Настоящее изобретение относится к генетическим конструкциям и, в частности, к рекомбинантным векторам, содержащим такие конструкции, а также к применениям данных конструкций и векторов в генотерапевтических способах лечения, предупреждения развития или облегчения нейродегенеративного нарушения, или лечения инсульта, или стимуляции регенерации и/или выживаемости нервов.
Нейродегенеративные заболевания представляют собой заболевания, которые в первую очередь поражают нейроны. Дегенеративный процесс может включать прогрессирующую потерю нейрональной структуры, прогрессирующую потерю нейрональной функции или прогрессирующую гибель нейронов. К категории нейродегенеративных заболеваний относят множество специфических нарушений. Болезнь Паркинсона представляет собой долгосрочное нейродегенеративное нарушение, которым, согласно оценкам, поражено приблизительно семь миллионов человек. Болезнь Хантингтона также представляет собой долгосрочное нейродегенеративное нарушение, и поэтому существует потребность в улучшенных средствах лечения болезни Паркинсона и болезни Хантингтона, и для таких пациентов может быть полезной стимуляция регенерации или выживаемости нервов.
К заболеванию двигательных нейронов относится любое нарушение, которое оказывает нейродегенеративное воздействие на двигательные нейроны. Сюда относятся амиотрофический латеральный склероз (ALS), первичный латеральный склероз (PLS), прогрессирующая мышечная атрофия (РМА), прогрессирующий бульбарный паралич (РВР), псевдобульбарный синдром или спинальные мышечные атрофии. Инсульт возникает, когда прерывается или уменьшается приток крови к головному мозгу, а плохой приток крови может привести к гибели клеток.
Болезнь Альцгеймера составляет приблизительно 60% всех случаев деменции, и, согласно оценкам, болезнью Альцгеймера страдают более 26 миллионов человек во всем мире [1]. Деменция включает в себя прогрессирующее снижение психической функции, что обычно включает нарушения памяти, речи и когнитивных процессов. Болезнь Альцгеймера может не только затрагивать самих пациентов, но и оказывает существенное влияние на миллионы людей, осуществляющих зачастую неоплачиваемый уход за лицами, которым необходим присмотр. Поскольку наибольшим фактором риска развития болезни Альцгеймера является возраст, имеет место существенное увеличение распространенности заболевания, так как увеличилась продолжительность дожития людей в пожилом возрасте [1]. Увеличение числа пациентов с болезнью Альцгеймера уже оказывает серьезное влияние на общемировые системы здравоохранения. Типичная патология, связанная с болезнью Альцгеймера, включает грубую атрофию головного мозга, истончение серого вещества в коре головного мозга, увеличенные желудочки, свидетельствующие об утрате нейронов, микроскопические внеклеточные амилоидные бляшки, содержащие бета-амилоидный пептид [Аβ], которые агрегируются в белковые агломераты, внутриклеточные нейрофибриллярные клубки, содержащие агрегированный тау-белок, и цереброваскулярный амилоид, т.е. амилоидный белок, окружающий кровеносные сосуды. При болезни Альцгеймера во многих областях головного мозга имеются амилоидные бляшки, появление которых обусловлено внеклеточными отложениями неправильно свернутого амилоидного β-пептида, и нейрофибриллярные клубки, состоящие из гиперфосфорилированного тау-белка, особенно в лобных, височных и теменных кортикальных слоях, гиппокампе и холинергическом ядре базального отдела переднего мозга. Эти участки головного мозга представляют собой ключевые области, участвующие в нейрональной схеме, которая играет важную роль в формировании кратковременной памяти. Отложение амилоидных бляшек происходит случайным образом во всем головном мозге, тогда как появление внутриклеточных нейрофибриллярных клубков, судя по всему, следует хорошо определенному паттерну [2], который обнаруживается первым в трансэнторинальной коре головного мозга. Затем наблюдается последовательное распространение нейрофибриллярных клубков в энторинальную кору, в области гиппокампа, а затем наружу в кору головного мозга. По результатам многочисленных исследований стало очевидно, что одно из наиболее ранних изменений при болезни Альцгеймера связано с утратой синапсов, что коррелирует со снижением умственных способностей [3], что в конечном итоге приводит к заметной утрате клеток во многих областях головного мозга. Следовательно, симптомы данного заболевания появляются совместно с медленным прогрессированием разрушения по всему головному мозгу, начиная с неспособности создать новые воспоминания, процесса, который зависит от гиппокампа.
Нейротрофический фактор головного мозга (BDNF) вместе с фактором роста нервов (NGF), нейротрофином-3 (NT-3) и нейротрофином-4/5 (NT-4/5), являются представителями нейротрофинового семейства трофических факторов [4-5]. Нейротрофины играют важную роль в развитии, выживании и функционировании широкого спектра нейронов как в периферической, так и в центральной нервной системе. Нейротрофины взаимодействуют с двумя рецепторами клеточной поверхности, низкоаффинными рецепторами р75NTR и представителями семейства высокоаффинных рецепторных тирозинкиназ (Trk) [4-5]. Фактор роста нервов (NGF) преимущественно связывает ТrkА, нейротрофический фактор головного мозга (BDNF), а нейротрофин-4/5 (NT4/5) связывается с тропомиозиновой рецепторной киназой В (ТrkВ), и нейротрофин-3 (NT-3) связывает ТrkС (и в меньшей степени ТrkА) [12-13].
Нейротрофический фактор головного мозга (BDNF) представляет собой белок, который характеризуется высоким уровнем экспрессии и широко распространен во всей центральной нервной системе, особенно в гиппокампе и коре головного мозга [6-7]. Было показано, что он играет важную роль в выживании и функционировании нейронов гиппокампа, коры головного мозга, холинергических и дофаминергических нейронов [8]. BDNF связан с рядом нарушений головного мозга, в том числе с болезнью Альцгеймера, болезнью Хантингтона, депрессией, шизофренией и синдромом Ретта. Было высказано предположение, что ранняя дисфункция памяти, наблюдаемая при болезни Альцгеймера, может быть связана с уровнями BDNF в гиппокампе, поскольку имеются публикации о значительном снижении уровней мРНК BDNF в гиппокампе при болезни Альцгеймера [9] и теменной коре головного мозга [10] и снижении уровней белка BDNF в энторинальной коре, гиппокампе, височной, лобной и теменной коре [11-16]. Тем не менее изменения в уровнях BDNF, по-видимому, обусловлены специфической даунрегуляцией определенных транскриптов BDNF. По результатам мета-анализа, также наблюдается значительное снижение уровней нейротрофинов в крови пациентов с болезнью Альцгеймера по сравнению со здоровыми субъектами [17]. Более того, было показано, что более низкая концентрация BDNF в спинномозговой жидкости является прогнозом прогрессирования от умеренного когнитивного нарушения (MCI) до болезни Альцгеймера [18].
По результатам ряда исследований виднол, что субъекты, у которых наблюдается полиморфизм Val66Met (при котором валин замещается метионином) про-домена BDNF, характеризуются связью с усилением прогрессирования болезни Альцгеймера [19], а также в этом могут быть задействованы и другие полиморфизмы BDNF. Утрата proBDNF, более крупной версии-предшественника BDNF и зрелого BDNF (mBDNF), происходит на ранней стадии заболевания (до отложения бляшек) и коррелирует с дефицитом памяти [20-21]. Эти данные убедительно свидетельствуют о связи между сниженными концентрациями BDNF, потерей синапсов и клеточной дисфункцией, которые лежат в основе когнитивного нарушения при болезни Альцгеймера. Также было показано, что BDNF индуцирует быстрое дефосфорилирование тау-белка в нейронах путем взаимодействий с рецептором ТrkВ и последующего увеличения передачи сигналов фосфоинозитол-3-киназы (PI3K) и протеинкиназы (Akt) [22-23]. Следовательно, снижения концентраций BDNF также могут способствовать гиперфосфорилированию тау-белка, патологическому признаку AD. Также заметен обратный эффект при увеличенном уровне тау-белка, что обуславливает снижение экспрессии BDNF у мышей [24]. По недавним данным также было выявлено значительное усиление нейротоксичности Ар при наличии про-домена нейротрофинов, включая BDNF [25].
При посмертном исследовании головного мозга пациента с болезнью Альцгеймера также были обнаружены изменения в нейронах, экспрессирующих рецептор mBDNF ТrkВ. Например, сообщалось об 47% уменьшении положительных по ТrkВ нейронов при посмертном исследовании головного мозга пациентов с болезнью Альцгеймера [26]. Это может быть связано либо с утратой нейронов, которые в норме экспрессируют данный рецептор, либо с биохимической даунрегуляцией экспрессии ТrkВ. Снижение ТrkВ также может быть усугублено увеличением уровней укороченных изоформ рецепторов TrkB-T1 и TrkB-Shc как во фронтальной, так и в височной доле коры головного мозга при болезни Альцгеймера, которые не обладают киназной активностью, необходимой для выживания нейронов [27]. Активация протеазы кальпаин под действием Аβ в культурах нейронов индуцирует снижение уровня ТrkВ [28] путем расщепления вблизи сайта стыковки рецептора Shc, что приводит к превращению полнофункциональных рецепторов в укороченную изоформу с дефектной киназной активностью. Результат превращения функциональных рецепторов ТrkВ в укороченную изоформу может затем играть роль приемника нейротрофина или доминантного отрицательного рецептора. На мышиной модели болезни Альцгеймера было обнаружено, что нокаут рецепторов ТrkВ усугубляет нарушения передачи сигналов и дефициты памяти, подобные тем, которые обнаруживаются при болезни Альцгеймера, не влияя на отложение Аβ [29]. Эти данные свидетельствуют, что утрата рецепторов ТrkВ и/или утрата активности вследствие сниженной выработки и секреции BDNF представляют собой важные элементы в возникновении симптомов и патофизиологии, подобных тем, которые имеют место при болезни Альцгеймера.
К другим важным механизмам, способствующим развитию нарушения передачи сигналов BDNF/TrkB в головном мозге при болезни Альцгеймера, относятся подавление путей с участием митоген-активируемой протеинкиназы (MAPK/ERK) и PI3K/Akt сублетальными концентрациями Аβ без негативного влияния на активацию TrkB-FL и фосфолипазы-γ (PLCγ) [30] и нарушение BDNF-индуцированного эндоцитоза ТrkВ. Воздействие олигомеров Аβ может нарушать эндоцитоз рецептора и последующую активацию Akt посредством опосредованного гликогенсинтазой-3β (GSK3β) фосфорилирования динамина-1 [31]. Кроме того, было показано, что олигомеры Аβ отрицательно влияют на BDNF-опосредованный ретроградный трафик ТrkВ [32] посредством нарушения работы убиквитиновой системы [33] и изменения гомеостаза кальция [34].
Такая общая картина характерна для значительного нарушения передачи сигналов с участием нейротрофических факторов при болезни Альцгеймера и, в частности, для системы BDNF. Дополнение или усиление передачи сигналов BDNF было исследовано на нескольких животных моделях болезни Альцгеймера. Например, инъекции BDNF уменьшают нарушения обучения у модельных крыс с болезнью Альцгеймера, индуцированной посредством Аβ [1-42] [35]. Инъекции нового слитого пептида, содержащего активный домен BDNF с кодируемым геномом ВИЧ трансактиватором транскрипции (ТАТ), который может проникать в головной мозг, значительно улучшали пространственную память с активацией пути TrkB/ERK1/2/Akt и восстановлением нескольких связанных с памятью белков у модельных животных [36]. Кроме того, было показано, что экспрессия BDNF с помощью лентивирусной генной терапии оказывает нейропротекторный эффект у трансгенных модельных мышей с болезнью Альцгеймера и у пожилых приматов, у которых наблюдается снижение когнитивных функций [37].
BDNF может вырабатываться в головном мозге и транспортироваться на периферию, где он может поддерживать нейроны и способствовать их выживанию [38-44]. При определенных условиях, таких как эксайтотоксичные поражения агонистами глутаматных рецепторов, такими как N-метил-D-аспартат, BDNF также может вырабатываться в периферических нейронах, хотя и с относительно низкими уровнями [45-46]. BDNF в норме вырабатывается в виде препрополипептида (т.е. preproBDNF), содержащего короткую сигнальную пептидную последовательность, которая способствует переносу всего полипептида в везикулы для высвобождения во внеклеточное пространство. Расщепление и удаление сигнального пептида превращает preproBDNF в proBDNF. Затем во внутриклеточном или внеклеточном пространстве отщепляется N-концевая последовательность proBDNF с образованием зрелого BDNF (mBDNF) [47]. Как pro-BDNF, так и mBDNF обладают биологической активностью в отношении pro-BDNF, преимущественно активируя рецепторы p75NTR, и более коротких mBDNF, активирующих рецепторы ТrkВ [48-50]. Активация p75NTR и рецепторов ТrkВ в сетчатке, например, оказывает противоположные эффекты на выживание ганглиозных клеток сетчатки (RGC), причем первый ответственен за апоптоз путем прямой активации p75NTR клеточного тела RGC [48-51] или опосредованно путем активации p75NTR на клетках Мюллера, таким образом стимулируя высвобождение фактора некроза опухоли-альфа (TNF-α), который дополнительно способствует утрате RGC [52].
На модельных животных с глаукомой было продемонстрировано, что после сдавливания нерва или повышенного IOР происходит переход от передачи нейротрофических сигналов mBDNF/TrkB к использованию путей pro-BDNF/p75NTR. Было продемонстрировано снижение уровней mBDNF и рецепторов ТrkВ в сетчатке [50, 53-54] одновременно с противоположным увеличением относительных уровней pro-BDNF [28] и рецепторов p75NTR [55]. Внесение mBDNF с помощью глазных инъекций рекомбинантного белка крысам с экспериментально повышенными уровнями IOР увеличивает выживаемость RGC-клеток по сравнению с необработанными глазами, подтверждая тем самым ключевую нейропротективную роль этого нейротрофина [42-44].
Таким образом, ввиду вышесказанного, существует потребность в улучшенной генной терапии для стимуляции регенерации или выживаемости нервов, для лечения, предупреждения развития или облегчения нейродегенеративного нарушения или инсульта.
Авторами настоящего изобретения была сконструирована новая генетическая конструкция, которая кодирует тирозинкиназный рецептор В (ТrkВ), и агонист рецептора ТrkВ под контролем одного промотора. Промотор данной конструкции можно использовать для обеспечения того, что агонист и рецептор будут экспрессироваться только в соответствующих нервных клетках и способствовать выживанию данных клеток.
Таким образом, согласно первому аспекту настоящего изобретения, предложена генетическая конструкция, содержащая промотор, функционально связанный с первой кодирующей последовательностью, которая кодирует тирозинкиназный рецептор В (ТrkВ), и второй кодирующей последовательностью, которая кодирует агонист рецептора ТrkВ, для применения при лечении, предупреждении развития или облегчения нейродегенеративного нарушения или инсульта.
Авторами настоящего изобретения было продемонстрировано в примерах, что можно объединить гены, которые кодируют как рецептор ТrkВ, так и его агонист, в одну генетическую конструкцию. Это было особенно сложно, учитывая их большие размеры, и нельзя было спрогнозировать, возможна ли их совместная экспрессия в физиологически полезных концентрациях. Преимущественно, с конструкцией по настоящему изобретению нет необходимости вводить инъекцией рекомбинантный белок, как описано в предшествующем уровне техники [56]. Кроме того, в предшествующем уровне техники все еще необходимо было производить регулярные инъекции белка, тогда как в случае конструкции по настоящему изобретению необходимо лишь однократное проведение генной терапии.
Предпочтительно, при применении рецептор ТrkВ активируется агонистом, тем самым способствуя выживанию нервных клеток. Генетическую конструкцию по настоящему изобретению предпочтительно применяют для лечения, предупреждения развития или облегчения нейродегенеративного нарушения, выбранного из группы, состоящей из болезни Александера, болезни Альпера, болезни Альцгеймера, латерального амиотрофического склероза (ALS), телеангиоэктатической атаксии, форм нейронального цероидлипофусциноза, болезни Баттена, губчатой энцефалопатии крупного рогатого скота (BSE), болезни Канавана, церебрального паралича, синдрома Коккейна, кортико-базальной дегенерации, болезни Крейтцфельда Якоба, лобно-височной лобарной дегенерации, болезни Гоше, болезни Хантингтона, ВИЧ-ассоциированного слабоумия, болезни Кеннеди, болезни Краббе, деменции с тельцами Леви, лизосомальных болезней накопления, нейроборрелиоза, болезни Мачадо-Джозефа, заболевания двигательных нейронов, множественной системной атрофии, рассеянного склероза, множественной сульфатазной недостаточности, муколипидозов, нарколепсии, болезни Нимана Пика типа С, болезни Нимана Пика, болезни Паркинсона, болезни Пелицеуса Мерцбахера, болезни Пика, болезни Помпе, первичного латерального склероза, прионных заболеваний, прогрессирующего надъядерного паралича, болезни Рефсума, болезни Сандхоффа, болезни Шилдера, подострой комбинированной дегенерации спинного мозга на фоне злокачественной анемии, болезни Шпильмейера Фогта Шегрена Баттена, спинально-церебеллярной атаксии, спинальной мышечной атрофии, болезни Стила-Ричардсона-Ольшевского, сухотки спинного мозга и болезни Тея-Сакса.
В соответствии с предпочтительным вариантом осуществления, генетическую конструкцию применяют для лечения, предупреждения развития или облегчения болезни Альцгеймера.
В соответствии с предпочтительным вариантом осуществления, генетическая конструкция предназначена для лечения, предупреждения развития или облегчения болезни Хантингтона.
В соответствии с предпочтительным вариантом осуществления, генетическая конструкция предназначена для лечения, предупреждения развития или облегчения болезни Паркинсона.
В соответствии с предпочтительным вариантом осуществления, генетическая конструкция предназначена для лечения, предупреждения развития или облегчения заболевания двигательных нейронов.
В соответствии с предпочтительным вариантом осуществления, генетическая конструкция предназначена для лечения, предупреждения развития или облегчения инсульта.
Генотерапевтическая конструкция может оказывать несколько благоприятных терапевтических эффектов для лечения нейродегенеративных нарушений, таких как болезнь Альцгеймера или инсульт. К благоприятным эффектам относятся терапевтическое восполнение сниженных концентраций mBDNF в головном мозге или восполнение других трофических факторов из семейства нейротрофинов. К другим благоприятным эффектам относятся восстановление уровней плотности рецепторов ТrkВ в нормальной ткани головного мозга. Возможность включения агониста в генетическую конструкцию, в которой не кодируется про-последовательность, например, не кодируется proBDNF, также позволяет восстановить баланс в сторону передачи сигналов типа mBDNF/TrkB и уйти от эффектов типа pro-BDNF/p75NTR. Более того, поскольку генную терапию можно применять для получения зрелой формы агониста, такого как mBDNF, без создания нейротрофина с про-доменом, будет значительно снижен риск усиления нейротоксичности Аβ, который мог бы возникнуть, если бы конструкция производила и высвобождала про-форму агониста, такую как proBDNF. Предпочтительно, с помощью конструкции по настоящему изобретению можно уменьшить фосфорилирование тау в нейронах (что является одним из патофизиологических признаков, ассоциируемых с головным мозгом при болезни Альцгеймера).
Следовательно, конструкцию по настоящему изобретению преимущественно можно применять для целенаправленного воздействия на нервные клетки с целью поддержания или усиления передачи сигналов ТrkВ в этих клетках. Таким образом, данную конструкцию можно применять для максимизации защиты от патофизиологических стрессоров и для стимуляции регенерации и/или выживаемости нервов. Более того, конструкцию можно применять для обеспечения долгосрочного лечения нейродегенеративных нарушений или инсультов благодаря экспрессии рецептора ТrkВ и агониста данного рецептора под контролем одного или нескольких промоторов. Следовательно, данная конструкция позволяет преодолеть необходимость применения нескольких альтернативных способов лечения, которые, даже в сочетании, обеспечивают временный терапевтический эффект. Более того, конструкция по настоящему изобретению является преимущественной, поскольку ее можно применять для значительного повышения чувствительности нервных клеток к агонистам рецептора ТrkВ вследствие локального увеличения уровня как рецептора ТrkВ, так и агониста данного рецептора.
Предпочтительно, генетическая конструкция по настоящему изобретению содержит кассету экспрессии, один вариант осуществления которой показан на фиг. 1. Как можно видеть на фиг. 1, конструкция содержит промотор, первую нуклеотидную последовательность, кодирующую рецептор ТrkВ, и вторую нуклеотидную последовательность, кодирующую зрелый нейротрофический белок зрелого головного мозга (mBDNF), который выполняет роль предпочтительного агониста рецептора ТrkВ. Тем не менее будет понятно, что можно применять другой агонист, как рассмотрено в настоящем документе. Также, как показано на фиг. 1, кассета экспрессии также включает в себя спейсерную последовательность 2А, последовательность, кодирующую посттранскрипционный регуляторный элемент вируса гепатита (WHPE), последовательность, кодирующую поли-А-хвост, и левую и правую последовательности инвертированных концевых повторов (ITR).
Так, генетическая конструкция предпочтительно содержит спейсерную последовательность, расположенную между первой и второй кодирующими последовательностями, при этом спейсерная последовательность кодирует пептидный спейсер, который может быть расщеплен или разрезан с образованием таким образом рецептора ТrkВ и агониста в форме отдельных молекул. В соответствии с вариантом осуществления, проиллюстрированном на фиг. 1, кодирующая последовательность для рецептора ТrkВ расположена 5' от кодирующей последовательности для агониста рецептора (BDNF) со спейсерной последовательностью между ними. Тем не менее в соответствии с другим вариантом осуществления, последовательность, кодирующая агонист рецептора, может быть расположена 5' от кодирующей последовательности для рецептора со спейсерной последовательностью между ними.
Предпочтительно генетическая конструкция содержит нуклеотидную последовательность, кодирующую посттранскрипционный регуляторный элемент вируса гепатита сурков (WHPE), который усиливает экспрессию двух трансгенов, т.е. рецептора ТrkВ и его агониста, которым предпочтительно является BDNF. Кодирующая последовательность WHPE предпочтительно расположена 3' от кодирующей последовательности трансгена.
Один вариант осуществления посттранскрипционного регуляторного элемента вируса гепатита сурков (WHPE) имеет длину 592 п.о., включая гамма-альфа-бета-элементы, и в настоящем документе указан под SEQ ID No: 57 и имеет следующий вид: 
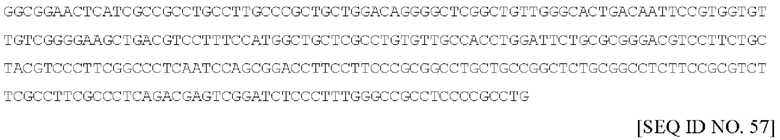
WHPE предпочтительно содержит последовательность нуклеиновой кислоты по существу как изложена под SEQ ID No: 57, или ее фрагмент или вариант.
Однако в соответствии с предпочтительным вариантом осуществления, применяют укороченный WHPE, длина которого составляет 247 п.о. из-за удаления бета-элемента, и который в настоящем документе указан под SEQ ID No: 58 и имеет следующий вид:

В последовательности укороченного WHPE, используемой в конструкции, преимущественно сохранено итого приблизительно 300 п.о. без отрицательного влияния на экспрессию трансгена. WHPE предпочтительно содержит последовательность нуклеиновой кислоты по существу как изложена под SEQ ID No: 58, или ее фрагмент или вариант.
Генетическая конструкция предпочтительно содержит нуклеотидную последовательность, кодирующую поли-А-хвост. Последовательность, кодирующая поли-А-хвост, предпочтительно расположена 3' от последовательности, кодирующей трансген, и предпочтительно 3' от последовательности, кодирующей WHPE.
Поли-А-хвост предпочтительно представляет собой последовательность поли-А вируса обезьян 40 длиной 224 п.о. Один вариант осуществления поли-А-хвоста в настоящем документе обозначен под SEQ ID No: 59 и имеет следующий вид:

Поли-А-хвост предпочтительно содержит последовательность нуклеиновой кислоты по существу, как изложена под SEQ ID No: 59, или ее фрагмент или вариант.
Генетическая конструкция предпочтительно содержит левую и/или правую последовательности инвертированных концевых повторов (ITR). Каждая ITR предпочтительно расположена на 5'- и/или 3'-конце конструкции.
Промотор в генетической конструкции по первому аспекту может представлять собой любую нуклеотидную последовательность, которая способна индуцировать связывание с РНК-полимеразой и транскрипцию первой и второй кодирующих последовательностей. В соответствии с одним вариантом осуществления, промотор в генетической конструкции по первому аспекту может представлять собой конститутивный промотор цитомелаловируса (CMV). Его считают неселективным как для нейронов, так и для глиальных клеток.
В соответствии с одним предпочтительным вариантом осуществления, промотор представляет собой промотор человеческого синапсина I (SYN I), для которого было показано, что он функционирует в головном мозге человека. Один вариант осуществления нуклеотидной последовательности длиной 469 нуклеотидов, кодирующей промотор человеческого синапсина I (SYN I), в настоящем документе обозначен под SEQ ID NO. 1 и имеет следующий вид:

Следовательно, промотор предпочтительно может содержать последовательность нуклеиновой кислоты, по существу как изложена под SEQ ID No: 1, или ее фрагмент или вариант.
В соответствии с другим предпочтительным вариантом осуществления, промотор представляет собой промотор CAG, для которого было показано, что он также функционирует в головном мозге человека. Промотор CAG предпочтительно содержит ранний энхансерный элемент цитомегаловируса, первый экзон и первый интрон гена бета-актина курицы и акцепторный сайт сплайсинга гена бета-глобина кролика. Один вариант осуществления нуклеотидной последовательности длиной 1733 нуклеотидов, кодирующей промотор CAG, в настоящем документе обозначен под SEQ ID NO.2 и имеет следующий вид: 

В соответствии с другим предпочтительным вариантом осуществления, промотор представляет собой укороченную форму промотора CAG, такую как 664-нуклеотидная форма промотора, которая обозначена в данном документе под SEQ ID NO.3 и имеет следующий вид:

В соответствии с еще одним предпочтительным вариантом осуществления, промотор представляет собой укороченную форму промотора CAG, такую как 584-нуклеотидная форма промотора, которая обозначена в данном документе под SEQ ID NO. 48 и имеет следующий вид: 
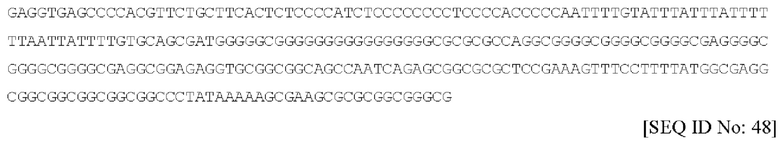
Следовательно, промотор предпочтительно содержит последовательность нуклеиновой кислоты по существу, как изложена под SEQ ID No: 2, 3 или 48, или ее фрагмент или вариант.
Многие бицистронные генные конструкции, представленные в научной литературе, имеют либо (i) встроенные двойные промоторы для раздельного управления экспрессией двух генов, либо (ii) в них использован участок внутренней посадки рибосомы (IRES) вируса энцефаломиокардита (EMCV), связывающий два транскрибируемых гена с одного промотора в рекомбинантных вирусных векторах [45-46]. Тем не менее, эффективность IRES-зависимой трансляции может варьировать в различных клетках и тканях, и в бицистронных векторах IRES-зависимая экспрессия второго гена может быть значительно ниже, чем кэп-зависимая экспрессия первого гена [47]. Более того, ограничение размера векторов rAAV (обычно <5 т.о.) будет препятствовать встраиванию крупных генных конструкций, таких как рецептор ТrkВ, вместе с BDNF с использованием двойных промоторов или линкеров IRES.
Следовательно, в соответствии с предпочтительным вариантом осуществления, генетическая конструкция предпочтительно содержит спейсерную последовательность, расположенную между первой и второй кодирующими последовательностями, при этом спейсерная последовательность кодирует пептидный спейсер, который может быть расщеплен с образованием таким образом рецептора ТrkВ и агониста в форме отдельных молекул. Спейсерная последовательность предпочтительно содержит и кодирует последовательность вирусного пептидного спейсера, более предпочтительно - последовательность вирусного пептидного спейсера 2А [47]. Последовательность пептида 2А предпочтительно соединяет первую кодирующую последовательность со второй кодирующей последовательностью. Это позволяет преодолеть для конструкции ограничения по размеру, которые возникают при экспрессии в различных векторах, и делает возможной экспрессию всех пептидов, кодируемых конструкцией по первому аспекту, под контролем одного промотора в форме единого белка.
Таким образом, после трансляции единого белка, содержащего последовательности ТrkВ, пептида 2А и агониста (предпочтительно BDNF), происходит расщепление последовательности вирусного пептида 2А по связи концевой глицин-пролин, тем самым освобождая два белка, т.е. ТrkВ и агонист (например, mBDNF). Генетическая конструкция сконструирована таким образом, что оставшаяся короткая N-концевая аминокислотная последовательность вирусного пептида 2А остается присоединенной к внутриклеточной части рецептора ТrkВ, тем самым устраняя риски иммуногенности и не оказывая отрицательного влияния на способность внутриклеточной передачи сигналов зрелого рецептора. Остаточная аминокислота пролин из С-концевой последовательности вирусного 2А остается присоединенной к N-концевому сигнальному пептиду агониста и в конечном итоге удаляется из белка-агониста после отщепления сигнальной последовательности от зрелого белка.
Авторами настоящего изобретения было создано два варианта осуществления спейсерной последовательности. Одним важным участком последовательности пептидного спейсера, который является общим для обоих вариантов осуществления, описанных в настоящем документе, является С-конец. Соответственно, последовательность пептидного спейсера предпочтительно содержит аминокислотную последовательность, обозначенную в настоящем документе под SEQ ID NO. 4, или ее фрагмент или вариант, которая имеет следующий вид:

Сайт расщепления или разрезания последовательности пептидного спейсера предпочтительно расположен между концевым глицином и концевым пролином в SEQ ID No:4.
В соответствии с первым предпочтительным вариантом осуществления, спейсерная последовательность содержит нуклеотидную последовательность, обозначаемую в настоящем документе под SEQ ID NO. 5, или ее фрагмент или вариант, которая имеет следующий вид:
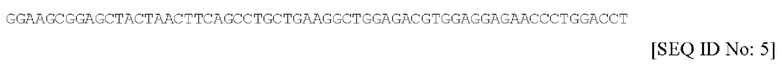
В соответствии с таким первым вариантом осуществления, последовательность пептидного спейсера содержит аминокислотную последовательность, обозначенную в настоящем документе под SEQ ID NO. 6, или ее фрагмент или вариант, которая имеет следующий вид:

В соответствии со вторым предпочтительным вариантом осуществления, спейсерная последовательность содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 7, или ее фрагмент или вариант, которая имеет следующий вид:
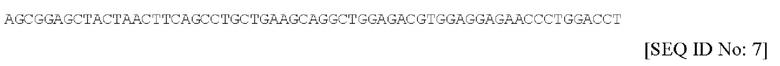
В соответствии с таким вторым вариантом осуществления, последовательность пептидного спейсера содержит аминокислотную последовательность, обозначенную в настоящем документе под SEQ ID NO. 8, или ее фрагмент или вариант, которая имеет следующий вид:

Авторами настоящего изобретения были тщательно проанализированы последовательности рецептора ТrkВ и были созданы несколько предпочтительных вариантов осуществления рецептора, который кодируется первой кодирующей последовательностью в генетической конструкции по первому аспекту.
В соответствии с одним предпочтительным вариантом осуществления, первая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую человеческую каноническую изоформу ТrkВ. Каноническая изоформа ТrkВ предпочтительно содержит аминокислотную последовательность (822 остатка), обозначаемую в настоящем документе под SEQ ID NO. 9, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с данным вариантом осуществления, первая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 10, или ее фрагмент или вариант, которая изложена ниже: 

В соответствии с другим предпочтительным вариантом осуществления, первая кодирующая последовательность содержит нуклеотидную последовательность, которая кодирует изоформу 4 ТrkВ. Изоформа 4 ТrkВ предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под SEQ ID NO. 11, или ее фрагмент или вариант, которая изложена ниже:


В соответствии с данным вариантом осуществления, первая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 12, или ее фрагмент или вариант, которая изложена ниже:


Авторами настоящего изобретения были приложены значительные изобретательские усилия при исследовании последовательности рецептора ТrkВ и было установлено, что ТrkВ содержит пять остатков тирозина (в положении 516, 701, 705, 706 и 816 в SEQ ID No: 9), которые обычно фосфорилируются после димеризации и аутофосфорилирования в присутствии димера BDNF. Проблема, связанная с фосфорилированием этих пяти остатков тирозина, заключается в том, что рецептор может быть легко дезактивирован фосфатазой, такой как фосфатаза Shp-2. Соответственно, для предупреждения фосфорилирования и последующей дезактивации рецептора in vivo один или несколько из этих ключевых тирозинов предпочтительно подвергают мутации (более предпочтительно - в глутаминовую кислоту) для имитации получаемого в результате фосфотирозина и получения рецептора, который остается активным в присутствии BDNF и который не может быть дезактивирован фосфатазой, такой как фосфатаза Shp-2. Такие мутантные формы ТrkВ нацелены на продуцирование активности рецептора ТrkВ, который остается активным в течение более длительных периодов или до интернализации данного рецептора.
В представленных ниже последовательностях ДНК и аминокислотных последовательностях проиллюстрированы положения этих пяти остатков тирозина (Y), которые были подвергнуты мутации в пять остатков глутаминовой кислоты (Е). Понятно, что в соответствии с вариантами осуществления настоящего изобретения, подвергнуты мутации в глутаминовую кислоту могут быть 1, 2, 3, 4 или 5 таких остатков. Также предусмотрены различные комбинации этих мутаций, например, только положения 516 и 701 или только положения 705, 706 и 816 и так далее.
Следовательно, в соответствии с другим предпочтительным вариантом осуществления, первая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую мутантную форму рецептора ТrkВ, причем один или несколько остатков тирозина в положении 516, 701, 705, 706 и/или 816 SEQ ID No: 9 подвергнуты модификации или мутации. Предпочтительно модификации подвергнуты по меньшей мере два, три или четыре остатка тирозина в положении 516, 701, 705, 706 и/или 816 SEQ ID No: 9. Наиболее предпочтительно модификации подвергнуты все пять остатков тирозина в положении 516, 701, 705, 706 и/или 816 SEQ ID No: 9.
Предпочтительно, каждый остаток тирозина модифицирован в другой аминокислотный остаток, более предпочтительно - в глутаминовую кислоту. Таким образом, мутантная форма рецептора ТrkВ предпочтительно содержит Y516Е, Y701E, Y705E, Y706E и/или Y816E.
Модифицированная форма рецептора ТrkВ предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под SEQ ID NO. 13, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с данным вариантом осуществления, первая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 14, или ее фрагмент или вариант, которая изложена ниже:


Будет понятно, что вторая кодирующая последовательность кодирует агонист рецептора ТrkВ, который предпочтительно является представителем нейротрофинового семейства трофических факторов. Агонист рецептора ТrkВ может быть представителем нейротрофинового семейства трофических факторов, у которых отсутствует пропоследовательность. Агонист рецептора ТrkВ может быть представителем нейротрофинового семейства трофических факторов в зрелой форме. Поэтому предпочтительные агонисты рецептора ТrkВ могут быть выбраны из группы агонистов, состоящей из нейротрофического фактора головного мозга (BDNF), фактора роста нервов (NGF), нейротрофина-3 (NT-3), нейротрофина-4 (NT-4) и нейротрофина-5 (NT-5) или их фрагментов. Предпочтительные агонисты рецептора ТrkВ могут быть выбраны из группы агонистов, состоящей из нейротрофического фактора головного мозга (BDNF) без пропоследовательности, фактора роста нервов (NGF) без пропоследовательности, нейротрофина-3 (NT-3) без пропоследовательности, нейротрофина-4 (NT-4) без пропоследовательности и нейротрофина-5 (NT-5) без пропоследовательности или их фрагментов. Предпочтительные агонисты рецептора ТrkВ могут быть выбраны из группы агонистов, состоящей из зрелого нейротрофического фактора головного мозга (BDNF), зрелого фактора роста нервов (NGF), зрелого нейротрофина-3 (NT-3), зрелого нейротрофина-4 (NT-4) и зрелого нейротрофина-5 (NT-5) или их фрагментов.
Нуклеотидные и аминокислотные последовательности каждого из этих агонистов будут известны специалисту в данной области техники. Тем не менее в качестве примера, аминокислотная последовательность одного варианта осуществления нейротрофина-4 (NT-4) является по существу такой как изложена под SEQ ID NO. 49 и имеющая следующий вид:

Кодирующая последовательность нуклеиновой кислоты данного варианта осуществления нейротрофина-4 (NT-4) является по существу такой, как изложена под SEQ ID NO. 50 и имеющая следующий вид:

Аминокислотная последовательность сигнального пептида для последовательности NT-4 является по существу такой, как изложена под SEQ ID NO. 51 и имеющая следующий вид:

Последовательность нуклеиновой кислоты данного сигнального пептида является по существу такой, как изложена под SEQ ID NO. 52 и имеющая следующий вид:
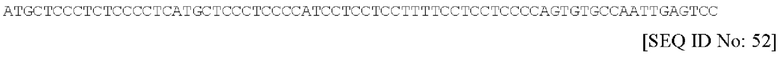
Аминокислотная последовательность пропептида для данной последовательности NT-4 является по существу такой, как изложена под SEQ ID NO. 53 и имеющая следующий вид:
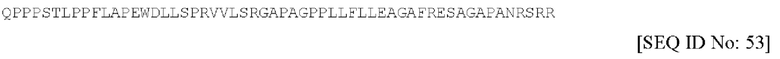
Последовательность нуклеиновой кислоты данного пропептида является по существу такой, как изложена под SEQ ID NO. 54 и имеющая следующий вид:
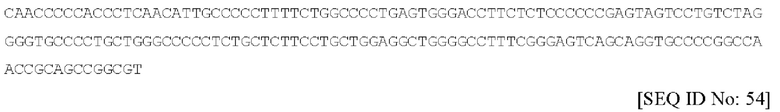
Аминокислотная последовательность зрелой белковой последовательности для данной последовательности NT-4 является по существу такой, как изложена под SEQ ID NO. 55 и имеющая следующий вид:

Кодирующая последовательность нуклеиновой кислоты данного зрелого белка NT-4 является по существу такой, как изложена под SEQ ID NO. 56 и имеющая следующий вид:

Следовательно, в соответствии с одним предпочтительным вариантом осуществления, вторая кодирующая последовательность кодирует нейротрофин-4 (NT-4), который может содержать аминокислотную последовательность по существу как изложена под SEQ ID NO: 49 или 55, или ее фрагмент или вариант. Так, вторая кодирующая последовательность может содержать нуклеотидную последовательность, по существу как изложена под SEQ ID No: 50 или 56, или ее фрагмент или вариант.
Однако к наиболее предпочтительным агонистам рецептора ТrkВ относятся препронейротрофический фактор головного мозга (pre-pro-BDNF), pro-BDNF или зрелый BDNF (mBDNF). BDNF изначально синтезируется в форме белка-предшественника preproBDNF рибосомами, которые находятся на поверхности эндоплазматического ретикулума. Существует по меньшей мере 17 известных сплайс-вариантов, кодируемых человеческим геном preproBDNF (ENSG00000176697). После попадания preproBDNF в шероховатый эндоплазматический ретикулум preproBDNF превращается в proBDNF путем отщепления сигнального пептида (т.е. «рrе» последовательности). ProBDNF превращается в mBDNF путем отщепления дополнительной N-концевой пептидной последовательности, которая присутствует на proBDNF. Как proBDNF, так и mBDNF затем секретируются во внеклеточное пространство, где они связываются с рецепторами на различных клетках и активируют эти рецепторы.
ProBDNF преимущественно связывается с рецептором p75NTR и активирует его, при этом рецептор при активации может индуцировать апоптоз у некоторых типов клеток.
Таким образом, в соответствии с одним предпочтительным вариантом осуществления, proBDNF является агонистом рецептора p75NTR. В соответствии с одним вариантом осуществления, proBDNF является каноническим proBDNF. Канонический proBDNF предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под SEQ ID NO. 15, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с данным вариантом осуществления, вторая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 16, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с другим вариантом осуществления, proBDNF представляет собой изоформу 2 proBDNF, которая предпочтительно содержит мутацию замены валина на метионин (подчеркнутая аминокислота). Изоформа 2 у proBDNF предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под SEQ ID NO. 17, или ее фрагмент или вариант, которая изложена ниже:

Тем не менее, в соответствии с одним вариантом осуществления, агонист не представляет собой proBDNF или его фрагмент или вариант, а вместо этого вторая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, которая кодирует зрелый BDNF. Зрелый BDNF (mBDNF) предпочтительно связывается и активирует ТrkВ, который при активации способствует выживанию нервных клеток. Таким образом, зрелый BDNF является наиболее предпочтительным агонистом ТrkВ. Конструкция по первому аспекту является преимущественной, поскольку, в отличие от других известных генетических конструкций, данная конструкция способна вырабатывать зрелый белок BDNF, который имеет правильную конформацию после свертывания.
Таким образом, в соответствии с одним предпочтительным вариантом осуществления, вторая кодирующая последовательность содержит нуклеотидную последовательность, которая кодирует зрелый BDNF. mBDNF является общим для всех 17 изоформ, кодируемых данным геном. Имеется 7 различных белковых последовательностей, пять из которых имеют удлиненные сигнальные последовательности относительно канонической формы, а одна имеет каноническую сигнальную последовательность, но содержит мутацию замены валина на метионин (которая является общей для изоформ 2, 4, 7, 8, 9, 10, 11, 12, 13, 14 и 16). Полагают, что мутация замены валина на метионин уменьшает высвобождение BDNF из клетки.
Зрелый BDNF предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под SEQ ID NO. 18, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с данным вариантом осуществления, вторая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 19, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с другим вариантом осуществления, агонист является представителем нейротрофинового семейства трофических факторов без пропоследовательности, но с сигнальным пептидом, конъюгированным с N-концом. Агонистом может быть любой представитель нейротрофинового семейства трофических факторов в зрелой форме и с сигнальным пептидом, конъюгированным с N-концом. Сигнальным пептидом может быть любой сигнальный пептид, который способствует правильному свертыванию или выработке агониста. В соответствии с предпочтительными вариантами осуществления, сигнальным пептидом может быть любой раскрываемый в настоящем документе сигнальный пептид.
В соответствии с еще одним предпочтительным вариантом осуществления, агонистом является mBDNF с сигнальным пептидом, конъюгированным с его N-концом. Как рассмотрено ниже, сигнальный пептид может быть каноническим сигнальным пептидом из preproBDNF, или сигнальным пептидом из IL-2, или новой сигнальной последовательностью, de novo созданной авторами настоящего изобретения.
Вторая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, кодирующую сигнальный пептид для агониста рецептора ТrkВ, наиболее предпочтительно - сигнальный пептид для BDNF. В соответствии с одним предпочтительным вариантом осуществления, нуклеотидная последовательность кодирует канонический сигнальный пептид для BDNF. В соответствии с данным вариантом осуществления, вторая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, которая кодирует сигнальный пептид, содержащий аминокислотную последовательность, обозначенную в настоящем документе под SEQ ID NO. 20, или ее фрагмент или вариант, которая изложена ниже:

В соответствии с данным вариантом осуществления, вторая кодирующая последовательность предпочтительно содержит нуклеотидную последовательность, обозначенную в настоящем документе под SEQ ID NO. 21, или ее фрагмент или вариант, которая изложена ниже:
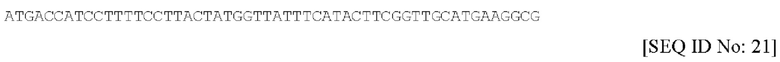
Авторами настоящего изобретения была создана серия удлиненных сигнальных пептидов. В соответствии с предпочтительными вариантами осуществления, нуклеотидная последовательность, кодирующая изоформу сигнального пептида для BDNF, выбрана из группы, состоящей из изоформ 2, 3, 6, 5 и 4. Ниже изложены последовательности нуклеиновой кислоты и аминокислотные последовательности для каждого из этих удлиненных сигнальных пептидов.
Изоформа 2

Изоформа 3 и 6
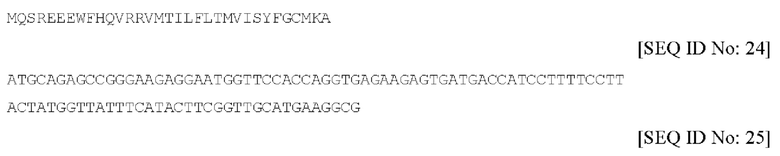
Изоформа 5

Изоформа 4

Следовательно, в соответствии с предпочтительными вариантами осуществления, вторая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую пептид сигнальной последовательности, обозначенную в настоящем документе под любым из SEQ ID NO. 23, 25, 27 или 29. Сигнальный пептид предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под любым из SEQ ID NO. 22, 24, 26 или 28.
Авторами настоящего изобретения также были созданы различные варианты новых сигнальных пептидов для агониста, предпочтительно BDNF. Эти сигнальные пептиды повышают уровень основности N-концевого участка (с добавленными остатками лизина (K) и аргинина (R)) и продолжающейся далее гидрофобной области (с добавлением остатков лейцина (L)), что увеличивает секрецию BDNF по сравнению с уровнями, наблюдаемыми с канонической сигнальной последовательностью дикого типа.
a) QTA003P (сигнальная последовательность IL-2)

b) QTA004P
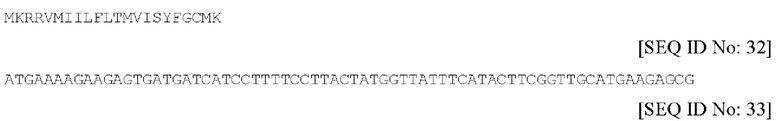
c) QTA009P (модифицированный IL-2)
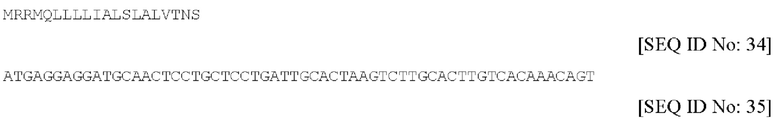
d) QTA010P

e) QTA0012P

f) QTA0013P

g) QTA0014P

i) QTA0015P

На фиг. 6 показаны нуклеотидные и аминокислотные последовательности для дополнительных предпочтительных вариантов осуществления сигнального пептида, применяемого в конструкции по настоящему изобретению для усиления секреции агониста, предпочтительно BDNF. Вторым остатком в сигнальном пептиде является треонин (Т), который предпочтительно заменен одним или несколькими основными остатками, такими как лизин (K) или аргинин (R). Следующий фрагмент из остатков в сигнальном пептиде, включающий изолейцин (I), лейцин (L), фенилаланин (F) и лейцин (L), предпочтительно заменен одним или несколькими гидрофобными остатками.
Следовательно, в соответствии с предпочтительными вариантами осуществления, вторая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую пептид сигнальной последовательности, обозначенную в настоящем документе под любым из SEQ ID NO. 31, 33, 35, 37, 39, 41, 43, 45, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 89, 91, 93, 95, 97, 99, 101 или 103. Сигнальный пептид предпочтительно содержит аминокислотную последовательность, обозначаемую в настоящем документе под любым из SEQ ID NO. 30, 32, 34, 36, 38, 40, 42, 44, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100 или 102.
Соответственно, будет понятно, что авторами настоящего изобретения была модифицирована последовательность гена BDNF путем удаления пропоследовательности, что также никогда не совершалось ранее, в результате чего был получен правильно свернутый зрелый BDNF, в сочетании с введением абсолютно новых сигнальных пептидов, которые значительно увеличивают выработку и высвобождение BDNF, чем когда-либо достигалось с эндогенной последовательностью.
Генетическая конструкция предпочтительно содержит левую и/или правую последовательности инвертированных концевых повторов (ITR). Каждая ITR предпочтительно расположена на 5'- и/или 3'-конце конструкции. ITR может быть специфичной для серотипа вируса (например, AAV или лентивируса) и может быть любой последовательностью, при условии, что она образует шпилечную петлю в своей вторичной структуре.
Последовательность ДНК из одного варианта осуществления (левая ITR из коммерчески доступной плазмиды AAV) ITR представлена в настоящем документе под SEQ ID No: 46 и имеет следующий вид:
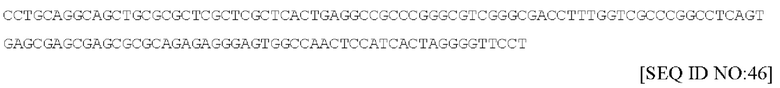
Последовательность ДНК из другого варианта осуществления (правая ITR из коммерчески доступной плазмиды AAV) ITR представлена в настоящем документе под SEQ ID No: 47 и имеет следующий вид:
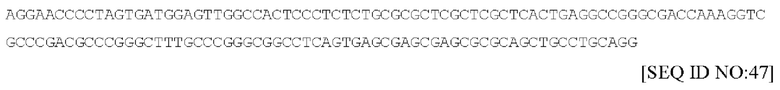
Исходя из вышеизложенного, специалисту в настоящей области техники будет понятна нуклеотидная последовательность из варианта осуществления конструкции по первому аспекту, а также аминокислотная последовательность кодируемого трансгена. Тем не менее с целью исключения двусмысленного толкования, кодирующая последовательность подвергнутой оптимизации кодона последовательности из 2940 п.о. для конструкции мышиный рецептор TrkB-вирусный пептид 2A-mBDNF, содержащаяся в плазмиде QTA020P (и векторе QTA020V), обозначена в настоящем документе под SEQ ID No: 107 и имеет следующий вид:


Кодирующая последовательность подвергнутой оптимизации кодона последовательности из 2943 п.о. для конструкции человеческий рецептор TrkB-вирусный пептид 2A-mBDNF, содержащаяся в плазмиде QTA029P (и векторе QTA029V), обозначена в настоящем документе под SEQ ID No: 108 и имеет следующий вид:


Следовательно, в соответствии с наиболее предпочтительным вариантом осуществления, конструкция содержит нуклеотидную последовательность по существу, как изложена под SEQ ID No: 107 или 108, или ее фрагмент или вариант.
Авторами настоящего изобретения была создана серия рекомбинантных экспрессирующих векторов, содержащих конструкцию по настоящему изобретению.
Так, согласно второму аспекту, настоящее изобретение относится к рекомбинантному вектору, содержащему генетическую конструкцию по первому аспекту, для применения при лечении, предупреждении развития или облегчении нейродегенеративного нарушения или инсульта.
Описываемые в настоящем документе конструкции и векторы экспрессии можно применять для стимуляции регенерации и выживаемости нервов. В соответствии с некоторыми вариантами осуществления, рекомбинантный вектор предназначен для лечения, предупреждения развития или облегчения болезни Альцгеймера, болезни Хантингтона, болезни Паркинсона, заболевания двигательных нейронов или инсульта. Описываемые в настоящем документе рекомбинантные векторы могут быть предназначены для любого описываемого в настоящем документе лечения или применения.
Рекомбинантный вектор может представлять собой рекомбинантный вектор AAV (rAAV). RAAV может быть природным вектором или вектором с гибридным AAV- серотипом. RAAV может быть AAV-1, AAV-2, AAV-3A, AAV-3B, AAV-4, AAV-5, AAV-6, AAV-7, AAV-8, AAV-9, AAV-10, и AAV-11. RAAV предпочтительно представляет собой rAAV серотипа 2.
Рекомбинантный AAV2 преимущественно вызывает минимальный иммунный ответ в организмах хозяев и обеспечивает долгосрочную экспрессию трансгена, которая может сохраняться в течение по меньшей мере одного года после введения вектора.
Используемый в настоящем документе термин «рекомбинантный AAV-вектор (rAAV-вектор)» означает полученную из AAV рекомбинантную нуклеиновую кислоту, содержащую по меньшей мере одну концевую повторяющуюся последовательность.
Предпочтительные варианты осуществления такого вектора показаны на фиг. 2-5.
Согласно третьему аспекту, настоящее изобретение относится к способу лечения, предупреждения развития или облегчения нейродегенеративного нарушения или инсульта у субъекта или для стимуляции регенерации и/или выживаемости нервов у субъекта, при этом способ предусматривает введение субъекту, нуждающемуся в таком лечении, терапевтически эффективного количества генетической конструкции по первому аспекту или рекомбинантного вектора по второму аспекту.
В соответствии с несколькими вариантами осуществления, способ может быть предназначен для лечения, предупреждения развития или облегчения болезни Альцгеймера, болезни Паркинсона, заболевания двигательных нейронов, болезни Хантингтона или любого другого раскрытого в настоящем документе нейродегенеративного нарушения.
Генетическая конструкция или рекомбинантный вектор по настоящему изобретению предпочтительно применяют в генотерапевтической методике. Агонист, кодируемый конструкцией или вектором, активирует ТrkВ, также кодируемый конструкцией/вектором, тем самым стимулируя выживаемость нейронов.
В соответствии с другим вариантом осуществления, такие конструкции и векторы можно применять для стимуляции регенерации и/или выживаемости нервов.
Будет понятно, что генетическую конструкцию по первому аспекту или рекомбинантный вектор по второму аспекту можно применять в лекарственном препарате, который можно применять в виде монотерапии (т.е. применение генетической конструкции по первому аспекту или вектора по второму аспекту настоящего изобретения) для лечения, облегчения или предупреждения развития нейродегенеративного нарушения или инсульта или для стимуляции регенерации и/или выживаемости нервов. Альтернативно, генетическую конструкцию или рекомбинантный вектор по настоящему изобретению можно применять в качестве дополнения или в сочетании с известными терапиями для лечения, облегчения или предупреждения развития нейродегенеративного нарушения или инсульта или для стимуляции регенерации и/или выживаемости нервов.
Генетическую конструкцию по настоящему изобретению или рекомбинантный вектор по настоящему изобретению можно объединить в композициях, имеющих множество различных форм, в зависимости, в частности, от способа, которым необходимо применять данную композицию. Таким образом, например, композиция может быть представлена в форме порошка, таблетки, капсулы, жидкости, мази, крема, геля, гидрогеля, аэрозоля, спрея, мицеллярного раствора, трансдермального пластыря, суспензии липосом или в любой другой подходящей форме, которую можно вводить нуждающемуся в лечении человеку или животному. Будет понятно, что наполнитель лекарственных препаратов по настоящему изобретению должен быть таким, который хорошо переносится субъектом, который его принимает.
Генетическую конструкцию или рекомбинантный вектор по настоящему изобретению также можно включить в устройство с замедленным или отсроченным высвобождением. Такие устройства можно, например, вставить на поверхности кожи или под кожу, и лекарственный препарат может высвобождаться в течение недель или даже месяцев. Устройство можно расположить по меньшей мере рядом с местом лечения. Такие устройства могут обладать особым преимуществом при необходимости долгосрочного лечения генетической конструкцией или рекомбинантным вектором, и при этом обычно необходимо частое их введение (например, по меньшей мере ежедневная инъекция).
В соответствии с предпочтительным вариантом осуществления, лекарственные препараты по настоящему изобретению можно вводить субъекту путем инъекции в кровоток, нерв или непосредственно в нуждающееся в лечении место. Например, лекарственный препарат способен проходить через гематоэнцефалический барьер. Инъекции могут быть внутривенными (струйной или инфузией), или подкожными (струйной или инфузией), или внутрикожными (струйной или инфузией).
Будет понятно, что необходимое количество генетической конструкции или рекомбинантного вектора определяют по их биологической активности и биодоступности, которая, в свою очередь, зависит от способа введения, физико-химических свойств генетической конструкции или рекомбинантного вектора и того, применяют ли их в качестве монотерапии или в комбинированной терапии. На частоту введения также будет влиять период полужизни циклического полипептида в организме подвергаемого лечению субъекта. Оптимальные дозы, подлежащие введению, могут быть определены специалистами в настоящей области техники и будут варьировать в зависимости от конкретной генетической конструкции или рекомбинантного вектора при применении, эффективности фармацевтической композиции, способа введения и запущенности или стадии развития нарушения. Дополнительные факторы, зависящие от конкретного субъекта, подвергаемого лечению, приведут к необходимости корректировать дозы, в том числе возраст субъекта, масса, пол, рацион и время введения.
Обычно для лечения, облегчения или предупреждения развития нейродегенеративного нарушения, болезни Альцгеймера, болезни Паркинсона, болезни Хантингтона, заболевания двигательных нейронов или инсульта можно применять суточную дозу от 0,001 мкг/кг массы тела до 10 мг/кг массы тела или от 0,01 мкг/кг массы тела до 1 мг/кг массы тела конструкции или вектора по настоящему изобретению, в зависимости от применяемой генетической конструкции или применяемого рекомбинантного вектора.
Генетическую конструкцию или рекомбинантный вектор можно вводить до, во время или после проявления нарушения. Суточные дозы можно вводить в виде однократного введения (например, однократной суточной инъекции или ингаляции назального спрея). Альтернативно, для генетической конструкции или рекомбинантного вектора может быть необходимо введение два или более раз в течение суток. В качестве примера, генетическую конструкцию или рекомбинантный вектор можно вводить в виде двух (или более, в зависимости от тяжести подвергаемого лечению нарушения) суточных доз, составляющих от 0,07 мкг до 700 мг (т.е. исходя из массы тела в 70 кг). Пациент, получающий лечение, может принимать первую дозу после пробуждения, а затем вторую дозу вечером (при двухдозовом режиме) или через 3- или 4-часовые промежутки после этого. Альтернативно, для введения оптимальных доз генетической конструкции или рекомбинантного вектора по настоящему изобретению пациенту без необходимости введения повторных доз можно применять устройство с замедленным высвобождением.
Для получения конкретных составов с генетической конструкцией или рекомбинантным вектором по настоящему изобретению и для обеспечения точных терапевтических режимов (таких как суточные дозы средств и частота их введения) можно использовать известные процедуры, такие как обычно используемые в фармацевтической промышленности (например, эксперименты in vivo, клинические испытания и т.д.). Авторы настоящего изобретения считают, что они первыми предложили генетическую конструкцию, кодирующую промотор, функционально связанный с кодирующими последовательностями рецептора ТrkВ и агониста рецептора ТrkВ.
Согласно четвертому аспекту, настоящее изобретение относится к фармацевтической композиции, содержащей генетическую конструкцию по первому аспекту или рекомбинантный вектор по второму аспекту и фармацевтически приемлемый наполнитель.
Согласно пятому аспекту, настоящее изобретение относится к способу получения фармацевтической композиции по четвертому аспекту, при этом способ предусматривает приведение в контакт генетической конструкции по первому аспекту или рекомбинантного вектора по второму аспекту с фармацевтически приемлемым наполнителем.
«Субъектом» может быть позвоночное, млекопитающее или домашнее животное. Следовательно, композиции и лекарственные препараты по настоящему изобретению можно применять для лечения любого млекопитающего, например, домашнего скота (например, лошади), домашних животных, или их можно применять в других ветеринарных целях. Однако наиболее предпочтительно субъектом является человек.
«Терапевтически эффективное количество» генетической конструкции, рекомбинантного вектора или фармацевтической композиции представляет собой любое количество, которое при введении субъекту представляет собой количество вышеупомянутого, которое необходимо для лечения нейродегенеративного нарушения, болезни Альцгеймера, болезни Паркинсона, болезни Хантингтона, заболевания двигательных нейронов, инсульта или получения требуемого эффекта, такого как стимуляция регенерации и/или выживаемости нервов.
Например, терапевтически эффективное количество применяемой генетической конструкции, рекомбинантного вектора или фармацевтической композиции может составлять от приблизительно 0,01 мг до приблизительно 800 мг и предпочтительно от приблизительно 0,01 мг до приблизительно 500 мг. Предпочтительно, чтобы количество генетической конструкции, рекомбинантного вектора или фармацевтической композиции представляло собой количества от приблизительно 0,1 мг до приблизительно 250 мг и наиболее предпочтительно - от приблизительно 0,1 мг до приблизительно 20 мг.
«Фармацевтически приемлемый наполнитель» в контексте настоящего документа представляет собой любое известное соединение или комбинацию известных соединений, которые известны специалистам в настоящей области техники как пригодные при составлении фармацевтических композиций.
В соответствии с одним вариантом осуществления, фармацевтически приемлемый наполнитель может быть твердым веществом, а композиция может быть представлена в форме порошка или таблетки. Твердый фармацевтически приемлемый наполнитель может включать одно или несколько веществ, которые также могут играть роль ароматизаторов, смазывающих веществ, солюбилизаторов, суспендирующих средств, красителей, заполнителей, веществ, способствующих скольжению, добавок для прессования, инертных связующих, подсластителей, консервантов, красителей, покрытий или веществ для улучшения распадаемости таблеток. Наполнитель также может представлять собой инкапсулирующий материал. В порошках наполнитель представляет собой тонкоизмельченное твердое вещество, которое находится в смеси с тонкоизмельченными активными средствами по настоящему изобретению. В таблетках действующее средство (например, генетическую конструкцию или рекомбинантный вектор по настоящему изобретению) можно смешать с наполнителем, имеющим необходимые свойства прессования, в подходящих пропорциях и спрессовать в требуемую форму и размер. Порошки и таблетки предпочтительно содержат до 99% действующих средств. Подходящие твердые наполнители включают, например, фосфат кальция, стеарат магния, тальк, сахара, лактозу, декстрин, крахмал, желатин, целлюлозу, поливинилпирролидин, легкоплавкие воски и ионообменные смолы. В соответствии с другим вариантом осуществления, фармацевтический наполнитель может быть гелем, а композиция может быть представлена в форме крема или тому подобного.
Тем не менее фармацевтический наполнитель может быть жидкостью, а фармацевтическая композиция представлена в форме раствора. Жидкие наполнители применять при приготовлении растворов, суспензий, эмульсий, сиропов, настоек и композиций под давлением. Генетическую конструкцию или рекомбинантный вектор по настоящему изобретению можно растворить или суспендировать в фармацевтически приемлемом жидком наполнителе, таком как вода, органический растворитель, их смесь или фармацевтически приемлемые масла или жиры. Жидкий наполнитель может содержать другие подходящие фармацевтические добавки, такие как солюбилизаторы, эмульгаторы, буферы, консерванты, подсластители, ароматизаторы, суспендирующие средства, загустители, пигменты, регуляторы вязкости, стабилизаторы или осморегуляторы. Подходящие примеры жидких наполнителей для перорального и парентерального введения включают воду (частично содержащую указанные выше добавки, например, производные целлюлозы, предпочтительно раствор карбоксиметилцеллюлозы натрия), спирты (в том числе одноатомные спирты и многоатомные спирты, например, гликоли) и их производные и масла (например, фракционированное кокосовое масло и арахисовое масло). В случае парентерального введения наполнитель также может представлять собой сложный маслянистый эфир, такой как этилолеат и изопропилмиристат. Стерильные жидкие наполнители пригодны в композициях в стерильных жидких формах для парентерального введения. Жидкий наполнитель для композиций под давлением может представлять собой галогенированный углеводород или другой фармацевтически приемлемый газ-вытеснитель.
Жидкие фармацевтические композиции, которые представляют собой стерильные растворы или суспензии, можно использовать, например, путем внутримышечной, интратекальной, эпидуральной, внутрибрюшинной, внутривенной и особенно подкожной инъекции. Генетическую конструкцию или рекомбинантный вектор можно приготовить в виде стерильной твердой композиции, которую можно растворить или суспендировать в момент введения с помощью стерильной воды, солевого раствора или другой подходящей стерильной инъекционной среды.
Генетическую конструкцию, рекомбинантный вектор и фармацевтическую композицию по настоящему изобретению можно вводить перорально в форме стерильного раствора или стерильной суспензии, которые содержат другие растворенные вещества или суспендирующие средства (например, достаточное количество солевого раствора или глюкозы для придания раствору изотоничности), соли желчных кислот, аравийскую камедь, желатин, сорбитанмоноолеат, полисорбат 80 (сложные олеатные эфиры сорбита и его ангидридов, сополимеризованных с этиленоксидом) и др. Генетическую конструкцию, рекомбинантный вектор или фармацевтическую композицию по настоящему изобретению также можно вводить перорально в форме либо жидкой, либо твердой композиции. Композиции, подходящие для перорального введения, включают твердые формы, такие как пилюли, капсулы, гранулы, таблетки и порошки, и жидкие формы, такие как растворы, сиропы, настойки и суспензии. Формы, подходящие для парентерального введения, включают стерильные растворы, эмульсии и суспензии.
Согласно дополнительному аспекту, настоящее изобретение относится к генетической конструкции по первому аспекту или рекомбинантному вектору по второму аспекту для применения при лечении, предупреждении развития или облегчении нарушения зрительного нерва или нарушения улиткового нерва или для стимуляции регенерации и/или выживаемости нервов; причем вторая кодирующая последовательность содержит последовательность, кодирующую зрелую форму трофического фактора из нейтрофинового семейства. Вторая кодирующая последовательность может содержать последовательность, кодирующую сигнальный пептид. Конструкция или вектор могут быть такими, что у агониста отсутствует пропоследовательность, но имеется сигнальный пептид. Сигнальный пептид может быть присоединен к N-концу и может улучшать секрецию, экспрессию или сворачивание агониста. Вторая кодирующая последовательность может содержать последовательность, кодирующую любой из зрелого фактора роста нервов (NGF), зрелого нейротрофина-3 (NT-3), зрелого нейротрофина-5 (NT-5) или их фрагменты или варианты.
Будет понятно, что настоящее изобретение относится к любой нуклеиновой кислоте или любому пептиду или варианту, их производному или аналогу, которые содержат по существу такие же аминокислотные последовательности или последовательности нуклеиновых кислот, что и любая из упомянутых в настоящем документе последовательностей, включая их варианты или фрагменты. Термины «по существу такая же аминокислотная/нуклеотидная/пептидная последовательность», «вариант» и «фрагмент» могут обозначать последовательность, которая имеет по меньшей мере 40% идентичности последовательности с аминокислотными/нуклеотидными/пептидными последовательностями любой из упоминаемых в настоящем документе последовательностей, например, 40% идентичности с последовательностью, указанной под SEQ ID No: 1-108, и так далее.
Также предусмотрены аминокислотные/полинуклеотидные/полипептидные последовательности с идентичностью последовательностей, превышающей 65%, более предпочтительно превышающей 70%, еще более предпочтительно превышающей 75% и еще более предпочтительно превышающей 80% с любой из упомянутых последовательностей. Аминокислотная/полинуклеотидная/полипептидная последовательность предпочтительно имеет по меньшей мере 85% идентичности с любой из упомянутых последовательностей, более предпочтительно по меньшей мере 90% идентичности, еще более предпочтительно по меньшей мере 92% идентичности, еще более предпочтительно по меньшей мере 95% идентичности, еще более предпочтительно по меньшей мере на 97% идентичности, еще более предпочтительно по меньшей мере 98% идентичности и наиболее предпочтительно по меньшей мере 99% идентичности с любой из упомянутых в настоящем документе последовательностей.
Специалисту в настоящей области техники будет известно, как рассчитать процент идентичности между двумя аминокислотными/полинуклеотидными/полипептидными последовательностями. Для расчета процента идентичности между двумя аминокислотными/полинуклеотидными/полипептидными последовательностями сначала необходимо подготовить выравнивание двух последовательностей, после чего рассчитать значение идентичности последовательностей. Процент идентичности для двух последовательностей может принимать различные значения в зависимости от: (i) способа, применяемого для выравнивания последовательностей, например, ClustalW, BLAST, FASTA, Смита-Уотермана (реализован в различных программах) или структурное выравнивание по результатам 3D-сравнения; и (ii) параметров, применяемых способом выравнивания, например, локальное или глобальное выравнивание, используемая матрица парных замен (например, BLOSUM62, РАМ250, Gonnet и т.д.) и штраф за введения гэпа, например, функциональная форма и константы.
После выполнения выравнивания есть много различных способов расчета процента идентичности между двумя последовательностями. Например, можно разделить количество идентификаторов на: (i) длину наиболее короткой последовательности; (ii) длину участка выравнивания; (iii) среднюю длину последовательности; (iv) количество положений без гэпов; или (v) количество эквивалентных положений, исключая выступающие части. Более того, понятно, что процент идентичности также сильно зависит от длины. Следовательно, чем короче пара последовательностей, тем выше идентичность последовательностей, которую можно ожидать получить без возможности ее прогноза.
Следовательно, понятно, что точное выравнивание белковых последовательностей или последовательностей ДНК является сложным процессом. Предпочтительным способом создания множественных выравниваний белков или ДНК по настоящему изобретению является популярная программа множественного выравнивания ClustalW (Thompson et al., 1994, Nucleic Acids Research, 22, 4673-4680; Thompson et al., 1997, Nucleic Acids Research, 24, 4876-4882). Подходящими параметрами для ClustalW могут быть следующие: для выравниваний ДНК: штраф за открытие гэпа (Gap Open Penalty)=15,0, штраф за продолжение гэпа (Gap Extension Penalty)=6,66 и тип матрицы (Matrix)=Identity; для выравниваний белков: штраф за открытие гэпа (Gap Open Penalty)=10,0, штраф за продолжение гэпа (Gap Extension Penalty)=0,2 и тип матрицы (Matrix)=Gonnet; для выравниваний ДНК и белков: ENDGAP=-1 и GAPDIST=4. Специалистам в данной области техники будет понятно, что для оптимального выравнивания последовательностей может быть необходимо изменить эти и другие параметры.
Предпочтительно, чтобы расчет процента идентичности между двумя аминокислотными/полинуклеотидными/полипептидными последовательностями затем можно было произвести из результатов такого выравнивания по формуле (N/T)*100, где N обозначает количество положений, в которых последовательности имеют идентичный остаток, а Т обозначает общее количество сравниваемых положений, включая гэпы, но исключая выступающие участки. Следовательно, наиболее предпочтительный способ расчета процента идентичности между двумя последовательностями предусматривает (i) подготовку выравнивания последовательностей с помощью программы ClustalW с использованием подходящего набора параметров, например, как изложено выше; и (ii) подстановку значений N и Т в следующую формулу: - идентичность последовательностей=(N/T)*100.
Специалистам в настоящей области техники известны и альтернативные способы выявления схожих последовательностей. Например, по существу подобная нуклеотидная последовательность будет кодироваться последовательностью, которая гибридизуется с последовательностями ДНК или их комплементарными цепями в жестких условиях. Под жесткими условиями подразумевают, что нуклеотид гибридизуется со связанной с фильтром ДНК или РНК в 3× хлориде натрия/цитрате натрия (SSC) при приблизительно 45°С с последующей по меньшей мере однократной промывкой в 0,2×SSC/0,1% SDS при приблизительно 20-65°С. Альтернативно, по существу подобный полипептид может отличаться по меньшей мере на 1, но менее чем на 5, 10, 20, 50 или 100 аминокислот от последовательностей, показанных, например, под SEQ ID NO: 3 и 5.
Из-за вырожденности генетического кода ясно, что любая описываемая в настоящем документе последовательность нуклеиновой кислоты можно подвергнуть вариациям или изменениям, практически не затрагивая последовательность кодируемого таким образом белка, с получением его функционального варианта. Подходящими вариантами нуклеотидных последовательностей являются варианты, имеющие последовательность, измененную путем замены разных кодонов, которые кодируют одну и ту же аминокислоту в последовательности, таким образом получая молчащее изменение. Другими подходящими вариантами являются варианты, имеющие гомологичные нуклеотидные последовательности, но содержащие всю последовательность или ее части, которые изменены путем замены различных кодонов, кодирующих аминокислоту с боковой цепью со схожими биофизическими свойствами, на аминокислоту, которая ее заменяет, с получением консервативного изменения. Например, к небольшим неполярным гидрофобным аминокислотам относятся глицин, аланин, лейцин, изолейцин, валин, пролин и метионин. К крупным неполярным гидрофобным аминокислотам относятся фенилаланин, триптофан и тирозин. К полярным нейтральным аминокислотам относятся серии, треонин, цистеин, аспарагин и глутамин. К положительно заряженным (основным) аминокислотам относятся лизин, аргинин и гистидин. К отрицательно заряженным (кислым) аминокислотам относятся аспарагиновая кислота и глутаминовая кислота. Поэтому будет понятно, какие аминокислоты можно заменить аминокислотой со схожими биофизическими свойствами, и специалисту в настоящей области техники будут известны нуклеотидные последовательности, кодирующие такие аминокислоты.
Все описываемые в настоящем документе признаки (в том числе любые пункты прилагаемой формулы изобретения, реферат и чертежи) и/или все стадии любого раскрываемого таким образом способа или процесса можно объединить с любым из вышеуказанных аспектов в любой комбинации, за исключением комбинаций, где по меньшей мере некоторые из таких признаков и/или стадий являются взаимоисключающими.
Для лучшего понимания настоящего изобретения и для демонстрации того, как могут быть реализованы варианты его осуществления, далее обратимся, в качестве примера, к прилагаемым чертежам, на которых:
фиг. 1 представляет собой схематическое изображение одного варианта осуществления генетической конструкции по настоящему изобретению;
фиг. 2 представляет собой схематический чертеж первого варианта рекомбинантного вектора по настоящему изобретению, известного как «плазмида QTA001PA», содержащая каноническую сигнальную последовательность (синим цветом) плюс proBDNF (красным цветом) и mBDNF (черным цветом). Сюда также относится последовательность -IRES-GFP- (голубым и фиолетовым цветом);
фиг. 3 представляет собой схематический чертеж второго варианта рекомбинантного вектора по настоящему изобретению, известного как «плазмида QTA002PA», без proBDNF (но который производит только mBDNF) и с той же сигнальной последовательностью (синим цветом), что и у QTA001PA. Сюда также относится последовательность -IRES-GFP- (голубым и фиолетовым цветом);
фиг. 4 представляет собой схематический чертеж третьего варианта рекомбинантного вектора по настоящему изобретению, известного как «плазмида QTA003PA», без proBDNF (но который производит только mBDNF) и с сигнальной последовательностью IL-2 (синим цветом). Сюда также относится последовательность -IRES-GFP (голубым и фиолетовым цветом);
фиг. 5 представляет собой схематический чертеж четвертого варианта рекомбинантного вектора по настоящему изобретению, известного как «плазмида QTA004PA», без proBDNF (но который производит только mBDNF) и с новой сигнальной последовательностью (синим цветом). Сюда также относится последовательность -IRES-GFP- (голубым и фиолетовым цветом);
на фиг. 6 показаны нуклеотидные и аминокислотные последовательности для различных вариантов осуществления сигнального пептида, применяемого в конструкции по настоящему изобретению. Вторым остатком является треонин (t), который может быть заменен одним или несколькими основными остатками, такими как лизин (K) или аргинин (R). Следующий фрагмент из остатков, включающий изолейцин (I), лейцин (L), фенилаланин (F) и лейцин (L), может быть заменен одним или несколькими гидрофобными остатками;
на фиг. 7 показано высвобождение BDNF из клеток НЕK293 с помощью специфичного ИФА через 24 часа после трансдукции плазмиды (4 мкг ДНК/лунка), содержащей гены, кодирующие mBDNF с различными сигнальными пептидными последовательностями и без кодирующей последовательности для удлиненного компонента proBDNF (данные показаны как среднее ±SEM для n=4);
на фиг. 8 показаны результаты вестерн-блоттинга клеточных концентраций BDNF-иммунореактивного материала (в произвольных единицах) в лизатах клеток НЕK293 через 24 часа после трансдукции плазмиды (данные представлены как среднее ±SEM для n=4);
на фиг. 9 показана BDNF-иммунореактивность в вестерн-блотах клеточных лизатов, где видны два бэнда с молекулярной массой (32 кДа и 14 кДа) при трансдукции клеток посредством QTA001PA, по сравнению только с одним бэндом массой 14 кДа при трансдукции посредством QTA002P, QTA003P и QTA004P;
на фиг. 10 показаны концентрации proBDNF в инкубационной среде НЕK293, измеренные с помощью специфичного ИФА через 24 часа после трансдукции плазмиды с помощью ИФА, селективного по proBDNF (данные показаны как среднее ±SEM для n=4);
на фиг. 11 показана экспрессия BDNF в лизате клеток НЕK293, осуществляемая плазмидами QTA002P (с эндогенной канонической сигнальной пептидной последовательностью) и QTA009P - QTA013P. Данные показаны как среднее + S.E.M. ** Р<0,01 в сравнении с QTA002P;
на фиг. 12 показана экспрессия BDNF в среде для инкубации клеток НЕK293, осуществляемая плазмидами QTA002P (с эндогенной канонической сигнальной пептидной последовательностью) и QTA009P - QTA013P. Данные показаны как среднее + S.E.M. ** Р<0,01 в сравнении с QTA002P;
на фиг. 13 показаны вестерн-блоты, полученные из клеток НЕK293 через 24 часа после их трансдукции плазмидами QTA015P (экспрессирующей BDNF и eGFP, отделенные спейсером IRES), QTA021P (экспрессирующей BDNF, за которым идет eGFP, отделенный функциональной последовательностью вирусного пептида 2А), QTA022P (экспрессирующей BDNF, за которым идет eGFP, отделенный нефункциональной последовательностью вирусного пептида 2А) и QTA023P (экспрессирующей eGFP, за которым идет последовательность, кодирующая BDNF, отделенный функциональной последовательностью вирусного пептида 2А). Данные представлены как BDNF-иммунореактивность (А), eGFP-иммунореактивность (В) и количество BDNF, высвобожденное из клеток НЕK293 в среду для инкубации (С). Данные представлены в виде среднего + S.E.M. плотности в бэндах;
на фиг. 14А показан вестерн-блот гомогенатов клеток НЕK293 через 48 часов после трансфекции вектором QTA020V, и виден эффективный процессинг участка, кодирующего большой предшественник, который включает рецептор ТrkВ и BDNF, разделенные пептидной последовательностью вирусного 2А. на фиг. 14В и 14С показано, что трансгенные белки, получаемые после расщепления вирусного пептида 2А, транспортировались в надлежащие внутриклеточные компартменты в клетках НЕK293 после процессинга (рецепторы ТrkВ на поверхность клетки, a BDNF в везикулы для хранения до высвобождения);
на фиг. 15А показана экспрессия рецептора ТrkВ, а на фиг. 15В показана экспрессия BDNF в гомогенате сетчатки мыши для вектора rAAV2, QTA020V. Данные представлены в виде среднего + S.E.M. плотности в вестерн-блоте гомогенатов сетчатки мыши. ** Р<0,01 в сравнении с интактными животными (без введения);
на фиг. 16 показана экспрессия трансгенов ТrkВ (А) и BDNF (В) в слое ганглиозных клеток сетчатки мыши, которая продемонстрирована с помощью иммуноцитохимии после инъекции QTA020V, rААУ2-вектора, содержащего последовательность, кодирующую рецептор ТrkВ и BDNF, разделенные последовательностью вирусного пептида 2А;
на фиг. 17 показано выживание ганглиозных клеток сетчатки (RGC) после сдавливания зрительного нерва (ONC) у мышей по сравнению с контрольными животными, получавшими вектор rAAV2-CAG-eGFP. Данные представлены в виде среднего + S.E.M. для средних количеств ганглиозных клеток сетчатки по всей сетчатке на одно животное по результатам подсчета Вrn3А-положительных клеток в плоских препаратах сетчатки. ***Р<0,001, *Р<0,05 по сравнению с контролями;
на фиг.18 показана экспрессия трансгенов BDNF (фиг. 18А) и ТrkВ (фиг. 18В) в гомогенатах недифференцированных человеческих клеток нейробластомы SH-SY5Y с помощью вестерн-блоттинга после трансфекции вирусными векторами rAAV2, которые не экспрессируют трансгены (нуль-вирус), экспрессируют только BDNF (QTA027V), только TrkB (QTA025V) и как BDNF, так и TrkB (QTA020V). На фиг. 18С показан уровень активированных фосфорилированных рецепторов TrkB в клетках SH-SY5Y в вестерн-блотах после трансфекции вирусными векторами «нуль-вектор», QTA020V, QTA025V или QTA027V. Было обнаружено, что только вектор QTA020V, который экспрессирует как BDNF, так и TrkB, значительно увеличивал активацию рецепторов TrkB по сравнению с нетрансфицированными клетками (**Р<0,01; дисперсионный анализ, после чего рассчитывали модифицированные по Бонферрони t-критерии для множественных сравнений). Данные показаны как среднее + S.E.M. для n=4 эксперимента;
на фиг. 19 показан уровень апоптозной клеточной гибели недифференцированных клеток SH-SY5Y в культуре после воздействия окислительного стресса, вызванного добавлением перекиси водорода (Н2О2 в количестве 0,1 мМ или 1,0 мМ) путем окрашивания с помощью TUNEL. Было обнаружено, что клетки, трансфицированные перед добавлением перекиси водорода rААV2-вектором QTA020V, который экспрессирует рецепторы как BDNF, так и ТrkВ, были значительно защищены от апоптоза по сравнению с необработанными клетками (**Р<0,01; дисперсионный анализ, после чего рассчитывали модифицированные по Бонферрони t-критерии для множественных сравнений). Данные показаны как среднее + S.E.M. для n=6-10; и
на фиг. 20 показаны репрезентативные иммуноцитохимические изображения зрительных нервов, полученных от мышей, трансгенных по мутантному человеческому тау-белку P301S, и окрашенных антителами, которые распознают фосфорилированный тау в положениях серии 396/серин 404 (PHF-1) или серии 202/серин 205 (АТ8). Мышам интравитреальной инъекцией вводили rААV2-вектор QTA020V (который экспрессирует рецепторы как mBDNF, так и TrkB) в возрасте 3 месяцев и умерщвляли через три недели с целью удаления зрительных нервов для иммуноцитохимии.
Примеры
В приведенных далее примерах продемонстрировано применение вариантов осуществления настоящего изобретения для стимуляции регенерации и/или выживаемости нервов. Идеи, выводимые из применений, способов и средств лечения, раскрытых в Примерах, также применимы к лечению нейродегенеративных нарушений и инсульта, как раскрыто в настоящем документе.
Способы и материалы
Молекулярное клонирование и плазмидные конструкции
Оптимизацию кодона последовательностей ДНК проводили с помощью онлайн-инструментария (http://www.idtdna.com/CodonOpt), а блоки ДНК были синтезированы компанией Integrated DNA technologies, Inc. (IDT; 9180 N. McCormick Boulevard, Скоки, Иллинойс 60076-2920, США) или компанией GenScript (860 Centennial Ave, Пискатауэй, Нью-Джерси 08854, США). Клонирование для получения мастер-плазмиды QTA001PA и последующих плазмид проводили с помощью стандартных методик молекулярной биологии и клонирования.
Наращивание количества плазмид и их очистка
ДНК-плазмиды наращивали в компетентных клетках SURE (Agilent Technologies; кат. №200238) в течение ночи с получением 2,29 мкг/мкл плазмиды после очистки с помощью набора Maxi-prep. Оставшиеся плазмиды наращивали в количестве до 500-мкг масштаба и качества трансдукции с минимальным присутствием эндотоксина.
Культивирование НЕK293 и трансдукция клеток плазмидной ДНК
Клетки НЕK293 (400000 клеток) культивировали в покрытых поли-L-лизином (10 мкг/мл, Sigma-Aldrich; кат. №Р1274) 6-луночных планшетах в 1,5 мл минимально обогащенной среде Дульбекко (DMEM), содержащей 10% фетальной бычьей сыворотки (FBS), 1% пенициллина и 1% стрептомицина (1% пен./стреп.) до 80% конфлюентности. Затем среду заменяли на 2 мл DMEM (без добавок). Спустя два-три часа в каждую лунку вносили 0,5 мл среды для трансфекции, содержащей 4 мкг плазмидной ДНК плюс 10 мкл липофектамина (4 мкл/мл; Thermo Fisher Scientific; кат. №12566014), в результате чего получали общий объем 2,5 мл в течение периода трансфекции и в ходе сбора супернатанта.
Культивирование SH-SY5Y и трансфекция клеток вирусными векторами rААV2 Клетки SH-SY5Y культивировали в 6-луночных планшетах (300000 клеток), 96-луночных планшетах (10000 клеток) или на стеклянных покровных стеклах размером 13 мм (100000 клеток), покрытых поли-L-лизином (10 мкг/мл, продукт компании Sigma №Р1274). Минимально обогащенную среду Дульбекко (DMEM), содержащую 10% фетальной бычьей сыворотки (FBS), 1% пенициллина и 1% стрептомицина (1% пен./стреп.), использовали для культивирования клеток до 80% конфлюентности при 37°С перед заменой на DMEM без добавок до проведения трансфекции. Использованные объемы DMEM представляли собой 6-луночные планшеты (2 мл), 96-луночные планшеты (100 мкл), покровные стекла (500 мкл). Разведенные в PBS векторы вносили непосредственно в культуральную среду в конечной концентрации 1,0×1010 (VP (вирусных частиц))/мл и инкубировали в течение 48 часов при температуре 37°С.
Индуцированная перекисью водорода гибель клеток SH-SY5Y и окрашивание с помощью TUNEL
Спустя 48 часов после трансфекции клеток SH-SY5Y среду заменяли на свежую DMEM (без добавок). Перекись водорода (Н2О2) (Thermo Fisher Scientific; продукт № ВР2633500, лот №1378087) разводили в профильтрованной воде (до концентрации 0,1 или 1,0 мМ) и вносили в равном объеме в лунки или планшеты на дополнительных 24 часа. Профильтрованная вода служила контрольным наполнителем. Покровные стекла дважды промывали в PBS и фиксировали в течение 30 минут в 4% параформальдегиде в 1 М фосфатно-солевом буферном растворе (PBS) при комнатной температуре. После еще трех промывок в PBS клетки блокировали и пермеабилизировали путем инкубации в 5% нормальной козьей сыворотке (NGS), 3% бычьем сывороточном альбумине (BSA) и 0,3% Тритоне Х-100 в PBS в течение 60 минут при комнатной температуре. Затем клетки инкубировали в течение ночи при 4°С с коммерческими кроличьими поликлональными антителами к TrkB (Abeam; продукт №ab33655, лот №GR232306-1, разведенный 1:500), кроличьими поликлональными антителами к BDNF (Santa Cruz Biotechnology Inc; продукт №sc-546; лот №C0915 в разведении 1:300) или p-Tyr515-TrkB (продукт компании Abcam, №ab109684, лот №GR92849-4, 1:750), разведенными в блокирующем растворе. Окрашивание проявляли с применением вторичных антител к кроличьим антителам, конъюгированных с красителем Alexa Fluor 488 (Life Technologies; продукт №A11034 в разведении 1:1000), в течение 2 часов при комнатной температуре. В ходе окрашивания с помощью TUNEL (Promega; продукт №G3250; лот №0000215719) клетки трижды промывали в PBS и погружали в уравновешивающий буфер TUNEL на 10 минут. Реакционную смесь TUNEL готовили в соответствии с протоколом производителя и добавляли ее к клеткам в количестве 100 мкл/покровное стекло на 1 час при 37°С. Реакцию останавливали путем инкубации в 1× стандартном цитратном растворе (SCS) в течение 15 минут. Ядра клеток докрашивали посредством 1 мкг/мл DAPI (Thermo Scientific; продукт №D1306 в соотношении 1:8000). Клетки дополнительно три раза промывали перед приготовлением препарата с реагентом FluorSave™ (Calbiochem/EMD Chemicals Inc., Гиббстаун, Нью-Джерси, США) перед визуализацией. Визуализацию производили с применением 20× объектива и эпифлуоресцентного микроскопа Leica DM6000 (Leica Microsystems, Вецлар, Германия).
Измерение BDNF с помощью ИФА
Количество BDNF, секретируемого из клеток НЕK293, измеряли в среде для культивирования клеток через 24 часа после трансфекции. Среду центрифугировали для удаления дебриса и измеряли с применением коммерческого набора для ИФА человеческого BDNF (Sigma-Aldrich, продукт №RAB0026). Концентрацию BDNF определяли путем сравнения образцов со свежеприготовленными стандартами BDNF.
Вестерн-блоттинт в отношении рецепторов BDNF и ТrkВ
Величину BDNF- и ТrkВ-иммунореактивности в клетках НЕK293 измеряли путем удаления инкубационной среды DMEM, промывки клетки в холодном фосфатно-солевом буферном растворе и добавлением в лунки 350 мкл свежеприготовленного лизирующего буфера (10 мл реагента Lysis-M+1 таблетка полного коктейля Mini ингибиторов протеаз, Roche; кат. №04719964001, + 100 мкл коктейля Halt ингибиторов фосфатаз (100×), Thermo Scientific; кат. №78428). После гомогенизации клеток белковую суспензию белка количественно оценивали с помощью анализа ВСА (набора для анализа белка ВСА Pierce, Thermo Scientific; кат. №23227). От 6 мкг до 15 мкг белка из клеточного лизата НЕK293/бэнд гнали вниз по Bis-Tris-гелю (12% NuPAGE Novex; кат. №NP0342BOX, Thermo Scientific) и изучали с помощью вестерн-блоттинга с применением первичных кроличьих поликлональных антител к BDNF (Santa Cruz Biotechnology Inc; продукт №sc-546; в разведении 1:500), кроличьих поликлональных антител к TrkB (Abeam; кат. №ab33655, используемых в разведении 1:2000) или антител к eGFP (продукт Abeam №ab-290, используемый в разведении 1:500), которые инкубировали в течение ночи. Первичные антитела визуализировали с помощью конъюгированных с HRP антител к кроличьим антителам (Vector Laboratories; кат. №PI-1000, в разведении 1:8000) и детектирования сигнала с помощью ECL Prime (Amersham, GE Healthcare, Великобритания) и системы для визуализации вестерн-блотов Alliance Western (UVItec Ltd, Кембридж, Великобритания). В случае вестерн-блотов мышиной сетчатки глаза от обработанных вектором животных гомогенизировали в 500 мкл свежеприготовленного лизирующего буфера (10 мл реагента Lysis-M+1 таблетка полного коктейля Mini ингибиторов протеаз, продукт Roche №04719964001+100 мкл коктейля Halt ингибиторов фосфатаз (100×), продукт Thermo Scientific №78428). Ткань разрушали в течение 1 минуты (Qiagen, продукт TissueRuptor №9001273), а затем выдерживали на льду в течение дополнительных 15 минут. Затем белок анализировали с помощью вестерн-блоттинга, как описано выше.
Иммуноцитохимия
Клетки НЕK293 (70000) высевали на 13 мм покрытые поли-L-лизином покровные стекла в 4-луночных планшетах и инкубировали в DMEM, содержащей 10% FBS и 1% пен./стреп., в 0,5 мл среды. После того как клетки выросли до 80% конфлюентности, среду заменяли на 0,4 мл DMEM (без добавок) на 2-3 часа, затем вносили дополнительные 0,1 мл среды для трансфекции (0,8 мкг плазмидной ДНК + 2 мкл липофектамина) так, чтобы конечный объем достигал 0,5 мл. Покровные стекла дважды промывали в PBS и фиксировали в течение 30 минут в 4% параформальдегиде в 1 М фосфатно-солевом буферном растворе (PBS) при комнатной температуре. После еще трех промывок в PBS клетки блокировали и пермеабилизировали путем инкубации в 5% нормальной козьей сыворотке (NGS), 3% бычьем сывороточном альбумине (BSA) и 0,3% Тритоне Х-100 в PBS в течение 60 минут при комнатной температуре. Затем клетки инкубировали в течение ночи при 4°С с коммерческими кроличьими поликлональными антителами к BDNF (Santa Cruz Biotechnology Inc; продукт №sc-546; в разведении 1:300) или TrkB (продукт компании Abcam, №ab33655, в разведении 1:500), разведенными в блокирующем растворе. Окрашивание проявляли с применением вторичных антител к кроличьим антителам, конъюгированных с красителем Alexa Fluor 647 (Invitrogen; продукт №А21248 в разведении 1:1000), в течение 2 часов при комнатной температуре. Ядра клеток также докрашивали посредством 1 мкг/мл DAPI (Thermo Scientific; продукт №D1306 в соотношении 1:8000). Клетки дополнительно три раза промывали перед приготовлением препарата с реагентом FluorSave™ (Calbiochem/EMD Chemicals Inc., Гиббстаун, Нью-Джерси, США) перед визуализацией. Визуализацию проводили с применением 20× объектива и эпифлуоресцентного микроскопа Leica DM6000 (Leica Microsystems, Вецлар, Германия) или конфокального микроскопа Leica SP5 (Leica Microsystems, Вецлар, Германия), оснащенного 63× объективом для масляной иммерсии, с 3× цифровым зумом и 0,5-0,8 последовательным интервалом сканирования по оси Z.
Для иммуноцитохимии структур сетчатки и зрительных нервов от обработанных контролем или обработанных вектором животных (через 3 или 4 недели после инъекции) аккуратно иссеченные глаза фиксировали в 4% параформальдегиде/0,1% PBS (рН 7,4) в течение ночи и дегидратировали в 30% сахарозе/0,1% PBS при 4°С (24 часа). Затем глаза погружали в силиконовые формы, содержащие смесь с оптимальной температурой для разрезания (OCT) (Sakura Finetek, Зутервауде, Нидерланды), и замораживали на сухом льду. 13-мкм срезы через дорсально-вентральную/верхне-нижнюю оси сетчатки или продольные срезы через зрительный нерв мышей P301S собирали на предметные стекла Superfrost plus (продукт VWR №631-0108) с использованием криостата Bright OTF 5000 (Bright Instruments, Хантингдон, Великобритания). Предметные стекла трижды промывали в PBS и пермеабилизировали в 5% нормальной козьей сыворотке (NGS), 3% бычьем сывороточном альбумине (BSA) и 0,3% Тритоне Х-100 в PBS в течение 60 минут при комнатной температуре. Затем предметные стекла инкубировали в течение ночи при 4°С с коммерческими кроличьими поликлональными антителами к BDNF (Santa Cruz Biotechnology Inc; продукт №sc-546, 1:300), TrkB (Abeam; продукт №ab33655, 1:500), тау-белок с Ser396/404 (PHF-1; получен в Кембридже, 1:500) или тау-белок с Ser202/205 (АТ8; продукт Invitrogen №MN1020, 1:500), разведенными в блокирующем растворе. Окрашивание проявляли с применением вторичных антител к кроличьим антителам, конъюгированных с красителем Alexa Fluor 647 (Invitrogen; продукт №А21248 в разведении 1:1000), в течение 2 часов при комнатной температуре. Ядра клеток сетчатки также докрашивали посредством 1 мкг/мл DAPI (Thermo Scientific; продукт №D1306 в соотношении 1:8000). Предметные стекла дополнительно три раза промывали перед приготовлением препарата с реагентом FluorSave™ (Calbiochem/EMD Chemicals Inc., Гиббстаун, Нью-Джерси, США) перед визуализацией. Визуализацию проводили с применением 20× объектива и эпифлуоресцентного микроскопа Leica DM6000 (Leica Microsystems, Вецлар, Германия) или конфокального микроскопа Leica SP5 (Leica Microsystems, Вецлар, Германия), оснащенного 63× объективом для масляной иммерсии, с 3× цифровым зумом и 0,5-0,8 последовательным интервалом сканирования по оси Z.
Интравитреальные инъекции
После 7-10-дневного периода акклиматизации мышей C57/BL.6 в возрасте 12 недель или P301S в возрасте 16 недель (Harlan labs, Байсестер, Великобритания) рандомизировали в различные группы исследования. Затем их анестезировали внутрибрюшинной инъекцией кетамина (50 мг/кг) и ксилазина (5 г/кг). В 1-й день исследования закапывали местные глазные капли 1% тетракаина. С помощью глазных капель 1% тропикамида достигали расширение зрачка. С помощью операционного микроскопа в склере производили пробное отверстие на частичную глубину с применением иглы 30-го калибра для облегчения проникновения в нижележащую склеру, сосудистую оболочку и сетчатку с помощью тонкой металлической микропипетки с диаметром наконечника 30 мкм и длиной наконечника 2,5 мм. Затем микропипетку соединяли со стеклянным 10-мкл шприцем (Hamilton Co., Рено, Северная Вирджиния) перед внесением 2 мкл суспензий векторов в пипетку в зависимости от группы. Действия выполняли с осторожностью, чтобы избежать проникновения в хрусталик или повреждения вортикозных вен при интравитреальной инъекции. Место инъекции отмечали на расстоянии примерно на 3 мм за верхневисочной каймой. Инъекции производили медленно в течение 1 минуты для обеспечения диффузии суспензии векторов. Правый глаз оставался нетронутым и служил в качестве внутреннего контралатерального контроля.
Сдавливание зрительного нерва (ONC)
Через три недели (21 день) после введения вектора мышей подвергали процедуре ONC, оставляли без обработки или производили холостое сдавливание. Под бинокулярным увеличением делали небольшой разрез с помощью пружинных ножниц в конъюнктиве, начиная с чуть ниже глазного яблока и временно вокруг глаза. Это обнажило заднюю сторону глазного яблока, позволяя визуализировать зрительный нерв. Обнаженный зрительный нерв сжимали на расстоянии примерно 1-3 мм от глазного яблока с помощью перекрещенных щипцов (Dumont №N7, кат. №RS-5027; Roboz) на 10 с, при этом единственное давление оказывалось от самосжимающего действия, прилагающего давление на нерв. Спустя 10 с зрительный нерв высвобождали, щипцы удаляли и глаз возвращали обратно на свое место. Через 7 дней после ONC животных умерщвляли. Оба глаза из каждой группы фиксировали, помещая орган в 4% параформальдегид/0,1% PBS (рН 7,4) на ночь. Затем готовили плоские препараты сетчатки после отсечения задней структуры глаза от роговицы и удаления хрусталика. После этого плоские препараты сетчатки фиксировали в течение 30 минут в 4% параформальдегиде/0,1% PBS и промывали в 0,5% Тритоне Х-100 в PBS. Сетчатки замораживали при -80°С в течение 10 минут для проникновения через ядерную мембрану и улучшения проникновения антител перед блокированием в 10% нормальной ослиной сыворотке (NDS), 2% бычьем сывороточном альбумине (BSA) и 2% Тритоне Х-100 в PBS в течение 60 минут при комнатной температуре. RGC контрастировали с помощью антител к Вrn3А (1:200 Santa Cruz, №sc-31984) и визуализировали под флуоресцентным микроскопом с применением 20× объектива и эпифлуоресцентного микроскопа Leica DM6000 (Leica Microsystems, Вецлар, Германия). Изображения с более высоким разрешением получали с применением конфокального микроскопа Leica SP5 (Leica Microsystems), оснащенного 40× объективом для масляной иммерсии, с 1,5× цифровым зумом и 0,5-0,8 последовательным интервалом сканирования по оси Z. Количество клеток RGC измеряли с помощью ImageJ с применением плагина для подсчета ядер на изображении Image-based Tool for Counting Nuclei (ITCN) и выражали в виде плотности RGC/mm2.
Конструкции и векторы
Авторами настоящего изобретения была создана генетическая конструкция, представленная на фиг. 1, которую можно применять для лечения субъекта, страдающего патологией зрительного нерва, такой как глаукома или патология улиткового нерва, или для стимуляции регенерации и/или выживаемости нервов. Конструкция была разработана для поддержания или увеличения плотности рецепторов TrkB на клеточной поверхности RGC и поддержания или увеличения передачи сигналов через путь с участием рецептора ТrkВ посредством сопутствующей выработки и локального высвобождения mBDNF. Конструкция содержит трансгены, кодирующие рецептор TrkB и его агонист, зрелый нейротрофический фактор головного мозга. Эти трансгены функционально связаны с одним промотором, который является либо промотором гена человеческого синапсина I (SYN I), либо промотором CAG. Преимущественно, конструкцию с фиг. 1 можно поместить в вектор rAAV2 без ограничения размера трансгенов, которые она кодирует. Это связано с тем, что конструкция ориентирована таким образом, чтобы первый трансген TrkB был связан с последовательностью, кодирующей вирусный пептид 2А, после которой шла последовательность, кодирующая сигнальный пептид BDNF, а затем последовательность, кодирующая зрелый белок. Эта ориентация также минимизирует риски иммуногенности, поскольку короткая N-концевая аминокислотная последовательность вирусного пептида 2А остается присоединенной к внутриклеточной части рецептора TrkB, а остаток аминокислоты пролин из С-концевой последовательности вирусного 2А остается присоединенной к N-концевому сигнальному пептиду BDNF и в конечном итоге удаляется из белка mBDNF после расщепления. Вектор можно поместить в фармакологически приемлемый буферный раствор, который можно вводить субъекту. На фиг. 2-5 показаны различные варианты осуществления векторов экспрессии. На фиг. 2 показан вектор, известный как «плазмида QTA001PA», содержащий каноническую сигнальную последовательность (синим цветом) (т.е. MMTILFLTMVISYFGCMKA [SEQ ID N0:20]) плюс proBDNF (красным цветом) и mBDNF (черным цветом). На фиг.3 показан вектор, известный как «плазмида QTA002P». Он не кодирует proBDNF, а вырабатывает только mBDNF и кодирует ту же сигнальную последовательность (синим цветом), что и QTA001PA. На фиг.4 показан вектор, известный как «плазмида QTA003P», который также не кодирует proBDNF, а вырабатывает только mBDNF. Вместо канонической сигнальной последовательности для mBDNF он содержит сигнальную последовательность IL-2 (синим цветом). Наконец, на фиг. 5 показан вектор, известный как «плазмида QTA004P». Он не кодирует proBDNF, а вырабатывает только mBDNF. Также он кодирует новую сигнальную последовательность (синим цветом), [SEQ ID NO: 32].
Авторами настоящего изобретения были созданы и изучены конструкция и вектор, относящиеся к идее генотерапии глаукомы, начинающиеся с такого элемента, как зрелый BDNF (mBDNF). Была четко продемонстрирована выработка и высвобождение mBDNF из клеток НЕK293 после липофектаминовой трансдукции плазмидой, которая содержит последовательность BDNF без области, кодирующей proBDNF (QTA002P, см. фиг. 3) (см. фиг. 7). MBDNF, высвобождаемый из клеток, представлял собой спрогнозированный мономер массой 14 кДа (по результатам измерения с помощью вестерн-блоттинга и коммерчески доступного антитела к BDNF), и не было никаких свидетельств наличия агрегатов белка, о которых сообщалось несколькими группами, предпринимавшими попытки получения промышленных количеств mBDNF с помощью дрожжевого и других клеточных подходов производства. Следовательно, mBDNF высвобождался в форме, которая могла позволить молекулам белка образовывать нековалентные димеры для активации рецепторов ТrkВ.
С помощью ИФА по BDNF (который не позволяет различить mBDNF и более крупный удлиненный белок proBDNF) авторами настоящего изобретения также было продемонстрировано, что возможна замена последовательности ДНК, кодирующей эндогенную каноническую сигнальную пептидную последовательность из 18 аминокислот (MTILFLTMVISYFGCMKA), на последовательность, кодирующую новый пептид (QTA004P - см. фиг. 5), и высвобождение эквивалентных уровней BDNF в среду для инкубации НЕK293 после липофектаминовой трансдукции клеток плазмидами, содержащими ген BDNF (см. фиг. 7).
Замена последовательности, кодирующей эндогенный сигнальный пептид, последовательностью, кодирующей сигнальный пептид интерлейкина-2 (QTA003P - см. фиг. 4), была менее эффективной при высвобождении BDNF из среды. Уровни BDNF, высвобождаемого в среду, в настоящее время составляют около 1-2 нМ, а концентрации этого агониста достаточны для максимальной активации специфических рецепторов TrkB (IC50 около 0,9 нМ). Уровни высвобождения BDNF примерно в 35 раз выше (876±87 нг/мл BDNF) с плазмидой QTA001PA (см. фиг. 2), которая содержит объединенные последовательности, кодирующие proBDNF и mBDNF, и которая также включает последовательность, кодирующую канонический сигнальный пептид из 18 аминокислот, по сравнению с плазмидами QTA002P (см. фиг. 3) и QTA004P (см. фиг. 5).
По результатам измерения остаточного BDNF в клетке путем количественного вестерн-блоттинга через 24 часа после липофектаминовой трансдукции плазмидой были выявлены более низкие концентрации остаточного BDNF с QTA001PA, чем с QTA002P и QTA004P (см. фиг. 8).
Кроме того, около половины BDNF-иммунореактивности в лизатах клеток, трансдуцированных QTA001PA, была представлена в форме proBDNF (бэнд с молекулярной массой на уровне 32 кДа), тогда как бэнд proBDNF отсутствовал в лизатах клеток, трансдуцированных QTA002P, QTA003P и QTA004P (см. фиг. 9), вероятно потому, что эти плазмиды не содержали последовательность, кодирующую удлиненный proBDNF.
С помощью ИФА, специфичного к proBDNF, авторам настоящего изобретения удалось продемонстрировать, что около 70 нг/мл (2,2 нМ или 3,5%) иммунореактивной молекулы BDNF, высвобожденной из клеток, трансдуцированных QTA001PA, были представлены в форме proBDNF, тогда как большинство (96,5% или 876 нг/мл / 63 нМ) высвобождалось в форме mBDNF (см. фиг. 10). ProBDNF-иммунореактивность в клетках, трансдуцированных QTA002P, QTA003P или QTA004P, которые не содержали последовательность, кодирующую удлиненный proBDNF, детектирована не была.
Соответственно, ясно, что все эти плазмиды могут вырабатывать белок mBDNF 14 кДа, но что количества mBDNF, высвобождаемые из клеток НЕK293, в значительной степени зависят от эффективности хранения белка и упаковки в секреторные везикулы. Следовательно, удлиненная форма белка, содержащая объединенные последовательности proBDNF и mBDNF, которая вырабатывается с плазмиды QTA001PA (фиг. 2), упаковывается в секреторные везикулы и высвобождается в среду для инкубации гораздо эффективнее, чем в случае с более мелкими последовательностями mBDNF, которые, судя по всему, накапливаются внутри клетки.
На фиг. 11 видно, что замена последовательности, кодирующей эндогенную каноническую сигнальную пептидную последовательность, представленной в плазмиде QTA002P, на новые последовательности, включенные в плазмиды QTA009P - QTA013P, увеличивала концентрацию BDNF в клетках НЕK293 через 24 часа после трансдукции плазмидами. На фиг. 12 продемонстрировано, что замена последовательности, кодирующей эндогенный канонический сигнальный пептид, включенной в плазмиду QTA002P, на новые последовательности (плазмиды QTA009P на QTA013P) увеличивала высвобождение BDNF (по результатам измерения с помощью ИФА) из клеток НЕK293, по результатам измерения через 24 часа после трансдукции плазмидами.
Как показано на фиг. 13, добавление последовательности вирусного пептида 2А приводило к эффективному процессингу последовательности, кодирующей большой белок-предшественник на два трансгена, eGFP и BDNF. На вестерн-блотах показаны клетки НЕK через 24 часа после их трансдукции плазмидами: (i) QTA015P (экспрессирующей BDNF и eGFP, отделенные спейсером IRES), (ii) QTA021P (экспрессирующей BDNF, за которым идет eGFP, отделенный функциональной последовательностью вирусного пептида 2А), (iii) QTA022P (экспрессирующей BDNF, за которым идет eGFP, отделенный нефункциональной последовательностью вирусного пептида 2А) и (iv) QTA023P (экспрессирующей eGFP, за которым идет последовательность, кодирующая BDNF, отделенный функциональной последовательностью вирусного пептида 2А).
Кодирующая последовательность QTA021P (плазмиды, содержащей подвергнутую оптимизации кодона последовательность mBDNF-вирусный пептид 2A-eGFP) в настоящем документе обозначена под SEQ ID No: 104 и имеет следующий вид:

Кодирующая последовательность QTA022P (плазмиды, содержащей подвергнутую оптимизации кодона последовательность mBDNF-нефункциональный вирусный пептид 2A-eGFP) в настоящем документе обозначена под SEQ ID No: 105 и имеет следующий вид:


Кодирующая последовательность QTA023P (плазмиды, содержащей подвергнутую оптимизации кодона последовательность eGFP-вирусный пептид 2A-mBDNF) в настоящем документе обозначена под SEQ ID No: 106 и имеет следующий вид:

На фиг. 14А показан вестерн-блот гомогенатов клеток НЕK293 через 48 часов после трансфекции вектором QTA020V. На нем виден эффективный процессинг области, кодирующей большой предшественник, которая включает рецептор TrkB и BDNF, разделенные последовательностью вирусного пептида 2А. Два трансгена, кодирующих TrkB- и mBDNF-иммунореактивные молекулы, находятся в пределах спрогнозированных правильных величин молекулярной массы. Следует отметить отсутствие окрашивания большого белка-предшественника выше бэнда рецептора TrkB, что свидетельствовало о почти полном или полном процессинге белка-предшественника в пяти повторностях. На фиг. 14В и 14С показано, что трансгенные белки, получаемые после расщепления вирусного пептида 2А, транспортировались в надлежащие внутриклеточные компартменты в клетках НЕK293 после процессинга (рецепторы TrkB на поверхность клетки, a BDNF в везикулы для хранения до высвобождения).
На фиг. 15 показано, что добавление последовательности, кодирующей вирусный пептид 2А и разделяющей две области, кодирующие рецептор TrkB и BDNF, приводило к эффективному процессингу на два трансгена в сетчатке мыши после интравитреальной инъекции rААV2-вектора QTA020V.
На фиг. 16 показана экспрессия трансгенов в слое ганглиозных клеток сетчатки мыши, которая продемонстрирована с помощью иммуноцитохимии после инъекции QTA020V, rААV2-вектора, содержащего последовательность, кодирующую рецептор TrkB и BDNF, разделенные последовательностью вирусного пептида 2А. Целевые ганглиозные клетки сетчатки окрашивали в красный цвет антителами к Вrn3А, а ядра клеток докрашивали в синий цвет с помощью DAPI для различения слоев сетчатки.
На фиг. 17 показано, что предварительная обработка QTA02oV (содержащей последовательность, кодирующую рецептор TrkB и BDNF, разделенные последовательностью вирусного пептида 2А) посредством интравитреальной инъекции (2 мкл 9×1012 векторных частиц/мл) придавала значительную нейропротекторную эффективность в отношении выживаемости ганглиозных клеток сетчатки после сдавливания зрительного нерва у мышей по сравнению с контрольными животными, обработанными вектором rAAV2-CAG-eGFP. Уровень нейропротекции вектором QTA020V также был выше, чем уровень, обеспечиваемый вектором, экспрессирующим только BDNF. Все три группы животных подвергали процедуре сдавливания зрительного нерва и через 7 дней после нанесенной травмы измеряли количество ганглиозных клеток сетчатки. Ганглиозные клетки сетчатки уменьшались в количестве на 71% в контролях (черные столбцы) по сравнению с животными, подвергнутыми холостому сдавливанию (данные не показаны).
Нейропротекторные эффекты конструкций
На фиг. 18 показана экспрессия трансгенов BDNF (см. фиг. 18А) и трансгенов TrkB (см. фиг. 18В) в гомогенатах недифференцированных человеческих клеток нейробластомы SH-SY5Y с помощью вестерн-блоттинга после трансфекции вирусными векторами rAAV2, которые не экспрессируют трансгены (нуль-вирус), экспрессируют только BDNF (QTA027V), только TrkB (QTA025V) и как BDNF, так и TrkB (QTA020V). Было четко видно, что достигались хорошие уровни экспрессии.
На фиг. 18С показан уровень активированных фосфорилированных рецепторов TrkB в клетках SH-SY5Y в вестерн-блотах после трансфекции вирусными векторами «нуль-вектор», QTA020V, QTA025V или QTA027V. Было обнаружено, что только вектор QTA020V, который экспрессирует как BDNF, так и TrkB, значительно увеличивал активацию рецепторов TrkB по сравнению с нетрансфицированными клетками. Таким образом, было продемонстрировано, что конструкции по настоящему изобретению эффективно экспрессировали оба трансгена и приводили к образованию активированных фосфорилированных рецепторов TrkB у клеток нейробластомы SH-SY5Y, что свидетельствовало о возможности лечения нейродегенеративных нарушений, таких как болезнь Альцгеймера или инсульт.
На фиг. 19 показан уровень апоптозной клеточной гибели недифференцированных клеток нейробластомы SH-SY5Y в культуре после воздействия окислительного стресса, вызванного добавлением перекиси водорода (Н2О2 в количестве 0,1 мМ или 1,0 мМ) путем окрашивания с помощью TUNEL. Неожиданно было обнаружено, что клетки, трансфицированные перед добавлением перекиси водорода rААV2-вектором QTA020V, который экспрессирует рецепторы как BDNF, так и TrkB, были значительно защищены от апоптоза по сравнению с необработанными клетками. Опять же эти данные подтверждали мнение, что конструкции по настоящему изобретению можно применять при лечении, предупреждении развития или облегчении нейродегенеративного нарушения или инсульта.
На фиг. 20 показаны репрезентативные иммуноцитохимические изображения зрительных нервов, полученных от мышей, трансгенных по мутантному человеческому тау-белку P301S, и окрашенных антителами, которые распознают фосфорилированный тау в положениях серии 396/серин 404 (PHF-1) или серии 202/серин 205 (АТ8).
У трансгенных мышей P301S к восьми месяцам развивалась потеря нейронов и атрофия головного мозга, преимущественно в гиппокампе, но распространявшаяся и на другие области головного мозга, в том числе гомогенетическую кору и энторинальную область коры головного мозга. У них развивались многочисленные нейрофибриллярные клубочковые включения в омогенетической коре, миндалевидном теле, гиппокампе, стволе головного мозга и в спинном мозге. Клубочковая патология сопровождалась микроглиозом и астроцитозом, но без образования амилоидных бляшек [56, 57, 58].
Мышей обрабатывали посредством интравитреальной инъекции QTA020V, который экспрессирует как рецепторы TrkB, так и BDNF в целевых ганглиозных клетках сетчатки и их аксонах. На изображениях, представленных на фиг.20, видно, что степень гиперфосфорилирования тау-белка с помощью PHF-1 и АТ-8 значительно снижалась в аксонах, которые составляют зрительный нерв. Из этих данных in vivo видно, что повышенная экспрессия TrkB и BDNF, получаемая с помощью конструкций по настоящему изобретению, может значимо уменьшать фосфорилирование тау-белка в нейронах, что является одним из патофизиологических признаков, связанных с головным мозгом, пораженным болезнью Альцгеймера.
Выводы
Понятно, что для болезни Альцгеймера не существует единой доклинической модели, которая обычно считается заменителем данного заболевания и на которой можно протестировать генную терапию с некоторой степенью прогнозируемости в отношении клинического исхода. Однако доступны модельные животные, у которых модификации, произведенные в их геноме, приводили к внедрению грызунам одного из определяющих генетических/нейрохимических или биохимических изменений, которые были выявлены у людей с данным заболеванием. К этим изменениям относят избыточную выработку Аβ и образование бляшек [59] гиперфосфорилированного тау-белка в телах и аксонах нейронов, которые, как полагают, опосредуют транспорт в аксонах [60] и снижение как BDNF, так и его родственного рецептора TrkB [11-14, 27].
Если за основу брать посмертную человеческую ткань и способность различных средств, которые могут успешно удалять бета-амилоид у обоих экспериментальных животных путем блокады фермента ВАСЕ-1, ответственного за его образование (верубецестат; Merck), или путем нейтрализации антителами (например, соленезумабом; Eli Lilly, и бапинейзумабом; Pfizer/J&J), то оба таких подхода не принесли существенного клинического полезного результата в клинических исследованиях III фазы. Следовательно, из широко описанных посмертных изменений в головном мозге человека, у которого диагностирована болезнь Альцгеймера, утрата передачи сигналов BDNF и присутствие нейрофибриллярных клубков, связанных с гиперфосфорилированным тау-белком, являются единственными непротестированными подходами к восстановлению или замедлению патофизиологических изменений, связанных с этим неврологическим состоянием.
Приложив значительные изобретательские усилия, авторы настоящего изобретения смогли решить задачу преодоления утраты передачи сигналов BDNF с помощью новой конструкции, которая одновременно способна как экспрессировать, так и активировать как рецепторы TrkB, так и BDNF, причем для обоих из них сообщалось, что их уровень снижается при данном заболевании (см. приведенные выше ссылки).
Поскольку BDNF имеет короткий период полужизни, регулярное введение рекомбинантного BDNF, для чего может требоваться несколько инъекций в сутки в головной мозг или посредством постоянной инфузии, клинически неосуществимо и, возможно, будет связано с даунрегуляцией рецепторов TrkB. Более того, авторы настоящего изобретения также продемонстрировали на фиг. 18С, что в клетках SHSY-5Y вектора rAAV2, экспрессирующего только рецепторы TrkB, недостаточно для значительного увеличения активности этого рецептора, если измерять по уровням окрашивания активного pY515-TrkB. Конструкции по настоящему изобретению были специально разработаны для размещения больших последовательностей, кодирующих как рецептор TrkB, так и BDNF, посредством ряда изобретательских стадий, включающих: (i) удаление последовательности, кодирующей pro-BDBF, (ii) введение новой последовательности, кодирующей сигнальный пептид для преодоления проблем, связанных с внутриклеточным транспортом и нормальным сворачиванием белка BDNF из-за пропуска важной последовательности Pro-BDNF, (iii) конструирование единого трансгена, содержащего последовательность, кодирующую вирусный пептид 2А, который облегчает трансляционный «пропуск» между рибосомальным продуцированием последовательностей TrkB и BDNF, и (iv) окончательно сокращенные последовательности WPRE и поли-А. Таким образом, авторами настоящего изобретения были представлены доказательства того, что новая конструкция, которая экспрессирует два трансгена, BDNF и его родственный рецептор TrkB, намного превосходит простую положительную регуляцию только лишь рецепторов TrkB. Авторами настоящего изобретения также было продемонстрировано, что новые генотерапевтические конструкции способны обеспечивать оптимальную активность, как было продемонстрировано ранее [56], но без необходимости дополнительных (регулярных) инъекций BDNF.
Основная цель авторов настоящего изобретения заключалась в том, чтобы разработать генотерапию, с помощью которой можно бороться с низкими уровнями передачи сигналов BDNF/TrkB, что четко продемонстрировано в приведенных примерах. Что было неожиданным, так это то, что новая генотерапевтическая конструкция способна значительно снижать плотность гиперфосфорилированного тау-белка (по результатам измерения с помощью двух антител, которые распознают несколько фосфорилированных остатков серина в цепи тау-белка), что видно на фиг. 20. Тау является убиквитарным белком, который встречается в головном мозге и других нервных тканях, таких как зрительный нерв. С помощью зрительного нерва в качестве модельной системы было обнаружено, что увеличение передачи сигналов BDNF в глазу снижает предполагаемый патологический уровень данной изоформы белка. Следовательно, способность активировать передачу сигналов BDNF/TrkB у линии трансгенных мышей P301S и проявление такого основательного снижения плотности фосфорилированного тау-белка были неожиданными.
Перечень литературы
1. Brookmeyer R, Johnson Е, Ziegler-Graham K, Arrighi НМ. Forecasting the global burden of Alzheimer's disease (2007). Alzheimers. Dement, vol. 3(3) PP: 186-191.
2. Вrааk Н, Braak Е. (1991). Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol., vol. 82(4), PP:239-259.
3. Dekosky ST, Scheff SW. (1990). Synapse loss in frontal cortex biopsies in Alzheimer's disease: correlation with cognitive severity. Ann. Neurol., vol. 27(5), PP:457-464.
4. Chao MV (2003). Neurotrophins and their receptors: A convergence point for many signalling pathways. Nature Rev. Neurosci. vol. 4, PP: 299-309.
5. Dawbarn D, Allen SJ, (2003). Neurotrophins and neurodegeneration. Neuropathol. Appl. Neurobiol., vol.29, PP: 211-230.
6. Phillips HS, Hains JM, Laramee GR, Rosenthal A, Winslow JW (1990). Widespread expression of BDNF but not NT3 by target areas of basal forebrain cholinergic neurons. Science, vol. 250(4978), PP: 290-294.
7. Wetmore C, Ernfors P, Persson H, Olson L, (1990). Localization of brain-derived neurotrophic factor mRNA to neurons in the brain by in situ hybridization. Exp. Neurol., vol. 109(2), PP: 141-152
8. Ghosh A, Carnahan J, Greenberg ME, (1994). Requirement for BDNF in activity-dependent survival of cortical neurons. Science, vol. 263(5153), PP: 1618 1623.
9. Phillips HS, Hains JM, Armanini M, Laramee GR, Johnson SA, Winslow JW, (1991). BDNF mRNA is decreased in the hippocampus of individuals with Alzheimer's disease. Neuron, vol. 7(5), PP: 695-702.
10. Holsinger RM, Schnarr J, Henry P, Castelo VT, Fahnestock M, (2000). Quantitation of BDNF mRNA in human parietal cortex by competitive reverse transcription-polymerase chain reaction: decreased levels in Alzheimer's disease. Brain Res. Mol. Brain Res., vol. 76(2), PP: 347-354.
11. Narisawa-Saito M, Wakabayashi K, Tsuji S, Takahashi H, Nawa H, (1996). Regional specificity of alterations in NGF, BDNF and NT-3 levels in Alzheimer's disease. Neuroreport, vol. 7(18), PP: 2925-2928.
12. Connor B, Young D, Yan Q, Faull R L, Synek B, Dragunow M, (1997). Brain-derived neurotrophic factor is reduced in Alzheimer's disease. Brain Res. Mol. Brain Res., vol. 49(1-2), PP: 71-81.
13. Ferrer I, Marin C, Rey M J, Ribalta T, Goutan E, Blanco R, Tolosa E, Marti E, (1999). BDNF and full-length and truncated TrkB expression in Alzheimer disease. Implications in therapeutic strategies. J. Neuropathol. Exp.Neurol., vol. 58(7), PP: 729-739.
14. Savaskan E, Muller-Spahn F, Olivieri G, Bruttel S, Often U, Rosenberg C, Hulette C, Hock C, (2000). Alterations in trk A, trk В and trk С receptor immunoreactivities in parietal cortex and cerebellum in Alzheimer's disease. Eur. Neurol., vol. 44(3), PP: 172 180.
15. Peng S, Wuu J, Mufson E J, Fahnestock M, (2005). Precursor form of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinical stages of Alzheimer's disease. J. Neurochem., vol. 93(6) PP: 1412 1421.
16. Michalski B, Fahnestock M, (2003). Pro-brain-derived neurotrophic factor is decreased in parietal cortex in Alzheimer's disease. Brain Res. Mol. Brain Res., vol. 111(1-2), PP: 148 154.
17. Qin XY, Cao C, Cawley NX, Liu TT, Yuan J, Loh YP, Cheng Y, (2016). Decreased peripheral brain-derived neurotrophic factor levels in Alzheimer's disease: A meta-analysis study (N=7277). Mol. Psychiatry, vol. 22, PP: 312-320.
18. Forlenza OV, Diniz BS, Teixeira AL, Radanovic M, Talib LL, Rocha NP, Gattaz WF, (2015). Lower Cerebrospinal Fluid Concentration of Brain-Derived Neurotrophic Factor Predicts Progression from Mild Cognitive Impairment to Alzheimer's Disease. Neuromolecular Med., vol. 17(3), PP: 326-332.
19. Feher A, Juhasz A, Rimanoczy A, Kalman J, Janka Z, (2009). Association between BDNF Val66Met polymorphism and Alzheimer disease, dementia with Lewy bodies, and Pick disease. Alzheimer Dis. Assoc. Disord., vol. 23(3) PP: 224-228.
20. Peng S, Wuu J, Mufson EJ, Fahnestock M, (2005). Precursor form of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinical stages of Alzheimer's disease. J. Neurochem., vol. 93, PP: 1412-1421;
21. Michalski B, Corrada MM, Kawas CH, Fahnestock M, (2015). Brain-derived neurotrophic factor and TrkB expression in the "oldest-old", the 90+study: Correlation with cognitive status and levels of soluble amyloid-β. Neurobiol. Aging, vol. 36, PP: 3130-3139.
22. Elliott E, Atlas R, Lange A, Ginzburg I, (2005). Brain-derived neurotrophic factor induces a rapid dephosphorylation of tau protein through a PI-3 kinase signalling mechanism. Eur. J. Neurosci., vol. 22, PP: 1081-1089.
23. Jiao S-S, Shen L-L, Zhu C, Bu X-L, Liu Y-H, Liu C-H, Yao X-Q, Zhang L-L, Zhou H-D, Walker DG, Tan J, Gortz J, Zhou X-F, Wang Y-J, (2016). Brain-derived neurotrophic factor protects against tau-related neurodegeneration of Alzheimer's disease. Transl. Psychiatry, vol. 6, e907.
24. Rosa E, Mahendram S, Ke YD, Ittner LM, Ginsberg SD, Fahnestock M, (2016). Tau downregulates BDNF expression in animal and cellular models of Alzheimer's disease. Neurobiol. Aging, vol. 48,PP: 135-142.
25. Lim JY, Reighard CP, Crowther DC, (2015). The pro-domains of neurotrophins, including BDNF, are linked to Alzheimer's disease through a toxic synergy with Aβ. Hum. Mol. Genet, vol., 24(14) PP:3929-3938.
26. Hashimoto R, Hirata Y, Asada T, Yamashita F, Nemoto K, Mori T, Moriguchi Y, Kunugi H, Arima K, Ohnishi T, (2009). Effect of the brain-derived neurotrophic factor and the apolipoprotein E polymorphisms on disease progression in preclinical Alzheimer's disease. Genes Brain Behav., vol. 8(1) PP: 43-52.
27. Allen S J, Wilcock G K, Dawbarn D, (1999). Profound and selective loss of catalytic TrkB immunoreactivity in Alzheimer's disease. Biochem. Biophys. Res. Commun., vol. 264(3) PP: 648 651.
28. Kemppainen S, Rantamaki T, Jeronimo-Santos A, Lavasseur G, Autio H, Karpova N, Karkkainen E, Staven S, Vicente Miranda H, Outeiro TF, Diogenes MJ, Laroche S, Davis S,  AM, Castren E, Tanila H, (2012). Impaired TrkB receptor signaling contributes to memory impairment in APP/PS1 mice. Neurobiol. Aging, vol. 33, e1123-e1139.
AM, Castren E, Tanila H, (2012). Impaired TrkB receptor signaling contributes to memory impairment in APP/PS1 mice. Neurobiol. Aging, vol. 33, e1123-e1139.
29. Devi L, Ohno M (2015) TrkB reduction exacerbates Alzheimer's disease-like signalling aberrations and memory deficits without affecting β-amyloidosis in 5XFAD mice. Transl. Psychiatry, vol. 5, e562.
30. Tong L, Balazs R, Thornton PL, Cotman CW, (2004). β-Amyloid peptide at sublethal concentrations downregulates brain-derived neurotrophic factor functions in cultured cortical neurons. J. Neurosci., vol. 24, PP:6799-6809.
31. Liu XH, Geng Z, Yan J, Li T, Chen Q, Zhang QY, Chen ZY, (2015). Blocking GSK3β-mediated dynaminl phosphorylation enhances BDNF-dependent TrkB endocytosis and the protective effects of BDNF in neuronal and mouse models of Alzheimer's disease. Neurobiol. Dis., vol. 74, PP: 377 391.
32. Poon WW, Blurton-Jones M, Tu CH, Feinberg LM, Chabrier MA, Harris JW, Jeon NL, Cotman CW, (2011). β-amyloid impairs axonal BDNF retrograde trafficking. Neurobiol. Aging, vol. 32, PP: 821-833.
33. Gong B, Cao Z, Zheng P, Vitolo OV, Liu S, Staniszewski A, Moolman D, Zhang H, Shelanski M, Arancio O, (2006). Ubiquitin hydrolase Uch-L1 rescues β-amyloid-induced decreases in synaptic function and contextual memory. Cell, vol. 126, PP: 775-788.
34. Gan KJ, Silverman MA, (2015). Dendritic and axonal mechanisms of Ca2+elevation impair BDNF transport in aβ oligomer-treated hippocampal neurons. Mol. Biol. Cell, vol. 26, PP: 1058 1071.
35. Zhang L, Fang Y, Lian Y, Chen Y, Wu T, Zheng Y, Zong H, Sun L, Zhang R, Wang Z, Xu Y, (2015). Brain-derived neurotrophic factor ameliorates learning deficits in a rat model of Alzheimer's disease induced by aβ1-42., PLoS One, vol. 10(4), e0122415.
36. Wu Y, Luo X, Liu X, Liu D, Wang X, Guo Z, Zhu L, Tian Q, Yang X, Wang JZ, (2015). Intraperitoneal Administration of a Novel TAT-BDNF Peptide Ameliorates Cognitive Impairments via Modulating Multiple Pathways in Two Alzheimer's Rodent Models. Sci. Rep., vol. 5, PP: 15032.
37. Nagahara AH, Merrill DA, Coppola G, Tsukada S, Schroeder BE, Shaked GM, Wang L, Blesch A, Kim A, Conner JM, Rockenstein E, Chao MV, Koo EH, Geschwind D, Masliah E, Chiba AA, Tuszynski MH, (2009). Neuroprotective effects of brain-derived neurotrophic factor in rodent and primate models of Alzheimer's disease. Nat. Med., vol. 15(3), PP: 331-337.
38. Mey J, Thanos S, (1993). Intravitreal injections of neurotrophic factors support the survival of axotomized retinal ganglion cells in adult rats in vivo. Brain Res., vol. 602, PP: 304-317.
39. Mansour-Rabaey S, Clarke DB, Wang Y-C, Bray GM, Aguayo AJ, (1994). Effects of ocular injury and administration of brain-derived neurotrophic factor on survival and regrowth of axotomized retinal ganglion cells. Proc. Natl. Acad. Sci. USA., vol. 91, PP: 1632-1636.
40. Peinado-Ramon P. Salvador M, Viuegas-Perez MP, Vidal-Sanz M, (1996). Effects of axotomy and intraocular administration of NT-4, NT-3 and brain-derived neurotrophic factor on the survival of adult rat retinal ganglion cells. A quantitative in vivo study. Invest Ophthalmol. Vis. Sci., vol. 37, PP: 489-500.
41. Di Polo A, Aigner LJ, Dunn RJ, Bray GM, Aguayo AJ, (1998). Prolonged delivery of brain-derived neurotrophic factor by adenovirus- infected  cells temporarily rescues injured retinal ganglion cells. Proc. Natl. Acad. Sci. USA., vol. 95, PP: 3978-3983.
cells temporarily rescues injured retinal ganglion cells. Proc. Natl. Acad. Sci. USA., vol. 95, PP: 3978-3983.
42. Klocker N, Kermer P, Weishaupt JH, Labes M, Ankerhold R,  , (2000). Brain-derived neurotrophic factor-mediated neuroprotection of adult rat retinal ganglion cells in vivo does not exclusively depend on phosphatidyl-inositol-3'-kinase/protein kinase В signaling. J. Neurosci., vol. 20, PP: 6962-6967.
, (2000). Brain-derived neurotrophic factor-mediated neuroprotection of adult rat retinal ganglion cells in vivo does not exclusively depend on phosphatidyl-inositol-3'-kinase/protein kinase В signaling. J. Neurosci., vol. 20, PP: 6962-6967.
43. Kо ML, Hu DN, Ritch R, Sharma SC, Chen CF, (2001). Patterns of retinal ganglion cell survival after brain-derived neurotrophic factor administration in hypertensive eyes of rats. Neurosci. Lett., vol. 305, PP: 139-142.
44. Chen H, Weber AJ, (2001). BDNF enhances retinal ganglion cell survival in cats with optic nerve damage. Invest Ophthamol. Vis. Sci., vol. 42, PP: 966-974.
45. Porez MTR, Caminos E, (1995). Expression of brain-derived neurotrophic factor and its functional receptor in neonatal and adult rat retina. Neurosci. Lett., vol. 183, PP: 96-99.
46. Vecino E, Ugarte M, Nash MS, Osborne NN. (1999). NMDA induces BDNF expression in the albino rat retina in vivo. Neuroreport, vol. 10, PP: 1103-1106.
47. Mowla SJ, Farhadi HF, Pareek S, Atwal JK, Morris SJ, Seidah NG, Murphy RA, (2001). Biosynthesis and post-translational processing of the precursor to brain-derived neurotrophic factor. J. Biol. Chem., vol. 276, PP: 12660-12666.
48. Gupta VK, You Y, Gupta VB, Klistorner A, Graham SL, (2013). TrkB receptor signalling: Implications in neurodegenerative, psychiatric and proliferative disorders. Int. J. Mol. Sci., vol.14, PP: 10122-10142
49. Teng HK, Teng KK, Lee R, Wright S, Tevar S, Almeida RD, Kermani P, Torkin R, Chen ZY, Lee FS, Kraemer RT, Nykjaer A, Hempstead BL, (2005). ProBDNF induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilin. J. Neurosci., vol. 25, PP: 5455-5463.
50. Wei Y, Zhang F, Zao J, Jiang X, Lu Q, Gao E, Wand N, (2012). Enhanced protein expression of proBDNF and proNGF in elevated intraocular pressure-induced rat retinal ischemia. Chin. Med. J., vol. 125, PP: 3875-3879.
51. Woo NH, Teng HK, Siao C-J, Chiaruttini C, Pang PT, Milner ТА, Hempstead BL,Lu B, (2005). Activation of p75NTR by proBDNF facilitates hippocampal long-term depression. Nature Neurosci., vol. 8, PP: 1069-1077.
52. Lebrun-Julien F, Bertrand MJ, De Backer O, Stellwagen D, Morales CR, Di Polo A, Barker PA, (2010). ProNGF induces TNFalpha-dependent death of retinal ganglion cells through a p75NTR non-cell-autonomous signaling pathway. Proc. Natl. Acad. Sci. USA., vol. 107, PP: 3817-3822.
53. Quigley HA, McKinnon SJ, Zack DJ, Pease ME, Kerrigan-Baumrind LA, Kerrigan DF, Mitchell RS, (2000). Retrograde axonal transport of BDNF in retinal ganglion cells is blocked by acute IOP elevation in rats. Invest. Ophthalmol. Vis. Sci., vol. 41, PP: 3460-3466.
54. Pease ME McKinnon SJ, Quigley HA, Kerrigan-Baumrind LA, Zack DJ, (2000). Obstructed axonal transport of BDNF and its receptor TRKB in experimental glaucoma. Invest. Ophthalmol. Vis. Sci., vol. 41, PP: 764-774.
55. Wei Y, Wang N, Lu Q, Zhang N, Zheng D, Li J, (2007). Enhanced protein expressions of sortilin and p75NTR in retina of rat following elevated intraocular pressure-induced retinal ischemia. Neurosci. Lett., vol. 429, PP: 169-174.
56. Cheng L, Sapieha P, Kittlerova P, Hauswith WW, Di Polo A, (2002). TrkB gene transfer protects retinal ganglion cells from axotomy-induced death in vivo. J. Neurosci., vol. 22(10), PP: 3977-3986.
57. YoshiyamaY, Higuchi M, Zhang B, Huang SM, Iwata N, Saido TC, Maeda J, Suhara T, Trojanowski TC, Lee VM, (2007). Synapse loss and microglial activation precede tangles in a P301S Tauopathy mouse model. Neuron, vol. 53, PP: 337-351.
58. Takeuchi H, Iba M, Inoue H, Higuchi M, Таkао K, Tsukita K, Karatsu Y, Iwamoto Y, Miyakama T, Suhara T, Trojanowski JQ, Lee VM, Takahashi R, (2011). P301S mutant human Tau transgenic mice manifest early symptoms of human tauopathies with dementia and altered sensorimotor gating. PLoS One, vol. 6(6), e21050.
59. Hardy J, Allsop D, (1991). Amyloid deposition as the central event in the aetiology of Alzheimer's disease, Trends Pharmacol. Sci., vol. 12(10), PP:383-388.
60. Mudher A, Lovestone S, (2002). Alzheimer's disease - do tauists and baptists finally shake hands? Trends Neurosci., vol. 25(1), PP: 22-26.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> КУЭТЕРА ЛИМИТЕД
<120> Генетическая конструкция для применения при лечении
нейродегенеративного нарушения или инсульта
<130> 83344PCT1
<160> 108
<170> PatentIn version 3.5
<210> 1
<211> 469
<212> DNA
<213> Homo sapiens
<400> 1
ctgcagaggg ccctgcgtat gagtgcaagt gggttttagg accaggatga ggcggggtgg 60
gggtgcctac ctgacgaccg accccgaccc actggacaag cacccaaccc ccattcccca 120
aattgcgcat cccctatcag agagggggag gggaaacagg atgcggcgag gcgcgtgcgc 180
actgccagct tcagcaccgc ggacagtgcc ttcgcccccg cctggcggcg cgcgccaccg 240
ccgcctcagc actgaaggcg cgctgacgtc actcgccggt cccccgcaaa ctccccttcc 300
cggccacctt ggtcgcgtcc gcgccgccgc cggcccagcc ggaccgcacc acgcgaggcg 360
cgagataggg gggcacgggc gcgaccatct gcgctgcggc gccggcgact cagcgctgcc 420
tcagtctgcg gtgggcagcg gaggagtcgt gtcgtgcctg agagcgcag 469
<210> 2
<211> 1733
<212> DNA
<213> Homo sapiens
<400> 2
ctcgacattg attattgact agttattaat agtaatcaat tacggggtca ttagttcata 60
gcccatatat ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc 120
ccaacgaccc ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag 180
ggactttcca ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac 240
atcaagtgta tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg 300
cctggcatta tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg 360
tattagtcat cgctattacc atggtcgagg tgagccccac gttctgcttc actctcccca 420
tctccccccc ctccccaccc ccaattttgt atttatttat tttttaatta ttttgtgcag 480
cgatgggggc gggggggggg ggggggcgcg cgccaggcgg ggcggggcgg ggcgaggggc 540
ggggcggggc gaggcggaga ggtgcggcgg cagccaatca gagcggcgcg ctccgaaagt 600
ttccttttat ggcgaggcgg cggcggcggc ggccctataa aaagcgaagc gcgcggcggg 660
cgggagtcgc tgcgcgctgc cttcgccccg tgccccgctc cgccgccgcc tcgcgccgcc 720
cgccccggct ctgactgacc gcgttactcc cacaggtgag cgggcgggac ggcccttctc 780
ctccgggctg taattagcgc ttggtttaat gacggcttgt ttcttttctg tggctgcgtg 840
aaagccttga ggggctccgg gagggccctt tgtgcggggg gagcggctcg gggggtgcgt 900
gcgtgtgtgt gtgcgtgggg agcgccgcgt gcggctccgc gctgcccggc ggctgtgagc 960
gctgcgggcg cggcgcgggg ctttgtgcgc tccgcagtgt gcgcgagggg agcgcggccg 1020
ggggcggtgc cccgcggtgc ggggggggct gcgaggggaa caaaggctgc gtgcggggtg 1080
tgtgcgtggg ggggtgagca gggggtgtgg gcgcgtcggt cgggctgcaa ccccccctgc 1140
acccccctcc ccgagttgct gagcacggcc cggcttcggg tgcggggctc cgtacggggc 1200
gtggcgcggg gctcgccgtg ccgggcgggg ggtggcggca ggtgggggtg ccgggcgggg 1260
cggggccgcc tcgggccggg gagggctcgg gggaggggcg cggcggcccc cggagcgccg 1320
gcggctgtcg aggcgcggcg agccgcagcc attgcctttt atggtaatcg tgcgagaggg 1380
cgcagggact tcctttgtcc caaatctgtg cggagccgaa atctgggagg cgccgccgca 1440
ccccctctag cgggcgcggg gcgaagcggt gcggcgccgg caggaaggaa atgggcgggg 1500
agggccttcg tgcgtcgccg cgccgccgtc cccttctccc tctccagcct cggggctgtc 1560
cgcgggggga cggctgcctt cgggggggac ggggcagggc ggggttcggc ttctggcgtg 1620
tgaccggcgg ctctagagcc tctgctaacc atgttcatgc cttcttcttt ttcctacagc 1680
tcctgggcaa cgtgctggtt attgtgctgt ctcatcattt tggcaaagaa ttg 1733
<210> 3
<211> 664
<212> DNA
<213> Homo sapiens
<400> 3
ctagatctga attcggtacc ctagttatta atagtaatca attacggggt cattagttca 60
tagcccatat atggagttcc gcgttacata acttacggta aatggcccgc ctggctgacc 120
gcccaacgac ccccgcccat tgacgtcaat aatgacgtat gttcccatag taacgccaat 180
agggactttc cattgacgtc aatgggtgga ctatttacgg taaactgccc acttggcagt 240
acatcaagtg tatcatatgc caagtacgcc ccctattgac gtcaatgacg gtaaatggcc 300
cgcctggcat tatgcccagt acatgacctt atgggacttt cctacttggc agtacatcta 360
cgtattagtc atcgctatta ccatggtcga ggtgagcccc acgttctgct tcactctccc 420
catctccccc ccctccccac ccccaatttt gtatttattt attttttaat tattttgtgc 480
agcgatgggg gcgggggggg ggggggggcg cgcgccaggc ggggcggggc ggggcgaggg 540
gcggggcggg gcgaggcgga gaggtgcggc ggcagccaat cagagcggcg cgctccgaaa 600
gtttcctttt atggcgaggc ggcggcggcg gcggccctat aaaaagcgaa gcgcgcggcg 660
ggcg 664
<210> 4
<211> 11
<212> PRT
<213> Homo sapiens
<400> 4
Gln Ala Gly Asp Val Glu Glu Asn Pro Gly Pro
1 5 10
<210> 5
<211> 63
<212> DNA
<213> Homo sapiens
<400> 5
ggaagcggag ctactaactt cagcctgctg aaggctggag acgtggagga gaaccctgga 60
cct 63
<210> 6
<211> 21
<212> PRT
<213> Homo sapiens
<400> 6
Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Gln Ala Gly Asp Val Glu
1 5 10 15
Glu Asn Pro Gly Pro
20
<210> 7
<211> 63
<212> DNA
<213> Homo sapiens
<400> 7
agcggagcta ctaacttcag cctgctgaag caggctggag acgtggagga gaaccctgga 60
cct 63
<210> 8
<211> 21
<212> PRT
<213> Homo sapiens
<400> 8
Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val Glu
1 5 10 15
Glu Asn Pro Gly Pro
20
<210> 9
<211> 822
<212> PRT
<213> Homo sapiens
<400> 9
Met Ser Ser Trp Ile Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Phe Cys Trp Leu Val Val Gly Phe Trp Arg Ala Ala Phe Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Phe Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
465 470 475 480
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
485 490 495
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
500 505 510
Asn Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
515 520 525
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
530 535 540
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
545 550 555 560
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
565 570 575
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
580 585 590
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
595 600 605
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
610 615 620
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
625 630 635 640
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
645 650 655
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
660 665 670
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
675 680 685
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr
690 695 700
Asp Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
705 710 715 720
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
725 730 735
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
740 745 750
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
755 760 765
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
770 775 780
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Met Arg Lys Asn Ile
785 790 795 800
Lys Gly Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
805 810 815
Tyr Leu Asp Ile Leu Gly
820
<210> 10
<211> 2466
<212> DNA
<213> Homo sapiens
<400> 10
atgtcgtcct ggataaggtg gcatggaccc gccatggcgc ggctctgggg cttctgctgg 60
ctggttgtgg gcttctggag ggccgctttc gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgctgtggt ggtgattgcg 1320
tctgtggtgg gattttgcct tttggtaatg ctgtttctgc ttaagttggc aagacactcc 1380
aagtttggca tgaaaggccc agcctccgtt atcagcaatg atgatgactc tgccagccca 1440
ctccatcaca tctccaatgg gagtaacact ccatcttctt cggaaggtgg cccagatgct 1500
gtcattattg gaatgaccaa gatccctgtc attgaaaatc cccagtactt tggcatcacc 1560
aacagtcagc tcaagccaga cacatttgtt cagcacatca agcgacataa cattgttctg 1620
aaaagggagc taggcgaagg agcctttgga aaagtgttcc tagctgaatg ctataacctc 1680
tgtcctgagc aggacaagat cttggtggca gtgaagaccc tgaaggatgc cagtgacaat 1740
gcacgcaagg acttccaccg tgaggccgag ctcctgacca acctccagca tgagcacatc 1800
gtcaagttct atggcgtctg cgtggagggc gaccccctca tcatggtctt tgagtacatg 1860
aagcatgggg acctcaacaa gttcctcagg gcacacggcc ctgatgccgt gctgatggct 1920
gagggcaacc cgcccacgga actgacgcag tcgcagatgc tgcatatagc ccagcagatc 1980
gccgcgggca tggtctacct ggcgtcccag cacttcgtgc accgcgattt ggccaccagg 2040
aactgcctgg tcggggagaa cttgctggtg aaaatcgggg actttgggat gtcccgggac 2100
gtgtacagca ctgactacta cagggtcggt ggccacacaa tgctgcccat tcgctggatg 2160
cctccagaga gcatcatgta caggaaattc acgacggaaa gcgacgtctg gagcctgggg 2220
gtcgtgttgt gggagatttt cacctatggc aaacagccct ggtaccagct gtcaaacaat 2280
gaggtgatag agtgtatcac tcagggccga gtcctgcagc gaccccgcac gtgcccccag 2340
gaggtgtatg agctgatgct ggggtgctgg cagcgagagc cccacatgag gaagaacatc 2400
aagggcatcc ataccctcct tcagaacttg gccaaggcat ctccggtcta cctggacatt 2460
ctaggc 2466
<210> 11
<211> 838
<212> PRT
<213> Homo sapiens
<400> 11
Met Ser Ser Trp Ile Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Phe Cys Trp Leu Val Val Gly Phe Trp Arg Ala Ala Phe Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Phe Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Asp Phe Ser Trp Phe Gly Phe Gly Lys Val Lys Ser Arg Gln Gly
465 470 475 480
Val Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
485 490 495
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
500 505 510
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
515 520 525
Asn Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
530 535 540
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
545 550 555 560
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
565 570 575
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
580 585 590
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
595 600 605
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
610 615 620
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
625 630 635 640
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
645 650 655
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
660 665 670
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
675 680 685
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
690 695 700
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr
705 710 715 720
Asp Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
725 730 735
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
740 745 750
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
755 760 765
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
770 775 780
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
785 790 795 800
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Met Arg Lys Asn Ile
805 810 815
Lys Gly Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
820 825 830
Tyr Leu Asp Ile Leu Gly
835
<210> 12
<211> 2514
<212> DNA
<213> Homo sapiens
<400> 12
atgtcgtcct ggataaggtg gcatggaccc gccatggcgc ggctctgggg cttctgctgg 60
ctggttgtgg gcttctggag ggccgctttc gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgctgtggt ggtgattgcg 1320
tctgtggtgg gattttgcct tttggtaatg ctgtttctgc ttaagttggc aagacactcc 1380
aagtttggca tgaaagattt ctcatggttt ggatttggga aagtaaaatc aagacaaggt 1440
gttggcccag cctccgttat cagcaatgat gatgactctg ccagcccact ccatcacatc 1500
tccaatggga gtaacactcc atcttcttcg gaaggtggcc cagatgctgt cattattgga 1560
atgaccaaga tccctgtcat tgaaaatccc cagtactttg gcatcaccaa cagtcagctc 1620
aagccagaca catttgttca gcacatcaag cgacataaca ttgttctgaa aagggagcta 1680
ggcgaaggag cctttggaaa agtgttccta gctgaatgct ataacctctg tcctgagcag 1740
gacaagatct tggtggcagt gaagaccctg aaggatgcca gtgacaatgc acgcaaggac 1800
ttccaccgtg aggccgagct cctgaccaac ctccagcatg agcacatcgt caagttctat 1860
ggcgtctgcg tggagggcga ccccctcatc atggtctttg agtacatgaa gcatggggac 1920
ctcaacaagt tcctcagggc acacggccct gatgccgtgc tgatggctga gggcaacccg 1980
cccacggaac tgacgcagtc gcagatgctg catatagccc agcagatcgc cgcgggcatg 2040
gtctacctgg cgtcccagca cttcgtgcac cgcgatttgg ccaccaggaa ctgcctggtc 2100
ggggagaact tgctggtgaa aatcggggac tttgggatgt cccgggacgt gtacagcact 2160
gactactaca gggtcggtgg ccacacaatg ctgcccattc gctggatgcc tccagagagc 2220
atcatgtaca ggaaattcac gacggaaagc gacgtctgga gcctgggggt cgtgttgtgg 2280
gagattttca cctatggcaa acagccctgg taccagctgt caaacaatga ggtgatagag 2340
tgtatcactc agggccgagt cctgcagcga ccccgcacgt gcccccagga ggtgtatgag 2400
ctgatgctgg ggtgctggca gcgagagccc cacatgagga agaacatcaa gggcatccat 2460
accctccttc agaacttggc caaggcatct ccggtctacc tggacattct aggc 2514
<210> 13
<211> 822
<212> PRT
<213> Homo sapiens
<400> 13
Met Ser Ser Trp Ile Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Phe Cys Trp Leu Val Val Gly Phe Trp Arg Ala Ala Phe Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Phe Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
465 470 475 480
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
485 490 495
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
500 505 510
Asn Pro Gln Glu Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
515 520 525
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
530 535 540
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
545 550 555 560
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
565 570 575
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
580 585 590
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
595 600 605
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
610 615 620
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
625 630 635 640
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
645 650 655
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
660 665 670
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
675 680 685
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Glu Ser Thr
690 695 700
Asp Glu Glu Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
705 710 715 720
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
725 730 735
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
740 745 750
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
755 760 765
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
770 775 780
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Met Arg Lys Asn Ile
785 790 795 800
Lys Gly Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
805 810 815
Glu Leu Asp Ile Leu Gly
820
<210> 14
<211> 2466
<212> DNA
<213> Homo sapiens
<400> 14
atgtcgtcct ggataaggtg gcatggaccc gccatggcgc ggctctgggg cttctgctgg 60
ctggttgtgg gcttctggag ggccgctttc gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgctgtggt ggtgattgcg 1320
tctgtggtgg gattttgcct tttggtaatg ctgtttctgc ttaagttggc aagacactcc 1380
aagtttggca tgaaaggccc agcctccgtt atcagcaatg atgatgactc tgccagccca 1440
ctccatcaca tctccaatgg gagtaacact ccatcttctt cggaaggtgg cccagatgct 1500
gtcattattg gaatgaccaa gatccctgtc attgaaaatc cccaggaatt tggcatcacc 1560
aacagtcagc tcaagccaga cacatttgtt cagcacatca agcgacataa cattgttctg 1620
aaaagggagc taggcgaagg agcctttgga aaagtgttcc tagctgaatg ctataacctc 1680
tgtcctgagc aggacaagat cttggtggca gtgaagaccc tgaaggatgc cagtgacaat 1740
gcacgcaagg acttccaccg tgaggccgag ctcctgacca acctccagca tgagcacatc 1800
gtcaagttct atggcgtctg cgtggagggc gaccccctca tcatggtctt tgagtacatg 1860
aagcatgggg acctcaacaa gttcctcagg gcacacggcc ctgatgccgt gctgatggct 1920
gagggcaacc cgcccacgga actgacgcag tcgcagatgc tgcatatagc ccagcagatc 1980
gccgcgggca tggtctacct ggcgtcccag cacttcgtgc accgcgattt ggccaccagg 2040
aactgcctgg tcggggagaa cttgctggtg aaaatcgggg actttgggat gtcccgggac 2100
gtggaaagca ctgacgaaga aagggtcggt ggccacacaa tgctgcccat tcgctggatg 2160
cctccagaga gcatcatgta caggaaattc acgacggaaa gcgacgtctg gagcctgggg 2220
gtcgtgttgt gggagatttt cacctatggc aaacagccct ggtaccagct gtcaaacaat 2280
gaggtgatag agtgtatcac tcagggccga gtcctgcagc gaccccgcac gtgcccccag 2340
gaggtgtatg agctgatgct ggggtgctgg cagcgagagc cccacatgag gaagaacatc 2400
aagggcatcc ataccctcct tcagaacttg gccaaggcat ctccggtcga actggacatt 2460
ctaggc 2466
<210> 15
<211> 229
<212> PRT
<213> Homo sapiens
<400> 15
Ala Pro Met Lys Glu Ala Asn Ile Arg Gly Gln Gly Gly Leu Ala Tyr
1 5 10 15
Pro Gly Val Arg Thr His Gly Thr Leu Glu Ser Val Asn Gly Pro Lys
20 25 30
Ala Gly Ser Arg Gly Leu Thr Ser Leu Ala Asp Thr Phe Glu His Val
35 40 45
Ile Glu Glu Leu Leu Asp Glu Asp Gln Lys Val Arg Pro Asn Glu Glu
50 55 60
Asn Asn Lys Asp Ala Asp Leu Tyr Thr Ser Arg Val Met Leu Ser Ser
65 70 75 80
Gln Val Pro Leu Glu Pro Pro Leu Leu Phe Leu Leu Glu Glu Tyr Lys
85 90 95
Asn Tyr Leu Asp Ala Ala Asn Met Ser Met Arg Val Arg Arg His Ser
100 105 110
Asp Pro Ala Arg Arg Gly Glu Leu Ser Val Cys Asp Ser Ile Ser Glu
115 120 125
Trp Val Thr Ala Ala Asp Lys Lys Thr Ala Val Asp Met Ser Gly Gly
130 135 140
Thr Val Thr Val Leu Glu Lys Val Pro Val Ser Lys Gly Gln Leu Lys
145 150 155 160
Gln Tyr Phe Tyr Glu Thr Lys Cys Asn Pro Met Gly Tyr Thr Lys Glu
165 170 175
Gly Cys Arg Gly Ile Asp Lys Arg His Trp Asn Ser Gln Cys Arg Thr
180 185 190
Thr Gln Ser Tyr Val Arg Ala Leu Thr Met Asp Ser Lys Lys Arg Ile
195 200 205
Gly Trp Arg Phe Ile Arg Ile Asp Thr Ser Cys Val Cys Thr Leu Thr
210 215 220
Ile Lys Arg Gly Arg
225
<210> 16
<211> 690
<212> DNA
<213> Homo sapiens
<400> 16
gcccccatga aagaagcaaa catccgagga caaggtggct tggcctaccc aggtgtgcgg 60
acccatggga ctctggagag cgtgaatggg cccaaggcag gttcaagagg cttgacatca 120
ttggctgaca ctttcgaaca cgtgatagaa gagctgttgg atgaggacca gaaagttcgg 180
cccaatgaag aaaacaataa ggacgcagac ttgtacacgt ccagggtgat gctcagtagt 240
caagtgcctt tggagcctcc tcttctcttt ctgctggagg aatacaaaaa ttacctagat 300
gctgcaaaca tgtccatgag ggtccggcgc cactctgacc ctgcccgccg aggggagctg 360
agcgtgtgtg acagtattag tgagtgggta acggcggcag acaaaaagac tgcagtggac 420
atgtcgggcg ggacggtcac agtccttgaa aaggtccctg tatcaaaagg ccaactgaag 480
caatacttct acgagaccaa gtgcaatccc atgggttaca caaaagaagg ctgcaggggc 540
atagacaaaa ggcattggaa ctcccagtgc cgaactaccc agtcgtacgt gcgggccctt 600
accatggata gcaaaaagag aattggctgg cgattcataa ggatagacac ttcttgtgta 660
tgtacattga ccattaaaag gggaagatag 690
<210> 17
<211> 229
<212> PRT
<213> Homo sapiens
<400> 17
Ala Pro Met Lys Glu Ala Asn Ile Arg Gly Gln Gly Gly Leu Ala Tyr
1 5 10 15
Pro Gly Val Arg Thr His Gly Thr Leu Glu Ser Val Asn Gly Pro Lys
20 25 30
Ala Gly Ser Arg Gly Leu Thr Ser Leu Ala Asp Thr Phe Glu His Met
35 40 45
Ile Glu Glu Leu Leu Asp Glu Asp Gln Lys Val Arg Pro Asn Glu Glu
50 55 60
Asn Asn Lys Asp Ala Asp Leu Tyr Thr Ser Arg Val Met Leu Ser Ser
65 70 75 80
Gln Val Pro Leu Glu Pro Pro Leu Leu Phe Leu Leu Glu Glu Tyr Lys
85 90 95
Asn Tyr Leu Asp Ala Ala Asn Met Ser Met Arg Val Arg Arg His Ser
100 105 110
Asp Pro Ala Arg Arg Gly Glu Leu Ser Val Cys Asp Ser Ile Ser Glu
115 120 125
Trp Val Thr Ala Ala Asp Lys Lys Thr Ala Val Asp Met Ser Gly Gly
130 135 140
Thr Val Thr Val Leu Glu Lys Val Pro Val Ser Lys Gly Gln Leu Lys
145 150 155 160
Gln Tyr Phe Tyr Glu Thr Lys Cys Asn Pro Met Gly Tyr Thr Lys Glu
165 170 175
Gly Cys Arg Gly Ile Asp Lys Arg His Trp Asn Ser Gln Cys Arg Thr
180 185 190
Thr Gln Ser Tyr Val Arg Ala Leu Thr Met Asp Ser Lys Lys Arg Ile
195 200 205
Gly Trp Arg Phe Ile Arg Ile Asp Thr Ser Cys Val Cys Thr Leu Thr
210 215 220
Ile Lys Arg Gly Arg
225
<210> 18
<211> 119
<212> PRT
<213> Homo sapiens
<400> 18
His Ser Asp Pro Ala Arg Arg Gly Glu Leu Ser Val Cys Asp Ser Ile
1 5 10 15
Ser Glu Trp Val Thr Ala Ala Asp Lys Lys Thr Ala Val Asp Met Ser
20 25 30
Gly Gly Thr Val Thr Val Leu Glu Lys Val Pro Val Ser Lys Gly Gln
35 40 45
Leu Lys Gln Tyr Phe Tyr Glu Thr Lys Cys Asn Pro Met Gly Tyr Thr
50 55 60
Lys Glu Gly Cys Arg Gly Ile Asp Lys Arg His Trp Asn Ser Gln Cys
65 70 75 80
Arg Thr Thr Gln Ser Tyr Val Arg Ala Leu Thr Met Asp Ser Lys Lys
85 90 95
Arg Ile Gly Trp Arg Phe Ile Arg Ile Asp Thr Ser Cys Val Cys Thr
100 105 110
Leu Thr Ile Lys Arg Gly Arg
115
<210> 19
<211> 744
<212> DNA
<213> Homo sapiens
<400> 19
atgaccatcc ttttccttac tatggttatt tcatactttg gttgcatgaa ggctgccccc 60
atgaaagaag caaacatccg aggacaaggt ggcttggcct acccaggtgt gcggacccat 120
gggactctgg agagcgtgaa tgggcccaag gcaggttcaa gaggcttgac atcattggct 180
gacactttcg aacacgtgat agaagagctg ttggatgagg accagaaagt tcggcccaat 240
gaagaaaaca ataaggacgc agacttgtac acgtccaggg tgatgctcag tagtcaagtg 300
cctttggagc ctcctcttct ctttctgctg gaggaataca aaaattacct agatgctgca 360
aacatgtcca tgagggtccg gcgccactct gaccctgccc gccgagggga gctgagcgtg 420
tgtgacagta ttagtgagtg ggtaacggcg gcagacaaaa agactgcagt ggacatgtcg 480
ggcgggacgg tcacagtcct tgaaaaggtc cctgtatcaa aaggccaact gaagcaatac 540
ttctacgaga ccaagtgcaa tcccatgggt tacacaaaag aaggctgcag gggcatagac 600
aaaaggcatt ggaactccca gtgccgaact acccagtcgt acgtgcgggc ccttaccatg 660
gatagcaaaa agagaattgg ctggcgattc ataaggatag acacttcttg tgtatgtaca 720
ttgaccatta aaaggggaag atag 744
<210> 20
<211> 18
<212> PRT
<213> Homo sapiens
<400> 20
Met Thr Ile Leu Phe Leu Thr Met Val Ile Ser Tyr Phe Gly Cys Met
1 5 10 15
Lys Ala
<210> 21
<211> 54
<212> DNA
<213> Homo sapiens
<400> 21
atgaccatcc ttttccttac tatggttatt tcatacttcg gttgcatgaa ggcg 54
<210> 22
<211> 26
<212> PRT
<213> Homo sapiens
<400> 22
Met Phe His Gln Val Arg Arg Val Met Thr Ile Leu Phe Leu Thr Met
1 5 10 15
Val Ile Ser Tyr Phe Gly Cys Met Lys Ala
20 25
<210> 23
<211> 78
<212> DNA
<213> Homo sapiens
<400> 23
atgttccacc aggtgagaag agtgatgacc atccttttcc ttactatggt tatttcatac 60
ttcggttgca tgaaggcg 78
<210> 24
<211> 33
<212> PRT
<213> Homo sapiens
<400> 24
Met Gln Ser Arg Glu Glu Glu Trp Phe His Gln Val Arg Arg Val Met
1 5 10 15
Thr Ile Leu Phe Leu Thr Met Val Ile Ser Tyr Phe Gly Cys Met Lys
20 25 30
Ala
<210> 25
<211> 99
<212> DNA
<213> Homo sapiens
<400> 25
atgcagagcc gggaagagga atggttccac caggtgagaa gagtgatgac catccttttc 60
cttactatgg ttatttcata cttcggttgc atgaaggcg 99
<210> 26
<211> 47
<212> PRT
<213> Homo sapiens
<400> 26
Met Leu Cys Ala Ile Ser Leu Cys Ala Arg Val Arg Lys Leu Arg Ser
1 5 10 15
Ala Gly Arg Cys Gly Lys Phe His Gln Val Arg Arg Val Met Thr Ile
20 25 30
Leu Phe Leu Thr Met Val Ile Ser Tyr Phe Gly Cys Met Lys Ala
35 40 45
<210> 27
<211> 141
<212> DNA
<213> Homo sapiens
<400> 27
atgctctgtg cgatttcatt gtgtgctcgc gttcgcaagc tccgtagtgc aggaaggtgc 60
gggaagttcc accaggtgag aagagtgatg accatccttt tccttactat ggttatttca 120
tacttcggtt gcatgaaggc g 141
<210> 28
<211> 100
<212> PRT
<213> Homo sapiens
<400> 28
Met Cys Gly Ala Thr Ser Phe Leu His Glu Cys Thr Arg Leu Ile Leu
1 5 10 15
Val Thr Thr Gln Asn Ala Glu Phe Leu Gln Lys Gly Leu Gln Val His
20 25 30
Thr Cys Phe Gly Val Tyr Pro His Ala Ser Val Trp His Asp Cys Ala
35 40 45
Ser Gln Lys Lys Gly Cys Ala Val Tyr Leu His Val Ser Val Glu Phe
50 55 60
Asn Lys Leu Ile Pro Glu Asn Gly Phe Ile Lys Phe His Gln Val Arg
65 70 75 80
Arg Val Met Thr Ile Leu Phe Leu Thr Met Val Ile Ser Tyr Phe Gly
85 90 95
Cys Met Lys Ala
100
<210> 29
<211> 300
<212> DNA
<213> Homo sapiens
<400> 29
atgtgtggag ccaccagttt tctccatgag tgcacaaggt taatccttgt tactactcag 60
aatgctgagt ttctacagaa agggttgcag gtccacacat gttttggcgt ctacccacac 120
gcttctgtat ggcatgactg tgcatcccag aagaagggct gtgctgtgta cctccacgtt 180
tcagtggaat ttaacaaact gatccctgaa aatggtttca taaagttcca ccaggtgaga 240
agagtgatga ccatcctttt ccttactatg gttatttcat acttcggttg catgaaggcg 300
<210> 30
<211> 20
<212> PRT
<213> Homo sapiens
<400> 30
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser
20
<210> 31
<211> 60
<212> DNA
<213> Homo sapiens
<400> 31
atgtacagga tgcaactcct gtcttgcatt gcactaagtc ttgcacttgt cacaaacagt 60
<210> 32
<211> 22
<212> PRT
<213> Homo sapiens
<400> 32
Met Lys Arg Arg Val Met Ile Ile Leu Phe Leu Thr Met Val Ile Ser
1 5 10 15
Tyr Phe Gly Cys Met Lys
20
<210> 33
<211> 70
<212> DNA
<213> Homo sapiens
<400> 33
atgaaaagaa gagtgatgat catccttttc cttactatgg ttatttcata cttcggttgc 60
atgaagagcg 70
<210> 34
<211> 20
<212> PRT
<213> Homo sapiens
<400> 34
Met Arg Arg Met Gln Leu Leu Leu Leu Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser
20
<210> 35
<211> 60
<212> DNA
<213> Homo sapiens
<400> 35
atgaggagga tgcaactcct gctcctgatt gcactaagtc ttgcacttgt cacaaacagt 60
<210> 36
<211> 21
<212> PRT
<213> Homo sapiens
<400> 36
Met Arg Arg Met Gln Leu Leu Leu Leu Thr Met Val Ile Ser Tyr Phe
1 5 10 15
Gly Cys Met Lys Ala
20
<210> 37
<211> 63
<212> DNA
<213> Homo sapiens
<400> 37
atgaggagga tgcaactcct gctcctgact atggttattt catacttcgg ttgcatgaag 60
gcg 63
<210> 38
<211> 18
<212> PRT
<213> Homo sapiens
<400> 38
Met Arg Ile Leu Leu Leu Thr Met Val Ile Ser Tyr Phe Gly Cys Met
1 5 10 15
Lys Ala
<210> 39
<211> 54
<212> PRT
<213> Homo sapiens
<400> 39
Ala Thr Gly Ala Gly Ala Ala Thr Cys Cys Thr Thr Cys Thr Thr Cys
1 5 10 15
Thr Thr Ala Cys Thr Ala Thr Gly Gly Thr Thr Ala Thr Thr Thr Cys
20 25 30
Ala Thr Ala Cys Thr Thr Cys Gly Gly Thr Thr Gly Cys Ala Thr Gly
35 40 45
Ala Ala Gly Gly Cys Gly
50
<210> 40
<211> 19
<212> PRT
<213> Homo sapiens
<400> 40
Met Arg Arg Ile Leu Phe Leu Thr Met Val Ile Ser Tyr Phe Gly Cys
1 5 10 15
Met Lys Ala
<210> 41
<211> 57
<212> DNA
<213> Homo sapiens
<400> 41
atgagaagaa tccttttcct tactatggtt atttcatact tcggttgcat gaaggcg 57
<210> 42
<211> 18
<212> PRT
<213> Homo sapiens
<400> 42
Met Arg Arg Phe Leu Phe Leu Leu Val Ile Ser Tyr Phe Gly Cys Met
1 5 10 15
Lys Ala
<210> 43
<211> 54
<212> DNA
<213> Homo sapiens
<400> 43
atgaggaggt tccttttcct tcttgttatt tcatacttcg gttgcatgaa ggcg 54
<210> 44
<211> 15
<212> PRT
<213> Homo sapiens
<400> 44
Met Arg Arg Phe Leu Phe Leu Leu Tyr Phe Gly Cys Met Lys Ala
1 5 10 15
<210> 45
<211> 45
<212> DNA
<213> Homo sapiens
<400> 45
atgaggaggt tccttttcct tctttacttc ggttgcatga aggcg 45
<210> 46
<211> 130
<212> DNA
<213> Homo sapiens
<400> 46
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcgtcg ggcgaccttt 60
ggtcgcccgg cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact 120
aggggttcct 130
<210> 47
<211> 141
<212> DNA
<213> Homo sapiens
<400> 47
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgcag ctgcctgcag g 141
<210> 48
<211> 584
<212> DNA
<213> Homo sapiens
<400> 48
gcgttacata acttacggta aatggcccgc ctggctgacc gcccaacgac ccccgcccat 60
tgacgtcaat aatgacgtat gttcccatag taacgccaat agggactttc cattgacgtc 120
aatgggtgga ctatttacgg taaactgccc acttggcagt acatcaagtg tatcatatgc 180
caagtacgcc ccctattgac gtcaatgacg gtaaatggcc cgcctggcat tatgcccagt 240
acatgacctt atgggacttt cctacttggc agtacatcta cgtattagtc atcgctatta 300
ccatggtcga ggtgagcccc acgttctgct tcactctccc catctccccc ccctccccac 360
ccccaatttt gtatttattt attttttaat tattttgtgc agcgatgggg gcgggggggg 420
ggggggggcg cgcgccaggc ggggcggggc ggggcgaggg gcggggcggg gcgaggcgga 480
gaggtgcggc ggcagccaat cagagcggcg cgctccgaaa gtttcctttt atggcgaggc 540
ggcggcggcg gcggccctat aaaaagcgaa gcgcgcggcg ggcg 584
<210> 49
<211> 210
<212> PRT
<213> Homo sapiens
<400> 49
Met Leu Pro Leu Pro Ser Cys Ser Leu Pro Ile Leu Leu Leu Phe Leu
1 5 10 15
Leu Pro Ser Val Pro Ile Glu Ser Gln Pro Pro Pro Ser Thr Leu Pro
20 25 30
Pro Phe Leu Ala Pro Glu Trp Asp Leu Leu Ser Pro Arg Val Val Leu
35 40 45
Ser Arg Gly Ala Pro Ala Gly Pro Pro Leu Leu Phe Leu Leu Glu Ala
50 55 60
Gly Ala Phe Arg Glu Ser Ala Gly Ala Pro Ala Asn Arg Ser Arg Arg
65 70 75 80
Gly Val Ser Glu Thr Ala Pro Ala Ser Arg Arg Gly Glu Leu Ala Val
85 90 95
Cys Asp Ala Val Ser Gly Trp Val Thr Asp Arg Arg Thr Ala Val Asp
100 105 110
Leu Arg Gly Arg Glu Val Glu Val Leu Gly Glu Val Pro Ala Ala Gly
115 120 125
Gly Ser Pro Leu Arg Gln Tyr Phe Phe Glu Thr Arg Cys Lys Ala Asp
130 135 140
Asn Ala Glu Glu Gly Gly Pro Gly Ala Gly Gly Gly Gly Cys Arg Gly
145 150 155 160
Val Asp Arg Arg His Trp Val Ser Glu Cys Lys Ala Lys Gln Ser Tyr
165 170 175
Val Arg Ala Leu Thr Ala Asp Ala Gln Gly Arg Val Gly Trp Arg Trp
180 185 190
Ile Arg Ile Asp Thr Ala Cys Val Cys Thr Leu Leu Ser Arg Thr Gly
195 200 205
Arg Ala
210
<210> 50
<211> 630
<212> DNA
<213> Homo sapiens
<400> 50
atgctccctc tcccctcatg ctccctcccc atcctcctcc ttttcctcct ccccagtgtg 60
ccaattgagt cccaaccccc accctcaaca ttgccccctt ttctggcccc tgagtgggac 120
cttctctccc cccgagtagt cctgtctagg ggtgcccctg ctgggccccc tctgctcttc 180
ctgctggagg ctggggcctt tcgggagtca gcaggtgccc cggccaaccg cagccggcgt 240
ggggtgagcg aaactgcacc agcgagtcgt cggggtgagc tggctgtgtg cgatgcagtc 300
agtggctggg tgacagaccg ccggaccgct gtggacttgc gtgggcgcga ggtggaggtg 360
ttgggcgagg tgcctgcagc tggcggcagt cccctccgcc agtacttctt tgaaacccgc 420
tgcaaggctg ataacgctga ggaaggtggc ccgggggcag gtggaggggg ctgccgggga 480
gtggacagga ggcactgggt atctgagtgc aaggccaagc agtcctatgt gcgggcattg 540
accgctgatg cccagggccg tgtgggctgg cgatggattc gaattgacac tgcctgcgtc 600
tgcacactcc tcagccggac tggccgggcc 630
<210> 51
<211> 24
<212> PRT
<213> Homo sapiens
<400> 51
Met Leu Pro Leu Pro Ser Cys Ser Leu Pro Ile Leu Leu Leu Phe Leu
1 5 10 15
Leu Pro Ser Val Pro Ile Glu Ser
20
<210> 52
<211> 72
<212> DNA
<213> Homo sapiens
<400> 52
atgctccctc tcccctcatg ctccctcccc atcctcctcc ttttcctcct ccccagtgtg 60
ccaattgagt cc 72
<210> 53
<211> 56
<212> PRT
<213> Homo sapiens
<400> 53
Gln Pro Pro Pro Ser Thr Leu Pro Pro Phe Leu Ala Pro Glu Trp Asp
1 5 10 15
Leu Leu Ser Pro Arg Val Val Leu Ser Arg Gly Ala Pro Ala Gly Pro
20 25 30
Pro Leu Leu Phe Leu Leu Glu Ala Gly Ala Phe Arg Glu Ser Ala Gly
35 40 45
Ala Pro Ala Asn Arg Ser Arg Arg
50 55
<210> 54
<211> 168
<212> DNA
<213> Homo sapiens
<400> 54
caacccccac cctcaacatt gccccctttt ctggcccctg agtgggacct tctctccccc 60
cgagtagtcc tgtctagggg tgcccctgct gggccccctc tgctcttcct gctggaggct 120
ggggcctttc gggagtcagc aggtgccccg gccaaccgca gccggcgt 168
<210> 55
<211> 130
<212> PRT
<213> Homo sapiens
<400> 55
Gly Val Ser Glu Thr Ala Pro Ala Ser Arg Arg Gly Glu Leu Ala Val
1 5 10 15
Cys Asp Ala Val Ser Gly Trp Val Thr Asp Arg Arg Thr Ala Val Asp
20 25 30
Leu Arg Gly Arg Glu Val Glu Val Leu Gly Glu Val Pro Ala Ala Gly
35 40 45
Gly Ser Pro Leu Arg Gln Tyr Phe Phe Glu Thr Arg Cys Lys Ala Asp
50 55 60
Asn Ala Glu Glu Gly Gly Pro Gly Ala Gly Gly Gly Gly Cys Arg Gly
65 70 75 80
Val Asp Arg Arg His Trp Val Ser Glu Cys Lys Ala Lys Gln Ser Tyr
85 90 95
Val Arg Ala Leu Thr Ala Asp Ala Gln Gly Arg Val Gly Trp Arg Trp
100 105 110
Ile Arg Ile Asp Thr Ala Cys Val Cys Thr Leu Leu Ser Arg Thr Gly
115 120 125
Arg Ala
130
<210> 56
<211> 390
<212> DNA
<213> Homo sapiens
<400> 56
ggggtgagcg aaactgcacc agcgagtcgt cggggtgagc tggctgtgtg cgatgcagtc 60
agtggctggg tgacagaccg ccggaccgct gtggacttgc gtgggcgcga ggtggaggtg 120
ttgggcgagg tgcctgcagc tggcggcagt cccctccgcc agtacttctt tgaaacccgc 180
tgcaaggctg ataacgctga ggaaggtggc ccgggggcag gtggaggggg ctgccgggga 240
gtggacagga ggcactgggt atctgagtgc aaggccaagc agtcctatgt gcgggcattg 300
accgctgatg cccagggccg tgtgggctgg cgatggattc gaattgacac tgcctgcgtc 360
tgcacactcc tcagccggac tggccgggcc 390
<210> 57
<211> 592
<212> DNA
<213> Homo sapiens
<400> 57
aatcaacctc tggattacaa aatttgtgaa agattgactg gtattcttaa ctatgttgct 60
ccttttacgc tatgtggata cgctgcttta atgcctttgt atcatgctat tgcttcccgt 120
atggctttca ttttctcctc cttgtataaa tcctggttgc tgtctcttta tgaggagttg 180
tggcccgttg tcaggcaacg tggcgtggtg tgcactgtgt ttgctgacgc aacccccact 240
ggttggggca ttgccaccac ctgtcagctc ctttccggga ctttcgcttt ccccctccct 300
attgccacgg cggaactcat cgccgcctgc cttgcccgct gctggacagg ggctcggctg 360
ttgggcactg acaattccgt ggtgttgtcg gggaagctga cgtcctttcc atggctgctc 420
gcctgtgttg ccacctggat tctgcgcggg acgtccttct gctacgtccc ttcggccctc 480
aatccagcgg accttccttc ccgcggcctg ctgccggctc tgcggcctct tccgcgtctt 540
cgccttcgcc ctcagacgag tcggatctcc ctttgggccg cctccccgcc tg 592
<210> 58
<211> 247
<212> DNA
<213> Homo sapiens
<400> 58
aatcaacctc tggattacaa aatttgtgaa agattgactg gtattcttaa ctatgttgct 60
ccttttacgc tatgtggata cgctgcttta atgcctttgt atcatgctat tgcttcccgt 120
atggctttca ttttctcctc cttgtataaa tcctggttag ttcttgccac ggcggaactc 180
atcgccgcct gccttgcccg ctgctggaca ggggctcggc tgttgggcac tgacaattcc 240
gtggtgt 247
<210> 59
<211> 224
<212> DNA
<213> Homo sapiens
<400> 59
agcagacatg ataagataca ttgatgagtt tggacaaacc acaactagaa tgcagtgaaa 60
aaaatgcttt atttgtgaaa tttgtgatgc tattgcttta tttgtaacca ttataagctg 120
caataaacaa gttaacaaca acaattgcat tcattttatg tttcaggttc agggggaggt 180
gtgggaggtt ttttaaagca agtaaaacct ctacaaatgt ggta 224
<210> 60
<211> 18
<212> PRT
<213> Homo sapiens
<400> 60
Met Thr Ile Leu Phe Leu Thr Met Val Ile Ser Tyr Phe Gly Cys Met
1 5 10 15
Lys Ala
<210> 61
<211> 54
<212> DNA
<213> Homo sapiens
<400> 61
atgaccatcc ttttccttac tatggttatt tcatacttcg gttgcatgaa ggcg 54
<210> 62
<211> 2
<212> PRT
<213> Homo sapiens
<400> 62
Met Arg
1
<210> 63
<211> 6
<212> DNA
<213> Homo sapiens
<400> 63
atgaga 6
<210> 64
<211> 3
<212> PRT
<213> Homo sapiens
<400> 64
Met Arg Arg
1
<210> 65
<211> 9
<212> DNA
<213> Homo sapiens
<400> 65
atgagaaga 9
<210> 66
<211> 4
<212> PRT
<213> Homo sapiens
<400> 66
Met Arg Arg Arg
1
<210> 67
<211> 12
<212> DNA
<213> Homo sapiens
<400> 67
atgagaagaa ga 12
<210> 68
<211> 2
<212> PRT
<213> Homo sapiens
<400> 68
Met Lys
1
<210> 69
<211> 6
<212> DNA
<213> Homo sapiens
<400> 69
atgaaa 6
<210> 70
<211> 3
<212> PRT
<213> Homo sapiens
<400> 70
Met Lys Lys
1
<210> 71
<211> 9
<212> DNA
<213> Homo sapiens
<400> 71
atgaaaaka 9
<210> 72
<211> 4
<212> PRT
<213> Homo sapiens
<400> 72
Met Lys Lys Lys
1
<210> 73
<211> 12
<212> DNA
<213> Homo sapiens
<400> 73
atgaaaaaaa aa 12
<210> 74
<211> 4
<212> PRT
<213> Homo sapiens
<400> 74
Met Lys Arg Arg
1
<210> 75
<211> 12
<212> DNA
<213> Homo sapiens
<400> 75
atgaaaagaa ga 12
<210> 76
<211> 4
<212> PRT
<213> Homo sapiens
<400> 76
Met Arg Lys Arg
1
<210> 77
<211> 12
<212> DNA
<213> Homo sapiens
<400> 77
atgagaaaaa ga 12
<210> 78
<211> 4
<212> PRT
<213> Homo sapiens
<400> 78
Met Arg Arg Lys
1
<210> 79
<211> 12
<212> DNA
<213> Homo sapiens
<400> 79
atgagaagaa aa 12
<210> 80
<211> 4
<212> PRT
<213> Homo sapiens
<400> 80
Met Lys Lys Arg
1
<210> 81
<211> 12
<212> DNA
<213> Homo sapiens
<400> 81
atgaaaaaaa ga 12
<210> 82
<211> 4
<212> PRT
<213> Homo sapiens
<400> 82
Phe Leu Phe Leu
1
<210> 83
<211> 12
<212> DNA
<213> Homo sapiens
<400> 83
ttccttttcc tt 12
<210> 84
<211> 4
<212> PRT
<213> Homo sapiens
<400> 84
Phe Phe Phe Leu
1
<210> 85
<211> 12
<212> DNA
<213> Homo sapiens
<400> 85
ttcttcttcc tt 12
<210> 86
<211> 4
<212> PRT
<213> Homo sapiens
<400> 86
Phe Ile Phe Leu
1
<210> 87
<211> 12
<212> DNA
<213> Homo sapiens
<400> 87
ttcatcttcc tt 12
<210> 88
<211> 4
<212> PRT
<213> Homo sapiens
<400> 88
Phe Ile Phe Ile
1
<210> 89
<211> 12
<212> DNA
<213> Homo sapiens
<400> 89
ttcatcttca tc 12
<210> 90
<211> 4
<212> PRT
<213> Homo sapiens
<400> 90
Phe Val Phe Ile
1
<210> 91
<211> 12
<212> DNA
<213> Homo sapiens
<400> 91
ttcgttttca tc 12
<210> 92
<211> 4
<212> PRT
<213> Homo sapiens
<400> 92
Phe Val Phe Val
1
<210> 93
<211> 12
<212> DNA
<213> Homo sapiens
<400> 93
ttcgttttcg tt 12
<210> 94
<211> 4
<212> PRT
<213> Homo sapiens
<400> 94
Phe Leu Phe Val
1
<210> 95
<211> 12
<212> DNA
<213> Homo sapiens
<400> 95
ttccttttcg tt 12
<210> 96
<211> 4
<212> PRT
<213> Homo sapiens
<400> 96
Phe Ile Phe Val
1
<210> 97
<211> 12
<212> DNA
<213> Homo sapiens
<400> 97
ttcatcttcg tt 12
<210> 98
<211> 4
<212> PRT
<213> Homo sapiens
<400> 98
Phe Phe Phe Ile
1
<210> 99
<211> 12
<212> DNA
<213> Homo sapiens
<400> 99
ttcttcttca tc 12
<210> 100
<211> 4
<212> PRT
<213> Homo sapiens
<400> 100
Phe Phe Phe Val
1
<210> 101
<211> 12
<212> DNA
<213> Homo sapiens
<400> 101
ttcttcttcg tt 12
<210> 102
<211> 5
<212> PRT
<213> Homo sapiens
<400> 102
Phe Ile Leu Phe Leu
1 5
<210> 103
<211> 15
<212> DNA
<213> Homo sapiens
<400> 103
ttcatccttt tcctt 15
<210> 104
<211> 1203
<212> DNA
<213> Homo sapiens
<400> 104
atgactatcc tgtttctgac aatggttatt agctatttcg gttgcatgaa ggctcacagt 60
gatcccgcac gccgcggaga acttagcgtg tgcgacagca tcagcgagtg ggtcaccgcc 120
gccgataaga agaccgctgt ggatatgtcc ggcgggaccg tcactgtact cgaaaaagtt 180
ccagtgagca aaggccaact gaaacaatat ttctatgaaa ctaagtgcaa ccccatgggg 240
tacaccaagg agggctgccg gggaatcgac aagagacact ggaattccca gtgccggacc 300
actcagagct acgtccgcgc cttgacgatg gattcaaaga agcgcatcgg atggcggttc 360
ataagaatcg acaccagttg tgtgtgcacg ctgacgataa aacgggggcg ggcccccgtg 420
aagcagaccc tgaactttga tttgctcaag ttggcggggg atgtggaaag caatcccggg 480
ccaatggtga gcaagggcga ggagctgttc accggcgttg tgccaatact ggttgagttg 540
gatggcgatg tcaacggaca caaatttagc gtaagcgggg agggagaggg cgacgccaca 600
tatggcaagc tgaccctgaa gttcatttgc acgaccggca aattgcccgt cccttggccc 660
acacttgtga cgaccctgac ttatggcgta cagtgcttca gcaggtaccc tgatcatatg 720
aagcaacacg acttctttaa gagtgccatg ccagagggat acgtccagga aagaaccata 780
ttcttcaaag atgatggaaa ttacaaaacc cgggcagagg tcaagtttga aggcgacacc 840
ctggtgaaca ggatcgaact caaaggcatc gatttcaaag aggacggaaa catcctcgga 900
cacaaactgg aatacaatta caacagccac aacgtctaca tcatggcaga taaacaaaag 960
aacggtatta aagtgaactt caagatccgg cacaacatcg aagacggctc cgtccagctt 1020
gccgaccact accagcaaaa taccccgatc ggcgacggcc ccgttctcct ccccgataat 1080
cactacctga gtacacagtc agccttgagc aaagacccta atgaaaagcg ggaccacatg 1140
gttttgctgg agttcgttac cgcagcgggt attacgctgg gtatggacga gctttacaag 1200
taa 1203
<210> 105
<211> 1203
<212> DNA
<213> Homo sapiens
<400> 105
atgactatcc tgtttctgac aatggttatt agctatttcg gttgcatgaa ggctcacagt 60
gatcccgcac gccgcggaga acttagcgtg tgcgacagca tcagcgagtg ggtcaccgcc 120
gccgataaga agaccgctgt ggatatgtcc ggcgggaccg tcactgtact cgaaaaagtt 180
ccagtgagca aaggccaact gaaacaatat ttctatgaaa ctaagtgcaa ccccatgggg 240
tacaccaagg agggctgccg gggaatcgac aagagacact ggaattccca gtgccggacc 300
actcagagct acgtccgcgc cttgacgatg gattcaaaga agcgcatcgg atggcggttc 360
ataagaatcg acaccagttg tgtgtgcacg ctgacgataa aacgggggcg ggcccctgtc 420
aaacaaaccc tcaattttga cttgctgaag cttgctgggg atgtcgagtc cgctgccgcg 480
gctatggtga gcaagggcga ggagctgttc accggcgttg tgccaatact ggttgagttg 540
gatggcgatg tcaacggaca caaatttagc gtaagcgggg agggagaggg cgacgccaca 600
tatggcaagc tgaccctgaa gttcatttgc acgaccggca aattgcccgt cccttggccc 660
acacttgtga cgaccctgac ttatggcgta cagtgcttca gcaggtaccc tgatcatatg 720
aagcaacacg acttctttaa gagtgccatg ccagagggat acgtccagga aagaaccata 780
ttcttcaaag atgatggaaa ttacaaaacc cgggcagagg tcaagtttga aggcgacacc 840
ctggtgaaca ggatcgaact caaaggcatc gatttcaaag aggacggaaa catcctcgga 900
cacaaactgg aatacaatta caacagccac aacgtctaca tcatggcaga taaacaaaag 960
aacggtatta aagtgaactt caagatccgg cacaacatcg aagacggctc cgtccagctt 1020
gccgaccact accagcaaaa taccccgatc ggcgacggcc ccgttctcct ccccgataat 1080
cactacctga gtacacagtc agccttgagc aaagacccta atgaaaagcg ggaccacatg 1140
gttttgctgg agttcgttac cgcagcgggt attacgctgg gtatggacga gctttacaag 1200
taa 1203
<210> 106
<211> 1203
<212> DNA
<213> Homo sapiens
<400> 106
atggtgagca agggcgagga gctgttcacc ggggtggtgc ccatcctggt cgagctggac 60
ggcgacgtaa acggccacaa gttcagcgtg tccggcgagg gcgagggcga tgccacctac 120
ggcaagctga ccctgaagtt catctgcacc accggcaagc tgcccgtgcc ctggcccacc 180
ctcgtgacca ccctgaccta cggcgtgcag tgcttcagcc gctaccccga ccacatgaag 240
cagcacgact tcttcaagtc cgccatgccc gaaggctacg tccaggagcg caccatcttc 300
ttcaaggacg acggcaacta caagacccgc gccgaggtga agttcgaggg cgacaccctg 360
gtgaaccgca tcgagctgaa gggcatcgac ttcaaggagg acggcaacat cctggggcac 420
aagctggagt acaactacaa cagccacaac gtctatatca tggccgacaa gcagaagaac 480
ggcatcaagg tgaacttcaa gatccgccac aacatcgagg acggcagcgt gcagctcgcc 540
gaccactacc agcagaacac ccccatcggc gacggccccg tgctgctgcc cgacaaccac 600
tacctgagca cccagtccgc cctgagcaag gaccccaacg agaagcgcga tcacatggtc 660
ctgctggagt tcgtgaccgc cgccgggatc actctcggca tggacgagct gtacaaggct 720
cccgttaaac aaactctgaa cttcgacctg ctgaagctgg ctggagacgt ggagtccaac 780
cctggaccta tgaccatcct tttccttact atggttattt catacttcgg ttgcatgaag 840
gcgcactccg accctgcccg ccgtggggag ctgagcgtgt gtgacagtat tagcgagtgg 900
gtcacagcgg cagataaaaa gactgcagtg gacatgtctg gcgggacggt cacagtccta 960
gagaaagtcc cggtatccaa aggccaactg aagcagtatt tctacgagac caagtgtaat 1020
cccatgggtt acaccaagga aggctgcagg ggcatagaca aaaggcactg gaactcgcaa 1080
tgccgaacta cccaatcgta tgttcgggcc cttactatgg atagcaaaaa gagaattggc 1140
tggcgattca taaggataga cacttcctgt gtatgtacac tgaccattaa aaggggaaga 1200
tag 1203
<210> 107
<211> 2940
<212> DNA
<213> Homo sapiens
<400> 107
atgagcccat ggctgaagtg gcacggacca gcaatggcaa gactgtgggg cctgtgcctg 60
ctggtgctgg gcttctggag agccagcctg gcctgtccaa cctcctgcaa gtgtagctcc 120
gccaggatct ggtgcacaga gccttctcca ggcatcgtgg cctttccccg cctggagcct 180
aacagcgtgg atcccgagaa tatcaccgag atcctgatcg ccaaccagaa gcggctggag 240
atcatcaatg aggacgatgt ggaggcctac gtgggcctga gaaacctgac aatcgtggac 300
tccggcctga agttcgtggc ctataaggcc tttctgaaga actctaatct gaggcacatc 360
aacttcaccc gcaataagct gacatctctg agccggagac actttcggca cctggatctg 420
tccgacctga tcctgaccgg caatccattc acatgctctt gtgacatcat gtggctgaag 480
accctgcagg agacaaagtc tagccccgat acccaggacc tgtactgtct gaacgagtcc 540
tctaagaata tgcctctggc caacctgcag atccctaatt gtggactgcc aagcgcccgg 600
ctggccgcac ctaacctgac agtggaggag ggcaagtccg tgacactgtc ctgttctgtg 660
ggcggcgatc ccctgcctac cctgtattgg gacgtgggca acctggtgtc taagcacatg 720
aatgagacct cccacacaca gggctctctg agaatcacaa atatcagctc cgacgatagc 780
ggcaagcaga tctcttgcgt ggcagagaac ctggtgggag aggatcagga cagcgtgaat 840
ctgaccgtgc acttcgcccc caccatcaca tttctggagt ctcctaccag cgatcaccac 900
tggtgcatcc ccttcacagt gcggggaaac ccaaagcccg ccctgcagtg gttttacaac 960
ggcgccatcc tgaatgagtc caagtatatc tgtaccaaga tccacgtgac caaccacaca 1020
gagtaccacg gctgcctgca gctggataat cccacccaca tgaacaatgg cgactacaca 1080
ctgatggcca agaacgagta tggcaaggac gagaggcaga tcagcgccca cttcatgggc 1140
cgccctggag tggattatga gaccaaccct aattacccag aggtgctgta tgaggactgg 1200
accacaccta ccgatatcgg cgacaccaca aacaagtcta atgagatccc aagcacagat 1260
gtggccgacc agtctaacag ggagcacctg agcgtgtacg cagtggtggt catcgcctcc 1320
gtggtgggct tctgcctgct ggtcatgctg ctgctgctga agctggcccg ccactctaag 1380
tttggcatga agggcccagc ctccgtgatc tctaatgacg atgacagcgc cagccccctg 1440
caccacatca gcaacggctc caatacccct tctagctccg agggcggccc agatgccgtg 1500
atcatcggca tgacaaagat ccccgtgatc gagaaccctc agtacttcgg catcaccaat 1560
tcccagctga agcctgacac atttgtgcag cacatcaagc ggcacaacat cgtgctgaag 1620
agggaactgg gagagggagc cttcggcaag gtgtttctgg ccgagtgcta taacctgtgc 1680
ccagagcagg ataagatcct ggtggccgtg aagaccctga aggatgccag cgacaacgcc 1740
cggaaggact tccacagaga ggccgagctg ctgacaaatc tgcagcacga gcacatcgtg 1800
aagttttacg gcgtgtgcgt ggagggcgac cctctgatca tggtgttcga gtatatgaag 1860
cacggcgatc tgaacaagtt tctgagagca cacggaccag atgccgtgct gatggcagag 1920
ggaaatcccc ctaccgagct gacacagtct cagatgctgc acattgcaca gcagattgca 1980
gcaggaatgg tgtacctggc cagccagcac ttcgtgcaca gggatctggc aaccagaaac 2040
tgcctggtgg gagagaatct gctggtgaag atcggcgact ttggcatgtc ccgggacgtg 2100
tactctaccg actactatag agtgggcggc cacacaatgc tgcccatcag gtggatgcca 2160
cccgagagca tcatgtatcg caagttcacc acagagtctg acgtgtggag cctgggcgtg 2220
gtgctgtggg agatctttac ctacggcaag cagccttggt atcagctgtc caacaatgaa 2280
gtgatcgagt gtattacaca gggacgcgtg ctgcagaggc cacgcacatg cccccaggag 2340
gtgtacgagc tgatgctggg ctgttggcag cgggagccac acaccagaaa gaacatcaag 2400
agcatccaca cactgctgca gaatctggcc aaggcctccc ccgtgtatct ggacatcctg 2460
ggcagcggag ctactaactt cagcctgctg aagcaggctg gagacgtgga ggagaaccct 2520
ggacctatga gaatccttct tcttactatg gttatttcat acttcggttg catgaaggcg 2580
cactccgacc ctgcccgccg tggggagctg agcgtgtgtg acagtattag cgagtgggtc 2640
acagcggcag ataaaaagac tgcagtggac atgtctggcg ggacggtcac agtcctagag 2700
aaagtcccgg tatccaaagg ccaactgaag cagtatttct acgagaccaa gtgtaatccc 2760
atgggttaca ccaaggaagg ctgcaggggc atagacaaaa ggcactggaa ctcgcaatgc 2820
cgaactaccc aatcgtatgt tcgggccctt actatggata gcaaaaagag aattggctgg 2880
cgattcataa ggatagacac ttcctgtgta tgtacactga ccattaaaag gggaagatag 2940
<210> 108
<211> 2943
<212> DNA
<213> Homo sapiens
<400> 108
atgtcatctt ggatccgctg gcacgggcca gcgatggccc gattgtgggg cttctgctgg 60
cttgttgtag gcttctggcg cgcggcgttc gcgtgtccga cctcttgcaa atgctcagca 120
agccgaattt ggtgctcaga ccctagtcca ggaattgttg cattcccccg actggaacca 180
aactccgtcg acccggagaa tataactgag atatttattg caaatcaaaa acgccttgaa 240
atcattaacg aggatgacgt ggaggcctac gttggtttga gaaatcttac tattgtcgac 300
tccggactta aatttgtagc tcataaagcc ttcctgaaga actctaatct gcagcacatt 360
aatttcacga gaaataagct gaccagcttg tcccggaagc atttccgcca tctcgacctg 420
agcgagctca tactggtcgg aaacccattt acgtgctcct gtgacatcat gtggatcaaa 480
actctgcaag aggcgaaaag tagtccggat acccaagacc tttactgtct taatgaaagc 540
tcaaaaaata tcccgctggc caacctgcag ataccgaact gcggacttcc tagtgcgaat 600
ttggctgccc caaatcttac cgtcgaagaa ggcaaatcaa tcacgctttc ttgttctgta 660
gctggagatc cagtgcctaa tatgtattgg gacgtgggta acctcgtctc aaaacatatg 720
aacgaaacga gccacaccca gggctctttg cggataacaa acatctcctc tgatgattct 780
ggaaagcaaa tcagttgcgt agctgaaaat ctggttggcg aagatcaaga ttcagtcaat 840
ctgacagtcc atttcgcccc aacgatcacc tttctggaga gcccaactag cgatcaccac 900
tggtgtattc cgtttacggt aaaaggaaat ccaaaacctg cactccaatg gttttataat 960
ggagccatct tgaatgaaag caaatatatc tgtactaaaa tccatgtgac gaatcacacc 1020
gagtatcacg ggtgtcttca attggataat ccaacccata tgaataatgg tgattatact 1080
ttgatagcga agaacgaata cggcaaagac gaaaagcaaa tatccgcaca tttcatgggt 1140
tggcctggca tcgacgacgg tgcgaacccg aactacccag atgttattta cgaggattat 1200
gggactgcgg caaacgacat tggcgacacc acaaaccgaa gcaacgagat accaagtact 1260
gacgtcactg acaaaacggg tcgagagcat ttgtctgttt acgccgttgt tgttatcgcc 1320
tcagttgtcg gattttgcct gttggtcatg cttttcctcc tgaagctcgc gcgacattcc 1380
aagtttggca tgaaggggcc agcaagtgtt atatccaatg atgatgatag cgcttctcca 1440
ttgcaccaca taagtaacgg ctcaaacacg ccgtcatcta gtgaaggtgg accagacgcg 1500
gtcattatag ggatgactaa aattcccgta atcgaaaacc ctcagtactt cggcataacc 1560
aacagtcagc ttaaacccga tactttcgtg cagcacatca aaaggcacaa catagtcctc 1620
aagcgcgaac tcggggaggg agccttcgga aaggtctttc ttgctgagtg ctataatttg 1680
tgtcctgagc aggataaaat tcttgtggct gtaaaaactc tcaaagatgc ttccgacaac 1740
gcacggaagg attttcatcg ggaggccgaa ctgttgacga atttgcagca cgagcatata 1800
gtaaagttct acggggtatg tgttgagggg gacccgttga ttatggtctt cgagtatatg 1860
aagcacgggg acctgaacaa atttttgcgc gcccatgggc ctgatgccgt ccttatggca 1920
gaagggaacc ctccaacaga actcacccag agtcagatgt tgcacatagc gcaacagatc 1980
gcggccggca tggtttacct ggccagtcaa cacttcgtgc atagagatct tgccactcgc 2040
aactgtttgg tcggggagaa ccttctggtt aagattggtg actttggtat gtcacgagat 2100
gtgtattcca ctgactatta cagagttggg ggtcatacaa tgcttcctat tcggtggatg 2160
ccccccgaat ccatcatgta cagaaagttc acgacagaga gtgatgtttg gagtctcggc 2220
gtggtgctct gggaaatttt cacatacgga aagcagccgt ggtatcaact tagcaacaat 2280
gaggtgatag agtgtattac acagggtcgg gtgttgcagc gccctcgaac gtgcccacaa 2340
gaagtatatg aacttatgct cgggtgctgg caaagagaac cacatatgag aaaaaatatc 2400
aaggggatac atacattgct tcagaacttg gccaaggcat cacccgtcta cctcgatata 2460
ctgggcagcg gagctactaa cttcagcctg ctgaagcagg ctggagacgt ggaggagaac 2520
cctggaccta tgagaatcct tcttcttact atggttattt catacttcgg ttgcatgaag 2580
gcgcactccg accctgcccg ccgtggggag ctgagcgtgt gtgacagtat tagcgagtgg 2640
gtcacagcgg cagataaaaa gactgcagtg gacatgtctg gcgggacggt cacagtccta 2700
gagaaagtcc cggtatccaa aggccaactg aagcagtatt tctacgagac caagtgtaat 2760
cccatgggtt acaccaagga aggctgcagg ggcatagaca aaaggcactg gaactcgcaa 2820
tgccgaacta cccaatcgta tgttcgggcc cttactatgg atagcaaaaa gagaattggc 2880
tggcgattca taaggataga cacttcctgt gtatgtacac tgaccattaa aaggggaaga 2940
tag 2943
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ | 2016 |
|
RU2734678C2 |
| НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ ЛОКУС TrkB | 2018 |
|
RU2800428C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| УНИВЕРСАЛЬНАЯ ВАКЦИНА НА ОСНОВЕ ОБЩИХ ОПУХОЛЕВЫХ НЕОАНТИГЕНОВ ДЛЯ ПРЕДУПРЕЖДЕНИЯ И ЛЕЧЕНИЯ ФОРМ РАКА С МИКРОСАТЕЛЛИТНОЙ НЕСТАБИЛЬНОСТЬЮ (MSI) | 2018 |
|
RU2792843C2 |
| НЕОАНТИГЕНЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2813924C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| ИММУНОДОМИНАНТНЫЕ БЕЛКИ И ФРАГМЕНТЫ ПРИ РАССЕЯННОМ СКЛЕРОЗЕ | 2019 |
|
RU2828590C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CTLA-4, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2756100C1 |
| ИСКУССТВЕННЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2763798C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ КОМПОЗИЦИЙ T-КЛЕТОК И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2835034C2 |
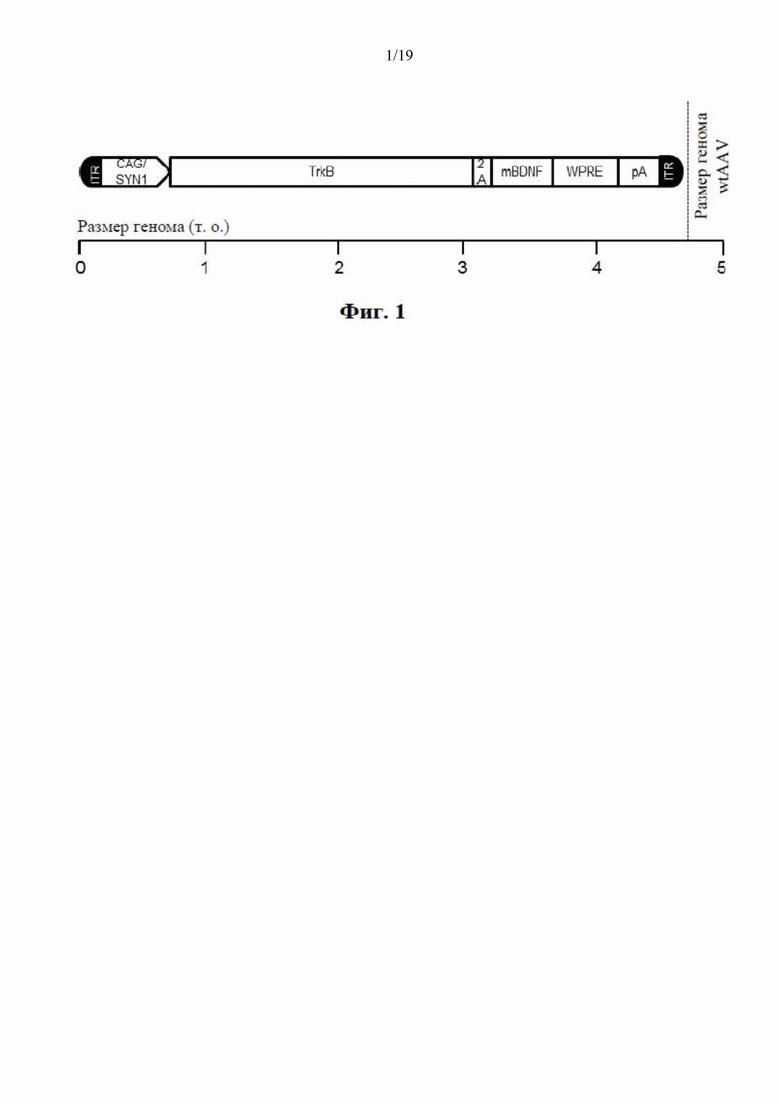
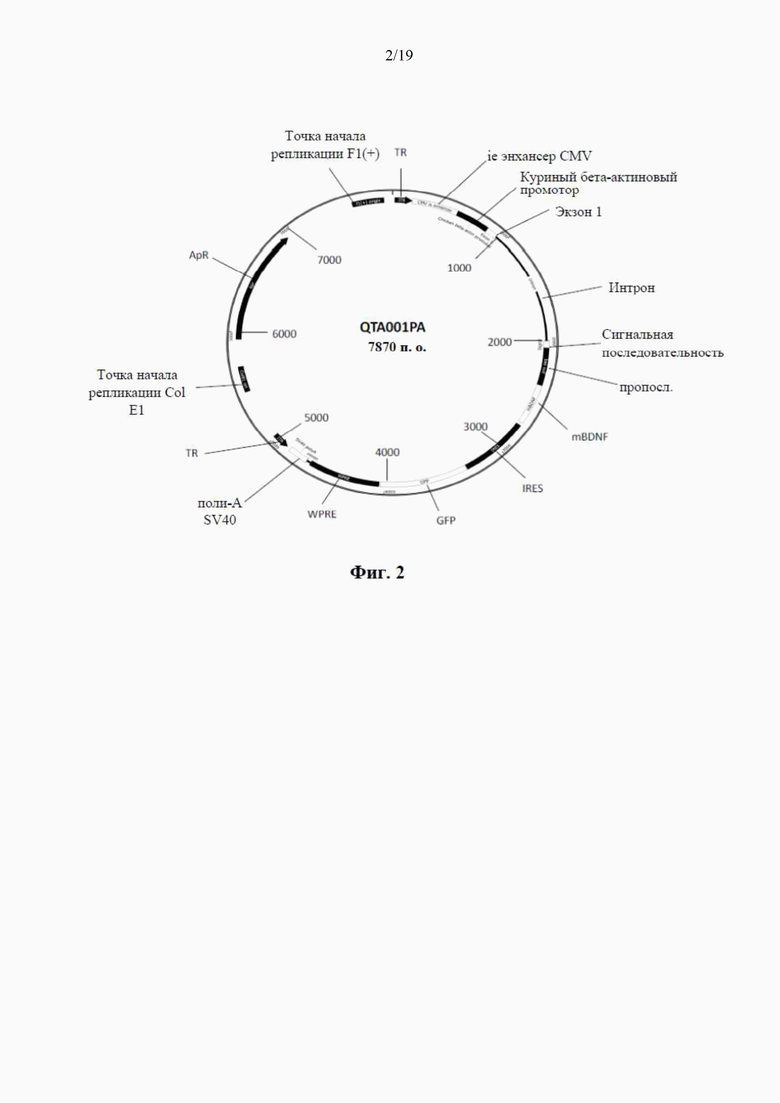
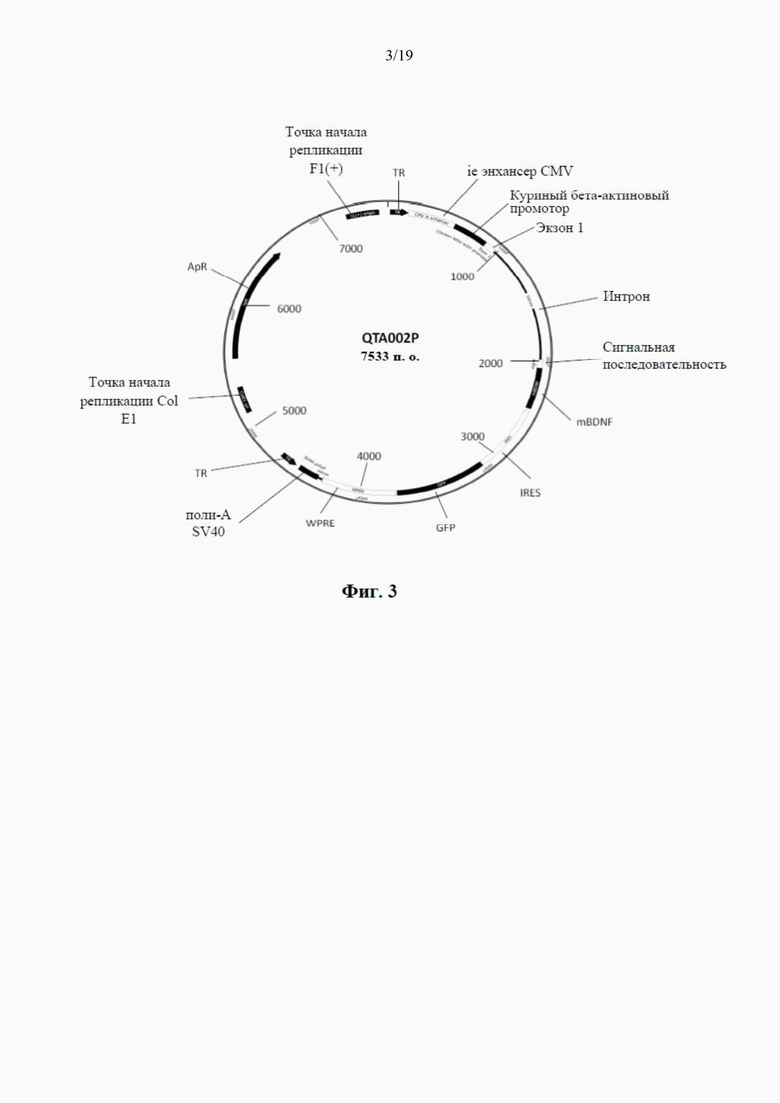
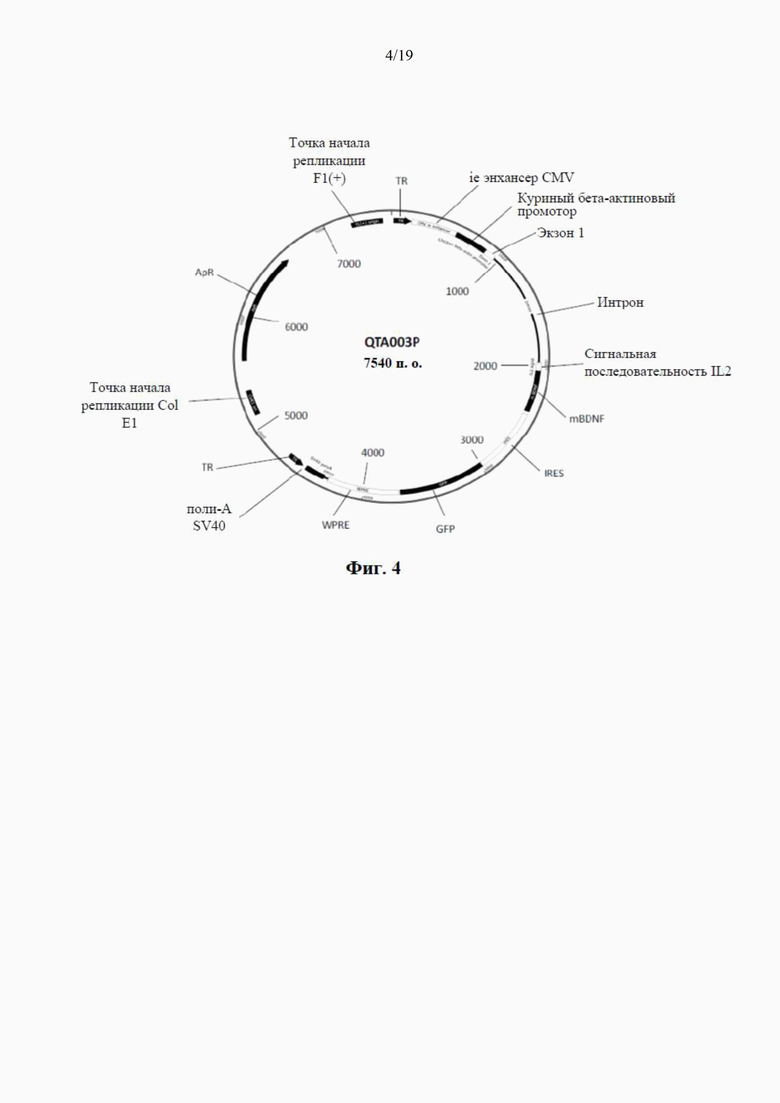
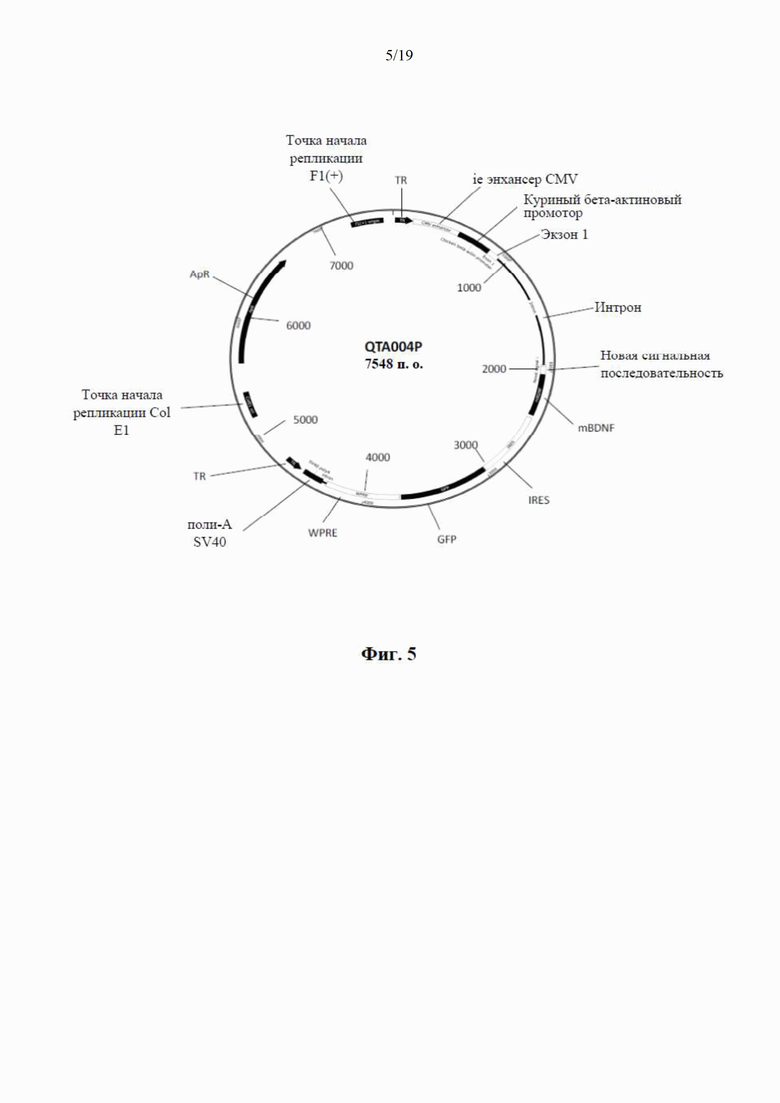

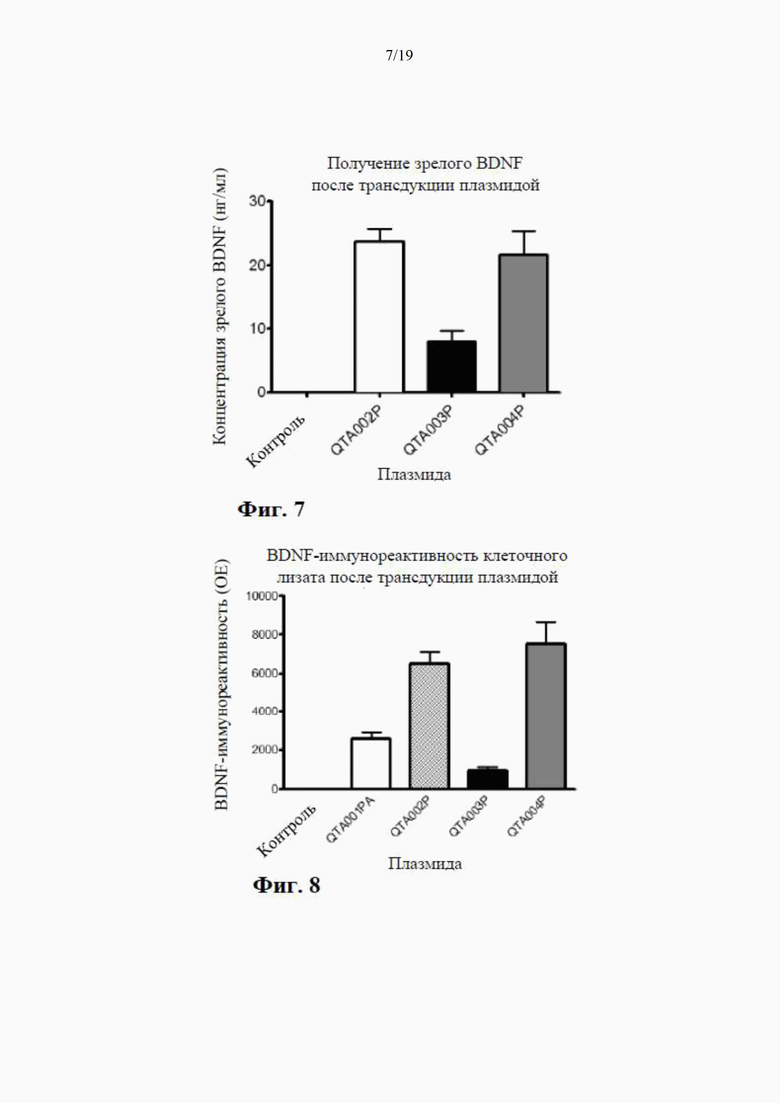
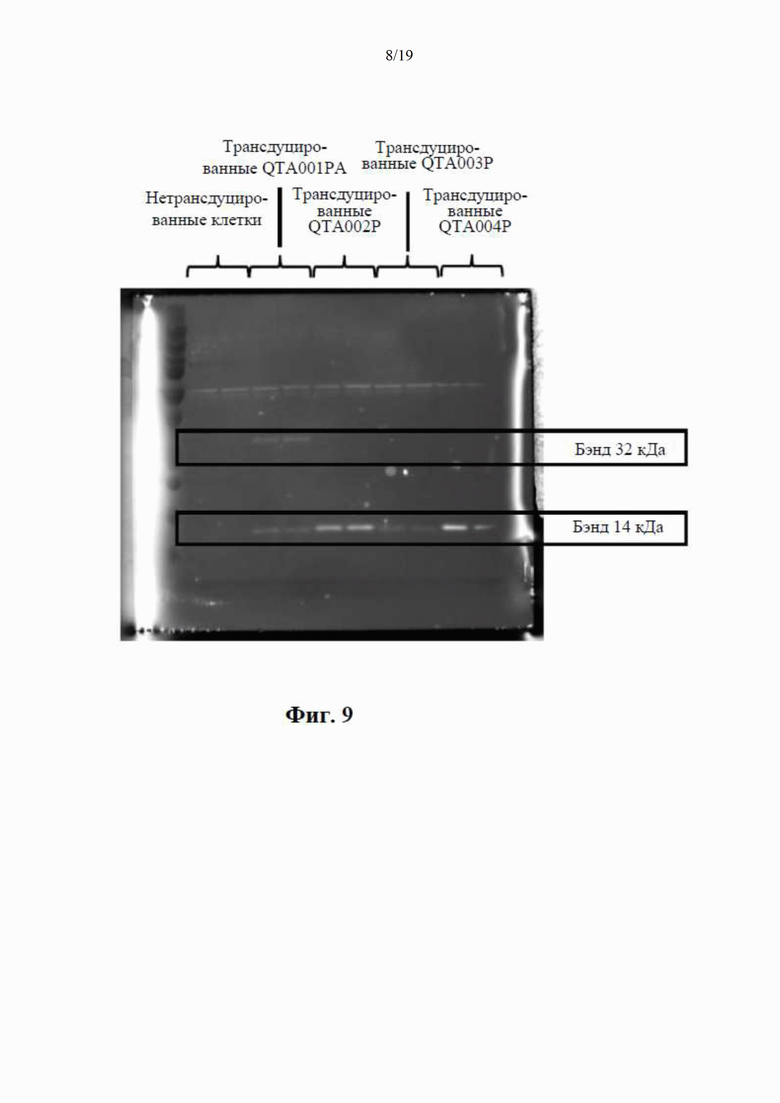
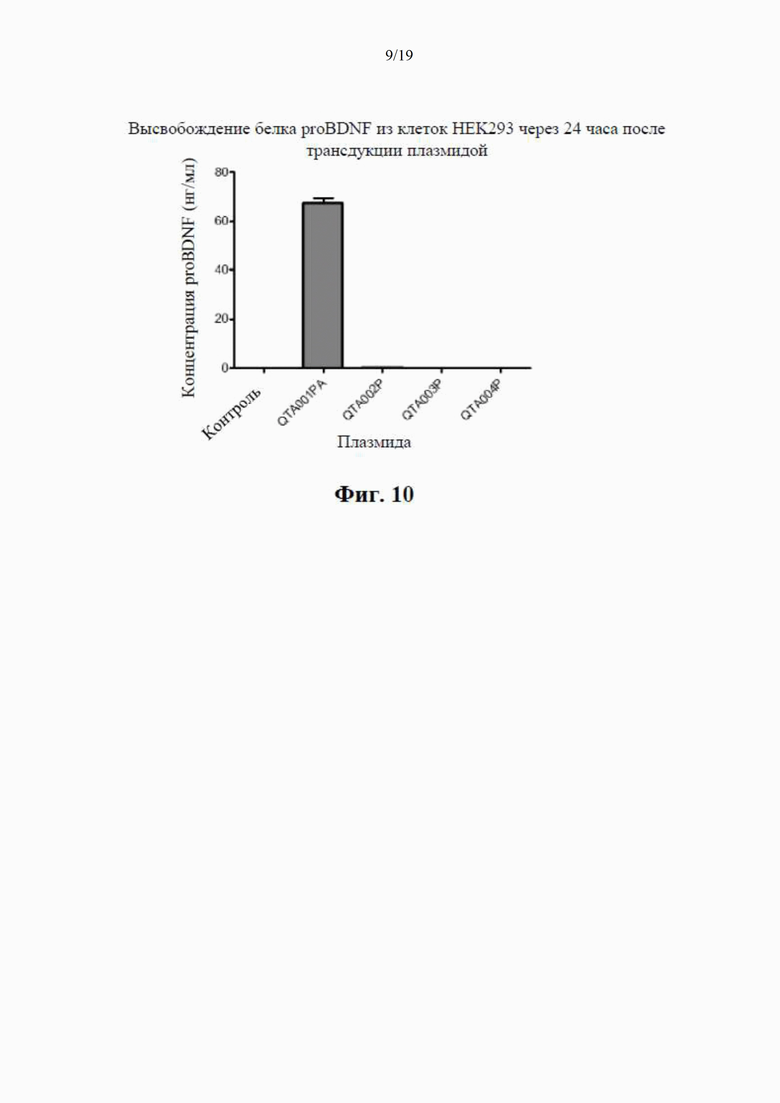
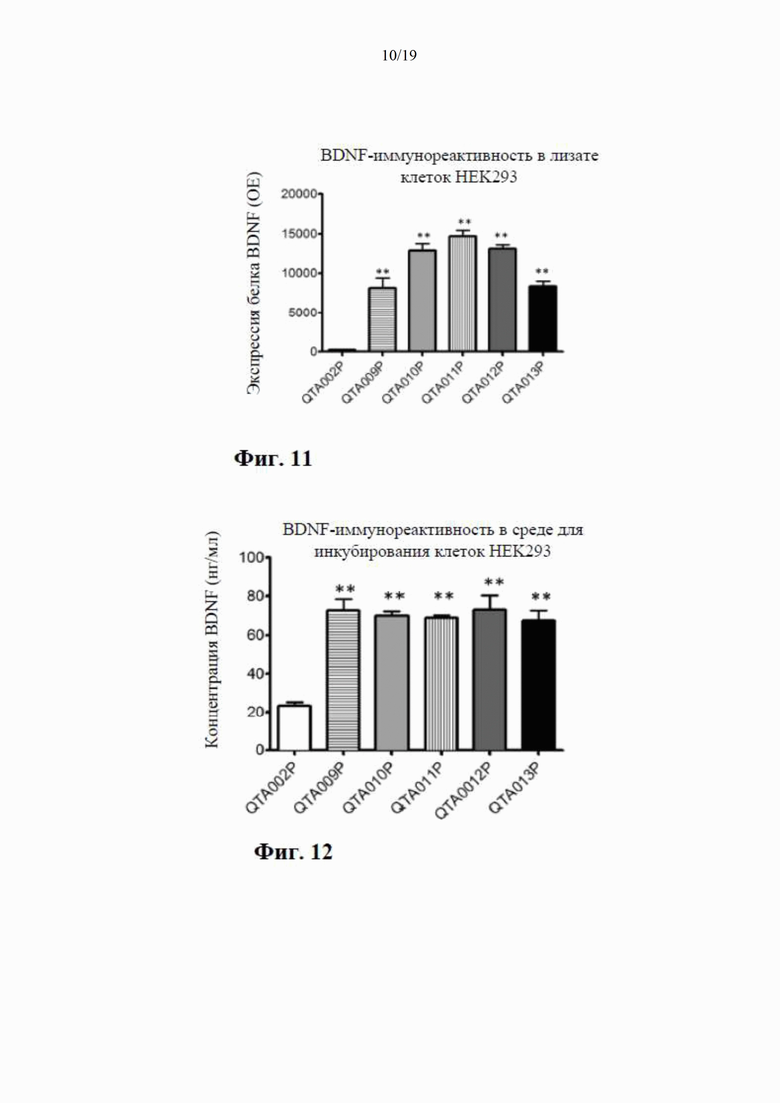
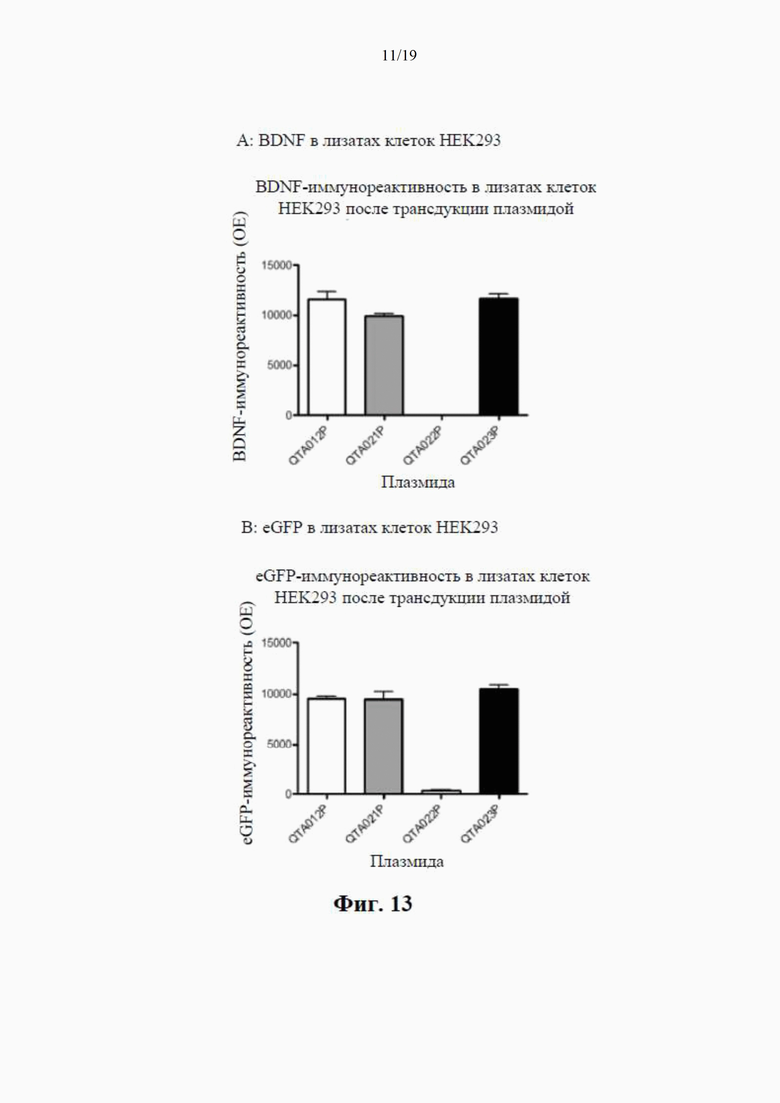
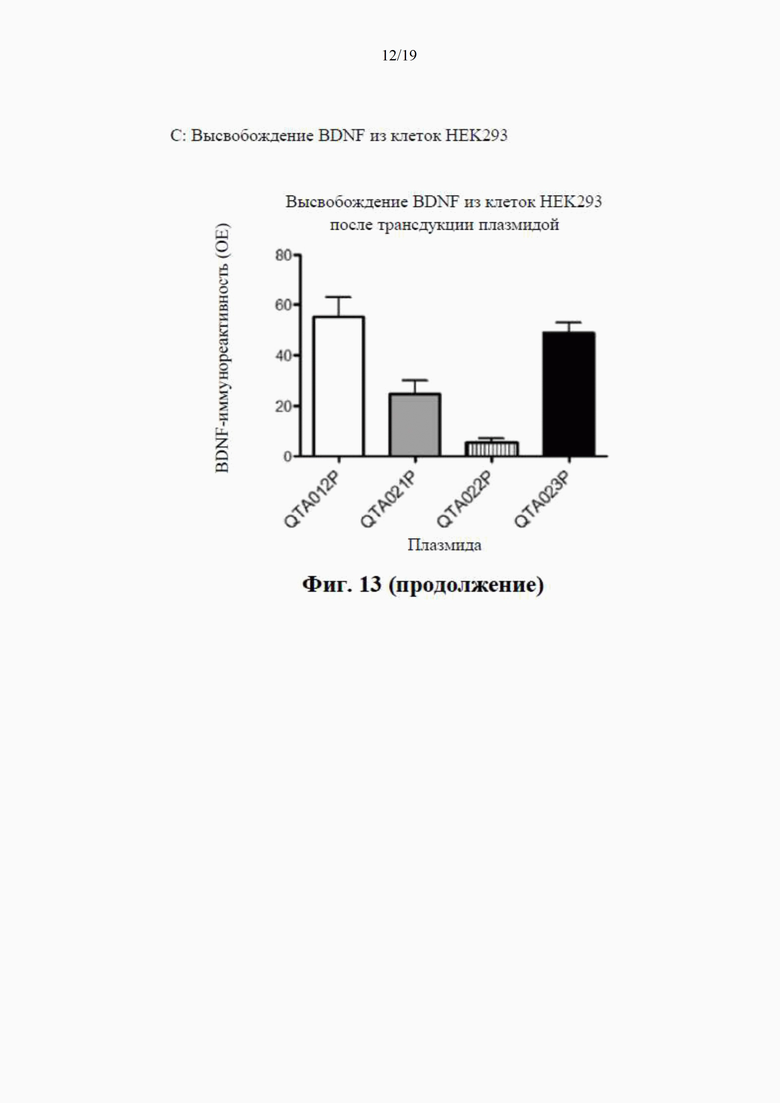
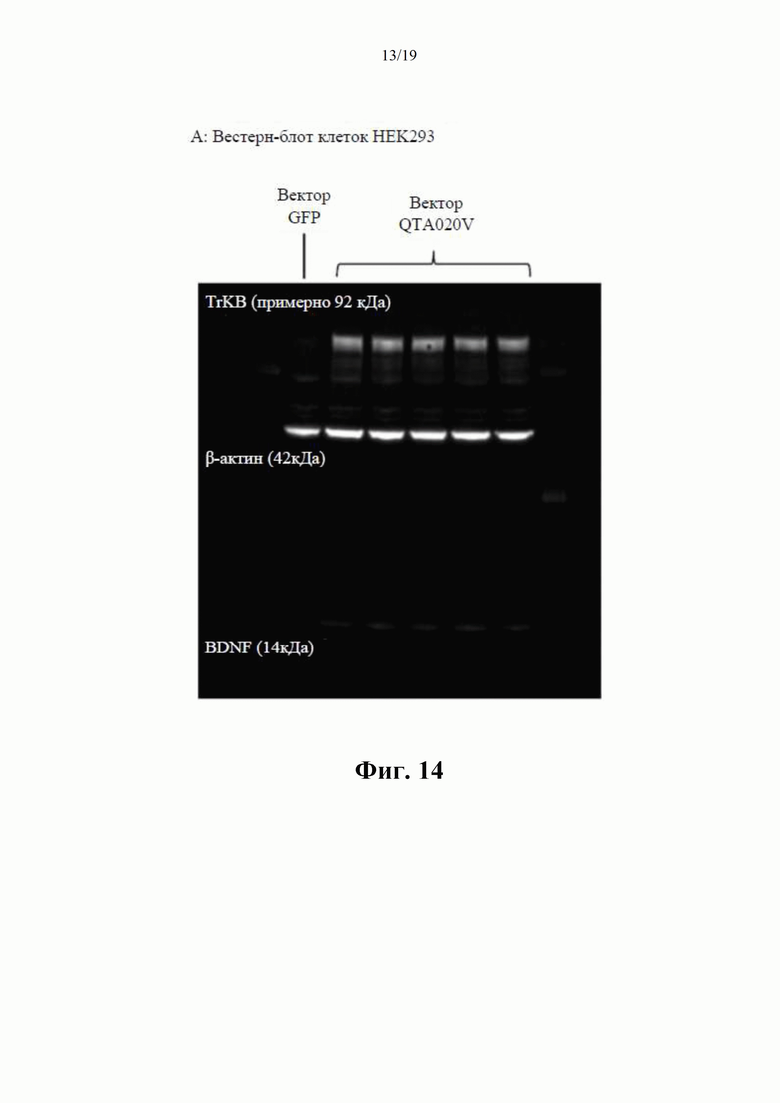
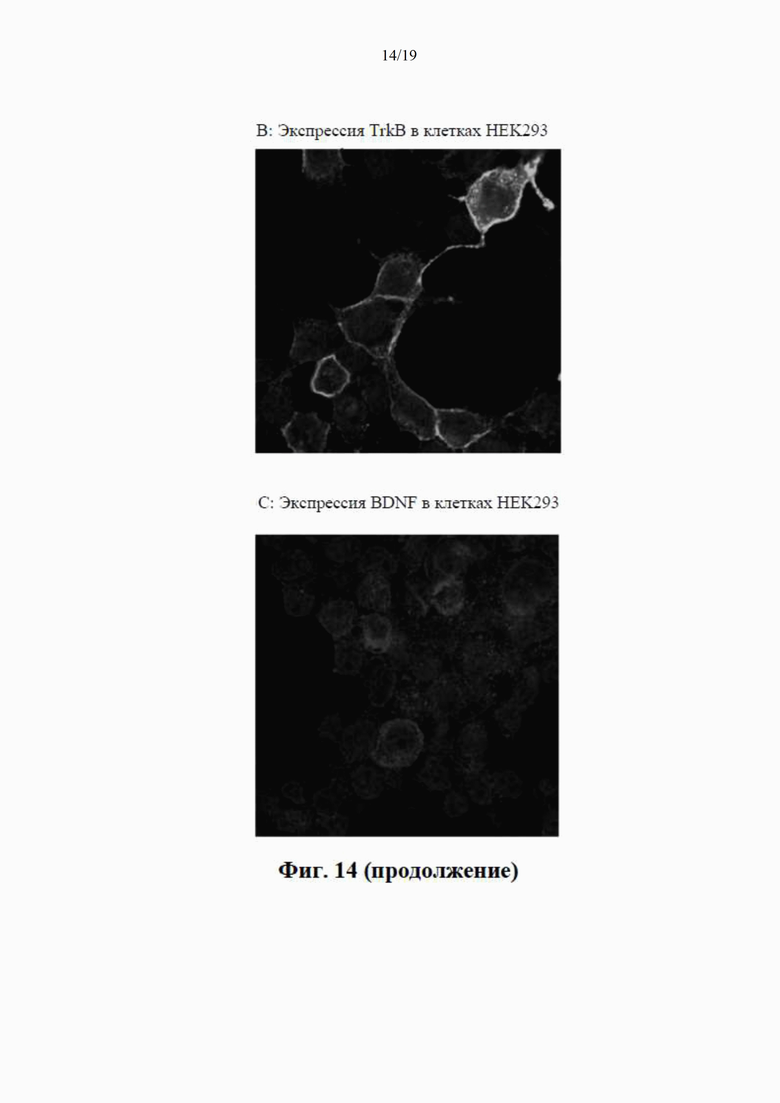
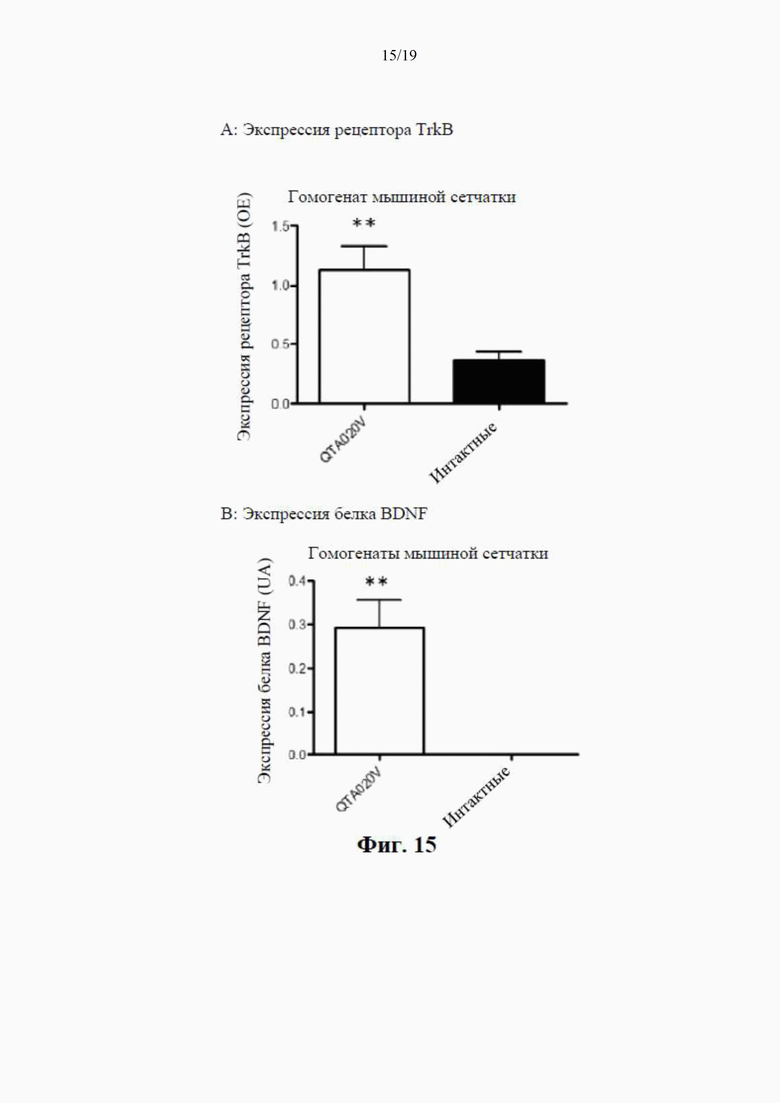
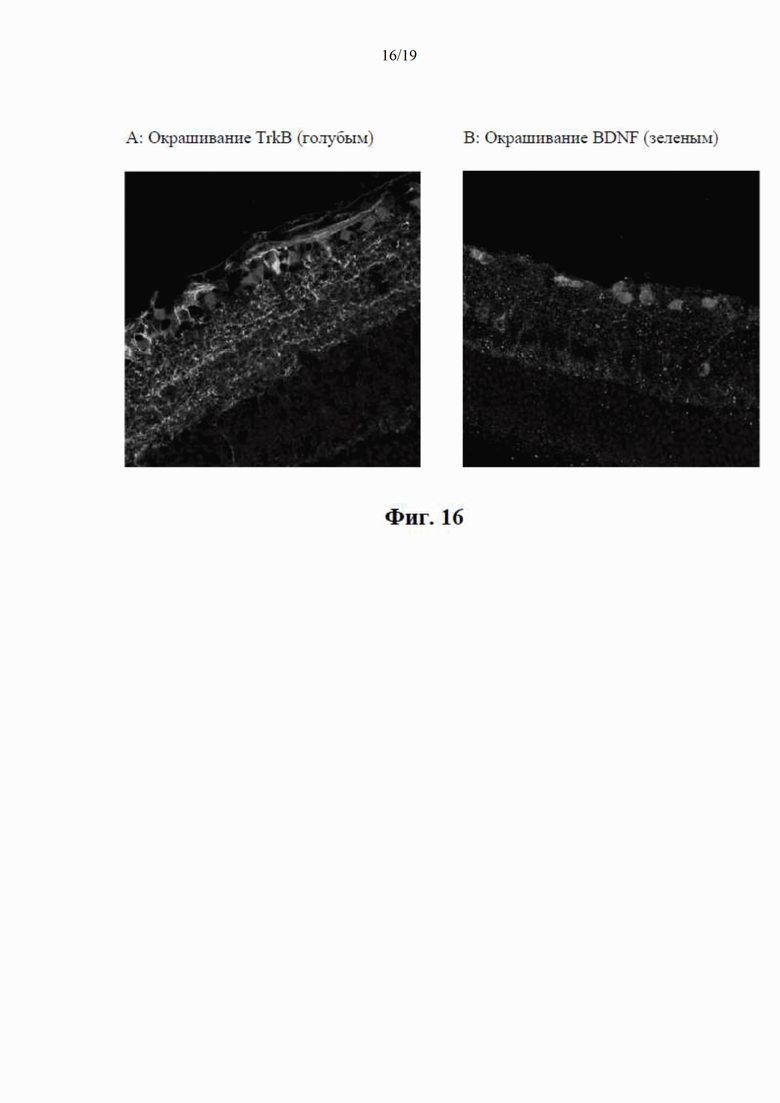
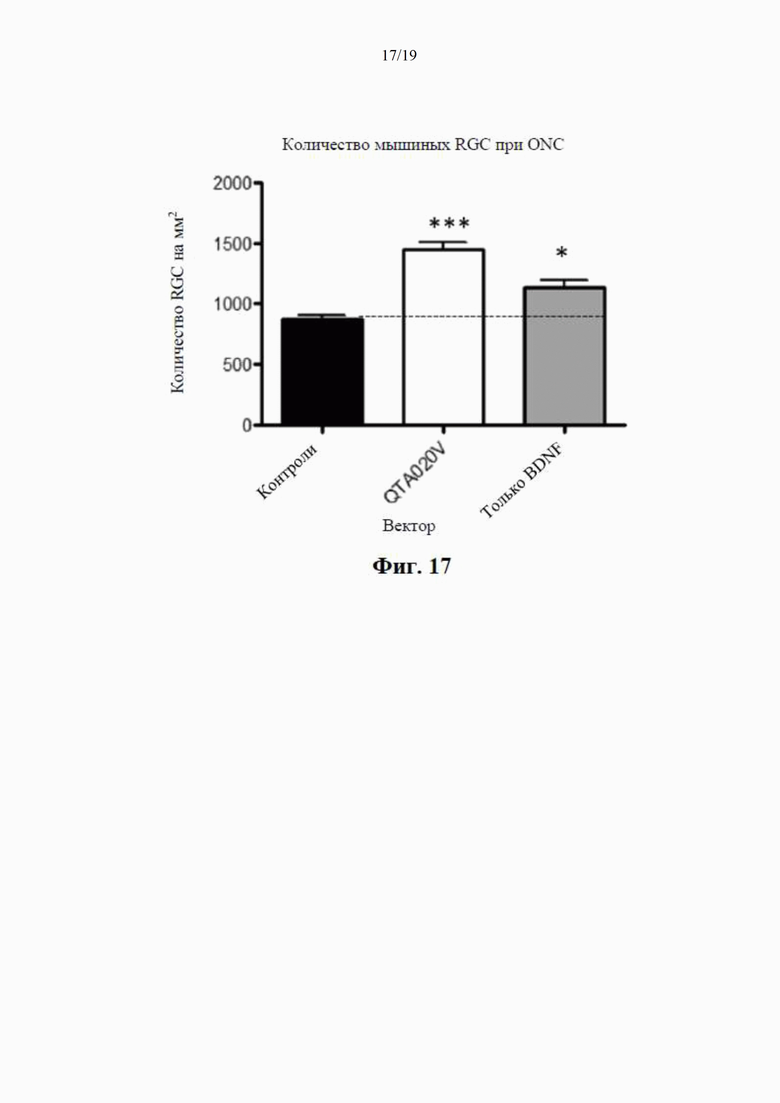
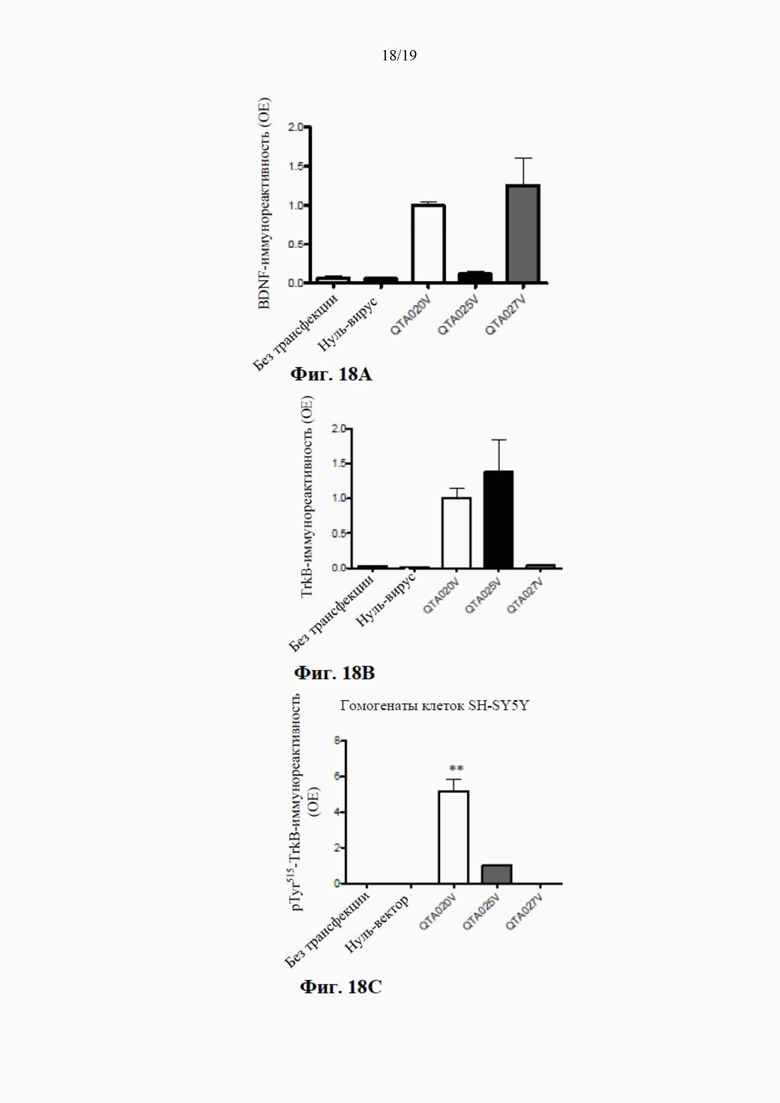
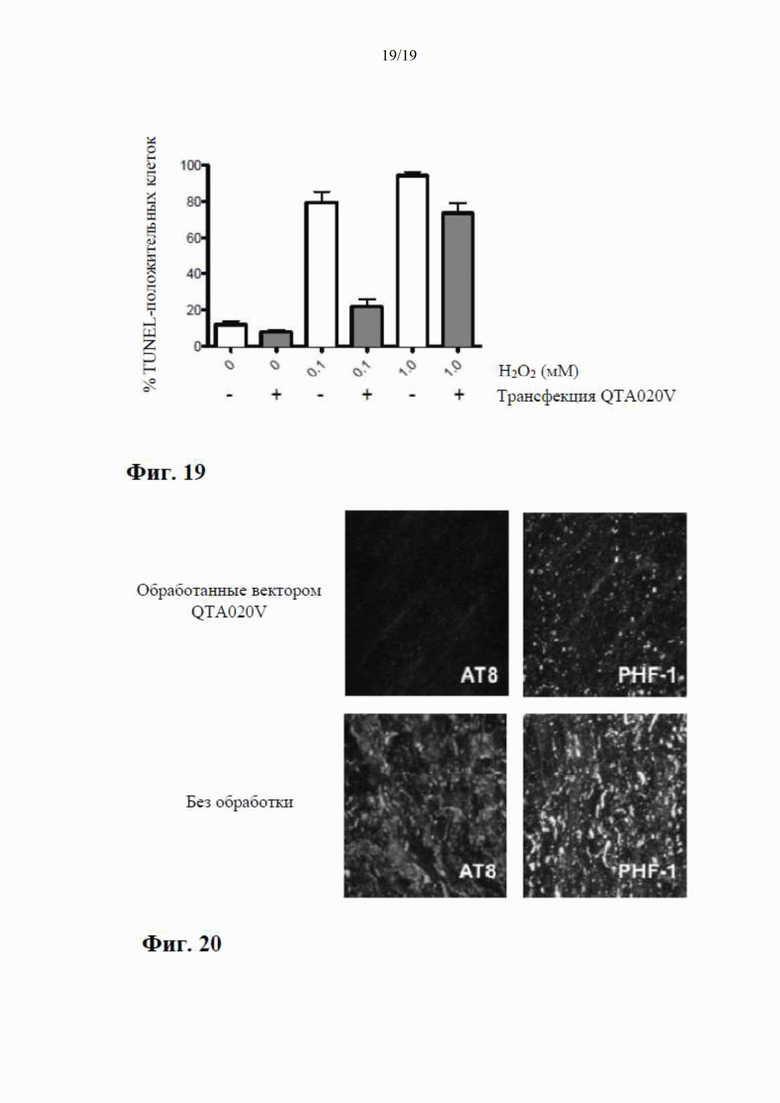
Изобретение относится к области биотехнологии, конкретно к рекомбинантному вектору, содержащему промотор, функционально связанный с последовательностью, которая кодирует тирозинкиназный рецептор В (TrkB), и последовательностью, которая кодирует агонист рецептора TrkB, и может быть использовано в медицине. Полученный вектор, где агонист рецептора TrkB представляет собой зрелый нейротрофический фактор головного мозга (BDNF) или зрелый нейротрофин-4 (NT-4), может быть эффективно использован в лечении, предупреждении или уменьшения интенсивности нейродегенеративного нарушения. 37 з.п. ф-лы, 20 ил., 1 пр.
1. Рекомбинантный вектор, содержащий генетическую конструкцию, содержащую промотор, функционально связанный с первой кодирующей последовательностью, которая кодирует тирозинкиназный рецептор В (TrkB), и второй кодирующей последовательностью, которая кодирует агонист рецептора TrkB, причем агонист представляет собой зрелый BDNF или зрелый NT-4, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую сигнальный пептид, который усиливает секрецию агониста рецептора TrkB, и причем генетическая конструкция содержит спейсерную последовательность, расположенную между первой и второй кодирующими последовательностями, при этом спейсерная последовательность кодирует пептидный спейсер, который выполнен с возможностью расщепления с образованием таким образом рецептора TrkB и агониста в виде отдельных молекул, для применения в лечении, предупреждении или уменьшения интенсивности нейродегенеративного нарушения.
2. Рекомбинантный вектор по п. 1, причем генетическая конструкция содержит нуклеотидную последовательность, кодирующую посттранскрипционный регуляторный элемент вируса гепатита сурков (WHPE), причем необязательно WHPE содержит последовательность нуклеиновой кислоты по существу как изложена в SEQ ID NO: 57 или 58, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 57 или 58.
3. Рекомбинантный вектор по п. 1 или 2, причем указанная конструкция содержит нуклеотидную последовательность, кодирующую поли-А-хвост, причем необязательно поли-А-хвост содержит последовательность нуклеиновой кислоты по существу как изложена в SEQ ID NO: 59, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 59.
4. Рекомбинантный вектор по любому из предыдущих пунктов, причем промотор представляет собой промотор гена человеческого синапсина I (SYN I), причем необязательно промотор содержит нуклеотидную последовательность по существу как изложена в SEQ ID NO: 1, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 1.
5. Рекомбинантный вектор по любому из пп. 1-3, причем промотор представляет собой промотор CAG, причем необязательно промотор содержит нуклеотидную последовательность по существу как изложена в SEQ ID NO: 2, 3 или 48, или ее фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 2, 3 или 48.
6. Рекомбинантный вектор по любому из предыдущих пунктов, причем спейсерная последовательность предпочтительно содержит и кодирует последовательность вирусного пептидного спейсера, более предпочтительно - последовательность вирусного пептидного спейсера 2А.
7. Рекомбинантный вектор по любому из предыдущих пунктов, причем последовательность пептидного спейсера содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 4, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 4.
8. Рекомбинантный вектор по любому из пп. 1-6, причем спейсерная последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 5, или ее фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 5.
9. Рекомбинантный вектор по любому из пп. 1-7, причем последовательность пептидного спейсера содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 6, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 6.
10. Рекомбинантный вектор по любому из пп. 1-6, причем спейсерная последовательность содержит нуклеотидную последовательность, которая изложена под SEQ ID NO: 7, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 7.
11. Рекомбинантный вектор по любому из пп. 1-7, причем последовательность пептидного спейсера содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 8, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 8.
12. Рекомбинантный вектор по любому из предыдущих пунктов, причем первая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую человеческую каноническую изоформу TrkB, причем каноническая изоформа TrkB содержит аминокислотную последовательность, которая изложена в SEQ ID NO. 9, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID No: 9.
13. Рекомбинантный вектор по любому из предыдущих пунктов, причем первая кодирующая последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 10, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 10.
14. Рекомбинантный вектор по любому из предыдущих пунктов, причем первая кодирующая последовательность содержит нуклеотидную последовательность, которая кодирует изоформу 4 TrkB.
15. Рекомбинантный вектор по п. 14, причем изоформа 4 TrkB содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 11, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 11.
16. Рекомбинантный вектор по любому из п. 14 или 15, причем первая кодирующая последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 12, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 12.
17. Рекомбинантный вектор по любому из предыдущих пунктов, причем первая кодирующая последовательность содержит нуклеотидную последовательность согласно SEQ ID NO: 9, причем один или несколько остатков тирозина в положении 516, 701, 705, 706 и/или 816 SEQ ID NO: 9 модифицированы в другой аминокислотный остаток.
18. Рекомбинантный вектор по п. 17, причем по меньшей мере два, три или четыре остатка тирозина в положении 516, 701, 705, 706 и/или 816 в SEQ ID NO: 9 модифицированы в другой аминокислотный остаток.
19. Рекомбинантный вектор по п. 18, причем все пять остатков тирозина в положении 516, 701, 705, 706 и/или 816 в SEQ ID NO: 9 модифицированы в другой аминокислотный остаток.
20. Рекомбинантный вектор по любому из пп. 17-19, причем каждый остаток тирозина модифицирован в глутаминовую кислоту.
21. Рекомбинантный вектор по любому из пп. 17-20, причем модифицированная форма рецептора TrkB содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 13, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 13.
22. Рекомбинантный вектор по п. 21, причем первая кодирующая последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 14, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 14.
23. Рекомбинантный вектор по любому из предыдущих пунктов, причем вторая кодирующая последовательность кодирует нейротрофин-4 (NT-4), который содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 49 или 55, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 49 или 55, и/или вторая кодирующая последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 50 или 56, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 50 или 56.
24. Рекомбинантный вектор по любому из предыдущих пунктов, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, которая кодирует зрелый BDNF.
25. Рекомбинантный вектор по п. 24, причем зрелый BDNF содержит аминокислотную последовательность, которая изложена в SEQ ID NO: 18, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 18.
26. Рекомбинантный вектор по п. 25, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 19, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 19.
27. Рекомбинантный вектор по любому из предыдущих пунктов, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую сигнальный пептид для агониста рецептора TrkB, наиболее предпочтительно сигнальный пептид для BDNF.
28. Рекомбинантный вектор по п. 27, причем нуклеотидная последовательность кодирует канонический сигнальный пептид для BDNF, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, которая кодирует сигнальный пептид, содержащий аминокислотную последовательность, которая изложена с SEQ ID NO: 20, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 20.
29. Рекомбинантный вектор по п. 27, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 21, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 21.
30. Рекомбинантный вектор по любому из предыдущих пунктов, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую последовательность сигнального пептида, которая изложена в любой из SEQ ID NO: 23, 25, 27 или 29, или причем сигнальный пептид содержит аминокислотную последовательность, которая изложена в любой из SEQ ID NO: 22, 24, 26 или 28.
31. Рекомбинантный вектор по любому из предыдущих пунктов, причем вторая кодирующая последовательность содержит нуклеотидную последовательность, кодирующую последовательность сигнального пептида, которая изложена в любой из SEQ ID NO: 31, 33, 35, 37, 39, 41, 43, 45, 61, 63, 65, 67, 69, 71, 73, 75, 77, 79, 81, 83, 85, 87, 89, 91, 93, 95, 97, 99, 101 или 103; или причем сигнальный пептид содержит аминокислотную последовательность, которая изложена в любой из SEQ ID NO: 30, 32, 34, 36, 38, 40, 42, 44, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80, 82, 84, 86, 88, 90, 92, 94, 96, 98, 100 или 102.
32. Рекомбинантный вектор по любому из предыдущих пунктов, причем конструкция содержит нуклеотидную последовательность, которая изложена в SEQ ID NO: 107 или 108, или фрагмент или вариант по меньшей мере с 65% идентичности последовательности с SEQ ID NO: 107 или 108.
33. Рекомбинантный вектор по любому из предыдущих пунктов, причем вектор представляет собой рекомбинантный AAV-вектор (rAAV-вектор).
34. Рекомбинантный вектор по п. 33, причем rAAV представляет собой AAV-1, AAV-2, AAV-3A, AAV-3B, AAV-4, AAV-5, AAV-6, AAV-7, AAV-8, AAV-9, AAV-10 или AAV-11.
35. Рекомбинантный вектор по п. 34, причем rAAV представляет собой rAAV серотипа 2.
36. Рекомбинантный вектор по любому из пп. 1-35, причем нейродегенеративное нарушение выбрано из группы, состоящей из болезни Александера, болезни Альпера, болезни Альцгеймера, латерального амиотрофического склероза (ALS), телеангиоэктатической атаксии, форм нейронального цероидлипофусциноза, болезни Баттена, губчатой энцефалопатии крупного рогатого скота (BSE), болезни Канавана, церебрального паралича, синдрома Коккейна, кортико-базальной дегенерации, болезни Крейтцфельда-Якоба, лобно-височной лобарной дегенерации, болезни Гоше, болезни Хантингтона, ВИЧ-ассоциированного слабоумия, болезни Кеннеди, болезни Краббе, деменции с тельцами Леви, лизосомальных болезней накопления, нейроборрелиоза, болезни Мачадо Джозефа, заболевания двигательных нейронов, множественной системной атрофии, рассеянного склероза, множественной сульфатазной недостаточности, муколипидозов, нарколепсии, болезни Нимана-Пика типа С, болезни Нимана-Пика, болезни Паркинсона, болезни Пелицеуса Мерцбахера, болезни Пика, болезни Помпе, первичного латерального склероза, прионных заболеваний, прогрессирующего надъядерного паралича, болезни Рефсума, болезни Сандхоффа, болезни Шилдера, подострой комбинированной дегенерации спинного мозга на фоне злокачественной анемии, болезни Шпильмейера Фогта Шегрена Баттена, спинально-церебеллярной атаксии, спинальной мышечной атрофии, болезни Стила-Ричардсона-Ольшевского, сухотки спинного мозга и болезни Тея Сакса.
37. Рекомбинантный вектор по любому из пп. 1-35, причем нейродегенеративное нарушение представляет собой болезнь Альцгеймера.
38. Рекомбинантный вектор по п. 37, причем в нейронах снижается фосфорилирование тау-белка.
| WO 2009120978 A2, 01.10.2009 | |||
| MARTIN K | |||
| R | |||
| G | |||
| et al., Gene therapy with brain-derived neurotrophic factor as a protection: retinal ganglion cells in a rat glaucoma model, Investigative ophthalmology and visual science, 2003, V | |||
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| РУДНИЦКАЯ Е | |||
| А | |||
| и др., Анализ вклада изменения нейротрофического обеспечения в развитие | |||

