Родственные заявки
Настоящая заявка связана с заявкой US 62/352482, поданной 20 июня 2016 г., содержание которой полностью включено в настоящий документ посредством ссылки.
Область изобретения
Настоящее изобретение относится к молекулам антител, связывающимся с лигандом-1 белка запрограммированной гибели клеток (PD-L1) и геном активации лимфоцитов 3 (LAG-3). Молекулы антител предпочтительно содержат сайт связывания антигена PD-L1 (антиген-связывающий сайт) на основе CDR и сайт связывания антигена LAG-3 на основе CDR, который может располагаться в двух или более структурных петлях CH3-домена молекулы антитела. Молекулы антител согласно настоящему изобретению находят применение, например, в лечении рака.
Уровень техники
Ген активации лимфоцитов-3 (LAG-3; CD223) является членом надсемейства Ig и характеризуется генетическим и структурным родством с CD4 (хотя лишь с 20% идентичностью последовательности). Как и CD4, LAG-3 связывается с молекулами MHC II класса, но с большим сродством, чем CD4 (KD = 60 нМ). LAG-3 экспрессируется на активированных Т-клетках, NK-клетках, pDC, B-клетках, γδ T-клетках и участвует в иммуносуппрессии, в частности, за счет устойчивой интенсивной экспрессии на некоторой доле регуляторных T-клеток (Treg) (Liang et al, 2008).
Ген LAG-3 располагается на 12 хромосоме человека, рядом с геном CD4, и включает 8 экзонов. Существует пять альтернативных транскриптов, два из которых образуют белковые продукты: полноразмерный трансмембранный белок и растворимую мономерную форму, полученную в результате альтернативного сплайсинга. Полноразмерный транскрипт кодирует белок из 525 аминокислот с молекулярной массой 70 кДа, обладающий функциональной активностью, в то время как растворимая форма, по-видимому, не связывает молекулы МНС II класса, и ее функция неизвестна. Полноразмерный белок LAG-3 человека характеризуется 93% идентичностью последовательности по отношению к LAG-3 Macaca fascicularis (яванского макака) и 70% идентичностью последовательности по отношению к LAG-3 Mus musculus (домовой мыши).
LAG-3 представляет собой трансмембранный белок с четырьмя внеклеточными Ig-подобными доменами (D1-D4) и цитоплазматическим фрагментом, ответственным за сигнальный путь LAG-3. Цитоплазматический домен содержит EP-мотив (глутаминовая кислота/пролин), связывающийся с LAG-3-ассоциированным белком (LAP), а также KIEELE-мотив, который считается необходимым для модуляции функции T-клеток за счет LAG-3. Сообщения о роли EP-мотива указывают, что он может отвечать за перенос LAG-3 к мембране T-клеток (Bae et al, 2014) или непосредственно за модуляцию дальнейшего сигнального пути STAT5 во время активации T-клеток (Durham et al, 2014), или, возможно, за оба указанных процесса.
Считается, что механизм иммуносуппрессии Т-клеток за счет LAG-3 осуществляется за счет перекрестного связывания LAG-3 на активированных Т-клетках, что приводит к ослаблению потока кальция и высвобождению ИЛ-2 при активации Т-клеток (Huard et al, 1997). На антиген-презентирующих клетках (АПК) связывание молекул ГКГС II LAG-3-положительными регуляторными Т-клетками вызывает снижение секреции ИЛ-12 и подавление функции CD86 (Liang et al, 2008), “вторичного сигнала” активации, что приводит к нечувствительности T-клеток из-за неправильной активации и/или пониженной презентации антигена АПК. Модели мышей с нокаутом LAG-3 жизнеспособны и характеризуются лишь легкой лимфогиперпролиферацией (Workman et al, 2003), что указывает на действие LAG-3 в качестве умеренного «тормоза» иммунной системы.
Предполагается, что это суппрессивное взаимодействие между LAG-3 и ГКГС II класса также происходит между Treg и CD4-положительными T-клетками (Sega et al, 2014). Treg подавляют иммунный ответ за счет высвобождения суппрессивных цитокинов (например, ИЛ-10 и ТФРβ), манипуляций с механизмом воспаления (например, аденозином, катаболизируемым CD73), регуляции созревания АПК или непосредственного взаимодействия между регуляторными Т-клетками и эффекторными T-клетками. Существуют доказательства, что Treg человека, положительные по ГКГС II класса, обладают улучшенными супрессивными функциями по сравнению с Treg, отрицательными по ГКГС II класса (Baecher-Allen et al, 2006), и активно подавляют иммунный ответ за счет непосредственного взаимодействия с LAG-3, экспрессируемым на эффекторных Т-клетках. Хотя LAG-3-отрицательные Treg могут подавлять пролиферацию обычных T-клеток, LAG-3-отрицательные CD4 и CD8 T-клетки устойчивы к иммуносуппрессии Treg. Описано, что данный процесс происходит между Т-клетками за счет процесса, известного как трогоцитоз (Sega et al, 2014), при котором Treg не только препятствуют созреванию АПК, но и получают ГКГС II класса, подавляя примированные LAG-3-положительные CD4 T-клетки.
Экспрессия LAG-3 также является маркером повторной антигенной стимуляции. При раке у T-клеток часто развивается “истощенный” фенотип с вовлечением экспрессии иммуносуппрессоров, например, PD-1, CTLA-4, TIM-3 и LAG-3 (Wherry et al, 2011), при котором клетки в целом не могут надлежащим образом пролиферировать и секретировать хемокины в ответ на антиген. Ингибирование этих иммуносуппрессоров понижает иммунный порог и (повторно) активирует надлежащий противоопухолевый ответ Т-клеток. В доклинических моделях это подтверждали с использованием антагонистических антител против LAG-3, CTLA-4 и PD-1, при котором наблюдали снижение опухолевой нагрузки. Считается, что ингибирование LAG-3 антагонистическими антителами реактивирует иммунный ответ в микроокружении опухоли, где экспрессия LAG-3 на CD4-положительных T-клетках и CD8- положительных T-клетках ассоциирована с истощенным фенотипом, а экспрессия LAG-3 на Treg ассоциирована с эффективностью иммуносуппрессии. Антитела, блокирующие LAG-3, усиливают пролиферацию эффекторных Т-клеток, продукцию цитокинов, цитотоксичность, а также снижают суппрессорную активность Treg, что приводит к ослаблению роста опухоли.
В опухолях человека повышенная экспрессия LAG-3 обнаружена на опухоль-инфильтрирующих лимфоцитах (ОИЛ) из светлоклеточных карцином и других опухолей человека, например, меланом и лимфом (Demeure et al, 2001; Wolchock et al, 2013). Важно отметить, что LAG-3 также коррелирует с дисфункцией T-клеток у пациентов с хронической вирусной инфекцией (Workman et al, 2005) и раком (Workman et al, 2003). Кроме того, LAG-3 выявлен в качестве поверхностного маркера опухоль-инфильтрирующих Treg при различных видах рака (Camisachi et al. 2010; Gandhi et al, 2006).
Ведется клиническая разработка моноклональных антител к LAG-3 человека для устранения иммуносуппрессии и потенциального усиления презентации антигенов при раке (плотных и гематологических злокачественных новообразованиях).
LAG-525 и IMP-701 (Novartis AG) представляют собой антитела человека против LAG-3; их испытания дошли до клинических исследований II и I фаз, соответственно, при раке почек (светлоклеточном раке); немелкоклеточном раке легких (НМРЛ); раке носоглотки; раке ободочной и прямой кишки; меланоме; раке желудка и аденокарциноме гастроэзофагеального перехода.
Антитело BMS-986016 против LAG-3 (Bristol-Myers Squibb Company) проходит клинические испытания I фазы при раке яичников; НМРЛ; раке ободочной и прямой кишки; раке шейки матки; меланоме; раке желудка; раке мочевого пузыря; плоскоклеточной карциноме головы и шеи; светлоклеточной карциноме, а также исследования II фазы при НМРЛ; рецидивирующем хроническом лимфолейкозе (ХЛЛ); стойком хроническом лимфолейкозе (ХЛЛ); меланоме; неходжкинской лимфоме; лимфоме Ходжкина; диффузной крупноклеточной B-клеточной лимфоме; медленно растущей лимфоме; лимфоме из клеток мантии; стойкой множественной миеломе; и рецидивирующей множественной миеломе в качестве как монотерапии, так и компонента комбинированной терапии.
Дополнительные антитела против LAG-3 также находятся на стадии доклинической разработки.
Белок-1 запрограммированной гибели клеток (PD-1) и его лиганды PD-L1 (CD274, B7-H1) и PD-L2 (B7-DC) обеспечивают передачу ингибиторных сигналов, регулирующих баланс между активацией Т-клеток, толерантностью и патологией иммунной системы. PD-L1 временно экспрессируется на всех иммунных клетках и некоторых опухолевых клетках.
PD-L1 представляет собой трансмембранный белок I типа, содержащий два Ig-подобных домена с внеклеточной областью, трансмембранный домен и короткий цитоплазматический домен. Цитоплазматический домен не содержит мотива для передачи сигнала, что указывает на отсутствие передачи сигнала от PD-L1 при взаимодействии этого лиганда с его рецептором. Его молекулярная масса - 40 кДа (290 аминокислот), его кодирует ген CD274 на хромосоме 19 мыши и хромосоме 9 человека, соответственно. PD-L1 является членом семейства белков B7 и характеризуется приблизительно 20% идентичностью аминокислотной последовательности по отношению к B7.1 и B7.2. PD-L1 человека характеризуется 70% и 93% идентичностью аминокислотной последовательности по отношению к ортологам PD-L1 мыши и яванского макака, соответственно.
PD-L1 связывается со своим рецептором PD-1 со сродством (KD), составляющим 770 нМ. PD-1 экспрессируется на активированных Т-клетках, B-клетках и миелоидных клетках и модулирует активацию или ингибирование клеточного иммунного ответа. Связывание PD-L1 с PD-1 обеспечивает передачу ингибиторного сигнала, снижение продукции цитокинов и пролиферации T-клеток. Кроме того, экспрессия PD-L1 клетками может опосредовать защиту от уничтожения цитотоксическими T-лимфоцитами (CTL) и представляет собой регуляторный механизм, ослабляющий хронические иммунные реакции во время вирусных инфекций. Рак как хроническое провоспалительное заболевание вызывает нарушение этого защитного пути иммунной системы за счет стимуляции экспрессии PD-L1, что приводит к избеганию иммунного ответа хозяина. В контексте активного иммунного ответа ИФНγ также стимулирует экспрессию PD-L1.
PD-L1 также опосредует иммуносуппрессию за счет взаимодействия с еще одним белком, B7.1 (также известным как CD80), блокирования его способности обеспечивать передачу одного из вторичных сигналов активации T-клеток посредством CD28. Значение этого специфического взаимодействия для устойчивости опухоли к иммунному ответу с точки зрения экспрессии PD-L1 на опухолевых клетках и его взаимодействия с B7.1 до настоящего времени неясно.
Экспрессия PD-L1 продемонстрирована в разнообразных плотных опухолях. 89 из 654 (14%) образцов, изученных в одном исследовании, охватывающем 19 опухолей из различных областей организма, являлись PD-L1-положительными (частота ≥5%). Максимальную частоту PD-L1-положительных клеток наблюдали при раке головы и шеи (17/54; 31%), шейки матки (10/34; 29%), неизвестного первичного происхождения (CUP; 8/29; 28%), мультиформной глиобластоме (GBM; 5/20; 25%), раке мочевого пузыря (8/37; 21%), пищевода (16/80; 20%), трижды негативном (TN) раке молочной железы (6/33; 18%) и гепатокарциноме (6/41; 15%) (Grosso et al, 2013). Показано, что опухоль-ассоциированная экспрессия PD-L1 придает опухоли устойчивость к иммунному ответу и, возможно, защищает опухолевые клетки от апоптоза, опосредованного T-клетками.
Терапевтические средства, адресно воздействующие на PD-L1, продемонстрировали превосходные результаты в исследованиях in vivo на мышах. На модели меланомы B16 у мышей лечение антагонистами PD-L1 в комбинации со стратегиями вакцинации GVAX или FVAX привело к значительному эффекту в отношении как выживаемости (30 дней для контрольных животных по сравнению с 52 днями при обработке PD-L1), так и процентной доле животных без опухолей (5%) по завершении исследования (Curran et al, 2010). Кроме того, терапевтическое средство на основе антагониста PD-L1 используется для исследования механизма иммуносуппрессии на модели мастомы P815 у мышей. Клетки P815, введенные в организм мыши, в норме инициируют мощный иммунный ответ, который приводит к их отторжению. При экспрессии PD-L1 на клетках P815 эти клетки избегают атаки иммунной системы, что, в свою очередь, нейтрализовать за счет введения антител против PD-L1 (Iwai et al, 2002). Очевидно, что адресное воздействие на ось PD-1/PD-L1 при иммуногенных злокачественных заболеваниях человека axis (Herbst et at, 2014) приводит к благоприятному действию на выживаемость за счет стимуляции противоракового иммунного ответа (Wolchock et al, 2013; Larkin et al, 2015).
Атезолизумаб (MPDL3280A, RG7466, TECENTRIQ) представляет собой гуманизированное антитело IgG1, связывающее PD-L1. Его применяют в клинических исследованиях в качестве единственного агента, а также в комбинации с другими биологическими и/или низкомолекулярными терапевтическими средствами для лечения плотных опухолей, в том числе рака ободочной и прямой кишки, рака молочной железы, немелкоклеточной карциномы легких, рака мочевого пузыря и почечноклеточной карциномы. Лечение с применением атезолизумаба приводит к частоте объективного ответа (ЧОР), составляющей 23% при НМРЛ, 36% при меланоме, 33% при раке мочевого пузыря, 14% при почечноклеточной карциноме и 13% при раке головы и шеи (Herbst et al, 2014; Powles et al, 2014).
Авелумаб (MSB0010718C) представляет собой полностью человеческое антитело IgG1, связывающееся с PD-L1, и проходит клинические исследования при ряде злокачественных заболеваний, в том числе раке мочевого пузыря, раке желудка, раке головы и шеи, мезотелиоме, немелкоклеточной карциноме легких, раке яичников, раке почек и карциноме из клеток Меркеля.
Дурвалумаб (MEDI4736) представляет собой антитело IgG1 человека, связывающееся с PD-L1 и тестируемое в клинических исследованиях отдельно или в комбинации с тремелимумабом при немелкоклеточном раке легких, плоскоклеточной карциноме головы и шеи, раке мочевого пузыря, раке поджелудочной железы и с другими биологическими и/или низкомолекулярными средствами в исследованиях дополнительных плотных злокачественных опухолей, например, раке желудка, меланоме и неоперабельной гепатоцеллюлярной карциноме.
В клинических исследованиях тестируются дополнительные антитела против PD-L1, в том числе BMS-936559 и другие антитела, проходящие доклинические исследования.
Вместе с тем, в настоящее время лишь небольшое количество терапевтических средств против LAG-3 проходят клинические испытания, и ни одно из них не утверждено для применения в терапевтических целях; таким образом, остается потребность в разработке дополнительных молекул, мишенью которых является LAG-3. Хотя некоторые терапевтические средства против PD-L1 разрабатываются в настоящее время, текущие данные показывают, что монотерапия антагонистами PD-L1 приводит к ответу у менее чем 50% раковых пациентов. Таким образом, в данной области техники остается потребность в дополнительных молекулах, мишенью которых может являться LAG-3 и/или PD-L1 и которые находят применение в лечении рака.
Сущность изобретения
Антитела против PD-1 и против PD-L1 преимущественно участвуют в нарушении иммунной толерантности и активации противоопухолевого иммунного ответа. LAG-3 экспрессируется на Т-клетках после активации, а также конститутивно экспрессируется на истощенных T-клетках, способствуя поддержанию этих клеток в состоянии суппрессии. Кроме того, показано, что блокада LAG-3 в комбинации с другими известными иммуносуппрессивными молекулами (т.е. PD-1, PD-L1) обеспечивает синергический усиленный иммунный ответ на моделях опухолей у мышей (Woo et al, 2012). Авторы настоящего изобретения предположили, что терапевтические средства, одновременно адресно воздействующие на оба эти пути, будут непосредственно влиять на механизмы, стимулирующие и поддерживающие истощение Т-клеток. Кроме того, авторы настоящего изобретения ожидают, что адресное воздействие на LAG-3 может подавлять презентацию антигена за счет действия LAG-3-экспрессирующих регуляторных T-клеток на АПК; в опубликованном исследовании документально подтверждено подавление CD86 (Grosso et al, 2013). Ожидается, что блокирование этого взаимодействия будет поддерживать презентацию антигена, в то время как блокирование сигнального пути PD-L1 будет нарушать толерантность, что будет приводить к значительному противоопухолевому ответу при одновременном ингибировании обоих путей.
Количество опубликованных данных о комбинациях антител против LAG-3 и PD-L1 ограничено, хотя имеется некоторое количество результатов доклинических исследований на сингенных моделях опухолевой и вирусной нагрузки у мышей. На модели миеломы у мышей комбинацию блокирующих антител против PD-L1 и LAG-3 вводили после облучения всего организма в низкой дозе и улучшения выживаемости до величины более 80% (Jing et al, 2015). Признаков системного или органоспецифического аутоиммунитета не наблюдали. Мыши с нокаутом LAG-3 и PD-1 демонстрировали значительно повышенную выживаемость при нескольких трансплантируемых опухолях и их ликвидацию (Woo et al, 2012).
Авторы настоящего изобретения предположили, что биспецифические антитела, связывающиеся как с LAG-3, так и с PD-L1, будут обладать рядом преимуществ по сравнению с комбинацией моноклональных антител против указанных антигенов, в том числе:
1. Направленная терапия
Активированные T-клетки экспрессируют LAG-3 в лимфатических узлах. Одна часть биспецифического антитела против LAG-3/PD-L1 адресно воздействует на примированные LAG-3-положительные T-клетки в лимфатических узлах, которые затм переносятся в область опухоли, осуществляя транспортировку биспецифического антитела. Попав в микроокружение опухоли, Т-клетки, несущие биспецифическое антитело, могут немедленно взаимодействовать с PD-L1 на опухолевых клетках посредством своей части против PD-L1 и блокировать его. Соответственно, все Т-клетки, переместившиеся в область опухоли, будут устойчивы к сигнальным путям LAG-3 и PD-L1/PD-1.
2. Образование мостиков
Примированные CD8-положительные T-клетки контактируют с опухолевыми антигенами в микроокружении опухоли, где они генерируют ответ, уничтожая опухолевые клетки в отсутствие суппрессивных сигналов. Ожидается, что биспецифические антитела будут превосходить комбинации отдельных терапевтических моноклональных антител с точки зрения поддержания и продления этого контакта между Т-клетками и опухолевыми клетками. Сила сигнала при активации Т-клеток имеет важное значение и может быть ключевым фактором в случае презентируемого антигена при злокачественных заболеваний (Engels et al, 2013); ожидается, что присутствие биспецифического антитела против LAG-3/PD-L1, связанного с мишенями на АПК или раковых клетках, будет увеличивать время, в течение которого Т-клетки успешно могут распознавать антиген и активироваться.
3. Локализация
В областях воспаления и текущего иммунного ответа экспрессия PD-L1 значительно усиливается из-за локализованного высвобождения ИФН-γ. Это верно для раковых клеток-мишеней, опухоль-ассоциированных макрофагов (ОАМ) или повторной стимуляции популяций Т-клеток. Ожидается, что биспецифическое антитело, обладающее антагонистическим действием по отношению к PD-L1 и LAG-3, будет локализваться и концентрироваться в областях с максимальной экспрессией PD-L1 в опухолях, что позволит их части против LAG-3 связываться и предотвращать LAG-3-опосредованную суппрессию T-клеток.
После интенсивного скрининга и программы созревания аффинности авторы настоящего изобретения смогли выявить десять партнеров по специфическому связыванию, содержащих сайт связывания, специфичный по отношению к LAG-3, в CH3-домене молекулы. Показано, что эти молекулы обладали высоким сродством к LAG-3 как человека, так и яванского макака. Ожидается, что высокое сродство к LAG-3 человека будет полезным в лечении, например, раковых опухолей, содержащих опухоль-инфильтрирующие лимфоциты (ОИЛ), экспрессирующие LAG-3, у пациентов- людей, в то время как высокое сродство к LAG-3 яванского макака, сопоставимое со сродством к LAG-3 человека, будет полезным при оценке свойств партнеров по специфическому связыванию на моделях заболеваний на основе яванского макака. Основанием для этого является то, что полученные результаты с большей вероятностью позволяют прогнозировать действие партнера по специфическому связыванию в организме пациентов-людей, чем для молекул с более высокой изменчивостью сродства по отношению к LAG-3 человека и яванского макака при тестировании на моделях яванского макака.
Кроме того, показано, что партнеры по специфическому связыванию обладали высокой активностью при анализе активации T-клеток, что, как ожидалось, является прогностическим показателем улучшенной эффективности у пациентов-людей за счет усиленного ингибирования LAG-3.
Авторы настоящего изобретения дополнительно объединили указанных партнеров по специфическому связыванию, содержащих сайт связывания, специфичный по отношению к LAG-3, в CH3-домене Fab-домена антитела, содержащем сайт связывания антигена PD-L1 на основе CDR, с целью получения молекул биспецифического антитела, содержащих сайты связывания как LAG-3, так и PD-L1, которые, как ожидается, будут обладать вышеописанными преимуществами. Авторы настоящего изобретения также получили суррогатные варианты этих молекул антител мышиного происхождения, связывающиеся с LAG-3 мыши и PD-L1 мыши; показано, что они могли в значительной степени ингибировать рост опухоли у сингенных моделей рака у мыши. В частности, применение указанных суррогатных молекул мышиного происхождения позволило продемонстрировать синергический эффект по отношению к суппрессии роста опухоли при введении молекулы антитела, содержащей сайты связывания как LAG-3, так и PD-L1, мышам в протестированных моделях. На основании аналогичного механизма действия LAG-3 и PD-L1 мыши и человека в окружении опухоли ожидается, что исследования на мышах, в которых продемонстрирована эффективность снижения опухолевой нагрузки, можно преобразовать в клинический терапевтический благоприятный эффект для пациентов-людей с раком. Кроме того, на основании этих результатов ожидается, что молекулы антител согласно настоящему изобретению будут демонстрировать лучший эффект в лечении рака у пациентов-людей, в частности, по отношению к подавлению роста опухоли, чем, например, введение двух отдельных молекул, связывающихся с LAG-3 и PD-L1, соответственно.
Таким образом, в первом аспекте настоящего изобретения предложена молекула антитела, связывающаяся как с PD-L1, так и с LAG-3. Конкретнее, эти антитела содержат:
(i) антиген-связывающий сайт на основе CDR для PD-L1; и
(ii) сайт связывания антигена LAG-3, располагающийся или сконструированный в двух или более структурных петлях CH3-домена молекулы антитела. Сайт связывания LAG-3 предпочтительно содержит аминокислотные последовательности WDEPWGED (SEQ ID NO: 1) и PYDRWVWPDE. (SEQ ID NO: 3).
Молекула антитела предпочтительно содержит аминокислотную последовательность, приведенную в SEQ ID NO: 1, в петле AB, и аминокислотную последовательность, приведенную в SEQ ID NO: 3, в петле EF CH3-домена.
Таким образом, в первом аспекте настоящего изобретения предложена молекула антитела, связывающаяся с лигандом-1 белка запрограммированной гибели клеток (PD-L1) и геном активации лимфоцитов 3 (LAG-3). Молекула антитела предпочтительно содержит (i) антиген-связывающий сайт на основе CDR для PD-L1; и (ii) сайт связывания антигена LAG-3, располагающийся в CH3-домене молекулы антитела.
Сайт связывания LAG-3 предпочтительно содержит аминокислотные последовательности WDEPWGED (SEQ ID NO: 1) и PYDRWVWPDE (SEQ ID NO: 3). Аминокислотная последовательность WDEPWGED предпочтительно располагается в первой структурной петле CH3-домена, а аминокислотная последовательность PYDRWVWPDE предпочтительно располагается во второй структурной петле CH3-домена.
Например, сайт связывания антигена LAG-3 может располагаться в области структурной петли CH3-домена молекулы антитела, причем указанная область структурной петли предпочтительно содержит две или более структурные петли, и сайт связывания LAG-3 предпочтительно содержит аминокислотные последовательности WDEPWGED (SEQ ID NO: 1) и PYDRWVWPDE (SEQ ID NO: 3).
В качестве дополнительного примера, сайт связывания антигена LAG-3 можно сконструировать в двух или более структурных петлях CH3-домена молекулы антитела, причем сайт связывания LAG-3 предпочтительно содержит аминокислотные последовательности WDEPWGED (SEQ ID NO: 1) и PYDRWVWPDE (SEQ ID NO: 3).
Как указано выше, последовательности сайта связывания LAG-3 предпочтительно располагаются в двух или более структурных петлях CH3-домена молекулы антитела. В предпочтительном варианте реализации сайт связывания антигена LAG-3 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 1, в петле AB, и аминокислотную последовательность, приведенную в SEQ ID NO: 3, в петле EF CH3-домена.
Аминокислотная последовательность, приведенная в SEQ ID NO: 1, предпочтительно располагается в области остатков 11-18 CH3-домена; и/или аминокислотная последовательность, приведенная в SEQ ID NO: 3, предпочтительно располагается в области остатков 92-101 CH3-домена; причем нумерация аминокислотных остатков соответствует схеме нумерации ImMunoGeneTics IMGT).
Сайт связывания антигена LAG-3 молекулы антитела может дополнительно содержать одну из следующих последовательностей, предпочтительно в петле CD CH3-домена молекулы антитела:
(i) SNGQPENNY (SEQ ID NOS 2, 8 и 18);
(ii) SNGQPEDNY (SEQ ID NO: 13);
(iii) SNGYPEIEF (SEQ ID NO: 23);
(iv) SNGIPEWNY (SEQ ID NO: 28);
(v) SNGYAEYNY (SEQ ID NO: 33);
(vi) SNGYKEENY (SEQ ID NO: 38);
(vii) SNGVPELNV (SEQ ID NO: 43); или
(viii) SNGYQEDNY (SEQ ID NO: 48).
Сайт связывания антигена LAG-3 молекулы антитела дополнительно предпочтительно содержит одну из следующих последовательностей, предпочтительно в петле CD CH3-домена молекулы антитела: аминокислотную последовательность, приведенную в SEQ ID NO: 2, 28 или 38, в петле CD CH3-домена. Более предпочтительно, сайт связывания антигена LAG-3 молекулы антитела содержит аминокислотную последовательность, приведенную в SEQ ID NO: 2, в петле CD CH3-домена
Аминокислотная последовательность, приведенная в SEQ ID NO: 2, 8, 13, 18, 23, 28, 33, 38, 43 или 48, предпочтительно располагается в области остатков 43-78 CH3-домена молекулы антитела, причем нумерация остатков соответствует схеме нумерации IMGT.
Последовательность CH3-домена молекулы антитела, за исключением последовательностей сайта связывания антигена LAG-3, не ограничена конкретным образом. CH3-домен предпочтительно представляет собой домен иммуноглобулина G человека, например, CH3-домен IgG1, IgG2, IgG3 или IgG4 человека, наиболее предпочтительно - CH3-домен IgG1 человека. Последовательности CH3-доменов IgG1, IgG2, IgG3, or IgG4 человека известны в данной области техники.
В предпочтительном варианте реализации молекула антитела содержит CH3-домен, приведенный в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50, более предпочтительно - CH3-домен, приведенный в SEQ ID NO: 5, 30 или 40, наиболее предпочтительно - CH3-домен, приведенный в SEQ ID NO: 5. В качестве альтернативы, молекула антитела может содержать CH3-домен с аминокислотной последовательностью, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50, предпочтительно SEQ ID NO: 5, 30 или 40, более предпочтительно - SEQ ID NO: 5.
Молекула антитела может дополнительно содержать CH2-домен. CH2-домен предпочтительно располагается на N-конце CH3-домена, как в случае молекулы IgG человека. CH2-домен молекулы антитела предпочтительно представляет собой CH2- домен IgG1, IgG2, IgG3 или IgG4 человека, более предпочтительно - CH2-домен IgG1 человека. Последовательности доменов IgG человека известны в данной области техники. В предпочтительном варианте реализации молекула антитела содержит CH2- домен IgG с последовательностью, приведенной в SEQ ID NO: 53 или SEQ ID NO: 54, или CH2-домен с аминокислотной последовательностью, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к SEQ ID NO: 53 или SEQ ID NO: 54.
В предпочтительном варианте реализации молекула антитела содержит последовательность, приведенную в SEQ ID NO: 6, 7, 11, 12, 16, 17, 21, 22, 26, 27, 31, 32, 36, 37, 41, 42, 46, 47, 51 или 52, или последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к последовательности, приведенной в SEQ ID NO: 6, 7, 11, 12, 16, 17, 21, 22, 26, 27, 31, 32, 36, 37, 41, 42, 46, 47, 51 или 52. Более предпочтительно, молекула антитела содержит последовательность, приведенную в SEQ ID NO: 6, 7, 31, 32, 41 или 42, или последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к последовательности, приведенной в SEQ ID NO: 6, 7, 31, 32, 41 или 42. Еще более предпочтительно, молекула антитела содержит последовательность, приведенную в SEQ ID NO: 6 или 7, или последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к последовательности, приведенной в SEQ ID NO: 6 или 7.
В предпочтительном варианте реализации молекула антитела представляет собой молекулу иммуноглобулина G человека, например, молекулу IgG1, IgG2, IgG3 или IgG4 человека, более предпочтительно - молекулу IgG1 человека. Последовательности молекул иммуноглобулина G человека известны в данной области техники, и внедрение CH3-домена или последовательности CH3-домена, описанных в настоящем документе, в такую молекулу не представит сложности для специалиста.
Молекула антитела предпочтительно содержит CDR VH- и/или VL-доменов, приведенные в SEQ ID NO: 92 и 93. Способы определения последовательности CDR в заданном VH- или VL-домене известны в данной области техники и включают системы нумерации Kabat и IMGT. Более предпочтительно, молекула антитела предпочтительно содержит одну или более, две или более, три или более, четыре или более, пять или более или все шесть областей, определяющих комплементарность, приведенных в SEQ ID NO 86-91. Молекула антитела предпочтительно содержит VH- и/или VL-домены, приведенные в SEQ ID NO: 92 и 93, соответственно.
В предпочтительном варианте реализации молекула антитела содержит последовательность тяжелой цепи, приведенную в любой из SEQ ID NO: 94-113, или тяжелую цепь с аминокислотной последовательностью, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к любой из SEQ ID NO: 94-113, при условии, что VH-домен последовательности тяжелой цепи остается неизменным. Более предпочтительно, молекула антитела содержит последовательность тяжелой цепи, приведенную в любой из SEQ ID NO: 94, 95, 104, 105, 108 и 109, или тяжелую цепь с аминокислотной последовательностью, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к любой из SEQ ID NO: 94, 95, 104, 105, 108 и 109, при условии, что VH-домен последовательности тяжелой цепи остается неизменным. Еще более предпочтительно, молекула антитела содержит последовательность тяжелой цепи, приведенную в SEQ ID NO: 94 или 95, или тяжелую цепь с аминокислотной последовательностью, обладающей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к SEQ ID NO: 94 или 95, при условии, что VH-домен последовательности тяжелой цепи остается неизменным.
В предпочтительном варианте реализации молекула антитела дополнительно или в качестве альтернативы содержит последовательность легкой цепи, приведенную в SEQ ID NO: 116, или легкую цепь с аминокислотной последовательностью, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичностью последовательности по отношению к SEQ ID NO: 116, при условии, что VL-домен последовательности легкой цепи остается неизменным.
Молекулы антитела предпочтительно может одновременно связываться с PD-L1 и LAG-3. PD-L1 и LAG-3 могут находиться, например, на двух различных клетках. Безотносительно к теоретическим представлениям считается, что результаты перекрестного связывания между клетками и интернализация PD-L1 и/или LAG-3 делают их недоступными для стимуляции.
Авторы настоящего изобретения показали, что молекула антитела, содержащая (i) антиген-связывающий сайт на основе CDR для PD-L1; и (ii) сайт связывания антигена LAG-3, располагающийся в CH3-домене молекулы антитела в соответствии с настоящим изобретением, FS18-7-9/84G09, оказалась способна опосредовать комплемент-зависимую цитотоксичность (КЗЦ) клеток, экспрессирующих PD-L1, но не клеток, экспрессирующих LAG-3, даже при наличии в образце смеси клеток, экспрессирующих LAG-3 и PD-L1. Ожидается, что это свойство может быть полезным там, где, как в случае с FS18-7-9/84G09, мишенью антиген-связывающего сайта на основе CDR является опухолевая клетка, а мишенью связывающего сайта, располагающегося в константном домене молекулы антитела, является иммунная клетка, так что иммунная клетка будет защищена от КЗЦ, опосредованной связыванием с молекулой антитела, в то время как опухолевая клетка будет подвергаться КЗЦ.
Таким образом, в дополнительном варианте реализации настоящее изобретение относится к молекуле антитела, связывающейся с опухолевым антигеном и антигеном иммунной клетки, причем молекула антитела содержит:
(i) антиген-связывающий сайт на основе CDR для опухолевого антигена; и
(ii) антиген-связывающий сайт для антигена иммунной клетки, располагающийся в константном домене,
предпочтительно CH3- или CH2-домене, более предпочтительно - в CH3-домене молекулы антитела,
причем молекула антитела не опосредует или не опосредует в значительной степени комплемент-зависимой цитотоксичности иммунной клетки, содержащей указанный антиген иммунной клетки, при связывании указанной иммунной клетки молекулой антитела.
Указанная молекула антитела предпочтительно опосредует комплемент-зависимую цитотоксичность опухолевой клетки, содержащей указанный опухолевый антиген, при связывании указанной опухолевой клетки молекулой антитела.
Способы измерения КЗЦ молекулы антитела известны в данной области техники и описаны в настоящем документе.
Авторы настоящего изобретения дополнительно показали, что молекула антитела, содержащая (i) антиген-связывающий сайт на основе CDR для PD-L1; и (ii) сайт связывания антигена LAG-3, располагающийся в CH3-домене молекулы антитела в соответствии с настоящим изобретением, FS18-7-9/84G09, оказалась способна опосредовать слабую антитело-зависимую клеточную цитотоксичность (АЗКЦ) клеток, экспрессирующих LAG-3, по сравнению с АЗКЦ клеток, экспрессирующих PD-L1. Аналогичным образом, ожидается, что это свойство может быть полезным там, где, как в случае с FS18-7-9/84G09, мишенью антиген-связывающего сайта на основе CDR является опухолевая клетка, а мишенью связывающего сайта, располагающегося в константном домене молекулы антитела, является иммунная клетка, так что иммунная клетка будет подвергаться более слабой АЗКЦ по сравнению с опухолевой клеткой, связываемой антителом.
Как описано в настоящем документе, в данной области техники известны мутации для снижения или устранения АЗКЦ-активности молекул антител. Одной из таких мутаций является мутация LALA, описанная в настоящем документе. Неожиданно обнаружилось, что FS18-7-9/84G09 обладает низкой АЗКЦ-активностью по отношению к клеткам, экспрессирующим LAG-3. Если полная нейтрализация АЗКЦ-активности не требуется, это может являться преимуществом.
Таким образом, в дополнительном варианте реализации настоящее изобретение относится к молекуле антитела, связывающейся с опухолевым антигеном и антигеном иммунной клетки, причем молекула антитела содержит:
(i) антиген-связывающий сайт на основе CDR для опухолевого антигена; и
(ii) антиген-связывающий сайт для антигена иммунной клетки, располагающийся в константном домене,
предпочтительно CH3- или CH2-домене, более предпочтительно - в CH3-домене молекулы антитела,
причем молекула антитела вызывает ослабленную АЗКЦ по отношению к иммунным клеткам, содержащим указанный антиген иммунной клетки, при связывании указанных иммунных клеток молекулой антитела, нежели по отношению к опухолевым клеткам, содержащим указанный опухолевый антиген, при связывании указанных опухолевых клеток молекулой антитела. Молекула антитела предпочтительно не опосредует или не опосредует в значительной степени АЗКЦ иммунной клетки, содержащей указанный антиген иммунной клетки, при связывании указанной иммунной клетки молекулой антитела. Молекула антитела также может не опосредовать или не опосредовать в значительной степени комплемент-зависимую цитотоксичность иммунной клетки, содержащей указанный антиген иммунной клетки, при связывании указанной иммунной клетки молекулой антитела, и/или может опосредовать комплемент-зависимую цитотоксичность опухолевой клетки, содержащей указанный опухолевый антиген, при связывании указанной опухолевой клети молекулой антитела.
Способы измерения АЗКЦ молекулы антитела известны в данной области техники и описаны в настоящем документе.
Различные опухолевые антигены и антигены иммунных клеток известны в данной области техники. Опухолевый антиген и антиген иммунной клетки предпочтительно являются антигенами поверхности клетки. Антиген иммунной клетки предпочтительно представляет собой антиген, присутствующий на опухоль-инфильтрирующих лимфоцитах.
Антиген-связывающий сайт для антигена иммунной клетки предпочтительно содержит одну или более из модификаций в одной или более из структурных петель константного домена молекулы антитела, например, в петле AB, CD или EF константного домена. Например, указанный сайт связывания может представлять сайт связывания LAG-3, описанный в настоящем документе.
Молекулу антитела согласно настоящему изобретению можно конъюгировать с модулятором иммунной системы, цитотоксической молекулой, радиоактивным изотопом или детектируемой меткой. Модулятор иммунной системы может представлять собой цитотоксическую молекулу, которая является цитокином.
В настоящем изобретении предложена нуклеиновая кислота, кодирующая молекулу антитела согласно настоящему изобретению, а также вектор, содержащий такую нуклеиновую кислоту.
Кроме того, предложена рекомбинантная клетка-хозяин, содержащая нуклеиновую кислоту или вектор согласно настоящему изобретению. Такую рекомбинантную клетку- хозяина можно применять для получения молекулы антитела. Таким образом, также предложен способ получения молекулы антитела согласно настоящему изобретению, причем указанный способ включает культивирование рекомбинантной клетки-хозяина в условиях для продуцирования молекулы антитела. Указанный способ может дополнительно включать этап выделения и/или очистки молекулы антитела.
Ожидается, что молекулы антител согласно настоящему изобретению найдут применение в терапии, в частности, терапии у людей, например, лечении рака. Таким образом, в настоящем изобретении также предложена фармацевтическая композиция, содержащая молекулу антитела согласно настоящему изобретению и фармацевтически приемлемое вспомогательное вещество.
В настоящем изобретении также предложена молекула антитела согласно настоящему изобретению для применения в способе лечения рака у пациента. Кроме того, предложен способ лечения рака у пациента, причем указанный способ включает введение пациенту терапевтически эффективного количества молекулы антитела согласно настоящему изобретению. Кроме того, предложено применение молекулы антитела согласно настоящему изобретению для применения в изготовлении лекарственного средства для лечения рака у пациента. Лечение может дополнительно включать введение пациенту противоопухолевой вакцины и/или химиотерапевтического агента.
Краткое описание чертежей
На фигуре 1: A показано выравнивание последовательностей девяти Fcab, выявленных после второго созревания аффинности, FS18-7-32; FS18-7-33; FS18-7-36; FS18-7-58; FS18-7-62; FS18-7-65; FS18-7-78; FS18-7-88; и FS18-7-95, против исходного Fcab, FS18-7-9. На фигуре B показана процентная идентичность последовательности каждого из этих Fcab по отношению к последовательности исходного Fcab, FS18-7-9.
На фигуре 2 показаны результаты анализа активации Т-клеток. Конкретно, на фигуре 2 показаны репрезентативные графики высвобождения ИЛ-2, которое является показателем активации T-клеток, для панели линий клеток, обработанных мАт2 FS18-7-9/84G09 (фигура 2A-C) или FS18-7-62/84G09 или FS18-7-78/84G09 (фигура 2D-F) и контрольными антителами. Фигура 2A: При анализе с LAG-3 и PD-L1 для активации требовалось ингибирование обеих мишеней (FS18-7-9/84G09LALA, FS18-7-9/4420LALA+84G09, 25F7+84G09LALA или 25F7+S1LALA). Фигура 2B: При анализе с LAG-3 для активации требовалось ингибирование только LAG-3 (FS18-7-9/84G09LALA, FS18-7-9/4420LALA+84G09, 25F7+84G09LALA, 25F7+S1LALA, 25F7, FS18-7-9/4420 LALA). Фигура 2С: При анализе с PD-L1 для активации требовалось ингибирование PD-L1 (FS18-7-9/84G09LALA, FS18-7-9/4420LALA+84G09, 25F7+84G09LALA или 25F7+S1LALA). Фигура 2D: При анализе с LAG-3 и PD-L1 для активации требовалось ингибирование обеих мишеней (FS18-7-62/84G09LALA, FS18-7-78/84G09LALA, FS18-7-62/4420LALA+84G09, FS18-7-78/4420LALA+84G09, 25F7+84G09LALA или 25F7+S1LALA). Фигура Е: При анализе только с LAG-3 для активации требовалось ингибирование LAG-3 (FS18-7-62/84G09LALA, FS18-7-62/4420LALA, FS18-7-78/84G09LALA, FS18-7-78/4420LALA, FS18-7-62/4420LALA+84G09, FS18-7-78/4420LALA+84G09, 25F7+84G09LALA или 25F7+S1LALA). Фигура 2F: При анализе только с PD-L1 для активации требовалось ингибирование PD-L1 (FS18-7-62/84G09LALA, FS18-7-78/84G09LALA, FS18-7-62/4420LALA+84G09, FS18-7-78/4420LALA+84G09, 25F7+84G09LALA или 25F7+S1LALA).
На фигуре 3 показано, что FS18-7-9/84G09 могло индуцировать активацию Т-клеток, определяемую по высвобождению ИЛ-2, в присутствии как cLAG-3 + cPD-L1, так и cLAG-3 или cPD-L1 по отдельности, что указывает на функциональную перекрестную реакционную способность с антигенами яванского макака.
На Фигуре 4 показан репрезентативный график анализа SEB. мАт2 LAG-3/PD-L1 и комбинация мАт2 LAG-3/4420 + 84G09LALA демонстрировали более выраженную активацию, чем только мАт 84G09LALA, в то время как мАт2 LAG-3/4420 или мАт 4420 не демонстрировали значимой активации.
На фигуре 5 показана масса конечных опухолей на 20 день на модели неразвившейся сингенной опухоли MC38. Мыши, обработанные мАт2 LAG-3/PD-L1 (FS18-29/S1), характеризовались конечными опухолями значительно меньшей массы по сравнению с мышами, обработанными комбинацией эталонных мАт C9B7W и S1.
На фигуре 6 показаны кривые роста в модели неразвившейся сингенной опухоли MC38. Мыши, обработанные мАт2 LAG-3/PD-L1 (FS18-29/S1), характеризовались опухолями меньшего размера по сравнению с мышами, обработанными комбинацией эталонных мАт C9B7W и S1 или только S1. мАт2 LAG-3/4420 и эталонное мАт против LAG-3 оказывали слабое влияние на рост опухоли.
На фигуре 7 показана масса конечных опухолей на 24 день на модели развившейся сингенной опухоли MC38. мАт2 против LAG-3/PD-L1 (FS18-29/S1) так же эффективно подавляло рост опухоли, как и комбинация эталонных антител C9B7W и S1. FS18-29/4420 само по себе не оказывало заметного влияния на рост опухоли, а S1 и C9B7W слегка влияли на итоговый рост опухоли.
На фигуре 8 показаны кривые роста опухоли в модели развившейся сингенной опухоли MC38. Объем опухолей у мышей, обработанных мАт2 LAG-3/PD-L1 (FS18-29/S1), был аналогичен объему опухолей у мышей, обработанных комбинацией эталонных мАт C9B7W и S1. Мыши, обработанные S1 или C9B7W по отдельности, демонстрировали опухоли промежуточного объема, в то время как обработка мАт2 LAG-3/4420 не оказывала влияния на рост опухоли.
На фигуре 9 показана масса конечных опухолей на 20 день на модели неразвившейся сингенной опухоли CT26. мАт2 LAG-3/PD-L1 (FS18-29/S1) подавляло рост опухоли в большей степени, чем комбинация эталонных антител C9B7W и S1.
На фигуре 10 показаны кривые роста опухоли в модели неразвившейся сингенной опухоли CT26. Продемонстрировано статистически значимое различие FS18-35/S1 и контрольного IgG при подавлении роста опухоли. Такого статистически значимого различия не наблюдали при использовании комбинации эталонных антител по сравнению с группой контрольного IgG.
На фигуре 11 показана масса конечных опухолей на 22 день на модели неразвившейся сингенной опухоли MC38, использовавшейся для сравнения влияния мутации LALA в мАт2 на ингибирование роста опухоли. Статистически значимые различия массы конечных опухолей у мышей, обработанных мАт2 с мутацией LALA и без нее, отсутствовали.
На фигуре 12 показаны кривые роста опухолей на модели неразвившейся сингенной опухоли MC38, использовавшейся для сравнения влияния мАт2 с мутацией LALA и без нее на ингибирование роста опухоли. Статистически значимые различия кривых роста опухоли у мышей, обработанных мАт2 с мутацией LALA и без нее, отсутствовали, однако имела место тенденция к увеличению ингибирования роста опухоли молекулами, содержавшими мутацию LALA.
На фигуре 13 показано действие обработки мАт2 на экспрессию LAG-3 Т-клетками. Показана экспрессия LAG-3 на CD8 (A), CD4 (B) и FoxP3 (C) опухоль-инфильтрирующих лимфоцитах (ОИЛ), обработанных мАт2 FS18-29/S1, FS18-29/4420, S1, FS18-29/4420 и S1, или контрольным антителом 4420, на 19 и 23 день после инокуляции опухоли, что соответствовало 3 и 7 дню после последнего введения мАт2/антитела, соответственно. Экспрессия LAG-3 снижалась после обработки мАт2 FS18-29/S1 на 19 и 23 день. Животные, получавшие комбинацию FS18-29/4420 и S1, также демонстрировали снижение экспрессии LAG-3, однако этот эффект был отсрочен до 23 дня, в то время как FS18-29/4420 и S1 при введении по отдельности приводили к незначительному снижению или отсутствию снижения экспрессии LAG-3.
На фигуре 14 показан процент лизиса клеток Raji, экспрессирующих PD-L1 и LAG-3, после обработки различными антителами/мАт2, при использовании анализа КЗЦ по высвобождению лактатдегидрогеназы (ЛДГ). На фигурах A и B показан КЗЦ-опосредованный лизис клеток Raji, экспрессирующих PD-L1 и LAG-3, соответственно. Клетки инкубировали с антителом 25F7 против LAG-3, антителом 84G09 против PD-L1, антителом 84G09 против PD-L1, содержащим мутацию LALA, антителом ритуксимаб против CD20, мАт2 FS18-7-9/84G09, мАт2 FS18-7-9/84G09, содержащим мутацию LALA, или ритуксимабом, содержащим мутацию LALA. Высвобождение ЛДГ измеряли через 4 ч после обработки комплементом детеныша кролика и выражали в виде процентной доли от полного лизиса. Концентрация антитела/мАт2 при обработке указана на оси X. Для возможности сравнения кривых для угла наклона всех кривых было задано общее значение.
На фигуре 15 показан процент мертвых клеток Raji, экспрессирующих PD-L1 и LAG-3, после различной обработки при использовании анализа КЗЦ на основе проточной цитометрии. Смесь клеток, экспрессирующих PD-L1 и LAG-3, меченых различными флуоресцентными метками, инкубировали с контрольным антителом 4420, антителом ритуксимаб (RIT) против CD20, комбинацией антитела 25F7 против LAG-3 и антитела 84G09 против PD-L1 или мАт2 FS18-7-9/84G09. Затем клетки обрабатывали комплементом детеныша кролика и окрашивали красителем, окрашивающим только мертвые клетки. Процентную долю мертвых клеток в двух популяциях клеток оценивали в виде процентной доли от общего количества клеток. Тип клеток (экспрессирующие PD-L1 или экспрессирующие LAG-3, что выявляли по различным флуоресцентным меткам), оцениваемый после каждой обработки, указан на фигуре 15 после названия соответствующей обработки, см., например, 4420 PDL1, что относится к оценке жизнеспособности клеток Raji, экспрессирующих PD-L1, после обработки контрольным антителом 4420. Концентрация антитела/мАт2 при обработке указана на оси X
На фигуре 16 показан процент лизиса (цитотоксичность) клеток Raji, экспрессирующих PD-L1 и LAG-3, после различной обработки при использовании анализа АЗКЦ по высвобождению лактатдегидрогеназы (ЛДГ). Клетки обрабатывали антителом ритуксимаб против CD20, антителом 84G09 против PD-L1, мАт2 FS18-7-9/84G09, ритуксимабом, содержащим мутацию LALA, мАт2 FS18-7-9/84G09, содержащим мутацию LALA, антителом 25F7 против LAG-3, антителом 84G09 против PD-L1, содержащим мутацию LALA, контрольным антителом 4420 или контрольным антителом 4420, содержащим мутацию LALA. Затем обработанные клетки совместно инкубировали с первичными NK-клетками и измеряли удельное высвобождение ЛДГ в виде процентной доли от полного лизиса клеток-мишеней. Концентрация антитела/мАт2 при обработке указана на оси X.
Подробное описание изобретения
Настоящее изобретение относится к молекулам антител, связывающимся как с PD-L1, так и с LAG-3. Конкретнее, молекулы антител согласно настоящему изобретению содержат сайт связывания антигена PD-L1 на основе CDR и сайт связывания антигена LAG-3, расположенный в константном домене молекулы антитела. Термины «PD-L1» и «LAG-3» могут относиться к PD-L1 и LAG-3 человека, PD-L1 и LAG-3 мыши и/или PD-L1 и LAG-3 яванского макака, если контекст не требует иного. Термины «PD-L1» и «LAG-3» предпочтительно относятся к PD-L1 и LAG-3 человека, если контекст не требует иного.
Термин "молекула антитела" описывает естественный либо полностью или частично синтезированный иммуноглобулин. Молекула антитела может представлять собой антитело человека или гуманизированное антитело.
Молекула антитела предпочтительно представляет собой молекулу моноклонального антитела. Примеры антител представляют собой изотипы иммуноглобулинов, например, иммуноглобулин G, и их изотипические подклассы, например, IgG1, IgG2, IgG3 и IgG4, а также их фрагменты.
Таким образом, в настоящем документе термин “молекула антитела” включает фрагменты антител, при условии, что такие фрагменты содержат сайт связывания антигена PD-L1 на основе CDR и сайт связывания антигена LAG-3, расположенный в константном домене, например, CH1-, CH2- или CH3-домене, предпочтительно CH3-домене молекулы антитела. Если контекст не требует иного, термин «молекула антитела» в настоящем документе эквивалентен термину «молекула антитела или ее фрагмент».
Можно взять моноклональные и другие антитела и использовать методики технологии рекомбинантных ДНК для получения других антител или химерных молекул, сохраняющих специфичность исходного антитела. Такие методики могут включать внедрение последовательностей CDR или вариабельных областей и/или константного домена, содержащего сайт связывания антигена LAG-3, в другой иммуноглобулин. Внедрение CDR одного иммуноглобулина в другой иммуноглобулин описано, например, в EP-A-184187, GB 2188638A или EP-A-239400. Аналогичные методики можно применить для последовательностей соответствующих константных доменов. В качестве альтернативы, гибридому или другую клетку, продуцирующую молекулу антитела, можно подвергать генетической мутации или другим изменениям, которые могут изменять или не изменять специфичность связывания полученных антител.
Поскольку антитела можно модифицировать различными способами, следует считать, что термин «молекула антитела» охватывает фрагменты антител, производные, функциональные эквиваленты и гомологи антител, в том числе любой полипептид, содержащий связывающий домен иммуноглобулина, природный или полностью или частично синтетический. Таким образом, данный термин включает химерные молекулы, содержащие связывающий домен иммуноглобулина или его эквивалент, присоединенный к другому полипептиду. Клонирование и экспрессия. химерных антител описаны в EP-A-0120694 и EP-A-0125023.
Примером фрагмента антител, содержащих как последовательности CDR, так и CH3-домен, является миниантитело, содержащее scFv, объединенный с CH3-доменом (Hu et al. (1996), Cancer Res., 56(13):3055-61).
Молекула антитела согласно настоящему изобретению связывается с PD-L1 и LAG-3. В данном контексте связывание может относиться к специфическому связыванию Термин "специфический" может относиться к ситуации, при которой молекула антитела не демонстрирует значительного связывания с молекулами, отличающимися от ее партнера(ов) по специфическому связыванию, в данном случае PD-L1 и LAG-3. Термин “специфический” также применим, если молекула антитела является специфичной по отношению к конкретным эпитопам, например, эпитопам на PD-L1 и LAG-3, которые несут ряд антигенов; в таком случае молекула антитела будет способна связываться с различными антигенами, несущими данный эпитоп.
LAG-3 обладает 40% идентичностью последовательности по отношению к CD4 - наиболее близкому к нему белку. Авторы настоящего изобретения протестировали Fcab FS18-7-9, который содержит аминокислотную последовательность, приведенную в SEQ ID NO: 1-3, на предмет связывания с CD4. Показано, что Fcab FS18-7-9 не связывался с CD4, что демонстрирует специфичность связывания данной молекулы с LAG-3. Таким образом, в предпочтительном варианте реализации сайт связывания LAG-3 молекулы антитела согласно настоящему изобретению не связывается или не демонстрирует значительного связывания с CD4.
Молекула антитела согласно настоящему изобретению предпочтительно содержит сайт связывания антигена LAG-3. Сайт связывания антигена LAG-3 располагается в константном домене молекулы антитела, например, CH1-, CH2-, CH3- или CH4-домене. Сайт связывания антигена LAG-3 предпочтительно располагается в CH3-домене молекулы антитела. Сайт связывания LAG-3 предпочтительно содержит аминокислотные последовательности WDEPWGED (SEQ ID NO: 1) и PYDRWVWPDE. (SEQ ID NO: 3). Эти последовательности присутствовали во всех основных клонах Fcab против LAG-3, выявленных авторами настоящего изобретения после интенсивной программы скрининга и исследования характеристик, описанной в разделе «Примеры».
Аминокислотные последовательности, приведенные в SEQ ID NO: 1 и 2, предпочтительно располагаются в структурных петлях константного домена молекулы антитела. Внедрение последовательностей в области структурных петель константных доменов антител с целью создания новых антиген-связывающих сайтов описано, например, в заявках WO 2006/072620 и WO 2009/132876.
Структурные петли константных доменов антител включают петли AB, CD и EF. Петли AB, CD и EF в CH3-домене располагаются в области остатков 11-18, 43-78 и 92-101 CH3-домена; причем нумерация аминокислотных остатков соответствует схеме нумерации ImMunoGeneTics (IMGT). Аминокислотная последовательность, приведенная в SEQ ID NO: 1, предпочтительно располагается в петле AB константного домена. Аминокислотная последовательность, приведенная в SEQ ID NO: 3, предпочтительно располагается в петле EF константного домена. Более предпочтительно, аминокислотная последовательность, приведенная в SEQ ID NO: 1, располагается в области остатков 11-18 CH3-домена; и/или аминокислотная последовательность, приведенная в SEQ ID NO: 3, предпочтительно располагается в области остатков 92-101 CH3-домена, причем нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Кроме того, молекула антитела предпочтительно содержит аминокислотную последовательность, приведенную в SEQ ID NO: 2, 8, 13, 18, 23, 28, 33, 38, 43 или 48, более предпочтительно - SEQ ID NO: 2, 28 или 38, еще более предпочтительно - SEQ ID NO: 2, в структурной петле константного домена молекулы антитела. Структурная петля предпочтительно представляет собой структурную петлю CD, а константный домен предпочтительно представляет собой CH3-домен. Аминокислотная последовательность, приведенная в SEQ ID NO: 2, 8, 13, 18, 23, 28, 33, 38, 43 или 48, предпочтительно располагается в области остатков 43-78 CH3-домена, причем нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Молекула антитела согласно настоящему изобретению дополнительно содержит остаток глутаминовой кислоты (E) в положении 36 и/или остаток тирозина (Y) в положении 85.2 CH3-домена (как показано на фигуре 1А),, причем нумерация аминокислотных остатков соответствует схеме нумерации IMGT. В частности, молекула антитела, содержащая область структурной петли CD, приведенную в SEQ ID NO: 8, предпочтительно содержит остаток глутаминовой кислоты (E) в положении 36 CH3-домена. Аналогичным образом, молекула антитела, содержащая область структурной петли, приведенную в SEQ ID NO: 18, предпочтительно содержит остаток тирозина (Y) в положении 85.2 CH3-домена.
В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит CH3-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50, предпочтительно CH3-домен с последовательностью, приведенной в SEQ ID NO: 5, 30 или 40, более предпочтительно- CH3 с последовательностью, приведенной в SEQ ID NO: 5.
Молекула антитела согласно настоящему изобретению может содержать CH3-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50, причем последовательность CH3-домена дополнительно содержит остаток лизина (K) на самом C-конце последовательности, приведенной в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50. Так, например, молекула антитела согласно настоящему изобретению может содержать CH3-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 5 с остатком лизина на самом C-конце последовательности, приведенной в SEQ ID NO: 5. Последовательность такого CH3-ломена будет представлять собой:
GQPREPQVYTLPPSWDEPWGEDVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDS DGSFFLYSKLTVPYDRWVWPDEFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 135)
Кроме того, молекула антитела согласно настоящему изобретению может содержать CH2-домен молекулы иммуноглобулина G, например, CH2-домен молекулы IgG1, IgG2, IgG3 или IgG4. Молекула антитела согласно настоящему изобретению предпочтительно содержит CH2-домен молекулы IgG1. CH2-домен может содержать последовательность, приведенную в SEQ ID NO: 53.
CH2-домен молекулы антитела может содержать мутацию, ослабляющую или устраняющую связывание CH2-домена с одним или более из Fcγ-рецепторов, например, FcγRI, FcγRIIa, FcγRIIb, FcγRIII, и/или с комплементом. CH2-домены IgG-доменов человека обычно связываются с Fcγ-рецепторами или комплементом, и авторы настоящего изобретения допускают, что пониженное связывание с Fcγ-рецепторами будет ослаблять антитело-зависимую клеточно-опосредованную цитотоксичность (АЗКЦ), а пониженное связывание с комплементом будет ослаблять комплемент-зависимую цитотоксичность (КЗЦ) молекулы антитела. Мутации для ослабления или устранения связывания CH2-домена с одним или более из Fcγ-рецепторов и комплементом известны и включают “LALA-мутацию”, описанную в работах Bruhns, et al. (2009) и Xu et al. (2000). Так, молекула антитела может содержать CH2-домен, причем указанный CH2-домен содержит остатки аланина в 4 и 5 положениях CH2-домена, причем нумерация соответствует схеме нумерации IMGT. Например, молекула антитела содержит CH2-домен IgG1, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 54.
Молекула антитела согласно настоящему изобретению содержит антиген- связывающий сайт на основе CDR для PD-L1. Термин “антиген-связывающий сайт на основе CDR” относится к антиген-связывающему сайту вариабельной области молекулы антитела, который состоит из шести CDR. Получение молекул антител против PD-L1 и определение последовательностей CDR таких молекул антител входят в возможности специалистов, и в данной области техники известно большое количество подходящих методик.
Молекула антитела согласно настоящему изобретению предпочтительно содержит HCDR3 антитела 84G09. Известно, что HCDR3 играет роль в определении специфичности молекулы антитела (Segal et al., (1974), PNAS, 71:4298-4302; Amit et al., (1986), Science, 233:747-753; Chothia et al., (1987), J. Mol. Biol., 196:901-917; Chothia et al., (1989), Nature, 342:877-883; Caton et al., (1990), J. Immunol., 144:1965-1968; Sharon et al., (1990a), PNAS, 87:4814-4817; Sharon et al., (1990b), J. Immunol., 144:4863-4869; Kabat et al., (1991b), J. Immunol., 147:1709-1719).
Молекула антитела может дополнительно содержать HCDR1, HCDR2, LCDR1, LCDR2 и/или LCDR3 антитела 84G09. Для специалиста не составит труда определить последовательности CDR из последовательностей VH- и VL-доменов антитела 84G09, показанных в SEQ ID NO 92 и 93, соответственно. Последовательности CDR можно определить, например, согласно схеме нумерации Kabat (Kabat, E.A. et al., (1991)) или IMGT.
Последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 антитела 84G09 согласно схеме нумерации IMGT приведены в SEQ ID NO 86, 87, 88, 89, 90 и 91, соответственно.
Последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 антитела 84G09 согласно Kabat приведены в SEQ ID NO 136, 137, 138, 139, 140 и 141, соответственно.
Кроме того, антитело может содержать VH- и/или VL-домен антитела 84G09. Последовательности VH- и VL-доменов антитела 84G09 показаны в SEQ ID NO 92 и 93, соответственно.
В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит (i) сайт связывания антигена PD-L1 на основе CDR, содержащий последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 антитела 84G09, и (ii) сайт связывания антигена LAG-3, расположенный в CH3-домене молекулы антитела, причем сайт связывания LAG-3 содержит аминокислотные последовательности, приведенные в SEQ ID NO 1 и 3, и аминокислотную последовательность, выбранную из группы, состоящей из: SEQ ID NO 2, 8, 13, 18, 23, 28, 33, 38, 43 и 48.
Более предпочтительно, молекула антитела согласно настоящему изобретению содержит (i) сайт связывания антигена PD-L1 на основе CDR, содержащий последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 антитела 84G09, и (ii) сайт связывания антигена LAG-3, расположенный в CH3-домене молекулы антитела, причем сайт связывания LAG-3 содержит аминокислотные последовательности, приведенные в SEQ ID NO 1 и 3, и аминокислотную последовательность, выбранную из группы, состоящей из: SEQ ID NO 2, 28 и 38.
Еще более предпочтительно, молекула антитела согласно настоящему изобретению содержит (i) сайт связывания антигена PD-L1 на основе CDR, содержащий последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 антитела 84G09, и (ii) сайт связывания антигена LAG-3, расположенный в CH3-домене молекулы антитела, причем сайт связывания LAG-3 содержит аминокислотные последовательности, приведенные в SEQ ID NO 1, 2 и 3.
В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит VH-домен и VL-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO 93 и 93, соответственно, и CH3-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50, предпочтительно CH3-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 5, 30 или 40, более предпочтительно - CH3-домен, содержащий или состоящий из последовательности, приведенной в SEQ ID NO: 5.
В дополнительном предпочтительном варианте реализации молекула антитела содержит тяжелую цепь, содержащую или состоящую из последовательности, приведенной в SEQ ID NO: 93-113, и легкую цепь, содержащую или состоящую из последовательности, приведенной в SEQ ID NO: 116. Более предпочтительно, молекула антитела содержит тяжелую цепь, содержащую или состоящую из последовательности, приведенной в SEQ ID NOS: 94, 95, 104, 105, 108 и 109, и легкую цепь, содержащую или состоящую из последовательности, приведенной в SEQ ID NO: 116. Наиболее предпочтительно, молекула антитела содержит тяжелую цепь, содержащую или состоящую из последовательности, приведенной в SEQ ID NO: 94 или 95, и легкую цепь, содержащую или состоящую из последовательности, приведенной в SEQ ID NO: 116.
Молекулы антител согласно настоящему изобретению также могут содержать варианты последовательности структурной петли, CH3-домена, CH2-домена, CH2- и CH3-домена, легкой цепи или тяжелой цепи, описанные в настоящем документе, при условии, что последовательности VL- и VH-доменов легкой и тяжелой цепи, соответственно, остаются неизменными. Подходящие варианты можно получить за счет способов изменения последовательностей или мутаций и скрининга. В предпочтительном варианте реализации молекула антитела, содержащая одну или более из вариантных последовательностей, сохраняет одну или более из функциональных характеристик исходной молекулы антитела, например, специфичность связывания и/или сродство связывания с LAG-3 и PD-L1. Например, молекула антитела, содержащая одну или более из вариантных последовательностей, предпочтительно связывается с LAG-3 с тем же или более высоким сродством, чем (исходная) молекула антитела. Исходная молекула антитела представляет собой молекулу антитела, не содержащую аминокислотных(ой) замен(ы), делеций(и) и/или инсерций(и), внедренных в вариантную молекулу антитела.
Например, молекула антитела согласно настоящему изобретению может содержать последовательность структурной петли, CH3-домена, CH2-домена, CH2- и CH3-домена, легкой цепи или тяжелой цепи, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8%, or по меньшей мере 99,9% идентичностью последовательности по отношению к последовательности структурной петли, CH3-домена, CH2-домена, CH2- и CH3-домена, легкой цепи или тяжелой цепи, описанной в настоящем документе, при условии, что последовательности VL- и VH-доменов легкой и тяжелой цепи, соответственно, остаются неизменными.
В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит последовательность CH3-домена, обладающую по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичностью последовательности по отношению к последовательности CH3-домена, приведенной в SEQ ID NO: 4, 5 или 135.
В дополнительном предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит последовательность CH3- и CH2-домена, обладающую по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичностью последовательности по отношению к последовательности CH2- и CH3-домена, приведенной в SEQ ID NO: 6 или 7.
Идентичность последовательности обычно определяют со ссылкой на алгоритм GAP (Wisconsin GCG package, Accelerys Inc, Сан-Диего, США). GAP использует алгоритм Needleman and Wunsch для выравнивания двух полных последовательностей, позволяющий максимизировать количество совпадений и минимизировать количество пробелов. В общем случае используют параметры по умолчанию, причем штраф за создание пробела = 12, а штраф за удлинение пробела = 4. Применение GAP может быть предпочтительным, однако можно применять и другие алгоритмы, например, BLAST (который использует способ из Altschul et al. (1990) J. MoI. Biol. 215: 405-410), FASTA (который использует способ из Pearson and Lipman (1988) PNAS USA 85: 2444-2448) или алгоритм Smith-Waterman (Smith and Waterman (1981) J. MoI Biol. 147: 195-197) или программу TBLASTN из Altschul et al. (1990) supra, в общем случае используя параметры по умолчанию. В частности, можно применять алгоритм psi-Blast (Nucl. Acids Res. (1997) 25 3389-3402).
Молекула антитела согласно настоящему изобретению также может содержать последовательность структурной петли, CH3-домена, CH2-домена, CH2- и CH3-домена, легкой цепи или тяжелой цепи, содержащую одну или более из модификаций аминокислотной последовательности (добавление, делецию замену и/или инсерцию аминокислотного остатка), предпочтительно 20 модификаций или менее, 15 модификаций или менее, 10 модификаций или менее, 5 модификаций или менее, 4 модификации или менее, 3 модификации или менее, 2 модификации или менее или 1 модификацию по сравнению с последовательностью структурной петли, CH3-домена, CH2-домена, CH2- и CH3-домена, легкой цепи или тяжелой цепи, описанной в настоящем документе, при условии, что последовательности VL- и VH-доменов легкой и тяжелой цепи, соответственно, остаются неизменными.. В частности, можно вносить модификации в одну или более из каркасных областей молекулы антитела за пределами последовательностей VH- и VL-доменов.
В предпочтительном варианте реализации молекула антитела может содержать последовательность CH3-домена, содержащую одну или более из модификаций аминокислотной последовательности (добавление, делецию замену и/или инсерцию аминокислотного остатка), предпочтительно 20 модификаций или менее, 15 модификаций или менее, 10 модификаций или менее, 5 модификаций или менее, 4 модификации или менее, 3 модификации или менее, 2 модификации или менее или 1 модификацию по сравнению с последовательностью CH3-домена, приведенной в SEQ ID NO: 4, 5 или 135.
В дополнительном предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит последовательность CH3- и CH2- домена, содержащую одну или более из модификаций аминокислотной последовательности (добавление, делецию замену и/или инсерцию аминокислотного остатка), предпочтительно 20 модификаций или менее, 15 модификаций или менее, 10 модификаций или менее, 5 модификаций или менее, 4 модификации или менее, 3 модификации или менее, 2 модификации или менее или 1 модификацию по сравнению с последовательностью CH2- и CH3-домена, приведенной в SEQ ID NO: 6 или 7.
Кроме того, в настоящем изобретении рассматривается молекула антитела, конкурирующая за связывание с LAG-3 и/или PD-L1 с молекулой антитела согласно настоящему изобретению или связывающаяся с тем же эпитопом LAG-3 и/или PD-L1, что и молекула антитела согласно настоящему изобретению, причем указанная молекула антитела содержит как сайт связывания антигена PD-L1 на основе CDR, так и сайт связывания антигена LAG-3, расположенный в CH3-домене молекулы антитела. В данной области техники известны способы определения конкуренции двух антител за антиген. Например, конкуренцию двух антител за связывание с антигеном можно определить с использованием BIAcore. Аналогичным образом, в данной области техники известны способы картирования эпитопа, связываемого антителом.
Молекула антитела согласно настоящему изобретению предпочтительно связывается с LAG-3 со сродством (KD), равным 1 x 10-9 M, или более высоким сродством. Например, молекула антитела согласно настоящему изобретению может связываться с LAG-3 со сродством (KD), равным 8 x 10-10 M, или более высоким сродством.
Fcab характеризуются меньшей поверхностью взаимодействия при связывании, чем моноклональные антитела, поскольку сайты связывания Fcab образуют сравнительно компактный фрагмент антитела с двумя сайтами связывания, расположенными в относительной близости. В противоположность этому, Fab-плечи типичного мАт разделены гибкой шарнирной областью. Два антиген-связывающих сайта Fcab также пространственно близки друг к другу по сравнению с антиген-связывающими сайтами типичного мАт. С учетом меньшей поверхности взаимодействия при связывании и пониженной гибкости двух сайтов связывания, способность Fcab против LAG-3 связываться с LAG-3 и ингибировать его со сродством и эффективностью, аналогичными сродству и эффективности эталонного моноклонального антитела, оказалась неожиданной.
Молекула антитела согласно настоящему изобретению предпочтительно связывается с PD-L1 со сродством (KD), равным 1 × 10-9 M или более высоким сродством.
Сродство связывания молекулы антитела с ее специфическим антигеном, например, LAG-3 или PD-L1, можно определить с помощью, например, поверхностного плазмонного резонанса. Сродство связывания молекулы антитела с ее специфическим антигеном, например, LAG-3 или PD-L1, экспрессируемым на поверхности клетки, можно определить с помощью проточной цитометрии.
Молекула антитела согласно настоящему изобретению предпочтительно может связываться с LAG-3 и PD-L1, экспрессируемыми на поверхности клетки. Указанная клетка предпочтительно является раковой клеткой.
Молекула антитела согласно настоящему изобретению предпочтительно может одновременно связываться с LAG-3 и PD-L1. В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению может одновременно связываться с LAG-3 и PD-L1, причем LAG-3 и PD-L1 экспрессируются на поверхности одиночной клетки или на поверхности двух отдельных клеток.
Молекула антитела согласно настоящему изобретению может связываться с LAG-3 человека, LAG-3 мыши и/или LAG-3 яванского макака. Молекула антитела согласно настоящему изобретению предпочтительно связывается с LAG-3 человека. Более предпочтительно, молекула антитела согласно настоящему изобретению связывается с LAG-3 человека и PD-L1 человека.
Молекула антитела согласно настоящему изобретению содержит (i) сайт связывания антигена PD-L1 на основе CDR и (ii) сайт связывания антигена LAG-3, расположенный в константном домене молекулы антитела. Таким образом, молекулы антител, не содержащие сайта связывания антигена LAG-3, расположенного в константном домене, например, CH3-домене молекулы антитела, не относятся к настоящему изобретению. Аналогичным образом, молекула, не содержащая антиген- связывающего сайта PD-L1 на основе CDR, не относится к настоящему изобретению.
Молекулу антитела согласно настоящему изобретению можно конъюгировать с терапевтическим агентом или детектируемой меткой. В этом случае молекулу антитела можно называть конъюгатом. Например, молекулу антитела можно конъюгировать с модулятором иммунной системы, цитотоксической молекулой, радиоактивным изотопом или детектируемой меткой. Модулятор иммунной системы или цитотоксическая молекула может представлять собой цитокин. Детектируемая метка может представлять собой радиоактивный изотоп, например, нетерапевтический радиоактивный изотоп.
Молекулу антитела можно конъюгировать с терапевтическим агентом или детектируемой меткой посредством пептидной связи или линкера, т.е. в составе гибридного полипептида, содержащего указанный терапевтический агент или детектируемую метку и молекулу антитела или компонент ее полипептидной цепи. Другие средства конъюгирования включают химическое конъюгирование, например, перекрестное связывание с использованием бифункционального реагента (например, с использованием руководства по выбору перекрестно сшивающих реагентов DOUBLE-REAGENTSTM Cross-linking Reagents Selection Guide, Pierce).
Таким образом, молекулу антитела и терапевтический агент или детектируемую метку можно присоединить непосредственно друг к другу, например, посредством любой подходящей химической связи, или посредством линкера, например, пептидного линкера.
Пептидный линкер может быть коротким (цепь из 2-20, предпочтительно 2-15 аминокислотных остатков). В данной области техники известны подходящие примеры последовательностей пептидных линкеров. Можно использовать один или более из различных линкеров. Длина линкера может составлять приблизительно 5 аминокислот.
Химическая связь может быть, например, ковалентной или ионной связью. Примеры ковалентных связей включают пептидные связи (амидные связи) и дисульфидные связи. Например, молекула антитела и терапевтический или диагностический агент могут быть ковалентно связаны. Например, пептидными связями (амидными связями). Таким образом, молекулу антитела и терапевтический или диагностический агент можно продуцировать (секретировать) в виде одноцепочечного полипептида.
В настоящем изобретении также предложены выделенные нуклеиновые кислоты, кодирующие молекулы антител согласно настоящему изобретению. Получение таких нуклеиновых кислот с использованием способов, хорошо известных в данной области техники, не составит затруднений для специалиста. Выделенную нуклеиновую кислоту можно применять для экспрессии молекулы антитела согласно настоящему изобретению, например, посредством экспрессии в клетке-хозяине бактерии, дрожжей, насекомого или млекопитающего. Предпочтительная клетка-хозяин является клеткой млекопитающего, например, клеткой CHO, HEK или NS0. Нуклеиновая кислота в общем случае может быть представлена в форме рекомбинантного экспрессирующего вектора.
Выделенная нуклеиновая кислота может, например, содержать последовательность, приведенную в SEQ ID NO: 142, 4, 9, 14, 19, 24, 29, 34, 39, 44 или 49, кодирующую CH3-домены FS18-7-9 (кодон-оптимизированная нуклеотидная последовательность CHO), FS18-7-9 (HEK293-экспрессируемая нуклеотидная последовательность), FS18-7-32, FS18-7-33, FS18-7-36, FS18-7-58, FS18-7-62, FS18-7-65, FS18-7-78, FS18-7-88 и FS18-7-95, соответственно.
В состав настоящего изобретения входят клетки-хозяева in vitro, содержащие такие нуклеиновые кислоты и векторы, а также их применение для экспрессии молекул антител согласно настоящему изобретению, которые впоследствии можно выделить из клеточной культуры и необязательно составить в виде фармацевтической композиции. Таким образом, в настоящем изобретении дополнительно предложен способ получения молекулы антитела согласно настоящему изобретению, включающий культивирование рекомбинантной клетки-хозяина согласно настоящему изобретению в условиях для продуцирования молекулы антитела. Способы культивирования подходящих клеток-хозяев, указанных выше, известны в данной области техники. Указанный способ может дополнительно включать выделение и/или очистку молекулы антитела. Указанный способ может также включать получение состава молекулы антитела в виде фармацевтической композиции, необязательно с фармацевтически приемлемым вспомогательным веществом или другим веществом, как описано ниже.
Известно, что PD-L1 экспрессируется на многих раковых клетках, в то время как экспрессия LAG-3 на раковых клетках носит более ограниченный характер. Оба указанные антигена экспрессируются на клетках иммунной системы. В частности, известно, что LAG-3 экспрессируется на истощенных T-клетках в окружении опухоли. Кроме того, авторы настоящего изобретения продемонстрировали, что применение молекулы антитела, связывающейся как с LAG-3, так и с PD-L1, эффективно подавляет рост опухолей на сингенных моделях у мышей, и что такие молекулы антител являются более эффективными, чем введение двух связывающих молекул, связывающихся с LAG-3 и PD-L1, соответственно.
Таким образом, молекулу антитела согласно настоящему изобретению можно применять в способе лечения рака у пациента. Пациент предпочтительно является пациентом-человеком.
Клетки рака, подлежащего лечению с применением молекулы антитела согласно настоящему изобретению, могут экспрессировать LAG-3, например, на своей поверхности. В одном варианте реализации может быть установлено, что клетки рака, подлежащего лечению, экспрессируют LAG-3, например, на своей поверхности. Например, показано, что B-клеточные лимфомы экспрессируют LAG-3 на поверхности своих клеток. Способы определения экспрессии антигена на поверхности клетки известны в данной области техники и включают, например, проточную цитометрию.
Ниже в примере 4 показано, что молекулы антител согласно настоящему изобретению можно применять для лечения опухолей с высоким уровнем LAG-3-экспрессирующих иммунных клеток, например, LAG-3-экспрессирующих ОИЛ у мышей. Таким образом, в качестве дополнения или альтернативы, раковые опухоли, подлежащие лечению с применением молекулы антитела согласно настоящему изобретению, могут содержать LAG-3-экспрессирующие иммунные клетки. LAG-3-экспрессирующие иммунные клетки, например, LAG-3-экспрессирующие ОИЛ, присутствуют между опухолевыми клетками при многих видах рака. В одном варианте реализации установлено, что раковые опухоли, подлежащие лечению с применением молекулы антитела согласно настоящему изобретению, содержат LAG-3-экспрессирующие иммунные клетки. Способы определения наличия LAG-3-экспрессирующих иммунных клеток в опухоли или на периферии опухоли известны в данной области техники.
Ниже в примере 4 также показано, что молекулы антител согласно настоящему изобретению можно применять для лечения опухолей, клетки которых экспрессируют PD-L1 на своей поверхности. Так, в качестве дополнения или альтернативы, клетки рака, подлежащего лечению с применением молекулы антитела согласно настоящему изобретению, могут экспрессировать PD-L1, например, на своей поверхности. В качестве дополнения или альтернативы, злокачественные опухоли, подлежащие лечению, могут содержать иммунные клетки, например, ОИЛ, которые экспрессируют PD-L1. Может быть установлено, что клетки рака, подлежащего лечению, экспрессируют PD-L1, например, на своей поверхности. В качестве дополнения или альтернативы, может быть установлено, что злокачественные опухоли, подлежащие лечению, содержат иммунные клетки, например, ОИЛ, которые экспрессируют PD-L1.
Ожидается, что экспрессия LAG-3 и PD-L1 на поверхности клеток позволит молекуле антитела связываться с LAG-3 и PD-L1, экспрессируемыми на поверхности иммунной клетки и/или раковой клетки. Считается, что это обеспечит направленную терапию, образование мостиков и локализацию раковых и иммунных клеток.
Рак, подлежащий лечению с применением молекулы антитела согласно настоящему изобретению, можно выбрать из группы, состоящей из лимфомы Ходжкина, неходжкинской лимфомы (например, диффузной крупноклеточной B-клеточной лимфомы, медленно растущей неходжкинской лимфомы, лимфомы из клеток мантии), рака яичников, рака предстательной железы, рака ободочной и прямой кишки, фибросаркомы, почечноклеточной карциномы, меланомы, рака поджелудочной железы, рака молочной железы, мультиформной глиобластомы, рака легких (например, немелкоклеточного рака легких), рака головы и шеи (например, плоскоклеточной карциномы головы и шеи), рака желудка, рака мочевого пузыря, рака шейки матки, рака матки, рака вульвы, рака яичек, рака полового члена, лейкоза (например, хронического лимфоцитарного лейкоза, миелолейкоза, острого лимфобластного лейкоза или хронического лимфобластного лейкоза), множественной миеломы, плоскоклеточного рака, рака яичек, рака пищевода (например, аденокарциномы гастроэзофагеального соединения), саркомы Капоши и лимфомы центральной нервной системы (ЦНС), гепатоцеллюлярной карциномы, рака носоглотки, карциномы из клеток Меркеля и мезотелиомы. Известно или предполагается, что опухоли этих видов рака экспрессируют PD-L1 на поверхности своих клеток и/или содержат иммунные клетки, например, ОИЛ, экспрессирующие PD-L1 и/или LAG-3.
Лечение почечноклеточной карциномы, рака легких (например, немелкоклеточного рака легких), рака носоглотки, рака ободочной и прямой кишки, меланомы, рака желудка, рака пищевода (например, аденокарциномы гастроэзофагеального соединения), рака яичников, рака шейки матки, рака мочевого пузыря, рака головы и шеи (например, плоскоклеточной карциномы головы и шеи), лейкоза (например, хронического лимфоцитарного лейкоза, лимфомы Ходжкина, неходжкинской лимфомы (например, диффузной крупноклеточной B-клеточной лимфомы, медленно растущей неходжкинской лимфомы, лимфомы из клеток мантии) и множественной миеломы с применением антител против LAG-3 исследуется в клинических исследованиях и демонстрирует перспективные результаты. Таким образом, рак, подлежащий лечению с применением молекул антител согласно настоящему изобретению, может представлять собой почечноклеточную карциному, рак легких (например, немелкоклеточный рак легких), рак носоглотки, рак ободочной и прямой кишки, меланому, рак желудка, рак пищевода (например, аденокарциному гастроэзофагеального соединения), рак яичников, рак шейки матки, рак мочевого пузыря, рак головы и шеи (например, плоскоклеточную карциному головы и шеи), лейкоз (например, хронический лимфоцитарный лейкоз, лимфому Ходжкина, неходжкинскую лимфому (например, диффузную крупноклеточную B-клеточную лимфому, медленно растущую неходжкинскую лимфому, лимфому из клеток мантии) или множественную миелому.
Лечение меланомы, рака ободочной и прямой кишки, рака молочной железы, рака мочевого пузыря, почечноклеточной карциномы, рака желудка, рака головы и шеи (например, плоскоклеточной карциномы головы и шеи), мезотелиомы, рака легких (например, немелкоклеточного рака легких), рака яичников, карциномы из клеток Меркеля, рака поджелудочной железы, меланомы и гепатоцеллюлярной карциномы с применением антител против PD-L1 также исследуется в клинических исследованиях и демонстрирует перспективные результаты. Таким образом, рак, подлежащий лечению с применением молекул антител согласно настоящему изобретению, может представлять собой меланому, рак ободочной и прямой кишки, рак молочной железы, рак мочевого пузыря, почечноклеточную карциному, рак желудка, рак головы и шеи (например, плоскоклеточную карциному головы и шеи), мезотелиому, рак легких (например, немелкоклеточный рак легких), рак яичников, карциному из клеток Меркеля, рак поджелудочной железы, меланому и гепатоцеллюлярную карциному.
Предпочтительными видами рака для лечения с применением молекул антител согласно настоящему изобретению являются рак легких (например, немелкоклеточный рак легких), рак мочевого пузыря, рак головы и шеи (плоскоклеточная карцинома головы и шеи), диффузная крупноклеточная В-клеточная лимфома, рак желудка, рак поджелудочной железы и гепатоцеллюлярная карцинома. Известно, что опухоли при этих видах рака содержат LAG-3-экспрессирующие иммунные клетки и экспрессируют PD-L1 на поверхности своих клеток или содержат иммунные клетки, экспрессирующие PD-L1.
Если заявка относится к конкретному виду рака, например, раку молочной железы, она относится к злокачественной трансформации соответствующей ткани, в данном случае - ткани молочной железы. Рак, происходящий от злокачественной трансформации другой ткани, например, ткани яичника, может приводить к появлению метастатических очагов в другой части тела, например, молочной железе, но из-за этого является не раком молочной железы, как указано в настоящем документе, а раком яичников.
Рак может быть первичным или вторичным раком. Таким образом, молекула антитела согласно настоящему изобретению может быть предназначена для применения в способе лечения рака у пациента, причем рак является первичной опухолью и//или метастазом опухоли.
Молекулы антител согласно настоящему изобретению предназначены для применения в способах лечения пациентов, предпочтительно пациентов-людей. Молекулы антител обычно следует вводить в форме фармацевтической композиции, которая может содержать по меньшей мере один компонент в дополнение к молекуле антитела, например, фармацевтически приемлемое вспомогательное вещество. Например, фармацевтическая композиция согласно настоящему изобретению может содержать, в дополнение к активному ингредиенту, фармацевтически приемлемое вспомогательное вещество, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области техники. Такие материалы должны быть нетоксичными и не должны мешать эффективности активного ингредиента. Точная природа носителя или другого материала зависит от пути введения, которое можно осуществлять, например, путем инъекции, например, внутривенной или подкожной инъекции. Молекулу антитела можно вводить внутривенно или подкожно.
Жидкие фармацевтические композиции в общем случае содержат жидкий носитель, например, воду, углеводородные, животные или растительные масла, минеральное масло или синтетическое масло. Можно включать физиологический раствор, раствор декстрозы или других сахаридов или гликоли, например, этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенного введения или инъекции в пораженную область молекула антитела или фармацевтическая композиция, содержащая молекулу антитела, предпочтительно представлена в форме парентерально приемлемого апирогенного водного раствора с подходящим pH, изотоничностью и стабильностью. Специалисты в данной области техники легко могут изготовить подходящие растворы, используя, например, изотонические носители, например, раствор хлорида натрия для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. При необходимости можно использовать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки. Специалистам в данной области техники известны многие способы изготовления фармацевтических составов. См., например, Robinson ed., Sustained and Controlled Release Drug Delivery Systems, Marcel Dekker, Inc., New York, 1978.
Композицию, содержащую молекулы антител согласно настоящему изобретению, можно вводить отдельно или в комбинации с другими терапевтическими средствами, одновременно или последовательно, или в виде комбинированного препарата с другим терапевтическим агентом или агентами, в зависимости от состояния, подвергаемого лечению. Например, молекулу антитела согласно настоящему изобретению можно вводить в комбинации с существующим терапевтическим агентом для заболевания, подлежащего лечению, например, рака, как указано выше. Например, молекулу антитела согласно настоящему изобретению можно вводить пациенту в комбинации со вторым противораковым терапевтическим средством, например, химиотерапевтическим средством, противоопухолевой вакциной (также называемой вакциной против рака), лучевой терапией, иммунотерапевтическим средством, онколитическим вирусом, терапевтическим средством на основе Т-клеток с химерным антигенным рецептором (CAR) или гормональным терапевтическим средством.
Ожидается, что молекула антитела согласно настоящему изобретению может действовать в качестве адьюванта при противораковой терапии, например, химиотерапии, противоопухолевой вакцинации или лучевой терапии. Безотносительно к теоретическим представлениям считается, что введение молекулы антитела пациенту в рамках химиотерапии, противоопухолевой вакцинации или лучевой терапии будет вызывать более сильный иммунный ответ против антигенов LAG-3 и PD-L1, ассоциированных с раком, чем ответ, достигаемый при применении только химиотерапии, противоопухолевой вакцинации или лучевой терапии. Например, показано, что терапевтические средства против LAG-3 характеризуются хорошей эффективностью в лечении вирусных патологических состояний у мышей (Blackburn SD, et al., 2009).
Таким образом, способ лечения рака у пациента может включать введение пациенту терапевтически эффективного количества молекулы антитела согласно настоящему изобретению в комбинации с химиотерапевтическим агентом, противоопухолевой вакциной, радионуклидом, иммунотерапевтическим агентом, онколитическими вирусами, CAR-T-клетками или агентом для гормональной терапии. Химиотерапевтический агент, противоопухолевая вакцина, радионуклид, иммунотерапевтический агент, онколитические вирусы, CAR-T-клетки или агент для гормональной терапии предпочтительно представляет собой химиотерапевтический агент, противоопухолевую вакцину, радионуклид, иммунотерапевтический агент, онколитические вирусы, CAR-T-клетки или агент для гормональной терапии рассматриваемого рака, т.е. химиотерапевтический агент, противоопухолевую вакцину, радионуклид, иммунотерапевтический агент, онколитические вирусы, CAR-T-клетки или агент для гормональной терапии, характеризующиеся продемонстрированной эффективностью в лечении рассматриваемого рака. Практикующий специалист способен выбрать подходящий химиотерапевтический агент, противоопухолевую вакцину, радионуклид, иммунотерапевтический агент, онколитические вирусы, CAR-T-клетки или агент для гормональной терапии, характеризующийся продемонстрированной эффективностью в лечении рассматриваемого рака.
Например, если способ включает введение молекулы антитела согласно настоящему изобретению пациенту в терапевтически эффективном количестве в комбинации с химиотерапевтическим агентом, указанный химиотерапевтический агент выбирают из группы, состоящей из таксанов, цитотоксических антибиотиков, ингибиторов тирозинкиназы, ингибиторов PARP, ингибиторов фермента B_RAF, алкилирующих агентов, аналогов платины, аналогов нуклеозидов, производных талидомида, противоопухолевых химиотерапевтических агентов и т.д. Таксаны включают доцетаксел, паклитаксел и паклитаксел, связанный с наночастицами альбумина; цитотоксические антибиотики включают актиномицин,блеомицин, антрациклины, доксорубицин и валрубицин; ингибиторы тирозинкиназы включают эрлотиниб, гефитиниб, акситиниб, PLX3397, иматиниб, кобиметиниб и траметиниб; ингибиторы PARP включают нирапариб; ингибиторы фермента B-Raf включают вемурафениб и дабрафениб; алкилирующие агенты включают дакарбазин, циклофосфамид, темозоломид; аналоги платины включают карбоплатин, цисплатин и оксаплатин; аналоги нуклеозидов включают гемцитабин и азацитидин; противоопухолевые средства включают флударабин. Другие химиотерапевтические агенты, подходящие для применения в настоящем изобретении, включают метотрексат, дефактиниб, энтиностат, пеметрексед, капецитабин, эрибулин, иринотекан, фторурацил и винбластин.
Стратегии вакцинации для лечения рака внедрены в клиническую практику и обсуждаются в научной литературе (например, Rosenberg, S. 2000 Development of Cancer Vaccines). Основные используемые стратегии заключаются в стимуляции иммунной системы к ответу на различные клеточные маркеры, экспрессируемые аутологичными или аллогенными раковыми клетками, с использованием указанных клеток при способе вакцинации как с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМКСФ), так и без него. ГМКСФ стимулирует сильный ответ при презентации антигена и действует в особенности эффективно при применении в рамках указанных стратегий.
Введение можно выполнять в "терапевтически эффективном количестве", которое является достаточным для оказания благоприятного действия на пациента. Такое благоприятное действие может представлять собой по меньшей мере улучшение по меньшей мере одного симптома. Такое «лечение» указанного заболевания относится к улучшению по меньшей мере одного симптома. Фактическое вводимое количество, скорость и период действия при введении зависят от природы и тяжести заболевания, подлежащего лечению, конкретного пациента, подлежащего лечению, клинического состояния отдельного пациента, причины расстройства, области доставки композиции, типа молекулы антитела, способа введения, схемы введения и других факторов, известных врачам-практикам. За назначение лечения, например, решение по дозировке и т.д., отвечают врачи общей практики и другие врачи; оно может зависеть от тяжести симптомов и / или прогрессирования заболевания, подлежащего лечению. Подходящие дозы молекул антител хорошо известны в данной области техники (Ledermann et al. (1991) Int. J. Cancer 47: 659-664; и Bagshawe et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4: 915-922). Можно использовать конкретные дозировки, указанные в настоящем документе или в Physician's Desk Reference (2003) как подходящие для вводимой молекулы антитела. Терапевтически эффективное количество или подходящую дозу молекулы антитела можно определить путем сравнения ее активности in vitro и in vivo на животной модели. Известны способы экстраполяции эффективных дозировок у мышей и других экспериментальных животных на человека. Точная доза зависит от ряда факторов, в том числе размера и местоположения области, подлежащей лечению, и точной природы молекулы антитела. Лечение можно повторить с ежедневными, два раза в неделю, еженедельными или ежемесячными интервалами по усмотрению врача. Терапевтические средства можно давать до и / или после операции, и можно вводить или наносить непосредственно на анатомическую область хирургического лечения.
Дополнительные аспекты и варианты реализации настоящего изобретения очевидны для специалистов в данной области техники в свете настоящего описания, включающего их экспериментальную реализацию.
Все документы, упомянутые в настоящем описании, полностью включены в настоящую заявку посредством ссылок.
Фразу "и / или", используемую в настоящем документе, следует рассматривать как специфическое описание каждой из двух указанных особенностей или компонентов друг с другом или по отдельности. Например, "А и / или В" следует рассматривать как специфическое описание каждого из (i) A, (ii) B и (iii) А и В, как если бы каждый из них по отдельности описывался в настоящем документе.
Если иное не следует из контекста, описания и определения особенностей, изложенных выше, не ограничиваются каким-либо конкретным аспектом или вариантом реализации настоящего изобретения и в равной степени применимы ко всем описанным аспектам и вариантам реализации.
Некоторые аспекты и варианты реализации настоящего изобретения будут проиллюстрированы посредством примеров со ссылкой на фигуры, описанные выше.
Примеры
Пример 1 - отбор и исследование характеристик молекул Fcab
1.1 Наивный отбор и созревание аффинности Fcab против LAG-3
1.1.1 Наивный отбор
Наивные библиотеки фагового дисплея, содержащие CH3-домен IgG1 человека (нумерация IMGT 1,4-130) с рандомизацией в пределах петель AB (остатки 14-18) и EF (остатки 92-101) использовали для отбора с использованием рекомбинантного Fc-меченого антигена LAG-3 (LAG-3 Fc) человека (R&D systems, 2319-L3-050). Отбор в библиотеках выполняли в три раунда с использованием антигена, иммобилизованного на гранулах белка А (Life Technologies, 10002D) или белка G (Life Technologies, 10004D). Результаты подвергали скринингу посредством твердофазного ИФА; молекулы, демонстрировавшие положительные результаты связывания, субклонировали и экспрессировали в виде растворимых Fcab (содержащих укороченную шарнирную область) в Pichia pastoris с использованием набора для экспрессии EasySelect Pichia Expression Kit (Life Technologies, K1740-01). Затем Fcab подвергали скринингу на предмет связывания с рекомбинантным LAG-3 Fc человека на Biacore 3000 (GE Healthcare). Вкратце, LAG-3 Fc (R&D systems, 2319-L3-050) присоединяли к чипу CM5 (GE Healthcare, BR-100012) при плотности 7200 РЕ с использованием реагентов для аминного связывания (GE Healthcare, BR-1000-50). Fcab разбавляли в буфере HBS-P (GE Healthcare, BR100368) и вводили в количестве 250 нМ, 500 нМ и 1000 нМ на 3 мин, а затем оставляли для диссоциации в буфере на 5 мин. Данные после вычитания эталонного контроля (проточная ячейка LAG-3 Fc 2 - холостая проточная ячейка) анализировали с использованием программного обеспечения BIAevaluation 3.2 с целью выявления связывания. Затем Fcab тестировали на предмет связывания с LAG-3-человека, экспрессируемым на клетках HEK (LAG-3 клонировали в векторе pcDNA5FRT [Life Technologies, V6010-20] [методологию см. в разделе 1.4.5]). Вкратце, клетки HEK 293, сверхэкспрессирующие LAG-3 человека, выращенные в среде DMEM (Life Technologies, 61965-026), содержащей 10% FBS (Life Technologies, 10270-1-6), 100 мг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мг/мл бластицидина (Melford Laboratories Ltd, B1105) и 1 мкг/мл доксициклина (Sigma, D9891), отделяли от флаконов для тканевых культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) и высевали в 96-луночные планшеты с V-образным дном, исходя из количества 2x105 клеток/лунку. Fcab инкубировали с клетками в концентрации 5 мкМ в объеме 100 мкл в течение 1 ч при 4°C. Планшеты промывали, вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098) разбавляли в соотношении 1:1000 PBS, добавляли в количестве 100 мкл к клеткам и инкубировали в течение 30 мин при 4°C. Планшеты промывали, клетки ресуспендировали в 100 мкл PBS, содержащего 1 мкг/мл ДАПИ (Biotium, 40043). Планшет считывали на цитометре BD FACSCanto II (BD Biosciences), данные анализировали с использованием FlowJoX. Затем Fcab экспрессировали в клетках млекопитающих путем трансформации с использованием липофектамина (Life Technologies, 11668-019) в клетки Flp-In T-Rex 293 (Life Technologies, R780-07). Fcab, связывающие LAG-3, тестировали на предмет ингибирования связывания ГКГС II класса человека на клетках A375 (ATCC, CRL-1619) с рекомбинантным LAG-3 Fc (с использованием методологии, описанной в примере 1.6). За три раунда фагового отбора выявили 54 уникальных последовательности Fcab; установили, что 12 из этих Fcab связывались с LAG-3 Fc при анализе на BIAcore и/или связывались с клетками HEK, экспрессирующими LAG-3. Три из отобранных Fcab также могли ингибировать взаимодействие LAG-3 с ГКГС II класса и были отобраны для созревания аффинности. Эти три Fcab получили названия FS18-3, FS18-7 и FS18-21.
1.1.2 Созревание аффинности
Первое созревание аффинности
Сконструировали шесть библиотек фагового дисплея для созревания аффинности, рандомизируя пять остатков в петле AB (остатки 14-18) и еще (остатки 92-94 и 97-98) или восемь (остатки 92-94 и 97-101) остатков в петле EF каждого из трех Fcab, выявленных с использованием наивного отбора, описанного выше.
Отбор в библиотеках для созревания аффинности выполняли с использованием рекомбинантного LAG-3 Fc человека (R&D systems, 2319-L3-050) и клеток HEK, экспрессирующих LAG-3 человека (как описано выше). Результаты подвергали скринингу посредством твердофазного ИФА; молекулы, демонстрировавшие положительные результаты связывания, субклонировали и экспрессировали в виде растворимых Fcab (содержащих укороченную шарнирную область) в клетках HEK Expi293 (Fcab клонировали в вектор pTT5 [Национальный совет Канады по исследованиям], трансфицированный с использованием набора для трансфекции ExpiFectamine 293 Transfection kit [Life Technologies, A14524] в клетки Expi293F [Life technologies, A14527]). Затем растворимые Fcab, экспрессируемые HEK, подвергали скринингу на предмет связывания с клетками, экспрессирующими LAG-3 человека, связывания с клетками, экспрессирующими LAG-3 яванского макака (методология согласно примеру 1.4.3), а также способности блокировать связывание ГКГС II класса с рекомбинантным LAG-3 Fc (методология согласно примеру 1.6). Блокирующие Fcab дополнительно тестировали на предмет определения способности обращать LAG-3-индуцированное ингибирование секреции ИЛ-2 в анализе активации T-клеток (методология согласно примеру 2.1). В шести библиотеках для созревания аффинности с использованием этих способов скрининга выявили 61 уникальный Fcab против LAG-3. Показано, что Fcab из линии FS18-7 после созревания аффинности обладали максимальным уровнем перекрестной реакционной способности по отношению к LAG-3 яванского макака. Три Fcab из этой линии с самым сильным связыванием с LAG-3 Fc яванского макака и максимальной активностью в анализе активации T-клеток (получившие названия FS18-7-7, FS18-7-9 и FS18-7-11) отобрали для дальнейшего созревания аффинности. Показано, что эти три Fcab также блокировали взаимодействие LAG-3 Fc с ГКГС II класса, экспрессируемым клетками.
Второе созревание аффинности
Пул трех Fcab (FS18-7-7, FS18-7-9 и FS18-7-11), полученных при первом созревании аффинности, использовали для дальнейшего создания библиотек для созревания аффинности Петлю CD интенсивно рандомизировали с использованием рандомизированных праймеров производства ELLA Biotech. Часть аминокислотных положений в петле CD (остатки 45,1-78) рандомизировали, используя эквимолярное распределение аминокислот, за исключением цистеина. Кроме того, выполнили ПЦР пониженной точности по всей последовательности CH3-домена с целью внедрения дополнительных мутаций, которые могли бы усилить связывание.
В фаге получили библиотеки для созревания аффинности, выполнили отбор с использованием биотинилированного рекомбинантного LAG-3 avi-Fc (BPS Bioscience, 71147) и клеток HEK hLAG-3 и скрининг на предмет связывания с рекомбинантным LAG-3 Fc (R&D systems, 2319-L3-050) посредством фагового твердофазного ИФА. 86 уникальных Fcab (содержащих укороченную шарнирную область) экспрессировали в клетках HEK293F. Отобранные Fcab также подвергли скринингу на предмет активности в анализе активации Т-клеток, как описано выше. Характеристики девяти Fcab с максимальной активностью в анализе активации Т-клеток, выявленных при втором созревании аффинности (FS18-7-32; FS18-7-33; FS18-7-36; FS18-7-58; FS18-7-62; FS18-7-65; FS18-7-78; FS18-7-88; и FS18-7-95), а также исходного клона Fcab, FS18-7-9, дополнительно исследовали, как онисано ниже. Выравнивание этих девяти Fcab по отношению к последовательности исходного клона Fcab, FS18-7-9, показано на фигуре 1А. На фигуре 1B подробно представлена процентная идентичность последовательности каждого из девяти клонов Fcab по отношению к исходному клону Fcab, FS18-7-9. Fcab, полученные при созревании аффинности двух других исходных клонов Fcab, FS18-7-7 и FS18-7-11, не являлись столь перспективными кандидатами, как молекулы, полученные при созревании аффинности FS18-7-9, и поэтому в дальнейшем не исследовались.
1.2 Отбор суррогатного Fcab, специфичного по отношению к LAG-3 мыши
Fcab FS18-7, отобранный с использованием вышеописанного протокола наивного отбора, использовали для получения фаговых библиотек для отбора с использованием LAG-3 мыши. Выполнили два раунда созревания аффинности, и после созревания аффинности отобрали клоны Fcab FS18-7-108-29 и FS18-7-108-35, демонстрировавшие специфическое связывание с высоким сродством с LAG-3 мыши. Подтвердили способность FS18-7-108-29 и FS18-7-108-35 ингибировать LAG-3 мыши в анализе активации Т-клеток. Картирование эпитопов с использованием Octet (Forteo Bio) показало, что Fcab против LAG-3 мыши конкурировали с Fcab против LAG-3 человека (отобранными после второго созревания аффинности, как описано выше) за связывание с LAG-3 человека. Между остатками Fcab против LAG-3 человека и против LAG-3 мыши имели место от 4 до 8 различий. Таким образом, ожидается, что Fcab против LAG-3 мыши представляют собой подходящие суррогаты для изучения связывания и функции Fcab против LAG-3 человека у мыши.
1.3 Конструирование и экспрессия имитационного мАт2
“Имитационные” мАт2, содержавшие основные Fcab против LAG-3 человека и против LAG-3 мыши, выявленные выше в разделах 1.1 и 1.2, получили для исследования характеристик этих Fcab в формате мАт2. Эти имитационные мАт2 получили из Fcab против LAG-3 и вариабельных областей антитела 4420 против FITC (подробную информацию см. в SEQ ID NO: 83, SEQ ID NO: 84 и SEQ ID NO: 85) (Bedzyk, W. D., et al. 1989 и Bedzyk, W. D., et al. 1989). Имитационное мАт2 получили как с мутацией LALA в CH2-домене тяжелой цепи (SEQ ID NO: 63, 65, 67, 69, 71, 73, 75, 77, 79 и 81), так и без нее (SEQ ID NO: 64, 66, 68, 70, 72, 74, 76, 78, 80 и 82) (подробную информацию см. ниже в разделе 1.5); они дополнительно содержали легкую цепь мАт 4420 против FITC (SEQ ID NO: 85). Имитационные мАт2 продуцировали путем временной экспрессии в клетках HEK293-6E и очищали с использованием колонок для отбора мАт с белком А mAb Select SuRe protein A.
1.4 Сродство связывания Fcab с LAG-3
1.4.1 Сродство связывания Fcab с LAG-3 человека при определении с помощью поверхностного плазмонного резонанса (ППР)
BIAcore T200 (GE Healthcare) использовали для измерения сродства Fcab против LAG-3 человека в формате имитационного мАт2 к LAG-3 человека. В проточной ячейке 4 на чипе сенсора CM5 (GE Healthcare, BR1005-30) иммобилизовали LAG-3-Fc человека (R&D Systems, 2319-L3-050), а в проточной ячейке 3 - буфер для эталонного сравнения с использованием набора для аминного связывания (GE Healthcare, BR-1000-50). LAG-3-Fc разбавляли до концентрации 5 мкг/мл ацетатом натрия, pH 5 (ForteoBio, 18-1069) и вводили при скорости потока 10 мкл/мин в течение 12 секунд с последующей деактивацией поверхности путем инъекции этаноламина в течение 420 с. Уровень иммобилизации составил 158 РЕ. Имитационное мАт2 (или контрольное мАт 25F7 против LAG-3 человека) разбавляли буфером HBS-P (GE Healthcare, BR-1003-68) в серии 2-кратных разведений с концентрации 4 мкг/мл. Контрольное мАт/имитационое мАт2 вводили со временем ассоциации, равным 240 секунд, со скоростью 30 мкл/мин, и временем диссоциации, равным 300 секунд, со скоростью 30 мкл/мин. Поверхность регенерировали с использованием 25 мМ NaOH в течение 30 секунд со скоростью 100 мкл/мин. После двойного вычитания показаний контрольного эталона данные анализировали с использованием программного обеспечения BIAevaluation 3.2, рассчитывая кинетические константы. Fcab в формате имитационных мАт2 обладали сродством к LAG-3 человека в диапазоне 0,8 - 1,1 нМ (таблица 1), что аналогично сродству эталонного мАт 25F7 против LAG-3 человека. Это оказалось неожиданным, поскольку Fcab характеризуются меньшей поверхностью взаимодействия при связывании, чем моноклональные антитела, поскольку сайты связывания Fcab образуют сравнительно компактный фрагмент антитела с двумя сайтами связывания, расположенными в относительной близости. В противоположность этому, Fab-плечи типичного мАТ разделены гибкой шарнирной областью. С учетом меньшей поверхности взаимодействия при связывании и ассоциированной с этим пониженной гибкости двух сайтов связывания в области Fc, способность Fcab против LAG-3 связываться с LAG-3 и ингибировать его со сродством и эффективностью, аналогичными сродству и эффективности эталонного антитела 25F7, оказалась неожиданной.
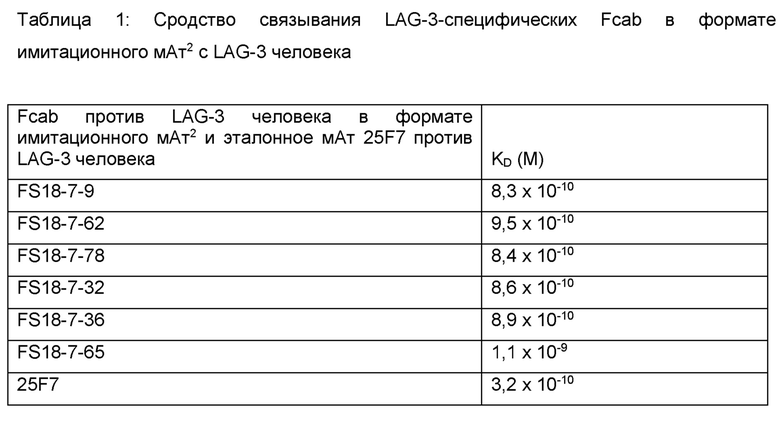
1.4.2 Сродство связывания суррогатного Fcab, специфичного по отношению к LAG-3 мыши, с LAG-3 мыши при определении с помощью ППР
BIAcore 3000 (GE Healthcare) использовали для измерения сродства суррогатных Fcab, специфичных по отношению к LAG-3 мыши, к LAG-3 мыши. Аминное связывание (набор для аминного связывания, GE Healthcare, BR-1000-50) использовали для иммобилизации mLAG-3 Fc (R&D Systems, 3328-L3-050), разбавленного 10 мМ ацетатом натрия, pH 5,0 (ForteBio, 18-1069), непосредственно на чипе CM5 (GE Healthcare, BR-1000-12). В проточной ячейке 1 иммобилизовали Fc мыши (SinoBiological, 51094-MNAH), а в проточной ячейке 2 - mLAG-3 Fc в количестве 950 РЕ. Fcab разбавляли буфером HBS-P (GE Healthcare, BR-1003-68) и вводили в различных концентрациях (четырехкратные разведения с концентрации 100 мМ) в течение 3 мин со скоростью 20 мкл/мин, а затем оставляли для диссоциации в буфере на 12 мин. Чип регенерировали инъекцией 10 мМ глицина, pH 2,5, в течение 30 с со скоростью 30 мкл/мин. После двойного вычитания показаний контрольного эталона данные анализировали с использованием программного обеспечения BIAevaluation 3.2, рассчитывая кинетические константы. Протестированные суррогатные Fcab связывались с LAG-3 мыши со сродством в области единиц наномоль, как представлено в таблице 2.

1.4.3 Сродство связывания Fcab с LAG-3 человека, экспрессируемым на клетках, при определении с помощью проточной цитометрии
Продукция линий клеток, сверхэкспрессирующих LAG-3
Для получения клеток DO11.10 (National Jewish Health), сверхэкспрессирующих LAG-3 человека, яванского макака или мыши, применяли методологию лентивирусной трансдукции с использованием системы упаковки Lenti-X HTX Packaging System (Clontech, № по каталогу 631249). Экспрессирующий вектор Lenti-X (pLVX) (Clontech, № по каталогу 631253), содержащий кДНК LAG-3 мыши (SEQ ID NO: 96), кДНК LAG-3 человека (SEQ ID NO: 95) или кДНК LAG-3 яванского макака (SEQ ID NO: 97), совместно трансфицировали с использованием смеси для упаковки Lenti-X HTX Packaging Mix в клетки линии Lenti-X 293T (Clontech, № по каталогу 632180) для получения вируса. Клетки линии DO11.10 трансдуцировали с использованием лентивирусных векторов, полученных с помощью системы упаковки Lenti-X HTX Packaging System.
Сродство Fcab против LAG-3 человека в формате имитационного мАт2 к клеткам, экспрессирующим LAG-3 человека (клеткам линии DO11.10, трансфицированным LAG-3 человека), измеряли с помощью проточной цитометрии. Разведения мАт2 и контрольного мАт (2 x конечная концентрация) получали в трех повторностях в 1 x DPBS (Gibco, 14190-094). Суспензии клеток DO11.10:LAG-3 получали в PBS+2%БСА (Sigma, A7906) и высевали из расчета 4 x 10-6 клеток/мл в объеме 50 мкл/лунку в 96-луночные планшеты с V-образным дном (Costar, 3897). 50 мкл разведений мАт2 или контрольного мАт (мАт 25F7 против LAG-3 человека) добавляли в лунки, содержащие клетки (конечный объем 100 мкл) и инкубировали при 4°C в течение 1 часа. Планшеты промывали, а затем добавляли вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098), разбавленное в соотношении 1:1000 PBS + 2% БСА, из расчета 100 мкл/лунку, и инкубировали в течение 30 мин при 4°C в темноте. Планшеты промывали и ресуспендировали в 100 мкл PBS, содержащего ДАПИ (Biotium, 40043) в концентрации 1 мкг/мл. Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Все протестированые Fcab в формате имитационных мАт2 и эталонное мАт 25F7 против LAG-3 человека связывали LAG-3 человека с аналогичным сродством (EC50) в диапазоне 1,2 - 2,1 нм, как представлено в таблице 3.


1.4.4 Сродство связывания Fcab с LAG-3 яванского макака, экспрессируемым на клетках, при определении с помощью проточной цитометрии
Сродство Fcab против LAG-3 человека в формате имитационного мАт2 к клеткам, экспрессирующим LAG-3 яванского макака (клеткам линии DO11.10, трансфицированным LAG-3 яванского макака), измеряли с помощью проточной цитометрии. Разведения мАт2 и контрольного мАт (2 x конечная концентрация) получали в трех повторностях в 1 x DPBS (Gibco, 14190-094). Суспензии клеток DO11.10:LAG-3 получали в PBS+2%БСА (Sigma, A7906) и высевали из расчета 4 x 10-6 клеток/мл в объеме 50 мкл/лунку в 96-луночные планшеты с V-образным дном (Costar, 3897). 50 мкл разведений мАт2 или контрольного мАт (мАт 25F7 против LAG-3 человека) добавляли в лунки, содержащие клетки (конечный объем 100 мкл) и инкубировали при 4°C в течение 1 часа. Планшеты промывали, а затем добавляли вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098), разбавленное в соотношении 1:1000 PBS + 2% БСА, из расчета 100 мкл/лунку, и инкубировали в течение 30 мин при 4°C в темноте. Планшеты промывали и ресуспендировали в 100 мкл PBS, содержащего ДАПИ (Biotium, 40043) в концентрации 1 мкг/мл. Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Протестированные Fcab в формате имитационных мАт2 связывались с LAG-3 яванского макака со сродством, равным 0,5-0,6 нМ, что указывало на возможность того, что токсикологические исследования у яванских макаков позволят прогнозировать эффекты, наблюдаемые у людей (см. таблицу 4). Эталонное мАт 25F7 против LAG-3 человека связывало LAG-3 яванского макака с 15- кратно худшим сродством (EC50) (таблица 4).
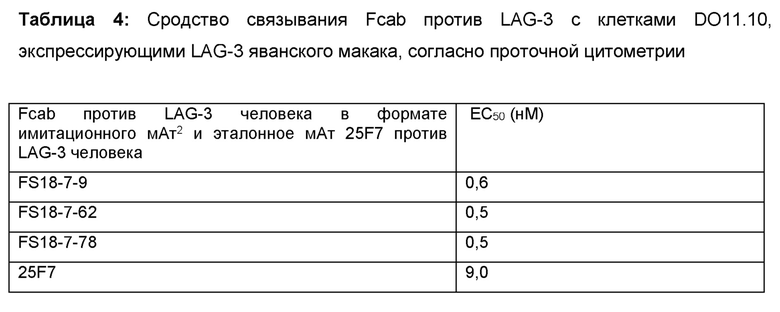
1.4.5 Сродство связывания суррогатных Fcab против LAG-3 мыши и Fcab против LAG-3 человека с LAG-3 мыши, экспрессируемым на клетках, при определении с помощью проточной цитометрии
Продукция клеток HEK, сверхэкспрессирующих mLAG-3
Последовательность LAG-3 мыши (SEQ ID NO: 96) субклонировали в вектор pcDNA5FRT (Life Technologies, V6010-20) с использованием рестриктазного гидролиза посредством KpnI (NEB, R0142) и NotI (NEB, R0146). Затем вектор трансформировали в клетки линии Flp-In T-REx 293 HEK (Life Technologies, R780-07) с использованием липофектамина 2000 (Life Technologies, 11668-019). Трансформированные клетки Flp-In T-REx 293 выращивали в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) в течение 3-4 недель до появления видимых колоний стабильно трансформированных клеток. Эти колонии амплифицировали в присутствии 1 мкг/мл доксициклина (Sigma, D9891) и тестировали на предмет экспрессии LAG-3 мыши с использованием PE-конъюгированного антитела против LAG-3 мыши (клон C9B7W, BD Biosciences, 552380).
Сродство суррогатных Fcab против LAG-3 мыши (содержавших укороченную шарнирную область; SEQ ID NO: 58) к LAG-3 мыши, экспрессируемому на клетках, определяли с помощью проточной цитометрии. Клетки HEK, экспрессирующие mLAG-3, выращенные в среде DMEM (Life Technologies, 61965-026), содержащей 10% FBS (Life Technologies, 10270-1-6), 100 мг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мг/мл бластицидина (Melford Laboratories Ltd, B1105) и 1 мкг/мл доксициклина (Sigma, D9891), отделяли от флаконов для тканевых культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) и высевали в 96-луночные планшеты с V-образным дном (Costar, 3897), исходя из количества 2x105 клеток/лунку. Планшеты центрифугировали при 1500 об/мин в течение 3 мин при 4°C для осаждения клеток. Серии разведений Fcab (или контрольного мАт) инкубировали с клетками в объеме 100 мкл в течение 1 ч при 4°C. Планшеты промывали, вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098 для Fcab или антитело против IgG крысы (H+L), конъюгат с Alexa Fluor 488, ThermoFisher, A-11006 для C9B7W) разбавляли PBS в соотношении 1:1000 и добавляли в количестве 100 мкл к клеткам на 30 мин при 4°C (планшеты держали в темноте). Затем планшеты промывали, клетки ресуспендировали в 100 мкл PBS, содержащего 1 мкг/мл ДАПИ (Biotium, 40043). Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Протестированные Fcab связывались с LAG-3 мыши с аналогичным сродством (см. таблицу 5). Эталонное мАт C9B7W против LAG-3 (2B Scientific, BE0174-50MG) связывало LAG-3 мыши с 17-кратно худшим сродством (EC50) по сравнению с Fcab (таблица 5).
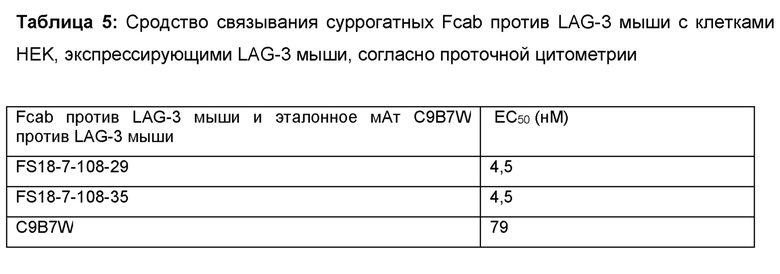
Сродство Fcab FS18-7-9 против LAG-3 человека в формате имитационного мАт2 к LAG-3 мыши, экспрессируемому клетками, определяли с помощью проточной цитометрии. Клетки HEK, экспрессирующие mLAG-3, выращенные в среде DMEM (Life Technologies, 61965-026), содержащей 10% FBS (Life Technologies, 10270-1-6), 100 мг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мг/мл бластицидина (Melford Laboratories Ltd, B1105) и 1 мкг/мл доксициклина (Sigma, D9891), отделяли от флаконов для тканевых культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) Клетки собирали центрифугированием при 1500 об/мин в течение 3 мин при 4°C, осаждая клетки, а затем ресуспендируя их в 1 x DPBS с последующим посевом в 96-луночные планшеты с V-образным дном (Costar, 3897) из расчета 1,2x105 клеток/лунку в объеме 30 мкл. Добавляли серии разведений мАт2 (или контрольного мАт) в объемном соотношении 1:1 и инкубировали их с клетками в течение 1 ч при 4°C. Планшеты промывали, вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098) разбавляли PBS в соотношении 1:1000 и добавляли в количестве 60 мкл к клеткам на 30 мин при 4°C (планшеты держали в темноте). Затем планшеты промывали, клетки ресуспендировали в 60 мкл PBS, содержащего 1 мкг/мл ДАПИ (Biotium, 40043). Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Fcab FS18-7-9 против LAG-3 человека в формате имитационного мАт2 связывался с LAG-3 мыши с EC50, равной 19 нМ, по сравнению с EC50 2,6 нМ для суррогатного Fcab FS18-7-9-108 против LAG-3 мыши (таблица 6). мАт 25F7 человека не демонстрировало детектируемого связывания с LAG-3 мыши, что указывало на наличие на LAG-3 другого эпитопа связывания для Fcab FS18-7-9 против LAG-3 человека по сравнению с эпитопом связывания для 25F7.

1.5 Сродство связывания Fcab с Fc-рецепторами
Известно, что внедрение мутации LALA в CH2-домен IgG1 человека ослабляет связывание с Fcγ-рецептором (Bruhns, P., et al. (2009); и Xu, D. et al. (2000)). Для подтверждения того, что мутация LALA снижала сродство связывания Fcab (в формате имитационного мАт2) с Fcγ-рецепторами, использовали BIAcore. Анализ связывания FcγR человека выполняли на приборе Biacore T200 (GE Healthcare) с использованием Fcab в формате имитационного мАт2. FcγR человека (R&D Systems, 1257-FC, 1330-CD, 1875-CD, 4325-FC) иммобилизовали с использованиям аминного связывания (набор для аминного связывания, GE Healthcare, BR-1000-50) на чипе Series S CM5 (GE Healthcare, BR-1005-30) при поверхностной плотности 370 РЕ для FcγRI, 264 РЕ для FcγRIII (FcγR человека с высоким сродством) и 500 РЕ для FcγRIIa и FcγRIIb (FcγR человека с низким сродством). Для каждого иммобилизованного чипа оставляли холостую проточную ячейку для вычитания фоновых значений. FcγR иммобилизовали в концентрации 5 мкг/мл в ацетате натрия, pH 5 (ForteBio, 18-1069), и вводили при скорости потока 10 мкл/мин в виде 15-секундных циклов до достижения требуемого уровня иммобилизации.
Для FcγRI и FcγRIII с высоким сродством 200 мкг/мл мАт или имитационных мАт2 пропускали в потоке через чип в течение 3 мин при скорости потока 30 мкл/мин с последующей диссоциацией в течение 5 мин. Проточный буфер представлял собой HBS-P (0,01 M HEPES, pH 7,4, 0,15 M NaCl, 0,005% об/об ПАВ P20, GE Healthcare, BR-1003-68). Для FcγRIIa и FcγRIIb с низким сродством концентрацию имитационного мАт2 увеличивали до 500 мкг/мл.
Положительный контроль представлял собой изотипическое IgG1-мАт дикого типа, которое сравнивали с контрольными мАт IgG1 LALA и моноклональными изотипическими IgG2- и IgG4-мАт к неспецифическим мишеням. Проточные ячейки регенерировали путем введения 10 мМ гидроксида натрия (VWR, 28244.262) при скорости потока 100 мкл/мин в течение 30 секунд. Анализ данных выполняли с помощью программного обеспечения BiaEvaluation версии 3.2 RC1 с учетом двойной поправки на на значение в холостой проточной ячейке (без иммобилизованного FcγR) и с вычитанием буферного цикла из тестируемых мАт2. Результаты показаны в таблице 7.

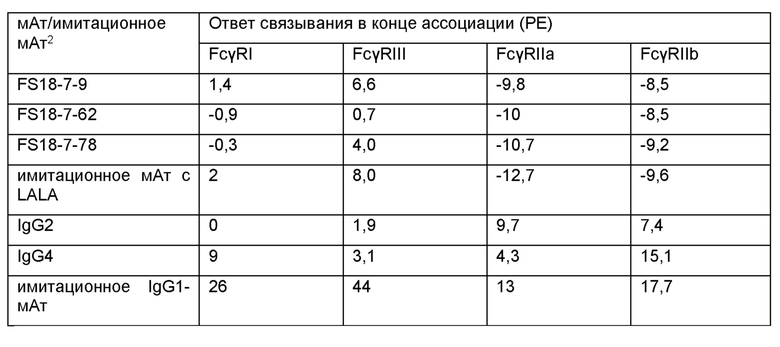
Все протестированные имитационные мАт2 (содержавшие мутацию LALA, приведенную выше) демонстрировали значительно ослабленное связывание с протестированными Fcγ-рецепторами по сравнению с контрольным антителом (имитационным IgG1-мАт) без мутации LALA, что указывало на ослабление связывания этих имитационных мАт2 с Fcγ-рецепторами за счет мутации LALA и, следовательно, ожидаемое снижение АЗКЦ-активности указанных мАт2.
1.6 Блокирование связывания ГКГС II класса с LAG-3
Способность Fcab (содержащих укороченную шарнирную область; SEQ ID NO: 58) блокировать взаимодействие между рекомбинантным LAG-3 Fc человека или мыши и ГКГС II класса человека исследовали, измеряя связывание LAG-3 Fc с клетками A375, клетками линии меланомы, экспрессирующими ГКГС II класса человека. Клетки A375 (ATCC, CRL-1619), выращенные в среде DMEM (Life Technologies, 61965-026), содержащей 10% FBS (Life Technologies, 10270-106-6), отделяли от флаконов для клеточных культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) и высевали в 96-луночные планшеты с V-образным дном (Costar, 3897), исходя из количества 2x105 клеток/лунку. Планшеты центрифугировали при 1500 об/мин в течение 3 мин при 4°C для осаждения клеток. Соответствующие концентрации Fcab или контрольного мАт инкубировали с 1 мкн/мл LAG-3 Fc (LAG-3-Fc человека, R&D Systems, 2319-L3-050 или LAG-3 Fc мыши, R&D Systems, 3328-L3-050) в 100 мкл DMEM, содержащей 10% FBS, в течение 1 ч при 4°C. Смесь LAG-3/Fcab добавляли к клеткам A375 и инкубировали в течение 1 ч при 4°C. Клетки промывали. Вторичное антитело (F(ab')2 козы против Fc человека, конъюгированное с Alexa Fluor 488, Jackson Immunoresearch, 109-546-098, или конъюгат антитела 488 козы против IgG (H+L) мыши, Life Technologies, A-1101) разбавляли PBS в соотношении 1:1000, и в количестве 100 мкл добавляли к клеткам на 30 мин при 4°C (планшеты держали в темноте). Клетки однократно промывали и ресуспендировали в 100 мкл PBS + 1 мкг/мл ДАПИ (Biotium, 40043). Планшеты считывали на цитометре BD FACSCanto II (BD Biosciences), данные анализировали с использованием программного обеспечения FlowJo.
Оба Fcab против LAG-3 мыши могли ингибировать взаимодействие ГКГС II человека с LAG-3 мыши, в то время как контрольное мАт против LAG-3 мыши (C9B7W, 2B Scientific, BE0174-50MG) такой способностью не обладало (см. таблицу 8).
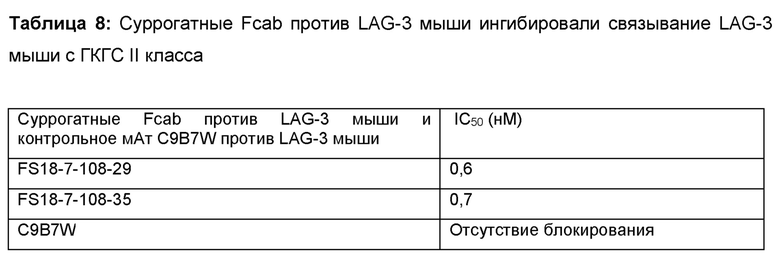
Протестированные Fcab против LAG-3 человека также могли ингибировать взаимодействие ГКГС II класса человека с LAG-3 человека с эффективностью, аналогичной эффективности контрольного мАт против LAG-3 человека (25F7).
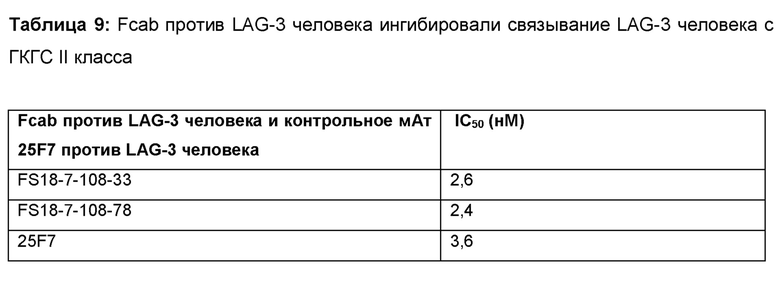
Пример 2: Получение и исследование характеристик молекул мАт и мАт2
2.1 Получение мАт 84G09
2.1.1 Получение ДНК-конструкта
Вставки ДНК, кодирующие вариабельные области тяжелой и легкой цепей 84G09, подвергли оптимизации кодонов для экспрессии в клетках животных и синтезировали с помощью DNA2.0 (Менло-Парк, штат Калифорния, США). Вставки в хозяйском векторе pJ-Amp-high субклонировали в экспрессирующих векторах pFS-hHC2.1-G1m17(z) LALA (тяжелая цепь IgG, содержащая мутацию LALA) или pFS-hHC2.1-G1m17(z) (тяжелая цепь IgG без мутации LALA) и pFShK1.0 (легкая каппа-цепь IgG) с использованием рестриктазного гидролиза посредством EcoRI и NheI.
Правильность клонирования проверяли посредством ПЦР колоний и последующего нуклеотидного секвенирования в сторонней организации (GATC Biotech).
2.1.2 Поддержание клеток
Клетки HEK293-6E (NRCC) субкультивировали в заранее подогретой среде F17 (Invitrogen, A13835-01) с добавлением 4 мм GlutaMAX-1 (Invitrogen, 35050-038), 0,1% плюроника F-68 (Invitrogen A13835-01) и 25 мкг/мл генетицина (Invitrogen, 10131-027). Клетки инкубировали при 37°C, 140 об/мин, 5% CO2 и субкультивировали из расчета 0,3x106 клеток/мл по схеме три и четыре дня.
2.1.3 Временная трансфекция
Клетки HEK293-6E временно трансфицировали с использованием PEIpro в концентрации 1 мг/мл (Polyplus, PPLU115). За 24 часа до трансфекции клетки высевали из расчета 0,8x106 клеток/мл в культуральную среду. На каждые 200 мл культуры клеток готовили смесь ДНК, смешивая 10 мл подогретой среды Opti-MEMI (Invitrogen, 11058-021), 100 мкг ДНК без эндотоксинов, кодирующей тяжелую цепь, и 100 мкг ДНК без эндотоксинов, кодирующей легкую цепь. Смесь ПЭИ готовили, смешивая 10 мл подогретой среды Opti-MEMI и 200 мкл PEIpro и перемешивая их на вортексе. Смесь ДНК быстро добавляли к перемешанной на вортексе смеси ПЭИ, перемешивали на вортексе в режиме 3 импульса в 1 с, инкубировали 3 мин при комнатной температуре и по каплям добавляли к клеткам. Через 48 часов после трансфекции в каждый флакон добавляли 20 мкл среды F17 с добавлением 0,5% триптона N1 (TekniScience Inc., 19553).
Через 6 дней после трансфекции клетки собирали центрифугированием при 4500 об/мин в течение 40 мин. Надосадочную жидкость фильтровали через 0,22-мкм полиэфирсульфоновый фильтровальный блок (Millipore, SCGPU01RE, SCGPU02RE, SCGPU05RE, SCGPU11RE) и хранили при +4°C до очистки.
2.1.4 Хроматография на белке A
Осветленную надосадочную жидкость очищали с использованием заранее упакованных 5-мл колонок HiTrap MabSelect SuRe (GE Healthcare, 11-0034-95) на ÄKTAexplorer или ÄKTAxpress. Вкратце, колонки уравновешивали 50 мМ трис-HCl, 250 мМ NaCl при pH 7,0, несвязавшийся материал отмывали тем же буфером при скорости потока 5 мл/мин. Продукты элюировали 10 мМ формиатом натрия, pH 3,0 при скорости потока 5 мл/мин. В элюированных образцах немедленно меняли буфер на PBS, pH 7,4 с использованием колонок PD-10 (GE Healthcare, 17-0851-01), заранее уравновешенных PBS, pH 7,4 в соответствии с рекомендациями производителя.
2.1.5 Измерение концентрации продукта посредством спектрометрии
Поглощение каждого очищенного продукта при 280 нм измеряли с использованием LabChip DS (PerkinElmer, 133089) с DropPlate 96 D+ (PerkinElmer, CLS135136). Концентрацию продукта рассчитывали с использованием коэффициента экстинкции (A280 для 1 мг/мл), рассчитанного с помощью программного обеспечения VectorNTI Advance v11.5.4 (Thermofisher Scientific, A13784)
2.1.6 Концентрация продукта
При необходимости очищенные фракции концентрировали с использованием блока для центробежной фильтрации Amicon Ultra-4 Centrifugal Filter Unit 30K (Millipore, UFC803024). После уравновешивания регенерированной целлюлозной мембраны Ultracel PBS, pH 7,4 посредством центрифугирования в течение 10 мин при 3000 об/мин образцы загружали в 4-мл блок и центрифугировали при 3000 об/мин до достижения желательной концентрации белка.
2.1.6 Стерилизация фильтрацией
Конечные образцы фильтровали с использованием заранее увлажненных шприцевых фильтров Millex-GV из ПВДФ (Millipore, SLGV013SL).
2.2 Получение мАт2 LAG-3/PD-L1 человека
Тяжелые цепи молекул мАт2 FS18-7-9/84G09 (SEQ ID NO 94 и 95), FS18-7-32/84G09 (SEQ ID NO 96 и 97), FS18-7-33/84G09 (SEQ ID NO 98 и 99), FS18-7-36/84G09 (SEQ ID NO 100 и 101), FS18-7-58/84G09 (SEQ ID NO 102 и 103), FS18-7-62/84G09 (SEQ ID NO 104 и 105), FS18-7-65/84G09 (SEQ ID NO 106 и 107), FS18-7-78/84G09 (SEQ ID NO 108 и 109), FS18-7-88/84G09 (SEQ ID NO 110 и 111) и FS18-7-95/84G09 (SEQ ID NO 112 и 113) получали путем замены CH3-доменов моноклональных антител 84G09 (с мутацией LALA и без нее) CH3-доменами Fcab FS18-7-9, FS18-7-32, FS18-7-33, FS18-7-36, FS18-7-58, FS18-7-62, FS18-7-65, FS18-7-78, FS18-7-88 и FS18-7-95, специфичных по отношению к LAG-3 человека, в пределах сайтов XhoI и BamHI, присутствующих в последовательности немодифицированного CH3-домена IgG1 человека. Тяжелую цепь мАт2 совместно трансфицировали с легкой цепью 84G09 (SEQ ID NO: 116), как описано для мАт 84G09 выше в разделе 2.1. Затем мАт2 экспрессировали и очищали, как описано для мАт 84G09 выше в разделе 2.1.
2.2 Получение мАт2 против LAG-3 человека/имитационного антигена
мАт против FITC (с мутацией LALA и без нее) получали, как описано для мАт 84G09 выше в разделе 2.1, с использованием тяжелых цепей (SEQ ID NO 83 и 84; с мутацией LALA и без нее) и легкой цепи (SEQ ID NO: 85) мАт 4420.
Тяжелые цепи молекул мАт2 FS18-7-9/4420 (SEQ ID NO 63 и 64), FS18-7-32/4420 (SEQ ID NO 65 и 66), FS18-7-33/4420 (SEQ ID NO 67 и 68, FS18-7-36/4420 (SEQ ID NO 69 и 70), FS18-7-58/4420 (SEQ ID NO 71 и 72), FS18-7-62/4420 (SEQ ID NO 73 и 74), FS18-7-65/4420 (SEQ ID NO 75 и 76), FS18-7-78/4420 (SEQ ID NO 77 и 78), FS18-7-88/4420 (SEQ ID NO 79 и 80) и FS18-7-95/4420 (SEQ ID NO 81 и 82) получали путем замены CH3-доменов моноклональных антител 4420 (с мутацией LALA и без нее) CH3-доменами Fcab FS18-7-9, FS18-7-32, FS18-7-33, FS18-7-36, FS18-7-58, FS18-7-62, FS18-7-65, FS18-7-78, FS18-7-88 и FS18-7-95, специфичных по отношению к LAG-3 человека, в пределах сайтов XhoI и BamHI, присутствующих в последовательности немодифицированного CH3-домена IgG1 человека. Тяжелые цепи мАт2 совместно трансфицировали с легкой цепью мАт 4420, как описано для мАт 84G09 выше в разделе 2.1. Затем белки экспрессировали и очищали, как описано для мАт 84G09 выше в разделе 2.1.
2.2 Получение мАт2 LAG-3/PD-L1 мыши
мАт против PD-L1 мыши (с мутацией LALA и без нее) получали, как описано для мАт 84G09 выше в разделе 2.1, с использованием тяжелой цепи (SEQ ID NO 122 и 123) и легкой цепи (SEQ ID NO: 119) мАт S1.
Тяжелую цепь молекул мАт2 FS18-7-108-29/S1 (SEQ ID NO 117 и 118) и FS18-7-108-35/S1 (SEQ ID NO 120 и 121) получали путем замены CH3-доменов моноклональных антител S1 (с мутацией LALA и без нее) CH3-доменами Fcab FS18-7-108-29 и FS18-7-108-35, специфичных по отношению к LAG-3 мыши, в пределах сайтов XhoI и BamHI, присутствующих в последовательности немодифицированного CH3-домена IgG1 человека. Тяжелую цепь мАт2 совместно трансфицировали с легкой цепью S1, как описано для мАт 84G09 выше в разделе 2.1. Затем белки экспрессировали и очищали, как описано для мАт 84G09 выше в разделе 2.1.
2.5 Сродство и кинетика связывания мАт2 по отношению к LAG-3 человека и PD-L1 человека
Белок L (Thermo, 21189) иммобилизовали на проточных ячейках 1 и 2 чипа CM5 серии S (GE Healthcare, BR-1005-30) посредством аминного присоединения (GE Healthcare, BR-1000-50) до поверхностной плотности 2000 РЕ, следуя инструкциям производителя для прибора BIAcore T200. Для связывания LAG-3 образцы мАт2 (все из которых содержали мутацию LALA) иммобилизовали только на проточной ячейке 2, и пропускали LAG-3Fc человека (R&D Systems, 2319-L3) в 4 концентрациях серии двукратных разведений, начиная с 0,5 нМ, через обе проточные ячейки 1 и 2 при скорости потока 30 мкл/мин. Время ассоциации составляло 3 мин, а время диссоциации - 6 мин. Подвижный буфер представлял собой HBS-P (GE Healthcare, BR-1003-68). Обе проточные ячейки регенерировали инъекцией 10 мМ гидроксида натрия (NaOH) при скорости потока 100 мкл/мин в течение 20 секунд. Данные анализировали при двойной поправке на холостую проточную ячейку.
Для связывания PD-L1 четыре концентрации серии двукратных разведений PD-L1 Fc (R&D Systems, 156-B7), начиная с 40 нМ, пропускали через мАт2, иммобилизованное на том же чипе с белком L. Все другие условия были такими же, как в эксперименте со связыванием LAG-3 (см. выше).
Кинетику связывания аппроксимировали моделью Лэнгмюра при соотношении 1:1, получая константы скорости ассоциации (ka) и диссоциации (kd) при связывании. Равновесные константы связывания (KD) рассчитывали путем деления скорости диссоциации на скорость ассоциации для каждого образца. Анализ данных выполняли с помощью программного обеспечения BiaEvaluation версии 3.2. Результаты показаны в таблицах 10 и 11.
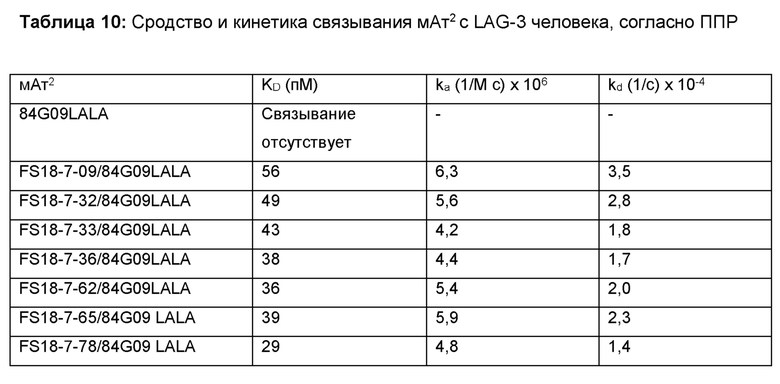
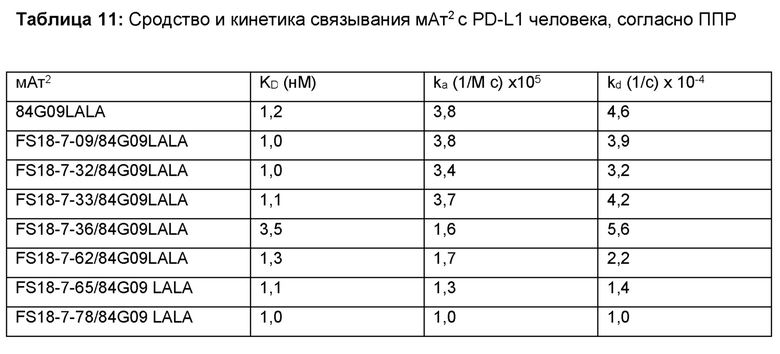
Сродство связывания по отношению к PD-L1 человека и LAG-3 человека было сопоставимо для всех протестированных мАт2. Сродство связывания мАт2 по отношению к PD-L1 человека было сопоставимо со сродством 84G09, что указывало на отсутствие влияния внедрения сайта связывания LAG-3 в CH3-домен на связывание PD-L1.
2.6 Одновременное связывание мАт2 с LAG-3 человека и PD-L1 человека
Способность мАт2 (FS18-7-09/84G09, FS18-7-32/84G09, FS18-7-33/84G09, FS18-7-36/84G09, FS18-7-62/84G09, FS18-7-65/84G09 и FS18-7-78/84G09, все из которых содержали мутацию LALA) одновременно связываться с LAG-3 и PD-L1 тестировали посредством ППР. PD-L1Fc человека (R&D Systems, 156-B7) иммобилизовали на 2 проточной ячейке чипа CM5 серии S (GE Healthcare, BR-1005-30) до поверхностной плотности 150 РЕ, следуя инструкциям производителя. Проточную ячейку 1 активировали и инактивировали без иммобилизации белка для вычитания фонового сигнала. Для каждого образца 10 мкг/мл мАт2 пропускали через проточные ячейки 1 и 2 при скорости потока 10 мкл/мин в течение 3 мин. Затем через обе проточные ячейки 1 и 2 пропускали 40 нМ LAG-3Fc (R&D Systems, 2319-L3) при скорости потока 10 мкл/мин в течение 3 мин. После каждого этапа связывания следовала диссоциация в течение 3 мин. Чип сенсора регенерировали после каждого цикла посредством 15-с инъекции 25 мМ NaOH при скорости потока 100 мкл/мин.
Все протестированные мАт2 могли одновременно связываться с LAG-3 и PD-L1. Исходное мАт 84G09 против PD-L1 связывается только с PD-L1.
2.7. Одновременное связывание суррогатного мАт2 с LAG-3 мыши и PD-L1 мыши
Способность двух суррогатных мАт2 мыши (FS18-7-108-29/S1 и FS18-7-108-35/S1, оба из которых содержали мутацию LALA) одновременно связываться с LAG-3 мыши и PD-L1 мыши протестировали посредством ППР на BIAcore 3000 (GE Healthcare). PD-L1Fc мыши (R&D Systems, 1019-B7-100) иммобилизовали на 4 проточной ячейке чипа CM5 до поверхностной плотности 830 РЕ, следуя инструкциям производителя. В 3 проточной ячейке иммобилизовали 820 РЕ Fc человека (R&D system, 110-HG) для вычитания фонового сигнала. Для каждого образца 50 мкг/мл мАт2 пропускали через проточные ячейки 1 и 2 при скорости потока 20 мкл/мин в течение 150 с. Затем через обе проточные ячейки 3 и 4 пропускали 50 нМ LAG-3Fc мыши (R&D Systems, 3328-L3-050) при скорости потока 20 мкл/мин в течение 150 с. После каждого этапа связывания следовала диссоциация в течение 3 мин. Чип сенсора регенерировали после каждого цикла с использованием 2 x 10 мкл 50 мМ NaOH. Оба протестированные мАт2 могли одновременно связываться с LAG-3 мыши и PD-L1 мыши и поэтому являлись подходящими суррогатами мАт2 LAG-3/PD-L1 человека.
2.8 Связывание рецепторов Fcγ человека мАт2, содержащими мутацию LALA
Рецепторы Fcγ человека иммобилизовали на чипе CM5 до поверхностной плотности приблизительно 200 РЕ для Fcγ RI (R&D Systems, 1257-FC) и Fcγ RIIIa (R&D Systems, 4325-FC) и приблизительно 500 РЕ для Fcγ RIIa (R&D Systems, 1330-CD) и Fcγ RIIb/c (R&D Systems, 1875-CD) согласно инструкции производителя для прибора BIAcore3000. Для Fcγ RI и Fcγ RIIIa 100 мкг/мл мАт или мАт2 пропускали через чип в течение 3 мин при скорости потока 10 мкл/мин с последующей диссоциацией в течение 5 мин. Подвижный буфер представлял собой PBS (Lonza, BE17-516F) + 0,05% (об/об) ПАВ P20 (GE Healthcare, BR-1000-54). Положительный контроль представлял собой IgG1 дикого типа - мАт 4420. Моноклональные мАт IgG2 и IgG4 по отношению к посторонним мишеням (20H4 и MOR7490) и IgG1 мыши (Sigma, P5305) включили в качестве эталонов. Регенерация не требовалась из-за быстрой диссоциации комплексов при связывании. Для тестирования Fcγ RIIa и Fcγ RIIb/c концентрацию мАт2 увеличили до 500 мкг/мл с целью компенсации более слабого связывания с этими двумя рецепторами. Результаты показаны в Таблице 12.
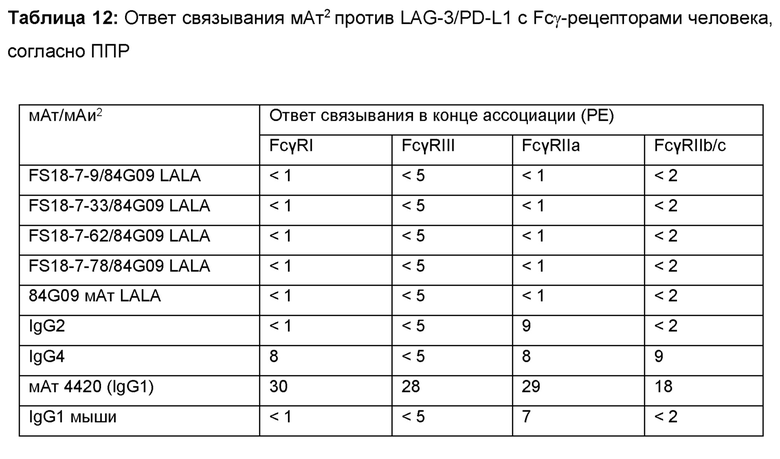
Как и ожидалось, вариант LALA, внедренный в мАт или мАт2, снижал способность этих молекул к связыванию рецепторов Fcγ человека.
2.9 Связывание мАт2 с клетками, экспрессирующими LAG-3 человека и яванского макака
Последовательность LAG-3 человека (SEQ ID NO: 126) или последовательность LAG-3 яванского макака (SEQ ID NO: 128) субклонировали в вектор pcDNA5FRT (Life Technologies, V6010-20) с использованием рестриктазного гидролиза посредством KpnI (NEB, R0142) и NotI (NEB, R0146). Затем вектор трансформировали в клетки линии Flp-In T-REx 293 HEK (Life Technologies, R780-07) с использованием липофектамина 2000 (Life Technologies, 11668-019). Трансформированные клетки Flp-In T-REx 293 выращивали в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) в течение 3-4 недель до появления видимых колоний стабильно трансформированных клеток. Эти колонии амплифицировали в присутствии 1 мкг/мл доксициклина (Sigma, D9891) и тестировали на предмет экспрессии LAG-3, которую подтвердили с помощью проточной цитометрии.
Сродство мАт2 (все из которых содержали мутацию LALA) к клеткам, экспрессировавшим LAG-3 человека или яванского макака, определяли с помощью проточной цитометрии. Клетки HEK, экспрессирующие LAG-3 человека или яванского макака, выращенные в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) и 1 мкг/мл доксициклина (Sigma, D9891), отделяли от флаконов для тканевых культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) и высевали в 96-луночные планшеты с V-образным дном (Costar, 3897) из расчета 2x105 клеток/лунку. Планшеты центрифугировали при 1500 об/мин в течение 3 мин при 4°C для осаждения клеток. Серии разведений мАт2 (или контрольного мАт) инкубировали с клетками в объеме 100 мкл в течение 1 ч при 4°C. Планшеты промывали, вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098) разбавляли PBS в соотношении 1:1000 и добавляли в количестве 100 мкл к клеткам на 30 мин при 4°C (планшеты держали в темноте). Планшеты промывали, а затем клетки ресуспендировали в 100 мкл PBS, содержащего 1 мкг/мл ДАПИ (Biotium, 40043). Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Планшеты считывали на цитометре BD FACSCanto II (BD Biosciences), данные анализировали с использованием программного обеспечения FlowJo. Результаты показаны в Таблице 13.
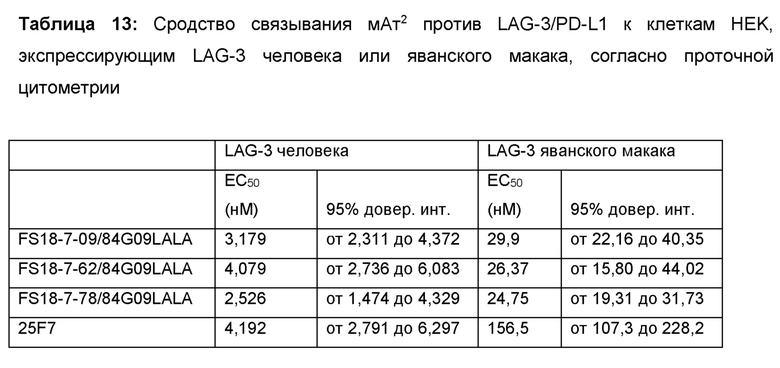
Результаты подтвердили связывание мАт2 с LAG-3 человека и яванского макака, экспрессируемым на клетках HEK. С точки зрения рассчитанных значений EC50, протестированные мАт2 демонстрировали лучшее или равное связывание с LAG-3 человека и по меньшей мере вдвое лучшее связывание с LAG-3 яванского макака по сравнению с контрольным антителом 25F7 против LAG-3. Перекрестной реакционной способности по отношению к другим белкам, экспрессируемым на поверхности клеток линии HEK, не наблюдали.
2.10 Связывание мАт2 с клетками, экспрессирующими PD-L1 человека и яванского макака
Последовательность PD-L1 человека (SEQ ID NO: 129) или последовательность PD-L1 яванского макака (SEQ ID NO: 131) субклонировали в вектор pcDNA5FRT (Life Technologies, V6010-20) с использованием рестриктазного гидролиза посредством KpnI (NEB, R0142) и NotI (NEB, R0146). Затем вектор трансформировали в клетки линии Flp-In T-REx 293 HEK (Life Technologies, R780-07) с использованием липофектамина 2000 (Life Technologies, 11668-019). Трансформированные клетки Flp-In T-REx 293 выращивали в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) в течение 3-4 недель до появления видимых колоний стабильно трансформированных клеток. Эти колонии амплифицировали в присутствии 1 мкг/мл доксициклина (Sigma, D9891) и подтверждали экспрессию LAG-3 с помощью проточной цитометрии.
Сродство связывания мАт2 (все из которых содержали мутацию LALA) с клетками, экспрессировавшими PD-L1 человека или яванского макака, или с исходными (нетрансформированными клетками) определяли с помощью проточной цитометрии. Клетки HEK, экспрессирующие PD-L1 человека или яванского макака, выращенные в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) и 1 мкг/мл доксициклина (Sigma, D9891), отделяли от флаконов для тканевых культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) и высевали в 96-луночные планшеты с V-образным дном (Costar, 3897) из расчета 2x105 клеток/лунку. Планшеты центрифугировали при 1500 об/мин в течение 3 мин при 4°C для осаждения клеток. Серии разведений мАт2 (или контрольного мАт) инкубировали с клетками в объеме 100 мкл в течение 1 ч при 4°C. Планшеты промывали, вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098) разбавляли PBS в соотношении 1:1000 и добавляли в количестве 100 мкл к клеткам на 30 мин при 4°C (планшеты держали в темноте). Планшеты промывали, а затем клетки ресуспендировали в 100 мкл PBS, содержащего 1 мкг/мл ДАПИ (Biotium, 40043). Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Планшеты считывали на цитометре BD FACSCanto II (BD Biosciences), данные анализировали с использованием программного обеспечения FlowJo. Результаты показаны в Таблице 14.

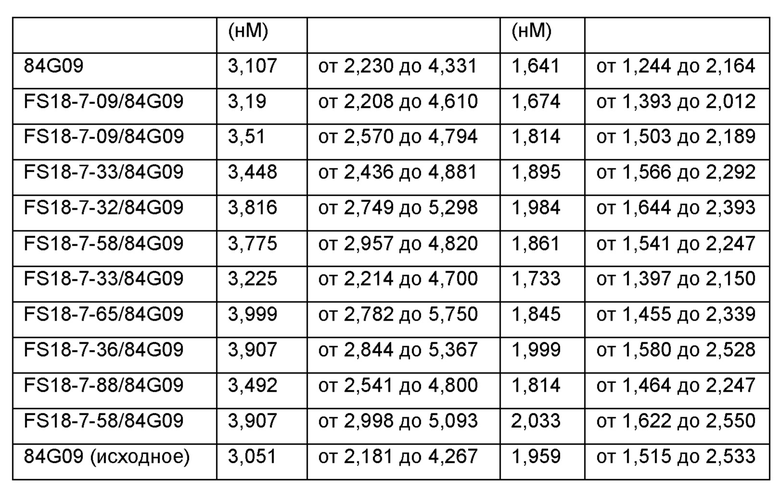
Все протестированные мАт2 LAG-3/PD-L1 связывались с PD-L1 человека и PD-L1 яванского макака с EC50, близким к аналогичному показателю мАт 84G09, что демонстрирует отсутствие влияния внедрения сайта связывания LAG-3 в CH3-домен мАт2 на сродство связывания с PD-L1.
2.11 Связывание суррогатного мАт2 мыши с клетками, экспрессирующими LAG-3 мыши или PD-L1 мыши
Последовательность LAG-3 мыши (SEQ ID NO: 127) или последовательность PD-L1 мыши (SEQ ID NO: 130) субклонировали в вектор pcDNA5FRT (Life Technologies, V6010-20) с использованием рестриктазного гидролиза посредством KpnI (NEB, R0142) и NotI (NEB, R0146). Затем вектор трансформировали в клетки линии Flp-In T-REx 293 HEK (Life Technologies, R780-07) с использованием липофектамина 2000 (Life Technologies, 11668-019). Трансформированные клетки Flp-In T-REx 293 выращивали в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) в течение 3-4 недель до появления видимых колоний стабильно трансформированных клеток. Эти колонии амплифицировали в присутствии 1 мкг/мл доксициклина (Sigma, D9891) и подтверждали экспрессию LAG-3 или PD-L1 с помощью проточной цитометрии.
Сродство связывания мАт2 (все из которых содержали мутации LALA) с клетками, экспрессировавшими LAG-3 мыши или PD-L1 мыши, определяли с помощью проточной цитометрии. Клетки HEK, экспрессирующие LAG-3 мыши или PD-L1 мыши, выращенные в среде DMEM (Life Technologies, 61965-026), содержавшей 10% FBS (Life Technologies, 10270-1-6), 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475), 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) и 1 мкг/мл доксициклина (Sigma, D9891), отделяли от флаконов для тканевых культур с использованием буфера для диссоциации клеток (Life Technologies, 13151-014) и высевали в 96-луночные планшеты с V-образным дном (Costar, 3897) из расчета 2x105 клеток/лунку. Планшеты центрифугировали при 1500 об/мин в течение 3 мин при 4°C для осаждения клеток. Серии разведений мАт2 (или контрольного мАт) инкубировали с клетками в объеме 60 мкл в течение 1 ч при 4°C. Планшеты промывали, вторичное антитело (антитело против Fc-488 человека, Jackson ImmunoResearch, 109-546-098 для Fcab или антитело против IgG крысы (H+L), конъюгат с Alexa Fluor 488, ThermoFisher, A-11006 для контрольного антитела C9B7W против LAG-3) разбавляли PBS в соотношении 1:1000 и добавляли в количестве 50 мкл к клеткам на 30 мин при 4°C (планшеты держали в темноте). Планшеты промывали, а затем клетки ресуспендировали в 50 мкл реагента для фиксации FACS Cell Fix (BD Bioscience, 340181) в течение 15 мин, затем промывали и ресуспендировали в 100 мкл PBS, содержащего 1 мкг/мл ДАПИ (Biotium, 40043). Планшеты считывали с помощью проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию на канале FITC (488 нм/530/30). Данные аппроксимировали логарифмической функцией (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. Планшеты считывали на цитометре BD FACSCanto II (BD Biosciences), данные анализировали с использованием программного обеспечения FlowJo. Результаты показаны в Таблице 15.
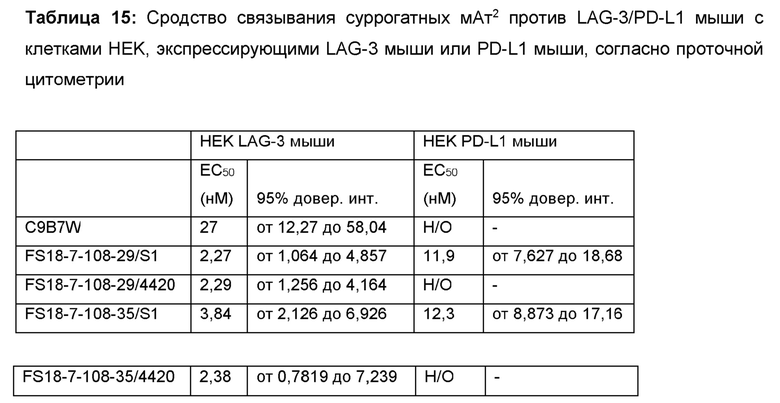
Суррогатные мАт2 могли связываться с LAG-3 мыши, экспрессируемым клетками, и с PD-L1 мыши, экспрессируемым клетками. Сродство связывания суррогатных мАт2 с LAG-3 мыши, экспрессируемым на клетках, было приблизительно аналогичным сродству мАт2 против LAG-3/PD-L1 человека с LAG-3 человека (2,3-3,8 нМ по сравнению с 2,5-4,2 нМ), а сродство связывания суррогатных мАт2 с PD-L1 мыши, экспрессируемым на клетках, до 3 раз превышало сродство мАт2 против LAG-3/PD-L1 человека с PD-L1 человека (11,9-12,3 нМ по сравнению с 3,1-4,0 нМ), демонстрируя, что эти мАт2 являлись подходящими суррогатами для мАт2 против LAG-3/PD-L1 человека для применения в исследованиях in vivo на мышах.
Пример 3: Активность молекул мАт2 в анализе активации Т-клеток и анализе SEB
3.1 Анализ активации Т-клеток
Анализ высвобождения ИЛ-2 на основе гибридомных линий Т-лимфоцитов DO11.10 OVA и В-лимфоцитов LK35.2 применили для функционального скрининга мАт2. Высвобождение ИЛ-2 является маркером активации Т-клеток. T-клетки, экспрессировавшие эндогенный PD-1 мыши, трансфицировали с пустым вектором (pLVX) или конструктом LAG-3 человека. B-клетки трансфицировали с пустым вектором (pLVX) или конструктом PD-L1 человека.
Три комбинации указанных четырех линий клеток параллельно использовали для тестирования активации Т-клеток посредством мАт2:
• DO11.10 pLVX + LK35.2 hPD-L1 для активности против PD-L1;
• DO11.10 hLAG-3 + LK35.2 pLVX для активности против LAG-3;
• DO11.10 hLAG-3 + LK35.2 hPD-L1 для одновременной активности против LAG-3/PD-L1.
Все мАт2 (все из которых содержали мутацию LALA) дважды тестировали в данном анализе активации Т-клеток. Перекрестную реакционную способность по отношению к LAG-3 и PD-L1 яванского макака тестировали при функциональном анализе активации Т-клеток с использованием клеток, сверхэкспрессирующих мишени яванского макака (cPD-L1 и cLAG-3).
Получение линий T клеток, сверхэкспрессирующих LAG-3
Для получения клеток DO11.10 (National Jewish Health), сверхэкспрессирующих LAG-3 человека, яванского макака или мыши, применяли методологию лентивирусной трансдукции с использованием системы упаковки Lenti-X HTX Packaging System (№ по каталогу 631249). Экспрессирующий вектор Lenti-X (pLVX) (№ по каталогу 631253), содержащий кДНК LAG-3 мыши (SEQ ID NO: 127), кДНК LAG-3 человека (SEQ ID NO: 126) или кДНК LAG-3 яванского макака (SEQ ID NO: 128), совместно трансфицировали с использованием смеси для упаковки Lenti-X HTX Packaging Mix в клетки линии Lenti- X 293T (№ по каталогу 632180) для получения вируса. Клетки линии DO11.10 трансдуцировали с использованием лентивирусных векторов, полученных с помощью системы упаковки Lenti-X HTX Packaging System.
Получение антиген-презентирующих клеток, сверхэкспрессирующих PD-L1
Для получения В-клеточной лимфомы LK35.2 (ATCC, HB-98), сверхэкспрессирующей PD-L1 человека, яванского макака или мыши, применяли методологию лентивирусной трансдукции с использованием системы упаковки Lenti-X HTX Packaging System (№ по каталогу 631249). Экспрессирующий вектор Lenti-X (pLVX) (№ по каталогу 631253), содержащий кДНК PD-L1 мыши (SEQ ID NO: 130), кДНК PD-L1 человека (SEQ ID NO: 129) или кДНК PD-L1 яванского макака (SEQ ID NO: 131), совместно трансфицировали с использованием смеси для упаковки Lenti-X HTX Packaging Mix в клетки линии Lenti-X 293T (№ по каталогу 632180) для получения вируса. Клетки линии LK35.2 трансдуцировали с использованием лентивирусных векторов, полученных с помощью системы упаковки Lenti-X HTX Packaging System.
Среда и пептид
Культуральная среда для клеток: DMEM (Gibco, 61965-026) 10% FBS (Gibco, 10270-106), 1 мМ пирувата натрия (Gibco, 11360-070), 1 мкг/мл пуромицина (Gibco, A11138-03)
Экспериментальная среда: полная культуральная среда для DO11.10 без пуромицина. Пептид OVA (молекулярная масса = 1773,9 Да): H-ISQAVHAAHAEINEAGR-OH (Pepscan)
Клетки:
• DO11.10 hLAG-3: Т-клеточная гибридома DO11.10, трансдуцированная лентивирусным вектором и сверхэкспрессирующая LAG-3 человека;
• DO11.10 pLVX: Т-клеточная гибридома DO11.10, трансдуцированная пустым лентивирусным вектором;
• DO11.10 cLAG-3: Т-клеточная гибридома DO11.10, трансдуцированная лентивирусным вектором и сверхэкспрессирующая LAG-3 яванского макака.
• LK 35.2 hPD-L1: В-клеточная гибридома, трансдуцированная лентивирусным вектором, содержащим hPD-L1, и сверхэкспрессирующая PD-L1 человека;
• LK 35.2 PLVX: B-клеточная гибридома, трансдуцированная пустым лентивирусным (pLVX) вектором;
• LK 35.2 cPD-L1: В-клеточная гибридома, трансдуцированная лентивирусным вектором и сверхэкспрессирующая PD-L1 яванского макака.
Клетки DO11.10 (клетки DO11.10 pLVX или клетки DO11.10 hLAG-3) в концентрации 0,3 x 106 клеток/мл смешивали в соотношении 1:1 с антителами в 3-кратной конечной концентрации. Антитела и клетки DO11.10 инкубировали при 37°C с 5% CO2 в течение 1 часа. Клетки LK 35.2 (как pLVX, так и PD-L1-pLVX) инкубировали в концентрации 3x105 клеток/мл экспериментальной среды с пептидом OVA в концентрации 1,5 мкМ в течение 30 мин. Клетки LK 35.2 + OVA добавляли к клеткам DO11.10/смеси для обработки в соотношении 1:2 в следующих комбинациях:
Функциональный скрининг с антигенами человека
DO11.10 pLVX + LK35.2 hPD-L1,
DO11.10 hLAG-3 + LK35.2 pLVX,
DO11.10 hLAG-3 + LK35.2 hPD-L1;
Скрининг перекрестной реакционной способности с антигенами яванского макака
DO11.10 pLVX + LK35.2 cPD-L1,
DO11.10 cLAG-3 + LK35.2 pLVX,
DO11.10 cLAG-3 + LK35.2 cPD-L1;
Клетки инкубировали при 37°C, 5% СО2 в течение 24 часов. Надосадочную жидкость собирали и анализировали с использованием набора для твердофазного ИФА ИЛ-2 мыши (eBioscience, 88-7024-88 или R&D systems, SM2000) согласно инструкциям производителя. Планшеты считывали при длине волны 450 нм с использованием планшет-ридера и программного обеспечения Gen5, BioTek. Значения поглощения при 570 нм вычитали из значений при 450 нм (поправка). Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации четырехпараметрической логистической моделью (программное обеспечение Gen5, BioTek). Строили график зависимости концентрации mIL-2 от десятичного логарифма концентрации Fcab или мАт, полученные кривые аппроксимировали уравнением логарифмической функции (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. В таблице 16 показаны значения EC50 и максимальное высвобождение ИЛ-2 для мАт2 и контрольных мАт, рассчитанные в процентах от контроля (84G09 + 25F7). На фигуре 2А-F показаны репрезентативные графики высвобождения ИЛ-2 для панели линий клеток, обработанных FS18-7-9/84G09, FS18-7-62/84G09 или FS18-7-78/84G09 и контрольными антителами. При анализе активации Т-клеток в присутствии обеих мишеней (DO11.10 hLAG-3 + LK35.2 hPD-L1) высвобождение ИЛ-2 индуцировалось только при ингибировании как LAG-3, так и PD-L1, например, при использовании мАт2 или комбинации антител против LAG-3 и PD-L1. Таким образом, мАт2 обладало преимуществом по сравнению с одиночными агентами по отдельности. В анализах в присутствии только LAG-3 (DO11.10 hLAG-3 + LK35.2 pvlx) или PD-L1 (DO11.10 pvlx + LK35.2 PD-L1) мАт2 показали аналогичную активность по ингибированию указанных мишеней по сравнению с одиночными агентами, демонстрируя, что мАт2 являлась единственной одиночной молекулой, способной приводить к активации Т-клеток в присутствии LAG-3, PD-L1 или LAG-3+PD-L1.
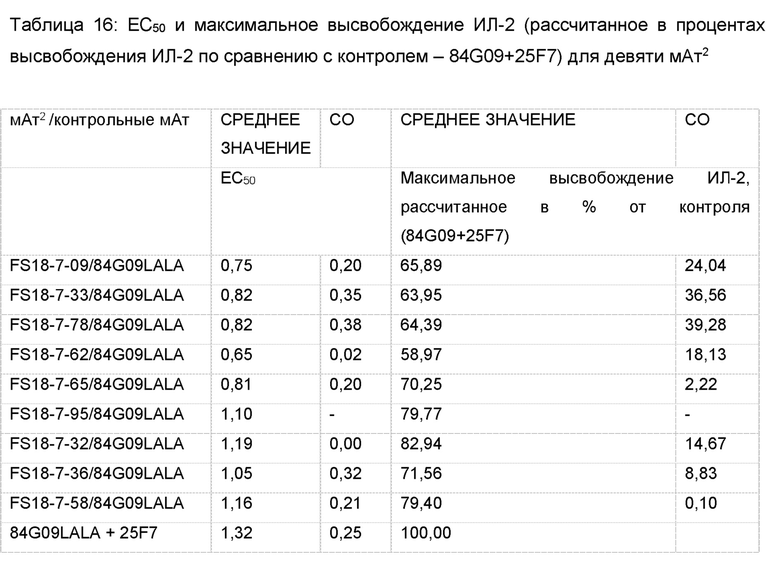
Одно из мАт2 (FS18-7-9/84G09, содержавшее мутацию LALA) тестировали на предмет функциональной перекрестной реакционной способности по отношению к антигенам яванского макака в анализе активации Т-клеток DO11.1-/LK35.2. На фигуре 3 A- C показаны результаты для панели из трех анализов на основе клеток. FS18-7-9/84G09 могло индуцировать активацию Т-клеток в присутствии как cLAG-3 + cPD-L1, так и cLAG-3 или cPD-L1 по отдельности, что указывало на функциональную перекрестную реакционную способность данного мАт2 по отношению к антигенам яванского макака и, следовательно, пригодность для применения в исследованиях безопасности на приматах.
3.2 Анализ стафилококкового энтеротоксина В
Три мАт2 (все из которых содержали мутацию LALA) тестировали в анализе стафилококкового энтеротоксина В (анализе SEB) на основе МПК человека. Стафилококковый энтеротоксин В является суперантигеном и связывается с молекулами ГКГС II класса на антиген-презентирующих клетках (АПК) и vβ-цепью T- клеточного рецептора (TCR), вызывая неспецифическую активацию T-клеток и высвобождение цитокинов. Требования к презентируемому антигену для наблюдения активации Т-клеток отсутствуют. При анализе SEB используют стимулированные клетки человека (МПК) с физиологическим уровнем ингибиторов ключевых компонентов иммунного ответа; его можно применять для подтверждения усиления активации Т-клеток за счет мАт2 у человека. Три мАт2 тестировали в системе SEB с клетками от четырех различных доноров.
Получение размноженных T-клеток
МПК выделяли из лейкоцитарного слоя посредством разделения в градиенте фиколла. CD4+ T-клетки выделяли с использованием набора для выделения Т-клеток человека CD4+ T Cell Isolation Kit (Miltenyi Biotec Ltd, 130-096-533) в соответствии с инструкцией производителя. Гранулы активатора Т-клеток человека Human T-Activator CD3/CD28 Dynabeads (Life technologies, 11131D) ресуспендирровали перемешиванием на вортексе. Гранулы переносили в стерильную 15-мл пробирку, и к промытым Dynabeads добавляли 10 мл среды RPMI (Life Technologies, 61870044) с 10% FBS (Life Technologies, 10270106) и 1x пенициллина-стрептомицина (Life Technologies, 15140122). Надосадочную жидкость выбрасывали. Необходимое количество CD4+ T- клеток в концентрации 1,0 x106 в среде RPMI с 10% FBS, 1x раствором пенициллина- стрептомицина и 50 МЕ/мл рекомбинантного ИЛ-2 человека (Peprotech, 200-02-50ug ) при соотношении гранул к клеткам 3:1 переносили во флакон T75 (Greiner Bio-one, 690195) и инкубировали при 37°C + 5% CO2. Через 3 дня клетки аккуратно ресуспендировали и подсчитывали. Плотность клеток поддерживали в диапазоне 0,8-1 x 106 клеток/мл, при необходимости добавляя свежую среду (RPMI-10% FBS + раствор пенициллина-стрептомицина 1X + 50 МЕ/мл rhuIL2). На 7 или 8 день гранулы CD3/28 удаляли, а CD4+ T-клетки оставляли на ночь в концентрации 1 x 106 клеток/мл свежей среды RPMI-10% FBS + раствор пенициллина-стрептомицина 1X с 10 МЕ/мл rhuIL2. Клетки до использования хранили в замороженном виде.
Получение MoiDC
Интактные моноциты выделяли из МПК человека с использованием набора для выделения моноцитов человека Pan Monocyte Isolation Kit (Miltenyi Biotec Ltd, 130-096-537) в соответствии с инструкцией производителя. Моноциты дифференцировали до iDC с использованием среды для дифференцировки Mo-DC человека (Miltenyi Biotec Ltd, 130-094-812) в соответствии с инструкцией производителя.
Анализ SEB
Размноженные T-клетки размораживали за день до эксперимента, промывали средой AIM (Gibco, 12055-091) и инкубировали при 37°C, 5% CO2 в течение ночи при концентрации 1x106 клеток/мл среды AIM. Концентрацию 2 мкМ каждого антитела/смеси готовили в DPBS (Gibco, 14190-169) и разбавляли средой в соотношении 1:10 (30 мкл + 270 мкл) до концентрации 200 нМ. В 96-луночном планшете получали последовательные разведения в соотношении 1:10 (30 мкл + 270 мкл экспериментальной среды; 2x конечная конц.). MoiDC размораживали, промывали средой AIM и смешивали с T-клетками от того же донора в соотношении 1:10 (5 мл iDC при концентрации 2x105 клеток/мл объединяли с 5 мл Т-клеток при концентрации 2x106 клеток/мл). 20 мкл SEB (Sigma, S4881) в концентрации 0,1 мкг/мл добавляли к 10 мл клеток. В круглодонном 96-луночном планшете добавляли 100 мкл смеси клетки/SEB к 100 мкл разведения антитела, получая соотношение 104 клеток iDC к 105 T-клеток с 0,1 нг/мл SEB в 200 мкл среды AIM на лунку при конечной концентрации антитела 100, 10, 1, 0,1, 0,01, 0,001 нМ. Клетки инкубировали при 37°С, 5% CO2 в течение 3 дней. Надосадочную жидкость собирали и немедленно анализировали с использованием набора для твердофазного ИФА ИФНγ человека (R&D Systems, PDIF50) или замораживали при -20°C для дальнейших анализов. Анализ выполняли в соответствии с инструкциями производителя набора с использованием надосадочной жидкости, разбавленной PBA (DPBS, 2% БСА (Sigma, A7906-100G)) в соотношении 1:30. Строили график зависимости концентрации ИФНγ человека от десятичного логарифма концентрации мАт2 или мАт, полученные кривые аппроксимировали уравнением логарифмической функции (для агонистического взаимодействия) в зависимости от ответа в программном обеспечении GraphPad Prism. В таблице 17 показаны значения EC50 и величина высвобождения ИФНγ в анализе SEB с клетками от четырех различных доноров клеток (доноров A-D). На фигуре 4 показан репрезентативный график анализа SEB с одним донором. Во всех шести анализах мАт2 LAG-3/PD-L1 и комбинация мАт2 LAG-3/FITC + 84G09LALA демонстрировали более выраженную активацию, чем только мАт 84G09LALA, в то время как мАт2 LAG-3/ FITC или мАт 4420 не демонстрировали значимой активации при анализе. Таким образом, результаты анализа SEB подтверждали результаты анализа DO11.10/LK35.2 в более физиологической системе.

Пример 4: Активность молекул мАт2 in vivo на моделях опухолей у мышей
3.2 Активность мАт2 в модели неразвившейся опухоли MC38
Сингенную модель опухоли MC38 использовали в данном эксперименте, поскольку известно, что опухоли MC38 экспрессируют PD-L1 на поверхности своих клеток и являются высокоиммуногенными, что приводит к усиленной экспрессии LAG-3 на иммунных клетках в опухоли и на периферии опухоли.
Суррогатное мАт2 мыши FS18-7-108-29/S1, содержащее мутацию LALA (SEQ ID NO: 117 и 119), называемое FS18-29/S1, протестировали на предмет активности in vivo с использованием сингенной модели роста опухоли MC38 у мышей. Способность мАт2 ингибировать рост опухоли сравнивали с действием LAG-3/имитационного мАт2 FS18-7-108-29/4420, содержащего мутацию LALA (SEQ ID NO: 132 и 85), называемого FS18-29/4420, эталонного мАт C9B7W против LAG-3 (2B scientific; № в каталоге BE0174-50MG), эталонного мАт S1 против PD-L1, содержащего мутацию LALA (SEQ ID NO: 122 и 119), и комбинации мАт C9B7W и S1.
Самок мышей C57BL/6 (Charles River) в возрасте 8-10 недель и массой по 20-25 г выдерживали перед началом исследования в течение одной недели. Всем животным вводили микрочипы и присваивали уникальные идентификаторы. Каждая группа включала 10 мышей. Клетки линии карциномы толстой кишки MC38 (S. Rosenberg, NIH) первоначально размножали, хранили и подвергали предварительному скринингу на предмет патогенных организмов с использованием IDEXX Bioresearch и протокола IMPACT I; продемонстрировано, что данная линия не содержала патогенных организмов. Клетки MC38 размораживали с температуры хранения -150°C и добавляли к 20 мл среды DMEM (Gibco, 61965-026) с 10% FCS (Gibco, 10270-106) во флаконах для культивирования тканей T175. Мышей анестезировали изофлураном (Abbott Laboratories), каждому животному подкожно вводили 2x106 клеток в левую подвзошную область. Через 7-8 дней после инокуляции клеток мышей, у которых отсутствовали опухоли в указанный момент, удаляли из исследования.
Все молекулы мАт2 и контрольные антитела анализировали в течение 24 часов перед введением, получая профиль эксклюзионной ВЭЖХ и проверяя их на предмет примесей. Антитела получали в конечной концентрации 10 мг/кг в PBS и в исследованиях комбинаций объединяли со вторым антителом. Молекулы мАт2 или контрольные антитела вводили мышам путем внутрибрюшинной (в/б) инъекции на 8, 11 и 14 день после инокуляции опухоли. В назначенную дату выполняли точные измерения опухолей и введение составов; в оставшееся время исследования мышей подвергали тщательному наблюдению. Измерение объема опухолей выполняли штангенциркулем, определяя самую длинную и самую короткую ось опухоли. Для расчета объема опухоли использовали следующую формулу:
L x (S2) / 2
где L = самая длинная ось; S = самая короткая ось
Исследование остановили на 20 день, когда опухолевую нагрузку считали близкой к заданному пределу. Всех мышей гуманно умертвили, опухоли хирургически удалили и взвесили.
Результаты показаны на фигурах 5 и 6. Мыши, обработанные мАт2 LAG-3/PD-L1 (FS18-29/S1), характеризовались конечными опухолями значительно меньшей массы по сравнению с мышами, обработанными комбинацией эталонных мАт C9B7W и S1. Конкретнее, статистический анализ конечной массы опухолей выполнили с использованием двустороннего t-критерия Стъюдента в программном пакете GraphPad Prism, продемонстрировав статистически значимое различие между введением FS18-29/S1 и введением комбинации эталонных антител C9B7W и S1 (p = 0,0125), указывавшее на то, что ингибирование LAG-3 и PD-L1 одной и той же молекулой приводило к синергическому действию на конечную массу опухолей по сравнению с ингибированием LAG-3 и PD-L1 отдельными молекулами.
Суррогатное мАт2 FS18-29/S1 также обладало выраженным действием на рост опухоли, предотвратив развитие 6 из 8 растущих опухолей MC38 и замедлив рост в остальных 2 случаях. Введение эталонных антител против LAG-3 и PD-L1 в комбинации замедлило рост опухоли у 7 животных, однако животные без опухолей отсутствовали.
FS18-29/4420 само по себе не оказывало выраженного действия на рост опухоли, что указывало на то, что для максимальной эффективности мАт2 требуется Fab против PD-L1. Эталонное антитело против LAG-3 мыши само по себе оказывало слабое влияние или не оказывало влияния на итоговый рост опухоли, в то время как эталонное антитело против PD-L1 мыши предотвратило развитие 1 из 7 опухолей в данной группе и оказывало некоторое общее влияние, замедляющее рост опухолей.
Сингенные модели мыши считаются подходящими системами на основе мышей для тестирования противоопухолевого действия ингибирования терапевтических мишеней и интенсивно используются для валидации разрабатываемых терапевтических средств медицинского назначения.
4.2 Активность молекул mAb2 на модели развившейся опухоли MC38
Суррогатное мАт2 мыши FS18-7-108-29/S1, содержащее мутацию LALA (SEQ ID NO: 117 и 119), называемое FS18-29/S1, протестировали на предмет активности in vivo на сингенной модели роста опухоли MC38 у мышей. Способность мАт2 ингибировать рост опухоли сравнивали с действием LAG-3/имитационного мАт2 FS18-7-108-29/4420, содержащего мутацию LALA (SEQ ID NO: 132 и 85), называемого FS18-29/4420, эталонного мАт C9B7W против LAG-3, эталонного мАт S1 против PD-L1, содержащего мутацию LALA (SEQ ID NO: 122 и 119), и комбинации C9B7W и S1.
Самок мышей C57BL/6 (Charles River) в возрасте 8-10 недель и массой по 20-25 г выдерживали перед началом исследования в течение одной недели. Всем животным вводили микрочипы и присваивали уникальные идентификаторы. Каждая группа включала 10 мышей. Клетки линии карциномы толстой кишки MC38 (S. Rosenberg, NIH) первоначально размножали, хранили и подвергали предварительному скринингу на предмет патогенных организмов с использованием IDEXX Bioresearch и протокола IMPACT I; продемонстрировано, что данная линия не содержала патогенных организмов. Клетки MC38 (приблизительно 3-5x106) размораживали с температуры хранения -150°C и добавляли к 20 мл среды DMEM (Gibco, 61965-026) с 10% FCS (Gibco, 10270-106) во флаконах для культивирования тканей T175. Мышей анестезировали изофлураном (Abbott Laboratories), каждой мыши подкожно вводили 2x106 клеток в объеме 100 мкл в левую подвзошную область. Через 7-8 дней после инокуляции клеток выполняли стандартный осмотр мышей на предмет состояния здоровья и роста опухолей, подходящего для начала исследования. Когда большинство мышей демонстрировали опухоли диаметром 5-10 мм, их сортировали и рандомизировали в исследуемые группы. Мышей, у которых отсутствовали опухоли соответствующего размера в указанный момент, удаляли из исследования.
Все молекулы мАт2 и контрольные антитела анализировали в течение 24 часов перед введением, получая профиль эксклюзионной ВЭЖХ и проверяя их на предмет примесей. Антитела получали в конечной концентрации 10 мг/кг в PBS и для исследований комбинаций объединяли со вторым антителом. Молекулы мАт2 или контрольные антитела вводили мышам путем в/б инъекции на 15, 18 и 21 день после инокуляции опухоли. У животных выполняли скрининг состояния здоровья под анестезией три раза в неделю в слепом режиме, выполняя при этом точное измерение опухолей. Измерение объема опухолей выполняли штангенциркулем, определяя самую длинную и самую короткую ось опухоли. Формула, использованная для расчета объема опухолей, приведена выше в разделе 4.1.
Исследование остановили на 24 день, когда опухолевую нагрузку считали близкой к заданному пределу. Всех мышей гуманно умертвили, опухоли хирургически удалили и взвесили. Результаты показаны на фигурах 7 и 8.
Эффективный контроль или подавление роста опухоли на сингенных моделях опухолей у мышей лучше всего выполнять путем терапевтического вмешательства на ранних стадиях (начиная с объема опухолей менее 40 мм3). Чем позже выполнено вмешательство, тем сложнее наблюдать положительные эффекты по отношению к росту опухоли, хотя, возможно, это в большей степени относится к ситуации в клинических условиях у человека.
FS18-29/S1 оказывало положительное влияние как при подавлении роста опухоли, так и при предотвращении развития карциномы толстой кишки MC38 у иммунокомпетентных мышей при его введении мышам C57BL/6 на ранних стадиях. При введении на более поздних стадиях (начиная с объема опухолей 50-125 мм3) FS18-29/S1 так же эффективно подавляло рост опухоли, как и комбинация эталонных антител. FS18-29/4420 само по себе не оказывало заметного влияния на рост опухоли, а S1 и C9B7W слегка влияли на итоговый рост опухоли.
3.2 Активность мАт2 в модели неразвившейся опухоли CT26
Сингенную модель опухоли CT26 использовали в данном эксперименте, поскольку известно, что опухоли CT26 экспрессируют PD-L1 на поверхности своих клеток и являются высокоиммуногенными, что приводит к усиленной экспрессии LAG-3 на иммунных клетках в опухоли и на периферии.
Суррогатное мАт2 мыши FS18-7-108-29/S1, содержащее мутацию LALA (SEQ ID NO: 117 и 119), называемого FS18-29/S1, и FS18-7-108-35/S1, содержащее мутацию LALA (SEQ ID NO: 120 и 119), называемое FS18-35/S1, протестировали на предмет активности in vivo на сингенной модели роста опухоли CT26 у мышей. Способность мАт2 ингибировать рост опухоли сравнивали с действием LAG-3/имитационного мАт2 FS18-7-108-29/4420, содержащего мутацию LALA (SEQ ID NO: 132 и 85), называемого FS18-29/4420, и FS18-7-108-35/4420, содержащего мутацию LALA (SEQ ID NO: 133 и 85), называемого FS18-35/4420, эталонного мАт C9B7W против LAG-3, эталонного мАт S1 против PD-L1, содержащего мутацию LALA (SEQ ID NO: 122 и 119), и комбинации C9B7W и S1.
Самок мышей BALB/с (Charles River) в возрасте 8-10 недель и массой по 20-25 г выдерживали перед началом исследования в течение одной недели. Всем животным вводили микрочипы и присваивали уникальные идентификаторы. Каждая группа включала 10 мышей. Клетки линии карциномы толстой кишки CT26 (ATCC, CRL-2638) первоначально размножали, хранили и подвергали предварительному скринингу на предмет патогенных организмов с использованием IDEXX Bioresearch и протокола IMPACT I; продемонстрировано, что данная линия не содержала патогенных организмов. Клетки CT26 (приблизительно 3-5x106) размораживали с температуры хранения -150°C и добавляли к 20 мл среды DMEM (Gibco, 61965-026) с 10% FCS (Gibco, 10270-106) во флаконах для культивирования тканей T175. Через 7-8 дней после инокуляции клеток выполняли стандартный осмотр мышей на предмет состояния здоровья и роста опухолей, подходящего для начала исследования. Когда большинство мышей демонстрировали опухоли диаметром 3-5 мм3, их сортировали и рандомизировали в исследуемые группы. Мышей, у которых отсутствовали опухоли в указанный момент, удаляли из исследования.
Все молекулы мАт2 и контрольные антитела анализировали в течение 24 часов перед введением, получая профиль эксклюзионной ВЭЖХ и проверяя их на предмет примесей. Антитела получали в конечной концентрации 10 мг/кг в PBS и в исследованиях комбинаций объединяли со вторым антителом. Молекулы мАт2 и контрольные антитела вводили мышам на 8, 11 и 14 день после инокуляции опухоли. У животных выполняли скрининг состояния здоровья, выполняя при этом точное измерение опухолей. Измерение объема опухолей выполняли штангенциркулем, определяя самую длинную и самую короткую ось опухоли. Формула, использованная для расчета объема опухолей, приведена выше в разделе 4.1.
Исследование остановили на 20 день, когда опухолевую нагрузку считали близкой к заданному пределу. Всех мышей гуманно умертвили, опухоли хирургически удалили и взвесили. Результаты показаны на фигурах 9 и 10.
Статистический анализ конечной массы опухолей выполняли с использованием двустороннего t-критерия Стъюдента в программном пакете GraphPad Prism. Статистический анализ кривых роста опухолей выполняли путем сравнения функции Growth Curves (кривые роста) из пакета статистического моделирования (Statistical Modeling), statmod (Elso et al., 2004 и Baldwin et al., 2007), доступного в проекте R для статистических расчетов (R Project for Statistical Computing).
Продемонстрировано статистически значимое различие между мАт2 FS18-35/S1 и контрольным IgG (нормальный рост) при подавлении роста опухоли. Такого статистически значимого различия не наблюдали при использовании комбинации эталонных антител или мАт2 FS18-29/S1 по сравнению с группой контрольного IgG или любой другой группой в данном исследовании.
Модель опухоли CT26 представляет собой модель агрессивной, быстро растущей опухоли, по своей природе склонной к развитию метастазов в кишечнике мышей, которая в итоге характеризуется крайне ограниченным терапевтическим окном.
Как ни странно, комбинация эталонных антител против LAG-3 и PD-L1 не подавляла рост опухоли в значительной степени по сравнению с группой контрольного IgG. В то же время в группе, обработанной FS18-35/S1, выявили значительное подавление роста по сравнению с группой контрольного IgG. Хотя FS18-29/S1 подавляло рост опухоли по сравнению с контрольным IgG, этот эффект не был статистически значимым. В данном исследовании показана вторая модель опухоли, в которой мАт2 против LAG-3/PD-L1 мыши продемонстрировали положительный эффект в отношении замедления роста опухоли по меньшей мере в той же степени, что и введение комбинации эталонных моноклональных антител.
4.4 Влияние мутации LALA на ингибирование роста опухоли в модели неразвившейся опухоли MC38
Два мАт2 (FS18-7-108-29/S1 LALA и FS18-7-108-29/S1) протестировали с целью оценки потенциальных различий противоопухолевой активности этих мАт2 с мутацией LALA в Fc-области или без нее. Суррогатное мАт2 мыши FS18-7-108-29/S1, называемое FS18-29/S1, содержащее (SEQ ID NO: 117 и 119) и не содержащее мутации LALA (SEQ ID NO: 118 и 119), протестировали на предмет активности in vivo с использованием сингенной модели роста опухоли MC38 у мышей. Способность мАт2 ингибировать рост опухоли сравнивали с действием LAG-3/имитационного мАт2 FS18-7-108-29/4420, содержащего (SEQ ID NO: 132 и 85) и не содержащего мутации LALA (SEQ ID NO: 134 и 85), называемого FS18-29/4420LALA и FS18-29/4420, и комбинации LAG-3/имитационного мАт2, содержащего и не содержащего мутации LALA, с эталонным мАт S1 против PD-L1, содержащим (SEQ ID NO: 122 и 119) и не содержащим мутации LALA (SEQ ID NO: 123 и 119).
Самок мышей C57BL/6 (Charles River) в возрасте 8-10 недель и массой по 20-25 г выдерживали перед началом исследования в течение одной недели. Всем животным вводили микрочипы и присваивали уникальные идентификаторы. Каждая группа включала 10 мышей. Клетки линии карциномы толстой кишки MC38 (S. Rosenberg, NIH) первоначально размножали, хранили и подвергали предварительному скринингу на предмет патогенных организмов с использованием IDEXX Bioresearch и протокола IMPACT I; продемонстрировано, что данная линия не содержала патогенных организмов. Клетки MC38 (приблизительно 3-5x106) размораживали с температуры хранения -150°C и добавляли к 20 мл среды DMEM (Gibco, 61965-026) с 10% FCS (Gibco, 10270-106) во флаконах для культивирования тканей T175. Мышей анестезировали изофлураном (Abbott Laboratories), каждой мыши подкожно вводили 2x106 клеток в объеме 100 мкл в левую подвзошную область. Мышам дали восстановиться под наблюдением; дату инокуляции отметили как день 0. Через 7-8 дней после инокуляции клеток выполняли стандартный осмотр мышей на предмет состояния здоровья и роста опухолей, подходящего для начала исследования. Мышей, у которых отсутствовали опухоли в указанный момент, удаляли из исследования.
Все молекулы мАт2 и контрольные антитела анализировали в течение 24 часов перед введением, получая профиль эксклюзионной ВЭЖХ и проверяя их на предмет примесей. Антитела получали в конечной концентрации 10 мг/кг в PBS и в исследованиях комбинаций объединяли со вторым антителом. Молекулы мАт2 или контрольные антитела вводили мышам путем в/б инъекции на 8, 11 и 14 день после инокуляции опухоли. У животных выполняли скрининг состояния здоровья под анестезией три раза в неделю в слепом режиме, выполняя при этом точное измерение опухолей. Измерение объема опухолей выполняли штангенциркулем, определяя самую длинную и самую короткую ось опухоли. Формула, использованная для расчета объема опухолей, приведена в разделе 4.1.
Всех мышей гуманно умертвили, опухоли хирургически удалили и взвесили. Результаты показаны на фигурах 11 и 12.
Данное исследование подтвердило, что наличие или отсутствие мутации LALA, устраняющей АЗКЦ-активность, не оказывало статистически значимого влияния на рост опухоли на модели карциномы толстой кишки MC38; в то же время мАт2, содержавшие мутацию LALA, имели тенденцию к усиленному подавлению роста опухоли. Как бы то ни было, основание для включения мутации, потенциально ингибирующей АЗКЦ-активность против T-клеток, экспрессирующих LAG-3 или PD-L1, подтверждено, поскольку мутация LALA не оказывала вредоносного влияния на противоопухолевую активность мАт2 против LAG-3/PD-L1. Имелись данные, указывающие на то, что включение мутации LALA было критично только для антитела против PD-L1.
В жанном исследовании также выполнена оценка возможности того, что мАт2 против LAG-3/PD-L1 обладает повышенной эффективностью по сравнению с введением отдельных антител (LAG-3 LALA + PD-L1 LALA). В данном случае значимые различия между двумя указанными группами отсутствовали. В обеих группах отмечено подавление роста на модели карциномы толстой кишки MC38.
4.5 Заключение
В целом вышеприведенные результаты ясно показывают наличие синергического эффекта по отношению к подавлению роста опухоли при введении молекулы мАт2, содержащей сайты связывания как LAG-3, так и PD-L1, мышам в протестированных моделях. На основании этих результатов ожидается, что молекулы антител согласно настоящему изобретению будут демонстрировать лучший эффект в лечении рака у пациентов-людей, в частности, по отношению к подавлению роста опухоли, чем введение двух отдельных молекул, связывающихся с LAG-3 и PD-L1, соответственно.
Пример 5: Действие мАт2 на экспрессию LAG-3 Т-клетками
Механизм, за счет которого суррогатное мАт2 мыши FS18-7-108-29/S1, содержащее мутацию LALA (SEQ ID NO: 117 и 119), называемое FS18-29/S1, снижало опухолевую нагрузку, протестировали на сингенной модели роста опухоли MC38, экспрессирующей овальбумин (MC38.OVA), у мышей. Влияние FS18-29/S1 сравнивали с влиянием LAG-3/имитационного мАт2 FS18-7-108-29/4420, содержащего мутацию LALA (SEQ ID NO: 132 и 85), называемого FS18-29/4420, эталонного мАт S1 против PD-L1, содержащего мутацию LALA (SEQ ID NO: 122 и 119), и комбинации FS18-29/4420 и S1.
В день имплантации культивированные клетки MC38.OVA собирали во время логарифмической фазы роста (конфлюэнтность ~75%) и ресуспендировали в PBS в концентрации 1 x 107 клеток/мл. Опухоли инициировали, вначале анестезировав каждое животное изофлураном, а затем подкожно имплантировав 1 x 106 клеток MC38.OVA (0,1 мл суспензии) в левую подвздошную область каждого тестируемого животного. Через одиннадцать дней после имплантации опухолевых клеток животных рандомизировали с использованием способа детерминированной рандомизации в пять групп с объемом отдельных опухолей от 32 до 62,5 мм3. Животным вводили 10 мг/кг антитела или мАт2 в 12, 14 и 16 день после инокуляции опухоли; опухоли собирали от трех животных/группу в 19 и 23 день после инокуляции опухоли. Для диссоциации опухолей использовали GentleMACSTM Dissociator, а затем клетки пропускали через сито для клеток с ячейками 70 μм для получения суспензии одиночных клеток. 1x106 клеток/лунку в 96-луночном планшете ресуспендировали в буфере для FACS с красителем для выявления жизнеспособности в соотношении 1:3000 и реагентом для блокирования Fc (антитело против CD16/32). Клетки для FACS-анализа окрашивали с использованием стандартной смеси, содержавшей меченые антитела против CD43, CD8a, CD4, FoxP3 и LAG-3. Для внутриклеточного окрашивания FoxP3 клетки фиксировали и пермеабилизировали перед окрашиванием с использованием антитела против FoxP3. Образцы анализировали на проточном цитометре Canto II с компенсационной матрицей и минимальным количеством 500000 подсчитываемых событий.
В данном эксперименте ОИЛ исследовали на предмет экспрессии LAG-3 после третьего введения антитела/мАт2, когда становилось очевидным различие роста опухолей при обработке контрольным антителом и мАт2, но до появления крупных различий в размере опухолей, которое могло исказить результаты. Обнаружено, что в указанный момент времени экспрессия LAG-3 на ОИЛ была существенно понижена у животных, обработанных мАт2 FS18-29/S1 или комбинацией FS18-29/4420 и S1. Конкретно, как показано на фигуре 13, экспрессия LAG-3 на CD8, CD4 и FoxP3 опухоль-инфильтрирующих лимфоцитах (ОИЛ) снижалась после обработки FS18-29/S1 на 19 и 23 день после инокуляции опухоли, что соответствовало 3 и 7 дню после последнего введения антитела/мАт2, соответственно. Снижение экспрессии LAG-3 было более выражено на 23 день, однако было очевидным уже на 19 день. Животные, получавшие комбинацию FS18-29/4420 и S1, также демонстрировали снижение экспрессии LAG-3 на ОИЛ, однако этот эффект был отсрочен до 23 дня, в то время как обработка FS18-29/4420 или S1 при введении по отдельности оказывала незначительное влияние или не оказывала влияния на экспрессию LAG-3 на ОИЛ.
Эти результаты показывают, что для снижения экспрессии LAG-3 ОИЛ требуется двойное ингибирование LAG-3 и PD-L1, поскольку это явление не наблюдалось у животных, получавших агенты против LAG-3 или PD-L1 по отдельности. Безотносительно к теоретическим представлениям считается, что двойная обработка против LAG-3 и против PD-L1 приводила к снижению экспрессии LAG-3 на ОИЛ, что ослабляло ингибирующее действие LAG-3 и позволяло ОИЛ преодолевать истощение. После активации ОИЛ они могут распознавать неоантигены, экспрессируемые опухолью, и вызывать ответ против нее, тем самым снижая опухолевую нагрузку. Таким образом, это считается механизмом, за счет которого обработка мАт2 против LAG3/PD-L1 приводила к снижению опухолевой нагрузки.
Пример 6: Антителозависимая клеточная цитотоксичность и комплемент- зависимая цитотоксичность мАт2
Антитела IgG1 обычно обладают эффекторными функциями за счет консервативных сайтов взаимодействия в константной области молекулы. Сюда входят антителозависимая клеточная цитотоксичность (АЗКЦ), опосредованная связыванием с FcγR, экспрессируемым на моноцитах/макрофагах, дендритных клетках, NK-клетках, нейтрофилах и других гранулоцитах, а также комплемент-зависимая цитотоксичность (КЗЦ), опосредованная индукцией каскада комплемента, инициируемой связыванием с компонентом C1q комплемента. Поскольку LAG-3 преимущественно экспрессируется на активированных Т-клетках, а PD-L1 экспрессируется на них, а также на опухолевых клетках в значительном количестве, исследовали способность мАт2 FS18-7-9/84G09 (SEQ ID NO 94 и 116) индуцировать АЗКЦ и КЗЦ.
Конкретнее, протестировали возможность индукции лизиса соответствующих клеток- мишеней за счет обработки клеток, экспрессирующих LAG-3 или PD-L1, антителом FS18-7-9/84G09 с последующим инкубированием с NK-клетками или комплементом. Кроме того, поскольку FS18-7-9/84G09 является биспецифическим антителом, также протестировали влияние связывания мишени с одним из сайтов специфического связывания на эффекторную функцию по отношению к клеткам, экспрессирующим мишень для другого сайта специфического связывания.
Информация об эффекторных функциях мАт2 FS18-7-9/84G09 полезна по ряду причин, в том числе из-за необходимости определить необходимость включения мутации, ослабляющей эффекторные функции, например, мутации LALA, в молекулу с целью защиты эффекторных Т-клеток, экспрессирующих LAG-3 и вовлеченных в уничтожение опухоли, от FS18-7-9/84G09-опосредованной АЗКЦ и/или КЗЦ.
6.1 Схема эксперимента
Для всех анализов использовали клетки Raji, рекомбинантно экспрессирующие LAG-3 или PD-L1, используя их эндогенную экспрессию CD20 в качестве контроля для адресного воздействия непатентованой версии ритуксимаба с целью демонстрации функциональной активности препаратов комплемента и NK-клеток независимо от рекомбинантной экспрессии белков мишеней. Экспрессию мишеней подтверждали перед этими экспериментами.
Для простейшего определения КЗЦ-активности против клеток, экспрессирующих LAG-3 или PD-L1, эту активность измеряли по высвобождению ЛДГ посредством преобразования субстрата в флуоресцентный краситель (CytoTox-ONE™ производства Promega). Чтобы определить, какие клетки-мишени в смеси клеток, содержащей как клетки, экспрессирующие LAG-3, так и клетки, экспрессирующие PD-L1, подвергались лизису, измеряли дифференциальную КЗЦ с помощью проточной цитометрии клеток-мишеней с различными флуоресцентными метками после инкубирования с мАт2/антителом, детектируя мертвые клетки с использованием флуоресцентного красителя, выводящегося из живых клеток.
Для определения АЗКЦ-активности против клеток, экспрессирующих LAG-3 или PD-L1, эту активность измеряли с использованием NK-клеток, выделенных из замороженных МПК, колориметрически измеряя высвобождение ЛДГ (CytoTox 96 производства Promega). Во всех этих исследованиях в качестве контроля использовали ритуксимаб в различных изотипах и Fc-конфигурациях. Надежный способ дифференциального измерения АЗКЦ неизвестен, поэтому дифференциальное измерение АЗКЦ- активности не выполняли.
Во всех экспериментах в качестве контролей использовали PD-L1-специфичное мАт (84G09) и LAG-3-специфичное мАт (25F7). Кроме того, использовали изотипический контрольный IgG (4420) в качестве отрицательного контроля или для получения фоновой КЗЦ-активности. Кроме того, в анализах КЗЦ и АЗКЦ выполняли сравнение LALA-версий соответствующих антител и мАт2 (за исключением 25F7) с версиями IgG1 дикого типа с целью определения влияния этой мутации на их эффекторные функции.
6.2 Материалы и способы
6.2.1 Анализы КЗЦ
Все антитела/мАт2, включая ритуксимаб, разбавляли, получая 10 последовательных разведений 1 к 2. Кроме того, подготавливали контрольные лунки, содержавшие IgG (4420 LALA) в максимальной используемой концентрации. Суспензии клеток Raji, рекомбинантно экспрессирующих LAG-3 или PD-L1, соответственно, получали в бессывороточной среде для анализа высвобождения ЛДГ и добавляли в равном объеме к подготовленным антителам/мАт2.
Для анализа КЗЦ на основе проточной цитометрии получали суспензии с концентрацией 5 x 107 клеток и ресуспендировали их в 0,5 мкМ CellTracker deep red (CellTracker™ Deep Red, Thermo Fisher, #C34565) или 5 мкМ CellTracker Green (CellTracker™ Green CMFDA (диацетат 5-хлорметилфлуоресцеина, Thermo Fisher, # C7025) с бессывороточной среде. После 30-мин инкубирования при 37°C клетки промывали бессывороточной средой и добавляли в лунки, содержащие подготовленное антитело/мАт2, или объединяли с другой дифференциально окрашенной линией клеток в равных объемах, а затем добавляли в лунки, содержащие подготовленное антитело/мАт2, как описано выше. Для обоих анализов после 30-мин инкубирования при условиях культивирования клеток лунки наполняли доверху равным объемом комплемента детеныша кролика, 10% в бессывороточной среде (Baby Rabbit Complement,TEBU-bio, #CL3441). Планшеты инкубировали в течение 4 часов в условиях культивирования клеток. Для анализа КЗЦ по высвобождению ЛДГ получали контрольные образцы со 100% лизисом путем добавления тритона X 100 в половину лунок, обработанных 4420 LALA, и выполняли анализ Cytotox согласно инструкциям производителя (CytotoxOne,Promega, G7891).
После считывания показаний сигнал контрольных образцов со 100% лизисом принимали за 100%, а сигналы лунок с образцами пересчитывали в процентах от этого уровня.
Для анализа КЗЦ на основе проточной цитометрии в конце периода инкубирования краситель для мертвых клеток (SYTOX® Blue Dead Cell Stain, Thermo Fisher, #S34857) разбавляли в соотношении 1 к 500 в PBS и заполняли лунки доверху равным объемом. Проточную цитометрию выполняли на проточном цитометре Cytoflex, настроенном на популяции интактных клеток на основе FSC и SSC и позволяющем определять процентную долю Sytox-положительных клеток (канал PB450) или как CellTracker™ Deep Red-положительных, так и CellTracker™ Green CMFDA- положительных популяций клеток.
6.2.2 Анализ АЗКЦ
АЗКЦ измеряли, как описано ранее (Broussas, Matthieu; Broyer, Lucile; and Goetsch, Liliane. (2013) Evaluation of Antibody-Dependent Cell Cytotoxicity Using Lactate Dehydrogenase (LDH) Measurement in Glycosylation Engineering of Biopharmaceuticals: Methods and Protocols, Methods in Molecular Biology. New York: Springer Science + Business Media. Volume 988, pp 305-317). Вкратце, клетки-мишени предварительно инкубировали с антителами, а затем добавляли первичные NK-клетки, выделенные из МПК человека (набор для выделения NK-клеток, Miltenyi Biotec, 130-092-657), в соотношении 20 к 1 на 4 ч. Анализ цитотоксичности выполняли в соответствии с инструкциями производителя (анализ цитотоксичности без использования радиоактивных меток CytoTox 96 Non-Radioactive Cytotoxicity Assay, Promega, G1780). % лизиса рассчитывали на основе 100% лизиса клеток-мишеней, учитывая спонтанный лизис эффекторных клеток и клеток-мишеней.
6.3 Результаты и выводы
6.3.1 Анализы КЗЦ
Клетки Raji, экспрессирующие PD-L1, подвергали воздействию антитела ритуксимаб против CD20, которое приводило к максимальному < 60% лизису при измерении КЗЦ по характерному высвобождению ЛДГ. Антитело 84G09 против PD-L1 (содержавшее F(ab)2-фрагмент мАт2 FS18-7-9/84G09) и FS18-7-9/84G09 в формате IgG1 продемонстрировали более высокие значения максимального лизиса и, кроме того, повышенную эффективность лизиса, причем рассчитанные полумаксимальные дозы были приблизительно в два раза меньше, чем доза ритуксимаба в формате IgG1. Это позволило продемонстрировать, что внедрение сайта связывания LAG-3 в антитело 84G09 не влияло на его специфическую активность к PD-L1 в отношении мощности или максимального ответа, поскольку оба эти показателя были очень близки при сравнении 84G09 и FS18-7-9/84G09. Внедрение мутации LALA привело к снижению максимального ответа для ритуксимаба, 84G09 и FS18-7-9/84G09, однако эффективность снизилась только для 84G09 и FS18-7-9/84G09. Как и ожидалось, антитело 25F7 против LAG-3 не влияло на жизнеспособность клеток Raji, экспрессирующих PD-L1, поскольку эти клетки не экспрессировали LAG-3 человека. Эти результаты показаны на фигуре 14A.
Для оценки КЗЦ клетки Raji, экспрессирующие LAG-3, подвергали воздействию антитела ритуксимаб против CD20, однако антитело 25F7 против LAG-3 демонстрировало еще лучшую эффективность, причем его рассчитанная полумаксимальная доза была приблизительно в два раза меньше, чем доза ритуксимаба. Ни одно из других антител (в том числе FS18-7-9/84G09) не демонстрировало КЗЦ-активности против клеток Raji, экспрессирующих LAG-3. Внедрение мутации LALA оказывало крайне ограниченное влияние на КЗЦ-активность ритуксимаба (фигура 14B).
6.3.2 Дифференциальный анализ КЗЦ
Авторы настоящего изобретения разработали дифференциальный анализ КЗЦ с использованием проточной цитометрии для распознавания лизируемых клеток, экспрессирующих мишени, при обработке FS18-7-9/84G09 или контрольными антителами. Этот анализ использовали для подтверждения результатов простейшего анализа КЗЦ по высвобождению ЛДГ, описанного выше. В отличие от IgG-изотипического контрольного антитела (4420), которое не влияло на процентную долю живых клеток, ритуксимаб опосредовал снижение количества живых клеток и увеличение количества мертвых клеток, причем как среди клеток, экспрессировавших PD-L1, так и среди клеток, экспрессировавших LAG-3. В то же время FS18-7-9/84G09 не влияло на клетки, экспрессирующие LAG-3, но крайне эффективно лизировало клетки, экспрессирующие PD-L1. Аналогичным образом, смесь LAG-3-специфического антитела 25F7 и PD-L1-специфического антитела 84G09 также демонстрировала снижение количества живых клеток в зависимости от дозы и обратное увеличение количества мертвых клеток; вместе с тем, максимальный лизис клеток, экспрессирующих LAG-3, составлял лишь 50% от общего количества клеток, однако достигался уже при концентрации приблизительно 1 нМ, что являлось минимальной дозой, позволяющей достичь максимального лизиса, среди всех протестированных антител. Это подтверждает предыдущие результаты, показавшие, что сайт связывания LAG-3 в CH3-домене FS18-7-9/84G09 не индуцирует КЗЦ-опосредованного лизиса клеток-мишеней, экспрессирующих LAG-3. Кроме того, данный эксперимент показал, что наличие клеток, экспрессирующих LAG-3, не влияло на КЗЦ-активность FS18-7-9/84G09 по отношению к клеткам, экспрессирующих PD-L1. Результаты показаны на фигуре 15.
6.3.3 Анализ АЗКЦ
Для анализа АЗКЦ клетки Raji, экспрессирующие PD-L1, подвергали воздействию антитела ритуксимаб против CD20, FS18-7-9/84G09 и 84G09, которые продемонстрировали очень близкую эффективность и приводили к максимальному лизису приблизительно 40% от общего количества клеток. Ритуксимаб и 84G09, содержащие мутацию LALA, не демонстрировали АЗКЦ-опосредованного лизиса, а FS18-7-9/84G09, содержащее мутацию LALA, демонстрировало крайне слабый лизис или отсутствие АЗКЦ-опосредованного лизиса клеток-мишеней, экспрессирующих PD-L1. LAG-3-специфическое антитело 25F7 и изотипический контроль 4420 с мутацией LALA и без нее не демонстрировали активности в данном анализе.
Эти результаты показывают, что внедрение сайта связывания LAG-3 в антитело 84G09 не влияло на его специфическую активность к PD-L1 в отношении мощности или максимального ответа, поскольку оба эти показателя были очень близки к показателям PD-L1-специфического антитела 84G09. Внедрение мутации LALA привело к устранению АЗКЦ-активности (фигура 16A).
Клетки Raji, экспрессирующие LAG-3, подвергали воздействию ритуксимаба и 25F7 с целью оценки АЗКЦ-опосредованного лизиса, что привело к максимальному лизису, составившему приблизительно 40%. FS18-7-9/84G09 также опосредовало лизис клеток, экспрессирующих LAG-3, за счет АЗКЦ, хотя и со значительно пониженной эффективностью, позволявшей достичь лишь 20% лизиса при концентрации 2,5 нМ. Внедрение мутации LALA полностью устраняло АЗКЦ-активность ритуксимаба и FS18-7-9/84G09. Как и ожидалось, 84G09 и изотипический контроль 4420 с мутацией LALA и без нее не демонстрировали АЗКЦ-активности в данном анализе, поскольку эти антитела не связываются с LAG-3 (фигура 16B).
Список последовательностей
Аминокислотные последовательности областей петель Fcab FS18-7-9
Петля AB FS18-7-9 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-9 - SNGQPENNY (SEQ ID NO: 2)
Петля EF FS18-7-9 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-9 (SEQ ID NO: 4)

CHO-кодон-оптимизированная нуклеотидная последовательность CH3-домена Fcab FS18-7-9 (SEQ ID NO: 142)

Аминокислотная последовательность CH3-домена Fcab FS18-7-9 (SEQ ID NO: 5)

Аминокислотная последовательность CH3-домена Fcab FS18-7-9, содержащая С-концевой остаток лизина (SEQ ID NO: 135)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-9, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 6)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-9 без мутации LALA (SEQ ID NO: 7)

Аминокислотные последовательности областей петель Fcab FS18-7-32
Петля AB FS18-7-32 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-32 - SNGQPENNY (SEQ ID NO: 8)
Петля EF FS18-7-32 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-32 (SEQ ID NO: 9)

Аминокислотная последовательность CH3-домена Fcab FS18-7-32 (SEQ ID NO: 10)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-32, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 11)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-32 без мутации LALA (SEQ ID NO: 12)

Аминокислотные последовательности областей петель Fcab FS18-7-33
Петля AB FS18-7-33 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-33 - SNGQPEDNY (SEQ ID NO: 13)
Петля EF FS18-7-33 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-33 (SEQ ID NO: 14)

Аминокислотная последовательность CH3-домена Fcab FS18-7-33 (SEQ ID NO: 15).

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-33, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 16)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-33 без мутации LALA (SEQ ID NO: 17)

Аминокислотные последовательности областей петель Fcab FS18-7-36
Петля AB FS18-7-36 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-36 - SNGQPENNY (SEQ ID NO: 18)
Петля EF FS18-7-36 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-36 (SEQ ID NO: 19)

Аминокислотная последовательность CH3-домена Fcab FS18-7-36 (SEQ ID NO: 20)

Аминокислотная последовательность CH2+CH3 CH2- и CH3-доменов Fcab FS18-7-36, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 21)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-36 без мутации LALA (SEQ ID NO: 22)

Аминокислотные последовательности областей петель Fcab FS18-7-58
Петля AB FS18-7-58 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-58 - SNGYPEIEF (SEQ ID NO: 23)
Петля EF FS18-7-58 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-58 (SEQ ID NO: 24)

Аминокислотная последовательность CH3-домена Fcab FS18-7-58 (SEQ ID NO: 25)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-58, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 26)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-58 без мутации LALA (SEQ ID NO: 27)

Аминокислотные последовательности областей петель Fcab FS18-7-62
Петля AB FS18-7-62 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-62 - SNGIPEWNY (SEQ ID NO: 28)
Петля EF FS18-7-62 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-62 (SEQ ID NO: 29)

Аминокислотная последовательность CH3-домена Fcab FS18-7-62 (SEQ ID NO: 30)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-62, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 31)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-62 без мутации LALA (SEQ ID NO: 32)

Аминокислотные последовательности областей петель Fcab FS18-7-65
Петля AB FS18-7-65 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-65 - SNGYAEYNY (SEQ ID NO: 33)
Петля EF FS18-7-65 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-65 (SEQ ID NO: 34)

Аминокислотная последовательность CH3-домена Fcab FS18-7-65 (SEQ ID NO: 35)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-65, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 36)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-65 без мутации LALA (SEQ ID NO: 37)

Аминокислотные последовательности областей петель Fcab FS18-7-78
Петля AB FS18-7-78 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-78 - SNGYKEENY (SEQ ID NO: 38)
Петля EF FS18-7-78 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-78 (SEQ ID NO: 39)

Аминокислотная последовательность CH3-домена Fcab FS18-7-78 (SEQ ID NO: 40)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-78, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 41)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-78 без мутации LALA (SEQ ID NO: 42)

Аминокислотные последовательности областей петель Fcab FS18-7-88
Петля AB FS18-7-88 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-88 - SNGVPELNV (SEQ ID NO: 43)
Петля EF FS18-7-88 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-88 (SEQ ID NO: 44)

Аминокислотная последовательность CH3-домена Fcab FS18-7-88 (SEQ ID NO: 45)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-88, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 46)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-88 без мутации LALA (SEQ ID NO: 47)

Аминокислотные последовательности областей петель Fcab FS18-7-95
Петля AB FS18-7-95 - WDEPWGED (SEQ ID NO: 1)
Петля CD FS18-7-95 - SNGYQEDNY (SEQ ID NO: 48)
Петля EF FS18-7-95 - PYDRWVWPDE (SEQ ID NO: 3)
Нуклеотидная последовательность CH3-домена Fcab FS18-7-95 (SEQ ID NO: 49)

Аминокислотная последовательность CH3-домена Fcab FS18-7-95 (SEQ ID NO: 50)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-95, содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 51)

Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-95 без мутации LALA (SEQ ID NO: 52)

Аминокислотная последовательность CH2-домена IgG1 человека дикого типа (SEQ ID NO: 53)

Аминокислотная последовательность CH2-домена IgG1 человека, содержащего «мутацию LALA» (выделено подчеркнутым шрифтом) (SEQ ID NO: 54)

Аминокислотная последовательность CH2- и CH3-доменов Fcab «дикого типа» без мутации LALA (SEQ ID NO: 55).

Аминокислотная последовательность CH2- и CH3-доменов Fcab «дикого типа», содержащих мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 56)

Аминокислотная последовательность шарнирной области IgG1 человека (SEQ ID NO: 57)

Аминокислотная последовательность укороченной шарнирной области IgG1 человека (SEQ ID NO: 58)

Аминокислотная последовательность Fcab FS18-7-108-29 против LAG-3 мыши, содержащего мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 59)
CH3-домен показан курсивом. Петли AB, CD и EF CH3-домена показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность Fcab FS18-7-108-29 против LAG-3 мыши без мутации LALA (SEQ ID NO: 60)
CH3-домен показан курсивом. Петли AB, CD и EF CH3-домена показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность Fcab FS18-7-108-35 против LAG-3 мыши, содержащего мутацию LALA (выделено подчеркнутым шрифтом) (SEQ ID NO: 61)
CH3-домен показан курсивом. Области петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность Fcab FS18-7-108-35 против LAG-3 мыши без мутации LALA (SEQ ID NO: 62)
CH3-домен показан курсивом. Области петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-9/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 63)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.


Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-9/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 64)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-32/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 65)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-32 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 66)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-33/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 67)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-33/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 68)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-36/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 69)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-36/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 70)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-58/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 71)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-58/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 72)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-62/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 73)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-62/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 74)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-65/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 75)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-65/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 76)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-78/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 77)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-78/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 78)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-88/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 79)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-88/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 80)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-95/4420 против LAG-3 человека/FITC, содержащей мутацию LALA (SEQ ID NO: 81)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-95/4420 против LAG-3 человека/FITC без мутации LALA (SEQ ID NO: 82)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт 4420 против FITC, содержащей мутацию LALA (SEQ ID NO: 83)
Положения CDR выделены подчеркиванием. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт 4420 против FITC без мутации LALA (SEQ ID NO: 84)
Положения CDR выделены подчеркиванием.

Аминокислотная последовательность легкой цепи мАт 4420 против FITC (SEQ ID NO: 85)
Положения CDR выделены подчеркиванием.

Аминокислотные последовательности CDR антитела 84G09 против PD-L1 (в соответствии с IMGT)

Аминокислотные последовательности CDR антитела 84G09 против PD-L1 (в соответствии с Kabat)

Аминокислотная последовательность VH-домена антитела 84G09 против PD-L1 (SEQ ID NO: 92)

Аминокислотная последовательность VL-домена антитела 84G09 против PD-L1 (SEQ ID NO: 93)
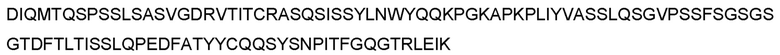
Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-9/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 94)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-9/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 95)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-32/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 96)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-32/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 97)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.


Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-33/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 98)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-33/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 99)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-36/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 100)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.


Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-36/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 101)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-58/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 102)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-58/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 103)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-62/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 104)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-62/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 105)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-65/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 106)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-65/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 107)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-78/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 108)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-78/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 109)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-88/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 110)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-88/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 111)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-95/84G09 против LAG-3/PD-L1 человека с мутацией LALA (SEQ ID NO: 112)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-95/84G09 против LAG-3/PD-L1 человека (SEQ ID NO: 113)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт 84G09 против PD-L1 человека с мутацией LALA (SEQ ID NO: 114)
Положения CDR выделены подчеркиванием. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт 84G09 против PD-L1 человека (SEQ ID NO: 115)
Положения CDR выделены подчеркиванием.

Аминокислотная последовательность легкой цепи мАт 84G09 против PD-L1 человека (SEQ ID NO: 116)
Положения CDR выделены подчеркиванием.

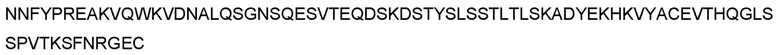
Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-29/S1 против LAG-3/PD-L1 мыши с мутацией LALA (SEQ ID NO: 117)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-29/S1 против LAG-3/PD-L1 мыши (SEQ ID NO: 118)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность легкой цепи мАт S1 против PD-L1 мыши (SEQ ID NO: 119) Положения CDR выделены подчеркиванием.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-35/S1 против LAG-3/PD-L1 мыши с мутацией LALA (SEQ ID NO: 120)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-35/S1 против LAG-3/PD-L1 мыши (SEQ ID NO: 121)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Аминокислотная последовательность тяжелой цепи контрольного мАт S1 против PD-L1 с мутацией LALA (SEQ ID NO: 122)
Положения CDR выделены подчеркиванием. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи контрольного мАт S1 против PD-L1 (SEQ ID NO: 123)
Положения CDR выделены подчеркиванием. Положение мутации LALA выделено полужирным шрифтом.

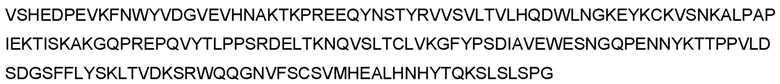
Аминокислотная последовательность тяжелой цепи контрольного мАт 25F7 против LAG-3 человека (SEQ ID NO: 124)
Положения CDR выделены подчеркиванием.

Аминокислотная последовательность легкой цепи контрольного мАт 25F7 против LAG-3 человека (SEQ ID NO: 125)
Положения CDR выделены подчеркиванием.

Аминокислотная последовательность LAG-3 человека (SEQ ID NO: 126)

Аминокислотная последовательность LAG-3 мыши (SEQ ID NO: 127)

Аминокислотная последовательность LAG-3 яванского макака (SEQ ID NO: 128)

Аминокислотная последовательность PD-L1 человека (SEQ ID NO: 129)

Аминокислотная последовательность PD-L1 мыши (SEQ ID NO: 130)

Аминокислотная последовательность PD-L1 яванского макака (SEQ ID NO: 131)

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-29/4420 против
LAG-3 мыши/FITC, содержащей мутацию LALA (SEQ ID NO: 132)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-35/4420 против
LAG-3 мыши/FITC, содержащей мутацию LALA (SEQ ID NO: 133)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом. Положение мутации LALA выделено полужирным шрифтом.

Аминокислотная последовательность тяжелой цепи мАт2 FS18-7-108-29/4420 против LAG-3 мыши/FITC, (SEQ ID NO: 134)
Положения CDR выделены подчеркиванием, а последовательности петель AB, CD и EF показаны полужирным подчеркнутым шрифтом.

Библиография
Все документы, упомянутые в настоящем описании, полностью включены в настоящую заявку посредством ссылок.
Bae J, Lee SJ, Park CG, Lee YS, Chun T. Trafficking of LAG-3 to the surface on activated T cells via its cytoplasmic domain and protein kinase C signaling. J Immunol. 193(6),3101-12 (2014).
Baecher-Allan C, Wolf E, Hafler DA. MHC class II expression identifies functionally distinct human regulatory T cells. J Immunol. 176(8),4622-31 (2006).
Camisaschi C, Casati C, Rini F, Perego M, De Filippo A, Triebel F, Parmiani G, Belli F, Rivoltini L, Castelli C. LAG-3 expression defines a subset of CD4(+)CD25(high)Foxp3(+) regulatory T cells that are expanded at tumor sites. J Immunol. 184(11),6545-51 (2010).
Curran MA, Montalvo W, Yagita H, Allison JP. PD-1 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumors. Proc Natl Acad Sci U S A. 107(9),4275-80 (2010).
Demeure, C. E., Wolfers, J., Martin-Garcia, N., Gaulard, P. & Triebel, F. T Lymphocytes infiltrating various tumour types express the MHC class II ligand lymphocyte activation gene-3 (LAG-3): role of LAG-3/MHC class II interactions in cell-cell contacts. European journal of cancer 37, 1709-1718 (2001).
Durham NM, Nirschl CJ, Jackson CM, Elias J, Kochel CM, Anders RA, Drake CG. Lymphocyte Activation Gene 3 (LAG-3) modulates the ability of CD4 T cells to be suppressed in vivo. PLoS One. 9(11), e109080 (2015).
Engels B, Engelhard VH, Sidney J, Sette A, Binder DC, Liu RB, Kranz DM, Meredith SC, Rowley DA, Schreiber H. Relapse or eradication of cancer is predicted by peptide-major histocompatibility complex affinity. Cancer Cell. 23(4),516-26 (2013).
Gandhi MK, Lambley E, Duraiswamy J, Dua U, Smith C, Elliott S, Gill D, Marlton P, Seymour J, Khanna R. Expression of LAG-3 by tumor-infiltrating lymphocytes is coincident with the suppression of latent membrane antigen-specific CD8+ T cell function in Hodgkin lymphoma patients. Blood 108(7),2280-9 (2006).
Grosso J, Inzunza D, Wu Q, et al. Programmed death-ligand 1 (PD-L1) expression in various tumor types. Journal for Immunotherapy of Cancer. 1(Suppl 1):P53. (2013).
Herbst RS, Soria JC, Kowanetz M, Fine GD, Hamid O, Gordon MS, Sosman JA, McDermott DF, Powderly JD, Gettinger SN, Kohrt HE, Horn L, Lawrence DP, Rost S, Leabman M, Xiao Y, Mokatrin A, Koeppen H, Hegde PS, Mellman I, Chen DS, Hodi FS. Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature 515(7528),563-7 (2014).
Huard B, Mastrangeli R, Prigent P,et al. Characterization of the major histocompatibility complex class II binding site on LAG-3 protein. Proc Natl Acad Sci USA 94,5744-9 (1997).
Iwai Y, Ishida M, Tanaka Y, Okazaki T, Honjo T, Minato N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc Natl Acad Sci U S A. 99(19),12293-7.
Jing W, Gershan JA, Weber J, Tlomak D, McOlash L, Sabatos-Peyton C, Johnson BD1. Combined immune checkpoint protein blockade and low dose whole body irradiation as immunotherapy for myeloma. J Immunother Cancer. 3(1):2 (2015).
Kabat, E.A. et al., In: Sequences of Proteins of Immunological Interest. NIH Publication, 91-3242 (1991).
Larkin J, Hodi FS, Wolchok JD. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N Engl J Med. 373(13),1270-1 (2015).
Huard B, Mastrangeli R, Prigent P,et al. Characterization of the major histocompatibility complex class II binding site on LAG-3 protein. Proc Natl Acad Sci USA 94,5744-9 (1997).
Powles T, Eder JP, Fine GD, Braiteh FS, Loriot Y, Cruz C, Bellmunt J, Burris HA, Petrylak DP, Teng SL, Shen X, Boyd Z, Hegde PS,Chen DS, Vogelzang NJ. MPDL3280A (anti-PD-L1) treatment leads to clinical activity in metastatic bladder cancer. Nature. 515(7528),558-62 (2014).
Sega EI, Leveson-Gower DB, Florek M, Schneidawind D, Luong RH, Negrin RS. Role of lymphocyte activation gene-3 (Lag-3) in conventional and regulatory T cell function in allogeneic transplantation.PLoS One. 9(1), e86551 (2014).
Wolchok J et al; Nivolumab plus ipilimumab in advanced melanoma. N Engl J Med. 369(2),122-33 (2013).
Woo SR, Turnis ME, Goldberg MV, Bankoti J, Selby M, Nirschl CJ, Bettini ML, Gravano DM, Vogel P, Liu CL, Tangsombatvisit S, Grosso JF, Netto G, Smeltzer MP, Chaux A, Utz PJ, Workman CJ, Pardoll DM, Korman AJ, Drake CG, Vignali DA. Immune inhibitory molecules LAG-3 and PD-1 synergistically regulate T cell function to promote tumoral immune escape. Cancer Res. 72(4),917-2 (2012).
Workman CJ, Vignali DA. Negative regulation of T cell homeostasis by lymphocyte activation gene-3 (CD223). J Immunol. 2005 Jan 15;174(2):688-95.
Workman CJ, Vignali DA. The CD4-related molecule, LAG-3 (CD223), regulates the expansion of activated T cells. Eur J Immunol. 2003 Apr;33(4):970-9.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> F-STAR DELTA LIMITED
<120> Партнеры по связыванию (2)
<130> TEK/CP7282171
<140> PCT/EP2017/065073
<141> 20.06.2017
<150> US 62/352482
<151> 20.06.2016
<160> 142
<170> Версия PatentIn 3.5
<210> 1
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля AB FS18-7-9
<400> 1
Trp Asp Glu Pro Trp Gly Glu Asp
1 5
<210> 2
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-9
<400> 2
Ser Asn Gly Gln Pro Glu Asn Asn Tyr
1 5
<210> 3
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля EF FS18-7-9
<400> 3
Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
1 5 10
<210> 4
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-9
<400> 4
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgaaat cgccgtggag 120
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 5
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-9
<400> 5
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 6
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-9,
содержащей мутацию LALA
<400> 6
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 7
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-9 без мутации LALA
<400> 7
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 8
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-32
<400> 8
Ser Asn Gly Gln Pro Glu Asn Asn Tyr
1 5
<210> 9
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-32
<400> 9
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgaaat cgccgtggag 120
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 10
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-32
<400> 10
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Glu Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 11
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-32,
содержащей мутацию LALA
<400> 11
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Glu Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 12
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-32 без мутации LALA
<400> 12
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Glu Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 13
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-33
<400> 13
Ser Asn Gly Gln Pro Glu Asp Asn Tyr
1 5
<210> 14
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-33
<400> 14
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggcagcc ggaggacaac tacaagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 15
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-33
<400> 15
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 16
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab FS18-7-33,
содержащей мутацию LALA
<400> 16
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asp Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 17
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-33 без мутации LALA
<400> 17
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asp Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 18
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-36
<400> 18
Ser Asn Gly Gln Pro Glu Asn Asn Tyr
1 5
<210> 19
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-36
<400> 19
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 180
gacggctcct acttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 20
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-36
<400> 20
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Tyr
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 21
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2+CH3 CH2- и CH3-доменов Fcab
FS18-7-36, содержащих мутацию LALA
<400> 21
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 22
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-36 без мутации LALA
<400> 22
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 23
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-58
<400> 23
Ser Asn Gly Tyr Pro Glu Ile Glu Phe
1 5
<210> 24
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-58
<400> 24
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggtatcc agaaatcgaa ttcaagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgcctt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 25
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-58
<400> 25
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Pro Glu
35 40 45
Ile Glu Phe Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 26
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-58, содержащей мутацию LALA
<400> 26
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Pro Glu Ile Glu
145 150 155 160
Phe Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 27
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-58 без мутации LALA
<400> 27
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Pro Glu Ile Glu
145 150 155 160
Phe Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 28
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-62
<400> 28
Ser Asn Gly Ile Pro Glu Trp Asn Tyr
1 5
<210> 29
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-62
<400> 29
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggatccc agaatggaac tataagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 30
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-62
<400> 30
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Ile Pro Glu
35 40 45
Trp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 31
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-62, содержащей мутацию LALA
<400> 31
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Ile Pro Glu Trp Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 32
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-62 без мутации LALA
<400> 32
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Ile Pro Glu Trp Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 33
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-65
<400> 33
Ser Asn Gly Tyr Ala Glu Tyr Asn Tyr
1 5
<210> 34
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-65
<400> 34
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggtatgc agaatataac tataagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 35
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-65
<400> 35
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Ala Glu
35 40 45
Tyr Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 36
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-65, содержащей мутацию LALA
<400> 36
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Ala Glu Tyr Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 37
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-65 без мутации LALA
<400> 37
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Ala Glu Tyr Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 38
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-78
<400> 38
Ser Asn Gly Tyr Lys Glu Glu Asn Tyr
1 5
<210> 39
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-78
<400> 39
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggtataa agaagaaaac tataagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 40
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-78
<400> 40
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Lys Glu
35 40 45
Glu Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 41
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-78, содержащей мутацию LALA
<400> 41
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Lys Glu Glu Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 42
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-78 без мутации LALA
<400> 42
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Lys Glu Glu Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 43
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-88
<400> 43
Ser Asn Gly Val Pro Glu Leu Asn Val
1 5
<210> 44
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-88
<400> 44
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atggggttcc agaactgaac gttaagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 45
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-88
<400> 45
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Val Pro Glu
35 40 45
Leu Asn Val Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 46
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-88, содержащей мутацию LALA
<400> 46
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Val Pro Glu Leu Asn
145 150 155 160
Val Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 47
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-88 без мутации LALA
<400> 47
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Val Pro Glu Leu Asn
145 150 155 160
Val Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 48
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD FS18-7-95
<400> 48
Ser Asn Gly Tyr Gln Glu Asp Asn Tyr
1 5
<210> 49
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность CH3-домена Fcab FS18-7-95
<400> 49
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cctgggatga gccgtggggt 60
gaagacgtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 120
tgggagagca atgggtatca ggaagataac tataagacca cgcctcccgt gctggactcc 180
gacggctcct tcttcctcta cagcaagctc accgtgccgt atgataggtg ggtttggccg 240
gatgagttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 300
ctctccctgt ctccgggt 318
<210> 50
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-95
<400> 50
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Gln Glu
35 40 45
Asp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 51
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-95, содержащей мутацию LALA
<400> 51
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Gln Glu Asp Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 52
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
FS18-7-95 без мутации LALA
<400> 52
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro
115 120 125
Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Tyr Gln Glu Asp Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 53
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2-домена IgG1 человека дикого типа
<400> 53
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 54
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2-домена IgG1 человека, содержащего
«мутацию LALA»
<400> 54
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 55
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab «дикого типа»
без мутации LALA
<400> 55
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 56
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH2- и CH3-доменов Fcab
«дикого типа», содержащих мутацию LALA
<400> 56
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly
210 215
<210> 57
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность шарнирной области IgG1 человека
<400> 57
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 58
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность укороченной шарнирной области
IgG1 человека
<400> 58
Thr Cys Pro Pro Cys Pro
1 5
<210> 59
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность Fcab FS18-7-108-29 против LAG-3 мыши,
содержащей мутацию LALA
<400> 59
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
100 105 110
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Val Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu Arg
180 185 190
Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 60
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность Fcab FS18-7-108-29 против LAG-3
мыши без мутации LALA
<400> 60
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
100 105 110
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Val Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu Arg
180 185 190
Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 61
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность Fcab FS18-7-108-35 против LAG-3 мыши,
содержащей мутацию LALA
<400> 61
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
100 105 110
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu Arg
180 185 190
Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 62
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность Fcab FS18-7-108-35 против LAG-3
мыши без мутации LALA
<400> 62
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
100 105 110
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu Arg
180 185 190
Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
210 215 220
<210> 63
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-9/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 63
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 64
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-9/4420 против LAG-3 человека/FITC без мутации LALA
<400> 64
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 65
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-32/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 65
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Glu Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 66
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-32 против LAG-3 человека/FITC без мутации LALA
<400> 66
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Glu Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 67
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-33/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 67
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 68
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-33/4420 против LAG-3 человека/FITC без мутации LALA
<400> 68
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 69
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-36/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 69
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 70
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-36/4420 против LAG-3 человека/FITC без мутации LALA
<400> 70
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 71
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-58/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 71
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Pro Glu Ile Glu Phe Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 72
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-58/4420 против LAG-3 человека/FITC без мутации LALA
<400> 72
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Pro Glu Ile Glu Phe Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 73
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-62/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 73
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Ile Pro Glu Trp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 74
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-62/4420 против LAG-3 человека/FITC без мутации LALA
<400> 74
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Ile Pro Glu Trp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
435 440 445
<210> 75
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-65/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 75
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Ala Glu Tyr Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 76
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-65/4420 против LAG-3 человека/FITC без мутации LALA
<400> 76
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Ala Glu Tyr Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 77
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-78/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 77
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Lys Glu Glu Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 78
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-78/4420 против LAG-3 человека/FITC без мутации LALA
<400> 78
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Lys Glu Glu Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 79
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-88/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 79
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Val Pro Glu Leu Asn Val Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 80
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-88/4420 против LAG-3 человека/FITC без мутации LALA
<400> 80
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Val Pro Glu Leu Asn Val Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 81
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-95/4420 против LAG-3 человека/FITC, содержащей мутацию LALA
<400> 81
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Gln Glu Asp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 82
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
FS18-7-95/4420 против LAG-3 человека/FITC без мутации LALA
<400> 82
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Tyr Gln Glu Asp Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp
405 410 415
Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 83
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт 4420 против FITC
содержащей мутацию LALA
<400> 83
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 84
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт 4420 против FITC
без мутации LALA
<400> 84
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 85
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность легкой цепи мАт 4420 против FITC
<400> 85
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Arg Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Val Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 86
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против PD-L1
(в соответствии с IMGT) HCDR1
<400> 86
Gly Phe Thr Phe Asp Asp Tyr Ala
1 5
<210> 87
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против PD-L1
(в соответствии с IMGT) HCDR2
<400> 87
Ile Ser Trp Lys Ser Asn Ile Ile
1 5
<210> 88
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против PD-L1
(в соответствии с IMGT) HCDR3
<400> 88
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro
1 5 10 15
<210> 89
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против PD-L1
(в соответствии с IMGT) LCDR1
<400> 89
Gln Ser Ile Ser Ser Tyr
1 5
<210> 90
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09
против PD-L1 (в соответствии с IMGT) LCDR2
<400> 90
Val Ala Ser
1
<210> 91
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против PD-L1
(в соответствии с IMGT) LCDR3
<400> 91
Gln Gln Ser Tyr Ser Asn Pro Ile Thr
1 5
<210> 92
<211> 122
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность VH-домена антитела 84G09 против PD-L1
<400> 92
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 93
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность VL-домена антитела 84G09 против PD-L1
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile
35 40 45
Tyr Val Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Ser Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Asn Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 94
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-9/84G09 с мутацией LALA
<400> 94
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 95
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-9/84G09
<400> 95
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 96
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3/PD-L1 человека FS18-7-32/84G09 с мутацией LALA
<400> 96
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Glu Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 97
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против LAG-3/PD-L1
человека FS18-7-32/84G09
<400> 97
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Glu Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 98
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против LAG-3/PD-L1
человека FS18-7-33/84G09 с мутацией LALA
<400> 98
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asp Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 99
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3/PD-L1 человека FS18-7-33/84G09
<400> 99
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asp Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 100
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-36/84G09 с мутацией LALA
<400> 100
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 101
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-36/84G09
<400> 101
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 102
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-58/84G09 с мутацией LALA
<400> 102
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Pro Glu Ile Glu Phe Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 103
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-58/84G09
<400> 103
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Pro Glu Ile Glu Phe Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 104
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-62/84G09 с мутацией LALA
<400> 104
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Ile Pro Glu Trp Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 105
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-62/84G09
<400> 105
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Ile Pro Glu Trp Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 106
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-65/84G09 с мутацией LALA
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Ala Glu Tyr Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 107
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-65/84G09
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Ala Glu Tyr Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 108
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-78/84G09 с мутацией LALA
<400> 108
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Lys Glu Glu Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 109
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-78/84G09
<400> 109
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Lys Glu Glu Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 110
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-88/84G09 с мутацией LALA
<400> 110
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Val Pro Glu Leu Asn Val Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 111
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-88/84G09
<400> 111
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Val Pro Glu Leu Asn Val Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 112
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-95/84G09 с мутацией LALA
<400> 112
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Gln Glu Asp Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 113
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2
против LAG-3/PD-L1 человека FS18-7-95/84G09
<400> 113
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Tyr Gln Glu Asp Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Pro Tyr Asp Arg Trp Val Trp Pro Asp Glu Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 114
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность мАт 84G09 против PD-L1 человека
с мутацией LALA,тяжелая цепь
<400> 114
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 115
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт 84G09
против PD-L1 человека
<400> 115
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 116
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность легкой цепи мАт 84G09 против
PD-L1 человека
<400> 116
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile
35 40 45
Tyr Val Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Ser Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Asn Pro Ile
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 117
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3/PD-L1 мыши FS18-7-108-29/S1 с мутацией LALA
<400> 117
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Val Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
435 440 445
<210> 118
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3/PD-L1 мыши FS18-7-108-29/S1
<400> 118
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Val Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 119
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность легкой цепи мАт S1 против
PD-L1 мыши
<400> 119
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Phe Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 120
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3/PD-L1 мыши FS18-7-108-35/S1 с мутацией LALA
<400> 120
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 121
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против LAG-3/PD-L1
мыши FS18-7-108-35/S1
<400> 121
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 122
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи контрольного мАт S1
против PD-L1 с мутацией LALA,
<400> 122
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 123
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи контрольного мАт S1
против
PD-L1
<400> 123
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 124
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность контрольного мАт 25F7 против
LAG-3 человека, тяжелая цепь
<400> 124
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Asp Tyr
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Arg Gly Ser Thr Asn Ser Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Leu Ser Leu Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Phe Gly Tyr Ser Asp Tyr Glu Tyr Asn Trp Phe Asp Pro Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 125
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность контрольного мАт 25F7 против
LAG-3 человека, легкая цепь
<400> 125
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Asn Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 126
<211> 525
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность LAG-3 человека
<400> 126
Met Trp Glu Ala Gln Phe Leu Gly Leu Leu Phe Leu Gln Pro Leu Trp
1 5 10 15
Val Ala Pro Val Lys Pro Leu Gln Pro Gly Ala Glu Val Pro Val Val
20 25 30
Trp Ala Gln Glu Gly Ala Pro Ala Gln Leu Pro Cys Ser Pro Thr Ile
35 40 45
Pro Leu Gln Asp Leu Ser Leu Leu Arg Arg Ala Gly Val Thr Trp Gln
50 55 60
His Gln Pro Asp Ser Gly Pro Pro Ala Ala Ala Pro Gly His Pro Leu
65 70 75 80
Ala Pro Gly Pro His Pro Ala Ala Pro Ser Ser Trp Gly Pro Arg Pro
85 90 95
Arg Arg Tyr Thr Val Leu Ser Val Gly Pro Gly Gly Leu Arg Ser Gly
100 105 110
Arg Leu Pro Leu Gln Pro Arg Val Gln Leu Asp Glu Arg Gly Arg Gln
115 120 125
Arg Gly Asp Phe Ser Leu Trp Leu Arg Pro Ala Arg Arg Ala Asp Ala
130 135 140
Gly Glu Tyr Arg Ala Ala Val His Leu Arg Asp Arg Ala Leu Ser Cys
145 150 155 160
Arg Leu Arg Leu Arg Leu Gly Gln Ala Ser Met Thr Ala Ser Pro Pro
165 170 175
Gly Ser Leu Arg Ala Ser Asp Trp Val Ile Leu Asn Cys Ser Phe Ser
180 185 190
Arg Pro Asp Arg Pro Ala Ser Val His Trp Phe Arg Asn Arg Gly Gln
195 200 205
Gly Arg Val Pro Val Arg Glu Ser Pro His His His Leu Ala Glu Ser
210 215 220
Phe Leu Phe Leu Pro Gln Val Ser Pro Met Asp Ser Gly Pro Trp Gly
225 230 235 240
Cys Ile Leu Thr Tyr Arg Asp Gly Phe Asn Val Ser Ile Met Tyr Asn
245 250 255
Leu Thr Val Leu Gly Leu Glu Pro Pro Thr Pro Leu Thr Val Tyr Ala
260 265 270
Gly Ala Gly Ser Arg Val Gly Leu Pro Cys Arg Leu Pro Ala Gly Val
275 280 285
Gly Thr Arg Ser Phe Leu Thr Ala Lys Trp Thr Pro Pro Gly Gly Gly
290 295 300
Pro Asp Leu Leu Val Thr Gly Asp Asn Gly Asp Phe Thr Leu Arg Leu
305 310 315 320
Glu Asp Val Ser Gln Ala Gln Ala Gly Thr Tyr Thr Cys His Ile His
325 330 335
Leu Gln Glu Gln Gln Leu Asn Ala Thr Val Thr Leu Ala Ile Ile Thr
340 345 350
Val Thr Pro Lys Ser Phe Gly Ser Pro Gly Ser Leu Gly Lys Leu Leu
355 360 365
Cys Glu Val Thr Pro Val Ser Gly Gln Glu Arg Phe Val Trp Ser Ser
370 375 380
Leu Asp Thr Pro Ser Gln Arg Ser Phe Ser Gly Pro Trp Leu Glu Ala
385 390 395 400
Gln Glu Ala Gln Leu Leu Ser Gln Pro Trp Gln Cys Gln Leu Tyr Gln
405 410 415
Gly Glu Arg Leu Leu Gly Ala Ala Val Tyr Phe Thr Glu Leu Ser Ser
420 425 430
Pro Gly Ala Gln Arg Ser Gly Arg Ala Pro Gly Ala Leu Pro Ala Gly
435 440 445
His Leu Leu Leu Phe Leu Ile Leu Gly Val Leu Ser Leu Leu Leu Leu
450 455 460
Val Thr Gly Ala Phe Gly Phe His Leu Trp Arg Arg Gln Trp Arg Pro
465 470 475 480
Arg Arg Phe Ser Ala Leu Glu Gln Gly Ile His Pro Pro Gln Ala Gln
485 490 495
Ser Lys Ile Glu Glu Leu Glu Gln Glu Pro Glu Pro Glu Pro Glu Pro
500 505 510
Glu Pro Glu Pro Glu Pro Glu Pro Glu Pro Glu Gln Leu
515 520 525
<210> 127
<211> 521
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность LAG-3 мыши
<400> 127
Met Arg Glu Asp Leu Leu Leu Gly Phe Leu Leu Leu Gly Leu Leu Trp
1 5 10 15
Glu Ala Pro Val Val Ser Ser Gly Pro Gly Lys Glu Leu Pro Val Val
20 25 30
Trp Ala Gln Glu Gly Ala Pro Val His Leu Pro Cys Ser Leu Lys Ser
35 40 45
Pro Asn Leu Asp Pro Asn Phe Leu Arg Arg Gly Gly Val Ile Trp Gln
50 55 60
His Gln Pro Asp Ser Gly Gln Pro Thr Pro Ile Pro Ala Leu Asp Leu
65 70 75 80
His Gln Gly Met Pro Ser Pro Arg Gln Pro Ala Pro Gly Arg Tyr Thr
85 90 95
Val Leu Ser Val Ala Pro Gly Gly Leu Arg Ser Gly Arg Gln Pro Leu
100 105 110
His Pro His Val Gln Leu Glu Glu Arg Gly Leu Gln Arg Gly Asp Phe
115 120 125
Ser Leu Trp Leu Arg Pro Ala Leu Arg Thr Asp Ala Gly Glu Tyr His
130 135 140
Ala Thr Val Arg Leu Pro Asn Arg Ala Leu Ser Cys Ser Leu Arg Leu
145 150 155 160
Arg Val Gly Gln Ala Ser Met Ile Ala Ser Pro Ser Gly Val Leu Lys
165 170 175
Leu Ser Asp Trp Val Leu Leu Asn Cys Ser Phe Ser Arg Pro Asp Arg
180 185 190
Pro Val Ser Val His Trp Phe Gln Gly Gln Asn Arg Val Pro Val Tyr
195 200 205
Asn Ser Pro Arg His Phe Leu Ala Glu Thr Phe Leu Leu Leu Pro Gln
210 215 220
Val Ser Pro Leu Asp Ser Gly Thr Trp Gly Cys Val Leu Thr Tyr Arg
225 230 235 240
Asp Gly Phe Asn Val Ser Ile Thr Tyr Asn Leu Lys Val Leu Gly Leu
245 250 255
Glu Pro Val Ala Pro Leu Thr Val Tyr Ala Ala Glu Gly Ser Arg Val
260 265 270
Glu Leu Pro Cys His Leu Pro Pro Gly Val Gly Thr Pro Ser Leu Leu
275 280 285
Ile Ala Lys Trp Thr Pro Pro Gly Gly Gly Pro Glu Leu Pro Val Ala
290 295 300
Gly Lys Ser Gly Asn Phe Thr Leu His Leu Glu Ala Val Gly Leu Ala
305 310 315 320
Gln Ala Gly Thr Tyr Thr Cys Ser Ile His Leu Gln Gly Gln Gln Leu
325 330 335
Asn Ala Thr Val Thr Leu Ala Val Ile Thr Val Thr Pro Lys Ser Phe
340 345 350
Gly Leu Pro Gly Ser Arg Gly Lys Leu Leu Cys Glu Val Thr Pro Ala
355 360 365
Ser Gly Lys Glu Arg Phe Val Trp Arg Pro Leu Asn Asn Leu Ser Arg
370 375 380
Ser Cys Pro Gly Pro Val Leu Glu Ile Gln Glu Ala Arg Leu Leu Ala
385 390 395 400
Glu Arg Trp Gln Cys Gln Leu Tyr Glu Gly Gln Arg Leu Leu Gly Ala
405 410 415
Thr Val Tyr Ala Ala Glu Ser Ser Ser Gly Ala His Ser Ala Arg Arg
420 425 430
Ile Ser Gly Asp Leu Lys Gly Gly His Leu Val Leu Val Leu Ile Leu
435 440 445
Gly Ala Leu Ser Leu Phe Leu Leu Val Ala Gly Ala Phe Gly Phe His
450 455 460
Trp Trp Arg Lys Gln Leu Leu Leu Arg Arg Phe Ser Ala Leu Glu His
465 470 475 480
Gly Ile Gln Pro Phe Pro Ala Gln Arg Lys Ile Glu Glu Leu Glu Arg
485 490 495
Glu Leu Glu Thr Glu Met Gly Gln Glu Pro Glu Pro Glu Pro Glu Pro
500 505 510
Gln Leu Glu Pro Glu Pro Arg Gln Leu
515 520
<210> 128
<211> 533
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность LAG-3 яванского макака
<400> 128
Met Trp Glu Ala Gln Phe Leu Gly Leu Leu Phe Leu Gln Pro Leu Trp
1 5 10 15
Val Ala Pro Val Lys Pro Pro Gln Pro Gly Ala Glu Ile Ser Val Val
20 25 30
Trp Ala Gln Glu Gly Ala Pro Ala Gln Leu Pro Cys Ser Pro Thr Ile
35 40 45
Pro Leu Gln Asp Leu Ser Leu Leu Arg Arg Ala Gly Val Thr Trp Gln
50 55 60
His Gln Pro Asp Ser Gly Pro Pro Ala Ala Ala Pro Gly His Pro Pro
65 70 75 80
Val Pro Gly His Arg Pro Ala Ala Pro Tyr Ser Trp Gly Pro Arg Pro
85 90 95
Arg Arg Tyr Thr Val Leu Ser Val Gly Pro Gly Gly Leu Arg Ser Gly
100 105 110
Arg Leu Pro Leu Gln Pro Arg Val Gln Leu Asp Glu Arg Gly Arg Gln
115 120 125
Arg Gly Asp Phe Ser Leu Trp Leu Arg Pro Ala Arg Arg Ala Asp Ala
130 135 140
Gly Glu Tyr Arg Ala Thr Val His Leu Arg Asp Arg Ala Leu Ser Cys
145 150 155 160
Arg Leu Arg Leu Arg Val Gly Gln Ala Ser Met Thr Ala Ser Pro Pro
165 170 175
Gly Ser Leu Arg Thr Ser Asp Trp Val Ile Leu Asn Cys Ser Phe Ser
180 185 190
Arg Pro Asp Arg Pro Ala Ser Val His Trp Phe Arg Ser Arg Gly Gln
195 200 205
Gly Arg Val Pro Val Gln Gly Ser Pro His His His Leu Ala Glu Ser
210 215 220
Phe Leu Phe Leu Pro His Val Gly Pro Met Asp Ser Gly Leu Trp Gly
225 230 235 240
Cys Ile Leu Thr Tyr Arg Asp Gly Phe Asn Val Ser Ile Met Tyr Asn
245 250 255
Leu Thr Val Leu Gly Leu Glu Pro Ala Thr Pro Leu Thr Val Tyr Ala
260 265 270
Gly Ala Gly Ser Arg Val Glu Leu Pro Cys Arg Leu Pro Pro Ala Val
275 280 285
Gly Thr Gln Ser Phe Leu Thr Ala Lys Trp Ala Pro Pro Gly Gly Gly
290 295 300
Pro Asp Leu Leu Val Ala Gly Asp Asn Gly Asp Phe Thr Leu Arg Leu
305 310 315 320
Glu Asp Val Ser Gln Ala Gln Ala Gly Thr Tyr Ile Cys His Ile Arg
325 330 335
Leu Gln Gly Gln Gln Leu Asn Ala Thr Val Thr Leu Ala Ile Ile Thr
340 345 350
Val Thr Pro Lys Ser Phe Gly Ser Pro Gly Ser Leu Gly Lys Leu Leu
355 360 365
Cys Glu Val Thr Pro Ala Ser Gly Gln Glu His Phe Val Trp Ser Pro
370 375 380
Leu Asn Thr Pro Ser Gln Arg Ser Phe Ser Gly Pro Trp Leu Glu Ala
385 390 395 400
Gln Glu Ala Gln Leu Leu Ser Gln Pro Trp Gln Cys Gln Leu His Gln
405 410 415
Gly Glu Arg Leu Leu Gly Ala Ala Val Tyr Phe Thr Glu Leu Ser Ser
420 425 430
Pro Gly Ala Gln Arg Ser Gly Arg Ala Pro Gly Ala Leu Arg Ala Gly
435 440 445
His Leu Pro Leu Phe Leu Ile Leu Gly Val Leu Phe Leu Leu Leu Leu
450 455 460
Val Thr Gly Ala Phe Gly Phe His Leu Trp Arg Arg Gln Trp Arg Pro
465 470 475 480
Arg Arg Phe Ser Ala Leu Glu Gln Gly Ile His Pro Pro Gln Ala Gln
485 490 495
Ser Lys Ile Glu Glu Leu Glu Gln Glu Pro Glu Leu Glu Pro Glu Pro
500 505 510
Glu Leu Glu Arg Glu Leu Gly Pro Glu Pro Glu Pro Gly Pro Glu Pro
515 520 525
Glu Pro Glu Gln Leu
530
<210> 129
<211> 290
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность PD-L1 человека
<400> 129
Met Arg Ile Phe Ala Val Phe Ile Phe Met Thr Tyr Trp His Leu Leu
1 5 10 15
Asn Ala Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu
35 40 45
Asp Leu Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile
50 55 60
Ile Gln Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser
65 70 75 80
Tyr Arg Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Arg Cys Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val
115 120 125
Lys Val Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val
130 135 140
Asp Pro Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr
145 150 155 160
Pro Lys Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser
165 170 175
Gly Lys Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn
180 185 190
Val Thr Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr
195 200 205
Cys Thr Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu
210 215 220
Val Ile Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr His
225 230 235 240
Leu Val Ile Leu Gly Ala Ile Leu Leu Cys Leu Gly Val Ala Leu Thr
245 250 255
Phe Ile Phe Arg Leu Arg Lys Gly Arg Met Met Asp Val Lys Lys Cys
260 265 270
Gly Ile Gln Asp Thr Asn Ser Lys Lys Gln Ser Asp Thr His Leu Glu
275 280 285
Glu Thr
290
<210> 130
<211> 290
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность PD-L1 мыши
<400> 130
Met Arg Ile Phe Ala Gly Ile Ile Phe Thr Ala Cys Cys His Leu Leu
1 5 10 15
Arg Ala Phe Thr Ile Thr Ala Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Val Thr Met Glu Cys Arg Phe Pro Val Glu Arg Glu Leu
35 40 45
Asp Leu Leu Ala Leu Val Val Tyr Trp Glu Lys Glu Asp Glu Gln Val
50 55 60
Ile Gln Phe Val Ala Gly Glu Glu Asp Leu Lys Pro Gln His Ser Asn
65 70 75 80
Phe Arg Gly Arg Ala Ser Leu Pro Lys Asp Gln Leu Leu Lys Gly Asn
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Cys Cys Ile Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu
115 120 125
Lys Val Asn Ala Pro Tyr Arg Lys Ile Asn Gln Arg Ile Ser Val Asp
130 135 140
Pro Ala Thr Ser Glu His Glu Leu Ile Cys Gln Ala Glu Gly Tyr Pro
145 150 155 160
Glu Ala Glu Val Ile Trp Thr Asn Ser Asp His Gln Pro Val Ser Gly
165 170 175
Lys Arg Ser Val Thr Thr Ser Arg Thr Glu Gly Met Leu Leu Asn Val
180 185 190
Thr Ser Ser Leu Arg Val Asn Ala Thr Ala Asn Asp Val Phe Tyr Cys
195 200 205
Thr Phe Trp Arg Ser Gln Pro Gly Gln Asn His Thr Ala Glu Leu Ile
210 215 220
Ile Pro Glu Leu Pro Ala Thr His Pro Pro Gln Asn Arg Thr His Trp
225 230 235 240
Val Leu Leu Gly Ser Ile Leu Leu Phe Leu Ile Val Val Ser Thr Val
245 250 255
Leu Leu Phe Leu Arg Lys Gln Val Arg Met Leu Asp Val Glu Lys Cys
260 265 270
Gly Val Glu Asp Thr Ser Ser Lys Asn Arg Asn Asp Thr Gln Phe Glu
275 280 285
Glu Thr
290
<210> 131
<211> 290
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность PD-L1 яванского макака
<400> 131
Met Arg Ile Phe Ala Val Phe Ile Phe Thr Ile Tyr Trp His Leu Leu
1 5 10 15
Asn Ala Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu
35 40 45
Asp Leu Thr Ser Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile
50 55 60
Ile Gln Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Asn
65 70 75 80
Tyr Arg Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn
85 90 95
Ala Ala Leu Arg Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Arg Cys Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val
115 120 125
Lys Val Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val
130 135 140
Asp Pro Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr
145 150 155 160
Pro Lys Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser
165 170 175
Gly Lys Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Leu Asn
180 185 190
Val Thr Ser Thr Leu Arg Ile Asn Thr Thr Ala Asn Glu Ile Phe Tyr
195 200 205
Cys Ile Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu
210 215 220
Val Ile Pro Glu Leu Pro Leu Ala Leu Pro Pro Asn Glu Arg Thr His
225 230 235 240
Leu Val Ile Leu Gly Ala Ile Phe Leu Leu Leu Gly Val Ala Leu Thr
245 250 255
Phe Ile Phe Tyr Leu Arg Lys Gly Arg Met Met Asp Met Lys Lys Cys
260 265 270
Gly Ile Arg Val Thr Asn Ser Lys Lys Gln Arg Asp Thr Gln Leu Glu
275 280 285
Glu Thr
290
<210> 132
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3 мыши/FITC FS18-7-108-29/4420, содержащей мутацию LALA
<400> 132
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Val Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 133
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3 мыши/FITC FS18-7-108-35/4420, содержащей мутацию LALA
<400> 133
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 134
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи мАт2 против
LAG-3 мыши/FITC FS18-7-108-29/4420
<400> 134
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Trp Asp Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Val Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Pro Phe Glu
405 410 415
Arg Trp Met Trp Pro Asp Glu Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 135
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность CH3-домена Fcab FS18-7-9,
содержащая C-концевой остаток лизина
<400> 135
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Trp Asp
1 5 10 15
Glu Pro Trp Gly Glu Asp Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Asp Arg Trp Val Trp Pro
65 70 75 80
Asp Glu Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
100 105
<210> 136
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09
против PD-L1 (в соответствии с Kabat) HCDR1
<400> 136
Asp Tyr Ala Met His
1 5
<210> 137
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против PD-L1
(в соответствии с Kabat) HCDR2
<400> 137
Gly Ile Ser Trp Lys Ser Asn Ile Ile Gly Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 138
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против
PD-L1 (в соответствии с Kabat) HCDR3
<400> 138
Asp Ile Thr Gly Ser Gly Ser Tyr Gly Trp Phe Asp Pro
1 5 10
<210> 139
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против
PD-L1 (в соответствии с Kabat) LCDR1
<400> 139
Arg Ala Ser Gln Ser Ile Ser Ser Tyr Leu Asn
1 5 10
<210> 140
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против
PD-L1 (в соответствии с Kabat) LCDR2
<400> 140
Val Ala Ser Ser Leu Gln Ser
1 5
<210> 141
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотные последовательности CDR антитела 84G09 против
PD-L1 (в соответствии с Kabat) LCDR3
<400> 141
Gln Gln Ser Tyr Ser Asn Pro Ile Thr
1 5
<210> 142
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CHO-кодон-оптимизированная нуклеотидная последовательность CH3-домена Fcab
FS18-7-9
<400> 142
ggccagcccc gggaacccca ggtgtacaca ctgcctccat cctgggatga gccctggggc 60
gaggatgtgt ctctgacctg tctcgtgaaa ggcttctacc cctccgatat cgccgtggaa 120
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggactcc 180
gacggctcat tcttcctgta cagcaagctg acagtgccct acgacagatg ggtgtggccc 240
gacgagttct cctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 300
ctgtccctga gccccggc 318
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ АНТИТЕЛ, КОТОРЫЕ СВЯЗЫВАЮТ CD137 И OX40 | 2019 |
|
RU2817602C2 |
| МОЛЕКУЛЫ АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТ PD-L1 и CD137 | 2019 |
|
RU2826084C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ МЕЗОТЕЛИН И CD137 | 2019 |
|
RU2815066C2 |
| ВЫСОКОАФФИННЫЕ АНТИ-PD-1 И АНТИ-LAG-3 АНТИТЕЛА И ПОЛУЧЕННЫЕ ИЗ НИХ БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ | 2019 |
|
RU2782381C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С МОДИФИЦИРОВАННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С FCRN И С СОХРАНЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С БЕЛКОМ А | 2015 |
|
RU2727639C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СПОСОБНОСТЯМИ СВЯЗЫВАТЬСЯ С FCRN | 2015 |
|
RU2730592C2 |
| АНТИТЕЛА К МЕЗОТЕЛИНУ | 2019 |
|
RU2837574C2 |
| СВЯЗЫВАЮЩИЙ ДОМЕН | 2019 |
|
RU2829934C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ НА ОСНОВЕ ПСЕВДО-FAB | 2019 |
|
RU2820254C2 |
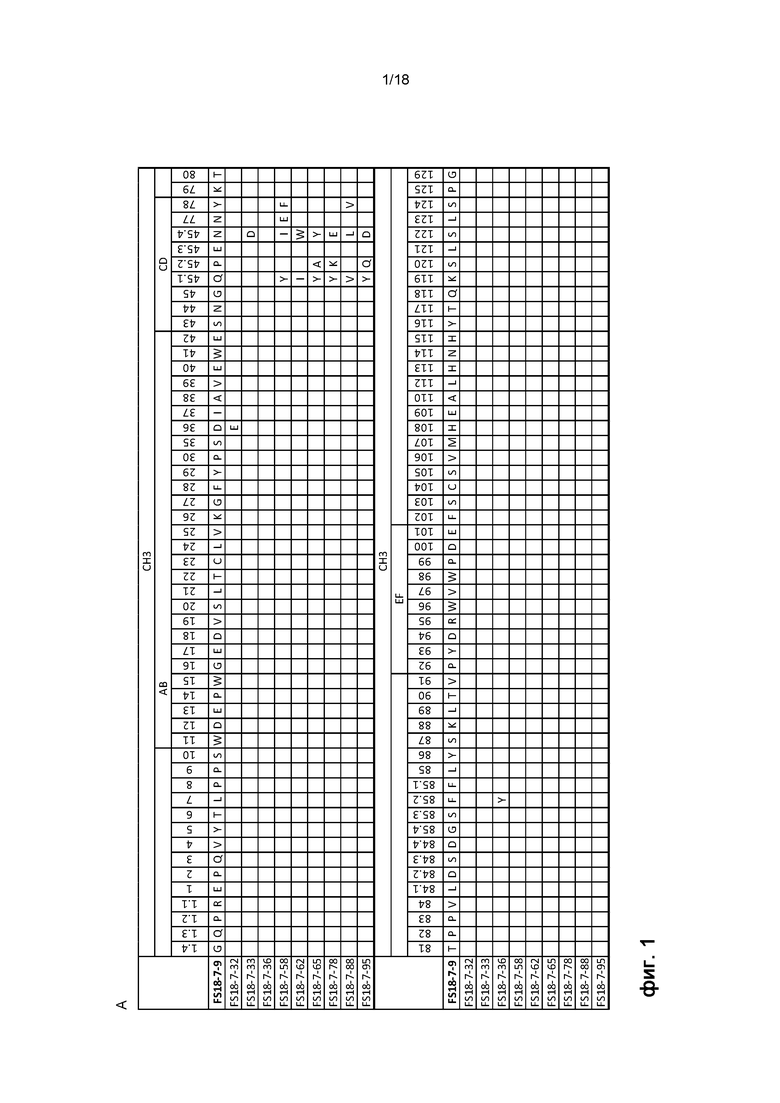

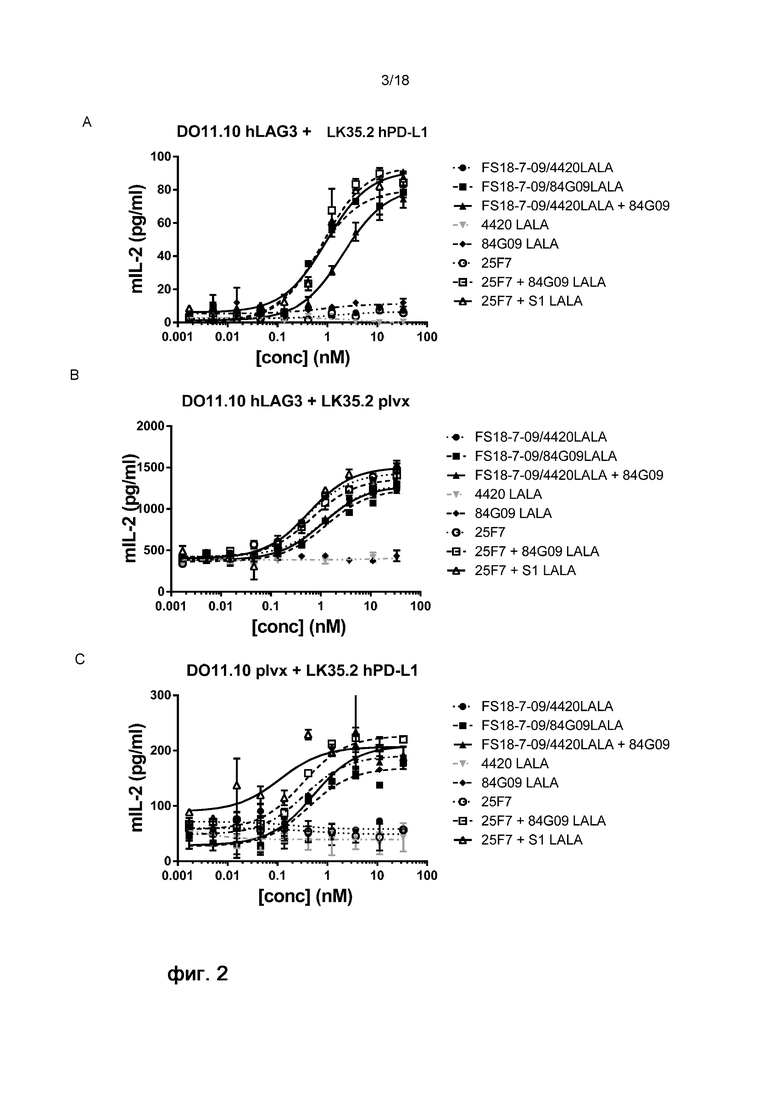
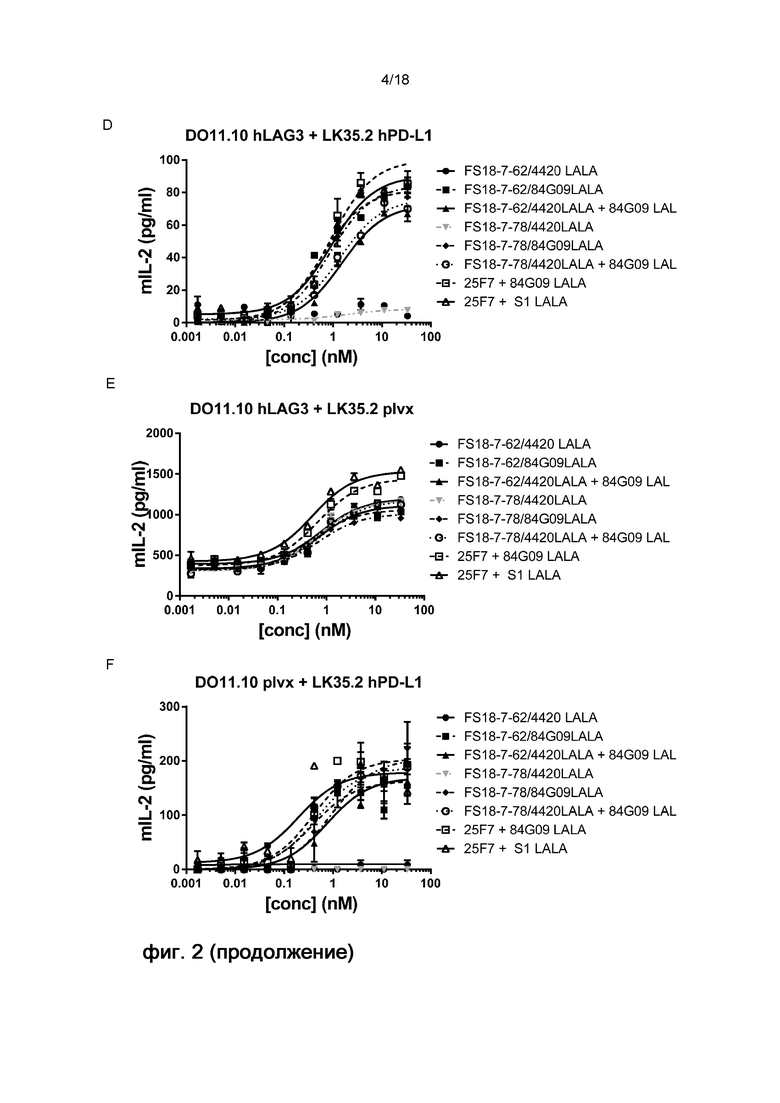
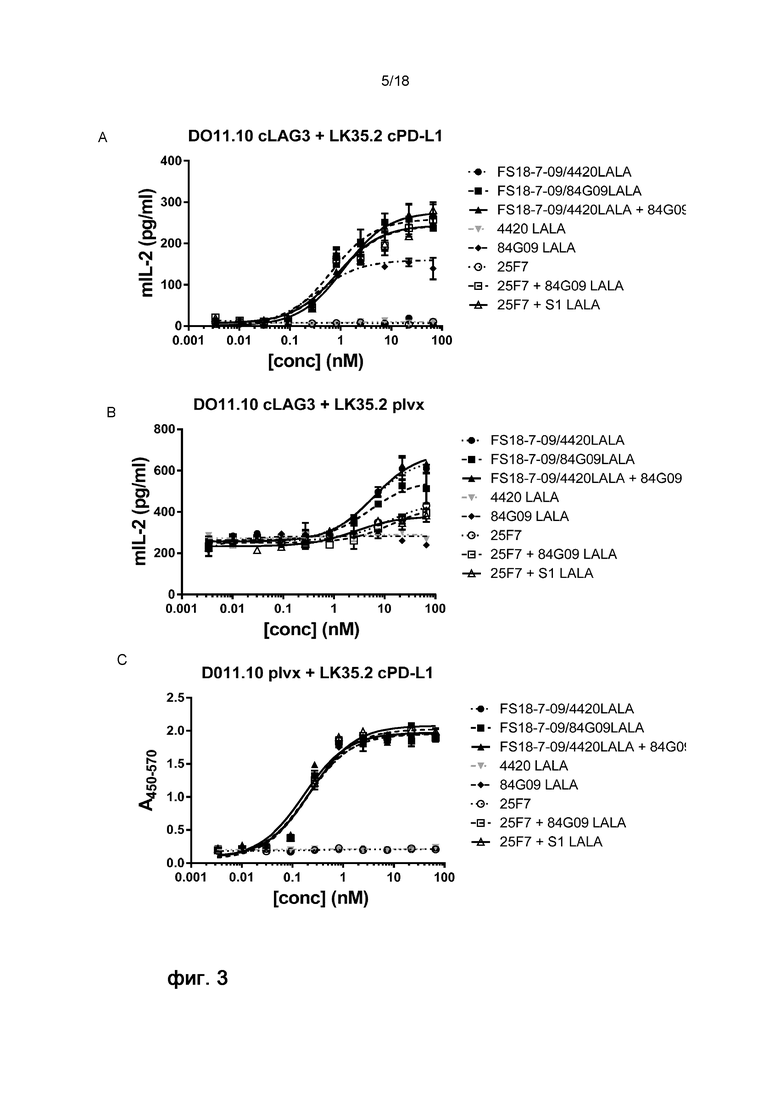
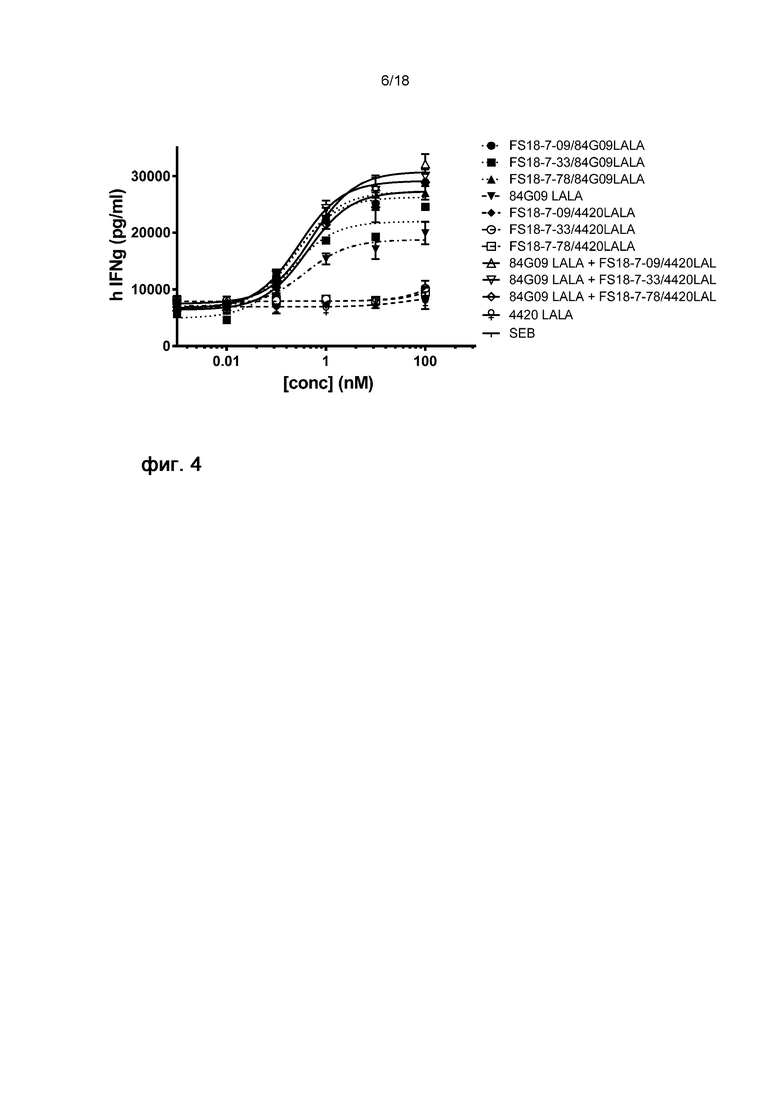
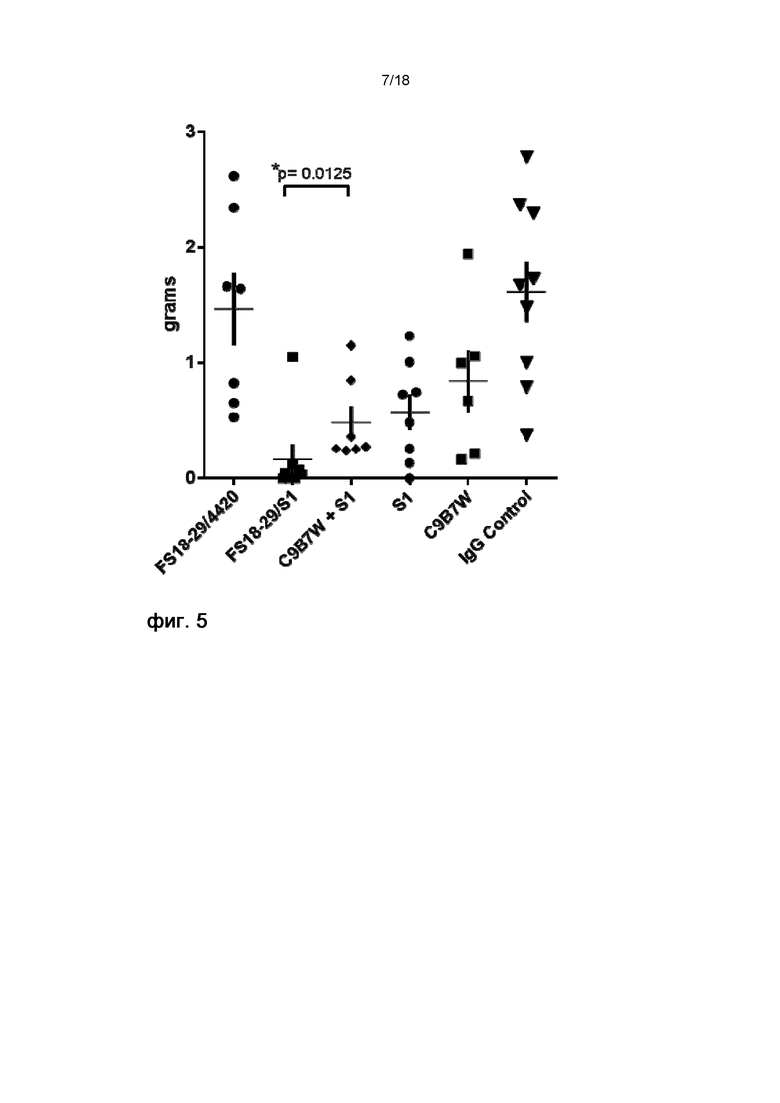
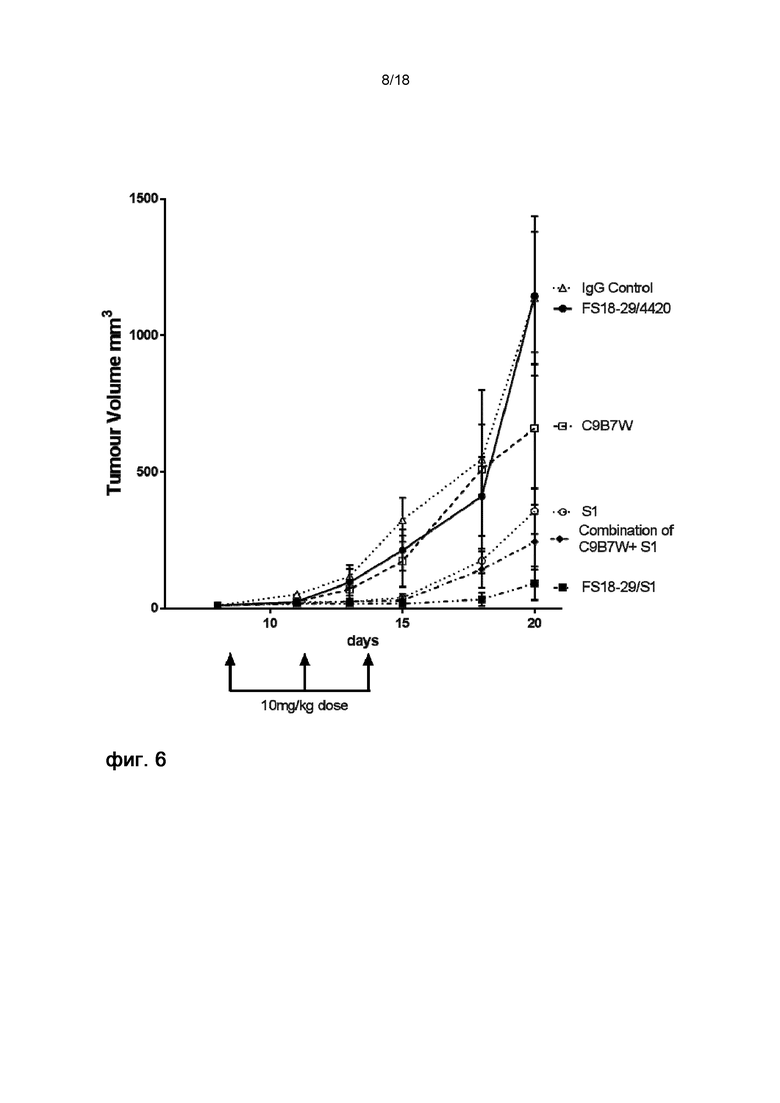
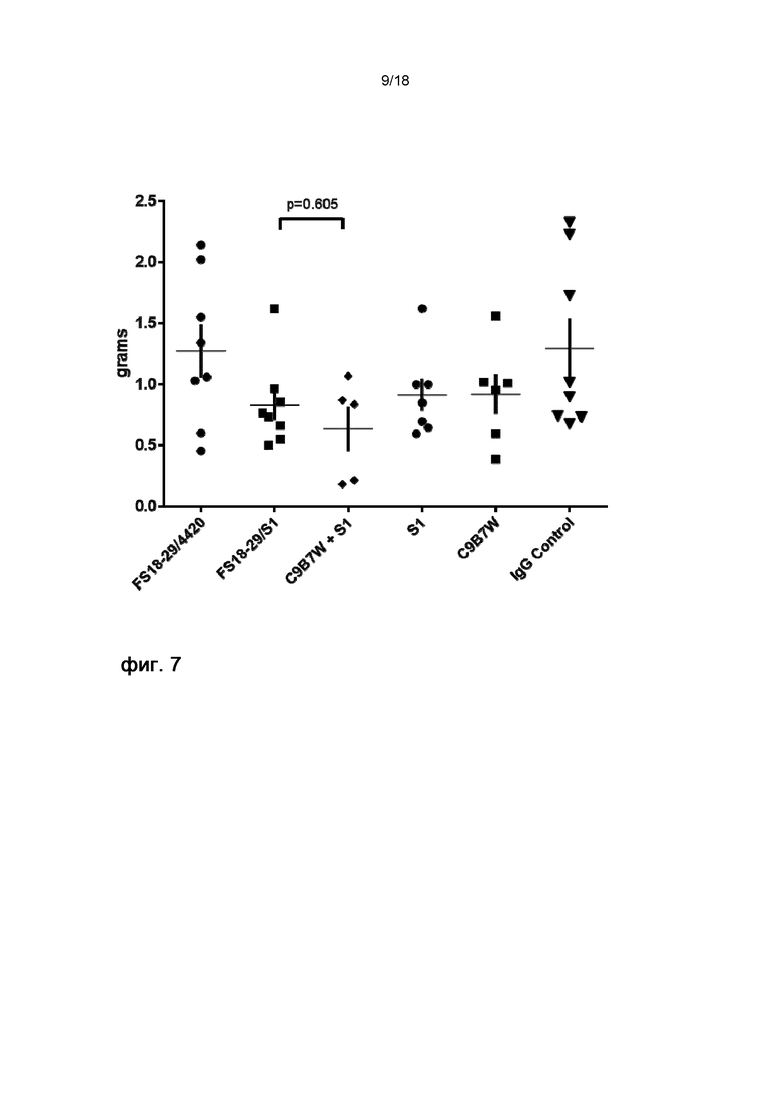
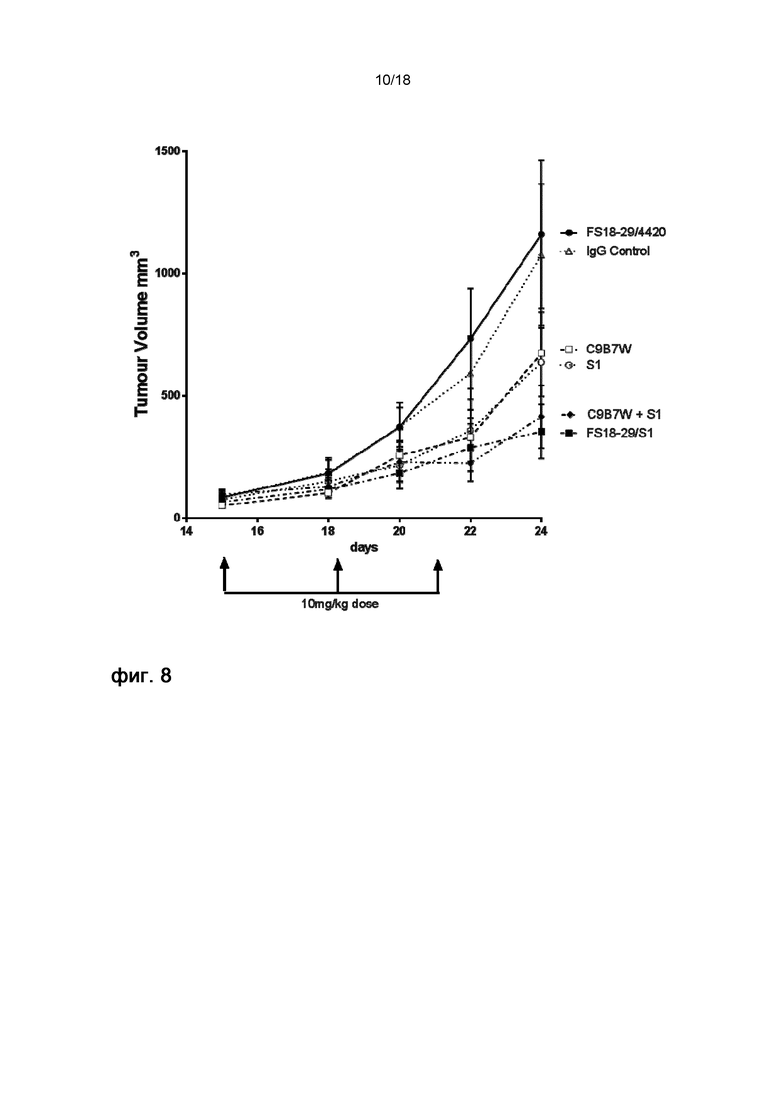
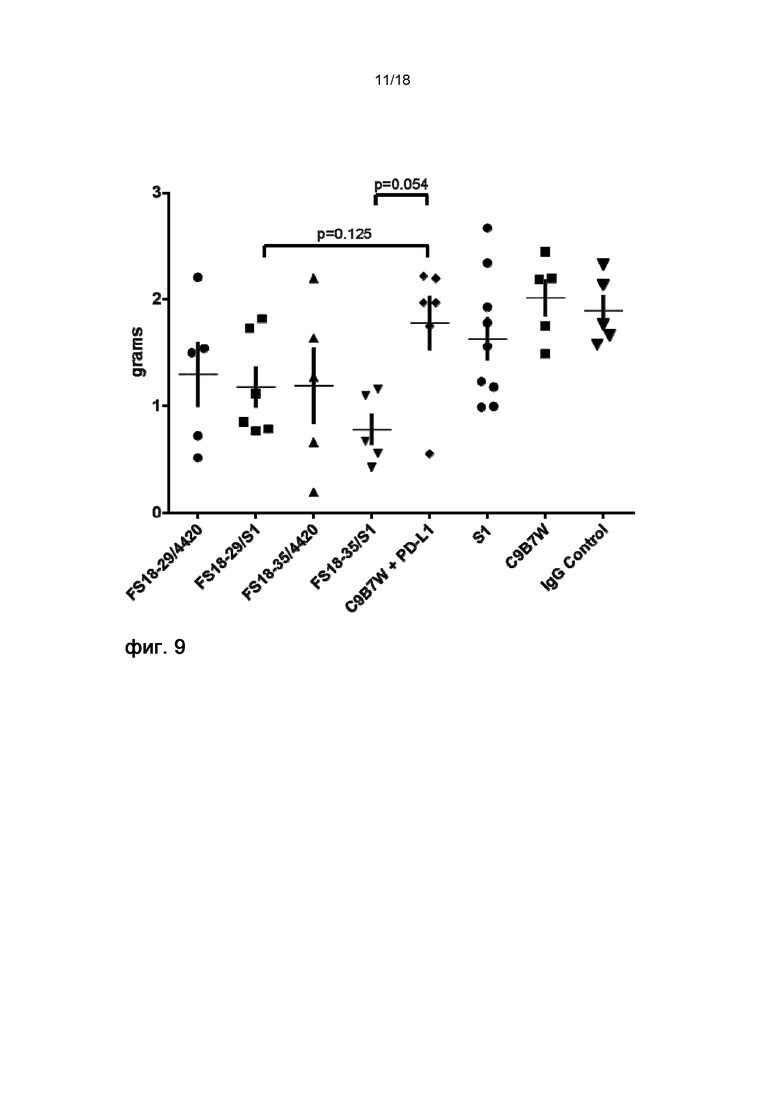
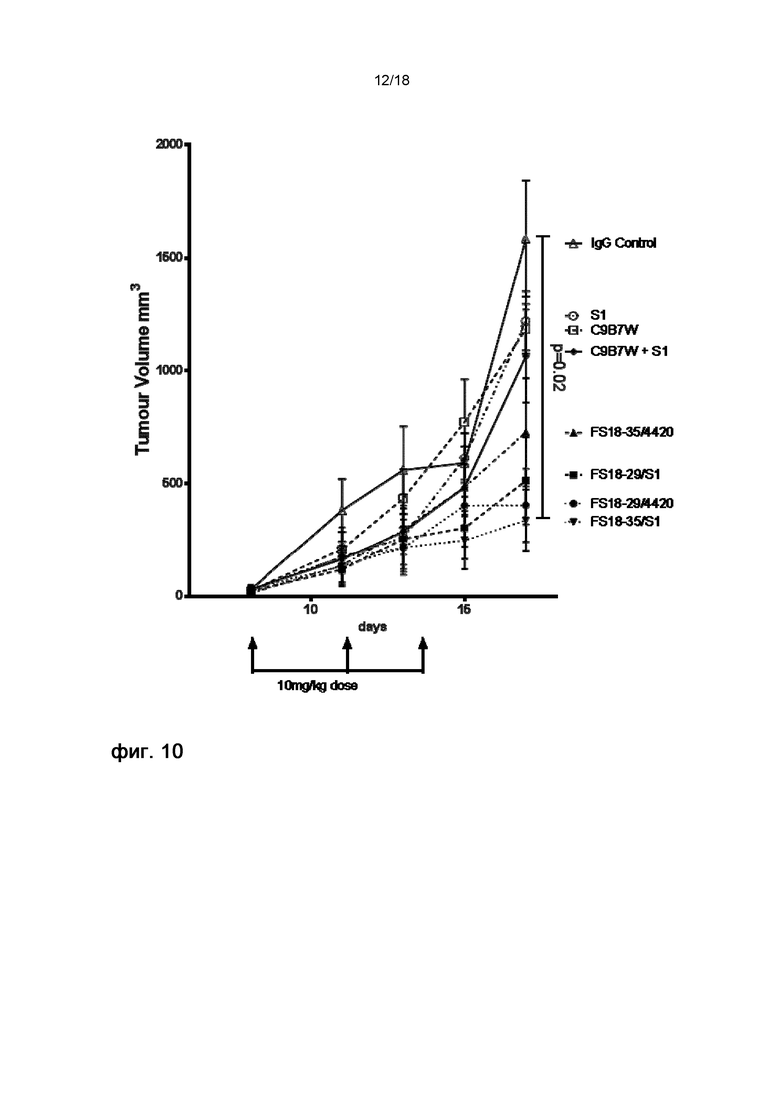
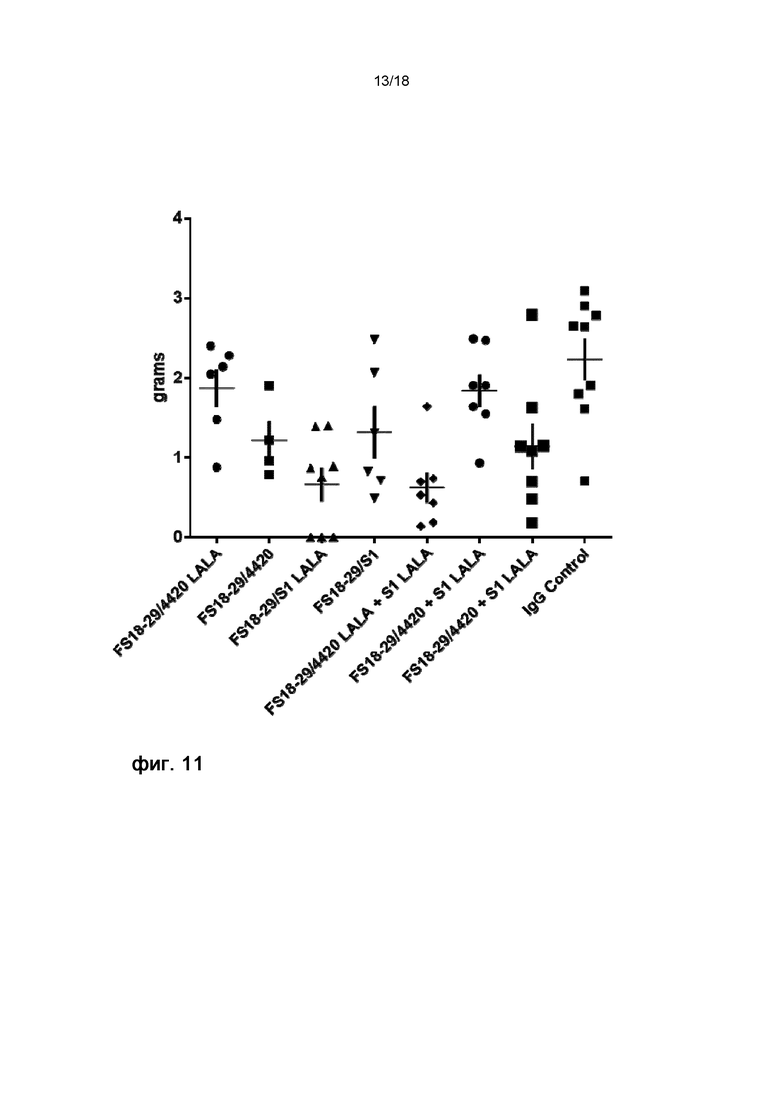
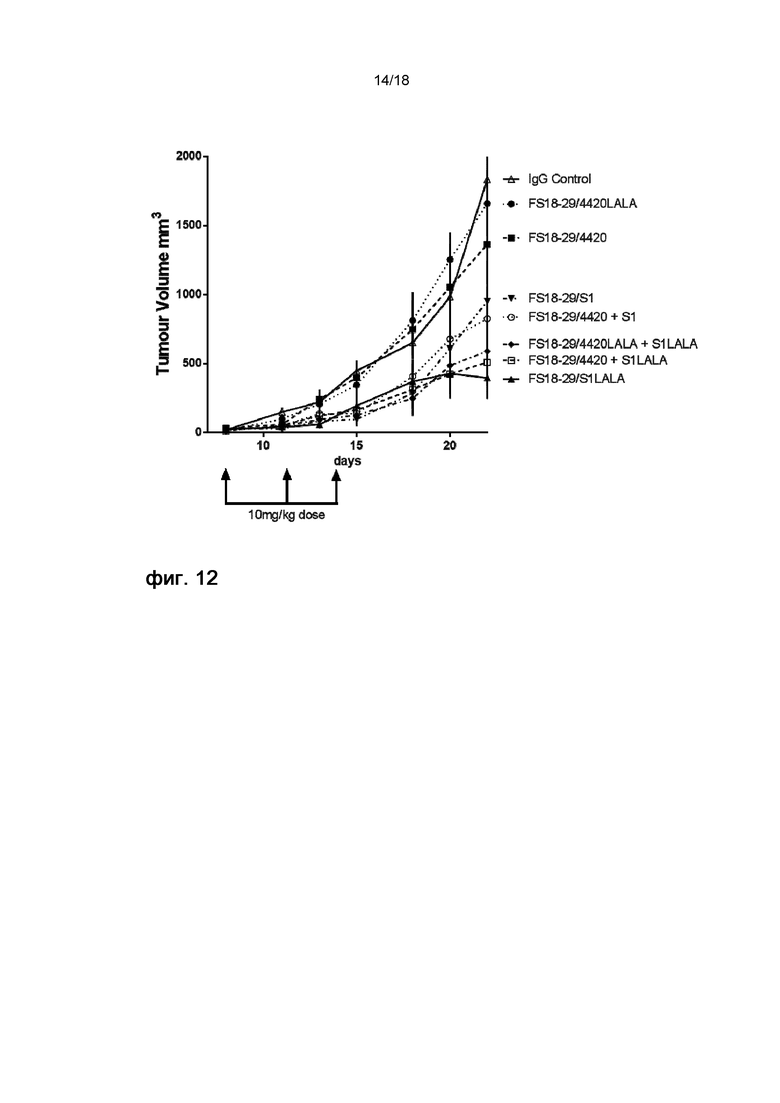
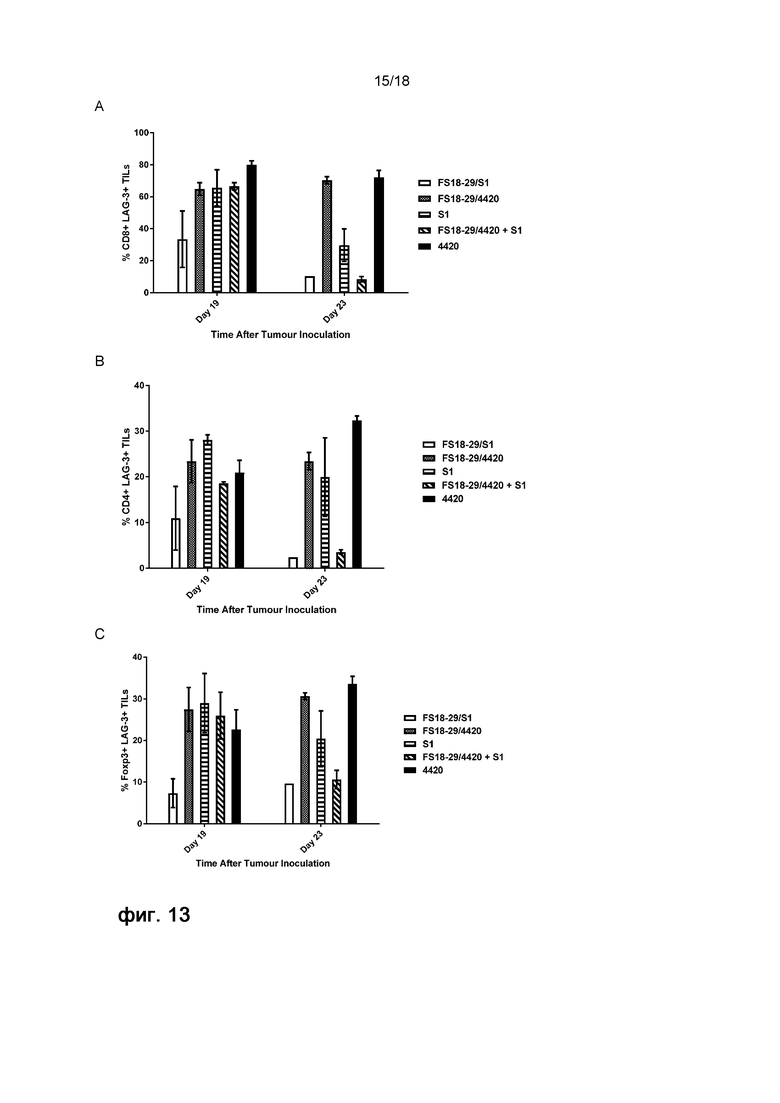
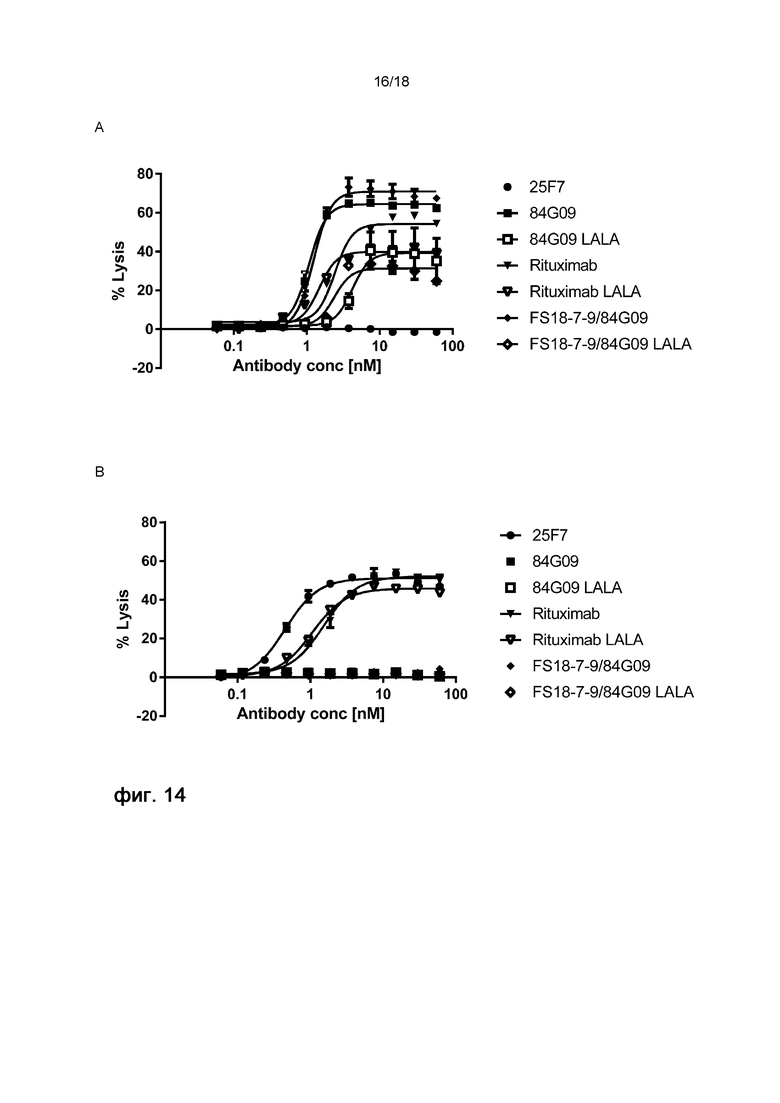
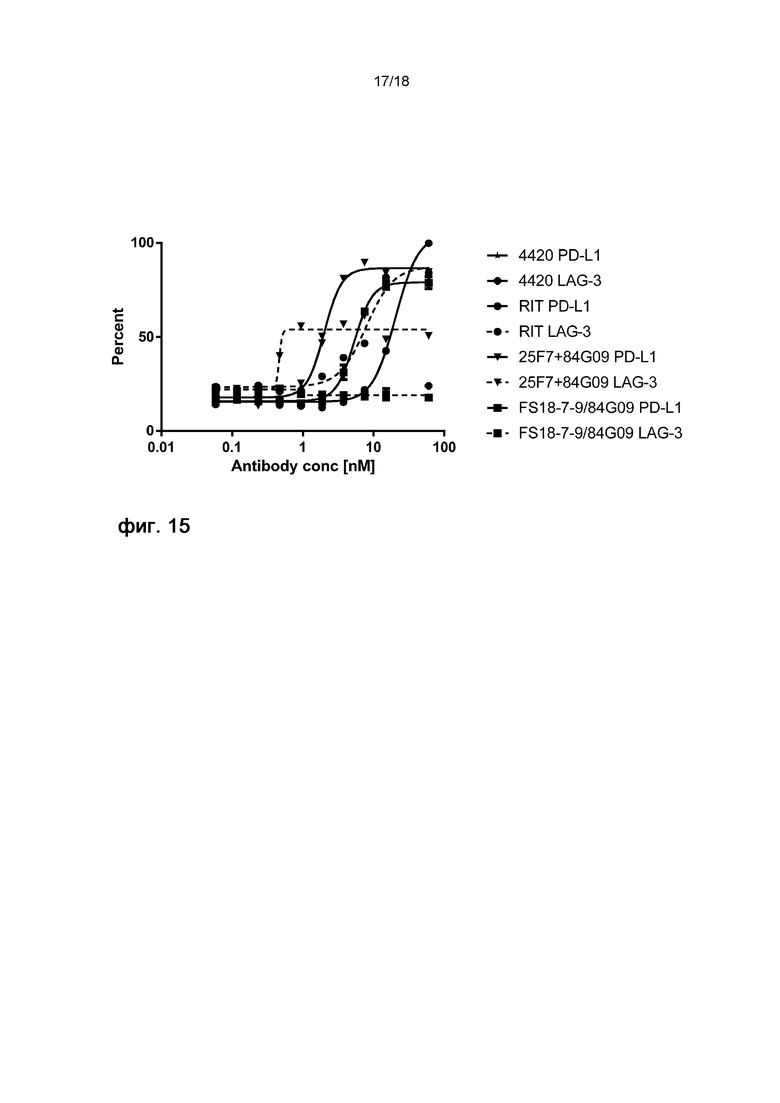
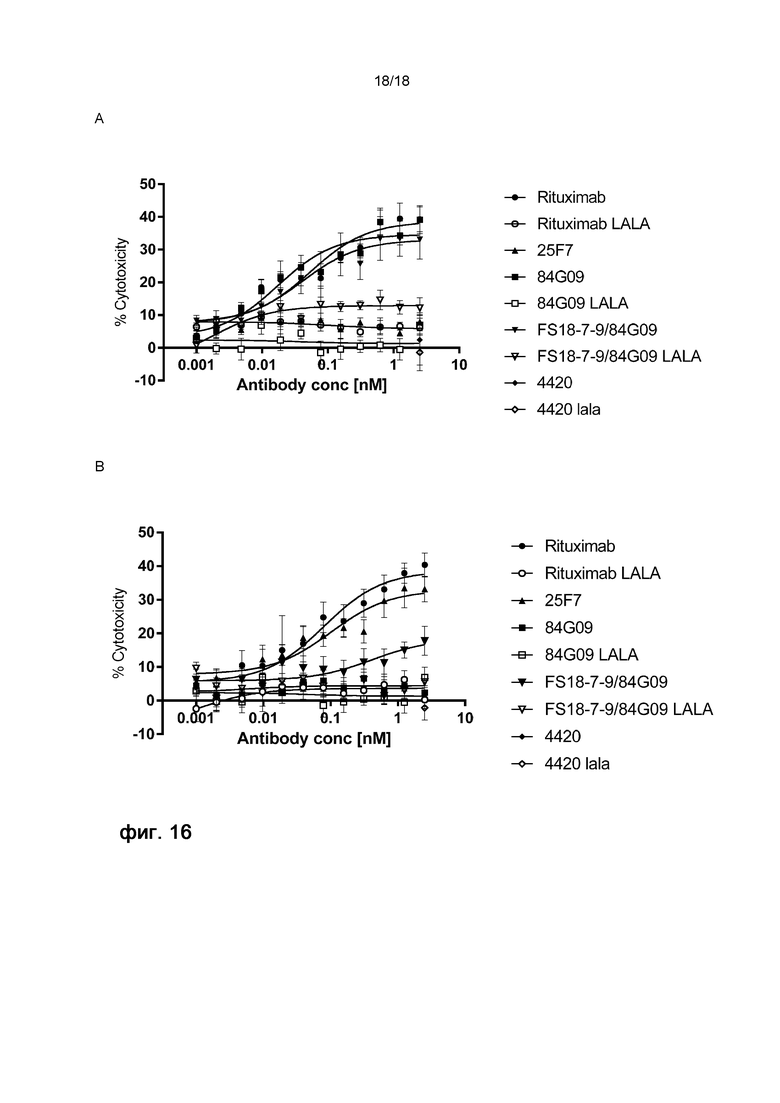
Изобретение относится к области биотехнологии. Предложена молекула антитела, связывающаяся с лигандом-1 белка запрограммированной гибели клеток (PD-L1), и геном активации лимфоцитов 3 (LAG-3). Молекула антитела содержит сайт связывания PD-L1 на основе CDR и сайт связывания LAG-3, который расположен в петлях AB и EF CH3-домена молекулы антитела. Также изобретение относится к кодирующей антитело нуклеиновой кислоте, экспрессирующему вектору и рекомбинантной клетке-хозяину, способу получения антитела. Кроме того, предложена фармацевтическая композиция, включающая указанное антитело. Молекулы антител согласно настоящему изобретению могут применяться в лечении рака. 7 н. и 29 з.п. ф-лы, 16 ил., 17 табл., 6 пр.
1. Молекула антитела, связывающаяся с лигандом-1 белка запрограммированной гибели клеток (PD-L1), и геном активации лимфоцитов 3 (LAG-3), причем указанная молекула антитела содержит:
(i) антигенсвязывающий сайт на основе областей, определяющих комплементарность (CDR), для PD-L1, содержащий HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 антитела против PD-L1; и
(ii) сайт связывания антигена LAG-3, расположенный в CH3-домене молекулы антитела, причем сайт связывания LAG-3 содержит аминокислотные последовательности WDEPWGED (SEQ ID NO: 1) и PYDRWVWPDE (SEQ ID NO: 3), и аминокислотная последовательность WDEPWGED располагается в петле AB CH3-домена, а аминокислотная последовательность PYDRWVWPDE располагается в петле EF CH3-домена.
2. Молекула антитела по п. 1, характеризующаяся тем, что указанный сайт связывания антигена LAG-3 дополнительно содержит одну из следующих последовательностей в петле CD CH3-домена:
(i) SNGQPENNY (SEQ ID NO: 2, 8 и 18);
(ii) SNGQPEDNY (SEQ ID NO: 13);
(iii) SNGYPEIEF (SEQ ID NO: 23);
(iv) SNGIPEWNY (SEQ ID NO: 28);
(v) SNGYAEYNY (SEQ ID NO: 33);
(vi) SNGYKEENY (SEQ ID NO: 38);
(vii) SNGVPELNV (SEQ ID NO: 43); или
(viii) SNGYQEDNY (SEQ ID NO: 48).
3. Молекула антитела по п. 2, характеризующаяся тем, что сайт связывания антигена LAG-3 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 2, 28 или 38, в петле CD CH3-домена.
4. Молекула антитела по п. 2, характеризующаяся тем, что сайт связывания антигена LAG-3 содержит аминокислотную последовательность, приведенную в SEQ ID NO: 2, в петле CD CH3-домена.
5. Молекула антитела по любому из пп. 1-4, характеризующаяся тем, что указанная молекула антитела представляет собой молекулу иммуноглобулина G, при этом необязательно указанная молекула антитела представляет собой молекулу IgG1, IgG2, IgG3 или IgG4 и далее необязательно указанная молекула антитела представляет собой молекулу IgG1.
6. Молекула антитела по любому из пп. 1-5, характеризующаяся тем, что указанная молекула антитела содержит CH3-домен, приведенный в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50.
7. Молекула антитела по п. 6, характеризующаяся тем, что указанная молекула антитела содержит CH3-домен, приведенный в SEQ ID NO: 5, 30 или 40.
8. Молекула антитела по п. 7, характеризующаяся тем, что указанная молекула антитела содержит CH3-домен, приведенный в SEQ ID NO: 5.
9. Молекула антитела по любому из пп. 1-8, характеризующаяся тем, что указанная молекула антитела содержит CH2-домен и указанный CH2-домен содержит последовательность, приведенную в SEQ ID NO: 53 или SEQ ID NO: 54.
10. Молекула антитела по любому из пп. 1-9, характеризующаяся тем, что указанная молекула антитела содержит последовательность, приведенную в SEQ ID NO: 6, 7, 11, 12, 16, 17, 21, 22, 26, 27, 31, 32, 36, 37, 41, 42, 46, 47, 51 или 52.
11. Молекула антитела по п. 10, характеризующаяся тем, что указанная молекула антитела содержит последовательность, приведенную в SEQ ID NO: 6, 7, 31, 32, 41 или 42.
12. Молекула антитела по п. 11, характеризующаяся тем, что указанная молекула антитела содержит последовательность, приведенную в SEQ ID NO: 6 или 7.
13. Молекула антитела по любому из пп. 1-12, характеризующаяся тем, что антигенсвязывающий сайт на основе CDR для PD-L1 содержит CDR, приведенные в SEQ ID NO: 86-91.
14. Молекула антитела по любому из пп. 1-12, характеризующаяся тем, что антигенсвязывающий сайт на основе CDR для PD-L1 содержит CDR, приведенные в SEQ ID NO: 136 - 141.
15. Молекула антитела по п. 13 или 14, характеризующаяся тем, что указанная молекула антитела содержит VH- и/или VL-домены, приведенные в SEQ ID NO: 92 и 93.
16. Молекула антитела по любому из пп. 13-15, характеризующаяся тем, что указанная молекула антитела содержит последовательность тяжелой цепи, приведенную в любой из SEQ ID NO: 94-113.
17. Молекула антитела по п. 16, характеризующаяся тем, что указанная молекула антитела содержит последовательность тяжелой цепи, приведенную в SEQ ID NO: 94, 95, 104, 105, 108 или 109.
18. Молекула антитела по п. 17, характеризующаяся тем, что указанная молекула антитела содержит последовательность тяжелой цепи, приведенную в SEQ ID NO: 94 или 95.
19. Молекула антитела по любому из пп. 13-18, характеризующаяся тем, что указанная молекула антитела содержит последовательность легкой цепи, приведенную в SEQ ID NO: 116.
20. Молекула антитела по п. 19, характеризующаяся тем, что указанная молекула антитела содержит последовательность тяжелой цепи, приведенную в SEQ ID NO: 94, и последовательность легкой цепи, приведенную в SEQ ID NO: 116.
21. Молекула антитела по любому из пп. 6-20, характеризующаяся тем, что последовательность CH3-домена молекулы антитела дополнительно содержит остаток лизина (K) на самом C-конце последовательности, приведенной в SEQ ID NO: 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50.
22. Молекула антитела по п. 21, характеризующаяся тем, что молекула антитела содержит CH3-домен, приведенный в последовательности SEQ ID NO: 135.
23. Молекула антитела по любому из пп. 1-22, характеризующаяся тем, что указанная молекула антитела способна одновременно связываться с PD-L1 и LAG-3.
24. Молекула антитела по п. 23, характеризующаяся тем, что PD-L1 и LAG-3 присутствуют на двух различных клетках.
25. Молекула антитела по любому из пп. 1-24, характеризующаяся тем, что указанная молекула антитела конъюгирована с модулятором иммунной системы, цитотоксической молекулой, радиоактивным изотопом или детектируемой меткой.
26. Молекула антитела по п. 25, характеризующаяся тем, что модулятор иммунной системы или цитотоксическая молекула представляет собой цитокин.
27. Нуклеиновая кислота, кодирующая молекулу антитела по любому из пп. 1-26.
28. Экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 27.
29. Рекомбинантная клетка-хозяин, содержащая нуклеиновую кислоту по п. 27 или вектор по п. 28, для экспрессии молекулы антитела по любому из пп. 1-26.
30. Способ получения молекулы антитела по любому из пп. 1-26, включающий культивирование рекомбинантной клетки-хозяина по п. 29 в условиях для продуцирования молекулы антитела.
31. Способ по п. 30, дополнительно включающий выделение и/или очистку молекулы антитела.
32. Фармацевтическая композиция для лечения рака у пациента, содержащая терапевтически эффективное количество молекулы антитела по любому из пп. 1-26 и фармацевтически приемлемое вспомогательное вещество.
33. Применение молекулы антитела по любому из пп. 1-26 для лечения рака у пациента.
34. Применение по п. 33, характеризующееся тем, что рак выбран из группы, состоящей из: лимфомы Ходжкина, неходжкинской лимфомы, рака яичников, рака предстательной железы, рака ободочной и прямой кишки, фибросаркомы, почечноклеточной карциномы, меланомы, рака поджелудочной железы, рака молочной железы, мультиформной глиобластомы, рака легких, рака головы и шеи, рака желудка, рака мочевого пузыря, рака шейки матки, рака матки, рака вульвы, рака яичек, рака полового члена, лейкоза, множественной миеломы, плоскоклеточного рака, рака яичек, рака пищевода, саркомы Капоши и лимфомы центральной нервной системы (ЦНС), гепатоцеллюлярной карциномы, карциномы из клеток Меркеля, рака носоглотки и мезотелиомы.
35. Применение по п. 33 или 34, характеризующееся тем, что лечение дополнительно включает введение противоопухолевой вакцины пациенту.
36. Применение по п. 33 или 34, характеризующееся тем, что указанное лечение дополнительно включает введение химиотерапевтического агента пациенту.
| WO 2015048312 A1, 02.04.2015 | |||
| WO 2010019570 A2, 18.02.2010 | |||
| JING W | |||
| et al | |||
| Combined immune checkpoint protein blockade and low dose whole body irradiation as immunotherapy for myeloma, Journal for ImmunoTherapy of Cancer, 2015, volume 3:2, pp.1-15 | |||
| КАРТРИДЖ ДЛЯ СТЕРИЛЬНОГО СМЕШИВАНИЯ ДВУХФАЗНОГО СОЕДИНЕНИЯ, В ЧАСТНОСТИ ДЛЯ ДВУХКОМПОНЕНТНЫХ АКРИЛОВЫХ ПОЛИМЕРОВ | 2006 |
|
RU2407487C2 |
| WOZNIAK-KNOPP G | |||
| et al | |||
| Introducing antigen-binding sites in structural | |||

