Изобретение относится к генетической конструкции (плазмиде), обеспечивающей синтез модифицированного белка - тканевого фактора (ТФ) человека, который, после выделения, очистки и релипидизации предназначен для использования в качестве одного из компонентов диагностического препарата, применяемого при постановке клинического лабораторного теста определения протромбинового времени (ПВ) и рекомбинантному штамму Е. coli - продуценту модифицированного ТФ.
Тканевой фактор - трансмембранный белок, выступающий в роли неферментативного кофактора и высокоаффинного клеточного рецептора фактора (ф) свертывания VII/VIIa [1-4]. Тканевой фактор стабилизирует каталитический сайт фVIIa на плазматической мембране и обеспечивает его взаимодействие с субстратами - фIX и фХ. Комплекс ТФ-фVIIa является первичным активатором ферментативного коагуляционного каскада внешнего пути свертывания крови [5, 6]. В норме ТФ не экспрессируется на поверхности клеток, которые контактируют с циркулирующей кровью [7]. При повреждении стенки кровеносного сосуда, вызванного травмой или воспалением, субэндотелиальный ТФ экспонируется в кровоток и связывается с фактором VII/VIIa, который циркулирует в виде фермента с минимальной (следовой) протеолитической активностью [8-11]. В составе комплекса ТФ-фVII/VIIa каталитическая эффективность фVIIa увеличивается на несколько порядков и инициирует свертывание крови путем протеолитической активации зимогенов факторов IX и X и превращения их в активные сериновые протеиназы -фIXa и фХа[12].
Ген ТФ человека локализован в хромосоме 1 (1р22-р21), насчитывает 12640 нуклеотидов и состоит из 6 экзонов. Открытая рамка считывания гена ТФ кодирует белок с молекулярной массой 33 кДа, состоящий из 295 аминокислот, включая сигнальную последовательность из 32 аминокислотных остатков (а.о.). В структуре ТФ выделяют 3 домена: внеклеточной (а.о. 1-219), трансмембранный (а.о. 220-242) и цитоплазматический (а.о. 243-263) [UniProtKB - PI3726 (TF_HUMAN) - https://www.uniprot.org/uniprot/P13726].
Пространственная структура внеклеточного участка представлена двумя Ig-подобными доменам [13, 14]. Внеклеточный домен ТФ взаимодействует с фVII/VIIa с образованием высокоактивного комплексного фермента. При этом максимальная каталитическая эффективность комплекса TФ-фVII/VIIa проявляется только в том случае, если он связан с фосфолипидной мембраной определенного состава [15-17]. В настоящее время общепризнано, что в качестве инициатора коагуляции ТФ без цитоплазматического домена функционально идентичен полноразмерному белку. По данным Fiore et al. [18] рекомбинантный ТФ (рТФ), лишенный и цитоплазматического и трансмембранного доменов не способен связываться с фосфолипидной мембраной, а образующийся комплекс внеклеточного домена с фVII проявляет очень низкую прокоагулянтную активность. Т.о., два из трех доменов ТФ - внеклеточный и трансмембранный - играют важную роль в процессе свертывания крови, тогда как цитоплазматический участок связан с сигнальными функциями [19].
В комплексе с фосфолипидами, ТФ образует тканевой тромбопластин (TP) - один из факторов гемостаза, участвующий во внешнем пути свертывания крови. В клинической практике препараты TP широко используются при постановке теста определения ПВ, для выявления нарушений в системе свертывания крови и при контроле за лечением антикоагулянтами непрямого действия [1, 5, 15, 20]. Разработка новых высокоактивных, стандартизированных и чувствительных препаратов TP является актуальной задачей биотехнологии, поскольку связана с перспективой повышения чувствительности и воспроизводимости клинических лабораторных исследований системы гемостаза человека.
Традиционно, для получения препаратов натурального TP используют плаценту человека, ткани легкого и мозга коровы или кролика. К недостаткам этих препаратов следует отнести возможность загрязнения активированными факторами свертывания крови и другими контаминантами, которые могут повлиять на интерпретацию полученных результатов. Другой серьезной проблемой является стандартизация реагентов для клинической диагностики, полученных из различных видов биологического сырья. Чтобы стандартизировать натуральные реагенты для определения ПВ, ВОЗ ввела систему расчета международного нормализованного отношения (MHO) [21]. MHO корректирует значения ПВ, полученные с использованием препаратов TP различного происхождения, путем сравнения с международным референтным препаратом TP, которому присваивается международный индекс чувствительности (МИЧ). Индекс - это мера чувствительности данного препарата TP, по отношению к международному референтному препарату. MHO рассчитывается как отношение ПВ пациента к нормальному значению ПВ, возведенное в МИЧ:
MHO=(ПВ пациента/нормальное значение ПВ)МИЧ
Начиная с 1983 года производители тромбопластинов для клинической лабораторной диагностики обязаны определять и указывать МИЧ для каждой партии ТР. Для высокочувствительных препаратов значение МИЧ приближается к 1,0.
Стандартизация - не единственная проблема, связанная с производством и использованием натуральных препаратов ТР. Тромбопластины изготовленные из тканей человека могут быть источниками ВИЧ-1 и других опасных вирусных инфекций. Кроме того препараты TP животного происхождения различаются по чувствительности к дефициту факторов свертывания и белкам, синтезируемым в условиях дефицита витамина К или в присутствие его антагонистов. Доступность высокоочищенных препаратов ТФ из натуральных источников в последние годы ограничена в связи с их высокой стоимостью [20, 22, 23].
Логичным решением, направленным на устранение этих недостатков, является производство диагностических препаратов TP с использованием генно-инженерного ТФ человека и искусственных липосом, оптимизированных по фосфолипидному составу [15, 24, 25]. Основным преимуществом препаратов рекомбинантного тромбопластина (рТР), производимых с использованием рТФ человека и комплекса синтетических фосфолипидов является то, что они не содержат примесей других факторов свертывания. Это обеспечивает высокую чувствительность препаратов рТР к дефициту факторов внешнего пути свертывания крови [20].
Полноразмерные варианты рТФ человека были синтезированы в системах экспрессии Escherichia coli [26], Baculovirus [27] и в клетках почек [26]. Внеклеточный домен рТФ получали с использованием генетически модифицированных клеток Е. coli [28,2 9], Saccharomyces cerevisiae [29] и Komaga-taella (Pichia) pastoris [30].
Поскольку свободный неклеточный домен ТФ (т.н. "растворимый" ТФ) не способен активировать фVII, на его основе не удается получить препараты TP, которые бы обладали достаточной прокоагулянтной активностью [31]. Поэтому очевидно, что для производства высокоактивных препаратов TP для клинико-диагностических исследований, молекула ТФ должна содержать трансмембранный домен. Наличие трансмембранного домена обеспечивает успешное инкорпорирование рекомбинантного ТФ (рТФ) в препараты искусственных липосом [25].
Основным недостатком существующих конструкций и продуцентов на основе Е. coli, является крайне низкий выход рекомбинантного ТФ. Это требует наработки большого количества биомассы, значительных затрат и существенно удорожает продукт.
В заявляемом техническом решении проведен инжиниринг структуры ТФ, в состав которого включен пептид Т7, представляющий лидерную последовательность (11 аминокислот) гена 10 бактериофага Т7. Присутствие этой последовательности в N- или С-концевой части белка способно значительно повысить выход целевого белка [32].
Техническим результатом заявленного изобретения является создание генетической конструкции "Т7 таг - модифицированный тканевой фактор человека", а на ее основе - рекомбинантного штамма Escherichia coli - продуцента рекомбинантного модифицированного ТФ человека, который обеспечивает эффективный синтез целевого белка.
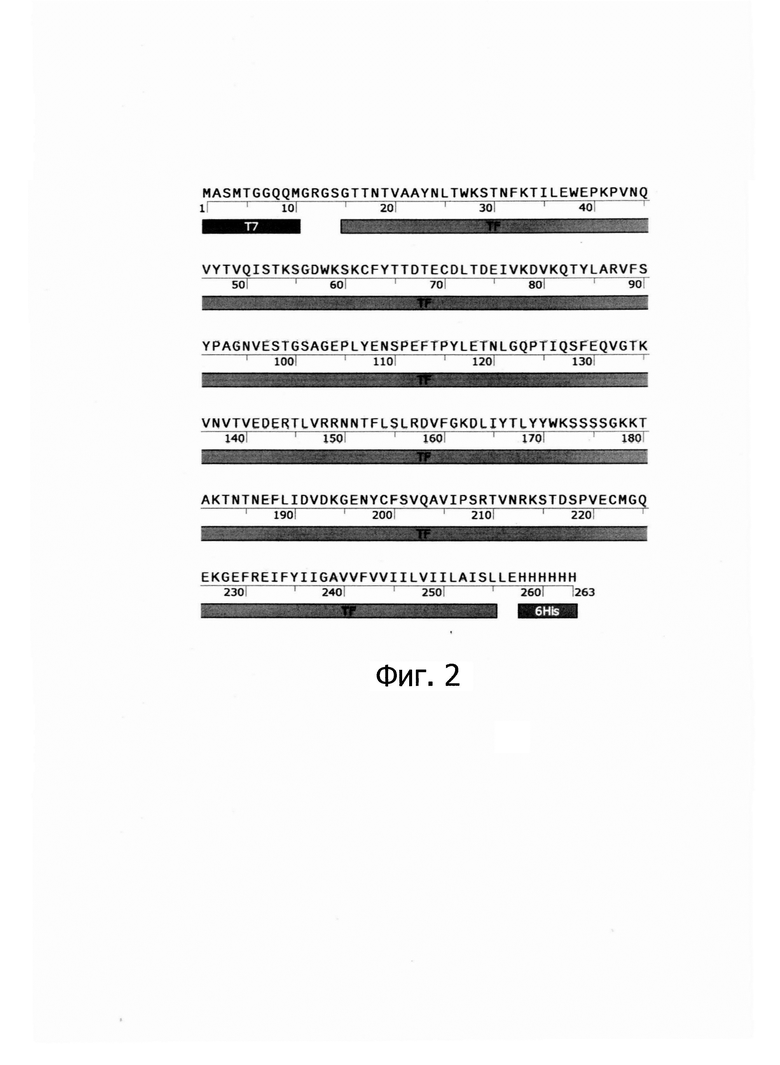
Указанный технический результат достигается созданием рекомбинантной плазмиды pET21-TF7 размером 6171 пар оснований (п.о.), характеризующейся физической и генетической картой, представленной на фиг. - 3, обеспечивающей синтез рекомбинантного ТФ человека размером 29,6 кДа в клетках Escherichia coli BL21(DE3), содержащей рекомбинантный ген ТФ человека, имеющей нуклеотидную последовательность SEQ ID NO: 1 от нук-леотида в положении 5205 п.о. до нуклеотида в положении 5996 п.о., кодирующей целевой рекомбинантный белок с аминокислотной последовательностью SEQ ID NO: 2, встроенный по сайтам рестрикции FauND I и CciNI в область полилинкера вектора рЕТ21а, содержащего промотор и терминатор РНК-полимеразы фага Т7.
Указанный технический результат достигается также созданием рекомбинантного штамма Escherichia coli BL21(DE3)/pET21-TF7 - продуцента рекомбинантного ТФ человека, полученного трансформацией рекомбинантной плазмидой pET21-TF7 по п. 1.
Заявляемый штамм Escherichia coli BL21(DE3)/pET21-TF7 - продуцент рекомбинантного ТФ человека, проявляющего, после включения в искусственные липосомы, прокоагулянтную активность и обеспечивающего активацию внешнего пути гемокоагуляционного каскада.
Для обеспечения высокого и стабильного выхода целевого продукта была проведена оптимизация кодонного состава выбранной последовательности ТФ человека для экспрессии в E. coli. В ходе теоретического дизайна проведена замена кодонов, используемых в клетках эукариот на кодоны, используемые прокариотами. В состав нуклеотидной последовательности гена на N-конец добавлена последовательность лидерного пептида Т7 (фиг. 1). Также было решено удалить из последовательности ТФ цитоплазматический домен, не влияющий на его прокоагуляционную активность. Оптимизированная последовательность была синтезирована.
В качестве вектора экспрессии выбрана плазмида рЕТ21а. После клонирования синтезированной последовательности в составе вектора, полученной рекомбинантной плазмидой была проведена трансформация бактериальных клеток Neb stable для наработки плазмидной ДНК.
В качестве продуцента выбран штамм E. coli BL21(DE3). Выбор штамма-реципиента обусловлен тем, что он несет ген РНК-полимеразы фага Т7, которая необходима для обеспечения эффективной транскрипции целевых генов, находящихся в векторной конструкции под контролем промотора фага Т7, а также тем, что данный штамм дефектен по синтезу протеаз, что существенно повышает выход синтезируемых гетерологичных белков.
При культивировании штамм E. coli BL21(DE3)/pET21-TF7, обеспечивает синтез рекомбинантного ТФ человека на уровне 10% от общего бактериального белка.
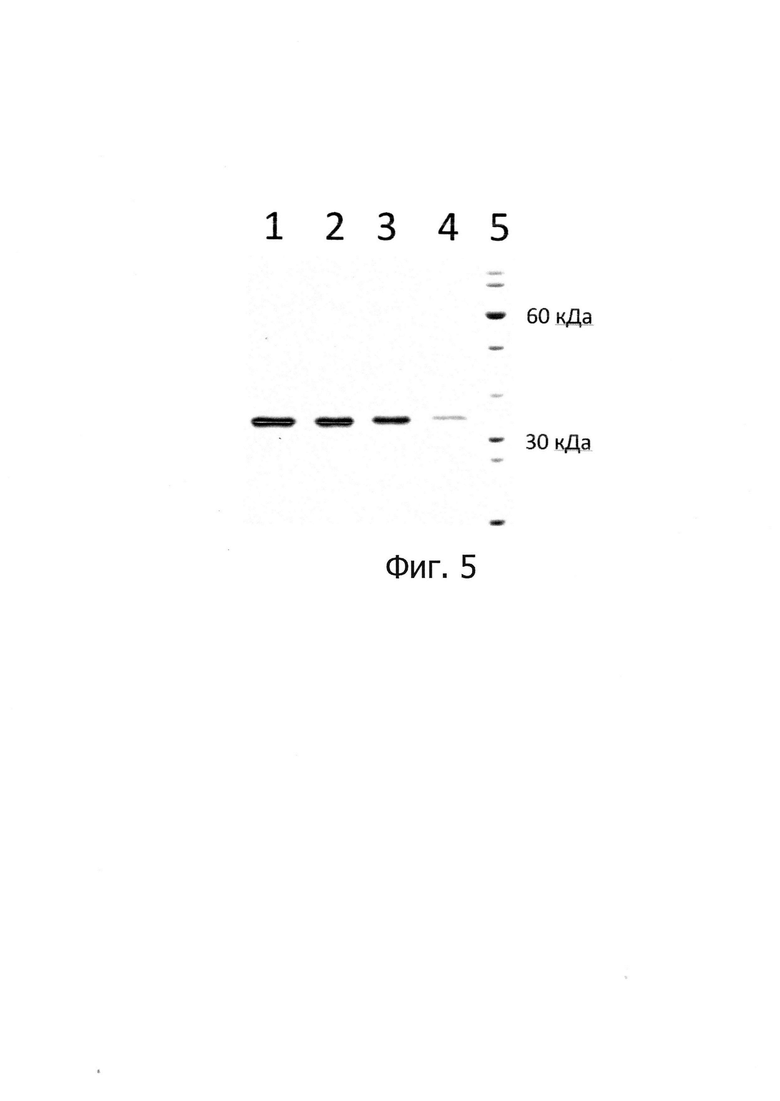
Изобретение иллюстрируется следующими графическими фигурами. На фиг. 1 приведена нуклеотидная последовательность рекомбинантного гена ТФ человека. На фиг. 2 представлена структура аминокислотной последовательности рекомбинантного модифицированного ТФ человека. На фиг. 3 изображена физическая и генетическая карта рекомбинантной плазмиды pET21-TF7, содержащая последовательность рекомбинантного модифицированного ТФ человека. На фиг. 4 представлена фотография электрофоретического разделения продуктов амплификации гена модифицированного ТФ человека в геле 1% агарозы. 1 - маркер молекулярной массы, 2, 3, 4 - продукт амплификации последовательности TF7. На фиг. 5 представлена фотография электрофоретического разделения препарата рекомбинантного модифицированного ТФ человека в ПААГ. 1, 2, 3, 4 - образец рекомбинантного модифицированного ТФ человека, 5 - маркер молекулярной массы.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам.
Пример 1. Синтез, встройка и анализ фрагмента ДНК, включающего кодирующую последовательность гена рекомбинантного тканевого фактора человека.
Заявляемая плазмида содержит следующие, существенные для ее функционирования и экспрессии целевого белка, структурные элементы:
- промотор бактериофага Т7, обеспечивающий эффективную транскрипцию контролируемой мРНК; нетранслируемую область терминации транскрипции бактериального оперона, терминатор РНК-полимеразы фага Т7, обеспечивающий эффективное окончание транскрипции;
- бактериальный оперон bla (AmpR), кодирующий белок бета-лактамазу, являющуюся селективным маркером для трансформации штаммов Е. coli;
- бактериальный участок инициации репликации типа ColE1 (ori), обеспечивающий репликацию плазмиды в штаммах Е. coli.
Выбранная нуклеотидная последовательность гена ТФ человека (NP_001984) после оптимизации структуры и кодонного состава была синтезирована на автоматическом синтезаторе ABI 3400 DNA/RNA Synthesizer. Синтезированный ген был клонирован в составе клонирующего вектора pGH («ДНК синтез», г. Москва). Для амплификации гена ТФ проводили ПЦР с использованием олигонуклеотидных праймеров, содержащих сайты гидролиза для последующей встройки продукта реакции в экспрессионный вектор: TF7-F (содержащий сайт FauNDI) и TF7-R (содержащий сайт CciNI) (таблица 1). Реакцию проводи с помощью PfuSE ДНК полимеразы (НПО "СибЭнзим", Россия) на ПЦР-амплификаторе «БИС» фирмы ООО "БИС-Н" (Россия). Реакционная смесь, объемом 50 мкл, содержащая 2 мкг ДНК (плазмиды pGH с геном тканевого фактора), 10 пкМ каждого праймера (67 мМ трис-HCl (рН 8,8), 15 мМ сульфата аммония, 2,5 мМ хлористого магния, 0,01% Твин-20, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. PfuSE ДНК полимеразы (НПО "СибЭнзим", Россия); реакцию осуществляли при следующих параметрах: 50 с - 96°С, 30 с - 60°С, 1 мин - 72°С (30 циклов).

После завершения амплификации компоненты реакционной смесь разделяли методом электрофореза в 1% агарозном геле и выявляли продукт реакции - единичный фрагмент размером 779 п.н. (Фиг 4). Фрагмент выделяли из геля с помощью набора «Gel Extraction Kit» фирмы Qiagen (Германия). Затем проводили гидролиз выделенного ПЦР-продукта и предварительно наработанного и очищенного вектора рЕТ21а эндонуклеазами рестрикции FauN-DI и CciNI («СибЭнзим», Россия) в условиях, рекомендованных производителем.
Плазмидную ДНК после ферментативного гидролиза фракционировали методом ЭФ в 1% геле агарозе. Рестрицированный вектор выделяли из агарозного геля с использованием набора «Gel Extraction Kit» фирмы Qiagen (Германия). Реакцию лигирования проводили с использованием ДНК-лигазы бактериофага Т4 («СибЭнзим», Россия). Реакция проводилась при +4°С в течение ночи. Полученной лигазной смесью трансформировали (как описано в примере 2) химически компетентные клетки E. coli штамм NEB ("New England Biolabs", Великобритания).
После трансформации клоны отбирали на селективной агаризованнной среде LB с антибиотиком ампициллином (50 мг/мл). Наличие вставки гена модифицированного тканевого фактора человека в рекомбинантной плазмиде проверяли при помощи анилитической ПЦР. Разделение продуктов амплификации проводили в 1%-м агарозном геле с последующим окрашиванием бромистым этидием (0,5 мкг/мл). Положительные колонии, вносили в 5 мл среды LB с ампициллином (50 мкг/мл) и растили в течение ночи на орбитальном шейкере при 37°С и 170 об/мин. Затем плазмидную ДНК выделяли из бактериальных клеток с помощью коммерческих наборов DNA mini kit фирмы «Qiagen» (Германия) согласно рекомендациям производителя. Нуклеотидную последовательность полученных плазмид подтверждали секвенированием методом Сэнгера в ЦКП «Геномика» СО РАН (Россия).
Секвенирование плазмидной ДНК положительных клонов в районе встройки гена модифицированного ТФ человека позволило отобрать клоны с отсутствием дефектов встраиваемого гена (вставки, делеции, замены), после чего из отобранных клонов была наработана и выделена плазмидная ДНК.
Пример 2. Получение рекомбинантного штамма Escherichia coli BL21(DE3)/pET21-TF7
С целью получения штамма-продуцента рекомбинантного ТФ человека была проведена трансформация клеток Е. coli BL21(DE3) сконструированной рекомбинантной плазмидой pET21-TF7 (Фиг 3), с использованием хлористого кальция.
Культуру клеток подращивали до ODλ600=0,5 единиц оптической плотности (е.о.п.), 1,5 мл суспензии центрифугировали при 6000 об/мин в течение 2 мин на центрифуге "Eppendorf 5415 R". Осадок клеток суспензировали в 700 мкл 0,2М CaCl2, инкубировали при 4°С в течение 15 мин, затем клетки осаждали центрифугированием 4000 об/мин 3 мин. Осадок клеток суспензировали в 100 мкл 0,2М CaCl2. К суспензии клеток добавляли 50-70 нг плазмидной ДНК, инкубировали 30 мин при 4°С, затем в течение 5 мин при 37°С. К суспензии клеток добавляли 1 мл среды LB и инкубировали при 37°С в течение 1 часа. Отбирали 100 мкл культуры и рассевали ее на чашку Петри с агаризованной средой с добавлением ампициллина до концентрации 100 мкг/мл. Остаток культуры осаждали центрифугированием при 4000 об/мин в течение 3 мин, отбирали супернатант, суспензировали в 100 мкл LB и высевали на чашку с агаризованной средой LB, содержащей 100 мкг/мл ампициллина. Чашки инкубировали в термостате при 37°С в течении ночи. На следующий день идентифицировали отдельные колонии BL21(DE3)/pET21-TF7.
Штамм E. coli BL21 (DE3), несущий рекомбинантную плазмиду рЕТ21-TF7, продуцент рекомбинантного ТФ человека, характеризуется следующими признаками: высокой физиологической стабильностью (снижение активности продуцируемого фермента после 50 генераций не превышает 20%) и высоким выходом активного продукта.
Клетки хорошо растут на стандартных питательных средах. Время генерации около 30 мин в жидкой LB-среде. На 2-2,5% питательном агаре "Difco" образуются круглые, гладкие, желтоватые колонии с ровными краями. При выращивании на жидких LB- и YT-средах образуется интенсивная ровная мутность.
Физиолого-биохимические признаки
Оптимальная температура культивирования - от 16 до 42°С, оптимум рН - 7,6. Источником азота служат органические соединения (в виде триптона, дрожжевого экстракта).
Существенным при использовании данных штаммов является их чувствительность к канамицину (до 25 мкг/мл). Проявляют устойчивость к ампициллину (до 100 мкг/мл), обусловленную наличием плазмид pET21-TF7.
Один из отобранных клонов обозначали как Escherichia coli BL21(DE3)/pET21-TF7, выращивали его в 5 мл жидкой среды LB, содержащей 100 мг/л ампициллина до ODλ600=5,0 е.о.п., добавляли к культуре равный объем стерильного 30% глицерина, разливали по 500 мкл в стерильные 1.5 мл пробирки, замораживали и хранили при - 70°С.
Пример 3. Наработка биомассы BL21(DE3)/pET21-TF7 и выделение рекомбинантного тканевого фактора человека
Индивидуальные колонии рекомбинантного штамма Е. coli, трансформированные плазмидой pET21-TF7, культивировали в течение ночи, на орбитальном шейкере, в среде LB, при 37°С и 180 об/мин. Инокулят, в соотношении 1:100, переносили в колбу Эрленмейера, содержащую среду LB и растили до оптической плотности (при λ=600 нм) равной 0,8. Добавляли индуктор (изопропил-β-D-1-тиогалактопиранозид), до конечной концентрации 0,1 мМ. Клетки дополнительно культивировали на шейкере в течение 12 часов при 25°С и 160 об/мин. Биомассу бактериальных клеток осаждали центрифугированием при 5000 G в течении 20 минут. Осадок растворяли, из расчета 20 мл на 1 грамм, в лизирующем буфере (8% сахароза, 50 мМ Трис, 20 мМ ЭДТА, 5% Triton X100 рН=8,0) с использованием ультразвукового гомогенизатора Soniprep 150 Plus ("MSE", Китай). Осадок, содержащий тельца включения, отделяли центрифугированием (20000 G в течение 25 мин, 4°С) и растворяли в базовом буфере (50 мМ KH2PO4, 150 мМ NaCl, рН=10,7), содержащем 8М мочевину.
После разделения все фракции были проанализированы в 12% ПААГ на наличие целевого белка. Результаты показали, что весь целевой белок находится в нерастворимой фракции и его процентное содержание от общего количества белков составляет порядка 10% (фиг 5).
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Grover, S.P. Tissue Factor An Essential Mediator of Hemostasis and Trigger of Thrombosis (Brief Review) / S.P. Grover, N. Mackman // Arterioscler. Thromb. Vasc. Biol.- 2018.- V. 38.- P. 709-725
2. Nemerson, Y. Tissue factor accelerates the activation of coagulation factor VII: the role of a bifunctional coagulation cofactor / Y. Nemerson, D. Repke // Thromb. Res.- 1985.- V. 40.- P. 351-358.
3. Butenas, S. Tissue factor in coagulation: Which? Where? When? /. Butenas, T. Orfeo, K.G. Mann // Arterioscler. Thromb. Vasc. Biol.- 2009.- V. 29.- P. 1989-1996.
4. Williams, J.C. Tissue factor in health and disease / J.C. Williams, N. Mackman // Front Biosci (Elite Ed).- 2012.- V.4.- P. 358-372.
5. Morrissey, J.H. Tissue factor: an enzyme cofactor and a true receptor / J.H. Morrissey // Thromb. Haemost.- 2001.- V. 86.- P. 66-74.
6. Mackman, N., Tilley R.E., Key N.S. Role of the extrinsic pathway of blood coagulation in hemostasis and thrombosis. Arterioscler. Thromb. Vase. Biol.- 2007.- V. 27.- P. 1687-1693.
7. Butenas, S. Tissue factor in thrombosis and hemorrhage / S. Butenas, T. Orfeo, K.E. Brummel-Ziedins, K.G. Mann // Surgery.- 2007.- V. 142 (4 Suppl.).- P. S2-S14.
8. Nemerson, Y. Tissue factor and hemostasis / Y. Nemerson // Blood.- 1988.- V. 71(1).- P. 1-8.
9. Edgington, T.S. The structural biology of expression and function of tissue factor /T.S. Edgington, N. Mackman, K. Brand, W. Ruf // Thromb. Haemost.- 1991.- V. 66(1).-P. 67-79.
10. Nakagaki, T. Kisiel Initiation of the extrinsic pathway of blood coagulation: evidence for the tissue factor dependent autoactivation of human coagulation factor VII / T. Nakagaki, D.C. Foster, K.L. Berkner, W. Kisiel // Biochemistry.- 1991.- V. 30.- 10819-10824.
11. Lawson, J.H. The evaluation of complex-dependent alterations in human factor VIIa / J.H. Lawson, S. Butenas, K.G. Mann // J. Biol.Chem.- 1992.- V. 267.-P. 4834-4843.
12. Komiyama, Y. Proteolytic activation of human factors IX and X by recombinant human factor VIIa: effects of calcium, phospholipids, and tissue factor / Y. Komiyama, A.H. Pedersen, W. Kisiel // Biochemistry.- 1990.- V. 29.- P. 9418-9425.
13. Harlos, K. Crystal structure of the extracellular region of human tissue factor / K. Harlos, D.M. Martin, D.P. O'Brien, E.Y. Jones, D.I. Stuart, I. Polikarpov, A. Miller, E.G. Tuddenham, C.W. Boys // Nature.- 1994.- V.370.- P. 662-666.
14. McVey, J. H. Review - The role of the tissue factor pathway in haemostasis and beyond /J. H. McVey // Curr. Opin. Hematol.- 2016.- V. 23 (5) P. 453-461.
15. Butenas, S. Comparison of natural and recombinant tissue factor proteins: new insights / S. Butenas // Biological Chemistry.- 2013.- V. 394 (7).- P. 819-829.
16. Boettcher, J.M. Backbone 1H, 13C and 15N resonance assignments of the extracellular domain of tissue factor / J.M. Boettcher, M.C. Clay, B.J. LaHood, J.H. Morrissey, C.M. Rienstra // Biomol. NMR Assign.- 2010.- V. 4.- P. 183-185.
17. Ohkubo, Y.Z. Dynamical view of membrane binding and complex formation of human factor VIIa and tissue factor / Y.Z. Ohkubo, J.H. Morrissey, E. Tajkhorshid // J. Thromb. Haemost- 2010.- V. 8.- P. 1044-1053.
18. Fiore, M.M. The biochemical basis for the apparent defect of soluble mutant tissue factor in enhancing the proteolytic activities of factor VIIa / M.M. Fiore, P.F. Neuenschwander, J.H. Morrissey // J. Biol. Chem.- 1994.- V. 269.- P. 143-149.
19. Schaffner, F. Tissue factor proangiogenic signaling in cancer progression / F. Schaffner, N. Yokota, W. Ruf // Thromb. Res.- 2012.- V. 129 (Suppl. 1).- S127-131.
20. Brucato, C.L. Expression of recombinant rabbit tissue factor in Pichia pastoris, and its application in a prothrombin time reagent./ C.L Brucato, C.A. Birr, P. Bruguera, J.A. Ruiz, 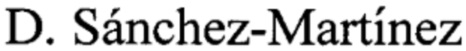 // Prot. Express. Purine- 2002.-V. 26 (3).- P. 386-393.
// Prot. Express. Purine- 2002.-V. 26 (3).- P. 386-393.
21. WHO Expert Commitee on Biological Standardization, Technical Report series 687, 33d Report, World Health Organization, Geneva, 1983, pp. 81-105.
22. Denson, K.W.E. Comparative studies of rabbit and human recombinant tissue factor reagents / K.W.E. Denson, S.W. Reed, M.E. Haddon, B. Woodhams, C. Brucato, J. Ruiz // Thromb. Res.- 1999.- V. 94.- P. 255-261.
23. Kovacs, M.J. Assessment of the validity of the INR system for patients with liver impairment / M.J. Kovacs, A. Wong, K. MacKinnon, K. Weir, M. Keeney, E. Boyle, M. Cruickshank // Thromb. Haemost.- 1994.- V. 71.- P. 727-730.
24. Kitchen, S. Two recombinant tissue factor reagents compared to conventional thromboplastins for determination of international normalised ratio: a thirty-three-laboratory collaborative study. The Steering Committee of the UK National External Quality Assessment Scheme for Blood Coagulation /S. Kitchen, I. Jennings, T.A. Woods, I.D. Walker, F.E. Preston // Thromb. Haemost.- 1996.- V. 76.-P. 372-376.
25. Smith, S.A. Rapid and efficient incorporation of tissue factor into liposomes. / S.A. Smith, J.H. Morrissey // J. Thromb. Haemost.- 2004.- V. 2.- P. 1155-1162.
26. Paborski, L.R. Purification of recombinant human tissue factor / L.R. Paborski, K.M. Tate, R.J. Harris, D.J. Yansura, L. Band, G. McCray, CM. Gorman, D.P. O_Brien, J.Y. Chang, J.R. Swartz, V.P. Fung, J.N. Thomas, G. Vehar // Biochemistry.- 1989.- V. 28.- P. 8072-8077.
27. Ruf, W. Two sites in the tissue factor extracellular domain mediate the recognition of the ligand factor VIIa / W. Ruf, T.S. Edgington // Proc. Natl. Acad. Sci. USA.- 1991.- V. 88.- P. 8430-8434.
28. Rezaie, A.R. Expression and purification of a soluble tissue factor fusion protein with an epitope for an unusual calciumdependent antibody / A.R. Rezaie, M.M. Fiore, P.F. Neuenschwander, C.T. Esmon, J.H. Morrissey // Protein Express. Purif.- 1992.- V. 3.- P. 453-460.
29. Stone, M.J. Recombinant soluble human tissue factor secreted by Saccharomyces cerevisiae and refolded from Escherichia coli inclusion bodies: glycosylation of mutants, activity, and physical characterization / M.J. Stone, R. Wolfram, D.J. Miles, T.S. Edgington, P.E. Wright // Biochem. J.- 1995.- V. 310.- P. 605-614.
30. A.J. Austin, C.E. Jones, G. Van Heeke, Production of human tissue factor using the Pichia pastoris expression system / A.J. Austin, C.E. Jones, G. Van Heeke // Protein Express. Purif-1998.- V. 13.- P. 136-142.
31. Prasad, R. Molecular determinants involved in differential behaviour between soluble tissue factor and full-length tissue factor towards factor VIIa / R. Prasad, P. Sen / Phys. Chem. Chem. Phys.- 2017.- V. 19.- P. 22230-22242
32. Studier, F.W. and Moffatt, B.A. (1986). Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol 189, 113-30.
--->
Перечень последовательностей
<110> федеральное государственное бюджетное образовательное учреждение высшего
образования «Алтайский государственный университет»
<120> Рекомбинантная плазмида pET21-TF7, обеспечивающая синтез модифицированного
тканевого фактора, и штамм Escherichia coli BL21(DE3) pET21-TF7 – продуцент
рекомбинантного тканевого фактора человека
<160> SEQ ID NO 1
<210> SEQ ID NO:1
<211> 792
<212> DNA
<213> Artificial Sequence
<220>
<223> Нуклеотидная последовательность гена TF7 рекомбинантного тканевого фактора
человека.
<400> 1
ATGGCTAGCATGACTGGTGGACAGCAAATGGGTCGCGGATCCGGCACCACCAATACCGTTGCAGCATATAATCTGACCTGG
AAAAGCACCAACTTTAAGACCATTCTGGAATGGGAACCGAAACCGGTTAATCAGGTTTATACCGTTCAGATTAGCACCAAA
AGCGGTGACTGGAAATCCAAATGTTTTTATACCACCGATACCGAATGTGATCTGACCGATGAAATTGTGAAAGATGTGAAA
CAGACCTATCTGGCACGTGTTTTTAGCTATCCGGCAGGTAATGTTGAAAGCACCGGTAGTGCCGGTGAACCGCTGTATGAA
AATAGTCCGGAATTTACCCCGTATCTGGAAACCAATCTGGGTCAGCCGACCATTCAGAGCTTTGAACAGGTTGGCACCAAA
GTTAATGTTACCGTTGAAGATGAACGTACCCTGGTTCGTCGTAATAACACCTTTCTGAGCCTGCGTGATGTTTTTGGTAAA
GATCTGATTTACACCCTGTATTATTGGAAGAGTAGCAGTAGCGGTAAGAAAACCGCAAAAACCAATACCAACGAGTTCCTG
ATTGATGTGGATAAAGGTGAGAACTATTGCTTTAGCGTTCAGGCAGTTATTCCGAGCCGTACCGTTAATCGTAAAAGCACC
GATTCACCGGTTGAATGTATGGGTCAAGAAAAAGGTGAATTTCGCGAGATCTTCTATATTATCGGTGCCGTTGTTTTTGTG
GTGATTATTCTGGTTATTATCCTGGCAATTAGCCTGCTGGAACATCATCACCATCACCATTGA (792
нуклеотида)
<210> SEQ ID NO:2
<211> 479
<212> DNA
<213> Artificial Sequence
<220>
<223> Аминокислотная последовательность рекомбинантного тканевого фактора
человека.
<400> 2
MASMTGGQQMGRGSGTTNTVAAYNLTWKSTNFKTILEWEPKPVNQVYTVQISTKSGDWKSKCFYTTDTECDLTDEIVKDVK
QTYLARVFSYPAGNVESTGSAGEPLYENSPEFTPYLETNLGQPTIQSFEQVGTKVNVTVEDERTLVRRNNTFLSLRDVFGK
DLIYTLYYWKSSSSGKKTAKTNTNEFLIDVDKGENYCFSVQAVIPSRTVNRKSTDSPVECMGQEKGEFREIFYIIGAVVFV
VIILVIILAISLLEHHHHHH
(263 аминокислотных остатка)
<---


Группа изобретений относится к рекомбинантной плазмиде, обеспечивающей синтез рекомбинантного модифицированного тканевого фактора человека, и рекомбинантному штамму, продуцирующему рекомбинантный тканевый фактор человека. Предложена рекомбинантная плазмида pET21-TF7 размером 6171 пар оснований (п.о.), содержая следующие структурные элементы: промотор бактериофага Т7; рекомбинантный ген, кодирующий модифицированный тканевой фактор человека; нетранслируемую область терминации транскрипции бактериального оперона; терминатор РНК-полимеразы фага Т7; последовательность bla (AmpR); последовательность ColE1 (ori). При этом рекомбинантный ген модифицированного тканевого фактора человека имеет нуклеотидную последовательность SEQ ID NO: 1 от нуклеотида в положении 5205 п.о. до нуклеотида в положении 5996 п.о., кодирующий целевой белок с полной аминокислотной последовательностью SEQ ID NO: 2, встроенный по сайтам рестрикции FauND I и CciNI в область полилинкера вектора рЕТ21а, содержащего промотор и терминатор РНК-полимеразы фага Т7. Указанная плазмида обеспечивает синтез рекомбинантного модифицированного тканевого фактора человека, размером 29,6 кДа в клетках Escherichia coli BL21(DE3). Предложен также рекомбинантный штамм Escherichia coli BL21(DE3)/pET21-TF7, продуцирующий рекомбинантный тканевый фактор человека и полученный трансформацией указанной рекомбинантной плазмидой pET21-TF7. Группа изобретений обеспечивает высокий и стабильный выход целевого продукта. 2 н.п. ф-лы, 5 ил., 1 табл., 3 пр.
1. Рекомбинантная плазмида pET21-TF7 размером 6171 пар оснований (п.о.), обеспечивающая синтез рекомбинантного модифицированного тканевого фактора человека, размером 29,6 кДа в клетках Escherichia coli BL21(DE3) и содержащая следующие структурные элементы: промотор бактериофага Т7; рекомбинантный ген, кодирующий модифицированный тканевой фактор человека; нетранслируемую область терминации транскрипции бактериального оперона; терминатор РНК-полимеразы фага Т7; последовательность bla (AmpR); последовательность ColE1 (ori); причем рекомбинантный ген модифицированного тканевого фактора человека имеет нуклеотидную последовательность SEQ ID NO: 1 от нуклеотида в положении 5205 п.о. до нуклеотида в положении 5996 п.о., кодирующий целевой белок с полной аминокислотной последовательностью SEQ ID NO: 2, встроенный по сайтам рестрикции FauND I и CciNI в область полилинкера вектора рЕТ21а, содержащего промотор и терминатор РНК-полимеразы фага Т7.
2. Рекомбинантный штамм Escherichia coli BL21(DE3)/pET21-TF7 - продуцент рекомбинантного тканевого фактора человека, полученный трансформацией рекомбинантной плазмидой pET21-TF7 по п. 1.
| ЩЕРБАКОВ Д.Н | |||
| и др | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| Всероссийская мультиконференция с международным участием "Биотехнология - медицине будущего", 29 июня - 2 июля 2019 г., г | |||
| Новосибирск, Россия, с | |||
| Переносное устройство для вырезания круглых отверстий в листах и т.п. работ | 1919 |
|
SU226A1 |
| ЭКСПРЕССИОННАЯ ПЛАЗМИДНАЯ ДНК p6E-tTF, КОДИРУЮЩАЯ ВНЕКЛЕТОЧНЫЙ И ТРАНСМЕМБРАННЫЙ ДОМЕНЫ ТКАНЕВОГО ФАКТОРА ЧЕЛОВЕКА И ШТАММ E.coli BL21[DE3]/p6E-tTF-ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ТКАНЕВОГО ФАКТОРА ЧЕЛОВЕКА | 2008 |
|
RU2426780C2 |
| STONE M.J | |||
| ET AL | |||

