Перекрестные ссылки на родственные заявки
По данной заявке испрашивается приоритет по предварительной патентной заявке США №61/553131, поданной 28 октября 2011 года; и предварительной патентной заявке США №61/711208, поданной 8 октября 2012 года, содержание каждой из которых включено в настоящий документ путем отсылки в полном объеме в отношении всех целей.
Ссылка на перечень последовательностей
Перечень последовательностей, записанный в файле 424859SEQLIST.txt, составляет 34 килобайта и был создан 24 октября 2012 года. Информация, содержащаяся в этом файле, включена в настоящий документ путем отсылки.
Предпосылки создания изобретения
Синуклеинопатии, также известные как заболевания с тельцами Леви (Lewy body diseases, LBD), характеризуются дегенерацией дофаминергической системы, нарушениями моторных функций, когнитивным расстройством и формированием телец Леви (LB), и/или нейритов Леви (McKeith et al., Neurology (1996) 47:1113-24). Синуклеинопатии включают болезнь Паркинсона (включая идиопатическую болезнь Паркинсона), болезнь с диффузными тельцами Леви (Diffuse Lewy Body Disease, DLBD), также известную как деменция с тельцами Леви (Dementia with Lewy Bodies, DLB), вариант болезни Альцгеймера с тельцами Леви (Lewy body variant of Alzheimer's disease, LBV), сочетание болезни Альцгеймера и болезни Паркинсона, идиопатическую ортостатическую гипотензию и множественную системную атрофию (MSA; например, оливопонтоцеребеллярную атрофию, стрионигральную дегенерацию и синдром Шая-Дрейджера). Некоторые немоторные признаки и симптомы считаются предвестниками синуклеинопатий в продромальной фазе заболеваний (то есть предсимптоматический, субклинический, предклинический или премоторный период). Такие ранние признаки включают, например, расстройство поведения в REM-стадии сна (RBD), утрату способности ощущать запахи и констатацию (Mahowald et al., Neurology (2010) 75:488-489). Заболевания с тельцами Леви продолжают оставаться общей причиной нарушений моторных функций и когнитивных расстройств у пожилых людей (Galasko et al., Arch. Neurol. (1994) 51:888-95).
Альфа-синуклеин является частью большого семейства белков, включая бета- и гамма-синуклеин, и синоретин. Альфа-синуклеин экспрессируется в нормальном состоянии, связанном с синапсами и, как полагают, играет роль в нервной пластичности, обучении и памяти. Некоторые исследования показали, что альфа-синуклеин играет центральную роль в патогенезе болезни Паркинсона (PD). Белок может агрегировать с образованием нерастворимых фибрилл в патологических условиях. Например, синуклеин накапливается в тельцах Леви (LB) (Spillantini et al., Nature (1997) 388:839-40; Takeda et al., J. Pathol. (1998) 152:367-72; Wakabayashi et al., Neurosci. Lett. (1997) 239:45-8). Мутации в гене альфа-синуклеина являются причиной редких семейных форм паркинсонизма (Kruger et al., Nature Gen. (1998) 18:106-8; Polymeropoulos, et al., Science (1997) 276:2045-7). Гиперэкспрессия альфа-синуклеина y трансгенных мышей (Masliah et al., Science (2000) 287:1265-9) и у мух вида Drosophila (Feany et al., Nature (2000) 404:394-8) имитирует некоторые патологические аспекты заболевания с тельцами Леви. Кроме того, было сделано предположение о том, что растворимые олигомеры синуклеина могут быть нейротоксическими (Conway KA, et al., Proc Natl Acad Sci USA (2000) 97:571-576; VollesMJ, Lansbury PT, Jr Biochemistry (2003) 42:7871-7878). Накопление альфа-синуклеина со сходными морфологическими и неврологическими изменениями в таких биологических видах и моделях животных, как человек, мыши и мухи, позволяет предположить, что эта молекула способствует развитию заболевания с тельцами Леви.
Краткое описание заявленного изобретения
Изобретение обеспечивает антитела, содержащие гуманизированную зрелую вариабельную область тяжелой цепи, содержащую три CDR по Kabat SEQ ID NO: 11, и являющуюся по меньшей мере на 90% идентичной SEQ ID NO: 11, и гуманизированную легкую цепь, содержащую три CDR по Kabat SEQ ID NO: 4, и являющуюся по меньшей мере на 90% идентичной SEQ ID NO: 4. В некоторых антителах зрелая вариабельная область тяжелой цепи, по меньшей мере, на 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO: 11, и зрелая вариабельная область легкой цепи, по меньшей мере, на 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO: 4. В некоторых антителах положение L36 (нумерация по Kabat) может быть занято Y или F, положение L83 (нумерация по Kabat) может быть занято F или L, положение Н73 (нумерация по Kabat) может быть занято N или D, и положение Н93 (нумерация по Kabat) может быть занято А или S. В некоторых таких антителах аминокислотная последовательность зрелой вариабельной области тяжелой цепи отличается от аминокислотной последовательности SEQ ID NO: 11, и аминокислотная последовательность зрелой вариабельной области легкой цепи отличается от аминокислотной последовательности SEQ ID NO: 4.
В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 8, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 3. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 8, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 4. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 8, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 5. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 9, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 3. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 9, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 4. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 9, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 5. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 10, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 3. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 10, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 4. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 10, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 5. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 11, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 3. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 11, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 4. В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 11, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 5.
Кроме того, изобретение обеспечивает антитело, содержащее гуманизированную тяжелую цепь, содержащую три CDR по Kabat SEQ ID NO: 11, и гуманизированную легкую цепь, содержащую по Kabat три CDR SEQ ID NO: 4, при условии, что положение L36 (нумерация по Kabat) занято F или Y, и/или положение L83 (нумерация по Kabat) занято L или F, и/или положение Н73 (нумерация по Kabat) занято D или N, и/или положение Н93 (нумерация по Kabat) занято S или А. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F и положение Н73 (нумерация по Kabat) занято D, и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 занято F. В некоторых таких антителах положение L83 занято L. В некоторых таких антителах положение Н73 занято D. В некоторых таких антителах положение Н93 занято А. В некоторых таких антителах положение L36 занято F и положение L83 занято L. В некоторых таких антителах положение L36 занято F и положение Н73 занято D. В некоторых таких антителах положение L36 занято F и положение Н93 занято А. В некоторых таких антителах положение L36 занято F, положение L83 занято L и положение Н73 занято D. В некоторых таких антителах положение L36 занято F, положение L83 занято L и положение Н93 занято А. В некоторых таких антителах положение L 36 занято F, положение L83 занято L, положение Н73 занято D и положение Н93 занято А. В некоторых таких антителах остатки в положениях L36, L83, Н73 и Н93 (нумерация по Kabat) заняты аминокислотами, как указано в Таблице 1, занято F, и положение Н73 (нумерация по Kabat) занято D, и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение Н73 (нумерация по Kabat) занято N. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L, положение Н73 (нумерация по Kabat) занято D, и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L и положение Н93 (нумерация по Kabat) занято S.
В любом из указанных выше антител зрелая вариабельная область тяжелой цепи может быть слита с константной областью тяжелой цепи, и зрелая константная область легкой цепи может быть слита с константной областью легкой цепи.
В любом из указанных выше антител константная область тяжелой цепи может представлять собой мутантную форму природной константной области человека, которая имеет пониженный уровень связывания с Fcγ-рецептором по сравнению с природной константной областью человека.
В любом из указанных выше антител константная область тяжелой цепи может быть изотипа IgG1 человека. В некоторых антителах аллотип представляет собой Glm3. В некоторых антителах аллотип представляет собой Glm1.
Кроме того, изобретение обеспечивает нуклеиновую кислоту, кодирующую любую из указанных выше зрелых вариабельных областей тяжелой цепи и/или любую указанную выше зрелую вариабельную область легкой цепи, например, SEQ ID NO: 15, 17, 18, 19 и 20.
Кроме того, изобретение обеспечивает клетку хозяина, содержащую вектор, содержащий любую из нуклеиновых кислот, описанных выше.
Кроме того, изобретение обеспечивает способ лечения пациента, имеющего или имеющего риск развития заболевания с тельцами Леви, включающий введение пациенту эффективного количества любого из указанных выше антител. В некоторых способах заболевание представляет собой болезнь Паркинсона. В некоторых способах ухудшение когнитивной функции у пациента является ингибированным. В некоторых способах количество нейритных и/или аксональных агрегатов альфа-синуклеина является уменьшенным. В некоторых способах нейритная дистрофия у пациента является уменьшенной. В некоторых способах синаптическая и/или дендритная плотность является сохраненной. В некоторых способах, способ сохраняет синаптофизин и/или МАР2 у пациента.
Кроме того, изобретение обеспечивает способ лечения пациента, имеющего или имеющего риск развития синуклеинопатии, включающий введение пациенту эффективного количества любого из указанных выше антител. В некоторых способах заболевание представляет собой болезнь Паркинсона. В некоторых способах, заболевание представляет собой расстройство поведения в REM-стадии сна (RBD). В некоторых способах, заболевание представляет собой деменцию с тельцами Леви (DLB) или множественную системную атрофию (MSA). В некоторых способах ухудшение когнитивной функции у пациента является ингибированным. В некоторых способах количество нейритных и/или аксональных агрегатов альфа-синуклеина является уменьшенным. В некоторых способах нейритная дистрофия у пациента является уменьшенной. В некоторых способах синаптическая и/или дендритная плотность является сохраненной. В некоторых способах, способ сохраняет синаптофизин и/или МАР2 у пациента.
Кроме того, изобретение обеспечивает способы обнаружения телец Леви у пациента, имеющего или имеющего риск развития заболевания с тельцами Леви, включающие введение пациенту эффективного количества любого из указанных выше антител, при этом антитело связывается с тельцами Леви и связанное антитело детектируется. В некоторых способах заболевание представляет собой болезнь Паркинсона. В некоторых способах заболевание представляет собой деменцию с тельцами Леви (DLB) или множественную системную атрофию (MSA). В некоторых способах антитело является меченым.
Кроме того, изобретение обеспечивает способ уменьшения формирования телец Леви у пациента, имеющего или имеющего риск развития заболевания с тельцами Леви, включающий введение пациенту эффективного количества любого из указанных выше антител. В некоторых способах заболевание представляет собой болезнь Паркинсона. В некоторых способах заболевание представляет собой деменцию с тельцами Леви (DLB) или множественную системную атрофию (MSA). В некоторых способах ухудшение когнитивной функции у пациента является ингибированным. В некоторых способах количество нейритных и/или аксональных агрегатов альфа-синуклеина является уменьшенным. В некоторых способах нейритная дистрофия у пациента является уменьшенной. В некоторых способах синаптическая и/или дендритная плотность является сохраненной. В некоторых способах, способ сохраняет синаптофизин и/или МАР2 у пациента.
Кроме того, изобретение обеспечивает способ ингибирования агрегации синуклеина, или очистки телец Леви или агрегатов синуклеина у пациента, имеющего или имеющего риск развития заболевания с тельцами Леви, включающий введение пациенту эффективного количества любого из указанных выше антител. В некоторых способах заболевание представляет собой болезнь Паркинсона. В некоторых способах заболевание представляет собой деменцию с тельцами Леви (DLB) или множественную системную атрофию (MSA). В некоторых способах ухудшение когнитивной функции у пациента является ингибированным. В некоторых способах количество нейритных и/или аксональных агрегатов альфа-синуклеина является уменьшенным. В некоторых способах нейритная дистрофия у пациента является уменьшенной. В некоторых способах синаптическая и/или дендритная плотность является сохраненной. В некоторых способах, способ сохраняет синаптофизин и/или МАР2 у пациента.
Кроме того, изобретение обеспечивает фармацевтическую композицию, содержащую любое из указанных выше антител.
Кроме того, изобретение обеспечивает способ получения антитела, включающий культивирование клеток, трансформированных нуклеиновыми кислотами, кодирующими тяжелые и легкие цепи антитела таким образом, что клетка секретирует антитело; и очистку антитела из клеточной культуральной среды; при этом антитело представляет собой любое из антител, описанных выше.
Кроме того, изобретение обеспечивает способ получения клеточной линии, продуцирующей антитело, включающий введение в клетки вектора, кодирующего тяжелые и легкие цепи антитела, и селектируемого маркера; размножение клеток в условиях, обеспечивающих отбор клеток, имеющих увеличенное число копий вектора; изолирование отдельных клеток из отобранной клетки; и консервацию клеток, клонированных из отдельной клетки, выбранной на основе выхода антитела; при этом антитело представляет собой любое из антител, описанных выше. Некоторые такие способы дополнительно включают размножение клеток в селективных условиях и скрининг на клеточные линии, природным образом экспрессирующие и секретирующие, по меньшей мере, 100 мг/л/106 клеток/24 ч.
Краткое описание фигур
Фиг. 1 показывает выравнивание аминокислотных последовательностей родительского мышиного mAb (называемого m9E4) со зрелой вариабельной областью тяжелой цепи гуманизированного 9Е4. 1791009Hu9E4VHFr (SEQ ID NO: 7) представляет собой человеческую акцепторную последовательность VH. CDR-участки согласно определению Kabat подчеркнуты и выделены жирным шрифтом.
Фиг. 2 показывает выравнивание аминокислотных последовательностей родительского мышиного mAb (называемого m9E4) со зрелой вариабельной областью легкой цепи гуманизированного 9Е4. 63102889Hu9E4VLFr (SEQ ID NO: 2) представляет собой человеческую акцепторную последовательность VL. CDR-участки согласно определению Kabat подчеркнуты и выделены жирным шрифтом.
Фиг. 3 показывает результаты воздействия пассивной иммунотерапии 9Е4 на характеристики памяти в пробной части теста в водном лабиринте Морриса.
Фиг. 4 показывает результаты воздействия пассивной иммунотерапии 9Е4 на скорость и ошибки в тесте на перекладине (round beam test).
Фиг. 5 показывает иммунопреципитацию различных вариантов гуманизированных антител 9Е4 в отношении его антигена, полученного из пораженной ткани. Ch9E4: химерное 9Е4; H1L3: Hu9E4VHvl- Hu9E4VLv3; H3L2: Hu9E4VHv3- Hu9E4VLv2; H3L3: Hu9E4VHv3- Hu9E4VLv3; N.S.=неспецифическое.
Фиг. 6 показывает Вестерн-блоттинг рекомбинантного человеческого суниклеина с мышиными химерными и гуманизированными антителами 9Е4.
Краткое описание последовательностей
SEQ ID NO: 1 представляет собой аминокислотную последовательность вариабельной области m9E4VL.
SEQ ID NO: 2 представляет собой аминокислотную последовательность вариабельной области 63102889Hu9E4VLFr.
SEQ ID NO: 3 представляет собой аминокислотную последовательность вариабельной области Hu9E4VLv1.
SEQ ID NO: 4 представляет собой аминокислотную последовательность вариабельной области Hu9E4VLv2.
SEQ ID NO: 5 представляет собой аминокислотную последовательность вариабельной области Hu9E4VLv3.
SEQ ID NO: 6 представляет собой аминокислотную последовательность вариабельной области m9E4VH.
SEQ ID NO: 7 представляет собой аминокислотную последовательность вариабельной области 1791009Hu9E4VHFr.
SEQ ID NO: 8 представляет собой аминокислотную последовательность вариабельной области Hu9E4VHv1.
SEQ ID NO: 9 представляет собой аминокислотную последовательность вариабельной области Hu9E4VHv2.
SEQ ID NO: 10 представляет собой аминокислотную последовательность вариабельной области Hu9E4VHv3.
SEQ ID NO: 11 представляет собой аминокислотную последовательность вариабельной области Hu9E4VHv4.
SEQ ID NO: 12 представляет собой аминокислотную последовательность природного человеческого альфа-синуклеина дикого типа.
SEQ ID NO: 13 представляет собой аминокислотную последовательность константной области легкой цепи гуманизированного 9Е4 с аргинином на N-конце.
SEQ ID NO: 14 представляет собой аминокислотную последовательность константной области тяжелой цепи гуманизированного 9Е4.
SEQ ID NO: 15 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VLv1.
SEQ ID NO: 16 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VLv2.
SEQ ID NO: 17 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VLv3.
SEQ ID NO: 18 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VHv1.
SEQ ID NO: 19 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VHv2.
SEQ ID NO: 20 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VHv3.
SEQ ID NO: 21 представляет собой нуклеотидную последовательность вариабельной области Hu9E4VHv4.
SEQ ID NO: 22 представляет собой аминокислотную последовательность сигнального пептида Hu9E4VL.
SEQ ID NO: 23 представляет собой нуклеотидную последовательность сигнального пептида Hu9E4VL.
SEQ ID NO: 24 представляет собой аминокислотную последовательность сигнального пептида Hu9E4VH.
SEQ ID NO: 25 представляет собой нуклеотидную последовательность сигнального пептида Hu9E4VH.
SEQ ID NO: 26 представляет собой консенсусную аминокислотную последовательность Hu9E4VL.
SEQ ID NO: 27 представляет собой консенсусную аминокислотную последовательность Hu9E4VH.
SEQ ID NO: 28 представляет собой аминокислотную последовательность константной области легкой цепи гуманизированного 9Е4 без аргинина на N-конце.
SEQ ID NO: 29 представляет собой аминокислотную последовательность варианта 3 легкой цепи гуманизированного 9Е4, содержащую (а) вариабельную область и (b) константную область с аргинином на N-конце.
SEQ ID NO: 30 представляет собой аминокислотную последовательность варианта 3 легкой цепи гуманизированного 9Е4, содержащую (а) вариабельную область и (b) константную область без аргинина на N-конце.
SEQ ID NO: 31 представляет собой аминокислотную последовательность варианта 3 тяжелой цепи гуманизированного 9Е4, содержащую вариабельную область и константную область.
SEQ ID NO: 32 представляет собой аминокислотную последовательность BIP версии константной области тяжелой цепи аллотипа Glm3 гуманизированного 9Е4.
Определения
Как правило, моноклональные антитела представлены в изолированной форме. Это означает, что антитело является в типичном случае чистым по меньшей мере на 50% масса/масса от белков и других макромолекул, образующихся в результате его выработки или очистки, но не исключает возможности комбинирования моноклонального антитела с избыточным количеством фармацевтически приемлемого носителя(ей) или другого наполнителя для облегчения его применения. В некоторых случаях моноклональные антитела являются, по меньшей мере, на 60%, 70%, 80%, 90%, 95 или 99% масса/масса чистыми от белков и других макромолекул, образующихся в результате выработки или очистки.
Специфическое связывание моноклонального антитела с его целевым антигеном означает аффинность, составляющую, по меньшей мере, 106, 107, 108, 109 или 1010 М-1. Специфическое связывание заметно выше по величине и отличается от неспецифического связывания, возникающего, по меньшей мере, с одной неродственной мишенью. Специфическое связывание может быть результатом образования связей между конкретными функциональными группами или конкретного пространственного соответствия (например, по типу «ключ-замок»), тогда как неспецифическое связывание обычно является результатом взаимодействия ван-дер-ваальсовых сил. Однако специфическое связывание не обязательно подразумевает, что моноклональное антитело связывается с одной и только одной мишенью.
Основным структурным элементом антитела является тетрамер, состоящий из субъединиц. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, каждая из которых имеет одну «легкую» цепь (примерно 25 кДа) и одну «тяжелую» цепь (примерно 50-70 кДа). Аминоконцевая часть каждой цепи содержит вариабельную область примерно из 100-110 или более аминокислот, ответственных главным образом за распознавание антигена. Эта вариабельная область первоначально экспрессируется связанной с отщепляемым сигнальным пептидом. Вариабельная область без сигнального пептида иногда называется зрелой вариабельной областью. Таким образом, например, зрелая вариабельная область легкой цепи означает вариабельную область легкой цепи без сигнального пептида легкой цепи. Карбокси-концевая часть каждой цепи определяет константную область, ответственную главным образом за эффекторную функцию.
Легкие цепи классифицируются как каппа- или лямбда-цепи. Тяжелые цепи классифицируются как гамма-, мю-, альфа-, дельта- или эпсилон-цепи, и определяют изотип антитела, такой как IgG, IgM, IgA, IgD и IgE, соответственно. В пределах легких и тяжелых цепей вариабельные и константные области связаны «J»-областью, состоящей из 12 или более аминокислот, при этом тяжелая цепь также включает «D»-область, состоящую примерно из 10 или более аминокислот. (Смотри, в общих чертах, Fundamental Immunology (Paul, W., ed., 2nd ed. Raven Press, N.Y., 1989, Ch. 7, полное содержание которого включено путем отсылки для всех целей).
Зрелые вариабельные области каждой пары легкой/тяжелой цепи образуют связывающий сайт антитела. Таким образом, интактное антитело имеет два связывающих сайта. За исключением бифункциональных или биспецифических антител эти два сайта связывания являются одинаковыми. Все цепи демонстрируют одинаковую общую структуру из относительно консервативных каркасных участков (FR), соединенных посредством трех гипервариабельных участков, также называемых участками, определяющими комплементарность, или CDR. CDR из двух цепей каждой пары соединяются посредством каркасных участков, обеспечивая связывание со специфическим эпитопом. От N-конца до С-конца легкая и тяжелая цепи содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Присвоение аминокислот каждому домену осуществляется в соответствии с нумерацией по Kabat, Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, MD, 1987 and 1991) или Chothia & Lesk, J. Mol. Biol. 196:901-917 (1987); Chothia et al., Nature 342:878-883 (1989). Кроме того, Kabat обеспечивает широко используемую систему нумерации (нумерацию по Kabat), в которой соответствующим остаткам между различными тяжелыми цепями или между различными легкими цепями присвоены одинаковые номера (например, Н83 означает положение 83 по нумерации Kabat в зрелой вариабельной области тяжелой цепи; аналогично, положение L36 означает положение 36 по нумерации Kabat в зрелой вариабельной области легкой цепи).
Термин «антитело» включает интактные антитела и их связывающие фрагменты. Как правило, фрагменты конкурируют с интактным антителом, из которого они получены, за специфическое связывание с мишенью, включая отдельные тяжелые цепи, легкие цепи Fab, Fab', F(ab')2, F(ab)c, диатела, Dabs, нанотела и Fv. Фрагменты могут быть получены методами рекомбинантных ДНК или путем ферментативного или химического расщепления интактных иммуноглобулинов. Термин «антитело» также относится к биспецифическому антителу и/или гуманизированному антителу. Биспецифическое или бифункциональное антитело представляет собой искусственное гибридное антитело, обладающее двумя различными парами тяжелой/легкой цепи и двумя различными сайтами связывания (смотри, например, Songsivilai and Lachmann, Clin. Exp. Immunol., 79:315-321 (1990); Kostelny et al., J. Immunol. 148:1547-53 (1992)). В некоторых биспецифических антителах две различные пары тяжелой/легкой цепи включают пару тяжелой цепи/легкой цепи гуманизированного 9Е4, и пару тяжелой цепи/легкой цепи, специфическую для эпитопа на альфа-синуклеине, отличного от эпитопа, связанного 9Е4. Гуманизированные антитела в целом описаны ниже в Разделе IV В.
Термин «эпитоп» относится к участку антигена, с которым связывается антитело. Эпитоп может быть сформирован из смежных или несмежными аминокислот, которые накладываются друг на друга при образовании третичной структуры одного или более белков. Эпитопы, сформированные смежными аминокислотами, обычно устойчивы при воздействии денатурирующих растворителей, в то время как эпитопы, образованные при формировании третичной белковой структуры, обычно разрушаются при обработке денатурирующими растворителями. Эпитоп обычно включает по меньшей мере 3 или более, чаще не менее 5 или 8-10 аминокислот в уникальной пространственной конформации. Методы определения пространственной конформации эпитопов включают, например, рентгеновскую кристаллографию и двумерный ядерный магнитный резонанс. Смотри, например, Epitope Mapping Protocols, in Methods in Molecular Biology, Vol.66, Glenn E. Morris, Ed. (1996).
Антитела, которые распознают одинаковые или перекрывающиеся эпитопы, могут быть идентифицированы с помощью обычного иммунного анализа, выявляющего способность одного антитела блокировать связывание другого антитела с антигеном-мишенью. Эпитоп антитела может быть также определен с помощью рентгеновской кристаллографии антитела, связанного с его антигеном, для идентификации контактных остатков. Альтернативно, два антитела обладают одним и тот же эпитопом, если все аминокислотные мутации в антигене, которые уменьшают или элиминируют связывание одного антитела, также уменьшают или элиминируют связывание другого антитела. Два антитела имеют перекрывающиеся эпитопы, если мутации некоторых аминокислот, которые уменьшают или элиминируют связывание одного антитела, уменьшают или элиминируют связывание другого антитела.
Конкуренцию между антителами определяют с помощью исследования, в котором тестируемое антитело ингибирует специфическое связывание контрольного антитела с известным антигеном (смотри, например, Junghans et al., Cancer Res. 50:1495, 1990). Тестируемое антитело конкурирует с контрольным антителом, если избыток тестируемого антитела (например, по меньшей мере, 2-кратный, 5-кратный, 10-кратный, 20-кратный или 100-кратный) ингибирует связывание контрольного антитела по меньшей мере на 50%, но предпочтительно на 75%, 90% или 99% по результатам измерения в тесте на конкурентное связывание. Антитела, идентифицированные с помощью методов конкурентного связывания (конкурирующие антитела), включают антитела, связывающиеся с тем же эпитопом, что и контрольное антитело, и антитела, связывающиеся со смежным эпитопом, расположенным проксимально по отношению к эпитопу, с которым связывается контрольное антитело, для поддержания стерического барьера.
Термин «пациент» относится к человеку или другому субъекту-млекопитающему, получающему профилактическое или терапевтическое лечение.
С целью классификации аминокислотных замен на консервативные и неконсервативные, аминокислоты сгруппированы следующим образом: Группа I (гидрофобные боковые цепи): met, ala, val, leu, ile; Группа II (нейтральные гидрофильные боковые цепи): cys, ser, thr; Группа III (кислые боковые цепи): asp, glu; Группа IV (основные боковые цепи): asn, gin, his, lys, arg; Группа V (остатки, влияющие на ориентацию цепи): gly, pro; и Группа VI (ароматические боковые цепи): trp, tyr, phe. Консервативные замены включают замены между аминокислотами одного класса. Неконсервативные замены представляют собой замены аминокислот одного класса на аминокислоты другого класса.
Процентные идентичности последовательностей определяют с помощью последовательностей антител, максимально выравненных с использованием системы нумерации Kabat. После выравнивания область представляющего интерес антитела (например, полной зрелой вариабельной области тяжелой или легкой цепи) сравнивают с той же областью контрольного антитела, процентная идентичность последовательностей между областями представляющего интерес и контрольного антитела представляет собой количество положений, занятых одинаковыми аминокислотами в областях как представляющего интерес, так и контрольного антитела, деленное на общее количество выровненных положений двух областей, без подсчета разрывов, умноженное на 100 для преобразования в процентное количество.
Композиции или способы, «содержащие» один или более указанных элементов, могут включать другие элементы, специально не указанные. Например, композиция, которая содержит антитело, может содержать антитело отдельно или в комбинации с другими ингредиентами.
Указание диапазона значений включает все целые значения внутри или в пределах указанного диапазона и все поддиапазоны, определенные целыми значениями внутри диапазона.
Если очевидным образом не вытекает из контекста, термин «примерно» охватывает значения в пределах SEM (стандартной ошибки среднего значения) от установленной величины.
Индивидуум имеет повышенный риск развития заболевания, если субъект имеет, по меньшей мере, один известный фактор риска (например, генетический, биохимический, семейный анамнез, воздействие ситуационных факторов), подвергающий индивидуумов с данным фактором риска статистически значимому большему риску развития заболевания по сравнению с индивидуумами без фактора риска.
Термин «симптом» относится к субъективному проявлению заболевания, такому как измененная походка, выявленному пациентом. «Признак» относится к объективному проявлению заболевания, наблюдаемому терапевтом.
Статистическая значимость означает р≤0.05.
«Когнитивная функция» относится к психическим процессам, таким как любой или все из внимания, памяти, воспроизведения и восприятия устной речи, разрешения проблем, и проявления интереса к окружающему и заботы о себе.
«Усиленная когнитивная функция» или «улучшенная когнитивная функция» относится к улучшению относительно исходного состояния, например, постановки диагноза или начала лечения. «Ухудшение когнитивной функции» относится к понижению функции относительно такого исходного состояния.
В системах экспериментальных животных, таких как крысы или мыши, когнитивную функцию можно оценить с помощью способов с использованием лабиринта, в котором субъекты используют пространственную информацию (например, водный лабиринт Морриса (Morris water maze), тест Барнеса с циркулярной платформой (Barnes circular maze), приподнятый радиальный лабиринт (elevated radial arm maze), Т-образный лабиринт (Τ maze) и другие), выработки условного рефлекса в связи с переживанием чувства страха, активного избегания, освещенного «открытого поля», измерения активности в темноте, приподнятого крестообразного лабиринта, двухкамерного экспериментального теста (two-compartment exploratory test) или теста принудительного плавания.
У человека когнитивную функцию можно оценить с применением одного или более из нескольких стандартизированных тестов. Примеры тестирования или оценки когнитивной функции были описаны (Ruoppila, 1 and Suutama, T. Scand. J. Soc. Med. Suppl. 53, 44-65, 1997) и включают стандартизированные психометрические тесты (например, шкалу памяти Векслера (Wechsler Memory Scale), шкалу интеллекта взрослых Векслера (Wechsler Adult Intelligence Scale), стандартные прогрессивные матрицы Равена (Raven's Standard Progressive Matrices), тест умственных способностей взрослых Шайе-Терстона (Schaie-Thurstone Adult Mental Abilities Test), нейропсихологические тесты (например, тест Лурии-Небраска (Luria-Nebraska)), самооценки метакогнитивных способностей (например, опросник функционирования мета-памяти (Metamemory Questionnaire)), тесты зрительно-пространственных фильтраций (например, фигур Поппелрейтера, распознавания часов, рисования и вычеркивания сот), тесты когнитивной фильтрации (например, краткая шкала оценки психического статуса Фольштейна (Folstein's Mini Mental State Test)) и тесты времени реагирования. Другие стандартные тесты для оценки когнитивных функций включают шкалу оценки когнитивных функций при болезни Альцгеймера (ADAS-cog); шкалу общего впечатления врача о происходящих изменениях, включая информацию лица, осуществляющего уход за больным (CIBIC-плюс); шкалу оценки повседневной деятельности при болезни Альцгеймера в ходе совместного исследования (ADCS-ADL); краткую шкалу оценки психического статуса (MMSE); психоневрологический опросник (NPI); клиническую рейтинговую шкалу деменции (CDR); Кембриджскую автоматизированную нейропсихологическую батарею тестов (CANTAB) или гериатрическую шкалу клинической оценки фирмы Сандоз (SCAG), тест Струпа (Stroop Test), тест на зрительно-моторную координацию (Trail Making), тест интеллекта Векслера (Wechsler Digit Span) и компьютерный тест на когнитивные функции CogState. Кроме того, когнитивную функцию можно оценить с использованием методов визуализации, таких как позитронно-эмиссионная томография (PET), функциональная магнитно-резонансная томография (fMRI), однофотонная эмиссионная компьютерная томография (SPECT) или любая другая технология изображения, которая позволяет измерить функцию мозга.
Подробное описание изобретения
I. Общие положения
Изобретение обеспечивает гуманизированные антитела 9Е4. Антитела являются полезными для лечения и диагностики заболевания с тельцами Леви.
II. Целевые молекулы
Природный человеческий альфа-синуклеин дикого типа представляет собой пептид из 140 аминокислот, имеющий следующую аминокислотную последовательность:

(Ueda et al., Proc. Natl. Acad. Sci. USA (1993) 90:11282-6); GenBank accession number: P37840). Белок имеет три идентифицированных домена, повторяющийся домен КТКЕ, охватывающий аминокислоты 1-61, NAC-домен (неамилоидный компонент, Non-amyloid component) расположенный между аминокислотами примерно 60-95, и кислый С-концевой домен, расположенный между аминокислотами примерно 98 и 140.
Если из контекста явно не следует иное, ссылка на альфа-синуклеин или его фрагменты включает природные человеческие аминокислотные последовательности дикого типа, указанные выше, и их человеческие аллельные варианты, в частности те, которые ассоциированы с заболеванием с тельцами Леви (например, Е46К, А30Р и А53Т, при этом первая буква обозначает аминокислоту в SEQ ID NO: 12, число обозначает положение кодона в SEQ ID NO: 12, и вторая буква представляет собой аминокислоту в аллельном варианте). Такие варианты могут необязательно быть представлены индивидуально или в любой комбинации в любом описанном ниже аспекте изобретения. Индуцированные мутации E83Q, A90V, А76Т, которые усиливают агрегацию альфа-синуклеина, также могут быть представлены индивидуально или в комбинации друг с другом, и/или аллельными вариантами Е46К, А30Р и А53Т человека.
III. Заболевания с тельцами Леви
Заболевание с тельцами Леви (LBD) характеризуются дегенерацией дофаминергической системы, нарушениями моторных функций, когнитивным расстройством и формированием телец Леви (LB) (McKeith et al., Neurology (1996) 47:1113-24). Тельца Леви представляют собой сферические белковые образования, выявляемые в нейронах. Их присутствие в мозге нарушает нормальное функционирование головного мозга, прерывая действие химических мессенджеров, включая ацетилхолин и дофамин. Заболевания с тельцами Леви включают болезнь Паркинсона (включая идиопатическую болезнь Пакринсона), болезнь с диффузными тельцами Леви (Diffuse Lewy Body Disease, DLBD), также известную как деменция с тельцами Леви (Dementia with Lewy Bodies, DLB), вариант болезни Альцгеймера с тельцами Леви (Lewy body variant of Alzheimer's disease, LBV), сочетание болезни Альцгеймера и болезни Паркинсона, и множественную системную атрофию (MSA; например, оливопонтоцеребеллярную атрофию, стрионигральную дегенерацию и синдром Шая-Дрейджера). DLBD имеет общие симптомы как болезни Альцгеймера, так и болезни Паркинсона. DLBD отличается от болезни Паркинсона, главным образом, локализацией телец Леви. В DLBD тельца Леви формируются преимущественно в коре головного мозга. В болезни Паркинсона они формируются в основном в черной субстанции. Другие заболевания с тельцами Леви включают идиопатическую ортостатическую гипотензию, дисфагию, связанную с тельцами Леви, сопутствующее LBD и наследственное LBD (например, мутации гена альфа-синуклеина, PARK3 и PARK4).
IV. Антитела по изобретению
А. Специфичность связывания и функциональные свойства
Гуманизированные антитела по изобретению специфически связываются с человеческим альфа-синуклеином. Аффинность некоторых гуманизированных антител (то есть константа ассоциации (Ка)) предпочтительно находится в пределах пятикратной или двукратной величины аффинности мышиного антитела 9Е4. Некоторые гуманизированные антитела обладают аффинностью, которая равна (в пределах экспериментальной ошибки) или больше, чем аффинность мышиного антитела 9Е4. Предпочтительные гуманизированные антитела связываются с одним и тем же эпитопом и/или конкурируют с мышиным антителом 9Е4 за связывание с человеческим альфа-синуклеином.
В некоторых антителах гуманизированное 9Е4 формирует одно плечо биспецифического антитела, другое плечо которого представляет собой антитело, которое связывается с рецептором, экспрессирующимся на поверхности гематоэнцефалического барьера, таким как рецептор инсулина, рецептор инсулиноподобного фактора роста (IGF), рецептор лептина или рецептор липопротеина, или предпочтительно рецептор трансферрина (Friden et al., PNAS 88:4771-4775, 1991; Friden et al., Science 259:373-377, 1993). Такое биспецифическое антитело может транспортироваться через гематоэнцефалический барьер посредством рецептор-опосредованного трансцитоза. Захват в головном мозге биспецифического антитела может быть дополнительно усилен путем конструирования биспецифического антитела для снижения его аффинности к рецептору гематоэнцефалического барьера. Пониженная аффинность к рецептору приводит к более широкому распространению в головном мозге (смотри, например, Atwal. et al. Sci. Trans. Med. 3, 84ra43, 2011; Yu et al. Sci. Trans. Med. 3, 84ra44,2011).
Иллюстративные биспецифические антитела могут также представлять собой (1) антитело с двойным вариабельным доменом (DVD-Ig), в котором каждая легкая цепь и тяжелая цепь содержит два вариабельных домена, связанных в тандеме посредством короткой пептидной связи (Wu et al., Generation and Characterization of a Dual Variable Domain Immunoglobulin (DVD-Ig™) Molecule, In: Antibody Engineering, Springer Berlin Heidelberg (2010)); (2) Tandab, который представляет собой слияние двух одноцепочечных диател, что приводит к образованию тетравалентного биспецифического антитела, которое имеет два участка связывания для каждого из целевых антигенов; (3) flexibody, которое представляет собой комбинацию scFvs с диателом, приводящую к образованию мультивалентной молекулы; (4) так называемую молекулу «dock and lock» на основе «домена димеризации и докинга» в протеинкиназе A, которая при применении к Fabs, может дать биспецифический трехвалентный связьшающий белок, состоящий из двух идентичных Fab-фрагментов, связанных с другим Fab-фрагментом; (5) так называемую молекулу Scorpion, содержащую, например, два scFvs, слитых с обоими концам Fc-области человека. Примеры платформ, полезных для получения биспецифических антител, включают, но без ограничения, BiTE (Micromet), DART (MacroGenics), Fcab и Mab2 (F-star), сконструированную форму Fc-домена IgG1 (Xencor) или DuoBody (на основе обмена Fab-фрагментов, Genmab).
B. Гуманизированные антитела
Гуманизированное антитело представляет собой полученное способами генетической инженерии антитело, в котором CDR из не-человеческого «донорного» антитела пересаживают в последовательности человеческого «акцепторного» антитела (смотри, например, Queen et al., US5,530,101 и 5,585,089; Winter et al., US 5,225,539, Carter, US 6,407,213, Adair, US 5,859,205 6,881,557, Foote, US 6,881,557). Последовательности акцепторного антитела могут представлять собой, например, последовательность зрелой вариабельной области антитела человека, композицию таких последовательностей, консенсусную последовательность, состоящую из последовательностей человеческого антитела (например, консенсусные последовательности вариабельной области легкой и тяжелой цепи по Kabat, 1991, supra) или последовательность вариабельной области зародышевой линии. Предпочтительной акцепторной последовательностью для тяжелой цепи является человеческая зрелая вариабельная область тяжелой цепи с NCBI accession code ААС50998 (GI: 1791009), или другая зрелая вариабельная область тяжелой цепи, полученная из зародышевой линии IGHV3-7'01 или IGHV3-7'02 (названия клонов V3-7 или VH3-11) (Glas et al., Clin Exp Immunol. 107:372-80, 1997), или последовательность зрелой вариабельной области тяжелой цепи, включающая одну из этих последовательностей зародышевой линии. В отношении легкой цепи, предпочтительная акцепторная последовательность представляет собой зрелую вариабельную область легкой цепи с NCBI accession code AAY33350 (GI:63102889) или другую последовательность зрелой легкой цепи, полученную из зародышевой линии IGKV1D-39 или IGKV1-39 (название клона O2 или O12) (Kramer et al., Eur J Immunol. 35:2131-45, 2005), или последовательность зрелой вариабельной области легкой цепи, включающую одну из этих последовательностей зародышевой линии. Таким образом, гуманизированное антитело по изобретению представляет собой антитело, имеющее три CDR легкой цепи и три CDR тяжелой цепи, как определено по Kabat, из донорного антитела 9Е4, и последовательности каркасных участков зрелой вариабельной области и константные области, в случае присутствия, полностью или в основном из последовательностей антитела человека. Аналогичным образом, гуманизированная тяжелая цепь представляет собой тяжелую цепь, имеющую три CDR тяжелой цепи, как определено по Kabat, из тяжелой цепи антитела 9Е4, и последовательность зрелой вариабельной тяжелой цепи и последовательность константной области тяжелой цепи, в случае присутствия, полностью или в основном из последовательности тяжелой цепи антитела человека. Аналогичным образом, гуманизированная легкая цепь представляет собой легкую цепь, имеющую три CDR легкой цепи, как определено по Kabat, из легкой цепи антитела 9Е4, и последовательность зрелой вариабельной легкой цепи и последовательность константной области легкой цепи, в случае присутствия, полностью и в основном из последовательности легкой цепи антитела человека. Последовательности каркасных участков зрелой вариабельной области цепи антитела или последовательность константной области цепи антитела происходят в основном из последовательности каркасных участков зрелой вариабельной области человека или последовательности человеческой константной области, соответственно, когда по меньшей мере 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% соответствующих остатков, согласно Kabat, являются идентичными.
Определенные аминокислоты из остатков человеческого каркасного участка зрелой вариабельной области могут быть выбраны для замещения исходя из их возможного влияния на CDR-конформацию и/или связывание с антигеном. Исследование таких возможных влияний проводят путем моделирования, оценки характеристик аминокислот в конкретных положениях или эмпирического наблюдения влияния замещения или мутагенеза конкретных аминокислот.
Например, если аминокислота в остатке каркасного участка зрелой вариабельной области мышей отличается от аминокислоты в выбранном остатке каркасного участка зрелой вариабельной области человека, то человеческая аминокислота каркасного участка может быть заменена эквивалентной аминокислотой каркасного участка из мышиного антитела в случае, если имеются основания ожидать, что данная аминокислота:
(1) нековалентно связывается непосредственно с антигеном,
(2) находится в непосредственной близости к CDR-участку,
(3) иным образом взаимодействует с CDR-участком (например, находится в пределах 6 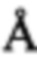 от CDR-участка)
от CDR-участка)
(4) опосредует взаимодействие между тяжелой и легкой цепями.
Изобретение обеспечивает гуманизированные формы мышиного антитела 9Е4, включая три иллюстративные гуманизированные зрелые вариабельные области легкой цепи (Hu9E4VLv1-v3; SEQ ID NOs:3-5) и четыре иллюстративные гуманизированные зрелые вариабельные области тяжелой цепи (Hu9E4VHv1-v4; SEQ ID NOs:8-11). SEQ ID NO: 4 включает три CDR по Kabat легкой цепи мышиного 9E4 и каркасные участки зрелой вариабельной области AAY33350. SEQ ID NOS. 3 и 5 включает обратные мутации, как показано в Таблице 2. SEQ ID NO. 11 включает три CDR по Kabat мышиного 9Е4 и каркасные участки зрелой вариабельной области ААС50998. SEQ ID NOs:8-10 включает обратные мутации, как показано в Таблице 3.
Изобретение обеспечивает варианты гуманизированного антитела 9Е4, в которых гуманизированная зрелая вариабельная область тяжелой цепи показывает по меньшей мере 90%, 95% или 99% идентичности с SEQ ID NOs:8-11, и гуманизированная зрелая вариабельная область легкой цепи показывает по меньшей мере 90, 95 или 99% идентичности последовательности с SEQ ID NOs:3-5, но в которых любое отклонение от указанной SEQ ID NO. возникает в каркасном участке зрелой вариабельной области скорее, чем CDR по Kabat. В некоторых таких антителах положение L36 занято Y или F, и/или положение L83 занято F или L, и/или положение Н73 занято N или D, и/или положение Н93 занято А или S (все положения здесь, так же как где-либо в другом месте в данном описании, представлены согласно нумерации Kabat). В некоторых таких антителах некоторые или все из обратных мутаций в Hu9E4VLv1-v3 и Hu9E4VHv1-v4 сохранены. Другими словами, одно или оба положения Н73 и Н93 тяжелой цепи заняты D и А, соответственно. Аналогичным образом, в некоторых антителах одно или оба положения L36 и L83 легкой цепи заняты F и L, соответственно. В некоторых антителах 1, 2, 3 или все четыре положения Н73, Н93, L36 и L83 занято/заняты D, A, F и L, соответственно. В некоторых антителах 0, 1 или 2 положения заменены в каркасном участке зрелой вариабельной области тяжелой цепи относительно SEQ ID NO: 11, и 0, 1 или 2 положения заменены в каркасном участке зрелой вариабельной области легкой цепи относительно SEQ ID NO: 4.
Некоторые антитела содержат гуманизированную тяжелую цепь, содержащую три CDR по Kabat SEQ ID NO: 11, и гуманизированную легкую цепь, содержащую три CDR по Kabat SEQ ID NO: 4, при условии, что положение L36 (нумерация по Kabat) занято F или Y и/или положение L83 (нумерация по Kabat) занято L или F, и/или положение Н73 (нумерация по Kabat) занято D или N, и/или положение Н93 (нумерация по Kabat) занято S или А. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F и положение L83 (нумерация по Kabat) занято L. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F и положение Н73 (нумерация по Kabat) занято D. В некоторых таких антителах положение L36 (Kabat numbering) занято F и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L и положение Н73 (нумерация по Kabat) занято D. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 занято F, положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение Н73 (нумерация по Kabat) занято D, и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L, положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L36 (нумерация по Kabat) занято F, положение L83 (нумерация по Kabat) занято L, положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L83 (нумерация по Kabat) занято L. В некоторых таких антителах положение L83 (нумерация по Kabat) занято L and положение Н73 (нумерация по Kabat) занято D. В некоторых таких антителах положение L83 (нумерация по Kabat) занято L и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L83 (нумерация по Kabat) занято L и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L83 (нумерация по Kabat) занято L, положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение L83 (нумерация по Kabat) занято L, положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение Н73 (нумерация по Kabat) занято D. В некоторых таких антителах положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение Н73 (нумерация по Kabat) занято D и положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение Н93 (нумерация по Kabat) занято S. В некоторых таких антителах положение Н93 (нумерация по Kabat) занято А. В некоторых таких антителах положение L36 занято Y, положение L83 занято F, положение Н73 занято N и положение Н93 занято S. Некоторые иллюстративные антитела с желательными остатками в положениях L36, L83, Н73 и Н93, и их комбинации перечислены в Таблице ниже:
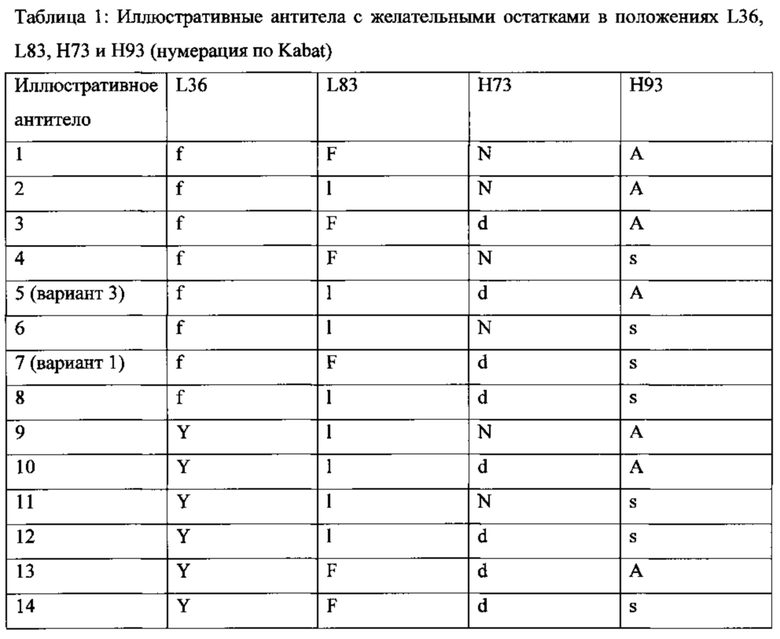

В некоторых антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 10. В некоторых антителах зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 5 или SEQ ID NO: 3. В некоторых таких антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 10, и зрелая вариабельная область легкой цепи содержит аминокислотную последовательность, обозначенную SEQ ID NO: 5 или SEQ ID NO: 3. В некоторых таких антителах зрелая вариабельная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 10, и зрелая вариабельная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 5.
Другие аминокислотные замены могут быть сделаны в каркасном участке зрелой вариабельной области, например, в остатках, не находящихся в контакте с CDR. Часто замены, проводимые в вариантных гуманизированных последовательностях, являются консервативными в отношении заменяемых аминокислот. В некоторых антителах замены в Hu9E4VLv1-v3 и Hu9E4VHv1-v4 (как консервативные, так и неконсервативные) не обладают существенным эффектом на аффинность связывания или эффективность полученного антитела в отношении Hu9E4VLv1-v3 и Hu9E4VHv1-v4, то есть его способность к связыванию человеческого альфа-синуклеина.
Варианты, как правило, отличаются от последовательностей зрелых вариабельных областей тяжелой и легкой цепи Hu9E4VLv1-v3 и Hu9E4VHv1-v4 незначительным числом (например, обычно не более чем 1, 2, 3, 5 или 10 в каркасном участке зрелой вариабельной области легкой цепи или тяжелой цепи, или обеих) замен, делеций или вставок.
С. Отбор константной области
Вариабельные области тяжелой и легкой цепи гуманизированных антител могут быть соединены, по меньшей мере, с частью человеческой константной области. Выбор константной области отчасти зависит от того, являются ли желательными антителозависимая клеточно-опосредованная цитотоксичность, антителозависимый клеточный фагоцитоз и/или комплементзависимая цитотоксичность. Например, изотипы IgG1 и IgG3 человека обладают комплементзависимой цитотоксичностью, а изотипы IgG2 и IgG4 человека не обладают. Человеческие IgG1 и IgG3 также индуцируют более сильные клеточно-опосредованные эффекторные функции, чем человеческие IgG2 и IgG4. Константные области легкой цепи могут представлять собой лямбда или каппа. Иллюстративная константная область человеческой легкой каппа-цепи имеет аминокислотную последовательность SEQ ID NO: 13. Некоторые такие константные области легкой цепи каппа могут быть кодированы нуклеотидной последовательностью. N-концевой аргинин SEQ ID NO: 13 может быть опущен, и в этом случае константная область легкой цепи каппа имеет аминокислотную последовательность SEQ ID NO: 28. Некоторые такие константные области легкой цепи каппа могут быть кодированы нуклеотидной последовательностью. Иллюстративная константная область тяжелой цепи IgG1 человека имеет аминокислотную последовательность SEQ ID NO: 14 (с С-концевым лизином или без него). Некоторые такие константные области тяжелой цепи могут быть кодированы нуклеотидной последовательностью. Антитела могут быть представлены в виде тетрамеров, содержащих две легкие и две тяжелые цепи, в виде отдельных тяжелых цепей, легких цепей, в виде Fab, Fab', F(ab')2 и Fv, или в виде одноцепочечных антител, в которых зрелые вариабельные области тяжелой и легкой цепи соединены с помощью спейсера.
Человеческие константные области показывают аллотипическую вариацию и изоаллотипическую вариацию между различными индивидуумами, то есть константные области могут различаться у разных индивидуумов в одном или нескольких полиморфных положениях. Изоаллотипы отличаются от аллотипов тем, что сыворотка, распознающая изоаллотип, связывается с неполиморфной областью одного или нескольких других изотипов. Таким образом, например, другая константная область тяжелой цепи является аллотипа Glm3 IgG1 и имеет аминокислотную последовательность SEQ ID NO: 32. Еще другая константная область тяжелой цепи имеет аминокислотную последовательность SEQ ID NO: 32 за исключением того, что она лишена С-концевого лизина.
Одна или несколько аминокислот на амино- или карбокси-концах легкой и/или тяжелой цепи, такие как С-концевой лизин тяжелой цепи, могут быть пропущены или дериватизированы в части или во всех молекулах. Замены могут быть проведены в константных областях для снижения или повышения эффекторной функции, такой как опосредуемая комплементом цитотоксичность или ADCC (смотри, например, Winter et al., патент США №5624821; Tso et al., патент США №5834597; и Lazar et al., Proc. Natl. Acad. Sci. USA 103:4005, 2006), или для продления времени полужизни у человека (смотри, например, Hinton et al., J. Biol. Chem. 279:6213, 2004). Иллюстративные замены включают Gln в положении 250 и/или Leu в положении 428 (нумерация по EU используется в этом параграфе для константной области) для увеличения времени полужизни антитела. Замена в любом или во всех положениях 234, 235, 236 и/или 237 снижает аффинность в отношении Fcγ-рецепторов, в частности рецептора FcγRI (смотри, например, US 6,624,821). Некоторые антитела имеют замену аланина в положениях 234, 235 и 237 человеческого IgG1 для снижения эффекторных функций. Необязательно, положения 234, 236 и/или 237 в человеческом IgG2 заменены аланином, и положение 235 глутамином (смотри, например, US 5,624,821).
D. Экспрессия рекомбинантных антител
Антитела могут быть получены путем рекомбинантной экспрессии. Нуклеиновые кислоты, кодирующие антитела, могут быть кодон-оптимизированы для экспрессии в желательном типе клетки (например, СНО или Sp2/0). Конструкции рекомбинантных нуклеиновых кислот обычно включают контролирующую экспрессию последовательность, функционально связанную с кодирующими последовательностями цепей антитела, включая природные или гетерологичные промоторные области. Регулирующие экспрессию последовательности могут представлять собой эукариотические промоторные системы, присутствующие в векторах, способных к трансформации или трансфекции эукариотических клеток-хозяев. После введения вектора в соответствующую клетку-хозяина, эту клетку выдерживают в условиях, подходящих для высокого уровня экспрессии нуклеотидных последовательностей, а затем осуществляют сбор и очистку перекрестнореагирующих антител. Вектор или векторы, кодирующие цепи антитела, могут также содержать селектируемый ген, такой как дигидрофолатредуктаза, для обеспечения амплификации числа копий нуклеиновых кислот, кодирующих цепи антитела.
Е. coli представляет собой прокариотического хозяина, особенно пригодного для экспрессии антител, в частности, фрагментов антител. Микробы, такие как дрожжи, также являются полезными для экспрессии. Сахаромицеты являются предпочтительными дрожжами-хозяевами с пригодными векторами, имеющими регулирующие экспрессию последовательности, точку начала репликации, стоп-кодоны и тому подобное по желанию. Типичные промотры включают 3-фосфоглицераткиназу и другие гликолитические ферменты. Индуцируемые дрожжевые промоторы включают, среди прочего, промоторы алкогольдегидрогеназы, изоцитохрома С и ферментов, ответственных за утилизацию мальтозы и галактозы.
Клетки млекопитающего могут быть использованы для экспрессии нуклеотидных сегментов, кодирующих иммуноглобулины или их фрагменты. Смотри Winnacker, From Genes to Clones, (VCH Publishers, NY, 1987). Специалистами был разработан ряд подходящих клеточных линий хозяев, способных экспрессировать интактные гетерологичные белки, и такими клеточными линиями являются клеточные линии СНО, различные клеточные линии COS, клетки HeLa, клетки НЕК293, L-клетки, и не вырабатывающие антитела клетки мышиной миеломы, включая Sp2/0 и NS0. Предпочтительно использование нечеловеческих клеток. Экспрессирующие векторы этих клеток могут включать последовательности, управляющие экспрессией, такие как точка начала репликации, промотор, энхансер (Queen et al., Immunol. Rev. 89:49 (1986)), и необходимые информационные сайты процессинга, такие как сайты связывания рибосомы, сайты сплайсинга РНК, сайты полиаденилирования, и последовательности терминатора транскрипции. Предпочтительными последовательностями, управляющими экспрессией, являются промоторы, полученные из генов иммуноглобулинов, цитомегаловируса, SV40, аденовируса, вируса бычьей папилломы и т.п. Смотри Со et al., J. Immunol. 148:1149 (1992).
После введения вектора(ов), кодирующего тяжелые и легкие цепи антитела в клеточную культуру, пулы клеток могут быть подвергнуты скринингу на продуктивность и качество продукта в бессывороточной среде. Пулы клеток с высокой продуктивностью затем могут быть подвергнуты одноклеточному клонированию на основе FACS для создания моноклональных линий. Удельные продуктивности более 50 пг или 100 пг на клетку в день, что соответствует титрам продукта больше, чем 7.5 г/л культуры, могут быть предпочтительными. Антитела, продуцируемые одноклеточными клонами, также могут быть тестированы на мутность, фильтрационные свойства, с помощью электрофореза в акриламидном геле (PAGE), изоэлектрического фокусирования (IEF), сканирования в УФ области, высокоэффективной эксклюзионной хроматографии (HP-SEC), углеводно-олигосахаридного картирования, масс-спектроскопии и анализа связывания, такого как ELISA или Biacore. Отобранный клон можно затем консервировать во множество флаконов и хранить в замороженном виде для последующего использования.
После экспрессии антитела могут быть очищены в соответствии со стандартными способами, известными в данной области, включая захват белком А, колоночную хроматографию (например, гидрофобное взаимодействие или ионный обмен), низкое значение рН для вирусной инактивации и тому подобное (смотри, в общих чертах, Scopes, Protein Purification (Springer-Verlag, NY, 1982)).
Методология для коммерческого производства антител включает оптимизацию кодона, отбор промоторов, элементов транскрипции и окончаний, бессывороточное одноклеточное клонирование, консервацию клеток, использование маркеров отбора для амплификации числа копий, СНО-окончание, сывороточное одноклеточное клонирование, улучшение титров белков (смотри, например, патенты США №№5786464, 6114148, 6063598, 7569339, заявки на патент США WO 2004/050884, WO 2008/012142, WO 2008/012142, WO 2005/019442, WO 2008/107388, и WO 2009/027471, и патент США №5888809).
V. Нуклеиновые кислоты
Изобретение дополнительно обеспечивает нуклеиновые кислоты, кодирующие любую из тяжелых и легких цепей, описанных выше. Как правило, нуклеиновые кислоты также кодируют сигнальный пептид, слитый со зрелыми тяжелыми и легкими цепями (например, сигнальные пептиды, имеющие аминокислотные последовательности SEQ ID NOS: 22 и 24, которые могут быть кодированы SEQ ID NOS: 23 и 25). Кодирующие последовательности на нуклеиновых кислотах могут быть функционально связанными с регуляторными последовательностями для обеспечения экспрессии кодирующих последовательностей, такими как промотор, энхансер, сайт связывания рибосомы, сигнал терминации транскрипции и тому подобное. Нуклеиновые кислоты, кодирующие тяжелые и легкие цепи, могут находиться в изолированной форме или могут быть клонированы в один или более векторов. Нуклеиновые кислоты могут быть синтезированы с помощью, например, твердофазного синтеза или полимеразной цепной реакции (PCR) с использованием перекрывающихся олигонуклеотидов. Нуклеиновые кислоты, кодирующие тяжелые и легкие цепи, могут быть соединены в виде одной непрерывной нуклеиновой кислоты, например, в пределах экспрессионного вектора, или могут быть раздельными, например, каждая клонирована в свой собственный экспрессионный вектор.
VI. Терапевтические применения
Изобретение обеспечивает несколько способов лечения или профилактического воздействия на заболевание с тельцами Леви у пациентов, страдающих или имеющих риск возникновения такого заболевания. Пациенты, подлежащие лечению, включают индивидуумов, которые имеют риск развития заболевания LBD, но не проявляют симптомов заболевания, а также пациентов, которые проявляют симптомы заболевания или ранние признаки синуклеинопатии, например, замедление EEG, нейропсихиатрические проявления (депрессия, деменция, галлюцинации, тревога, апатия, ангедония), изменения вегетативной нервной системы (ортостатическая гипотензия, дисфункция мочевого пузыря, запор, недержание кала, повышенное слюноотделение, дисфагия, сексуальная дисфункция, изменения мозгового кровообращения), сенсорные изменения (искажение обонятельных ощущений, боли, цвета), нарушения сна (нарушение поведения во время сна (RBD) в фазе REM, синдром беспокойных ног/периодических движений конечностями, гиперсомния, инсомния) и разные другие признаки и симптомы (утомляемость, диплопия, расфокусированное зрение, снижение/набор массы тела). Таким образом, способы по настоящему изобретению можно применять профилактически к индивидуумам, у которых существует известный генетический риск развития LBD. К таким индивидуумам относятся индивидуумы, которые имеют родственников, испытывающих данное заболевание, и индивидуумы, у которых был определен риск его развития по анализу генетических или биохимических маркеров. Генетические маркеры риска в отношении болезни Паркинсона (PD) включают мутации в генах альфа-синуклеина или Parkin, UCHLI и CYP2D6; в частности мутации в 30 и 53 положениях гена альфа-синуклеина. Индивидуумы, страдающие болезнью Паркинсона, могут быть распознаны по ее клиническим проявлениям, включая тремор покоя, мышечную ригидность, брадикинезию и постуральную неустойчивость.
Пациенты, у которых не обнаруживаются симптомы заболевания, могут начинать проходить лечение в любом возрасте (например, с 10-, 20-, 30-летнего возраста). Однако, как правило, необязательно начинать лечение до тех пор, пока пациент не достигнет 40-, 50-, 60- или 70-летнего возраста. Обычно курс лечения включает многократное дозирование на протяжении определенного периода времени. За курсом лечения можно наблюдать путем анализа антитела или ответов активированных Т-клеток или В-клеток на терапевтический агент (например, процессированную форму пептида альфа-синуклеина) со временем. В случае падения ответа вводят бустерную дозу.
Антитела можно применять для лечения или профилактического воздействия на заболевание с клетками Леви у пациентов путем введения в условиях, которые вызывают положительный терапевтический ответ у пациента (например, снижение нейритных и/или аксональных агрегатов альфа-синуклеина, уменьшение нейритной дистрофии, улучшение когнитивной функции и/или регрессию, лечение или предупреждение ухудшения когнитивных способностей) у пациента. В некоторых способах участки нейритной дистрофии в нейропиле неокортекса и/или базальные ганглии могут быть снижены в среднем, по меньшей мере, на 10%, 20%, 30% или 40% у прошедших лечение пациентов по сравнению с контрольной популяцией.
Когнитивное нарушение, прогрессирующее ухудшение когнитивной функции, изменения в морфологии головного мозга и изменения мозгового кровообращения обычно наблюдаются у пациентов, страдающих или имеющих риск развития заболевания с тельцами Леви. Введение антител по настоящему изобретению может ингибировать или замедлять ухудшение функции у таких пациентов.
Изобретение также обеспечивает способы сохранения или увеличения синаптической плотности и/или дендритной плотности. Индекс изменений синалтической или дендритной плотности может быть измерен с помощью маркеров образования синапса (синаптофизин) и/или дендритов (МАР2). В некоторых способах синаптическая или дендритная плотность может быть восстановлена до уровня синаптической или дендритной плотности здорового субъекта. В некоторых способах средний уровень синаптической или дендритной плотности у прошедших лечение пациентов может быть повышен на 5%, 10%, 15%, 20%, 25%, 30% или более по сравнению с популяцией не прошедших лечение контрольных пациентов.
VII. Фармацевтические композиции и способы лечения
В профилактических применениях антитело или агент для индуцирования антитела, или содержащую его фармацевтическую композицию вводят пациенту, восприимчивому или иным образом подверженному риску развития заболевания в режиме (доза, частота и способ введения), эффективном для снижения риска заболевания, уменьшения остроты или задержки появления, по меньшей мере, одного признака или симптома заболевания. В некоторых профилактических применениях режим является эффективным для ингибирования или замедления накопления альфа-синуклеина и процессированных фрагментов в головном мозге, и/или ингибирования или задержки его токсических эффектов, и/или ингибирования/или замедления развития поведенческих дефицитов. В терапевтических применениях антитело или агент для индуцирования антитела вводят пациенту, у которого подозревается или уже имеется заболевание с тельцами Леви, в режиме (доза, частота и способ введения), эффективном для того, чтобы ослабить или, по меньшей мере, ингибировать дальнейшее ухудшение, по меньшей мере, одного признака или симптома заболевания. В некоторых терапевтических применениях режим является эффективным для того, чтобы уменьшить или, по меньшей мере, ингибировать дальнейшее повышение уровней альфа-синуклеина и процессированных фрагментов, связанных токсичностей и/или поведенческих дефицитов.
Режим считается терапевтически или профилактически эффективным, если прошедший индивидуальное лечение пациент достигает результата, более благоприятного, чем средний результат в контрольной популяции сопоставимых пациентов, не получавших лечение способами по изобретению, или если более благоприятный результат продемонстрирован у прошедших лечение пациентов по сравнению с контрольными пациентами в контролируемом клиническом испытании (например, фаза II, фаза II/III или фаза III испытания) при уровне p<0.05 или 0.01, или даже 0.001.
Эффективные дозы могут изменяться в зависимости от многих различных факторов, включая способы введения, целевой участок, физиологическое состояние пациента, включая тип заболевания с тельцами Леви, является ли пациент носителем аполипопротеина Ε (АроЕ), является ли пациент человеком или животным, прием других лекарственных средства, и является ли лечение профилактическим или терапевтическим.
Иллюстративный диапазон доз антител составляют от примерно 0.01 до 5 мг/кг и чаще от 0.1 до 3 мг/кг или 0.15-2 мг/кг, или 0.15-1.5 мг/кг массы тела пациента. Антитело можно вводить в таких дозах ежедневно, через день, еженедельно, один раз в две недели, один раз в месяц, один раз в 3 месяца или по любой другой схеме, установленной с помощью эмпирического анализа. Иллюстративное лечение предусматривает введение многократных доз в течение продолжительного периода, например, по меньшей мере, шести месяцев. Дополнительные иллюстративные режимы лечения предусматривают введение один раз каждые две недели или один раз в месяц, или один раз каждые от 3 до 6 месяцев.
Антитела можно вводить периферическим путем (то есть способом, при котором введенное или индуцированное антитело пересекает гематоэнцефалитический барьер для достижения целевого участка в мозге). Способы введения включают местное, внутривенное, пероральное, подкожное, внутриартериальное, внутричерепное, интратекальное, интраперитонеальное, интраназальное или внутримышечное введение. Некоторые способы введения антител представляют собой внутривенное и подкожное введение. Этот тип введения наиболее часто выполняют в мышцы плеча или ноги. В некоторых способах агенты инъецируют непосредственно в конкретную ткань, где накоплены отложения, например, интракраниальная инъекция.
Фармацевтические композиции для парентерального введения могут быть стерильными и по существу изотоническими, и изготовленными в соответствии со всеми правилами Good Manufacturing Practice (GMP). Фармацевтические композиции могут быть обеспечены в стандартной лекарственной форме (то есть дозе для однократного введения). Фармацевтические композиции могут быть формулированы с использованием одного или нескольких физиологически приемлемых носителей, разбавителей, вспомогательных веществ или добавок. Формулирование зависит от выбранного способа введения. Для инъекции антитела могут быть формулированы в водных растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хэнка, раствор Рингера или физиологический солевой раствор, или ацетатный буфер (для снижения дискомфорта в участке введения). Раствор может содержать вспомогательные вещества, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно, антитела могут быть в лиофилизированной форме для совмещения с подходящим носителем, например, стерильной апирогенной водой, перед использованием.
Режимы по настоящему изобретению можно применять в комбинации с другим агентом, эффективным для лечения или профилактики заболевания, по поводу которого проводится лечение. Например, в случае болезни Паркинсона, иммунотерапия против альфа-синуклеина WO/2008/103472, леводопа, агонисты допамина, ингибиторы СОМТ, ингибиторы МАО-В, амантадин или антихолинергические средства можно применять в комбинации с режимами по настоящему изобретению.
VIII. Другие применения
Антитела, описанные выше, можно применять для выявления альфа-синуклеина в контексте клинической диагностики или лечения, или в исследовании. Антитела можно также применять в качестве реагентов для лабораторных исследований по выявлению клеток, несущих альфа-синуклеин, и их ответа на различные раздражители. В таких применениях моноклональные антитела могут быть мечены флуоресцентными молекулами, спин-мечеными молекулами, ферментами или радиоизотопами, и могут быть представлены в виде набора со всеми необходимыми реагентами для проведения анализа на альфа-синуклеин. Антитела также можно применять для очистки альфа-синуклеина, например, посредством аффинной хроматографии.
Антитела можно применять для обнаружения телец Леви (LB) у пациента. Такие способы являются полезными для диагностики или подтверждения диагноза болезни Паркинсона (PD) или другого заболевания, связанного с присутствием телец Леви (LB) в головном мозге, или восприимчивости к таким заболеваниям. Например, способы можно применять у пациента, проявляющего симптомы деменции. В случае, если пациент имеет тельца Леви (LB), то пациент, по-видимому, страдает заболеванием с тельцами Леви, таким как болезнь Паркинсона. Способы также можно применять у пациентов, не обнаруживающих симптомов заболевания. Присутствие телец Леви или других аномальных отложений альфа-синуклеина указывает на восприимчивость к будущему заболеванию с клиническими проявлениями. Способы являются также полезными для наблюдения за прогрессированием заболевания и/или ответом на лечение у пациентов, у которых было ранее диагностировано заболевание с тельцами Леви.
Способы можно выполнять путем введения антитела и затем детекции антитела после его связывания. При желании, элиминирующий ответ можно избежать путем использования фрагмента антитела, лишенного полноразмерной константной области, такого как Fab. В некоторых способах такое же антитело может служить в качестве реагента как лечения, так и для диагностики.
Для диагностики (например, визуализации in vivo) антитела могут быть введены путем внутривенной инъекции в организм пациента, или непосредственно в головной мозг путем внутричерепной инъекции, или путем сверления отверстия в черепе. Доза реагента должна находиться в таких же диапазонах, как и для способов лечения. Как правило, антитело является меченым, хотя в некоторых способах антитело является немеченым и вторичный агент для мечения используется для связывания с антителом. Выбор метки зависит от способа детекции. Например, флуоресцентная метка является пригодной для обнаружения оптическими методами. Использование парамагнитных меток является пригодным для томографического обнаружения без хирургического вмешательства. Радиоактивные метки также могут быть детектированы с использованием позитронно-эмиссионной томографии (PET) или однофотонной эмиссионной компьютерной томографии (SPECT).
Диагностику выполняют путем сравнения числа, размера и/или интенсивности меченых локусов с соответствующими исходными значениями. Исходные значения могут представлять средние уровни у популяции не имеющих заболевания индивидуумов. Исходные значения также могут представлять предшествующие уровни, определенные у этого же пациента. Например, исходные значения могут быть определены у пациента до начала лечения, и после этого произведено сравнение измеренных значений с исходными значениями. Снижение значений относительно исходных значений указывает на положительный ответ на лечение.
Антитела можно применять для создания анти-идиотипических антител (смотри, например, Greenspan & Bona, FASEB J. 7(5):437-444, 1989; и Nissinoff, J. Immunol. 147:2429-2438, 1991). Такие анти-идиотипические антитела можно использовать в фармакокинетических, фармакодинамических исследованиях, исследованиях биораспределения, а также в исследованиях клинических ответов на человеческие античеловеческие антитела (НАНА) у индивидуумов, прошедших лечение антителами. Например, анти-идиотипические антитела специфически связываются с вариабельной областью гуманизированных антител 9Е4 и, следовательно, могут быть использованы для детекции гуманизированных антител 9Е4 в фармакокинетических исследованиях и содействия количественной оценке ответов на человеческие античеловеческие антитела (НАНА) у прошедших лечение индивидуумов.
Все патенты, веб-сайты, другие публикации, учетные номера и тому подобное, цитируемые выше или ниже, включены путем отсылки в полном объеме для всех целей в такой степени, как если бы каждый из этих документов был конкретно и индивидуально указан для включения путем отсылки. Если различные версии последовательности связаны с номером доступа в разное время, то подразумевается версия, связанная с номером доступа на действительную дату подачи данной заявки. Действительная дата подачи заявки означает более раннюю из действительной даты подачи или даты подачи приоритетной заявки, ссылающейся на номер доступа, при необходимости. Аналогично, если различные версии публикации, веб-сайта или тому подобное опубликованы в разное время, то подразумевается версия, опубликованная самой последней на действительную дату подачи заявки, если не указано иное. Любой признак, стадия, элемент, вариант осуществления или аспект изобретения может быть использован в комбинации с любым другим, если специально не указано иное. Несмотря на то, что настоящее изобретение описано достаточно подробно при помощи иллюстрации и примера для прояснения и понимания, будет очевидно, что некоторые изменения и модификации могут быть произведены в пределах объема прилагаемой формулы изобретения.
Примеры
Пример I. Дизайн гуманизированных антител 9Е4
Исходной точкой или донорным антителом для гуманизирования является мышиное антитело 9Е4, продуцируемое гибридомой, имеющей АТСС Accession No. РТА-8221, и описанное в заявке на патент США №11/710248 (номер публикации US 2009/0208487). Вариабельный каппа (Vκ) 9Е4 принадлежит мышиной подгруппе 1 по Kabat, которая соответствует человеческой подгруппе 1 по Kabat. Вариабельная область тяжелой цепи (Vh) 9Е4 принадлежит мышиной подгруппе 3d по Kabat, которая соответствует человеческой подгруппе 3 по Kabat (Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition. NIH Publication No. 91-3242, 1991). В данном Примере используется нумерация по Kabat.
CDR-L1 из 17 остатков принадлежит каноническому классу 3, CDR-L2 из 7 остатков принадлежит классу 1, и CDR-L3 из 9 остатков принадлежит классу 1 в Vk (Martin & Thornton, J Mol Biol. 263:800-15, 1996). CDR-H1 из 5 остатков принадлежит классу 1, и CDR-H2 из 17 остатков принадлежит классу 2 (Martin & Thornton, J Mol Biol. 263:800-15,1996). CDR-H3 не имеет канонических классов, но петля из 7 остатков возможно имеет изогнутое основание согласно правилам Shirai et al. (FEBS Lett. 455:188-97,1999).
Поиск по белковым последовательностям проводят в базе данных PDB (Deshpande et al., Nucleic Acids Res. 33: D233-7, 2005) для нахождения структур, которые будут обеспечивать примерную структурную модель 9Е4. Кристаллическая структура димерного антитела Х836 (код в PDB 3МВХ) (Teplyakov et al., Mol Immunol., 47(14):2422-6, 2010) выбрана для структуры Vκ, так как она имеет хорошее разрешение (1.6А) и общее сходство последовательности с Vκ 9Е4, удерживая такие же канонические структуры для петель. 1Н3Р (Pizarro et al., FEBS Lett. 509:463-8, 2001) использовали для структуры Vh. 1Н3Р имеет хорошее общее сходство последовательности и приемлемое разрешение (2.6А), но также имеет CDR-H3 такой же длины с изогнутым основанием. К тому же, CDRs-H1 и Н2 имеют такие же канонические структуры, как Vh 9Е4. Для моделирования структуры использовали программы Deep View/Swiss-Pdv Viewer 3.7 (SP5) (Guex and Peitsch, Electrophoresis 18: 2714-2723,1997).
Поиск по базе данных «non-redundant protein sequence» из NCBI обеспечивает отбор пригодных человеческих каркасных участков, в которые прививают мышиные CDR. Для Vκ была выбрана человеческая легкая цепь каппа с NCBI accession code AAY33350 (GI:63102889) (Kramer et al., Eur J Immunol. 35:2131-45, 2005). Она имеет такие же канонические классы для CDR-L2 и L3, как 9Е4, и принадлежит человеческой зародышевой линии IGKV1D-39 или IGKV1-39 (название клона O2 или O12) согласно конвенции IMGT. Она является членом человеческой подгруппы каппа 1 по Kabat. Для Vh была выбрана тяжелая цепь Ig человека с NCBI accession code ААС50998 (GI:1791009) (Glas et al., Clin Exp Immunol. 107:372-80, 1997), снова с такими же каноническими классами, как 9Е4, и принадлежащая человеческой зародышевой линии IGHV3-7'01 или IGHV3-7'02 (названия клонов V3-7 или VH3-11). Она является членом человеческой подгруппы 3 тяжелой цепи по Kabat.
Следующие положения, различающиеся между человеческими акцепторными и мышиными донорными каркасными участками вариабельной области, были идентифицированы в качестве кандидатов для обратной мутации. Н73 находится на границе антигенсвязывающего сайта и взаимодействует с CDR-H2. Н93 представляет собой остаток поверхности раздела, который лежит под петлями CDR-H1 и Н3. L36 представляет собой остаток поверхности раздела Vκ/Vh. L83 находится в непосредственной близости от константной области. L83 в 9Е4 представляет собой лейцин, тогда как в человеческой каркасной области L83 представляет собой аминокислоту фенилаланин.
Три гуманизированные тяжелые цепи и три гуманизированные легкие цепи получали путем введения обратных мутаций при различных перестановках этих положений (Фиг. 1А, В, выравнивание последовательности, и Таблицы 2-3).



Версия 1>9E4Vκ
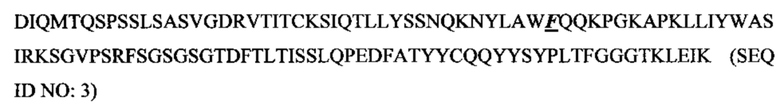
Версия 2>9E4Vκ
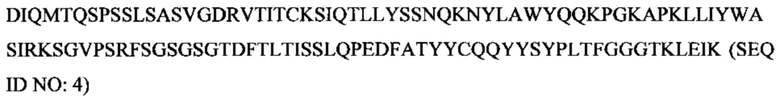
Версия 3>9E4Vκ
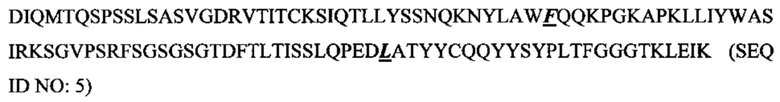
Версия 1>9E4vh

Версия 2>9E4vh

Версия 3>9E4vh

Нумерация по Kabat для AAY33350 легкой цепи и ААС50998 тяжелой цепи перечислены ниже:
Нумерация по Kabat для AAY33350 легкой цепи:
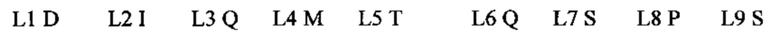

Нумерация по Kabat для ААС50998 тяжелой цепи:

Нумерация по Kabat других вариабельных областей тяжелых и легких цепей может быть определена путем выравнивания с соответствующими остатками, которым присвоен такой же номер, или с использованием коммерчески доступного программного обеспечения.


Пример II. Пассивная иммунизация антителами к альфа-синуклеину
Целью данного исследования является определение эффективности антител к альфа-синуклеину в in vitro и in vivo исследованиях, а также поведенческие анализы. Авторы использовали трансгенных по альфа-синуклеину (Линия 61), нокаутных по альфа-синуклеину и дикого типа самок мышей в возрасте 3-4 месяцев на момент начала эксперимента, и n=14/группа. Тестируемые антитела включали 9Е4 (IgG1, эпитоп: аминокислоты 118-126 альфа-синуклеина), 5С1 (IgG1, эпитоп: аминокислоты 118-126 альфа-синуклеина, с-линкер), 5D12, IgG2 (SN118-126), 1Н7, IgG1 (SN 91-99) и контрольное антитело IgG1 27-1. Мышам вводили дозу 10 мг/кг в течение 5 месяцев, в общем количестве 21 инъекций. К тому же, животных инъецировали лентивирусом (LV), экспрессирующим человеческий альфа-синуклеин (масса) путем одностороннего введения человеческого альфа-синуклеина (масса) в гиппокамп.
Считываемые антитела включают антитела, полученные от Chemicon (эпитоп: полноразмерный альфа-синуклеин), Millipore (эпитоп: полноразмерный альфа-синуклеин) и Neotope, ELADW 105 (эпитоп: аминокислоты 121-124 полноразмерного альфа-синуклеина).
Конечные показатели: Титры антител измеряют во время фазы развития. Поведенческие анализы включают тест водного лабиринта Морриса (MWW) и тест на горизонтальной перекладине. Тест на круглой перекладине представляет собой тестирование моторного баланса, координации и походки, который выполняется с использованием двух перекладин различного диаметра. Перекладина А имеет больший диаметр (более легкая, рассматриваемая как тренировочная перекладина), и Перекладина D имеет меньший диаметр (более трудная, рассматриваемая как испытательная перекладина). Данные представленные как «ошибки» (число промахов/10 см) и «скорость» (время, затраченное на прохождение 10 см/сек). Выполнение в водном лабиринте проводили на 10 неделе и в конце испытания. Производили следующие нейропатологические измерения: агрегация альфа-синуклеина, синаптофизин и МАР2. Производили следующие биохимические измерения: альфа-синуклеин, PSD95, синаптофизин. Выбранное множественное мечение и конфокальное мечение проводили с использованием синаптических, нейрональных и глиальных маркеров.
Результаты показали, что все антитела, за исключением 5D12, вызывали значительное уменьшение накопления альфа-синуклеина и сохранение синаптических и дендритных плотностей, а также положительные результаты выполнения MWM. Антитело 9Е4 является эффективным в in vitro и in vivo исследованиях, а также поведенческих анализах. Результаты считывания указывают на то, что антитело может уменьшать нейритные/аксональные агрегаты альфа-синуклеина.
Поведенческие результаты: Антитело 9Е4 улучшило характеристики выполнения теста в водном лабиринте у мышей, трансгенных по альфа-синуклеину (Фигуры 3-4). Напротив, антитело 5D12 не улучшило характеристики выполнения теста в водном лабиринте у мышей, трансгенных по альфа-синуклеину (Фигура 4). Антитела 9Е4 и 1Н7 улучшили характеристики выполнения теста на перекладине, измеренные по скорости и ошибкам, тогда как антитела 5D12 и 5С1 не улучшили (Фигура 4).
Нейропатологические результаты: Антитела 9Е4, 1Н7 и 5С1 уменьшили ELADW-105 положительную нейритную дистрофию, тогда как антитело 5D12 не уменьшило. У мышей, трансгенных по альфа-синуклеину, антитело 9Е4 уменьшило площадь нейропиля на 43% в неокортексе и на 40% в базальных ганглиях по сравнению с контролем. Антитело 9Е4 также сохранило синаптофизин и МАР2 в неокортексе и базальных ганглиях.
Пример III. Иммунопреципитация
Иммунопреципитацию выполняли для тестирования аффинности связывания различных вариантов гуманизированных антител 9Е4 в отношении его антигена, полученного из пораженной ткани (Фигура 5). 150 мкг растворимых в Tris лизатов головного мозга с деменцией с тельцами Леви иммунопреципитировали с помощью 5 мкг каждого указанного антитела с использованием магнитных частиц с иммобилизованным белком G (Protein G magnetic beads, New England Biolabs). Образцы промывали 5 раз PBS/350 мМ NaCl/0.5% NP-40, нагревали до кипения и полученные образцы анализировали с помощью SDS-PAGE. После блоттинга мембраны инкубировали с Ab5038 (Millipore), поликлональным антителом для детекции общего синуклеина. Эксперимент повторяли три раза для подтверждения достоверности.
Пример IV. Вестерн-блоттинг
Вестерн-блоттинг рекомбинантного человеческого синуклеина с мышиными химерными и гуманизированными антителами 9Е4 показан на Фигуре 6. Кривые разведения антител практически аналогичны для мышиных химерных и гуманизированных 9Е4 антител. Все антитела выявили полосу при 28 KDa и вторую полосу при 49 KDa. Полоса 49 KDa вероятно представляет собой мультимер синуклеина.
Указанные количества рекомбинантного бактериально-экспрессированного человеческого синуклеина дикого типа анализировали с помощью SDS-PAGE, и проводили блоттинг с идентичными количествами указанной формы 9Е4. После промывания наносили подходящие козьи поликлональные антитела, конъюгированные с флуорофором IRDye-800, и блот промывали. Времена воздействия были идентичны для различных антител.
Информация о депонировании
Следующую гибридому депонировали в Американской коллекции типовых культур «American Type Culture Collection)) (АТСС, P.O. Box 1549, Manassas, VA 20108) согласно положениям Будапештского договора на указанную дату. Этот депонированный образец будет храниться в уполномоченном банке депозитария и будет заменен в случае мутации, нежизнеспособности или разрушения в течение, по меньшей мере, пяти лет после получения депозитарием последнего запроса на выдачу образца в течение, по меньшей мере, тридцати лет после даты депонирования или в течение законной длительности связанного патента, в зависимости от того, какой из этих периодов будет длиннее. Все ограничения в доступе общества к этим клеточным линиям будут окончательно устранены после выдачи патента на основе заявки.

--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> САЛДАНХА, Хосе
НИЙДЖАР, Тарлочан С.
<120> ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, КОТОРЫЕ РАСПОЗНАЮТ АЛЬФА-СИНУКЛЕИН
<130> 057450/424859
<150> 61/553,131
<151> 2011-10-28
<160> 32
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 1
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 2
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 3
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 4
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 4
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 5
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 6
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 6
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Ser Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Glu Asp Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ser Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210> 7
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 7
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ser Asp Met Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 8
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 9
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 10
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 11
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 11
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 12
<211> 140
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 12
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 13
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 13
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 14
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 14
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Val Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 15
<211> 339
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 15
gacatccaga tgacccagtc cccctcctcc ctgtccgcct ccgtgggcga ccgcgtgacc 60
atcacctgca agtccatcca gaccctgctg tactcctcca accagaagaa ctacctggcc 120
tggttccagc agaagcccgg caaggccccc aagctgctga tctactgggc ctccatccgc 180
aagtccggcg tgccctcccg cttctccggc tccggctccg gcaccgactt caccctgacc 240
atctcctccc tgcagcccga ggacttcgcc acctactact gccagcagta ctactcctac 300
cccctgacct tcggcggcgg caccaagctg gagatcaag 339
<210> 16
<211> 339
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 16
gacatccaga tgacccagtc cccctcctcc ctgtccgcct ccgtgggcga ccgcgtgacc 60
atcacctgca agtccatcca gaccctgctg tactcctcca accagaagaa ctacctggcc 120
tggtaccagc agaagcccgg caaggccccc aagctgctga tctactgggc ctccatccgc 180
aagtccggcg tgccctcccg cttctccggc tccggctccg gcaccgactt caccctgacc 240
atctcctccc tgcagcccga ggacttcgcc acctactact gccagcagta ctactcctac 300
cccctgacct tcggcggcgg caccaagctg gagatcaag 339
<210> 17
<211> 339
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 17
gacatccaga tgacccagtc cccctcctcc ctgtccgcct ccgtgggcga ccgcgtgacc 60
atcacctgca agtccatcca gaccctgctg tactcctcca accagaagaa ctacctggcc 120
tggttccagc agaagcccgg caaggccccc aagctgctga tctactgggc ctccatccgc 180
aagtccggcg tgccctcccg cttctccggc tccggctccg gcaccgactt caccctgacc 240
atctcctccc tgcagcccga ggacctggcc acctactact gccagcagta ctactcctac 300
cccctgacct tcggcggcgg caccaagctg gagatcaag 339
<210> 18
<211> 348
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 18
gaggtgcagc tggtggagtc cggcggcggc ctggtgcagc ccggcggctc cctgcgcctg 60
tcctgcgccg cctccggctt caccttctcc aactacggca tgtcctgggt gcgccaggcc 120
cccggcaagg gcctggagtg ggtggcctcc atctcctccg gcggcggctc cacctactac 180
cccgacaacg tgaagggccg cttcaccatc tcccgcgacg acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg cgccgaggac accgccgtgt actactgctc ccgcggcggc 300
gccggcatcg actactgggg ccagggcacc ctggtgaccg tgtcctcc 348
<210> 19
<211> 348
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 19
gaggtgcagc tggtggagtc cggcggcggc ctggtgcagc ccggcggctc cctgcgcctg 60
tcctgcgccg cctccggctt caccttctcc aactacggca tgtcctgggt gcgccaggcc 120
cccggcaagg gcctggagtg ggtggcctcc atctcctccg gcggcggctc cacctactac 180
cccgacaacg tgaagggccg cttcaccatc tcccgcgaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg cgccgaggac accgccgtgt actactgctc ccgcggcggc 300
gccggcatcg actactgggg ccagggcacc ctggtgaccg tgtcctcc 348
<210> 20
<211> 348
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 20
gaggtgcagc tggtggagtc cggcggcggc ctggtgcagc ccggcggctc cctgcgcctg 60
tcctgcgccg cctccggctt caccttctcc aactacggca tgtcctgggt gcgccaggcc 120
cccggcaagg gcctggagtg ggtggcctcc atctcctccg gcggcggctc cacctactac 180
cccgacaacg tgaagggccg cttcaccatc tcccgcgacg acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg cgccgaggac accgccgtgt actactgcgc ccgcggcggc 300
gccggcatcg actactgggg ccagggcacc ctggtgaccg tgtcctcc 348
<210> 21
<211> 348
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 21
gaggtgcagc tggtggagtc cggcggcggc ctggtgcagc ccggcggctc cctgcgcctg 60
tcctgcgccg cctccggctt caccttctcc aactacggca tgtcctgggt gcgccaggcc 120
cccggcaagg gcctggagtg ggtggcctcc atctcctccg gcggcggctc cacctactac 180
cccgacaacg tgaagggccg cttcaccatc tcccgcgaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg cgccgaggac accgccgtgt actactgcgc ccgcggcggc 300
gccggcatcg actactgggg ccagggcacc ctggtgaccg tgtcctcc 348
<210> 22
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 22
Met Asp Met Arg Val Pro Ala Gln Leu Leu Gly Leu Leu Met Leu Trp
1 5 10 15
Val Ser Gly Ser Ser Gly
20
<210> 23
<211> 66
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 23
atggacatgc gcgtgcccgc ccagctgctg ggcctgctga tgctgtgggt gtccggctcc 60
tccggc 66
<210> 24
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 24
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys
<210> 25
<211> 57
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 25
atggagttcg gcctgtcctg gctgttcctg gtggccatcc tgaagggcgt gcagtgc 57
<210> 26
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<220>
<221> VARIANT
<222> 42
<223> Xaa = Y or F
<220>
<221> VARIANT
<222> 89
<223> Xaa = F or L
<400> 26
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Xaa Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Xaa Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 27
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<220>
<221> VARIANT
<222> 74
<223> Xaa = N or D
<220>
<221> VARIANT
<222> 97
<223> Xaa = A or S
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Xaa Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Xaa Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 28
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 28
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 29
<211> 220
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 29
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 30
<211> 219
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ile Gln Thr Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Ile Arg Lys Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Leu Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 31
<211> 446
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 31
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Ser Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Ala Gly Ile Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Val Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 32
<211> 330
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthesized
<400> 32
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВА, ПУТИ ПРИМЕНЕНИЯ И СПОСОБЫ ЛЕЧЕНИЯ СИНУКЛЕОПАТИИ | 2016 |
|
RU2765303C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| Моноклональные антитела к альфа-синуклеину для предотвращения агрегации тау-белка | 2017 |
|
RU2760334C2 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2831724C2 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2729416C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ БЕТА-АМИЛОИДУ/ЧЕЛОВЕЧЕСКОМУ РЕЦЕПТОРУ ТРАНСФЕРРИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2730682C1 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ЛЕЙКОТОКСИНОВ STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2805969C2 |

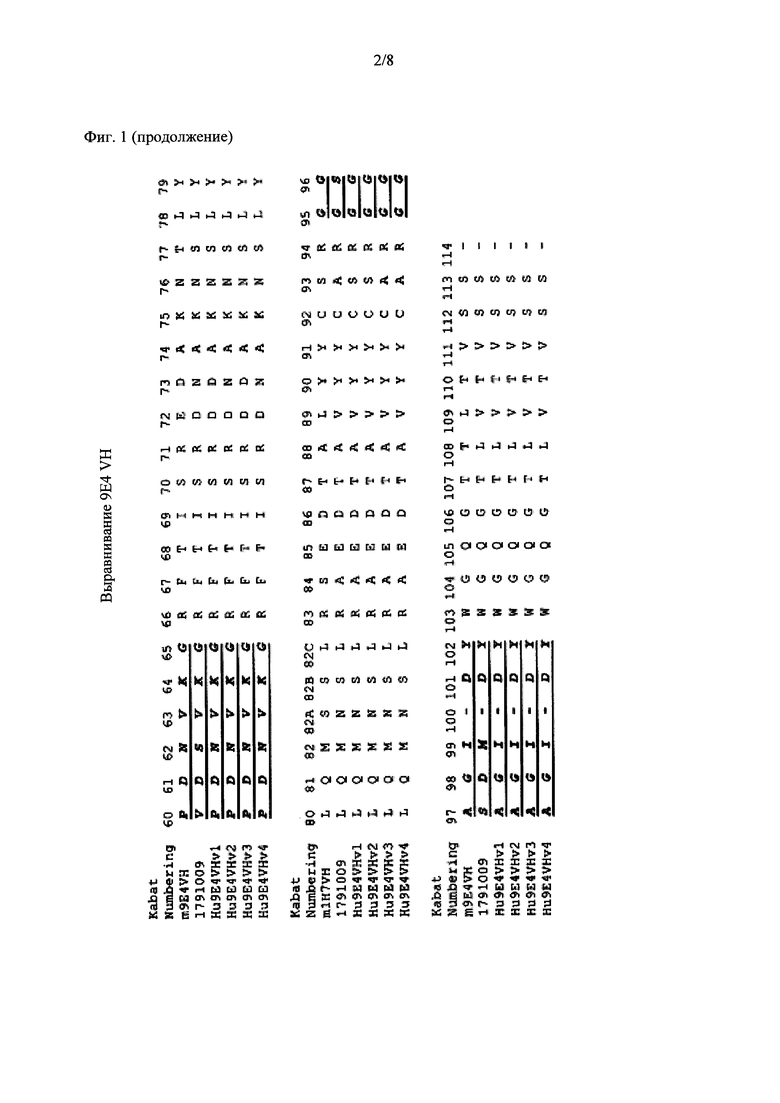
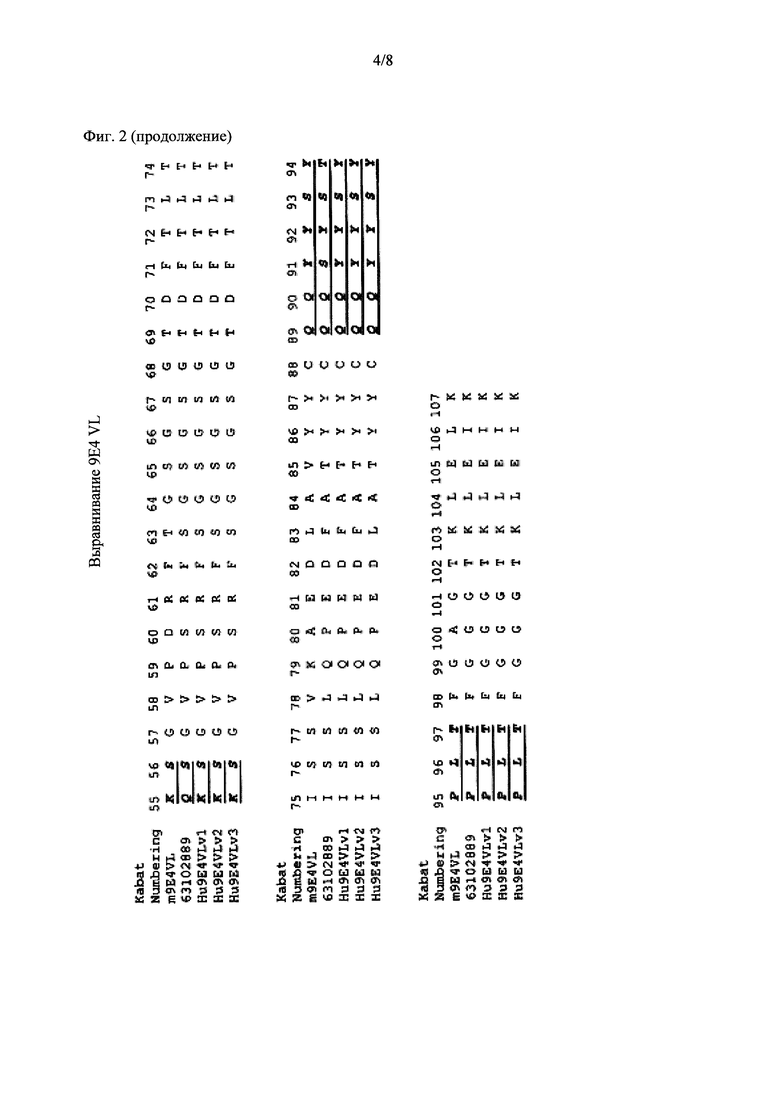

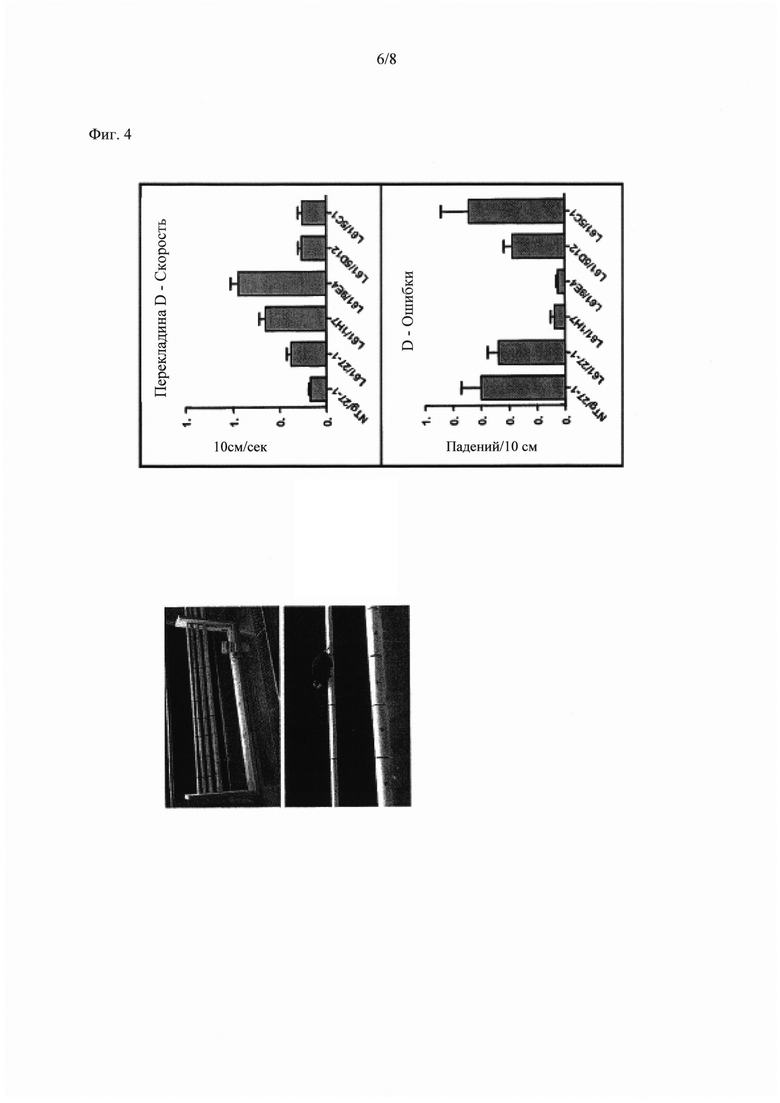
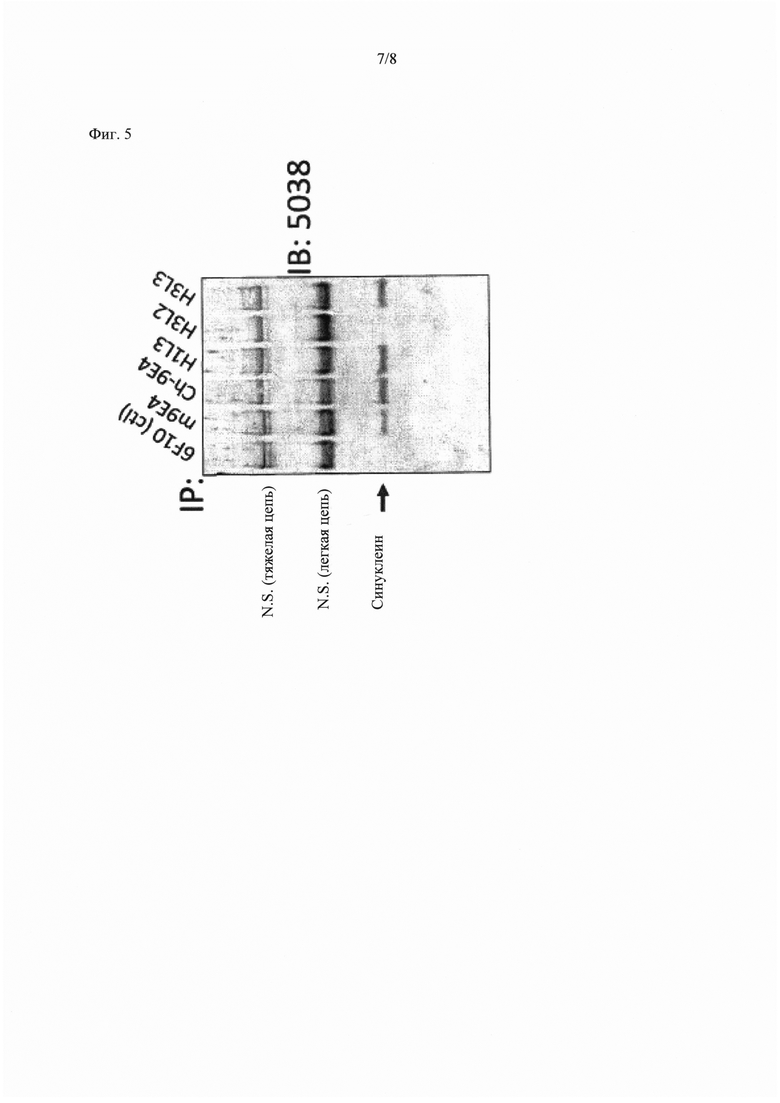
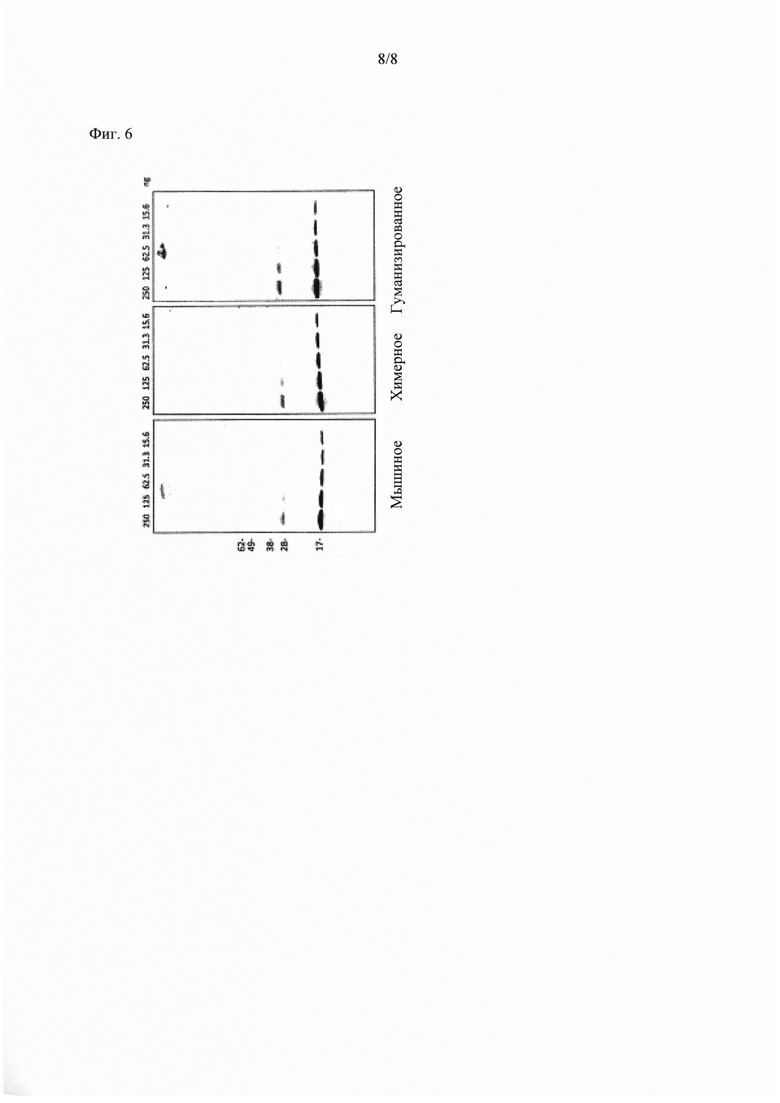
Изобретение относится к области биохимии, в частности к антителу, которое специфически связывается с человеческим альфа-синуклеином. Также раскрыты нуклеиновая кислота, кодирующая указанное антитело; клетка-хозяин для получения указанного антитела; фармацевтическая композиция, содержащая указанное антитело. Раскрыты способы лечения с помощью указанного антитела; обнаружения телец Леви и уменьшения формирования телец Леви, с помощью указанного антитела; ингибирования образования агрегатов синуклеина с помощью указанного антитела; способ получения указанного антитела и указанной клетки. Изобретение позволяет эффективно лечить заболевания, ассоциированные с человеческим альфа-синуклеином. 12 н. и 26 з.п. ф-лы, 4 пр., 4 табл., 6 ил.
1. Антитело, которое специфически связывается с человеческим альфа-синуклеином, содержащее гуманизированную тяжелую цепь, содержащую три CDR по Kabat последовательности SEQ ID NO: 11, и гуманизированную легкую цепь, содержащую три CDR последовательности SEQ ID NO: 4, при условии, что положение L36 (нумерация по Kabat) занято F, и/или положение L83 (нумерация по Kabat) занято L, и/или положение Н73 (нумерация по Kabat) занято D, и/или положение Н93 (нумерация по Kabat) занято S.
2. Антитело по п. 1, в котором положение L36 (нумерация по Kabat) занято F и положение Н73 (нумерация по Kabat) занято D, и положение Н93 (нумерация по Kabat) занято S.
3. Антитело по п. 1, в котором положение L36 (нумерация по Kabat) занято F и положение Н93 (нумерации по Kabat)-занято S.
4. Антитело по п. 1, в котором положение L36 (нумерация по Kabat) занято F и положение Н73 (нумерация по Kabat) занято D.
5. Антитело по п. 1, в котором положение Н73 (нумерация по Kabat) занято D и положение H93 (нумерация по Kabat) занято S.
6. Антитело по п. 1, в котором положение Н93 (нумерация по Kabat) занято S.
7. Антитело по п. 1, в котором положение Н73 (нумерация по Kabat) занято D.
8. Антитело по п. 1, в котором положение L36 (нумерация по Kabat) занято F и положение L83 (нумерация по Kabat) занято L, и положение Н73 (нумерация по Kabat) занято D, и положение Н93 (нумерация по Kabat) занято S.
9. Антитело по п. 1, в котором положение L36 (нумерация по Kabat) занято F и положение L83 (нумерация по Kabat) занято L, и положение Н93 (нумерация по Kabat) занято S.
10. Антитело по п. 1, в котором положение L36 (нумерация по Kabat) занято F и положение L83 (нумерация по Kabat) занято L, и положение Н73 (нумерация по Kabat) занято D.
11. Антитело по любому из пп. 1-10, в котором зрелая вариабельная область тяжелой цепи является слитой с константной областью тяжелой цепи, и зрелая вариабельная область легкой цепи является слитой с константной областью легкой цепи.
12. Антитело по п. 11, в котором константная область тяжелой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 14, при условии, что С-концевой лизиновый остаток может быть опущен.
13. Антитело по п. 11, в котором константная область легкой цепи имеет аминокислотную последовательность, обозначенную SEQ ID NO: 13.
14. Антитело по п. 11, в котором легкая цепь имеет константную область, обозначенную SEQ ID NO: 28.
15. Антитело по пп. 11, 13, 14, в котором константная область тяжелой цепи представляет собой мутантную форму природной человеческой константной области, которая имеет уменьшенное связывание с Fcγ-рецептором по сравнению с природной человеческой константной областью.
16. Антитело по пп. 11, 13, 14, в котором константная область тяжелой цепи имеет изотип IgGl человека.
17. Нуклеиновая кислота, кодирующая зрелую вариабельную область тяжелой цепи и зрелую вариабельную область легкой цепи, как определено по любому предшествующему пункту.
18. Нуклеиновая кислота по п. 17, имеющая последовательность, содержащую любую из SEQ ID NO: 15, 17, 18, 19 и 20.
19. Нуклеиновая кислота, кодирующая зрелую вариабельную область тяжелой цепи, как определено по любому из пп. 1-16.
20. Нуклеиновая кислота, кодирующая зрелую вариабельную область легкой цепи, как определено по любому из пп. 1-16.
21. Клетка-хозяин для получения антитела по любому из пп. 1-16, содержащая вектор, кодирующий зрелую вариабельную область тяжелой цепи и зрелую вариабельную область легкой цепи антитела; или содержащая вектор, кодирующий зрелую вариабельную область тяжелой цепи, и вектор, кодирующий зрелую вариабельную область легкой цепи антитела.
22. Способ лечения пациента, имеющего синуклеинопатию или имеющего риск развития синуклеинопатии, включающий введение пациенту антитела по любому из пп. 1-16 в эффективном режиме.
23. Способ по п. 22, в котором пациент имеет нарушение сна в REM-фазе (RBD).
24. Способ обнаружения телец Леви у пациента, имеющего заболевание с тельцами Леви или имеющего риск возникновения заболевания с тельцами Леви, включающий введение пациенту эффективного количества антитела, как определено по любому из пп. 1-16, при этом антитело связывается с тельцами Леви; и детекцию связанного антитела у пациента.
25. Способ по п. 24, в котором антитело является меченым.
26. Способ уменьшения формирования телец Леви у пациента, имеющего заболевание с тельцами Леви или имеющего риск возникновения заболевания с тельцами Леви, включающий введение пациенту эффективного количества антитела, как определено по любому из пп. 1-16.
27. Способ ингибирования образования агрегатов синуклеина или уменьшения телец Леви, или агрегатов синуклеина у пациента, имеющего заболевание с тельцами Леви или имеющего риск возникновения заболевания с тельцами Леви, включающий введение пациенту эффективного количества антитела, как определено по любому из пп. 1-16.
28. Способ по любому из пп. 22, 24-27, в котором заболевание представляет собой болезнь Паркинсона.
29. Способ по любому из пп. 22, 24-27, в котором заболевание представляет собой множественную системную атрофию (MSA) или деменцию с тельцами Леви (DLB).
30. Фармацевтическая композиция для лечения или профилактики заболевания с тельцами Леви, содержащая эффективное количество антитела по любому из пп. 1-16 и фармацевтически приемлемый носитель.
31. Способ получения антитела, включающий культивирование клеток-хозяев по п. 21 таким образом, что клетка секретирует антитело; и очистку антитела из клеточной культуральной среды; при этом антитело определено по любому из пп. 1-16.
32. Способ получения линии клеток-хозяев по п. 21, включающий введение вектора, кодирующего тяжелую и легкую цепи антитела, и селектируемого маркера в клетки; размножение клеток в условиях для отбора клеток, имеющих увеличенное число копий вектора; выделение единичных клеток из отобранной клетки; и консервацию клеток, клонированных из единичной клетки, отобранной на основе выхода антитела, при этом антитело представляет собой антитело, определенное по любому из пп. 1-16.
33. Способ по п. 32, дополнительно включающий размножение клеток в селективных условиях и скрининг на клеточные линии, природным образом экспрессирующие и секретирующие по меньшей мере 100 мг/л/106 клеток/24 ч.
34. Способ по любому из пп. 22, 26-27, в котором ухудшение когнитивной функции у пациента является ингибированным.
35. Способ по любому из пп. 22, 26-27, в котором нейритные и/или аксональные альфа- синуклеиновые агрегаты являются уменьшенными.
36. Способ по любому из пп. 22, 26-27, в котором нейритная дистрофия у пациента является уменьшенной.
37. Способ по любому из пп. 22, 26-27, в котором синаптическая и/или дендритная плотность является сохраненной.
38. Способ по любому из пп. 22, 26-27, при этом способ сохраняет синаптофизин и/или МАР2 у пациента.
| Способ очистки хлороформа | 1927 |
|
SU10469A1 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЯ | 2002 |
|
RU2358733C2 |
| Ударное приспособление для разгонки рельсовых зазоров | 1927 |
|
SU6630A1 |
| WO 2004065419 A1, 05.08.2004 | |||
| WO 2007011907 A2, 25.01.2007. | |||

