ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к новому антителу и фрагменту этого антитела, которые специфически связываются с TIGIT, а также к композиции, включающей это антитело или его фрагмент. Кроме того, настоящее изобретение относится к нуклеиновой кислоте, кодирующей это антитело или его фрагмент, клетке-хозяину, содержащей эту нуклеиновую кислоту, и ее использованию. Кроме того, настоящее изобретение относится к терапевтическому или диагностическому применению этого антитела или его фрагмента.
УРОВЕНЬ ТЕХНИКИ
TIGIT (T-клеточный иммунный рецептор, включающий домены иммуноглобулина (Ig) и ITIM, также известный как WUCAM, Vstm3 или Vsig9) был впервые открыт в качестве члена семейства CD28 в ходе сравнений методами биоинформатики. TIGIT - это коингибирующий рецептор, который экспрессируется на поверхностной мембране различных иммунных клеток (природных киллеров (NK-клеток), активированных T-лимфоцитов, T-клеток памяти, регуляторных T-клеток, фолликулярных T-хелперов и т. д.) и является членом суперсемейства иммуноглобулинов. Полагают, что молекулы TIGIT могут осуществлять регуляцию иммунной системы за счет трех механизмов: 1) TIGIT конкурирует с костимулирующим рецептором CD226 за связывание с общим лигандом CD155/CD112, экспрессирующимся на поверхности дендритных или опухолевых клеток, и передает сигналы ингибирования клеткам, экспрессирующим TIGIT, тем самым подавляя активацию этих клеток; 2) TIGIT взаимодействует непосредственно с костимулирующим рецептором CD226, препятствуя гомодимеризации CD226, блокируя его сигнал активации, передаваемый вниз по цепи передачи сигнала; 3) после того как TIGIT связывается с CD155, который экспрессируется дендритными клетками, внутриклеточная последовательность ITIM передает сигналы ингибирования, в результате чего экспрессируются иммуносупрессивные цитокины, подавляющие активацию иммунной системы. TIGIT может играть определенную роль в самых разных клетках, составляющих микроокружение опухоли. В числе таких клеток могут быть опухоль-инфильтрирующие CD8+ Т-лимфоциты, регуляторные T-клетки, а также NK-клетки. В целом, из целого ряда исследований следует, что иммуносупрессивное действие TIGIT опосредуется первым из перечисленных механизмов.
Показано, что связывание антител с рецептором TIGIT человека может быть использовано для лечения онкологических заболеваний. См., например, WO 2006/124667. Antibody blockade of PD-L1 and TIGIT can increase the CD8+ T cell-mediated tumor rejection in a synergistic manner in a mouse model. Grogan et al. (2014) J. Immunol. 192(1) Suppl. 203.15; Johnston et al. (2014) Cancer Cell 26:1-15. Сходные результаты получены на животных с экспериментальной моделью меланомы. Inozume et al. (2014) J. Invest. Dermatol. 134:S121-Abstract 693.
Та роль, которую TIGIT играет в иммунном ответе, позволяет считать его привлекательной мишенью для иммунотерапии опухолей. Соответственно, существует потребность в разработке новых антител к TIGIT, в частности TIGIT-связывающих антител, способных блокировать CD155/CD112, и, более конкретно, человеческих антител к TIGIT, а также комбинированных препаратов на их основе, для лечения заболеваний, в частности для лечения онкологических заболеваний.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Предметом настоящего изобретения являются антитело к TIGIT, кодирующий это антитело ген, и их использование. С использованием методов генной инженерии и дрожжевого дисплея изобретатель в ходе скрининга отобрал полностью человеческое антитело к рецептору TIGIT человека из библиотеки человеческих антител, представленной на поверхности клеток дрожжей, а затем с использованием аффинного созревания получил высокоаффинное антитело к рецептору TIGIT человека. Молекула полностью гуманизированного антитела по настоящему изобретению способна эффективно блокировать связывание TIGIT с его лигандом CD155, ослаблять или элиминировать сигналы ингибирования, передаваемые клеткам, увеличивать продукцию IL-2, а также подавлять опухолевый рост в случае применения in vivo, причем эффект подавления опухоли выражен особенно сильно в случае применения этого антитела в комбинации с антителом к PD-1. Таким образом, антитело по настоящему изобретению может быть использовано для различных целей, включая, помимо прочего, усиление иммунного ответа, подавление опухолевого роста, противодействие инфекциям и выявление белков TIGIT.
Предметом настоящего изобретения являются новое полностью гуманизированное антитело, связывающееся с рецептором TIGIT человека, и его антиген-связывающий фрагмент.
В некоторых вариантах осуществления изобретения антитело к TIGIT по настоящему изобретению обладает одним или несколькими из следующих свойств:
(i) способность связываться с TIGIT человека с высокой аффинностью;
(ii) способность давать перекрестную иммунологическую реакцию с TIGIT обезьяны и/или мыши;
(iii) эффективное связывание с TIGIT на клеточной поверхности;
(iv) блокирование связывания TIGIT с его лигандом CD155;
(v) ослабление ингибирующего действия связывания CD155 с TIGIT в отношении IL-2-зависимого нисходящего сигнального пути;
(vi) увеличение продукции IL-2 T-лимфоцитами;
(vii) противоопухолевая активность, например подавление опухолевого роста;
(viii) способность оказывать более выраженное подавляющее действие в отношении опухоли в комбинации с антителом к PD-1, например усиленное ингибирование роста опухоли.
В некоторых вариантах осуществления изобретения предметом настоящего изобретения являются антитело или его фрагмент, которые связываются с TIGIT, в том числе: последовательности HCDR1, HCDR2 и HCDR3 одной из вариабельных областей тяжелой цепи, представленные в SEQ ID NO: 84-103, и (или) последовательности LCDR1, LCDR2 и LCDR3 вариабельных областей легкой цепи, представленные в SEQ ID NO: 104-110, либо комбинация CDR-последовательностей, описанных выше.
В некоторых вариантах осуществления изобретения предметом настоящего изобретения также являются антитело к TIGIT или его антиген-связывающий фрагмент. Антитело к TIGIT и иллюстративное (типовое) антитело (например, антитело, сочетающее в себе комбинацию последовательностей антитела VH и VL, перечисленных в Таблице B) по настоящему изобретению связываются с одним и тем же эпитопом или с перекрывающимися эпитопами и (или) конкурируют за связывание с TIGIT, и (или) антитело к TIGIT подавляет (например, конкурентно ингибирует) иллюстративное антитело по настоящему изобретению.
В некоторых вариантах осуществления изобретения предметом настоящего изобретения является нуклеиновая кислота, кодирующая антитело или его антиген-связывающий фрагмент, раскрытая ниже, вектор, содержащий нуклеиновую кислоту, и клетка-хозяин, содержащая этот вектор.
В некоторых вариантах осуществления изобретения предметом настоящего изобретения является метод получения антитела или его антиген-связывающего фрагмента, раскрытый ниже.
В некоторых вариантах осуществления изобретения предметом настоящего изобретения является иммуноконъюгат, фармацевтическая композиция и комбинированный препарат, включающие антитело по настоящему изобретению.
Предметом настоящего изобретения также является способ блокирования связывания TIGIT с CD155 (например, с CD155, экспрессированным на поверхности дендритной клетки или опухолевой клетки) у субъекта, применяющего антитело по настоящему изобретению. В некоторых вариантах осуществления изобретения сигналы ингибирования, опосредованные TIGIT, ослабляются или элиминируются в клетках (в частности, в T-лимфоцитах и NK-клетках), экспрессирующих TIGIT, с тем чтобы вызвать активацию T-лимфоцитов и NK-клеток. В некоторых вариантах осуществления изобретения за счет предлагаемого метода увеличивается продукция IL-2 T-лимфоцитами. В некоторых вариантах осуществления изобретения за счет предлагаемого метода снижается опосредованная CD155 передача сигналов ингибирования в клетках (например, в дендритных клетках), экспрессирующих CD155, в результате чего уменьшается экспрессия иммуносупрессивных цитокинов. В некоторых вариантах осуществления изобретения за счет предлагаемого метода происходит активация иммунной системы. Соответственно, предметом настоящего изобретения также является способ профилактики или лечения опухолевых заболеваний или инфекции с использованием антитела по настоящему изобретению.
Настоящее изобретение также относится к способу обнаружения TIGIT в образце.
Настоящее изобретение дополнительно проиллюстрировано следующими рисунками и конкретными примерами. Однако эти рисунки и конкретные примеры осуществления изобретения не следует воспринимать как ограничивающие объем изобретения, и в объем правовой охраны, определяемый формулой изобретения, также будут включены любые модификации, легко осуществимые специалистами в данной области техники и отвечающие сути настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
РИС. 1A-F иллюстрируют связывание антител, экспрессированных дрожжами, с TIGIT человека, экспрессированным на поверхности клеток CHO, до и после аффинного созревания.
РИС. 2 иллюстрирует связывание 7 антител-кандидатов, экспрессированных клетками CHO, с TIGIT человека, экспрессированным на поверхности клеток CHO.
РИС. 3A-F иллюстрируют способность антител, экспрессированных дрожжами, блокировать связывание с CD155 до и после аффинного созревания.
РИС. 4 иллюстрирует способность 7 антител-кандидатов, экспрессированных клетками CHO, блокировать связывание с CD155.
РИС. 5A-E иллюстрируют результаты анализа биологической активности, лежащей в основе механизма действия (МД), для антител, экспрессированных дрожжами, до и после аффинного созревания.
РИС. 6 иллюстрирует результаты анализа биологической активности, лежащей в основе МД, для 7 антител-кандидатов, экспрессированных клетками CHO.
РИС. 7 иллюстрирует результаты фармакодинамических исследований молекул-кандидатов ADI-30293 и ADI-30278, которые вводились по отдельности (в дозе 10 мг/кг) мышам линии MC38 с внедренным (knock-in) геном рецептора TIGIT человека.
РИС. 8A-B иллюстрируют результаты фармакодинамических исследований молекул-кандидатов ADI-30293 и ADI-30278, которые вводились (в дозе 10+1 мг/кг) в комбинации с антителом к PD1 (антитело C, WO 2017133540A1) мышам линии MC38 с внедренным (knock-in) геном рецептора TIGIT человека.
РИС. 9 иллюстрирует типовую последовательность рецептора TIGIT человека, использованную в примерах по настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
Если не указано иное, все технические и научные термины, используемые в этом документе, имеют общепринятый смысл, придаваемый им специалистами в данной области техники. В целях настоящего изобретения, ниже даны определения следующих терминов.
Термин «примерно», используемый в комбинации с численным значением, предназначен для охвата диапазона численных значений от нижней границы, отличающейся от указанного численного значения на 5% в меньшую сторону, до верхней границы, отличающейся от указанного численного значения на 5% в большую сторону.
Термин «и (или)» следует понимать, как относящийся к любому из вариантов или к любым двум или большему числу вариантов.
Используемые в этом документе термины «содержать» или «включать» следует понимать как включение элементов, целых чисел или этапов, но не как исключение каких-либо других элементов, целых чисел или этапов. Используемые в этом документе термины «содержать» или «включать», если не указано иное, также относятся к ситуации, когда целое состоит из описанных элементов, целых чисел или этапов. Например, если сказано, что «включена» вариабельная область антитела с определенной последовательностью, также подразумевается, что включена вариабельная область антитела, состоящая из определенной последовательности.
Используемый в этом документе термин «антитело» относится к полипептиду, включающему по меньшей мере одну вариабельную область легкой цепи иммуноглобулина или вариабельную область тяжелой цепи иммуноглобулина, которая специфически узнает антиген и связывается с ним. Термин «антитело» охватывает различные структуры антител, включая, среди прочего, моноклональные антитела, поликлональные антитела, одноцепочечные или полицепочечные антитела, моноспецифические или полиспецифические антитела (например, биспецифические антитела), полностью человеческие или химерные, или гуманизированные антитела, а также полноразмерные антитела и фрагменты антител, при условии, что они обладают требуемой антиген-связывающей активностью.
Специалистам в данной области техники должно быть понятно, что «цельное антитело» (в этом документе термин используется на равных основаниях с терминами «полноразмерное антитело», «полное антитело» и «интактное антитело») включает по крайней мере две тяжелых цепи (H) и две легких цепи (L). Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (в этом документе используется сокращение VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из 3 доменов: CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (в этом документе используется сокращение VL) и константной области легкой цепи. Константнакя область легкой цепи состоит из домена CL. Вариабельные области - это домены тяжелых или легких цепей антител, которые участвуют в связывании антител с их антигенами. Константные области не принимают непосредственного участия в связывании антител с антигенами, однако выполняют ряд эффекторных функций. Легкие цепи антитела могут быть отнесены к одному из двух типов, каппа (κ) и лямбда (λ), в зависимости от аминокислотной последовательности их константного домена. В зависимости от аминокислотной последовательности константного участка тяжелой цепи, тяжелая цепь антитела может быть отнесена к одному из 5 основных типов, а именно IgA, IgD, IgE, IgG и IgM; некоторые типы тяжелых цепей могут быть дополнительно разделены на подтипы, например IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные участки тяжелой цепи, соответствующие 5 различным типам антител, называются α, δ, ε, γ и μ соответственно. Термин «изотип» относится к типу антител, который определяется константным участком тяжелой цепи антитела. См., например, издание Fundamental Immunology, Ch.7 (Paul, w. Eds., 2nd edition, Raven Press, N.Y. (1989)), которое включено в настоящий документ путем ссылки целиком и безоговорочно.
Термин «антиген-связывающая часть» (в этом документе термин используется на равных основаниях с терминами «фрагмент антитела» и «антиген-связывающий фрагмент») антитела относится к неполной молекуле антитела, включающей часть интактного антитела для связывания с антигеном, с которым связывается интактное антитело. Специалистам в данной области техники должно быть понятно, что антиген-связывающая часть антитела, как правило, включает в себя аминокислотные остатки из «участка, определяющего комплементарность» или «CDR». Антиген-связывающий фрагмент может быть получен с использованием технологии рекомбинантных ДНК либо путем ферментативного или химического расщепления интактного антитела. Антиген-связывающие фрагменты включают, в числе прочего, Fab, scFab, Fab', F(ab')2, Fab'-SH, Fv, одноцепочечный Fv, диатело, триатело, тетратело, минитело и однодоменное антитело (sdAb). Более подробные описания фрагментов антител см. в Fundamental Immunology, W.E. Paul eds., Raven Press, N.Y. (1993); Shao Rongguang et al. (eds.), Antibody Drug Research and Application, People's Medical Publishing House (2013); Hollinger et al., PNAS USA 90: 6444-6448 (1993); Hudson et al., Nat. Med. 9:129-134 (2003).
Термины «человеческое антитело» и «полностью гуманизированное антитело» в этом документе используются на равных основаниях и относятся к антителу, включающему вариабельные области, в которых каркасные области и CDR-участки происходят из последовательностей человеческих иммуноглобулинов в зародышевой конфигурации. Кроме того, если антитело включает константные области, то константные области также происходят из последовательностей человеческих иммуноглобулинов в зародышевой конфигурации. Человеческое антитело, раскрытое в этом документе, может включать аминокислотные последовательности (например, мутации, введенные путем случайного или сайт-специфического мутагенеза in vitro либо в ходе соматического мутагенеза in vivo), которые отсутствуют в последовательностях человеческого иммуноглобулина в зародышевой конфигурации, например в CDR, и, в частности, в CDR3. Однако термин «человеческое антитело», используемый в этом документе, не включает антитела, в которых CDR-последовательности происходят из зародышевой линии других видов млекопитающих (например, мышей), пересаженных на человеческие каркасные последовательности.
В настоящем документе термин «рекомбинантное человеческое антитело» включает любые человеческие антитела, которые получены, экспрессированы, произведены или выделены с использованием технологии рекомбинантных ДНК, например: (а) антитела, выделенные из трансгенных или трансхромосомных животных (например, мышей) с использованием генов человеческих иммуноглобулинов или из гибридом, полученных из генов человеческих иммуноглобулинов; (б) антитела, выделенные из клеток-хозяев (например, трансфектом), трансформированных для экспрессии человеческих антител; (в) антитела, выделенные из рекомбинантных или комбинаторных библиотек человеческих антител (например, библиотек на основе дрожжевого дисплея); (г) антитела, которые получены, экспрессированы, произведены или выделены любым другим способом, включая сплайсинг генов человеческих иммуноглобулинов с получением других последовательностей ДНК. Эти рекомбинантные человеческие антитела имеют вариабельные области, в которых каркасные области и CDR-участки происходят из последовательностей человеческих иммуноглобулинов в зародышевой конфигурации. Однако в определенных вариантах осуществления изобретения рекомбинантные человеческие антитела могут быть подвергнуты мутагенезу in vitro (или соматическому мутагенезу in vivo в случае трансгенных животных с использованием последовательности Ig человека), и аминокислотные последовательности VH- и VL-областей полученных таким образом рекомбинантных антител, хотя они и происходят из «зародышевых» VH- и VL-последовательностей человека, не встречаются в библиотеке человеческих антител, основанной на природных «зародышевых» последовательностях.
Термин «химерное антитело» относится к антителу, в котором последовательности вариабельной области происходят из одного вида, а последовательности константной области происходят из другого вида, например антитело, в котором последовательности вариабельной области происходят из мышиного антитела, а последовательности константной области происходят из человеческого антитела.
Термин «гуманизированное антитело» относится к антителу, в котором CDR-последовательности, происходящие из млекопитающего другого вида, например мыши, соединены с каркасными человеческими последовательностями. В каркасные человеческие последовательности могут быть внесены дополнительные модификации.
«Изолированное» антитело - это антитело, которое отделено от компонентов его природной среды. В некоторых вариантах осуществления изобретения антитело очищено до степени чистоты выше 95% или 99%, как определено, например, методом электрофореза (например, ДНС-ПААГ, изоэлектрическим фокусированием (ИЭФ), капиллярным электрофорезом) или хроматографии (например, ионообменной или обращенно-фазовой ВЭЖХ). Обзор методов определения чистоты антител приведен, например, в работе Flatman, S. et al., J.Chrom.B 848 (2007) 79-87.
Термин «эпитоп» означает участок антигена, с которым связывается антитело. Эпитопы могут быть образованы как аминокислотами, составляющими непрерывную последовательность, так и аминокислотами, не составляющими непрерывную последовательность, но сближенными в пространстве при сворачивании белка в третичную структуру.
В этом документе TIGIT относится к «T-клеточному иммунному рецептору, включающему домены Ig и ITIM». Этот термин также включает варианты, изотипы, гомологи и видовые гомологи TIGIT. Термин «TIGIT человека» относится к последовательности TIGIT человека. Одна специфическая последовательность TIGIT человека представлена в SEQ ID NO: 208. В некоторых вариантах осуществления изобретения аминокислотная последовательность TIGIT человека по меньшей мере на 95%, и даже по меньшей мере на 96%, 97%, 98% или 99%, идентична аминокислотной последовательности TIGIT человека, представленной в SEQ ID NO: 208. Белки TIGIT также могут включать фрагменты TIGIT, такие как фрагменты, содержащие внеклеточные домены, например фрагменты, сохраняющие способность связываться с любым из антител, раскрытых в настоящем документе. В этом документе CD155, также известный как PVR (полиовирусный рецептор), PVS, HVED, CD155, NECL5, TAGE4 и Necl-5, взаимодействует с TIGIT, индуцируя сигналы иммуносупрессии.
Термин «специфически связывается с» означает, что антитело избирательно или предпочтительно связывается с антигеном. Если антитело связывается с TIGIT человека с KD примерно 5×10-7 M или меньше, примерно 1×10-7 M или меньше, примерно 5×10-8 M или меньше, примерно 1×10-8 M или меньше, примерно 5×10-9 M или меньше, как определено методом оптической интерферометрии, то это антитело является антителом, «специфически связывающимся с TIGIT человека». Однако антитело, специфически связывающееся с TIGIT человека, может проявлять перекрестную реактивность с белками TIGIT из других видов. Например, в некоторых вариантах осуществления изобретения антитело, специфически связывающееся с TIGIT человека, может перекрестно реагировать с белками TIGIT из видов, отличных от человека. В других вариантах осуществления изобретения антитело, специфически связывающееся с TIGIT человека, может быть полностью специфично по отношению к TIGIT человека и не проявлять перекрестную реактивность с другими видами либо проявлять перекрестную реактивность только с TIGIT из определенных видов.
В настоящем документе термин «перекрестная реактивность» относится к способности антитела связываться с TIGIT из разных видов. Например, описанное в этом документе антитело, связывающееся с TIGIT человека, может также связываться с TIGIT из других видов (например, с TIGIT обезьяны и (или) мыши). Метод определения перекрестной реактивности включает метод, описанный в примерах, а также стандартные методы анализа, известные специалистам в данной области техники, такие как биологическая оптическая интерферометрия или поточная цитометрия.
Термины «аффинность» или «аффинность связывания» относятся к аффинности связывания, присущей данной паре молекул, которая отражает особенности взаимодействия между двумя молекулами, связывающимися друг с другом. Аффинность молекулы X по отношению к партнеру Y в общем случае может быть охарактеризована равновесной константой диссоциации (KD), представляющей собой отношение константы скорости диссоциации (kdis) и константы скорости ассоциации (kon). Аффинность может быть измерена обычными методами, известными специалистам в данной области техники. Одним из методов измерения аффинности является кинетический анализ связывания ForteBio, описанный в этом документе.
В случае IgG-антитела термин «высокоаффинное» означает, что антитело связывается с антигеном-мишенью с KD 1×10-7 M или меньше, в предпочтительном варианте 5×10-8 M или меньше, в более предпочтительном примерно 1×10-8 M или меньше, в наиболее предпочтительном примерно 5×10-9 M или меньше. Однако «высокоаффинное» связывание может пониматься по-разному в зависимости от изотипа антитела. Например, для антитела изотипа IgM «высокоаффинное» означает, что антитело имеет KD 1×10-6 M или меньше, в предпочтительном варианте 1×10-7 M или меньше, в более предпочтительном примерно 1×10-8 M или меньше.
Термин «антитело, конкурирующее за связывание» с антигеном (например, TIGIT) с референтным антителом означает, что антитело блокирует связывание референтного антитела с антигеном (например, TIGIT) на 50% или больше в анализе конкурентного связывания и, наоборот, референтное антитело блокирует связывание данного антитела с антигеном (например, TIGIT) на 50% или больше в анализе конкурентного связывания. Примеры анализов конкурентного связывания описаны в издании "Antibodies", Harbor and Lane (Cold Spring Harbor Press, Cold Spring Harbor, NY). Антитело, конкурирующее за связывание, и референтное антитело могут связываться с одной и той же эпитопной областью, например с одним и тем же эпитопом, соседними эпитопами или перекрывающимися эпитопами.
Под антителом, которое ингибирует (например, конкурентно ингибирует) связывание референтного антитела с его антигеном, понимается антитело, которое ингибирует связывание референтного антитела с его антигеном на 50%, 60%, 70%, 80%, 90%, 95% или больше. И, наоборот, референтное антитело ингибирует связывание данного антитела с его антигеном на 50%, 60%, 70%, 80%, 90%, 95% или больше. Связывание антитела с антигеном может быть охарактеризовано аффинностью (например, равновесной константой диссоциации). Методы определения аффинности известны специалистам в данной области техники.
Под антителом, демонстрирующим ту же или аналогичную аффинность связывания и (или) специфичность, что и референтное антитело, понимается антитело, способное демонстрировать аффинность связывания и (или) специфичность, составляющие по меньшей мере 50%, 60%, 70%, 80%, 90%, 95% или более процентов от таковых референтного антитела. Это может быть установлено любым методом, используемым для определения аффинности связывания и (или) специфичности и известным специалистам в данной области техники.
Термин «Fc-участок» используется в данном документе для определения C-концевого участка тяжелой цепи иммуноглобулина, включающего по крайней мере часть константной области. Этот термин включает Fc-участок нативной последовательности и вариантный Fc-участок. В одном из вариантов осуществления изобретения Fc-участок тяжелой цепи человеческого иммуноглобулина начинается от остатка Cys226 или Pro230 тяжелой цепи и заканчивается карбоксильным концом. Однако C-концевой лизин (Lys447) Fc-участка может присутствовать или отсутствовать. Если не указано иное, в этом документе нумерация аминокислотных остатков в Fc-участке или константной области основана на системе нумерации EU, также известной как индекс EU, которая описана в издании Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th edition, Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242.
Термин «вариант» в отношении к антителу в данном документе относится к антителу, включающему целевую область антитела, аминокислоты в которой изменены по сравнению с референтным антителом за счет по крайней мере одной модификации, например 1-30, 1-20 или 1-10 модификаций, например путем замен, делеций и (или) вставок 1, 2, 3, 4 или 5 аминокислот, и при этом вариант в существенной степени сохраняет по крайней мере одно биологическое свойство (например, способность связываться с антигеном) молекулы антитела до внесения изменений. Целевая область антитела может представлять собой полноразмерное антитело или вариабельную область тяжелой цепи, или вариабельную область легкой цепи, или их комбинацию, либо одну или несколько CDR-участков тяжелой цепи, или одну или несколько CDR-участков легкой цепи, или их комбинацию. В данном документе область антитела, аминокислоты в которой изменены по сравнению с областью референтного антитела, также называется «вариантом» области референтного антитела.
В данном документе термин «идентичность последовательности» относится к степени идентичности последовательностей нуклеотидов или аминокислот в окне сравнения. «Процент идентичности последовательности» может быть рассчитан с использованием следующих этапов: сравнение двух оптимально сопоставленных последовательностей в окне сравнения; определение числа положений, в которых идентичные остатки нуклеотидов (напр., A, T, C, G и I) или аминокислот (напр., Ala, Pro, Ser, Thr, Gly, Val, Leu, Ile, Phe, Tyr, Trp, Lys, Arg, His, Asp, Glu, Asn, Gln, Cys и Met) встречаются в обеих последовательностях, с получением числа соответствующих (правильно спаренных) положений; деление числа правильно спаренных положений на общее число положений в окне сравнения (то есть размер окна); и умножение результата на 100 для получения процента идентичности последовательности. Оптимальное сопоставление для определения процента идентичности последовательности может быть достигнуто различными методами, известными специалистам в данной области техники, например с использованием общедоступных компьютерных программ, таких как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить надлежащие параметры сопоставления последовательностей, включая алгоритм, необходимый для оптимального сопоставления сравниваемых последовательностей, будь то полноразмерных или целевых.
В применении к последовательностям антител, процент идентичности аминокислотной последовательности в данном документе определяется путем оптимального сопоставления последовательности антитела-кандидата с последовательностью референтного антитела, и, в предпочтительном варианте осуществления изобретения, оптимальное сопоставление выполняется в соответствии со схемой нумерации Kabat. В данном документе, если не задано окно сравнения (то есть целевая область антитела, подлежащая сравнению), то сопоставление выполняется с полной последовательностью референтного антитела. В некоторых вариантах осуществления изобретения в том, что касается антител, идентичность последовательности может быть обеспечена для вариабельной области тяжелой цепи и (или) вариабельной области легкой цепи, либо процент идентичности последовательности может относиться только к каркасным областям, в то время как последовательности соответствующих CDR-участков остаются идентичными на 100%.
Аналогичным образом, в отношении последовательностей антител, антитело-кандидат с целевой областью, аминокислоты в которой изменены по сравнению с референтным антителом, может быть определено на основании сопоставления.
В данном документе термин «консервативная замена» относится к изменению аминокислоты, в результате которого аминокислота заменяется на химически сходную аминокислоту. Модификации аминокислот, например замены, могут быть введены в антитело по настоящему изобретению стандартными методами, известными специалистам в данной области техники, например сайт-направленным мутагенезом или ПЦР-опосредованным мутагенезом.
Таблицы консервативных замен, в которых представлены функционально сходные аминокислоты, хорошо известны специалистам в данной области техники. В предпочтительном аспекте, консервативно замещенные остатки выбраны из Таблицы A консервативных замен, приведенной ниже, и, предпочтительно, являются предпочтительными консервативно замещенными остатками, представленными в Таблице A.
Таблица A
Все аспекты настоящего изобретения подробно обсуждаются в следующих разделах.
I. Антитело к TIGIT по настоящему изобретению
Одним аспектом настоящего изобретения является антитело или его антиген-связывающий фрагмент, в частности полностью гуманизированное антитело или его антиген-связывающий фрагмент, которые специфически связываются с TIGIT, предпочтительно с человеческим белком TIGIT (напр., с последовательностью TIGIT человека, представленной в SEQ ID NO: 208). В некоторых вариантах осуществления изобретения антиген-связывающий фрагмент антитела, раскрытого в настоящем документе, представляет собой фрагмент антитела, выбранный из следующего перечня: Fab, Fab', Fab'-SH, Fv, одноцепочечных антител, таких как scFv, (Fab')2-фрагментов, однодоменных антител, диател (dAb) или линейных антител.
Биологические свойства антител, обеспечивающие их преимущества
В некоторых вариантах осуществления изобретения антитело к TIGIT или его антиген-связывающий фрагмент, раскрытые в настоящем документе, связываются с TIGIT человека с высокой аффинностью, например с равновесной константой диссоциации (KD) меньше 100×10-9 M, меньше или равной примерно 50×10-9 M, в предпочтительном варианте меньше или равной примерно 1-30×10-9 M, и более предпочтительно - примерно 1×10-9 M, в еще более предпочтительном варианте примерно 1-10×10-10 M. В предпочтительном варианте, KD определяют методом биологической оптической интерферометрии (например, методом анализа аффинности Fortebio). В некоторых вариантах осуществления изобретения KD определяют путем измерения аффинности моновалентного связывания Fab-фрагмента (например, Fab-фрагмента, экспрессируемого дрожжами) антитела с TIGIT человека. В предпочтительном варианте KD для моновалентного взаимодействия составляет 1-100×10-10 M, в более предпочтительном варианте 1-50×10-10 M, в еще более предпочтительном варианте 1-10×10-10 M или 2-5×10-10 M. В других вариантах осуществления изобретения KD определяют путем измерения аффинности моновалентного связывания интактного антитела (например, интактного антитела, экспрессируемого клетками CHO) с TIGIT человека. В предпочтительном варианте KD для моновалентного взаимодействия составляет 1-50×10-10 M, в более предпочтительном варианте 1-30 × 10-10 M или 1-10 × 10-10 M.
В некоторых вариантах осуществления изобретения антитело к TIGIT или его антиген-связывающий фрагмент, раскрытые в данном документе, обладают перекрестной реактивностью с белком TIGIT обезьяны. В некоторых вариантах осуществления изобретения антитело связывается с TIGIT обезьяны с высокой аффинностью, причем KD (например, определенная путем измерения аффинности моновалентного связывания интактного антитела с TIGIT обезьяны) составляет примерно 0,1-100×10-9 M, в более предпочтительном варианте 0,1-50×10-9 M или 1-30×10-10 M. В некоторых вариантах осуществления изобретения антитело обладает перекрестной реактивностью с белком TIGIT мыши, причем KD (например, определенная путем измерения аффинности моновалентного связывания интактного антитела с TIGIT мыши) составляет примерно 1-100×10-9 M, например 1-10×10-7 M или 1-10×10-8 M, или 1-10×10-9 M. В других вариантах осуществления изобретения антитело к TIGIT по настоящему изобретению не обладает перекрестной реактивностью с TIGIT мыши.
В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, связываются с TIGIT, экспрессируемым на поверхности клеток, с высокой аффинностью. В одном варианте осуществления изобретения клетками, экспрессирующими TIGIT человека на своей поверхности, являются клетки CHO. В предпочтительном варианте значение EC50, характеризующее связывание антитела с клетками, экспрессирующими TIGIT человека, измеряют методом поточной цитометрии (например, FACS). В некоторых вариантах осуществления изобретения антителом является интактное антитело, экспрессируемое дрожжами, с EC50 меньше, чем примерно 10 нМ, например 0,1-1 нМ, в предпочтительном варианте меньше или равно примерно 1 нМ, и более предпочтительно - примерно 0,2-0,9 нМ, например 0,9 нМ, 0,6 нМ или 0,4 нМ. В некоторых вариантах осуществления изобретения антителом является интактное антитело, экспрессируемое клетками CHO, с EC50 меньше, чем примерно 10 нМ, например 0,1-1 нМ, в предпочтительном варианте примерно 0,1-0,3 нМ, например примерно 0,3 нМ, 0,2 нМ или 0,1 нМ.
В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, подавляют соответствующую активность TIGIT. В некоторых вариантах осуществления данного изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, блокируют связывание TIGIT с его лигандом CD155. В предпочтительном варианте способность антитела блокировать связывание TIGIT человека (TIGIT, экспрессированного на клетках) с человеческим CD155 (например, IC50) измеряют методом поточной цитометрии (например, FACS). В некоторых вариантах осуществления изобретения антителом является интактное антитело, экспрессируемое дрожжами, с IC50 меньше, чем примерно 10 нМ, например 0,1-2 нМ, в предпочтительном варианте примерно 0,1-1,0 нМ, например 0,8 нМ, 0,6 нМ, 0,4 нМ или 0,2 нМ. В других вариантах осуществления изобретения антителом является интактное антитело, экспрессируемое клетками CHO, с EC50 меньше чем примерно 1 нМ, например 0,1-0,5 нМ, в предпочтительном варианте примерно 0,3 нМ, 0,2 нМ или 0,1 нМ.
В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, ослабляют или элиминируют сигналы ингибирования, вызываемые связыванием TIGIT с CD155. В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, ослабляют или элиминируют опосредованную TIGIT передачу сигналов ингибирования в клетке (в частности, в Т-лимфоците), экспрессирующей TIGIT. В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, индуцируют экспрессию в Т-лимфоцитах генов, расположенных ниже промоторов IL-2, а в некоторых вариантах увеличивают продукцию IL-2 в Т-лимфоцитах. В некоторых вариантах осуществления изобретения способность антитела ослаблять или элиминировать сигналы ингибирования, вызываемые связыванием TIGIT с CD155 (например, EC50), определяют с помощью анализа с флуоресцентным репортерным белком (например, анализ, основанный на МД, по примеру 5). В некоторых вариантах осуществления изобретения антителом является интактное антитело, экспрессируемое дрожжами, с EC50 в предпочтительном варианте меньше, чем примерно 10 нМ, например 0,1-5 нМ, и более предпочтительно - примерно 0,1-3,0 нМ, например примерно 2,0 нМ, 1,5 нМ, 1,0 нМ или 0,5 нМ. В других вариантах осуществления изобретения антителом является интактное антитело, экспрессируемое клетками CHO, с EC50 в предпочтительном варианте меньше, чем 5 нМ, например примерно 0,1-3,0 нМ, например примерно 1,6 нМ, 1,2 нМ, 1,0 нМ или примерно 0,18-0,5 нМ.
В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, подавляют рост опухоли, включающей инфильтрирующие лимфоциты, которые экспрессируют TIGIT человека. В одном варианте осуществления изобретения опухолью является опухоль желудочно-кишечного тракта, предпочтительно колоректальный рак. Например, в экспериментальной модели трансплантированной опухоли in vivo, например у мышей линии MC38, подавляется рост рака толстой кишки. В некоторых вариантах осуществления изобретения комбинированная терапия антителом по настоящему изобретению и антителом к PD-1 обеспечивает гораздо более значительный противоопухолевый эффект, чем каждое из антител по отдельности.
В предпочтительном варианте антитело или его антиген-связывающий фрагмент, раскрытые в настоящем документе, обладают по крайней мере одним, предпочтительно по крайней мере двумя, в более предпочтительном варианте по крайней мере тремя, четырьмя или пятью, а в еще более предпочтительном варианте - всеми вышеперечисленными свойствами.
CDR-участки антител
«участок, определяющий комплементарность», «CDR-участок» или «CDR» (эти термины в настоящем документе используются на равных основаниях с термином «гипервариабельная область» или «HVR») - это образуемая аминокислотами область в вариабельной области антитела, которая в основном отвечает за связывание с эпитопом антигена. CDR-участки тяжелых и легких цепей, как правило, обозначаются как CDR1, CDR2 и CDR3, причем нумерация увеличивается в направлении от N-конца. CDR-участки, расположенные в вариабельной области тяжелой цепи антитела, обозначаются как HCDR1, HCDR2 и HCDR3, в то время как CDR-участки, расположенные в вариабельной области легкой цепи антитела, обозначаются как LCDR1, LCDR2 и LCDR3.
Комбинации последовательностей VH и VL некоторых иллюстративных антител, раскрытых в данном документе, перечислены в Таблице B ниже:
Специалистам в данной области техники известны различные схемы (методы) идентификации последовательности CDR для данной аминокислотной последовательности VH или VL. Например, для идентификации участков, определяющих комплементарность (CDR), чаще всего используют метод Kabat, основанный на вариабельности последовательностей (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). Метод Chothia основан на расположении структурных петель (Chothia and Lesk, J. mol. biol. 196:901-917 (1987)). HVR-области, определяемые методом AbM, представляют собой компромисс между HVR, определяемыми по Kabat, и структурными петлями Chothia, и используются в программном средстве моделирования антител Oxford Molecular's AbM. Определение «контактных» HVR-областей основано на анализе имеющихся сложных кристаллических структур. В соответствии с различными схемами определения CDR-участков, остатки, входящие в каждую HVR/CDR, для этих HVR-областей определены следующим образом.
HVR-области также могут представлять собой последовательности HVR, определяемые следующими положениями остатков согласно системе нумерации Kabat:
положения 24-36 или 24-34 (LCDR1), положения 46-56 или 50-56 (LCDR2) и положения 89-97 или 89-96 (LCDR3) в VL; положения 26-35 или 27-35B (HCDR1), положения 50-65 или 49-65 (HCDR2) и положения 93-102, 94-102 или 95-102 (HCDR3) в VH.
В одном варианте осуществления изобретения HVR-области антитела, раскрытого в данном документе, представляют собой последовательности HVR, определяемые следующими положениями остатков согласно системе нумерации Kabat:
положения 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в VL, и положения 27-35B (HCDR1), 50-65 (HCDR2) и 93-102 (HCDR3) в VH.
В одном варианте осуществления изобретения HVR-области антитела, раскрытого в данном документе, представляют собой последовательности HVR, определяемые следующими положениями остатков согласно системе нумерации Kabat:
положения 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в VL, и положения 26-35B (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3) в VH.
HVR-области также могут быть определены на основе положений остатков согласно системе нумерации Kabat, соответствующих референтной последовательности CDR (например, любой из раскрытых в данном документе иллюстративных CDR).
Если не указано иное, термины «CDR», «последовательность CDR», «HVR» или «последовательность HVR», используемые в настоящем документе, включают последовательности CDR или HVR, определенные любым из описанных выше способов.
Если не указано иное, в настоящем документе положения остатков вариабельной области антитела (включая остатки вариабельной области тяжелой цепи и остатки вариабельной области легкой цепи) в этом изобретении пронумерованы в соответствии с системой нумерации Kabat (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)).
В одном предпочтительном варианте осуществления изобретения последовательности CDR, раскрытые в данном документе, показаны в Таблице 2, где HCDR1 представляет собой последовательность CDR, определенную методом AbM, а остальные CDR представляют собой последовательности CDR, определенные по методу Kabat.
В другом предпочтительном варианте осуществления изобретения последовательности CDR, раскрытые в данном документе, показаны в Таблице 1.
Комбинации некоторых иллюстративных последовательностей CDR, раскрытых в данном документе, перечислены в Таблице C ниже:
Комбинации некоторых иллюстративных последовательностей CDR, раскрытых в данном документе, перечислены в Таблице D ниже:
Антитела, различающиеся по специфичности (то есть имеющие разные участки связывания для разных антигенов), различаются и CDR-участками. Однако, несмотря на то что разные антитела различаются по CDR, в связывании с антигеном непосредственно участвует лишь ограниченное число аминокислотных остатков внутри CDR. С использованием по крайней мере двух из описанных методов (Kabat, Chothia, AbM и метод контакта) может быть определена наименьшая перекрывающаяся область, что позволяет определить «минимальную связывающую единицу» для связывания антигена. Минимальной связывающей единицей может быть субфрагмент CDR. Специалистам в данной области техники будет понятно, что остатки, входящие в остальные последовательности CDR, могут быть определены на основе знания структуры антитела и сворачивания белка. Поэтому любые варианты CDR, приведенные в этом документе, также учитываются в этом изобретении. Например, в одном из вариантов CDR аминокислотные остатки в минимальной связывающей единице могут оставаться неизменными, в то время как другие остатки CDR, определенные по Kabat или Chothia, могут быть замещены консервативными остатками аминокислот.
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению содержит по крайней мере одну, две, три, четыре, пять или шесть CDR, которые идентичны, или являются вариантами, соответствующим CDR внутри последовательностей вариабельной области любого из антител, перечисленных в Таблице B. В некоторых вариантах осуществления изобретения антитело по настоящему изобретению содержит по крайней мере одну, две или три HCDR, которые идентичны, или являются вариантами, соответствующим CDR тяжелой цепи внутри последовательностей вариабельной области любого из антител, перечисленных в Таблице B. В некоторых вариантах осуществления изобретения антитело по настоящему изобретению содержит по крайней мере одну, две или три LCDR, которые идентичны, или являются вариантами, соответствующим CDR легкой цепи внутри последовательностей вариабельной области любого из антител, перечисленных в Таблице B. В настоящем документе термин «соответствующие CDR» относится к CDR, которые расположены внутри аминокислотных последовательностей вариабельной области антитела-кандидата таким образом, что их расположение наиболее сходно с расположением CDR в референтном антителе после оптимального сопоставления. В настоящем документе вариантом CDR является CDR, который был модифицирован путем замены, делеции и (или) вставки по крайней мере одной аминокислоты, например одной или двух, или трех аминокислот, и при этом антиген-связывающая молекула, содержащая вариант CDR, в существенной степени сохраняет биологические свойства антиген-связывающей молекулы, содержащей немодифицированные CDR, например сохраняет по крайней мере 60%, 70%, 80%, 90% или 100% биологической активности (например, антиген-связывающей способности). Следует понимать, что каждый CDR может быть модифицирован независимо или в комбинации. В предпочтительном варианте, аминокислотная модификация представляет собой замену аминокислоты, в частности замену консервативной аминокислоты, например замену одной из консервативных аминокислот, перечисленных в Таблице A. В некоторых вариантах осуществления изобретения аминокислотные замены предпочтительно осуществляются в положениях аминокислот, соответствующих остаткам X консенсусных последовательностей CDR (например, представленные в SEQ ID NO: 5, 10,15, 21, 24, 30, 33, 37, 40, 47, 52, 56, 62, 65, 68, 74, 77 и 80), приведенных в этом документе.
Кроме того, специалистам в данной области техники известно, что область CDR3, которая является независимой от областей CDR1 и (или) CDR2, может сама по себе определять специфичность связывания антитела с его антигеном. Также на основе консенсусной последовательности CDR3 может быть получено множество других антител с той же специфичностью связывания. См., например, патенты США № 6,951,646; 6,914,128; 6,090,382; 6,818,216; 6,156,313; 6,827,925; 5,833,943; 5,762,905 и 5,760,185. Все эти патенты включены в настоящий документ путем ссылки.
Так, в одном из вариантов осуществления изобретения антитело по настоящему изобретению включает последовательность CDR3 из вариабельной области тяжелой и (или) легкой цепи любого из антител, приведенных в Таблице B, причем антитело способно специфически связываться с TIGIT человека. В еще одном варианте осуществления изобретения антитело может дополнительно включать CDR2 из вариабельной области тяжелой и (или) легкой цепи того же антитела или CDR2 из вариабельной области тяжелой и (или) легкой цепи другого антитела к TIGIT. В еще одном варианте осуществления изобретения антитело может дополнительно включать CDR1 из вариабельной области тяжелой и (или) легкой цепи того же антитела или CDR1 из вариабельной области тяжелой и (или) легкой цепи другого антитела к TIGIT. Активность этих антител, включая способность связываться с TIGIT человека, способность блокировать связывание TIGIT с молекулами CD155 и (или) способность подавлять рост опухоли, может быть охарактеризована аналитическими методами, описанными в этом документе.
В еще одном аспекте, учитывая что антиген-связывающая специфичность зависит главным образом от областей CDR1, CDR2 и CDR3, в некоторых вариантах осуществления изобретения последовательности VH CDR 1, 2 и 3, и последовательности VL CDR 1, 2 и 3 могут «комбинироваться и объединяться в пары» (например, CDR из разных антител, которые связываются с одним и тем же антигеном TIGIT, могут комбинироваться и объединяться в пары, и каждое антитело предпочтительно включает VH CDR 1, 2 и 3, и VL CDR 1, 2 и 3) для получения других молекул по настоящему изобретению, которые связываются с TIGIT. Связывание таких «скомбинированных и объединенных в пары» антител с TIGIT может быть изучено методами анализа связывания, которые известны специалистам в данной области техники (например, ELISA, SET и Biacore), а также другими методами анализа, описанными в примерах. В случае когда последовательности VH CDR комбинируются и объединяются в пары, последовательности CDR1, CDR2 и (или) CDR3 из конкретной последовательности VH в предпочтительном варианте замещаются структурно сходными последовательностями CDR. Сходным образом, когда последовательности VL CDR комбинируются и объединяются в пары, последовательности CDR1, CDR2 и (или) CDR3 из конкретной последовательности VL в предпочтительном варианте замещаются структурно сходными последовательностями CDR. CDR могут «комбинироваться и объединяться в пары» для антител по настоящему изобретению, показанных в Таблице 3. Кроме того, специалистам в данной области техники должно быть понятно, что антитела по настоящему изобретению могут быть также получены путем замены структурно сходных последовательностей CDR, раскрытых в этом документе, на одну или несколько последовательностей VH CDR и (или) VL CDR из других отличающихся антител.
Так, в некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают вариабельную область тяжелой цепи, содержащую определяющий комплементарность участок 3 тяжелой цепи (HCDR3), причем HCDR3:
(i) идентична HCDR3 вариабельной области тяжелой цепи какого-либо из антител, перечисленных в Таблице B;
(ii) идентична какой-либо из последовательностей HCDR3, перечисленных в Таблице C или D; или
(iii) включает по крайней мере 1 (предпочтительно 1-2, а более предпочтительно 1) аминокислотную модификацию (предпочтительно замену, а более предпочтительно консервативную замену) по сравнению с HCDR3 по пункту (i) или пункту (ii).
В некоторых вариантах осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем определяющие комплементарность участки 3 тяжелой цепи (HCDR3) и определяющие комплементарность участки 3 легкой цепи (LCDR3) антитела:
(i) идентичны последовательностям HCDR3 и LCDR3 вариабельной области тяжелой и легкой цепи какого-либо из антител, перечисленных в Таблице B;
(ii) идентичны последовательностям HCDR3 и LCDR3, перечисленных в Таблице C или D, в любой комбинации; или
(iii) включают всего по крайней мере 1 (предпочтительно 1-2, а более предпочтительно 1) аминокислотную модификацию (предпочтительно замену, а более предпочтительно консервативную замену) по сравнению с HCDR3 и LCDR3 по пункту (i) или пункту (ii).
В одном варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают вариабельную область тяжелой цепи (VH), причем VH включает:
(i) последовательности HCDR1, HCDR2 и HCDR3, содержащиеся в последовательности VH какого-либо из антител, перечисленных в Таблице B;
(ii) последовательности HCDR1, HCDR2 и HCDR3, перечисленные в Таблице C или D, в любой комбинации; или
(iii) последовательности, имеющие всего по крайней мере одну и не более 5, 4, 3, 2 или 1 аминокислотных модификаций (предпочтительно замен аминокислот, предпочтительно консервативных замен) в трех CDR по сравнению с последовательностями по пункту (i) или пункту (ii).
В другом варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают вариабельную область легкой цепи (VL), причем VL включает:
(i) последовательности LCDR1, LCDR2 и LCDR3, содержащиеся в последовательности VL какого-либо из антител, перечисленных в Таблице B;
(ii) последовательности LCDR1, LCDR2 и LCDR3, перечисленные в Таблице C или D, в любой комбинации; или
(iii) последовательности, имеющие всего по крайней мере одну и не более 5, 4, 3, 2 или 1 аминокислотных модификаций (предпочтительно замен аминокислот, предпочтительно консервативных замен) в трех CDR по сравнению с последовательностями по пункту (i) или пункту (ii).
В другом варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем антитело включает:
(i) шесть последовательностей CDR, содержащихся в последовательностях VH и VL какого-либо из антител, перечисленных в Таблице B;
(ii) шесть последовательностей CDR, перечисленных в Таблице C или D, в любой комбинации; или
(iii) последовательности, имеющие всего по крайней мере одну и не более 10, 5, 4, 3, 2 или 1 аминокислотных модификаций (предпочтительно замен аминокислот, предпочтительно консервативных замен) в шести CDR по сравнению с последовательностями по пункту (i) или пункту (ii).
В одном варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в данном документе, включают:
(i) последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, представленные в SEQ ID NO: 84, 85, 86 или 87 и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, представленные в SEQ ID NO: 104;
(ii) последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, представленные в SEQ ID NO: 88, 89 или 90 и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, представленные в SEQ ID NO: 105 или 106;
(iii) последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, представленные в SEQ ID NO: 91, 92 или 93 и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, представленные в SEQ ID NO: 107;
(iv) последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, представленные в SEQ ID NO: 94, 95, 96 или 97 и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, представленные в SEQ ID NO: 108;
(v) последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, представленные в SEQ ID NO: 98, 99 или 100 и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, представленные в SEQ ID NO: 109; или
(vi) последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, представленные в SEQ ID NO: 101, 102 или 103 и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, представленные в SEQ ID NO: 110.
В предпочтительном варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают три определяющие комплементарность участка из вариабельной области тяжелой цепи (HCDR) и три определяющие комплементарность участка из вариабельной области легкой цепи (LCDR), где
(i) HCDR1 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 1-5 или 178-181, HCDR2 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 6-10, HCDR3 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 11-15 или 182-185, LCDR1 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 16, LCDR2 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 17, а LCDR3 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 18;
(ii) HCDR1 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 19-21 или 186-187, HCDR2 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 22-24, HCDR3 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 25 и 188, LCDR1 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 26, LCDR2 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 27, а LCDR3 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 28-30;
(iii) HCDR1 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 31-33 или 189-190, HCDR2 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 34-37, HCDR3 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 38-40 или 191-192, LCDR1 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 41, LCDR2 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 42, а LCDR3 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 43;
(iv) HCDR1 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 44-47 или 193-195, HCDR2 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 48-52, HCDR3 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 53-56 или 196-198, LCDR1 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 57, LCDR2 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 58, а LCDR3 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 59;
(v) HCDR1 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 60-62 или 199-200, HCDR2 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 6 или 63-65, HCDR3 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 66-68 или 201-202, LCDR1 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 69, LCDR2 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 17, а LCDR3 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 70; или
(vi) HCDR1 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 71-74 или 203-205, HCDR2 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 22 или 75-77, HCDR3 содержит или состоит из аминокислотной последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 78-80 или 206-207, LCDR1 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 81, LCDR2 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 82, а LCDR3 содержит или состоит из аминокислотной последовательности, представленной в SEQ ID NO: 83.
В одном предпочтительном варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают 6 последовательностей CDR в одной из комбинаций, перечисленных в Таблице C.
В другом предпочтительном варианте осуществления изобретения антитело или его антиген-связывающий фрагмент, раскрытые в этом документе, включают 6 последовательностей CDR в одной из комбинаций, перечисленных в Таблице D.
Вариабельные области антител
«Вариабельная область» или «вариабельный участок» - это домен тяжелой или легкой цепи антитела, который участвует в связывании антитела с его антигеном. В вариабельной области тяжелой цепи (VH) и в вариабельной области легкой цепи (VL) можно дополнительно выделить гипервариабельные области (HVR, также известные как участки, определяющие комплементарность (CDR)) с более консервативными областями (каркасными областями (FR)), расположенными между ними. Каждый VH- или VL-домен состоит из трех CDR и четырех FR, которые располагаются в следующем порядке в направлении от N-конца к C-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В некоторых случаях антиген-связывающая специфичность может обеспечиваться одним единственным VH- или VL-доменом. Кроме того, антитела, связывающиеся с конкретными антигенами, могут быть выделены в ходе скрининга библиотек VL- или VH-доменов, определяющих комплементарность, благодаря способности VL- или VH-доменов антител связываться с антигенами (см, например, Portolano, S. et al., J. Immunol. 150 (1993) 880-887; Clackson, T. et al., Nature 352 (1991) 624-628).
Специалистам в данной области техники известно, что один или более остатков в одной или обеих вариабельных областях (VH и (или) VL) могут быть модифицированы, например могут быть модифицированы остатки в одном или более CDR-участке и (или) в одной или более каркасных областей, в частности путем замены консервативных остатков, и модифицированное таким образом антитело в существенной степени сохраняет по крайней мере одно биологическое свойство (например, способность связываться с антигеном) молекулы антитела до внесения изменений. Например, остатки в CDR-участках могут быть изменены за счет мутаций для улучшения одной или более характеристик антитела, относящихся к его связыванию с антигеном (например, аффинности). Антиген-связывающие характеристики или другие функциональные характеристики мутированного антитела могут изучаться методами анализа in vitro или in vivo. В предпочтительном варианте осуществляют консервативные замены. В предпочтительном варианте в CDR-участке вводят не более 1, 2, 3, 4 или 5 модификаций остатков. Кроме того, могут быть изменены за счет мутаций остатки в каркасных областях, например для улучшения характеристик антитела. Например, один или несколько остатков в каркасных областях могут быть заменены на соответствующие остатки «зародышевой» последовательности за счет «обратных мутаций».
Еще одним методом модификации вариабельной области антитела, известным специалистам в данной области техники, является «пересадка CDR». Поскольку последовательности CDR отвечают за большинство взаимодействий «антиген-антитело», может быть сконструирован вариант рекомбинантного антитела, имитирующий свойства известных антител. В таком варианте антитела последовательности CDR из известных антител «пересажены» на каркасные области других антител с отличающимися свойствами. Соответственно, в одном варианте осуществления изобретения настоящее изобретение относится к антителу к TIGIT или его антиген-связывающему фрагменту, где антитело или его фрагмент включает последовательности CDR из вариабельных областей тяжелых и легких цепей одного из антител, указанных в Таблице B, и другие последовательности каркасных областей. Последовательность каркасной области для замены может быть получена из общедоступных баз данных ДНК, включающих последовательности генов антител зародышевой линии, или из опубликованных последовательностей антител к TIGIT. Например, ДНК зародышевой линии, кодирующая гены вариабельных областей тяжелой и легкой цепи, может быть получена из базы данных GenBank. Последовательности белков антител могут быть сопоставлены с белковыми последовательностями в базе данных с использованием инструментов для анализа сходства последовательностей, таких как Gapped BLAST. В предпочтительном варианте последовательность каркасной области для замены обладает структурным сходством с каркасной последовательностью антитела по настоящему изобретению, выбранной для изменения, например с каркасной последовательностью, имеющей идентичность последовательности по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или более.
В еще одном варианте осуществления изобретения последовательности VH и VL из иллюстративного антитела по настоящему изобретению (одного из антител, приведенных в Таблице B) и других отличных от него антител к TIGIT (предпочтительно другого антитела, приведенного в Таблице B) могут «комбинироваться и объединяться в пары» для получения других антител по настоящему изобретению, связывающихся с TIGIT. В случае комбинирования и объединения в пары таких цепей предпочтительно, чтобы последовательность VH из определенной пары VH/VL заменялась на структурно сходную последовательность VH. Сходным образом, последовательность VL из определенной пары VH/VL в предпочтительном варианте заменяется на структурно сходную последовательность VL. Связывание таких «скомбинированных и объединенных в пары» антител с TIGIT может быть изучено методами анализа связывания, которые известны специалистам в данной области техники (например, ELISA, а также другими методами анализа, описанными в примерах).
Так, в одном варианте осуществления изобретения антитело по настоящему изобретению включает или состоит из последовательности вариабельной области тяжелой цепи (VH) какого-либо из антител, перечисленных в Таблице B. В другом варианте осуществления изобретения антитело по настоящему изобретению включает вариант последовательности VH.
В еще одном варианте осуществления изобретения антитело по настоящему изобретению включает или состоит из последовательности вариабельной области легкой цепи (VL) какого-либо из антител, перечисленных в Таблице B. В другом варианте осуществления изобретения антитело по настоящему изобретению включает вариант последовательности VL.
В еще одном варианте осуществления изобретения антитело по настоящему изобретению включает:
(i) последовательность VH, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 84, 85, 86 или 87, или ее вариант, и (или) последовательность VL, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 104 или ее вариант;
(ii) последовательность VH, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 88, 89 или 90, или ее вариант, и (или) последовательность VL, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 105 или 106, или ее вариант;
(iii) последовательность VH, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 91, 92 или 93, или ее вариант, и (или) последовательность VL, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 107 или ее вариант;
(iv) последовательность VH, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 94, 95, 96 или 97, или ее вариант, и (или) последовательность VL, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 108 или ее вариант;
(v) последовательность VH, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 98, 99 или 100, или ее вариант, и (или) последовательность VL, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 109 или ее вариант; или
(vi) последовательность VH, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 101, 102 или 103, или ее вариант, и (или) последовательность VL, содержащую аминокислотную последовательность, представленную в SEQ ID NO: 110 или ее вариант.
В одном варианте осуществления изобретения в том, что касается аминокислотной последовательности, вариант последовательности VH идентичен референтной последовательности VH (предпочтительно при сравнении с полноразмерной последовательностью или с областями CDR1, CDR2 и CDR3) по меньшей мере на 80%, 85%, 90%, 92%, 95%, 97%, 98%, 99% или более. В одном варианте осуществления изобретения в том, что касается аминокислотной последовательности, вариант последовательности VH включает не менее одной и не более 30, 10, 5, 4, 3, 2, 1 или 0 аминокислотных модификаций (предпочтительно замен аминокислот, а более предпочтительно консервативных замен) по сравнению с референтной последовательностью VH (предпочтительно при сравнении с полноразмерной последовательностью или с областями CDR1, CDR2 и CDR3). В предпочтительном варианте отличия в последовательностях не встречаются в CDR.
В предпочтительном варианте осуществления изобретения в том, что касается аминокислотной последовательности, вариант последовательности VL идентичен референтной последовательности VL (предпочтительно при сравнении с полноразмерной последовательностью или с областями CDR1, CDR2 и CDR3) по меньшей мере на 80%, 85%, 90%, 92%, 95%, 97%, 98%, 99% или более. В предпочтительном варианте осуществления изобретения в том, что касается аминокислотной последовательности, вариант последовательности VL включает не менее одной и не более 30, 10, 5, 4, 3, 2, 1 или 0 аминокислотных модификаций (предпочтительно замен аминокислот, а более предпочтительно консервативных замен) по сравнению с референтной последовательностью VH (предпочтительно при сравнении с полноразмерной последовательностью или с областями CDR1, CDR2 и CDR3). В предпочтительном варианте отличия в последовательностях не встречаются в CDR.
В предпочтительном варианте осуществления изобретения антитело по настоящему изобретению включает или состоит из пары последовательностей VH/VL из вариабельных областей тяжелой и легкой цепи какого-либо из антител, перечисленных в Таблице B. Предметом настоящего изобретения также является вариант антитела, например вариант, имеющий идентичность не менее 95-99% или содержащий не более 10 аминокислотных модификаций при сравнении с VH, VL или VH и VL.
В любом из вышеуказанных вариантов осуществления изобретения, предпочтительно в том, что касается одной или более CDR (предпочтительно всех трех CDR), вариабельная область тяжелой цепи варианта антитела включает не более 10, предпочтительно не более 5 (например, 3, 2, 1 или 0) аминокислотных модификаций (предпочтительно замен аминокислот, а более предпочтительно консервативных замен) по сравнению с референтным антителом.
В любом из вышеуказанных вариантов осуществления изобретения, предпочтительно в том, что касается одной или более CDR (предпочтительно всех трех CDR), вариабельная область легкой цепи (VL) варианта антитела включает не более 10, предпочтительно не более 5 (например, 3, 2, 1 или 0) аминокислотных модификаций (предпочтительно замен аминокислот, а более предпочтительно консервативных замен) по сравнению с референтным антителом.
Тяжелые и легкие цепи антител
В некоторых вариантах осуществления изобретения антитело по настоящему изобретению включает Fc-участок тяжелой цепи, например Fc-участок иммуноглобулинов с изотипами IgG1, IgG2 или IgG4. В одном варианте осуществления изобретения антитело по настоящему изобретению включает IgG4-Fc-участок, в котором имеется мутация с заменой серина на пролин (S228P) в положении аминокислоты 228 (нумерация EU). В еще одном предпочтительном варианте осуществления изобретения антитело по настоящему изобретению включает Fc-участок IgG4-PAA. В Fc-участке IgG4-PAA имеется мутация с заменой серина на пролин (S228P) в положении 228, мутация с заменой фенилаланина на аланин в положении 234 (нумерация EU) и мутация с заменой лейцина на аланин в положении 235 (нумерация EU). Мутация S228P - это мутация в шарнирном участке «опухолевой» константной области, которая способна уменьшить или устранить гетерогенность дисульфидного мостика между тяжелыми цепями. Мутации F234A и L235A могут дополнительно ослабить эффекторную функцию человеческого иммуноглобулина с изотипом IgG4 (эффекторная функция которого уже снижена). В некоторых вариантах осуществления изобретения антитело по настоящему изобретению включает Fc-участок IgG4-PAA с тяжелой цепью, с C-конца которой удален лизин (des-Lys). В некоторых вариантах осуществления изобретения антитело по настоящему изобретению включает константную область легкой цепи каппа (κ), например константную область человеческой легкой цепи κ.
В еще одном предпочтительном варианте осуществления изобретения Fc-участок включает аминокислотную последовательность, представленную в SEQ ID NO: 177 или аминокислотную последовательность, включающую не менее одной, двух или трех, но не более 20, 10 или 5 аминокислотных модификаций по сравнению с аминокислотной последовательностью, представленной в SEQ ID NO: 177, или последовательность идентична аминокислотной последовательности, представленной в SEQ ID NO: 177 по меньшей мере на 95-99%.
В предпочтительном варианте осуществления изобретения антитело по настоящему изобретению включает константную область легкой цепи. В предпочтительном варианте осуществления изобретения константная область легкой цепи представляет собой константную область легкой цепи κ человека. В еще одном предпочтительном варианте осуществления изобретения константная область легкой цепи включает аминокислотную последовательность, представленную в SEQ ID NO: 209 или аминокислотную последовательность, содержащую не менее одной, двух или трех, но не более 20, 10 или 5 аминокислотных модификаций по сравнению с аминокислотной последовательностью, представленной в SEQ ID NO: 209, или аминокислотная последовательность идентична аминокислотной последовательности, представленной в SEQ ID NO: 209 по меньшей мере на 95-99%.
В некоторых предпочтительных вариантах осуществления изобретения антитело по настоящему изобретению включает тяжелую цепь, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 111-130, или аминокислотную последовательность, содержащую не менее одной, двух или трех, но не более 20, 10 или 5 аминокислотных модификаций по сравнению с указанными последовательностями, или аминокислотная последовательность идентична указанным последовательностям по меньшей мере на 80%, 85%, 90%, 92%, 95%, 97%, 98%, 99% или более. Предпочтительно аминокислотные модификации не встречаются в CDR, а более предпочтительно аминокислотные модификации не встречаются в вариабельных областях.
В некоторых предпочтительных вариантах осуществления изобретения антитело по настоящему изобретению включает легкую цепь, содержащую аминокислотную последовательность, выбранную из SEQ ID NO: 137-143, или аминокислотную последовательность, содержащую не менее одной, двух или трех, но не более 20, 10 или 5 аминокислотных модификаций по сравнению с указанными последовательностями, или аминокислотная последовательность идентична указанным последовательностям по меньшей мере на 80%, 85%, 90%, 92%, 95%, 97%, 98%, 99% или более. Предпочтительно аминокислотные модификации не встречаются в CDR, а более предпочтительно аминокислотные модификации не встречаются в вариабельных областях.
В предпочтительном варианте осуществления изобретения антитело по настоящему изобретению включает последовательность тяжелой цепи и (или) последовательность легкой цепи, выбранные из:
(a) последовательность тяжелой цепи, включающая аминокислотную последовательность, выбранную из SEQ ID NO: 111-114, или ее вариант, и (или) последовательность легкой цепи, включающая аминокислотную последовательность, представленную в SEQ ID NO: 137 или ее вариант;
(b) последовательность тяжелой цепи, включающая аминокислотную последовательность, выбранную из SEQ ID NO: 115-117 или ее вариант, и (или) последовательность легкой цепи, включающая аминокислотную последовательность, представленную в SEQ ID NO: 138 или 139, или ее вариант;
(c) последовательность тяжелой цепи, включающая аминокислотную последовательность, выбранную из SEQ ID NO: 118-120 или ее вариант, и (или) последовательность легкой цепи, включающая аминокислотную последовательность, представленную в SEQ ID NO: 140 или ее вариант;
(d) последовательность тяжелой цепи, включающая аминокислотную последовательность, выбранную из SEQ ID NO: 121-124, или ее вариант, и (или) последовательность легкой цепи, включающая аминокислотную последовательность, представленную в SEQ ID NO: 141 или ее вариант;
(e) последовательность тяжелой цепи, включающая аминокислотную последовательность, выбранную из SEQ ID NO: 125-127, или ее вариант, и (или) последовательность легкой цепи, включающая аминокислотную последовательность, представленную в SEQ ID NO: 142 или ее вариант; и
(f) последовательность тяжелой цепи, включающая аминокислотную последовательность, выбранную из SEQ ID NO: 128-130 или ее вариант, и (или) последовательность легкой цепи, включающая аминокислотную последовательность, представленную в SEQ ID NO: 143 или ее вариант,
где вариант включает аминокислотную последовательность, содержащую не менее одной, двух или трех, но не более 20, 10 или 5 аминокислотных модификаций или имеющую идентичность не менее 80%, 85%, 90%, 92%, 95%, 97%, 98%, 99% или более процентов при сравнении с соответствующей референтной последовательностью. Предпочтительно аминокислотные модификации не встречаются в CDR, а более предпочтительно аминокислотные модификации не встречаются в вариабельных областях.
В одном варианте осуществления изобретения модификации остатков осуществлены в константной области антитела, например для изменения свойств антитела, таких как эффекторная функция.
В некоторых вариантах осуществления изобретения тяжелая и (или) легкая цепь антитела к TIGIT или его фрагмента, раскрытых в этом документе, дополнительно включают сигнальную пептидную последовательность, например METDTLLLWVLLLWVPGSTG.
Последовательности иллюстративных антител
Предметом настоящего изобретения является полностью гуманизированное антитело, специфически связывающееся с TIGIT (например, TIGIT человека), выделенное и охарактеризованное как описано в примерах. Последовательности VH и VL из вариабельных областей иллюстративных антител, раскрытых в этом документе, перечислены в Таблице 3 ниже. Иллюстративные последовательности CDR антител перечислены в Таблицах 1 и 2 ниже. В перечне последовательностей показаны аминокислотные последовательности тяжелых и легких цепей иллюстративных антител, раскрытых в этом документе, и кодирующие нуклеотидные последовательности вариабельных областей (VH и VL) иллюстративных антител, раскрытых в этом документе.
Варианты антител
Одним из аспектов настоящего изобретения является любое из антител, раскрытых в этом документе, в частности варианты иллюстративных антител, перечисленных в Таблице B. В одном варианте осуществления изобретения вариант антитела сохраняет не менее 60%, 70%, 80%, 90% или 100% биологической активности (например, антиген-связывающей активности) антитела до внесения модификаций. В некоторых вариантах осуществления изобретения модификация не приводит к потере способности варианта антитела связываться с антигеном, но необязательно может влиять на его свойства, например увеличивать аффинность связывания или другие эффекторные функции.
Должно быть понятно, что вариабельные области тяжелых или легких цепей или все CDR-участки антитела могут быть модифицированы независимо или в комбинации. В некоторых вариантах осуществления изобретения в один или более, или во все три CDR-участка тяжелой цепи внесено не более 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных модификаций. Предпочтительно аминокислотная модификация относится к замене аминокислоты, предпочтительно консервативной замене. В некоторых вариантах осуществления изобретения в один или более, или во все три CDR-участка легкой цепи внесено не более 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных модификаций. В некоторых вариантах осуществления изобретения в один или более, или во все шесть CDR-участков внесено не более 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных модификаций. Предпочтительно аминокислотная модификация относится к замене аминокислоты, предпочтительно консервативной замене. В некоторых вариантах осуществления изобретения вариант антитела имеет аминокислотную последовательность, которая по меньшей мере на 80%, 85%, 90%, 95%, 99% или более процентов идентична целевой области последовательности референтного антитела. Например, в одном варианте осуществления изобретения антитело по настоящему изобретению по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более процентов идентично референтному антителу (например, одному из антител, перечисленных в Таблице 3) в отношении трех CDR-участков тяжелой цепи. В одном варианте осуществления изобретения антитело по настоящему изобретению по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более процентов идентично референтному антителу (например, одному из антител, перечисленных в Таблице 3) в отношении трех CDR-участков легкой цепи. В другом варианте осуществления изобретения антитело по настоящему изобретению по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более процентов идентично референтному антителу (например, одному из антител, перечисленных в Таблице 3) в отношении шести CDR-участков. В еще одном варианте осуществления изобретения антитело по настоящему изобретению по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более процентов идентично референтному антителу (например, одному из антител, перечисленных в Таблице 3) в отношении вариабельной области тяжелой цепи. В еще одном варианте осуществления изобретения антитело по настоящему изобретению по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более процентов идентично референтному антителу (например, одному из антител, перечисленных в Таблице 3) в отношении вариабельной области легкой цепи. В еще одном варианте осуществления изобретения антитело по настоящему изобретению по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более процентов идентично референтному антителу (например, одному из антител, перечисленных в Таблице 3) в отношении вариабельных областей тяжелых и (или) легких цепей.
Кроме того, модификации могут быть внесены в Fc-участок антитела. Модификации в Fc-участке могут вноситься независимо или в комбинации с модификациями в каркасных областях и (или) CDR, описанными выше. Fc-участок может быть модифицирован, например, с целью изменения одной или более функций антитела, например периода полувыведения из сыворотки, связывания комплемента, связывания с Fc-рецептором и (или) антиген-зависимой цитотоксичности. Кроме того, антитело по настоящему изобретению может быть химически модифицировано (например, связано с ПЭГ) или может быть изменен его профиль гликозилирования.
В определенных вариантах осуществления изобретения Fc-участок может включать Fc-участок, имеющий одну или более аминокислотных замен, увеличивающих антителозависимую клеточную цитотоксичность (АЗКЦ), например замен в положениях 298, 333 и (или) 334 (нумерация остатков EU) Fc-участка. В некоторых вариантах осуществления изобретения Fc-участок также может быть модифицирован с целью изменения (то есть увеличения или уменьшения) связывания C1q и (или) комплементзависимой цитотоксичности (КЗЦ) (см., например, US 6,194,551, WO 99/51642 и Idusogie, E. E. et al., J. Immunol. 164 (2000) 4178-4184).
В других вариантах осуществления изобретения Fc-участок может быть модифицирован с целью увеличения или уменьшения степени его гликозилирования и (или) изменения его профиля гликозилирования. Удобным способом добавления или удаления сайтов гликозилирования в Fc-участке является добавление или удаление одного или более сайтов гликозилирования путем изменения аминокислотной последовательности. Например, для удаления одного или более сайтов гликозилирования и тем самым исключения гликозилирования в этих сайтах могут быть заменены одна или более аминокислот. Могут быть получены антитела с измененным профилем гликозилирования, например низко-фукозилированные или нефукозилированные антитела с пониженным содержанием фукозильных остатков, или антитела с повышенным содержанием структур с бисектным GlcNac. Показано, что такие измененные профили гликозилирования могут увеличивать АЗКЦ антител. В настоящем документе также рассматриваются варианты антител, имеющие по крайней мере один галактозный остаток в олигосахариде, связанном с Fc-участком. В вариантах антител может быть усилена функция КЗЦ.
В определенных вариантах осуществления изобретения настоящее изобретение также касается вариантов антител, обладающих некоторыми, но не всеми эффекторными функциями, что делает их желательными кандидатами для некоторых сфер применения, в которых важен период полувыведения антител in vivo, в то время как другие эффекторные функции (например, способность связывать комплемент или АЗКЦ) не нужны или вредны. Например, Fc-участок может содержать мутацию, которая элиминирует или снижает эффекторные функции, в частности Fc-участок человеческого IgG1 может иметь мутации P329G и (или) L234A и L235A, либо Fc-участок человеческого IgG4 может иметь мутации P329G и (или) S228P и L235E.
В определенных вариантах осуществления изобретения настоящее изобретение может касаться получения антител, модифицированных методами цистеиновой инженерии, таких как «сульфо-МАт», в которых один или более остатков антитела заменены на цистеиновые остатки. Например, ряд цистеиновых остатков в шарнирной области антитела могут быть изменены, например для облегчения сборки легких и тяжелых цепей, или для увеличения или уменьшения стабильности антитела. См., например, патент США № 5677425.
В определенных вариантах осуществления изобретения антитела, представленные в этом документе, могут быть дополнительно модифицированы с целью введения в них других небелковых элементов. Подходящие элементы для модификации антител включают в числе прочего водорастворимые полимеры. Примером водорастворимого полимера является в числе прочего полиэтиленгликоль (ПЭГ), который вводят, например, для увеличения периода полувыведения антитела (например, из сыворотки крови). Методы ПЭГилирования белков известны специалистам в данной области техники и могут быть применены к антителам по настоящему изобретению. См., например, EP 0154316 и EP 0401384.
II. Полинуклеотиды, векторы и организмы-хозяева
Предметом настоящего изобретения является нуклеиновая кислота, кодирующая любое из вышеописанных антител к TIGIT или его фрагментов, а также вектор, содержащий такую нуклеиновую кислоту. В одном варианте осуществления изобретения вектор является экспрессионным вектором. Кроме того, предметом изобретения является клетка-хозяин, содержащая такую нуклеиновую кислоту или вектор. В одном варианте осуществления изобретения клетка-хозяин является эукариотической. В другом варианте осуществления изобретения клетка-хозяин выбрана из клетки дрожжей и клетки млекопитающих (например, клетки 293 или клетки CHO). В еще одном варианте осуществления изобретения клетка-хозяин является прокариотической.
Одним из аспектов настоящего изобретения является нуклеиновая кислота, кодирующая любое из вышеописанных антител к TIGIT или его фрагментов. Нуклеиновая кислота может включать нуклеиновую кислоту, кодирующую аминокислотные последовательности вариабельных областей легких цепей и (или) вариабельных областей тяжелых цепей антител, или нуклеиновую кислоту, кодирующую аминокислотные последовательности легких цепей и (или) тяжелых цепей антител. Иллюстративные последовательности нуклеиновых кислот, кодирующих вариабельные области тяжелых цепей, включают последовательности нуклеиновых кислот, которые по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям нуклеиновых кислот, выбранных из SEQ ID NO: 150-169, или включают последовательности нуклеиновых кислот, выбранные из SEQ ID NO: 150-169. Иллюстративные последовательности нуклеиновых кислот, кодирующих вариабельные области легких цепей, включают последовательности нуклеиновых кислот, которые по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям нуклеиновых кислот, выбранных из SEQ ID NO: 170-176, или включают последовательности нуклеиновых кислот, выбранные из SEQ ID NO: 170-176. Полипептиды, кодируемые полинуклеотидами, могут демонстрировать способность связываться с антигеном (связываться с TIGIT) в случае их экспрессии в подходящем экспрессионном векторе.
Предметом настоящего изобретения также является полинуклеотид, кодирующий по крайне мере одну CDR, а, как правило, все три CDR из последовательности тяжелой цепи (VH) или последовательности легкой цепи (VL) описанных выше антител, связывающихся с TIGIT. В некоторых других вариантах осуществления изобретения полинуклеотид кодирует полную или практически полную последовательность вариабельной области тяжелых цепей и (или) легких цепей описанных выше антител, связывающихся с TIGIT.
Специалистам в данной области техники будет понятно, что аминокислотная последовательность каждого антитела или полипептида может кодироваться различными нуклеотидными последовательностями, что обусловлено вырожденностью кодона.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота, кодирующая антитело по настоящему изобретению, дополнительно включает нуклеотидную последовательность, кодирующую Fc-участок тяжелой цепи, например последовательность Fc-участка, представленную в SEQ ID NO: 177, или последовательность, практически идентичную ей.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота, кодирующая антитело по настоящему изобретению, дополнительно включает нуклеотидную последовательность, кодирующую последовательность константной области легкой цепи, например последовательность, представленную в SEQ ID NO: 209, или последовательность, практически идентичную ей.
С использованием методов, известных специалистам в данной области техники, полинукеотидные последовательности могут быть получены путем твердофазного синтеза ДНК de novo или путем ПЦР-мутагенеза существующих последовательностей (например, VH-кодирующей последовательности ДНК, представленную в SEQ ID NO: 150-169 или VL-кодирующей последовательности ДНК, представленной в SEQ ID NO: 170-176), кодирующих антитела, связывающиеся с TIGIT, или их антиген-связывающие фрагменты.
В одном варианте осуществления изобретения имеется один или более векторов, содержащих нуклеиновую кислоту по настоящему изобретению. В одном варианте осуществления изобретения вектор является экспрессионным вектором, например эукариотическим экспрессионным вектором. Вектор включает в числе прочего вирус, плазмиду, космиду, фаг лямбда или искусственную хромосому дрожжей (YAC). В предпочтительном варианте осуществления изобретения экспрессионным вектором по настоящему изобретению является экспрессионный вектор pTT5.
В одном варианте осуществления изобретения имеется клетка-хозяин, содержащая вектор. Подходящая клетка-хозяин для клонирования или экспрессии вектора, кодирующего антитело, включает прокариотическую или эукариотическую клетку, описанную в этом документе. Например, антитела могут быть произведены в бактериях, в частности когда не требуются гликозилирование или эффекторные функции, опосредованные Fc-участком. Экспрессия фрагментов антител и полипептидов в бактериях описаны, например, в патентах США № 5648237, 5789199 и 5840523, а также в издании Charlton, Methods in Molecular Biology, Vol. 248 (B. K. C. Lo, ed., Humana Press, Totowa, NJ, 2003), стр. 245-254, где описана экспрессия фрагментов антител в E. coli. После экспрессии антитела в растворимой фракции могут быть выделены из массы бактериальных клеток и дополнительно очищены.
В одном варианте осуществления изобретения клетка-хозяин является эукариотической. В другом варианте осуществления изобретения клетка-хозяин выбрана из клетки дрожжей, клетки млекопитающих и других клеток, пригодных для получения антитела или его антиген-связывающего фрагмента. Например, подходящими хозяевами для клонирования или экспрессии векторов, кодирующих антитела, являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи. Например, штаммы грибов или дрожжей, в которых путь гликозилирования «гуманизирован», позволяют производить антитела с частично или полностью человеческим профилем гликозилирования. См. Gerngross, Nat. Biotech. 22: 1409-1414 (2004), а также Li et al., Nat. Biotech. 24:210-215 (2006). Клетки-хозяева, пригодные для экспрессии гликозилированных антител, также могут происходить из многоклеточных организмов (беспозвоночных и позвоночных). Клетки позвоночных организмов также могут использоваться в качестве хозяев. Например, может быть использована линия клеток млекопитающих, модифицированная для роста в суспензии. Примерами полезных линий клеток-хозяев млекопитающих являются линия клеток CV1 из почки обезьяны, трансформированных вирусом SV40 (COS-7), линия клеток из почки человеческого эмбриона (клетки 293 HEK или 293, как описано, например в Graham et al., J. Gen Virol. 36:59 (1977)) и тому подобное. Другими полезными линиями клеток-хозяев млекопитающих являются клетки яичников китайского хомячка (CHO), включая клетки DHFR- CHO (Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 216 (1980)), и линии клеток миеломы, такие как Y0, NS0 и Sp2/0. Обзоры, посвященные определенным линиям клеток-хозяев млекопитающих, пригодных для продукции антител, приведены, например, в Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B. K. C. Lo, ed., Humana Press, Totowa, NJ), стр. 255-268 (2003).
III. Получение антител
В одном варианте осуществления изобретения имеется метод получения антитела к TIGIT, где этот метод включает культивирование клеток-хозяев, содержащих кодирующую антитело нуклеиновую кислоту в условиях, пригодных для экспрессии антитела, как описано выше, и, необязательно, выделение антитела из клеток-хозяев (или среды культивирования клеток-хозяев). Для получения антитела к TIGIT по методу рекомбинантных ДНК нуклеиновую кислоту, кодирующую антитело (например, антитело, описанное выше), выделяют и вставляют в один или более векторов для дальнейшего клонирования и (или) экспрессии в клетках-хозяевах. Такая нуклеиновая кислота может быть легко выделена и секвенирована с использованием обычных процедур (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи антител).
IV. Анализ
Антитела к TIGIT, являющиеся предметом настоящего изобретения, могут быть идентифицированы, подвергнуты скринингу или охарактеризованы с точки зрения их физических/химических свойств и (или) биологической активности с помощью различных методов анализа, известных специалистам в данной области техники.
В одном аспекте изобретения антитела по настоящему изобретению подвергают анализу на предмет антиген-связывающей активности. Например, способность связываться с TIGIT человека может быть определена такими методами, известными специалистам в данной области техники, как твердофазный иммуноанализ (ELISA), вестерн-блоттинг и тому подобное, либо иллюстративными методами, раскрытыми в примерах, приведенных в настоящем документе. Например, может быть проведен анализ методом поточной цитометрии, где антитела реагируют с линией клеток, экспрессирующих TIGIT человека, например с клетками CHO, трансфицированными для экспрессии TIGIT человека на клеточной поверхности. Метод поточной цитометрии можно использовать и для других клеток, включая T-клетки, экспрессирующие природный TIGIT. В альтернативном варианте для определения связывания антител, в том числе кинетики связывания (например, KD), может быть использован метод биологической оптической интерферометрии с применением рекомбинантных белков TIGIT. Например, в некоторых вариантах осуществления изобретения используется адаптированный метод анализа аффинности Fortebio.
В другом аспекте изобретения для идентификации антитела, конкурирующего за связывание с TIGIT с каким-либо из антител, раскрытых в этом документе, может быть использован конкурентный анализ. В определенных вариантах осуществления изобретения такое конкурирующее антитело связывается с тем же или перекрывающимся эпитопом (например, линейным или конформационным эпитопом), что и какое-либо из антител к TIGIT, раскрытых в этом документе. Иллюстративный метод определения положения эпитопа, с которым связывается антитело, подробно описан в издании Morris (1996) "Epitope Mapping Protocols", Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ).
Предметом настоящего изобретения также является метод определения антител к TIGIT, обладающих биологической активностью. Биологическая активность может включать, например, связывание с TIGIT (например, TIGIT человека), блокирование связывания TIGIT (например, TIGIT человека) с молекулами CD155, блокирование опосредованной TIGIT передачи сигналов ингибирования, увеличение продукции IL-2 в T-клетках и (или) подавление роста опухоли. Например, способность антител подавлять рост опухоли проверяют в модели супрессии опухоли in vivo (см., например, Пример 6). Антитела, обладающие такой биологической активностью in vivo и (или) in vitro, также описаны в этом документе.
Специалистам в данной области техники должно быть понятно, что любой из упомянутых выше методов анализа может быть осуществлен с использованием иммуноконъюгатов или полиспецифических антител по настоящему изобретению вместо антител к TIGIT и в дополнение к ним.
V. Полиспецифические антитела
Еще одним аспектом настоящего изобретения является полиспецифическая (в том числе биспецифическая) молекула антитела, которая специфически связывается с TIGIT, предпочтительно TIGIT человека. В одном варианте осуществления изобретения антитела (или их антиген-связывающие фрагменты) по настоящему изобретению, когда идет речь о полиспецифических антителах, специфически связываются с TIGIT в качестве первого антигена. В еще одном варианте осуществления изобретения полиспецифические антитела дополнительно обладают способностью специфически связываться со вторым антигеном, либо, в еще одном варианте осуществления изобретения, обладают способностью специфически связываться со вторым и третьим антигеном. В еще одном варианте осуществления изобретения полиспецифические антитела являются биспецифическими антителами.
В одном варианте осуществления изобретения специфичность связывания зависит от «связывающего участка» или «антиген-связывающего участка» антитела (области молекулы антитела, которая фактически связывается с антигеном). В предпочтительном варианте осуществления изобретения антиген-связывающий участок образован парой VH/VL, состоящей из вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH) антитела. Так, в одном варианте осуществления изобретения «полиспецифические» антитела содержат по крайней мере два антиген-связывающих участка, каждый из которых может связываться с разными эпитопами одного и того же антигена или с разными эпитопами разных антигенов.
Полиспецифические антитела и методы их получения описаны, например, в патентах WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 и WO 2010/145793.
VI. Иммуноконъюгаты
Еще одним аспектом настоящего изобретения является иммуноконъюгат, полученный за счет конъюгирования антител по настоящему изобретению с гетерологичной молекулой. В одном варианте осуществления изобретения иммуноконъюгат получают конъюгированием антител (или их антиген-связывающих фрагментов) по настоящему изобретению с терапевтическим или диагностическим агентом. В некоторых вариантах осуществления изобретения антитела по настоящему изобретению могут быть конъюгированы с гетерологичной молекулой в форме полноразмерных антител или фрагментов антител. Например, антитела конъюгируют в форме Fab-фрагментов, Fab'-фрагментов, F(ab)'2-фрагментов, одноцепочечных scFab-антител, одноцепочечных scFvs или других фрагментов.
В других вариантах осуществления изобретения антитела по настоящему изобретению конъюгируют с терапевтической молекулой. Для ковалентного связывания антител с терапевтической молекулой могут быть использованы линкеры. Подходящие линкеры включают химические или пептидные линкеры.
В других вариантах осуществления изобретения антитела по настоящему изобретению могут быть конъюгированы с диагностическим или подлежащим обнаружению агентом. Такие конъюгаты могут использоваться как часть метода, применяемого в клиническом исследовании (например, для определения эффективности той или иной терапии), для мониторинга или прогнозирования начала, развития, прогрессирования и (или) тяжести заболевания или нарушения. Такая диагностика или детекция могут быть осуществлены за счет связывания антител с агентом, подлежащим обнаружению.
VII. Фармацевтические композиции и фармацевтические препараты
Предметом настоящего изобретения также является композиция (включая фармацевтическую композицию или фармацевтический препарат), включающая антитела к TIGIT или их иммуноконъюгаты, или полиспецифические антитела, а также композиция, включающая полинуклеотид, кодирующий антитела к TIGIT или их иммуноконъюгаты, или полиспецифические антитела. Такие композиции опционально могут дополнительно включать подходящие фармацевтические вспомогательные вещества, например фармацевтический носитель, фармацевтическое вспомогательное вещество и тому подобные компоненты, известные специалистам в данной области техники, включая буферы.
В одном варианте осуществления изобретения композиция дополнительно включает второй терапевтический агент. Второй терапевтический агент может быть выбран из группы, включающей в числе прочего антитела к PD-1 и антитела к PD-L1. В предпочтительном варианте второй терапевтический агент представляет собой антагонист PD-1, в частности антитело к PD-1.
Фармацевтическим носителем, применимым к настоящему изобретению, может быть стерильная жидкость, такая как вода или масло, включая жидкости, происходящие из нефти, животных, растений или полученные синтетическим путем, такие как арахисовое масло, соевое масло, вазелиновое масло, кунжутное масло и т.п. В случае если фармацевтическую композицию вводят внутривенно, предпочтительным носителем является вода. В качестве жидких носителей, в частности для инъекционных растворов, также могут использоваться солевые растворы, водная декстроза и растворы глицерина. Подходящие фармацевтические вспомогательные вещества включают крахмал, глюкозу, лактозу, сахарозу, желатин, ржаной солод, рис, муку, мел, силикагель, натрия стеарат, глицеролмоностеарат, тальк, натрия хлорид, сухое обезжиренное молоко, глицерин, пропилен, гликоль, воду, этанол и т. п. Использование и сферы применения вспомогательных веществ описаны в Handbook of Pharmaceutical Excipients, 5-е изд., R. C. Rowe, P. J. Seskey and S. C. Owen, Pharmaceutical Press, London, Chicago. Композиция может дополнительно включать небольшое количество смачивающих агентов или эмульсификаторов, либо, при желании, буфер для поддержания pH. Композиция может быть изготовлена в форме раствора, суспензии, эмульсии, таблетки, пилюли, капсулы, порошка, препарата с пролонгированным высвобождением и тому подобное. Пероральные препараты могут включать стандартный носитель, например маннитол, лактозу, крахмал, магния стеарат и сахарин фармацевтической степени чистоты.
Фармацевтический препарат по настоящему изобретению может быть получен путем смешивания антител к TIGIT, иммуноконъюгатов или полиспецифических антител по настоящему изобретению желаемой степени чистоты с одним или более опциональных фармацевтических вспомогательных веществ, предпочтительно в форме лиофилизированного препарата или водного раствора (Remington's Pharmaceutical Sciences, 16-е изд., Osol, A. eds. (1980)).
Иллюстративный препарат лиофилизированного антитела описан в патенте США № 6,267,958. Водный препарат антител включает препараты, описанные в патентах США № 6,171,586 и WO 2006/044908, а последний препарат включает гистидин-ацетатный буфер.
Фармацевтическая композиция или препарат по настоящему изобретению могут дополнительно включать один или более других активных ингредиентов, требуемых для конкретного показания, подлежащего лечению, и в предпочтительном варианте активные ингредиенты обладают взаимно дополняющими свойствами, не влияющими друг на друга неблагоприятным образом. Например, может оказаться желательным дополнительно включить другие активные ингредиенты с противоопухолевой активностью, такие как антагонисты, связывающиеся с PD-1, или антагонисты, связывающиеся с PD-L1, например антитело к PD-1 или антитело к PD-L1. Активные ингредиенты комбинируются надлежащим образом в количествах, являющихся эффективными для поставленной цели.
Может быть получен препарат с пролонгированным высвобождением. Примером препарата с пролонгированным высвобождением является полупроницаемая матрица из твердого гидрофобного полимера, включающая антитело. Матрица изготовлена в форме профилированного изделия, например пленки или микрокапсулы.
Другие компоненты фармацевтического препарата описаны в патенте WO 2015/153513.
VIII. Комбинированные препараты
Одним из аспектов настоящего изобретения также является комбинированный препарат, включающий антитела или их антиген-связывающие фрагменты, биспецифические антитела или иммуноконъюгаты по настоящему изобретению и один или более терапевтических агентов (например, химиотерапевтический агент, другие антитела, цитотоксический агент, вакцину, антиинфекционный активный агент). Комбинированный препарат по настоящему изобретению может быть использован в терапевтическом методе, раскрытом в этом документе.
В некоторых вариантах осуществления изобретения имеется комбинированный препарат, в котором другие терапевтические агенты относятся, например, к такому терапевтическому агенту, как антитело, которое обладает способностью стимулировать иммунный ответ и тем самым дополнительно усиливать, стимулировать или увеличивать иммунный ответ у субъекта. В некоторых вариантах осуществления изобретения другие антитела относятся, например, к антителу к PD-1 или к антителу к PD-L1.
В некоторых вариантах осуществления изобретения комбинированный препарат используется для профилактики или лечения опухолевого заболевания. В некоторых вариантах осуществления изобретения опухоль представляет собой злокачественное новообразование, например злокачественное новообразование желудочно-кишечного тракта (например, рак желудка, рак прямой кишки, рак толстой кишки и колоректальный рак) или злокачественное новообразование кожи (например, злокачественная меланома). В некоторых вариантах осуществления изобретения комбинированный препарат используется для профилактики или лечения инфекции, например бактериальной инфекции, вирусной инфекции, грибковой инфекции, протозойной инфекции и тому подобное.
IX. Метод терапии и использование антител
В настоящем документе термины «индивидуум» и «субъект» могут использоваться взаимозаменяемым образом и относятся к млекопитающему. Млекопитающие включают в числе прочего домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, человекообразных и не человекообразных приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В частности, субъектом является человек.
В настоящем документе термин «лечение» относится к клиническому вмешательству с целью изменить естественное течение заболевания у субъекта, подвергаемого лечению. Желательные терапевтические эффекты включают в числе прочего предотвращение появления или рецидива заболеваний, облегчение симптомов, улучшение любых прямых или косвенных исходов заболеваний, предотвращение метастазов, замедление прогрессирования заболевания, улучшение или облегчения состояния и улучшение прогноза.
В одном из аспектов настоящее изобретение относится к методу усиления иммунного ответа организма субъекта, где данный метод включает введение субъекту эффективного количества любого из антител к TIGIT или их фрагментов, описанных в этом документе, иммуноконъюгата или полиспецифического антитела, включающих антитела или их фрагменты, или фармацевтической композиции. В некоторых вариантах осуществления изобретения антитела к TIGIT или их антиген-связывающие фрагменты по настоящему изобретению вводят субъекту с опухолью для стимуляции противоопухолевого иммунного ответа. В других вариантах осуществления изобретения антитела или их антиген-связывающие фрагменты по настоящему изобретению вводят субъекту с инфекцией для стимуляции иммунного ответа против инфекции.
В другом аспекте настоящее изобретение относится к методу лечения опухолевого заболевания, например злокачественного новообразования, у субъекта, где данный метод включает введение субъекту эффективного количества любого из антител к TIGIT или их фрагментов, описанных в этом документе, иммуноконъюгата или полиспецифического антитела, включающих антитела или их фрагменты, или фармацевтической композиции. Злокачественное новообразование может быть ранней, промежуточной или поздней стадии, или метастатическим злокачественным новообразованием.
Показано, что TIGIT в высокой степени экспрессирован на поверхности человеческих T-клеток CD8+, инфильтрирующих опухоль. В некоторых вариантах осуществления изобретения метод по настоящему изобретению применяют для лечения онкологического заболевания, в частности солидной опухоли, инфильтрированной опухоль-инфильтрирующими лимфоцитами, экспрессирующими TIGIT.
В одном варианте осуществления изобретения онкологическим заболеванием является злокачественное новообразование желудочно-кишечного тракта, например рак толстой кишки.
В некоторых вариантах осуществления изобретения опухоль или опухолевая клетка могут быть выбраны из колоректального новообразования, новообразования яичников, новообразования поджелудочной железы, новообразования легкого, новообразования печени, новообразования молочной железы, новообразования почки, новообразования простаты, новообразования желудочно-кишечного тракта, меланомы, новообразования шейки матки, новообразования мочевого пузыря, глиобластомы, а также новообразования головы и шеи. В некоторых вариантах осуществления изобретения злокачественное новообразование может быть выбрано из колоректального рака, рака яичников, рака поджелудочной железы, рака легких, рака печени, рака молочной железы, рака почек, рака простаты, злокачественного новообразования желудочно-кишечного тракта, меланомы, рака шейки матки, рака мочевого пузыря, глиобластомы, а также злокачественного новообразования головы и шеи.
В некоторых вариантах осуществления изобретения имеется метод лечения опухолевого заболевания (например, злокачественного новообразования), где данный метод включает введение субъекту антител к TIGIT по настоящему изобретению и антагонистических антител к PD-1. В некоторых вариантах осуществления изобретения метод применяют для лечения опухолевого заболевания, при котором опухоль коэкспрессирует TIGIT и PD-1, например меланомы, коэкспрессирующей TIGIT и PD-1 (Chauvin et al. (2015) J. Clin. Invest. 125:2046), немелкоклеточного рака легких (НМРЛ) и почечно-клеточной карциномы (ПКК).
В другом аспекте настоящее изобретение относится к методу лечения инфекционного заболевания, например хронической инфекции, у субъекта, где данный метод включает введение субъекту эффективного количества любого из антител к TIGIT или их фрагментов, описанных в этом документе, иммуноконъюгата или полиспецифического антитела, включающих антитела или их фрагменты, или фармацевтической композиции. В одном варианте осуществления изобретения инфекция является вирусной инфекцией.
В одном варианте осуществления изобретения инфекционное заболевание возникает в результате вирусной инфекции. Некоторыми примерами патогенных вирусов являются вирусы гепатита (A, B и C), вирусы гриппа (A, B и C), ВИЧ, герпетические вирусы (например, VZV, HSV-1, HAV-6, HSV-II, CMV и вирус Эпштейна-Барр), аденовирусы, флавивирусы, эховирусы, риновирусы, вирусы Коксаки, коронавирусы, респираторно-синцитиальные вирусы, вирусы эпидемического паротита, ротавирусы, вирусы кори, вирусы краснухи, парвовирусы, вирусы осповакцины, вирусы Т-клеточного лейкоза человека (HTLV), вирусы денге, папиллярные карциномы, вирусы моллюска, полиовирусы, вирусы бешенства, вирусы JC и вирусы энцефалита, передаваемые членистоногими.
В некоторых вариантах осуществления изобретения метод, описанный в настоящем документе, дополнительно включает применение к субъекту одного или более терапевтических воздействий в комбинации друг с другом (например, терапевтического метода и (или) других терапевтических агентов). В некоторых вариантах осуществления изобретения терапевтический метод включает хирургическое лечение и (или) лучевую терапию.
В некоторых вариантах осуществления изобретения T-клеточный ответ может быть стимулирован комбинацией антител к TIGIT по настоящему изобретению и одного или более терапевтических агентов. В некоторых вариантах осуществления изобретения, помимо введения антител по настоящему изобретению, метод по настоящему изобретению дополнительно включает введение по крайней мере одного дополнительного антитела с иммуностимулирующей активностью, например антитела к PD-1 и антитела к PD-L1, которые могут быть, например, полностью гуманизированными, химерными или гуманизированными.
В других вариантах осуществления изобретения другими терапевтическими агентами, которые могут быть использованы в комбинации с антителами по настоящему изобретению, являются антагонисты, связывающиеся с PD-1, или антагонисты, связывающиеся с PD-L1. Альтернативные названия «PD-1» включают CD279 и SLEB2. Альтернативные названия «PD-L1» включают B7-H1, B7-4, CD274 и B7-H. В некоторых вариантах осуществления изобретения PD-1 и PD-L1 являются человеческим PD-1 и человеческим PD-L1 соответственно. В некоторых вариантах осуществления изобретения антагонистом, связывающимся с PD-1, является молекула, ингибирующая связывание PD-1 с его лигандом/партнером по связыванию. В одном конкретном аспекте лигандом/партнером по связыванию PD-1 является PD-L1. В другом варианте осуществления изобретения антагонистом, связывающимся с PD-L1, является молекула, ингибирующая связывание PD-L1 с его партнером по связыванию. В одном конкретном аспекте партнером по связыванию PD-L1 является PD-1. Антагонистом может быть антитело, антиген-связывающий фрагмент, иммуноадгезин, слитый белок или олигопептид. В некоторых вариантах осуществления изобретения антагонистом, связывающимся с PD-1, является антитело к PD-1 (например, человеческое антитело, гуманизированное антитело или химерное антитело). В некоторых вариантах осуществления изобретения антитело к PD-1 выбрано из такого списка: IBI308 (моноклональное антитело синтилумаб, WO2017/025016A1), MDX-1106 (ниволумаб, OPDIVO), Merck 3475 (MK-3475, пембролизумаб, KEYTRUDA) и CT-011 (пидилизумаб). В некоторых вариантах осуществления изобретения антителом к PD-1 является MDX-1106. В некоторых вариантах осуществления изобретения антителом к PD-1 является ниволумаб (номер в реестре CAS: 946414-94-4). В предпочтительном варианте осуществления изобретения антителом к PD-1 является «Антитело C», описанное в этом документе.
В некоторых других вариантах осуществления изобретения антитела к TIGIT или их фрагменты, по отдельности или в комбинации с антагонистом, связывающимся с PD-1/PD-L1, могут применяться в комбинации с одним или более терапевтическим воздействием, например терапевтическими методами и (или) другими терапевтическими агентами. В некоторых вариантах осуществления изобретения терапевтические методы включают хирургическое вмешательство (например, резекцию опухоли), лучевую терапию (например, дистанционную лучевую терапию, включая трехмерную конформную лучевую терапию, при которой специальным образом определяется зона облучения), частичное облучение (например, облучение, направленное на заранее выбранную мишень или орган), фокусированное облучение и тому подобное.
В некоторых вариантах осуществления изобретения антитела к TIGIT или их антиген-связывающие фрагменты по настоящему изобретению могут применяться в комбинации с химиотерапией или химиотерапевтическим агентом. В некоторых вариантах осуществления изобретения антитела к TIGIT или их антиген-связывающие фрагменты по настоящему изобретению могут применяться в комбинации с лучевой терапией или радиотерапевтическим агентом. В некоторых вариантах осуществления изобретения антитела к TIGIT или их антиген-связывающие фрагменты по настоящему изобретению могут применяться в комбинации с таргетной терапией или таргетным терапевтическим агентом. В некоторых вариантах осуществления изобретения антитела к TIGIT или их антиген-связывающие фрагменты по настоящему изобретению могут применяться в комбинации с иммунотерапией или иммунотерапевтическим агентом, например моноклональным антителом.
Антитело по настоящему изобретению (фармацевтическая композиция или иммуноконъюгат, включающие те же или любые другие терапевтические агенты) могут применяться любым подходящим способом, например путем парентерального введения, внутрилегочного введения, интраназального введения и внутриочагового введения, если требуется местное лечение. Парентеральное введение включает внутримышечное, внутривенное, внутриартериальное, интраперитонеальное или подкожное введение. Лекарственные препараты могут вводиться любым подходящим способом, в частности инъекцией, например внутривенной или подкожной инъекцией, что в определенной степени зависит от того, является лечение краткосрочным или длительным. Изобретение охватывает различные схемы введения, включая в числе прочего однократное введение или многократное введение, болюсные инъекции и дробные инфузии в различные моменты времени.
В случае применения для профилактики и лечения заболеваний надлежащая доза антител по настоящему изобретению (при его использовании в отдельности или в комбинации с одним или более терапевтическими агентами) будет зависеть от типа заболевания, подлежащего лечению, типа антител, тяжести и прогрессирования заболевания, цели применения (профилактического или терапевтического), ранее применявшейся терапии, клинического анамнеза пациента, ответа на антитело и мнения лечащего врача. Приемлемым вариантом применения антитела у пациента является однократное введение или серия из нескольких введений.
В методах, описанных выше, композиция, полиспецифическое антитело или иммуноконъюгат по настоящему изобретению могут применяться вместо антитела или его антиген-связывающего фрагмента, описанных в этом документе. В альтернативном варианте, в этих методах композиция, полиспецифическое антитело или иммуноконъюгат по настоящему изобретению также могут применяться после применения антитела или его антиген-связывающего фрагмента, описанных в этом документе.
Еще одним аспектом настоящего изобретения является использование антитела к TIGIT, композиции, иммуноконъюгата и полиспецифического антитела по настоящему изобретению для получения лекарственных препаратов, применяемых в рамках методов, описанных выше (например, для лечения).
X. Методы и композиции для диагностики и детекции (обнаружения)
Еще один аспект настоящего изобретения относится к методу и набору для обнаружения TIGIT в образце, где данный метод включает следующие этапы: (a) приведение образца в контакт с антителом или антиген-связывающим фрагментом, или иммуноконъюгатом, раскрытых в этом документе; и (b) обнаружение комплекса, образованного между антителом или его антиген-связывающим фрагментом, или иммуноконъюгатом и белком TIGIT. В некоторых вариантах осуществления изобретения образец получен от пациента с онкологическим заболеванием, например от пациента с раком кожи. Детекция может осуществляться in vitro или in vivo.
Термин «детекция (обнаружение)», используемый в данном документе, включает количественное или качественное определение, а иллюстративные варианты детекции могут включать детекцию с помощью иммуногистохимического исследования, иммуноцитохимического исследования, поточной цитометрии (например, FACS), магнитных шариков, связанных с молекулами антитела, твердофазного иммуноанализа (ELISA) и методов ПЦР (например, ПЦР-ОТ). В определенных вариантах осуществления изобретения биологический образец представляет собой кровь, сыворотку или другой жидкий образец биологического происхождения. В определенных вариантах осуществления изобретения биологический образец включает клетки или ткани. В некоторых вариантах осуществления изобретения биологический образец происходит из пролиферативного или злокачественного поражения. В определенных вариантах осуществления изобретения TIGIT, подлежащий обнаружению, представляет собой TIGIT человека.
В одном варианте осуществления изобретения антитело к TIGIT используют для отбора субъекта, подходящего для лечения антителом к TIGIT, например TIGIT является биомаркером для отбора субъекта. В одном варианте осуществления изобретения антитело по настоящему изобретению может быть использовано для диагностики злокачественных новообразований и опухолей, например для оценки (например, мониторинга) лечения или прогрессирования, диагностики и (или) определения стадии заболевания (например, гиперпролиферативного или злокачественного заболевания), описанного в этом документе, у субъекта.
В определенных вариантах осуществления изобретения имеется меченое антитело к TIGIT. Метка включает в числе прочего метку или группу (например, флуоресцентную метку, хромофорную метку, электронноплотную метку, хемилюминесцентную метку и радиоактивную метку), которая может быть обнаружена непосредственно, а также группу, которая может быть обнаружена опосредованно, в частности фермент или лиганд, которые могут быть обнаружены, например, с помощью ферментативной реакции или молекулярного взаимодействия. Примерами меток являются в числе прочего радиоизотопы 32P, 14C, 125I, 3H и 131I, флуорофоры (например, хелаты редкоземельных элементов или флуоресцеин) и их производные, родамин и его производные, дансил, умбеллиферон, люцифераза (например, люцифераза светлячка и бактериальная люцифераза (патент США № 4,737,456)), флуоресцеин, 2,3-дигидрофталазиндион, пероксидаза хрена (ПХ), щелочная фосфатаза, β-галактозидаза, глюкоамилаза, лизоцим, карбогидратоксидаза (например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфатдегидрогеназа), гетероциклическая оксидаза (например, уриказа и ксантиноксидаза), ферменты, окисляющие предшественники красителей в присутствии перекиси водорода (например, ПХ, лактопероксидаза или микропероксидаза), биотин/авидин, спиновые метки, фаговые метки, стабильные свободные радикалы и т.п.
Для того чтобы облегчить понимание настоящего изобретения, ниже приведены следующие примеры. Эти примеры не предназначены для уменьшения объема правовой охраны настоящего изобретения и не должны интерпретироваться как таковые.
Таблица 1. Последовательности CDR иллюстративных антител по настоящему изобретению
NO:
NO:
NO:
NO:
NO:
NO:
(где X1 выбран из T, L, R, G и остатков, обеспечивающих их консервативную замену, а X2 выбран из S, N, G и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из N, D и остатков, обеспечивающих их консервативную замену, X2 выбран из S, R и остатков, обеспечивающих их консервативную замену, X3 выбран из Y, F и остатков, обеспечивающих их консервативную замену, а X4 выбран из Q, R и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из Y, T и остатков, обеспечивающих их консервативную замену, а X2 выбран из M, T и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из S, G и остатков, обеспечивающих их консервативную замену, а X2 выбран из R, Q и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из Y, R и остатков, обеспечивающих их консервативную замену, а X2 выбран из S, G и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из F, H и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из F, I и остатков, обеспечивающих их консервативную замену, X2 выбран из S, G и остатков, обеспечивающих их консервативную замену, а X3 выбран из S, G и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из делеции, S и остатков, обеспечивающих его консервативную замену, X2 выбран из G, S и остатков, обеспечивающих их консервативную замену, X3 выбран из S, G и остатков, обеспечивающих их консервативную замену, а X4 выбран из Y, N, H и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из A, G и остатков, обеспечивающих их консервативную замену, а X2 выбран из D, R и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из S, G и остатков, обеспечивающих их консервативную замену, X2 выбран из S, P и остатков, обеспечивающих их консервативную замену, а X3 выбран из A, G и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из A, S и остатков, обеспечивающих их консервативную замену, X2 выбран из G, A и остатков, обеспечивающих их консервативную замену, X3 выбран из S, R и остатков, обеспечивающих их консервативную замену, X4 выбран из Y, W и остатков, обеспечивающих их консервативную замену, а X5 выбран из Y, H и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из D, H и остатков, обеспечивающих их консервативную замену, X2 выбран из S, Y и остатков, обеспечивающих их консервативную замену, а X3 выбран из S, T и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из S, E и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из N, S и остатков, обеспечивающих их консервативную замену, X2 выбран из G, A и остатков, обеспечивающих их консервативную замену, а X3 выбран из S, K и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из A, R и остатков, обеспечивающих их консервативную замену, а X2 выбран из A, L и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из A, S и остатков, обеспечивающих их консервативную замену, а X2 выбран из S, L, V и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из Y, F, W и остатков, обеспечивающих их консервативную замену, а X2 выбран из L, F и остатков, обеспечивающих их консервативную замену)
(где X1 выбран из G, T и остатков, обеспечивающих их консервативную замену, а X2 выбран из T, G и остатков, обеспечивающих их консервативную замену)
Таблица 2. Последовательности CDR иллюстративных антител по настоящему изобретению
Таблица 3. Последовательности VH и VL вариабельных областей иллюстративных антител по настоящему изобретению
Таблица 4. Номера SEQ ID родственных последовательностей всех антител, являющихся предметом настоящего изобретения
Примечание. HC была образована слиянием последовательности VH с IgG4-PAA частью последовательности, представленной в SEQ ID NO: 177, а LC была образована слиянием последовательности VL с Cκ-цепью последовательности, представленной в SEQ ID NO: 209.
20 иллюстративных антител (ADI-27238, ADI-30263, ADI-30267, ADI-30268, ADI-27243, ADI-30302, ADI-30336, ADI-27278, ADI-30293, ADI-30296, ADI-27291, ADI-30283, ADI-30286, ADI-30288, ADI-27297, ADI-30272, ADI-30278, ADI-27301, ADI-30306 и ADI-30311), включенных в примеры по настоящему изобретению, описанные ниже, аминокислотные последовательности CDR, вариабельные области легких и тяжелых цепей, легкие и тяжелые цепи этих антител, а также соответствующие нуклеотидные последовательности приведены в Таблицах 1-3 и в перечне последовательностей настоящей заявки. Номера соответствующих последовательностей приведены в Таблице 4.
Примеры
Пример 1. Получение антител
Скрининг полностью гуманизированных антител к TIGIT с использованием дрожжевого дисплея
Дрожжевые библиотеки антител (Adimab) были амплифицированы методами, известными специалистам в данной области техники (описаны в патентах WO 2009036379, WO 2010105256 и WO 2012009568), с разнообразием каждой библиотеки 1×109. Вкратце, в первых двух раундах скрининга клетки сортировали на магнитных шариках с использованием системы MACS, выпускаемой компанией Miltenyi. Сначала клетки дрожжей (примерно 1×1010 клеток/библиотеку) из библиотек инкубировали в буфере FACS (фосфатный буфер, содержащий 0,1% бычьего сывороточного альбумина и 100 нМ человеческих антигенов TIGIT, меченых биотином (Acro Biosystems, TIT-H52H3)) в течение 15 мин при комнатной температуре. Клетки один раз промывали 50 мл предварительно охлажденного буфера FACS и ресуспендировали в 40 мл того же буфера, после чего добавляли 500 мкл микрошариков со стрептавидином (Miltenyi LS) и инкубировали при температуре 4°C в течение 15 мин. Смесь центрифугировали при 1000 об/мин в течение 5 мин. После отделения надосадочной жидкости клетки ресуспендировали в 5 мл буфера FACS. Полученную клеточную суспензию наносили на колонку Miltenyi LS. После нанесения колонку три раза промывали буфером FACS, каждый раз по 3 мл. Колонку Miltenyi LS извлекали из магнитного поля и элюировали 5 мл ростовой среды. Элюированные клетки дрожжей собирали и инкубировали в течение ночи при температуре 37°C.
Следующий раунд сортировки осуществляли с использованием поточного цитометра; при этом примерно 1×108 клеток дрожжей, отобранных с помощью системы MACS, три раза промывали буфером FACS и инкубировали совместно с антигенами TIGIT, мечеными биотином в низкой концентрации (100-1 нМ) при комнатной температуре. Надосадочную жидкость сливали. Клетки два раза промывали буфером FACS и смешивали с LC-FITC (меченое FITC антитело к легкой цепи каппа человеческого иммуноглобулина, Southern Biotech) (разведение 1:100) и SA-633 (стрептавидин-633, Molecular Probes) (разведение 1:500) или SA-PE (стрептавидин-PE, Sigma) (разведение 1:50), после чего смесь инкубировали при температуре 4°C в течение 15 мин. Клетки дважды элюировали предварительно охлажденным буфером FACS, ресуспедировали в 0,4 мл буфера и переносили в пробирку для разделения с фильтром. Клетки сортировали с использованием прибора FACS ARIA (BD Biosciences).
Клетки дрожжей, экспрессирующие антитела к TIGIT, которые были получены в ходе скрининга, индуцировали путем встряхивания при температуре 30°C в течение 48 ч для экспрессии антител к TIGIT. После индукции клетки дрожжей отделяли центрифугированием при 1300 об/мин в течение 10 мин и собирали надосадочную жидкость. Перед сбором антитела к TIGIT, содержащиеся в надосадочной жидкости, очищали с помощью Протеина A и элюировали с использованием ацетатного буфера с pH 2,0. Чистота антител составляла более 95%. Антитела расщепляли папаином и очищали на смоле KappaSelect (GE Healthcare) для получения соответствующих Fab-фрагментов. В результате такого скрининга были получены антитела к TIGIT (ADI-27301, ADI-27238, ADI-27278, ADI-27243, ADI-27297 и ADI-27291).
Аффинная оптимизация антител к TIGIT человека
Для получения антител к TIGIT человека с более высокой аффинностью антитела (ADI-27301, ADI-27238, ADI-27278, ADI-27243, ADI-27297 и ADI-27291) оптимизировали следующими методами.
Скрининг VHmut
Данный метод использовали для введения мутаций в области тяжелых цепей антител с помощью стандартной ПЦР с неправильным спариванием оснований. Вероятность неправильного спаривания оснований в ходе ПЦР увеличивали примерно до 0,01 п.н. путем добавления 1 мкМ аналогов оснований dPTP и 8-оксо-dGTP с высоким потенциалом образования мутаций.
Образованные продукты ПЦР с неправильным спариванием оснований вводили в вектор, содержащий константную область тяжелой цепи, с помощью гомологичной рекомбинации. Этим методом была получена вторичная библиотека с емкостью 1×107 в жестких условиях скрининга, включавшего титры к антигенам TIGIT, конкуренцию с немеченым антигеном и конкуренцию с родительскими (исходными) антителами. С использованием FACS были успешно осуществлены три раунда скрининга.
Скрининг CDRH1/CDRH2
Гены CDRH3 дочерних антител, полученных методом VHmut, использовали для конструирования пула генов CDRH1/CDRH2 с разнообразием 1×108 и осуществляли 3 раунда скрининга генов. В первом раунде скрининга использовали систему MACS, а во втором и третьем раундах - FACS. Конъюгаты антиген-антитело подвергали жесткому скринингу («под давлением») для отбора антител с максимальной аффинностью.
В результате вышеописанного процесса аффинного созревания 6 родительских антител были получены 14 моноклональных антител к TIGIT человека с улучшенной аффинностью: ADI-30263/ADI-30267/ADI-30268 (из ADI-27238), ADI-30302/ADI-30336 (из ADI-27243), ADI-30293/ADI-30296 (из ADI-27278), ADI-30283/ADI-30286/ADI-30288 (из ADI-27291), ADI-30272/ADI-30278 (из ADI-27297) и ADI-30306/ADI-30311 (из ADI-27301).
Клетки дрожжей, экспрессирующие антитела к TIGIT, которые были получены в ходе скрининга, индуцировали путем встряхивания при температуре 30°C в течение 48 ч для экспрессии антител к TIGIT. После индукции клетки дрожжей отделяли центрифугированием при 1300 об/мин в течение 10 мин и собирали надосадочную жидкость. Перед сбором антитела к TIGIT, содержащиеся в надосадочной жидкости, очищали с помощью Протеина A и элюировали с использованием ацетатного буфера с pH 2,0. Чистота антител составляла более 95%. Антитела расщепляли папаином и очищали на смоле KappaSelect (GE Healthcare) для получения соответствующих Fab-фрагментов.
Экспрессия и очистка антител
Информация о последовательностях и номера последовательностей антител к TIGIT человека, являющихся предметом настоящего изобретения, представлены в Таблицах 1-9; при этом как родительские антитела, так и дочерние антитела после аффинного созревания были экспрессированы и очищены в дрожжах методом, описанным выше.
Следующие референтные антитела, использованные в примерах, были экспрессированы и очищены в клетках HEK293: 22G2, антитело к TIGIT человека компании BMS, которое было транзитно экспрессировано в клетках HEK293, причем последовательности вариабельных областей легких и тяжелых цепей были идентичны таковым антитела «22G2», описанного в патентной заявке № WO 2016/106302A1; 31C6, антитело к TIGIT человека компании Merck, которое было транзитно экспрессировано в клетках HEK293, причем последовательности вариабельных областей легких и тяжелых цепей были идентичны таковым антитела «31C6», описанного в патентной заявке № WO 2016/028656A1; 10A7 и 1F4, антитела к TIGIT человека компании Genentech, которые были транзитно экспрессированы в клетках HEK293, причем последовательности вариабельных областей легких и тяжелых цепей были идентичны таковым антител «10A7» и «1F4» в патентной заявке № WO2015009856A2 соответственно. Константные области 4 референтных антител соответствовали последовательностям IgG4 дикого типа.
Для транзитной экспрессии антитела в клетках HEK293 использовали вектор pTT5. Тяжелые и легкие цепи антитела сначала клонировали в отдельных векторах pTT5. Векторы pTT5, несущие тяжелые и легкие цепи молекулы антитела, переносили в клетки HEK293 методом химической трансфекции. Культивируемые клетки HEK293 транзитно трансфицировали с помощью реагента для химической трансфекции ПЭИ (приобретенного у Polysciences) по схеме, предоставленной производителем. Плазмидные ДНК и реагент для трансфекции готовили в ламинарном шкафу, после чего в две пробирки для центрифугирования на 50 мл переносили среду F17 (Gibco) (объем составлял 1/5 от объема, требуемого для трансфекции). Отфильтрованные плазмиды (130 мкг / 100 мл) добавляли в одну пробирку, а отфильтрованный ПЭИ (1 г/л, Polysciences) (массовое отношение «плазмида:ПЭИ» = 1:3) добавляли в другую пробирку. Содержимое каждой пробирки тщательно перемешивали в течение 5 мин, затем обе смеси объединяли и аккуратно перемешивали 20 раз, после чего полученную смесь оставляли в покое на 15-30 мин (не больше 30 мин). Смесь ДНК/ПЭИ аккуратно добавляли к клеткам HEK293 и тщательно перемешивали. Клетки культивировали при 37°C, 8% CO2 в течение 7 дней, каждые 48 ч добавляя свежую среду. Через семь дней или после того, как процент жизнеспособных клеток на фоне непрерывного культивирования достигал ≤60%, смесь центрифугировали при 13 000 об/мин в течение 20 мин. Отбирали надосадочную жидкость и очищали антитела на Протеине A, в результате чего получали антитела с чистотой более 95%.
Кроме того, интактные антитела (ADI-30268, ADI-30278, ADI-30286, ADI-30288, ADI-30293, ADI-30306 и ADI-30336) экспрессировали и очищали в клетках CHO.
Экспрессия и очистка в клетках CHO: линии клеток CHO, экспрессирующих антитела, получали с использованием линии HD-BIOP1 (GS Null CHO-K1) компании Horizon в соответствии с инструкцией производителя. Последовательности ДНК тяжелых и легких цепей молекулы антитела сначала вставляли в плазмиды pD2531 компании ATUM. Затем сконструированные плазмиды переносили в клетки CHO с помощью электротрансфекции, а продукцию антител обнаруживали методом ForteBio, который позволял определить эффективность трансфекции через 24 ч после нее. Трансфицированные клетки подвергали жесткому скринингу («под давлением») для получения пула клеток с высоким уровнем экспрессии антител. Этот пул клеток затем амплифицировали для экспрессии больших количеств антител. С клеток собирали надосадочную жидкость и очищали антитела с помощью Протеина A и гель-фильтрации, в результате чего получали антитела с чистотой более 95%.
Пример 2. Анализ аффинности антител
Равновесные константы диссоциации (KD), характеризующие связывание 20 иллюстративных антител по настоящему изобретению с TIGIT человека, измеряли методом биологической оптической интерферометрии (ForteBio).
Анализ аффинности методом ForteBio проводили, как описано ранее (Estep, P, et al., High throughput solution based measurement of antibody-antigen affinity and epitope binning. MAbs, 2013.5(2): p. 270-8).
Измерение аффинности моновалентного связывания Fab-фрагмента антитела-кандидата с TIGIT-FC: датчик уравновешивали оффлайн в буфере для анализа в течение 20 мин, после чего осуществляли детекцию онлайн в течение 120 с для установления исходного уровня. На датчик AHQ (ForteBio) наносили TIGIT человека -FC для проведения анализа аффинности по методу ForteBio. Датчик с нанесенными на него антигенами помещали в раствор, содержащий 100 нМ Fab-фрагмента, до достижения плато, после чего переносили его как минимум на 2 мин в буфер для анализа, в котором происходила диссоциация, для измерения скорости диссоциации. Анализ кинетики выполняли с использованием модели связывания 1:1.
Измерение аффинности моновалентного связывания интактных антител-кандидатов с TIGIT-his человека (ACRO, TIT-H52H3), мыши (ACRO, TIT-M52E6) и обезьяны (ACRO, TIT-C5223): датчик уравновешивали оффлайн в буфере для анализа в течение 20 мин, после чего осуществляли детекцию онлайн в течение 120 с для установления исходного уровня. На датчик AHQ (ForteBio) наносили очищенные антитела до толщины 1 нанометр для проведения анализа аффинности по методу ForteBio. Датчик с нанесенными на него антителами помещали в раствор, содержащий 100 нМ антигенов TIGIT-his, до достижения плато, после чего переносили его как минимум на 2 мин в буфер для анализа, в котором происходила диссоциация, для измерения скорости диссоциации. Анализ кинетики выполняли с использованием модели связывания 1:1.
В экспериментах с использованием анализов, описанных выше, были определены значения KD для 20 антител-кандидатов (Fab), экспрессированных в дрожжах; эти значения приведены в Таблице 5.
Таблица 5. Значения KD для моновалентного связывания 20 антител-кандидатов (Fab), экспрессированных в дрожжах, с TIGIT человека-FC
В экспериментах с использованием анализов, описанных выше, были определены значения KD для моновалентного связывания 7 интактных антител-кандидатов, экспрессированных клетками CHO, с TIGIT человека, мыши и обезьяны; эти значения приведены в Таблице 6.
Таблица 6. Значения KD для моновалентного связывания 7 антител-кандидатов, экспрессированных клетками CHO, и референтных антител с TIGIT человека, мыши и обезьяны
Из приведенных выше таблиц можно видеть, что: (1) после аффинного созревания каждое дочернее антитело имеет существенно более высокую аффинность связывания с TIGIT человека, чем родительское антитело; (2) после аффинного созревания аффинность связывания 7 отобранных антител, экспрессированных в клетках CHO, с TIGIT человека была примерно такой же или выше, чем у референтных антител; все 7 антител-кандидатов могут перекрестно реагировать с TIGIT обезьяны; при этом 4 (ADI-30268/ADI-30286/ADI-30288/ADI-30293) из 7 антител-кандидатов могут перекрестно реагировать с TIGIT мыши.
Пример 3. Связывание антител к TIGIT с TIGIT человека, экспрессированным на поверхности клеток
Способность 20 указанных выше иллюстративных антител по настоящему изобретению связываться с TIGIT человека, экспрессированным на поверхности клеток CHO, измеряли с помощью поточной цитометрии. Связывающую способность различных антител определяли, сравнивая кривые их связывания с TIGIT человека, экспрессированным на поверхности клеток CHO.
В частности, для экспрессии родительских и дочерних антител после аффинного созревания использовали клетки дрожжей. После сбора антител (с чистотой примерно 70%) проводили эксперименты по связыванию этих антител с TIGIT человека, экспрессированным на поверхности клеток CHO. Эксперименты проводили следующим образом: (1) клетки CHOS трансфицировали плазмидами, несущими TIGIT человека, сконструированными на основе pCHO1.0, а затем проводили скрининг с метотрексатом и пуромицином для отбора пула стабильных клеток. (2) Антитела-кандидаты в различных концентрациях инкубировали с пулом стабильных клеток при температуре 4°C в течение 30 мин. После трех промывок фосфатно-солевым буфером (ФСБ) клетки инкубировали с мышиными вторыми антителами к человеческим иммуноглобулинам, меченными фикоэритрином (PE), в течение 30 мин при температуре 4°C. После трех дополнительных промывок ФСБ клетки ресуспендировали в 100 мкл ФСБ. (3) С помощью поточной цитометрии определяли медианные значения флуоресценции для клеток в двух каналах. Значения EC50 и максимальные значения для разных кривых сравнивали с использованием графиков, построенных в координатах «десятичный логарифм концентрации антител» (по оси абсцисс) - «медиана флуоресценции для двух каналов» (по оси ординат). Результаты показаны в следующей таблице и на РИС. 1 (EC50: нМ).
Из приведенных выше результатов экспериментов можно сделать следующие выводы: (1) после аффинного созревания каждое дочернее антитело имеет более высокую способность связываться с TIGIT человека, экспрессированным на поверхности клеток CHO, чем родительское антитело (о чем говорит более высокое плато на кривой связывания); (2) способность дочерних антител после аффинного созревания связываться с TIGIT человека, экспрессированным на поверхности клеток CHO, выше, чем для двух антител от компании Genentech (10A7 и 1F4), и сопоставима с таковой для антитела 22G2 от компании BMS и антитела 31C6 от компании Merck.
Кроме того, 7 антител, отобранных после аффинного созревания, были экспрессированы и очищены с использованием системы CHO для получения антител высокой степени чистоты. Был проведен эксперимент по связыванию антител высокой степени чистоты с TIGIT человека, экспрессированным на поверхности клеток CHO. В экспериментах, проведенных, как описано выше, антитела ADI-30268, ADI-30336, ADI-30293, ADI-30286, ADI-30288, ADI-30278 и ADI-30306 связывались с TIGIT человека, гиперэкспрессированным на поверхности клеток CHO, со значениями EC50 0,1392 нМ, 0,1300 нМ, 0,2288 нМ, 0,1758 нМ, 0,1860 нМ, 0,1321 нМ и 0,1999 нМ соответственно, причем их способность связываться с TIGIT человека, гиперэкспрессированным на поверхности клеток CHO, была выше таковой для референтных антител 22G2 и 1F4 (значения EC50 составляют 0,7249 нМ и 0,6073 нМ соответственно) и сопоставима с таковой для референтного антитела 31C6 (значение EC50 составляет 0,1846 нМ). Хотя значение EC50 для референтного антитела 10A7 (0,1949 нМ) сопоставимо с таковыми для 7 антител-кандидатов, из кривой связывания этого референтного антитела видно, что плато на этой кривой находится ниже, чем плато на кривых 7 антител-кандидатов. Как правило, это свидетельствует о более быстрой диссоциации комплекса антиген-антитело (см. РИС. 2). Поэтому связывающая способность 7 антител-кандидатов с TIGIT человека, экспрессированным на поверхности клеток CHO, признана более высокой, чем у антитела 10A7.
Пример 4. Блокирование антителом к TIGIT связывания TIGIT с его лигандом CD155
Способность антител-кандидатов блокировать лиганды определяли методом поточной цитометрии. Такое определение выполняли следующим образом: к полученному пулу клеток CHOS, стабильно гиперэкспрессирующих TIGIT человека, добавляли в концентрации 50 нМ человеческий белок CD155 (ACRO, TIT-H5253-1MG) c Fc-фрагментом мышиного IgG2a, а также антитела в различных концентрациях, после чего инкубировали клетки в течение 30 мин при температуре 4°C. После трех промывок ФСБ молекулы-лиганды CD155, оставшиеся на клетках, прокрашивали козьими антителами к Fc-участку мышиного иммуноглобулина (в концентрации 1%) и вторыми антителами, меченными аллофикоцианином (APC) (Biolegend, 405308). После еще трех промывок ФСБ измеряли медианную флуоресценцию в соответствующем канале (C6, канал 4) с помощью поточного цитометра. Для каждого антитела строили график зависимости медианной флуоресценции (ось ординат) от десятичного логарифма концентрации антитела в нМ (ось абсцисс). Способность различных антител блокировать связывание TIGIT с CD155 оценивали по значениям IC50, которые определяли из построенных графиков.
В экспериментах по оценке способности различных антител-кандидатов до и после аффинного созревания блокировать связывание TIGIT с CD155 в качестве отрицательного контроля использовали IgG1, а в качестве положительных контролей - антитела 10A7, 22G2 и 31C6. РИС. 3 и приведенная ниже таблица демонстрируют способность антител, экспрессированных в дрожжах, до и после аффинного созревания блокировать связывание TIGIT с CD155 (IC50: нМ).
Из результатов проведенных экспериментов можно сделать вывод о том, что большинство дочерних антител после аффинного созревания обладали более высокой способностью блокировать связывание TIGIT с CD155, чем их родительские антитела, были сопоставимы по блокирующей способности с референтными антителами 22G2 и 31C6 и превосходили референтное антитело 10A7.
Способность блокировать связывание TIGIT с CD155 также определили для 7 антител высокой степени чистоты, экспрессированных в клетках CHO. В экспериментах, проведенных, как описано выше, в качестве отрицательного контроля использовали IgG4, а в качестве положительных контролей - антитела 22G2, 31C6 и 10A7. Из результатов проведенных экспериментов можно сделать вывод о том, что значения IC50 для антител ADI-30268, ADI-30336 и ADI-30278 составляют 0,1402 нМ, 0,1219 нМ и 0,1164 нМ соответственно, то есть ниже таковых для референтных антител 22G2 (0,6069 нМ), 31C6 (0,1659 нМ) и 10A7 (0,3252 нМ), из чего следует, что три названных антитела обладают более высокой способностью блокировать связывание лиганда (CD155), чем референтные антитела. Четыре других антитела-кандидата (ADI-30293, ADI-30286, ADI-30288 и ADI-30306) характеризуются значениями IC50 0,2510 нМ, 0,1893 нМ, 0,2238 нМ и 0,2420 нМ соответственно; все эти значения ниже значений для референтных антител 22G2 и 10A7, но выше такового для референтного антитела 31C6, из чего следует, что эти 4 антитела обладают более высокой способностью блокировать связывание лиганда (CD155), чем референтные антитела 22G2 и 10A7, но более низкой, чем референтное антитело 31C6. Результаты показаны в следующей таблице и на РИС. 4 (IC50: нМ).
Пример 5. Анализ биологической активности антител-кандидатов, лежащей в основе механизма их действия (МД)
Все антитела к TIGIT за счет блокирования связывания TIGIT с CD155 способны ослаблять эффект ингибирования в отношении нисходящего IL-2-зависимого сигнального пути. В этом примере линия клеток для анализа биологической активности, лежащей в основе МД (J2201), от компании Promega была использована для выявления экспрессии генов репортерных флуоресцентных белков, отражающей активацию IL-2-зависимого сигнального пути, и тем самым для выявления способности антител подавлять связывание TIGIT с CD155. Анализ проводили согласно инструкциям производителя, компании Promega. Процедура анализа состояла в следующем: клетки Jurkat, стабильно экспрессирующие TIGIT человека и гены репортерного белка люциферазы под контролем промоторов IL-2, клетки CHO-K1, стабильно экспрессирующие человеческий CD155 и элементы активации T-лимфоцитов, и антитела к TIGIT человека в различных концентрациях совместно инкубировали в CO2-инкубаторе при температуре 37°C в течение 6 ч, после чего измеряли интенсивность флуоресценции.
Результаты проведенных экспериментов показали, что дочерние антитела после аффинного созревания (непосредственно экспрессированные дрожжами, с чистотой примерно 70%) обладали существенно более высокой биологической активностью in vitro, чем их родительские антитела, и в большинстве случаев достигали уровня биологической активности, сопоставимого с таковым для двух референтных антител 22G2 и 31C6. Результаты показаны в следующей таблице и на РИС. 5 (EC50: нМ).
Анализ биологической активности, лежащей в основе МД, был проведен и для 7 высокоочищенных антител, экспрессированных клетками CHO. В анализе, описанном выше, в качестве отрицательного контроля использовали IgG4, а в качестве положительного контроля - антитело 31C6. Из результатов проведенных экспериментов можно сделать вывод о том, что антитело-кандидат ADI-30278 характеризуется значением EC50 0,1856 нМ, которое ниже соответствующих значений для других антител-кандидатов и для референтного антитела 31C6 (0,3424 нМ), что говорит о более высокой биологической активности in vitro; антитела ADI-30268, ADI-30286, ADI-30288 и ADI-30306 характеризуются значениями EC50 0,5065 нМ, 0,3963 нМ, 0,3844 нМ и 0,3984 нМ соответственно, то есть обладают примерно такой же биологической активностью, что и референтное антитело 31C6; антитела ADI-30336 и ADI-30293 характеризуются значениями EC50 1,274 нМ и 1,624 нМ соответственно, что говорит о более низкой биологической активности. Результаты показаны в следующей таблице и на РИС. 6 (EC50: нМ).
Пример 6. Исследование эффективности in vivo (на животных)
В этом исследовании изучали противоопухолевую активность антител к TIGIT в экспериментальной модели опухоли на мышах линии MC38 с внедренным (knock-in) геном TIGIT человека.
Изучали фармакодинамику антител-кандидатов ADI-30293 и ADI-30278, которые вводились по отдельности (в дозе 10 мг/кг) мышам линии MC38 с внедренным геном рецептора TIGIT человека. Клетки рака толстой кишки мыши MC38 (OBiO Technology (Shanghai) Co., Ltd., HYC0116) пересаживали самкам трансгенных мышей с внедренным геном TIGIT (Biocytogen Beijing Co., Ltd.). Мышам из каждой группы подкожно вводили антитела в дни 6, 10, 13 и 17 после пересадки. В дни 6, 10, 13, 17, 21 и 25 после пересадки измеряли объем опухоли, после чего мышей усыпляли. Результаты показаны на РИС. 7.
Из результатов, показанных на РИС. 7, видно, что моноклональные антитела к TIGIT ADI-30293 и ADI-30278 по настоящему изобретению при их применении в отдельности в определенной степени подавляют рост опухоли по сравнению с Ig4, который служил отрицательным контролем. По способности подавлять рост опухоли два антитела-кандидата в случае их применения в отдельности статистически значимо не отличались от антитела 31C6, которое служило положительным контролем.
Дополнительно изучали фармакодинамику антител-кандидатов ADI-30293 и ADI-30278, которые вводились (в дозе 10 + 1 мг/кг) в комбинации с антителом к PD1 (антитело C, WO2017133540A1) мышам линии MC38 с внедренным (knock-in) геном TIGIT человека.
Клетки рака толстой кишки мыши MC38 (OBiO Technology (Shanghai) Co., Ltd., HYC0116) культивировали в среде DMEM. 1×106 клеток MC38 в 0,2 мл основной суспензионной среды DMEM пересаживали на правый бок самкам трансгенных мышей с внедренным геном TIGIT (SHANGHAI MODEL ORGANISMS). Два раза в неделю в ходе исследования измеряли объем опухоли и вес тела. На 8-й день после трансплантации опухолевых клеток измеряли длину и ширину опухоли штангенциркулем. Объем опухоли рассчитывали по следующей формуле: ширина2 × длина/2 (мм3). Мышей со средним объемом опухоли примерно 71 мм3 случайным образом распределяли по группам (по 6 мышей в каждой) и усыпляли после достижения конечной точки для определения объема опухоли или когда мыши теряли более 20% веса тела. В дни 8, 12, 15 и 19 после трансплантации мышам из каждой группы подкожно вводили человеческий IgG (10 мг/кг), антитело C (1 мг/кг) и антитела к TIGIT или их изотипы (10 мг/кг) соответственно. Средний объем опухоли в конечной точке определяли в день 29 после трансплантации клеток, после чего мышей усыпляли. Результаты показаны на РИС. 8.
Из результатов, показанных на РИС. 8, видно, что антитело к TIGIT (ADI-30293), применяемое в комбинации с антителом к PD1 (антитело C), сильнее подавляет рост опухоли, чем каждое из антител в отдельности; при этом подавляющий эффект комбинации антитела ADI-30293 и антитела к PD1 превосходит эффект комбинации антитела 31C6 и антитела к PD1. Сходным образом, эффект комбинации антитела ADI-30278 и антитела к PD1 превосходил эффект комбинации антитела 31C6 и антитела к PD1 (антитела C).
В этом исследовании статистически значимого изменения веса тела мышей в день 29 после трансплантации клеток ни в одной из групп выявлено не было.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Innovent Biologics (Suzhou) Co., Ltd.
<120> Антитело к TIGIT и его использование
<130> PF190269PCT
<160> 209
<170> Патент редакции 3.3
<210> 1
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 1 ADI-27238 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 1
Tyr Thr Phe Thr Ser Tyr Tyr Met Ser
1 5
<210> 2
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 2 ADI-30263 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 2
Tyr Thr Phe Leu Ser Tyr Tyr Met Asn
1 5
<210> 3
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 3 ADI-30267 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 3
Tyr Thr Phe Arg Ser Tyr Tyr Met Ser
1 5
<210> 4
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 4 ADI-30268 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 4
Tyr Thr Phe Gly Ser Tyr Tyr Met Gly
1 5
<210> 5
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 5, консенсусная последовательность
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из T, L, R, G, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, N, G, и ее консервативно замещенным остатком
<400> 5
Tyr Thr Phe Xaa Ser Tyr Tyr Met Xaa
1 5
<210> 6
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 6 ADI-27238, ADI-27297 HCDR2
KABAT, пронумерованные положения H50-65
<400> 6
Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 7
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 7 ADI-30263 HCDR2 KABAT, H50-65
<400> 7
Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 8
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 8 ADI-30267 HCDR2 KABAT, H50-65
<400> 8
Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Phe Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 9
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 9 ADI-30268 HCDR2 KABAT, H50-65
<400> 9
Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Tyr Ala Arg Lys Phe Gln
1 5 10 15
Gly
<210> 10
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 10, консенсусная последовательность
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из N, D, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, R, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из Y, F, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из Q, R, и ее консервативно замещенным остатком
<400> 10
Ile Ile Xaa Pro Ser Gly Gly Xaa Thr Ser Xaa Ala Xaa Lys Phe Gln
1 5 10 15
Gly
<210> 11
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 11 ADI-27238 HCDR3 Kabat, H93-102
<400> 11
Ala Arg Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val
1 5 10 15
<210> 12
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 12 ADI-30263 HCDR3 Kabat, H93-102
<400> 12
Ala Arg Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Thr Asp Val
1 5 10 15
<210> 13
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 13 ADI-30267 HCDR3 Kabat, H93-102
<400> 13
Ala Arg Ala Arg Tyr Pro Glu Ser Trp Pro Tyr Gly Met Asp Val
1 5 10 15
<210> 14
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 14 ADI-30268 HCDR3 Kabat, H93-102
<400> 14
Ala Arg Ala Arg Thr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val
1 5 10 15
<210> 15
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 15, консенсусная последовательность
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из Y, T, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (13)..(13)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из M, T, и ее консервативно замещенным остатком
<400> 15
Ala Arg Ala Arg Xaa Pro Ser Ser Trp Pro Tyr Gly Xaa Asp Val
1 5 10 15
<210> 16
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 16 ADI-27238, ADI-30263, ADI-30267,
ADI-30268 LCDR1 Kabat, L24-34
<400> 16
Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asn
1 5 10
<210> 17
<211> 7
<212> PRT
<220>
<223> Последовательность антитела 17 ADI-27238, ADI-30263, ADI-30267,
ADI-30268, ADI-27297, ADI-30272, ADI-30272 LCDR2 Kabat, L50-56
<400> 17
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 18
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 18 ADI-27238, ADI-30263, ADI-30267,
ADI-30268 LCDR3 Kabat, L89-97
<400> 18
Gln Gln Ser Leu Leu Thr Pro Phe Thr
1 5
<210> 19
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 19 ADI-27243, ADI-30336 HCDR1
KABAT, пронумерованные положения H27-35B
<400> 19
Gly Ser Ile Ser Ser Ser Arg Tyr Tyr Trp Gly
1 5 10
<210> 20
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 20 ADI-30302 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 20
Gly Ser Ile Gly Ser Ser Gln Tyr Tyr Trp Gly
1 5 10
<210> 21
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 21, консенсусная последовательность
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, G, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из R, Q, и ее консервативно замещенным остатком
<400> 21
Gly Ser Ile Xaa Ser Ser Xaa Tyr Tyr Trp Gly
1 5 10
<210> 22
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 22 ADI-27243, ADI-30336, ADI-27301 HCDR2 KABAT, H50-65
<400> 22
Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 23
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 23 ADI-30302 HCDR2 KABAT, H50-65
<400> 23
Ser Ile Tyr Arg Ser Gly Gly Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 24
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 24, консенсусная последовательность
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из Y, R, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, G, и ее консервативно замещенным остатком
<400> 24
Ser Ile Tyr Xaa Ser Gly Xaa Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 25
<211> 14
<212> PRT
<220>
<223> Последовательность антитела 25 ADI-27243, ADI-30302, ADI-30336
HCDR3 Kabat, H93-102
<400> 25
Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His
1 5 10
<210> 26
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 26 ADI-27243, ADI-30302, ADI-30336
LCDR1 Kabat, L24-34
<400> 26
Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala
1 5 10
<210> 27
<211> 7
<212> PRT
<220>
<223> Последовательность антитела 27 ADI-27243, ADI-30302, ADI-30336
LCDR2 Kabat, L50-56
<400> 27
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 28
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 28 ADI-27243, ADI-30302 LCDR3 Kabat,
L89-97
<400> 28
Gln Gln Arg Phe Ala Phe Pro Tyr Thr
1 5
<210> 29
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 29 ADI-30336 LCDR3 Kabat, L89-97
<400> 29
Gln Gln Arg Phe Ala His Pro Tyr Thr
1 5
<210> 30
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 30, консенсусная последовательность
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из F, H, и ее консервативно замещенным остатком
<400> 30
Gln Gln Arg Phe Ala Xaa Pro Tyr Thr
1 5
<210> 31
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 31 ADI-27278, ADI-30293 HCDR1
KABAT, пронумерованные положения H27-35B
<400> 31
Phe Thr Phe Ser Ser Tyr Ser Met Asn
1 5
<210> 32
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 32 ADI-30296 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 32
Phe Thr Ile Gly Gly Tyr Ser Met Asn
1 5
<210> 33
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 33, консенсусная последовательность
<220>
<221> misc_feature
<222> (3)..(5)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 3 выбрана из F, I и их консервативно замещенных остатков,
Xaa в положении 4 выбрана из S, G и их консервативно замещенных остатков,
а Xaa в положении 5 выбрана из S, G и их консервативно замещенных остатков
<400> 33
Phe Thr Xaa Xaa Xaa Tyr Ser Met Asn
1 5
<210> 34
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 34 ADI-27278 HCDR2 KABAT, H50-65
<400> 34
Tyr Ile Ser Gly Ser Ser Ser Thr Ile Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 35
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 35 ADI-30293 HCDR2 KABAT, H50-65
<400> 35
Tyr Ile Ser Ser Ser Gly Thr Ile Asn Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 36
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 36 ADI-30296 HCDR2 KABAT, H50-65
<400> 36
Tyr Ile Ser Ser Ser Ser Thr Ile His Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 37
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 37, консенсусная последовательность
<220>
<221> misc_feature
<222> (3)..(4)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 3 выбрана из делеции, S и ее консервативно замещенного остатка,
а Xaa в положении 4 выбрана из G, S и их консервативно замещенных остатков
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, G, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из Y, N, H, и ее консервативно замещенным остатком
<400> 37
Tyr Ile Xaa Xaa Ser Ser Xaa Thr Ile Xaa Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 38
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 38 ADI-27278, ADI-30293 HCDR3 Kabat, H93-102
<400> 38
Ala Arg His Arg Ile Ala Asp Ser Pro Ser Arg Ala Phe Asp Ile
1 5 10 15
<210> 39
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 39 ADI-30296 HCDR3 Kabat, H93-102
<400> 39
Ala Arg His Arg Ile Gly Arg Ser Pro Ser Arg Ala Phe Asp Ile
1 5 10 15
<210> 40
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 40, консенсусная последовательность
<220>
<221> misc_feature
<222> (6)..(7)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 6 выбрана из A, G и их консервативно замещенных остатков,
а Xaa в положении 7 выбрана из D, R и их консервативно замещенных остатков
<400> 40
Ala Arg His Arg Ile Xaa Xaa Ser Pro Ser Arg Ala Phe Asp Ile
1 5 10 15
<210> 41
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 41 ADI-27278, ADI-30293, ADI-30296
LCDR1-Kabat, L24-34
<400> 41
Lys Ser Ser Gln Ser Val Leu Phe Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 42
<211> 7
<212> PRT
<220>
<223> Последовательность антитела 42 ADI-27278, ADI-30293, ADI-30296
LCDR2-Kabat, L50-56
<400> 42
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 43
<211> 8
<212> PRT
<220>
<223> Последовательность антитела 43 ADI-27278, ADI-30293, ADI-30296
LCDR3 Kabat, L89-97
<400> 43
Gln Gln Ser Tyr Phe Phe Pro Thr
1 5
<210> 44
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 44 ADI-27291 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 44
Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5
<210> 45
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 45 ADI-30283 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 45
Phe Thr Phe Ser Pro Tyr Gly Met Ser
1 5
<210> 46
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 46 ADI-30286, ADI-30288 HCDR1
KABAT, пронумерованные положения H27-35B
<400> 46
Phe Thr Phe Gly Pro Tyr Gly Met Ser
1 5
<210> 47
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 47, консенсусная последовательность
<220>
<221> misc_feature
<222> (4)..(5)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 4 выбрана из S, G и их консервативно замещенных остатков,
а Xaa в положении 5 выбрана из S, P и их консервативно замещенных остатков
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из A, G, и ее консервативно замещенным остатком
<400> 47
Phe Thr Phe Xaa Xaa Tyr Xaa Met Ser
1 5
<210> 48
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 48 ADI-27291 HCDR2 KABAT, H50-65
<400> 48
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 49
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 49 ADI-30283 HCDR2 KABAT, H50-65
<400> 49
Ser Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 50
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 50 ADI-30286 HCDR2 KABAT, H50-65
<400> 50
Ala Ile Ser Gly Ser Gly Ala Ser Thr Trp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 51
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 51 ADI-30288 HCDR2 KABAT, H50-65
<400> 51
Ala Ile Ser Gly Ser Gly Ala Ser Thr Trp His Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 52
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 52, консенсусная последовательность
<220>
<221> misc_feature
<222> (1)..(1)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из A, S, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (7)..(8)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 7 выбрана из G, A и их консервативно замещенных остатков,
а Xaa в положении 8 выбрана из S, R и их консервативно замещенных остатков
<220>
<221> misc_feature
<222> (10)..(11)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 10 выбрана из Y, W и их консервативно замещенных остатков,
а Xaa в положении 11 выбрана из Y, H и их консервативно замещенных остатков
<400> 52
Xaa Ile Ser Gly Ser Gly Xaa Xaa Thr Xaa Xaa Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 53
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 53 ADI-27291, ADI-30283 HCDR3 Kabat, H93-102
<400> 53
Ala Lys Asp Pro Gly Thr Asp Ser Ser Gly Tyr Tyr Tyr Ile Trp Arg
1 5 10 15
Tyr
<210> 54
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 54 ADI-30286 HCDR3 Kabat, H93-102
<400> 54
Ala Lys Asp Pro Gly Thr His Tyr Ser Gly Tyr Tyr Tyr Ile Trp Arg
1 5 10 15
Tyr
<210> 55
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 55 ADI-30288 HCDR3 Kabat, H93-102
<400> 55
Ala Lys Asp Pro Gly Thr Asp Ser Thr Gly Tyr Tyr Tyr Ile Trp Arg
1 5 10 15
Tyr
<210> 56
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 56, консенсусная последовательность
<220>
<221> misc_feature
<222> (7)..(9)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 7 выбрана из D, H и их консервативно замещенных остатков,
Xaa в положении 8 выбрана из S, Y и их консервативно замещенных остатков,
а Xaa в положении 9 выбрана из S, T и их консервативно замещенных остатков
<400> 56
Ala Lys Asp Pro Gly Thr Xaa Xaa Xaa Gly Tyr Tyr Tyr Ile Trp Arg
1 5 10 15
Tyr
<210> 57
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 57 ADI-27291, ADI-30296, ADI-30286,
ADI-30288 LCDR1 Kabat, L24-34
<400> 57
Arg Ala Ser Gln Ser Val Ser Ser Asn Leu Ala
1 5 10
<210> 58
<211> 7
<212> PRT
<220>
<223> Последовательность антитела 58 ADI-27291, ADI-30283, ADI-30286,
ADI-30288 LCDR2 Kabat, L50-56
<400> 58
Ser Ala Ser Thr Arg Ala Thr
1 5
<210> 59
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 59 ADI-27291, ADI-30283, ADI-30286,
ADI-30288 LCDR3 Kabat, L89-97
<400> 59
Gln Gln Tyr Val Pro His Pro Pro Phe Thr
1 5 10
<210> 60
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 60 ADI-27297 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 60
Tyr Thr Phe Thr Ser Tyr Tyr Met His
1 5
<210> 61
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 61 ADI-30272, ADI-30278 HCDR1
KABAT, пронумерованные положения H27-35B
<400> 61
Tyr Thr Phe Thr Glu Tyr Tyr Met His
1 5
<210> 62
<211> 9
<212> PRT
<220>
<220>
<221> misc_feature
<222> (5)..(5)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, E, и ее консервативно замещенным остатком
<400> 62
Tyr Thr Phe Thr Xaa Tyr Tyr Met His
1 5
<210> 63
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 63 ADI-30272 HCDR2 KABAT, H50-65
<400> 63
Ile Ile Ser Pro Ser Ala Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 64
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 64 ADI-30278 HCDR2 KABAT, H50-65
<400> 64
Ile Ile Ser Pro Ser Ala Gly Ser Thr Lys Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 65
<211> 17
<212> PRT
<220>
<223> Последовательность антитела 65, консенсусная последовательность
<220>
<221> misc_feature
<222> (3)..(3)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из N, S, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из G, A, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, K, и ее консервативно замещенным остатком
<400> 65
Ile Ile Xaa Pro Ser Xaa Gly Ser Thr Xaa Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 66
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 66 ADI-27297, ADI-30272 HCDR3 Kabat, H93-102
<400> 66
Ala Arg Asp His Asp Ile Ala Ala Ala Gly Arg Leu Ala Asp Tyr
1 5 10 15
<210> 67
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 67 ADI-30278 HCDR3 Kabat, H93-102
<400> 67
Ala Arg Asp His Asp Ile Arg Leu Ala Gly Arg Leu Ala Asp Tyr
1 5 10 15
<210> 68
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 68, консенсусная последовательность
<220>
<221> misc_feature
<222> (7)..(8)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 7 выбрана из A, R и их консервативно замещенных остатков,
а Xaa в положении 8 выбрана из A, L и их консервативно замещенных остатков
<400> 68
Ala Arg Asp His Asp Ile Xaa Xaa Ala Gly Arg Leu Ala Asp Tyr
1 5 10 15
<210> 69
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 69 ADI-27297, ADI-30272, ADI-30278
LCDR1-Kabat, L24-34
<400> 69
Arg Ala Ser Gln Gly Ile Ser Ser Trp Leu Ala
1 5 10
<210> 70
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 70 ADI-27297, ADI-30272, ADI-30278
LCDR3 Kabat, L89-97
<400> 70
Gln Gln Ala Val Ile Leu Pro Ile Thr
1 5
<210> 71
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 71 ADI-27301 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 71
Gly Ser Ile Ser Ser Ser Ser Tyr Tyr Trp Gly
1 5 10
<210> 72
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 72 ADI-30306 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 72
Gly Ser Ile Ser Ser Ser Leu Tyr Tyr Trp Gly
1 5 10
<210> 73
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 73 ADI-30311 HCDR1 KABAT,
пронумерованные положения H27-35B
<400> 73
Gly Ser Ile Ala Ser Ser Val Tyr Tyr Trp Gly
1 5 10
<210> 74
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 74, консенсусная последовательность
<220>
<221> misc_feature
<222> (4)..(4)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из A, S, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из S, L, V, и ее консервативно замещенным остатком
<400> 74
Gly Ser Ile Xaa Ser Ser Xaa Tyr Tyr Trp Gly
1 5 10
<210> 75
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 75 ADI-30306 HCDR2 KABAT, H50-65
<400> 75
Ser Ile Tyr Tyr Ser Gly Ser Thr Phe Tyr Asn Pro Ser Phe Lys Ser
1 5 10 15
<210> 76
<211> 16
<212> PRT
<220>
<223> Последовательность антитела 76 ADI-30311 HCDR2 KABAT, H50-65
<400> 76
Ser Ile Tyr Tyr Ser Gly Ser Thr Trp Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 77
<211> 16
<212> PRT
<220>
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из Y, F, W, и ее консервативно замещенным остатком
<220>
<221> misc_feature
<222> (14)..(14)
<223> Xaa может быть любой природной аминокислотой, предпочтительно
выбранной из L, F, и ее консервативно замещенным остатком
<400> 77
Ser Ile Tyr Tyr Ser Gly Ser Thr Xaa Tyr Asn Pro Ser Xaa Lys Ser
1 5 10 15
<210> 78
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 78 ADI-27301, ADI-30306 HCDR3 Kabat, H93-102
<400> 78
Ala Arg Glu Ala Gly Arg Gly Thr Thr Gly Leu Leu Phe Asp Tyr
1 5 10 15
<210> 79
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 79 ADI-30311 HCDR3 Kabat, H93-102
<400> 79
Ala Arg Glu Ala Gly Arg Thr Gly Thr Gly Leu Leu Phe Asp Tyr
1 5 10 15
<210> 80
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 80, консенсусная последовательность
<220>
<221> misc_feature
<222> (7)..(8)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa в положении 7 выбрана из G, T и их консервативно замещенных остатков,
а Xaa в положении 8 выбрана из T, G и их консервативно замещенных остатков
<400> 80
Ala Arg Glu Ala Gly Arg Xaa Xaa Thr Gly Leu Leu Phe Asp Tyr
1 5 10 15
<210> 81
<211> 11
<212> PRT
<220>
<223> Последовательность антитела 81 ADI-27301, ADI-30306, ADI-30311
LCDR1 Kabat, L24-34
<400> 81
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 82
<211> 7
<212> PRT
<220>
<223> Последовательность антитела 82 ADI-27301, ADI-30306, ADI-30311
LCDR2-Kabat, L50-56
<400> 82
Lys Ala Ser Ser Leu Glu Ser
1 5
<210> 83
<211> 9
<212> PRT
<220>
<223> Последовательность антитела 83 ADI-27301, ADI-30306, ADI-30311
LCDR3 Kabat, L89-97
<400> 83
Gln Gln Tyr Gly Ile Leu Pro Arg Thr
1 5
<210> 84
<211> 122
<212> PRT
<220>
<400> 84
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 85
<211> 122
<212> PRT
<220>
<400> 85
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Leu Ser Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Thr Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 86
<211> 122
<212> PRT
<220>
<400> 86
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Arg Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Phe Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Tyr Pro Glu Ser Trp Pro Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 87
<211> 122
<212> PRT
<220>
<400> 87
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Gly Ser Tyr
20 25 30
Tyr Met Gly Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Tyr Ala Arg Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Thr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 88
<211> 122
<212> PRT
<220>
<400> 88
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Arg Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 89
<211> 122
<212> PRT
<220>
<400> 89
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Gly Ser Ser
20 25 30
Gln Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Arg Ser Gly Gly Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 90
<211> 122
<212> PRT
<220>
<400> 90
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Arg Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 91
<211> 122
<212> PRT
<220>
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Gly Ser Ser Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Arg Ile Ala Asp Ser Pro Ser Arg Ala Phe Asp Ile Trp
100 105 110
Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 92
<211> 121
<212> PRT
<213> искусственная последовательность
<220>
<400> 92
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Thr Ile Asn Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Arg Ile Ala Asp Ser Pro Ser Arg Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 93
<211> 121
<212> PRT
<220>
<400> 93
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Ile Gly Gly Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Thr Ile His Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Arg Ile Gly Arg Ser Pro Ser Arg Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 94
<211> 124
<212> PRT
<220>
<400> 94
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr Asp Ser Ser Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 95
<211> 124
<212> PRT
<220>
<400> 95
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr Asp Ser Ser Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 96
<211> 124
<212> PRT
<220>
<400> 96
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Pro Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Ala Ser Thr Trp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr His Tyr Ser Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 97
<211> 124
<212> PRT
<220>
<400> 97
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Pro Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Ala Ser Thr Trp His Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr Asp Ser Thr Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 98
<211> 122
<212> PRT
<220>
<400> 98
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Asp Ile Ala Ala Ala Gly Arg Leu Ala Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 99
<211> 122
<212> PRT
<220>
<400> 99
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Ser Pro Ser Ala Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Asp Ile Ala Ala Ala Gly Arg Leu Ala Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 100
<211> 122
<212> PRT
<220>
<400> 100
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Ser Pro Ser Ala Gly Ser Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Asp Ile Arg Leu Ala Gly Arg Leu Ala Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 101
<211> 123
<212> PRT
<220>
<400> 101
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Ala Gly Arg Gly Thr Thr Gly Leu Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 102
<211> 123
<212> PRT
<220>
<400> 102
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Leu Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Phe Tyr Asn Pro Ser
50 55 60
Phe Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Ala Gly Arg Gly Thr Thr Gly Leu Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 103
<211> 123
<212> PRT
<220>
<400> 103
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ala Ser Ser
20 25 30
Val Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Trp Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Ala Gly Arg Thr Gly Thr Gly Leu Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 104
<211> 107
<212> PRT
<220>
<223> Последовательность антитела 104 ADI-27238, ADI-30263, ADI-30267,
<400> 104
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Leu Leu Thr Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 105
<211> 107
<212> PRT
<220>
<400> 105
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Phe Ala Phe Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 106
<211> 107
<212> PRT
<220>
<400> 106
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Phe Ala His Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 107
<211> 112
<212> PRT
<220>
<400> 107
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Phe Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ser Tyr Phe Phe Pro Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 108
<211> 108
<212> PRT
<220>
<223> Последовательность антитела 108 ADI-27291, ADI-30283, ADI-30286,
<400> 108
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Val Pro His Pro Pro
85 90 95
Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 109
<211> 107
<212> PRT
<213> искусственная последовательность
<220>
<400> 109
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Ser Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Val Ile Leu Pro Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 110
<211> 107
<212> PRT
<220>
<400> 110
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ile Leu Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 111
<211> 448
<212> PRT
<220>
<400> 111
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 112
<211> 448
<212> PRT
<220>
<400> 112
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Leu Ser Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Thr Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 113
<211> 448
<212> PRT
<213> искусственная последовательность
<220>
<400> 113
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Arg Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Phe Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Tyr Pro Glu Ser Trp Pro Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 114
<211> 448
<212> PRT
<220>
<400> 114
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Gly Ser Tyr
20 25 30
Tyr Met Gly Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Arg Thr Ser Tyr Ala Arg Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Arg Thr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 115
<211> 448
<212> PRT
<220>
<400> 115
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Arg Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 116
<211> 448
<212> PRT
<220>
<400> 116
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Gly Ser Ser
20 25 30
Gln Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Arg Ser Gly Gly Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 117
<211> 448
<212> PRT
<220>
<400> 117
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Arg Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 118
<211> 448
<212> PRT
<220>
<400> 118
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Gly Ser Ser Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Arg Ile Ala Asp Ser Pro Ser Arg Ala Phe Asp Ile Trp
100 105 110
Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 119
<211> 447
<212> PRT
<220>
<400> 119
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Thr Ile Asn Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Arg Ile Ala Asp Ser Pro Ser Arg Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 120
<211> 447
<212> PRT
<220>
<400> 120
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Ile Gly Gly Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Thr Ile His Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg His Arg Ile Gly Arg Ser Pro Ser Arg Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 121
<211> 450
<212> PRT
<220>
<400> 121
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr Asp Ser Ser Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly
450
<210> 122
<211> 450
<212> PRT
<220>
<400> 122
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr Asp Ser Ser Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly
450
<210> 123
<211> 450
<212> PRT
<220>
<400> 123
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Pro Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Ala Ser Thr Trp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr His Tyr Ser Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly
450
<210> 124
<211> 450
<212> PRT
<220>
<400> 124
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Gly Pro Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Ala Ser Thr Trp His Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Gly Thr Asp Ser Thr Gly Tyr Tyr Tyr Ile Trp Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
130 135 140
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
195 200 205
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
210 215 220
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
260 265 270
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Leu Gly
450
<210> 125
<211> 448
<212> PRT
<220>
<400> 125
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Asp Ile Ala Ala Ala Gly Arg Leu Ala Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 126
<211> 448
<212> PRT
<220>
<400> 126
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Ser Pro Ser Ala Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Asp Ile Ala Ala Ala Gly Arg Leu Ala Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 127
<211> 448
<212> PRT
<220>
<400> 127
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Ser Pro Ser Ala Gly Ser Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp His Asp Ile Arg Leu Ala Gly Arg Leu Ala Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 128
<211> 449
<212> PRT
<220>
<400> 128
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Ala Gly Arg Gly Thr Thr Gly Leu Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly
<210> 129
<211> 449
<212> PRT
<220>
<400> 129
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Leu Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Phe Tyr Asn Pro Ser
50 55 60
Phe Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Ala Gly Arg Gly Thr Thr Gly Leu Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly
<210> 130
<211> 449
<212> PRT
<220>
<400> 130
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ala Ser Ser
20 25 30
Val Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Trp Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Ala Gly Arg Thr Gly Thr Gly Leu Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
260 265 270
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
435 440 445
Gly
<210> 131
<211> 441
<212> PRT
<220>
<400> 131
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Thr Gln Pro Gly Lys
1 5 10 15
Ser Leu Lys Leu Ser Cys Glu Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Thr Met His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Ser Gly Ser Gly Ile Val Phe Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Leu Leu Phe
65 70 75 80
Leu Gln Met Asn Asp Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Pro Leu Gly His Asn Thr Phe Asp Ser Trp Gly Gln Gly
100 105 110
Thr Leu Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
115 120 125
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
210 215 220
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
225 230 235 240
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
245 250 255
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
260 265 270
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
275 280 285
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
290 295 300
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
305 310 315 320
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
325 330 335
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
340 345 350
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
355 360 365
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
370 375 380
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
385 390 395 400
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
405 410 415
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
420 425 430
Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440
<210> 132
<211> 457
<212> PRT
<220>
<400> 132
Gln Val His Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Ile Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Tyr Tyr Val Ser Gly Asn Tyr Tyr Asn Val Asp Tyr
100 105 110
Tyr Phe Phe Gly Val Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
130 135 140
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455
<210> 133
<211> 446
<212> PRT
<220>
<400> 133
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Val Met His Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asp Pro Tyr Asn Asp Gly Ala Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Pro Tyr Gly Trp Tyr Phe Asp Val Trp Gly Ala Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 134
<211> 446
<212> PRT
<220>
<400> 134
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Thr
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly His
20 25 30
Leu Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Ile Pro Tyr Asn Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Asp Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ser Arg Gly Leu Arg Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val
260 265 270
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 135
<211> 467
<212> PRT
<220>
<400> 135
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Arg Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Thr Ser Arg Asp Asp Ser Lys Asn
85 90 95
Ala Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Gly Pro Gly Trp Tyr Ala Ala Asp Val Trp
115 120 125
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
225 230 235 240
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly
465
<210> 136
<211> 465
<212> PRT
<220>
<400> 136
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Arg Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Thr Ser Arg Asp Asp Ser Lys Asn
85 90 95
Ala Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Gly Pro Gly Trp Tyr Ala Ala Asp Val Trp
115 120 125
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
225 230 235 240
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
275 280 285
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
450 455 460
Lys
465
<210> 137
<211> 214
<212> PRT
<220>
<223> Последовательность антитела 137 ADI-27238, ADI-30263, ADI-30267,
<400> 137
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Leu Leu Thr Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 138
<211> 214
<212> PRT
<220>
<400> 138
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Phe Ala Phe Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 139
<211> 214
<212> PRT
<220>
<400> 139
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Phe Ala His Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 140
<211> 219
<212> PRT
<220>
<400> 140
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Phe Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ser Tyr Phe Phe Pro Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 141
<211> 215
<212> PRT
<220>
<223> Последовательность антитела 141 ADI-27291, ADI-30283, ADI-30286,
<400> 141
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Val Pro His Pro Pro
85 90 95
Phe Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 142
<211> 214
<212> PRT
<220>
<400> 142
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Ser Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Val Ile Leu Pro Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 143
<211> 214
<212> PRT
<220>
<400> 143
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ile Leu Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 144
<211> 220
<212> PRT
<220>
<400> 144
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Lys Ser Ser Gln Ser Leu Tyr Tyr Ser
20 25 30
Gly Val Lys Glu Asn Leu Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Tyr Ala Ser Ile Arg Phe Thr Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr
65 70 75 80
Ile Thr Ser Val Gln Ala Glu Asp Met Gly Gln Tyr Phe Cys Gln Gln
85 90 95
Gly Ile Asn Asn Pro Leu Thr Phe Gly Asp Gly Thr Lys Leu Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 145
<211> 214
<212> PRT
<220>
<400> 145
Gly Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Asp Val Ile Asn Tyr
20 25 30
Leu Ala Trp Cys Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln His Tyr Ser Pro Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 146
<211> 214
<212> PRT
<220>
<400> 146
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu His Ile Tyr Ser Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val
35 40 45
Tyr Asn Ala Lys Thr Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Phe Ser Leu Lys Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Thr Tyr Tyr Cys Gln His His Phe Gly Ser Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Thr Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 147
<211> 219
<212> PRT
<220>
<400> 147
Asp Val Val Leu Thr Gln Thr Pro Leu Ser Leu Ser Val Ser Phe Gly
1 5 10 15
Asp Gln Val Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Asn Ser
20 25 30
Tyr Gly Asn Thr Phe Leu Ser Trp Tyr Leu His Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Phe Gly Ile Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Thr Ile Lys Pro Glu Asp Leu Gly Met Tyr Tyr Cys Leu Gln Gly
85 90 95
Thr His Gln Pro Pro Thr Phe Gly Pro Gly Thr Lys Leu Glu Val Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 148
<211> 236
<212> PRT
<220>
<400> 148
Met Asp Phe Gln Val Gln Ile Ile Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
35 40 45
Gln Ser Ile Ser Ser Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ala Asp Leu Pro Ala Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 149
<211> 236
<212> PRT
<220>
<400> 149
Met Asp Phe Gln Val Gln Ile Ile Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Met Ser Arg Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
35 40 45
Gln Ser Ile Ser Ser Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
100 105 110
Ala Asp Leu Pro Ala Phe Ala Phe Gly Gly Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 150
<211> 366
<212> ДНК
<220>
<400> 150
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttcacc agctactata tgtcatgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atcaacccta gtggtggtag cacaagctac 180
gcacagaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagggctaga 300
tacccatcaa gctggccata cggaatggac gtatggggcc agggaacaac tgtcaccgtc 360
tcctca 366
<210> 151
<211> 366
<212> ДНК
<220>
<400> 151
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata cacattcctg agctactata tgaattgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atcgatccta gtggtggtag gacaagctac 180
gcacagaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagggctaga 300
tacccatcaa gctggccata cggaacggat gtatggggcc agggaacaac tgtcaccgtc 360
tcctca 366
<210> 152
<211> 366
<212> ДНК
<220>
<400> 152
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttccgg agctactata tgagttgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atcgatccta gtggtggtcg tacaagcttc 180
gcacaaaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagggctaga 300
tacccagagt cttggccata cggaatggac gtatggggcc agggaacaac tgtcaccgtc 360
tcctca 366
<210> 153
<211> 366
<212> ДНК
<220>
<400> 153
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttcggg agctactata tggggtgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atcgacccta gtggtggtag gacaagttac 180
gcacggaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagggctcgt 300
acgccatcaa gctggccata cggaatggac gtatggggcc agggaacaac tgtcaccgtc 360
tcctca 366
<210> 154
<211> 366
<212> ДНК
<220>
<400> 154
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtagtcgct actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggagtatct attatagtgg gagcacctac 180
tacaacccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagttctgt gaccgccgca gacacggcgg tgtactactg cgccagagat 300
gggttgtacc acaccccaga atacttccaa cactggggac agggtacatt ggtcaccgtc 360
tcctca 366
<210> 155
<211> 366
<212> ДНК
<220>
<400> 155
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcggg agtagtcagt actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggagtatct atcgtagtgg ggggacctac 180
tacaacccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagttctgt gaccgccgca gacacggcgg tgtactactg cgccagagat 300
gggttgtacc acaccccaga atacttccaa cactggggac agggtacatt ggtcaccgtc 360
tcctca 366
<210> 156
<211> 366
<212> ДНК
<220>
<400> 156
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtagtcgct actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggagtatct attatagtgg gagcacctac 180
tacaacccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagttctgt gaccgccgca gacacggcgg tgtactactg cgccagagat 300
gggttgtacc acaccccaga atacttccaa cactggggac agggtacatt ggtcaccgtc 360
tcctca 366
<210> 157
<211> 366
<212> ДНК
<220>
<400> 157
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtttcatac attagtggaa gtagtagtac catatactac 180
gcagactctg tgaagggccg attcaccatc tccagagaca atgccaagaa ctcactgtat 240
ctgcaaatga acagcctgag agccgaggac acggcggtgt actactgcgc cagacaccgc 300
atagcagact caccaagcag agccttcgat atatggggtc agggtacaat ggtcaccgtc 360
tcctca 366
<210> 158
<211> 363
<212> ДНК
<220>
<400> 158
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg gactggagtg ggtttcatac attagtagta gtgggaccat aaattacgca 180
gactctgtga agggccgatt caccatctcc agagacaatg ccaagaactc actgtatctg 240
caaatgaaca gcctgagagc cgaggacacg gcggtgtact actgcgccag acaccgcata 300
gcagactcac caagcagagc cttcgatata tggggtcagg gtacaatggt caccgtctcc 360
tca 363
<210> 159
<211> 363
<212> ДНК
<220>
<400> 159
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccatcggg ggttatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtttcatac attagtagta gtagtaccat acattacgca 180
gactctgtga agggccgatt caccatctcc agagacaatg ccaagaactc actgtatctg 240
caaatgaaca gcctgagagc tgaggacacg gcggtgtact actgcgccag acaccgcata 300
gggcgttcac caagcagagc cttcgatata tggggtcagg gtacaatggt caccgtctcc 360
tca 363
<210> 160
<211> 372
<212> ДНК
<220>
<400> 160
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggcggtgt actactgcgc caaggacccc 300
ggaacagaca gcagcggcta ctactacata tggcgatact ggggacaggg tacattggtc 360
accgtctcct ca 372
<210> 161
<211> 372
<212> ДНК
<220>
<400> 161
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc ccgtatggga tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcaagt attagtggaa gtggtggtag gacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggcggtgt actactgcgc caaggacccc 300
ggaacagaca gcagcggcta ctactacata tggcgatact ggggacaggg tacattggtc 360
accgtctcct ca 372
<210> 162
<211> 372
<212> ДНК
<220>
<400> 162
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttggc ccgtatggta tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggaa gtggtgcgag cacatggtac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca actccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggcggtgt actactgcgc caaggacccc 300
ggaacacatt atagcggcta ctactacata tggcgatact ggggacaggg tacattggtc 360
accgtctcct ca 372
<210> 163
<211> 372
<212> ДНК
<220>
<400> 163
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttggc ccgtatggta tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggaa gtggtgcgag cacatggcac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggcggtgt actactgcgc caaggacccc 300
ggaacagaca gcactgggta ctactacata tggcgatact ggggacaggg tacattggtc 360
accgtctcct ca 372
<210> 164
<211> 366
<212> ДНК
<220>
<400> 164
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttcacc agctactata tgcactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atcaacccta gtggtggtag cacaagctac 180
gcacagaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagagaccac 300
gacatagcag cagcaggaag attggctgat tactggggac agggtacatt ggtcaccgtc 360
tcctca 366
<210> 165
<211> 366
<212> ДНК
<220>
<400> 165
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttcacg gagtactata tgcactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atctcgccta gtgcgggtag cacaagctac 180
gcacagaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagagaccac 300
gacatagcag cagcaggaag attggctgat tactggggac agggtacatt ggtcaccgtc 360
tcctca 366
<210> 166
<211> 366
<212> ДНК
<220>
<400> 166
caggtgcagc tggtgcagtc tggggctgag gtgaagaagc ctggggcctc agtgaaggtt 60
tcctgcaagg catctggata caccttcacg gagtactata tgcactgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggaata atctcgccta gtgcgggtag cacaaagtac 180
gcacagaagt tccagggcag agtcaccatg accagggaca cgtccacgag cacagtctac 240
atggagctga gcagcctgag atctgaggac acggcggtgt actactgcgc aagagaccac 300
gacatacgtc tggcaggaag attggctgat tactggggac agggtacatt ggtcaccgtc 360
tcctca 366
<210> 167
<211> 369
<212> ДНК
<220>
<400> 167
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtagtagtt actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggagtatct attatagtgg gagcacctac 180
tacaacccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagttctgt gaccgccgca gacacggcgg tgtactactg cgccagagaa 300
gccggacgcg gcaccactgg tctcttgttc gactactggg gacagggtac attggtcacc 360
gtctcctca 369
<210> 168
<211> 369
<212> ДНК
<220>
<400> 168
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcagc agtagtcttt actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggtctatct attatagtgg gagcaccttt 180
tacaacccgt ccttcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagttctgt gaccgccgca gacacggcgg tgtactactg cgccagagaa 300
gccggacgcg gcaccactgg tctcttgttc gactactggg gacagggtac attggtcacc 360
gtctcctca 369
<210> 169
<211> 369
<212> ДНК
<220>
<400> 169
cagctgcagc tgcaggagtc gggcccagga ctggtgaagc cttcggagac cctgtccctc 60
acctgcactg tctctggtgg ctccatcgcg agtagtgttt actactgggg ctggatccgc 120
cagcccccag ggaaggggct ggagtggatt gggagtatct attatagtgg gagcacctgg 180
tacaacccgt ccctcaagag tcgagtcacc atatccgtag acacgtccaa gaaccagttc 240
tccctgaagc tgagttctgt gaccgccgca gacacggcgg tgtactactg cgccagagaa 300
gccggacgca ctgggactgg tctcttgttc gactactggg gacagggtac attggtcacc 360
gtctcctca 369
<210> 170
<211> 321
<212> ДНК
<220>
<223> Последовательность антитела 170 ADI-27238, ADI-30263, ADI-30267,
<400> 170
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattaac agctatttaa attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcagcaa agcctcctca ctcctttcac ttttggcgga 300
gggaccaagg ttgagatcaa a 321
<210> 171
<211> 321
<212> ДНК
<220>
<400> 171
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agctacttag cctggtacca acagaaacct 120
ggccaggctc ccaggctcct catctatgat gcatccaaca gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg cagtttatta ctgtcagcag agattcgcct tcccttacac ttttggcgga 300
gggaccaagg ttgagatcaa a 321
<210> 172
<211> 321
<212> ДНК
<220>
<400> 172
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agctacttag cctggtacca acagaaacct 120
ggccaggctc ccaggctcct catctatgat gcatccaaca gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg cagtttatta ctgtcagcag agattcgccc atccttacac ttttggcgga 300
gggaccaagg ttgagatcaa a 321
<210> 173
<211> 336
<212> ДНК
<220>
<223> Последовательность антитела 173 ADI-27278, ADI-30293,
<400> 173
gacatcgtga tgacccagtc tccagactcc ctggctgtgt ctctgggcga gagggccacc 60
atcaactgca agtccagcca gagtgtttta ttcagctcca acaataagaa ctacttagct 120
tggtaccagc agaaaccagg acagcctcct aagctgctca tttactgggc atctacccgg 180
gaatccgggg tccctgaccg attcagtggc agcgggtctg ggacagattt cactctcacc 240
atcagcagcc tgcaggctga agatgtggca gtttattact gtcagcagtc ctacttcttc 300
cctacttttg gcggagggac caaggttgag atcaaa 336
<210> 174
<211> 324
<212> ДНК
<213> искусственная последовательность
<220>
<223> Последовательность антитела 174 ADI-27291, ADI-30283, ADI-30286,
<400> 174
gaaatagtga tgacgcagtc tccagccacc ctgtctgtgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gagtgttagc agcaacttag cctggtacca gcagaaacct 120
ggccaggctc ccaggctcct catctatagc gcatccacca gggccactgg tatcccagcc 180
aggttcagtg gcagtgggtc tgggacagag ttcactctca ccatcagcag cctgcagtct 240
gaagattttg cagtttatta ctgtcagcag tacgtccccc accctccttt cacttttggc 300
ggagggacca aggttgagat caaa 324
<210> 175
<211> 321
<212> ДНК
<220>
<400> 175
gacatccaga tgacccagtc tccatcttcc gtgtctgcat ctgtaggaga cagagtcacc 60
atcacttgtc gggcgagtca gggtattagc agctggttag cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctccgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagattttg caacttatta ctgtcagcag gcagtcatcc tccctatcac ttttggcgga 300
gggaccaagg ttgagatcaa a 321
<210> 176
<211> 321
<212> ДНК
<220>
<400> 176
gacatccaga tgacccagtc tccttccacc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gagtattagt agctggttgg cctggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctataaa gcctccagtt tggaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagaa ttcactctca ccatcagcag cctgcagcct 240
gatgattttg caacttatta ctgccagcag tacggcatcc tccctaggac ttttggcgga 300
gggaccaagg ttgagatcaa a 321
<210> 177
<211> 327
<212> PRT
<220>
<220>
<221> misc_feature
<222> (327)..(327)
<223> Xaa может быть любой природной аминокислотой, и, предпочтительно,
Xaa является лизином или делетирована
<400> 177
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Xaa
325
<210> 178
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 178 ADI-27238 HCDR1-AbM
<400> 178
Gly Tyr Thr Phe Thr Ser Tyr Tyr Met Ser
1 5 10
<210> 179
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 179 ADI-30263 HCDR1-AbM
<400> 179
Gly Tyr Thr Phe Leu Ser Tyr Tyr Met Asn
1 5 10
<210> 180
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 180 ADI-30267 HCDR1-AbM
<400> 180
Gly Tyr Thr Phe Arg Ser Tyr Tyr Met Ser
1 5 10
<210> 181
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 181 ADI-30268 HCDR1-AbM
<400> 181
Gly Tyr Thr Phe Gly Ser Tyr Tyr Met Gly
1 5 10
<210> 182
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 182 ADI-27238 HCDR3-Kabat
<400> 182
Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val
1 5 10
<210> 183
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 183 ADI-30263 HCDR3-Kabat
<400> 183
Ala Arg Tyr Pro Ser Ser Trp Pro Tyr Gly Thr Asp Val
1 5 10
<210> 184
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 184 ADI-30267 HCDR3-Kabat
<400> 184
Ala Arg Tyr Pro Glu Ser Trp Pro Tyr Gly Met Asp Val
1 5 10
<210> 185
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 185 ADI-30268 HCDR3-Kabat
<400> 185
Ala Arg Thr Pro Ser Ser Trp Pro Tyr Gly Met Asp Val
1 5 10
<210> 186
<211> 12
<212> PRT
<220>
<223> Последовательность антитела 186 ADI-27243, ADI-30336 HCDR1-AbM
<400> 186
Gly Gly Ser Ile Ser Ser Ser Arg Tyr Tyr Trp Gly
1 5 10
<210> 187
<211> 12
<212> PRT
<213> искусственная последовательность
<220>
<223> Последовательность антитела 187 ADI-30302 HCDR1-AbM
<400> 187
Gly Gly Ser Ile Gly Ser Ser Gln Tyr Tyr Trp Gly
1 5 10
<210> 188
<211> 12
<212> PRT
<220>
<223> Последовательность антитела 188 ADI-27243, ADI-30302, ADI-30336
HCDR3-Kabat
<400> 188
Asp Gly Leu Tyr His Thr Pro Glu Tyr Phe Gln His
1 5 10
<210> 189
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 189 ADI-27278, ADI-30293 HCDR1-AbM
<400> 189
Gly Phe Thr Phe Ser Ser Tyr Ser Met Asn
1 5 10
<210> 190
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 190 ADI-30296 HCDR1-AbM
<400> 190
Gly Phe Thr Ile Gly Gly Tyr Ser Met Asn
1 5 10
<210> 191
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 191 ADI-27278, ADI-30293 HCDR3-Kabat
<400> 191
His Arg Ile Ala Asp Ser Pro Ser Arg Ala Phe Asp Ile
1 5 10
<210> 192
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 192 ADI-30296 HCDR3-Kabat
<400> 192
His Arg Ile Gly Arg Ser Pro Ser Arg Ala Phe Asp Ile
1 5 10
<210> 193
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 193 ADI-27291 HCDR1-AbM
<400> 193
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5 10
<210> 194
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 194 ADI-30283 HCDR1-AbM
<400> 194
Gly Phe Thr Phe Ser Pro Tyr Gly Met Ser
1 5 10
<210> 195
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 195 ADI-30286, ADI-30288 HCDR1-AbM
<400> 195
Gly Phe Thr Phe Gly Pro Tyr Gly Met Ser
1 5 10
<210> 196
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 196 ADI-27291, ADI-30283 HCDR3-Kabat
<400> 196
Asp Pro Gly Thr Asp Ser Ser Gly Tyr Tyr Tyr Ile Trp Arg Tyr
1 5 10 15
<210> 197
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 197 ADI-30286 HCDR3-Kabat
<400> 197
Asp Pro Gly Thr His Tyr Ser Gly Tyr Tyr Tyr Ile Trp Arg Tyr
1 5 10 15
<210> 198
<211> 15
<212> PRT
<220>
<223> Последовательность антитела 198 ADI-30288 HCDR3-Kabat
<400> 198
Asp Pro Gly Thr Asp Ser Thr Gly Tyr Tyr Tyr Ile Trp Arg Tyr
1 5 10 15
<210> 199
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 199 ADI-27297 HCDR1-AbM
<400> 199
Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His
1 5 10
<210> 200
<211> 10
<212> PRT
<220>
<223> Последовательность антитела 200 ADI-30272, ADI-30278 HCDR1-AbM
<400> 200
Gly Tyr Thr Phe Thr Glu Tyr Tyr Met His
1 5 10
<210> 201
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 201 ADI-27297, ADI-30272 HCDR3-Kabat
<400> 201
Asp His Asp Ile Ala Ala Ala Gly Arg Leu Ala Asp Tyr
1 5 10
<210> 202
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 202 ADI-30278 HCDR3-Kabat
<400> 202
Asp His Asp Ile Arg Leu Ala Gly Arg Leu Ala Asp Tyr
1 5 10
<210> 203
<211> 12
<212> PRT
<220>
<223> Последовательность антитела 203 ADI-27301 HCDR1-AbM
<400> 203
Gly Gly Ser Ile Ser Ser Ser Ser Tyr Tyr Trp Gly
1 5 10
<210> 204
<211> 12
<212> PRT
<220>
<223> Последовательность антитела 204 ADI-30306 HCDR1-AbM
<400> 204
Gly Gly Ser Ile Ser Ser Ser Leu Tyr Tyr Trp Gly
1 5 10
<210> 205
<211> 12
<212> PRT
<220>
<223> Последовательность антитела 205 ADI-30311 HCDR1-AbM
<400> 205
Gly Gly Ser Ile Ala Ser Ser Val Tyr Tyr Trp Gly
1 5 10
<210> 206
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 206 ADI-27301, ADI-30306 HCDR3-Kabat
<400> 206
Glu Ala Gly Arg Gly Thr Thr Gly Leu Leu Phe Asp Tyr
1 5 10
<210> 207
<211> 13
<212> PRT
<220>
<223> Последовательность антитела 207 ADI-30311 HCDR3-Kabat
<400> 207
Glu Ala Gly Arg Thr Gly Thr Gly Leu Leu Phe Asp Tyr
1 5 10
<210> 208
<211> 244
<212> PRT
<220>
<223> Последовательность 208, последовательность TIGIT человека
<400> 208
Met Arg Trp Cys Leu Leu Leu Ile Trp Ala Gln Gly Leu Arg Gln Ala
1 5 10 15
Pro Leu Ala Ser Gly Met Met Thr Gly Thr Ile Glu Thr Thr Gly Asn
20 25 30
Ile Ser Ala Glu Lys Gly Gly Ser Ile Ile Leu Gln Cys His Leu Ser
35 40 45
Ser Thr Thr Ala Gln Val Thr Gln Val Asn Trp Glu Gln Gln Asp Gln
50 55 60
Leu Leu Ala Ile Cys Asn Ala Asp Leu Gly Trp His Ile Ser Pro Ser
65 70 75 80
Phe Lys Asp Arg Val Ala Pro Gly Pro Gly Leu Gly Leu Thr Leu Gln
85 90 95
Ser Leu Thr Val Asn Asp Thr Gly Glu Tyr Phe Cys Ile Tyr His Thr
100 105 110
Tyr Pro Asp Gly Thr Tyr Thr Gly Arg Ile Phe Leu Glu Val Leu Glu
115 120 125
Ser Ser Val Ala Glu His Gly Ala Arg Phe Gln Ile Pro Leu Leu Gly
130 135 140
Ala Met Ala Ala Thr Leu Val Val Ile Cys Thr Ala Val Ile Val Val
145 150 155 160
Val Ala Leu Thr Arg Lys Lys Lys Ala Leu Arg Ile His Ser Val Glu
165 170 175
Gly Asp Leu Arg Arg Lys Ser Ala Gly Gln Glu Glu Trp Ser Pro Ser
180 185 190
Ala Pro Ser Pro Pro Gly Ser Cys Val Gln Ala Glu Ala Ala Pro Ala
195 200 205
Gly Leu Cys Gly Glu Gln Arg Gly Glu Asp Cys Ala Glu Leu His Asp
210 215 220
Tyr Phe Asn Val Leu Ser Tyr Arg Ser Leu Gly Asn Cys Ser Phe Phe
225 230 235 240
Thr Glu Thr Gly
<210> 209
<211> 107
<212> PRT
<220>
<400> 209
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СКОНСТРУИРОВАННЫЕ PH-ЗАВИСИМЫЕ АНТИТЕЛА К CD3, А ТАКЖЕ СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2832079C2 |
| СТАБИЛЬНЫЕ СОСТАВЫ АНТИТЕЛ ПРОТИВ TIGIT, ОТДЕЛЬНО И В КОМБИНАЦИИ С АНТИТЕЛАМИ ПРОТИВ РЕЦЕПТОРА 1 ПРОГРАММИРУЕМОЙ СМЕРТИ (PD-1), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2820576C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ ПРОТИВ TIGIT И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2776714C2 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732591C2 |
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2817838C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| ХИМЕРНЫЕ БЕЛКИ НА ОСНОВЕ TIGIT И LIGHT | 2018 |
|
RU2775490C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА CAIX, ANO1, МЕЗОТЕЛИН, TROP2, СEA ИЛИ КЛАУДИН-18.2 | 2018 |
|
RU2792671C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА NKG2D, CD16 И TROP2 | 2018 |
|
RU2820629C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ CD33, NKG2D И CD16 | 2018 |
|
RU2820603C2 |
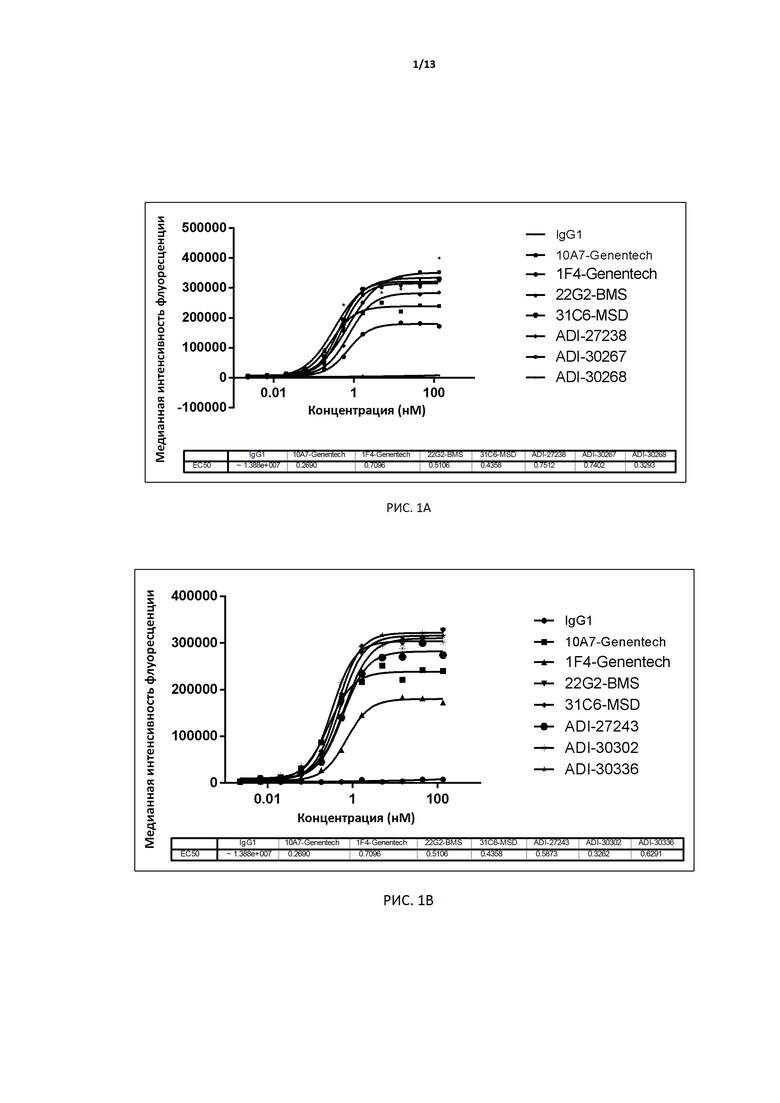
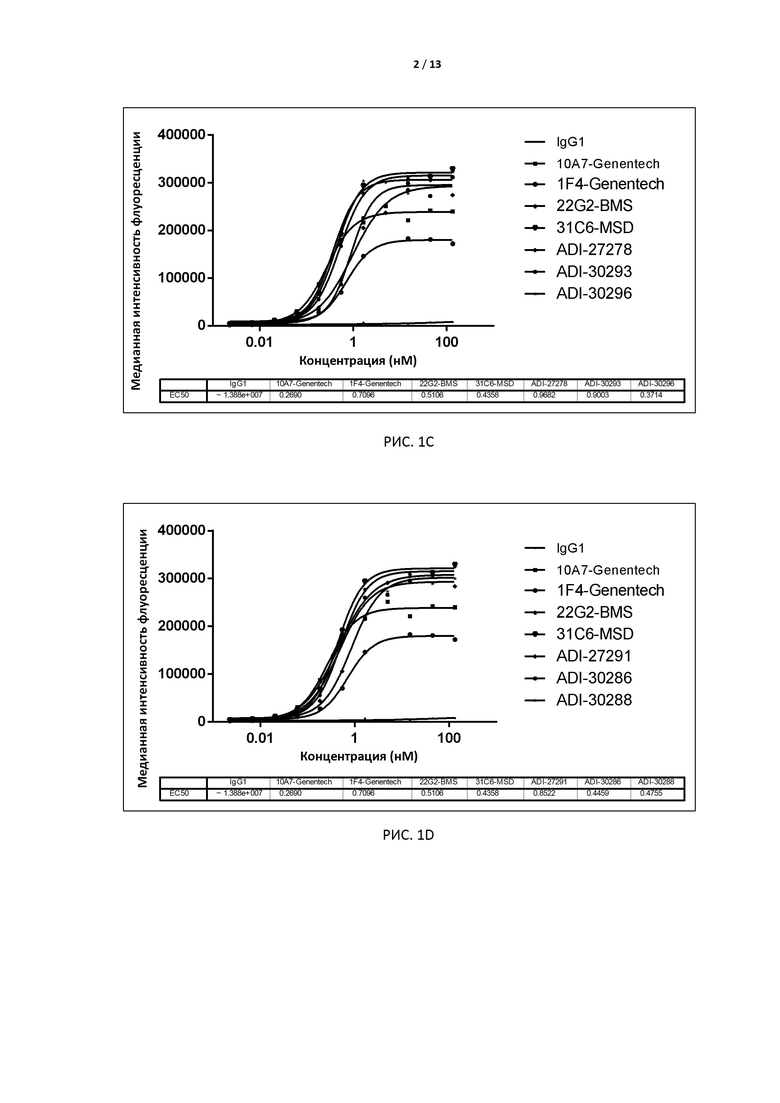
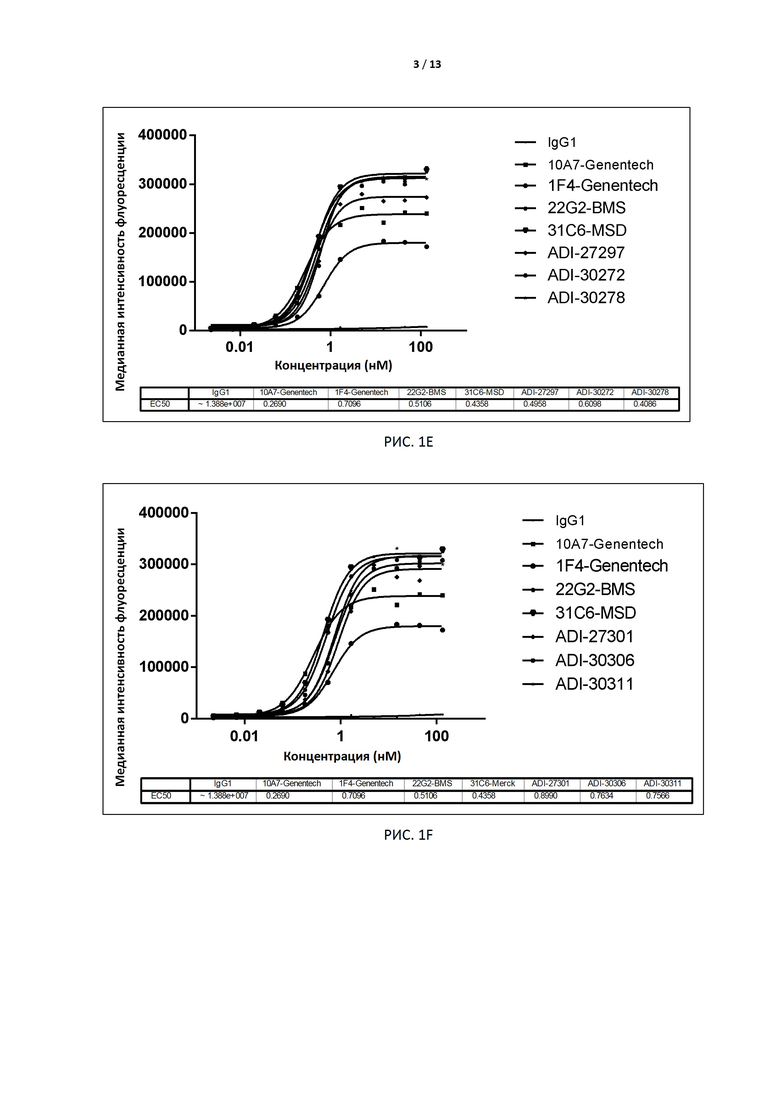
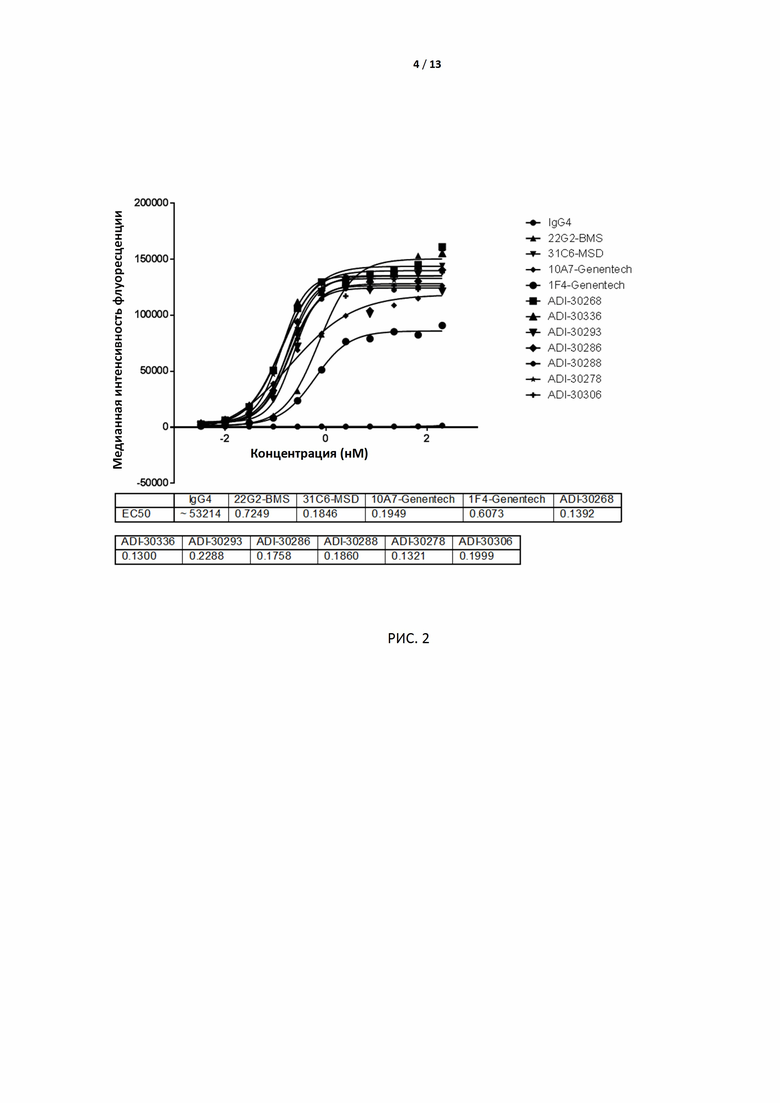
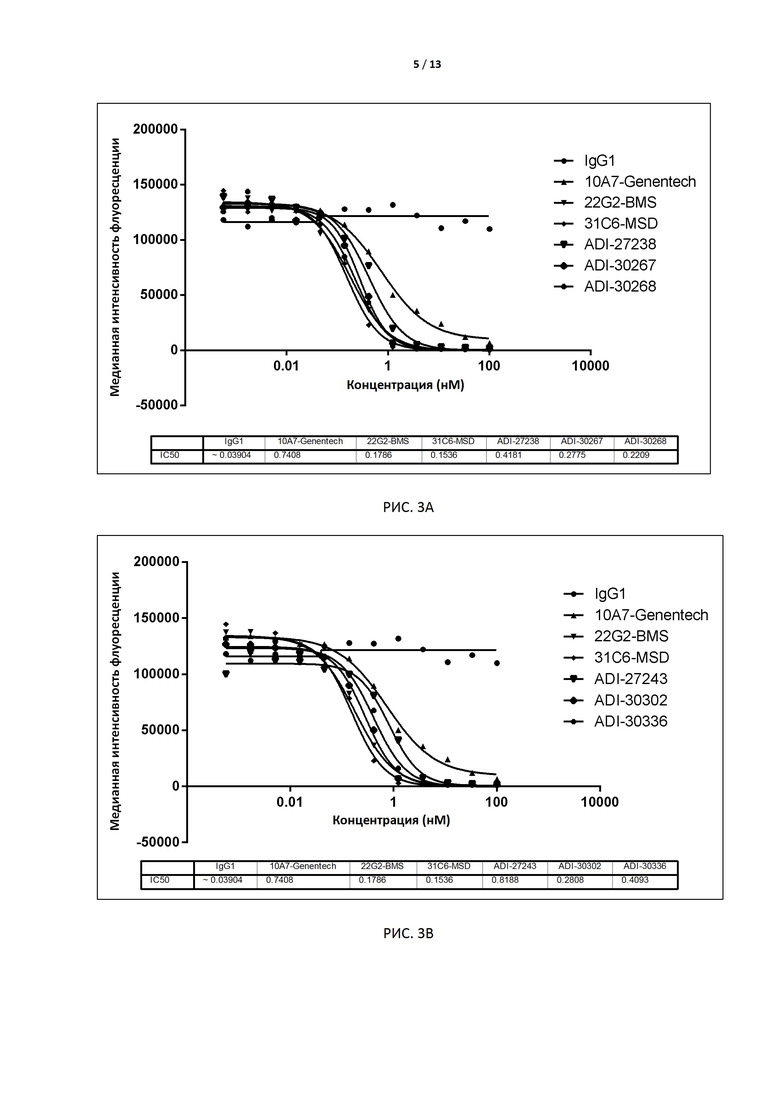
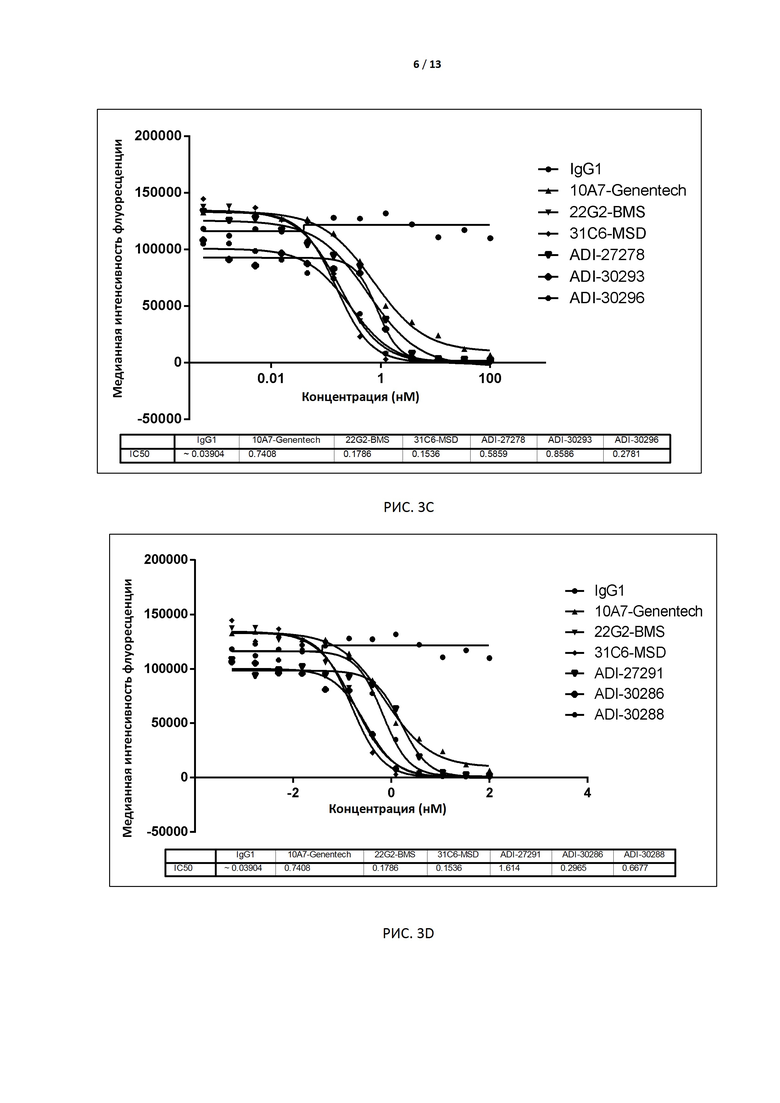
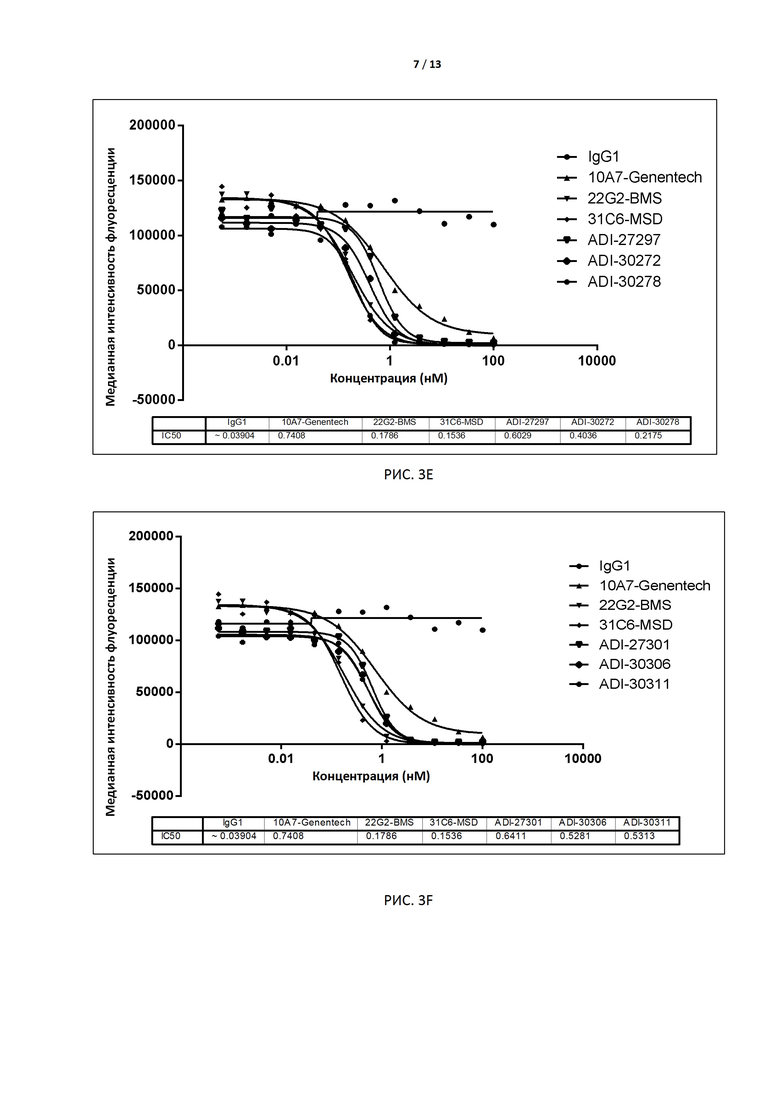
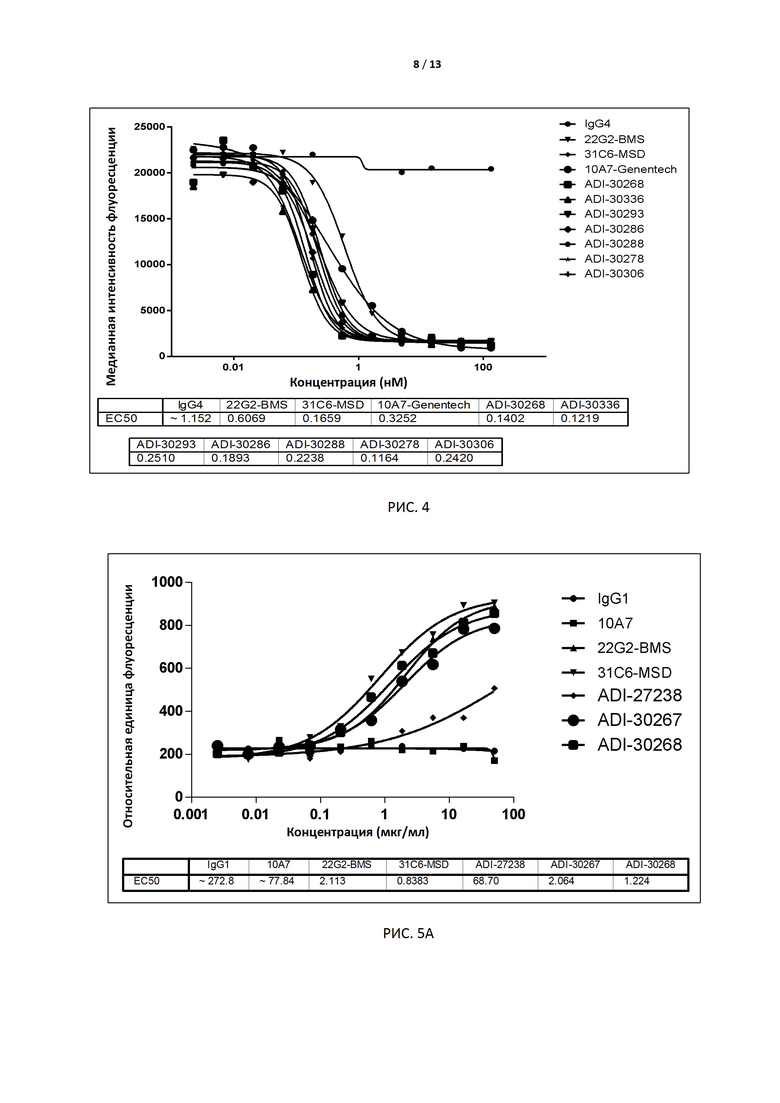
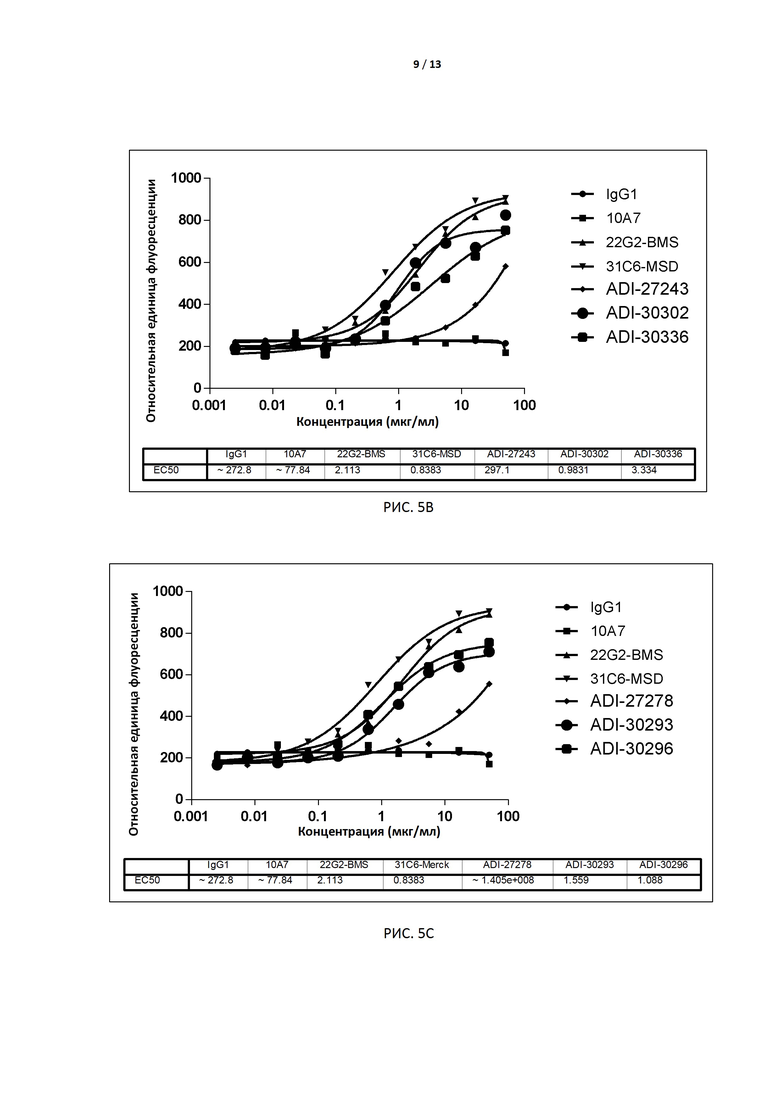
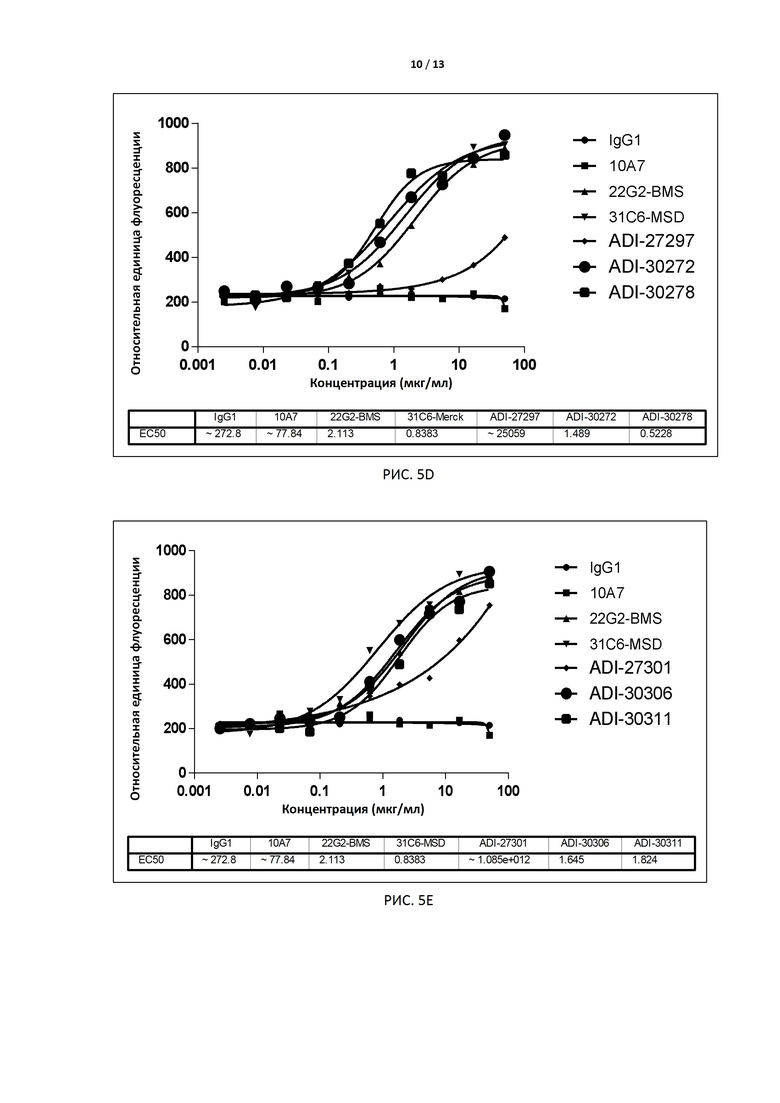

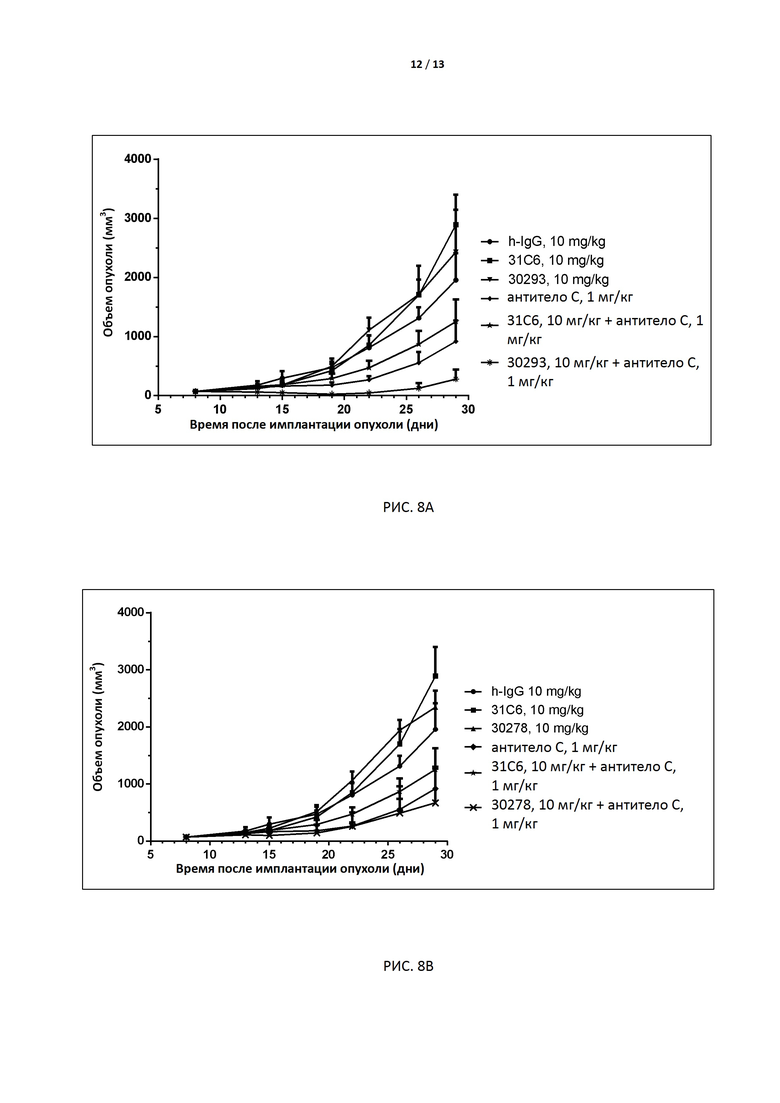

Настоящее изобретение относится к биотехнологии, в частности к новому антителу и фрагменту этого антитела, которые специфически связываются с TIGIT, а также к композиции, включающей это антитело или его фрагмент. Кроме того, настоящее изобретение относится к нуклеиновой кислоте, кодирующей это антитело или его фрагмент, клетке-хозяину, содержащей эту нуклеиновую кислоту, и их использованию. Кроме того, настоящее изобретение относится к терапевтическому или диагностическому применению этого антитела или его фрагмента. 13 н. и 8 з.п. ф-лы, 9 ил., 16 табл., 6 пр.
1. Антитело или его антиген-связывающий фрагмент, которые связываются с TIGIT, содержащие три определяющие комплементарность участка тяжелой цепи (HCDR) и три определяющие комплементарность участка легкой цепи (LCDR), где:
(a) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 1, 2, 3, 4 или 5, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 6, 7, 8, 9 или 10, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 11, 12, 13, 14 или 15, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 16, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 17, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 18;
(b) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 19, 20 или 21, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, 23 или 24, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 25, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 26, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 27, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 28, 29 или 30;
(c) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 31, 32 или 33, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 34, 35, 36 или 37, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 38, 39 или 40, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 41, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 42, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 43;
(d) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 44, 45, 46 или 47, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 48, 49, 50, 51 или 52, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 53, 54, 55 или 56, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 57, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 58, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 59; или
(e) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 60, 61 или 62, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 6, 63, 64 или 65, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 66, 67 или 68, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 69, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 17, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 70;
(f) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 71, 72, 73 или 74, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, 75, 76 или 77, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 78, 79 или 80, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 81, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 82, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 83;
(g) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 178, 179, 180 или 181, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 6, 7, 8 или 9, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 182, 183, 184 или 185, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 16, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 17, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 18;
(h) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 186 или 187, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 22 или 23, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 188, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 26, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 27, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 28 или 29;
(i) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 189 или 190, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 34, 35 или 36, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 191 или 192, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 41, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 42, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 43;
(j) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 193, 194 или 195, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 48, 49, 50 или 51, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 196, 197 или 198, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 57, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 58, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 59;
(k) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 199 или 200, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 6, 63 или 64, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 201 или 202, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 69, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 17, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 70; или
(l) HCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 203, 204 или 205, HCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, 75 или 76, HCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 206 или 207, LCDR1 содержит аминокислотную последовательность, представленную в SEQ ID NO: 81, LCDR2 содержит аминокислотную последовательность, представленную в SEQ ID NO: 82, а LCDR3 содержит аминокислотную последовательность, представленную в SEQ ID NO: 83.
2. Антитело или его антиген-связывающий фрагмент по п. 1, содержащие:
(i) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 84, 85, 86 или 87, и (или) вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 104;
(ii) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 88, 89 или 90, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 105 или 106;
(iii) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 91, 92 или 93, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 107;
(iv) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 94, 95, 96 или 97, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 108;
(v) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 98, 99 или 100, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 109; или
(vi) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 101, 102 или 103, и/или вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая по меньшей мере на 80%, 85% или 90% идентична аминокислотной последовательности, представленной в SEQ ID NO: 110.
3. Антитело или его антиген-связывающий фрагмент по любому из пп. 1–2, содержащий вариабельную область тяжелой цепи и вариабельную область легкой цепи, выбранные из:
(i) вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 84, 85, 86 или 87, и вариабельной области легкой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 104;
(ii) вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 88, 89 или 90, и вариабельной области легкой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 105 или 106;
(iii) вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 91, 92 или 93, и вариабельной области легкой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 107;
(iv) вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 94, 95, 96 или 97, и вариабельной области легкой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 108;
(v) вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 98, 99 или 100, и вариабельной области легкой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 109; или
(vi) вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 101, 102 или 103, и вариабельной области легкой цепи, содержащей аминокислотную последовательность, представленную в SEQ ID NO: 110.
4. Антитело или его антиген-связывающий фрагмент по любому из предшествующих пунктов, где антитело является антителом изотипа IgG1, IgG2 или IgG4, либо его антиген-связывающим фрагментом, предпочтительно Fc-участком IgG4, содержащим мутации S228P, F234A и L235A.
5. Антитело или его антиген-связывающий фрагмент по любому из предшествующих пунктов, где антитело является полностью гуманизированным антителом, гуманизированным антителом или химерным антителом.
6. Антитело или его антиген-связывающий фрагмент по любому из предшествующих пунктов, где антиген-связывающий фрагмент является фрагментом антитела, выбранным из: Fab, Fab', Fab'-SH, Fv, одноцепочечных антител, таких как scFv, (Fab')2-фрагментов, однодоменных антител, диател (dAb) или линейных антител.
7. Антитело или его антиген-связывающий фрагмент по любому из предшествующих пунктов, где антитело обладает одним или более из следующих свойств:
(i) способность связываться с TIGIT человека с высокой аффинностью;
(ii) способность давать перекрестную иммунологическую реакцию с TIGIT обезьяны и/или мыши;
(iii) эффективное связывание с TIGIT на клеточной поверхности;
(iv) блокирование связывания TIGIT с его лигандом CD155;
(v) ослабление ингибирующего действия связывания CD155 с TIGIT в отношении IL-2-зависимого нисходящего сигнального пути;
(vi) увеличение продукции IL-2 T-лимфоцитами;
(vii) противоопухолевая активность, например, подавление опухолевого роста;
(viii) более выраженное подавляющее действие в отношении опухоли в комбинации с антителом к PD-1, например, усиленное ингибирование роста опухоли.
8. Выделенная нуклеиновая кислота, кодирующая антитело к TIGIT или его антиген-связывающий фрагмент по любому из предшествующих пунктов.
9. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 8.
10. Клетка-хозяин для экспрессии антитела к TIGIT или его антиген-связывающего фрагмента, содержащая нуклеиновую кислоту по пункту 8 или экспрессионный вектор по п. 9.
11. Способ получения антитела к TIGIT или его антиген-связывающего фрагмента, включающий культивирование клетки-хозяина по пункту 10 в условиях, подходящих для экспрессии нуклеиновой кислоты, кодирующей антитело к TIGIT или его антиген-связывающий фрагмент по любому из пп. 1–7, и, необязательно, выделение антитела или его антиген-связывающего фрагмента, где необязательно указанный способ дополнительно включает выделение антитела к TIGIT или его антиген-связывающего фрагмента из клетки-хозяина.
12. Иммуноконъюгат для усиления иммунного ответа у субъекта, содержащий антитело или его антиген-связывающий фрагмент по любому из пп. 1–7, конъюгированные с терапевтическим агентом.
13. Иммуноконъюгат для обнаружения TIGIT в образце, содержащий антитело или его антиген-связывающий фрагмент по любому из пп. 1-7, конъюгированные с диагностическим агентом.
14. Полиспецифическое антитело для усиления иммунного ответа у субъекта, включающее антитело или его антиген-связывающий фрагмент по любому из пп. 1–7, где полиспецифическое антитело предпочтительно является биспецифическим антителом.
15. Фармацевтическая композиция для усиления иммунного ответа у субъекта, содержащая антитело или его антиген-связывающий фрагмент по любому из пп. 1–7, и фармацевтическое вспомогательное вещество.
16. Фармацевтическая композиция по п. 15, содержащая второй терапевтический агент, где второй терапевтический агент предпочтительно выбран из антитела к PD-1 или антитела к PD-L1.
17. Способ профилактики или лечения опухоли у субъекта, включающий введение субъекту эффективного количества антитела к TIGIT или его антиген-связывающего фрагмента по любому из пп. 1–7 или фармацевтической композиции по п. 15 или 16.
18. Способ по п. 17, где опухоль является злокачественным новообразованием желудочно-кишечного тракта, таким как рак толстой кишки.
19. Способ профилактики или лечения инфекционного заболевания у субъекта, включающий введение субъекту эффективного количества антитела к TIGIT или его антиген-связывающего фрагмента по любому из пп. 1–7 или фармацевтической композиции по п. 15 или 16.
20. Способ блокирования связывания TIGIT с CD155 с целью ослабления или элиминации иммуносупрессивного действия TIGIT у субъекта, включающий введение субъекту эффективного количества антитела к TIGIT или его антиген-связывающего фрагмента по любому из пп. 1–7 или фармацевтической композиции по п. 15 или 16.
21. Способ обнаружения TIGIT в образце, включающий:
(a) приведение образца в контакт с антителом или его антиген-связывающим фрагментом по любому из пп. 1–7; и
(b) обнаружение комплекса, образованного антителом или его антиген-связывающим фрагментом и TIGIT, где антитело содержит обнаруживаемую метку.
| WO 2017152088 A1 08.09.2017 | |||
| US 9713641 B2 25.07.2017 | |||
| АНТИ-TIGIT АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732591C2 |

