Изобретение относится к биологии и медицине, а именно, к генной инженерии. Предлагаемый репрессор может быть использован для научных экспериментов в области фундаментальной клеточной биологии, биомедицинских исследований при моделировании наследственных заболеваний, а также в терапии наследственных заболеваний. Настоящее изобретение не предназначено для модификации генетической целостности клеток зародышевой линии человека.
В настоящее время известно несколько технологий контроля экспрессии целевого гена в клетках млекопитающих. Широко используется РНК-интерференция (RNAi). RNAi, однако, имеет ряд ограничений, например, неполный нокдаун экспрессии и большое количество нецелевых мишеней (Krueger U, Bergauer Т, Kaufmann В, Wolter I, Pilk S, Heider-Fabian M, Kirch S, Artz-Oppitz C, Isselhorst M, Konrad J, Insights into effective RNAi gained from large-scale siRNA validation screening, Oligonucleotides, 2007; 17(2): 237-2S0.; Sigoillot FD, Lyman S, Huckins JF, Adamson B, Chung E, Quattrochi B, King RW, A bioinformatics method identifies prominent off-targeted transcripts in RNAi screens," Nat Methods. 2012; 9(4): 363-6).
Более специфичными являются технологии на основе ДНК-связывающих белков с доменами цинковых пальцев или эффекторы, подобные активаторам транскрипции (TALE), слитые с доменами-репрессорами транскрипции, например, KRAB (Margolin JF1, Friedman JR, Meyer WK, Vissing H, Thiesen HJ, Rauscher FJ 3 rd., Kriippel-associated boxes are potent transcriptional repression domains, Proc Natl Acad Sci USA. 1994; 91(10): 4509-13.).
Однако для каждой отдельной мишени требуется создание дизайна и последовательности гена отдельного белка, что является достаточно долгой и весьма трудозатратной процедурой (Gaj Т, Gersbach СА, Barbas CF 3 rd., ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering, Trends Biotechnol. 2013; 31(7): 397-405; Joung JK, Sander JD, "TALENs: a widely applicable technology for targeted genome editing," Nat Rev Mol Cell Biol. 2013; 14(1): 49-55).
Современная технология редактирования генома с помощью системы CRISPR/Cas9 (Jinek М, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E, A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity, Science. 2012; 337(6096): 816-21; Cho SW, Kim S, Kim JM, Kim JS, Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease, Nat Biotechnol. 2013; 31(3): 230-2) может быть трансформирована для получения универсальных искусственных репрессоров. Ключевыми компонентами системы являются РНК-зависимая ДНК эндонуклеаза Cas9 и направляющая РНК. Направляющая РНК состоит из конститутивной структурной части, распознаваемой Cas9 и геном-специфичной вариабельной части, которая нацеливает Cas9 на специфичный участок генома.
Введение двух мутаций D10A и Н840А в последовательность гена Cas9 лишает белок Cas9 способности разрезать ДНК, однако сохраняет его способность связывать направляющую РНК и распознавать целевой локус генома (Qi LS, Larson МН, Gilbert LA, Doudna JA, Weissman JS, Arkin AP, Lim WA, Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression, Cell. 2013; 152(5): 1173-83). Такой белок получил название dCas9. Изменить специфичность dCas9 белка очень легко путем изменения вариабельной части направляющей РНК. Будучи направленным на сайт инициации транскрипции, dCas9 способен подавлять экспрессию гена (Qi LS, Larson MH, Gilbert LA, Doudna JA, Weissman JS, Arkin AP, Lim WA, Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression, Cell. 2013; 152(5): 1173-83).
Однако эффективность этого способа контроля экспрессии гена невысокая, так как он требует знания точного положения старта транскрипции и не эффективен в случае, если у гена несколько стартов транскрипции.
Эффективность ингибирования транскрипции в свое время была повышена путем добавления к dCas9 Kruppel-ассоциированного домена репрессии (KRAB) (Gilbert LA, Larson MH, Morsut L, Liu Z, Brar GA, Torres SE, Stern-Ginossar N, Brandman O, Whitehead EH, Doudna JA, Lim WA, Weissman JS, Qi LS, CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes, Cell. 2013; 154(2): 442-51; Gilbert LA, Horlbeck MA, Adamson B, Villalta JE, Chen Y, Whitehead EH, Guimaraes C, Panning B, Ploegh HL, Bassik MC, Qi LS, Kampmann M, Weissman JS, Genome-Scale CRISPR-Mediated Control of Gene Repression and Activation, Cell. 2014; 159(3): 647-61; Thakore PI, Black JB, Hilton IB, Gersbach CA, Editing the epigenome: technologies for programmable transcription and epigenetic modulation, Nat Methods. 2016; 13(2): 127-37) или гистон-метил трансферазы Ezh2 (O'Geen H., Bates SL., Carter SS. Nisson KA, Halmai J, Fink KD, Rhie SK, Farnham PJ, Segal DJ. Ezh2-dCas9 and KRAB-dCas9 enable engineering of epigenetic memory in a context-dependent manner. Epigenetics & Chromatin. 2019; 12, 26).
Тем не менее, для достаточно большого числа генов эффективность таких искусственных репрессоров с одним доменом до сих пор не высока, что стимулирует разработку новых эффективных репрессоров транскрипции.
На момент подачи заявки из открытых источников было известно о следующих аналогах.
Известны способы применения CRISPR/Cas9 систем для редактирования паттернов экспрессии генов человека, в том числе применение белка dCas9, слитого с доменами активации транскрипции VP64 или репрессии транскрипции KRAB или SID4X (RU 2725502, С2).
Известны также способы применения CRISPR/Cas9 систем для целенаправленного воздействия и терапии в клетках печени, в том числе применения белка dCas9, слитого с доменами активации транскрипции VP64 или репрессии транскрипции KRAB или SID4X (RU 2716420, С2).
Кроме того, известны различные способы и составы лечения болезни Хантингтона с применением CRISPR/Cas9 систем, в то числе с применением белка dCas9, слитого с такими репрессорами транскрипции как ДНК-метилтрансфераза 3А (DNMT3A) или метил-CpG-связывающий белок (МеСР) (RU 22639277, С2).
Однако, как было уже упомянуто ранее, подобные репрессорные системы первого поколения с одним доменом репрессии не обеспечивают достаточную эффективность ингибирования транскрипции для ряда генов.
В качестве прототипа нами выбран искусственный репрессор, представляющий собой химерный белок dCas9-KRAB-MeCP2, в котором к KRAB домену добавлен метил-CpG-связывающий белок 2 (МеСР2). (Yeo NC, Chavez A, Lance-Byrne A, Chan Y, Menn D, Milanova D, Kuo C-C, Guo X, Sharma S, Tung A, Cecchi RJ, Tuttle M, Pradhan S, Lim ET, Davidsohn N, Ebrahimkhani MR, Collins JJ, Lewis NE, Kiani S, Church GM. An enhanced CRISPR repressor for targeted mammalian gene regulation. Nat Methods. 2018; 15, 611-616).
Однако этот репрессор не эффективен в случае тех генов, где не эффективен репрессор dCas9-KRAB (O'Geen Н., Bates SL., Carter SS. Nisson KA, Halmai J, Fink KD, Rhie SK, Farnham PJ, Segal DJ. Ezh2-dCas9 and KRAB-dCas9 enable engineering of epigenetic memory in a context-dependent manner. Epigenetics & Chromatin. 2019; 12, 26).
Проблемой, на решение которой направлено данное изобретение, является разработка высокоэффективного универсального репрессора транскрипции генов в клетках человека, исключая модификацию генетической целостности клеток зародышевой линии человека.
Технический результат, достигаемый при использовании предлагаемого репрессора, заключается в обеспечении эффективной репрессии транскрипции гена за счет одновременного действия на него факторов репрессии KRAB и МеСР2 и CpG-метилирования посредством ДНК-метилтрансферазы DNMT3a-L.
Благодаря наличию в составе предлагаемого репрессора ДНК-метилтрансферазы, он может использоваться не только для изменения экспрессии различных генов, но и для метилирования регуляторных участков генов, что также подавляет экспрессию генов. Универсальность предполагаемого репрессора также обеспечивается за счет изменения последовательности направляющей РНК; модификация самого репрессора не требуется. При использовании нескольких направляющих РНК предлагаемый репрессор может быть использован для одновременного подавления экспрессии нескольких генов.
Сущность изобретения заключается в следующем.
Предложен эпигеномный репрессор транскрипции гена или генов в клетках человека, исключая клетки зародышевой линии человека, представляющий собой белок dCas9, кодируемый последовательностью SEQ ID NO: 1, соединенный линкером 1, кодируемым последовательностью SEQ ID NO: 2, с белком DNMT3a-L, кодируемым последовательностью SEQ ID NO: 3, который соединен линкером 2, кодируемым последовательностью SEQ ID NO: 4, с репрессорным доменом KRAB, кодируемым последовательностью SEQ ID NO: 5, который соединен линкером 3, кодируемым последовательностью SEQ ID NO: 6, с белком МеСР2, кодируемым последовательностью SEQ ID NO: 7.
Белок dCas9 - это белок Cas9, содержащий две мутации D10A и Н840А
и лишенный эндонуклеазной активности.
Белок DNMT3a-L - это химерный белок мышиной ДНК-метилтрансферазы DNMT3a, состоящий из двух слитых субъединиц 3а и 3L.
Репрессорный домен KRAB - это Kruppel-ассоциированный домен репрессии человека.
Белок МеСР2 - это метил-CpG-связывающий белок 2 человека.
Заявлена универсальная система высокоэффективной репрессии транскрипции целевых генов на основе химерного белка - искусственного регулятора транскрипции, состоящего из белка dCas9, белка DNMT3a-L, репрессорного домена KRAB и белка МеСР2, связанных тремя оригинальными линкерами последовательно между собой.
Готовая последовательность гена, кодирующего созданный химерный белок-репрессор, состоит из последовательности SEQ ID NO: 1, кодирующей кодон-оптимизированный для экспрессии в клетках человека белок - dCas9 (содержит 1368 а.о), соединенной линкером 1, кодируемым последовательностью SEQ ID NO: 2, с последовательностью SEQ ID NO: 3, кодирующей химерный белок мышиной ДНК-метилтрансферазы - DNMT3a-L, состоящий из двух слитых субъединиц 3а и 3L (содержит 564 а.о.), соединенной линкером 2, кодируемым последовательностью SEQ ID NO: 4, с последовательностью SEQ ID NO: 5, кодирующей Kruppel-ассоциированный репрессорный домен человека - KRAB (содержит 65 а.о.), и соединенной линкером 3, кодируемым последовательностью SEQ ID NO: 6, с последовательностью SEQ ID NO: 7, кодирующей метил-CpG-связывающий белок 2 человека - МеСР2 (содержит 298 а.о.).
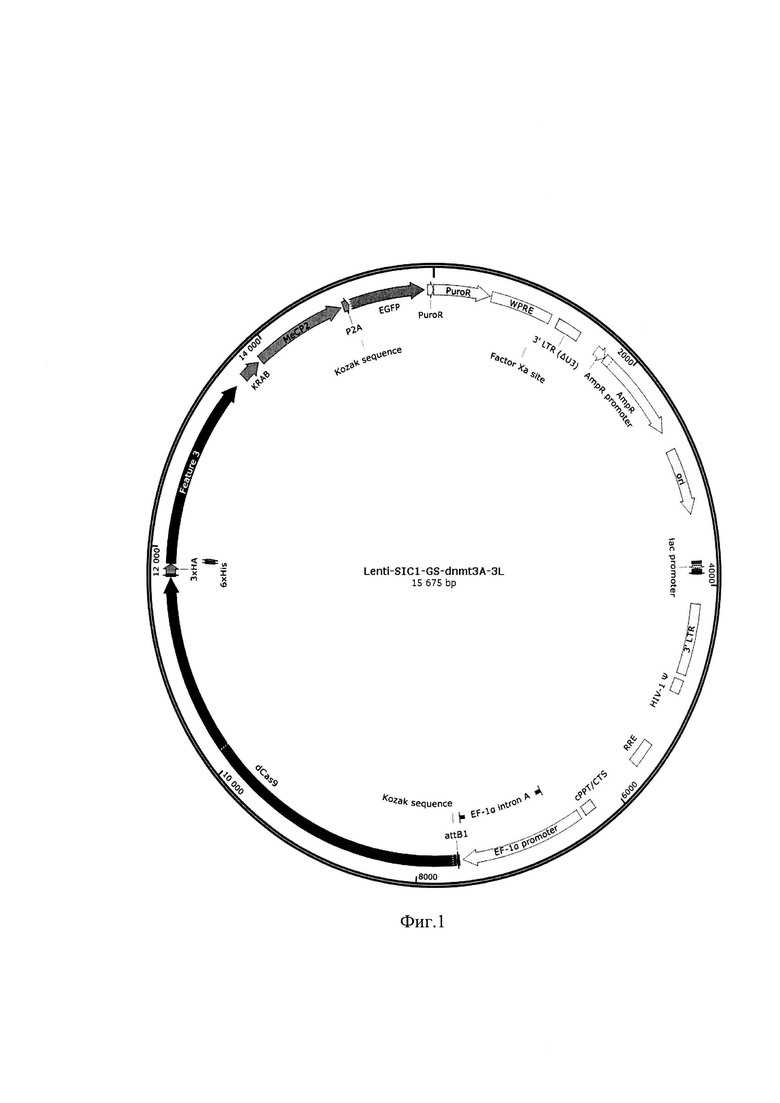
В настоящий момент данный ген клонирован в лентивирусный вектор Lenti-SIC1-GS-dnmt3A-3L (фиг. 1), обеспечивающий экспрессию искусственного репрессора транскрипции, который содержит также репортерный зеленый флуоресцентный белок eGFP и селективный маркер пуромицин-N-ацетилтранферазу. Вектор имеет размер 15675 п.н.
Искусственный репрессор может быть доставлен в клетку в виде плазмидного вектора, в виде лентивирусного вектора или готового белка, наработанного и очищенного из штамма-продуцента.
Направляющая РНК может быть доставлена в клетку в виде плазмидного вектора pU6-gRNA, в виде лентивирусного вектора, в виде готовой РНК, созданной при помощи химического синтеза или Т7-транскрипции.
Репрессор и направляющая РНК могут быть доставлены в клетку при помощи липофекции, кальций-фосфатной преципитации, электропорации, микроинъекции.
После доставки всех компонентов в клетку, репрессорный комплекс с помощью направляющей РНК связывается с регуляторной областью целевого гена и подавляет его экспрессию, привлекая другие внутриклеточные репрессоры транскрипции, а также производя CpG-метилирование.
Для доказательства эффективности предлагаемого искусственного репрессора мы продемонстрировали его работу на примере метилирования энхансера гена эпоксидгидролазы (ЕРНХ2) и подавления экспрессии гена допа-декарбоксилазы (DDC) в нейрональной линии клеток SH-S5Y5 (Biedler J.L., Helson L., Spengler B.A. Morphology and growth, tumorigenicity, and cytogenetics of human neuroblastoma cells in continuous culture // Cancer research. - 1973. - T. 33. - №. 11. - C. 2643-2652).
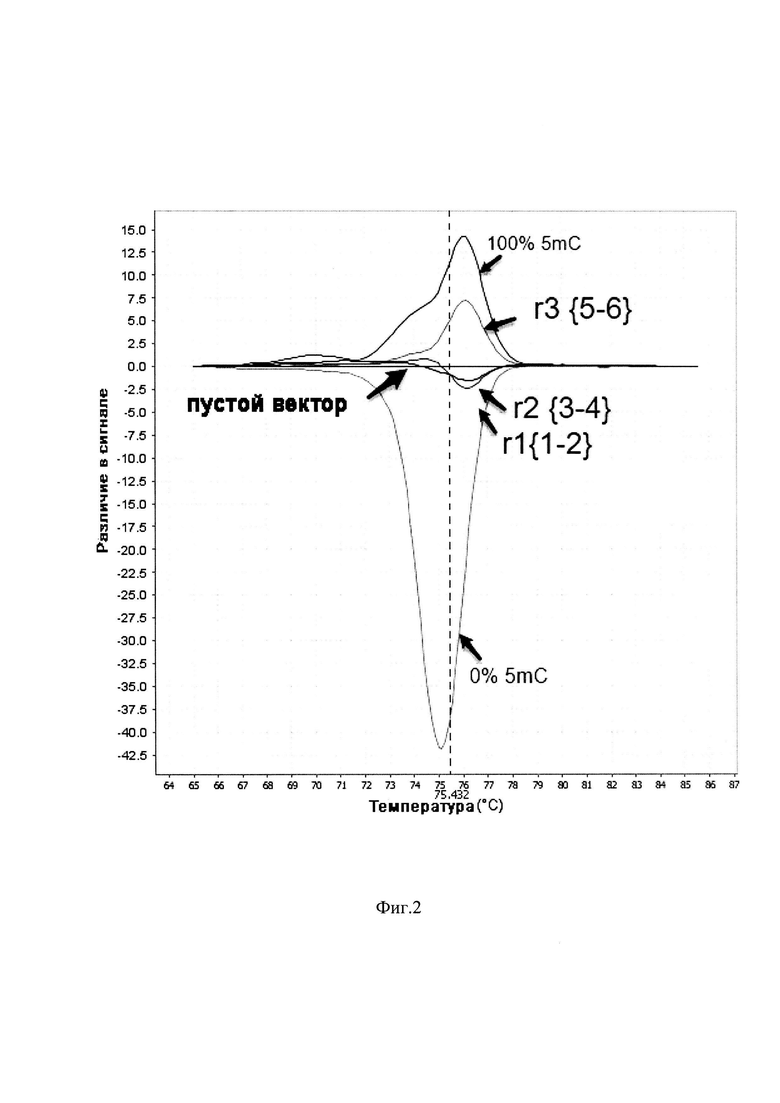
Внутри энхансера EPHX2 мы выбрали три участка-мишени для связывания репрессора, к каждому из которых подобрали по две последовательности направляющей РНК. На фиг. 2 представлены результаты MS-HRM анализа (Wojdacz Т.K., Dobrovic A. Methylation-sensitive high resolution melting (MS-HRM): a new approach for sensitive and high-throughput assessment of methylation // Nucleic acids research. - 2007. - T. 35. - №. 6. - C. e41) этого энхансера. Представлен график первой производной от графика плавления фрагментов ДНК. Кривые r1, r2 и r3 демонстрируют уровень метилирования первого, второго и третьего участков-мишеней, соответственно. Кривые 100% 5 mC и 0% 5 mC демонстрируют контроли гиперметилирования и гипометилирования, соответственно. Видно, что при заражении пустым вектором, не несущим последовательности направляющих РНК, уровень метилирования не изменяется. В то же время при заражении вектором, кодирующим направляющие РНК, уровень метилирования энхансера в третьем участке r3 значительно возрастает по сравнению с пустым вектором, r1 и r2, где направляюще РНК были написаны на другие участки энхансера.
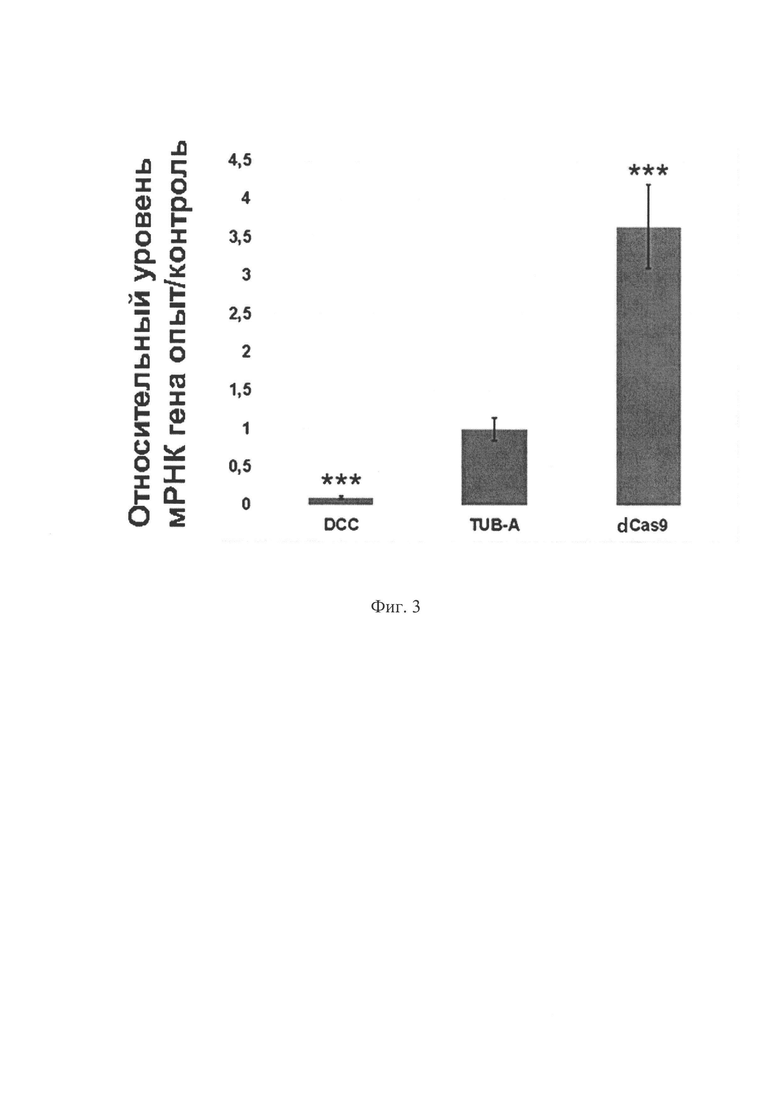
На фиг. 3 представлены результаты ПЦР в реальном времени в виде относительных уровней экспрессии генов DDC, Tub-A и dCas9. В качестве опытных образцов взяты клетки, экспрессирующие искусственный репрессор транскрипции, в качестве контрольных - клетки без репрессора. Разброс данных представлен стандартным отклонением от среднего (n=3). ***р<0.001. Статистическая значимость рассчитана с помощью двустороннего критерия Стьюдента для сравнения двух независимых средних. Сравнивали значения в парах DCC/Tub-A, Cas9/Tub-A. Поскольку с введением в клетки гена репрессора уровень экспрессии гена Tub-A не меняется, его относительный уровень экспрессии равен единице и принят в качестве контроля.
На фиг. 3 видно, что в опытных клетках уровень мРНК dCas9 значительно повышен, что свидетельствует об эффективной экспрессии химерного репрессора на основе dCas9; понижение уровня экспрессии гена-мишени DCC свидетельствует о репрессии транскрипции данного гена вследствие работы искусственного репрессора.
Следует понимать, что данное описание и фигуры служат только для иллюстрации осуществления изобретения, и не ограничивают объема настоящего изобретения.
--->
Перечень последовательностей
<110> Федеральное государственное автономное образовательное учреждение
высшего образования «Российский национальный исследовательский медицинский
университет имени Н.И. Пирогова» Министерства здравоохранения Российской
Федерации (Federalnoe gosudarstvennoe autonomnoe obrazovatelnoe uchrezhdenie
vysshego obrazovaniya «Rossijskij natsionalnyj issledovatelskij meditsinskij
universitet imeni N.I. Pirogova» Ministerstva zdravookhraneniya Rossijskoj
Federatsii)
<120> Универсальный эпигеномный репрессор транскрипции целевых генов в
клетках человека
<160> 7
<210> SEQ ID NO: 1
<211> 4104
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая белок dCas9
<400> 1
atggacaaga agtactccat tgggctcgct atcggtacca acagcgtcgg ctgggccgtc 60
attacggacg agtacaaggt gccgagcaaa aaattcaaag ttctgggcaa taccgatcgc 120
cacagcataa agaagaacct cattggagcc ctcctgttcg actccgggga gacggccgaa 180
gccacgcggc tcaaaagaac agcacggcgc agatataccc gcagaaagaa tcggatctgc 240
tacctgcagg agatctttag taatgagatg gctaaggtgg atgactcttt cttccatagg 300
ctggaggagt cctttttggt ggaggaggat aaaaagcacg agcgccaccc aatctttggc 360
aatatcgtgg acgaggtggc gtaccatgaa aagtacccaa ccatatatca tctgaggaag 420
aagctggtag acagtactga taaggctgac ttgcggttga tctatctcgc gctggcgcac 480
atgatcaaat ttcggggaca cttcctcatc gagggggacc tgaacccaga caacagcgat 540
gtcgacaaac tctttatcca actggttcag acttacaatc agcttttcga ggagaacccg 600
atcaacgcat ccggcgttga cgccaaagca atcctgagcg ctaggctgtc caaatcccgg 660
cggctcgaaa acctcatcgc acagctccct ggggagaaga agaacggcct gtttggtaat 720
cttatcgccc tgtcactcgg gctgaccccc aactttaaat ctaacttcga cctggccgaa 780
gatgccaagc tgcaactgag caaagacacc tacgatgatg atctcgacaa tctgctggcc 840
cagatcggcg accagtacgc agaccttttt ttggcggcaa agaacctgtc agacgccatt 900
ctgctgagtg atattctgcg agtgaacacg gagatcacca aagctccgct gagcgctagt 960
atgatcaagc gctatgatga gcaccaccaa gacttgactt tgctgaaggc ccttgtcaga 1020
cagcaactgc ctgagaagta caaggaaatt ttcttcgatc agtctaaaaa tggctacgcc 1080
ggatacattg acggcggagc aagccaggag gaattttaca aatttattaa gcccatcttg 1140
gaaaaaatgg acggcaccga ggagctgctg gtaaagctga acagagaaga tctgttgcgc 1200
aaacagcgca ctttcgacaa tggaagcatc ccccaccaga ttcacctggg cgaactgcac 1260
gctatcctca ggcggcaaga ggatttctac ccctttttga aagataacag ggaaaagatt 1320
gagaaaatcc tcacatttcg gataccctac tatgtaggcc ccctcgctcg gggaaattcc 1380
agattcgcgt ggatgactcg caaatcagaa gagaccatca ctccctggaa cttcgaggaa 1440
gtcgtggata agggggcctc tgcccagtcc ttcatcgaaa ggatgactaa ctttgataaa 1500
aatctgccta acgaaaaggt gcttcctaaa cactctctgc tgtacgagta cttcacagtt 1560
tataacgagc tcaccaaggt caaatacgtc acagaaggga tgagaaagcc agcattcctg 1620
tctggagagc agaagaaagc tatcgtggac ctcctcttca agacgaaccg gaaagttacc 1680
gtgaaacagc tcaaagaaga ctatttcaaa aagattgaat gtttcgactc tgttgaaatc 1740
agcggagtgg aggatcgctt caacgcatcc ctgggaacgt atcacgatct cctgaaaatc 1800
attaaagaca aggacttcct ggacaatgag gagaacgagg acattcttga ggacattgtc 1860
ctcaccctta cgttgtttga agatagggag atgattgaag aacgcttgaa aacttacgct 1920
catctcttcg acgacaaagt catgaaacag ctcaagagac gccgatatac aggatggggg 1980
cggctgtcaa gaaaactgat caatggcatc cgagacaagc agagtggaaa gacaatcctg 2040
gattttctta agtccgatgg atttgccaac cggaacttca tgcagttgat ccatgatgac 2100
tctctcacct ttaaggagga catccagaaa gcacaagttt ctggccaggg ggacagtctt 2160
cacgagcaca tcgctaatct tgcaggtagc ccagctatca aaaagggaat actgcagacc 2220
gttaaggtcg tggatgaact cgtcaaagta atgggaaggc ataagcccga gaatatcgtt 2280
atcgagatgg cccgagagaa ccaaactacc cagaagggac agaagaacag tagggaaagg 2340
atgaagagga ttgaagaggg tataaaagaa ctggggtccc aaatccttaa ggaacaccca 2400
gttgaaaaca cccagcttca gaatgagaag ctctacctgt actacctgca gaacggcagg 2460
gacatgtacg tggatcagga actggacatc aaccggttgt ccgactacga cgtggatgct 2520
atcgtgcccc aaagctttct caaagatgat tctattgata ataaagtgtt gacaagatcc 2580
gataaaaata gagggaagag tgataacgtc ccctcagaag aagttgtcaa gaaaatgaaa 2640
aattattggc ggcagctgct gaacgccaaa ctgatcacac aacggaagtt cgataatctg 2700
actaaggctg aacgaggtgg cctgtctgag ttggataaag ccggcttcat caaaaggcag 2760
cttgttgaga cacgccagat caccaagcac gtggcccaaa ttctcgattc acgcatgaac 2820
accaagtacg atgaaaatga caaactgatt cgagaggtga aagttattac tctgaagtct 2880
aagctggtct cagatttcag aaaggacttt cagttttata aggtgagaga gatcaacaat 2940
taccaccatg cgcatgatgc ctacctgaat gcagtggtag gcactgcact tatcaaaaaa 3000
tatcccaagc tggaatctga atttgtttac ggagactata aagtgtacga tgttaggaaa 3060
atgatcgcaa agtctgagca ggaaataggc aaggccaccg ctaagtactt cttttacagc 3120
aatattatga attttttcaa gaccgagatt acactggcca atggagagat tcggaagcga 3180
ccacttatcg aaacaaacgg agaaacagga gaaatcgtgt gggacaaggg tagggatttc 3240
gcgacagtcc gcaaggtcct gtccatgccg caggtgaaca tcgttaaaaa gaccgaagta 3300
cagaccggag gcttctccaa ggaaagtatc ctcccgaaaa ggaacagcga caagctgatc 3360
gcacgcaaaa aagattggga ccccaagaaa tacggcggat tcgattctcc tacagtcgct 3420
tacagtgtac tggttgtggc caaagtggag aaagggaagt ctaaaaaact caaaagcgtc 3480
aaggaactgc tgggcatcac aatcatggag cgatccagct tcgagaaaaa ccccatcgac 3540
tttctcgaag cgaaaggata taaagaggtc aaaaaagacc tcatcattaa gctgcccaag 3600
tactctctct ttgagcttga aaacggccgg aaacgaatgc tcgctagtgc gggcgagctg 3660
cagaaaggta acgagctggc actgccctct aaatacgtta atttcttgta tctggccagc 3720
cactatgaaa agctcaaagg gtctcccgaa gataatgagc agaagcagct gttcgtggaa 3780
caacacaaac actaccttga tgagatcatc gagcaaataa gcgagttctc caaaagagtg 3840
atcctcgccg acgctaacct cgataaggtg ctttctgctt acaataagca cagggataag 3900
cccatcaggg agcaggcaga aaacattatc cacttgttta ctctgaccaa cttgggcgcg 3960
cctgcagcct tcaagtactt cgacaccacc atagacagaa agcggtacac ctctacaaag 4020
gaggtcctgg acgccacact gattcatcag tcaattacgg ggctctatga aacaagaatc 4080
gacctctctc agctcggtgg agac 4104
<210> SEQ ID NO: 2
<211> 189
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая линкер между белком dCas9 и белком
DNMT3a-L
<400 >2
ggcaccggcg ggcccaagaa gaagaggaag gtatacccat acgatgttcc tgactatgcg 60
ggctatccct atgacgtccc ggactatgca ggatcgtatc cttatgacgt tccagattac 120
gctgGATCCA TGGGCAGCAG CCATCATCAT CAtcatcaca gcagcggcct ggtgccgcgc 180
ggcagccat 189
<210> SEQ ID NO: 3
<211> 1632
<212>ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая белок DNMT3a-L
<400> 3
atgaaccatg accaggaatt tgacccccca aaggtttacc cacctgtgcc agctgagaag 60
aggaagccca tccgcgtgct gtctctcttt gatgggattg ctacagggct cctggtgctg 120
aaggacctgg gcatccaagt ggaccgctac attgcctccg aggtgtgtga ggactccatc 180
acggtgggca tggtgcggca ccagggaaag atcatgtacg tcggggacgt ccgcagcgtc 240
acacagaagc atatccagga gtggggccca ttcgacctgg tgattggagg cagtccctgc 300
aatgacctct ccattgtcaa ccctgcccgc aagggacttt atgagggtac tggccgcctc 360
ttctttgagt tctaccgcct cctgcatgat gcgcggccca aggagggaga tgatcgcccc 420
ttcttctggc tctttgagaa tgtggtggcc atgggcgtta gtgacaagag ggacatctcg 280
cgatttcttg agtctaaccc cgtgatgatt gacgccaaag aagtgtctgc tgcacacagg 540
gcccgttact tctggggtaa ccttcctggc atgaacaggc ctttggcatc cactgtgaat 600
gataagctgg agctgcaaga gtgtctggag cacggcagaa tagccaagtt cagcaaagtg 660
aggaccatta ccaccaggtc aaactctata aagcagggca aagaccagca tttccccgtc 720
ttcatgaacg agaaggagga catcctgtgg tgcactgaaa tggaaagggt gtttggcttc 780
cccgtccact acacagacgt ctccaacatg agccgcttgg cgaggcagag actgctgggc 840
cgatcgtgga gcgtgccggt catccgccac ctcttcgctc cgctgaagga atattttgct 900
tgtgtgtcta gcggcaatag taacgctaac agccgcgggc cgagcttcag cagcggcctg 960
gtgccgttaa gcttgcgcgg cagccatatg ggccctatgg agatatacaa gacagtgtct 1020
gcatggaaga gacagccagt gcgggtactg agccttttta gaaatattga taaagtacta 1080
aagagtttgg gctttttaga aagcggttct ggttctgggg gaggaacgct gaagtacgtg 1140
gaagatgtca caaatgtcgt gaggagagac gtggagaaat ggggcccctt tgacctggtg 1200
tacggctcga cgcagcccct aggcagctcc tgtgatcgct gtcccggctg gtacatgttc 1260
cagttccacc ggatcctgca gtatgcgctg cctcgccagg agagtcagcg gcccttcttc 1320
tggatattca tggacaatct gctgctgact gaggatgacc aagagacaac tacccgcttc 1380
cttcagacag aggctgtgac cctccaggat gtccgtggca gggactacca gaatgctatg 1440
cgggtgtgga gcaacattcc agggctgaag agcaagcatg cgcccctgac cccaaaggaa 1500
gaagagtatc tgcaagccca agtcagaagc aggagcaagc tggacgcccc gaaagttgac 1560
ctcctggtga agaactgcct tctcccgctg agagagtact tcaagtattt ttctCAAAAC 1620
TCACTTCCTC TT 1632
<210> SEQ ID NO: 4
<211> 102
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая линкер между белком DNMT3a-L и
реперссорным доменом KRAB
<400> 4
GGTGgatccg ctggaggagg tggaagcgga ggaggaggaa gcggaggagg aggtagcgga 60
cctaagaaaa agaggaaggt gcgatccctt acggcatggt cg 102
<210> SEQ ID NO: 5
<211> 195
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая репрессорный домен KRAB
<400> 5
agaacactgg ttacgttcaa ggacgtgttt gtggacttta cacgtgagga gtggaaattg 60
ctggatactg cgcaacaaat tgtgtatcga aatgtcatgc ttgagaatta caagaacctc 120
gtcagtctcg gataccagtt gacgaaaccg gatgtgatcc ttaggctcga aaagggggaa 180
gaaccttggc tggta 195
<210> SEQ ID NO: 6
<211> 63
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая линкер между репрессорным доменом KRAB
и белком MeCP2
<400> 6
tcgggaggtg gttcgggtgg ctctggatca agcccaaaga agaaacggaa ggtggaagcc 60
tca 63
<210> SEQ ID NO: 7
<211> 861
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность, кодирующая белок MeCP2
<400> 7
gtgcaggtga aaagggtgct ggaaaaatcc cccggcaaac tcctcgtgaa gatgcccttc 60
caggcttccc ctggcggaaa aggtgaaggg ggtggcgcaa ccacatctgc ccaggtcatg 120
gtcatcaagc gacctggaag gaaaagaaag gccgaggctg accctcaggc cattccaaag 180
aaacggggac gcaagccagg gtccgtggtc gcagctgcag cagctgaggc taagaaaaag 240
gcagtgaagg aaagctccat ccgcagtgtg caggagactg tcctgcccat caagaagagg 300
aaaactaggg agaccgtgtc catcgaggtc aaagaagtgg tcaagcccct gctcgtgtcc 360
accctgggcg aaaaatctgg aaaggggctc aaaacatgca agtcacctgg acggaaaagc 420
aaggagtcta gtccaaaggg gcgctcaagc tccgcttcta gtccccctaa aaaggaacac 480
catcaccatc accatcacgc cgagtctcct aaggctccta tgccactgct cccaccacct 540
ccaccacctg agccacagtc aagcgaagat cccatcagcc cacccgagcc tcaggatctg 600
tcctctagta tttgcaaaga ggaaaagatg cccagagcag gcagcctgga gagtgatggc 660
tgtccaaaag aacccgccaa gacccagcct atggtggcag ccgctgcaac taccaccaca 720
accacaacta ccacagtggc cgaaaaatac aagcatcgcg gcgagggcga acgaaaggac 780
attgtgtcaa gctccatgcc cagacctaac cgggaggaac cagtcgatag taggacaccc 840
gtgactgaga gagtctcatg a 861
<---
Изобретение относится к области биотехнологии, в частности к эпигеномному репрессору транскрипции гена или генов в клетках человека, исключая клетки зародышевой линии человека, представляющему собой белок dCas9, соединенный линкером с белком DNMT3a-L, который соединен линкером с репрессорным доменом KRAB, который соединен линкером с белком МеСР2. Изобретение эффективно для научных экспериментов в области фундаментальной клеточной биологии, биомедицинских исследований при моделировании наследственных заболеваний, а также в терапии наследственных заболеваний. 3 ил.
Эпигеномный репрессор транскрипции гена или генов в клетках человека, исключая клетки зародышевой линии человека, представляющий собой белок dCas9, кодируемый последовательностью SEQ ID NO: 1, соединенный линкером 1, кодируемым последовательностью SEQ ID NO: 2, с белком DNMT3a-L, кодируемым последовательностью SEQ ID NO: 3, который соединен линкером 2, кодируемым последовательностью SEQ ID NO: 4, с репрессорным доменом KRAB, кодируемым последовательностью SEQ ID NO: 5, который соединен линкером 3, кодируемым последовательностью SEQ ID NO: 6, с белком МеСР2, кодируемым последовательностью SEQ ID NO: 7.
| NAN CHER YEO et al., An enhanced CRISPR repressor for targeted mammalian gene regulation, Nature Method, 2018, Vol.15, pp.611-616 | |||
| HENRIETTE O’GEEN et al., Ezh2-dCas9 and KRAB-dCas9 enable engineering of epigenetic memory in a context-dependent manner, Epigenetics & Chromatin, 2019, Vol.12, N.26 | |||
| СПОСОБЫ И СОСТАВЫ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2013 |
|
RU2639277C2 |
| ДОСТАВКА И ПРИМЕНЕНИЕ СИСТЕМ CRISPR-CAS, ВЕКТОРОВ И КОМПОЗИЦИЙ ДЛЯ ЦЕЛЕНАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ И ТЕРАПИИ В ПЕЧЕНИ | 2014 |
|
RU2716420C2 |

