Область техники, к которой относится изобретение:
В рамках настоящего изобретения предложены выделенные клетки-естественные киллеры (NK-клетки) человека в криоконсервированном состоянии, которые были предварительно нагружены перед замораживанием конструкцией антитела, указанная конструкция антитела содержит по меньшей мере первый связывающий домен, связывающийся с антигеном рецептора на клеточной поверхности иммунологической эффекторной клетки, в особенности, с тем, который экспрессируется на NK-клетках, и второй связывающий домен, связывающийся с антигеном клеточной поверхности на клеточной поверхности клетки-мишени.
Уровень техники
Клетки-естественные киллеры (NK-клетки) представляют собой мощные цитотоксические иммунные эффекторные клетки врожденной иммунной системы. Они способны распознавать и уничтожать опухолевые клетки, а также клетки, инфицированные вирусами или бактериями. NK-клетки могут вызывать антиген-независимый иммунный ответ против злокачественных клеток. Более того, в дополнение к NK-клеткам, существуют другие иммунологические эффекторные клетки, например моноциты, макрофаги, нейтрофилы и т.д., которые способны убивать опухолевые клетки или клетки, инфицированные вирусами или бактериями. В целом это понимают как врожденный иммунитет.
Растущее число научных отчетов и клинических исследований демонстрирует многообещающие и сильные противоопухолевые эффекты при применении иммунотерапии на основе NK-клеток. В настоящее время исследуют различные подходы с целью увеличения количества и функции NK-клеток. В одном из указанных подходов используют конструкции би- или мультиспецифичных антител с целью повышения специфичности эндогенных NK-клеток путем перекрестного связывания их с соответствующими клетками-мишенями. Чтобы найти применение такой синергии, адоптивную клеточную терапию в сочетании с терапией на основе антител комбинируют при различных показаниях. Различные источники NK-клеток для адоптивного переноса находятся в стадии оценки и включают алло генные гаплоидентичные NK-клетки, которые подвергли краткосрочным или долгосрочным активации или размножению, NK-клетки пуповины, которые также выращивают/активируют в определенных условиях, клеточные линии NK-клеток или полученные из стволовых клеток/дифференцированные NK-клетки. Все источники теоретически также могут быть применены для создания NK-клеток с химерным антигенным рецептором (CAR-NK-клеток), которые получены за счет нового типа генетической модификации ленти- или ретровирусами с получением стабильных антиген-специфичных NK-клеток. Первые примеры включают подход на основе CD19-CAR-NK-клеток, который в настоящее время тестируют на пациентах с неходжкинской лимфомой (НХЛ, NHL).
Однако при комбинировании конструкций антител с любым из вышеуказанных источников NK-клеток для адоптивного переноса всегда необходимо получить клетки в месте применения, то есть в больнице, в которой осуществляют терапию соответствующего пациента, и, впоследствии, осуществить совместное введение указанному пациенту требуемой дозы соответствующих конструкций антител. Помимо того факта, что это может быть логистической проблемой для большинства больниц, следует далее отметить, что все комплексы работ, требуемые для подготовки клеток к введению, должны выполняться в соответствии со строго контролируемыми протоколами, чтобы исключить любые проблемы, которые можно предотвратить, что увеличивает такой риск.
Соответственно, существует потребность в средствах и способах для упрощения такой терапии, чтобы обеспечить ее применимость для большей группы пациентов, особенно в неспециализированных больницах или медицинских центрах.
На Фигурах показано:
Фигура 1: 4-часовой анализ цитотоксичности с высвобождением кальцеина на клетках-мишенях MM.1S (А) или DK-MG (В), NK-клетки в качестве эффекторных клеток с указанным соотношением Е:Т, которые были предварительно нагружены 10 мкг/мл указанных антител и отмыты (с) или не отмыты (без) перед одним циклом замораживания/размораживания. В качестве контроля те же самые NK-клетки не были предварительно нагружены, но отмыты и подвергнуты одному циклу замораживания/размораживания, и указанное антитело было добавлено в анализ цитотоксичности (свежие; сплошные серые ромбы).
Фигура 2: Связывание различных антител к CD16 с вариантами антигена CD16 в виде покрытия, проанализированное с помощью иммуноферментного анализа (ИФА (ELISA)). 96-луночные планшеты для ИФА (ELISA) покрывали рекомбинантными вариантами антигена, показанными в легенде к панели А. Различные антитела, показанные в (А)-(D), серийно разводили и инкубировали на планшетах в указанном диапазоне концентраций. Связанные антитела выявляли с помощью анти-His-HRP в (А)-(С) или Белка L-HRP. Реакции субстрата тетраметилбензидина, ТМБ (ТМВ) измеряли при 450 нм (Ext (450 нм)). Кривые связывания были подобраны с применением четырехпараметрической логистической (4PL) модели кривой зависимости логарифма (агониста) от ответа - переменный наклон в Graphpad Prism.
Фигура 3: Реактивность различных антител scFv к CD16 или контрольных scFv или моноклональных антител с рекомбинантными вариантами антигена CD16 или контрольным антигеном рецептора эпидермального фактора роста, РЭФР (EGFR), экспрессируемым на клетках яичника китайского хомячка (СНО) с трансмембранным доменом РЭФР (EGFR) (ТМ) или гликозилфосфатидилинозитол (ГФИ (CPI))-якорем.
Фигура 4: Выравнивание последовательностей внеклеточных доменов CD16A человека и яванского макака и вариантов CD16B человека, применяемых в качестве рекомбинантных антигенов в экспериментах, обобщенных в Таблице 3. Вариации в последовательности внеклеточного домена (ECD) CD16A яванского макака выделены серым цветом. Для выравнивания применяли инструмент множественного выравнивания последовательностей CLUSTAL О (1.2.4).
Фигура 5: Связывание различных антител к CD16 с вариантами антигена CD16 после разделения с помощью электрофорез белков в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-ΠΑΑΓ (SDS-PAGE)) и вестерн-блоттинга. 2 мкг рекомбинантного белка антигена на дорожку загружали, разделяли с помощью SDS-ΠΑΑΓ (SDS-PAGE), переносили на мембрану PVDF и инкубировали с указанными первичными и вторичными антителами. Сигналы от колориметрического проявления с субстратом 3,3'-диаминобензидином (ДАБ (DAB)) показывают отчетливое распознавание вариантов антигена CD16A и CD16B.
Фигура 6: Конкурентное связывание с CD16A различных антител scFv против CD16 с моноклональными антителами 3G8 в ИФА (ELISA). Планшеты, покрытые антигеном CD16A (48R, 158V)-mFc.67, инкубировали со смесью 1 нМ моноклональных антител 3G8 и серийных разведений различных scFv против CD16 в указанном диапазоне концентраций. CD16A-связанный 3G8 обнаруживали с помощью антитела козы против IgG мыши (H+L)-HRPO. Реакции субстрата тетраметилбензидина (ТМБ (ТМВ)) измеряли при 450 нм (Ext (450 нм)). Кривые связывания были подобраны с применением четырехпараметрической логистической (4PL) модели кривой зависимости логарифма(агониста) от ответа - переменный наклон в Graphpad Prism.
Фигура 7: Титрование scFv_CD16-1, содержащего человеческие домены Fv из клона CD16-1 (A), scFv_CD16-2, содержащего человеческие домены Fv против CD16 из клона CD16-2 (В), и scFv_3G8, содержащего мышиные домены Fv от моноклональных антител 3G8 (С) на первичных NK-клетках человека при 37°С в присутствии или в отсутствие 10 мг/мл поликлонального человеческого IgG.
Фигура 8: 4-часовой анализ цитотоксичности с высвобождением кальцеина на клетках-мишенях COLO 205 (А) или KARPAS-299 (В), NK-клетки в качестве эффекторных клеток при указанном соотношении Е:Т, которые были предварительно нагружены 10 мкг/мл указанных антител и заморожены при -80°С. В качестве контроля аликвоты тех же NK-клеток не были предварительно нагружены (без антител), но также были подвергнуты одному циклу замораживания/размораживания, и указанные антитела (scFv-IgAb_75 EpCAM/NKG2D или тандемные диатела CD30/CD16A) были сразу добавлены в концентрации 10 мкг/мл в анализ цитотоксичности. На графике нанесены среднее и стандартное отклонение значений лизиса в двух проворностях. Exp.No.: RBL 1464.
Определения
Термин «конструкция антитела» относится к молекуле, в которой структура и/или функция основана/основаны на структуре и/или функции антитела, например, полноразмерной или целой молекулы иммуноглобулина и/или является/являются взятыми из доменов вариабельной тяжелой цепи (VH) и/или вариабельной легкой цепи (VL) антитела или его фрагмента. Следовательно, конструкция антитела способна связываться со своей специфичной мишенью или антигеном. Кроме того, связывающий домен конструкции антитела, определенный в контексте настоящего изобретения, включает минимальные структурные требования для антитела, которые позволяют связываться с мишенью. Это минимальное требование может, например, быть определено по присутствию по меньшей мере трех участков, определяющих комплементарность (CDR) легкой цепи (т.е. CDR1, CDR2 и CDR3 участка VL) и/или трех CDR тяжелой цепи (т.е. CDR1, CDR2 и CDR3 участка VH), предпочтительно всех шести CDR. Альтернативный подход к определению минимальных структурных требований к антителу - это определение эпитопа антитела в структуре специфичной мишени, соответственно, белкового домена белка-мишени, составляющего участок эпитопа (кластер эпитопа), или по отношению к специфичному антителу, конкурирующему с эпитопом определенного антитела. Антитела, на которых основаны конструкции, определенные в контексте настоящего изобретения, содержат, например, моноклональные, рекомбинантные, химерные, деиммунизированные, гуманизированные и человеческие антитела (антитела человека).
Связывающий домен конструкции антитела, определенного в контексте настоящего изобретения, может, например, включать упомянутые выше группы CDR. Предпочтительно, эти CDR включены в каркасный участок вариабельного участка легкой цепи (VL) антитела и вариабельного участка тяжелой цепи (VH) антитела; однако они не обязательно должны включать и то, и другое. Фрагменты Fd, например, имеют два участка VH и часто сохраняют некоторую антиген-связывающую функцию интактного антиген-связывающего домена. Дополнительные примеры формата фрагментов антител, вариантов антител или связывающих доменов содержат (1) фрагмент Fab, моновалентный фрагмент, имеющий домены VL, VH, CL и СН1; (2) фрагмент F (ab')2, бивалентный фрагмент, имеющий два фрагмента Fab, связанных дисульфидным мостиком в шарнирном участке; (3) фрагмент Fd, имеющий два домена VH и СН1; (4) фрагмент Fv, имеющий домены VL и VH одного плеча антитела, (5) фрагмент dAb (Ward et al., (1989) Nature 341: 544-546), который имеет домен VH; (6) выделенный участок, определяющий комплементарность (CDR), и (7) одноцепочечный вариабельный фрагмент (scFv), последний является предпочтительным (например, полученный из библиотеки scFV).
Кроме того, в рамках определения «связывающий домен» или «домен, который связывает» находятся фрагменты полноразмерных антител, такие как VH, VHH, VL, (s)dAb, Fv, Fd, Fab, Fab', F (ab')2 или «r IgG» («полуантитело»). Конструкции антител, как определено в контексте изобретения, могут также включать модифицированные фрагменты антител, также называемые вариантами антител, такие как scFv, ди-scFv или би(c)-scFv, scFv-Fc, scFv-зиппер, scFab, Fab2, Fab3, диатела, одноцепочечные диатела, тандемные диатела (Tandab's), тандемные ди-scFv, тандемные три-scFv, «мультитела», такие как триатела или тетратела, и однодоменные антитела, такие как нанотела или антитела с одиночным вариабельным доменом, включающие только один вариабельный домен, которым может быть VHH, VH или VL, который специфично связывается с антигеном или эпитопом независимо от других V-участков или доменов.
В настоящем документе термины «одноцепочечный Fv», «одноцепочечные антитела» или «scFv» относятся к фрагментам антител с одноцепочечной полипептидной цепью, которые включают вариабельные участки и тяжелой, и легкой цепей, но не имеют константных участков. Обычно одноцепочечное антитело дополнительно включает полипептидный линкер между доменами VH и VL, который позволяет ему образовывать желаемую структуру, которая допускает связывание антигена. Одноцепочечные антитела подробно обсуждаются Plueckthun в The Pharmacology of Monoclonal Antibodies, т.113, Rosenburg и Moore ред. Springer-Verlag, Нью-Йорк, с. 269-315 (1994). Известны различные способы получения одноцепочечных антител, включая методы, описанные в патентах США №№4694778 и 5260203; публикации международной патентной заявки №WO 88/01649; Bird (1988) Science 242: 423-442; Huston et al. (1988) Proc. Natl. Acad. Sci. США 85: 5879-5883; Ward et al. (1989) Nature 334: 54454; Skerra et al. (1988) Science 242: 1038-1041. В специфичных вариантах осуществления одноцепочечные антитела также могут быть биспецифичными, мультиспецифичными, человеческими и/или гуманизированными и/или синтетическими.
Кроме того, определение термина «конструкция антитела» охватываетт моновалентные, двухвалентные и поливалентные/мультивалентные конструкции и, таким образом, биспецифичные конструкции, специфично связывающиеся только с двумя антигенными структурами, а также полиспецифичные/мультиспецифичные конструкции, которые специфично связывают более чем две антигенные структуры, например три, четыре или более через отдельные связывающие домены. Более того, определение термина «конструкция антитела» включает молекулы, состоящие только из одной полипептидной цепи, а также молекулы, состоящие из более чем одной полипептидной цепи, причем эти цепи могут быть как идентичными (гомодимеры, гомотримеры или гомоолигомеры), так и разными (гетеродимер, гетеротример или гетероолигомер). Примеры идентифицированных выше антител и их вариантов или производных описаны, среди прочего, в Harlow and Lane, Antibodies a lab manual, CSHL Press (1988) и Using Antibodies: a lab manual, CSHL Press (1999), Kontermann and Dubel, Antibody Engineering, Springer, 2-е изд. 2010 и Little, Recombinant Antibodies for Immunotherapy, Cambridge University Press, 2009.
Термин «валентный» обозначает наличие определенного количества антигенсвязывающих доменов в антигенсвязывающем белке. Природный IgG имеет два антиген-связывающих домена и является бивалентным. Антигенсвязывающие белки, как определено в контексте изобретения, являются по меньшей мере трехвалентными. Примеры тетра-, пента- и гексавалентных антиген-связывающих белков описаны в настоящем документе.
В настоящем документе термин «биспецифичная» относится к конструкции антитела, которая является «по меньшей мере биспецифичной», т.е. он включает по меньшей мере первый связывающий домен и второй связывающий домен, причем первый связывающий домен связывается с одним антигеном или мишенью (здесь: рецептор NK-клеток, например CD16a), и второй связывающий домен связывается с другим антигеном или мишенью (здесь: поверхностный антиген клетки-мишени). Соответственно, конструкции антител, как определено в контексте изобретения, включают специфичности по меньшей мере к двум различным антигенам или мишеням. Например, первый домен предпочтительно связывается с внеклеточным эпитопом рецептора NK-клетки одного или более видов, выбранных из человека, видов макак и видов грызунов.
Термин «рецептор NK-клетки», используемый в контексте изобретения, определяет белки и белковые комплексы на поверхности NK-клеток. Таким образом, этот термин определяет молекулы клеточной поверхности, которые характерны для NK-клеток, но необязательно экспрессируются исключительно на поверхности NK-клеток, но также и на других клетках, таких как макрофаги или Т-клетки. Примеры рецепторов NK-клеток включают, без ограничения перечисленными, CD16a, CD16b, NKp46 и NKG2D.
«CD16A» относится к активирующему рецептору CD16A, также известному как FCγRIIIA, экспрессируемому на клеточной поверхности NK-клеток. CD16A - это активирующий рецептор, запускающий цитотоксическую активность NK-клеток. Аффинность антител к CD16A напрямую коррелирует с их способностью запускать активацию NK-клеток, таким образом, более высокая аффинность к CD16A снижает дозу антител, требуемую для активации. Антиген-связывающий сайт антиген-связывающего белка связывается с CD16A, но не с CD16B. Например, антиген-связывающий сайт, содержащий вариабельные домены тяжелой (VH) и легкой (VL) цепи, связывающиеся с CD16A, но не связывающиеся CCD16B, может быть обеспечен антигенсвязывающим сайтом, который специфично связывается с эпитопом CD16A, который включает аминокислотные остатки С-концевой последовательности SFFPPGYQ (SEQ ID NO: 172) и/или остатки G130 и/или Y141 CD16A(SEQ ID NO: 23)), которые не присутствуют в CD16B.
«CD16B» относится к рецептору CD16B, также известному как FCγRIIIB, экспрессируемому на нейтрофилах и эозинофилах. Рецептор является ГФИ (GPI)-заякоренным, и предполагается, что он не запускает какую-либо цитотоксическую активность CD16B-положительных иммунных клеток.
«NKp46» относится к рецептору, активирующему цитотоксичность, который может способствовать повышению эффективности активированных клеток-естественных киллеров (NK) клеток по опосредованию лизиса опухолевых клеток. Он также известен как NCR1 или CD335.
«NKG2D» относится к активирующему и костимулирующему рецептору, вовлеченному в иммунный надзор за связыванием с различными лигандами, индуцируемыми клеточным стрессом, отображаемыми на поверхности аутологичных опухолевых клеток и инфицированных вирусом клеток. Обеспечивает и стимулирующий, и костимулирующий врожденные иммунные ответы на активированные киллерные (NK) клетки, приводя к цитотоксической активности. Действует как костимулирующий рецептор для Т-клеточного рецептора (TCR) в CD8(+) Т-клеточно-опосредованных адаптивных иммунных ответах за счет усиления активации Т-клеток. Стимулирует опосредованное перфорином элиминирование лиганд-экспрессирующих опухолевых клеток. Он также известен как лектин-подобный рецептор киллерных клеток К1, KLRK1 или CD314.
Термин «поверхностный антиген клетки-мишени» относится к антигенной структуре, экспрессируемой клеткой и присутствующей на клеточной поверхности, так что она доступна для конструкции антитела, которая описана в данном документе. Это может быть белок, предпочтительно внеклеточная часть белка, пептид, который представлен на клеточной поверхности в контексте главного комплекса гистосовместимости, ГКГ (МНС) (включая HLA-A2, HLA-A11, HLA-A24, HLA-B44, HLA-C4) или углеводная структура, предпочтительно углеводная структура белка, такого как гликопротеин. Это предпочтительно ассоциированный с опухолью или ограниченный опухолью антиген.
Термин «биспецифичная конструкция антитела», как определено в контексте изобретения, также охватывает мультиспецифичные конструкции антител, такие как триспецифичные конструкции антител, последние из которых содержат три связывающих домена, или конструкции, имеющие более трех (например, четыре, пять...) специфичностей. Примеры конструкций би- или мультиспецифичных антител обеспечены, например, в WO 2006/125668, WO 2015/158636, WO 2017/064221, РСТ/ЕР2019/056516, PCT/IB2019/053040 и Ellwanger et al. (MAbs. 2019 Jul; 11 (5):899-918).
Учитывая, что конструкции антител, как определено в контексте изобретения, являются (по меньшей мере) биспецифичными, они не встречаются в природе и заметно отличаются от продуктов природного происхождения. Конструкция «биспецифичного» антитела или иммуноглобулин, следовательно, представляет собой искусственное гибридное антитело или иммуноглобулин, имеющее по меньшей мере два различных сайта связывания с разными специфичностями. Конструкции биспецифичных антител могут быть созданы различными способами, содержащими слияние гибридом или связывание фрагментов Fab'. См, например, Songsivilai & Lachmann, Clin. Exp.Immunol. 79:315-321 (1990).
Эти по крайней мере два связывающих домена и вариабельные домены (VH/VL) конструкции антитела согласно настоящему изобретению могут включать или не включать пептидные линкеры (спейсерные пептиды). Термин «пептидный линкер» включает в соответствии с настоящим изобретением аминокислотную последовательность, согласно которой аминокислотные последовательности одного (вариабельного и/или связывающего) домена и другого (вариабельного и/или связывающего) домена конструкции антитела, определенного в настоящем документе, являются связаны друг с другом. Пептидные линкеры также могут быть применены для слияния третьего домена с другими доменами конструкции антитела, определенного в настоящем документе. Существенной технической особенностью такого пептидного линкера является то, что он не включает какую-либо полимеризационную активность.
Конструкции антител, как определено в контексте изобретения, предпочтительно представляют собой «конструкции антител, генерируемые in vitro». Этот термин относится к конструкции антитела в соответствии с приведенным выше определением, где весь или часть вариабельного участка (например, по меньшей мере, один CDR) генерируется в результате неиммунного отбора клеток, например, фагового дисплея in vitro, белкового чипа или любого другого способа, в котором последовательности-кандидаты могут быть протестированы на их способность связываться с антигеном. Таким образом, этот термин предпочтительно исключает последовательности, созданные исключительно в результате геномной перестройки в иммунной клетке животного. «Рекомбинантное антитело» - это антитело, полученное с применением технологии рекомбинантной ДНК или генной инженерии.
В настоящем документе термин «моноклональное антитело» (mAb) или конструкция моноклонального антитела относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, включенные в популяцию, идентичны, за исключением возможных естественно происходящих мутаций и/или посттрансляционных модификаций (например, изомеризации, амидирования), которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, будучи направленными против одного антигенного сайта или детерминанты антигена, в отличие от общепринятых (поликлональных) препаратов антител, которые обычно содержат различные антитела, направленные против различных детерминант (или эпитопов). Вдобавок к своей специфичности, преимущество моноклональных антител в том, что они синтезируются культурой гибридомы и, следовательно, не контаминированы другими иммуноглобулинами. Модификатор «моноклональное» указывает на характер антитела как полученного из по существу гомогенной популяции антител, и его не следует истолковывать как требующий получения антитела каким-либо особенным способом.
Для получения моноклональных антител можно применять любую технику, обеспечивающую получение антител, продуцируемых культурами непрерывных клеточных линий. Например, моноклональные антитела, которые будут применяться, могут быть получены гибридомным способом, впервые описанным Koehler et al., Nature, 256: 495 (1975), или могут быть сделаны способами рекомбинации ДНК (см., например, патент США №4816567). Примеры дополнительных техник получения человеческих моноклональных антител содержат технику триомы, технику гибридомы В-клеток человека (Kozbor, Immunology Today 4 (1983), 72) и технику EBV-гибридомы (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc. (1985), 77-96).
Гибридомы затем можно подвергнуть скринингу с применением стандартных способов, таких как иммуноферментный анализ, ИФА (ELISA) и анализ поверхностного плазмонного резонанса (BIACORE™), чтобы идентифицировать одну или более гибридом, которые продуцируют антитело, которое специфично связывается с определенным антигеном. Любая форма соответствующего антигена может быть применена в качестве иммуногена, например рекомбинантный антиген, встречающиеся в природе формы, любые его варианты или фрагменты, а также его антигенный пептид.
Поверхностный плазмонный резонанс, используемый в системе BIAcore, можно применять для увеличения эффективности фаговых антител, которые связываются с эпитопом поверхностного антигена клетки-мишени (Schier, Human Antibodies Hybridomas 7 (1996), 97-105; Malmborg, J. Immunol. Methods 183 (1995), 7-13). Другой иллюстративный способ производства моноклональных антител содержит скрининг библиотек экспрессии белков, например библиотек фагового дисплея или рибосомного дисплея. Фаговый дисплей описан, например, в Ladner et al., патент США №5223409; Smith (1985) Science 228:1315-1317, Clackson et al, Nature, 352: 624-628 (1991) и Marks et al., J. Mol. Biol., 222: 581-597 (1991).
Вдобавок к применению дисплейных библиотек, соответствующий антиген можно применять для иммунизации животного, не являющегося человеком, например грызуна (такого как мышь, хомяк, кролик или крыса). В одном варианте осуществления животное, не являющееся человеком, содержит по меньшей мере часть гена иммуноглобулина человека. Например, можно сконструировать линии мышей, дефицитных по продукции мышиных антител, с большими фрагментами локусов человеческого Ig (иммуноглобулина). Применяя гибридомную технологию, можно получить и выбрать антиген-специфичные моноклональные антитела, происходящие от генов с желаемой специфичностью. См., например, XENOMOUSE™, Green et al. (1994) Nature Genetics 7:13-21, США 2003-0070185, WO 96/34096, и WO 96/33735.
Моноклональное антитело также может быть получено от животного, не являющегося человеком, а затем модифицировано, например, гуманизировано, деиммунизировано, превращено в химерное и т.д., с использованием методов рекомбинации ДНК, известных в данной области. Примеры конструкций модифицированных антител содержат гуманизированные варианты нечеловеческих (не относящихся к человеку) антител, антитела с «созревшей аффинностью» (см., например, Hawkins et al. J. Mol. Biol. 254, 889-896 (1992) и Lowman et al., Biochemistry 30, 10832-10837 (1991)) и мутанты антител с измененной (-ыми) эффекторной (-ыми) функцией (-ями) (см., например, патент США 5648260, Kontermann and Dubel (2010), loc. cit. и Little (2009), loc. cit).
В иммунологии созревание аффинности - это процесс, при котором В-клетки продуцируют антитела с повышенной аффинностью к антигену в ходе иммунного ответа. При многократном воздействии одного и того же антигена хозяин будет продуцировать антитела со все большей аффинностью. Как и в природном прототипе, созревание аффинности in vitro основано на принципах мутации и отбора. Созревание аффинности in vitro успешно применялось для оптимизации антител, конструкций антител и фрагментов антител. Случайные мутации внутри CDR вносятся с применением радиации, химических мутагенов или подверженной ошибкам полимеразной цепной реакции, ПЦР (PCR). Кроме того, генетическое разнообразие может быть увеличено путем перестановки цепей. Два или три раунда мутации и отбора с применением дисплейных способов, таких, как фаговый дисплей, обычно приводят к получению фрагментов антител с аффинностями в низком наномолярном диапазоне.
Предпочтительный тип заместительной вариации аминокислот в конструкциях антител подразумевает замену одного или более остатков гипервариабельного участка родительского антитела (например, гуманизированного или человеческого антитела). Как правило, итоговый (-е) вариант (-ы), выбранный (-е) для дальнейшей разработки, будет (-ут) иметь улучшенные биологические свойства по сравнению с родительским антителом, из которого они были получены. Подходящий способ создания таких заместительных вариантов подразумевает созревание аффинности с применением фагового дисплея. Вкратце, несколько сайтов гипервариабельного участка (например, 6-7 сайтов) мутируют для создания всех возможных аминокислотных замен на каждом сайте. Полученные таким образом варианты антител отображаются моновалентным образом из частиц нитчатого фага в виде слияния с продуктом гена III М13, упакованным в каждой частице. Отображенные фагом варианты подвергаются скринингу на их биологическую активность (например, аффинность связывания), как описано в настоящем документе. Чтобы идентифицировать сайты гипервариабельного участка-кандидата для модификации, можно выполнить мутагенез с аланиновым сканированием для идентификации остатков гипервариабельного участка, вносящих значительный вклад в связывание антигена. Альтернативно или дополнительно может быть полезно проанализировать кристаллическую структуру комплекса антиген-антитело для определения точек контакта между связывающим доменом и, например, поверхностным антигеном клетки-мишени человека. Такие контактные остатки и соседние остатки являются кандидатами на замену в соответствии с техниками, разработанными в настоящем документе. Когда такие варианты созданы, панель вариантов подвергается скринингу, как описано в данном документе, и антитела с превосходящими свойствами в одном или более соответствующих анализах могут быть выбраны для дальнейшей разработки.
Моноклональные антитела и конструкции антител, согласно настоящему изобретению, в частности, содержат «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, происходящих от определенного вида или принадлежащих к определенному классу или подклассу антител, в то время как остальная часть цепи (-ей) идентична или гомологична соответствующим последовательностям в антителах, происходящих от другого вида или принадлежащих к другому классу или подклассу антител, а также фрагментам таких антител, при условии, что они проявляют желаемую биологическую активность (патент США № 4816567; Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984)). Представляющие интерес химерные антитела содержат «примитизированные» антитела, включающие антиген-связывающие последовательности вариабельного домена, полученные от приматов, не являющихся человеком (например, обезьяны Старого Света, широконосые обезьяны и т.д.), и последовательностей константных участков человека. Описаны различные подходы к производству химерных антител. См., например, Morrison et al., Proc. Natl. Acad. Sci U.S. A. 81:6851, 1985; Takeda et al., Nature 314:452, 1985, Cabilly et al., патент США №4816567; Boss et al., патент США №4816397; Tanaguchiet al., ЕР 0171496; ЕР 0173494; и GB 2177096.
Антитело, конструкция антитела, фрагмент антитела или вариант антитела также могут быть модифицированы путем специфичного удаления эпитопов Т-клеток человека (способ, называемый «деиммунизация») способами, описанными, например, в WO 98/52976 или WO 00/34317. Вкратце, вариабельные домены тяжелой и легкой цепи антитела можно анализировать на наличие пептидов, которые связываются с главным комплексом гистосовместимости, ГКГС (МНС) II класса; эти пептиды представляют собой потенциальные Т-клеточные эпитопы (как определено в WO 98/52976 и WO 00/34317). Для обнаружения потенциальных эпитопов Т-клеток может быть задействован подход компьютерного моделирования, называемый «пептидное протягивание» и, кроме того, в базе данных пептидов, связывающих человеческий ГКГС (МНС) II класса, можно искать мотивы, присутствующие в последовательностях VH и VL, как описано в WO 98/52976 и WO 00/34317. Эти мотивы связываются с любым из 18 основных аллотипов DR ГКГС (МНС) II класса и, таким образом, составляют потенциальные эпитопы Т-клеток. Обнаруженные потенциальные Т-клеточные эпитопы могут быть элиминированы путем замены небольшого количества аминокислотных остатков в вариабельных доменах или, предпочтительно, путем замены одной аминокислоты. Обычно производятся консервативные замены. Часто, но не исключительно, может применяться аминокислота, общая для положения в последовательностях антител человеческой зародышевой линии. Последовательности человеческой зародышевой линии описаны, например, в Tomlinson, et al. (1992) J. Mol. Biol. 227:776-798; Cook, G.P. et al. (1995) Immunol. Today Vol. 16 (5): 237-242; и Tomlinson et al. (1995) EMBO J. 14: 14:4628-4638. Каталог V BASE обеспечивает обширный каталог последовательностей вариабельных участков человеческого иммуноглобулина (составлен Tomlinson, LA. et. al. MRC Center for Protein Engineering, Кембридж, Великобритания). Эти последовательности можно применять в качестве источника человеческих последовательностей, например, для каркасных участков и CDR. Также можно применять консенсусные каркасные участки человека, например, как описано в патенте США №6300064.
«Гуманизированные» антитела, конструкции антител, их варианты или фрагменты (такие как Fv, Fab, Fab', F (ab')2 или другие антиген-связывающие подпоследовательности антител) представляют собой антитела или иммуноглобулины преимущественно человеческих последовательностей, которые содержат минимальную (-ые) последовательность (-и), полученную (-ые) из нечеловеческого иммуноглобулина. По большей части гуманизированные антитела являются человеческими иммуноглобулинами (реципиентные антитела), в которых остатки гипервариабельного участка (также CDR) реципиента заменены остатками гипервариабельного участка нечеловеческого (например, грызуна) вида (донорское антитело), такого как мышь, крыса, хомяк или кролик, имеющими желаемую специфичность, аффинность и емкость. В некоторых случаях остатки каркасного участка (FR) Fv иммуноглобулина человека заменяются соответствующими остатками нечеловеческого происхождения. Кроме того, в контексте настоящего документа «гуманизированные антитела» могут также включать остатки, которых нет ни в антителе реципиента, ни в антителе донора. Эти модификации сделаны для дальнейшего усовершенствования и оптимизации производительности антител. Гуманизированное антитело может также включать по меньшей мере часть константного участка иммуноглобулина (Fc), обычно иммуноглобулина человека. Подробнее см. Jones et al., Nature, 321: 522-525 (1986); Reichmann et al., Nature, 332: 323-329 (1988); и Presta, Curr. Op.Struct. Biol., 2: 593- 596 (1992).
Гуманизированные антитела или их фрагменты могут быть получены путем замены последовательностей вариабельного домена Fv, которые непосредственно не подразумевают связывания антигена эквивалентными последовательностями человеческих вариабельных доменов Fv. Иллюстративные способы получения гуманизированных антител или их фрагментов обеспечены Morrison (1985) Science 229:1202-1207; по Oi et al. (1986) BioTechniques 4:214; и по US 5585089; US 5693761; US 5693762; US 5859205; и US 6407213. Эти способы содержат выделение, манипулирование и экспрессию последовательностей нуклеиновых кислот, которые кодируют все или часть вариабельных доменов Fv иммуноглобулина из, по меньшей мере, одной тяжелой или легкой цепи. Такие нуклеиновые кислоты могут быть получены из гибридомы, продуцирующей антитело против заранее определенной мишени, как описано выше, а также из других источников. Рекомбинантная ДНК, кодирующая молекулу гуманизированного антитела, может быть затем клонирована в соответствующий вектор экспрессии.
Гуманизированные антитела также могут быть получены с применением трансгенных животных, таких как мыши, которые экспрессируют человеческие гены тяжелой и легкой цепей, но неспособны экспрессировать гены тяжелой и легкой цепей эндогенного мышиного иммуноглобулина. Winter описывает иллюстративный способ CDR-привития, который можно применять для получения описанных здесь гуманизированных антител (патент США №5225539). Все CDR конкретного человеческого антитела могут быть заменены, по меньшей мере, частью нечеловеческих CDR, или только некоторые из CDR могут быть заменены нечеловеческими CDR. Необходимо только заменить количество CDR, требуемое для связывания гуманизированного антитела с заранее определенным антигеном.
Гуманизированное антитело может быть оптимизировано путем внедрения консервативных замен, замен консенсусной последовательности, замен зародышевой линии и/или обратных мутаций. Такие измененные молекулы иммуноглобулина могут быть созданы любым из нескольких способов, известных в данной области (например, Teng et al., Proc. Natl. Acad. Sci. U.S.A., 80: 7308-7312, 1983; Kozbor ei al., Immunology Today, 4: 7279, 1983; Olsson et al., Meth. Enzymol., 92: 3-16, 1982, и ЕР 239 400).
Термин «человеческое антитело/антитело человека», «конструкция человеческого антитела» и «связывающий домен человека» содержит антитела, конструкции антител и связывающие домены, имеющие участки антитела, такие как вариабельные и константные участки или домены, которые по существу соответствуют последовательностям иммуноглобулина человеческой зародышевой линии, известным в данной области, содержащим, например, описанные Kabat et al. (1991) (loc. cit.). Человеческие антитела, конструкции антител или связывающие домены, как определено в контексте изобретения, могут содержать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов человеческой зародышевой линии (например, мутации, внесенные случайным или сайт-специфичным мутагенезом in vitro или соматической мутацией in vivo), например, в CDR и, в частности, в CDR3. Человеческие антитела, конструкции антител или связывающие домены могут иметь по меньшей мере одно, два, три, четыре, пять или более положений, замененных аминокислотным остатком, который не кодируется последовательностью иммуноглобулина зародышевой линии человека. Однако определение человеческих антител, конструкций антител и связывающих доменов в настоящем документе также предполагает «полностью человеческие антитела», которые содержат только неискусственно и/или генетически измененные человеческие последовательности антител, поскольку они могут быть получены с применением технологий или систем, таких как Xenomouse. Предпочтительно, «полностью человеческое антитело» не содержит аминокислотные остатки, не кодируемые последовательностями иммуноглобулина человеческой зародышевой линии.
В некоторых вариантах осуществления конструкции антител, определенные в данном документе, представляют собой «выделенные» или «по существу чистые» конструкции антител. «Выделенная» или «по существу чистая» при применении для описания описанных здесь конструкций антител означает конструкцию антитела, которая была идентифицирован, отделен и/или извлечен из компонента его производственной среды. Предпочтительно конструкция антитела свободна или по существу свободна от связи со всеми другими компонентами из среды, в которой ее получали. Контаминирующие компоненты его производственной среды, такие как те, которые являются продуктом получения рекомбинантных трансфицированных клеток, являются материалами, которые обычно вмешиваются в диагностическое или терапевтическое применение полипептида, и могут содержать ферменты, гормоны и другие белковые или небелковые растворенные вещества. Конструкции антител могут, например, составлять по меньшей мере около 5% или по меньшей мере около 50% по весу от общего белка в данном образце. Понятно, что выделенный белок может составлять от 5% до 99,9% по весу от общего содержания белка, в зависимости от обстоятельств. Полипептид может быть создан в значительно более высокой концентрации за счет применения индуцибельного промотора или промотора высокой экспрессии, так что он будет получен в повышенных уровнях концентрации. Это определение содержит получение конструкции антитела в широком спектре организмов и/или клеток-хозяев, известных в данной области. В предпочтительных вариантах осуществления конструкция антитела будет очищена (1) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с применением секвенатора с вращающейся чашей, или (2) до гомогенности с помощью SDS-ΠΑΑΓ (SDS-PAGE) в невосстанавливающих или восстанавливающих условиях с применением окрашивания кумасси синим или, предпочтительно, серебром. Обычно, однако, выделенная конструкция антитела будет получена, по меньшей мере, за один этап очистки.
Термин «связывающий домен» характеризует в связи с настоящим изобретением домен, который (специфично) связывается с/взаимодействует с/распознает данный целевой эпитоп или заданный сайт-мишень на целевых молекулах (антигенах), например антиген рецептора NK-клеток, например CD16, и поверхностный антиген клетки-мишени соответственно. Структура и функция первого связывающего домена (распознавание, например, CD16), и предпочтительно также структура и/или функция второго связывающего домена (распознавание поверхностного антигена клетки-мишени) основана/основаны на структуре и/или функции антитела, например полноразмерной или цельной молекулы иммуноглобулина и/или взята/взяты из доменов вариабельной тяжелой цепи (VH) и/или вариабельной легкой цепи (VL) антитела или его фрагмента. Предпочтительно первый связывающий домен характеризуется наличием трех CDR легкой цепи (т.е. CDR1, CDR2 и CDR3 участка VL) и/или трех CDR тяжелой цепи (т.е. CDR1, CDR2 и CDR3 участка VH). Второй связывающий домен предпочтительно также включает минимальные структурные требования антитела, которые позволяют связываться с мишенью. Более предпочтительно, второй связывающий домен включает по меньшей мере три CDR легкой цепи (т.е. CDR1, CDR2 и CDR3 участка VL) и/или три CDR тяжелой цепи (т.е. CDR1, CDR2 и CDR3 участка VH). Предполагается, что первый и/или второй связывающий домен продуцируется или может быть получен скорее способами фагового дисплея или скрининга библиотеки, чем путем привития последовательностей CDR из ранее существовавшего (моноклонального) антитела в скелет.
В соответствии с настоящим изобретением связывающие домены находятся в форме одного или более полипептидов. Такие полипептиды могут содержать белковые части и небелковые части (например, химические линкеры или химические перекрестно связывающие агенты, такие как глутаральдегид). Белки (определение содержит их фрагменты, предпочтительно биологически активные фрагменты и пептиды, обычно имеющие менее 30 аминокислот) включают две или более аминокислот, связанных друг с другом ковалентной пептидной связью (что приводит к образованию цепи аминокислот).
В настоящем документе термин «полипептид» описывает группу молекул, которые обычно состоят из более чем 30 аминокислот. Полипептиды, далее, могут образовывать мультимеры, такие какдимеры, тримеры и более высокие олигомеры, т.е. состоящие из более чем одной полипептидной молекулы. Молекулы полипептидов, образующие такие димеры, тримеры и т.д., могут быть идентичными или неидентичными. Соответствующие структуры более высокого порядка таких мультимеров, следовательно, называются гомо- или гетеродимерами, гомо- или гетеротримерами и т.д. Примером гетеромультимера является молекула антитела, которая в своей природной форме состоит из двух идентичных легких полипептидных цепей и двух идентичных тяжелых полипептидных цепей. Термины «пептид», «полипептид» и «белок» также относятся к естественно модифицированным пептидам/полипептидам/белкам, причем на модификацию влияют, например, посттрансляционные модификации, такие как гликозилирование, ацетилирование, фосфорилирование и т.п. «Пептид», «полипептид» или «белок», когда они упоминаются в настоящем документе, также могут быть химически модифицированы, например пегилированы. Такие модификации хорошо известны в данной области и описаны в настоящем документе ниже.
Предпочтительно связывающий домен, который связывается с антигеном рецептора NK-клеток, например CD16 и/или связывающий домен, который связывается с поверхностным антигеном клетки-мишени, является/являются человеческими связывающими доменами. Антитела и конструкции антител, включающие по меньшей мере один из группы, состоящей из связывающих доменов человека, минуют некоторые проблемы, связанные с антителами или конструкциями антител, которые обладают нечеловеческими, такими как принадлежащие грызунам (например, мышиным, крысам, хомякам или кроликам) вариабельными и/или константными участками. Присутствие таких белков, происходящих от грызунов, может привести к быстрому удалению антител или конструкций антител или может привести к выработке пациентом иммунного ответа против антитела или конструкции антитела. Чтобы избежать применения антител или конструкций антител, происходящих от грызунов, можно генерировать человеческие или полностью человеческие антитела/конструкции антител путем внедрения функции человеческого антитела грызуну, так что грызун продуцирует полностью человеческие антитела.
Способность клонировать и реконструировать человеческие локусы размером млн п. о. в искусственных дрожжевых хромосомах (YAC) и вносить их в мышиную зародышевую линию обеспечивает мощный подход к объяснению функциональных компонентов очень больших или грубо картированных локусов, а также к созданию полезных моделей заболеваний человека. Кроме того, применение такой технологии для замены мышиных локусов их человеческими эквивалентами может обеспечить уникальное проникновение в механизмы экспрессии и регуляции продуктов генов человека во время развития, их связь с другими системами и их вовлечение в индукцию и прогрессирование заболевания.
Важным практическим воздействием такой стратегии является «гуманизация» мышиной гуморальной иммунной системы. Внедрение локусов человеческого иммуноглобулина (Ig) мышам, у которых были инактивированы эндогенные гены Ig, дает возможность изучить механизмы, лежащие в основе запрограммированной экспрессии и сборки антител, а также их роль в развитии В-клеток. Кроме того, такая стратегия могла бы обеспечить идеальный источник для производства полностью человеческих моноклональных антител (mAb) - важную веху на пути к реализации терапии антителами при заболеваниях человека. Ожидается, что полностью человеческие антитела или конструкции антител минимизируют иммуногенные и аллергические ответы, присущие мышиным или производным от мыши моноклональным антителам, и, таким образом, увеличат эффективность и безопасность вводимых антител/конструкций антител. Можно ожидать, что применение полностью человеческих антител или конструкций антител обеспечит существенное преимущество при лечении хронических и рецидивирующих заболеваний человека, таких как воспаление, аутоиммунитет и рак, которые требуют повторных введений соединения.
Один из подходов к этой цели состоял в том, чтобы сконструировать линии мышей, дефицитных по продуцированию мышиных антител, с большими фрагментами локусов человеческого Ig в ожидании того, что такие мыши будут продуцировать большой репертуар человеческих антител в отсутствие мышиных антител. Большие фрагменты человеческого Ig сохранят большое разнообразие вариабельных генов, а также правильную регуляцию продуцирования и экспрессии антител. Используя механизм мышей для диверсификации и отбора антител и отсутствие иммунологической толерантности к человеческим белкам, репертуар репродуцированных человеческих антител в этих линиях мышей должен давать высокоаффинные антитела против любого интересующего антигена, понятие содержит и человеческие антигены. Применяя гибридомную технологию, антиген-специфичные человеческие mAb с желаемой специфичностью можно легко получить и отобрать. Эта общая стратегия была продемонстрирована в связи с созданием первых линий мышей XenoMouse (см. Green et al. Nature Genetics 7: 13-21 (1994)). Линии XenoMouse были сконструированы с использованием искусственных дрожжевых хромосом (YAC), содержащих фрагменты конфигурации зародышевой линии размером 245 т.п.о. и 190 т.п.о. локуса человеческой тяжелой цепи и локуса легкой цепи каппа соответственно, которые вмещали сердцевинные последовательности вариабельного и константного участка. YAC, содержащие человеческий Ig, оказались совместимыми с мышиной системой как в отношении реаранжировки, так и в отношении экспрессии антител и были способны к замещению инактивированных генов мышиного Ig. Это было продемонстрировано их способностью индуцировать развитие В-клеток, продуцировать репертуар полностью человеческих антител как у взрослого человека, и генерировать антиген-специфичные человеческие моноклональные антитела. Эти результаты также заставляют предполагать, что внедрение больших частей локусов человеческого Ig, содержащих большее количество V-генов, дополнительных регуляторных элементов и константных участков человеческого Ig, может повторять, по существу, полный репертуар, характерный для человеческого гуморального ответа на инфекцию и иммунизацию. Работа Green et al. недавно была расширена до внедрения более чем примерно 80% репертуара человеческих антител путем внедрения фрагментов YAC размером млн п.о. конфигурации зародышевой линии локусов человеческой тяжелой цепи и локусов легкой цепи каппа, соответственно. См. Mendez et al. Nature Genetics 15: 146-156 (1997) и заявка на патент США сер. №08/759620.
Производство мышей XenoMouse дополнительно обсуждается и описывается в патентных заявках США сер. №07/466008, сер. №07/610515, сер. №07/919297, сер. №07/922649, сер. №08/031801, сер. №08/1 12848, сер. №08/234145, сер. №08/376279, сер. №08/430938, сер. №08/464584, сер. №08/464582, сер. №08/463191, сер. №08/462837, сер. №08/486853, сер. №08/486857, сер. №08/486859, сер. №08/462513, сер. №08/724752 и сер. №08/759620; и патенты США №№6162963; 6150584; 61 14598; 6075181 и 5939598 и патенты Японии №№3 068 180 В2, 3 068 506 В2 и 3 068 507 В2. См. также Mendez et al. Nature Genetics 15:146-156 (1997) и Green and Jakobovits J. Exp.Med. 188:483-495(1998), EP0 463 151 B1, WO 94/02602, WO 96/34096, WO 98/24893, WO 00/76310, и WO 03/47336.
В альтернативном подходе другие, в том числе GenPharm International, Inc., использовали «минилокусный» подход. В минилокусном подходе экзогенный локус Ig имитируется путем включения частей (индивидуальных генов) из локуса Ig. Таким образом, один или более генов VH, один или более генов DH, один или более генов JH, константный участок мю и второй константный участок (предпочтительно константный участок гамма) формируются в конструкцию для вставки в животное. Этот подход описан в патенте США №5545807, принадлежащем Surani et al. и патентах США. №№5545806; 5625825; 5625126; 5633425; 5661016; 5770429; 5789650; 5814318; 5877397; 5874299; и 6255458 каждый из которых принадлежит Lonberg and Kay, патенты США №№5591669 и 6023010, принадлежащие Krimpenfort and Berns, патенты США №№5612205; 5721367; и 5789215, принадлежащие Berns et al., и патент США №5643763, принадлежащие Choi and Dunn, и GenPharm International, патентные заявки США сер. №07/574748, сер. №07/575962, сер. №07/810279, сер. №07/853408, сер. №07/904068, сер. №07/990860, сер. №08/053131, сер. №08/096762, сер. №08/155301, сер. №08/161739, сер. №08/165699, сер. №08/209741. См. также ЕР 0546073 В1, WO 92/03918, WO 92/22645, WO 92/22647, WO 92/22670, WO 93/12227, WO 94/00569, WO 94/25585, WO 96/14436, WO 97/ 13852 и WO 98/24884 и патент США №5981175. См., далее, Taylor et al. (1992), Chen et al. (1993), Tuaillon et al. (1993), Choi et al. (1993), Lonberg et al. (1994), Taylor et al. (1994), и Tuaillon et al. (1995), Fishwild et al. (1996).
Kirin также продемонстрировал создание человеческих антител посредством мышей, которым путем слияния микроклеток были внесены большие части хромосом или целые хромосомы. См. европейские патентные заявки №№773288 и 843961. Xenerex Biosciences разрабатывает технологию потенциального создания человеческих антител. В этой технологии мышей с тяжелым комбинированным иммунодефицитом, ТКИД (SCID) восстанавливают лимфатическими клетками человека, например В- и/или Т-клетками. Затем мышей иммунизируют антигеном, и они могут генерировать иммунный ответ против антигена. См. патенты США №№5476996; 5698767; и 5958765.
Ответы с участием антител человека к Ig мыши (НАМА) привели промышленность к тому, чтобы получить химерные или иным образом гуманизированные антитела. Однако ожидается, что будут наблюдаться определенные ответы с участием антител человека против против химерных антител (НАСА), особенно при хроническом или многодозовом использовании антитела. Таким образом, было бы желательно обеспечить конструкции антител, включающие человеческий связывающий домен против поверхностного антигена клетки-мишени и человеческий связывающий домен против CD16, чтобы ослабить опасения по поводу и/или эффекты НАМА- или НАСА-ответа.
Термины "(специфично) связывается с", "(специфично) распознает", "(специфично) направлено" и "(специфично) реагирует с" означают в соответствии с данным изобретением, что связывающий домен взаимодействует или специфично взаимодействует с данным эпитопом или данным сайтом-мишенью на молекулах-мишенях (антигенах), здесь: рецептор NK-клеток, например CD16a, и поверхностный антиген клетки-мишени, соответственно.
Термин «эпитоп» относится к сайту антигена, с которым связывающий домен, такой как антитело или иммуноглобулин, или производное, фрагмент или вариант антитела или иммуноглобулина, специфично связывается. «Эпитоп» является антигенным, и поэтому к термину эпитоп в настоящем документе иногда также относится «антигенная структура» или «антигенная детерминанта». Таким образом, связывающий домен является «сайтом, с которым взаимодействует антиген». Также подразумевается, что указанное связывание/взаимодействие определяет «специфичное распознавание».
«Эпитопы» могут быть образованы как заменимыми аминокислотами, так и незаменимыми аминокислотами, наложенными друг на друга посредством третичной укладки белка. «Линейный эпитоп» представляет собой эпитоп, в котором первичная аминокислотная последовательность включает распознаваемый эпитоп. Линейный эпитоп обычно содержит по меньшей мере 3 или по меньшей мере 4, а чаще по меньшей мере 5 или по меньшей мере 6 или по меньшей мере 7, например, от около 8 до около 10 аминокислот в уникальной последовательности.
«Конформационный эпитоп», в отличие от линейного эпитопа, представляет собой эпитоп, в котором первичная последовательность аминокислот, которые включает эпитоп, не является единственным определяющим компонентом распознаваемого эпитопа (например, эпитоп, в котором первичная последовательность аминокислот не обязательно распознается связывающим доменом). Обычно конформационный эпитоп включает большее количество аминокислот по сравнению с линейным эпитопом. Что касается распознавания конформационных эпитопов, связывающий домен распознает трехмерную структуру антигена, предпочтительно пептида или белка или его фрагмента (в контексте настоящего изобретения антигенная структура для одного из связывающих доменов включена в белок поверхностного антигена клетки-мишени). Например, когда молекула белка укладывается для формирования трехмерной структуры, определенные аминокислоты и/или полипептидный остов, формирующий (-е) конформационный эпитоп, сопоставляются, позволяя антителу распознавать эпитоп. Способы определения конформации эпитопов содержат, без ограничения перечисленными, рентгеновскую кристаллографию, спектроскопию двумерного ядерного магнитного резонанса, 2D-ЯМР (2D-NMR) и сайт-направленное спиновое мечение и спектроскопию электронного парамагнитного резонанса, ЭПР (EPR).
Взаимодействие между связывающим доменом и эпитопом или участком, включающим эпитоп, подразумевает, что связывающий домен проявляет заметную аффинность к эпитопу/участку, включающему эпитоп, на конкретном белке или антигене (здесь: рецептор NK-клеток, например CD16a, и поверхностный антиген клетки-мишени, соответственно) и, как правило, не проявляет значительной реактивности с белками или антигенами, кроме рецептора NK-клеток, например CD16a, и поверхностного антигена клетки-мишени. «Заметная аффинность» содержит связывание с аффинностью примерно 106 Μ (кД) или сильнее. Предпочтительно, связывание считается специфичным, когда аффинность связывания составляет около от 10-12 до 10-8 М, от 10-12 до 10-9 М, от 10-12 до 10-10 М, от 10-11 до 10-8 М, предпочтительно около от 10-11 до 10-9 М. Может ли связывающий домен специфично реагировать с или связываться с мишенью, можно легко проверить, среди прочего, путем сравнения реакции указанного связывающего домена с белком-мишенью или антигеном с реакцией указанного связывания домена с белками или антигенами, отличными от рецептора NK-клеток, например CD16a, и поверхностного антигена клетки-мишени. Предпочтительно, связывающий домен, как определено в контексте изобретения, по сути или по существу не связывается с белками или антигенами, отличными от рецептора NK-клетки, например CD16a и поверхностного антигена клетки-мишени (т.е. первый связывающий домен не способен связываться с белками, отличными от рецептора NK-клетки, например CD16a, и второй связывающий домен не способен связываться с белками, отличными от поверхностного антигена клетки-мишени).
Термин «по сути/по существу не связывается» или «не способен к связыванию» означает, что связывающий домен по настоящему изобретению не связывает белок или антиген, отличный от рецептора NK-клетки, например CD16a, и поверхностного антигена клетки-мишени, т.е. не проявляет реактивности более 30%, предпочтительно не более 20%, более предпочтительно не более 10%, особенно предпочтительно не более 9%, 8%, 7%, 6% или 5% с белками или антигенами, отличными от рецептора NK-клетки, например CD16a и поверхностного антигена клетки-мишени, каковым образом связывание с рецептором NK-клетки, например CD16a, и поверхностным антигеном клетки-мишени соответственно установлено равным 100%.
Полагают, что на специфичное связывание влияют специфичные мотивы в аминокислотной последовательности связывающего домена и антигена. Таким образом, связывание достигается как результат их первичной, вторичной и/или третичной структуры, а также как результат вторичных модификаций указанных структур. Специфичное взаимодействие сайта взаимодействия антигена с его специфичным антигеном может привести к простому связыванию указанного сайта с антигеном. Более того, специфичное взаимодействие сайта взаимодействия антигена с его специфичным антигеном может альтернативно или дополнительно привести к инициированию сигнала, например, за счет индукции изменения конформации антигена, олигомеризации антигена и т.д.
Термин «вариабельный» относится к частям доменов антитела или иммуноглобулина, которые проявляют вариабельность своей последовательности и которые подразумевают определение специфичности и аффинности связывания конкретного антитела (то есть «вариабельного (-ых) домена (-ов)»). Спаривание вариабельной тяжелой цепи (VH) и вариабельной легкой цепи (VL) вместе образует единый антиген-связывающий сайт.
Вариабельность неравномерно распределена по вариабельным доменам антител; она сконцентрирована в субдоменах каждого из вариабельных участков тяжелой и легкой цепи. Эти субдомены называются «гипервариабельными участками» или «участками, определяющими комплементарность» (CDR). Более консервативные (т.е. негипервариабельные) части вариабельных доменов называются «каркасными» участками (FRM или FR) и обеспечивают скелет для шести CDR в трехмерном пространстве для формирования антигенсвязывающей поверхности. Каждый из вариабельных доменов встречающихся в природе тяжелых и легких цепей включает четыре участка FRM (FR1, FR2, FR3 и FR4), в основном принимающих конфигурацию β-листа, соединенных тремя гипервариабельными участками, которые формируют петли, соединяющие, и в некоторых случаях образующие часть, структуры β-листа. Гипервариабельные участки в каждой цепи удерживаются вместе в непосредственной близости посредством FRM и вместе с гипервариабельными участками из другой цепи вносят вклад в формирование антигенсвязывающего сайта (см. Kabat et al., loc. cit.).
Термины «CDR» и его множественное число «CDR» относятся к участкам, определяющим комплементарность, три из которых выполняют связывающую роль в вариабельном участке легкой цепи (CDR-L1, CDR-L2 и CDR-L3), и три выполняют связывающую роль в вариабельном участке тяжелой цепи (CDR-H1, CDR-H2 и CDR-H3). CDR содержат большинство остатков, ответственных за специфичные взаимодействия антитела с антигеном, и, следовательно, вносят вклад в функциональную активность молекулы антитела: они являются основными детерминантами специфичности антигена.
Точные дефиниционные границы и длины CDR подлежат различным системам классификации и нумерации. Таким образом, CDR могут упоминаться согласно Kabat, Chothia, контактным или любым другим граничным определениям, содержа описанную в настоящем документе систему нумерации. Несмотря на разные границы, каждая из этих систем имеет некоторую степень сообщения в том, что составляет так называемые «гипервариабельные участки» внутри вариабельных последовательностей. Определения CDR в соответствии с этими системами могут, следовательно, различаться по длине и граничным областям по отношению к прилежащему каркасному участку. См., например, Kabat (подход, основанный на межвидовой изменчивости последовательностей), Chothia (подход, основанный на кристаллографических исследованиях комплексов антиген-антитело) и/или MacCallum (Kabat et al., loc. cit; Chothia et al., J Mol. Biol, 1987, 196: 901-917 и MacCallum et al., J. Mol. Biol, 1996, 262: 732). Еще одним стандартом для характеристики сайта связывания антигена является определение AbM, применяемое программой моделирования антител Oxford Molecular's AbM. См., например, Анализ последовательности и структуры белка вариабельных доменов антитела. В: Antibody Engineering Lab Manual (Ed.: Duebel, S. и Kontermann, R., Springer-Verlag, Heidelberg). В той мере, в какой два метода идентификации остатков определяют перекрывающиеся, но не идентичные участки, они могут быть объединены для определения гибридного CDR. Однако предпочтительна нумерация по так называемой системе Kabat.
Обычно CDR формируют петлевую структуру, которую можно классифицировать как каноническую структуру. Термин «каноническая структура» относится к конформации основной цепи, которую принимают антиген-связывающие (CDR) петли. Сравнительными структурными исследованиями было обнаружено, что пять из шести антиген-связывающих петель имеют только ограниченный репертуар доступных конформаций. Каждую каноническую структуру можно охарактеризовать торсионными углами полипептидного остова. Соответствующие петли между антителами могут, следовательно, иметь очень похожие трехмерные структуры, несмотря на высокую вариабельность аминокислотной последовательности в большинстве частей петель (Chothia и Lesk, J. Mol. Biol., 1987, 196: 901; Chothia et al., Nature, 1989, 342: 877; Martin and Thornton, J. Mol. Biol, 1996, 263: 800). Кроме того, существует взаимосвязь между принятой структурой петли и окружающими ее аминокислотными последовательностями. Конформация конкретного канонического класса определяется длиной петли и аминокислотными остатками, находящимися в ключевых положениях внутри петли, а также внутри консервативного каркаса (т.е. вне петли). Отнесение к конкретному каноническому классу, следовательно, может быть выполнено на основе присутствия этих ключевых аминокислотных остатков.
Термин «каноническая структура» может также содержать соображения относительно линейной последовательности антитела, например, как каталогизировано Kabat (Kabat et al., Ioc. cit.). Схема (система) нумерации по Kabat является широко принятым стандартом для последовательной нумерации аминокислотных остатков вариабельного домена антитела и является предпочтительной схемой, задействованной в настоящем изобретении, как также упоминается везде в настоящем документе. Дополнительные структурные соображения также могут быть применены для определения канонической структуры антитела. Например, те различия, которые не полностью отражены в нумерации Kabat, могут быть описаны системой нумерации Chothia et al. и/или выявленные другими методами, например кристаллографией и двух- или трехмерным компьютерным моделированием. Соответственно, данная последовательность антитела может быть помещена в канонический класс, который позволяет, среди прочего, идентифицировать соответствующие последовательности шасси (например, на основании желания сделать библиотеку содержащей множество канонических структур). Нумерация по Kabat аминокислотных последовательностей антител и структурные соображения, как описано Chothia et al., Ioc. cit. и их значение для конструирования канонических аспектов структуры антитела описаны в литературе. Структуры субъединиц и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны в данной области. Для обзора структуры антител см. Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, ред. Harlow et al., 1988. Глобальным референтным источником в иммуноинформатике является база данных трехмерной (3D) структуры Международной информационной системы ImMunoGenetics (IMGT) (Ehrenmann et al., 2010, Nucleic Acids Res., 38, D301-307). Структурные данные базы данных трехмерной структуры IMGT (IMGT/3Dstructure-DB) извлекаются из Банка данных белков (PDB) и аннотируются в соответствии с концепциями классификации IMGT с применением внутренних инструментов. Таким образом, IMGT/3Dstructure-DB обеспечивает ближайшие гены и аллели, которые экспрессируются в аминокислотных последовательностях трехмерных структур, путем выравнивания этих последовательностей со справочным каталогом IMGT. Этот каталог вмещает, для рецепторов антигенов, аминокислотные последовательности доменов, кодируемых константными генами, а также трансляцию вариабельной зародышевой линии и присоединяющихся генов. Участки CDR наших аминокислотных последовательностей предпочтительно определяли с применением базы данных IMGT/3Dstructure.
CDR3 легкой цепи и, особенно, CDR3 тяжелой цепи могут составлять наиболее важные детерминанты связывания антигена в вариабельных участках легкой и тяжелой цепи. В некоторых конструкциях антител CDR3 тяжелой цепи, по-видимому, составляет основную площадь контакта между антигеном и антителом. Схемы отбора in vitro, в которых варьируется только CDR3, можно применять для изменения связывающих свойств антитела или определения того, какие остатки вносят вклад в связывание антигена. Следовательно, CDR3 обычно является самым большим источником молекулярного разнообразия в сайте связывания антитела. Например, НЗ может состоять всего из двух аминокислотных остатков или из более чем 26 аминокислот.
В классическом полно размерном антителе или иммуноглобулине каждая легкая (L) цепь связана с тяжелой (Н) цепью одной ковалентной дисульфидной связью, в то время как две Η-цепи связаны друг с другом одной или более дисульфидными связями в зависимости от изотипа Η-цепи. Домен СН, наиболее проксимальный к VH, обычно обозначается как СН1. Константные («С») домены не подразумевают непосредственного участия в связывании антигена, но проявляют различные эффекторные функции, такие как антителозависимая, клеточно-опосредованная цитотоксичность и активация комплемента. Участок Fc антитела включен в константные домены тяжелой цепи и, например, способен взаимодействовать с рецепторами Fc, расположенными на клеточной поверхности.
Последовательность генов антител после сборки и соматической мутации сильно различается, и оценочно эти разнообразные гены кодируют 1010 различных молекул антител (Immunoglobulin Genes, 2-е изд., ред. Jonio et al., Academic Press, San Diego, CA, 1995). Соответственно, иммунная система обеспечивает репертуар иммуноглобулинов. Термин «репертуар» относится по меньшей мере к одному из группы, состоящей из нуклеотидных последовательностей, полностью или частично полученных из по меньшей мере одной последовательности, кодирующей по меньшей мере один иммуноглобулин. Последовательность (-и) может (-гут) быть получена (-ы) путем перестройки in vivo сегментов V, D и J тяжелых цепей и сегментов V и J легких цепей. Альтернативно, последовательность (-и) может (-гут) быть получена (-ы) из клетки, в ответ на которую происходит перестройка, например, стимуляция in vitro. Альтернативно, часть или вся последовательность (-и) может (-гут) быть получена (-ы) сплайсингом ДНК, синтезом нуклеотидов, мутагенезом и другими методами, см., например, патент США 5565332. Репертуар может содержать только одну последовательность или может содержать множество последовательностей, содержа те, которые входят в генетически диверсифицированную коллекцию.
Конструкция антитела, определенная в контексте изобретения, может также включать дополнительные домены, которые, например, полезны при выделении молекулы или относятся к адаптированному фармакокинетическому профилю молекулы. Домены, полезные для выделения конструкции антитела, могут быть выбраны из пептидных мотивов или вторично внесенных фрагментов, которые могут быть захвачены способом выделения, например колонкой для выделения. Неограничивающие варианты осуществления таких дополнительных доменов включают пептидные мотивы, известные как Мус-метка, НАТ-метка, НА-метка, ТАР-метка, GST-метка, хитин-связывающий домен (CBD-метка), мальтозо-связывающий белок (МВР-метка), Flag-метка, Strep-метка и их варианты (например, Strepll-метка) и His-метка. Все описанные в настоящем документе конструкции антител, характеризующиеся идентифицированными CDR, могут включать домен His-метка, который обычно известен как повтор последовательных остатков гистидина (His) в аминокислотной последовательности молекулы, предпочтительно пяти, а более предпочтительно шести остатков His (гекса-гистидин). His-метка может быть расположена, например, на N- или С-конце конструкции антитела, предпочтительно она расположена на С-конце. Наиболее предпочтительно гекса-гистидиновая метка (НННННН) (SEQ ID NO: 25) связана посредством пептидной связи с С-концом конструкции антитела согласно изобретению. Кроме того, система конъюгата полилактид-ко-гликолид полиэтиленгликоль - полилактид-ко-гликолид, ПЛГА-ПЭГ-ПЛГА (PLGA-PEG-PLGA) может быть объединена с полигистидиновой меткой для приложения с замедленным высвобождением и улучшения фармакокинетического профиля.
Также обдуманы модификации аминокислотной последовательности описанных в настоящем документе конструкций антител. Например, может быть желательно улучшить аффинность связывания и/или другие биологические свойства конструкции антитела. Варианты аминокислотных последовательностей конструкций антител получают путем внесения соответствующих нуклеотидных изменений в нуклеиновую кислоту конструкций антител или путем пептидного синтеза. Все описанные ниже модификации аминокислотной последовательности должны приводить к конструкции антитела, которая все же сохраняет желаемую биологическую активность (связывание с рецептором NK-клетки, например, CD16a, и поверхностным антигеном клетки-мишени) немодифицированной родительской молекулы.
Термин «аминокислота» или «аминокислотный остаток» обычно относится к аминокислоте, имеющей определение, принятое в данной области, такой как аминокислота, выбранная из группы, состоящей из: аланина (Ala или А); аргинина (Arg или R); аспарагина (Asn или N); аспарагиновой кислоты (Asp или D); цистеина (Cys или С); глутамина (Gin или Q); глутаминовой кислоты (Glu или Е); глицина (Gly или G); гистидина (His или Н); изолейцина (Не или I): лейцина (Leu или L); лизина (Lys или K); метионина (Met или М); фенилаланина (Phe или F); пролина (Pro или Р); серина (Ser или S); треонина (Thr или Т); триптофана (Trp или W); тирозина (Tyr или Y); и валина (Val или V), хотя при желании можно применять модифицированные, синтетические или редкие аминокислоты. Обычно аминокислоты могут быть сгруппированы как имеющие неполярную боковую цепь (например, Ala, Cys, Не, Leu, Met, Phe, Pro, Val); отрицательно заряженную боковую цепь (например, Asp, Glu); положительно заряженную боковую цепь (например, Arg, His, Lys); или незаряженную полярную боковую цепь (например, Asn, Cys, Gin, Gly, His, Met, Phe, Ser, Thr, Trp и Tyr).
Аминокислотные модификации содержат, например, удаления и/или вставки, и/или замены остатков в аминокислотных последовательностях конструкций антител. Любая комбинация удаления, вставки и замены делается для получения конечной конструкции при обеспечении условия, что конечная конструкция обладает желаемыми характеристиками. Изменения аминокислот также могут изменять посттрансляционные процессы конструкций антител, такие как изменение количества или положения сайтов гликозилирования.
Например, 1, 2, 3, 4, 5 или 6 аминокислот могут быть вставлены, заменены или удалены в каждый из CDR (конечно, в зависимости от их длины), в то время как 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 25 аминокислот могут быть вставлены, заменены или удалены в каждом из FRob. Предпочтительно, вставки аминокислотной последовательности в конструкцию антитела содержат амино- и/или карбоксиконцевые слияния в диапазоне длины от 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 остатков до полипептидов, содержащих сто или более остатков, а также вставки внутрь последовательности одного или нескольких аминокислотных остатков. Соответствующие модификации также могут быть выполнены в третьем домене конструкции антитела, определенном в контексте изобретения. Вставочный вариант конструкции антитела, определенный в контексте изобретения, содержит слияние с N-концом или с С-концом конструкции антитела фермента или слияние с полипептидом.
Сайты, представляющие наибольший интерес для замещающего мутагенеза, содержат (без ограничения перечисленными) CDR тяжелой и/или легкой цепи, особенно гипервариабельные участки, но изменения FR в тяжелой и/или легкой цепи также рассматриваются. Замены предпочтительно представляют собой консервативные замены, как описано в настоящем документе. Предпочтительно, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот могут быть заменены в CDR, в то время как 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 25 аминокислот могут быть заменены в каркасных участках (FR), в зависимости от длины CDR или FR. Например, если последовательность CDR охватывает 6 аминокислот, предполагается, что одна, две или три из этих аминокислот заменены. Аналогичным образом, если последовательность CDR охватывает 15 аминокислот, предполагается, что одна, две, три, четыре, пять или шесть из этих аминокислот заменены.
Полезный способ идентификации определенных остатков или участков конструкций антител, которые являются предпочтительными местами для мутагенеза, называется «аланин-сканирующий мутагенез», какописано Cunningham и Wells в Science, 244:1081-1085 (1989). Здесь остаток или группа остатков-мишеней в конструкции антитела идентифицируются (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяются нейтральной или отрицательно заряженной аминокислотой (наиболее предпочтительно аланином или полиаланином), чтобы влиять на взаимодействие аминокислот с эпитопом.
Те положения аминокислот, которые демонстрируют функциональную чувствительность к заменам, затем усовершенствуют путем внесения дополнительных или других вариантов в, или для, сайты (-ов) замены. Таким образом, хотя сайт или участок для внесения вариации аминокислотной последовательности предопределены, природа мутации как таковая не требует предварительного определения. Например, чтобы проанализировать или оптимизировать эффект мутации в данном сайте, можно провести сканирование аланином или случайный мутагенез в целевом кодоне или участке, и экспрессированные варианты конструкции антитела подвергаются скринингу на предмет оптимальной комбинации желаемой активности. Техники создания заменяющих мутаций в заранее определенных сайтах ДНК, имеющих известную последовательность, хорошо известны, например, мутагенез с помощью праймера М13 и мутагенез с помощью ПЦР. Скрининг мутантов проводится с применением анализов активности связывания антигена, такого как рецептор NK-клеток, например CD16a, и связывания поверхностного антигена клетки-мишени.
Обычно, если аминокислоты замещены в одной или более или всех CDR тяжелой и/или легкой цепи, предпочтительно, чтобы полученная таким образом «замещенная» последовательность была по меньшей мере на 60% или 65%, еще более предпочтительно на 70% или 75%, даже более предпочтительно на 80% или 85% и особенно предпочтительно на 90% или 95% идентична «оригинальной» последовательности CDR. Это означает, что от длины CDR зависит то, в какой степени она идентична «замещенной» последовательности. Например, CDR, имеющий 5 аминокислот, предпочтительно на 80% идентичен его замещенной последовательности, чтобы иметь по меньшей мере одну аминокислоту замещенной. Соответственно, CDR конструкции антитела могут иметь различные степени идентичности с их замещенными последовательностями, например, CDRL1 может иметь 80%, тогда как CDRL3 может иметь 90%.
Предпочтительные замены (или замещения) представляют собой консервативные замены. Однако любая замена (содержа при этом неконсервативную замену или одну или более «иллюстративных замен», перечисленных в Таблице 3 ниже) предусматривается до тех пор, пока конструкция антитела сохраняет свою способность связываться с рецептором NK-клетки, например CD16a, через первый домен, и с поверхностным антигеном клетки-мишени через второй домен и/или его CDR идентичны замещенной последовательности (по меньшей мере на 60% или 65%, более предпочтительно на 70% или 75%, еще более предпочтительно на 80%% или 85%, и особенно предпочтительно на 90% или 95% идентичны «оригинальной» последовательности CDR).
Консервативные замены показаны в Таблице 1 под заголовком «предпочтительные замены». Если такие замены приводят к изменению биологической активности, тогда более существенные изменения, названные «иллюстративными заменами» в Таблице 1 или, как дополнительно описано ниже в соотношении с классами аминокислот, могут быть внесены, и продукты будут подвергаться скринингу на предмет желаемой характеристики.
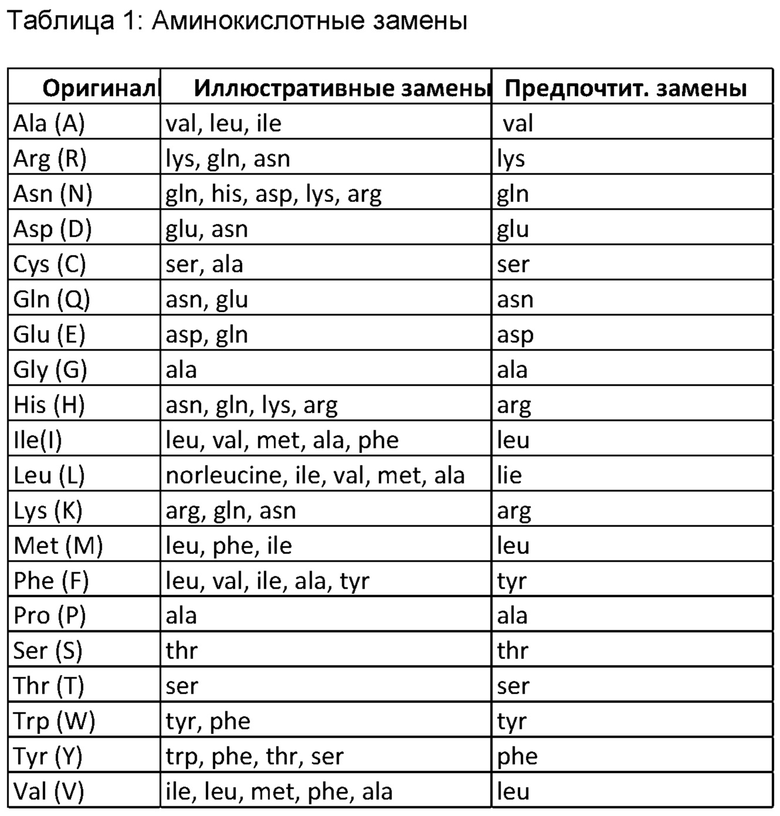
Существенные модификации биологических свойств конструкции антитела по настоящему изобретению достигаются путем выбора замен, которые значительно различаются по своему влиянию на поддержание (а) структуры полипептидного остова в области замены, например, в виде листа или спиральной конформации, (b) заряда или гидрофобности молекулы в целевом сайте, или (с) массива боковой цепи. Встречающиеся в природе остатки делятся на группы на основе общих свойств боковой цепи: (1) гидрофобные: норлейцин, met, ala, val, leu, ile; (2) нейтральные гидрофильные: cys, ser, thr, asn, gin; (3) кислые: asp, glu; (4) основные: his, lys, arg; (5) остатки, влияющие на ориентацию цепи: gly, pro; и (6) ароматические: trp, tyr, phe.
Неконсервативные замены повлекут за собой замену члена одного из этих классов на другой класс. Любой остаток цистеина, в отношении которого не подразумевается, что он участвует в поддержании надлежащей конформации конструкции антитела, может быть заменен, как правило, серином для улучшения окислительной стабильности молекулы и предотвращения аберрантного перекрестного связывания. Напротив, цистеиновая (-ые) связь (-и) может (-гут) быть добавлена (-ы) к антителу для повышения его стабильности (особенно когда антитело представляет собой фрагмент антитела, такой как фрагмент Fv).
Для аминокислотных последовательностей идентичность и/или сходство последовательностей определяют с применением стандартных методик, известных в данной области, содержа, без ограничения перечисленными, алгоритм идентичности локальных последовательностей Smith и Waterman, 1981, Adv. Appl. Math. 2: 482, алгоритм выравнивания идентичности последовательностей Needleman и Wunsch, 1970, J. Mol. Biol. 48: 443, способ поиска сходства Pearson и Lipman, 1988, Proc. Nat. Acad. Sci. USA 85: 2444, компьютеризированные варианты реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Drive, Madison, Wis.), программу последовательностей Best Fit, описанная Devereux et al., 1984, Nucl. Acid Res. 12: 387-395, желательно с применением настроек по умолчанию или путем проверки. Предпочтительно процент идентичности рассчитывается FastDB на основе следующих параметров: штраф за несовпадение, составляющий 1; штраф за гэп, составляющий 1; штраф за длину гэпа, составляющий 0,33; и штраф за присоединение, составляющий 30, "Current Methods in Sequence Comparment and Analysis", Macromolecule Sequencing and Synthesis, Selected Methods and Applications, c. 127-149 (1988), Alan R. Liss, Inc.
Примером полезного алгоритма является PILEUP. PILEUP создает множественное выравнивание последовательностей из группы связанных последовательностей с применением прогрессивных попарных выравниваний. Он также может построить дерево, показывающее отношения кластеризации, применяемые для создания выравнивания. PILEUP применяет упрощение способа прогрессивного выравнивания Feng & Doolittle, 1987, J. Mol. Evol. 35: 351-360; способ аналогичен описанному Higgins и Sharp, 1989, CABIOS 5: 151-153. Полезные параметры PILEUP содержат вес гэпа по умолчанию, составляющий 3,00, вес длины гэпа по умолчанию, составляющий 0,10, и концевые гэпы с приданным весом.
Другим примером полезного алгоритма является алгоритм BLAST (Средство поиска основного локального выравнивания), описанный в: Altschul et al., 1990, J. Mol. Biol. 215: 403-410; Altschul et al., 1997, Nucleic Acids Res. 25: 3389-3402; и Karin et al., 1993, Proc. Natl. Acad. Sci. U.S.A. 90: 5873-5787. Особенно полезной программой BLAST является программа WU-BLAST-2, которая была получена от Altschul et al., 1996, Methods in Enzymology 266: 460-480. WU-BLAST-2 использует несколько параметров поиска, в большинстве из которых установлены значения по умолчанию. Регулируемые параметры устанавливаются со следующими значениями: охват перекрытия = 1, фракция перекрытия = 0,125, порог слова (Т)=II. Параметры HSP S и HSP S2 являются динамическими значениями и устанавливаются самой программой в зависимости от композиции конкретной последовательности и композиции конкретной базы данных, по которой выполняется поиск интересующей последовательности; однако значения можно отрегулировать для увеличения чувствительности.
Дополнительным полезным алгоритмом является BLAST с гэпами, как сообщается Altschul et al., 1993, Nucl. Acids Res. 25: 3389-3402. BLAST с гэпами применяет очки за замену BLOSUM-62; пороговый параметр Τ установлен на 9; способ двух попаданий для запуска расширений без гэпов штрафует за длину гэпов, составляющую k, в размере 10+k; для Xu установлено значение 16, а для Xg установлено значение 40 для этапа поиска в базе данных и 67 для этапа вывода алгоритмов. Выравнивания с гэпами запускаются счетом, соответствующим около 22 битам.
Как правило, аминокислотная гомология, сходство или идентичность между индивидуальными вариантными CDR или последовательностями VH/VL составляет по меньшей мере 60% от последовательностей, указанных в данном документе, и более типично с предпочтительным увеличением гомологии или идентичности по меньшей мере на 65% или 70%, более предпочтительно по меньшей мере на 75% или 80%, еще более предпочтительно по меньшей мере на 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% и почти на 91%. Аналогичным образом, «процент (%) идентичности последовательности нуклеиновой кислоты» по отношению к последовательности нуклеиновой кислоты связывающих белков, идентифицированных в настоящем документе, определяется как процент нуклеотидных остатков в последовательности-кандидате, которые идентичны нуклеотидным остаткам в кодирующей последовательности конструкции антитела. В специфичном способе используется модуль BLASTN (Медленное сравнение с целью поиска всех сходных последовательностей) программы WU-BLAST-2, настроенный на параметры по умолчанию, с охватом перекрытия и фракцией перекрытия, установленными на 1 и 0,125, соответственно.
Как правило, гомология, сходство или идентичность последовательностей нуклеиновых кислот между нуклеотидными последовательностями, кодирующими индивидуальные вариантные CDR или последовательности VH/VL, и нуклеотидными последовательностями, указанными в данном документе, составляют по меньшей мере 60%, и более типично с предпочтительно увеличивающейся гомологией или идентичностью по меньшей мере 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% и почти 100%. Таким образом, «вариантный CDR» или «вариантный участок VH/VL» является участком с указанной гомологией, сходством или идентичностью с родительским CDR/VH/VL, определенным в контексте изобретения, и разделяет биологическую функцию, включая, без ограничения перечисленным, по меньшей мере 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% специфичности и/или активности родительского CDR или VH/VL.
В одном варианте осуществления процент идентичности с человеческой зародышевой линией конструкции антитела в соответствии с изобретением составляет более 70% или более 75%, более предпочтительно более 80% или более 85%, даже более предпочтительно более 90% и наиболее предпочтительно более 91%, более 92%, более 93%, более 94%, более 95% или даже более 96%. Идентичность продуктам генов зародышевой линии человеческих антител считается важным признаком снижения риска вызывания терапевтическими белками иммунного ответа против лекарственного средства у пациента во время лечения. Hwang & Foote ("Immunogenicity of engineered antibodies"; Methods 36 (2005) 3-10) демонстрируют, что уменьшение нечеловеческих частей конструкций лекарственных антител приводит к снижению риска индукции антител к лекарствам у пациентов во время лечения. Путем сравнения исчерпывающего количества клинически оцененных лекарств на основе антител и соответствующих данных об иммуногенности показана тенденция, согласно которой гуманизация V-участков антител делает белок менее иммуногенным (в среднем 5,1% пациентов), чем антитела, несущие неизмененные нечеловеческие V-участки (в среднем 23,59% пациентов). Более высокая степень идентичности человеческим последовательностям, следовательно, желательна для белковых терапевтических средств на основе V-участка в форме конструкций антител. Для этой цели определения идентичности зародышевой линии V-участки VL могут быть выровнены с аминокислотными последовательностями V-сегментов и J-сегментов человеческой зародышевой линии (http://vbase.mrc-cpe.cam.ac.uk/) с применением программного обеспечения Vector NTI и аминокислотной последовательности, рассчитанной путем деления идентичных аминокислотных остатков на общее количество аминокислотных остатков VL в процентах. То же самое может быть сделано для сегментов VH (http://vbase.mrc-cpe.cam.ac.uk/) за исключением того, что VH CDR3 может быть исключен из-за его высокого разнообразия и отсутствия существующих партнеров для выравнивания VH CDR3 человеческой зародышевой линии. Затем можно применять рекомбинантные техники для увеличения идентичности последовательности к генам антител человеческой зародышевой линии.
Термин «миеломная клетка» означает злокачественную (раковую) плазматическую клетку, возникающую из плазматической клетки в костном мозге в результате неопластической трансформации. При миеломе злокачественные плазматические клетки продуцируют большое количество аномальных антител, которые не способны бороться с инфекцией. Эти аномальные антитела представляют собой так называемый моноклональный белок или М-белок, который функционирует как опухолевый маркер миеломы. Клетка миеломы имеет фенотип CD19 -/CD38+/CD138+/BCMA+. Следовательно, CD38, CD138 и ВСМА представляют собой антигены, экспрессируемые на миеломной клетке. Также здесь содержатся злокачественные фенотипы в линии В-клеток, которые положительны по CD19/CD20/CD22/BCMA и другим антигенам (это должно содержать фенотипы, которые классически не понимаются как плазматические клетки, но могут развиваться из В-клеток памяти или пре-плазматической клеточной линии).
Термин «РЭФР» («EGFR») относится к рецептору эпидермального фактора роста (EGFR; ErbB-1; HER1) у человека, содержа все изоформы или варианты, описанные с активацией, мутациями и вовлеченные в патофизиологические процессы. Связывающий сайт антигена РЭФР (EGFR) распознает эпитоп во внеклеточном домене РЭФР (EGFR). В некоторых вариантах осуществления антиген-связывающий сайт специфично связывается с РЭФР (EGFR) человека и яванского макака. Рецептор эпидермального фактора роста, РЭФР (EGFR) является членом семейства HER рецепторных тирозинкиназ и состоит из четырех членов: EGFR (ErbB1/HER1), HER2/neu (ErbB2), HER3 (ErbB3) и HER4 (ErbB4). Стимуляция рецептора посредством связывания лиганда (например, EGF, TGFa, HB-EGF, нейрегулины, бетацеллулин, амфирегулин) активирует внутреннюю рецепторную тирозинкиназу во внутриклеточном домене через фосфорилирование тирозина и способствует гомо- или гетеродимеризации рецептора с членами семейства HER. Эти внутриклеточные фосфотирозины служат в качестве стыковочных сайтов для различных адаптерных белков или ферментов, включая SHC, GRB2, PLCg и PI(3)K/Akt, которые одновременно инициируют множество сигнальных каскадов, которые влияют на пролиферацию клеток, ангиогенез, устойчивость к апоптозу, инвазию и метастазирование.
СИФ (MFI) означает среднюю интенсивность флуоресценции.
Подробное описание изобретения:
Таким образом, в первом аспекте настоящее изобретение обеспечивает выделенные человеческие NK-клетки в криоконсервированном состоянии, предварительно нагруженные перед замораживанием конструкцией антитела, указанная конструкция антитела включает по меньшей мере первый связывающий домен, связывающийся с антигеном рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки и второй связывающий домен, связывающий с клеточной поверхностью антиген на клеточной поверхности клетки-мишени.
Более того, в другом аспекте настоящее изобретение обеспечивает способ восстановления/получения таких жизнеспособных человеческих NK-клеток, предварительно нагруженных такой конструкцией антитела, из криоконсервированного состояния.
На сегодняшний день готовые продукты (аллогенные) сильно отстают от индивидуализированных (аутологичных) подходов. Хотя аутологичный перенос клеток валидирован клинически, производство клеточных продуктов требуется наладить для каждого отдельного пациента. Индивидуализированное производства требует от 7 до 14 дней для выделения, размножения и получения клеток, несмотря на то, что используются различные стратегии оптимизации. Одним из ограничений является то, что для некоторых пациентов аутологичное размножение их иммунных клеток не дает достаточного количества клеток. Кроме того, было описано, что иммунные эффекторные клетки больного раком функционально ослаблены из-за иммуносупрессии и измененных фенотипов. Следовательно, вариабельность между пациентами отрицательно влияет как на индивидуализированное производство, так и на клиническую эффективность.
В отличие от аутологичных подходов, готовые аллогенные продукты предлагают уникальные преимущества: такие продукты могут быть задействованы по отношению к пациенту сразу же, то есть сразу после размораживания, что значительно сокращает время цепи снабжения/логистики и увеличивает доступность продукта вне одного централизованного госпиталя. Большие партии аллогенных иммунных эффекторных клеток могут быть произведены в централизованной производственной точке, что не только снижает производственные затраты, но также увеличивает однородность продукта иммунных эффекторных клеток, обеспечивая соблюдение высоких стандартов качества. Такие централизованные предприятия могут также позволить отбор подходящих здоровых доноров для применения наиболее функциональной популяции эффекторных клеток для производственного процесса.
Как уже указывалось выше, согласно первому варианту осуществления настоящего изобретения обеспечены выделенные человеческие NK-клетки в криоконсервированном состоянии, предварительно нагруженные перед замораживанием конструкцией антитела, указанная конструкция антитела содержит по меньшей мере первый связывающий домен, связывающийся с антигеном рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки, и второй связывающий домен, связывающий с клеточной поверхностью антиген на клеточной поверхности клетки-мишени, который может происходить из любого из вышеуказанных источников.
NK-клетки, применяемые в контексте изобретения, представляют собой, например, выделенные из мононуклеарных клеток периферической крови, МКПК (РВМС) здоровых доноров, характеризующиеся различными фенотипами (например, адаптивные клетки памяти NK100), выделенные из ткани пуповины или плаценты, NK-клетки, полученные из индуцированных плюрипотентных стволовых клеток (полученные из ИПСК (iPSC)) (например, FT500 или FT516)) или NK-клеточных линий. Примеры подходящих линий NK-клеток включают, без ограничения перечисленными, клетки NK-92 (АТСС® CRL-2407™), клетки NK-YS (Tsuchiyama et al. 1998, Blood) или клетки NK-YT (Teshigawara et al., J. Immunol., 1985). Также в группу подходящих NK-клеток входят NK-клетки с химерным антигенным рецептором (CAR), которые могут быть получены из любого из вышеуказанных источников.
Термин «ИПСК» (iPSC) в контексте изобретения определяет индуцированные плюрипотентные стволовые клетки, которые можно применять для создания дифференцированных и специализированных типов клеток (например, NK-клеток) с использованием принципов генной инженерии, определенных средств лечения с помощью малых молекул и размножения клеток.
В некоторых вариантах осуществления изобретения необходимо размножить и/или активировать NK-клетки до стадии предварительной нагрузки клеток указанной конструкцией антитела. Протоколы такого размножения известны в данной области, например, из WO2017/042393, WO2015/132415, US20110014162, US20120308986A1, US9062287B2, US8026097B2, US9260696B2; US20170119865.
В соответствии с изобретением NK-клетки можно культивировать перед инкубацией с конструкцией антитела. Такой этап клеточной культуры может быть необходим для размножения и/или активации соответствующих клеток. Типичные протоколы размножения могут включать отбор клеток CD34+, деплецию клеток CD3+ и инкубацию с питающими клетками, несущими 4-1BBL и/или трансмембранный IL-21, и, необязательно, дополнительную стимуляцию лимфопролиферативными цитокинами, такими как IL-2. В некоторых вариантах осуществления партия NK-клеток, предварительно нагруженных конструкцией антитела, включает 106 и 1012 клеток. Количество предварительно инкубированных клеток, которые могут быть введены пациенту, может находиться в диапазоне от 1×105 до 1×108 NK-клеток на кг веса тела.
В контексте изобретения «клетки в криоконсервированном состоянии» представляют собой клетки, которые были консервированы путем охлаждения до температуры ниже нуля (градусов Цельсия). Криоконсервированные клетки могут или могут не быть консервированы в присутствии криопротекторного агента. Криопротекторный агент, такой как диметилсульфоксид, ДМСО (DMSO), глицерин, этиленгликоль или пропиленгликоль, представляет собой вещество, которое защищает клетки от повреждений, связанных охранением при температуре ниже нуля и/или замораживании, например, от повреждения клеточной мембраны из-за образования кристаллов льда.
Термин «предварительно нагруженный перед замораживанием» относится к предварительной инкубации клеток перед охлаждением до температуры ниже нуля с идентифицированной конструкцией антитела.
Следует отметить, что первый связывающий домен описанной конструкции антитела связывает антиген рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки с этим рецептором таким образом, который позволяет поддерживать связывание на клеточной поверхности даже во время и после того, как клетка находится в криоконсервированном состоянии. Как показано в прилагаемом примере 1, конструкции антител, имеющие первый связывающий домен, специфичный для CD16 (а или b), связываются со специфичным эпитопом, отличным от связывающего домена IgG FcRγIII. Предполагается, что связывание с таким специфичным эпитопом позволяет поддерживать связывание даже во время процесса охлаждения NK-клеток согласно одному аспекту изобретения до температуры ниже нуля и последующего восстановления клеток для введения субъекту, испытывающему в этом потребность. Подобные эпитопы существуют также для других антигенов рецептора NK-клеток на клеточной поверхности иммунологических эффекторных клеток, например NKp46 или NKG2D.
Согласно одному варианту осуществления изобретения выделенные человеческие NK-клетки были выделены из ткани пуповины или МКПК (РВМС) от здоровых доноров.
Кроме того, в одном варианте осуществления выделенные человеческие NK-клетки согласно одному аспекту изобретения консервированы в криорастворе. Криораствор в контексте изобретения включает, по меньшей мере, базальную среду для культивирования клеток и криозащитный агент. Более того, криораствор может дополнительно включать изотонический раствор, такой как Plasma-lyte, и/или сывороточный альбумин.
Неограничивающие примеры криозащитных агентов были обеспечены в данном документе выше. Определенными вариантами осуществления криозащитных агентов в контексте изобретения являются ДМСО (DMSO) и глицерин. Концентрация криозащитного агента может находиться в диапазоне от 1% до 30% (об/об), В некоторых вариантах осуществления в диапазоне от 5% до 25% (об/об), в других вариантах осуществления в диапазоне от 10% до 25% (об/об), также в некоторых вариантах осуществления в концентрации, составляющей 20% (об/об). Термин «базальная среда для культивирования клеток» относится в контексте изобретения к жидкой питательной среде для клеточной культуры, которая подходит для клеточной культуры клеток млекопитающих. Примеры таких «базальных среда для культивирования клеток» хорошо известны в данной области. Концентрация изотонического раствора может находиться в диапазоне от 1% до 60% (об/об), в некоторых вариантах осуществления в диапазоне от 10% до 50% (об/об), в других вариантах осуществления в диапазоне от 20% до 45% (об/об) и в одном варианте осуществления в концентрации, составляющей 40% (об/об). Сывороточный альбумин может быть человеческим или (фетальным) бычьим (предпочтительно человеческим) сывороточным альбумином, который был выделен от доноров или получен рекомбинантно. Концентрация альбумина в криорастворе может находиться в диапазоне от 1% до 40% (об/об), в некоторых вариантах осуществления в диапазоне от 5% до 25% (об/об), также в некоторых вариантах осуществления в диапазоне от 10% до 20% (об/об). Пример криораствора (замораживающей среды) может включать базальную среду, 40% Plasma-lyte, человеческий сывороточный альбумин, ЧСА (HSA) и 10% диметилсульфоксид, ДМСО (DMSO). Альтернативно можно применять фетальную бычью сыворотку, ФБС (FBS) или глицерин для замены ЧСА (HSA) и/или ДМСО (DMSO) соответственно. Наиболее часто применяемым криопротектором является ДМСО (DMSO). В силу разных причин ДМСО (DSMO) может быть заменен альтернативными соединениями, такими как глицерин, этиленгликоль или пропиленгликоль.
Согласно определенному варианту осуществления NK-клетки одного аспекта изобретения были предварительно нагружены в растворе, содержащем конструкцию антитела в концентрации по меньшей мере 5 нМ. В определенных вариантах осуществления концентрация составляет по меньшей мере 10 нМ, более того, в определенных вариантах осуществления концентрация составляет по меньшей мере 25 нМ, также в определенных вариантах осуществления концентрация составляет по меньшей мере 50 нМ, в определенных вариантах осуществления концентрация составляет по меньшей мере 75 нМ, 90 нМ, 100 нМ, 125 нМ или 150 нМ.
Антиген рецептора NK-клетки, с которым связывается первый связывающий домен конструкции антитела на поверхности выделенных человеческих NK-клеток согласно одному аспекту изобретения выбран из группы, состоящей из CD16a, CD16b, NKp46, NKG2D и CD16a+CD16b.
Определенные варианты осуществления рецепторов NK-клеток были определены в настоящем документе выше. Первый связывающий домен указанной конструкции антитела связывается, таким образом, либо специфично с CD16a, CD16b, либо с CD16a и CD16b. Более того, первый связывающий домен может специфично связываться с NKp46 или NKG2D.
Антиген клеточной поверхности на клеточной поверхности клетки-мишени, с которой связывается второй связывающий домен конструкции антитела, выбран из группы, состоящей из CD19, CD20, CD22, CD30, CD33, CD52, CD70, CD74, CD79b, CD123, CLL1, ВСМА, FCRH5, EGFR, EGFRvIII, HER2, GD2.
Эти антигены клеточной поверхности на поверхности клеток-мишеней связаны со специфичными патологическими образованиями. CD30 является антигеном клеточной поверхности, характерным для злокачественных клеток лимфомы Ходжкина. CD19, CD20, CD22, CD70, CD74 и CD79b являются антигенами клеточной поверхности, характерными для злокачественных клеток неходжкинских лимфом (диффузная крупно-В-клеточная лимфома (DLBCL), мантийноклеточная лимфома (MCL), фолликулярная лимфома (FL), Т- клеточные лимфомы (как периферические, так и кожные, содержа трансформированный грибовидный микоз/синдром Сезари TMF/SS и анапластическую крупноклеточную лимфому (ALCL)). CD52, CD33, CD123, CLL1 являются антигенами клеточной поверхности, характерными для злокачественных клеток при лейкозах (хронический лимфоцитарный лейкоз (CLL), острый лимфобластный лейкоз (ALL), острый миелоидный лейкоз (AML)). ВСМА, FCRH5 являются антигенами клеточной поверхности, характерными для злокачественных клеток при множественной миеломе. РЭФР (EGFR), HER2, GD2 являются антигенами клеточной поверхности, характерными для солидного рака (трижды отрицательный рак молочной железы (TNBC), рак молочной железы (ВС), коло ректальный рак (CRC), немелкоклеточная карцинома (NSCLC), мелкоклеточная карцинома (SCLC), также известная как «мелкоклеточный рак легкого» или «овсяноклеточная карцинома»), рак простаты (PC), глиобластома (также известная как мультиформная глиобластома (GBM)).
В одном варианте осуществления человеческих NK-клеток согласно одному аспекту изобретения конструкция антитела включает в первом связывающем домене три CDR тяжелой цепи и три CDR легкой цепи, выбранные из группы, состоящей из:
(a) CDR-H1, указанного под SEQ ID NO: 29, CDR-H2, указанного под SEQ ID NO: 30, CDR-H3, указанного под SEQ ID NO: 31, CDR-L1, указанного под SEQ ID NO: 32, CDR-L2, указанного под SEQ ID NO: 33, CDR-L3, указанного под SEQ ID NO: 34;
(b) CDR-H1, указанного под SEQ ID NO: 40, CDR-H2, указанного под SEQ ID NO: 41, CDR-H3, указанного под SEQ ID NO: 42, CDR-L1, указанного под SEQ ID NO: 43, CDR-L2, указанного под SEQ ID NO: 44, CDR-L3, указанного под SEQ ID NO: 45;
(c) CDR-H1, указанного под SEQ ID NO: 51, CDR-H2, указанного под SEQ ID NO: 52, CDR-H3, указанного под SEQ ID NO: 53, CDR-L1, указанного под SEQ ID NO: 54, CDR-L2, указанного под SEQ ID NO: 55, CDR-L3, указанного под SEQ ID NO: 56;
(d) CDR-H1, указанного под SEQ ID NO: 62, CDR-H2, указанного под SEQ ID NO: 63, CDR-H3, указанного под SEQ ID NO: 64, CDR-L1, указанного под SEQ ID NO: 65, CDR-L2, указанного под SEQ ID NO: 66, CDR-L3, указанного под SEQ ID NO: 67;
(e) CDR-H1, указанного под SEQ ID NO: 73, CDR-H2, указанного под SEQ ID NO: 74, CDR-H3, указанного под SEQ ID NO: 75, CDR-L1, указанного под SEQ ID NO: 76, CDR-L2, указанного под SEQ ID NO: 77, CDR-L3, указанного под SEQ ID NO: 78;
(f) CDR-H1, указанного под SEQ ID NO: 84, CDR-H2, указанного под SEQ ID NO: 85, CDR-H3, указанного под SEQ ID NO: 86, CDR-L1, указанного под SEQ ID NO: 87, CDR-L2, указанного под SEQ ID NO: 88, CDR-L3, указанного под SEQ ID NO: 89; и
(g) CDR-H1, указанного под SEQ ID NO: 95, CDR-H2, указанного под SEQ ID NO: 96, CDR-H3, указанного под SEQ ID NO: 97, CDR-L1, указанного под SEQ ID NO: 98, CDR-L2, указанного под SEQ ID NO: 99, CDR-L3, указанного под SEQ ID NO: 100.
Также в одном варианте осуществления выделенных человеческих NK-клеток в соответствии с одним аспектом изобретения конструкция антитела включает в первом связывающем домене пары VH- и VL-цепей, имеющих последовательность, указанную в парах последовательностей, выбранных из группы, состоящей из SEQ ID NO: 35 и SEQ ID NO: 36, SEQ ID NO: 46 и SEQ ID NO: 47, SEQ ID NO: 57 и SEQ ID NO: 58, SEQ ID NO: 68 и SEQ ID NO: 69, SEQ ID NO: 79 и SEQ ID NO: 80, SEQ ID NO: 90 и SEQ ID NO: 91, и SEQ ID NO: 101 и SEQ ID NO: 102.
В варианте осуществления выделенных человеческих NK-клеток согласно одному аспекту изобретения конструкция антитела включает во втором связывающем домене три CDR тяжелой цепи и три CDR легкой цепи, выбранные из группы, состоящей из:
(a) CDR-H1, указанного под SEQ ID NO: 106, CDR-H2, указанного под SEQ ID NO: 107, CDR-H3, указанного под SEQ ID NO: 108, CDR-L1, указанного под SEQ ID NO: 109, CDR-L2, указанного под SEQ ID NO: 110, CDR-L3, указанного под SEQ ID NO: 111;
(b) CDR-H1, указанного под SEQ ID NO: 106, CDR-H2, указанного под SEQ ID NO: 107, CDR-H3, указанного под SEQ ID NO: 108, CDR-L1, указанного под SEQ ID NO: 109, CDR-L2, указанного под SEQ ID NO: 110, CDR-L3, указанного под SEQ ID NO: 111;
(c) CDR-H1, указанного под SEQ ID NO: 117, CDR-H2, указанного под SEQ ID NO: 118, CDR-H3, указанного под SEQ ID NO: 119, CDR-L1, указанного под SEQ ID NO: 120, CDR-L2, указанного под SEQ ID NO: 121, CDR-L3, указанного под SEQ ID NO: 122.
Кроме того, в одном варианте осуществления выделенных человеческих NK-клеток в соответствии с одним аспектом изобретения конструкция антитела включает во втором связывающем домене пары VH- и VL-цепей, имеющих последовательность, указанную в парах последовательностей, выбранных из группы, состоящей из SEQ ID NO: 112 и SEQ ID NO: 113, SEQ ID NO: 123 и SEQ ID NO: 124, и SEQ ID NO: 134 и SEQ ID NO: 135.
В одном варианте осуществления выделенной NK-клетки человека согласно одному аспекту изобретения конструкция антитела включает последовательность белка, указанную в SEQ ID NOs: 161-171.
В альтернативном варианте осуществления согласно настоящему изобретению предложен способ получения криоконсервированных предварительно нагруженных человеческих NK-клеток в криоконсервированном состоянии, указанный способ включает
(i) инкубирование NK-клеток с конструкцией антитела, указанная конструкция антитела включает, по меньшей мере, первый связывающий домен, связывающийся с антигеном рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки, и второй связывающий домен, связывающий с клеточной поверхностью антиген на клеточной поверхности клетки-мишени; и
(ii) замораживание NK-клеток.
Кроме того, согласно настоящему изобретению предложен такой способ, в котором NK-клетки выделяют из ткани пуповины, ИПСК (iPSC) или МКПК (РВМС) от здоровых доноров.
Как описано выше, NK-клетки, указанные в способе согласно одному аспекту изобретения, были предварительно нагружены в растворе, содержащем конструкцию антитела в концентрации по меньшей мере 5 нМ. В некоторых вариантах осуществления концентрация составляет по меньшей мере 10 нМ, в некоторых вариантах осуществления в концентрации по меньшей мере 25 нМ, по меньшей мере 50 нМ или по меньшей мере 75 нМ. В некоторых вариантах осуществления концентрация составляет по меньшей мере 90 нМ, 100 нМ, 125 нМ или 150 нМ.
В одном варианте осуществления способа согласно одному аспекту изобретения антиген рецептора NK-клетки, с которым связывается первый связывающий домен конструкции антитела, выбран из группы, состоящей из CD16a, CD16b, NKp46, NKG2D и CD16a+CD16b.
Кроме того, согласно определенным вариантам осуществления, антиген клеточной поверхности на клеточной поверхности клетки-мишени, с которой связывается второй связывающий домен конструкции антитела, выбран из группы, состоящей из CD19, CD20, CD22, CD30, CD33, CD52, CD70, CD74, CD79b, CD123, CLL1, ВСМА, FCRH5, EGFR, HER2, GD2.
В соответствии со способом согласно одному аспекту изобретения для получения криоконсервированных предварительно нагруженных человеческих NK-клеток этап замораживания NK-клеток выполняется с применением среды для замораживания/криораствора. Некоторые композиции и соединения замораживающей среды/криораствора в соответствии с изобретением были описаны в настоящем документе выше.
В некоторых вариантах этого способа NK-клетки выделяют из ткани пуповины, ИПСК (iPSC) или МКПК (РВМС) от здоровых доноров.
Также для этого способа в одном аспекте изобретения предусмотрено, что NK-клетки предварительно нагружают в растворе, включающем конструкцию антитела в концентрации по меньшей мере 5 нМ.
Более того, в соответствии с изобретением находится то, что антиген рецептора NK-клетки, с которым связывается первый связывающий домен конструкции антитела, выбран из группы, состоящей из CD16a, CD16b, NKp46, NKG2D и CD16a+CD16b.
Также для этого способа в одном аспекте изобретения предусмотрено, что антиген клеточной поверхности на клеточной поверхности клетки-мишени, с которой связывается второй связывающий домен конструкции антитела, выбран из группы, состоящей из CD19, CD22, CD30, CD33, CD52, CD70, CD74, CD79b, CD123, CLL1, ВСМА, FCRH5, EGFR, HER2, GD2.
Согласно одному варианту осуществления этого способа в одном аспекте изобретения этап замораживания NK-клеток выполняется с применением среды для замораживания, которая вмещает, по меньшей мере, базальную среду для культивирования клеток и криозащитный агент.
В одном аспекте способа по изобретению конструкция антитела, применяемая в способе, представляет собой одну из конструкций антитела, описанных в настоящем документе выше. В некоторых вариантах осуществления способа изобретения указанная конструкция антитела включает последовательность белка, указанную в SEQ ID NOs: 161-171.
В альтернативном варианте осуществления согласно настоящему изобретению предложен способ восстановления/получения жизнеспособных предварительно нагруженных человеческих NK-клеток из человеческих NK-клеток согласно одному аспекту изобретения в криоконсервированном состоянии, описанном в настоящем документе выше, или полученных согласно способу согласно одному аспекту изобретения для получения криоконсервированных предварительно нагруженных человеческих NK-клеток в криоконсервированном состоянии для введения указанных клеток субъекту, испытывающему в этом потребность, указанный способ включает этап восстановления/получения клеток для введения пациенту путем размораживания.
Размораживание замороженных клеток может быть выполнено хорошо известными способами. Общие процедуры размораживания подразумевают быстрое перемещение замороженных клеток в водяную баню с температурой 37°С и осторожное перемешивание до полного размораживания активированных NK-клеток. После размораживания размороженные NK-клетки можно получить для введения пациенту путем добавления, предпочтительно по каплям, фармацевтически приемлемых носителей при комнатной температуре, например, для разбавления концентрации среды для замораживания и/или промывки ранее активированных NK-клеток. Как показано в настоящем документе, популяции клеток по настоящему изобретению сохраняют свое активированное состояние в комбинации с конструкцией антитела после такой консервации. Подготовка клеток к введению после консервирования зависит от способа консервирования. Например, клетки могут быть получены для введения после консервации клеточной культурой и/или замораживанием одним или более способами: промывка клеток, ресуспендирование клеток в фармацевтически приемлемом носителе и/или помещением клеток в подходящее устройство для доставки, например, шприц. В одном варианте осуществления клетки готовят для введения после криоконсервации путем размораживания, например, осторожным перемешиванием на водяной бане при 37°С, а затем помещают клетки в подходящее устройство для доставки, например, шприц. Неожиданно, эти размороженные популяции клеток могут применяться в клинике почти сразу по мере потребности, например, без реактивации или других обширных и/или длительных манипуляций.
Соответственно, хотя и не необходимо, клетки, полученные для введения после криоконсервации, могут быть дополнительно приготовлены путем добавления, предпочтительно по каплям, фармацевтически приемлемого носителя при комнатной температуре, например, для разбавления концентрации и/или отмывания клеток от среды для замораживания. В одном варианте фармацевтически приемлемый носитель по существу не содержит активирующего агента.
В другом альтернативном варианте осуществления согласно настоящему изобретению предложена фармацевтическая композиция, включающая человеческие NK-клетки, которые были восстановлены из человеческих NK-клеток в криоконсервированном состоянии в соответствии с изобретением или получены способом в соответствии с изобретением.
Некоторые варианты осуществления обеспечивают фармацевтические композиции, включающие предварительно нагруженные NK-клетки, определенные в контексте изобретения, и дополнительно один или более вспомогательных веществ, таких какте, которые иллюстративно описаны в этом разделе и в любых других местах настоящего документа. Вспомогательные вещества могут применяться в изобретении в этом отношении для широкого круга целей, таких как регулировка физических, химических или биологических свойств препаратов, таких как регулировка вязкости, и или способы одного аспекта изобретения для повышения эффективности и или для стабилизации таких препаратов и способов от деградации и порчи вследствие, например, стрессов, возникающих во время производства, транспортировки, хранения, подготовки перед применением, введения и после этого.
В некоторых вариантах осуществления фармацевтическая композиция может вмещать материалы препаратов для целей модификации, поддержания или консервации, например, рН, осмолярности, вязкости, прозрачности, цвета, изотоничности, запаха, стерильности, стабильности, скорости растворения или высвобождения, адсорбции или проникновения в композицию (см. REMINGTON'S PHARMACEUTICAL SCIENCES, 18" Edition, (AR Genrmo, ed.), 1990, Mack Publishing Company). В таких вариантах осуществления подходящие материалы препаратов могут содержать, без ограничения перечисленными:
• аминокислоты, такие как глицин, аланин, глутамин, аспарагин, треонин, пролин, 2-фенилаланин, содержа заряженные аминокислоты, предпочтительно лизин, ацетат лизина, аргинин, глутамат и/или гистидин
• противомикробные, такие как антибактериальные и противогрибковые агенты
• антиоксиданты, такие как аскорбиновая кислота, метионин, сульфит натрия или гидросульфит натрия;
• буферы, буферные системы и буферные агенты, которые применяются для поддержания композиции при физиологическом рН или немного более низком рН; примерами буферов являются борат, бикарбонат,
• Трис-HCl, цитраты, фосфаты или другие органические кислоты, сукцинат, фосфат и гистидин; например, Трис-буфер с рН около 7,0-8,5;
• неводные растворители, такие как пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и сложные органические эфиры для инъекций, такие как этилолеат;
• водные носители, содержа воду, спиртовые/водные растворы, эмульсии или суспензии, содержа физиологический раствор и забуференные среды;
• биоразлагаемые полимеры, такие как полиэфиры;
• объемообразующие агенты, такие как маннит или глицин;
• хелатирующие агенты, такие как этилендиаминтетрауксусная кислота, ЭДТА (EDTA);
• изотонические и замедляющие абсорбцию агенты;
• комплексообразователи, такие как кофеин, поливинилпирролидон, бета-циклодекстрин или гидроксипропил-бета-циклодекстрин)
• наполнители;
• моносахариды; дисахариды; и другие углеводы (такие как глюкоза, манноза или декстрины); углеводы могут быть невосстанавливающими сахарами, предпочтительно трегалозой, сахарозой, октасульфатом, сорбитолом или ксилитом;
• (низкомолекулярные) белки, полипептиды или белковые носители, такие как человеческий или бычий сывороточный альбумин, желатин или иммуноглобулины, предпочтительно человеческого происхождения;
• красители и ароматизаторы;
• серосодержащие восстановители, такие как глутатион, тиоктовая кислота, тиогликолят натрия, тиоглицерин, альфа-монотиоглицерин и тиосульфат натрия
• разбавители;
• эмульгаторы;
• гидрофильные полимеры, такие как поливинилпирролидон)
• солеобразующие противоионы, такие как натрий;
• консерванты, такие как противомикробные средства, антиоксиданты, хелатирующие агенты, инертные газы и тому подобное; примерами являются: бензалкония хлорид, бензойная кислота, салициловая кислота, тиомерсал, фенилэтиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота или пероксид водорода);
• комплексы металлов, такие как комплексы Zn-белок;
• растворители и сорастворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль);
• сахара и сахарные спирты, такие как трегалоза, сахароза, октасульфат, маннит, сорбитол или ксилит, стахиоза, манноза, сорбоза, ксилоза, рибоза, миоинизитоза, галактоза, лактитол, рибитол, миоинозитол, галактитол, глицерин, циклитолы (например, инозитол), полиэтиленгликоль; и многоатомные сахарные спирты;
• суспендирующие агенты;
• поверхностно-активные вещества или смачивающие агенты, такие как плюроники, ПЭГ, эфиры сорбитана, полисорбаты, такие как полисорбат 20, полисорбат, тритон, трометамин, лецитин, холестерин, тилоксапал; поверхностно-активные вещества могут быть детергентами, предпочтительно с молекулярной массой более 1,2 кД и/или полиэфиром, предпочтительно с молекулярной массой более 3 кД; неограничивающими примерами предпочтительных детергентов являются Твин 20, Твин 40, Твин 60, Твин 80 и Твин 85; неограничивающими примерами предпочтительных полиэфиров являются ПЭГ 3000, ПЭГ 3350, ПЭГ 4000 и ПЭГ 5000;
• агенты, повышающие стабильность, такие как сахароза или сорбитол;
• агенты, повышающие тоничность, такие как галогениды щелочных металлов, предпочтительно хлорид натрия или калия, маннитол сорбитол;
• основы для парентеральной доставки, содержащие раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия, лактат Рингера или жирные масла;
• основы для внутривенной доставки, содержащие пополнители жидкости и питательных веществ, пополнители электролитов (например, на основе декстрозы Рингера).
Для специалистов в данной области очевидно, что различные составляющие фармацевтической композиции (например, перечисленные выше) могут иметь разные эффекты, например, и аминокислота может действовать как буфер, стабилизатор и/или антиоксидант; маннит может действовать как наполнитель и/или агент, повышающий тоничность; хлорид натрия может действовать как основа для доставки и/или агент, повышающий тоничность; и т.д.
В некоторых вариантах осуществления оптимальная фармацевтическая композиция будет определена специалистом в данной области в зависимости, например, от предполагаемого способа введения, формата доставки и желаемой дозировки. См., например, REMINGTON'S PHARMACEUTICAL SCIENCES, выше. Например, подходящей основой или носителем может быть вода для инъекций, физиологический раствор или искусственная спинномозговая жидкость, возможно, с добавлением других материалов, обычно используемых в композициях для парентерального введения. Нейтральный забуференный физиологический раствор или физиологический раствор, смешанный с сывороточным альбумином, являются дополнительными иллюстративными основами.
В одном варианте осуществления фармацевтической композиции согласно одному аспекту изобретения композиция вводится пациенту внутривенно.
Способы и протоколы внутривенного (iv) введения фармацевтических композиций, описанных здесь, хорошо известны в данной области.
В одном аспекте фармацевтической композиции по изобретению указанная фармацевтическая композиция применяется для предотвращения, лечения или облегчения заболевания, выбранного из пролиферативного заболевания, опухолевого заболевания или иммунологического расстройства. Предпочтительно указанное опухолевое заболевание представляет собой злокачественное заболевание, предпочтительно рак.
В одном варианте осуществления фармацевтической композиции по изобретению идентифицированное злокачественное заболевание выбрано из группы, состоящей из лимфомы Ходжкина, неходжкинской лимфомы, лейкоза, множественной миеломы и солидных опухолей.
Кроме того, в одном варианте осуществления согласно настоящему изобретению предложен способ лечения или облегчения заболевания, указанный способ включает этап введения субъекту, испытывающему в этом потребность, предварительно нагруженных человеческих NK-клеток, которые были восстановлены из человеческих NK-клеток в криоконсервированном состоянии в соответствии с изобретением, или получены способом в соответствии с аспектами изобретения для восстановления/получения жизнеспособных предварительно нагруженных человеческих NK-клеток из криоконсервированного состояния.
В соответствии с одним аспектом способа лечения или облегчения по изобретению предварительно нагруженные человеческие NK-клетки вводятся пациенту внутривенно.
В одном варианте осуществления указанного способа лечения или облегчения субъект страдает пролиферативным заболеванием, опухолевым заболеванием или иммунологическим расстройством.
Предпочтительно, чтобы указанное опухолевое заболевание представляло собой злокачественное заболевание, предпочтительно рак.
В одном варианте осуществления указанного способа для лечения или облегчения заболевания упомянутое злокачественное заболевание выбрано из группы, состоящей из лимфомы Ходжкина, неходжкинской лимфомы, лейкоза, множественной миеломы и солидных опухолей.
Следует отметить, что в настоящем документе формы единственного числа содержат и ссылки на множественное число, если контекст явно не указывает иное. Таким образом, например, ссылка на «реагент» содержит один или более таких различных реагентов, и ссылка на «способ» содержит ссылку на эквивалентные этапы и способы, известные рядовым специалистам в данной области, которые могут быть модифицированы или заменены на способы, описанные в настоящем документе.
Если не указано иное, термин «по меньшей мере», предшествующий серии элементов, следует понимать как относящийся к каждому элементу в серии. Специалисты в данной области поймут или смогут установить, используя не более чем рутинное экспериментирование, многие эквиваленты специфичных вариантов осуществления изобретения, описанных в данном документе. Предполагается, что такие эквиваленты охватываются настоящим изобретением.
Термин «и/или», где бы он ни применялся в настоящем документе, содержит значение «и», «или» и «все или любая другая комбинация элементов, связанных указанным термином».
Термин «около» или «примерно» в контексте настоящего описания означает в пределах 20%, предпочтительно в пределах 10%, и более предпочтительно в пределах 5% от заданного значения или диапазона. Однако оно содержит и конкретное число, например, около 20 включает 20.
Термин «менее чем» или «более чем» содержит конкретное число. Например, менее 20 означает менее или равно. Аналогично, более или больше означает более или равно, или больше или равно, соответственно.
В этом описании и формуле изобретения, если контекст не требует иного, слово «включать» и варианты, такие как «включает» и «включающий», будут пониматься как подразумевающие включение указанного целого или этапа или группы целых или этапов, но не исключение любого другого целого или этапа или группы целых или этапа. При применении в данном документе термин «включающий» может быть заменен термином «вмещающий» или «содержащий», а иногда при применении в настоящем документе термином «имеющий».
При применении в настоящем документе термин «состоящий из» исключает любой элемент, этап или ингредиент, не указанные в элементе формулы изобретения. При применении в настоящем документе термин «по сути состоящий из» не исключает материалы или этапы, которые вещественно не влияют на основные и новые характеристики формулы изобретения.
В каждом случае в настоящем документе любой из терминов «включающий», «по сути состоящий из» и «состоящий из» может быть заменен любым из двух других терминов.
Следует понимать, что это изобретение не ограничивается конкретной методологией, протоколами, материалами, реагентами и веществами и т.д., описанными в настоящем документе, и как таковое может варьироваться. Применяемая в настоящем документе терминология предназначена только для описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения, которое определяется исключительно формулой изобретения.
Все публикации и патенты, цитируемые по всему тексту настоящего описания (включая все патенты, патентные заявки, научные публикации, описания производителя, инструкции и т.д.), выше или ниже, включены в настоящий документ посредством ссылки во всей своей полноте. Ничто в настоящем документе не должно толковаться как признание того, что изобретение не имеет права датировать такое раскрытие задним числом на основании предшествующего изобретения. В той степени, в которой материал, включенный посредством ссылки, противоречит или не согласуется с этим описанием, данное описание заменяет любой такой материал.
Примеры конструкций антител, которые можно применять в соответствии с данным изобретением, описаны в MAbs. 2019 Jul; 11(5):899-918.
Лучшее понимание настоящего изобретения и его преимуществ будет получено из следующих примеров, предлагаемых только для иллюстративных целей. Примеры никоим образом не предназначены для ограничения объема настоящего изобретения.
Пример 1: Оценка опосредованной антителами цитотоксичности NK-клетками после предварительной нагрузки и замораживания/размораживания.
Культура клеточных линий
MM.1S (Американская коллекция типовых культур, АКТК (АТСС), каталожный №: CRL2974) и DK-MG (Немецкая коллекция микроорганизмов и клеточных культур, НКМКК (DSMZ), каталожный №: АСС277) культивировали в среде RPMI 1640 с добавлением 10% инактивированной нагреванием фетальной телячьей сыворотки, ФТС (FCS), 2 мМ L-глутамина и 100 МЕ/мл пенициллина G натрия и 100 мкг/мл стрептомицина сульфата. Все клеточные линии культивировали в стандартных условиях и субкультивпровали в соответствии с рекомендациями поставщика при 37°С в увлажненной атмосфере с 5% CO2.
Выделение МКПК (РВМС) из лейкоцитарной пленки и обогащение человеческих NK-клеток
МКПК (РВМС) выделяли из лейкоцитарной пленки (Немецкий Красный Крест, Мангейм, Германия) центрифугированием в градиенте плотности. Образцы лейкоцитарной пленки разбавляли двукратным или трехкратным объемом фосфатно-солевого буфера, ФСБ (PBS) (Invitrogen, каталожный №: 14190-169), наносили на подушку из Lymphoprep (Stem Cell Technologies, каталожный №: 07861) и центрифугировали при 800 × g в течение 25 мин при комнатной температуре без тормоза. МКПК (РВМС), расположенные на границе раздела сред, забирали и промывали 3 раза ФСБ (PBS) перед тем, как их культивировали в полной среде RPMI 1640 (среда RPMI 1640 с добавкой 10% инактивированной нагреванием ФТС (FCS), 2 мМ L-глутамина и 100 МЕ/мл пенициллина G натрия и 100 мкг/мл стрептомицина сульфата (все компоненты от Invitrogen)) в течение ночи без стимуляции. Для обогащения NK-клеток МКПК (РВМС) собирали из ночных культур и применяли для одного раунда отрицательного отбора с применением набора EasySep™ Human N K Cell Enrichment Kit (Stem Cell Technologies, каталожный №: 17055) для иммуномагнитного выделения нетронутых человеческих NK-клеток и Big Easy EasySep™ Magnet (Stem Cell Technologies, каталожный №: 18001) в соответствии с инструкциями производителя.
Предварительная нагрузка NK-клеток и замораживание/размораживание
Аликвоты обогащенных первичных человеческих NK-клеток ресуспендировали в полной среде RPMI 1640, содержащей 10 мкг/мл указанных конструкций антител в общем объеме 0,5 мл, и инкубировали в течение 30 мин при комнатной температуре. Как указано, половину аликвот дважды промывали 5 мл полной среды RPMI 1640 и затем ресуспендировали в 0,5 мл полной среды RPMI 1640, а вторую половину аликвот не промывали перед тем, как добавка ФТС (FCS) с 20% ДМСО (DMSO) (Sigma) была добавлена. Затем клеточные суспензии переносили в криопробирки 2 мл (Greiner bio-one) и замораживали при -80°С в крио 1°С морозильном контейнере (NaIgene) более чем на 12 часов.
4-часовой анализ цитотоксичности с высвобождением кальцеина
Для анализов цитотоксичности с высвобождением кальцеина указанные клетки-мишени собирали из культур, промывали средой RPMI 1640 без ФТС (FCS) и метили 10 мкМ кальцеина AM (Invitrogen/Molecular Probes, каталожный №: С3100МР) в течение 30 минут в среде RPMI без ФТС (FCS) при температуре 37°С. После осторожной отмывки меченые клетки ресуспендировали в полной среде RPMI 1640 до плотности 1×105/мл, а затем аликвоты 1×104 клеток-мишеней высевали в отдельные лунки 96-луночного круглодонного микропланшета. Параллельно, NK-клетки размораживали из -80°С путем инкубации криопробирок на водяной бане при 37°С. Когда были видны последние кристаллы льда, клеточную суспензию разбавляли полной средой RPMI 1640, центрифугировали и один раз промывали перед добавлением клеток к клеткам-мишеням при указанном соотношении эффектор-мишень (Е:Т) с добавлением или без добавления свежего антитела, как указано, в общем объеме 200 мкл в дубликатах. Спонтанное высвобождение определяли инкубацией клеток-мишеней в отсутствие эффекторных клеток и без антител, и максимальное высвобождение индуцировали добавлением 1% Тритона Х-100 (конечная концентрация, Roth), и также измеряли четырежды на каждом планшете.
После центрифугирования в течение 2 мин при 200 g анализ инкубировали в течение 4 ч при 37°С в увлажненной атмосфере с 5% СО2. 100 мкл супернатанта клеточной культуры собирали из каждой лунки после дополнительного центрифугирования в течение 5 минут при 500 g, и флуоресценцию высвобожденного кальцеина измеряли при 520 нм с применением флуоресцентного мультипланшетного ридера (Victor 3 или EnSight, Perkin Elmer). На основе измеренных количеств был рассчитан специфичный лизис клеток по следующей формуле: [флуоресценция (образец) - флуоресценция (спонтанная)]/[флуоресценция (максимальная) - флуоресценция (спонтанная)] × 100%. Флуоресценция (спонтанная) представляет собой количество флуоресценции от клеток-мишеней в отсутствие эффекторных клеток и антител, а флуоресценция (максимальная) представляет общий лизис клеток, индуцированный добавлением Тритона Х-100. Величины лизиса анализировали, и среднее значение и стандартное отклонение строили с применением программного обеспечения GraphPad Prism (GraphPad Prism версии 7.04 для Windows, GraphPad Software, La Jolla California USA).
Результаты показаны на Фигуре 1.
Пример 2: Анализ связывания антител к CD16 с различными вариантами антигена
FcgRIII (CD16), низкоаффинный рецептор к части Fc IgG, существует в двух изоформах, кодируемых двумя почти идентичными генами: FCGR3A (CD16A) и FCGR3B (CD16B), что приводит ктканеспецифичной экспрессии альтернативных заякоренных в мембране изоформ (Ravetch и Perussia, 1989, J Exp Med.):
- FcgRIIIA (CD16A) экспрессируется как трансмембранный белок на NK-клетках, макрофагах и подгруппах моноцитов и Т-клеток
- FcgRIIIB (CD16B) экспрессируется гликозилфосфатидилинозитол, ГФИ (GPI) -заякоренным нейтрофилами и после стимуляции гамма-интерфероном эозинофилами.
Аллельные варианты/полиморфизмы и CD16A, и CD16B хорошо известны (обобщены в Таблице 2).
Аллотипические варианты CD16B различаются пятью аминокислотами ECD (Таблица 2). Два из них делают возможным дополнительное гликозилирование, приводя к трем различным вариантам ГФИ (GPI) - заякоренных гранулоцитарных антигенов FcγRIIIB HNA-1a, -1b и -1с (ранее называвшихся NA1, NA2 и SH).
Есть лишь несколько различий в аминокислотных последовательностях CD16A и В. Их различия ограничиваются двумя альтернативными аминокислотами в зрелом ECD (Таблица 2). Дополнительные различия заключаются в несопоставимых режимах заякоривания CD16A и CD16B к мембране. С-концевая пропептидная последовательность CD16B, которая отщепляется в зрелой ГФИ (GPI) - заякоренной форме, высоко гомологична последовательности, соединяющей ECD CD16A с его трансмембранным доменом (Таблица 2).
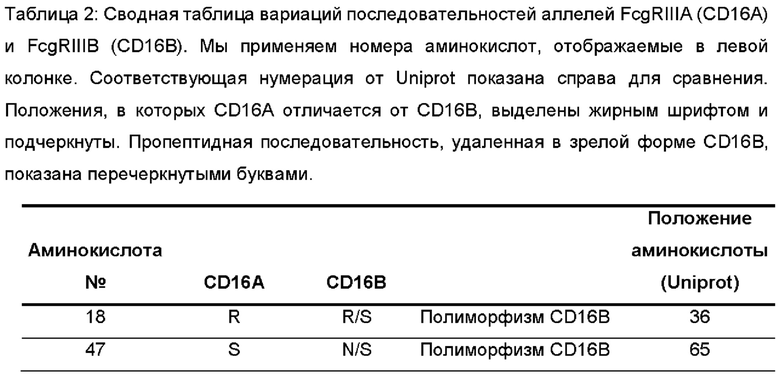
Выделение CD16A-специфичного антитела «LSIV21», лишенного связывания с CD16B, было описано ранее (Reusch et al., 2014, MAbs).
Чтобы исследовать, какая из вариаций последовательностей CD16A/CD16B определяет связывающий сайт для наших CD16A-селективных антител, мы создали и охарактеризовали набор растворимых рекомбинантных вариантов CD16A- и CD16B-антигенов.
Рекомбинантный белок ECDCD16B (NA1) экспрессировался либо в виде зрелой формы (без пропептида) (CD16Bmat), либо в виде варианта, содержащего последовательность пропептида (CD16Bpr). Вдобавок, были сконструированы, экспрессированы и протестированы в анализах связывания мутанты ECD CD16B, в которых CD16B-специфичные аминокислоты были заменены соответствующими остатками CD16A: CD16BD129G, CD16BH140Y, CD16BS185F. Все эти варианты антигенов были слиты посредством глицин-серинового линкера с растворимым мономерным Fc (Ying et al., 2012, J. Biol. Chem.), экспрессированы в яичник китайского хомяка (СНО) и очищены от супернатантов клеточных культур.
Антитело scFv, включающее вариабельные последовательности тяжелой и легкой цепи опубликованного CD16A- и CD16B-реактивного антитела 3G8, распознает все протестированные варианты антигенов CD16A и CD16B (Фигура 2, Таблица 3). Различные антитела, содержащие вариабельные последовательности тяжелой и легкой цепей «LSIV21» или «Р2С47» с улучшенной аффинностью (в форме моновалентных scFv или бивалентно связывающихся тетравалентных биспецифичных тандемных днательных антител), распознавали CD16A, но не зрелые или содержащие пропептид антигены CD16B, сливающиеся с ECD. Однако если гистидин (Н) в положении 140 в ECD CD16B мутировал в тирозин (Y) - аминокислоту, присутствующую в соответствующем положении в CD16A, - эта мутантная форма CD16B (CD16BH140Y) хорошо распознавалась антителами, содержащими вариабельные домены «LSIV21» или «Р2С47» (Фигура 2, Таблица 3).
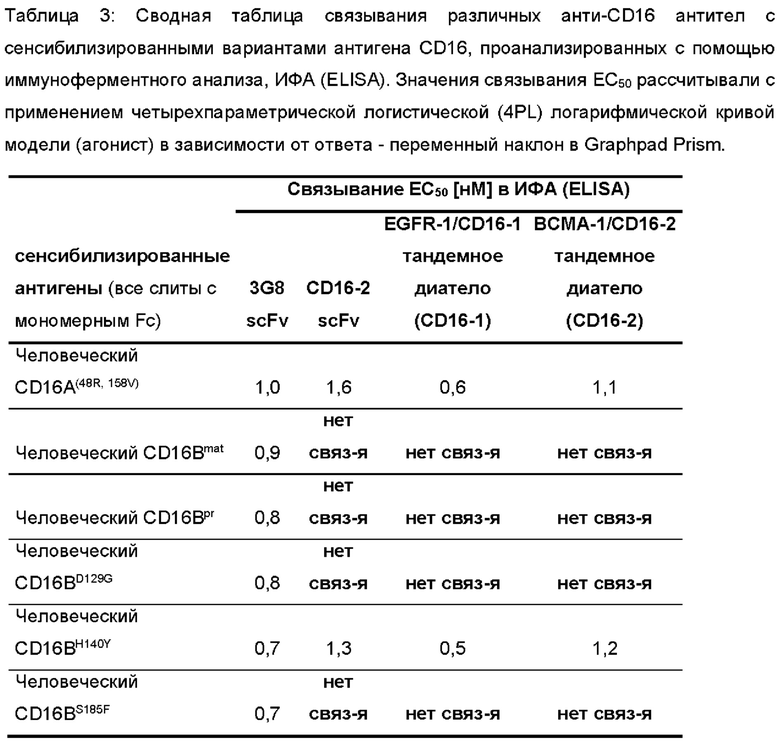
Дополнительную аффинность созревших антител scFv к CD16 анализировали аналогичным образом с помощью ИФА (ELISA). Результаты показывают, что, за исключением scFv «CD16-5», которые демонстрируют слабую перекрестную реактивность с CD16B, другие антитела scFv против CD16 с созревшей аффинностью поддерживают свойство избирательного связывания CD16A родительского антитела. Они не связываются с CD16B, если только CD16B не вмещает ранее описанную аминокислотную замену CD16BH140Y (Таблица 4). Все антитела против CD16 демонстрируют очень хорошую перекрестную реактивность с CD16A яванского макака (Таблица 4), несмотря на то, что последовательность ECD CD16A яванского макака отличается в общей сложности на 16 аминокислот от человеческого CD16A (см. выравнивание на Фигуре 4).
Важность тирозина (Y) в положении 140 в ECD CD16 для CD16A-специфичного распознавания нашими антителами также была подтверждена с применением рекомбинантных клеток яичника китайского хомяка (СНО) с трансгенной экспрессией различных вариантов антигена CD16 на клеточной поверхности. Последовательности ECD были либо слиты с трансмембранным доменом человеческого РЭФР (EGFR) (Wang et al., 2011, Blood 118 (5) 1255), либо заякорены через ГФИ (GPI) с использованием эндогенной последовательности человеческого CD16B с полноразмерной пропептидной последовательностью для посттрансляционного процессинга и липидирования для GPI-заякоривания.
Антитело scFv, включающее вариабельные последовательности тяжелой и легкой цепи опубликованного CD16A- и CD16B-реактивного антитела 3G8, распознает все варианты антигена CD16An CD16B, экспрессируемые на клеточной поверхности (Фигура 3), но не рекомбинантный РЭФР (EGFR), экспрессируемый в качестве контроля специфичности. Антитела scFv, содержащие вариабельные последовательности тяжелой и легкой цепей «LSIV21» или «Р2С47» с улучшенной аффинностью, распознают варианты CD16A, экспрессируемые на поверхности клеток яичника китайского хомяка (СНО), но не зрелые или содержащие пропептиды ECD CD16B (NA1). Однако, если гистидин (Н) в положении 140 в ECD мембранно-заякоренного CD16B клеток яичника китайского хомяка (СНО) мутировал в тирозин (Y) - аминокислоту, присутствующую в соответствующем положении в CD16A - эта мутантная форма CD16B (CD16BH140Y) хорошо распознавалась антителами scFv, содержащими вариабельные домены «LSIV21» или «Р2С47» (Фигура 3).
Варианты антигенов CD16A и CD16BH140Y, но не антигена CD16B, также распознавались после SDS-ΠΑΑΓ (SDS-PAGE) и вестерн-блоттинга, когда образцы обрабатывались в невосстанавливающих условиях антителами, содержащими «LSIV21», «Р2С47» или вариабельные домены с созревшей аффинностью (Фиг. 5), тогда как антитело 3G8 распознает все варианты антигенов CD16A и CD16B. Когда образцы редуцировались перед загрузкой в гели SDS-ΠΑΑΓ (SDS-PAGE), на вестерн-блоттинге не получали сигналов (данные не показаны), что свидетельствует о нелинейной последовательности, но скорее о трехмерной структуре в качестве связывающего эпитопа. В случае распознавания «LSIV21», «Р2С47» или вариантами этих антител с созревшей аффинностью, трехмерная структура эпитопа, вероятно, определяется тирозином в положении 140 CD16.
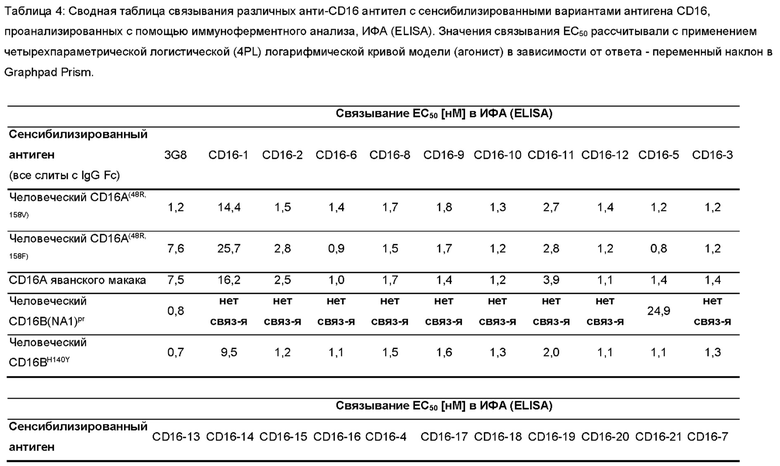
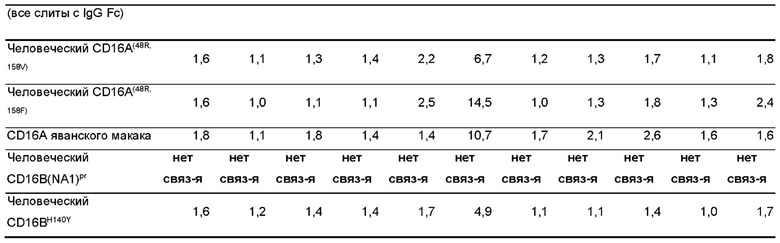
Связывающий сайт IgG FcgRIII представляет собой прерывистую область связывания на мембранном проксимальном внеклеточном домене рецептора. Ключевые аминокислоты для низкоаффинного взаимодействия FcgRIIIA (CD16A) с Fc-частью IgG были ранее идентифицированы и описаны (например, Ahmed et al., 2016). Остатками CD16A, подразумевающими взаимодействие с доменом СН2 частей Fc, являются К120, Y132, Н134, Н135 и К161. Хотя сообщалось, что антитело 3G8 связывает CD16A и CD16B и блокирует связывание IgG (Tamm and Schmidt, 1996), мы не наблюдали блокирования связывания наших антител к CD16 с CD16A посредством IgG. Чтобы поддержать гипотезу о том, что связывающий сайт наших специфичных антител к CD16A отличается от связывающего сайта IgG на FcgRIIIA, мы провели конкурентный ИФА (ELISA). Хотя связывание моноклональных антител 3G8 с CD16A блокируется возрастающими концентрациями антитела scFv, содержащего вариабельные домены 3G8, другие антитела scFv против CD16 не блокируют или не вытесняют моноклональные антитела 3G8 на CD16A и связываются независимо (Фигура 6). Поскольку известно, что связывающий сайт 3G8 блокирует связывание IgG, мы пришли к выводу, что связывающий сайт наших антител к CD16 отличается от связывающего сайта IgG.
Анализ связывания ИФА (ELISA)
96-луночные планшеты для ИФА (ELISA) (Immuno MaxiSorp; Nunc) покрывали на ночь при 4°С рекомбинантно полученными гибридными белками вариантов антигена внеклеточного домена (ECD) CD16, слитыми с Fc человеческого IgG1 или мономерным mFc.67 (Ying et al, 2012, J. Biol. Chem.) в 100 мМ карбонатно-бикарбонатного буфера в концентрации 1,5 или 3 мкг/мл. После этапа блокирования 3% (мас./об) сухого обезжиренного молока (Merck), растворенного в ФСБ (PBS), серийные разведения различных антител в ФСБ (PBS), содержащем 0,3% (мас./об) сухого обезжиренного молока, инкубировали на планшетах в течение 1,5 ч при комнатной температуре. После трехкратного отмывания 300 мкл на лунку ФСБ (PBS), содержащего 0,1% (об/об) Твин 20, планшеты инкубировали с детектирующими антителами в течение 1 ч при комнатной температуре. Для детекции аналитов, меченных His, применяли Penta-HIS-HRP (Qiagen) при разведении 1:3000. Для обнаружения четырехвалентных биспецифичных антител тандемных диател BCMA/CD16, Т763, применяли конъюгат Protein L-HRP (Pierce) при разведении 1:20000 в течение 1 часа при комнатной температуре. После трехкратного отмывания 300 мкл на лунку ФСБ (PBS), содержащего 0,1% (об/об) Твин 20, планшеты инкубировали с субстратом ТМБ (ТМВ) (Seramun) до тех пор, пока не стало ясно видимым проявление цвета. Реакцию останавливали добавлением 100 мкл на лунку 0,5 Μ H2SO4. Поглощение измеряли при 450 нм с применением многометочного планшетного ридера (Victor, Perkin Elmer). Значения абсорбции были построены и проанализированы с применением нелинейной регрессии, сигмоидальной зависимости доза-ответ (переменный наклон), аппроксимации методом наименьших квадратов (обычный) с помощью GraphPad Prism версии 6.07 (GraphPad Software, La Jolla California USA).
Анализ связывания после SDS-ΠΑΑΓ (SDS-PAGE) и вестерн-блоттинга
Очищенные рекомбинантные варианты антигена CD16 (человеческий CD16A48R, 158V" mFc.67, человеческий CD16B(NA1)pr-mFc.67 и huCD16B(NA1)prH140Y-mFc.67 разводили в ФСБ (PBS) до 0,5 мкг/мл, смешивая с равным объем невосстанавливающего буфера для образцов и 8 мкл смеси загружали в лунки с 4-20% гелями Criterion TGX Precast Gels (Biorad). Гели прогоняли при 300 В в 1х TGS-буфере (Biorad) в течение 22 минут, и общий белок отображали с применением системы документирования гелей Biorad Molecular Imager Gel Documentation system. Белок переносили вестерн-блоттингом на миди-мембраны PVDF Midi Membranes (Biorad) с использованием системы Trans-Blot Turbo system (Biorad). Мембраны инкубировали в течение 30 минут с 3% (мас/об) сухим обезжиренным молоком (Merck), растворенным в трис-забуференном физиологическом растворе, ТЗФР (TBS) для блокирования неспецифичных связывающих сайтов, а затем инкубировали с разведениями первичных антител: антитела KCD16 разбавляли в ТЗФР (TBS), содержащем 3% (мас/об)) сухого обезжиренного молока до концентрации 4 мкг/мл и инкубировали на мембранах в течение 1 часа при комнатной температуре. После промывания ТЗФРТ (TBST) (ТЗФР (TBS), содержащий 0,1% (об/об) Твин 20) и два раза с ТЗФР (TBS), мембраны инкубировали с конъюгатами для обнаружения вторичных антител: Мембраны, инкубированные санти-CD16A, содержащими антитела scFv-IgAb, инкубировали с Protein L-HRP (Pierce), 1: 10000, разведенным в ТЗФР (TBS), содержащем 3% (мас/об) сухого обезжиренного молока, мембраны, инкубированные с антителами против CD16 scFv, инкубировали с анти-Penta-His-HRP (QIAGEN) 1:3000, разведенным в ТЗФР (TBS), содержащем 3% (мас/об) сухого обезжиренного молока, в течение 1 часа при комнатной температуре. После отмывки ТЗФРТ (TBST) и двухкратной отмывки ТЗФР (TBS) колориметрическое проявление начинали добавлением свежеприготовленной смеси 0,66 мг/мл ДАБ (DAB), 0,02% CoCl2, 0,015% Н202 в ТЗФР (TBS) и инкубировали на мембранах до проявления цвета. Реакцию останавливали промыванием мембран водой. Мембраны высушивали и фотографировали.
Конкурентный ИФА (ELISA) CD16A
96-луночные планшеты для ИФА (ELISA) (Immuno MaxiSorp; Nunc) покрывали на ночь при 4°С рекомбинантно полученными гибридными белками ECD CD16A (48R, 158V), слитыми с мономерным mFc.67 (Ying et al, 2012, J. Biol. Chem.) в 100 мМ карбонатно-б и карбонатного буфера в концентрации 1,5 мкг/мл. После этапа блокирования 3% (мас./об) сухого обезжиренного молока (Merck), растворенного в ФСБ (PBS), серийные разведения различных scFv-антител в ФСБ (PBS), содержащем 0,3% (мас./об) сухого обезжиренного молока, смешивали с 1 нМ анти-CD16 mAb 3G8 (BD Bioscience) и совместно инкубировали на планшетах в течение 1,5 ч при комнатной температуре. После трехкратного отмывания 300 мкл на лунку ФСБ (PBS), содержащего 0,1% (об/об) Твин 20, планшеты инкубировали с детектирующими антителами в течение 1 ч при комнатной температуре. Для обнаружения CD16A-связанного 3G8 применяли козий антимышиный IgG (H+L)-HRPO (Dianova 115-035-003) при разведении 1:10000. После трехкратного отмывания 300 мкл на лунку ФСБ (PBS), содержащего 0,1% (об/об) Твин 20, планшеты инкубировали с субстратом ТМБ (ТМВ) (Seramun) до тех пор, пока не стало ясно видимым проявление цвета. Реакцию останавливали добавлением 100 мкл на лунку 0,5 Μ H2S04. Поглощение измеряли при 450 нм с применением многометочного планшетного ридера (Victor, Perkin Elmer). Значения абсорбции были построены и проанализированы с применением нелинейной регрессии, сигмоидальной зависимости доза-ответ (переменный наклон), аппроксимации методом наименьших квадратов (обычный) с помощью GraphPad Prism версии 6.07 (GraphPad Software, La Jolla California USA).
Пример 3: Связывание анти-CD16A scFv с первичными NK-клетками человека при 37°С в присутствии или в отсутствие 10 мг/мл поликлонального человеческого IgG
Способы:
Выделение МКПК (РВМС) из лейкоцитарной пленки и обогащение человеческих NK-клеток
МКПК (РВМС) выделяли из лейкоцитарной пленки (Немецкий Красный Крест, Мангейм, Германия) центрифугированием в градиенте плотности. Образцы лейкоцитарной пленки разбавляли двукратным или трехкратным объемом фосфатно-солевого буфера, ФСБ (PBS) (Invitrogen, каталожный №: 14190-169), наносили на подушку из Lymphoprep (Stem Cell Technologies, каталожный №: 07861) и центрифугировали при 800 × g в течение 25 мин при комнатной температуре без тормоза. МКПК (РВМС), расположенные на границе раздела сред, забирали и промывали 3 раза ФСБ (PBS) перед тем, как их культивировали в полной среде RPMI 1640 (среда RPMI 1640 с добавкой 10% инактивированной нагреванием ФТС (FCS), 2 мМ L-глутамина и 100 МЕ/мл пенициллина G натрия и 100 мкг/мл стрептомицина сульфата (все компоненты от Invitrogen)) в течение ночи без стимуляции. Для обогащения NK-клеток МКПК (РВМС) собирали из ночных культур и применяли для одного раунда отрицательного отбора с применением набора EasySep™ Human NKCell Enrichment Kit (Stem Cell Technologies, каталожный №: 17055) для иммуномагнитного выделения нетронутых человеческих NK-клеток и Big Easy EasySep™ Magnet (Stem Cell Technologies, каталожный №: 18001) в соответствии с инструкциями производителя.
Анализы связывания клеток и анализы проточной цитометрии
Аликвоты 0,2-1 × 106 обогащенных человеческих NK-клеток инкубировали со 100 мкл серийных разведений указанных конструкций scFv в буфере флуоресцентно активируемой клеточной сортировки, ФАКС (FACS) (PBS, Invitrogen, каталожный №: 14190-169), содержащем 2% инактивированную нагреванием ФТС (FCS) (Invitrogen, каталожный №: 10270-106), 0,1% азид натрия (Roth, Карлсруэ, Германия, каталожный №: А1430.0100) в присутствии 10 мкг/мл поликлонального человеческого IgG (Gammanorm, Octapharma) или соответствующего буфера в течение 45 мин при 37°С. После повторной отмывки буфером для ФАКС (FACS) связанные с клетками антитела выявляли с помощью 10 мкг/мл анти-His моноклональных антител 13/45/31-2 (Dianova, Гамбург, Германия, каталожный №: DIA910-1MG) с последующим введением 15 мкг/мл флуоресцеин изотиоцианат, ФИТЦ (FITC) - конъюгированного козьего антимышиного IgG (Dianova, каталожный №: 115-095-062). После последнего этапа окрашивания клетки снова отмывали и ресуспендировали в 0,2 мл буфера для ФАКС (FACS), содержащего 2 мкг/мл йодида пропидия (PI) (Sigma, каталожный №: Р4170), чтобы исключить мертвые клетки. Флуоресценцию 2-5 × 103 живых клеток измеряли с применением проточного цитометра Millipore Guava EasyCyte (Merck Millipore, Швальбах, Германия) или цитометра CytoFlex (Beckman Coulter, Крефельд, Германия) и определяли среднюю интенсивность флуоресценции образцов клеток. После вычитания значений интенсивности флуоресценции клеток, окрашенных только вторичными и третичными реагентами, эти значения применяли для анализа нелинейной регрессии с применением программного обеспечения GraphPad Prism (GraphPad Prism версии 7.04 для Windows, GraphPad Software, La Jolla California USA). Для расчета KD применяли уравнение для связывания с одним сайтом (гипербола).
Результаты
ScFv против CD16, содержащие домены Fv человека из клона CD16-1, scFv, содержащие домены Fv человека против CD16 из клона CD16-2, и scFv_3G8, содержащие мышиные домены Fv из моноклональных антител 3G8, титровали на первичных NK-клетках человека при 37°С в присутствии или отсутствии 10 мг/мл поликлонального человеческого IgG. Соответствующие результаты изображены на Фигуре 7.
Пример 4: Оценка опосредованной антителами цитотоксичности NK-клеток после предварительной нагрузки антителами против ЕрСАМ и CD30 и замораживания/размораживания.
Способы:
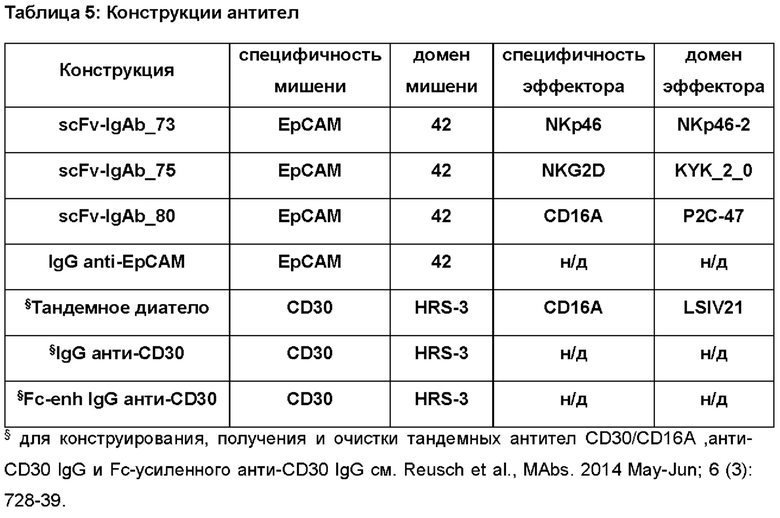
Культура клеточных линий
Клетки COLO 205 (любезно предоставлены доктором G. Moldenhauer, German Cancer Research Center, Гейдельберг, Германия) и клетки KARPAS-299 (DSMZ, каталожный №: АСС31) культивировали в среде RPMI 1640 с добавкой 10% инактивированной нагреванием ФТС (FCS), 2 мМ L-глутамина и 100 МЕ/мл пенициллина G натрия и 100 мкг/мл стрептомицина сульфата. Все клеточные линии культивировали в стандартных условиях и субкультивировали в соответствии с рекомендациями поставщика при 37°С в увлажненной атмосфере с 5% СО2.
Выделение МКПК (РВМС) из лейкоцитарной пленки и обогащение человеческих NK-клеток
МКПК (РВМС) выделяли из лейкоцитарной пленки (Немецкий Красный Крест, Мангейм, Германия) центрифугированием в градиенте плотности. Образцы лейкоцитарной пленки разбавляли двукратным или трехкратным объемом фосфатно-солевого буфера, ФСБ (PBS) (Invitrogen, каталожный №: 14190-169), наносили на подушку из Lymphoprep (Stem Cell Technologies, каталожный №: 07861) и центрифугировали при 800 × g в течение 25 мин при комнатной температуре без тормоза. МКПК (РВМС), расположенные на границе раздела сред, забирали и промывали 3 раза ФСБ (PBS) перед тем, как их культивировали в полной среде RPMI 1640 (среда RPMI 1640 с добавкой 10% инактивированной нагреванием ФТС (FCS), 2 мМ L-глутамина и 100 МЕ/мл пенициллина G натрия и 100 мкг/мл стрептомицина сульфата (все компоненты от Invitrogen)) в течение ночи без стимуляции. Для обогащения NK-клеток МКПК (РВМС) собирали из ночных культур и применяли для одного раунда отрицательного отбора с применением набора EasySep™ Human NK Cell Enrichment Kit (Stem Cell Technologies, каталожный №: 17055) для иммуномагнитного выделения нетронутых человеческих NK-клеток и Big Easy EasySep™ Magnet (Stem Cell Technologies, каталожный №: 18001) в соответствии с инструкциями производителя.
Предварительная нагрузка NK-клеток и замораживание/размораживание
Аликвоты обогащенных первичных NK-клеток человека ресуспендировали в полной среде RPMI 1640, содержащей 10 мкг/мл указанных конструкций антител в общем объеме 0,5 мл, и инкубировали в течение 30 мин при комнатной температуре. После добавления 0,5 мл ФТС (FCS) с добавкой 20% ДМСО (DMSO) (Sigma) клеточные суспензии затем переносили в криопробирки 2 мл (Greiner bio-one) и замораживали при -80°С в крио 1°С морозильном контейнере (NaIgene) более чем на 12 часов.
4-часовой анализ цитотоксичности с высвобождением кальцеина
Для анализов цитотоксичности с высвобождением кальцеина указанные клетки-мишени собирали из культур, промывали средой RPMI 1640 без ФТС (FCS) и метили 10 мкМ кальцеина AM (Invitrogen/Molecular Probes, каталожный №: С3100МР) в течение 30 минут в среде RPMI без ФТС (FCS) при температуре 37°С. После осторожной отмывки меченые клетки ресуспендировали в полной среде RPMI 1640 до плотности 1×105/мл, а затем аликвоты 1×104 клеток-мишеней высевали в отдельные лунки 96-луночного круглодонного микропланшета. Параллельно, NK-клетки размораживали из -80°С путем инкубации криопробирок на водяной бане при 37°С. Когда были видны последние кристаллы льда, клеточную суспензию разбавляли полной средой RPMI 1640, центрифугировали и один раз промывали перед добавлением клеток к клеткам-мишеням при указанном соотношении эффектор-мишень (Е:Т) с добавлением или без добавления свежего антитела, как указано, в общем объеме 200 мкл в дубликатах. Спонтанное высвобождение определяли инкубацией клеток-мишеней в отсутствие эффекторных клеток и без антител, и максимальное высвобождение индуцировали добавлением 1% Тритона Х-100 (конечная концентрация, Roth), и также измеряли четырежды на каждом планшете.
После центрифугирования в течение 2 мин при 200 g анализ инкубировали в течение 4 ч при 37°С в увлажненной атмосфере с 5% CO2. 100 мкл супернатанта клеточной культуры собирали из каждой лунки после дополнительного центрифугирования в течение 5 минут при 500 g, и флуоресценцию высвобожденного кальцеина измеряли при 520 нм с применением флуоресцентного мультипланшетного ридера (Victor 3 или EnSight, Perkin Elmer). На основе измеренных количеств был рассчитан специфичный лизис клеток по следующей формуле: [флуоресценция (образец) - флуоресценция (спонтанная)]/[флуоресценция (максимальная) - флуоресценция (спонтанная)] × 100%. Флуоресценция (спонтанная) представляет собой количество флуоресценции от клеток-мишеней в отсутствие эффекторных клеток и антител, а флуоресценция (максимальная) представляет общий лизис клеток, индуцированный добавлением Тритона Х-100. Величины лизиса анализировали, и среднее значение и стандартное отклонение строили с применением программного обеспечения GraphPad Prism (GraphPad Prism версии 7.04 для Windows, GraphPad Software, La Jolla California USA).
Результаты:
Первичные NK-клетки человека, которые были предварительно нагружены конструкциями антител против ЕрСАМ, такими как scFv-IgAb_73 EpCAM/NKp46, scFv-IgAb_75 EpCAM/NKG2D, scFv-IgAb_80 EpCAM/CD16A и IgG против ЕрСАМ, индуцировали специфичный лизис ЕрСАМ-положительных клеток COLO 205, но не ЕрСАМ-отрицательных клеток KARPAS-299. Среди биспецифичных конструкций против ЕрСАМ, NK-клетки, предварительно нагруженные scFv-IgAb_80 EpCAM/CD16A, проявляли наивысшую эффективность в лизисе клеток COLO 205, а NK-клетки, нагруженные scFv-IgAb_75 EpCAM/NKG2D, имели самую низкую эффективность. Ни один из препаратов NK-клеток, предварительно нагруженных конструкциями анти-CD30 антител, не опосредовал существенный лизис COLO 205 (Фигура 8А и Таблица 6).
Напротив, NK-клетки, которые были предварительно нагружены тандемными диателами CD30/CD16A или Fc-усиленным анти-CD30 IgG, опосредовали лизис CD30-положительных клеток KARPAS-299, но не CD30-отрицательных клеток COLO 205. NK-клетки, предварительно нагруженные анти-CD30 IgG дикого типа, не опосредовали лизис KARPAS-299, что позволяет предположить, что этот анти-CD30 IgG дикого типа слишком быстро диссоциировал от NK-клеток во время замораживания, размораживания и отмывки, и что количество оставшегося IgG было слишком низким, чтобы опосредовать существенный лизис клеток-мишеней (Фигура 8 В и Таблица 6).
Интересно, что NK-клетки, которые были предварительно нагружены 10 мкг/мл scFv-IgAb_75 EpCAM/NKG2D или тандемными диателами CD30/CD16A, заморожены, разморожены и отмыты, индуцировали такой же лизис клеток COLO 205 или KARPAS-299 соответственно, как NK-клетки, которые не были предварительно нагружены, но к ним в анализе цитотоксичности было добавлено 10 мкг/мл свежих scFv-IgAb_75 EpCAM/NKG2D или тандемных диател CD30/CD16A.
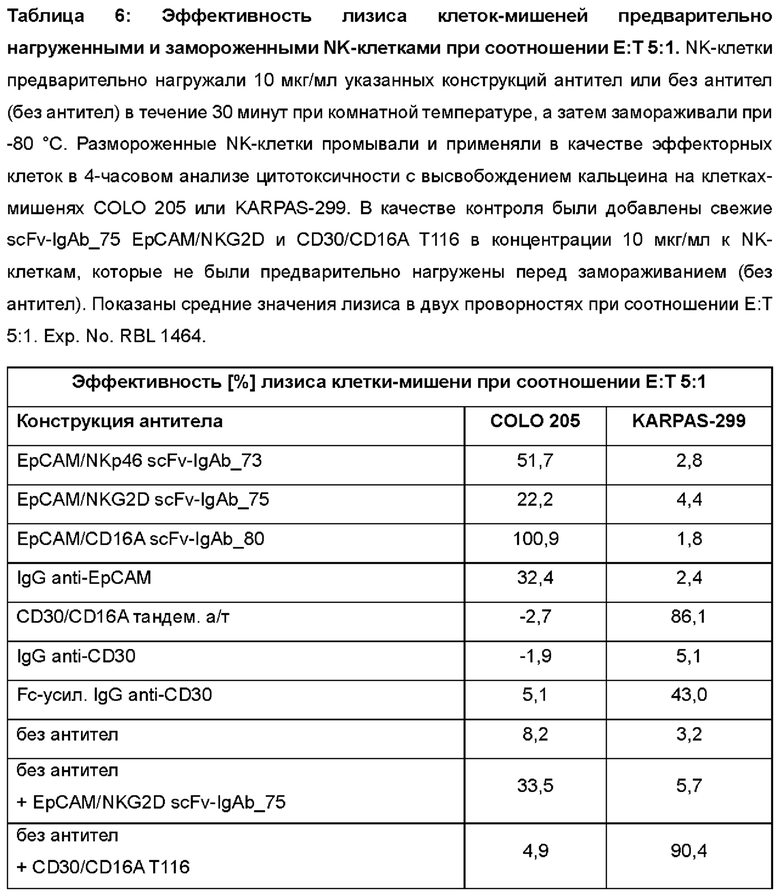






































--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Affimed GmbH
<120> Криоконсервированные натуральные киллерные клетки,
предварительно нагруженные конструктом антитела
<130> A 3319PCT
<150> EP18191031.6
<151> 2018-08-27
<160> 178
<170> BiSSAP 1.3.6
<210> 1
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 1
Gly Gly Ser Gly Gly Ser
1. 5
<210> 2
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 2
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
1. 5 10 15
Gly Ser
<210> 3
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 3
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
1. 5 10 15
Gly Ser Gly Gly Ser
20
<210> 4
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 4
Gly Gly Ser Gly Gly Ser Gly Gly Ser
1. 5
<210> 5
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 5
Gly Gly Gly Gly Ser
1. 5
<210> 6
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 6
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10
<210> 7
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 7
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1. 5 10 15
Gly Gly Gly Ser
20
<210> 8
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 8
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1. 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 9
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> шарнир
<400> 9
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1. 5 10 15
<210> 10
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> шарнир
<400> 10
Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1. 5 10
<210> 11
<211> 328
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный домен тяжелой цепи
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
65 70 75 80
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
85 90 95
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
100 105 110
Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
115 120 125
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
130 135 140
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
145 150 155 160
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
165 170 175
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
180 185 190
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
195 200 205
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
210 215 220
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
225 230 235 240
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
245 250 255
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
260 265 270
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
275 280 285
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
290 295 300
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
305 310 315 320
Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 12
<211> 328
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> константный домен тяжелой цепи
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
65 70 75 80
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
85 90 95
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
100 105 110
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
115 120 125
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
130 135 140
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
145 150 155 160
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
165 170 175
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
180 185 190
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
195 200 205
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
210 215 220
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
225 230 235 240
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
245 250 255
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
260 265 270
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
275 280 285
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
290 295 300
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
305 310 315 320
Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 13
<211> 106
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> константный домен легкой цепи
<400> 13
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1. 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 14
<211> 107
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> константный домен легкой цепи
<400> 14
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1. 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 15
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный домен CH2-CH3
<400> 15
Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1. 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 16
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный домен CH2-CH3
<400> 16
Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1. 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Lys Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Val Pro Val Leu Asp Ser Asp Gly Ser Phe Arg Leu
165 170 175
Ala Ser Tyr Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro
210 215
<210> 17
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный домен ch2-ch3
<400> 17
Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1. 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Tyr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 18
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> константный домен ch2-ch3
<400> 18
Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1. 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Thr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 19
<211> 98
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> домен ch1
<400> 19
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 20
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> домен cl
<400> 20
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1. 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Ser Ser
100 105
<210> 21
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> домен cl
<400> 21
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1. 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Ser
100 105
<210> 22
<211> 216
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> домен ch2-ch3
<400> 22
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1. 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 23
<211> 192
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> CD16a
<400> 23
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
1. 5 10 15
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
20 25 30
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
35 40 45
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
50 55 60
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
65 70 75 80
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
85 90 95
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
100 105 110
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
115 120 125
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
130 135 140
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Phe
145 150 155 160
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
165 170 175
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
180 185 190
<210> 24
<211> 192
<212> БЕЛОК
<213> Яванский макак
<220>
<223> CD16a
<400> 24
Gly Met Arg Ala Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
1. 5 10 15
Gln Trp Tyr Arg Val Leu Glu Lys Asp Arg Val Thr Leu Lys Cys Gln
20 25 30
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Arg Trp Phe His Asn Glu
35 40 45
Ser Leu Ile Ser Ser Gln Thr Ser Ser Tyr Phe Ile Ala Ala Ala Arg
50 55 60
Val Asn Asn Ser Gly Glu Tyr Arg Cys Gln Thr Ser Leu Ser Thr Leu
65 70 75 80
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
85 90 95
Ala Pro Arg Trp Val Phe Lys Glu Glu Glu Ser Ile His Leu Arg Cys
100 105 110
His Ser Trp Lys Asn Thr Leu Leu His Lys Val Thr Tyr Leu Gln Asn
115 120 125
Gly Lys Gly Arg Lys Tyr Phe His Gln Asn Ser Asp Phe Tyr Ile Pro
130 135 140
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Ile
145 150 155 160
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
165 170 175
Asp Leu Ala Val Ser Ser Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
180 185 190
<210> 25
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептидная метка
<400> 25
His His His His His His
1. 5
<210> 26
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> пептидная метка
<400> 26
Glu Pro Glu Ala
1
<210> 27
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 27
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 28
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 28
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 29
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 29
Gly Tyr Thr Phe Thr Ser Tyr Tyr
1. 5
<210> 30
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 30
Ile Asn Pro Ser Gly Gly Ser Thr
1. 5
<210> 31
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 31
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 32
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 32
Asn Ile Gly Ser Lys Asn
1. 5
<210> 33
<400> 33
000
<210> 34
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 34
Gln Val Trp Asp Asn Tyr Ser Val Leu
1. 5
<210> 35
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 35
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 36
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 36
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 37
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 37
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
130 135 140
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys
145 150 155 160
Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg
165 170 175
Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 38
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 38
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 39
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 39
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 40
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 40
Gly Tyr Thr Phe Thr Ser Tyr Tyr
1. 5
<210> 41
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 41
Ile Glu Pro Met Tyr Gly Ser Thr
1. 5
<210> 42
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 42
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 43
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 43
Asn Ile Gly Ser Lys Asn
1. 5
<210> 44
<400> 44
000
<210> 45
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 45
Gln Val Trp Asp Asn Tyr Ser Val Leu
1. 5
<210> 46
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 46
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 47
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 47
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 48
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 48
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
130 135 140
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys
145 150 155 160
Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg
165 170 175
Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 49
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 49
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 50
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 50
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 51
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 51
Gly Tyr Thr Phe Thr Asn Tyr Tyr
1. 5
<210> 52
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 52
Ile Asn Pro Gly Gly Gly Ser Thr
1. 5
<210> 53
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 53
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 54
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 54
Asn Ile Gly Ser Gln Ser
1. 5
<210> 55
<400> 55
000
<210> 56
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 56
Gln Val Trp Asp Asn Tyr Ser Val Val
1. 5
<210> 57
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 57
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asn Pro Gly Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 58
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 58
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly His Asn Ile Gly Ser Gln Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 59
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 59
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asn Pro Gly Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
130 135 140
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys
145 150 155 160
Gly Gly His Asn Ile Gly Ser Gln Ser Val His Trp Tyr Gln Gln Lys
165 170 175
Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gln Asp Ser Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Asn Tyr Ser Val Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 60
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 60
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Asn Pro Gly Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 61
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 61
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly His Asn Ile Gly Ser Gln Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 62
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 62
Gly Tyr Ser Phe Ser Asp Phe Tyr
1. 5
<210> 63
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 63
Ile Asn Pro Gly Gly Ala Ser Thr
1. 5
<210> 64
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 64
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 65
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 65
Asn Ile Gly Arg Gln Ser
1. 5
<210> 66
<400> 66
000
<210> 67
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 67
Gln Val Trp Asp Asn Tyr Thr Val Val
1. 5
<210> 68
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 68
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Ser Asp Phe
20 25 30
Tyr Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Gly Gly Ala Ser Thr Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 69
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 69
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Tyr Asn Ile Gly Arg Gln Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Thr Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 70
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 70
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Ser Asp Phe
20 25 30
Tyr Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Gly Gly Ala Ser Thr Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
130 135 140
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys
145 150 155 160
Gly Gly Tyr Asn Ile Gly Arg Gln Ser Val His Trp Tyr Gln Gln Lys
165 170 175
Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gln Asp Ser Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Asn Tyr Thr Val Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 71
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 71
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Ser Asp Phe
20 25 30
Tyr Ile Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Gly Gly Ala Ser Thr Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 72
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 72
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Tyr Asn Ile Gly Arg Gln Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Thr Val Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 73
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 73
Gly Tyr Thr Phe Ser Ser Tyr Tyr
1. 5
<210> 74
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 74
Ile Glu Pro Arg Gly Val Arg Ile
1. 5
<210> 75
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 75
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 76
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 76
Asn Ile Gly Ser Thr Asn
1. 5
<210> 77
<400> 77
000
<210> 78
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 78
Gln Val Trp Asp Asn Tyr Ser Val Gln
1. 5
<210> 79
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 79
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Glu Pro Arg Gly Val Arg Ile Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 80
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 80
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly His Asn Ile Gly Ser Thr Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Gln
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 81
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 81
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Glu Pro Arg Gly Val Arg Ile Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
130 135 140
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys
145 150 155 160
Gly Gly His Asn Ile Gly Ser Thr Asn Val His Trp Tyr Gln Gln Lys
165 170 175
Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gln Asp Ser Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Asn Tyr Ser Val Gln Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 82
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 82
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Glu Pro Arg Gly Val Arg Ile Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 83
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 83
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly His Asn Ile Gly Ser Thr Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Gln
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 84
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 84
Gly Tyr Thr Phe Thr Asn Tyr Tyr
1. 5
<210> 85
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 85
Ile Asn Pro Ser Gly Gly Val Thr
1. 5
<210> 86
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 86
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 87
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 87
Asn Ile Gly Ser Lys Ser
1. 5
<210> 88
<400> 88
000
<210> 89
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 89
Gln Val Trp Asp Asp Tyr Ile Val Leu
1. 5
<210> 90
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 90
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Val Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 91
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 91
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asp Tyr Ile Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 92
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 92
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Val Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
130 135 140
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys
145 150 155 160
Gly Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys
165 170 175
Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gln Asp Lys Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Asp Tyr Ile Val Leu Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 93
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 93
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Val Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 94
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 94
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asp Tyr Ile Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 95
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 95
Gly Tyr Thr Phe Thr Asn Tyr Tyr
1. 5
<210> 96
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 96
Ile Glu Pro Asp Gly Gly Arg Arg
1. 5
<210> 97
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 97
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
1. 5 10
<210> 98
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 98
Gln Gly Val Ser Gly Asp
1. 5
<210> 99
<400> 99
000
<210> 100
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 100
Gln Gln Trp Asp Asn Tyr Ser Val Thr
1. 5
<210> 101
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 101
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Glu Pro Asp Gly Gly Arg Arg Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 102
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 102
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Gly His Gln Gly Val Ser Gly Asp
20 25 30
Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gln Ala Asn Lys Arg Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Asp Asn Tyr Ser Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 103
<211> 256
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 103
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Glu Pro Asp Gly Gly Arg Arg Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Ile Val Leu Thr Gln
130 135 140
Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser
145 150 155 160
Cys Arg Gly His Gln Gly Val Ser Gly Asp Val His Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Gln Ala Asn Lys Arg
180 185 190
Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu
195 200 205
Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr
210 215 220
Tyr Cys Gln Gln Trp Asp Asn Tyr Ser Val Thr Phe Gly Gln Gly Thr
225 230 235 240
Lys Val Glu Ile Lys Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 104
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 104
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Glu Pro Asp Gly Gly Arg Arg Thr Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 105
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 105
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Gly His Gln Gly Val Ser Gly Asp
20 25 30
Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gln Ala Asn Lys Arg Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Asp Asn Tyr Ser Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 106
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 106
Gly Phe Thr Phe Ser Asn Tyr Asp
1. 5
<210> 107
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 107
Ile Ser Thr Arg Gly Asp Ile Thr
1. 5
<210> 108
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 108
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr
1. 5 10
<210> 109
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 109
Glu Asp Ile Tyr Asn Gly
1. 5
<210> 110
<400> 110
000
<210> 111
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 111
Ala Gly Pro His Lys Tyr Pro Leu Thr
1. 5
<210> 112
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 112
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 113
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 113
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr Asn Gly
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Glu Ala Thr Tyr Tyr Cys Ala Gly Pro His Lys Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 114
<211> 254
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 114
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
145 150 155 160
Ala Ser Glu Asp Ile Tyr Asn Gly Leu Ala Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp
180 185 190
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Glu Ala Thr Tyr Tyr Cys
210 215 220
Ala Gly Pro His Lys Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Ala Ala Ala Gly Ser His His His His His His
245 250
<210> 115
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 115
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 116
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 116
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr Asn Gly
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Glu Ala Thr Tyr Tyr Cys Ala Gly Pro His Lys Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 117
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 117
Gly Phe Thr Phe Ser Asn Phe Asp
1. 5
<210> 118
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 118
Ile Thr Thr Gly Gly Gly Asp Thr
1. 5
<210> 119
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 119
Val Arg His Gly Tyr Tyr Asp Gly Tyr His Leu Phe Asp Tyr Trp Gly
1. 5 10 15
<210> 120
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 120
Gln Gly Ile Ser Asn Asn
1. 5
<210> 121
<400> 121
000
<210> 122
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 122
Gln Gln Phe Thr Ser Leu Pro Tyr Thr
1. 5
<210> 123
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 123
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Ser Ile Thr Thr Gly Gly Gly Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg His Gly Tyr Tyr Asp Gly Tyr His Leu Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 124
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Asn Gln Gly Ile Ser Asn Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile
35 40 45
Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Thr Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 125
<211> 254
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 125
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Ser Ile Thr Thr Gly Gly Gly Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg His Gly Tyr Tyr Asp Gly Tyr His Leu Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
145 150 155 160
Ala Asn Gln Gly Ile Ser Asn Asn Leu Asn Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Pro Leu Ile Tyr Tyr Thr Ser Asn Leu Gln Ser
180 185 190
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
210 215 220
Gln Gln Phe Thr Ser Leu Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu
225 230 235 240
Glu Ile Lys Ala Ala Ala Gly Ser His His His His His His
245 250
<210> 126
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 126
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Phe
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Ser Ile Thr Thr Gly Gly Gly Asp Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg His Gly Tyr Tyr Asp Gly Tyr His Leu Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 127
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 127
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Asn Gln Gly Ile Ser Asn Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile
35 40 45
Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Thr Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 128
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 128
Gly Ser Val Ser Ser Gly Ser Tyr Tyr
1. 5
<210> 129
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 129
Ile Tyr Tyr Ser Gly Ser Thr
1. 5
<210> 130
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 130
Ala Arg Asn Pro Ile Ser Ile Pro Ala Phe Asp Ile
1. 5 10
<210> 131
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 131
Asn Ile Gly Ser Lys Ser
1. 5
<210> 132
<400> 132
000
<210> 133
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 133
Gln Val Trp Asp Thr Ser Ser Asp His Val Leu
1. 5 10
<210> 134
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 134
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Ser Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asn Pro Ile Ser Ile Pro Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 135
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 135
Gln Pro Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 136
<211> 257
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 136
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Ser Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asn Pro Ile Ser Ile Pro Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Pro Val Leu Thr Gln
130 135 140
Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Arg Ile Thr Cys
145 150 155 160
Gly Gly Asn Asn Ile Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys
165 170 175
Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp Ser Asp Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Val Trp Asp Thr Ser Ser Asp His Val Leu Phe Gly Gly Gly
225 230 235 240
Thr Lys Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His
245 250 255
His
<210> 137
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 137
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Ser Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asn Pro Ile Ser Ile Pro Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 138
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 138
Gln Pro Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 139
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 139
Gly Tyr Met Phe Ser Thr Phe Trp
1. 5
<210> 140
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 140
Ile Tyr Pro Gly Asp Ser Asn Thr
1. 5
<210> 141
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 141
Ala Lys Ile Arg Asp Gly Tyr Ser Tyr Asp Ala Phe Asp Leu
1. 5 10
<210> 142
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 142
Ser Ser Asp Val Gly Gly Tyr Asn Tyr
1. 5
<210> 143
<400> 143
000
<210> 144
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 144
Ser Ser Tyr Thr Ser Ser Ser Thr Leu Ala Tyr Val
1. 5 10
<210> 145
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 145
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Ala Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Arg Ile Ser Cys Arg Ala Ser Gly Tyr Met Phe Ser Thr Phe
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Tyr Pro Gly Asp Ser Asn Thr Ile Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Asn Thr Thr Tyr
65 70 75 80
Leu Gln Trp Ser Gly Leu Lys Ala Ser Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Lys Ile Arg Asp Gly Tyr Ser Tyr Asp Ala Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 146
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 146
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1. 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Leu Ala Tyr Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 147
<211> 262
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 147
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Ala Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Arg Ile Ser Cys Arg Ala Ser Gly Tyr Met Phe Ser Thr Phe
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Tyr Pro Gly Asp Ser Asn Thr Ile Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Asn Thr Thr Tyr
65 70 75 80
Leu Gln Trp Ser Gly Leu Lys Ala Ser Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Lys Ile Arg Asp Gly Tyr Ser Tyr Asp Ala Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Ser Ala Leu Thr
130 135 140
Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln Ser Ile Thr Ile Ser
145 150 155 160
Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr Asn Tyr Val Ser Trp
165 170 175
Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile Tyr Asp Val
180 185 190
Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe Ser Gly Ser Lys Ser
195 200 205
Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala Glu Asp Glu
210 215 220
Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser Ser Thr Leu Ala Tyr
225 230 235 240
Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Ala Ala Ala Gly Ser
245 250 255
His His His His His His
260
<210> 148
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 148
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Ala Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Arg Ile Ser Cys Arg Ala Ser Gly Tyr Met Phe Ser Thr Phe
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Tyr Pro Gly Asp Ser Asn Thr Ile Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Asn Thr Thr Tyr
65 70 75 80
Leu Gln Trp Ser Gly Leu Lys Ala Ser Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Lys Ile Arg Asp Gly Tyr Ser Tyr Asp Ala Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 149
<211> 218
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 149
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1. 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Leu Ala Tyr Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 150
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 150
Gly Phe Thr Phe Ser Ser Tyr Ala
1. 5
<210> 151
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 151
Ile Ser Tyr Asp Gly Ser Asn Lys
1. 5
<210> 152
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 152
Ala Arg Asp Gly Gly Tyr Tyr His Tyr Gly Leu Asp Val
1. 5 10
<210> 153
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 153
Lys Leu Gly Asp Lys Tyr
1. 5
<210> 154
<400> 154
000
<210> 155
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> участок, определяющий комплементарность (cdr)
<400> 155
Gln Ala Trp Asp Ser Ser Thr Gly Val
1. 5
<210> 156
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 156
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Tyr His Tyr Gly Leu Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 157
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> вариабельный домен
<400> 157
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1. 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Leu Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Arg Lys Pro Gly Gln Ser Pro Val Leu Leu Ile Tyr
35 40 45
Gln Asp Ala Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser Thr Gly Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 158
<211> 255
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> scFv
<400> 158
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Tyr His Tyr Gly Leu Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Glu Leu Thr Gln
130 135 140
Pro Pro Ser Val Ser Val Ser Pro Gly Gln Thr Ala Ser Ile Thr Cys
145 150 155 160
Ser Gly Asp Lys Leu Gly Asp Lys Tyr Ala Ser Trp Tyr Gln Arg Lys
165 170 175
Pro Gly Gln Ser Pro Val Leu Leu Ile Tyr Gln Asp Ala Lys Arg Pro
180 185 190
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
195 200 205
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
210 215 220
Cys Gln Ala Trp Asp Ser Ser Thr Gly Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
245 250 255
<210> 159
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VH
<400> 159
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Gly Tyr Tyr His Tyr Gly Leu Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val
210 215
<210> 160
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fab VL
<400> 160
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1. 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Leu Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Arg Lys Pro Gly Gln Ser Pro Val Leu Leu Ile Tyr
35 40 45
Gln Asp Ala Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser Thr Gly Val
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 161
<211> 489
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> конструкт тандемного диатела
<400> 161
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1. 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Ser Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asn Pro Ile Ser Ile Pro Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly
130 135 140
Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn
145 150 155 160
Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile
165 170 175
Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
180 185 190
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala
195 200 205
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val
210 215 220
Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly
225 230 235 240
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
245 250 255
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
260 265 270
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
275 280 285
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
290 295 300
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
305 310 315 320
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
325 330 335
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
340 345 350
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
355 360 365
Gly Ser Gln Pro Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro
370 375 380
Gly Lys Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys
385 390 395 400
Ser Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val
405 410 415
Ile Tyr Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser
420 425 430
Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu
435 440 445
Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser
450 455 460
Asp His Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ala
465 470 475 480
Ala Gly Ser His His His His His His
485
<210> 162
<211> 705
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-IgAb
<400> 162
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1. 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln
450 455 460
Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr Leu Ser Leu Thr
465 470 475 480
Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly Ser Tyr Tyr Trp Ser
485 490 495
Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile Gly Tyr Ile
500 505 510
Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser Arg Val
515 520 525
Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu Lys Leu Ser
530 535 540
Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asn Pro
545 550 555 560
Ile Ser Ile Pro Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr
565 570 575
Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
580 585 590
Gly Ser Gly Gly Ser Gln Pro Val Leu Thr Gln Pro Pro Ser Val Ser
595 600 605
Val Ala Pro Gly Lys Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile
610 615 620
Gly Ser Lys Ser Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
625 630 635 640
Val Leu Val Ile Tyr Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu
645 650 655
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
660 665 670
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
675 680 685
Thr Ser Ser Asp His Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val
690 695 700
Leu
705
<210> 163
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-IgAb
<400> 163
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 164
<211> 481
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> конструкт тандемного диатела
<400> 164
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser
130 135 140
Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile
145 150 155 160
Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro
165 170 175
Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu
180 185 190
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
195 200 205
Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
210 215 220
Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
225 230 235 240
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
245 250 255
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
260 265 270
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
275 280 285
Trp Met Gly Ala Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln
290 295 300
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
305 310 315 320
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
325 330 335
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
340 345 350
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser
355 360 365
Gly Gly Ser Gly Gly Ser Ala Ile Gln Met Thr Gln Ser Pro Ser Ser
370 375 380
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
385 390 395 400
Glu Asp Ile Tyr Asn Gly Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
405 410 415
Ala Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val
420 425 430
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr
435 440 445
Ile Ser Ser Leu Gln Pro Glu Asp Glu Ala Thr Tyr Tyr Cys Ala Gly
450 455 460
Pro His Lys Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
465 470 475 480
Lys
<210> 165
<211> 727
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-IgAb
<400> 165
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
465 470 475 480
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
485 490 495
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
500 505 510
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
515 520 525
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
530 535 540
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
545 550 555 560
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
565 570 575
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser
580 585 590
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln
595 600 605
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser
610 615 620
Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr
625 630 635 640
Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
645 650 655
Ala Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln
660 665 670
Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met
675 680 685
Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala
690 695 700
Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly
705 710 715 720
Thr Leu Val Thr Val Ser Ser
725
<210> 166
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-IgAb
<400> 166
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile Tyr Asn Gly
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Glu Ala Thr Tyr Tyr Cys Ala Gly Pro His Lys Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 167
<211> 729
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь KiH-scDb-Fc
<400> 167
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly
1. 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Thr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Tyr Val Leu
225 230 235 240
Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile
245 250 255
Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln
260 265 270
Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys
275 280 285
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn
290 295 300
Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp
305 310 315 320
Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly
325 330 335
Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gln Val Gln Leu
340 345 350
Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val
355 360 365
Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp
370 375 380
Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ala Ile Glu
385 390 395 400
Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val
405 410 415
Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser
420 425 430
Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser
435 440 445
Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val
450 455 460
Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
465 470 475 480
Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val
485 490 495
Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn
500 505 510
Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser
515 520 525
Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro
530 535 540
Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile
545 550 555 560
Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp
565 570 575
Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
580 585 590
Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu
595 600 605
Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly
610 615 620
Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly
625 630 635 640
Gln Gly Leu Glu Trp Met Gly Ala Ile Glu Pro Met Tyr Gly Ser Thr
645 650 655
Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr
660 665 670
Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp
675 680 685
Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe
690 695 700
Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala
705 710 715 720
Ala Gly Ser His His His His His His
725
<210> 168
<211> 469
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь KiH-scDb-Fc
<400> 168
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
145 150 155 160
Ala Ser Glu Asp Ile Tyr Asn Gly Leu Ala Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp
180 185 190
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Glu Ala Thr Tyr Tyr Cys
210 215 220
Ala Gly Pro His Lys Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
370 375 380
Gln Val Ser Leu Tyr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly
465
<210> 169
<211> 721
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь Db-Fc
<400> 169
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1. 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser
100 105 110
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
115 120 125
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
130 135 140
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
145 150 155 160
Gly Ala Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe
165 170 175
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
180 185 190
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
195 200 205
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
210 215 220
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
225 230 235 240
Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe
245 250 255
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
465 470 475 480
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
485 490 495
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr Asp Met
500 505 510
Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser
515 520 525
Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val Lys Gly
530 535 540
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
545 550 555 560
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
565 570 575
Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly Gln Gly
580 585 590
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
595 600 605
Ser Gly Gly Gly Gly Ser Ala Ile Gln Met Thr Gln Ser Pro Ser Ser
610 615 620
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
625 630 635 640
Glu Asp Ile Tyr Asn Gly Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
645 650 655
Ala Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val
660 665 670
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr
675 680 685
Ile Ser Ser Leu Gln Pro Glu Asp Glu Ala Thr Tyr Tyr Cys Ala Gly
690 695 700
Pro His Lys Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
705 710 715 720
Lys
<210> 170
<211> 960
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scDb-mFc
<400> 170
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
145 150 155 160
Ala Ser Glu Asp Ile Tyr Asn Gly Leu Ala Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp
180 185 190
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Glu Ala Thr Tyr Tyr Cys
210 215 220
Ala Gly Pro His Lys Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Pro Glu
245 250 255
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Lys Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Ser Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Val Pro Val Leu Asp Ser Asp Gly Ser Phe Arg Leu Ala Ser Tyr
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Tyr
465 470 475 480
Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala
485 490 495
Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp
500 505 510
Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp
515 520 525
Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser
530 535 540
Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu
545 550 555 560
Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly
565 570 575
Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gln Val
580 585 590
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu
595 600 605
Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met
610 615 620
His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ala
625 630 635 640
Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly
645 650 655
Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu
660 665 670
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
675 680 685
Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr
690 695 700
Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
705 710 715 720
Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser
725 730 735
Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly
740 745 750
His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly
755 760 765
Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly
770 775 780
Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu
785 790 795 800
Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln
805 810 815
Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr
820 825 830
Val Leu Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly
835 840 845
Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala
850 855 860
Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala
865 870 875 880
Pro Gly Gln Gly Leu Glu Trp Met Gly Ala Ile Glu Pro Met Tyr Gly
885 890 895
Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg
900 905 910
Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser
915 920 925
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr
930 935 940
Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
945 950 955 960
<210> 171
<211> 960
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь Bi-scFv-Fc
<400> 171
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Asp Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Arg Gly Asp Ile Thr Ser Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asp Tyr Tyr Thr Asp Tyr Met Gly Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Ala Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg
145 150 155 160
Ala Ser Glu Asp Ile Tyr Asn Gly Leu Ala Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp
180 185 190
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Glu Ala Thr Tyr Tyr Cys
210 215 220
Ala Gly Pro His Lys Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Val
225 230 235 240
Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Pro Glu
245 250 255
Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Lys Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
370 375 380
Gln Val Ser Leu Ser Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Val Pro Val Leu Asp Ser Asp Gly Ser Phe Arg Leu Ala Ser Tyr
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Tyr
465 470 475 480
Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala
485 490 495
Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp
500 505 510
Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp
515 520 525
Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser
530 535 540
Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu
545 550 555 560
Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly
565 570 575
Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gln Val
580 585 590
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu
595 600 605
Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met
610 615 620
His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ala
625 630 635 640
Ile Glu Pro Met Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly
645 650 655
Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu
660 665 670
Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
675 680 685
Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr
690 695 700
Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
705 710 715 720
Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser
725 730 735
Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly
740 745 750
His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly
755 760 765
Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly
770 775 780
Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu
785 790 795 800
Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln
805 810 815
Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr
820 825 830
Val Leu Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly
835 840 845
Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala
850 855 860
Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala
865 870 875 880
Pro Gly Gln Gly Leu Glu Trp Met Gly Ala Ile Glu Pro Met Tyr Gly
885 890 895
Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg
900 905 910
Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser
915 920 925
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr
930 935 940
Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
945 950 955 960
<210> 172
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 172
Ser Phe Phe Pro Pro Gly Tyr Gln
1. 5
<210> 173
<211> 725
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-Ig
<400> 173
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Thr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ser Ser Gly Trp Trp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly
435 440 445
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr
465 470 475 480
Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly Glu Thr Val Thr Ile
485 490 495
Thr Cys Arg Val Ser Glu Asn Ile Tyr Ser Tyr Leu Ala Trp Tyr Gln
500 505 510
Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val Tyr Asn Ala Lys Thr
515 520 525
Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
530 535 540
Gln Phe Ser Leu Lys Ile Asn Ser Leu Gln Pro Glu Asp Phe Gly Ser
545 550 555 560
Tyr Tyr Cys Gln His His Tyr Gly Thr Pro Trp Thr Phe Gly Gly Gly
565 570 575
Thr Lys Leu Glu Ile Lys Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
580 585 590
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Gln
595 600 605
Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln Ser Leu Ser Leu Thr
610 615 620
Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Asp Tyr Ala Trp Asn Trp
625 630 635 640
Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp Met Gly Tyr Ile Thr
645 650 655
Tyr Ser Gly Ser Thr Ser Tyr Asn Pro Ser Leu Glu Ser Arg Ile Ser
660 665 670
Ile Thr Arg Asp Thr Ser Thr Asn Gln Phe Phe Leu Gln Leu Asn Ser
675 680 685
Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys Ala Arg Gly Gly Tyr
690 695 700
Tyr Gly Ser Ser Trp Gly Val Phe Ala Tyr Trp Gly Gln Gly Thr Leu
705 710 715 720
Val Thr Val Ser Ala
725
<210> 174
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-Ig
<400> 174
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1. 5 10 15
Arg Val Ile Ile Ser Cys Ser Gly Ser His Ser Asn Ile Gly Ser Asn
20 25 30
Asn Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asn Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ser Asp Tyr Phe Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 175
<211> 727
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-Ig
<400> 175
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Thr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ser Ser Gly Trp Trp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly
435 440 445
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ser Ala Leu Thr
465 470 475 480
Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln Ser Ile Thr Ile Ser
485 490 495
Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn Ala Val Asn Trp Tyr
500 505 510
Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Asp Asp
515 520 525
Leu Leu Pro Ser Gly Val Ser Asp Arg Phe Ser Gly Ser Lys Ser Gly
530 535 540
Thr Ser Ala Phe Leu Ala Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala
545 550 555 560
Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu Asn Gly Pro Val Phe
565 570 575
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly
580 585 590
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val
595 600 605
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu
610 615 620
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly Met
625 630 635 640
His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Phe
645 650 655
Ile Arg Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys Gly
660 665 670
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
675 680 685
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
690 695 700
Asp Arg Gly Leu Gly Asp Gly Thr Tyr Phe Asp Tyr Trp Gly Gln Gly
705 710 715 720
Thr Thr Val Thr Val Ser Ser
725
<210> 176
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-Ig
<400> 176
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1. 5 10 15
Arg Val Ile Ile Ser Cys Ser Gly Ser His Ser Asn Ile Gly Ser Asn
20 25 30
Asn Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asn Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ser Asp Tyr Phe Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 177
<211> 722
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-Ig
<400> 177
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Thr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ser Ser Gly Trp Trp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Ala Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly
435 440 445
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
450 455 460
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Tyr Val Leu Thr
465 470 475 480
Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser
485 490 495
Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln
500 505 510
Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg
515 520 525
Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr
530 535 540
Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr
545 550 555 560
Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr
565 570 575
Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
580 585 590
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln
595 600 605
Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys
610 615 620
Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg
625 630 635 640
Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ala Ile Glu Pro Met
645 650 655
Tyr Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met
660 665 670
Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu
675 680 685
Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr
690 695 700
Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
705 710 715 720
Ser Ser
<210> 178
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> полипептидная цепь scFv-Ig
<400> 178
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1. 5 10 15
Arg Val Ile Ile Ser Cys Ser Gly Ser His Ser Asn Ile Gly Ser Asn
20 25 30
Asn Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asn Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ser Asp Tyr Phe Cys Gly Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-LILRB1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2801535C1 |
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, ЭКСПРЕССИОННАЯ КАССЕТА (ВАРИАНТЫ), ПЛАЗМИДА (ВАРИАНТЫ), КЛЕТКА-ХОЗЯИН, ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, НАБОР, СПОСОБ ЛЕЧЕНИЯ БОЛЬНОГО, ПОДВЕРЖЕННОГО РИСКУ ИЛИ СТРАДАЮЩЕГО ОТ ИНФЕКЦИИ E.COLI, И СПОСОБ ДИАГНОСТИКИ ОПРЕДЕЛЕНИЯ ИНФЕКЦИЙ E.COLI | 2014 |
|
RU2724530C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ ДОМЕНЫ ПРОТИВ TRBC1 | 2018 |
|
RU2791327C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ ПРОТИВ TIGIT И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2776714C2 |
| АМАТОКСИНОВЫЕ КОНЪЮГАТЫ АНТИТЕЛА С ЛЕКАРСТВЕННЫМ СРЕДСТВОМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826004C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| МОЛЕКУЛЫ АНТИ-SIGLEC-9 АНТИТЕЛА | 2020 |
|
RU2833330C1 |
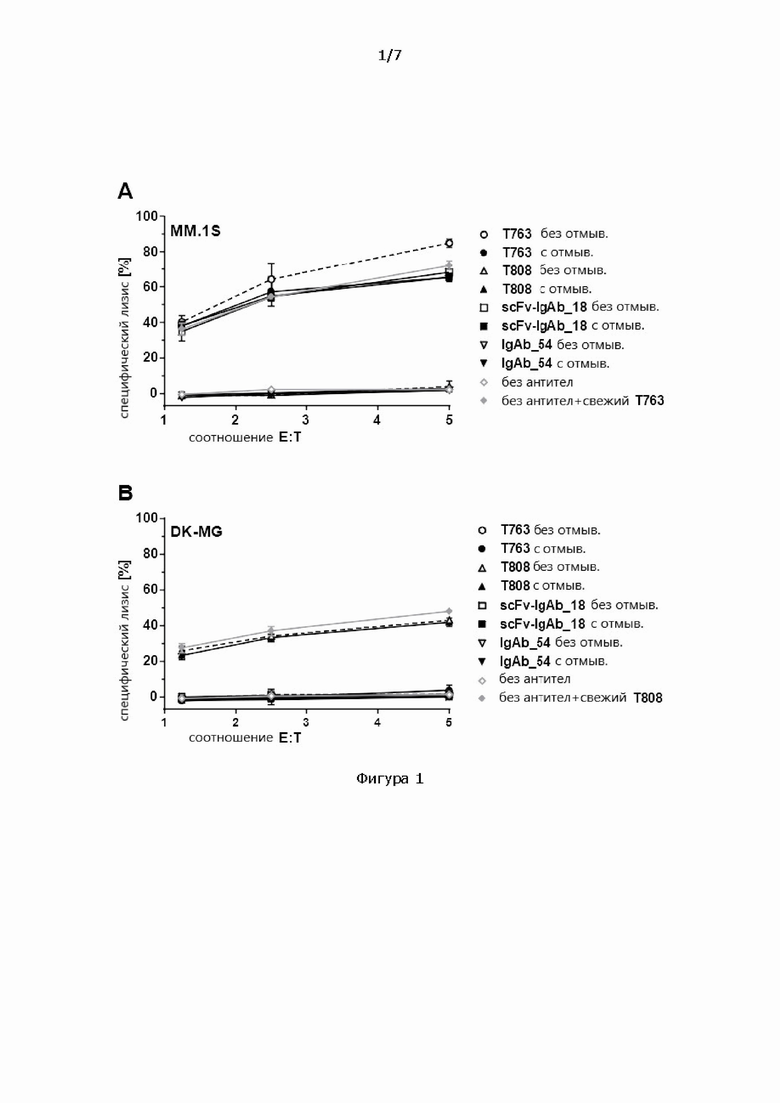
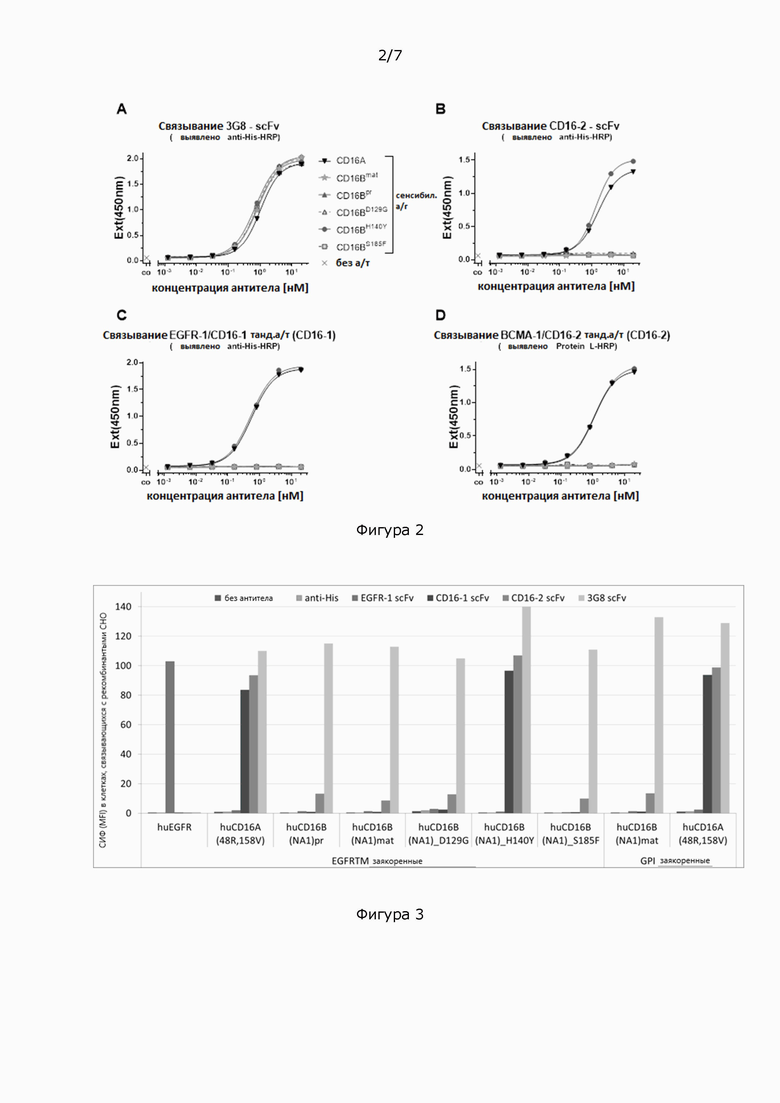


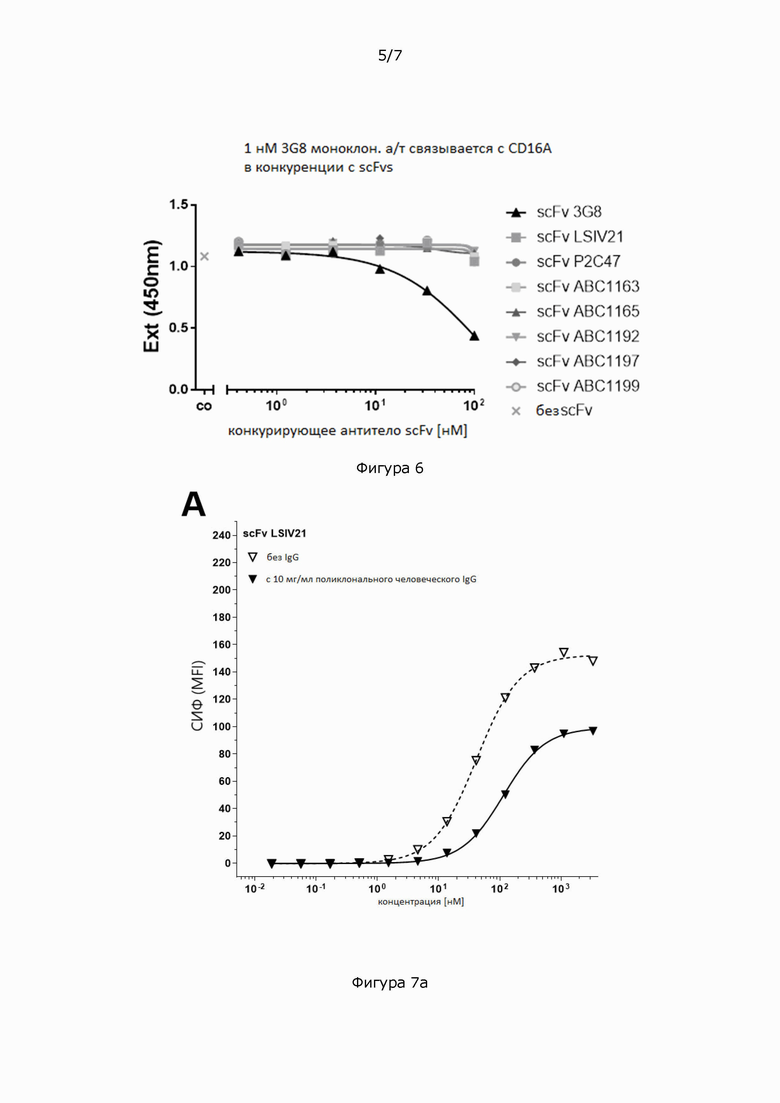
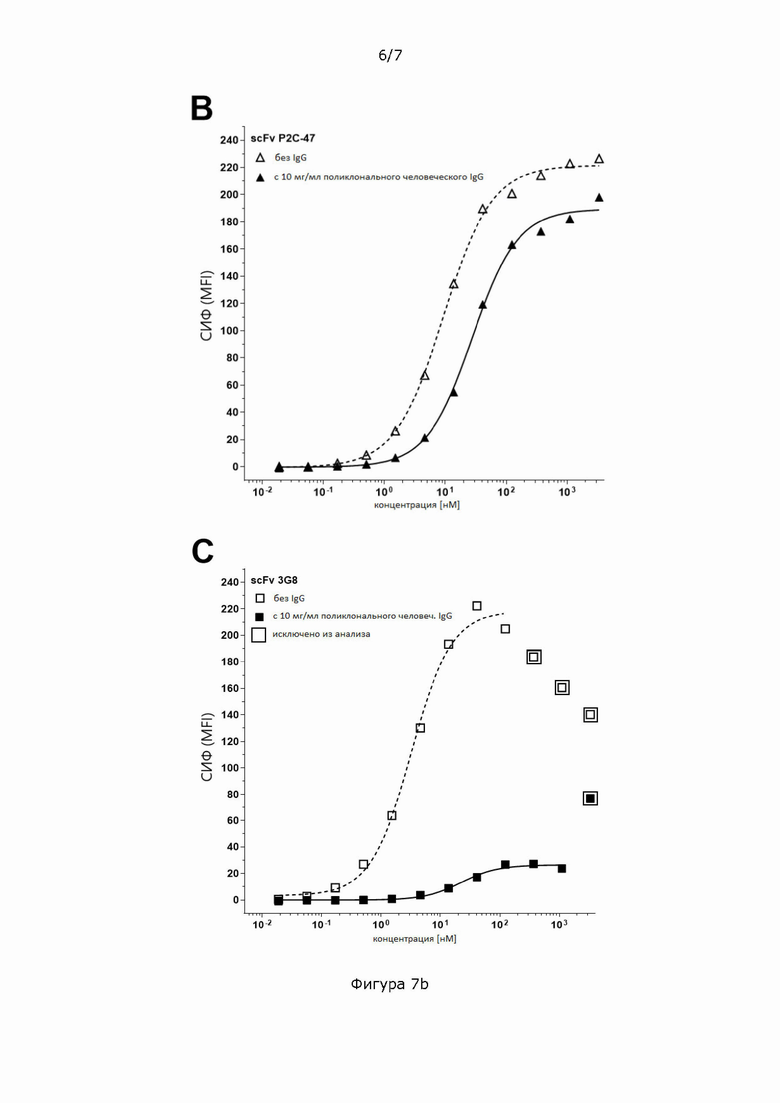
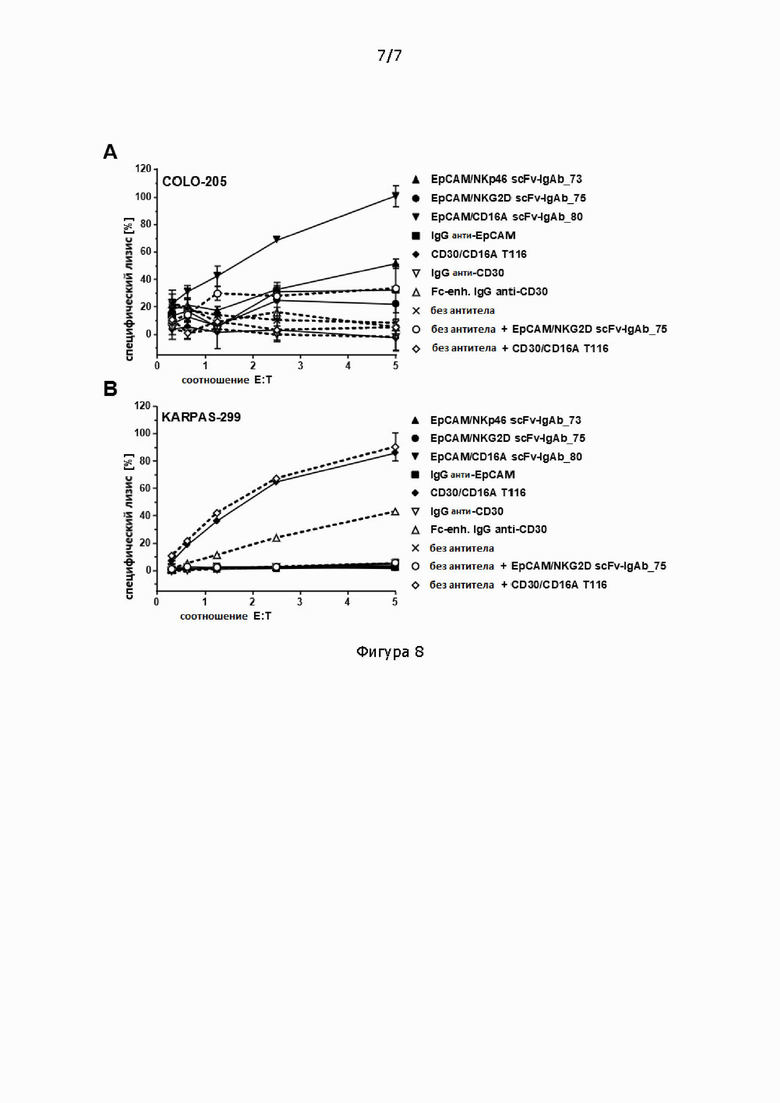
Изобретение относится к области биотехнологии, в частности к выделенной клетке-естественному киллеру (NK-клетке) человека в криоконсервированном состоянии. Указанная клетка предназначена для получения NK-клетки, способной проявлять цитотоксическое действие в отношении клетки-мишени, и предварительно нагружена перед замораживанием конструкцией антитела, содержащей первый связывающий домен, связывающийся с антигеном рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки, и второй связывающий домен, связывающийся с антигеном клеточной поверхности на клеточной поверхности клетки-мишени. Указанные первый и второй связывающие домены содержат три CDR легкой цепи и три CDR тяжелой цепи. Кроме того, настоящее изобретение также относится к способам получения NK-клеток, фармацевтической композиции и способу лечения опухолевого заболевания. Настоящее изобретение позволяет поддерживать конструкцию антитела в связанном с поверхностью клетки состоянии даже во время и после нахождения NK-клетки в криоконсервированном состоянии. 5 н. и 24 з.п. ф-лы, 9 ил., 6 табл., 4 пр.
1. Выделенная клетка-естественный киллер (NK-клетка) человека в криоконсервированном состоянии для получения NK-клетки, способной проявлять цитотоксическое действие в отношении клетки-мишени, предварительно нагруженная перед замораживанием конструкцией антитела, указанная конструкция антитела содержит по меньшей мере первый связывающий домен, связывающийся с аффинностью по меньшей мере 106 М с антигеном рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки, указанный первый связывающий домен содержит три CDR легкой цепи и три CDR тяжелой цепи, и второй связывающий домен, связывающийся с антигеном клеточной поверхности на клеточной поверхности клетки-мишени, указанный второй связывающий домен содержит три CDR легкой цепи и три CDR тяжелой цепи.
2. Выделенная NK-клетка человека согласно п. 1, отличающаяся тем, что указанная NK-клетка выделена из ткани пуповины или плаценты, индуцированных плюрипотентных стволовых клеток (iPSC) или мононуклеарных клеток периферической крови (РВМС) от здоровых доноров.
3. Выделенная NK-клетка человека согласно п. 1 или 2, отличающаяся тем, что указанная NK-клетка консервирована в криорастворе.
4. Выделенная NK-клетка человека согласно любому из пп. 1-3, отличающаяся тем, что указанная NK-клетка предварительно нагружена в растворе, содержащем указанную конструкцию антитела в концентрации по меньшей мере 5 нМ.
5. Выделенная NK-клетка человека согласно любому из пп. 1-4, отличающаяся тем, что указанный антиген рецептора NK-клеток, с которым связывается указанный первый связывающий домен конструкции антитела, выбран из группы, состоящей из CD16a, CD16b, NKp46, NKG2D и CD16a + CD16b.
6. Выделенная NK-клетка человека согласно любому из пп. 1-5, отличающаяся тем, что указанный антиген клеточной поверхности на клеточной поверхности клетки-мишени, с которым связывается указанный второй связывающий домен конструкции антитела, выбран из группы, состоящей из CD19, CD20, CD22, CD30, CD33, CD52, CD70, CD74, CD79b, CD123, CLL1, BCMA, FCRH5, EGFR, EGFRvIII, HER2 и GD2.
7. Выделенная NK-клетка человека согласно любому из пп. 1-6, отличающаяся тем, что указанная конструкция антитела содержит первый связывающий домен, связывающийся с CD16a, и второй связывающий домен, связывающийся с антигеном, выбранным из группы, состоящей из CD19, CD20, CD22, CD30, CD33, CD52, CD70, CD74, CD79b, CD123, CLL1, BCMA, FCRH5, EGFR, EGFRvIII, HER2 и GD2.
8. Выделенная NK-клетка человека согласно п. 7, отличающаяся тем, что указанная конструкция антитела содержит в указанном первом связывающем домене три CDR тяжелой цепи и три CDR легкой цепи, выбранные из группы, состоящей из:
(a) CDR-H1, указанного под SEQ ID NO: 29, CDR-H2, указанного под SEQ ID NO: 30, CDR-H3, указанного под SEQ ID NO: 31, CDR-L1, указанного под SEQ ID NO: 32, CDR-L2, указанного под SEQ ID NO: 33, CDR-L3, указанного под SEQ ID NO: 34;
(b) CDR-H1, указанного под SEQ ID NO: 40, CDR-H2, указанного под SEQ ID NO: 41, CDR-H3, указанного под SEQ ID NO: 42, CDR-L1, указанного под SEQ ID NO: 43, CDR-L2, указанного под SEQ ID NO: 44, CDR-L3, указанного под SEQ ID NO: 45;
(c) CDR-H1, указанного под SEQ ID NO: 51, CDR-H2, указанного под SEQ ID NO: 52, CDR-H3, указанного под SEQ ID NO: 53, CDR-L1, указанного под SEQ ID NO: 54, CDR-L2, указанного под SEQ ID NO: 55, CDR-L3, указанного под SEQ ID NO: 56;
(d) CDR-H1, указанного под SEQ ID NO: 62, CDR-H2, указанного под SEQ ID NO: 63, CDR-H3, указанного под SEQ ID NO: 64, CDR-L1, указанного под SEQ ID NO: 65, CDR-L2, указанного под SEQ ID NO: 66, CDR-L3, указанного под SEQ ID NO: 67;
(e) CDR-H1, указанного под SEQ ID NO: 73, CDR-H2, указанного под SEQ ID NO: 74, CDR-H3, указанного под SEQ ID NO: 75, CDR-L1, указанного под SEQ ID NO: 76, CDR-L2, указанного под SEQ ID NO: 77, CDR-L3, указанного под SEQ ID NO: 78;
(f) CDR-H1, указанного под SEQ ID NO: 84, CDR-H2, указанного под SEQ ID NO: 85, CDR-H3, указанного под SEQ ID NO: 86, CDR-L1, указанного под SEQ ID NO: 87, CDR-L2, указанного под SEQ ID NO: 88, CDR-L3, указанного под SEQ ID NO: 89; и
(g) CDR-H1, указанного под SEQ ID NO: 95, CDR-H2, указанного под SEQ ID NO: 96, CDR-H3, указанного под SEQ ID NO: 97, CDR-L1, указанного под SEQ ID NO: 98, CDR-L2, указанного под SEQ ID NO: 99, CDR-L3, указанного под SEQ ID NO: 100.
9. Выделенная NK-клетка человека согласно п. 8, отличающаяся тем, что указанная конструкция антитела содержит в указанном первом связывающем домене пары VH- и VL-цепей, имеющих последовательность, указанную в парах последовательностей, выбранных из группы, состоящей из SEQ ID NO: 35 и SEQ ID NO: 36, SEQ ID NO: 46 и SEQ ID NO: 47, SEQ ID NO: 57 и SEQ ID NO: 58, SEQ ID NO: 68 и SEQ ID NO: 69, SEQ ID NO: 79 и SEQ ID NO: 80, SEQ ID NO: 90 и SEQ ID NO: 91 и SEQ ID NO: 101 и SEQ ID NO: 102.
10. Выделенная NK-клетка человека согласно любому из пп. 7-9, отличающаяся тем, что указанная конструкция антитела содержит в указанном втором связывающем домене три CDR тяжелой цепи и три CDR легкой цепи, выбранные из группы, состоящей из:
(a) CDR-H1, указанного под SEQ ID NO: 106, CDR-H2, указанного под SEQ ID NO: 107, CDR-H3, указанного под SEQ ID NO: 108, CDR-L1, указанного под SEQ ID NO: 109, CDR-L2, указанного под SEQ ID NO: 110, CDR-L3, указанного под SEQ ID NO: 111;
(b) CDR-H1, указанного под SEQ ID NO: 128, CDR-H2, указанного под SEQ ID NO: 129, CDR-H3, указанного под SEQ ID NO: 130, CDR-L1, указанного под SEQ ID NO: 131, CDR-L2, указанного под SEQ ID NO: 132, CDR-L3, указанного под SEQ ID NO: 133;
(c) CDR-H1, указанного под SEQ ID NO: 117, CDR-H2, указанного под SEQ ID NO: 118, CDR-H3, указанного под SEQ ID NO: 119, CDR-L1, указанного под SEQ ID NO: 120, CDR-L2, указанного под SEQ ID NO: 121, CDR-L3, указанного под SEQ ID NO: 122.
11. Выделенная NK-клетка человека согласно любому из пп. 7-10, отличающаяся тем, что указанная конструкция антитела содержит в указанном втором связывающем домене пары VH- и VL-цепей, имеющих последовательность, указанную в парах последовательностей, выбранных из группы, состоящей из SEQ ID NO: 112 и SEQ ID NO: 113, SEQ ID NO: 123 и SEQ ID NO: 124, и SEQ ID NO: 134 и SEQ ID NO: 135.
12. Выделенная NK-клетка человека согласно любому из пп. 7-11, отличающаяся тем, что указанная конструкция антитела содержит последовательность белка, указанную под SEQ ID NO: 161-171.
13. Способ получения криоконсервированной предварительно нагруженной NK-клетки человека, указанный способ включает
(i) инкубирование NK-клетки с конструкцией антитела в базальной среде для культивирования клеток, причем указанная конструкция антитела находится в концентрации по меньшей мере 5 нМ и содержит по меньшей мере первый связывающий домен, связывающийся с аффинностью по меньшей мере 106 М с антигеном рецептора NK-клетки на клеточной поверхности иммунологической эффекторной клетки, указанный первый связывающий домен содержит три CDR легкой цепи и три CDR тяжелой цепи, и второй связывающий домен, связывающийся с антигеном клеточной поверхности на клеточной поверхности клетки-мишени, указанный второй связывающий домен содержит три CDR легкой цепи и три CDR тяжелой цепи; и
(ii) замораживание указанной NK-клетки путем охлаждения до температуры ниже нуля в среде для замораживания, которая содержит базальную среду для культивирования клеток, криозащитный агент и, необязательно, изотонический раствор.
14. Способ по п. 13, отличающийся тем, что указанная NK-клетка выделена из ткани пуповины, iPSC или РВМС от здоровых доноров.
15. Способ по пп. 13 и 14, отличающийся тем, что криопротекторный агент находится в концентрации от 1% до 30% (об./об.).
16. Способ согласно любому из пп. 13-15, отличающийся тем, что указанный антиген рецептора NK-клетки, с которым связывается указанный первый связывающий домен конструкции антитела, выбран из группы, состоящей из CD16a, CD16b, NKp46, NKG2D и CD16a + CD16b.
17. Способ согласно любому из пп. 13-16, отличающийся тем, что указанный антиген клеточной поверхности на клеточной поверхности клетки-мишени, с которым связывается указанный второй связывающий домен конструкции антитела, выбран из группы, состоящей из CD19, CD22, CD30, CD33, CD52, CD70, CD74, CD79b, CD123, CLL1, BCMA, FCRH5, EGFR, HER2, GD2.
18. Способ согласно любому из пп. 13-17, отличающийся тем, что среда для замораживания содержит изотонический раствор в диапазоне от 1% до 60% (об./об.).
19. Способ согласно любому из пп. 13-18, отличающийся тем, что указанная конструкция антитела содержит последовательность белка, указанную под SEQ ID NO: 161-171.
20. Способ получения жизнеспособной предварительно нагруженной NK-клетки человека из NK-клетки человека в криоконсервированном состоянии согласно любому из пп. 1-12 для введения указанной клетки субъекту, нуждающемуся в этом, указанный способ характеризуется тем, что он включает стадию получения указанной клетки для введения пациенту путем размораживания NK клетки по любому из пп. 1-12.
21. Фармацевтическая композиция для введения NK-клетки человека пациенту, содержащая NK-клетку человека, которая получена из NK-клетки человека в криоконсервированном состоянии согласно любому из пп. 1-12, и по меньшей мере одно фармацевтическое вспомогательное вещество.
22. Фармацевтическая композиция согласно п. 21, которую вводят пациенту внутривенно.
23. Фармацевтическая композиция согласно п. 21 или 22 для применения для предотвращения, лечения или облегчения заболевания, выбранного из пролиферативного заболевания, опухолевого заболевания или иммунологического расстройства.
24. Фармацевтическая композиция согласно п. 23, отличающаяся тем, что указанное опухолевое заболевание представляет собой злокачественное заболевание, предпочтительно рак.
25. Фармацевтическая композиция согласно п. 24, отличающаяся тем, что указанное злокачественное заболевание выбрано из группы, состоящей из лимфомы Ходжкина, неходжкинской лимфомы, лейкоза, множественной миеломы и солидных опухолей.
26. Способ лечения опухолевого заболевания, указанный способ включает стадию введения субъекту, нуждающемуся в этом, предварительно нагруженной NK-клетки человека, которая была получена из NK-клетки человека в криоконсервированном состоянии согласно любому из пп. 6-12.
27. Способ согласно п. 26, отличающийся тем, что указанную предварительно нагруженную NK-клетку человека вводят пациенту внутривенно.
28. Способ согласно п. 26 или 27, отличающийся тем, что указанное опухолевое заболевание представляет собой злокачественное заболевание, предпочтительно рак.
29. Способ согласно п. 28, отличающийся тем, что указанное злокачественное заболевание выбрано из группы, состоящей из лимфомы Ходжкина, неходжкинской лимфомы, лейкоза, множественной миеломы и солидных опухолей.
| REUSCH U | |||
| et al., A novel tetravalent bispecific TandAb (CD30/CD16A) efficiently recruits NK cells for the lysis of CD30 + tumor cells, MABS, 2014, vol | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| НЕФТЯНАЯ РЕГЕНЕРАТИВНАЯ ПЕЧЬ | 1921 |
|
SU727A1 |
| WO 2011004201 A1, 13.01.2011 | |||
| АНТИ-CD 16 СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2006 |
|
RU2491294C2 |
| СПОСОБ КРИОКОНСЕРВАЦИИ КЛЕТОК, ИСКУССТВЕННЫЕ КЛЕТОЧНЫЕ КОНСТРУКЦИИ ИЛИ ТРЕХМЕРНЫЕ СЛОЖНЫЕ КОМПЛЕКСЫ ТКАНЕЙ | 2010 |
|
RU2573307C2 |
| S | |||
| PASLEY et al., Natural killer-92 cells maintain cytotoxic activity after long-term | |||

