Область изобретения
Настоящее изобретение относится к молекуле, являющейся, по меньшей мере, биспецифической для антигенов CD45 и CD79, составу, содержащему указанную молекулу, и применению любого из них в лечении. Настоящее изобретение также относится к способам получения указанных молекул и указанных составов. В независимом аспекте настоящее изобретение также относится к новым последовательностям и фрагментам антител, представленным в настоящем описании.
Уровень техники
Биологические механизмы in vivo представляют собой крайне сложные каскады сигналов, которые тяжело подвергнуть деконволюции и понять. Примером такой передачи сигнала является сигнал, необходимый для активации B-клеток. B-клеточный антигенный рецептор (BCR) состоит из мембранных молекул иммуноглобулина (mIg) и ассоциированных гетеродимеров Igα/Igβ (CD79a/CD79b) (α/β). Субъединицы mIg связывают антиген, что приводит к агрегации рецептора, в то время как субъединицы α/β передают сигнал внутрь клетки. Агрегация BCR быстро активирует киназы семейства Src Lyn, Blk и Fyn, а также тирозинкиназы Syk и Btk. Это инициирует образование "сигналосомы", состоящей из BCR, указанных выше тирозинкиназ, адаптерных белков, таких как CD19 и BLNK, и сигнальных ферментов, таких как PLCγ2, PI3K и Vav.
Сигналы, происходящие из сигналосомы, активируют множество каскадов передачи сигнала, включающих киназы, ГТФазы и факторы транскрипции. Это приводит к изменениям метаболизма клеток, экспрессии генов и организации цитоскелета. Сложность передачи сигнала BCR делает возможным множество различных исходов, включая выживание, толерантность (анергию) или апоптоз, пролиферацию и дифференцировку в антитело-продуцирующие клетки или B-клетки памяти. Исход ответа определяется стадией созревания клетки, природой антигена, величиной и длительностью передачи сигнала BCR и сигналами других рецепторов, таких как CD40, рецептор ИЛ-21 и BAFF-R.
Многие другие трансмембранные белки, некоторые из которых являются рецепторами, модулируют конкретные элементы передачи сигнала BCR. Некоторые из них включают CD45, CD19, CD22, PIR-B и FcγRIIB1 (CD32). Величина и длительность передачи сигнала BCR ограничена петлями отрицательной обратной связи, включая те, которые включают путь Lyn/CD22/SHP-1, путь Cbp/Csk, SHIP, Cbl, Dok-1, Dok-3, FcγRIIB1, PIR-B и интернализацию BCR.
In vivo B-клетки часто активируются антигенпрезентирующими клетками, захватывающими антигены и презентирующими их на своей клеточной поверхности. Для активации B-клеток такими мембрано-ассоциированными антигенами необходима BCR-индуцированная реорганизация цитоскелета.
Аутореактивные B-клетки отвечают за продукцию патогенных аутоантител, напрямую или косвенно вызывающих или усиливающих аутоиммунные состояния. Истощение CD20-положительных B-клеток используют для успешного лечения ряда аутоиммунных состояний, и, таким образом, окончательно установлено, что B-клетки играют важную роль в вызывании или поддержании ряда аутоиммунных заболеваний. Хотя истощение B-клеток является успешным терапевтическим вариантом, также существуют доказательства того, что контроль роста B-клеток и статуса активации также могут являться эффективным способом модуляции функции B-клеток. Таким образом, желательными являются альтернативные стратегии, не приводящие к истощению B-клеток и придающие гибкость контролю B-клеток без длительной супрессии B-клеточного иммунитета, которая, как показано, ассоциирована с некоторыми побочными эффектами. Кроме того, не все B-клеточные ответы или активности являются вредными, и доказательства позволяют предполагать, что поддержание популяций регуляторных B-клеток может являться протективным. Такой подход должен являться эффективным при заболеваниях, сопровождающихся аномальной функцией B-клеток, вызываемой неправильной или избыточной передачи сигнала BcR. Неограничивающие примеры включают воспаление, аутоиммунность и злокачественные новообразования. Конкретный интерес представляют заболевания, имеющие прямую потребность в передаче сигнала BcR или ингибировании или стимуляции гуморальных иммунных ответов.
В значительной степени ожидают, что биспецифические антитела будут играть основную роль в следующем поколении биотерапевтических средств (D. Holmes, Nature Rev Drug Disc Nov 2011:10; 798). Они обладают потенциалом превосходящей, длительной эффективности широкого спектра у более высокой доли пациентов. Этого можно достигать посредством одновременного совместного вовлечения различных антигенов в пределах общего пути заболевания, таким образом снижая избыточность; или посредством таргетинга антигенов из независимых путей для обеспечения аддитивного или синергического действия.
К настоящему времени стратегии ингибирования функции B-клеток без истощения B-клеток сфокусированы на использовании природного механизма регуляции с помощью CD32b (FcgRIIB). Они включают биспецифические антитела против CD79b/CD32b (Veri et al., Arthritis & Rheumatism 2010 62 1933-1943), CD19/CD32b (Karnell et al., J.Immunol 2014 192 1430-1490) и антитело против CD19 с Fc с усиленным связыванием CD32b (Chu et al., Arthritis & Rheumatology 2014 66 1153-1164).
В значительной степени ожидают, что биспецифические антитела будут играть основную роль в следующем поколении биотерапевтических средств (D. Holmes, Nature Rev Drug Disc Nov 2011:10; 798). Они обладают потенциалом превосходящей, длительной эффективности широкого спектра у более высокой доли пациентов. Этого можно достигать посредством одновременного совместного вовлечения различных антигенов в пределах общего пути заболевания, таким образом снижая избыточность; или посредством таргетинга антигенов из независимых путей для обеспечения аддитивного или синергического действия.
К настоящему времени стратегии ингибирования функции B-клеток без истощения B-клеток сфокусированы на использовании природного механизма регуляции с помощью CD32b (FcgRIIB). Они включают биспецифические антитела против CD79b/CD32b (Veri et al., Arthritis & Rheumatism 2010 62 1933-1943), CD19/CD32b (Karnell et al., J.Immunol 2014 192 1430-1490) и антитело против CD19 с Fc с усиленным связыванием CD32b (Chu et al., Arthritis & Rheumatology 2014 66 1153-1164).
Совместное лигирование Fc-рецептора гамма IIb (CD32b) с B-клеточным рецептором, по-видимому, естественным образом регулирует передачу сигнала, в частности, когда антиген связан с антителом в небольших иммунных комплексах. Затем CD32b рекрутирует фосфатазы SHP-1 и SHIP-1, противодействующие активации BcR. Хотя этот природный механизм регуляции может контролировать функцию B-клеток, нарушение функции CD32b, вызванное изменением белковой последовательности CD32b, может приводить к аутоиммунному заболеванию, и при аутоиммунном заболевании этот рецептор может подвергаться отрицательной регуляции, например, как в случае SLE. Таким образом, желательными являются альтернативные пути блокирования активности B-клеток, т.к. они представляют альтернативные, неприродные, пути регуляции функции BcR. Эти альтернативные механизмы вероятно являются особенно важными, если природные механизмы являются дисфункциональными при указанном заболевании.
Биспецифические антитела облегчают доступность новых биологических способов и средств, таких как:
1) перекрестное сшивание рецепторов на клетке, при необходимости,
2) индуцирование клеточно-опосредованных эффектов,
3) локализация цитокина в клетке для регуляции передачи сигнала или локального блокирования функции цитокинов,
4) одновременное вовлечение множества эпитопов для получения "новой активности", повышения функции или специфичности, которые не проявляет отдельное моноклональное антитело или, фактически, смеси несвязанных антител ("поли-моноклональных"), включая смеси против разных антигенов.
В нормальных физиологических условиях после связывания антигена CD45 исключается из комплекса BcR. Авторы настоящего изобретения неожиданно обнаруживали, что, используя биспецифическое антитело для соединения BcR (CD79) с молекулой CD45, можно ингибировать передачу сигнала BCR. Таким образом, физически связывая BcR с CD45 с использованием биспецифического антитела, авторы настоящего изобретения неожиданно обнаруживали, что можно ингибировать активацию B-клеток.
Таким образом, авторы настоящего изобретения идентифицировали синергическую функцию молекул, являющихся, по меньшей мере, биспецифическими для CD45 и CD79. По-видимому, эта функция является детектируемой, главным образом, когда связывающие области с комбинацией специфичностей предоставляют в биспецифическом (полиспецифическом) формате в отличие от их предоставления в виде смеси, например, моноклональных антител или их связывающих фрагментов.
Таким образом, полиспецифические молекулы по изобретению применимы в контроле аномальных функций B-клеток, ассоциированных с конкретными заболеваниями, такими как аутоиммунные заболевания и злокачественные новообразования.
Сущность изобретения
Таким образом, настоящее изобретение относится к полиспецифической молекуле, содержащей связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79.
Комбинация по настоящему изобретению в биспецифическом формате демонстрирует интересную биологическую активность в функциональных анализах in vitro, например, ингибирование передачи сигнала B-клеток, измеряемое с помощью любого из следующего: ингибирования фосфорилирования Akt S473, ингибирования фосфорилирования P38 и PLCγ2, ингибирования Y759 IkB в дополнение к ингибированию экспрессии CD86, CD71 и/или CD40 на B-клетках. Тот же уровень активности не является очевидным для отдельных компонентов в отдельности или компонентов, предоставленных в смеси. Однако, активность является очевидной при предоставлении биспецифической конструкции со специфичностью для CD45 и CD79.
Ингибирование конкретных функций B-клеток, наблюдаемое в этих анализах, свидетельствует о том, что полиспецифическую молекулу по изобретению, содержащую связывающий домен, специфичный для CD45, и связывающий домен, специфичный для CD79, можно использовать для изменения функции B-клеток и, например, для получения терапевтического средства, альтернативного истощению B-клеток.
Передача сигнала через B-клеточный рецептор является критической функцией B-клетки и требованием для антигенспецифической активации B-клеток. Передача сигнала BcR является критической с ранних стадий развития B-клеток до активации и развития ответов B-клеток памяти. B-клеточный рецептор состоит из поверхностной молекулы иммуноглобулина (Ig), ассоциированной с гетеродимерным комплексом CD79a и CD79b. Считают, что, когда поверхностный Ig распознает антиген, это приводит к кластеризации комплекса CD79a/b, что приводит к нижележащей активации промежуточного каскада передачи сигнала, включающего киназы семейства Src, а также тирозинкиназы Syk и Btk. Затем этот сигнальный комплекс может рекрутировать адаптерные белки, такие как CD19 и BLNK, и приводить к активации PLCγ2 и PI3K, которые, в свою очередь, могут активировать дополнительные нижележащие пути, такие как те, которые контролируют рост, выживаемость и дифференцировку B-клеток. Это сигнальный комплекс может дополнительно регулироваться другими вторыми сигналами посредством передачи сигнала через BAFF-R, ИЛ-21R и CD40 и также может регулироваться другими сигнальными молекулами, такими как, среди прочего, CD19, CD21, CD83, CD22, CD32b и CD45. После распознавания антигена посредством BcR одним из первых активируемых ответов является положительная регуляция поверхностных рецепторов, таких как костимуляторные молекулы CD80 и CD86. Эти молекулы связываются с соответствующими рецепторами на T-клетках, передающими дополнительные сигналы выживаемости и активации, делающие возможной выживаемость и экспансию T-клеток, распознающих антиген в случае MHC класса II. Этот ответ дополнительно амплифицируется способностью B-клеток презентировать антиген в случае MHC класса II обратно T-клетке, высвобождающей такие факторы, как ИЛ-2 и ИЛ-21. Эти цитокины, в свою очередь, значительно повышают количество B-клеток.
Кроме того, ингибирование передачи сигнала через B-клеточный рецептор может приводить к ингибированию нижележащих функций. Одним из таких исходов может являться ингибирование костимуляторных молекул, таких как CD86 (или их сниженная экспрессия), которое будет приводить к ингибированию функции, выживаемости и дифференцировки T-клеток.
Таким образом, ингибирование передачи сигнала через B-клеточный рецептор может являться полезным в контроле аномальных функций B-клеток, ассоциированных с аутоиммунностью и злокачественными новообразованиями. Передача сигнала через B-клеточный рецептор необходима для пролиферации, дифференцировки, презентации антигена и высвобождения цитокинов B-клетками при аутоиммунном заболевании. Таким образом, ингибирование активности BcR может регулировать функции B-клеток, таких как секреция иммуноглобулинов, активация T-клеток и контроль неправильной активности B-клеток, ассоциированной, например, с аутоиммунными состояниями. Кроме того, существует несколько B-клеточных лейкозов и лимфом, в случае которых передача сигнала через B-клеточный рецептор необходима для выживаемости и роста, которые можно контролировать с помощью ингибиторов активации B-клеточного рецептора.
В одном из вариантов осуществления каждый связывающий домен или связывающие домены полиспецифических молекул по настоящему изобретению независимо содержат один или два (например, два) вариабельных домена антитела, специфичных для соответствующего антигена (такого как CD45 или CD79, или дополнительного антигена, если молекула является, по меньшей мере, триспецифической).
Как применяют в настоящем описании, термин "CD79" относится к комплексу, состоящему из CD79a и CD79b. Таким образом, антитела, связывающиеся с CD79, могут связываться с CD79a и/или CD79b. Как применяют в настоящем описании, термин "связывается с CD79a и/или CD79b" относится к: специфичным для CD79a, специфичным для CD79b, специфичным для CD79a и b (т.е. распознает эпитоп на CD79a, а также распознает эпитоп на CD79b, т.е. является панспецифическим) или является специфичным для комплекса CD79a и CD79b (т.е. распознает эпитоп, образующийся при взаимодействии CD79a и CD79b в форме комплекса).
В одном из вариантов осуществления антитело или его связывающий фрагмент, используемое в молекулах по настоящему изобретению, является специфичным для CD79a.
В одном из вариантов осуществления антитело или его связывающий фрагмент, используемое в молекулах по настоящему изобретению, является специфичным для CD79b.
В одном из вариантов осуществления антитело или его связывающий фрагмент, используемое в молекулах по настоящему изобретению, является специфичным для комплекса CD79, т.е. оно распознает эпитоп, присутствующий в комплексе, и является специфичным для него, например, эпитоп, включающий взаимодействие между CD79a и CD79b.
В одном из вариантов осуществления, даже если связывающий домен является специфичным для CD79a или CD79b, следует понимать, что связывающий домен все равно будет связываться с CD79a или CD79b в форме комплекса.
Если в связывающем домене или в каждом связывающем домене есть две вариабельные области, то две вариабельные области, как правило, будут действовать совместно, приводя к специфичности для соответствующего антигена, например, они являются когнатной парой или аффинно созревают, приводя к достаточной аффинности таким образом, что домен является специфичным для конкретного антигена. Как правило, они являются парой вариабельных областей тяжелой и легкой цепи (парой VH/VL).
В одном из вариантов осуществления молекула по настоящему изобретению является биспецифической.
В одном из вариантов осуществления молекула по настоящему изобретению является триспецифической, например, если третий связывающий домен является специфичным для сывороточного альбумина, например, сывороточного альбумина человека.
В одном из вариантов осуществления молекула по настоящему изобретению является моноспецифической для CD79 и CD45, т.е. молекула содержит только один связывающий домен, связывающийся с CD79, и один связывающий домен, связывающийся с CD45.
В одном из вариантов осуществления полиспецифическая молекула по настоящему изобретению является одноцепочечной.
В одном из вариантов осуществления полиспецифическая молекула по настоящему изобретению содержит тяжелую цепь, а также легкую цепь. В одном примере, как применяют в настоящем описании, спаривание тяжелой и легкой цепи не обозначают как димер, в частности, если в одном из вариантов осуществления молекула по настоящему изобретению не содержит мультимеры, такие как димеры антитела, единицу/фрагмент или компоненты.
В одном из аспектов настоящее изобретение относится к молекуле полиспецифического антитела, содержащей или состоящей из:
a) полипептидной цепи формулы (I):
VH-CH1-X-(V1)p;
b) полипептидной цепи формулы (II):
VL-CL-Y-(V2)q;
где:
VH представляет собой вариабельный домен тяжелой цепи;
CH1 представляет собой домен константной области тяжелой цепи, например, ее домен 1;
X представляет собой связь или линкер, например, аминокислотный линкер;
Y представляет собой связь или линкер, например, аминокислотный линкер;
V1 представляет собой dab, scFv, dsscFv или dsFv;
VL представляет собой вариабельный домен, например, вариабельный домен легкой цепи;
CL представляет собой домен константной области, например, домен константной области легкой цепи, такой как Ckappa;
V2 представляет собой dab, scFv, dsscFv или dsFv;
p является 0 или 1;
q является 0 или 1; и
если p является 1, q является 0 или 1, и если q является 1, p является 0 или 1, т.е. p и q оба не представляют собой 0.
В одном из вариантов осуществления молекула содержит не более одного участка связывания для CD45 и не более одного участка связывания для CD79.
Указанный выше формат является особенно пригодным для скрининга комбинаций вариабельных областей, например, в более длительных анализах и терапевтического использования.
В одном из вариантов осуществления q является 0, и p является 1.
В одном из вариантов осуществления q является 1, и p является 1.
В одном из вариантов осуществления V1 представляет собой dab, и V2 представляет собой dab, и вместе они образуют единый связывающий домен совместно действующей пары вариабельных областей, такой как когнатная пара VH/VL.
В одном из вариантов осуществления VH и VL являются специфичными для CD79, например, CD79a или CD79b.
В одном из вариантов осуществления V1 является специфичным для CD79, например, CD79a.
В одном из вариантов осуществления V2 является специфичным для CD79, например, CD79a.
В одном из вариантов осуществления V1 является специфичным для CD79, например, CD79b.
В одном из вариантов осуществления V2 является специфичным для CD79, например, CD79b.
В одном из вариантов осуществления V1 и V2 вместе (например, в виде одного связывающего домена) являются специфичными для CD79, например, CD79a или CD79b.
В одном из вариантов осуществления VH и VL являются специфичными для CD45.
В одном из вариантов осуществления V1 является специфичным для CD45.
В одном из вариантов осуществления V2 является специфичным для CD45.
В одном из вариантов осуществления V1 и V2 вместе (например, в виде одного связывающего домена) являются специфичными для CD45.
В одном из вариантов осуществления молекула по настоящему изобретению является или содержит слитый белок.
В одном из вариантов осуществления настоящее изобретение относится к полиспецифической молекуле по настоящему изобретению, являющейся биспецифическим белковым комплексом, имеющим формулу A-X:Y-B, где:
A-X является первым слитым белком;
Y-B является вторым слитым белком;
X:Y является гетеродимерным соединением;
A содержит первый связывающий домен, специфичный для CD45, CD79a, CD79b или комплекса CD79a и b;
B содержит второй связывающий домен, специфичный для CD45, CD79a, CD79b или комплекса CD79a и b;
X является первым партнером по связыванию из связывающейся пары;
Y является вторым партнером по связыванию из связывающейся пары; и
: представляет собой взаимодействие (такое как связывающее взаимодействие) между X и Y, и
где по меньшей мере один из A или B является специфичным для CD45, а другой является специфичным для CD79a, CD79b или их комплексной формы.
Указанный выше формат является удобным, т.к. он обеспечивает быстрый и эффективный путь сборки биспецифических форматов, которые, например, можно подвергать тестированию in vitro в функциональных анализах. Это может облегчать выбор предпочтительной пары вариабельных областей, которые затем можно встраивать в альтернативный, терапевтический формат полиспецифического антитела.
Не будучи связанными какой-либо теорией, различные пермутации вариабельных областей, специфичных для CD45, в комбинации с диапазоном вариабельных областей, специфичных для CD79, могут обеспечивать доступ к различным нюансам биологической функции.
Изобретение также относится к новым антителам против CD45, например, для применения в полиспецифических молекулах по настоящему изобретению или для встраивания в любой другой подходящий формат антитела.
Изобретение также относится к новым антителам против CD79, например, для применения в полиспецифических молекулах по настоящему изобретению или для встраивания в любой другой подходящий формат антитела.
Краткое описание чертежей
Фигура 1 является гистограммой относительной активности ингибирования фосфорилированного Akt в случае биспецифических и бивалентных комбинаций антител со специфичностью для CD45 и CD79b.
Фигура 2 является гистограммой относительной активности ингибирования фосфорилированного PLCg2 в случае биспецифических и бивалентных комбинаций антител со специфичностью для CD45 и CD79b.
Фигура 3 является графиком, на котором показано титрование эффекта биспецифической комбинации CD45 и CD79b в отношении экспрессии CD86 на стимулированных антителом против IgM B-клеток.
Фигура 4 является графиком ингибирования фосфорилированного PLCg2 в случае биспецифических белков со специфичностью для CD45 и CD79b с разными V-областями.
Фигура 5 является выдержкой из Chan and Carter Reviews Immunology vol 10, May 2010, 301, где показаны конкретные форматы антител.
На фигуре 6 показаны данные о перекрестных специфичностях сетки антигенов. Антиген 2=CD79b и антиген 4=CD45. Значения представляют собой процентную долю ингибирования (отрицательное значение для активации) фосфорилирования Syk и среднее значение для множества оцениваемых комбинаций V-областей.
На фигуре 7 показаны данные о перекрестных специфичностях сетки антигенов. Антиген 2=CD79b и антиген 4=CD45. Значения представляют собой процентную долю ингибирования (отрицательное значение для активации) PLCγ2 и среднее значение для множества оцениваемых комбинаций V-областей.
На фигуре 8 показаны данные о перекрестных специфичностях сетки антигенов. Антиген 2=CD79b и антиген 4=CD45. Значения представляют собой процентную долю ингибирования (отрицательное значение для активации) AKT и среднее значение для множества оцениваемых комбинаций V-областей.
На фигуре 9 показана процентная доля ингибирования фосфорилирования Syk, PLCγ2 и AKT для каждой комбинации V-областей в случае специфичности Fab-X для CD79b в комбинации со специфичностью Fab-Y для CD45.
На фигуре 10 показана процентная доля ингибирования фосфорилирования Syk, PLCγ2 и AKT для каждой комбинации V-областей в случае специфичности Fab-Y для CD79b в комбинации со специфичностью Fab-X для CD45.
На фигура 11 и 12 показано ингибирование PLCγ2 (+/-SD) очищенным CD79b-CD45 (транзиторно экспрессируемым) на стимулируемых IgM B-клетках донора 129 и 130.
На фигуре 13 и 14 показано ингибирование p38 (+/-SD) очищенным CD79b-CD45 (транзиторно экспрессируемым) на стимулируемых IgM B-клетках донора 129 и 130.
На фигура 15 и 16 показано ингибирование Akt (+/-SD) очищенным CD79b-CD45 (транзиторно экспрессируемым) на стимулируемых IgM B-клетках донора 129 и 130.
На фигуре 17 показано ингибирование продукции IgG против столбнячного токсина из PBMC, культивируемых с различными полиспецифическими молекулами.
Подробное описание изобретения
Как применяют в настоящем описании, термин "полиспецифическая молекула" относится к молекуле со способностью специфически связываться по меньшей мере с двумя отдельными антигенами, например, разными антигенами. В одном из вариантов осуществления полиспецифическая молекула является биспецифической, триспецифической или тетраспецифической молекулой, в частности, биспецифической молекулой или триспецифической молекулой.
Таким образом, в одном из аспектов настоящее изобретение относится к молекуле подходящего формата, специфичного, по меньшей мере, для CD45 и CD79a, и применению антител/фрагментов или их комбинаций, специфичных для CD45 и CD79a, в полиспецифической молекуле, такой как биспецифический формат или триспецифический формат.
Таким образом, в одном из аспектов настоящее изобретение относится к молекуле подходящего формата, специфичного, по меньшей мере, для CD45 и CD79b, и применению антител/фрагментов или их комбинаций, специфичных для CD45 и CD79b, в полиспецифической молекуле, такой как биспецифический формат или триспецифический формат.
Таким образом, в одном из аспектов настоящее изобретение относится к молекуле подходящего формата, специфичной, по меньшей мере, для комплекса CD45 и CD79a/b, и применению антител/фрагментов или их комбинаций, специфичных для комплекса CD45 и CD79a/b, в полиспецифической молекуле, такой как биспецифический формат или триспецифический формат.
В одном из вариантов осуществления молекула по настоящему изобретению является триспецифической, например, где третий связывающий домен способен повышать время полужизни молекулы, например, посредством связывания сывороточного белка-носителя.
В плазме есть множество белков, и они включают тироксин-связывающий белок, транстиретин, α1-кислый гликопротеин, трансферрин, фибриноген и альбумин или фрагмент любого из них (Bartalena & Robbins, 1993, Clinics in Lab. Med. 13:583-598; Bree et al., 1986, Clin. Pharmacokin. 11:336-342; Gitlin et al. 1964, J. Clin. Invest. 10:1938-1951; Peters, 1985, Adv Protein Chem. 37:161-245; Waldeman & Strober, 1969, Progr. Allergy, 13:1-110). В одном примере третий связывающий домен является специфичным для сывороточного альбумина, например, сывороточного альбумина человека.
Полиспецифические форматы молекул
В этой области известны примеры подходящих полиспецифических молекул, например, как описано в обзоре ʺThe coming of Age of Engineered Multivalent Antibodies, Nunez-Prado et al Drug Discovery Today Vol 20 Number 5 Mar 2015, page 588-594, D. Holmes, Nature Rev Drug Disc Nov 2011:10; 798, Chan and Carter, Nature Reviews Immunology vol 10, May 2010, 301, включенном в настоящее описание в качестве ссылки.
В одном из вариантов осуществления полиспецифические форматы включают форматы, известные в этой области и представленные в настоящем описании, такие как те, где формат молекулы выбран из группы, содержащей или состоящей из:
• Тандемного sdAb, тандемного sdAb-sdAb (трех sdAb);
• (scFv)2 (также обозначаемого как тандемный scFv), scFv-dsFv, dsscFv-dsFv (dsFv)2;
• Диатела, ds-диатела, ди-ds-диатела,
• Sc-диатела, dssc-диатела, ди-dssc-диатела;
• Антитела Dart, т.е. VL1-линкер-VH2-линкер и VH1-линкер-VL2, где C-концы VH1 и VH2 соединены дисульфидной связью;
• BiTE®, dsBiTE, ди-dsBiTE;
• Ди-диатела (см. Nunez-Prado et al, в частности, молекулу № 25 на фиг. 1), ds-ди-диатела, ди-ds-ди-диатела;
• Триатела, ds-триатела, ди-ds-триатела, три-ds-триатела;
• Тетрател, ds-тетратела, ди-ds-тетратела, три-ds-тетратела, тетра-ds-тетратела;
• tandAb (см. Nunez-Prado et al, в частности, молекулу № 22 на фиг. 1); ds-tandab, ди-ds-tandab, три-ds-tandab, тетра-ds-tandab;
• [sc(Fv)2]2, (см. Nunez-Prado et al, в частности, молекулу № 22 на фиг. 1), ds[sc(Fv)2]2, ди-ds[sc(Fv)2]2, три-ds[sc(Fv)2]2, тетра-ds[sc(Fv)2]2;
• Пентатела (см. Nunez-Prado et al, в частности, молекулу № 27 на фиг. 1);
• Fab-scFv (также обозначаемого как битело), FabʹscFv, FabdsscFv (или BYbe), FabʹdsscFv;
• тритела, ds-тритела, ди-ds-тритела (также обозначаемого как Fab-ди-dsscFv или TrYbe или Fab-(dsscFv)2), Fabʹди-dsscFv;
• Fabdab, FabFv, Fabʹdab, FabʹFv;
• Fa-один линкер-Fv (также обозначаемого в настоящем описании как FabdsFv, как описано в WO2014/096390), Fabʹ-один линкер-Fv (также обозначаемого в настоящем описании как FabʹdsFv);
• FabscFv-один линкер-Fv, FabʹscFv-один линкер-Fv;
• FabdsscFv-один линкер-Fv, FabʹdsscFv-один линкер-Fv;
• FvFabFv, FvFabʹFv, dsFvFabFv, dsFvFabʹFv, FvFabdsFv, FvFabʹdsFv, dsFvFabdsFv, dsFvFabʹdsFv,
• FabFvFv, FabʹFvFv, FabdsFvFv, FabʹdsFvFv, FabFvdsFv, FabʹFvdsFv, FabdsFvdsFv, FabʹdsFvdsFv,
• diFab, diFabʹ, включая химически конъюгированный diFabʹ,
• (FabscFv)2, (Fab)2scFvdsFv, (Fab)2dsscFvdsFv, (FabdscFv)2,
• (FabʹscFv)2, (Fabʹ)2scFvdsFv, (Fabʹ)2dsscFvdsFv, (FabʹdscFv)2,
• VHHCK (см. Nunez-Prado et al, в частности, молекулу № 6 на фиг. 1);
• минитела, ds-минитела, ди-ds-минитела,
• миниантитела (ZIP) [см. Nunez-Prado et al, в частности, молекулу № 7 на фиг. 1], ds-миниантитела (ZIP) и ди-ds-миниантитела (ZIP);
• триби-минитела [см. Nunez-Prado et al, в частности, молекулу № 15 на фиг. 1], ds-триби-минитела, ди-ds-триби-минитела, три-ds-триби-минитело;
• диатела-CH3, ds-диатела-CH3, ди-ds-диатела-CH3, sc-диатела-CH3, dssc-диатела-CH3, ди-dssc-диатела-CH3,
• тандемного scFv-CH3, тандемного dsscFv-CH3, тандемного ди-dsscFv-CH3, тандемного три-dsscFv-CH3, тандемного тетра-dsscFv-CH3,
• scFv-Fc (также обозначаемого в настоящем описании как (scFvCH2CH3)2), как описано в WO2008/012543, и их одноцепочечной версии, dsscFvscFv-Fc, dsscFv-Fc (также обозначаемого в настоящем описании как (dsscFvCH2CH3)2), scFv-dsFv-Fc, dsscFv-dsFv-Fc, dsFv-Fc (также обозначаемого в настоящем описании как (dsFvCH2CH3)2),
• молекулы типа "скорпион" (Trubion), т.е. связывающего домена, линкера-CH2CH3 связывающего домена, как описано в патенте США № 8409577;
• SMIP (Trubion) т.е. (scFv-CH2CH3)2;
• (dsFvCH2CH3)2, тандемного scFv-Fc, тандемного dsscFvscFv-Fc, тандемного dsscFv-Fc,
• scFv-Fc-scFv, dsscFv-Fc-scFv, scFv-Fc-dsscFv,
• диатела-Fc, ds-диатела-Fc, ди-ds-диатела-Fc, триатела-Fc, ds-триатела-Fc, ди-ds-триатела-Fc, три-ds-триатела-Fc, тетратела-Fc, ds-тетратела-Fc, ди-ds-тетратела-Fc, три-ds-тетратела-Fc, тетра-ds-тетратела-Fc, ds-тетратела-Fc, ди-ds-тетратела-Fc, три-ds-тетратела-Fc, тетра-ds-тетратела-Fc, sc-диатела-Fc, dssc-диатела, ди-dssc-диатела;
• би- или трифункционального антитела, например, с разными вариабельными областями тяжелой цепи и общими легкими цепями, например, формат биспецифического антитела Merus (Biclonics®) с общими легкими цепями с фиксированной последовательностью и разными тяжелыми цепями (включая разные CDR) и сконструированным доменом CH3 для регуляции димеризации разных тяжелых цепей,
• дуотела (т.е. антитела, где одна полноразмерная цепь в антителе имеет специфичность, отличающуюся от другой полноразмерной цепи в антителе);
• полноразмерного антитела, где для получения биспецифического формата используют обмен плеча Fab;
• би- или трифункционального антитела, где полноразмерное антитело имеет общую тяжелую цепь и разные легкие цепи, также обозначаемого как каппа/лямбда-тело или κ/λ-тело, см., например, WO2012/023053, включенный в настоящее описание в качестве ссылки;
• антитела, включающего один, два, три или четыре Ig-scFv с C-конца тяжелой или легкой цепи, один, два, три или четыре scFv-Ig с N-конца тяжелой или легкой цепи, один линкер Ig-Fv, один, два, три или четыре Ig-dsscFv с C-конца тяжелой или легкой цепи (с одной, двумя, тремя или четырьмя дисульфидными связями);
• антитела, включающего один, два, три или четыре Ig-dsscFv с N-конца тяжелой или легкой цепи (с одной, двумя, тремя или четырьмя дисульфидными связями),
• Ig-один линкер-Fv (см. PCT/EP2015/064450),
• Ig-dab, dab-Ig, scFv-Ig, V-Ig, Ig-V,
• scFabFvFc, scFabdsFvFc (версия scFavFv с одним линкером), (FabFvFc)2, (FabdsFvFc)2, scFabʹFvFc, scFabʹdsFvFc, (FabʹFvFc)2, (FabʹdsFvFc)2 и
• DVDIg, более подробно описываемые ниже.
В одном из вариантов осуществления форматы полиспецифической молекулы включают форматы, известные в этой области и представленные в настоящем описании, например, где формат молекулы выбран из группы, содержащей или состоящей из: диатела, sc-диатела, триатела, тритела, тетрател, тандемного scFv, FabFv, FabʹFv, FabdsFv, Fab-scFv, Fab-dsscFv, Fab-(dsscFv)2, diFab, diFabʹ, тандемного scFv-Fc, scFv-Fc-scFv, sc-диатела-Fc, sc-диатела-CH3, Ig-scFv, scFv-Ig, V-Ig, Ig-V, дуотела и DVDIg, более подробно описываемых ниже.
В одном из вариантов осуществления молекула полиспецифического антитела по настоящему изобретению не содержит домен Fc т.е. не содержит домен CH2 и CH3, например, молекула выбрана из группы, содержащей тандемный scFv, scFv-dsFv, dsscFv-dsFv ди-dsFv, диатело, ds-диатело, ди-ds-диатело, sc-диатело (также обозначаемое как (scFv)2), dssc-диатело, триатело, ds-триатело, ди-ds-триатело, три-ds-триатело, тетратела, ds-тетратело, ди-ds-тетратело, три-ds-тетратело, тетра-ds-тетратело, тритело, ds-тритело, ди-ds-тритело, Fabdab, FabFv, Fabʹdab, FabʹFv, Fab-один линкер-Fv (как описано в WO2014/096390), Fabʹ-один линкер-Fv, FabdsFv, FabʹdsFv, Fab-scFv (также обозначаемое как битело), FabʹscFv, FabdsscFv, FabʹdsscFv, Fabʹди-dsscFv, Fabʹди-dsscFv, FabscFv-один линкер-Fv, FabʹscFv-один линкер-Fv, FabdsscFvs-один линкер-Fv, FabʹdsscFv-один линкер-Fv, FvFabFv, FvFabʹFv, dsFvFabFv, dsFvFabʹFv, FvFabdsFv, FvFabʹdsFv, dsFvFabdsFv, dsFvFabʹdsFv, FabFvFv, FabʹFvFv, FabdsFvFv, FabʹdsFvFv, FabFvdsFv, FabʹFvdsFv, FabdsFvdsFv, FabʹdsFvdsFv, diFab, diFabʹ, включая химически конъюгированное diFabʹ, (FabscFv)2, (Fab)2scFvdsFv, (Fab)2dsscFvdsFv, (FabdscFv)2, минитело, ds-минитело, ди-ds-минитело, диатело-CH3, ds-диатело-CH3, ди-ds-диатело-CH3, sc-диатело-CH3, dssc-диатело-CH3, ди-dssc-диатело-CH3, тандемное scFv-CH3, тандемное dsscFv-CH3, тандемное ди-dsscFv-CH3, тандемное три-dsscFv-CH3 и тандемное тетра-dsscFv-CH3.
В одном из вариантов осуществления молекула по настоящему изобретению не содержит домен Fc.
В одном из вариантов осуществления молекула по настоящему изобретению содержит измененный домен Fc, как представлено в настоящем описании ниже.
Как применяют в настоящем описании, термин "домен Fc", в основном, относится к -(CH2CH3)2, если контекст четко не указывает на иное.
В одном из вариантов осуществления молекула по настоящему изобретению не содержит -CH2CH3-фрагмент.
В одном из вариантов осуществления молекула по настоящему изобретению не содержит домен CH2.
В одном из вариантов осуществления молекула по настоящему изобретению не содержит домен CH3.
Как применяют в настоящем описании, термин "молекула" используют в его биохимическом смысле для обозначения группы атомов, образующих органическую, в частности, белковоподобную массу, включающую комплекс, подходящий для обращения как с отдельной молекулой в соответствующих условиях после образования комплекса, например, комплекса, образованного двумя или более полипептидными цепями.
Термины "молекула" и "конструкция" в настоящем описании используют взаимозаменяемо, если контекст не указывает на иное. Хотя термин "конструкция" можно использовать чаще для обозначения молекулы полинуклеотида, а термин "молекула" можно использовать чаще для обозначения частицы, главным образом, содержащей аминокислотную последовательность.
Как применяют в настоящем описании, термин "специфичность" (или "специфичный") относится к случаю, когда партнеры по взаимодействию распознают только друг друга или имеют значительно более высокую аффинность друг для друга по сравнению с не-партнерами, например, по меньшей мере в 2, 3, 4, 5, 6, 7, 8, 9, 10 раз более высокую аффинность, чем, например, фоновый уровень связывания или связывание с другим неродственным белком.
Как применяют в настоящем описании, термин "связывающий домен" относится к связывающей области, как правило, полипептиду, способному связываться с антигеном-мишенью, например, с достаточной аффинностью для характеризации домена как специфичного для антигена.
В полиспецифических молекулах по настоящему изобретению можно использовать любые подходящие связывающие домены. Их можно получать из любого подходящего источника.
В одном из вариантов осуществления в связывающей домене молекул по настоящему изобретению используют биосовместимую структуру каркаса, и такие структуры основаны на белковых каркасах или скелетах, иных, чем иммуноглобулиновые домены. Например, можно использовать те, которые основаны на фибронектине, анкирине, липокалине, неокарциностатине, цитохроме b, CP1 с цинковыми пальцами, PST1, биспирали, LACI-D1, Z-домене и доменах тендрамисата (см., например, Nygren and Uhlen, 1997, Current Opinion in Structural Biology, 7, 463-469).
Как применяют в настоящем описании, термин "полиспецифические молекулы" также могут включать связывающие средства на основе биологических каркасов, включая аднектины, аффитела, дарпины, филомеры, авимеры, аптамеры, антикалины, тетранектины, микротела, аффилины и домены Куница.
Полиспецифическая молекула по настоящему изобретению, как правило, является молекулой полиспецифического антитела, т.е. по меньшей мере один или несколько связывающих доменов полиспецифической молекулы получают из антитела или его фрагмента.
Если связывающий домен получают из антитела, "связывающий домен или участок", как применяют в настоящем описании, является частью антитела, контактирующей с антигеном. В одном из вариантов осуществления связывающий домен содержит по меньшей мере один вариабельный домен или его производное, например, пару вариабельных доменов или их производных, таких как когнатная пара вариабельных доменов или их производных. Как правило, она является парой VH/VL.
Вариабельные области (также обозначаемые в настоящем описании как вариабельные домены), как правило, содержат 3 CDR и подходящий каркас. В одном из вариантов осуществления связывающий домен содержит две вариабельные области, вариабельную область легкой цепи и вариабельную область тяжелой цепи, и вместе эти элементы вносят свой вклад в специфичность связывающего взаимодействия антитела или связывающего фрагмента.
Как применяют в настоящем описании, термин "когнатная пара" относится к паре вариабельных доменов (или их производному, такому как их гуманизированная версия) тяжелой и легкой цепи, выделенной из хозяина в виде предварительно образовавшейся пары. Это определение не включает вариабельные домены, выделенные из библиотеки, где исходная пара из хозяина не сохраняется. Когнатные пары могут являться предпочтительными, т.к. зачастую они являются аффинно зрелыми в хозяине и, таким образом, могут иметь более высокую аффинность к антигену, к которому они являются специфичными, чем комбинация пар вариабельных доменов, выбранных из библиотеки, такой как фаговая библиотека.
Как применяют в настоящем описании, термин "производное природного домена" предназначен для обозначения того, одна, две, три, четыре или пять аминокислот в природной последовательности подвергнуты замене или делеции, например, для оптимизации свойств домена, такой как устранение нежелательных свойств, но с сохранением характерных признаков домена. Примерами модификаций являются удаление участков гликозилирования, GPI-якорей или экспонируемых в направлении растворителя лизинов. Эти модификации можно осуществлять посредством замены важных аминокислотных остатков с использованием консервативной аминокислотной замены.
Другая модификация в CDR может включать, например, замену одного или нескольких цистеинов, например, остатком серина. Asn может являться субстратом дезаминирования, и это свойство можно снижать посредством замены Asn и/или соседней аминокислоты альтернативной аминокислотой, такой как консервативная замена. Аминокислоту Asp в CDR можно подвергать изомеризации. Последнее можно минимизировать посредством замены Asp и/или соседней аминокислоты альтернативной аминокислотой, например, консервативной замены.
Гуманизированные версии вариабельной области также являются его производным в контексте настоящего описания. Гуманизация может включать замену не принадлежащего человеку каркаса каркасом человека и, необязательно, обратную мутацию одного или нескольких остатков в "донорные остатки". Как применяют в настоящем описании, термин "донорные остатки" относится к остаткам, обнаруживаемым в исходной вариабельной области, выделенной из хозяина, в частности, к замене указанной аминокислоты в каркасе человека аминокислотой в соответствующем участке в донорном каркасе.
В одном из вариантов осуществления связывающий домен или каждый связывающий домен является частью (включен или встроен в) антитела или фрагмента антитела.
В одном из вариантов осуществления связывающие домены в молекулах по настоящему изобретению находятся в молекулах иммуноглобулина/антитела.
Как применяют в настоящем описании, "молекула антитела" включает антитела и их связывающие фрагменты.
Как применяют в настоящем описании, в одном из вариантов осуществления термин "антитело" относится к молекуле иммуноглобулина, способной специфически связываться с антигеном-мишенью, таким как углевод, полинуклеотид, липид, полипептид, пептид и т.д., с помощью по меньшей мере одного участка распознавания антигена (также обозначаемого в настоящем описании как участок связывания или связывающий домен), локализованного в вариабельной области молекулы иммуноглобулина.
Как применяют в настоящем описании, термин "фрагменты антител" относится к связывающим фрагментам антител, включающим, в качестве неограничивающих примеров, Fab, модифицированный Fab, Fabʹ, модифицированный Fabʹ, F(abʹ)2, Fv, однодоменные антитела, scFv, Fv, би-, три- или тетравалентные антитела, бис-scFv, диатела, триатела, тетратела и эпитоп-связывающие фрагменты любого из указанного выше (см., например, Holliger and Hudson, 2005, Nature Biotech. 23(9):1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217).
Как применяют в настоящем описании, термин "связывающий фрагмент" относится к фрагменту, способному связываться с пептидом- или антигеном-мишенью с аффинностью, достаточной для характеризации фрагмента как специфичного для пептида или антигена.
Способы получения и производства эти фрагментов антител хорошо известны в этой области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216:165-181). Другие фрагменты антител для использования в настоящем изобретении включают Fab- и Fab'-фрагменты, описываемые в WO05/003169, WO05/003170 и WO05/003171. Мультивалентные антитела могут иметь множество специфичностей, например, могут являться биспецифическими или моноспецифическими (см., например, WO92/22853, WO05/113605, WO2009/040562 и WO2010/035012).
Как применяют в настоящем описании, термин "Fab-фрагмент" относится к фрагменту антитела, содержащему фрагмент легкой цепи, содержащий домен VL (вариабельный домен легкой цепи) и константный домен легкой цепи (CL), и домен VH (вариабельный домен тяжелой цепи) и первый константный домен (CH1) тяжелой цепи.
Термин "Fv" относится к двум вариабельным доменам, например, действующим совместно вариабельным доменам, таким как когнатная пара или аффинно зрелые вариабельные домены, т.е. пара VH и VL.
Как применяют в настоящем описании, действующие совместно вариабельные домены являются вариабельными доменами, дополняющими друг друга и/или участвующими в связывании антигена, делая Fv (пару VH/VL) специфичным для интересующего антигена.
Как применяют в настоящем описании, термин "однодоменное антитело" (также обозначаемое в настоящем описании как dab и sdAb) относится к фрагменту антитела, состоящему из одного мономерного вариабельного домена антитела. Примеры однодоменных антител включают VH или VL или VHH.
Как применяют в настоящем описании, термин "тандемный sdAb" относится к двухдоменным антителам, соединенным линкером, например, пептидным линкером, в частности, когда доменные антитела имеют специфичность к разным антигенам.
Как применяют в настоящем описании, термин "тандемный sdAb-sdAb" относится к трехдоменным антителам, соединенным серией из двух линкеров, например, пептидных линкеров, в частности, когда доменные антитела имеют специфичность к разным антигенам.
Как применяют в настоящем описании, термин "dsFv" относится к Fv с дисульфидной связью внутри вариабельной области. dsFv может являться компонентом более крупной молекулы, например, один из вариабельных доменов можно соединять, например, аминокислотным линкером с другим фрагментом/компонентом антитела.
Как применяют в настоящем описании, термин "(dsFv)2" относится к dsFv с одним доменом, соединенным, например, пептидным линкером или дисульфидной связью (например, между C-концами двух VH) с доменом во втором dsFv, формат схож с (scFv)2, описываемым ниже, но каждая пара вариабельных областей содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "компонент" относится к структурному элементу или части полиспецифической молекулы по настоящему изобретению, в частности, если компонент является фрагментом антитела, таким как scFv, Fab или другой фрагмент, в частности, как представлено в настоящем описании.
Как применяют в настоящем описании, термин "одноцепочечный Fv" или, сокращенно "scFv", относится к фрагменту антитела, содержащему домены антитела VH и VL, соединенные (например, пептидным линкером) с образованием единой полипептидной цепи. В этом формате отсутствуют константные области тяжелой и легкой цепи.
Как применяют в настоящем описании, термин "dsscFv" относится к scFv с дисульфидной связью внутри вариабельной области.
Как применяют в настоящем описании, термин "тандемный scFv" (также обозначаемый в настоящем описании как discFv или (scFv)2)) относится к двум scFv, соединенным одним линкером таким образом, что есть один линкер между Fv, например, как показано на фигуре 5b.
Как применяют в настоящем описании, термин "тандемный dsscFv" (также обозначаемый в настоящем описании как scFvdsscFv или dsscFvscFv) относится к двум scFv, соединенный одним линкером таким образом, что есть один линкер между Fv, например, как показано на фигуре 5b, и где один из scFv имеет дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "тандемный ди-dsscFv" (также обозначаемый в настоящем описании как ди-dsscFv) относится к двум scFv, соединенным одним линкером таким образом, что есть один линкер между Fv, например, как показано на фигуре 5b, и где каждый scFv содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, scFv-dsFv является scFv, соединенным, например, пептидным линкером, с FV-доменом, состоящим из двух вариабельных доменов, соединенных дисульфидной связью с образованием dsFv. В этом формате VH или VL scFv можно соединять с VH или VL dsFv.
Как применяют в настоящем описании dsscFv-dsFv является dsscFv, соединенным, например, пептидным линкером, с FV-доменом, состоящим из двух вариабельных доменов, соединенных дисульфидной связью с образованием dsFv. В этом формате VH или VL dsscFv можно соединять с VH или VL dsFv.
Как применяют в настоящем описании, термин "диатело" относится к двум парам VH/VL Fv и дополнительной паре VH/VL, имеющим два линкера между Fv, таким образом, что VH первого Fv соединяют с VL второго Fv и VL первого Fv соединяют с VH второго Fv.
Как применяют в настоящем описании, термин "ds-диатело" относится к диателу, содержащему дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "ди-ds-диатело" относится к диателу, содержащему две дисульфидные связи внутри вариабельной области, т.е. одну ds между каждой парой вариабельных областей.
Как применяют в настоящем описании, термин "sc-диатело" относится к диателу, содержащему линкер внутри Fv, таким образом, что молекула содержит три линкера и образует два нормальных scFv, например, VH1-линкер-VL1-линкер-VH2-линкер-VL2.
Как применяют в настоящем описании, термин "dssc-диатело" относится к sc-диателу с дисульфидной связью внутри вариабельной области.
Как применяют в настоящем описании, термин "ди-dssc-диатело" относится к sc-диателу с дисульфидной связью внутри вариабельной области между каждой парой вариабельных областей.
Как применяют в настоящем описании, термин "Dart" относится к VL1-линкер-VH2-линкер и VH1-линкер-VL2, где C-конец VH1 и VH2 соединяют дисульфидной связью (Paul A. Moore et al Blood, 2011; 117(17):4542-4551).
Как применяют в настоящем описании, термин "Bite®" относится к молекуле, содержащей две пары вариабельных доменов в следующем формате; домен из пары 1 (например, VH1), соединенный линкером с доменом из пары 2 (например, VH2 или VL2), при этом указанный второй домен соединен линкером с дополнительным доменом из пары 1 (например, VL1), в свою очередь, соединенным с оставшимся доменом из пары два (т.е. VL2 или VH2).
Термин "ди-диатело" см. в Nunez-Prado et al, в частности, молекулу № 25 на фиг. 1.
Как применяют в настоящем описании, термин "ds-ди-диатело" является ди-диателом с дисульфидной связью внутри вариабельной области.
Как применяют в настоящем описании, термин "ди-ds-ди-диатело" является ди-диателом с дисульфидной связью внутри вариабельной области между каждой парой вариабельных областей.
Как применяют в настоящем описании, термин "триатело" относится к формату, схожему с диателом, содержащему три Fv и три линкера между Fv.
Как применяют в настоящем описании, термин "ds-триатело" относится к триателу, содержащему дисульфидную связь внутри вариабельной области между одной из пар вариабельных доменов.
Как применяют в настоящем описании, термин "ди-ds-триатело" относится к триателу, содержащему две дисульфидные связи внутри вариабельной области, т.е. одну ds между каждой из двух пар вариабельных доменов.
Как применяют в настоящем описании, термин "три-ds-триатело" относится к триателу, содержащему три дисульфидные связи внутри вариабельной области, т.е. одну ds между каждой парой вариабельных областей.
Как применяют в настоящем описании, термин "тетратело" относится к формату, схожему с диателом, содержащему четыре Fv и четыре линкера между Fv.
Как применяют в настоящем описании, термин "ds-тетратело" относится к тетрателу, содержащему дисульфидную связь внутри вариабельной области между одной из пар вариабельных доменов.
Как применяют в настоящем описании, термин "ди-ds-тетратело" относится к тетрателу, содержащему две дисульфидные связи внутри вариабельной области, т.е. одну ds между каждой из двух пар вариабельных доменов.
Как применяют в настоящем описании, термин "три-ds-тетратело" относится к тетрателу, содержащему три дисульфидные связи внутри вариабельной области т.е. одну ds между каждой из трех пар вариабельных областей.
Как применяют в настоящем описании, термин "тетра-ds-тетратело" относится к тетрателу, содержащему четыре дисульфидные связи внутри вариабельной области т.е. одну ds между каждым вариабельным доменом.
Как применяют в настоящем описании, термин "тритело" (также обозначаемый как Fab(scFv)2) относится к Fab-фрагменту с первым scFv, присоединенным к C-концу легкой цепи, и вторым scFv, присоединенным к C-концу тяжелой цепи.
Как применяют в настоящем описании, термин "ds-тритело" относится к трителу, содержащему dsscFv в одном из двух положений.
Как применяют в настоящем описании, термин "ди-ds-тритело" или "TrYbe" относится к трителу, содержащему два dsscFv.
Как применяют в настоящем описании, термин "dsFab" относится к Fab с дисульфидной связью внутри вариабельной области.
Как применяют в настоящем описании, термин "dsFabʹ" относится к Fabʹ с дисульфидной связью внутри вариабельной области.
scFab является одноцепочечным Fab-фрагментом.
scFabʹ является одноцепочечным Fab'-фрагментом.
dsscFab является dsFab в одноцепочечной форме.
dsscFabʹ является dsFabʹ в одноцепочечной форме.
Как применяют в настоящем описании, термин "Fabdab" относится к Fab-фрагменту с доменным антителом, присоединенным к его тяжелой или легкой цепи, необязательно, с помощью линкера.
Как применяют в настоящем описании, термин "Fabʹdab" относится к Fab'-фрагменту с доменным антителом, присоединенным к его тяжелой или легкой цепи, необязательно, с помощью линкера.
Как применяют в настоящем описании, термин "FabFv" относится к Fab-фрагменту с дополнительной вариабельной областью, присоединенной к C-концу каждого из следующего, CH1 тяжелой цепи и CL легкой цепи, см., например, WO2009/040562. Формат можно получать в его пегилированной версии, см., например, WO2011/061492.
Как применяют в настоящем описании, термин "FabʹFv" схож с FabFv, где Fab-часть заменяют Fabʹ. Формат можно получать в его пегилированной версии.
Как применяют в настоящем описании, термин "FabdsFv" относится к FabFv, где дисульфидная связь внутри Fv стабилизирует присоединенные C-концевые вариабельные области, см., например, WO2010/035012. Формат можно получать в его пегилированной версии.
Как применяют в настоящем описании, термин "Fab-один линкер-Fv" и "Fabʹ-один линкер" относится к Fab- или Fab'-фрагменту, соединенному с вариабельным доменом, например, пептидным линкером, и указанный вариабельный домен соединяют со вторым вариабельным доменом с помощью дисульфидной связи внутри вариабельного домена, таким образом, получая dsFv, см., например, WO2014/096390.
Как применяют в настоящем описании, термин "Fab-scFv" (также обозначаемый как битело) является Fab-молекулой с scFv, присоединенным к C-концу легкой или тяжелой цепи, необязательно, с помощью линкера.
Как применяют в настоящем описании, Fabʹ-scFv является Fabʹ-молекулой с scFv, присоединенным к C-концу легкой или тяжелой цепи, необязательно, с помощью линкера.
Как применяют в настоящем описании, FabdsscFv или BYbe является FabscFv с дисульфидной связью между вариабельными областями одноцепочечного Fv.
Как применяют в настоящем описании, FabʹdsscFv является FabʹscFv с дисульфидной связью между вариабельными областями одноцепочечного Fv.
Как применяют в настоящем описании, термин "FabscFv-dab" относится к Fab с scFv, присоединенным к C-концу одной цепи, и доменным антителом, присоединенным к C-концу другой цепи.
Как применяют в настоящем описании, термин "FabʹscFv-dab" относится к Fabʹ с scFv, присоединенным к C-концу одной цепи, и доменным антителом, присоединенным к C-концу другой цепи.
Как применяют в настоящем описании, термин "FabdsscFv-dab" относится к Fab с dsscFv, присоединенным к C-концу одной цепи, и доменным антителом, присоединенным к C-концу другой цепи.
Как применяют в настоящем описании, термин "FabʹdsscFv-dab" относится к Fabʹ с dsscFv, присоединенным к C-концу одной цепи, и доменным антителом, присоединенным к C-концу другой цепи.
Как применяют в настоящем описании, термин "FabscFv-один линкер-Fv" относится к "Fab-один линкер-Fv", где домен Fv соединяют с тяжелой или легкой цепью Fab, scFv соединяют с другой цепью Fab и домены Fv соединяют дисульфидной связью внутри вариабельной области.
Как применяют в настоящем описании, "FabdsscFv-один линкер-Fv" относится к "FabscFv-один линкер-Fv", где scFv содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabʹscFv-один линкер-Fv" относится к "Fabʹ-один линкер-Fv", где домен Fv соединяют с тяжелой или легкой цепью Fab, scFv соединяют с другой цепью Fab и домены Fv соединяют дисульфидной связью внутри вариабельной области.
Как применяют в настоящем описании, термин "FabʹdsscFv-один линкер-Fv" относится к "FabʹscFv-один линкер-Fv", где scFv содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FvFabFv" относится к Fab с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fab, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи.
Как применяют в настоящем описании, термин "FvFabʹFv" относится к Fabʹ с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fabʹ, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи.
Как применяют в настоящем описании, термин "dsFvFabFv" относится к Fab с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fab, где первый Fv содержит дисульфидную связь внутри вариабельной области, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи.
Как применяют в настоящем описании, термин "FvFabdsFv" относится к Fab с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fab, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи, и где второй Fv содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "dsFvFabʹFv" относится к Fabʹ с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fabʹ, где первый Fv содержит дисульфидную связь внутри вариабельной области, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи.
Как применяют в настоящем описании, термин "FvFabʹdsFv" относится к Fabʹ с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fabʹ, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи, и где второй Fv содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "dsFvFabdsFv" относится к Fab с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fab, где первый Fv содержит дисульфидную связь внутри вариабельной области, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи, и где второй Fv также содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "dsFvFabʹdsFv" относится к Fabʹ с доменами первого Fv, присоединенного к N-концу тяжелой и легкой цепи Fabʹ, где первый Fv содержит дисульфидную связь внутри вариабельной области, и доменами второго Fv, присоединенного к C-концу тяжелой и легкой цепи, и где второй Fv также содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabFvFv" относится к Fab-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, см., например, WO2011/086091.
Как применяют в настоящем описании, термин "FabʹFvFv" относится к Fab'-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, см., например, WO2011/086091.
Как применяют в настоящем описании, термин "FabdsFvFv" относится к Fab-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, см., например, WO2011/086091, где первая пара Fv, соединенная напрямую с C-концом, содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabʹdsFvFv" относится к Fab'-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, см., например, WO2011/086091, где первая пара Fv, соединенная напрямую с C-концом, содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabFvdsFv" относится к Fab-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, где вторая пара Fv на "C"-конце молекулы содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabʹFvdsFv" относится к Fab'-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, где вторая пара Fv на "C"-конце молекулы содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabdsFvdsFv" относится к Fab-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, где первая и вторая пара Fv содержат дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "FabʹdsFvdsFv" относится к Fab'-фрагменту с двумя парами Fv, присоединенными серией к C-концу тяжелой и легкой цепи, где первая и вторая пара Fv содержат дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "ди-Fab" относится к двум Fab-молекулам, соединенным C-концами своих тяжелых цепей.
Как применяют в настоящем описании, термин "ди-Fabʹ" относится к двум Fabʹ-молекулам, соединенным одной или несколькими дисульфидными связями в своей шарнирной области.
Молекулы ди-Fab и ди-Fabʹ включают их химически конъюгированные формы.
Как применяют в настоящем описании, термин "(FabscFv)2" относится к молекуле ди-Fab с двумя scFv, присоединенными к ней, например, присоединенными к C-концу тяжелой или легкой цепи, такой как тяжелая цепь.
Как применяют в настоящем описании, термин (FabʹscFv)2 относится к молекуле ди-Fabʹ с двумя scFv, присоединенными к ней, например, присоединенными к C-концу тяжелой или легкой цепи, такой как тяжелая цепь.
Как применяют в настоящем описании, термин "(Fab)2scFvdsFv" относится к diFab с присоединенными scFv и dsFv, например, одним из каждого C-конца тяжелой цепи.
Как применяют в настоящем описании, термин "(Fabʹ)2scFvdsFv" относится к diFabʹ с присоединенными scFv и dsFv, например, одним из каждого C-конца тяжелой цепи.
Как применяют в настоящем описании, термин "(Fab)2dsscFvdsFv", относится к diFab с присоединенными dsscFv и dsFv, например, из C-конца тяжелой цепи.
(Fabʹ)2dsscFvdsFv как применяют в настоящем описании относится к diFabʹ с присоединенными dsscFv и dsFv, например, из C-конца тяжелой цепи.
Как применяют в настоящем описании, термин "минитело" относится к (VL/VH-CH3)2.
Как применяют в настоящем описании, термин "ds-минитело" относится к (VL/VH-CH3)2, где одна VL/VH содержит дисульфидную связь внутри вариабельной области.
Как применяют в настоящем описании, термин "ди-ds-минитело" относится к (dsFv-CH3)2.
Как применяют в настоящем описании, термин "scFv-Fc" относится к scFv, присоединенному к N-концу домена CH2, например, с помощью шарнирной области, фрагмента константной области- (CH2CH3), таким образом, что молекула имеет 2 связывающих домена.
Как применяют в настоящем описании, термин "dsscFv-Fc" относится к dsscFv, присоединенному к N-концу домена CH2, и scFv, присоединенный к N-концу второго домена CH2, например, с помощью шарнирной области, фрагмента константной области -(CH2CH3)2, таким образом, что молекула имеет 2 связывающих домена.
Как применяют в настоящем описании, термин "ди-dsscFv-Fc" относится к scFv, присоединенному к N-концу домена CH2, например, с помощью шарнирной области, фрагмента константной области -(CH2CH3)2, таким образом, что молекула имеет 2 связывающих домена.
Как применяют в настоящем описании, термин "тандемный scFv-Fc" относится к двум тандемным scFv, где каждый присоединен серией к N-концу домена CH2, например, с помощью шарнирной области, фрагмента константной области -(CH2CH3), таким образом, что молекула имеет 4 связывающих домена.
Как применяют в настоящем описании, sc-диатело-Fc является двумя sc-диателами, где каждое присоединено к N-концу домена CH2, например, с помощью шарнирной области, фрагмента константной области -CH2CH3.
Как применяют в настоящем описании, термин "scFv-Fc-scFv" относится к четырем scFv, где один из каждого присоединен к N-концу и C-концу и тяжелой, и легкой цепи фрагмента -CH2CH3.
Как применяют в настоящем описании, термин "sc-диатело-CH3" относится к двум молекулам sc-диателам, где каждая соединена, например, с помощью шарнирной области, с доменом CH3.
Каппа/лямбда-телоʹ или ʹκ/λ-тело является форматом нормального IgG с двумя тяжелыми цепями и двумя легкими цепями, где две легкие цепи отличаются друг от друга, одна является легкой лямбда-цепью (VL CL), а другая является легкой каппа-цепью (VK-CK). Тяжелая цепь является идентичной, даже в отношении CDR, как описано в WO2012/023053.
Как применяют в настоящем описании, IgG-scFv является полноразмерным антителом с scFv на C-конце каждой из тяжелых цепей или каждой из легких цепей.
Как применяют в настоящем описании, scFv-IgG является полноразмерным антителом с scFv на N-конце каждой из тяжелых цепей или каждой из легких цепей.
Как применяют в настоящем описании, V-IgG является полноразмерным антителом с вариабельным доменом на N-конце каждой из тяжелых цепей или каждой из легких цепей.
Как применяют в настоящем описании, IgG-V является полноразмерным антителом с вариабельным доменом на C-конце каждой из тяжелых цепей или каждой из легких цепей.
DVD-Ig (также известный как IgG с двойным V-доменом) является полноразмерным антителом с 4 дополнительными вариабельными доменами, один на N-конце каждой из тяжелых цепей или каждой из легких цепей.
Как применяют в настоящем описании дуотело или "обмен плеча Fab" является биспецифическим форматом антитела IgG, где совпадающие и комплементарные сконструированные замены аминокислот в константных доменах (как правило, CH3) двух разных моноклональных антител после смешивания приводят к образованию гетеродимеров. Пара тяжелая:легкая цепь из первого антитела в результате конструирования остатков предпочтительно будет связываться с парой тяжелая:легкая цепь второго антитела. См., например, WO2008/119353, WO2011/131746 и WO2013/060867.
Если одна или несколько пар вариабельных областей в полиспецифической молекуле по настоящему изобретению содержат дисульфидную связь между VH и VL, она может находиться в любом подходящем положении, например, между двумя остатками, указанными ниже (если контекст не указывает на иное, в представленном ниже списке используют нумерацию по Kabat). Если сделана ссылка на нумерацию по Kabat, соответствующей ссылкой является Kabat et al., 1987, в Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA.
В одном из вариантов осуществления дисульфидная связь находится в положении, выбранном из группы, содержащей:
VH37+VL95C см., например, Protein Science 6, 781-788 Zhu et al (1997);
VH44+VL100 см., например; Biochemistry 33 5451-5459 Reiter et al (1994); или Journal of Biological Chemistry Vol. 269 No. 28 pp.18327-18331 Reiter et al (1994); или Protein Engineering, vol.10 no.12 pp.1453-1459 Rajagopal et al (1997);
VH44+VL105 см., например, J Biochem. 118, 825-831 Luo et al (1995);
VH45+VL87 см., например, Protein Science 6, 781-788 Zhu et al (1997);
VH55+VL101 см., например, FEBS Letters 377 135-139 Young et al (1995);
VH100+VL50 см., например, Biochemistry 29 1362-1367 Glockshuber et al (1990);
VH100b+VL49;
VH98+VL 46 см., например, Protein Science 6, 781-788 Zhu et al (1997);
VH101+VL46;
VH105+VL43 см., например; Proc. Natl. Acad. Sci. USA Vol. 90 pp.7538-7542 Brinkmann et al (1993); или Proteins 19, 35-47 Jung et al (1994),
VH106+VL57 см., например, FEBS Letters 377 135-139 Young et al (1995),
и положение, соответствующее ему в паре вариабельных областей, локализованной в молекуле.
В одном из вариантов осуществления дисульфидная связь образуется между положениями VH44 и VL100.
Как применяют в настоящем описании, термин "моноспецифический" относится к способности связываться с антигеном-мишенью лишь однократно. Таким образом, в одном из вариантов осуществления полиспецифические молекулы по настоящему изобретению являются моноспецифическими для каждого антигена.
Таким образом, в одном из вариантов осуществления связывающие домены полиспецифических молекул по настоящему изобретению являются моноспецифическими. Это является предпочтительным при некотором терапевтическом применении, т.к. молекулы по настоящему изобретению неспособны поперчено сшивать антиген посредством связывания антигена-мишени неоднократно. Таким образом, в одном из вариантов осуществления биспецифические или полиспецифические молекулы по настоящему изобретению неспособны к поперечной сшивке посредством связывания одной и той же мишени дважды в двух разных участках, например, на одной и той же клетке или на двух разных клетках.
Перекрестное сшивание, в частности, в отношении CD79b на одной и той же клетке или различных клетках может генерировать сигналы in vivo, например, стимулирующие активность антигена-мишени.
В другом варианте осуществления, например, если молекулы по настоящему изобретению содержат по меньшей мере три связывающих домена, то два или три связывающих домена (например, антитела, фрагменты или комбинация антитела и фрагмента) могут иметь разные антигенные специфичности, например, связывание с двумя или тремя разными антигенами-мишенями.
В одном примере полиспецифические молекулы по настоящему изобретению содержат не более одного связывающего домена для CD22 и не более одного связывающего домена для CD79. Каждый связывающий домен является моноспецифическим.
В одном из вариантов осуществления каждое антитело или фрагмент антитела, используемое в полиспецифических молекулах по настоящему изобретению, является моновалентным.
Таким образом в одном из вариантов осуществления связывающие домены полиспецифических молекул по настоящему изобретению являются моновалентными.
Таким образом в одном из вариантов осуществления связывающие домены полиспецифических молекул по настоящему изобретению являются моновалентными и моноспецифическими.
В одном из вариантов осуществления полиспецифическая молекула по настоящему изобретению состоит из двух или более моноспецифических, моновалентных связывающих доменов, таких как Fab, Fabʹ, scFv, VH, VL, VHH, Fv, dsFv, комбинированных или связанных любым подходящим образом для конструирования полиспецифической молекулы, например, как представлено в настоящем описании выше.
Константные области
Домены константной области антитела полиспецифической молекулы по настоящему изобретению, при их наличии, можно выбирать с учетом предполагаемой функции молекулы полиспецифического антитела и, в частности, эффекторных функций, которые могут быть необходимыми. Например, домены константной области могут являться доменами IgA, IgD, IgE, IgG или IgM человека. В частности, можно использовать домены константной области IgG человека, особенно, изотипов IgG1 и IgG3, если молекула антитела предназначена для терапевтического применения, и необходимы эффекторные функции антитела. Альтернативно, можно использовать изотипы IgG2 и IgG4, если молекула антитела предназначена для терапевтических целей, и эффекторные функции антитела не являются необходимыми.
Следует понимать, что также можно использовать варианты последовательности этих доменов константной области. Например, можно использовать молекулы IgG4, в которых серин в положении 241 заменяют пролином, как описано в Angal et al., 1993, Molecular Immunology, 1993, 30:105-108. Таким образом, в варианте осуществления, где антитело является антителом IgG4, антитело может включать мутацию S241P.
В одном из вариантов осуществления тяжелая цепь антитела содержит домен CH1, и легкая цепь антитела содержит домен CL, каппа или лямбда.
В одном из вариантов осуществления тяжелая цепь антитела содержит домен CH1, домен CH2 и домен CH3, и легкая цепь антитела содержит домен CL, каппа или лямбда.
Четыре изотипа IgG человека связываются с активирующими рецепторами Fcγ (FcγRI, FcγRIIa, FcγRIIIa), ингибиторным рецептором FcγRIIb и первым компонентом комплемента (C1q) с различными аффинностями, что приводит к очень разным эффекторным функциям (Bruhns P. et al., 2009. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood. 113(16):3716-25), также см. Jeffrey B. Stavenhagen, et al. Cancer Research 2007 Sep 15; 67(18):8882-90.
Связывание IgG с FcγR или C1q зависит от остатков, локализованных в шарнирной области и домене CH2. Две области домена CH2 являются критичными для связывания FcγR и C1q и имеют уникальные последовательности в IgG2 и IgG4. Показано, что замены остатков IgG1 или IgG2 человека в положениях 233-236 и остатков IgG4 в положениях 327, 330 и 331 значительно снижают ADCC и CDC (Armour KL. et al., 1999. Recombinant human IgG molecules lacking Fcgamma receptor I binding and monocyte triggering activities. Eur J Immunol. 29(8):2613-24 and Shields RL. et al., 2001. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J Biol Chem. 276(9):6591-604). Кроме того, Idusogie et al. показали, что замена аланина в разных положениях, включая K322, значительно снижала активацию комплемента (Idusogie EE. et al., 2000. Mapping of the C1q binding site on rituxan, a chimeric antibody with a human IgG1 Fc. J Immunol. 164(8):4178-84). Аналогично, показано, что мутации в домене CH2 IgG2A мыши снижают связывание с FcγRI и C1q (Steurer W. et al., 1995. Ex vivo coating of islet cell allografts with murine CTLA4/Fc promotes graft tolerance. J Immunol. 155(3):1165-74).
В одном из вариантов осуществления используемая Fc-область является мутантной, в частности, с мутацией, представленной в настоящем описании. В одном из вариантов осуществления мутация предназначена для устранения связывания и/или эффекторной функции.
В одном из вариантов осуществления мутация Fc выбрана из группы, содержащей мутацию для устранения связывания Fc-области, мутацию для повышения или устранения эффекторной функции, мутацию для повышения времени полужизни и их комбинацию.
Некоторые антитела, селективно связывающиеся с FcRn при pH 6,0, но не pH 7,4, демонстрируют большее время полужизни во множестве моделей на животных. Показано, что несколько мутаций, локализованных на границе между доменами CH2 и CH3, таких как T250Q/M428L (Hinton PR. et al., 2004. Engineered human IgG antibodies with longer serum half-lives in primates. J Biol Chem. 279(8):6213-6) и M252Y/S254T/T256E+H433K/N434F (Vaccaro C. et al., 2005. Engineering the Fc region of immunoglobulin G to modulate in vivo antibody levels. Nat Biotechnol. 23(10):1283-8), повышают аффинность связывания с FcRn и время полужизни IgG1 in vivo.
Однако, не всегда существует прямая взаимосвязь между повышенным связыванием FcRn и улучшенным временем полужизни (Datta-Mannan A. et al., 2007. Humanized IgG1 Variants with Differential Binding Properties to the Neonatal Fc Receptor: Relationship to Pharmacokinetics in Mice and Primates. Drug Metab. Dispos. 35: 86-94).
Подкласс IgG4 демонстрирует сниженное связывание Fc-рецептора (FcγRIIIa), антитела других подклассов IgG, как правило, демонстрируют сильное связывание. Сниженного связывания рецептора в этих других подтипах IgG можно достигать посредством изменения, например, замены одной или нескольких аминокислот, выбранных из группы, содержащей Pro238, Aps265, Asp270, Asn270 (утрата углевода в Fc), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435.
В одном из вариантов осуществления молекула по настоящему изобретению имеет Fc подкласса IgG, например, IgG1, IgG2 или IgG3, где Fc является мутантным в одном, двух или всех следующих положениях S228, L234 и/или D265.
В одном из вариантов осуществления мутации в Fc-области независимо выбраны из S228P, L234A, L235A, L235A, L235E и их комбинаций.
Желательным может являться снижение или повышение эффекторной функции Fc-области. Необходимы антитела, направленно воздействующие на молекулы поверхности клетки, в частности, иммунных клеток, устраняющие эффекторные функции. В некоторых вариантах осуществления, например, для лечения аутоиммунных заболеваний, желательным может являться повышенное связывание Fc на иммунных клетках посредством повышения отрицательного связывания Fc-рецептора (FcgRIIb или CD32b), см. Stavenhagen JB, et al Advances in Enzyme Regulation 2007 December 3 и Veri MC, et al. Arthritis Rheum, 2010 Mar 30;62(7):1933-43. И наоборот, в случае антител, предназначенных для онкологического применения, повышение эффекторных функций может улучшать терапевтическую активность.
Получено множество мутаций в домене CH2 IgG1 человека, и их эффект в отношении ADCC и CDC тестировали in vitro (Idusogie EE. et al., 2001. Engineered antibodies with increased activity to recruit complement. J Immunol. 166(4):2571-5). Примечательно, что сообщали о том, что замена аланина в положении 333 повышает и ADCC, и CDC. Lazar et al. описывали тройного мутанта (S239D/I332E/A330L) с более высокой аффинностью к FcγRIIIa и более низкой аффинностью к FcγRIIb, что приводит к повышенной ADCC (Lazar GA. et al., 2006. Engineered antibody Fc variants with enhanced effector function. PNAS 103(11): 4005-4010). Те же мутации использовали для получения антитела с повышенной ADCC (Ryan MC. et al., 2007. Antibody targeting of B-cell maturation antigen on malignant plasma cells. Mol. Cancer Ther., 6: 3009-3018). Richards et al. исследовали немного другого тройного мутанта (S239D/I332E/G236A) с улучшенной аффинностью к FcγRIIIa и соотношением FcγRIIa/FcγRIIb, опосредующим повышенный фагоцитоз клеток-мишеней макрофагами (Richards JO et al 2008. Optimization of antibody binding to Fcgamma RIIa enhances macrophage phagocytosis of tumor cells. Mol Cancer Ther. 7(8):2517-27).
В результате отсутствия эффекторных функций, антитела IgG4 представляют собой подходящий подкласс IgG для блокирования рецептора без истощения клеток. Молекулы IgG4 могут заменять полу-молекулы в динамическом процессе под названием "замена плеча Fab". Это явление может происходить между терапевтическими антителами и эндогенным IgG4. Показано, что мутация S228P предотвращает этот процесс рекомбинации, делая возможным дизайн менее непредсказуемых терапевтических антител IgG4 (Labrijn AF. et al., 2009. Therapeutic IgG4 antibodies engage in Fab-arm exchange with endogenous human IgG4 in vivo. Nat Biotechnol. 27(8):767-71). Эту технологию можно использовать для получения молекул биспецифического антитела.
Специалисту в этой области также будет понятно, что антитела могут подвергаться множеству посттрансляционных модификаций. Тип и степень этих модификаций часто зависят от линии клеток-хозяев, используемой для экспрессии антитела, а также условий культивирования. Такие модификации могут включать варианты гликозилирования, окисления метионина, образование дикетопиперазина, изомеризацию аспартата и дезамидирование аспарагина. Частой модификацией является утрата карбокси-концевого основного остатка (такого как лизин или аргинин) в результате действия карбоксипептидаз (как описано в Harris, RJ. Journal of Chromatography 705:129-134, 1995). Таким образом, C-концевой лизин тяжелой цепи антитела может отсутствовать.
Аффинность
Полиспецифические молекулы по настоящему изобретению содержат связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79a и/или CD79b.
В одном из вариантов осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным для CD45.
В одном из вариантов осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным для CD79a.
В одном из вариантов осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным для CD79b.
В одном из вариантов осуществления связывающий домен, используемый в молекулах по настоящему изобретению, является специфичным для комплекса CD79, т.е. он распознает эпитоп, присутствующий в комплексе и специфичный для него, например, эпитоп, включающий взаимодействие между CD79a и CD79b.
CD45 (также известный как PTPRC) является известным белком. CD45 является членом белкового семейства тирозинфосфатаз (PTP). PTP известны как сигнальные молекулы, регулирующие множество клеточных процессов, включая рост клеток, дифференцировку, митотический цикл и онкогенную трансформацию. Эта PTP содержит внеклеточный домен, единственный трансмембранный сегмент и два тандемных внутрицитоплазматических каталитических домена и, таким образом, принадлежит к рецепторным PTP. Существуют различные изоформы CD45: CD45RA, CD45RB, CD45RC, CD45RAB, CD45RAC, CD45RBC, CD45RO, CD45R (ABC). CD45RA локализуется на наивных T-клетках, а CD45RO локализуется на T-клетках памяти. Варианты сплайсинга изоформ A, B и C CD45 дифференциально экспрессируются на B-клетках человека. CD45 является членом белкового семейства тирозинфосфатаз (PTP): его внутриклеточная (COOH-концевая) область содержит два каталитических домена PTP, а внеклеточная область является крайне изменчивой в результате альтернативного сплайсинга экзонов 4, 5, и 6 (обозначаемых A, B, и C, соответственно) и разных уровней гликозилирования. Определяемые изоформы CD45 являются специфичными в отношении типа клеток, созревания и статуса активации. В основном, длинная форма белка (A, B или C) экспрессируется на наивных или неактивированных B-клетках, и зрелая или укороченная форма CD45 (RO) экспрессируется на активированных или зрелых B-клетках/B-клетках памяти.
Последовательность человека доступна в UniProt под регистрационным номером P08575 и представлена в настоящем описании в SEQ ID NO: 143 или как аминокислоты 24-1304 SEQ ID NO: 143 без сигнального пептида. Мышиная версия представлена в UniProt под регистрационным номером P06800. Настоящее изобретение относится ко всем формам CD45 из любого вида. В одном из вариантов осуществления CD45 относится к человеческой форме белка и его природным вариантам и изоформам.
В одном из вариантов осуществления аффинность связывающего домена к CD45 в молекуле по настоящему изобретению составляет приблизительно 100 нМ или более, например, приблизительно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пМ, 250 пМ, 200 пМ, 100 пМ или более, в частности, аффинность связывания составляет 50 пМ или более.
Связывающий домен для CD79 может связываться с CD79a и/или CD79b.
CD79a (также известный как иммуноглобулин альфа и альфа-цепь белка, ассоциированного с комплексом B-клеточного антигенного рецептора) является известным белком. Экспрессия CD79a ограничена B-лимфоцитами. Последовательность человека доступна в UniProt под регистрационным номером P11912 (SEQ ID NO: и аминокислоты 33-226 SEQ ID NO: 141 без сигнальной последовательности). Мышиная версия доступна в UniProt под регистрационным номером 11911. Настоящее изобретение относится ко всем формам CD79a любого вида, в частности, человека, и любым его природным вариантам. В одном из вариантов осуществления CD79a относится к человеческой форме белка.
CD79b (также известный как иммуноглобулин бета и кластер дифференцировки 79B) является известным белком. Экспрессия CD79b ограничена B-лимфоцитами. Последовательность человека доступна в UniProt под регистрационным номером P40259 (SEQ ID NO: 142 и аминокислоты 29-229 SEQ ID NO: 142 без сигнальной последовательности). Мышиная версия доступна в UniProt под регистрационным номером P15530. Настоящее изобретение относится ко всем формам CD79b любого вида, в частности, человека, и любым его природным вариантам. В одном из вариантов осуществления CD79b относится к человеческой форме белка.
В одном из вариантов осуществления связывающий домен, специфичный для CD79, связывается с CD79a.
В одном из вариантов осуществления связывающий домен, специфичный для CD79, связывается с CD79b.
В одном из вариантов осуществления связывающий домен, специфичный для CD79, связывается с комплексом CD79a и CD79b.
В одном из вариантов осуществления аффинность связывающего домена к CD79 в молекуле по настоящему изобретению составляет приблизительно 100нМ или более, например, приблизительно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пМ, 250 пМ, 200 пМ, 100 пМ или более, в частности, аффинность связывания составляет 50 пМ или более.
В одном из вариантов осуществления аффинность связывающего домена к CD79a в молекуле по настоящему изобретению составляет приблизительно 100 нМ или более, например, приблизительно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пМ, 250 пМ, 200 пМ, 100 пМ или более, в частности, аффинность связывания составляет 50 пМ или более.
В одном из вариантов осуществления аффинность связывающего домена к CD79b в молекуле по настоящему изобретению составляет приблизительно 100 нМ или более, например, приблизительно 50 нМ, 20 нМ, 10 нМ, 1 нМ, 500 пМ, 250 пМ, 200 пМ, 100 пМ или более, в частности, аффинность связывания составляет 50 пМ или более.
Следует понимать, что аффинность связывающего домена к CD45 может являться той же или отличаться от аффинности связывающего домена к CD79.
В одном из вариантов осуществления молекулы полиспецифического антитела по настоящему изобретению или компоненты антитела/фрагмента обрабатывают для достижения улучшенной аффинности к антигену-мишени или антигенам. Такие варианты можно получать рядом способов созревания аффинности, включая мутагенез CDR (Yang et al., J. Mol. Biol., 254, 392-403, 1995), перестановку цепей (Marks et al., Bio/Technology, 10, 779-783, 1992), использование мутаторных штаммов E. coli (Low et al J. Mol. Biol., 250, 359-368, 1996), ДНК-перестановку (Patten et al Curr. Opin. Biotechnol., 8, 724-733, 1997), фаговый дисплей (Thompson et al., J. Mol. Biol., 256, 77-88, 1996) и ПЦР с имитацией полового размножения (Crameri et al Nature, 391, 288-291, 1998). В Vaughan et al (выше) описывают эти способы созревания аффинности.
Антитела и их получение
Связывающие домены для использования в настоящем изобретении можно получать любым подходящим известным в этой области способом, например, можно получать CDR из не принадлежащих человеку антител, включая коммерчески доступные антитела, и пересаживать на каркасы человека или, альтернативно, химерные антитела можно получать с использованием не принадлежащих человеку вариабельных областей и константных областей человека и т.д.
Как правило, связывающие домены для использования в настоящем изобретении являются связывающими доменами, полученными из антител, связывающихся с выбранным антигеном, таких как антитела, связывающиеся с CD45, CD79a и/или CD79b.
В этой области известны примеры антител против CD45 и CD79, и их можно использовать напрямую в молекулах по настоящему изобретению или подвергать скринингу на пригодность с использованием способов, представленных в настоящем описании, и затем, при необходимости, модифицировать, например, гуманизировать, способами, представленными в настоящем описании. В этой области описывают терапевтические антитела против CD45 и против CD79, например, антитела против CD45, описываемые в US2011/0076270, антитела против CD79b, описываемые в WO2014/011521 и WO2015/021089.
Примеры антител против CD45 включают моноклональное вантитело крысы YTH54, YTH25.4, моноклональное антитело мыши от Miltenyi клон 5B1 и клон 30F11, моноклональное антитело крысы YAML568, от BD Bioscience моноклональный клон мыши 2D1, кат. № 347460, от Novus моноклональное антитело мыши 5D3A3, кат. № NBP2-37293, моноклональное HI30 мыши, кат. № NBP1-79127, моноклональное антитело мыши 4A8A4C7A2, кат. № NBP1-47428, моноклональное антитело мыши 2B11, кат. № NBP2-32934, моноклональное антитело крысы YTH24.5, кат. № NB100-63828, моноклональное антитело кролика Y321, кат. № NB110-55701, моноклональное антитело мыши PD7/26/16, кат. № NB120-875, от Santa Cruz моноклональное антитело мыши из клона B8, кат. № sc-28369, моноклональное антитело мыши из клона F10-89-4, кат. № sc-52490, моноклональное антитело кролика из клона H-230, кат. № sc-25590, моноклональное антитело козы из клона N-19, кат. № sc-1123, моноклональное антитело мыши из клона OX1, кат. № sc-53045, моноклональное антитело крысы (T29/33), кат. № sc-18901, моноклональное антитело крысы (YAML 501.4), кат. № sc65344, моноклональное антитело крысы (YTH80.103), кат. № sc-59071, моноклональное антитело мыши (351C5), кат. № sc-53201, моноклональное антитело мыши (35-Z6), кат. № sc-1178, моноклональное антитело мыши (158-4D3), кат. № sc-52386, моноклональное антитело мыши против CD45RO (UCH-L1), кат. № sc-1183, моноклональное антитело мыши против CD45RO (2Q1392), кат. № sc-70712.
Антитела против CD45 также описывают в WO2005/026210, WO02/072832 и WO2003/048327, включенных в настоящее описание в качестве ссылки.
Коммерчески доступные антитела против CD79a включают моноклональное антитело мыши LS-B4504 (LSBio) из клона HM57, моноклональное антитело мыши LS-B8330, моноклональное антитело мыши LS-C44954, моноклональное антитело кролика LS-B9093, моноклональное антитело мыши LS-B8513 из клона JCB117, моноклональное антитело кролика LS-C210607 из клона SP18, моноклональное антитело мыши LS-C175441 из клона 5E2, моноклональное антитело мыши LS-C338670 из клона 3D3, моноклональное антитело мыши LS-C88120 из клона HM47/A9, моноклональное антитело мыши LS-C191714, моноклональное антитело мыши LS-C87592, моноклональное антитело мыши LS-C44955, моноклональное антитело мыши LS-C95934, моноклональное антитело мыши LS-C121584, моноклональное антитело мыши LS-C121585, моноклональное антитело мыши LS-C204347, моноклональное антитело мыши LS-C88122, моноклональное антитело мыши Abcam ab3121 [HM47/A9], моноклональное антитело кролика ab79414 и моноклональное антитело кролика ab133483.
Коммерчески доступные антитела против CD79b включают моноклональное антитело мыши Abcam ab33295, моноклональное антитело крысы ab23826, моноклональное антитело мыши ab103422, моноклональное антитело кролика ab134103, моноклональное антитело кролика ab134147 и моноклональное антитело кролика ab183343.
Некоторые коммерчески доступные антитела могут являться полезными средствами в обнаружении терапевтических антител.
Специалист в этой области может получать антитела для использования в полиспецифических молекулах по изобретению любым подходящим известным в этой области способом.
Антигенные полипептиды для использования в получении антител, например, для использования для иммунизации хозяина или для использования в пэннинге, таком как фаговый дисплей, можно получать способами, хорошо известными в этой области, из генетически сконструированных клеток-хозяев, содержащих системы экспрессии, или их можно выделять из природных биологических источников. В настоящей заявке термин "полипептиды" включает пептиды, полипептиды и белки. Их используют взаимозаменяемо, если не указано иначе. Антигенный полипептид в некоторых случаях может являться частью более крупного белка, такого как слитый белок например, слитый с аффинной меткой или т.п. В одном из вариантов осуществления хозяина можно иммунизировать с использованием клетки, трансфицированной с использованием соответствующего белка или полипептида, например, котрансфицированной с использованием CD79a и CD79b.
Можно получать антитела против антигенного полипептида, где необходима иммунизация животного, посредством введения полипептидов животному, предпочтительно - не относящемуся к человеку животному, хорошо известными и общепринятыми способами, см., например, Handbook of Experimental Immunology, D. M. Weir (ed.), Vol 4, Blackwell Scientific Publishers, Oxford, England, 1986. Можно иммунизировать многих теплокровных животных, таких как кролики, мыши, крысы, овцы, коровы, верблюжьи или свиньи. Однако, наиболее подходящими, как правило, являются мыши, кролики, свиньи и крысы.
Моноклональные антитела можно получать любым известным в этой области способом, таким как гибридомный способ (Kohler & Milstein, 1975, Nature, 256:495-497), триомный способ, способ с использованием гибридомы B-клеток человека (Kozbor et al 1983, Immunology Today, 4:72) и EBV-гибридомный способ (Cole et al Monoclonal Antibodies and Cancer Therapy, pp77-96, Alan R Liss, Inc., 1985).
Антитела также можно получать способами получения антител из выбранных лимфоцитов посредством клонирования и экспрессии кДНК вариабельной области иммуноглобулина, полученной из отдельных лимфоцитов, выбранных для продукции специфических антител, например, способами, описываемыми в Babcook, J. et al 1996, Proc. Natl. Acad. Sci. USA 93(15):7843-7848l; WO92/02551; WO2004/051268 и WO2004/106377.
Антитела для использования в настоящем изобретении также можно получать с использованием различных известных в этой области способов фагового дисплея, и они включают описываемые Brinkman et al. (в J. Immunol. Methods, 1995, 182: 41-50), Ames et al. (J. Immunol. Methods, 1995, 184:177-186), Kettleborough et al. (Eur. J. Immunol. 1994, 24:952-958), Persic et al. (Gene, 1997 187 9-18), Burton et al. (Advances in Immunology, 1994, 57:191-280) и WO90/02809; WO91/10737; WO92/01047; WO92/18619; WO93/11236; WO95/15982; WO95/20401; и US 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743; 5969108 и WO20011/30305.
В одном примере полиспецифические молекулы по настоящему изобретению являются полностью человеческими, в частности, один или несколько вариабельных доменов являются полностью человеческими.
Полностью человеческие молекулы являются молекулами, в которых все вариабельные области и константные области (при их наличии) тяжелых и легких цепей имеют человеческое происхождение или являются, по существу, идентичными последовательностям человеческого происхождения, необязательно из одного и того же антитела. Примеры полностью человеческих антител могут включать антитела, получаемые, например, способами фагового дисплея, описываемыми выше, и антитела, продуцируемые мышами, в которых гены вариабельной и, необязательно, константной области иммуноглобулина мыши заменяют соответствующими генами человека, например, как описано в общих терминах в EP0546073, US5545806, US5569825, US5625126, US5633425, US5661016, US5770429, EP0438474 и EP0463151.
В одном примере связывающие домены полиспецифических молекул по настоящему изобретению гуманизируют.
Как применяют в настоящем описании, термин "гуманизированный" (что включает антитела с пересаженными CDR) относится к молекулам, имеющим одну или несколько определяющих комплементарность областей (CDR) из не являющихся человеком видов и каркасную область из молекулы иммуноглобулина человека (см., например, US 5585089; WO91/09967). Следует понимать, что необходимым может являться лишь перенос определяющих специфичность остатков CDR, а не полной CDR (см., например, Kashmiri et al., 2005, Methods, 36, 25-34). Гуманизированные антитела, необязательно, могут дополнительно содержать один или несколько каркасных остатков, полученных из не являющихся человеком видов, из которых получали CDR.
Как применяют в настоящем описании, термин "гуманизированная молекула антитела" относится к молекуле антитела, где тяжелая и/или легкая цепь содержит одну или несколько CDR (включая, при желании, одну или несколько модифицированных CDR) из донорного антитела (например, моноклонального антитела мыши), пересаженные на каркас вариабельной области тяжелой и/или легкой цепи акцепторного антитела (например, антитела человека). Обзор см. в Vaughan et al, Nature Biotechnology, 16, 535-539, 1998. В одном из вариантов осуществления вместо переноса полной CDR на каркас антитела переносят лишь один или несколько определяющих специфичность остатков человека из любой из CDR, представленных в настоящем описании выше (см., например, Kashmiri et al., 2005, Methods, 36, 25-34). В одном из вариантов осуществления на каркас антитела человека переносят лишь определяющие специфичность остатки из одной или нескольких CDR, представленных в настоящем описании выше. В другом варианте осуществления на каркас антитела человека переносят лишь определяющие специфичность остатки из каждой из CDR, представленных в настоящем описании выше.
Если пересаживают CDR или определяющие специфичность остатки, можно использовать любую подходящую акцепторную последовательность каркаса вариабельной области с учетом класса/типа донорного антитела, из которого получают CDR, включая каркасные области мыши, примата и человека. Соответственно, гуманизированное антитело по настоящему изобретению имеет вариабельный домен, содержащий акцепторные каркасные области человека, а также одну или несколько CDR, представленных в настоящем описании.
Примерами каркасов человека, которые можно использовать в настоящем изобретении, являются KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (Kabat et al, выше). Например, можно использовать KOL и NEWM для тяжелой цепи, можно использовать REI для легкой цепи и можно использовать EU, LAY и POM для тяжелой цепи и легкой цепи. Альтернативно, можно использовать последовательности зародышевой линии человека; они доступны на: http://www2.mrc-lmb.cam.ac.uk/vbase/list2.php.
В гуманизированной молекуле антитела по настоящему изобретению акцепторные тяжелые и легкие цепи не обязательно необходимо получать из одного антитела, и, при желании, они могут содержать композитные цепи, имеющие каркасные области, полученные из различных цепей.
Каркасные области не обязательно должны иметь точно ту же последовательность, что и в акцепторном антителе. Например, нетипичные остатки можно заменять остатками, чаще встречающимися в этом классе или типе акцепторной цепи. Альтернативно, выбранные остатки в акцепторных каркасных областях можно заменять таким образом, что они соответствуют остатку, обнаруживаемому в том же положении в донорном антителе (см. Reichmann et al 1998, Nature, 332, 323-324). Такие изменения необходимо сводить к минимуму, необходимому для достижения аффинности донорного антитела. Способ выбора остатков в акцепторных каркасных областях, замена которых может быть необходимой, приведен в WO91/09967.
Производные каркасов могут иметь 1, 2, 3 или 4 аминокислот, замененных альтернативной аминокислотой, например, донорным остатком.
Донорные остатки являются остатками из донорного антитела, т.е. антитела, из которого исходно получали CDR, в частности, переносят остаток в соответствующем положении из донорной последовательности. Донорные остатки можно заменять подходящим остатком, полученным из рецепторного каркаса человека (акцепторные остатки).
Остатки в вариабельных доменах антитела общепринято нумеруют по системе, разработанной Kabat et al. Эта система изложена в Kabat et al., 1987, in Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA (ниже обозначаемой как "Kabat et al. (выше)"). Эту систему нумерации используют в настоящем описании кроме случаев, когда указано иное.
Обозначение остатков по Kabat не всегда напрямую соответствует линейной нумерации аминокислотных остатков. Конкретная линейная аминокислотная последовательность может содержать меньше аминокислот или дополнительные аминокислоты относительно точной нумерации по Kabat, соответствующие укорачиванию или инсерции в структурном компоненте, является ли он каркасом или определяющей комплементарность областью (CDR), основной структуре вариабельного домена. Для указанного антитела корректную нумерацию остатков по Kabat можно определять посредством выравнивания гомологичных остатков в последовательности антитела со "стандартной" пронумерованной по Kabat последовательностью.
CDR вариабельного домена тяжелой цепи локализованы в остатках 31-35 (CDR-H1), остатках 50-65 (CDR-H2) и остатках 95-102 (CDR-H3) согласно системе нумерации по Kabat. Однако, согласно Chothia (Chothia, C. и Lesk, A.M. J. Mol. Biol., 196, 901-917 (1987)) петля, эквивалентная CDR-H1, распространяется от остатка 26 до остатка 32. Таким образом, если не указано иначе, как применяют в настоящем описании, термин "CDR-H1" предназначен для обозначения остатков от 26 до 35, как описывают посредством комбинирования системы нумерации по Kabat и топологического определения петель по Chothia.
CDR вариабельного домена легкой цепи локализованы в остатках 24-34 (CDR-L1), остатках 50-56 (CDR-L2) и остатках 89-97 (CDR-L3) согласно системе нумерации по Kabat.
В одном примере настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD79, содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 78, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 79, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 80.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD79, содержащую 3 CDR тяжелой цепи, SEQ ID NO: 88 в случае CDRH1, SEQ ID NO: 89 в случае CDRH2 и SEQ ID NO: 90 в случае CDRH3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область легкой цепи, специфичную для CD79, содержащую 3 CDR легкой цепи, SEQ ID NO: 75 в случае CDRL1, SEQ ID NO: 76 в случае CDRL2 и SEQ ID NO: 77 в случае CDRL3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область легкой цепи, специфичную для CD79, содержащую 3 CDR легкой цепи, SEQ ID NO: 85 в случае CDRL1, SEQ ID NO: 86 в случае CDRL2 и SEQ ID NO: 87 в случае CDRL3.
В одном примере настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD79, содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 78, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 79, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 80, и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 75, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 76 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 77.
В одном примере настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD79, содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 88, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 89, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 90, и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 85, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 86 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 87.
В одном из вариантов осуществления полиспецифическая молекула по настоящему изобретению содержит связывающий домен, специфичный для CD45, содержащий 3 CDR тяжелой цепи, выбранные из группы, содержащей SEQ ID NO: 98, 99, 100, 108, 109, 110, 118, 119, 120, 128, 129 и 130.
В одном из вариантов осуществления полиспецифическая молекула по настоящему изобретению содержит связывающий домен, специфичный для CD45, содержащий 3 CDR легкой цепи, выбранные из группы, содержащей SEQ ID NO: 95, 97, 97, 105, 106, 107, 115, 116, 117, 125, 126 и 127.
В одном из вариантов осуществления полиспецифическая молекула по настоящему изобретению содержит связывающий домен, специфичный для CD45, содержащий 3 CDR тяжелой цепи, выбранные из группы содержащей SEQ ID NO: 98, 99, 100, 108, 109, 110, 118, 119, 120, 128, 129, 130, и 3 CDR легкой цепи, выбранные из группы, содержащей SEQ ID NO: 95, 97, 97, 105, 106, 107, 115, 116, 117, 125, 126 и 127.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD45, содержащую 3 CDR тяжелой цепи, SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD45, содержащую 3 CDR тяжелой цепи, SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD45, содержащую 3 CDR тяжелой цепи, SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область тяжелой цепи (VH), специфичную для CD45, содержащую 3 CDR тяжелой цепи, SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область легкой цепи, специфичную для CD45, содержащую 3 CDR легкой цепи, SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область легкой цепи, специфичную для CD45, содержащую 3 CDR легкой цепи, SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область легкой цепи, специфичную для CD45, содержащую 3 CDR легкой цепи, SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3.
В одном из вариантов осуществления настоящее изобретение относится к связывающему домену, содержащему вариабельную область легкой цепи, специфичную для CD45, содержащую 3 CDR легкой цепи, SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для CD45, содержащему вариабельную область тяжелой цепи (VH), содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 98, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 99, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 100, и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 95, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 96 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 97.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для CD45, содержащему вариабельную область тяжелой цепи (VH), содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 108, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 109, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 110, и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 105, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 106 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 107.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для CD45, содержащему вариабельную область тяжелой цепи (VH), содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 118, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 119, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 120, и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 115, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 116 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 117.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для CD45, содержащему вариабельную область тяжелой цепи (VH), содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 128, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 129, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 130 и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 125, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 126 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 127.
В одном примере настоящее изобретение относится к полиспецифической молекуле, содержащей связывающий домен, специфичный для антигена CD79, и связывающий домен, специфичный для антигена CD45, где эта каждая пара связывающих доменов содержит 6 CDR из пары антител против CD79 и CD45, например, выбранные из следующих пар антител; 4447 и 4122, 4447 и 4129, 4447 и 4131, 4447 и 4133, 4450 и 4122, 4450 и 4129, 4450 и 4131 и 4447 и 4133.
Последовательности этих антител против CD79 (антитела 4447 и антитела 4450), включая последовательности VH, VL и CDR, представлены в настоящем описании ниже. Последовательности этих антител против CD45 (антител 4122, 4129, 4131 и 4133), включая последовательности VH, VL и CDR, представлены в настоящем описании ниже, и их можно комбинировать как связывающие домены в молекулах по настоящему изобретению.
В одном из вариантов осуществления настоящее изобретение относится к последовательности антитела, представленной в настоящем описании.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для альбумина, содержащему вариабельную область тяжелой цепи (VH), содержащую три CDR, где CDR H1 имеет последовательность, приведенную в SEQ ID NO: 131, CDR H2 имеет последовательность, приведенную в SEQ ID NO: 132, и CDR H3 имеет последовательность, приведенную в SEQ ID NO: 133, и вариабельную область легкой цепи (VL), содержащую три CDR, где CDR L1 имеет последовательность, приведенную в SEQ ID NO: 134, CDR L2 имеет последовательность, приведенную в SEQ ID NO: 135 и CDR L3 имеет последовательность, приведенную в SEQ ID NO: 136.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для альбумина, содержащему вариабельную область тяжелой цепи (VH), имеющую последовательность, приведенную в SEQ ID NO: 137, и вариабельную область легкой цепи (VL), имеющую последовательность, приведенную в SEQ ID NO: 139.
В одном примере настоящее изобретение относится к связывающему домену, специфичному для альбумина, содержащему вариабельную область тяжелой цепи (VH), имеющую последовательность, приведенную в SEQ ID NO: 138, и вариабельную область легкой цепи (VL), имеющую последовательность, приведенную в SEQ ID NO: 140.
В одном примере связывающие домены гуманизируют.
В одном примере одну или несколько CDR, представленных в настоящем описании, можно модифицировать для удаления нежелательных остатков или участков, таких как остатки цистеина или участки изомеризации аспарагиновой кислоты (D) или участки дезамидирования аспарагина (N). В одном примере участок дезамидирования аспарагина можно удалять из одной или нескольких CDR посредством мутации остатка аспарагина (N) и/или соседнего остатка в любую другую подходящую аминокислоту. В одном примере участок дезамидирования аспарагина, такой как NG или NS, может являться мутантным, например, в NA или NT.
В одном примере участок изомеризации аспарагиновой кислоты можно удалять из одной или нескольких CDR посредством мутации остатка аспарагиновой кислоты (D) и/или соседнего остатка в любую другую подходящую аминокислоту. В одном примере участок изомеризации аспарагиновой кислоты, такой как DG или DS, может являться мутантным, например, в EG, DA или DT.
Например, один или несколько остатков цистеина в любой из CDR можно заменять другой аминокислотой, такой как серин.
В одном примере участок N-гликозилирования, такой как NLS можно удалять посредством мутации остатка аспарагина (N) в любую другую подходящую аминокислоту, например, в SLS или QLS. В одном примере участок N-гликозилирования, такой как NLS, можно удалять посредством мутации остатка серина (S) в любой другой остаток, за исключением треонина (T).
Специалист в этой области способен тестировать варианты CDR или гуманизированные последовательности в любом подходящем анализе, таком как анализы, представленные в настоящем описании, для подтверждения поддержания активности.
Специфическое связывание с антигеном можно тестировать с использованием любого подходящего анализа, включая, например, способы ELISA или поверхностного плазмонного резонанса, такого как BIAcore, где можно измерять связывание с антигеном (CD45 и/или CD79). В таких анализах можно использовать выделенный природный или рекомбинантный CD45 или CD79 (или b) или подходящий слитый белок/полипептид. В одном примере связывание измеряют с использованием рекомбинантного CD45 (SEQ ID NO: 143 или аминокислот 23-1304 SEQ ID NO: 143) или CD79, такого как последовательность, приведенная в SEQ ID NO: 141 и SEQ ID NO: 142 и аминокислоты 33-226 SEQ ID NO: 141 и аминокислоты 29-229 SEQ ID NO: 142, например, с помощью поверхностного плазмонного резонанса, такого как BIAcore. Альтернативно, белки могут экспрессироваться на клетке, такой как клетка HEK, и аффинность измеряют с использованием определения аффинности на основе проточной цитометрии.
Последовательности антитела, представленные в настоящем описании, можно использовать для идентификации дополнительных антител и, таким образом, связывающих доменов, пригодных для использования в полиспецифических молекулах по настоящему изобретению. Антитела, перекрестно блокирующие связывание молекулы антитела по настоящему изобретению против CD79, в частности, молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 73, и последовательность легкой цепи, приведенную в SEQ ID NO: 71, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 83, и последовательность легкой цепи, приведенную в SEQ ID NO: 81, аналогично, могут являться применимыми в связывании CD79 и, таким образом, аналогично, применимыми в полиспецифических молекулах по настоящему изобретению. Таким образом, настоящее изобретение также относится к полиспецифической молекуле, содержащей связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79b, где связывающий домен для CD79b перекрестно блокирует связывание любой из молекул антител, представленных в настоящем описании выше, с CD79 и/или перекрестно блокирован от связывания CD79 любым из этих антител. В одном из вариантов осуществления такое антитело связывается с тем же эпитопом, что и антитело, представленное в настоящем описании выше.
Аналогично, антитела, перекрестно блокирующие связывание молекулы антитела по настоящему изобретению с CD45, в частности, молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 93, и последовательность легкой цепи, приведенную в SEQ ID NO: 91, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 103, и последовательность легкой цепи, приведенную в SEQ ID NO: 101, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 113, и последовательность легкой цепи, приведенную в SEQ ID NO: 111, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 123, и последовательность легкой цепи, приведенную в SEQ ID NO: 121, или молекулы антитела, содержащей последовательность тяжелой цепи, приведенную в SEQ ID NO: 133, и последовательность легкой цепи, приведенную в SEQ ID NO: 131, аналогично, могут являться применимыми в связывании CD45 и, таким образом, аналогично, применимыми в полиспецифических молекулах по настоящему изобретению. Таким образом, настоящее изобретение также относится к полиспецифической молекуле, содержащей связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79, где связывающий домен для CD45 перекрестно блокирует связывание любой из молекул антител, представленных в настоящем описании выше, с CD45 и/или перекрестно блокирован от связывания CD45 любым из этих антител. В одном из вариантов осуществления такое антитело связывается с тем же эпитопом, что и антитело, представленное в настоящем описании выше. В другом варианте осуществления перекрестно блокирующее антитело связывается с эпитопом, граничащим и/или перекрывающимся с эпитопом, связываемым антителом, представленным в настоящем описании выше.
В другом варианте осуществления перекрестно блокирующее, нейтрализующее антитело связывается с эпитопом, граничащим и/или перекрывающимся с эпитопом, связываемым антителом, представленным в настоящем описании выше.
Перекрестно блокирующие антитела можно идентифицировать любым подходящим способом, известным в этой области, например, с использованием конкурентного ELISA или анализа BIAcore, где связывание перекрестно блокирующего антитела с антигеном (CD45 и/или CD79) предотвращает связывание антитела по настоящему изобретению или наоборот. В таких анализах перекрестного блокирования можно использовать выделенный природный или рекомбинантный CD45 или CD79 (a и/или b) или подходящий слитый белок/полипептид. В одном примере связывание и перекрестно блокирование измеряют с использованием рекомбинантного CD45 (SEQ ID NO: 143) или CD79.
Альтернативно или дополнительно, антитела по этому аспекту изобретения можно перекрестно блокировать от связывания с антигеном (CD45 или CD79) с помощью связывающего домена, представленного в настоящем описании, например, содержащего CDR, полученные из последовательности вариабельной области тяжелой цепи и последовательности легкой цепи, приведенных в SEQ ID NO: 71 и 73, 81 и 83, 91 и 93, 101 и 103, 111 и 113, и 121 и 123. Таким образом, настоящее изобретение также относится к полиспецифической молекуле, содержащей связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79b, где связывающий домен для CD79b перекрестно блокирует связывание любой из молекул антител, представленных в настоящем описании выше, с CD79b и/или перекрестно блокирован от связывания CD79 любым из этих антител более чем на 80%, например, более чем на 85%, например, более чем на 90%, в частности, более чем на 95% и, необязательно, где связывающий домен для CD45 перекрестно блокирует связывание любой из молекул антител, представленных в настоящем описании выше, с CD45 и/или перекрестно блокирован от связывания CD45 любым из этих антител более чем на 80%, например, более чем на 85%, например, более чем на 90%, в частности, более чем на 95%.
В одном из аспектов настоящее изобретение относится к полиспецифической молекуле антитела, содержащей или состоящей из:
a) полипептидной цепи формулы (I):
VH-CH1-X-(V1)p;
b) полипептидной цепи формулы (II):
VL-CL-Y-(V2)q;
где:
VH представляет собой вариабельный домен тяжелой цепи;
CH1 представляет собой домен константной области тяжелой цепи, например, ее домен 1;
X представляет собой связь или линкер, например, аминокислотный линкер;
Y представляет собой связь или линкер, например, аминокислотный линкер;
V1 представляет собой dab, scFv, dsscFv или dsFv;
VL представляет собой вариабельный домен, например, вариабельный домен легкой цепи;
CL представляет собой домен из константной области, например, домен константной области легкой цепи, такой как C-каппа;
V2 представляет собой dab, scFv, dsscFv или dsFv;
p является 0 или 1;
q является 0 или 1; и
если p является 1, q является 0 или 1, и если q является 1, p является 0 или 1, т.е. p и q не представляют собой 0.
В одном из вариантов осуществления молекула полиспецифического антитела содержит не более одного связывающего домена для CD45 и не более одного связывающего домена для CD79.
В одном из вариантов осуществления q является 0, и p является 1.
В одном из вариантов осуществления q является 1, и p является 1.
В одном из вариантов осуществления V1 является dab, и V2 является dab, и вместе они образуют единый связывающий домен из действующей совместно пары вариабельных областей, такой как когнатная пара VH/VL, необязательно, соединенная дисульфидной связью.
В одном из вариантов осуществления VH и VL являются специфичными для CD79, например, CD79a или CD79b.
В одном из вариантов осуществления V1 является специфичным для CD79, например, CD79a или CD79b.
В одном из вариантов осуществления V2 является специфичным для CD79, например, CD79a или CD79b.
В одном из вариантов осуществления V1 и V2 вместе (например, в виде связывающего домена) являются специфичными для CD79, например, CD79a или CD79b, и VH и VL являются специфичными для CD45.
В одном из вариантов осуществления V1 является специфичным для CD45.
В одном из вариантов осуществления V2 является специфичным для CD45.
В одном из вариантов осуществления V1 и V2 вместе (например, в виде одного связывающего домена) являются специфичными для CD45, и VH и VL являются специфичными для CD79.
В одном из вариантов осуществления V1 является специфичным для CD45, V2 является специфичным для альбумина, и VH и VL являются специфичными для CD79.
В одном из вариантов осуществления V1 является специфичным для альбумина, V2 является специфичным для CD45, и VH и VL являются специфичными для CD79.
В одном из вариантов осуществления V1 является специфичным для CD79, V2 является специфичным для альбумина, и VH и VL являются специфичными для CD45.
В одном из вариантов осуществления V1 является специфичным для альбумина, V2 является специфичным для CD79, и VH и VL являются специфичными для CD45.
В одном из вариантов осуществления V1 является dsscFv, специфичным для CD45, V2 является dsscFv, специфичным для альбумина, и VH и VL являются специфичными для CD79.
В одном из вариантов осуществления V1 является dsscFv, специфичным для альбумина, V2 является dscFv, специфичным для CD45, и VH и VL являются специфичными для CD79.
В одном из вариантов осуществления V1 является dsscFv, специфичным для CD79, V2 является dsscFv, специфичным для альбумина, и VH и VL являются специфичными для CD45.
В одном из вариантов осуществления V1 является dsscFv, специфичным для альбумина, V2 является dsscFv, специфичным для CD79, и VH и VL являются специфичными для CD45.
Каждая из V1, V2, VH и VL в указанных выше конструкциях может представлять собой связывающий домен и включать любую из последовательностей, представленных в настоящем описании.
X и Y представляют собой любой подходящий линкер, например, X и Y могут являться SGGGGSGGGGS (SEQ ID NO: 17).
В одном из вариантов осуществления, если V1 и/или V2 являются dab, dsFv или a dsscFv, дисульфидная связь между вариабельными доменами VH и VL из V1 и/или V2 образуется между положениями VH44 и VL100.
Настоящее изобретение также относится к новым полипептидным последовательностям, представленным в настоящем описании, и последовательностям, по меньшей мере на 80% схожим или идентичным им, например, на 85% или более, например, на 90% или более, в частности, на 95% или более схожим или идентичным.
Как применяют в настоящем описании, термин "идентичность" свидетельствует о том, что в любом конкретном положении в выровненных последовательностях аминокислотный остаток является идентичным между последовательностями. Как применяют в настоящем описании, термин "сходство" свидетельствует о том, что в любом конкретном положении в выровненных последовательностях аминокислотный остаток имеет схожий тип между последовательностями. Например, лейцин можно заменять изолейцином или валином. Другие аминокислоты, которые часто можно заменять друг другом, включают, в качестве неограничивающих примеров:
- фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи);
- лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи);
- аспартат и глутамат (аминокислоты, имеющие кислые боковые цепи);
- аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и
- цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи).
Степени идентичности и схожести легко можно вычислять и (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, программное обеспечение BLAST™, доступное в NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215:403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3:266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266:131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25:3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7:649-656)).
В частности, в одном из аспектов настоящее изобретение относится к антителам против CD45 и CD79, представленным в настоящем описании, в любом подходящем формате антитела.
Таким образом, в одном из аспектов настоящее изобретение относится к антителам против CD45 или их фрагментам, содержащим один или несколько связывающих доменов, представленных в настоящем описании выше, содержащим CDR, представленные в настоящем описании и в SEQ ID NO: 95, 96, 97, 98, 99 и 100 (антитело 4122) или 105, 106, 107, 108, 109 и 110 (антитело 4129) или 115, 116, 117, 118, 119 и 120 (антитело 4131) или 125, 126, 127, 128, 129 и 130 (антитело 4133). Кроме того, настоящее изобретение относится к антителам против CD79 или их фрагментам, содержащим один или несколько связывающих доменов, представленных в настоящем описании выше, содержащих CDR, представленные в настоящем описании и в SEQ ID NO: 75, 76, 77, 78, 79 и 80 (антитело 4447) или SEQ ID NO: 85, 86, 87, 88, 89 и 90 (антитело 4450).
Указанные CDR можно встраивать в любой подходящий каркас антитела и в любой подходящий формат антитела. Такие антитела включают целые антитела и функционально активные фрагменты или его производные, которые могут являться, в качестве неограничивающих примеров, моноклональными, гуманизированными, полностью человеческими или химерными антителами. Таким образом, такие антитела могут содержать полную молекулу антитела, содержащую полноразмерные тяжелые и легкие цепи, или ее фрагмент и могут являться, в качестве неограничивающих примеров, Fab, модифицированным Fab, Fabʹ, F(abʹ)2, Fv, однодоменными антителами, scFv, би-, три- или тетравалентными антителами, бис-scFv, диателами, триателами, тетрателами и эпитоп-связывающими фрагментами любого из указанного выше (см., например, Holliger and Hudson, 2005, Nature Biotech. 23(9):1126-1136; Adair and Lawson, 2005, Drug Design Reviews - Online 2(3), 209-217). Способы получения и производства эти фрагментов антител хорошо известны в этой области (см., например, Verma et al., 1998, Journal of Immunological Methods, 216, 165-181). Поливалентные антитела могут иметь множество специфичностей или могут являться моноспецифическими (см., например, WO 92/22853 и WO05/113605). Следует понимать, что этот аспект изобретения также относится к вариантам этих антител против CD45 и CD79, включая гуманизированные версии и модифицированные версии, включая те, в которых аминокислоты подвергают мутации в CDR для удаления одного или нескольких участков изомеризации, дезамидирования, гликозилирования или остатков цистеина, как представлено в настоящем описании выше.
Линкеры
Руководство в настоящем описании в отношении линкеров в одном контексте в равной степени можно использовать в отношении линкеров в других контекстах, если используют линкер, как в случае любой полиспецифической молекулы по настоящему изобретению.
В одном из вариантов осуществления линкер, используемый в молекуле по настоящему изобретению, является аминокислотным линкером длиной 50 остатков или менее, например, выбранным из последовательности, приведенной в последовательностях 5-70.
Таблица 1. Последовательности линкера в шарнирной области
Таблица 2. Последовательности гибкого линкера
(S) является необязательным в последовательностях 17-20.
Примеры жестких линкеров включают пептидные последовательности GAPAPAAPAPA (SEQ ID NO: 69), PPPP (SEQ ID NO: 70) и PPP.
Другие линкеры представлены в таблице 3:
ЭФФЕКТОРНЫЕ МОЛЕКУЛЫ
При желании полиспецифическую молекулу для использования в настоящем изобретении можно конъюгировать с одной или несколькими эффекторными молекулами. Следует понимать, что эффекторная молекула может содержать одну эффекторную молекулу или две или более таких молекул, соединенных таким образом, что получают единый остаток, который можно присоединять к полиспецифическим молекулам по настоящему изобретению. Если желательным является получение антитела или полиспецифической молекулы по настоящему изобретению, соединенной с эффекторной молекулой, ее можно получать стандартными химическими способами или способами рекомбинантной ДНК, в которых фрагмент антитела соединяют с эффекторной молекулой напрямую или с помощью связывающего средства. Способы конъюгации таких эффекторных молекул с антителами хорошо известны в этой области (см. Hellstrom et al., Controlled Drug Delivery, 2nd Ed., Robinson et al., eds., 1987, pp. 623-53; Thorpe et al., 1982, Immunol. Rev., 62:119-58 и Dubowchik et al., 1999, Pharmacology and Therapeutics, 83, 67-123). Конкретные химические способы включают, например, способы, описываемые в WO93/06231, WO92/22583, WO89/00195, WO89/01476 и WO03/031581. Альтернативно, если эффекторная молекула является белком или полипептидом, связывание можно осуществлять способами рекомбинантной ДНК, например, как описано в WO86/01533 и EP0392745.
В одном из вариантов осуществления полиспецифические молекулы по настоящему изобретению могут содержать эффекторную молекулу.
Как применяют в настоящем описании, термин "эффекторная молекула" включает, например, антинеопластические средства, лекарственные средства, токсины, биологически активные белки, например, ферменты, другое антитело или фрагменты антител, синтетические или природные полимеры, нуклеиновые кислоты и их фрагменты, например, ДНК, РНК и их фрагменты, радионуклиды, в частности, радиоактивный йод, радиоактивные изотопы, хелатообразующие металлы, наночастицы и репортерные группы, такие как флуоресцентные соединения или соединения, которые можно определять посредством ЯМР- или ЭПР-спектроскопии.
Примеры эффекторных молекул могут включать цитотоксины или цитотоксические средства, включая любое средство, вредное для клеток (например, убивающее их). Примеры включают комбрестатины, доластатины, эпотилоны, стауроспорин, майтанзиноиды, спонгистатины, ризоксин, галихондрины, роридины, гемиастерлины, таксол, цитохалазин B, грамицидин D, бромистый этидий, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, и пуромицин и их аналоги или гомологи.
Эффекторные молекулы также включают, в качестве неограничивающих примеров, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил декарбазин), алкилирующие средства (например, мехлоретамин, тиотепа хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромоманнит, стрептозотоцин, митомицин C, и цис-дихлордиамин платины (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее известный как дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее известный как актиномицин), блеомицин, митрамицин, антрамицин (AMC), калихимицины или дуокармицины) и антимитотические средства (например, винкристин и винбластин).
Другие эффекторные молекулы могут включать хелатообразующие радионуклиды, такие как 111In и 90Y, Lu177, висмут-213, калифорний-252, иридий-192 и вольфрам-188/рений-188; или лекарственные средства, такие как, в качестве неограничивающих примеров, алкилфосфохолины, ингибиторы топоизомеразы I, таксоиды и сурамин.
Другие эффекторные молекулы включают белки, пептиды и ферменты. Интересующие ферменты включают, в качестве неограничивающих примеров, протеолитические ферменты, гидролазы, лиазы, изомеразы, трансферазы. Интересующие белки, полипептиды и пептиды включают, в качестве неограничивающих примеров, иммуноглобулины, токсины, такие как абрин, рицин A, псевдомонадный экзотоксин или дифтерийный токсин, белок, такой как инсулин, фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, фактор роста тромбоцитов или тканевый активатор плазминогена, тромботическое средство или антиангиогенное средство, например, ангиостатин или эндостатин, или модификатор биологического ответа, такой как лимфокин, интерлейкин-1 (ИЛ-1), интерлейкин-2 (ИЛ-2), гранулоцитарно-макрофагальный колониестимулирующий фактор (ГМ-КСФ), гранулоцитарный колониестимулирующий фактор (Г-КСФ), фактор роста нервов (NGF) или другой фактор роста и иммуноглобулины.
Другие эффекторные молекулы могут включать детектируемые вещества, применимые, например, в диагностике. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радионуклиды, позитронно-активные металлы (для использования в позитронно-эмиссионной томографии) и ионы нерадиоактивных парамагнитных металлов. В целом, см. патент США № 4741900 на предмет ионов металлов, которые можно конъюгировать с антителами для использования в качестве диагностических средств. Подходящие ферменты включают пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; подходящие простетические группы включают стрептавидин, авидин и биотин; подходящие флуоресцентные материалы включают умбеллиферон, флуоресцеин, флуоресцеинизотиоцианат, родамин, дихлортриазиниламинфлуоресцеин, дансил хлорид и фикоэритрин; подходящие люминесцентные материалы включают люминол; подходящие биолюминесцентные материалы включают люциферазу, люциферин и экворин; и подходящие радионуклиды включают 125I, 131I, 111In и 99Tc.
В другом примере эффекторная молекула может повышать время полужизни антитела in vivo, и/или снижать иммуногенность антитела и/или повышать доставку антитела через эпителиальный барьер в иммунную систему. Примеры подходящих эффекторных молекул этого типа включают полимеры, альбумин, альбумин-связывающие белки или альбумин-связывающие соединения, такие, как описывают в WO05/117984.
Если эффекторная молекула является полимером, он, в основном, может являться синтетическим или природным полимером, например, необязательно замещенным неразветвленным или разветвленным полиалкиленовым, полиалкениленовым или полиоксиалкиленовым полимером или разветвленным или неразветвленным полисахаридом, например, гомо- или гетерополисахаридом.
Конкретные необязательные заместители, которые могут присутствовать в указанных выше синтетических полимерах, включают один или несколько гидрокси-, метильных или метоксигрупп.
Конкретные примеры синтетических полимеров включают необязательно замещенные неразветвленные или разветвленные полиэтиленгликоль, полипропиленгликоль, поливиниловый спирт или его производные, в частности, необязательно замещенный полиэтиленгликоль, такой как метоксиполиэтиленгликоль или его производные.
Функциональные анализы и форматы скрининга
Как правило, подходящие связывающие домены для использования в настоящем изобретении можно идентифицировать посредством тестирования одной или нескольких пар связывающих доменов в функциональном анализе. Например, полиспецифическую молекулу, содержащую связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79a и/или CD79b, можно тестировать в одном или нескольких функциональных анализах.
Как применяют в настоящем описании, термин "функциональный анализ" относится к анализу, который можно использовать для определения одного или нескольких желаемых свойств или активностей белковых комплексов, комплексов антител или смеси антител, подвергаемых воздействию условий анализа. Подходящие функциональные анализы могут являться анализами связывания, анализами апоптоза, анализами антителозависимой клеточной цитотоксичности (ADCC), анализами комплементзависимой цитотоксичности (CDC), анализами ингибирования роста клеток или пролиферации (цитостатического эффекта), анализами гибели клеток (цитотоксического эффекта), анализами передачи сигнала в клетках, анализами продукции цитокинов, анализами продукции антител и переключения изотипа и анализами дифференцировки клеток.
Эффективность полиспецифических антител по настоящему изобретению можно сравнивать с отдельными антителами или смесями антител (или фрагментов) в таких моделях способами, как правило, известными специалисту в этой области.
Функциональные анализы можно повторять множество раз по мере необходимости для повышения достоверности результатов. Для определения статистически значимых результатов и, таким образом, идентификации полиспецифических молекул с биологическими функциями можно использовать различные статистические тесты, известные специалисту в этой области.
Примеры подходящих функциональных анализов описывают в примерах в настоящем описании, и они включают измерение способности полиспецифической молекулы по настоящему изобретению ингибировать активацию B-клеток после стимуляции с использованием антитела против IgM, что измеряют посредством определения ингибирования маркеров активации B-клеток, таких как экспрессия фосфорилированного Akt, экспрессия фосфорилированного P38, передача сигнала PLCγ, экспрессия CD40, экспрессия CD71 и/или экспрессия CD86.
При разработке функционального анализа для скрининга специалист в этой области может устанавливать подходящее пороговое значение, идентифицированную активность свыше которого считают "совпадением". При использовании нескольких функциональных анализов пороговое значение для каждого анализа можно устанавливать на подходящем уровне для определения контролируемого коэффициента совпадений. В одном примере коэффициент совпадений может составлять 3-5%. В одном примере набором критериев при поиске пар связывающих доменов, ингибирующих функцию B-клеток, может являться по меньшей мере 30% ингибирования, по меньшей мере два считывания фосфорилирования в анализе активации B-клеток.
В одном примере полиспецифическая молекула по настоящему изобретению имеет IC50 менее 5 нМ для ингибирования экспрессии CD86 в B-клетках, стимулируемых антителом против IgM.
В одном из вариантов осуществления для тестирования молекул по настоящему изобретению можно использовать анализы in vivo, такие как модели на животных, включая модели опухолей мыши, модели аутоиммунного заболевания, модели грызунов или приматов, инфицированных вирусом или бактериями, и т.п.
Пример подходящего формата для скрининга и обнаружения связывающих доменов представлен в настоящем описании ниже.
В скрининге для идентификации связывающих доменов для использования в настоящем изобретении можно использовать биспецифический белковый комплекс.
Как применяют в настоящем описании, термин "биспецифический белковый комплекс" относится к молекуле, содержащей два белка (A и B, обозначаемые в настоящем описании как биспецифические компоненты), удерживаемые вместе посредством гетеродимерного соединения. В одном из вариантов осуществления один или оба белка имеют связывающий домен, например, один или оба белка являются антителами или их фрагментами.
Как правило, биспецифический белковый комплекс имеет формулу A-X:Y-B, где:
A-X является первым слитым белком;
Y-B является вторым слитым белком;
X:Y является гетеродимерным соединением;
A содержит первый связывающий домен;
B содержит второй связывающий домен;
X является первым партнером по связыванию из связывающейся пары;
Y является вторым партнером по связыванию из связывающейся пары; и
: представляет собой взаимодействие (такое как связывающее взаимодействие) между X и Y.
Как применяют в настоящем описании, "слитые белки" содержат белковый компонент, например, A или B, слитые с другой молекулой, например, партнером по связыванию X или Y (при необходимости). В варианте осуществления слитый белок является трансляционным белком, экспрессируемым рекомбинантными способами с генетической конструкции, например, экспрессируемым в хозяине с конструкции ДНК.
Функцией соединения X:Y является удержание белков A и B вблизи друг друга таким образом, что можно реализовывать синергическую функцию A и B.
Как применяют в настоящем описании, термин "гетеродимерное соединение" относится к соединению, содержащему два разных партнера по связыванию X и Y, образующих взаимодействие (такое как связывание) друг с другом, имеющее общую аффинность, достаточную для удержания двух партнеров по связыванию вместе. В одном из вариантов осуществления X и/или Y не подходят для образования гомодимеров.
Термины "гетеродимерно соединенные" и "гетеродимерное соединение" используют взаимозаменяемо в настоящем описании.
Как применяют в настоящем описании, В одном из вариантов осуществления термин "неподходящий для образования гомодимеров" относится к образованию гетеродимеров X-Y, более предпочтительных, например, стабильных, например, термодинамически стабильных и/или физически стабильных (например, о чем свидетельствует отсутствие агрегации), после образования.
В одном из вариантов осуществления взаимодействие X-Y является более предпочтительным, чем взаимодействие X-X или Y-Y. Это снижает образование гомодимеров X-X или Y-Y при смешивании слитых белков A-X и B-Y. Это также делает удаление гомодимеров относительно простым, например, за один этап очистки, такой как колоночная хроматография, получают, по существу, чистые слитые белки и/или биспецифические белковые комплексы по настоящему изобретению.
В одном из вариантов осуществления этап очистки осуществляют после экспрессии слитого белка. Таким образом, в одном из вариантов осуществления перед смешиванием in vitro слитые белки получают в, по существе, чистой форме. Как применяют в настоящем описании, термин "по существу, чистая форма" относится к случаю, когда слитый белок имеет содержание мономера в диапазоне от 85 до 100%, например, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 или 100%.
В одном из вариантов осуществления очистка не является необходимой после образования биспецифического белкового комплекса.
В одном из вариантов осуществления соотношение слитых белков, используемых на этапе смешивания in vitro в способе по настоящему изобретению, представляет собой A-X к B-Y от 0,8:1 до 3:1, такое как 1,5: 1 или 2:1.
В одном из вариантов осуществления соотношение слитых белков, используемых на этапе смешивания in vitro в способе по настоящему изобретению представляет собой B-Y к A-X от 0,8:1 до 3:1, такое как 1,5: 1 или 2:1.
В одном из вариантов осуществления соотношение составляет 1:1.
В одном из вариантов осуществления один (или по меньшей мере один) из партнеров по связыванию неспособен образовывать гомодимер, например, аминокислотную последовательность партнера по связыванию подвергают мутации для устранения или минимизации образования гомодимеров.
В одном из вариантов осуществления оба партнера по связыванию неспособны образовывать гомодимер, например, один из партнеров по связыванию является пептидом, а другой партнер по связыванию является VHH, специфичным для указанного пептида.
В одном из вариантов осуществления scFv, используемый в молекулах по настоящему изобретению, неспособен образовывать гомодимер.
Как применяют в настоящем описании, термин "неспособный образовывать гомодимеры" относится к низкой или нулевой способности образовывать гомодимеры. Как применяют в настоящем описании, термин "низкий" относится к 5% или менее, например, 4, 3, 2, 1, 0,5% агрегатов или менее.
Небольшие количества агрегата в слитых белках или остаточные количества в гетеродимерно соединенном биспецифическом белковом комплексе, как правило, имеют минимальный эффект в отношении способа по настоящему изобретению.
В одном из вариантов осуществления ":" представляет собой связывающее взаимодействие, например, на основе сил притяжения, таких как ван-дер-ваальсовы силы, таких как водородные связи и электростатические взаимодействия, например, на основе специфичности антитела к антигену, такому как пептид.
В одном из вариантов осуществления ":" представляет собой ковалентную связь, образующуюся при специфическом химическом взаимодействии, таком как клик-химия.
В одном из вариантов осуществления ":" не является ковалентной связью.
Как применяют в настоящем описании, термин "образуют комплекс" относится к взаимодействию, включая связывающие взаимодействия или химическую реакцию, являющуюся достаточно специфичной и сильной, если компоненты слитого белка A-X и B-Y приводят в контакт в соответствующих условиях таким образом, что собирается комплекс, и слитые белки удерживаются вместе.
Как применяют в настоящем описании, термин "удерживаемые вместе" относится к удержанию компонентов (слитых белков) вблизи друг друга, таким образом, что после связывания с комплексом можно обращаться так, как если бы он являлся одной молекулой, и он во многих случаях ведет себя и действует как единая молекула. В одном из вариантов осуществления удержание делает комплекс пригодным для использования в способе, представленном в настоящем описании, т.е. пригодном для использования по меньшей мере в одном функциональном скрининге.
В одном из вариантов осуществления связывающее взаимодействие является обратимым.
Как применяют в настоящем описании, термин "специфичность" в отношении к X и Y относится к случаю, когда партнеры по связыванию X и Y во взаимодействии распознают только друг друга или имеют значительно более высокую аффинность друг для друга по сравнению с не-партнерами, например, по меньшей мере в 2, 3, 4, 5, 6, 7, 8, 9, 10 раз более высокую аффинность.
В одном из вариантов осуществления связывающее взаимодействие между X и Y имеет низкую константу диссоциации.
Примеры низкой константы диссоциации включают 1-9×10-2 с-1 или менее, например, 1-9×10-3 с-1, 1-9×10-4 с-1, 1-9×10-5 с-1, 1-9×10-6 с-1 или 1-9×10-7 с-1. Особенно подходящие константы диссоциации включают 1×10-4 с-1 или менее, например, 1×10-5 с-1, 1×10-6 с-1 или 1×10-7 с-1.
Не будучи связанными какой-либо теорией, считают, что низкая константа диссоциации (также обозначаемая как скорость обратной реакции) позволяет молекулам быть достаточно стабильными, чтобы сделать биспецифический белковый комплекс применимым, в частности, в анализах для функционального скрининга.
В одном из вариантов осуществления аффинность X и Y друг для друга составляет 5 нМ или более, например, 4 нМ, 3 нМ, 2 нМ, 1 нМ или более.
В одном из вариантов осуществления аффинность X и Y друг для друга составляет 900 пМ или более, например, 800, 700, 600, 500, 400, 300, 200, 100 или 50 пМ или более.
В другом варианте осуществления аффинность X и Y друг для друга составляет 10 пМ или более, например, 9, 8, 7, 6 или 5 пМ.
Аффинность является значением, вычисляемым из скорости прямой и обратной реакций взаимодействия. Как применяют в настоящем описании, термин "аффинность" относится к силе общей суммы нековалентных взаимодействий между одним участком связывания молекулы (например, антитела) и его партнером по связыванию (например, пептидом). Аффинность молекулы для ее партнера по связыванию, как правило, можно представлять посредством равновесной константы диссоциации (KD). Аффинность можно измерять общепринятыми известными в этой области способами, включая представленные в настоящем описании, такие как способы поверхностного плазмонного резонанса, в частности, BIAcore.
В одном из вариантов осуществления множество биспецифических белковых комплексов по настоящему изобретению тестируют параллельно или, по существу, одновременно.
Как применяют в настоящем описании, термин "одновременно" относится к случаю, когда образцы/молекулы/комплексы анализируют в одном анализе, например, за один "раз".
В одном из вариантов осуществления одновременно относится к одновременному анализу, в котором исходящий сигнал анализируют, по существу, одновременно с помощью инструмента. Для этого сигнала может потребоваться деконволюция для интерпретации полученных результатов.
Преимущественно, тестирование множества биспецифических белковых комплексов делает возможным более эффективный скрининг большого количества биспецифических белковых комплексов и идентификацию новых и интересных взаимосвязей. Значительно отличающиеся вариабельные области к интересующим антигенам-мишеням CD45 и CD79 могут обеспечить доступ к тонким нюансам биологической функции.
В одном из вариантов осуществления множество биспецифических белковых комплексов тестируют с использованием мультиплекса, как определено выше, и подвергают их одному или нескольким функциональным анализам.
Как применяют в настоящем описании, термин "биологическая функция" относится к активности, являющейся природной или целевой для биологической молекулы, подвергаемой тестированию, например, природной активностью клетки, белка или т.п. В идеале наличие функции можно тестировать с использованием функционального анализа in vitro, включая анализы с использованием живых клеток млекопитающих. Как применяют в настоящем описании, термин "природная функция" включает аномальную функцию, такую как функции, ассоциированные со злокачественными новообразованиями.
Как применяют в настоящем описании, термин "соответствующий "биологический компаратор"" относится к подходящей молекуле для оценки активности в том же анализе, который используют для биспецифического белкового комплекса, для определения того, присутствует ли какое-либо изменение или новая активность или функция. Подходящие компараторы для A-X:Y-B могут включать очищенный белок (включая рекомбинантные белки) в природной форме или присутствовать в том же формате, что и биспецифические, т.е. если A и B являются одной и той же молекулой, такой как A-X:Y-A или B-X:Y-B. Альтернативно, в качестве компаратора можно использовать слитый белок A-X или B-Y в некомплексированной форме. Альтернативно, можно использовать множество компараторов различных форматов (в частности, как представлено в настоящем описании). Специалист в этой области способен идентифицировать и включать подходящий контроль/компаратор с учетом общих сведений или информации, обнаруживаемой в литературе.
Как применяют в настоящем описании, термин "синергическая функция" относится к биологической активности, не наблюдаемой или более высокой, чем наблюдаемая, если первый и второй белки из биспецифического белкового комплекса по настоящему изобретению не используют совместно, например, активности, наблюдаемой только в биспецифической форме. Таким образом, термин "синергический" включает новую биологическую функцию.
Настоящее изобретение относится к молекуле со специфичностью, по меньшей мере, к CD45 и CD79 с новой биологической функцией.
Как применяют в настоящем описании, термин "новая биологическая функция" относится к функции, не очевидной или отсутствующей до тех пор, пока две или более синергических молекулы [белок A и белок B] не объединяют (в биспецифическом или ином виде), или ранее неидентифицированной функции.
Как применяют в настоящем описании, термин "более высокий" относится к повышению активности, включая повышение относительно нуля, т.е. некоторой активности в биспецифической форме, где отдельный некомплексированный биспецифический компонент или компоненты не имеют активности в соответствующем функциональном анализе, также обозначаемой в настоящем описании как новая активность или новая биологическая функция. Как применяют в настоящем описании, термин "более высокий" также включает большую, чем аддитивная, функцию в биспецифической форме в соответствующем функциональном анализе по сравнению с отдельными некомплексированными биспецифическими компонентами или бивалентными связывающими доменами, например, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 200, 300% или большее повышение соответствующей активности.
В одном из вариантов осуществления новая синергическая функция является более высокой ингибиторной активностью.
В одном из вариантов осуществления молекула полиспецифического антитела по настоящему изобретению имеет более высокую ингибиторную активность, чем сумма активности бивалентного связывающего домена для CD45 и бивалентного связывающего домена для CD79a, представленных в отдельности или в смеси.
В одном из вариантов осуществления по меньшей мере один из первого партнера по связыванию, X, и второго партнера по связыванию, Y, из связывающейся пары независимо выбран из пептида и белка; например, первый партнер по связыванию или второй партнер по связыванию является пептидом.
Подходящие пептиды включают группу, содержащую GCN4, Fos/Jun (Fos человека и мыши имеют регистрационный номер Uniprot P01100 и P01101, соответственно, и jun человека и мыши имеют регистрационный номер Uniprot P05412 и P05627, соответственно), гемагглютинин вируса гриппа человека (HA), полигистидин (His), зеленый флуоресцентный белок (GFP) и FLAG. Другие пептиды также включены в качестве пригодных для использования в настоящем изобретении, и особенно подходящими пептидами являются аффинные метки для очистки белка, т.к. такие пептиды имеют склонность к связыванию с высокой аффинностью с соответствующими партнерами по связыванию.
Как применяют в настоящем описании, термин "пептид" относится к короткому полимеру из аминокислот, соединенных пептидными связями, где пептид содержит диапазон от 2 до 100 аминокислот, например, от 5 до 99, например, от 6 до 98, от 7 до 97 или от 8 до 96. В одном из вариантов осуществления пептид, используемый в настоящем изобретении, представляет собой аминокислотную последовательность из 50 аминокислотных остатков или менее, например, 40, 30, 10 или менее. Пептиды, используемые в настоящем изобретении, имеют достаточную длину, чтобы соответствовать цели, например, если пептид является линкером, он должен быть достаточно длинным, чтобы позволить фрагменту, который он соединяет, осуществлять свою биологическую функцию; альтернативно, если пептид является партнером по связыванию, он должен быть способен специфически связываться с другой молекулой, такой как антитело.
В одном из вариантов осуществления другой партнер по связыванию из связывающейся пары (альтернативный первый или второй партнер по связыванию) является белком.
Как применяют в настоящем описании, термин "белок" относится к аминокислотной последовательности из 100 аминокислот или более. Как применяют в настоящем описании, в одном из вариантов осуществления термин "белок" относится к аминокислотной последовательности с вторичной или третичной структурой.
В одном из вариантов осуществления первый белок, A, и/или второй белок, B, из биспецифического белкового комплекса является антителом или фрагментом антитела. Такой биспецифический белковый комплекс можно обозначать как биспецифический комплекс антитела.
В одном из вариантов осуществления каждое антитело или фрагмент, используемый в биспецифическом комплексе антитела по настоящему изобретению, содержит один участок связывания.
Полноразмерное антитело или фрагмент антитела, используемое в слитых белках (A-X или B-Y), может являться моноспецифическим, мультивалентным или биспецифическим.
Преимущественно, использование двух биспецифических антител или фрагментов антител позволяет молекулам по настоящему изобретению, таким как биспецифический комплекс антитела, представленный в настоящем описании, потенциально являться специфичными для до 4 различных антигенов (т.е. комплекс может являться тетраспецифическим). Это позволяет исследовать эффекты типа авидности.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как первый слитый белок A-X, являются моноспецифическим антителом или фрагментом антитела, например, Fab, Fabʹ, scFv или т.п., и, в частности, являются специфичными для CD45.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как второй слитый белок B-Y, является моноспецифическим антителом или фрагментом антитела, например, Fab, Fabʹ, scFv или т.п., и, в частности, являются специфичными для CD79a и/или CD79b.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как второй слитый белок B-Y, являются мультивалентными, т.е. имеют два или более связывающих домена.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как первый слитый белок A-X, являются моновалентными, и антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как второй слитый белок B-X, являются моновалентными.
Таким образом, в одном из вариантов осуществления связывающие домены полиспецифических молекул по настоящему изобретению являются моновалентными.
Таким образом, в одном из вариантов осуществления связывающие домены полиспецифических молекул по настоящему изобретению являются моновалентными и моноспецифическими.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как первый слитый белок A-X, являются моновалентными, и антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как второй слитый белок B-Y, являются мультивалентными.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как первый слитый белок A-X, являются мультивалентными, и антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как второй слитый белок B-Y, являются моновалентными.
В одном из вариантов осуществления антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как первый слитый белок A-X, являются мультивалентными, и антитело или фрагмент антитела, используемое в молекулах по настоящему изобретению, или его компоненты, такие как второй слитый белок B-Y, являются мультивалентными.
В одном из вариантов осуществления первое антитело, второе антитело или первое и второе антитело из молекул по настоящему изобретению или его компоненты, такие как биспецифический комплекс антитела, может представлять собой формат IgG, например, антитело против CD45 и/или CD79 можно получать в формате IgG.
В одном из вариантов осуществления фрагмент антитела выбран из группы, состоящей из: фрагмента антитела (Fab), одноцепочечного вариабельного фрагмента (scFv) и однодоменного антитела (sdAb), такого как scFv, используют в первом (A-X) или втором слитом белке (B-Y). Преимущественно, небольшой размер scFv может облегчать правильный фолдинг биспецифических комплексов антитела.
В одном из вариантов осуществления первое (A), второе антитело/фрагмент (B) или первое и второе антитело/фрагмент из биспецифического комплекса антитела по настоящему изобретению могут являться Fab.
В одном из вариантов осуществления первое, второе антитело/фрагмент или первое и второе антитело/фрагмент из биспецифического комплекса антитела по настоящему изобретению являются VHH.
Как применяют в отношении биспецифического комплекса по настоящему изобретению, термин "слитый белок" относится к белку, например, антителу или фрагменту антитела, соединенному с партнером по связыванию.
Для удобства биспецифические белковые комплексы по настоящему изобретению обозначают в настоящем описании как A-X:Y-B. Однако эта номенклатура не предназначена для ограничения дизайна слитого белка A-X и B-Y, т.к. эксперименты авторов настоящего изобретения показывают, что партнеры по связыванию X и Y могут являться противоположными, т.е. A-Y и B-X, без неблагоприятного влияния на способ. Таким образом, A и B и X и Y представляет собой условные обозначения, приведенные для облегчения объяснения технологии по настоящему изобретению.
Как применяют в настоящем описании, термин "соединенный" относится к соединению напрямую или косвенно с помощью линкера, такого как пептидный линкер, примеры которого описаны ниже. Прямое соединение включает слияние (например, пептидную связь) или химическую конъюгацию.
Как применяют в настоящем описании, термин "партнер по связыванию" относится к одному компоненту связывающейся пары.
В одном из вариантов осуществления аффинность партнеров по связыванию является высокой, 5 нМ или более, такой 900, 800, 700, 600, 500, 400, 300 пМ или более.
Как применяют в настоящем описании, термин "связывающаяся пара" относится к двум партнерам по связыванию, специфически связывающимся друг с другом. Примеры связывающейся пары включают пептид и антитело или связывающий фрагмент, специфичный для него, или фермент и лиганд, или фермент и ингибитор этого фермента.
В одном из вариантов осуществления первый партнер по связыванию (X) выбран из группы, содержащей: полноразмерное антитело, Fab, Fabʹ, scFv, пептид и sdAb, где примеры sdAb включают VH или VL или VHH.
В одном из вариантов осуществления второй партнер (Y) выбран из группы, содержащей: полноразмерное антитело, Fab, Fabʹ, scFv, пептид и sdAb, где примеры sdAb включают VH или VL или VHH.
В одном из вариантов осуществления, где A является антителом или его фрагментом, первого партнера по связыванию (X) соединяют с C-концом тяжелой или легкой цепи первого антитела или фрагмента антитела, например, первого партнера по связыванию соединяют с C-концом тяжелой цепи первого антитела или фрагмента антитела.
В другом варианте осуществления, где B является антителом или его фрагментом, второго партнера по связыванию (Y) соединяют с C-концом тяжелой или легкой цепи второго антитела или фрагмента антитела, например, второго партнер по связыванию соединяют с C-концом тяжелой цепи второго антитела или фрагмента антитела.
В одном из вариантов осуществления X соединяют с C-концом тяжелой цепи антитела или фрагмента (белка A) и Y соединяют с C-концом антитела или фрагмента (белка B).
В одном из вариантов осуществления X соединяют с помощью линкера (такого как ASGGGG или ASGGGGSG) с C-концом тяжелой цепи антитела или фрагмента (белка A) и Y соединяют с помощью линкера (такого как ASGGGG или ASGGGGSG) с C-концом антитела или фрагмента (белка B).
В одном из вариантов осуществления первый или второй партнер по связыванию (X или Y) является пептидом.
Примеры подходящей связывающейся пары могут включать GCN4 (SEQ ID NO: 1) или его вариант и 52SR4 (SEQ ID NO: 3) или его вариант, являющийся scFv, специфичным для GCN4.
В одном из вариантов осуществления первый партнер по связыванию (а именно X) является GCN4 (например, как представлено в SEQ ID NO: 1) или его вариантом (например, без гистидиновой метки), и второй партнер по связыванию (а именно Y) является scFv, специфичным для GCN4 (например, как представлено в SEQ ID NO: 3) или его вариантом.
В одном из вариантов осуществления первый партнер по связыванию (а именно X) является sFv, специфичным для GCN4 (например, как представлено в SEQ ID NO: 3), или его вариантом, и второй партнер по связыванию (а именно Y) является GCN4 (например, как представлено в SEQ ID NO: 1) или его вариантом.
Варианты GCN4 включают аминокислотную последовательность по меньшей мере с 80%, 85%, 90%, 91%, 92%, 93%, 94% 95%, 96%, 97% или 98% или 99% идентичности по отношению к SEQ ID NO: 1. Варианты GCN4 также включают аминокислотную последовательность, имеющую по меньшей мере 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности по отношению к последовательности, кодируемой нуклеотидной последовательностью SEQ ID NO: 2 или нуклеотидной последовательностью, гибридизующейся с SEQ ID NO: 2 в строгих условиях.
Подходящим scFv, специфичный для GCN4, является 52SR4 (SEQ ID NO: 3) или его вариант. Варианты 52SR4 включают аминокислотную последовательность по меньшей мере с 80%, или 85%, или 90%, или 95%, или 98% или 99% идентичности по отношению к SEQ ID NO: 3. Варианты 52SR4 также включают аминокислотную последовательность, имеющую по меньшей мере 80%, или 85%, или 90%, или 95%, или 98%, или 99% идентичности по отношению к последовательности, кодируемой нуклеотидной последовательностью SEQ ID NO: 4 или нуклеотидной последовательностью, гибридизующейся с SEQ ID NO: 2 в строгих условиях.
Авторы настоящего изобретения обнаруживали, что одноцепочечное антитело 52SR4 и пептид GCN4 являются связывающейся парой, пригодной для использования в биспецифических белковых комплексах по настоящему изобретению.
Альтернативно, в качествен X и Y можно использовать любое подходящее антитело/фрагмент и антиген (такой как пептид).
В одном из вариантов осуществления первый партнер по связыванию (X) и второй партнер по связыванию(Y) являются белком.
В одном из вариантов осуществления первый партнер по связыванию (X) является ферментом или его активным фрагментом, и второй партнер по связыванию (Y) является лигандом, или наоборот.
В одном из вариантов осуществления первый партнер по связыванию (X) является ферментом или его активным фрагментом, и второй партнер по связыванию (Y) является ингибитором этого фермента, или наоборот.
Как применяют в настоящем описании, термин "активный фрагмент" относится к аминокислотному фрагменту, являющемуся меньшим, чем целая аминокислотная последовательность молекулы, и сохраняющему, по существу, ту же биологическую активность или соответствующую биологическую активность, например, активность более 50%, такую как 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%.
В другом варианте осуществления первый партнер по связыванию X является глутатионом (GSH), и второй партнер по связыванию Y является глутатион-S-трансферазой (GST), или наоборот.
В другом варианте осуществления X является Fos, и Y является Jun, или наоборот.
В другом варианте осуществления X является His, и Y является антителом против His, или наоборот.
В другом варианте осуществления связывающаяся пара является кальмодулин-связывающим пептидом, и Y является кальмодулином, или наоборот.
В другом варианте осуществления X является мальтоза-связывающим белком, и Y является антителом против мальтоза-связывающего белка или его фрагментом, или наоборот.
Другие комбинации фермент-лиганд также предусмотрены для использования в партнерах по связыванию. Кроме того, подходящими являются аффинные метки, известные в этой области для очистки белка, т.к. они имеют склонность к связыванию с высокой аффинностью с соответствующими партнерами по связыванию.
Степени идентичности и сходства легко можно вычислять (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, программное обеспечение BLAST™, доступное в NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215:403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3:266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266:131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25:3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7:649-656).
В одном из вариантов осуществления первый или второй партнер по связыванию (X или Y) является белком или пептидом.
В одном из вариантов осуществления первый и второй слитые белки содержат один или несколько пептидных линкеров. Линкеры можно встраивать в разные участки в слитых белках. Например, можно встраивать линкер между партнером по связыванию и присоединенным к нему белком.
В одном из вариантов осуществления линкер является пептидным линкером.
Как применяют в настоящем описании, термин "пептидный линкер" относится к пептиду с аминокислотными последовательностями. Диапазон подходящих пептидных линкеров известен специалисту в этой области.
В одном из вариантов осуществления пептидный линкер может иметь синтетическое происхождение, т.е. его можно получать химическими способами синтеза.
В одном из вариантов осуществления партнеры по связыванию из биспецифических белковых комплексов соединяют с соответствующими белками с помощью пептидных линкеров.
В одном из вариантов осуществления слитые белки представляет собой трансляционное слияние, т.е. слитый белок, экспрессирующийся в клетках-хозяевах, содержащих генетическую конструкцию, с которой экспрессируется слитый белок.
В одном из вариантов осуществления слитый белок получают посредством конъюгации A с X или B с Y, необязательно, с помощью пептидного линкера.
В одном из вариантов осуществления пептидный линкер составляет 50 аминокислот в длину или менее, например, 20 аминокислот или менее.
Как правило, более эффективной может являться рекомбинантная экспрессия слитого белка, и, таким образом, предпочтительной может являться прямая пептидная связь или пептидный линкер, который может экспрессироваться клеткой-хозяином.
В одном из аспектов настоящее изобретение относится к способу получения биспецифического белкового комплекса по настоящему изобретению, включающему этапы:
(a) получения первого слитого белка (A-X), содержащего связывающий домен, специфичный для CD45 или CD79 и/или CD79b (A), соединенный с первым партнером по связыванию (X) из связывающейся пары;
(b) получения второго слитого белка (B-Y), содержащего связывающий домен, специфичный для CD45 или CD79a и/или CD79b (B), соединенный со вторым партнером по связыванию (Y) из связывающейся пары;
где, по меньшей мере, первый слитый белок или второй слитый белок содержат связывающий домен, специфичный для CD45, и оставшийся слитый белок содержит связывающий домен, специфичный для CD79a и/или CD79b, и
(c) смешивания первого (A-X) и второго слитых белков (B-Y), полученных на этапе a) и b).
В частности, гетеродимерно соединенный биспецифический белковый комплекс получают посредством смешивания A-X и B-Y in vitro. Таким образом, в одном из вариантов осуществления способ включает этап смешивания in vitro с приведением A-X и B-Y в контакт.
Таким образом, как правило, слитые белки A-X и B-Y не коэкспрессируются в одной клетке. Это является предпочтительным, т.к. это позволяет, например, 100 слитых белков A-X и 100 слитых белков A-Y раздельно экспрессировать и, необязательно, очищать, и с помощью последующего смешивания 200 слитых белков в различных пермутациях можно получать 10000 гетеродимерно соединенных биспецифических белковых комплексов.
В отличие от этого, в способах предшествующего уровня техники необходима коэкспрессия биспецифических молекул и, таким образом, в случае 10000 комплексов необходимо 10000 трансфекций, экспрессий и очисток.
Партнеры по связыванию X и Y имеют аффинность друг для друга и действуют в качестве биологического эквивалента velcro® или "застежки" и "магнита" и удерживают комплекс вместе. Преимущественно, это означает, что слитые белки A-X и Y-B легко можно собирать в биспецифический белковый комплекс просто посредством смешивания слитых белков. Таким образом, биспецифический белковый комплекс по настоящему изобретению имеет модульную структуру, позволяющую легко собирать два разных белка для получения больших панелей пермутаций биспецифических белковых комплексов с различными комбинациями антигенных специфичностей, например, в виде сетки. Это делает возможным эффективный и систематический скрининг большого количества биспецифических белковых комплексов для детекции аддитивной, синергической или новой биологической функции.
Учитывая, что X и Y являются специфичными друг для друга, это значительно снижает способность образовывать гомодимеры. В настоящем описании X и Y в совокупности обозначают как связывающуюся пару или партнеров по связыванию. В одном из вариантов осуществления X не имеет высокую аффинность к другим X. В одном из вариантов осуществления Y не имеет высокую аффинность к другим Y. Преимущественно, если X и Y не образуют гомодимеры, это предотвращает образование нежелательных моноспецифических белковых комплексов, повышает выход желаемых биспецифических белковых комплексов и устраняет потребность в трудоемких этапах очистки для удаления моноспецифических белковых комплексов.
Этой быстрой сборки биспецифических белковых комплексов, уровня выхода и/или чистоты нельзя достигать способами предшествующего уровня техники, в частности, для способов предшествующего уровня техники, как правило, необходимы этапы интенсивной очистки.
Преимущественно, компоненты X и Y делают возможной быструю и легкую сборку мультиплекса, содержащего биспецифические белковые комплексы, полученные посредством различных пермутаций слитых белков.
В одном из вариантов осуществления белки A и B являются антителами или фрагментами антител. Если антитело или фрагменты антител удерживаются вместе в виде комплекса с помощью X и Y, они образуют биспецифический комплекс антитела.
Смешивание, как правило, осуществляют в условиях, в которых X и Y могут взаимодействовать. В одном из вариантов осуществления слитые белки инкубируют в средах для культивирования клеток в условиях культивирования клеток, например, слитые белки инкубируют в течение 90 минут в условиях 37°C/5%CO2.
В одном из вариантов осуществления слитые белки по настоящему изобретению смешивают в водной среде, например, один слитый белок может связываться с твердой поверхностью, такой как бусина или планшет, а другой слитый белок можно включать в водный раствор/суспензию. Твердая фаза позволяет легко вымывать избыточные компоненты и реагенты. В одном из вариантов осуществления слитый белок не присоединяют к твердой фазе, а просто смешивают в жидкости/растворе/среде.
Преимущественно, способ по настоящему изобретению можно использовать для получения комплексов, образующихся между гетерогенными парами (т.е. между первым слитым белком [A-X] и вторым слитым белком [B-Y]), где минимизируют взаимодействия между гомогенными парами (т.е. между двумя первыми слитыми белками [A-X] или двумя вторыми слитыми белками [B-Y]). Таким образом, способ по настоящему изобретению позволяет получать большие количества биспецифических белковых комплексов с минимальной контаминацией гомодимерными комплексами или без нее. Этот уровень чистоты и выход невозможны при использовании способов предшествующего уровня техники.
В одном из вариантов осуществления для образующихся комплексов не требуются дополнительные этапы очистки.
В одном из вариантов осуществления для образующихся комплексов необходим один этап очистки, например, колоночной хроматографии.
В одном из вариантов осуществления способ дополнительно включает по меньшей мере один этап очистки, например, после экспрессии слитого белка по настоящему изобретению.
Как применяют в настоящем описании, термин "функциональный анализ" относится к анализу, который можно использовать для определения одного или нескольких желаемых свойств или активностей белковых комплексов, комплексов антител или смеси антител, подвергаемых воздействию условий анализа. Подходящие функциональные анализы могут являться анализами связывания, анализами апоптоза, анализами антителозависимой клеточной цитотоксичности (ADCC), анализами комплементзависимой цитотоксичности (CDC), анализами ингибирования роста или пролиферации клеток (цитостатического эффекта), анализами гибели клеток (цитотоксического эффекта), анализами передачи сигнала в клетки, анализами продукции цитокинов, анализами продукции антител и переключения изотипа и анализами дифференцировки клеток. В одном из вариантов осуществления для тестирования молекул по настоящему изобретению можно использовать анализы in vivo, такие как модели на животных, включая модели опухолей мыши, модели аутоиммунного заболевания, модели грызунов или приматов, инфицированных вирусом или бактериями, и т.п.
Что касается биспецифических комплексов антитела, эффективность биспецифических комплексов антител по настоящему изобретению можно сравнивать с отдельными антителами или смесями антител (или фрагментов) в таких моделях способами, как правило, известными специалисту в этой области.
Функциональные анализы можно повторять множество раз по мере необходимости с использованием различных образцов конкретного биспецифического комплекса антитела или без них для повышения достоверности результатов. Для идентификации статистически значимых результатов можно использовать различные статистические тесты, известные специалисту в этой области, и, таким образом, идентифицировать биспецифические комплексы антител с биологическими функциями, и в частности, для идентификации оптимальных пар вариабельных областей для использования в полиспецифической молекуле по настоящему изобретению.
Композиции и медицинское применение
В одном из аспектов настоящее изобретение относится к молекуле по настоящему изобретению или компоненту, такому как слитый белок, гетеродимерно соединенному биспецифическому белковому комплексу, композиции, содержащей молекулу по изобретению, включая слитый белок или указанный биспецифический белковый комплекс, мультиплексу, чипу, библиотеке, как определено в настоящем описании.
В одном из вариантов осуществления молекулы по настоящему изобретению, например, антитело, представленное в настоящем описании, полиспецифическая молекула и/или биспецифические белковые комплексы, подходят для терапевтического применения и могут представлять собой новые терапевтические средства для лечения заболеваний. Таким образом, в дополнительном аспекте настоящее изобретение относится к молекуле по настоящему изобретению, например, биспецифическому белковому комплексу, как описано выше, для применения в терапии. Молекулы по настоящему изобретению, включая полиспецифические молекулы и биспецифические белковые комплексы, представленные в настоящем описании, подходят для лечения спектра заболеваний, таких как злокачественное новообразование.
Молекулы по настоящему изобретению, включая полиспецифические молекулы и биспецифические белковые комплексы, представленные в настоящем описании, также особенно подходят для ингибирования функции B-клеток для контроля иммунных и аутоиммунных реакций при различных аутоиммунных заболеваниях.
Таким образом, настоящее изобретение относится к способу лечения заболевания у пациента, включающему введение терапевтически эффективного количества молекулы по настоящему изобретению, например, полиспецифической молекулы или биспецифического белкового комплекса по настоящему изобретению.
В одном из аспектов настоящее изобретение относится к фармацевтической композиции, содержащей одну или несколько молекул по настоящему изобретению, например, полиспецифическую молекулу по настоящему изобретению.
В композицию можно включать различные компоненты, включая фармацевтически приемлемые носители, эксципиенты и/или дилюенты. Композиция, необязательно, может содержать дополнительные молекулы, способные изменять характеристики популяции полиспецифических молекул по изобретению, таким образом, например, снижая, стабилизируя, замедляя, модулируя и/или активируя функцию антител. Композиция может находиться в твердой или жидкой форме и может находиться, помимо прочего, в форме порошка, таблетки, раствора или аэрозоля.
Настоящее изобретение также относится к фармацевтической или диагностической композиции, содержащей молекулу антитела или полиспецифическую молекулу по настоящему изобретению в комбинации с одним или несколькими фармацевтически приемлемыми эксципиентами, дилюентами или носителями. Таким образом, настоящее изобретение относится к применению полиспецифической молекулы по изобретению для применения в лечении и производстве лекарственного средства для лечения патологического состояния или нарушения.
Патологические состояния
Патологическое состояние или нарушение, например, можно выбирать из группы, состоящей из инфекций (вирусных, бактериальных, грибковых и паразитарных), эндотоксического шока, ассоциированного с инфекцией, артрита, такого как ревматоидный артрит, астмы, такой как тяжелая астма, хронической обструктивной болезни легких (COPD), воспалительного заболевания органов малого таза, болезни Альцгеймера, воспалительного заболевания кишечника, болезни Крона, язвенного колита, болезни Пейрони, целиакии, заболевания желчного пузыря, пилонидальной болезни, перитонита, псориаза, васкулита, послеоперационных спаек, инсульта, диабета типа I, болезни Лайма, менингоэнцефалита, аутоиммунного увеита, иммуно-опосредованных воспалительных нарушений центральной и периферической нервной системы, таких как рассеянный склероз, системной красной волчанки (такой как системная красная волчанка) и синдрома Гийена-Барре, атопического дерматита, аутоиммунного гепатита, фиброзирующего альвеолита, болезни Грейвса, IgA-нефропатии, идиопатической тромбоцитопенической пурпура, болезни Меньера, пемфигоида, первичного биллиарного цирроза, саркоидоза, склеродермии, гранулематоза Вегенера, других аутоиммунных нарушений, панкреатита, травмы (хирургического вмешательства), реакции "трансплантат против хозяина", отторжения трансплантата, заболевания сердца, включая ишемические заболевания, такие как инфаркт миокарда, а также атеросклероза, внутрисосудистой коагуляции, резорбции кости, остеопороза, остеоартрита, периодонтита, гипохлоргидрии и злокачественного новообразования, включая рак молочной железы, рак легких, рак желудка, рак яичников, печеночноклеточный рак, рак толстого кишечника, рак поджелудочной железы, рак пищевода, рак головы и шеи, рак почек, в частности, почечноклеточную карциному, рак предстательной железы, рак печени, меланому, саркому, миелому, нейробластому, плацентарную хориокарциному, рак шейки матки и рак щитовидной железы, и их метастазирующие формы.
В одном из вариантов осуществления нарушение является злокачественным новообразованием, например, лейкозом, включая лимфоцитарный лейкоз, такой как острый лимфобластный лейкоз или хронический лимфоцитарный лейкоз; или миелогенный лейкоз, такой как острый миелогенный лейкоз или хронический миелогенный лейкоз.
В одном из вариантов осуществления аутоиммунное заболевание включает: острый диссеминированный энцефаломиелит (adem), острый некротизирующий геморрагический лейкоэнцефалит, болезнь Аддисона, недостаточность надпочечников, гипокортицизм, очаговую алопецию, амилоидоз, анкилозирующий спондилит, спондилоартрит, анкилозирующий спондилит, анти-GBM/анти-TBM-нефрит, антифосфолипидный синдром (aps), аутоиммунную ангиоэдему, аутоиммунную апластическую анемию, аутоиммунную дистонию, аутоиммунный гепатит, аутоиммунную гиперлипидемию, аутоиммунный иммунодефицит, аутоиммунное заболевание внутреннего уха (AIED), аутоиммунный лимфопролиферативный синдром (ALPS), синдром Канале-Смита, аутоиммунный миокардит, аутоиммунный оофорит, аутоиммунный панкреатит (AIP), аутоиммунные полигландулярные синдромы (типов I, II и III), аутоиммунную ретинопатию (AR), аутоиммунную тромбоцитопеническую пурпуру (АТФ), аутоиммунное заболевание щитовидной железы, аутоиммунную крапивницу, аксональные/нейрональные невропатии, болезнь Бало, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, болезнь Кастлемена, целиакию, болезнь Шагаса, хроническую воспалительную демиелинизирующую полиневропатию (CIDP), хронический рецидивирующий мультифокальный остеомиелит (CRMO), синдром Чарга-Стросса, рубцующийся пемфигоид/доброкачественный слизистый пемфигоид (CP), болезнь Крона, воспалительное заболевание кишечника, колит, энтерит, илеит, синдром Когана, болезнь холодовых агглютининов, врожденную блокаду сердца, миокардит, вызванный вирусом Коксаки, CREST-синдром, криоглобулинемию, демиелинизирующие невропатии, герпетиформный дерматит, болезнь Дюринга, дерматомиозит, диабет типа I, дискоидную красную волчанку (DLE), синдром Дресслера, эндометриоз, буллезный эпидермолиз (EB) и EB Acquisita (EBA), эозинофильный гастроэнтерит, эзофагит, эозинофильный фасцит, синдром Шульмана, нодозную эритему, экспериментальный аллергический энцефаломиелит, синдром Эванса, фиброзирующий альвеолит, гигантоклеточный артериит (височный артериит), гигантоклеточный миокардит, гломерулонефрит (непролиферативный: фокальный сегментарный гломерулосклероз и мембранозный гломерулонефрит; пролиферативный: IgA-нефропатию), синдром Гудпасчера, гранулематоз с полиангиитом (GPA) (ранее известный гранулематоз Вегенера), болезнь Грейвса, синдром Гийена-Барре, синдром Миллера-Фишера, острую моторную аксональную невропатию, острую моторно-сенсорную аксональную невропатию, острую панавтономную невропатию, стволовой энцефалит Бикерстафа, энцефалит Хашимото, тиреоидит Хашимото, гемолитическую анемию, пурпуру Шенлейн-Геноха, герпес беременных, гипогаммаглобулинемию, идиопатический легочный фиброз, идиопатическую тромбоцитопеническую пурпуру (ITP), IgA-нефропатию (IGAN), синдром Бергера, синфарингитический гломерулонефрит, IgA-пемфигоид, IgG4-связанное склерозирующее заболевание, иммуно-регулируемое бесплодие, миозит с тельцами включения, инсулинозависимый сахарный диабет, интерстициальный цистит, синдром Исаакса, нейромиотонию, ювенильный артрит, ювенильный миозит, синдром Кавасаки, синдром Ламберта-Итона, лейкоцитокластический васкулит, красный плоский лишай, склерозирующий лишай, деревянистый конъюнктивит, линейный IgA-дерматоз (LAD), пемфигоид, системную красную волчанку (SLE), болезнь Лайма, болезнь Меньера, микроскопический полиангиит (MPA), смешанную болезнь соединительной ткани (MCTD), моноклональную гаммапатию, язву Мурена, болезнь Мухи-Габерманна, рассеянный склероз, миастению, миозит, нарколепсию, оптикомиелит (болезнь Девика), нейромиотонию, синдром Исаакса (приобретенный, паранеопластический, наследственный), нейтропению, глазной рубцующийся пемфигоид, неврит зрительного нерва, оофорит, опсоклонус-миоклонус-синдром, орхит, мигрирующий артрит, PANDAS (детские аутоиммунные нервно-психические расстройства, ассоциированные со стрептококковыми инфекциями), паранеопластический аутоиммунный полиорганный синдром (PAMS), паранепластическую дегенерацию мозжечка, паранеопластический пемфигоид (PNP), пароксизмальную ночную гемоглобинурию (PNH), синдром Парри-Ромберга, синдром Персонейдж-Тернера, Pars planitis (периферический увеит), пемфигоид беременных (PG), обыкновенную пузырчатку (PV), эксфолиативный пемфигоид (PF), периферическую невропатию, перивенозный энцефаломиелит, пернициозную анемию, POEMS-синдром, нодозный полиартериит (PAN), ревматическую полимиалгию, полимиозит, постинфарктный синдром миокарда, постперикардиотомический синдром, прогестероновый дерматит, первичный биллиарный цирроз, синдром Ано, первичный склерозирующий холангит (PSC), склерозирующий холангит, псориаз, псориатический артрит, гангренозную пиодермию, истинную эритроцитарную аплазию, энцефалит Расмуссена, хронический фокальный энцефалит (CFE), синдром Рейно, реактивный артрит, синдром Рейтера, рековерин-ассоциированную ретинопатию (RAR), рефлекторную симпатическую дистрофию, синдром Рейтера, рецидивирующий полихондрит, синдром беспокойных ног, ретроперитонеальный фиброз, ревматическую лихорадку, ревматоидный артрит, саркоидоз, синдром Шмидта, склерит, склеродермию, системный склероз, синдром Шегрена, аутоиммунные поражения спермы и яичек, синдром скованного человека, подострый бактериальный эндокардит (SBE), синдром Сусака, симпатическую офтальмию, артериит Такаясу, височный артериит/гигантоклеточный артериит, облитерирующий тромбангиит, болезнь Бюргера, тромбоцитопеническую пурпуру (TTP), синдром Толосы-Ханта, поперечный миелит, язвенный колит, недифференцированное заболевание соединительной ткани (UCTD), увеит, ревматическую полимиалгию, артериит Такаясу, височный артериит, болезнь Бюргера, кожный васкулит, болезнь Кавасаки, нодозный полиартериит, болезнь Бехчета, синдром Чарга-Стросса, кожный васкулит, пурпура Шенлейн-Геноха, микроскопический полиангиит, гранулематоз Вегенера, васкулит гольфиста, везикулобуллезный дерматоз, витилиго, гранулематоз Вегенера (в настоящее время обозначаемый как гранулематоз с полиангиитом (GPA).
В одном из вариантов осуществления аутоиммунное заболевание выбрано из группы, содержащей или состоящей из: ANCA-васкулита, IgA-нефропатии (Бергера), обыкновенной пузырчатки/буллезного пемфигоида, ITP, первичного биллиарного цирроза, аутоиммунного тиреоидита (болезнь Грейвса), болезни Хашимото, волчаночного нефрита, мембранозного гломерулонефрита (или мембранозной нефропатии), APS, миастении, оптикомиелита, первичного синдрома Шегрена, аутоиммунной нейтропении, аутоиммунного панкреатита, дерматомиозита, аутоиммунного увеита, аутоиммунной ретинопатии, болезни Бехчета, IPF, системного склероза, фиброза печени, аутоиммунного гепатита, первичного склерозирующего холангита, витилиго, синдрома Гудпасчера, легочно-альвеолярного протеиноза, хронической аутоиммунной крапивницы, псориаза, ревматоидного артрита, псориатического артрита, аксиального спондилоартрита, трансплантации (включая GvHD), астмы, COPD, гигантоклеточного артериита, рефрактерный аутоиммунных цитопений, синдрома Эванса (аутоиммунной гемолитической анемии), диабета типа I, саркоидоза, полимиозита, язвенного колита, болезни Крона, целиакии, макроглобулинемии Вальденстрема, фокального сегментарного гломерулосклероза, хронической болезни Лайма (боррелиоза), красного плоского лишая, синдрома скованного человека, дилатационной кардиомиопатии, аутоиммунного (лимфоцитарного) оофорита, буллезного эпидермолиза Acquisita, аутоиммунного атрофического гастрита, пернициозной анемии, атопического дерматита, атеросклероза, рассеянного склероза, энцефалита Расмуссена, синдрома Гийена Барре и приобретенной нейромиотонии, инсульта.
В одном из вариантов осуществления нарушение является злокачественным новообразованием, например, лейкозом, например, лимфоцитарным лейкозом, таким как острый лимфобластный лейкоз или хронический лимфоцитарный лейкоз; или миелогенным лейкозом, таким как острый миелогенный лейкоз или хронический миелогенный лейкоз; или лимфомой, такой как диффузная крупноклеточная В-клеточная лимфома, или лимфома Ходжкина, или неходжкинская лимфома.
Настоящее изобретение также относится к фармацевтической или диагностической композиции, содержащей молекулу по настоящему изобретению, такую как полиспецифическая молекула, представленная в настоящем описании, в комбинации с одним или несколькими из фармацевтически приемлемого эксципиента, дилюента или носителя. Таким образом, настоящее изобретение относится к применению молекулы по настоящему изобретению, такой как полиспецифическая молекула, как представлено в настоящем описании, для применения в лечении и производстве лекарственного средства.
Композицию, как правило, будут поставлять как часть стерильной фармацевтической композиции, которая, как правило, будет включать фармацевтически приемлемый носитель. Фармацевтическая композиция по настоящему изобретению может дополнительно содержать фармацевтически приемлемый адъювант.
Настоящее изобретение также относится к способу получения фармацевтической или диагностической композиции, включающему добавление и смешивание полиспецифической молекулы по настоящему изобретению с одним или несколькими из фармацевтически приемлемого эксципиента, дилюента или носителя.
Как применяют в настоящем описании, термин "фармацевтически приемлемый эксципиент" относится к фармацевтически приемлемому носителю, раствору или добавке для состава для повышения желаемых характеристик композиций по настоящему изобретению. Эксципиенты хорошо известны в этой области и включают буферы (например, цитратный буфер, фосфатный буфер, ацетатный буфер и бикарбонатный буфер), аминокислоты, мочевину, спирты, аскорбиновую кислоту, фосфолипиды, белки (например, сывороточный альбумин), ЭДТА, хлорид натрия, липосомы, маннит, сорбит и глицерин. Растворы или суспензии можно инкапсулировать в липосомы и биодеградирующие микросферы. Как правило, состав будут получать, по существу, стерильной форме с использованием стерильных способов производства.
Они могут включать получение и стерилизацию фильтрацией забуференного раствора растворителя, используемого для состава, асептической суспензии антитела в стерильном забуференном растворе растворителя, и диспергирование состава в стерильных сосудах способами, известными специалистам в этой области.
Фармацевтически приемлемый носитель не должен сам по себе индуцировать продукцию антител, вредных для индивидуума, которому вводят композицию, и не должен являться токсичным. Подходящие носители могут являться крупными, медленно метаболизируемыми макромолекулами, такими как белки, полипептиды, липосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, coполимеры аминокислот и неактивные вирусные частицы.
Можно использовать фармацевтически приемлемые соли, например, соли неорганических кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, или соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты.
Фармацевтически приемлемые носители в терапевтических композициях могут дополнительно содержать жидкости, такие как вода, физиологический раствор, глицерин и этанол. Такие носители позволяют составлять фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, суспензий для переваривания пациентом.
Молекулы по настоящему изобретению, такие как полиспецифическая молекула, представленная в настоящем описании, можно доставлять в форме дисперсии в растворителе, например, в форме раствора или суспензии. Ее можно суспендировать в соответствующем физиологическом растворе, например, физиологическом растворе, фармакологически приемлемом растворителе или забуференном растворе. Известные в этой области забуференные растворы могут содержать от 0,05 мг до 0,15 мг диэдетата натрия, от 8,0 мг до 9,0 мг NaCl, от 0,15 мг до 0,25 мг полисорбата, от 0,25 мг до 0,30 мг безводной лимонной кислоты и от 0,45 мг до 0,55 мг цитрата натрия на 1 мл воды для достижения pH приблизительно от 4,0 до 5,0. Как указано выше, суспензию можно получать, например, из лиофилизированного антитела.
Всестороннее обсуждение фармацевтически приемлемых носителей доступно в Remington's Pharmaceutical Sciences (Mack Publishing Company, N.J. 1991).
Как применяют в настоящем описании, термин "терапевтически эффективное количество" относится к количеству терапевтического средства, необходимому для лечения, улучшения или профилактики целевого заболевания или состояния или для проявления определяемого терапевтического или профилактического эффекта. В случае любого антитела терапевтически эффективное количество можно оценивать исходно в анализах культур клеток или на моделях на животных, как правило, на грызунах, кроликах, собаках, свиньях или приматах. Модель на животных также можно использовать для определения подходящего диапазона концентраций и пути введения. Затем такую информацию можно использовать для определения применимых доз и путей введения у людей.
Точное терапевтически эффективное количество для человека будет зависеть от тяжести заболевания, общего состояния здоровья индивидуума, возраста, массы и пола индивидуума, питания, времени и частоты введения, комбинаций лекарственных средств, чувствительности реакций и толерантности/ответа на терапию. Это количество можно определять посредством рутинного экспериментирования, и оно находится в пределах суждения клинициста. Как правило, терапевтически эффективное количество будет составлять от 0,01 мг/кг до 50 мг/кг, например, от 0,1 мг/кг до 20 мг/кг. Альтернативно, доза может составлять от 1 до 500 мг в сутки, например, от 10 до 100, 200, 300 или 400 мг в сутки. Фармацевтические композиции может быть удобным составлять в форме однократных доз, содержащих заранее определенное количество активного средства по изобретению.
Композиции можно вводить пациенту в отдельности или можно вводить в комбинации (например, одновременно, последовательно или раздельно) с другими средствами, лекарственными средствами или гормонами.
Доза, в которой вводят полиспецифическую молекулу по настоящему изобретению, зависит от природы состояния, подлежащего лечению, степени присутствующего воспаления и того, используют ли молекулу антитела профилактически или для лечения существующего состояния.
Частота введения доз будет зависеть от времени полужизни полиспецифической молекулы и длительности ее эффекта. Если полиспецифическая молекула имеет короткое время полужизни (например, от 2 до 10 часов), необходимым может являться введение одной или нескольких доз в сутки. Альтернативно, если полиспецифическая молекула имеет большое время полужизни (например, от 2 до 15 дней), необходимым может являться введение дозы один раз в сутки, один раз в неделю или даже один раз каждые 1 или 2 месяца.
В настоящем изобретении pH конечного состава несхож со значением изоэлектрической точки полиспецифической молекулы, в случае, если pH состава составляет 7, то подходящей может являться pI 8-9 или выше. Не будучи связанными какой-либо теорией, считают, что, в конечном итоге, посредством этого можно получать конечный состав с улучшенной стабильностью, например, антитело или фрагмент остается в растворе.
Фармацевтические композиции по настоящему изобретению можно вводить любым количеством путей, включая, в качестве неограничивающих примеров, пероральный, внутривенный, внутримышечный, внутриартериальный, интрамедуллярный, интратекальный, внутрижелудочковый, трансдермальный, чрескожный (например, см. WO98/20734), подкожный, интраперитонеальный, интраназальный, энтеральный, местный, сублингвальный, внутривлагалищный или ректальный пути. Для введения фармацевтических композиций по изобретению также можно использовать безыгольные инжекторы.
Прямую доставку композиций, как правило, будут осуществлять посредством инъекции, подкожно, интраперитонеально, внутривенно или внутримышечно или доставлять в интерстициальное пространство ткани. Композиции также можно вводить в конкретную интересующую ткань. Режим введения доз может представлять собой режим однократных доз или режим многократных доз.
Если продукт предназначен для инъекции или инфузии, его можно получать в форме суспензии, раствора или эмульсии в масляном или водном наполнителе, и он может содержать средства для составления, такие как суспендирующие средства, консервант, стабилизирующие и/или диспергирующие средства. Альтернативно, полиспецифическая молекула может находится в сухой форме для восстановления перед использованием с помощью соответствующей стерильной жидкости. Если композиция предназначена для введения путем с использованием желудочно-кишечного тракта, композиция должна содержать средства, защищающие антитело от деградации, но высвобождающие биспецифический белковый комплекс после его абсорбции из желудочно-кишечного тракта.
Можно получать небулизируемый состав по настоящему изобретению, например, в виде единиц с однократными дозами (например, запаянных пластиковых контейнеров или сосудов), упакованных в фольгу. Каждый сосуд содержит однократную дозу в объеме, например, 2 мл, растворителя/раствора буфера.
Как применяют в настоящем описании, термин "вариант" относится к пептиду или белку, содержащему по меньшей мере одно изменение аминокислотной последовательности или нуклеотидной последовательности по сравнению с аминокислотной или нуклеотидной последовательностью соответствующего пептида или белка дикого типа. Вариант может иметь по меньшей мере 80%, или 85%, или 90%, или 95%, или 98% или 99% идентичности последовательности в отношении соответствующего пептида или белка дикого типа. Однако, вариант может иметь менее 80% идентичности последовательности, при условии, что вариант проявляет, по существу, функцию, схожую с соответствующим пептидом или белком дикого типа.
В одном из вариантов осуществления конструкция по настоящему изобретению является, по меньшей мере, триспецифической. В этом случае дополнительная специфичность может быть направлена на любой интересующий антиген, например, антигены, для повышения времени полужизни, такой как альбумин или неонатальный Fc-рецептор (FcRn); антигены для эффекторной функции, такие как активирующие или ингибирующие Fc-рецепторы или костимуляторные молекулы; антигены для таргетинга тканей или клеток; или антигены для облегчения перехода через гематоэнцефалический барьер (BBB), такие как рецептор трансферрина или LRP1.
Настоящее изобретение также относится к композициям, таким как фармацевтические композиции, содержащие указанные новые форматы с конкретной специфичностью к антигену.
В дополнительном аспекте настоящее изобретение включает применение форматов и композиций в лечении.
Настоящее изобретение также относится к способу получения фармацевтической или диагностической композиции, включающему добавление и смешивание молекулы антитела по настоящему изобретению с одним или несколькими из фармацевтически приемлемого эксципиента, дилюента или носителя.
Молекула антитела может являться единственным активным ингредиентом в фармацевтической или диагностической композиции или ей могут сопутствовать другие активные ингредиенты, включая другие ингредиенты-антитела или ингредиенты, не являющиеся антителами, такими как стероиды или другие молекулы лекарственных средств.
Соответственно, фармацевтические композиции содержат терапевтически эффективное количество антитела по изобретению. Как применяют в настоящем описании, термин "терапевтически эффективное количество" относится к количеству терапевтического средства, необходимому для лечения, улучшения или профилактики целевого заболевания или состояния или для проявления определяемого терапевтического или профилактического эффекта. В случае любого антитела терапевтически эффективное количество можно оценивать исходно в анализах культур клеток или на моделях на животных, как правило, на грызунах, кроликах, собаках, свиньях или приматах. Модель на животных также можно использовать для определения соответствующего диапазона концентраций и пути введения. Затем такую информацию можно использовать для определения применимых доз и путей введения у людей.
Точное терапевтически эффективное количество для человека будет зависеть от тяжести заболевания, общего состояния здоровья индивидуума, возраста, массы и пола индивидуума, питания, времени и частоты введения, комбинаций лекарственных средств, чувствительностей реакций и толерантности/ответа на терапию. Это количество можно определять посредством рутинного экспериментирования, и оно находится в пределах суждения клинициста. Как правило, терапевтически эффективное количество будет составлять от 0,01 мг/кг до 500 мг/кг, например, от 0,1 мг/кг до 200 мг/кг, такое как 100мг/кг.
Фармацевтические композиции может быть удобным составлять в форме однократных доз, содержащих заранее определенное количество активного средства по изобретению на дозу.
Композиции можно вводить пациенту в отдельности или можно вводить в комбинации (например, одновременно, последовательно или раздельно) с другими средствами, лекарственными средствами или гормонами.
Как применяют в настоящем описании, термин "средства" относится к веществу, которая при введении имеет физиологический эффект.
Как применяют в настоящем описании, термин "лекарственное средство" относится к химическому веществу, которое в терапевтической дозе имеет соответствующий физиологический эффект.
В одном из вариантов осуществления антитела или фрагменты по настоящему изобретению используют с иммуносупрессорной терапией, такой как стероид, в частности, преднизон.
В одном из вариантов осуществления антитела или фрагменты по настоящему изобретению используют с ритуксимабом или другими B-клеточными терапевтическими средствами.
В одном из вариантов осуществления антитела или фрагменты по настоящему изобретению используют с любым B-клеточным или T-клеточным модулирующим средством или иммуномодулятором. Примеры включают метотрексат, микофенолат и азатиоприн.
Доза, в которой вводят молекулу антитела по настоящему изобретению, зависит от природы состояния, подлежащего лечению, степени присутствующего воспаления и того, используют ли молекулу антитела профилактически или для лечения существующего состояния.
Частота введения дозы будет зависеть от времени полужизни молекулы антитела и длительности его эффекта. Если молекула антитела имеет короткое время полужизни (например, от 2 до 10 часов), необходимым может являться введение одной или нескольких доз в сутки. Альтернативно, если молекула антитело имеет большое время полужизни (например, от 2 до 15 дней) и/или длительный фармакодинамический (PD) профиль, необходимым может являться введение дозы лишь один раз в сутки, один раз в неделю или даже один раз в 1 или 2 месяца.
В одном из вариантов осуществления дозу вводят раз в две недели, т.е. дважды в месяц.
В одном из вариантов осуществления делают интервалы между введениями доз, чтобы позволить ответам против лекарственного средства (в этом случае против антитела) ослабнуть перед введением следующей дозы.
Как применяют в настоящем описании, термин "время полужизни" предназначен для обозначения длительности нахождения молекулы в кровотоке, например, в сыворотке/плазме.
Как применяют в настоящем описании, термин "фармакодинамика" относится к профилю и, в частности, длительности биологического действия молекулы по настоящему изобретению.
Фармацевтически приемлемый носитель сам по себе не должен индуцировать продукцию антител, вредных для индивидуума, которому вводят композицию, и не должен являться токсичным. Подходящие носители могут являться крупными, медленно метаболизирующимися макромолекулами, такими как белки, полипептиды, лимпосомы, полисахариды, полимолочные кислоты, полигликолевые кислоты, полимерные аминокислоты, coполимеры аминокислот и неактивные вирусные частицы.
Можно использовать фармацевтически приемлемые соли, например, соли неорганических кислот, такие как гидрохлориды, гидробромиды, фосфаты и сульфаты, или соли органических кислот, такие как ацетаты, пропионаты, малонаты и бензоаты.
Фармацевтически приемлемые носители в терапевтических композициях могут дополнительно содержать жидкости, такие как вода, физиологический раствор, глицерин и этанол. Кроме того, в таких композициях могут присутствовать вспомогательные средства, такие как увлажнители или эмульгаторы или pH-буферные вещества. Такие носители позволяют составлять фармацевтические композиции в виде таблеток, пилюль, драже, капсул, жидкостей, гелей, сиропов, суспензий для переваривания пациентом.
Подходящие формы для введения включают формы, подходящие для парентерального введения, например, посредством инъекции или инфузии, например, посредством болюсной инъекции или непрерывной инфузии. Если продукт предназначен для инъекции или инфузии, его также можно получать в форме суспензии, раствора или эмульсии в масляном или водном наполнителе, и он может содержать средства для составления, такие как суспендирующие средства, консерванты, стабилизирующие и/или диспергирующие средства. Альтернативно, молекула антитело может находиться в сухой форме для восстановления перед использованием с помощью подходящей стерильной жидкости.
После составления композиции по изобретению можно вводить напрямую индивидууму. Индивидуумы, подлежащие лечению, могут являться животными. Однако в одном или нескольких вариантах осуществления композиции адаптируют для введения людям.
Соответственно, в составах по настоящему изобретению pH конечного состава несхож со значением изоэлектрической точки антитела или фрагмента, например, если pI белка находится в диапазоне 8-9 или выше, то подходящем может являться pH состава 7. Не будучи связанными какой-либо теорией считают, что посредством этого в конечном итоге можно получать конечный состав с улучшенной стабильностью, например, антитело или фрагмент остается в растворе.
В одном примере фармацевтический состав при pH в диапазоне от 4,0 до 7,0 содержит: от 1 до 200 мг/мл молекулы антитела по настоящему изобретению, от 1 до 100 мМ буфера, от 0,001 до 1% поверхностно-активного вещества, a) от 10 до 500 мМ стабилизатора, b) от 10 до 500 мМ стабилизатора и от 5 до 500 мМ регулятора тоничности, или c) от 5 до 500 мМ регулятора тоничности.
Фармацевтические композиции по настоящему изобретению можно вводить любым количеством путей включая, в качестве неограничивающих примеров, пероральный, внутривенный, внутримышечный, внутриартериальный, интрамедуллярный, интратекальный, внутрижелудочковый, трансдермальный, чрескожный (например, см. WO98/20734), подкожный, интраперитонеальный, интраназальный, энтеральный, местный, сублингвальный, внутривлагалищный или ректальный пути. Для введения фармацевтических композиций по изобретению также можно использовать безыгольные инжекторы. Как правило, терапевтические композиции можно получать в инъецируемой форме, в виде жидких растворов или суспензий. Также можно получать твердые формы, подходящие для растворения или суспендирования в жидких наполнителях перед инъекцией.
Прямую доставку композиций, как правило, будут осуществлять посредством инъекции, подкожно, интраперитонеально, внутривенно или внутримышечно или доставлять в интерстициальное пространство ткани. Композиции также можно вводить в очаг поражения. Режим введения доз может представлять собой режим однократных доз или режим многократных доз.
Следует понимать, что активный ингредиент в композиции будет молекулой антитела. В связи с этим, он будет восприимчив к деградации в желудочно-кишечном тракте. Таким образом, если композиция подлежит введению путем с использованием желудочно-кишечного тракта, композиция должна будет содержать средства, защищающие антитело от деградации, но высвобождающие антитело после абсорбции из желудочно-кишечного тракта.
Всестороннее обсуждение фармацевтически приемлемых носителей доступно в Remington's Pharmaceutical Sciences (Mack Publishing Company, N.J. 1991).
В одном из вариантов осуществления состав получают в виде состава для местного введения, включая ингаляцию.
Подходящие ингалируемые препараты включают ингалируемые порошки, дозированные аэрозоли, содержащие газы-пропелленты, или ингалируемые растворы, не содержащие газы-пропелленты. Ингалируемые порошки по настоящему изобретению, содержащие активное вещество могут состоять исключительно из указанных выше активных веществ или смеси указанных выше активных веществ с физиологически приемлемым эксципиентом.
Эти ингалируемые порошки могут включать моносахариды (например, глюкозу или арабинозу), дисахариды (например, лактозу, сахарозу, мальтозу), олиго- и полисахариды (например, декстраны), полиспирты (например, сорбит, маннит, ксилит), соли (например, хлорид натрия, карбонат кальция) или их смеси друг с другом. Моно- или дисахариды используют соответствующим образом, лактозу или глюкозу, в частности, в качестве неограничивающих примеров, используют в форме их гидратов.
В случае частиц для накопления в легких необходим размер частиц менее 10 мкм, такой как 1-9 мкм, например, от 1 до 5 мкм. Размер частиц активного ингредиента (такого как антитело или фрагмент) представляет особую важность.
Газы-пропеленты, которые можно использовать для получения ингалируемых аэрозолей, известны в этой области. Подходящие газы-пропеленты выбирают из углеводородов, таких как n-пропан, n-бутан или изобутан, и галогенуглеводородов, таких как хлорированные и/или фторированные производные метана, этана, пропана, бутана, циклопропана или циклобутана. Указанные выше газы-пропеленты можно использовать в отдельности или в смесях.
Особенно подходящими газами-пропелентами являются галогенированные производные алканов, выбранные из TG 11, TG 12, TG 134a и TG227. Из указанных выше галогенированных углеводородов, особенно подходят TG134a (1,1,1,2-тетрафторэтана) и TG227 (1,1,1,2,3,3,3-гептафторпропана) и их смеси.
Ингалируемые аэрозоли, содержащие газ-пропелент, также могут содержать другие ингредиенты, такие как сорастворители, стабилизаторы, поверхностно-активные средства, антиоксиданты, смазочные средства и средства для коррекции pH. Все эти ингредиенты известны в этой области.
Ингалируемые аэрозоли по изобретению, содержащие газ-пропелент, могут содержать до 5% по массе активного вещества. Аэрозоли по изобретению содержат, например, о 0,002 до 5% по массе, от 0,01 до 3% по массе, от 0,015 до 2% по массе, от 0,1 до 2% по массе, от 0,5 до 2% по массе или от 0,5 до 1% по массе активного ингредиента.
Альтернативно, местное введение в легкие также можно осуществлять посредством введения жидкого раствора или состава суспензии, например, с использованием устройства, такого как небулайзер, например, небулайзер, соединенный с компрессором (например, небулайзер Pari LC-Jet Plus(R), соединенный с компрессором Pari Master(R), производимый Pari Respiratory Equipment, Inc., Richmond, Va.).
Антитело или полиспецифическую молекулу по изобретению можно доставлять в форме дисперсии в растворителе, например, в форме раствора или суспензии. Его можно суспендировать в подходящем физиологическом растворе, например, физиологическом растворе или другом фармакологически приемлемом растворителе или забуференном растворе. Известные в этой области забуференные растворы могут содержать от 0,05 мг до 0,15 мг диэдетата натрия, от 8,0 мг до 9,0 мг NaCl, от 0,15 мг до 0,25 мг полисорбата, от 0,25 мг до 0,30 мг безводной лимонной кислоты и от 0,45 мг до 0,55 мг цитрата натрия на 1 мл воды таким образом, что достигают pH приблизительно от 4,0 до 5,0. В суспензии можно использовать, например, лиофилизированное антитело.
Терапевтические составы суспензий или растворов также могут содержать один или несколько эксципиентов. Эксципиенты хорошо известны в этой области и включают буферы (например, цитратный буфер, фосфатный буфер, ацетатный буфер и бикарбонатный буфер), аминокислоты, мочевину, спирты, аскорбиновую кислоту, фосфолипиды, белки (например, сывороточный альбумин), ЭДТА, хлорид натрия, липосомы, маннит, сорбит, и глицерин. Растворы или суспензии можно инкапсулировать в липосомах или биодеградирующих микросферах. Как правило, состав будут получать в, по существу, стерильной форме с использованием стерильных способов производства.
Они могут включать получение и стерилизацию фильтрацией забуференного растворителя/раствора, используемого для состава, асептической суспензии антитела в стерильном забуференном растворе растворителя, и диспергирование состава в стерильных сосудах способами, известными специалистам в этой области.
Можно получать небулизируемый состав по настоящему изобретению, например, в виде единиц с однократными дозами (например, запаянных пластиковых контейнеров или сосудов), упакованных в фольгу. Каждый сосуд содержит однократную дозу в объеме, например, 2 мл, растворителя/раствора буфера.
Антитела, представленные в настоящем описании, могут подходить для доставки посредством распыления.
Также предполагают, что антитело по настоящему изобретению можно вводить с использованием генотерапии. Для ее осуществления последовательности ДНК, кодирующие тяжелые и легкие цепи молекулы антитела под контролем подходящих компонентов ДНК вводят пациенту таким образом, что цепи антитела экспрессируются с последовательностей ДНК и собираются in situ.
В одном из вариантов осуществления молекулу по настоящему изобретению, такую как биспецифический белковый комплекс, представленный в настоящем описании, можно использовать для функционального изменения активности интересующего антигена или антигенов. Например, биспецифический белковый комплекс может нейтрализовать, являться антагонистом или агонистом активности указанного антигена или антигенов, прямо или косвенно.
Настоящее изобретение также относится к набору, содержащему молекулу по настоящему изобретению или его компонент. В одном из вариантов осуществления набор содержит:
a) один или несколько слитых белков (A-X), содержащих первое антитело или фрагмент антитела (A), специфичное для CD45 или CD79a и/или CD79b, соединенное с первым партнером по связыванию из связывающейся пары (X); и
b) один или несколько слитых белков (B-Y), содержащих второе антитело или фрагмент антитела (B), специфичное для CD45 или CD79a и/или CD79b, соединенное со вторым партнером по связыванию из связывающейся пары (Y), где последний является специфичным для первого партнера по связыванию;
например, где первый партнер по связыванию (X) является пептидом или полипептидом, и второй партнер по связыванию (Y) является антителом или фрагментом антитела, специфичным к нему;
где Y, второй партнер по связыванию, является специфичным для первого партнера по связыванию X, и второй партнер по связыванию является, например, антителом или фрагментом антитела, специфичным к нему; и при специфическом взаимодействии (таком как связывающее взаимодействие) двух партнеров по связыванию образуется гетеродимерное соединение, физически удерживающее два слитых белка из a) и b) вместе с образованием биспецифического белкового комплекса; и
где по меньшей мере один из A или B является специфичным для CD45, а другой является специфичным для CD79a и/или CD79b, и
слитые белки находятся в комплексированной или некомплексированной форме.
Преимущественно, набор может содержать биспецифические белковые комплексы по настоящему изобретению или может содержать слитые белки, находящиеся в комплексированной или некомплексированной форме. В первом случае биспецифические белковые комплексы готовы к использованию "по умолчанию", что обеспечивает удобство и простоту использования, в то время как в последнем случае биспецифические белковые комплексы можно собирать в соответствии с требованиями пользователя посредством комбинирования различных слитых белков.
В другом варианте осуществления набор дополнительно содержит инструкции по использованию.
В еще одном варианте осуществления набор дополнительно содержит один или несколько реагентов для осуществления одного или нескольких функциональных анализов.
В одном из вариантов осуществления молекулы по настоящему изобретению, включая слитые белки, биспецифические белковые комплексы или композиции, содержащие их, предоставляют для применения в качестве лабораторного реагента.
Дополнительные аспекты
В дополнительном аспекте настоящее изобретение относится к нуклеотидной последовательности, например, последовательности ДНК, кодирующей конструкцию, как представлено в настоящем описании, включая полиспецифическую молекулу или слитый белок, как определено выше.
В одном из вариантов осуществления настоящее изобретение относится к нуклеотидной последовательности, например, последовательности ДНК, кодирующей конструкцию, как представлено в настоящем описании, включая полиспецифическую молекулу или биспецифический белковый комплекс или антитело по настоящему изобретению.
Настоящее изобретение также относится к вектору, содержащему нуклеотидную последовательность, как определено выше.
Как применяют в настоящем описании, термин "вектор" относится к молекуле нуклеиновой кислоты, способной переносить другую нуклеиновую кислоту, с которой ее связывают. Примером вектора является "плазмида", являющаяся замкнутой кольцевой двухцепочечной ДНК, в которую можно лигировать дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, где дополнительные сегменты ДНК можно лигировать в вирусный геном. Конкретные векторы способны к автономной репликации в клетке-хозяине, в которую их встраивают (например, бактериальные векторы, имеющие бактериальный участок начала репликации и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) можно встраивать в геном клетки-хозяина, где они впоследствии могут реплицироваться вместе с геномом хозяина. В настоящем описании, термины "плазмида" и "вектор" могут быть использованы взаимозаменяемо, т.к. плазмида является наиболее общеупотребительной формой вектора.
Общие способы, которыми можно конструировать векторы, способы трансфекция и способы культивирования хорошо известны специалистам в этой области. В связи с этим дают ссылку на "Current Protocols in Molecular biology", 1999, F. M. Ausubel (ed), Wiley Interscience, New York и руководство Maniatis, изданное Cold Spring Harbor Publishing.
Как применяют в настоящем описании, термин "селективный маркер" относится к белку, экспрессия которого позволяет идентифицировать клетки, трансформированные или трансфицированные с использованием вектора, содержащего маркерный ген. В этой области известен широкий диапазон селективных маркеров. Например, как правило, ген селективного маркера придает резистентность к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую встраивают вектор. Селективный маркер также может являться визуально идентифицируемым маркером, таким как, например, флуоресцентный маркер. Примеры флуоресцентных маркеров включают родамин, FITC, TRITC, Alexa Fluor и различные их конъюгаты.
Настоящее изобретение также относится к клетке-хозяину, содержащей один или несколько клонирующих или экспрессирующих векторов, содержащих одну или несколько последовательностей ДНК, кодирующих антитело по настоящему изобретению. Можно использовать любую подходящую систему клетка-хозяин/вектор для экспрессии последовательностей ДНК, кодирующих молекулу антитела по настоящему изобретению. Можно использовать бактериальные, например, E. coli, и другие микробные системы или также можно использовать эукариотические системы экспрессии в клетках-хозяевах, например, млекопитающих. Подходящие клетки-хозяева млекопитающих включают CHO, миеломные или гибридомные клетки.
Настоящее изобретение также относится к способу получения молекулы по настоящему изобретению или ее компонента, включающему культивирование клетки-хозяина, содержащей вектор по настоящему изобретению, в условиях, подходящих для экспрессии белка с ДНК, кодирующей молекулу по настоящему изобретению, и выделение молекулы.
Молекулы по настоящему изобретению, включая биспецифические белковые комплексы, представленные в настоящем описании, можно использовать в наборах для диагностики/детекции. Наборы могут содержать, например, биспецифические комплексы антител, являющиеся специфичными для двух антигенов, оба из которых присутствуют на одном и том же типе клеток, и где положительную диагностику можно исключать, только если оба антигена успешно определены. Используя молекулу по настоящему изобретению, такую как биспецифические комплексы антител, представленные в настоящем описании, а не два отдельных антитела или фрагмента антител в некомплексированной форме, можно значительно повышать специфичность детекции.
В одном из вариантов осуществления молекулы по настоящему изобретению, такие как биспецифические комплексы антител, фиксируют на твердой поверхности. Твердая поверхность, например, может являться чипом или планшетом для ELISA.
Кроме того, настоящее изобретение относится к применению молекулы по настоящему изобретению, например, биспецифического белкового комплекса, представленного в настоящем описании, для детекции наличия первого и второго пептида в образце, при этом указанные молекулы используют в качестве средств детекции.
Молекулы по настоящему изобретению, такие как биспецифические комплексы антител, представленные в настоящем описании, например, можно конъюгировать с флуоресцентным маркером, облегчающим детекцию связавшихся комплексов антитело-антиген. Такие биспецифические комплексы антител можно использовать для иммунофлуоресцентной микроскопии. Альтернативно, биспецифические комплексы антитела также можно использовать для вестерн-блоттинга или ELISA.
В одном из вариантов осуществления настоящее изобретение относится к способу очистки молекулы по настоящему изобретению или ее компонента.
В одном из вариантов осуществления настоящее изобретение относится к способу очистки молекулы по настоящему изобретению или ее компонента, включающему этапы: осуществления анионообменной хроматографии в режиме без связывания таким образом, что примеси остаются на колонке, и антитело сохраняют в несвязавшейся фракции. Этап, например, можно осуществлять при pH приблизительно 6-8.
Способ может дополнительно включать исходный этап захвата с использованием катионообменной хроматографии, осуществляемый, например, при pH приблизительно от 4 до 5.
Способ может дополнительно включать дополнительные этапы хроматографии для обеспечения того, что относящиеся к продукту и способу примеси соответствующим образом отделяют от потока продукта.
Способ очистки также может включать один или несколько этапов ультрафильтрации, такие как этап концентрирования и диафильтрации.
Как применяют выше, термин "очищенная форма" предназначен для обозначения по меньшей мере 90% чистоты, например, 91, 92, 93, 94, 95, 96, 97, 98, 99% масс./масс. или более чистоты.
Что касается настоящего описания, термин "содержащий" следует интерпретировать как "включающий".
Аспекты настоящего изобретения, содержащие конкретные элементы, также должны относиться к альтернативным вариантам осуществления, "состоящим" или "по существу, состоящим" из соответствующих элементов.
Конкретно процитированные варианты осуществления можно использовать в настоящем описании как основу для письменного отказа от пункта формулы.
Все ссылки, приведенные в настоящем описании, конкретно включены в настоящее описание в качестве ссылок.
Подзаголовки в настоящем описании используют для облегчения структурирования описания, и они не предназначены для использования в качестве ограничения значения технических терминов в настоящем описании.
Последовательности по настоящему изобретению представлены в настоящем описании ниже.
Последовательности GCN4(7P14P)
ASGGGRMKQLEPKVEELLPKNYHLENEVARLKKLVGERHHHHHH SEQ ID NO: 1, где аминокислоты, выделенные полужирным шрифтом, являются необязательными
GCTAGCGGAGGCGGAAGAATGAAACAACTTGAACCCAAGGTTGAAGAATTGCTTCCGAAAAATTATCACTTGGAAAATGAGGTTGCCAGATTAAAGAAATTAGTTGGCGAACGCCATCACCATCACCATCAC - SEQ ID NO: 2
Последовательность 52SR4 ds scFv
DAVVTQESALTSSPGETVTLTCRSSTGAVTTSNYASWVQEKPDHLFTGLIGGTNNRAPGVPARFSGSLIGDKAALTITGAQTEDEAIYFCVLWYSDHWVFGCGTKLTVLGGGGGSGGGGSGGGGSGGGGSDVQLQQSGPGLVAPSQSLSITCTVSGFLLTDYGVNWVRQSPGKCLEWLGVIWGDGITDYNSALKSRLSVTKDNSKSQVFLKMNSLQSGDSARYYCVTGLFDYWGQGTTLTVSSAAAHHHHHHEQKLISEEDL - SEQ ID NO: 3
GATGCGGTGGTGACCCAGGAAAGCGCGCTGACCAGCAGCCCGGGCGAAACCGTGACCCTGACCTGCCGCAGCAGCACCGGCGCGGTGACCACCAGCAACTATGCGAGCTGGGTGCAGGAAAAACCGGATCATCTGTTTACCGGCCTGATTGGCGGCACCAACAACCGCGCGCCGGGCGTGCCGGCGCGCTTTAGCGGCAGCCTGATTGGCGATAAAGCGGCGCTGACCATTACCGGCGCGCAGACCGAAGATGAAGCGATTTATTTTTGCGTGCTGTGGTATAGCGACCATTGGGTGTTTGGCTGCGGCACCAAACTGACCGTGCTGGGTGGAGGCGGTGGCTCAGGCGGAGGTGGCTCAGGCGGTGGCGGGTCTGGCGGCGGCGGCAGCGATGTGCAGCTGCAGCAGAGCGGCCCGGGCCTGGTGGCGCCGAGCCAGAGCCTGAGCATTACCTGCACCGTGAGCGGCTTTCTCCTGACCGATTATGGCGTGAACTGGGTGCGCCAGAGCCCGGGCAAATGCCTGGAATGGCTGGGCGTGATTTGGGGCGATGGCATTACCGATTATAACAGCGCGCTGAAAAGCCGCCTGAGCGTGACCAAAGATAACAGCAAAAGCCAGGTGTTTCTGAAAATGAACAGCCTGCAGAGCGGCGATAGCGCGCGCTATTATTGCGTGACCGGCCTGTTTGATTATTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGCGGCCGCCCATCACCATCACCATCACGAACAGAAACTGATTAGCGAAGAAGATCTGTAATAG - SEQ ID NO: 4
Антитела против CD79b
Ab 4447
Область VL Ab 4447 кролика - SEQ ID NO: 71
AQVLTQTPSP VSAPVGGTVT INCQASQSVV SGNYLAWLQQ KPGQPPKQLI HSASTLASGV SSRFSGSGSG TQFTLTISGV QCEDAATYYC LGEFSCSSHD CNAFGGGTEV VVK
Область VL Ab 4447 кролика - SEQ ID NO: 72
gcccaagtgc tgacccagac tccgtcccct gtgtctgcac ctgtgggagg cacagtcacc atcaattgcc aggccagtca gagtgttgtt agtggcaatt acctagcctg gcttcagcag aaaccagggc agcctcccaa gcaactgatc cattctgcat ccactctggc atctggggtc tcatcgcggt tcagcggcag tggatctggg acacaattca ctctcaccat cagcggcgtg cagtgtgaag atgctgccac ttactactgt ctaggcgaat ttagttgtag tagtcatgat tgtaatgctt tcggcggagg gaccgaggtg gtggtcaaa
Область VH Ab 4447 кролика - SEQ ID NO: 73
QSLEESGGRL VTPGTPLTLT CTVSGFSLSN YAVSWVRQAP GEGLEWIGII YIETGTTWYA NWAKGRFTIS KTSTTVDLTI TSPSTEDTAT YFCAREPYEP YDDSNIYYGM DPWGPGTLVT VSS
Область VH Ab 4447 кролика - SEQ ID NO: 74
cagtcgctgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc tgcaccgtct ctggattctc cctcagtaac tatgcagtaa gctgggtccg ccaggctcca ggggagggac tggaatggat cgggatcatt tatattgaaa ctggtaccac atggtacgcg aactgggcga aaggccgatt caccatctcc aaaacctcga ccacggtgga tctgacaatc accagtccgt caaccgagga cacggccacc tatttctgtg ccagagaacc ttatgaacct tatgatgata gtaatattta ctacggcatg gacccctggg gcccaggcac cctcgtcacc gtctcgagt
CDRL1 - SEQ ID NO: 75 QASQSVVSGNYLA
CDRL2 - SEQ ID NO: 76 SASTLAS
CDRL3 - SEQ ID NO: 77 LGEFSCSSHDCNA
CDRH1 - SEQ ID NO: 78 GFSLSNYAVS
CDRH2 - SEQ ID NO: 79 IIYIETGTTWYANWAKG
CDRH3 - SEQ ID NO: 80 EPYEPYDDSNIYYGMDP
Настоящее изобретение также относится к производному SEQ ID NO: 77, где один или оба цистеина заменяют другой аминокислотой, например, серином, в частности, где первый Cys заменяют серином, а второй Cys оставляют неизменным, или первый цистеин оставляет неизменным, а второй цистеин заменяют серином, или где оба цистеина заменяют серином.
Ab 4450
Область VL Ab 4450 кролика - SEQ ID NO: 81
AIDMTQTPSP VSAAVGGTVT INCQSSQSIY NNNDLAWYQQ KPGQPPKLLI YEASKLASGV PSRFKGSGSG TQFTLTISGV QCDDAATYYC QGGGSGGDGI AFGGGTKVVV E
Область VL Ab 4450 кролика - SEQ ID NO: 82
gccattgata tgacccagac tccatccccc gtgtctgcag ctgtgggagg cacagtcacc atcaattgcc agtccagtca gagtatttat aataataatg acttagcctg gtatcagcag aaaccagggc agcctcccaa gctcctgatc tacgaagcat ccaaactggc atctggggtc ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagtggcgtg cagtgtgatg atgctgccac ttactactgt cagggcggtg gtagtggtgg tgatggcatt gctttcggcg gagggaccaa ggtggtcgtc gaa
Область VH Ab 4450 кролика - SEQ ID NO: 83
QSVEESGGRL VTPGAPLTLT CTVSGFSLNN YVMVWVRQAP GKGLEWIGII YVSGNAYYAS WAKGRFTISR TSTTVDLKVT SLTTEDTATY FCARDAGHSD VDVLDIWGPG TLVTVSS
Область VH Ab 4450 кролика - SEQ ID NO: 84
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg gggcacccct gacactcacc tgcacagtct ctggattctc cctcaataac tatgtaatgg tctgggtccg ccaggctcca gggaaggggc tggaatggat cggaatcatt tatgttagtg gtaatgcata ctacgcgagc tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaagtgacc agtctgacaa ccgaggacac ggccacctat ttctgtgcca gagatgctgg tcatagtgat gtcgatgttt tggatatttg gggcccgggc accctcgtca ccgtctcgag t
CDRL1 - SEQ ID NO: 85 QSSQSIYNNNDLA
CDRL2 - SEQ ID NO: 86 EASKLAS
CDRL3 - SEQ ID NO: 87 QGGGSGGDGIA
CDRH1 - SEQ ID NO: 88 GFSLNNYVMV
CDRH2 - SEQ ID NO: 89 IIYVSGNAYYASWAKG
CDRH3 - SEQ ID NO: 90 DAGHSDVDVLDI
Настоящее изобретение также относится к производному SEQ ID NO: 87, где по меньшей мере одну из аминокислот в мотиве DG заменяют другой аминокислотой, например, мотив подвергают мутации в EG, DA или DS.
Антитела против CD45
Ab 4122
Область VL Ab 4122 кролика - SEQ ID NO: 91
DIVMTQTPAS VSEPVGGTVT IMCQASQSIS NWLAWYQQKP GQPPKLLIYQ ASKLASGVPS RFKGSGSGTE YTLTISDLEC ADAATYYCQS YYDSGSNVFF AFGGGTKVVV E
Область VL Ab 4122 кролика - SEQ ID NO: 92
gacattgtga tgacccagac tccagcctcc gtgtctgaac ctgtgggagg cacagtcacc atcatgtgcc aggccagtca gagcattagc aattggttag cctggtatca acagaaacca gggcagcctc ccaagctcct gatctaccag gcatccaaac tggcatctgg ggtcccatcg cggttcaaag gcagtggatc tgggacagag tacactctca ccatcagcga cctggagtgt gccgatgctg ccacttacta ctgtcaaagc tattatgata gtggtagtaa tgtttttttt gctttcggcg gagggaccaa ggtggtggtc gaa
Область VH Ab 4122 кролика - SEQ ID NO: 93
LSLEESGGDL VKPGASLTLT CTASGFSFSA GYWICWVRQA PGKGLEWIAC TYAGRSGSTY YANWVNGRFT IPKTSSTTVT LQMTSLSGAD TASYFCARGN AGVAVGALWG PGTLVTVSS
Область VH Ab 4122 кролика - SEQ ID NO: 94
ctgtcgttgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc tgcacagcct ctggattctc cttcagtgcc ggctattgga tatgttgggt ccgccaggct ccagggaagg ggctggagtg gatcgcatgc acttatgctg gtcgtagtgg tagcacttac tacgcgaact gggtgaatgg ccgattcacc atccccaaaa cctcgtcgac cacggtgact ctgcaaatga ccagtctgtc aggcgcggac acggccagct atttctgtgc gagaggtaat gctggtgttg ctgttggtgc cttgtggggc ccaggcaccc tggtcaccgt ctcgagt
CDRL1 - SEQ ID NO: 95 QASQSISNWLA
CDRL2 - SEQ ID NO: 96 QASKLAS
CDRL3 - SEQ ID NO: 97 QSYYDSGSNVFFA
CDRH1 - SEQ ID NO: 98 GFSFSAGYWIC
CDRH2 - SEQ ID NO: 99 CTYAGRSGSTYYANWVNG
CDRH3 - SEQ ID NO: 100 GNAGVAVGAL
Настоящее изобретение также относится к производному SEQ ID NO: 97, где по меньшей мере одну из аминокислот в мотиве DS заменяют другой аминокислотой, например, мотив подвергают мутации в DA или DT.
Настоящее изобретение также относится к производному SEQ ID NO: 98, где цистеин заменяют другой аминокислотой, например, серином.
Настоящее изобретение также относится к производному SEQ ID NO: 99, где цистеин заменяют другой аминокислотой, например, серином.
Настоящее изобретение также относится к производному SEQ ID NO: 99, где по меньшей мере одну из аминокислот в мотиве NG заменяют другой аминокислотой, например, мотив подвергают мутации в NA, NS или NT.
Ab 4129
Область VL Ab 4129 кролика - SEQ ID NO: 101
DIVMTQTPAS VEAAVGGTVT INCQASQSIS SWLSWYQQKP GQPPKLLIYG ASNLASGVPS RFSGSGSGTQ FSLTISDLEC ADAATYYCQS YYDSGSSVFF NFGGGTKVVV K
Область VL Ab 4129 кролика - SEQ ID NO: 102
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc atcaattgcc aagccagtca gagcattagc agttggttat cctggtatca gcagaaacca gggcagcctc ccaagctcct gatctatggt gcatccaatc tggcatctgg ggtcccatca cggttcagcg gcagtggatc tgggacacag ttcagtctca ccatcagcga cctggagtgt gccgatgctg ccacttacta ctgtcaaagc tattatgata gtggtagtag tgtttttttt aatttcggcg gagggaccaa ggtggtcgtc aaa
Область VH Ab 4129 кролика - SEQ ID NO: 103
QSLEESGGDL VKPGASLTLT CTASGFSFSA GYWICWVRQA PGKGLEWIAC IYAGSSGSTY YASWAKGRFT IPKTSSTTVT LQMTSLTGAD TATYFCARGN AGVAVGALWG PGTLVTVSS
Область VH Ab 4129 кролика - SEQ ID NO: 104
cagtcgttgg aggagtccgg gggagacctg gttaagcctg gggcatccct gacactcacc tgcacagcct ctggattctc cttcagtgcc ggctattgga tatgttgggt ccgccaggct ccagggaagg ggctggagtg gatcgcatgc atttatgctg gtagtagtgg tagcacttac tacgcgagct gggcgaaagg ccgattcacc atccccaaaa cctcgtcgac cacggtgact ctgcaaatga ccagtctgac aggcgcggac acggccacct atttctgtgc gagaggtaat gctggtgttg ctgttggtgc cttgtggggc ccaggcaccc tcgtcaccgt ctcgagt
CDRL1 - SEQ ID NO: 105 QASQSISSWLS
CDRL2 - SEQ ID NO: 106 GASNLAS
CDRL3 - SEQ ID NO: 107 QSYYDSGSSVFFN
CDRH1 - SEQ ID NO: 108 GFSFSAGYWIC
CDRH2 - SEQ ID NO: 109 CIYAGSSGSTYYASWAKG
CDRH3 - SEQ ID NO: 110 GNAGVAVGAL
Настоящее изобретение также относится к производному SEQ ID NO: 107, где по меньшей мере одну из аминокислот в мотиве DS заменяют другой аминокислотой, например, мотив подвергают мутации в DA или DT.
Настоящее изобретение также относится к производному SEQ ID NO: 108, где цистеин заменяют другой аминокислотой, например, серином.
Настоящее изобретение также относится к производному SEQ ID NO: 109, где цистеин заменяют другой аминокислотой, например, серином.
Ab 4131
Область VL Ab 4131 кролика - SEQ ID NO: 111
DIVMTQTPAS VSEPVGGSVT IKCQASQSFY NLLAWYQQKP GQPPKLLIYD ASDLASGVPS RFKGSGSGTD FTLTISDLEC ADAAAYYCQS ADGSSYAFGG GTEVVVK
Область VL Ab 4131 кролика - SEQ ID NO: 112
gacattgtga tgacccagac tccagcctcc gtgtctgaac ctgtgggagg ctcagtcacc atcaagtgcc aggccagtca gagcttttac aacctcttag cctggtatca gcagaaacca gggcagcctc ccaaactcct gatctatgat gcatccgatc tggcatctgg ggtcccatcg cggttcaaag gcagtggatc tgggactgat ttcactctca ccatcagcga cctggagtgt gccgatgctg ccgcttacta ctgtcaaagt gctgatggta gtagttacgc tttcggcgga gggaccgagg tggtcgtcaa a
Область VH Ab 4131 кролика - SEQ ID NO: 113
QEQLEESGGG LVKPEGSLTL TCTASGVSFS SSYWIYWVRQ APGKGLEWIA CIYTGSSGST YYASWAKGRF TVSETSSTTV TLQMTSLTAA DTATYFCARA SAWTYGMDLW GPGTLVTVSS
Область VH Ab 4131 кролика - SEQ ID NO: 114
caggagcaat tggaggagtc cgggggaggc ctggtcaagc ctgagggatc cctgacactc acctgcacag cctctggagt ctccttcagt agcagctatt ggatatactg ggtccgccag gctccaggga aggggctgga gtggatcgca tgcatttata ctggtagtag tggtagcact tactacgcga gctgggcgaa aggccgattc accgtctccg aaacctcgtc gaccacggtg actctgcaaa tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagagca agcgcttgga cctacggcat ggacctctgg ggcccgggca ccctcgtcac cgtctcgagt
CDRL1 - SEQ ID NO: 115 QASQSFYNLLA
CDRL2 - SEQ ID NO: 116 DASDLAS
CDRL3 - SEQ ID NO: 117 QSADGSSYA
CDRH1 - SEQ ID NO: 118 GVSFSSSYWIY
CDRH2 - SEQ ID NO: 119 CIYTGSSGSTYYASWAKG
CDRH3 - SEQ ID NO: 120 ASAWTYGMDL
Настоящее изобретение также относится к производному SEQ ID NO: 117, где, по меньшей мере, одну из аминокислот в мотиве DS заменяют другой аминокислотой, например, мотив подвергают мутации в DA или DT.
Настоящее изобретение также относится к производному SEQ ID NO: 119, где цистеин заменяют другой аминокислотой, например, серином.
Ab 4133
Область VL Ab 4133 кролика - SEQ ID NO: 121
AQVLTQTPSP VSAVVGGTVS ISCQASQSVY NNNNLSWYQQ KPGQPPKLLI YDASKLASGV PSRFKGSGSG TQFTLTISGV QCDDAATYYC LGGYYSSGWY FAFGGGTKVV VK
Область VL Ab 4133 кролика - SEQ ID NO: 122
gcgcaagtgc tgacccagac tccatctccc gtgtctgcag ttgtgggagg cacagtcagc atcagttgcc aggccagtca gagtgtttat aataacaaca acttatcctg gtatcagcag aaaccagggc agcctcccaa gctcttgatc tacgatgcat ccaaattggc atctggggtc ccatcccggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagcggcgtg cagtgtgacg atgctgccac ttactactgt ctaggcggtt attatagtag tggttggtat tttgctttcg gcggagggac caaggtggtg gtcaaa
Область VH Ab 4133 кролика - SEQ ID NO: 123
QEQLVESGGG LVQPEGSLTL TCTASGFSFS GNYYMCWVRQ APGKGLEWIG CLYTGSSGST YYASWAKGRF TISKTSSTTV TLQMTSLTAA DTATYFCARD LGYEIDGYGG LWGQGTLVTV SS
Область VH Ab 4133 кролика - SEQ ID NO: 124
caggagcagc tggtggagtc cgggggaggc ctggtccagc ctgagggatc cctgacacta acctgcacag cttctggatt ctccttcagt ggcaactact acatgtgctg ggtccgccag gctccaggga aggggctgga gtggatcgga tgcctttata ctggtagtag tggtagcaca tattacgcga gctgggcgaa aggccgattc accatctcca aaacctcgtc gaccacggtg actctgcaaa tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagagat ctaggttatg aaattgatgg ttatgggggc ttgtggggcc agggcaccct cgtcaccgtc tcgagt
CDRL1 - SEQ ID NO: 125 QASQSVYNNNNLS
CDRL2 - SEQ ID NO: 126 DASKLAS
CDRL3 - SEQ ID NO: 127 LGGYYSSGWYFA
CDRH1 - SEQ ID NO: 128 GFSFSGNYYMC
CDRH2 - SEQ ID NO: 129 CLYTGSSGSTYYASWAKG
CDRH3 - SEQ ID NO: 130 DLGYEIDGYGGL
Настоящее изобретение также относится к производному SEQ ID NO: 125, где удаляют участок гликозилирования NLS, например, подвергают мутации в SLS или QLS.
Настоящее изобретение также относится к производному SEQ ID NO: 128, где цистеин заменяют другой аминокислотой, например, серином.
Настоящее изобретение также относится к производному SEQ ID NO: 130, где по меньшей мере одну из аминокислот в мотиве DG заменяют другой аминокислотой, например, мотив подвергают мутации в EG, DA или DS.
Антитела, связывающие сывороточный альбумин
CDRH1 dAbH1 - SEQ ID NO: 131 Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
CDRH2 dAbH1 - SEQ ID NO: 132 Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
CDRH3 dAbH1 - SEQ ID NO: 133 Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu
CDRL1 dAbL1 - SEQ ID NO: 134 Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
CDRL2 dAbL1 - SEQ ID NO: 135 Glu Ala Ser Lys Leu Thr Ser
CDRL3 dAbL1 - SEQ ID NO: 136 Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
Вариабельный домен тяжелой цепи антитела против альбумина (без ds) - SEQ ID NO: 137
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
Вариабельный домен тяжелой цепи антитела против альбумина (ds) - SEQ ID NO: 138
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
Вариабельный домен легкой цепи антитела против альбумина (без ds) - SEQ ID NO: 139
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
Вариабельный домен легкой цепи антитела против альбумина (ds) - SEQ ID NO: 140
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
CD79a человека - SEQ ID NO: 141
MPGGPGVLQA LPATIFLLFL LSAVYLGPGC QALWMHKVPA SLMVSLGEDA HFQCPHNSSN NANVTWWRVL HGNYTWPPEF LGPGEDPNGT LIIQNVNKSH GGIYVCRVQE GNESYQQSCG TYLRVRQPPP RPFLDMGEGT KNRIITAEGI ILLFCAVVPG TLLLFRKRWQ NEKLGLDAGD EYEDENLYEG LNLDDCSMYE DISRGLQGTY QDVGSLNIGD VQLEKP
CD79b человека - SEQ ID NO: 142
MARLALSPVP SHWMVALLLL LSAEPVPAAR SEDRYRNPKG SACSRIWQSP RFIARKRGFT VKMHCYMNSA SGNVSWLWKQ EMDENPQQLK LEKGRMEESQ NESLATLTIQ GIRFEDNGIY FCQQKCNNTS EVYQGCGTEL RVMGFSTLAQ LKQRNTLKDG IIMIQTLLII LFIIVPIFLL LDKDDSKAGM EEDHTYEGLD IDQTATYEDI VTLRTGEVKW SVGEHPGQE
CD45 человека - SEQ ID NO: 143
MYLWLKLLAF GFAFLDTEVF VTGQSPTPSP TGLTTAKMPS VPLSSDPLPT HTTAFSPAST FERENDFSET TTSLSPDNTS TQVSPDSLDN ASAFNTTGVS SVQTPHLPTH ADSQTPSAGT DTQTFSGSAA NAKLNPTPGS NAISDVPGER STASTFPTDP VSPLTTTLSL AHHSSAALPA RTSNTTITAN TSDAYLNASE TTTLSPSGSA VISTTTIATT PSKPTCDEKY ANITVDYLYN KETKLFTAKL NVNENVECGN NTCTNNEVHN LTECKNASVS ISHNSCTAPD KTLILDVPPG VEKFQLHDCT QVEKADTTIC LKWKNIETFT CDTQNITYRF QCGNMIFDNK EIKLENLEPE HEYKCDSEIL YNNHKFTNAS KIIKTDFGSP GEPQIIFCRS EAAHQGVITW NPPQRSFHNF TLCYIKETEK DCLNLDKNLI KYDLQNLKPY TKYVLSLHAY IIAKVQRNGS AAMCHFTTKS APPSQVWNMT VSMTSDNSMH VKCRPPRDRN GPHERYHLEV EAGNTLVRNE SHKNCDFRVK DLQYSTDYTF KAYFHNGDYP GEPFILHHST SYNSKALIAF LAFLIIVTSI ALLVVLYKIY DLHKKRSCNL DEQQELVERD DEKQLMNVEP IHADILLETY KRKIADEGRL FLAEFQSIPR VFSKFPIKEA RKPFNQNKNR YVDILPYDYN RVELSEINGD AGSNYINASY IDGFKEPRKY IAAQGPRDET VDDFWRMIWE QKATVIVMVT RCEEGNRNKC AEYWPSMEEG TRAFGDVVVK INQHKRCPDY IIQKLNIVNK KEKATGREVT HIQFTSWPDH GVPEDPHLLL KLRRRVNAFS NFFSGPIVVH CSAGVGRTGT YIGIDAMLEG LEAENKVDVY GYVVKLRRQR CLMVQVEAQY ILIHQALVEY NQFGETEVNL SELHPYLHNM KKRDPPSEPS PLEAEFQRLP SYRSWRTQHI GNQEENKSKN RNSNVIPYDY NRVPLKHELE MSKESEHDSD ESSDDDSDSE EPSKYINASF IMSYWKPEVM IAAQGPLKET IGDFWQMIFQ RKVKVIVMLT ELKHGDQEIC AQYWGEGKQT YGDIEVDLKD TDKSSTYTLR VFELRHSKRK DSRTVYQYQY TNWSVEQLPA EPKELISMIQ VVKQKLPQKN SSEGNKHHKS TPLLIHCRDG SQQTGIFCAL LNLLESAETE EVVDIFQVVK ALRKARPGMV STFEQYQFLY DVIASTYPAQ NGQVKKNNHQ EDKIEFDNEV DKVKQDANCV NPLGAPEKLP EAKEQAEGSE PTSGTEGPEH SVNGPASPAL NQGS
Ссылки
1. Ribosome display efficiently selects and evolves high-affinity antibodies in vitro from immune libraries. Hanes J, Jermutus L, Weber-Bornhauser S, Bosshard HR, Plückthun A. (1998) Proc. Natl. Acad. Sci. U.S.A. 95, 14130-14135
2. Directed in Vitro Evolution and Crystallographic Analysis of a Peptide-binding Single Chain Antibody Fragment (scFv) with Low Picomolar Affinity. Zhand C, Spinelli S, Luginbuhl B, Amstutz P, Cambillau C, Pluckthun A. (2004) J. Biol. Chem. 279, 18870-18877
3. Antigen recognition by conformational selection. Berger C, Weber-Bornhauser S, Eggenberger Y, Hanes J, Pluckthun A, Bosshard H. R. (1999) F.E.B.S. Letters 450, 149-153
ПРИМЕРЫ
Как применяют в примерах, с помощью термина "Fab-Kd-Fab" описывают биспецифический белковый комплекс, имеющий формулу A-X:Y-B, где:
A-X является первым слитым белком;
Y-B является вторым слитым белком;
X:Y является гетеродимерным соединением;
A содержит Fab-фрагмент, специфичный для антигена, такого как CD45 или CD79;
B содержит Fab-фрагмент, специфичный для антигена, такого как CD45 или CD79;
X является первым партнером по связыванию из связывающейся пары, таким как scFv;
Y является вторым партнером по связыванию из связывающейся пары, таким как пептид; и
: представляет собой взаимодействие (такое как связывающее взаимодействие) между X и Y.
Пример 1 - Получение Fab-A (Fab-scFv [A-X]) и Fab-B (Fab-пептид [B-Y]) для функциональных анализов
Стратегии клонирования
ДНК вариабельной области антитела получали посредством ПЦР или генетического синтеза последовательности ДНК, фланкирующей участки распознавания рестрикционных ферментов. Эти участки соответствовали HindIII и XhoI в случае вариабельных областей тяжелых цепей, и HindIII и BsiWI - в случае вариабельных областей легких цепей. Это делает вариабельную область тяжелой цепи доступной для лигирования в два вектора тяжелой цепи (pNAFH с FabB-Y и pNAFH с FabA-Xds [стабилизированные дисульфидной связью]), т.к. они имеют комплементарные участки рестрикции. С помощью них лигируют вариабельную область выше (или в 5ʹ-направлении) константных областей мыши и пептида Y (GCN4) или scFv X (52SR4), получая целую рамку считывания. Легкие цепи клонировали в стандартные собственные векторы с константной областью каппа-цепи мыши (pMmCK или pMmCK S171C), в которых снова использовали те же комплементарные участки рестрикции. Используют вектор pMmCK S171C, если вариабельную область выделяют из кролика. Клонированные объекты подтверждали посредством секвенирования с использованием праймеров, фланкирующих целую открытую рамку считывания.
Культивирование CHOS
Суспензию клеток CHOS предварительно адаптировали для сред CDCHO (Invitrogen), дополненных 2 мМ (100X) Glutamax. Клетки поддерживали в фазе логарифмического роста, перемешивали при 140 об./мин. в шейкере-инкубаторе (Kuner AG, Birsfelden, Switzerland) и культивировали при 37°C при 8% CO2.
Трансфекция посредством электропорации
Перед трансфекцией количества и жизнеспособность клеток определяли с использованием цитометра CEDEX (Innovatis AG. Bielefeld, Germany), необходимое количество клеток (2×108 клеток/мл) переносили в конические центрифужные пробирки и центрифугировали при 1400 об./мин. в течение 10 минут. Осажденные клетки ресуспендировали в стерильном сбалансированном солевом растворе Эрла и центрифугировали при 1400 об./мин. еще в течение 10 минут. Супернатант выбрасывали и осадки ресуспендировали до желаемой плотности клеток.
ДНК вектора в конечной концентрации 400 мкг для смеси 2×108 клетки/мл и 800 мкл пипетировали в кюветах (Biorad) и электропорировали с использованием собственной системы электропорации.
Трансфицированные клетки переносили непосредственно в 1×3л колбы Эрленмайера, содержащие среды ProCHO 5, обогащенные 2 мМ Glutamax и раствор антимитотических антибиотиков (100-кратный) (1 в 500) и клетки культивировали в шейкере-инкубаторе Kuhner при 37°C, 5% CO2 и встряхивании при 140 об./мин. Добавку 2 г/л ASF (AJINOMOTO) добавляли через 24 ч. после трансфекции и снижали температуру до 32°C для культивирования еще в течение 13 дней. На четвертый день в культуру добавляли 3 мМ бутирата натрия (натриевой соли n-масляной кислоты, Sigma B-5887).
В день 14 культуры переносили в пробирки и отделяли супернатант от клеток после центрифугирования в течение 30 минут при 4000 об./мин. Оставшиеся супернатанты дополнительно фильтровали через фильтр 0,22 мкм SARTO BRAN P Millipore, а затем фильтры 0,22 мкм Gamma gold. Конечные уровни экспрессии определяли посредством ВЭЖХ с протеином G.
Крупномасштабная очистка (1,0 л)
Fab-A и Fab-B очищали посредством аффинного захвата с использованием систем AKTA Xpress и заранее упакованных никелевых колонок HisTrap Excel (GE Healthcare). Супернатанты культур стерилизовали фильтрацией с помощью фильтра 0,22 мкм и корректировали pH до нейтрального, при необходимости, с использованием слабой кислоты или основания до нагрузки на колонки. Использовали этап вторичной промывки, включающий 15-25 мМ имидазола, для вытеснения каких-либо слабо связавшихся белков клеток-хозяев/неспецифических His-связывающих средств из никелевой смолы. Элюцию осуществляли с использованием 10 мМ фосфата натрия, pH 7,4+1M NaCl+250 мМ имидазола и собирали 2 мл фракций. Один объем колонки при элюции системы задерживали в течение 10 минут для сужения пика элюции и, таким образом, снижения общего объема элюции. Наиболее чистые фракции объединяли, заменяли буфер PBS (Sigma), pH 7,4, и фильтровали с помощью фильтров 0,22 мкм. Конечные совокупности анализировали посредством сканирования A280, SE-ВЭЖХ (способа G3000), SDS-PAGE (в восстанавительных и невосстанавительных условиях) и на эндотоксин с использованием системы PTS Endosafe.
Пример 2 - Биспецифический комплекс Fab против CD45/Fab против CD79, но не смесь Fab против CD45 и CD79 или бивалентный комплекс Fab против CD79, ингибирует передачу сигнала Akt
PBMC человека, полученные из роторов при тромбоцитаферезе, хранили в виде замороженных аликвот. Перед осуществлением анализа клетки размораживали, промывали в DMEM (Life Technologies) и позволяли им акклиматизироваться в условиях 37°C/5% CO2. В течение этого периода сетки биспецифических или бивалентных антител получали посредством разведения эквимолярных (200 нМ) количеств Fabʹ-A (Fab-scFv) и Fab-B (Fab-пептид) или Fab-A (Fab-пептид) и Fab-B (Fab-пептид) с антигенной специфичностью к белкам поверхности клетки CD45 и CD79b в DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Эта сетка представлена в таблице 4.
Таблица 4: Сетка биспецифических и бивалентных комбинаций антител со специфичностью к CD45 и CD79b.
Fab A
где X является scFv (52SR4), и Y является пептидом (GCN4).
FabA-X и FabB-Y или Fab-A-Y и Fab-B-Y инкубировали совместно в течение 90 минут (в условиях 37°C/5% CO2) перед смешиванием с 2,5×105 PBMC в 96-луночных планшетах с V-образным дном. Затем PBMC и биспецифические или бивалентные комбинации инкубировали совместно еще в течение 90 минут. После этого B-клетки активировали посредством добавления 200 нМ F(abʹ)2 козы против IgM человека (Southern Biotechnology) в течение 8 минут при 37°C. Затем реакцию передачи сигнала останавливали добавлением равного объема буфера Cytofix (BD Biosciences). Затем планшеты оставляли при комнатной температуре в течение 15 минут перед центрифугированием при 500×g в течение 5 минут. Избыточный супернатант выбрасывали из клеточного осадка, который ресуспендировали в проточном буфере (PBS+1%BSA+0,01%NaN3) и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут перед двукратным промыванием в проточном буфере. Затем клетки окрашивали с использованием флуоресцентно меченого антитела против CD20 (BD Biosciences) и флуоресцентно меченого антитела против фосфорилированного Akt, распознающего модифицированный остаток серина в положении 473 в белке. Затем планшеты ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и ресуспендировали в 25 мкл проточного буфера. Клеточную экспрессию CD20 и Akt измеряли с использованием проточного цитометра Intellicyt HTFC™.
Используя пакет программ для анализа данных Forecyt™ (Intellicyt), B-клетки идентифицировали отдельно от других популяций клеток и вычисляли среднее геометрическое уровней Akt для каждой лунки. Затем все данные выражали в виде процентной доли ингибирования максимального ответа (только против IgM) минус фоновый уровень (только клетки). Относительный эффект комбинаций CD45 и CD79b представлен в таблице 5 (↓=ингибирование, ↑=стимуляция и ↔=без общего эффекта).
Таблица 5: Таблица относительной активности ингибирования фосфорилированного Akt в случае биспецифических бивалентных комбинаций антител со специфичностью к CD45 и CD79b.
Fab A
где X является scFv (52SR4), и Y является пептидом (GCN4).
Эти данные также представлены в форме гистограммы (фигура 1): данные представляют собой средние значения, и планки погрешностей представляют собой 95%-ные достоверные интервалы. Данные свидетельствуют о том, что биспецифическая комбинация против CD45 и CD79b может ингибировать экспрессию фосфорилированного Akt в B-клетках, стимулированных антителом против IgM, в то время как комбинация CD79b-Y с CD79b-Y, являющаяся смесью, которая не может образовывать биспецифическое средство, - нет.
Пример 3 - Биспецифический комплекс Fab против CD45/Fab против CD79, но не смесь Fab против CD45 и CD79 или бивалентный комплекс Fabʹ против CD79, ингибирует передачу сигнала PLCγ2
PBMC человека, полученные из роторов при тромбоцитаферезе, хранили в виде замороженных аликвот. Перед осуществлением анализа клетки размораживали, промывали в DMEM (Life Technologies) и позволяли им акклиматизироваться в условиях 37°C/5% CO2. В течение этого периода сетки биспецифических или бивалентных антител получали посредством разведения эквимолярных (200 нМ) количеств Fab-a (Fab-scFv [A-X]) и Fabʹ-B (Fab-пептид [B-Y]) или Fab-A (Fab-пептид) и Fab-B (Fab-пептид) с антигенной специфичностью к белкам поверхности клетки CD45 и CD79b в DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Эта сетка представлена в таблице 4.
FabʹA-X и FabʹB-Y или Fab-A-Y и Fab-B-Y инкубировали совместно в течение 90 минут (в условиях 37°C/5% CO2) перед смешиванием с 2,5×105 PBMC в 96-луночных планшетах с V-образным дном. Затем PBMC и биспецифические или бивалентные комбинации инкубировали совместно еще в течение 90 минут. После этого B-клетки активировали посредством добавления 200 нМ F(abʹ)2 козы против IgM человека (Southern Biotechnology) в течение 8 минут при 37°C. Затем реакцию передачи сигнала останавливали добавлением равного объема буфера Cytofix (BD Biosciences). Затем планшеты оставляли при комнатной температуре в течение 15 минут перед центрифугированием при 500×g в течение 5 минут. Избыточный супернатант выбрасывали из клеточного осадка, который ресуспендировали в проточном буфере и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут перед двукратным промыванием в проточном буфере.
Затем клетки окрашивали с использованием флуоресцентно меченого антитела против CD20 (BD Biosciences) и флуоресцентно меченого антитела против фосфорилированного PLCγ2, распознающего модифицированный остаток тирозина в положении 759 в белке. Затем планшеты ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и ресуспендировали в 25 мкл проточного буфера. Клеточную экспрессию CD20 и PLCg2 измеряли с использованием проточного цитометра Intellicyt HTFC™.
Используя пакет программ для анализа данных Forecyt™ (Intellicyt), B-клетки идентифицировали отдельно от других популяций клеток и геометрическое среднее уровней PLCγ2 вычисляли для каждой лунки. Затем все данные выражали в виде процентной доли ингибирования максимального ответа (только против IgM) минус фоновый уровень (только клетки). Относительный эффект комбинаций CD45 и CD79b представлен в таблице 6 (↓=ингибирование, ↑=стимуляция и ↔=без общего эффекта).
Таблица 6: Таблица относительной активности ингибирования фосфорилированного PLCg2 в случае биспецифических бивалентных комбинаций антител со специфичностью к CD45 и CD79b.
Fab A
где X является scFv и Y является пептидом.
Эти данные также представлены в форме гистограммы (фигура 2), данные представляют собой средние значения, и планки погрешностей представляют собой 95%-ные достоверные интервалы. Данные свидетельствуют о том, что биспецифическая комбинация против CD45 и CD79b ингибирует экспрессию фосфорилированного PLCγ2 в B-клетках, стимулированных антителом против IgM, в то время как комбинация CD79b-Y с CD79b-Y, являющаяся смесью, которая не может образовывать биспецифическое средство, - нет.
Пример 4 - Биспецифический комплекс против CD45 и CD79b может значительно ингибировать экспрессию CD86 на B-клетках
PBMC человека, полученные из роторов при тромбоцитаферезе, хранили в виде замороженных аликвот. Перед осуществлением анализа клетки размораживали, промывали в DMEM (Life Technologies) и позволяли им акклиматизироваться в условиях 37°C/5% CO2. В течение этого периода биспецифические комбинации получали посредством разведения эквимолярных (500 нМ) количеств Fab-X (Fab-scFv) и Fab-Y (Fab-пептид) с антигенной специфичностью к белкам поверхности клетки CD45 и CD79b в DMEM, содержащей 10% телячьей сыворотки и 2 мМ глутамина. Затем эти комбинации разводили при 8 постепенных разведениях 1 к 2,5 для титрования дозы для этой комбинации. Fab-X и Fab-Y инкубировали совместно в течение 90 минут (в условиях 37°C/5% CO2) перед добавлением 2,5×105 PBMC в 96-луночных планшетах с V-образным дном. Затем PBMC добавляли к комбинациям Fabʹ-X и Fabʹ-Y и инкубировали совместно еще в течение 90 минут. После этого B-клетки активировали посредством добавления 200 нМ F(abʹ)2 козы против IgM человека (Southern Biotechnology) в течение 24 часов при 37°C. Чтобы сделать возможной детекцию поверхностных маркеров активации клеток планшеты помещали на лед и однократно промывали в ледяном проточном буфере (PBS+1% BSA+0,01% NaN3). Затем клетки окрашивали флуоресцентно меченым антителом против CD19 (BD Biosciences) и флуоресцентно меченым антителом против CD86 и инкубировали на льду в течение 1 часа в темноте. После этого планшеты промывали еще два раза и ресуспендировали а 25 мкл проточного буфера. Клеточную экспрессию CD19 и CD86 измеряли с использованием проточного цитометра Intellicyt HTFC™. Используя пакет программ для анализа данных Forecyt™ (Intellicyt), B-клетки идентифицировали отдельно от других популяций клеток и геометрическое среднее уровней CD86 вычисляли для каждой лунки. Затем все данные выражали в виде процентной доли ингибирования максимального ответа (только против IgM) минус фоновый уровень (только клетки). Как видно на фигуре 3, титрование комбинации CD45-X/CD79b-Y способно ингибировать индуцированную антителом против IgM экспрессию CD86 на B-клетках через 24 часа. IC50, как экстраполировали с использованием аппроксимации 4-параметрической логистической кривой с использованием Graphpad Prism 6, составляла 4,7нМ (данные представляют собой средние значения, и планки погрешностей представляют собой стандартные отклонения).
Пример 5 - Ингибиторный эффект биспецифического белка против CD45 и CD79b можно воспроизводить с использованием различных V-областей антитела
Иммунизация: ДНК, кодирующую антигены CD79a и CD79b и CD45, получали посредством генетического синтеза или из коммерческих источников и клонировали в экспрессирующий вектор с сильным конститутивным промотором. Затем с помощью плазмидной ДНК трансфицировали фибробласты Rab-9 кролика (ATCC® CRL-1414™) с использованием собственной системы электропорации. В случае иммунизаций CD79 котрансфицировали CD79a и CD79b. Через двадцать четыре часа клетки проверяли на экспрессию антигена посредством проточной цитометрии и замораживали в аликвотах в жидком азоте до момента использования. Иммунизацию проводили с использованием до 6 антигенов на кролика посредством коэкспрессии на одной и той же клетке или получения смесей отдельно или многократно трансфицированных клеток. Кроликов иммунизировали 3 дозами клеток.
Обнаружение антител: Культуры B-клеток получали способом, схожим со способом, описываемым Zubler et al. (1985). В кратком изложении, B-клетки, полученные из селезенки или PBMC иммунизированных кроликов, культивировали при плотности приблизительно 2000-5000 клеток на лунку в меченых штрих-кодами 96-луночных планшетах для культивирования тканей с 200 мкл/лунку среды RPMI 1640 (Gibco BRL), дополненной 10% FCS (PAA laboratories ltd), 2% HEPES (Sigma Aldrich), 1% L-глутамина (Gibco BRL), 1% раствора пенициллина/стрептомицина (Gibco BRL), 0,1% β-меркаптоэтанола (Gibco BRL), 3% супернатанта активированной культуры спленоцитов и гамма-облученных мутантных клеток тимомы мыши EL4 (5×104/лунку) в течение семи дней при 37°C в атмосфере 5% CO2.
Наличие антиген-специфичных антител в супернатантах культур B-клеток определяли с использованием гомогенного анализа связывания на основе флуоресценции с использованием клеток HEK293, котрансфицированных с использованием CD79a и CD79b или CD45. Скрининг включал перенос 10 мкл супернатанта из меченых штрих-кодами 96-луночных планшетов для культивирования тканей в меченые штрих-кодами 384-луночные планшеты с черными стенками, содержащие клетки HEK293, трансфицированные с использованием антигена-мишени (приблизительно 3000 клетки/лунку) с помощью устройства для обработки жидкостей Matrix Platemate. Связывание выявляли с использованием Fcγ-специфичного конъюгата антитела козы против IgG кролика с Cy-5 (Jackson). Планшеты считывали с помощью системы детекции клеток Applied Biosystems 8200.
После первичного скрининга положительные супернатанты объединяли на 96-луночных меченых штрих-кодами мастер-планшетах с использованием робота для выбора совпадений Aviso Onyx и B-клетки в планшетах для клеточных культур замораживали при -80°C. Затем мастер-планшеты подвергали скринингу в гомогенном анализе связывания на основе флуоресценции с использованием клеток HEK293, трансфицированных с использованием антигенов CD79a и CD79b или CD45, или бус Superavidin™ (Bangs Laboratories), покрытых рекомбинантным белком CD45 в качестве источника антигена. Это осуществляли для определения антигенной специфичности для каждой лунки.
Чтобы сделать возможным выделение генов вариабельных областей антитела из набора интересующих лунок, осуществляли этап деконволюции, позволяющий идентифицировать антиген-специфичные B-клетки в указанной лунке, содержащей гетерогенную популяцию B-клеток. Этого достигали способом фокусов флуоресценции (Clargo et al., 2014. Mabs 2014 Jan 1: 6(1) 143-159; EP1570267B1). В кратком изложении, иммуноглобулин-секретирующие B-клетки из положительной лунки смешивали с клетками HEK293, трансфицированными с использованием антигена-мишени, или стрептавидиновых бус (New England Biolabs), покрытых биотинилированным антигеном-мишенью и конечным разведением 1:1200 Fcγ-фрагмент-специфичного конъюгата антитела козы против кролика с FITC (Jackson). После статической инкубации при 37°C в течение 1 часа антиген-специфичные B-клетки можно идентифицировать благодаря наличию флуоресцентного ореола, окружающего эту B-клетку. Затем ряд этих отдельных B-клеточных клонов, идентифицированных с использованием микроскопа Olympus, выбирали с помощью микроманипулятора Eppendorf и помещали в пробирку для ПЦР. Способ фокусов флуоресценции также использовали для идентификации антиген-специфичных B-клеток из гетерогенной популяции B-клеток напрямую из костного мозга иммунизированных кроликов.
Гены вариабельной области антитела выделяли из отдельных клеток посредством ПЦР с обратной транскрипцией (RT) с использованием праймеров, специфичных для вариабельных областей тяжелых и легких цепей. Осуществляли два раунда ПЦР с вторичной гнездовой ПЦР, посредством которой встраивают участки рестрикции на 3ʹ- и 5'-концы, позволяющие клонировать вариабельную область в векторы млекопитающих, экспрессирующие Fab-X и Fab-Y мыши (VH) или каппа-цепь (VL) мыши. С помощью конструкций тяжелых и легких цепей для векторов, экспрессирующих Fab-X и Fab-Y, котрансфицировали клетки HEK-293 с использованием Fectin 293 (Life Technologies) или клетки Expi293 с использованием Expifectamine (Life Technologies) и рекомбинантное антитело экспрессировали в 6-луночных планшетах для культивирования тканей в объеме 5 мл. После 5-7 дней экспрессии собирали супернатанты. Супернатанты тестировали в гомогенном анализе связывания на основе флуоресценции с использованием клеток HEK293, трансфицированных с использованием антигена, и бус Superavidin™ (Bangs Laboratories), покрытых рекомбинантным белком или трансфицированными с использованием антигена клетками HEK. Это осуществляли для подтверждения специфичности клонированных антител.
Маломасштабное получение Fab A-X и Fab B-Y (маломасштабная (50 мл) трансфекция Expi293)
Клетки Expi293 общепринято субклонировали в экспрессионной среде Expi293™ до конечной концентрации 0,5×106 жизнеспособных клеток/мл и инкубировали на орбитальном шейкере (Multitron, Infors HT) при 120 об./мин., 8% CO2 и 37°C.
В день трансфекции жизнеспособность клеток и концентрацию измеряли с использованием автоматизированного цитометра (Vi-CELL, Beckman Coulter). Для достижения конечной концентрации клеток 2,5×106 жизнеспособных клеток/мл соответствующий объем клеточной суспензии добавляли в стерильную колбу Эрленмайера емкостью 250 мл и доводили до объема 42,5 мл посредством добавления свежей, предварительно нагретой экспрессионной среды Expi293™ для каждой трансфекции в объеме 50 мл.
Для получения комплексов липид-ДНК для каждой трансфекции всего 50 мкг плазмидной ДНК тяжелой цепи и легкой цепи разводили в среде Opti-MEM® I (LifeTechnologies) до общего объема 2,5 мл и 135 мкл реагента ExpiFectamine™ 293 (LifeTechnologies) разводили в среде Opti-MEM® I до общего объема 2,5 мл. Все разведения осторожно смешивали и инкубировали в течение не более 5 минут при комнатной температуре перед добавлением раствора ДНК к соответствующему разведению реагента ExpiFectamine™ 293 для достижения общего объема 5 мл. Смеси ДНК-реагент ExpiFectamine™ 293 осторожно смешивали и инкубировали в течение 20-30 минут при комнатной температуре, чтобы позволить образоваться комплексам ДНК-реагент ExpiFectamine™ 293.
После завершения инкубации комплекса ДНК-реагент ExpiFectamine™ 293 5 мл комплекса ДНК-реагент ExpiFectamine™ 293 добавляли в каждую встряхиваемую колбу. Встряхиваемые колбы инкубировали на орбитальном шейкере (Multitron, Infors HT) при 120 об./мин., 8% CO2 и 37°C.
Через приблизительно 16-18 часов после трансфекции 250 мкл усилителя трансфекции 1 ExpiFectamine™ 293 (LifeTechnologies) и 2,5 мл усилителя трансфекции 2 ExpiFectamine™ 293 (LifeTechnologies) добавляли в каждую встряхиваемую колбу.
Культуры клеток собирали через 7 дней после трансфекции. Клетки переносили в центрифужные пробирки емкостью 50 мл (Falcon) и центрифугировали в течение 30 мин при 4000 об./мин. с последующей стерилизацией фильтрацией через фильтры 0,22 мкм Stericup (Merck Millipore). Очищенные и стерилизованные фильтрацией супернатанты хранили при 4°C. Конечные уровни экспрессии определяли посредством ВЭЖХ с протеином G.
Маломасштабная (50 мл) очистка: Fab-X и Fab-Y очищали раздельно посредством аффинного захвата с использованием маломасштабной вакуумной системы очистки. В кратком изложении, 50 мл супернатантов культур стерилизовали фильтрацией через фильтры 0,22 мкм перед добавлением 500 мкл бус Ni-сефарозы (GE Healthcare). Затем смесь супернатанта с бусами переворачивали в течение приблизительно часа перед удалением супернатанта с помощью вакуума. Затем бусы промывали промывочным раствором 1 (50 мМ фосфата натрия, 1 M NaCl, pH 6,2) и промывочным раствором 2 (0,5 M NaCl). Элюцию осуществляли с использованием 50 мМ ацетата натрия, pH 4,0+1M NaCl. Буфер в элюированных фракциях заменяли PBS (Sigma), pH 7,4, и фильтровали через фильтры 0,22 мкм. Конечные совокупности анализировали посредством сканирования A280, SE-UPLC (способа BEH200), электрофореза в ПААГ в присутствие SDS (в восстановительных и невосстановительных условиях) и анализировали на эндотоксин с использованием системы PTS Endosafe.
PBMC человека, полученные из роторов при тромбоцитаферезе, хранили в виде замороженных аликвот. Перед осуществлением анализа клетки размораживали, промывали в RPMI 1640 (Life Technologies) и позволяли им акклиматизироваться в условиях 37°C/5% CO2. В течение этого периода комбинации биспецифических, бивалентных средств или смесей антител получали посредством разведения эквимолярных (200 нМ) количеств Fabʹ-X (Fab-scFv) и Fabʹ-Y (Fab-пептид) с антигенной специфичностью к белкам поверхности клетки CD45 и CD79b в RPMI 1640, содержащей 10% эмбриональной телячьей сыворотки, 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 2 мМ L-глутамина. Эти комбинации 3 разных Fab-Y против CD79b и 2 разных Fab-X против CD45 представлены в таблице 7.
Таблица 7: Сетка биспецифических белков со специфичностью к CD45 и CD79b.
Fab A
VR4248
где X является scFv (52SR4) и Y является пептидом (GCN4).
FabA-X и FabB-Y инкубировали совместно в течение 60 минут (в условиях 37°C/5% CO2) перед смешиванием с 2,5×105 PBMC в 96-луночных планшетах с V-образным дном. Затем PBMC и комбинации FabA-X и/или FabB-Y инкубировали совместно еще в течение 90 минут. После этого B-клетки активировали посредством добавления 12,5 мкг/мл F(abʹ)2 козы против IgM человека (Southern Biotechnology) в течение 10 минут при 37°C. Затем реакцию передачи сигнала останавливали добавлением равного объема буфера Cytofix (BD Biosciences). Затем планшеты оставляли при комнатной температуре в течение 15 минут перед центрифугированием при 500×g в течение 5 минут. Избыточный супернатант выбрасывали из клеточного осадка, который ресуспендировали в проточном буфере (PBS+1% BSA+0,1% NaN3+2 мМ ЭДТА) и промывали еще раз. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут перед двукратным промыванием в проточном буфере.
Затем клетки окрашивали с использованием флуоресцентно меченого антитела против CD20 (BD Biosciences) и антитела против фосфорилированного PLCγ2, распознающего модифицированный остаток тирозина в положении 759. Затем планшеты ресуспендировали и инкубировали в течение 1 часа при комнатной температуре в темноте. После этого планшеты промывали еще два раза и ресуспендировали в 40 мкл проточного буфера. Клеточную экспрессию CD20 и PLCγ2 измеряли с использованием проточного цитометра Intellicyt HTFC™.
Используя пакет программ для анализа данных Forecyt™ (Intellicyt), B-клетки идентифицировали отдельно от других популяций клеток и геометрическое среднее уровней PLCγ2 вычисляли для каждой лунки. Затем все данные выражали в виде процентной доли ингибирования максимального ответа (только против IgM) минус фоновый уровень (только клетки).
Как можно видеть на фигуре 4, данные свидетельствуют о том, что комбинация CD45 с CD79b с разными V-областями антитела может ингибировать экспрессию фосфорилированного PLCγ2 в B-клетках, стимулированных антителом против IgM.
Пример 6: Скрининг сетки больших панелей гетеродимерно соединенных белковых комплексов для идентификации новых мишеней биспецифического антитела
Введение: После успешной проверки биспецифического формата и способа скрининга в приведенных выше примерах скрининг распространяли на большее количество пар антигенов. Получали панель пар вариабельных областей (V) антитела к 23 разным антигенам, экспрессирующимся на B-клетках. Используя формат Fab-Kd-Fab [т.е. A-X:Y-B, где A и B являются Fab-фрагментами] получали сетку гетеродимерно соединенных белковых комплексов, представляющую собой множество комбинаций V-областей из каждой из 315 разных комбинаций пар антигенов. Эти комбинации подвергали скринингу на их способность модулировать передачу сигнала BCR (B-клеточного рецептора) в высокопроизводительном анализе посредством проточной цитометрии для выбора новых пар мишеней для воздействия с использованием биспецифического антитела.
Иммунизация: ДНК, кодирующую выбранные антигены, получали посредством генетического синтеза или из коммерческих источников и клонировали в экспрессирующий вектор с сильным конститутивным промотором. Затем с помощью плазмидной ДНК трансфицировали фибробласты кролика Rab-9 (ATCC® CRL-1414™) с использованием собственной системы электропорации. Через двадцать четыре часа клетки проверяли на экспрессию антигена посредством проточной цитометрии и замораживали в аликвотах в жидком азоте до использования. Иммунизацию проводили с использованием до 6 антигенов на кролика посредством коэкспрессии на одной и той же клетке или получения смесей отдельно или многократно трансфицированных клеток. Кроликов иммунизировали 3 дозами клеток.
Обнаружение антител: Культуры B-клеток получали способом, схожим со способом, описываемым Zubler et al. (1985). В кратком изложении, B-клетки, полученные из селезенки или PBMC иммунизированных кроликов, культивировали при плотности приблизительно 2000-5000 клеток на лунку в меченых штрих-кодами 96-луночных планшетах для культивирования тканей с 200 мкл/лунку среды RPMI 1640 (Gibco BRL), дополненной 10% FCS (PAA laboratories ltd), 2% HEPES (Sigma Aldrich), 1% L-глутамина (Gibco BRL), 1% раствора пенициллина/стрептомицина (Gibco BRL), 0,1% β-меркаптоэтанола (Gibco BRL), 3% супернатанта активированной культуры спленоцитов и гамма-облученных мутантных клеток тимомы мыши EL4 (5×104/лунку) в течение семи дней при 37°C в атмосфере 5% CO2.
Наличие антиген-специфичных антител в супернатантах культур B-клеток определяли с использованием гомогенного анализа связывания на основе флуоресценции с использованием клеток HEK293, котрансфицированных с использованием антигенов, которыми иммунизировали кроликов. Скрининг включал перенос 10 мкл супернатанта из меченых штрих-кодами 96-луночных планшетов для культивирования тканей в меченые штрих-кодами 384-луночные планшеты с черными стенками, содержащие клетки HEK293, трансфицированные с использованием антигена-мишени (приблизительно 3000 клетки/лунку) с помощью устройства для обработки жидкостей Matrix Platemate. Связывание выявляли с использованием Fcγ-специфичного конъюгата антитела козы против IgG кролика с Cy-5 (Jackson). Планшеты считывали с помощью системы детекции клеток Applied Biosystems 8200.
После первичного скрининга положительные супернатанты объединяли на 96-луночных меченых штрих-кодами мастер-планшетах с использованием робота для выбора совпадений Aviso Onyx и B-клетки в планшетах для клеточных культур замораживали при -80°C. Затем мастер-планшеты подвергали скринингу в гомогенном анализе связывания на основе флуоресценции с использованием клеток HEK293, трансфицированных с использованием антигенов раздельно, и бус Superavidin™ (Bangs Laboratories), покрытых рекомбинантным белком в качестве источника антигена. Это осуществляли для определения антигенной специфичности для каждой лунки.
Чтобы сделать возможным выделение генов вариабельных областей антитела из набора интересующих лунок, осуществляли этап деконволюции, позволяющий идентифицировать антиген-специфичные B-клетки в указанной лунке, содержащей гетерогенную популяцию B-клеток. Этого достигали способом фокусов флуоресценции (Clargo et al., 2014. Mabs 2014 Jan 1: 6(1) 143-159; EP1570267B1). В кратком изложении, иммуноглобулин-секретирующие B-клетки из положительной лунки смешивали с клетками HEK293, трансфицированными с использованием антигена-мишени, или стрептавидиновых бус (New England Biolabs), покрытых биотинилированным антигеном-мишенью и конечным разведением 1:1200 Fcγ-фрагмент-специфичного конъюгата антитела козы против кролика с FITC (Jackson). После статической инкубации при 37°C в течение 1 часа антиген-специфичные B-клетки можно идентифицировать благодаря наличию флуоресцентного ореола, окружающего эту B-клетку. Затем ряд этих отдельных B-клеточных клонов, идентифицированных с использованием микроскопа Olympus, выбирали с помощью микроманипулятора Eppendorf и помещали в пробирку для ПЦР. Способ фокусов флуоресценции также использовали для идентификации антиген-специфичных B-клеток из гетерогенной популяции B-клеток напрямую из костного мозга иммунизированных кроликов.
Гены вариабельной области антитела выделяли из отдельных клеток посредством ПЦР с обратной транскрипцией (RT) с использованием праймеров, специфичных для вариабельных областей тяжелых и легких цепей. Осуществляли два раунда ПЦР с вторичной гнездовой ПЦР, посредством которой встраивают участки рестрикции на 3ʹ- и 5'-концы, позволяющие клонировать вариабельную область в векторы млекопитающих, экспрессирующие Fab-X и Fab-Y мыши (VH) или каппа-цепь (VL) мыши. С помощью конструкций тяжелых и легких цепей для векторов, экспрессирующих Fab-X и Fab-Y, котрансфицировали клетки HEK-293 с использованием Fectin 293 (Life Technologies) или клетки Expi293 с использованием Expifectamine (Life Technologies) и рекомбинантное антитело экспрессировали в 6-луночных планшетах для культивирования тканей в объеме 5 мл. После 5-7 дней экспрессии собирали супернатанты. Супернатанты тестировали в гомогенном анализе связывания на основе флуоресценции с использованием клеток HEK293, трансфицированных с использованием антигена, и бус Superavidin™ (Bangs Laboratories), покрытых рекомбинантным белком или трансфицированными с использованием антигена клетками HEK. Это осуществляли для подтверждения специфичности клонированных антител.
Маломасштабное получение Fab A-X и Fab B-Y (маломасштабная (50 мл) трансфекция Expi293)
Клетки Expi293 общепринято субклонировали в экспрессионной среде Expi293™ до конечной концентрации 0,5×106 жизнеспособных клеток/мл и инкубировали на орбитальном шейкере (Multitron, Infors HT) при 120 об./мин., 8% CO2 и 37°C.
В день трансфекции жизнеспособность клеток и концентрацию измеряли с использованием автоматизированного цитометра (Vi-CELL, Beckman Coulter). Для достижения конечной концентрации клеток 2,5×106 жизнеспособных клеток/мл соответствующий объем клеточной суспензии добавляли в стерильную колбу Эрленмайера емкостью 250 мл и доводили до объема 42,5 мл посредством добавления свежей, предварительно нагретой экспрессионной среды Expi293™ для каждой трансфекции в объеме 50 мл.
Для получения комплексов липид-ДНК для каждой трансфекции всего 50 мкг плазмидной ДНК тяжелой цепи и легкой цепи разводили в среде Opti-MEM® I (LifeTechnologies) до общего объема 2,5 мл и 135 мкл реагента ExpiFectamine™ 293 (LifeTechnologies) разводили в среде Opti-MEM® I до общего объема 2,5 мл. Все разведения осторожно смешивали и инкубировали в течение не более 5 минут при комнатной температуре перед добавлением раствора ДНК к соответствующему разведению реагента ExpiFectamine™ 293 для достижения общего объема 5 мл. Смеси ДНК-реагент ExpiFectamine™ 293 осторожно смешивали и инкубировали в течение 20-30 минут при комнатной температуре, чтобы позволить образоваться комплексам ДНК-реагент ExpiFectamine™ 293.
После завершения инкубации комплекса ДНК-реагент ExpiFectamine™ 293 5 мл комплекса ДНК-реагент ExpiFectamine™ 293 добавляли в каждую встряхиваемую колбу. Встряхиваемые колбы инкубировали на орбитальном шейкере (Multitron, Infors HT) при 120 об./мин., 8% CO2 и 37°C.
Через приблизительно 16-18 часов после трансфекции 250 мкл усилителя трансфекции 1 ExpiFectamine™ 293 (LifeTechnologies) и 2,5 мл усилителя трансфекции 2 ExpiFectamine™ 293 (LifeTechnologies) добавляли в каждую встряхиваемую колбу.
Культуры клеток собирали через 7 дней после трансфекции. Клетки переносили в центрифужные пробирки емкостью 50 мл (Falcon) и центрифугировали в течение 30 мин при 4000 об./мин. с последующей стерилизацией фильтрацией через фильтры 0,22 мкм Stericup (Merck Millipore). Очищенные и стерилизованные фильтрацией супернатанты хранили при 4°C. Конечные уровни экспрессии определяли посредством ВЭЖХ с протеином G.
Маломасштабная (50 мл) очистка: Fab-X и Fab-Y очищали раздельно посредством аффинного захвата с использованием маломасштабной вакуумной системы очистки. В кратком изложении, 50 мл супернатантов культур стерилизовали фильтрацией через фильтры 0,22 мкм перед добавлением 500 мкл бус Ni-сефарозы (GE Healthcare). Затем смесь супернатанта с бусами переворачивали в течение приблизительно часа перед удалением супернатанта с помощью вакуума. Затем бусы промывали промывочным раствором 1 (50 мМ фосфата натрия, 1 M NaCl, pH 6,2) и промывочным раствором 2 (0,5 M NaCl). Элюцию осуществляли с использованием 50 мМ ацетата натрия, pH 4,0+1M NaCl. Буфер в элюированных фракциях заменяли PBS (Sigma), pH 7,4, и фильтровали через фильтры 0,22 мкм. Конечные совокупности анализировали посредством сканирования A280, SE-UPLC (способа BEH200), электрофореза в ПААГ в присутствие SDS (в восстановительных и невосстановительных условиях) и анализировали на эндотоксин с использованием системы PTS Endosafe.
Скрининговые анализы
PBMC донора быстро размораживали с использованием водяной бани, установленной на 37°C, и осторожно переносили в пробирку Falcon емкостью 50 мл. Затем их по каплям разводили до 5 Мл в аналитических средах для минимизации осмотического шока. Затем клетки осторожно разводили до 20 мл перед добавлением конечных сред для разведения для достижения объема 50 мл. Затем клетки центрифугировали при 500×g в течение 5 минут перед удалением супернатанта и ресуспендирования клеток в 1 мл сред. Затем клетки подсчитывали и разводили до 1,66×106 клеток/мл перед распределением по 30 мкл на лунку в планшете с TC V-образным дном, достигая конечной концентрации для анализа 5,0×104 клеток/лунку. Затем планшет для культивирования клеток хранили закрытым в инкубаторе при 37°C, 5% CO2 до того момента, когда они понадобятся, оставляя их минимум на 1 час в покое.
Реагенты Fab-X и Fab-Y смешивают в эквимолярном соотношении при 5-кратной конечной концентрации для анализа в средах для анализа и инкубировали в течение 90 мин при 37°C, 5% CO2. Образцы получали в 96-луночнуночном полипропиленовом планшете с U-образным дном и закрывали их на время инкубации.
10 мкл 5-кратной смеси Fab-KD-Fab добавляли в соответствующие тестовые лунки, содержащие клетки, и смешивали посредством встряхивания при 1000 об./мин. в течение 30 сек перед инкубацией в течение 90 мин при 37°C, 5% CO2.
Затем клетки стимулировали 10 мкл антитела против IgM человека. Конечную концентрацию стимула для анализа варьировали в зависимости от считываний в панели при анализе, три смеси антител A, B и C (подробно описаны ниже) стимулировали при конечной концентрации для анализа 50 мкг/мл (смесь A и C) или 25 мкг/мл (смесь B). Затем планшеты для анализа осторожно перемешивали при 1000 об./мин. в течение 30 сек перед инкубацией при 37°C, 5% CO2 в течение 5 мин (смесь антител A и C) или 2 мин (смесь антител B). Анализ останавливали добавлением 150 мкл ледяного BD CytoFix во все лунки и инкубировали в течение 15 мин при RT. Затем фиксированные клетки центрифугировали при 500×g в течение 5 мин для осаждения клеток и удаления супернатанта с использованием устройства для промывания планшетов BioTek ELx405. Осадок ресуспендировали посредством перемешивания планшета на центрифуге типа вортекс при 2400 об./мин. в течение 30 сек. Затем клетки пермеабилизовали при 4°C добавлением 100 мкл ледяного буфера для пермеабилизации клеток III BD в течение 30 мин. Затем клетки промывали в 100 мкл буфера для FACS и центрифугировали при 500×g в течение 5 мин. Супернатант снова удаляли с помощью ELx405 перед его использованием для быстрого распределения 200 мкл буфера для FACS для отмывания любого остатка буфера для пермеабилизации. Клетки снова центрифугировали при 500×g и удаляли супернатант переворачиванием. На предшествующем этапе центрифугирования смесь антитела получали в буфере для FACS и защищали от света. Затем клетки ресуспендировали посредством перемешивания на центрифуге типа вортекс (2400 об./мин., 30 сек) перед добавлением 20 мкл смеси антитела во все лунки и планшет встряхивали в течение 30 сек при 1000 об./мин. Затем клетки инкубировали в течением 60 мин при RT в темноте.
Затем клетки дважды промывали 200 мкл буфера для FACS с центрифугированием при 500×g и удаляли супернатант после каждого этапа. В конечном итоге, клетки ресуспендировали посредством перемешивания на центрифуге типа вортекс в течение 30 сек при 2400 об./мин. перед добавлением конечных 20 мкл буфера для FACS. Затем планшеты считывали с помощью устройства Intellicyt HTFC/iQue.
Буфер для FACS=PBS+1% BSA+0,05% NaN3+2 мМ ЭДТА
Смесь антител A=1:2 CD20 PerCp-Cy5,5 (BD Biosciences)+1:5 PLCγ2 AF88+1:10 Akt AF647+1:50 ERK1/2 PE (разведенная в буфере для FACS).
Смесь антител B=1:2 CD20 PerCp-Cy5,5 (BD Biosciences)+1:5 Syk PE+1:5 BLNK AF647 (разведенная в буфере для FACS)
Смесь антител C=1:5 CD20 PerCp-Cy5,5 (Biolegend)+1:5 PLCγ2 AF488+1:10 Akt AF647+1:5 Syk PE (разведенная в буфере для FACS)
Комбинации Fab-X+Fab-Y подвергали скринингу с использованием смеси антител A и B или C в отдельности. Все скрининги осуществляли на клетках из роторов для тромбоцитафереза от 2 разных доноров крови. Данные регистрировали и оценивали с использованием коммерчески доступного программного обеспечения. Всего 2500 комбинаций Fab-X+Fab-Y подвергали скринингу в отношении 315 различных комбинаций антигенов.
Результаты
Вычисляли процентную долю ингибирования индукции фосфорилирования белков каскада передачи сигнала BCR каждой комбинацией Fab-Kd-Fab [т.е. A-X:Y-B, где A и B являются Fab-фрагментами], в этом примере, в котором искали новые комбинации антигенов, ингибирующих функцию B-клеток, устанавливали такие критерии положительной комбинаций, как по меньшей мере 30% ингибирование по меньшей мере двух считываний фосфорилирования по меньшей мере одной комбинацией V-областей. В соответствии с этим пороговым значением необходимым критериям соответствовали 11 новых комбинаций пар антигенов из 315 исследуемых. Это представляет собой 3,5% коэффициент совпадений, что свидетельствует о важности скрининга больших количеств комбинаций для обнаружения тех, которые имеют желаемую активность, и том, насколько редкой является активность комбинации CD79b и CD45.
На фигурах 6-8 представлены данные для сеток перекрестных специфичностей антигена. Значения представляют собой процентную долю ингибирования (отрицательное значение для активации) фосфорилирования Syk, PLCγ2 и AKT, соответственно, и среднее значение для множества оцениваемых комбинаций V-областей. Тестировали 315 различных комбинаций антигенов, и, как можно видеть, эффект в отношении передачи сигнала BCR различными комбинациями антител значительно варьировался от сильного ингибирования, например, антигена 2 (CD79b) в Fab-X в комбинации с антигеном 4 (CD45) в Fab-Y (70,4% ингибирования фосфорилированного Syk, фигура 6) до активации, например, антигеном 6 в X и антигеном 11 в Y (минус 118,10% фосфорилированного Syk, фигура 6).
Каждая точка данных, представляющая собой средние значения %, показанные на фигурах 6-8, представлена для комбинации антигена 2 (CD79b) в Fab-X и антигена 4 (CD45) в Fab-Y на фигуре 9. В этом случае оценивали 10 различных комбинаций различных V областей антитела. Та же комбинация антигенов, но в альтернативной ориентации, т.е. антиген 2 (CD79b) в Fab-Y и антиген 4 (CD45) в Fab-X, представлена на фигуре 10. В этом случае оценивали 6 различных комбинаций различных V-областей антитела. И снова, все V-области демонстрировали ингибирование, но этим способом можно идентифицировать и выбрать оптимальные комбинации V-областей.
Пример 7 - Скрининг транзиторно экспрессируемых V-областей к антигену CD45 в виде Fab-X с использованием очищенного Fab-Y против CD79b в гетеродимерно соединенных белковых комплексов для селекции оптимальных V-областей антитела против CD45
Введение: Новые V-области к CD45, ингибирующие передачу сигнала B-клетками, в виде биспецифического антитела в комбинации с CD79b-специфичными V-областями идентифицировали с использованием скрининга сетки гетеродимерно соединенных белковых комплексов. V-области CD45 транзиторно экспрессировали как Fab-X и комбинировали с очищенным Fab-Y против CD79b. Измеряли ингибирование активации передачи сигнала B-клетками для селекции наиболее активных V-областей против CD45 и против CD79b.
Получение антиген-экспрессирующих клеток и иммунизацию кроликов осуществляли так же, как описано в примере 6.
Обнаружение антител: Культуры B-клеток получали так же, как описано в примере 6.
Скрининг антиген-специфичных антител в супернатантах культур B-клеток и этап деконволюции для идентификации антиген-специфичных B-клеток определяли так же, как описано в примере 6.
Гены вариабельной области антитела выделяли из отдельных клеток посредством ПЦР с обратной транскрипцией (RT) с использованием праймеров, специфичных для вариабельных областей тяжелых и легких цепей. Осуществляли два раунда ПЦР с вторичной гнездовой ПЦР, посредством которой встраивают участки рестрикции на 3ʹ- и 5'-концы, позволяющие клонировать вариабельную область в векторы млекопитающих, экспрессирующие Fab-X и Fab-Y мыши (VH) или каппа-цепь (VL) мыши. Затем с помощью этих векторов котрансфицировали клетки HEK-293 с использованием 293Fectin (Life Technologies) или клетки Expi293 с использованием Expifectamine (Life Technologies) и оставляли для экспрессии в течение 6 дней. Супернатанты тестировали в гомогенном анализе связывания на основе флуоресценции с использованием клеток HEK293, трансфицированных с использованием антигена, и бус Superavidin™ (Bangs Laboratories), покрытых рекомбинантным белком или трансфицированными с использованием антигена клетками HEK. Это осуществляли для подтверждения специфичности клонированных антител.
В дополнение к супернатантам с транзиторно экспрессируемыми Fab-X, тем же способом с помощью неродственной контрольной ДНК получали ложные супернатанты отрицательного контроля.
Уровни экспрессии Fab-X определяли посредством ВЭЖХ с протеином G.
Получение очищенных Fab-X и Fab-Y: Очищенные Fab-X и Fab-Y получали с использованием того же способа, который описан в примере 6.
Анализ PHosFlow: CD79b-специфичный Fab-Y и CD45-специфичный Fab-X, очищенный или транзиторно экспрессированный в супернатанте, инкубировали совместно в течение 60 минут (в условиях 37°C и 5% CO2) при эквимолярной концентрации 200 нМ и 90 нМ. Также включали ложный супернатант без примесей. В лунки 96-луночных планшетов с V-образным дном добавляли 5,0×104 PBMC, к которым добавляли титрованные комбинации Fab-X и Fab-Y или ложный супернатант. Затем комбинации и клетки инкубировали совместно еще в течение 90 минут. После этого B-клетки активировали посредством добавления 25 мкг/мл F(abʹ)2 козы против IgM человека (Southern Biotechnology) в течение 15 минут при 37°C и 5% CO2. Затем реакцию передачи сигнала останавливали добавлением равного объема буфера Cytofix (BD Biosciences). Затем планшеты оставляли при комнатной температуре в течение 15 минут перед центрифугированием при 500×g в течение 5 минут. Избыточный супернатант выбрасывали из клеточного осадка, который ресуспендировали в буфере для FACS (PBS+1% BSA+0,01% NaN3+2 мМ ЭДТА) и еще раз промывали. Затем клетки ресуспендировали в ледяном буфере Perm Buffer III (BD Biosciences) в течение 30 минут перед двукратным промыванием в проточном буфере.
Затем клетки окрашивали, как описано в примере 6, за исключением того, что вместо 3 разных смесей антител, использовали только одну смесь с теми же концентрациями для анализа и условиями инкубации, которые описаны для смеси антител A в примере 6.
Смесь антител=1:3 CD20 PerCp-Cy5,5+1:5 PLCγ2 AF88+1:10 Akt AF647+1:5 p38 MAPK PE (разбавленная в буфере для FACS).
Результаты
Как видно на фигурах 11-16, данные свидетельствуют о том, что комбинация различных транзиторно экспрессируемых V-областей к антигену CD45 в Fab-X с 2 разными очищенными V-областями к антигену CD79b (VR447 и VR4450) в Fab-Y может ингибировать активацию B-клеток (что измеряли по ингибированию PLCγ2, p38 и Akt) до различных уровней, и, таким образом, скрининг в биспецифическом формате облегчает селекцию оптимальных комбинаций V-областей. Комбинации с транзиторно экспрессируемым Fab-X сравнивают с референсной комбинацией с очищенным Fab-X против CD45 (VR4122).
Пример 8 - Эффект совместного таргетинга антигена CD79b и антигена CD45 в отношении функции B-клеток памяти с использованием молекулярно связанных биспецифических Bybe с добавлением антитела против альбумина или без него
Введение: Для проверки того, что таргетинг CD79b/CD45 имеет функциональный эффект в отношении B-клеток при длительном культивировании измеряли продукцию IgG B-клетками в смешанной культуре PBMC. Измерение специфических антител для повторного воздействия антигена столбнячного токсина обеспечивает считывание функции B-клеток памяти.
Специфичности к антигену CD79b (VR4447) и специфичности к антигену CD45 (VR4248 и VR4133) достигали в формате BYbe с добавлением фрагмента против альбумина (VR0645) или без него. Фрагмент антитела против альбумина подвергали слиянию с легкой цепью Fab против антигена CD45 из формата BYbe, как описано в примере 8.
Описание конструкций, используемых в этих экспериментах.
Способы
Очистка BYbe с дополнительной специфичностью антитела против альбумина или без нее
Форматы BYbe (Fab-dsscFv [scFv без C-конца тяжелой цепи Fab]) и BYbe против альбумина (Fab-2xdsscFv [scFv без C-конца тяжелой цепи и легкой цепи Fab]) очищали следующим образом. Очищенные супернатанты культур клеток из стандартным систем экспрессии expiHEK или CHO стерилизовали фильтрацией через фильтры 0,22 мкм. Профильтрованные супернатанты нагружали в количестве 2 мл/мин на 50-мл колонки GammabindPlus SepHarose XK26 (GE Healthcare), уравновешенные PBS при pH 7,4 (Sigma Aldrich Chemicals). После нагрузки колонки промывали PBS, pH 7,4, а затем элюировали 0,1 M глицином/HCl, pH 2,7. После элюции определяли поглощение при 280 нм, собирали пики элюции, а затем нейтрализовали 1/25 объема 2 M Трис/HCl, pH 8,5. Нейтрализованные образцы концентрировали с использованием концентраторов Amicon Ultra-15 с мембраной с отсечкой по молекулярной массе 10 кДа или 30 кДа и центрифугирования при 4000×g с использованием ротора со свободно подвешенными стаканами. Концентрированные образцы наносили на колонку XK16/60 или XK26/60 Superdex 200 (GE Healthcare), уравновешенную PBS, pH 7,4. Колонки обрабатывали с помощью изократического градиента PBS, pH 7,4, при 1 мл/мин или 2,6 мл/мин соответственно. Фракции собирали и анализировали с помощью эксклюзионной хроматографии в геле TSK G3000SWXL; колонку 5 мкм, 7,8 × 300 мм обрабатывали с помощью изократического градиента 0,2 M фосфата, pH 7,0, при 1 мл/мин, с детекцией поглощения при 280 нм. Выбранные мономерные фракции объединяли и концентрировали до >1 мг/мл с использованием концентратора Amicon Ultra-15 с мембраной с отсечкой по молекулярной массе 10 кДа или 30 кДа и центрифугированием при 4000×g с использованием ротора со свободно подвешенными стаканами. Анализировали конечные образцы; на концентрацию - с помощью сканирующего УФ-спектрофотометра A280 (Cary 50Bio); на % мономера - посредством эксклюзионной хроматографии в геле TSK G3000SWXL; колонке 5 мкм, 7,8×300 мм, обрабатываемой с помощью изократического градиента 0,2 M фосфат, pH 7,0, при 1 мл/мин, с детекцией поглощения при 280 нм; посредством электрофореза в ПААГ в присутствие SDS в восстановительных и невосстановительных условиях, осуществляемом в 4-20%-ных гелях с Трис-глицином 1,5 мм (Novex) при 50 мА (на гель) в течение 53 минут; и на эндотоксин с помощью портативной тест-системы Charles Riverʹs EndoSafe® с тестовыми картриджами Limulus Amebocyte Lysate (LAL).
Активация B-клеток и измерение специфичных для столбнячного токсина IgG
PBMC человека стимулировали 500 нг/мл CD40L, 1 мкг/мл CpG и 50 нг/мл ИЛ-21 в средах 1640 с 10% эмбриональной телячьей сывороткой и 2 мМ Glutamax (среда R10) в течение 6 дней. Добавляли конструкции очищенного белка в конечной концентрации 100 нМ в день 0 и оставляли в среде для культивирования на время анализа. Через 6 дней собирали супернатанты и определяли количество специфичных для столбнячного токсина IgG с помощью ELISA. В кратком изложении, планшеты для ELISA Maxisorp с половинным объемом лунок (Nunc) покрывали 10 мкг/мл столбнячного токсина в PBS в течение ночи при 4°C. Затем планшеты блокировали в 5% молоке в PBS, содержащем 0,05% Tween 20, в течение 2 часов. Супернатанты разводили, а затем добавляли в течение 2 часов при комнатной температуре. Планшеты промывали PBS-0,05% Tween20 и связавшееся со столбнячным токсином антитело определяли с использованием конъюгированного с пероксидазой антитела козы против IgG(H+L) человека в количестве 1 мкг/мл в 5% молоке в PBS/0,05% Tween 20. Планшеты проявляли с использованием раствора субстрата TMB (KPL) и измеряли поглощение при 450 нМ с использованием спектрофотометра для чтения микропланшетов Synergy 2 (Biotek). Данные экспортировали в Excel и вычисляли процентную долю ингибирования относительно клеток, культивируемых без тестовых антител. Затем данные импортировали в Graphpad Prism® и строили гистограммы.
На фигуре 17 показано ингибирование продукции IgG против столбнячного токсина PBMC, культивируемыми с VR4447/VR4248 BYbe, VR4447/VR4133 BYbe, VR4447/VR4248/VR645 BYbe/альбумин и VR4447/VR4133/VR645 BYbe/альбумин. Данные представлены для отдельного донора.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCB Pharma
<120> МОЛЕКУЛЫ СО СПЕЦИФИЧНОСТЬЮ К CD45 И CD79
<130> G0225_WO
<150> GB1412659.3
<151> 2014-07-16
<160> 143
<170> PatentIn version 3.5
<210> 1
<211> 44
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность GCN4(7P14P)
<400> 1
Ala Ser Gly Gly Gly Arg Met Lys Gln Leu Glu Pro Lys Val Glu Glu
1. 5 10 15
Leu Leu Pro Lys Asn Tyr His Leu Glu Asn Glu Val Ala Arg Leu Lys
20 25 30
Lys Leu Val Gly Glu Arg His His His His His His
35 40
<210> 2
<211> 132
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность GCN4(7P14P)
<400> 2
gctagcggag gcggaagaat gaaacaactt gaacccaagg ttgaagaatt gcttccgaaa 60
aattatcact tggaaaatga ggttgccaga ttaaagaaat tagttggcga acgccatcac 120
catcaccatc ac 132
<210> 3
<211> 262
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность 52SR4 ds scFv
<400> 3
Asp Ala Val Val Thr Gln Glu Ser Ala Leu Thr Ser Ser Pro Gly Glu
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Ser Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Asn Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Val Leu Trp Tyr Ser Asp
85 90 95
His Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
115 120 125
Gly Ser Asp Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Ala Pro
130 135 140
Ser Gln Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Leu Leu Thr
145 150 155 160
Asp Tyr Gly Val Asn Trp Val Arg Gln Ser Pro Gly Lys Cys Leu Glu
165 170 175
Trp Leu Gly Val Ile Trp Gly Asp Gly Ile Thr Asp Tyr Asn Ser Ala
180 185 190
Leu Lys Ser Arg Leu Ser Val Thr Lys Asp Asn Ser Lys Ser Gln Val
195 200 205
Phe Leu Lys Met Asn Ser Leu Gln Ser Gly Asp Ser Ala Arg Tyr Tyr
210 215 220
Cys Val Thr Gly Leu Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
225 230 235 240
Val Ser Ser Ala Ala Ala His His His His His His Glu Gln Lys Leu
245 250 255
Ile Ser Glu Glu Asp Leu
260
<210> 4
<211> 792
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность 52SR4 ds scFv
<400> 4
gatgcggtgg tgacccagga aagcgcgctg accagcagcc cgggcgaaac cgtgaccctg 60
acctgccgca gcagcaccgg cgcggtgacc accagcaact atgcgagctg ggtgcaggaa 120
aaaccggatc atctgtttac cggcctgatt ggcggcacca acaaccgcgc gccgggcgtg 180
ccggcgcgct ttagcggcag cctgattggc gataaagcgg cgctgaccat taccggcgcg 240
cagaccgaag atgaagcgat ttatttttgc gtgctgtggt atagcgacca ttgggtgttt 300
ggctgcggca ccaaactgac cgtgctgggt ggaggcggtg gctcaggcgg aggtggctca 360
ggcggtggcg ggtctggcgg cggcggcagc gatgtgcagc tgcagcagag cggcccgggc 420
ctggtggcgc cgagccagag cctgagcatt acctgcaccg tgagcggctt tctcctgacc 480
gattatggcg tgaactgggt gcgccagagc ccgggcaaat gcctggaatg gctgggcgtg 540
atttggggcg atggcattac cgattataac agcgcgctga aaagccgcct gagcgtgacc 600
aaagataaca gcaaaagcca ggtgtttctg aaaatgaaca gcctgcagag cggcgatagc 660
gcgcgctatt attgcgtgac cggcctgttt gattattggg gccagggcac caccctgacc 720
gtgagcagcg cggccgccca tcaccatcac catcacgaac agaaactgat tagcgaagaa 780
gatctgtaat ag 792
<210> 5
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 5
Asp Lys Thr His Thr Cys Ala Ala
1. 5
<210> 6
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 6
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1. 5 10
<210> 7
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 7
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Thr Cys Pro Pro Cys
1. 5 10 15
Pro Ala
<210> 8
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 8
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Thr Cys Pro Pro Cys
1. 5 10 15
Pro Ala Thr Cys Pro Pro Cys Pro Ala
20 25
<210> 9
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 9
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Gly Lys Pro Thr Leu
1. 5 10 15
Tyr Asn Ser Leu Val Met Ser Asp Thr Ala Gly Thr Cys Tyr
20 25 30
<210> 10
<211> 31
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 10
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Gly Lys Pro Thr His
1. 5 10 15
Val Asn Val Ser Val Val Met Ala Glu Val Asp Gly Thr Cys Tyr
20 25 30
<210> 11
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 11
Asp Lys Thr His Thr Cys Cys Val Glu Cys Pro Pro Cys Pro Ala
1. 5 10 15
<210> 12
<211> 26
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 12
Asp Lys Thr His Thr Cys Pro Arg Cys Pro Glu Pro Lys Ser Cys Asp
1. 5 10 15
Thr Pro Pro Pro Cys Pro Arg Cys Pro Ala
20 25
<210> 13
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Линкер в шарнирной области
<400> 13
Asp Lys Thr His Thr Cys Pro Ser Cys Pro Ala
1. 5 10
<210> 14
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 14
Ser Gly Gly Gly Gly Ser Glu
1. 5
<210> 15
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 15
Asp Lys Thr His Thr Ser
1. 5
<210> 16
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 16
Ser Gly Gly Gly Gly Ser
1. 5
<210> 17
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 17
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10
<210> 18
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 18
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10 15
<210> 19
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 19
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10 15
Gly Gly Gly Gly Ser
20
<210> 20
<211> 26
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 20
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10 15
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 21
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 21
Ala Ala Ala Gly Ser Gly Gly Ala Ser Ala Ser
1. 5 10
<210> 22
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может являться встречающейся в природе аминокислотой
<400> 22
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
1. 5 10 15
<210> 23
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может являться встречающейся в природе аминокислотой
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa может являться встречающейся в природе аминокислотой
<400> 23
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1. 5 10 15
Gly Ala Ser Ala Ser
20
<210> 24
<211> 26
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может являться встречающейся в природе аминокислотой
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa может являться встречающейся в природе аминокислотой
<220>
<221> misc_feature
<222> (17)..(17)
<223> Xaa может являться встречающейся в природе аминокислотой
<400> 24
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1. 5 10 15
Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
20 25
<210> 25
<211> 31
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может являться встречающейся в природе аминокислотой
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa может являться встречающейся в природе аминокислотой
<220>
<221> misc_feature
<222> (17)..(17)
<223> Xaa может являться встречающейся в природе аминокислотой
<220>
<221> misc_feature
<222> (22)..(22)
<223> Xaa может являться встречающейся в природе аминокислотой
<400> 25
Ala Ala Ala Gly Ser Gly Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser
1. 5 10 15
Xaa Gly Gly Gly Ser Xaa Gly Gly Gly Ser Gly Ala Ser Ala Ser
20 25 30
<210> 26
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может являться встречающейся в природе аминокислотой
<400> 26
Ala Ala Ala Gly Ser Gly Xaa Ser Gly Ala Ser Ala Ser
1. 5 10
<210> 27
<211> 28
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 27
Pro Gly Gly Asn Arg Gly Thr Thr Thr Thr Arg Arg Pro Ala Thr Thr
1. 5 10 15
Thr Gly Ser Ser Pro Gly Pro Thr Gln Ser His Tyr
20 25
<210> 28
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 28
Ala Thr Thr Thr Gly Ser Ser Pro Gly Pro Thr
1. 5 10
<210> 29
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 29
Ala Thr Thr Thr Gly Ser
1. 5
<210> 30
<211> 0
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 30
000
<210> 31
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 31
Glu Pro Ser Gly Pro Ile Ser Thr Ile Asn Ser Pro Pro Ser Lys Glu
1. 5 10 15
Ser His Lys Ser Pro
20
<210> 32
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 32
Gly Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
1. 5 10 15
<210> 33
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 33
Gly Gly Gly Gly Ile Ala Pro Ser Met Val Gly Gly Gly Gly Ser
1. 5 10 15
<210> 34
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 34
Gly Gly Gly Gly Lys Val Glu Gly Ala Gly Gly Gly Gly Gly Ser
1. 5 10 15
<210> 35
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 35
Gly Gly Gly Gly Ser Met Lys Ser His Asp Gly Gly Gly Gly Ser
1. 5 10 15
<210> 36
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 36
Gly Gly Gly Gly Asn Leu Ile Thr Ile Val Gly Gly Gly Gly Ser
1. 5 10 15
<210> 37
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 37
Gly Gly Gly Gly Val Val Pro Ser Leu Pro Gly Gly Gly Gly Ser
1. 5 10 15
<210> 38
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 38
Gly Gly Glu Lys Ser Ile Pro Gly Gly Gly Gly Ser
1. 5 10
<210> 39
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 39
Arg Pro Leu Ser Tyr Arg Pro Pro Phe Pro Phe Gly Phe Pro Ser Val
1. 5 10 15
Arg Pro
<210> 40
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 40
Tyr Pro Arg Ser Ile Tyr Ile Arg Arg Arg His Pro Ser Pro Ser Leu
1. 5 10 15
Thr Thr
<210> 41
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 41
Thr Pro Ser His Leu Ser His Ile Leu Pro Ser Phe Gly Leu Pro Thr
1. 5 10 15
Phe Asn
<210> 42
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 42
Arg Pro Val Ser Pro Phe Thr Phe Pro Arg Leu Ser Asn Ser Trp Leu
1. 5 10 15
Pro Ala
<210> 43
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 43
Ser Pro Ala Ala His Phe Pro Arg Ser Ile Pro Arg Pro Gly Pro Ile
1. 5 10 15
Arg Thr
<210> 44
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 44
Ala Pro Gly Pro Ser Ala Pro Ser His Arg Ser Leu Pro Ser Arg Ala
1. 5 10 15
Phe Gly
<210> 45
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 45
Pro Arg Asn Ser Ile His Phe Leu His Pro Leu Leu Val Ala Pro Leu
1. 5 10 15
Gly Ala
<210> 46
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 46
Met Pro Ser Leu Ser Gly Val Leu Gln Val Arg Tyr Leu Ser Pro Pro
1. 5 10 15
Asp Leu
<210> 47
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 47
Ser Pro Gln Tyr Pro Ser Pro Leu Thr Leu Thr Leu Pro Pro His Pro
1. 5 10 15
Ser Leu
<210> 48
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 48
Asn Pro Ser Leu Asn Pro Pro Ser Tyr Leu His Arg Ala Pro Ser Arg
1. 5 10 15
Ile Ser
<210> 49
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 49
Leu Pro Trp Arg Thr Ser Leu Leu Pro Ser Leu Pro Leu Arg Arg Arg
1. 5 10 15
Pro
<210> 50
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 50
Pro Pro Leu Phe Ala Lys Gly Pro Val Gly Leu Leu Ser Arg Ser Phe
1. 5 10 15
Pro Pro
<210> 51
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 51
Val Pro Pro Ala Pro Val Val Ser Leu Arg Ser Ala His Ala Arg Pro
1. 5 10 15
Pro Tyr
<210> 52
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 52
Leu Arg Pro Thr Pro Pro Arg Val Arg Ser Tyr Thr Cys Cys Pro Thr
1. 5 10 15
Pro
<210> 53
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 53
Pro Asn Val Ala His Val Leu Pro Leu Leu Thr Val Pro Trp Asp Asn
1. 5 10 15
Leu Arg
<210> 54
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Гибкий линкер
<400> 54
Cys Asn Pro Leu Leu Pro Leu Cys Ala Arg Ser Pro Ala Val Arg Thr
1. 5 10 15
Phe Pro
<210> 55
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 55
Asp Leu Cys Leu Arg Asp Trp Gly Cys Leu Trp
1. 5 10
<210> 56
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 56
Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1. 5 10
<210> 57
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 57
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Gly Asp
1. 5 10 15
<210> 58
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 58
Gln Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1. 5 10 15
Glu Asp Asp Glu
20
<210> 59
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 59
Gln Gly Leu Ile Gly Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1. 5 10 15
Gly Arg Ser Val
20
<210> 60
<211> 21
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 60
Gln Gly Leu Ile Gly Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp
1. 5 10 15
Gly Arg Ser Val Lys
20
<210> 61
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 61
Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp Asp
1. 5 10 15
<210> 62
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 62
Arg Leu Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu
1. 5 10 15
Asp Asp
<210> 63
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 63
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp Asp
1. 5 10 15
<210> 64
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 64
Met Glu Asp Ile Cys Leu Pro Arg Trp Gly Cys Leu Trp Glu Asp
1. 5 10 15
<210> 65
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 65
Arg Leu Met Glu Asp Ile Cys Leu Ala Arg Trp Gly Cys Leu Trp Glu
1. 5 10 15
Asp Asp
<210> 66
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 66
Glu Val Arg Ser Phe Cys Thr Arg Trp Pro Ala Glu Lys Ser Cys Lys
1. 5 10 15
Pro Leu Arg Gly
20
<210> 67
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 67
Arg Ala Pro Glu Ser Phe Val Cys Tyr Trp Glu Thr Ile Cys Phe Glu
1. 5 10 15
Arg Ser Glu Gln
20
<210> 68
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Альбумин-связывающий пептид
<400> 68
Glu Met Cys Tyr Phe Pro Gly Ile Cys Trp Met
1. 5 10
<210> 69
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Жесткие линкеры
<400> 69
Gly Ala Pro Ala Pro Ala Ala Pro Ala Pro Ala
1. 5 10
<210> 70
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Жесткий линкер
<400> 70
Pro Pro Pro Pro
1
<210> 71
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4447 кролика
<400> 71
Ala Gln Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Pro Val Gly
1. 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Val Ser Gly
20 25 30
Asn Tyr Leu Ala Trp Leu Gln Gln Lys Pro Gly Gln Pro Pro Lys Gln
35 40 45
Leu Ile His Ser Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe
50 55 60
Ser Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Glu Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Glu Phe Ser Cys
85 90 95
Ser Ser His Asp Cys Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val
100 105 110
Lys
<210> 72
<211> 339
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4447 кролика
<400> 72
gcccaagtgc tgacccagac tccgtcccct gtgtctgcac ctgtgggagg cacagtcacc 60
atcaattgcc aggccagtca gagtgttgtt agtggcaatt acctagcctg gcttcagcag 120
aaaccagggc agcctcccaa gcaactgatc cattctgcat ccactctggc atctggggtc 180
tcatcgcggt tcagcggcag tggatctggg acacaattca ctctcaccat cagcggcgtg 240
cagtgtgaag atgctgccac ttactactgt ctaggcgaat ttagttgtag tagtcatgat 300
tgtaatgctt tcggcggagg gaccgaggtg gtggtcaaa 339
<210> 73
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4447 кролика
<400> 73
Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1. 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Asn Tyr Ala
20 25 30
Val Ser Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Tyr Ile Glu Thr Gly Thr Thr Trp Tyr Ala Asn Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Thr Ile
65 70 75 80
Thr Ser Pro Ser Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Glu
85 90 95
Pro Tyr Glu Pro Tyr Asp Asp Ser Asn Ile Tyr Tyr Gly Met Asp Pro
100 105 110
Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 74
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4447 кролика
<400> 74
cagtcgctgg aggagtccgg gggtcgcctg gtcacgcctg ggacacccct gacactcacc 60
tgcaccgtct ctggattctc cctcagtaac tatgcagtaa gctgggtccg ccaggctcca 120
ggggagggac tggaatggat cgggatcatt tatattgaaa ctggtaccac atggtacgcg 180
aactgggcga aaggccgatt caccatctcc aaaacctcga ccacggtgga tctgacaatc 240
accagtccgt caaccgagga cacggccacc tatttctgtg ccagagaacc ttatgaacct 300
tatgatgata gtaatattta ctacggcatg gacccctggg gcccaggcac cctcgtcacc 360
gtctcgagt 369
<210> 75
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 75
Gln Ala Ser Gln Ser Val Val Ser Gly Asn Tyr Leu Ala
1. 5 10
<210> 76
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 76
Ser Ala Ser Thr Leu Ala Ser
1. 5
<210> 77
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 77
Leu Gly Glu Phe Ser Cys Ser Ser His Asp Cys Asn Ala
1. 5 10
<210> 78
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 78
Gly Phe Ser Leu Ser Asn Tyr Ala Val Ser
1. 5 10
<210> 79
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 79
Ile Ile Tyr Ile Glu Thr Gly Thr Thr Trp Tyr Ala Asn Trp Ala Lys
1. 5 10 15
Gly
<210> 80
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 80
Glu Pro Tyr Glu Pro Tyr Asp Asp Ser Asn Ile Tyr Tyr Gly Met Asp
1. 5 10 15
Pro
<210> 81
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4450 кролика
<400> 81
Ala Ile Asp Met Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1. 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Ile Tyr Asn Asn
20 25 30
Asn Asp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Glu Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Gly Gly Ser Gly
85 90 95
Gly Asp Gly Ile Ala Phe Gly Gly Gly Thr Lys Val Val Val Glu
100 105 110
<210> 82
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4450 кролика
<400> 82
gccattgata tgacccagac tccatccccc gtgtctgcag ctgtgggagg cacagtcacc 60
atcaattgcc agtccagtca gagtatttat aataataatg acttagcctg gtatcagcag 120
aaaccagggc agcctcccaa gctcctgatc tacgaagcat ccaaactggc atctggggtc 180
ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagtggcgtg 240
cagtgtgatg atgctgccac ttactactgt cagggcggtg gtagtggtgg tgatggcatt 300
gctttcggcg gagggaccaa ggtggtcgtc gaa 333
<210> 83
<211> 117
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4450 кролика
<400> 83
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Ala Pro
1. 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Asn Asn Tyr Val
20 25 30
Met Val Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
35 40 45
Ile Ile Tyr Val Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Val Thr
65 70 75 80
Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Asp Ala
85 90 95
Gly His Ser Asp Val Asp Val Leu Asp Ile Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 84
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4450 кролика
<400> 84
cagtcggtgg aggagtccgg gggtcgcctg gtcacgcctg gggcacccct gacactcacc 60
tgcacagtct ctggattctc cctcaataac tatgtaatgg tctgggtccg ccaggctcca 120
gggaaggggc tggaatggat cggaatcatt tatgttagtg gtaatgcata ctacgcgagc 180
tgggcaaaag gccgattcac catctccaga acctcgacca cggtggatct gaaagtgacc 240
agtctgacaa ccgaggacac ggccacctat ttctgtgcca gagatgctgg tcatagtgat 300
gtcgatgttt tggatatttg gggcccgggc accctcgtca ccgtctcgag t 351
<210> 85
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 85
Gln Ser Ser Gln Ser Ile Tyr Asn Asn Asn Asp Leu Ala
1. 5 10
<210> 86
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 86
Glu Ala Ser Lys Leu Ala Ser
1. 5
<210> 87
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 87
Gln Gly Gly Gly Ser Gly Gly Asp Gly Ile Ala
1. 5 10
<210> 88
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 88
Gly Phe Ser Leu Asn Asn Tyr Val Met Val
1. 5 10
<210> 89
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 89
Ile Ile Tyr Val Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys Gly
1. 5 10 15
<210> 90
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 90
Asp Ala Gly His Ser Asp Val Asp Val Leu Asp Ile
1. 5 10
<210> 91
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4122 кролика
<400> 91
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Ser Glu Pro Val Gly
1. 5 10 15
Gly Thr Val Thr Ile Met Cys Gln Ala Ser Gln Ser Ile Ser Asn Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Glu Tyr Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Tyr Tyr Asp Ser Gly Ser
85 90 95
Asn Val Phe Phe Ala Phe Gly Gly Gly Thr Lys Val Val Val Glu
100 105 110
<210> 92
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4122 кролика
<400> 92
gacattgtga tgacccagac tccagcctcc gtgtctgaac ctgtgggagg cacagtcacc 60
atcatgtgcc aggccagtca gagcattagc aattggttag cctggtatca acagaaacca 120
gggcagcctc ccaagctcct gatctaccag gcatccaaac tggcatctgg ggtcccatcg 180
cggttcaaag gcagtggatc tgggacagag tacactctca ccatcagcga cctggagtgt 240
gccgatgctg ccacttacta ctgtcaaagc tattatgata gtggtagtaa tgtttttttt 300
gctttcggcg gagggaccaa ggtggtggtc gaa 333
<210> 93
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4122 кролика
<400> 93
Leu Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1. 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Ala Gly Tyr
20 25 30
Trp Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Thr Tyr Ala Gly Arg Ser Gly Ser Thr Tyr Tyr Ala Asn Trp
50 55 60
Val Asn Gly Arg Phe Thr Ile Pro Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Gln Met Thr Ser Leu Ser Gly Ala Asp Thr Ala Ser Tyr Phe Cys
85 90 95
Ala Arg Gly Asn Ala Gly Val Ala Val Gly Ala Leu Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 94
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4122 кролика
<400> 94
ctgtcgttgg aggagtccgg gggagacctg gtcaagcctg gggcatccct gacactcacc 60
tgcacagcct ctggattctc cttcagtgcc ggctattgga tatgttgggt ccgccaggct 120
ccagggaagg ggctggagtg gatcgcatgc acttatgctg gtcgtagtgg tagcacttac 180
tacgcgaact gggtgaatgg ccgattcacc atccccaaaa cctcgtcgac cacggtgact 240
ctgcaaatga ccagtctgtc aggcgcggac acggccagct atttctgtgc gagaggtaat 300
gctggtgttg ctgttggtgc cttgtggggc ccaggcaccc tggtcaccgt ctcgagt 357
<210> 95
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 95
Gln Ala Ser Gln Ser Ile Ser Asn Trp Leu Ala
1. 5 10
<210> 96
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 96
Gln Ala Ser Lys Leu Ala Ser
1. 5
<210> 97
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 97
Gln Ser Tyr Tyr Asp Ser Gly Ser Asn Val Phe Phe Ala
1. 5 10
<210> 98
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 98
Gly Phe Ser Phe Ser Ala Gly Tyr Trp Ile Cys
1. 5 10
<210> 99
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 99
Cys Thr Tyr Ala Gly Arg Ser Gly Ser Thr Tyr Tyr Ala Asn Trp Val
1. 5 10 15
Asn Gly
<210> 100
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 100
Gly Asn Ala Gly Val Ala Val Gly Ala Leu
1. 5 10
<210> 101
<211> 111
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4129 кролика
<400> 101
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Glu Ala Ala Val Gly
1. 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Ser Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Thr Tyr Tyr Cys Gln Ser Tyr Tyr Asp Ser Gly Ser
85 90 95
Ser Val Phe Phe Asn Phe Gly Gly Gly Thr Lys Val Val Val Lys
100 105 110
<210> 102
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4129 кролика
<400> 102
gacattgtga tgacccagac tccagcctcc gtggaggcag ctgtgggagg cacagtcacc 60
atcaattgcc aagccagtca gagcattagc agttggttat cctggtatca gcagaaacca 120
gggcagcctc ccaagctcct gatctatggt gcatccaatc tggcatctgg ggtcccatca 180
cggttcagcg gcagtggatc tgggacacag ttcagtctca ccatcagcga cctggagtgt 240
gccgatgctg ccacttacta ctgtcaaagc tattatgata gtggtagtag tgtttttttt 300
aatttcggcg gagggaccaa ggtggtcgtc aaa 333
<210> 103
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4129 кролика
<400> 103
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1. 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Ala Gly Tyr
20 25 30
Trp Ile Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp
50 55 60
Ala Lys Gly Arg Phe Thr Ile Pro Lys Thr Ser Ser Thr Thr Val Thr
65 70 75 80
Leu Gln Met Thr Ser Leu Thr Gly Ala Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Gly Asn Ala Gly Val Ala Val Gly Ala Leu Trp Gly Pro Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 104
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4129 кролика
<400> 104
cagtcgttgg aggagtccgg gggagacctg gttaagcctg gggcatccct gacactcacc 60
tgcacagcct ctggattctc cttcagtgcc ggctattgga tatgttgggt ccgccaggct 120
ccagggaagg ggctggagtg gatcgcatgc atttatgctg gtagtagtgg tagcacttac 180
tacgcgagct gggcgaaagg ccgattcacc atccccaaaa cctcgtcgac cacggtgact 240
ctgcaaatga ccagtctgac aggcgcggac acggccacct atttctgtgc gagaggtaat 300
gctggtgttg ctgttggtgc cttgtggggc ccaggcaccc tcgtcaccgt ctcgagt 357
<210> 105
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 105
Gln Ala Ser Gln Ser Ile Ser Ser Trp Leu Ser
1. 5 10
<210> 106
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 106
Gly Ala Ser Asn Leu Ala Ser
1. 5
<210> 107
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 107
Gln Ser Tyr Tyr Asp Ser Gly Ser Ser Val Phe Phe Asn
1. 5 10
<210> 108
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 108
Gly Phe Ser Phe Ser Ala Gly Tyr Trp Ile Cys
1. 5 10
<210> 109
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 109
Cys Ile Tyr Ala Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala
1. 5 10 15
Lys Gly
<210> 110
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 110
Gly Asn Ala Gly Val Ala Val Gly Ala Leu
1. 5 10
<210> 111
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4131 кролика
<400> 111
Asp Ile Val Met Thr Gln Thr Pro Ala Ser Val Ser Glu Pro Val Gly
1. 5 10 15
Gly Ser Val Thr Ile Lys Cys Gln Ala Ser Gln Ser Phe Tyr Asn Leu
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asp Leu Ala Ser Gly Val Pro Ser Arg Phe Lys Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asp Leu Glu Cys
65 70 75 80
Ala Asp Ala Ala Ala Tyr Tyr Cys Gln Ser Ala Asp Gly Ser Ser Tyr
85 90 95
Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105
<210> 112
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4131 кролика
<400> 112
gacattgtga tgacccagac tccagcctcc gtgtctgaac ctgtgggagg ctcagtcacc 60
atcaagtgcc aggccagtca gagcttttac aacctcttag cctggtatca gcagaaacca 120
gggcagcctc ccaaactcct gatctatgat gcatccgatc tggcatctgg ggtcccatcg 180
cggttcaaag gcagtggatc tgggactgat ttcactctca ccatcagcga cctggagtgt 240
gccgatgctg ccgcttacta ctgtcaaagt gctgatggta gtagttacgc tttcggcgga 300
gggaccgagg tggtcgtcaa a 321
<210> 113
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4131 кролика
<400> 113
Gln Glu Gln Leu Glu Glu Ser Gly Gly Gly Leu Val Lys Pro Glu Gly
1. 5 10 15
Ser Leu Thr Leu Thr Cys Thr Ala Ser Gly Val Ser Phe Ser Ser Ser
20 25 30
Tyr Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Ala Cys Ile Tyr Thr Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Val Ser Glu Thr Ser Ser Thr Thr Val
65 70 75 80
Thr Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe
85 90 95
Cys Ala Arg Ala Ser Ala Trp Thr Tyr Gly Met Asp Leu Trp Gly Pro
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 114
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4131 кролика
<400> 114
caggagcaat tggaggagtc cgggggaggc ctggtcaagc ctgagggatc cctgacactc 60
acctgcacag cctctggagt ctccttcagt agcagctatt ggatatactg ggtccgccag 120
gctccaggga aggggctgga gtggatcgca tgcatttata ctggtagtag tggtagcact 180
tactacgcga gctgggcgaa aggccgattc accgtctccg aaacctcgtc gaccacggtg 240
actctgcaaa tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagagca 300
agcgcttgga cctacggcat ggacctctgg ggcccgggca ccctcgtcac cgtctcgagt 360
<210> 115
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 115
Gln Ala Ser Gln Ser Phe Tyr Asn Leu Leu Ala
1. 5 10
<210> 116
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 116
Asp Ala Ser Asp Leu Ala Ser
1. 5
<210> 117
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 117
Gln Ser Ala Asp Gly Ser Ser Tyr Ala
1. 5
<210> 118
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 118
Gly Val Ser Phe Ser Ser Ser Tyr Trp Ile Tyr
1. 5 10
<210> 119
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 119
Cys Ile Tyr Thr Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala
1. 5 10 15
Lys Gly
<210> 120
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 120
Ala Ser Ala Trp Thr Tyr Gly Met Asp Leu
1. 5 10
<210> 121
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4133 кролика
<400> 121
Ala Gln Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Val Val Gly
1. 5 10 15
Gly Thr Val Ser Ile Ser Cys Gln Ala Ser Gln Ser Val Tyr Asn Asn
20 25 30
Asn Asn Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Asp Ala Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Gly Tyr Tyr Ser
85 90 95
Ser Gly Trp Tyr Phe Ala Phe Gly Gly Gly Thr Lys Val Val Val Lys
100 105 110
<210> 122
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VL-область Ab 4133 кролика
<400> 122
gcgcaagtgc tgacccagac tccatctccc gtgtctgcag ttgtgggagg cacagtcagc 60
atcagttgcc aggccagtca gagtgtttat aataacaaca acttatcctg gtatcagcag 120
aaaccagggc agcctcccaa gctcttgatc tacgatgcat ccaaattggc atctggggtc 180
ccatcccggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagcggcgtg 240
cagtgtgacg atgctgccac ttactactgt ctaggcggtt attatagtag tggttggtat 300
tttgctttcg gcggagggac caaggtggtg gtcaaa 336
<210> 123
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4133 кролика
<400> 123
Gln Glu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Glu Gly
1. 5 10 15
Ser Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Gly Asn
20 25 30
Tyr Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Cys Leu Tyr Thr Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val
65 70 75 80
Thr Leu Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe
85 90 95
Cys Ala Arg Asp Leu Gly Tyr Glu Ile Asp Gly Tyr Gly Gly Leu Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VH-область Ab 4133 кролика
<400> 124
caggagcagc tggtggagtc cgggggaggc ctggtccagc ctgagggatc cctgacacta 60
acctgcacag cttctggatt ctccttcagt ggcaactact acatgtgctg ggtccgccag 120
gctccaggga aggggctgga gtggatcgga tgcctttata ctggtagtag tggtagcaca 180
tattacgcga gctgggcgaa aggccgattc accatctcca aaacctcgtc gaccacggtg 240
actctgcaaa tgaccagtct gacagccgcg gacacggcca cctatttctg tgcgagagat 300
ctaggttatg aaattgatgg ttatgggggc ttgtggggcc agggcaccct cgtcaccgtc 360
tcgagt 366
<210> 125
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1
<400> 125
Gln Ala Ser Gln Ser Val Tyr Asn Asn Asn Asn Leu Ser
1. 5 10
<210> 126
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2
<400> 126
Asp Ala Ser Lys Leu Ala Ser
1. 5
<210> 127
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3
<400> 127
Leu Gly Gly Tyr Tyr Ser Ser Gly Trp Tyr Phe Ala
1. 5 10
<210> 128
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1
<400> 128
Gly Phe Ser Phe Ser Gly Asn Tyr Tyr Met Cys
1. 5 10
<210> 129
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2
<400> 129
Cys Leu Tyr Thr Gly Ser Ser Gly Ser Thr Tyr Tyr Ala Ser Trp Ala
1. 5 10 15
Lys Gly
<210> 130
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3
<400> 130
Asp Leu Gly Tyr Glu Ile Asp Gly Tyr Gly Gly Leu
1. 5 10
<210> 131
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH1 dAbH1
<400> 131
Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
1. 5 10
<210> 132
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH2 dAbH1
<400> 132
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
1. 5 10 15
<210> 133
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRH3 dAbH1
<400> 133
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu
1. 5 10
<210> 134
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL1 dAbL1
<400> 134
Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
1. 5 10
<210> 135
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL2 dAbL1
<400> 135
Glu Ala Ser Lys Leu Thr Ser
1. 5
<210> 136
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> CDRL3 dAbL1
<400> 136
Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
1. 5 10
<210> 137
<211> 121
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи антитела без альбумина (без ds)
<400> 137
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 138
<211> 121
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи антитела без альбумина (ds)
<400> 138
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 139
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи антитела без альбумина (без ds)
<400> 139
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
100 105 110
<210> 140
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи антитела без альбумина (ds)
<400> 140
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Cys Gly Thr Tyr Val Glu Ile Lys Arg Thr
100 105 110
<210> 141
<211> 226
<212> PRT
<213> Искусственная последовательность
<220>
<223> CD79a человека
<400> 141
Met Pro Gly Gly Pro Gly Val Leu Gln Ala Leu Pro Ala Thr Ile Phe
1. 5 10 15
Leu Leu Phe Leu Leu Ser Ala Val Tyr Leu Gly Pro Gly Cys Gln Ala
20 25 30
Leu Trp Met His Lys Val Pro Ala Ser Leu Met Val Ser Leu Gly Glu
35 40 45
Asp Ala His Phe Gln Cys Pro His Asn Ser Ser Asn Asn Ala Asn Val
50 55 60
Thr Trp Trp Arg Val Leu His Gly Asn Tyr Thr Trp Pro Pro Glu Phe
65 70 75 80
Leu Gly Pro Gly Glu Asp Pro Asn Gly Thr Leu Ile Ile Gln Asn Val
85 90 95
Asn Lys Ser His Gly Gly Ile Tyr Val Cys Arg Val Gln Glu Gly Asn
100 105 110
Glu Ser Tyr Gln Gln Ser Cys Gly Thr Tyr Leu Arg Val Arg Gln Pro
115 120 125
Pro Pro Arg Pro Phe Leu Asp Met Gly Glu Gly Thr Lys Asn Arg Ile
130 135 140
Ile Thr Ala Glu Gly Ile Ile Leu Leu Phe Cys Ala Val Val Pro Gly
145 150 155 160
Thr Leu Leu Leu Phe Arg Lys Arg Trp Gln Asn Glu Lys Leu Gly Leu
165 170 175
Asp Ala Gly Asp Glu Tyr Glu Asp Glu Asn Leu Tyr Glu Gly Leu Asn
180 185 190
Leu Asp Asp Cys Ser Met Tyr Glu Asp Ile Ser Arg Gly Leu Gln Gly
195 200 205
Thr Tyr Gln Asp Val Gly Ser Leu Asn Ile Gly Asp Val Gln Leu Glu
210 215 220
Lys Pro
225
<210> 142
<211> 229
<212> PRT
<213> Искусственная последовательность
<220>
<223> CD79b человека
<400> 142
Met Ala Arg Leu Ala Leu Ser Pro Val Pro Ser His Trp Met Val Ala
1. 5 10 15
Leu Leu Leu Leu Leu Ser Ala Glu Pro Val Pro Ala Ala Arg Ser Glu
20 25 30
Asp Arg Tyr Arg Asn Pro Lys Gly Ser Ala Cys Ser Arg Ile Trp Gln
35 40 45
Ser Pro Arg Phe Ile Ala Arg Lys Arg Gly Phe Thr Val Lys Met His
50 55 60
Cys Tyr Met Asn Ser Ala Ser Gly Asn Val Ser Trp Leu Trp Lys Gln
65 70 75 80
Glu Met Asp Glu Asn Pro Gln Gln Leu Lys Leu Glu Lys Gly Arg Met
85 90 95
Glu Glu Ser Gln Asn Glu Ser Leu Ala Thr Leu Thr Ile Gln Gly Ile
100 105 110
Arg Phe Glu Asp Asn Gly Ile Tyr Phe Cys Gln Gln Lys Cys Asn Asn
115 120 125
Thr Ser Glu Val Tyr Gln Gly Cys Gly Thr Glu Leu Arg Val Met Gly
130 135 140
Phe Ser Thr Leu Ala Gln Leu Lys Gln Arg Asn Thr Leu Lys Asp Gly
145 150 155 160
Ile Ile Met Ile Gln Thr Leu Leu Ile Ile Leu Phe Ile Ile Val Pro
165 170 175
Ile Phe Leu Leu Leu Asp Lys Asp Asp Ser Lys Ala Gly Met Glu Glu
180 185 190
Asp His Thr Tyr Glu Gly Leu Asp Ile Asp Gln Thr Ala Thr Tyr Glu
195 200 205
Asp Ile Val Thr Leu Arg Thr Gly Glu Val Lys Trp Ser Val Gly Glu
210 215 220
His Pro Gly Gln Glu
225
<210> 143
<211> 1304
<212> PRT
<213> Искусственная последовательность
<220>
<223> CD45 человека
<400> 143
Met Tyr Leu Trp Leu Lys Leu Leu Ala Phe Gly Phe Ala Phe Leu Asp
1. 5 10 15
Thr Glu Val Phe Val Thr Gly Gln Ser Pro Thr Pro Ser Pro Thr Gly
20 25 30
Leu Thr Thr Ala Lys Met Pro Ser Val Pro Leu Ser Ser Asp Pro Leu
35 40 45
Pro Thr His Thr Thr Ala Phe Ser Pro Ala Ser Thr Phe Glu Arg Glu
50 55 60
Asn Asp Phe Ser Glu Thr Thr Thr Ser Leu Ser Pro Asp Asn Thr Ser
65 70 75 80
Thr Gln Val Ser Pro Asp Ser Leu Asp Asn Ala Ser Ala Phe Asn Thr
85 90 95
Thr Gly Val Ser Ser Val Gln Thr Pro His Leu Pro Thr His Ala Asp
100 105 110
Ser Gln Thr Pro Ser Ala Gly Thr Asp Thr Gln Thr Phe Ser Gly Ser
115 120 125
Ala Ala Asn Ala Lys Leu Asn Pro Thr Pro Gly Ser Asn Ala Ile Ser
130 135 140
Asp Val Pro Gly Glu Arg Ser Thr Ala Ser Thr Phe Pro Thr Asp Pro
145 150 155 160
Val Ser Pro Leu Thr Thr Thr Leu Ser Leu Ala His His Ser Ser Ala
165 170 175
Ala Leu Pro Ala Arg Thr Ser Asn Thr Thr Ile Thr Ala Asn Thr Ser
180 185 190
Asp Ala Tyr Leu Asn Ala Ser Glu Thr Thr Thr Leu Ser Pro Ser Gly
195 200 205
Ser Ala Val Ile Ser Thr Thr Thr Ile Ala Thr Thr Pro Ser Lys Pro
210 215 220
Thr Cys Asp Glu Lys Tyr Ala Asn Ile Thr Val Asp Tyr Leu Tyr Asn
225 230 235 240
Lys Glu Thr Lys Leu Phe Thr Ala Lys Leu Asn Val Asn Glu Asn Val
245 250 255
Glu Cys Gly Asn Asn Thr Cys Thr Asn Asn Glu Val His Asn Leu Thr
260 265 270
Glu Cys Lys Asn Ala Ser Val Ser Ile Ser His Asn Ser Cys Thr Ala
275 280 285
Pro Asp Lys Thr Leu Ile Leu Asp Val Pro Pro Gly Val Glu Lys Phe
290 295 300
Gln Leu His Asp Cys Thr Gln Val Glu Lys Ala Asp Thr Thr Ile Cys
305 310 315 320
Leu Lys Trp Lys Asn Ile Glu Thr Phe Thr Cys Asp Thr Gln Asn Ile
325 330 335
Thr Tyr Arg Phe Gln Cys Gly Asn Met Ile Phe Asp Asn Lys Glu Ile
340 345 350
Lys Leu Glu Asn Leu Glu Pro Glu His Glu Tyr Lys Cys Asp Ser Glu
355 360 365
Ile Leu Tyr Asn Asn His Lys Phe Thr Asn Ala Ser Lys Ile Ile Lys
370 375 380
Thr Asp Phe Gly Ser Pro Gly Glu Pro Gln Ile Ile Phe Cys Arg Ser
385 390 395 400
Glu Ala Ala His Gln Gly Val Ile Thr Trp Asn Pro Pro Gln Arg Ser
405 410 415
Phe His Asn Phe Thr Leu Cys Tyr Ile Lys Glu Thr Glu Lys Asp Cys
420 425 430
Leu Asn Leu Asp Lys Asn Leu Ile Lys Tyr Asp Leu Gln Asn Leu Lys
435 440 445
Pro Tyr Thr Lys Tyr Val Leu Ser Leu His Ala Tyr Ile Ile Ala Lys
450 455 460
Val Gln Arg Asn Gly Ser Ala Ala Met Cys His Phe Thr Thr Lys Ser
465 470 475 480
Ala Pro Pro Ser Gln Val Trp Asn Met Thr Val Ser Met Thr Ser Asp
485 490 495
Asn Ser Met His Val Lys Cys Arg Pro Pro Arg Asp Arg Asn Gly Pro
500 505 510
His Glu Arg Tyr His Leu Glu Val Glu Ala Gly Asn Thr Leu Val Arg
515 520 525
Asn Glu Ser His Lys Asn Cys Asp Phe Arg Val Lys Asp Leu Gln Tyr
530 535 540
Ser Thr Asp Tyr Thr Phe Lys Ala Tyr Phe His Asn Gly Asp Tyr Pro
545 550 555 560
Gly Glu Pro Phe Ile Leu His His Ser Thr Ser Tyr Asn Ser Lys Ala
565 570 575
Leu Ile Ala Phe Leu Ala Phe Leu Ile Ile Val Thr Ser Ile Ala Leu
580 585 590
Leu Val Val Leu Tyr Lys Ile Tyr Asp Leu His Lys Lys Arg Ser Cys
595 600 605
Asn Leu Asp Glu Gln Gln Glu Leu Val Glu Arg Asp Asp Glu Lys Gln
610 615 620
Leu Met Asn Val Glu Pro Ile His Ala Asp Ile Leu Leu Glu Thr Tyr
625 630 635 640
Lys Arg Lys Ile Ala Asp Glu Gly Arg Leu Phe Leu Ala Glu Phe Gln
645 650 655
Ser Ile Pro Arg Val Phe Ser Lys Phe Pro Ile Lys Glu Ala Arg Lys
660 665 670
Pro Phe Asn Gln Asn Lys Asn Arg Tyr Val Asp Ile Leu Pro Tyr Asp
675 680 685
Tyr Asn Arg Val Glu Leu Ser Glu Ile Asn Gly Asp Ala Gly Ser Asn
690 695 700
Tyr Ile Asn Ala Ser Tyr Ile Asp Gly Phe Lys Glu Pro Arg Lys Tyr
705 710 715 720
Ile Ala Ala Gln Gly Pro Arg Asp Glu Thr Val Asp Asp Phe Trp Arg
725 730 735
Met Ile Trp Glu Gln Lys Ala Thr Val Ile Val Met Val Thr Arg Cys
740 745 750
Glu Glu Gly Asn Arg Asn Lys Cys Ala Glu Tyr Trp Pro Ser Met Glu
755 760 765
Glu Gly Thr Arg Ala Phe Gly Asp Val Val Val Lys Ile Asn Gln His
770 775 780
Lys Arg Cys Pro Asp Tyr Ile Ile Gln Lys Leu Asn Ile Val Asn Lys
785 790 795 800
Lys Glu Lys Ala Thr Gly Arg Glu Val Thr His Ile Gln Phe Thr Ser
805 810 815
Trp Pro Asp His Gly Val Pro Glu Asp Pro His Leu Leu Leu Lys Leu
820 825 830
Arg Arg Arg Val Asn Ala Phe Ser Asn Phe Phe Ser Gly Pro Ile Val
835 840 845
Val His Cys Ser Ala Gly Val Gly Arg Thr Gly Thr Tyr Ile Gly Ile
850 855 860
Asp Ala Met Leu Glu Gly Leu Glu Ala Glu Asn Lys Val Asp Val Tyr
865 870 875 880
Gly Tyr Val Val Lys Leu Arg Arg Gln Arg Cys Leu Met Val Gln Val
885 890 895
Glu Ala Gln Tyr Ile Leu Ile His Gln Ala Leu Val Glu Tyr Asn Gln
900 905 910
Phe Gly Glu Thr Glu Val Asn Leu Ser Glu Leu His Pro Tyr Leu His
915 920 925
Asn Met Lys Lys Arg Asp Pro Pro Ser Glu Pro Ser Pro Leu Glu Ala
930 935 940
Glu Phe Gln Arg Leu Pro Ser Tyr Arg Ser Trp Arg Thr Gln His Ile
945 950 955 960
Gly Asn Gln Glu Glu Asn Lys Ser Lys Asn Arg Asn Ser Asn Val Ile
965 970 975
Pro Tyr Asp Tyr Asn Arg Val Pro Leu Lys His Glu Leu Glu Met Ser
980 985 990
Lys Glu Ser Glu His Asp Ser Asp Glu Ser Ser Asp Asp Asp Ser Asp
995 1000 1005
Ser Glu Glu Pro Ser Lys Tyr Ile Asn Ala Ser Phe Ile Met Ser
1010 1015 1020
Tyr Trp Lys Pro Glu Val Met Ile Ala Ala Gln Gly Pro Leu Lys
1025 1030 1035
Glu Thr Ile Gly Asp Phe Trp Gln Met Ile Phe Gln Arg Lys Val
1040 1045 1050
Lys Val Ile Val Met Leu Thr Glu Leu Lys His Gly Asp Gln Glu
1055 1060 1065
Ile Cys Ala Gln Tyr Trp Gly Glu Gly Lys Gln Thr Tyr Gly Asp
1070 1075 1080
Ile Glu Val Asp Leu Lys Asp Thr Asp Lys Ser Ser Thr Tyr Thr
1085 1090 1095
Leu Arg Val Phe Glu Leu Arg His Ser Lys Arg Lys Asp Ser Arg
1100 1105 1110
Thr Val Tyr Gln Tyr Gln Tyr Thr Asn Trp Ser Val Glu Gln Leu
1115 1120 1125
Pro Ala Glu Pro Lys Glu Leu Ile Ser Met Ile Gln Val Val Lys
1130 1135 1140
Gln Lys Leu Pro Gln Lys Asn Ser Ser Glu Gly Asn Lys His His
1145 1150 1155
Lys Ser Thr Pro Leu Leu Ile His Cys Arg Asp Gly Ser Gln Gln
1160 1165 1170
Thr Gly Ile Phe Cys Ala Leu Leu Asn Leu Leu Glu Ser Ala Glu
1175 1180 1185
Thr Glu Glu Val Val Asp Ile Phe Gln Val Val Lys Ala Leu Arg
1190 1195 1200
Lys Ala Arg Pro Gly Met Val Ser Thr Phe Glu Gln Tyr Gln Phe
1205 1210 1215
Leu Tyr Asp Val Ile Ala Ser Thr Tyr Pro Ala Gln Asn Gly Gln
1220 1225 1230
Val Lys Lys Asn Asn His Gln Glu Asp Lys Ile Glu Phe Asp Asn
1235 1240 1245
Glu Val Asp Lys Val Lys Gln Asp Ala Asn Cys Val Asn Pro Leu
1250 1255 1260
Gly Ala Pro Glu Lys Leu Pro Glu Ala Lys Glu Gln Ala Glu Gly
1265 1270 1275
Ser Glu Pro Thr Ser Gly Thr Glu Gly Pro Glu His Ser Val Asn
1280 1285 1290
Gly Pro Ala Ser Pro Ala Leu Asn Gln Gly Ser
1295 1300
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К LY6G6D И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2818569C1 |
| Антагонистические антитела, которые связываются с человеческими TGFβ1, TGFβ2 и TGFβ3, и их применение при лечении фиброза легких | 2017 |
|
RU2746926C2 |
| МОЛЕКУЛЫ СО СПЕЦИФИЧНОСТЬЮ В ОТНОШЕНИИ CD79 И CD22 | 2015 |
|
RU2743590C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |
| АНТИ-αvβ8 АНТИТЕЛА И КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812478C2 |
| Композиции и способы лечения и предупреждения инфекций, вызванных Staphylococcus aureus | 2015 |
|
RU2764981C1 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ НА ОСНОВЕ АКТИВИРУЮЩИХ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРОТИВ CD3 И ФОЛАТНОГО РЕЦЕПТОРА 1 (FOLR1) И АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С ОСЬЮ PD-1 | 2015 |
|
RU2753902C2 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |

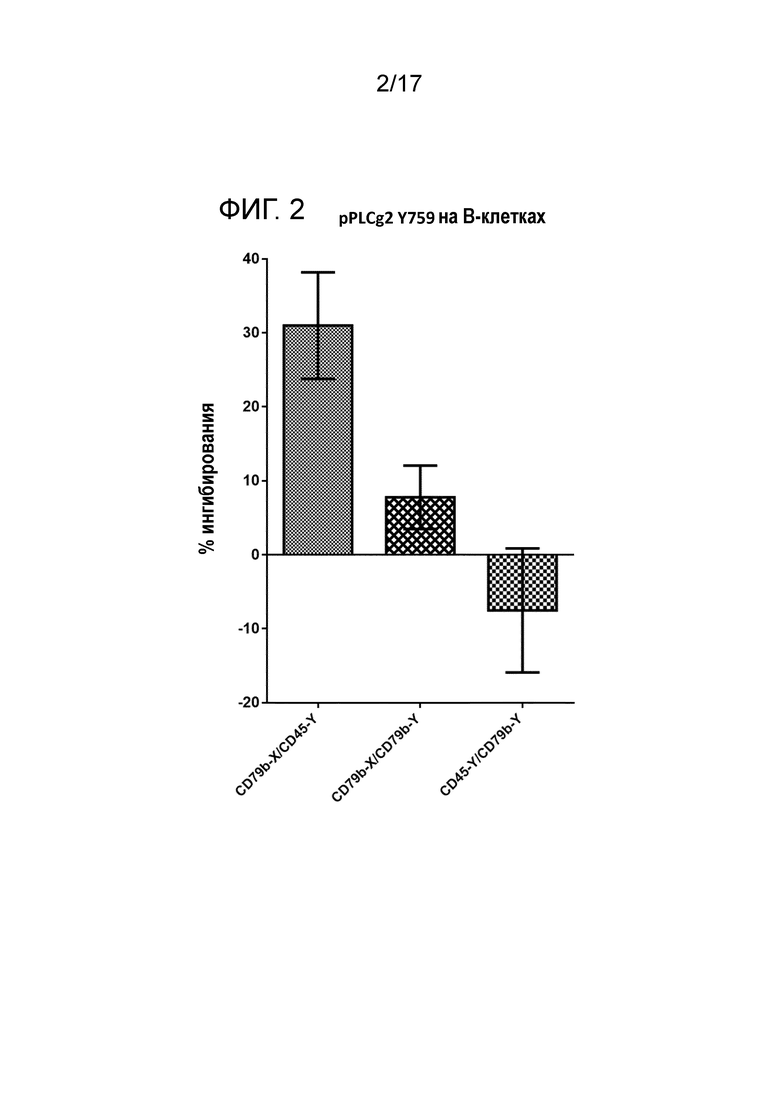





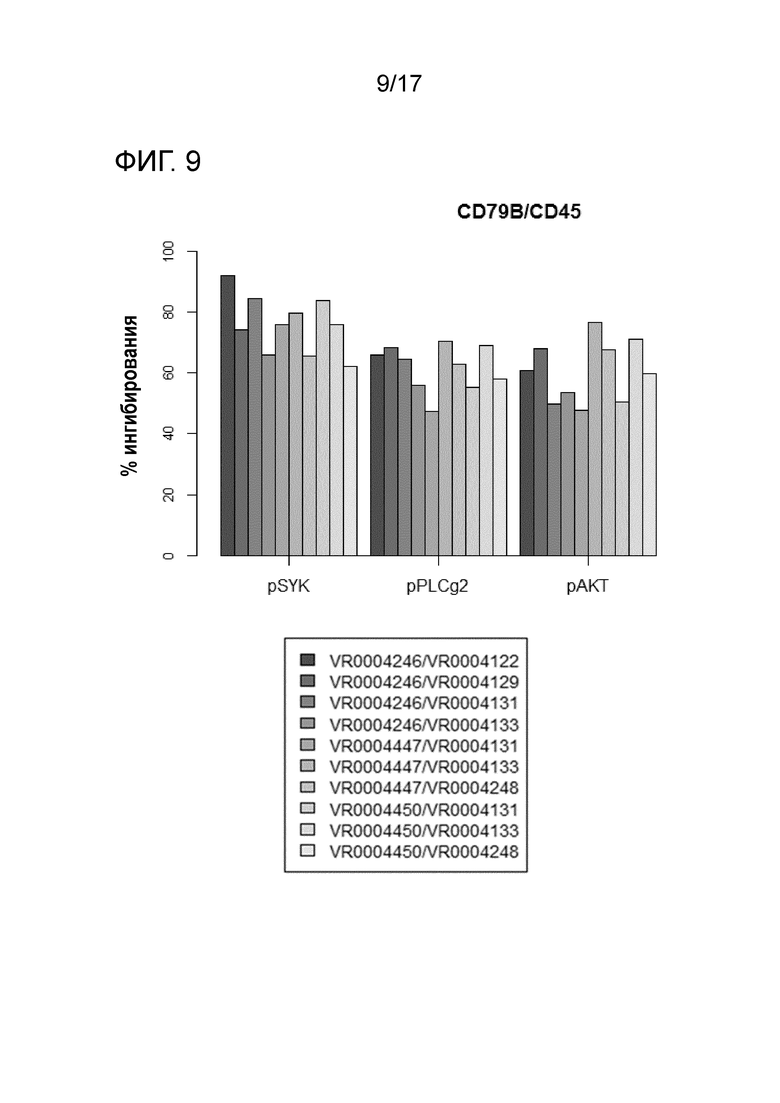
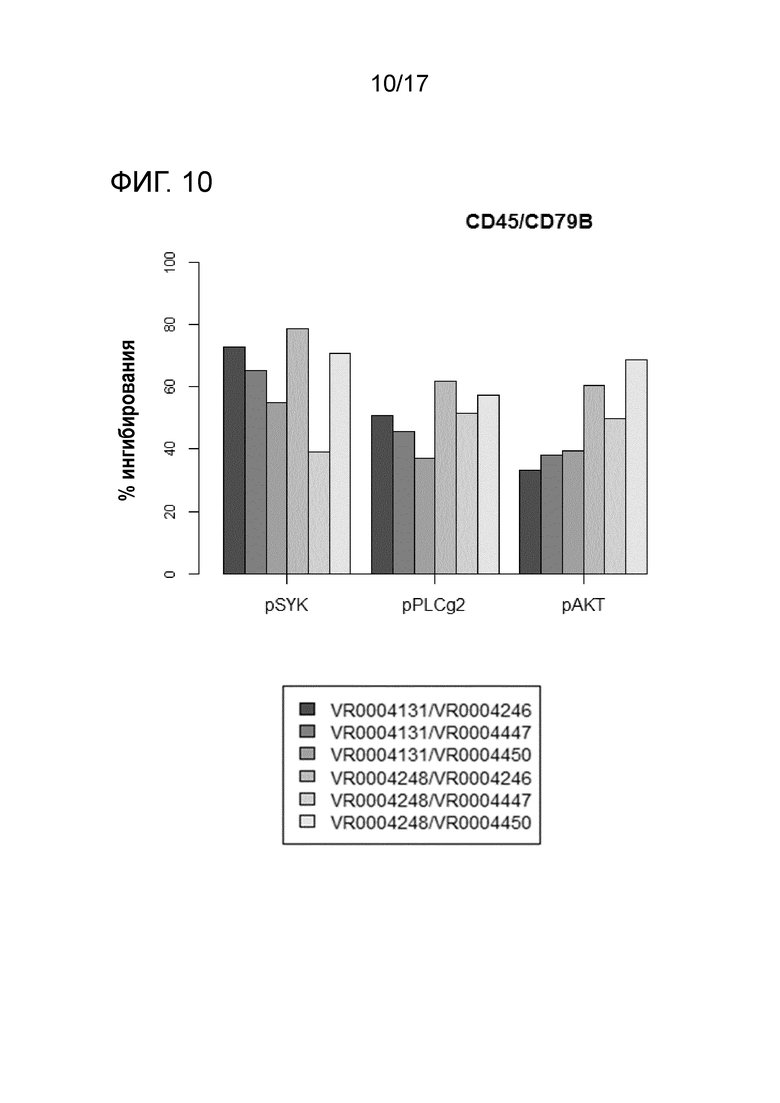


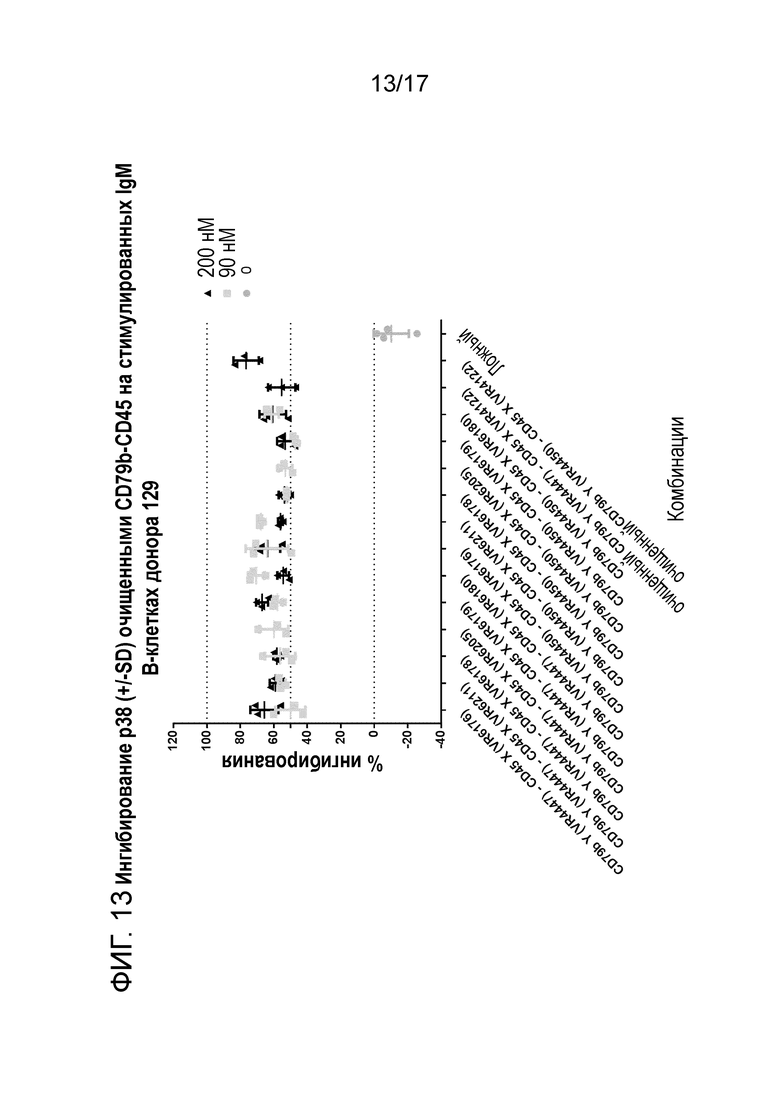
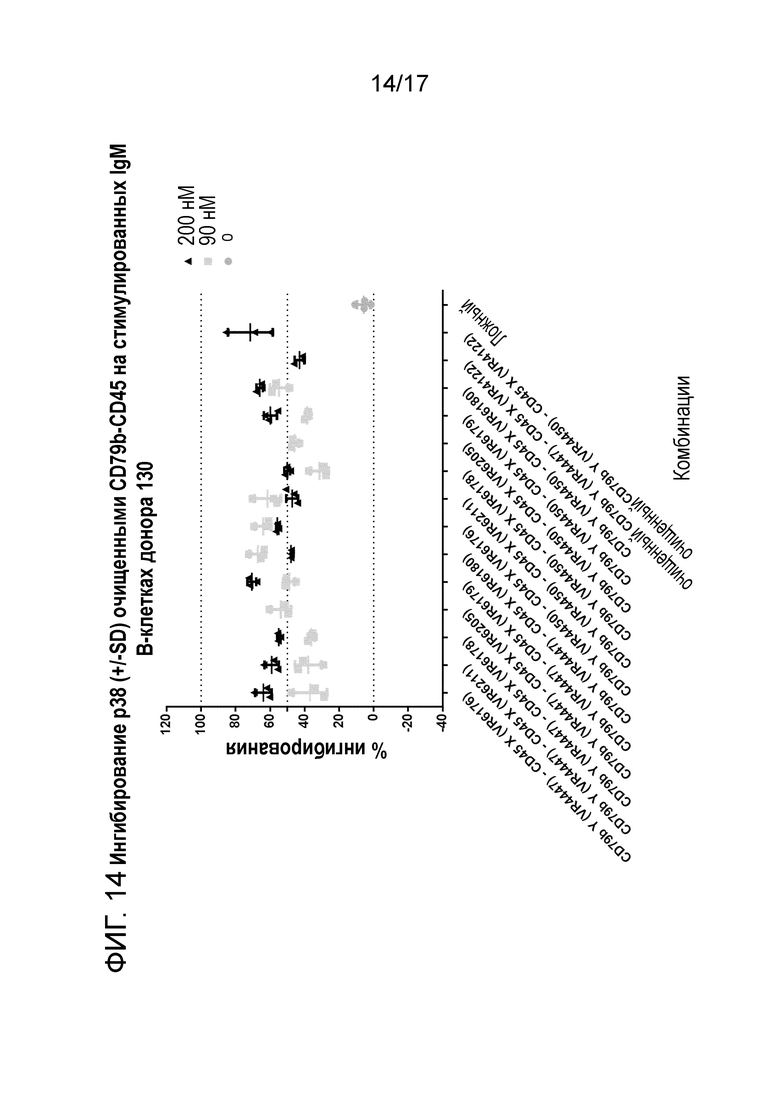
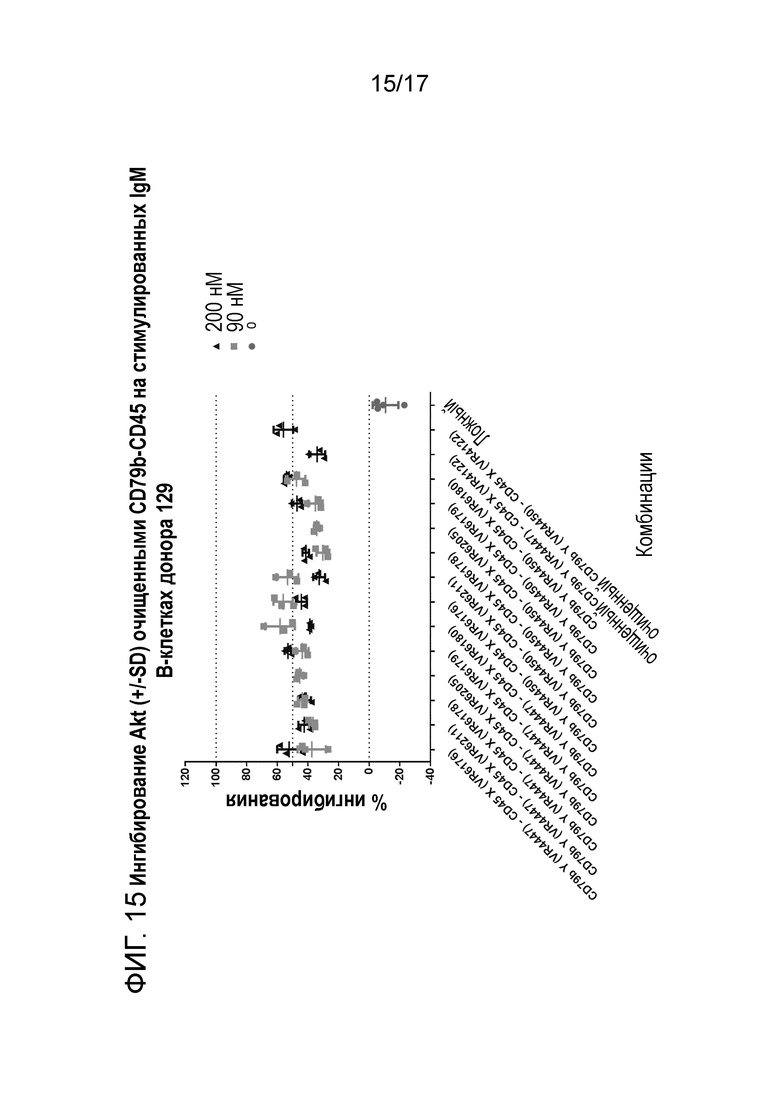
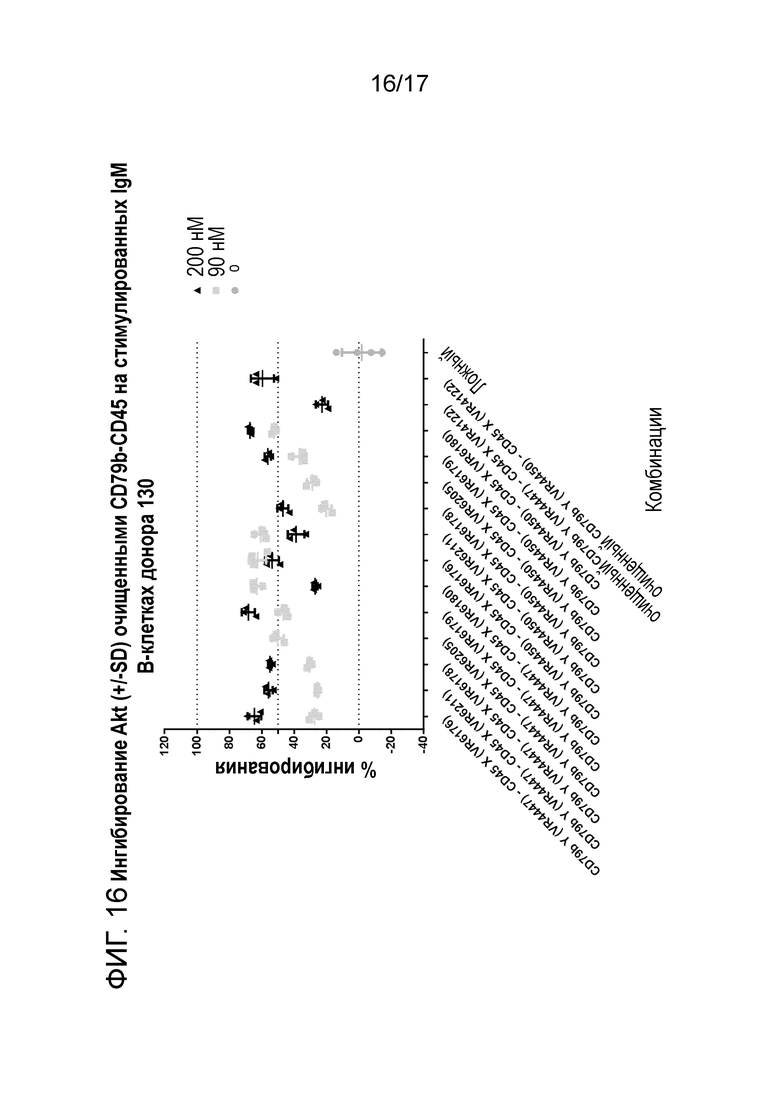
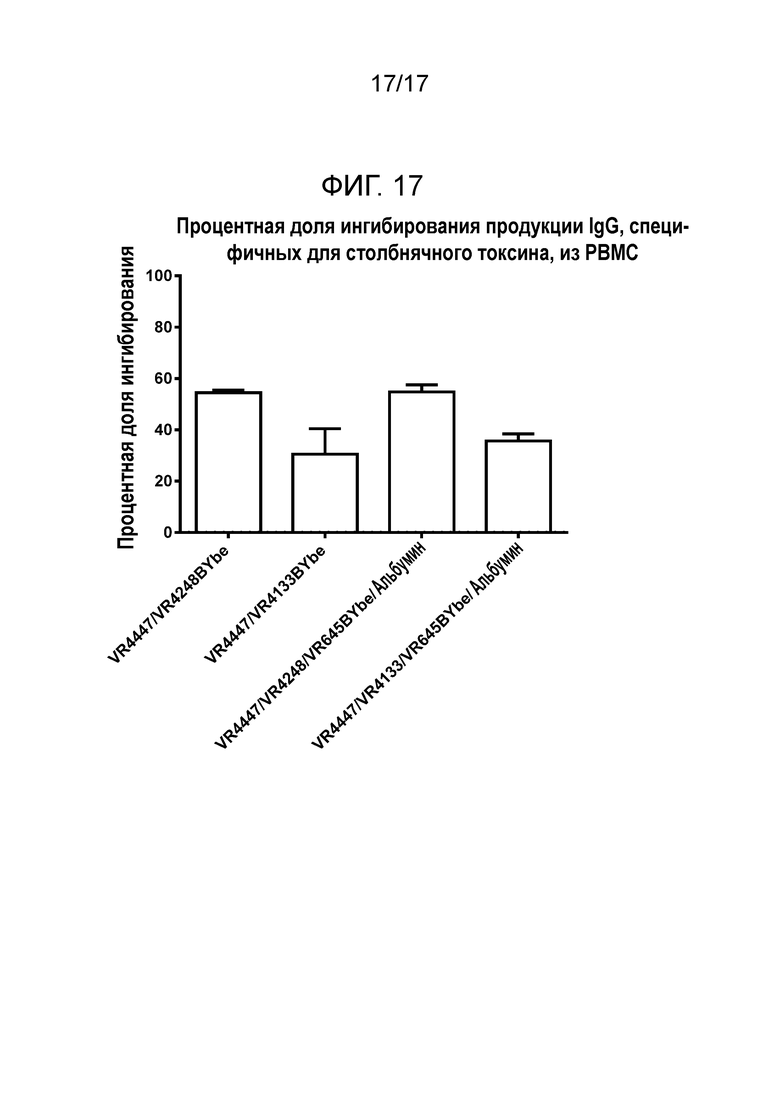
Изобретение относится к области биохимии, в частности к молекуле, специфической для антигенов CD45 и CD79b, где указанная молекула содержит связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79b. Изобретение также относится к молекуле, специфической для антигенов CD45, CD79b и сывороточного альбумина, где указанная молекула содержит связывающий домен, специфичный для антигена CD45, связывающий домен, специфичный для антигена CD79b, и связывающий домен, специфичный для сывороточного альбумина. Изобретение эффективно в контроле аномальных функций B-клеток, ассоциированных с заболеваниями, такими как аутоиммунные заболевания и злокачественные новообразования. 2 н. и 9 з.п. ф-лы, 17 ил., 7 табл., 8 пр.
1. Молекула, специфическая для антигенов CD45 и CD79b, где указанная молекула содержит связывающий домен, специфичный для антигена CD45, и связывающий домен, специфичный для антигена CD79b, где связывающий домен, являющийся специфичным для CD45, содержит:
3 CDR тяжелой цепи, имеющие последовательность, приведенную в SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3;
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; и
где домен, являющийся специфичным для CD79, содержит:
3 CDR тяжелой цепи, содержащие SEQ ID NO: 78 в случае CDRH1, SEQ ID NO: 79 в случае CDRH2 и SEQ ID NO: 80 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 75 в случае CDRL1, SEQ ID NO: 76 в случае CDRL2 и SEQ ID NO: 77 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 78 в случае CDRH1, SEQ ID NO: 79 в случае CDRH2 и SEQ ID NO: 80 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 85 в случае CDRL1, SEQ ID NO: 86 в случае CDRL2 и SEQ ID NO: 87 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 88 в случае CDRH1, SEQ ID NO: 89 в случае CDRH2 и SEQ ID NO: 90 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 85 в случае CDRL1, SEQ ID NO: 86 в случае CDRL2 и SEQ ID NO: 87 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 88 в случае CDRH1, SEQ ID NO: 89 в случае CDRH2 и SEQ ID NO: 90 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 75 в случае CDRL1, SEQ ID NO: 76 в случае CDRL2 и SEQ ID NO: 77 в случае CDRL3.
2. Молекула по п. 1, где связывающий домен или связывающие домены содержат вариабельную область антитела, специфичную для соответствующего антигена.
3. Молекула по п. 1 или 2, где каждый связывающий домен содержит два вариабельных домена антитела.
4. Молекула по п. 3, где два вариабельных домена антитела являются парой VH/VL.
5. Молекула по любому из пп. 1-4, где молекула является слитым белком.
6. Молекула по любому из пп. 1-3, где формат молекулы выбран из диатела, sc-диатела, триатела, тандемного scFv, FabFv, Fab'Fv, FabdsFv, Fab-scFv, Fab-dsscFv, Fab-(dsscFv)2 ди-Fab, ди-Fab', тритела, тандемного scFv-Fc, scFv-Fc-scFv, sc-диатело-Fc, sc-диатело-CH3, Ig-scFv, scFv-Ig, V-Ig, Ig-V, дуотела и DVD-Ig.
7. Молекула по любому из пп. 1-6, где каждый связывающий домен является моноспецифическим.
8. Молекула по любому из пп. 1-7, где связывающий домен, являющийся специфичным для CD45, и связывающий домен, являющийся специфичным для CD79b, независимо выбран из Fab, scFv, Fv, dsFv и dsscFv.
9. Молекула по любому из пп. 1-8, в которой связывающие домены гуманизируют.
10. Молекула, специфическая для антигенов CD45, CD79b и сывороточного альбумина, где указанная молекула содержит связывающий домен, специфичный для антигена CD45, связывающий домен, специфичный для антигена CD79b, и связывающий домен, специфичный для сывороточного альбумина, где связывающий домен, являющийся специфичным для CD45, содержит:
3 CDR тяжелой цепи, имеющие последовательность, приведенную в CDR SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 98 в случае CDRH1, SEQ ID NO: 99 в случае CDRH2 и SEQ ID NO: 100 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3;
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 108 в случае CDRH1, SEQ ID NO: 109 в случае CDRH2 и SEQ ID NO: 110 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 118 в случае CDRH1, SEQ ID NO: 119 в случае CDRH2 и SEQ ID NO: 120 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 125 в случае CDRL1, SEQ ID NO: 126 в случае CDRL2 и SEQ ID NO: 127 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 95 в случае CDRL1, SEQ ID NO: 96 в случае CDRL2 и SEQ ID NO: 97 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 105 в случае CDRL1, SEQ ID NO: 106 в случае CDRL2 и SEQ ID NO: 107 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 128 в случае CDRH1, SEQ ID NO: 129 в случае CDRH2 и SEQ ID NO: 130 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 115 в случае CDRL1, SEQ ID NO: 116 в случае CDRL2 и SEQ ID NO: 117 в случае CDRL3; и
где связывающий домен, являющийся специфичным для CD79, содержит:
3 CDR тяжелой цепи, содержащие SEQ ID NO: 78 в случае CDRH1, SEQ ID NO: 79 в случае CDRH2 и SEQ ID NO: 80 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 75 в случае CDRL1, SEQ ID NO: 76 в случае CDRL2 и SEQ ID NO: 77 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 78 в случае CDRH1, SEQ ID NO: 79 в случае CDRH2 и SEQ ID NO: 80 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 85 в случае CDRL1, SEQ ID NO: 86 в случае CDRL2 и SEQ ID NO: 87 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 88 в случае CDRH1, SEQ ID NO: 89 в случае CDRH2 и SEQ ID NO: 90 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 85 в случае CDRL1, SEQ ID NO: 86 в случае CDRL2 и SEQ ID NO: 87 в случае CDRL3; или
3 CDR тяжелой цепи, содержащие SEQ ID NO: 88 в случае CDRH1, SEQ ID NO: 89 в случае CDRH2 и SEQ ID NO: 90 в случае CDRH3, и 3 CDR легкой цепи, содержащие SEQ ID NO: 75 в случае CDRL1, SEQ ID NO: 76 в случае CDRL2 и SEQ ID NO: 77 в случае CDRL3.
11. Молекула по п. 10, где сывороточный альбумин представляет собой сывороточный альбумин человека.
| US 2014099254 A1, 10.04.2014 | |||
| MARK S | |||
| DENNIS et al | |||
| Albumin Binding as a General Strategy for Improving the Pharmacokinetics of Proteins, The Journal of Biological Chemistry, 2002, Vol.277, pp.35035-35043 | |||
| SCHOONJANS R et al | |||
| A new model for intermediate molecular weight recombinant bispecific and trispecific antibodies by efficient |

