протяжении от нескольких лет до нескольких десятилетий, генотерапия может являться идеальной для показаний RP посредством конститутивной экспрессии RdCVF в сетчатке. Для острых неотложных показаний, таких как отслоение сетчатки, белковая терапия может обеспечивать преимущество. Это представляет собой использование рекомбинантного белка RdCVF для защиты фоторецепторов от гибели перед тем, как сетчатку можно будет присоединить обратно к задней поверхности глаза, т.е., к пигментному эпителию сетчатки и хороидным слоям. К сожалению, ученые в данной области столкнулись с значительными сложностями для эффективной экспрессии и секреции белка RdCVF, особенно короткой формы RdCVF. См., например, Публикацию патента США No. 20110034546, параграф [0004].
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к слитому белку, содержащему первую N-концевую сигнальную пептидную последовательность, вторую пептидную последовательность на С-конце относительно сигнальной пептидной последовательности, и третью пептидную последовательность на С-конце относительно второй пептидной последовательности; где одна из второй пептидной последовательности или третьей пептидной последовательности представляет собой последовательность RdCVF-короткого пептида, а другая представляет собой гидрофильную пептидную последовательность. После трансляции сигнальный пептид отщепляется в эндоплазматическом ретикулуме, оставляя слитый белок, содержащий вторую пептидную последовательность и третью пептидную последовательность минус сигнальный пептид. Таким образом, это изобретение также относится к слитому белку, содержащему вторую пептидную последовательность и третью пептидную последовательность на С-конце относительно второй пептидной последовательности; где одна из второй пептидной последовательности и третьей пептидной последовательности представляет собой последовательность RdCVF-короткого пептида, и другая представляет собой гидрофильную пептидную последовательность. Настоящее изобретение также относится к нуклеиновым кислотам и к экспрессирующим векторам, кодирующим слитый белок, к клеткам, содержащим нуклеиновую кислоту или вектор экспрессии, так же, как к способам лечения и применениям слитого белка, нуклеиновой кислоты и экспрессирующего вектора. Настоящее изобретение также относится к способу получения слитого белка, включающему культивирование клеток по настоящему изобретению в условиях, позволяющих экспрессию, и секрецию кодированного слитого белка, и выделение слитого белка из культуры клеток.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
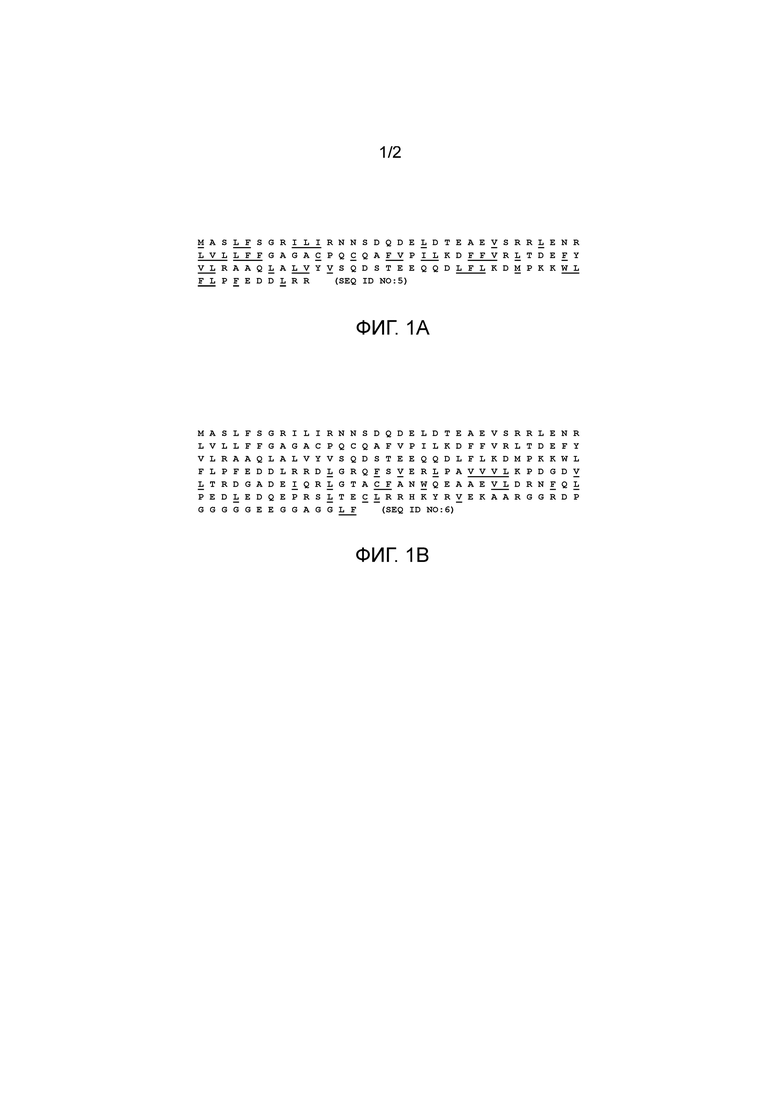
На фигуре 1A показана аминокислотная последовательность короткой формы происходящего из палочек фактора жизнеспособности колбочек человека. Аминокислотный состав короткого RdCVF человека является высоко гидрофобным. 38,5% аминокислот являются гидрофобными в коротком RdCVF. Подчеркнутые аминокислоты являются гидрофобными.
На фигуре 1B показана аминокислотная последовательность длинной формы происходящего из палочек фактора жизнеспособности колбочек человека. 25% аминокислот являются гидрофобными на C-конце длинного RdCVF (Подчеркнутые аминокислоты являются гидрофобными на C-конце длинной формы RdCVF).
Фигура 2. Слитый белок между коротким RdCVF и гидрофильным доменом.
На фигуре 2 показано схематическое представление двух слитых белков с коротким RdCVF человека. Короткий RdCVF человека слит с гидрофильным доменом с N-конца или с C-конца. Слитый белок имеет сигнальный пептид для облегчения секреции из клеток.
Фигура 3. Экспрессия и секреция новых слитых белков между альбумином человека и коротким RdCVF человека.
На фигуре 3 показан анализ Вестерн-блоттинга слитых белков короткого RdCVF человека и альбумина человека.
Дорожка 1: 30 мкл культуральной среды от клеток 293 человека, трансдуцированных с использованием AAV-GFP, вектора AAV, кодирующего GFP, в качестве контроля;
Дорожка 2: 30 мкл культуральной среды от клеток 293 человека, трансдуцированных с использованием AAV-ALB-RdCVFS, вектора AAV, кодирующего короткую форму RdCVF человека, слитую с альбумином человека на N-конце короткого RdCVF;
Дорожка 3: 30 мкл культуральной среды из клеток 293 человека, трансдуцированных с использованием AAV-RdCVFS-ALB, вектора AAV, кодирующего короткую форму RdCVF человека, слитую с альбумином человека на C-конце короткого RdCVF.
Числа слева показывают маркер молекулярной массы в кДа.
КРАТКОЕ ОПИСАНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1. Аминокислотная последовательность слитого белка между альбумином человека и коротким RdCVF, с альбумином человека на N-конце слитого белка (ALB-RdCVFS)
SEQ ID NO:2. Нуклеотидная последовательность, кодирующая короткий RdCVF человека, слитый с альбумином человека на N-конце (ALB-RdCVFS)
SEQ ID NO:3. Аминокислотная последовательность слитого белка между альбумином человека и коротким RdCVF, с альбумином человека на C-конце слитого белка (RdCVFS-ALB). Присутствует спейсер из четырех аминокислот между RdCVFS и альбумином человека. Первые 21 аминокислота от N-конца представляют собой сигнальную последовательность из Igk мыши. Присутствует спейсер (линкер) из 14 аминокислот между сигнальной последовательностью и RdCVFS.
SEQ ID NO:4. Нуклеотидная последовательность, кодирующая короткий RdCVF человека, слитый с альбумином человека на C-конце (RdCVFS-ALB)
SEQ ID NO:5. Аминокислотная последовательность короткой формы происходящего из палочек фактора жизнеспособности колбочек человека.
SEQ ID NO:6. Аминокислотная последовательность длинной формы происходящего из палочек фактора жизнеспособности колбочек человека.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Без намерения быть связанными теорией, сложность в экспрессии и секреции короткой формы RdCVF потенциально может быть обусловлена его высоко гидрофобным аминокислотным составом. Тщательный анализ аминокислотного состава как короткой формы, так и длинной формы белков RdCVF человека выявил, что короткая форма белка RdCVF является чрезвычайно гидрофобной (Фигуры 1A и 1B). Сорок две аминокислоты из 109 (38,5%) являются гидрофобными аминокислотами в коротком RdCVF. Присутствует один фрагмент из шести гидрофобных аминокислот, один фрагмент из четырех гидрофобных аминокислот и два фрагмента из трех гидрофобных аминокислот каждый. Существует высокая вероятность того, что высокий процент гидрофобных аминокислот в составе делает короткий RdCVF очень сложным для эффективной экспрессии и секреции in vitro и in vivo из клеток млекопитающих, поскольку он с большей вероятностью прилипает к липидным мембранам посредством гидрофобно-гидрофобных взаимодействий. Интересно, что несмотря на то, что N-концевые 109 аминокислот в длинной форме RdCVF являются идентичными полному короткому RdCVF, C-концевые 103 аминокислоты длинного RdCVF не являются гидрофобными, где только 25% аминокислот являются гидрофобными (26 из 103). Наиболее длинный фрагмент из гидрофобных аминокислот на этом C-конце длинного RdCVF имеет длину только четыре аминокислоты. Не присутствует фрагментов из трех гидрофобных аминокислот. Относительно гидрофильный характер C-конца длинного RdCVF может играть важную роль в уменьшении общей гидрофобности длинного RdCVF.
В одном из вариантов осуществления слитого белка по настоящему изобретению, вторая пептидная последовательность представляет собой последовательность RdCVF-короткого пептида, и третья пептидная последовательность представляет собой гидрофильную пептидную последовательность. В другом варианте осуществления вторая пептидная последовательность представляет собой гидрофильную пептидную последовательность, и третья пептидная последовательность представляет собой последовательность RdCVF-короткого пептида.
Должна присутствовать N-концевая сигнальная пептидная последовательность, которая может облегчать секрецию слитого белка из клеток, экспрессирующих этот слитый белок. Можно использовать любой сигнальный пептид, позволяющий секрецию белка из клеток млекопитающих. В следующих вариантах осуществления, сигнальная пептидная последовательность выбрана из группы, состоящей из сигнальной пептидной последовательности Igk и сигнальной пептидной последовательности альбумина. В более конкретных вариантах осуществления сигнальная пептидная последовательность IgK представляет собой сигнальную пептидную последовательность IgK мыши, и сигнальная пептидная последовательность альбумина представляет собой сигнальную пептидную последовательность альбумина человека.
В предпочтительном варианте осуществления настоящего изобретения последовательность RdCVF-короткого пептида представляет собой последовательность RdCVF-короткого пептида человека. Примеры подходящих последовательностей RdCVF-короткого пептида включают последовательность RdCVF1-короткого пептида, последовательность RdCVF2-короткого пептида и последовательность RdCVF-короткого пептида, которая отличается от соответствующей последовательности дикого типа, например, одной или несколькими консервативными аминокислотными заменами.
По настоящему изобретению, можно использовать любую гидрофильную пептидную последовательность, при условии, что она уменьшает индекс гидрофобности слитого белка по сравнению с последовательностью RdCVF-короткого пептида без гидрофильной пептидной последовательности. В одном варианте осуществления настоящего изобретения слитый белок, включая любую сигнальную пептидную последовательность, если она присутствует, имеет индекс гидрофобности менее чем минус 0,20, более предпочтительно, менее чем минус 0,30.
Предпочтительно, гидрофильная пептидная последовательность не является иммуногенной для человека, не оказывает какого-либо другого отрицательного эффекта на физиологию сетчатки человека или нормальную функцию сетчатки и не влияет на биологическую функцию коротких RdCVF. Гидрофильная пептидная последовательность может представлять собой гидрофильный белок, гидрофильный домен белка, гидрофильный олигопептид или гидрофильный полипептид. Более одного гидрофильного домена могут являться кандидатами в качестве гидрофильного партнера по слиянию с коротким RdCVF. В одном варианте осуществления настоящего изобретения гидрофильная пептидная последовательность представляет собой альбумин, например альбумин человека.
В соответствии со слитым белком по настоящему изобретению, необязательно, может присутствовать спейсер из одной или нескольких аминокислот между первой пептидной последовательностью и второй пептидной последовательностью, между второй пептидной последовательностью и третьей пептидной последовательностью, или в обоих случаях. В одном варианте осуществления настоящего изобретения не присутствует спейсера между первой пептидной последовательностью и второй пептидной последовательностью, или иными словами, первая пептидная последовательность является ковалентно связанной с второй пептидной последовательностью посредством одной пептидной связи. В другом варианте осуществления присутствует спейсер между первой пептидной последовательностью и второй пептидной последовательностью. В другом варианте осуществления настоящего изобретения не присутствует спейсера между второй пептидной последовательностью и третьей пептидной последовательностью, или иными словами, вторая пептидная последовательность является ковалентно связанной с третьей пептидной последовательностью посредством одной пептидной связи. В другом варианте осуществления присутствует спейсер между второй пептидной последовательностью и третьей пептидной последовательностью. В принципе, не существует ограничения размера спейсеров. В одном варианте осуществления настоящего изобретения спейсер между первой и второй пептидными последовательностями имеет от двух до четырнадцати аминокислот. В другом варианте осуществления спейсер между второй и третьей пептидными последовательностями имеет от двух до четырнадцати аминокислот, более конкретно, между двумя и четырьмя аминокислотами.
Один вариант осуществления кодирующей последовательности слитого белка по настоящему изобретению дополнительно содержит сигнал полиаденилирования на С-конце относительно кодирующей последовательности для третьей пептидной последовательности. Сигнал полиаденилирования (поли-A) может представлять собой любой поли-A.
В одном варианте осуществления слитого белка по настоящему изобретению, первая пептидная последовательность представляет собой сигнальную последовательность альбумина человека, вторая пептидная последовательность представляет собой альбумин человека, и третья пептидная последовательность представляет собой последовательность RdCVF-короткого пептида. В более конкретном варианте осуществления слитый белок имеет последовательность (SEQ ID NO:1) или аминокислоты 25-717 из (SEQ ID NO:1). В другом варианте осуществления слитого белка по настоящему изобретению первая пептидная последовательность представляет собой сигнальную последовательность Igk, вторая пептидная последовательность представляет собой последовательность RdCVF-короткого пептида, и третья пептидная последовательность представляет собой альбумин человека. В более конкретном варианте осуществления слитый белок имеет последовательность (SEQ ID NO:3) или аминокислоты 22-732 из (SEQ ID NO:3).
В одном варианте осуществления слитого белка по настоящему изобретению, одна, две или все из сигнальной пептидной последовательности, последовательности RdCVF-короткого пептида и гидрофильной пептидной последовательности отличаются от соответствующей последовательности дикого типа. В более конкретном варианте осуществления различия включают одну или несколько консервативных аминокислотных замен. И еще в более конкретном варианте осуществления аминокислотные замены ограничены одной или несколькими консервативными аминокислотными заменами.
Слитый белок по настоящему изобретению может являться гликозилированным или негликозилированным. Говоря в общем, гликозилирование обеспечивает преимущество для стабильности и растворимости белка. В одном варианте осуществления слитый белок, экспрессированный в клетках, трансдуцированных экспрессирующим вектором по настоящему изобретению, является гликозилированным и является гликозилированным после секреции из клеток.
В одном варианте осуществления нуклеиновая кислота по настоящему изобретению представляет собой ДНК. В другом варианте осуществления кодирующую последовательность для одной, двух или всех из сигнальной пептидной последовательности, последовательности RdCVF-короткого пептида и гидрофильной пептидной последовательности перекодируют по сравнению с соответствующей последовательностью дикого типа. В более конкретном варианте осуществления перекодируют кодирующую последовательность для последовательности RdCVF-короткого пептида. Нуклеиновая кислота по настоящему изобретению может, необязательно, содержать один или несколько интронов между первой и второй пептидными последовательностями, между второй и третьей пептидными последовательностями или в другом месте. В одном варианте осуществления настоящего изобретения нуклеиновая кислота кодирует слитый белок, имеющий последовательность (SEQ ID NO:1), например, нуклеиновая кислота, имеющая последовательность (SEQ ID NO:2). В другом варианте осуществления нуклеиновая кислота кодирует слитый белок, имеющий последовательность (SEQ ID NO:3), например, нуклеиновая кислота, имеющая последовательность (SEQ ID NO:4). Можно легко предполагать другие последовательности нуклеиновой кислоты с учетом вырожденности генетического кода.
Один вариант осуществления настоящего изобретения представляет собой вектор экспрессии, содержащий нуклеиновую кислоту, описанную выше, функционально связанную с контрольной последовательностью, например, промотором. Промотор, управляющий слитым белком RdCVFS, может представлять собой любой промотор и не является ограниченным промотором CMV. Когда присутствует интрон между промотором и кодирующей последовательностью слитого белка, можно использовать любой пригодный и общепринятый интрон. Например, интрон β-глобина является пригодным.
В экспериментах два слитых белка получали посредством слияния короткого RdCVF человека с гидрофильным доменом (см. схему на фигуре 2). Один представлял собой короткий RdCVF человека, слитый с альбумином человека на N-конце, с сигнальным пептидом альбумина на N-конце слитого белка (SEQ ID NO:1). Экспрессирующую конструкцию для экспрессии и секреции этого слитого белка конструировали в контексте вектора AAV, обозначенного rAAV-ALB-RdCVFS. Этот вектор кодировал оптимизированный по кодонному составу (перекодированный) короткий RdCVF человека, слитый с альбумином человека на N-конце (SED ID NO:2). Слитый белок назван ALB-RdCVFS. Кодирующую последовательность для сигнального пептида альбумина человека включали также в экспрессирующую слитый белок конструкцию выше кодирующей последовательности для слитого белка. Экспрессирующая конструкция дополнительно содержала промотор CMV и интрон, связанный с кодирующей последовательностью для слитого белка. Экспрессирующая конструкция дополнительно содержала сигнал полиаденилирования на C-конце кодирующей последовательности слитого белка. Полную экспрессирующую кассету клонировали в экспрессирующую плазмиду AAV, и плазмиду подвергали секвенированию ДНК для подтверждения целостности экспрессирующей конструкции.
Другой иллюстративный слитый белок представлял собой короткий RdCVF человека, слитый с альбумином человека на C-конце, с сигнальным пептидом Igk мыши на N-конце слитого белка (SEQ ID NO:3). Сконструирована экспрессирующая конструкция для экспрессии и секреции этого слитого белка в контексте вектора AAV, обозначенного rAAV-RdCVFS-ALB. Этот вектор кодировал оптимизированный по кодонному составу (перекодированный) короткий RdCVF человека, слитый с альбумином человека на C-конце (SEQ ID NO:4). Слитый белок назван RdCVFS-ALB. Кодирующую последовательность для модифицированного сигнального пептида Igk мыши также включали в экспрессирующую конструкцию для слитого белка выше кодирующей последовательности слитого белка. Экспрессирующая конструкция дополнительно содержала промотор CMV и интрон, связанный с кодирующей последовательностью для слитого белка. Экспрессирующая конструкция дополнительно содержала сигнал полиаденилирования в кодирующей последовательности на C-конце слитого белка. Полную экспрессирующую кассету клонировали в экспрессирующую плазмиду AAV, и плазмиду подвергали секвенированию ДНК для подтверждения целостности экспрессирующей конструкции.
Всего существует 20 природных аминокислот. Некоторые из них являются гидрофобными, и некоторые из них являются гидрофильными. Индекс гидрофобности аминокислоты представляет собой количество, представляющее гидрофобные или гидрофильные свойства ее боковой цепи. Чем больше составляет количество, тем более гидрофобной является аминокислота (Таблица 1). Наиболее гидрофобными аминокислотами являются изолейцин (4,5) и валин (4,2). Наиболее гидрофильными являются аргинин (-4,5) и лизин (-3,9). Несмотря на то что гидрофобный или гидрофильный характер белка зависит от того, из каких аминокислот он состоит, так же, как от вторичной, третичной и четвертичной структуры, индекс гидрофобности белка может иметь прогностическую ценность для гидрофобности белка.
Таблица 1. Индекс гидрофобности аминокислот
Ala: 1,800
Arg: -4,500
Asn: -3,500
Asp: -3,500
Cys: 2,500
Gln: -3,500
Glu: -3,500
Gly: -0,400
His: -3,200
Ile: 4,500
Leu: 3,800
Lys: -3,900
Met: 1,900
Phe: 2,800
Pro: -1,600
Ser: -0,800
Thr: -0,700
Trp: -0,900
Tyr: -1,300
Val: 4,200
Публично доступную программу GPMAW использовали для расчета индекса гидрофобности для короткого RdCVF, и двух слитых белков из короткого RdCVF и альбумина. Индекс гидрофобности для нативной короткой формы RdCVF человека (RdCVFS) составляет -0,12. После слияния альбумина человека с N-концом RdCVFS, полученный слитый белок, ALB-RdCVFS, имел индекс гидрофобности, уменьшенный от -0,12 до -0,32 (включая сигнальный пептид), уменьшение индекса гидрофобности до 266,7%. После слияния альбумина человека с C-концом RdCVFS, полученный слитый белок, RdCVFS-ALB имел индекс гидрофобности, уменьшенный от -0,12 до -0,33 (включая сигнальный пептид), уменьшение индекса гидрофобности до 275%. Очень сильное уменьшение индекса гидрофобности может вносить вклад в эффективную экспрессию и секрецию слитых белков.
Кроме того, кодирующую последовательность для слитого белка клонировали в экспрессирующую конструкцию AAV-2, данные, включенные в настоящее описание, ясно показывают, что этот новый слитый белок эффективно экспрессировался, как детектировано посредством Вестерн-блоттинга со специфическими для RdCVF антителами (Фигура 3). Партнер по слиянию для короткого RdCVF не является ограниченным альбумином человека. Любой гидрофильный белок или гидрофильный домен белка, или гидрофильный пептид можно использовать для получения слитого белка с коротким RdCVF для уменьшения гидрофобности белка. Предпочтительный гидрофильный домен является неиммуногенным для человека.
Белок можно кодировать и доставлять посредством вектора для генотерапии, такого как AAV (аденоассоциированный вирусный) вектор, лентивирусный вектор, синтетический вектор, аденовирусный вектор, ретровирусный вектор, голая ДНК, наночастицы и т.д. Альтернативно, слитый белок можно доставлять в сетчатку в форме рекомбинантного белка. Гидрофильный слитый домен не должен оказывать какого-либо отрицательного эффекта на физиологию сетчатки человека или нормальную функцию сетчатки и не должен влиять на биологическую функцию коротких RdCVF. Потенциально, более одного гидрофильного домена могут являться кандидатами в качестве гидрофильного партнера по слиянию с коротким RdCVF. В примерах, альбумин человека использовали в качестве гидрофильного домена, чтобы служить партнером по слиянию для короткого RdCVF человека.
Это изобретение относится к фармацевтической композиции, содержащей: (i) компонент, выбранный из группы, состоящей из слитого белка, нуклеиновой кислоты, экспрессирующего вектора или клетки по настоящему изобретению; и (ii) фармацевтически приемлемый носитель.
Это изобретение относится к способу лечения состояния, поддающегося такому лечению, у пациента-млекопитающего, где лечение включает введение пациенту эффективного количества белка, нуклеиновой кислоты, вектора, клетки или фармацевтической композиции, описанной в настоящем документе, таким образом, лечение состояния у пациента. В вариантах осуществления настоящего изобретения состояние выбрано из группы, состоящей из дистрофии сетчатки, болезни Штаргардта, пигментного ретинита, сухой связанной с возрастом дегенерации желтого пятна (сухой AMD), географической атрофии (поздней стадии сухой AMD), влажной связанной с возрастом дегенерации желтого пятна (влажной AMD), глаукомы с внутриглазной гипертензией или без нее, диабетической ретинопатии, синдрома Барде-Бидля, синдрома Бессена-Корнцвейга, болезни Беста, хороидемы, гиратной атрофии, врожденного амавроза, болезни Рефсума, синдрома Ушера, связанного с щитовидной железой заболевания глаз, болезни Грэйва, заболевания, ассоциированного с клетками пигментированного эпителия сетчатки, заболевания переднего сегмента глаза, заболевания хрусталика/катаракт, повреждения глазного яблока, увеита, болезни Альцгеймера, болезни Гентингтона, болезни Паркинсона и заболевания органов обоняния. Можно использовать любой общепринятый способ введения, например, инъекцию в глаз, внутривенную инъекцию или другое системное введение. В одном варианте осуществления способа по настоящему изобретению состояние представляет собой офтальмологическое состояние, и введение выбрано из группы, состоящей из субретинальной инъекции и инъекции в стекловидное тело. В более конкретном варианте осуществления пациент представляет собой пациента-человека.
Это изобретение относится к способу защиты фоторецепторных клеток глаза у пациента, включающему введение в глаз пациента эффективного количества слитого белка, нуклеиновой кислоты, вектора, клеток или фармацевтической композиции по настоящему изобретению, таким образом, защиту фоторецепторных клеток глаза у пациента. В более конкретном варианте осуществления введение выбрано из группы, состоящей из субретинальной инъекции и инъекции в стекловидное тело. В более конкретном варианте осуществления пациент представляет собой пациента-человека.
В настоящее время изобретение описано со ссылкой на следующие примеры. Эти примеры представлены только с целью иллюстрации, и изобретение не следует рассматривать как ограниченное этими примерами, но вместо этого следует рассматривать, как включающее все без исключения варианты, которые становятся очевидными в результате объяснений, представленных в настоящем описании.
В то время как конкретные варианты осуществления изобретения описаны в настоящем документе с целью описания, специалисту в данной области понятно, что можно вносить различные изменения в детали без отклонения от изобретения, как описано в прилагаемой формуле изобретения.
Полное содержание всех публикаций, патентов и патентных заявок, упомянутых в этом описании, приведено в настоящем описании в такой же степени, как если бы было конкретно и индивидуально указано, что содержание каждой индивидуальной публикации, патента или патентной заявки приведено в настоящем документе в качестве ссылки. Также приведена в качестве ссылки любая дополнительная информация, опубликованная вместе с любой из вышеупомянутых публикаций, патентов и патентных заявок. Например, некоторые статьи в журналах опубликованы вместе с дополнительной информацией, которая, как правило, доступна онлайн.
ПРИМЕРЫ
ПРИМЕР 1: Получение векторов rAAV, кодирующих слитые белки короткой формы происходящего из палочек фактора жизнеспособности колбочек человека и альбумина
Клонирование плазмиды
кДНК оптимизированной по кодонному составу короткой формы белка RdCVF человека, слитой с альбумином человека на N-конце, синтезировали посредством GENEART® (компании, оказывающей платные услуги) и клонировали в аденоассоциированную вирусную векторную плазмиду pAAV-MCS (Cell Biolabs, San Diego, CA), получая плазмиду pAAV-ALB-RdCVFS. Сигнальный пептид из альбумина человека также включали выше кодирующей последовательности слитого белка RdCVF и альбумина.
Плазмида pAAV-ALB-RdCVFS содержит следующие признаки между AAV-ITR:
промотор CMV - интрон β-глобина - сигнальная последовательность - альбумин - короткий RdCVF - поли-A
Полученная аминокислотная последовательность для этого слитого белка RdCVF и альбумина показана в SEQ ID NO:1.
кДНК оптимизированной по кодонному составу короткой формы белка RdCVF человека, слитой с альбумином человека на C- конце, синтезировали посредством GENEART® (компании, оказывающей платные услуги) и клонировали в аденоассоциированную вирусную векторную плазмиду pAAV-MCS (Cell Biolabs, San Diego, CA), получая плазмиду pAAV-RdCVFS-ALB. Этот сигнальный пептид из Igk мыши также включали выше кодирующей последовательности слитого белка RdCVF и альбумина.
Плазмида pAAV-RdCVFS-ALB содержит следующие признаки между AAV-ITR:
промотор CMV - интрон β-глобина - сигнальная последовательность - короткий RdCVF - альбумин - поли-A
Полученная аминокислотная последовательность для этого слитого белка RdCVF и альбумина показана в SEQ ID NO:3.
Получение и очистка рекомбинантных векторов AAV-ALB-RdCVFS и AAV-RdCVFS-ALB
Плазмидами pAAV-ALB-RdCVFS или pAAV-RdCVFS-ALB, pHELPER (Cell BioLabs, каталожный No. 340202) и pRC2 (Cell BioLabs, каталожный No. 340201) трансформировали компетентные клетки бактерий DH10B (Invitrogen, каталожный No. 18297-010) и увеличивали масштаб с использованием наборов для выделения плазмидной ДНК Qiagen EndoFree Plasmid Maxi Kit или EndoFree Plasmid Mega Kit, в соответствии с инструкциями производителя. Концентрации плазмиды определяли с использованием спектрофотометра Beckman DU-600. Идентичность каждой плазмиды подтверждали посредством анализа рестрикционного расщепления и секвенирования ДНК.
Для получения вектора rAAV-ALB-RdCVFS или rAAV-RdCVFS-ALB, клетки 293AAV (Cell BioLabs, каталожный No. AAV-100) рассевали при 4 миллионов клеток на 15 см чашку в cDMEM (DMEM, дополненную 10% FBS, 1% глутамином, 1% заменимых аминокислот и 1% пенициллином/стрептомицином). На следующие сутки среду заменяли на 25 мл свежей cDMEM. Через два часа проводили трансфекцию. Воду (57,4 мл) смешивали с 1,3 мг pHELPER, 650 мкг pRC2, 650 мкг pAAV-ALB-RdCVFS или pAAV-RdCVFS-ALB и 8,1 мл 2 M CaCl2 (смесь вода/плазмида/CaCl2). Объем 12,5 мл 2xHBS (Lonza, Sku:RR07005) переносили в каждую из пяти 50 мл конических пробирок. При встряхивании, 12,5 мл смеси вода/плазмида/CaCl2 медленно добавляли в каждую из конических пробирок, содержащих 2xHBS. После 5-минутной инкубации, 2,5 мл суспензии добавляли в каждую культуральную чашку, содержащую клетки 293AAV.
На следующие сутки среду заменяли на 25 мл новой среды cDMEM на чашку. Через двое суток клетки собирали с использованием скребка для клеток, и смесь клетки/среда переносили в 250 мл конические пробирки. Образцы центрифугировали при 3000 об/мин в течение 15 минут при 4°C, супернатант отбрасывали, и осадки клеток ресуспендировали в 110 мл DMEM. Образцы ресуспендированных клеток переносили по аликвотам (30 мл) в 50 мл конические пробирки, и стадию замораживания/размораживания/замораживания проводили с использованием бани с этанолом/сухим льдом и водяной бани при 37°C. Пробирки сохраняли при -80°C до дальнейшей переработки материала. Такой же способ использовали для получения контрольного вектора rAAV-GFP, заменяя плазмиду pAAV-ALB-RdCVFS на плазмиду pAAV-GFP (Cell BioLabs, каталожный No. AAV-400).
Для очистки вектора rAAV-ALB-RdCVFS, четыре 50 мл конических пробирки, содержащие вектор, со стадии замораживания/размораживания/замораживания размораживали при 37°C в водяной бане. Сорок микролитров BENZONASE® (нуклеазы) (Sigma, каталожный No. E8263-25kU) добавляли в каждую пробирку, которую затем инкубировали при 37°C в течение 30 минут. Пробирки центрифугировали в течение 10 минут при 3000 об./мин, и супернатанты переносили в 500 мл бутыль. Добавляли шесть миллилитров раствора 10% дезоксихолата натрия (8,2 г в 82 мл воды). Образец кратко перемешивали и инкубировали при 37°C в течение 30 минут. Суспензию фильтровали с использованием 5 мкм фильтров. Затем проводили другую стадию фильтрации с использованием 0,8 мкм фильтра. Подготавливали колонку с гепарин-агарозой (8 мл) (Sigma, каталожный No. H6508-25 мл), и колонку уравновешивали с использованием 48 мл фосфатно-солевого буфера (PBS) (Invitrogen, каталожный No. 10010-049). Фильтрованный лизат клеток наносили на колонку, и колонку промывали с использованием 40 мл буфера для промывки (20 мл 5 M NaCl, 980 мл PBS). Вектор элюировали с использованием 15 мл буфера для элюции (80 мл 5 M NaCl, 920 мл PBS) и собирали в новую 50 мл коническую пробирку.
Вектор концентрировали посредством фильтрации центрифугированием. Установку для фильтрации центрифугированием AMICON ULTRA-15 (Millipore, каталожный No. UFC910024) промывали один раз с использованием PBS, и элюированный образец добавляли в устройство. Центрифугирование проводили в центрифуге Beckman Allegro 6KR при 2200 об/мин, 22°C, пока образец не концентрировали до объема 1-2 мл. Добавляли объем 15 мл PBS, и центрифугирование повторяли, пока объем образца не достигал ≤ 1 мл. Очищенный вектор собирали, и стенки фильтра промывали с использованием 100 мкл PBS. Образец перемешивали, и аликвоты вектора по 30 мкл хранили при 80°C в 600 мкл конических пробирках до использования.
Этот процесс повторяли для очистки векторов rAAV-RdCVFS-ALB и rAAV-GFP.
ПРИМЕР 2: Экспрессия и секреция слитых белков короткого RdCVF человека и альбумина человека, опосредованные векторами rAAV
Анализ Вестерн-блоттинга с использованием SDS-PAGE в 4-20% геле использовали для детекции экспрессии слитого белка RdCVF и альбумина с использованием стандартных способов. В качестве контроля, объем 5 мкл белкового стандарта для Вестерн-блоттинга MAGICMARK™ XP (Invitrogen, каталожный No. LC5602) добавляли во внешние лунки. По 30 мкл культуральной среды от трансдуцированных каждым вектором rAAV клеток 293 человека, смешанной с буфером для образцов белка, добавляли в каждую лунку. Проводили разделение в геле при 200 В, пока краситель не достигал низа геля. Анализ Вестерн-блоттинга проводили с использованием набора для анализа Вестерн-блоттинга VECTASTAIN® ABC-Amp от Vector Laboratories, в соответствии с модифицированным вариантом инструкций производителя. SDS-PAGE уравновешивали в буфере для переноса в течение 20 мин, и белки, разделенные посредством SDS-PAGE, переносили на нитроцеллюлозную мембрану с использованием ячейки для полусухого переноса Trans Blot Semi-Dry Transfer Cell при 20 В в течение 40 минут. После завершения переноса, мембрану блокировали в 200 мл 1x раствора казеина с осторожным покачиванием на качающейся платформе в течение по меньшей мере двух часов при комнатной температуре (RT) или при 4°C в течение ночи. Мембрану инкубировали с 50 мл 1x раствора казеина, содержащего антитело кролика, специфическое для белка RdCVF (первичное антитело, полученное из Covance (Denver, PA), разведенное 1:2000, с осторожным покачиванием при 4°C в течение ночи или 1 часа при комнатной температуре. Мембрану промывали с использованием 30 мл 1x раствора казеина 4 раза в течение 5 минут каждый при RT с осторожным покачиванием. Мембрану инкубировали с 30 мл биотинилированного антитела козы против IgG кролика (вторичного антитела), разведенного 1:10000 в 1x растворе казеина, в течение 1 часа при RT с осторожным покачиванием. Мембрану промывали в 30 мл 1x раствора казеина 3 раза по 5 минут каждый при RT с осторожным покачиванием. Мембрану инкубировали в течение 45 минут в Vectastain ABC-AmP в 50 мл 1x казеина, содержащего 100 мкл реагента A и 100 мкл реагента B. Мембрану промывали в 30 мл 1X раствора казеина 3 раза по 5 минут каждый при RT с осторожным покачиванием.
Мембрану инкубировали в Трис, pH 9,5. Хемилюминесцентный сигнал получали с использованием 6 мл субстрата DUOLOX (Vector Laboratories, каталожный No. SK 6605) и экспонировали мембрану с рентгеновской пленкой KODAK BIOMAX MS (Kodak Carestream Health, каталожный No. 8572786) в кассете для пленки в течение от 10 секунд до 5 минут, с последущей проявкой пленки с использованием раствора для проявки KODAK (Kodak GBX, каталожный No. 1900984) и раствора для фиксации KODAK.
Как показано на фигуре 3, культуральная среда от клеток 293 человека, трансдуцированных как rAAV-ALB-RdCVFS, так и rAAV-RdCVFS-ALB, содержала полосу с молекулярной массой приблизительно 80 кДа, вступающую в специфическую реакцию с антителом кролика против RdCVF. Эту полосу не детектировали в культуральной среде от клеток, трансдуцированных контрольным вектором rAAV-GFP. Данные позволяют предполагать, что оба вектора rAAV-ALB-RdCVFS и rAAV-RdCVFS-ALB опосредовали экспрессию и секрецию в клетках человека слитого белка короткого RdCVF человека и альбумина.
Векторы rAAV-ALB-RdCVFS или rAAV-RdCVFS-ALB можно использовать для внутриглазного введения для лечения заболеваний, перечисленных выше. Конкретно, векторы можно доставлять посредством субретинальной инъекции или инъекции в стекловидное тело.
Обобщение
Рекомбинантный вектор AAV, кодирующий оптимизированный по кодонному составу (перекодированный) короткий RdCVF человека, слитый с кодирующей последовательностью слитого белка с альбумином человека на N-конце, являлся способным опосредовать экспрессию слитого белка в клетках человека и секрецию слитого белка из клеток человека. Рекомбинантный вектор AAV, кодирующий оптимизированный по кодонному составу короткий RdCVF человека, слитый со слитым белком с альбумином человека на C-конце, являлся способным опосредовать экспрессию слитого белка в клетках человека и секрецию слитого белка из клеток человека.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ВЕЛЛСТАТ ОФТЭЛМИКС КОРПОРЭЙШН
<120> СЛИТЫЙ БЕЛОК КОРОТКОЙ ФОРМЫ ФАКТОРА ЖИЗНЕСПОСОБНОСТИ КОЛБОЧЕК,
ПОЛУЧЕННОГО ИЗ ПАЛОЧЕК, И ГИДРОФИЛЬНОГО ПЕПТИДА
<130> WOC-019PCT
<150> US 62/406,552
<151> 2016-10-11
<160> 6
<170> PatentIn версии 3.5
<210> 1
<211> 717
<212> Белок
<213> Homo sapiens
<220>
<221> Сигнальная последовательность
<222> (1)..(24)
<223> Сигнальная последовательность из альбумина человека
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (1)..(609)
<223> Альбумин человека (включая N-концевой сигнальный пептид)
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (25)..(609)
<223> Альбумин человека (исключая N-концевой сигнальный пептид)
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (610)..(717)
<223> RdCVFS
<400> 1
Met Lys Trp Val Thr Phe Ile Ser Leu Leu Phe Leu Phe Ser Ser Ala
1 5 10 15
Tyr Ser Arg Gly Val Phe Arg Arg Asp Ala His Lys Ser Glu Val Ala
20 25 30
His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu
35 40 45
Ile Ala Phe Ala Gln Tyr Leu Gln Gln Cys Pro Phe Glu Asp His Val
50 55 60
Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp
65 70 75 80
Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp
85 90 95
Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala
100 105 110
Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln
115 120 125
His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val
130 135 140
Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys
145 150 155 160
Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro
165 170 175
Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys
180 185 190
Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu
195 200 205
Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys
210 215 220
Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val
225 230 235 240
Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser
245 250 255
Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly
260 265 270
Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile
275 280 285
Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu
290 295 300
Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp
305 310 315 320
Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser
325 330 335
Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly
340 345 350
Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val
355 360 365
Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys
370 375 380
Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu
385 390 395 400
Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys
405 410 415
Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu
420 425 430
Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val
435 440 445
Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His
450 455 460
Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val
465 470 475 480
Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg
485 490 495
Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe
500 505 510
Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala
515 520 525
Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu
530 535 540
Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys
545 550 555 560
Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala
565 570 575
Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe
580 585 590
Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly
595 600 605
Leu Ala Ser Leu Phe Ser Gly Arg Ile Leu Ile Arg Asn Asn Ser Asp
610 615 620
Gln Asp Glu Leu Asp Thr Glu Ala Glu Val Ser Arg Arg Leu Glu Asn
625 630 635 640
Arg Leu Val Leu Leu Phe Phe Gly Ala Gly Ala Cys Pro Gln Cys Gln
645 650 655
Ala Phe Val Pro Ile Leu Lys Asp Phe Phe Val Arg Leu Thr Asp Glu
660 665 670
Phe Tyr Val Leu Arg Ala Ala Gln Leu Ala Leu Val Tyr Val Ser Gln
675 680 685
Asp Ser Thr Glu Glu Gln Gln Asp Leu Phe Leu Lys Asp Met Pro Lys
690 695 700
Lys Trp Leu Phe Leu Pro Phe Glu Asp Asp Leu Arg Arg
705 710 715
<210> 2
<211> 2154
<212> ДНК
<213> Homo sapiens
<400> 2
atgaagtggg taacctttat ttcccttctt tttctcttta gctcggctta ttccaggggt 60
gtgtttcgtc gagatgcaca caagagtgag gttgctcatc ggtttaaaga tttgggagaa 120
gaaaatttca aagccttggt gttgattgcc tttgctcagt atcttcagca gtgtccattt 180
gaagatcatg taaaattagt gaatgaagta actgaatttg caaaaacatg tgttgctgat 240
gagtcagctg aaaattgtga caaatcactt catacccttt ttggagacaa attatgcaca 300
gttgcaactc ttcgtgaaac ctatggtgaa atggctgact gctgtgcaaa acaagaacct 360
gagagaaatg aatgcttctt gcaacacaaa gatgacaacc caaacctccc ccgattggtg 420
agaccagagg ttgatgtgat gtgcactgct tttcatgaca atgaagagac atttttgaaa 480
aaatacttat atgaaattgc cagaagacat ccttactttt atgccccgga actccttttc 540
tttgctaaaa ggtataaagc tgcttttaca gaatgttgcc aagctgctga taaagctgcc 600
tgcctgttgc caaagctcga tgaacttcgg gatgaaggga aggcttcgtc tgccaaacag 660
agactcaagt gtgccagtct ccaaaaattt ggagaaagag ctttcaaagc atgggcagta 720
gctcgcctga gccagagatt tcccaaagct gagtttgcag aagtttccaa gttagtgaca 780
gatcttacca aagtccacac ggaatgctgc catggagatc tgcttgaatg tgctgatgac 840
agggcggacc ttgccaagta tatctgtgaa aatcaagatt cgatctccag taaactgaag 900
gaatgctgtg aaaaacctct gttggaaaaa tcccactgca ttgccgaagt ggaaaatgat 960
gagatgcctg ctgacttgcc ttcattagct gctgattttg ttgaaagtaa ggatgtttgc 1020
aaaaactatg ctgaggcaaa ggatgtcttc ctgggcatgt ttttgtatga atatgcaaga 1080
aggcatcctg attactctgt cgtgctgctg ctgagacttg ccaagacata tgaaaccact 1140
ctagagaagt gctgtgccgc tgcagatcct catgaatgct atgccaaagt gttcgatgaa 1200
tttaaacctc ttgtggaaga gcctcagaat ttaatcaaac aaaattgtga gctttttgag 1260
cagcttggag agtacaaatt ccagaatgcg ctattagttc gttacaccaa gaaagtaccc 1320
caagtgtcaa ctccaactct tgtagaggtc tcaagaaacc taggaaaagt gggcagcaaa 1380
tgttgtaaac atcctgaagc aaaaagaatg ccctgtgcag aagactatct atccgtggtc 1440
ctgaaccagt tatgtgtgtt gcatgagaaa acgccagtaa gtgacagagt caccaaatgc 1500
tgcacagaat ccttggtgaa caggcgacca tgcttttcag ctctggaagt cgatgaaaca 1560
tacgttccca aagagtttaa tgctgaaaca ttcaccttcc atgcagatat atgcacactt 1620
tctgagaagg agagacaaat caagaaacaa actgcacttg ttgagctcgt gaaacacaag 1680
cccaaggcaa caaaagagca actgaaagct gttatggatg atttcgcagc ttttgtagag 1740
aagtgctgca aggctgacga taaggagacc tgctttgccg aggagggtaa aaaacttgtt 1800
gctgcaagtc aagctgcctt aggcttagcc agcctgttca gcggccggat cctgatcagg 1860
aacaacagcg accaggacga gctggacacc gaggccgaag tgagcaggag gctggagaac 1920
agactggtgc tgctgttctt tggcgccgga gcctgccctc agtgccaggc cttcgtgccc 1980
atcctgaagg atttctttgt gcggctgacc gacgagttct acgtgctgag agccgcccag 2040
ctggccctgg tgtatgtgag ccaggacagc accgaggagc agcaggacct gttcctgaag 2100
gacatgccca agaagtggct gttcctgccc ttcgaggacg acctgcggag atga 2154
<210> 3
<211> 732
<212> Белок
<213> Искусственная последовательность
<220>
<223> N-концевая сигнальная последовательность из IgK мыши, за которой
следует спейсер, за которым следует RdCVFS, за которым следует
спейсер, за которым следует альбумин человека на C-конце.
<220>
<221> Сигнальная последовательность
<222> (1)..(21)
<223> Сигнальная последовательность из Igk мыши.
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (22)..(35)
<223> Спейсер
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (36)..(143)
<223> RdCVFS
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (144)..(147)
<223> Спейсер
<220>
<221> НЕОПРЕДЕЛЕННЫЙ ПРИЗНАК
<222> (148)..(732)
<223> Альбумин человека
<400> 3
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ala Ala Gln Pro Ala Arg Arg Ala Val Arg Ser
20 25 30
Leu Val Pro Ala Ser Leu Phe Ser Gly Arg Ile Leu Ile Arg Asn Asn
35 40 45
Ser Asp Gln Asp Glu Leu Asp Thr Glu Ala Glu Val Ser Arg Arg Leu
50 55 60
Glu Asn Arg Leu Val Leu Leu Phe Phe Gly Ala Gly Ala Cys Pro Gln
65 70 75 80
Cys Gln Ala Phe Val Pro Ile Leu Lys Asp Phe Phe Val Arg Leu Thr
85 90 95
Asp Glu Phe Tyr Val Leu Arg Ala Ala Gln Leu Ala Leu Val Tyr Val
100 105 110
Ser Gln Asp Ser Thr Glu Glu Gln Gln Asp Leu Phe Leu Lys Asp Met
115 120 125
Pro Lys Lys Trp Leu Phe Leu Pro Phe Glu Asp Asp Leu Arg Arg Gly
130 135 140
Arg Asp Ala Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp
145 150 155 160
Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln
165 170 175
Tyr Leu Gln Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu
180 185 190
Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn
195 200 205
Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val
210 215 220
Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys
225 230 235 240
Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn
245 250 255
Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr
260 265 270
Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu
275 280 285
Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe
290 295 300
Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp
305 310 315 320
Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly
325 330 335
Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys
340 345 350
Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln
355 360 365
Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp
370 375 380
Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys
385 390 395 400
Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp
405 410 415
Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu
420 425 430
Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp
435 440 445
Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys
450 455 460
Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu
465 470 475 480
Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu
485 490 495
Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp
500 505 510
Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val
515 520 525
Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln
530 535 540
Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys
545 550 555 560
Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn
565 570 575
Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg
580 585 590
Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys
595 600 605
Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys
610 615 620
Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val
625 630 635 640
Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe
645 650 655
His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys
660 665 670
Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys
675 680 685
Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys
690 695 700
Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys
705 710 715 720
Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
725 730
<210> 4
<211> 2199
<212> ДНК
<213> Искусственная последовательность
<220>
<223> N-концевая сигнальная последовательность из IgK мыши, за которой
следует спейсер, за которым следует RdCVFS, за которым следует
спейсер, за которым следует альбумин человека на C-конце.
<400> 4
atggagacag acacactcct gctatgggta ctgctgctct gggttccagg ttccactggt 60
gacgcggccc agccggccag gcgcgccgta cgaagcttgg tacccgccag cctgttcagc 120
ggccggatcc tgatcaggaa caacagcgac caggacgagc tggacaccga ggccgaagtg 180
agcaggaggc tggagaacag actggtgctg ctgttctttg gcgccggagc ctgccctcag 240
tgccaggcct tcgtgcccat cctgaaggat ttctttgtgc ggctgaccga cgagttctac 300
gtgctgagag ccgcccagct ggccctggtg tatgtgagcc aggacagcac cgaggagcag 360
caggacctgt tcctgaagga catgcccaag aagtggctgt tcctgccctt cgaggacgac 420
ctgcggagag gacgagatgc agatgcacac aagagtgagg ttgctcatcg gtttaaagat 480
ttgggagaag aaaatttcaa agccttggtg ttgattgcct ttgctcagta tcttcagcag 540
tgtccatttg aagatcatgt aaaattagtg aatgaagtaa ctgaatttgc aaaaacatgt 600
gttgctgatg agtcagctga aaattgtgac aaatcacttc ataccctttt tggagacaaa 660
ttatgcacag ttgcaactct tcgtgaaacc tatggtgaaa tggctgactg ctgtgcaaaa 720
caagaacctg agagaaatga atgcttcttg caacacaaag atgacaaccc aaacctcccc 780
cgattggtga gaccagaggt tgatgtgatg tgcactgctt ttcatgacaa tgaagagaca 840
tttttgaaaa aatacttata tgaaattgcc agaagacatc cttactttta tgccccggaa 900
ctccttttct ttgctaaaag gtataaagct gcttttacag aatgttgcca agctgctgat 960
aaagctgcct gcctgttgcc aaagctcgat gaacttcggg atgaagggaa ggcttcgtct 1020
gccaaacaga gactcaagtg tgccagtctc caaaaatttg gagaaagagc tttcaaagca 1080
tgggcagtag ctcgcctgag ccagagattt cccaaagctg agtttgcaga agtttccaag 1140
ttagtgacag atcttaccaa agtccacacg gaatgctgcc atggagatct gcttgaatgt 1200
gctgatgaca gggcggacct tgccaagtat atctgtgaaa atcaagattc gatctccagt 1260
aaactgaagg aatgctgtga aaaacctctg ttggaaaaat cccactgcat tgccgaagtg 1320
gaaaatgatg agatgcctgc tgacttgcct tcattagctg ctgattttgt tgaaagtaag 1380
gatgtttgca aaaactatgc tgaggcaaag gatgtcttcc tgggcatgtt tttgtatgaa 1440
tatgcaagaa ggcatcctga ttactctgtc gtgctgctgc tgagacttgc caagacatat 1500
gaaaccactc tagagaagtg ctgtgccgct gcagatcctc atgaatgcta tgccaaagtg 1560
ttcgatgaat ttaaacctct tgtggaagag cctcagaatt taatcaaaca aaattgtgag 1620
ctttttgagc agcttggaga gtacaaattc cagaatgcgc tattagttcg ttacaccaag 1680
aaagtacccc aagtgtcaac tccaactctt gtagaggtct caagaaacct aggaaaagtg 1740
ggcagcaaat gttgtaaaca tcctgaagca aaaagaatgc cctgtgcaga agactatcta 1800
tccgtggtcc tgaaccagtt atgtgtgttg catgagaaaa cgccagtaag tgacagagtc 1860
accaaatgct gcacagaatc cttggtgaac aggcgaccat gcttttcagc tctggaagtc 1920
gatgaaacat acgttcccaa agagtttaat gctgaaacat tcaccttcca tgcagatata 1980
tgcacacttt ctgagaagga gagacaaatc aagaaacaaa ctgcacttgt tgagctcgtg 2040
aaacacaagc ccaaggcaac aaaagagcaa ctgaaagctg ttatggatga tttcgcagct 2100
tttgtagaga agtgctgcaa ggctgacgat aaggagacct gctttgccga ggagggtaaa 2160
aaacttgttg ctgcaagtca agctgcctta ggcttataa 2199
<210> 5
<211> 109
<212> Белок
<213> Homo sapiens
<400> 5
Met Ala Ser Leu Phe Ser Gly Arg Ile Leu Ile Arg Asn Asn Ser Asp
1 5 10 15
Gln Asp Glu Leu Asp Thr Glu Ala Glu Val Ser Arg Arg Leu Glu Asn
20 25 30
Arg Leu Val Leu Leu Phe Phe Gly Ala Gly Ala Cys Pro Gln Cys Gln
35 40 45
Ala Phe Val Pro Ile Leu Lys Asp Phe Phe Val Arg Leu Thr Asp Glu
50 55 60
Phe Tyr Val Leu Arg Ala Ala Gln Leu Ala Leu Val Tyr Val Ser Gln
65 70 75 80
Asp Ser Thr Glu Glu Gln Gln Asp Leu Phe Leu Lys Asp Met Pro Lys
85 90 95
Lys Trp Leu Phe Leu Pro Phe Glu Asp Asp Leu Arg Arg
100 105
<210> 6
<211> 212
<212> Белок
<213> Homo sapiens
<400> 6
Met Ala Ser Leu Phe Ser Gly Arg Ile Leu Ile Arg Asn Asn Ser Asp
1 5 10 15
Gln Asp Glu Leu Asp Thr Glu Ala Glu Val Ser Arg Arg Leu Glu Asn
20 25 30
Arg Leu Val Leu Leu Phe Phe Gly Ala Gly Ala Cys Pro Gln Cys Gln
35 40 45
Ala Phe Val Pro Ile Leu Lys Asp Phe Phe Val Arg Leu Thr Asp Glu
50 55 60
Phe Tyr Val Leu Arg Ala Ala Gln Leu Ala Leu Val Tyr Val Ser Gln
65 70 75 80
Asp Ser Thr Glu Glu Gln Gln Asp Leu Phe Leu Lys Asp Met Pro Lys
85 90 95
Lys Trp Leu Phe Leu Pro Phe Glu Asp Asp Leu Arg Arg Asp Leu Gly
100 105 110
Arg Gln Phe Ser Val Glu Arg Leu Pro Ala Val Val Val Leu Lys Pro
115 120 125
Asp Gly Asp Val Leu Thr Arg Asp Gly Ala Asp Glu Ile Gln Arg Leu
130 135 140
Gly Thr Ala Cys Phe Ala Asn Trp Gln Glu Ala Ala Glu Val Leu Asp
145 150 155 160
Arg Asn Phe Gln Leu Pro Glu Asp Leu Glu Asp Gln Glu Pro Arg Ser
165 170 175
Leu Thr Glu Cys Leu Arg Arg His Lys Tyr Arg Val Glu Lys Ala Ala
180 185 190
Arg Gly Gly Arg Asp Pro Gly Gly Gly Gly Gly Glu Glu Gly Gly Ala
195 200 205
Gly Gly Leu Phe
210
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ IMMTAC, СВЯЗЫВАЮЩИЕ РЕСТРИКТИРОВАННЫЙ ПО HLA-A*02 ПЕПТИД, С УВЕЛИЧЕННЫМ ПЕРИОДОМ ПОЛУВЫВЕДЕНИЯ | 2020 |
|
RU2833530C2 |
| РЕКОМБИНАНТНЫЕ БЕЛКИ С ДОМЕНАМИ CCN И СЛИТЫЕ БЕЛКИ | 2020 |
|
RU2825102C2 |
| МНОГОФУНКЦИОНАЛЬНЫЕ СЛИТЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2815388C2 |
| ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2722374C1 |
| УСОВЕРШЕНСТВОВАННЫЕ СВЯЗЫВАЮЩИЕСЯ С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ ВЕЩЕСТВА | 2018 |
|
RU2789495C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2698655C2 |
| НЕДЕСТРУКТИВНАЯ ГЕННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ MMA | 2018 |
|
RU2820602C2 |
| Слитый белок человеческого фактора свертывания IХ (FIX), способ его получения и применения | 2017 |
|
RU2736339C1 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2728861C1 |
| ГЕННАЯ ТЕРАПИЯ ГИПОФОСФАТЕМИЧЕСКИХ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ФАКТОРОМ РОСТА ФИБРОБЛАСТОВ 23 | 2020 |
|
RU2815545C2 |

Изобретение относится к области биотехнологии, конкретно к рекомбинантным белкам на основе RdCVF-подобного тиоредоксину белка, специфически экспрессируемого палочковыми фоторецепторными клетками в сетчатке, и может быть использовано в медицине для защиты фоторецепторных клеток глаза от гибели. Предложен слитый белок, содержащий последовательность RdCVF-короткого пептида и последовательность альбумина человека на С-конце относительно последовательность RdCVF-короткого пептида. Также слитый белок может содержать N-концевую сигнальную пептидную последовательность Igk. Изобретение обеспечивает экспрессию слитого белка на основе короткой формы пептида RdCVF человека в клетках человека и секрецию указанного слитого белка из клеток человека. 7 н. и 10 з.п. ф-лы, 3 ил., 1 табл., 2 пр.
1. Слитый белок для защиты фоторецепторов, содержащий:
(a) первую N-концевую сигнальную пептидную последовательность,
вторую пептидную последовательность на С-конце относительно сигнальной пептидной последовательности, и
третью пептидную последовательность на С-конце относительно второй пептидной последовательности; или
(b) вторую пептидную последовательность, и
третью пептидную последовательность на С-конце относительно второй пептидной последовательности;
где первая пептидная последовательность представляет собой сигнальную последовательность Igk,
вторая пептидная последовательность представляет собой последовательность RdCVF-короткого пептида и
третья пептидная последовательность представляет собой альбумин человека; и
где слитый белок имеет последовательность SEQ ID NO:3 или аминокислоты 22-732 SEQ ID NO:3.
2. Нуклеиновая кислота, кодирующая слитый белок по п.1.
3. Нуклеиновая кислота по п.2, кодирующая слитый белок с последовательностью SEQ ID NO:3.
4. Нуклеиновая кислота по п.3, имеющая последовательность SEQ ID NO:4.
5. Вектор экспрессии, содержащий нуклеиновую кислоту по п.2, функционально связанную с промотором.
6. Вектор экспрессии по п.5, где промотор представляет собой промотор CMV.
7. Вектор экспрессии по п.5, где вектор экспрессии представляет собой плазмиду.
8. Вектор экспрессии по п.7, где вектор экспрессии представляет собой экспрессионную плазмиду AAV.
9. Вектор экспрессии по п.5, где вектор экспрессии представляет собой вирусный вектор.
10. Вектор экспрессии по п.9, где вирусный вектор выбран из группы, состоящей из вектора AAV, лентивирусного вектора, ретровирусного вектора, аденовирусного вектора и синтетического вирусного вектора.
11. Клетка для получения слитого белка по п.1, где клетка содержит вектор экспрессии по любому из пп.5-10.
12. Фармацевтическая композиция для защиты фоторецепторных клеток глаза от гибели, содержащая:
(i) компонент, выбранный из группы, состоящей из слитого белка по п.1, и вектора экспрессии по любому из пп.5-10; и
(ii) фармацевтически приемлемый носитель.
13. Способ защиты фоторецепторных клеток глаза от гибели у пациента, включающий введение в глаз пациенту эффективного количества слитого белка по п.1, вектора экспрессии по любому из пп.5-10 или фармацевтической композиции по п.12, таким образом защищая фоторецепторные клетки глаза у пациента.
14. Способ по п.13, где введение выбрано из группы, состоящей из субретинальной инъекции и инъекции в стекловидное тело.
15. Способ по п.13 или 14, где пациентом является человек.
16. Способ по любому из пп.13-15, где пациент представляет собой млекопитающее, страдающее глазным заболеванием, выбранным из группы, состоящей из дистрофии сетчатки, болезни Штаргардта, пигментного ретинита, сухой связанной с возрастом дегенерации желтого пятна (сухой AMD), географической атрофии (поздней стадии сухой AMD), влажной связанной с возрастом дегенерации желтого пятна (влажной AMD), глаукомы с внутриглазной гипертензией или без нее, диабетической ретинопатии, синдрома Барде-Бидля, синдрома Бессена-Корнцвейга, болезни Беста, хороидемы, гиратной атрофии, врожденного амавроза, болезни Рефсума, синдрома Ушера, связанного со щитовидной железой заболевания глаз, болезни Грэйва, заболевания, ассоциированного с клетками пигментированного эпителия сетчатки, заболевания переднего сегмента глаза, заболевания хрусталика/катаракт, повреждения глазного яблока, увеита.
17. Способ получения слитого белка по п.1, включающий культивирование клетки по п.11,
где клетка содержит вектор экспрессии, вектор экспрессии содержит нуклеиновую кислоту, кодирующую слитый белок с последовательностью SEQ ID NO:3, и нуклеиновая кислота функционально связана с контрольной последовательностью,
в условиях, обеспечивающих экспрессию и секрецию кодируемого слитого белка, и выделение слитого белка из клеточной культуры.
| US 2014328821 A1, 06.11.2014 | |||
| ПРОТЕИН, СВЯЗАННЫЙ С ЗАБОЛЕВАНИЕМ | 2002 |
|
RU2384586C2 |
| LEVEILLARD TH | |||
| et al., Identification and characterization of rod-derived cone viability factor, NATURE GENETICS, 2004, v | |||
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| US 2005054570 A1, 10.03.2005 | |||
| WO 2009146183 A1, 03.12.2009 | |||
| ORLANDO M., Modification of proteins and low molecular weight substances with | |||

