Изобретение относится к области биотехнологии, а именно к созданию и способу получения ферментных препаратов (ФП), содержащих высокоактивные ферменты по отношению к целлюлозосодержащему сырью. Изобретение может быть использовано при производстве технических сахаров из целлюлозосодержащих материалов для их дальнейшей биоконверсии в продукты микробиологического синтеза.
Возобновляемая растительная биомасса и биополимеры, входящие в состав растительной клеточной стенки (целлюлоза, гемицеллюлозы и лигнин), представляют собой основную часть органического материала на Земле и являются практически неисчерпаемым источником сырья и энергии [Гусаков А.В., Синицын А.П. Деполимеризация природных биополимеров. Ферментативный гидролиз целлюлозы. Обзор. «Химия биомассы: биотоплива и биопластики» под ред. С.Д. Варфоломеева. М.: Научный мир, 2017, с.65-99]. Ферментативный гидролиз растительных полимеров позволяет получить С6- (в основном глюкоза) и С5-сахара (ксилоза, арабиноза и др.), т.н. технические сахара, которые могут являться субстратом для таких востребованных продуктов микробиологического синтеза как: биоспирты, органические кислоты, аминокислоты, диолы, фураны, сложные эфиры, алкены, алканы, акрилаты, полимеры, смолы, кормовые продукты [Синицын А.П., Синицына О.А. // Успехи биологической химии. 2021. Т. 61. С. 347-414. doi:10.1134/S0006297921140121].
Технические сахара являются продуктами гидролиза биополимеров растительной биомассы, осуществляемого целлюлазным комплексом ферментов, в который входят три ключевые целлюлазы – экзо-1,4-β-глюканазы (целлобиогидролазы, ЦБГ, КФ 3.2.1.91), эндо-1,4-β-глюканазы (эндоглюканазы, ЭГ, КФ 3.2.1.4) и β-глюкозидазы (БГ, КФ 3.2.1.74), осуществляющие конверсию целлюлозы до глюкозы, целлобиозы и целлоолигосахаридов. К вспомогательным ферментам, усиливающим действие целлюлаз, относятся литические полисахаридмонооксигеназы (ПМО, КФ 1.14.99.54), катализирующие окислительную деструкцию основных цепей целлюлозы, способствующую аморфизации её кристаллических зон [Moon M, Lee JP, Park GW, Lee JS, Park HJ, Min K. Lytic polysaccharide monooxygenase (LPMO)-derived saccharification of lignocellulosic biomass. Bioresour Technol. 2022. 359:127501. doi: 10.1016/j.biortech.2022.127501].
Недавно, в статье [М.В. Семенова, А.В. Гусаков, В.Д. Телицин, В.Ю. Матыс, Т.В. Бубнова, В.А. Немашкалов, А.М. Рожкова, А.П. Синицын, Новый ферментный препарат, содержащий полисахаридмонооксигеназу и β-глюкозидазу –синергетические добавки к целлюлазам, 2022, Прикл. биохим. и микроб, Т.58, №4, стр. 366-373] нами было показано, что совместное воздействие полисахаридмонооксигеназы и β-глюкозидазы в составе ферментного препарата (ФП) ПМО/БГ на основе штамма Penicillium verruculosum B1-537 (DniaD) приводит к увеличению эффективности гидролиза целлюлозосодержащих материалов от 20 до 100% при условии добавки ФП ПМО/БГ к основному комплексу B1-537 в количестве 10-20% от общего белка, используемого в реакции гидролиза. Низкая эффективность использования ФП ПМО/БГ как индивидуального ферментного препарата объясняется собственной низкой авицеллазной активностью, характеризующей целлобиогидролазу I, которая гидролизует кристалличные участки целлюлозы. Эффект снижения авицеллазной активности в рекомбинантных штаммах на основе штамма P. verruculosum B1-537 (DniaD) и, соответственно, в ферментных препаратах, полученных на их основе, наблюдался нами и ранее [Чулкин А.М., Кислицин В.Ю., Зоров И.Н., Синицын А.П., Рожкова А.М. Определение копийности целевых генов карбогидраз в рекомбинантных штаммах гриба Penicillium verruculosum, 2019, Биотехнология, Т.35, №5, стр. 51-57].
Для решения данной проблемы нами был получен новый штамм-реципиент P. verruculosum ΔniaDΔtacA [Чулкин А.М., Кислицин В.Ю., Зоров И.Н., Шашков И.А. Рожкова А.М. Влияние нокаута транскрипционного фактора TacA на транскрипцию и экспрессию гена cbhI в штамме мицелиального гриба Penicillium verruculosum // Вестн. Моск. ун-та. Сер. 2. 2023, Химия. Т. 64. № 2. С. 121–129], в котором проведен нокаут транскрипционного фактора TacA отвечающего за репрессию гена cbh1, что позволяет сохранить уровень авицеллазной активности в реципиентном штамме (депонирован в коллекцию ВКМ как Penicillium verruculosum B6 (ВКМ F-5023D)).
Таким образом, техническая задача, на решение которой направлено предлагаемое изобретение, состоит в получении штамма мицелиального гриба P. verruculosum B6/PMO-BG-25, характеризуюшегося наличием одновременно ферментативных активностей - гомологичной литической полисахаридмонооксигеназы (ПМО, КФ 1.14.99.54) и целлобиогидролазы I (ЦБГI, КФ 3.2.1.91), а также гетерологичной β-глюкозидазы (БГ, КФ 3.2.1.74) гриба A. niger.
Прототипами предлагаемого изобретения могут являться патенты RU 2574206 C1, RU 2646136 C2, RU 2653429 C1, RU 2696036 C1, RU 2810538 C2, RU 2711578 C1, RU 2550044 C2, RU 2532840 C2. Однако во всех приведенных изобретениях в качестве реципиентного штамма использовался мицелиальный гриб-ауксотроф P. verruculosum 537, не имеющий нокаута гена tacA, кодирующего негативный фактор транскрипции TacA.
В изобретении RU 2361918 C1 описывается способ получения ферментного препарата комплекса целлюлаз, ксиланазы и ксилоглюканазы для гидролиза целлюлозы и гемицеллюлозы, получающегося в результате культивирования штамма Penicillium verruculosum BKM F-3972D. Однако описанный способ отличается от предложенного способа культивирования штамма P. verruculosum B6/PMO-BG-25 по параметрам состава ферментационный среды и времени культивирования штамма.
Технический результат от предлагаемого изобретения состоит в сокращении времени культивирования рекомбинантного штамма P. verruculosum B6/PMO-BG-25, а также увеличении выхода глюкозы и восстанавливающих сахаров при ферментативном гидролизе целлюлозосодержащих материалов вследствие применения ферментного препарата, полученного на основе рекомбинантного штамма P. verruculosum B6/PMO-BG-25, за счет комплексного воздействия ПМО, ЦБГI и БГ.
Сущность изобретения заключается в получении рекомбинантного штамма гриба P. verruculosum B6/PMO-BG-25 путем одновременной трансформации плазмидами, содержащими гены lpmo1 и bgl1, кодирующими ПМО P. verruculosum и БГ A. niger соответственно, с последующим получением комплексного ферментного препарата для гидролиза целлюлозосодержащих материалов.
Изобретение реализуется следующим образом:
- Штамм P. verruculosum B6/PMO-BG-25 получают из исходного штамма P. Verruculosum B6 (ВКМ F-5023D) путем котрансформации плазмидами pLPMOI, pCBHI-BGL и pSTA10 с последующей селекцией на агаризованной среде с 10 мМ NaNO3. Получение плазмиды pLPMOI было описано в [Semenova M.V., Gusakov A.V., Volkov P.V., Matys V.Yu., Nemashkalov V.A., Telitsin V.D., Rozhkova A.M., Sinitsyn A.P. Enhancement of the enzymatic cellulose saccharification by Penicillium verruculosum multienzyme cocktails containing homologously overexpressed lytic polysaccharide monooxygenase, Molecular Biology Reports (2019) 46:2363–2370]. Получение плазмиды pCBHI-BGL было описано в [Dotsenko G.S., Gusakov A.V., Rozhkova A.M., Korotkova O.G., Sinitsyn A.P. Heterologous b-glucosidase in a fungal cellulase system: Comparisonof different methods for development of multienzyme cocktails, Process Biochemistry 50 (2015) 1258–1263]. Генетические элементы плазмид, промотор и терминатор гена cbh1 описаны в патенте RU 2378372 ранее.
Способ получения ферментного препарата для гидролиза целлюлозосодержащих материалов предусматривает глубинное культивирование штамма P. verruculosum B6/PMO-BG-25 в течение 96 ч на среде, содержащей микрокристаллическую целлюлозу с последующей распылительной сушкой культуральной жидкости.
Изобретение позволяет получить ФП с высокой активностью гомологичной литической полисахаридмонооксигеназы P. verruculosum и гетерологичной β-глюкозидазы A. niger. Использование штамма P. verruculosum B6 (ВКМ F-5023D) в качестве реципиента для получения P. verruculosum B6/PMO-BG-25 позволяет также сохранить ферментативную активность целлобиогидролазы I в ФП B6/PMO-BG-25.
Применение нового ФП B6/PMO-BG-25 позволяет увеличить эффективность гидролиза целлюлозосодержащих материалов, что характеризуется повышенным выходом восстанавливающих сахаров и глюкозы через 48 ч гидролиза.
Культурально-морфологические и микроскопические особенности штамма P. verruculosum B6/PMO-BG-25:
Растет на агаризованных средах (среда Чапека с дрожжевым автолизатом, Мальц-агар, глюкозо-картофельный агар, сусло-агар) при Т 26-30°С в течение 7-10 суток, рН 4.5-5.0.
На среде Чапека с дрожжевым экстрактом при культивировании гриба при 25°С на 7 сутки колонии достигают 24-30 мм в диаметре, складчатые, поверхность сильно радиально плотная, тонкая, ростовая зона врастает в агар, имеет ширину 1.5-2.0 мм. Мицелий светло-желтоватый, шерстистый, центр колонии выпуклый, конидиогенез слабый, серо-зеленоватого оттенка. Экссудата и растворимого пигмента нет. Обратная сторона светлая. При температуре 37°С колонии диаметром 5 мм, мицелий светлый, конидиообразования нет. При температуре 5°С роста нет.
При росте на Мальц-агаре диаметр колонии 23-24 мм, поверхность сильно радиально складчатая, плотная, тонкая, ростовая зона врастает в агар, шириной 1,5-2,0 мм. Мицелий белый, шерстистый, прижатый, конидиогенез очень слабый, практически отсутствует. Эксудата и растворимого пигмента нет. Обратная сторона светлая.
При микроскопировании штамм имеет конидиеносцы двухярусные, терминальные, бивертициллятные, гладкие длиной около 150 мкм, шириной 2-3мкм. Метулы расходящиеся размером 10-13 × 2.5-3.0 мкм, фиалиды ампуллиформные размером 7-8 × 2.8-3.0 мкм. Конидии округлые, шероховатые размером 3.0-3.5 мкм.
При культивировании в глубинных условиях с использованием растворимых субстратов (глюкоза, фруктоза, лактоза) образуется рыхлый разветвленный мицелий со слабой пеллетизацией, удельная начальная скорость роста мицелия составляла 0,35 ч-1, в конце культивирования 0,1 ч-1.
Физиолого-биохимические признаки штамма:
Мезофилен. Оптимальная температура роста мицелия 32°С (29-34°С), оптимум для образования целлюлаз 28°С (26-29°С). Оптимальные значения рН роста и секреции целлюлаз 3,5-5,0. Рост мицелия наблюдается и при рН 2,5, но при этом наблюдается очень слабое образование целлюлаз и других карбогидраз.
Является прототрофом. Способен быстро ассимилировать глюкозу, глицерин, галактозу, ксилозу, D-маннозу, D-маннит, трегалозу, сорбозу и сорбит, медленнее - D-ксилозу, L- и D-арабинозу, L-рамнозу и рибозу. Слабо ассимилирует: D-глюкозамин, дезоксирибозу, дезоксигалактозу, 2-дезокси-D-глюкозу и 5-тио-D-глюкозу.
Использует неорганический и органический азот, хорошо ассимилирует аммонийную и нитратную форму азота.
Штамм P. verruculosum B6/PMO-BG-25 отличается от своих предшественников одновременным биосинтезом гомологичной литической полисахаридмонооксигеназы и гетерологичной β-глюкозидазы A. niger. На 96 ч культивирования ферментативная активность по п-нитрофенил-β-D-глюкопиранозиду составляет 140 Ед/мл, по 2,6 –диметоксифенолу – 0,6 Ед/мл, по микрокристаллической целлюлозе – 10 Ед/мл.
Методы определения активности
β-глюкозидазную активность измеряли, проводя гидролиз 0,5 мг/мл п-нитрофенил-β-D-глюкопиранозида (пНФГ) при рН 5,0 (0,05 М ацетатный буфер) и 40°С в течение 10 минут. Реакцию останавливают 1 М Nа2СО3, после чего измеряли оптическую плотность при 400 нм. За единицу бета-глюкозидазной активности принимали такое количество фермента, которое в течение 1 минуты при рН 5,0 и 40°С приводит к образованию 1 микромоля п-нитрофенола.
Целлобиогидролазную (авицелазную) активность измеряли, проводя гидролиз МКЦ (5 мг/мл) при рН 5,0 (0,1 М ацетатный буфер) и 50°С в течение 60 минут. После этого определяли в реакционной смеси концентрацию восстанавливающих сахаров (ВС) методом Нельсона-Сомоджи. За единицу целлобиогидролазной активности принимали такое количество фермента, которое в течение 1 минут при температуре 50°С и рН 5,0 при гидролизе суспензии МКЦ концентрацией 5 мг/мл освобождает количество ВС, эквивалентных 1 микромолю глюкозы, и определяемых методом Сомоджи-Нельсона.
Эндоглюканазную (КМЦ-азную) активность измеряли, проводя гидролиз водорастворимой Na-соли карбоксиметилцеллюлозы (5 мг/мл) при рН 5,0 (0,1 М ацетатный буфер) и 50°С в течение 10 минут. За единицу эндоглюканазной активности принимали такое количество фермента, которое в течение 1 минуты при температуре 50°С и рН 5,0 при гидролизе раствора КМЦ концентрацией 5 мг/мл приводило к образованию количества ВС, эквивалентного 1 микромолю глюкозы, определяемых методом Сомоджи-Нельсона [А.П.Синицын, А.В.Гусаков, И.М.Черноглазов. Биоконверсия лигноцеллюлозных материалов, Учебное пособие, М.: Изд-во МГУ, 1995, с.144-156].
Полисахаримонооксигеназную активность определяли с использованием 2,6-диметоксифенола (2,6-ДМФ) в качестве модельного субстрата и H2O2 в качестве второго субстрата при рН 7.5 и 30°С согласно методике, описанной в работе [Breslmayr E., Hanzek M., Hanrahan A., Leitner C.,Kittl R., Santek B., Oostenbrink C., Ludwig R. // Biotechnol.Biofuels. 2018. V. 11. P. 79]. Концентрацию хромогенного продукта рассчитывали, используя коэффициент экстинкции 53200 М-1 см-1.
Содержание белка в ФП определяли методом Лоури, используя БСА в качестве стандарта [Peterson GL. Review of the Folin phenol protein quantitation method of Lowry,Rosebrough, Farr and Randall. Anal Biochem 1979;100:201–20.].
Новый ФП B6/PMO-BG-25, полученный на основе КЖ рекомбинантного штамма P. verruculosum B6/PMO-BG-25 будет востребован в процессах ферментативной обработки предобработанных целлюлозосодержащих материалов за счет повышенной эффективности ферментативного гидролиза, обусловленного наличием литической полисахаридмонооксигеназы и β-глюкозидазы, действующих совместно с базовой целлобиогидролазой I штамма P. verruculosum B6 (ВКМ F-5023D).
Изобретение характеризуется следующими примерами.
Пример 1. Получение рекомбинантного штамма P. verruculosum B6/PMO-BG-25
Реципиентный штамм P. verruculosum B6 (ВКМ F-5023D) одновременно трансформируют плазмидами pLPMOI и pCBHI-BGL, совместно с плазмидой pSTA10 в соотношении 3:1 (мкг каждой ДНК) по стандартной методике [Sambrook, J., and Russell, D.W. (2001) Molecular cloning:a laboratory manual, Cold Spring Harbor Laboratory Press, N.Y.; A.Y. Aleksenko, N.A. Makarova, I.V. Nikolaev, A.J. Clutterbuck, Integrative and replicative transformation of Penicillium canescens with a heterologous nitrate-reductase gene, Curr. Genet. 28 (1995) 474-478]. В результате трансформации получают более 50 рекомбинантных штаммов (клонов) серии P. verruculosum B6/PMO-BG, выросших на агаризованной среде Чапека с добавкой селективного агента - 10мМ NaNO3. В результате первичного скрининга при культивировании в 100 мл стандартной питательной среды в колбах Эрленмейра отбирают клоны, обладающие высокой полисахаридмонооксигеназной, β-глюкозидазной и целлобиогидролазной (авицеллазной) активностями. Стандартная среда культивирования имеет следующий состав (г/л):
МКЦ - 40,
пшеничные отруби - 10,
кукурузный экстракт - 25,
дрожжевой экстракт - 10,
KH2PO4 - 15,
(NH4)2SO4 - 5,
MgSO4×7H2O - 0,3,
CaCl2×2H2O - 0,3.
Условия культивирования: 250 об/мин, 144 ч, 32°С, рН в начале культивирования 5.5 (далее рН не контролируется).
Полисахаридмонооксигеназная активность в отобранных при культивировании клонах серии B6/PMO-BG варьирует от 0,2 до 0,6 ед/мл, β-глюкозидазная от 100 до 140 ед/мл, целлобиогидролазная (авицеллазная) от 9 до 11 ед/мл. Для дальнейших испытаний был отобран клон B6/PMO-BG-25, отличающийся наибольшими целевыми активностями по сравнению с реципиентным штаммом P. verruculosum B6 (ВКМ F-5023D).
Пример 2. Культивирование штамма P. verruculosum B6/PMO-BG-25 в ферментере, получение сухого ФП B6/PMO-BG-25
Культивирование штамма P.verruculosum B6/PMO-BG-25 проводят в ферментере объемом 3 л (КФ-108, «Проинтех», Россия), оснащенном барботером для подачи воздуха в аппарат и турбинной мешалкой на стандартной среде следующего состава (г/л):
МКЦ - 40,
пшеничные отруби - 10,
кукурузный экстракт - 25,
дрожжевой экстракт - 10,
KH2PO4 - 15,
(NH4)2SO4 - 5,
MgSO4×7H2O - 0,3,
CaCl2×2H2O - 0,3.
Культивирование проводят в течение 144 ч, при рН 4,5-5,0 и 32°С.
Параллельно, в аналогичных условиях, проводят культивирование реципиентного штамма P. verruculosum B6 (ВКМ F-5023D) и исходного штамма P. verruculosum PV2007 (ВКМ F-3972D), являющегося предшественником P. verruculosum B6 (ВКМ F-5023D).
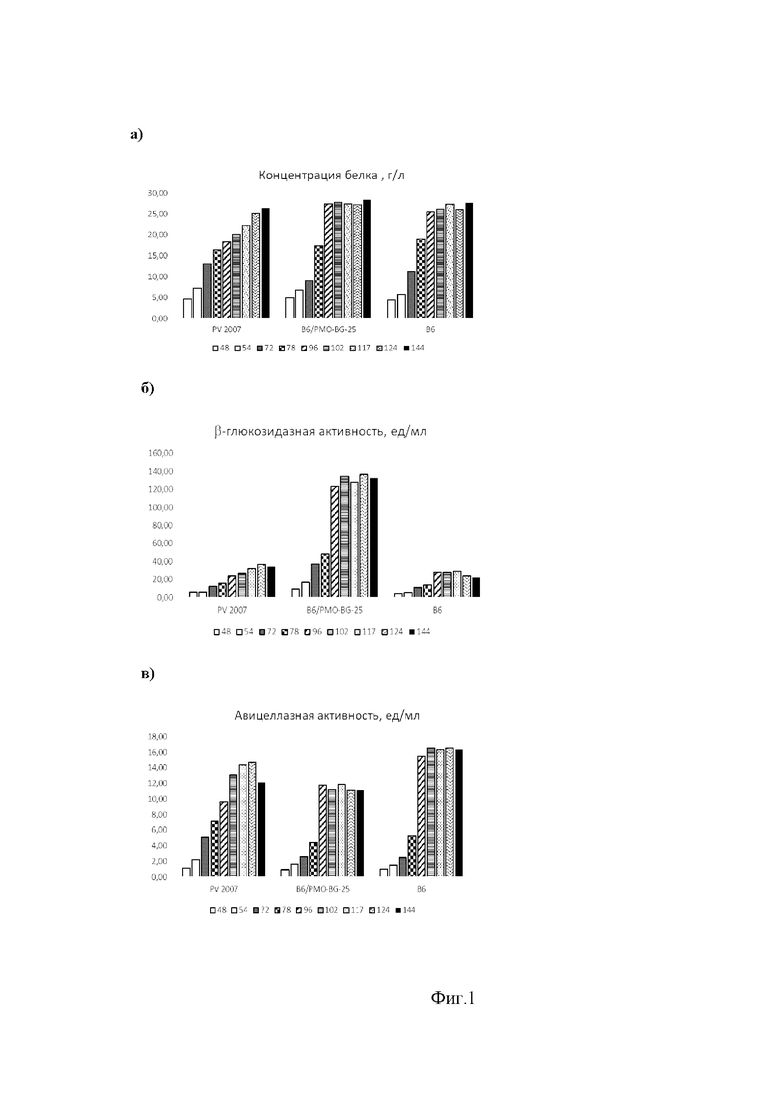
Образцы КЖ отбирают, начиная с 48 ч культивирования (48, 54, 72, 78, 96, 102, 117, 124 и 144 ч), центрифугируют и определяют концентрацию белка (Фиг. 1, а) и β-глюкозидазную активность (Фиг. 1, б). Из Фиг. 1 следует, что максимальная концентрация секретируемого белка при культивировании штамма P. verruculosum B6/PMO-BG-25 достигается уже к 96 ч культивирования (также как и для реципиентного штамма P. verruculosum B6, но не для исходного штамма P. verruculosum PV2007), однако секретируемый комплекс штамма P.verruculosum B6/PMO-BG-25 содержит β-глюкозидазу, уровень которой также не меняется, начиная с 96 ч культивирования. Примечательно, что и уровень авицеллазной активности в КЖ штамма P.verruculosum B6/PMO-BG-25 увеличивается после 96 ч незначительно (Фиг. 1, в).
По окончании ферментации грибную биомассу удаляют путем центрифугирования (4000 об/мин в течение 20 мин на центрифуге Avanti JXN-26, «Beckman coulter», США), свободную от клеток КЖ концентрируют с помощью ультрафильтрации (с пределом отсечения 10 кДа), ультраконцентрат сушат на распылительной сушилке (Buchi MiniSpray Dryer B-290, «BUCHI Labortechnik», Швейцария, Твх=135°С, Твых=55-65°С, степень аспирации=70%, скорость потока 0,5 л КЖ в час) с получением сухого ФП, который представляет собой светло-бежевый легко растворимый в водной среде порошок.
Таким образом, получают сухой ФП B6/PMO-BG-25 (отбирая КЖ после 96 ч культивирования). Параллельно в качестве контролей получают ФП PV2007 и ФП В6 (отбирая КЖ после 144 ч культивирования). Ферментативные активности ФП показаны в Табл. 1.
Таблица 1- Ферментативные активности и концентрация белка в ФП PV2007, ФП B6 и ФП B6-PMO-BG-25, ед/г белка
Пример 3. Определение восстанавливающих сахаров (ВС) и глюкозы после проведения гидролиза целлюлозосодержащих субстратов
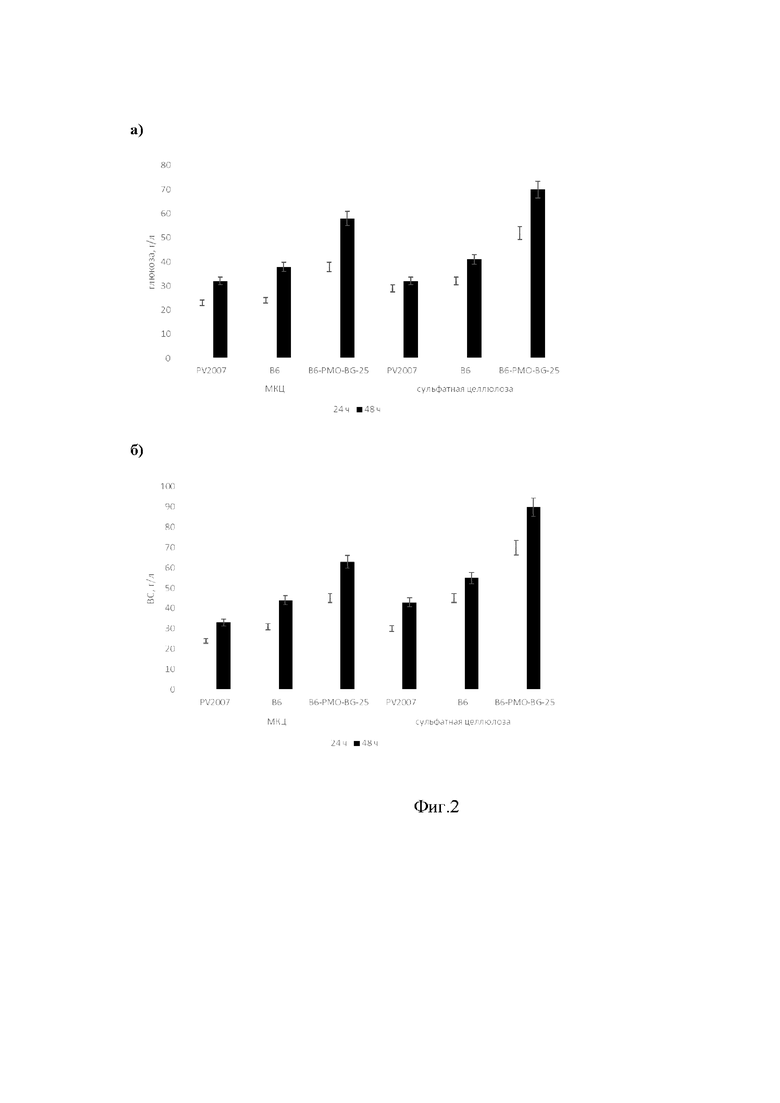
Эксперимент проводили в термостатируемой при заданной температуре ячейке (50°С), помещенной в шейкер “INNOVA 40 Thermo Shaker” (США). Концентрация субстрата (либо МКЦ, либо сульфатная целлюлоза) в реакционной смеси составляла 100 г/л (в пересчете на сухое вещество). Реакцию проводили в 0.1 М ацетатном буфере при перемешивании (250 об/мин). Гидролиз проводили под действием ФП (либо PV2007, либо В6, либо В6/PMO-BG-25). Конечная концентрация белка в реакционной смеси составляла 10 мг/г сухого субстрата, объем реакционной смеси – 20 мл. Гидролиз проводили в течение 2 суток при 50°С. Реакционная ячейка представляла собой пластиковый сосуд с крышкой объемом 50 мл, для обеспечения дополнительного перемешивания реакционной смеси в ячейку помещали металлический цилиндр (d=7 мм, h=10 мм) из нержавеющей стали. Через определенные промежутки времени из реакционной смеси отбирали пробы (по 0.5 мл), центрифугировали 3 мин при 10000 g и измеряли в супернатанте концентрацию восстанавливающих сахаров (ВС) методом Шомоди–Нельсона [Синицын А.П., Гусаков А.В., Черноглазов В.А. Биоконверсия лигноцеллюлозных материалов. – М.: МГУ, 1995. – 144 с] и глюкозы глюкозооксидазным методом [Полыгалина Г.В., Чередниченко В.С., Римарева Л.В. Определение активности ферментов. М.: ДеЛиПринт, 2003. С. 147–169.]. Выход ВС и глюкозы представлен на Фиг. 2.
Как следует из результатов эксперимента гидролитическая способность ФП В6/PMO-BG-25 превосходит гидролитическую способность контролей как в случае гидролиза МКЦ, так и при гидролизе сульфатной целлюлозы. Выход ВС при гидролизе МКЦ под действием ФП В6/PMO-BG-25 был в 1,5 и 2 раза выше, чем под действием ФП В6 и ФП PV2007 соответственно. При гидролизе сульфатной целлюлозы под действием ФП В6/PMO-BG-25 выход ВС был в 1, 7 и 2,2 раза выше, чем в соответствующих контролях.
Увеличение гидролитической способности ФП В6/PMO-BG-25 объясняется наличием дополнительной гетерологичной БГ и гомологичной ПМО, а также собственным высоким уровнем авицеллазной активности, что приводит к сбалансированности целлюлолитического комплекса ферментов, секретируемого новым штаммом P. verruculosum В6/PMO-BG-25.
Таким образом, технический результат от предлагаемого изобретения состоит в сокращении времени культивирования рекомбинантного штамма P. verruculosum B6/PMO-BG-25 до 96 ч, а также увеличении выхода глюкозы и восстанавливающих сахаров при ферментативном гидролизе целлюлозосодержащих материалов вследствие применения ферментного препарата, полученного на основе рекомбинантного штамма P. verruculosum B6/PMO-BG-25, за счет комплексного воздействия ПМО, ЦБГI и БГ.
На Фиг. 1 приведена динамика изменения концентрации белка (а), β-глюкозидазной (б) и авицеллазной активности (с) в культуральной жидкости исходного штамма P. verruculosum PV2007 (ВКМ F-3972D), рекомбинантного штамма P. verruculosum B6/PMO-BG-25 и реципиентного штамма P. verruculosum B6.
На Фиг. 2 приведена концентрация глюкозы (а) и восстанавливающих сахаров (б) в гидролизатах микрокристаллической целлюлозы и сульфатной целлюлозы после 24 и 48 ч гидролиза под действием ФП PV2007, ФП B6 и ФП B6-PMO-BG-25.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКАЯ ФЬЮЖН-КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ МУЛЬТИФЕРМЕНТНОГО КОМПЛЕКСА КАРБОГИДРАЗ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА Penicillium verruculosum, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ШТАММА ГРИБА Penicillium verruculosum И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА НА ЕГО ОСНОВЕ | 2013 |
|
RU2550044C2 |
| НОВЫЙ РЕКОМБИНАНТНЫЙ ШТАММ (ВАРИАНТЫ) МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM И ФЕРМЕНТНЫЙ ПРЕПАРАТ (ВАРИАНТЫ), ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ГИДРОЛИЗА ПЛОДОВО-ЯГОДНОГО СЫРЬЯ, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2014 |
|
RU2574206C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ КОМПЛЕКСА ФЕРМЕНТОВ ЭНДОГЛЮКАНАЗ И КСИЛАНАЗ В КЛЕТКАХ ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ НА ЕГО ОСНОВЕ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ КОРМОПРОИЗВОДСТВА | 2017 |
|
RU2653429C1 |
| ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM - ПРОДУЦЕНТ КОМПЛЕКСА ЦЕЛЛЮЛАЗ, КСИЛАНАЗЫ И КСИЛОГЛЮКАНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА КОМПЛЕКСА ЦЕЛЛЮЛАЗ, КСИЛАНАЗЫ И КСИЛОГЛЮКАНАЗЫ ДЛЯ ГИДРОЛИЗА ЦЕЛЛЮЛОЗЫ И ГЕМИЦЕЛЛЮЛОЗЫ | 2008 |
|
RU2361918C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-ГЛЮКАНАЗЫ II И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2022 |
|
RU2810538C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM ( ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА С ЕГО ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2015 |
|
RU2646136C2 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ЕЕ-105 ПРОДУЦЕНТ КОМПЛЕКСА ВЫСОКОЭФФЕКТИВНЫХ ЭНДОГЛЮКАНАЗ И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ КОРМОВОЙ ДОБАВКИ В ЗЕРНОВЫХ КОРМАХ | 2018 |
|
RU2696036C1 |
| ШТАММ ГРИБА PENICILLIUM CANESCENS mtCBHI ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ ЦЕЛЛОБИОГИДРОЛАЗЫ I И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ БИОКОНВЕРСИИ ВОЗОБНОВЛЯЕМОГО РАСТИТЕЛЬНОГО СЫРЬЯ В САХАРА | 2019 |
|
RU2741078C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM ПРОДУЦЕНТ КОМПЛЕКСА ФИТАЗЫ А И ЭНДО-1,4-β-КСИЛАНАЗЫ Е И ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ ДОБАВКИ В КОРМАХ | 2023 |
|
RU2819918C1 |
| ШТАММ ГРИБА Penicillium verruculosum B10 EGII ПРОДУЦЕНТ ЭНДО-1.3/1.4-β-ГЛЮКАНАЗЫ, ЦЕЛЛЮЛАЗЫ, β-ГЛЮКОЗИДАЗЫ И КСИЛАНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА | 2012 |
|
RU2532840C2 |
Изобретение относится к биотехнологии. Предложены рекомбинантный штамм Penicillium verruculosum B6/PMO-BG-25, предназначенный для продукции гомологичной литической полисахаридмонооксигеназы, гомологичной целлобиогидролазы I и гетерологичной β-глюкозидазы Aspergillus niger, а также способ получения ферментного препарата B6/PMO-BG-25 с использованием указанного рекомбинантного штамма. Изобретение обеспечивает сокращение времени культивирования указанного рекомбинантного штамма, а также увеличение выхода глюкозы и восстанавливающих сахаров при ферментативном гидролизе целлюлозосодержащих материалов при использовании указанного ферментного препарата. 2 н.п. ф-лы, 2 ил., 1 табл., 3 пр.
1. Рекомбинантный штамм Penicillium verruculosum B6/PMO-BG-25, предназначенный для продукции гомологичной литической полисахаридмонооксигеназы, гомологичной целлобиогидролазы I и гетерологичной β-глюкозидазы Aspergillus niger, полученный путем котрансформации штамма Penicillium verruculosum ΔniaDΔtacA плазмидами pLPMOI, содержащей ген lpmo1, кодирующий полисахаридмонооксигеназу P. verruculosum, и pCBHI-BGL, содержащей ген bgl1, кодирующий β-глюкозидазу A. niger, совместно с плазмидой pSTA10.
2. Способ получения ферментного препарата B6/PMO-BG-25, полученного на основе рекомбинантного штамма Penicillium verruculosum B6/PMO-BG-25 по п.1, при выращивании на ферментационной среде, обеспечивающий получение ферментного препарата комплексного действия, включающего гомологичную литическую полисахаридмонооксигеназу, гомологичную целлобиогидролазу I и гетерологичную β-глюкозидазу Aspergillus niger, и состоящий в культивировании рекомбинантного штамма по п.1 при 32°С в течение 96 ч, на ферментационной среде, состоящей из (г/л):
| СЕМЕНОВА М.В | |||
| и др | |||
| Новый ферментный препарат, содержащий полисахаридмонооксигеназу и β-глюкозидазу - синергетические добавки к целлюлазам | |||
| ПРИКЛАДНАЯ БИОХИМИЯ И МИКРОБИОЛОГИЯ, 2022, т | |||
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
| Саморазгружающаяся железнодорожная платформа | 1921 |
|
SU366A1 |
| ЧУЛКИН А.М | |||
| и др | |||
| Влияние нокаута транскрипционного фактора TacA на транскрипцию и экспрессию гена cbhI в штамме мицелиального гриба | |||

