ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка испрашивает приоритет. по предварительной заявке США № 62/872425, поданной 10 июля 2019 года, которая включена в настоящий документ путем ссылки.
ЗАЯВЛЕНИЕ О СПИСКЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Список последовательностей, связанный с этой заявкой, предоставляется в текстовом формате вместо бумажной копии и таким образом включен в данное описание посредством ссылки. Названием текстового файла, содержащего список последовательностей, является 72245_SEQ_Final_2020-07-08.txt. Текстовый файл имеет размер 145 килобайт, был создан 8 июля 2020 года и подан через EFS-Web вместе с подачей спецификации изобретения.
УРОВЕНЬ ТЕХНИКИ
[0003] Мышечные заболевания человека включают как относительно редкие состояния, таких как мышечные дистрофии, так и распространенные сопутствующие состоянии, такие как связанная с диабетом и возрастная мышечная слабость (саркопения). В настоящее время не существует эффективного лечения для многих заболеваний мышц, включая наиболее распространенную и тяжелую форму мышечной дистрофии, известную как мышечная дистрофия Дюшенна (DMD). Краткий обзор основных признаков DMD показан схематически на фиг. 1. Так как дистрофин, ген, ответственный за DMD, был открыт в 1987 году, исследователи пытались разработать методики доставки гена дистрофина в мышцы и за счет этого ослабить или даже вылечить заболевание. Однако основным недостатком этого подхода в случае DMD является огромный размер гена дистрофина дикого типа, составляющий приблизительно 14 т.п.о. у зрелого транскрипта, который является слишком большим для вставки в вирусные векторы, доступные для доставки генов в ткани. Для иллюстрации, наиболее широко используемый в генной терапии вирусный вектор представляет собой аденоассоциированный вирус (AAV), общая вместимость которого составляет всего 5 т.п.о. Следовательно, чтобы обеспечить возможность доставки терапевтического дистрофинового трансгена, ген дистрофина должен быть усечен до «микродистрофинов», которые составляют приблизительно одну треть от полноразмерного дистрофина и обеспечивают только частичное улучшение функции мышц in vivo у мышей mdx (DMD). Другие основные проблемы, связанные с вирусными векторами, включают отсутствие специфичности в таргетинге вектора к желаемому типу клеток и потенциальный иммунный ответ, связанный с доставкой вируса. Возможность активации иммунного ответа является существенным ограничением в случае подходов с использованием AAV, так как она функционально ограничивает терапевтическую доставку всего одним введением. Весьма вероятно, что этого будет недостаточно для лечения дегенеративных заболеваний мышц, таких как DMD, которые могут требовать вмешательства в течение нескольких лет или даже десятилетий. Кроме того, многие люди уже встречались с некоторыми серотипами AAV, что исключает у них применение генной терапии с использованием АAV.
[0004] Соответственно, несмотря на успехи в области понимания патологии мышечных заболеваний и создания терапевтических вирусных векторов, сохраняется потребность в неиммуногенных векторных платформах, которые могут специфично доставлять терапевтическую полезную нагрузку в мышечные клетки без ограничений по размеру или типу полезной нагрузки. Данное изобретение решает эти и связанные потребности.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0005] Это краткое изложение сущности изобретения приведено для представления подборки концепций в сокращенном виде, которые дополнительно описаны ниже в подробном описании. Это краткое изложение не предназначено для определения ключевых признаков заявленного объекта изобретения, а также не предназначено для использования в качестве помощи при определении объема заявленного изобретения.
[0006] В одном из аспектов настоящее изобретение относится к композиции, содержащей связывающий домен, например, антитело, или его фрагмент или производное, которое специфично связывается с белком-переносчиком глюкозы 4 («GLUT4»), и терапевтическую полезную нагрузку, конъюгированную со связывающим доменом.
[0007] В другом аспекте настоящее изобретение относится к способу таргетинга терапевтической полезной нагрузки для доставки в клетку, которая экспрессирует GLUT4, причем способ включает контактирование клетки с композицией по изобретению. Способ может быть осуществлен in vitro. В альтернативном варианте способ может представлять собой способ in vivo лечения заболевания или состояния, связанного с клетками, экспрессирующими GLUT4.
[0008] В другом аспекте настоящее изобретение относится к способу лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин, включающему введение субъекту терапевтически эффективного количества композиции по изобретению. Композиция в этом аспекте способа содержит связывающий домен, например, антитело, или его фрагмент или производное, которое специфично связывается с белком-переносчиком глюкозы 4 («GLUT4»), и терапевтическую полезную нагрузку, конъюгированную с антителом, или его фрагментом или производным. Терапевтическая полезная нагрузка содержит нуклеиновую кислоту с промоторной последовательностью, функционально связанную с последовательностью, кодирующей белок дистрофин или его функциональный фрагмент.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0009] Фигура 1 представляет собой схематичный обзор мышечной дистрофии Дюшенна (DMD).
[0010] Фигура 2 представляет собой схематичный обзор аспектов невирусной, мышечно-таргетированной платформы доставки генов, раскрытой в данном документе.
[0011] Фигура 3 представляет собой схематическое изображение общей структуры репрезентативной платформы композиций по изобретению, которые использовались в клеточных экспериментах in vitro, более подробно описанных ниже.
[0012] Фигура 4 иллюстрирует результаты Вестерн-блоттинга из эксперимента 1, описанного ниже, показывающего экспрессию nNOS и НА-тэга. GAPDH использовали в качестве контроля нанесения белка. Показано, что инкубация платформы с мышечными клетками в течение всего 15 мин приводит к экспрессии nNOS.
[0013] Фигура 5 иллюстрирует результаты Вестерн-блоттинга из эксперимента 2, описанного ниже, показывая экспрессию nNOS. GAPDH использовали в качестве контроля нанесения белка. В этом эксперименте проверяли влияние различных концентраций плазмидной ДНК (pDNA) и различных соотношений DC-chol:pDNA.
[0014] Фигура 6 иллюстрирует результаты Вестерн-блоттинга из эксперимента 3, описанного ниже, показывая экспрессию полноразмерного дистрофина с использованием антитела 331 (антитела к С-концевой области белка) при возрастающих концентрациях вводимого инсулина. GAPDH использовали в качестве контроля нанесения белка. Результаты показали, что добавление инсулина до 5 нМ повышает экспрессию дистрофина.
[0015] Фигура 7 иллюстрирует результаты Вестерн-блоттинга из эксперимента 4, описанного ниже, показывая экспрессию полноразмерного дистрофина с использованием антитела 331 (антитела к С-концевой области белка). GAPDH использовали в качестве контроля нанесения белка. Как показано, экспрессия дистрофина выше в клетках, сверхэкспрессирующих (+) GLUT4 (mdx GLUT4), и при увеличении количества альбумина, добавленного при экспрессии.
[0016] Фигура 8 представляет собой схематическое изображение общей структуры и компонентов другого аспекта композиции по изобретению, которая была предназначена для экспериментов in vivo, более подробно описанных ниже.
[0017] Фигура 9A-B иллюстрирует результаты Вестерн-блоттинга из эксперимента 5, описанного ниже, показывающего экспрессию nNOS и НА-тэга после введения in vivo платформы без добавления анти-GLUT4 антитела. GAPDH использовали в качестве контроля нанесения белка. Повышение экспрессии как nNOS, так и НА-тэга наблюдалось в икроножной (9A) и передней большеберцовой (9В) мышцах у получавшей инъекцию мыши по сравнению с не получавшей инъекцию мышью mdx Con. Экспрессия nNOS наблюдалась как в получивших инъекцию (+) мышцах, так и в не получивших инъекцию (-) мышцах, что указывает на то, что композиция способна выходить из получивших инъекцию мышц и поступать в системное кровообращение, чтобы в конечном итоге трансфецировать мышцы в противоположной (не получавшей инъекцию) конечности.
[0018] Фигура 10A-B. (10A) Окрашивание Кумасси, демонстрирующее успешную очистку двух клонов анти-GLUT4 моноклональных антител (mAb) с использованием колонок с A/G-белком. Окрашивание кумасси каждого клона показано в невосстановительных (NR) или восстановительных (R) условиях; (10В) графическая иллюстрация связывания клонов mAb с нативным белком GLUT4 с инсулином и без инсулина. Mdx-миобласты, сверхэкспрессирующие GLUT4, инкубировали с клонами мышиных mAb 1 и 2 (при различных разведениях исходного раствора 1 мг/мл), в присутствии или без 100 нМ инсулина, в течение 20 мин при 37°С с последующей инкубацией с конъюгированным с HRP вторичным антителом в течение того же времени. Связывание с антителами (поглощение) измеряли с помощью колориметрического анализа. Повышенная абсорбционная способность показывает, что оба клона связываются с нативным белком GLUT4 in vitro, при этом Клон 2 обладает большей аффинностью связывания, чем Клон 1. Для обоих клонов инсулин увеличивает поглощение за счет переноса большего количества GLUT4 в плазматическую мембрану. В качестве контроля показано поглощение для только вторичного антитела (пунктирная линия).
[0019] Фигура 11 иллюстрирует результаты Вестерн-блоттинга из эксперимента 6, описанного ниже, показывающего экспрессию НА-тэга после введения in vivo (внутримышечной инъекции в переднюю большеберцовую и икроножную мышцы) композиции платформы без добавления («без mAb») или с добавлением («mAb») анти-GLUT4 антитела. GAPDH использовали в качестве контроля нанесения белка. Многократное увеличение экспрессии HA-тэга наблюдалось в обеих мышцах мышей, обработанных композицией платформы, конъюгированной с mAb.
[0020] Фигура 12A-B иллюстрирует результаты эксперимента 7, показывающие экспрессию α-синтрофина у мышей mdx4CV после введения in vivo (внутримышечной (в/м) инъекции в переднюю большеберцовую и икроножную мышцы или внутривенной (в/в) инъекции через ретроорбитальный синус) композиции платформы, содержащей анти-GLUT4 антитело (см. фиг. 8). ДНК-конструкция α-синтрофина содержала последовательность пальмитоилирования RAS для таргетинга белка на поверхностную мембрану мышечной клетки (сарколемму). (12А) Результаты Вестерн-блоттинга показывают значительно более высокую экспрессию α-синтрофина в мышцах мышей как с в/м, так и с в/в инъекций, по сравнению с не получавшей инъекции контрольной мышью mdx4CV. GAPDH использовали в качестве контроля нанесения белка. (12B) Иммуноокрашивание поперечных срезов мышц, показывающее множество мышечных волокон с локализованным на поверхности мембраны окрашенным α-синтрофином у получавших в/м или в/в инъкцию мышей, что неочевидно у не получавших инъекцию контрольных мышей. Следует обратить внимание, что очень яркие области окрашивания в некоторых мышечных волокнах у мыши с в/м инъекцией, скорее всего, представляют собой нервно-мышечные соединения, которые имеют большую площадь поверхности мембраны для экспрессии α-синтрофина.
[0021] Фигура 13A-B иллюстрирует результаты Вестерн-блоттинга в эксперименте 8, показывающие экспрессию nNOS у мышей mdx4CV после введения in vivo платформы, содержащей конструкцию nNOS, описанную на фиг. 11. (13A) В этом эксперименте двум мышам в/м вводили композицию в икроножную мышцу одной задней конечности, и двум мышам вводили в/в через ретроорбитальный синус. Вестерн-блот показывает, что как в/м, так и в/м инъекции привели к значительному увеличению экспрессии nNOS в икроножной мышце по сравнению с двумя икроножными мышцами контрольной мыши mdx4CV, не получавшей инъекции. В соответствии с предыдущими экспериментами, в/м инъекция приводила к увеличению экспрессии nNOS в мышцах как получавшей инъекцию (в/м+), так и противоположной (в/м-) конечности, что указывает на системное распространение платформы после в/м введения. Кроме того, уровень гомолога дистрофина, атрофина, также был значительно увеличен в мышцах мышей, которым платформу вводили с помощью инъекции. GAPDH использовали в качестве контроля нанесения белка. (13B) Трем мышам mdx внутривенно вводили различное количество плазмиды nNOS (20, 60 или 120 мкг). Как показал Вестерн-блоттинг, уровни экспрессии как nNOS, так и HA-тэга постепенно повышаются по мере увеличения дозы плазмиды. GAPDH использовали в качестве контроля нанесения белка.
[0022] Фигура 14 иллюстрирует результаты из эксперимента 9, где экспрессию полноразмерного дистрофина в мышцах измеряли после системной доставки композиции невирусной платформы (см. фиг. 8). Двум мышам mdx4CV в/в вводили различные дозы платформы, содержащей 50 или 100 мкг конструкции CK8-дистрофина. Как показал Вестерн-блоттинг, обе дозы демонстрируют экспрессию полноразмерного дистрофина в трех мышцах задних конечностей (четырехглавой, икроножной и передней большеберцовой) с уровнем, составляющим приблизительно от 0,2 до 0,25% от показателей для мыши дикого типа. Общий белок, вносимый в лунку WT (мышь дикого типа), был такой же, как и во всех других лунках, но состоял из 0,25% белка мыши дикого типа (WT), смешанного с 99,75% белка мыши mdx4CV Con. Слабые полосы у контрольной мыши mdx4CV без инъекций указывают на очень небольшое количество ревертантных волокон, которые экспрессируют низкий уровень дистрофина у мышей mdx.
[0023] Фигура 15 представляет собой результаты эксперимента 9, показывающие экспрессию полноразмерного дистрофина в сердечной мышце после системной доставки состава платформы (см. фиг.8). Двум мышам mdx4CV в/в вводили платформу, содержащей конструкцию либо CK8-дистрофина, либо CMV-дистрофина. Как показано с помощью Вестерн-блоттинга, обе конструкции обеспечивали экспрессию полноразмерного дистрофина в сердце, причем плазмида CMV-дистрофина давала более высокий уровень, чем конструкция с CK8. При этом уровень экспрессии в сердечной мышце также составлял приблизительно 0,1-0,2% от экспрессии у мыши дикого типа (WT). Общий белок, вносимый в лунку WT, был такой же, как и во всех других лунках, но состоял из 0,25% белка мыши дикого типа (WT), смешанного с 99,75% белка мыши mdx4CV Con. В качестве мембранно-специфичного контроля нанесения использовали кавеолин-3.
[0024] На фигуре 16A-B продемонстрированы результаты эксперимента 10, показывающего анализ отслеживания частиц композиции невирусной платформы (см. фиг. 8), содержащей плазмиду СК8-полноразмерного дистрофина. (16A) Изображение частиц платформы, идентифицированных и отслеженных программным обеспечением Nanosight ns300 для анализа размера. (16B) График распределения частиц, показывающий размер частиц в зависимости от концентрации частиц. Значения представляют собой среднее значение ±SEM для 4 отдельных входных данных из одного образца.
[0025] Фигура 17 иллюстрирует результаты эксперимента 11, в котором анализ ELISA был разработан для измерения аффинности связывания 6 моноклональных антител, полученных против пептидной последовательности мышиного GLUT4 (GRQGPGGPDSI; SEQ ID NO:4). Как показано, все антитела демонстрировали дозозависимое увеличение поглощения, указывающее на повышенное связывание с пептидом мышиного GLUT4.
[0026] На фигуре 18 продемонстрированы результаты эксперимента 11 анализа ELISA, в котором проверяли аффинность связывания 7 моноклональных антител, полученных против пептидной последовательности GLUT4 человека (GRQGPEGPSSI; SEQ ID NO:2). Как показано, все антитела демонстрировали дозозависимое увеличение поглощения, указывающее на повышенное связывание с рекомбинантным человеческим белком GLUT4.
[0027] На фигуре 19 показаны результаты эксперимента 11, в котором анализ ELISA был разработан для проверки того, могут ли антитела, полученные к последовательности GLUT4 человека (GRQGPEGPSSI; SEQ ID NO:2), связываться с последовательностью пептида GLUT4 мыши (GRQGPGGPDSI; SEQ ID №:4). Как показано, клоны антител, полученные к последовательности белка человека (5E3, 8G2, 12E11, 1A7), связывали мышиный пептид намного слабее, чем антитела, полученные к последовательности мышиного белка (22F6 и 21E5), которые достигли насыщения связывания при относительно низкой концентрации (приблизительно 2 нг/мл).
[0028] На фигуре 20 продемонстрированы результаты анализа ELISA из эксперимента 11, в котором проверяли, могут ли антитела, полученные к последовательности мышиного GLUT4 (GRQGPGGPDSI; SEQ ID NO:4), связываться с последовательностью GLUT4 человека (GRQGPEGPSSI; SEQ ID NO:2). Как показано, антитела, полученные к последовательности мышиного белка (22F6 и 21E5), очень слабо связывались с антигеном, в то время как антитела к последовательности белка человека связывались сильно (5E3, 8G2).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0029] Это изобретение основано на разработке авторами невирусных векторных платформ для специфичного таргетинга терапевтических полезных нагрузок, как генетических, так и фармакологических, в ткани скелетных и сердечных мышц. Вкратце, конструкция платформы, показанная на фиг. 2, была ориентирована на доставку большего количества полезной нагрузки, например, полноразмерного транскрипта гена дистрофина, для лечения заболеваний, связанных с нарушением экспрессии дистрофина или дисфункцией продукта гена.
[0030] Платформа использует специфичную и инсулинозависимую экспрессию белка-переносчика глюкозы 4 (GLUT4) в мышцах и другом ограниченном наборе тканей. GLUT4 кодируется геном SLC2A4, который преимущественно экспрессируется в поперечнополосатых мышцах (скелетных и сердечных) и жировой ткани, но также экспрессируется в определенных областях головного мозга, включая гиппокамп и мозжечок. GLUT4 является членом семейства белков, переносящих глюкозу. Переносчики глюкозы представляют собой интегральные мембранные белки, содержащие 12 трансмембранных спиральных доменов. GLUT4 на высоком уровне экспрессируется в мышечной и жировой ткани, в то время как другие члены семейства переносчиков глюкозы специфичны для других тканей. Например, GLUT3, высокоаффинный переносчик глюкозы, является основным переносчиком глюкозы в нейронах. Без активации, GLUT4 в основном находится во внутриклеточных везикулах, но перемещается к плазматической мембране и сливается с ней при введении инсулина, тем самым опосредуя инсулинозависимое поглощение глюкозы. После связывания инсулина с рецептором инсулина активируется сигнальный путь, ведущий к транслокации GLUT4 в плазматическую мембрану. Этот процесс происходит в течение 15 минут после связывания инсулина. Как только концентрация инсулина снижается, со временем GLUT4 интернализуется в везикулах и либо проходит через эндосомальную систему, либо возвращается обратно в плазматическую мембрану.
[0031] Описанная векторная платформа использует это двунаправленное опосредованное везикулами перемещение GLUT4 к плазматической мембране, а затем обратно внутрь клетки, чтобы обеспечить стратегию доставки терапевтических средств в клетки и ткани, экспрессирующие GLUT4. Как более подробно описано ниже, впервые были разработаны новые антитела, как кроличьи поликлональные, так и мышиные моноклональные, которые связываются с внеклеточным доменом мышиного белка GLUT4. С использованием различных методов конъюгации антител было получено несколько структур полезной нагрузки/вектора. Был проведен ряд экспериментов in vitro и in vivo для оценки специфичности, пропускной способности и других полезных аспектов этих репрезентативных вариантов осуществления векторной платформы. Например, было продемонстрировано, что эффективно нацеленный терапевтический трансген, в том числе плазмидная ДНК размером до 19 т.п.о., содержащая полноразмерный ген дистрофина, может поглощаться и экспрессироваться мышечными клетками. Плазмидная ДНК была успешно транспортирована в клетку после интернализации GLUT4, и была инициирована экспрессия гена дистрофина. Анализы в конечном итоге демонстрируют полезность платформы для лечения мышечных заболеваний, которые до сих пор трудно поддавались лечению. Платформа является не только гибкой, что позволяет использовать различные типы терапевтической полезной нагрузки, но и не требует вирусных компонентов. Соответственно, платформа допускает многократное введение в течение длительного времени, в том числе в течение нескольких лет, для лечения хронических и дегенеративных заболеваний мышц без риска иммуногенности.
[0032] В соответствии с вышеизложенным в описании предложена композиция, содержащая связывающий домен, который специфично связывается с белком-переносчиком глюкозы 4 («GLUT4»). Композиция дополнительно содержит терапевтическую полезную нагрузку, конъюгированную со связывающим доменом.
Связывающий домен
[0033] В некоторых вариантах осуществления связывающий домен представляет собой или содержит антитело, его фрагмент или производное. В таких вариантах осуществления терапевтическая полезная нагрузка может быть прямо или непрямо конъюгирована с антителом, его фрагментом или производным.
[0034] Термин «антитело» используется в настоящем документе в самом широком смысле и охватывает различные структуры антител, полученные из любого млекопитающего, продуцирующего антитела (например, мыши, крысы, кролика и примата, включая человека), и которые специфично связываются с целевым антигеном. Фрагмент антитела конкретно относится к интактной части или субдомену исходного антитела, который все еще сохраняет антигенсвязывающую способность. Производное антитела относится к молекуле, которая включает одно или несколько антител или фрагментов антител. Как правило, имеется по меньшей мере некоторая дополнительная модификация в структуре антитела или его фрагмента или в представлении или конфигурации антитела или его фрагмента. Примеры антител по настоящему изобретению включают поликлональные, моноклональные и рекомбинантные антитела. Примеры антител или производных антител по настоящему изобретению также включают полиспецифичные антитела (например, биспецифичные антитела); гуманизированные антитела; мышиные антитела; химерные, мышь-человек, мышь-примат, примат-
[0035] Как указано, фрагмент антитела представляет собой часть или субдомен, полученный из полноразмерного антитела или относящийся к нему, предпочтительно включающий его определяющие комплементарность области (CDR), антигенсвязывающие области или вариабельные области, а производные антитела относятся к дальнейшей структурной модификации или комбинациям в полученной молекуле. Иллюстративные примеры фрагментов или производных антител, охватываемых настоящим изобретением, включают Fab, Fab’, F(ab)2, F(ab’)2 и Fv-фрагменты, диантитела, молекулы одноцепочечных антител, VHH-фрагменты, VNAR-фрагменты, полиспецифичные антитела, образованные фрагментами антител, нанотела и т.п. Например, типовое производное одноцепочечного антитела, охватываемое настоящим описанием, представляет собой «одноцепочечный Fv» или «scFv» фрагмент антитела, который содержит домены VH и VL антитела, где эти домены присутствуют в одной полипептидной цепи. Полипептид Fv может дополнительно содержать полипептидный линкер между доменами VH и VL, который позволяет scFv образовывать желаемую структуру для связывания антигена. Еще одно иллюстративное одноцепочечное антитело, охватываемое настоящим изобретением, представляет собой одноцепочечный Fab-фрагмент (scFab).
[0036] Как указано, антитела могут быть дополнительно модифицированы для создания производных, подходящих для различных целей. Например, «химерное антитело» представляет собой рекомбинантный белок, который содержит домены из разных источников. Например, вариабельные домены и определяющие комплементарность области (CDR) могут быть получены из антител животных (например, грызунов), в то время как остальная часть молекулы антитела получена из человеческого антитела. «Гуманизированное антитело» представляет собой химерное антитело, содержащее минимальную последовательность, соответствующую специфичным определяющим комплементарность областям, полученным из иммуноглобулина животного, перенесенного в каркас антитела человека. Гуманизированные антитела обычно представляют собой рекомбинантные белки, в которых только определяющие комплементарность области (CDR) антитела получены из антитела животного. Любое из этих антител или их фрагментов или производных охвачено настоящим изобретением.
[0037] Фрагменты и производные антител, распознающие специфичное эпитопы, могут быть получены любым способом, известным специалистам в данной области. Например, Fab- и F(ab’)2-фрагменты по настоящему изобретению могут быть получены протеолитическим расщеплением молекул иммуноглобулина с использованием таких ферментов, как папаин (для получения Fab-фрагментов) или пепсин (для получения F(ab’)2-фрагментов). F(ab’)2-фрагменты содержат вариабельную область, константную область легкой цепи и домен CHI тяжелой цепи. Кроме того, антитела, или их фрагменты или производные по настоящему изобретению также могут быть получены с использованием различных методов фагового дисплея, известных в данной области. Наконец, антитела, или их фрагменты или производные могут быть получены рекомбинантно в соответствии с известными методами.
[0038] Специалисту-практику будет очевидно, что связывающий домен может содержать антигенсвязывающие молекулы, отличные от домена на основе антител, такие как пептидные антитела, антигенсвязывающие каркасы (например, DARPin, белки с повторами HEAT, белки с повторами ARM, белки с тетратрикопептидными повторами и другие каркасы на основе природных белков с повторами и т. д. [см., например, Boersma and Pluckthun, Curr. Opin. Biotechnol. 22:849-857, 2011, и цитируемые в них ссылки, включенные в настоящий документ путем ссылки]), которые включают функциональный связывающий домен или его антигенсвязывающий фрагмент.
[0039] В некоторых вариантах осуществления связывающий домен (например, антитело, или его фрагмент или производное) является биотинилированным. Как более подробно описано ниже, биотинилирование облегчает некоторые варианты сборки/конъюгации композиции.
[0040] Используемый в настоящем документе термин «специфично связываться» или его варианты относятся к способности связывающего домена (например, антитела или его фрагмента или производного) связываться с представляющим интерес антигеном (например, GLUT4), без значительного связывания с другими молекулами в стандартных условиях, известных в данной области. Связывающий домен может связываться с другими пептидами, полипептидами или белками, но с более низкой аффинностью, определяемой, например, с помощью иммунологических анализов, BIAcore или других методов анализа, известных в данной области. Однако предпочтительно, чтобы связывающий домен практически не реагировал перекрестно с другими антигенами.
[0041] В некоторых вариантах осуществления GLUT4-связывающий домен композиции имеет аффинность связывания в диапазоне, характеризуемом константой диссоциации (Kd) от примерно 50 нМ (более низкая аффинность связывания) до примерно 0,001 нМ (более высокая аффинность связывания). Например, связывающий домен имеет аффинность связывания с белком GLUT, характеризующуюся (Kd) примерно 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 5 нМ, 1 нМ, 0,75 нМ, 0,5 нМ, 0,1 нМ, 0,05 нМ, 0,01 нМ, 0,005 нМ и 0,001 нМ или даже меньше. Типичные диапазоны (Kd), характеризующие аффинность связывания домена для таргетинга в клетки с антигеном, характерным для интересующего типа клеток, включают от примерно 30 нМ до примерно 10 нМ, от примерно 20 нМ до примерно 1 нМ, от примерно 10 нМ до примерно 0,1 нМ, от примерно 0,5 нМ до примерно 0,05 нМ и от примерно 0,1 нМ до примерно 0,001 нМ или даже ниже, или любой субдиапазон в них.
Мишень для связывания
[0042] Как указано выше, связывающий домен (например, антитело или его фрагмент или производное) специфично связывается с белком-переносчиком глюкозы 4 (GLUT4). GLUT4 кодируется геном SLC2A4 (или его гомологами), который преимущественно экспрессируется в поперечнополосатых мышцах (скелетных и сердечных) и жировой ткани, но также экспрессируется в определенных областях головного мозга, включая гиппокамп и мозжечок. Это изобретение охватывает GLUT4 любого млекопитающего, такого как человек, мышь, крыса, кошка, собака, лошадь и т. д. В некоторых вариантах осуществления связывающий домен специфично связывается с внеклеточным доменом белка GLUT4.
[0043] В некоторых вариантах осуществления связывающий домен специфично связывается с доменом первой внеклеточной петли GLUT4. В некоторых вариантах осуществления домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:2, которая является субдоменом домена первой внеклеточной петли GLUT4 человека. Например, иллюстративный домен GLUT4, рассматриваемый в настоящем описании, включает домены, представленные в последовательностях с номером доступа Genbank M91463.1 (человек), номером доступа Genbank AB008453.1 (мышь), номером доступа Genbank NM_001159327.1 (собака) и номером доступа Genbank L36125.1 (крыса), каждая из которых полностью включена в настоящий документ посредством ссылки. В некоторых вариантах осуществления домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:1, которая представляет собой репрезентативный домен первой внеклеточной петли GLUT4 человека. В некоторых вариантах осуществления связывающий домен специфично связывается с субдоменом домена первой внеклеточной петли GLUT4, который содержит последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно на 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:2.
[0044] В некоторых вариантах осуществления домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:4, которая является субдоменом домена первой внеклеточной петли мышиного GLUT4. В некоторых вариантах осуществления домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:3, которая представляет собой репрезентативный домен первой внеклеточной петли мышиного GLUT4. В некоторых вариантах осуществления связывающий домен специфично связывается с субдоменом домена первой внеклеточной петли GLUT4, который содержит последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98%) с последовательностью, представленной в SEQ ID NO:4.
Полезная нагрузка
[0045] Терапевтическая полезная нагрузка может представлять собой любую полезную нагрузку, способную индуцировать изменение в клетке-мишени, которая, по крайней мере временно, экспрессирует GLUT4 на своей поверхности. Изменение может иметь терапевтический эффект, если оно происходит in vivo. Типичные, неограничивающие терапевтические полезные нагрузки могут содержать нуклеиновую кислоту, белок или пептид, липид, низкомолекулярный фармацевтический препарат и/или радиоизотоп.
[0046] Например, в некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, такую как ДНК (включая кДНК), мРНК, киРНК, кшРНК и нРНК (направляющая РНК). Молекулы нРНК можно использовать в методах для редактирования ДНК (например, методах CRISPR/Cas9) и редактирования РНК.
[0047] В некоторых вариантах осуществления нуклеиновая кислота содержит последовательность (например, открытую рамку считывания), кодирующую белок, который оказывает терапевтическое действие на клетку-мишень. В некоторых вариантах осуществления нуклеиновая кислота содержит открытую рамку считывания, функционально связанную с промоторной последовательностью для обеспечения экспрессии открытой рамки считывания в клетке-мишени. Эту открытую рамку считывания можно назвать трансгеном. Термин «промотор» относится к регуляторной нуклеотидной последовательности, которая может активировать транскрипцию (экспрессию) трансгена и/или его сплайсированные варианты. Промотор обычно расположен выше гена, но может быть расположен и в других областях, прилегающих к гену, или даже внутри гена. Промотор обычно содержит сайты связывания РНК-полимеразы и одного или нескольких факторов транскрипции, которые участвуют в сборке транскрипционного комплекса. Используемый в настоящем документе термин «функционально связанный» означает, что промотор и кодирующая нуклеиновая кислота сконфигурированы и расположены относительно друг друга таким образом, что промотор может активировать транскрипцию кодирующей нуклеиновой кислоты с помощью транскрипционного аппарата клетки. Промотор может быть конститутивным или индуцибельным. Конститутивные промоторы можно определить на основании характеристик клетки-мишени и конкретных факторов транскрипции, доступных в цитозоле. Специалист в данной области может выбрать подходящий промотор в зависимости от предполагаемого хозяина, поскольку различные промоторы известны и широко используются в данной области. Примеры неограничивающих промоторов включают промотор мышечной креатинкиназы (CK) (номер доступа Genbank AF188002.1) и промотор скелетного актина человека (номер доступа Genbank NG_006672.1), каждый из которых включен в настоящий документ путем ссылки. В то время как усовершенствования, предусмотренные в настоящем документе, позволяют использовать полноразмерные промоторы, укороченные промоторы, которые сохраняют некоторую активность, по-прежнему охватываются настоящим изобретением. Типичные, не ограничивающие примеры укороченных промоторов СК, которые сохраняют активность, описаны в Hauser MA, et al., Analysis of muscle creatine kinase regulatory elements in recombinant adenoviral vectors, Mol Ther. 2000l 2(1):16-25, и в Brennan KJ and Hardeman EC, Quantitative analysis of the human alpha-skeletal actin gene in transgenic mice, J Biol Chem. 1993 268(1):719-725, каждая из которых полностью включена в настоящий документ посредством ссылки.
[0048] Терапевтическая полезная нагрузка нуклеиновой кислоты может иметь любой размер и любую конфигурацию, такую как линейная форма, плазмидная форма (например, кольцевая форма) или миникольцевая форма. Как показано ниже, было продемонстрировано, что композиции по настоящему изобретению способны доставлять очень большие трансгены для успешной экспрессии в клетках-мишенях. Таким образом, полезная нагрузка нуклеиновой кислоты не ограничена в отношении размера, что практически запрещает использование типичных вирусных векторов. Таким образом, линейные формы нуклеиновых кислот могут включать короткие олигонуклеотиды или более длинные трансгены, без ограничений. Как понятно специалистам-практикам, миникольца представляют собой производные кольцевых плазмид меньшего размера, в которых отсутствует большинство или все компоненты прокариотического вектора. Миникольца могут функционировать как носители трансгена с тем преимуществом, что они менее подвержены деградации из-за меньшего размера и/или отсутствия прокариотической последовательности.
[0049] Как указано выше, нуклеиновая кислота может представлять собой трансген, сконфигурированный таким образом, чтобы он содержал последовательность, кодирующую любой желаемый белок или фрагмент белка, экспрессируемый в клетке-мишени. Трансген может кодировать любой белок, который желательно экспрессировать в клетке. В некоторых вариантах осуществления трансген кодирует белок, который оказывает благотворное влияние на клетку по сравнению с мутантной эндогенной версией, присутствующей в клетке-мишени. Например, как описано выше, мышечная дистрофия Дюшенна (DMD) представляет собой тяжелое сцепленное с полом заболевание, возникающее из-за аберрантного белка дистрофина (или аберрантной экспрессии белка дистрофина). Ген, кодирующий дистрофин, называемый геном «DMD», является одним из самых длинных генов человека, охватывающим около 2,3 миллионов пар оснований (0,08% генома человека) в локусе Xp21 с примерно 79 экзонами. Первичный транскрипт белка дикого типа составляет около 21 тысяч пар оснований, а зрелая мРНК - около 14 тысяч пар оснований. Как описано ниже, авторы изобретения установили, что векторная платформа по изобретению может доставлять такие длинные трансгены в клетки GLUT4+ (например, мышечные клетки) для успешного получения экспрессии. Соответственно, типичный и не ограничивающий пример полезной нагрузки нуклеиновой кислоты включает кодирующую нуклеотидную последовательность, кодирующую белок дистрофин, или его функциональный фрагмент, функционально связанную с промоторной последовательностью. В некоторых вариантах осуществления кодируемый белок дистрофин представляет собой полноразмерный белок дистрофин человека или по существу полноразмерный белок дистрофин человека. Типичная полноразмерная белковая последовательность дистрофина человека представлена в SEQ ID NO:5. В некоторых вариантах осуществления полезная нагрузка нуклеиновой кислоты кодирует дистрофин с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно а 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:5.
[0050] Термин «практически полноразмерный» охватывает варианты осуществления, в которых кодируется значительная, но неполная версия дистрофина, как, например, кодируется по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по по меньшей мере примерно 96%, по меньшей мере примерно 98% или по меньшей мере примерно 99% полноразмерного белка дистрофина (например, с идентичностью последовательности по меньшей мере 80% с SEQ ID NO:5, описанной выше), обеспечивая видимое фенотипическое улучшение в клетке-мишени.
[0051] Также было показано, что белки «микродистрофины», вводимые или экспрессируемые в клетке с аберрантным или мутантным геном DMD, могут обеспечивать по меньшей мере некоторый эффект улучшения, компенсирующий эндогенную мутацию в гене DMD. Не ограничиваясь какой-либо конкретной теорией, считается, что многие укороченные версии дистрофина обладают улучшенной функциональностью по сравнению с мутированной или аберрантной экспрессией дистрофина у субъектов с DMD и другими заболеваниями, связанными с дистрофином. Соответственно, доставка или индукция экспрессии этих укороченных или микродистрофиновых белков в мышечной клетке может начать восстановление относительно здорового или улучшенного фенотипа. Соответственно, в некоторых вариантах осуществления полезная нагрузка нуклеиновой кислоты может кодировать функциональный фрагмент белка дистрофина, такой как любой микродистрофин, известный в данной области. Например, в статье Ramos et al., 2019 Molecular Therapies 27: 623-635, включенной в настоящий документ путем ссылки, описаны несколько миниатюрных конструкций микродистрофина, которые оптимизированы для повышения эффективности. Иллюстративные и не ограничивающие области и домены белка дистрофина, которые, как было показано, обладают характеристиками, способствующими функциональности конструкций микродистрофина, включают: палочковидный домен с по меньшей мере 4 SR-доменами, α-синтрофин-связывающий домен в SR16-17, актин-связывающий домен и богатый цистеином домен для связывания β-дистрогликана.
[0052] В некоторых вариантах осуществления полноразмерный белок дистрофин имеет аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:5, которая представляет собой предсказанную последовательность полноразмерного белка дистрофина человека. Последовательность имеет номер доступа Genbank AAA53189.1 и полностью включена в настоящий документ посредством ссылки. В других вариантах осуществления полноразмерный белок дистрофин имеет аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:6, которая представляет собой предсказанную последовательность полноразмерного белка дистрофина мыши. Последовательность имеет номер доступа Genbank NP_031894.1 и полностью включена в настоящий документ посредством ссылки. В других вариантах осуществления полноразмерный белок дистрофин человека имеет аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO: 7, которая представляет собой предсказанную последовательность полноразмерного белка дистрофина крысы. Последовательность имеет номер доступа Genbank XP_017457396.1 и полностью включена в настоящий документ посредством ссылки. В других вариантах осуществления полноразмерный белок дистрофин человека имеет аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO: 8, которая представляет собой предсказанную последовательность полноразмерного белка дистрофина кролика. Последовательность имеет номер доступа Genbank XP_017205220.1 и полностью включена в настоящий документ посредством ссылки. Как указано выше, терапевтическая полезная нагрузка нуклеиновой кислоты в этих вариантах осуществления может содержать полноразмерную последовательность белка дистрофина, по существу полноразмерную последовательность дистрофина или быть ее функциональным фрагментом, который сохраняет способность обеспечивать по меньшей мере некоторый терапевтический эффект в отношении восстановления улучшенного фенотипа.
[0053] Как указано выше, полезная нагрузка нуклеиновой кислоты может включать линейные нуклеиновые кислоты, такие как короткие олигомеры. В некоторых вариантах осуществления нуклеиновая кислота не является трансгеном, кодирующим белок, а скорее изменяет эндогенную экспрессию и трансляцию белков в клетке-мишени. Например, в некоторых терапевтических подходах к лечению геномных заболеваний, таких как DMD, вводят олигонуклеотиды для пропуска экзонов в пре-мРНК, которые содержат мутацию, приводящую к сдвигу рамки считывания. Вводимые олигонуклеотиды (например, фосфодиамидат-морфолино олигомеры) сконфигурированы для последовательность-специфичной гибридизации с мутированной пре-мРНК и стерически препятствуют связыванию сплайсосомы в местах соединения мутантного экзона в кодирующей молекуле пре-мРНК. В результате сплайсосома вырезает не только интроны, но и экзон, содержащий мутацию сдвига рамки считывания. Полученная зрелая мРНК кодирует слегка укороченный белок, лишенный мутантного экзона и сохраняющий рамку считывания по всем оставшимся интронов. Укороченный белок сохраняет по меньшей мере некоторые характеристики белка дикого типа по сравнению с версией, содержащей сдвиг рамки считывания. Этот подход был реализован для DMD. Однако такие подходы требуют очень большого количества олигонуклеотидов из-за высокой скорости деградации после введения. См., например, Verhaart IEC and Aartsma-Rus A, Therapeutic developments for Duchenne muscular dystrophy, Nat Rev Neurol. 2019 Jul;15(7):373-386, полностью включенную в настоящий документ посредством ссылки. Полезная нагрузка нуклеиновой кислоты по настоящему изобретению включает такие терапевтические олигомеры. Описанная платформа обеспечивает преимущество специфичного таргетинга олигомеров в клетку-мишень, что приводит к эффективному поглощению клеткой. Таким образом, раскрытая платформа для таргетинга не требует таких высоких доз олигонуклеотидов и обеспечивает более высокую вероятность доставки и поглощения клетками-мишенями.
[0054] В других вариантах осуществления полезная нагрузка нуклеиновой кислоты по настоящему изобретению может быть сконфигурирована так, чтобы стимулировать РНК-интерференцию. Например, в некоторых вариантах осуществления полезная нагрузка нуклеиновой кислоты представляет собой или содержит молекулы короткой интерферирующей РНК (киРНК) или короткой шпилечной РНК (кшРНК) или нуклеиновой кислоты, кодирующей молекулы киРНК или кшРНК. Последовательности таких молекул нуклеиновых кислот могут быть легко сконфигурированы в соответствии с навыками и знаниями в данной области, чтобы привести к связыванию и последующей деградации транскриптов мРНК, что приведет к нокдауну любого желаемого белка-мишени в клетке.
[0055] В любом варианте полезной нагрузки нуклеиновой кислоты, но особенно когда полезная нагрузка нуклеиновой кислоты довольно велика (например, для длинных нуклеиновых кислот, кодирующих дистрофин или его большие участки), композиция может также содержать дополнительные компоненты для облегчения эффективного свертывания или упаковки (например, конденсации) нуклеиновой кислоты. Дополнительная упаковка может быть полезна для линейных (например, олиго или длинных последовательностей), плазмидных и миникольцевых вариантов полезной нагрузки нуклеиновой кислоты, описанных выше, без ограничений. Например, в одном варианте осуществления композиция дополнительно содержит гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой. Гистоновый белок (белки) может способствовать свертыванию полезной нагрузки нуклеиновой кислоты для эффективной упаковки и переноса. В некоторых вариантах осуществления гистоновый белок или по меньшей мере часть множества гистоновых белков биотинилируют для облегчения конъюгации, как более подробно описано ниже.
[0056] В других вариантах осуществления композиция дополнительно содержит множество катионных фрагментов, связанных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота. Не ограничиваясь какой-либо конкретной теорией, считается, что катионные молекулы потенциально могут конденсировать ДНК, которая представляет собой полимерную анионную молекулу. Таким образом, пептиды, содержащие множество положительно заряженных аминокислот, пригодны для конденсации полезной нагрузки полимерной нуклеиновой кислоты. Примеры катионных аминокислот включают лизин (Lys, K), гистидин (His, H) и аргинин (Arg, R). Соответственно, в некоторых вариантах осуществления композиция дополнительно содержит один или несколько катионных пептидов, где катионные пептиды содержат множество катионных аминокислот, например, выбранных из His, Arg и Lys, возможно, в комбинации. Пептид обычно имеет общий положительный заряд. В некоторых вариантах осуществления пептид содержит одну или несколько пептидных фрагментов поли-L-Lys («PLL»). Дополнительные не ограничивающие катионные молекулы или фрагменты, которые могут быть включены в композицию для конденсации полезной нагрузки нуклеиновой кислоты, включают линейный полиэтиленимин («LPEI»), разветвленный полиэтиленимин («BPEI»), хитозан, спермидин и спермин, полиамидоамин («РАМАМ»), поли(2-диметиламиноэтилметакрилат) («PDMAEMA»), поли(бета-аминоэфиры) («PBAE»), поли{N-[N-(2-аминоэтил)-2-аминоэтил]аспартамид («PAsp(DET)»), поли(2-аминоэтилэтиленфосфат («PPEEA») и т.п.
[0057] В некоторых вариантах осуществления по меньшей мере часть катионных фрагментов, таких как PLL или другие катионные молекулы, указанные выше, биотинилируют для облегчения конъюгации, как более подробно описано ниже.
Конъюгация
[0058] Связывающий домен, например, антитело или его фрагменты или производные, можно конъюгировать с терапевтической полезной нагрузкой любым известным способом. Конъюгация может быть ковалентной или ионной.
[0059] В некоторых иллюстративных неограничивающих вариантах осуществления конъюгация основана на взаимодействии между биотином и связывающимся партнером, таким как авидин, нейтравидин и стрептавидин, для обеспечения очень высокой аффинности и специфичной конъюгации составляющих частей композиции.
[0060] Например, в одном варианте осуществления связывающий домен (например, антитело или его фрагмент, или производное) и сворачивающий или конденсирующий ДНК элемент (например, гистон или катионный пептид) являются биотинилированными. Полезная нагрузка нуклеиновой кислоты связана со сворачивающим или конденсирующим ДНК элементом (например, сворачивается вокруг сворачивающего элемента или конденсируется с его помощью). Благодаря взаимодействию нескольких биотинилированных компонентов с одной и той же молекулой нейтравидина, связывающий домен и сворачивающий или конденсирующий элемент и ассоциированная полезная нагрузка нуклеиновой кислоты конъюгируются вместе с высокой аффинностью.
[0061] В некоторых дополнительных вариантах осуществления может быть агрегировано множество связывающих доменов и полезной нагрузки нуклеиновой кислоты (с ассоциированным сворачивающими или конденсирующими ДНК элементами). Этого можно достичь, просто в результате связывания нескольких биотинилированных элементов с одним и тем же связывающим партнером на основе авидина. В качестве альтернативы может быть несколько биотинилированных сворачивающих или конденсирующих ДНК элементов, связанных с одной и той же полезной нагрузкой нуклеиновой кислоты, каждый из которых связан с другим связывающим партнером на основе авидина, который, в свою очередь, связан с другим связывающим доменом. Этой агрегации можно дополнительно способствовать путем включения в состав одной или нескольких частиц, таких как липосомы, которые могут связываться с несколькими нуклеотидными полезными нагрузками. Например, как показано на фиг. 3, несколько полезных нагрузок в виде плазмид связаны с катионной структурой липосомы. Каждая плазмида, в свою очередь, связана с множеством гистоновых белков, часть которых биотинилирована. Биотинилированные гистоны связываются с биотинилированными связывающими доменами (например, антителами против GLUT4) в результате связывания с нейтравидином. Получение такой иллюстративной композиции более подробно описано ниже.
[0062] Соответственно, в некоторых вариантах осуществления раскрытая композиция может дополнительно содержать катионную частицу, такую как липосома, которая может дополнительно ассоциироваться и/или агрегировать с полезной нагрузкой нуклеиновой кислоты. Катионные липосомы известны и могут быть легко сконструированы специалистами в данной области для облегчения надлежащей агрегации и/или ассоциации с полезной нагрузкой нуклеиновой кислоты. В некоторых иллюстративных неограничивающих вариантах осуществления липосома может содержать один или несколько липидов, выбранных из DC-холестерин HCl,3b-[N-(N',N'-диметиламиноэтан)-карамоил]холестерин гидрохлорида («DC-Chol»), 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламина («DOPE»), 1,2-ди-О-октадеценил-3-триметиламмония («DOTMA»), N-[1-(2,3-диолеилокси)пропил]-N, N,N-триметиламмонийпропана («DOTAP»), диметилдиоктадециламмоний бромида («DDAB»), 1,2-димиристоил-3-триметиламмоний-пропана («DMTAP»), 1,2-диолеоил-3-диметиламмоний-пропана («DODAP»), гемисукцинатхолестерина («CHEMS») и холестерина («CHOL»), 2,3-диолеилокси-N-[2(сперминкарбоксамидо)этил]-N, N-диметил-1-пропанаминия («DOSPA»), диоктадециламидоглицилспермина («DOGS»), диолеоилфосфатидилхолина («DOPC») и т.п. В некоторых вариантах осуществления липосома дополнительно содержит полиэтиленгликоль («PEG»).
[0063] В другом иллюстративном варианте осуществления, показанном на фиг. 8, полезная нагрузка плазмидной ДНК связана с множеством катионных фрагментов PLL, которые являются биотинилированными. Биотинилированные фрагменты PLL связываются со связывающими доменами (например, антителами против GLUT4) путем взаимной ассоциации с одним и тем же связывающим партнером нейтравидином.
[0064] В дополнительных вариантах осуществления композиция дополнительно содержит альбумин, который может способствовать усиленной экспрессии полезной нагрузки нуклеиновой кислоты. Как более подробно описано ниже, недавние данные показали, что альбумин может усиливать экспрессию белка с доставляемых ДНК/липосомных векторов. Среди других причин считается, что альбумин усиливает доставку композиции к клеткам-мишеням, включая усиление исключения из эндосомальной утилизации и проникновения в ядро полезной нагрузки нуклеиновой кислоты. Отрицательно заряженный альбумин может связываться с положительно заряженными липосомами, связанными с полезной нагрузкой нуклеиновыми кислотами. В других вариантах осуществления, где для конъюгации используются связывающие партнеры на основе биотина и авидина, альбумин может быть биотинилирован. Таким образом, положительно заряженная липосома не является обязательным условием эффективности альбумина. Это показано на фиг. 8.
[0065] В одном аспекте изобретение относится к композициям, описанным в настоящем документе, для применения при лечении заболевания, при этом заболевание поддается лечению путем доставки полезной нагрузки в клетку, которая экспрессирует GLUT4. Подходящие заболевания, охватываемые данным аспектом, в дополнение к другим элементам термина «лечение», используемого в данном документе, более подробно описаны ниже. В некоторых вариантах осуществления применение предполагается для лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин. В некоторых вариантах осуществления композиции, описанные в настоящем документе, предназначены для лечения мышечной дистрофии Дюшенна («DMD») или мышечной дистрофии Беккера («BMD»), которые более подробно описаны выше и ниже. В этих вариантах осуществления полезная нагрузка композиций по изобретению может содержать нуклеиновую кислоту, кодирующую полноразмерный белок дистрофин (например, дикого типа), или его функциональный фрагмент (например, около 80%, 85%, 90%, 95%, 98%), или его вариант, аминокислотная последовательность которого по меньшей мере примерно на 90%, 95% или 98% идентична последовательности белка дикого типа. В некоторых вариантах осуществления изменение идентичности последовательностей по сравнению с последовательностью дикого типа является результатом консервативных замен, которые существенно не влияют на функцию белка. В некоторых вариантах осуществления применение может дополнительно включать введение субъекту инсулина для увеличения доступной мишени GLUT4 и таргетинга полезной нагрузки.
Способы
[0066] Настоящее изобретение также охватывает способы использования и изготовления раскрытых композиций.
[0067] В одном аспекте относится к способу таргетинга полезной нагрузки (например, терапевтической полезной нагрузки) для доставки в клетку, которая экспрессирует GLUT4. Способ включает контакт клетки с любым вариантом раскрытой композиции, например, как указано выше.
[0068] В некоторых вариантах осуществления способ дополнительно включает контакт клетки с инсулином до контакта клетки с композицией. Как описано выше, GLUT4 в высокой степени экспрессируется в поперечнополосатых мышцах и жировой ткани, а также в клетках определенных областей мозга, включая гиппокамп и мозжечок. GLUT4 в основном находится во внутриклеточных везикулах хранения GLUT4 (GSV). Однако после связывания инсулина с рецептором инсулина (в чувствительных к инсулину клетках, включая скелетные мышцы, сердце, жировые клетки и некоторые нейроны) активируется сигнальный путь, ведущий к транслокации GLUT4 к плазматической мембране. См., например, Klip, A, et al., 30 sweet years of GLUT4. 2019, REV119.008351, полностью включенную в настоящий документ посредством ссылки. Таким образом, в некоторых вариантах осуществления инсулин контактирует с клеткой по меньшей мере около 10 минут (например, около 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 80, 90, 120 минут или больше) до контакта клетки с композицией.
[0069] В некоторых вариантах осуществления клетка представляет собой клетку скелетной мышцы, клетку сердечной мышцы, клетку гладкой мышцы, клетку жировой ткани, клетку гиппокампа или клетку мозжечка.
[0070] Как указано выше, полезная нагрузка (например, терапевтическая полезная нагрузка) может представлять собой любую композицию, которая улучшает или изменяет фенотип клетки-мишени. В некоторых вариантах осуществления терапевтическая полезная нагрузка представляет собой нуклеиновую кислоту, такую как трансген, которая обеспечивает экспрессию желаемого белка в клетке. Иллюстративный вариант осуществления представляет собой трансген, кодирующий дистрофин или его функциональный фрагмент для обеспечения экспрессии белка дистрофина в клетке. Соответственно, в некоторых вариантах осуществления способ характеризуется как способ экспрессии или увеличения экспрессии функционального белка дистрофина в клетке или увеличения экспрессии функционального фрагмента белка дистрофина в клетке. Увеличение может быть любым уровнем экспрессии от начальной точки отсутствия экспрессии до осуществления способа. В некоторых вариантах осуществления клетка может иметь остаточную (то есть обнаруживаемую) экспрессию белка дистрофина или его функционального фрагмента. В некоторых вариантах осуществления клетка может экспрессировать белок дистрофин или его функциональный фрагмент на уровне, который составляет по меньшей мере примерно 0,01%, 0,025%, 0,05%, 0,1%, 1%, 2,5%, 5%, 10%, 25%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 100% или более по сравнению с аналогичной клеткой (например, из того же источника ткани), которая имеет интактный ген дикого типа, кодирующий дистрофин. В некоторых вариантах осуществления клетка представляет собой мышечную клетку.
[0071] В других вариантах осуществления клетка представляет собой раковую клетку, экспрессирующую GLUT4 на поверхности. Неограничивающие примеры таких раковых клеток включают клетки меланомы, клетки рака предстательной железы, раковые клетки мышечной ткани, такие как клетки рабдомиосаркомы, и клетки рака молочной железы. В таких вариантах осуществления терапевтическая полезная нагрузка может представлять собой любую противораковую терапевтическую нагрузку, такую как радиоизотоп или другой токсин.
[0072] Способ можно осуществлять in vitro. В качестве альтернативы способ может представлять собой метод in vivo, направленный на лечение заболевания, связанного с дисфункцией клеток, экспрессирующих GLUT4, конститутивно или временно, на поверхности. Способ in vivo включает введение эффективного количества раскрытой композиции нуждающемуся в этом субъекту.
[0073] Используемый в настоящем документе термин «лечить» относится к медикаментозному контролю заболевания, нарушения или состояния (например, мышечной дистрофии, такой как мышечная дистрофия Дюшенна (DMD) или мышечная дистрофия Беккера (BMD) или рак) у субъекта (например, человека или другого млекопитающего, такое как примат, лошадь, собака, мышь, крыса, морская свинка, кролик и т.п.). Лечение может включать любые признаки успеха в лечении или облегчение заболевания или состояния (например, DMD, BMD или рака), включая любой параметр, такой как ослабление, ремиссия, ослабление симптомов или улучшение переносимости заболевания или состояния пациентом, замедление скорости дегенерации или ухудшения, или менее тяжелая дегенерация. В неограничивающих примерах термин «лечить» в контексте DMD может включать замедление или ингибирование прогрессирования потери мышечной массы или слабости; стабилизацию или увеличение мышечной силы, мышечного контроля или двигательных навыков; снижение мышечной усталости; стабилизацию или улучшение функции диафрагмы; стабилизацию или улучшение сердечного выброса и т.д. В неограничивающих примерах термин «лечить» в контексте рака может включать замедление или ингибирование скорости роста опухоли или снижение вероятности рецидива по сравнению с отсутствием лечения. В некоторых вариантах осуществления лечение включает достижение некоторой поддающейся обнаружению степени гибели раковых клеток у пациента. Лечение или облегчение симптомов может основываться на объективных или субъективных параметрах, включая результаты осмотра врачом. Соответственно, термин «лечение» включает введение композиций по настоящему изобретению для облегчения или остановки или ингибирования развития симптомов или состояний, связанных с заболеванием или состоянием (например, DMD, BMD или рака). Термин «терапевтический эффект» относится к облегчению, уменьшению или устранению заболевания или состояния, симптомов заболевания или состояния или побочных эффектов заболевания или состояния у субъекта. Термин «терапевтически эффективный» относится к количеству композиции, которое приводит к терапевтическому эффекту и может быть легко определено.
[0074] Как указано выше, способ может включать введение субъекту терапевтически эффективного количества раскрытой композиции. Как указано, полезная нагрузка нуклеиновой кислоты обычно сконфигурирована для экспрессии в клетке-мишени, например, функционально связана с соответствующим промотором. Эффективное количество представляет собой количество, которое дает результаты в благоприятном изменении фенотипа. Преимущество наличия промотора, специфичного для клетки, например, промотора, специфичного для мышц, заключается в том, что он будет придавать дополнительную специфичность по сравнению с уже обеспечиваемым таргетингом связывающего домена на GLUT4.
[0075] В некоторых вариантах осуществления способ лечения сочетается или координируется с другими терапевтическими стратегиями. В этом комбинаторном аспекте предусмотрена любая другая терапевтическая стратегия для лечения состояний, связанных с мутированной или аберрантной экспрессией дистрофина. В контексте рака, экспрессирующего GLUT4, другой стратегией противораковой терапии может быть противораковая иммунотерапия, в которой используются иммуномодулирующие композиции (например, антитела, иммунные клетки, цитокины и т.д.), которые могут усиливать собственный иммунный ответ субъекта против раковой мишени. Такая иммунная терапия включает адоптивную терапию иммунными клетками, включая Т-клетки CAR, терапию ингибиторами иммунных контрольных точек, противораковые вакцины и т.п.
[0076] Способы лечения могут включать композицию по изобретению для устранения ряда состояний, связанных с клетками, экспрессирующими GLUT4. Например, к заболеваниям, которые можно лечить введением композиции по изобретению, относятся заболевания скелетных мышц, такие как: мышечные дистрофии (миотоническая дистрофия (болезнь Штейнерта), мышечная дистрофия Дюшенна, мышечная дистрофия Беккера, поясно-конечностные мышечные дистрофии, плечелопаточно-лицевая мышечная дистрофия врожденная мышечная дистрофия, окулофарингеальная мышечная дистрофия, дистальная мышечная дистрофия и мышечная дистрофия Эмери-Дрейфуса), другие миопатии, такие как воспалительные миопатии (например, полимиозит, дерматомиозит и миозит с включениями), миопатии, обусловленные эндокринными нарушениями (гипертиреоидные и гипотиреоидные миопатии), заболевания, вызывающие мышечную атрофию, такие как раковая кахексия и саркопения, врожденная миотония, врожденная парамиотония, центроядерная миопатия, немалиновая миопатия, миотубулярная миопатия, периодический паралич, миопатии RYR-1 и воспалительные миопатии (например, полимиозит, дерматомиозит и миозит с включенными тельцами). Предлагаемый способ может быть также применен для лечения заболеваний двигательных нейронов (например, бокового амиотрофического склероза (ALS), инфантильной прогрессирующей спинальной мышечной атрофии (1-го типа - болезни Верднига-Гоффмана), промежуточной спинальной мышечной атрофии (2-го тип), ювенильной спинальной мышечной атрофии (3-го типа - болезни Кугельберга-Веландера), спинальной мышечной атрофии взрослых (4-го типа) и спинально-бульбарной мышечной атрофии (болезни Кеннеди)), заболеваний нервно-мышечного синапса, таких как миастения, синдрома Ламберта-Итона (миастенического) и врожденных миастенических синдромов, заболеваний периферических нервов, таких как болезнь Шарко-Мари-Тута, атаксия Фридрейха и болезнь Держерина-Сотта, кардиомиопатий, метаболических заболеваний, поражающих мышцы, например, дефицита фосфорилазы (болезни Мак-Ардла), дефицита кислой мальтазы (болезни Помпе), дефицита фосфофруктокиназы (болезни Таруи), дефицита фермента амило-1,6-глюкозидазы (болезни Кори или Форбса), митохондриальной миопатии, дефицита карнитина, дефицита карнитин-пальмитил-трансферазы, дефицита фосфоглицераткиназы, дефицита фосфоглицератмутазы, дефицита лактатдегидрогеназы и дефицита миоаденилатдезаминазы), а также заболеваний, поражающих жировую ткань, таких как диабет, болезненное ожирение, врожденная генерализованная липодистрофия (врожденная липодистрофия Берардинелли-Сейпа), семейная парциальная липодистрофия, семейный дефицит липопротеинлипазы, болезнь накопления нейтральных липидов и синдром Чанарина-Дорфмана. Поскольку GLUT4 экспрессируется в гиппокампе и мозжечке, заболевания головного мозга, включая болезнь Альцгеймера, атаксию, телеангиэктазию, NBIA (нейродегенерация с накоплением железа в головном мозге), синдром Фара, синдром Барта и болезнь Паркинсона, можно лечить с использованием композиций по изобретению. Наконец, поскольку GLUT4 сверхэкспрессируется при некоторых видах рака, раскрытые композиции можно использовать для таргетинга и лечения злокачественной меланомы и рака предстательной железы, а также рака, поражающего мышцы, такого как рабдомиосаркома.
[0077] В конкретном аспекте изобретение относится к способу лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин. Способ включает введение субъекту терапевтически эффективного количества композиции, содержащей:
связывающий домен, который специфично связывает белок-транспортер глюкозы 4 («GLUT4»), и
терапевтическую полезную нагрузку, конъюгированную со связывающим доменом, при этом терапевтическая полезная нагрузка содержит нуклеиновую кислоту с промоторной последовательностью, функционально связанной с последовательностью, кодирующей белок дистрофин или его функциональный фрагмент.
[0078] В некоторых вариантах осуществления субъект имеет или является генетическим носителем мышечной дистрофии Дюшенна («DMD»). Как описано выше, мышечная дистрофия Дюшенна (DMD) представляет собой тяжелое сцепленное с полом заболевание, возникающее из-за аберрантного белка дистрофина (или аберрантной экспрессии белка дистрофина). Будучи сцепленным с полом заболеванием, DMD в основном поражает мужчин и обычно проявляется сначала потерей мышечной ткани ног и таза, а затем рук. Случаи могут прогрессировать до того, что субъект испытывает затруднения в дыхании и плохой сердечный выброс. У пострадавших субъектов в конечном итоге значительно сокращается продолжительность жизни из-за обширных потерь и деградации мышечной ткани. В некоторых вариантах осуществления субъект имеет или является генетическим носителем мышечной дистрофии Беккера («BMD»). Мышечная дистрофия Беккера («BMD») связана с DMD тем, что она также возникает в результате мутаций в гене дистрофина. Мышечная дистрофия Беккера («BMD»), однако, имеет более мягкое течение симптомов по сравнению с DMD, вероятно, потому, что уровень остаточной функции дистрофина у субъектов с BMD относительно более высокий по сравнению с субъектами с DMD из-за локализации мутации в гене дистрофина.
[0079] Примеры элементов композиции более подробно описаны выше и применимы к данному аспекту. В некоторых вариантах осуществления связывающий домен представляет собой антитело, его фрагмент или производное, которое специфично связывается с GLUT4. Такие антитела или их фрагменты, или производные более подробно описаны выше. В некоторых вариантах осуществления антитело, его фрагмент или производное специфично связывается с первым внеклеточным доменом GLUT4. Домен и конкретные эпитопы в нем более подробно описаны выше. В некоторых вариантах осуществления связывающий домен специфичо связывается с доменом первой внеклеточной петли GLUT4. В некоторых вариантах осуществления домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:2, которая является субдоменом домена первой внеклеточной петли GLUT4 человека. В некоторых вариантах осуществления домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью последовательности по меньшей мере примерно 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно на 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98% и 100%) с последовательностью, представленной в SEQ ID NO:1, которая представляет собой репрезентативный домен первой внеклеточной петли GLUT4 человека. В некоторых вариантах осуществления связывающий домен специфично связывается с субдоменом домена первой внеклеточной петли GLUT4, который содержит последовательность с идентичностью последовательности по меньшей мере 80% (например, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 92%, по меньшей мере примерно 94%, по меньшей мере примерно 96%, по меньшей мере примерно 98%) с последовательностью, представленной в SEQ ID NO:2.
[0080] В некоторых вариантах осуществления связывающий домен представляет собой моноклональное антитело, которое связывается с GLUT4, как описано выше. В некоторых вариантах осуществления связывающий домен представляет собой фрагмент антитела или производное антитела. Фрагмент антитела или производное антитела может представлять собой или содержать одноцепочечное антитело, Fab-фрагмент, F(ab)2-фрагмент, VHH-фрагмент, VNAR-фрагмент или нанотело. Одноцепочечное антитело может представлять собой одноцепочечный вариабельный фрагмент (scFv) или одноцепочечный Fab-фрагмент (scFab).
[0081] В некоторых вариантах осуществления антитело, его фрагмент или производное является биотинилированным.
[0082] Нуклеиновая кислота в этом аспекте способа может представлять собой любую полезную нагрузку нуклеиновой кислоты, как описано выше. Например, нуклеиновая кислота имеет линейную форму, форму плазмиды или форму миникольца.
[0083] В некоторых вариантах осуществления композиция дополнительно содержит гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой. В некоторых вариантах осуществления гистоновый белок или по меньшей мере часть множества гистоновых белков биотинилированы.
[0084] В некоторых вариантах осуществления композиция дополнительно содержит липосому, связанную с нуклеиновой кислотой. Элементы липосом более подробно описаны выше и применимы к этому аспекту изобретения. В некоторых вариантах осуществления липосома содержит один или несколько липидов, выбранных из DC-холестерин HCl,3b-[N-(N',N'-диметиламиноэтан)-карамоил]холестерин гидрохлорида («DC-Chol»), 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламина («DOPE»), 1,2-ди-О-октадеценил-3-триметиламмония («DOTMA»), N-[1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмонийпропана («DOTAP»), диметилдиоктадециламмоний бромида («DDAB»), 1,2-димиристоил-3-триметиламмоний-пропана («DMTAP»), 1,2-диолеоил-3-диметиламмоний-пропана («DODAP»), гемисукцинатхолестерина («CHEMS») и холестерина («CHOL»), 2,3-диолеилокси-N-[2(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминия («DOSPA»), диоктадециламидоглицилспермина («DOGS»), диолеоилфосфатидилхолина («DOPC») и т.п. В некоторых вариантах осуществления липосома дополнительно содержит полиэтиленгликоль («PEG»).
[0085] В некоторых вариантах осуществления композиция содержит множество катионных пептидных фрагментов, связанных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота. В некоторых вариантах осуществления катионные фрагменты представляют собой или включают поли-L-лизин («PLL»), линейный полиэтиленимин («LPEI»), разветвленный полиэтиленимин («BPEI»), хитозан, спермидин и спермин, полиамидоамин («PAMAM»), поли(2-диметиламиноэтилметакрилат) («PDMAEMA»), поли(бета-аминоэфир)ы («PBAE»), поли{N-[N-(2-аминоэтил)-2-аминоэтил]аспартамид («PAsp(DET)»), поли(2-аминоэтилэтиленфосфат («PPEEA») и т.п. В некоторых вариантах осуществления по меньшей мере часть множества катионных пептидных фрагментов является биотинилированной.
[0086] В некоторых вариантах осуществления композиция дополнительно содержит альбумин. В некоторых вариантах осуществления по меньшей мере часть альбумина является биотинилированной.
[0087] В некоторых вариантах осуществления антитело, его фрагмент или производное биотинилировано, и композиция дополнительно содержит нейтравидин.
[0088] В некоторых вариантах осуществления кодируемый белок дистрофин представляет собой по существу полноразмерный белок дистрофин человека. В некоторых вариантах осуществления полноразмерный белок дистрофин человека имеет аминокислотную последовательность, по меньшей мере на 80% идентичную последовательности, представленной в SEQ ID NO:5.
[0089] Субъектом может быть млекопитающее, такое как человек, другой примат, крыса, мышь, кролик, кошка, собака и т.п. В некоторых вариантах осуществления субъектом является человек.
Состав и введение
[0090] Настоящее изобретение также охватывает составы, подходящие для способов введения для применения в терапевтических целях in vivo у субъектов (например, млекопитающих с DMD, BMD, раком или состояниями, описанными выше). Согласно навыкам и знаниям, общепринятым в данной области, композиции по изобретению могут быть изготовлены с соответствующими носителями и неактивными связующими веществами и т.п. для введения в конкретные клетки-мишени. Поскольку композиции содержат связывающие домены, которые придают специфичность к клеткам-мишеням, композиции могут быть составлены для прямого или системного введения в соответствии с навыками и знаниями в данной области.
Иллюстративные анти-GLUT4 антитела.
[0091] Известны анти-GLUT4 антитела, которые связываются с внеклеточным доменом GLUT4 человека (см., например, Tucker et al, Proc. Natl. Acad. Sci. 115:E4990-E4999, опубликовано онлайн 16 мая 2018 г.). В другом аспекте изобретение относится к моноклональным антителам, которые связывают пептид GLUT4 человека GRQGPEGPSSI (SEQ ID NO:2). Таким образом, в некоторых вариантах осуществления настоящее изобретение относится к анти-GLUT4 антителу, или его фрагменту или производному, содержащему вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL). В некоторых аспектах VH содержит CDR1, содержащую последовательность (D/E)Y(S/T)(I/M)H, CDR2, содержащую последовательность WINTE(S/T)G(D/E)X1(T/S)YADDFKG, и CDR3, содержащую последовательность RX2X3Y; и VL содержит CDR1, содержащую последовательность (R/K)(A/S)SQS(L/V)X4X5(N/S), CDR2, содержащую последовательность (V/A)SNR(F/Y)(S/T), и CDR3, содержащую последовательность QDX6X7X8PT. В некоторых вариантах осуществления X1 представляет собой P или T; X2 представляет собой A, F, S или G; X3 представляет собой A, D, E или G; X4 представляет собой S, V, N или Т; X5 представляет собой N, H, R, K или T; X6 представляет собой R, Y, S, T или K; X7 представляет собой H, N, E, T, Y или S; или X8 представляет собой V, S, L или I.
[0092] Соответственно, в некоторых вариантах осуществления анти-GLUT4 антитело, или его фрагмент или производное содержит по меньшей мере одну, две, три, четыре, пять или шесть CDR, выбранных из (а) VH-CDR1, содержащей последовательность (D/E)Y(S/T)(I/M)H; (b) VH-CDR2, содержащей последовательность WINTE(S/T)G(D/E)X1(T/S)YADDFKG; (c) VH-CDR3, содержащей последовательность RX2X3Y; (d) VL-CDR1, содержащей последовательность (R/K)(A/S)SQS(L/V)X4X5(N/S); (e) VL-CDR2, содержащей последовательность (V/A)SNR(F/Y)(S/T); и (f) VL-CDR3, содержащей последовательность QDX6X7X8PT. В некоторых вариантах осуществления X1 представляет собой P или T; X2 представляет собой A, F, S или G; X3 представляет собой A, D, E или G; X4 представляет собой S, V, N или Т; X5 представляет собой N, H, R, K или T; X6 представляет собой R, Y, S, T или K; X7 представляет собой H, N, E, T, Y или S; или X8 представляет собой V, S, L или I.
[0093] В некоторых вариантах осуществления изобретения анти-GLUT4 антитело, или его фрагмент или производное содержит по меньшей мере одну, две, три, четыре, пять или шесть CDR, выбранных из (а) VH-CDR1, имеющей последовательность, выбранную из SEQ ID NO:10-17; (b) VH-CDR2, имеющей последовательность, выбранную из SEQ ID NO:18-25; (c) VH-CDR3, имеющей последовательность, выбранную из LDF и SEQ ID NO:27-33; (d) VL-CDR1, имеющей последовательность, выбранную из SEQ ID NO:34-41; (e) VL-CDR2, имеющей последовательность, выбранную из SEQ ID NO:42-49; и (f) VL-CDR3, имеющей последовательность, выбранную из SEQ ID NO:50-57.
[0094] В некоторых вариантах осуществления настоящее изобретение относится к анти-GLUT4 антителу, или его фрагменту или производному, содержащему вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL). В некоторых вариантах осуществления VH содержит VH-CDR1, имеющую последовательность, выбранную из SEQ ID NO:10-17, VH-CDR2, имеющую последовательность, выбранную из SEQ ID NO:18-25, и VH-CDR3, имеющую последовательность, выбранную из LDF и SEQ ID NO:27-33; и VL содержит VL-CDR1, имеющую последовательность, выбранную из SEQ ID NO:34-41, VL-CDR2, имеющую последовательность, выбранную из SEQ ID NO:42-49, и VL-CDR3, имеющую последовательность, выбранную из SEQ ID NO:50-57.
[0095] В некоторых вариантах осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:10, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:18, и VH-CDR3, имеющую последовательность LDF; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:34, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:42, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:50.
[0096] В некоторых вариантах осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:11, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:19, и VH-CDR3, имеющую последовательность приведенную в SEQ ID NO:27; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:35, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:43, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:51.
[0097] В некоторых вариантах осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:12, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:20, и VH-CDR3, имеющую последовательность приведенную в SEQ ID NO:28; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:36, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:44, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:52.
[0098] В некоторых вариантах осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:13, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:21, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:29; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:37, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:45, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:53.
[0099] В некоторых вариантах осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:14, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:22, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:30; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:38, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:46, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:54.
[0100] В некоторых вариантах осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:15, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:23, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:31; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:39, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:47, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:55.
[0101] В другом варианте осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:16, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:24, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:32; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:40, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:48, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:56.
[0102] В еще одном варианте осуществления изобретения VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:17, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:25, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:33; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:41, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:49, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:57.
[0103] В некоторых вариантах осуществления замены, вставки или делеции могут присутствовать в одной или нескольких CDR без существенного снижения способности антитела связывать антиген. Например, в CDR могут быть внесены консервативные изменения (например, консервативные замены, предусмотренные в настоящем документе), которые существенно не снижают аффинность связывания. В некоторых вариантах осуществления каждая представленная выше CDR либо не изменена, либо содержит одну, две, три или четыре аминокислотные замены.
[0104] В некоторых вариантах осуществления анти-GLUT4 антитело, или его фрагмент или производное дополнительно содержит CDR по предшествующим абзацам в этом разделе, и последовательность каркасных областей, имеющую идентичность по меньшей мере 90% или по меньшей мере 95% с каркасной последовательностью иммуноглобулина человека. В некоторых вариантах осуществления каждая из каркасной области 1 (FR1), каркасной области 2 (FR2), каркасной области 3 (FR3) и каркасной области 4 (FR4) имеет по меньшей мере 90% идентичности или по меньшей мере 95% идентичности соответствующей последовательности FR1, FR2, FR3 или FR4 человека.
[0105] Антитело, описанное выше, которое связывает пептид GLUT4 человека с аминокислотной последовательностью GRQGPEGPSSI (SEQ ID NO:2), можно использовать для доставки полезной нагрузки, как описано в любом из вариантов осуществления, подробно описанных в настоящем документе.
Иллюстративные варианты осуществления
[0106] Иллюстративные варианты осуществления включают, но не ограничиваются, следующее:
[0107] В некоторых вариантах осуществления настоящее изобретение относится к композиции, содержащей антитело, или его фрагмент или производное, которое специфично связывается с белком-переносчиком глюкозы 4 («GLUT4»), и терапевтическую полезную нагрузку, конъюгированную с антителом, или его фрагментом или производным. В одном варианте осуществления антитело, или его фрагмент или производное, специфично связывается с доменом первой внеклеточной петли GLUT4. В некоторых аспектах домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью по меньшей мере 80% с последовательностью INAPQKVIEQSYNETWLGRQGPEGPSSIPPGTLTTL (SEQ ID NO:1). В одном аспекте домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью по меньшей мере 80% с последовательностью GRQGPEGPSSI (SEQ ID NO:2). В некоторых аспектах антитело, или его фрагмент или производное, специфично связывается с субдоменом первого внеклеточного домена GLUT4, который содержит аминокислотную последовательность с идентичностью по меньшей мере 80% с последовательностью GRQGPGGPDSI (SEQ ID NO:4). В одном аспекте, антитело, или его фрагмент или производное, специфично связывается с субдоменом первого внеклеточного домена GLUT4, который содержит последовательность с идентичностью по меньшей мере 80% с последовательностью GRQGPEGPSSI (SEQ ID NO:2).
[0108] В некоторых вариантах осуществления антитело представляет собой моноклональное антитело. В одном варианте фрагмент антитела или производное антитела представляет собой или содержит одноцепочечное антитело, Fab-фрагмент, F(ab)2-фрагмент, VHH-фрагмент, VNAR-фрагмент или нанотело. В некоторых аспектах одноцепочечное антитело может представлять собой одноцепочечный вариабельный фрагмент (scFv) или одноцепочечный Fab-фрагмент (scFab).
[0109] В некоторых вариантах осуществления антитело или его фрагмент или производное биотинилировано.
[0110] В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, белок или пептид, липид, низкомолекулярный фармацевтический препарат или радиоизотоп. В одном варианте осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, где нуклеиновая кислота выбрана из ДНК, мРНК, киРНК и кшРНК. В одном варианте осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, где нуклеиновая кислота содержит открытую рамку считывания, функционально связанную с промоторной последовательностью. В одном варианте осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту в линейной форме, нуклеиновую кислоту в плазмидной форме или нуклеиновую кислоту в форме миникольца.
[0111] В некоторых вариантах осуществления композиция дополнительно содержит гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой. В одном аспекте гистоновый белок или по меньшей мере часть множества гистоновых белков биотинилирована.
[0112] В некоторых вариантах осуществления композиция дополнительно содержит липосому, связанную с нуклеиновой кислотой. В одном аспекте липосома содержит один или несколько липидов, выбранных из DC-холестерин HCl,3b-[N-(N',N'-диметиламиноэтан)-карамоил]холестерин гидрохлорида («DC-Chol»), 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламина («DOPE»), 1,2-ди-О-октадеценил-3-триметиламмония («DOTMA»), N-[1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмонийпропана («DOTAP»), диметилдиоктадециламмоний бромида («DDAB»), 1,2-димиристоил-3-триметиламмоний-пропана («DMTAP»), 1,2-диолеоил-3-диметиламмоний-пропана («DODAP»), гемисукцинатхолестерина («CHEMS») и холестерина («CHOL»), 2,3-диолеилокси-N-[2(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминия («DOSPA»), диоктадециламидоглицилспермина («DOGS»), диолеоилфосфатидилхолина («DOPC») и т.п. В одном аспекте осуществления липосома дополнительно содержит полиэтиленгликоль («PEG»).
[0113] В некоторых вариантах осуществления композиция дополнительно содержит множество катионных пептидных фрагментов, ассоциированных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота. В одном варианте осуществления катионные пептидные фрагменты представляют собой или содержат поли-L-лизин («PLL»), линейный полиэтиленимин («LPEI»), разветвленный полиэтиленимин («BPEI»), хитозан, спермидин и спермин, полиамидоамин («PAMAM»), поли(2-диметиламиноэтилметакрилат) («PDMAEMA»), поли(бета-аминоэфир)ы («PBAE»), поли{N-[N-(2-аминоэтил)-2-аминоэтил]аспартамид («PAsp(DET)»), поли(2-аминоэтилэтиленфосфат («PPEEA») и т.п. В одном аспекте по меньшей мере часть множества катионных пептидных фрагментов является биотинилированной.
[0114] В некоторых вариантах осуществления композиция по настоящему изобретению дополнительно содержит альбумин. В одном варианте осуществления по меньшей мере часть альбумина биотинилирована.
[0115] В некоторых вариантах осуществления антитело, или его фрагмент или производное биотинилировано, и композиция дополнительно содержит нейтравидин.
[0116] В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту с промоторной последовательностью, функционально связанную с последовательностью, кодирующей белок дистрофин или его функциональный фрагмент. В одном варианте осуществления белок дистрофин представляет собой по существу полноразмерный белок дистрофин человека. В одном аспекте полноразмерный белок дистрофин человека имеет аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% с последовательностью, представленной в SEQ ID NO: 5.
[0117] Изобретение дополнительно относится к способу таргетинга терапевтической полезной нагрузки для доставки в клетку, которая экспрессирует GLUT4, причем способ включает контакт клетки с композицией. В некоторых вариантах осуществления способ дополнительно включает контакт клетки с инсулином перед контактом клетки с композицией. В одном варианте осуществления клетка представляет собой клетку скелетной мышцы, клетку сердечной мышцы, клетку гладкой мускулатуры, клетку жировой ткани, клетку гиппокампа или клетку мозжечка. В одном аспекте клетка представляет собой клетку меланомы, клетку рака предстательной железы, раковую клетку мышечной ткани, такую как клетка рабдомиосаркомы, или клетку рака молочной железы.
[0118] В другом аспекте настоящее изобретение включает способ лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин, включающий в себя введение субъекту терапевтически эффективного количества композиции, содержащей: антитело, или его фрагмент или производное, которое специфично связывается с белком-переносчиком глюкозы 4 («GLUT4»), и терапевтическую полезную нагрузку, конъюгированную с антителом, или его фрагментом или производным, где терапевтическая полезная нагрузка содержит нуклеиновую кислоту с промоторной последовательностью, функционально связанной с последовательностью, кодирующей белок дистрофин или его функциональный фрагмент. В некоторых вариантах осуществления субъект имеет или представляет собой генетического носителя мышечной дистрофии Дюшенна («DMD») или мышечную дистрофию Беккера («BMD»).
[0119] В некоторых вариантах осуществления антитело, или его фрагмент или производное, используемое в способе лечения заболевания или состояния, описанного в предыдущих двух абзацах, специфично связывается с доменом первой внеклеточной петли GLUT4. В одном из вариантов осуществления изобретения домен первой внеклеточной петли GLUT4 содержит аминокислотную последовательность с идентичностью по меньшей мере 80% с последовательностью INAPQKVIEQSYNETWLGRQGPEGPSSIPPGTLTTL (SEQ ID NO:1). В некоторых аспектах антитело, или его фрагмент или производное, специфично связывается с субдоменом первого внеклеточного домена GLUT4, который содержит последовательность с идентичностью по меньшей мере 80% с последовательностью GRQGPEGPSSI (SEQ ID NO: 2). В некоторых вариантах осуществления антитело представляет собой моноклональное антитело. В одном варианте осуществления изобретения фрагмент антитела или производное антитела представляет собой или содержит одноцепочечное антитело, Fab-фрагмент, F(ab)2-фрагмент, VHH-фрагмент, VNAR-фрагмент или нанотело. В некоторых аспектах одноцепочечное антитело может представлять собой одноцепочечный вариабельный фрагмент (scFv) или одноцепочечный Fab-фрагмент (scFab). В некоторых вариантах осуществления антитело, или его фрагмент или производное, биотинилировано. В некоторых вариантах осуществления изобретения нуклеиновая кислота находится в линейной форме, плазмидной форме или миникольцевой форме. В одном варианте осуществления композиция дополнительно содержит гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой. В одном аспекте гистоновый белок или по меньшей мере часть множества гистоновых белков биотинилированы. В некоторых вариантах осуществления композиция дополнительно содержит липосому, связанную с нуклеиновой кислотой. В одном варианте осуществления липосома содержит один или несколько липидов, выбранных из DC-холестерин HCl,3b-[N-(N',N'-диметиламиноэтан)-карамоил]холестерин гидрохлорида («DC-Chol»), 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламина («DOPE»), 1,2-ди-О-октадеценил-3-триметиламмония («DOTMA»), N-[1-(2,3-диолеилокси)пропил]-N,N,N-триметиламмонийпропана («DOTAP»), диметилдиоктадециламмоний бромида («DDAB»), 1,2-димиристоил-3-триметиламмоний-пропана («DMTAP»), 1,2-диолеоил-3-диметиламмоний-пропана («DODAP»), гемисукцинатхолестерина («CHEMS») и холестерина («CHOL»), 2,3-диолеилокси-N-[2(сперминкарбоксамидо)этил]-N,N-диметил-1-пропанаминия («DOSPA»), диоктадециламидоглицилспермина («DOGS»), диолеоилфосфатидилхолина («DOPC») и т.п. В одном аспекте осуществления липосома дополнительно содержит полиэтиленгликоль («PEG»). В некоторых вариантах осуществления композиция дополнительно содержит множество катионных пептидных фрагментов, ассоциированных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота. В одном варианте осуществления катионные пептидные фрагменты представляют собой или содержат поли-L-лизин («PLL»), линейный полиэтиленимин («LPEI»), разветвленный полиэтиленимин («BPEI»), хитозан, спермидин и спермин, полиамидоамин («PAMAM»), поли(2-диметиламиноэтилметакрилат) («PDMAEMA»), поли(бета-аминоэфир)ы («PBAE»), поли{N-[N-(2-аминоэтил)-2-аминоэтил]аспартамид («PAsp(DET)»), поли(2-аминоэтилэтиленфосфат («PPEEA») и т.п. В одном аспекте по меньшей мере часть множества катионных пептидных фрагментов является биотинилированной. В некоторых вариантах осуществления композиция дополнительно содержит альбумин. В одном варианте осуществления по меньшей мере часть альбумина биотинилирована. В некоторых вариантах осуществления антитело, или его фрагмент или производное, биотинилировано, и композиция дополнительно содержит нейтравидин. В некоторых вариантах осуществления кодируемый белок дистрофин представляет собой по существу полноразмерный белок дистрофин человека. В одном варианте осуществления полноразмерный белок дистрофин человека имеет аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% с последовательностью (SEQ ID NO: 5). В некоторых вариантах осуществления субъект представляет собой человека.
[0120] В еще одном аспекте настоящее изобретение относится к анти-GLUT4 антителу, или его фрагменту или производному, содержащему вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где: (а) VH содержит CDR1, содержащую последовательность (D/E)Y(S/T)(I/M)H, CDR2, содержащую последовательность WINTE(S/T)G(D/E)X1(T/S)YADDFKG, и CDR3, содержащую последовательность RX2X3Y; и (b) VL содержит CDR1, содержащую последовательность (R/K)(A/S)SQS(L/V)X4X5(N/S), CDR2, содержащую последовательность (V/A)SNR(F/Y)(S/T), и CDR3, содержащую последовательность QDX6X7X8PT. В некоторых вариантах осуществления X1 представляет собой P или T; X2 представляет собой A, F, S или G; X3 представляет собой A, D, E или G; X4 представляет собой S, V, N или Т; X5 представляет собой N, H, R, K или T; X6 представляет собой R, Y, S, T или K; X7 представляет собой H, N, E, T, Y или S; или X8 представляет собой V, S, L или I.
[0121] В еще одном аспекте настоящее изобретение включает анти-GLUT4 антитело, или его фрагмент или производное, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где: (а) VH содержит VH-CDR1, имеющую последовательность, выбранную из SEQ ID NO:10-17, VH-CDR2, имеющую последовательность, выбранную из SEQ ID NO:18-25, и VH-CDR3, имеющую последовательность, выбранную из LDF и SEQ ID NO:27-33; и (b) VL содержит VL-CDR1, имеющую последовательность, выбранную из SEQ ID NO:34-41, VL-CDR2, имеющую последовательность, выбранную из SEQ ID NO:42-49, и VL-CDR3, имеющую последовательность, выбранную из SEQ ID NO:50-57.
[0122] В некоторых вариантах осуществления изобретения анти-GLUT4 антитело или его фрагмент или производное, содержит вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где: (а) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:10, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:18, и VH-CDR3, имеющую последовательность LDF; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:34, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:42, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:50; (b) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:11, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:19, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:27, и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:35, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:43, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO: 51; (c) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:12, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:20, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:28, и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:36, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:44, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:52; (d) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:13, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:21, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:29, и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:37, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:45, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:53; (e) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:14, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:22, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:30, и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:38, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:46, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:54; (f) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:15, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:23, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:31; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:39, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:47, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:55; (g) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:16, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:24, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:32, и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:40, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:48, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:56; или (h) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:17, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:25, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:33, и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:41, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:49, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:57.
[0123] В некоторых вариантах осуществления изобретение относится к композиции, содержащей антитело, или его фрагмент или производное по любому из вариантов осуществления, описанных выше в трех предыдущих абзацах, и терапевтическую полезную нагрузку, конъюгированную с антителом, или его фрагментом или производным. В некоторых вариантах осуществления фрагмент антитела или производное антитела представляет собой или содержит одноцепочечное антитело, Fab-фрагмент, F(ab)2-фрагмент, VHH-фрагмент, VNAR-фрагмент или нанотело. В некоторых аспектах одноцепочечное антитело может представлять собой одноцепочечный вариабельный фрагмент (scFv) или одноцепочечный Fab-фрагмент (scFab). В одном аспекте одноцепочечное антитело представляет собой одноцепочечный вариабельный фрагмент (scFv), или одноцепочечный Fab-фрагмент (scFv). В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, белок или пептид, липид, низкомолекулярный фармацевтический препарат или радиоизотоп. В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, где нуклеиновая кислота выбрана из ДНК, мРНК, киРНК и кшРНК. В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту, где нуклеиновая кислота содержит открытую рамку считывания, функционально связанную с промоторной последовательностью. В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту в линейной форме, нуклеиновую кислоту в плазмидной форме или нуклеиновую кислоту в миникольцевой форме. В некоторых вариантах осуществления композиция дополнительно содержит гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой. В некоторых вариантах осуществления композиция дополнительно содержит липосому, связанную с нуклеиновой кислотой. В некоторых вариантах осуществления композиция дополнительно содержит множество катионных пептидных фрагментов, ассоциированных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота. В некоторых вариантах осуществления терапевтическая полезная нагрузка содержит нуклеиновую кислоту с промоторной последовательностью, функционально связанную с последовательностью, кодирующей белок дистрофин или его функциональный фрагмент. В одном варианте осуществления белок дистрофин представляет собой по существу полноразмерный белок дистрофин человека, имеющий аминокислотную последовательность с идентичностью последовательности по меньшей мере 80% с последовательностью, представленной в SEQ ID NO:5.
[0124] В некоторых вариантах осуществления настоящее изобретение относится к способу таргетинга терапевтической полезной нагрузки для доставки в клетку, которая экспрессирует GLUT44, причем способ включает контакт клетки с композицией, содержащей анти-GLUT4 антитело, или его фрагмент или производное, описанные в предыдущем абзаце. В некоторых вариантах осуществления способ дополнительно включает контакт клетки с инсулином перед контактом клетки с композицией. В одном варианте осуществления клетка представляет собой клетку скелетной мышцы, клетку сердечной мышцы, клетку гладкой мускулатуры, клетку жировой ткани, клетку гиппокампа или клетку мозжечка. В одном варианте осуществления клетка представляет собой клетку меланомы, клетку рака предстательной железы, раковую клетку мышечной ткани, такую как клетка рабдомиосаркомы, или клетку рака молочной железы. В некоторых вариантах осуществления субъект представляет собой человека.
[0125] В некоторых вариантах осуществления изобретение относится к способу лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин, включающему в себя введение субъекту терапевтически эффективного количества композиции, содержащей антитело по любому из вышеуказанных вариантов осуществления. В некоторых вариантах осуществления субъект имеет или представляет собой генетического носителя мышечной дистрофии Дюшенна («DMD») или мышечной дистрофии Беккера(«BMD»). В некоторых вариантах осуществления кодируемый белок дистрофин имеет идентичность последовательности по меньшей мере 80% с SEQ ID NO:5. В некоторых вариантах осуществления субъект представляет собой человека.
Общие определения
[0126] Если специально не указано в данном документе, все термины, используемые в данном документе, имеют то же значение, что и для специалистов в данной области. Для определений и терминов в данной области практикующие специалисты могут обратиться к Ausubel, F.M., et al. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, New York (2010), Coligan, J.E., et al. (eds.), Current Protocols in Immunology, John Wiley & Sons, New York (2010), Mirzaei, H. and Carrasco, M. (eds.), Modern Proteomics - Sample Preparation, Analysis and Practical Applications in Advances in Experimental Medicine and Biology, Springer International Publishing, 2016, и Comai, L, et al., (eds.), Proteomic: Methods and Protocols in Methods in Molecular Biology, Springer International Publishing, 2017.
[0127] Для удобства, некоторые термины, используемые в этом описании и/или формуле изобретения, представлены в данном документе. Определения приведены для помощи в описании конкретных вариантов осуществления и не предназначены для ограничения заявленного изобретения, поскольку объем изобретения ограничен только формулой изобретения.
[0128] Использование термина «или» в формуле изобретения используется для обозначения «и/или», если явно не указано, что он относится к только альтернативам, или альтернативные варианты являются взаимоисключающими, хотя изобретение поддерживает определение, которое относится только к альтернативам и «и/или».
[0129] Слова в единственном числе при использовании в сочетании со словом «содержащий» в формуле изобретения или в описании означают один или несколько, если специально не указано иное.
[0130] Если контекст явно не требует иного, во всем описании и формуле изобретения, слова «содержать», «содержащий» и т.п. должны быть истолкованы во включительном смысле, в отличие от исключающего или исчерпывающего смысла, что должно означать «включающий, но не ограниченный этим». Слова, используемые в единственном или множественном числе, также включают множественное и единственное число, соответственно. Слово «примерно» указывает число в диапазоне минорного изменения выше или ниже установленного эталонного числа. Например, в пределах 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% выше или ниже указанного эталонного числа.
[0131] В настоящем документе термин «полипептид» или «белок» относится к полимеру, в котором мономеры представляют собой аминокислотные остатки, которые соединены вместе амидные связи. Когда аминокислоты представляют собой альфа-аминокислоты, можно использовать либо L-оптический изомер, либо D-оптический изомер, причем L-изомеры являются предпочтительными. Термин полипептид или белок, используемый в настоящем документе, охватывает любую аминокислотную последовательность и включает модифицированные последовательности, такие как гликопротеины. Термин «полипептид» конкретно предназначен для охвата встречающихся в природе белков, а также тех, которые получены рекомбинантно или синтетически.
[0132] Специалисту понятно, что отдельные замены, делеции или добавления в пептиде, полипептиде или белковой последовательности, которые изменяют, добавляют или удаляют одну аминокислоту или часть аминокислот в последовательности, являются «консервативно модифицированным вариантом», где изменение приводит к замене аминокислоты химически подобной аминокислотой. Консервативные аминокислотные замены, обеспечивающие функционально аналогичные аминокислоты, хорошо известны специалисту в данной области. Следующие шесть групп являются примерами аминокислот, которые считаются консервативными заменами друг для друга:
(1) аланин (А), серин (S), треонин (Т),
(2) аспарагиновая кислота (D), глутаминовая кислота (Е),
(3) аспарагин (N), глутамин (Q),
(4) аргинин (R), лизин (K),
(5) изолейцин (I), лейцин (L), метионин (М), валин (V), и
(6) фенилаланин (F), тирозин (Y), триптофан (W).
[0133] Ссылка на идентичность последовательностей указывает степень схожести двух полимерных последовательностей, таких как белковые последовательности. Определение идентичности последовательностей может быть легко осуществлено специалистами в данной области с использованием принятых алгоритмов и/или методик. Идентичность последовательностей обычно определяют путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где часть последовательности пептида или полинуклеотида в окне сравнения может содержать добавления или делеции (например, разрывы) по сравнению с эталонной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания двух последовательностей. Процент рассчитывают путем определения количества положений, в которых идентичный аминокислотный остаток или нуклеотидное основание встречаются в обеих последовательностях, получая количество совпадающих положений, деления количества совпадающих положений на общее число положений в окне сравнения и умножения результата на 100 для получения процента идентичности последовательностей. Для осуществления таких сравнений легко доступны различные выполняемые в программах алгоритмы, такие как BLAST N или BLAST P.
[0134] Раскрыты материалы, композиции и компоненты, которые могут быть использованы для, могут быть использованы в сочетании с, могут быть использованы в приготовлении для, или представляют собой продукты раскрытых способов и композиций. Понятно, что, когда комбинации, подмножества, взаимодействия, группы и т.д. из этих материалов раскрыты, то специально предполагается каждая из различных индивидуальных и сборных комбинаций, даже несмотря на то, что конкретные ссылки на каждую и все одиночную комбинацию и перестановку этих соединений могут быть явно не раскрыты. Эта концепция применяется ко всем аспектам этого изобретения, включая, но не ограничиваясь этим, стадии в описанных способах. Таким образом, конкретные элементы любых вышеуказанных вариантов осуществления могут быть объединены или замещенны на элементы в других вариантах осуществления. Например, если существует множество дополнительных стадий, которые могут быть выполнены, следует понимать, что каждая из этих дополнительных стадий может быть выполнена с помощью любых конкретных стадий способа или комбинации стадий способа раскрытых способов, и что каждая такая комбинация или выборка комбинаций конкретно предусмотрена и должна рассматриваться как раскрытая. Кроме того, понятно, что варианты осуществления, описанные в данном документе, могут быть реализованы с использованием любого подходящего материала, такого как те, которые описаны в другом месте в данном документе или известные в данной области техники.
[0135] Публикации, цитируемые в данном документе, и предмет изобретения, из-за которого они указаны, включены в настоящий документ посредством ссылки во всей их полноте.
ПРИМЕРЫ
[0136] Ниже приведено описание ряда иллюстративных анализов, проводимых на репрезентативных вариантах осуществления композиций мышечно-специфичной векторной платформы для определения полезности конструкции платформы в отношении эффективной доставки терапевтических полезных нагрузок в мышечные и другие ткани.
Введение
[0137] В невирусной векторной платформе по изобретению используется анти-GLUT4 антитело (или другие связывающие агенты (например, фрагмент антитела, пептиды, рецепторы и т.д.)), способное специфично связывать внеклеточный домен GLUT4. Первая внеклеточная петля является наиболее крупной, наиболее доступной и, вероятно, иммуногенной областью для внеклеточного таргетинга GLUT4. Этот домен содержит аминокислоты 45-80 в последовательности GLUT4 мыши: inapqkvieqsynatwlgrqgpggpdsipqgtlttl (SEQ ID NO: 3). Антитела получали против пептида с аминокислотами 61-72 в данной последовательности: grqgpggpdsi (SEQ ID NO: 4). Аналогичная область в последовательности белка человека, grqgpegpssi (SEQ ID NO: 2) имеет 83,3% гомологии с последовательностью мышиного белка. Предсказано, что эта последовательность не имеет какой-либо вторичной структуры и гипотетически была предложена для получения антител, которые могли связываться с этим доменом нативного белка GLUT4.
[0138] С использованием этой платформы, анти-GLUT4 антитело (или его фрагмент) может быть затем конъюгировано с терапевтическим геном или другим лекарственным средством с использованием широкого спектра материалов, таких как белки, пептиды, липиды, липосомы и т.д., для инкапсулирования терапевтического агента, оптимизации его стабильности и биодоступности in vivo, и усиления исключения из эндосомальной утилизации и доставки в ядро (для нуклеиновых кислот) после проникновения в клетку-мишень. Как продемонстрировано в нижеследующих экспериментах, размер ДНК не является ограничивающим фактором, предоставляя возможность доставки больших генов, таких как полноразмерный дистрофин (приблизительно 14 т.п.о.), и также использования сильных мышечноспецифичных промоторов для активации экспрессии гена.
Эксперименты на культуре клеток in vitro
[0139] Для начальных экспериментов по культивированию клеток in vitro использовали систему биотин-нейтравидин для конъюгации нацеливающего антитела с катионной липосомой, содержащей терапевтическую плазмиду. Этот вариант осуществления векторной платформы для экспериментов in vitro показан схематически на фиг. 3. Сначала, плазмиду смешивают с гистоном H1 (H-H1), часть которого биотинилирована. H-H1 представляет собой положительно заряженный ядерный белок, который связывается с отрицательно заряженной ДНК и частично конденсирует ее. Далее, добавляют биотинилированное анти-GLUT4 антитело, которое также биотинилировано. После чего, негликозилированный авидидин (дегликозилированный авидидин), высокоспецифичный биотинсвязывающий белок с минимальным неспецифичным связыванием по сравнению с авидином, добавляют для связывания биотинилированных белков. Далее, белки и ДНК смешивают с катионными липосомами, содержащими два липида, DC-Chol(DC-холестерин HCl,3b-[N-(N’,N’-диметиламиноэтан)-карамоил] холестерина гидрохлорид) и DOPE (1,2-диолеоил-sn-глицеро-3-фосфоэтаноламин). Молярное соотношение DC-Chol:DOPE составляет 30:70. Каждая из четырех стадий инкубации, описанных выше, проводится при комнатной температуре в течение 30 минут.
[0140] В первоначальных экспериментах для доказательства концепции, эту конъюгированную структуру оценивали по тому, может ли она проникать в мышечные клетки in vitro, и экспрессируется ли в клетках белок, кодируемый плазмидой ДНК. В этих экспериментах использовали коммерчески доступную клеточную линию крысиных миобластов (L6-GLUT4myc), которая сверхэкспрессирует GLUT4, содержащий myc-тэг (аминокислотную последовательность AEEQKLISEEDLLK; SEQ ID NO: 9), вставленную в первую внеклеточную петлю между аминокислотами 66 и 67. Для начальных экспериментов с использованием этой клеточной линии биотинилировали коммерческое антитело, которое специфично связывается с myc-тэгом.
Эксперимент 1
[0141] Клетки L6-GLUT4myc в 6-луночных планшетах трансфицировали структурой, показанной на фиг. 3. В этом эксперименте использовали плазмиду, содержащую модифицированную конструкцию нейронной синтазы оксида азота (NOS-M), которая включает С-концевой НА-тэг для обнаружения и последовательность пальмитоилирования RAS для таргетинга на плазматическую мембрану (Rebolledo DL, et al. (2016). Sarcolemmal targeting of nNOSmu improves contractile function of mdx muscle. Hum Mol Genet 25, 158-166). Эта плазмида управляется промотором скелетного актина человека (HSA) и имеет длину приблизительно 11 т.п.о. В этом эксперименте для каждой лунки использовали 10 мкг плазмиды NOS-М. Основные задачи этого эксперимента заключались в проверке того, достаточной для связывания антитела с GLUT4myc, и определения того, может ли степень связывания усиливаться инсулином. После инкубации клетки промывали и затем собирали через 3 дня для анализа экспрессии nNOS и НА-тэга методом Вестерн-блоттинга. Как показано на фиг. 4, как минимум 15-минутная инкубация была достаточной для получения экспрессии nNOS и HA, а уровни белка после инкубации в течение 30 и 40 мин были одинаковыми. Важно отметить, что для каждого временного интервала 100 нМ инсулин заметно увеличивал экспрессию белка, что указывает на то, что связывание и проникновение структуры в клетку зависело от перемещения GLUT4 из цитозоля на мембрану.
Эксперимент 2
[0142] Для второго эксперимента были поставлены две основные задачи. Первая задача заключалась в исследовании влияния различного количества плазмидной ДНК на поглощение клеткой и экспрессию. Второй задачей являлось определение оптимального количества ДНК относительно липосом, определяемого соотношением DC-Chol к ДНК (мас./мас.). Как показано на фиг. 5, из всех концентраций ДНК, соотношение DC-Chol и ДНК, равное 2,0, давало самую высокую экспрессию nNOS в соответствии с предшествующими исследованиями. В пересчете на ДНК, 5 мкг было достаточным для получения высокой экспрессии nNOS, тогда как уровень экспрессии снижался при 20 мкг, в частности, из-за более высокого соотношения DC-Chol и ДНК. Таким образом, в будущих экспериментах на клеточных культурах использовали от 5 до 10 мкг ДНК при соотношении DC-Chol и ДНК 2,0.
Эксперимент 3
[0143] В этом эксперименте миобласты L6-GLUT4myc инкубировали с той же структурой, показанной на фиг. 3, за исключением того, что плазмида NOS-M была заменена полноразмерной плазмидой, кодирующей дистрофин. Эта конструкция находится под контролем широко известного промотора CAG и имеет приблизительно 20 т.п.о. в длину (Farruggio AP, et al. (2017), Biotechnol J 12(4), 1600477). Двумя основными задачами этого эксперимента были: (1) проверка того, может ли очень большой ген дистрофина проникать в целевую мышечную клетку и экспрессироваться с использованием этой нацеливающей платформы, и (2) проверка эффекта широкого диапазона концентраций инсулина на таргетинг и инициацию экспрессии в мышечных клетках. Диапазон инсулина начинался с низкого уровня, соответствующего уровню в крови человека. В этом эксперименте клетки инкубировали с вектором в течение 15 мин, промывали и затем собирали через 3 дня. Как показано на фиг. 6, полноразмерный дистрофин экспрессировался на высоком уровне, и уровень дистрофина был выше в клетках, обработанных инсулином. Важно отметить, что при низких концентрациях инсулина, обычно присутствующих в крови человека (0,5 и 1,0 нМ), происходит увеличение экспрессии дистрофина по сравнению с клетками без инсулина. Эти результаты поддерживают гипотезу от том, что усиление или индукция транслокации GLUT4 к плазматической мембране инсулином приводит к большему связыванию и поглощению платформы и последующей экспрессии дистрофина.
Эксперимент 4
[0144] Были получены как поликлональные, так и мышиные моноклональные антитела против пептидной последовательности (GRQGPGGPDSI; SEQ ID NO:4), взятой из первого внеклеточной петли мышиного GLUT4. Эти антитела хорошо связываются с пептидом в анализе ELISA. См. например, таблицу 1, в которой приведены результаты тестов ELISA для 20 отобранных мышиных моноклональных антител.
[0145] Таблица 1. Данные анализа ELISA, показывающие уровни поглощения при различных разведениях для 20 мышиных моноклональных антител против пептидной последовательности GRQGPGGPDSI (SEQ ID NO:4). Все клеточные линии, продуцирующие антитела, хорошо связываются с пептидом.
[0146] В этом эксперименте поликлональные кроличьи антитела сначала очищали с помощью аффинной очистки на пептиде. Далее проверяли способность антител связываться с нативным GLUT4 и опосредовать поглощение полноразмерного дистрофина мышечными клетками, отрицательными по дистрофину (mdx-H2N) (Morgan et al., 1994). Несмотря на то, что эти клетки экспрессируют некоторое количество эндогенного GLUT4, было желательно определить, будет ли увеличение экспрессии GLUT4 дополнительно улучшать поглощение и экспрессию вектора по изобретению. Поэтому, клетки mdx-H2K трансфицировали плазмидой, содержащей ген GLUT4 мыши (mdx-H2K-GLUT4), и с помощью антибактериального отбора была получена поликлональная клеточная линия, которая имеет уровень экспрессии GLUT4 выше примерно в два-три раза. С использованием этих дистрофин-отрицательных клеток было желательно оценить, приводит ли повышенная экспрессия GLUT4 к большему связыванию и поглощению невирусной векторной платформы, и, следовательно, к усилению экспрессии дистрофина.
[0147] Другая основная задача этого эксперимента заключалась в проверке того, может ли альбумин повышать экспрессию гена. Основой для этой гипотезы послужили недавние данные о том, что покрытие ДНК/липосомальных векторов сывороточным альбумином человека может усиливать экспрессию репортерного белка люциферазы в клетках легких и селезенки мышей. Поскольку альбумин отрицательно заряжен при нейтральном рН, он может связываться с положительно заряженными липосомами, вместе с плазмидной ДНК. Результаты этого эксперимента выявляют ряд важных фактов (фиг. 7). Во-первых, экспрессия полноразмерного дистрофина была определена и измерена, указывая на то, что поликлональные антитела кролика были способны эффективно связываться с GLUT4 и доставлять плазмиду внутрь мышечных клеток в ядро. Во-вторых, экспрессия дистрофина была значительно выше в клетках mdx-H2K-GLUT4 по сравнению с обычными клетками mdx-H2K. В-третьих, увеличение концентраций альбумина повышает экспрессию дистрофина в обеих мышечных клеточных линиях.
Эксперименты in vivo
[0148] ДНК/липосомные векторы для введения in vivo могут иметь потенциальные проблемы вследствие относительно больших размеров этих структур (обычно≥200 нм). Таким образом, была разработана нелипосомальная альтернативная структура (фиг. 8) для улучшения применения in vivo. При этом плазмидная ДНК в высокой степени конденсируется положительно заряженным поли-L-лизином (PLL) в структуры, которые могут иметь диаметр от 30 до 60 нм для плазмиды 20 т.п.о. (Fink TL, et al. (2006)). Размер плазмиды до 20 т.п.о. не ограничивает эффективный перенос генов в клетках легких in vivo с использованием упакованных наночастиц ДНК. Gene Ther 13, 1048-1051). PLL является биотинилированным, что дает возможность последующего связывания с нейтравидином. Затем добавляют биотинилированное мышиное моноклональное анти-GLUT4 антитело.
[0149] Наконец, добавляют биотинилированный рекомбинантный сывороточный альбумин человека. Утверждается, что альбумин способствует in vivo введению этой структуры по ряду причин. (1) Альбумин пересекает плотный эндотелиальный барьер мышечных капилляров за счет трансцитоза. Это обеспечивает механизм для перемещения альбумина и, следовательно, невирусной векторной платформы по изобретению из кровотока в мышечное внеклеточное пространство. (2) Известно, что альбумин предпочтительно накапливается во внеклеточном пространстве скелетных и сердечных мышц. (3) Альбумин может входить в мышечные клетки за счет эндоцитоза. Это может иметь особое значение в дистрофических мышцах, которые имеют повышенную проницаемость мембраны и накапливают альбумин внутриклеточно. (4) Было показано, что альбумин способствует усилению исключения из эндосомальной утилизации и доставке в ядро плазмидной ДНК. (5) Альбумин является очень стабильным и имеет длительный период полужизни in vivo 19 дней. (6) Альбумин с очень низкой вероятностью будет вызывать любые побочные эффекты или иммунные ответы, учитывая, что он является распространенным сывороточным белком. Это позволит осуществлять многократное введение терапевтической полезной нагрузки через невирусную векторную платформу.
Эксперимент 5
[0150] В этом первом эксперименте in vivo внутримышечные (в/м) инъекции сначала выполняли с использованием структуры, показанной на фиг. 8, без анти-GLUT4 антитела для определения фонового эффекта невирусной векторной платформы без нацеливающего антитела. Инъекции проводили в две мышцы (переднюю большеберцовую и икроножную мышцы) одной конечности 8-недельных мышей mdx, общепринятой животной модели DMD. Структура содержала плазмиду NOS-M, описанную в эксперименте 1. Интересно, что через три дня, как передняя большеберцовая, так и икроножная мышцы экспрессировали NOS-M, как было показано как с помощью антител к nNOS, так и с помощью антител к НА-тэгу (фиг. 9А-В). Предположительно, более проницаемые мышечные волокна у mdx-мышей позволяют некоторое поглощение альбумин-содержащей структуры. По сравнению с получившими инъекции mdx-мышами, не получившие инъекций mdx-мыши, имели эндогенную экспрессию nNOS, но не имели экспрессии НА-тэга. Не получавшая инъекций мышь дикого типа (WT) имела более высокую эндогенную экспрессию nNOS, чем mdx-мышь, но также не имела экспрессию НА-тэга. Другое очень интересное открытие состоит в том, что для обеих мышц, но, в особенности, для передней большеберцовой мышцы, наблюдалась экспрессия NOS-M (НА-тэг) как в получившей инъекцию, так и в не получившей инъекцию конечности (см. фиг. 9A-B). Это указывает на то, что структура способна выйти в системный кровоток из получивших инъекцию мышц, а затем проникнуть в не получавшие инъекцию мышцы другой конечности.
Эксперимент 6
[0151] В следующем эксперименте была проверена гипотеза о том, что конъюгация моноклональных мышиных анти-GLUT4 антител со структурой будет значительно усиливать проникновение платформы по изобретению в мышцы mdx-мышей. Сначала были получены и очищены два мышиных моноклональных антитела. Корректную молекулярную массу и чистоту антител проверяли в восстановительных и невосстановительных условиях с помощью окрашивания Кумасси полиакриламидного геля (фиг. 10А). Затем проверяли аффинность связывания каждого антитела in vitro с использованием линии мышечных клеток mdx-H2K-GLUT4. Конкретно, клетки инкубировали с каждым антителом при различных разведениях (от исходной концентрации1 мг/мл) в течение 20 мин при 37°С с или без 100 нМ инсулина. Клетки промывали, инкубировали с вторичным меченным HRP антителом и снова промывали. Колориметрический анализ использовали для количественного определения относительного количества связанного антитела с нативным белком GLUT4 путем измерения поглощения с помощью спектрофотометра. Как показано на фиг. 10В, оба антитела, Клон 1 и Клон 2, связывались с нативным белком GLUT4 дозозависимым образом. Во всех разведениях Клон 2 имел более высокое поглощение, чем Клон 1. Важно, что для обоих антител инсулин (100 нМ) увеличивал поглощение (см. фиг. 10В), что указывает на большее связывание, когда большее количество GLUT4 переносится в плазматическую мембрану посредством инсулинового сигнального пути.
[0152] Далее антитело Клона 2 биотинилировали и встраивали в невирусную векторную платформу путем связывания с нейтравидином. В качестве контроля использовали также невирусную векторную платформу без антитела (описанную в эксперименте 5). Двум mdx-мышей вводили с помощью в/м инъекции невирусную векторную платформу с или без конъюгированного антитела. Аналогично, инъекции проводили в переднюю большеберцовую и икроножную мышцы одной конечности. Через шесть дней после инъекций мышцы собирали и использовали для Вестерн-блот-анализа. Как показано на фиг. 11, уровень экспрессии nNOS, измеренный по НА-тэгу, был значительно (в несколько раз) выше как в передней большеберцовой, так и в икроножной мышцах мыши, получившей инъекцию платформы, содержащей mAb Клона 1. Эти данные свидетельствуют о том, что анти-GLUT4 антитело сильно повышает таргетинг и проникновению невирусной векторной платформы в скелетные мышцы.
Эксперимент 7
[0153] Затем проверяли, может ли внутривенное (в/в) введение нацеливающей невирусной платформы привести к системной доставке и экспрессии в мышцах белков, кодирующих гены различных размеров. Для проверки этой гипотезы использовали три гена различного размера (эксперименты 7-9). В этом эксперименте использовали наименьший из этих генов. Этот ген представляет собой α-синтрофин, с размером плазмиды приблизительно 5,5 т.п.о. В этом эксперименте мышам mdx4CV вводили с помощью в/м или в/в инъекции невирусную платформу, описанную на фиг. 11, содержащую ген α-синтрофина, который имеет последовательность пальмитоилирования RAS на С-конце для таргетинга в сарколемму. Таргетинг α-синтропина в сарколемму в нормальной мышце происходит путем взаимодействия с дистрофином, отсутствующим в мышцах мышей mdx4CV. Как показано на фиг. 12, как в/м, так и в/в доставка приводит к значительному увеличению экспрессии α-синтропина в мышце через 12 дней после инъекции по сравнению с не получавший инъекцию контрольной мышцей, как подтверждено Вестерн-блоттингом (фиг. 12A) и иммуноокрашиванием (фиг.12В) поперечных срезов мышц.
Эксперимент 8
[0154] Вторым геном был выбран nNOS, как описано на фиг. 11, с размером плазмиды приблизительно 10,5 т.п.о. Вновь мышам mdx4CV снова проводили либо в/м, либо в/в инъекции. Конкретно, двум мышам вводили с помощью в/м инъекции в икроножную мышцу одной задней конечности, а двум мышам вводили с помощью в/в инъекции в ретроорбитальный синус. Как показано на фиг. 13А, экспрессия nNOS была значительно выше (приблизительно на 50%) в мышцах мышей после в/м и в/в инъекций по сравнению с не получавшей инъекции контрольной мышцей. В данном эксперименте также измеряли экспрессию утрофина (гомолога дистрофина) с помощью Вестерн-блоттинга, где было показано значительное увеличение у мышей, которым вводили с помощью инъекции конструкцию nNOS (см. фиг. 13A). В другом эксперименте исследовали влияние дозы плазмиды nNOS. Для этого mdx-мышам вводили в/в платформу, содержащую различные дозы плазмиды nNOS (20, 60 или 120 мкг). Как показано на фиг. 13, наблюдалось прогрессирующее увеличение экспрессии nNOS в передней большеберцовой мышце по мере увеличения дозы плазмиды nNOS, что было измерено как по экспрессии nNOS, так и НА-тэга.
Эксперимент 9
[0155] Используемый для проверки третий и наибольший исследуемый ген представлял собой полноразмерный мышиный дистрофин. При этом использовали две плазмидные конструкции, каждая из которых содержала полноразмерный дистрофин, но с различными промоторами и плазмидными каркасами. Первая конструкция содержала укороченный мышечноспецифичный промотор креатин-киназы 8 (CK8) (PMID: 21266958, PMID: 19298131). Эта конструкция имела размер 15,5 т.п.о. Вторая конструкция содержала повсеместно распространенный промотор цитомегаловируса (CMV). Эта конструкция имела размер 17,5 т.п.о. Как показано на фиг. 14, мыши mdx4CV, получившие в/в инъекцию CK8-конструкции, демонстировала экспрессию полноразмерного дистрофина в ряде мышц задних конечностей. Уровни экспрессии были намного выше, чем у не получавших инъекции мышей mdx4CV, образцы от которых имели слабые полосы на Вестерн-блоте из-за присутствия очень малого количества ревертантных волокон, которые экспрессируют дистрофин (PMID: 1635838). Кроме того, в этом эксперименте для Вестерн-блоттинга использовали обогащенный мембранами лизат мышц, который, вероятно, позволяет обнаружение даже очень низкого уровня дистрофина скелетных мышц из ревертантных волокон. Сравнение с мышью WT показало уровни дистрофина у получивших инъекции мышей в пределах от -0,2 до 0,5% для различных мышц. Далее мышам mdx4CV вводили с помощью в/в инъекции платформу, содержащую либо CK8-, либо CMV-конструкцию для измерения экспрессии дистрофина в сердце. Важно, что, как показано на фиг. 15, сердечные мышцы у получивших инъекцию мышей показали четкую экспрессию полноразмерного дистрофина по сравнению с не получавшим инъекции контролем. Уровень экспрессии дистрофина был выше с использованием CMV-конструкции по сравнению с CK8-конструкцией, при значениях от 0,1 до 0,2% от уровня белка у мыши WT. Взятые вместе, данные из экспериментов 8 и 9 показывают, что композиция невирусной платформы (см. фиг. 8), содержащая большой ген, такой как полноразмерный дистрофин, эффективно поглощается и экспрессируется как скелетными, так и сердечными мышцами.
Эксперимент 10
[0156] В этом эксперименте проводили анализ отслеживания частиц для определения распределения размеров частиц, полученных с использованием невирусной платформы по изобретению. В этом эксперименте композиция платформы (см. фиг. 8) содержала плазмиду для экспрессии полноразмерного дистрофина под контролем CK8-промотора. Анализ отслеживания частиц проводили с использованием стандартных процедур для NanoSight ns300 (Malvern Panalytical), что позволяет достоверно и быстро измерять распределение частиц по размеру в диапазоне от 0,01 до 1,0 мкм. Образец платформы разводили 1:100 деионизированной водой, и затем проводили 4 отдельных измерения распределения частиц по размерам. Как показано на фиг. 20, график распределения размера частиц указывает, что модальный (пиковый) размер частиц составлял 171 нм. В целом, средний размер частиц составлял 236 нм ±4,4 и. Анализ данных показал, что 50% частиц имели размер менее 213 нм, а 90% частиц имели размер менее 344 нм. Средний размер частиц по изобретению является очень сходным со средним размером частиц, содержащих нейтравидин и киРНК (237 нм), которые были эффективными для подавления экспрессии генов in vivo у мышей (PMID: 28026957). Таким образом, данные, представленные в этом эксперименте, показывают, что даже с крупной плазмидной ДНК, такой как плазмида, кодирующая полноразмерный дистрофин (15,5 т.п.о.), невирусная платформа образует мелкие частицы, которые могут эффективно поглощаться и экспрессироваться мышцами in vivo (см. фиг. 14 и 15).
Эксперимент 11
[0157] В этом эксперименте получали ряд моноклональных антител против GLUT4 и оценивали аффинность связывания каждого антитела. Мышиные моноклональные антитела против как пептида GLUT4 мыши (GRQGPGGPDSI; SEQ ID NO:4), так и пептида GLUT4 человека (GRQGPPGPSSI; SEQ ID NO: 2) были изготовлены в компании Genscript (NJ, USA). Было получено всего 14 уникальных клонов; 6 против последовательности GLUT4 мыши и 8 против последовательности GLUT4 человека. Каждую линию клеток гибридомы выращивали в культуре в течение нескольких недель, и моноклональные антитела очищали с использованием A/G-латексных шариков. Аффинность связывания каждого моноклонального антитела измеряли с помощью ELISA. Для антител против последовательности GLUT4 мыши ELISA-планшет покрывали пептидом (SEQ ID NO: 4). Для антител против последовательностей GLUT4 человека, рекомбинантный белок GLUT4 человека использовали для покрытия планшета для ELISA. В обоих случаях каждую лунку планшета для ELISA покрывали 100 нг антигена в течение ночи при 4°С после блокирования, моноклональные антитела добавляли на 1 ч при комнатной температуре. После промывки добавляли вторичное конъюгированное с HRP анти-мышиное антитело на 1 ч при комнатной температуре. После дальнейшей промывки добавляли хромогенный субстрат для HRP (1-STEP™ Ultra TMB-ELISA substrate solution; Thermo Fisher Scientific) на 10-20 минут. Наконец, реакцию останавливали добавлением 2М серной кислоты и измеряли поглощение при 450 нм на планшетном ридере. Как показано на фиг. 16, все 6 моноклональных антител демонстрировали зависимое от концентрации увеличение поглощения. Наилучшими связывающими клонами были 25D4, а затем 22F6 и 21Е5. Остальные три клона продемонстрировали аналогичные, более низкие кривые связывания (см. фиг. 16). Для моноклональных антител, специфичных к последовательности GLUT человека, использовали такой же способ, за исключением того, что планшет покрывали рекомбинантным белком GLUT4 человека. Как показано на фиг. 17, 7 моноклональных антител демонстрировали дозозависимое увеличение поглощения, причем клоны 5E3 и 8G2 имели наилучшую аффинность связывания, и клоны 2В9 и 4G2 показывали наиболее слабую аффинность связывания. Один клон, 15G2, невозможно было проанализировать из-за недостаточной продукции антител этой линией гибридомных клеток.
[0158] Для проверки того, могут ли антитела против мышиной последовательности связываться с белком GLUT4 человека, или наоборот, могут ли антитела против последовательности из белка человека связываться с последовательностью пептида GLUT4 мыши, были проведены два дополнительных анализа ELISA. Как показано на фиг. 18, два антитела, которые прочно связываются с последовательностью мыши (22F6 и 21Е5), не связываются эффективно с белком GLUT4 человека, в то время как антитела, полученные против последовательности пептида GLUT4 человека (5E3 и 8G2), показали сильное связывание. Как показано на фиг. 19, в обратном эксперименте антитела, полученные против последовательности GLUT4 человека, связывались с различной степенью аффинности с пептидом GLUT4 мыши (8G2 являлись наилучшими, тогда как 12Е11 не связывались), но все четверо этих антител связывались намного хуже, чем антитела, полученные против мышиной последовательности (22F6 и 21E5). Таким образом, несмотря на 83,3% гомологии между последовательностями мышиного и человеческого пептида, полученные антитела связываются значительно сильнее с видоспецифичной последовательностью GLUT4, против которой они были получены.
Эксперимент 12
[0159] Для каждого из полученных 14 уникальных моноклональных антител, 6 против последовательности GLUT4 мыши и 8 против последовательности GLUT4 человека, в компании GenScript (NJ, USA) были отсеквенированы вариабельные области тяжелой и легкой цепей следующим способом. Общую РНК выделяли из клеток гибридомы (с использованием реагента TRIzol®). Затем тотальную РНК обратно транскрибировали в кДНК, используя либо изотип-специфичные антисмысловые праймеры, либо универсальные праймеры (с помощью набора PrimeScript™ 1st Strand cDNA Synthesis Kit). Фрагменты тяжелой цепи и легкой цепи антител амплифицировали по стандартной рабочей процедуре (SOP) быстрой амплификации концов кДНК (RACE) от GenScript. Амплифицированные фрагменты антител клонировали в стандартный вектор для клонирования по отдельности. ПЦР на колониях проводили для скрининга клонов со вставками правильного размера, и были получены последовательности для каждого антитела. В таблице 2 приведены определяющие комплементарность области (CDR) для тяжелой и легкой цепей для 6 моноклональных антител, полученных против пептида GLUT4 мыши (GRQGPGGPDSI; SEQ ID NO:4). Также показаны предсказанные консенсусные последовательности для каждой CDR. В таблице 3 приведены определяющие комплементарность области (CDR) для тяжелой и легкой цепей для 8 моноклональных антител, полученных против пептида GLUT4 человека (GRQGPPGPSSI; SEQ ID NO:2). Также показаны прогнозируемые консенсусные последовательности для каждой CDR. Интересно, что два клона (2В9 и 4G7), которые наиболее слабо связываются с белком GLUT4 человека в анализе ELISA (см. фиг. 17), не содержат аргинина (R), найденного в консенсусной последовательности CDR3VH у других 6 клонов, что позволяет предположить, что этот остаток может иметь важную роль в активности связывания антигена. Следует также отметить, что клон 15G2 не был представлен в анализе ELISA (см. фиг. 17), поскольку это антитело не росло хорошо при культивировании в лаборатории авторов изобретения.
[0160] В таблице 2 приведены результаты эксперимента 12, в котором получены аминокислотные последовательности определяющих комплементарность областей (CDR) для тяжелой цепи (VH) и легкой цепи (VL) 6 клонов моноклональных мышиных антител против последовательности пептида GRQGPGGPDSI мышиного GLUT4 (SEQ ID NO: 4). Также ниже таблицы 2 приведены иллюстративные консенсусные последовательности для каждой CDR.
Консенсусные последовательности CDR
CDR1 (VH): T Y G M S/T
CDR2 (VH): W I N T Y/F/S S G L/V P T Y X X D F K G
CDR3 (VH): P I H/T X V V A/P X D Y
CDR1 (VL): K/R S S Q S L L/V Y X5-6 Y L A/H
CDR2 (VL): K/W A/V S N/T R E/F S
CDR3 (VL): Q Q S/Y F/R/Y H/S/T X P Y/W T
[0161] В таблице 3 приведены результаты эксперимента 12, в котором получены аминокислотные последовательности, определяющих комплементарность областей (CDR), для тяжелой цепи (VH) и легкой цепи (VL) 8 клонов моноклональных антител против последовательности пептида GRQGPEGPSSI GLUT4 человека (SEQ ID NO:2). Также ниже таблица 3 приведены иллюстративные консенсусные последовательности для каждой CDR.
Консенсусные последовательности CDR
CDR1 (VH): D/E Y S/T I/M H
CDR2 (VH): W I N T E S/T G D/E X T/S Y A D D F K G
CDR3 (VH): R X X Y
CDR1 (VL): R/K A/S S Q S L/V X X N/S
CDR2 (VL): V/A S N R F/Y S/T
CDR3 (VL): Q D X X X P W T
Заключение
[0162] Показано, что невирусная векторная платформа по изобретению может эффективно и специфично доставлять терапевтические полезные нагрузки в ткани, которые экспрессируют GLUT4, включая, но не ограничиваясь ими, скелетные мышцы, сердечные мышцы, жировую ткань, гладкие мышцы, определенные области головного мозга (например, гиппокамп и мозжечок) и определенные раковые клетки (например, меланоциты). Терапевтические средства, которые могут быть доставлены в эти ткани с помощью платформы по изобретению, включают нуклеиновые кислоты (например, плазмидную ДНК, миникольцевую ДНК, мРНК, киРНК, кшРНК, олигонуклеотиды, конструкции CRISPR/Cas9), пептиды, липиды и лекарственные средства. Платформа не ограничена конкретными размерами полезной нагрузки, а скорее может успешно доставлять различные крупные нуклеотидные конструкции, обеспечивая поглощение и экспрессию кодируемых больших трансгенов. Эффекты и эффективность таргетинга и поглощения дополнительно усиливаются индуцируемой инсулином транслокацией GLUT4 в плазматическую мембрану клетки и введением альбумина в состав векторной платформы. Терапевтические композиции, включающие невирусную векторную платформу по изобретению, могут быть доставлены в качестве монотерапии или как часть комбинированной терапии.
[0163] Хотя были проиллюстрированы и описаны иллюстративные варианты осуществления, следует понимать, что в них могут быть внесены различные изменения, не выходящие за рамки сущности и объема изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> University of Washington
Whitehead, Nicholas P.
Froehner, Stanley C.
<120> НЕВИРУСНЫЙ ИММУНОТАРГЕТИНГ
<130> UWOTL-1-72245
<150> US 62/872,425
<151> 2019-07-10
<160> 89
<170> PatentIn version 3.5
<210> 1
<211> 36
<212> Белок
<213> Homo sapiens
<400> 1
Ile Asn Ala Pro Gln Lys Val Ile Glu Gln Ser Tyr Asn Glu Thr Trp
1 5 10 15
Leu Gly Arg Gln Gly Pro Glu Gly Pro Ser Ser Ile Pro Pro Gly Thr
20 25 30
Leu Thr Thr Leu
35
<210> 2
<211> 11
<212> Белок
<213> Homo sapiens
<400> 2
Gly Arg Gln Gly Pro Glu Gly Pro Ser Ser Ile
1 5 10
<210> 3
<211> 36
<212> Белок
<213> Mus musculus
<400> 3
Ile Asn Ala Pro Gln Lys Val Ile Glu Gln Ser Tyr Asn Ala Thr Trp
1 5 10 15
Leu Gly Arg Gln Gly Pro Gly Gly Pro Asp Ser Ile Pro Gln Gly Thr
20 25 30
Leu Thr Thr Leu
35
<210> 4
<211> 11
<212> Белок
<213> Mus musculus
<400> 4
Gly Arg Gln Gly Pro Gly Gly Pro Asp Ser Ile
1 5 10
<210> 5
<211> 3685
<212> Белок
<213> Homo sapiens
<400> 5
Met Leu Trp Trp Glu Glu Val Glu Asp Cys Tyr Glu Arg Glu Asp Val
1 5 10 15
Gln Lys Lys Thr Phe Thr Lys Trp Val Asn Ala Gln Phe Ser Lys Phe
20 25 30
Gly Lys Gln His Ile Glu Asn Leu Phe Ser Asp Leu Gln Asp Gly Arg
35 40 45
Arg Leu Leu Asp Leu Leu Glu Gly Leu Thr Gly Gln Lys Leu Pro Lys
50 55 60
Glu Lys Gly Ser Thr Arg Val His Ala Leu Asn Asn Val Asn Lys Ala
65 70 75 80
Leu Arg Val Leu Gln Asn Asn Asn Val Asp Leu Val Asn Ile Gly Ser
85 90 95
Thr Asp Ile Val Asp Gly Asn His Lys Leu Thr Leu Gly Leu Ile Trp
100 105 110
Asn Ile Ile Leu His Trp Gln Val Lys Asn Val Met Lys Asn Ile Met
115 120 125
Ala Gly Leu Gln Gln Thr Asn Ser Glu Lys Ile Leu Leu Ser Trp Val
130 135 140
Arg Gln Ser Thr Arg Asn Tyr Pro Gln Val Asn Val Ile Asn Phe Thr
145 150 155 160
Thr Ser Trp Ser Asp Gly Leu Ala Leu Asn Ala Leu Ile His Ser His
165 170 175
Arg Pro Asp Leu Phe Asp Trp Asn Ser Val Val Cys Gln Gln Ser Ala
180 185 190
Thr Gln Arg Leu Glu His Ala Phe Asn Ile Ala Arg Tyr Gln Leu Gly
195 200 205
Ile Glu Lys Leu Leu Asp Pro Glu Asp Val Asp Thr Thr Tyr Pro Asp
210 215 220
Lys Lys Ser Ile Leu Met Tyr Ile Thr Ser Leu Phe Gln Val Leu Pro
225 230 235 240
Gln Gln Val Ser Ile Glu Ala Ile Gln Glu Val Glu Met Leu Pro Arg
245 250 255
Pro Pro Lys Val Thr Lys Glu Glu His Phe Gln Leu His His Gln Met
260 265 270
His Tyr Ser Gln Gln Ile Thr Val Ser Leu Ala Gln Gly Tyr Glu Arg
275 280 285
Thr Ser Ser Pro Lys Pro Arg Phe Lys Ser Tyr Ala Tyr Thr Gln Ala
290 295 300
Ala Tyr Val Thr Thr Ser Asp Pro Thr Arg Ser Pro Phe Pro Ser Gln
305 310 315 320
His Leu Glu Ala Pro Glu Asp Lys Ser Phe Gly Ser Ser Leu Met Glu
325 330 335
Ser Glu Val Asn Leu Asp Arg Tyr Gln Thr Ala Leu Glu Glu Val Leu
340 345 350
Ser Trp Leu Leu Ser Ala Glu Asp Thr Leu Gln Ala Gln Gly Glu Ile
355 360 365
Ser Asn Asp Val Glu Val Val Lys Asp Gln Phe His Thr His Glu Gly
370 375 380
Tyr Met Met Asp Leu Thr Ala His Gln Gly Arg Val Gly Asn Ile Leu
385 390 395 400
Gln Leu Gly Ser Lys Leu Ile Gly Thr Gly Lys Leu Ser Glu Asp Glu
405 410 415
Glu Thr Glu Val Gln Glu Gln Met Asn Leu Leu Asn Ser Arg Trp Glu
420 425 430
Cys Leu Arg Val Ala Ser Met Glu Lys Gln Ser Asn Leu His Arg Val
435 440 445
Leu Met Asp Leu Gln Asn Gln Lys Leu Lys Glu Leu Asn Asp Trp Leu
450 455 460
Thr Lys Thr Glu Glu Arg Thr Arg Lys Met Glu Glu Glu Pro Leu Gly
465 470 475 480
Pro Asp Leu Glu Asp Leu Lys Arg Gln Val Gln Gln His Lys Val Leu
485 490 495
Gln Glu Asp Leu Glu Gln Glu Gln Val Arg Val Asn Ser Leu Thr His
500 505 510
Met Val Val Val Val Asp Glu Ser Ser Gly Asp His Ala Thr Ala Ala
515 520 525
Leu Glu Glu Gln Leu Lys Val Leu Gly Asp Arg Trp Ala Asn Ile Cys
530 535 540
Arg Trp Thr Glu Asp Arg Trp Val Leu Leu Gln Asp Ile Leu Leu Lys
545 550 555 560
Trp Gln Arg Leu Thr Glu Glu Gln Cys Leu Phe Ser Ala Trp Leu Ser
565 570 575
Glu Lys Glu Asp Ala Val Asn Lys Ile His Thr Thr Gly Phe Lys Asp
580 585 590
Gln Asn Glu Met Leu Ser Ser Leu Gln Lys Leu Ala Val Leu Lys Ala
595 600 605
Asp Leu Glu Lys Lys Lys Gln Ser Met Gly Lys Leu Tyr Ser Leu Lys
610 615 620
Gln Asp Leu Leu Ser Thr Leu Lys Asn Lys Ser Val Thr Gln Lys Thr
625 630 635 640
Glu Ala Trp Leu Asp Asn Phe Ala Arg Cys Trp Asp Asn Leu Val Gln
645 650 655
Lys Leu Glu Lys Ser Thr Ala Gln Ile Ser Gln Ala Val Thr Thr Thr
660 665 670
Gln Pro Ser Leu Thr Gln Thr Thr Val Met Glu Thr Val Thr Thr Val
675 680 685
Thr Thr Arg Glu Gln Ile Leu Val Lys His Ala Gln Glu Glu Leu Pro
690 695 700
Pro Pro Pro Pro Gln Lys Lys Arg Gln Ile Thr Val Asp Ser Glu Ile
705 710 715 720
Arg Lys Arg Leu Asp Val Asp Ile Thr Glu Leu His Ser Trp Ile Thr
725 730 735
Arg Ser Glu Ala Val Leu Gln Ser Pro Glu Phe Ala Ile Phe Arg Lys
740 745 750
Glu Gly Asn Phe Ser Asp Leu Lys Glu Lys Val Asn Ala Ile Glu Arg
755 760 765
Glu Lys Ala Glu Lys Phe Arg Lys Leu Gln Asp Ala Ser Arg Ser Ala
770 775 780
Gln Ala Leu Val Glu Gln Met Val Asn Glu Gly Val Asn Ala Asp Ser
785 790 795 800
Ile Lys Gln Ala Ser Glu Gln Leu Asn Ser Arg Trp Ile Glu Phe Cys
805 810 815
Gln Leu Leu Ser Glu Arg Leu Asn Trp Leu Glu Tyr Gln Asn Asn Ile
820 825 830
Ile Ala Phe Tyr Asn Gln Leu Gln Gln Leu Glu Gln Met Thr Thr Thr
835 840 845
Ala Glu Asn Trp Leu Lys Ile Gln Pro Thr Thr Pro Ser Glu Pro Thr
850 855 860
Ala Ile Lys Ser Gln Leu Lys Ile Cys Lys Asp Glu Val Asn Arg Leu
865 870 875 880
Ser Gly Leu Gln Pro Gln Ile Glu Arg Leu Lys Ile Gln Ser Ile Ala
885 890 895
Leu Lys Glu Lys Gly Gln Gly Pro Met Phe Leu Asp Ala Asp Phe Val
900 905 910
Ala Phe Thr Asn His Phe Lys Gln Val Phe Ser Asp Val Gln Ala Arg
915 920 925
Glu Lys Glu Leu Gln Thr Ile Phe Asp Thr Leu Pro Pro Met Arg Tyr
930 935 940
Gln Glu Thr Met Ser Ala Ile Arg Thr Trp Val Gln Gln Ser Glu Thr
945 950 955 960
Lys Leu Ser Ile Pro Gln Leu Ser Val Thr Asp Tyr Glu Ile Met Glu
965 970 975
Gln Arg Leu Gly Glu Leu Gln Ala Leu Gln Ser Ser Leu Gln Glu Gln
980 985 990
Gln Ser Gly Leu Tyr Tyr Leu Ser Thr Thr Val Lys Glu Met Ser Lys
995 1000 1005
Lys Ala Pro Ser Glu Ile Ser Arg Lys Tyr Gln Ser Glu Phe Glu
1010 1015 1020
Glu Ile Glu Gly Arg Trp Lys Lys Leu Ser Ser Gln Leu Val Glu
1025 1030 1035
His Cys Gln Lys Leu Glu Glu Gln Met Asn Lys Leu Arg Lys Ile
1040 1045 1050
Gln Asn His Ile Gln Thr Leu Lys Lys Trp Met Ala Glu Val Asp
1055 1060 1065
Val Phe Leu Lys Glu Glu Trp Pro Ala Leu Gly Asp Ser Glu Ile
1070 1075 1080
Leu Lys Lys Gln Leu Lys Gln Cys Arg Leu Leu Val Ser Asp Ile
1085 1090 1095
Gln Thr Ile Gln Pro Ser Leu Asn Ser Val Asn Glu Gly Gly Gln
1100 1105 1110
Lys Ile Lys Asn Glu Ala Glu Pro Glu Phe Ala Ser Arg Leu Glu
1115 1120 1125
Thr Glu Leu Lys Glu Leu Asn Thr Gln Trp Asp His Met Cys Gln
1130 1135 1140
Gln Val Tyr Ala Arg Lys Glu Ala Leu Lys Gly Gly Leu Glu Lys
1145 1150 1155
Thr Val Ser Leu Gln Lys Asp Leu Ser Glu Met His Glu Trp Met
1160 1165 1170
Thr Gln Ala Glu Glu Glu Tyr Leu Glu Arg Asp Phe Glu Tyr Lys
1175 1180 1185
Thr Pro Asp Glu Leu Gln Lys Ala Val Glu Glu Met Lys Arg Ala
1190 1195 1200
Lys Glu Glu Ala Gln Gln Lys Glu Ala Lys Val Lys Leu Leu Thr
1205 1210 1215
Glu Ser Val Asn Ser Val Ile Ala Gln Ala Pro Pro Val Ala Gln
1220 1225 1230
Glu Ala Leu Lys Lys Glu Leu Glu Thr Leu Thr Thr Asn Tyr Gln
1235 1240 1245
Trp Leu Cys Thr Arg Leu Asn Gly Lys Cys Lys Thr Leu Glu Glu
1250 1255 1260
Val Trp Ala Cys Trp His Glu Leu Leu Ser Tyr Leu Glu Lys Ala
1265 1270 1275
Asn Lys Trp Leu Asn Glu Val Glu Phe Lys Leu Lys Thr Thr Glu
1280 1285 1290
Asn Ile Pro Gly Gly Ala Glu Glu Ile Ser Glu Val Leu Asp Ser
1295 1300 1305
Leu Glu Asn Leu Met Arg His Ser Glu Asp Asn Pro Asn Gln Ile
1310 1315 1320
Arg Ile Leu Ala Gln Thr Leu Thr Asp Gly Gly Val Met Asp Glu
1325 1330 1335
Leu Ile Asn Glu Glu Leu Glu Thr Phe Asn Ser Arg Trp Arg Glu
1340 1345 1350
Leu His Glu Glu Ala Val Arg Arg Gln Lys Leu Leu Glu Gln Ser
1355 1360 1365
Ile Gln Ser Ala Gln Glu Thr Glu Lys Ser Leu His Leu Ile Gln
1370 1375 1380
Glu Ser Leu Thr Phe Ile Asp Lys Gln Leu Ala Ala Tyr Ile Ala
1385 1390 1395
Asp Lys Val Asp Ala Ala Gln Met Pro Gln Glu Ala Gln Lys Ile
1400 1405 1410
Gln Ser Asp Leu Thr Ser His Glu Ile Ser Leu Glu Glu Met Lys
1415 1420 1425
Lys His Asn Gln Gly Lys Glu Ala Ala Gln Arg Val Leu Ser Gln
1430 1435 1440
Ile Asp Val Ala Gln Lys Lys Leu Gln Asp Val Ser Met Lys Phe
1445 1450 1455
Arg Leu Phe Gln Lys Pro Ala Asn Phe Glu Leu Arg Leu Gln Glu
1460 1465 1470
Ser Lys Met Ile Leu Asp Glu Val Lys Met His Leu Pro Ala Leu
1475 1480 1485
Glu Thr Lys Ser Val Glu Gln Glu Val Val Gln Ser Gln Leu Asn
1490 1495 1500
His Cys Val Asn Leu Tyr Lys Ser Leu Ser Glu Val Lys Ser Glu
1505 1510 1515
Val Glu Met Val Ile Lys Thr Gly Arg Gln Ile Val Gln Lys Lys
1520 1525 1530
Gln Thr Glu Asn Pro Lys Glu Leu Asp Glu Arg Val Thr Ala Leu
1535 1540 1545
Lys Leu His Tyr Asn Glu Leu Gly Ala Lys Val Thr Glu Arg Lys
1550 1555 1560
Gln Gln Leu Glu Lys Cys Leu Lys Leu Ser Arg Lys Met Arg Lys
1565 1570 1575
Glu Met Asn Val Leu Thr Glu Trp Leu Ala Ala Thr Asp Met Glu
1580 1585 1590
Leu Thr Lys Arg Ser Ala Val Glu Gly Met Pro Ser Asn Leu Asp
1595 1600 1605
Ser Glu Val Ala Trp Gly Lys Ala Thr Gln Lys Glu Ile Glu Lys
1610 1615 1620
Gln Lys Val His Leu Lys Ser Ile Thr Glu Val Gly Glu Ala Leu
1625 1630 1635
Lys Thr Val Leu Gly Lys Lys Glu Thr Leu Val Glu Asp Lys Leu
1640 1645 1650
Ser Leu Leu Asn Ser Asn Trp Ile Ala Val Thr Ser Arg Ala Glu
1655 1660 1665
Glu Trp Leu Asn Leu Leu Leu Glu Tyr Gln Lys His Met Glu Thr
1670 1675 1680
Phe Asp Gln Asn Val Asp His Ile Thr Lys Trp Ile Ile Gln Ala
1685 1690 1695
Asp Thr Leu Leu Asp Glu Ser Glu Lys Lys Lys Pro Gln Gln Lys
1700 1705 1710
Glu Asp Val Leu Lys Arg Leu Lys Ala Glu Leu Asn Asp Ile Arg
1715 1720 1725
Pro Lys Val Asp Ser Thr Arg Asp Gln Ala Ala Asn Leu Met Ala
1730 1735 1740
Asn Arg Gly Asp His Cys Arg Lys Leu Val Glu Pro Gln Ile Ser
1745 1750 1755
Glu Leu Asn His Arg Phe Ala Ala Ile Ser His Arg Ile Lys Thr
1760 1765 1770
Gly Lys Ala Ser Ile Pro Leu Lys Glu Leu Glu Gln Phe Asn Ser
1775 1780 1785
Asp Ile Gln Lys Leu Leu Glu Pro Leu Glu Ala Glu Ile Gln Gln
1790 1795 1800
Gly Val Asn Leu Lys Glu Glu Asp Phe Asn Lys Asp Met Asn Glu
1805 1810 1815
Asp Asn Glu Gly Thr Val Lys Glu Leu Leu Gln Arg Gly Asp Asn
1820 1825 1830
Leu Gln Gln Arg Ile Thr Asp Glu Arg Lys Arg Glu Glu Ile Lys
1835 1840 1845
Ile Lys Gln Gln Leu Leu Gln Thr Lys His Asn Ala Leu Lys Asp
1850 1855 1860
Leu Arg Ser Gln Arg Arg Lys Lys Ala Leu Glu Ile Ser His Gln
1865 1870 1875
Trp Tyr Gln Tyr Lys Arg Gln Ala Asp Asp Leu Leu Lys Cys Leu
1880 1885 1890
Asp Asp Ile Glu Lys Lys Leu Ala Ser Leu Pro Glu Pro Arg Asp
1895 1900 1905
Glu Arg Lys Ile Lys Glu Ile Asp Arg Glu Leu Gln Lys Lys Lys
1910 1915 1920
Glu Glu Leu Asn Ala Val Arg Arg Gln Ala Glu Gly Leu Ser Glu
1925 1930 1935
Asp Gly Ala Ala Met Ala Val Glu Pro Thr Gln Ile Gln Leu Ser
1940 1945 1950
Lys Arg Trp Arg Glu Ile Glu Ser Lys Phe Ala Gln Phe Arg Arg
1955 1960 1965
Leu Asn Phe Ala Gln Ile His Thr Val Arg Glu Glu Thr Met Met
1970 1975 1980
Val Met Thr Glu Asp Met Pro Leu Glu Ile Ser Tyr Val Pro Ser
1985 1990 1995
Thr Tyr Leu Thr Glu Ile Thr His Val Ser Gln Ala Leu Leu Glu
2000 2005 2010
Val Glu Gln Leu Leu Asn Ala Pro Asp Leu Cys Ala Lys Asp Phe
2015 2020 2025
Glu Asp Leu Phe Lys Gln Glu Glu Ser Leu Lys Asn Ile Lys Asp
2030 2035 2040
Ser Leu Gln Gln Ser Ser Gly Arg Ile Asp Ile Ile His Ser Lys
2045 2050 2055
Lys Thr Ala Ala Leu Gln Ser Ala Thr Pro Val Glu Arg Val Lys
2060 2065 2070
Leu Gln Glu Ala Leu Ser Gln Leu Asp Phe Gln Trp Glu Lys Val
2075 2080 2085
Asn Lys Met Tyr Lys Asp Arg Gln Gly Arg Phe Asp Arg Ser Val
2090 2095 2100
Glu Lys Trp Arg Arg Phe His Tyr Asp Ile Lys Ile Phe Asn Gln
2105 2110 2115
Trp Leu Thr Glu Ala Glu Gln Phe Leu Arg Lys Thr Gln Ile Pro
2120 2125 2130
Glu Asn Trp Glu His Ala Lys Tyr Lys Trp Tyr Leu Lys Glu Leu
2135 2140 2145
Gln Asp Gly Ile Gly Gln Arg Gln Thr Val Val Arg Thr Leu Asn
2150 2155 2160
Ala Thr Gly Glu Glu Ile Ile Gln Gln Ser Ser Lys Thr Asp Ala
2165 2170 2175
Ser Ile Leu Gln Glu Lys Leu Gly Ser Leu Asn Leu Arg Trp Gln
2180 2185 2190
Glu Val Cys Lys Gln Leu Ser Asp Arg Lys Lys Arg Leu Glu Glu
2195 2200 2205
Gln Lys Asn Ile Leu Ser Glu Phe Gln Arg Asp Leu Asn Glu Phe
2210 2215 2220
Val Leu Trp Leu Glu Glu Ala Asp Asn Ile Ala Ser Ile Pro Leu
2225 2230 2235
Glu Pro Gly Lys Glu Gln Gln Leu Lys Glu Lys Leu Glu Gln Val
2240 2245 2250
Lys Leu Leu Val Glu Glu Leu Pro Leu Arg Gln Gly Ile Leu Lys
2255 2260 2265
Gln Leu Asn Glu Thr Gly Gly Pro Val Leu Val Ser Ala Pro Ile
2270 2275 2280
Ser Pro Glu Glu Gln Asp Lys Leu Glu Asn Lys Leu Lys Gln Thr
2285 2290 2295
Asn Leu Gln Trp Ile Lys Val Ser Arg Ala Leu Pro Glu Lys Gln
2300 2305 2310
Gly Glu Ile Glu Ala Gln Ile Lys Asp Leu Gly Gln Leu Glu Lys
2315 2320 2325
Lys Leu Glu Asp Leu Glu Glu Gln Leu Asn His Leu Leu Leu Trp
2330 2335 2340
Leu Ser Pro Ile Arg Asn Gln Leu Glu Ile Tyr Asn Gln Pro Asn
2345 2350 2355
Gln Glu Gly Pro Phe Asp Val Gln Glu Thr Glu Ile Ala Val Gln
2360 2365 2370
Ala Lys Gln Pro Asp Val Glu Glu Ile Leu Ser Lys Gly Gln His
2375 2380 2385
Leu Tyr Lys Glu Lys Pro Ala Thr Gln Pro Val Lys Arg Lys Leu
2390 2395 2400
Glu Asp Leu Ser Ser Glu Trp Lys Ala Val Asn Arg Leu Leu Gln
2405 2410 2415
Glu Leu Arg Ala Lys Gln Pro Asp Leu Ala Pro Gly Leu Thr Thr
2420 2425 2430
Ile Gly Ala Ser Pro Thr Gln Thr Val Thr Leu Val Thr Gln Pro
2435 2440 2445
Val Val Thr Lys Glu Thr Ala Ile Ser Lys Leu Glu Met Pro Ser
2450 2455 2460
Ser Leu Met Leu Glu Val Pro Ala Leu Ala Asp Phe Asn Arg Ala
2465 2470 2475
Trp Thr Glu Leu Thr Asp Trp Leu Ser Leu Leu Asp Gln Val Ile
2480 2485 2490
Lys Ser Gln Arg Val Met Val Gly Asp Leu Glu Asp Ile Asn Glu
2495 2500 2505
Met Ile Ile Lys Gln Lys Ala Thr Met Gln Asp Leu Glu Gln Arg
2510 2515 2520
Arg Pro Gln Leu Glu Glu Leu Ile Thr Ala Ala Gln Asn Leu Lys
2525 2530 2535
Asn Lys Thr Ser Asn Gln Glu Ala Arg Thr Ile Ile Thr Asp Arg
2540 2545 2550
Ile Glu Arg Ile Gln Asn Gln Trp Asp Glu Val Gln Glu His Leu
2555 2560 2565
Gln Asn Arg Arg Gln Gln Leu Asn Glu Met Leu Lys Asp Ser Thr
2570 2575 2580
Gln Trp Leu Glu Ala Lys Glu Glu Ala Glu Gln Val Leu Gly Gln
2585 2590 2595
Ala Arg Ala Lys Leu Glu Ser Trp Lys Glu Gly Pro Tyr Thr Val
2600 2605 2610
Asp Ala Ile Gln Lys Lys Ile Thr Glu Thr Lys Gln Leu Ala Lys
2615 2620 2625
Asp Leu Arg Gln Trp Gln Thr Asn Val Asp Val Ala Asn Asp Leu
2630 2635 2640
Ala Leu Lys Leu Leu Arg Asp Tyr Ser Ala Asp Asp Thr Arg Lys
2645 2650 2655
Val His Met Ile Thr Glu Asn Ile Asn Ala Ser Trp Arg Ser Ile
2660 2665 2670
His Lys Arg Val Ser Glu Arg Glu Ala Ala Leu Glu Glu Thr His
2675 2680 2685
Arg Leu Leu Gln Gln Phe Pro Leu Asp Leu Glu Lys Phe Leu Ala
2690 2695 2700
Trp Leu Thr Glu Ala Glu Thr Thr Ala Asn Val Leu Gln Asp Ala
2705 2710 2715
Thr Arg Lys Glu Arg Leu Leu Glu Asp Ser Lys Gly Val Lys Glu
2720 2725 2730
Leu Met Lys Gln Trp Gln Asp Leu Gln Gly Glu Ile Glu Ala His
2735 2740 2745
Thr Asp Val Tyr His Asn Leu Asp Glu Asn Ser Gln Lys Ile Leu
2750 2755 2760
Arg Ser Leu Glu Gly Ser Asp Asp Ala Val Leu Leu Gln Arg Arg
2765 2770 2775
Leu Asp Asn Met Asn Phe Lys Trp Ser Glu Leu Arg Lys Lys Ser
2780 2785 2790
Leu Asn Ile Arg Ser His Leu Glu Ala Ser Ser Asp Gln Trp Lys
2795 2800 2805
Arg Leu His Leu Ser Leu Gln Glu Leu Leu Val Trp Leu Gln Leu
2810 2815 2820
Lys Asp Asp Glu Leu Ser Arg Gln Ala Pro Ile Gly Gly Asp Phe
2825 2830 2835
Pro Ala Val Gln Lys Gln Asn Asp Val His Arg Ala Phe Lys Arg
2840 2845 2850
Glu Leu Lys Thr Lys Glu Pro Val Ile Met Ser Thr Leu Glu Thr
2855 2860 2865
Val Arg Ile Phe Leu Thr Glu Gln Pro Leu Glu Gly Leu Glu Lys
2870 2875 2880
Leu Tyr Gln Glu Pro Arg Glu Leu Pro Pro Glu Glu Arg Ala Gln
2885 2890 2895
Asn Val Thr Arg Leu Leu Arg Lys Gln Ala Glu Glu Val Asn Thr
2900 2905 2910
Glu Trp Glu Lys Leu Asn Leu His Ser Ala Asp Trp Gln Arg Lys
2915 2920 2925
Ile Asp Glu Thr Leu Glu Arg Leu Gln Glu Leu Gln Glu Ala Thr
2930 2935 2940
Asp Glu Leu Asp Leu Lys Leu Arg Gln Ala Glu Val Ile Lys Gly
2945 2950 2955
Ser Trp Gln Pro Val Gly Asp Leu Leu Ile Asp Ser Leu Gln Asp
2960 2965 2970
His Leu Glu Lys Val Lys Ala Leu Arg Gly Glu Ile Ala Pro Leu
2975 2980 2985
Lys Glu Asn Val Ser His Val Asn Asp Leu Ala Arg Gln Leu Thr
2990 2995 3000
Thr Leu Gly Ile Gln Leu Ser Pro Tyr Asn Leu Ser Thr Leu Glu
3005 3010 3015
Asp Leu Asn Thr Arg Trp Lys Leu Leu Gln Val Ala Val Glu Asp
3020 3025 3030
Arg Val Arg Gln Leu His Glu Ala His Arg Asp Phe Gly Pro Ala
3035 3040 3045
Ser Gln His Phe Leu Ser Thr Ser Val Gln Gly Pro Trp Glu Arg
3050 3055 3060
Ala Ile Ser Pro Asn Lys Val Pro Tyr Tyr Ile Asn His Glu Thr
3065 3070 3075
Gln Thr Thr Cys Trp Asp His Pro Lys Met Thr Glu Leu Tyr Gln
3080 3085 3090
Ser Leu Ala Asp Leu Asn Asn Val Arg Phe Ser Ala Tyr Arg Thr
3095 3100 3105
Ala Met Lys Leu Arg Arg Leu Gln Lys Ala Leu Cys Leu Asp Leu
3110 3115 3120
Leu Ser Leu Ser Ala Ala Cys Asp Ala Leu Asp Gln His Asn Leu
3125 3130 3135
Lys Gln Asn Asp Gln Pro Met Asp Ile Leu Gln Ile Ile Asn Cys
3140 3145 3150
Leu Thr Thr Ile Tyr Asp Arg Leu Glu Gln Glu His Asn Asn Leu
3155 3160 3165
Val Asn Val Pro Leu Cys Val Asp Met Cys Leu Asn Trp Leu Leu
3170 3175 3180
Asn Val Tyr Asp Thr Gly Arg Thr Gly Arg Ile Arg Val Leu Ser
3185 3190 3195
Phe Lys Thr Gly Ile Ile Ser Leu Cys Lys Ala His Leu Glu Asp
3200 3205 3210
Lys Tyr Arg Tyr Leu Phe Lys Gln Val Ala Ser Ser Thr Gly Phe
3215 3220 3225
Cys Asp Gln Arg Arg Leu Gly Leu Leu Leu His Asp Ser Ile Gln
3230 3235 3240
Ile Pro Arg Gln Leu Gly Glu Val Ala Ser Phe Gly Gly Ser Asn
3245 3250 3255
Ile Glu Pro Ser Val Arg Ser Cys Phe Gln Phe Ala Asn Asn Lys
3260 3265 3270
Pro Glu Ile Glu Ala Ala Leu Phe Leu Asp Trp Met Arg Leu Glu
3275 3280 3285
Pro Gln Ser Met Val Trp Leu Pro Val Leu His Arg Val Ala Ala
3290 3295 3300
Ala Glu Thr Ala Lys His Gln Ala Lys Cys Asn Ile Cys Lys Glu
3305 3310 3315
Cys Pro Ile Ile Gly Phe Arg Tyr Arg Ser Leu Lys His Phe Asn
3320 3325 3330
Tyr Asp Ile Cys Gln Ser Cys Phe Phe Ser Gly Arg Val Ala Lys
3335 3340 3345
Gly His Lys Met His Tyr Pro Met Val Glu Tyr Cys Thr Pro Thr
3350 3355 3360
Thr Ser Gly Glu Asp Val Arg Asp Phe Ala Lys Val Leu Lys Asn
3365 3370 3375
Lys Phe Arg Thr Lys Arg Tyr Phe Ala Lys His Pro Arg Met Gly
3380 3385 3390
Tyr Leu Pro Val Gln Thr Val Leu Glu Gly Asp Asn Met Glu Thr
3395 3400 3405
Pro Val Thr Leu Ile Asn Phe Trp Pro Val Asp Ser Ala Pro Ala
3410 3415 3420
Ser Ser Pro Gln Leu Ser His Asp Asp Thr His Ser Arg Ile Glu
3425 3430 3435
His Tyr Ala Ser Arg Leu Ala Glu Met Glu Asn Ser Asn Gly Ser
3440 3445 3450
Tyr Leu Asn Asp Ser Ile Ser Pro Asn Glu Ser Ile Asp Asp Glu
3455 3460 3465
His Leu Leu Ile Gln His Tyr Cys Gln Ser Leu Asn Gln Asp Ser
3470 3475 3480
Pro Leu Ser Gln Pro Arg Ser Pro Ala Gln Ile Leu Ile Ser Leu
3485 3490 3495
Glu Ser Glu Glu Arg Gly Glu Leu Glu Arg Ile Leu Ala Asp Leu
3500 3505 3510
Glu Glu Glu Asn Arg Asn Leu Gln Ala Glu Tyr Asp Arg Leu Lys
3515 3520 3525
Gln Gln His Glu His Lys Gly Leu Ser Pro Leu Pro Ser Pro Pro
3530 3535 3540
Glu Met Met Pro Thr Ser Pro Gln Ser Pro Arg Asp Ala Glu Leu
3545 3550 3555
Ile Ala Glu Ala Lys Leu Leu Arg Gln His Lys Gly Arg Leu Glu
3560 3565 3570
Ala Arg Met Gln Ile Leu Glu Asp His Asn Lys Gln Leu Glu Ser
3575 3580 3585
Gln Leu His Arg Leu Arg Gln Leu Leu Glu Gln Pro Gln Ala Glu
3590 3595 3600
Ala Lys Val Asn Gly Thr Thr Val Ser Ser Pro Ser Thr Ser Leu
3605 3610 3615
Gln Arg Ser Asp Ser Ser Gln Pro Met Leu Leu Arg Val Val Gly
3620 3625 3630
Ser Gln Thr Ser Asp Ser Met Gly Glu Glu Asp Leu Leu Ser Pro
3635 3640 3645
Pro Gln Asp Thr Ser Thr Gly Leu Glu Glu Val Met Glu Gln Leu
3650 3655 3660
Asn Asn Ser Phe Pro Ser Ser Arg Gly Arg Asn Thr Pro Gly Lys
3665 3670 3675
Pro Met Arg Glu Asp Thr Met
3680 3685
<210> 6
<211> 3678
<212> Белок
<213> Mus musculus
<400> 6
Met Leu Trp Trp Glu Glu Val Glu Asp Cys Tyr Glu Arg Glu Asp Val
1 5 10 15
Gln Lys Lys Thr Phe Thr Lys Trp Ile Asn Ala Gln Phe Ser Lys Phe
20 25 30
Gly Lys Gln His Ile Asp Asn Leu Phe Ser Asp Leu Gln Asp Gly Lys
35 40 45
Arg Leu Leu Asp Leu Leu Glu Gly Leu Thr Gly Gln Lys Leu Pro Lys
50 55 60
Glu Lys Gly Ser Thr Arg Val His Ala Leu Asn Asn Val Asn Lys Ala
65 70 75 80
Leu Arg Val Leu Gln Lys Asn Asn Val Asp Leu Val Asn Ile Gly Ser
85 90 95
Thr Asp Ile Val Asp Gly Asn His Lys Leu Thr Leu Gly Leu Ile Trp
100 105 110
Asn Ile Ile Leu His Trp Gln Val Lys Asn Val Met Lys Thr Ile Met
115 120 125
Ala Gly Leu Gln Gln Thr Asn Ser Glu Lys Ile Leu Leu Ser Trp Val
130 135 140
Arg Gln Ser Thr Arg Asn Tyr Pro Gln Val Asn Val Ile Asn Phe Thr
145 150 155 160
Ser Ser Trp Ser Asp Gly Leu Ala Leu Asn Ala Leu Ile His Ser His
165 170 175
Arg Pro Asp Leu Phe Asp Trp Asn Ser Val Val Ser Gln His Ser Ala
180 185 190
Thr Gln Arg Leu Glu His Ala Phe Asn Ile Ala Lys Cys Gln Leu Gly
195 200 205
Ile Glu Lys Leu Leu Asp Pro Glu Asp Val Ala Thr Thr Tyr Pro Asp
210 215 220
Lys Lys Ser Ile Leu Met Tyr Ile Thr Ser Leu Phe Gln Val Leu Pro
225 230 235 240
Gln Gln Val Ser Ile Glu Ala Ile Gln Glu Val Glu Met Leu Pro Arg
245 250 255
Thr Ser Ser Lys Val Thr Arg Glu Glu His Phe Gln Leu His His Gln
260 265 270
Met His Tyr Ser Gln Gln Ile Thr Val Ser Leu Ala Gln Gly Tyr Glu
275 280 285
Gln Thr Ser Ser Ser Pro Lys Pro Arg Phe Lys Ser Tyr Ala Phe Thr
290 295 300
Gln Ala Ala Tyr Val Ala Thr Ser Asp Ser Thr Gln Ser Pro Tyr Pro
305 310 315 320
Ser Gln His Leu Glu Ala Pro Arg Asp Lys Ser Leu Asp Ser Ser Leu
325 330 335
Met Glu Thr Glu Val Asn Leu Asp Ser Tyr Gln Thr Ala Leu Glu Glu
340 345 350
Val Leu Ser Trp Leu Leu Ser Ala Glu Asp Thr Leu Arg Ala Gln Gly
355 360 365
Glu Ile Ser Asn Asp Val Glu Glu Val Lys Glu Gln Phe His Ala His
370 375 380
Glu Gly Phe Met Met Asp Leu Thr Ser His Gln Gly Leu Val Gly Asn
385 390 395 400
Val Leu Gln Leu Gly Ser Gln Leu Val Gly Lys Gly Lys Leu Ser Glu
405 410 415
Asp Glu Glu Ala Glu Val Gln Glu Gln Met Asn Leu Leu Asn Ser Arg
420 425 430
Trp Glu Cys Leu Arg Val Ala Ser Met Glu Lys Gln Ser Lys Leu His
435 440 445
Lys Val Leu Met Asp Leu Gln Asn Gln Lys Leu Lys Glu Leu Asp Asp
450 455 460
Trp Leu Thr Lys Thr Glu Glu Arg Thr Lys Lys Met Glu Glu Glu Pro
465 470 475 480
Phe Gly Pro Asp Leu Glu Asp Leu Lys Cys Gln Val Gln Gln His Lys
485 490 495
Val Leu Gln Glu Asp Leu Glu Gln Glu Gln Val Arg Val Asn Ser Leu
500 505 510
Thr His Met Val Val Val Val Asp Glu Ser Ser Gly Asp His Ala Thr
515 520 525
Ala Ala Leu Glu Glu Gln Leu Lys Val Leu Gly Asp Arg Trp Ala Asn
530 535 540
Ile Cys Arg Trp Thr Glu Asp Arg Trp Ile Val Leu Gln Asp Ile Leu
545 550 555 560
Leu Lys Trp Gln His Phe Thr Glu Glu Gln Cys Leu Phe Ser Thr Trp
565 570 575
Leu Ser Glu Lys Glu Asp Ala Met Lys Asn Ile Gln Thr Ser Gly Phe
580 585 590
Lys Asp Gln Asn Glu Met Met Ser Ser Leu His Lys Ile Ser Thr Leu
595 600 605
Lys Ile Asp Leu Glu Lys Lys Lys Pro Thr Met Glu Lys Leu Ser Ser
610 615 620
Leu Asn Gln Asp Leu Leu Ser Ala Leu Lys Asn Lys Ser Val Thr Gln
625 630 635 640
Lys Met Glu Ile Trp Met Glu Asn Phe Ala Gln Arg Trp Asp Asn Leu
645 650 655
Thr Gln Lys Leu Glu Lys Ser Ser Ala Gln Ile Ser Gln Ala Val Thr
660 665 670
Thr Thr Gln Pro Ser Leu Thr Gln Thr Thr Val Met Glu Thr Val Thr
675 680 685
Met Val Thr Thr Arg Glu Gln Ile Met Val Lys His Ala Gln Glu Glu
690 695 700
Leu Pro Pro Pro Pro Pro Gln Lys Lys Arg Gln Ile Thr Val Asp Ser
705 710 715 720
Glu Leu Arg Lys Arg Leu Asp Val Asp Ile Thr Glu Leu His Ser Trp
725 730 735
Ile Thr Arg Ser Glu Ala Val Leu Gln Ser Ser Glu Phe Ala Val Tyr
740 745 750
Arg Lys Glu Gly Asn Ile Ser Asp Leu Gln Glu Lys Val Asn Ala Ile
755 760 765
Ala Arg Glu Lys Ala Glu Lys Phe Arg Lys Leu Gln Asp Ala Ser Arg
770 775 780
Ser Ala Gln Ala Leu Val Glu Gln Met Ala Asn Glu Gly Val Asn Ala
785 790 795 800
Glu Ser Ile Arg Gln Ala Ser Glu Gln Leu Asn Ser Arg Trp Thr Glu
805 810 815
Phe Cys Gln Leu Leu Ser Glu Arg Val Asn Trp Leu Glu Tyr Gln Thr
820 825 830
Asn Ile Ile Thr Phe Tyr Asn Gln Leu Gln Gln Leu Glu Gln Met Thr
835 840 845
Thr Thr Ala Glu Asn Leu Leu Lys Thr Gln Ser Thr Thr Leu Ser Glu
850 855 860
Pro Thr Ala Ile Lys Ser Gln Leu Lys Ile Cys Lys Asp Glu Val Asn
865 870 875 880
Arg Leu Ser Ala Leu Gln Pro Gln Ile Glu Gln Leu Lys Ile Gln Ser
885 890 895
Leu Gln Leu Lys Glu Lys Gly Gln Gly Pro Met Phe Leu Asp Ala Asp
900 905 910
Phe Val Ala Phe Thr Asn His Phe Asn His Ile Phe Asp Gly Val Arg
915 920 925
Ala Lys Glu Lys Glu Leu Gln Thr Ile Phe Asp Thr Leu Pro Pro Met
930 935 940
Arg Tyr Gln Glu Thr Met Ser Ser Ile Arg Thr Trp Ile Gln Gln Ser
945 950 955 960
Glu Ser Lys Leu Ser Val Pro Tyr Leu Ser Val Thr Glu Tyr Glu Ile
965 970 975
Met Glu Glu Arg Leu Gly Lys Leu Gln Ala Leu Gln Ser Ser Leu Lys
980 985 990
Glu Gln Gln Asn Gly Phe Asn Tyr Leu Ser Asp Thr Val Lys Glu Met
995 1000 1005
Ala Lys Lys Ala Pro Ser Glu Ile Cys Gln Lys Tyr Leu Ser Glu
1010 1015 1020
Phe Glu Glu Ile Glu Gly His Trp Lys Lys Leu Ser Ser Gln Leu
1025 1030 1035
Val Glu Ser Cys Gln Lys Leu Glu Glu His Met Asn Lys Leu Arg
1040 1045 1050
Lys Phe Gln Asn His Ile Lys Thr Leu Gln Lys Trp Met Ala Glu
1055 1060 1065
Val Asp Val Phe Leu Lys Glu Glu Trp Pro Ala Leu Gly Asp Ala
1070 1075 1080
Glu Ile Leu Lys Lys Gln Leu Lys Gln Cys Arg Leu Leu Val Gly
1085 1090 1095
Asp Ile Gln Thr Ile Gln Pro Ser Leu Asn Ser Val Asn Glu Gly
1100 1105 1110
Gly Gln Lys Ile Lys Ser Glu Ala Glu Leu Glu Phe Ala Ser Arg
1115 1120 1125
Leu Glu Thr Glu Leu Arg Glu Leu Asn Thr Gln Trp Asp His Ile
1130 1135 1140
Cys Arg Gln Val Tyr Thr Arg Lys Glu Ala Leu Lys Ala Gly Leu
1145 1150 1155
Asp Lys Thr Val Ser Leu Gln Lys Asp Leu Ser Glu Met His Glu
1160 1165 1170
Trp Met Thr Gln Ala Glu Glu Glu Tyr Leu Glu Arg Asp Phe Glu
1175 1180 1185
Tyr Lys Thr Pro Asp Glu Leu Gln Thr Ala Val Glu Glu Met Lys
1190 1195 1200
Arg Ala Lys Glu Glu Ala Leu Gln Lys Glu Thr Lys Val Lys Leu
1205 1210 1215
Leu Thr Glu Thr Val Asn Ser Val Ile Ala His Ala Pro Pro Ser
1220 1225 1230
Ala Gln Glu Ala Leu Lys Lys Glu Leu Glu Thr Leu Thr Thr Asn
1235 1240 1245
Tyr Gln Trp Leu Cys Thr Arg Leu Asn Gly Lys Cys Lys Thr Leu
1250 1255 1260
Glu Glu Val Trp Ala Cys Trp His Glu Leu Leu Ser Tyr Leu Glu
1265 1270 1275
Lys Ala Asn Lys Trp Leu Asn Glu Val Glu Leu Lys Leu Lys Thr
1280 1285 1290
Met Glu Asn Val Pro Ala Gly Pro Glu Glu Ile Thr Glu Val Leu
1295 1300 1305
Glu Ser Leu Glu Asn Leu Met His His Ser Glu Glu Asn Pro Asn
1310 1315 1320
Gln Ile Arg Leu Leu Ala Gln Thr Leu Thr Asp Gly Gly Val Met
1325 1330 1335
Asp Glu Leu Ile Asn Glu Glu Leu Glu Thr Phe Asn Ser Arg Trp
1340 1345 1350
Arg Glu Leu His Glu Glu Ala Val Arg Lys Gln Lys Leu Leu Glu
1355 1360 1365
Gln Ser Ile Gln Ser Ala Gln Glu Ile Glu Lys Ser Leu His Leu
1370 1375 1380
Ile Gln Glu Ser Leu Glu Phe Ile Asp Lys Gln Leu Ala Ala Tyr
1385 1390 1395
Ile Thr Asp Lys Val Asp Ala Ala Gln Met Pro Gln Glu Ala Gln
1400 1405 1410
Lys Ile Gln Ser Asp Leu Thr Ser His Glu Ile Ser Leu Glu Glu
1415 1420 1425
Met Lys Lys His Asn Gln Gly Lys Asp Ala Asn Gln Arg Val Leu
1430 1435 1440
Ser Gln Ile Asp Val Ala Gln Lys Lys Leu Gln Asp Val Ser Met
1445 1450 1455
Lys Phe Arg Leu Phe Gln Lys Pro Ala Asn Phe Glu Gln Arg Leu
1460 1465 1470
Glu Glu Ser Lys Met Ile Leu Asp Glu Val Lys Met His Leu Pro
1475 1480 1485
Ala Leu Glu Thr Lys Ser Val Glu Gln Glu Val Ile Gln Ser Gln
1490 1495 1500
Leu Ser His Cys Val Asn Leu Tyr Lys Ser Leu Ser Glu Val Lys
1505 1510 1515
Ser Glu Val Glu Met Val Ile Lys Thr Gly Arg Gln Ile Val Gln
1520 1525 1530
Lys Lys Gln Thr Glu Asn Pro Lys Glu Leu Asp Glu Arg Val Thr
1535 1540 1545
Ala Leu Lys Leu His Tyr Asn Glu Leu Gly Ala Lys Val Thr Glu
1550 1555 1560
Arg Lys Gln Gln Leu Glu Lys Cys Leu Lys Leu Ser Arg Lys Met
1565 1570 1575
Arg Lys Glu Met Asn Val Leu Thr Glu Trp Leu Ala Ala Thr Asp
1580 1585 1590
Thr Glu Leu Thr Lys Arg Ser Ala Val Glu Gly Met Pro Ser Asn
1595 1600 1605
Leu Asp Ser Glu Val Ala Trp Gly Lys Ala Thr Gln Lys Glu Ile
1610 1615 1620
Glu Lys Gln Lys Ala His Leu Lys Ser Val Thr Glu Leu Gly Glu
1625 1630 1635
Ser Leu Lys Met Val Leu Gly Lys Lys Glu Thr Leu Val Glu Asp
1640 1645 1650
Lys Leu Ser Leu Leu Asn Ser Asn Trp Ile Ala Val Thr Ser Arg
1655 1660 1665
Val Glu Glu Trp Leu Asn Leu Leu Leu Glu Tyr Gln Lys His Met
1670 1675 1680
Glu Thr Phe Asp Gln Asn Ile Glu Gln Ile Thr Lys Trp Ile Ile
1685 1690 1695
His Ala Asp Glu Leu Leu Asp Glu Ser Glu Lys Lys Lys Pro Gln
1700 1705 1710
Gln Lys Glu Asp Ile Leu Lys Arg Leu Lys Ala Glu Met Asn Asp
1715 1720 1725
Met Arg Pro Lys Val Asp Ser Thr Arg Asp Gln Ala Ala Lys Leu
1730 1735 1740
Met Ala Asn Arg Gly Asp His Cys Arg Lys Val Val Glu Pro Gln
1745 1750 1755
Ile Ser Glu Leu Asn Arg Arg Phe Ala Ala Ile Ser His Arg Ile
1760 1765 1770
Lys Thr Gly Lys Ala Ser Ile Pro Leu Lys Glu Leu Glu Gln Phe
1775 1780 1785
Asn Ser Asp Ile Gln Lys Leu Leu Glu Pro Leu Glu Ala Glu Ile
1790 1795 1800
Gln Gln Gly Val Asn Leu Lys Glu Glu Asp Phe Asn Lys Asp Met
1805 1810 1815
Ser Glu Asp Asn Glu Gly Thr Val Asn Glu Leu Leu Gln Arg Gly
1820 1825 1830
Asp Asn Leu Gln Gln Arg Ile Thr Asp Glu Arg Lys Arg Glu Glu
1835 1840 1845
Ile Lys Ile Lys Gln Gln Leu Leu Gln Thr Lys His Asn Ala Leu
1850 1855 1860
Lys Asp Leu Arg Ser Gln Arg Arg Lys Lys Ala Leu Glu Ile Ser
1865 1870 1875
His Gln Trp Tyr Gln Tyr Lys Arg Gln Ala Asp Asp Leu Leu Lys
1880 1885 1890
Cys Leu Asp Glu Ile Glu Lys Lys Leu Ala Ser Leu Pro Glu Pro
1895 1900 1905
Arg Asp Glu Arg Lys Leu Lys Glu Ile Asp Arg Glu Leu Gln Lys
1910 1915 1920
Lys Lys Glu Glu Leu Asn Ala Val Arg Arg Gln Ala Glu Gly Leu
1925 1930 1935
Ser Glu Asn Gly Ala Ala Met Ala Val Glu Pro Thr Gln Ile Gln
1940 1945 1950
Leu Ser Lys Arg Trp Arg Gln Ile Glu Ser Asn Phe Ala Gln Phe
1955 1960 1965
Arg Arg Leu Asn Phe Ala Gln Ile His Thr Leu His Glu Glu Thr
1970 1975 1980
Met Val Val Thr Thr Glu Asp Met Pro Leu Asp Val Ser Tyr Val
1985 1990 1995
Pro Ser Thr Tyr Leu Thr Glu Ile Ser His Ile Leu Gln Ala Leu
2000 2005 2010
Ser Glu Val Asp His Leu Leu Asn Thr Pro Glu Leu Cys Ala Lys
2015 2020 2025
Asp Phe Glu Asp Leu Phe Lys Gln Glu Glu Ser Leu Lys Asn Ile
2030 2035 2040
Lys Asp Asn Leu Gln Gln Ile Ser Gly Arg Ile Asp Ile Ile His
2045 2050 2055
Lys Lys Lys Thr Ala Ala Leu Gln Ser Ala Thr Ser Met Glu Lys
2060 2065 2070
Val Lys Val Gln Glu Ala Val Ala Gln Met Asp Phe Gln Gly Glu
2075 2080 2085
Lys Leu His Arg Met Tyr Lys Glu Arg Gln Gly Arg Phe Asp Arg
2090 2095 2100
Ser Val Glu Lys Trp Arg His Phe His Tyr Asp Met Lys Val Phe
2105 2110 2115
Asn Gln Trp Leu Asn Glu Val Glu Gln Phe Phe Lys Lys Thr Gln
2120 2125 2130
Asn Pro Glu Asn Trp Glu His Ala Lys Tyr Lys Trp Tyr Leu Lys
2135 2140 2145
Glu Leu Gln Asp Gly Ile Gly Gln Arg Gln Ala Val Val Arg Thr
2150 2155 2160
Leu Asn Ala Thr Gly Glu Glu Ile Ile Gln Gln Ser Ser Lys Thr
2165 2170 2175
Asp Val Asn Ile Leu Gln Glu Lys Leu Gly Ser Leu Ser Leu Arg
2180 2185 2190
Trp His Asp Ile Cys Lys Glu Leu Ala Glu Arg Arg Lys Arg Ile
2195 2200 2205
Glu Glu Gln Lys Asn Val Leu Ser Glu Phe Gln Arg Asp Leu Asn
2210 2215 2220
Glu Phe Val Leu Trp Leu Glu Glu Ala Asp Asn Ile Ala Ile Thr
2225 2230 2235
Pro Leu Gly Asp Glu Gln Gln Leu Lys Glu Gln Leu Glu Gln Val
2240 2245 2250
Lys Leu Leu Ala Glu Glu Leu Pro Leu Arg Gln Gly Ile Leu Lys
2255 2260 2265
Gln Leu Asn Glu Thr Gly Gly Ala Val Leu Val Ser Ala Pro Ile
2270 2275 2280
Arg Pro Glu Glu Gln Asp Lys Leu Glu Lys Lys Leu Lys Gln Thr
2285 2290 2295
Asn Leu Gln Trp Ile Lys Val Ser Arg Ala Leu Pro Glu Lys Gln
2300 2305 2310
Gly Glu Leu Glu Val His Leu Lys Asp Phe Arg Gln Leu Glu Glu
2315 2320 2325
Gln Leu Asp His Leu Leu Leu Trp Leu Ser Pro Ile Arg Asn Gln
2330 2335 2340
Leu Glu Ile Tyr Asn Gln Pro Ser Gln Ala Gly Pro Phe Asp Ile
2345 2350 2355
Lys Glu Ile Glu Val Thr Val His Gly Lys Gln Ala Asp Val Glu
2360 2365 2370
Arg Leu Leu Ser Lys Gly Gln His Leu Tyr Lys Glu Lys Pro Ser
2375 2380 2385
Thr Gln Pro Val Lys Arg Lys Leu Glu Asp Leu Arg Ser Glu Trp
2390 2395 2400
Glu Ala Val Asn His Leu Leu Arg Glu Leu Arg Thr Lys Gln Pro
2405 2410 2415
Asp Arg Ala Pro Gly Leu Ser Thr Thr Gly Ala Ser Ala Ser Gln
2420 2425 2430
Thr Val Thr Leu Val Thr Gln Ser Val Val Thr Lys Glu Thr Val
2435 2440 2445
Ile Ser Lys Leu Glu Met Pro Ser Ser Leu Leu Leu Glu Val Pro
2450 2455 2460
Ala Leu Ala Asp Phe Asn Arg Ala Trp Thr Glu Leu Thr Asp Trp
2465 2470 2475
Leu Ser Leu Leu Asp Arg Val Ile Lys Ser Gln Arg Val Met Val
2480 2485 2490
Gly Asp Leu Glu Asp Ile Asn Glu Met Ile Ile Lys Gln Lys Ala
2495 2500 2505
Thr Leu Gln Asp Leu Glu Gln Arg Arg Pro Gln Leu Glu Glu Leu
2510 2515 2520
Ile Thr Ala Ala Gln Asn Leu Lys Asn Lys Thr Ser Asn Gln Glu
2525 2530 2535
Ala Arg Thr Ile Ile Thr Asp Arg Ile Glu Arg Ile Gln Ile Gln
2540 2545 2550
Trp Asp Glu Val Gln Glu Gln Leu Gln Asn Arg Arg Gln Gln Leu
2555 2560 2565
Asn Glu Met Leu Lys Asp Ser Thr Gln Trp Leu Glu Ala Lys Glu
2570 2575 2580
Glu Ala Glu Gln Val Ile Gly Gln Val Arg Gly Lys Leu Asp Ser
2585 2590 2595
Trp Lys Glu Gly Pro His Thr Val Asp Ala Ile Gln Lys Lys Ile
2600 2605 2610
Thr Glu Thr Lys Gln Leu Ala Lys Asp Leu Arg Gln Arg Gln Ile
2615 2620 2625
Ser Val Asp Val Ala Asn Asp Leu Ala Leu Lys Leu Leu Arg Asp
2630 2635 2640
Tyr Ser Ala Asp Asp Thr Arg Lys Val His Met Ile Thr Glu Asn
2645 2650 2655
Ile Asn Thr Ser Trp Gly Asn Ile His Lys Arg Val Ser Glu Gln
2660 2665 2670
Glu Ala Ala Leu Glu Glu Thr His Arg Leu Leu Gln Gln Phe Pro
2675 2680 2685
Leu Asp Leu Glu Lys Phe Leu Ser Trp Ile Thr Glu Ala Glu Thr
2690 2695 2700
Thr Ala Asn Val Leu Gln Asp Ala Ser Arg Lys Glu Lys Leu Leu
2705 2710 2715
Glu Asp Ser Arg Gly Val Arg Glu Leu Met Lys Pro Trp Gln Asp
2720 2725 2730
Leu Gln Gly Glu Ile Glu Thr His Thr Asp Ile Tyr His Asn Leu
2735 2740 2745
Asp Glu Asn Gly Gln Lys Ile Leu Arg Ser Leu Glu Gly Ser Asp
2750 2755 2760
Glu Ala Pro Leu Leu Gln Arg Arg Leu Asp Asn Met Asn Phe Lys
2765 2770 2775
Trp Ser Glu Leu Gln Lys Lys Ser Leu Asn Ile Arg Ser His Leu
2780 2785 2790
Glu Ala Ser Ser Asp Gln Trp Lys Arg Leu His Leu Ser Leu Gln
2795 2800 2805
Glu Leu Leu Val Trp Leu Gln Leu Lys Asp Asp Glu Leu Ser Arg
2810 2815 2820
Gln Ala Pro Ile Gly Gly Asp Phe Pro Ala Val Gln Lys Gln Asn
2825 2830 2835
Asp Ile His Arg Ala Phe Lys Arg Glu Leu Lys Thr Lys Glu Pro
2840 2845 2850
Val Ile Met Ser Thr Leu Glu Thr Val Arg Ile Phe Leu Thr Glu
2855 2860 2865
Gln Pro Leu Glu Gly Leu Glu Lys Leu Tyr Gln Glu Pro Arg Glu
2870 2875 2880
Leu Pro Pro Glu Glu Arg Ala Gln Asn Val Thr Arg Leu Leu Arg
2885 2890 2895
Lys Gln Ala Glu Glu Val Asn Ala Glu Trp Asp Lys Leu Asn Leu
2900 2905 2910
Arg Ser Ala Asp Trp Gln Arg Lys Ile Asp Glu Ala Leu Glu Arg
2915 2920 2925
Leu Gln Glu Leu Gln Glu Ala Ala Asp Glu Leu Asp Leu Lys Leu
2930 2935 2940
Arg Gln Ala Glu Val Ile Lys Gly Ser Trp Gln Pro Val Gly Asp
2945 2950 2955
Leu Leu Ile Asp Ser Leu Gln Asp His Leu Glu Lys Val Lys Ala
2960 2965 2970
Leu Arg Gly Glu Ile Ala Pro Leu Lys Glu Asn Val Asn Arg Val
2975 2980 2985
Asn Asp Leu Ala His Gln Leu Thr Thr Leu Gly Ile Gln Leu Ser
2990 2995 3000
Pro Tyr Asn Leu Ser Thr Leu Glu Asp Leu Asn Thr Arg Trp Arg
3005 3010 3015
Leu Leu Gln Val Ala Val Glu Asp Arg Val Arg Gln Leu His Glu
3020 3025 3030
Ala His Arg Asp Phe Gly Pro Ala Ser Gln His Phe Leu Ser Thr
3035 3040 3045
Ser Val Gln Gly Pro Trp Glu Arg Ala Ile Ser Pro Asn Lys Val
3050 3055 3060
Pro Tyr Tyr Ile Asn His Glu Thr Gln Thr Thr Cys Trp Asp His
3065 3070 3075
Pro Lys Met Thr Glu Leu Tyr Gln Ser Leu Ala Asp Leu Asn Asn
3080 3085 3090
Val Arg Phe Ser Ala Tyr Arg Thr Ala Met Lys Leu Arg Arg Leu
3095 3100 3105
Gln Lys Ala Leu Cys Leu Asp Leu Leu Ser Leu Ser Ala Ala Cys
3110 3115 3120
Asp Ala Leu Asp Gln His Asn Leu Lys Gln Asn Asp Gln Pro Met
3125 3130 3135
Asp Ile Leu Gln Ile Ile Asn Cys Leu Thr Thr Ile Tyr Asp Arg
3140 3145 3150
Leu Glu Gln Glu His Asn Asn Leu Val Asn Val Pro Leu Cys Val
3155 3160 3165
Asp Met Cys Leu Asn Trp Leu Leu Asn Val Tyr Asp Thr Gly Arg
3170 3175 3180
Thr Gly Arg Ile Arg Val Leu Ser Phe Lys Thr Gly Ile Ile Ser
3185 3190 3195
Leu Cys Lys Ala His Leu Glu Asp Lys Tyr Arg Tyr Leu Phe Lys
3200 3205 3210
Gln Val Ala Ser Ser Thr Gly Phe Cys Asp Gln Arg Arg Leu Gly
3215 3220 3225
Leu Leu Leu His Asp Ser Ile Gln Ile Pro Arg Gln Leu Gly Glu
3230 3235 3240
Val Ala Ser Phe Gly Gly Ser Asn Ile Glu Pro Ser Val Arg Ser
3245 3250 3255
Cys Phe Gln Phe Ala Asn Asn Lys Pro Glu Ile Glu Ala Ala Leu
3260 3265 3270
Phe Leu Asp Trp Met Arg Leu Glu Pro Gln Ser Met Val Trp Leu
3275 3280 3285
Pro Val Leu His Arg Val Ala Ala Ala Glu Thr Ala Lys His Gln
3290 3295 3300
Ala Lys Cys Asn Ile Cys Lys Glu Cys Pro Ile Ile Gly Phe Arg
3305 3310 3315
Tyr Arg Ser Leu Lys His Phe Asn Tyr Asp Ile Cys Gln Ser Cys
3320 3325 3330
Phe Phe Ser Gly Arg Val Ala Lys Gly His Lys Met His Tyr Pro
3335 3340 3345
Met Val Glu Tyr Cys Thr Pro Thr Thr Ser Gly Glu Asp Val Arg
3350 3355 3360
Asp Phe Ala Lys Val Leu Lys Asn Lys Phe Arg Thr Lys Arg Tyr
3365 3370 3375
Phe Ala Lys His Pro Arg Met Gly Tyr Leu Pro Val Gln Thr Val
3380 3385 3390
Leu Glu Gly Asp Asn Met Glu Thr Pro Val Thr Leu Ile Asn Phe
3395 3400 3405
Trp Pro Val Asp Ser Ala Pro Ala Ser Ser Pro Gln Leu Ser His
3410 3415 3420
Asp Asp Thr His Ser Arg Ile Glu His Tyr Ala Ser Arg Leu Ala
3425 3430 3435
Glu Met Glu Asn Ser Asn Gly Ser Tyr Leu Asn Asp Ser Ile Ser
3440 3445 3450
Pro Asn Glu Ser Ile Asp Asp Glu His Leu Leu Ile Gln His Tyr
3455 3460 3465
Cys Gln Ser Leu Asn Gln Asp Ser Pro Leu Ser Gln Pro Arg Ser
3470 3475 3480
Pro Ala Gln Ile Leu Ile Ser Leu Glu Ser Glu Glu Arg Gly Glu
3485 3490 3495
Leu Glu Arg Ile Leu Ala Asp Leu Glu Glu Glu Asn Arg Asn Leu
3500 3505 3510
Gln Ala Glu Tyr Asp Arg Leu Lys Gln Gln His Glu His Lys Gly
3515 3520 3525
Leu Ser Pro Leu Pro Ser Pro Pro Glu Met Met Pro Thr Ser Pro
3530 3535 3540
Gln Ser Pro Arg Asp Ala Glu Leu Ile Ala Glu Ala Lys Leu Leu
3545 3550 3555
Arg Gln His Lys Gly Arg Leu Glu Ala Arg Met Gln Ile Leu Glu
3560 3565 3570
Asp His Asn Lys Gln Leu Glu Ser Gln Leu His Arg Leu Arg Gln
3575 3580 3585
Leu Leu Glu Gln Pro Gln Ala Glu Ala Lys Val Asn Gly Thr Thr
3590 3595 3600
Val Ser Ser Pro Ser Thr Ser Leu Gln Arg Ser Asp Ser Ser Gln
3605 3610 3615
Pro Met Leu Leu Arg Val Val Gly Ser Gln Thr Ser Glu Ser Met
3620 3625 3630
Gly Glu Glu Asp Leu Leu Ser Pro Pro Gln Asp Thr Ser Thr Gly
3635 3640 3645
Leu Glu Glu Val Met Glu Gln Leu Asn Asn Ser Phe Pro Ser Ser
3650 3655 3660
Arg Gly Arg Asn Ala Pro Gly Lys Pro Met Arg Glu Asp Thr Met
3665 3670 3675
<210> 7
<211> 3699
<212> Белок
<213> Rattus norvegicus
<400> 7
Met Glu Asp Glu Arg Glu Asp Val Gln Lys Lys Thr Phe Thr Lys Trp
1 5 10 15
Ile Asn Ala Gln Phe Ser Lys Phe Gly Lys Gln His Ile Asp Asn Leu
20 25 30
Phe Ser Asp Leu Gln Asp Gly Lys Arg Leu Leu Asp Leu Leu Glu Gly
35 40 45
Leu Thr Gly Gln Lys Leu Pro Lys Glu Lys Gly Ser Thr Arg Val His
50 55 60
Ala Leu Asn Asn Val Asn Lys Ala Leu Gln Val Leu Gln Lys Asn Asn
65 70 75 80
Val Asp Leu Val Asn Ile Gly Ser Thr Asp Ile Val Asp Gly Asn His
85 90 95
Lys Leu Thr Leu Gly Leu Ile Trp Asn Ile Ile Leu His Trp Gln Val
100 105 110
Lys Asn Val Met Lys Thr Ile Met Ala Gly Leu Gln Gln Thr Asn Ser
115 120 125
Glu Lys Ile Leu Leu Ser Trp Val Arg Glu Ser Thr Arg Asn Tyr Pro
130 135 140
Gln Val Asn Val Leu Asn Phe Thr Ser Ser Trp Ser Asp Gly Leu Ala
145 150 155 160
Leu Asn Ala Leu Ile His Ser His Arg Pro Asp Leu Phe Asp Trp Asn
165 170 175
Ser Val Val Ser Gln Gln Ser Ala Thr Gln Arg Leu Glu His Ala Phe
180 185 190
Asn Ile Ala Lys Cys Gln Leu Gly Ile Glu Lys Leu Leu Asp Pro Glu
195 200 205
Asp Val Ala Thr Thr Tyr Pro Asp Lys Lys Ser Ile Leu Met Tyr Ile
210 215 220
Thr Ser Leu Phe Gln Val Leu Pro Gln Gln Val Ser Ile Glu Ala Ile
225 230 235 240
Arg Glu Val Glu Met Leu Pro Arg Pro Ser Lys Val Thr Arg Glu Glu
245 250 255
His Phe Gln Leu His His Gln Met His Tyr Ser Gln Gln Ile Thr Val
260 265 270
Ser Leu Ala Gln Gly Tyr Glu Gln Thr Ser Ser Ser Pro Lys Pro Arg
275 280 285
Phe Lys Ser Tyr Ala Phe Thr Gln Ala Ala Tyr Val Ala Thr Ser Asp
290 295 300
Ser Ser Gln Ser Pro Tyr Pro Ser Gln His Leu Glu Ala Pro Gly Ser
305 310 315 320
Lys Ser Phe Gly Ser Ser Leu Ile Glu Thr Glu Val Asn Leu Asp Ser
325 330 335
Tyr Gln Thr Ala Leu Glu Glu Val Leu Ser Trp Leu Leu Ser Ala Glu
340 345 350
Asp Thr Leu Arg Ala Gln Gly Glu Ile Ser Lys Asp Val Glu Glu Val
355 360 365
Lys Glu Gln Phe His Ala His Glu Gly Phe Met Met Asp Leu Thr Ser
370 375 380
His Gln Gly Leu Val Gly Asn Val Leu Gln Leu Gly Ser Arg Leu Val
385 390 395 400
Gly Lys Gly Lys Leu Thr Glu Asp Glu Glu Thr Glu Val Gln Glu Gln
405 410 415
Met Asn Leu Leu Asn Ser Arg Trp Glu Cys Leu Arg Val Ala Ser Met
420 425 430
Glu Lys Gln Ser Asn Leu His Lys Val Leu Met Asp Leu Gln Asn Gln
435 440 445
Lys Leu Lys Glu Leu Asp Asp Trp Leu Thr Lys Thr Glu Glu Arg Thr
450 455 460
Lys Lys Met Glu Glu Glu Pro Leu Gly Pro Asp Leu Glu Asp Leu Lys
465 470 475 480
Cys Gln Val Gln Gln His Lys Val Leu Gln Glu Asp Leu Glu Gln Glu
485 490 495
Gln Val Arg Val Asn Ser Leu Thr His Met Val Val Val Val Asp Glu
500 505 510
Ser Ser Gly Asp His Ala Thr Ala Ala Leu Glu Glu Gln Leu Lys Val
515 520 525
Leu Gly Asp Arg Trp Ala Asn Ile Cys Lys Trp Thr Glu Asp Arg Trp
530 535 540
Ile Leu Leu Gln Asp Ile Leu Leu Lys Trp Gln Arg Phe Thr Glu Glu
545 550 555 560
Gln Cys Leu Phe Ser Lys Trp Leu Ser Glu Lys Glu Asp Ala Met Lys
565 570 575
Asn Ile Gln Thr Ser Gly Phe Glu Asp Gln Asn Glu Met Val Ser Ser
580 585 590
Leu Gln Asn Ile Ser Ala Leu Lys Ile Asp Leu Glu Lys Lys Lys Gln
595 600 605
Ser Met Glu Lys Leu Ser Ser Leu Asn Gln Asp Leu Leu Ser Ala Leu
610 615 620
Lys Asn Lys Ser Val Thr Gln Lys Met Glu Met Trp Met Glu Asn Phe
625 630 635 640
Ala Gln Arg Trp Asp Asn Leu Thr Gln Lys Leu Glu Lys Ser Ser Ala
645 650 655
Gln Ile Ser Gln Ala Val Thr Thr Thr Gln Pro Ser Leu Thr Gln Thr
660 665 670
Thr Val Met Glu Thr Val Thr Met Val Thr Thr Arg Glu Gln Ile Met
675 680 685
Val Lys His Ala Gln Glu Glu Leu Pro Pro Pro Pro Pro Gln Lys Lys
690 695 700
Arg Gln Ile Thr Val Asp Ser Glu Ile Arg Lys Arg Leu Asp Val Asp
705 710 715 720
Ile Thr Glu Leu His Ser Trp Ile Thr Arg Ser Glu Ala Val Leu Gln
725 730 735
Ser Ser Glu Phe Ala Val Tyr Arg Lys Glu Gly Asn Ile Ser Asp Leu
740 745 750
Lys Glu Lys Val Asn Ala Ile Ala Arg Glu Lys Ala Glu Lys Phe Arg
755 760 765
Lys Leu Gln Asp Ala Ser Arg Ser Ala Gln Ala Leu Val Glu Gln Met
770 775 780
Val Asn Glu Gly Val Asn Ala Glu Ser Ile Arg Gln Ala Ser Glu Gln
785 790 795 800
Leu Asn Ser Arg Trp Thr Glu Phe Cys Gln Leu Leu Ser Glu Arg Val
805 810 815
Asn Trp Leu Glu Tyr Gln Asn Asn Ile Ile Thr Phe Tyr Asn Gln Leu
820 825 830
Gln Gln Leu Glu Gln Met Thr Thr Thr Ala Glu Asn Leu Leu Lys Thr
835 840 845
Gln Pro Thr Thr Leu Ser Glu Pro Thr Ala Ile Lys Ser Gln Leu Lys
850 855 860
Ile Cys Lys Asp Glu Val Asn Arg Leu Ser Ala Leu Gln Pro Gln Ile
865 870 875 880
Glu Arg Leu Lys Ile Gln Ser Leu Thr Leu Lys Glu Lys Gly Gln Gly
885 890 895
Pro Met Phe Leu Asp Ala Asp Phe Val Ala Phe Thr Asn His Phe Asn
900 905 910
Tyr Val Phe Asp Gly Val Arg Ala Arg Glu Lys Glu Leu Gln Thr Ile
915 920 925
Phe Asp Thr Leu Pro Pro Met Arg Tyr Gln Glu Thr Met Ser Ser Ile
930 935 940
Arg Thr Trp Ile Gln Gln Ser Glu Asn Lys Leu Ser Ile Pro His Leu
945 950 955 960
Ser Val Thr Glu Tyr Glu Ile Met Glu Glu Arg Leu Gly Lys Leu Gln
965 970 975
Ala Leu Gln Ser Ser Leu Lys Glu Gln Gln Asn Gly Phe Asn Tyr Leu
980 985 990
Asn Ala Thr Val Lys Glu Ile Ala Lys Lys Ala Pro Ser Glu Ile Ser
995 1000 1005
Gln Lys Tyr Gln Ser Glu Phe Glu Glu Val Glu Gly Arg Trp Lys
1010 1015 1020
Lys Leu Ser Thr Gln Leu Val Glu His Cys Gln Lys Leu Glu Glu
1025 1030 1035
His Met Asn Lys Leu Arg Lys Phe Gln Asn His Lys Lys Thr Leu
1040 1045 1050
Gln Lys Trp Met Ala Glu Val Asp Val Phe Leu Lys Glu Glu Trp
1055 1060 1065
Pro Ala Leu Gly Asp Ala Glu Ile Leu Lys Lys Gln Leu Lys Gln
1070 1075 1080
Cys Arg Leu Leu Val Gly Asp Ile Gln Thr Ile Gln Pro Ser Leu
1085 1090 1095
Asn Ser Val Asn Glu Gly Gly Gln Lys Ile Lys Ser Glu Ala Glu
1100 1105 1110
Phe Glu Phe Ala Ser Arg Leu Glu Lys Glu Leu Lys Glu Leu Asn
1115 1120 1125
Thr Gln Trp Asp His Ile Cys Arg Gln Val Tyr Thr Arg Lys Glu
1130 1135 1140
Ala Leu Lys Ala Gly Leu Asp Lys Thr Val Ser Leu Gln Lys Asp
1145 1150 1155
Leu Ser Glu Met His Glu Trp Met Thr Gln Ala Glu Glu Glu Tyr
1160 1165 1170
Leu Glu Arg Asp Phe Glu Tyr Lys Thr Pro Asp Glu Leu Gln Thr
1175 1180 1185
Ala Val Glu Glu Met Lys Arg Ala Lys Glu Glu Ala Leu Gln Lys
1190 1195 1200
Glu Ala Lys Val Lys Leu Leu Thr Glu Thr Val Asn Ser Val Ile
1205 1210 1215
Ser Gln Ala Pro Pro Ala Ala Gln Glu Ala Leu Lys Lys Glu Leu
1220 1225 1230
Glu Thr Leu Thr Thr Asn Tyr Gln Trp Leu Cys Thr Arg Leu Asn
1235 1240 1245
Gly Lys Cys Lys Thr Leu Glu Glu Val Trp Ala Cys Trp His Glu
1250 1255 1260
Leu Leu Ser Tyr Leu Glu Lys Ala Asn Lys Trp Leu Asn Glu Val
1265 1270 1275
Glu Leu Lys Leu Lys Ala Thr Glu Asn Val Pro Ala Gly Ala Glu
1280 1285 1290
Glu Ile Thr Glu Val Leu Glu Ser Leu Glu Asn Leu Met His His
1295 1300 1305
Ser Glu Glu Asn Pro Asn Gln Ile Arg Leu Leu Ala Gln Thr Leu
1310 1315 1320
Thr Asp Gly Gly Val Met Asp Glu Leu Ile Asn Glu Glu Leu Glu
1325 1330 1335
Thr Phe Asn Ser Arg Trp Arg Glu Leu His Glu Glu Ala Val Arg
1340 1345 1350
Lys Gln Lys Leu Leu Glu Gln Ser Ile Gln Ser Ala Gln Glu Ile
1355 1360 1365
Glu Lys Ser Leu His Leu Ile Gln Glu Ser Leu Glu Phe Ile Asp
1370 1375 1380
Lys Gln Leu Ala Ala Tyr Ile Ala Asp Lys Val Asp Ala Ala Gln
1385 1390 1395
Met Pro Gln Glu Ala Gln Lys Ile Gln Ser Asp Leu Thr Ser His
1400 1405 1410
Glu Ile Ser Leu Glu Glu Met Lys Lys His Asn Gln Gly Lys Asp
1415 1420 1425
Ala Asn Gln Arg Val Leu Ser Gln Ile Asp Val Ala Gln Lys Lys
1430 1435 1440
Leu Gln Asp Val Ser Ile Lys Phe Arg Leu Phe Gln Lys Pro Ala
1445 1450 1455
Asn Phe Glu Gln Arg Leu Glu Glu Ser Lys Met Ile Leu Asp Glu
1460 1465 1470
Val Lys Met His Leu Pro Ala Leu Glu Thr Lys Ser Val Glu Gln
1475 1480 1485
Glu Val Val Gln Ser Gln Leu Ser His Cys Val Asn Leu Tyr Lys
1490 1495 1500
Ser Leu Ser Glu Val Lys Ser Glu Val Glu Met Val Ile Lys Thr
1505 1510 1515
Gly Arg Gln Ile Val Gln Lys Lys Gln Thr Glu Asn Pro Lys Glu
1520 1525 1530
Leu Asp Glu Arg Val Thr Ala Leu Lys Leu His Tyr Asn Glu Leu
1535 1540 1545
Gly Ala Lys Val Thr Glu Arg Lys Gln Gln Leu Glu Lys Cys Leu
1550 1555 1560
Lys Leu Ser Arg Lys Met Arg Lys Glu Met Asn Val Leu Thr Glu
1565 1570 1575
Trp Leu Ala Ala Thr Asp Thr Glu Leu Thr Lys Arg Ser Ala Val
1580 1585 1590
Glu Gly Met Pro Ser Asn Leu Asp Ser Glu Val Ala Trp Gly Lys
1595 1600 1605
Ala Thr Gln Lys Glu Ile Glu Lys Gln Lys Ala His Leu Lys Ser
1610 1615 1620
Val Thr Glu Leu Gly Asp Ser Leu Lys Thr Val Leu Gly Lys Lys
1625 1630 1635
Glu Thr Leu Val Glu Asp Lys Leu Thr Leu Leu Asn Ser Asn Trp
1640 1645 1650
Ile Ala Val Thr Ser Arg Val Glu Glu Trp Leu Asn Leu Leu Leu
1655 1660 1665
Glu Tyr Gln Lys His Met Glu Ser Phe Asp Gln Asn Val Glu His
1670 1675 1680
Ile Thr Lys Trp Ile Ile His Thr Asp Glu Leu Leu Asp Glu Ser
1685 1690 1695
Glu Lys Arg Lys Pro Gln Gln Lys Glu Asp Ile Leu Lys Arg Leu
1700 1705 1710
Lys Ala Glu Met Asn Asp Ile Arg Pro Lys Val Asp Ala Thr Arg
1715 1720 1725
Asp Gln Ala Ala Lys Leu Met Ala Asn Arg Gly Asp Tyr Cys Arg
1730 1735 1740
Lys Ile Val Glu Pro Gln Ile Ser Glu Leu Asn Arg Arg Phe Ala
1745 1750 1755
Ala Ile Ser His Arg Ile Lys Thr Gly Lys Ala Ser Ile Pro Leu
1760 1765 1770
Lys Glu Leu Glu Gln Phe Asn Ser Asp Ile Gln Lys Leu Leu Glu
1775 1780 1785
Pro Leu Glu Ala Glu Ile Gln Gln Gly Val Asn Leu Lys Glu Glu
1790 1795 1800
Asp Phe Asn Lys Asp Met Ser Glu Asp Asn Glu Gly Thr Val Asn
1805 1810 1815
Glu Leu Leu Gln Arg Gly Asp Asn Leu Gln Gln Arg Ile Thr Asp
1820 1825 1830
Glu Arg Lys Arg Glu Glu Ile Lys Leu Lys Gln Gln Leu Leu Gln
1835 1840 1845
Thr Lys His Asn Ala Leu Lys Asp Leu Arg Ser Gln Arg Arg Lys
1850 1855 1860
Lys Ala Leu Glu Ile Ser His Gln Trp Tyr Gln Tyr Lys Ser Gln
1865 1870 1875
Ala Asp Asp Leu Leu Lys Cys Leu Asp Glu Ile Glu Lys Lys Leu
1880 1885 1890
Ala Ser Leu Pro Glu Pro Arg Asp Glu Arg Lys Ile Lys Glu Ile
1895 1900 1905
Asp Arg Glu Leu Gln Lys Lys Lys Glu Glu Leu Asn Ala Val Arg
1910 1915 1920
Arg Gln Ala Glu Ser Leu Ser Glu Asn Gly Ala Ala Met Ala Val
1925 1930 1935
Glu Pro Thr Gln Ile Gln Leu Ser Lys Arg Trp Arg Glu Ile Glu
1940 1945 1950
Ser Asn Phe Ala Gln Phe Arg Arg Leu Asn Phe Ala Gln Ile His
1955 1960 1965
Thr Leu His Glu Glu Thr Met Val Val Thr Thr Glu Asp Met Pro
1970 1975 1980
Leu Asp Val Ser Tyr Val Pro Ser Thr Tyr Leu Thr Glu Ile Ser
1985 1990 1995
His Ile Leu Gln Ala Leu Ser Glu Val Glu Gln Leu Leu Asn Ala
2000 2005 2010
Pro Glu Leu Asn Ala Lys Asp Phe Glu Asp Leu Phe Lys Gln Glu
2015 2020 2025
Glu Ser Leu Lys Asn Ile Lys Glu Asn Leu Gln Gln Ile Ser Gly
2030 2035 2040
Arg Ile Asp Val Ile His Lys Lys Lys Thr Ala Ala Leu Gln Ser
2045 2050 2055
Ala Thr Pro Met Glu Arg Val Lys Leu Gln Glu Ala Val Ser Gln
2060 2065 2070
Met Asp Phe His Trp Glu Lys Leu Asn Arg Met Tyr Lys Glu Arg
2075 2080 2085
Gln Gly Arg Phe Asp Arg Ser Val Glu Lys Trp Arg His Phe His
2090 2095 2100
Tyr Asp Met Lys Val Phe Asn Gln Trp Leu Asn Asp Val Glu Gln
2105 2110 2115
Phe Phe Lys Lys Thr Gln Asn Pro Glu Asn Trp Glu His Ala Lys
2120 2125 2130
Tyr Lys Trp Tyr Leu Lys Glu Leu Gln Asp Gly Ile Gly Gln Arg
2135 2140 2145
Gln Ala Val Val Arg Thr Leu Asn Ala Thr Gly Glu Glu Ile Ile
2150 2155 2160
Gln Gln Ser Ser Lys Thr Asp Ala Asn Ile Leu Gln Glu Lys Leu
2165 2170 2175
Gly Ser Leu Ser Leu Arg Trp His Glu Val Cys Lys Glu Leu Ala
2180 2185 2190
Glu Arg Arg Lys Arg Val Glu Glu Gln Lys Asn Val Phe Ser Glu
2195 2200 2205
Phe Gln Arg Asp Leu Asn Glu Phe Val Ser Trp Leu Glu Glu Ala
2210 2215 2220
Asp Asn Ile Ala Thr Thr Pro Pro Gly Asp Glu Glu Gln Leu Lys
2225 2230 2235
Glu Lys Leu Glu Gln Val Lys Leu Leu Thr Glu Glu Leu Pro Leu
2240 2245 2250
Arg Gln Gly Ile Leu Lys Gln Leu Asn Glu Thr Gly Gly Ala Val
2255 2260 2265
Leu Val Ser Ala Pro Ile Arg Pro Glu Glu Gln Asp Lys Leu Glu
2270 2275 2280
Lys Lys Leu Lys Gln Thr Asn Leu Gln Trp Ile Lys Val Ser Arg
2285 2290 2295
Ala Leu Pro Glu Lys Gln Gly Glu Leu Glu Val His Ile Lys Asp
2300 2305 2310
Phe Arg Gln Phe Glu Glu Gln Leu Asp His Leu Leu Leu Trp Leu
2315 2320 2325
Ser Pro Ile Arg Asn Gln Leu Glu Ile Tyr Asn Gln Pro Ser Gln
2330 2335 2340
Pro Gly Pro Phe Asp Leu Lys Glu Thr Glu Val Thr Val Gln Ala
2345 2350 2355
Lys Gln Pro Asp Val Glu Arg Leu Leu Ser Lys Gly Gln His Leu
2360 2365 2370
Tyr Lys Glu Lys Pro Ser Thr Gln Pro Val Lys Arg Lys Leu Glu
2375 2380 2385
Asp Leu Arg Ser Glu Trp Glu Ala Val Asn His Leu Leu Trp Glu
2390 2395 2400
Leu Arg Thr Lys Gln Pro Asp Arg Ala Pro Gly Leu Ser Thr Thr
2405 2410 2415
Gly Ala Ser Ala Ser Gln Thr Val Thr Val Val Thr Gln Pro Val
2420 2425 2430
Asp Thr Lys Glu Thr Val Ile Ser Lys Leu Glu Met Pro Ser Ser
2435 2440 2445
Leu Leu Leu Glu Val Pro Ala Leu Ala Asp Phe Asn Arg Ala Trp
2450 2455 2460
Thr Glu Leu Thr Asp Trp Leu Ser Leu Leu Asp Arg Val Ile Lys
2465 2470 2475
Ser Gln Arg Val Met Val Gly Asp Leu Glu Asp Ile Asn Glu Met
2480 2485 2490
Ile Ile Lys Gln Lys Leu Val Glu Ser Met Met Asn Ser His Leu
2495 2500 2505
Gln Gly Ala Thr Leu Gln Asp Leu Glu Gln Arg Arg Pro Gln Leu
2510 2515 2520
Glu Glu Leu Ile Thr Ala Ala Gln Asn Leu Lys Asn Lys Thr Ser
2525 2530 2535
Asn Gln Glu Ala Arg Thr Ile Ile Thr Asp Arg Ile Glu Arg Ile
2540 2545 2550
Gln Ile Gln Trp Asp Glu Val Gln Glu Gln Leu Gln Asn Arg Arg
2555 2560 2565
Gln Gln Leu Asn Glu Met Leu Lys Asp Ser Thr Gln Trp Leu Glu
2570 2575 2580
Ala Lys Glu Glu Ala Glu Gln Val Ile Gly Gln Ala Arg Gly Lys
2585 2590 2595
Leu Asp Ser Trp Lys Glu Gly Pro His Thr Met Asp Ala Ile Gln
2600 2605 2610
Lys Lys Ile Thr Glu Thr Lys Gln Leu Ala Lys Asp Leu Arg Gln
2615 2620 2625
Arg Gln Ile Asn Val Asp Val Ala Asn Asp Leu Ala Leu Lys Leu
2630 2635 2640
Leu Arg Asp Tyr Ser Ala Asp Asp Thr Arg Lys Val His Met Ile
2645 2650 2655
Thr Glu Asn Ile Asn Thr Ser Trp Gly Asn Ile Leu Lys Arg Val
2660 2665 2670
Ser Glu Arg Glu Ala Ala Leu Glu Glu Thr Gln Arg Leu Leu Gln
2675 2680 2685
Gln Phe Pro Leu Asp Leu Glu Lys Phe Leu Ala Trp Ile Thr Glu
2690 2695 2700
Ala Glu Thr Thr Ala Asn Val Leu Gln Asp Ala Ser Arg Lys Glu
2705 2710 2715
Lys Leu Leu Glu Asp Ser Arg Gly Val Arg Glu Leu Met Lys Pro
2720 2725 2730
Trp Gln Asp Leu Gln Gly Glu Ile Glu Ala His Thr Asp Ile Tyr
2735 2740 2745
His Asn Leu Asp Glu Asn Gly Gln Lys Ile Leu Arg Ser Leu Glu
2750 2755 2760
Gly Ser Asp Glu Ala Pro Leu Leu Gln Arg Arg Leu Asp Asn Met
2765 2770 2775
Asn Phe Lys Trp Ser Glu Leu Arg Lys Lys Ser Leu Asn Ile Arg
2780 2785 2790
Ser His Leu Glu Val Ser Ser Asp Gln Trp Lys Arg Leu His Leu
2795 2800 2805
Ser Leu Gln Glu Leu Leu Val Trp Leu Gln Leu Lys Asp Asp Glu
2810 2815 2820
Leu Ser Arg Gln Ala Pro Ile Gly Gly Asp Phe Pro Ala Val Gln
2825 2830 2835
Lys Gln Asn Asp Val His Arg Ala Phe Lys Arg Glu Leu Lys Thr
2840 2845 2850
Lys Glu Pro Val Ile Met Ser Thr Leu Glu Thr Val Arg Ile Phe
2855 2860 2865
Leu Thr Glu Gln Pro Leu Glu Gly Leu Glu Lys Leu Tyr Gln Glu
2870 2875 2880
Pro Arg Glu Leu Pro Pro Glu Glu Arg Ala Gln Asn Val Thr Arg
2885 2890 2895
Leu Leu Arg Lys Gln Ala Glu Glu Val Asn Thr Glu Trp Asp Lys
2900 2905 2910
Leu Asn Leu His Ser Ala Asp Trp Gln Arg Lys Ile Asp Glu Ala
2915 2920 2925
Leu Glu Arg Leu Gln Glu Leu Gln Glu Ala Ala Asp Glu Leu Asp
2930 2935 2940
Leu Lys Leu Arg Gln Ala Glu Val Ile Lys Gly Ser Trp Gln Pro
2945 2950 2955
Val Gly Asp Leu Leu Ile Asp Ser Leu Gln Asp His Leu Glu Lys
2960 2965 2970
Val Lys Ala Leu Arg Gly Glu Ile Ala Pro Leu Lys Glu Asn Val
2975 2980 2985
Asn His Val Asn Asp Leu Ala His His Leu Thr Thr Leu Gly Ile
2990 2995 3000
Gln Leu Ser Pro Tyr Asn Leu Ser Ile Leu Glu Asp Leu Asn Thr
3005 3010 3015
Arg Trp Arg Leu Leu Gln Val Ala Val Glu Asp Arg Val Arg Gln
3020 3025 3030
Leu His Glu Ala His Arg Asp Phe Gly Pro Ala Ser Gln His Phe
3035 3040 3045
Leu Ser Thr Ser Val Gln Gly Pro Trp Glu Arg Ala Ile Ser Pro
3050 3055 3060
Asn Lys Val Pro Tyr Tyr Ile Asn His Glu Thr Gln Thr Thr Cys
3065 3070 3075
Trp Asp His Pro Lys Met Thr Glu Leu Tyr Gln Ser Leu Ala Asp
3080 3085 3090
Leu Asn Asn Val Arg Phe Ser Ala Tyr Arg Thr Ala Met Lys Leu
3095 3100 3105
Arg Arg Leu Gln Lys Ala Leu Cys Leu Asp Leu Leu Ser Leu Ser
3110 3115 3120
Ala Ala Cys Asp Ala Leu Asp Gln His Asn Leu Lys Gln Asn Asp
3125 3130 3135
Gln Pro Met Asp Ile Leu Gln Ile Ile Asn Cys Leu Thr Thr Ile
3140 3145 3150
Tyr Asp Arg Leu Glu Gln Glu His Asn Asn Leu Val Asn Val Pro
3155 3160 3165
Leu Cys Val Asp Met Cys Leu Asn Trp Leu Leu Asn Val Tyr Asp
3170 3175 3180
Thr Gly Arg Thr Gly Arg Ile Arg Val Leu Ser Phe Lys Thr Gly
3185 3190 3195
Ile Ile Ser Leu Cys Lys Ala His Leu Glu Asp Lys Tyr Arg Tyr
3200 3205 3210
Leu Phe Lys Gln Val Ala Ser Ser Thr Gly Phe Cys Asp Gln Arg
3215 3220 3225
Arg Leu Gly Leu Leu Leu His Asp Ser Ile Gln Ile Pro Arg Gln
3230 3235 3240
Leu Gly Glu Val Ala Ser Phe Gly Gly Ser Asn Ile Glu Pro Ser
3245 3250 3255
Val Arg Ser Cys Phe Gln Phe Ala Asn Asn Lys Pro Glu Ile Glu
3260 3265 3270
Ala Ala Leu Phe Leu Asp Trp Met Arg Leu Glu Pro Gln Ser Met
3275 3280 3285
Val Trp Leu Pro Val Leu His Arg Val Ala Ala Ala Glu Thr Ala
3290 3295 3300
Lys His Gln Ala Lys Cys Asn Ile Cys Lys Glu Cys Pro Ile Ile
3305 3310 3315
Gly Phe Arg Tyr Arg Ser Leu Lys His Phe Asn Tyr Asp Ile Cys
3320 3325 3330
Gln Ser Cys Phe Phe Ser Gly Arg Val Ala Lys Gly His Lys Met
3335 3340 3345
His Tyr Pro Met Val Glu Tyr Cys Thr Pro Thr Thr Ser Gly Glu
3350 3355 3360
Asp Val Arg Asp Phe Ala Lys Val Leu Lys Asn Lys Phe Arg Thr
3365 3370 3375
Lys Arg Tyr Phe Ala Lys His Pro Arg Met Gly Tyr Leu Pro Val
3380 3385 3390
Gln Thr Val Leu Glu Gly Asp Asn Met Glu Thr Pro Val Thr Leu
3395 3400 3405
Ile Asn Phe Trp Pro Val Asp Ser Ala Pro Ala Ser Ser Pro Gln
3410 3415 3420
Leu Ser His Asp Asp Thr His Ser Arg Ile Glu His Tyr Ala Ser
3425 3430 3435
Arg Leu Ala Glu Met Glu Asn Ser Asn Gly Ser Tyr Leu Asn Asp
3440 3445 3450
Ser Ile Ser Pro Asn Glu Ser Ile Asp Asp Glu His Leu Leu Ile
3455 3460 3465
Gln His Tyr Cys Gln Ser Leu Asn Gln Asp Ser Pro Leu Ser Gln
3470 3475 3480
Pro Arg Ser Pro Ala Gln Ile Leu Ile Ser Leu Glu Ser Glu Glu
3485 3490 3495
Arg Gly Glu Leu Glu Arg Ile Leu Ala Asp Leu Glu Glu Glu Asn
3500 3505 3510
Arg Asn Leu Gln Ala Glu Tyr Asp Arg Leu Lys Gln Gln His Glu
3515 3520 3525
His Lys Gly Leu Ser Pro Leu Pro Ser Pro Pro Glu Met Met Pro
3530 3535 3540
Thr Ser Pro Gln Ser Pro Arg Asp Ala Glu Leu Ile Ala Glu Ala
3545 3550 3555
Lys Leu Leu Arg Gln His Lys Gly Arg Leu Glu Ala Arg Met Gln
3560 3565 3570
Ile Leu Glu Asp His Asn Lys Gln Leu Glu Ser Gln Leu His Arg
3575 3580 3585
Leu Arg Gln Leu Leu Glu Gln Pro Gln Ala Glu Ala Lys Val Asn
3590 3595 3600
Gly Thr Thr Val Ser Ser Pro Ser Thr Ser Leu Gln Arg Ser Asp
3605 3610 3615
Ser Ser Gln Pro Met Leu Leu Arg Val Val Gly Ser Gln Thr Ser
3620 3625 3630
Glu Ser Met Gly Glu Glu Asp Leu Leu Ser Pro Pro Gln Asp Thr
3635 3640 3645
Ser Thr Gly Leu Glu Glu Val Met Glu Gln Leu Asn Asn Ser Phe
3650 3655 3660
Pro Ser Ser Arg Gly His Asn Val Gly Ser Leu Phe His Met Ala
3665 3670 3675
Asp Asp Leu Gly Arg Ala Met Glu Ser Leu Val Ser Val Met Ile
3680 3685 3690
Asp Glu Glu Gly Ala Glu
3695
<210> 8
<211> 3677
<212> Белок
<213> Oryctolagus cuniculus
<220>
<221> прочие признаки
<222> (2570)..(2570)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> прочие признаки
<222> (2597)..(2597)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 8
Met Leu Trp Trp Glu Glu Val Glu Asp Cys Tyr Glu Arg Glu Asp Val
1 5 10 15
Gln Lys Lys Thr Phe Thr Lys Trp Ile Asn Ala Gln Phe Ser Lys Phe
20 25 30
Gly Lys Gln His Ile Glu Asn Leu Phe Ser Asp Leu Gln Asp Gly Arg
35 40 45
Arg Leu Leu Asp Leu Leu Glu Gly Leu Thr Gly Gln Gln Leu Pro Lys
50 55 60
Glu Lys Gly Ser Thr Arg Val His Ala Leu Asn Asn Val Asn Lys Ala
65 70 75 80
Leu Arg Val Leu Gln Lys Asn Asn Val Asp Leu Val Asn Ile Gly Ser
85 90 95
Thr Asp Ile Val Asp Gly Asn His Lys Leu Thr Leu Gly Leu Ile Trp
100 105 110
Asn Ile Ile Leu His Trp Gln Val Lys Asn Val Met Lys Asn Ile Met
115 120 125
Ala Gly Leu Gln Gln Thr Asn Ser Glu Lys Ile Leu Leu Ser Trp Val
130 135 140
Arg Gln Ser Thr Arg Asn Tyr Ser Gln Val Asn Val Ile Asn Phe Thr
145 150 155 160
Thr Ser Trp Ser Asp Gly Leu Ala Leu Asn Ala Leu Ile His Ser His
165 170 175
Arg Pro Asp Leu Phe Asp Trp Asn Ser Val Val Cys Gln Gln Ser Ala
180 185 190
Thr Gln Arg Leu Glu His Ala Phe Asn Ile Ala Lys Cys Gln Leu Gly
195 200 205
Ile Glu Lys Leu Leu Asp Pro Glu Asp Val Ala Thr Thr Tyr Pro Asp
210 215 220
Lys Lys Ser Ile Leu Met Tyr Ile Thr Ser Leu Phe Gln Val Leu Pro
225 230 235 240
Gln Gln Val Ser Ile Glu Ala Ile Gln Glu Val Glu Leu Leu Pro Arg
245 250 255
Pro Ser Lys Val Thr Arg Glu Glu His Phe Gln Leu His His Gln Met
260 265 270
His Tyr Ser Gln Gln Ile Thr Val Ser Leu Ala Gln Gly Tyr Glu Arg
275 280 285
Thr Ser Ser Pro Lys Pro Arg Phe Lys Ser Tyr Ala Tyr Thr Gln Ala
290 295 300
Ala Tyr Val Ser Thr Ser Asp Pro Thr Arg Ser Pro Phe Pro Ser Gln
305 310 315 320
His Leu Glu Ala Pro Gly Asp Lys Ser Phe Gly Ser Ser Leu Met Glu
325 330 335
Thr Glu Val Ser Leu Asp Ser Tyr Gln Thr Ala Leu Glu Glu Val Leu
340 345 350
Ser Trp Leu Leu Ser Ala Glu Asp Thr Leu Gln Ala Gln Lys Glu Ile
355 360 365
Ser Ser Asp Val Glu Glu Val Lys Glu Gln Phe His Thr His Glu Gly
370 375 380
Tyr Met Met Asp Leu Thr Ser His Gln Gly Arg Val Gly Asn Val Leu
385 390 395 400
Gln Leu Gly Ser Gln Leu Ile Arg Thr Gly Lys Leu Ser Glu Glu Glu
405 410 415
Glu Thr Glu Val Gln Glu Gln Met Asn Leu Leu Asn Ser Arg Trp Glu
420 425 430
Cys Leu Arg Val Ala Ser Met Glu Lys Gln Ser Asn Leu His Lys Val
435 440 445
Leu Met Asp Leu Gln Asn Gln Lys Leu Lys Glu Leu Asn Asp Trp Leu
450 455 460
Thr Lys Thr Glu Glu Arg Thr Arg Lys Met Glu Glu Glu Pro Leu Gly
465 470 475 480
Pro Asp Leu Glu Asp Leu Lys Arg Gln Val Gln Gln His Lys Val Leu
485 490 495
Gln Glu Asp Leu Glu Gln Glu Gln Val Arg Val Asn Ser Leu Thr His
500 505 510
Met Val Val Val Val Asp Glu Ser Ser Gly Glu His Ala Thr Ala Ala
515 520 525
Leu Glu Glu Gln Leu Lys Val Leu Gly Asp Arg Trp Ala Asn Ile Cys
530 535 540
Arg Trp Thr Glu Asp Arg Trp Val Leu Leu Gln Asp Ile Leu Leu Lys
545 550 555 560
Trp Gln Arg Phe Thr Glu Glu Gln Cys Leu Phe Ser Thr Trp Leu Ser
565 570 575
Glu Lys Glu Asp Ala Val Asn Lys Ile His Thr Thr Gly Phe Lys Asp
580 585 590
Gln Asn Glu Met Leu Ser Ser Leu His Lys Leu Thr Val Leu Lys Thr
595 600 605
Asp Leu Glu Lys Lys Lys Gln Ser Met Asp Lys Leu Ser Ser Leu Asn
610 615 620
Gln Ala Leu Leu Ser Thr Leu Lys Asn Lys Ser Val Thr Gln Lys Met
625 630 635 640
Glu Ala Trp Leu Glu Asn Phe Ala Leu Arg Trp Asp Asn Leu Val Gln
645 650 655
Lys Leu Glu Lys Ser Ser Ala Gln Ile Ser Gln Ala Val Thr Thr Thr
660 665 670
Gln Pro Ser Leu Thr Gln Thr Thr Val Met Glu Thr Val Thr Met Val
675 680 685
Thr Thr Arg Glu Gln Ile Leu Val Lys His Ala Gln Glu Glu Leu Pro
690 695 700
Pro Pro Pro Pro Gln Lys Lys Arg Gln Ile Ile Val Asp Ser Glu Ile
705 710 715 720
Arg Lys Arg Leu Asp Val Asp Ile Thr Glu Leu His Ser Trp Ile Thr
725 730 735
Arg Ser Glu Ala Val Leu Gln Ser Pro Glu Phe Ala Ile Tyr Arg Lys
740 745 750
Glu Gly Asn Phe Ser Asp Leu Lys Glu Lys Val Asn Ala Ile Glu Arg
755 760 765
Glu Lys Ala Glu Lys Phe Arg Lys Leu Gln Asp Ala Ser Arg Ser Ala
770 775 780
Gln Ala Leu Val Glu Gln Met Val Asn Glu Gly Val Asn Ala Asp Ser
785 790 795 800
Ile Lys Gln Ala Leu Glu Gln Leu Asn Ser Arg Trp Ile Glu Phe Cys
805 810 815
Gln Leu Leu Ser Glu Arg Leu His Trp Leu Glu Tyr Gln Asn Ser Ile
820 825 830
Ile Thr Phe Tyr Asn Gln Leu Gln Gln Leu Glu Gln Met Ile Thr Thr
835 840 845
Ala Glu Asn Trp Leu Lys Thr Gln Pro Thr Thr Ala Ser Glu Pro Thr
850 855 860
Ala Ile Lys Ser Gln Leu Lys Met Cys Lys Asp Glu Val Asn Arg Leu
865 870 875 880
Ser Ala Leu Gln Pro Gln Ile Glu Arg Leu Lys Ile Gln Ser Thr Ala
885 890 895
Leu Lys Glu Lys Gly Gln Gly Pro Met Phe Leu Asp Ala Asp Phe Val
900 905 910
Ala Phe Thr Asn His Phe Asn Gln Val Phe Ala Asp Ala Gln Ala Arg
915 920 925
Glu Lys Glu Leu Gln Thr Ile Phe Asp Thr Leu Pro Pro Thr Arg Tyr
930 935 940
Gln Glu Thr Ile Ser Thr Ile Arg Thr Trp Ile Gln Gln Ser Glu Pro
945 950 955 960
Lys Leu Ser Ile Pro Gln Leu Ser Val Thr Glu Tyr Glu Ile Met Glu
965 970 975
Gln Arg Leu Gly Glu Leu Gln Ala Leu Gln Ser Ser Leu Gln Glu Gln
980 985 990
Gln Ser Gly Leu Thr Tyr Leu Ser Thr Thr Val Lys Glu Met Ala Lys
995 1000 1005
Lys Ala Pro Ser Glu Ile Ser Arg Lys Tyr Gln Ser Glu Phe Glu
1010 1015 1020
Glu Ile Glu Gly His Trp Lys Lys Leu Ser Tyr Gln Leu Val Asp
1025 1030 1035
His Cys Gln Lys Leu Glu Glu Gln Met Asn Lys Leu Arg Lys Ile
1040 1045 1050
Gln Asn His Ile Lys Thr Leu Lys Lys Trp Met Ala Glu Val Asp
1055 1060 1065
Val Phe Leu Lys Glu Glu Trp Pro Ala Leu Gly Asp Ser Glu Ile
1070 1075 1080
Leu Lys Lys Gln Leu Lys Gln Cys Arg Leu Leu Val Asn Asp Ile
1085 1090 1095
Gln Thr Ile Gln Pro Ser Leu Asn Ser Val Asn Glu Ser Gly Gln
1100 1105 1110
Lys Ile Lys Asn Glu Ala Glu Pro Glu Phe Ala Ser Arg Leu Glu
1115 1120 1125
Thr Glu Leu Arg Glu Leu Asn Ser Gln Trp Asp His Met Cys Arg
1130 1135 1140
Gln Val Tyr Thr Arg Lys Asp Ala Leu Lys Ala Gly Leu Asp Lys
1145 1150 1155
Thr Leu Ser Leu Gln Lys Asp Leu Ser Glu Met His Glu Trp Met
1160 1165 1170
Thr Gln Ala Glu Glu Glu Tyr Leu Glu Arg Asp Phe Glu Tyr Lys
1175 1180 1185
Thr Pro Asp Glu Leu Gln Thr Ala Val Glu Glu Met Lys Arg Ala
1190 1195 1200
Lys Glu Glu Ala Gln Gln Lys Glu Ser Lys Val Lys Leu Leu Thr
1205 1210 1215
Glu Ser Val Asn Ser Val Ile Ala Gln Ala Pro Pro Ala Ala Gln
1220 1225 1230
Glu Ala Leu Lys Lys Glu Leu Asp Thr Leu Thr Thr Asn Tyr Gln
1235 1240 1245
Trp Leu Cys Thr Arg Leu Asn Gly Lys Cys Lys Thr Leu Glu Glu
1250 1255 1260
Val Trp Ala Cys Trp His Glu Leu Leu Ser Tyr Leu Glu Lys Ala
1265 1270 1275
Asn Lys Trp Leu Asn Glu Val Glu Val Lys Leu Lys Thr Thr Glu
1280 1285 1290
Thr Leu Pro Gly Gly Ala Glu Glu Ile Ser Glu Val Leu Asp Ser
1295 1300 1305
Leu Glu Asn Leu Met Gln His Ser Glu Asp Asn Pro Asn Gln Ile
1310 1315 1320
Arg Ile Leu Ala Gln Thr Leu Thr Asp Gly Gly Val Met Asp Glu
1325 1330 1335
Leu Ile Asn Glu Glu Leu Glu Thr Phe Asn Ser Arg Trp Arg Glu
1340 1345 1350
Leu His Glu Glu Ala Val Arg Arg Gln Lys Leu Leu Glu Gln Ser
1355 1360 1365
Ile Gln Ser Ala Gln Glu Ile Glu Lys Ser Leu His Leu Ile Gln
1370 1375 1380
Glu Ser Leu Ala Phe Ile Asp Lys Gln Leu Ala Ala Tyr Ile Ala
1385 1390 1395
Asp Lys Val Asp Ala Ala Gln Met Pro Gln Glu Ala Gln Lys Ile
1400 1405 1410
Gln Ser Asp Leu Thr Ser His Glu Ile Ser Leu Glu Glu Met Lys
1415 1420 1425
Lys His Asn Gln Gly Lys Glu Ala Ala Gln Arg Val Leu Ser Gln
1430 1435 1440
Ile Asp Val Ala Gln Lys Lys Leu Gln Asp Val Ser Met Lys Phe
1445 1450 1455
Arg Leu Phe Gln Lys Pro Ala Asn Phe Glu Gln Arg Leu Glu Glu
1460 1465 1470
Ser Lys Met Ile Leu Asp Glu Val Lys Met His Leu Pro Ala Leu
1475 1480 1485
Glu Thr Lys Ser Val Glu Gln Glu Val Val Gln Ser Gln Leu Asn
1490 1495 1500
His Cys Val Asn Leu Tyr Lys Ser Leu Ser Glu Val Lys Ser Glu
1505 1510 1515
Val Glu Met Val Ile Lys Thr Gly Arg Gln Ile Val Gln Lys Lys
1520 1525 1530
Gln Thr Glu Asn Pro Lys Glu Leu Asp Glu Arg Val Thr Ala Leu
1535 1540 1545
Lys Leu His Tyr Asn Glu Leu Gly Ala Lys Val Thr Glu Arg Lys
1550 1555 1560
Gln Gln Leu Glu Lys Cys Leu Lys Leu Ser Arg Lys Met Arg Lys
1565 1570 1575
Glu Met Asn Ala Leu Thr Glu Trp Leu Ala Ala Thr Asp Met Glu
1580 1585 1590
Leu Thr Lys Arg Ser Ala Val Glu Gly Met Pro Ser Asn Leu Asp
1595 1600 1605
Ala Glu Ile Ala Trp Gly Lys Ala Thr Gln Lys Glu Val Glu Lys
1610 1615 1620
Gln Lys Ala His Leu Lys Ser Val Ile Glu Leu Gly Glu Ala Leu
1625 1630 1635
Lys Thr Val Leu Gly Lys Lys Glu Thr Leu Val Glu Asp Lys Leu
1640 1645 1650
Ser Leu Leu Asn Ser Asn Trp Val Ala Val Thr Ser Arg Ala Glu
1655 1660 1665
Glu Trp Leu Asn Leu Leu Leu Glu Tyr Gln Lys His Met Glu Thr
1670 1675 1680
Phe Asp Gln Asn Val Asp His Ile Thr Lys Trp Ile Ile Gln Ala
1685 1690 1695
Asp Thr Leu Leu Asp Glu Ser Glu Lys Lys Lys Pro Gln Gln Lys
1700 1705 1710
Glu Asp Val Leu Lys Arg Leu Lys Ala Glu Met Asn Asp Ile Arg
1715 1720 1725
Pro Lys Val Asp Ser Ile Arg Asp Gln Ala Ala Asn Leu Met Ala
1730 1735 1740
Asn Arg Gly Asp His Cys Arg Lys Val Val Glu Pro Lys Ile Ser
1745 1750 1755
Glu Leu Asn His Arg Phe Ala Ala Ile Ser His Arg Ile Lys Thr
1760 1765 1770
Gly Lys Ala Ser Ile Pro Leu Lys Glu Leu Glu Gln Phe Asn Ser
1775 1780 1785
Asp Ile Gln Lys Leu Leu Glu Pro Leu Glu Ala Glu Ile Gln Gln
1790 1795 1800
Gly Val Asn Leu Lys Glu Glu Asp Phe Asn Lys Asp Met Ser Glu
1805 1810 1815
Asp Asn Glu Gly Thr Val Lys Glu Leu Leu Gln Arg Gly Asp Asn
1820 1825 1830
Leu His Glu Arg Ile Thr Asp Glu Arg Lys Arg Glu Glu Ile Lys
1835 1840 1845
Ile Lys Gln Gln Leu Leu Gln Thr Lys His Asn Ala Leu Lys Asp
1850 1855 1860
Leu Arg Ser Gln Arg Arg Lys Lys Ala Leu Glu Ile Ser His Gln
1865 1870 1875
Trp Tyr Gln Tyr Lys Arg Gln Ala Asp Asp Leu Leu Lys Cys Leu
1880 1885 1890
Asp Asp Ile Glu Lys Lys Leu Ala Ser Leu Pro Asp Pro Lys Asp
1895 1900 1905
Glu Arg Lys Ile Lys Glu Ile Asp Arg Glu Leu Gln Lys Lys Lys
1910 1915 1920
Glu Glu Leu Asn Ala Val His Arg Gln Ala Glu Gly Leu Ser Glu
1925 1930 1935
Asp Gly Ala Ala Met Ala Val Glu Pro Ile Gln Ile Gln Leu Ser
1940 1945 1950
Lys Arg Trp Arg Glu Ile Glu Ser Lys Phe Ala Gln Phe Arg Arg
1955 1960 1965
Leu Asn Phe Ala Gln Ile His Thr Val Arg Glu Glu Thr Met Val
1970 1975 1980
Val Met Thr Glu Asp Met Pro Leu Glu Ile Ser Tyr Val Pro Ser
1985 1990 1995
Thr Tyr Leu Thr Glu Ile Thr His Val Ser Gln Ala Leu Ser Glu
2000 2005 2010
Val Asp Gln Leu Leu Asn Ala Pro Asp Leu Ser Ala Lys Asp Phe
2015 2020 2025
Glu Asp Leu Phe Lys Gln Glu Glu Ser Leu Lys Asn Ile Lys Glu
2030 2035 2040
Cys Met Gln Gln Ile Ser Gly Arg Ile Asp Val Ile His Asn Lys
2045 2050 2055
Lys Ala Ala Ala Leu Gln Ser Ala Thr Ala Ala Glu Arg Val Lys
2060 2065 2070
Leu Gln Glu Ala Leu Ala Gln Leu Asp Ser Gln Trp Glu Lys Val
2075 2080 2085
Asn Lys Met Tyr Lys Asp Arg Gln Gly Arg Phe Asp Arg Ser Val
2090 2095 2100
Glu Lys Trp Arg Arg Phe His Tyr Asp Met Lys Ile Phe Asn Gln
2105 2110 2115
Trp Leu Thr Glu Ala Glu Gln Phe Leu Lys Lys Thr Gln Ile Pro
2120 2125 2130
Glu Asn Trp Glu His Ala Lys Tyr Lys Trp Tyr Leu Lys Glu Leu
2135 2140 2145
Gln Asp Gly Ile Gly Gln Arg Gln Thr Val Val Arg Thr Leu Asn
2150 2155 2160
Thr Thr Gly Glu Glu Ile Ile Gln Gln Ser Ser Lys Thr Asp Ala
2165 2170 2175
Asn Ile Leu Gln Glu Lys Leu Gly Asn Leu Asn Leu Arg Trp Gln
2180 2185 2190
Glu Val Cys Lys Gln Leu Ala Glu Arg Arg Lys Arg Leu Glu Glu
2195 2200 2205
Gln Lys Asn Ile Leu Ser Glu Phe Gln Arg Asp Leu Asn Glu Phe
2210 2215 2220
Val Leu Trp Leu Glu Glu Ala Asp Asn Ile Thr Ser Ile Pro Leu
2225 2230 2235
Glu Pro Gly Asn Glu Gln Gln Leu Lys Glu Lys Leu Glu Gln Val
2240 2245 2250
Lys Leu Leu Ala Glu Glu Leu Pro Leu Arg Gln Gly Ile Leu Lys
2255 2260 2265
Gln Leu Asn Glu Ala Gly Gly Thr Ala Leu Val Ser Ala Pro Ile
2270 2275 2280
Ser Pro Glu Glu Gln Asp Lys Leu Glu Asn Lys Leu Lys Gln Thr
2285 2290 2295
Asn Leu Gln Trp Ile Lys Val Ser Lys Ala Leu Pro Glu Lys Gln
2300 2305 2310
Glu Glu Ile Val Ala His Val Lys Glu Leu Gly Gln Leu Glu Glu
2315 2320 2325
Gln Leu Asn His Leu Leu Leu Trp Leu Thr Pro Ile Arg Asn Gln
2330 2335 2340
Leu Glu Ile Tyr Asn Gln Pro Asn Gln Thr Gly Pro Phe Asp Ile
2345 2350 2355
Lys Glu Thr Glu Val Ala Val Gln Ala Lys Gln Pro Asp Val Glu
2360 2365 2370
Arg Ile Leu Ser Lys Gly Gln His Leu Tyr Lys Glu Lys Pro Ala
2375 2380 2385
Thr Gln Pro Val Lys Arg Lys Leu Glu Asp Leu Ser Ser Glu Trp
2390 2395 2400
Lys Val Val Asn Tyr Leu Leu Gln Glu Leu Arg Ala Lys Arg Pro
2405 2410 2415
Asp Leu Pro Pro Gly Pro Ala Thr Ile Gly Ala Ser Pro Ser Gln
2420 2425 2430
Thr Val Thr Leu Val Thr Gln Thr Val Val Thr Lys Glu Ile Ser
2435 2440 2445
Leu Ser Glu Leu Glu Met Pro Ser Ser Leu Leu Leu Glu Val Pro
2450 2455 2460
Ala Leu Ala Asp Phe Asn Arg Ala Trp Thr Glu Leu Thr Asp Trp
2465 2470 2475
Leu Ser Leu Leu Asp Arg Val Leu Lys Ser Gln Gln Val Ile Val
2480 2485 2490
Gly Asp Leu Glu Asp Ile Asn Glu Met Ile Ile Lys Gln Lys Ala
2495 2500 2505
Thr Leu Gln Glu Leu Glu Gln Arg Arg Pro Gln Leu Glu Glu Leu
2510 2515 2520
Ile Thr Ala Ala Gln Asn Leu Lys Asn Lys Thr Ser Asn Gln Glu
2525 2530 2535
Ala Arg Thr Ile Ile Thr Asp Arg Ile Glu Arg Ile Gln Asn Gln
2540 2545 2550
Trp Asp Glu Val Gln Glu His Leu Gln Asn Arg Ser Ser Ser Leu
2555 2560 2565
Asn Xaa Met Leu Lys Asp Ser Thr Gln Trp Leu Glu Ala Lys Glu
2570 2575 2580
Glu Leu Ser Ser Leu Gly Gln Ala Arg Ala Lys Leu Ser Xaa Met
2585 2590 2595
Glu Glu Gly Pro Tyr Pro Met Asn Ala Leu Gln Lys Lys Ile Glu
2600 2605 2610
Glu Thr Lys Gln Leu Ala Lys Asp Leu Arg Gln Trp Gln Ile Asn
2615 2620 2625
Val Asp Val Ala Asn Asp Leu Ala Leu Lys Leu Leu Arg Asp Tyr
2630 2635 2640
Ser Ala Asp Asp Thr Arg Lys Val His Met Ile Thr Glu Asn Ile
2645 2650 2655
Asn Ala Ser Trp Gly Ser Ile His Lys Arg Val Asn Glu Arg Glu
2660 2665 2670
Ala Thr Leu Glu Glu Ala Tyr Arg Leu Leu Gln Gln Phe Pro Leu
2675 2680 2685
Asp Leu Glu Lys Phe Leu Ala Trp Leu Thr Glu Ala Glu Thr Thr
2690 2695 2700
Ala Asn Val Leu Gln Asp Ala Thr His Lys Glu Arg Leu Pro Glu
2705 2710 2715
Asp Ser Lys Gly Val Arg Glu Leu Met Lys Gln Trp Gln Asp Leu
2720 2725 2730
Gln Gly Glu Ile Glu Ala His Thr Asp Ile Tyr His Asn Leu Asp
2735 2740 2745
Glu Asn Gly Gln Lys Ile Leu Arg Ser Leu Glu Gly Ser Asp Glu
2750 2755 2760
Ala Val Leu Leu Gln Arg Arg Leu Asp Asn Met Asn Phe Lys Trp
2765 2770 2775
Ser Glu Leu Arg Lys Lys Ser Leu Asn Ile Arg Ser His Leu Glu
2780 2785 2790
Ala Ser Ser Asp Gln Trp Lys Arg Leu His Leu Ser Leu Gln Glu
2795 2800 2805
Leu Leu Val Trp Leu Gln Leu Lys Asp Asp Glu Leu Asn Arg Gln
2810 2815 2820
Ala Pro Val Gly Gly Asp Phe Pro Ala Val Gln Lys Gln Asn Asp
2825 2830 2835
Val His Arg Ala Phe Lys Arg Glu Leu Lys Thr Lys Glu Pro Val
2840 2845 2850
Ile Met Ser Thr Leu Glu Thr Val Arg Ile Phe Leu Thr Glu Gln
2855 2860 2865
Pro Leu Gly Gly Leu Glu Lys Leu Tyr Gln Glu Pro Arg Glu Leu
2870 2875 2880
Pro Pro Asp Glu Arg Ala Gln Asn Val Thr Arg Leu Leu Arg Lys
2885 2890 2895
Gln Ala Glu Glu Val Asn Ala Glu Trp Glu Lys Leu Asn Leu His
2900 2905 2910
Ser Thr Asp Trp Gln Arg Lys Leu Asp Glu Ala Leu Glu Arg Leu
2915 2920 2925
Gln Glu Leu Gln Glu Ala Thr Asp Glu Leu Asp Leu Lys Leu Arg
2930 2935 2940
Gln Ala Glu Val Ile Lys Gly Ser Trp Gln Pro Val Gly Asp Leu
2945 2950 2955
Leu Ile Asp Ser Leu Gln Asp His Leu Glu Lys Val Lys Ala Leu
2960 2965 2970
Arg Gly Glu Ile Ala Pro Leu Lys Glu Asn Val Ser His Val Asn
2975 2980 2985
Asp Leu Gly Arg Gln Leu Thr Thr Leu Gly Ile Gln Leu Ser Pro
2990 2995 3000
Tyr Asn Leu Ser Thr Leu Glu Asp Leu Asn Thr Arg Trp Lys Leu
3005 3010 3015
Leu Gln Val Ala Val Glu Asp Arg Ile Arg Gln Leu His Glu Ala
3020 3025 3030
His Arg Asp Phe Gly Pro Ala Ser Gln His Phe Leu Ser Thr Ser
3035 3040 3045
Val Gln Gly Pro Trp Glu Arg Ala Ile Ser Pro Asn Lys Val Pro
3050 3055 3060
Tyr Tyr Ile Asn His Glu Thr Gln Thr Thr Cys Trp Asp His Pro
3065 3070 3075
Lys Met Thr Glu Leu Tyr Gln Ser Leu Ala Asp Leu Asn Asn Val
3080 3085 3090
Arg Phe Ser Ala Tyr Arg Thr Ala Met Lys Leu Arg Arg Leu Gln
3095 3100 3105
Lys Ala Leu Cys Leu Asp Leu Leu Thr Leu Ser Ala Ala Cys Asp
3110 3115 3120
Ala Leu Asp Gln His Asn Leu Lys Gln Asn Asp Gln Pro Met Asp
3125 3130 3135
Ile Leu Gln Ile Ile Asn Cys Leu Thr Thr Val Tyr Asp Arg Leu
3140 3145 3150
Glu Gln Glu His Asn Asn Leu Val Asn Val Pro Leu Cys Val Asp
3155 3160 3165
Met Cys Leu Asn Trp Leu Leu Asn Val Tyr Asp Thr Gly Arg Thr
3170 3175 3180
Gly Arg Ile Arg Val Leu Ser Phe Lys Thr Gly Ile Ile Ser Leu
3185 3190 3195
Cys Lys Ala His Leu Glu Asp Lys Tyr Arg Tyr Leu Phe Lys Gln
3200 3205 3210
Val Ala Ser Ser Thr Gly Phe Cys Asp Gln Arg Arg Leu Gly Leu
3215 3220 3225
Leu Leu His Asp Ser Ile Gln Ile Pro Arg Gln Leu Gly Glu Val
3230 3235 3240
Ala Ser Phe Gly Gly Ser Asn Ile Glu Pro Ser Val Arg Ser Cys
3245 3250 3255
Phe Gln Phe Ala Asn Asn Lys Pro Glu Ile Glu Ala Ala Leu Phe
3260 3265 3270
Leu Asp Trp Met Arg Leu Glu Pro Gln Ser Met Val Trp Leu Pro
3275 3280 3285
Val Leu His Arg Val Ala Ala Ala Glu Thr Ala Lys His Gln Ala
3290 3295 3300
Lys Cys Asn Ile Cys Lys Glu Cys Pro Ile Ile Gly Phe Arg Tyr
3305 3310 3315
Arg Ser Leu Lys His Phe Asn Tyr Asp Ile Cys Gln Ser Cys Phe
3320 3325 3330
Phe Ser Gly Arg Val Ala Lys Gly His Lys Met His Tyr Pro Met
3335 3340 3345
Val Glu Tyr Cys Thr Pro Thr Thr Ser Gly Glu Asp Val Arg Asp
3350 3355 3360
Phe Ala Lys Val Leu Lys Asn Lys Phe Arg Thr Lys Arg Tyr Phe
3365 3370 3375
Ala Lys His Pro Arg Met Gly Tyr Leu Pro Val Gln Thr Val Leu
3380 3385 3390
Glu Gly Asp Asn Met Glu Thr Pro Val Thr Leu Ile Asn Phe Trp
3395 3400 3405
Pro Val Asp Ser Ala Pro Ala Ser Ser Pro Gln Leu Ser His Asp
3410 3415 3420
Asp Thr His Ser Arg Ile Glu His Tyr Ala Ser Arg Leu Ala Glu
3425 3430 3435
Met Glu Asn Ser Asn Gly Ser Tyr Leu Asn Asp Ser Ile Ser Pro
3440 3445 3450
Asn Glu Ser Ile Asp Asp Glu His Leu Leu Ile Gln His Tyr Cys
3455 3460 3465
Gln Ser Leu Asn Gln Asp Ser Pro Leu Ser Gln Pro Arg Ser Pro
3470 3475 3480
Ala Gln Ile Leu Ile Ser Leu Glu Ser Glu Glu Arg Gly Glu Leu
3485 3490 3495
Glu Arg Ile Leu Ala Asp Leu Glu Glu Glu Asn Arg Asn Leu Gln
3500 3505 3510
Ala Glu Tyr Asp Arg Leu Lys Gln Gln His Glu His Lys Gly Leu
3515 3520 3525
Ser Pro Leu Pro Ser Pro Pro Glu Met Met Pro Thr Ser Pro Gln
3530 3535 3540
Ser Pro Arg Asp Ala Glu Leu Ile Ala Glu Ala Lys Leu Leu Arg
3545 3550 3555
Gln His Lys Gly Arg Leu Glu Ala Arg Met Gln Ile Leu Glu Asp
3560 3565 3570
His Asn Lys Gln Leu Glu Ser Gln Leu His Arg Leu Arg Gln Leu
3575 3580 3585
Leu Glu Gln Pro Gln Ala Glu Ala Lys Val Asn Gly Thr Thr Val
3590 3595 3600
Ser Ser Pro Ser Thr Ser Leu Gln Arg Ser Asp Ser Ser Gln Pro
3605 3610 3615
Met Leu Leu Arg Val Val Gly Ser Gln Thr Ser Glu Ser Met Gly
3620 3625 3630
Glu Glu Asp Leu Pro Ser Pro Pro Gln Asp Thr Ser Thr Gly Leu
3635 3640 3645
Glu Glu Val Met Glu Gln Leu Asn Asn Ser Phe Pro Ser Ser Arg
3650 3655 3660
Gly Arg Asn Thr Pro Gly Lys Pro Met Arg Glu Asp Thr Met
3665 3670 3675
<210> 9
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 9
Ala Glu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Leu Lys
1 5 10
<210> 10
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 10
Asp Asp Ala Met His
1 5
<210> 11
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 11
Asp Tyr Ser Met His
1 5
<210> 12
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 12
Glu Tyr Thr Ile His
1 5
<210> 13
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 13
Asp Tyr Ser Met His
1 5
<210> 14
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 14
Asp Phe Ser Met His
1 5
<210> 15
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 15
Ala Tyr Ser Met His
1 5
<210> 16
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 16
Asp Tyr Ser Met His
1 5
<210> 17
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 17
Asp Tyr Ser Met His
1 5
<210> 18
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 18
Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 19
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 19
Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 20
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 20
Gly Ile Asn Cys Asn Ser Gly Gly Thr Ser Tyr Thr Gln Lys Phe Lys
1 5 10 15
Asp
<210> 21
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 21
Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 22
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 22
Trp Ile Asn Thr Glu Thr Gly Asp Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 23
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 23
Trp Ile Asn Thr Glu Thr Gly Glu Ala Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 24
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 24
Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 25
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 25
Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 26
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> misc_feature
<222> (1)..(1)
<223> Xaa can be any naturally occurring amino acid
<400> 26
Xaa Leu Asp Phe
1
<210> 27
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 27
Gly Ala Ser Gly Tyr
1 5
<210> 28
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 28
Arg Phe Phe Tyr Gly Ser Ser Gln Phe Ala Tyr
1 5 10
<210> 29
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 29
Arg Gly Glu Phe
1
<210> 30
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 30
Arg Gly Asp Tyr
1
<210> 31
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 31
Arg Gly Asp Tyr
1
<210> 32
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 32
Arg Gly Asp Tyr
1
<210> 33
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 33
Gly Arg Ser Ala Tyr
1 5
<210> 34
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 34
Arg Ser Ser Gln Ser Leu Val His Ser Asn Gly Lys Thr Tyr Leu His
1 5 10 15
<210> 35
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 35
Lys Ala Ser Gln Ser Val Ser Lys Asn Val Ala
1 5 10
<210> 36
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 36
Arg Ala Ser Lys Ser Val Thr Thr Ser Gly Tyr Ser Tyr Met His
1 5 10 15
<210> 37
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 37
Lys Ala Ser Gln Ser Val Asn Lys Asn Val Ala
1 5 10
<210> 38
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 38
Arg Ser Ser Gln Ser Leu Val His Ser Asn Arg Asn Ile Tyr Leu His
1 5 10 15
<210> 39
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 39
Arg Ser Ser Gln Ser Leu Val Arg Ser Asn Arg Asn Ile Tyr Leu His
1 5 10 15
<210> 40
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 40
Arg Ser Ser Gln Ser Leu Val His Asn Asn Arg Asn Ile Tyr Leu His
1 5 10 15
<210> 41
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 41
Lys Ala Ser Gln Thr Val Ser Asn Asn Val Ala
1 5 10
<210> 42
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 42
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 43
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 43
Ser Ser Ser Lys Arg Tyr Ser
1 5
<210> 44
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 44
Leu Ala Ser Asn Leu Glu Ser
1 5
<210> 45
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 45
Ser Ala Ser Lys Arg Tyr Thr
1 5
<210> 46
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 46
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 47
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 47
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 48
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 48
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 49
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 49
Ser Ala Ser Asn Arg Tyr Thr
1 5
<210> 50
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 50
Ser Gln Ser Thr His Val Pro Trp Thr
1 5
<210> 51
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 51
His Gln Asp Tyr Asn Ser Pro Trp Thr
1 5
<210> 52
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 52
Gln His Ser Arg Glu Leu Pro Leu Thr
1 5
<210> 53
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 53
Leu Gln Asp Tyr Thr Ser Pro Trp Thr
1 5
<210> 54
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 54
Ser Gln Asp Lys Tyr Ile Pro Trp Thr
1 5
<210> 55
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 55
Ser Gln Asp Ser His Ile Pro Trp Thr
1 5
<210> 56
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 56
Ser Gln Asp Arg Tyr Ile Pro Trp Thr
1 5
<210> 57
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 57
Gln Gln Asp Tyr Ser Ser Pro Trp Thr
1 5
<210> 58
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 58
Thr Tyr Gly Met Ser
1 5
<210> 59
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 59
Thr Tyr Gly Met Thr
1 5
<210> 60
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 60
Trp Ile Asn Thr Ser Ser Gly Val Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 61
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 61
Trp Ile Asn Thr Tyr Ser Gly Val Pro Thr Tyr Ala Asn Asp Phe Lys
1 5 10 15
Gly
<210> 62
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 62
Trp Ile Asn Thr Tyr Ser Gly Leu Pro Thr Tyr Thr Asn Asp Phe Lys
1 5 10 15
Gly
<210> 63
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 63
Trp Ile Asn Thr Tyr Ser Gly Leu Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 64
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 64
Trp Ile Asn Thr Tyr Ser Gly Val Pro Thr Tyr Ala Asp Asp Phe Lys
1 5 10 15
Gly
<210> 65
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 65
Trp Ile Asn Thr Phe Ser Gly Val Pro Thr Tyr Thr Asp Asp Phe Lys
1 5 10 15
Gly
<210> 66
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 66
Pro Ile His Met Val Val Ala Glu Asp Tyr
1 5 10
<210> 67
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 67
Pro Ile His Lys Val Val Ala Glu Asp Tyr
1 5 10
<210> 68
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 68
Pro Ile Thr Thr Val Val Pro Phe Asp Tyr
1 5 10
<210> 69
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 69
Pro Ile Thr Thr Val Val Pro Phe Asp Tyr
1 5 10
<210> 70
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 70
Pro Ile Thr Lys Val Val Ala Gly Asp Phe
1 5 10
<210> 71
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 71
Pro Ile Thr Thr Val Val Pro Phe Asp Tyr
1 5 10
<210> 72
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 72
Lys Ser Ser Gln Ile Leu Leu Tyr Gly Arg Asn Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 73
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 73
Arg Ser Ser Gln Ser Leu Leu Tyr Ser Gly Asn Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 74
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 74
Arg Ser Ser Gln Ser Leu Val Tyr Thr Tyr Gly Asn Thr Tyr Leu His
1 5 10 15
<210> 75
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 75
Arg Ser Ser Gln Ser Leu Val Tyr Asn Tyr Gly Asn Thr Tyr Leu His
1 5 10 15
<210> 76
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 76
Lys Ser Ser Gln Ser Leu Leu Tyr Arg Ser Asn Gln Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 77
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 77
Arg Ser Ser Gln Ser Leu Val Tyr Ser Thr Gly Asn Thr Tyr Leu His
1 5 10 15
<210> 78
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 78
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 79
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 79
Trp Ala Ser Thr Trp Glu Ser
1 5
<210> 80
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 80
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 81
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 81
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 82
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 82
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 83
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 83
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 84
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 84
Gln Gln Tyr Tyr Ser Lys Pro Tyr Thr
1 5
<210> 85
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 85
Gln Gln Tyr Tyr Thr Phe Pro Tyr Thr
1 5
<210> 86
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 86
Ser Gln Ser Arg His Ser Pro Trp Thr
1 5
<210> 87
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 87
Ser Gln Ser Arg His Ser Pro Trp Thr
1 5
<210> 88
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 88
Gln Gln Tyr Phe Ser Ser Pro Tyr Thr
1 5
<210> 89
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 89
Ser Gln Ser Arg His Gly Pro Trp Thr
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Композиции и способы активации интегринов | 2020 |
|
RU2836262C1 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
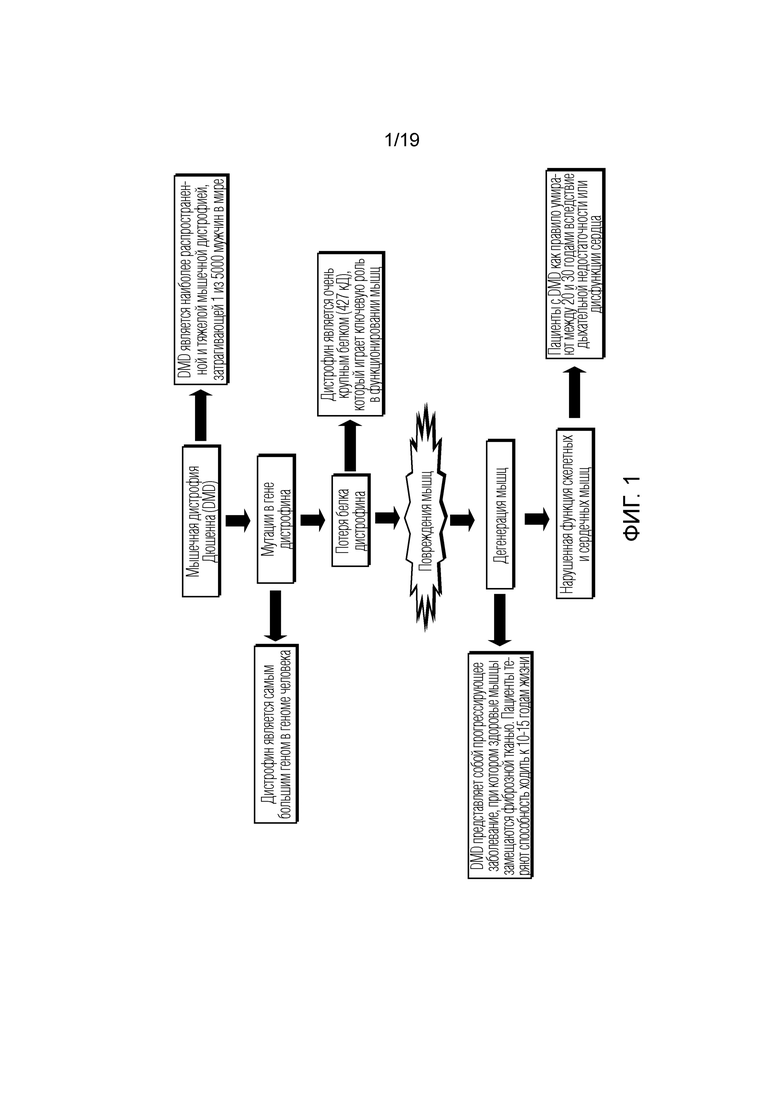
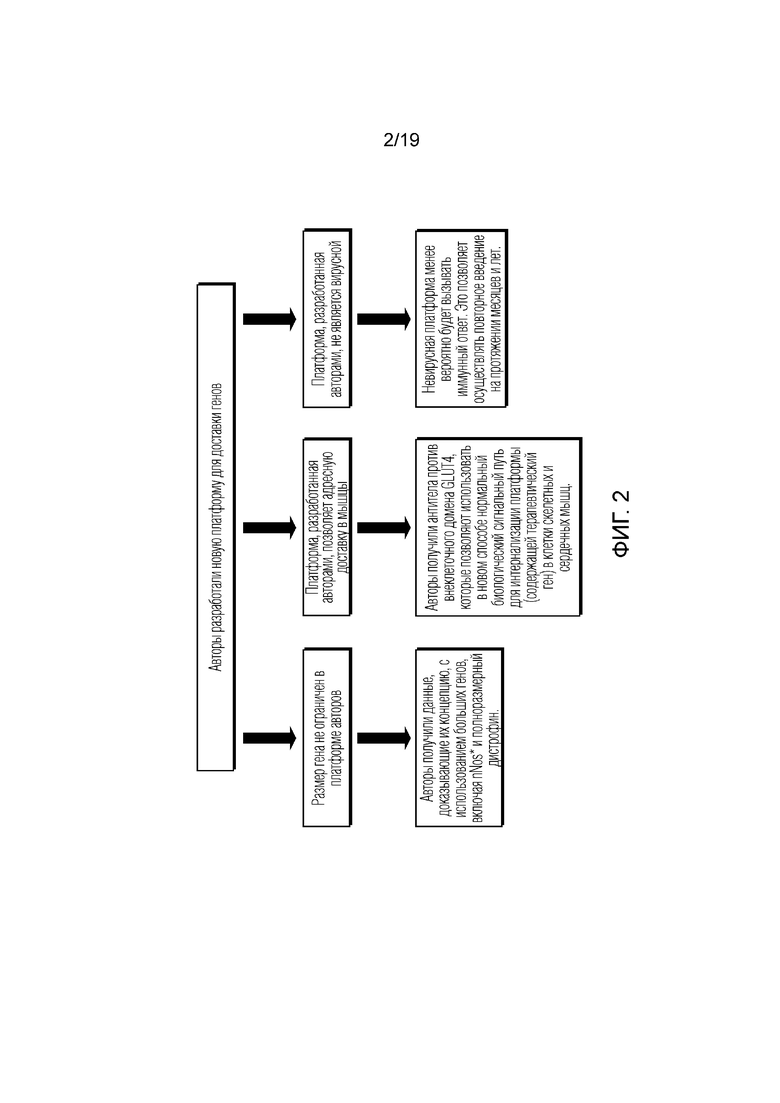
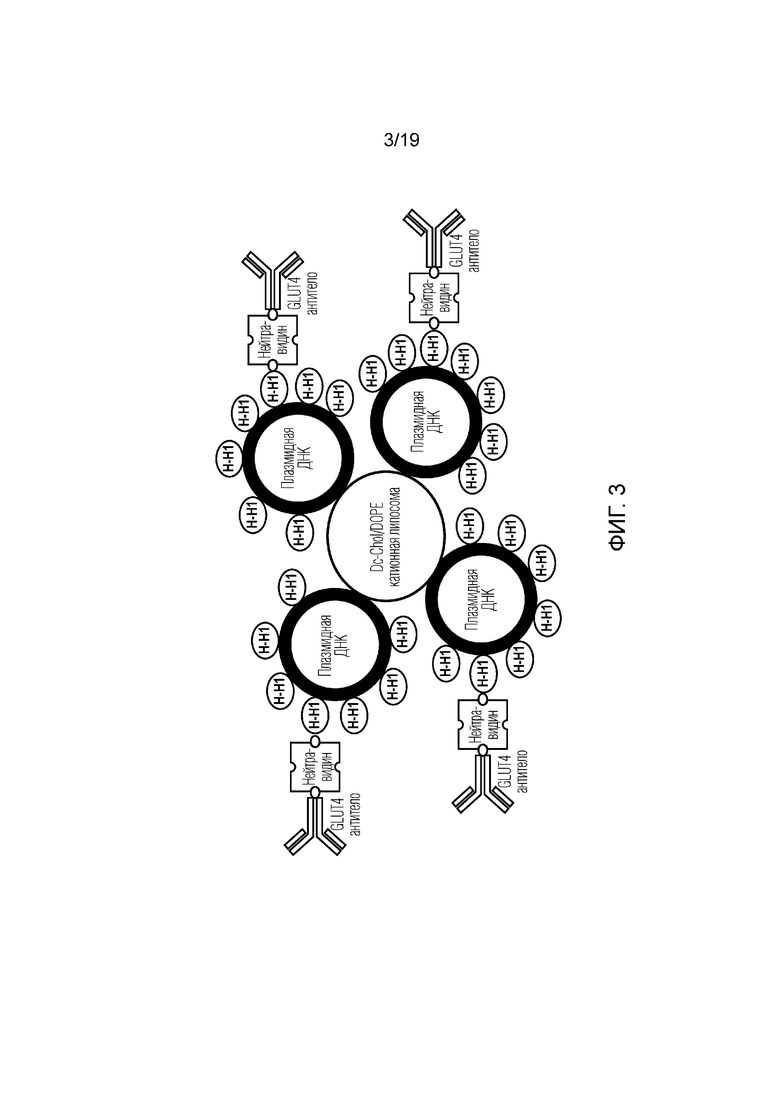
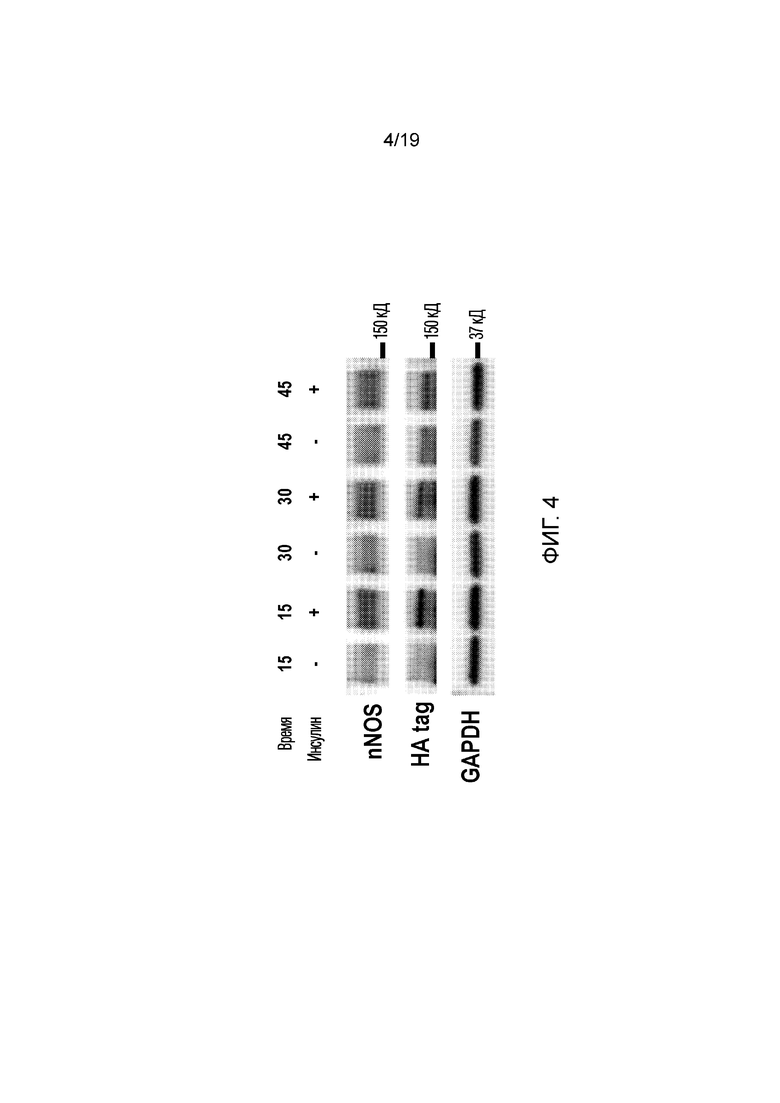
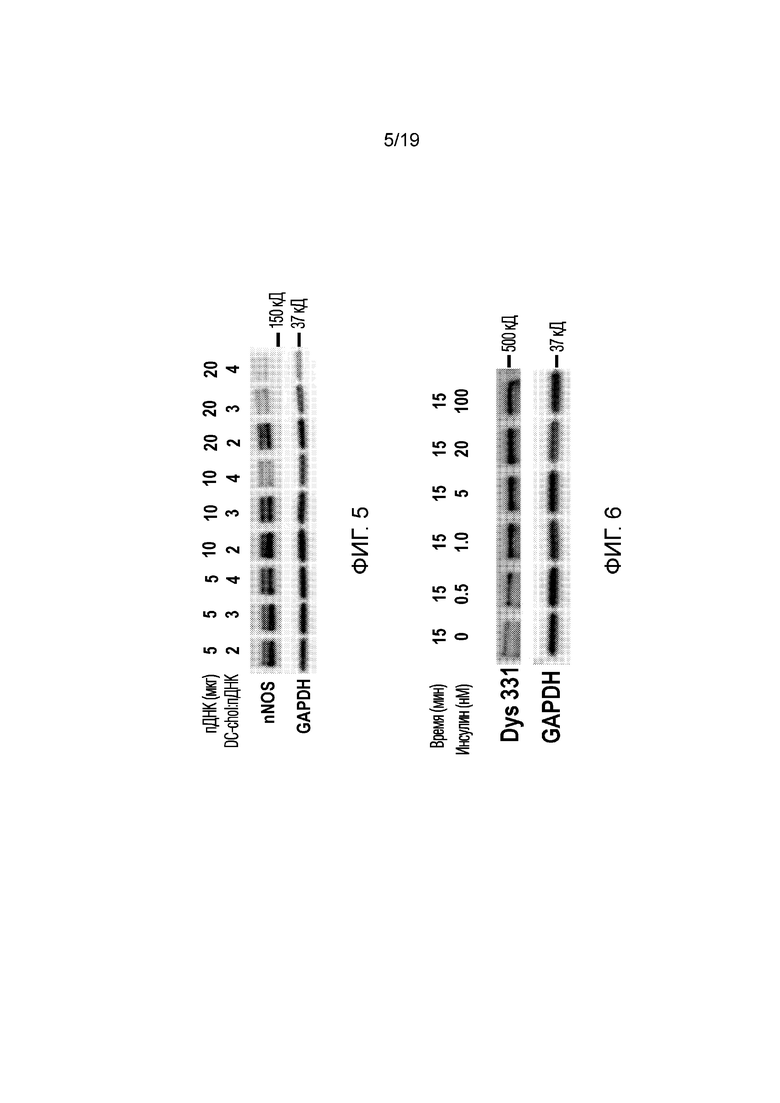
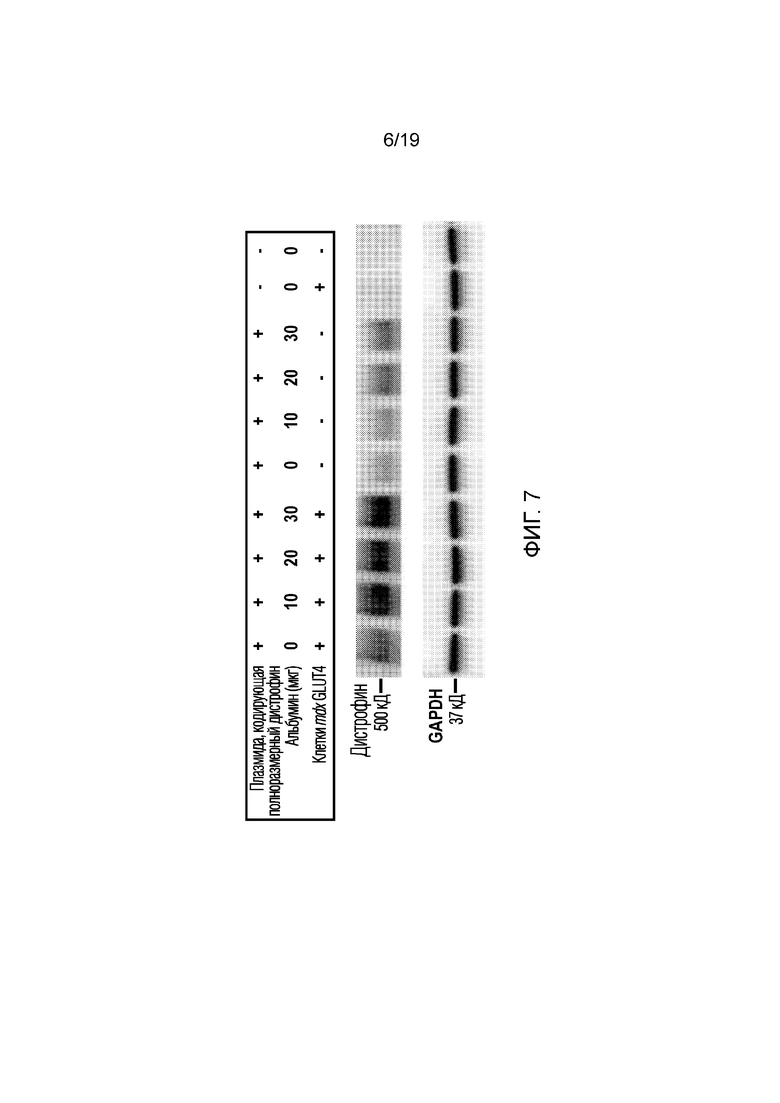
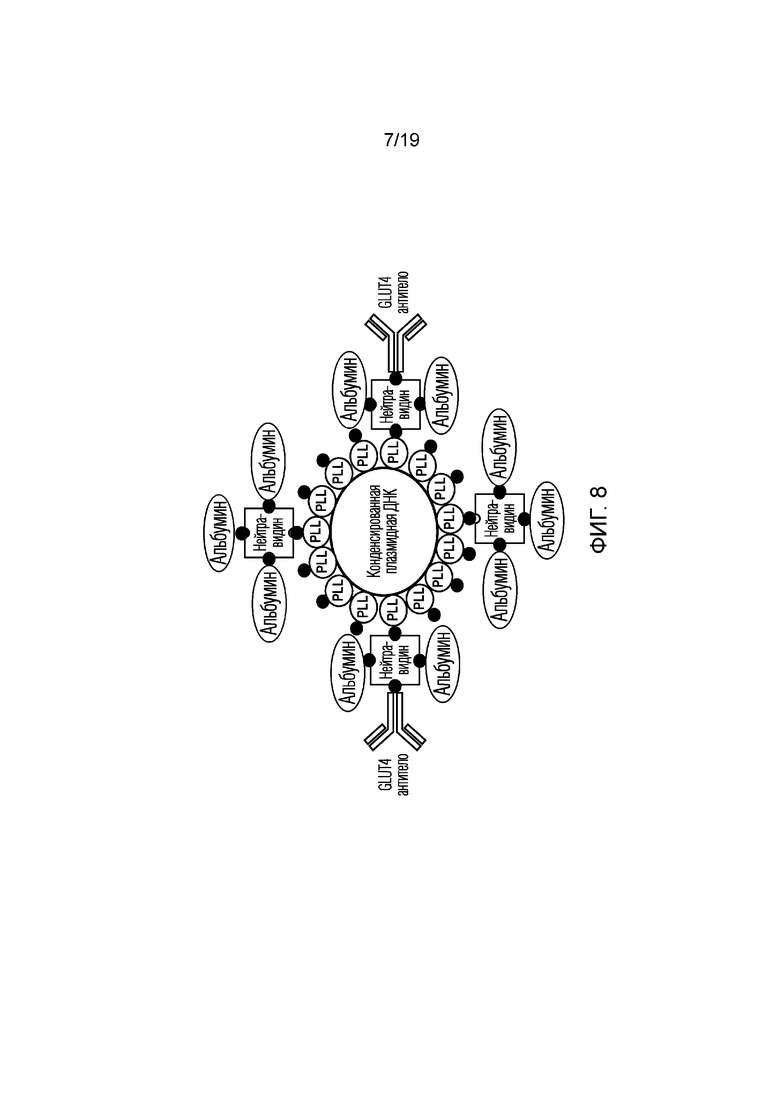
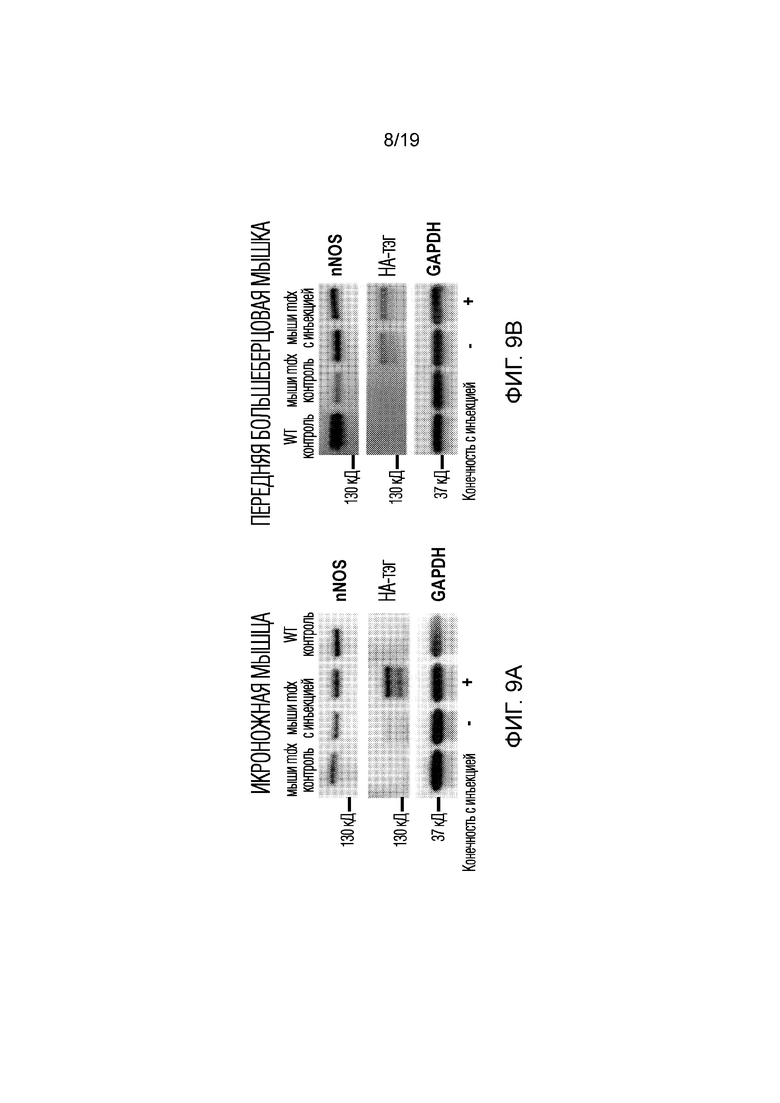
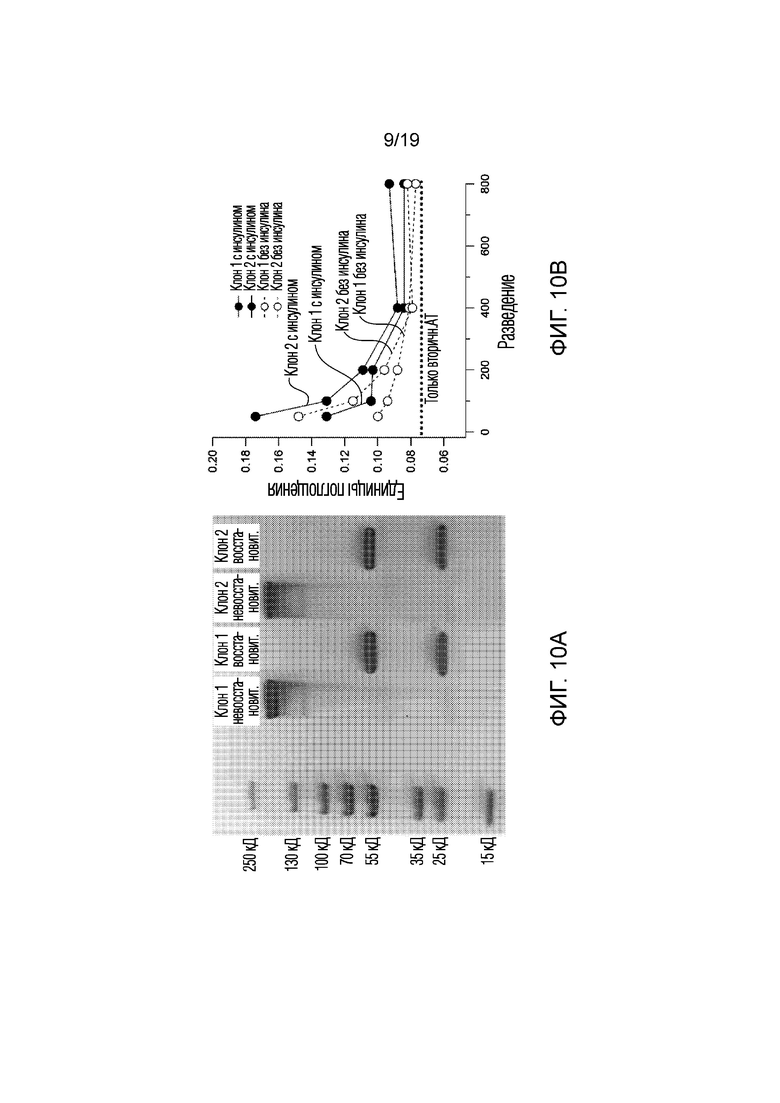
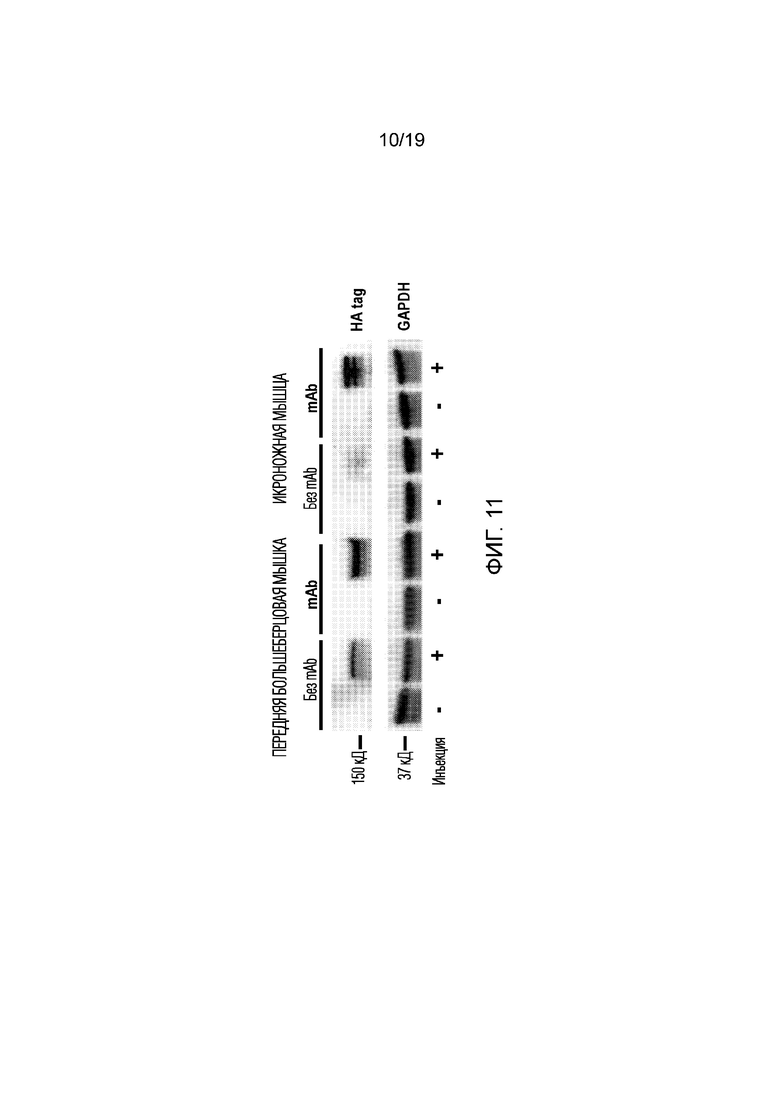

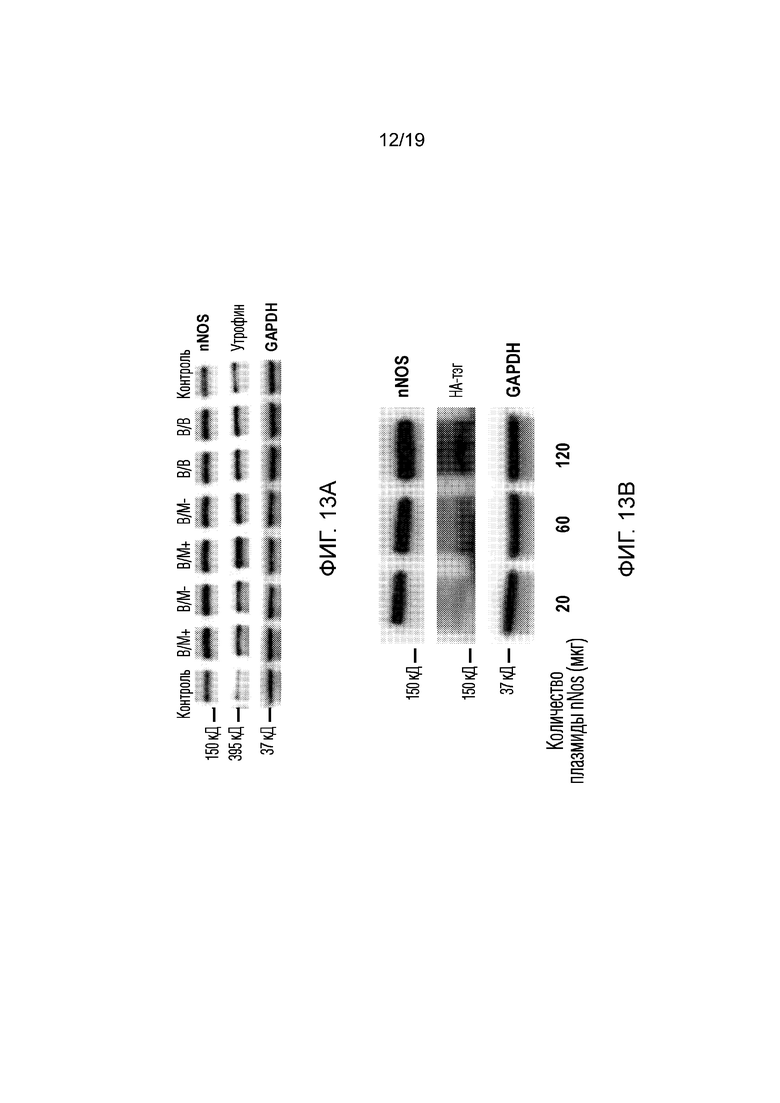
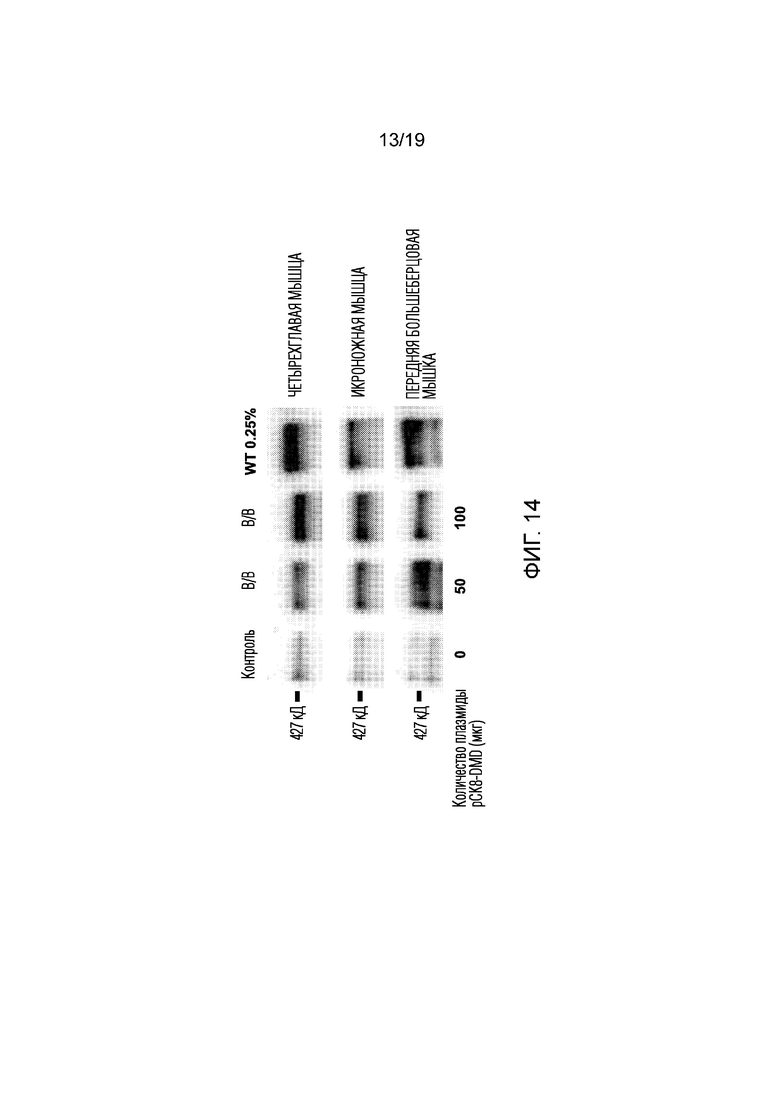
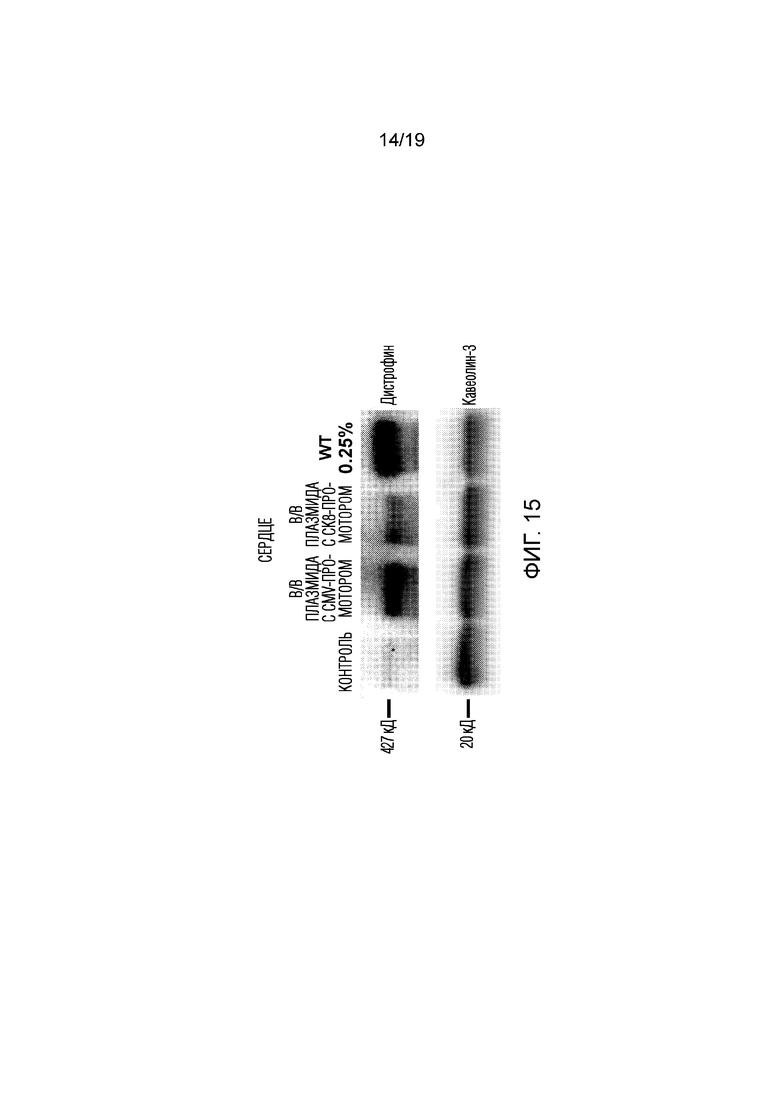
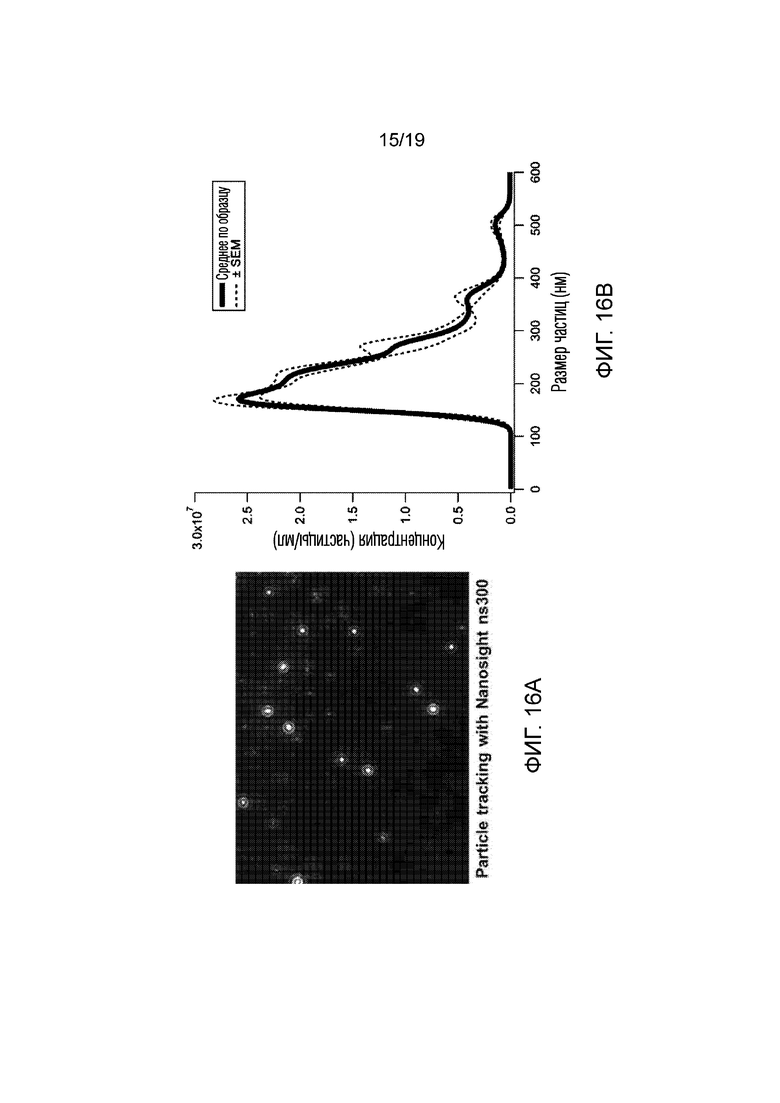
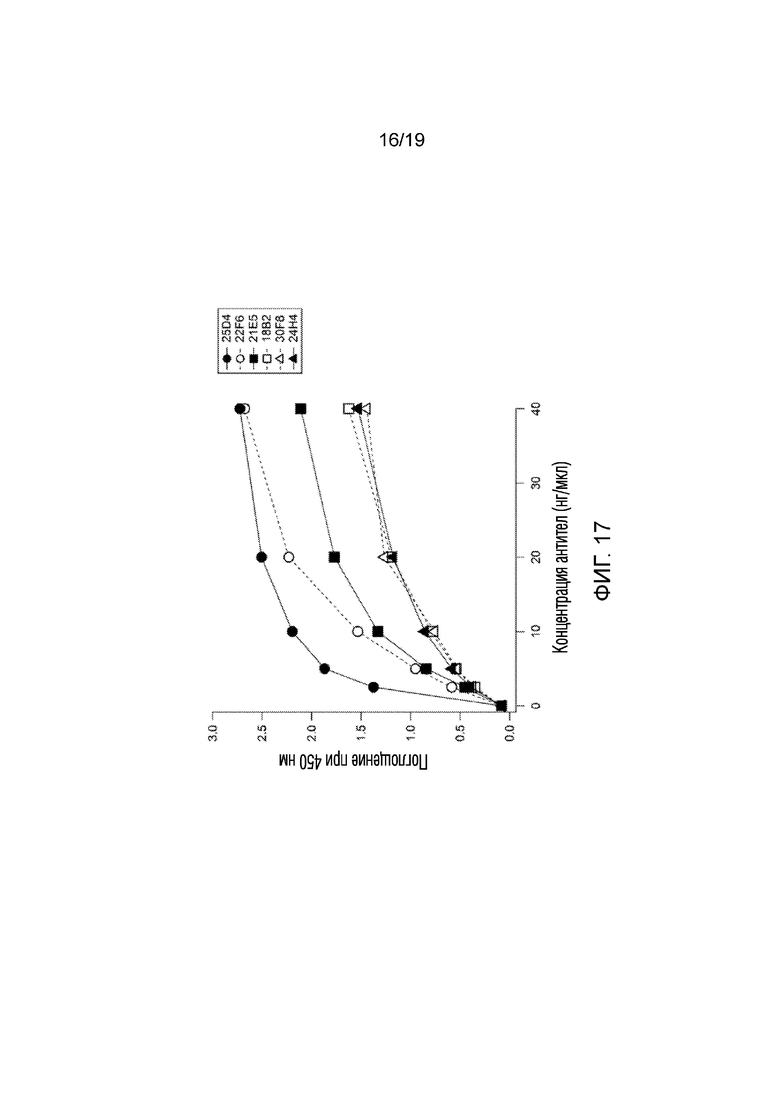
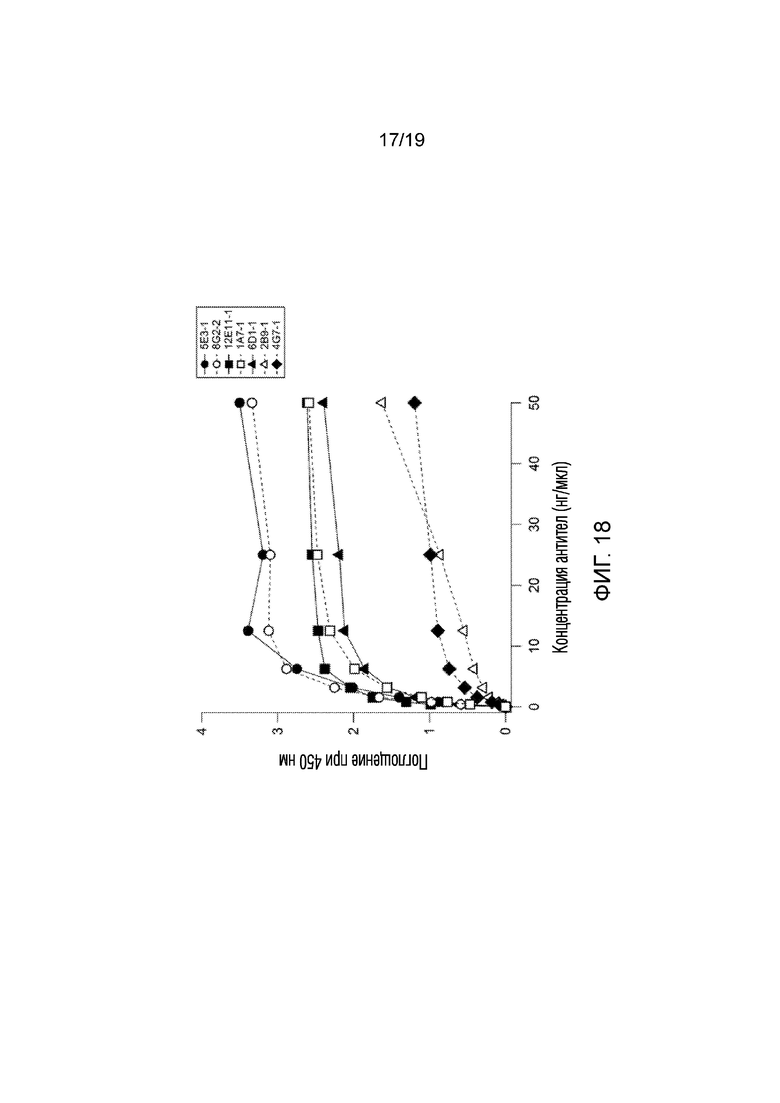
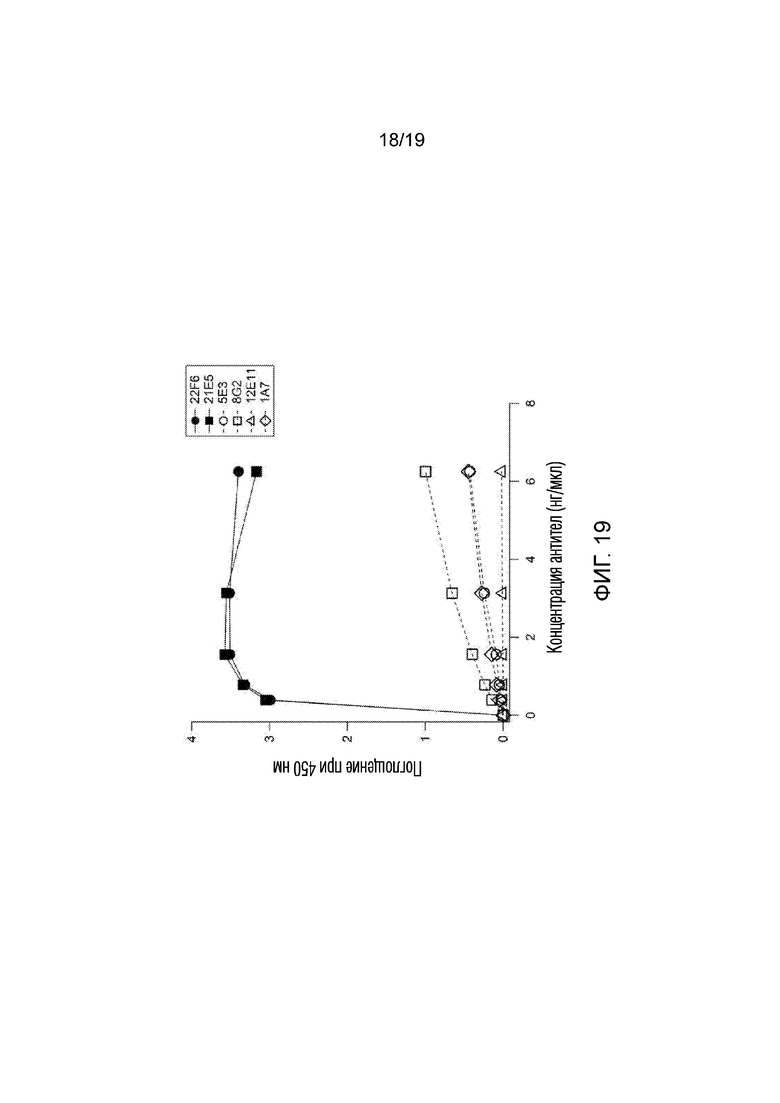
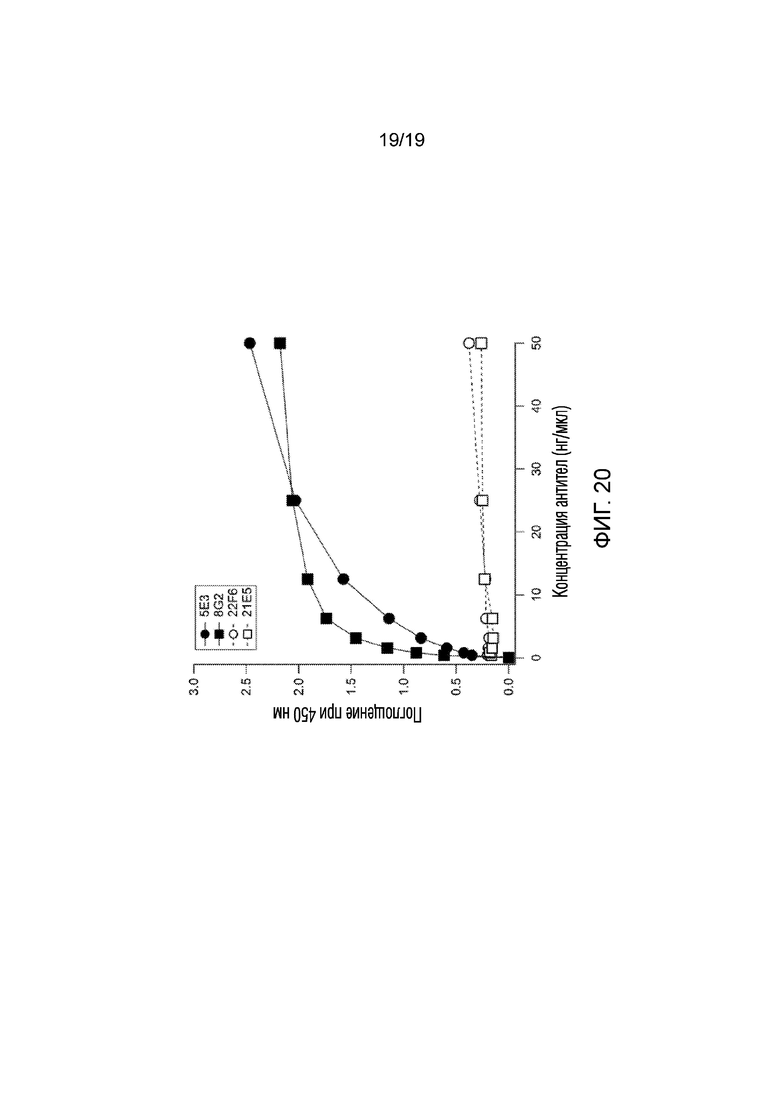
Изобретение относится к области биотехнологии. Предложены антитело или его связывающий фрагмент, которые специфично связываются с белком-переносчиком глюкозы 4 (GLUT4). Также изобретение относится к композициям и способам доставки полезной нагрузки в клетки или ткани, которые экспрессируют GLUT4. Композиции содержат указанные антитело или его связывающий фрагмент, конъюгированные с терапевтической полезной нагрузкой. Полезная нагрузка содержит нуклеиновую кислоту, кодирующую функциональный белок дистрофин или его функциональный фрагмент. Композиции по изобретению применимы для способов лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин. Изобретение обеспечивает невирусные векторные платформы, которые могут специфично доставлять терапевтическую полезную нагрузку в мышечные клетки без ограничений по размеру полезной нагрузки. 4 н. и 23 з.п. ф-лы, 25 ил., 3 табл.
1. Анти-GLUT4 антитело или его связывающий фрагмент, содержащие вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где:
а) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:10, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:18, и VH-CDR3, имеющую последовательность LDF; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:34, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:42, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:50;
b) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:11, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:19, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:27; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:35, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:43, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:51;
c) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:12, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:20, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:28; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:36, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:44, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:52;
d) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:14, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:22, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:30; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:38, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:46, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:54;
e) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:15, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:23, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:31; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:39, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:47, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:55;
f) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:16, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:24, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:32; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:40, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:48, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:56; или
g) VH содержит VH-CDR1, имеющую последовательность, приведенную в SEQ ID NO:17, VH-CDR2, имеющую последовательность, приведенную в SEQ ID NO:25, и VH-CDR3, имеющую последовательность, приведенную в SEQ ID NO:33; и VL содержит VL-CDR1, имеющую последовательность, приведенную в SEQ ID NO:41, VL-CDR2, имеющую последовательность, приведенную в SEQ ID NO:49, и VL-CDR3, имеющую последовательность, приведенную в SEQ ID NO:57.
2. Композиция для доставки терапевтической полезной нагрузки в клетку, которая экспрессирует GLUT4, где композиция содержит антитело или его связывающий фрагмент по п. 1, конъюгированные с терапевтической полезной нагрузкой.
3. Композиция по п. 2, где антитело или его связывающий фрагмент представляют собой или содержат одноцепочечное антитело, Fab-фрагмент, F(ab)2-фрагмент, VHH-фрагмент, VNAR-фрагмент или нанотело.
4. Композиция по п. 3, где одноцепочечное антитело представляет собой одноцепочечный вариабельный фрагмент (scFv) или одноцепочечный Fab-фрагмент (scFab).
5. Композиция по п. 2, где терапевтическая полезная нагрузка содержит нуклеиновую кислоту, белок или пептид, липид, низкомолекулярный фармацевтический препарат или радиоизотоп.
6. Композиция по п. 5, где терапевтическая полезная нагрузка содержит нуклеиновую кислоту, при этом нуклеиновая кислота выбрана из ДНК, мРНК, киРНК и кшРНК.
7. Композиция по п. 5, где терапевтическая полезная нагрузка содержит нуклеиновую кислоту, при этом нуклеиновая кислота содержит открытую рамку считывания, функционально связанную с промоторной последовательностью.
8. Композиция по п. 5, где терапевтическая полезная нагрузка содержит нуклеиновую кислоту в линейной форме, нуклеиновую кислоту в форме плазмиды или нуклеиновую кислоту в форме мини-кольца.
9. Композиция по п. 5, дополнительно содержащая гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой.
10. Композиция по п. 8, дополнительно содержащая липосому, связанную с нуклеиновой кислотой.
11. Композиция по п. 8, дополнительно содержащая множество катионных пептидных фрагментов, связанных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота.
12. Композиция по п. 2, где терапевтическая полезная нагрузка содержит нуклеиновую кислоту с промоторной последовательностью, функционально связанной с последовательностью, кодирующей белок дистрофин или его функциональный фрагмент.
13. Композиция по п. 12, где белок дистрофин представляет собой по существу полноразмерный белок дистрофин человека, имеющий аминокислотную последовательность, по меньшей мере на 80% идентичную последовательности, представленной в SEQ ID NO:5.
14. Способ таргетинга терапевтической полезной нагрузки для доставки в клетку, которая экспрессирует GLUT4, включающий приведение клетки в контакт с композицией по любому из пп. 2-13.
15. Способ по п. 14, дополнительно включающий контакт клетки в контакт с инсулином до контакта клетки с композицией.
16. Способ по п. 14, где клетка представляет собой клетку скелетной мышцы, клетку сердечной мышцы, клетку гладкой мышцы, клетку жировой ткани, клетку гиппокампа или клетку мозжечка.
17. Способ по п. 14, где клетка представляет собой клетку меланомы, клетку рака простаты, клетку рака мышечной ткани, такую как клетка рабдомиосаркомы, или клетку рака молочной железы.
18. Способ лечения заболевания или состояния у субъекта с генетической мутацией в гене, кодирующем белок дистрофин, включающий введение субъекту терапевтически эффективного количества композиции по п. 12.
19. Способ по п. 18, где субъект имеет или является генетическим носителем мышечной дистрофии Дюшенна («DMD») или мышечной дистрофии Беккера («BMD»).
20. Способ по п. 18, где антитело или его связывающий фрагмент представляет собой или содержит одноцепочечное антитело, Fab-фрагмент, F(ab)2-фрагмент, VHH-фрагмент, VNAR-фрагмент или нанотело.
21. Способ по п. 20, где одноцепочечное антитело представляет собой одноцепочечный вариабельный фрагмент (scFv) или одноцепочечный Fab-фрагмент (scFab).
22. Способ по п. 18, где нуклеиновая кислота находится в линейной форме, плазмидной форме или форме мини-кольца.
23. Способ по п. 22, где композиция дополнительно содержит гистоновый белок или множество гистоновых белков, связанных с нуклеиновой кислотой.
24. Способ по п. 22, где композиция дополнительно содержит липосому, связанную с нуклеиновой кислотой.
25. Способ по п. 18, дополнительно включающий множество катионных пептидных фрагментов, связанных с нуклеиновой кислотой, в результате чего образуется конденсированная нуклеиновая кислота.
26. Способ по п. 18, где кодируемый белок дистрофин имеет идентичности последовательности по меньшей мере 80% с SEQ ID NO:5.
27. Способ по п. 18, где субъектом является человек.
| TUCKER D.F | |||
| et al., Isolation of state-dependent monoclonal antibodies against the 12-transmembrane domain glucose transporter 4 using virus-like particles" PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES, 2018, vol | |||
| Ударно-долбежная врубовая машина | 1921 |
|
SU115A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| LAURITZEN HANS P | |||
| M | |||
| M., SCHERTZER JONATHAN D., Measuring GLUT4 translocation in mature muscle | |||

