Изобретение относится к области биотехнологии, а именно, к созданию и производству ферментных препаратов (ФП), содержащих высокоактивные и термостабильные ферменты. Изобретение может быть использовано в биотопливной индустрии.
Ускорение научно-технического прогресса, рост мировой экономики стимулируют потребление традиционных источников энергии, что в конечном счете приведет к их дефициту. По данным Министерства энергетики США, потребление всех видов энергоносителей в мире (нефтепродуктов, угля, природного газа, электроэнергии и альтернативного топлива из возобновляемых источников энергии) с 1992 по 2009 увеличилось на 40%, при этом мировое потребление нефти и нефтепродуктов выросло почти на треть [Коротких А.А. // Биотопливная индустрия США в новом веке // США и Канада: экономика, политика, культура. 2015. Вып. 5. С 103-118]. В Российской Федерации производство тепловой энергии только за период с 2013 по 2017 увеличилось на 18,14% по данным Министерства энергетики РФ [Министерство энергетики РФ. Открытые данные // https://minenergo.gov.ru/opendata].
В условиях дальнейшей интенсификации экономической деятельности и роста мировых цен на энергоносители, замена импортной нефти и других видов минерального топлива на «чистую энергию» с преобладанием возобновляемых энергетических источников - одна из острейших глобальных и региональных проблем.
Растительная биомасса является основным источником органической материи на Земле. Общие запасы растительной биомассы составляют ≈1 трлн т, а ежегодный прирост биомассы в мире составляет ≈50 млрд τ [Bungay H.R. //Energy: the biomass options. Wiley and Sons, New York. 1981. Bioprocessing of renewable resources to commodity bioproducts, First edition / Edited by V.S. Bisaria and A. Kondo. Wiley and Sons, New York. 2014. Sticklen M. // Curr. Opin. Biotechnol. 2006. Vol. 17. P. 315-319]. Современные технологии позволяют перерабатывать растительное сырье в биотопливо и другие разнообразные химические соединения, являющиеся коммерчески востребованными продуктами [From the Sugar Platform to biofuels and biochemical. Final report for the European Commission Directorate-General Energy № ENER/C2/423-2012/SI2.673791, April 2015].
Целлюлоза, являющаяся основным компонентом, растительной биомассы может быть конвертирована в глюкозу и другие сбраживаемые сахара с помощью ферментативного гидролиза. Полученные таким образом сахара являются незаменимым сырьем для микробиологических процессов получения топлива (этанола, бутанола и др.), этилена, органических и аминокислот, полимеров, кормового белка и многих других полезных продуктов.
Глубокая биоконверсия растительного целлюлозосодержащего сырья в простые сахара является ключевой стадией и осуществляется под действием ферментного комплекса. Для эффективной конверсии целлюлозы используются ферментные комплексы целлюлаз, полученные с использованием различных грибных штаммов-продуцентов (микроскопические грибы родов Trichoderma, Penicillium, Aspergillus, Humicola и др.). В состав таких ферментных комплексов входят в различном соотношении ключевые ферменты, участвующие в гидролизе целлюлозы, -эндоглюканазы (ЭГ), осуществляющие деструкцию целлюлозы до олигосахаридов, целлобиогидролазы (ЦБГ), превращающие целлюлозу и олигосахариды в целлобиозу, и β-глюкозидазы (БГ), гидролизующие целлобиозу до глюкозы [Гусаков А.В., Синицын А.П. Деполимеризация природных биополимеров. Ферментативный гидролиз целлюлозы. Обзор. «Химия биомассы: биотоплива и биопластики» под ред. С.Д.Варфоломеева, изд. Научный мир, Москва, 2017, с. 65-99, ISBN 978-5-91522-451-2].
Качественный и количественный состав ферментного комплекса, а также активность и стабильность входящих в него ферментов, определяют эффективность гидролиза целлюлозосодержащего сырья. Изменение и оптимизация состава комплекса целлюлаз позволяет получать ферментные смеси с увеличенной биокаталитической способностью. Помимо этого белковая инженерия целлюлаз позволяет увеличить каталитическую активность и термостабильность индивидуальных ферментов, что открывает новые возможности увеличения общей активности и стабильности конечной ферментной смеси [Illanes Α., Cauerhff Α., Wilson L., Castro G.R. // Bioresour. Technol. 2012. Vol. 115. P. 48-57.].
Мицелиальный гриб Penicillium verruculosum является перспективным продуцентом гидролитического комплекса ферментов карбогидраз и секретирует целлюлазный комплекс высокоактивных ферментов [Gusakov Α. V, Sinitsyn А.Р. Cellulases from Penicillium species for producing fuel from biomass. Review. Biofuels (2012) 3(4), 463-477], что позволяет использовать его в качестве основы для получения мультиферментных комплексов для биодеградации целлюлозосодержащих материалов.
Основным компонентом комплекса ферментов Penicillium verruculosum является целлобиогидролаза I (Се17А), ее количество в комплексе составляет почти 50% от общего секретируемого белка. Показано, что ферменты 7-ой семьи гликозид-гидролаз (GH7) играют ключевую роль в процессе конверсии биомассы [Ilmen М., Saloheimo Α., Onnela Μ. L., and Penttila Μ. Ε. (1997)
Regulation of cellulase gene expression in the filamentous fungus Trichoderma reesei. Appl. Environ. Microbiol. 63, 1298-1306], а целлобиогидролаза I является основным ограничивающим скорость деградации биомассы ферментом [Payne С.М., Knott В. С, Mayes Η. В., Hans son Η., Himmel Μ. Ε., Sandgren Μ., Stahlberg J., and Beckham G T. (2015) Fungal cellulases. Chem. Rev. 115, 1308-1448; Suominen P. L., Mantyla A. L., Karhunen Т., Hakola S., and Nevalainen H. (1993) High-frequency one-step gene replacement in Trichoderma reesei. 2. Effects of deletions of individual cellulase genes. Mol. Gen. Genet. 241, 523-530]. Однако, термостабильность целлобиогидролазы I мезофильного гриба P.verruculosum ограничивается 50°C, что не позволяет использовать ее в процессах с повышенной температурой, например, ферментативном гидролизе древесины. Это свойство ЦБП оказывает негативное влияние на эффективность промышленной ферментативной конверсии вследствие фундаментальной зависимости скорости реакции от температуры.
Таким образом, улучшение технических характеристик целлобиогидролазы I Penicillium verruculosum, в частности, ее термостабильности является актуальной и перспективной задачей.
Мицелиальный гриб Penicillium canescens является продуцентом внеклеточных ферментов. По сравнению с другими грибами данный штамм имеет ряд преимуществ: обладает более высокой скоростью роста и синтеза внеклеточных ферментов; штамм выращивается на простой и дешевой среде со свекловичным жомом; процесс ферментации легко масштабируется; получен штамм-реципиент с ауксотрофным признаком селекции; отработана система экспрессии и трансформации штамма экзогенной ДНК [Патент РФ 2001 124 653. Вавилова Ε. Α., Винецкий Ю. П., Николаев И. В., Серебряный В. Α., Чулкин A.M., Синицын А. П., Синицына О.А., Окунев О.Н., Черноглазое В.М.// Новая трансформационная система для экспрессии генов грибов под контролем регуляторных систем мщелиального гриба Penicillium canescnes//Дата публикации заявки: 27.07.2003]. Это позволяет использовать данный гриб как основу для получения рекомбинантных штаммов -продуцентов ферментов для практического применения в различных областях промышленности.
Одним из современных подходов к повышению термостабильности является биоинженерия ферментов с использованием комбинации методов компьютерного моделирования и сайт-направленного мутагенеза. Данный подход был успешно применен к увеличению термостабильности ксиланазы А P.canescens [Denisenko Υ.Α., Gusakov A.V., Rozhkova A.M., Osipov D.O., Zorov I.N., Matys V.Y., Uporov I.V., Sinitsyn A.P. Site-directed mutagenesis of GH10 xylanase A from Penicillium canescens for determining factors affecting the enzyme thermostability, International Journal of Biological Macromolecules, 2017, v.104, p.665-671]. Аналогичный подход был применен и к модификации ЦБГ I P. verruculosum.
Таким образом, создание нового штамма-продуцента термостабильной модифицированной ЦБГ I P. verruculosum и получение ФП на его основе, который будет эффективен в составе ферментных смесей, использующихся для биодеградации растительной биомассы, является важной и актуальной задачей современной биотехнологии.
Техническая задача, на решение которой направлено данное изобретение, состоит в введении 5-и мутаций в ген cel7A, кодирующий модифицированную форму ЦБГ I, и получении сухого ФП на основе нового рекомбинантного штамма P. canescens mtCBHI (ВКМ F-4850D), являющегося продуцентом гетерологичной мутантной ЦБГ I P. verruculosum, относящаяся к 7-й семье гликозид-гидролаз (КФ 3.2.1.176, мол. масса 55 кДа) для применения в биотопливной индустрии для осуществления эффективного процесса биоконверсии целлюлозосодержащей биомассы в технические сахара.
Технический результат от предлагаемого изобретения состоит в сокращении времени ферментативного гидролиза целлюлозосодержащей биомассы за счет способности основного действующего фермента ЦБГ I эффективно гидролизовать целлюлозу при повышенной температуре в следствии увеличения термостабильности фермента.
Сущность изобретения заключается в получении рекомбинантного штамма гриба P.canescens путем трансформации его генетической конструкцией, содержащей модифицированный ген, кодирующий мутантную целлобиогидролазу I P.verruculosum, с последующим получением ферментного препарата термостабильной ЦБП на его основе.
Изобретение реализуется следующим образом:
- Штамм P. canescens mtCBHI (ВКМ F-4850D) получен путем трансформации ауксотрофного штамма P. canescens RN-3-11 экспрессионной плазмидой pCBHI-mt5 с последующей селекцией на агаризованной среде с 10 мМ NaNC>3 и отбором наиболее перспективного клона по критерии каталитической активности и термостабильности по отношению к МКЦ.
- Способ получения ФП предусматривает глубинное культивирование штамма-продуцента P. canescens mtCBHI (ВКМ F-4850D) на среде, содержащей свекловичный жом с последующей распылительной сушкой КЖ и доведении ферментативной активности препарата до 0,5 ед/мг белка по МКЦ.
Изобретение позволяет получить ФП с высокой активностью целевой целлобиогидролазы I с повышенной термостабильностью. Применение нового ФП позволит увеличить эффективность гидролиза целлюлозы и сократит время ферментативной обработки.
Культурально-морфологические и микроскопические особенности штамма P. canescens mtCBHI (ВКМ F-4850D):
Растет на агаризованных средах (среда Чапека с дрожжевым автолизатом, Мальц-агар, глюкозо-картофельный агар, сусло-агар) при Т=26-30°С в течение 7-10 суток, рН 4.5-5.0.
На среде Чапека с дрожжевым экстрактом при культивировании гриба при 25°С на 7 сутки колонии достигают 24-30 мм в диаметре, складчатые, поверхность сильно радиально складчатая, плотная, тонкая, ростовая зона врастает в агар, имеет ширину 1.5-2.0 мм. Мицелий светло-желтоватый, шерстистый, центр колонии выпуклый, конидиогенез слабый, серо-зеленоватого оттенка. Экссудата и растворимого пигмента нет. Обратная сторона светлая, в центре колонии - палево-оранжевая. При температуре 37°С колонии диаметром 5 мм, мицелий светлый, конидиообразования нет.При температуре 5°С роста нет.
При росте на Мальц-агаре диаметр колонии 23-24 мм, поверхность сильно радиально складчатая, плотная, тонкая, ростовая зона врастает в агар, шириной 1,5-2,0 мм. Мицелий белый, шерстистый, прижатый, конидиогенез очень слабый, практически отсутствует. Экссудата и растворимого пигмента нет.Обратная сторона светлая.
При микроскопировании штамм имеет конидиеносцы двухярусные, терминальные, бивертициллятные, гладкие длиной около 150 мкм, шириной 2-3 мкм. Метулы расходящиеся размером 10-13×2,5-3,0 мкм, фиалиды ампуллиформные размером 7-8×2,8-3,0 мкм. Конидии округлые, шероховатые размером 3,0-3,5 мкм.
При культивировании в глубинных условиях с использованием растворимых субстратов (глюкоза, фруктоза, лактоза) образуется рыхлый разветвленный мицелий со слабой пеллетизацией, удельная начальная скорость роста мицелия составляла 0,35 ч-1, в конце культивирования 0,1 ч-1.
Физиолого-биохимические признаки штамма:
Мезофилен. Оптимальная температура роста мицелия 32°С (29-34°С), оптимум для образования целлюлаз 28°С (26-29°С). Оптимальные значения рН роста и секреции целлюлаз 3,5-5,0. Рост мицелия наблюдается и при рН 2,5, но при этом наблюдается очень слабое образование целлюлаз и других карбогидраз.
Резистентность к нистатину хорошая. При поверхностном культивировании устойчив к концентрации до 0,5 мкг/мл, при концентрации 2,5 мкг/мл рост подавляется. При добавлении в среду дигитонина (3,5-4,0 мкг/мл) или бенгальского розового (30-50 мкг/мл) размер колоний уменьшается.
Является прототрофом. Способен быстро ассимилировать глюкозу, глицерин, галактозу, D-маннозу, D-маннит, трегалозу, сорбозу и сорбит, медленнее - D-ксилозу, L- и D-арабинозу, L-рамнозу и рибозу. Слабо ассимилирует: D-глюкозамин, дезоксирибозу, дезоксигалактозу, 2-дезокси-D-глюкозу и 5-тио-В-глюкозу.
Использует неорганический и органический азот, хорошо ассимилирует нитратную и аммонийную формы азота.
Образует ферментные системы, позволяющие расти на соответствующих комплексных субстратах: целлюлозе, крахмале, ксилане, бета-глюкане.
Штамм P. canescens mtCBHI (ВКМ F-4850D) отличается наличием продукции термостабильной гетерологичной мутантной ЦБГ I с увеличенной в 2,5 раза - термостабильностью по отношению к МКЦ при 65°С по сравнению с нативной ЦБГ I.
В предлагаемом изобретении метод определения активности целлобиогидролазы I основан на измерении скорости образования восстанавливающих сахаров (ВС) методом Шомоди-Нельсона при гидролизе полисахаридного субстрата (МКЦ). За единицу активности принимается такое количество фермента, которое приводит к образованию 1 мкмоль ВС в минуту при рН 5,0 и 40°С [Синицын А.П., Гусаков А.В., Черноглазое В.А. Биоконверсия лигноцеллюлозных материалов. - М.: МГУ, 1995. - 144 с]. Активность по п-НФ-Р-О-лактозиду определяли по скорости образования n-нитрофенола при гидролизе пНФЛ, за единицу активности принимали количество фермента, необходимое для образования 1 мкмоль «-нитрофенола в течение 1 минуты при рН 5,0 и 40°С [Синицын А.П., Черноглазое В.М., Гусаков А.В. //Итоги науки и техники, сер. Биотехнология. - М.: ВИНИТИ, 1993. Т. 25, С.30-37].
Новый ФП, mtCBHI-5, полученный на основе P. canescens mtCBHI (ВКМ F-4850D) обладает улучшенными эксплуатационными характеристиками за счет введения 5-ти мутаций в ген се17А, кодирующий мутантную форму ЦБГ I, имеющую высокую экзо-Р-1,4-глюканазную активность и увеличенную термостабильность. Это является преимуществом данного ФП mtCBHI-5.
Новый ФП mtCBHI-5, полученный на основе КЖ мутантного штамма P. canescens mtCBHI (ВКМ F-4850D), продуцента термостабильной высокоактивной ЦБГ I, будет иметь высокую рентабельность применения в процессах ферментативной обработки лигноцеллюлозной биомассы за счет возможности проведения гидролиза при повышенной температуре.
Возможность использования изобретения иллюстрируется примерами, которые не ограничивают объем и сущность притязаний, связанных с ними.
Изобретение характеризуется следующими примерами.
Пример 1. Белковая инженерия ЦБГ I и введение точечных мутаций в ген сеНА.
На основании биоинформатического анализа пространственной структуры белка ЦБП Ρ.verruculosum, построенной на основе пространственных структур кристаллов целлобиогидролазы из Trichoderma reesei (анаморф Hypocrea jecorina, PDB ID: 50A5 и 4C4C) был выбран неструктурированный участок полипептидной цепи (с Т301 по N380, на Фиг 1 данная область обведена красным цветом). Методом сетевого анализа ограничений (CNA, Constraint Network Analysis, http://cpclab.uni-duesseldorf.de/cna) в структуре ЦБП на участке 301-380 были определены 5 «слабых» точек, т.е. 5 положений а/к в которых средний индекс жесткости, Ri, минимален. Такими точками являлись Т291, V290, Е302, V312, G342. Далее, методом молекулярной динамики в каждой «слабой» точке были обсчитаны варианты замен нативной аминокислоты на 19 других. Таким образом, принимая во внимание изменений параметра жесткости и изменения энергии стабилизации, AAG, наиболее перспективными заменами являлись Т291Р, V290F, E302L, V213W, G342R.
Для введения мутаций в ген се17А, были использованы олигонуклеотиды, приведенные в Таблице 1.
Участок полинуклеотидной цепи, содержащий пять обозначенных выше мутаций был синтезирован отдельно в ЗАО «Евроген». Структура мутированного участка гена представлен на Фиг. 2.
Амплификация ПЦР-продуктов проводилась в термоциклере MyCycler (BioRad, США). В качестве матрицы была взята либо геномная ДНК штамма P.verruculosum, предварительно выделенная из 2-х дневного мицелия гриба с помощью набора "DNeasy Plant Mini Kit» (QIAGEN, Германия), либо плазмида pCBHImt с мутированным учаском гена cbh7A. Получение полноразмерного гена проводилась в два этапа методом модифицированной гнездовой ПЦР. На первом этапе были получены отдельные фрагменты с использованием форвардных олигонуклеотидов OVERLAP 1-FWD, OVERLAP2-FWD или CBHI B1-UPLIC и реверсных OVERLAP 1-REV, OVERLAP2-REV или CBHII_Bl-LOWERLIC. Полученные фрагменты были выделены из 1% агарозного геля с использованием набора GelExtraction KIT (QIAGEN, Германия) и смешаны в качестве матриц для получения полноразмерного гена се17А ПЦР-методом с использованием крайних праймеров CBHI_B1-UPLIC и CBHII_Bl-LOWERLIC, которые содержали сайты независимого лигирования для последующего лигирования полноразмерного гена с мутацией в вектор pXEG. Полноразмерный ПЦР-πηοπνκτ Гпазмеп 1578 п. оЛ и панее полученный линеапизованный вектсю pXEG были обработаны Т4 ДНК полимеразой (Thermo Fisher Scientific Inc., Waltham, MA, USA) в присутствии dATP и dTTP (Thermo Fisher Scientific Inc., Waltham, MA, USA), соответственно. Условия проведения реакции были следующими: 22° - 30 мин, 80° - 30 мин. Обработанные продукты были лигированы в вектор pXEG в соотношении 50 нг вектора с 150 нг вставки. Затем смесь инкубировали в течение 1 часа при 22°С без добавления лигазы, после этого такой смесью были трансформированы бактериальные компетентные клетки штамма E.coli MachI (Invitrogene, Carlsbad, CA, USA) по стандартному протоколу трансформации [J. Sambrook, D. Russell, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, New York, 2001]. Таким образом, были получены бактериальные колонии, способные расти на среде с ампициллином, благодаря гену bla в составе полученной плазмидной конструкции.
Скрининг полученных клонов осуществляли с помощью ПЦР. Для этого использовали клетки E.coli в качестве матрицы, BIOTAQ™ Red DNA Polymerase (Bioline Reagents Ltd, UK) и концевые олигонуклеотиды CBHI_B1-UPLIC и CBHI_Bl-LOWERLIC. Реакцию проводили при следующих условиях: 1,5 мин при 95°С (первичная денатурация); 30 последующих циклов - 1 мин при 95°С (денатурация в цикле), 2 мин при 50°С, 1 мин при 72°С (синтез); 10 мин при 72°С, 10 мин при 4°С.Полученные продукты были исследованы с помощью 1%-ного агарозного гель-электрофореза в буфере ТВЕ. Было показано, что эффективность клонирования составила 80%. Таким образом была получена плазмида pCBHI-mt5, содержащая ген cell А с 5-ю мутациями.
Секвенирование плазимиды показало наличие искомых мутаций в полинуклеотидной последовательности при отсутствии инсерций, делеций или дополнительных мутаций в остальной части гена cell А.
Пример 2. Получение рекомбинантного штамма P. canescens mtCBHI (ВКМ F-4850D) и ферментного препарата mtCBHI-5 на его основе.
Плазмида pCBHI-mt5, полученная в Примере 1 была трансформирована в реципиентный штамм P.canescens RN-3-11-7 (ΔniaD) совместно с плазмидой pSTA10 в соотношении 5:1 (мкг каждой ДНК) по стандартной методике [Sambrook, J., and Russell, D.W. (2001) Molecular cloning:a laboratory manual, Cold Spring Harbor Laboratory Press, N.Y.; A.Y. Aleksenko, N.A. Makarova, I.V. Nikolaev, A.J. Clutterbuck, Integrative and replicative transformation of Penicillium canescens with a heterologous nitrate-reductase gene, Curr. Genet. 28 (1995) 474-478]. В результате трансформации было получено более 40 рекомбинантных штаммов. Трансформанты были культивированы в колбах на стандартной среде культивирования следующего состава (г/л): свекловичный жом - 30, пептон - 50, КН2РО4- 25, рН 4,5. В результате первичного скрининга по критерию наличия полосы, размером 55 к Да, состоящего в проведении электрофореза культуральных жидкостей, секретируемых рекомбинантными штаммами (данные не показаны), был отобран штамм P. canescens mtCBHI с наибольшей ферментативной активностью по кристаллической целлюлозе (МКЦ).
Культивирование штамма P. canescens mtCBHI проводили в ферментере объемом 3 л КФ-108 («Проинтех», Россия), оснащенном барботером для подачи воздуха в аппарат и турбинной мешалкой на среде 1 следующего состава:
Среда 1 (г/л):
Культивирование проводили 144 ч, при рН не ниже 5,2 и 30°С.
По окончании ферментации грибную биомассу удаляли путем центрифугирования (4000 об/мин в течение 20 мин на центрифуге Avanti JXN-26, «Весктап coulter», США), свободную от клеток КЖ концентрировали с помощью ультрафильтрации (с пределом отсечения 15 кДа), ультраконцентрат сушили на распылительной сушилке (Buchi MiniSpray Dryer В-290, условия: ТВХ=135°С, ТВЫХ=55-65°С, степень аспирации 70%, скорость потока 0,5 л КЖ в час) с получением сухих ФП, которые представляли собой бежевый рассыпчатый порошок легко растворимый в водной среде. Таким образом был получен сухой ФП mtCBHI-5, активность которого представлена таблице 2. Ферментные препараты mtCBHI-5 и RN-3-11-7 (ферментный препарат, полученный аналогичным способом на основе реципиентного штамма) обладали базовой ксиланазной активностью в пределах. Ферментный препарат mtCBHI-5 обладал также повышенной активностью по микрокристаллической целлюлозе, что свидетельствует о дополнительной экспрессии привнесенной целлобиогидролазы I.
Далее был проведен масс-спектрометрический (МС) анализ образцов целевых белков для подтверждения аминокислотных замен. Для проведения МС анализа были использованы образцы целевого белка. МС анализ показал присутствие мутаций E302L, V213W, G342R. Мутации Т291Р и V290F не попадали в зоны фрагментирования белка после трипсинолиза, однако молекулярная масса мутантного белка соответствовала ожидаемым 55 кДа, из чего был сделан вывод о присутствии мутаций Т291Р и V290F, также как и доказанных мутаций E302L, V213W, G342R.
Пример 3. Изучение термостабильности модифицированной ЦБГ I P.verruculosum, содержащей мутации Т291Р, V290F, E302L, V213W, G342R.
Для изучения термостабильности модифицированной ЦБГ I целевой фермент был выделен из ФП mtCBHI-5 в гомогенном виде в три стадии по приведенной ниже схеме с использованием хроматографа АКТА purifier (GE Health Care, Швеция):
1. Гель-проникающая хроматография (колонка BioGel Р4, буфер 0,02 Μ Bis-Tris/HCl; рН 6.80) для обессоливания раствора ФП mtCBHI-5.
2. Фракционирование обессоленного ФП mtCBHI-5 с помощью анионообменной хроматографии (колонка Sourse 15Q 10 мл, элюент 0,01 Μ Bis-Tris/HCl; рН 6.8) в градиенте NaCl от 0 до 0,4 М.
3. Доочистка фракций, содержащих мутантную ЦБГ I, с помощью гидрофобной хроматографии (колонка Sourse 15Iso 1 мл, элюент 50 mM NaAc; рН 5.0) в градиенте сульфата аммония от 1,4 до 0 М. Таким образом, была выделена в гомогенном виде мутантная форма
ЦБГ I из ФП mtCBHI-5, полученного на основе рекомбинантного штамма P. canescens mtCBHI.
Для оценки термостабильности мутантной ЦБГ I проводили гидролиз МКЦ (микрокристаллической целлюлозы) при различных температурах. Эксперимент проводили в пробирках на 2 мл в термошейкере при 50, 65 и 75°С. Концентрация субстрата в реакционной смеси составляла 10 г/л (в пересчете на сухое вещество), реакция проводилась в 0.1 Μ ацетатном буфере рН 5,0, при перемешивании 1000 об/мин. Гидролиз проводили в течение двух суток. Для обеспечения дополнительного перемешивания реакционной смеси в пробирки помещали маленькую магнитную мешалку (п=5 мм, с1=2 мм). Гомогенный фермент добавлялся в пересчете на белок - 0,5 мг белка на 1 г сухого вещества субстрата. В качестве контроля использовали гомогенный нативный фермент ЦБГ I P. verruculosum, выделенный из штамма P.canescens ЦБП по аналогичной схеме.
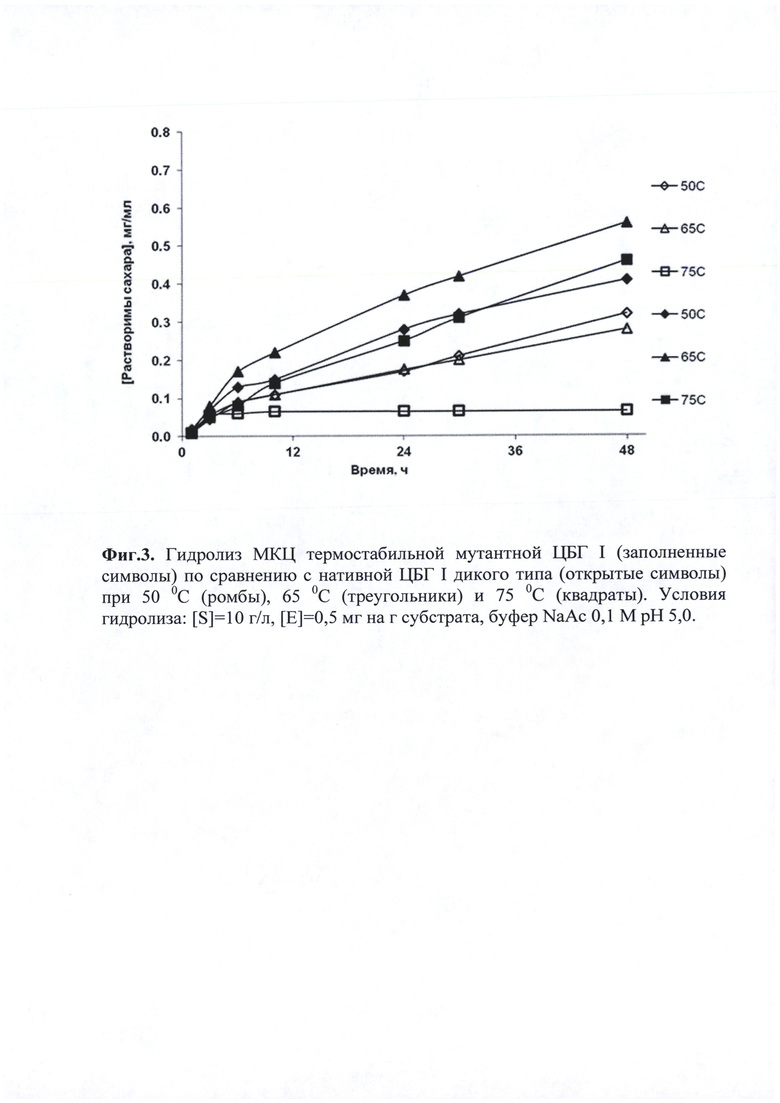
Через определенные промежутки времени из реакционной смеси отбирали пробы и определяли концентрацию сахаров методом ВЭЖХ. На фиг. 3 представлен график зависимости концентрации сахаров от времени при различных температурах.
Мутантная форма ЦБГ I оказалась более активна и обеспечивала более высокий выход сахаров при температурах 50°, 65° и 75°С, чем нативная ЦБГ I дикого типа в том же диапазоне температур. Для нативной ЦБГ I дикого типа выход сахаров был наиболее высокий при температуре 50°С, выход сахаров при 65°С был немного ниже, при 75°С фермент был практически неактивен. Мутантная форма ЦБГ I сохранила относительно высокую активность даже при 75°С, а наибольший выход сахаров наблюдался при 65°С.Можно заключить, что при 65°С фермент проявлял наибольшую активность.
Таким образом, гомогенная мутантная ЦБГ I, содержащая 5 аминокислотных замен, обладает увеличенной термостабильностью при высоких температурах по сравнению с ферментом дикого типа. Выход сахаров при гидролизе МКЦ после 48 часов при 65°С под действием модифицированной ЦБГ I был в 2 раза выше по сравнению с действием фермента дикого типа.
Пример 4. Определение восстанавливающих сахаров (ВС) после проведения гидролиза растительных субстратов.
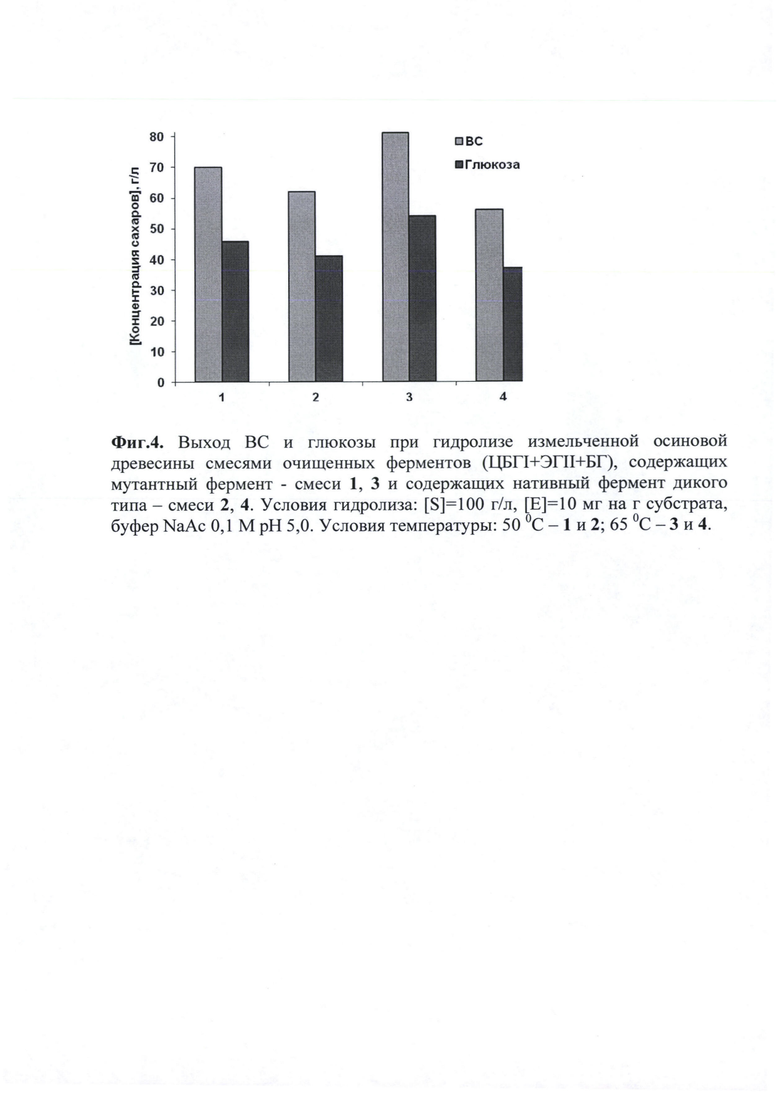
Эксперимент проводили в термостатируемой при заданной температуре ячейке (50°С и 65°С), помещенной в шейкер 'TNNOVA 40 Thermo Shaker" (США). Концентрация субстрата в реакционной смеси составляла 100 г/л (в пересчете на сухое вещество). Реакцию проводили в 0.1 Μ ацетатном буфере при перемешивании (250 об/мин). Гидролиз проводили смесями очищенных ферментов. Смеси состояли из ЦБГ I (мутантная форма и нативная в качестве контроля), ЭГ II (эндо-1,4-β-глюканаза II P.verruculosum) и БГ (β-глюкозидаза A.niger). Ферменты дозировали по концентрации белка, в соотношении ЦБП: ЭГП как 8:2. Конечная концентрация белка в реакционной смеси составляла 10 мг/г сухого субстрата, объем реакционной смеси - 20 мл. β-Глюкозидаза A.niger была дозирована по активности из расчета40 ед. активности на 1 г сухого субстрата. Гидролиз проводили в течение 2 суток. В качестве контроля вместо мутантной ЦБП в реакционную смесь добавляли соответствующее количество нативной ЦБП дикого типа, выделенной из штамма P.canescens ЦБП по схеме из Примера 3. Реакционная ячейка представляла собой пластиковый сосуд с крышкой объемом 50 мл, для обеспечения дополнительного перемешивания реакционной смеси в ячейку помещали металлический цилиндр (d=7 мм, h=10 мм) из нержавеющей стали. Через определенные промежутки времени из реакционной смеси отбирали пробы (по 0.5 мл), центрифугировали 3 мин при 10000 g и измеряли в супернатанте концентрацию восстанавливающих сахаров (ВС) методом Шомоди-Нельсона [Синицын А.П., Гусаков А.В., Черноглазое В.А. Биоконверсия лигноцеллюлозных материалов. - М.: МГУ, 1995. - 144 с] и глюкозы глюкозооксидазным методом [Полыгалина Г.В., Чередниченко B.C., Римарева Л.В. Определение активности ферментов. М.: ДеЛиПринт, 2003. С.147-169.]. Выход ВС и глюкозы представлен на фиг. 4.
Гидролитическая способность смесей 2 и 4, содержащих нативную форму ЦБГ I по отношению к осиновой древесине была ниже, чем у смесей 1 и 3, содержащих мутантную форму ЦБГ I. Причем выход ВС и глюкозы при 65°С смеси 4 был меньше, чем при 50°С (смесь 2).
Гидролитическая способность смеси 1 при 50°С была выше, чем у контрольной смеси в аналогичных условиях. Выход ВС и глюкозы составил 70 и 46 г/л, соответственно, что на 13% больше, чем у контроля.
При повышении температуры существенно увеличилась гидролитическая способность смеси 3 (65°С). Выход ВС и глюкозы составил 81 и 54 г/л, соответственно, что на 45% больше, чем у контроля.
Таким образом, при увеличении термостабильности у мутантной формы ЦБП существенно увеличилась ее гидролитическая способность.




| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM MX-73 ПРОДУЦЕНТ МОДИФИЦИРОВАННОЙ КСИЛАНАЗЫ Е С ПОВЫШЕННОЙ ТЕРМОСТАБИЛЬНОСТЬЮ, ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В ПИЩЕВОЙ И КОРМОВОЙ ПРОМЫШЛЕННОСТИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2018 |
|
RU2711578C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-КСИЛАНАЗЫ Е И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2023 |
|
RU2819918C1 |
| НОВЫЙ РЕКОМБИНАНТНЫЙ ШТАММ (ВАРИАНТЫ) МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM И ФЕРМЕНТНЫЙ ПРЕПАРАТ (ВАРИАНТЫ), ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ГИДРОЛИЗА ПЛОДОВО-ЯГОДНОГО СЫРЬЯ, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2014 |
|
RU2574206C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-ГЛЮКАНАЗЫ II И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2022 |
|
RU2810538C2 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM B6/PMO-BG-25 И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА β-ГЛЮКОЗИДАЗЫ, ЦЕЛЛОБИОГИДРОЛАЗЫ И ПОЛИСАХАРИДМОНООКСИГЕНАЗЫ НА ЕГО ОСНОВЕ ДЛЯ БИОКОНВЕРСИИ ЦЕЛЛЮЛОЗОСОДЕРЖАЩИХ МАТЕРИАЛОВ | 2024 |
|
RU2836771C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ КОМПЛЕКСА ФЕРМЕНТОВ ЭНДОГЛЮКАНАЗ И КСИЛАНАЗ В КЛЕТКАХ ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ НА ЕГО ОСНОВЕ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ КОРМОПРОИЗВОДСТВА | 2017 |
|
RU2653429C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS CL14, ПРОДУЦИРУЮЩИЙ КОМПОНЕНТ ЦЕЛЛЛЮЛОСОМЫ CLOSTRIDIUM THERMOCELLUM, И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2016 |
|
RU2646132C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ЕЕ-105 ПРОДУЦЕНТ КОМПЛЕКСА ВЫСОКОЭФФЕКТИВНЫХ ЭНДОГЛЮКАНАЗ И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ КОРМОВОЙ ДОБАВКИ В ЗЕРНОВЫХ КОРМАХ | 2018 |
|
RU2696036C1 |
| ГЕНЕТИЧЕСКАЯ ФЬЮЖН-КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ МУЛЬТИФЕРМЕНТНОГО КОМПЛЕКСА КАРБОГИДРАЗ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА Penicillium verruculosum, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ШТАММА ГРИБА Penicillium verruculosum И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА НА ЕГО ОСНОВЕ | 2013 |
|
RU2550044C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM ( ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА С ЕГО ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2015 |
|
RU2646136C2 |

Изобретение относится к области биотехнологии, а именно к созданию и производству ферментных препаратов (ФП), содержащих высокоактивные и термостабильные ферменты. Задача, на решение которой направлено данное изобретение, состоит в введении 5-и мутаций в ген сеl7А, кодирующий модифицированную форму ЦБГ I, и получении сухого ФП на основе нового рекомбинантного штамма P. canescens mtCBHI (ВКМ F-4850D), являющегося продуцентом гетерологичной мутантной ЦБГ I P. verruculosum, относящейся к 7-й семье гликозид-гидролаз (КФ 3.2.1.176, мол. масса 55 кДа) для применения в биотопливной индустрии для осуществления эффективного процесса биоконверсии целлюлозосодержащей биомассы в технические сахара. 3 н.п. ф-лы, 4 ил., 2 табл., 4 пр.
1. Гетерологичный ген cel7 А гриба Penicillium verruculosum, содержащий мутации в целевой кодирующей последовательности для обеспечения аминокислотных замен в положениях Т291Р, V290F, E302L, V213W, G342R белка целлобиогидролазы I с увеличенной термостабильностью и повышенной гидролитической активностью при 65°С.
2. Рекомбинантный штамм Penicillium canescens mtCBHI (ВКМ F-4850D), полученный путем трансформации клеток гриба Penicillium canescens RN3-11-7 генетической конструкцией с геном cel7A по п. 1 и предназначенный для секреции гетерологичной термостабильной целлобиогидролазы I Penicillium verruculosum.
3. Ферментный препарат mtCBHI-5, полученный на основе рекомбинантного штамма P. canescens mtCBHI и характеризующийся активностью по микрокристаллической целлюлозе и n-НФ-лактозиду - 240 и 124 ед на 1 г ферментного препарата mtCBHI-5.
| ГУСАКОВ А.В., СИНИЦЫН А.П | |||
| Деполимеризация природных биополимеров | |||
| Ферментативный гидролиз целлюлозы | |||
| Обзор | |||
| "Химия биомассы: биотоплива и биопластики" под ред | |||
| С.Д.Варфоломеева, изд | |||
| Научный мир, Москва, 2017, с | |||
| Разборное приспособление для накатки на рельсы сошедших с них колес подвижного состава | 1920 |
|
SU65A1 |
| DENISENKO Υ.Α., et al | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

