ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к молекуле, которая является по меньшей мере биспецифической в отношении антигенов CD22 и CD79, к препарату, содержащему указанную молекулу, и к применению любого из перечисленного в лечении. Настоящее изобретение также относится к способам получения указанных молекул и указанных препаратов. В независимом аспекте изобретение также относится к новым последовательностям антител и фрагментов, описанных в настоящем документе.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Биологические механизмы in vivo представляют собой чрезвычайно сложные каскады сигналов, которые с трудом поддаются выяснению и пониманию. Примерами таких сигналов являются сигналы, необходимые для активации B-клеток. B-клеточный рецептор антигена (BCR) состоит из молекул мембранных иммуноглобулинов (mIg) и ассоциированных гетеродимеров (α/β) Igα/Igβ (CD79a/CD79b). Субъединицы mIg связывают антиген, что приводит к агрегации рецепторов, в то время как α/β субъединицы передают сигналы внутрь клетки. Агрегация BCR быстро активирует киназы Lyn, Blk и Fyn семейства Src, а также тирозинкиназы Syk и Btk. Этот процесс инициирует образование «сигналосомы», состоящей из BCR, вышеупомянутых тирозинкиназ, адаптерных белков, таких как CD19 и BLNK, и сигнальных ферментов, таких как PLCγ2, PI3K и Vav.
Сигналы, исходящие из сигналосомы, активируют несколько сигнальных каскадов, включающих киназы, ГТФазы и факторы транскрипции. Это приводит к изменениям в клеточном метаболизме, экспрессии генов и организации цитоскелета. Сложность сигнализации BCR допускает множество различных исходов, включая выживание, толерантность (анергию) или апоптоз, пролиферацию и дифференциацию в клетки, продуцирующие антитела, или B-клетки памяти. Результат ответа определяется состоянием зрелости клетки, природой антигена, амплитудой и продолжительностью сигнализации BCR, а также сигналами от других рецепторов, таких как рецептор CD40, IL-21 и BAFF-R. Многие другие трансмембранные белки, некоторые из которых являются рецепторами, моделируют определенные элементы сигнализации BCR. Некоторые из них включают CD45, CD19, CD22, PIR-B и FcγRIIB1 (CD32). Амплитуда и продолжительность сигнализации BCR ограничены отрицательными петлями обратной связи, в том числе теми, в которые вовлечены путь Lyn/CD22/SHP-1, путь Cbp/Csk, SHIP, Cbl, Dok-1, Dok-3, FcγRIIB1, PIR-B и интернализация BCR. In vivo B-клетки часто активируются антигенпредставляющими клетками, которые захватывают антигены и экспонируют их на своей клеточной поверхности. Активация B-клеток такими ассоциированными с мембраной антигенами требует индуцируемой BCR реорганизации цитоскелета.
Аутореактивные B-клетки отвечают за продуцирование патогенных аутоантител, которые способны либо прямо, либо косвенно вызывать или обострять аутоиммунные состояния. Истощение пула CD20-положительных B-клеток было использовано для успешного лечения ряда аутоиммунных состояний и, таким образом, было окончательно установлено, что В-клетки играют важную роль в возникновении или поддержании целого ряда аутоиммунных заболеваний. Хотя истощение пула B-клеток явилось успешным методом лечения, существуют также доказательства того, что контроль роста и статуса активации B-клеток также может быть эффективным способом модуляции функции B-клеток. Таким образом, существует потребность в альтернативных стратегиях, не требующих истощения пула B-клеток и предлагающих гибкость контролирования B-клеток без долгосрочного подавления B-клеточного иммунитета, что, как показано, связано с некоторыми побочными эффектами. Кроме того, не все B-клеточные ответы или активности являются вредными, и данные свидетельствуют о том, что поддержание популяций регуляторных B-клеток может играть защитную роль. Такой подход может быть эффективным в случае заболеваний, при которых имеет место аномальная функция B-клеток, вызываемая неадекватной или чрезмерной сигнализацией BcR. Примеры включают, но не ограничиваются ими, воспаление, аутоиммунные заболевания и рак. Особый интерес представляют заболевания, при которых существует либо прямая необходимость в сигнализации BcR, либо необходимость в ингибировании или стимуляции гуморальных иммунных ответов.
Ожидается, что биспецифические антитела будут играть главную роль среди биотерапевтических средств следующего поколения (D. Holmes, Nature Rev Drug Disc Nov 2011, 10, 798). Они обладают потенциалом обеспечения превосходной, долговременной, широкого профиля эффективности у большого числа пациентов. Это может быть достигнуто либо путем одновременного совместного вовлечения различных антигенов в рамках общего пути заболевания, с соответствующим уменьшением избыточности; либо путем таргетирования антигенов из независимых путей для обеспечения аддитивного или синергического эффекта.
Современные стратегии ингибирования B-клеточной функции без истощения пула B-клеток сфокусированы на использовании природного механизма регуляции с помощью CD32b (FcgRIIB). Сюда относятся биспецифические антитела к CD79b/CD32b (Veri et al., Arthritis & Rheumatism 2010, 62, 1933-1943), CD19/CD32b (Karnell et al., J.Immunol 2014, 192, 1480-1490) и антитело к CD19 с областью Fc, отличающейся усиленным связыванием CD32b (Chu et al., Arthritis & Rheumatology 2014 66 1153-1164).
Связывание Fc-гамма рецептора IIb (CD32b) с B-клеточным рецептором естественным образом регулирует сигнализацию, в частности, когда антиген связан с антителом в небольших иммунных комплексах. Затем CD32b рекрутирует фосфатазы SHP-1 и SHIP-1, которые препятствуют активации BcR. Хотя этот природный регуляторный механизм может контролировать функцию B-клеток, нарушение функции CD32b, вызываемое изменениями в белковой последовательности CD32b, может приводить к аутоиммунному заболеванию, и при аутоиммунном заболевании может иметь место понижающая регуляция этого рецептора - например, как в случае SLE. Таким образом, существует потребность в альтернативных путях блокирования активности B-клеток, предлагающих альтернативные, неприродные пути регуляции функции BcR. Эти альтернативные механизмы, вероятно, будут наиболее важны в случае, когда природные механизмы не работают при конкретном заболевании.
Биспецифические антитела облегчают доступ к новым биологическим возможностям, таким как:
1) сшивка рецепторов с поверхностью клетки, в случае необходимости,
2) индукция опосредованных клетками эффектов,
3) локализация цитокина на клетке для регуляции сигнализации или локального блокирования функции цитокина,
4) вовлечение одновременно нескольких эпитопов для создания «новой активности», усиления функции или специфичности, что может не проявляться при использовании моноклонального антитела или фактической смеси несвязанных антител («поли-моноклональных антител»), включая смеси, направленные на разные антигены.
Авторы настоящего изобретения неожиданно обнаружили, что за счет использования биспецифического антитела для связывания BcR (CD79) с молекулой отрицательной регуляции CD22, которая в нормальных физиологических условиях была бы исключена из комплекса, сигнализация BcR может быть подавлена. CD22 отвечает за регуляцию тонизирующей сигнализации через BcR в отсутствие связывания антигена. Однако при связывании антигена CD22 обычно исключается из комплекса BcR. Авторы изобретения обнаружили, что путем физического связывания BcR с сигнальной молекулой CD22 за счет использования биспецифического антитела активацию в B-клетках можно ингибировать.
Таким образом, авторы настоящего изобретения выявили синергетическую функцию для молекул, которые являются по меньшей мере биспецифическими в отношении CD22 и CD79. Эта функция, судя по всему, поддается обнаружению, главным образом, когда связывающие области с комбинацией специфичностей предоставлены в биспецифическом (мультиспецифическом) формате, в отличие от того, когда они предоставлены просто в виде смеси, например, моноклональных антител или их связывающих фрагментов.
Мультиспецифические молекулы по изобретению, таким образом, являются полезными для контролирования аберрантных функций B-клеток, ассоциированных с некоторыми заболеваниями, такими как аутоиммунные заболевания и рак.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, предложена мультиспецифическая молекула, содержащая связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79.
Сочетание по настоящему изобретению в биспецифическом формате демонстрирует интересную биологическую активность в функциональных анализах in vitro, например, ингибирование B-клеточной сигнализации, что измеряют одним из следующих методов, позволяющих определять: ингибирование фосфорилирования Akt S473, ингибирование фосфорилирования P38 и PLCγ2 Y759, ингибирование IκB, в дополнение к ингибированию экспрессии CD86, CD71 и/или CD40 на B-клетках. Такой же уровень активности не наблюдается в случае индивидуальных компонентов, используемых самостоятельно, или в случае компонентов, предоставленных в смеси. Однако активность является очевидной при использовании биспецифического конструкта со специфичностью в отношении CD22 и CD79b.
Ингибирование, наблюдаемое в этих анализах, свидетельствует о том, что мультиспецифическая молекула по изобретению, содержащая связывающий домен, специфичный в отношении CD22, и связывающий домен, специфичный в отношении CD79, может быть использована для изменения функции B-клеток и создания терапевтической альтернативы истощению пула B-клеток.
Сигнализация B-клеточного рецептора является крайне важной функцией B-клетки и необходимым условием для антиген-специфической активации B-клеток. Сигнализация BcR является крайне важной с ранних стадий развития B-клеток и далее для активации и развития B-клеточных ответов памяти. B-клеточный рецептор состоит из молекулы поверхностного иммуноглобулина (Ig), ассоциированной с гетеродимерным комплексом из CD79a и CD79b. Когда поверхностный Ig узнает антиген, считается, что это приводит к кластеризации комплекса CD79a/b, результатом чего является понижающая активация непосредственного сигнального каскада, включающего киназы семейства Src, а также тирозинкиназы Syk и Btk. Этот сигнальный комплекс затем может рекрутировать адаптерные белки, такие как CD19 и BLNK, и это приводит к активации PLCγ2 и PI3K, что, в свою очередь, может активировать другие последующие пути, такие как те, которые контролируют рост, выживание и дифференциацию B-клеток. Этот сигнальный комплекс может дополнительно регулироваться другими вторыми сигналами за счет сигнализации через BAFF-R, IL-21R и CD40 и также может регулироваться другими сигнальными молекулами, такими как CD19, CD21, CD83, CD22, CD32b и CD45, в числе прочих. При узнавании антигена BcR одним из первых активированных ответов является повышающая регуляция поверхностных рецепторов, таких как костимулирующие молекулы CD80 и CD86. Эти молекулы связываются с соответствующими рецепторами на T-клетках, которые обеспечивают дополнительные сигналы выживания и активации, позволяющие выживать и размножаться T-клеткам, которые узнают антиген в контексте молекул MHC класса II. Этот ответ дополнительно усиливается за счет способности B-клеток представлять антиген в контексте молекул MHC класса II обратно T-клетке, которая высвобождает такие факторы, как IL-2 и IL-21. Эти цитокины, в свою очередь, способствуют значительному увеличению числа B-клеток. Таким образом, понижающая регуляция CD86 на поверхности клеток может быть показателем ингибирования B-клеточной сигнализации.
Кроме того, ингибирование сигнализации B-клеточного рецептора может приводить к ингибированию последующих функций. Одним из таких результатов могло бы быть ингибирование костимулирующих молекул, таких как CD86 (или снижение их экспрессии), что будет приводить к ингибированию функции, выживания и дифференциации T-клеток.
Таким образом, ингибирование сигнализации B-клеточного рецептора может быть полезным для контролирования аберрантных функций B-клеток, ассоциированных с аутоиммунными заболеваниями и раком. Сигнализация B-клеточного рецептора необходима для B-клеточной пролиферации, дифференциации, представления антигена и секреции цитокинов при аутоиммунном заболевании. Таким образом, ингибирование активности BcR может регулировать функции B-клеток, такие как секреция иммуноглобулинов, активация T-клеток и контроль неадекватной активности B-клеток, ассоциированной, например, с аутоиммунными состояниями. Кроме того, существуют некоторые формы B-клеточного лейкоза и лимфомы, при которых необходима сигнализация B-клеточного рецептора для выживания и роста клеток, это можно контролировать с помощью ингибиторов активации B-клеточных рецепторов.
В одном варианте осуществления связывающий домен или связывающие домены мультиспецифических молекул по настоящему изобретению, каждый независимо, содержат один или два (например, два) вариабельных домена антитела, специфичных в отношении релевантного антигена (такого как CD22 или CD79, или другого антигена, если молекула является по меньшей мере триспецифической).
При использовании в настоящем документе «CD79» означает комплекс, состоящий из CD79a и CD79b. Соответственно, антитела или связывающие домены, которые связывают CD79, могут связывать CD79a и/или CD79b. При использовании в настоящем документе «связывание CD79a и/или CD79b» означает связывание, специфичное в отношении CD79a, специфичное в отношении CD79b, специфичное как в отношении CD79a, так и в отношении CD79b (то есть, одно и то же антитело или связывающий домен узнает эпитоп на CD79a и также узнает эпитоп на CD79b, то есть, является полиспецифическим), или специфичное в отношении комплекса из CD79a и CD79b (то есть узнает эпитоп, образованный в результате взаимодействия CD79a и CD79b в форме комплекса, и оно способно отличать комплекс от отдельных компонентов).
В одном варианте осуществления антитело, или его связывающий фрагмент, используемое в молекулах по настоящему изобретению, является специфичным в отношении CD79a.
В одном варианте осуществления антитело, или его связывающий фрагмент, используемое в молекулах по настоящему изобретению, является специфичным в отношении CD79b.
В одном варианте осуществления антитело, или его связывающий фрагмент, используемое в молекулах по настоящему изобретению, является специфичным в отношении комплекса CD79, то есть оно узнает эпитоп, присутствующий в комплексе, и является специфичным в отношении него, например, эпитоп, включающий участок соприкосновения между CD79a и CD79b.
В одном варианте осуществления, даже если связывающий домен является специфичным в отношении CD79a или CD79b, следует понимать, что связывающий домен предпочтительно все еще будет связывать CD79a или CD79b, находящиеся в форме комплекса, поскольку два белка естественным образом совместно экспрессируются на клеточной поверхности.
Если в связывающем домене и/или в каждом связывающем домене имеются две вариабельные области, тогда две вариабельные области, как правило, будут действовать совместно, обеспечивая специфичность в отношении релевантного антигена, например, они являются когнатной парой или парой с созревшей аффинностью для обеспечения надлежащей аффинности таким образом, что домен является специфичным в отношении конкретного антигена. Как правило, они представляют собой пару вариабельных областей тяжелой и легкой цепей (пару VH/VL).
В одном варианте осуществления молекула по настоящему изобретению является биспецифической.
В одном варианте осуществления молекула по настоящему изобретению является триспецифический.
В одном варианте осуществления молекула по настоящему изобретению является моноспецифической в отношении CD79 и моноспецифической в отношении CD22, то есть, молекула содержит только один связывающий домен, который связывает CD79, и один связывающий домен, который связывает CD22.
В одном варианте осуществления мультиспецифическая молекула по настоящему изобретению является одноцепочечной.
В одном варианте осуществления мультиспецифическая молекула по настоящему изобретению содержит тяжелую цепь, а также легкую цепь. В одном примере, приведенном в настоящем документе, спаренную тяжелую и легкую цепь не называют димером, в частности, когда в одном варианте осуществления молекула по настоящему изобретению не содержит мультимеры, например, димеры, антитела, части/фрагмента или компонентов.
В одном аспекте предложена мультиспецифическая молекула антитела, содержащая или состоящая из:
a) полипептидной цепи формулы (I):
VH-CH1-X-(V1)p;
b) полипептидной цепи формулы (II):
VL-CL-Y-(V2)q;
где:
VH представляет собой вариабельный домен тяжелой цепи;
CH1 представляет собой домен константной области тяжелой цепи, например, домен 1 из нее;
X представляет собой связь или линкер, например, аминокислотный линкер;
Y представляет собой связь или линкер, например, аминокислотный линкер;
V1 представляет собой dab, scFv, dsscFv или dsFv;
VL представляет собой вариабельный домен, например, вариабельный домен легкой цепи;
CL представляет собой домен из константной области, например, домен константной области легкой цепи, такой как Cкаппа;
V2 представляет собой dab, scFv, dsscFv или dsFv;
p равно 0 или 1;
q равно 0 или 1; и
если p равно 1, q равно 0 или 1, и если q равно 1, p равно 0 или 1, то есть, p и q оба не равны 0.
В одном варианте осуществления молекула содержит не более одного связывающего домена для CD22 и не более одного связывающего домена для CD79.
Вышеуказанный формат является особенно полезным для скрининга сочетаний вариабельных областей, например, в долгосрочных анализах и для терапевтического применения.
В одном варианте осуществления q равно 0 и p равно 1.
В одном варианте осуществления q равно 1 и p равно 1.
В одном варианте осуществления V1 представляет собой dab и V2 представляет собой dab, и вместе они образуют один связывающий домен из кооперативной пары вариабельных областей, такой как когнатная пара VH/VL.
В одном варианте осуществления VH и VL являются специфичными в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления V1 является специфичной в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления V2 является специфичной в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления V1 и V2 вместе (например, в виде одного связывающего домена) являются специфичными в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления VH и VL являются специфичными в отношении CD22.
В одном варианте осуществления V1 является специфичной в отношении CD22.
В одном варианте осуществления V2 является специфичной в отношении CD22.
В одном варианте осуществления V1 и V2 вместе (например, в виде одного связывающего домена) являются специфичными в отношении CD22.
В одном варианте осуществления молекула по настоящему изобретению представляет собой или содержит слитый белок.
В одном варианте осуществления предложена мультиспецифическая молекула по настоящему изобретению, которая представляет собой биспецифический белковый комплекс, имеющий формулу A-X:Y-B, где:
A-X представляет собой первый слитый белок;
Y-B представляет собой второй слитый белок;
X:Y представляет собой гетеродимерное сплетение;
A содержит первый связывающий домен, специфичный в отношении CD22 или CD79;
B содержит второй связывающий домен, специфичный в отношении CD22 или CD79;
X представляет собой первый партнер по связыванию связывающейся пары;
Y представляет собой второй партнер по связыванию связывающейся пары; и
: обозначает взаимодействие (такое как взаимодействие связывания) между X и Y, и
при этом по меньшей мере один из A или B является специфичным в отношении CD22 и другой является специфичным в отношении CD79.
Вышеуказанный формат является удобным форматом, поскольку он обеспечивает быстрый и эффективный путь сборки биспецифических форматов, которые, например, можно тестировать in vitro в функциональных анализах. Это может облегчать выбор предпочтительной пары вариабельных областей, которые впоследствии можно включать в альтернативный формат терапевтического мультиспецифического антитела. Без привязки к конкретной теории, различные пермутации вариабельных областей, специфичных в отношении CD22, в сочетании с диапазоном вариабельных областей, специфичных в отношении CD79, могут открывать доступ к различным нюансам биологической функции.
По изобретению также предложены новые антитела к CD22 для использования в мультиспецифических молекулах по настоящему изобретению или для включения в любой другой подходящий формат антитела.
По изобретению также предложены новые антитела к CD79 для использования в мультиспецифических молекулах по настоящему изобретению или для включения в любой другой подходящий формат антитела.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 представляет собой гистограмму относительной эффективности ингибирования фосфорилирования Akt для биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b.
Фигура 2 представляет собой гистограмму относительной эффективности ингибирования фосфорилирования PLCγ2 для биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b.
Фигура 3 представляет собой гистограмму относительной эффективности ингибирования экспрессии CD86 для биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b.
Фигура 4 представляет собой гистограмму относительной эффективности ингибирования фосфорилирования Akt для биспецифических, двухвалентных сочетаний или смесей антител со специфичностью в отношении CD22 и CD79b.
Фигура 5 представляет собой гистограмму относительной эффективности ингибирования фосфорилирования PLCγ2 для биспецифических, двухвалентных сочетаний или смесей антител со специфичностью в отношении CD22 и CD79b.
Фигура 6 представляет собой график, показывающий титрование эффекта биспецифического сочетания для CD22 и CD79b на общие уровни IκB в стимулированных анти-IgM B-клетках.
Фигура 7 представляет собой график, показывающий титрование эффекта биспецифического сочетания для CD22 и CD79b на экспрессию CD86 на стимулированных анти-IgM B-клетках.
Фигура 8 представляет собой график ингибирования фосфорилирования PLCγ2 для биспецифических белков со специфичностью в отношении CD22 и CD79b и с разными V-областями.
Фигура 9 представляет собой выдержку из статьи Chan and Carter, Nature Reviews Immunology vol 10, May 2010, 301, демонстрирующую некоторые форматы антител.
Фигура 10 представляет собой таблицу, показывающую данные для сетки перекрестных специфичностей в отношении антигенов. Антиген 2=CD79b и антиген 3=CD22. Значения соответствуют выраженному в процентах ингибированию (отрицательные значения для активации) фосфорилирования Syk и представляют собой среднее значение для нескольких оцененных сочетаний V-областей.
Фигура 11 представляет собой таблицу, показывающую данные для сетки перекрестных специфичностей в отношении антигенов. Антиген 2=CD79b и антиген 3=CD22. Значения соответствуют выраженному в процентах ингибированию (отрицательные значения для активации) фосфорилирования PLCγ2 и представляют собой среднее значение для нескольких оцененных сочетаний V-областей.
Фигура 12 представляет собой таблицу, показывающую данные для сетки перекрестных специфичностей в отношении антигенов. Антиген 2=CD79b и антиген 3=CD22. Значения соответствуют выраженному в процентах ингибированию (отрицательные значения для активации) фосфорилирования AKT и представляют собой среднее значение для нескольких оцененных сочетаний V-областей.
Фигура 13 представляет собой график, показывающий выраженное в процентах ингибирование фосфорилирования Syk, PLCγ2 и AKT для каждого сочетания V-областей со специфичностью в отношении CD79b в Fab-X в сочетании со специфичностью в отношении CD22 в Fab-Y.
Фигура 14 представляет собой график, показывающий выраженное в процентах ингибирование фосфорилирования Syk, PLCγ2 и AKT для каждого сочетания V-областей со специфичностью в отношении CD22 в Fab-X в сочетании со специфичностью в отношении CD79b в Fab-Y.
Фигура 15 демонстрирует данные для выраженного в процентах ингибирования индуцированного анти-IgM фосфорилирования PLCγ2 в B-клетках за счет специфичных в отношении CD79b и CD22 Fab-Kd-Fab или BYbe.
Фигура 16 демонстрирует данные для выраженного в процентах ингибирования индуцированного анти-IgM фосфорилирования P38 в B-клетках за счет специфичных в отношении CD79b и CD22 Fab-Kd-Fab или BYbe.
Фигура 17 демонстрирует данные для выраженного в процентах ингибирования индуцированного анти-IgM фосфорилирования Akt в B-клетках за счет специфичных в отношении CD79b и CD22 Fab-Kd-Fab или BYbe.
Фигура 18 демонстрирует данные для выраженного в процентах ингибирования индуцированной анти-IgM экспрессии CD71 на B-клетках за счет специфичных в отношении CD79b и CD22 Fab-Kd-Fab или BYbe.
Фигура 19 демонстрирует данные для выраженного в процентах ингибирования индуцированной анти-IgM экспрессии CD40 на B-клетках за счет специфичных в отношении CD79b и CD22 Fab-Kd-Fab или BYbe.
Фигура 20 демонстрирует данные для выраженного в процентах ингибирования индуцированной анти-IgM экспрессии CD86 на B-клетках за счет специфичных в отношении CD79b и CD22 Fab-Kd-Fab или BYbe.
Фигура 21 показывает ингибирование экспрессии CD27 на B-клетках за счет специфичных в отношении CD79b и CD22 VR4447/VR4126 BYbe и VR4447/VR4126/VR645 BYbe/альбумин.
Фигура 22 показывает ингибирование экспрессии CD71 на B-клетках за счет специфичных в отношении CD79b и CD22 VR4447/VR4126 BYbe и VR4447/VR4126/VR645 BYbe/альбумин.
Фигура 23 показывает ингибирование экспрессии CD86 на B-клетках за счет специфичных в отношении CD79b и CD22 VR4447/VR4126 BYbe и VR4447/VR4126/VR645 BYbe/альбумин.
Фигура 24 показывает ингибирование экспрессии CD27 на B-клетках за счет специфичных в отношении CD79b и CD22 VR4447/VR4130 BYbe и VR4447/VR4130/VR645 BYbe/альбумин.
Фигура 25 показывает ингибирование экспрессии CD71 на B-клетках за счет специфичных в отношении CD79b и CD22 VR4447/VR4130 BYbe и VR4447/VR4130/VR645 BYbe/альбумин.
Фигура 26 показывает ингибирование экспрессии CD86 на B-клетках за счет специфичных в отношении CD79b и CD22 VR4447/VR4130 BYbe и VR4447/VR4130/VR645 BYbe/альбумин.
Фигура 27 показывает ингибирование продуцирования IgG против токсоида столбняка клетками МКПК, культивируемыми с VR4447/VR4126 BYbe, VR4447/VR4127 BYbe и VR4447/VR4130 BYbe. Данные представляют собой объединенные данные от 3 доноров.
Фигура 28 показывает ингибирование продуцирования IgG против токсоида столбняка очищенными B-клетками, культивируемыми с VR4447/VR4126 BYbe, VR4447/VR4127 BYbe и VR4447/VR4130 BYbe. Данные представляют собой объединенные данные от 2 доноров.
Фигура 29 показывает ингибирование продуцирования IgG против токсоида столбняка либо МКПК, либо очищенными B-клетками, культивируемыми с VR4447/VR4126 BYbe, VR4447/VR4127 BYbe, VR4447/VR4130 BYbe, VR4447/VR4126/VR645, BYbe/альбумин и VR4447/VR4130/VR645 BYbe/альбумин. Представлены данные от одного донора.
Фигура 30 показывает базовые уровни фосфорилирования в не стимулированных B-клетках в образцах от 12 здоровых доноров и 12 пациентов с SLE.
Фигура 31 показывает эффект специфичных в отношении CD79b+CD22 VR4447/VR4130 BYbe на индуцированное анти-IgM фосфорилирование NFκB в B-клетках от 12 здоровых доноров (ЗД) и 12 пациентов с SLE.
Фигура 32 показывает эффект специфичных в отношении CD79b+CD22 VR4447/VR4130 BYbe на индуцированное анти-IgM фосфорилирование Akt в B-клетках от 12 здоровых доноров (ЗД) и 12 пациентов с SLE.
Фигура 33 показывает эффект специфичных в отношении CD79b+CD22 VR4447/VR4130 BYbe на индуцированное анти-IgM фосфорилирование Syk в B-клетках от 12 здоровых доноров (ЗД) и 12 пациентов с SLE.
Фигура 34 показывает эффект специфичных в отношении CD79b+CD22 VR4447/VR4130 BYbe на индуцированное анти-IgM фосфорилирование Erk 1 и 2 в B-клетках от 12 здоровых доноров (ЗД) и 12 пациентов с SLE.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Используемый в настоящем документе термин «мультиспецифическая молекула» означает молекулу, способную специфически связывать по меньшей мере два определенных антигена, например, разные антигены. В одном варианте осуществления мультиспецифическая молекула представляет собой биспецифическую, триспецифическую или тетраспецифическую молекулу, в частности, биспецифическую или триспецифическую молекулу.
В одном аспекте изобретение относится к молекуле соответствующего формата, специфичной в отношении по меньшей мере CD22 и CD79a, и к использованию антител/фрагментов или их сочетаний, специфичных в отношении CD22 и CD79a, в мультиспецифической молекуле, например, молекуле биспецифического или триспецифического формата.
В одном аспекте изобретение относится к молекуле соответствующего формата, специфичной в отношении по меньшей мере CD22 и CD79a, и к использованию антител/фрагментов или их сочетаний, специфичных в отношении CD22 и CD79a, в мультиспецифической молекуле, например, молекуле биспецифического или триспецифического формата.
В одном аспекте изобретение относится к молекуле соответствующего формата, специфичной в отношении по меньшей мере комплекса CD22 и CD79a/b, и к использованию антител/фрагментов или их сочетаний, специфичных в отношении комплекса CD22 и CD79a/b, в мультиспецифической молекуле, например, молекуле биспецифического или триспецифического формата.
В одном варианте осуществления молекула по настоящему изобретению является триспецифической, например, в случае, когда третий связывающий домен способен продлевать время полувыведения молекулы, например, за счет связывания сывороточного белка-носителя.
Различные белки существуют в плазме и включают тироксин-связывающий белок, транстиретин, α1 кислый гликопротеин, трансферрин, фибриноген и альбумин, либо фрагмент любого из них (Bartalena & Robbins, 1993, Clinics in Lab. Med. 13: 583-598; Bree et al., 1986, Clin. Pharmacokin. 11: 336-342; Gitlin et al. 1964, J. Clin. Invest. 10: 1938-1951; Peters, 1985, Adv Protein Chem. 37: 161-245; Waldeman & Strober, 1969, Progr. Allergy, 13: 1-110). В одном примере третий связывающий домен специфичен в отношении сывороточного альбумина, например, человеческого сывороточного альбумина.
ФОРМАТЫ МУЛЬТИСПЕЦИФИЧЕСКИХ МОЛЕКУЛ
Примеры подходящих мультиспецифических молекул известны в данной области, например, те, которые описаны в обзорах «The coming of Age of Engineered Multivalent Antibodies», Nunez-Prado et al., Drug Discovery Today Vol 20 Number 5 Mar 2015, page 588-594, D. Holmes, Nature Rev Drug Disc Nov 2011: 10; 798, Chan and Carter, Nature Reviews Immunology vol 10, May 2010, 301, содержание которых включено в настоящий документ посредством ссылки.
В одном варианте осуществления мультиспецифические форматы включают те, которые известны в данной области и которые описаны в настоящем документе, например, формат молекул выбирают из группы, включающей или состоящей из:
- тандем-sdAb, тандем-sdAb-sdAb (три sdAb);
- (scFv)2 (также называемых тандем-scFv), scFv-dsFv, dsscFv-dsFv (dsFv)2;
- диатела, ds-диатела, dids-диатела,
- sc-диатела, dssc-диатела, didssc-диатела;
- дарт-антитела, то есть, VL1 линкер VH2 линкер и VH1 линкер VL2, где C-концы VH1 и VH2 соединены дисульфидной связью;
- BiTE®, ds-BiTE, dids-BiTE;
- di-диатела (смотри статью Nunez-Prado et al., в частности, молекулу номер 25 на фигуре 1 в статье), dsdi-диатела, didsdi-диатела;
- триатела, ds-триатела, dids-триатела, trids-триатела;
- тетратела, ds-тетратела, dids-тетратела, trids-тетратела, tetrads-тетратела;
- tandab (смотри статью Nunez-Prado et al., в частности, молекулу номер 22 на фигуре 1 в статье); ds-tandab, dids-tandab, trids-tandab, tetrads-tandab;
- [sc(Fv)2]2, (смотри статью Nunez-Prado et al., в частности молекулу номер 22 на фигуре 1 в статье), ds[sc(Fv)2]2, dids[sc(Fv)2]2, trids[sc(Fv)2]2, tetrads[sc(Fv)2]2;
- пентатела (смотри статью Nunez-Prado et al., в частности молекулу номер 27 на фигуре 1 в статье);
- Fab-scFv (также называемого бителом), Fab'scFv, FabdsscFv (или BYbe), Fab'dsscFv;
- тритела, ds-тритела, dids-тритела (также называемого FabdidsscFv или TrYbe или Fab-(dsscFv)2), Fab'didsscFv;
- Fabdab, FabFv, Fab'dab, Fab'Fv;
- FabFv с одним линкером (также называемого в настоящем документе FabdsFv, раскрытого в WO 2014/096390), Fab'Fv с одним линкером (также называемого в настоящем документе Fab'dsFv);
- FabscFvFv с одним линкером, Fab'scFvFv с одним линкером;
- FabdsscFvFv с одним линкером, Fab'dsscFvFv с одним линкером;
- FvFabFv, FvFab'Fv, dsFvFabFv, dsFvFab'Fv, FvFabdsFv, FvFab'dsFv, dsFvFabdsFv, dsFvFab'dsFv,
- FabFvFv, Fab'FvFv, FabdsFvFv, Fab'dsFvFv, FabFvdsFv, Fab'FvdsFv, FabdsFvdsFv, Fab'dsFvdsFv,
- diFab, diFab', включая химически конъюгированные diFab',
- (FabscFv)2, (Fab)2scFvdsFv, (Fab)2dsscFvdsFv, (FabdscFv)2,
- (Fab'scFv)2, (Fab')2scFvdsFv, (Fab')2dsscFvdsFv, (Fab'dscFv)2,
- VHHCK (смотри статью Nunez-Prado et al., в частности молекулу номер 6 на фигуре 1 в статье);
- минитела, ds-минитела, dids-минитела,
- миниантитела (ZIP) [смотри статью Nunez-Prado et al., в частности молекулу номер 7 на фигуре 1 в статье], ds-миниантитела (ZIP) и dids-миниантитела (ZIP);
- tribi-минитела [смотри статью Nunez-Prado et al., в частности молекулу номер 15 на фигуре 1 в статье], dstribi-минитела, didstribi-минитела, tridstribi-минитела;
- диатела-CH3, ds-диатела-CH3, dids-диатела-CH3, sc-диатела-CH3, dssc-диатела-CH3, didssc-диатела-CH3,
- тандем-scFv-CH3, тандем-dsscFv-CH3, тандем-didsscFv-CH3, тандем-tridsscFv-CH3, тандем-tetradsscFv-CH3,
- scFv-Fc (также называемого в настоящем документе (scFvCH2CH3)2), описанного в WO 2008/012543, и его одноцепочечного варианта, dsscFvscFv-Fc, dsscFv-Fc (также называемого в настоящем документе (dsscFvCH2CH3)2), scFv-dsFv-Fc, dsscFv-dsFv-Fc, dsFv-Fc (также называемого в настоящем документе (dsFvCH2CH3)2),
- молекулы-скорпиона (Trubion) то есть, связывающий домен, линкер-CH2CH3 связывающий домен, как описано в US8409577;
- SMIP (Trubion), то есть, (scFv-CH2CH3)2;
- (dsFvCH2CH3)2, тандем-scFv-Fc, тандем-dsscFvscFv-Fc, тандем-dsscFv-Fc,
- scFv-Fc-scFv, dsscFv-Fc-scFv, scFv-Fc-dsscFv,
- диатела-Fc, ds-диатела-Fc, dids-диатела-Fc, триатела-Fc, ds-триатела-Fc, dids-триатела-Fc, trids-триатела-Fc, тетратела-Fc, ds-тетратела-Fc, dids-тетратела-Fc, trids-тетратела-Fc, tetrads-тетратела-Fc, ds-тетратела-Fc, dids-тетратела-Fc, trids-тетратела-Fc, tetrads-тетратела-Fc, sc-диатела-Fc, dssc-диатела, didssc-диатела;
- би- или трифункционального антитела, например, с разными вариабельными областями тяжелой цепи и общими легкими цепями, например, формата Merus биспецифического антитела (Biclonics®) с общими легкими цепями неизменной последовательности и разными тяжелыми цепями (включая разные CDR) и сконструированным CH3-доменом для управления димеризацией разных тяжелых цепей,
- дуотела (то есть, когда одна полноразмерная цепь в антителе имеет иную специфичность, чем другая полноразмерная цепь в антителе);
- полноразмерного антитела, в котором была проведена замена плеча Fab для создания биспецифического формата;
- би- или трифункционального антитела, когда полноразмерное антитело имеет общую тяжелую цепь и разные легкие цепи, также называемого «каппа/лямбда телом» или «κ/λ-телом», смотри WO 2012/023053;
- Ig-scFv одна, две, три или четыре с C-конца тяжелой или легкой цепи, scFv-Ig одна, две, три или четыре с N-конца тяжелой или легкой цепи, Ig-Fv с одним линкером, Ig-dsscFv одна, две, три или четыре с C-конца тяжелой или легкой цепи (с одной, двумя, тремя или четырьмя дисульфидными связями);
- Ig-dsscFv одна, две, три или четыре с N-конца тяжелой или легкой цепи (с одной, двумя, тремя или четырьмя дисульфидными связями),
- Ig-Fv с одним линкером (смотри PCT/EP2015/064450),
- Ig-dab, dab-Ig, scFv-Ig, V-Ig, Ig-V,
- scFabFvFc, scFabdsFvFc (вариант scFavFv с одним линкером), (FabFvFc)2, (FabdsFvFc)2, scFab'FvFc, scFab'dsFvFc, (Fab'FvFc)2, (Fab'dsFvFc)2 и
- DVDIg, которые обсуждаются более подробно ниже.
В одном варианте осуществления форматы мультиспецифической молекулы включают те, которые известны в данной области и которые описаны в настоящем документе, например формат молекулы, выбранный из группы, включающей или состоящей из: диатела, sc-диатела, триатела, тритела, тетратела, тандем-scFv, FabFv, Fab'Fv, FabdsFv, Fab-scFv, Fab-dsscFv, Fab-(dsscFv)2, diFab, diFab', тандем-scFv-Fc, scFv-Fc-scFv, sc-диатела-Fc, sc-диатела-CH3, Ig-scFv, scFv-Ig, V-Ig, Ig-V, дуотела и DVDIg, которые обсуждаются более подробно ниже.
В одном варианте осуществления мультиспецифическая молекула антитела по настоящему изобретению не содержит домен Fc, то есть, не содержит домен CH2 и CH3, например, молекула выбрана из группы, включающей тандем-scFv, scFv-dsFv, dsscFv-dsFv, didsFv, диатело, ds-диатело, dids-диатело, sc-диатело (также называемое (scFv)2), dssc-диатело, триатело, ds-триатело, dids-триатело, trids-триатело, тетратело, ds-тетратело, dids-тетратело, trids-тетратело, tetrads-тетратело, тритело, ds-тритело, dids-тритело, Fabdab, FabFv, Fab'dab, Fab'Fv, FabFv с одним линкером (описанные в WO 2014/096390), Fab'Fv с одним линкером, FabdsFv, Fab'dsFv, Fab-scFv (также называемое бителом), Fab'scFv, FabdsscFv, Fab'dsscFv, FabdidsscFv, Fab'didsscFv, FabscFvFv с одним линкером, Fab'scFvFv с одним линкером, FabdsscFvsFv с одним линкером, Fab'dsscFvFv с одним линкером, FvFabFv, FvFab'Fv, dsFvFabFv, dsFvFab'Fv, FvFabdsFv, FvFab'dsFv, dsFvFabdsFv, dsFvFab'dsFv, FabFvFv, Fab'FvFv, FabdsFvFv, Fab'dsFvFv, FabFvdsFv, Fab'FvdsFv, FabdsFvdsFv, Fab'dsFvdsFv, diFab, diFab' включая химически конъюгированные diFab', (FabscFv)2, (Fab)2scFvdsFv, (Fab)2dsscFvdsFv, (FabdscFv)2, минитело, ds-минитело, dids-минитело, диатело-CH3, ds-диатело-CH3, dids-диатело-CH3, sc-диатело-CH3, dssc-диатело-CH3, didssc-диатело-CH3, тандем-scFv-CH3, тандем-dsscFv-CH3, тандем-didsscFv-CH3, тандем-tridsscFv-CH3 и тандем-tetradsscFv-CH3.
В одном варианте осуществления молекула по настоящему изобретению не содержит домен Fc.
В одном варианте осуществления молекула по настоящему изобретению содержит измененный домен Fc, описанный ниже в настоящем документе.
Используемый в настоящем документе термин «домен Fc», как правило, означает -(CH2CH3)2, если из контекста явно не следует иное.
В одном варианте осуществления молекула по настоящему изобретению не содержит фрагмент -CH2CH3.
В одном варианте осуществления молекула по настоящему изобретению не содержит домен CH2.
В одном варианте осуществления молекула по настоящему изобретению не содержит домен CH3.
В настоящем документе термин «молекула» используют в биохимическом смысле для обозначения группы атомов, образующих органическую, в частности, белковую массу, которая включает комплекс, с которым можно обращаться как с единым целым в соответствующих условиях, когда комплекс образовался, например, комплекс, образованный двумя или более полипептидными цепями.
Термины «молекула» и «конструкт» в настоящем документе используют взаимозаменяемо, если из контекста не следует иное. Хотя термин «конструкт» может быть использован чаще для обозначения полинуклеотидной молекулы, а термин «молекула» может быть использован чаще для обозначения структурной единицы, главным образом представляющей собой аминокислотную последовательность.
Используемый в настоящем документе термин «специфичность» (или «специфический») относится к ситуации, когда партнеры при взаимодействии узнают только друг друга или имеют значительно более высокую аффинность друг в отношении друга, чем в отношении тех, которые не являются партнерами, например, по меньшей мере в 2, 3, 4, 5, 6, 7, 8, 9, 10 раз более высокую аффинность, чем, например, для фонового уровня связывания или связывания другого постороннего белка.
Используемый в настоящем документе термин «связывающий домен» означает связывающую область, как правило, полипептид, способный связывать целевой антиген, например, с достаточной аффинностью, чтобы охарактеризовать домен как специфический в отношении антигена.
Любые подходящие связывающие домены можно использовать в мультиспецифических молекулах по настоящему изобретению. Они могут быть получены из любого подходящего источника.
В одном варианте осуществления используют биосовместимую каркасную структуру в связывающем домене молекул по настоящему изобретению, и такие структуры основаны на белковых каркасах или скелетах, отличных от доменов иммуноглобулина. Например, можно использовать структуры на основе фибронектина, анкирина, липокалина, неокарциностатина, цитохрома b, цинкового пальца CP1, PST1, двойной спирали, LACI-D1, Z-домена и доменов тендамистата (смотри, например, Nygren and Uhlen, 1997, Current Opinion in Structural Biology, 7, 463-469).
Используемый в настоящем документе термин «мультиспецифические молекулы» может также включать связующие вещества на основе биологических каркасов, в том числе аднектины, аффитела, дарпины, филомеры, авимеры, аптамеры, антикалины, тетранектины, микротела, аффилины и домены Кунитца.
Мультиспецифическая молекула по настоящему изобретению, как правило, представляет собой мультиспецифическую молекулу антитела, то есть, по меньшей мере один или более из связывающих доменов мультиспецифической молекулы происходят из антитела или его фрагмента.
Если связывающий домен происходит из антитела, используемый в настоящем документе термин «связывающий домен или сайт» означает часть антитела, которая контактирует с антигеном. В одном варианте осуществления связывающий домен содержит по меньшей мере один вариабельный домен или его производное, например, пару вариабельных доменов или их производных, такую как когнатная пара вариабельных доменов или их производных. Как правило, эта пара представляет собой пару VH/VL.
Вариабельные области (также называемые в настоящем документе вариабельными доменами), как правило, содержат 3 области CDR и соответствующий каркас. В одном варианте осуществления связывающий домен содержит две вариабельные области, вариабельную область легкой цепи и вариабельную область тяжелой цепи, и в совокупности эти элементы вносят вклад в специфичность взаимодействия связывания антитела или связывающего фрагмента.
Используемый в настоящем документе термин «когнатная пара» означает пару тяжелой и легкой цепей вариабельных доменов (или их производных, например, их гуманизированных вариантов), выделенную из хозяина в виде предварительно образовавшейся пары. Это определение не включает вариабельные домены, выделенные из библиотеки, когда исходные пары хозяина не сохраняются. Когнатные пары могут быть полезными, поскольку они часто проходят процесс созревания аффинности в хозяине и, таким образом, могут иметь более высокую аффинность в отношении антигена, в отношении которого они специфичны, чем сочетание пар вариабельных доменов, выбранных из библиотеки, такой как фаговая библиотека.
Используемый в настоящем документе термин «производное природного домена» относится к ситуации, когда одна, две, три, четыре или пять аминокислот в природной последовательности были заменены или делетированы, например, для оптимизации свойств домена с тем, чтобы удалить нежелательные свойства, однако при этом характерная особенность(и) домена сохраняется (сохраняются). Примерами модификаций являются модификации для удаления сайтов гликозилирования, GPI якорей или экспонированных для растворителя остатков лизина. Такие модификации можно осуществлять, заменяя соответствующие аминокислотные остатки путем консервативной аминокислотной замены.
Модификация в областях CDR может, например, включать замену одного или более остатков цистеина, например, остатками серина. Asn может быть субстратом для дезаминирования и эта предрасположенность может быть снята путем замены Asn и/или соседней аминокислоты на альтернативную аминокислоту, например, путем консервативной замены. Аминокислота Asp в областях CDR может быть подвержена изомеризации. Это можно сводить к минимуму путем замены Asp или соседней аминокислоты на альтернативную аминокислоту, например, путем консервативной замены.
Гуманизированные варианты вариабельной области также являются ее производными в контексте настоящей спецификации. Гуманизация может включать замену не принадлежащей человеку каркасной области на человеческую каркасную область и, необязательно, обратную мутацию одного или более остатков в «донорские остатки». Используемый в настоящем документе термин «донорские остатки» означает остатки, присутствующие в исходной вариабельной области хозяина, в частности, речь идет о замене конкретной аминокислоты в человеческой каркасной области аминокислотой, находящейся в соответствующем положении в донорской каркасной области.
В одном варианте осуществления связывающий домен или каждый связывающий домен представляет собой часть (включенную или встроенную в) антитела или фрагмент антитела.
В одном варианте осуществления связывающие домены в молекулах по настоящему изобретению находятся в молекулах иммуноглобулина/антитела.
Используемый в настоящем документе термин «молекула антитела» включает антитела и их связывающие фрагменты.
В одном варианте осуществления используемый в настоящем документе термин «антитело» означает молекулу иммуноглобулина, способную к специфическому связыванию с целевым антигеном, таким как углевод, полинуклеотид, липид, полипептид, пептид и так далее, через по меньшей мере один сайт узнавания антигена (также называемый в настоящем документе сайтом связывания или связывающим доменом), расположенный в вариабельной области молекулы иммуноглобулина.
Используемый в настоящем документе термин «фрагменты антитела» означают связывающие фрагменты антитела, включая, но без ограничения, Fab, модифицированные Fab, Fab', модифицированные Fab', F(ab')2, Fv, однодоменные антитела, scFv, Fv, двух-, трех- или четырехвалентные антитела, bis-scFv, диатела, триатела, тетратела и эпитопсвязывающие фрагменты любых из вышеперечисленных (смотри, например, Holliger and Hudson, 2005, Nature Biotech. 23(9): 1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217).
Используемый в настоящем документе термин «связывающий фрагмент» означает фрагмент, способный связывать целевой пептид или антиген с аффинностью, достаточной, чтобы охарактеризовать фрагмент как специфический в отношении пептида или антигена.
Способы создания и производства таких фрагментов антитела хорошо известны в данной области (смотри, например, Verma et al., 1998, Journal of Immunological Methods, 216: 165-181). Другие фрагменты антитела для использования по настоящему изобретению включают Fab и Fab'-фрагменты, описанные в WO 05/003169, WO 05/003170 и WO 05/003171. Мультивалентные антитела могут иметь несколько специфичностей, например, могут быть биспецифическими, или могут быть моноспецифическими (смотри, например, WO 92/22853, WO 05/113605, WO 2009/040562 и WO 2010/035012).
Используемый в настоящем документе термин «Fab-фрагмент» означает фрагмент антитела, содержащий фрагмент легкой цепи, включающий домен VL (вариабельный домен легкой цепи) и константный домен легкой цепи (CL), а также домен VH (вариабельный домен тяжелой цепи) и первый константный домен (CH1) тяжелой цепи.
Термин «Fv» означает два вариабельных домена, например, кооперативные вариабельные домены, такие как когнатная пара или вариабельный домены с созревшей аффинностью, то есть, пара VH и VL.
Термин «кооперативные вариабельный домены» при использовании в настоящем документе означает вариабельные домены, которые дополняют друг друга и/или оба участвуют в связывании антигена, придавая Fv (паре VH/VL) специфичность в отношении конкретного антигена.
Используемый в настоящем документе термин «однодоменное антитело» (другое название в настоящем документе dab и sdAb) означает фрагмент антитела, состоящий из одного мономерного вариабельного домена антитела. Примеры однодоменных антител включают VH или VL или VHH.
Используемый в настоящем документе термин «тандем-sdAb» означает двудоменные антитела, связанные с помощью линкера, например, пептидного линкера, в частности, когда домены антитела обладают специфичностью в отношении разных антигенов.
Используемый в настоящем документе термин «тандем-sdAb-sdAb» означает трехдоменные антитела, связанные последовательно с помощью двух линкеров, например, пептидных линкеров, в частности, когда домены антитела обладают специфичностью в отношении разных антигенов.
Используемый в настоящем документе термин «dsFv» означает Fv с дисульфидной связью внутри вариабельной области. dsFv может быть компонентом большей по размеру молекулы, например, один из вариабельных доменов может быть связан, например, через аминокислотный линкер, с другим фрагментом/компонентом антитела.
Используемый в настоящем документе термин «(dsFv)2» означает dsFv, в которых один домен связан, например, через пептидный линкер или дисульфидную связь (например, между C-концами двух VH), с доменом во второй dsFv, формат напоминает формат (scFv)2, описанный ниже, но каждая пара вариабельных областей содержит дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «компонент» означает строительный блок или часть мультиспецифической молекулы по настоящему изобретению, в частности, представляющий собой фрагмент антитела, такой как scFv, Fab или другой фрагмент, в частности, описанный в настоящем документе.
Используемый в настоящем документе термин «одноцепочечная Fv», или сокращенно «scFv», означает фрагмент антитела, содержащий домены VH и VL антитела, связанные (например, с помощью пептидного линкера), с образованием одной полипептидной цепи. Константные области тяжелой и легкой цепи опущены в данном формате.
Используемый в настоящем документе термин «dsscFv» означает scFv с дисульфидной связью внутри вариабельной области.
Используемый в настоящем документе термин «тандем-scFv» (другое название в настоящем документе discFv или (scFv)2) означает две scFv, связанные с помощью одного линкера таким образом, что имеется один линкер между Fv, например, как показано на фигуре 9b.
Используемый в настоящем документе термин «тандем-dsscFv) (другое название в настоящем документе scFvdsscFv или dsscFvscFv) означает две scFv, связанные с помощью одного линкера таким образом, что имеется один линкер между Fv, например, как показано на фигуре 9b, и при этом одна из scFv имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «тандем-didsscFv (другое название в настоящем документе didsscFv) означает две scFv, связанные с помощью одного линкера таким образом, что имеется один линкер между Fv, например, как показано на фигуре 9b, и при этом каждая scFv имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «scFv-dsFv» означает scFv, связанную, например, с помощью пептидного линкера, с доменом Fv, который состоит из двух вариабельных доменов, связанных дисульфидной связью, с образованием dsFv. В данном формате VH или VL из scFv могут быть связаны с VH или VL из dsFv.
Используемый в настоящем документе термин «dsscFv-dsFv» означает dsscFv, связанную, например, с помощью пептидного линкера, с доменом Fv, который состоит из двух вариабельных доменов, связанных дисульфидной связью, с образованием dsFv. В данном формате VH или VL из dsscFv могут быть связаны с VH или VL из dsFv.
Используемый в настоящем документе термин «диатело» означает две пары VH/VL Fv, которые имеют два линкера между Fv таким образом, что VH из первой Fv связана с VL из второй Fv и VL из первой Fv связана с VH из второй Fv.
Используемый в настоящем документе термин «ds-диатело» означает диатело, содержащее дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «dids-диатело» означает диатело, содержащее две дисульфидные связи внутри вариабельной области, то есть, одна ds между каждой парой вариабельных областей.
Используемый в настоящем документе термин «sc-диатело» означает диатело, содержащее линкер внутри Fv таким образом, что молекула содержит три линкера и образует две нормальные scFv, например, VH1 линкер VL1 линкер VH2 линкер VL2.
Используемый в настоящем документе термин «dssc-диатело» означает sc-диатело с дисульфидной связью внутри вариабельной области.
Используемый в настоящем документе термин «didssc-диатело» означает sc-диатело с дисульфидной связью внутри вариабельной области между каждой парой вариабельных областей.
Используемый в настоящем документе термин «дарт» означает VL1 линкер VH2 линкер и VH1 линкер VL2, где C-концы VH1 и VH2 связаны дисульфидной связью, Paul A. Moore et al. Blood, 2011; 117(17): 4542-4551.
Используемый в настоящем документе термин «Bite®» означает молекулу, содержащую две пары вариабельных доменов в следующем формате; домен из пары 1 (например, VH1) связан с помощью линкера с доменом из пары 2 (например, VH2 или VL2), при этом указанный второй домен связан с помощью линкера со следующим доменом из пары 1 (например, VL1), в свою очередь связанным с оставшимся доменом из пары два (то есть, VL2 или VH2).
«Di-диатело» - смотри статью Nunez-Prado et al., в частности, молекулу номер 25 на фигуре 1 в статье.
Используемый в настоящем документе термин «dsdi-диатело» означает di-диатело с дисульфидной связью внутри вариабельной области.
Используемый в настоящем документе термин «didsdi-диатело» означает di-диатело с дисульфидной связью внутри вариабельной области между каждой парой вариабельных областей.
Используемый в настоящем документе термин «триатело» означает формат, аналогичный формату диатела, с содержанием трех Fv и трех линкеров между Fv.
Используемый в настоящем документе термин «ds-триатело» означает триатело, имеющее дисульфидную связь внутри вариабельной области между одной из пар вариабельных доменов.
Используемый в настоящем документе термин «dids-триатело» означает триатело, имеющее две дисульфидные связи внутри вариабельной области, то есть, одну ds между каждой из двух пар вариабельных доменов.
Используемый в настоящем документе термин «trids-триатело» означает триатело, имеющее три дисульфидные связи внутри вариабельной области, то есть, одну ds между каждой парой вариабельных областей.
Используемый в настоящем документе термин «тетратело» означает формат, аналогичный формату диатела, с содержанием четырех Fv и четырех линкеров между Fv.
Используемый в настоящем документе термин «ds-тетратело» означает тетратело, имеющее дисульфидную связь внутри вариабельной области между одной из пар вариабельных доменов.
Используемый в настоящем документе термин «dids-тетратело» означает тетратело, имеющее две дисульфидные связи внутри вариабельной области, то есть, одну ds между каждой из двух пар вариабельных доменов.
Используемый в настоящем документе термин «trids-тетратело» означает тетратело, имеющее три дисульфидные связи внутри вариабельной области, то есть, одну ds между каждой из трех пар вариабельных областей.
Используемый в настоящем документе термин «tetrads-тетратело» означает тетратело, имеющее четыре дисульфидные связи внутри вариабельной области, то есть, одну ds между каждым вариабельным доменом.
Используемый в настоящем документе термин «тритело» (другое название в настоящем документе Fab(scFv)2) означает Fab-фрагмент с первой scFv, присоединенной к C-концу легкой цепи, и второй scFv, присоединенной к C-концу тяжелой цепи.
Используемый в настоящем документе термин «ds-тритело» означает тритело, содержащее dsscFv в одном из двух положений.
Используемый в настоящем документе термин «dids-тритело» или «TrYbe» означает тритело, содержащее две dsscFv.
Используемый в настоящем документе термин «dsFab» означает Fab с дисульфидной связью внутри вариабельной области.
Используемый в настоящем документе термин «dsFab'» означает Fab' с дисульфидной связью внутри вариабельной области.
«scFab» означает одноцепочечный Fab-фрагмент.
«scFab'» означает одноцепочечный Fab'-фрагмент.
«dsscFab» означает dsFab в виде одной цепи.
«dsscFab'» означает dsFab' в виде одной цепи.
Используемый в настоящем документе термин «Fabdab» означает Fab-фрагмент с доменом антитела, присоединенным к тяжелой или легкой цепи антитела, необязательно, через линкер.
Используемый в настоящем документе термин «Fab'dab» означает Fab'-фрагмент с доменом антитела, присоединенным к тяжелой или легкой цепи антитела, необязательно, через линкер.
Используемый в настоящем документе термин «FabFv» означает Fab-фрагмент с дополнительной вариабельной областью, присоединенной к C-концу каждого из следующего: CH1 тяжелой цепи и CL легкой цепи, смотри, например, WO 2009/040562. Формат может быть предоставлен в виде его пегилированного варианта, смотри, например, WO 2011/061492,
Используемый в настоящем документе термин «Fab'Fv» является аналогичным термину «FabFv», при этом Fab-часть заменена на Fab'. Формат может быть предоставлен в виде его пегилированного варианта.
Используемый в настоящем документе термин «FabdsFv» означает FabFv, при этом дисульфидная связь внутри Fv стабилизирует присоединенные C-концевые вариабельные области, смотри, например, WO 2010/035012. Формат может быть предоставлен в виде его пегилированного варианта.
Используемые в настоящем документе термины «FabFv с одним линкером» и «Fab'Fv с одним линкером» означают Fab или Fab'-фрагмент, связанный с вариабельным доменом, например, с помощью пептидного линкера, и указанный вариабельный домен связан со вторым вариабельным доменом с помощью дисульфидной связи внутри вариабельного домена, тем самым образуя dsFv, смотри, например, WO 2014/096390.
Используемый в настоящем документе термин «Fab-scFv» (другое название битело) означает молекулу Fab с scFv, присоединенной к C-концу легкой или тяжелой цепи, необязательно, с помощью линкера.
Используемый в настоящем документе термин «Fab'-scFv» означает молекулу Fab' с scFv, присоединенной к C-концу легкой или тяжелой цепи, необязательно, с помощью линкера.
Используемый в настоящем документе термин «FabdsscFv» или «BYbe» означает Fab-scFv с дисульфидной связью между вариабельными областями одноцепочечной Fv.
Используемый в настоящем документе термин «Fab'dsscFv» означает Fab'scFv с дисульфидной связью между вариабельными областями одноцепочечной Fv.
Используемый в настоящем документе термин «FabscFv-dab» означает Fab с scFv, присоединенной к C-концу одной цепи и доменом антитела, присоединенным к C-концу другой цепи.
Используемый в настоящем документе термин «Fab'scFv-dab» означает Fab' с scFv, присоединенной к C-концу одной цепи и доменом антитела, присоединенным к C-концу другой цепи.
Используемый в настоящем документе термин «FabdsscFv-dab» означает Fab с dsscFv, присоединенной к C-концу одной цепи и доменом антитела, присоединенным к C-концу другой цепи.
Используемый в настоящем документе термин «Fab'dsscFv-dab» означает Fab' с dsscFv, присоединенной к C-концу одной цепи и доменом антитела, присоединенным к C-концу другой цепи.
Используемый в настоящем документе термин «FabscFvFv с одним линкером» означает FabFv с одним линкером, когда домен Fv связан с тяжелой или легкой цепью Fab и scFv связана с другой цепью Fab, и домены Fv связаны дисульфидной связью внутри вариабельной области.
Используемый в настоящем документе термин «FabdsscFvFv с одним линкером» означает FabscFvFv с одним линкером, когда scFv имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «Fab'scFvFv с одним линкером» означает Fab'Fv с одним линкером, когда домен Fv связан с тяжелой или легкой цепью Fab и scFv связана с другой цепью Fab, и домены Fv связаны дисульфидной связью внутри вариабельной области.
Используемый в настоящем документе термин «Fab'dsscFvFv с одним линкером» означает Fab'scFvFv с одним линкером, когда scFv имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «FvFabFv» означает Fab с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи.
Используемый в настоящем документе термин «FvFab'Fv» означает Fab' с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab', и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи.
Используемый в настоящем документе термин «dsFvFabFv» означает Fab с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab, при этом первая Fv имеет дисульфидную связь внутри вариабельной области, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи.
Используемый в настоящем документе термин «FvFabdsFv» означает Fab с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи, и при этом вторая Fv имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «dsFvFab'Fv» означает Fab' с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab', при этом первая Fv имеет дисульфидную связь внутри вариабельной области, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи.
Используемый в настоящем документе термин «FvFab'dsFv» означает Fab' с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи Fab', и при этом вторая Fv имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «dsFvFabdsFv» означает Fab с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab, при этом первая Fv имеет дисульфидную связь внутри вариабельной области, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи, и при этом вторая Fv также имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «dsFvFab'dsFv» означает Fab' с доменами первой Fv, присоединенными к N-концу тяжелой и легкой цепи Fab', при этом первая Fv имеет дисульфидную связь внутри вариабельной области, и доменами второй Fv, присоединенными к C-концу тяжелой и легкой цепи, и при этом вторая Fv также имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «FabFvFv» означает Fab-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, смотри, например, WO 2011/086091.
Используемый в настоящем документе термин «Fab'FvFv» означает Fab'-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, смотри, например, WO 2011/086091.
Используемый в настоящем документе термин «FabdsFvFv» означает Fab-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, смотри, например, WO 2011/086091, при этом первая пара Fv, присоединенная непосредственно к C-концу, имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «Fab'dsFvFv» означает Fab'-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, смотри, например, WO 2011/086091, при этом первая пара Fv, присоединенная непосредственно к C-концу, имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «FabFvdsFv» означает Fab-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, при этом вторая пара Fv на «C»-конце молекулы имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «Fab'FvdsFv» означает Fab'-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, при этом вторая пара Fv на «C»-конце молекулы имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «FabdsFvdsFv» означает Fab-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, при этом первая и вторая пара Fv имеют дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «Fab'dsFvdsFv» означает Fab'-фрагмент с двумя парами Fv, присоединенными последовательно к C-концу тяжелой и легкой цепи, при этом первая и вторая пара Fv имеют дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «diFab» означает две молекулы Fab, связанные через их C-концы тяжелых цепей.
Используемый в настоящем документе термин «diFab'» означает две молекулы Fab', связанные с помощью одной или более дисульфидных связей в их шарнирной области.
Молекулы diFab и diFab' включают их химически конъюгированные формы.
Используемый в настоящем документе термин (FabscFv)2 означает молекулу diFab с двумя scFv, присоединенными к ней, например, присоединенными к C-концу тяжелой или легкой цепи, например, тяжелой цепи.
Используемый в настоящем документе термин «(Fab'scFv)2» означает молекулу diFab' с двумя scFv, присоединенными к ней, например, присоединенными к C-концу тяжелой или легкой цепи, например, тяжелой цепи.
Используемый в настоящем документе термин «(Fab)2scFvdsFv» означает diFab с областями scFv и dsFv, присоединенными, например, по одной с каждого C-конца тяжелой цепи.
Используемый в настоящем документе термин «(Fab')2scFvdsFv» означает diFab' с областями scFv и dsFv, присоединенными, например, по одной с каждого C-конца тяжелой цепи.
Используемый в настоящем документе термин «(Fab)2dsscFvdsFv» означает diFab с областями dsscFv и dsFv, присоединенными, например, с C-конца тяжелой цепи.
Используемый в настоящем документе термин «(Fab')2dsscFvdsFv» означает diFab' с областями dsscFv и dsFv, присоединенными, например, с C-конца тяжелой цепи.
Используемый в настоящем документе термин «минитело» означает (VL/VH-CH3)2.
Используемый в настоящем документе термин «ds-минитело» означает (VL/VH-CH3)2, где один комплекс VL/VH имеет дисульфидную связь внутри вариабельной области.
Используемый в настоящем документе термин «dids-минитело» означает (dsFv-CH3)2.
Используемый в настоящем документе термин «каппа/лямбда тело» или «κ/λ-тело» означает формат нормального IgG с двумя тяжелыми цепями и двумя легкими цепями, при этом две легкие цепи отличаются друг от друга, одна представляет собой легкую цепь лямбда (VL-CL) и другая представляет собой легкую цепь каппа (VK-CK). Тяжелые цепи являются идентичными, даже в областях CDR, как описано в WO 2012/023053.
Используемый в настоящем документе термин «scFv-Fc» означает scFv, присоединенную, например, с помощью шарнира, к N-концу домена CH2 фрагмента константной области -(CH2CH3), так что молекула имеет 2 связывающих домена.
Используемый в настоящем документе термин «dsscFv-Fc» означает dsscFv, присоединенную, например, с помощью шарнира, к N-концу домена CH2, и scFv, присоединенную к N-концу второго домена CH2 фрагмента константной области -(CH2CH3)2, так что молекула имеет 2 связывающих домена.
Используемый в настоящем документе термин «didsscFv-Fc» означает scFv, присоединенную, например, с помощью шарнира, к N-концу домена CH2 фрагмента константной области -(CH2CH3)2, так что молекула имеет 2 связывающих домена.
Используемый в настоящем документе термин «тандем-scFv-Fc» означает две тандем-scFv, при этом каждая присоединена последовательно, например, с помощью шарнира, к N-концу домена CH2 фрагмента константной области -(CH2CH3), так что молекула имеет 4 связывающих домена.
Используемый в настоящем документе термин «sc-диатело-Fc» означает два sc-диатела, при этом каждое из них присоединено, например, с помощью шарнира, к N-концу домена CH2 фрагмента константной области -CH2CH3.
Используемый в настоящем документе термин «scFv-Fc-scFv» означает четыре scFv, при этом каждая из них присоединена к N-концу и C-концу как тяжелой, так и легкой цепи фрагмента -CH2CH3.
Используемый в настоящем документе термин «sc-диатело-CH3» означает две молекулы sc-диатела, каждая из которых связана, например, с помощью шарнира, с доменом CH3.
Используемый в настоящем документе термин «IgG-scFv» означает полноразмерное антитело с scFv на C-конце каждой из тяжелых цепей или каждой из легких цепей.
Используемый в настоящем документе термин «scFv-IgG» означает полноразмерное антитело с scFv на N-конце каждой из тяжелых цепей или каждой из легких цепей.
Используемый в настоящем документе термин «V-IgG» означает полноразмерное антитело с вариабельным доменом на N-конце каждой из тяжелых цепей или каждой из легких цепей.
Используемый в настоящем документе термин «IgG-V» означает полноразмерное антитело с вариабельным доменом на C-конце каждой из тяжелых цепей или каждой из легких цепей.
Используемый в настоящем документе термин «DVD-Ig» (другое название IgG с двойным V-доменом) означает полноразмерное антитело с 4 дополнительными вариабельными доменами, по одному на N-конце каждой из тяжелых и легких цепей.
Используемый в настоящем документе термин «дуотело» или «антитело с заменой плеча Fab» означает формат биспецифического IgG антитела, в котором подобранные и комплементарно разработанные аминокислотные замены в константных доменах (как правило, CH3) двух различных моноклональных антител приводят, при смешивании, к образованию гетеродимеров. Пара тяжелая:легкая цепь из первого антитела в результате инженерии остатков будет предпочтительно связываться с парой тяжелая:легкая цепь второго антитела. Смотри, например, WO 2008/119353, WO 2011/131746 и WO 2013/060867.
Если одна или более пар вариабельных областей в мультиспецифической молекуле антитела содержат дисульфидную связь между VH и VL, это может быть в любом подходящем положении, например, между двумя из остатков, перечисленных ниже (если из контекста не следует иное, в приведенном ниже списке использована нумерация Kabat). При любом упоминании нумерации Kabat соответствующим литературным источником является Kabat et al., 1987, в «Sequences of Proteins of Immunological Interest», US Department of Health and Human Services, NIH, USA.
В одном варианте осуществления дисульфидная связь находится в положении, выбранном из группы, включающей:
- VH37+VL95C смотри, например, Protein Science 6, 781-788 Zhu et al. (1997);
- VH44+VL100 смотри, например; Biochemistry 33, 5451-5459 Reiter et al. (1994); или Journal of Biological Chemistry Vol. 269 No. 28 pp. 18327-18331 Reiter et al. (1994); или Protein Engineering, vol.10 no. 12 pp. 1453-1459 Rajagopal et al. (1997);
- VH44+VL105 смотри, например, J Biochem. 118, 825-831 Luo et al. (1995);
- VH45+VL87 смотри, например, Protein Science 6, 781-788 Zhu et al. (1997);
- VH55+VL101 смотри, например, FEBS Letters 377, 135-139 Young et al. (1995);
- VH100+VL50 смотри, например, Biochemistry 29, 1362-1367 Glockshuber et al. (1990);
- VH100b+VL49;
- VH98+VL46 смотри, например, Protein Science 6, 781-788 Zhu et al. (1997);
- VH101+VL46;
- VH105+VL43 смотри, например; Proc. Natl. Acad. Sci. USA Vol. 90 pp. 7538-7542 Brinkmann et al. (1993); или Proteins 19, 35-47 Jung et al. (1994),
- VH106+VL57 смотри, например, FEBS Letters 377, 135-139 Young et al. (1995),
и соответствующем положении в паре вариабельной области, расположенной в молекуле.
В одном варианте осуществления дисульфидная связь образована между положениями VH44 и VL100.
Используемый в настоящем документе термин «моноспецифические» означает способность связывать целевой антиген только один раз.
Таким образом, в одном варианте осуществления мультиспецифические молекулы по настоящему изобретению являются моноспецифическими в отношении каждого антигена.
Таким образом, в одном варианте осуществления связывающие домены мультиспецифических молекул по настоящему изобретению являются моноспецифическими. Это является преимуществом в некоторых терапевтических вариантах применения, поскольку молекулы по изобретению не способны к перекрестному связыванию антигена за счет связывания целевого антигена более одного раза. Таким образом, в одном варианте осуществления биспецифические или мультиспецифические молекулы по настоящему изобретению не способны к перекрестному связыванию за счет связывания одной и той же мишени дважды в двух разных местах, например, на одной и той же клетке или на двух разных клетках.
Перекрестное связывание, в частности, применительно к CD79b, на одной и той же клетке или разных клетках может генерировать сигналы in vivo, например, которые стимулируют активность целевого антигена.
В одном примере мультиспецифические молекулы по настоящему изобретению содержат не более одного связывающего домена для CD22 и не более одного связывающего домена для CD79. Каждый связывающий домен является моноспецифическим.
Таким образом, в одном примере мультиспецифическая молекула является одновалентной в отношении CD22 и одновалентной в отношении CD79.
В одном варианте осуществления каждое антитело или фрагмент антитела, которые используют в мультиспецифических молекулах по настоящему изобретению, является одновалентным.
Таким образом, в одном варианте осуществления связывающие домены мультиспецифических молекул по настоящему изобретению являются одновалентными.
Таким образом, в одном варианте осуществления связывающие домены мультиспецифических молекул по настоящему изобретению являются одновалентными и моноспецифическими.
В одном варианте осуществления мультиспецифическая молекула по настоящему изобретению состоит из двух или более моноспецифических, одновалентных связывающих доменов, таких как Fab, Fab', scFv, VH, VL, VHH, Fv, dsFv, объединенных или связанных любым подходящим способом для конструирования мультиспецифической молекулы, например, как описано выше в настоящем документе.
В другом варианте осуществления, например, когда молекулы по изобретению содержат по меньшей мере три связывающих домена, тогда два или три связывающих домена (например, антитела, фрагменты или сочетание антитела и фрагмента) могут иметь разные антигенные специфичности, например, связываются с тремя разными целевыми антигенами.
КОНСТАНТНЫЕ ОБЛАСТИ
Домены константной области антитела в мультиспецифической молекуле по настоящему изобретению, при наличии, могут быть выбраны с учетом предполагаемой функции мультиспецифической молекулы антитела, и в частности, эффекторных функций, которые могут быть необходимы. Например, домены константной области могут представлять собой домены IgA, IgD, IgE, IgG или IgM человека. В частности, домены константной области IgG человека, особенно изотипов IgG1 и IgG3, можно использовать, когда молекула антитела предназначена для терапевтического применения и необходимы эффекторные функции антитела. Альтернативно, можно использовать изотипы IgG2 и IgG4, когда молекула антитела предназначена для терапевтических целей и эффекторные функции антитела не нужны.
Следует понимать, что также можно использовать варианты последовательностей этих доменов константной области. Например, можно использовать молекулы IgG4, в которых остаток серина в положении 241 заменен на остаток пролина, как описано в статье Angal et al., 1993, Molecular Immunology, 1993, 30: 105-108. Соответственно, в варианте осуществления, в котором антитело представляет собой антитело IgG4, антитело может содержать мутацию S241P.
В одном варианте осуществления тяжелая цепь антитела содержит домен CH1, и легкая цепь антитела содержит домен CL, либо каппа, либо лямбда.
В одном варианте осуществления тяжелая цепь антитела содержит домен CH1, домен CH2 и домен CH3, и легкая цепь антитела содержит домен CL, либо каппа, либо лямбда.
Человеческие IgG четырех изотипов связывают активирующие Fcγ рецепторы (FcγRI, FcγRIIa, FcγRIIIa), ингибирующий FcγRIIb рецептор и первый компонент комплемента (C1q) с разными аффинностями, результатом чего являются очень разные эффекторные функции (Bruhns P. et al., 2009. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood. 113(16): 3716-25), смотри также Jeffrey B. Stavenhagen, et al. Cancer Research 2007 Sep 15; 67(18): 8882-90.
Связывание IgG с рецепторами FcγR или C1q зависит от остатков, расположенных в шарнирной области и домене CH2. Две области домена CH2 являются критичными для связывания FcγR и C1q и имеют уникальные последовательности в IgG2 и IgG4. Показано, что замены в IgG1 человека на остатки IgG2 в положениях 233-236 и остатки IgG4 в положениях 327, 330 и 331 приводят к сильному снижению показателей ADCC и CDC (Armour K.L. et al., 1999. Recombinant human IgG molecules lacking Fcgamma receptor I binding and monocyte triggering activities. Eur J Immunol. 29(8): 2613-24 и Shields R.L. et al., 2001. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem. 276(9): 6591-604). Кроме того, в статье Idusogie et al. продемонстрировано, что замена остатков аланина в разных положениях, включая K322, приводит к значительному снижению активации комплемента (Idusogie E.E. et al., 2000. Mapping of the C1q binding site on rituxan, a chimeric antibody with a human IgG1 Fc. J Immunol. 164(8): 4178-84). Аналогично, показано, что мутации в домене CH2 мышиного IgG2A приводят к уменьшению связывания с FcγRI и C1q (Steurer W. et al., 1995. Ex vivo coating of islet cell allografts with murine CTLA4/Fc promotes graft tolerance. J Immunol. 155(3): 1165-74).
В одном варианте осуществления используемая Fc-область является мутантной, в частности, содержит мутацию, описанную в настоящем документе. В одном варианте осуществления мутация введена для устранения связывающей и/или эффекторной функции.
В одном варианте осуществления мутацию Fc выбирают из группы, включающей мутацию для устранения связывания Fc-области, мутацию для усиления или устранения эффекторной функции, мутацию для увеличения времени полувыведения, а также их сочетание.
Некоторые антитела, которые избирательно связывают FcRn при pH 6,0, но не при pH 7,4, имеют более продолжительное время полувыведения в различных животных моделях. Некоторые мутации, находящиеся на границе раздела между доменами CH2 и CH3, например, T250Q/M428L (Hinton P.R. et al., 2004. Engineered human IgG antibodies with longer serum half-lives in primates. J Biol Chem. 279(8): 6213-6) и M252Y/S254T/T256E+H433K/N434F (Vaccaro C. et al., 2005. Engineering the Fc region of immunoglobulin G to modulate in vivo antibody levels. Nat Biotechnol. 23(10): 1283-8), как показано, приводят к увеличению аффинности связывания с FcRn и времени полувыведения IgG1 in vivo.
Однако не всегда существует прямая связь между усилением связывания FcRn и увеличением времени полувыведения (Datta-Mannan A. et al., 2007. Humanized IgG1 Variants with Differential Binding Properties to the Neonatal Fc Receptor: Relationship to Pharmacokinetics in Mice and Primates. Drug Metab. Dispos. 35: 86-94).
Подкласс IgG4 характеризуется пониженным связыванием Fc рецептора (FcγRIIIa), антитела других подклассов IgG, как правило, демонстрируют сильное связывание. Ослабления связывания с рецептором в случае этих других подтипов IgG можно добиться путем изменения, например, замены одной или более аминокислот, выбранных из группы, включающей Pro238, Aps265, Asp270, Asn270 (потеря углевода Fc), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435.
В одном варианте осуществления молекула по настоящему изобретению имеет Fc подкласса IgG, например, IgG1, IgG2 или IgG3, при этом Fc имеет мутацию в одном, двух или всех из следующих положений: S228, L234 и/или D265.
В одном варианте осуществления мутации в Fc-области независимо выбирают из S228P, L234A, L235A, L235A, L235E и их сочетаний.
Может быть желательным либо ослаблять, либо усиливать эффекторную функцию Fc-области. У антител, направленных на молекулы клеточной поверхности, особенно находящиеся на иммунных клетках, необходимо устранять эффекторные функции. В некоторых вариантах осуществления, например, для лечения аутоиммунных заболеваний, может быть желательным увеличение связывания Fc на иммунных клетках за счет увеличения связывания с ингибиторным Fc рецептором (FcgRIIb или CD32b), смотри Stavenhagen J.B, et al. Advances in Enzyme Regulation 2007 December 3 и Veri M.C., et al. Arthritis Rheum, 2010 Mar 30; 62(7): 1933-43. Напротив, в случае антител, предназначенных для использования в онкологии, усиление эффекторных функций может повышать терапевтическую активность.
Многочисленные мутации были введены в домен CH2 человеческого IgG1 и их влияние на ADCC и CDC было протестировано in vitro (Idusogie E.E. et al., 2001. Engineered antibodies with increased activity to recruit complement. J Immunol. 166(4): 2571-5). Примечательно, что замена остатка аланина в положении 333, как сообщалось, приводит к усилению как ADCC, так и CDC. В статье Lazar et al. описан тройной мутант (S239D/I332E/A330L) с повышенной аффинностью в отношении FcγRIIIa и пониженной аффинностью в отношении FcγRIIb, что приводит к усилению ADCC (Lazar G.A. et al., 2006, Engineered antibody Fc variants with enhanced effector function. PNAS 103(11): 4005-4010). Те же мутации были использованы для создания антитела с усиленной ADCC (Ryan M.C. et al., 2007, Antibody targeting of B-cell maturation antigen on malignant plasma cells. Mol. Cancer Ther., 6: 3009-3018). В статье Richards et al. описан немного отличающийся тройной мутант (S239D/I332E/G236A) с повышенной аффинностью в отношении FcγRIIIa и соотношением FcγRIIa/FcγRIIb, способствующим усиленному фагоцитозу клеток-мишеней макрофагами (Richards J.O. et al. 2008, Optimization of antibody binding to Fcgamma RIIa enhances macrophage phagocytosis of tumor cells. Mol Cancer Ther, 7(8): 2517-27).
Из-за отсутствия у них эффекторных функций антитела IgG4 являются подходящим подклассом IgG для блокирования рецепторов без истощения пула клеток. У молекул IgG4 может происходить обмен половины молекулы в динамическом процессе, называемом заменой плеча Fab. Это явление может иметь место между терапевтическими антителами и эндогенными IgG4. Показано, что мутация S228P предотвращает этот процесс рекомбинации, что позволяет конструировать более предсказуемые терапевтические антитела IgG4 (Labrijn A.F. et al., 2009. Therapeutic IgG4 antibodies engage in Fab-arm exchange with endogenous human IgG4 in vivo. Nat Biotechnol. 27(8): 767-71). Данную технологию можно использовать для создания биспецифических молекул антител.
Специалисту в данной области также будет понятно, что антитела могут подвергаться различным посттрансляционным модификациям. Тип и степень данных модификаций часто зависит от линии клетки-хозяина, используемой для экспрессии антитела, а также от условий культивирования. Такие модификации могут включать вариации в гликозилировании, окислении метионина, образовании дикетопиперазина, изомеризации аспартата и дезамидировании аспарагина. Частой модификацией является удаление карбокси-концевого основного остатка (такого как лизин или аргинин) в результате действия карбоксипептидаз (как описано в статье Harris, R.J. Journal of Chromatography 705: 129-134, 1995). Соответственно, C-концевой остаток лизина в тяжелой цепи антитела может отсутствовать.
АФФИННОСТЬ
Мультиспецифические молекулы по настоящему изобретению содержат связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79a и/или CD79b.
В одном варианте осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным в отношении CD22.
В одном варианте осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным в отношении CD79a.
В одном варианте осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным в отношении CD79b.
В одном варианте осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным в отношении комплекса CD79, то есть, он узнает эпитоп, присутствующий в комплексе, и является специфичным в отношении него, например, эпитоп, включающий участок взаимодействия между CD79a и CD79b.
CD22 (также известный как кластер дифференцировки-22) является известным белком. CD22 представляет собой ингибирующий корецептор B-клеточного рецептора (BCR) и играет очень важную роль в установлении пороговых значений сигнализации для активации B-клеток. Последовательность белка человека доступна под регистрационным номером UniProt P20273 (SEQ ID NO: 161, и без сигнального пептида аминокислоты 20-847 в SEQ ID NO: 161). Мышиный вариант белка имеет регистрационный номер UniProt P35329. Настоящее изобретение относится ко всем формам CD22 из любых видов, в частности, к белку человека и его природным вариантам. В одном варианте осуществления CD22 означает белок человека.
В одном варианте осуществления аффинность связывающего домена в отношении CD22 в молекуле по настоящему изобретению составляет примерно 100 нМ или является более сильной, например, составляет примерно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пM, 250 пM, 200 пM, 100 пM или является более сильной, в частности, аффинность связывания составляет 50 пM или является более сильной.
Связывающий домен для CD79 может связываться с CD79a и/или CD79b.
CD79a (также известный как альфа цепь белка, ассоциированного с комплексом иммуноглобулина альфа и B-клеточного антигенного рецептора) является известным белком. Экспрессия CD79a ограничена B-лимфоцитами. Последовательность белка человека доступна под регистрационным номером UniProt P11912 (SEQ ID NO: 162, и без сигнальной последовательности аминокислоты 33-226 в SEQ ID NO: 162). Мышиный вариант белка имеет регистрационный номер UniProt 11911. Настоящее изобретение относится ко всем формам CD79a из любых видов, в частности, к белку человека и его природным вариантам. В одном варианте осуществления CD79a означает белок человека.
CD79b (также известный как иммуноглобулин-ассоциированный бета и кластер дифференцировки 79B) является известным белком. Экспрессия CD79b ограничена B-лимфоцитами. Последовательность белка человека доступна под регистрационным номером UniProt P40259 (SEQ ID NO: 163, и без сигнальной последовательности аминокислоты 29-229 в SEQ ID NO: 163). Мышиный вариант белка имеет регистрационный номер UniProt P15530. Настоящее изобретение относится ко всем формам CD79b из любых видов, в частности, к белку человека и его природным вариантам. В одном варианте осуществления CD79b означает белок человека.
В одном варианте осуществления связывающий домен, специфичный в отношении CD79, связывает CD79a.
В одном варианте осуществления связывающий домен, специфичный в отношении CD79, связывает CD79b.
В одном варианте осуществления связывающий домен, специфичный в отношении CD79, связывает комплекс CD79a и CD79b.
В одном варианте осуществления аффинность связывающего домена для CD79 в молекуле по настоящему изобретению составляет примерно 100 нМ или является более сильной, например, составляет примерно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пM, 250 пM, 200 пM, 100 пM или является более сильной, в частности, аффинность связывания составляет 50 пM или является более сильной.
В одном варианте осуществления аффинность связывающего домена для CD79a в молекуле по настоящему изобретению составляет примерно 100 нМ или является более сильной, например, составляет примерно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пM, 250 пM, 200 пM, 100 пM или является более сильной, в частности, аффинность связывания составляет 50 пM или является более сильной.
В одном варианте осуществления аффинность связывающего домена для CD79b в молекуле по настоящему изобретению составляет примерно 100 нМ или является более сильной, например, составляет примерно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пM, 250 пM, 200 пM, 100 пM или является более сильной, в частности, аффинность связывания составляет 50 пM или является более сильной.
Следует понимать, что аффинность связывающего домена для CD22 может быть такой же или отличающейся от аффинности связывающего домена для CD79.
В одном варианте осуществления мультиспецифические молекулы антитела по настоящему изобретению или их фрагменты/компоненты подвергают обработке в целях повышения аффинности в отношении целевого антигена или антигенов. Такие варианты могут быть получены с использованием целого ряда протоколов созревания аффинности, включая введение мутаций в области CDR (Yang et al., J. Mol. Biol., 254, 392-403, 1995), перетасовку цепей (Marks et al., Bio/Technology, 10, 779-783, 1992), использование штаммов-мутаторов E. coli (Low et al., J. Mol. Biol., 250, 359-368, 1996), перетасовку ДНК (Patten et al., Curr. Opin. Biotechnol., 8, 724-733, 1997), фаговый дисплей (Thompson et al., J. Mol. Biol., 256, 77-88, 1996) и ПЦР с имитацией полового размножения (Crameri et al., Nature, 391, 288-291, 1998). В статье Vaughan et al. (выше) описаны данные способы созревания аффинности.
АНТИТЕЛА И ИХ ПОЛУЧЕНИЕ
Связывающие домены для использования по настоящему изобретению можно получать любым подходящим способом, известным в данной области, например, области CDR могут быть взяты из антител, отличных от человеческих, включая коммерчески доступные антитела, и привиты на каркасы антитела человека или, альтернативно, могут быть получены химерные антитела с вариабельными областями, отличными от человеческих, и константными областями антитела человека, и так далее.
Как правило, связывающие домены для использования по настоящему изобретению представляют собой связывающие домены, полученные из антител, которые связывают выбранный антиген, таких как антитела, которые связывают CD22, CD79a и/или CD79b.
Примеры антител к CD22 и CD79 известны в данной области и их можно использовать непосредственно в молекулах по настоящему изобретению или подвергать скринингу на пригодность с использованием способов, описанных в настоящем документе, а затем модифицировать, при необходимости, например, гуманизировать с использованием способов, описанных в настоящем документе. Примеры антител к CD22, имеющих клиническое применение, включают эпратузумаб и инотузумаб. В данной области описаны и другие терапевтические антитела, например, анти-CD22 антитела, раскрытые в US2003202975 и WO 14011520, анти-CD79b антитела, раскрытые в WO 14011521 и WO 15021089. Анти-CD22 антитела, отличные от человеческих, включают кроличье моноклональное антитело LS-C2210357 (LSBio) из клона SP104, мышиное моноклональное антитело LS-C174778 из клона 4C3, мышиное моноклональное антитело LS-C4802, мышиное моноклональное антитело LS-B9996 из клона 1B1, мышиное моноклональное антитело LS-C340404 из клона 2E6, мышиное моноклональное антитело LS-C312263, мышиное моноклональное антитело LS-C152867, мышиное моноклональное антитело LS-C87523, мышиное моноклональное антитело LS-C134333 из клона FRB4, мышиное моноклональное антитело LS-C134336, мышиное моноклональное антитело LS-C40961 из клона HIB22, мышиное моноклональное антитело LS-C134332, следующие антитела от компании Santa Cruz Biotechnology: sc-271579, sc-377304, sc-7032, sc-18909, sc-7932, sc-7323, sc-7307, sc-7031, sc-20053, sc-189000, sc-136440, sc-136507, sc-53031, sc-73363, sc-53032, кроличье моноклональное антитело Ab33859 (EP498Y) от компании Abcam, мышиное моноклональное антитело AA 1-687 с каталожным номером ABIN1999423, мышиное моноклональное антитело от компании Biolegend workshop с номером V CD22.14 из клона HIB22.
Коммерчески доступные анти-CD79a антитела включают мышиное моноклональное антитело LS-B4504 (LSBio) из клона HM57, мышиное моноклональное антитело LS-B8330, мышиное моноклональное антитело LS-C44954, кроличье моноклональное антитело LS-B9093, мышиное моноклональное антитело LS-B8513 из клона JCB117, кроличье моноклональное антитело LS-C210607 из клона SP18, мышиное моноклональное антитело LS-C175441 из клона 5E2, мышиное моноклональное антитело LS-C338670 из клона 3D3, мышиное моноклональное антитело LS-C88120 из клона HM47/A9, мышиное моноклональное антитело LS-C191714, мышиное моноклональное антитело LS-C87592, мышиное моноклональное антитело LS-C44955, мышиное моноклональное антитело LS-C95934, мышиное моноклональное антитело LS-C121584, мышиное моноклональное антитело LS-C121585, мышиное моноклональное антитело LS-C204347, мышиное моноклональное антитело LS-C88122, мышиное моноклональное антитело ab3121 [HM47/A9] от компании Abcam, кроличье моноклональное антитело ab79414 и кроличье моноклональное антитело ab133483.
Коммерчески доступные антитела к CD79b включают мышиное моноклональное антитело ab33295 от компании Abcam, крысиное моноклональное антитело ab23826, мышиное моноклональное антитело ab103422, кроличье моноклональное антитело ab134103, кроличье моноклональное антитело ab134147 и кроличье моноклональное антитело ab183343.
Такие коммерчески доступные антитела могут быть полезными инструментами в разработке следующих терапевтических антител.
Квалифицированный специалист может получать антитела для использования в мультиспецифических молекулах по изобретению любым подходящим методом, известным в данной области.
Антигенные полипептиды, используемые для получения антител, например, используемые для иммунизации хозяина или используемые для пэннинга, например, в фаговом дисплее, можно получать методами, хорошо известными в данной области, из генетически модифицированных клеток-хозяев, содержащих экспрессионные системы, или их можно извлекать из природных биологических источников. В настоящей заявке термин «полипептиды» включает пептиды, полипептиды и белки. Их используют взаимозаменяемо, если не указано иначе. Антигенный полипептид в некоторых случаях может быть частью большего белка, такого как слитый белок, например, слит с аффинной меткой или тому подобным. В одном варианте осуществления хозяина можно иммунизировать клеткой, такой как фибробласт, трансфицированной соответствующим белком или полипептидом, например, совместно трансфицированной CD79a и CD79b.
Антитела, вырабатываемые против антигенного полипептида, в случае, когда необходима иммунизация животного, можно получать путем введения полипептидов животному, предпочтительно животному, отличному от человека, с использованием хорошо известных и обычных протоколов, смотри, например, сборник «Handbook of Experimental Immunology», D.M. Weir (ed.), Vol 4, Blackwell Scientific Publishers, Oxford, England, 1986. Можно иммунизировать многих теплокровных животных, таких как кролики, мыши, крысы, овцы, коровы, верблюды или свиньи. Однако мыши, кролики, свиньи и крысы, как правило, являются наиболее подходящими.
Моноклональные антитела можно получать любым методом, известным в данной области, таким как метод гибридом (Kohler & Milstein, 1975, Nature, 256: 495-497), метод триом, метод гибридом с B-клетками человека (Kozbor et al., 1983, Immunology Today, 4: 72) и методом EBV-гибридом (Cole et al., Monoclonal Antibodies and Cancer Therapy, pp. 77-96, Alan R. Liss, Inc., 1985).
Антитела также можно получать методом одиночных лимфоцитов путем клонирования и экспрессии кДНК вариабельных областей иммуноглобулина, полученных из одиночных лимфоцитов, выбранных для продуцирования специфических антител, например, методами, описанными в статье Babcook, J. et al., 1996, Proc. Natl. Acad. Sci. USA 93(15): 7843-7848l; WO 92/02551; WO 2004/051268 и WO 2004/106377.
Антитела для использования по настоящему изобретению также можно получать с использованием различных методов фагового дисплея, известных в данной области, включая те, которые описаны в статьях Brinkman et al. (в J. Immunol. Methods, 1995, 182: 41-50), Ames et al. (J. Immunol. Methods, 1995, 184: 177-186), Kettleborough et al. (Eur. J. Immunol. 1994, 24: 952-958), Persic et al. (Gene, 1997, 187: 9-18), Burton et al. (Advances in Immunology, 1994, 57: 191-280), а также WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401 и US 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743; 5969108 и WO 20011/30305.
В одном примере мультиспецифические молекулы по настоящему изобретению являются полностью человеческими, в частности, один или более из вариабельных доменов являются полностью человеческими.
Полностью человеческие молекулы представляют собой такие молекулы, в которых все вариабельные области и константные области (при наличии) как тяжелой, так и легкой цепей являются человеческими, или по существу идентичными последовательностям из антитела человека, необязательно из одного и того же антитела. Примеры полностью человеческих антител могут включать антитела, полученные, например, методами фагового дисплея, описанными выше, и вырабатываемые мышами антитела, в которых мышиные гены вариабельной и, необязательно, константной области иммуноглобулина были заменены их человеческими аналогами, например, как описано в общих чертах в EP0546073, US5545806, US5569825, US5625126, US5633425, US5661016, US5770429, EP 0438474 и EP0463151.
В одном примере связывающие домены мультиспецифических молекул по изобретению являются гуманизированными.
Используемый в настоящем документе термин «гуманизированные» (который включает CDR-привитые антитела) означает молекулы, имеющие одну или более определяющих комплементарность областей (CDR) из видов, отличных от человека, и каркасную область из молекулы человеческого иммуноглобулина (смотри, например, US5585089; WO 91/09967). Следует понимать, что может быть необходимо переносить только определяющие специфичность остатки из областей CDR, а не полностью CDR (смотри, например, Kashmiri et al., 2005, Methods, 36, 25-34). Гуманизированные антитела, необязательно, могут дополнительно содержать один или более каркасных остатков из видов, отличных от человека, из которых получены области CDR.
Используемый в настоящем документе термин «молекула гуманизированного антитела» означает молекулу антитела, в которой тяжелая и/или легкая цепь содержит одну или более областей CDR (включая, при необходимости, одну или более модифицированных областей CDR) из донорского антитела (например, мышиного моноклонального антитела), привитые на каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, человеческого антитела). Для обзора смотри статью Vaughan et al., Nature Biotechnology, 16, 535-539, 1998. В одном варианте осуществления вместо переноса всей области CDR на каркас антитела человека переносят только один или более определяющих специфичность остатков из любой из областей CDR, описанных выше в настоящем документе (смотри, например, Kashmiri et al., 2005, Methods, 36, 25-34). В одном варианте осуществления на каркас антитела человека переносят только определяющие специфичность остатки из одной или более областей CDR, описанных выше в настоящем документе. В другом варианте осуществления на каркас антитела человека переносят только определяющие специфичность остатки из каждой из областей CDR, описанных выше в настоящем документе.
Если прививают области CDR или определяющие специфичность остатки, можно использовать любую подходящую акцепторную каркасную последовательность вариабельной области, с учетом класса/типа донорского антитела, из которого получены области CDR, включая каркасные области иммуноглобулинов мыши, приматов и человека. Предпочтительно, гуманизированное антитело в соответствии с настоящим изобретением имеет вариабельный домен, содержащий акцепторные каркасные области иммуноглобулина человека, а также одну или более областей CDR, описанных в настоящем документе.
Примерами каркасов иммуноглобулинов человека, которые можно использовать по настоящему изобретению, являются KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (Kabat et al., выше). Например, KOL и NEWM можно использовать для тяжелой цепи, REI можно использовать для легкой цепи и EU, LAY и POM можно использовать как для тяжелой цепи, так и для легкой цепи. Альтернативно, можно использовать последовательности зародышевой линии иммуноглобулинов человека; они доступны на сайте: http://www2.mrc-lmb.cam.ac.uk/vbase/list2.php.
В молекуле гуманизированного антитела по настоящему изобретению акцепторные тяжелая и легкая цепи необязательно должны быть получены из одного и того же антитела и, при необходимости, могут представлять собой составные цепи, имеющие каркасные области, полученные из разных цепей.
Каркасные области необязательно должны иметь точно такую же последовательность, как у акцепторного антитела. Например, необычные остатки могут быть заменены на более часто встречающиеся остатки для такого класса или типа акцепторной цепи. Альтернативно, выборочные остатки в акцепторных каркасных областях могут быть изменены таким образом, чтобы они соответствовали остатку, находящемуся в том же положении в донорском антителе (смотри Reichmann et al., 1998, Nature, 332, 323-324). Такие изменения должны быть сведены к минимуму, необходимому для восстановления аффинности донорского антитела. Протокол для выбора остатков в акцепторных каркасных областях, которые может потребоваться заменять, приведен в WO 91/09967.
В производных каркасных областях 1, 2, 3 или 4 аминокислоты могут быть заменены на альтернативную аминокислоту, например, на донорский остаток.
Донорские остатки представляют собой остатки из донорского антитела, то есть, антитела, из которого были исходно получены области CDR, в частности, выбирают остаток в соответствующем положении из донорской последовательности. Донорские остатки могут быть заменены соответствующим остатком, происходящим из акцепторной каркасной последовательности иммуноглобулина человека (акцепторные остатки).
Остатки в вариабельных доменах антитела, как правило, нумеруют в соответствии с системой, разработанной Kabat с соавторами. Данная система изложена в публикации Kabat et al., 1987, в Sequences of Proteins of Immunological Interest, US Department of Health и Human Services, NIH, USA (далее по тексту «Kabat et al. (выше)»). В настоящей спецификации используют данную систему нумерации, если не указано иначе.
Обозначение остатков по Kabat не всегда прямо соответствует линейной нумерации аминокислотных остатков. Фактическая линейная аминокислотная последовательность может содержать меньше или больше аминокислот, чем в четкой нумерации Kabat, отражая укорочение или вставку в структурный компонент, либо каркас, либо определяющую комплементарность область (CDR), базовой структуры вариабельного домена. Правильную нумерацию остатков по Kabat можно определять для конкретного антитела путем выравнивания гомологичных остатков в последовательности антитела со «стандартной» нумерованной по Kabat последовательностью.
Области CDR вариабельного домена тяжелой цепи расположены на остатках 31-35 (CDR-H1), остатках 50-65 (CDR-H2) и остатках 95-102 (CDR-H3) в соответствии с системой нумерации Kabat. Однако в соответствии с системой Chothia (Chothia, C. and Lesk, A.M. J. Mol. Biol., 196, 901-917 (1987)) петля, эквивалентная CDR-H1, продолжается от остатка 26 до остатка 32. Таким образом, если не указано иначе, используемый в настоящем документе термин «CDR-H1» должен означать остатки 26-35, при использовании сочетания системы нумерации Kabat и топологического определения петли Chothia.
Области CDR вариабельного домена легкой цепи расположены на остатках 24-34 (CDR-L1), остатках 50-56 (CDR-L2) и остатках 89-97 (CDR-L3) в соответствии с системой нумерации Kabat.
В одном примере предложен связывающий домен, содержащий вариабельную область тяжелой цепи (например, VH), специфичную в отношении CD79, которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 78, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 79, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 80.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (например, VH), специфичную в отношении CD79, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 88 для CDR-H1, SEQ ID NO: 89 для CDR-H2 и SEQ ID NO: 90 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи (например, VL), специфичную в отношении CD79, которая содержит 3 области CDR легкой цепи SEQ ID NO: 75 для CDR-L1, SEQ ID NO: 76 для CDR-L2 и SEQ ID NO: 77 для CDR-L3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи (например, VL), специфичную в отношении CD79, которая содержит 3 области CDR легкой цепи SEQ ID NO: 85 для CDR-L1, SEQ ID NO: 86 для CDR-L2 и SEQ ID NO: 87 для CDR-L3.
В одном примере предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD79, которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 78, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 79, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 80, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 75, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 76, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 77.
В одном примере предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD79, которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 88, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 89, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 90, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 85, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 86, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 87.
В одном варианте осуществления мультиспецифическая молекула по настоящему изобретению содержит связывающий домен, специфичный в отношении CD22, который содержит 3 области CDR тяжелой цепи, выбранные из группы, включающей SEQ ID NO: 98, 99, 100, 108, 109, 110, 118, 119, 120, 128, 129, 130, 138, 139, 140, 148, 149 и 150.
В одном варианте осуществления мультиспецифическая молекула по настоящему изобретению содержит связывающий домен, специфичный в отношении CD22, который содержит 3 области CDR легкой цепи, выбранные из группы, включающей SEQ ID NO: 95, 97, 97, 105, 106, 107, 115, 116, 117, 125, 126, 127, 136, 137, 138, 145, 146 и 147.
В одном варианте осуществления мультиспецифическая молекула по настоящему изобретению содержит связывающий домен, специфичный в отношении CD22, который содержит 3 области CDR тяжелой цепи, выбранные из группы, включающей SEQ ID NO: 98, 99, 100, 108, 109, 110, 118, 119, 120, 128, 129, 130, 138, 139, 140, 148, 149 и 150, и 3 области CDR легкой цепи, выбранные из группы, включающей SEQ ID NO: 95, 97, 97, 105, 106, 107, 115, 116, 117, 125, 126, 127, 136, 137, 138, 145, 146 и 147.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD22, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 98 для CDR-H1, SEQ ID NO: 99 для CDR-H2 и SEQ ID NO: 100 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD22, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 108 для CDR-H1, SEQ ID NO: 109 для CDR-H2 и SEQ ID NO: 110 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD22, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 118 для CDR-H1, SEQ ID NO: 119 для CDR-H2 и SEQ ID NO: 120 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD22, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 128 для CDR-H1, SEQ ID NO: 129 для CDR-H2 и SEQ ID NO: 130 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD22, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 138 для CDR-H1, SEQ ID NO: 139 для CDR-H2 и SEQ ID NO: 140 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область тяжелой цепи (VH), специфичную в отношении CD22, которая содержит 3 области CDR тяжелой цепи SEQ ID NO: 148 для CDR-H1, SEQ ID NO: 149 для CDR-H2 и SEQ ID NO: 150 для CDR-H3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи, специфичную в отношении CD22, которая содержит 3 области CDR легкой цепи SEQ ID NO: 95 для CDR-L1, SEQ ID NO: 96 для CDR-L2 и SEQ ID NO: 97 для CDR-L3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи, специфичную в отношении CD22, которая содержит 3 области CDR легкой цепи SEQ ID NO: 105 для CDR-L1, SEQ ID NO: 106 для CDR-L2 и SEQ ID NO: 107 для CDR-L3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи, специфичную в отношении CD22, которая содержит 3 области CDR легкой цепи SEQ ID NO: 115 для CDR-L1, SEQ ID NO: 116 для CDR-L2 и SEQ ID NO: 117 для CDR-L3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи, специфичную в отношении CD22, которая содержит 3 области CDR легкой цепи SEQ ID NO: 125 для CDR-L1, SEQ ID NO: 126 для CDR-L2 и SEQ ID NO: 127 для CDR-L3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи, специфичную в отношении CD22, которая содержит 3 области CDR легкой цепи SEQ ID NO: 135 для CDR-L1, SEQ ID NO: 136 для CDR-L2 и SEQ ID NO: 137 для CDR-L3.
В одном варианте осуществления предложен связывающий домен, содержащий вариабельную область легкой цепи, специфичную в отношении CD22, которая содержит 3 области CDR легкой цепи SEQ ID NO: 145 для CDR-L1, SEQ ID NO: 146 для CDR-L2 и SEQ ID NO: 147 для CDR-L3.
В одном примере предложен связывающий домен, специфичный в отношении CD22, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 98, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 99, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 100, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 95, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 96, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 97.
В одном примере предложен связывающий домен, специфичный в отношении CD22, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 108, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 109, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 110, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 105, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 106, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 107.
В одном примере предложен связывающий домен, специфичный в отношении CD22, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 118, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 119, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 120, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 115, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 116, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 117.
В одном примере предложен связывающий домен, специфичный в отношении CD22, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 128, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 129, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 130, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 125, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 126, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 127.
В одном примере предложен связывающий домен, специфичный в отношении CD22, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 138, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 139, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 140, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 135, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 136, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 137.
В одном примере предложен связывающий домен, специфичный в отношении CD22, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 148, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 149, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 150, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 145, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 146, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 147.
В одном примере по настоящему изобретению предложена мультиспецифическая молекула, содержащая связывающий домен, специфичный в отношении антигена CD79, и связывающий домен, специфичный в отношении антигена CD22, при этом данная пара связывающих доменов содержит 6 областей CDR от пары антител к CD79 и к CD22, причем указанную пару антител выбирают из следующего списка пар антител к CD79 и к CD22: 4447 и 4120, 4447 и 4126, 4447 и 4127, 4447 и 4128, 4447 и 4130, 4447 и 4132, 4450 и 4120, 4450 и 4126, 4450 и 4127, 4450 и 4128, 4450 и 4130, а также 4450 и 4132.
Последовательности этих антител к CD79 (антитела 4447 и антитела 4450), включая последовательности VH, VL и CDR, приведены ниже в настоящем документе. Последовательности этих антител к CD22 (антител 4120, 4126, 4127, 4128, 4130, 4132), включая последовательности VH, VL и CDR, приведены ниже в настоящем документе и могут быть скомбинированы в качестве связывающих доменов в молекулах по настоящему изобретению.
В одном варианте осуществления изобретение относится к последовательности антитела, раскрытой в настоящем документе.
В одном примере предложен связывающий домен, специфичный в отношении альбумина, содержащий вариабельную область тяжелой цепи (VH), которая содержит три области CDR, при этом CDR-H1 имеет последовательность, приведенную в SEQ ID NO: 151, CDR-H2 имеет последовательность, приведенную в SEQ ID NO: 152, и CDR-H3 имеет последовательность, приведенную в SEQ ID NO: 153, и вариабельную область легкой цепи (VL), которая содержит три области CDR, при этом CDR-L1 имеет последовательность, приведенную в SEQ ID NO: 154, CDR-L2 имеет последовательность, приведенную в SEQ ID NO: 155, и CDR-L3 имеет последовательность, приведенную в SEQ ID NO: 156.
В одном примере предложен связывающий домен, специфичный в отношении альбумина, содержащий вариабельную область тяжелой цепи (VH), имеющую последовательность, приведенную в SEQ ID NO: 157, и вариабельную область легкой цепи (VL), имеющую последовательность, приведенную в SEQ ID NO: 159.
В одном примере предложен связывающий домен, специфичный в отношении альбумина, содержащий вариабельную область тяжелой цепи (VH), имеющую последовательность, приведенную в SEQ ID NO: 158, и вариабельную область легкой цепи (VL), имеющую последовательность, приведенную в SEQ ID NO: 160.
В одном примере связывающие домены являются гуманизированными.
В одном примере одна или более областей CDR, предложенных в настоящем документе, могут быть модифицированы для удаления нежелательных остатков или сайтов, например, остатков цистеина или сайтов изомеризации аспарагиновой кислоты (D), или сайтов дезамидирования аспарагина (N).
Например, один или более остатков цистеина в любой из областей CDR могут быть заменены на любую другую аминокислоту, например, серин.
В одном примере сайт дезамидирования аспарагина может быть удален из одной или более областей CDR за счет мутации остатка аспарагина (N) и/или соседнего остатка на любую другую подходящую аминокислоту. В одном примере сайт дезамидирования аспарагина, такой как NG или NS, может быть изменен мутацией, например, на NA или NT.
В одном примере сайт изомеризации аспарагиновой кислоты может быть удален из одной или более областей CDR за счет мутации остатка аспарагиновой кислоты (D) и/или соседнего остатка на любую другую подходящую аминокислоту. В одном примере сайт изомеризации аспарагиновой кислоты, такой как DG или DS, может быть изменен мутацией, например, на EG, DA или DT.
В одном примере сайт N-гликозилирования, такой как NLS, может быть удален за счет мутации остатка аспарагина (N) на любую другую подходящую аминокислоту, например, на SLS или QLS. В одном примере сайт N-гликозилирования, такой как NLS, может быть удален за счет мутации остатка серина (S) на любой другой остаток, за исключением треонина (T).
Квалифицированный специалист может тестировать варианты областей CDR или гуманизированные последовательности в любом подходящем анализе, таком как те, которые описаны в настоящем документе, для подтверждения сохранения активности.
Специфическое связывание с антигеном можно тестировать в любом подходящем анализе, включая, например, методы ELISA или поверхностного плазмонного резонанса, например, BIAcore, в которых можно измерять связывание с антигеном (CD22 или CD79). В таких анализах можно использовать выделенные природные или рекомбинантные белки CD22 или CD79 (a и/или b), или соответствующий слитый белок/полипептид. В одном примере связывание измеряют с использованием рекомбинантного CD22 (например, последовательности, приведенной в SEQ ID NO: 161 или аминокислотах 20-847 из SEQ ID NO: 161) или CD79 (например, последовательности, приведенной в SEQ ID NO: 162 и SEQ ID NO: 163 и аминокислотах 33-226 из SEQ ID NO: 162, и аминокислотах 29-229 из SEQ ID NO: 163), например, методом поверхностного плазмонного резонанса, таким как BIAcore. Альтернативно, белки могут быть экспрессированы на клетке, такой как клетка HEK, и аффинность можно измерять методом определения аффинности на основе проточной цитометрии.
Последовательности антител, предложенные по настоящему изобретению, можно использовать для идентификации дополнительных антител и, следовательно, связывающих доменов, подходящих для использования в мультиспецифических молекулах по настоящему изобретению. Антитела, которые перекрестно блокируют связывание молекулы антитела по настоящему изобретению с CD79, в частности, молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 73, и последовательность легкой цепи, приведенную в SEQ ID NO: 71, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 83, и последовательность легкой цепи, приведенную в SEQ ID NO: 81, могут быть так же полезны для связывания CD79 и, следовательно, так же полезны в мультиспецифических молекулах по настоящему изобретению. Соответственно, настоящее изобретение также относится к мультиспецифической молекуле, содержащей связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79b, в случае, когда связывающий домен для CD79b перекрестно блокирует связывание любой из молекул антител, описанных выше в настоящем документе, с CD79 и/или перекрестно блокирован от связывания CD79 любым из данных антител. В одном варианте осуществления такое антитело связывается с тем же эпитопом, что и антитело, описанное выше в настоящем документе. В другом варианте осуществления перекрестно блокирующее антитело связывается с эпитопом, который граничит и/или перекрывается с эпитопом, который связывает антитело, описанное выше в настоящем документе.
Аналогично, антитела, которые перекрестно блокируют связывание молекулы антитела по настоящему изобретению с CD22, в частности, молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 93, и последовательность легкой цепи, приведенную в SEQ ID NO: 91, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 103, и последовательность легкой цепи, приведенную в SEQ ID NO: 101, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 113, и последовательность легкой цепи, приведенную в SEQ ID NO: 111, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 123, и последовательность легкой цепи, приведенную в SEQ ID NO: 121, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 133, и последовательность легкой цепи, приведенную в SEQ ID NO: 131, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 143, и последовательность легкой цепи, приведенную в SEQ ID NO: 141, могут быть так же полезны для связывания CD22 и, следовательно, так же полезны в мультиспецифических молекулах по настоящему изобретению. Соответственно, настоящее изобретение также относится к мультиспецифической молекуле, содержащей связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79, в случае, когда связывающий домен для CD22 перекрестно блокирует связывание любой из молекул антител, описанных выше в настоящем документе, с CD22 и/или перекрестно блокирован от связывания CD22 любым из данных антител. В одном варианте осуществления такое антитело связывается с тем же эпитопом, что и антитело, описанное выше в настоящем документе. В другом варианте осуществления перекрестно блокирующее антитело связывается с эпитопом, который граничит и/или перекрывается с эпитопом, который связывает антитело, описанное выше в настоящем документе.
Перекрестно блокирующие антитела можно идентифицировать любым подходящим методом в данной области, например, в конкурентном анализе ELISA или анализе BIAcore, в которых связывание перекрестно блокирующего антитела с антигеном (CD22 и/или CD79) предотвращает связывание антитела по настоящему изобретению, или наоборот. В таких анализах перекрестного блокирования можно использовать экспрессированный в клетке, выделенный природный, либо рекомбинантный CD22 или CD79 (a и/или b), или соответствующий слитый белок/полипептид. В одном примере связывание и перекрестное блокирование измеряют с использованием рекомбинантного CD22, либо его подходящего фрагмента или природного варианта (например, последовательности, приведенной в SEQ ID NO: 161, или последовательности, приведенной в аминокислотах 20-847 из SEQ ID NO: 161), либо CD79, например, последовательности, приведенной в SEQ ID NO: 162, или последовательности, приведенной в аминокислотах 33-226 из SEQ ID NO: 162 (CD79a), и/или последовательности, приведенной в SEQ ID NO: 163, или последовательности, приведенной в аминокислотах 29-229 из SEQ ID NO: 163.
Альтернативно или дополнительно, антитела по данному аспекту изобретения могут быть перекрестно блокированы от связывания с антигеном (CD22 или CD79) связывающим доменом, раскрытым в настоящем документе, например, содержащим области CDR из вариабельной последовательности тяжелой цепи и последовательности легкой цепи, приведенные в SEQ ID NO: 71 и 73, 81 и 83, 91 и 93, 101 и 103, 111 и 113, 121 и 123, 131 и 133, а также 141 и 143. Таким образом, также предложена мультиспецифическая молекула, содержащая связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79b, в случае, когда связывающий домен для CD79b перекрестно блокирует связывание любой из молекул антител, описанных выше в настоящем документе, с CD79b и/или перекрестно блокирован от связывания CD79b любым из данных антител на более чем 80%, например, на более чем 85%, например, на более чем 90%, в частности, на более чем 95%, и необязательно, в случае, когда связывающий домен для CD22 перекрестно блокирует связывание любой из молекул антител, описанных выше в настоящем документе, с CD22 и/или перекрестно блокирован от связывания CD22 любым из данных антител на более чем 80%, например, на более чем 85%, например, на более чем 90%, в частности, на более чем 95%.
В одном аспекте предложена мультиспецифическая молекула антитела, содержащая или состоящая из:
a) полипептидной цепи формулы (I):
VH-CH1-X-(V1)p;
b) полипептидной цепи формулы (II):
VL-CL-Y-(V2)q;
где:
VH представляет собой вариабельный домен тяжелой цепи;
CH1 представляет собой домен константной области тяжелой цепи, например, домен 1 из нее;
X представляет собой связь или линкер, например, аминокислотный линкер;
Y представляет собой связь или линкер, например, аминокислотный линкер;
V1 представляет собой dab, scFv, dsscFv или dsFv;
VL представляет собой вариабельный домен, например, вариабельный домен легкой цепи;
CL представляет собой домен из константной области, например, домен константной области легкой цепи, такой как Cкаппа;
V2 представляет собой dab, scFv, dsscFv или dsFv;
p равно 0 или 1;
q равно 0 или 1; и
если p равно 1, q равно 0 или 1, и если q равно 1, p равно 0 или 1, то есть, p и q оба не равны 0.
В одном варианте осуществления мультиспецифическая молекула антитела содержит не более одного связывающего домена для CD22 и не более одного связывающего домена для CD79.
В одном варианте осуществления q равно 0 и p равно 1.
В одном варианте осуществления q равно 1 и p равно 1.
В одном варианте осуществления V1 представляет собой dab и V2 представляет собой dab, и вместе они образуют один связывающий домен из кооперативной пары вариабельных областей, такой как когнатная пара VH/VL, необязательно связанных дисульфидной связью.
В одном варианте осуществления VH и VL являются специфичными в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления V1 является специфичной в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления V2 является специфичной в отношении CD79, например, CD79a или CD79b.
В одном варианте осуществления V1 и V2 вместе (например, в виде связывающего домена) являются специфичными в отношении CD79, например, CD79a или CD79b, и VH и VL являются специфичными в отношении CD22.
В одном варианте осуществления V1 является специфичной в отношении CD22.
В одном варианте осуществления V2 является специфичной в отношении CD22.
В одном варианте осуществления V1 и V2 вместе (например, в виде одного связывающего домена) являются специфичными в отношении CD22, и VH и VL являются специфичными в отношении CD79.
В одном варианте осуществления V1 является специфичной в отношении CD22, V2 является специфичной в отношении альбумина, и VH и VL являются специфичными в отношении CD79.
В одном варианте осуществления V1 является специфичной в отношении альбумина, V2 является специфичной в отношении CD22, и VH и VL являются специфичными в отношении CD79.
В одном варианте осуществления V1 является специфичной в отношении CD79, V2 является специфичной в отношении альбумина, и VH и VL являются специфичными в отношении CD22.
В одном варианте осуществления V1 является специфичной в отношении альбумина, V2 является специфичной в отношении CD79, и VH и VL являются специфичными в отношении CD22.
В одном варианте осуществления V1 представляет собой dsscFv, специфичную в отношении CD22, V2 представляет собой dsscFv, специфичную в отношении альбумина, и VH и VL являются специфичными в отношении CD79.
В одном варианте осуществления V1 представляет собой dsscFv, специфичную в отношении альбумина, V2 представляет собой dscFv, специфичную в отношении CD22, и VH и VL являются специфичными в отношении CD79.
В одном варианте осуществления V1 представляет собой dsscFv, специфичную в отношении CD79, V2 представляет собой dsscFv, специфичную в отношении альбумина, и VH и VL являются специфичными в отношении CD22.
В одном варианте осуществления V1 представляет собой dsscFv, специфичную в отношении альбумина, V2 представляет собой dsscFv, специфичную в отношении CD79, и VH и VL являются специфичными в отношении CD22.
Каждый из V1, V2, VH и VL в конструктах, описанных выше, может представлять собой связывающий домен и включать любую из последовательностей, приведенных в настоящем документе.
X и Y представляют собой любой подходящий линкер, например, X и Y могут представлять собой SGGGGSGGGGS (SEQ ID NO: 17).
В одном варианте осуществления, когда V1 и/или V2 представляют собой dab, dsFv или dsscFv, дисульфидная связь между вариабельными доменами VH и VL в V1 и/или V2 образована между положениями VH44 и VL100.
Настоящее изобретение также относится к новым полипептидным последовательностям, раскрытым в настоящем документе, и последовательностям, которые по меньшей мере на 80% аналогичны или идентичны им, например, имеют 85% или более, например, 90% или более, в частности, 95% или более сходства или идентичности.
Используемый в настоящем документе термин «идентичность» означает, что в любом конкретном положении в выровненных последовательностях находятся идентичные аминокислотные остатки. Используемый в настоящем документе термин «сходство» означает, что в любом конкретном положении в выровненных последовательностях находятся аминокислотные остатки сходного типа. Например, лейцин может быть заменен на изолейцин или валин. Другие аминокислоты, которые часто могут быть заменены друг на друга, включают, но не ограничиваются ими:
- фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи);
- лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи);
- аспартат и глутамат (аминокислоты, имеющие кислые боковые цепи);
- аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и
- цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи).
Степень идентичности и сходства можно легко рассчитывать (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, программа BLAST™, доступная от NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215: 403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3: 266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266: 131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25: 3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7: 649-656).
В частности, в одном аспекте настоящее изобретение относится к антителам против CD22 и CD79, описанным в настоящем документе, в любом подходящем формате антител.
Соответственно, в одном аспекте настоящее изобретение относится к анти-CD22 антителам или их фрагментам, содержащим один или более из связывающих доменов, описанных выше в настоящем документе, содержащих области CDR, приведенные в настоящем документе и в SEQ ID NOS: 95, 96, 97, 98, 99 и 100 (антитело 4120) или 105, 106, 107, 108, 109 и 110 (антитело 4126), или 115, 116, 117, 118, 119 и 120 (антитело 4127), или 125, 126, 127, 128, 129 и 130 (антитело 4128), или 135, 136, 137, 138, 139 и 140 (антитело 4130), или 145, 146, 147, 148, 149 и 150 (антитело 4132). Также предложены анти-CD79 антитела или их фрагменты, содержащие один или более из связывающих доменов, описанных выше в настоящем документе, содержащих области CDR, приведенные в настоящем документе и в SEQ ID NOS: 75, 76, 77, 78, 79 и 80 (антитело 4447) или SEQ ID NOs 85, 86, 87, 88, 89 и 90 (антитело 4450).
Указанные области CDR могут быть включены в любой подходящий каркас антитела и в любой подходящий формат антитела. Такие антитела включают целые антитела, а также их функционально активные фрагменты или производные, которые могут представлять собой, но не ограничиваются ими, моноклональные, гуманизированные, полностью человеческие или химерные антитела. Соответственно, такие антитела могут включать целую молекулу антитела, имеющую полноразмерную тяжелую и легкую цепи, или ее фрагмент и могут представлять собой, но не ограничиваются ими, Fab, модифицированные Fab, Fab', F(ab')2, Fv, однодоменные антитела, scFv, двух-, трех- или четырехвалентные антитела, bis-scFv, диатела, триатела, тетратела и эпитопсвязывающие фрагменты любых из вышеперечисленных (смотри, например, Holliger and Hudson, 2005, Nature Biotech. 23(9): 1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217). Способы создания и производства таких фрагментов антител хорошо известны в данной области (смотри, например, Verma et al., 1998, Journal of Immunological Methods, 216: 165-181). Мультивалентные антитела могут иметь несколько специфичностей или могут быть моноспецифическими (смотри, например, WO 92/22853 и WO 05/113605). Следует понимать, что данный аспект изобретения также относится к вариантам таких антител против CD22 и CD79, включая гуманизированные варианты и модифицированные варианты, в том числе те, в которых аминокислоты были мутированы в областях CDR для удаления одного или более из сайтов изомеризации, дезамидирования, гликозилирования или остатков цистеина, как описано выше в настоящем документе.
ЛИНКЕРЫ
В настоящем документе определение линкеров в одном контексте может быть в равной степени применено к линкерам в других контекстах, связанных с использованием линкеров, например, в любой мультиспецифической молекуле по настоящему изобретению.
В одном варианте осуществления линкер, используемый в молекуле по настоящему изобретению, представляет собой аминокислотный линкер, имеющий 50 или менее остатков в длину, например, выбранный из последовательностей 5-70.
Таблица 1. Последовательности шарнирных линкеров
Таблица 2. Последовательности гибких линкеров
(S) является необязательным остатком в последовательностях 17-20.
Примеры жестких линкеров включают пептидные последовательности GAPAPAAPAPA (SEQ ID NO: 69), PPPP (SEQ ID NO: 70) и PPP.
Другие линкеры приведены в Таблице 3:
ЭФФЕКТОРНЫЕ МОЛЕКУЛЫ
При необходимости, мультиспецифическая молекула, предназначенная для использования по настоящему изобретению, может быть конъюгирована с одной или более эффекторными молекулами. Следует понимать, что эффекторная молекула может включать одну эффекторную молекулу, либо две или более таких молекул, связанных таким образом, что они образуют единый фрагмент, который может быть присоединен к мультиспецифическим молекулам по настоящему изобретению. Если необходимо получать антитело или мультиспецифическую молекулу по настоящему изобретению, связанную с эффекторной молекулой, этот конструкт можно создавать стандартными химическими методами или методами рекомбинантной ДНК, в которых фрагмент антитела связывают либо напрямую, либо через сшивающий агент с эффекторной молекулой. Методы конъюгирования таких эффекторных молекул с антителами хорошо известны в данной области (смотри, Hellstrom et al., Controlled Drug Delivery, 2е издание, Robinson et al., eds., 1987, pp. 623-53; Thorpe et al., 1982, Immunol. Rev., 62: 119-58 и Dubowchik et al., 1999, Pharmacology and Therapeutics, 83, 67-123). Конкретные химические методы включают, например, те, которые описаны в WO 93/06231, WO 92/22583, WO 89/00195, WO 89/01476 и WO 03/031581. Альтернативно, если эффекторная молекула представляет собой белок или полипептид, связывание можно осуществлять с использованием методов рекомбинантной ДНК, например, как описано в WO 86/01533 и EP0392745.
В одном варианте осуществления мультиспецифические молекулы по настоящему изобретению могут содержать эффекторную молекулу.
Используемый в настоящем документе термин «эффекторная молекула» охватывает, например, антинеопластические средства, лекарственные средства, токсины, биологически активные белки, например, ферменты, другое антитело или фрагменты антитела, синтетические или природные полимеры, нуклеиновые кислоты и их фрагменты, например, ДНК, РНК и их фрагменты, радионуклиды, в частности, радиоактивный йод, радиоизотопы, хелатные металлы, наночастицы и репортерные группы, такие как флуоресцентные соединения или соединения, которые могут быть обнаружены методами ЯМР или ЭПР-спектроскопии.
Примеры эффекторных молекул могут включать цитотоксины или цитотоксические средства, в том числе любое средство, которое отрицательно влияет на (например, уничтожает) клетки. Примеры включают комбрестатины, доластатины, эпотилоны, стауроспорин, майтанзиноиды, спонгистатины, ризоксин, халихондрины, роридины, гемиастерлины, таксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрацин-дион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, а также их аналоги или гомологи.
Эффекторные молекулы также включают, но не ограничиваются ими, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), алкилирующие средства (например, мехлорэтамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиамин платины (II) (DDP) цисплатин), антрациклины (например, даунорубицин (бывший дауномицин) и доксорубицин), антибиотики (например, дактиномицин (бывший актиномицин), блеомицин, митрамицин, антрамицин (AMC), калихеамицины или дуокармицины), а также антимитотические средства (например, винкристин и винбластин).
Другие эффекторные молекулы могут включать хелатные радионуклиды, такие как 111In и 90Y, Lu177, висмут213, калифорний252, иридий192 и вольфрам188/рений188; или лекарственные средства, такие как, но без ограничения, алкилфосфохолины, ингибиторы топоизомеразы I, таксоиды и сурамин.
Другие эффекторные молекулы включают белки, пептиды и ферменты. Представляющие интерес ферменты включают, но не ограничиваются ими, протеолитические ферменты, гидролазы, лиазы, изомеразы, трансферазы. Представляющие интерес белки, полипептиды и пептиды включают, но не ограничиваются ими, иммуноглобулины, токсины, такие как абрин, рицин A, экзотоксин псевдомонад или дифтерийный токсин, белок, такой как инсулин, фактор некроза опухолей, α-интерферон, β-интерферон, фактор роста нервов, тромбоцитарный фактор роста или тканевой активатор плазминогена, антитромботическое средство или антиангиогенное средство, например, ангиостатин или эндостатин, или модификатор биологического ответа, такой как лимфокин, интерлейкин-1 (IL-1), интерлейкин-2 (IL-2), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), гранулоцитарный колониестимулирующий фактор (G-CSF), фактор роста нервов (NGF) или другой фактор роста и иммуноглобулины.
Другие эффекторные молекулы могут включать поддающиеся обнаружению вещества, полезные, например, для диагностики. Примеры поддающихся обнаружению веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радионуклиды, позитрон-излучающие металлы (для использования в позитронно-эмиссионной томографии) и не радиоактивные парамагнитные ионы металлов. Смотри, в основном, патент США № 4741900 для информации, относящейся к ионам металлов, которые можно конъюгировать с антителами для использования в качестве диагностических средств. Подходящие ферменты включают пероксидазу хрена, щелочную фосфатазу, бета галактозидазу или ацетилхолинэстеразу; подходящие простетические группы включают стрептавидин, авидин и биотин; подходящие флуоресцентные материалы включают умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид и фикоэритрин; подходящие люминесцентные материалы включают люминол; подходящие биолюминесцентные материалы включают люциферазу, люциферин и экворин; и подходящие радионуклиды включают 125I, 131I, 111In и 99Tc.
В другом примере эффекторная молекула может увеличивать время полувыведения антитела in vivo и/или уменьшать иммуногенность антитела, и/или увеличивать доставку антитела через эпителиальный барьер к иммунной системе. Примеры подходящих эффекторных молекул такого типа включают полимеры, альбумин, альбумин-связывающие белки или альбумин-связывающие соединения, такие как те, которые описаны в WO 05/117984.
Если эффекторная молекула представляет собой полимер, она, как правило, может представлять собой синтетический или природный полимер, например, полиалкилен с необязательно замещенной прямой или разветвленной цепью, полиалкениленовый или полиоксиалкиленовый полимер, или разветвленный или неразветвленный полисахарид, например, гомо- или гетерополисахарид.
Конкретные возможные заместители, которые могут присутствовать на вышеупомянутых синтетических полимерах, включают одну или более гидрокси, метил или метокси групп.
Конкретные примеры синтетических полимеров включают поли(этиленгликоль) с необязательно замещенной прямой или разветвленной цепью, поли(пропиленгликоль), поли(виниловый спирт) или их производные, в частности, необязательно замещенный поли(этиленгликоль), например, метоксиполи(этиленгликоль) или его производные.
ФУНКЦИОНАЛЬНЫЕ АНАЛИЗЫ И ФОРМАТЫ СКРИНИНГА
Как правило, подходящие связывающие домены для использования по настоящему изобретению можно идентифицировать путем тестирования одной или более пар связывающих доменов в функциональном анализе. Например, мультиспецифическую молекулу, содержащую связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79a и/или CD79b, можно тестировать в одном или более функциональных анализах.
Используемый в настоящем документе термин «функциональный анализ» означает анализ, который можно использовать для определения одного или более желательных свойств или активностей белковых комплексов, антительных комплексов или смеси антител, подвергнутых воздействию условий анализа. Подходящие функциональные анализы могут представлять собой анализы на связывание, анализы на апоптоз, анализы на антителозависимую клеточную цитотоксичность (ADCC), анализы на комплементзависимую цитотоксичность (CDC), анализы на ингибирование клеточного роста или пролиферации (цитостатический эффект), анализы на уничтожение клеток (цитотоксический эффект), анализы на клеточную сигнализацию, анализы на продуцирование цитокинов, анализы на продуцирование антител и переключение изотипов, а также анализы на клеточную дифференциацию.
Эффективность мультиспецифических антител по настоящему изобретению можно сравнивать с эффективностью отдельных антител или смесей антител (или фрагментов) в таких моделях методами, как правило, известными специалисту в данной области.
Функциональные анализы можно выполнять столько раз, сколько потребуется, для повышения надежности полученных результатов. Можно применять различные статистические критерии, известные специалистам, для получения статистически значимых результатов и, таким образом, для идентификации мультиспецифических молекул с биологическими функциями.
Примеры подходящих функциональных анализов приведены в разделе «Примеры» настоящего документа и включают определение способности мультиспецифической молекулы по настоящему изобретению к ингибированию активации B-клеток после стимуляции анти-IgM, измеряемой по обнаруживаемому ингибированию маркеров активации B-клеток, таких как экспрессия фосфорилированного Akt, экспрессия фосфорилированного P38, сигнализация PLCγ, экспрессия CD40, экспрессия CD71 и/или экспрессия CD86.
При разработке функционального анализа для скрининга специалист может устанавливать соответствующие пороговые значения, свыше которых попытка обнаружения активности считается «успехом». При использовании более одного функционального анализа пороговые значения для каждого анализа могут быть установлены на соответствующем уровне для достижения контролируемого процента успеха. В одном примере процент успеха может составлять 3-5%. В одном примере критерием, установленным для поиска пар связывающих доменов, которые ингибируют функцию B-клеток, может являться по меньшей мере 30% ингибирование по меньшей мере двух показателей фосфорилирования, как описано выше и в примерах настоящего документа, в анализе на активацию B-клеток.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 1 нМ для ингибирования фосфорилирования P38 в стимулированных анти-IgM B-клетках, предпочтительно IC50 менее 0,5 нМ. В одном примере IC50 мультиспецифической молекулы в данном анализе составляет менее 0,05 нМ.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 1 нМ для ингибирования фосфорилирования Akt в стимулированных анти-IgM B-клетках, предпочтительно IC50 менее 0,1 нМ. В одном примере IC50 мультиспецифической молекулы в данном анализе составляет менее 0,05 нМ.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 1 нМ для ингибирования фосфорилирования PLCγ2 в стимулированных анти-IgM B-клетках, предпочтительно IC50 менее 0,8 нМ. В одном примере IC50 мультиспецифической молекулы в данном анализе составляет менее 0,05 нМ.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 5 нМ для ингибирования экспрессии CD71 в стимулированных анти-IgM B-клетках, предпочтительно IC50 менее 3 нМ. В одном примере IC50 мультиспецифической молекулы в данном анализе составляет менее 0,5 нМ.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 5 нМ для ингибирования экспрессии CD40 в стимулированных анти-IgM B-клетках. В одном примере IC50 мультиспецифической молекулы в данном анализе составляет менее 0,5 нМ.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 5 нМ для ингибирования экспрессии CD86 в стимулированных анти-IgM B-клетках, предпочтительно IC50 менее 2 нМ. В одном примере IC50 мультиспецифической молекулы в данном анализе составляет менее 0,5 нМ.
В одном примере мультиспецифическая молекула по настоящему изобретению имеет IC50 менее 5 нМ для ингибирования экспрессии CD71, CD40 и CD86 в стимулированных анти-IgM B-клетках и/или IC50 менее 1 нМ для ингибирования фосфорилирования PLCγ2, P38 и AKT в стимулированных анти-IgM B-клетках.
В одном варианте осуществления можно использовать in vivo анализы, такие как животные модели, включая модели опухолей мышей, модели аутоиммунных заболеваний, модели на грызунах или приматах, инфицированных вирусами или бактериями, и тому подобное, для тестирования молекул по настоящему изобретению.
Примеры подходящего формата для скрининга и выявления связывающих доменов описаны ниже в настоящем документе.
В скрининге с целью выявления связывающих доменов для использования по настоящему изобретению можно использовать биспецифический белковый комплекс.
Используемый в настоящем документе термин «биспецифический белковый комплекс» означает молекулу, содержащую два белка (A и B, называемые в настоящем документе биспецифическими компонентами), которые удерживаются вместе за счет гетеродимерного сплетения. В одном варианте осуществления один или оба из белков имеют связывающий домен, например, один или оба из белков являются антителами или их фрагментами.
Как правило, биспецифический белковый комплекс имеет формулу A-X:Y-B, где:
A-X представляет собой первый слитый белок;
Y-B представляет собой второй слитый белок;
X:Y представляет собой гетеродимерное сплетение;
A содержит первый связывающий домен;
B содержит второй связывающий домен;
X представляет собой первый партнер по связыванию связывающейся пары;
Y представляет собой второй партнер по связыванию связывающейся пары; и
: обозначает взаимодействие (такое как взаимодействие связывания) между X и Y.
«Слитые белки», используемые по настоящему изобретению, включают белковый компонент, например, A или B, слитый с другим фрагментом, например, партнером по связыванию X или Y (в зависимости от обстоятельств). В одном варианте осуществления слитый белок представляет собой транслированный белок, экспрессированный рекомбинантными методами с генетического конструкта, например, экспрессированный в хозяине с ДНК конструкта.
Функция сплетения X:Y заключается в удержании белков A и B в непосредственной близости друг к другу таким образом, что синергетическая функция A и B может быть реализована.
Используемый в настоящем документе термин «гетеродимерное сплетение» означает сплетение, заключающее в себе два разных партнера по связыванию X и Y, которые вступают друг с другом во взаимодействие (например, связывание), общая аффинность которого является достаточной для удержания вместе двух партнеров по связыванию. В одном варианте осуществления X и/или Y непригодны для образования гомодимеров.
Термины «гетеродимерно сплетенные» и «гетеродимерное сплетение» в настоящем документе используют взаимозаменяемо.
В одном варианте осуществления используемая в настоящем документе фраза «непригодны для образования гомодимеров» означает, что образование гетеродимеров X-Y имеет преимущества, например, стабильность, такую как термодинамическая стабильность и/или физическая стабильность (например, подтвержденная отсутствием агрегации), образованных гетеродимеров.
В одном варианте осуществления взаимодействие X-Y является более эффективным, чем взаимодействие X-X или Y-Y. Это уменьшает образование гомодимеров X-X или Y-Y при смешивании слитых белков A-X и B-Y. Это также делает удаление гомодимеров относительно простой процедурой, например, одна стадия очистки, такая как хроматография на колонке, позволяет получать по существу чистые слитые белки и/или биспецифические белковые комплексы по настоящему изобретению.
В одном варианте осуществления стадию очистки используют после экспрессии слитого белка. Таким образом, в одном варианте осуществления перед in vitro смешиванием слитый белок(белки) предоставляют в по существу чистой форме. Фраза «по существу чистая форма» при использовании в настоящем документе означает, что слитый белок на 85-100%, например, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100% представляет собой мономер.
В одном варианте осуществления после образования биспецифического белкового комплекса очистка не требуется.
В одном варианте осуществления соотношение слитых белков A-X и B-Y, используемых на стадии in vitro смешивания настоящего способа, составляет от 0,8:1 до 3:1, например, 1,5:1 или 2:1.
В одном варианте осуществления соотношение слитых белков B-Y и A-X, используемых на стадии in vitro смешивания настоящего способа составляет от 0,8:1 до 3:1, например, 1,5:1 или 2:1.
В одном варианте осуществления соотношение составляет 1:1.
В одном варианте осуществления один (или по меньшей мере один) из партнеров по связыванию не способен к образованию гомодимера, например, аминокислотная последовательность партнера по связыванию мутирована для устранения или сведения к минимуму образования гомодимеров.
В одном варианте осуществления оба из партнеров по связыванию не способны к образованию гомодимеров, например, один из партнеров по связыванию представляет собой пептид и другой партнер по связыванию представляет собой VHH, специфичную в отношении указанного пептида.
В одном варианте осуществления scFv, используемая в молекулах по настоящему изобретению, не способна к образованию гомодимеров.
Фраза «неспособность к образованию гомодимеров» при использовании в настоящем документе означает низкую или нулевую предрасположенность к образованию гомодимеров. При использовании в настоящем документе «низкая» предрасположенность означает наличие 5% или менее, например, 4, 3, 2, 1, 0,5% или менее агрегатов.
Небольшое количество агрегатов в слитых белках или остаточное количество в гетеродимерно сплетенном биспецифическом белковом комплексе, как правило, оказывает минимальный эффект на способ по настоящему изобретению.
В одном варианте осуществления «:» обозначает взаимодействие связывания, например, основанное на силах притяжения, таких как силы Ван-дер-Ваальса, например, водородных связях и электростатических взаимодействиях, например, на основе специфичности антитела в отношении антигена, такого как пептид.
В одном варианте осуществления «:» представляет собой ковалентную связь, образованную в результате специфического химического взаимодействия, такого как клик-химия.
В одном варианте осуществления «:» не является ковалентной связью.
Используемый в настоящем документе термин «образование комплекса» означает взаимодействие, включая взаимодействия связывания или химическую реакцию, которое является достаточно специфическим и сильным, когда слитые белковые компоненты A-X и B-Y вступают в контакт при соответствующих условиях, чтобы комплекс образовывался и слитые белки удерживались вместе.
При использовании в настоящем документе фраза «удерживаются вместе» означает удержание компонентов (слитых белков) в непосредственной близости друг от друга таким образом, что после связывания с комплексом можно обращаться так, как будто он является единой молекулой, и во многих случаях он ведет себя и действует, как единая молекула. В одном варианте осуществления такое удержание делает комплекс подходящим для использования в способе, раскрытом в настоящем документе, то есть, подходящим для использования по меньшей мере в одном функциональном скрининге.
В одном варианте осуществления взаимодействие связывания является обратимым.
При использовании в настоящем документе «специфичность» применительно к X и Y означает, что партнеры по связыванию X и Y при взаимодействии узнают только друг друга или имеют гораздо более высокую аффинность друг в отношении друга в сравнении с молекулами, не являющимися партнерами, например, по меньшей мере в 2, 3, 4, 5, 6, 7, 8, 9, 10 раз более высокую аффинность.
В одном варианте осуществления взаимодействие связывания между X и Y имеет низкую константу диссоциации.
Примеры низкой константы диссоциации включают 1-9×10-2с-1 или менее, например, 1-9×10-3с-1, 1-9×10-4с-1, 1-9×10-5с-1, 1-9×10-6с-1 или 1-9×10-7с-1. В частности, подходящие константы диссоциации включают 1×10-4с-1 или менее, например, 1×10-5с-1, 1×10-6с-1 или 1×10-7с-1.
Без привязки к конкретной теории, считается, что низкая константа диссоциации (также называемая скоростью диссоциации) позволяет молекулам оставаться достаточно стабильными, чтобы биспецифический белковый комплекс можно было использовать, в частности, в функциональных анализах скрининга.
В одном варианте осуществления аффинность X и Y друг в отношении друга составляет 5 нМ или является более сильной, например, составляет 4 нМ, 3 нМ, 2 нМ, 1 нМ или является более сильной.
В одном варианте осуществления аффинность X и Y друг в отношении друга составляет 900 пM или является более сильной, например, составляет 800, 700, 600, 500, 400, 300, 200, 100 или 50 пM или является более сильной.
В другом варианте осуществления аффинность X и Y друг в отношении друга составляет 10 пM или является более сильной, например, составляет 9, 8, 7, 6 или 5 пM.
Аффинность является величиной, рассчитываемой на основании скорости образования и распада взаимодействия. Используемый в настоящем документе термин «аффинность» означает силу суммарно всех нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнером по связыванию (например, пептидом). Аффинность молекулы в отношении ее партнера по связыванию, как правило, может быть выражена в виде равновесной константы диссоциации (KD). Аффинность можно измерять общепринятыми методами, известными в данной области, включая те, которые описаны в настоящем документе, такие как методы поверхностного плазмонного резонанса, в частности, BIAcore.
В одном варианте осуществления несколько биспецифических белковых комплексов по настоящему изобретению тестируют параллельно или практически одновременно.
При использовании в настоящем документе «одновременно» означает, что образцы/молекулы/комплексы анализируют в одном и том же анализе, например, в одном и том же «прогоне».
В одном варианте осуществления «одновременно» относится к параллельному анализу, в котором выходной сигнал анализируется прибором практически в одно и то же время. Для интерпретации полученных результатов данному сигналу может потребоваться конволюция.
Тестирование нескольких биспецифических белковых комплексов имеет то преимущество, что позволяет проводить более эффективный скрининг большого количества биспецифических белковых комплексов и идентифицировать новые и интересные взаимодействия. Очевидно, что разные вариабельные области для целевых антигенов интересующих белков CD22 и CD79 могут обеспечить доступ к тонким нюансам биологической функции.
В одном варианте осуществления несколько биспецифических белковых комплексов тестируют с использованием мультиплекса, описанного выше, и подвергают их одному или более функциональным анализам.
Используемый в настоящем документе термин «биологическая функция» означает активность, которая является естественной или планируемой для тестируемого биологического объекта, например, природная активность клетки, белка или тому подобного. В идеале, наличие функции можно тестировать с использованием функционального анализа in vitro, включая анализы, в которых используют живые клетки млекопитающих. При использовании в настоящем документе «естественная» функция включает аберрантную функцию, такую как функции, ассоциированные с раком.
Релевантный «биологический компаратор» при использовании в настоящем документе означает соответствующий объект для оценки активности, используемый в том же анализе, который используют для биспецифического белкового комплекса, с целью установления наличия каким-либо образом измененной, либо новой активности или функции. Соответствующие компараторы для A-X:Y-B могут включать очищенный белок (в том числе рекомбинантные белки) в естественной форме или представленный в том же формате, что и биспецифический комплекс, то есть где A и B представляют собой те же соединения, например, A-X:Y-A или B-X:Y-B. Альтернативно, в качестве компаратора можно использовать слитый белок A-X или B-Y не в форме комплекса. Альтернативно, можно использовать несколько компараторов в разных форматах (в частности, как описано в настоящем документе). Специалист в данной области способен идентифицировать и включать в анализ соответствующий контроль/компаратор на основании общеизвестных фактов или информации, полученной из литературных источников.
Используемый в настоящем документе термин «синергетическая функция» означает биологическую активность, которая отсутствует или выражена в гораздо меньшей степени, когда первый и второй белки биспецифического белкового комплекса по настоящему изобретению не связаны вместе, например, активность, которую наблюдают только в случае биспецифической формы. Таким образом, термин «синергетическая» включает новую биологическую функцию.
Настоящее изобретение относится к молекуле со специфичностью по меньшей мере к CD22 и CD79, обладающей новой биологической функцией.
Используемый в настоящем документе термин «новая биологическая функция» означает функцию, которая не является очевидной или отсутствует до тех пор, пока два или более синергетических соединений [белок A и белок B] не объединяются (в виде биспецифического соединения или иным образом), либо ранее не идентифицированную функцию.
При использовании в настоящем документе выражение «более высокая» означает возрастание активности, включая возрастание от нуля, то есть, появление некоторой активности у биспецифического соединения в случае, когда у отдельного не входящего в комплекс биспецифического компонента или компонентов активность отсутствует в соответствующем функциональном анализе, в этом случае в настоящем документе также используют выражение «новая активность» или «новая биологическая функция». При использовании в настоящем документе выражение «более высокая» также охватывает большую, чем аддитивная, функцию биспецифического соединения в соответствующем функциональном анализе по сравнению с функцией отдельных не входящих в комплекс биспецифических компонентов или двухвалентных связывающих доменов, например, возросшую на 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300% активность в соответствующем анализе.
В одном варианте осуществления новая синергетическая функция представляет собой более высокую ингибирующую активность.
В одном варианте осуществления мультиспецифическая молекула антитела по настоящему изобретению имеет более высокую ингибирующую активность, чем суммарная активность двухвалентного связывающего домена для CD22 и двухвалентного связывающего домена для CD79a, отдельно или в смеси.
В одном варианте осуществления по меньшей мере один из первого партнера по связыванию, X, и второго партнера по связыванию, Y, связывающейся пары независимо выбран из пептида и белка; например, первый партнер по связыванию или второй партнер по связыванию представляет собой пептид.
Подходящие пептиды включают группу, включающую GCN4, Fos/Jun (человеческий и мышиный Fos имеют номера Uniprot P01100 и P01101, соответственно, и человеческий и мышиный jun имеют номера Uniprot P05412 и P05627, соответственно), гемагглютинин вируса гриппа человека (HA), полигистидин (His), зеленый флуоресцентный белок (GFP) и FLAG. Другие пептиды также рассматриваются как подходящие для использования по настоящему изобретению и особенно подходящими пептидами являются аффинные метки для очистки белков, поскольку такие пептиды, как правило, связываются с высокой аффинностью с их соответствующими партнерами по связыванию.
Используемый в настоящем документе термин «пептид» означает короткий полимер из аминокислот, связанных пептидными связями, при этом пептид содержит от 2 до 100 аминокислот, например, 5-99, например, 6-98, 7-97 или 8-96 аминокислот. В одном варианте осуществления пептид, используемый по настоящему изобретению, представляет собой аминокислотную последовательность из 50 или менее аминокислотных остатков, например, 40, 30, 10 или менее аминокислотных остатков. Пептиды, используемые по настоящему изобретению, имеют достаточную длину для соответствующих целей, например, если пептид представляет собой линкер, он должен быть достаточно длинным, чтобы позволять фрагменту, который он связывает, выполнять его биологическую функцию; альтернативно, если пептид является партнером по связыванию, он должен быть способен специфически связываться с другим соединением, таким как антитело.
В одном варианте осуществления другой партнер по связыванию связывающейся пары (альтернативно, первый или второй партнер по связыванию) представляет собой белок.
Используемый в настоящем документе термин «белок» означает аминокислотную последовательность из 100 или более аминокислот. В одном варианте осуществления используемый в настоящем документе термин «белок» означает аминокислотную последовательность со вторичной или третичной структурой.
В одном варианте осуществления первый белок, A, и/или второй белок, B, биспецифического белкового комплекса представляет собой антитело или фрагмент антитела. Такой биспецифический белковый комплекс можно называть биспецифическим антительным комплексом.
В одном варианте осуществления каждое антитело или фрагмент, которые используют в биспецифическом антительном комплексе по изобретению, содержит один сайт связывания.
Полноразмерное антитело или фрагмент антитела, которые используют в слитых белках (A-X или B-Y), могут быть моноспецифическими, мультивалентными или биспецифическими.
Предпочтительно, использование двух биспецифических антител или фрагментов антител позволяет молекулам по настоящему изобретению, таким как биспецифический антительный комплекс, описанный в настоящем документе, потенциально быть специфичными в отношении, в общей сложности, 4 разных антигенов (то есть комплекс может быть тетраспецифическим). Это позволяет исследовать эффекты авидности.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как первый слитый белок A-X, представляют собой моноспецифическое антитело или фрагмент антитела, например, Fab, Fab', scFv или тому подобное, и в частности, являются специфичными в отношении CD22.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как второй слитый белок B-Y, представляют собой моноспецифическое антитело или фрагмент антитела, например, Fab, Fab', scFv или тому подобное, и в частности, являются специфичными в отношении CD79a и/или CD79b.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как второй слитый белок B-Y, являются мультивалентными, то есть имеют два или более связывающих доменов.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как первый слитый белок A-X, являются одновалентными, и антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как второй слитый белок B-X, являются одновалентными.
Таким образом, в одном варианте осуществления связывающие домены мультиспецифических молекул по настоящему изобретению являются одновалентными.
Таким образом, в одном варианте осуществления связывающие домены мультиспецифических молекул по настоящему изобретению являются одновалентными и моноспецифическими.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как первый слитый белок A-X, являются одновалентными, и антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как второй слитый белок B-X, являются мультивалентными.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как первый слитый белок A-X, являются мультивалентными, и антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как второй слитый белок B-X, являются одновалентными.
В одном варианте осуществления антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как первый слитый белок A-X, являются мультивалентными, и антитело или фрагмент антитела, используемые в молекулах по настоящему изобретению или их компонентах, таких как второй слитый белок B-X, являются мультивалентными.
В одном варианте осуществления первое антитело, второе антитело или и первое, и второе антитело молекул по настоящему изобретению или их компонентов, например, биспецифического антительного комплекса, могут иметь формат IgG, например, анти-CD22 и/или анти-CD79 антитело может быть предоставлено в формате IgG.
В одном варианте осуществления фрагмент антитела выбирают из группы, состоящей из: антигенсвязывающего (Fab) фрагмента, одноцепочечного вариабельного фрагмента (scFv) и однодоменного антитела (sdAb), например, scFv используют в первом (A-X) или втором слитом белке (B-Y). Предпочтительно, небольшой размер scFv может облегчать правильное сворачивание биспецифических антительных комплексов.
В одном варианте осуществления первое (A), второе антитело/фрагмент (B) или как первое, так и второе антитело/фрагмент биспецифического антительного комплекса по настоящему изобретению может представлять собой Fab.
В одном варианте осуществления первое, второе антитело/фрагмент или как первое, так и второе антитело/фрагмент биспецифического антительного комплекса по настоящему изобретению представляет/представляют собой VHH.
Термин «слитый белок», используемый в контексте биспецифического комплекса по настоящему изобретению, означает белок, например, антитело или фрагмент антитела, присоединенный к партнеру по связыванию.
В настоящем документе биспецифические белковые комплексы по настоящему изобретению для удобства называют A-X:Y-B. Однако данное название не должно ограничивать то, как обозначены слитые белки A-X и B-Y, поскольку эксперименты авторов изобретения свидетельствуют о том, что партнеры по связыванию X и Y могут меняться местами, то есть, A-Y и B-X, без отрицательного влияния на данный способ. Таким образом, A и B, а также X и Y являются номинальными обозначениями, предназначенными для облегчения объяснения настоящей технологии.
Используемый в настоящем документе термин «присоединенные» означает связанные или объединенные прямо, либо косвенно через линкер, такой как пептидный линкер, примеры которого обсуждаются ниже. Прямое связывание включает слияние (например, пептидную связь) или химическую конъюгацию.
Используемый в настоящем документе термин «партнер по связыванию» относится к одному компоненту связывающейся пары.
В одном варианте осуществления аффинность партнеров по связыванию является высокой, составляет 5 нМ или является более сильной, например, составляет 900, 800, 700, 600, 500, 400, 300 пM или является более сильной.
Используемый в настоящем документе термин «связывающаяся пара» относится к двум партнерам по связыванию, которые специфически связываются друг с другом. Примеры связывающейся пары включают пептид и специфичное в отношении него антитело или связывающий фрагмент, или фермент и лиганд, или фермент и ингибитор этого фермента.
В одном варианте осуществления первый партнер по связыванию (X) выбран из группы, включающей: полноразмерное антитело, Fab, Fab', scFv, пептид и sdAb, при этом примеры sdAb включают VH или VL, или VHH.
В одном варианте осуществления второй партнер (Y) выбран из группы, включающей: полноразмерное антитело, Fab, Fab', scFv, пептид и sdAb, при этом примеры sdAb включают VH или VL, или VHH.
В одном варианте осуществления, если A представляет собой антитело или его фрагмент, первый партнер по связыванию (X) присоединен к C-концу тяжелой или легкой цепи первого антитела или фрагмента антитела, например, первый партнер по связыванию присоединен к C-концу тяжелой цепи первого антитела или фрагмента антитела.
В другом варианте осуществления, если B представляет собой антитело или его фрагмент, второй партнер по связыванию (Y) присоединен к C-концу тяжелой или легкой цепи второго антитела или фрагмента антитела, например, второй партнер по связыванию присоединен к C-концу тяжелой цепи второго антитела или фрагмента антитела.
В одном варианте осуществления X присоединен к C-концу тяжелой цепи антитела или фрагмента (белка A) и Y присоединен к C-концу антитела или фрагмента (белка B).
В одном варианте осуществления X присоединен через линкер (такой как ASGGGG или ASGGGGSG) к C-концу тяжелой цепи антитела или фрагмента (белка A) и Y присоединен через линкер (такой как ASGGGG или ASGGGGSG) к C-концу антитела или фрагмента (белка B).
В одном варианте осуществления первый или второй партнер по связыванию (X или Y) представляет собой пептид.
Примеры подходящей связывающейся пары могут включать последовательность GCN4 (SEQ ID NO: 1) или ее вариант и последовательность 52SR4 (SEQ ID NO: 3) или ее вариант, который представляет собой scFv, специфичную в отношении GCN4.
В одном варианте осуществления первый партнер по связыванию (условно X) представляет собой последовательность GCN4 (например, приведенную в SEQ ID NO: 1) или ее вариант (например, без His-метки) и второй партнер по связыванию (условно Y) представляет собой scFv, специфичную в отношении последовательности GCN4 (например, приведенную в SEQ ID NO: 3), или ее вариант.
В одном вариант осуществления первый партнер по связыванию (условно X) представляет собой sFv, специфичную в отношении последовательности GCN4 (например, приведенную в SEQ ID NO: 3), или ее вариант и второй партнер по связыванию (условно Y) представляет собой последовательность GCN4 (например, приведенную в SEQ ID NO: 1), или ее вариант.
Варианты GCN4 включают аминокислотную последовательность, имеющую по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94% 95%, 96%, 97% или 98%, или 99% идентичности с SEQ ID NO: 1. Варианты GCN4 также включают аминокислотную последовательность, имеющую по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, или 99% идентичности с последовательностью, кодируемой нуклеотидной последовательностью SEQ ID NO: 2, или нуклеотидной последовательностью, которая гибридизуется с SEQ ID NO: 2 в строгих условиях.
Подходящей scFv, специфичной в отношении GCN4, является 52SR4 (SEQ ID NO: 3) или ее вариант. Варианты 52SR4 включают аминокислотную последовательность, имеющую по меньшей мере 80% или 85%, или 90%, или 95%, или 98%, или 99% идентичности с SEQ ID NO: 3. Варианты 52SR4 также включают аминокислотную последовательность, имеющую по меньшей мере 80% или 85%, или 90%, или 95%, или 98%, или 99% идентичности с последовательностью, кодируемой нуклеотидной последовательностью SEQ ID NO: 4, или нуклеотидной последовательностью, которая гибридизуется с SEQ ID NO: 2 в строгих условиях.
Авторы настоящего изобретения обнаружили, что одноцепочечное антитело 52SR4 и пептид GCN4 представляют собой связывающуюся пару, подходящую для использования в биспецифических белковых комплексах по настоящему изобретению.
Альтернативно, любое подходящее антитело/фрагмент и антиген (такой как пептид) можно использовать в качестве X и Y.
В одном варианте осуществления первый партнер по связыванию (X) и второй партнер по связыванию (Y) представляют собой белок.
В одном варианте осуществления первый партнер по связыванию (X) представляет собой фермент или его активный фрагмент и второй партнер по связыванию (Y) представляет собой лиганд, или наоборот.
В одном варианте осуществления первый партнер по связыванию (X) представляет собой фермент или его активный фрагмент и второй партнер по связыванию (Y) представляет собой ингибитор этого фермента, или наоборот.
Используемый в настоящем документе термин «активный фрагмент» означает аминокислотный фрагмент, который по размеру меньше целой аминокислотной последовательности и сохраняет практически такую же биологическую активность или вполне достаточную биологическую активность, например, более 50% активности, например, 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%.
В другом варианте осуществления первый партнер по связыванию X представляет собой глутатион (GSH) и второй партнер по связыванию Y представляет собой глутатион-S-трансферазу (GST), или наоборот.
В другом варианте осуществления X представляет собой Fos и Y представляет собой Jun, или наоборот.
В другом варианте осуществления X представляет собой His и Y представляет собой анти-His антитело, или наоборот.
В другом варианте осуществления X представляет собой кальмодулин-связывающий пептид и Y представляет собой кальмодулин, или наоборот.
В другом варианте осуществления X представляет собой мальтоза-связывающий белок и Y представляет собой антитело против мальтоза-связывающего белка, либо его фрагмент, или наоборот.
Другие сочетания фермент-лиганд также предусмотрены для использования в качестве партнеров по связыванию. Подходящими также являются аффинные метки, известные в данной области, предназначенные для очистки белков, поскольку они, как правило, связываются с высокой аффинностью с их соответствующими партнерами по связыванию.
В одном варианте осуществления первый или второй партнер по связыванию (X или Y) представляет собой белок или пептид.
В одном варианте осуществления первый и второй слитые белки содержат один или более пептидных линкеров. Линкеры могут быть включены в разные участки слитых белков. Например, линкер может быть включен между партнером по связыванию и белком, присоединенным к нему.
В одном варианте осуществления, линкер представляет собой пептидный линкер.
Используемый в настоящем документе термин «пептидный линкер» означает пептид с аминокислотной последовательностью. Ассортимент подходящих пептидных линкеров известен специалистам в данной области.
В одном варианте осуществления пептидный линкер может иметь синтетическое происхождение, то есть, получен химическими методами синтеза.
В одном варианте осуществления партнеры по связыванию в биспецифических белковых комплексах связаны со своими соответствующими белками через пептидные линкеры.
В одном варианте осуществления слитые белки являются трансляционными слитыми продуктами, то есть, слитый белок экспрессируется в клетках-хозяевах, содержащих генетический конструкт, с которого экспрессируется слитый белок.
В одном варианте осуществления слитый белок получен путем конъюгации A с X или B с Y, необязательно, через пептидный линкер.
В одном варианте осуществления пептидный линкер имеет 50 или менее аминокислот в длину, например, 20 или менее аминокислот.
Как правило, более эффективным способом является рекомбинантная экспрессия белка и, таким образом, непосредственная пептидная связь или пептидный линкер, который может быть экспрессирован клеткой-хозяином, может быть предпочтительным.
В одном аспекте предложен способ продуцирования биспецифического белкового комплекса по настоящему изобретению, включающий этапы:
(a) продуцирования первого слитого белка (A-X), содержащего связывающий домен, специфичный в отношении CD22 или CD79a, и/или CD79b (A), присоединенный к первому партнеру по связыванию (X) связывающейся пары;
(b) продуцирования второго слитого белка (B-Y), содержащего связывающий домен, специфичный в отношении CD22 или CD79a, и/или CD79b (A), присоединенный ко второму партнеру по связыванию (Y) связывающейся пары;
при этом по меньшей мере первый слитый белок или второй слитый белок содержит связывающий домен, специфичный в отношении CD22, и оставшийся слитый белок содержит связывающий домен, специфичный в отношении CD79a и/или CD79b, и
(c) смешивания первого (A-X) и второго (B-Y) слитых белков, полученных на этапах a) и b).
В частности, гетеродимерно сплетенный биспецифический белковый комплекс получают путем смешивания A-X и B-Y in vitro. Таким образом, в одном варианте осуществления способ включает этап смешивания in vitro, приводящий к установлению контакта между A-X и B-Y.
Таким образом, как правило, слитые белки A-X и B-Y не экспрессируются совместно в одной и той же клетке. Это является предпочтительным, поскольку позволяет, например, 100 слитых белков A-X и 100 слитых белков A-Y экспрессировать отдельно и, необязательно, очищать, и последующее смешивание 200 слитых белков в различных пермутациях позволяет получать 10000 гетеродимерно сплетенных биспецифических белковых комплексов.
Напротив, в способах предшествующего уровня техники необходима совместная экспрессия биспецифических конструкций и, таким образом, для получения 10000 комплексов необходимо проводить 10000 трансфекций, экспрессий и очисток.
Партнеры по связыванию X и Y имеют сродство друг к другу и действуют как биологический эквивалент велкро® или стержня и магнита, удерживая компоненты комплекса вместе. Предпочтительно, это означает, что слитые белки A-X и Y-B могут быть с легкостью собраны в биспецифический белковый комплекс путем простого смешивания слитых белков. Таким образом, биспецифический белковый комплекс по настоящему изобретению имеет модульную структуру, которая позволяет легко собирать вместе два разных белка для получения больших панелей пермутаций биспецифических белковых комплексов с разными сочетаниями антигенных специфичностей, например, подобно сетке. Это позволяет проводить эффективный и систематический скрининг большого количества биспецифических белковых комплексов для обнаружения аддитивных, синергетических или новых биологических функций.
В условиях, когда X и Y являются специфичными друг в отношении друга, значительно снижается возможность образования гомодимеров. В настоящем документе X и Y собирательно называют связывающейся парой или партнерами по связыванию. В одном варианте осуществления X не обладает высокой аффинностью к другим X. В одном варианте осуществления Y не обладает высокой аффинностью к другим Y. То, что X и Y не образуют гомодимеры, является преимуществом, это предотвращает образование нежелательных моноспецифических белковых комплексов, повышает выход желаемых биспецифических белковых комплексов и устраняет необходимость в обременительных стадиях очистки для удаления моноспецифических белковых комплексов.
Такая быстрая сборка биспецифических белковых комплексов, уровень выхода и/или чистоты не могут быть эффективно достигнуты способами предшествующего уровня техники, в частности, в способах предшествующего уровня техники, как правило, необходимы масштабные стадии очистки.
Преимуществом является то, что компоненты X и Y позволяют быстро и легко собирать мультиплекс, включающий биспецифические белковые комплексы, полученные из разных пермутаций слитых белков.
В одном варианте осуществления белки A и B представляют собой антитела или фрагменты антител. Если антитела или фрагменты антител удерживаются вместе в виде комплекса через X и Y, то образуется биспецифический антительный комплекс.
Смешивание, как правило, выполняют в условиях, в которых X и Y могут взаимодействовать. В одном варианте осуществления слитые белки инкубируют в среде для культивирования клеток в условиях, подходящих для культивирования клеток, например, слитые белки инкубируют в течение 90 минут в условиях 37°C/5%CO2.
В одном варианте осуществления слитые белки по настоящему изобретению смешивают в водной среде, например, один слитый белок может быть связан с твердой поверхностью, такой как гранула или планшет, а другой слитый белок может быть внесен в водном растворе/суспензии. Использование твердой фазы позволяет быстро отмывать избыток компонентов и реагентов. В одном варианте осуществления ни один из слитых белков не связан с твердой фазой и их просто смешивают в жидкости/растворе/среде.
Преимуществом является то, что способ по настоящему изобретению можно использовать для получения комплексов, образованных между гетерогенными парами (то есть, между первым слитым белком [A-X] и вторым слитым белком [B-Y]), при этом взаимодействия между гомогенными парами (то есть, между двумя первыми слитыми белками [A-X] или двумя вторыми слитыми белками [B-Y]) сведены к минимуму. Таким образом, настоящий способ позволяет получать большие количества биспецифических белковых комплексов с минимальными примесями, или совсем без, гомодимерных комплексов. Такой уровень чистоты и выхода невозможен при использовании способов предшествующего уровня техники.
В одном варианте осуществления образовавшиеся комплексы не требуют дополнительных стадий очистки.
В одном варианте осуществления образовавшиеся комплексы требуют одну стадию очистки, например, хроматографию на колонке.
В одном варианте осуществления способ дополнительно включает по меньшей мере одну стадию очистки, например, после экспрессии слитого белка по настоящему изобретению.
Используемый в настоящем документе термин «функциональный анализ» означает анализ, который можно использовать для определения одного или более желательных свойств или активностей белковых комплексов, антительных комплексов или смеси антител, подвергнутых воздействию условий анализа. Подходящие функциональные анализы могут представлять собой анализы на связывание, анализы на апоптоз, анализы на антителозависимую клеточную цитотоксичность (ADCC), анализы на комплементзависимую цитотоксичность (CDC), анализы на ингибирование клеточного роста или пролиферации (цитостатический эффект), анализы на уничтожение клеток (цитотоксический эффект), анализы на клеточную сигнализацию, анализы на продуцирование цитокинов, анализы на продуцирование антител и переключение изотипов, а также анализы на клеточную дифференциацию. В одном варианте осуществления можно использовать in vivo анализы, такие как животные модели, включая модели опухолей мышей, модели аутоиммунных заболеваний, модели на грызунах или приматах, инфицированных вирусами или бактериями, и тому подобное, для тестирования молекул по настоящему изобретению.
В контексте биспецифических антительных комплексов эффективность биспецифических антительных комплексов по настоящему изобретению можно сравнивать с эффективностью антител или смесей антител (или фрагментов) в таких моделях методами, как правило, известными специалисту в данной области.
Функциональные анализы можно выполнять столько раз, сколько потребуется, с использованием или без использования разных образцов конкретного биспецифического антительного комплекса для повышения надежности результатов. Можно применять различные статистические критерии, известные специалистам, для получения статистически значимых результатов и, таким образом, для идентификации биспецифических антительных комплексов с биологическими функциями и, в частности, для идентификации оптимальных пар вариабельных областей для использования в мультиспецифической молекуле по настоящему изобретению.
КОМПОЗИЦИИ И МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ
В одном аспекте предложена молекула по настоящему изобретению или компонент, такой как слитый белок, гетеродимерно сплетенный биспецифический белковый комплекс, композиция, содержащая молекулу по изобретению, в том числе слитый белок или указанный биспецифический белковый комплекс, мультиплекс, матрица, библиотека, описанные в настоящем документе.
В одном варианте осуществления молекулы по настоящему изобретению, например, антитело, описанное в настоящем документе, мультиспецифическая молекула и биспецифический белковый комплекс, подходят для терапевтического применения и могут являться новыми терапевтическими средствами для лечения заболеваний. Таким образом, в следующем аспекте предложена молекула по настоящему изобретению, например, биспецифический белковый комплекс, описанный выше, для использования в терапии. Молекулы по настоящему изобретению, включая биспецифические белковые комплексы, описанные в настоящем документе, подходят для лечения ряда заболеваний, таких как рак.
Молекулы по настоящему изобретению, включая мультиспецифические молекулы и биспецифические белковые комплексы, описанные в настоящем документе, также, в частности, подходят для ингибирования функции B-клеток с целью контролирования иммунных и аутоиммунных реакций при различных аутоиммунных заболеваниях.
Таким образом, настоящее изобретение относится к способу лечения заболевания у пациента, включающему введение терапевтически эффективного количества молекулы по настоящему изобретению, например, мультиспецифической молекулы или биспецифического белкового комплекса по настоящему изобретению.
В одном аспекте предложена фармацевтическая композиция, содержащая одну или более молекул по настоящему изобретению, например, мультиспецифических молекул по настоящему изобретению.
Множество различных компонентов можно включать в композицию, в том числе фармацевтически приемлемые носители, эксципиенты и/или разбавители. Композиция может, необязательно, содержать дополнительные молекулы, способные изменять характеристики группы мультиспецифических молекул по изобретению, тем самым, например, снижая, стабилизируя, замедляя, модулируя и/или активируя функцию антител. Композиция может быть в твердой или жидкой форме и, в частности, может быть в форме порошка, таблетки, раствора или аэрозоля.
Настоящее изобретение также относится к фармацевтической или диагностической композиции, содержащей молекулу антитела или мультиспецифическую молекулу по настоящему изобретению в сочетании с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя. Соответственно, предложено применение мультиспецифической молекулы по изобретению в лечении и в производстве лекарственного средства для лечения патологического состояния или заболевания.
ПАТОЛОГИЧЕСКИЕ СОСТОЯНИЯ
Патологическое состояние или заболевание может быть, например, выбрано из группы, состоящей из инфекций (вирусных, бактериальных, грибковых и паразитарных), эндотоксинового шока, ассоциированного с инфекцией, артрита, например, ревматоидного артрита, астмы, например, тяжелой формы астмы, хронического обструктивного заболевания легких (COPD), воспаления тазовых органов, болезни Альцгеймера, воспалительного заболевания кишечника, болезни Крона, язвенного колита, болезни Пейрони, целиакии, заболевания желчного пузыря, пилонидальной болезни, перитонита, псориаза, васкулита, хирургических спаек, инсульта, диабета I типа, болезни Лайма, менингоэнцефалита, аутоиммунного увеита, опосредованных иммунными реакциями воспалительных заболеваний центральной и периферической нервной системы, таких как рассеянный склероз, волчанка (например, системная красная волчанка) и синдром Гийена-Барре, атопического дерматита, аутоиммунного гепатита, фиброзирующего альвеолита, болезни Грэйвса, IgA нефропатии, идиопатической тромбоцитопенической пурпуры, болезни Меньера, пемфигуса, первичного билиарного цирроза, саркоидоза, склеродермии, гранулематоза Вегенера, других аутоиммунных заболеваний, панкреатита, травмы (хирургической), реакции «трансплантат против хозяина», отторжения трансплантата, заболеваний сердца, включая ишемические заболевания, такие как инфаркт миокарда, а также атеросклероза, внутрисосудистой коагуляции, костной резорбции, остеопороза, остеоартрита, периодонтита, гипохлоргидрии и рака, включая рак молочной железы, рак легкого, рак желудка, рак яичника, печеночно-клеточный рак, рак толстого кишечника, рак поджелудочной железы, рак пищевода, рак головы и шеи, а также рак почки, в частности, почечноклеточный рак, рак предстательной железы, рак печени, меланому, саркому, миелому, нейробластому, плацентарную хориокарциному, рак шейки матки и рак щитовидной железы, а также их метастатические формы.
В одном варианте осуществления заболевание представляет собой рак, например, лейкоз, включая лимфоцитарный лейкоз, такой как острый лимфобластный лейкоз или хронический лимфоцитарный лейкоз; или миелогенный лейкоз, такой как острый миелогенный лейкоз или хронический миелогенный лейкоз.
В одном варианте осуществления аутоиммунное заболевание включает: острый диссеминированный энцефаломиелит (adem), острый некротизирующий геморрагический лейкоэнцефалит, болезнь Аддисона, недостаточность надпочечников, гипокортизолизм, очаговую алопецию, амилоидоз, анкилозирующий спондилит, спондилоартрит, болезнь Штрюмпелля-Мари, анти-GBM/анти-TBM нефрит, антифосфолипидный синдром (aps), аутоиммунный ангиоотек, аутоиммунную апластическую анемию, аутоиммунную вегетативную дистонию, аутоиммунный гепатит, аутоиммунную гиперлипидемию, аутоиммунный иммунодефицит, аутоиммунную болезнь внутреннего уха (AIED), аутоиммунный лимфопролиферативный синдром (ALPS), синдром Канали-Смита, аутоиммунный миокардит, аутоиммунный оофорит, аутоиммунный панкреатит (AIP), аутоиммунные полигландулярные синдромы (I, II и III типов), аутоиммунную ретинопатию (AR), аутоиммунную тромбоцитопеническую пурпуру (ATP), аутоиммунное заболевание щитовидной железы, аутоиммунную крапивницу, аксональные/нейрональные невропатии, болезнь Бало, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, болезнь Кастлемана, целиакию, болезнь Шагаса, хроническую воспалительную демиелинизирующую полиневропатию (CIDP), хронический рецидивирующий мультифокальной остеомиелит (CRMO), синдром Черджа-Стросс, рубцовый пемфигоид/доброкачественный пемфигоид слизистых оболочек (CP), болезнь Крона, воспалительное заболевание кишечника, колит, энтерит, илеит, синдром Когана, болезнь холодовых агглютининов, врожденную блокаду сердца, миокардит Коксаки, болезнь CREST, криоглобулинемию, демиелинизирующие невропатии, герпетиформный дерматит, болезнь Дюринга, дерматомиозит, диабет I типа, дискоидную красную волчанку (DLE), синдром Дресслера, эндометриоз, буллезный эпидермолиз (EB) и приобретенный буллезный эпидермолиз (EBA), эозинофильный гастроэнтерит, эзофагит, эозинофильный фасциит, синдром Шульмана, узловатую эритему, экспериментальный аллергический энцефаломиелит, синдром Эванса, фиброзирующий альвеолит, гигантоклеточный артериит (височный артериит), гигантоклеточный миокардит, гломерулонефрит (не пролиферативный: фокальный сегментный гломерулосклероз и мембранный гломерулонефрит; пролиферативный: IgA нефропатию), синдром Гудпасчера, гранулематоз с полиангиитом (GPA) (ранее называемый гранулематозом Вегенера), болезнь Грэйвса, синдром Гийена-Барре, синдром Миллера-Фишера, острую моторную аксональную невропатию, острую моторно-сенсорную аксональную невропатию, острую параавтономную невропатию, стволовой энцефалит Бикерстаффа, энцефалит Хашимото, треоидит Хашимото, гемолитическую анемию, пурпуру Геноха-Шенлейна, герпес беременных, гипогаммаглобулинемию, идиопатический пульмонарный фиброз, идиопатическую тромбоцитопеническую пурпуру (ITP), IgA нефропатию (IGAN), синдром Бергера, синфарингитный гломерулонефрит, IgA пемфигус, IgG4-связанную склерозирующую болезнь, иммунное бесплодие, миозит с тельцами включения, инсулинозависимый сахарный диабет, интерстициальный цистит, синдром Исаака, нейромиотонию, ювенильный артрит, ювенильный миозит, синдром Кавасаки, синдром Ламберта-Итона, лейкоцитокластический васкулит, плоский лишай, склерозирующий лишай, деревянистый конъюнктивит, линейный IgA дерматоз (LAD), пемфигоид, волчанку (SLE), болезнь Лайма, болезнь Меньера, микроскопический полиангиит (MPA), смешанное заболевание соединительной ткани (MCTD), моноклональную гаммапатию, язву Мурена, болезнь Муха-Хаберманн, рассеянный склероз, миастению, миозит, нарколепсию, оптиконевромиелит (болезнь Девика), невромиотонию, синдром Исаака (приобретенный, паранеопластический, наследственный), нейтропению, глазной рубцовый пемфигоид, неврит зрительного нерва, оофорит, опсо-миоклональный синдром, орхит, палиндромный ревматизм, pandas (педиатрические аутоиммунные психоневрологические расстройства, вызванные стрептококком), паранеопластический аутоиммунный полиорганный синдром (PAMS), паранеопластическую мозжечковую дегенерацию, паранеопластический пемфигус (PNP), пароксизмальную ночную гемоглобинурию (PNH), синдром Парри-Ромберга, синдром Персонейджа-Тернера, парспланит (периферический увеит), пемфигоид беременных (PG), обыкновенную пузырчатку (PV), листовидную пузырчатку (PF), периферическую невропатию, перивенозный энцефаломиелит, злокачественную анемию, POEMS-синдром, узелковый полиартериит (PAN), ревматическую полимиалгию, полимиозит, постинфарктный синдром, постперикардиотомный синдром, прогестероновый дерматит, первичный билиарный цирроз, синдром Гано, первичный склерозирующий холангит (PSC), склерозирующий холангит, псориаз, псориатический артрит, гангренозную пиодермию, истинную эритроцитарную аплазию, энцефалит Расмуссена, хронический очаговый энцефалит (CFE), феномен Рейно, реактивный артрит, синдром Рейтера, рековерин-ассоциированную ретинопатию (RAR), рефлекторную симпатическую дистрофию, синдром Рейтера, рецидивирующий полихондрит, синдром беспокойных ног, забрюшинный фиброз, ревматическую атаку, ревматоидный артрит, саркоидоз, синдром Шмидта, склерит, склеродермию, системную склеродермию, синдром Шегрена, аутоиммунитет к антигенам спермы и яичек, синдром жесткого человека, подострый бактериальный эндокардит (SBE), синдром Сусака, симпатическую офтальмию, артериит Такаясу, височный артериит/гигантоклеточный артериит, облитерирующий тромбангиит, болезнь Бергера, тромбоцитопеническую пурпуру (TTP), синдром Толоса-Ханта, поперечный миелит, язвенный колит, недифференцированное заболевание соединительной ткани (UCTD), увеит, ревматическую полимиалгию, артериит Такаясу, височный артериит, болезнь Бергера, кожный васкулит, болезнь Кавасаки, узелковый полиартериит, синдром Бехчета, синдром Черджа-Стросс, кожный васкулит, пурпуру Геноха-Шенлейна, микроскопический полиангиит, гранулематоз Вегенера, васкулит гольфиста, везикулобуллезный дерматоз, витилиго, гранулематоз Вегенера (современное название гранулематоз с полиангиитом (GPA).
В одном варианте осуществления аутоиммунное заболевание выбирают из группы, включающей или состоящей из: ANCA васкулита, IgA нефропатии (Бергера), обыкновенной пузырчатки/буллезного пемфигоида, ITP, первичного билиарного цирроза, аутоиммунного тиреоидита (болезни Грэйвса), болезни Хашимото, волчанкового нефрита, мембранного гломерулонефрита (или мембранной нефропатии), APS, миастении, оптиконевромиелита, первичного сидрома Шегрена, аутоиммунной нейтропении, аутоиммунного панкреатита, дерматомиозита, аутоиммунного увеита, аутоиммунной ретинопатии, болезни Бехчета, IPF, системной склеродермии, фиброза печени, аутоиммунного гепатита, первичного склерозирующего холангита, витилиго, синдрома Гудпасчера, легочного альвеолярного протеиноза, хронической аутоиммунной крапивницы, псориаза, ревматоидного артрита, псориатического артрита, осевого спондилоартрита, трансплантации (включая реакции «трансплантат против хозяина»), астмы, COPD, гигантоклеточного артериита, рефрактерной аутоиммунной цитопении, синдрома Эванса (аутоиммунной гемолитической анемии), диабета I типа, саркоидоза, полимиозита, язвенного колита, болезни Крона, целиакии, макроглобулинемии Вальденстрема, фокального сегментного гломерулосклероза, хронической болезни Лайма (клещевого боррелиоза), плоского лишая, синдрома жесткого человека, дилатационной кардиомиопатии, аутоиммунного (лимфоцитарного) оофорита, приобретенного буллезного эпидермолиза, аутоиммунного атрофического гастрита, злокачественной анемии, атопического дерматита, атеросклероза, рассеянного склероза, энцефалита Расмуссена, синдрома Гийена-Барре, приобретенной нейромиотонии, инсульта.
В одном варианте осуществления заболевание представляет собой рак, например, лейкоз, например, лимфоцитарный лейкоз, такой как острый лимфобластный лейкоз или хронический лимфоцитарный лейкоз; или миелогенный лейкоз, такой как острый миелогенный лейкоз или хронический миелогенный лейкоз; или лимфому, такую как диффузная крупноклеточная B-клеточная лимфома, либо лимфома Ходжкина или неходжкинская лимфома.
Настоящее изобретение также относится к фармацевтической или диагностической композиции, содержащей молекулу по настоящему изобретению, такую как мультиспецифическая молекула, описанная в настоящем документе, в сочетании с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя. Соответственно, предложено применение молекулы по настоящему изобретению, такой как мультиспецифическая молекула, описанная в настоящем документе, для лечения и для производства лекарственного средства.
Композиция, как правило, будет предоставлена в виде части стерильной фармацевтической композиции, которая обычно включает фармацевтически приемлемый носитель. Кроме того, фармацевтическая композиция по настоящему изобретению может содержать фармацевтически приемлемый адъювант.
Настоящее изобретение также относится к способу получения фармацевтической или диагностической композиции, включающему добавление и смешивание мультиспецифической молекулы по настоящему изобретению с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя.
Используемый в настоящем документе термин «фармацевтически приемлемый эксципиент» означает фармацевтически приемлемый носитель, раствор или добавку к препарату для усиления желательных характеристик композиций по настоящему изобретению. Эксципиенты хорошо известны в данной области и включают буферы (например, цитратный буфер, фосфатный буфер, ацетатный буфер и бикарбонатный буфер), аминокислоты, мочевину, спирты, аскорбиновую кислоту, фосфолипиды, белки (например, сывороточный альбумин), ЭДТА, хлорид натрия, липосомы, маннит, сорбит и глицерин. Растворы или суспензии могут быть инкапсулированы в липосомы или биоразлагаемые микросферы. Препарат, как правило, будет предоставлен в практически стерильной форме при использовании производственных процессов в стерильных условиях.
Сюда может входить получение и стерилизация фильтрованием буферного растворителя, используемого для препарата, асептическое суспендирование антитела в стерильном буферном растворителе и распределение препарата в стерильные емкости методами, известными специалистам в данной области.
Фармацевтически приемлемый носитель сам по себе не должен вызывать продуцирование антител, вредных для индивидуума, получающего композицию, и не должен быть токсичным. Подходящими носителями могут быть крупные, медленно метаболизируемые макромолекулы, такие как белки, полипептиды, липосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы.
Можно использовать фармацевтически приемлемые соли, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, или соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты.
Кроме того, фармацевтически приемлемые носители в терапевтических композициях могут содержать такие жидкости как вода, солевой раствор, глицерин и этанол. Такие носители позволяют формулировать фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, кашиц и суспензий для приема внутрь пациентом.
Молекулы по изобретению, такие как мультиспецифическая молекула, описанная в настоящем документе, можно доставлять диспергированными в растворителе, например, в форме раствора или суспензии. Их можно суспендировать в соответствующем физиологическом растворе, например, физиологическом солевом растворе, фармакологически приемлемом растворителе или буферном растворе. Буферные растворы, известные в данной области, могут содержать от 0,05 мг до 0,15 мг динатрия эдетата, от 8,0 мг до 9,0 мг NaCl, от 0,15 мг до 0,25 мг полисорбата, от 0,25 мг до 0,30 мг безводной лимонной кислоты и от 0,45 мг до 0,55 мг цитрата натрия на 1 мл воды, так, чтобы было достигнуто значение pH примерно 4,0-5,0. Как упомянуто выше, суспензия может быть получена, например, из лиофилизированного антитела.
Подробное описание фармацевтически приемлемых носителей приведено в сборнике Remington's Pharmaceutical Sciences (Mack Publishing Company, N.J. 1991).
Используемый в настоящем документе термин «терапевтически эффективное количество» означает количество лекарственного средства, необходимое для лечения, облегчения или предотвращения целевого заболевания или состояния, или для оказания поддающегося определению терапевтического или превентивного эффекта. Для любого антитела терапевтически эффективное количество можно оценивать сначала либо в анализах на культуре клеток, либо в животных моделях, как правило, на грызунах, кроликах собаках, свиньях или приматах. Животную модель также можно использовать для определения подходящего диапазона концентраций и пути введения. Такую информацию затем можно использовать для определения полезных доз и путей введения для человека.
Точное терапевтически эффективное количество для субъекта-человека будет зависеть от тяжести заболевания, общего состояния здоровья субъекта, возраста, массы тела и пола субъекта, его диеты, времени и частоты введения, сочетания(ий) лекарственных средств, чувствительности реакции и толерантности/ответа на терапию. Это количество может быть определено путем обычного экспериментирования и остается на усмотрение лечащего врача. Как правило, терапевтически эффективное количество будет составлять от 0,01 мг/кг до 50 мг/кг, например, от 0,1 мг/кг до 20 мг/кг. Альтернативно, доза может составлять 1-500 мг в сутки, например, 10-100, 200, 300 или 400 мг в сутки. Фармацевтические композиции для удобства могут быть предоставлены в стандартных лекарственных формах, содержащих заранее определенное количество активного средства по изобретению.
Композиции можно вводить пациенту индивидуально или можно вводить в сочетании (например, одновременно, последовательно или раздельно) с другими средствами, лекарственными средствами или гормонами.
Доза, в которой вводят мультиспецифическую молекулу по настоящему изобретению, зависит от природы состояния, подвергаемого лечению, степени имеющегося воспаления, а также от того, используют ли молекулу антитела профилактически или для лечения существующего состояния.
Частота введения доз будет зависеть от времени полувыведения мультиспецифической молекулы и продолжительности ее эффекта. Если мультиспецифическая молекула имеет короткое время полувыведения (например, от 2 до 10 часов), может потребоваться вводить одну или более доз в сутки. Альтернативно, если мультиспецифическая молекула имеет длительное время полувыведения (например, от 2 до 15 дней), может потребоваться вводить дозу только один раз в сутки, один раз в неделю или даже один раз каждые 1 или 2 месяца.
В настоящем изобретении значение pH конечного препарата не соответствует значению изоэлектрической точки мультиспецифической молекулы, так что, если pH препарата равно 7, тогда величина pI, составляющая 8-9 или выше, может быть подходящей. Без привязки к конкретной теории, считается, что это в конечном счете может придавать препарату повышенную стабильность, например, антитело или его фрагмент остается в растворе.
Фармацевтические композиции по данному изобретению можно вводить самыми разными способами, включая, но без ограничения, пероральный, внутривенный, внутримышечный, внутриартериальный, интрамедулярный, интратекальный, интравентрикулярный, трансдермальный, чрескожный (например, смотри WO 98/20734), подкожный, внутрибрюшинный, интраназальный, энтеральный, топический, подъязычный, интравагинальный и ректальный пути введения. Безыгольные шприцы также можно использовать для введения фармацевтических композиций по изобретению.
Прямая доставка композиций, как правило, будет осуществляться путем инъекции, подкожно, внутрибрюшинно, внутривенно или внутримышечно, или будет производиться в интерстициальное пространство ткани. Композиции также можно вводить в определенную интересующую ткань. Режим дозирования может быть режимом однократной дозы или режимом многократных доз.
Когда препарат предназначен для инъекции или инфузии, он может иметь форму суспензии, раствора или эмульсии в масляной или водной среде и может содержать вспомогательные вещества, такие как суспендирующее средство, консервант, стабилизирующее и/или диспергирующее средство. Альтернативно, мультиспецифическая молекула может находиться в сухой форме, которую восстанавливают перед использованием в соответствующей стерильной жидкости. Если композицию предстоит вводить через желудочно-кишечный тракт, композиция должна будет содержать средства, защищающие антитело от деградации, но которые будут высвобождать биспецифический белковый комплекс после того, как он будет абсорбирован из желудочно-кишечного тракта.
Можно предоставлять препарат по настоящему изобретению в распыляемой форме, например, в виде стандартной дозы (например, в запаянных пластиковых контейнерах или флаконах), упакованной в конверт из фольги. Каждый флакон содержит стандартную дозу в растворителе/буферном растворе объемом, например, 2 мл.
Используемый в настоящем документе термин «вариант» относится к пептиду или белку, который имеет по меньшей мере одно изменение в аминокислотной последовательности или нуклеотидной последовательности по сравнению с аминокислотной или нуклеотидной последовательностью соответствующего пептида или белка дикого типа. Вариант может иметь по меньшей мере 80% или 85%, или 90%, или 95%, или 98% или 99% идентичности последовательности с соответствующим пептидом или белком дикого типа. Однако вариант может иметь менее 80% идентичности последовательности, при условии, что вариант обладает функцией, по существу аналогичной функции соответствующего пептида или белка дикого типа.
В одном варианте осуществления конструкт по настоящему изобретению является по меньшей мере триспецифическим. В данной ситуации дополнительная специфичность может быть направлена на любой интересующий антиген, например, антигены, способствующие продлению времени полувыведения, такие как альбумин или неонатальный Fc-рецептор (FcRn); антигены для эффекторной функции, например, активирующие или ингибирующие Fc-рецепторы или костимулирующие молекулы; антигены, нацеленные на ткани или клетки; или антигены, способствующие проникновению за гематоэнцефалический барьер (ГЭБ), такие как рецептор трансферрина или LRP1.
Изобретение также относится к композициям, таким как фармацевтические композиции, содержащим указанные новые форматы с конкретной антигенной специфичностью.
В следующем аспекте изобретение относится к применению форматов и композиций в лечении.
Настоящее изобретение также относится к способу получения фармацевтической или диагностической композиции, включающему добавление и смешивание молекулы антитела или мультиспецифической молекулы по настоящему изобретению с одним или более из фармацевтически приемлемого эксципиента, разбавителя или носителя.
Молекула антитела или мультиспецифическая молекула может быть единственным активным ингредиентом в фармацевтической или диагностической композиции, либо может находиться в сочетании с другими активными ингредиентами, включая другие ингредиенты-антитела или не являющиеся антителами ингредиенты, такие как стероиды или другие молекулы лекарственного средства.
Фармацевтические композиции, соответственно, содержат терапевтически эффективное количество антитела по изобретению. Используемый в настоящем документе термин «терапевтически эффективное количество» означает количество лекарственного средства, необходимое для лечения, облегчения или предотвращения целевого заболевания или состояния, или для оказания поддающегося определению терапевтического или превентивного эффекта. Для любого антитела терапевтически эффективное количество можно оценивать сначала либо в анализах на культуре клеток, либо в животных моделях, как правило, на грызунах, кроликах собаках, свиньях или приматах. Животную модель также можно использовать для определения подходящего диапазона концентраций и пути введения. Такую информацию затем можно использовать для определения полезных доз и путей введения для человека.
Точное терапевтически эффективное количество для субъекта-человека будет зависеть от тяжести заболевания, общего состояния здоровья субъекта, возраста, массы тела и пола субъекта, его диеты, времени и частоты введения, сочетания(ий) лекарственных средств, чувствительности реакции и толерантности/ответа на терапию. Это количество может быть определено путем обычного экспериментирования и остается на усмотрение лечащего врача. Как правило, терапевтически эффективное количество будет составлять от 0,01 мг/кг до 500 мг/кг, например, от 0,1 мг/кг до 200 мг/кг, например, 100 мг/кг.
Фармацевтические композиции для удобства могут быть предоставлены в стандартных лекарственных формах, содержащих заранее определенное количество активного средства по изобретению.
Композиции можно вводить пациенту индивидуально или можно вводить в сочетании (например, одновременно, последовательно или раздельно) с другими средствами, лекарственными средствами или гормонами.
Используемый в настоящем документе термин «средство» означает соединение, которое при введении проявляет физиологический эффект.
Используемый в настоящем документе термин «лекарственное средство» означает химическое соединение, которое в терапевтической дозе проявляет соответствующий физиологический эффект.
В одном варианте осуществления антитела или фрагменты по настоящему изобретению используют совместно с терапией иммунодепрессантами, такими как стероиды, в частности, преднизон.
В одном варианте осуществления антитела или фрагменты по настоящему изобретению используют в сочетании с ритуксимабом или другими видами терапии, направленными против B-клеток.
В одном варианте осуществления антитела или фрагменты по настоящему изобретению используют в сочетании с любым модулирующим средством для B-клеток или T-клеток, или иммуномодулятором. Примеры включают метотрексат, микофенолат и азатиоприн.
Доза, в которой вводят молекулу антитела по настоящему изобретению, зависит от природы состояния, подвергаемого лечению, степени имеющегося воспаления, а также от того, используют ли молекулу антитела профилактически или для лечения существующего состояния. Частота введения доз будет зависеть от времени полувыведения молекулы антитела и продолжительности ее эффекта. Если молекула антитела имеет короткое время полувыведения (например, от 2 до 10 часов), может потребоваться вводить одну или более доз в сутки. Альтернативно, если молекула антитела имеет длительное время полувыведения (например, от 2 до 15 дней) и/или более продолжительный профиль фармакодинамики (ФД), может потребоваться вводить дозу только один раз в сутки, один раз в неделю или даже один раз каждые 1 или 2 месяца.
В одном варианте осуществления дозу вводят раз в две недели, то есть дважды в месяц.
В одном варианте осуществления между дозами выдерживают интервалы, давая возможность реакциям против лекарственного средства (в данном случае анти-антителам) ослабевать до введения следующей дозы.
Используемый в настоящем документе термин «время полувыведения» означает продолжительность нахождения молекулы в системе циркуляции, например, в сыворотке/плазме.
Используемый в настоящем документе термин «фармакодинамика» означает профиль и, в частности, продолжительность биологического действия молекулы по настоящему изобретению.
Фармацевтически приемлемый носитель сам по себе не должен вызывать продуцирование антител, вредных для индивидуума, получающего композицию, и не должен быть токсичным. Подходящими носителями могут быть крупные, медленно метаболизируемые макромолекулы, такие как белки, полипептиды, липосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, сополимеры аминокислот и неактивные вирусные частицы.
Можно использовать фармацевтически приемлемые соли, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, или соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты.
Кроме того, фармацевтически приемлемые носители в терапевтических композициях могут содержать такие жидкости как вода, солевой раствор, глицерин и этанол. Кроме того, в таких композициях могут присутствовать вспомогательные вещества, такие как увлажняющие средства или эмульгаторы, или поддерживающие pH буферные вещества. Такие носители позволяют формулировать фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, кашиц и суспензий для приема внутрь пациентом.
Подходящие формы для введения включают формы, подходящие для парентерального введения, например, путем инъекции или инфузии, например, путем болюсной инъекции или непрерывной инфузии. Когда препарат предназначен для инъекции или инфузии, он может иметь форму суспензии, раствора или эмульсии в масляной или водной среде и может содержать вспомогательные вещества, такие как суспендирующее средство, консервант, стабилизирующее и/или диспергирующее средство. Альтернативно, молекула антитела может находиться в сухой форме, которую восстанавливают перед использованием в соответствующей стерильной жидкости.
Сформулированные композиции по изобретению можно вводить непосредственно субъекту. Субъекты, получающие лечение, могут быть животными. Однако в одном или более вариантах осуществления композиции адаптированы для введения субъектам-людям.
В препаратах по настоящему изобретению значение pH конечного препарата не соответствует значению изоэлектрической точки антитела или фрагмента, например, если значение pI белка находится в диапазоне 8-9 или является более высоким, тогда значение pH 7 для препарата может быть подходящим. Без привязки к конкретной теории, считается, что это в конечном счете может придавать препарату повышенную стабильность, например, антитело или его фрагмент остается в растворе.
В одном примере фармацевтический препарат, имеющий значение pH в диапазоне 4,0-7,0, содержит: 1-200 мг/мл молекулы антитела по настоящему изобретению, 1-100 мМ буфера, 0,001-1% сурфактанта, a) 10-500 мМ стабилизатора, b) 10-500 мМ стабилизатора и 5-500 мМ средства для поддержания тоничности, или c) 5-500 мМ средства для поддержания тоничности.
Фармацевтические композиции по данному изобретению можно вводить самыми разными способами, включая, но без ограничения, пероральный, внутривенный, внутримышечный, внутриартериальный, интрамедулярный, интратекальный, интравентрикулярный, трансдермальный, чрескожный (например, смотри WO 98/20734), подкожный, внутрибрюшинный, интраназальный, энтеральный, топический, подъязычный, интравагинальный или ректальный пути введения. Безыгольные шприцы также можно использовать для введения фармацевтических композиций по изобретению. Как правило, терапевтические композиции можно изготавливать в виде инъекционных препаратов, либо в виде жидких растворов, либо в виде суспензий. Также можно изготавливать твердые формы, подходящие для растворения или суспендирования в жидких средах перед инъекцией.
Прямая доставка композиций, как правило, будет осуществляться путем инъекции, подкожно, внутрибрюшинно, внутривенно или внутримышечно, или будет производиться в интерстициальное пространство ткани. Композиции также можно вводить в лезию. Режим дозирования может быть режимом однократной дозы или режимом многократных доз.
Следует понимать, что активным ингредиентом в композиции будет молекула антитела. В силу этого, она будет подвержена деградации в желудочно-кишечном тракте. Таким образом, если композицию предстоит вводить через желудочно-кишечный тракт, композиция должна будет содержать средства, защищающие антитело от деградации, но которые будут высвобождать антитело после того, как оно будет абсорбировано из желудочно-кишечного тракта.
Подробное описание фармацевтически приемлемых носителей приведено в сборнике Remington's Pharmaceutical Sciences (Mack Publishing Company, N.J. 1991).
В одном варианте осуществления препарат предоставляют в виде препарата для топического введения, включая ингаляцию.
Подходящие ингаляционные препараты включают ингаляционные порошки, дозируемые аэрозоли, содержащие газ-пропеллент, или ингаляционные растворы, не содержащие газ-пропеллент. Ингаляционные порошки по изобретению, содержащие активное вещество, могут состоять только из вышеупомянутых активных веществ или из смеси вышеупомянутых активных веществ с физиологически приемлемым эксципиентом.
Эти ингаляционные порошки могут включать моносахариды (например, глюкозу или арабинозу), дисахариды (например, лактозу, сахарозу, мальтозу), олиго- и полисахариды (например, декстраны), полиспирты (например, сорбит, маннит, ксилит), соли (например, хлорид натрия, карбонат кальция) или смеси этих веществ друг с другом. Соответственно, можно использовать моно- или дисахариды, используя лактозу или глюкозу предпочтительно, но не исключительно, в форме их гидратов.
Частицы для осаждения в легких должны иметь размер частиц менее 10 микрон, например, 1-9 микрон, например, от 1 до 5 мкм. Размер частиц активного ингредиента (такого как антитело или его фрагмент) имеет первостепенное значение.
Газы-пропелленты, которые можно использовать для изготовления ингаляционных аэрозолей, известны в данной области. Соответствующие пропелленты выбирают из углеводородов, таких как н-пропан, н-бутан или изобутан, и галогеноуглеводородов, таких как хлорированные и/или фторированные производные метана, этана, пропана, бутана, циклопропана или циклобутана. Вышеупомянутые газы-пропелленты можно использовать самостоятельно или в виде их смесей.
Особенно подходящими газами-пропеллентами являются галогенированные алкановые производные, выбранные из TG11, TG12, TG134a и TG227. Из вышеуказанных галогенированных углеводородов TG134a (1,1,1,2-тетрафторэтан) и TG227 (1,1,1,2,3,3,3-гептафторпропан), а также их смеси являются особенно подходящими.
Содержащие газ-пропеллент ингаляционные аэрозоли также могут содержать другие ингредиенты, такие как coрастворители, стабилизаторы, поверхностно-активные средства (сурфактанты), антиоксиданты, смазывающие средства и средства для корректировки pH. Все эти ингредиенты известны в данной области.
Содержащие газ-пропеллент ингаляционные аэрозоли по изобретению могут содержать до 5% по массе активного вещества. Аэрозоли по изобретению содержат, например, 0,002-5% по массе, 0,01-3% по массе, 0,015-2% по массе, 0,1-2% по массе, 0,5-2% по массе или 0,5-1% по массе активного ингредиента.
Альтернативно, топическое введение в легкие также можно осуществлять путем введения жидкого раствора или суспензионного препарата, например, с использованием такого устройства, как небулайзер, например, небулайзер, соединенный с компрессором (например, небулайзер Pari LC-Jet Plus® соединенный с компрессором Pari Master® производства компании Pari Respiratory Equipment, Inc., Richmond, Va.).
Антитело или мультиспецифическую молекулу по изобретению можно доставлять диспергированными в растворителе, например, в форме раствора или суспензии. Их можно суспендировать в подходящем физиологическом растворе, например, солевом растворе или другом фармакологически приемлемом растворителе или буферном растворе. Буферные растворы, известные в данной области, могут содержать от 0,05 мг до 0,15 мг динатрия эдетата, от 8,0 мг до 9,0 мг NaCl, от 0,15 мг до 0,25 мг полисорбата, от 0,25 мг до 0,30 мг безводной лимонной кислоты и от 0,45 мг до 0,55 мг цитрата натрия на 1 мл воды так, чтобы было достигнуто значение pH примерно 4,0-5,0. В суспензии можно использовать, например, лиофилизированное антитело.
Терапевтические препараты в виде суспензий или растворов также могут содержать один или более эксципиентов. Эксципиенты хорошо известны в данной области и включают буферы (например, цитратный буфер, фосфатный буфер, ацетатный буфер и бикарбонатный буфер), аминокислоты, мочевину, спирты, аскорбиновую кислоту, фосфолипиды, белки (например, сывороточный альбумин), ЭДТА, хлорид натрия, липосомы, маннит, сорбит и глицерин. Растворы или суспензии могут быть инкапсулированы в липосомы или биоразлагаемые микросферы. Препарат, как правило, будет предоставлен в практически стерильной форме при использовании производственных процессов в стерильных условиях.
Сюда может входить получение и стерилизация фильтрованием буферного растворителя/раствора, используемого для препарата, асептическое суспендирование антитела в стерильном буферном растворителе и распределение препарата в стерильные емкости методами, известными специалистам в данной области.
Препарат по настоящему изобретению в распыляемой форме можно предоставлять, например, в виде стандартных доз (например, в запаянных пластиковых контейнерах или флаконах), упакованных в конверты из фольги. Каждый флакон содержит стандартную дозу в растворителе/буферном растворе объемом, например, 2 мл.
Антитела, раскрытые в настоящем документе, могут быть подходящими для доставки путем распыления.
Кроме того, предусмотрено, что антитело по настоящему изобретению можно вводить с использованием методов генной терапии. Для этого последовательности ДНК, кодирующие тяжелую и легкую цепи молекулы антитела под контролем соответствующих компонентов ДНК, вводят пациенту так, чтобы цепи антитела экспрессировались с последовательностей ДНК и сборка происходила in situ.
В одном варианте осуществления молекулу по настоящему изобретению, такую как биспецифический белковый комплекс, описанный в настоящем документе, можно использовать для функционального изменения активности интересующего антигена или антигенов. Например, биспецифический белковый комплекс может нейтрализовать, противодействовать или способствовать активности указанного антигена или антигенов, прямо или косвенно.
Настоящее изобретение также относится к набору, включающему молекулу по настоящему изобретению или ее компонент. В одном варианте осуществления набор включает:
a) один или более слитых белков (A-X), содержащих первое антитело или фрагмент антитела (A), специфичный в отношении CD22 или CD79a, и/или CD79b, присоединенный к первому партнеру по связыванию связывающейся пары (X); и
b) один или более слитых белков (B-Y), содержащих второе антитело или фрагмент антитела (B), специфичный в отношении CD22 или CD79a, и/или CD79b, присоединенный ко второму партнеру по связыванию связывающейся пары (Y), при этом последний является специфичным в отношении первого партнера по связыванию; например, когда первый партнер по связыванию (X) представляет собой пептид или полипептид и второй партнер по связыванию (Y) представляет собой специфичное в отношении него антитело или фрагмент антитела;
при этом второй партнер по связыванию Y является специфичным в отношении первого партнера по связыванию X, и второй партнер по связыванию представляет собой, например, специфичное в отношении него антитело или фрагмент антитела; и при специфическом взаимодействии (таком как взаимодействие связывания) двух партнеров по связыванию образуется гетеродимерное сплетение, которое физически сближает два слитых белка из пунктов a) и b), с образованием биспецифического белкового комплекса; и
при этом по меньшей мере один из A или B является специфичным в отношении CD22 и другой является специфичным в отношении CD79a и/или CD79b, и
слитый белок (белки) находится/находятся в форме комплекса или не в форме комплекса.
Предпочтительно, набор может включать биспецифические белковые комплексы по настоящему изобретению или может включать слитые белки, которые находятся в форме комплекса или не в форме комплекса. В первом случае биспецифические белковые комплексы готовы для использования «сразу из упаковки», что обеспечивает удобство и легкость применения, при этом в последнем случае биспецифические белковые комплексы могут быть собраны в соответствии с потребностями пользователя путем объединения разных слитых белков.
В другом варианте осуществления набор дополнительно включает инструкции по применению.
В другом варианте осуществления набор дополнительно включает один или более реагентов для выполнения одного или более функциональных анализов.
В одном варианте осуществления молекулы по настоящему изобретению, включая слитые белки, биспецифические белковые комплексы или композиции, содержащие их, предоставляют для использования в качестве лабораторного реагента.
ДОПОЛНИТЕЛЬНЫЕ АСПЕКТЫ
В следующем аспекте предложена нуклеотидная последовательность, например, последовательность ДНК, кодирующая конструкт, описанный в настоящем документе, включающий мультиспецифическую молекулу или слитый белок, определенный выше.
В одном варианте осуществления предложена нуклеотидная последовательность, например, последовательность ДНК, кодирующая конструкт, описанный в настоящем документе, включающий мультиспецифическую молекулу или биспецифический белковый комплекс, или антитело по настоящему изобретению.
Настоящее изобретение также относится к вектору, содержащему нуклеотидную последовательность, определенную выше.
Используемый в настоящем документе термин «вектор» означает молекулу нуклеиновой кислоты, способную переносить другую нуклеиновую кислоту, с которой она связана. Примером вектора является «плазмида», которая представляет собой кольцевую двухцепочечную петлю ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, не эписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина, где они впоследствии реплицируются вместе с геномом хозяина. В настоящей спецификации термины «плазмида» и «вектор» могут быть использованы взаимозаменяемо, поскольку плазмида является наиболее широко используемой формой вектора.
Общие методы, с помощью которых векторы могут быть сконструированы, методы трансфекции и методы культивирования клеток хорошо известны специалистам в данной области. В этом отношении приводится ссылка на сборник Current Protocols in Molecular Biology, 1999, F.M. Ausubel (ed.), Wiley Interscience, New York и Maniatis Manual, издательства Cold Spring Harbor Publishing.
Используемый в настоящем документе термин «селективный маркер» означает белок, экспрессия которого позволяет идентифицировать клетки, которые были трансформированы или трансфицированы вектором, содержащим ген маркера. Широкий спектр селективных маркеров известен в данной области. Например, как правило, ген селективного маркера придает устойчивость к лекарственным средствам, таким как G 418, гигромицин или метотрексат, клетке-хозяину, в которую данный вектор был введен. Селективный маркер также может представлять собой визуально определяемый маркер, например, такой как флуоресцентный маркер. Примеры флуоресцентных маркеров включают родамин, FITC, TRITC, Alexa Fluors и различные конъюгаты с ними.
Также предложена клетка-хозяин, содержащая один или более клонирующих или экспрессионных векторов, содержащих одну или более последовательностей ДНК, кодирующих антитело по настоящему изобретению. Любую подходящую систему клетка-хозяин/вектор можно использовать для экспрессии последовательностей ДНК, кодирующих молекулу антитела по настоящему изобретению. Можно использовать бактериальную, например, E. coli, и другие микробные системы, или также можно использовать эукариотические системы экспрессии, например, в клетке-хозяине млекопитающих. Подходящие клетки-хозяева млекопитающих включают клетки CHO, миеломы или гибридомы.
Настоящее изобретение также относится к способу получения молекулы по настоящему изобретению или ее компонента, включающему культивирование клетки-хозяина, содержащей вектор по настоящему изобретению, в условиях, подходящих для экспрессии белка с ДНК, кодирующей молекулу по настоящему изобретению, и выделение данной молекулы.
Молекулы по настоящему изобретению, включая биспецифические белковые комплексы, описанные в настоящем документе, можно использовать в наборах для диагностики/обнаружения. Наборы могут, например, включать биспецифические антительные комплексы, которые специфичны в отношении двух антигенов, оба из которых присутствуют на клетках одного и того же типа, и при этом диагноз может быть положительным только в случае успешного обнаружения обоих антигенов. За счет использования молекулы по настоящему изобретению, такой как биспецифические антительные комплексы, описанные в настоящем документе, а не двух отдельных антител или фрагментов антител не в форме комплекса, специфичность обнаружения может быть значительно повышена.
В одном варианте осуществления молекулы по настоящему изобретению, такие как биспецифические антительные комплексы, закреплены на твердой поверхности. Твердая поверхность может представлять собой, например, чип, или планшет для ELISA.
Кроме того, предложено применение молекулы по настоящему изобретению, например, биспецифического белкового комплекса, описанного в настоящем документе, для обнаружения в образце присутствия первого и второго пептида, при этом указанные молекулы используют в качестве средств обнаружения.
Молекулы по настоящему изобретению, такие как биспецифические антительные комплексы, описанные в настоящем документе, могут, например, быть конъюгированы с флуоресцентным маркером, который облегчает обнаружение связанных комплексов антитело-антиген. Такие биспецифические антительные комплексы можно использовать для иммунофлуоресцентной микроскопии. Альтернативно, биспецифические антительные комплексы также можно использовать для вестерн-блоттинга или ELISA.
В одном варианте осуществления предложен способ очистки молекулы по настоящему изобретению или ее компонента.
В одном варианте осуществления предложен способ очистки молекулы по настоящему изобретению или ее компонента, включающий стадии: проведения анионообменной хроматографии в режиме «без связывания» так, что примеси удерживаются на колонке и антитело остается в не связавшейся фракции. Данную стадию можно, например, выполнять при pH примерно 6-8.
Способ может дополнительно включать начальную стадию захвата в процессе катионообменной хроматографии, которую выполняют, например, при pH примерно 4-5.
Кроме того, способ может включать дополнительную стадию(и) хроматографии для гарантии того, что связанные с продуктом или способом примеси надлежащим образом удалены из потока продукта.
Способ очистки также может включать одну или более стадий ультрафильтрации, такие как стадия концентрирования и диафильтрации.
Использованный выше термин «очищенная форма» означает форму, имеющую чистоту по меньшей мере 90%, например, 91, 92, 93, 94, 95, 96, 97, 98, 99% по массе, или более высокую.
Последовательности по изобретению приведены ниже в настоящем документе.
Последовательности
Последовательности GCN4(7P14P)
ASGGGRMKQLEPKVEELLPKNYHLENEVARLKKLVGERHHHHHH SEQ ID NO: 1,
в которой аминокислоты, выделенные жирным шрифтом, являются необязательными.
GCTAGCGGAGGCGGAAGAATGAAACAACTTGAACCCAAGGTTGAAGAATTGCTTCCGAAAAATTATCACTTGGAAAATGAGGTTGCCAGATTAAAGAAATTAGTTGGCGAACGCCATCACCATCACCATCAC SEQ ID NO: 2
Последовательность dsscFv 52SR4
DAVVTQESALTSSPGETVTLTCRSSTGAVTTSNYASWVQEKPDHLFTGLIGGTNN RAPGVPARFSGSLIGDKAALTITGAQTEDEAIYFCVLWYSDHWVFGCGTKLTVLGGGG GSGGGGSGGGGSGGGGSDVQLQQSGPGLVAPSQSLSITCTVSGFLLTDYGVNWVRQSP GKCLEWLGVIWGDGITDYNSALKSRLSVTKDNSKSQVFLKMNSLQSGDSARYYCVTGL FDYWGQGTTLTVSSAAAHHHHHHEQKLISEEDL- SEQ ID NO: 3.
GATGCGGTGGTGACCCAGGAAAGCGCGCTGACCAGCAGCCCGGGCGAAACCGTGACCCTGACCTGCCGCAGCAGCACCGGCGCGGTGACCACCAGCAACTATGCGAGCTGGGTGCAGGAAAAACCGGATCATCTGTTTACCGGCCTGATTGGCGGCACCAACAACCGCGCGCCGGGCGTGCCGGCGCGCTTTAGCGGCAGCCTGATTGGCGATAAAGCGGCGCTGACCATTACCGGCGCGCAGACCGAAGATGAAGCGATTTATTTTTGCGTGCTGTGGTATAGCGACCATTGGGTGTTTGGCTGCGGCACCAAACTGACCGTGCTGGGTGGAGGCGGTGGCTCAGGCGGAGGTGGCTCAGGCGGTGGCGGGTCTGGCGGCGGCGGCAGCGATGTGCAGCTGCAGCAGAGCGGCCCGGGCCTGGTGGCGCCGAGCCAGAGCCTGAGCATTACCTGCACCGTGAGCGGCTTTCTCCTGACCGATTATGGCGTGAACTGGGTGCGCCAGAGCCCGGGCAAATGCCTGGAATGGCTGGGCGTGATTTGGGGCGATGGCATTACCGATTATAACAGCGCGCTGAAAAGCCGCCTGAGCGTGACCAAAGATAACAGCAAAAGCCAGGTGTTTCTGAAAATGAACAGCCTGCAGAGCGGCGATAGCGCGCGCTATTATTGCGTGACCGGCCTGTTTGATTATTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGCGGCCGCCCATCACCATCACCATCACGAACAGAAACTGATTAGCGAAGAAGATCTGTAATAG SEQ ID NO: 4.
Антитела к CD79b
Aт 4447
Область VL кроличьего Aт 4447 SEQ ID NO: 71
AQVLTQTPSP VSAPVGGTVT INCQASQSVV SGNYLAWLQQ KPGQPPKQLI HSASTLASGV SSRFSGSGSG TQFTLTISGV QCEDAATYYC LGEFSCSSHD CNAFGGGTEV VVK
Область VL кроличьего Aт 4447 SEQ ID NO: 72
gcccaagtgc tgacccagac tccgtcccct gtgtctgcac ctgtgggagg cacagtcacc atcaattgcc aggccagtca gagtgttgtt agtggcaatt acctagcctg gcttcagcag aaaccagggc agcctcccaa gcaactgatc cattctgcat ccactctggc atctggggtc tcatcgcggt tcagcggcag tggatctggg acacaattca ctctcaccat cagcggcgtg cagtgtgaag atgctgccac ttactactgt ctaggcgaat ttagttgtag tagtcatgat tgtaatgctt tcggcggagg gaccgaggtg gtggtcaaa
Область VH кроличьего Aт 4447 SEQ ID NO: 73
QSLEESGGRL VTPGTPLTLT CTVSGFSLSN YAVSWVRQAP GEGLEWIGII YIETGTTWYA NWAKGRFTIS KTSTTVDLTI TSPSTЭДТАT YFCAREPYEP YDDSNIYYGM DPWGPGTLVT VSS
Область VH кроличьего Aт 4447 SEQ ID NO: 74
cagtcgctgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc tgcaccgtct ctggattctc cctcagtaac tatgcagtaa gctgggtccg ccaggctcca ggggagggac tggaatggat cgggatcatt tatattgaaa ctggtaccac atggtacgcg aactgggcga aaggccgatt caccatctcc aaaacctcga ccacggtgga tctgacaatc accagtccgt caaccgagga cacggccacc tatttctgtg ccagagaacc ttatgaacct tatgatgata gtaatattta ctacggcatg gacccctggg gcccaggcac cctcgtcacc gtctcgagt
CDRL1 SEQ ID NO: 75 QASQSVVSGNYLA
CDRL2 SEQ ID NO: 76 SASTLAS
CDRL3 SEQ ID NO: 77 LGEFSCSSHDCNA
CDRH1 SEQ ID NO: 78 GFSLSNYAVS
CDRH2 SEQ ID NO: 79 IIYIETGTTWYANWAKG
CDRH3 SEQ ID NO: 80 EPYEPYDDSNIYYGMDP.
Изобретение также относится к производному SEQ ID NO: 77, в котором один или оба остатка цистеина заменены другой аминокислотой, например, серином, в частности, когда первый остаток cys заменен остатком серина и второй остаток cys оставлен без изменения, или первый остаток cys оставлен без изменения и второй остаток цистеина заменен остатком серина, или когда оба остатка цистеина заменены остатками серина.
Aт 4450
Область VL кроличьего Aт 4450 SEQ ID NO: 81
AIDMTQTPSP VSAAVGGTVT INCQSSQSIY NNNDLAWYQQ KPGQPPKLLI YEASKLASGV PSRFKGSGSG TQFTLTISGV QCDDAATYYC QGGGSGGDGI AFGGGTKVVV E
Область VL кроличьего Aт 4450 SEQ ID NO: 82
gccattgata tgacccagac tccatccccc gtgtctgcag ctgtgggagg cacagtcacc atcaattgcc agtccagtca gagtatttat aataataatg acttagcctg gtatcagcag aaaccagggc agcctcccaa gctcctgatc tacgaagcat ccaaactggc atctggggtc ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagtggcgtg cagtgtgatg atgctgccac ttactactgt cagggcggtg gtagtggtgg tgatggcatt gctttcggcg gagggaccaa ggtggtcgtc gaa
Область VH кроличьего Aт 4450 SEQ ID NO: 83
QSVEESGGRL VTPGAPLTLT CTVSGFSLNN YVMVWVRQAP GKGLEWIGII YVSGNAYYAS WAKGRFTISR TSTTVDLKVT SLTTЭДТАTY FCARDAGHSD VDVLDIWGPG TLVTVSS
Область VH кроличьего Aт 4450 SEQ ID NO: 84
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg gggcacccct gacactcacc tgcacagtct ctggattctc cctcaataac tatgtaatgg tctgggtccg ccaggctcca gggaaggggc tggaatggat cggaatcatt tatgttagtg gtaatgcata ctacgcgagc tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaagtgacc agtctgacaa ccgaggacac ggccacctat ttctgtgcca gagatgctgg tcatagtgat gtcgatgttt tggatatttg gggcccgggc accctcgtca ccgtctcgag t
CDRL1 SEQ ID NO: 85 QSSQSIYNNNDLA
CDRL2 SEQ ID NO: 86 EASKLAS
CDRL3 SEQ ID NO: 87 QGGGSGGDGIA
CDRH1 SEQ ID NO: 88 GFSLNNYVMV
CDRH2 SEQ ID NO: 89 IIYVSGNAYYASWAKG
CDRH3 SEQ ID NO: 90 DAGHSDVDVLDI.
Изобретение также относится к производному SEQ ID NO: 87, в котором по меньшей мере одна из аминокислот в мотиве DG заменена другой аминокислотой, например, мотив мутирован на EG, DA или DS.
Антитела к CD22
Aт 4120
Область VL кроличьего Aт 4120 SEQ ID NO: 91
AFELSQTPAS VEAAVGGTVT IKCQASQSIS TALAWYQQKP GQRPKLLIYG ASTLASGVSS RFKGSGSGTE FTLTISDLEC ADAATYYCQS YYGTSSGGSW AFGGGTKVVV K
Область VL кроличьего Aт 4120 SEQ ID NO: 92
gcattcgaat tgagccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc atcaagtgcc aggccagtca gagcattagc actgcattag cctggtatca gcagaaacca gggcagcgtc ccaagctcct gatctatggt gcatccactc tggcatctgg ggtctcatcg cggttcaaag gcagtggatc tgggacagag ttcactctca ccatcagcga cctggagtgt gccgatgctg ccacttacta ctgtcaaagc tattatggta cgagtagtgg tggttcttgg gctttcggcg gagggaccaa ggtggtcgtc aaa
Область VH кроличьего Aт 4120 SEQ ID NO: 93
QSLEESGGDL VKPGASLTLT CTASGFSFSS SYYMCWVRQS PGKGLEWIAC IYTGSSGDTY YASWAKGRFT ISKTSSTTVS LQMTSLTAAD TATYFCARGP YVGYGYDLQY LYLWGPGTLV TVSS
Область VH кроличьего Aт 4120 SEQ ID NO: 94
cagtcattgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc tgcacagcct ctggattctc cttcagtagt agctactaca tgtgctgggt ccgccagtct ccagggaagg ggctggagtg gatcgcatgc atttatactg gtagtagtgg tgacacttac tacgcgagct gggcgaaagg ccgattcacc atctccaaaa cctcgtcgac cacggtgtct ctgcaaatga ccagtctgac agccgcggac acggccactt atttctgtgc gagagggcct tatgttggtt atggttatga tcttcaatac ttgtacttgt ggggcccggg gaccctcgtc accgtctcga gt
CDRL1 SEQ ID NO: 95 QASQSISTALA
CDRL2 SEQ ID NO: 96 GASTLAS
CDRL3 SEQ ID NO: 97 QSYYGTSSGGSWA
CDRH1 SEQ ID NO: 98 GFSFSSSYYMC
CDRH2 SEQ ID NO: 99 CIYTGSSGDTYYASWAKG
CDRH3 SEQ ID NO: 100 GPYVGYGYDLQYLYL.
Изобретение также относится к производному SEQ ID NO: 98, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 99, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Aт 4126
Область VL кроличьего Aт 4126 SEQ ID NO: 101
DIVMTQTPAS VEAAVGGTVT IKCQASQNIG SGLAWYQQKP GQPPKLLIYY ASTLASGVPS RFKGSGSGTQ FTLTISDLEC ADAATYYCQS HDYSSVRSYG NAFGGGTEVV VK
Область VL кроличьего Aт 4126 SEQ ID NO: 102
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc atcaagtgcc aggccagtca gaacattggt agtggtttag cctggtatca gcagaaacca gggcagcctc ccaagctcct gatctattat gcatccactc tggcatctgg ggtcccatca aggttcaaag gcagtggatc tgggacacag ttcactctca ccatcagcga cctggagtgt gccgacgctg ccacttacta ctgtcaaagt catgattata gtagtgttcg gagttacggt aatgctttcg gcggagggac cgaggtggtg gtcaaa
Область VH кроличьего Aт 4126 SEQ ID NO: 103
QQHLEESGGG LVKPGGTLTL TCKASGIDFS SYYYMCWVRQ APGKGLEWVA CIDPASSGTT YYATWAKGRF TISKTSSTTV TLQMTSLTAA DTATYFCARA YGSGGSGYIG CYFDLWGQGT LVTVSS
Область VH кроличьего Aт 4126 SEQ ID NO: 104
cagcagcacc tggaggagtc cgggggaggc ctggtcaagc ctggaggaac cctgacactc acctgcaaag cctctggaat cgacttcagt agctactact acatgtgctg ggtccgccag gctccaggga aggggctgga gtgggtcgcg tgcattgatc ctgctagtag tggtactact tactacgcga cctgggcgaa aggccgattc accatctcca aaacctcgtc gaccacggtg actctgcaaa tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagggca tatggtagtg ggggtagtgg ttatataggg tgctactttg acttgtgggg ccaaggcacc ctcgtcaccg tctcgagt
CDRL1 SEQ ID NO: 105 QASQNIGSGLA
CDRL2 SEQ ID NO: 106 YASTLAS
CDRL3 SEQ ID NO: 107 QSHDYSSVRSYGNA
CDRH1 SEQ ID NO: 108 GIDFSSYYYMC
CDRH2 SEQ ID NO: 109 CIDPASSGTTYYATWAKG
CDRH3 SEQ ID NO: 110 AYGSGGSGYIGCYFDL.
Изобретение также относится к производному SEQ ID NO: 108, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 109, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 110, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Aт 4127
Область VL кроличьего Aт 4127 SEQ ID NO: 111
AIVMTQTPSS KSVPMGGTVT INCQASQSVY GNNELSWYQQ KPGQPPKLLI YLASRLASGV PSRFSGSGSG TQFTLTISGV QCDDAATYYC AGYKSDSDDG TTFGGGTKVV VE
Область VL кроличьего Aт 4127 SEQ ID NO: 112
gccatcgtga tgacccagac tccatcttcc aagtctgtcc ctatgggagg cacagtcacc atcaactgcc aggccagtca gagtgtttat ggtaataacg aattatcctg gtatcagcag aaaccagggc agcctcccaa gctcctgatc tatttggcat ccaggctggc atcgggggtc ccatcgcggt ttagcggcag tggatctggg acacagttca ctctcaccat cagcggcgtg cagtgtgacg atgctgccac ttactactgt gcaggctata aaagtgatag tgatgatggc actactttcg gcggagggac caaggtggtg gtcgaa
Область VH кроличьего Aт 4127 SEQ ID NO: 113
QQLEESGGDL VKPGASLTLT CTASGFSFSN LYYMCWVRQA PGKGLELIGC IDISSSGSTY YASWAKGRFT ISKTSSTTVT LQMTSLTAAD TATYFCARDY YSSDWGVRFN LWGQGTLVTV SS
Область VH кроличьего Aт 4127 SEQ ID NO: 114
cagcagctgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc tgcacagcct ctggattctc cttcagtaat ctctattaca tgtgttgggt ccgccaggct ccagggaagg ggctggagtt gatcggatgc attgatatta gcagtagtgg tagcacttac tacgcgagct gggcgaaagg ccgattcacc atctccaaaa cctcgtcgac cacggtgact ctgcagatga ccagtctgac agccgcggac acggccacct atttctgtgc gagagattac tattctagtg actggggtgt tagatttaac ttgtggggcc agggcaccct cgtcaccgtc tcgagt
CDRL1 SEQ ID NO: 115 QASQSVYGNNELS
CDRL2 SEQ ID NO: 116 LASRLAS
CDRL3 SEQ ID NO: 117 AGYKSDSDDGTT
CDRH1 SEQ ID NO: 118 GFSFSNLYYMC
CDRH2 SEQ ID NO: 119 CIDISSSGSTYYASWAKG
CDRH3 SEQ ID NO: 120 DYYSSDWGVRFNL.
Изобретение также относится к производному SEQ ID NO: 117, в котором независимо выполнены следующие мутации, например, DS изменено на EA, DA или DT и DG изменено на EG, DA или DS. В одном варианте осуществления последовательности DS: DG представляют собой DS, EG; DS, DA; DS, DS; EA, DG; EA, EG; EA, DA; EA, DS; DA, DG; DA, EG; DA, DA; DA, DS; DT, DG; DT, EG; DT, DA; а также DT, DS.
Изобретение также относится к производному SEQ ID NO: 118, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 119, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Aт 4128
Область VL кроличьего Aт 4128 SEQ ID NO: 121
DIVMTQTPAS VEAAVGGTVT IKCQASESIS NYLSWFQQKP GQPPKLLIYA SSKLSSGVPS RFKGDRSGTE YTLTISDLEC ADAATYYCQI YYSASGSRDW TFGGGTKVVV E
Область VL кроличьего Aт 4128 SEQ ID NO: 122
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc atcaagtgcc aggccagtga aagcattagc aactacttat cctggtttca gcagaaacca gggcagcctc ccaagctcct gatctatgct tcatccaaac tgtcatctgg ggtcccatcg cggttcaaag gcgatagatc tgggacagag tacactctca ccatcagcga cctggagtgt gccgatgctg ccacttacta ctgtcaaatc tattattcgg ctagtggcag tcgtgattgg actttcggcg gagggaccaa ggtggtcgtc gaa
Область VH кроличьего Aт 4128 SEQ ID NO: 123
QSLEESGGDL VQPEGSLTLT CKGSGLDFSS YWICWVRQAP GKGLEWIACI VTGSSDNTYY ASWAKGRFTI SKTSSTTVTL QMTSLTAADT ATYFCARGGG AGYSGAFDLW GQGTLVTVSS
Область VH кроличьего Aт 4128 SEQ ID NO: 124
cagtcgttgg aggagtccgg gggagacctg gtccagcctg agggatccct gacactcacc tgcaaaggct ccgggttaga cttcagtagc tactggatat gctgggtccg ccaggctcca gggaaggggc tggagtggat cgcatgcatt gttactggta gtagtgataa cacttactac gcgagctggg cgaaaggccg attcaccatc tccaaaacct cgtcgaccac ggtgactctg caaatgacca gtctgacagc cgcggacacg gccacctatt tctgtgcgag aggtggtggt gctggttata gtggtgcctt tgacttgtgg ggccaaggga ccctcgtcac cgtctcgagt
CDRL1 SEQ ID NO: 125 QASESISNYLS
CDRL2 SEQ ID NO: 126 ASSKLSS
CDRL3 SEQ ID NO: 127 QIYYSASGSRDWT
CDRH1 SEQ ID NO: 128 GLDFSSYWIC
CDRH2 SEQ ID NO: 129 CIVTGSSDNTYYASWAKG
CDRH3 SEQ ID NO: 130 GGGAGYSGAFDL.
Изобретение также относится к производному SEQ ID NO: 128, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 129, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Aт 4130
Область VL кроличьего Aт 4130 SEQ ID NO: 131
AAVLTQTPSP VSAAVGGTVS ISCQSSQSVY NTKDLAWYQQ KPGQPPKLLI YGTSTLASGV SSRFSGSGSG TEFTLTISDL ECDDAATYYC QGGFSSSDLN VFGGGTKVVV K
Область VL кроличьего Aт 4130 SEQ ID NO: 132
gccgccgtgc tgacccagac tccatctccc gtgtctgcag ctgtgggagg cacagtcagc atcagttgcc agtccagtca gagtgtttat aatacaaagg acttagcctg gtatcagcag aaaccagggc agcctcccaa gctcctgatc tatggtacat ccactctggc atctggggtc tcatcacggt tcagcggcag tggatctggg acagagttca ctctcaccat cagcgacctg gagtgtgacg atgctgccac ttattactgt caaggcggtt ttagtagtag tgatttgaat gttttcggcg gagggaccaa ggtggtggtc aaa
Область VH кроличьего Aт 4130 SEQ ID NO: 133
QQQLEESGGD LVRPEGSLTL TCTASGFDFS GGYDISWVRQ APGKGLEWIG CIYGGINSVT DYASWAKGRV TISKTSSTTV TLQMTSLTAA DTATYFCARD VSNSDHYTRL DLWGQGTLVT VSS
Область VH кроличьего Aт 4130 SEQ ID NO: 134
cagcagcagc tggaggagtc cgggggagac ctggtcaggc ctgagggatc cctgacactc acctgcacag cctctggatt cgacttcagt ggcggctacg acatttcctg ggtccgccag gctccaggga aggggctgga gtggatcgga tgcatttatg gtggtatcaa tagtgtcact gactacgcga gctgggcgaa aggccgagtc accatctcca aaacctcgtc gaccacggtg actctgcaga tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagagat gttagtaata gcgatcatta tactcggttg gatctctggg gccaaggcac cctggtcacc gtctcgagt
CDRL1 SEQ ID NO: 135 QSSQSVYNTKDLA
CDRL2 SEQ ID NO: 136 GTSTLAS
CDRL3 SEQ ID NO: 137 QGGFSSSDLNV
CDRH1 SEQ ID NO: 138 GFDFSGGYDIS
CDRH2 SEQ ID NO: 139 CIYGGINSVTDYASWAKG
CDRH3 SEQ ID NO: 140 DVSNSDHYTRLDL.
Изобретение также относится к производному SEQ ID NO: 139, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 139, в котором мотив NS изменен, например, на NA или NT.
Изобретение также относится к производному SEQ ID NO: 140, в котором мотив NS изменен, например, на NA или NT.
Aт 4132
Область VL кроличьего Aт 4132 SEQ ID NO: 141
DIVMTQTPAS VEAAVGGTVT IKCQASETIS SRLAWYQQKL GQPPKLLIYS ASTLASGVPS RFKGSGSGTE YTLTISGVQC ADAATYYCQG YYYSSGSDYG FGGGTKVVVK
Область VL кроличьего Aт 4132 SEQ ID NO: 142
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc atcaagtgcc aggccagtga gaccattagt agtagattag cctggtatca gcagaagcta gggcagcctc ccaaactcct gatctattct gcatccactc tggcgtctgg ggtcccatcg cggttcaaag gcagtggatc tgggacagag tacactctca ccatcagcgg cgtgcagtgt gccgatgctg ccacttatta ctgtcaaggc tattattata gtagtggtag tgattatggt ttcggcggag ggaccaaggt ggtcgtcaaa
Область VH кроличьего Aт 4132 SEQ ID NO: 143
QSLEESGGDL VKPGASLTLT CTASGFSFSS SYWICWVRQA PGKGLEWSGC INSGTGGTAY ASWAKGRFTI SNSSSTTVTL QMTSLTAADT ATYFCAREWV SGYYKDAFDL WGQGTLVTVS S
Область VH кроличьего Aт 4132 SEQ ID NO: 144
cagtcgttgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc tgcacagcct ctggattctc cttcagtagc agctactgga tatgctgggt ccgccaggct ccagggaagg ggctggagtg gagcggatgc attaatagtg gtactggtgg cactgcctac gcgagctggg cgaaaggccg attcaccatc tccaattcct cgtcgaccac ggtgactctt caaatgacca gtctgacagc cgcggacacg gccacctatt tctgtgcgag agaatgggtt agtggttatt ataaagatgc ttttgatctc tggggccagg gcaccctggt caccgtctcg agt
CDRL1 SEQ ID NO: 145 QASETISSRLA
CDRL2 SEQ ID NO: 146 SASTLAS
CDRL3 SEQ ID NO: 147 QGYYYSSGSDYG
CDRH1 SEQ ID NO: 148 GFSFSSSYWIC
CDRH2 SEQ ID NO: 149 CINSGTGGTAYASWAKG
CDRH3 SEQ ID NO: 150 EWVSGYYKDAFDL.
Изобретение также относится к производному SEQ ID NO: 148, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 149, в котором остаток цистеина заменен другой аминокислотой, например, серином.
Изобретение также относится к производному SEQ ID NO: 149, в котором мотив NS изменен, например, на NA или NT.
Антитела, связывающие сывороточный альбумин
Вариабельный домен тяжелой цепи антитела против альбумина (без ds) SEQ ID NO: 157
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
Вариабельный домен тяжелой цепи антитела против альбумина (ds) SEQ ID NO: 158
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
Вариабельный домен легкой цепи антитела против альбумина (без ds) SEQ ID NO: 159
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
Вариабельный домен легкой цепи антитела против альбумина (ds) SEQ ID NO: 160
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
CD22 человека SEQ ID NO: 161
MHLLGPWLLL LVLEYLAFSD SSKWVFEHPE TLYAWEGACV WIPCTYRALD GDLESFILFH NPEYNKNTSK FDGTRLYEST KDGKVPSEQK RVQFLGDKNK NCTLSIHPVH LNDSGQLGLR MESKTEKWME RIHLNVSERP FPPHIQLPPE IQESQEVTLT CLLNFSCYGY PIQLQWLLEG VPMRQAAVTS TSLTIKSVFT RSELKFSPQW SHHGKIVTCQ LQDADGKFLS NDTVQLNVKH TPKLEIKVTP SDAIVREGDS VTMTCEVSSS NPEYTTVSWL KDGTSLKKQN TFTLNLREVT KDQSGKYCCQ VSNDVGPGRS EEVFLQVQYA PEPSTVQILH SPAVEGSQVE FLCMSLANPL PTNYTWYHNG KEMQGRTEEK VHIPKILPWH AGTYSCVAEN ILGTGQRGPG AELDVQYPPK KVTTVIQNPM PIREGDTVTL SCNYNSSNPS VTRYEWKPHG AWEEPSLGVL KIQNVGWDNT TIACAACNSW CSWASPVALN VQYAPRDVRV RKIKPLSEIH SGNSVSLQCD FSSSHPKEVQ FFWEKNGRLL GKESQLNFDS ISPEDAGSYS CWVNNSIGQT ASKAWTLEVL YAPRRLRVSM SPGDQVMEGK SATLTCESDA NPPVSHYTWF DWNNQSLPYH SQKLRLEPVK VQHSGAYWCQ GTNSVGKGRS PLSTLTVYYS PETIGRRVAV GLGSCLAILI LAICGLKLQR RWKRTQSQQG LQENSSGQSF FVRNKKVRRA PLSEGPHSLG CYNPMMEDGI SYTTLRFPEM NIPRTGDAES SEMQRPPPDC DDTVTYSALH KRQVGDYENV IPDFPEDEGI HYSELIQFGV GERPQAQENV DYVILKH
CD79a человека SEQ ID NO: 162
MPGGPGVLQA LPATIFLLFL LSAVYLGPGC QALWMHKVPA SLMVSLGEDA HFQCPHNSSN NANVTWWRVL HGNYTWPPEF LGPGEDPNGT LIIQNVNKSH GGIYVCRVQE GNESYQQSCG TYLRVRQPPP RPFLDMGEGT KNRIITAEGI ILLFCAVVPG TLLLFRKRWQ NEKLGLDAGD EYEDENLYEG LNLDDCSMYE DISRGLQGTY QDVGSLNIGD VQLEKP
CD79b человека SEQ ID NO: 163
MARLALSPVP SHWMVALLLL LSAEPVPAAR SEDRYRNPKG SACSRIWQSP RFIARKRGFT VKMHCYMNSA SGNVSWLWKQ EMDENPQQLK LEKGRMEESQ NESLATLTIQ GIRFEDNGIY FCQQKCNNTS EVYQGCGTEL RVMGFSTLAQ LKQRNTLKDG IIMIQTLLII LFIIVPIFLL LDKDDSKAGM EEDHTYEGLD IDQTATYEDI VTLRTGEVKW SVGEHPGQE.
В контексте данной спецификации «содержащий» следует понимать, как «включающий».
Аспекты изобретения, включающие определенные элементы, также должны относиться к альтернативным вариантам осуществления, «состоящим» или «состоящим по существу» из соответствующих элементов.
Непосредственно изложенные варианты осуществления могут быть использованы в настоящем документе в качестве основы для отказа от пунктов формулы изобретения.
Все литературные источники, приведенные в настоящем документе, специально включены в него посредством ссылки.
Список литературы
1. Ribosome display efficiently selects and evolves high-affinity antibodies in vitro from immune libraries. Hanes J., Jermutus L., Weber-Bornhauser S., Bosshard H.R., Plückthun A. (1998) Proc. Natl. Acad. Sci. U.S.A. 95, 14130-14135.
2. Directed in Vitro Evolution and Crystallographic Analysis of a Peptide-binding Single Chain Antibody Fragment (scFv) with Low Picomolar Affinity. Zhand C., Spinelli S., Luginbuhl B., Amstutz P., Cambillau C., Pluckthun A. (2004) J. Biol. Chem. 279, 18870-18877.
3. Antigen recognition by conformational selection. Berger C., Weber-Bornhauser S., Eggenberger Y., Hanes J., Pluckthun A., Bosshard H.R. (1999) F.E.B.S. Letters 450, 149-153.
ПРИМЕРЫ
Термин «Fab-Kd-Fab», используемый в примерах, означает биспецифический белковый комплекс, имеющий формулу A-X:Y-B, где:
A-X представляет собой первый слитый белок;
Y-B представляет собой второй слитый белок;
X:Y представляет собой гетеродимерное сплетение;
A включает Fab-фрагмент, специфичный в отношении антигена, такого как CD22 или CD79;
B включает Fab-фрагмент, специфичный в отношении антигена, такого как CD22 или CD79;
X является первым партнером по связыванию связывающейся пары, например, scFv;
Y является вторым партнером по связыванию связывающейся пары, например, пептидом; и
: означает взаимодействие (такое как взаимодействие связывания) между X и Y.
Пример 1 - Получение Fab'-A (Fab-scFv [A-X]) и Fab'-B (Fab-пептид [B-Y) для функциональных анализов
Стратегия клонирования
ДНК вариабельной области антитела получали путем ПЦР или синтеза гена, фланкируя последовательность ДНК сайтами для ферментов рестрикции. Этими сайтами были HindIII и XhoI для вариабельных тяжелых цепей и HindIII и BsiWI для вариабельных легких цепей. Это создает возможность лигирования вариабельной области тяжелой цепи в два вектора для тяжелой цепи (pNAFH с FabB-Y и pNAFH с FabA-Xds [стабилизированные дисульфидными связями]), поскольку они имеют комплементарные сайты рестрикции. При этом вариабельная область лигируется выше (или 5') относительно константных областей мышиного антитела и пептида Y (GCN4) или scFv X (52SR4), создавая целую рамку считывания. Легкие цепи клонировали в стандартные векторы мышиных константных областей каппа (pMmCK или pMmCK S171C) собственного изготовления, в которых опять-таки использовали те же комплементарные сайты рестрикции. Вектор pMmCK S171C использовали, если вариабельная область была получена от кролика. События клонирования подтверждали секвенированием с использованием праймеров, фланкирующих полную открытую рамку считывания.
Культивирование CHO-S
Суспензию клеток CHO-S предварительно адаптировали к среде CDCHO (Invitrogen), содержащей 2 мМ (100x) глютамакса. Клетки поддерживали в логарифмической фазе роста с вращением при 140 об/мин в инкубаторе с шейкером (Kuner AG, Birsfelden, Switzerland) и культивировали при температуре 37°C в атмосфере 8% CO2.
Трансфекция электропорацией
Перед трансфекцией определяли число и жизнеспособность клеток с использованием устройства для счета клеток CEDEX (Innovatis AG. Bielefeld, Germany) и необходимое количество клеток (2×108 клеток/мл) переносили в конические центрифужные пробирки и центрифугировали при 1400 об/мин в течение 10 минут. Осажденные клетки ресуспендировали в стерильном сбалансированном солевом растворе Эрла и вновь центрифугировали при 1400 об/мин в течение 10 минут. Супернатант отбрасывали и осадки ресуспендировали до нужной плотности клеток.
Смешивали векторную ДНК в конечной концентрации 400 мкг для 2×108 клеток/мл, 800 мкл переносили пипеткой в кюветы (Biorad) и проводили электропорацию с использованием системы для электропорации собственного изготовления.
Трансфицированные клетки переносили непосредственно в 1X3 л колбы Эрленмейера, содержащие среду ProCHO 5, обогащенную 2 мМ глютамаксом и раствором антибиотика-антимикотика (100X) (1 в 500), и клетки культивировали в инкубаторе с шейкером Kuhner в условиях 37°C, 5% CO2 со встряхиванием при 140 об/мин. Питательную добавку 2 г/л ASF (AJINOMOTO) добавляли через 24 часа после трансфекции и температуру снижали до 37°C на следующие 13 дней культивирования. В день четвертый в культуру добавляли 3 мМ бутирата натрия (натриевая соль н-масляной кислоты, Sigma B-5887).
В день 14 культуры переносили в пробирки и супернатант отделяли от клеток центрифугированием в течение 30 минут при 4000 об/мин. Оставшиеся супернатанты дополнительно фильтровали через 0,22-мкм мембраны SARTO BRAN P Millipore с последующим фильтрованием через 0,22-мкм золотые фильтры Gamma. Конечные уровни экспрессии определяли методом ВЭЖХ с белком G.
Крупномасштабная (1,0 л) очистка
Fab-A и Fab-B очищали методом аффинного захвата с использованием систем AKTA Xpress и предварительно заполненных никелевых колонок HisTrap Excel (GE Healthcare). Культуральные супернатанты стерильно фильтровали через 0,22-мкм фильтры и pH доводили до нейтрального значения, при необходимости, слабой кислотой или основанием перед нанесением на колонки. Стадию вторичной промывки раствором, содержащим 15-25 мМ имидазола, использовали для вытеснения любых слабо связанных белков клеток-хозяев/неспецифически связанных His-соединений с никелевой смолы. Элюцию выполняли 10 мМ фосфатом натрия, pH 7,4+1M NaCl+250 мМ имидазола, и собирали 2-мл фракции. Один объем колонки при элюции системы удерживали в течение 10 минут для сжатия элюируемого пика и последующего уменьшения общего элюированного объема. Наиболее чистые фракции объединяли, буфер заменяли на PBS (Sigma), pH 7,4, и фильтровали через 0,22-мкм фильтр. Конечные пулы анализировали сканированием при A280, эксклюзионной ВЭЖХ (метод G3000), SDS-ПААГ (в восстанавливающих и не восстанавливающих условиях), а также на эндотоксины с использованием системы PTS Endosafe.
Пример 2 - Использование Fab'-A (Fab-scFv [A-X]) и Fab'-b (Fab-пептид [B-Y]) в формате гетеродимерно сплетенного биспецифического белкового комплекса для демонстрации того, что биспецифические, но не двухвалентные сочетания для CD79/CD22 ингибируют сигнализацию Akt
Человеческие МКПК, полученные из конусов при аферезе тромбоцитов, замораживали аликвотами. Перед проведением анализа клетки размораживали, промывали в DMEM (Life Technologies) и оставляли для акклиматизации в условиях 37°C/5% CO2. В это время готовили сетки из биспецифических или двухвалентных антител путем разбавления эквимолярных количеств (200 нМ) Fab'-A (Fab-scFv) и Fab'-B (Fab-пептид) с антигенной специфичностью в отношении белков клеточной поверхности CD22 и CD79b в среде DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Данная сетка показана в таблице 4.
Таблица 4: Возможная сетка биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b
Fab A
где X представляет собой scFv (52SR4) и Y представляет собой пептид (GCN4).
Fab'A-X и Fab'B-Y инкубировали вместе в течение 90 минут (в условиях 37°C/5% CO2), а затем смешивали с 2,5×105 МКПК в 96-луночных планшетах с V-образным дном. Затем МКПК плюс биспецифические или двухвалентные сочетания инкубировали вместе в течение следующих 90 минут. После этого B-клетки активировали добавлением 200 нМ F(ab')2 козы против IgM человека (Southern Biotechnology) на 8 минут при температуре 37°C. Реакцию сигнализации затем останавливали добавлением равного объема буфера Cytofix (BD Biosciences). После этого планшеты оставляли при комнатной температуре на 15 минут, с последующим центрифугированием при 500 g в течение 5 минут. Отбрасывали лишний супернатант над осадком клеток, который ресуспендировали в проточном буфере (PBS+1% БСА+0,01% NaN3) и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут, с последующим двукратным промыванием в проточном буфере.
Вслед за этим клетки окрашивали флуоресцентно меченым анти-CD20 антителом (BD Biosciences) и флуоресцентно меченым анти-фосфо-Akt антителом, которое узнает модифицированный остаток серина в положении 473 на белке. Затем клетки в планшетах ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 25 мкл проточного буфера. Измеряли экспрессию в клетках CD20 и Akt с помощью проточного цитометра Intellicyt HTFC™.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни Akt. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки). Относительный эффект сочетаний для CD22 и CD79b представлен в таблице 5 (↓=ингибирование, ↑=стимуляция и ↔=отсутствие общего эффекта).
Таблица 5: Таблица относительной эффективности ингибирования фосфорилирования Akt для биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b
где X представляет собой scFv (52SR4) и Y представляет собой пептид (GCN4).
Эти данные также представлены в виде гистограммы (фигура 1): данные представляют собой средние значения и планки погрешностей соответствуют 95% доверительным интервалам. Данные показывают, что сочетания для CD22 с CD79b могут ингибировать экспрессию фосфо-Akt в B-клетках, стимулированных анти-IgM. Напротив, в случае сочетания для CD22 с CD22 наблюдали повышенные уровни экспрессии фосфо-Akt.
Пример 3 - Использование Fab'-A (Fab-scFv [A-X]) и Fab'-b (Fab-пептид [B-Y]) в формате гетеродимерно сплетенного биспецифического белкового комплекса для демонстрации того, что биспецифические, но не двухвалентные сочетания для CD79/CD22 ингибируют сигнализацию PLCγ2
Человеческие МКПК, полученные из конусов при аферезе тромбоцитов, замораживали аликвотами. Перед проведением анализа клетки размораживали, промывали в DMEM (Life Technologies) и оставляли для акклиматизации в условиях 37°C/5% CO2. В это время готовили сетки из биспецифических или двухвалентных антител путем разбавления эквимолярных количеств (200 нМ) Fab'-A (Fab-scFv) и Fab'-B (Fab-пептид) с антигенной специфичностью в отношении белков клеточной поверхности CD22 и CD79b в среде DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Данная сетка показана в таблице 4.
Fab'A-X и Fab'B-Y инкубировали вместе в течение 90 минут (в условиях 37°C/5% CO2), а затем смешивали с 2,5×105 МКПК в 96-луночных планшетах с V-образным дном. Затем МКПК плюс биспецифические или двухвалентные сочетания инкубировали вместе в течение следующих 90 минут. После этого B-клетки активировали добавлением 200 нМ F(ab')2 козы против IgM человека (Southern Biotechnology) на 8 минут при температуре 37°C. Реакцию сигнализации затем останавливали добавлением равного объема буфера Cytofix (BD Biosciences). После этого планшеты оставляли при комнатной температуре на 15 минут, с последующим центрифугированием при 500 g в течение 5 минут. Отбрасывали лишний супернатант над осадком клеток, который ресуспендировали в проточном буфере и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут, с последующим двукратным промыванием в проточном буфере.
Вслед за этим клетки окрашивали флуоресцентно меченым анти-CD20 антителом (BD Biosciences) и флуоресцентно меченым анти-фосфо-PLCγ2 антителом, которое узнает модифицированный остаток тирозина в положении 759 на белке. Затем клетки в планшетах ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 25 мкл проточного буфера. Измеряли экспрессию в клетках CD20 и PLCγ2 с помощью проточного цитометра Intellicyt HTFC™.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни PLCγ2. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки). Относительный эффект сочетаний для CD22 и CD79b представлен в таблице 6 (↓=ингибирование, ↑=стимуляция и ↔=отсутствие общего эффекта).
Таблица 6: Таблица относительной эффективности ингибирования фосфорилирования PLCγ2 для биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b
где X представляет собой scFv и Y представляет собой пептид.
Эти данные также представлены в виде гистограммы (фигура 2): данные представляют собой средние значения и планки погрешностей соответствуют 95% доверительным интервалам. Данные показывают, что все сочетания для CD22 с CD79b и CD79b с CD79b могут ингибировать экспрессию фосфо-PLCγ2 в B-клетках, стимулированных анти-IgM. Напротив, в случае сочетания для CD22 с CD22 наблюдали повышенные уровни экспрессии фосфо-PLCγ2.
Пример 4 - Использование Fab'-A (Fab-scFv [A-X]) и Fab'-b (Fab-пептид [B-Y]) в формате гетеродимерно сплетенного биспецифического белкового комплекса для демонстрации того, что биспецифические сочетания для CD79/CD22 ингибируют экспрессию CD86
Человеческие МКПК, полученные из конусов при аферезе тромбоцитов, замораживали аликвотами. Перед проведением анализа клетки размораживали, промывали в DMEM (Life Technologies) и оставляли для акклиматизации в условиях 37°C/5% CO2. В это время готовили сетки из биспецифических или двухвалентных антител путем разбавления эквимолярных количеств (200 нМ) Fab'-A (Fab-scFv) и Fab'-B (Fab-пептид) с антигенной специфичностью в отношении белков клеточной поверхности CD22 и CD79b в среде DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Данная сетка показана в таблице 4.
Fab'A-X и Fab'B-Y инкубировали вместе в течение 90 минут (в условиях 37°C/5% CO2), а затем смешивали с 2,5×105 МКПК в 96-луночных планшетах с V-образным дном. Затем МКПК плюс биспецифические или двухвалентные сочетания инкубировали вместе в течение следующих 90 минут. После этого B-клетки активировали добавлением 200 нМ F(ab')2 козы против IgM человека (Southern Biotechnology) на 24 минуты при температуре 37°C. Затем планшеты помещали на лед и промывали один раз ледяным проточным буфером (PBS+1% БСА+0,01% NaN3). Вслед за этим клетки окрашивали флуоресцентно меченым анти-CD19 антителом (BD Biosciences) и флуоресцентно меченым анти-CD86 антителом, и инкубировали на льду в течение 1 часа в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 25 мкл проточного буфера. Измеряли экспрессию в клетках CD19 и CD86 с помощью проточного цитометра Intellicyt HTFC™.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни CD86. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки). Относительный эффект сочетаний для CD22 и CD79b представлен в таблице 7 (↓=ингибирование, ↑=стимуляция и ↔=отсутствие общего эффекта).
Таблица 7: Таблица относительной эффективности ингибирования экспрессии CD86 B-клетками для биспецифических и двухвалентных сочетаний антител со специфичностью в отношении CD22 и CD79b
где X представляет собой scFv (52SR4) и Y представляет собой пептид (GCN4).
Эти данные также представлены в виде гистограммы (фигура 3), данные представляют собой средние значения и планки погрешностей соответствуют 95% доверительным интервалам. Данные показывают, что все сочетания для CD22 с CD79b и CD79b с CD79b могут ингибировать экспрессию CD86 на B-клетках, стимулированных анти-IgM. Напротив, в случае сочетания для CD22 с CD22 наблюдали повышенные уровни экспрессии CD86.
Пример 5 - Ингибирующий эффект сочетаний для CD22 и CD79b может быть воспроизведен, только если антитела собраны в биспецифической ориентации
Человеческие МКПК, полученные из конусов при аферезе тромбоцитов, замораживали аликвотами. Перед проведением анализа клетки размораживали, промывали в DMEM (Life Technologies) и оставляли для акклиматизации в условиях 37°C/5% CO2. В это время готовили сочетания биспецифических, двухвалентных или смесей антител путем разбавления эквимолярных количеств (200 нМ) Fab'-A (Fab-scFv) и Fab'-B (Fab-пептид) с антигенной специфичностью в отношении белков клеточной поверхности CD22 и CD79b в среде DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Эти сочетания представлены в таблице 8. Для эксперимента с построением кривой титрования эти сочетания затем разбавляли 8-ступенчато с шагом разведения 1:2,5, выполняя титрование дозы для данного сочетания.
Таблица 8: Сетка биспецифических, двухвалентных или смесей антител со специфичностью в отношении CD22 и CD79b
где X представляет собой scFv (52SR4) и Y представляет собой пептид (GCN4).
Fab'A-X и/или Fab'B-Y инкубировали вместе в течение 90 минут (в условиях 37°C/5% CO2), а затем смешивали с 2,5×105 МКПК в 96-луночных планшетах с V-образным дном. Затем МКПК плюс сочетания Fab'A-X и/или Fab'B-Y инкубировали вместе в течение следующих 90 минут. После этого B-клетки активировали добавлением 200 нМ F(ab')2 козы против IgM человека (Southern Biotechnology) на 8 минут при температуре 37°C. Реакцию сигнализации затем останавливали добавлением равного объема буфера Cytofix (BD Biosciences). После этого планшеты оставляли при комнатной температуре на 15 минут, с последующим центрифугированием при 500 g в течение 5 минут. Отбрасывали лишний супернатант над осадком клеток, который ресуспендировали в проточном буфере (PBS+1% БСА+0,01% NaN3) и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут, с последующим двукратным промыванием в проточном буфере.
Вслед за этим клетки окрашивали флуоресцентно мечеными анти-CD20 антителом (BD Biosciences), анти-фосфо-Akt антителом, которое узнает модифицированный остаток серина в положении 473, и анти-фосфо-PLCγ2 антителом, которое узнает модифицированный остаток тирозина в положении 759. Затем клетки в планшетах ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 25 мкл проточного буфера. Измеряли экспрессию в клетках CD20, Akt и PLCγ2 с помощью проточного цитометра Intellicyt HTFC™.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни Akt и PLCγ2. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки). На фигурах 4 и 5 показано, что только биспецифическое сочетание антител для CD22 и CD79b, но не смеси антител для CD22 и CD79b ингибирует экспрессию фосфорилированных Akt и PLCγ2 (данные представляют собой среднее значение и планки погрешностей соответствуют 95% доверительным интервалам).
Для подтверждения ингибирования, наблюдаемого в случае биспецифического сочетания для CD22 и CD79b, данное сочетание, наряду со смесью антител для CD22 и CD79b, титровали и в B-клетках измеряли ингибирование общего внутриклеточного IkB (показатель сигнализации) и CD86 (маркер активации через 24 часа).
Как можно видеть на фигуре 6, сочетание для CD22-X/CD79b-Y, но не сочетание для CD22-X/CD79b-X, было способно ингибировать активацию сигнала NF-κB после стимуляции анти-IgM, что было измерено по уровню суммарного белка IkB. Значение IC50, экстраполированное путем построения 4-параметрической логистической кривой с использованием Graphpad Prism 6, составляло 7,5 нМ (данные представляют собой средние значения и планки погрешностей соответствуют стандартному отклонению). Кроме того, при титровании сочетание для CD22-X/CD79b-Y, но не сочетание для CD22-X/CD79b-X, было способно ингибировать индуцированную анти-IgM экспрессию CD86 на B-клетках через 24 часа (смотри фигуру 7).
Человеческие МКПК, полученные из конусов при аферезе тромбоцитов, замораживали аликвотами. Перед проведением анализа клетки размораживали, промывали в DMEM (Life Technologies) и оставляли для акклиматизации в условиях 37°C/5% CO2. В это время готовили биспецифические сочетания путем разбавления эквимолярных количеств (500 нМ) Fab'-X (Fab-scFv) и Fab'-Y (Fab-пептид) с антигенной специфичностью в отношении белков клеточной поверхности CD22 и CD79b в среде DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Эти сочетания затем разбавляли 8-ступенчато с шагом разведения 1:2,5, выполняя титрование дозы для данного сочетания. Fab'-X и Fab'-Y инкубировали вместе в течение 90 минут (в условиях 37 градусов C/5%CO2), с последующим добавлением 2,5×105 МКПК в 96-луночные планшеты с V-образным дном. МКПК затем добавляли к сочетаниям Fab'-X и Fab'-Y и инкубировали вместе в течение следующих 90 минут. После этого B-клетки активировали добавлением 200 нМ F(ab')2 козы против IgM человека (Southern Biotechnology) на 24 часа при температуре 37 градусов C. Для обнаружения маркеров активации на клеточной поверхности планшеты помещали на лед и промывали один раз ледяным проточным буфером (PBS+1% БСА+0,01% NaN3). Затем клетки окрашивали флуоресцентно меченым анти-CD19 антителом (BD Biosciences) и флуоресцентно меченым анти-CD86 антителом и инкубировали на льду в течение 1 часа в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 25 мкл проточного буфера. Измеряли экспрессию в клетках CD19 и CD86 с помощью проточного цитометра Intellicyt HTFC™. С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни CD86. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки). Как можно видеть на фигуре 7, при титровании сочетание для CD22-X/CD79b-Y было способно ингибировать индуцированную анти-IgM экспрессию CD86 на B-клетках через 24 часа. Значение IC50, экстраполированное путем построения 4-параметрической логистической кривой с использованием Graphpad Prism 6, составляло 10,3 нМ (данные представляют собой средние значения и планки погрешностей соответствуют стандартному отклонению).
Пример 6 - Ингибирующий эффект биспецифического белка для CD22 и CD79b может быть воспроизведен с другими V-областями антитела
Иммунизация: ДНК, кодирующую выбранные антигены, получали путем синтеза генов или из коммерческих источников и клонировали в экспрессионный вектор с сильным конститутивным промотором. Плазмидную ДНК затем трансфицировали в кроличьи фибробласты Rab-9 (ATCC® CRL-1414™) с использованием системы электропорации собственного изготовления. Через двадцать четыре часа клетки проверяли на экспрессию антигенов методом проточной цитометрии и замораживали аликвотами в жидком азоте до использования. До 6 антигенов были использованы для иммунизации каждого кролика либо за счет совместной экспрессии на одной и той же клетке, либо за счет создания смесей из однократно или множественно трансфицированных клеток. Кроликов иммунизировали 3 дозами клеток.
Обнаружение антител: B-клеточные культуры получали с использованием метода, аналогичного тому, который описан в статье Zubler et al. (1985). Вкратце, B-клетки, полученные из селезенки или МКПК от иммунизированных кроликов, культивировали при плотности примерно 2000-5000 клеток на лунку в 96-луночных культуральных планшетах с нанесенными штрих-кодами, содержащих по 200 мкл/лунке среду RPMI 1640 (Gibco BRL) с добавлением 10% ЭТС (PAA laboratories ltd), 2% HEPES (Sigma Aldrich), 1% L-глутамина (Gibco BRL), 1% раствора пенициллина/стрептомицина (Gibco BRL), 0,1% β-меркаптоэтанола (Gibco BRL), 3% культурального супернатанта активированных спленоцитов и гамма-облученных мутантных клеток EL4 мышиной тимомы (5×104/лунку), в течение семи дней при температуре 37°C в атмосфере 5% CO2.
Присутствие антиген-специфических антител в культуральных супернатантах B-клеток определяли с использованием гомогенного анализа связывания на основе флуоресценции, используя клетки HEK293, совместно трансфицированные антигенами, которыми были иммунизированы кролики. Скрининг включал перенос 10 мкл супернатанта из 96-луночных культуральных планшетов с нанесенными штрихкодами в закодированные штрихкодами 384-луночные аналитические планшеты с темными стенками, содержащие клетки HEK293, трансфицированные целевым антигеном (примерно 3000 клеток/лунку), с использованием устройства для манипуляций с жидкостями Matrix Platemate. Связывание обнаруживали при помощи Fcγ-специфичных антител козы против IgG кролика, конъюгированных с Cy-5 (Jackson). Планшеты просчитывали на устройстве на обнаружения клеток Applied Biosystems 8200.
После первичного скрининга положительные супернатанты объединяли в 96-луночных закодированных штрихкодами мастер-планшетах с использованием робота Aviso Onyx, отбирающего положительные образцы, и B-клетки в культуральных планшетах замораживали при -80°C. Затем проводили скрининг мастер-планшетов в гомогенном анализе связывания на основе флуоресценции, используя отдельно клетки HEK293, трансфицированные антигенами, и гранулы Superavidin™ (Bangs Laboratories), покрытые рекомбинантным белком, в качестве источника антигена. Это было сделано для определения антигенной специфичности для каждой лунки.
Чтобы иметь возможность извлечь гены вариабельной области антител из набора интересующих лунок, проводили этап деконволюции для идентификации антиген-специфических B-клеток в конкретной лунке, содержащей гетерогенную популяцию B-клеток. Это выполняли методом флуоресцентных очагов (Clargo et al., 2014, Mabs 2014 Jan 1: 6(1) 143-159; EP1570267B1). Вкратце, иммуноглобулин-секретирующие B-клетки из положительной лунки смешивали либо с клетками HEK293, трансфицированными целевым антигеном, либо с гранулами Superavidin™ (New England Biolabs), покрытыми биотинилированным целевым антигеном, и со специфическими в отношении Fcγ-фрагмента антителами козы против иммуноглобулинов кролика, конъюгированными с FITC, в конечном разведении 1:1200 (Jackson). После статической инкубации при температуре 37°C в течение 1 часа антиген-специфические B-клетки могли быть идентифицированы благодаря наличию флуоресцентного гало, окружающего такую B-клетку. Ряд таких отдельных B-клеточных клонов, идентифицированных при помощи микроскопа Olympus, затем отбирали микроманипулятором Эппендорф и помещали в пробирку для ПЦР. Метод флуоресцентных очагов также использовали для идентификации антиген-специфических B-клеток в гетерогенной популяции B-клеток непосредственно из костного мозга иммунизированных кроликов.
Гены вариабельной области антител получали из отдельных клеток методом ПЦР с обратной транскрипцией (ОТ-ПЦР) с использованием праймеров, специфичных для вариабельных областей тяжелой и легкой цепей. Проводили два раунда ПЦР с вложенной вторичной ПЦР, включающей сайты рестрикции на 3' и 5'-концах, позволяющие клонировать вариабельную область в экспрессионные векторы млекопитающих для мышиных Fab-X и Fab-Y (VH) или мышиной каппа (VL). Конструкты тяжелой и легкой цепей для экспрессионных векторов Fab-X и Fab-Y были совместно трансфицированы в клетки HEK-293 с использованием Fectin 293 (Life Technologies) или в клетки Expi293 с использованием Expifectamine (Life Technologies), и рекомбинантное антитело было экспрессировано в 6-луночных культуральных планшетах в объеме 5 мл. Через 5-7 дней экспрессии супернатанты собирали. Супернатанты тестировали в гомогенном анализе связывания на основе флуоресценции, используя клетки HEK293, трансфицированные антигеном, и гранулы Superavidin™ (Bangs Laboratories), покрытые рекомбинантным белком или трансфицированными антигеном клетками HEK. Данный эксперимент выполняли для подтверждения специфичности клонированных антител.
Мелкомасштабное производство Fab A-X и Fab B-Y (мелкомасштабная (50 мл) трансфекция Expi293)
Клетки Expi293 обычным образом субкультивировали в среде для экспрессии Expi293™ до конечной концентрации 0,5×106 жизнеспособных клеток/мл и инкубировали в инкубаторе с орбитальным шейкером (Multitron, Infors HT) при 120 об/мин в условиях 8% CO2 и 37°C.
В день трансфекции жизнеспособность и концентрацию клеток определяли с использованием автоматизированного устройства для счета клеток (Vi-CELL, Beckman Coulter). Для достижения конечной концентрации клеток 2,5×106 жизнеспособных клеток/мл соответствующий объем клеточной суспензии добавляли в стерильную 250-мл встряхиваемую колбу Эрленмейера и доводили объем до 42,5 мл добавлением свежей, предварительно подогретой среды Expi293™ для каждой трансфекции в объеме 50 мл.
Для получения комплексов липид-ДНК для каждой трансфекции в общей сложности 50 мкг плазмидной ДНК тяжелой цепи и легкой цепи разбавляли в среде Opti-MEM® I (LifeTechnologies) до общего объема 2,5 мл, и 135 мкл реагента Expifectamine™ 293 (LifeTechnologies) разбавляли в среде Opti-MEM® I до общего объема 2,5 мл. Все разведенные растворы осторожно перемешивали и инкубировали не более 5 минут при комнатной температуре перед тем, как каждый раствор ДНК добавляли к соответствующему разбавленному реагенту Expifectamine™ 293, получая общий объем 5 мл. Смеси ДНК с реагентом Expifectamine™ 293 осторожно перемешивали и инкубировали в течение 20-30 минут при комнатной температуре, давая возможность образовываться комплексам ДНК-Expifectamine™ 293.
После завершения инкубации комплекса ДНК-Expifectamine™ 293, 5 мл комплекса ДНК-Expifectamine™ 293 добавляли в каждую встряхиваемую колбу. Встряхиваемые колбы инкубировали в инкубаторе с орбитальным шейкером (Multitron, Infors HT) при 120 об/мин в условиях 8% CO2 и 37°C.
Через примерно 16-18 часов после трансфекции 250 мкл Expifectamine™ 293 усилителя трансфекции 1 (LifeTechnologies) и 2,5 мл Expifectamine™ 293 усилителя трансфекции 2 (LifeTechnologies) добавляли в каждую встряхиваемую колбу.
Клеточные культуры собирали через 7 дней после трансфекции. Клетки переносили в 50-мл центрифужные пробирки (Falcon) и центрифугировали в течение 30 минут при 4000 об/мин, с последующим стерильным фильтрованием через 0,22-мкм фильтр Stericup (Merck Millipore). Осветленные и стерильно профильтрованные супернатанты хранили при температуре 4°C. Конечные уровни экспрессии определяли методом ВЭЖХ с белком G.
Мелкомасштабная (50 мл) очистка: Как Fab-X, так и Fab-Y очищали отдельно методом аффинного захвата с использованием мелкомасштабной вакуумной системы очистки. Вкратце, 50 мл культуральных супернатантов стерильно фильтровали через 0,22-мкм фильтры, с последующим добавлением 500 мкл гранул Ni Sepharose (GE Healthcare). Смесь супернатантов и гранул затем обрабатывали во вращающемся барабане в течение примерно часа, с последующим удалением супернатанта вакуумной аспирацией. Затем гранулы промывали промывочной средой 1 (50 мМ фосфат натрия, 1 M NaCl, pH 6,2) и промывочной средой 2 (0,5 M NaCl). Элюцию выполняли 50 мМ ацетатом натрия, pH 4,0+1 M NaCl. В элюированных фракциях проводили замену буфера на PBS (Sigma), pH 7,4, с последующим фильтрованием через 0,22-мкм фильтр. Конечные пулы анализировали сканированием при A280, эксклюзионной ВЭЖХ (метод BEH200), SDS-ПААГ (в восстанавливающих и не восстанавливающих условиях), а также на эндотоксины с использованием системы PTS Endosafe.
Человеческие МКПК, полученные из конусов при аферезе тромбоцитов, замораживали аликвотами. Перед проведением анализа клетки размораживали, промывали в RPMI 1640 (Life Technologies) и оставляли для акклиматизации в условиях 37°C/5% CO2. В это время готовили сочетания биспецифических, двухвалентных или смесей антител путем разбавления эквимолярных количеств (200 нМ) Fab'-X (Fab-scFv) и/или Fab'-Y (Fab-пептид) с антигенной специфичностью в отношении белков клеточной поверхности CD22 и CD79b в среде RPMI 1640, содержащей 10% эмбриональной бычьей сыворотки, 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 2 мМ глутамина. Данные сочетания 3 разных Fab-Y для CD79b и 3 разных Fab-X для CD22 представлены в таблице 9.
Таблица 9: Сетка биспецифических белков со специфичностью в отношении CD22 и CD79b
где X представляет собой scFv (52SR4) и Y представляет собой пептид (GCN4).
Fab'A-X и Fab'B-Y инкубировали вместе в течение 60 минут (в условиях 37°C/5% CO2), а затем смешивали с 2,5×105 МКПК в 96-луночных планшетах с V-образным дном. Затем МКПК плюс сочетания Fab'A-X и/или Fab'B-Y инкубировали вместе в течение следующих 90 минут. После этого B-клетки активировали добавлением 12,5 мкг/мл F(ab')2 козы против IgM человека (Southern Biotechnology) на 10 минут при температуре 37°C. Реакцию сигнализации затем останавливали добавлением равного объема буфера Cytofix (BD Biosciences). После этого планшеты оставляли при комнатной температуре на 15 минут, с последующим центрифугированием при 500 g в течение 5 минут. Отбрасывали лишний супернатант над осадком клеток, который ресуспендировали в проточном буфере (PBS+1% БСА+0,01% NaN3+2 мМ ЭДТА) и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут, с последующим двукратным промыванием в проточном буфере.
Вслед за этим клетки окрашивали флуоресцентно мечеными анти-CD20 антителом (BD Biosciences) и анти-фосфо-PLCγ2 антителом, которое узнает модифицированный остаток тирозина в положении 759. Затем клетки в планшетах ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 40 мкл проточного буфера. Измеряли экспрессию в клетках CD20 и PLCγ2 с помощью проточного цитометра Intellicyt HTFC™.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни PLCγ2. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки).
Как можно видеть на фигуре 8, данные свидетельствуют о том, что сочетание для CD22 с CD79b при использовании совершенно разных V-областей антитела может ингибировать экспрессию фосфо-PLCγ2 в B-клетках, стимулированных анти-IgM.
Пример 7: Сканирование в формате сетки больших панелей гетеродимерно сплетенных белковых комплексов для выявления новых мишеней биспецифических антител
Введение: После успешной валидации биспецифического формата и метода скрининга в более ранних примерах, скрининг был использован для большего числа антигенных пар. Была получена панель пар вариабельных (V) областей антитела к 23 разным антигенам, экспрессируемым на B-клетках. С использованием формата Fab-Kd-Fab [то есть, A-X:Y-B, где A и B представляют собой Fab-фрагменты] была создана сетка гетеродимерно сплетенных белковых комплексов, представляющая множественные сочетания V-областей каждого из 315 сочетаний разных антигенных пар. Проводили скрининг этих сочетаний на их способность модулировать сигнализацию BCR (B-клеточного рецептора) методом проточной цитометрии с высокой пропускной способностью для выбора новых пар мишеней для взаимодействия с биспецифическим антителом.
Антитела выделяли, как описано в примере 6.
Анализы скрининга
Донорские МКПК быстро размораживали с использованием водяной бани, установленной на 37°C, и осторожно переносили в 50-мл пробирку Фалькона. Затем их разбавляли, добавляя среду для анализа до объема 5 мл по каплям, чтобы свести к минимуму осмотический шок. Затем клетки осторожно разбавляли до 20 мл, с последующим окончательным добавлением разбавителя до объема 50 мл. После этого клетки центрифугировали при 500 g в течение 5 минут, с последующим удалением супернатанта и ресуспендированием клеток в 1 мл среды. Затем клетки подсчитывали и разбавляли до концентрации 1,66×106 клеток/мл, с последующим внесением по 30 мкл на лунку в TC планшет с V-образным дном, что составляло конечную концентрацию в анализе 5,0×104 клеток/лунку. Планшет с клетками затем хранили накрытым в инкубаторе при температуре 37°C и 5% CO2 до использования, оставляя их в состоянии покоя минимум на 1 час.
Реагенты Fab-X и Fab-Y смешивали в эквимолярном соотношении при 5x конечной аналитической концентрации в среде для анализа и инкубировали в течение 90 минут в условиях 37°C, 5% CO2. Образцы готовили в 96-луночном полипропиленовом планшете с U-образным дном и накрывали в процессе инкубации.
10 мкл 5x смеси Fab-KD-Fab добавляли в соответствующие анализируемые лунки, содержащие клетки, и смешивали встряхиванием при 1000 об/мин в течение 30 секунд, с последующей инкубацией в течение 90 мин в условиях 37°C, 5% CO2.
Затем клетки стимулировали 10 мкл антител против IgM человека. Конечные концентрации стимула в анализе варьировались в зависимости от результатов анализа панелей, три коктейля из антител A, B и C (описаны ниже) стимулировали при конечной концентрации в анализе либо 50 мкг/мл (коктейли A и C), либо 25 мкг/мл (коктейль B). Содержимое аналитических планшетов затем осторожно перемешивали при 1000 об/мин в течение 30 секунд, с последующей инкубацией в условиях 37°C, 5% CO2 в течение 5 минут (коктейли антител A и C) или 2 минут (коктейль антител B). Анализ останавливали добавлением 150 мкл ледяного реагента BD CytoFix во все лунки и инкубировали в течение 15 минут при RT. Фиксированные клетки затем центрифугировали при 500 g в течение 5 минут для осаждения клеток и возможности удаления супернатанта при помощи устройства для промывки планшетов BioTek ELx405. Осадок ресуспендировали встряхиванием планшета на вортексе при 2400 об/мин в течение 30 секунд. После этого клетки пермеабилизировали при температуре 4°C добавлением 100 мкл ледяного буфера пермеабилизации BD Cell Permeabilisation Buffer III на 30 минут. Затем клетки промывали в 100 мкл FACS буфера и центрифугировали при 500 g в течение 5 минут. Супернатант вновь удаляли при помощи устройства ELx405, с последующим использованием его для быстрого внесения 200 мкл FACS буфера для промывания от любых остаточных количеств буфера пермеабилизации. Клетки вновь центрифугировали при 500 g и супернатант удаляли переворачиванием планшетов. Во время предыдущей стадии центрифугирования готовили коктейль антител в FACS буфере и хранили его, защищая от света. Затем клетки ресуспендировали встряхиванием планшета на вортексе (2400 об/мин, 30 секунд), с последующим добавлением 20 мкл коктейля антител во все лунки, после чего планшет встряхивали в течение 30 секунд при 1000 об/мин. Затем клетки инкубировали в течение 60 минут при RT в темноте.
Вслед за этим клетки промывали дважды в 200 мкл FACS буфера с центрифугированием при 500 g и удалением супернатанта после каждого этапа. В конечном итоге клетки ресуспендировали встряхиванием планшета на вортексе в течение 30 секунд при 2400 об/мин, с последующим добавлением последних 20 мкл FACS буфера. После этого планшет(ы) считывали на приборе Intellicyt HTFC/iQue.
FACS буфер=PBS+1% БСА+0,05% NaN3+2 мМ ЭДТА
Коктейль антител A=1:2 CD20 PerCp-Cy5,5 (BD Biosciences)+1:5 PLCγ2 AF88+1:10 Akt AF647+1:50 ERK1/2 PE (разбавление в FACS буфере).
Коктейль антител B=1:2 CD20 PerCp-Cy5,5 (BD Biosciences)+1:5 Syk PE+1:5 BLNK AF647 (разбавление в FACS буфере).
Коктейль антител C=1:5 CD20 PerCp-Cy5,5 (Biolegend)+1:5 PLCγ2 AF488+1:10 Akt AF647+1:5 Syk PE (разбавление в FACS буфере).
Проводили скрининг сочетаний Fab-X+Fab-Y либо с коктейлем антител A и B, либо только с C. Весь скрининг проводили на клетках, собранных из конусов (аферез), от 2 разных доноров крови. Данные собирали и оценивали с использованием коммерчески доступного программного обеспечения. Был проведен скрининг в общей сложности 2500 сочетаний Fab-X+Fab-Y к 315 разным сочетаниям антигенов.
Результаты
Был рассчитан процент ингибирования индукции фосфорилирования белков сигнального каскада BcR каждым из сочетаний Fab-Kd-Fab [то есть, A-X:Y-B, где A и B представляют собой Fab-фрагменты], в данном примере при поиске новых сочетаний антигенов, которые ингибируют функцию B-клеток, критерием для признания сочетания положительным являлось по меньшей мере 30% ингибирование по меньшей мере двух показателей фосфорилирования по меньшей мере сочетанием V-областей. С учетом таких пороговых показателей, 11 новых сочетаний пар антигенов из изученных 315 соответствовали необходимым критериям. Это соответствует коэффициенту успеха 3,5%, демонстрируя важность скрининга большого числа сочетаний для обнаружения тех, которые обладают желаемой активностью, и то, насколько редкой является активность сочетания для CC79b и CD22.
На фигурах 10-12 представлены данные для перекрестных специфичностей сетки антигенов. Значения соответствуют процентам ингибирования (отрицательное значение для активации) фосфорилирования Syk, PLCγ2 и AKT, соответственно, и представляют собой средние значения для нескольких оцененных сочетаний V-областей. Было протестировано 315 разных сочетаний антигенов и, как можно видеть, эффект разных сочетаний антител на сигнализацию BcR варьировался значительно от сильного ингибирования, например, антиген 2 (CD79b) на Fab-X в сочетании с антигеном 3 (CD22) на Fab-Y (69,66% ингибирование фосфо-Syk) и антиген 2 (CD79b) на Fab-Y в сочетании с антигеном 3 (CD22) на Fab-X (52,32% ингибирование фосфо-Syk), представлено на фигуре 11), до активации например, антиген 6 на X и антиген 11 на Y (минус 118,10% фосфо-Syk, фигура 11).
Каждая точка данных, представляющая средний % значений, представленных на фигурах 10-12, показана для антигена 2 (CD79b) на Fab-X и антигена 3 (CD22) на Fab-Y на фигуре 13. В этом случае оценивали 23 разных сочетания разных V-областей антитела. То же сочетание антигенов, но в альтернативной ориентации, то есть, антиген 2 (CD79b) на Fab-Y и антиген 3 (CD22) на Fab-X, показано на фигуре 14. В этом случае оценивали 9 разных сочетаний разных V-областей антитела. Все V-области демонстрировали ингибирование, однако преимуществом является то, что данный способ также можно использовать при выборе оптимальных сочетаний V-областей.
Пример 8 Сравнение активности совместного таргетирования антигена CD79b плюс антигена CD22 в скринируемом формате Fab-Kd-Fab и молекулярно связанном биспецифическом формате BYbe
Введение: Для проверки того, что активность в отношении целевой пары CD79b/CD22, обнаруженная в случае гетеродимерно сплетенного скринируемого комплекса Fab-Kd-Fab, может транслироваться в аналогичную желаемую активность в случае альтернативного терапевтического молекулярно связанного формата, специфичность в отношении антигена CD79b (VR4447) и специфичность в отношении антигена CD22 (VR4130) объединяли в формате BYbe. Данный формат BYbe состоит из V-областей против антигена CD22 (VR4130) в виде стабилизированной дисульфидными связями (ds) одноцепочечной (sc)-Fv, слитых с тяжелой цепью Fab против антигена CD79b (VR4447) через линкер SGGGGSGGGGS (SEQ ID NO: 17).
Методы:
Очистку BYbes для функционального скрининга проводили следующим образом:
Форматы для функционального скрининга BYbe (Fab-dsscFv [scFv с C-конца тяжелой цепи Fab]) очищали следующим образом. Осветленные супернатанты клеточных культур после стандартной экспрессии клеток expiHEK или CHO стерильно фильтровали через 0,22-мкм фильтры. Профильтрованные супернатанты наносили со скоростью 2 мл/мин на 50-мл колонки Gammabind Plus Sepharose XK26 (GE Healthcare), уравновешенные в PBS pH 7,4 (Sigma Aldrich Chemicals). После нанесения колонки промывали PBS pH 7,4, а затем элюировали 0,1 M раствором глицин/HCl, pH 2,7. Элюирование отслеживали по поглощению при 280 нм, элюируемый пик собирали и затем нейтрализовали 1/25 объема 2 M Tris/HCl, pH 8,5. Нейтрализованные образцы концентрировали с использованием устройств для концентрирования Amicon Ultra-15 с мембраной, имеющей порог отсечения по молекулярной массе 10 кДа (BYbes), и центрифугировали при 4000 x g в роторе со свободно подвешенными стаканами. Концентрированные образцы наносили на колонку либо XK16/60, либо XK26/60 Superdex 200 (GE Healthcare), уравновешенную в PBS, pH 7,4. Колонки промывали изократическим градиентом PBS, pH 7,4, со скоростью либо 1 мл/мин, либо 2,6 мл/мин, соответственно. Фракции собирали и анализировали методом эксклюзионной хроматографии на TSK геле G3000SWXL; колонку 5 мкм, 7,8 X 300 мм промывали изократическим градиентом 0,2 M фосфата, pH 7,0 со скоростью 1 мл/мин, с детекцией по поглощению при 280 нм. Выбранные фракции мономеров объединяли и концентрировали до концентрации >1 мг/мл с использованием устройства для концентрирования Amicon Ultra-15 с мембраной, имеющей порог отсечения по молекулярной массе 10 кДа, и центрифугировали при 4000 x g в роторе со свободно подвешенными стаканами. Конечные образцы анализировали: на концентрацию при помощи сканирующего в УФ-видимой области спектрофотометра (Cary 50Bio) при A280; на % мономера методом эксклюзионной хроматографии на TSK геле G3000SWXL; колонку 5 мкм, 7,8 X 300 мм промывали изократическим градиентом 0,2 M фосфата, pH 7,0, со скоростью 1 мл/мин, с детекцией по поглощению при 280 нм; методом SDS-ПААГ в восстанавливающих и не восстанавливающих условиях в 1,5-мм 4-20% гелях с Tris-глицином (Novex) при 50 мА (на гель) в течение 53 минут; и на эндотоксин при помощи переносной тест-системы EndoSafe® от компании Charles River, используя картриджи для тестирования с лизатом амебоцитов мечехвоста (LAL).
Функциональные анализы
Анализ на маркеры активации: Специфичные в отношении антигена CD79b Fab'-Y и специфичные в отношении антигена CD22 Fab'-X инкубировали вместе в течение 60 минут (в условиях 37°C и 5% CO2) в эквимолярных концентрациях. Сочетания титровали, начиная с молярности 100 нМ, в серийных разведениях 1:4. Специфичные в отношении антигенов CD79b и CD22 BYbe также титровали, начиная с молярности 100 нМ, в серийных разведениях 1:4. В лунки 96-луночных планшетов с V-образным дном вносили 1,5×105 МКПК, к ним добавляли титрованные сочетания Fab'-X и Fab'-Y или титрованные BYbe. Сочетания Fab'-X и Fab'-Y или BYbe инкубировали с клетками в течение следующих 90 минут. После этого B-клетки активировали добавлением 25 мкг/мл F(ab')2 козы против IgM человека (Southern Biotechnology) на 24 часа в условиях 37°C плюс 5% CO2.
В лунки добавляли 100 мкл ледяного FACS буфера (PBS+1% БСА+0,1% NaN3+2 мМ ЭДТА), планшеты запечатывали и засыпали мокрым льдом примерно на 15 минут, с последующим центрифугированием при 500 x g в течение 5 минут при температуре 4°C. Лишний супернатант с клеточных осадков отбрасывали и планшеты встряхивали при 2000 об/мин в течение 30 секунд.
Затем клетки окрашивали коктейлем из флуоресцентно меченых анти-CD19, анти-CD20 и анти-CD71, анти-CD40 и анти-CD86 антител (BD Biosciences). Планшеты быстро встряхивали и инкубировали в течение 1 часа на мокром льду в темноте. После этого планшеты промывали дважды и клетки ресуспендировли в 20 мкл FACS буфера. Измеряли экспрессию в клетках CD19, CD20 и CD71, CD40 и CD86 с использованием проточного цитометра Intellicyt iQUE®.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни CD71, CD40 и CD86. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки).
Анализ PhosFlow: Специфичные в отношении антигена CD79b Fab'-Y и специфичные в отношении антигена CD22 Fab'-X инкубировали вместе в течение 60 минут (в условиях 37°C и 5% CO2) в эквимолярных концентрациях. Сочетания титровали, начиная с молярности 100 нМ, в серийных разведениях 1:4. Специфичные в отношении антигена CD79b и антигена CD22 BYbe также титровали, начиная с молярности 100 нМ, в серийных разведениях 1:4. В лунки 96-луночных планшетов с V-образным дном вносили 5,0 x104 МКПК, к ним добавляли титрованные сочетания Fab'-X и Fab'-Y или титрованные BYbe. Сочетания Fab'-X и Fab'-Y или BYbe инкубировали с клетками в течение следующих 90 минут. После этого B-клетки активировали добавлением 25 мкг/мл F(ab')2 козы против IgM человека (Southern Biotechnology) на 15 минут в условиях 37°C плюс 5% CO2. Реакцию сигнализации затем останавливали добавлением равного объема буфера Cytofix (BD Biosciences). После этого планшеты оставляли при комнатной температуре на 15 минут, с последующим центрифугированием при 500 g в течение 5 минут. Отбрасывали лишний супернатант над осадком клеток, который ресуспендировали в FACS буфере (PBS+1% БСА+0,01% NaN3+2 мМ ЭДТА) и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) на 30 минут, с последующим двукратным промыванием в проточном буфере.
Затем клетки окрашивали флуоресцентно мечеными анти-CD20 антителом (BD Biosciences) и антителами против фосфорилированного PLCγ2, антителами против фосфорилированного Akt и антителами против фосфорилированного p38 (BD Biosciences). Затем клетки в планшетах ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и клетки ресуспендировали в 20 мкл FACS буфера. Измеряли экспрессию в клетках CD20 и фосфо-PLCγ2, фосфо-Akt и фосфо-p38 с использованием проточного цитометра Intellicyt iQUE®.
С использованием пакета программ для анализа данных Forecyt™ (Intellicyt) B-клетки были вычленены из других популяций клеток и для каждой лунки были рассчитаны геометрические средние уровни PLCγ2, Akt и p38. Затем все данные были выражены в виде процента ингибирования максимального ответа (только анти-IgM) за вычетом уровня фона (только клетки).
Результаты
Анализ PhosFlow: Данные на фигуре 15 показывают, что при таргетировании антигена CD79b и антигена CD22 любой из форматов Fab-Kd-Fab или BYbe может ингибировать фосфорилирование PLCγ2 в B-клетках, стимулированных анти-IgM. Данные на фигуре 16 показывают, что при таргетировании антигена CD79b и антигена CD22 любой из форматов Fab-Kd-Fab или BYbe может ингибировать фосфорилирование P38 в B-клетках, стимулированных анти-IgM. Данные на фигуре 17 показывают, что при таргетировании антигена CD79b и антигена CD22 любой из форматов Fab-Kd-Fab или BYbe может ингибировать фосфорилирование Akt в B-клетках, стимулированных анти-IgM.
Анализ на маркеры активации: Как можно видеть на фигуре 18, данные показывают, что при таргетировании антигена CD79b и антигена CD22 любой из форматов Fab-Kd-Fab или BYbe может ингибировать экспрессию CD71 на B-клетках, стимулированных анти-IgM. Данные на фигуре 19 показывают, что при таргетировании антигена CD79b и антигена CD22 любой из форматов Fab-Kd-Fab или BYbe может ингибировать экспрессию CD40 на B-клетках, стимулированных анти-IgM. Данные на фигуре 20 показывают, что при таргетировании антигена CD79b и антигена CD22 любой из форматов Fab-Kd-Fab или BYbe может ингибировать экспрессию CD86 на B-клетках, стимулированных анти-IgM.
Пример 9 - Сравнение активности совместного таргетирования антигена CD79b плюс антигена CD22 в формате молекулярно связанного биспецифического Bybe с активностью в случае добавления связывающего домена, направленного против альбумина, с целью продления времени полувыведения in vivo
Введение: Для проверки того, что активность в отношении целевой пары CD79b/CD22, обнаруженная в случае гетеродимерно сплетенного скринируемого комплекса Fab-Kd-Fab, может транслироваться в аналогичную желаемую активность в случае потенциального терапевтического молекулярно связанного формата с продленным периодом полувыведения in vivo за счет дополнительной направленности против альбумина, проводили слияние направленного против альбумина фрагмента антитела с легкой цепью Fab против антигена CD22 в формате BYbe, описанном в примере 8, через линкер, имеющий последовательность SGGGGSGGGGS (SEQ ID NO: 17). Специфичность в отношении антигена CD79b (VR4447) и специфичность в отношении антигена CD22 (VR4130 и VR4126) объединяли в формате Bybe с добавлением и без добавления фрагмента, направленного против альбумина (VR0645).
Описание конструктов, использованных в данном эксперименте.
Методы
Очистка BYbe с/без дополнительной специфичности против альбумина
Форматы BYbe (Fab-dsscFv [scFv от C-конца тяжелой цепи Fab]) и BYbe с фрагментом против альбумина (Fab-2xdsscFv [scFv от C-конца тяжелой цепи и легкой цепи Fab]) очищали следующим образом. Осветленные супернатанты клеточных культур после стандартной экспрессии клеток expiHEK или CHO стерильно фильтровали через 0,22-мкм фильтры. Профильтрованные супернатанты наносили со скоростью 2 мл/мин на 50-мл колонки Gammabind Plus Sepharose XK26 (GE Healthcare), уравновешенные в PBS, pH 7,4 (Sigma Aldrich Chemicals). После нанесения колонки промывали PBS, pH 7,4, а затем элюировали 0,1 M раствором глицин/HCl, pH 2,7. Элюирование отслеживали по поглощению при 280 нм, элюируемый пик собирали и затем нейтрализовали 1/25 объема 2 M Tris/HCl, pH 8,5. Нейтрализованные образцы концентрировали с использованием устройств для концентрирования Amicon Ultra-15 с мембраной, имеющей порог отсечения по молекулярной массе 10 кДа или 30 кДа, и центрифугировали при 4000 x g в роторе со свободно подвешенными стаканами. Концентрированные образцы наносили на колонку либо XK16/60, либо XK26/60 Superdex 200 (GE Healthcare), уравновешенную в PBS, pH 7,4. Колонки промывали изократическим градиентом PBS, pH 7,4, со скоростью либо 1 мл/мин, либо 2,6 мл/мин, соответственно. Фракции собирали и анализировали методом эксклюзионной хроматографии на TSK геле G3000SWXL; колонку 5 мкм, 7,8 X 300 мм промывали изократическим градиентом 0,2 M фосфата, pH 7,0, со скоростью 1 мл/мин, с детекцией по поглощению при 280 нм. Выбранные фракции мономеров объединяли и концентрировали до концентрации >1 мг/мл с использованием устройства для концентрирования Amicon Ultra-15 с мембраной, имеющей порог отсечения по молекулярной массе 10 кДа или 30 кДа, и центрифугировали при 4000 x g в роторе со свободно подвешенными стаканами. Конечные образцы анализировали: на концентрацию при помощи сканирующего в УФ-видимой области спектрофотометра (Cary 50Bio) при A280; на % мономера методом эксклюзионной хроматографии на TSK геле G3000SWXL; колонку 5 мкм, 7,8 X 300 мм промывали изократическим градиентом 0,2 M фосфата, pH 7,0, со скоростью 1 мл/мин, с детекцией по поглощению при 280 нм; методом SDS-ПААГ в восстанавливающих и не восстанавливающих условиях в 1,5-мм 4-20% гелях с Tris-глицином (Novex) при 50 мА (на гель) в течение 53 минут; и на эндотоксин при помощи переносной тест-системы EndoSafe® от компании Charles River, используя картриджи для тестирования с лизатом амебоцитов мечехвоста (LAL).
100 нМ очищенного белка каждого конструкта преинкубировали с человеческими МКПК, полученными от пяти отдельных доноров, в течение 60 минут в условиях 37 градусов C/5%CO2 в среде RMPI 1640 плюс 10% эмбриональной бычьей сыворотки и 2 мМ глютамакса (среда R10). Через 60 минут клетки стимулировали 25 мкг/мл анти-IgM антителом козы, предназначенным для стимуляции только B-клеток. Через 24 часа планшеты помещали на лед для остановки любой дальнейшей активации клеток, с последующим промыванием один раз ледяным буфером для проточной цитометрии (PBS+1% БСА+0,01% NaN3). Весь супернатант удаляли и осадки клеток ресуспендировали. Клетки помещали на лед и добавляли коктейль из анти-CD19, анти-CD20, анти-CD27, анти-CD71 и анти-CD86 антител. Клетки инкубировали в течение 60 минут, с последующим двукратным промыванием в буфере для проточной цитометрии. Данные по связыванию анти-CD27, анти-CD71 и анти-CD86 с положительными по CD19/CD20 B-клетками получали с использованием проточного цитометра iQUE с высокой пропускной способностью. Программное обеспечение Forecyt использовали для построения гистограмм и выведения геометрических средних показателей интенсивности для связывания анти-CD27, анти-CD71 и анти-CD86 антител с B-клетками. Эти данные были импортированы в Excel и значения процента ингибирования были рассчитаны для каждого сочетания. Затем данные были импортированы в Graphpad Prism и диаграммы размаха («ящик с усами») строили для каждого сочетания со средним значением, указанным знаком «+».
На фигуре 21 показано ингибирование экспрессии CD27 на B-клетках, индуцированное VR4447/VR4126 BYbe и VR4447/VR4126/VR645 BYbe/альбумин. Для пяти протестированных доноров оба конструкта показали неизменно сходные уровни ингибирования индуцированной анти-IgM экспрессии CD27. На фигуре 22 показано ингибирование экспрессии CD71 на B-клетках, индуцированное VR4447/VR4126 BYbe и VR4447/VR4126/VR645 BYbe/альбумин. Для пяти доноров оба конструкта показали неизменно сходные уровни ингибирования индуцированной анти-IgM экспрессии CD71. На фигуре 23 показано ингибирование экспрессии CD86 на B-клетках, индуцированное VR4447/VR4126 BYbe и VR4447/VR4126/VR645 BYbe/альбумин. Для пяти доноров оба конструкта показали неизменно сходные уровни ингибирования индуцированной анти-IgM экспрессии CD86.
На фигуре 24 показано ингибирование экспрессии CD27 на B-клетках, индуцированное VR4447/VR4130 BYbe и VR4447/VR4130/VR645 BYbe/альбумин. Для пяти протестированных доноров оба конструкта показали неизменно сходные уровни ингибирования индуцированной анти-IgM экспрессии CD27. На фигуре 25 показано ингибирование экспрессии CD71 на B-клетках, индуцированное VR4447/VR4130 BYbe и VR4447/VR4130/VR645 BYbe/альбумин. Для пяти доноров оба конструкта показали неизменно сходные уровни ингибирования индуцированной анти-IgM экспрессии CD71. На фигуре 26 показано ингибирование экспрессии CD86 на B-клетках, индуцированное VR4447/VR4130 BYbe и VR4447/VR4130/VR645 BYbe/альбумин. Для пяти доноров оба конструкта показали неизменно сходные уровни ингибирования индуцированной анти-IgM экспрессии CD86.
Пример 10 - Эффект совместного таргетирования антигена CD79b плюс антигена CD22 на функцию B-клеток памяти с использованием молекулярно связанных биспецифических Bybe с добавлением или без добавления фрагмента против альбумина
Введение: Для оценки того, оказывает ли таргетирование CD79b/CD22 функциональный эффект на B-клетки в долгосрочной культуре, измеряли продуцирование IgG B-клетками, культивируемыми в изоляции или в смешанной культуре МКПК. Количественное определение специфических антител к сенсибилизирующему антигену токсоида столбняка является показателем функции B-клеток памяти.
Специфичность в отношении антигена CD79b (VR4447) и специфичность в отношении антигена CD22 (VR4126, VR4127 и VR4130) объединяли в формате Bybe с добавлением и без добавления фрагмента, направленного против альбумина (VR0645). Фрагмент антитела против альбумина был слит с легкой цепью Fab для антигена CD22 в формате BYbe, как описано в примере 8, через линкер, имеющий последовательность SGGGGSGGGGS (SEQ ID NO: 17).
Описание конструктов, использованных в данном эксперименте
Методы
Очистка BYbe с/без дополнительной специфичности против альбумина
Форматы BYbe (Fab-dsscFv [scFv от C-конца тяжелой цепи Fab]) и BYbe с фрагментом против альбумина (Fab-2xdsscFv [scFvs от C-конца тяжелой цепи и легкой цепи Fab]) очищали, как описано в примере 9.
Активация B-клеток и определение специфических IgG в отношении токсоида столбняка
Человеческие МКПК или очищенные B-клетки, полученные от 3 отдельных доноров, стимулировали 500 нг/мл CD40L, 1 мкг/мл CpG и 50 нг/мл IL-21 в среде 1640 плюс 10% эмбриональной бычьей сыворотки и 2 мМ глютамакса (среда R10) в течение 6 дней. Конструкты очищенного белка добавляли в конечной концентрации 100 нМ в день 0 и оставляли в культуральной среде на всем протяжении анализа. Через 6 дней супернатанты собирали и количество IgG, специфичных в отношении токсоида столбняка, определяли методом ELISA. Вкратце, лунки планшетов для ELISA Maxisorp с половинчатыми лунками (Nunc) покрывали раствором токсоида столбняка в PBS с концентрацией 10 мкг/мл в течение ночи при температуре 4°C. Затем планшеты блокировали 5% раствором молока в PBS, содержащем 0,05% Tween 20, в течение 2 часов. Супернатанты разбавляли и затем добавляли в лунки на 2 часа при комнатной температуре. Планшеты промывали PBS-0,05% Tween 20 и связавшееся с токсоидом столбняка антитело обнаруживали с использованием конъюгированных с пероксидазой антител козы против IgG (H+L) человека, разбавленных до концентрации 1 мкг/мл в 5% растворе молока в PBS-0,05% Tween 20. Развитие окраски в планшетах вызывали добавлением раствора субстрата TMB (KPL) и поглощение измеряли при 450 нМ на микропланшетном ридере Synergy 2 (Biotek). Данные экспортировали в Excel и процент ингибирования рассчитывали относительно клеток, культивируемых без тестируемых антител. Затем данные импортировали в Graphpad Prism® и использовали для построения гистограмм.
На фигуре 27 показано ингибирование продуцирования IgG против токсоида столбняка в клетках МКПК, культивируемых с VR4447/VR4126 BYbe, VR4447/VR4127 BYbe и VR4447/VR4130 BYbe. Данные представляют собой объединенные данные для 3 доноров.
На фигуре 28 показано ингибирование продуцирования IgG против токсоида столбняка в очищенных B-клетках, культивируемых с VR4447/VR4126 BYbe, VR4447/VR4127 BYbe и VR4447/VR4130 BYbe. Данные представляют собой объединенные данные для 2 доноров.
На фигуре 29 показано ингибирование продуцирования IgG против токсоида столбняка или в МКПК, или в очищенных B-клетках, культивируемых с VR4447/VR4126 BYbe, VR4447/VR4127 BYbe, VR4447/VR4130 BYbe, VR4447/VR4126/VR645 BYbe/альбумин и VR4447/VR4130/VR645 BYbe/альбумин. Приведены данные для одного донора.
Пример 11 - Нарушение регуляции сигнализации BcR в B-клетках пациента с SLE и эффект совместного таргетирования антигена CD79b плюс антигена CD22 на функцию B-клеток при SLE
Введение: Для оценки того, можно ли использовать сочетание для CD79b/CD22 в лечении людей с аутоиммунными заболеваниями, авторы изобретения использовали B-клетки от пациентов с системной красной волчанкой (SLE) в качестве модельной системы. Тестировали влияние сочетания для CD79b/CD22 (VR4447/VR4130) на статус активации сигнальных белков, которые, как известно, вовлечены в функцию B-клеток, но их регуляция нарушена у пациентов с SLE, в сравнении со здоровыми добровольцами. В данном эксперименте B-клетки от 12 пациентов с SLE и 12 здоровых добровольцев сравнивали в отношении эффекта, который оказывало совместное таргетирование CD79b и CD22 на статус активации белков.
Методы:
Анализ PhosFlow: Во всех анализах использовали 2×105 МКПК на лунку.
В обработанных образцах специфические в отношении антигена CD79b и антигена CD22 BYbe тестировали в концентрации 100 нМ. МКПК как от здоровых добровольцев, так и от пациентов с SLE, преинкубировали с BYbe в течение 90 минут при температуре 37°C. В необработанных образцах BYbe просто не добавляли во время этого периода инкубации. После этого клетки активировали 25 мкг/мл F(ab')2 козы против IgM человека (Southern Biotechnology) в течение 10 минут в условиях 37°C плюс 5% CO2 и реакцию останавливали добавлением фиксатора (Cytofix - BD Biosciences). В не стимулированных образцах антитела против IgM человека просто не добавляли во время этого периода инкубации. Через 15 минут при комнатной температуре клетки осаждали (500 x g в течение 5 минут) и затем ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences), с последующим двукратным промыванием в проточном буфере (PBS+1% БСА+0,01% NaN3+2 мМ ЭДТА). Затем клетки окрашивали анти-CD20, антителами против фосфорилированного (p) NF-κB, анти-pSyk, анти-pAtk и анти-pErk 1&2 антителами и инкубировали при комнатной температуре в темноте в течение одного часа. В завершение планшеты промывали дважды в проточном буфере, с последующим измерением на проточном цитометре iQUE (Intellicyt). Затем рассчитывали геометрические средние значения (средняя интенсивность флуоресценции, СИФ) для экспрессии pNF-κB, pSyk, pAkt и pErk 1&2 в B-клетках и выражали результаты в графической форме.
Результаты:
На фигуре 30 показано, что базовый уровень фосфорилирования NF-κB, Syk, Akt и Erk 1&2 (в отсутствие стимуляции и обработки) повышен в B-клетках у пациентов с SLE по сравнению с таковым у здоровых добровольцев.
На фигурах 31-34 показано, что BYbe для CD79/CD22 может в равной степени ингибировать pNF-κB, pSyk, pAkt и pErk 1&2 у здоровых добровольцев и у пациентов с SLE.
Выводы:
Полученные данные свидетельствуют о том, что B-клетки от пациентов с SLE являются активированными до какой-либо стимуляции in vitro, в отличие от клеток здоровых добровольцев. При стимуляции клеток через B-клеточный рецептор как у здоровых добровольцев, так и у пациентов с SLE, наблюдали повышение уровней активации по сравнению с фоновым сигналом. Как у здоровых добровольцев, так и у пациентов с SLE, этот сигнал в значительной степени был блокирован сочетанием для CD79b/CD22. Эти данные указывают на то, что сочетание для CD79b/CD22 может ингибировать B-клетки как у здоровых добровольцев, так и у людей, страдающих аутоиммунным заболеванием, свидетельствуя о том, что этот путь имеет принципиальное значение для активации B-клеток.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCB Pharma
<120> МОЛЕКУЛЫ СО СПЕЦИФИЧНОСТЬЮ ДЛЯ CD79 и CD22
<130> G0224_WO
<150> GB1412658.5
<151> 2014-07-16
<160> 163
<170> PatentIn version 3.5
<210> 1
<211> 44
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность GCN4(7P14P)
<400> 1
Ala Ser Gly Gly Gly Arg Met Lys Gln Leu Glu Pro Lys Val Glu Glu
1 5 10 15
Leu Leu Pro Lys Asn Tyr His Leu Glu Asn Glu Val Ala Arg Leu Lys
20 25 30
Lys Leu Val Gly Glu Arg His His His His His His
35 40
<210> 2
<211> 132
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность GCN4(7P14P)
<400> 2
gctagcggag gcggaagaat gaaacaactt gaacccaagg ttgaagaatt gcttccgaaa 60
aattatcact tggaaaatga ggttgccaga ttaaagaaat tagttggcga acgccatcac 120
catcaccatc ac 132
<210> 3
<211> 262
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность 52SR4 dsscFv
<400> 3
Asp Ala Val Val Thr Gln Glu Ser Ala Leu Thr Ser Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Ser Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Asn Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Val Leu Trp Tyr Ser Asp
85 90 95
His Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Ser Asp Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Ala Pro
130 135 140
Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Leu Leu Thr
145 150 155 160
Asp Tyr Gly Val Asn Trp Val Arg Gln Ser Pro Gly Lys Cys Leu Glu
165 170 175
Trp Leu Gly Val Ile Trp Gly Asp Gly Ile Thr Asp Tyr Asn Ser Ala
180 185 190
Leu Lys Ser Arg Leu Ser Val Thr Lys Asp Asn Ser Lys Ser Gln Val
195 200 205
Phe Leu Lys Met Asn Ser Leu Gln Ser Gly Asp Ser Ala Arg Tyr Tyr
210 215 220
Cys Val Thr Gly Leu Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
225 230 235 240
Val Ser Ser Ala Ala Ala His His His His His His Glu Gln Lys Leu
245 250 255
Ile Ser Glu Glu Asp Leu
260
<210> 4
<211> 792
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность 52SR4 dsscFv
<400> 4
gatgcggtgg tgacccagga aagcgcgctg accagcagcc cgggcgaaac cgtgaccctg 60
acctgccgca gcagcaccgg cgcggtgacc accagcaact atgcgagctg ggtgcaggaa 120
aaaccggatc atctgtttac cggcctgatt ggcggcacca acaaccgcgc gccgggcgtg 180
ccggcgcgct ttagcggcag cctgattggc gataaagcgg cgctgaccat taccggcgcg 240
cagaccgaag atgaagcgat ttatttttgc gtgctgtggt atagcgacca ttgggtgttt 300
ggctgcggca ccaaactgac cgtgctgggt ggaggcggtg gctcaggcgg aggtggctca 360
ggcggtggcg ggtctggcgg cggcggcagc gatgtgcagc tgcagcagag cggcccgggc 420
ctggtggcgc cgagccagag cctgagcatt acctgcaccg tgagcggctt tctcctgacc 480
gattatggcg tgaactgggt gcgccagagc ccgggcaaat gcctggaatg gctgggcgtg 540
atttggggcg atggcattac cgattataac agcgcgctga aaagccgcct gagcgtgacc 600
aaagataaca gcaaaagcca ggtgtttctg aaaatgaaca gcctgcagag cggcgatagc 660
gcgcgctatt attgcgtgac cggcctgttt gattattggg gccagggcac caccctgacc 720
gtgagcagcg cggccgccca tcaccatcac catcacgaac agaaactgat tagcgaagaa 780
gatctgtaat ag 792
<210> 5
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 5
Asp Lys Thr His Thr Cys Ala Ala
1 5
<210> 6
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 6
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10
<210> 7
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 7
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Thr Cys Pro Pro Cys
1 5 10 15
Pro Ala
<210> 8
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 8
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Thr Cys Pro Pro Cys
1 5 10 15
Pro Ala Thr Cys Pro Pro Cys Pro Ala
20 25
<210> 9
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 9
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Gly Lys Pro Thr Leu
1 5 10 15
Tyr Asn Ser Leu Val Met Ser Asp Thr Ala Gly Thr Cys Tyr
20 25 30
<210> 10
<211> 31
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 10
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Gly Lys Pro Thr His
1 5 10 15
Val Asn Val Ser Val Val Met Ala Glu Val Asp Gly Thr Cys Tyr
20 25 30
<210> 11
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 11
Asp Lys Thr His Thr Cys Cys Val Glu Cys Pro Pro Cys Pro Ala
1 5 10 15
<210> 12
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 12
Asp Lys Thr His Thr Cys Pro Arg Cys Pro Glu Pro Lys Ser Cys Asp
1 5 10 15
Thr Pro Pro Pro Cys Pro Arg Cys Pro Ala
20 25
<210> 13
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Шарнирный линкер
<400> 13
Asp Lys Thr His Thr Cys Pro Ser Cys Pro Ala
1 5 10
<210> 14
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 14
Ser Gly Gly Gly Gly Ser Glu
1 5
<210> 15
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 15
Asp Lys Thr His Thr Ser
1 5
<210> 16
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 16
Ser Gly Gly Gly Gly Ser
1 5
<210> 17
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 17
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 18
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 18
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 19
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 19
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Gly Ser
20
<210> 20
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 20
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 21
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 21
Ala Ala Ala Gly Ser Gly Gly Ala Ser Ala Ser
1 5 10
<210> 22
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> разное
<222> (7)..(7)
<223> Xaa может быть природной аминокислотой
<400> 22
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
1 5 10 15
<210> 23
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> разное
<222> (7)..(7)
<223> Xaa может быть природной аминокислотой
<220>
<221> разное
<222> (12)..(12)
<223> Xaa может быть природной аминокислотой
<400> 23
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1 5 10 15
Gly Ala Ser Ala Ser
20
<210> 24
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> разное
<222> (7)..(7)
<223> Xaa может быть природной аминокислотой
<220>
<221> разное
<222> (12)..(12)
<223> Xaa может быть природной аминокислотой
<220>
<221> разное
<222> (17)..(17)
<223> Xaa может быть природной аминокислотой
<400> 24
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1 5 10 15
Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
20 25
<210> 25
<211> 31
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> разное
<222> (7)..(7)
<223> Xaa может быть природной аминокислотой
<220>
<221> разное
<222> (12)..(12)
<223> Xaa может быть природной аминокислотой
<220>
<221> разное
<222> (17)..(17)
<223> Xaa может быть природной аминокислотой
<220>
<221> разное
<222> (22)..(22)
<223> Xaa может быть природной аминокислотой
<400> 25
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1 5 10 15
Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
20 25 30
<210> 26
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> разное
<222> (7)..(7)
<223> Xaa может быть природной аминокислотой
<400> 26
Ala Ala Ala Gly Ser Gly Xaa Ser Gly Ala Ser Ala Ser
1 5 10
<210> 27
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 27
Pro Gly Gly Asn Arg Gly Thr Thr Thr Thr Arg Arg Pro Ala Thr Thr
1 5 10 15
Thr Gly Ser Ser Pro Gly Pro Thr Gln Ser His Tyr
20 25
<210> 28
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 28
Ala Thr Thr Thr Gly Ser Ser Pro Gly Pro Thr
1 5 10
<210> 29
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 29
Ala Thr Thr Thr Gly Ser
1 5
<210> 30
<211> 0
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 30
000
<210> 31
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 31
Glu Pro Ser Gly Pro Ile Ser Thr Ile Asn Ser Pro Pro Ser Lys Glu
1 5 10 15
Ser His Lys Ser Pro
20
<210> 32
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 32
Gly Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
1 5 10 15
<210> 33
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 33
Gly Gly Gly Gly Ile Ala Pro Ser Met Val Gly Gly Gly Gly Ser
1 5 10 15
<210> 34
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 34
Gly Gly Gly Gly Lys Val Glu Gly Ala Gly Gly Gly Gly Gly Ser
1 5 10 15
<210> 35
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 35
Gly Gly Gly Gly Ser Met Lys Ser His Asp Gly Gly Gly Gly Ser
1 5 10 15
<210> 36
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 36
Gly Gly Gly Gly Asn Leu Ile Thr Ile Val Gly Gly Gly Gly Ser
1 5 10 15
<210> 37
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 37
Gly Gly Gly Gly Val Val Pro Ser Leu Pro Gly Gly Gly Gly Ser
1 5 10 15
<210> 38
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 38
Gly Gly Glu Lys Ser Ile Pro Gly Gly Gly Gly Ser
1 5 10
<210> 39
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 39
Arg Pro Leu Ser Tyr Arg Pro Pro Phe Pro Phe Gly Phe Pro Ser Val
1 5 10 15
Arg Pro
<210> 40
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 40
Tyr Pro Arg Ser Ile Tyr Ile Arg Arg Arg His Pro Ser Pro Ser Leu
1 5 10 15
Thr Thr
<210> 41
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 41
Thr Pro Ser His Leu Ser His Ile Leu Pro Ser Phe Gly Leu Pro Thr
1 5 10 15
Phe Asn
<210> 42
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 42
Arg Pro Val Ser Pro Phe Thr Phe Pro Arg Leu Ser Asn Ser Trp Leu
1 5 10 15
Pro Ala
<210> 43
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 43
Ser Pro Ala Ala His Phe Pro Arg Ser Ile Pro Arg Pro Gly Pro Ile
1 5 10 15
Arg Thr
<210> 44
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 44
Ala Pro Gly Pro Ser Ala Pro Ser His Arg Ser Leu Pro Ser Arg Ala
1 5 10 15
Phe Gly
<210> 45
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 45
Pro Arg Asn Ser Ile His Phe Leu His Pro Leu Leu Val Ala Pro Leu
1 5 10 15
Gly Ala
<210> 46
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 46
Met Pro Ser Leu Ser Gly Val Leu Gln Val Arg Tyr Leu Ser Pro Pro
1 5 10 15
Asp Leu
<210> 47
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 47
Ser Pro Gln Tyr Pro Ser Pro Leu Thr Leu Thr Leu Pro Pro His Pro
1 5 10 15
Ser Leu
<210> 48
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 48
Asn Pro Ser Leu Asn Pro Pro Ser Tyr Leu His Arg Ala Pro Ser Arg
1 5 10 15
Ile Ser
<210> 49
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 49
Leu Pro Trp Arg Thr Ser Leu Leu Pro Ser Leu Pro Leu Arg Arg Arg
1 5 10 15
Pro
<210> 50
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 50
Pro Pro Leu Phe Ala Lys Gly Pro Val Gly Leu Leu Ser Arg Ser Phe
1 5 10 15
Pro Pro
<210> 51
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 51
Val Pro Pro Ala Pro Val Val Ser Leu Arg Ser Ala His Ala Arg Pro
1 5 10 15
Pro Tyr
<210> 52
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 52
Leu Arg Pro Thr Pro Pro Arg Val Arg Ser Tyr Thr Cys Cys Pro Thr
1 5 10 15
Pro
<210> 53
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 53
Pro Asn Val Ala His Val Leu Pro Leu Leu Thr Val Pro Trp Asp Asn
1 5 10 15
Leu Arg
<210> 54
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 54
Cys Asn Pro Leu Leu Pro Leu Cys Ala Arg Ser Pro Ala Val Arg Thr
1 5 10 15
Phe Pro
<210> 55
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 55
Asp Leu Cys Leu Arg Asp Trp Gly Cys Leu Trp
1 5 10
<210> 56
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 56
Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10
<210> 57
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 57
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Gly Asp
1 5 10 15
<210> 58
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 58
Gln Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Glu Asp Asp Glu
20
<210> 59
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 59
Gln Gly Leu Ile Gly Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Gly Arg Ser Val
20
<210> 60
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 60
Gln Gly Leu Ile Gly Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1 5 10 15
Gly Arg Ser Val Lys
20
<210> 61
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 61
Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp Asp
1 5 10 15
<210> 62
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 62
Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu
1 5 10 15
Asp Asp
<210> 63
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 63
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp Asp
1 5 10 15
<210> 64
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 64
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp
1 5 10 15
<210> 65
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 65
Arg Leu Met Glu Asp Ile Cys Leu Ala Arg Trp Gly Cys Leu Trp Glu
1 5 10 15
Asp Asp
<210> 66
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 66
Glu Val Arg Ser Phe Cys Thr Arg Trp Pro Ala Glu Lys Ser Cys Lys
1 5 10 15
Pro Leu Arg Gly
20
<210> 67
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 67
Arg Ala Pro Glu Ser Phe Val Cys Tyr Trp Glu Thr Ile Cys Phe Glu
1 5 10 15
Arg Ser Glu Gln
20
<210> 68
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептид, связывающий альбумин
<400> 68
Glu Met Cys Tyr Phe Pro Gly Ile Cys Trp Met
1 5 10
<210> 69
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> жесткие линкеры
<400> 69
Gly Ala Pro Ala Pro Ala Ala Pro Ala Pro Ala
1 5 10
<210> 70
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> жесткий линкер
<400> 70
Pro Pro Pro Pro
1
<210> 71
<211> 113
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4447
<400> 71
Ala Gln Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Pro Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Val Ser Gly
20 25 30
Asn Tyr Leu Ala Trp Leu Gln Gln Lys Pro Gly Gln Pro Pro Lys Gln
35 40 45
Leu Ile His Ser Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Glu Phe Ser Cys
85 90 95
Ser Ser His Asp Cys Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val
100 105 110
Lys
<210> 72
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4447
<400> 72
gcccaagtgc tgacccagac tccgtcccct gtgtctgcac ctgtgggagg cacagtcacc 60
atcaattgcc aggccagtca gagtgttgtt agtggcaatt acctagcctg gcttcagcag 120
aaaccagggc agcctcccaa gcaactgatc cattctgcat ccactctggc atctggggtc 180
tcatcgcggt tcagcggcag tggatctggg acacaattca ctctcaccat cagcggcgtg 240
cagtgtgaag atgctgccac ttactactgt ctaggcgaat ttagttgtag tagtcatgat 300
tgtaatgctt tcggcggagg gaccgaggtg gtggtcaaa 339
<210> 73
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4447
<400> 73
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Asn Tyr Ala
20 25 30
Val Ser Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Tyr Ile Glu Thr Gly Thr Thr Trp Tyr Ala Asn Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Thr Ile
65 70 75 80
Thr Ser Pro Ser Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Glu
85 90 95
Pro Tyr Glu Pro Tyr Asp Asp Ser Asn Ile Tyr Tyr Gly Met Asp Pro
100 105 110
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 74
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4447
<400> 74
cagtcgctgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcaccgtct ctggattctc cctcagtaac tatgcagtaa gctgggtccg ccaggctcca 120
ggggagggac tggaatggat cgggatcatt tatattgaaa ctggtaccac atggtacgcg 180
aactgggcga aaggccgatt caccatctcc aaaacctcga ccacggtgga tctgacaatc 240
accagtccgt caaccgagga cacggccacc tatttctgtg ccagagaacc ttatgaacct 300
tatgatgata gtaatattta ctacggcatg gacccctggg gcccaggcac cctcgtcacc 360
gtctcgagt 369
<210> 75
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 75
Gln Ala Ser Gln Ser Val Val Ser Gly Asn Tyr Leu Ala
1 5 10
<210> 76
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 76
Ser Ala Ser Thr Leu Ala Ser
1 5
<210> 77
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 77
Leu Gly Glu Phe Ser Cys Ser Ser His Asp Cys Asn Ala
1 5 10
<210> 78
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 78
Gly Phe Ser Leu Ser Asn Tyr Ala Val Ser
1 5 10
<210> 79
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 79
Ile Ile Tyr Ile Glu Thr Gly Thr Thr Trp Tyr Ala Asn Trp Ala Lys
1 5 10 15
Gly
<210> 80
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 80
Glu Pro Tyr Glu Pro Tyr Asp Asp Ser Asn Ile Tyr Tyr Gly Met Asp
1 5 10 15
Pro
<210> 81
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4450
<400> 81
Ala Ile Asp Met Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Ile Tyr Asn Asn
20 25 30
Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Glu Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Gly Ser Gly
85 90 95
Gly Asp Gly Ile Ala Phe Gly Gly Gly Thr Lys Val Val Val Glu
100 105 110
<210> 82
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4450
<400> 82
gccattgata tgacccagac tccatccccc gtgtctgcag ctgtgggagg cacagtcacc 60
atcaattgcc agtccagtca gagtatttat aataataatg acttagcctg gtatcagcag 120
aaaccagggc agcctcccaa gctcctgatc tacgaagcat ccaaactggc atctggggtc 180
ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagtggcgtg 240
cagtgtgatg atgctgccac ttactactgt cagggcggtg gtagtggtgg tgatggcatt 300
gctttcggcg gagggaccaa ggtggtcgtc gaa 333
<210> 83
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4450
<400> 83
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Ala Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Asn Asn Tyr Val
20 25 30
Met Val Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Tyr Val Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Val Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp Ala
85 90 95
Gly His Ser Asp Val Asp Val Leu Asp Ile Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 84
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4450
<400> 84
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg gggcacccct gacactcacc 60
tgcacagtct ctggattctc cctcaataac tatgtaatgg tctgggtccg ccaggctcca 120
gggaaggggc tggaatggat cggaatcatt tatgttagtg gtaatgcata ctacgcgagc 180
tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaagtgacc 240
agtctgacaa ccgaggacac ggccacctat ttctgtgcca gagatgctgg tcatagtgat 300
gtcgatgttt tggatatttg gggcccgggc accctcgtca ccgtctcgag t 351
<210> 85
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 85
Gln Ser Ser Gln Ser Ile Tyr Asn Asn Asn Asp Leu Ala
1 5 10
<210> 86
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 86
Glu Ala Ser Lys Leu Ala Ser
1 5
<210> 87
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 87
Gln Gly Gly Gly Ser Gly Gly Asp Gly Ile Ala
1 5 10
<210> 88
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 88
Gly Phe Ser Leu Asn Asn Tyr Val Met Val
1 5 10
<210> 89
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 89
Ile Ile Tyr Val Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 90
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 90
Asp Ala Gly His Ser Asp Val Asp Val Leu Asp Ile
1 5 10
<210> 91
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4120
<400> 91
Ala Phe Glu Leu Ser Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Ile Ser Thr Ala
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Arg Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Tyr Tyr Gly Thr Ser Ser
85 90 95
Gly Gly Ser Trp Ala Phe Gly Gly Gly Thr Lys Val Val Val Lys
100 105 110
<210> 92
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4120
<400> 92
gcattcgaat tgagccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc 60
atcaagtgcc aggccagtca gagcattagc actgcattag cctggtatca gcagaaacca 120
gggcagcgtc ccaagctcct gatctatggt gcatccactc tggcatctgg ggtctcatcg 180
cggttcaaag gcagtggatc tgggacagag ttcactctca ccatcagcga cctggagtgt 240
gccgatgctg ccacttacta ctgtcaaagc tattatggta cgagtagtgg tggttcttgg 300
gctttcggcg gagggaccaa ggtggtcgtc aaa 333
<210> 93
<211> 124
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4120
<400> 93
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Ser Ser Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Ile Tyr Thr Gly Ser Ser Gly Asp Thr Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Ser
65 70 75 80
Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Gly Pro Tyr Val Gly Tyr Gly Tyr Asp Leu Gln Tyr Leu Tyr
100 105 110
Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 94
<211> 372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4120
<400> 94
cagtcattgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc 60
tgcacagcct ctggattctc cttcagtagt agctactaca tgtgctgggt ccgccagtct 120
ccagggaagg ggctggagtg gatcgcatgc atttatactg gtagtagtgg tgacacttac 180
tacgcgagct gggcgaaagg ccgattcacc atctccaaaa cctcgtcgac cacggtgtct 240
ctgcaaatga ccagtctgac agccgcggac acggccactt atttctgtgc gagagggcct 300
tatgttggtt atggttatga tcttcaatac ttgtacttgt ggggcccggg gaccctcgtc 360
accgtctcga gt 372
<210> 95
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 95
Gln Ala Ser Gln Ser Ile Ser Thr Ala Leu Ala
1 5 10
<210> 96
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 96
Gly Ala Ser Thr Leu Ala Ser
1 5
<210> 97
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 97
Gln Ser Tyr Tyr Gly Thr Ser Ser Gly Gly Ser Trp Ala
1 5 10
<210> 98
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 98
Gly Phe Ser Phe Ser Ser Ser Tyr Tyr Met Cys
1 5 10
<210> 99
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 99
Cys Ile Tyr Thr Gly Ser Ser Gly Asp Thr Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 100
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 100
Gly Pro Tyr Val Gly Tyr Gly Tyr Asp Leu Gln Tyr Leu Tyr Leu
1 5 10 15
<210> 101
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4126
<400> 101
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Gln Asn Ile Gly Ser Gly
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ser His Asp Tyr Ser Ser Val
85 90 95
Arg Ser Tyr Gly Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 102
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4126
<400> 102
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc 60
atcaagtgcc aggccagtca gaacattggt agtggtttag cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctattat gcatccactc tggcatctgg ggtcccatca 180
aggttcaaag gcagtggatc tgggacacag ttcactctca ccatcagcga cctggagtgt 240
gccgacgctg ccacttacta ctgtcaaagt catgattata gtagtgttcg gagttacggt 300
aatgctttcg gcggagggac cgaggtggtg gtcaaa 336
<210> 103
<211> 126
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4126
<400> 103
Gln Gln His Leu Glu Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Thr Leu Thr Leu Thr Cys Lys Ala Ser Gly Ile Asp Phe Ser Ser Tyr
20 25 30
Tyr Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Val Ala Cys Ile Asp Pro Ala Ser Ser Gly Thr Thr Tyr Tyr Ala Thr
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val
65 70 75 80
Thr Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe
85 90 95
Cys Ala Arg Ala Tyr Gly Ser Gly Gly Ser Gly Tyr Ile Gly Cys Tyr
100 105 110
Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 104
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4126
<400> 104
cagcagcacc tggaggagtc cgggggaggc ctggtcaagc ctggaggaac cctgacactc 60
acctgcaaag cctctggaat cgacttcagt agctactact acatgtgctg ggtccgccag 120
gctccaggga aggggctgga gtgggtcgcg tgcattgatc ctgctagtag tggtactact 180
tactacgcga cctgggcgaa aggccgattc accatctcca aaacctcgtc gaccacggtg 240
actctgcaaa tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagggca 300
tatggtagtg ggggtagtgg ttatataggg tgctactttg acttgtgggg ccaaggcacc 360
ctcgtcaccg tctcgagt 378
<210> 105
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 105
Gln Ala Ser Gln Asn Ile Gly Ser Gly Leu Ala
1 5 10
<210> 106
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 106
Tyr Ala Ser Thr Leu Ala Ser
1 5
<210> 107
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 107
Gln Ser His Asp Tyr Ser Ser Val Arg Ser Tyr Gly Asn Ala
1 5 10
<210> 108
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 108
Gly Ile Asp Phe Ser Ser Tyr Tyr Tyr Met Cys
1 5 10
<210> 109
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 109
Cys Ile Asp Pro Ala Ser Ser Gly Thr Thr Tyr Tyr Ala Thr Trp Ala
1 5 10 15
Lys Gly
<210> 110
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 110
Ala Tyr Gly Ser Gly Gly Ser Gly Tyr Ile Gly Cys Tyr Phe Asp Leu
1 5 10 15
<210> 111
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4127
<400> 111
Ala Ile Val Met Thr Gln Thr Pro Ser Ser Lys Ser Val Pro Met Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Gly Asn
20 25 30
Asn Glu Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Ala Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Asp
85 90 95
Ser Asp Asp Gly Thr Thr Phe Gly Gly Gly Thr Lys Val Val Val Glu
100 105 110
<210> 112
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4127
<400> 112
gccatcgtga tgacccagac tccatcttcc aagtctgtcc ctatgggagg cacagtcacc 60
atcaactgcc aggccagtca gagtgtttat ggtaataacg aattatcctg gtatcagcag 120
aaaccagggc agcctcccaa gctcctgatc tatttggcat ccaggctggc atcgggggtc 180
ccatcgcggt ttagcggcag tggatctggg acacagttca ctctcaccat cagcggcgtg 240
cagtgtgacg atgctgccac ttactactgt gcaggctata aaagtgatag tgatgatggc 300
actactttcg gcggagggac caaggtggtg gtcgaa 336
<210> 113
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4127
<400> 113
Gln Gln Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Asn Leu Tyr
20 25 30
Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Leu Ile
35 40 45
Gly Cys Ile Asp Ile Ser Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Asp Tyr Tyr Ser Ser Asp Trp Gly Val Arg Phe Asn Leu Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 114
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4127
<400> 114
cagcagctgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc 60
tgcacagcct ctggattctc cttcagtaat ctctattaca tgtgttgggt ccgccaggct 120
ccagggaagg ggctggagtt gatcggatgc attgatatta gcagtagtgg tagcacttac 180
tacgcgagct gggcgaaagg ccgattcacc atctccaaaa cctcgtcgac cacggtgact 240
ctgcagatga ccagtctgac agccgcggac acggccacct atttctgtgc gagagattac 300
tattctagtg actggggtgt tagatttaac ttgtggggcc agggcaccct cgtcaccgtc 360
tcgagt 366
<210> 115
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 115
Gln Ala Ser Gln Ser Val Tyr Gly Asn Asn Glu Leu Ser
1 5 10
<210> 116
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 116
Leu Ala Ser Arg Leu Ala Ser
1 5
<210> 117
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 117
Ala Gly Tyr Lys Ser Asp Ser Asp Asp Gly Thr Thr
1 5 10
<210> 118
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 118
Gly Phe Ser Phe Ser Asn Leu Tyr Tyr Met Cys
1 5 10
<210> 119
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 119
Cys Ile Asp Ile Ser Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 120
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 120
Asp Tyr Tyr Ser Ser Asp Trp Gly Val Arg Phe Asn Leu
1 5 10
<210> 121
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-облсть кроличьего Ат 4128
<400> 121
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Glu Ser Ile Ser Asn Tyr
20 25 30
Leu Ser Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Ser Lys Leu Ser Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Asp Arg Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ile Tyr Tyr Ser Ala Ser Gly
85 90 95
Ser Arg Asp Trp Thr Phe Gly Gly Gly Thr Lys Val Val Val Glu
100 105 110
<210> 122
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4128
<400> 122
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc 60
atcaagtgcc aggccagtga aagcattagc aactacttat cctggtttca gcagaaacca 120
gggcagcctc ccaagctcct gatctatgct tcatccaaac tgtcatctgg ggtcccatcg 180
cggttcaaag gcgatagatc tgggacagag tacactctca ccatcagcga cctggagtgt 240
gccgatgctg ccacttacta ctgtcaaatc tattattcgg ctagtggcag tcgtgattgg 300
actttcggcg gagggaccaa ggtggtcgtc gaa 333
<210> 123
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4128
<400> 123
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Gln Pro Glu Gly Ser
1 5 10 15
Leu Thr Leu Thr Cys Lys Gly Ser Gly Leu Asp Phe Ser Ser Tyr Trp
20 25 30
Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Ala
35 40 45
Cys Ile Val Thr Gly Ser Ser Asp Asn Thr Tyr Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr Leu
65 70 75 80
Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Gly Gly Gly Ala Gly Tyr Ser Gly Ala Phe Asp Leu Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4128
<400> 124
cagtcgttgg aggagtccgg gggagacctg gtccagcctg agggatccct gacactcacc 60
tgcaaaggct ccgggttaga cttcagtagc tactggatat gctgggtccg ccaggctcca 120
gggaaggggc tggagtggat cgcatgcatt gttactggta gtagtgataa cacttactac 180
gcgagctggg cgaaaggccg attcaccatc tccaaaacct cgtcgaccac ggtgactctg 240
caaatgacca gtctgacagc cgcggacacg gccacctatt tctgtgcgag aggtggtggt 300
gctggttata gtggtgcctt tgacttgtgg ggccaaggga ccctcgtcac cgtctcgagt 360
<210> 125
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 125
Gln Ala Ser Glu Ser Ile Ser Asn Tyr Leu Ser
1 5 10
<210> 126
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 126
Ala Ser Ser Lys Leu Ser Ser
1 5
<210> 127
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 127
Gln Ile Tyr Tyr Ser Ala Ser Gly Ser Arg Asp Trp Thr
1 5 10
<210> 128
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 128
Gly Leu Asp Phe Ser Ser Tyr Trp Ile Cys
1 5 10
<210> 129
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 129
Cys Ile Val Thr Gly Ser Ser Asp Asn Thr Tyr Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 130
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 130
Gly Gly Gly Ala Gly Tyr Ser Gly Ala Phe Asp Leu
1 5 10
<210> 131
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4130
<400> 131
Ala Ala Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Ser Ile Ser Cys Gln Ser Ser Gln Ser Val Tyr Asn Thr
20 25 30
Lys Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Thr Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Asp Leu
65 70 75 80
Glu Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Phe Ser Ser
85 90 95
Ser Asp Leu Asn Val Phe Gly Gly Gly Thr Lys Val Val Val Lys
100 105 110
<210> 132
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4130
<400> 132
gccgccgtgc tgacccagac tccatctccc gtgtctgcag ctgtgggagg cacagtcagc 60
atcagttgcc agtccagtca gagtgtttat aatacaaagg acttagcctg gtatcagcag 120
aaaccagggc agcctcccaa gctcctgatc tatggtacat ccactctggc atctggggtc 180
tcatcacggt tcagcggcag tggatctggg acagagttca ctctcaccat cagcgacctg 240
gagtgtgacg atgctgccac ttattactgt caaggcggtt ttagtagtag tgatttgaat 300
gttttcggcg gagggaccaa ggtggtggtc aaa 333
<210> 133
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4130
<400> 133
Gln Gln Gln Leu Glu Glu Ser Gly Gly Asp Leu Val Arg Pro Glu Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Asp Phe Ser Gly Gly
20 25 30
Tyr Asp Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Cys Ile Tyr Gly Gly Ile Asn Ser Val Thr Asp Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Val Thr Ile Ser Lys Thr Ser Ser Thr Thr Val
65 70 75 80
Thr Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe
85 90 95
Cys Ala Arg Asp Val Ser Asn Ser Asp His Tyr Thr Arg Leu Asp Leu
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 134
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4130
<400> 134
cagcagcagc tggaggagtc cgggggagac ctggtcaggc ctgagggatc cctgacactc 60
acctgcacag cctctggatt cgacttcagt ggcggctacg acatttcctg ggtccgccag 120
gctccaggga aggggctgga gtggatcgga tgcatttatg gtggtatcaa tagtgtcact 180
gactacgcga gctgggcgaa aggccgagtc accatctcca aaacctcgtc gaccacggtg 240
actctgcaga tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagagat 300
gttagtaata gcgatcatta tactcggttg gatctctggg gccaaggcac cctggtcacc 360
gtctcgagt 369
<210> 135
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 135
Gln Ser Ser Gln Ser Val Tyr Asn Thr Lys Asp Leu Ala
1 5 10
<210> 136
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 136
Gly Thr Ser Thr Leu Ala Ser
1 5
<210> 137
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 137
Gln Gly Gly Phe Ser Ser Ser Asp Leu Asn Val
1 5 10
<210> 138
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 138
Gly Phe Asp Phe Ser Gly Gly Tyr Asp Ile Ser
1 5 10
<210> 139
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 139
Cys Ile Tyr Gly Gly Ile Asn Ser Val Thr Asp Tyr Ala Ser Trp Ala
1 5 10 15
Lys Gly
<210> 140
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 140
Asp Val Ser Asn Ser Asp His Tyr Thr Arg Leu Asp Leu
1 5 10
<210> 141
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4132
<400> 141
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Lys Cys Gln Ala Ser Glu Thr Ile Ser Ser Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Leu Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Gly Val Gln Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Tyr Tyr Tyr Ser Ser Gly
85 90 95
Ser Asp Tyr Gly Phe Gly Gly Gly Thr Lys Val Val Val Lys
100 105 110
<210> 142
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область кроличьего Ат 4132
<400> 142
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc 60
atcaagtgcc aggccagtga gaccattagt agtagattag cctggtatca gcagaagcta 120
gggcagcctc ccaaactcct gatctattct gcatccactc tggcgtctgg ggtcccatcg 180
cggttcaaag gcagtggatc tgggacagag tacactctca ccatcagcgg cgtgcagtgt 240
gccgatgctg ccacttatta ctgtcaaggc tattattata gtagtggtag tgattatggt 300
ttcggcggag ggaccaaggt ggtcgtcaaa 330
<210> 143
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4132
<400> 143
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Ser Ser Tyr
20 25 30
Trp Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ser
35 40 45
Gly Cys Ile Asn Ser Gly Thr Gly Gly Thr Ala Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Asn Ser Ser Ser Thr Thr Val Thr Leu
65 70 75 80
Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Glu Trp Val Ser Gly Tyr Tyr Lys Asp Ala Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 144
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область кроличьего Ат 4132
<400> 144
cagtcgttgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc 60
tgcacagcct ctggattctc cttcagtagc agctactgga tatgctgggt ccgccaggct 120
ccagggaagg ggctggagtg gagcggatgc attaatagtg gtactggtgg cactgcctac 180
gcgagctggg cgaaaggccg attcaccatc tccaattcct cgtcgaccac ggtgactctt 240
caaatgacca gtctgacagc cgcggacacg gccacctatt tctgtgcgag agaatgggtt 300
agtggttatt ataaagatgc ttttgatctc tggggccagg gcaccctggt caccgtctcg 360
agt 363
<210> 145
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 145
Gln Ala Ser Glu Thr Ile Ser Ser Arg Leu Ala
1 5 10
<210> 146
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 146
Ser Ala Ser Thr Leu Ala Ser
1 5
<210> 147
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 147
Gln Gly Tyr Tyr Tyr Ser Ser Gly Ser Asp Tyr Gly
1 5 10
<210> 148
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 148
Gly Phe Ser Phe Ser Ser Ser Tyr Trp Ile Cys
1 5 10
<210> 149
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 149
Cys Ile Asn Ser Gly Thr Gly Gly Thr Ala Tyr Ala Ser Trp Ala Lys
1 5 10 15
Gly
<210> 150
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 150
Glu Trp Val Ser Gly Tyr Tyr Lys Asp Ala Phe Asp Leu
1 5 10
<210> 151
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH1 dAbH1
<400> 151
Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
1 5 10
<210> 152
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH2 dAbH1
<400> 152
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
1 5 10 15
<210> 153
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRH3 dAbH1
<400> 153
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu
1 5 10
<210> 154
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL1 dAbL1
<400> 154
Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
1 5 10
<210> 155
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL2 dAbL1
<400> 155
Glu Ala Ser Lys Leu Thr Ser
1 5
<210> 156
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDRL3 dAbL1
<400> 156
Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
1 5 10
<210> 157
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи антитела против альбумина (без ds)
<400> 157
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 158
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи антитела против альбумина (ds)
<400> 158
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 159
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи антитела против альбумина (без ds)
<400> 159
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
100 105 110
<210> 160
<211> 128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи антитела против альбумина (ds)
<400> 160
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Gly Lys Pro Thr His
1 5 10 15
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
20 25 30
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
35 40 45
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
50 55 60
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
65 70 75 80
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
85 90 95
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
100 105 110
Ser Asp Thr Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
<210> 161
<211> 847
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CD22 человека
<400> 161
Met His Leu Leu Gly Pro Trp Leu Leu Leu Leu Val Leu Glu Tyr Leu
1 5 10 15
Ala Phe Ser Asp Ser Ser Lys Trp Val Phe Glu His Pro Glu Thr Leu
20 25 30
Tyr Ala Trp Glu Gly Ala Cys Val Trp Ile Pro Cys Thr Tyr Arg Ala
35 40 45
Leu Asp Gly Asp Leu Glu Ser Phe Ile Leu Phe His Asn Pro Glu Tyr
50 55 60
Asn Lys Asn Thr Ser Lys Phe Asp Gly Thr Arg Leu Tyr Glu Ser Thr
65 70 75 80
Lys Asp Gly Lys Val Pro Ser Glu Gln Lys Arg Val Gln Phe Leu Gly
85 90 95
Asp Lys Asn Lys Asn Cys Thr Leu Ser Ile His Pro Val His Leu Asn
100 105 110
Asp Ser Gly Gln Leu Gly Leu Arg Met Glu Ser Lys Thr Glu Lys Trp
115 120 125
Met Glu Arg Ile His Leu Asn Val Ser Glu Arg Pro Phe Pro Pro His
130 135 140
Ile Gln Leu Pro Pro Glu Ile Gln Glu Ser Gln Glu Val Thr Leu Thr
145 150 155 160
Cys Leu Leu Asn Phe Ser Cys Tyr Gly Tyr Pro Ile Gln Leu Gln Trp
165 170 175
Leu Leu Glu Gly Val Pro Met Arg Gln Ala Ala Val Thr Ser Thr Ser
180 185 190
Leu Thr Ile Lys Ser Val Phe Thr Arg Ser Glu Leu Lys Phe Ser Pro
195 200 205
Gln Trp Ser His His Gly Lys Ile Val Thr Cys Gln Leu Gln Asp Ala
210 215 220
Asp Gly Lys Phe Leu Ser Asn Asp Thr Val Gln Leu Asn Val Lys His
225 230 235 240
Thr Pro Lys Leu Glu Ile Lys Val Thr Pro Ser Asp Ala Ile Val Arg
245 250 255
Glu Gly Asp Ser Val Thr Met Thr Cys Glu Val Ser Ser Ser Asn Pro
260 265 270
Glu Tyr Thr Thr Val Ser Trp Leu Lys Asp Gly Thr Ser Leu Lys Lys
275 280 285
Gln Asn Thr Phe Thr Leu Asn Leu Arg Glu Val Thr Lys Asp Gln Ser
290 295 300
Gly Lys Tyr Cys Cys Gln Val Ser Asn Asp Val Gly Pro Gly Arg Ser
305 310 315 320
Glu Glu Val Phe Leu Gln Val Gln Tyr Ala Pro Glu Pro Ser Thr Val
325 330 335
Gln Ile Leu His Ser Pro Ala Val Glu Gly Ser Gln Val Glu Phe Leu
340 345 350
Cys Met Ser Leu Ala Asn Pro Leu Pro Thr Asn Tyr Thr Trp Tyr His
355 360 365
Asn Gly Lys Glu Met Gln Gly Arg Thr Glu Glu Lys Val His Ile Pro
370 375 380
Lys Ile Leu Pro Trp His Ala Gly Thr Tyr Ser Cys Val Ala Glu Asn
385 390 395 400
Ile Leu Gly Thr Gly Gln Arg Gly Pro Gly Ala Glu Leu Asp Val Gln
405 410 415
Tyr Pro Pro Lys Lys Val Thr Thr Val Ile Gln Asn Pro Met Pro Ile
420 425 430
Arg Glu Gly Asp Thr Val Thr Leu Ser Cys Asn Tyr Asn Ser Ser Asn
435 440 445
Pro Ser Val Thr Arg Tyr Glu Trp Lys Pro His Gly Ala Trp Glu Glu
450 455 460
Pro Ser Leu Gly Val Leu Lys Ile Gln Asn Val Gly Trp Asp Asn Thr
465 470 475 480
Thr Ile Ala Cys Ala Ala Cys Asn Ser Trp Cys Ser Trp Ala Ser Pro
485 490 495
Val Ala Leu Asn Val Gln Tyr Ala Pro Arg Asp Val Arg Val Arg Lys
500 505 510
Ile Lys Pro Leu Ser Glu Ile His Ser Gly Asn Ser Val Ser Leu Gln
515 520 525
Cys Asp Phe Ser Ser Ser His Pro Lys Glu Val Gln Phe Phe Trp Glu
530 535 540
Lys Asn Gly Arg Leu Leu Gly Lys Glu Ser Gln Leu Asn Phe Asp Ser
545 550 555 560
Ile Ser Pro Glu Asp Ala Gly Ser Tyr Ser Cys Trp Val Asn Asn Ser
565 570 575
Ile Gly Gln Thr Ala Ser Lys Ala Trp Thr Leu Glu Val Leu Tyr Ala
580 585 590
Pro Arg Arg Leu Arg Val Ser Met Ser Pro Gly Asp Gln Val Met Glu
595 600 605
Gly Lys Ser Ala Thr Leu Thr Cys Glu Ser Asp Ala Asn Pro Pro Val
610 615 620
Ser His Tyr Thr Trp Phe Asp Trp Asn Asn Gln Ser Leu Pro Tyr His
625 630 635 640
Ser Gln Lys Leu Arg Leu Glu Pro Val Lys Val Gln His Ser Gly Ala
645 650 655
Tyr Trp Cys Gln Gly Thr Asn Ser Val Gly Lys Gly Arg Ser Pro Leu
660 665 670
Ser Thr Leu Thr Val Tyr Tyr Ser Pro Glu Thr Ile Gly Arg Arg Val
675 680 685
Ala Val Gly Leu Gly Ser Cys Leu Ala Ile Leu Ile Leu Ala Ile Cys
690 695 700
Gly Leu Lys Leu Gln Arg Arg Trp Lys Arg Thr Gln Ser Gln Gln Gly
705 710 715 720
Leu Gln Glu Asn Ser Ser Gly Gln Ser Phe Phe Val Arg Asn Lys Lys
725 730 735
Val Arg Arg Ala Pro Leu Ser Glu Gly Pro His Ser Leu Gly Cys Tyr
740 745 750
Asn Pro Met Met Glu Asp Gly Ile Ser Tyr Thr Thr Leu Arg Phe Pro
755 760 765
Glu Met Asn Ile Pro Arg Thr Gly Asp Ala Glu Ser Ser Glu Met Gln
770 775 780
Arg Pro Pro Pro Asp Cys Asp Asp Thr Val Thr Tyr Ser Ala Leu His
785 790 795 800
Lys Arg Gln Val Gly Asp Tyr Glu Asn Val Ile Pro Asp Phe Pro Glu
805 810 815
Asp Glu Gly Ile His Tyr Ser Glu Leu Ile Gln Phe Gly Val Gly Glu
820 825 830
Arg Pro Gln Ala Gln Glu Asn Val Asp Tyr Val Ile Leu Lys His
835 840 845
<210> 162
<211> 226
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CD79a человека
<400> 162
Met Pro Gly Gly Pro Gly Val Leu Gln Ala Leu Pro Ala Thr Ile Phe
1 5 10 15
Leu Leu Phe Leu Leu Ser Ala Val Tyr Leu Gly Pro Gly Cys Gln Ala
20 25 30
Leu Trp Met His Lys Val Pro Ala Ser Leu Met Val Ser Leu Gly Glu
35 40 45
Asp Ala His Phe Gln Cys Pro His Asn Ser Ser Asn Asn Ala Asn Val
50 55 60
Thr Trp Trp Arg Val Leu His Gly Asn Tyr Thr Trp Pro Pro Glu Phe
65 70 75 80
Leu Gly Pro Gly Glu Asp Pro Asn Gly Thr Leu Ile Ile Gln Asn Val
85 90 95
Asn Lys Ser His Gly Gly Ile Tyr Val Cys Arg Val Gln Glu Gly Asn
100 105 110
Glu Ser Tyr Gln Gln Ser Cys Gly Thr Tyr Leu Arg Val Arg Gln Pro
115 120 125
Pro Pro Arg Pro Phe Leu Asp Met Gly Glu Gly Thr Lys Asn Arg Ile
130 135 140
Ile Thr Ala Glu Gly Ile Ile Leu Leu Phe Cys Ala Val Val Pro Gly
145 150 155 160
Thr Leu Leu Leu Phe Arg Lys Arg Trp Gln Asn Glu Lys Leu Gly Leu
165 170 175
Asp Ala Gly Asp Glu Tyr Glu Asp Glu Asn Leu Tyr Glu Gly Leu Asn
180 185 190
Leu Asp Asp Cys Ser Met Tyr Glu Asp Ile Ser Arg Gly Leu Gln Gly
195 200 205
Thr Tyr Gln Asp Val Gly Ser Leu Asn Ile Gly Asp Val Gln Leu Glu
210 215 220
Lys Pro
225
<210> 163
<211> 229
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CD79b человека
<400> 163
Met Ala Arg Leu Ala Leu Ser Pro Val Pro Ser His Trp Met Val Ala
1 5 10 15
Leu Leu Leu Leu Leu Ser Ala Glu Pro Val Pro Ala Ala Arg Ser Glu
20 25 30
Asp Arg Tyr Arg Asn Pro Lys Gly Ser Ala Cys Ser Arg Ile Trp Gln
35 40 45
Ser Pro Arg Phe Ile Ala Arg Lys Arg Gly Phe Thr Val Lys Met His
50 55 60
Cys Tyr Met Asn Ser Ala Ser Gly Asn Val Ser Trp Leu Trp Lys Gln
65 70 75 80
Glu Met Asp Glu Asn Pro Gln Gln Leu Lys Leu Glu Lys Gly Arg Met
85 90 95
Glu Glu Ser Gln Asn Glu Ser Leu Ala Thr Leu Thr Ile Gln Gly Ile
100 105 110
Arg Phe Glu Asp Asn Gly Ile Tyr Phe Cys Gln Gln Lys Cys Asn Asn
115 120 125
Thr Ser Glu Val Tyr Gln Gly Cys Gly Thr Glu Leu Arg Val Met Gly
130 135 140
Phe Ser Thr Leu Ala Gln Leu Lys Gln Arg Asn Thr Leu Lys Asp Gly
145 150 155 160
Ile Ile Met Ile Gln Thr Leu Leu Ile Ile Leu Phe Ile Ile Val Pro
165 170 175
Ile Phe Leu Leu Leu Asp Lys Asp Asp Ser Lys Ala Gly Met Glu Glu
180 185 190
Asp His Thr Tyr Glu Gly Leu Asp Ile Asp Gln Thr Ala Thr Tyr Glu
195 200 205
Asp Ile Val Thr Leu Arg Thr Gly Glu Val Lys Trp Ser Val Gly Glu
210 215 220
His Pro Gly Gln Glu
225
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ СО СПЕЦИФИЧНОСТЬЮ К CD45 И CD79 | 2015 |
|
RU2747980C2 |
| АНТИТЕЛО ПРОТИВ CD79B, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2817143C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2826270C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |
| КРИОКОНСЕРВИРОВАННЫЕ КЛЕТКИ-ЕСТЕСТВЕННЫЕ КИЛЛЕРЫ, ПРЕДВАРИТЕЛЬНО НАГРУЖЕННЫЕ КОНСТРУКЦИЕЙ АНТИТЕЛА | 2019 |
|
RU2819927C2 |
| АНТИ-CD79b АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2791984C2 |
| СПОСОБЫ ПРИМЕНЕНИЯ CD32B X CD79B-СВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРИ ЛЕЧЕНИИ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ И РАССТРОЙСТВ | 2017 |
|
RU2772434C2 |
| АНТИТЕЛА ПРОТИВ CD79b, КОНЪЮГАТЫ С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2805251C2 |
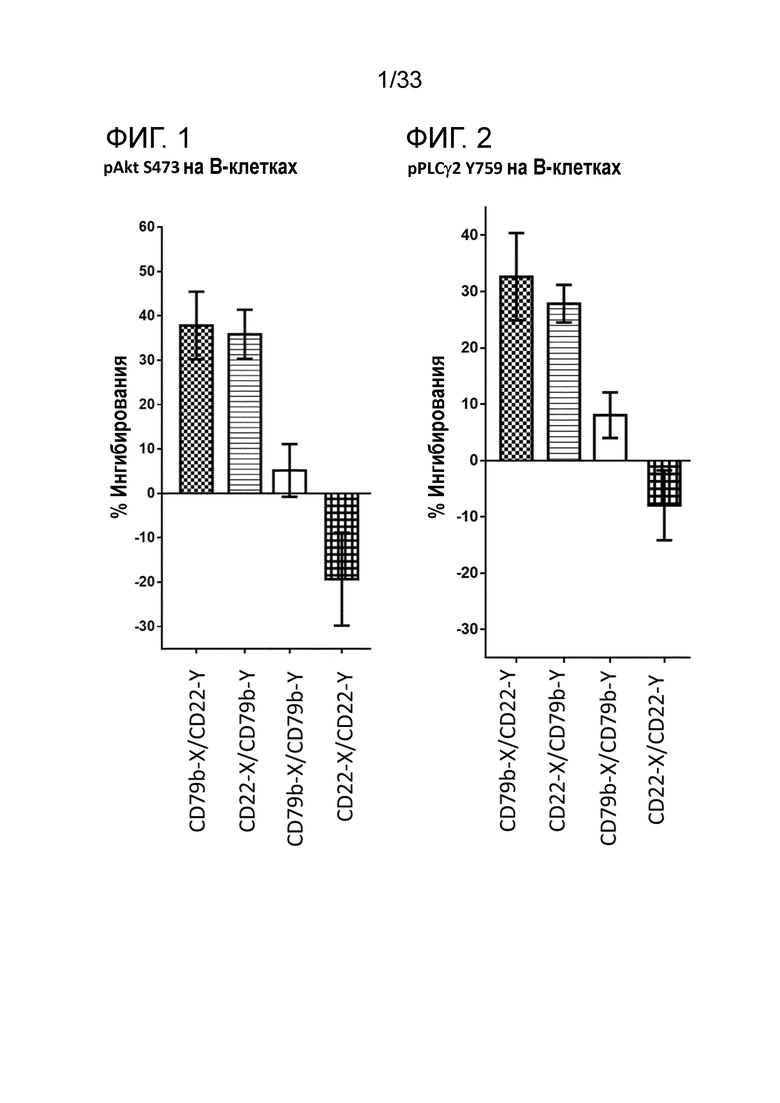
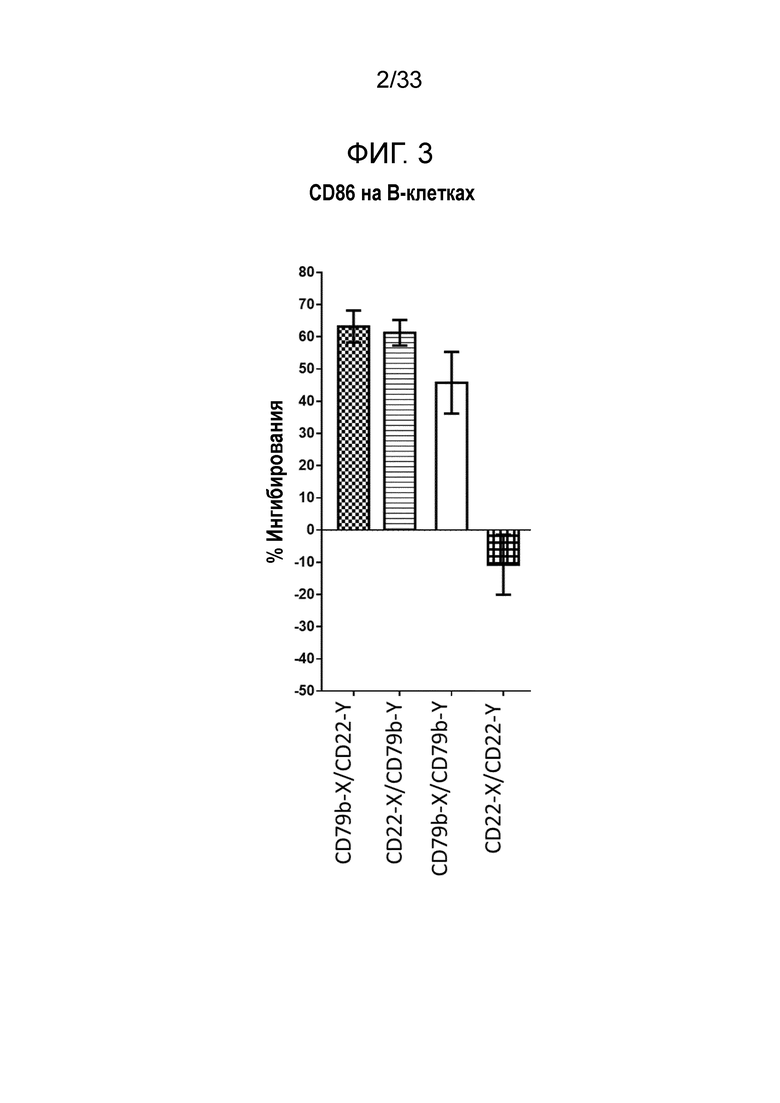

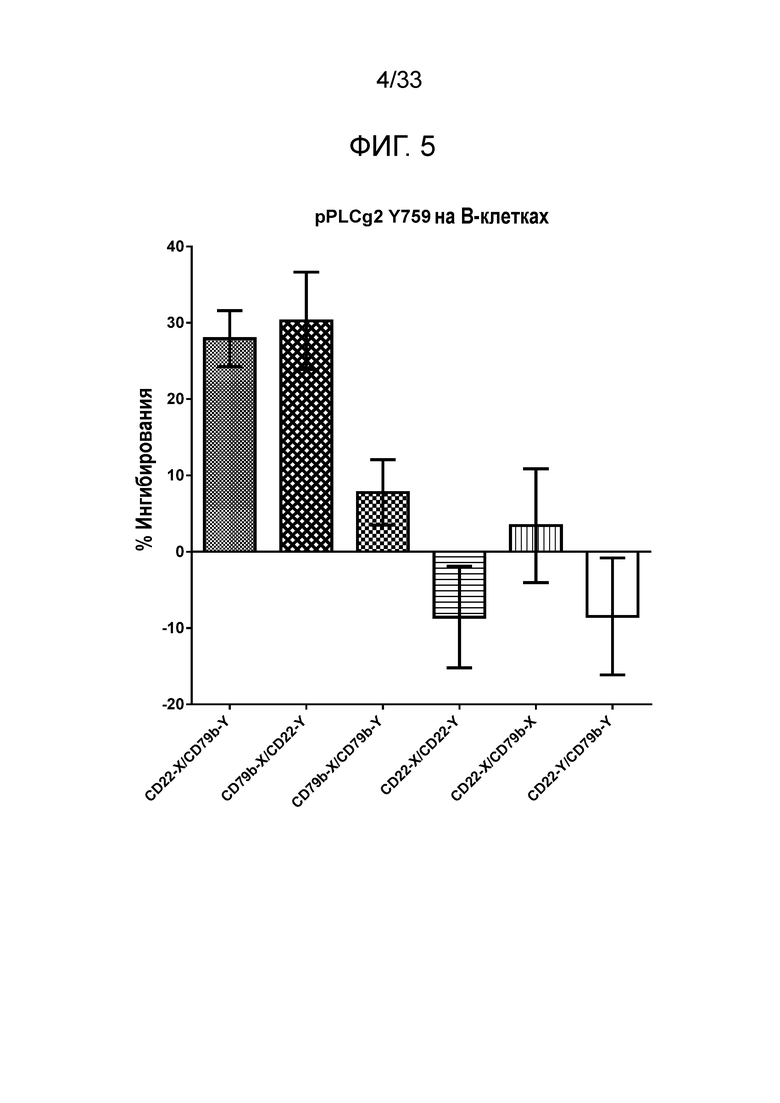
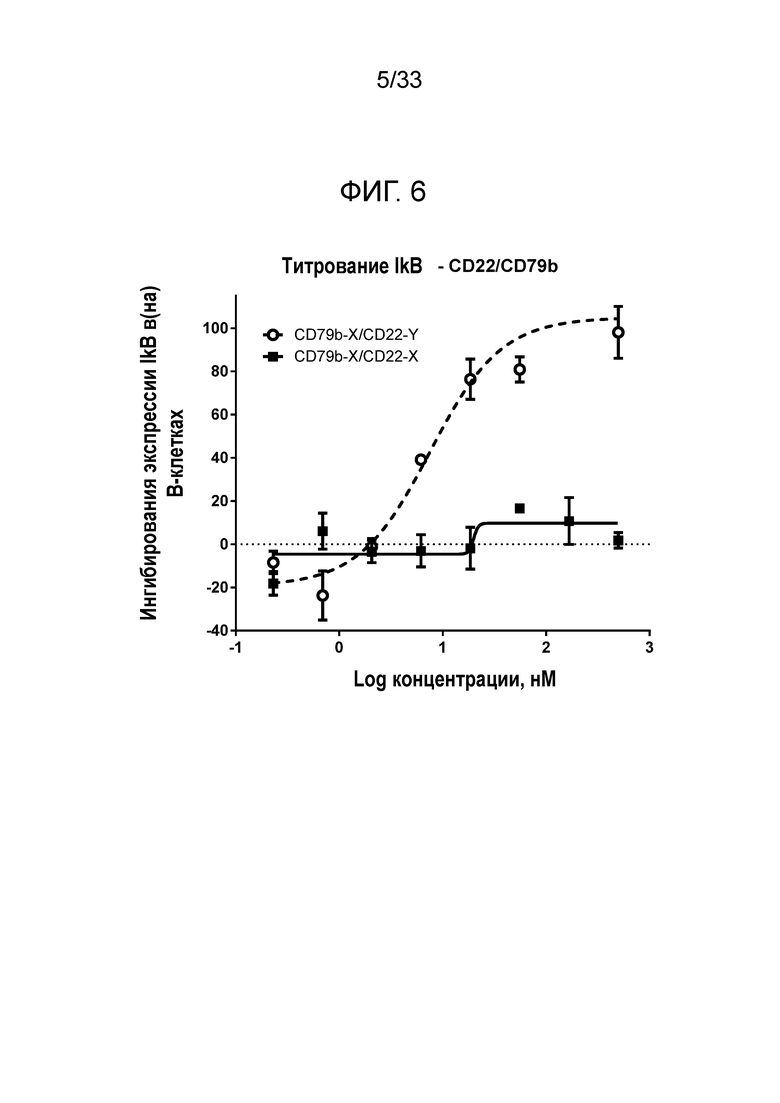
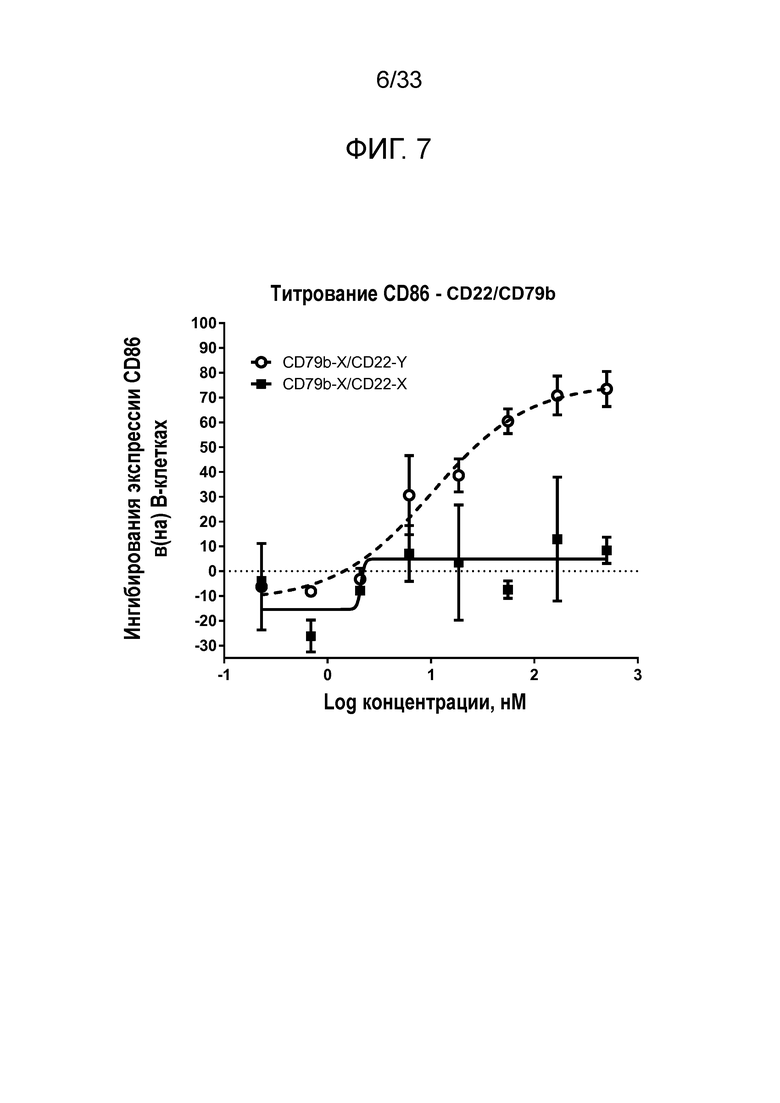
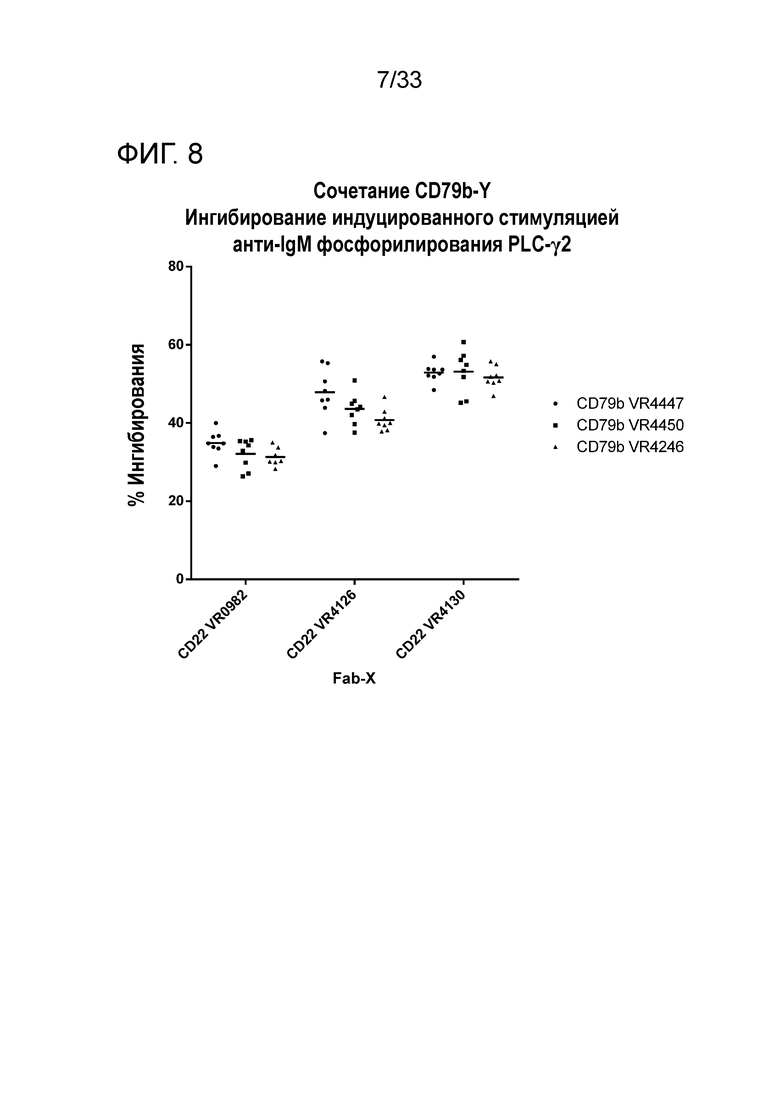






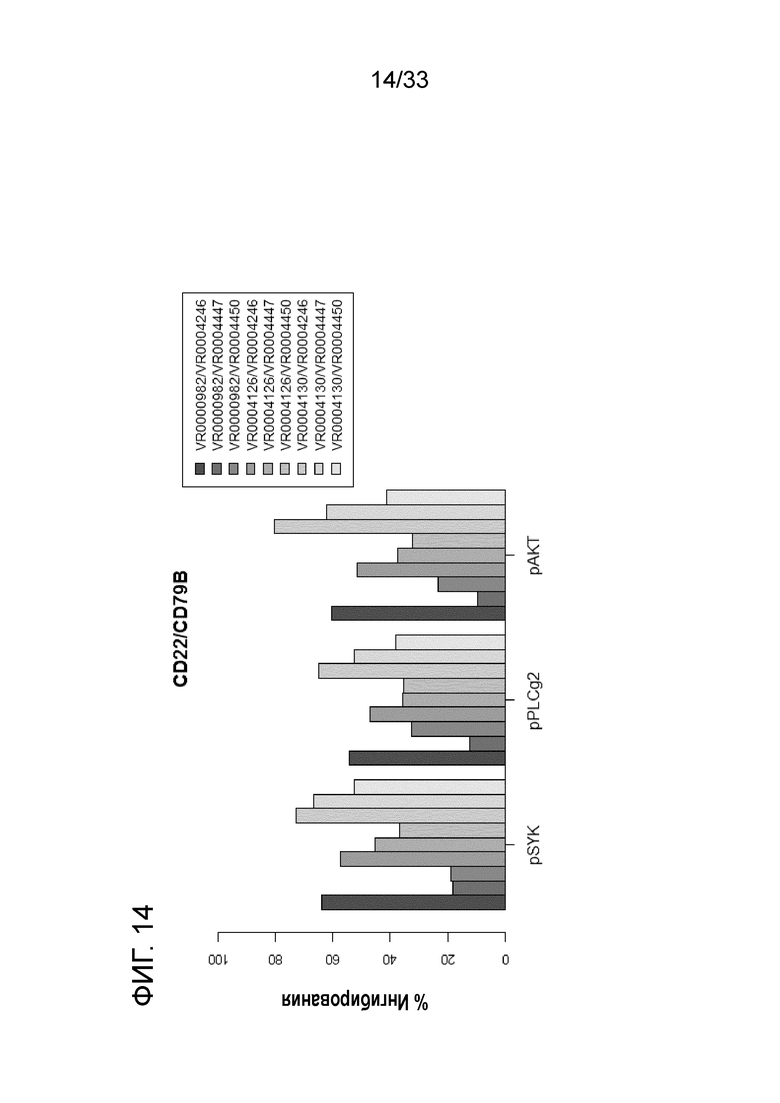
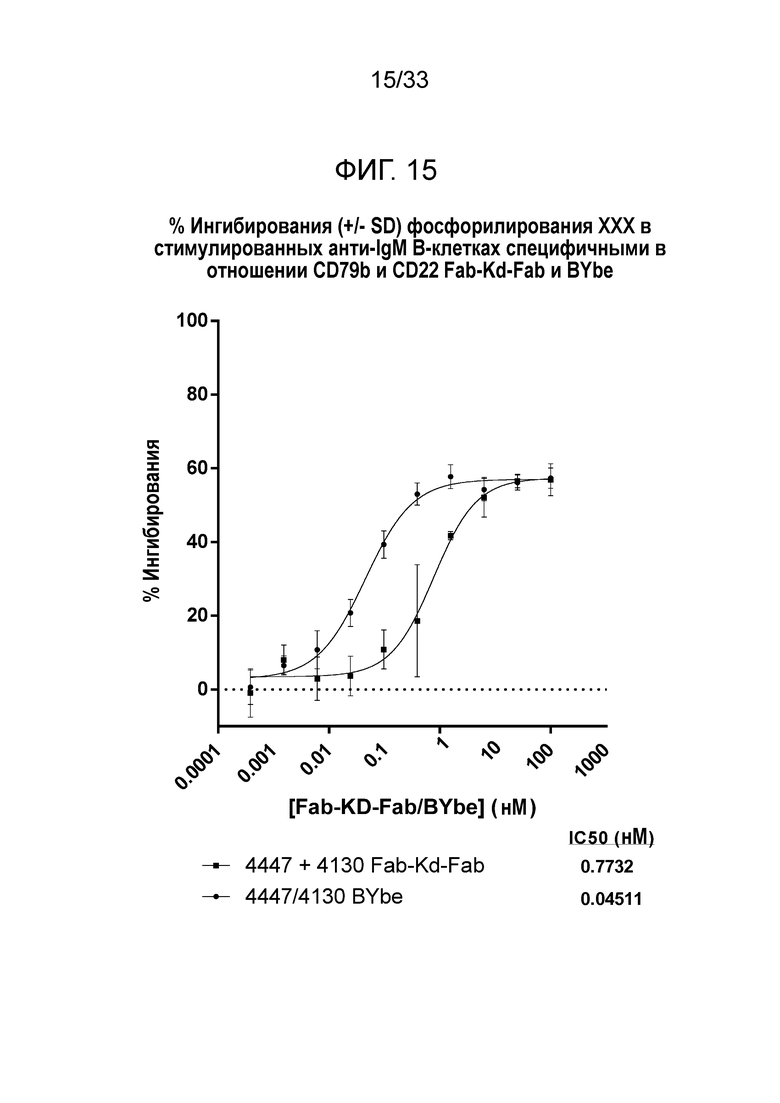
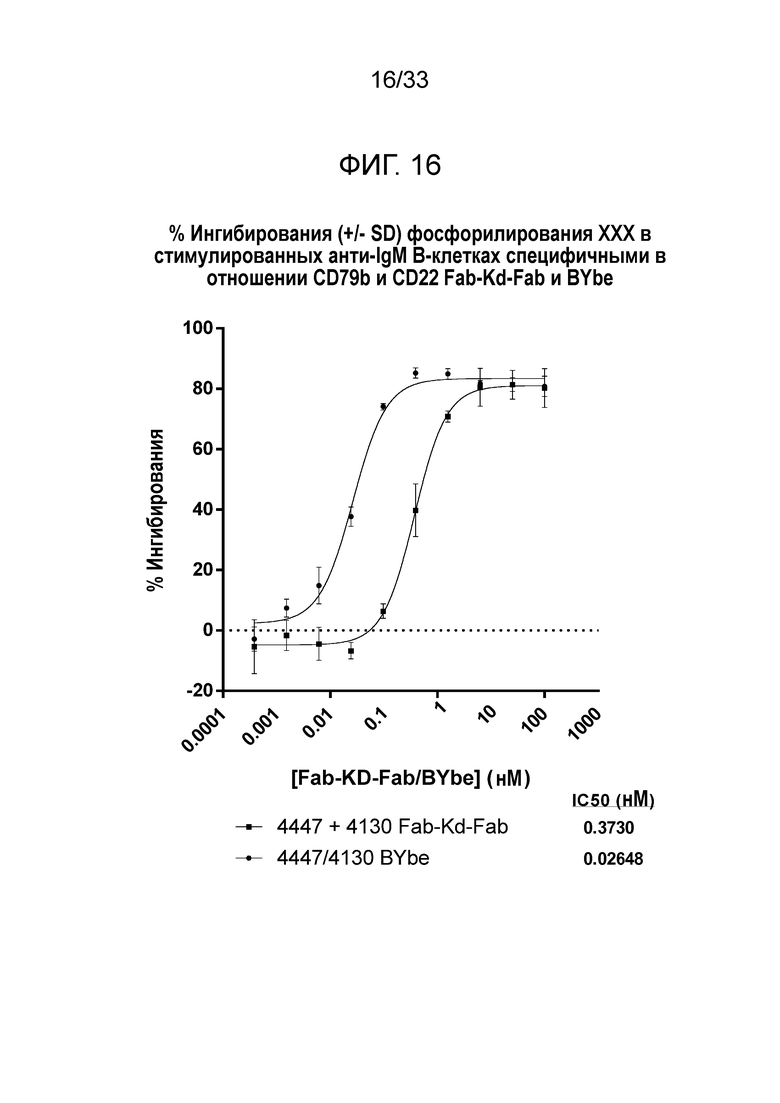
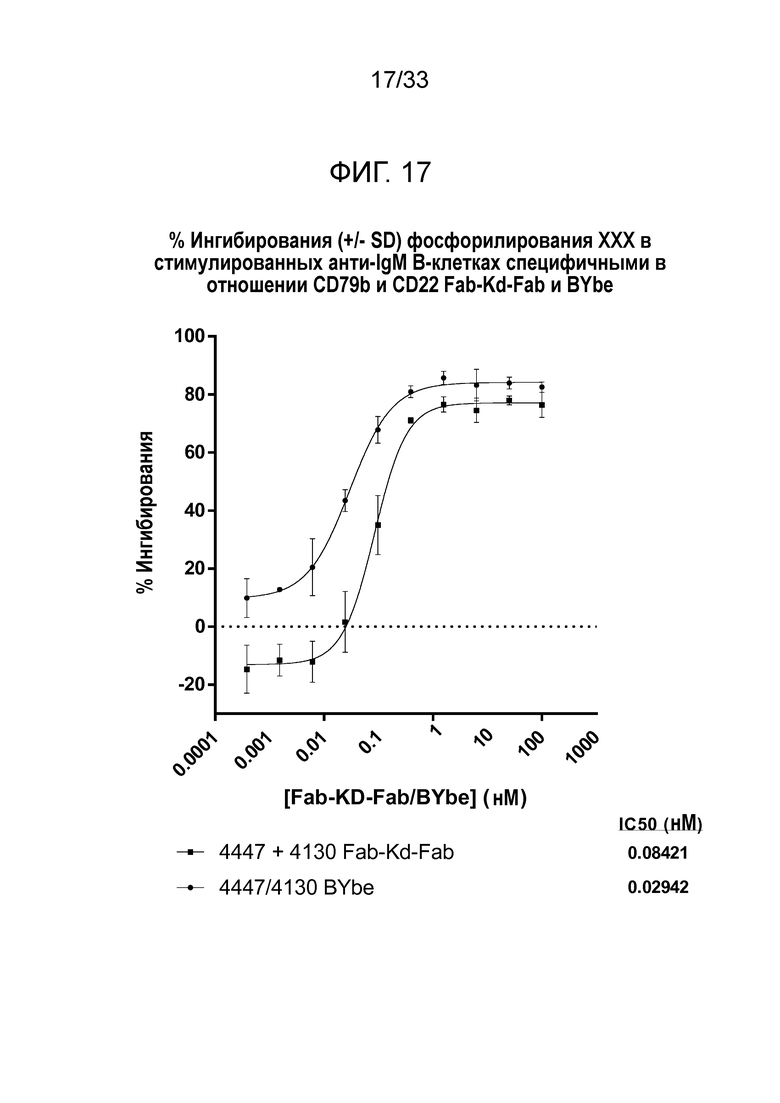


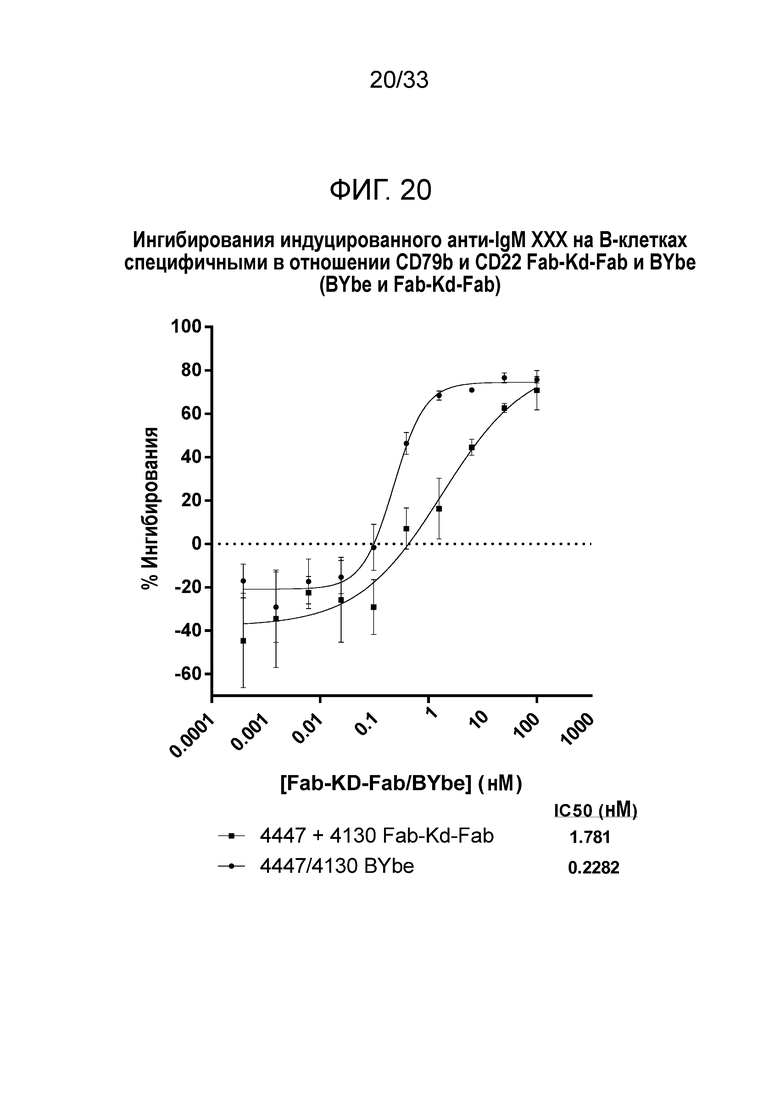
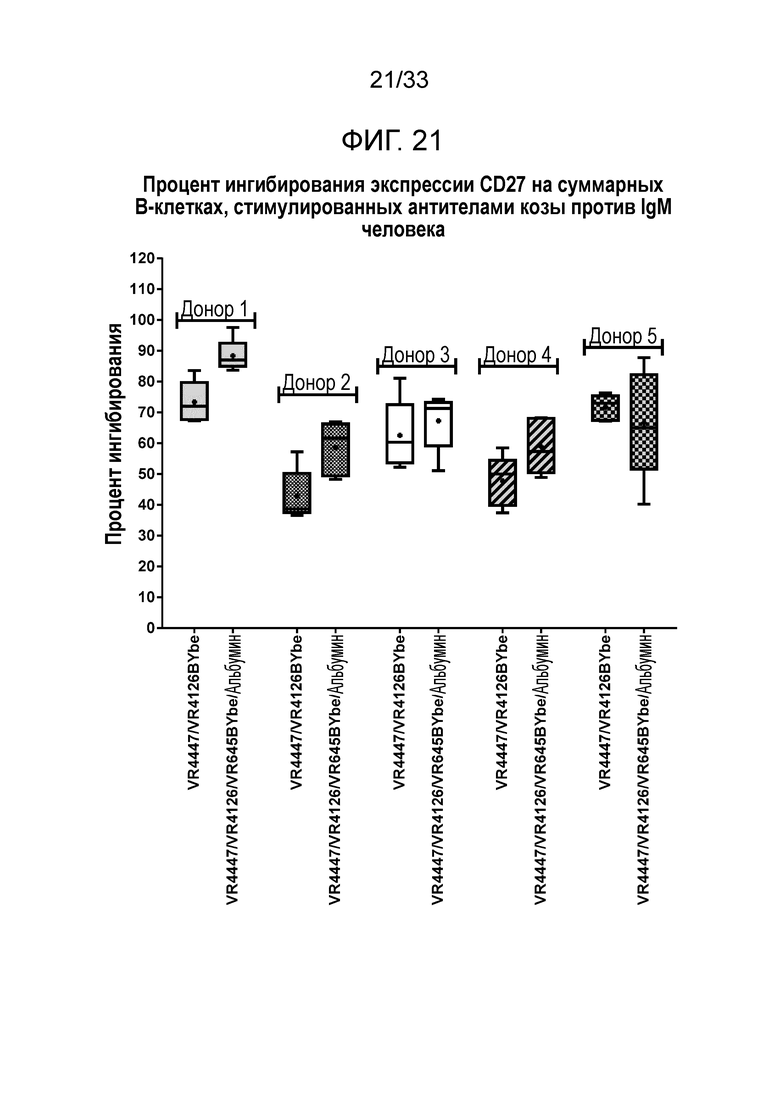
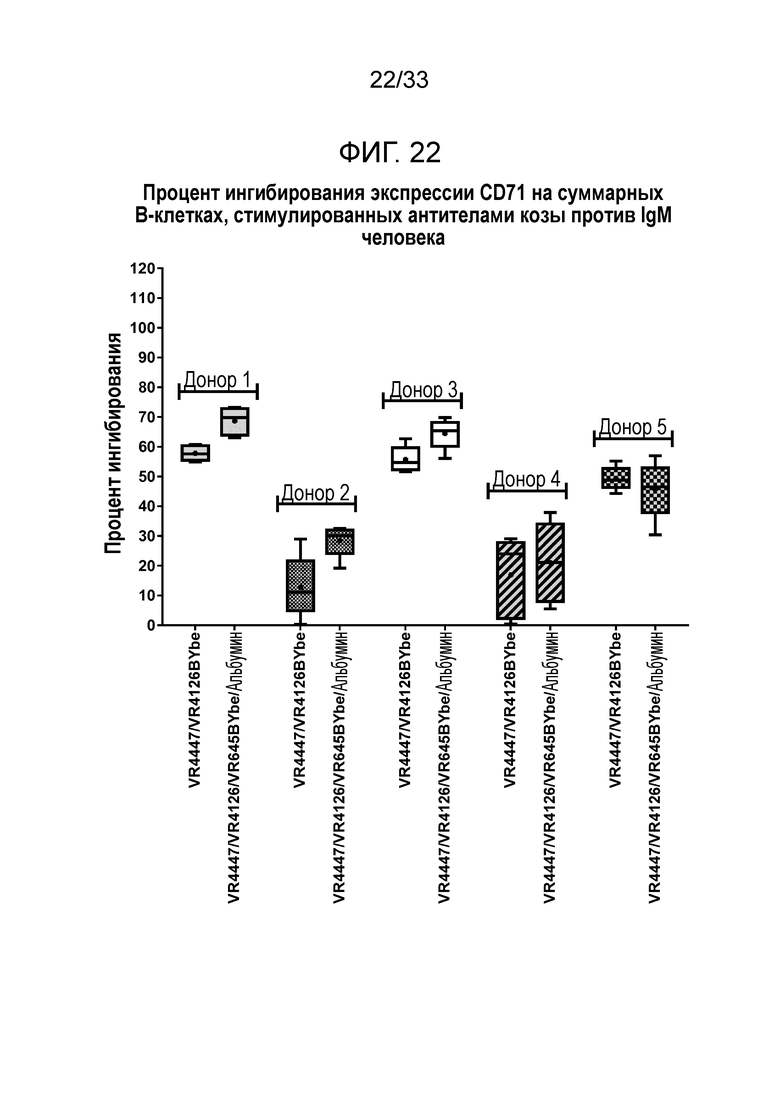
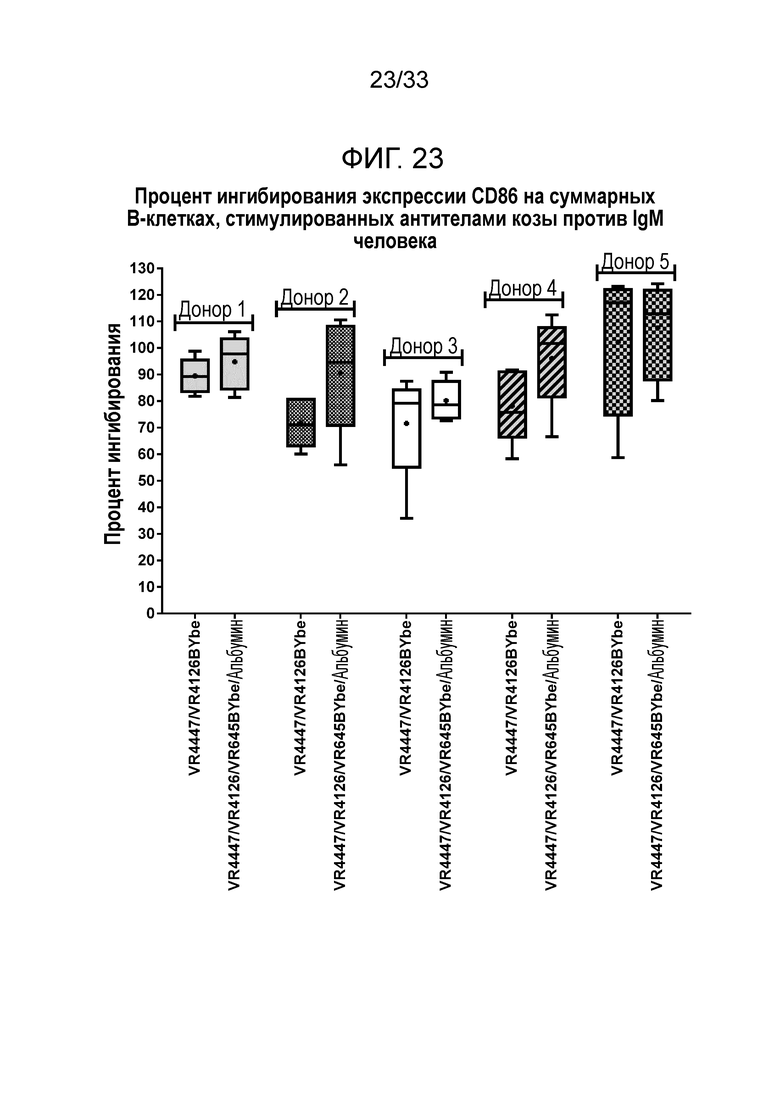
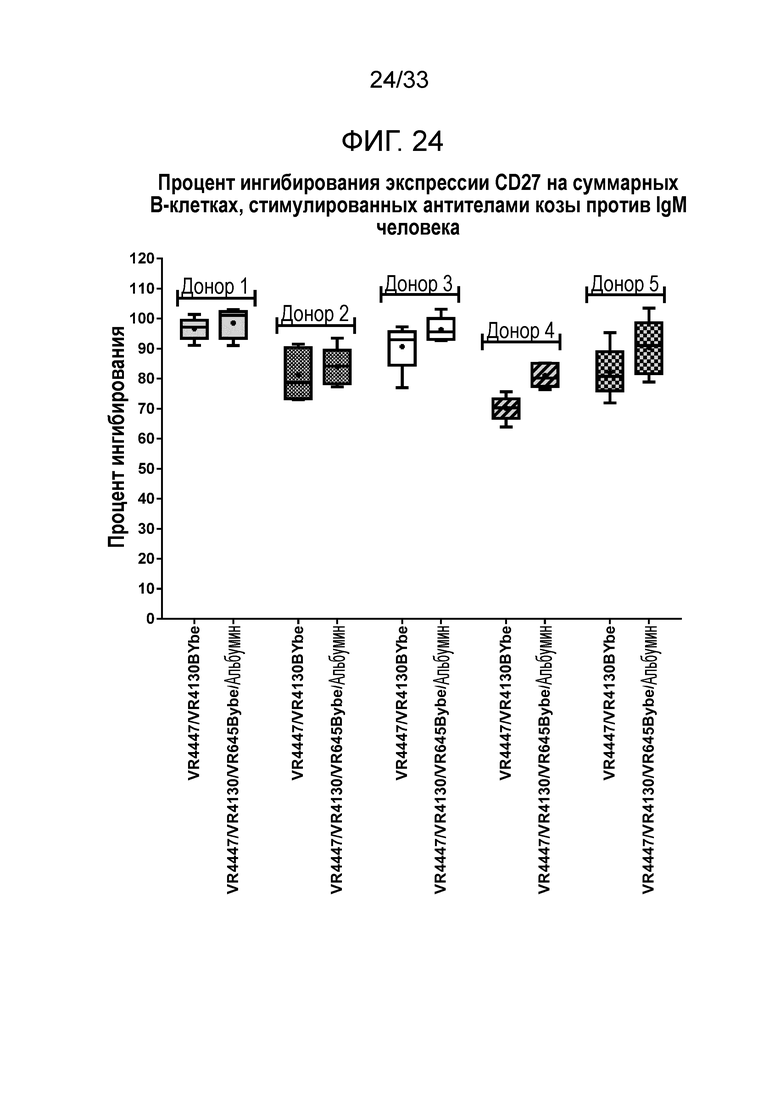
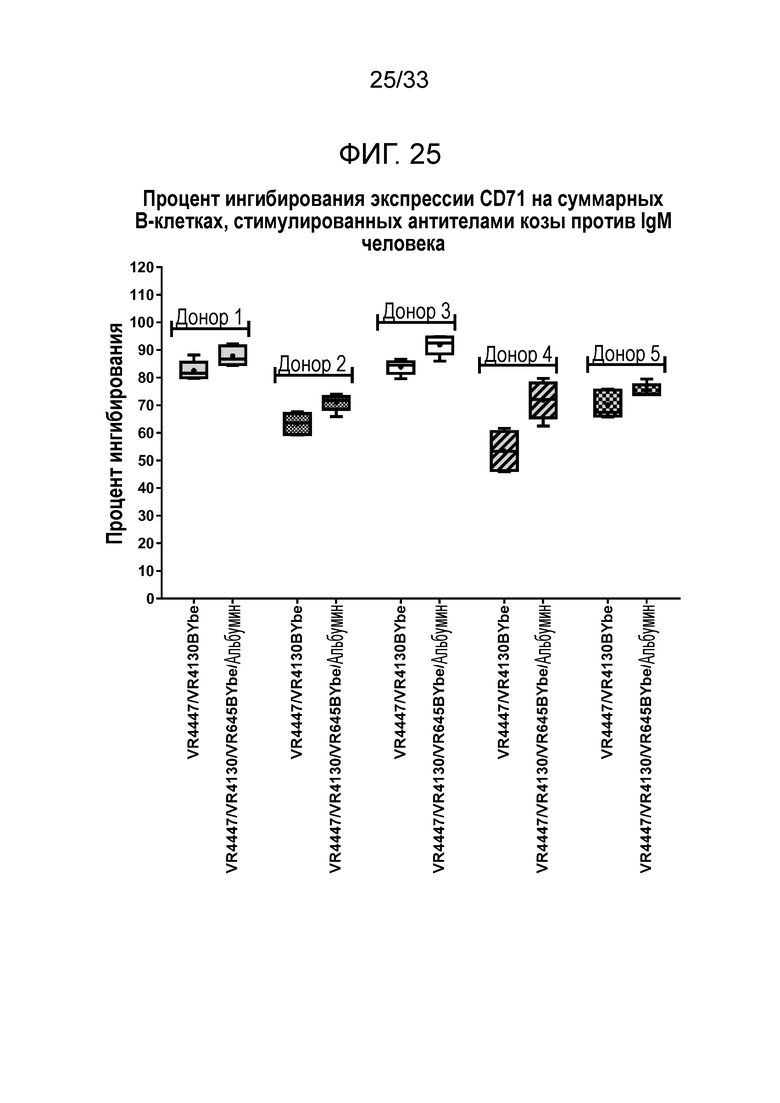
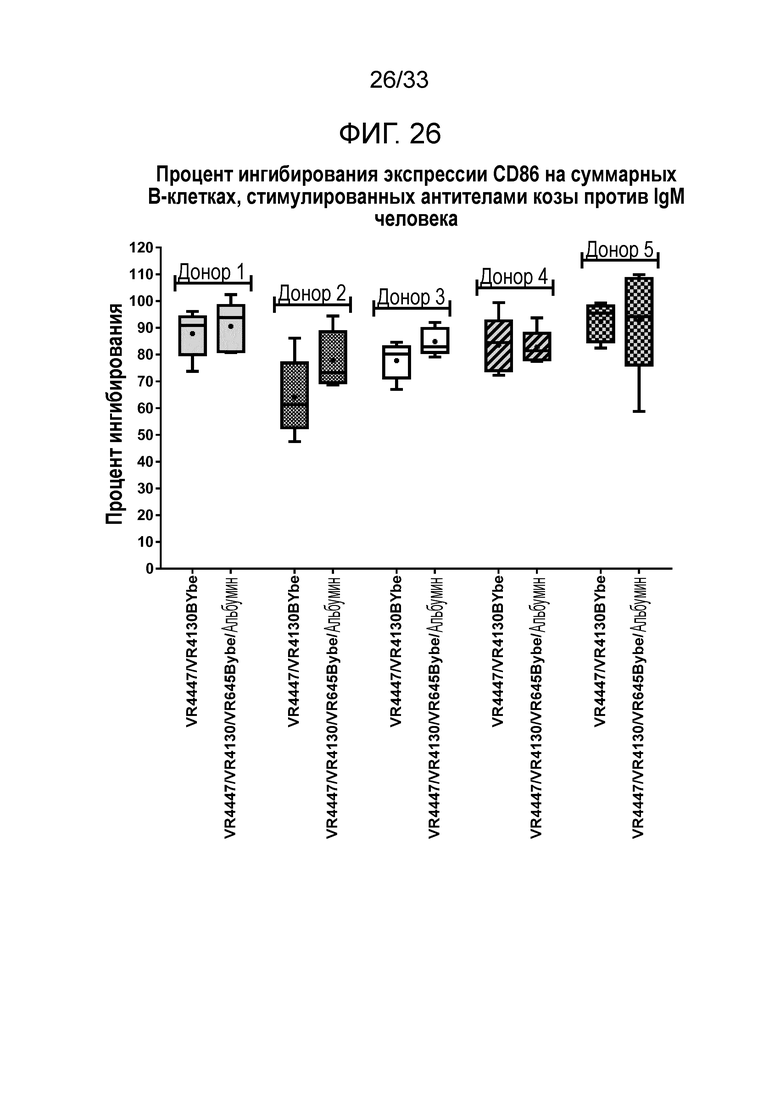
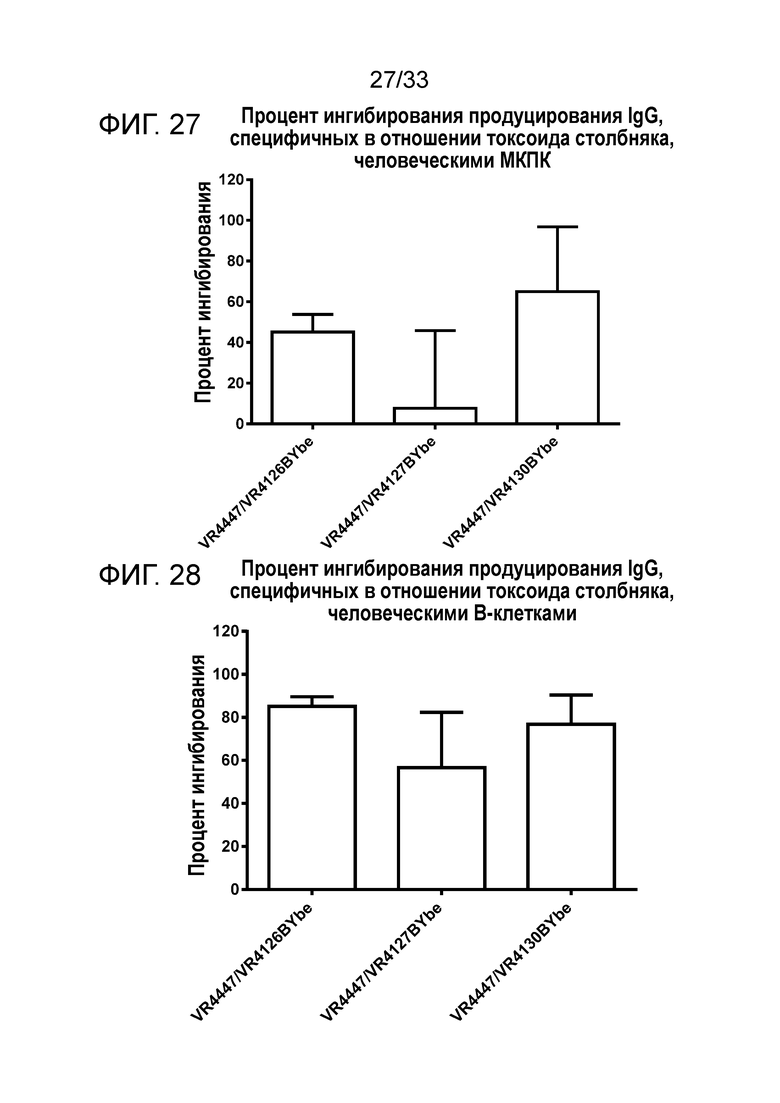
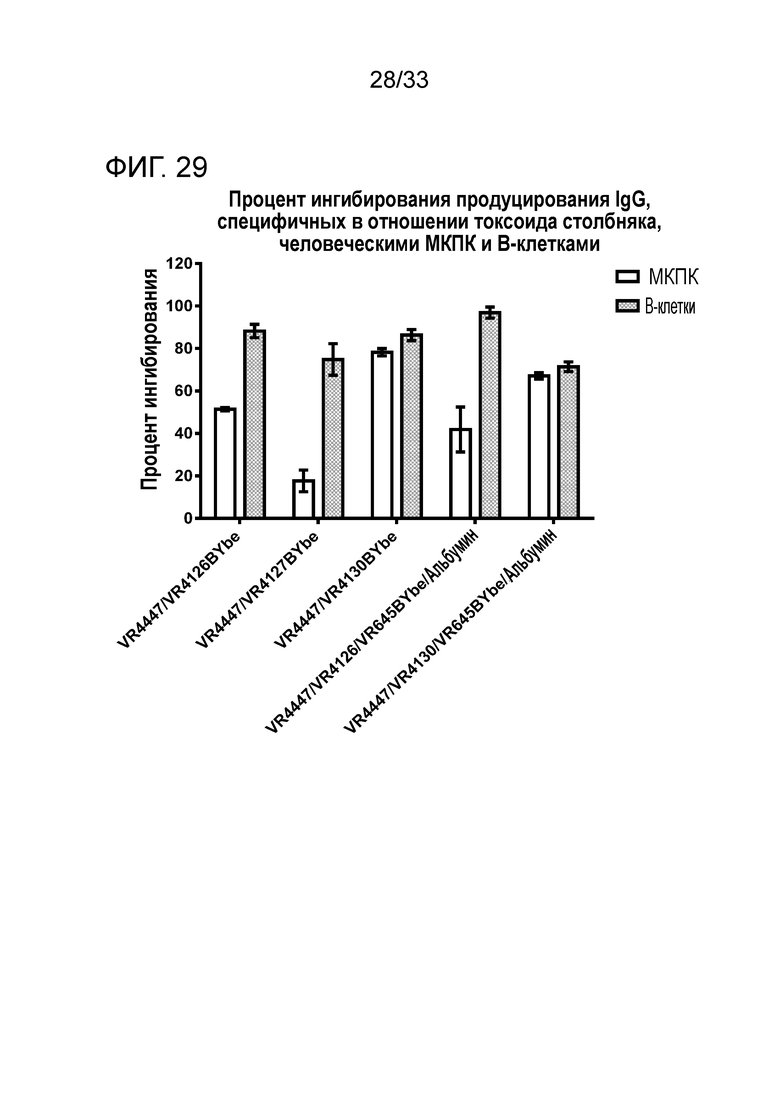
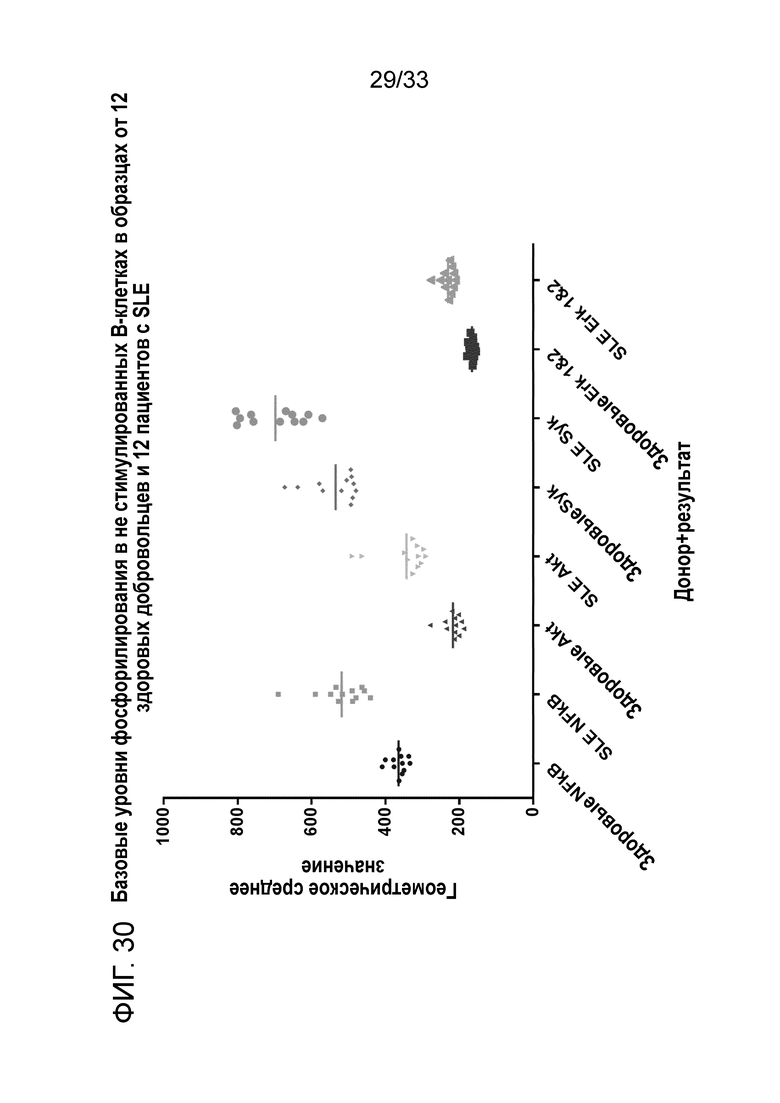
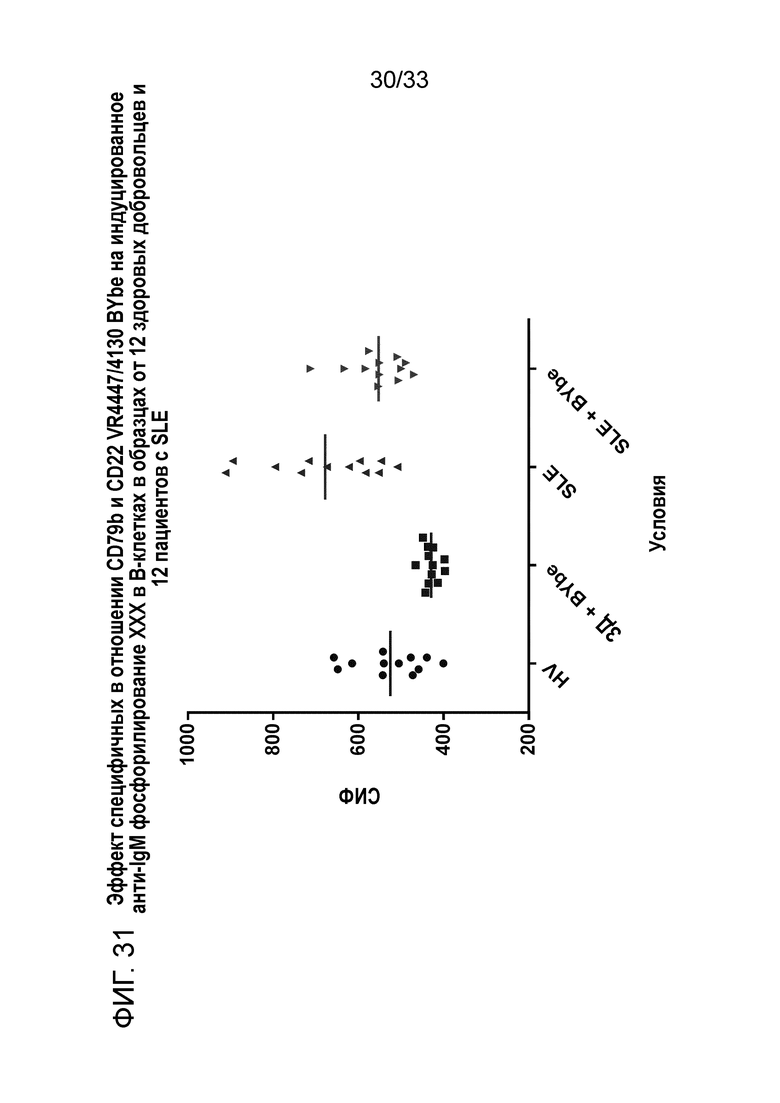

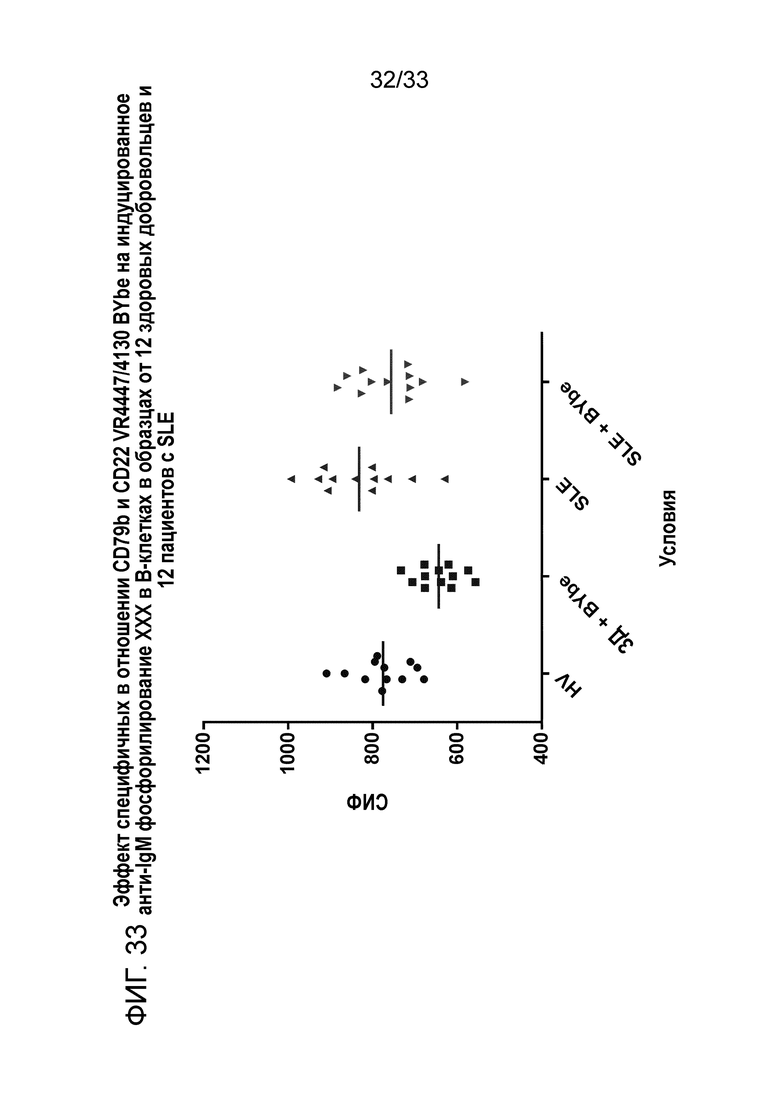
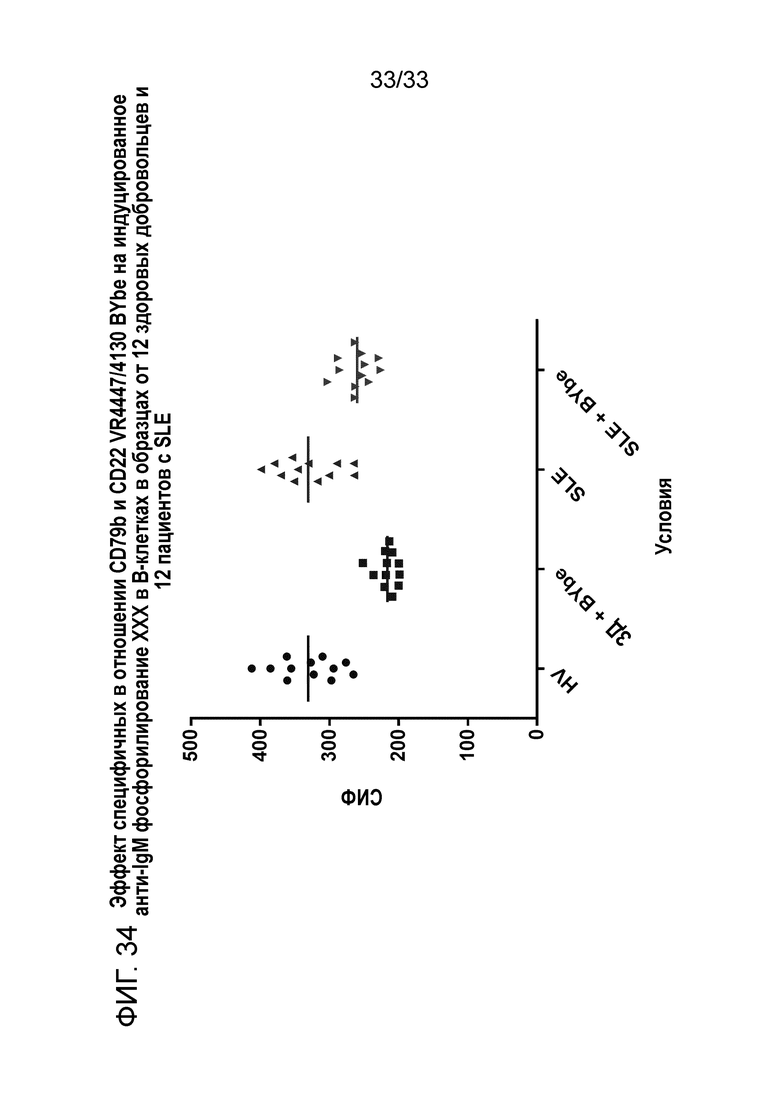
Изобретение относится к биотехнологии, в частности к мультиспецифической молекуле, содержащей связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79b. Изобретение может быть эффективно использовано в лечении аутоиммунного заболевания. 4 н. и 5 з.п. ф-лы, 9 табл., 11 пр., 34 ил.
1. Молекула, специфичная в отношении антигенов CD22 и CD79b, содержащая связывающий домен, специфичный в отношении антигена CD22, и связывающий домен, специфичный в отношении антигена CD79b,
где указанный связывающий домен, специфичный в отношении CD22, содержит
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 128 для CDRH1, SEQ ID NO: 129 для CDRH2 и SEQ ID NO: 130 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 125 для CDRL1, SEQ ID NO: 126 для CDRL2 и SEQ ID NO: 127 для CDRL3; или
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 138 для CDRH1, SEQ ID NO: 139 для CDRH2 и SEQ ID NO: 140 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 135 для CDRL1, SEQ ID NO: 136 для CDRL2 и SEQ ID NO: 137 для CDRL3, и
где указанный связывающий домен, специфичный в отношении CD79b, содержит
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 78 для CDRH1, SEQ ID NO: 79 для CDRH2 и SEQ ID NO: 80 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 75 для CDRL1, SEQ ID NO: 76 для CDRL2 и SEQ ID NO: 77 для CDRL3; или
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 88 для CDRH1, SEQ ID NO: 89 для CDRH2 и SEQ ID NO: 90 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 85 для CDRL1, SEQ ID NO: 86 для CDRL2 и SEQ ID NO: 87 для CDRL3.
2. Молекула по п. 1, где формат молекулы выбирают из диатела, sc-диатела, триатела, тандем-scFv, FabFv, Fab'Fv, FabdsFv, Fab-scFv, Fab-dsscFv, Fab-(dsscFv)2 diFab, diFab', тритела, тандем-scFv-Fc, scFv-Fc-scFv, sc-диатела-Fc, sc-диатела-CH3, Ig-scFv, scFv-Ig, V-Ig, Ig-V, дуотела и DVD-Ig.
3. Молекула по п. 1 или 2, где каждый связывающий домен является моноспецифическим.
4. Молекула по любому из пп. 1-3, содержащая не более одного связывающего домена, который является специфичным в отношении CD22, и не более одного связывающего домена, который является специфичным в отношении CD79b.
5. Молекула по любому из пп. 1-4, в которой связывающие домены являются гуманизированными.
6. Молекула, специфичная в отношении антигенов CD22, CD79b и сывороточного альбумина, содержащая связывающий домен, специфичный в отношении антигена CD22, связывающий домен, специфичный в отношении антигена CD79b, и связывающий домен, специфичный в отношении сывороточного альбумина,
где указанный связывающий домен, специфичный в отношении CD22, содержит
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 128 для CDRH1, SEQ ID NO: 129 для CDRH2 и SEQ ID NO: 130 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 125 для CDRL1, SEQ ID NO: 126 для CDRL2 и SEQ ID NO: 127 для CDRL3; или
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 138 для CDRH1, SEQ ID NO: 139 для CDRH2 и SEQ ID NO: 140 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 135 для CDRL1, SEQ ID NO: 136 для CDRL2 и SEQ ID NO: 137 для CDRL3, и
где указанный связывающий домен, специфичный в отношении CD79b, содержит
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 78 для CDRH1, SEQ ID NO: 79 для CDRH2 и SEQ ID NO: 80 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 75 для CDRL1, SEQ ID NO: 76 для CDRL2 и SEQ ID NO: 77 для CDRL3; или
3 области CDR тяжелой цепи, имеющие последовательности, приведенные в SEQ ID NO: 88 для CDRH1, SEQ ID NO: 89 для CDRH2 и SEQ ID NO: 90 для CDRH3, и 3 области CDR легкой цепи, имеющие последовательности, приведенные в SEQ ID NO: 85 для CDRL1, SEQ ID NO: 86 для CDRL2 и SEQ ID NO: 87 для CDRL3.
7. Молекула по п. 6, где указанный сывороточный альбумин представляет собой человеческий сывороточный альбумин.
8. Нуклеиновая кислота, кодирующая молекулу по любому из пп. 1-7.
9. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 8.
| WO 2007146968 A2, 21.12.2007 | |||
| WO 2014011519 A1, 16.01.2014 | |||
| УСТРОЙСТВО ДЛЯ ТРАНСПОРТИРОВКИ КОНВЕРТОВ И ОТНОСЯЩИЕСЯ К НЕМУ СПОСОБЫ | 2009 |
|
RU2514410C2 |

